ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Изобретение относится к рекомбинантному микроорганизму Saccharomyces sp., продуцирующему молочную кислоту, и к способу получения молочной кислоты из культуральной среды, содержащей данный микроорганизм, путем его культивирования.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
В общем, молочная кислота представляет собой важную органическую кислоту с широким диапазоном применений, включая пищевые добавки, такие как пищевые консерванты, ароматизаторы или подкислители и т.д., и она широко используется в промышленных целях, как, например, в косметике, химии, производстве металлов, электроники, тканей, окрашивании тканей, в фармацевтической промышленности и т.д. Кроме того, молочная кислота представляет собой важный ингредиент полимолочной кислоты - одной из биодеградируемых пластмасс, и, таким образом, спрос на молочную кислоту значительно увеличивается. Она также используется в качестве важного вещества для получения многих химических соединений, включающих полимолочную кислоту, ацетальдегид, полипропиленгликоль, акриловую кислоту, 2,3-пентатион и т.д. В частности, молочная кислота D-типа является основным ингредиентом при получении стереокомплексной PLA (полимолочная кислота), которая является оптическим изомером, необходимым для получения высокоустойчивой к нагреванию PLA.
В частности, способ получения молочной кислоты включает традиционный химический синтез и биологический процесс брожения. При получении молочной кислоты посредством химического синтеза она получается в форме рацемической смеси, состоящей из 50% молочной кислоты D-типа и 50% молочной кислоты L-типа, данное соотношение в составе сложно контролировать, и, таким образом, получающаяся посредством него полимолочная кислота может становиться аморфным полимером, имеющим низкую температуру плавления, накладывая посредством этого ограничения на разработку ее применения. С другой стороны, биологический процесс брожения обеспечивает селективную продукцию молочной кислоты D-типа или молочной кислоты L-типа, в зависимости от используемого штамма. Таким образом, последний является коммерчески предпочтительным, так как возможно получать конкретную изоформу молочной кислоты.
Тем временем предпринимали попытки для того, чтобы улучшить продуктивность в отношении молочной кислоты посредством различных генетических манипуляций с использованием микроорганизма Saccharomyces sp., имеющего способность к продукции D-молочной кислоты, путем введения гена фермента для превращения в молочную кислоту D-типа. В частности, предпринимали попытки для повышения уровня продукции молочной кислоты посредством усиления активности дегидрогеназы молочной кислоты (LDH) при уменьшении активностей пируватдекарбоксилазы (PDC), альдегиддегидрогеназы (ALD) и/или ацетил-КоА-синтетазы (ACS) (публикации заявок на патент США №2012 - 021421, 2010 - 0248233 и 2006 - 0148050). Однако общая продуктивность брожения была низкой из-за слабого роста клеток штамма, продуцирующего молочную кислоту.
Соответственно, авторы настоящего изобретения предприняли интенсивные попытки для того, чтобы получить микроорганизм, имеющий улучшенную продуктивность в отношении молочной кислоты, с эффективным ростом клеток при уменьшении активности PDC. В результате подтвердили, что штаммы, у которых контролировались активности изотипов PDC и были увеличенными активности альдегиддегидрогеназы и ацетил-КоА-синтетазы, были способны увеличивать выход продукции молочной кислоты и облегчать рост клеток штаммов, улучшая, посредством этого, общую продуктивность молочнокислого брожения, и это составило настоящее изобретение.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
ТЕХНИЧЕСКАЯ ПРОБЛЕМА
Целью настоящего изобретения является предложение микроорганизма Saccharomyces sp., имеющего улучшенную продуктивность в отношении молочной кислоты.
Другой целью настоящего изобретения является предложение способа получения молочной кислоты с использованием микроорганизма Saccharomyces sp.
ТЕХНИЧЕСКОЕ РЕШЕНИЕ
В первом аспекте настоящего изобретения для достижения описанных выше целей предложен микроорганизм Saccharomyces sp., имеющий улучшенную продуктивность в отношении молочной кислоты, где данный микроорганизм мутирован так, что (а) активность пируватдекарбоксилазы снижается по сравнению с активностью не мутантного штамма, продуцирующего молочную кислоту; и (б) активности альдегиддегидрогеназы и ацетил-КоА-синтетазы улучшаются по сравнению с активностями не мутантного штамма, продуцирующего молочную кислоту.
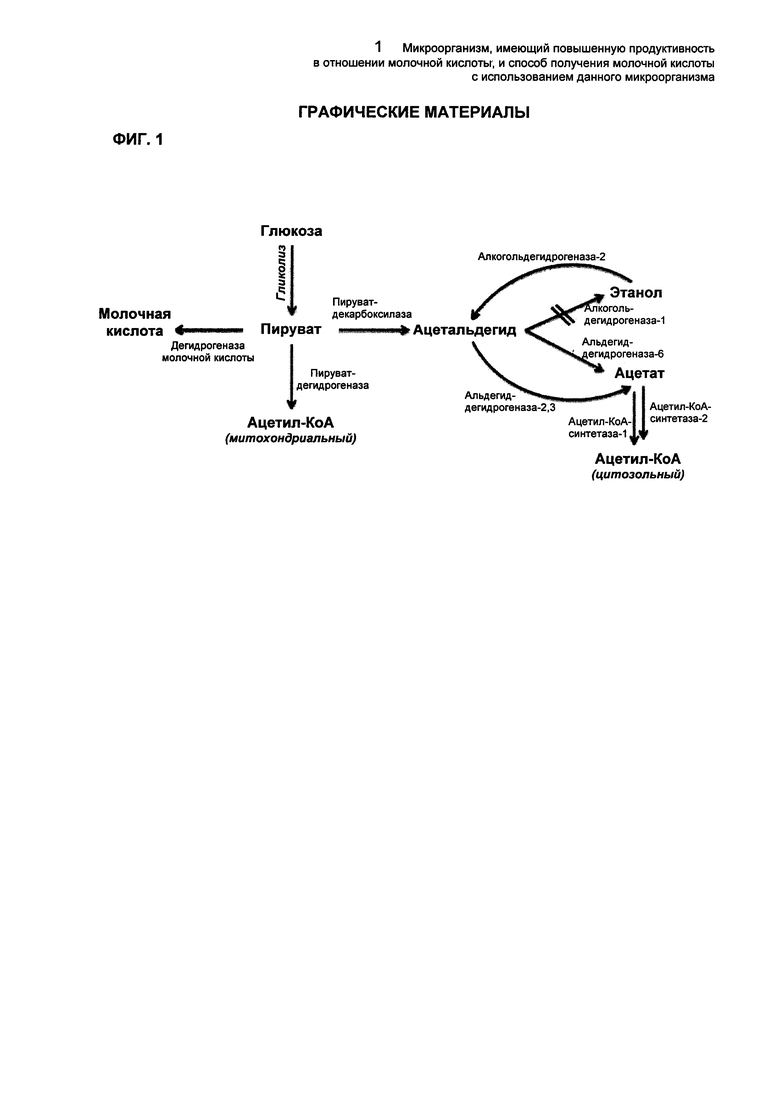
В общем, микроорганизм Saccharomyces sp., продуцирующий молочную кислоту, продуцирует ее посредством дегидрогеназы молочной кислоты (LDH) с использованием пирувата в качестве субстрата. Путь брожения с образованием этанола и путь образования ацетил-КоА, типичные метаболические пути, использующие пируват в качестве общего субстрата, были заблокированы. Уменьшение активности PDC может быть полезным при получении молочной кислоты и повышении ее выхода, однако при достижении определенного уровня уменьшения образовалось недостаточное количество цитозольного ацетил-КоА, что, в свою очередь, блокировало рост клеток, и, таким образом, не достигалось нормальное брожение. Соответственно, авторы настоящего изобретения разработали микроорганизм Saccharomyces sp., имеющий улучшенную продуктивность в отношении молочной кислоты путем улучшения общей продуктивности молочнокислого брожения, в котором поддерживалась скорость роста микроорганизма с улучшенным выходом продуктивности молочной кислоты путем регуляции пути ацетил-КоА на минимальном уровне.
Используемый здесь термин «пируватдекарбоксилаза» (PDC) относится к белку, имеющему активность, способную опосредовать реакцию, ответственную за образование карбоновой кислоты и ацетальдегида из пирувата, но не ограничивается каким-либо его производным или изотипом, имеющим такую же активность. Было известно, что данный белок участвует в стадии спиртового брожения и, главным образом, присутствует в дрожжах и растениях. Пируватдекарбоксилаза по настоящему изобретению может по своей природе присутствовать в микроорганизме Saccharomyces sp. или может представлять собой PDC1, PDC5 и/или PDC6 или конкретно PDC1, PDC5 и/или PDC6 Saccharomyces cerevisiae, но не ограничивается ими. Белок может включать его любые варианты или аналоги при условии, что они являются биологически идентичными и имеют активности, соответствующие этому белку. Аминокислотные последовательности данного белка могут быть получены из известной базы данных и т.д., например GenBank NCBI и т.д., но не ограничиваясь ей. В частности, PDC1 может состоять из аминокислотной последовательности SEQ ID NO: 71, PDC5 - из аминокислотной последовательности SEQ ID NO: 72, и PDC6 - из аминокислотной последовательности SEQ ID NO: 73. Белок может включать аминокислотные последовательности, имеющие гомологию более чем 70%, конкретно более чем 80%, более конкретно более чем 90% и даже более конкретно более чем 95% с каждой из перечисленных выше аминокислотных последовательностей. В настоящее изобретение также может быть включен любой вариант из перечисленных выше последовательностей, кодирующий те же самые аминокислотные последовательности, который образуется в результате вырожденности генетического кода.
Используемый здесь термин «гомология» относится к степени сходства между множеством нуклеотидных последовательностей или аминокислотных последовательностей и представляет собой единицу, представляющую последовательность, имеющую такие же последовательности, что и аминокислотные последовательности или нуклеотидные последовательности по настоящему изобретению, с равной или большей вероятностью, чем приведенная выше вероятность. Такую гомологию можно определять посредством сравнения двух данных последовательностей невооруженным глазом, но скорее, ее можно измерять с использованием программы сравнения последовательностей, которая легко доступна, которая интерпретирует степень гомологии путем размещения бок о бок последовательностей, подлежащих сравнению. Программы сравнения последовательностей, известные в данной области, включают FASTP, BLAST, BLAST2, PSIBLAST и программу, включающую CLUSTAL W и т.д.
Были приведены многочисленные примеры относительно получения молочной кислоты путем обеспечения дефекта в PDC1, в которых продемонстрирована большая активность продукции молочной кислоты (Appl Microbiol Biotechnol. 2009, 82(5): 883-90). В данном случае, поскольку PDC6 редко экспрессируется, реальная активность PDC, по-видимому, обусловлена экспрессией гена PDC5. Согласно данным сообщениям дефект одной PDC1 не мешает росту клеток штамма дикого типа, и также может сохраняться примерно 60-70% активности PDC по сравнению с активностью дикого типа, и, таким образом, в данном штамме не наблюдали значимого фенотипического изменения (J Bacteriol. 1990, 172(2): 678-685).
В качестве альтернативы, можно получить штамм, имеющий одновременный двойной дефект в генах и PDC1, и PDC5, которые демонстрируют большие активности PDC в дрожжах. В таком случае молочнокислое брожение может быть проведено с использованием такого источника сахара, как гликоген, в отсутствие косубстрата, такого как уксусная кислота или этанол. Однако это приводило к уменьшению скорости роста штамма дрожжей из-за быстрого уменьшения активности PDC, посредством этого уменьшая продуктивность молочнокислого брожения (Biosci Biotechnol Biochem. 2006, 70(5): 1148-1153).
Тем временем, для того чтобы максимизировать уровень молочной кислоты, образующейся посредством пути LDH, которая конкурирует с PDC за пируват, можно получить штамм с одновременным тройным дефектом в PDC1, PDC5 и PDC6. В данном случае выход молочнокислого брожения может быть максимизирован, но метаболические емкости этанола и уксусной кислоты из-за катаболитной репрессии, индуцированной в присутствии глюкозы, могут быть дополнительно ингибированы, посредством этого уменьшая рост клеток и, в конечном счете, приводя к уменьшению продуктивности брожения (Curr Genet. 2003, 43(3): 139-160).
В частности, уменьшение активности пируватдекарбоксилазы (PDC) по настоящему изобретению может 1) инактивировать активность PDC1 и уменьшать активность PDC5; или 2) уменьшать активность PDC1 и инактивировать активность PDC5.
В типичном воплощении настоящего изобретения на основе штамма Saccharomyces cerevisiae, в котором была инактивирована активность PDC1, получали четыре разных штамма, которые включают штамм с пониженной активностью PDC5 посредством замены промотора гена PDC5, штамм, в котором вызван дефект в гене PDC5 посредством восстановления активности PDC1, штамм, в котором вызван двойной дефект в генах PDC1 и PDC5, и штамм, в котором вызван тройной дефект в генах PDC1, PDC5 и PDC6. Среди шаммов, полученных таким образом, было показано, что штамм, имеющий тройной дефект генов, редко демонстрирует рост клеток.
Используемый здесь термин «альдегиддегидрогеназа (ALD)» относится к белку, имеющему, главным образом, активность продуцирования уксусной кислоты из ацетальдегида в качестве белка, имеющего активность продуцирования карбоновой кислоты или ацильной группы путем окисления альдегида, но не ограничивается в настоящем изобретении ее производным или изотипом, имеющим такую же активность. Альдегиддегидрогеназа по настоящему изобретению может происходить из микроорганизма Saccharomyces sp. или может представлять собой ALD2 и/или ALD3. В частности, белок может представлять собой ALD2 и/или ALD3 Saccharomyces cerevisiae, но не ограничивается ими, и может включать любой их вариант или аналог при условии, что они являются биологически идентичными и имеют активности, соответствующие данному белку. Аминокислотные последовательности данного белка могут быть получены из базы данных и т.д., известной в данной области, например GenBank NCBI и т.д., но не ограничиваясь ей. В частности, ALD2 может состоять из аминокислотной последовательности SEQ ID NO: 74, и ALD3 может состоять из аминокислотной последовательности SEQ ID NO: 75. Данный белок может включать аминокислотные последовательности, имеющие гомологию более чем 70%, конкретно более чем 80%, более конкретно более чем 90% и даже более конкретно более чем 95% в отношении аминокислотных последовательностей. В настоящее изобретение также может быть включен любой вариант последовательностей, кодирующий идентичные аминокислотные последовательности, который возникает из-за вырожденности генетического кода.
Используемый здесь термин «ацетил-КоА-синтетаза (ACS)» относится к белку, имеющему активность катализирования тиоэтерификации уксусной кислоты и КоА в сочетании с реакцией распада АТФ, но не ограничивается в настоящем изобретении производным или изотипом, имеющим такую же активность. Было известно, что данный белок присутствует в микроорганизмах, растениях и животных и т.д. Ацетил-КоА-синтетаза по настоящему изобретению может происходить из микроорганизма Saccharomyces sp. или может представлять собой ACS1. В частности, данный белок может представлять собой ACS1 Saccharomyces cerevisiae, но не ограничивается им, и может включать его любой вариант или аналог, при условии, что они являются биологически идентичными и имеют активности, соответствующие данному белку. Аминокислотные последовательности данного белка могут быть получены из известной базы данных и т.д., например GenBank NCBI и т.д., но не ограничиваясь ей. В частности, ACS1 может состоять из аминокислотной последовательности SEQ ID NO: 76 и может включать аминокислотные последовательности, имеющие гомологию более чем 70%, конкретно более чем 80%, более конкретно более чем 90% и даже более конкретно более чем 95% в отношении аминокислотной последовательности. В настоящее изобретение также может быть включен мутант белка с последовательностью, кодирующей идентичные аминокислотные последовательности, который возникает из-за вырожденности генетического кода.
В типичном воплощении настоящего изобретения штаммы, в которых активности ALD2 и ACS или активности ALD3 и ACS были увеличены, получали на основе штамма, имеющего сниженную активность PDC по сравнению с активностью немутированного микроорганизма. В частности, штаммы, имеющие повышенные активности ALD и ACS, были получены на основе штамма, имеющего инактивированную PDC1 посредством дефекта PDC1 и пониженную активность PDC5 посредством замены промотора гена PDC5 промотором, имеющим низкую способность к экспрессии. Более конкретно, были получены штаммы микроорганизма Saccharomyces sp., в котором активность PDC1 была инактивирована, активность PDC5 была снижена, активность по меньшей мере одного фермента, выбранного из группы, состоящей из ALD2 и ALD3, была повышена, и активность ACS1 была повышена. Соответственно, подтвердили, что скорость роста штаммов, скорость продукции D-молочной кислоты и ее выход были значительно улучшены.
Термин «инактивация» активности фермента по настоящему изобретению относится к способу инактивирования активностей фермента, включающему любой способ, который ингибирует экспрессию фермента или обеспечивает экспрессию фермента, не способного демонстрировать его исходные активности. Данный способ может включать частичную делецию гена или полную делецию гена, вызванную гомологичной рекомбинацией, ингибирование экспрессии фермента, вызванное вставкой гена чужеродного происхождения в релевантный ген, ингибирование экспрессии фермента, вызванное заменой или модификацией последовательности промотора гена фермента или мутацию до неактивного фермента, имеющего потерю его исходных функций, вызванную заменой или модификацией фермента и т.д., но не ограничиваясь ими.
Используемый здесь термин «снижение» ферментативной активности относится к способу уменьшения активности фермента, включающему любой способ уменьшения уровня экспрессии фермента или уменьшения активности фермента, который экспрессируется. Данный способ может включать уменьшение экспрессии, вызванное заменой или модификацией последовательности промотора гена фермента, или мутацию до фермента, имеющего пониженную активность, вызванную заменой или модификацией фермента и т.д., но не ограничиваясь ими.
Используемый здесь термин «увеличение» ферментативной активности относится к вставке плазмиды, содержащей гены фермента, увеличению числа копий гена, кодирующего фермент, на хромосоме или увеличению активности фермента, вызванному заменой или модификацией, или к мутации последовательности промотора гена фермента и т.д., но не ограничивается ими.
Используемый здесь термин «дрожжевой микроорганизм» относится к микроорганизму, принадлежащему к Eumycetes, который размножается почкованием, но не ограничивается им, при условии, что он участвует в любом пути продукции молочной кислоты, пути продукции спирта и/или пути продукции ацетил-КоА. Дрожжевой микроорганизм может классифицироваться на Saccharomyces sp., Pichia sp., Candida sp. и Saccharomycopsis sp., в зависимости от формы дрожжей, и, в частности, в настоящем изобретении может применяться Saccharomyces sp., который включает разные виды. Конкретно микроорганизм может быть выбран из группы, состоящей из Saccharomyces bayanus, Saccharomyces boulardii, Saccharomyces bulderi, Saccharomyces cariocanus, Saccharomyces cariocus, Saccharomyces cerevisiae, Saccharomyces chevalieri, Saccharomyces dairenensis, Saccharomyces ellipsoideus, Saccharomyces eubayanus, Saccharomyces exiguus, Saccharomyces florentinus, Saccharomyces kluyveri, Saccharomyces martiniae, Saccharomyces monacensis, Saccharomyces norbensis, Saccharomyces paradoxus, Saccharomyces pastorianus, Saccharomyces spencerorum, Saccharomyces turicensis, Saccharomyces unisporus, Saccharomyces uvarum и Saccharomyces zonatus, и, более конкретно, он может представлять собой Saccharomyces cerevisiae.
Путем получения микроорганизма, имеющего пониженную активность PDC и улучшенные активности ALD и ACS, на основе Saccharomyces cerevisiae, типичного примера Saccharomyces sp., подтвердили значительное увеличение продукции молочной кислоты.
Микроорганизм по настоящему изобретению может включать алкогольдегидрогеназу (ADH), которая подлежит дальнейшей инактивации.
Используемый здесь термин «алкогольдегидрогеназа» относится к белку, имеющему активность катализирования обратной реакции, ответственной за продукцию альдегида или кетона путем удаления водорода от спирта, но не ограничивается производным или изотипом, имеющим такую же активность. Алкогольдегидрогеназа по настоящему изобретению может происходить из Saccharomyces sp. или может представлять собой ADH1. В частности, данный белок может представлять собой ADH1 Saccharomyces cerevisiae, но не ограничивается им, и может включать его любой вариант или аналог при условии, что они являются биологически идентичными и имеют активности, соответствующие данному белку. Аминокислотные последовательности данного белка могут быть получены из известной базы данных и т.д., например GenBank NCBI и т.д., но не ограничиваясь ей. В частности, ADH1 может состоять из аминокислотной последовательности SEQ ID NO: 77 и может включать аминокислотные последовательности, имеющие гомологию более чем 70%, конкретно более чем 80%, более конкретно более чем 90% и даже более конкретно более чем 95% по отношению к данной аминокислотной последовательности. В настоящее изобретение также может быть включен мутант белка с последовательностью, кодирующей идентичные аминокислотные последовательности, который возникает из-за вырожденности генетического кода.
Микроорганизм по настоящему изобретению может включать дегидрогеназу D-молочной кислоты (DLD), которая дополнительно инактивирована.
Используемый здесь термин «дегидрогеназа D-молочной кислоты» относится к белку, имеющему активность образования пирувата путем ангидризации D-молочной кислоты, но не ограничивается изотипом, имеющим такую же активность. Дегидрогеназа D-молочной кислоты по настоящему изобретению может происходить из Saccharomyces sp. В частности, данный белок может представлять собой DLD1 Saccharomyces cerevisiae, но не ограничивается им, и может включать его любой вариант или аналог при условии, что они являются биологически идентичными и имеют активности, соответствующие данному белку. Аминокислотные последовательности данного белка могут быть получены из известной базы данных и т.д., например GenBank NCBI, но не ограничиваясь ей. В частности, DLD1 может состоять из аминокислотной последовательности SEQ ID NO: 78 и может включать аминокислотные последовательности, имеющие гомологию более чем 70%, конкретно более чем 80%, более конкретно более чем 90% и даже более конкретно более чем 95% по отношению к данной аминокислотной последовательности. В настоящее изобретение также может быть включен любой вариант последовательности, кодирующей идентичные аминокислотные последовательности, который возникает из-за вырожденности генетического кода.
В настоящем изобретении штаммы, имеющие дефект в ADH1, ферменте, участвующем в пути спиртового брожения, использующем альдегид в качестве субстрата, который далее продуцируется из пирувата, и дефект в DLD1, ферменте, который расщепляет продуцированную молочную кислоту, использовали для точного измерения изменений эффективности молочнокислого брожения клеток согласно регуляции пути образования уксусной кислоты. В типичном воплощении настоящего изобретения штаммы, в которых регулировались активности PDC, ALD и ACS, демонстрировали значительное увеличение продуктивности молочнокислого брожения. Результаты обобщены в Таблице 12.
В другом аспекте в настоящем изобретении предложен способ получения молочной кислоты с использованием микроорганизма по настоящему изобретению.
Конкретно в типичном воплощении настоящего изобретения в настоящем изобретении предложен способ получения молочной кислоты, включающий культивирование микроорганизма по настоящему изобретению и сбор молочной кислоты из культуральной среды, содержащей данный микроорганизм.
Культивирование можно осуществлять с использованием подходящей среды и условий культивирования, известных в данной области. В соответствии с используемыми штаммами, способ культивирования может быть легко скорректирован обычным специалистом в данной области. Примеры способов культивирования включают культивирование периодического типа, непрерывного типа и с подпиткой, но не ограничиваются ими. Среды, используемые в способе культивирования, должны подходящим образом удовлетворять требованиям конкретного штамма.
Среда, используемая в настоящем изобретении, содержит сахарозу или глюкозу в качестве главного источника углерода, и в качестве источника углерода также можно использовать мелассу, содержащую высокую концентрацию сахарозы. Можно использовать другие источники углерода в адекватном количестве разными способами. В качестве источника азота можно использовать источники органического азота, включающие пептон, дрожжевой экстракт, мясной экстракт, солодовый экстракт, жидкий кукурузный экстракт и экстракты сои и пшеницы, и источники неорганического азота, включающие азот, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Эти источники азота можно использовать либо одиночно, либо в комбинации. В среду можно добавлять источники фосфора, такие как дигидрофосфат калия, гидрофосфат дикалия или соответствующие натрийсодержащие соли. Кроме того, среда может содержать соли металлов, такие как сульфат магния и сульфат железа. Кроме того, среда может быть дополнена аминокислотами, витаминами и подходящими предшественниками. Данные среды или предшественники можно добавлять в культуры периодическим или непрерывным способом.
На протяжении процесса культивирования можно подходящим образом добавлять такие соединения, как гидроксид аммония, гидроксид калия, фосфорную кислоту и серную кислоту, для того, чтобы подводить рН культуры. Кроме того, можно добавлять пеногаситель, такой как сложный эфир полигликоля с жирной кислотой, для того, чтобы ингибировать образование пены в культуре. Кроме того, для поддержания культуры в аэробном состоянии в культуру можно инъецировать кислород или кислородсодержащий газ, и для поддержания культуры в анаэробном и микроаэробном состоянии в культуру можно инъецировать газообразные азот, водород или диоксид углерода без инъецирования какого-либо газа.
Может поддерживаться температура культуры от 20 до 40°С, конкретно от 25 до 35°С и более конкретно 30°С. Культивирование можно продолжать, пока не получается желаемое количество желаемого вещества, конкретно, в течение 10-100 часов.
Молочную кислоту, продуцируемую в способах культивирования по настоящему изобретению, можно отбирать из культуральной среды подходящим способом, известным в данной области, в зависимости от способа культивирования, например периодического типа, непрерывного типа или с подпиткой.
ПОЛЕЗНЫЕ ЭФФЕКТЫ
Настоящее изобретение относится к применению микроорганизма, имеющего улучшенную продуктивность молочнокислого брожения, путем контролирования активностей изотипов PDC и увеличения активностей альдегиддегидрогеназы (ALD) и ацетил-КоА-синтетазы (ACS). Следовательно, его можно широко использовать в промышленном производстве молочной кислоты посредством брожения.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
ФИГ. 1 представляет собой схематическую диаграмму, иллюстрирующую связь между путем продукции молочной кислоты микроорганизма Saccharomyces sp., путем спиртового брожения и путем продукции ацетил-КоА.
ЛУЧШИЙ СПОСОБ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Далее настоящее изобретение будет подробно описано с использованием сопровождающих типичных воплощений. Однако раскрытые здесь типичные воплощения предназначены лишь для иллюстративных целей и не должны истолковываться как ограничивающие объем настоящего изобретения.
Пример 1: получение штамма, продуцирующего молочную кислоту
Для получения штаммов, продуцирующих молочную кислоту, Saccharomyces cerevisiae CEN.PK2-1D, репрезентативные дрожжи дикого типа, приобретенные у EUROSCARF, подвергали генетической манипуляции.
В частности, в качестве базового штамма использовали штамм, где алкогольдегидрогеназа 1 (ADH1) и пируватдекарбоксилаза 1 (PDC1) были дефектными для минимизации потери пирувата в пути синтеза спирта и дегидрогеназа D-молочной кислоты 1 (DLD1) была дефектной для блокирования пути распада молочной кислоты D-типа.
DLD1 не является решающим фактором, который может иметь прямое влияние на улучшение роста, но она была известна в качестве главного фермента, способного превращать D-молочную кислоту до пирувата с использованием NAD+ (никотинамидадениндинуклеотид) в качестве дегидрогеназы D-молочной кислоты. Соответственно, последующий штамм конструировали на основе штамма, имеющего генные дефекты в DLD1, ферменте, который потребляет продуцированную им молочную кислоту, для сравнения полной продуктивности брожения дрожжей, продуцирующих молочную кислоту D-типа, которые планируется получить в настоящем изобретении. В результате сравнивали продуктивность брожения.
В настоящем изобретении для генных манипуляций использовали обычное молекулярное клонирование.
Во-первых, для того чтобы удалить гены ADH1 и PDC1 дрожжевых штаммов, проводили эксперимент со ссылкой на содержание, раскрытое в ссылке: Lee ТН, et al. (J. Microbiol. Biotechnol. (2006), 16(6), 979-982), используя плазмиды pWAL100 и pWBR100. Каждую вставку, которую вводили в векторы на основе плазмид, получали с использованием подходящих праймеров (соответствующих нуклеотидным последовательностям SEQ ID NO: 1-8) посредством ПЦР (полимеразная цепная реакция).
Кроме того, для делеции гена DLD1 посредством двойного кроссинговера вводили HIS3, который представляет собой маркерный ген, и делали его дефектным. Используемые здесь фрагменты ДНК получали с использованием праймеров, соответствующих нуклеотидным последовательностям SEQ ID NO: 9 и 10. Праймеры, используемые в манипуляциях с генами, обобщены в Таблице 1, приведенной ниже.


Дегидрогеназу D-молочной кислоты (D-LDH), специфично требующуюся для образования D-молочной кислоты, вводили на основе штамма, имеющего дефекты в трех генах, таких KaKADHI, PDC1 и DLD1.
D-LDH затем клонировали в вектор, имеющий сайты рестрикционных ферментов Xhol и Spel на 5' и 3' концах соответственно, для того чтобы IdhD, происходящий из Lactobacillus plantarum (Lb. plantarum), был включен между промотором TEF1, происходящим из S. cerevisiae, и терминатором CYC1. В частности, эту вставку получали двойным расщеплением Sacl/Pvull, и вектор был расщеплен с образованием тупых концов посредством нуклеазы Mungbean из фрагмента ДНК, который дважды расщепляли из p-δ-neo в BamHl/Notl. Наконец, вектор обрабатывали Sacl для того, чтобы получить, посредством этого, вектор, имеющий липкий конец Sacl и тупой конец, полученный посредством BamHl.
Конструирование вектора pTL573 завершали посредством лигирования полученного вектора со вставкой. Плазмида pTL573 содержит ген IdhD, происходящий из Lb. plantarum, и его конструировали так, что он может включать случайную вставку многих копий генов в частичный домен δ-последовательности среди ретротранспозируемого элемента штамма CEN.PK2-1D pdc1Δ adh1Δ dld1Δ S. cerevisiae. Для множественной вставки соответствующего гена конструировали фрагменты ДНК, способные индуцировать одиночный кроссинговер на δ-последовательности, путем расщепления плазмиды pTL_573 с использованием Sall. Путем введения фрагментов ДНК в родительский штамм посредством трансформации из чашки с YPD (1% дрожжевой экстракт, 2% бактопептон и 2% глюкоза) получали многочисленные колонии при максимальной концентрации G418 5 мг/мл. Наконец, подтверждали, что в штамме, полученном таким образом, D-LDH, происходящая из Lb. plantarum, подвергалась многократной вставке с целью обеспечения способности продуцировать D-молочную кислоту, и его обозначили штамм СС02-0064.
Пример 2: получение мутантных штаммов, имеющих пониженную активность PDC5
Мутантный штамм, имеющий замененный промотор PDC5, получали на основе штамма СС02-0064, полученного в Примере 1. На протяжении данного процесса осуществляли процессы получения кассеты и селекции штамма согласно способу, раскрытому в Lee Т. Н. et al. (Development of reusable split URA3-marked knockout vectors for budding yeast, Saccharomyces cerevisiae. J Microbiol Biotechnol, 2006, 16:979-982).
Конкретно, всего получали пять новых штаммов путем замены промотора PDC5 штамма СС02-0064 промоторами SCO1, SCO2, ACS1, IDP2 и FBA1 соответственно и затем получали кассеты с заменой промотора, используя праймеры, соответствующие нуклеотидным последовательностям SEQ ID NO: 11-36.
Праймеры, используемые при замене промотора, обобщены в Таблице 2, приведенной ниже.


Полученные таким образом новые штаммы обозначали СС02-0167, СС02-0168, СС02-0169, СС02-0170 и СС02-0174 соответственно. Соответствующие штаммы и их генетические признаки обобщены в Таблице 3, приведенной ниже.

Пример 3: оценка молочнокислого брожения для мутантных штаммов, имеющих пониженную активность PDC5
Оценку молочнокислого брожения проводили для штаммов с мутацией PDC5, полученных в Примере 2. Для этого готовили специфическую среду для оценки молочнокислого брожения.
В частности, для того чтобы получить синтетические сложные среды (среды SC), лимитирующую среду для дрожжей, 0,67% дрожжевых азотистых оснований без аминокислот, служащих в качестве основы, смешивали со средой с выпадением аминокислот (Sigma) согласно протоколу изготовителя и, при необходимости, добавляли аминокислоты, которые были исключены из основы. Кроме того, в образующуюся среду добавляли 380 мг/л лейцина и добавляли урацил, триптофан и гистидин в концентрации 76 мг/л соответственно. Также добавляли 8% глюкозы в качестве источника углерода и 1% СаСО3 в качестве нейтрализующего агента. Полученную таким образом среду использовали для оценки молочнокислого брожения дрожжевых штаммов.
Среди штаммов с мутацией промотора PDC5, полученных в Примере 2, мутантные штаммы с заменой на более слабый промотор, чем исходный промотор PDC5, не могли расти, тогда как мутантные штаммы с заменой на более сильный промотор демонстрировали улучшенный рост. В частности, мутантные штаммы с заменой на промоторы SCO1, SCO2, IDP2 или ACS1, которые представляют собой более слабые промоторы, чем промотор PDC5, не могли расти, оставляя штаммы, промотор которых был заменен промотором FBA1, единственными штаммами, подлежащими оценке. Результат оценки молочнокислого брожения для штаммов СС02-0064 и СС02-0174, который был измеримым, обобщен в Таблице 4, приведенной ниже.
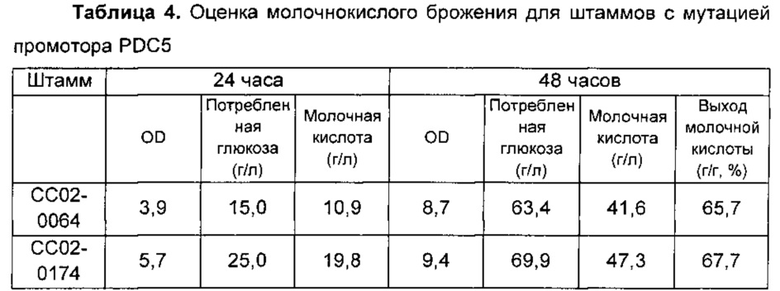
Выход молочной кислоты рассчитывали следующим образом:
Выход молочной кислоты (г/г, %)=Молочная кислота (г/л) / Потребленная глюкоза (г/л) (%).
Как показано в оценке, приведенной выше, подтвердили, что на протяжении пути, стимулирующего продукцию ацетил-КоА, штамм, в котором промотор PDC5 дикого типа был заменен промотором FBA1, демонстрировал улучшенную скорость роста клеток и их продуктивность в отношении молочной кислоты по сравнению с таковыми исходного штамма (СС02-0064). Однако при сравнении результата образцов, отобранных в 24 часа и 48 часов соответственно, подтвердили, что улучшения скорости роста клеток и их продуктивности в отношении молочной кислоты согласно указанному времени продолжали уменьшаться посредством простого усиления активности одной PDC без усиления активностей ALD и ACS, которые участвуют в последующем пути продукции ацетил-КоА. В примере по настоящему изобретению улучшение потребления глюкозы посредством усиления активности PDC составляло 10,3%, и максимальная концентрация продуцированной молочной кислоты составляла 47,3 г/л. Соответственно, общее улучшение продуктивности в отношении молочной кислоты составляло 13,7%.
Пример 4: получение штамма, имеющего дефект гена PDC5
Помимо штамма, имеющего дефект гена PDC1 и пониженную активность PDC5, полученного в Примере 2, получали штамм, имеющий дефект в гене PDC5 и пониженную активность PDC1, для того, чтобы подтвердить, посредством этого, был ли ослаблен в соответствующем штамме путь PDC.
В частности, с целью получения дефекта гена PDC5 использовали праймеры, соответствующие нуклеотидным последовательностям SEQ ID NO: 37-40, для получения кассеты с дефектом гена PDC5 на основе штамма СС02-0064. Дефектный штамм получали тем же самым способом, который описан в литературе Примера 1. Праймеры, использованные в Примере 4, обобщены в Таблице 5 ниже.

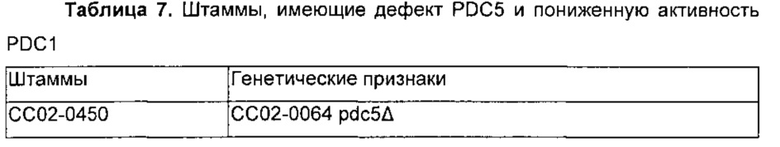
Полученный таким образом штамм, имеющий дефект гена PDC5, был обозначен СС02-0450 (СС02-0064, pdc5Δ).
Пример 5: получение штаммов с мутацией промотора PDC1 на основе штамма, имеющего дефект PDC5
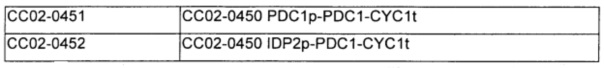
Штамм, имеющий замену промотора PDC1, получали на основе штамма СС02-0450, полученного в Примере 4. Для этого штамм СС02-0451 (СС02-0450, PDC1p-PDC1), в котором был восстановлен дефект гена PDC1, получали для того, чтобы он служил в качестве группы сравнения, и штамм СС02-0452 (СС02-0450, IDP1p-PDC1), имеющий пониженную активность PDC1, получали для того, чтобы он служил в качестве экспериментальной группы.
Каждый штамм получали таким образом, чтобы в штамм были включены векторы PDC1p-PDC1-CYC1t и pRS406-IDP2p-PDC1-CYC1, которые были сконструированы путем клонирования намеченной кассеты гена в вектор pRS406 без точки начала репликации у дрожжей.
В частности, проводили ПЦР (полимеразная цепная реакция) с использованием праймеров, имеющих нуклеотидные последовательности SEQ ID NO: 41 и 42, с хромосомной ДНК дрожжей, служащей в качестве матрицы, для получения, посредством этого, продукта, включающего ген PDC1. Затем получали последовательность терминатора CYC1 с использованием праймеров, имеющих нуклеотидные последовательности SEQ ID NO: 43 и 44. Кроме того, получали фрагменты ДНК, соединяющие PDC1 и терминатор CYC1, посредством ПЦР с использованием праймеров, соответствующих нуклеотидным последовательностям SEQ ID NO: 41 и 44, с последовательностями PDC1 и терминатора CYC1 соответственно, служащими в качестве матрицы. Плазмидный вектор pRS406-PDC1-CYC1t получали путем обработки фрагментов ДНК терминатора PDC1-CYC1 и вектора pRS406 рестрикционными ферментами Spel и Xhol с их последующим лигированием. Тем временем, для введения домена промотора в плазмидные векторы, полученные таким образом, плазмидные векторы, в которые были включены промоторы PDC1 и IDP2 соответственно, получали слиянием праймеров, имеющих нуклеотидные последовательности SEQ ID NO: 45 и 46, и 47 и 48 соответственно, посредством ПЦР с использованием хромосомной ДНК в качестве матрицы. Фрагменты ДНК, включающие каждый промотор и плазмиду pRS406-PDC1-CYC1t, расщепляли и лигировали для получения, посредством этого, плазмидных векторов pRS406-PDC1p-PDC1-CYC1t и pRS406-IDP2p-PDC1-CYC1t соответственно, которые представляют собой плазмиду, требующуюся для вставки в дрожжевую хромосому, сконструированную так, что экспрессия гена контролируется промотором PDC1 и промотором IDP2.
Праймеры, используемые в Примере 5, обобщены в Таблице 6.
Два полученных таким образом плазмидных вектора расщепляли Stul соответственно и немедленно вставляли в штаммы. Конечные штаммы обозначали СС02-0451(СС02-0450, PDC1p-PDC1) и СС02-0452(СС02-0450, IDP2p-PDC1) соответственно. Полученные таким образом штаммы и их генетические признаки обобщены в Таблице 7.
Пример 6: получение штаммов, имеющих двойные или тройные дефекты в генах PDC
Из генов семейства PDC намеревались получить штаммы, имеющие одиночный дефект в гене PDC1, двойной дефект в генах PDC1 и PDC5 и тройной дефект в генах PDC1, PDC5 и PDC6. Штамм СС02-0064, полученный в Примере 1, использовали в качестве штамма, имеющего одиночный дефект в гене PDC1. Кассету для дефекта PDC5 получали с использованием праймеров, соответствующих нуклеотидным последовательностям SEQ ID NO: 49-56, и вставляли в СС02-0064 для получения штамма, имеющего двойные дефекты в генах PDC1 и PDC5. Затем полученный таким образом штамм обозначали СС02-0256. Кроме того, получали штамм, имеющий тройной дефект в генах PDC1, PDC5 и PDC6, на основе штамма, имеющего двойные дефекты в генах PDC1 и PDC5, с использованием праймеров, соответствующих нуклеотидной последовательности SEQ ID NO: 57-64, и обозначали СС02-0257.
Получение кассеты с дефектом и процесс отбора штаммов проводили тем же самым способом, который описан в литературе, раскрытой в Примере 1. Праймеры, использованные в Примере 6, обобщены в Таблице 8.


Полученные таким образом штаммы и их генетические признаки обобщены в Таблице 9.

Пример 7: получение штаммов, сверхэкспрессирующих ALD и ACS1
Для получения штаммов, сверхэкспрессирующих ALD и ACS1, получали плазмиды, сверхэкспрессирующие ALD2, ALD3 и ACS1.
В частности, получали открытую рамку считывания (ORF) ALD2 с использованием праймеров, соответствующих нуклеотидным последовательностям SEQ ID NO: 65 и 66, получали ORF ALD3 с использованием праймеров, соответствующих нуклеотидным последовательностям SEQ ID NO: 67 и 68, и получали ORF ACS1 с использованием праймеров, соответствующих нуклеотидным последовательностям SEQ ID NO: 69 и 70. Кроме того, p415ADH-ALD2, p415ADH-ALD3, p414ADH-ACS1 и p416ADH-ACS1, которые представляют собой рекомбинантные векторы на основе плазмид p414ADH, p415ADH и p416ADH, получали рестрикционными ферментами Spel, Xnol или EcoRl. Праймеры, использованные в Примере 7, обобщены в Таблице 10, приведенной ниже.

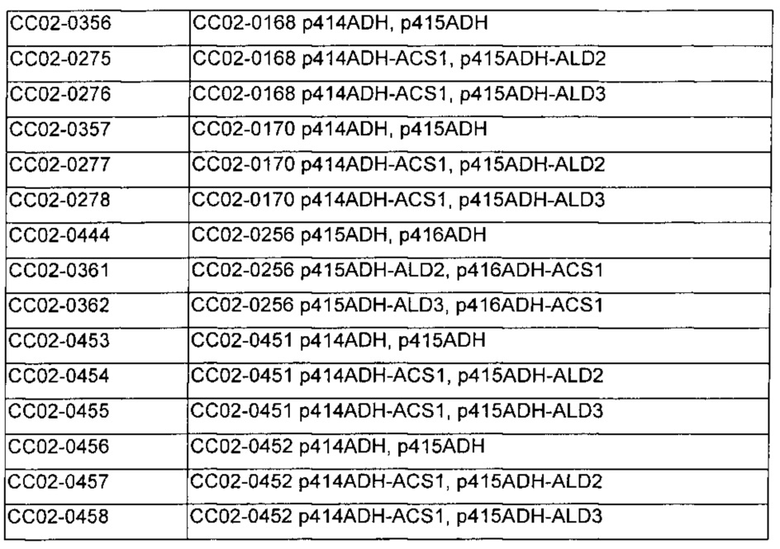
Полученные таким образом рекомбинантные плазмиды вводили в штаммы, включающие СС02-0064, СС02-0168, СС02-0170, СС02-0256, СС02-0257, СС02-0451 и СС02-0452, посредством трансформации дрожжей комбинацией p415ADH-ALD2, p414ADH-ACS1, комбинацией p415ADH-ALD3, p414ADH-ACS1, комбинацией p415ADH-ALD2, p416ADH-ACS1 или комбинацией p415ADH-ALD3, p416ADH-ACS1. Однако не получали трансформанта в штамме СС02-0257, имеющем тройные дефекты в генах PDC, в котором не демонстрировалась активность PDC.
Полученные таким образом штаммы и их генетические признаки обобщены в Таблице 11.

Пример 8: оценка молочнокислого брожения для дрожжевых штаммов
Проводили оценку способности к молочнокислому брожению для штаммов, сверхэкспрессирующих ALD и ACS1, полученных в Примере 7.
В частности, дрожжи инокулировали в каждый флакон, содержащий 25 мл среды, полученной в Примере 3, для оценки молочнокислого брожения и культивировали при аэробных условиях при 30°С в течение 71 часа. Анализировали количество молочной кислоты D-типа, присутствующей в ферментационном бульоне, и проводили ферментативный анализ (уксусная кислота, R-Biopharm, Германия) для определения количества присутствующей в нем уксусной кислоты.
Результаты приведенных выше экспериментов обобщены в Таблице 12, приведенной ниже.
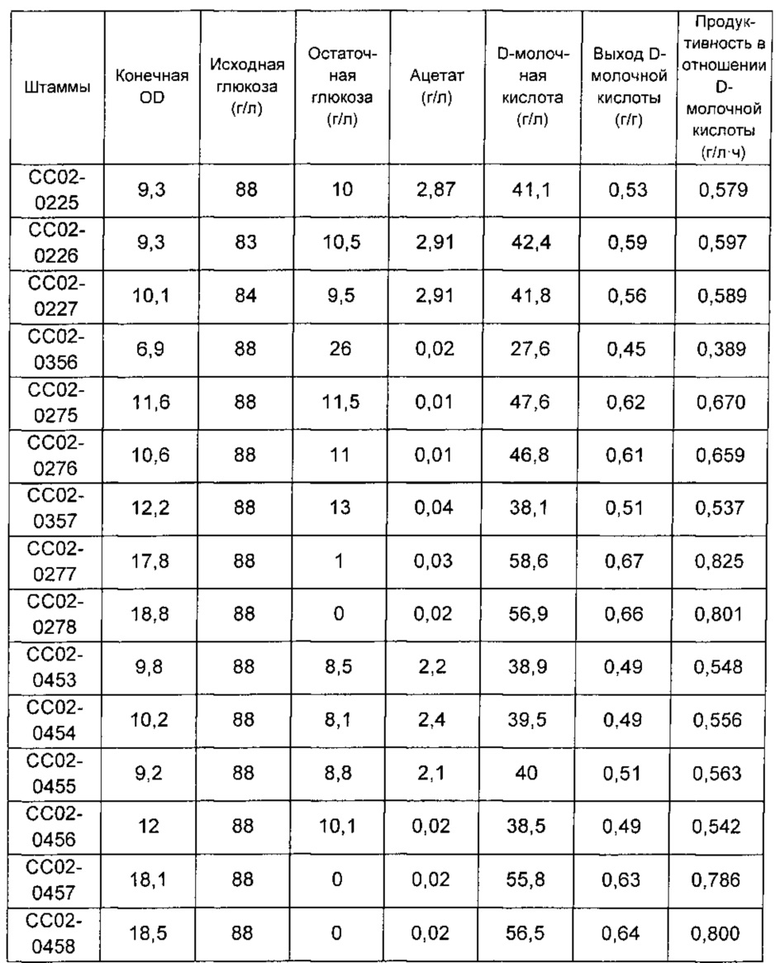
Таблица 12. Оценка скорости роста, молочнокислого брожения, побочных продуктов, выхода продукции и т.д. для штаммов, сверхэкспрессирующих ALD и ACS.
Выход D-молочной кислоты рассчитывали следующим образом:
Выход D-молочной кислоты (г/г)=D-молочная кислота (г/л) / (Исходная глюкоза - Остаточная глюкоза) (г/л).
Как подтверждено в Таблице 12, штаммы, имеющие пониженную посредством промотора IDP2 или промотора SC02 активность PDC5, имели сильное уменьшение накопления ацетата - побочного продукта, т.е. было подтверждено малое выявление ацетата по сравнению со штаммом с нормальной активностью PDC5. В таком случае конечная клеточная концентрация штаммов, в которых активности ALD и ACS не были увеличены, имела тенденцию к снижению согласно заменам промотора PDC5. С другой стороны, штаммы, в которых экспрессия PDC5 была снижена и активности ALD и ACS были повышены, демонстрировали увеличение конечной клеточной концентрации. Соответственно, было подтверждено улучшение роста клеток. В частности, штаммы СС02-0277 и СС02-0278, имеющие повышенные активности ALD и ACS, полученные на основе штамма СС02-0170, где промотор PDC5 был заменен IDP2, демонстрировали улучшенные скорость роста, концентрацию продуцированной D-молочной кислоты, ее выход и продуктивность брожения по мере увеличения активностей ALD и ACS.
В заключение, штамм, в котором промотор PDC5 был заменен на слабо экспрессирующийся промотор IDP2, имел уменьшение накопления ацетата, и конечная OD (оптическая плотность) была в 1,3 раза выше по сравнению со штаммом, демонстрирующим нормальную экспрессию PDC5. Кроме того, подтвердили, что при соэкспрессии ALD и ACS под контролем промотора ADH1 потребление глюкозы и его скорость увеличивались и, в конечном счете, процентный выход возрастал от 56% или 59% до 66% или 67%, соответственно, демонстрируя улучшенный выход.
В частности, посредством сравнения двух видов промоторов, применяемых для слабой экспрессии PDC5, подтверждали, что продуктивность в отношении молочной кислоты улучшалась у обоих штаммов, имеющих промотор SC02 и промотор IDP2, соответственно. Однако штамм, имеющий промотор IDP2, может считаться самой оптимизированной формой штамма в терминах общей клеточной концентрации, потребления глюкозы и его скорости.
Пример 9: оценка молочнокислого брожения для дрожжевого штамма, имеющего двойные дефекты в генах PDC1 и PDC5, и увеличение активностей ALD и ACS
Поскольку эффекты на рост клеток и улучшение выхода, возникающие в результате пониженной активности PDC5, были ясно подтверждены, предпринимали оценку для определения эффектов дополнительного дефекта гена PDC в продукции молочной кислоты. Способ оценки для каждого штамма был идентичен способу, описанному в Примере 8, и культивирование проводили в течение 74 часов.
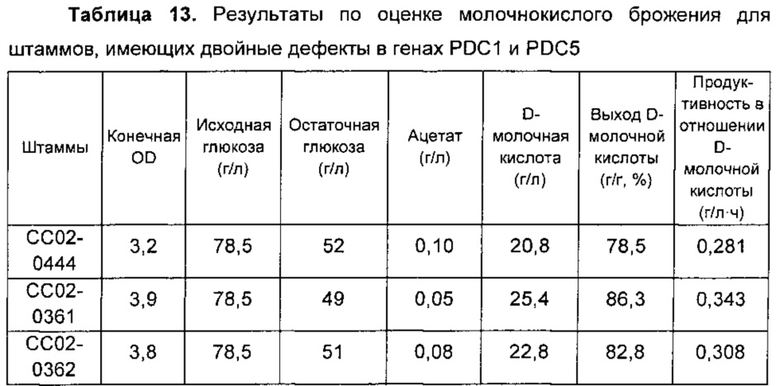
Полученные таким образом результаты эксперимента обобщены в Таблице 13, приведенной ниже.
Выход D-молочной кислоты рассчитывали следующим образом:
Выход D-молочной кислоты (г/л)=D-молочная кислота (г/л) / (Исходная глюкоза - Остаточная глюкоза) (г/л) (%).
Как подтверждено в Таблице 13, концентрация ацетата явно снижалась в штаммах, имеющих двойные дефекты в генах PDC1 и PDC5, однако также наблюдали уменьшение концентрации продуцируемой D-молочной кислоты из-за уменьшения роста клеток и потребления глюкозы. Кроме того, штамм, в котором путь PDC практически инактивирован, что происходило из-за двойных дефектов в генах PDC1 и PDC5, не имел какого-либо улучшения в росте клеток, потреблении глюкозы и его продуктивности, несмотря на то что данный штамм демонстрировал повышенные активности ALD и ACS.
Пример 10: оценка молочнокислого брожения для штаммов, в которых путь PDC ослаблен, с использованием сахарозы
С целью оценки брожения с использованием сахарозы использовали штаммы дрожжей, продуцирующих молочную кислоту, с ослабленным путем PDC - штаммы, идентичные оцениваемым в Примере 8 и 9, для подтверждения эффекта на продукцию молочной кислоты. Для этого в качестве источника углерода вместо глюкозы использовали сахарозу. Способ оценки осуществляли так же, как и в Примере 8.
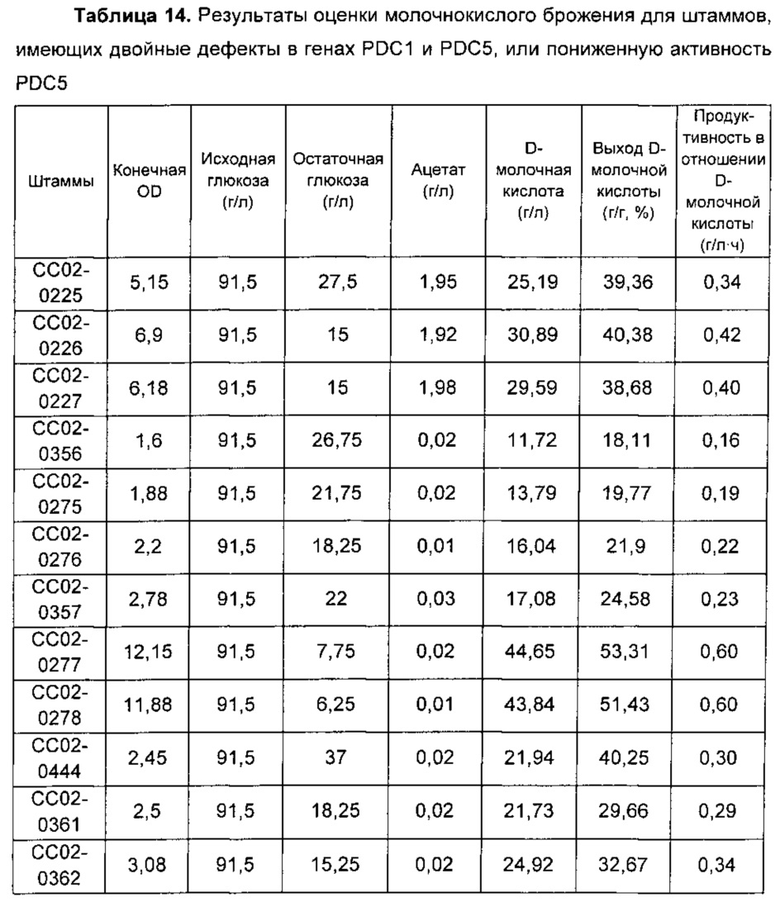
Полученные таким образом результаты экспериментов обобщены в Таблице 14, приведенной ниже.
Выход D-молочной кислоты рассчитывали следующим образом:
Выход D-молочной кислоты (г/г)=D-молочная кислота (г/л) / (Исходная глюкоза - Остаточная глюкоза) (г/л) (%).
Применение сахарозы вместо глюкозы для штаммов, используемых так же, как и в Примерах 8 и 9, обеспечивало улучшенные рост и выход брожения путем увеличения активностей ALD и ACS в штаммах, где ослаблен путь PDC, демонстрируя такую же картину результатов, что и у штаммов в Примере 8, где в качестве источника углерода использовали глюкозу. Соответственно, настоящее изобретение подтверждает то, что улучшенные выход брожения и рост из-за пониженной активности PDC и повышенных активностей ALD и ACS, которые были подтверждены в настоящем изобретении, не ограничиваются типом используемого сахара.
Для обобщения приведенных выше результатов подтвердили то, что при мутировании штаммов таким образом, что ослабевал путь PDC и улучшались активности ALD и ACS по сравнению с таковыми у немутировавших штаммов, продукция молочной кислоты возрастала и одновременно поддерживалась их скорость роста.
Из приведенного выше специалист в области, к которой относится настоящее изобретение, сможет понять, что настоящее изобретение может быть воплощено в других конкретных формах без модификации технических идей или существенных характеристик настоящего изобретения. В данном отношении раскрытые здесь типичные воплощения служат только для иллюстративных целей и не должны истолковываться как ограничивающие объем настоящего изобретения. С другой стороны, подразумевается то, что настоящее изобретение охватывает не только типичные воплощения, но также разные альтернативы, модификации, эквиваленты и другие воплощения, которые могут быть включены в пределы сущности и объема настоящего изобретения, как определено приложенной формулой изобретения.



























Изобретение относится к области биохимии, генной инженерии и биотехнологии, в частности к модифицированному микроорганизму Saccharomyces cerevisiae, имеющему повышенную продуктивность в отношении молочной кислоты. Настоящий модифицированный микроорганизм характеризуется тем, что в нем снижена активность пируватдекарбоксилазы, усилены активности альдегиддегидрогеназы и ацетил-КоА-синтетазы и в него введена дегидрогеназа молочной кислоты. В результате указанных модификаций этот микроорганизм способен продуцировать молочную кислоту с высоким выходом. Изобретение также относится к способу получения молочной кислоты. Настоящий способ предусматривает культивирование указанного микроорганизма и выделение из культуральной среды молочной кислоты, продуцируемой при культивировании этого микроорганизма. Настоящее изобретение позволяет получать молочную кислоту с высоким выходом. 2 н. и 4 з.п. ф-лы, 1 ил., 14 табл., 10 пр.
1. Модифицированный микроорганизм Saccharomyces cerevisiae, имеющий повышенную продуктивность в отношении молочной кислоты, который модифицирован так, что:
(а) активность пируватдекарбоксилазы (PDC) микроорганизма снижена по сравнению с таковой у немодифицированного штамма, продуцирующего молочную кислоту;
(б) активности альдегиддегидрогеназы (ALD) и ацетил-КоА-синтетазы (ACS) микроорганизма усилены по сравнению с таковыми у немодифицированного штамма, продуцирующего молочную кислоту; и
(в) введена дегидрогеназа молочной кислоты (LDH).
2. Микроорганизм по п. 1, в котором пируватдекарбоксилаза представляет собой по меньшей мере одну пируватдекарбоксилазу, выбранную из группы, состоящей из PDC1, PDC5 и PDC6.
3. Микроорганизм по п. 2, который модифицирован для того, чтобы:
1) инактивировать активность PDC1 и снизить активность PDC5 или
2) снизить активность PDC1 и инактивировать активность PDC5.
4. Микроорганизм по п. 1, в котором альдегиддегидрогеназа представляет собой по меньшей мере одну альдегиддегидрогеназу, выбранную из группы, состоящей из ALD2 и ALD3, и ацетил-КоА-синтетаза представляет собой ACS1.
5. Микроорганизм по п. 1, в котором дополнительно инактивирована алкогольдегидрогеназа (ADH).
6. Способ получения молочной кислоты, включающий:
а) культивирование микроорганизма по любому из пп. 1-5 в культуральной среде и
б) выделение молочной кислоты из культуральной среды со стадии а).
| US 8071357 B2, 06.12.2011 | |||
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА МОЛОЧНОЙ КИСЛОТЫ И РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ SCHIZOSACCHAROMYCES POMBE ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2004 |
|
RU2268304C1 |
| СПОСОБ ПРЕОБРАЗОВАНИЯ ТЕПЛОВОЙ ЭНЕРГИИ НАГРЕТОЙ ВОДЫ В КИНЕТИЧЕСКУЮ ЭНЕРГИЮ РЕАКТИВНОЙ СТРУИ И УСТРОЙСТВО ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2005 |
|
RU2281881C1 |
| US 20100062505 A1, 11.03.2010. | |||

