Область техники
Данное изобретение относится к области получения антител, в частности, настоящее изобретение относится к мутантному полностью человеческому антителу против HER2, к кодирующему гену и к их применению.
Описание предшествующего уровня техники
Рак молочной железы представляет собой самое распространенное злокачественное заболевание у женщин во всем мире. Семейство белков HER регулирует рост и развитие нормальной молочной железы, избыточная экспрессия HER2 связана с раком молочной железы. Трастузумаб, известный под торговым названием герцептин (Herceptin) является первым человеческим моноклональным антителом для применения в качестве лекарственного средства при лечении HER2-положительного метастатического рака молочной железы (HER2-рецептор 2 типа эпидермального фактора роста человека). Хотя герцептин стал стандартным терапевтическим средством для лечения HER2-положительного злокачественного заболевания, 40% пациентов не отвечают на терапию при помощи этого лекарственного средства. Более того, серьезной проблемой при таком терапевтическом лечении является резистентность (устойчивость) к указанному лекарству. Полагают, что избыток фактора роста и интерференция среди внутриклеточных путей сигнальной трансдукции являются основными причинами, которые способствуют возникновению резистентности к лекарству у пациентов с раком молочной железы. Пертузумаб, созданный недавно в компании Genentech Corporation (USA) в сотрудничестве с Roche Corporation, представляет собой новое человеческое антитело против HER2. В отличие от герцептина пертузумаб нацелен на эпитоп антигена, который расположен во внеклеточной области II рецептора HER2. Результаты клинических экспериментальных исследований показали, что применение одного пертузумаба дает слабый противоопухолевый терапевтический эффект. Однако исследования показали, что применение пертузумаба в комбинации с герцептином может полностью блокировать сигнальную трансдукцию HER благодаря комплементации их механизмов действия, что приводит к более эффективному ингибированию роста опухолевых клеток.
Гликозилирование белка представляет собой процесс, при котором цепь полисахарида добавляется в белок в эндоплазматическом ретикулюме с образованием олигосахаридной цепи. Гликозилирование белка является сайт-специфическим и фермент-направленным. В зависимости от вида связывания с белковым фрагментом модификация белка путем гликозилирования может быть или О-связанной, или N-связанной, при этом консервативный сайт N-гликозилирования представляет собой Asn-X-Thr/Ser, где X является любой аминокислотой, отличающейся от Pro. Существует консервативный N-связанный сайт гликозилирования Asn297 в области СН2 сегмента Fc тяжелой цепи человеческого IgG. Fc полисахаридная цепь является существенной для оптимального связывания антитела с различными рецепторами, эффективного удаления патогенов антителом, а также для регулирования клинических свойств терапевтического антитела. Модификация путем N-гликозилирования Fab человеческого IgG может оказывать очевидное ускоряющее или ингибирующее действие на функцию связывания антитела с антигеном. Незначительные изменения в положении, где происходит модификация гликозилированием, может приводить к полностью отличающемуся эффекту при последующем процессинге полисахаридной цепи и активности связывания антитела с антигеном.
Данный заявитель ранее получил полностью человеческое моноклональное антитело GB235-019 против человеческого HER2 (Her-2/neu) путем применения технологии скрининга фаговых библиотек полностью человеческого scFV-фрагментов и технологии экспрессии рекомбинантных белков (можно дать отсылку на заявку CN №201410705404.0). Моноклональное антитело может снизить вероятность протекания реакции при трансфузии и иммуногенность, повысить безопасность лекарств и представляет лучший фармакокинетический профиль. Более того, GB235-019 может быть использовано в комбинации с другими терапевтическими агентами против HER2-положительных опухолей для лечения HER2-положительных опухолей.
Сущность изобретения
Настоящее изобретение предусматривает мутантное полностью человеческое антитело GB235-019 против HER2, где аминокислотная последовательность вариабельной области тяжелой цепи и такая же аминокислотная последовательность вариабельной области легкой цепи мутантного антитела соответственно представлены SEQ ID NO: 10, SEQ ID NO: 2; SEQ ID NO: 11, SEQ ID NO: 2; или SEQ ID NO: 12, SEQ ID NO: 2.
Согласно одному из вариантов мутантное полностью человеческое антитело к HER2 GB235-019 по данному изобретению находится в виде Fab, Fab', F(ab') 2, Fv или scFv. Обозначения Fab, Fab', F(ab') 2, Fv или scFv имеют значения, общеизвестные в уровне техники.
Согласно одному из вариантов мутантное полностью человеческое антитело GB235-019 против HER2 по изобретению, описанное выше, может также содержать константную область тяжелой цепи и константную область легкой цепи человеческого IgG. В соответствии с конкретным вариантом человеческий IgG представляет собой IgG1. Согласно конкретному варианту аминокислотная последовательность константную область тяжелой цепи человеческого IgG представляет собой SEQ ID NO: 5, и аминокислотная последовательность константной области легкой цепи человеческого IgG представляет собой SEQ ID NO: 6.
Настоящее изобретение предусматривает нуклеотидную последовательность, кодирующую мутантное полностью человеческое антитело к HER2 GB235-019 по изобретению.
Согласно конкретному варианту нуклеотидная последовательность, кодирующая вариабельную область тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 10, представляет собой SEQ ID NO: 13, нуклеотидная последовательность, кодирующая вариабельную область тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 11, представляет собой SEQ ID NO: 14, нуклеотидная последовательность, кодирующая вариабельную область тяжелой цепи, имеющую аминокислотную последовательность SEQ ID NO: 12, представляет собой SEQ ID NO: 15, и нуклеотидная последовательность, кодирующая вариабельную область легкой цепи, имеющую аминокислотную последовательность SEQ ID NO: 2, представляет собой SEQ ID NO: 4.
Согласно конкретному варианту, когда мутантное полностью человеческое антитело к HER2 GB235-019 по изобретению является полноразмерным антителом, нуклеотидная последовательность по изобретению, кодирующая константную область тяжелой цепи представляет собой SEQ ID NO: 7, и нуклеотидная последовательность по изобретению, кодирующая константную область легкой цепи представляет собой SEQ ID NO: 8.
Настоящее изобретение предусматривает экспрессионный вектор, в котором нуклеотидная последовательность по изобретению функционально связана с последовательностью, контролирующей экспрессию, экспрессионного вектора.
Согласно конкретным вариантам экспрессионный вектор представляет собой вектор pGEM-T или вектор 293.
Настоящее изобретение предусматривает также клетки, которые содержат экспрессионный вектор по изобретению. Клетки могут быть или прокариотными, или эукариотными. Согласно конкретным вариантам клетки могут быть клетками млекопитающих, такими как клетки FreeStyle 293F.
Настоящее изобретение предусматривает фармацевтическую композицию, содержащую мутантное полностью человеческое антитело GB235-019 против HER2 по изобретению и фармацевтически приемлемый носитель.
Настоящее изобретение предусматривает комбинированное лекарственное средство, содержащее мутантное полностью человеческое антитело GB235-019 против HER2 по изобретению и дополнительный HER2-положительный противоопухолевый терапевтический агент (агенты), причем HER2-положительный противоопухолевый терапевтический агент (агенты), является герцептином и/или пертузумабом. Комбинированное лекарственное средство может вводиться субъекту в следующих количествах: 0.001-500 мг/кг мутантного антитела GB235-019+0.001-500 мг/кг герцептина и/или пертузумаба; 0.001-300 мг/кг мутантного антитела GB235-019+0.001-300 мг/кг герцептина и/или пертузумаба; 0.001-200 мг/кг мутантного антитела GB235-019+0.001-200 мг/кг герцептина и/или пертузумаба; 0.01-200 мг/кг мутантного антитела GB235-019+0.001-200 мг/кг герцептина и/или пертузумаба; 0.01-100 мг/кг мутантного антитела GB235-019+0.001-100 мг/кг герцептина и/или пертузумаба; 0.1-90 мг/кг мутантного антитела GB235-019+0.001-90 мг/кг герцептина и/или пертузумаба; 0.1-70 мг/кг мутантного антитела GB235-019+0.001 - герцептина и/или пертузумаба; 0.1-60 мг/кг мутантного антитела GB235-019+0.1-60 мг/кг мутантного антитела герцептина и/или пертузумаба; 0.1-50 мг/кг мутантного антитела GB235-019+0.1-50 мг/кг герцептина и/или пертузумаба; 0.1-40 мг/кг мутантного антитела GB235-019+0.1-40 мг/кг герцептина и/или пертузумаба; 1-40 мутантного антитела GB235-019+1-40 мг/кг герцептина и/или пертузумаба. Мутантное полностью человеческое антитело к HER2 GB235-019 по изобретению можно вводить субъекту в отдельности или одновременно с дополнительным HER2-положительным противоопухолевым средством (средствами). Путь введения может быть общепринятым в области применения антител.
Настоящее изобретение предусматривает также набор, содержащий мутантное полностью человеческое антитело GB235-019 к HER2 по изобретению. Этот набор можно использовать для детектирования белка HER2 в образце. Набор может также содержать дополнительные компоненты, обычно используемые в наборах для детектирования HER2 в данной области.
Настоящее изобретение предусматривает также применение мутантного полностью человеческого антитела GB235-019 к HER2 по изобретению для приготовления лекарственного средства, пригодного для лечения HER2-положительной опухоли, слабо положительной опухоли или отрицательной опухоли у субъекта.
Термин "HER2-положительная опухоль" означает, что если результат IHC (иммуногистохимии) является результатом с тремя плюсами (+++), то есть более 30% опухолевых клеток дает полное и сильное окрашивание цитоплазматической мембраны, тогда опухоль считается HER2-положительной; и если получается результат с двумя плюсами (++), то есть, по меньшей мере 10% опухолевых клеток являются окрашенными в цитоплазматической мембране в степени от слабой до умеренной, тогда затем осуществляют FISH (флуоресцентную гибридизацию in-situ) или CISH (хромогенную гибридизацию in-situ), и если получается положительный результат (происходит генная амплификация), тогда может быть поставлен диагноз - HER2-положительная опухоль. Предпочтительно, диагноз о наличии HER2-положительной опухоли является результатом, полученным при использовании набора для детектирования (набора для детектирования методами IHC, FISH или CISH), одобренного Китайским Управлением по контролю за пищевыми продуктами и лекарствами. Практикующий врач хорошо знает, как можно определить, является ли опухоль HER2-положительной.
Термин "HER2-слабо положительная опухоль" означает, что, если результат анализа имеет два плюса (++), то есть, по меньшей мере 10% опухолевых клеток являются окрашенными в цитоплазматической мембране в степени от слабой до умеренной, тогда затем осуществляют FISH или CISH, и, если не происходит генной амплификации, тогда может быть поставлен диагноз - HER2-слабо положительная опухоль. Соответственно, термин "HER2-отрицательная опухоль" означает, что, если получен результат с одним плюсом (+) или 0, тогда опухоль считается HER2-отрицательной.
HER2-положительная опухоль может выбрана из HER2-положительного рака молочной железы, рака желудка, рака легкого, немелкоклеточного рака легких, рака кости, рака поджелудочной железы, рака кожи, рака головы и шеи, меланомы кожи или внутриглазной меланомы, рака матки, рака яичника, рака прямой кишки, рака заднего прохода, рака толстой кишки, рака фаллопиевой маточной трубы, рака эндометрия, рака шейки матки, рака влагалища, рака наружных половых органов (вульвы), болезни Ходжкина, рака пищевода, рака тонкой кишки, рака эндокринной системы, рака щитовидной железы, рака паращитовидной железы, рака надпочечной железы, рака мягких тканей, рака мочеточника, рака полового члена, рака простаты, рака мочевого пузыря, рака почки или уретры, почечно-клеточного рака, рака почечной лоханки, мезотелиомы, печеночно-клеточного рака, рака желчного пузыря, хронического или острого лейкоза, рака лимфатической системы, рака центральной нервной системы (ЦНС), рака позвоночника, нейроглиомы ствола головного мозга, многоформной глиобластомы, астроцитомы, нейролеммомы, эпендимомы, медуллобластомы, менингиомы, сквамозно-клеточной карциномы и аденомы гипофиза.
Субъектом, предпочтительно, является человек.
Краткое описание чертежей
На Фигуре 1А представлена диаграмма структуры тяжелой цепи экспрессионного вектора (293-VH-CH) рекомбинантного полноразмерного антитела GB235-019 против человеческого HER2; и на Фигуре 1В показана диаграмма структуры легкой цепи экспрессионного вектора (293-VL-CL) рекомбинантного полноразмерного антитела GB235-019 против человеческого HER2. Фрагмент гена сигнального пептида, содержащий 5'-конец рестрикционного сайта EcoRI, фрагмент гена вариабельной области тяжелой цепи (VH) и фрагмент гена константной области тяжелой цепи (СН), включающий терминирующий кодон TGA и 3'-конец рестрикционного сайта BamH I был получен методом ПЦР (PCR) с использованием соответствующих темплатов и праймеров, соответственно (подробности описаны в Примере 5), и три фрагмента были связаны при помощи метода перекрывающей ПЦР для получения фрагмента гена тяжелой цепи полноразмерного антитела GB235-019. Используя такой же подход, получали фрагмент гена легкой цепи полноразмерного антитела GB235-019, содержащий сигнальный пептид, вариабельную область легкой цепи (VL) и константную область легкой цепи (CL). Фрагмент гена тяжелой цепи и фрагмент гена легкой цепи соответственно клонировали в вектор pGEM-T, используя липкий конец, образованный после расщепления при действии рестриктаз EcoR I и BamH I.
На Фигуре 2 показана схема электрофореза SDS-PAGE (с додецилсульфонатом натрия) рекомбинантного полноразмерного антитела GB235-019 против человеческого HER2. Очищенное антитело GB235-019 и контрольный образец герцептина подвергали электрофорезу в 10% геле полиакриламида в условиях восстановления при помощи 50 мМ дитиотреитола. Результаты показывают, что и антитело GB235-019 и герцептин дают две полосы с молекулярной массой 50 кДа и 25 кДа, которые соответствуют тяжелой цепи и легкой цепи антител.
На Фигурах 3А и 3В показаны результаты анализа GB235-019 с пониженной молекулярной массой. Антитело GB235-019 анализировали с использованием хроматографа Waters H-Class Bio для хроматографии ультравысокого разрешения в условиях восстановления дитиотреитолом, и исходные сигналы масс-спекроскопии деконволютировали с использованием компьютерной программы PROMASS для получения соответствующего значения молекулярной массы. Теоретическая молекулярная масса тяжелой цепи (Fc, содержащего форму гликана G0F) была рассчитана с использованием программы GPMAW6.0, она была равна 50416.7 Да, а теоретическая молекулярная масса легкой цепи была равна 23120.8 Да. Результат, показанный на Фигуре 3А, показывает, что найденная молекулярная масса легкой цепи антитела GB235-019 сопоставима с теоретической (вычисленной) молекулярной массой, что свидетельствует об отсутствии гликозилирования в легкой цепи. Результат, показанный на Фигуре 3В, показывает, что найденная молекулярная масса тяжелой цепи антитела GB235-019 значительно отличалась от теоретической молекулярной массы (>1500 Da). С помощью выравнивании с теоретической последовательностью было установлено, что кроме Fc области теоретический сайт N-гликозилирования (Asn-Thr-Ser) имеет Fab каркасный фрагмент.
На Фигуре 4 показана схема электрофореза SDS-PAGE (с додецилсульфонатом натрия) рекомбинантного мутантного антитела GB235-019. Очищенные мутантные антитела GB235-019N73D, GB235-019N73Q и GB235-019S75A и контрольный образец герцептина подвергали электрофорезу в 10% геле полиакриламида в условиях восстановления при помощи 50 мМ дитиотреитола. Результаты показывают, что мутантные антитела GB235-019 и антитело герцептин все давали две полосы, соответствующие молекулярной массе равной 50 кДа и 25 кДа, которые соответствовали тяжелой и легкой цепям антител.
Фигуры 5А, 5В и Fig. 5С показывают результаты определения пониженной молекулярной массы мутантных антител GB235-019. Мутантные антитела GB235-019N73D, GB235-019N73Q и GB235-019S75A анализировали с использованием хроматографа Waters H-Class Bio для хроматографии ультравысокого разрешения в условиях восстановления дитиотреитолом, и исходные сигналы масс-спекроскопии деконволютировали с использованием компьютерной программы PROMASS для получения соответствующего значения молекулярной массы. Теоретическая молекулярная масса тяжелой цепи (формы Fc, содержащего гликан G0F) была рассчитана с использованием программы GPMAW6.0, она была равна 50416.7 Да, а теоретическая молекулярная масса легкой цепи была равна 50416.7 Да. Фигура 5А показывает, что полученная молекулярная масса легкой цепи антитела GB235-019N73D соответствовала теоретической молекулярной массе, что свидетельствует об отсутствии гликозилирования в легкой цепи. Фигуры 5В и Fig. 5С показывают, что полученная молекулярная масса тяжелой цепи мутантных антител GB235-019N73Q и GB235-019S75A соответствовала теоретической молекулярной массе (с разницей<1 Да), что свидетельствует о том, что сайт N-гликозилирования в Fab каркасной области был удален.
Фигура 6 показывает связывание рекомбинантных полноразмерных мутантных антител GB235-019 против человеческого HER2 с человеческим HER2 антигеном. Планшет ELISA был покрыт антигеном человеческого HER2, антитела GB235-019WT, GB235-019N73D, GB235-019N73Q, GB235-019S75A, герцептин и пертузумаб в различных концентрациях связывались с молекулами антигена на планшете, и затем проводили детектирование связанных антител при помощи HRP-меченого антитела козы против Fc IgG человека. Результат, показанный на Фигуре 6, свидетельствует о том, что антитело GB235-019WT, а также мутантные антитела GB235-019N73D, GB235-019N73Q и GB235-019S75A способны специфически связываться с HER2 антигеном человека.
Фигура 7А показывает in vitro ингибирующее действие рекомбинантных полноразмерных мутантных антител GB235-019 против HER2 человека на пролиферацию клеток ВТ-474. HER2-положительные клетки ВТ-474 рака молочной железы, экспрессирующие высокий уровень P-HER2, инкубировали в течение 6 дней в полной культуральной среде, дополненной херегулином-α. Клетки обрабатывали GB235-019WT и раздельно вводили мутантные антитела GB235-019WT, включая GB235-019N73D, GB235-019N73Q и GB235-019S75A, а также герцептин, каждый в различных концентрациях. Жизнеспособность клеток определяли, используя краситель Alarmar Blue. Результаты показывают, что добавление херегулина-α в полную среду вызвало пролиферацию клеток ВТ-474. Клетки ВТ-474 стали нечувствительными (резистентными) к герцептину, введенному отдельно, действие антитела GB235-019WT и мутантных антител GB235-019N73D, GB235-019N73Q и GB235-019S75A, введенных по отдельности, не было значительным, когда их использовали в отдельности.
Фигура 7В показывает результаты in vitro эксперимента, показывающие, что резистентность клеток ВТ-474 к герцептину, вызванная херегулином-α, становилась обратимой под действием рекомбинантных полноразмерных антител против HER2 человека. HER2-положительные раковые клетки ВТ-474 молочной железы, экспрессирующие высокий уровень P-HER2, инкубировали в течение 6 дней в полной культуральной среде, дополненной херегулином-α. Затем клетки обрабатывали путем совместного введения антитела GB235-019WT и мутантных антител, включающих GB235-019N73D, GB235-019N73Q и GB235-019S75A, каждого в комбинации с герцептином. Жизнеспособность клеток определяли, используя краситель Alarmar Blue. Результаты показывают, что добавление херегулина-α в полную культуральную среду индуцировало пролиферацию клеток ВТ-474. Клетки ВТ-474 становились резистентными к герцептину, вводимому в отдельности, в то время, как комбинированное введение каждого из антитела GB235-019WT и мутантных антител GB235-019N73D, GB235-019N73Q и GB235-019S75A в комбинации с герцептином ингибировало индуцированную херегулином-α пролиферацию, существенно ниже уровня, который был перед индукцией херегулином-α, в зависимости от концентрации. Действие мутантных антител GB235-019N73D, GB235-019N73Q и GB235-019S75A было эквивалентно действию антитела GB235-019WT.
Фигура 8 отражает ингибирующее действие рекомбинантных полноразмерных антител против человеческого HER2 на сигнальную трансдукцию в клетках рака молочной железы линии ВТ-474. HER2-положительные клетки рака молочной железы линии ВТ-474, экспрессирующие высокие уровни P-HER2, помещали в культуральную среду с низким содержанием сыворотки, содержащую 0.1% фетальной бычьей сыворотки, на 24 ч. Затем добавляли по 20 мкг/мл каждого из антител GB235-019WT и мутантного антитела GB235-019N73D, и 20 мкг/мл герцептина и 20 мкг/мл пертузумаба в отдельности. После обработки клеток линии ВТ-474 антителами в течение 6 ч для индуцирования в течение 10 мин добавляли херегулин-α с конечной концентрацией равной 100 нг/мл, в это время отбирали образцы. Проводили иммуноблоттинг собранного клеточного лизата и лизата цельной крови и детектировали фосфорилированные HER3, Akt и ERK, используя соответствующие антитела. Результаты, представленные на Фигуре 8, показывают, что по сравнению с контрольной группой без добавления херегулина-α, херегулин-α вызывал позитивную регуляцию фосфорилирования HER3 в ВТ-474 клетках. Введенные в отдельности антитела GB235-019WT и GB235-019N73D в значительной степени ингибировали позитивную регуляцию фосфорилирования HER3 в клетках ВТ-474, индуцированную херегулином-α и полностью отменяли позитивную регуляцию фосфорилирования HER3, индуцированную херегулином-α. Мутантное антитело GB235-019N73D вызывало такой же эффект, что и антитело GB235-019WT. Пертузумаб также полностью ингибировал позитивную регуляцию фосфорилирования HER3 в клетках ВТ-474, индуцированную херегулином-α, и герцептин также в значительной степени ингибировал позитивную регуляцию фосфорилирования HER3 в клетках ВТ-474, индуцированную герегулином-α.
Отдельно введенное антитело GB235-019WT и мутантное антитело GB235-019N73D не ингибировали позитивную регуляцию фосфорилирования Akt, вызванную херегулином-α. Отдельно введенный пертузумаб в значительной степени ингибировал позитивную регуляцию фосфорилирования Akt, вызванную херегулином-α. Отдельно введенное мутантное антитело GB235-019N73D и антитело GB235-019WT в значительной степени ингибировали позитивную регуляцию фосфорилирования ERK1/2, индуцированную херегулином-α. Мутантное антитело GB235-019N73D вызывало такой же эффект, что и антитело GB235-019WT.
На Фигуре 9 показано действие GB235-019 рекомбинантных полноразмерных антител против человеческого HER2 на сигнальную трансдукцию в раковых клетках MCF 7 молочной железы. HER2-отрицательные клетки MCF 7 клетки рака молочной железы, экспрессирующие низкий уровень HER2 и высокий уровень HER3, но не экспрессирующие P-HER2 и P-HER3, помещали в культуральную среду с низким содержанием сыворотки, содержащую 0.1% фетальной бычьей сыворотки, на 24 ч. Затем добавляли по 20 мкг/мл каждого из антител GB235-019WT и мутантного антитела GB235-019N73D, GB235-019N73Q и GB235-019S75A и 20 мкг/мл герцептина и 20 мкг/мл пертузумаба в отдельности. После обработки клеток линии MCF 7 антителами в течение 6 ч для индуцирования в течение 10 мин добавляли херегулин-α с конечной концентрацией равной 100 нг/мл, в это время отбирали образцы. Проводили иммуноблоттинг собранного клеточного лизата и лизата цельной крови и детектировали фосфорилированные HER3, Akt и ERK, используя соответствующие антитела. Результаты, показанные на Фигуре 9, свидетельствуют, что по сравнению с контрольной группой без добавления херегулина-α, херегулин-α вызывал позитивную регуляцию фосфорилирования HER3 в MCF 7 клетках. Введенные в отдельности антитела GB235-019WT и мутантные антитела GB235-019N73D, GB235-019N73Q, GB235-019S75A в значительной степени ингибировали позитивную регуляцию фосфорилирования HER3 в клетках MCF 7, индуцированную херегулином-α и полностью отменяли позитивную регуляцию фосфорилирования HER3, индуцированную херегулином-α. Введенные в отдельности герцептин и пертузумаб также в значительной степени ингибировали позитивную регуляцию фосфорилирования HER3 в клетках MCF 7, индуцированную херегулином-α. Отдельно введенные антитело GB235-019WT и мутантные антитела GB235-019N73D, GB235-019N73Q, GB235-019S75A в значительной степени ингибировали положительную регуляцию фосфорилирования Akt, индуцированную херегулином-α, а введенный в отдельности пертузумаб полностью отменял положительную регуляцию фосфорилирования Akt, индуцированную херегулином-α. Введенные отдеотер мутантные антитела GB235-019N73D и GB235-019N73Q в значительной степени ингибировали положительную регуляцию фосфорилирования ERK1/2 в клетках MCF 7, индуцированную херегулином-α, и отдельно введенное антитело GB235-019WT и мутантное антитело GB235-019S75A слабо ингибировали регуляцию фосфорилирования ERK1/2, индуцированную херегулином-α. Введенный в отдельности пертузумаб полностью отменял положительную регуляцию фосфорилирования ERK1/2, индуцированную херегулином-α.
На Фиг. 10 показана эксклюзионная хроматограмма трех мутантных антител GB235-019N73D, GB235-019N73Q и GB235-019S75A. Имеются небольшие количества полимеров и фрагментов, хотя их содержание мало. Осуществляли эксклюзионную хроматографию (гель-фильтрацию) (SEC-HPLC) 30 мкг каждого из трех мутантных антител для определения соответствующей степени их чистоты. Содержание (чистота) основного пика антитела GB235-019N73D составляет 88.3%, содержание (чистота) основного пика антитела GB235-019N73Q составляет 89.7% и содержание (чистота) основного пика антитела GB235-019S75A составляет 93.1%.
На Фиг. 11 представлены электрофореграммы, полученные в результате капиллярного изоэлектрического фокусирования трех мутантных антител GB235-019N73D, GB235-019N73Q и GB235-019S75A. Осуществляли капиллярное изоэлектрическое фокусирование (iCIEF) трех мутантных антител для определения изоэлектрической точки (pI) соответствующих основных пиков и степени чистоты изомеров. Экспериментальные значения для изоэлектрических точек трех мутантных антител находятся в интервале от около 9.4 до 9.6. Изоэлектрическая точка GB235-019N73D была примерно на 0.1 ниже изоэлектрической точки GB235-019N73Q и GB235-019S75A, а чистота его основного пика была наивысшей.
На Фиг. 12 представлены электрофореграммы капиллярного электрофореза трех мутантных антител GB235-019N73D, GB235-019N73Q и GB235-019S75A в редуцирующих условиях. Осуществляли капиллярный электрофорез (rCE-SDS) трех мутантных антител для определения их относительной чистоты (суммы показателей для легкой цепи и тяжелой цепи). Все три мутантных антитела содержали небольшое количество низкомолекулярных примесей и высокомолекулярных примесей. Антитело GB235-019N73D содержало наименьшее количество примесей по сравнению с другими антителами, и степень его чистоты относительно суммы показателей для легкой цепи и тяжелой цепи (LC+HC) была наивысшей.
На Фиг. 13 представлены диаграммы, построенные по результатам дифференциальной сканирующей калориметрии для трех мутантных антител GB235-019N73D, GB235-019N73Q и GB235-019S75A. Три мутантных антитела исследовали методом дифференциальной сканирующей калориметрии (DSC) для определения tсоответствующей величины Tm (температуры фазового перехода, которая означает, что 50% биологических молекул являются при этой температуре неструктурированными). Температуры Tm1 указанных трех молекул отличались. Температура Tm1 антитела GB235-019N73Q была самой низкой по сравнению с температурами двух других антител, тогда Tm1 антитела GB235-019N73Q была близка к Tm1 антитела GB235-019S75A. Чем выше значение Tm, тем выше термическая устойчивость. Разница между Tm1 указывает на то, что в известной мере мутация оказывает определенное влияние на СН2 домен. GB235-019N73D и GB235-019S75A обладают повышенной термической устойчивостью по сравнению с GB235-019N73Q.
Осуществление изобретения
Технические решения, предусмотренные данным изобретением, будут далее проиллюстрированы следующими конкретными примерами, которые не ограничивают настоящее изобретение.
Пример 1. Скрининг клонов, специфически связывающихся с человеческим HER2-Fc из фаговой библиотеки полностью человеческих одноцепочечных scFV антител
Используя специфичность связывания антиген-антитело в методе ELISA, антиген слитого (химерного, гибридного) белка человеческий HER2 (внеклеточный домен)-Fc (сокращенно hHER2-Fc) иммобилизовали на планшете ELISA, и фаги, специфически связывающиеся с иммобилизованным антигеном, отмывали и разделяли, используя пэннинг. Антиген HER2-Fc (от Sino Biological Inc., номер по каталогу: 10004-Н02Н) разводили в PBS (0.01 М Na2HPO4⋅12H2O+0.002 М KH2PO4+0.14 М NaCl+0.002 М KCl, рН=8.6) до концентрации 5 мкг/мл, добавляли в планшет ELISA в количестве 100 мкл/лунка и иммобилизовали в течение ночи при 4°С. Планшет 4 раза отмывали в PBST (буфер PBS, содержащий 0.05% Tween 20), затем в лунки добавляли 5% BSA (от Amresco Inc., USA, номер по каталогу: 0332-100g, раствор в PBS) в количестве 300 мкл/лунка и блокировали при 37°С в течение 1 часа. Планшет снова дважды отмывали в PBST. Суспензию полностью человеческого антитела из фаговой библиотеки scFv антител (созданной в компании EUREKA (Beijing) Biotechnology Ltd. путем связывания генов вариабельных областей из клеток лимфатических узлов группы здоровых людей с искусственно синтезированным геном CDR3 тяжелой цепи), содержащую 7×1010 независимых клонов, добавляли в планшет ELISA в количестве 100 мкл/лунка и инкубировали при 37°С в течение 2 часов. По окончании инкубации суспензию фага в лунках планшета ELISA удаляли аспирацией. Затем в каждую лунку добавляли PBST, 300 мкл/лунка, и через содержимое лунок пропускали воздух в течение 5 минут для удаления фагов, не связанных специфически с иммобилизованным антигеном. Добавляли элюент 0.2 М глицин-HCl (рН=2.2), содержащий 0.1%BSA (от Amresco Inc., USA, номер по каталогу: 0332-100g, раствор в PBS), и инкубировали при комнатной температуре в течение 10 минут. Затем через содержимое лунок пропускали воздух для элюирования фагов, которые были специфически связаны с иммобилизованным антигеном. Суспензию элюированных фагов нейтрализовали буфером 1 М Tris-HCl (рН 9.1). Элюированные фаги добавляли к 1 мл бактерий TG1 (Lucigen Inc., USA, номер по каталогу 60500-0) в log-фазе (как указывает величина OD600 примерно от 0.3-0.4), и полученный бактериальный раствор оставляли стоять при 37°С в течение 1 часа для инфицирования. Из 10 мкл инфицированного бактериального раствора готовили 10-кратные серийные разведения и 10-кратные, 100-кратные и 1000-кратные разведения засевали и считали. 90 мкл инфицированного бактериального раствора хранили при -80°С в глицерине с конечной концентрацией глицерина 10%. Весь остальной инфицированный бактериальный раствор помещали на сплошной планшет 2×YT-A 150 мм (17 г/л триптона, 10 г/л дрожжевого экстракта, 5 г/л хлорида натрия, 15 г/л агар-агара и 100 мкг/мл ампициллина) и культивировали при 37°С в течение ночи. 5 мл среды 2×YT-А-10% глицерин добавляли в планшет 150 мм и оставляли на ночь. Планшет осторожно очищали (скребли) стерильным шпателем до тех пор, пока на нем не исчезли какие-либо остатки бактериального раствора. Во втором раунде амплификации соответствующее количество счищенного с планшета бактериального раствора добавляли к 5 мл жидкой питательной среды 2×YT-AMP-глюкоза (17 г/л триптона, 10 г/л дрожжевого экстракта, 5 г/л хлорида натрия, 2% глюкозы и 100 мкг/мл ампициллина) (желательно, чтобы значение OD600 было примерно от 0.05 до 0.1), и культивировали при 37°С, 200 об/мин до log-фазы (OD600 примерно от 0.3 до 0.4). Затем добавляли хелперный фаг M13K07 (от NEB Inc., USA, номер по каталогу: N0315S) в количестве, в 20 раз превышающем общее количество бактерий, для инфицирования, которое проводили при 37°С в течение 1 часа. После инфицирования осадок бактерий, собранный центрифугированием при 1500 g в течение 5 минут, ресуспендировали в среде 2×YT-AMP-Kana (17 г/л триптона, 10 г/л дрожжевого экстракта, 5 г/л хлорида натрия, 50 мкг/мл канамицина и 100 мкг/мл ампициллина) и культивировали при 30°С, 200 об/мин в течение ночи до завершения амплификации и получения рекомбинантных фагов. Второй раунд пэннинга и третий раунд амплификации и пэннинга проводили таким же способом. Колонии бактерий пересадили на 5 мл жидкой питательной среды 2×YT-AMP-глюкоза (17 г/л триптона, 10 г/л дрожжевого экстракта, 5 г/л хлорида натрия, 2% глюкозы и 100 мкг/мл ампициллина) и культивировали при температуре 37°С 200 об/мин в течение ночи. Плазмиды извлекали с использованием набора для извлечения плазмид (от Qiagen Inc., USA, номер по каталогу: 12943), идентифицировали секвенированием и хранили при -80°С.
Пример 2. Определение иммунореактивности фагов, специфически связывающихся с человеческим HER2-Fc, с использованием твердофазного иммуноферментного анализа (ELISA, твердофазный ИФА)
Используя твердофазный иммуноферментный анализ (ELISA), определяли также иммунореактивность фагов, которые специфически связываются с человеческим HER2-Fc, полученных в Примере 1. Человеческий HER2-Fc антиген (приобретен в Sino Biological Inc., номер по каталогу: 10004-Н02Н) разводили в PBS (рН=8.6) до концентрации 2 мкг/мл, добавляли в планшет ELISA в количестве 100 мкл/лунку и иммобилизовали при 4°С в течение ночи. Планшет отмывали в PBST четыре раза, затем добавляли 5% BSA (приобретен в Amresco, USA, номер по каталогу: 0332-100g, раствор в PBS) в количестве 300 мл/лунку и осуществляли блокирование при 37°С в течение 1 часа. Планшет снова отмывали дважды в PBST, затем добавляли суспензии фаговых клонов, 100 мкл/лунку, инкубировали при 37°С в течение 2 часов. Планшет отмывали в PBST четыре раза. Добавляли HRP-меченные анти-M13K07 фаговые антитела (GE Inc., USA, номер по каталогу: 27-9421-01, разведение 1:5000 в PBST, 100 мкл/лунку) и инкубировали при комнатной температуре в течение 1 часа. Планшет отмывали в PBST четыре раза, затем добавляли однокомпонентный раствор субстрата 3,3',5,5'-тетраметилбензидина (приобретен в Tiangen Co., Ltd., номер по каталогу: РА107-01), 100 мкл/лунку, и инкубировали при комнатной температуре в течение 15 минут для визуализации. Добавляли стоп-реагент (1 М серную кислоту) по 50 мкл/лунку и считывали оптическую плотность при длине волны 450/570 нм на многофункциональном планшетном ридере ELISA (Bio-Rad, Model 680 Micro reader, USA).
Результаты показывают, что после трех раундов повторяющегося скрининга было получено в общей сложности 1312 фаговых клонов scFV антител, которые могут связываться с человеческим HER2-Fc антигеном, среди которых 499 клона scFV антител могут специфически связываться с человеческим HER2-Fc антигеном. Секвенирование ДНК показало, что у 102 scFvs из этих клонов отличались как последовательности ДНК, так и аминокислотные последовательности (как показано в Таблице 1).

Пример 3. Детектирование межвидовой перекрестной реактивности и межмолекулярной перекрестной реактивности у членов семейства HER в 102 человеческих HER2-Fc-специфических scFvs методом ELISA
Межвидовую перекрестную реактивность и межмолекулярную перекрестную реактивность у членов семейства HER в 102 человеческих HER2-Fc-специфических scFvs детектировали методом ELISA. При этом следовали методике, описанной в Примере 2, за исключением того, что человеческий HER2-Fc антиген был заменен на обезьяний антиген HER2-Fc (приобретен в Sino Biological Inc., номер по каталогу: 90295-С02Н), мышиный HER2-Fc (приобретен в Sino Biological Inc., номер по каталогу: 50714-М02Н), человеческий HER1-Fc (приобретен в Sino Biological Inc., номер по каталогу: 10001-Н02Н), человеческий HER3-Fc (приобретен в Sino Biological Inc., номер по каталогу: 10201-Н05Н) и человеческий HER4-Fc (приобретен в Sino Biological Inc., номер по каталогу: 10363-Н02Н). Соответствующие антигены разводили в PBS (рН=8.6) до концентрации 2 мкг/мл, добавляли в планшет ELISA, по 100 мкл/лунку, и иммобилизовали при 4°С в течение ночи. Планшет четыре раза отмывали в PBST, затем добавляли 5% BSA (приобретен в Amresco, USA, номер по каталогу: 0332-100g, раствор в PBS), по 300 мкл/лунку, и блокировали при 37°С в течение 1 часа. Планшет снова дважды отмывали в PBST, затем добавляли суспензии 102 фаговых клонов ScFv, по 100 мкл/лунку, и инкубировали при 37°С в течение 2 часов. Планшет четыре раза отмывали в PBST. Добавляли HRP-меченные анти-M13K07 фаговые антитела (приобретен в GE Inc., USA, номер по каталогу: 27-9421-01, разведение 1:5000 в PBST, 100 мкл/лунку) и инкубировали при комнатной температуре в течение 1 часа. Планшет отмывали в PBST четыре раза, затем добавляли однокомпонентный раствор субстрата 3,3',5,5'-тетраметилбензидина (приобретен в Tiangen Co., Ltd., номер по каталогу: РА107-01), 100 мкл/лунку, и инкубировали при комнатной температуре в течение 15 минут для визуализации. Добавляли стоп-реагент (1 М серную кислоту) по 50 мкл/лунку и считывали оптическую плотность при длине волны 450/570 нм на многофункциональном планшетном ридере ELISA (Bio-Rad, Model 680 Micro reader, USA).
Результаты показали, что 96 фаговых клонов ScFv обладают перекрестной реактивностью к обезьяньему HER2-Fc, а 20 клонов обладают перекрестной реактивностью к мышиному HER2-Fc. Ни один из 102 клонов не обладал перекрестной реактивностью к человеческому HER1-Fc, человеческому HER3-Fc человеческому HER4-Fc (как показано в Таблице 2).


Пример 4. Классификация 102 фаговых клонов ScFv по аффинности
Классификацию 102 фаговых клонов ScFv антител по аффинности осуществляли методом ELISA. Исходя из человеческого HER2-Fc антигена с концентрацией 25 мкг/мл, готовили 10-кратные разведения в буфере PBS, получали 8 разведений в градиенте концентрации. Разведения инкубировали соответственно со 102 фаговыми клонами ScFv антител при комнатной температуре в течение 4 часов до достижения равновесия. Затем полученные смеси помещали на планшет ELISA, на котором предварительно иммобилизовали человеческий HER2-Fc антиген с концентрацией 2 мкг/мл (рН=8.6 PBS, 4°С в течение ночи, 100 мкл/лунку), и планшет ELISA блокировали с использованием 5% BSA (приобретен в Amresco Inc., USA, номер по каталогу: 0332-100g, в растворе PBS) для связывания неиммобилизованных ScFv антител. Добавляли HRP-меченные анти-М13 фаговые антитела (приобретен в GE Inc., USA, номер по каталогу: 27-9421-01, разведение 1:5000 в PBST, 100 мкл/лунку) и детектирование осуществляли методом, аналогичным методу, описанному в Примере 2. Классификацию 102 положительных клонов по аффинности проводили по значению IC50 (чем ниже значение IC50, тем выше аффинность).
Результаты выявили интервал распределения значений IC50 102 фаговых клонов ScFv антител, из которых 4 клона обладали аффинностью, превышающей аффинность герцептина.

Пример 5. Конструирование эукариотического вектора для экспрессии GB235-019 рекомбинантного полноразмерного антитела IgG1 изотипа
Эукариотический экспрессионный вектор для рекомбинантного полноразмерного антитела GB235-019 IgG1 изотипа (последовательность рекомбинантного полноразмерного антитела клона 019 была обозначена как GB235-019) конструировали при использовании последовательностей 102 фаговых клонов ScFv фрагментов антител. Нуклеотидная последовательность одноцепочечного антитела клона WG1-019, полученная скринингом фаговой библиотеки на основе полностью человеческих ScFv (клон с последовательностью одноцепочечного антитела, полученный при использовании фаговой библиотеки ScFv, был обозначен как WG1-019), представляла собой последовательность SEQ ID NO: 9, содержащую вариабельную область тяжелой цепи, имеющую нуклеотидные последовательности SEQ ID NO: 3 (которая кодирует аминокислотную последовательность SEQ ID NO: 1) и SEQ ID NO: 4 (которая кодирует аминокислотную последовательность SEQ ID NO: 2), соответственно. Сигнальный пептид имел аминокислотную последовательность  и аминокислотную последовательность
и аминокислотную последовательность 
 (которые синтезированы в Shanghai Generay Biotech Со, Ltd).
(которые синтезированы в Shanghai Generay Biotech Со, Ltd).
Рекомбинантное полноразмерное антитело GB235-019 имеет константную область тяжелой цепи и константную область легкой цепи, которые, соответственно, имеют нуклеотидные последовательности SEQ ID NO: 7 (которая кодирует аминокислотную последовательность SEQ ID NO: 5) и SEQ ID NO: 8 (которая кодирует аминокислотную последовательность SEQ ID NO: 6) (синтезированные в Shanghai Generay Biotech Со, Ltd).
Ниже представлены праймеры, предназначенные для конструирования эукариотического экспрессионного вектора для тяжелой цепи и легкой цепи рекомбинантного полноразмерного антитела GB235-019 IgG1 изотипа:

Амплификацией методом PCR (ПЦР) с использованием синтезированной последовательности сигнального пептида в качестве матрицы и 1-1 и 2-3 в качестве праймеров был получен генный фрагмент, содержащий сайт расщепления рестриктазой EcoR I, его обозначили как "SPL-GB235-019"; амплификацией методом PCR (ПЦР) с использованием синтезированной последовательности вариабельной области легкой цепи SEQ ID NO: 4 в качестве матрицы и 3-3 и 4-4 в качестве праймеров был получен генный фрагмент для вариабельной области легкой цепи и обозначен как "VL-GB235-019"; и амплификацией методом PCR (ПЦР) с использованием синтезированной последовательности константной области легкой цепи SEQ ID NO: 8 в качестве матрицы и 5-2 и 6-1 в качестве праймеров был получен генный фрагмент для константной области легкой цепи, содержащий стоп-кодон TGA и сайт рестрикции BamH I, и обозначен как "CL-GB235-019". Амплификацией методом ПЦР с перекрывающимися ДНК (Higuchi R, et al. A general method of in vitro preparation and specific mutagenesis of DNA fragments: study of protein and DNA interactions. Nucleic Acids Research, 1988, 16(15):7351-67) с использованием генных фрагментов SPL-GB235-019, VL-GB235-019 и CL-GB235-019 в качестве матриц и 1-1 и 6-1 в качестве праймеров был получен фрагмент полноразмерного гена легкой цепи антитела GB235-019.
Аналогично, амплификацией методом ПЦР (полимеразной цепной реакции) с использованием последовательности сигнального пептида в качестве матрицы и 1-1 и 7-7 в качестве праймеров был получен генный фрагмент, содержащий сайт расщепления рестриктазой EcoR I, его обозначили как "SPH-GB235-019"; амплификацией методом ПЦР с использованием синтезированной последовательности вариабельной области тяжелой цепи SEQ ID NO: 3 в качестве матрицы и 8-7 и 9-1 в качестве праймеров получен генный фрагмент для вариабельной области тяжелой цепи и обозначен как "VH-GB235-019", и амплификацией методом ПЦР с использованием синтезированной последовательности константной области тяжелой цепи SEQ ID NO: 7 в качестве матрицы и 10-1 и 11-1 в качестве праймеров был получен генный фрагмент для константной области тяжелой цепи, содержащий стоп-кодон TGA и сайт рестрикции BamH I, и обозначен как "CH-GB235-019". Амплификацией методом ПЦР с перекрывающимися ДНК с использованием генных фрагментов SPH-GB235-019, VH-GB235-019 и CH-GB235-019 в качестве матриц и 1-1 и 11-1 в качестве праймеров был получен фрагмент полноразмерного гена тяжелой цепи антитела GB235-019.
Вышеуказанные фрагменты полноразмерных генов тяжелой цепи и легкой цепи клонировали в вектор pGEM-T (приобретен в Promega Inc., USA, номер по каталогу: А3600), так чтобы 5'-конец генных фрагментов содержал сайт рестрикции EcoR I и 3'-конец содержал стоп-кодон TGA и сайт рестрикции BamH I. После секвенирования ДНК осуществляли двойное расщепление корректных, по данным секвенирования, клонов с применением EcoR I (приобретен в NEB Inc., USA, номер по каталогу: R0101S) и BamH I (приобретен в NEB Inc., USA, номер по каталогу: R0136S) при 37°С в течение 4 часов для регенерации соответствующих генных фрагментов. Фрагмент полноразмерного гена тяжелой цепи антитела и фрагмент полноразмерного гена легкой цепи антитела, полученные в результате вышеуказанного расщепления, клонировали в вектор 293 (приобретен в Invitrogen Inc., USA, номер по каталогу: K8300-01). После идентификации с помощью секвенирования ДНК получали клоны, содержащие корректно сконструированный эукариотический вектор экспрессии тяжелой цепи полноразмерного антитела или эукариотический вектор экспрессии легкой цепи полноразмерного антитела.
На Фиг. 1А схематически изображена структура вектора экспрессии тяжелой цепи рекомбинантного полноразмерного антитела к человеческому HER2 (293-VH-СН); и на Фиг. 1В схематически изображена структура вектора экспрессии легкой цепи рекомбинантного полноразмерного антитела к человеческому HER2 (293-VL-CL).
Пример 6. Экспрессия при транзиторной трансфекции антитела GB235-019 в эукариотических клетках и его очистка
Для экспрессии рекомбинантного вектора для антитела GB235-019, сконструированного в Примере 5, можно использовать метод котрансфекции FreeStyle 293F клеток (приобретены в Invitrogen, USA, номер по каталогу: R790-07). За 24 часа до трансфекции клетки FreeStyle 293F пересевали при плотности 6×105 клеток/мл и культивировали на термостатированной качалке в таких условиях (135 об/мин, 37°С, 8% СО2), чтобы на день трансфекции плотность клеток (определенная методом подсчета гемоцитов на планшете) составляла 1.2-1.5×106 клеток/мл. Клетки разводили в среде FreeStyle 293 (приобретены в Invitrogen Inc., USA, номер по каталогу: 12338-018) до плотности 1×106 клеток/мл. Для обеспечения наилучшей трансфекции жизнеспособность клеток (определенная методом окрашивания трипановым синим) должна быть выше 95%.
Реагент для трансфекции FreeStyle Max Reagent (полученный в Invitrogen Inc., USA, номер по каталогу: 16447-500) осторожно переворачивали (на 180°) 4 раза для получения однородной смеси. 315 мкг каждого из плазмидных векторов для экспрессии тяжелой цепи и легкой цепи, соответственно, добавляли в культуральный (питательный) раствор для трансфекции OptiPRO SFM (приобретен в Invitrogen Inc., USA, номер по каталогу: 12309-050). Объем дополняли до 10 мл раствором для трансфекции OptiPRO SFM и смесь перемешивали до однородного состояния. В другой центрифужной пробирке 625 мкл реагента FreeStyle Max Reagent разводили до 10 мл в растворе OptiPRO SFM. Пробирку осторожно переворачивали для однородного (гомогенного) смешения. Разбавленную смесь с плазмидой и разбавленный реагент FreeStyle Max Reagent перемешивали до однородного состояния и инкубировали при комнатной температуре в течение 15 минут. Полученный смешанный раствор (20 мл) медленно добавляли в качалочную колбу, содержащую 500 мл среды FreeStyle 293F (полученную в Invitrogen Inc., USA, номер по каталогу: 12338-018). Для культивирования качалочную колбу помещали на термостатированную качалку на 7 дней (135 об/мин, 37°С, 8% СО2). Культуру центрифугировали на рефрижераторной центрифуге при 9000 об/мин в течение 20 минут и супернатант собирали для последующей очистки белка.
Вышеуказанный супернатант клеток FreeStyle 293F, содержащий антитело GB235-019, центрифугировали. Антитело IgG1 изотипа помещали на колонку с Протеином А (приобретен в GE Healthcare Bio-Sciences Inc., USA, номер по каталогу: 17-5080-02) и элюировали, используя в качестве элюента 50 мМ буфер лимонная кислота-цитрат натрия (рН 3.3). Элюат собирали (0.5 мл) и нейтрализовали (до нейтрального рН), добавляя 100 мкл 1 М буфера трис(гидроксиметил)аминометан-соляная кислота (Tris-HCl) (рН 11.0). Затем проводили диализ элюента против фосфатного буферного раствора PBS (0.01 М Na2HPO4⋅12Н2О+0.002 М KH2PO4+0.14 М NaCl+0.002 М KCl, рН=7.2) с использованием диализной мембраны 10K (приобретена в Shanghai Generay Biotech Со, Ltd, номер по каталогу: M1915), и содержание белка определяли при OD280 нм. Полученный раствор фильтровали через фильтр 0.22 мкм (приобретен в Millipore Inc., Germany, номер по каталогу: GVHP01300) для стерилизации и фильтрат хранили при -80°С. После очистки проверяли чистоту антитела GB235-019 с помощью электрофореза в 10%-ном полиакриламидном геле в редуцирующих условиях в присутствии дитиотреитола с конечной концентрацией 50 мкМ.
Результаты, представленные на Фиг. 2, показывают, что в полностью редуцирующих условиях (условиях полного восстановления) антитело GB235-019 проявлялось в виде двух полос (пятен) с молекулярной массой 50 кДа и 25 кДа, соответственно, которые представляют собой полосы для тяжелой цепи и легкой цепи антитела (в качестве положительного контроля служил герцептин приобретен в Roche Corp.). Эти результаты показывают, что сконструированное антитело GB235-019 имело корректную структуру и его молекулярная масса соответствовала теоретическому (расчетному) значению.
Пример 7. Анализ антитела GB235-019 с пониженной молекулярной массой
К 10 мкг антитела GB235-019 добавляли дитиотреитол с конечной концентрацией 20 мМ и смесь инкубировали на водяной бане при 37°С в течение 30 минут, чтобы разорвать все межцепные дисульфидные связи. Разделенные легкую цепь и тяжелую цепь анализировали с помощью обращенно-фазовой хроматографии в сочетании с масс-спектрометрией. В исследовании использовали высокоэффективный жидкостной хроматограф Waters Н-Class Bio (Waters Inc., USA) с хроматографической колонкой: PLRP-S 300  , 3.0 мкм, 2.1×150 мм (приобретена в Agilent Inc., USA, номер по каталогу: 1912-3301); подвижная фаза: А (вода), В (ацетонитрил) и С (1% трифторуксусная кислота (TFA)), градиент приобретен в 35% В на 4ой минуте до 42% В на 20ой минуте, а содержание фазы С поддерживалось на уровне 10%, при этом скорость потока составляла 0.3 мл/мин, а нагрузка (нанесенный образец) составляла 20 мкг. Применяли масс-спектрометр Thermo LTQ-Orbitrap Discovery (Thermo Fisher Inc., USA), напряжение при впрыскивании пробы 3.7 кВ, цилиндрическая линза 230 В, температура в капилляре 300°С, разрешение 30000, и интервал отношения массы к заряду приобретен в 1000 до 3000. Теоретическая молекулярная масса тяжелой цепи (форма: Fc содержит G0F гликан), вычисленная с помощью программы GPMAW6.0, составляет 50416.7 Да, а теоретическая (вычисленная) молекулярная масса легкой цепи составляет 23120.8 Да. Обратную свертку (деконволюцию) исходных сигналов, получаемых масс-спектрометрией, проводили с использованием программы PROMASS, получая соответствующую молекулярную массу.
, 3.0 мкм, 2.1×150 мм (приобретена в Agilent Inc., USA, номер по каталогу: 1912-3301); подвижная фаза: А (вода), В (ацетонитрил) и С (1% трифторуксусная кислота (TFA)), градиент приобретен в 35% В на 4ой минуте до 42% В на 20ой минуте, а содержание фазы С поддерживалось на уровне 10%, при этом скорость потока составляла 0.3 мл/мин, а нагрузка (нанесенный образец) составляла 20 мкг. Применяли масс-спектрометр Thermo LTQ-Orbitrap Discovery (Thermo Fisher Inc., USA), напряжение при впрыскивании пробы 3.7 кВ, цилиндрическая линза 230 В, температура в капилляре 300°С, разрешение 30000, и интервал отношения массы к заряду приобретен в 1000 до 3000. Теоретическая молекулярная масса тяжелой цепи (форма: Fc содержит G0F гликан), вычисленная с помощью программы GPMAW6.0, составляет 50416.7 Да, а теоретическая (вычисленная) молекулярная масса легкой цепи составляет 23120.8 Да. Обратную свертку (деконволюцию) исходных сигналов, получаемых масс-спектрометрией, проводили с использованием программы PROMASS, получая соответствующую молекулярную массу.
Результат, представленный на Фиг. 3А, показывает, что найденная молекулярная масса легкой цепи антитела GB235-019 сопоставима с вычисленной (теоретической) молекулярной массой, что указывает на отсутствие гликозилирования в легкой цепи. Результат, представленный на Фиг. 3В, показывает, что найденная молекулярная масса тяжелой цепи антитела GB235-019 значительно отличается от вычисленной молекулярной массы (>1500 Da). С помощью выравнивания с теоретической последовательностью было найдено, что помимо Fc области, Fab каркасный фрагмент также содержал теоретический (расчетный) сайт N-гликозилирования (Asn-Thr-Ser), что приводит к увеличению молекулярной массы.
Пример 8. Конструирование эукариотических векторов для экспрессии GB235-019 с мутантной знаковой (signature) последовательностью N-связанного гликана на Fab конце тяжелой цепи.
Консервативный сайт N-гликозилирования представлял собой Asn-X-Thr/Ser, где X означал любую аминокислоту, отличную приобретен в Pro. N-связанный гликан был связан с остатком Asn в характеристической последовательности Asn-X-Ser/Thr (Imperiali В, O'Connor SE. Effect of N-linked glycosylation on glycopeptide and glycoprotein structure. Curr. Opin. Chem. Biol. 3 (6): 643-649). Сайт Asn73 в Fab каркасном фрагменте 3 полноразмерного антитела GB235-019, полученного в Примере 5, являлся сайтом N-гликозилирования. Консервативный сайт N-гликозилирования удаляли посредством мутации уникальной последовательности Asn-X-Ser/Thr (Walsh G. Biopharmaceutical benchmarks-2003. Nat Biotechnol, 2003, 21:865-870). Выравнивание с использованием открытых баз данных IgBLAST выявило присутствие комбинации Asp-Thr-Ser в гене зародышевой линии. Как показано с помощью выравнивания нуклеотидных последовательностей, кодон "ААС", соответствующий Asn, мог мутировать в "GAC", тем самым Asn заменяется на Asp. Как Asn, так и Gln представляют собой аминокислоты амидного типа, и они являются консервативными заменами друг для друга, поскольку Gln содержит на один метальный радикал в группе боковой цепи больше, чем Asn. Аналогично, может осуществляться мутация кодона "ААС", соответствующего Asn, в "CAG", для замены Asn на Gln. Выравнивание с использованием баз данных IgBLAST выявило присутствие Ala в положении Ser в гене зародышевой линии. Выравнивание с нуклеотидной последовательностью зародышевой линии показывает, что может осуществляться мутация кодона "ТСС", соответствующего Ser, в "GCC" для замены Ser на Ala.
Векторы для экспрессии тяжелой цепи мутантных антител для GB235-019 Fab фрагмента с мутантной знаковой (signature) последовательностью N-связанного гликана создавали точковой мутацией, с использованием эукариотического вектора для экспрессии тяжелой цепи полноразмерного антитела, полученного в Примере 5, в качестве матрицы (Kunkel, Т. A, et al. "Rapid and efficient site-specific mutagenesis without phenotypic selection. Proc. Natl. Acad. Sci, 1985(82):488-492). Были разработаны три схемы мутаций, которые представляли собой мутацию Asn73 (N73) в тяжелой цепи антитела GB235-019 в Asp73 (D73), мутацию Asn73 (N73) в тяжелой цепи антитела GB235-019 в Gln73 (Q73) и мутацию Ser75 (S75) в тяжелой цепи антитела GB235-019 в Ala75 (А75), соответственно. Были созданы праймеры для конструирования вектора экспрессии тяжелой цепи для GB235-019 антитела с вышеуказанными точковыми мутациями в знаковой (signature) последовательности N-связанного гликана на Fab конце GB235-019 антитела, эти праймеры выглядят следующим образом:
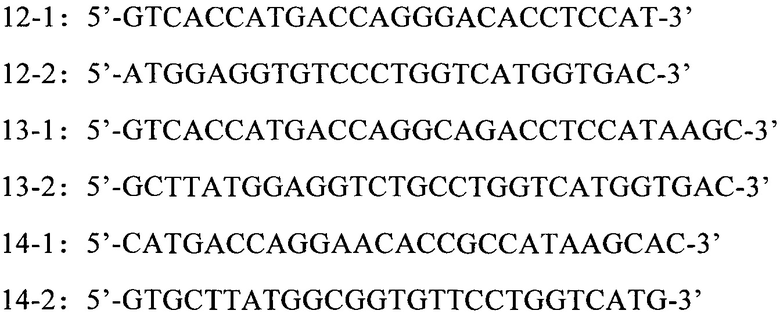
С использованием экспрессионного вектора 293-VH-CH, полученного в Примере 5, в качестве матрицы и 12-1 и 12-2 в качестве праймеров амплификацией методом PCR (ПЦР) был получен продукт ПЦР. Для расщепления продукта ПЦР 2 мкл Dpn I (приобретен в компании NEB Inc., USA, номер по каталогу: 1235А) добавляли к 20 мкл продукта ПЦР и выдерживали при 37°С в течение 1 часа. Продукт ПЦР очищали с применением набора для очистки продукта ПЦР (приобретен в компании Axygen Inc., USA, номер по каталогу: AP-PCR-50). DH5α Е. coli компетентные клетки (полученные в Tiangen Co., Ltd., номер по каталогу: СВ101) трансформировали очищенным продуктом ПЦР методом теплового шока (42°С, 90 секунд). После идентификации с помощью секвенирования ДНК получали эукариотический вектор для экспрессии тяжелой цепи мутантного антитела, который был обозначен "293-VH-CH-N73D" (который содержал вариабельную область тяжелой цепи, имеющую аминокислотную последовательность и нуклеотидную последовательность SEQ ID NO: 10 и SEQ ID NO: 13, соответственно). Аналогично, 13-1 и 13-2 использовали в качестве праймеров для получения продукта ПЦР амплификации методом ПЦР. 2 мкл Dpn I (приобретен в компании NEB Inc., USA, номер по каталогу: 1235А) добавляли к 20 мкл продукта ПЦР для расщепления при 37°С в течение 1 часа. Продукт ПЦР очищали с применением набора для очистки продукта ПЦР (приобретен в компании Axygen Inc., USA, номер по каталогу: АР-PCR-50). DH5α Е. coli компетентные клетки (полученные в Tiangen Co., Ltd., номер по каталогу: СВ101) трансформировали очищенным продуктом ПЦР методом теплового шока (42°С, 90 секунд). После идентификации с помощью секвенирования ДНК получали эукариотический вектор для экспрессии тяжелой цепи мутантного антитела, который был обозначен "293-VH-CH-N73Q" (который содержал вариабельную область тяжелой цепи, имеющую аминокислотную последовательность и нуклеотидную последовательность SEQ ID NO: 11 и SEQ ID NO: 14, соответственно). И подобным же образом 14-1 и 14-2 использовали в качестве праймеров для получения продукта ПЦР амплификацией методом ПЦР. 2 мкл Dpn I (приобретен в компании NEB Inc., USA, номер по каталогу: 1235А) добавляли к 20 мкл продукта ПЦР для расщепления при 37°С в течение 1 часа. Продукт ПЦР очищали с применением набора для очистки продукта ПЦР (приобретен в компании Axygen Inc., USA, номер по каталогу: AP-PCR-50). DH5α Е. coli компетентные клетки (полученные в Tiangen Co., Ltd., номер по каталогу: СВ101) трансформировали очищенным продуктом ПЦР методом теплового шока (42°С, 90 секунд). После идентификации с помощью секвенирования ДНК получали эукариотический вектор для экспрессии тяжелой цепи мутантного антитела, который был обозначен "293-VH-CH-S75A" (который содержал вариабельную область тяжелой цепи, имеющую аминокислотную последовательность и нуклеотидную последовательность SEQ ID NO: 12 и SEQ ID NO: 15, соответственно).
Пример 9. Экспрессия при транзиторной трансфекции GB235-019 мутантных антител в эукариотических клетках и их очистка.
Экспрессию рекомбинантных векторов для мутантных антител, созданных в Примере 8, осуществляли по методике, описанной в Примере 6. 293-VH-CH-N73D, 293-VH-CH-N73Q и 293-VH-CH-S75A применяли, соответственно, с 293-VL-CL для котрансфекции клеток FreeStyle 293F (полученных в Invitrogen, USA, номер по каталогу: R790-07).
Реагент для трансфекции FreeStyle Max Reagent (полученный в Invitrogen Inc., USA, номер по каталогу: 16447-500) осторожно переворачивали (на 180°) 4 раза для получения однородной смеси. 315 мкг каждой из экспрессионных плазмид для экспрессии тяжелой цепи и легкой цепи, соответственно, добавляли в культуральный (питательный) раствор для трансфекции OptiPRO SFM (полученный приобретен в Invitrogen Inc., USA, номер по каталогу: 12309-050). Объем доводили до 10 мл с помощью OptiPRO SFM и смесь перемешивали до однородного состояния. В другой центрифужной пробирке 625 мкл реагента FreeStyle Max Reagent доводили до 10 мл с помощью OptiPRO SFM. Пробирку осторожно переворачивали для гомогенного смешения. Разбавленную смесь с плазмидой и разбавленный реагент FreeStyle Мах Reagent перемешивали до гомогенного состояния и инкубировали при комнатной температуре в течение 15 минут. Полученный смешанный раствор (20 мл) медленно добавляли в качалочную колбу, содержащую 500 мл среды FreeStyle 293F (полученную в Invitrogen Inc., USA, номер по каталогу: 12338-018). Для культивирования качалочную колбу помещали на термостатированную качалку на 7 дней (135 об/мин, 37°С, 8% СО2). Культуру центрифугировали на рефрижераторной центрифуге при 9000 об/мин в течение 20 минут и супернатант собирали для последующей очистки белка. Способ очистки мутантных антител был такой же, как и в Примере 6. Полученные мутантные антитела фильтровали через фильтр 0.22 мкм (приобретен в Millipore Inc., номер по каталогу: GVHP01300) для стерилизации и фильтрат хранили при -80°С. Мутантные антитела, полученные в результате очистки, были обозначены как GB235-019N73D, GB235-019N73Q и GB235-019S75A. Чистоту и молекулярную массу мутантных антител проверяли, используя электрофорез в 10-ном полиакриламидном геле в восстанавливающих (редуцирующих) условиях в присутствии дитиотреитола с конечной концентрацией 50 мМ.
Результаты, представленные на Фиг. 4, показывают, что в условиях полного (исчерпывающего) восстановления (в полностью редуцирующих условиях) каждое из антител GB235-019N73D, GB235-019N73Q и GB235-019S75A имеет две полосы с молекулярной массой 50 кДа и 25 кДа, соответственно, для тяжелой цепи и для легкой цепи соответствующих антител (в качестве положительного контроля служил герцептин, полученный в Roche Corp.). Эти результаты показывают, что сконструированные антитела GB235-019N73D, GB235-019N73Q и GB235-019S75A имели корректные структуры и молекулярные массы, согласующиеся с вычисленным (теоретическим) значением.
Пример 10. Анализ рекомбинантных полноразмерных GB235-019 мутантных антител с пониженной молекулярной массой
Способ анализа мутантных антител GB235-019N73D, GB235-019N73Q и GB235-019S75A с пониженной молекулярной массой, полученных в Примере 9, был идентичен способу, описанному в Примере 7. К 10 мкг каждого из GB235-019 мутантных антител GB235-019N73D, GB235-019N73Q и GB235-019S75A, добавляли дитиотреитол с конечной концентрацией 20 мкМ и мутантные антитела инкубировали на водяной бане при температуре 37°С в течение 30 минут, чтобы разорвать межцепные дисульфидные связи. Разделенные легкую цепь и тяжелую цепь отдельно анализировали с помощью обращенно-фазовой хроматографии с масс-спектрометрией. В исследовании использовали высокоэффективный жидкостной хроматограф Waters Н-Class Bio (Waters Inc., USA) с хроматографической колонкой: PLRP-S 300  , 3.0 мкм, 2.1×150 мм (приобретен в Agilent Inc., USA, номер по каталогу: 1912-3301); подвижная фаза: А (вода), В (ацетонитрил) и С (1% TFA), градиент от 35% В на 4ой минуте до 42% В на 20ой минуте, а содержание фазы С поддерживается на уровне 10%, при этом скорость потока составляла 0.3 мл/мин, а нагрузка (нанесенный образец) составляла 20 мкг. Применяли масс-спектрометр Thermo LTQ-Orbitrap Discovery (Thermo Fisher Inc., USA), напряжение при впрыскивании пробы 3.7 кВ, цилиндрическая линза 230 В, температура в капилляре 300°С, разрешение 30000, и интервал отношения массы к заряду равен от 1000 до 3000. Теоретическая молекулярная масса тяжелых цепей (форма: Fc содержит G0F гликан) трех мутантных антител, вычисленная с помощью программы GPMAW6.0, составляет 50400.7 Да для GB235-019S75A, 50417.7 Да для GB235-019N73D и 50430.7 Да для GB235-019 N73Q. Обратную свертку исходных сигналов, получаемых масс-спектрометрией, проводили с использованием программы PROMASS, получая соответствующие молекулярные массы.
, 3.0 мкм, 2.1×150 мм (приобретен в Agilent Inc., USA, номер по каталогу: 1912-3301); подвижная фаза: А (вода), В (ацетонитрил) и С (1% TFA), градиент от 35% В на 4ой минуте до 42% В на 20ой минуте, а содержание фазы С поддерживается на уровне 10%, при этом скорость потока составляла 0.3 мл/мин, а нагрузка (нанесенный образец) составляла 20 мкг. Применяли масс-спектрометр Thermo LTQ-Orbitrap Discovery (Thermo Fisher Inc., USA), напряжение при впрыскивании пробы 3.7 кВ, цилиндрическая линза 230 В, температура в капилляре 300°С, разрешение 30000, и интервал отношения массы к заряду равен от 1000 до 3000. Теоретическая молекулярная масса тяжелых цепей (форма: Fc содержит G0F гликан) трех мутантных антител, вычисленная с помощью программы GPMAW6.0, составляет 50400.7 Да для GB235-019S75A, 50417.7 Да для GB235-019N73D и 50430.7 Да для GB235-019 N73Q. Обратную свертку исходных сигналов, получаемых масс-спектрометрией, проводили с использованием программы PROMASS, получая соответствующие молекулярные массы.
На Фиг. 5А показано, что полученная молекулярная масса тяжелой цепи GB235-019N73D соответствует вычисленной (теоретической) молекулярной массе, что указывает на отсутствие гликозилирования в тяжелой цепи. На Фиг. 5В и Фиг. 5С показано, что полученная молекулярная масса тяжелой цепи мутантных антител GB235-019N73Q и GB235-019S75A точно соответствует теоретической молекулярной массе (с разницей<1 Da), это означает, что сайт N-гликозилирования в Fab каркасной области удален.
В пределах СН2 области в Fc сегменте тяжелой цепи человеческого IgG имеется консервативный N-связанный сайт гликозилирования Asn297. Полисахаридная цепь, связанная по Asn297, может сохранять четвертичную структуру антитела и термическую устойчивость Fc сегмента и может регулировать антителозависимую клеточную цитотоксичность (ADCC), комплементзависимую клеточную цитотоксичность (CDC) и период полужизни, соответственно, за счет эффекта связывания молекулы IgG с FcRs, Clq и FcRn.
Модификация фрагмента Fab человеческого IgG с помощью N-гликозилирования может оказывать явное промотирующее или ингибирующее действие на функцию связывания антитела с антигеном. Минорные изменения в положении, в котором происходит модификация за счет гликозилирования, могут оказывать абсолютно отличное воздействие на последующую обработку полисахаридной цепи и активность связывания антитела с антигеном, и вызывать сложности в контроле качества в процессе получения антител. Мутантные антитела GB235-019N73D, GB235-019N73Q и GB235-019S75A были получены с помощью точковой мутации в антителе GB235-019 дикого типа. В трех мутантных антителах был изменен специфический сайт для N-связанного гликана (Asn-Thr-Ser) Fab тяжелой цепи в антителе GB235-019 дикого типа. Определение пониженной молекулярной массы трех мутантных антител выявило отсутствие гликозилирования в Fab тяжелой цепи, что может быть полезным для контроля качества в процессе получения. Также проводили анализ биологической активности и физико-химический анализ этих трех мутантных антител.
Пример 11. Идентификация иммунологической активности рекомбинантного полноразмерного антитела GB235-019 дикого типа и мутантных антител
Способность GB235-019 антитела дикого типа (GB235-019WT) и мутантных антител GB235-019N73D, GB235-019N73Q и GB235-019S75A связываться с человеческим HER2 антигеном оценивали, используя ELISA анализ связывания в соответствии с описанной ниже методикой. Человеческий HER2 антиген (полученный в Sino Biological Inc., номер по каталогу: 10004-Н08Н) разводили до концентрации 1 мкг/мл в буфере PBS и добавляли в планшет ELISA по 100 мкл/лунку и сенсибилизировали при 4°С в течение ночи. Планшет отмывали в PBST четыре раза, затем добавляли 5% BSA (приобретен в Amresco, USA, номер по каталогу: 0332-100 г, раствор PBS) 300 мкл/лунку для осуществления блокады при комнатной температуре в течение 1 часа. После четырехкратной отмывки планшета в PBST делали пятикратные разведения каждого из антител: антитела GB235-019WT, мутантных антител GB235-019N73D, GB235-019N73Q и GB235-019S75A, и пертузумаба (приобретен в Roche Corp.) и герцептина (приобретен в Roche Corp.), соответственно, начиная с концентрации 5 мкг/мл, получали 7 разведений в градиенте концентрации. Каждое из разведений помещали в планшет ELISA, 100 мкл/лунку, и инкубировали при комнатной температуре в течение 1 часа. Планшет отмывали четыре раза в PBST, затем меченное HRP антитело козы к Fc IgG человека (приобретен в CalBiochem Inc., USA, номер по каталогу: АР113А-K) разводили в PBS буфере в соотношении 1:10000, добавляли в планшет ELISA, по 100 мкл/лунку, и проводили инкубацию при комнатной температуре в течение 1 часа. Планшет отмывали четыре раза в PBST, затем добавляли раствор субстрата 3,3',5,5'-тетраметилбензидина (приобретен в Tiangen Co., Ltd., номер по каталогу: РА107-01) в количестве 100 мкл/лунку и инкубировали в течение 15 минут при комнатной температуре для визуализации. Добавляли стоп-реагент (1 М серную кислоту) по 50 мкл/лунку и считывали оптическую плотность при длине волны 450/630 нм на М5 многофункциональном планшетном ридере ELISA (Molecular Devices Inc., USA).
Результаты, представленные на Фиг. 6, показали, что мутантные антитела GB235-019N73D, GB235-019N73Q и GB235-019S75A обладают способностью специфически связываться с человеческим HER2 антигеном в зависимости от концентрации и до насыщения. Мутантные антитела GB235-019N73D, GB235-019N73Q и GB235-019S75A обладают активностью связывания, аналогичной активности связывания с антителом GB235-019WT, без существенных различий.
Пример 12. Рекомбинантные полноразмерные GB235-019 мутантные антитела ингибировали in vitro активность пролиферации опухолевых клеток ВТ-474 рака молочной железы
В клетках ВТ-474 рака молочной железы наблюдаются умеренные уровни экспрессии HER2 и HER3 и высокие уровни экспрессии P-HER2, но эти клетки не экспрессируют P-HER3 (Richard М. Neve. Acollection of breast cancer cell lines for the study of functionally distinct cancer subtypes. CANCER CELL, 2006, 515-527). Как говорится выше в данном описании, клетки ВТ-474 рака молочной железы представляют собой HER2-положительные опухолевые клетки. В анализе ингибирования пролиферации в полной культуральной среде, дополненной херегулином-α (приобретен в R&D Inc., USA, номер по каталогу: 296-HR), клетки ВТ-474 в log-фазе культивировали в 96-луночном культуральном планшете при плотности 5000 клеток/лунку в RPMI1640 полной культуральной среде (приобретена в Invitrogen Inc., USA, номер по каталогу: А10491), содержащей 10% фетальной бычьей сыворотки (приобретена в Invitrogen Inc., USA, номер по каталогу: 10099-141) при 37°С в 5% СО2 в течение 24 часов. Анализ ингибирования проводили, вводя антитело GB235-019WT и мутантные антитела GB235-019N73D, GB235-019N73Q и GB235-019S75A как отдельно, так и в комбинации с герцептином. В отдельной группе экспериментов добавляли, соответственно, антитело GB235-019WT и мутантные антитела GB235-019N73D, GB235-019N73Q и GB235-019S75A и пертузумаб и герцептин (с конечной рабочей концентрацией 75, 18.8, 4.7, 1.2, 0.29, 0.07, 0.018, 0.005, 0.0011 и 0 мкг/мл); и в группе с комбинированным введением каждое из антител GB235-019WT, GB235-019N73D, GB235-019N73Q, GB235-019S75A и пертузумаб вводили в каждой из вышеуказанных доз в комбинации с герцептином. После обработки антителом в течение 2 часов, как указано выше, добавляли раствор херегулина-α с конечной рабочей концентрацией 100 нг/мл, были также включены контрольные лунки без добавления раствора херегулина-α (для холостого опыта). Культивирование в планшете проводили при 37°С в 5% СО2 в течение 6 дней. Для оценки жизнеспособности клеток ВТ-474 добавляли реагент AlamarBlue (приобретен в Invitrogen Inc., USA, номер по каталогу: DAL1100) и значение флуоресценции считывали при длине волны 544/590 нм на М5 многофункциональном ELISA планшетном ридере (Molecular Devices Inc., USA).
Результаты анализа показали, что рекомбинантные полноразмерные мутантные антитела к HER2 GB235-019N73D, GB235-019N73Q и GB235-019S75A отменяли, in vitro, резистентность к герцептину HER2-положительных ВТ-474 клеток, индуцированную херегулином-α. Результат, представленный на Фиг. 7А, показал, что добавление херегулина-α в полную культуральную среду индуцировало пролиферацию ВТ-474 клеток. Клетки ВТ-474 становились нечувствительными к вводимому отдельно герцептину, и мутантные антитела GB235-019N73D, GB235-019N73Q и GB235-019S75A в равной степени не вызывали ингибирующего эффекта при раздельном введении, аналогично эффекту антитела GB235-019WT. Результат, представленный на Фиг. 7В, показал, что мутантные антитела GB235-019N73D, GB235-019N73Q и GB235-019S75A ингибировали индуцированную херегулином-α пролиферацию, когда каждое из них вводили в комбинации с герцептином. Совместное (комбинированное) введение (применение) каждого из трех мутантных антител с герцептином не только ингибировало пролиферацию клеток ВТ-474, индуцированную херегулином-α, но также заметно ингибировало до уровня ниже уровня, который был перед индукцией с помощью херегулина-α, в зависимости от концентрации. Эффект, вызываемый мутантными антителами GB235-019N73D, GB235-019N73Q и GB235-019S75A был аналогичен эффекту антитела GB235-019WT, без существенных различий.
Пример 13. In vitro ингибирование сигнальной трансдукции в клетках ВТ-474 рака молочной железы с помощью рекомбинантных полноразмерных GB235-019 антител к человеческому HER2
Клетки ВТ-474 в log-фазе культивировали в 6-луночном культуральном планшете при плотности 1.8×105 клеток/лунку в RPMI1640 полной культуральной среде (приобретена в Invitrogen Inc., USA, номер по каталогу: А10491), содержащей 10% фетальной бычьей сыворотки (приобретена в Invitrogen Inc., USA, номер по каталогу: 10099-141), при 37°С в 5% СО2 в течение 24 часов. На следующий день среду отбрасывали и вместо нее для инкубации еще в течение 24 часов использовали культуральную среду с низким содержанием сыворотки, содержащую 0.1% фетальной бычьей сыворотки (приобретен в Invitrogen Inc., USA, номер по каталогу: 10099-141).
Затем по отдельности вводили по 20 мкг/мл GB235-019WT и мутантного GB235-019N73D, герцептина и пертузумаба. После обработки клеток ВТ-474 антителами в течение 6 часов добавляли херегулин-α (приобретен в R&D Inc., USA, номер по каталогу: 296-HR) с конечной концентрацией 100 нг/мл для индукции в течение 15 минут, также имелись лунки для холостого контрольного опыта, в которые не добавляли херегулин-α. Планшет отмывали предварительно охлажденным PBS при 4°С, для прекращения реакции. Затем в лунки планшета добавляли 120 мкл LDS (приобретен в Invitrogen Inc., USA, номер по каталогу: NP0007) и планшет помещали на лед. Клеточный лизат сразу же быстро собирали и хранили при -80°С для последующего использования.
Проводили вестерн-блоттинг собранного клеточного лизата в восстанавливающих условиях в присутствии дитиотреитола (приобретен в Sangon Inc., номер по каталогу: D0281) с конечной концентрацией 50 мМ, чтобы определить воздействие антител на фосфорилирование HER-3, Akt и ERK1/2 в клетках SK-BR-3, индуцированное херегулином-α (полученным в R&D Inc., номер по каталогу: 296-HR). Вестерн-блоттинг (иммуноблот) осуществляли следующим образом. Белки на геле после электрофореза методом электрофоретического переноса (300 мА, 80 минут) переносили на NC мембрану (приобретена в Pall Inc., USA, номер по каталогу: S80209) и блокировали 5% сухим обезжиренным молоком (приобретено в Sangon Inc., номер по каталогу: NB0669). Затем добавляли, соответственно, кроличье первичное антитело к P-HER3 Y1289 (приобретено в Cell Signaling Technology Inc., USA, номер по каталогу: 8017) в разведении 1:1000, кроличье первичное антитело к HER3 (приобретено в Cell Signaling Technology Inc., USA, номер по каталогу: 12708) в разведении 1:1000, кроличье первичное антитело к Р-AktS473 (приобретено в Cell Signaling Technology Inc., USA, номер по каталогу: 4060) в разведении 1:1000, кроличье первичное антитело к Akt (приобретено в Cell Signaling Technology Inc., USA, номер по каталогу: 4691) в разведении 1:1000, кроличье первичное антитело к Р-ERK1/2 (приобретено в Cell Signaling Technology Inc., USA, номер по каталогу: 4370) в разведении 1:500, кроличье первичное антитело к ERK1/2 (приобретено в Cell Signaling Technology Inc., USA, номер по каталогу: 4695) в разведении 1:1000 и кроличье первичное антитело GAPDH (приобретено в Cell Signaling Technology Inc., USA, номер по каталогу: 5174) в разведении 1:5000 и инкубировали при 4°С в течение ночи. NC мембрану трижды отмывали в 1×TBST, затем добавляли HRP-меченное антитело козы к кроличьему иммуноглобулину (приобретен в MERCK Inc., USA, номер по каталогу: 401315) в разведении 1:10000. NC мембрану трижды отмывали в 1×TBST, затем добавляли ECL (приобретен в PerkinElmer Inc., USA, номер по каталогу: NEL104001EA) для визуализации. Пленку (приобретена в Kodak Inc., номер по каталогу: FF057) экспонировали, регистрируя сигнал.
Раковые клетки молочной железы ВТ-474 экспрессируют высокие уровни Р-HER2, но не экспрессируют P-HER3, и являются клетками линии, чувствительной герцептину. Результаты, представленные на Фиг. 8, показывают, что херегулин-α вызывал позитивную регуляцию фосфорилирования HER3 в ВТ-474 клетках по сравнению с контрольной группой без добавления херегулина-α. Отдельно введенное антитело GB235-019WT и мутантное антитело GB235-019N73D в значительной степени ингибировали позитивную регуляцию фосфорилирования HER3 в клетках ВТ-474, индуцированную херегулином-α, и полностью отменяли позитивную регуляцию фосфорилирования HER3, индуцированную херегулином-α, мутантное антитело GB235-019N73D вызывало такой же эффект, что и антитело GB235-019WT. Пертузумаб также полностью ингибировал позитивную регуляцию фосфорилирования HER3 в клетках ВТ-474, индуцированную херегулином-α, и герцептин также в значительной степени ингибировал позитивную регуляцию фосфорилирования HER3 в клетках ВТ-474, индуцированную херегулином-α.
Отдельно введенное антитело GB235-019WT и мутантное антитело GB235-019N73D не ингибировали позитивную регуляцию фосфорилирования Akt, вызванную херегулином-α. Отдельно введенный пертузумаб в значительной степени ингибировал позитивную регуляцию фосфорилирования Akt, вызванную херегулином-α. Отдельно введенное мутантное антитело GB235-019N73D и антитело GB235-019WT в значительной степени ингибировали позитивную регуляцию фосфорилирования ERK1/2, индуцированную херегулином-α, мутантное антитело GB235-019N73D вызывало такой же эффект, что и антитело GB235-019WT.
Пример 14. In vitro ингибирование сигнальной трансдукции с помощью рекомбинантных полноразмерных мутантных антител к HER2 человека в раковых клетках MCF7 молочной железы
Раковые клетки MCF 7 молочной железы экспрессируют HER2 с низкими уровнями и HER3 с высокими уровнями, но не экспрессируют P-HER2 и P-HER3 (Richard М. Neve. A collection of breast cancer cell lines for the study of functionally distinct cancer subtypes. CANCER CELL, 2006: 515-527). Согласно определению, приведенному выше, в данной заявке клетки MCF 7 рака молочной железы представляют собой HER2-отрицательные опухолевые клетки. Клетки MCF 7 в log-фазе культивировали в 6-луночном культуральном планшете при плотности 1.8×105 клеток/лунку в полной среде RPMI1640 (приобретена в Invitrogen Inc., USA, номер по каталогу: А10491), содержащей 10% фетальной бычьей сыворотки (приобретен B Invitrogen Inc., USA, номер по каталогу: 10099-141) в течение 24 часов. На следующий день среду отбрасывали и вместо нее применяли культуральную среду с низким содержанием сыворотки, содержащую 0.1% фетальной бычьей сыворотки, для выращивания клеток на минимальной среде в течение 24 часов. Клетки MCF 7 обрабатывали, добавляя 20 мкг/мл каждого из антител: антитела GB235-019WT и мутантных антител GB235-019N73D, GB235-019N73Q и GB235-019S75A, и пертузумаба, и герцептина, по отдельности или в комбинации в культуральной среде RPMI, содержащей 0.1% фетальной бычьей сыворотки, в течение 6 часов. Затем добавляли херегулин-α (приобретен в R&D Inc., USA, номер по каталогу: 296-HR) с конечной рабочей концентрацией 100 нг/мл для индукции в течение 10 минут, при этом были также включены контрольные лунки без добавления раствора херегулина-α (для холостого опыта). Планшет отмывали в предварительно охлажденном PBS при 4°С, чтобы остановить реакцию. Затем в лунки планшета добавляли 120 мкл LDS (приобретен в Invitrogen Inc., USA, номер по каталогу: NP0007) и планшет помещали на лед. Клеточный лизат сразу же быстро собирали и хранили при -80°С для последующего использования.
Проводили вестерн-блоттинг собранного клеточного лизата в восстанавливающих условиях в присутствии дитиотреитола с конечной концентрацией 50 мМ, чтобы определить действие антител на фосфорилирование HER-3 в клетках MCF 7, индуцированное херегулином-α, действие антител на фосфорилирование Akt и ERK1/2 после HER3. Вестерн-блоттинг осуществляли по той же методике, что и в Примере 13.
На Фиг. 9 показано ингибирование сигнальной трансдукции в HER2-отрицательных раковых клетках MCF 7 молочной железы GB235-019 мутантными антителами. Результаты показали, что херегулин-α индуцировал позитивную регуляцию (активацию) фосфорилирования HER3 в MCF 7 клетках по сравнению с контрольной группой без добавления херегулина-α. Каждое из вводимых отдельно антител: антитело GB235-019WT и мутантные антитела GB235-019N73D, GB235-019N73Q и GB235-019S75A - заметно ингибировало позитивную регуляцию фосфорилирования HER3 в клетках MCF 7, индуцированную херегулином-α; антитело GB235-019N73D полностью отменяло позитивную регуляцию фосфорилирования HER3, индуцированную херегулином-α; отдельно введенный герцептин также заметно ингибировал позитивную регуляцию фосфорилирования HER3 в клетках MCF 7, индуцированную херегулином-α; и пертузумаб полностью отменял позитивную регуляцию фосфорилирования HER3, индуцированную херегулином-α. Каждое из отдельно введенных антител - антитело GB235-019WT и мутантные антитела GB235-019N73D, GB235-019N73Q и GB235-019S75A - заметно ингибировало позитивную регуляцию фосфорилирования Akt в клетках MCF 7, индуцированную херегулином-α; и пертузумаб полностью отменял позитивную регуляцию фосфорилирования Akt, индуцированную херегулином-α. Каждое из отдельно введенных антител - антитело GB235-019WT и мутантные антитела GB235-019N73D, GB235-019N73Q и GB235-019S75A - заметно ингибировало позитивную регуляцию фосфорилирования ERK1/2 в клетках MCF 7, индуцированную херегулином-α; и пертузумаб полностью отменял позитивную регуляцию фосфорилирования ERK1/2, индуцированную херегулином-α. Следовательно, антитела по настоящему изобретению также можно применять для лечения HER2-отрицательных опухолей.
Пример 15. Анализ мутантного GB235-019 антитела методом гель-фильтрации (эксклюзионной хроматографии)
Мутантные GB235-019 антитела, GB235-019N73D, GB235-019N73Q и GB235-019S75A, хроматографировали методом эксклюзионной хроматографии (ситовой, гель-хроматографии, гель-фильтрации) (SEC-HPLC) для определения их чистоты. Ниже даются условия эксперимента: жидкостной хроматограф Waters 2695 (Waters Inc., USA); хроматографические колонки TSKgel G3000SWXL, расположенные последовательно (2 колонки) (приобретены в Waters Inc., USA); подвижная фаза 0.1 М фосфатный буфер, 0.1 М хлорид натрия, рН 7.0, 1.0 мл/мин, изократический режим в течение 30 минут; нагрузка 30 каждого из антител; и длина волны детектора 280 нм.
На Фиг. 10 показана эксклюзионная хроматограмма трех мутантных антител. Имеются небольшие количества полимеров и фрагментов, хотя их содержание мало. В Таблице 4 представлены результаты проверки чистоты соответствующих антител по данным эксклюзионной хроматографии (гель-фильтрации). Содержание (чистота) главного пика антитела GB235-019N73D составляет 88.3%, содержание (чистота) главного пика антитела GB235-019N73Q составляет 89.7% и содержание (чистота) главного пика антитела GB235-019S75A составляет 93.1%, содержание (чистота) главного пика для всех трех мутантных антител превышает 85%.
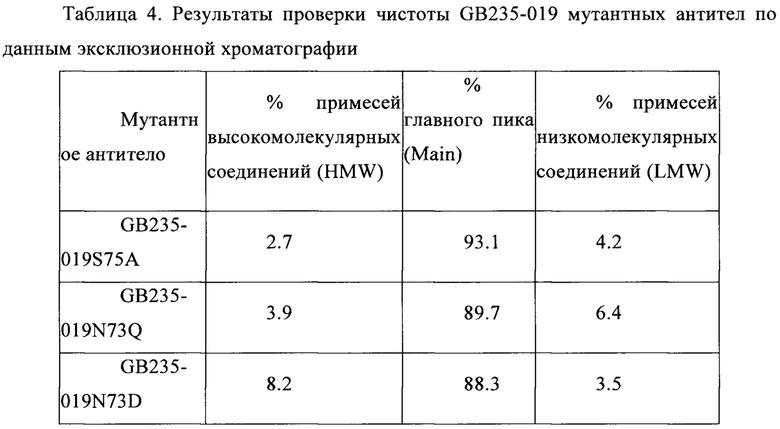
Пример 16. Визуализация GB235-019 мутантных антител методом капиллярного изоэлектрического фокусирования
Для определения изоэлектрической точки (pI) соответствующих главных пиков и чистоты зарядовых изомеров проводили анализ мутантных антител, соответственно, GB235-019N73D, GB235-019N73Q и GB235-019S75A методом капиллярного изоэлектрического фокусирования (iCIEF, КИЭФ). Ниже даются условия эксперимента: прибор для капиллярного изоэлектрического фокусирования Protein Simple iCE280 (Protein Simple Inc., USA); iCIEF картридж (приобретен в Protein Simple Inc., USA); раствор амфотерного электролита готовили, смешивая 12 мкл Pharmalyte 3-10 (приобретен в GE Inc., USA, номер по каталогу: 17045601), 0.5 мкл pI Marker 5.85 (приобретен в GE Inc., USA, номер по каталогу: 17-0472-01), 0.5 мкл pI Marker 9.77 (приобретен в GE Inc., USA, номер по каталогу: 17-0473-01), 2 мкл 500 мМ L-аргинина (приобретен в Sangon Inc., номер по каталогу: AB0205-100g), 2 мкл 200 мМ иминодиуксусной кислоты (приобретена в Sangon Inc., номер по каталогу: IB0530-100g), 70 мкл 1% метилцеллюлозы (приобретена в Sangon Inc., номер по каталогу: MB0616-250g) и 113 мкл 5.3 М мочевины (приобретена в Sangon Inc., номер по каталогу: UB0148-500g). Раствор для нанесения на колонку готовили, смешивая 180 мкл раствора амфотерного электролита с 20 мкл раствора белка с концентрацией 2.5 мг/мл. Систему отбора проб предварительно фокусировали при 1,500 В 1 минуту и в течение 10 минут фокусировали при 3,000 В. Паттерны фокусировки фиксировали, используя камеру CCD с длиной волны детектора 280 нм.
На Фиг. 11 представлены электрофореграммы, полученные в результате капиллярного изоэлектрического фокусирования трех мутантных антител на системе для капиллярного электрофореза. В Таблице 5 представлены данные для изоэлектрических точек соответствующих главных пиков и чистоты зарядовых изомеров трех мутантных антител. Экспериментальные значения для изоэлектрических точек трех мутантных антител находятся в интервале от около 9.4 до 9.6. Изоэлектрическая точка GB235-019N73D примерно на 0.1 ниже изоэлектрической точки GB235-019N73Q и GB235-019S75A, а степень чистоты его главного пика наивысшая по сравнению с GB235-019N73Q и GB235-019S75A.
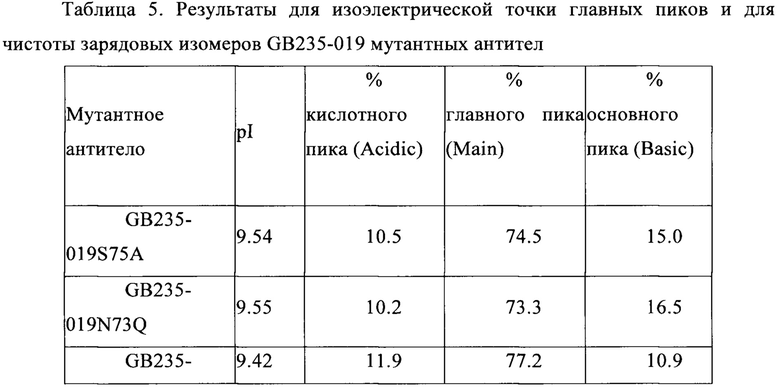

Пример 17. Анализ GB235-019 мутантных антител методом капиллярного гель-электрофореза
Для определения относительной (степени) чистоты (процентного содержания от суммы легкой цепи и тяжелой цепи) мутантных антител GB235-019N73D, GB235-019N73Q и GB235-019S75A, соответственно, проводили анализ методом капиллярного гель-электрофореза в редуцирующих (с восстанавливающим агентом) условиях (rCE-SDS). Ниже приводятся условия эксперимента. Для анализа использовали аналитический набор SDS-MW Analysis Kit (приобретен в Beckman Inc., USA, номер по каталогу: 390953) и непокрытую капиллярную трубку (приобретена в Micro solv Inc.). 100 мМ раствор Tris-HCl, рН 9.0, 1% SDS (приобретен в Sangon Inc., номер по каталогу: SB0485-100g) смешивали с β-меткаптоэтанолом (приобретен в Sigma Inc., USA, номер по каталогу: М6250) в соотношении 55:5. 60 мкл этого смешанного раствора смешивали с 40 мкл раствора образца с концентрацией 2.5 мг/мл и инкубировали на водяной бане при 70°С в течение 10 минут. Нанесение образца осуществляли при 5 кВ в течение 20 секунд, а разделение проводили при 15 кВ в течение 30 минут. Использовали UV (УФ)-детектор, установлена длина волны 214 нм.
На Фиг. 12 представлены электрофореграммы капиллярного электрофореза трех мутантных антител в редуцирующих условиях. В Таблице 6 приводятся результаты, полученные для (степени) чистоты по отношению к сумме легкой цепи и тяжелой цепи трех мутантных антител. Все три мутантных антитела содержали небольшое количество низкомолекулярных примесей и высокомолекулярных примесей. Антитело GB235-019N73D содержало наименьшее количество примесей по сравнению с другими антителами, и степень его чистоты относительно суммы легкой цепи и тяжелой цепи (LC+HC) была наивысшей.


Пример 18. Анализ GB235-019 мутантных антител методом дифференциальной сканирующей калориметрии
Для определения значения соответствующей Tm (температуры фазового перехода, означающей, что при этой температуре 50% биологических молекул являются неструктурированными) проводили анализ мутантных антител GB235-019N73D, GB235-019N73Q и GB235-019S75A, соответственно, методом дифференциальной сканирующей калориметрии (DSC). Условия эксперимента указаны далее. Использовали дифференциальный сканирующий калориметр MicroCal VP-Capillary DSC (GE Inc., USA) с объемом измерительной ячейки 130 мкл. Три мутантных антитела, соответственно, разводили в PBS (0.01 М Na2HPO4⋅12Н2О+0.002 М KH2PO4+0.14 М NaCl+0.002 М KCl, рН=8.6) до концентрации 0.9 мг/мл. PBS использовали в качестве контроля (холостой опыт), устанавливали изменение температуры от 30°С до 95°С, и скорость сканирования составляла 60°С/час.
На Фиг. 13 представлены диаграммы, построенные по результатам дифференциальной сканирующей калориметрии для трех GB235-019 мутантных антител. Температуры Tm1 указанных трех молекул отличались. Температура Tm1 антитела GB235-019N73Q была самой низкой по сравнению с температурами двух других антител, тогда Tm1 антитела GB235-019N73Q была близка к Tm1 антитела GB235-019S75A. Чем выше значение Tm, тем выше термическая устойчивость. Разница между Tm1 указывает на то, что в известной мере мутация оказывает определенное влияние на СН2 домен. GB235-019N73D и GB235-019S75A обладают повышенной термической устойчивостью по сравнению с GB235-019N73Q.
Ранее мы получили полностью человеческое моноклональное антитело GB235-019 к человеческому HER2 (Her-2/neu) скринингом фаговой библиотеки полностью человеческих scFV антител и экспрессией с использованием методов рекомбинантной ДНК (генная инженерия) (можно сослаться на китайскую заявку на патент №201410705404.0). Антитело GB235-019 может связываться с HER2 человека, HER2 обезьяны и мышиным HER2 в зависимости от концентрации и до насыщения, но оно не может связываться с человеческими антигенами HER1, HER3 и HER4. Введение GB235-019 в комбинации с герцептином может отменить резистентность клеток ВТ-474 к герцептину, индуцированную лигандом к HER3 херегулином-α, а отдельное введение GB235-019 ингибирует позитивную регуляцию фосфорилирования HER3 в ВТ-474 клетках, индуцированное херегулином-α, лигандом к HER3. Также введение антитела GB235-019 в комбинации с герцептином может отменить обусловленную лигандом к HER3, херегулином-α, резистентность клеток SK-BR-3 к герцептину, так же, как и к пертузумабу. Введение антитела GB235-019 в комбинации с герцептином может в значительной степени ингибировать рост ксенотрансплантата рака молочной железы человека (KPL-4) у мышей. Asn73 в Fab фрагменте каркасной области 3 антитела GB235-019 является сайтом N-гликозилирования, и благодаря определению антитела с пониженной молекулярной массой было обнаружено, что на GB235-019 Fab присутствует сложная полисахаридная цепь. Модификация человеческого IgG Fab посредством N-гликозилирования может оказывать явное промотирующее или ингибирующее действие на функцию связывания антитела с антигеном. Минорные изменения в положении, в котором происходит модификация посредством гликозилирования, могут оказывать абсолютно отличное воздействие на последующую обработку полисахаридной цепи и активность связывания антитела с антигеном, и вызывать сложности в контроле качества в процессе получения антител. Три мутантных антитела, GB235-019N73D, GB235-019N73Q и GB235-019S75A, были получены после введения точковой мутации в антитело GB235-019 дикого типа. В трех мутантных антителах была изменена знаковая (signature) последовательность для N-связанного гликана (Asn-Thr-Ser) Fab тяжелой цепи в антителе GB235-019 дикого типа. Обнаружение антитела с пониженной молекулярной массой подтвердило, что три мутантных антитела не являются гликозилированными в Fab фрагменте тяжелой цепи.
Идентификация иммунологической активности показала, что мутантные антитела GB235-019N73D, GB235-019N73Q и GB235-019S75A обладают способностью специфически связываться с человеческим антигеном HER2 в зависимости от концентрации и до насыщения (полное связывание). Эта способность мутантных антител GB235-019N73D, GB235-019N73Q и GB235-019S75A к связыванию аналогична способности к связыванию антитела GB235-019WT, без существенных различий. Введение каждого из мутантных антител, GB235-019N73D, GB235-019N73Q и GB235-019S75A, в комбинации с герцептином ингибировало пролиферацию, индуцированную херегулином-α. Введение каждого из трех мутантных антител в комбинации с герцептином не только ингибировало пролиферацию клеток ВТ-474, индуцированную херегулином-α, но также заметно ингибировало до уровня ниже уровня, который был перед индукцией с помощью херегулина-α, в зависимости от концентрации. Эффект, вызываемый мутантными антителами GB235-019N73D, GB235-019N73Q и GB235-019S75A, был аналогичен эффекту антитела GB235-019WT, без существенных различий. Отдельное введение мутантного антитела GB235-019N73D в значительной степени ингибировало позитивную регуляцию фосфорилирования HER3 в клетках ВТ-474, индуцированную херегулином-α, эффект мутантного антитела GB235-019N73D был аналогичен эффекту антитела GB235-019WT. Отдельное введение каждого из мутантных антител, GB235-019N73D и GB235-019WT, в значительной степени ингибировало позитивную регуляцию фосфорилирования ERK1/2 в клетках ВТ-474, индуцированную херегулином-α, при этом эффект мутантного антитела GB235-019N73D был аналогичен эффекту антитела GB235-019WT. Отдельное введение каждого из антител: антитела GB235-019WT и мутантных антител GB235-019N73D, GB235-019N73Q и GB235-019S75A, в заметной степени ингибировало позитивную регуляцию фосфорилирования HER3 в клетках MCF 7, индуцированную херегулином-α, а антитело GB235-019N73D полностью отменяло позитивную регуляцию фосфорилирования HER3 в клетках MCF 7, индуцированную херегулином-α. Отдельное введение каждого из антител: антитела GB235-019WT и мутантных антител GB235-019N73D, GB235-019N73Q и GB235-019S75A, заметно ингибировало позитивную регуляцию фосфорилирования Akt в клетках MCF 7, индуцированную херегулином-α. Отдельное введение каждого из антител: антитела GB235-019WT и мутантных антител GB235-019N73D, GB235-019N73Q и GB235-019S75A, заметно ингибировало позитивную регуляцию фосфорилирования ERK1/2 в клетках MCF 7, индуцированную херегулином-α, что также является подтверждением биологической активности этих трех мутантных антител. Суммируя вышесказанное, можно сделать вывод, что антитела по изобретению можно применять не только для лечения HER2-положительных опухолей, но также и для лечения HER2-отрицательных опухолей.
Мы анализировали физические и химические свойства мутантных антител GB235-019N73D, GB235-019N73Q и GB235-019S75A. Анализ мутантных антител методом эксклюзионной хроматографии (гель-фильтрации) показал, что чистота главного пика антитела GB235-019N73D составляет 88.3%, чистота главного пика антитела GB235-019N73Q составляет 89.7% и чистота главного пика антитела GB235-019S75A составляет 93.1%, т.е. чистота главного пика для всех трех мутантных антител выше 85%. Анализ антител методом капиллярного изоэлектрического фокусирования (iCIEF, КИЭФ) позволил определить изоэлектрическую точку (pI) соответствующих главных пиков и чистоту зарядовых изомеров трех мутантных антител. Экспериментально определенные значения изоэлектрических точек находятся в интервале примерно от 9.4 до 9.6. Изоэлектрическая точка GB235-019N73D примерно на 0.1 ниже изоэлектрической точки GB235-019N73Q и GB235-019S75A, а степень чистоты его главного пика наивысшая по сравнению с GB235-019N73Q и GB235-019S75A. Анализ методом капиллярного гель-электрофореза в редуцирующих условиях (rCE-SDS) позволил определить относительную чистоту (процентное содержание от суммы легкой цепи и тяжелой цепи) этих трех мутантных антител. Все три мутантных антитела содержали небольшие количества низкомолекулярных примесей и высокомолекулярных примесей. Антитело GB235-019N73D содержало наименьшее количество примесей по сравнению с другими антителами, и степень его чистоты относительно суммы легкой цепи и тяжелой цепи (LC+HC) была наивысшей. Анализ методом дифференциальной сканирующей калориметрии (DSC) позволил определить значение соответствующей Tm (температуры фазового перехода, означающей, что при этой температуре 50% биологических молекул являются неструктурированными) трех мутантных антител. Температуры Tm1 указанных трех молекул отличались. Температура Tm1 антитела GB235-019N73Q была самой низкой по сравнению с температурами двух других антител, тогда Tm1 антитела GB235-019N73Q была близка к Tm1 антитела GB235-019S75A. Чем выше значение Tm, тем выше термическая устойчивость. Разница между Tm1 указывает на то, что в известной мере мутация оказывает определенное влияние на СН2 домен. Антитела GB235-019N73D и GB235-019S75A обладают повышенной термической устойчивостью по сравнению с GB235-019N73Q.
| название | год | авторы | номер документа |
|---|---|---|---|
| СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СПЕЦИФИЧНЫЕ ПО ОТНОШЕНИЮ К HER3, И ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2620068C2 |
| КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ АНТИТЕЛО, КОТОРОЕ СВЯЗЫВАЕТСЯ С ДОМЕНОМ II HER2, И ЕГО КИСЛЫЕ ВАРИАНТЫ | 2009 |
|
RU2543664C2 |
| НЕКОНКУРЕНТНЫЕ В ОТНОШЕНИИ НЕЙРЕГУЛИНА АЛЛОСТЕРИЧЕСКИЕ АНТИТЕЛА ПРОТИВ ЧЕЛОВЕЧЕСКОГО HER3 И ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2704228C2 |
| АНТИ-НЕR2 АНТИТЕЛО И ЕГО КОНЪЮГАТ | 2014 |
|
RU2656161C1 |
| ВВЕДЕНИЕ ФИКСИРОВАННЫХ ДОЗ HER-АНТИТЕЛ | 2005 |
|
RU2438705C2 |
| КОМПОЗИЦИЯ АНТИТЕЛ К HER2 | 2005 |
|
RU2361880C2 |
| АНТИТЕЛА, СПОСОБНЫЕ СПЕЦИФИЧЕСКИ СВЯЗЫВАТЬСЯ С HER2 | 2014 |
|
RU2628094C2 |
| КОМПОЗИЦИИ АНТИТЕЛ | 2005 |
|
RU2426554C2 |
| МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, АНАЛОГИ АНТИТЕЛ, КОМПОЗИЦИИ И СПОСОБЫ | 2010 |
|
RU2580038C2 |
| ИДЕНТИФИКАЦИЯ ПАЦИЕНТОВ, НУЖДАЮЩИХСЯ В СОВМЕСТНОЙ ТЕРАПИИ С ИСПОЛЬЗОВАНИЕМ ИНГИБИТОРА PD-L1 | 2013 |
|
RU2692773C2 |









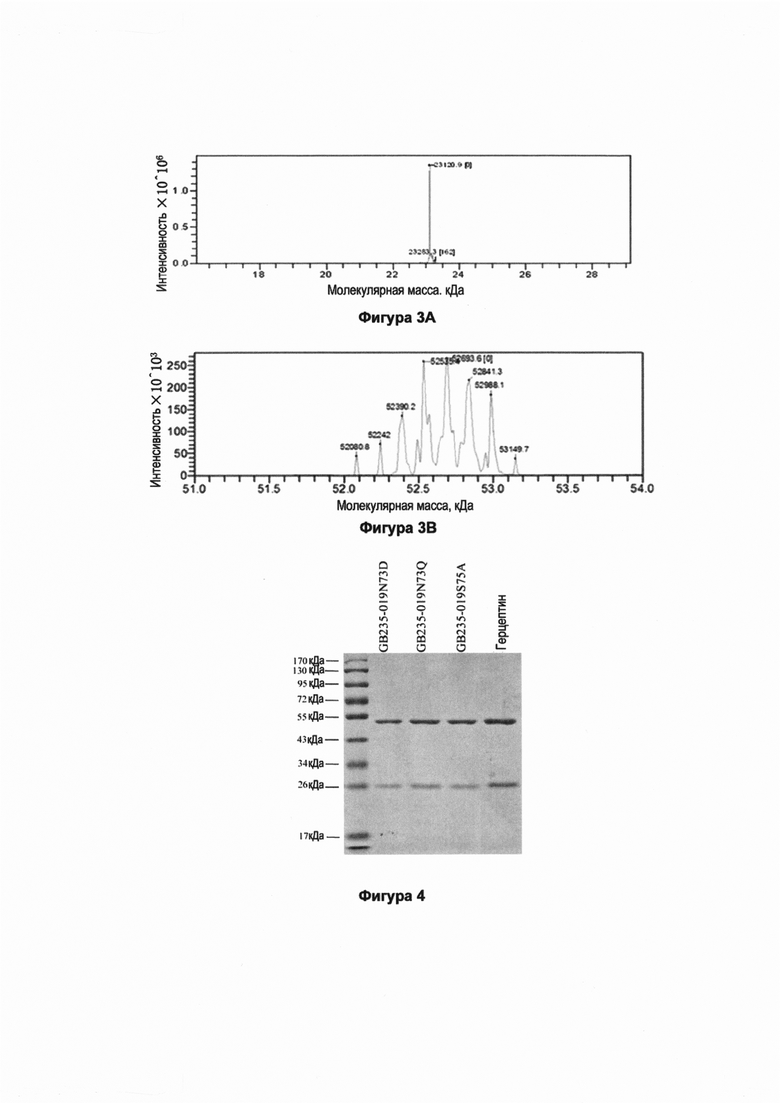
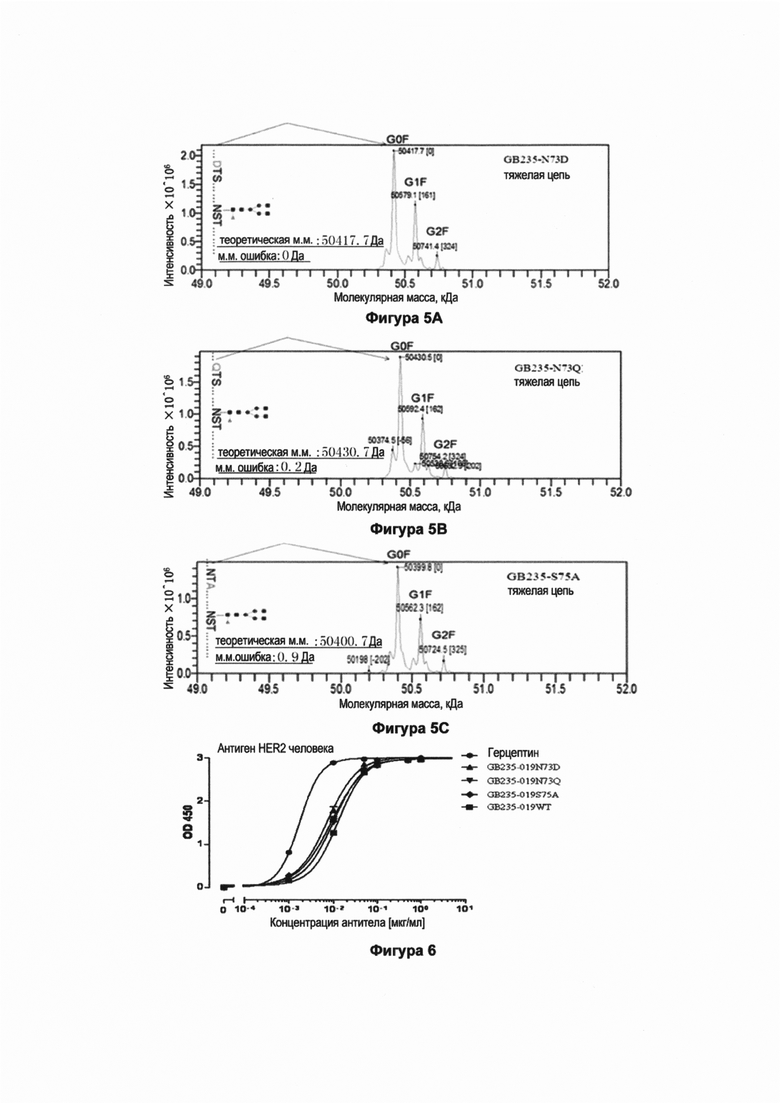
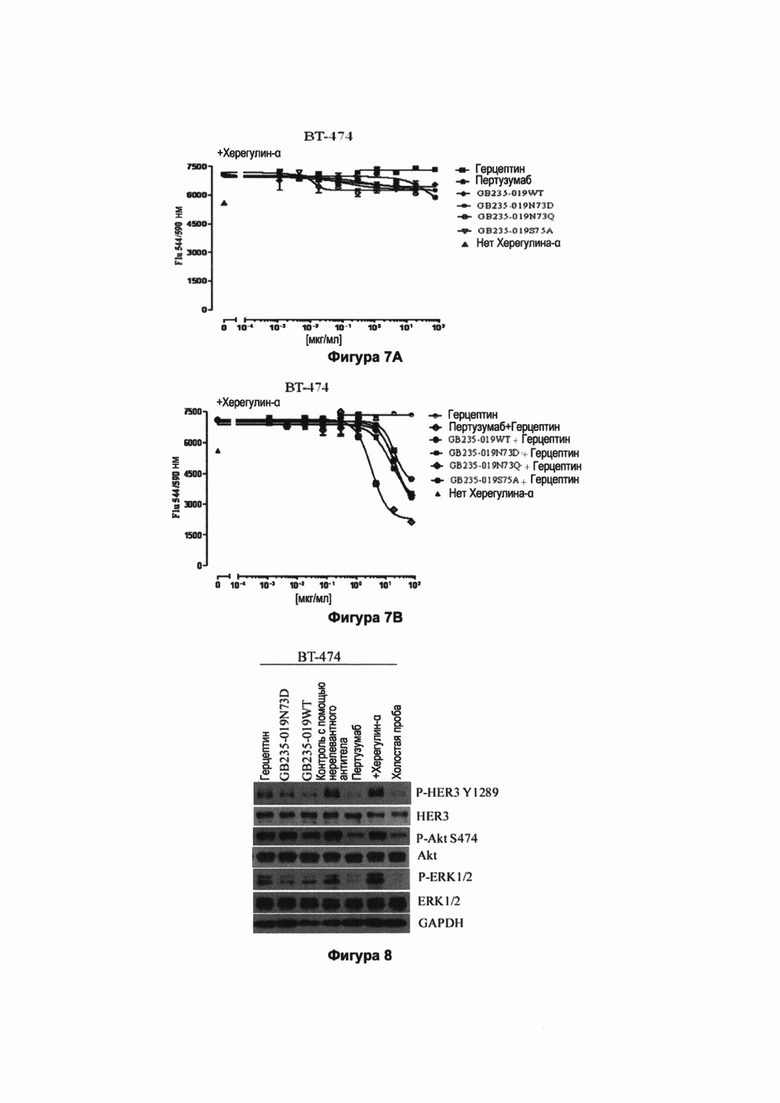
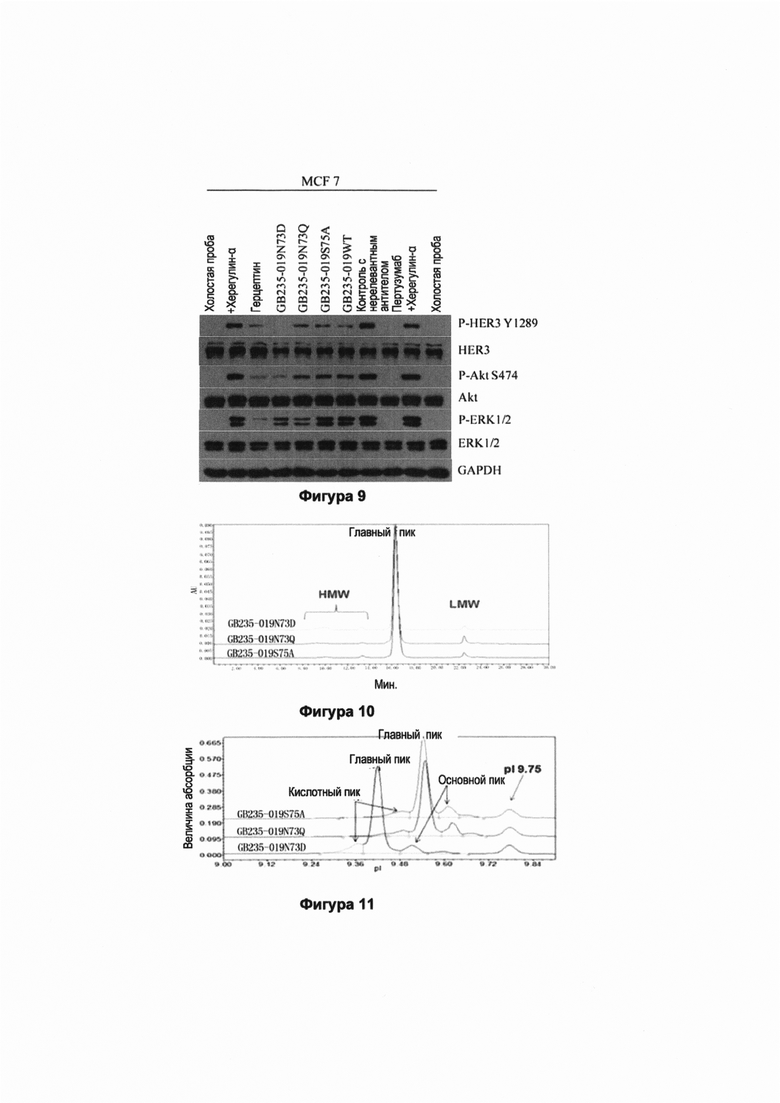
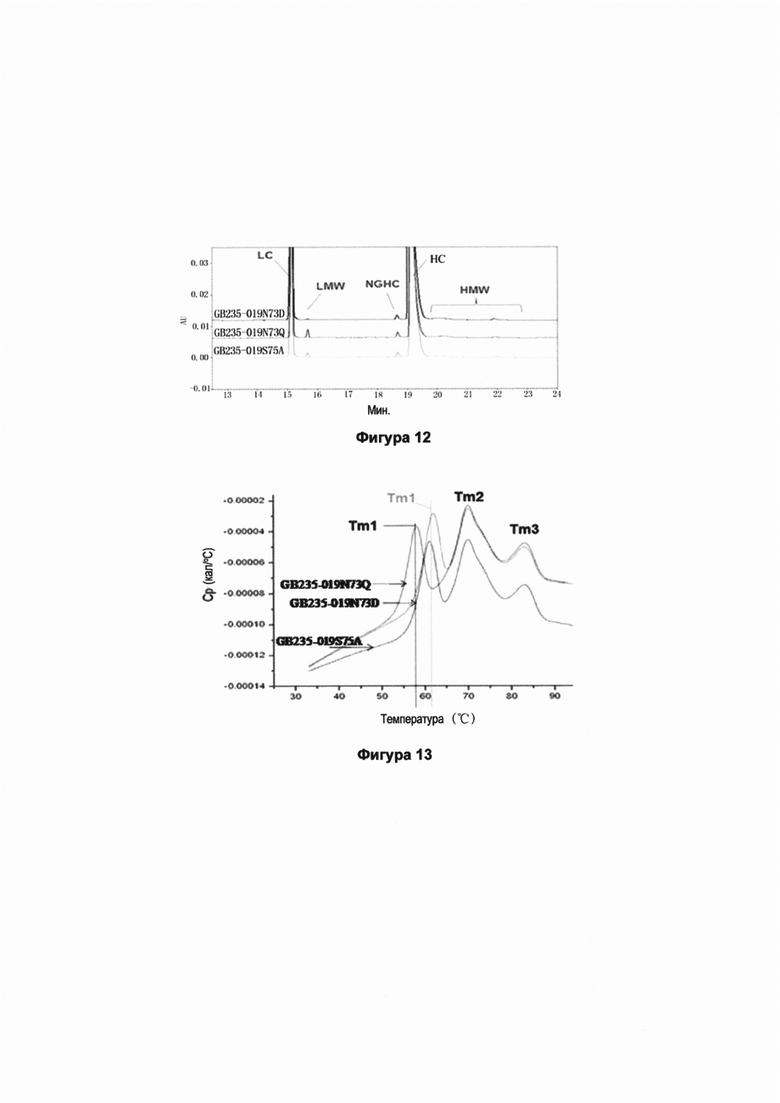
Изобретение относится к биохимии. Описано мутантное полностью человеческое антитело против HER2, при этом аминокислотная последовательность вариабельной области тяжелой цепи и аминокислотная последовательность вариабельной области легкой цепи антитела представляют собой SEQ ID NO: 10 и SEQ ID NO: 2; SEQ ID NO: 11 и SEQ ID NO: 2; или SEQ ID NO: 12 и SEQ ID NO: 2, соответственно. Также представлены нуклеиновая кислота, кодирующая описанное антитело. Вектор, содержащий такую кислоту. Композиция, содержащая описанное антитело. Изобретение может использоваться в комбинации с дополнительными терапевтическими агентами против HER2-положительных опухолей для лечения HER2-положительной опухоли, слабо положительной опухоли и отрицательной опухоли. 6 н. и 3 з.п. ф-лы, 13 ил., 6 табл., 18 пр.
1. Полностью гуманизированное антитело, специфически связывающееся с HER2, в котором аминокислотная последовательность вариабельной области тяжелой цепи и аминокислотная последовательность вариабельной области легкой цепи антитела представляют собой SEQ ID NO: 10 и SEQ ID NO: 2; SEQ ID NO: 11 и SEQ ID NO: 2; или SEQ ID NO: 12 и SEQ ID NO: 2, соответственно.
2. Антитело по п. 1, которое находится в виде Fab, Fab', F(ab')2, Fv или scFv.
3. Антитело по п. 1, которое содержит константную область тяжелой цепи и константную область легкой цепи человеческого IgG; предпочтительно, человеческий IgG представляет собой IgG1, и, наиболее предпочтительно, аминокислотная последовательность константной области тяжелой цепи человеческого IgG представляет собой SEQ ID NO: 5 и аминокислотная последовательность константной области легкой цепи человеческого IgG представляет собой SEQ ID NO: 6.
4. Нуклеиновая кислота, кодирующая полностью гуманизированное антитело по любому из пп. 1-3, в которой, предпочтительно, нуклеиновая кислота, кодирующая вариабельную область тяжелой цепи, представляет собой SEQ ID NO: 13, SEQ ID NO: 14 или SEQ ID NO: 15, и нуклеиновая кислота, кодирующая вариабельную область легкой цепи, представляет собой SEQ ID NO: 4.
5. Экспрессионный вектор, включающий нуклеиновую кислоту по п. 4, причем указанная нуклеиновая кислота функционально связана с последовательностью, контролирующей экспрессию указанного экспрессионного вектора; и указанный экспрессионный вектор представляет собой вектор pGEM-T или вектор 293.
6. Фармацевтическая композиция для лечения рака, содержащая в эффективном терапевтическом количестве полностью гуманизированное антитело, специфически связывающееся с HER2, по любому из пп. 1-3, и фармацевтически приемлемый носитель.
7. Комбинированное лекарственное средство для лечения рака, содержащее в эффективном терапевтическом количестве полностью гуманизированное антитело, специфически связывающееся с HER2, по любому из пп. 1-3 и дополнительный (-ые) терапевтический (-ие) агент (-ы) против HER2-положительных опухолей, причем указанный дополнительный (-ые) терапевтический (-ие) агент (-ы) против HER2-положительных опухолей является герцептином или пертузумабом.
8. Применение полностью гуманизированного антитела, специфически связывающегося с HER2 по любому из пп. 1-3 для приготовления лекарственного средства для лечения HER2-положительной опухоли, слабо положительной опухоли и отрицательной опухоли у субъекта, причем, предпочтительно, указанная HER2-положительная опухоль выбрана из HER2-положительного рака молочной железы, рака желудка, рака легкого, немелкоклеточного рака легких, рака кости, рака поджелудочной железы, рака кожи, рака головы и шеи, меланомы кожи или внутриглазной меланомы, рака матки, рака яичника, рака прямой кишки, рака заднего прохода, рака толстой кишки, рака фаллопиевой маточной трубы, рака эндометрия, рака шейки матки, рака влагалища, рака наружных половых органов (вульвы), болезни Ходжкина, рака пищевода, рака тонкой кишки, рака эндокринной системы, рака щитовидной железы, рака паращитовидной железы, рака надпочечной железы, рака мягких тканей, рака мочеточника, рака полового члена, рака простаты, рака мочевого пузыря, рака почки или уретры, почечно-клеточного рака, рака почечной лоханки, мезотелиомы, печеночно-клеточного рака, рака желчного пузыря, хронического или острого лейкоза, рака лимфатической системы, рака центральной нервной системы (ЦНС), рака позвоночника, нейроглиомы ствола головного мозга, многоформной глиобластомы, астроцитомы, нейролеммомы, эпендимомы, медуллобластомы, менингиомы, сквамозно-клеточной карциномы и аденомы гипофиза.
9. Применение по п. 9, где указанный субъект является человеком, имеющим HER2-положительную опухоль, слабо положительную опухоль и отрицательную опухоль.
| CN 103153339 А, 12.06.2013 | |||
| КОМПОЗИЦИЯ АНТИТЕЛ К HER2 | 2005 |
|
RU2361880C2 |

