Область техники
Изобретение относится к биотехнологии и касается микробиологического способа получения молочной кислоты с использованием дрожжей Schizosaccharomyces pombe.
Уровень техники
Молочная кислота традиционно используется во многих областях промышленности, однако в последнее десятилетие спрос на нее значительно возрос в связи с развитием «зеленых» технологий, в частности - получением биоразлагаемого полимера молочной кислоты полилактида, занимающего лидирующее положение по объемам выпуска среди биопластиков и находящего все большее применение.
Известно, что для производства полилактида с улучшенными физическими свойствами необходимы два оптически активных энантиомера молочной кислоты: L- и D-молочная кислота. Различное соотношение оптически чистых изомеров позволяет улучшить термохимические свойства, скорость биоразложения и другие характеристики полилактида.
В связи с этим разработка эффективных способов производства L- и. D-формы молочной кислоты имеет коммерческое значение и является актуальной задачей.
Получение молочной кислоты микробиологическим способом из возобновляемых ресурсов является экономически выгодной и экологически чистой альтернативой нефтехимическому синтезу. В качестве штаммов-продуцентов в промышленности чаще всего используют молочнокислые бактерии и генно-модифицированные дрожжи.
Природные продуценты молочной кислоты (молочнокислые бактерии) характеризуются высокими продуктивностью, скоростью синтеза и конверсией, однако главным их недостатком является чувствительность к низким значениям рН и высоким концентрациям молочной кислоты в культуральной жидкости (КЖ). Это приводит к значительным экономическим затратам для поддержания нейтральных значений рН в течение процесса и усложняет очистку конечного продукта.
В качестве альтернативы бактериальным штаммам в производстве широко используют штаммы дрожжей, способные к росту и брожению в условиях повышенной кислотности.
Дрожжевые штаммы-продуценты молочной кислоты получают путем экспрессии в дрожжевых клетках гетерологичных генов лактатдегидрогеназы (L-LDH или D-LDH), в основном из молочнокислых бактерий. При этом в качестве реципиента используют разные виды дрожжей.
Штамм Saccharomyces cerevisiae YPH499/dPdA3-34/DLDH/1-18 [Biotechnol. Bioeng. 2017; 114: 2075-2084] содержит в составе хромосомы дополнительные гены, усиливающие гликолитический путь, и несколько копий гена D-лактатдегидрогеназы из Lactobacillus mesenteroides. Штамм продуцирует D-молочную кислоту в количестве 60 г/л в колбах на богатой среде с мелом.
Штамм Saccharomyces cerevisiae JHY5330 [Appl. Microbiol. Biotechnol. 2016; 100:2737-2748], содержащий в составе хромосомы несколько копий гена D-лактатдегидрогеназы из Lactobacillus mesenteroides, а также делеции генов JEN1, DLD1, PDC1, ADH1, GPD1 и GPD2, продуцирует D-молочную кислоту в количестве 112 г/л в колбах на среде с дрожжевым экстрактом, пептоном и мелом.
Недостатками данных работ являются использование относительно дорогих сред и необходимость поддержания нейтральных значений рНмелом в процессе культивирования, что требует в дальнейшем дополнительной очистки продукта.
Делящиеся дрожжи Schizosaccharomyces pombe обладают высокой устойчивостью к низким значениям рН и, в частности, к высоким концентрациям молочной кислоты [RU 2268304], вследствие чего также могут быть использованы в качестве реципиента для получения штаммов-продуцентов органических кислот.
Традиционно дрожжи S. pombe использовались для быстрого сбраживания Сахаров в этанол. Таким образом, для конструирования продуцента молочной кислоты на базе дрожжей S. pombe необходимо создать путь биосинтеза молочной кислоты из пирувата путем интеграции гетерологичных лактатдегидрогеназ и перенаправить поток углерода с синтеза этанола на биосинтез целевого продукта.
Одним из подходов, используемым для ослабления потока пирувата, идущего на синтез этанола, является инактивация генов PDC, которые кодируют пируватдекарбоксилазы, ответственные за конверсию пирувата в ацетальдегид (интермедиат для образования этанола).
В дрожжах S. pombe выявлено 4 гена PDC: PDC1 (SPAC13A11.06), PDC2 (SPAClF8.07c), PDC3 (SPAC186.09) и PDC4 (SPAC3G9.11c). Как правило, для конструирования штаммов-продуцентов молочной кислоты на этих дрожжах используют штаммы с делециями генов PDC1 и/или PDC2.
Так штамм Schizosaccharomyces pombe ВКПМ Y-4822 [RU 2752896], содержащий в составе хромосомы делецию гена PDC1 и гены трех различных гетерологичных Z-лактатдегидрогеназ, продуцирует L-молочную кислоту в пробирке в количестве 80 г/л при кислых значениях рН=2.7.
Известен штамм Schizosaccharomyces pombe ATR5-LA2 [Metab Eng Commun. 2017; 5:60-67], содержащий в составе хромосомы делеции генов PDC1, PDC2, L-LDH и ADH, интеграцию в хромосому гетерологичных генов MhpF и EutE для прямой конверсии ацетальдегида в ацетил-КоА, а также 2 копии гена D-лактатдегидрогеназы из Lactobacillus plantarum в локусы GPD2 и Adh8. Штамм S. pombe ATR5-LA2 продуцирует D-молочную кислоту в количестве 25 г/л на минимальной среде с добавлением урацила, аденина и лейцина в пробирках.
Для определения оптимальной комбинации делеций генов PDC в дрожжах S. pombe в экспериментах in vitro была измерена суммарная пируватдекарбоксилазная активность различных PDC-мутантов. Вместе с ростовыми характеристиками эти данные позволяют выбрать комбинацию делеций, подходящую для дальнейшего конструирования продуцента молочной кислоты. Показано, что минимальное значение суммарной пируватдекарбоксилазной активности наблюдается у мутанта, с делециями генов PDC1, PDC3 и PDC4.
Полученные результаты позволяют предположить, что мутантный штамм дрожжей Schizosaccharomyces pombe, содержащий в составе хромосомы указанные делеций, перспективен для использования в качестве реципиента для конструирования продуцентов молочной кислоты.
Технической проблемой, на решение которой направлено заявляемое изобретение, является расширение арсенала рекомбинантных микроорганизмов - реципиентов для конструирования продуцентов молочной кислоты и расширение арсенала микроорганизмов - продуцентов молочной кислоты.
Раскрытие сущности изобретения
Техническим результатом заявляемого изобретения является получение мутантного штамма дрожжей Schizosaccharomyces pombe.
Техническим результатом заявляемого изобретения является поучение трансформанта дрожжей Schizosaccharomyces pombe с делетированными генами URA4, PDC1, PDC3 и PDC4.
Техническим результатом заявляемого изобретения является получение рекомбинантного штамма дрожжей Schizosaccharomyces pombe Sp-D3 ВКПМ Y-5151.
Технический результат достигается тем, что предложен мутантный штамм дрожжей Schizosaccharomyces pombe Y-4539, имеющий генотип Δurа4 Δpdc1 Δpdc3 Δpdc4, в качестве реципиента для конструирования продуцентов молочной кислоты.
Технический результат достигается тем, что предложен трансформант дрожжей Schizosaccharomyces pombe с делетированными генами URA4, PDC1, PDC3 и PDC4, несущий в составе хромосомной ДНК гены гетерологичных D-лактатдегидрогеназ из Lactobacillus plantarum и Lactobacillus delbruecki - продуцент D-молочной кислоты.
Технический результат достигается тем, что предложен штамм дрожжей Schizosaccharomyces pombe ВКПМ Y-5151, имеющий генотип Δpdc1 Δpdc3 Δpdc4 pHsp9-LDHplant URA4 pADH-LDHdelbr - продуцент D-молочной кислоты.
Краткое описание чертежей
Изобретение проиллюстрировано следующими фигурами графического изображения:
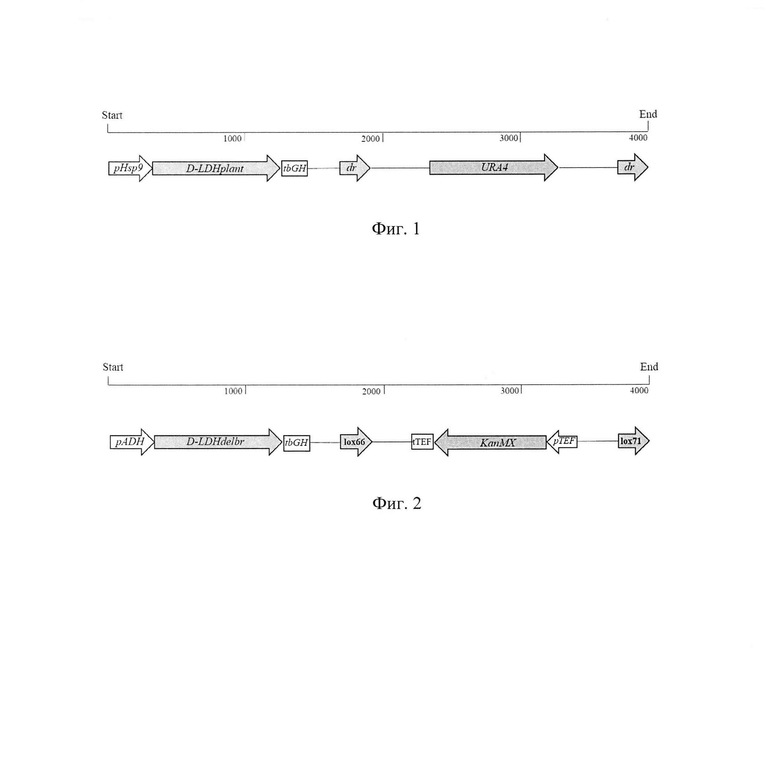
Фиг. 1. Кассета для трансформации, содержащая ген URA4 и ген D-лактатдегидрогеназы из Lactobacillus plantarum.
Фиг. 2. Кассета для трансформации, содержащая ген KanMX и ген D-лактатдегидрогеназы из Lactobacillus delbruecki.
Осуществление изобретения
Штамм Schizosaccharomyces pombe Sp-1.4 ВКПМ Y-4539 получен в результате делеционной мутации, а именно, путем делетирования изхромосомной ДНК родительского штамма S. pombe Y-285 генов URA4, PDC1, PDC3 и PDC4.
Штамм Schizosaccharomyces pombe Sp-D3 ВКПМ Y-5151 получен путем интеграции в состав хромосомной ДНК штамма S. pombe Y-4539 кассеты, содержащей ген URA4 и ген D-лактатдегидрогеназы из Lactobacillus plantarum, кассеты, содержащей ген KanMX и ген D-лактатдегидрогеназы из Lactobacillus delbruecki, и делетирования гена KanMX.
Пример 1 Конструирование мутанта дрожжей Schizosaccharomyces pombe - реципиента для конструирования продуцентов молочной кислоты
1.1. Делетирование гена URA4 из хромосомной ДНК штамма S. pombe Y-285
Готовят компетентные клетки штамма S. pombe Y-285, которые трансформируют методом электропорации линейной плазмидой, содержащей дрожжевой селективный маркер KanMX и объединенные методом ПЦР последовательности Up- и Dn-, расположенные выше и ниже гена URA4 с регуляторными элементами.
Селекцию трансформантов ведут на агаризованной среде YES состава (мас. %): дрожжевой экстракт - 0.5; глюкоза - 3.0; агар - 2; остальное - вода, с генетицином в количестве 50 мкг/мл в течение 5 суток при температуре 30°С.
Отбирают трансформант Т1 с заданным местом инсерции линейной плазмиды в Up-плечо гена URA4.
Делетирование генов URA4 и KanMX из хромосомной ДНК трансформанта Т1 проводят на минеральной агаризованной среде PMG [https://dornsife.usc.edu/pombenet/media/] с урацилом (200 мкг/мл) с добавлением 5-фтороротовой кислоты (1 мас. %) по методу [https://dornsife.usc.edu/pombenet/drugs/]. Делетирование гена URA4 и селективного маркера КапМХ происходит по прямым повторам, образовавшимся в результате встраивания плазмиды, по механизмугомологичной рекомбинации. Из отдельных колоний выделяют хромосомную ДНК и анализируют на наличие делеций генов URA4 и KanMX методом ПЦР. Отбирают мутант Sp-1.1, который содержит конструкцию Δura4.
1.2. Делетирование гена PDC1 из хромосомной ДНК мутанта S. pombe Sp-1.1
Компетентные клетки штамма S. pombe Sp-1.1 трансформируют методом электропорации линейной плазмидой, содержащей дрожжевой селективный маркер URA4 и объединенные методом ПЦР последовательности Up- и Dn-, расположенные выше и ниже структурной части гена PDC1.
Селекцию трансформантов ведут на агаризованной среде PMG в течение 5 суток при температуре 30°С.
Отбирают трансформант Т2 с заданным местом инсерции линейной плазмиды в Up-плечо гена PDC1.
Далее проводят делетирование генов URA4 и PDC1 из хромосомной ДНК трансформанта Т2, как описано в п. 1.1. Из отдельных колоний выделяют хромосомную ДНК и анализируют на наличие делеций генов URA4 и PDC1 методом ПЦР. Отбирают мутант S. pombe Sp-1.2, который содержит конструкцию Δura4 Δpdc1.
1.3. Делетирование гена PDC3 из хромосомной ДНК штамма S. pombe Sp-1.2
Компетентные клетки штамма S. pombe Sp-1.2 трансформируют методом электропорации линейной плазмидой, содержащей дрожжевой селективный маркер URA4 и объединенные методом ПЦР последовательности Up- и Dn-, расположенные выше и ниже структурной части гена PDC3.
Селекцию трансформантов ведут на агаризованной среде PMG в течение 5 суток при температуре 30°С.
Отбирают трансформант Т3 с заданным местом инсерции линейной плазмиды в Up-плечо гена PDC3.
Далее проводят делетирование генов URA4 и PDC3 из хромосомной ДНК трансформанта Т3, как описано в п. 1.1. Из отдельных колоний выделяют хромосомную ДНК и анализируют на наличие делеций генов URA4 и PDC3 методом ПЦР. Отбирают мутант S. pombe Sp-1.3, который содержит конструкцию Δura4 Δpdc1 Δpdc3.
1.4. Делетирование гена PDC4 из хромосомной ДНК штамма S. pombe Sp-1.3
Компетентные клетки штамма S. pombe Sp-1.3 трансформируют методом электропорации линейной плазмидой, содержащей дрожжевой селективный маркер URA4 и объединенные методом ПЦР последовательности Up- и Dn-, расположенные выше и ниже структурной части гена PDC4.
Селекцию трансформантов ведут на агаризованной среде PMG в течение 5 суток при температуре 30°С.
Отбирают трансформант Т4 с заданным местом инсерции линейной плазмиды в Up-плечо гена PDC4.
Далее проводят делетирование генов URA4 и PDC4 из хромосомной ДНК трансформанта Т4, как описано в п. 1.1. Из отдельных колоний выделяют хромосомную ДНК и анализируют на наличие делеций генов URA4 и PDC4 методом ПЦР. Отбирают мутант S. pombe Sp-1.4, который содержит конструкцию Δura4 Δpdc1 Δpdc3 Δpdc4.
Пример 2. Штамм дрожжей Schizosaccharomyces pombe Y-4539
Полученный мутант, в котором делетированы ген урацила URA4 и гены пируватдекарбоксилаз PDC1, PDC3 и PDC4, депонирован в Биоресурсном центре Всероссийская Коллекция Промышленных Микроорганизмов (БРЦ ВКПМ) НИЦ «Курчатовский институт» - ГосНИИгенетика (117545 Москва, 1-ый Дорожный проезд, д.1) как штамм Schizosaccharomyces pombe ВКПМ Y-4539.
Штамм характеризуется следующими признаками:
Культурально-морфологические признаки
При культивировании при температуре 30°С в течение 72 ч на агаризованной среде PMG с урацилом (200 мкг/мл) культура образует колонии белого или слегка кремоватого оттенка, округлые, выпуклые, с ровным краем.
При культивировании в среде YES в течение 2-х суток клетки имеют палочковидную форму и размеры 3-4 мкм в диаметре и 7-14 мкм в длину. Осадок формируется.
Вегетативное размножение - деление клетки пополам.
Физиолого-биохимические признаки
Культура способна сбраживать глюкозу, сахарозу, мальтозу и раффинозу. Не способна к брожению галактозы, лактозы и мелибиозы. Ассимилирует в качестве единственного источника углерода сахарозу, мальтозу, раффинозу. Не ассимилирует галактозу, целлобиозу, трегалозу, лактозу, ксилозу, арабинозу, рибозу, рамнозу, эритрит, рибитол, маннит, крахмал, янтарную, лимонную кислоты, инозит. Культура не ассимилирует нитраты, не способна расти на среде без витаминов и урацила.
Оптимальные условия для поддержания штамма
Температура 30°С, рН 6, агаризованная дрожжевая среда YES.
Штамм хранится в лиофилизированном виде или в среде YES с глицерином (2 мас. %) при -70°С.
Полученный мутантный штамм может быть использован в качестве реципиента для конструирования продуцентов молочной кислоты.
Пример 3. Конструирование трансформанта дрожжей Schizosaccharomyces pombe - продуцента L- молочной кислоты
Введение гена L-LDH L-лактатдегидрогеназы из Lactobacillus acidophilus в состав хромосомной ДНК штамма Y-4539 осуществляют путем трансформации кассетой, содержащей ген L-LDH из Lactobacillus acidophilus, транскрипция которого находится под контролем промотора pHsp9 итерминатора транскрипции tbGH, и дрожжевой селективный маркер - ген урацила URA4, фланкированный прямыми повторами dr и обеспечивающий прототрофность по урацилу.
Селекцию трансформантов ведут на агаризованной среде PMG в течение 5 суток при температуре 30°С.
Отбирают трансформанты, содержащие конструкцию Δpdc1 Δpdc3 Δpdc4 pHsp9-LDHaci URA4, и культивируют их в ферментационной среде для определения продукции L-молочной кислоты. Результаты представлены в табл. 1.
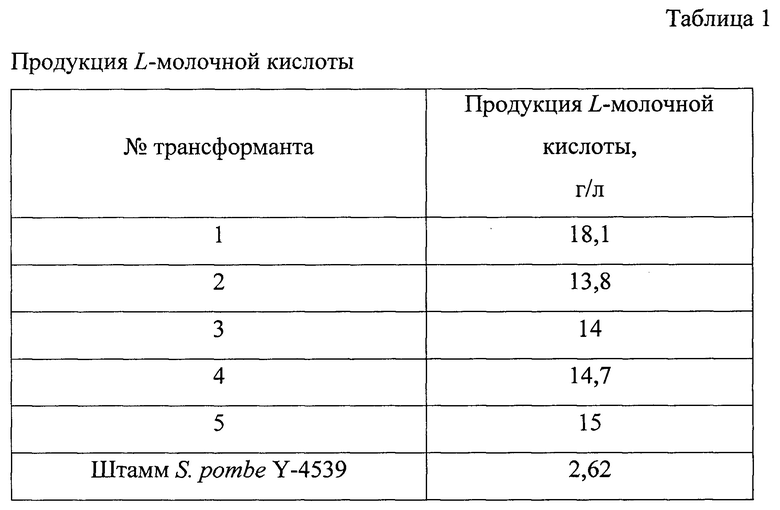
Полученные результаты показывают, что штамм S. pombe Y-4539 может быть использован в качестве реципиента для конструирования продуцентов L- молочной кислоты.
Пример 4. Конструирование трансформанта дрожжей Schizosaccharomyces pombe -продуцента D-молочной кислоты
4.1. Введение гена D-LDH D-лактатдегидрогеназы из Lactobacillus plantarum в состав хромосомной ДНК штамма S. pombe Y-4539
Полученные результаты показывают, что штамм S. pombe Y-4539 может быть использован в качестве реципиента для конструирования продуцентов L- молочной кислоты.
Пример 4. Конструирование трансформанта дрожжей Schizosaccharomyces pombe -продуцента D-молочной кислоты
4.1. Введение гена D-LDH D-лактатдегидрогеназы из Lactobacillus plantarum в состав хромосомной ДНК штамма S. pombe Y-4539
Введение гена D-LDH D-лактатдегидрогеназы из Lactobacillus plantarum в состав хромосомной ДНК штамма S. pombe Y-4539 осуществляют путем трансформации кассетой, содержащей ген D-LDH из Lactobacillus plantarum, транскрипция которого находится под контролем промотора pHsp9 и терминатора транскрипции tbGH, и дрожжевой селективный маркер - ген урацила URA4, фланкированный прямыми повторами dr и обеспечивающий прототрофность по урацилу (фиг. 1).
Селекцию трансформантов ведут на агаризованной среде PMG в течение 5 суток при температуре 30°С.
Отбирают трансформанты с инсерцией кассеты, содержащей ген D-LDH из Lactobacillus plantarum, в хромосомную ДНК S. pombe.
Трансформанты культивируют в ферментационной среде для определения продукции D-молочной кислоты.
Посевную культуру выращивают в пробирках (50 мл) с 3 мл жидкой питательной среды YPD состава (мас. %): дрожжевой экстракт - 1.0; пептон -1.5; глюкоза - 2.0; остальное - вода, при 30°С в течение 24 ч на качалке (250 об/мин). Культивирование проводят при 30°С на качалке (250 об/мин) в ферментационной среде ФС состава (мас. %): кукурузный экстракт - 2, калий-фосфатный буфер с рН 6.0-5, остальное - вода, с добавлением глюкозы (20 мас. %) в пробирках (50 мл) с рабочим объемом 5 мл. Посевной материал вносят в ферментационную среду в соотношении 1/10 об. Ферментацию продолжают в течение 72 часов. Концентрацию D-молочной кислоты в КЖ определяют согласно методу ВЭЖХ [Acta Biotechnol. 1990;10 (5): 459-468]. По результатам ферментации отбирают наиболее продуктивный трансформант S. pombe Sp-Dl, который содержит конструкцию Δpdc1 Δpdc3 Δpdc4 pHsp9-LDHplant URA4.
4.2. Введение гена D-LDH D-лактатдегидрогеназы из Lactobacillus delbruecki в состав хромосомной ДНК трансформанта S. pombe Sp-D1
Введение гена D-LDH лактатдегидрогеназы из Lactobacillus delbruecki в состав хромосомной ДНК трансформанта S. pombe Sp-D1 осуществляютпутем трансформации кассетой (фиг. 2), содержащей ген D-LDH из Lactobacillus delbruecki, транскрипция которого находится под контролем промотора pADH и терминатора транскрипции tbGH. В качестве селективного маркера используют фланкированный сайтами lox66 и lox71 ген устойчивости к генетицину KanMX, транскрипция которого находится под контролем промотора pTEF и терминатора транскрипции tTEF.
Селекцию трансформантов ведут на агаризованной среде YES с генетицином в количестве 50 мкг/мл в течение 5 суток при температуре 30°С.
Отбирают трансформанты с инсерцией кассеты, содержащей ген D-LDH из Lactobacillus delbruecki, в хромосомную ДНК S. pombe.
Отбор трансформантов осуществляют как описано в п. 4.1. Отобран наиболее продуктивный трансформант Sp-D2, который содержит конструкцию Δpdc1 Δpdc3 Δpdc4 pHsp9-LDHplant URA4 pADH-LDHdelbr KanMX.
4.3. Делетирование селективного маркера KanMX по lox-сайтам из хромосомной ДНК трансформанта S. pombe Sp-D2
Делетирование селективного маркера KanMX из хромосомной ДНК трансформанта S. pombe Sp-D2 осуществляют в результате рекомбинации по lox-сайтам с использованием Cre-рекомбиназы. [Biosci Biotechnol Biochem 2004;68(3): 545-550; Yeast 2006;23(11): 813-823].
Трансформант Sp-D2 трансформируют плазмидой, содержащей ген cre-рекомбиназы под контролем промотора nmt1 с селективным маркером Hph, обеспечивающим устойчивость к гигромицину. Трансформанты отбирают на агаризованной среде YES с гигромицином (50 мкг/мл). Отдельную колонию засевают в пробирку с жидкой минеральной средой PMG. Пробирку инкубируют на качалке (250 об/мин) при температуре 30° в течение 24 ч. Культуру высевают на чашки с агаризованной средой PMG с 5FOA. Через 10 суток отельные колонии с помощью репликатора переносят на агаризованные среды PMG, PMG с урацилом (200 мкг/мл), YES, YES с генетицином (50 мкг/мл) и YES с гигромицином (50 мкг/мл). В результатеотбирают колонию, чувствительную к генетицину и гигромицину. Трансформант назван Sp-D3 и содержит конструкцию Δpdc1 Δpdc3 Δpdc4 pHsp9-LDHplant URA4 pADH-LDHdelbr.
Пример 5. Штамм дрожжей Schizosaccharomyces pombe Y-5151 -продуцент D-молочной кислоты
Отобранный трансформант Sp-D3, в котором делетированы гены пируватдекарбоксилаз PDC1, PDC3 и PDC4, а в хромосомную ДНК интегрированы гены D-лактатдегидрогеназ D-LDH из Lactobacillus plantarum и Lactobacillus delbruecki. депонирован в Биоресурсном центре Всероссийская Коллекция Промышленных Микроорганизмов (БРЦ ВКПМ) НИЦ «Курчатовский институт» - ГосНИИгенетика (117545 Москва, 1-ый Дорожный проезд, д. 1) как штамм Schizosaccharomyces pombe ВКПМ Y-5151.
Штамм способен продуцировать D-молочную кислоту.
Штамм характеризуется следующими признаками.
Культурально-морфологические признаки
При культивировании при температуре 30°С в течение 72 ч на агаризованной среде PMG культура образует колонии белого или слегка кремоватого оттенка, округлые, выпуклые, с ровным краем.
При культивировании в жидкой среде YES в течение 2-х суток клетки имеют палочковидную форму и размеры 3-4 мкм в диаметре и 7-14 мкм в длину. Осадок формируется.
Вегетативное размножение - деление клетки пополам.
Физиолого-биохимические признаки
Культура способна сбраживать глюкозу, сахарозу, мальтозу и раффинозу. Не способна к брожению галактозы, лактозы и мелибиозы. Ассимилирует в качестве единственного источника углерода сахарозу, мальтозу, раффинозу. Не ассимилирует галактозу, целлобиозу, трегалозу, лактозу, ксилозу, арабинозу, рибозу, рамнозу, эритрит, рибитол, маннит, крахмал, янтарную, лимонную кислоты, инозит. Культура не ассимилирует нитраты, не способна расти на среде без витаминов.
Оптимальные условия для поддержания штамма
Температура 30°С, рН 6, агаризованная дрожжевая среда YES.
Штамм хранится в лиофилизированном виде или в жидкой среде YES с глицерином (2 мас. %) при -70°С.
Заявляемый штамм Schizosaccharomyces pombe Sp-D3 сохраняет способность к образованию D-молочной кислоты после 10 последовательных пересевов на полноценной среде.
Пример 6. Культивирование заявляемого штамма в пробирке
Штамм Schizosaccharomyces pombe Sp-D3 культивируют в ферментационной среде для определения уровня продукции D-молочной кислоты. В качестве контроля используют исходный штамм Schizosaccharomyces pombe Y-4539.
Культивирование проводят в среде ФС при 30°С на качалке (250 об/мин) в пробирках 50 мл с рабочим объемом 5 мл в течение 72 часов. Концентрацию D-молочной кислоты в КЖ определяют согласно методу ВЭЖХ.
У исходного штамма Schizosaccharomyces pombe Y-4539 продукция D-молочной кислоты отсутствует. Продукция D-молочной кислоты у штамма Schizosaccharomyces pombe Sp-D3 составляет 70 г/л, что почти в 3 раза превышает результат, описанный в литературе для дрожжей Schizosaccharomyces pombe, [Metab Eng Commun. 2017; 5:60-67].
Таким образом, сконструирован высокопродуктивный прототрофный штамм Schizosaccharomyces pombe Sp-D3 ВКПМ Y-5151, способный продуцировать D-молочную кислоту при кислых значениях рН на небогатой среде с кукурузным экстрактом (отходе крахмало-паточного и глюкозного производства). Полученный штамм не обладает антибиотикорезистентностью.
| название | год | авторы | номер документа |
|---|---|---|---|
| Штамм дрожжей Schizosaccharomyces pombe, продуцирующий L-молочную кислоту, содержащий в составе хромосомы гены трех различных гетерологичных лактатдегидрогеназ | 2020 |
|
RU2752896C1 |
| Трансформант дрожжей Schizosaccharomyces pombe, продуцирующий молочную кислоту (варианты), способ его получения (варианты), способ микробиологического синтеза молочной кислоты с использованием такого трансформанта | 2015 |
|
RU2614233C1 |
| Штамм Schizosaccharomyces pombe - продуцент молочной кислоты | 2016 |
|
RU2650669C1 |
| Способ модификации дрожжей Schizosaccharomyces pombe с помощью Cre-lox системы бактериофага Р1, трансформант, полученный таким способом | 2016 |
|
RU2652877C9 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА МОЛОЧНОЙ КИСЛОТЫ И РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ SCHIZOSACCHAROMYCES POMBE ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2004 |
|
RU2268304C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Schizosaccharomyces pombe - ПРОДУЦЕНТ МОЛОЧНОЙ КИСЛОТЫ | 2013 |
|
RU2539092C1 |
| ПОЛИПЕПТИД, ОБЛАДАЮЩИЙ АКТИВНОСТЬЮ D-ЛАКТАТДЕГИДРОГЕНАЗЫ, ПОЛИНУКЛЕОТИД, КОДИРУЮЩИЙ ЭТОТ ПОЛИПЕПТИД, И СПОСОБ ПОЛУЧЕНИЯ D-МОЛОЧНОЙ КИСЛОТЫ | 2010 |
|
RU2553563C2 |
| Трансформант дрожжей Yarrowia lipolytica, продуцирующий кантаксантин | 2023 |
|
RU2827562C1 |
| Интегративная кассета для экспрессии генов и ее использование при конструировании трансформанта дрожжей Komagataella mondaviorum | 2023 |
|
RU2836330C1 |
| Микроорганизм, имеющий повышенную продуктивность в отношении молочной кислоты, и способ получения молочной кислоты с использованием данного микроорганизма | 2015 |
|
RU2636467C2 |
Изобретение относится к области биотехнологии. Предложен мутантный штамм Schizosaccharomyces pombe ВКПМ Y-4539 с делетированными генами URA4, PDC1, PDC3 и PDC4 в качестве реципиента для конструирования продуцентов молочной кислоты. Также предложены трансформант дрожжей Schizosaccharomyces pombe и штамм дрожжей Schizosaccharomyces pombe ВКПМ Y-5151, являющиеся продуцентами D-молочной кислоты и имеющие делетированные гены PDC1, PDC3, PDC4 и введенные ген D-LDH D-лактатдегидрогеназы из Lactobacillus plantarum, транскрипция которого находится под контролем промотора pHsp9, ген D-LDH D-лактатдегидрогеназы из Lactobacillus delbruecki, транскрипция которого находится под контролем промотора pADH, и ген урацила URA4. Изобретение обеспечивает расширение арсенала рекомбинантных микроорганизмов-реципиентов для конструирования продуцентов молочной кислоты и расширение арсенала микроорганизмов-продуцентов молочной кислоты. 3 н.п. ф-лы, 2 ил., 1 табл., 6 пр.
1. Мутантный штамм дрожжей Schizosaccharomyces pombe ВКПМ Y-4539, имеющий генотип Δura4 Δpdc1 Δpdc3 Δpdc4, в качестве реципиента для конструирования продуцентов молочной кислоты.
2. Трансформант дрожжей Schizosaccharomyces pombe, имеющий делетированные гены PDC1, PDC3, PDC4 и введенный ген D-LDH D-лактатдегидрогеназы из Lactobacillus plantarum, транскрипция которого находится под контролем промотора pHsp9, введенный ген D-LDH D-лактатдегидрогеназы из Lactobacillus delbruecki, транскрипция которого находится под контролем промотора pADH, и введенный ген урацила URA4 -продуцент D-молочной кислоты.
3. Штамм дрожжей Schizosaccharomyces pombe ВКПМ Y-5151, содержащий делетированные гены PDC1, PDC3, PDC4, введенный ген D-LDH D-лактатдегидрогеназы из Lactobacillus plantarum, транскрипция которого находится под контролем промотора pHsp9, введенный ген D-LDH D-лактатдегидрогеназы из Lactobacillus delbruecki, транскрипция которого находится под контролем промотора pADH, и введенный ген урацила URA4 -продуцент D-молочной кислоты.

