ОБЛАСТЬ ПРИМЕНЕНИЯ
Настоящее изобретение относится к биотехнологии, биохимии и медицинской химии. Оно обеспечивает возможность использования ω-амидазы человека (Nit2) в качестве потенциальной мишени для создания новых средств противоопухолевой терапии.
Уровень техники
Из уровня техники известен ряд патентов, раскрывающих способы получения ω-амидазы человека (Nit2) и других ферментов, сходных по структурной организации и/или субстратной специфичности, с использованием рекомбинантных продуцентов, а также способы применения этих ферментов в медицине и конструирования их ингибиторов в качестве кандидатных терапевтических средств.
В патенте RU 2642323 [Дерябина Ю.И., Исакова Е.П., Красников Б.Ф., Антипов А.Н., Белякова А.В., Бирюкова Ю.К., патент RU 2642323, опубл. 24.01.2018] раскрыто получение рекомбинантного штамма Escherichia coli BL21 (DE3; pQE-Nit2) - продуцента ω-амидазы человека (Nit2). Указанный штамм получен путем трансформации штамма Escherichia coli BL21 (DE3) плазмидной конструкцией pQE-Nit2, которая находится под контролем промотора фага Т5. Он предназначен для получения целевого фермента при культивировании на питательных средах на основе пептона, дрожжевого экстракта и глюкозы. Особенностью гена Nit2 в составе конструкции pQE-Nit2 является оптимизированный для экспрессии в Е. coli кодоновый состав, а также наличие искусственно введенной аффинной метки 6His-Tar непосредственно после инициаторного кодона ATG (Met), обеспечивающего возможность одностадийной очистки продукта методом металлоаффинной хроматографии на Ni-NTA-агарозе. Описанное изобретение позволяет получить ω-амидазу человека с повышенным выходом.
В заявке на изобретение RU 2015146605 [Эпова Е.Ю., Дерябина Ю.И., Исакова Е.П., Красников Б.Ф., Антипов А.Н., Секова В.Ю., Бирюкова Ю.К., Гусева М.А., Белякова А.В., патент RU 2015146605, опубл. 03.05.2017] описывается штамм Yarrowia lipolytica, получаемый путем трансформации коллекционного штамма Y. lipolytica W29 конструкцией pUVGT2, предназначенный для получения глутаминтрансаминазы К человека при культивировании на питательных средах на основе пептона, дрожжевого экстракта и глюкозы.
В патенте на изобретение RU 2375055 [Перзиновский С., Радзкий Р.П., Биенко М., патент RU 2375055, опубл. 10.12.2009] предложено применение α-кетоглутаровой кислоты или ее фармацевтически приемлемой соли для изготовления фармацевтического препарата или пищевой или кормовой добавки для стимулирования продукции ЛВП у позвоночных животных, включая человека и соответствующий способ стимулирования продукции ЛВП α-кетоглутаровой кислотой (или ее солью) и фармацевтически приемлемых физических смесей α-кетоглутаровой кислоты или ее фармацевтически приемлемой соли и по меньшей мере одной аминокислоты. Показано статистически значимое повышение концентрации ЛВП при снижении общего холестерина, ЛНП и триглицеридов.
Патент US 2002115698 [Newcomb R., Newocmb М., патент US 2002115698, опубл. 17.09.2002] описывает способ снижения концентрации глутамина путем селективного ингибирование глутаминазы бис-тиадиазолом.
В патенте на изобретение WO 2008100977 [О Dasse., Parrott J.A., Putman D., Adam J., патент WO 2008100977, опубл. 21.08.2008] описывается фармакологическое ингибирование активности амид-гидролазы жирных кислот (FAAH), приводящее к повышению уровня амидов жирных кислот в крови. В данном патенте раскрыты способы получения сложных эфиров алкилкарбаминовой кислоты и способы их применения.
Изобретение, раскрытое в патенте ЕА 014853 В1 [Стрекстра X., патент ЕА 014853 В1, опубл. 28.02.2011], заключается в описании способа получения пищевого продукта с пониженным содержанием акриламида, включающий стадию тепловой обработки с последующим добавлением фермента - амидазы к промежуточной форме пищевого продукта. Изобретение также относится к изолированным амидазам из Aspergillus niger и рекомбинантным амидазам, кодирующим их полинуклеотидам, векторам и рекомбинантным клеткам-хозяевам. Применение данного изобретения - амидазы - позволяет снизить уровень акриламида в пищевом продукте или продукте питания.
Изобретение, раскрываемое в патенте RU 2260051 [Касиваги Т., Симба Н., Исикава К., Сузуки Е., Хираяма К., Йокояма К., патент RU 2260051, опубл. 10.09.2005] относится к способу конструирования и получения мутантных трансглутаминаз на основе пространственной структуры трансглутаминазы, полученной из Streptoverticillium mobaraense (MTG), и полученной таким образом мутантной MTG. Настоящее изобретение относится также к способу модификации MTG на основе пространственной структуры и трансглутаминазе, обладающей усиленной этим способом реакционной способностью по отношению к субстрату. Данное изобретение позволяет предсказать сайт связывания MTG с субстратом на основе пространственной структуры, определенной с помощью рентгеноструктурного анализа кристаллов MTG, и сконструировать мутантные трансглутаминазы путем замены, вставки или делеции аминокислотных остатков, расположенных в субстрат-связывающем сайте трансглутаминазы.
В патенте на изобретение US 2010016207 [Wurtman R.J., Wang L., патент US 2010016207, опубл. 21.01.2010] описываются способы повышения уровня ГАМК в головном мозге, способы лечения болезни Альцгеймера, болезни Хантингтона и ее симптомов, бессонницы, когнитивных нарушений или нарушений памяти. Так же описываются способы лечения первичных или метастатических опухолей головного мозга, депрессии (например, биполярная депрессия) путем введения глутамина, его соли или богатого глутамином пептида.
В патенте US 5514579 [Patrick J. O'Hara, Francis J. Grant, Paul О. Sheppard, патент US 5514579, опубл. 07.05.1996] описаны методы идентификации и клонирования трансглутаминазы простаты и плаценты человека. Трансглутаминазы, кодируемые геномом человека, описанные в данном изобретении, полезны, среди прочего, для терапевтического заживления ран, закрытия кожных трансплантатов, стабилизации пищевых препаратов и маркеров для идентификации агентов, которые действуют как агонисты или антагонисты клеточного апоптоза.
Настоящее изобретение WO 2016170447 [Rickert М, Strop Р., патент WO 2016170447, опубл. 27.10.2016] описывает способы получения растворимых микробных трансглутаминаз с использованием рекомбинантных продуцентов. Данный патент описывает искусственно сконструированную микробную трансглутаминазу, содержащую одну или несколько замен в области про-домена для модуляции взаимодействия между про-доменом и ферментным доменом трансглутаминазы. Изобретение описывает также особенности векторов и микроорганизмов-хозяев, имеющие значение для получения целевого фермента с высоким выходом.
В патенте JP 2012246294 [Jalan R., Jalan K.N., патент JP 2012246294, опубл. 22.10.2014] раскрыто лекарственное средство для профилактики или лечения гипераммониемии. Описанное в патенте средство содержит орнитин, фенилацетат, либо фенилбутират, и используется для предотвращения или лечения гипераммониемии, отека мозга и уменьшения фагоцитоза нейтрофилов. Лечение пациентов проводят на ранней стадии заболевания, чтобы предотвратить или отсрочить наступление печеночной декомпенсации и, как следствие, возникновение печеночной энцефалопатии. В качестве альтернативы предлагается метод лечения печеночной энцефалопатии путем снижения концентрации аммиака в крови и поддержания функции нейтрофилов.
Изобретение, описанное в патенте TW 201339166 [Szillat Н., Leeuw Т., Lorenz М, патент TW 201339166, опубл. 01.10.2013], раскрывает структуру замещенных изотиазоло[5,4-b]пиридин-2-карбоксамидов. Описанные в данном патенте соединения эффективны в качестве ингибиторов трансглутаминаз, в частности трансглутаминазы 2, благодаря чему подходят для лечения ряда заболеваний человека, например, остеоартрита и других дегенеративных заболеваний суставов. Кроме того, в изобретении описываются способы получения таких соединений, и применение фармацевтических препаратов на их основе.
Патент US 20030144227 [Chin-Hsiang Chien, Taipei; Chun-Hung Lin, патент US 20030144227, опубл. 31.07.2003] раскрывает новые способы применения гена Nit2 для ингибирования роста опухолевых клеток. В частности, изобретение обеспечивает фармацевтические композиции для ингибирования роста опухолевых клеток, содержащие ген Nit2 и белок Nit2 соответственно.
В ходе анализа уровня техники по теме «Методы получения рекомбинантных продуцентов ферментов, родственных омега-амидазы человека (Nit2), и создания их ингибиторов» собраны аналитические данные о действующей охранной документации. Проведенный анализ по теме позволил выявить 15 наиболее близких по тематике патентов, из которых 10 являются действующими. Среди них присутствует патент RU 2642323 С2 [Дерябина Ю.И., Исакова Е.П., Красников Б.Ф., Антипов А.Н., Белякова А.В., Бирюкова Ю.К.,
Наиболее близким к заявляемому является патент RU 2642323, опубл. 24.01.2018], раскрывающий способ получения рекомбинантного продуцента омега-амидазы человека Nit2 на основе Е. coli.
Однако, в описании изобретения и формуле отсутствуют показатели выхода целевого рекомбинантного белка (в единицах активности или массы продукта) в исходной биомассе и на стадиях очистки.
Раскрытие изобретения
Изобретение раскрывает искусственно сконструированную последовательность фрагмента гена фермента ω-амидазы человека SEQ_ID# 1 длиной 81 п.н. вблизи сайта инициации трансляции, позволяющую повысить выход продукта при культивировании рекомбинантного штамма-продуцента на основе Е. coli гена Nit2. Использование варианта гена Nit2, содержащего раскрываемую последовательность, облегчает наработку и очистку рекомбинантной ω-амидазы человека, необходимой для тестирования ее специфических ингибиторов - потенциальных средств противоопухолевой терапии.
Описание графических изображений
Фиг. 1. Карта плазмидной конструкции pQE-Nit22, предназначенной для получения ω-амидазы человека Nit2 в клетках Е. coli. Конструкция содержит искусственный ген Nit2 с оптимизированным для экспрессии в Е. coli кодоновым составом в составе вектора pQE30 (Qiagen). Плазмида содержит промотор и терминатор транскрипции бактериофага Т5 и маркер устойчивости к ампициллину BLA.
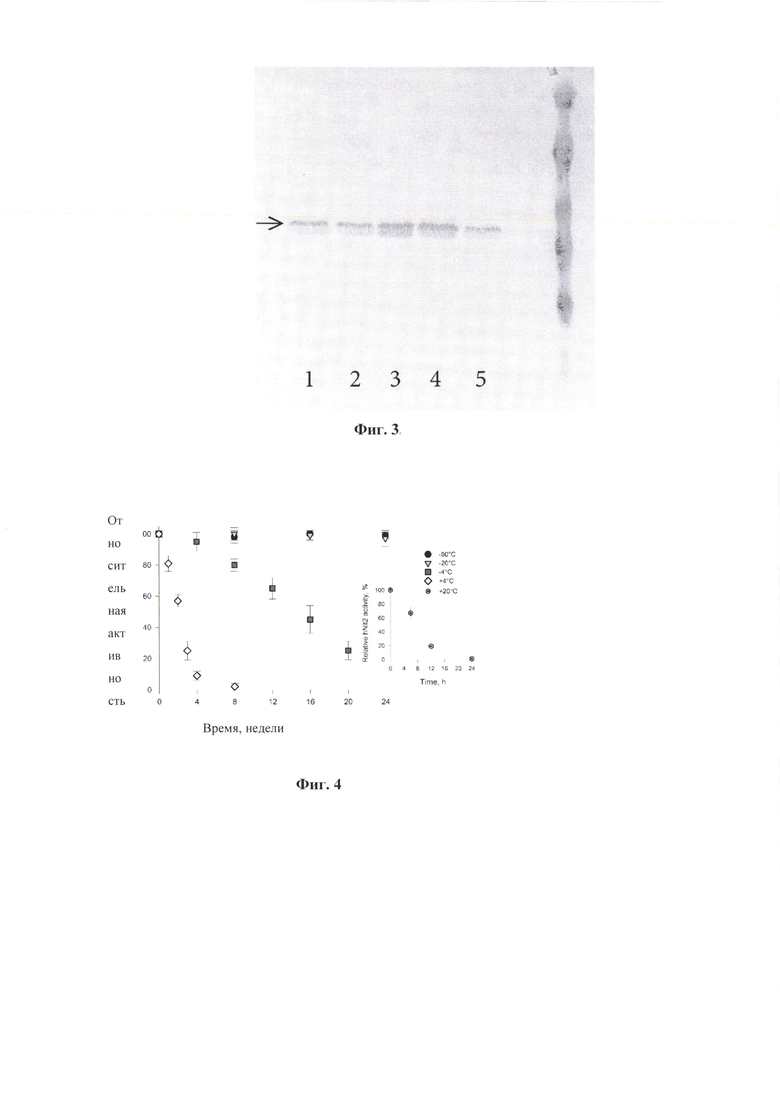
Фиг. 2. Электрофоретический анализ белковых фракций штамма BL21 (DE3; pQE-Nit22), полученных по ходу очистки целевого продукта Nit2 человека в 12,5% денатурирующем полиакриламидном геле после окрашивания в 1% Кумасси R-250.
На дорожки геля нанесены образцы:
L - стандарт молекулярных масс (14,4, 18,4, 25, 35, 45, 66 кДа);
1 - грубый лизат Е. coli BL21 (DE3, pQE-Nit22);
2 - растворимая фракция грубого лизата Е. coli BL21 (DE3, pQE-Nit22);
3 - растворимая фракция грубого лизата Е. coli BL21 (DE3, pQE-Nit22) после дробного фракционирования 50% и 70% сульфатом аммония;
4 - Фракция 1 после элюции с Ni-NTA-агарозы (V=0,5 мл);
5 - Фракция 2 после элюции с Ni-NTA-агарозы (V=0,5 мл);
6 - Фракция 3 после элюции с Ni-NTA-агарозы (V=0,5 мл);
7 - Фракция 4 после элюции с Ni-NTA-агарозы (V=0,5 мл);
L - стандарт молекулярных масс (14,4, 18,4, 25, 35, 45, 66 кДа).
Фиг. 3. Электрофоретический анализ белковых фракций штамма BL21 (DE3; pQE-Nit22), полученных по ходу очистки целевого продукта Nit2 человека в 12,5% денатурирующем полиакриламидном геле после полусухого электропереноса на мембрану PVDF (Immobilon, Millipore) и окрашивания разведенными в 1000 раз мышиными моноклональными антителами к аффинной метке 6His (GE HealthCate, USA) и разведенными в 3000 раз поликлональными антителами козы против IgG мыши, конъюгированными с пероксидазой хрена (ООО Биалекса, Россия). В качестве хромогенного субстрата на пероксидазную активность использовали 3,3'-диаминобензидин в концентрации 20 мкг/мл и Н2О2 в концентрации 0,01%.
На дорожки геля нанесены:
3 - растворимая фракция грубого лизата Е. coli BL21 (DE3, pQE-Nit22) после дробного фракционирования 50% и 70% сульфатом аммония;
4 - Фракция 1 после элюции с Ni-NTA-агарозы (V=0,5 мл);
5 - Фракция 2 после элюции с Ni-NTA-агарозы (V=0,5 мл);
6 - Фракция 3 после элюции с Ni-NTA-агарозы (V=0,5 мл);
7 - Фракция 4 после элюции с Ni-NTA-агарозы (V=0,5 мл);
L - стандарт молекулярных масс 120, 90, 50, 34, 26 и 20 кДа (ThermoFisher Scientific, cat. # 26612).
Фиг. 4. Относительная активность (%) очищенного hNit2 при хранении в различных температурных условиях (n=3).
Осуществление изобретения
1. Генно-инженерное конструирование
Для получения генно-инженерной конструкции pQE-Nit22, используют конструкцию pQE-Nit2, описанную раннее в патенте RU 2642323 и синтетические дезоксирибонуклеотиды, содержащие 3'-фосфорилированные концы:
производства ООО «Синтол» (Москва, Россия). По 30 пмоль каждого олигонуклеотида и 0,4 мкг плазмиды pQE-Nit2, предварительно расщепленной эндонуклеазами рестрикции BamHI и SphI с последующей очисткой на кремнеземном сорбенте, смешивают в пробирке, добавляют 3 мкл Т4-лигазного буфера и доводят до общего объема 30 мкл водой. Смесь нагревают при 70°С в течение 5 мин и медленно охлаждают до комнатной температуры в течение 10 минут. Затем добавляют 5 ед. ДНК-лигазы Т4 (Thermo Fisher Scientific, США) и инкубируют смесь при +4°С в течение 14 часов.
Лигазную смесь используют для трансформации штамма TG1 Е. coli, как описано в [Sambrook, J., Fritch, Е., and Maniatis, Т. (1989) Molecular Cloning: A Laboratory Manual, 2nd Edn., Vol. 3, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY]. Трансформанты отбирают на среде LB с добавлением 100 мкг/мл ампициллина. Подлинность структуры плазмиды подтверждают секвенированием с использованием стандартных праймеров pQE (промотор Т5 и терминатор Т5), рекомендованных производителем (Qiagen).
2. Поддержание штаммов E. coli
Культивирование штамма Е. coli BL21 (DE3) проводят на жидкой среде LB при температуре +28-30°С при максимально возможной аэрации (не менее 250 оборотов в минуту) в течение 12-24 часов. Культуру клеток Е. coli BL21 (DE3) инкубируют в течение ночи, отбирают аликвоту 30 мкл и переносят в 3 мл свежей среды LB. Культуру выдерживают при +30 С и максимально возможной аэрации в течение 2-3 ч до достижения оптической плотности А600 ~ 0,3. В стерильных условиях отбирают по 1 мл культуры в три стерильные 1,5-мл пробирки типа Эппендорф и держат на льду в течение 5 мин. Клетки осаждают центрифугированием при 800 g в течение 2 мин, ресуспендируют в 750 мкл 70 мМ CaCl2 и выдерживают на льду в течение 30-40 мин. Затем клетки осаждают центрифугированием при 800 g в течение 2 мин и ресуспендируют в 100 мкл 70 мМ CaCl2. Полученная суспензия должна быть использована для трансформации в течение 24 ч после приготовления.
Для выращивания и поддержания штамма используют следующие среды:
• бульон LB: 1% триптон (Difco); 0,5% дрожжевой экстракт (Difco); 1% NaCl (Лаверна);
• агар LB: бульон LB; 1,5% бактоагар (Difco);
• бульон 2YT: 1% триптон (Difco); 1% дрожжевой экстракт (Difco); 0,5% NaCl (Лаверна);
• Минимальная среда: 10 г/л минимальные соли М9 (Gibco); 1,5% бактоагар (Difco); 0,4% глюкоза (Уфавита); витамин В1 (тиамин) в конечной концентрации 100 мкг/мл (Уфавита).
3. Трансформация плазмидной конструкцией Е. coli BL21 (DE3, pQE-Nit22) и поддержание плазмидного продуцента
Перед трансформацией готовят чашки Петри диаметром 90 мм с агаризованной средой LB с добавлением 100 мкг/мл ампициллина и выдерживают 1-3 дня при комнатной температуре (избегая попадания прямых солнечных лучей) для удаления избыточной влаги. Трансформацию бактериальных клеток проводят по оригинальному методу Mandel и Higa, модифицированному Sambrook et al. [Sambrook, J., Fritch, E., and Maniatis, T. (1989) Molecular Cloning: A Laboratory Manual, 2nd Edn., Vol. 3, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY]. Засевают 0,1 мкг очищенной ДНК pQE-Nit22 и 3×107 бактериальных клеток на чашку Петри. Чашки помещают на 14-30 ч в термостат при температуре +28-30°С. Выросшие колонии используют в качестве посевного материала.
В каждую чашку Петри добавляют 6 мл свежей среды LB. Биомассу бактерий тщательно суспендируют в жидкой среде шпателем Дригальского в стерильных условиях и переносят в полипропиленовую пробирку типа Falcon. Свежую суспензию немедленно используют для инокуляции колб Эрленмейера объемом 750 мл, содержащих 30 мл соответствующей жидкой среды с добавлением 100 мкг/мл ампициллина. Исходное содержание бактериальных клеток на момент инокуляции должно быть не менее 3×107 КОЕ на мл. Колбы Эрленмейера инкубируют на микробиологическом шейкере Biosan (Латвия) при +28-30°С в течение 16-18 ч со скоростью вращения 200 об/мин.
4. Культивирование и первичная переработка биомассы штамма Е. coli BL21 (DE3, pQENit2)
После завершения ферментации колбы Эрленмейера охлаждают при +4°С в течение 30 мин. Затем культуру помещают в пробирки объемом 50 мл с завинчивающейся крышкой и центрифугируют при 4400 g в течение 20 мин. Супернатант полностью удаляют. В каждую пробирку собирают 100 мл культуры путем двукратного осаждения аликвот по 50 мл. Биомассу ресуспендируют в 25 мл ледяного стерильного физиологического раствора, осаждают в тех же условиях, что и супернатант.
Промытую биомассу ресуспендируют в 10 мл буфера, содержащего 10 мМ Трис-HCl, 10 мМ ЭДТА, рН 8,2, путем встряхивания и гомогенизации на ультразвуковом диспергаторе Soniprep 150 MSE (Япония) 5 сериями по 1 мин каждая при мощности 100 Вт. В ходе обработки ультразвуком используют ледяную баню для предотвращения нагрева лизата. Гомогенат центрифугируют при 12000 g в течение 1 ч, супернатант удаляют. Осадок суспендируют в 10 мМ Трис-HCl, рН 8,2, 10 мМ ЭДТА, 100 мМ NaCl, 0,1% Triton Х-100 и подвергают повторной обработке ультразвуком в тех же условиях. Гомогенат центрифугируют при 12000 g в течение 1 ч и супернатант отбрасывают. Осадок в каждой пробирке суспендируют в 25 мл 10 мМ Трис-HCl, 10 мМ ЭДТА, рН 8,2 и подвергают повторной обработке ультразвуком в тех же условиях. Гомогенат центрифугируют при 12000 g в течение 1 ч и супернатант декантируют в чистую колбу. Объем осветленного супернатанта тщательно измеряют и медленно добавляют сульфат аммония в соотношении 377 г на 1 л растворимой фракции клеточного лизата (до ±50% насыщения от насыщения при +20°С) при +4°С при перемешивании на магнитной мешалке. После полного растворения соли лизат инкубируют при +4°С в течение ночи, а затем центрифугируют при +4°С и 12000 g в течение 1 ч. Осадок отбрасывают, а супернатант сливают в чистую колбу. Объем супернатанта тщательно измеряют и медленно добавляют сульфат аммония в соотношении 150 г на 1 л лизата (до насыщения 70% при +20°С) при +4°С при перемешивании на магнитной мешалке. После полной солюбилизации лизат инкубируют при +4°С в течение ночи и центрифугируют при +4°С и 12000 g в течение 1 ч. Супернатант отбрасывают, и осадок растворяют в буфере 50 мМ фосфат натрия, рН 8,0, имидазол 10 мМ в соотношение 10 мл на 1 г осадка.
Солюбилизированную фракцию в количестве 30-35 мл помещают в целлофановый мешок и подвергают диализу против 0,5 л буфера (50 мМ Na3PO4, 10 мМ имидазол, рН 8,0). Диализ проводят в течение 14 ч при +4 С
Полученный раствор осветляют центрифугированием при 12000 g в течение 1 ч, осадок отбрасывают. Супернатант наносят на колонку Ni-NTA с 10 мл сорбента TALON Metal Affinity Resin (Clontech, США, Кат. №635501) и промывают 50 мл буфера (50 мМ Na3PO4, 10 мМ имидазол, рН 8,0) и 50 мл буфера (50 мМ Na3PO4, 50 мМ имидазол, рН 8,0). Элюцию проводят в 5 мл буфера (50 мМ Na3PO4, 250 мМ имидазол, рН 8,0). Фракции, которые обладают ω-амидазной активностью, объединяют и фильтруют через систему фильтрации центробежного концентратора Vivaspin-20 с отсечкой 100 кДа (Sartorius, Германия, № по каталогу VS2021, 100000 MWCO PES). Полученный фильтрат концентрируют до объема 1,5-2 мл с помощью системы фильтрации центробежного концентратора Vivaspin-20 с отсечкой 30 кДа (Sartorius, Германия, каталожный № VS2021, 30000 MWCO PES). Остаточный имидазол удаляют путем повторного суспендирования концентрата в 20 мл 10 мМ пирофосфатного (PPi) буфера с добавлением 2 мМ ДТТ, рН 8,5 с последующим концентрированием суспензии до объема 1,5-2 мл с использованием Vivaspin с отсечкой 10 кДа. Очищенный фермент разделяют на аликвоты по 100 мкл и хранят при -80°С.
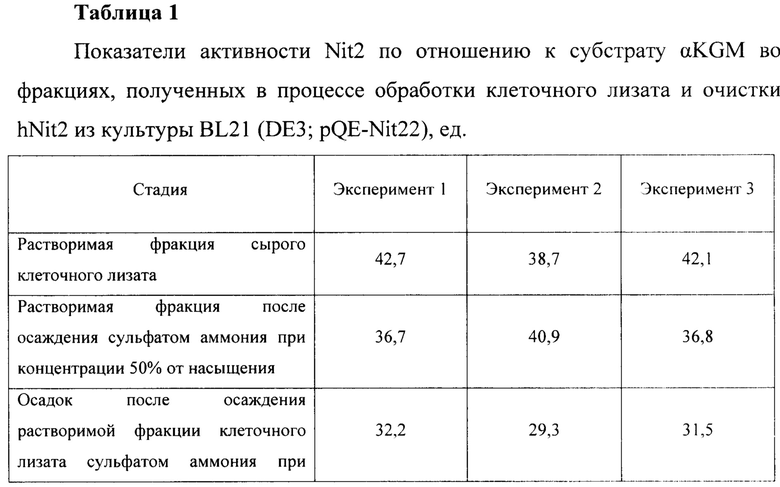
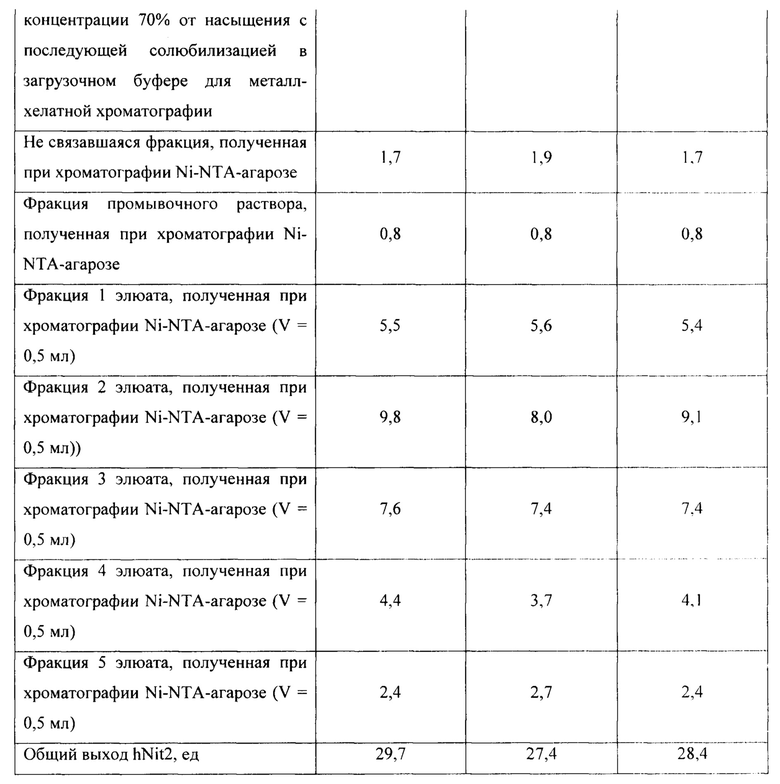
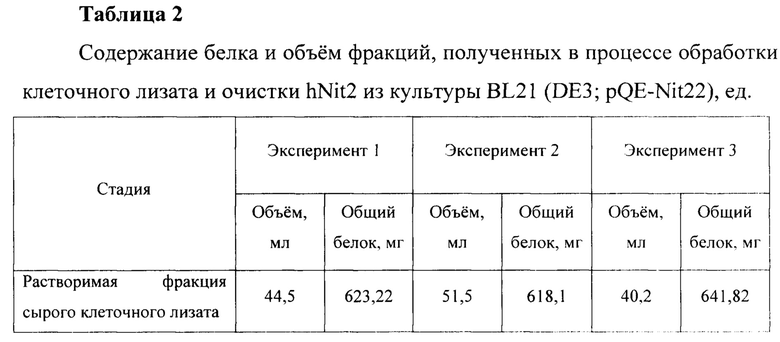
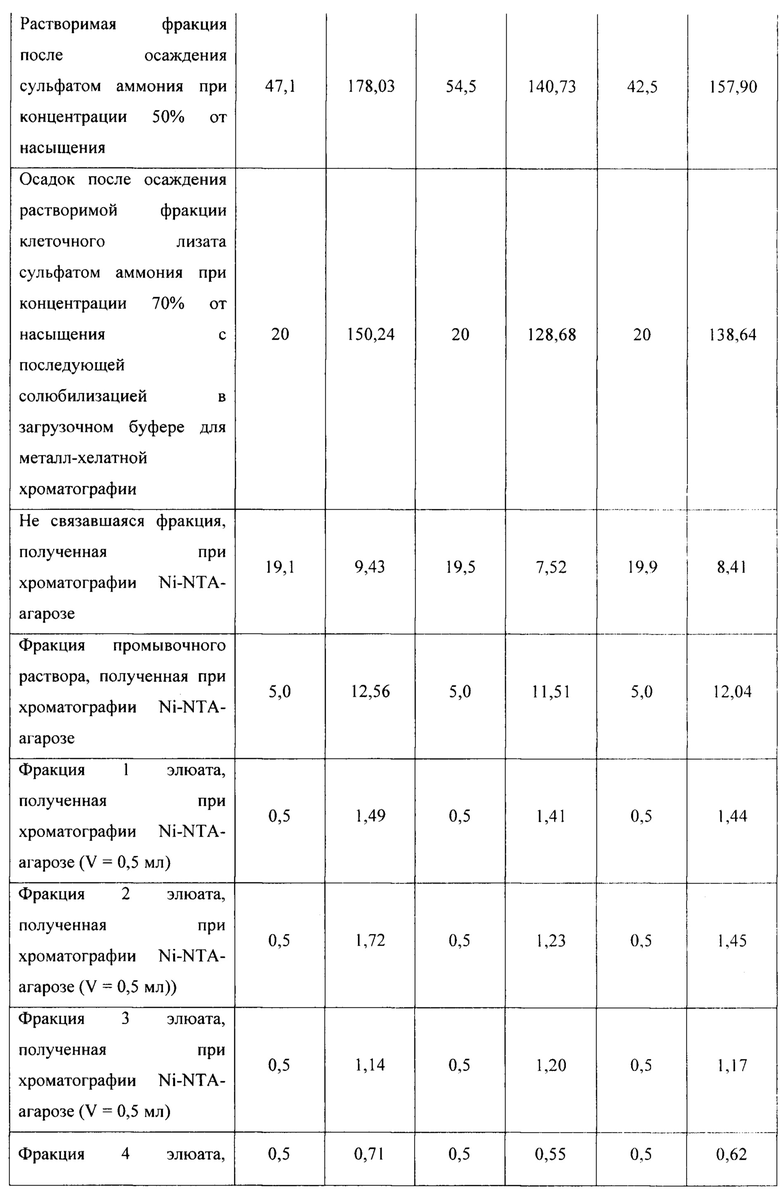
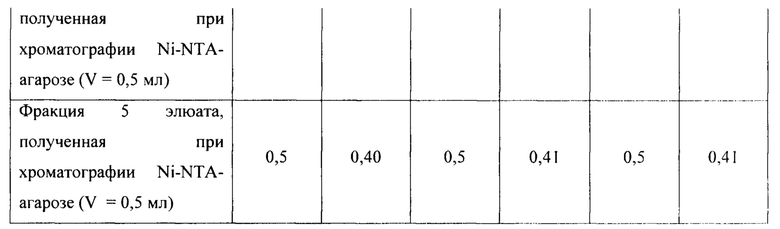
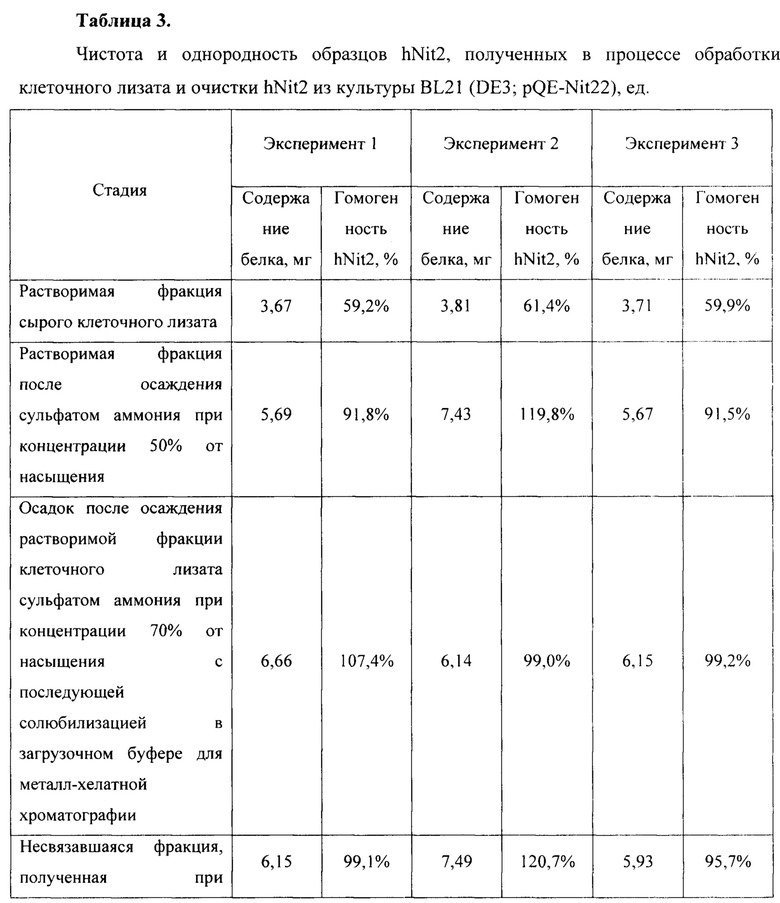
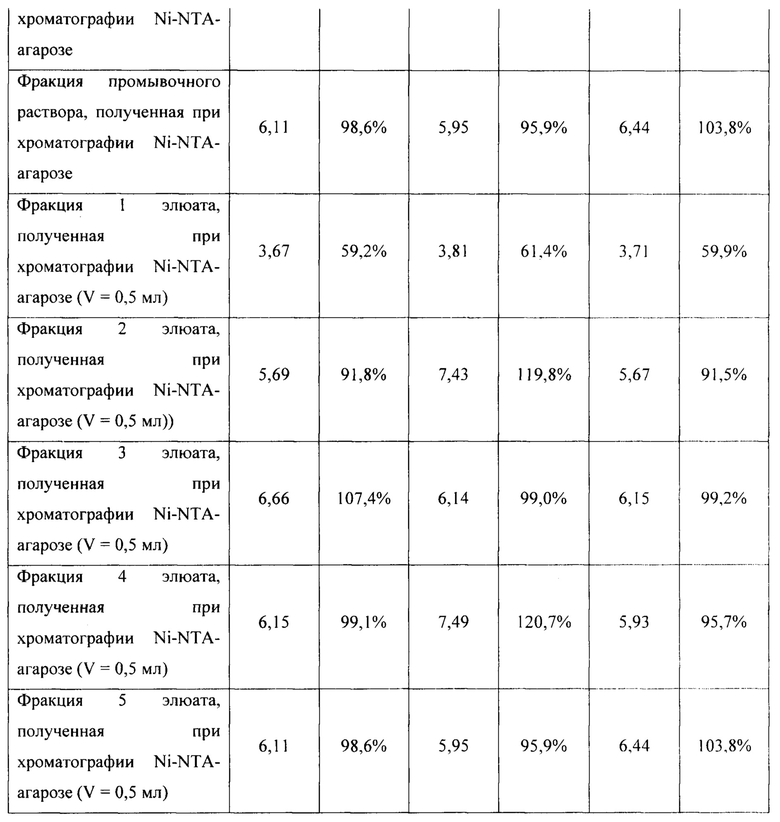
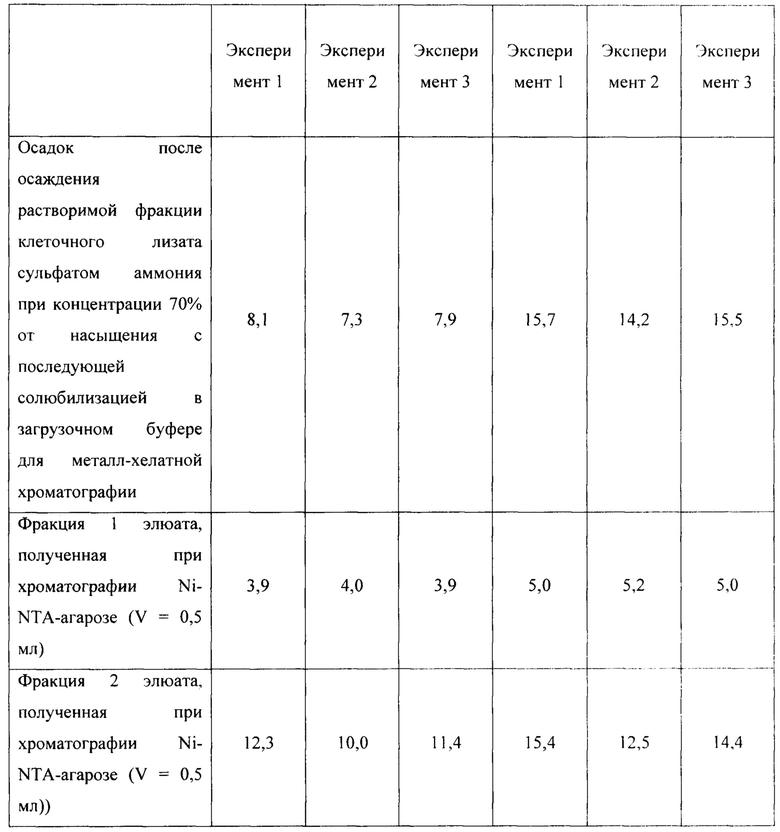
Для оценки эффективности нового продуцента hNit2, полученного из конструкции pQE-Nit22, культивирование соответствующих штаммов повторяют в трех повторностях. Каждое культивирование выполняют в 12 колбах Эрленмейера (общий объем питательной среды 360 мл). Выход на каждой стадии обработки культуры и очистки Nit2 показан в таблицах 1-3.
Определение концентрации белка проводят с использованием модифицированного метода Лоури с бицинхониновой кислотой, как описано в [Smith PK, Krohn RI, Hermanson GT, Mallia AK, Gartner FH, Provenzano MD, et al. Measurement of protein using bicinchoninic acid. Anal Biochem. 1985 Oct; 150 (1): 76-85].
5. Определение ω-амидазной активности во фракциях биомассы
Реакцию проводят в 96-луночных виниловых плоскодонных планшетах Costar (каталожный №2595).
Для измерения ω-амидазной активности с использованием αKGM в качестве субстрата используют спектрофотометрический анализ конечной точки [Krasnikov BF, Nostramo R, Pinto JT, Cooper AJ. Assay and purification of omega-amidase/Nit2, a ubiquitously expressed putative tumor suppressor, that catalyzes the deamidation of the alpha-keto acid analogues of glutamine and asparagine. Anal. Biochem. 2009; 391(2): 144-150. doi: 10.1016/j.ab.2009.05.025]. Реакционная смесь (общий объем 50 мкл) содержит: 100 мМ Трис-HCl буфер (рН 8,5), 5 мМ ДТТ и 5 мкл источника фермента. Началом реакции служит добавление 5 мМ αKGM. Реакцию останавливают после инкубации образцов в течение 5-15 мин при +37°С добавлением 20 мкл 5 мМ DNPD в 2 М HCl. После дополнительной инкубации в течение 5 мин при +37°С в каждую лунку вносят 130 мкл 1 М NaOH и считывают абсорбцию при 430 нм по сравнению с контролем, содержащим αKGM, но без фермента, в течение 5 мин. Коэффициент экстинкции продукта реакции α-кетоглутарата в DNPD при 430 нм в представленных условиях составляет 16000 М-1×см-1. При этом инкубация с DNPD не должна длиться более 5 мин, поскольку в кислых условиях происходит медленное дезамидирование αKGM в α-кетоглутарат.
Для измерения активности α-амидазы с использованием α-сукцинамата в качестве субстрата используют следующие исходные растворы: 500 мМ α-сукцинамат натрия, рН 7,5; 1 М Трис-HCl, 1 М гидроксиламин натрия, 50 мМ ДТТ. Все растворы фильтруют через шприцевую насадку для стерильной фильтрации с размером пор 0,22 мкм. Все исходные растворы хранят при -20°С. Реакционная смесь на лунку (50 мкл) содержит: 50 мМ α-сукцинамат - 5 мкл; 100 мМ Трис-HCl - 5 мкл; 100 мМ гидроксиламин - 5 мкл; 5 мМ ДТТ - 5 мкл; деионизированная вода 20 мкл; образец фермента - 10 мкл.
Контрольный образец содержит все компоненты реакционной смеси, но вместо фермента добавляют 20 мкл воды. Реакцию проводят в течение 30-60 мин при 37°С и останавливали добавлением 150 мкл стоп-раствора (0,37 М FeCl3, 0,67 М HCl и 0,2 М трихлоруксусная кислота). А535 измеряют на планшетном спектрофотометре Uniplan (Пикон, Россия). Молярный коэффициент экстинкции α-сукцинилгидроксамата с Fe3+ равен 850 М-1×см-1.
Общий выход белка и удельную активность ω-амидазы измеряют на каждой стадии, как описано в Таблицах 1-3. Содержание белка и его чистоту оценивают визуально с использованием данных денатурирующего дискового электрофореза по Лаэммли.
Общий выход рекомбинантного hNit2 после всех стадий, усредненный для трех повторностей, составляет 85,5 ед., что соответствует 13,8 мг фермента с молярной активностью 6,3 ед./мг.
Сравнение активности фракций, полученных в процессе обработки клеточного лизата и очистки hNit2 от BL21 (DE3; pQE-Nit22) по отношению к αKGM и сукцинамату, показано в таблице 4.

6. Электрофорез белков в денатурирующих условиях и иммунноблоттинг
Электрофорез белков в денатурирующих условиях проводят, используя следующие растворы: электродный буфер - Трис 25 мМ, рН 8,6; глицин 92 мМ; 0,1% SDS; верхний (концентрирующий) гелевый буфер (4×) - 50 мМ Трис-HCl; 0,4% SDS; нижний (разделяющий) гель-буфер (4×) - 150 мМ Трис-HCl, рН 8,8; буфер Лаэммли с мочевиной - Трис (свободное основание) 25 мМ, 192 мМ глицин; рН 8,6; 1% SDS, 7 М мочевина, 50 мМ ДТТ, 0,001% бромфеноловый синий. Электрофорез проводят при 80 В на стадии концентрирования и 250 В на стадии разделения. Длина геля 5 см.
После электрофореза гель окрашивают 1% Кумасси R-250, разбавленным 10% уксусной кислотой, и обесцвечивают в 10% уксусной кислоте.
Полусухой электроблоттинг выделенных белков из ПААГ проводят на мембране из PVDF (Immobilon, Millipore, США) в камере Mini-Protean 3 Cell (Bio-Rad, США) в течение 1,5 ч и плотности тока 0,8 мА/см2 при 20°С. Мембрану окрашивают 1%-ным Понсо красным, растворенным в 10%-ной уксусной кислоте. Для определения положения полос используют маркеры молекулярных масс 120, 90, 50, 34, 26 и 20 кДа (ThermoFisher Scientific, каталожный №26612).
Мембрану обесцвечивают 10-кратной промывкой в воде, а неспецифическую связывающую способность блокируют инкубацией в течение 4-20 ч в 0,1% растворе BSA в PBS (10 мМ Na3PO4, рН 7,2, 100 мМ NaCl). Связывание первичных мышиных моноклональных антител против аффинной метки 6His (GE HealthCare, США) проводят в PBS с добавлением 0,03% Tween-20. Мембрану трижды промывают PBS с добавлением 0,03% Tween-20. Промытый фильтр инкубируют в течение 2 ч при осторожном перемешивании в растворе вторичных козьих антител против мышиного IgG, конъюгированных с пероксидазой хрена (ООО «Биалекса», Москва), разведенных в соотношении 1:3000 в PBS с добавлением 0,03% Tween-20. Мембрану трижды промывают PBS с добавлением 0,03% Tween-20. Хромогенный тест на пероксидазную активность проводят в растворе 3,3'-диаминобензидина 20 мкг/мл и 0,01% H2O2. Реакцию останавливают инкубацией в 10%-ной серной кислоте с последующей промывкой деионизированной водой. Обработанную таким образом мембрану высушивают на воздухе.
7. Оценка стабильности hNit2 при различных температурных условиях хранения
На основе метода измерения конечной активности ω-амидазы определяют зависимость стабильности очищенной hNit2 от температуры в условиях кратковременного и длительного хранения (Фиг. 4). Ко испытуемым фракциям hNit2, полученным после хроматографии на Ni-NTA-агарозе, добавляют ДТТ до 2 мМ и смесь ингибиторов протеиназ в разведении 1:100. Образцы помещают на хранение при комнатной температуре, +4°С, -80°С и -20°С. Замороженные образцы размораживают однократно через 1 неделю, 1, 2, 3, 4, 5 и 6 месяцев. Путем определения активности в отношении αKGM устанавливают, что образцы, хранящиеся при -20°С, сохраняют >95% в течение 1 месяца, а затем теряют по 15-18% активности каждый месяц. Образцы, хранящиеся при +4°С, после первой недели утрачивают 19-22% активности, а через 4 недели активность в них не определяется. Хранение образцов при комнатной температуре (+19-25°С) приводит к полной потере активности в течение 24 ч.
Устанавливают, что: 1) после трех последовательных циклов замораживания-оттаивания образцы теряют до 75-85% своей исходной активности и 2) присутствие глицерина влияло на конечную точку определения оптической плотности, увеличивая фоновый сигнал до неизмеримых значений (данные не показаны).
Сравнение раскрываемой последовательности SEQ_ID#1 (в составе плазмиды pQE-NIT22) с соответствующей ей последовательностью из состава плазмиды-прототипа pQE-NIT2

Полная последовательность плазмиды pQE-NIT22
Последовательность SEQ_ID#1 выделена фиолетовым, сайты рестриктаз, использованные для ее стыковки с плазмидой-прототипом выделены красным


--->
<110> Federal state budgetary institution “Center of strategic planning and
management of medical and biological risks to the health” of Russian federal
medical and biological agency”
<120> Recombinant producer of human omega-amidase Nit2
<160> SEQ ID NO 1
<210> SEO ID NO:1
<211> 81 bp
<212> DNA
<213> artificial sequence (optimized for expression in E. coli)
<220> A novel producer of human omega-amidase (Nit2) in Escherichia coli
<221>
<400> 1
GGA TCC CGC ACC AGC TTT CGT CTG GCG CTG ATT CAG CTG CAG ATT AGC 48
AGT ATT AAA AGC GAT AAC GTC ACC CGC GCA TGC 81
<110> Federal state budgetary institution “Center of strategic planning
and management of medical and biological risks to the health” of Russian
federal medical and biological agency”
<120> Recombinant producer of human omega-amidase Nit2
<160> SEQ ID NO 2
<210> SEO ID NO:2
<211> 27 aa
<212> DNA
<213> artificial sequence (optimized for expression in E. coli)
<220> A novel producer of human omega-amidase (Nit2) in Escherichia coli
<221>
<400> 2
Gly Ser Arg Thr Ser Phe Arg Leu Ala Leu Ile Gln Leu Gln Ile Ser 16
Ser Ile Lis Ser Asp Asn Val Thr Arg Ala Cys 27
<---
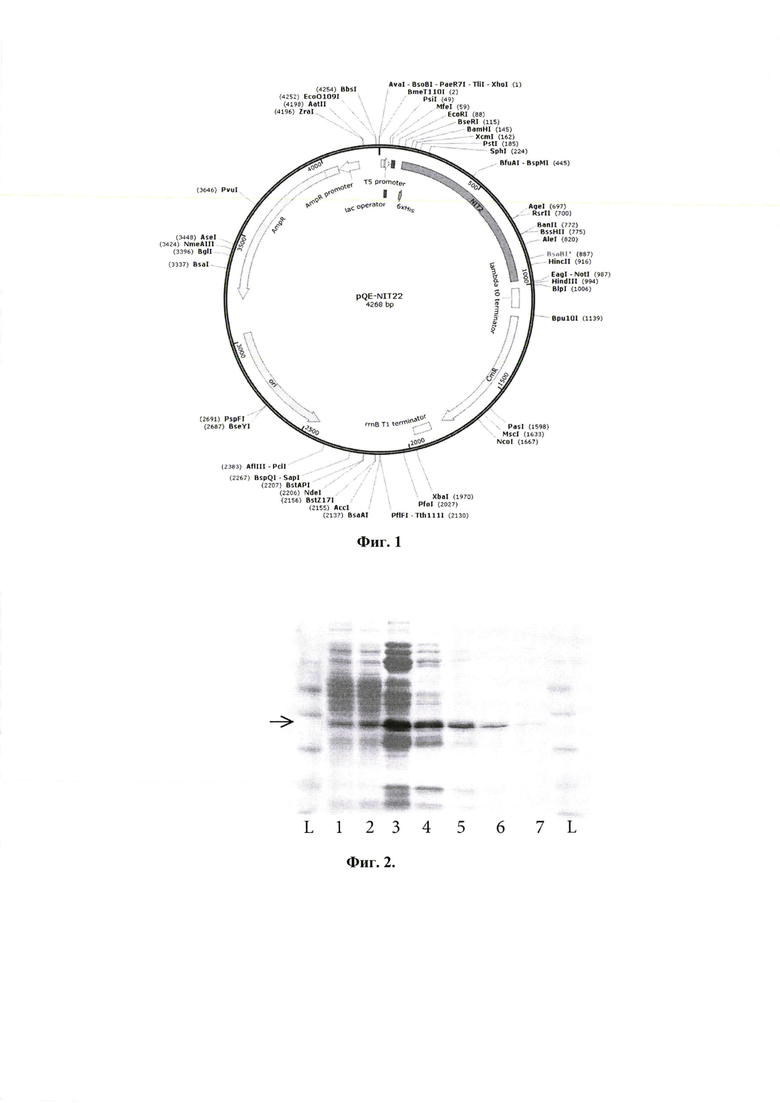
Изобретение относится к штамму Escherichia coli BL21(DE3), предназначенному для получения ω-амидазы человека. Предложен штамм Escherichia coli BL21(DE3), трансформированный экспрессионной плазмидной конструкцией pQE-Nit22, представленной на фиг. 1. При этом в составе экспрессионной плазмидной конструкции pQE-Nit22 81 нуклеотид кодирующей области гена Nit2, ограниченный сайтами рестриктаз BamHI и SphI, соответствует SEQ ID#1. Изобретение позволяет повысить выход продукта при культивировании на питательных средах на основе пептона, дрожжевого экстракта и глюкозы. 4 ил., 4 табл.
Штамм Escherichia coli BL21(DE3), предназначенный для получения ω-амидазы человека при культивировании на питательных средах на основе пептона, дрожжевого экстракта и глюкозы и трансформированный экспрессионной плазмидной конструкцией pQE-Nit22, представленной на фиг. 1, при этом 81 нуклеотид кодирующей области гена Nit2, ограниченный сайтами рестриктаз BamHI и SphI в составе экспрессионной плазмидной конструкции pQE-Nit22, соответствует SEQ ID#1.
| Рекомбинантный продуцент омега-амидазы человека Nit2 на основе Escherichia coli | 2016 |
|
RU2642323C2 |
| COOPER A.J.L | |||
| ET AL | |||
| ω-Amidase: an underappreciated, but important enzyme in L-glutamine and L-asparagine metabolism; relevance to sulfur and nitrogen metabolism, tumor biology and hyperammonemic diseases | |||
| Приспособление для автоматической односторонней разгрузки железнодорожных платформ | 1921 |
|
SU48A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Способ очистки нефти и нефтяных продуктов и уничтожения их флюоресценции | 1921 |
|
SU31A1 |

