Область техники, к которой относится изобретение
Изобретение относится к области биоинженерии, в частности к разработке метода получения α-амидазы человека (Nit2) – ключевого фермента метаболизма азота в мозге человека. Это фермент катализирует гидролиз α-кетоглутарамата с образованием α-кетоглутарата. Получаемый рекомбинантный фермент может быть использован, в частности, при разработке ингибиторов, представляющих собой потенциальные средства медикаментозного лечения печеночной энцефалопатии.
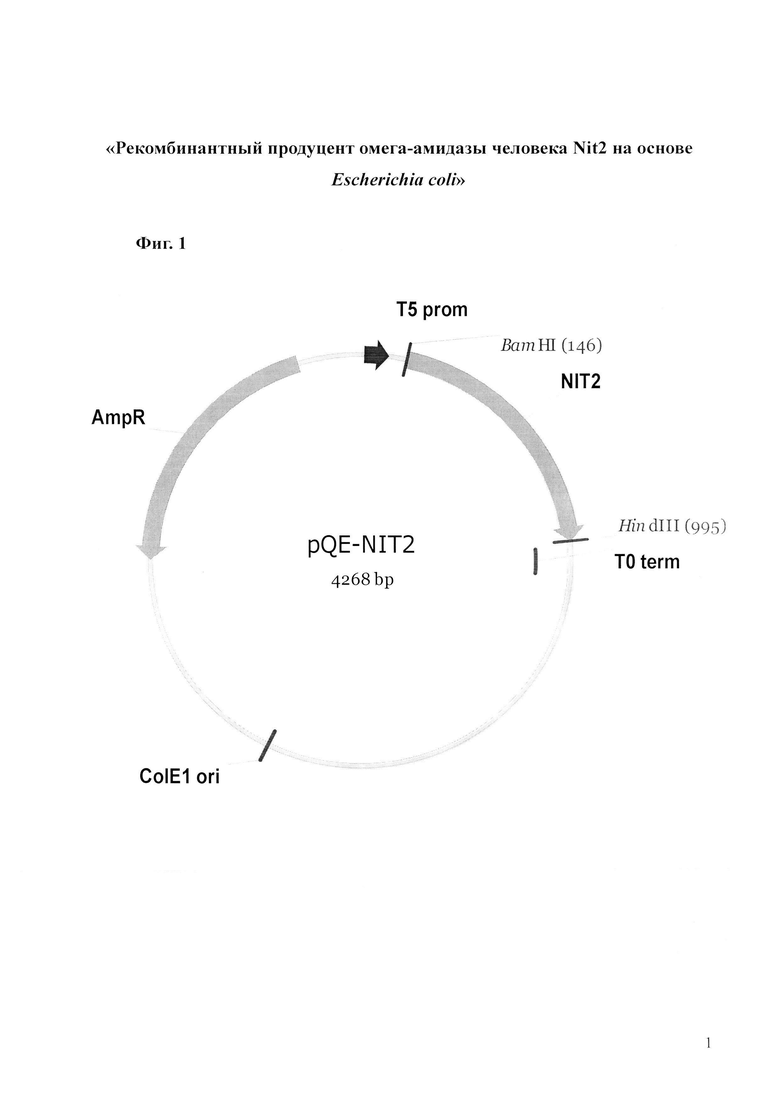
Предметом изобретения является последовательность генетической конструкции pQE-Nit2 (находится под контролем промотора фага Т5), а также способ ее введения в штаммы Escherichia coli и определения его ферментативной активности. Особенностью гена Nit2 в составе конструкции pQE-Nit2 является оптимизированный кодоновый состав, а также наличие искусственно введенной аффинной метки 6His-таг непосредственно после инициаторного кодона ATG (Met), обеспечивающего возможность одностадийной очистки продукта методом металлоаффинной хроматографии на Ni-NTA-агарозе. Оптимизация кодонового состава гена Nit2 проведена таким образом, чтобы вблизи стартовой точки трансляции (первые тридцать кодонов) находилось несколько кодонов, редко встречающихся в природных открытых рамках считывания генома E.coli, а остальные приходились на кодоны с умеренной встречаемостью. На остальном протяжении открытой рамки считывания использовались только кодоны, наиболее часто употребляемые в открытых рамках считывания генома E.coli.
Уровень техники
Выбор E.coli в качестве объекта для получения Nit2 обусловлен тем, что в геноме этого организма не содержится собственного гена ω-амидазы, хотя такой ген есть у большинства биологических видов организмов, в том числе у высших растений, водорослей, животных дрожжеподобных и плесневых грибов и бактерий рода Rhizobium. Энзиматическая цепочка в составе глутаминтрансаминазы и ω-амидазы, первоначально описанная в различных тканях млекопитающих [Jaisson et al Biochimie. 2009; 91(9): 1066-10671], недавно обнаружена в растениях (томат, кукуруза) и бактериях (Bacillus subtilis) [Ellens K.V., Richardson L.G.L., Frelin O., Collins J., Leite Ribeira C., Hsieh Y., Mullen R.T., Nanson A.D. Evidence that glutamine transaminase and omega-amidase potentially act in tandem to close the methionine salvage cycle in bacteria and plants. Phytochemistry, V. 3, 2015, pp.160–169]. Таким образом, число природных объектов, лишенных собственной активности Nit2, может оказаться ограниченным, причем только небольшая доля среди них допускает создание рекомбинантных продуцентов.
Характеризуя уровень техники в области создания продуцентов ω-амидазы Nit2 человека и шире, амидаз животных, необходимо указать на публикацию Jaisson et al. [Biochimie. 2009; 91(9): 1066-10671 «Молекулярная идентификация омега-амидазы Nit2, фермента, функционального связанного с глутаминтрнасаминазой, в качестве потенциального онкосупрессора»]. В публикации описано конструирование двух рекомбинантных продуцентов на основе E.coli с использованием кДНК генов мыши, идентифицированных по известной аминокислотной последовательности, как представителей семейства нитрилазы – NIT1 и NIT2. Авторы очистили соответствующие рекомбинантные продукты экспрессии и показали, что продукт гена NIT2 способен гидролизовать α-кетоглутарамат и другие известные субстраты α-амидазы, а продукт гена NIT1 не обладает такой способностью. Анализируя назначение гена NIT2 млекопитающих, этом авторы акцентируются на функциональной сцепленности ω-амидазы и глутаминтрнасаминазы. Они отмечают, что в геноме Bacillus subtilis (организм, не родственный человеку в таксономическом отношении) непосредственно рядом с геном глутаминтрансаминазы mtnV имеется ген, продукт которого по последовательности может быть идентифицирован как «углерод-азот-гидролаза», т.е. потенциально может оказаться ω-амидазой.
В работе Krasnikov et al. [Anal. Biochem. 2009; 391(2):144-150 «Исследование активности и выделение омега амидазы/Nit2, повсеместно распространенного онкосупрессора, катализирующего дезаминирование альфа-кетокислот - аналогов глутамина и аспарагина»] сообщается, что ω-амидазой (омега-амидодикарбоксилатаминогидролазой, EC 3.5.1.3) называется фермент из цитозоля клеток печени крысы, обладающий активностями амидазы, трансамидазы и эстеразы. Достаточно высокий уровень активности ω-амидазы в отношении α-кетоглутарамата и α-кетосукцинамата наблюдается в клетках млекопитающих, включая опухолевые линии, в растениях, бактериях и грибах. Однако количественные данные о назначении и физиологически обусловленной регуляции активности этого фермента до настоящего времени остаются недостаточными, что во многом обусловлено недоступностью коммерческих препаратов α-кетоглутарамата. Авторы разработали препаративный метод получения этого соединения и колориметрический метод определения ω-амидазы с использованием 96-луночных плоскодонных полистироловых планшетов. Они предложили также метод определения активности ω-амидазы с использованием в качестве субстрата коммерчески доступного сукцинамата. Продукт этой реакции - сукцинил гидроксамат - в кислой среде дает окрашенный комплекс с хлоридом железа, который может быть легко измерен колориметрическим методом. Характеризуя специфичность своего метода определения активности, авторы отмечают, что измеряя соотношение активности фракций клеточного лизата на различных стадиях фракционирования (в конечном итоге фермент был обогащен в 9600 раз по сравнению с исходным материалом), они наблюдали неизменное соотношение активностей по двум субстратам: α-кетоглутарамату и сукцинамату. Актуальность своей работы авторы, в первую очередь, аргументируют перспективностью использования ω-амидазы в качестве онкосупрессора.
В работе Chien et al. [Biol. Chem. 2012; 287(31): 25715-25726 «Особенности структуры каталитического центра Nit2/ω-амидазы человека: кинетические исследования и симуляция молекулярной динамики»] сообщается, что ω-амидаза человека hNit2 наряду с гидролизом α-кетоглутарамата и α-кетосукцинамата способна осуществлять реакцию трансаминирования между глутамином и α-кето-γ-метиолбутиратом, промежуточным метаболитом синтеза метионина. Таким образом, hNit2 может иметь отношение к включению серы в метаболиты цикла Кребса. С целью исследовать возможности каталитического действия hNit2 авторы провели симуляцию фермент-субстратного взаимодействия фермента дикого типа и его гипотетических мутантных вариантов: E43A, K112A и C153A, а также варианта с делецией петли, расположенной между остатками 116 и 128. Авторы считают, что их данные вносят существенный вклад в создание ингибиторов ω-амидаз – потенциальных лекарственных препаратов для лечения рака и гипераммониемии. Появление данной работы свидетельствует о недоступности системы рекомбинантной продукции ω-амидазы человека Nit2, в результате чего экспериментальные данные исследования мутантных вариантов фермента пришлось заменить теоретическими расчетами.
Заключая анализ уровня техники в области создания продуцентов ω-амидазы NIT2 человека, необходимо отметить, что компания US Biologicals (США) коммерчески распространяет этот фермент в варианте, имеющем на N-конце аффинную метку «6His-таг», получаемый в клетках рекомбинантного штамма E.coli. Известен также набор для определения ω-амидазы человека NIT2(NIT2) с помощью иммуноферментного анализа производства компании Cusabio Biotech (каталожный номер CSB-EL015823HU), а также аналогичный набор того же производителя для определения ω-амидазы быка (CSB-EL015823BO) и мыши производства Antibody Research Corporation (#789848). Таким образом, рекомбинантные продуценты ω-амидазы NIT2 человека на основе E.coli существуют, однако, они не описаны в патентной литературе и других открытых источниках.
Анализируя уровень техники в области промышленного применения амидаз человека, необходимо отметить патент [Dasse et al, WO 2008100977 A2, опубл. 21.08.2008], в котором излагается результат отбора соединений, обладающих активностью ингибитора амидазы жирных кислот (FAAH). Авторы этого патента отмечают, что для всех известных амидаз вне зависимости от типа субстратной специфичности характерно наличие консервативного участка последовательности, обогащенного глицином, серином и аланином. В последовательности амидазы жирных кислот человека этот маркерный участок располагается между адресами 215 и 257. В работах [Chebrou H., et al., Study of the amidase signature group.Biochim Biophys Acta 1996; 1298: 285-293; Kobayashi M., et al. Identification of active sites in amidase: evolutionary relationship between amide bond- and peptide bond-cleaving enzymes. Proc Natl Acad Sci USA 1997; 94: 11986- 11991] описывается еще 17 единичных аминокислот, участвующих в формировании активного центра амидаз семейства FAAH, удобных для идентификации генов амидаз в базах неаннотированных последовательностей. В работе [Cravatt B. F., et al. Molecular characterization of an enzyme that degrades neuromodulatory fatty-acid amides. Nature 1996; 384: 83-87] обнаружено, что белки семейства FAAH вне зависимости от происхождения имеют одинаковую длину последовательности: 579 а.о.
Практический интерес к FAAH в качестве мишени для фармацевтического воздействия (создания ингибиторов) обусловлено высокой биологической активностью целого ряда амидов жирных кислот, разлагаемых FAAH. К этой группе относится прежде всего, анандамид, а также N-пальмитоил этаноламид, N-олеоил этаноламид, олеамид, 2-арахидоноилглицерин [Deutsch et al. 2003 Prostaglandins Leukot Essent Fatty Acids 66:201; and Cravatt and Lichtman 2003 Current Opinion in Chemical Biology 7:469]. Подавление активности FAAH должно приводиь к повышению уровня накопления анандамида и других амидов жирных кислот. Накопление этих соединений, в свою очередь, должно приводить к увеличению болевого порога. Таким образом, ингибиторы FAAH рассматриваются как потенциальные анестетики. Они перспективны также в качестве средства воздействия на канабиоидные рецепторы при разработке успокаивающих препаратов, регуляторов аппетита, средств для лечения расстройств сердечно-сосудистой системы.
Известен целый ряд природных и неприродных лигандов канабиоидных рецепторов. Среди них наиболее активно изучается анандамид – арахидоноилэтаноламид [Devane et al Science 1992; 258: 1946-1949; Piomelli, D. Nat Rev. Neurosci. 2003; 4: 873-884]. Существуют доказательства того, что этот медиатор, распространяющийся через мембраны, может высвобождаться во внешнюю среду при стимуляции определенных типов нейронов [Di Marzo et al Nature, 1994;372:686-691; Giuffrida et al Nat. Neurosci. 1999, 2:358-363], вызывая мощную активацию канабиоидных рецепторов [Devane et al Science 1992;258:1946-1949]. Однако анандамид быстро элиминируется за счет двухступенчатого механизма, включающего его импорт в клетку и гидролиз внутриклеточной амидазой [Beltramo et al Science 1997; 277:1094-1097; Di Marzo et al Nature 1994; 372:686-691; Hillard et al. J. Lipid Res. 1997; 38:2383-2398]. Будучи компонентами мембраны, в которую интегрирован канабиоидный рецептор (например, CB1), и не реагируя непосредственно с его лиганд-связывающим центром, анадамид и 2-арахидоноилглицерид (2-AG) способны приводит его в постоянно активированное состояние, снижая тем самым болевой порог [Meng et al, Nature 1998;Sep 24;395(6700):381-3]. При этом анандамид, действуя в качестве агониста CB1 в экспериментах на мышах, проявляет активность, сравнимую с синтетическими канабиоидами.
FAAH эффективно расщепляет амиды жирных кислот, в том числе N-ацилэтаноламины (NAE) и первичные амиды жирных кислот (FAPA). В группу NAE входит анандамид (AEA), пальмитоилэтаноламид (PEA) олеоилэтаноламид (OEA). В группу FAPA входит 9-Z-окатдеценамид и олеамид [McKinney MK, Cravatt BF. 2005. Annu Rev Biochem 74:411-32].
Мутантные линии мышей, искусственно лишенные генов FAAH, обладают резко сниженной способностью к гидролизу АЕА и других амидов жирных кислот, благодаря чему у них повышена активность канабиоидных рецепторов. В конечном итоге это внешне проявляется в резком снижении у таких животных болевого порога [Cravatt, et al. 2001. Proc Nat Acad Sci USA 98: 9371-9376]. Эти данные свидетельствуют о том, что ингибиторы FAAH способны увеличивать физиологическую активность EAE in vivo. В то же время они не приводят к эффектам, характерным для системной активации канабиоидных рецепторов, как это наблюдается при введении Δ9-THC (активный компонент марихуаны) и других активаторов канабиоидных рецепторов.
Известен патент [Unkefer et al, WO 2011106794 A1, опубл. 01.09.2011] «Ускорение роста растений путем модулирования экспрессии омега-амидазы». Пользуясь гомологией с геном ω-амидазы человека (NP_064587.1). В нем, в частности, описана идентификация генов ω-амидазы у ряда организмов самых различных таксономических групп: модельного растения Arabidopsis thaliana – номера доступа в базе данных NCBI GenBank AY075592.1, NP445766, винограда Vitis vinifera (XP_002279687.1), кукурузы Zea mays (ACN3091 1.1), тополя Populus trichocarpa (XP_002309478.1), североамериканской пихты Picea sitchensis (ABK22312.1), риса Oryza sativa (NP_001049134.1), сорго Sorghum bicolor (Genbank accession XP_002468410.1), клещевины Ricinus communis (XP_0025161 16.1), мха Physcomitrella patens subsp.patens (XP_001766085.1), плауна Selaginella moellendorffii (Плаунок Меллендорфа) (Genbank accession XP_002969787.1), люцерны Medicago truncatula (ACJ85250.1), одноклеточных зеленых водорослей Chlorella variabilis (EFN54567.1), Micromonas pusilla (XP_003064056.1) и Chlamydomonas reinhardtii (Genbank accession XP_001690839.1), микроскопической колониальной зеленой водоросли Volvox carteri (XP_002948137.1), морской бурой водоросли Ectocarpus siliculosus (CBJ25483.1), морской диатомовой водоросли Phaeodactylum tricornutum (XP_002183613.1), делящихся дрожжей Schizosaccharomyces pombe (NP_594154.1), плесневого гриба-дейтеромицета Aspergillus oryzae RIB40 (XP_001819629.1), нитчатого аскомицета Neurospora crassa OR74A (XP_960906.1), грамотрицательных бактерий Rhizobium leguminosarum (YP_769862.1), Rhizobium etli (YP_471237.1), Rhizobium leguminosarum (YP_002977603.1), Bradyrhizobium sp.(YP_001202760.1) и оомицета Phytophthora infestans T30-4 (XP_002999170.1).
Раскрытие изобретения
1) Фрагмент, кодирующий ген NIT2 человека, соответствующий последовательности SEQ ID NO 1, получают с помощью полуферментативного метода синтеза в виде продукта 859 п.н. Продукт очищают и подвергают рестрикции по сайтам BamHI и HindIII. Проводят клонирование в вектор pQE30, линеаризованный по сайтам BamHI и HindIII. Проводят трансформацию E.coli TG1,
отбирают колонии, содержащие целевой фрагмент при рестрикции BamHI и HindIII, получая конструкцию pQE-Nit2.
2) Полученная конструкция pQE-Nit2 при введении в штамм E.coli BL21(DE3) обеспечивает получение трансформантов, обладающих воспроизводимо определяемой активностью Nit2 в клеточных лизатах, зависимой от условий культивирования рекомбинантного штамма.
Краткое описание графических изображений:
Фиг.1. Схема функциональных элементов конструкций pQE-Nit2 (SEQ ID NO 2), предназначенной для экспрессии в клетках E.coli синтетического гена NIT2 человека. Конструкции содержат промотор и терминатор фага T5, синтетический ген NIT2 с оптимизированным для E.coli кодоновым составом, маркер лекарственной устойчивости к ампициллину. Поддержание конструкции в E.coli осуществляется за счет элементов: репликона плазмиды pMC16 и гена β-лактамазы (детерминанта устойчивости к ампициллину).
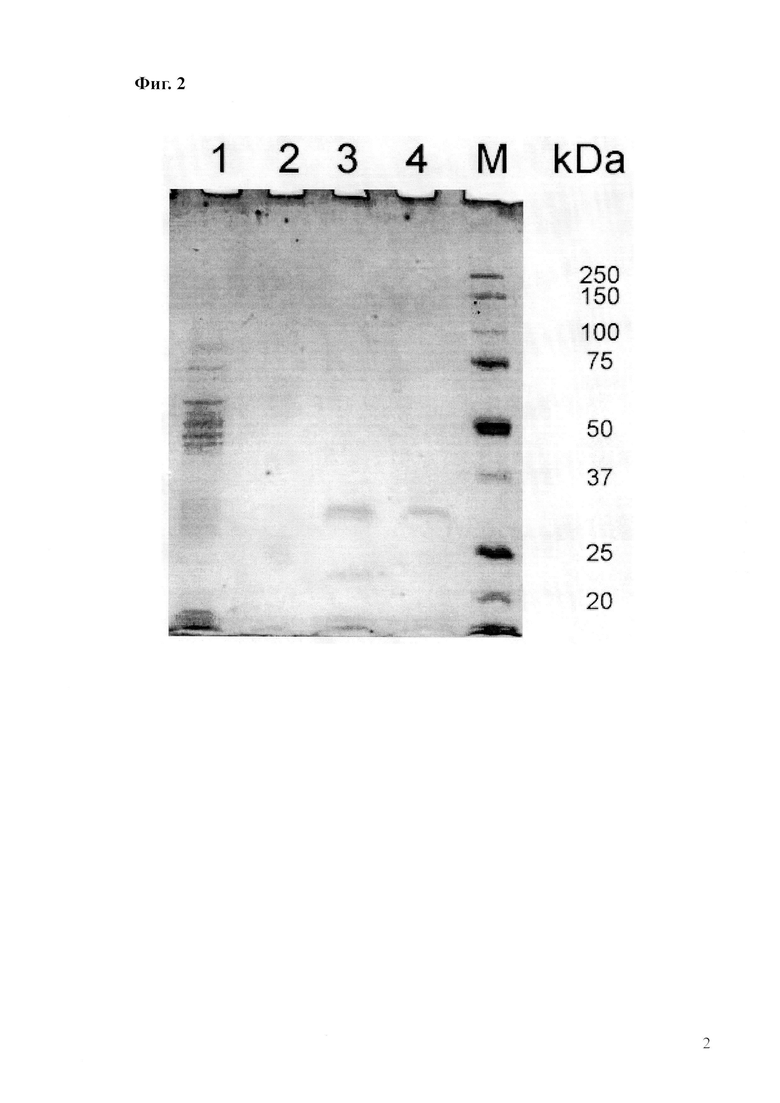
Фиг.2. Результаты электрофоретического анализа препарата ω-амидазы, очищенной из растворимой фракции гомогената рекомбинантных клеток E.coli BL21(DE3, pQE-Nit2). Электрофореграмма в 12,5%-ном денатурирующем полиакриламидном геле окрашена в растворе 1%-ного Кумасси R-250 и обесцвечена четырехкратным промыванием в горячей в 10%-ной уксусной кислоте.
На дорожки геля нанесены:
1 – растворимая клеточная фракция лизата рекомбинантного штамма E.coli BL21(DE3, pQE-Nit2) до хроматографической очистки (общее содержание белка в образце, проанализированном методом электрофореза - 50 мкг); 2 – пермеат элюат с сорбента Ni-NTA, полученный при нанесении на колонку хроматографического буфера №3 с последующей фильтрацией через патрон-концентратор с пределом пропускания 100 кДа (20 мкг); 3 – концентрат, полученный при сжатии объема целевого препарата в фильтрующем патроне-концентраторе с пределом пропускания 10 кДа (20 мкг); 4 – концентрат, полученный при очистке целевого препарата от низкомолекулярной примеси в фильтрующем патроне-концентраторе с пределом пропускания 25 кДа; М – маркер молекулярных масс (20 мкг).
Осуществление изобретения:
1. Генно-инженерное конструирование
Фрагмент ДНК длиной 859 п.н. в составе кодирующей области гена NIT2 синтезируют с полуферментативным способом в соответствии с последовательность SEQ ID NO 1. 1 мкг препарата ДНК гена NIT2 используют для получения конструкции pQE-Nit2. Продукт подвергают рестрикции по сайтам BamHI и HindIII, очищают и используют для лигирования с ДНК вектора pQE30 (Quiagen), линеаризованного по сайтам BamHI и HindIII.
2. Поддержание штаммов E.coli
Для выращивания и поддержания штамма используются следующие среды:
• бульон LB: 1% триптон (Difco); 0,5% дрожжевой экстракт (Difco); 1% NaCl (Лаверна)
• агар LB: бульон LB; 1,5% бактоагар (Difco)
• бульон 2YT: 1% триптон (Difco); 1% дрожжевой экстракт (Difco); 0,5% NaCl (Лаверна)
• Минимальная среда: 10 г/л минимальные соли M9 (Gibco); 1,5% бактоагар (Difco); 0,4% глюкоза (Уфавита); витамин В1 (тиамин) в конечной концентрации 100 мкг/мл (Уфавита)
3. Трансформация плазмидной конструкцией E.coli BL21(DE3, pQE-Nit2) и поддержание плазмидного продуцента
Трансформация бактерий проводится по модифицированному методу Манделя и Хиги с использованием 70 мМ CaCl2 [Sambrook et al., 1989]. Отбор трансформированных клонов, несущих плазмиды, проводился на селективных средах, содержащих соответствующие антибиотики: ампициллин 100 мкг/мл.
Для экспрессии конструкций на базе вектора pQE30 и производных векторов с промотором фага Т5 используется штамм E.coli BL21(DE3).
Для получения инокулята в ходе ферментации целесообразно использовать смыв биомассы рекомбинантного штамма со свежих чашек, имеющих возраст не более 30 ч с момента высева трансформантов на агаризованную селективную среду. Селективная среда представляет собой LB агар с 0,4% глюкозой и 100 мг/л ампициллина. Культивирование рекомбинантных штаммов на агаризованной среде целесообразно проводить при температуре не более 30°С.
Образовавшиеся колонии смыть с чашки диаметром 90 мм 6 мл свежей среды LB и использовать полученный смыв для засева не более двух колб Эрленмейера объемом 750 мл, содержащих по 30 мл соответствующей жидкой среды с 100 мг/л ампициллина.
4. Культивирование и первичная переработка биомассы штамма E.coli BL21(DE3, pQE-Nit2)
Для культивирования штаммов, продуцирующих белок в растворимой форме, использовать жидкую среду LB. Культивирование ведется в течение 12-24 ч при температуре 20-28ºC при максимально возможной аэрации (не менее 250 об/мин). Клетки собрать центрифугированием в течение 20 мин при 4400 g. Биомассу смешать с 5 объемами буфера (10 мМ трис-HCl, 10 мM ЭДТА, pH 8,0), гомогенизировать с помощью встряхивателя Vortex и дезинтегрировать трехкратной обработкой на ультразвуковом диспергаторе Soniprep 150 MSE (Япония) в присутствии фрагментов льда, не допуская замерзания биомассы и ее нагревания выше 20°С. Нерастворимую клеточную фракцию отделить центрифугированием в течение 60 мин при 12000 g.
Для удаления из клеточного лизата коллоидных примесей и нуклеиновых кислот применяется дробное осаждение сульфатом аммония по нижеописанной методике.
К растворимой фракции добавить сульфат аммония из расчета 1,2 г на 10 мл исходного раствора, после чего инкубировали раствор при +4ºС в течение 2 ч или более и отделить нерастворимую фракцию центрифугированием в течение 60 мин при 12000 g. Осадок отбросить, а к надосадочной жидкости добавить сульфат аммония из расчета 2 г на 10 мл жидкости, затем инкубировать раствор при +4ºС в течение 2 ч или более и отделить нерастворимую фракцию центрифугированием в течение 60 мин при 12000 g. Надосадочную жидкость удалить, а осадок растворить в 15-20 мл деионизованной воды или любого удобного буфера, обеспечивающего нативность целевого белка. При необходимости для удаления следов сульфата аммония раствор белка диализовать против деионизованной воды или буфера.
5. Определение активности Nit2 в лизатах рекомбинантного штамма E.coli BL21(DE3, pQE-Nit2)
5.1 Подготовка образцов к измерению. Стандартизация и нормировка клеточных экстрактов (гомогенатов)
В случае проведения измерений на готовых ферментных препаратах (гомогенатах) первым этапом определения является разделение водорастворимой и водонерастворимой фракций и определение концентрации белка в каждой из них по методу Лоури. Для этого исследуемый препарат встряхивают на вортексе в течение 30 с, не допуская нагревания, автоматической пипеткой Gilson отбирают аликвоту 1-100 мкл, помещают в полипропиленовую пробирку объемом 1,5 мл и доводят объем до 100 мкл забуференным физиологическим раствором (NaCl 100 мМ, фосфат натрия 10 мМ, рН 7,2). Для отбора аликвот объемом менее 10 мкл используют автоматическую пипетку с номинальным объемом 0,5-10 мкл. Аликвоту встряхивают на вортексе в течение 30 с, не допуская нагревания, и центрифугируют на настольной центрифуге при 10 тыс. g в течение 5 мин. Отбирают супернатант и переносят в чистую пробирку объемом 1,5 мкл. В пробирку с осадком вносят 100 мкл охлажденного на льду забуференным физиологическим раствором и гомогенизируют, встряхивая на вортексе в течение 20 с.
Из пробирок с растворимой и нерастворимой клеточной фракциями делают серию и 2-4 разведений с шагом 10 раз, перенося аликвоты объемом 10 мкл в полипропиленовые пробирки или лунки 96-луночного иммунологического планшета, содержащие по 100 мкл забуференного физиологического раствора. Для определения используют 10 мкл каждого разведения.
В случае определения активности ω-амидазы в биомассе рекомбинантного продуцента отбирают аликвоту ~500 мг влажного осадка биомассы свежей культуры, осажденной низкоскоростным центрифугированием, помещают в полипропиленовую пробирку с завинчивающейся крышкой типа Falcon oбъемом 15 мл, суспендируют в 5 мл охлажденного забуференного физиологического раствора (NaCl 100 мМ, фосфат натрия 10 мМ, рН 7,2) и добавляют 300 мг стеклянных шариков диаметром 0,5 мм. Биомассу гомогенизируют, встряхивая смесь на лабораторном ручном вортексе 5 раз по 1 мин. В промежутках между встряхиванием пробирки охлаждают на ледяной бане в течение 5-10 мин. Лизат осветляют центрифугированием в течение 30 мин при 12000 g, в полученном супернатанте определяют общий белок по модифицированному методу Лоури с использованием бицинхонинового реагента.
Для определения общего белка по модифицированному методу Лоури используют следующие стандартные растворы:
• Реагент А – 25 мМ ВСА-Na2, 0,2 М Na2CO3, 9 мМ KNaC4H4O6, 100 мМ NaOH, 113 мМ NaHCO3 (pH 11,25);
• Реагент В – 250 мМ CuSO4.
Для построения калибровочного графика используют серию разведений бычьего сывороточного альбумина (Promega), поставляемого в виде раствора с концентрацией 10 мг/мл.
Для определения общего содержания белка в анализируемом растворе получают рабочий раствор смешением реагента А и реагента В в соотношении 50:1. В микробиологический планшет вносят 10 мкл исследуемого раствора белка. Затем добавляют 200 мкл рабочего раствора. Перемешивают смесь в течение 30 с на шейкере и инкубируют при 37ºС в течение 1 ч. Измеряют коэффициент поглощения раствора на спектрофотометре при длине волны 595 нм. Результат считается достоверным при условии, что величина оптической плотности находится в диапазоне от 0,05 до 0,25 ОЕ. Концентрацию анализируемого раствора белка определяют по калибровочному графику.
При работе с очищенным ферментом со степенью гомогенности 20-100% для определения специфической активности отбирают аликвоты, содержащие 0,1-2 мкг растворимого или нерастворимого общего белка. При работе с грубыми лизатами количество белка увеличивают пропорционально ожидаемому содержанию в нем целевого фермента (ω-амидазы).
5.2 Приготовление реакционной смеси
Реакционная смесь для определения активности ω-амидазы в свободноклеточных экстрактах содержит:
5 мМ αKГM,
5 мМ DTT,
100 мМ Трис-HCl буфера (pH 8.5),
Аликвота клеточного гомогената объемом 50 мкл.
Максимальный объем измерения – 1 мл, минимальный объем измерения – 20 мкл (при использовании 96-луночных планшетов)
Реакционная смесь для определения активности коммерческого ферментного препарата ω-амидазы (OriGene TP310660, Recombinant protein of human nitrilase family, member 2 (NIT2) содержит:
5 мМ αKGM,
5 мМ DTT,
100 мМ Трис-HCl буфера (pH 8.5),
Аликвота препарата ω-амидазы, содержащая 0,5-1,0 единиц ферментативной активности в 1 мл.
Максимальный объем измерения – 1 мл, минимальный объем измерения – 20 мкл (при использовании 96-луночных планшетов)
5.3 Проведение измерений. Определение ω-амидазы с использованием αKГM в качестве субстрата на 96-луночных планшетах
Метод было определения ω-амидазы не использует солей ртути и основан на кинетическом принципе, при котором концентрация α-кетоглутарата, образующегося из αKГM, в реальном времени измеряется с помощью восстановительного аминирования в присутствии NADH, аммония и глутаматдегидрогеназы. При этом непрерывно регистрировалось падение абсорбции NADH при 340 нм (ε=6 220×M-1×cм-1).
Этот метод был адаптирован к анализу с использованием 96-луночных планшетов. После 5-минутной инкубации реакционной смеси, содержащей ω-амидазу, при 37°C реакцию останавливают добавлением 0,02 мл 5 мМ 2,4-динитрофенилгидразина в 2 M HCl. После 5-минутной инкубации в кислой среде при 37°C в реакционную смесь нейтрализуют добавлением 0,13 мл 1 М NaOH и измеряют абсорбцию при λ=430 нм в течение 5 мин (ε430 комплекса α-кетоглутарат-2,4- динитрофенилгидразона в этих условиях составляет 16000 M-1×cм-1; инкубация с 2,4-динитрофенилгидразином не должна продолжаться более 5 мин, та как при этом происходит медленное дезаминирование αKГM с образованием α-кетоглутарата - реакция ускоряется в кислой среде). Чтобы замедлить фоновую абсорбцию, объемы реакционной смеси и 2,4-динитрофенилгидразина уменьшают до 20 мкл и 10 мкл соответственно.
При проведении измерений с очищенной ω-амидазой и коммерческим препаратом ω-амидазы за контроль принимают реакционную смесь без добавления фермента. При измерении неочищенных гомогенатов за контроль принимают реакционную смесь, в которую добавлены гомогенат и 200 мМ глицилглицина. Глицилглицин является ингибитором ω-амидазы при pH>8,5 [Cooper 1977, J. Biol. Chem. 252: 2032–2038]. Такой контроль компенсирует мелкие карбонилсодержащие молекулы в неочищенных гомогенатах, которые могут реагировать с 2,4-динитрофенилгидразином.
При измерении αKГM смесь показывает относительно высокую активность ω-амидазы, поэтому инкубацию проводят в течение 5 или 10 мин при 37°C перед добавлением 2,4-динитрофенилгидразина. Однако для менее концентрированных очищенных ферментных препаратов образование α-кетоглутарата линейно возрастает в течение более длительного времени (до 90 мин).
Расчет активности фермента выполняют, занося полученные данные в таблицу (Табл. 1). За единицу ферментативной активности принимают количество фермента, катализируемого образование 1 мкмоль α-кетоглутарата в 1 мин при 37 °С (рН 9,0).
Таблица 1. Активность ω-амидазы в составе растворимой фракции гомогената рекомбинантных клеток E.coli BL21(DE3, pQE-Nit2)
6. Очистка ω-амидазы из растворимой фракции гомогената рекомбинантных клеток E.coli BL21(DE3, pQE-Nit2) методом металлоаффинной хроматографии
Для металлоаффинной хроматографии используют следующие стандартные растворы:
• 10× фосфатный буфер– 500 мM NaH2PO4×2H2O, 3 M NaCl, 4 M NaOH (рН 8,0);
• Раствор имидазола – 1 M имидазол, 2 мМ ЭДТА;
• Буфер 1 – 1× фосфатный буфер, 10 мМ раствор имидазола;
• Буфер 2 – 1× фосфатный буфер, 20 мМ раствор имидазола;
• Буфер 3 – 1× фосфатный буфер, 250 мМ раствор имидазола.
Все операции проводят при комнатной температуре. Колонку с Ni-NTA-агарозой с полным объемом сорбента 1,5 мл (Qiagen) уравновешивают 10 мл буфера 1. Растворимую клеточную фракцию гомогената рекомбинантных клеток E.coli BL21(DE3, pQE-Nit2) осветляют центрифугированием при 50 тыс.g в течение 1 ч и наносят на колонку в объеме 10-20 мл. Колонку промывают 20 мл буфера 2 и элюируют продукт буфером 3 в виде одной или более фракций объемом 1 мл каждая. Во фракциях определяют поглощение при λ=280 нм и по результатам готовят образцы для электрофореза в полиакриламидном геле в денатурирующих условиях (Фиг.1). Фракции, содержащие наиболее гомогенный целевой белок, объединяют и подвергают дополнительно очистке и концентрированию на центрифужных патронах-концентраторах производства Millipore.
Вначале проводят очистку элюата с сорбента Ni-NTA от высокомолекулярных примесей с помощью фильтрующего патрона-концентратора с пределом пропускания 100 кДа, собирая фракцию пермеата. Затем собранную фракцию пермеата концентрируют в 5 раз с помощью фильтрующего патрона-концентратора с пределом пропускания 10 кДа, собирают фракцию концентрата. Наконец, с целью удаления низкомолекулярной примеси проводят фракционирование на фильтрующем патроне-концентраторе с пределом пропускания 25 кДа. При этом процедуру повторяют три раза, каждый раз доводя объем концентрируемого препарата до исходного 1× фосфатным буфером, и сжимая его центрифугированием в пять раз. В результате достигается очистка целевого белка от избытка имидазола. В заключение определяют общее содержание белка в отобранных по ходу очистки фракций по модифицированному методу Лоури с использованием бицинхонинового реагента.
Специфическую ферментативную активность очищенного препарата определяют по методике, описанной выше. Она составляет 0,21 ед/мг белка.


Настоящее изобретение относится к биохимии и генетической инженерии, в частности к рекомбинантному штамму Escherichia coli BL21(DE3; pQE-Nit2). Указанный штамм получен путём трансформации штамма Escherichia coli BL21(DE3) плазмидной конструкцией pQE-Nit2, которая находится под контролем промотора фага Т5, и предназначен для получения ω-амидазы человека (Nit2) при культивировании на питательных средах на основе пептона, дрожжевого экстракта и глюкозы. Особенностью гена Nit2 в составе конструкции pQE-Nit2 является оптимизированный кодоновый состав, а также наличие искусственно введенной аффинной метки 6His-таг непосредственно после инициаторного кодона ATG (Met), обеспечивающего возможность одностадийной очистки продукта методом металлоаффинной хроматографии на Ni-NTA-агарозе. Настоящее изобретение позволяет получить ω-амидазу человека с повышенным выходом. 2 ил., 1 табл., 6 пр.
Штамм Escherichia coli BL21(DE3; pQE-Nit2), предназначенный для получения ω-амидазы человека при культивировании на питательных средах на основе пептона, дрожжевого экстракта и глюкозы, который получен путём трансформации штамма Escherichia coli BL21(DE3) плазмидной конструкцией pQE-Nit2, характеризующейся нуклеотидной последовательностью SEQ ID NO:2.
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pSVH0106, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ Gl7ACA-АЦИЛАЗЫ В КЛЕТКАХ Escherichia coli, И РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli BL21(DE3)/pSVH0106-ПРОДУЦЕНТ Gl7ACA-АЦИЛАЗЫ | 2005 |
|
RU2300566C2 |
| KRASNIKOV BORIS F., et al., Assay and purification of ω-amidase/Nit2, a ubiquitously expressed putative tumor suppressor that catalyzes the deamidation of the α-keto acid analogues of glutamine and asparagine, Anal Biochem., 2009, 391(2), pp.144-150 | |||
| ПАВЛОВА Ю.А., и др., Сравнительный анализ последовательностей генов амидаз почвенных актинобактерий рода Rhodococcus, Известия Самарского научного центра Российской академии наук, 2011, Т.13, No.5(3), с.272-276 | |||
| JEONG HAEYOUNG, et al., Complete genome sequence of Escherichia coli strain BL21, Genome Announcements, 19.03.2015, Vol.3, Issue 2, e00134-15. | |||

