ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Область техники, к которой относится изобретение
Настоящая заявка относится к микроорганизму из рода Saccharomyces, продуцирующему молочную кислоту, и к способу продуцирования молочной кислоты с его использованием.
Предшествующий уровень техники
Молочная кислота имеет широкий диапазон применений в промышленности, в том числе в производстве пищевых продуктов, лекарственных препаратов, косметики и т.д. Недавно благодаря применению молочной кислоты в качестве мономера для полимолочной кислоты потребность в ней значительно возросла. Способы продуцирования молочной кислоты включают традиционный химический синтез и биологический ферментационный процесс, в котором в качестве субстратов используют углеводы, и этот способ в последнее время стал предпочтительным.
Как правило, у продуцирующих молочную кислоту дрожжевых микроорганизмов лактатдегидрогеназа (LDH) и пируватдекарбоксилаза (PDC) конкурируют за использование пирувата в качестве субстрата. В связи с этим для того, чтобы получить максимальный выход пирувата, необходимо свести к минимуму функции PDC для повышения продукции молочной кислоты (LA) лактатдегидрогеназой (LDH). У дрожжей Saccharomyces cerevisiae PDC присутствует в трех различных изоферментных формах, т.е. PDC1, PDC5 и PDC6. Поэтому для максимальной продукции молочной кислоты можно использовать способ получения штамма с одновременной тройной делецией генов PDC1, PDC5 и PDC6. Однако, хотя инактивация активности PDC может повысить выход молочной кислоты, она также имеет недостатки, заключающиеся в снижении производительности и в том, что штаммы не могут расти равномерно (Европейский патент № ЕР 2041264), что создает трудности для получения желаемого количества молочной кислоты. Кроме того, в отличие от прокариот, таких как бактерии, у дрожжей, например рода Saccharomyces, невозможно точно прогнозировать, могут ли различные клеточные органеллы и различные органические системы продуцировать желаемое количество молочной кислоты в результате простых манипуляций с генами.
Описание изобретения
Техническая задача изобретения
Авторы настоящего изобретения предприняли попытку разработки способа для повышения как выхода продукции, так и количества продуцируемой молочной кислоты, с сохранением равномерного роста продуцирующего молочную кислоту микроорганизма, и в результате было подтверждено, что усиление биохимического пути восполнения ацетил-КоА и биохимического пути восполнения пирувата из оксалоацетата (ОАА) приводит к увеличению количества продуцируемой молочной кислоты и, таким образом, выполнили настоящую заявку на изобретение.
Техническое решение
Задача настоящего изобретения состоит в обеспечении микроорганизма рода Saccharomyces, продуцирующего молочную кислоту.
В задачу настоящей заявки также входит обеспечение способа продуцирования молочной кислоты с использованием этого микроорганизма.
Благоприятные эффекты
Продуцирующий молочную кислоту модифицированный штамм в соответствии с настоящей заявкой, в котором инактивирована PDC активность, введена чужеродная активность ACL и усилен путь биосинтеза пирувата, обладает превосходным выходом ферментации молочной кислоты и продуктивностью, сравнимой с традиционными штаммами, за счет сведения к минимуму ферментации продуцирования спирта и блокирования биохимического пути разложения молочной кислоты. Таким образом был усилен рост продуцирующего молочную кислоту штамма, и модифицированный продуцирующий молочную кислоту штамм в соответствии с настоящей заявкой может быть широко использован для повышения производительности производства различных продуктов, получаемых с использованием молочной кислоты в качестве исходного материала. Полученную таким путем молочную кислоту можно поставлять в качестве исходного материала для различных видов продуктов.
Краткое описание графических материалов
На ФИГ. 1 представлен биохимический путь усиления биосинтеза пирувата посредством сверхэкспрессии РЕР-карбоксикиназы (PCK 1, ЕС 4.1.1.49) и пируваткиназы (PYK 2, ЕС 2.7.1.40), присутствующих в дрожжевом микроорганизме, и показана схематическая диаграмма, иллюстрирующая стратегию улучшения продуктивности синтеза молочной кислоты путем введения чужеродной ACL (АТР (аденозинтрифосфат)-цитратлиазы) и усиления биохимических путей PCK и PYK в соответствии с настоящей заявкой.
На ФИГ. 2 представлен биохимический путь усиления биосинтеза пирувата посредством усиления малатдегидрогеназы 2 (MDH2, ЕС 1.1.1.37) и цитозольного яблочного фермента 1 (МАЕ1, ЕС 1.1.1.38), присутствующих в дрожжевом микроорганизме, и показана схематическая диаграмма, иллюстрирующая стратегию улучшения продуктивности синтеза молочной кислоты путем введения чужеродной ACL и посредством биохимических путей MDH и цитозольного МАЕ в соответствии с настоящей заявкой.
Лучший путь осуществления изобретения
Для решения вышеуказанной задачи в одном аспекте в настоящей заявке предложен микроорганизм рода Saccharomyces, продуцирующий молочную кислоту, модифицированный путем инактивации активности пируватдекарбоксилазы (PDC) по сравнению с ее эндогенной активностью, введения активности АТР-цитратлиазы (ACL) и усиления пути биосинтеза пирувата по сравнению с эндогенным путем его биосинтеза.
Продуцирующий молочную кислоту микроорганизм рода Saccharomyces может представлять собой микроорганизм, в котором повышен выход ферментации молочной кислоты, и/или усилен рост микроорганизма рода Saccharomyces (увеличение роста микроорганизма-продуцента), и/или повышена продуктивность синтеза молочной кислоты по сравнению с немодифицированными штаммами, в которые не введена активность ACL и/или не усилен путь биосинтеза пирувата относительно эндогенного пути его биосинтеза.
Используемый в настоящем документе термин «молочная кислота (LA)» относится к органической кислоте, представленной формулой С2Н4ОНСООН. При получении такой молочной кислоты путем химического синтеза молочная кислота образуется в форме рацемической смеси, в которой молочная кислота D-типа и молочная кислота L-типа смешаны в соотношении 50/50, и соотношение их в композиции невозможно контролировать. Поэтому при получении полимолочной кислоты молочная кислота образует аморфный полимер с низкой точкой плавления, и поэтому имеются многочисленные ограничения при разработке его применений. В отличие от этого, при получении молочной кислоты биологическим ферментационным способом с использованием микроорганизма молочная кислота D-типа и молочная кислота L-типа могут продуцироваться селективно в зависимости от используемых бактерий или вводимой в них лактатдегидрогеназы (LDH).
Используемый в настоящем документе термин «продуцирующий молочную кислоту микроорганизм» в настоящей заявке относится к штамму микроорганизма, который продуцирует молочную кислоту, может преобразовывать сахар в молочную кислоту и, например, может включать в себя любой дрожжевой микроорганизм без каких-либо ограничений, если он включает в себя путь синтеза молочной кислоты и путь синтеза ацетил-СоА по настоящей заявке.
В соответствии с их формами дрожжевые микроорганизмы можно классифицировать на род Saccharomyces, род Pichia, род Candida и род Saccharomycopsis, и конкретно в настоящей заявке можно использовать микроорганизм рода Saccharomyces, включая его различные виды, если этот микроорганизм способен продуцировать молочную кислоту. В частности, микроорганизм Saccharomyces sp. может быть выбран из группы, состоящей из Saccharomyces bayanus, Saccharomyces boulardii, Saccharomyces bulderi, Saccharomyces cariocanus, Saccharomyces cariocus, Saccharomyces cerevisiae, Saccharomyces chevalieri, Saccharomyces dairenensis, Saccharomyces ellipsoideus, Saccharomyces eubayanus, Saccharomyces exiguus, Saccharomyces florentinus, Saccharomyces kluyveri, Saccharomyces martiniae, Saccharomyces monacensis, Saccharomyces norbensis, Saccharomyces paradoxus, Saccharomyces pastorianus, Saccharomyces spencerorum, Saccharomyces turicensis, Saccharomyces unisporus, Saccharomyces uvarum и Saccharomyces zonatus, и более конкретно Saccharomyces cerevisiae.
Продуцирующий молочную кислоту микроорганизм рода Saccharomyces по настоящей заявке может представлять собой микроорганизм, модифицированный таким образом, что активность пируватдекарбоксилазы (PDC) инактивирована по сравнению с ее эндогенной активностью, введена активность АТР-цитратлиазы и усилен биохимический путь биосинтеза пирувата по сравнению с эндогенным биохимическим путем его биосинтеза.
В частности, продуцирующий молочную кислоту микроорганизм Saccharomyces sp. может представлять собой микроорганизм, модифицированный для (1) инактивации активности пируватдекарбоксилазы (PDC) по сравнению с ее эндогенной активностью, (2) введения активности АТР-цитратлиазы и (3) усиления пути биосинтеза пирувата по сравнению с эндогенным путем его биосинтеза, и дополнительно модифицированный для (4) введения активности лактатдегидрогеназы (LDH), (5) ослабления или инактивирования активности алкогольдегидрогеназы 1 по сравнению с ее эндогенной активностью, (6) ослабления или инактивирования активности пируватдекарбоксилазы 1 по сравнению с ее эндогенной активностью и/или (7) ослабления или инактивирования активности D-лактатдегидрогеназы 1 по сравнению с ее эндогенной активностью.
Кроме того, микроорганизм по настоящей заявке может быть дополнительно модифицирован для (1) инактивирования активности алкогольдегидрогеназы 1 (ADH1) по сравнению с ее эндогенной активностью; (2) инактивирования активности пируватдекарбоксилазы 1 (PDC1) по сравнению с ее эндогенной активностью; и (3) инактивирования активности D-лактатдегидрогеназы 1 (DLD1) по сравнению с ее эндогенной активностью.
Используемый в настоящем документе термин «пируватдекарбоксилаза (PDC)», который можно использовать взаимозаменяемо с ферментом, катализирующим декарбоксилирование пирувата, относится к ферменту, который превращает пируват в ацетальдегид и диоксид углерода (СО2). Пируватдекарбоксилаза представляет собой фермент, вовлеченный в процесс ферментации в анаэробных условиях, происходящий у дрожжей, в частности, Saccharomyces sp., и представляет собой фермент, который продуцирует этанол посредством ферментирования. Как правило, PDC у Saccharomyces sp. присутствует в трех различных изоферментных формах, т.е. PDC1, PDC5 и PDC6. Последовательности белка и гена PDC могут быть получены из известной базы данных, такой как GenBank NCBI (Национальный центр биотехнологической информации), но не ограничиваясь ею. В частности, что касается этого фермента, то PDC1 может представлять собой белок, представленный аминокислотной последовательностью SEQ ID NO: 39, PDC5 может представлять собой белок, представленный аминокислотной последовательностью SEQ ID NO: 41, и PDC6 может представлять собой белок, представленный аминокислотной последовательностью SEQ ID NO: 43, но может быть включена без ограничений любая аминокислотная последовательность, обладающая активностью PDC. Кроме того, гены, кодирующие PDC1, PDC5 и PDC6, могут быть конкретно представлены нуклеотидными последовательностями, например SEQ ID NO: 40, 42 и 44, соответственно, но может быть без ограничений включена любая нуклеотидная последовательность, которая может кодировать этот фермент.
Используемый в настоящем документе термин «АТР-цитратлиаза (ACL, ЕС 2.3.3.8)» относится к ферменту, который преобразует цитрат в оксалоацетат (ОАА) и ацетил-СоА, и о котором известно, что он присутствует у высших организмов и некоторых дрожжей (ATP-citrate lyase: A mini-review, Biochemical and Biophysical Research Communications, 422, (2012), 1-2).
Схема реакции приведена ниже:
Лимонная кислота + АТР + СоА + Н2О → ОАА + ацетил-СоА + A + Pi
Ацетил-СоА является незаменимым ферментом для роста микроорганизмов, и его значимость подчеркивается в недавно опубликованных литературных источниках. В качестве репрезентативного примера можно привести сообщение об исследовании по улучшению продуктивности в эукариотическом организме, способном продуцировать 1,3-бутандиол (1,3-BDO), путем обеспечения цитозольного ацетил-СоА посредством неприродного биохимического пути (публикация международной заявки на патент № WO 2013/036764).
В этом отношении создана возможность обеспечения ацетил-СоА, незаменимого для роста штамма, в котором активность PDC ослаблена или удалена, путем введения экзогенной ACL, в результате чего этот микроорганизм становится способным расти независимо от активности PDC. Последовательности белка и гена могут быть получены из известной базы данных, например GenBank NCBI, но не ограничиваясь ею. В частности, АТР-цитратлиаза может иметь аминокислотную последовательность SEQ ID NO: 29, но может быть включена без ограничений любая белковая последовательность, обладающая этой ферментативной активностью. Кроме того, ген, кодирующий ACL, может быть конкретно представлен нуклеотидной последовательностью SEQ ID NO: 30, но может быть включена без ограничений любая последовательность, кодирующая этот фермент.
Используемый в настоящем документе термин «путь биосинтеза пирувата», относящийся к пути биосинтеза, который может обеспечивать пируват у микроорганизма рода Saccharomyces, может представлять собой путь восполнения от ОАА до пирувата. Конкретные примеры представлены на ФИГ. 1 и 2.
В иллюстративном воплощении изобретения путь биосинтеза пирувата может быть осуществлен посредством модификации активностей либо фосфоенолпируваткарбоксикиназы 1 (PCK1), либо пируваткиназы 2 (PYK2), либо обоих ферментов для усиления их активностей по сравнению с их эндогенными активностями.
Альтернативно путь биосинтеза пирувата может быть осуществлен посредством модификации активностей либо малатдегидрогеназы 2 (MDH2), либо цитозольного яблочного фермента 1 (цитозольного МАЕ1), либо обоих ферментов для усиления их активностей по сравнению с их эндогенными активностями.
Используемый в настоящем документе термин «фосфоенолпируваткарбоксикиназа 1 (PCK1)» относится к ферменту, катализирующему превращение ОАА в фосфоенолпируват (PEP). PCK1 представляет собой фермент, необходимый для глюконеогенеза для превращения ОАА в PEP в дрожжевом микроорганизме, и известно, что в присутствии глюкозы происходит ингибирование его экспрессии (Differential post-transcriptional regulation of yeast mRNAs in response to high and low glucose concentrations. Mol Microbiol 35 (3): 553-65 (2000)).
Используемый в настоящем документе термин «пируваткиназа 2 (PYK2)» относится к ферменту, катализирующему продуцирование пирувата и АТР посредством доставки фосфатной группы от PEP к ADP. PYK2 представляет собой фермент заключительной стадии гликолиза у дрожжевого микроорганизма, и также известно, что в присутствии глюкозы происходит его ингибирование (Characterization of a glucose-repressed pyruvate kinase (Pyk2p) in Saccharomyces cerevisiae that is catalytically insensitive to fructose-1,6-bisphosphate, J Bacteriol. 1997 May; 179 (9): 2987-93).
Каждая из последовательностей белка и гена может быть получена из известной базы данных, например GenBank NCBI и т.д., но не ограничиваясь ею. PCK1 может иметь аминокислотную последовательность SEQ ID NO: 31, но может быть включена без ограничений любая белковая последовательность, обладающая этой ферментативной активностью. Кроме того, ген, кодирующий PCK1, может быть конкретно представлен нуклеотидной последовательностью SEQ ID NO: 32, но может быть включена без ограничений любая последовательность, кодирующая этот фермент. PYK2 может иметь аминокислотную последовательность SEQ ID NO: 33, но может быть включена без ограничений любая белковая последовательность, обладающая этой ферментативной активностью. Кроме того, в качестве конкретного примера, ген, кодирующий PYK2, может быть представлен нуклеотидной последовательностью SEQ ID NO: 34, но может быть включена без ограничений любая последовательность, которая может кодировать этот фермент.
Используемый в настоящем документе термин «малатдегидрогеназа 2 (MDH2)» относится к обратимому ферменту, превращающему ОАА в малат. MDH2 представляет собой фермент, первично локализованный в цитозоле.
Используемый в настоящем документе термин «цитозольный яблочный фермент 1 (МАЕ1)» относится к ферменту, модифицированному для локализации в цитозоле путем удаления из МАЕ1 последовательности митохондриального нацеливания, который представляет собой фермент замещения малата пируватом. Фермент МАЕ1 представляет собой белок, исходно локализованный в митохондриях, и он преобразует малат, являющийся промежуточным веществом в цикле трикарбоновых кислот (ТСА), в пируват в митохондриях (Metabolic Engineering, 6 (2004), 352-363). Каждая из последовательностей белка и гена может быть получена из известной базы данных, например GenBank NCBI и т.д., но не ограничиваясь ею. MDH2 может иметь аминокислотную последовательность SEQ ID NO: 35, но может быть включена без ограничений любая белковая последовательность, обладающая этой ферментативной активностью. Кроме того, в качестве конкретного примера, ген, кодирующий MDH2, может быть представлен нуклеотидной последовательностью SEQ ID NO: 36, но может быть включена без ограничений любая последовательность, которая может кодировать этот фермент. МАЕ1 может иметь аминокислотную последовательность SEQ ID NO: 37, но может быть включена без ограничений любая белковая последовательность, обладающая этой ферментативной активностью. Кроме того, для того, чтобы МАЕ1 присутствовал в цитозоле, МАЕ1 может иметь аминокислотную последовательность, соответствующую последовательности SEQ ID NO: 37, в которой аминокислотные остатки в положениях от начала до положения 30 удалены (т.е. аминокислотную последовательность, из которой удалена последовательность SEQ ID NO: 51, представляющая собой последовательность митохондриального нацеливания), и последовательность представлена SEQ ID NO: 52. Кроме того, в конкретном примере ген, кодирующий МАЕ1, может быть представлен нуклеотидной последовательностью SEQ ID NO: 38, но может быть включена без ограничений любая последовательность, которая может кодировать этот фермент.
Используемый в настоящем документе термин «лактатдегидрогеназа (LDH)» относится к ферменту, который может катализировать преобразование лактата в пируват и обратно, и последовательности гена и белка могут быть получены из известной базы данных, например GenBank NCBI и т.д., но не ограничиваясь ею. LDH может иметь аминокислотную последовательность SEQ ID NO: 49, но может быть включена без ограничений любая белковая последовательность, обладающая этой ферментативной активностью. Кроме того, ген, кодирующий LDH, может быть представлен нуклеотидной последовательностью SEQ ID NO: 50, но может быть включена без ограничений любая последовательность, которая может кодировать этот фермент.
Каждый из описанных выше ферментов может без ограничений включать помимо аминокислотных последовательностей, представленных SEQ ID NO, любую аминокислотную последовательность, имеющую гомологию 70% или выше, в частности, 80% или выше, более конкретно 90% или выше, еще более конкретно 95% или выше, еще более конкретно 98% или выше и еще более конкретно 99% или выше, с каждой из перечисленных выше аминокислотных последовательностей, до тех пор, пока фермент проявляет практически такое же или соответствующее действие каждого из этих ферментов. Кроме того, очевидно, что в объем настоящей заявки может входить любой модифицированный фермент, который имеет описанную выше гомологию и действие, соответствующее каждому ферменту, хотя такой фермент может иметь аминокислотную последовательность с частичной делецией, модификацией, заменой или добавлением.
Кроме того, гены, кодирующие каждый из описанных выше ферментов, могут также включать без ограничений в дополнение к нуклеотидным последовательностям, представленным SEQ ID NO, любую последовательность гена, кодирующего ферменты, которая имеет гомологию 70% или выше, в частности, 80% или выше, более конкретно 90% или выше, еще более конкретно 95% или выше, еще более конкретно 98% или выше и еще более конкретно 99% или выше, с каждой из перечисленных выше нуклеотидных последовательностей до тех пока эта последовательность кодирует фермент, обладающий по существу таким же или соответствующим действием каждого из этих ферментов. Кроме того, очевидно, что в объем настоящей заявки может входить любая нуклеотидная последовательность, которая имеет описанную выше гомологию, хотя эта последовательность может иметь частичную делецию, модификацию, замену или добавление.
Используемый в настоящем документе термин «гомология» относится к идентичности, выраженной в процентах, между двумя полинуклеотидными или полипептидными группировками. Соответствие последовательности от одной группировки до другой можно определить методом, известным в данной области техники. Например, гомологию можно определить путем прямого выравнивания информации о последовательности (например, параметров, таких как балл, идентичность и подобие) на двух полинуклеотидных молекулах или двух полипептидных молекулах с использованием компьютерной программы (например, BLAST 2.0), которая легкодоступна и способна осуществлять выравнивание информации последовательности. Кроме того, гомологию можно определить путем гибридизации полинуклеотидов в условиях образования стабильной двойной нити в гомологичных областях и расщепления гибридизованной нити нуклеазой однонитевой специфичности для определения размера расщепленных фрагментов.
Используемый в настоящем документе термин «эндогенная активность» относится к условиям, где микроорганизм характеризуется естественным состоянием ферментов или уровнем активации ферментов до модификации соответствующих ферментов.
Используемый в настоящем документе термин «активность фермента модифицирована для инактивации по сравнению с эндогенной активностью» относится к состоянию, при котором ген совсем не экспрессируется по сравнению с геном нативного штамма или штамма до модификации, или даже ген экспрессируется, но активность отсутствует или снижена.
Указанное выше снижение активности представляет собой положение, которое включает случай снижения активности самого фермента вследствие модификации гена, кодирующего этот фермент, и т.д. по сравнению с эндогенной активностью, которой исходно обладал микроорганизм, случай, где общий уровень активности фермента в клетке ниже по сравнению с уровнем штамма дикого типа или штамма до модификации, а также комбинацию этих случаев.
Инактивация фермента может быть достигнута различными способами, известными в данной области техники. Примеры этих способов могут включать способ замены гена, кодирующего фермент, на хромосоме геном, мутированным для снижения ферментативной активности, в том числе случай, где активность фермента удалена; способ введения модификации в регуляторную последовательность экспрессии гена, кодирующего фермент, на хромосоме; способ замены регуляторной последовательности экспрессии гена, кодирующего фермент, последовательностью, обладающей слабой активностью или не обладающей активностью; способ делеции всего гена, кодирующего фермент, или его участка на хромосоме; способ введения антисмыслового олигонуклеотида (например антисмысловой РНК), который комплементарно связывается с транскриптом гена на хромосоме, ингибируя таким образом трансляцию с мРНК в фермент; способ искусственного включения последовательности, комплементарной последовательности Шайна-Дальгарно (SD), выше последовательности Шайна-Дальгарно гена, кодирующего фермент, образующей вторичную структуру, в результате чего присоединение рибосомы к этой последовательности становится невозможным; способ вставки промотора в 3' конец открытой рамки считывания (ORF) для индукции обратной транскрипции (обратно-транскрипционное конструирование (reverse transcription engineering, RTE) и т.д., а также их комбинацию, но не ограничиваясь ими.
В частности, способ делеции всего гена, кодирующего фермент, или его участка может быть выполнен путем замены полинуклеотида, кодирующего эндогенный целевой белок, на хромосоме полинуклеотидом или геном-маркером, имеющим частичную делецию в нуклеиново-кислотной последовательности, с использованием вектора для встраивания в хромосому в пределах данного штамма. В иллюстративном воплощении способа делеции всего гена или его участка можно использовать способ делеции гена посредством гомологичной рекомбинации.
Используемый в настоящем документе термин «участок» может изменяться в зависимости от видов полинуклеотидов и может конкретно относиться к участку длиной от 1 до 300, более конкретно от 1 до 100 и еще более конкретно от 1 до 50, но без особых ограничений.
Используемый в настоящем документе термин «гомологичная рекомбинация» относится к генетической рекомбинации, идущей посредством кроссинговера в генетических локусах, обладающих взаимной гомологией.
В частности, регуляторная последовательность экспрессии может быть модифицирована путем индукции модификации регуляторной последовательности экспрессии в результате делеции, вставки, неконсервативной или консервативной замены или их комбинации в нуклеиново-кислотной последовательности регуляторной последовательности экспрессии; либо в результате замены промотора более слабым промотором и т.д. Регуляторная последовательность экспрессии может включать промотор, последовательность оператора, последовательность, кодирующую участок связывания рибосомы, и последовательности, контролирующие терминацию транскрипции и трансляции.
Кроме того, последовательность гена на хромосоме может быть модифицирована путем индукции модификации последовательности в результате делеции, вставки, неконсервативной или консервативной замены или их комбинации в последовательности гена для снижения ферментативной активности; или посредством замены последовательностью гена, которая была усовершенствована для ослабления активности, или последовательностью гена, которая была усовершенствована для отсутствия активности.
Используемый в настоящем документе термин «усиление активности по сравнению с эндогенной активностью» относится к повышению внутриклеточной активности белка (или фермента) у микроорганизма в результате модификации этого белка для улучшения внутриклеточной активности по сравнению с активностью белка, которой он обладает в естественном состоянии. «Усиление» может включать в себя проектирование повышенного действия по сравнению с исходной функцией за счет повышения активности самого белка (или фермента) и может быть выполнено по меньшей мере одним способом, выбранным из группы, состоящей из способа увеличения числа копий полинуклеотида, кодирующего белок (или фермент), способа введения модификации в регуляторную последовательность гена, кодирующего белок (или фермент), способа замены регуляторной последовательности гена, кодирующего белок (или фермент), на хромосоме последовательностью, обладающей более сильной активностью, способа замены гена, кодирующего белок (или фермент), геном, мутированным для повышения активности белка (или фермента), и способа введения модификации в ген, кодирующий белок (или фермент), на хромосоме для усиления активности белка (или фермента), но может включать в себя без ограничений любой известный способ, с использованием которого можно усилить активность белка (или фермента) по сравнению с его эндогенной активностью или усилить введенную активность.
Используемый в настоящем документе термин «введение активности белка (или фермента)» относится к обеспечению активности конкретного белка (или фермента) в микроорганизме, не обладающем активностью этого конкретного белка (или фермента); или к повышению внутриклеточной активности конкретного белка (или фермента) в микроорганизме, не обладающем активностью конкретного белка (или фермента), посредством модификации этого микроорганизма для дополнительного улучшения внутриклеточной активности белка (или фермента) после обеспечения активности этого конкретного белка (или фермента) в микроорганизме.
«Введение активности белка (или фермента)» может быть выполнено различными способами, известными в данной области техники, например: способом встраивания полинуклеотида, включающего нуклеотидную последовательность, кодирующую белок (или фермент), в хромосому; способом увеличения числа копий полинуклеотида таким способом, как введение полинуклеотида в микроорганизм посредством его встраивания в векторную систему; способом введения промотора, способного к проявлению улучшенной активности, или введения белка (или фермента) с модификацией в промоторе в участок, находящийся выше нуклеотидной последовательности, кодирующей белок (или фермент); способом введения варианта нуклеотидной последовательности, кодирующей белок (или фермент); и т.д., но может включать в себя без ограничений любой известный способ, с использованием которого можно ввести активность белка (или фермента).
В описанном выше способе увеличение числа копий полинуклеотида можно осуществить в такой форме, чтобы полинуклеотид был функционально связан с вектором, или путем вставки полинуклеотида в хромосому клетки-хозяина, хотя способ конкретно не ограничен этим. В частности, увеличение числа копий полинуклеотида можно осуществить путем введения в клетку-хозяина вектора, который способен к репликации и функционированию независимо от клетки-хозяина, и полинуклеотид, кодирующий белок по настоящей заявке, функционально связан с ним; либо можно осуществить путем введения в клетку-хозяина вектора, который способен к встраиванию полинуклеотида в хромосому клетки-хозяина, и полинуклеотид функционально связан с ним.
Вектор представляет собой конструкцию ДНК, включающую в себя последовательность полинуклеотида, кодирующего целевой белок, которая функционально связана с соответствующей регуляторной последовательностью для обеспечения экспрессии целевого пептида в клетке-хозяине. Регуляторная последовательность включает в себя промотор, способный к инициации транскрипции, любую последовательность оператора для регуляции транскрипции, последовательность, кодирующую соответствующий домен связывания рибосомы мРНК, и последовательность, регулирующую терминацию транскрипции и трансляции. После того, как вектор трансформирован в соответствующую клетку-хозяина, он может реплицироваться или функционировать независимо от генома клетки-хозяина или может быть интегрирован в сам геном клетки-хозяина.
Для экспрессионного вектора дрожжей можно использовать как интегративную дрожжевую плазмиду (YIp), так и экстрахромосомный плазмидный вектор. Экстрахромосомный плазмидный вектор может включать в себя эписомную дрожжевую плазмиду (YEp), репликативную дрожжевую плазмиду (YRp) и центромерную дрожжевую плазмиду (YCp). Кроме того, в качестве экспрессионных векторов в соответствии с настоящей заявкой можно также использовать искусственные хромосомы дрожжей (artificial yeast chromosomes, YAC). Например, векторы для использования в настоящей заявке могут включать pESC-HIS, pESC-LEU, pESC-TRP, pESC-URA, Gateway pYES-DEST52, pAO815, pGAPZ A, pGAPZ B, pGAPZ C, pGAPα A, pGAPα B, pGAPα C, pPIC3.5K, pPIC6 A, pPIC6 В, pPIC6 С, pPIC6α А, pPIC6α В, pPIC6α C, pPIC9K, pYC2/CT, вектор дрожжевого дисплея pYD1, pYES2, pYES2/CT, pYES2/NT A, pYES2/NT B, pYES2/NT С, pYES2/CT, pYES2.1, pYES-DEST52, pTEF1/Zeo, pFLD1, PichiaPink™, p427-TEF, p417-CYC, pGAL-MF, p427-TEF, p417-CYC, PTEF-MF, pBY011, pSGP47, pSGP46, pSGP36, pSGP40, ZM552, pAG303GAL-ccdB, pAG414GAL-ccdB, pAS404, pBridge, pGAD-GH, pGAD T7, pGBK T7, pHIS-2, pOBD2, pRS408, pRS410, pRS418, pRS420, pRS428, микронную ДНК дрожжей формы A, pRS403, pRS404, pRS405, pRS406, pYJ403, pYJ404, pYJ405 и pYJ406, но не ограничены ими.
Более конкретно дрожжевой вектор может представлять собой дрожжевую репликативную плазмиду, включающую в себя ориджин репликации (ori) и кассету устойчивости к антибиотикам, которую можно размножать и отбирать в Е. coli. Как правило, экспрессионные векторы могут включать в себя экспрессионную конструкцию промотор-ген-последовательность терминации транскрипции.
Например, когда клетки-хозяева представляют собой дрожжи, промоторы, которые можно использовать в экспрессионной конструкции, могут включать промотор TEF1, промотор TEF2, промотор GAL10, промотор GAL1, промотор ADH1, промотор ADH2, промотор PHO5, промотор GAL1-10, промотор TDH3 (промотор GPD), промотор TDH2, промотор TDH1, промотор PGK1, промотор PYK2, промотор ENO1, промотор ENO2 и промотор TPI1, но без ограничений ими.
Последовательности терминации транскрипции, которые можно использовать в экспрессионной конструкции, могут включать терминатор ADH1, терминатор CYC1, терминатор GAL10, терминатор PGK1, терминатор PHO5, терминатор ENO1, терминатор ENO2 и терминатор TPI1, но без ограничений ими.
Кроме того, полинуклеотид, кодирующий эндогенный целевой белок, может быть заменен модифицированным полинуклеотидом внутри хромосомы посредством вектора для вставки в хромосому внутри клетки-хозяина. Альтернативно полинуклеотид, кодирующий чужеродный целевой белок, для введения в хромосому может быть заменен модифицированным полинуклеотидом. Вставку полинуклеотида в хромосому можно осуществить любым способом, известным в данной области техники, например, посредством гомологичной рекомбинации. Поскольку вектор по настоящей заявке может быть встроен в хромосому посредством гомологичной рекомбинации, дополнительно может быть включен селективный маркер для подтверждения вставки в хромосому. Селективный маркер используют для селекции трансформированной клетки, т.е. для подтверждения вставки целевого полинуклеотида, и можно использовать маркеры, способные обеспечивать селективные фенотипы, такие как устойчивость к лекарственным средствам, потребность в питательных веществах, устойчивость к цитотоксическим агентам и экспрессия поверхностных белков. В условиях обработки селективными агентами только клетки, способные к экспрессии селективных маркеров, могут выжить или экспрессировать другие фенотипические признаки и, таким образом, можно отбирать трансформированные клетки.
Используемый в настоящем документе термин «трансформация» относится к процессу введения в клетку-хозяина вектора, включающего в себя полинуклеотид, кодирующий целевой белок и, следовательно, обеспечивающего экспрессию полинуклеотида, кодирующего белок, в клетке-хозяине. Для трансформированного полинуклеотида не имеет значения, встроен ли он в хромосому клетки-хозяина и локализован в ней или локализован вне хромосомы, если он может экспрессироваться в клетке-хозяине. Кроме того, полинуклеотид включает ДНК и РНК, которые кодируют целевой белок. Полинуклеотид может быть встроен в любой форме, если он может быть введен в клетку-хозяина и экспрессироваться в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, представляющей собой генную конструкцию, включающую в себя все существенные элементы, требующиеся для самостоятельной экспрессии. Экспрессионная кассета традиционно может включать в себя промотор, функционально связанный с полинуклеотидом, сигнал терминации транскрипции, домен связывания рибосомы и сигнал терминации трансляции. Экспрессионная кассета может находиться в форме экспрессионного вектора, способного к самостоятельной репликации. Кроме того, полинуклеотид можно вводить в клетку-хозяина самого по себе и функционально связанным с последовательностью, существенной для его экспрессии в клетке-хозяине.
Кроме того, используемый в настоящем документе термин «функционально связанный» относится к функциональному соединению между последовательностью промотора, которая инициирует и опосредует транскрипцию полинуклеотида, кодирующего целевой белок по настоящей заявке, и описанной выше последовательностью целевого гена.
Способ трансформации вектором по настоящей заявке может включать любой способ, позволяющий вводить нуклеиновые кислоты в клетки, и трансформацию можно осуществлять путем подбора соответствующей методики, известной в данной области техники, в зависимости от клетки-хозяина. Например, способ может включать электропорацию, кальций-фосфатную (CaPO4) преципитацию, кальций-хлоридную (CaCl2) преципитацию, микроинъекцию, способ на основе полиэтиленгликоля (PEG), способ на основе DEAE (диэтиламиноэтил)-декстрана, катионно-липосомного кальция и способ на основе ацетата лития/DMSO (диметилсульфоксид) и т.д., но без ограничений ими.
В частности, используемая клетка-хозяин должна обладать высокой эффективностью введения ДНК и высокой экспрессионной эффективностью введенной ДНК, и для целей настоящей заявки клетка-хозяин может представлять собой микроорганизм рода Saccharomyces.
Затем введение модификации в регуляторную последовательность экспрессии для повышения экспрессии полинуклеотида, хотя оно конкретно этим не ограничено, можно осуществлять путем индукции модификации в нуклеиново-кислотной последовательности в результате делеции, вставки, консервативной замены или неконсервативной замены или их комбинации с целью дополнительного усиления активности регуляторной последовательности экспрессии; или путем замены полинуклеотидной последовательности нуклеиново-кислотной последовательностью, обладающей усиленной активностью. Регуляторная последовательность экспрессии, хотя конкретно этим не ограничена, может включать промотор, последовательность оператора, последовательность, кодирующую домен связывания рибосомы и последовательность регуляции терминации транскрипции и трансляции и т.д.
С участком, расположенным выше экспрессионной единицы полинуклеотида, вместо исходного промотора может быть связан сильный экзогенный промотор.
Как правило, введение или усиление активности белка может приводить к повышению активности или концентрации соответствующего белка относительно активности или концентрации белка дикого типа или белка в штамме микроорганизма от по меньшей мере 10%, 25%, 50%, 75%, 100%, 150%, 200%, 300%, 400% или 500% до максимальных значений 1000% или 2000%.
В другом аспекте в настоящей заявке предложен способ получения молочной кислоты, включающий а) культивирование в питательной среде нового микроорганизма рода Saccharomyces, продуцирующего молочную кислоту, модифицированного для инактивации активности пируватдекарбоксилазы (PDC) по сравнению с ее эндогенной активностью, для введения активности АТР-цитратлиазы (ACL) и для усиления пути биосинтеза пирувата по сравнению с эндогенным путем его биосинтеза; и б) выделение молочной кислоты из культивируемого микроорганизма и культуры.
Микроорганизм рода Saccharomyces, продуцирующий молочную кислоту, является таким, как описано выше.
Используемый в настоящем документе термин «культивирование» относится к выращиванию микроорганизма в соответствующей искусственно подобранной окружающей среде. В настоящей заявке культивирование с использованием микроорганизма рода Saccharomyces можно выполнять любым соответствующим способом, хорошо известным в данной области техники. В частности, культивирование можно выполнять непрерывно в периодическом процессе, периодическом процессе с подпиткой или в многократном периодическом процессе с подпиткой, но без ограничений ими.
Среды, используемые для культивирования микроорганизма по настоящей заявке, и другие условия культивирования конкретно не ограничены, но можно использовать любую среду, применяемую для традиционного культивирования микроорганизма рода Saccharomyces. В частности, микроорганизм по настоящей заявке можно культивировать в традиционной среде, содержащей приемлемые источники углерода, источники азота, источники фосфора, неорганические соединения, аминокислоты и/или витамины и т.д., в аэробных условиях при регулировании температуры, рН и т.д.
В качестве примера источников углерода можно использовать сахарозу или глюкозу, а можно использовать мелассы, содержащие большое количество сахарозы, и необходимое количество различных других видов источников углерода.
Примеры источников азота могут включать органические источники азота, такие как пептон, дрожжевой экстракт, мясной экстракт, солодовый экстракт, кукурузный сироп и соевая мука; и неорганические источники азота, такие как мочевина, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Эти источники азота можно использовать отдельно или в комбинации. В описанной выше среде в качестве источников фосфора могут содержаться дигидрофосфат калия, двухзамещенный фосфат калия и соответствующие натрийсодержащие соли. Дополнительно могут содержаться соли металлов, такие как сульфат магния или сульфат железа. Кроме того, могут содержаться аминокислоты, витамины и соответствующие предшественники. Эти среды или предшественники можно добавлять в культуру в процессе периодического культивирования или в процессе непрерывного культивирования.
В период культивирования рН культуры можно регулировать добавлением в культуру соединения, такого как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота и серная кислота, соответствующим образом. Кроме того, в период культивирования для предотвращения образования пены можно добавлять пеногаситель, такой как сложный эфир жирной кислоты и полигликоля. Дополнительно для поддержания аэробного состояния культуры в культуру можно вводить кислород или кислородсодержащий газ, а для поддержания анаэробного и микроаэрофильного состояния культуры можно вводить газ азот, водород или диоксид углерода без введения воздуха.
Температура культуры обычно может составлять от 20°С до 40°С, конкретно от 25°С до 35°С и более конкретно 30°С, но может изменяться без ограничений в зависимости от желаемых целей. Дополнительно культивирование можно продолжать до того момента, когда может быть получено желаемое количество продукта, и, в частности, от 10 часов до 100 часов, но без ограничений этим.
Способ получения молочной кислоты по настоящей заявке может включать выделение молочной кислоты из культивируемого микроорганизма или культуры. Способ выделения молочной кислоты из культивируемого микроорганизма или культуры можно выполнять соответствующим способом, известным в данной области техники, например посредством центрифугирования, фильтрования, анионообменной хроматографии, кристаллизации, высокоэффективной жидкостной хроматографии (HPLC) и т.д., но не ограничиваясь ими.
Выделение может включать в себя процесс очистки.
Способы осуществления изобретения
Далее настоящая заявка подробно описана сопроводительными иллюстративными воплощениями. Тем не менее раскрытые в настоящем документе иллюстративные воплощения предназначены только для иллюстративных целей и их не следует истолковывать как ограничивающие объем настоящей заявки.
Пример 1. Получение штамма, продуцирующего молочную кислоту
Для получения репрезентативного продуцирующего молочную кислоту штамма для использования в настоящей заявке, Saccharomyces cerevisiae CEN. PK2-1D, который является представителем штаммов дрожжей среди штаммов дрожжей дикого типа, полученных от компании Euroscarf, подвергали серии генетических манипуляций.
В частности, делетировали алкогольдегидрогеназу 1 (ADH1) и пируватдекарбоксилазу 1 (PDC1) для сведения к минимуму ферментации с образованием спирта, при этом для блокирования пути разложения молочной кислоты в качестве базового штамма по настоящей заявке использовали штамм с делецией D-лактатдегидрогеназы 1 (DLD1).
DLD1 не является фактором, непосредственно влияющим на улучшение роста, но DLD1, представляющий собой дегидрогеназу молочной кислоты D-типа, известен как основной фермент, который превращает молочную кислоту в пируват, используя NAD+. Соответственно, последующие штаммы получали на основе штамма с делецией гена DLD1, который представляет собой фермент, расходующий молочную кислоту, и сравнивали продуктивность молочной кислоты.
В настоящей заявке генетические манипуляции были выполнены с использованием общих методов молекулярного клонирования. Сначала проводили эксперименты по делетированию генов ADH1 и PDC1 ферментов с помощью плазмид pWAL100 и pWBR100, основываясь на описании, приведенном в публикации (Lee ТН, et al., J. Microbiol. Biotechnol. (2006), 16 (6), 979-982). Каждая вставка, встроенная в каждую плазмиду, была получена путем полимеразной цепной реакции (PCR) с использованием праймеров, соответствующих каждой вставке (SEQ ID NO: 1-SEQ ID NO: 8).
PCR проводили, используя геномную ДНК штаммов дрожжей дикого типа в качестве матрицы. Для делеции ADH1 PCR проводили с использованием праймеров SEQ ID NO: 1 и SEQ ID NO: 2, и полученный в результате продукт клонировали в pWAL100 с использованием ферментов рестрикции BamHI и NcoI. Следующую PCR проводили с использованием праймеров SEQ ID NO: 3 и SEQ ID NO: 4, и полученный в результате продукт клонировали в pWBR100 с использованием ферментов рестрикции BamHI и NcoI. PCR проводили путем денатурации при 95°С в течение 5 мин, отжига при 53°С в течение 1 мин и полимеризации при 72°С в течение 1 мин 30 с.
Для получения делеции PDC1 PCR проводили с использованием праймеров SEQ ID NO: 5 и SEQ ID NO: 6, и полученный в результате продукт клонировали в pWAL100 с использованием ферментов рестрикции BamHI и NcoI. Следующую PCR проводили с использованием праймеров SEQ ID NO: 7 и SEQ ID NO: 8, и полученный в результате продукт клонировали в pWBR100 с использованием ферментов рестрикции BamHI и NcoI. PCR проводили путем денатурации при 95°С в течение 5 мин, отжига при 53°С в течение 1 мин и полимеризации при 72°С в течение 1 мин 30 с.
Дополнительно для делеции гена DLD1 маркерный ген HIS3 был делетирован в результате введения посредством двойного кроссинговера. Используемые при этом фрагменты ДНК были получены путем проведения PCR с использованием геномной ДНК штамма дрожжей дикого типа вместе с праймерами SEQ ID NO: 9 и SEQ ID NO: 10. PCR проводили путем денатурации при 95°С в течение 5 мин, отжига при 53°С в течение 1 мин и полимеризации при 72°С в течение 1 мин 30 с.
Сводные данные праймеров, используемых при манипуляциях с генами, представлены в Таблице 1 ниже.
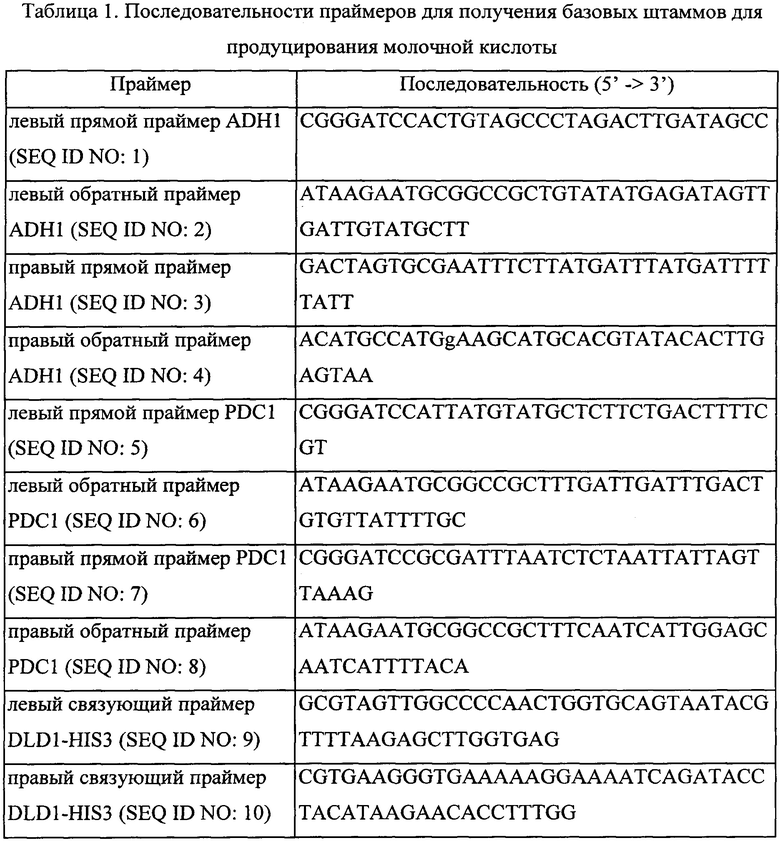
На основе штаммов, имеющих делеции трех генов (ADH1, PDC1, и DLD1), была введена D-лактатдегидрогеназа (D-LDH) для продуцирования молочной кислоты. 5'-Конец и 3'-конец гена ldhD, имеющего происхождение из Lb. plantarum, были соответственно встроены в вектор p413TEF1 так, что ген ldhD мог быть включен в вектор между промотором TEF1 и терминатором CYC1, имеющими происхождение от S. cerevisiae, в котором вставка была получена путем двойного расщепления SaxI/PvuII. В векторе были созданы тупые концы с использованием нуклеазы золотистой фасоли на фрагментах ДНК, образовавшихся в результате двойного расщепления BamHI/NotI вектора р-δ-neo, и снова обрабатывали SacI, в результате чего был получен участок вектора, имеющий липкий конец SacI и тупой конец BamHI.
Полученные таким путем вектор и вставку лигировали с получением вектора pTL573, который был назван вектором pTL573. Плазмида pTL573 содержит ген ldhD, имеющий происхождение от Lb. plantarum и сконструированный так, что множественные копии могут быть случайным образом встроены в δ-последовательность, которая составляет часть участка ретротранспозона S. cerevisiae CEN штамма PK2-1D pdc1 adh1 dld1. Для множественной вставки соответствующего гена плазмиду pTL573 расщепляли SacI с получением фрагмента ДНК, который может индуцировать однократный кроссинговер в δ-последовательности. Полученный в результате вектор вводили в родительский штамм путем трансфекции, и в среде YPD (1% дрожжевого экстракта, 2% бакто-пептона и 2% глюкозы) при максимальной концентрации 5 мг/мл G418 были получены многочисленные колонии. Было подтверждено, что в полученный таким образом штамм встроены многочисленные копии D-LDH, имеющей происхождение от Lb. plantarum, что обеспечивает продуктивность молочной кислоты D, и этот штамм был обозначен СС02-0064.
Пример 2. Получение штаммов со сниженным титром или инактивацией PDC
Штамм со сниженным титром или с инактивацией PDC был получен путем получения штамма с делецией PDC5, т.е. изофермента PDC, в штамме СС02-0064, представляющем собой базовый штамм, полученный в примере 1, и штамма с делецией обоих генов PDC5 и PDC6 в штамме СС02-0064.
В частности, для делеции гена дрожжей использовали плазмиды pWAL100 и pWBR100 (J. Microbiol. Biotechnol., (2006) 16 (6), 979-982).
Для делеции PDC5 проводили PCR с использованием праймеров SEQ ID NO: 11 и SEQ ID NO: 12, и полученный в результате продукт клонировали в pWAL100 с использованием ферментов рестрикции BamHI и NotI. Следующую PCR проводили с использованием праймеров SEQ ID NO: 13 и SEQ ID NO: 14, и полученный в результате продукт клонировали в pWBR100 с использованием ферментов рестрикции SpeI и NcoI. PCR проводили путем денатурации при 95°С в течение 5 мин, отжига при 53°С в течение 1 мин и полимеризации при 72°С в течение 1 мин 30 с.
Для делеции PDC6 PCR проводили с использованием праймеров SEQ ID NO: 15 и SEQ ID NO: 16, и полученный в результате продукт клонировали в pWAL100 с использованием ферментов рестрикции BamHI и NotI. Следующую PCR проводили с использованием праймеров SEQ ID NO: 17 и SEQ ID NO: 18, и полученный в результате продукт клонировали в pWBR100 с использованием ферментов рестрикции SpeI и NcoI. PCR проводили путем денатурации при 95°С в течение 5 мин, отжига при 53°С в течение 1 мин и полимеризации при 72°С в течение 1 мин 30 с.

Полученные выше новые штаммы были названы СС02-0256 и СС02-0553 соответственно, и сводные данные генетических признаков этих новых штаммов приведены в таблице 3 ниже.

Пример 3. Оценка продуктивности молочной кислоты штаммов со сниженной или инактивированной активностью PDC
Среда, используемая для оценки штаммов, представляла собой синтетическую комплексную среду (SC). Для приготовления среды смесь с исключением аминокислот (Sigma) смешивали с 0,67% дрожжевой азотной основой без аминокислот, которую использовали в качестве основы, в соответствии с протоколом производителя, и не включенные в смесь аминокислоты добавляли по мере необходимости. Лейцин добавляли до концентрации 380 мг/л, а урацил, триптофан и гистидин добавляли до концентрации 76 мг/л, соответственно, и в смесь добавляли глюкозу (8%) в качестве источника углерода и 1% СаСО3 в качестве нейтрализующего агента. Приготовленную таким образом среду использовали для оценки ферментации молочной кислоты в штаммах дрожжей.
В качестве условий для оценки способности штаммов к ферментации молочной кислоты среду, приготовленную для оценки ферментации молочной кислоты, делили на аликвоты в количестве 25 мл на каждую колбу и засевали каждым из штаммов дрожжей, культивировали при 30°С в аэробных условиях в течение 48 часов и проводили количественный анализ содержания молочной кислоты в ферментационной жидкости методом высокоэффективной жидкостной хроматографии (HPLC).
Сводные результаты экспериментов представлены в Таблице 4 ниже.
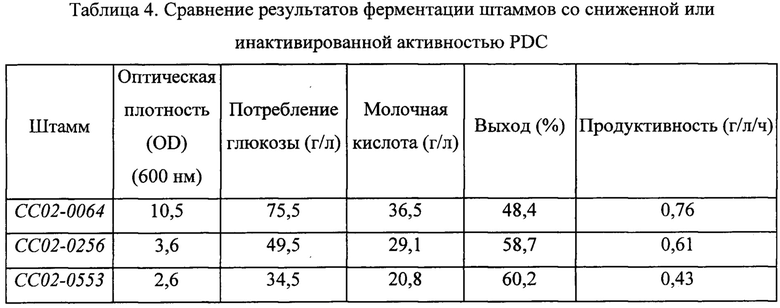
В результате, как можно видеть из основании данных, приведенных в Таблице 4 выше, по мере снижения активности PDC выход повышался, но продуктивность снижалась.
Пример 4. Введение АТР-цитратлиазы (ACL) на основе штаммов с инактивированной PDC и получение штаммов с усиленной активностью фосфоенолпируваткарбоксикиназы 1 (PCK1) и пируваткиназы 2 (PYK2)
(1) Получение вектора для введения чужеродной ACL в штамм, продуцирующий молочную кислоту
Был получен рекомбинантный вектор для введения чужеродного фермента ACL и одновременной сверхэкспрессии PCK1 и PYK2, одного из путей биосинтеза пирувата.
Для чужеродной ACL использовали ген, имеющий происхождение от млекопитающего Mus musculus, и соответствующий ген был подтвержден в базе данных Национального центра биотехнологической информации (NCBI) (номер доступа NP_001186225).
Конкретно, этот ген был синтезирован с использованием аминокислотной последовательности SEQ ID NO: 29 (или аминокислотной последовательности SEQ ID NO: 30), вектор был получен с использованием промотора GPD на основе pRS415, вектора для экспрессии генов для дрожжей, и вектор со вставкой гена был получен и назван p415GPDpro-ACL.
(2) Получение вектора, содержащего PCK1 и PYK2 с усиленной активностью, для усиления пути биосинтеза пирувата
Был получен рекомбинантный вектор для одновременной сверхэкспрессии PCK1 и PYK2 для усиления пути биосинтеза пирувата.
PYK2 представляет собой ген, присутствующий в дрожжевом микроорганизме, и он может быть представлен SEQ ID NO: 33. PCR проводили с использованием геномной ДНК S. cerevisiae в качестве матрицы вместе с праймерами SEQ ID NO: 19 и 20, и из продуктов реакции получали фрагменты гена PYK2. PCR проводили путем денатурации при 95°С в течение 5 мин, отжига при 53°С в течение 1 мин и полимеризации при 72°С в течение 1 мин 30 с. Клонирование выполняли с использованием фрагментов гена и ферментов рестрикции в дрожжевом экспрессионном векторе, происходящем из pRS416, т.е. SpeI, XhoI, и сверхэкспрессию осуществляли с использованием промотора TEF1. Соответствующий рекомбинантный вектор был назван pRS416-TEF1pro-PYK2.
PYK1 также представляет собой ген, присутствующий в дрожжевом микроорганизме, и он может быть представлен SEQ ID NO: 31. PCR проводили с использованием геномной ДНК S. cerevisiae в качестве матрицы вместе с праймерами SEQ ID NO: 23 и 24, и из продуктов реакции получали фрагменты гена PYK1. PCR проводили путем денатурации при 95°С в течение 5 мин, отжига при 53°С в течение 1 мин и полимеризации при 72°С в течение 1 мин 30 с. PCR в таких же условиях проводили для получения фрагмента PCK1 с использованием геномной ДНК S. cerevisiae в качестве матрицы вместе с праймерами SEQ ID NO: 21 и 22, так что PYK1 может быть экспрессирован с использованием промотора TEF2, и был получен промотор TEF2. Затем для одновременной экспрессии PCK1 и PYK2 в одном рекомбинантном векторе, рекомбинантный вектор pRS416-TEF1pro-PYK2, полученный, как описано выше, расщепляли XhoI и одновременно клонировали фрагмент промотора TEF2 и фрагмент PCK1 с использованием набора реагентов для клонирования In-Fusion (Clontech). Наконец, был получен единый рекомбинантный вектор, который может сверхэкспрессировать PCK1 и PYK2 с промотором TEF2 и промотором TEF1, соответственно, и он был назван pRS416-TEF1pro-PYK2-TEF2pro-PCK1. Сводные данные праймеров, использованных при получении вектора для сверхэкспрессии PCK1 и PYK2, приведены в Таблице 5 ниже.


(3) Введение чужеродной ACL в продуцирующие молочную кислоту штаммы с инактивированной PDC и получение штаммов с усиленной активностью PCK1 и PYK2
Вводили чужеродную ACL, полученную в примере 4-(1) на основе штамма СС02-0553, полученного в примере 2, и вектор для одновременной сверхэкспрессии PCK1/PYK2, полученный в примере 4-(2), встраивали путем трансфекции.
Трансфекцию выполняли, используя способ, включающий обработку штамма СС02-0553, выращенного в среде YPD (1% дрожжевого экстракта, 2% бактопептона и 2% глюкозы) в течение 18 часов, раствором, содержащим 0,1 М ацетата лития, 0,01 М трис-HCl и 0,001 М EDTA (этилендиаминотетрауксусная кислота, далее в настоящем документе буфер LiAc/TE) и обрабатывали нагреванием вместе с буфером LiAc/TE, содержащим 40% PEG (полиэтиленгликоль), при 42°С в течение 15 минут для встраивания рекомбинантного вектора. Полученные таким образом штаммы были названы СС02-0652 и СС02-0553 соответственно, и сводные данные их генетических признаков приведены в Таблице 6 ниже.

Пример 5. Введение ACL на основе инактивированной PDC и оценка ферментации в штамме с усиленной активностью PCK1 и PYK2
Для оценки ACL-PCK1-PYK2-усиленных штаммов способность к ферментации молочной кислоты оценивали в штаммах с инактивированной активностью PDC, полученных в примере 4-(3), таким же путем, как описано в примере 3. Сводные результаты представлены в Таблице 7 ниже.
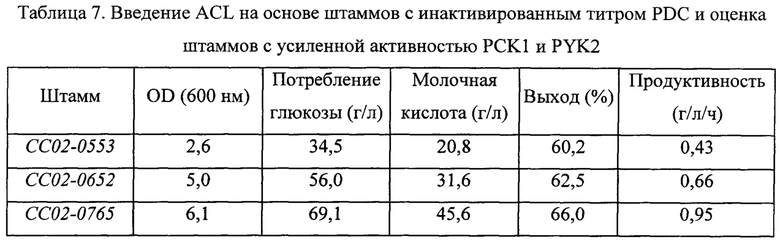
В результате, как можно видеть на основании Таблицы 7 выше, штаммы, в которые была введена чужеродная ACL и были усилены активности PCK1 и PYK2, проявляли повышение значений OD600, которые характеризуют бактериальный рост, относительно штамма с инактивированной PDC, на 130%; увеличение количества потребляемой глюкозы за тот же период на 100%; и улучшение выхода ферментации молочной кислоты на 10%. Дополнительно эти штаммы продемонстрировали конечное улучшение продуктивности молочной кислоты на 120%. На основании результата ферментации штамма СС02-0652 было подтверждено, что введение чужеродной ACL может усиливать рост дрожжевых микроорганизмов за счет продуцирования ацетил-СоА новым путем продуцирования. Дополнительно было подтверждено, что введение чужеродной ACL может привести не только к усилению роста, но также к повышению продуктивности.
Кроме того, на основании результата ферментации, полученного для штамма СС02-0765, подтвердили, что продуктивность ферментации молочной кислоты можно дополнительно повысить за счет усиления биосинтеза пирувата и, таким образом, было подтверждено, что способ получения молочной кислоты путем стратегии, предложенной в настоящей заявке, применяя продуцирование ацетил-СоА посредством нового пути и усиления биосинтеза пирувата, представляет собой способ, позволяющий не только повысить выход ферментации молочной кислоты и усилить рост данного микроорганизма, но также повысить продуктивность ферментации молочной кислоты, в отличие от существующей технологии.
Соответственно, штамм СС02-0765 был депонирован в Корейском центре культур микроорганизмов (Korean Culture Center of Microorganisms, KCCM) 28 ноября 2014 г. с номером доступа KCCM11616P в соответствии с Будапештским договором.
Пример 6. Введение ACL на основе PDC-инактивированных штаммов и получение штаммов с усиленной активностью малатдегидрогеназы 2 (MDH2) и цитозольного яблочного фермента 1 (МАЕ1)
На основании результата примера 5 было подтверждено, что стратегия продуцирования ацетил-СоА посредством нового биохимического пути и усиления биосинтеза пирувата представляет собой эффективный способ повышения выхода ферментации молочной кислоты, усиления роста микроорганизма, обладающего продуктивностью, и повышения продуктивности ферментации молочной кислоты и, таким образом, авторы настоящего изобретения предприняли попытку подтвердить, что усиление биосинтеза пирувата с использованием других генов может обладать подобными эффектами.
(1) Получение вектора с повышенной активностью MDH2 и цитозольного МАЕ1
Поскольку ОАА, продуцируемый в результате введения чужеродной ACL, может быть биосинтезирован в пируват посредством другого биохимического пути, авторы настоящего изобретения предприняли попытку осуществления сверхэкспрессии MDH2, изначально локализованного в цитозоле, и сверхэкспрессии МАЕ1, представляющего собой фермент, локализованный в митохондриях, путем изменения его локализации на цитозоль. С этой целью был получен рекомбинантный вектор.
MDH2 представляет собой ген, присутствующий в дрожжевых микроорганизмах, и он может быть представлен аминокислотной последовательностью SEQ ID NO: 35. Фрагмент гена MDH2 был получен с помощью PCR, которую проводили с использованием геномной ДНК S. cerevisiae в качестве матрицы вместе с праймерами SEQ ID NO: 25 и 26. PCR проводили путем денатурации при 95°С в течение 5 мин, отжига при 53°С в течение 1 мин и полимеризации при 72°С в течение 1 мин 30 с. Полученный таким образом фрагмент гена MDH2 клонировали на основе вектора pRS414 после его расщепления ферментами рестрикции SpeI и XhoI, в котором ген MDH2 был расположен так, чтобы происходила его сверхэкспрессия с использованием промотора TEF1. Полученный таким образом рекомбинантный вектор был назван pRS414-TEF1pro-MDH2.
МАЕ1 представляет собой ген, который изначально присутствует в митохондриях дрожжевых микроорганизмов и может быть представлен аминокислотной последовательностью SEQ ID NO: 37. Для экспрессии гена МАЕ1 в цитозоле ген МАЕ1 был клонирован с исключением последовательности нацеливания в митохондрии (представленной аминокислотной последовательностью SEQ ID NO: 51), которая состоит из последовательности длиной 90 нуклеотидов от стартового кодона гена МАЕ1. PCR проводили с использованием геномной ДНК S. cerevisiae в качестве матрицы вместе с праймерами SEQ ID NO: 27 и 28. PCR проводили путем денатурации при 95°С в течение 5 мин, отжига при 53°С в течение 1 мин и полимеризации при 72°С в течение 2 мин. Полученные таким образом фрагменты клонировали на основе вектора pRS416 после его расщепления ферментами рестрикции SpeI и XmaI. Праймер SEQ ID NO: 27 был получен аналогичным образом с получением нуклеотидных последовательностей, начинающихся с положения 91, с целью получения последовательности 90 нуклеотидов, начиная со стартового кодона ОРС МАЕ1. Дополнительно была предпринята попытка сверхэкспрессии цитозольного фермента МАЕ1 с использованием промотора TEF1. Дополнительно полученный таким образом рекомбинантный вектор был назван pRS416-TEF1pro-cytosolic МАЕ1. Сводные данные праймеров, использованных для получения рекомбинантных векторов, т.е. pRS414-TEF1pro-MDH2 и pRS416-TEF1pro-cytosolic МАЕ1, приведены в Таблице 8 ниже.

(2) Получение штаммов с усиленной активностью MDH2 и цитозольного MAE1 в продуцирующем молочную кислоту штамме с инактивированной PDC
Вектор с введенной чужеродной ACL, полученный в примере 4-(1), и вектор сверхэкспрессии MDH2 и сверхэкспрессии цитозольного МАЕ1, полученные в примере 6-(1), клонировали путем трансфекции на основе штамма СС02-0553, полученного в примере 2. Трансфекцию проводили, используя способ, раскрытый в примере 4-(3). Полученный таким образом штамм был назван СС02-0821, и сводные данные генетических признаков этого штамма приведены в Таблице 9 ниже.
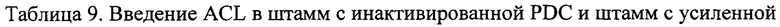

Пример 7. Введение ACL на основе штаммов с инактивированной PDC и оценка ферментации в штаммах с усиленной активностью MDH2 и цитозольного МАЕ1
Для оценки ACL-MDH2-цитозольный МАЕ1-усиленных штаммов оценивали способность к ферментации молочной кислоты в штаммах с инактивированной PDC, полученных в примере 6, таким же путем, как описано в примере 2. Сводные результаты представлены в Таблице 10 ниже.
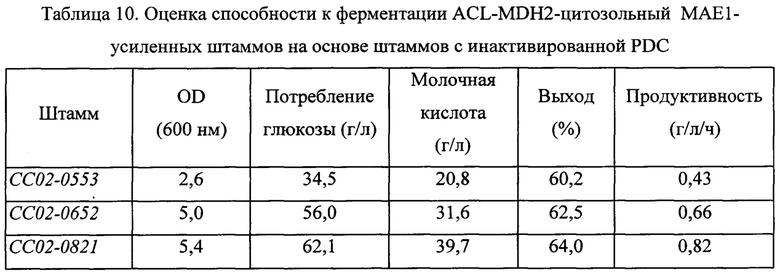
Как можно видеть на основании приведенной выше Таблицы 10, в результате оценки способности к ферментации для штамма СС02-0821, в котором биосинтез пирувата был усилен в результате сверхэкспрессии MDH2 и цитозольного МАЕ1, также было показано повышение значения OD600, которое характеризует бактериальный рост, относительно штамма с удаленной активностью PDC, на 110%; увеличение количества потребляемой глюкозы за тот же период на 80%; и улучшение выхода ферментации молочной кислоты на 6%, как и в случае штамма СС02-0765. Дополнительно для этих штаммов показано конечное улучшение продуктивности молочной кислоты на 90%. На основании описанных выше результатов было подтверждено, что не только введение чужеродной ACL для улучшения продуцирования молочной кислоты в дрожжевом микроорганизме, но также усиление путей биосинтеза пирувата посредством различных путей также эффективно для дополнительного улучшения продуктивности молочной кислоты.
Пример 8. Введение ACL в штаммах с пониженной активностью PDC и получение штаммов с повышенной активностью PCK1 и PYK2 и штаммов с повышенной активностью MDH2 и цитозольного МАЕ1
Как вывод из результатов, полученных в примерах, было подтверждено, что штаммы, в которых титр PDC был удален, могли повышать продуктивность ферментации молочной кислоты в результате введения чужеродной ACL и усиления пути биосинтеза пирувата. В связи с этим авторы настоящего изобретения предприняли попытку подтвердить, что такой же результат может быть получен для штаммов, в которых титр PDC снижен, а также для штаммов, в которых титр PDC удален.
Рекомбинантные векторы, т.е. pRS415-GPDpro-ACL и pRS416-TEF1pro-PYK2-TEF2p-PCK1, полученные в примере 4, были встроены на основе штамма СС02-0256, представляющего собой штамм с пониженной активностью PDC, полученный в примере 2, таким же путем, как в примере 4-(3). Полученные таким образом штаммы были названы СС02-0819 и СС02-0820 соответственно. Рекомбинантные векторы, т.е. pRS415-GPDpro-ACL, полученный в примере 4, и pRS414-TEF1pro-MDH2 и pRS416-TEF1pro-cytosolic МАЕ1, полученные в примере 6-(1), были встроены на основе штамма СС02-0256, представляющего собой штамм с пониженной активностью PDC, полученный в примере 2, таким же путем, как в примере 4-(3). Полученный таким образом штамм был назван СС02-0831, и сводные данные генетических признаков этого штамма приведены в Таблице 11 ниже.

Пример 9. Введение ACL в штаммы с пониженной активностью PDC и оценка ферментации в штаммах с усиленной активностью PCK1 и PYK2
Способность к ферментации молочной кислоты штаммов СС02-0819 и СС02-0820, полученных в примере 8, оценивали параллельно со штаммом СС02-0256, представляющим собой контрольную группу, таким же путем, как в примере 3. Сводные результаты экспериментов представлены в Таблице 12 ниже.
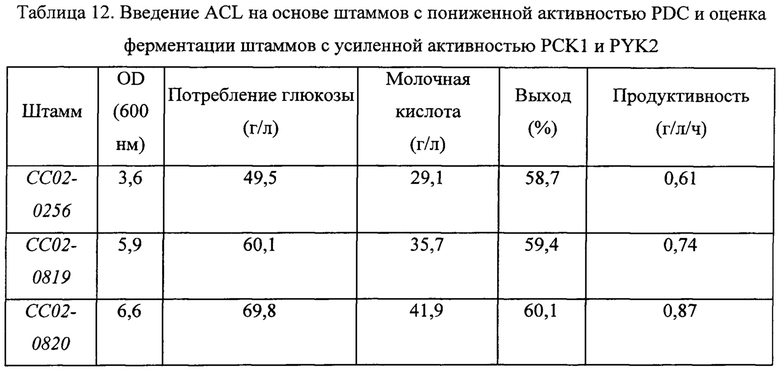
В результате, как можно видеть на основании приведенной выше Таблицы 12, штамм СС02-0256 не показал значительного увеличения эффектов в результате введения чужеродной ACL и усиления пути биосинтеза пирувата, в отличие от штамма СС02-0553, но наблюдалось улучшение выхода ферментации молочной кислоты, увеличение бактериального роста микроорганизмов, обладающих продуктивностью, а также повышение продуктивности ферментации молочной кислоты.
Штамм СС02-0820, в который была введена чужеродная ACL и в котором была усилена активность PCK1 и PYK2, демонстрировал повышение значения OD600, которое характеризует бактериальный рост, относительно штамма со сниженным титром PDC, на 80%; увеличение количества потребляемой глюкозы за тот же период на 40%; и улучшение выхода ферментации молочной кислоты на 2%. Дополнительно продуктивность молочной кислоты была, в конечном счете, повышена на 40%. Штамм СС02-0820 демонстрировал повышение всех значений: OD, количества потребляемой глюкозы за тот же период и выхода ферментации молочной кислоты - по сравнению со штаммом СС02-0819, в который была только введена чужеродная ACL, и показал повышение продуктивности на 20%.
Пример 10: Введение ACL в штаммы с пониженной активностью PDC и сравнительная оценка ферментации в штаммах с усиленной активностью MDH2 и цитозольного МАЕ1
Штамм СС02-0831, полученный в примере 8, оценивали параллельно со штаммами СС02-0256 и СС02-0819, представляющими собой контрольные группы, таким же путем, как в примере 3. Сводные результаты экспериментов представлены в Таблице 13 ниже.
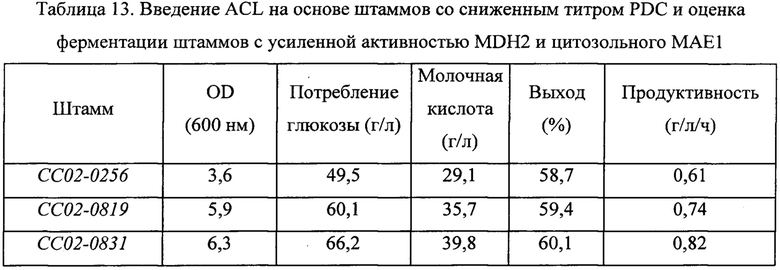
В результате, как можно видеть на основании Таблицы 13 выше, штамм СС02-0831, в который была введена чужеродная ACL и в котором была усилена активность MDH2 и цитозольного МАЕ1, демонстрировал повышение значения OD600 относительно значения для родительского штамма, СС02-0256, на 80%; увеличение количества потребляемой глюкозы за тот же период на 30%; и улучшение выхода ферментации молочной кислоты на 2%. Дополнительно продуктивность молочной кислоты была, в конечном счете, повышена на 30%. Штамм СС02-0831 демонстрировал повышение всех значений: OD, количества потребляемой глюкозы за тот же период и выхода ферментации молочной кислоты - по сравнению со штаммом СС02-0819, в который была только введена чужеродная ACL, и демонстрировал повышение продуктивности на 20%.
Описанные выше результаты подтверждают, что способ продуцирования молочной кислоты путем стратегии, в которой применяют продуцирование ацетил-СоА новым биохимическим путем и усиленный биосинтез пирувата, представляет собой способ, позволяющий, в отличие от существующей технологии, не только повысить выход ферментации молочной кислоты и усилить рост микроорганизма, обладающего продуктивностью, но также повысить продуктивность ферментации молочной кислоты. В частности, результаты подтверждают, что микроорганизм, который был получен в результате стратегии применения продуцирования ацетил-СоА новым биохимическим путем за счет введения чужеродной ACL и усиленного биосинтеза пирувата, позволяет не только повысить выход ферментации молочной кислоты и усилить рост микроорганизма, обладающего продуктивностью, но также повысить продуктивность ферментации молочной кислоты, в результате чего можно получить превосходный микроорганизм, продуцирующий молочную кислоту.
На основании описанного выше специалист в области техники, к которой относится настоящая заявка, сможет понять, что настоящая заявка может быть применена в других конкретных формах без модификации технических концепций или существенных характеристик настоящей заявки. В связи с этим раскрытые в настоящем документе иллюстративные воплощения предназначены только для иллюстративных целей, и их не следует истолковывать как ограничивающие объем настоящей заявки. Напротив, предусмотрено, что настоящая заявка включает в себя не только иллюстративные воплощения, но также различные альтернативы, модификации, эквиваленты и другие воплощения, которые могут быть включены в сущность и объем настоящей заявки, определенные в прилагаемой формуле изобретения.
Группа изобретений относится к микроорганизму рода Saccharomyces, обладающему способностью продуцировать молочную кислоту по сравнению с немодифицированным микроорганизмом, и к способу продуцирования молочной кислоты с его использованием. Микроорганизм рода Saccharomyces модифицирован так, что у него инактивирована активность пируватдекарбоксилазы (PDC) по сравнению с ее эндогенной активностью, введена активность АТР-цитратлиазы (ACL) и усилен путь биосинтеза пирувата по сравнению с эндогенным путем его биосинтеза. Способ получения молочной кислоты включает культивирование указанного микроорганизма рода Saccharomyces в среде и выделение молочной кислоты из культивируемого микроорганизма или из среды. Группа изобретений обеспечивает увеличение количества продуцируемой молочной кислоты. 2 н. и 10 з.п. ф-лы, 2 ил., 13 табл., 10 пр.
1. Микроорганизм рода Saccharomyces, обладающий повышенной продуктивностью по молочной кислоте по сравнению с немодифицированным микроорганизмом, модифицированный для обладания инактивированной активностью пируватдекарбоксилазы (PDC) по сравнению с ее эндогенной активностью, для введения активности АТP-цитратлиазы (ACL) и для усиления пути биосинтеза пирувата по сравнению с его эндогенным путем биосинтеза.
2. Микроорганизм по п. 1, где усиление пути биосинтеза пирувата достигнуто путем усиления активности фосфоенолпируваткарбоксикиназы 1 (PCK1), пируваткиназы 2 (PYK2) или активности обоих ферментов по сравнению с их эндогенными активностями.
3. Микроорганизм по п. 1, где усиление пути биосинтеза пирувата достигнуто путем усиления активности малатдегидрогеназы 2 (MDH2), цитозольного яблочного фермента 1 (цитозольного MAE1) или активности обоих ферментов по сравнению с их эндогенными активностями.
4. Микроорганизм по п. 1, где пируватдекарбоксилаза представляет собой фермент, представленный по меньшей мере одной аминокислотной последовательностью, выбранной из группы, состоящей из аминокислотных последовательностей SEQ ID NO:39, 41 и 43.
5. Микроорганизм по п. 1, где АТP-цитратлиаза представляет собой фермент, представленный аминокислотной последовательностью SEQ ID NO: 29.
6. Микроорганизм по п. 2, где фосфоенолпируваткарбоксикиназа 1 представляет собой фермент, представленный аминокислотной последовательностью SEQ ID NO: 31, а пируваткиназа 2 представляет собой фермент, представленный аминокислотной последовательностью SEQ ID NO: 33.
7. Микроорганизм по п. 3, где малатдегидрогеназа 2 представляет собой фермент, представленный аминокислотной последовательностью SEQ ID NO: 35, а цитозольный яблочный фермент 1 представляет собой фермент, представленный аминокислотной последовательностью SEQ ID NO: 37 или SEQ ID NO: 52.
8. Микроорганизм по п. 1, дополнительно модифицированный для введения активности лактатдегидрогеназы (LDH).
9. Микроорганизм по п. 8, где лактатдегидрогеназа представляет собой фермент, представленный аминокислотной последовательностью SEQ ID NO: 49.
10. Микроорганизм по п. 1, где микроорганизм дополнительно модифицирован для:
(1) инактивации активности алкогольдегидрогеназы 1 (ADH1) по сравнению с ее эндогенной активностью;
(2) инактивации активности пируватдекарбоксилазы 1 (PDC1) по сравнению с ее эндогенной активностью; и
(3) инактивации активности D-лактатдегидрогеназы 1 (DLD1) по сравнению с ее эндогенной активностью.
11. Микроорганизм по п. 1, где микроорганизм рода Saccharomyces представляет собой Saccharomyces cerevisiae.
12. Способ получения молочной кислоты, включающий:
a) культивирование микроорганизма рода Saccharomyces по любому из пп. 1-11 в среде; и
б) выделение молочной кислоты из культивируемого микроорганизма или из среды.
| ISHIDA N | |||
| ET AL | |||
| "The Effect of Pyruvate Decarboxylase Gene Knockout in Saccharomyces Cerevisiae on L-lactic Acid Production", Bioscience, Biotechnology, and Biochemistry, 2006, vol | |||
| Деревянный торцевой шкив | 1922 |
|
SU70A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| LIAN J | |||
| ET AL., "Recent Advances in Biosynthesis of Fatty Acids Derived Products in Saccharomyces Cerevisiae via Enhanced Supply of | |||

