Изобретение относится к области медицины, к клеточным технологиям и регенеративной медицине и может быть использовано для стимуляции дифференцировки панкреатических предшественников бета-клеток в продуцирующие и секретирующие инсулин панкреатические бета-клетки при инсулинзависимом сахарном диабете.
В 2014 г. уровень заболеваемости диабетом составил 9% среди взрослого населения 18 лет и старше [1]. Согласно прогнозам ВОЗ, к 2030 г. число человек, страдающих диабетом, возрастет до 552 миллионов [2]. Из этого числа доля пациентов с инсулинзависимым сахарным диабетом составит около 10% [3]. Главной причиной инсулинзависимого сахарного диабета выступает разрушение продуцирующих и секретирующих инсулин бета-клеток поджелудочной железы и снижение продукции или недостаточная секреция инсулина этими клетками. Это может быть вызвано вирусной инфекцией или аутоиммунной патологией, возникающей вследствие вирусной инфекции, не исключают и наследственный фактор в развитии инсулинзависимого сахарного диабета [4].
Основными средствами лечения гипергликемии у пациентов, страдающих инсулинзависимым сахарным диабетом, является ежедневное введение инсулиновых препаратов, бигуанид-содержащих средств; препаратов, содержащих сульфонилмочевину, тиазолидиндион-содержащих средств и т.п., с регулярным контролем уровня глюкозы крови. Тем не менее, инсулинотерапия и назначение других лекарственных средств не восстанавливают нормальную физиологическую регуляцию уровня глюкозы крови. Лечение не устраняет риск опасных гипогликемических состояний, остается высокий риск острых и отдаленных осложнений [5]. Оказывается не эффективной трансплантация поджелудочной железы и клеток островковой ее части. Единственным возможным средством для лечения пациентов с инсулинзависимым диабетом представляется устранение основной причины заболевания - катастрофического дефицита бета-клеток. Исключительность бета-клеток состоит в том, что только эта популяция клеток естественным образом способна осуществлять оценку уровня сахаров в крови и зависимую от уровня глюкозы продукцию и секрецию инсулина.
Физиологическим подходом восстановления популяции продуцирующих и секретирующих инсулин панкреатических бета-клеток при инсулинзависимом сахарном диабете с минимальными побочными эффектами, без использования иммунодепрессантов и риска возникновения неопластического процесса может выступать стимуляция дифференцировки панкреатических предшественников бета-клеток в продуцирующие и секретирующие инсулин бета-клетки (неогенез бета-клеток). Специалистам известен глюкагон-подобный пептид-1 (7-36) амид (Glucagon-like peptide-1 (7-36)amide - GLP-1), представляющий собой полипептид (гормон), вырабатываемый L клетками желудочно-кишечного тракта в ответ на прием пищи [6]. Мишенью для GLP-1 выступает принадлежащий к семейству G-протеиновых рецепторов GPP-1-рецептор (GPP-1R), экспрессирующийся на бета-клетках поджелудочной железы [6]. На животных моделях GLP-1 стимулировал дифференцировку предшественников бета-клеток в инсулин продуцирующие бета-клетки в естественных условиях и при диабете 1 типа [7].
Данное средство является наиболее близким к заявляемому по достигаемому результату и выбрано в качестве прототипа.
Недостатком данного средства является кратковременность действия фермента, время полураспада для GLP-1 составляет 2 мин [6]. Быстрая деградация GLP-1 ферментами дипептидил пептидазой-IV (dipeptidyl peptidase-IV - DPP-IV) и нейтральной эндопептидазой 24.11 (neutral endopeptidase 24.11 - NEP 24.11) и высокий клиренс являются основными препятствиями на пути создания на основе глюкагон-подобного пептида-1 потенциального агента для снижения глюкозы [8-10].
Одна из возможностей повысить устойчивость к действию ферментов биологически активных молекул - это пегилирование. Ковалентно присоединенная молекула полиэтиленгликоля экранирует сайты расщепления и таким образом увеличивает стабильность биологически активной молекулы, снижает почечный клиренс. Известен устойчивый к энзиматическому воздействию конъюгированный с полиэтиленгликолем (polyethylene glycol-PEG) GLP-1 - (7-36)NH2 [11]. Молекула PEG ковалентно монозамещает N-концевую позицию Lys34 - Lys-PEG/GLP-1. Lys-PEG/GLP-1 значительно снижает высокий уровень глюкозы в крови во время орального глюкозотолерантного теста и устойчива к действию DPP-IV. Глюкозогликемический эффект Lys-PEG/GLP-1 связан с дополнительной секрецией инсулина бета-клетками. Модификация Lys-PEG/GLP-1 может потенциально лечить сахарный диабет 2 типа. Недостатком Lys-PEG/GLP-1 является невозможность стимулировать дифференцировку панкреатических предшественников бета-клеток в продуцирующие и секретирующие инсулин панкреатические бета-клетки и таким образом снижать уровень сахара в крови при инсулинзависимом сахарном диабете [11].
Задачей, решаемой данным изобретением, является создание нового эффективного средства для стимуляции дифференцировки панкреатических предшественников бета-клеток в продуцирующие и секретирующие инсулин бета-клетки при инсулинзависимом сахарном диабете на основе пегилированной формы глюкагон-подобного пептида 1.
Поставленная задача достигается созданием средства для стимуляции дифференцировки панкреатических предшественников бета-клеток в продуцирующие и секретирующие инсулин бета-клетки при инсулинзависимом сахарном диабете, содержащего пегилированную форму глюкагон-подобного пептида 1 (GLP-1(7-37)), отличающегося заменой в полипептидной последовательности глюкагон-подобного пептида 1 аминокислоты Ala8 на аминокислоту Gly8 и осуществлением ковалентной связи молекулы полиэтиленгликоля с молекулой глюкагон-подобного пептида 1 через аминокислоты His7, Lys26, Lys34.
Новым в предлагаемом способе является использование для снижения сахара и повышения уровня инсулина в крови при инсулинзависимом сахарном диабете устойчивой к действию протеолитических ферментов пегилированной формы глюкагон-подобного пептида 1 с заменой в полипептидной последовательности глюкагон-подобного пептида 1 аминокислоты Ala8 на аминокислоту Gly8 и осуществление ковалентной связи молекулы полиэтиленгликоля с молекулой глюкагон-подобного пептида 1 через аминокислоты His7, Lys26, Lys34.
Поскольку настоящее изобретение описывает средство, которое стимулирует дифференцировку или регенерацию панкреатических бета-клеток, продуцирующих и секретирующих инсулин, то преимущество настоящего изобретения заключается в том, что оно позволяет устранить главную причину гипергликемии, то есть дефицит панкреатических бета-клеток и недостаточное продуцирование или секрецию инсулина в этих клетках.
Отличительные признаки проявили в заявляемой совокупности новые свойства, явным образом не вытекающие из уровня техники в данной области и неочевидные для специалиста. Идентичной совокупности признаков не обнаружено в проанализированной патентной и научно-медицинской литературе.
Исходя из вышеизложенного, заявляемое изобретение соответствует критериям патентоспособности изобретения «Новизна», «Изобретательский уровень» и «Промышленная применимость».
Предлагаемое действие пегилированной формы глюкагон-подобного пептида изучено в экспериментах на 200 мышах-самцах линии C57B1/6 в возрасте 4 недель. Животные поступили из отдела экспериментальных биологических моделей НИИФиРМ имени Е.Д. Гольдберга (ветеринарное удостоверение имеется). Содержание животных и экспериментальный дизайн одобрены Этическим комитетом НИИФиРМ имени Е.Д. Гольдберга и соответствуют международным правилам, принятым Европейской Конвенцией по защите позвоночных животных, используемых для экспериментальных и иных научных целей.
С целью исключения сезонных колебаний изучаемых показателей все эксперименты проведены в осенне-зимний период. Забор материала осуществляют в утренние часы. Мышей умерщвляют передозировкой СО2. Количество животных в каждой группе не менее 10 особей.
Изобретение будет понятно из следующего описания и приложенной к нему фиг. 1.
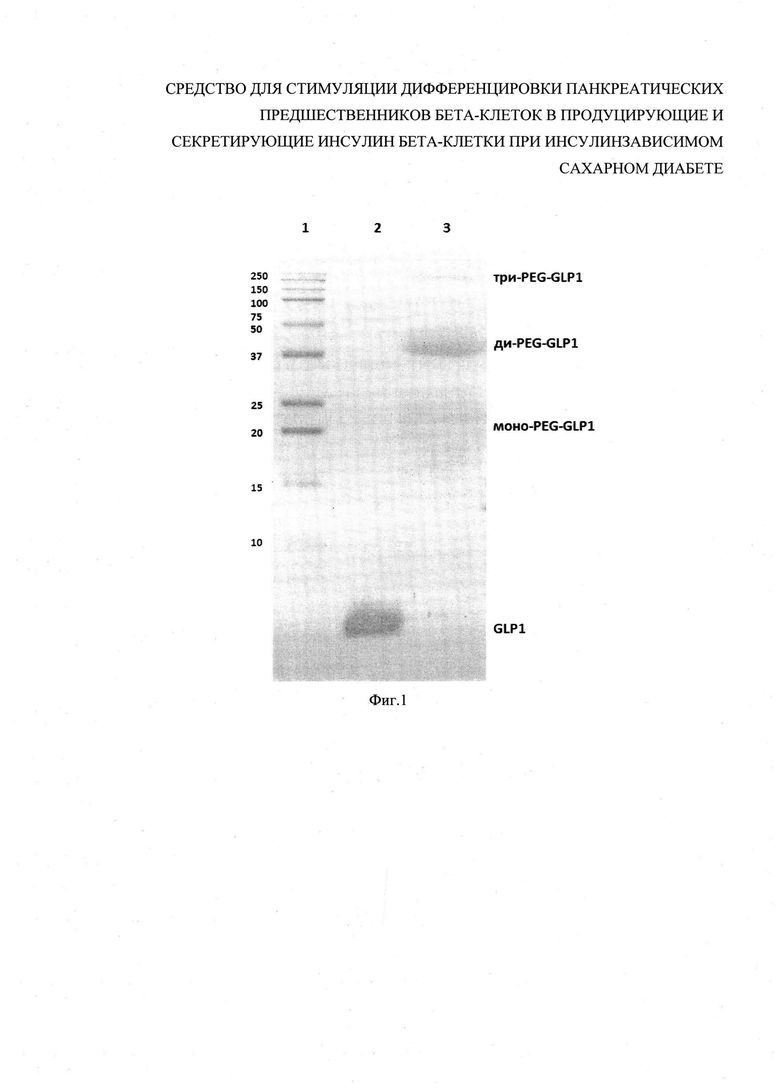
На фиг. 1 изображен молекулярно-массовый состав пегилированной формы аналога глюкагон-подобного пептида 1 (GLP-1): 1 - маркеры молекулярных масс [кДа]; 2 - контроль (непегилированный GLP-1); 3 - пегилированная форма аналога GLP-1.
Изобретение осуществляют следующим образом.
Препарат представляет собой глюкагон-подобный пептид 1 (GLP-1(7-37)) с аминокислотными заменами аминокислоты Ala8 на аминокислоту Gly8, ковалентно связанный с молекулой полиэтиленгликоля (ПЭГ).
Полипептид, используемый в изобретении, представляет собой последовательность:
His7-Gly8-Glu9-Gly10-Thr11-Phe12-Thr13-Ser14-Asp15-Val16-Ser17-Ser18-Tyr19-Leu20-Glu21-Gly22-Gln23-Ala24-Ala25-Lys26-Glu27-Phe28-Ile29-Ala30-Trp31-Leu32-Val33-Lys34-Gly35-Arg36-Gly37
Полипептид получен методами пептидного синтеза или экспрессии рекомбинантных ДНК в организме-продуценте. Очистка полипептида проведена комплексом стандартных методов, включающих ионообменную, обращенно-фазовую и эксклюзионную хроматографию.
Полиэтиленгликоль, используемый в изобретении, имеет молекулярную массу от 500 до 100000 дальтон, может быть линейным или разветвленным, активированным функциональной группой сукцинимида.
Ковалентная связь полиэтиленгликоля с молекулой GLP-1 осуществлена через одну из аминокислот His7, Lys26, Lys34.
Полученная таким образом молекула пегилированного GLP-1 обладает характерными для GLP-1 противодиабетическими свойствами, отличается от нативной высокой устойчивостью к протеиназам и имеет сниженный почечный клиренс. Период полувыведения пегилированных производных GLP-1 не менее 18 часов.
Формула:
α-methoxy-ω-{[1-(Nδ1-His7)GLP1]carbonyl}poly(oxyethylene)
α-methoxy-ω-{[1-(Nα-His7)GLP1]carbamoyl}poly(oxyethylene)
α-methoxy-ω-[((Nε-Lys26)GLP1)carbamoyl]poly(oxyethylene)
α-methoxy-ω-[((Nε-Lys34)GLP1)carbamoyl] poly(oxyethylene)
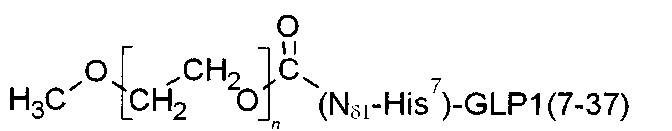
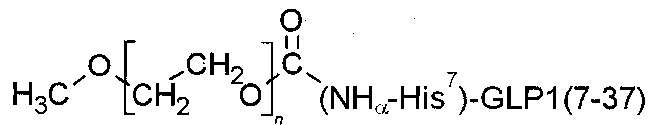
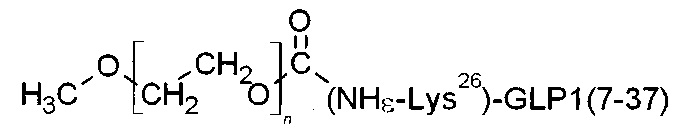
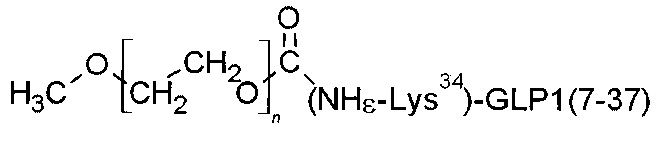
У лабораторного животного (мыши-самцы линии C57B1/6) диабет 1 типа моделируют стрептозотоцином («Sigma», USA). Стрептозотоцин - противоопухолевое средство алкилирующего действия из группы производных нитрозомочевины. Механизм противоопухолевого действия полностью не изучен, но, вероятно, обусловлен образованием метилкарбониевых ионов, которые вызывают алкилирование или связываются с различными внутриклеточными структурами, включая нуклеиновые кислоты. Образует поперечные сшивки между нитями ДНК, что и приводит к ингибированию ее синтеза [12]. Оказывает также гипергликемический эффект, вызывая необратимое повреждение бета-клеток поджелудочной железы [13, 14]. Известна способность стрептозотоцина индуцировать инсулинзависимый сахарный диабет в эксперименте [7]. Стрептозотоцин вводят после 4-часовой голодовки животных внутрибрюшинно в дозе 40 мг/кг в 250 мкл фосфатного буфера 1 раз в сутки в течение 5 дней. Контролируется модель инсулинзависимого сахарного диабета по уровню глюкозы в крови при помощи глюкометра (Accu-Chek Performa Nanu ("Roche Diagnostes GmbH", Germany) и по количеству инсулин-продуцирующих клеток в поджелудочной железе [15]. Животным контрольной группы внутрибрюшинно вводят 250 мкл фосфатного буфера. За 0-й день эксперимента принимают последнее введение стрептозотоцина.
Лабораторным животным с диабетом глюкагон-подобный пептид 1 (GLP-1) или пегилированный аналог глюкагон-подобного пептида 1 GLP-1 (pegGLP-1) вводят внутрижелудочно в дозе 0,02 мг/мышь в 0,1 мл фосфатного буфера один раз в день (с 9 ч до 9.15 ч) курсом в течение 4 суток (с 24-х по 27-е сутки эксперимента).
У интактных мышей линии C57B1/6 исследуют уровень GLP-1 в сыворотке крови до и через 2, 5, 15, 30 и 60 мин, 2, 4, 8 и 12 ч после внутрижелудочного введения GLP-1 или pegGLP-1. У мышей-самцов линии C57B1/6 с диабетом на 27 день эксперимента исследуют уровень GLP-1 в сыворотке крови через 4 ч после внутрижелудочного введения GLP-1 или pegGLP-1. Уровень GLP-1 в сыворотке оценивают иммуноферментным методом в соответствии с протоколами производителя (Cusabio Biotech CO., LTD, Китай). При оценке специфической активности in vitro концентрация GLP-1 или pegGLP-1 в культуральной среде составила 10-7 М.
На 4, 14, 21, 27 и 29 сутки эксперимента проводят оценку уровня глюкозы в крови у мышей линии C57B1/6 с диабетом. Уровень глюкозы в крови определяют при помощи глюкометра (Accu-Chek Performa Nanu ("Roche Diagnostes GmbH", Germany). Измерение исходного уровня глюкозы в крови у животных проводят после 16-часовой депривации корма, все последующие измерения уровня глюкозы также производят после 16-часовой депривации еды.
На 27 сутки эксперимента проводят глюкозотолерантный тест. На первом этапе проводят забор образца крови для оценки исходного уровня глюкозы у экспериментальных животных. Через 1 ч после этого производят внутривенную инъекцию глюкозы (D-глюкоза, Sigma-Aldrich, США) в хвостовую вену в дозе 2 г/кг. Первый забор крови для исследования уровня глюкозы проводят через 30 мин после инъекции глюкозы, далее исследование содержания глюкозы производят с интервалом в 30 мин, продолжительность исследования составит 2,5 ч. Доза глюкозы 2 г/кг была выбрана в силу того обстоятельства, что, согласно данным литературы, ответ организма животных на низкие дозы глюкозы при ее внутривенном введении слабый, концентрация инсулина возрастает только в первую минуту и редко сохраняется повышенной в течение первых 5 мин наблюдения [16].
На 27 и 29 сутки после введения стрептозотоцина иммуноферментным методом определяют уровень инсулина в соответствии с протоколами производителя (Cusabio Biotech CO., LTD, Китай).
На 27 сутки оценивают количество предшественников бета-клеток и бета-клеток в поджелудочной железе и функциональную активность предшественников бета-клеток в поджелудочной железе [7]. Из поджелудочной железы выделяют неприлипающие мононуклеары. Неприлипающие мононуклеары разделяют на 3 группы. У клеток 1 группы цитометрически изучают экспрессию CD45, TER119, CD117 (c-kit), CD49f, FLK-1, CD 133. Для анализа используют прибор FACS Canto II с программным обеспечением FACS Diva («Becton Dickinson», США). У мононуклеаров 2 подгруппы прижизненным окрашиванием дитизоном выявляют инсулин-продуцирующие клетки (Pancreatic Cell DTZ Detection Assay, США). У мононуклеаров 3 группы изучают потенциал к самоподдержанию при длительном культивировании. Для этого мононуклеары в количестве 1×106 вносят в культуральный флакон («ТРР Techno Plastic Products AG», Швейцария) и инкубируют в стандартных условиях (37°C, 5% СО2, 100% влажность воздуха) в течение 30 дней в среде, содержащей 45% DMEM («Sigma», США), 45% DMEM/F-12 («Sigma», США), 10% инактивированной фетальной бычьей сыворотки («Sigma», США), 5 мг/л ITS+3 (смесь рекомбинантного человеческого инсулина, трансферина и селинита Na) («Sigma», США), 1×10-7 M дексаметозона («Sigma», США), 10 ммоль/л никотиновой кислоты («Sigma», США), 5 ммоль/л фосфатного буфера (HEPES) («Sigma», США) раствор антибиотиков 100 ед./мл пенициллина и 100 мкг/мл стрептомицина («Sigma», США), 2 ммоль/л L-глутамина («Sigma», США), β-меркаптоэтанола 50 ммоль/л («Thermoscientific», США). Эпидермальный фактор роста 4 нг/мл («Sigma», США) и фактор роста гепатоцитов 10 нг/мл («Sigma», США) добавляют через 24 часа от начала культивирования. Каждые 3-4 дня проводят смену среды. По окончании цикла культивирования супернатанты от мононуклеаров собирают и методом иммуноферментного анализа исследуют уровень свободного инсулина.
Полученные в результате культивирования мононуклеаров 3 группы клеток разделяют на две подгруппы. У клеток 1 подгруппы оценивают экспрессию CD45, TER119, CD117 (c-kit), CD49f, FLK-1, CD133. У 0,5×106 клеток 2 подгруппы с использованием глюкагон-подобного пептида GLP-1 (7-37) («Sigma», США) изучают дифференцировку в инсулин-продуцирующие клетки мультипотентных прогениторных клеток и олигопотентных предшественников бета-клеток в присутствии GLP-1 или пегилированного аналога GLP-1. По окончании 30 дней у клеток 2 подгруппы прижизненным окрашиванием дитизоном выявляют инсулин-продуцирующие клетки и изучают секрецию инсулина. Оценивают базальный уровень секреции (при концентрации глюкозы в фосфатном буфере 3 ммоль/л) и секреторную активность в условиях повышения концентрации глюкозы до 20 ммоль/л у 1×105 мононуклеаров [17].
Статистическую обработку полученных результатов проводят методами вариационной статистики с использованием пакета статистической обработки данных SPSS 12,0. Вычисляют среднее арифметическое (М), ошибку среднего арифметического (m), значение вероятности (Р). Различие двух сравниваемых величин считают достоверным в том случае, если вероятность их тождества была меньше 5% (Р<0,05). Используя выборочные коэффициенты асимметрии и эксцесса, оценивают степень приближения закона распределения исследуемого признака к нормальному. В случаях нормального распределения признаков для статистической оценки применяют параметрический t-критерий Стьюдента. При больших отклонениях распределений признака от нормального вида для независимых выборок используют непараметрический критерий U-критерий Уилкоксона. Для выявления достоверности различий качественных показателей используют критерий углового преобразования Фишера.
Пример 1. Получение пегилированной формы аналога глюкагон-подобного пептида
5 мг глюкагон-подобного пептида (GLP-1(7-37)) (Sigma-Aldrich) растворяют в 10 мл 20 мМ фосфатного буфера рН 7,0, прибавляют 100 мг коммерческого пегилирующего агента (линейный метокси-ПЭГ с молекулярной массой 12 кДа, активированный сукцинимидом). Через 3 часа реакцию останавливают внесением 2 мл 2 М глицина. Молекулярно-массовый состав изучают с помощью электрофореза в полиакриламидном геле с SDS по стандартной методике (фиг. 1). В результате проведения пегилирования весь GLP-1 расходуется на образование моно-, ди- и трипегилированных форм, свободный GLP-1 не выявляется (табл.1).
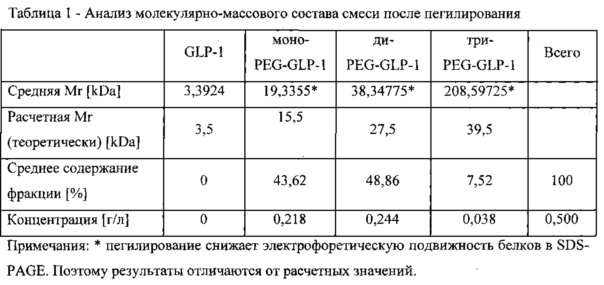
Выход монопегилированного GLP-1 составляет 43,6%. Учитывая, что в данных условиях исходный GLP-1 был полностью пропегилирован, а ди- и трипегилированные молекулы теряют активность, можно ожидать, что биологическая активность смеси обусловлена активностью монопегилированной формы аналога GLP-1 (пегилированной формы аналога GLP-1 - pegGLP-1).
Пример 2. Исследование концентрации GLP-1 в сыворотке крови у мышей С57B1/6 в естественных условиях и с диабетом
Однократное внутрижелудочное введение GLP-1 не влияет на концентрацию GLP-1 в сыворотке крови у мышей на протяжении всего периода наблюдения (Табл. 2). После однократного внутрижелудочного введения pegGLP-1 отмечается достоверное повышение уровня сывороточного GLP-1 относительно мышей интактного контроля и мышей, леченных GLP-1, на 60 мин, 2 ч, 4 ч, 8 ч, 12 ч, 16 ч, 18 ч эксперимента. Максимальные значения показателя регистрируются через 4-8 ч после назначения pegGLP-1. В настоящем исследовании период полувыведения (Т1/2) pegGLP-1 составляет 8 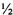 ч. Среднее время удерживания в организме (MRT) составляет 15 ч.
ч. Среднее время удерживания в организме (MRT) составляет 15 ч.
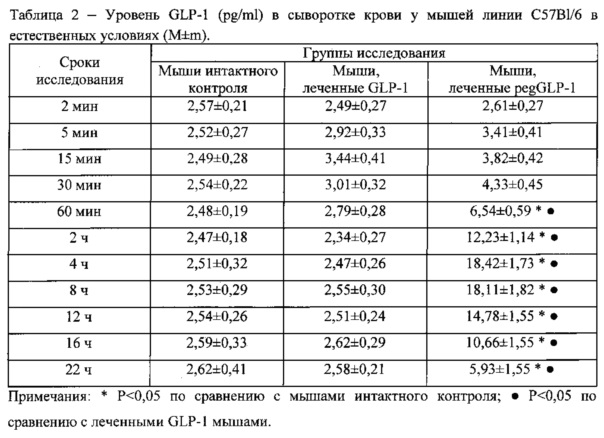
По результатам ИФА GLP-1 в сыворотке крови у мышей в естественных условиях концентрацию сывороточного GLP-1 у мышей с диабетом исследуют через 4 ч после внутрижелудочного введения GLP-1 или pegGLP-1. Как видно из Таблицы 2, для мышей с диабетом характерно снижение уровня GLP-1. Курсовое назначение GLP-1 не оказывает влияние на концентрацию сывороточного GLP-1 у диабетических мышей. Между тем, после курсового введения pegGLP-1 уровень GLP-1 у мышей с диабетом резко повышается и значительно превосходит таковой не только у нелеченых диабетических мышей, но и у мышей интактного контроля (Табл. 3).
Пример 3. Исследование концентрации глюкозы в сыворотке и глюкозотолерантный тест у мышей C57B1/6 при диабете
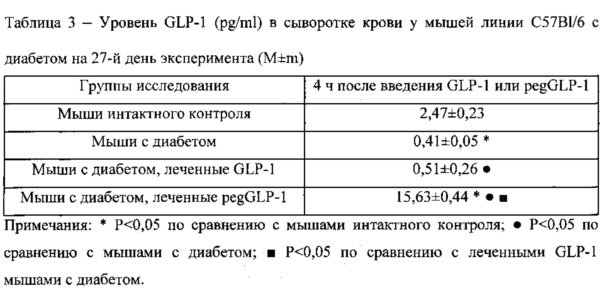
Моделирование стрептозотоцин-индуцированного диабета приводит к стойкому увеличению уровня глюкозы в крови у мышей, начиная с 4-х суток по 29-е сутки эксперимента (Табл. 3). Максимальные значения показателя выявлены на 21, 27 и 29-е сутки. При курсовом введении GLP-1 наблюдается тенденция к снижению уровня глюкозы у диабетических мышей на 27 сутки эксперимента. pegGLP-1 оказывает выраженное гипогликемическое действие. Так, на 27 и 29-е сутки регистрируется снижение концентрации глюкозы в 2,1 и 1,79 раза соответственно по сравнению с группой нелеченых мышей с диабетом (Табл. 4).
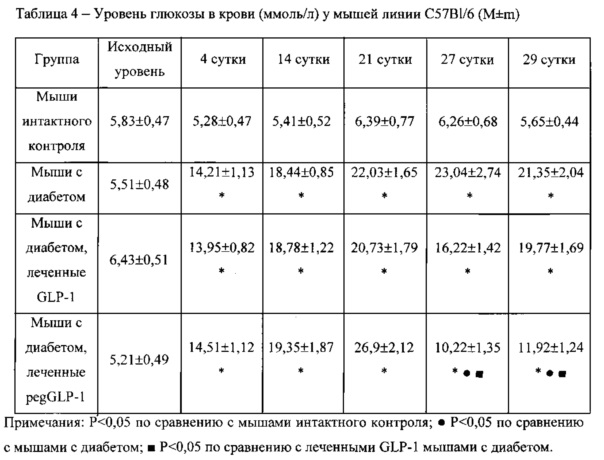
Критерием выраженности нарушения метаболизма глюкозы является глюкозотолерантный тест (ГТТ). У животных интактного контроля и у мышей с диабетом проведен ГТТ. В ответ на внутривенную инъекцию глюкозы (доза 2 г/кг) у интактных животных регистрировалось достоверное повышение уровня глюкозы в крови на протяжении 60 мин (Табл. 5). Далее на 90, 120, 150 мин наблюдения отмечается поступательное снижение концентрации глюкозы. Как было уже продемонстрировано выше, у животных с диабетом отмечается гипергликемия, на 27-е сутки эксперимента концентрация глюкозы в крови максимальная (Табл. 4). У мышей с диабетом ГТТ проводился на 27-е сутки. Высокая концентрация глюкозы у диабетических мышей сохранялась с 30 мин по 150 мин проведения ГТТ (Табл. 5). Курсовое введение GLP-1 не влияет на результаты ГТТ у мышей с диабетом. В свою очередь назначение pegGLP-1 вызывает значительное снижение уровня глюкозы на 120 мин и 150 мин наблюдения.
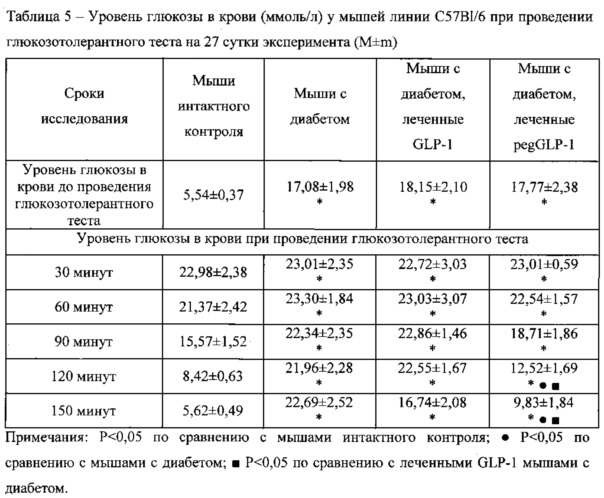
Пример 4. Исследование концентрации инсулина в сыворотке крови у мышей С57В1/6 при диабете 1 типа
Моделирование диабета вызывает резкое снижение концентрации инсулина в сыворотке крови у мышей на 27 и 29 сутки эксперимента (Табл. 6).
Курсовое введение GLP-1 и pegGLP-1 повышает уровень инсулина в сыворотке крови у мышей с диабетом на 27 сутки. Через 2 суток после курса лечения - 29 сутки эксперимента, у леченных pegGLP-1 диабетических мышей концентрация инсулина в сыворотке остается высокой и превышает показатель у нелеченых мышей с диабетом в 3 раза. Между тем, в группе мышей с диабетом, леченных GLP-1, уровень инсулина, напротив, снижается до уровня диабетического контроля.

Пример 5. Исследование содержания предшественников бета-клеток и бета-клеток в поджелудочной железе у мышей C57B1/6 при диабете 1 типа
5-кратное введение стрептозотоцина вызывает изменения предшественников бета-клеток и инсулин-продуцирующих бета-клеток в поджелудочной железе на 27-е сутки эксперимента, свойственные для диабета 1 типа. Так, окрашивание дитизоном демонстрирует разрушение панкреатических инсулин-продуцирующих клеток. По сравнению с животными интактного контроля их количество уменьшается в 5,2 раза (Р<0,05). По данным цитометрических исследований в поджелудочной железе у мышей с диабетом 1 типа значительно снижается количество клеток с фенотипом CD45- TER119- CD133+ CD49flow (олигопотентные предшественники бета-клеток) по сравнению с интактным контролем (Табл. 7).
Внутрижелудочное введение GLP-1 не влияет на содержание олигопотентных предшественников бета-клеток в поджелудочной железе мышей с диабетом 1 типа (табл.7). Оставалось низким и количество дитизон-положительных инсулин-продуцирующих клеток: по сравнению с животными интактного контроля их количество уменьшается в 5,3 раза (Р<0,05). В отличие от GLP-1 внутрижелудочное введение pegGLP-1 более чем в 1,87 раза повышает количество олигопотентных предшественников бета-клеток в поджелудочной железе мышей с диабетом 1 типа, их количество превышает интактный контроль на 30% (Р<0,05). Между тем прирост дитизон-положительных инсулин-продуцирующих клеток у мышей с диабетом 1 типа, леченных pegGLP-1, не столь интенсивен как олигопотентных предшественников бета-клеток: их количество составляет 75% от интактного контроля.
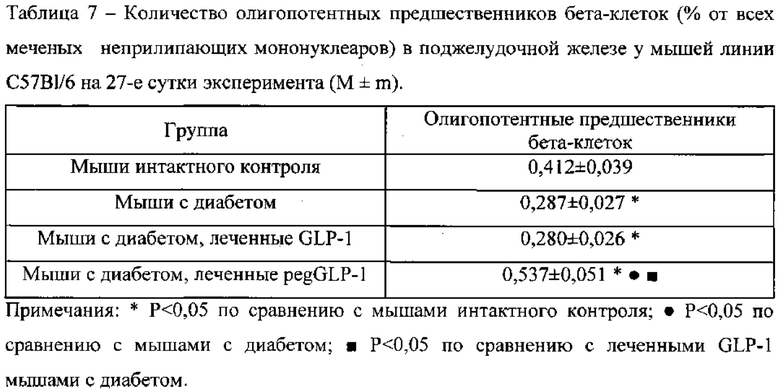
Пример 6. Исследование самоподдержания панкреатических предшественников бета-клеток in vitro
На 27-е сутки эксперимента выделяют мононуклеары из поджелудочной железы у мышей интактного контроля, мышей с диабетом и мышей с диабетом, леченных GLP-1 или pegGLP-1. Далее обогащают общую фракцию мононуклеаров клетками с иммунофенотипом мультипотентных предшественников бета-клеток (CD45- TER119- cKit-1- Flk-1-) и олигопотентных предшественников бета-клеток (CD45- TER119- CD133- CD49flow), и в количестве 1×105 вносят в культуральный флакон и проводят культивирование в течение 30 суток. Из результатов цитометрических исследований следует, что панкреатические олигопотентные предшественники бета-клеток демонстрируют различную способность к самоподдержанию in vitro. Так, содержание клеток, экспрессирующих маркеры CD45, TER119, CD117 (c-kit), CD49f, FLK-1, CD133, в культуре интактного контроля на 10% превышает тканевые значения, в культуре диабетического контроля - на 29%, в культуре клеток диабетических мышей, леченных GLP-1, - на 28% (Табл. 8).
При культивировании панкреатических клеток диабетических мышей, леченных pegGLP-1, обогащение предшественниками бета-клеток в образцах происходит более интенсивно (+82% по сравнению с тканевыми значениями), чем в образцах интактного контроля, диабетического контроля и группы с диабетом, леченным GLP-1. Прижизненное окрашивание дитизоном мононуклеаров, полученных в результате культивирования, не выявило инсулин-продуцирующих клеток в образцах интактного контроля и клеток диабетических мышей. В супернатантах не определяется инсулин.
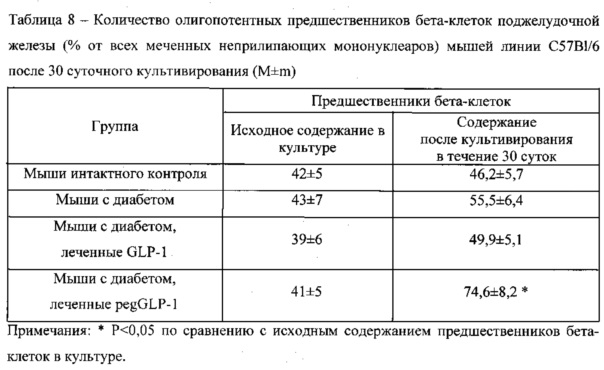
Пример 7. Исследование дифференцировки предшественников бета-клеток в продуцирующие и секретирующие инсулин бета-клетки in vitro
У полученных в результате культивирования мультипотентных предшественников бета-клеток (CD45-TER119-cKit-1-Flk-1-) и олигопотентных предшественников бета-клеток (CD45-TER119-CD133+CD49flow) изучают способность дифференцироваться в инсулин-продуцирующие клетки (дитизон-положительные) в присутствии GLP-1 или pegGLP-1 in vitro. Концентрация GLP-1 или pegGLP-1 в культуре составляла 10-7 М. Забор мононуклеаров поджелудочной железы проводится на 27-е сутки эксперимента. Полученные данные указывают на то, что по окончании цикла дифференцировки предшественников бета-клеток в соответствующей среде без GLP-1 или pegGLP-1 (спонтанная дифференцировка) появляются единичные дитизон-положительные клетки в группе интактного и диабетического контроля. При внесении в культуру интактных предшественников бета-клеток GLP-1 или pegGLP-1 наблюдается прирост дитизон-положительных клеток до n=100 и n=87 соответственно. В культуре предшественников бета-клеток диабетического контроля прирост дитизон-положительных клеток после внесения в культуру препаратов GLP-1 или pegGLP-1 возрастает более значительно: на 105 клеток выявлено n=329 и n=284 дитизон-положительных клеток соответственно.
В супернатантах дитизон-положительных клеток, полученных в результате дифференцировки предшественников, ИФА оценивают уровень инсулина. В культуральной среде моделируют базальный уровень глюкозы (концентрация глюкозы 3 ммоль/л) и глюкозную нагрузку (концентрация глюкозы 20 ммоль/л). Из результатов, представленных в Таблице 9, следует, что инсулин определяется только в супернатантах от дитизон-положительных клеток, полученных в результате GLP-1 или pegGLP-1-индуцированной дифференцировки предшественников бета-клеток. Обращает на себя внимание, во-первых, тот факт, что уровень инсулина при моделировании глюкозной нагрузки значительно превосходит таковой в условиях базального уровня глюкозы. Во-вторых, то обстоятельство, что уровень глюкозы в группе диабетического контроля значительно превосходит уровень глюкозы в интактном контроле.
Таким образом, предлагаемая пегилированная форма аналога GLP-1 значительно более устойчива к действию протеолитических ферментов по сравнению с GLP-1 за счет экранирования полиэтиленгликолем сайтов расщепления, биологическая активность средства не уступает активности GLP-1 in vitro (инициирует дифференцировку панкреатических предшественников бета-клеток в продуцирующие и секретирующие инсулин бета-клетки) и превосходит активности GLP-1; in vivo (гипогликемический эффект, глюкозотолерантный тест, содержание предшественников бета-клеток и бета-клеток в поджелудочной железе). Гипогликемический эффект пегилированной формы аналога GLP-1 регистрируется после окончания курса лечения.
ЛИТЕРАТУРА
1. Global status report on noncommunicable diseases 2014. Geneva, World Health Organization, 2012.
2. Whiting DR, Guariguata L, Weil С, Shaw J. IDF diabetes atlas: global estimates of the prevalence of diabetes for 2011 and 2030. Diabetes research and clinical practice. 2011; 94(3): 311-321. DOI: http://dx.doi.org/10.1016/j.diabres.2011.10.029.
3. Stanekzai J, Isenovic ER, Mousa SA. Treatment options for diabetes: Potential role of stem cells. Diabetes Research and Clinical Practice. 2012; 98(3): 361-368. DOI: http://dx.doi.org/10.1016/j.diabres.2012.09.010.
4. Пеллегрини С., Сорди В., Пьемонта Л. Замещение β-клеток поджелудочной железы при сахарном диабете. Сахарный диабет. 2013; (3). Т 1-20. DOI: http://dx.doi.org/10.14341/2072-0351-812.
5. Van Belle TL, Coppieters KT, Von Herrath MG. Type 1 Diabetes: Etiology, Immunology, and Therapeutic Strategies. Physiological Reviews. 2011; 91(1): 79-118. DOI: http://dx.doi.org/10.1152/physrev.00003.2010.
6. Kieffer TJ, Habener JF. The glucagon-like peptides. Endocr Rev. 1999; 20: 876-913.
7. Скурихин Е.Г., Ермакова H.H., Хмелевская Е.С., Першина О.В., Крупин В.А., Ермолаева Л.А., Дыгай A.M. Дифференцировка стволовых и прогениторных β-клеток поджелудочной железы в инсулин-секретирующие клетки у мышей при сахарном диабете. Бюл. эксперим. биол. и медицины. 2013. Т. 156. №12. - С. 681-686.
8. Hansen L., Deacon CF., Orskov С. et al. Glucagon-like pep-tide-1-(7-36)amide is transformed to glucagon-like peptide-1-(9-36)amide by dipeptidyl peptidase IV in the capillaries supplying the L cells of the porcine intestine. Endocrinology. 1999. 140. P. 5356-5363.
9. Kieffer TJ, Mcintosh CH, Pederson RA. Degradation of glucose-dependent insulinotropic polypeptide and truncated glucagon-like peptide 1 in vitro and in vivo by dipeptidyl peptidase IV. Endocrinology. 1995. 136: 3585-3596.
10. Plamboeck A, Hoist JJ, Carr RD, Deacon CF. Neutral endopeptidase 24.11 and dipeptidyl peptidase IV are both mediators of the degradation of glucagon-like peptide 1 in the anaesthetised pig. Diabetologia. 2005. 48: 1882-1890.
11. Lee YS, Youn S-H, Lee Y, Byun K, Lee C. PEGylated glucagon-like peptide-1 displays preserved effects on insulin release in isolated pancreatic islets and improved biological activity in db/db mice. Diabetologia. 2006. 49: 1608-1611 DOI 10.1007/s00125-006-0234-3.
12. Справочник лекарственных средств Vidal, 2014. 1600 с.
13. Jinzi Wu, Liang-Jun Yan Streptozotocin-induced type 1 diabetes in rodents as a model for studying mitochondrial mechanisms of diabetic β cell glucotoxicity. Diabetes, Metabolic Syndrome and Obesity: Targets and Therapy, 2015. V.8. P. 181-188.
14. Huang SW, Taylor GE. Immune insulitis and antibodies to nucleic acids induced with streptozotocin in mice. Clin. exp. Immunol. 1981. 43, P. 425-429.
15. Akira S, Masahide Y, Hiroshi Y. et al., Identification of insulinproducing cells derived 107. from embryonic stem cells by zinc-chelating dithizone. Stem Cells 2002, 20: 284-292.
16. Pacini G,  M, В. Reappraisal of the intravenous glucose tolerance index for a simple assessment of insulin sensitivity in mice. American Journal of Physiology. 2009. Vol.296. №5, R1316-1324.
M, В. Reappraisal of the intravenous glucose tolerance index for a simple assessment of insulin sensitivity in mice. American Journal of Physiology. 2009. Vol.296. №5, R1316-1324.
17. Okuno M, Minami K, Okumachi A, et al. Generation of insulin-secreting cells from pancreatic acinar cells of animal models of type 1 diabetes. Am J Physiol Endocrinol Metab. 2007. Jan; 292(l): E158-65.
Настоящее изобретение относится к биохимии и медицине, в частности к средству для стимуляции дифференцировки панкреатических предшественников бета-клеток в продуцирующие и секретирующие инсулин бета-клетки при инсулинзависимом сахарном диабете. Указанное средство представляет собой пегилированную форму глюкагон-подобного пептида 1 GLP-1 (7-37). Настоящее изобретение позволяет расширить арсенал средств для лечения инсулинзависимого сахарного диабета за счет способности настоящего средства стимулировать дифференцировку панкреатических предшественников бета-клеток в продуцирующие и секретирующие инсулин бета-клетки. 1 ил., 9 табл., 7 пр.
Средство для стимуляции дифференцировки панкреатических предшественников бета-клеток в продуцирующие и секретирующие инсулин бета-клетки при инсулинзависимом сахарном диабете, представляющее собой пегилированную форму глюкагон-подобного пептида 1 (GLP-1 (7-37)).
| SELISA FABIO et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Способ окисления боковых цепей ароматических углеводородов и их производных в кислоты и альдегиды | 1921 |
|
SU58A1 |
| СПОСОБ И СИСТЕМА АВТОРИЗАЦИИ ВЕБ-САЙТА В ВЕБ-БРАУЗЕРЕ | 2018 |
|
RU2718480C2 |
| STROHL WILLIAM R | |||
| Fusion proteins for half-life extension of biologics as a strategy to make biobetters, BioDrugs., 2015, 29(4), pp | |||
| Кузнечный горн | 1921 |
|
SU215A1 |
| RU 2014130726 A, 20.02.2016. | |||

