ПЕРЕКРЕСТНАЯ ССЫЛКА НА СМЕЖНУЮ ЗАЯВКУ
Настоящая заявка испрашивает преимущество предварительной заявки на патент США № 61/657160, поданной 8 июня 2012 г., содержание которой полностью включено в настоящий документ путем ссылки для любых целей.
ОБЛАСТЬ ПРИМЕНЕНИЯ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к области дифференцировки клеток. Более конкретно, настоящее изобретение раскрывает применение эфриновых лигандов и сфингозин-1-фосфата как регуляторов дифференцировки плюрипотентных стволовых клеток в эндокринные клетки.
УРОВЕНЬ ТЕХНИКИ
Последние достижения в области заместительной клеточной терапии для лечения сахарного диабета 1 типа и нехватка островков Лангерганса для трансплантации заставили обратить внимание на разработку источников инсулин-продуцирующих клеток, или β-клеток, подходящих для трансплантации. Одним из подходов является формирование функциональных β-клеток из плюрипотентных стволовых клеток, таких как, например, эмбриональные стволовые клетки.
При эмбриональном развитии позвоночных плюрипотентные клетки дают начало группе клеток, формирующих три зародышевых листка (эктодерму, мезодерму и энтодерму) в ходе процесса, именуемого гаструляцией. Ткани, такие как ткань щитовидной железы, вилочковой железы, поджелудочной железы, кишечника и печени, будут развиваться из энтодермы через промежуточную стадию. Промежуточной стадией данного процесса является формирование дефинитивной энтодермы. Клетки дефинитивной энтодермы экспрессируют ряд маркеров, таких как HNF3-бета, GATA4, MIXL1, CXCR4 и SOX17.
К концу гаструляции энтодерма разделяется на передний и задний отделы, которые могут быть распознаны по экспрессии ряда факторов, однозначно выделяющих переднюю, среднюю и заднюю области энтодермы. Например, Hhex и Sox2 идентифицируют переднюю область, в то время как Cdx1, 2 и 4 идентифицируют заднюю половину энтодермы.
Миграция ткани энтодермы приближает энтодерму к различным мезодермальным тканям, которые способствуют регионализации кишечной трубки. Это достигается за счет целого ряда секретируемых факторов, таких как FGF, Wnt, TGF-B, ретиноевая кислота (RA), лиганды BMP и их антагонисты. Например, FGF4 и BMP способствуют экспрессии Cdx2 в предполагаемой энтодерме задней кишки и подавляют экспрессию передних генов Hhex и SOX2 (Development 2000 г., 127:1563-1567). Было также продемонстрировано, что сигнализация WNT действует параллельно с сигнализацией FGF, способствуя развитию задней кишки и препятствуя зачаткам передней кишки (Development 2007 г., 134:2207-2217). Наконец, секретируемая мезенхимой ретиноевая кислота регулирует границу между передней и задней кишкой (Curr Biol 2002 г., 12:1215-1220).
Уровень экспрессии специфических факторов транскрипции может использоваться для определения типа ткани. Во время преобразования дефинитивной энтодермы в примитивную кишечную трубку кишечная трубка становится разделенной на широкие домены, которые можно наблюдать на молекулярном уровне с помощью ограниченных паттернов экспрессии генов. Например, регионализованный домен поджелудочной железы в кишечной трубке демонстрирует очень высокую экспрессию PDX-1 и очень низкую экспрессию CDX2 и SOX2. Аналогичным образом, наличие высоких уровней Foxe1 свидетельствует о ткани пищевода; в легочной ткани высок уровень экспрессии NKX2.1; в ткани желудка высок уровень экспрессии SOX2/Odd1 (OSR1); уровень экспрессии PROX1/Hhex/AFP высок в ткани печени; SOX17 имеет высокий уровень экспрессии в тканях желчного тракта; уровень экспрессии PDX1, NKX6.1/PTf1a и NKX2.2 высок в ткани поджелудочной железы; а уровень экспрессии CDX2 высок в ткани кишечника. Приведенная выше сводка взята из Dev Dyn 2009 г., 238:29-42 и Annu Rev Cell Dev Biol 2009 г., 25:221-251.
Формирование поджелудочной железы происходит в результате дифференцировки дефинитивной энтодермы в панкреатическую энтодерму (Annu Rev Cell Dev Biol 2009 г., 25:221-251; Dev Dyn 2009 г., 238:29-42). Дорсальный и вентральный домены поджелудочной железы формируются из эпителия передней кишки. Передняя кишка также дает начало формированию пищевода, трахеи, легких, щитовидной железы, желудка, печени, поджелудочной железы и системы желчных протоков.
Клетки панкреатической энтодермы экспрессируют панкреодуоденальный, содержащий гомеобокс ген PDX1. При отсутствии PDX1 развитие поджелудочной железы не идет дальше формирования вентрального и дорсального зачатков. Таким образом, экспрессия PDX1 характеризует критическую стадию органогенеза поджелудочной железы. Зрелая поджелудочная железа содержит, помимо других типов клеток, экзокринную ткань и эндокринную ткань. Экзокринная и эндокринная ткани образуются при дифференцировке панкреатической энтодермы.
D'Amour et al. описывают производство обогащенных культур дефинитивной энтодермы, полученной из эмбриональных стволовых (ЭС) клеток человека в присутствии высокой концентрации активина и низкой концентрации сыворотки (Nature Biotechnol 2005 г., 23:1534-1541; патент США № 7,704,738). Трансплантация этих клеток под капсулу почки у мышей приводила к дифференцировке в более зрелые клетки с характеристиками ткани энтодермы (патент США № 7,704,738). Клетки дефинитивной энтодермы, полученные из эмбриональных стволовых клеток человека, могут быть далее дифференцированы в PDX1-положительные клетки после добавления FGF-10 и ретиноевой кислоты (патентная публикация США № 2005/0266554 A1). Последующая трансплантация таких клеток-предшественников панкреатических клеток в жировое тело иммунодефицитных мышей привела к образованию функциональных панкреатических эндокринных клеток с последующей 3-4-месячной стадией созревания (патент США № 7993920 и патент США № 7534608).
Fisk et al. сообщают о системе получения клеток панкреатических островков из эмбриональных стволовых клеток человека (патент США № 7033831). В данном случае процесс дифференцировки был разделен на три стадии. Эмбриональные стволовые клетки человека сначала были дифференцированы в энтодерму при помощи комбинации бутирата натрия и активина A (патент США № 7326572). Затем клетки культивировали с антагонистами BMP, такими как Noggin, в комбинации с EGF или бета-целлюлином для генерирования PDX1-положительных клеток. Окончательную дифференцировку индуцировали никотинамидом.
Низкомолекулярные ингибиторы также применяли для индуцирования клеток-предшественников панкреатических эндокринных клеток. Например, низкомолекулярные ингибиторы рецепторов TGF-B и рецепторов BMP (Development 2011 г., 138:861-871; Diabetes 2011 г., 60:239-247) использовали для значительного увеличения количества панкреатических эндокринных клеток. Кроме того, для генерирования клеток дефинитивной энтодермы или клеток-предшественников панкреатических клеток также применяли низкомолекулярные активаторы (Curr Opin Cell Biol 2009 г., 21:727-732; Nature Chem Biol 2009 г., 5:258-265).
Несмотря на большие успехи, достигнутые в оптимизации протоколов по получению панкреатических клеток из плюрипотентных стволовых клеток человека, существует необходимость разработки протоколов для получения функциональных эндокринных клеток и, в частности, бета-клеток. В данной работе мы демонстрируем, что класс эфринов и сфингозин-1-фосфат или агонисты рецептора сфингозина повышают эффективность получения эндокринных клеток и ускоряют кластеризацию эндокринных гормонов и эндокринных клеток-предшественников.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В одном из вариантов осуществления настоящее изобретение относится к способу повышения экспрессии инсулина и NKX6.1 путем культивирования популяции клеток панкреатической энтодермы в среде, содержащей эфрин А4 или эфрин А3. В некоторых вариантах осуществления популяция клеток панкреатической энтодермы по существу не экспрессирует CDX2 или SOX2. В некоторых вариантах осуществления популяцию клеток панкреатической энтодермы получают путем поэтапной дифференцировки плюрипотентных клеток. В некоторых вариантах осуществления плюрипотентные клетки являются эмбриональными плюрипотентными клетками человека.
В одном из вариантов осуществления настоящее изобретение относится к способу повышения экспрессии соматостатина при подавлении экспрессии инсулина, глюкагона и грелина путем культивирования клеток панкреатической энтодермы в среде, содержащей активин A или активин C. В некоторых вариантах осуществления популяция клеток панкреатической энтодермы, обработанных активином A или активином С, экспрессирует больше соматостатина, чем популяция клеток панкреатической энтодермы, не обработанных активином A или активином C. В некоторых вариантах осуществления экспрессия инсулина подавлена в популяции клеток панкреатической энтодермы, обработанных активином А или активином С, по сравнению с экспрессией инсулина в популяции клеток панкреатической энтодермы, не обработанных активином А или активином С. В некоторых вариантах осуществления экспрессия глюкагона подавлена в популяции клеток панкреатической энтодермы, обработанных активином А или активином С, по сравнению с экспрессией глюкагона в популяции клеток панкреатической энтодермы, не обработанных активином А или активином С. В некоторых вариантах осуществления экспрессия грелина подавлена в популяции клеток панкреатической энтодермы, обработанных активином А или активином С, по сравнению с экспрессией грелина в популяции клеток панкреатической энтодермы, не обработанных активином А или активином С. В некоторых вариантах осуществления клетки панкреатической энтодермы по существу не экспрессируют CDX2 или SOX2. В некоторых вариантах осуществления клетки панкреатической энтодермы, обработанные активином А или активином С, получают путем поэтапной дифференцировки плюрипотентных клеток. В некоторых вариантах осуществления плюрипотентные клетки, из которых получают клетки панкреатической энтодермы, являются эмбриональными плюрипотентными клетками человека.
В одном из вариантов осуществления настоящее изобретение относится к способу повышения экспрессии NKX6.1 путем обработки клеток панкреатической энтодермы средой, содержащей семафорин 3А или эпиген. В некоторых вариантах осуществления популяция клеток панкреатической энтодермы, обработанных средой, содержащей семафорин 3А или эпиген, экспрессирует повышенное количество NKX6.1 по сравнению с клетками панкреатической энтодермы, не обработанных средой, содержащей семафорин 3А или эпиген. В некоторых вариантах осуществления уровень экспрессии таких гормонов, как инсулин, глюкагон и грелин в клетках панкреатической энтодермы, обработанных средой, содержащей семафорин 3А или эпиген, остается таким же, как и в клетках панкреатической энтодермы, не обработанных средой, содержащей семафорин 3А или эпиген. В некоторых вариантах осуществления клетки панкреатической энтодермы по существу не экспрессируют CDX2 или SOX2. В некоторых вариантах осуществления клетки панкреатической энтодермы, обработанные средой, содержащей семафорин 3А или эпиген, получают путем поэтапной дифференцировки плюрипотентных клеток. В некоторых вариантах осуществления плюрипотентные клетки, из которых получают клетки панкреатической энтодермы, являются эмбриональными плюрипотентными клетками человека.
В некоторых вариантах осуществления настоящее изобретение относится к способу поэтапной дифференцировки плюрипотентных клеток, содержащему культивирование клеток панкреатической энтодермы в среде, содержащей эфрин А4, эфрин А3, активин А, активин С, семафорин 3А или эпиген. В некоторых вариантах осуществления клетки панкреатической энтодермы культивируют в среде, содержащей эфрин А4 или эфрин А3. В некоторых вариантах осуществления клетки панкреатической энтодермы культивируют в среде, содержащей активин А или активин С. В некоторых вариантах осуществления клетки панкреатической энтодермы культивируют в среде, содержащей семафорин 3А или эпиген. В некоторых вариантах осуществления плюрипотентные клетки, из которых получают клетки панкреатической энтодермы, являются эмбриональными плюрипотентными клетками человека.
В одном из вариантов осуществления настоящее изобретение относится к способу индукции экспрессии эндокринных кластеров путем обработки панкреатических эндокринных клеток агонистом рецептора сфингозина-1. В некоторых вариантах осуществления для обработки панкреатических эндокринных клеток в качестве агониста рецептора сфингозина-1 используют сфингозин-1-фосфат (S1P).
Также в качестве вариантов осуществления настоящего изобретения рассматриваются клетки, полученные способами настоящего изобретения и способами с использованием клеток настоящего изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
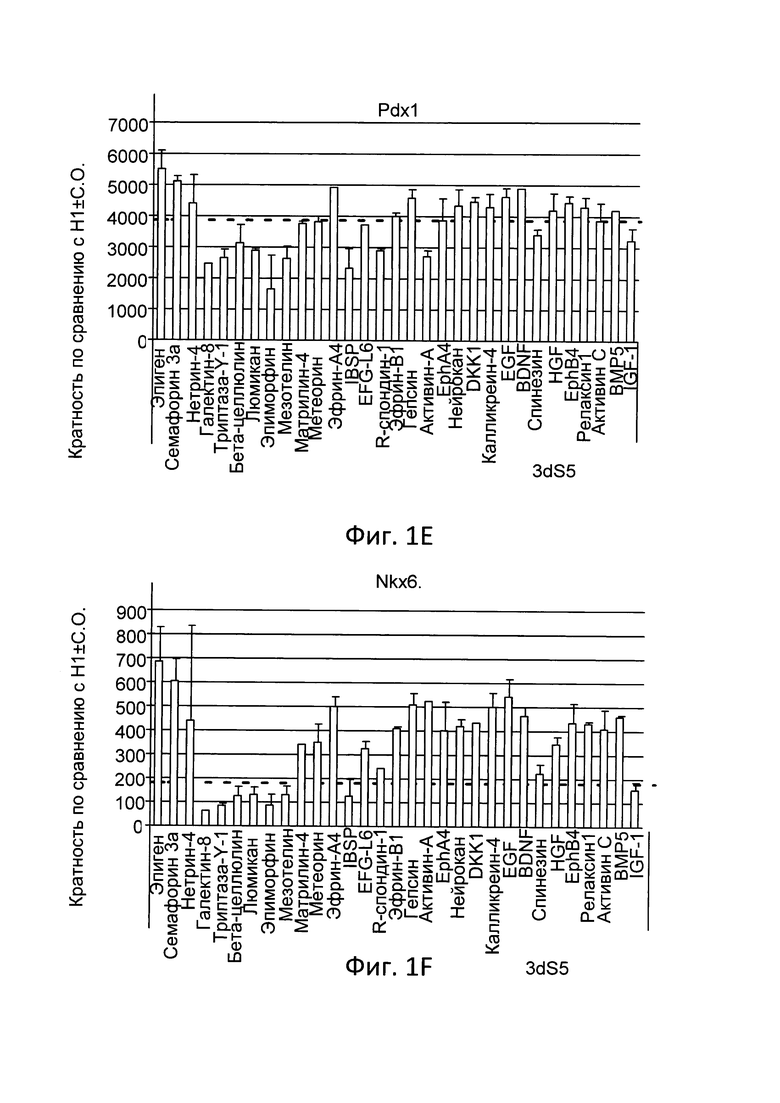
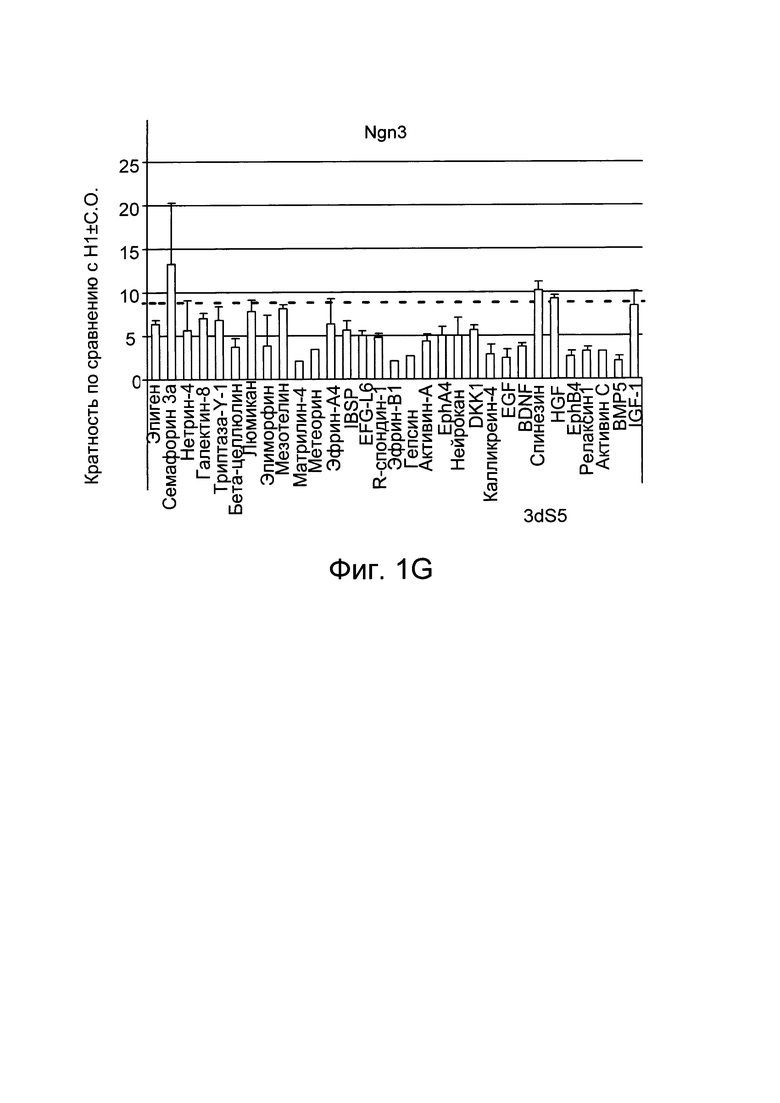
На Фиг. 1A-1G представлены данные анализов ПЦР в реальном времени по экспрессии следующих генов в клетках линии эмбриональных стволовых клеток человека H1, дифференцированных, как описано в примере 1: инсулин (Фиг. 1A), соматостатин (Фиг. 1B), грелин (Фиг. 1C), глюкагон (Фиг. 1D), PDX-1 (Фиг. 1E), NKX6.1 (Фиг. 1F) и NGN3 (Фиг. 1G).
На Фиг. 2A-2C представлены изображения клеток после иммуноокрашивания на инсулин. Фиг. 2A - контроль; Фиг. 2B - клетки, обработанные раствором эфрина А3, 50 нг/мл; и Фиг. 2C - клетки, обработанные раствором эфрина А3, 100 нг/мл, как описано в примере 2.
На Фиг. 3A-3C представлены изображения клеток после иммуноокрашивания на инсулин. Фиг. 3A - контроль; Фиг. 3B - клетки, обработанные раствором эфрина А4, 50 нг/мл; и Фиг. 3C - клетки, обработанные раствором эфрина А4, 100 нг/мл, как описано в примере 2.
На Фиг. 4A-4D представлены фазово-контрастные изображения клеток культуры S6, которые обработали сфингозин-1-фосфатом (S1P) и сфотографировали на 1-й день (Фиг. 4A), 7-й день (Фиг. 4B) и при двух различных увеличениях на 10-й день (Фиг. 4C и Фиг. 4D). Эти изображения показывают, что на 7-й день наблюдались четкие признаки кластеризации эндокринных клеток, а на 10-й день кластеры отделялись друг от друга тонким слоем панкреатического энтодермального эпителия.
На Фиг. 5A-5D представлены изображения клеток, обработанных S1P и иммуноокрашенных на Hb9 (Фиг. 5A) и NKX6.1 (Фиг. 5B) или иммуноокрашенных на инсулин (Фиг. 5C) и Hb9 (Фиг. 5D).
На Фиг. 6A и Фиг. 6B представлены фазово-контрастные изображения различного увеличения клеток, обработанных 10 мкM S1P и собранных через три дня после начала стадии 6. На Фиг. 6C и Фиг. 6D представлены изображения клеток после иммуноокрашивания на NKX2.2. Фиг. 6C - клетки контроля; Фиг. 6D - клетки, обработанные S1P.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Для четкости описания, а не для ограничения изобретения подробное описание изобретения разделено на следующие подразделы, описывающие или иллюстрирующие определенные особенности, варианты осуществления или области применения настоящего изобретения.
Определения
Стволовые клетки представляют собой недифференцированные клетки, определяемые как обладающие способностью на одноклеточном уровне к самообновлению и дифференциации. Стволовые клетки могут производить клетки-потомки, включая самообновляющиеся прогениторные клетки, необновляющиеся прогениторные клетки и окончательно дифференцированные клетки. Стволовые клетки также характеризуются своей способностью дифференцироваться in vitro в функциональные клетки различных клеточных линий дифференцировки из множества зародышевых листков (энтодермы, мезодермы и эктодермы). Стволовые клетки также дают начало тканям множества зародышевых листков после трансплантации и вносят значительный вклад в образование большинства, если не всех тканей после инъекции в бластоцисты.
По потенциалу развития стволовые клетки разделяют на: (1) тотипотентные, т.е. способные преобразоваться в любой из эмбриональных и внеэмбриональных типов клеток; (2) плюрипотентные, т.е. способные преобразоваться во все типы эмбриональных клеток; (3) мультипотентные, т.е. способные преобразоваться в подмножество клеточных линий дифференцировки, но в рамках одной ткани, органа или физиологической системы (например, гемопоэтические стволовые клетки (ГСК) могут порождать клетки-потомки, которые включают в себя ГСК (самообновление), ограниченные клетками крови олигопотентные прогениторные клетки и все типы клеток и клеточные элементы (например, тромбоциты), являющиеся стандартными компонентами крови); (4) олигопотентные, т.е. способные преобразоваться в более ограниченное подмножество клеточных линий дифференцировки, чем мультипотентные стволовые клетки; и (5) унипотентные, т.е. способные преобразоваться в единственную клеточную линию дифференцировки (например, сперматогенные стволовые клетки).
Дифференцировка представляет собой процесс, при помощи которого неспециализированная («некоммитированная») или менее специализированная клетка приобретает свойства специализированной клетки, например нервной или мышечной клетки. Дифференцированная клетка или клетка, индуцированная дифференциацией, представляет собой клетку, занявшую более специализированное («коммитированное») положение в линии дифференцировки клетки. Термин «коммитированный» в применении к процессу дифференцировки относится к клетке, которая прошла по пути дифференцировки до той стадии, на которой при нормальных условиях она продолжит дифференцироваться в клетку заданного типа или подмножества типов клеток и при этом не может в нормальных условиях дифференцироваться в клетку другого типа или вернуться к клетке менее дифференцированного типа. Термин «дедифференцировка» относится к процессу, посредством которого клетка возвращается на менее специализированную (или коммитированную) стадию клеточной линии дифференцировки. Используемый в настоящей заявке термин «линия дифференцировки клетки» определяет наследственность клетки, то есть определяет, из какой клетки произошла данная клетка и каким клеткам она может дать начало. В линии дифференцировки клетка помещается в наследственную схему развития и дифференцировки. Маркером, специфичным для линии дифференцировки, называется характерная особенность, специфически ассоциированная с фенотипом клеток конкретной линии дифференцировки, которая может использоваться для оценки дифференцировки некоммитированных клеток в клетки данной линии дифференцировки.
Используемый в настоящей заявке термин «маркеры» означает молекулы нуклеиновых кислот или полипептидов с дифференциальной экспрессией в интересующих клетках. В данном контексте под дифференциальной экспрессией понимают повышенный уровень положительного маркера и пониженный уровень отрицательного маркера по сравнению с недифференцированной клеткой. Поддающийся обнаружению уровень маркерной нуклеиновой кислоты или полипептида в интересующих клетках оказывается значительно выше или ниже по сравнению с другими клетками, что позволяет выявить интересующую клетку и отличить ее от других клеток с помощью любого из множества известных в данной области способов.
В настоящем документе клетка «положительна по» заданному маркеру или «положительна», если заданный маркер обнаруживается в клетке. Аналогичным образом, клетка «отрицательна по» заданному маркеру или «отрицательна», если заданный маркер не обнаруживается в клетке.
В настоящем документе термины «плотность клеток» и «плотность посева» применяются взаимозаменяемо и относятся к числу клеток, высеваемых на единицу площади поверхности твердого или полутвердого плоского или криволинейного субстрата.
В настоящем документе термины «стадия 1» и «S1» применяются взаимозаменяемо для обозначения клеток, экспрессирующих маркеры, характерные для дефинитивной энтодермы (ДЭ).
В настоящем документе термин «дефинитивная энтодерма» относится к клеткам, которые несут в себе характеристики клеток, появившихся из эпибласта во время гаструляции, и которые формируют желудочно-кишечный тракт и его производные. Клетки дефинитивной энтодермы экспрессируют по меньшей мере один из следующих маркеров: HNF3-бета, GATA4, SOX17, CXCR4, Cerberus, OTX2, goosecoid, C-Kit, CD99 и MIXL1.
Термин «кишечная трубка» в настоящем документе относится к клеткам, полученным из дефинитивной энтодермы, экспрессирующим по меньшей мере один из следующих маркеров: HNF3-бета, HNF1-бета или HNF4-альфа. Клетки кишечной трубки могут дать начало всем энтодермальным органам, таким как легкие, печень, поджелудочная железа, желудок и кишечник.
Используемые в настоящем документе взаимозаменяемые термины «стадия 2» и «S2» обозначают клетки, экспрессирующие маркеры, характерные для примитивной кишечной трубки.
Термин «энтодерма передней кишки» относится к клеткам энтодермы, которые дают начало пищеводу, легким, желудку, печени, поджелудочной железе, желчному пузырю и части двенадцатиперстной кишки.
Термин «задняя часть передней кишки» относится к клеткам энтодермы, которые могут дать начало задней части желудка, поджелудочной железе, печени и части двенадцатиперстной кишки.
Термин «энтодерма средней кишки» относится к клеткам энтодермы, которые могут дать начало кишечнику, частям двенадцатиперстной кишки, аппендиксу и восходящей ободочной кишке.
Термин «энтодерма задней кишки» относится к клеткам энтодермы, которые могут дать начало дистальной трети поперечной ободочной кишки, нисходящей ободочной кишки, сигмовидной кишки и прямой кишки.
Термины «стадия 3» и «S3» используются взаимозаменяемо для обозначения клеток, экспрессирующих маркеры, характерные для энтодермы передней кишки. Используемый в настоящем документе термин «клетки, экспрессирующие маркеры, характерные для линии передней кишки» относится к клеткам, экспрессирующим по меньшей мере один из следующих маркеров: PDX-1, FOXA2, CDX2, SOX2 и HNF4-альфа.
В настоящем документе термины «стадия 4» и «S4» используются взаимозаменяемо для обозначения клеток, экспрессирующих маркеры, характерные для клеток-предшественников панкреатических клеток из передней кишки. Используемый в настоящем документе термин «клетки, экспрессирующие маркеры, характерные для линии клетки-предшественника панкреатической клетки из передней кишки» относится к клеткам, экспрессирующим по меньшей мере один из следующих маркеров: PDX-1, NKX6.1, HNF6, FOXA2, PTF1a, Prox1 и HNF4-альфа.
В настоящем документе термины «стадия 5» и «S5» используются взаимозаменяемо для обозначения клеток, экспрессирующих маркеры, характерные для клеток панкреатической энтодермы и клеток-предшественников панкреатических эндокринных клеток. Используемый в настоящем документе термин «клетки, экспрессирующие маркеры, характерные для линии панкреатической энтодермы» относится к клеткам, экспрессирующим по меньшей мере один из следующих маркеров: PDX1, NKX6.1, HNF1-бета, PTF1-альфа, HNF6, HNF4-альфа, SOX9, HB9 или PROX1. Клетки, экспрессирующие маркеры, характерные для линии панкреатической энтодермы, по существу не экспрессируют CDX2 или SOX2.
Используемые в настоящей заявке термины «панкреатическая эндокринная клетка», или «клетка, экспрессирующая гормон поджелудочной железы», или «клетка, экспрессирующая маркеры, характерные для линии панкреатических эндокринных клеток», или «клетки стадии 6», или «клетки S6» используются взаимозаменяемо и относятся к клетке, способной экспрессировать по меньшей мере один из следующих гормонов: инсулин, глюкагон, соматостатин, грелин и панкреатический полипептид.
Термин «панкреатические инсулин-положительные клетки» относится к популяции эндокринных клеток, экспрессирующих инсулин, HB9, NKX2.2 и NKX6.1.
Термин «клетка-предшественник панкреатической эндокринной клетки» или «прогениторная клетка панкреатической эндокринной клетки» относится к клеткам панкреатической энтодермы, способным стать клеткой, экспрессирующей гормон поджелудочной железы. Такая клетка может экспрессировать по меньшей мере один из следующих маркеров: NGN3, NKX2.2, NeuroD, ISL-1, Pax4, Pax6 или ARX.
В настоящем документе взаимозаменяемо применяются выражения «d1», «d 1» и «день 1»; «d2», «d 2» и «день 2»; «d3», «d 3» и «день 3» и так далее. Эти комбинации цифр и букв относятся к конкретному дню инкубации на различных стадиях в процессе поэтапного протокола дифференцировки настоящей заявки.
Термины «глюкоза» и «D-глюкоза» используются в настоящем документе взаимозаменяемо и относятся к декстрозе, сахару, широко встречающемуся в природе.
В настоящем документе термины «NeuroD» и «NeuroD1» используются взаимозаменяемо для обозначения белка, экспрессируемого в прогениторных клетках панкреатических эндокринных клеток, и кодирующего его гена.
В настоящем документе взаимозаменяемо используются выражения «LDN» и «LDN-193189», обозначающие ингибитор рецептора BMP, поставляемый компанией Stemgent, штат Калифорния, США.
Выделение, размножение и культивирование плюрипотентных стволовых клеток
Плюрипотентные стволовые клетки могут экспрессировать один или более стадиеспецифических эмбриональных антигенов (SSEA) 3 и 4, а также маркеры, обнаруживаемые с использованием антител, обозначаемых как Tra-1-60 и Tra-1-81 (Thomson et al., 1998 г. Science 282:1145, 1147). Дифференцировка плюрипотентных стволовых клеток in vitro приводит к потере экспрессии SSEA-4, Tra-1-60 и Tra-1-81. Недифференцированные плюрипотентные стволовые клетки, как правило, имеют щелочнофосфатазную активность, которую можно обнаружить путем обработки клеток 4%-ным параформальдегидом с последующим проявлением Vector Red в качестве субстрата, как описано производителем (Vector Laboratories, штат Калифорния, США). Недифференцированные плюрипотентные стволовые клетки также, как правило, экспрессируют OCT4 и TERT, обнаруживаемые с помощью ОТ-ПЦР.
Другим желательным фенотипическим свойством выращенных плюрипотентных стволовых клеток является потенциал дифференцировки в клетки всех трех зародышевых листков: в энтодермальные, мезодермальные и эктодермальные ткани. Плюрипотентность стволовых клеток может быть подтверждена, например, путем инъекции клеток мышам с тяжелым комбинированным иммунодефицитом (SCID), обработкой всех образующихся тератом 4%-ным раствором параформальдегида, а затем гистологическим исследованием на наличие типов клеток из трех зародышевых листков. В качестве альтернативы плюрипотентность можно определить по созданию эмбриоидных телец и анализа их на предмет присутствия маркеров, ассоциирующихся с тремя зародышевыми листками.
Выращенные линии плюрипотентных стволовых клеток можно кариотипировать с применением стандартного способа окрашивания с использованием красителя Гимза (G-banding) и сравнения с опубликованными кариотипами соответствующих видов приматов. Желательно получить клетки, имеющие «нормальный кариотип», т.е. эуплоидные клетки, в которых все человеческие хромосомы присутствуют и не имеют видимых изменений. Плюрипотентные клетки можно легко размножить в культуре путем использования различных питательных слоев или с помощью сосудов, покрытых матриксными белками. Альтернативно для стандартного размножения клеток можно использовать химически определенные поверхности в комбинации со средами определенного состава, такими как среды mTeSR® 1 (StemCell Technologies, г. Ванкувер, Канада). Плюрипотентные клетки можно легко снять с культуральных планшетов при помощи ферментативной, механической обработки или с использованием различных кальцийхелатирующих агентов, таких как EDTA (этилендиаминтетрауксусная кислота). Альтернативно плюрипотентные клетки можно размножать в суспензии в отсутствие каких-либо матриксных белков или питательного слоя.
Источники плюрипотентных стволовых клеток
Типы возможных для применения плюрипотентных стволовых клеток могут включать в себя устойчивые линии плюрипотентных клеток, полученных из тканей, образованных после беременности, в том числе преэмбриональной ткани (такой как, например, бластоцист), эмбриональной ткани или ткани плода, взятой в любой момент во время беременности, как правило, но не обязательно перед сроком приблизительно от 10 до 12 недель беременности. Неограничивающими примерами являются устойчивые линии эмбриональных стволовых клеток человека (ЭСК) или эмбриональные зародышевые клетки человека, такие как, например, эмбриональные стволовые клетки человека линий H1, H7 и H9 (WiCell Research Institute, г. Мэдисон, штат Висконсин, США). Также соответствуют целям настоящего изобретения клетки, взятые из популяции плюрипотентных стволовых клеток, уже культивированных в отсутствие питающих клеток. Также соответствуют целям настоящего изобретения индуцибельные плюрипотентные клетки (IPS) или перепрограммированные плюрипотентные клетки, которые могут быть получены из взрослых соматических клеток с помощью принудительной экспрессии ряда факторов, относящихся к плюрипотентным транскрипционным факторам, таким как OCT4, NANOG, Sox2, KLF4 и ZFP42 (Annu Rev Genomics Hum Genet 2011 г., 12:165-185). Эмбриональные стволовые клетки человека, которые используются в способах настоящего изобретения, также могут быть получены, как описано Thomson et al. (патент США № 5843780; Science, 1998 г., 282:1145-1147; Curr Top Dev Biol 1998 г., 38:133-165; Proc Natl Acad Sci U.S.A. 1995 г., 92:7844-7848).
Формирование клеток, экспрессирующих маркеры, характерные для линии панкреатической энтодермы, из плюрипотентных стволовых клеток
Характеристики плюрипотентных стволовых клеток хорошо известны специалистам в данной области, и продолжается выявление дополнительных характеристик плюрипотентных стволовых клеток. Маркеры плюрипотентных стволовых клеток включают в себя, например, экспрессию одного или более из следующих маркеров: ABCG2, cripto, FOXD3, коннексин 43, коннексин 45, OCT4, SOX2, NANOG, hTERT, UTF1, ZFP42, SSEA-3, SSEA-4, Tra 1-60, Tra 1-81.
Соответствующие целям настоящего изобретения плюрипотентные стволовые клетки включают в себя, например, человеческие эмбриональные стволовые клетки линии H9 (код NIH: WA09), человеческие эмбриональные стволовые клетки линии H1 (код NIH: WA01), человеческие эмбриональные стволовые клетки линии H7 (код NIH: WA07) и человеческие эмбриональные стволовые клетки линии SA002 (Cellartis, Швеция). Также соответствуют целям настоящего изобретения клетки, экспрессирующие по меньшей мере один из следующих маркеров, характерных для плюрипотентных клеток: ABCG2, cripto, CD9, FOXD3, коннексин 43, коннексин 45, OCT4, SOX2, NANOG, hTERT, UTF1, ZFP42, SSEA-3, SSEA-4, Tra 1-60 и Tra 1-81.
Маркеры, характерные для линии дефинитивной энтодермы, выбираются из группы, состоящей из SOX17, GATA4, HNF3-бета, GSC, CER1, Nodal, FGF8, Brachyury, Mix-подобного гомеобоксного белка, FGF4, CD48, эомезодермина (EOMES), DKK4, FGF17, GATA6, CXCR4, C-Kit, CD99 и OTX2. Целям настоящего изобретения соответствуют клетки с экспрессией по меньшей мере одного из маркеров, характерных для линии сформированной энтодермы. В одном из аспектов настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии сформированной энтодермы, представляет собой клетку-предшественник первичной полоски. В другом аспекте настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии сформированной энтодермы, представляет собой мезэнтодермальную клетку. В другом аспекте настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии сформированной энтодермы, представляет собой клетку сформированной энтодермы.
Маркеры, характерные для линии панкреатической энтодермы, выбираются из группы, содержащей PDX1, NKX6.1, HNF1-бета, PTF1-альфа, HNF6, HNF4-альфа, SOX9, HB9 и PROX1. Целям настоящего изобретения соответствуют клетки с экспрессией по крайней мере одного из маркеров, характерных для линии панкреатической энтодермы. В одном из аспектов настоящего изобретения клетки, экспрессирующие маркеры, характерные для линии панкреатической энтодермы, представляют собой клетки панкреатической энтодермы, в которых экспрессия PDX-1 и NKX6.1 по существу превышает экспрессию CDX2 и SOX2.
Маркеры, характерные для линии панкреатических эндокринных клеток, выбираются из группы, состоящей из NGN3, NEUROD, ISL1, PDX1, NKX6.1, PAX4, ARX, NKX2.2 и PAX6. В одном варианте осуществления панкреатическая эндокринная клетка способна к экспрессии по меньшей мере одного из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид. Целям настоящего изобретения соответствуют клетки, экспрессирующие по меньшей мере один из маркеров, характерных для линии панкреатических эндокринных клеток. В одном из аспектов настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии панкреатических эндокринных клеток, представляет собой панкреатическую эндокринную клетку. Панкреатическая эндокринная клетка может представлять собой панкреатическую клетку, экспрессирующую гормоны. Альтернативно панкреатическая эндокринная клетка может представлять собой панкреатическую клетку, секретирующую гормоны.
Панкреатические эндокринные клетки настоящего изобретения представляют собой клетки, экспрессирующие маркеры, характерные для клеточной линии дифференцировки. Клетка с экспрессией маркеров, характерных для клеточной линии дифференцировки клеток, экспрессирует PDX1 и по меньшей мере один из следующих транскрипционных факторов: NKX2.2, NKX6.1, NEUROD, ISL1, HNF3-бета, MAFA, PAX4 и PAX6. В одном аспекте настоящего изобретения клетка с экспрессией маркеров, характерных для клеточной линии дифференцировки клеток, представляет собой клетку.
В одном из вариантов осуществления настоящее изобретение относится к способу повышения экспрессии инсулина и NKX6.1 путем культивирования клеток на стадии 5 в среде, содержащей эфрин А4 или эфрин А3. В некоторых вариантах осуществления экспрессия инсулина и NKX6.1 в популяции клеток повышается по меньшей мере в 2 раза по сравнению с экспрессией инсулина и NKX6.1 в популяции необработанных клеток. В некоторых вариантах осуществления популяция клеток на стадии 5 по существу не экспрессирует CDX2 или SOX2. В некоторых вариантах осуществления популяцию клеток на стадии 5 получают путем поэтапной дифференцировки плюрипотентных клеток. В некоторых вариантах осуществления плюрипотентные клетки представляют собой эмбриональные плюрипотентные клетки человека.
В одном из вариантов осуществления настоящее изобретение относится к способу повышения экспрессии соматостатина при подавлении экспрессии инсулина, глюкагона и грелина путем культивирования клеток на стадии 5 в среде, содержащей активин A или активин C. В некоторых вариантах осуществления обработанная популяция клеток экспрессирует по меньшей мере в два раза больше соматостатина, чем необработанные культуры. В некоторых вариантах осуществления экспрессия инсулина подавлена приблизительно в два раза по сравнению с экспрессией инсулина в необработанных культурах. В некоторых вариантах осуществления экспрессия глюкагона подавлена приблизительно до 1/10 от уровня экспрессии глюкагона в необработанных культурах. В некоторых вариантах осуществления экспрессия грелина подавлена приблизительно до 1/3 от уровня экспрессии грелина в необработанных культурах. В некоторых вариантах осуществления клетки на стадии 5 по существу не экспрессируют CDX2 или SOX2. В некоторых вариантах осуществления клетки на стадии 5 получают путем поэтапной дифференцировки плюрипотентных клеток. В некоторых вариантах осуществления плюрипотентные клетки представляют собой эмбриональные плюрипотентные клетки человека.
В одном из вариантов осуществления настоящее изобретение относится к способу повышения экспрессии NKX6.1 путем обработки клеток на стадии 5 средой, содержащей семафорин 3А или эпиген. В некоторых вариантах осуществления обработанная популяция клеток экспрессирует по меньшей мере в два раза больше NKX6.1, чем необработанные культуры. В некоторых вариантах осуществления уровень экспрессии гормонов в обработанных клетках остается тем же, что и в необработанных культурах. В некоторых вариантах осуществления клетки на стадии 5 по существу не экспрессируют CDX2 или SOX2. В некоторых вариантах осуществления клетки на стадии 5 получают путем поэтапной дифференцировки плюрипотентных клеток. В некоторых вариантах осуществления плюрипотентные клетки представляют собой эмбриональные плюрипотентные клетки человека.
В некоторых вариантах осуществления настоящее изобретение относится к способу поэтапной дифференцировки плюрипотентных клеток, содержащему культивирование клеток на стадии 5 в среде, содержащей эфрин А4, эфрин А3, активин А, активин С, семафорин 3А или эпиген. В некоторых вариантах осуществления клетки на стадии 5 культивировали в среде, содержащей эфрин А4 или эфрин А3. В некоторых вариантах осуществления клетки на стадии 5 культивировали в среде, содержащей активин А или активин С. В некоторых вариантах осуществления клетки на стадии 5 культивировали в среде, содержащей семафорин 3А или эпиген. В некоторых вариантах осуществления плюрипотентные стволовые клетки представляют собой эмбриональные плюрипотентные стволовые клетки человека.
В одном из вариантов осуществления настоящее изобретение относится к способу индукции экспрессии инсулина, содержащему культивирование клеток панкреатической энтодермы с эфриновым лигандом. В некоторых вариантах осуществления эфриновый лиганд выбирается из эфрина А3 или эфрина А4. В некоторых вариантах осуществления культивирование клеток панкреатической энтодермы с эфриновым лигандом усиливает экспрессию инсулина и NKX6.1. В некоторых вариантах осуществления культивирование клеток панкреатической энтодермы с эфриновым лигандом повышает экспрессию инсулина и NKX6.1 в клетках панкреатической энтодермы по меньшей мере в 2 раза по сравнению с экспрессией инсулина и NKX6.1 в необработанных клетках панкреатической энтодермы. В некоторых вариантах осуществления клетки панкреатической энтодермы по существу не экспрессируют CDX2 или SOX2. В некоторых вариантах осуществления клетки панкреатической энтодермы получают путем поэтапной дифференцировки плюрипотентных стволовых клеток. В некоторых вариантах осуществления плюрипотентные стволовые клетки, используемые в рамках способов настоящего изобретения, являются плюрипотентными стволовыми клетками человека.
В одном из вариантов осуществления настоящее изобретение касается клеток, экспрессирующих инсулин и NKX6.1, полученных способами настоящего изобретения.
В одном из вариантов осуществления настоящее изобретение относится к способу индукции образования эндокринных кластеров, содержащему культивирование клеток панкреатической энтодермы с агонистом рецептора сфингозина-1. В некоторых вариантах осуществления клетки панкреатической энтодермы получают путем поэтапной дифференцировки плюрипотентных стволовых клеток. В некоторых вариантах осуществления плюрипотентные стволовые клетки представляют собой эмбриональные плюрипотентные стволовые клетки человека.
Цитируемые в настоящем документе публикации полностью включаются в настоящий документ путем ссылки. Настоящее изобретение дополнительно иллюстрируется, среди прочего, следующими примерами.
ПРИМЕРЫ
Пример 1
Выявление эфрина А4 как мощного индуктора экспрессии инсулина
Данный пример выполнили для понимания роли различных белков в генерации культур панкреатической энтодермы/эндокринных культур из дифференцировки эмбриональных стволовых клеток человека.
Эмбриональные стволовые клетки человека линии H1 (hESC H1, пассаж 40) рассеивали как одиночные клетки в концентрации 1×105 клеток/см2 на MATRIGEL™ (разведение 1:30; BD Biosciences, штат Нью-Джерси) с покрытием чашек средами mTeSR®1 (StemCell Technologies, г. Ванкувер, Канада) с добавлением 10 мкМ Y27632 (Rock inhibitor, номер по каталогу Y0503, SigmaAldrich, г. Сент-Луис, штат Миссури). Спустя сорок восемь часов после посева культуры промыли в неполном ФСБ (фосфатно-солевом буферном растворе без Mg или Ca). Культуры дифференцировали в линии панкреатической энтодермы/панкреатических эндокринных клеток следующим образом:
a) стадия 1 (дефинитивная энтодерма (ДЭ) - 3 дня): клетки культивировали один день в среде стадии 1: среда MCDB-131 (№ по каталогу 10372-019, Invitrogen, г. Карлсбад, штат Калифорния) с добавлением 0,1%-ного не содержащего жирных кислот БСА (№ по каталогу 68700, Proliant, г. Энкени, штат Айова), 0,0012 г/мл бикарбоната натрия (№ по каталогу S3187, SigmaAldrich, г. Сент-Луис, штат Миссури), 1X GlutaMax™ (№ по каталогу 35050-079 Invitrogen), 4,5 мM D-глюкозы (№ по каталогу G8769, SigmaAldrich), 100 нг/мл GDF8 (R&D Systems, г. Миннеаполис, штат Миннесота) и 1 мкМ композиции MCX (ингибитор GSK3B, 14-проп-2-ен-1-ил-3,5,7,14,17,23,27-гептаазотетрацикло [19.3.1.1~2,6~0,1~8,12~]гептакоза-1(25),2(27),3,5,8(26),9,11,21,23-нонаен-16-он, публикация заявки на патент США № 2010-0015711; полностью включенной в настоящий документ путем ссылки). Затем клетки дополнительно культивировали один день в среде MCDB-131 с добавлением 0,1%-ного не содержащего жирных кислот БСА, 0,0012 г/мл бикарбоната натрия, 1X GlutaMax™, 4,5 мM D-глюкозы, 100 нг/мл GDF8 и 0,1 мкМ композиции MCX. Затем клетки дополнительно культивировали один день в среде MCDB-131 с добавлением 0,1%-ного не содержащего жирных кислот БСА, 0,0012 г/мл бикарбоната натрия, 1X GlutaMax™, 4,5 мM D-глюкозы и 100 нг/мл GDF8, затем
b) стадия 2 (первичная кишечная трубка - 2 дня): в течение двух дней клетки обрабатывали средой MCDB-131 с добавлением 0,1%-ного не содержащего жирных кислот БСА; 0,0012 г/мл бикарбоната натрия; 1X GlutaMax™; 4,5 мМ D-глюкозы; 0,25 мM аскорбиновой кислоты (Sigma, г. Сент-Луис, штат Миссури) и 25 нг/мл FGF7 (R&D Systems, г. Миннеаполис, штат Миннесота), затем
c) стадия 3 (передняя кишка - 2 дня): клетки обрабатывали средой MCDB-131 с добавлением 1:200 раствора ITS-X (Invitrogen); 4,5 мМ глюкозы; 1X GlutaMax™; 0,0017 г/мл бикарбоната натрия; 2%-ного не содержащего жирных кислот БСА; 0,25 мкM SANT-1 (Sigma, г. Сент-Луис, штат Миссури); 10 нг/мл активина A (R&D Systems); 1 мкM ретиноевой кислоты (RA; Sigma); 25 нг/мл FGF7; 0,25 мМ аскорбиновой кислоты; 200 нM TPB (активатор PKC; № по каталогу 565740; EMD Chemicals, г. Джибстаун, штат Нью-Джерси); 10 мкМ форсколина (FSK, Sigma) и 100 нM LDN (ингибитор рецептора BMP; № по каталогу 04-0019; Stemgent; г. Сан-Диего, штат Калифорния) в течение дня 1. На 2-й день клетки обрабатывали средой MCDB-131 с добавлением 1:200 раствора ITS-X; 4,5 мМ глюкозы; 1X GlutaMax™; 0,0017 г/мл бикарбоната натрия; 2%-ного не содержащего жирных кислот БСА; 0,25 мкМ SANT-1; 10 нг/мл активина; 1 мкM ретиноевой кислоты; 25 нг/мл FGF7; 0,25 мM аскорбиновой кислоты, 200 нM TPB, 10 мкM форсколина и 10 нM LDN, затем
d) стадия 4 (клетка-предшественник панкреатической клетки из передней кишки - 2 дня): клетки обрабатывали средой MCDB-131 с добавлением 1:200 раствора ITS-X; 4,5 мМ глюкозы; 1X GlutaMax™; 0,0015 г/мл бикарбоната натрия; 2%-ного не содержащего жирных кислот БСА; 0,25 мкМ SANT-1; 50 нM ретиноевой кислоты; 50 нM LDN-193189; 10 мкМ форсколина; 0,25 мМ аскорбиновой кислоты; и 100 нM TPB в течение двух дней, затем
e) стадия 5 (панкреатическая энтодерма/эндокринная часть поджелудочной железы - 3 дня): клетки стадии 4 обрабатывали средой MCDB-131 с добавлением 1:200 раствора ITS-X; 20 мМ глюкозы; 1X GlutaMax™; 0,0015 г/мл бикарбоната натрия; 2%-ного не содержащего жирных кислот БСА; 0,25 мкМ SANT-1; 50 нM ретиноевой кислоты; 10 мкМ форсколина; 0,25 нM аскорбиновой кислоты в течение трех дней, с добавлением 100 нM ингибитора ALk5 SD-208 (описан в работе Molecular Pharmacology, 2007 г., 72:152-161) только на дни 2-3.
В день 1 стадии 5 указанные ниже в таблице I факторы вносили в среду и по завершении S5 (день 3 стадии 5) выделяли мРНК для ПЦР-анализа генов, характерных для панкреатической энтодермы/эндокринной части поджелудочной железы. В качестве контроля культуры обрабатывали только средой S5, описанной выше. Тотальную РНК экстрагировали с помощью набора RNeasy Mini Kit (Qiagen, г. Валенсия, штат Калифорния) и обратно транскрибировали с помощью набора High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, г. Фостер-Сити, штат Калифорния) в соответствии с инструкциями изготовителя. кДНК амплифицировали с помощью систем Taqman Universal Master Mix и Taqman Gene Expression Assays, которые предварительно наносили на специализированные чипы Taqman Arrays (Applied Biosystems). Данные анализировали с помощью программного обеспечения Sequence Detection Software (Applied Biosystems) и нормировали по недифференцированным эмбриональным стволовым клеткам человека (hES) с применением способа ΔΔCt. Все праймеры приобретали в Applied Biosystems.
Список факторов, протестированных на стадии 5 в примере 1
(продолжение)
На Фиг. 1А-1G представлены данные ПЦР в реальном времени при анализе экспрессии следующих генов в эмбриональных стволовых клетках человека линии Н1, дифференцированных до стадии 5, как описано в примере 1, и в присутствии факторов, перечисленных в таблице I: инсулин (Фиг. 1A), соматостатин (Фиг. 1B), грелин (Фиг. 1C), глюкагон (Фиг. 1D), PDX-1 (Фиг. 1E), NKX6.1 (Фиг. 1F) и NGN3 (Фиг. 1G).
Как показано на Фиг. 1, эфрин-A4 увеличивает экспрессию мРНК NKX6.1 и инсулина по сравнению с контрольными культурами (Фиг. 1F), при этом минимально влияя на экспрессию PDX-1 (Фиг. 1E) и NGN3 (Фиг. 1G). Такие факторы, как активин A и активин С, значительно увеличивают экспрессию соматостатина (Фиг. 1B), при этом подавляя экспрессию инсулина (Фиг. 1A), глюкагона (Фиг. 1D) и грелина (Фиг. 1C). Более того, такие факторы, как семафорин 3А и эпиген, повышают экспрессию NKX6.1, при этом не влияя на экспрессию гормонов по сравнению с необработанными культурами. Пунктирной линией на графике Фиг. 1 отмечен средний уровень экспрессии различных маркеров в контрольных культурах.
Пример 2
Подтверждение воздействия эфринов на экспрессию инсулина на стадии 5
Этот пример описывает подтверждение эффектов, выявленных в примере 1. В частности, эффект добавления эфрина A3 или эфрина A4 на стадии 5 в указанном ниже протоколе.
Эмбриональные стволовые клетки человека линии H1 (hESC H1, пассаж 40) рассеивали как одиночные клетки при концентрации 1×105 клеток/см2 на чашки, покрытые MATRIGEL™ (разведение 1:30; BD Biosciences, штат Нью-Джерси) в среде mTeSR®1 с добавлением 10 мкM Y27632. Спустя сорок восемь часов после посева культуры промыли в неполном ФСБ (фосфатно-солевом буферном растворе без Mg или Ca). Культуры дифференцировали в линии панкреатической энтодермы/панкреатических эндокринных клеток следующим образом:
a) стадия 1 (дефинитивная энтодерма (ДЭ) - 3 дня): клетки культивировали в течение одного дня в среде для стадии 1 (см. пример 1 выше). Затем клетки дополнительно культивировали один день в среде MCDB-131 с добавлением 0,1%-ного не содержащего жирных кислот БСА, 0,0012 г/мл бикарбоната натрия, 1X GlutaMax™, 4,5 мM D-глюкозы, 100 нг/мл GDF8 и 0,1 мкМ композиции MCX. Затем клетки дополнительно культивировали один день в среде MCDB-131 с добавлением 0,1%-ного не содержащего жирных кислот БСА, 0,0012 г/мл бикарбоната натрия, 1X GlutaMax™, 4,5 мM D-глюкозы и 100 нг/мл GDF8, затем
b) стадия 2 (первичная кишечная трубка - 2 дня): в течение двух дней клетки обрабатывали средой MCDB-131 с добавлением 0,1% не содержащего жирных кислот БСА; 0,0012 г/мл бикарбоната натрия; 1X GlutaMax™; 4,5 мМ D-глюкозы; 0,25 мM аскорбиновой кислоты (Sigma, штат Миссури) и 25 нг/мл FGF7 (R&D Systems, штат Миннесота), затем
c) стадия 3 (передняя кишка - 2 дня): клетки обрабатывали средой MCDB-131 с добавлением 1:200 раствора ITS-X (Invitrogen, штат Калифорния); 4,5 мМ глюкозы; 1X GlutaMax™; 0,0017 г/мл бикарбоната натрия; 2%-ного не содержащего жирных кислот БСА; 0,25 мкM SANT-1 (Sigma, штат Миссури); 10 нг/мл активина А (R & D Systems, штат Миннесота); 1 мкM ретиноевой кислоты (Sigma, штат Миссури); 25 нг/мл FGF7; 0,25 мМ аскорбиновой кислоты; 200 нM TPB (активатор PKC; № по каталогу 565740; EMD Chemicals, г. Джибстаун, штат Нью-Джерси); 10 мкM форсколина и 100 нM LDN (ингибитор рецептора BMP; № по каталогу 04-0019; Stemgent) в день 1. На 2-й день клетки обрабатывали средой MCDB-131 с добавлением 1:200 раствора ITS-X; 4,5 мМ глюкозы; 1X GlutaMax™; 0,0017 г/мл бикарбоната натрия; 2%-ного не содержащего жирных кислот БСА; 0,25 мкМ SANT-1; 10 нг/мл активина А; 1 мкM ретиноевой кислоты; 25 нг/мл FGF7; 0,25 мМ аскорбиновой кислоты, 200 нM TPB, 10 мкM форсколина и 10 нM LDN, затем
d) стадия 4 (клетка-предшественник панкреатической клетки из передней кишки - 2 дня): клетки обрабатывали средой MCDB-131 с добавлением 1:200 раствора ITS-X; 4,5 мМ глюкозы; 1X GlutaMax™; 0,0015 г/мл бикарбоната натрия; 2%-ного не содержащего жирных кислот БСА; 0,25 мкМ SANT-1; 50 нM ретиноевой кислоты; 50 нM LDN-193189; 10 мкМ форсколина; 0,25 мМ аскорбиновой кислоты; и 100 нM TPB в течение двух дней, затем
e) стадия 5 (панкреатическая энтодерма/эндокринная часть поджелудочной железы - 3 дня): Клетки стадии 4 обрабатывали средой MCDB-131 с добавлением 1:200 раствора ITS-X; 4,5 мМ глюкозы; 1X GlutaMax™; 0,0015 г/мл бикарбоната натрия; 2%-ного не содержащего жирных кислот БСА; 0,25 мкМ SANT-1; 50 нM ретиноевой кислоты; 10 мкМ форсколина; 0,25 мМ аскорбиновой кислоты; 100 нМ ингибитора ALk5 (только в течение дней 2 и 3) (SD-208, описан в Molecular Pharmacology, 2007 г., 72:152-161) и +/-0-100 нг/мл эфрина A3 или эфрина A4 (R & D Systems, штат Миннесота) в течение трех дней.
В конце стадии 5 контрольные и обработанные эфрином культуры фиксировали и окрашивали для определения экспрессии инсулина (использовали антитела против инсулина, полученные в морской свинке, Millipore; г. Кембридж, штат Массачусетс). На Фиг. 2 представлены изображения клеток, иммуноокрашенных на инсулин. Фиг. 2A - клетки контроля; Фиг. 2B - клетки, обработанные 50 нг/мл эфрина A3; Фиг. 2C - клетки, обработанные 100 нг/мл эфрина A3. На Фиг. 3 представлены изображения клеток, иммуноокрашенных на инсулин. Фиг. 3A - клетки контроля; Фиг. 3B - клетки, обработанные 50 нг/мл эфрина A4; Фиг. 3C -клетки, обработанные 100 нг/мл эфрина А4. Представленные данные демонстрируют, что добавление как эфрина A3, так и эфрина A4 на стадии 5 значительно усиливает экспрессию инсулина, что согласуется с данными примера 1.
Пример 3
Добавление сфингозин-1-фосфата на стадии 6 значительно ускоряет образование клеточных кластеров, содержащих эндокринные гормоны
Этот пример описывает последовательность образования эндокринных кластеров на стадии 6 и влияние сфингозин-1-фосфата на ускорение образования насыщенных эндокринных кластеров.
Эмбриональные стволовые клетки человека линии H1 (hESC H1, пассаж 40) рассеивали как одиночные клетки при концентрации 1×105 клеток/см2 на чашки, покрытые MATRIGEL™ (разведение 1:30; BD Biosciences, штат Нью-Джерси), в среде mTeSR®1 (StemCell Technologies, г. Ванкувер, Канада) с добавлением 10 мкM Y27632. Спустя сорок восемь часов после посева культуры промыли в неполном ФСБ (фосфатно-солевом буферном растворе без Mg или Ca). Культуры дифференцировали в линии панкреатической энтодермы/панкреатических эндокринных клеток следующим образом:
a) стадия 1 (дефинитивная энтодерма (ДЭ) - 3 дня): клетки культивировали в течение одного дня в среде для стадии 1 (см. пример 1 выше). Затем клетки дополнительно культивировали один день в среде MCDB-131 с добавлением 0,1%-ного не содержащего жирных кислот БСА, 0,0012 г/мл бикарбоната натрия, 1X GlutaMax™, 4,5 мM D-глюкозы, 100 нг/мл GDF8 и 0,1 мкМ композиции MCX. Затем клетки дополнительно культивировали один день в среде MCDB-131 с добавлением 0,1%-ного не содержащего жирных кислот БСА, 0,0012 г/мл бикарбоната натрия, 1X GlutaMax™, 4,5 мM D-глюкозы и 100 нг/мл GDF8, затем
b) стадия 2 (первичная кишечная трубка - 2 дня): в течение двух дней клетки обрабатывали средой MCDB-131 с добавлением 0,1%-ного не содержащего жирных кислот БСА; 0,0012 г/мл бикарбоната натрия; 1X GlutaMax™; 4,5 мМ D-глюкозы; 0,25 мM аскорбиновой кислоты (Sigma, штат Миссури) и 25 нг/мл FGF7 (R & D Systems, штат Миннесота), затем
c) стадия 3 (передняя кишка - 2 дня): клетки обрабатывали средой MCDB-131 с добавлением 1:200 раствора ITS-X (Invitrogen, штат Калифорния); 4,5 мМ глюкозы; 1X GlutaMax™; 0,0017 г/мл бикарбоната натрия; 2%-ного не содержащего жирных кислот БСА; 0,25 мкM SANT-1 (Sigma, штат Миссури); 10 нг/мл активина А (R& D Systems, штат Миннесота); 1 мкM ретиноевой кислоты (Sigma, штат Миссури); 25 нг/мл FGF7; 0,25 мМ аскорбиновой кислоты; 200 нM TPB (активатор PKC; № по каталогу 565740; EMD Chemicals, г. Джибстаун, штат Нью-Джерси); 10 мкM форсколина (FSK, Sigma, штат Миссури) и 100 нM LDN (ингибитор рецептора BMP; № по каталогу 04-0019; Stemgent, штат Калифорния) в течение дня 1. На 2-й день клетки обрабатывали средой MCDB-131 с добавлением 1:200 раствора ITS-X; 4,5 мМ глюкозы; 1X GlutaMax™; 0,0017 г/мл бикарбоната натрия; 2%-ного не содержащего жирных кислот БСА; 0,25 мкМ SANT-1; 10 нг/мл активина А; 1 мкM ретиноевой кислоты; 25 нг/мл FGF7; 0,25 мM аскорбиновой кислоты, 200 нM TPB, и 10 нM LDN, затем
d) стадия 4 (клетка-предшественник панкреатической клетки из передней кишки - 2 дня): клетки обрабатывали средой MCDB-131 с добавлением 1:200 раствора ITS-X; 4,5 мМ глюкозы; 1X GlutaMax™; 0,0015 г/мл бикарбоната натрия; 2%-ного не содержащего жирных кислот БСА; 0,25 мкМ SANT-1; 50 нM ретиноевой кислоты; 50 нM LDN-193189; 10 мкМ форсколина; 0,25 мМ аскорбиновой кислоты; 2 нг/мл FGF7; 1 нг/мл AA; и 100 нM TPB в течение двух дней, затем
e) стадия 5 (панкреатическая энтодерма/эндокринная часть поджелудочной железы - 3 дня): клетки стадии 4 обрабатывали средой MCDB-131 с добавлением 1:200 раствора ITS-X; 15 мМ глюкозы; 1X GlutaMax™; 0,0015 г/мл бикарбоната натрия; 2%-ного не содержащего жирных кислот БСА; 0,25 мкМ SANT-1; 50 нM ретиноевой кислоты; 10 мкМ форсколина; 0,25 мМ аскорбиновой кислоты; и 1 нг/мл FGF7 на протяжении трех дней; с добавлением 100 нM ингибитора ALk5 SD-208 только в дни 2-3, затем
f) стадия 6 (эндокринная часть поджелудочной железы - 3-10 дней): клетки стадии 5 обрабатывали средой MCDB-131 с добавлением 1:200 раствора ITS-X; 15 мМ глюкозы; 1X GlutaMax™; 0,0015 г/мл бикарбоната натрия; 2%-ного не содержащего жирных кислот БСА; 0,25 мкМ SANT-1; 50 нM ретиноевой кислоты; 0,25 мМ аскорбиновой кислоты; на протяжении 3-10 дней. В некоторые культуры в течение трех дней добавляли 10 мкM сфингозин-1-фосфат (Sigma, штат Миссури).
На Фиг. 4A-4D представлены фазово-контрастные изображения клеток культуры S6, обработанных сфингозин-1-фосфатом (S1P) и сфотографированных на 1-й день (Фиг. 4A), 7-й день (Фиг. 4B) и при двух различных увеличениях на 10-й день (Фиг. 4C и Фиг. 4D). Эти изображения показывают, что на 7-й день наблюдались четкие признаки кластеризации эндокринных клеток, а на 10-й день кластеры отделялись друг от друга тонким слоем панкреатического энтодермального эпителия.
На Фиг. 5A-5D представлены изображения клеток, иммуноокрашенных на Hb9 (Фиг. 5A) и NKX6.1 (Фиг. 5B) или иммуноокрашенных на инсулин (Фиг. 5C) и Hb9 (Фиг. 5D). Фиг. 5A и Фиг. 5B показывают, что эндокринные кластеры обогащены Hb9, в то время как панкреатический эпителий, окружающий кластеры, обогащен NKX6.1. Некоторые клетки в кластерах, обогащенных Hb9, также содержали NKX6.1. Кластеры были обогащены инсулином и Hb9, что показано на Фиг. 5C и Фиг. 5D. Такое морфологическое изменение сильно напоминает развитие поджелудочной железы, когда эпителий, обогащенный NKX6.1+ и PDX-1+, дает начало эндокринным кластерам. В каждом случае пару фотографий получали, используя разные фильтры для съемки одной и той же области клеток.
На Фиг. 6A и Фиг. 6B представлены полученные при разном увеличении фазово-контрастные изображения клеток, обработанных 10 мкM сфингозин-1-фосфата (S1P) и собранных через три дня после начала стадии 6. Эти фотографии показывают, что эндокринные кластеры возникают всего лишь через 3 дня после начала стадии 6. Это приблизительно на 7 дней опережает формирование кластеров в контрольных культурах.
На Фиг. 6C и Фиг. 6D представлены фотографии контрольных клеток (Фиг. 6C) и клеток, обработанных S1P (Фиг. 6D), после иммуноокрашивания на NKX2.2. По сравнению с контрольными культурами, где NKX2.2-положительные клетки равномерно распространялись по культуре (Фиг. 6D), в обработанных S1P культурах NKX2.2-положительные клетки были сосредоточены в эндокринных кластерах (Фиг. 6C).








Изобретение относится к области клеточной биологии, конкретно к получению клеток, экспрессирующих инсулин и индукции экспрессии инсулина в клетках панкреатической энтодермы. Способ включает дифференцировку плюрипотентных стволовых клеток, выбранных из группы, состоящей из клеточных линий Н1, Н7, SA002, индуцибельных плюрипотентных клеток и перепрограммированных плюрипотентных стволовых клеток, в клетки дефинитивной эндодермы, дифференцировку клеток дефинитивной энтодермы в клетки первичной кишечной трубки, дифференцировку клеток первичной кишечной трубки в клетки передней кишки, дифференцировку клеток передней кишки в клетки-предшественники панкреатических клеток, дифференцировку клеток-предшественников панкреатических клеток в клетки панкреатической эндодермы и культивирование клеток панкреатической энтодермы с эфриновым лигандом. Изобретение позволяет повысить эффективность и направленность дифференцировки. 2 н. и 15 з.п. ф-лы, 1 табл., 6 ил., 3 пр.
1. Способ индукции экспрессии инсулина в клетках панкреатической энтодермы, экспрессирующих гормоны, включающий культивирование клеток панкреатической энтодермы с эфриновым лигандом, где клетки панкреатической энтодермы получены из плюрипотентных стволовых клеток, выбранных из группы, состоящей из: клеток H1, клеток H7, клеток H9, клеток SA002, индуцибельных плюрипотентных клеток и перепрограммированных плюрипотентных стволовых клеток.
2. Способ по п. 1, в котором культивирование клеток панкреатической энтодермы с эфриновым лигандом также усиливает экспрессию NKX6.1.
3. Способ по п. 2, в котором культивирование клеток панкреатической энтодермы с эфриновым лигандом усиливает экспрессию инсулина и NKX6.1 в клетках панкреатической энтодермы по сравнению с экспрессией инсулина и NKX6.1 в необработанных клетках панкреатической энтодермы.
4. Способ по п. 3, в котором клетки панкреатической энтодермы по существу не экспрессируют CDX2 или SOX2.
5. Способ по п. 4, в котором клетки панкреатической энтодермы экспрессируют приблизительно менее 10% CDX2 или SOX2.
6. Способ по любому из пп.1-5, в котором эфриновым лигандом является эфрин A3 или эфрин A4.
7. Способ по п. 6, в котором эфриновым лигандом является эфрин A3.
8. Способ по п. 6, в котором эфриновым лигандом является эфрин A4.
9. Способ по п. 6, в котором клетки панкреатической энтодермы получают путем поэтапной дифференцировки плюрипотентных стволовых клеток.
10. Способ по п. 9, в котором плюрипотентные стволовые клетки представляют собой эмбриональные плюрипотентные стволовые клетки человека.
11. Способ получения клеток, экспрессирующих инсулин, включающий:
(a) дифференцировку плюрипотентных стволовых клеток в клетки эндодермы;
(b) дифференцировку клеток дефинитивной эндодермы в клетки первичной кишечной трубки;
(c) дифференцировку клеток первичной кишечной трубки в клетки передней кишки;
(d) дифференцировку клеток передней кишки в клетки-предшественники панкреатических клеток;
(e) дифференцировку клеток-предшественников панкреатических клеток в клетки панкреатической эндодермы; и
(f) культивирование клеток панкреатической эндодермы с эфриновым лигандом с получением таким образом клеток, экспрессирующих инсулин,
где плюрипотентные стволовые клетки выбраны из группы, состоящей из: H1, клеток H7, клеток H9, клеток SA002, индуцибельных плюрипотентных клеток и перепрограммированных плюрипотентных стволовых клеток.
12. Способ по п. 11, где способ усиливает экспрессию инсулина и NKX6.1.
13. Способ по п. 11, в котором клетки со стадии (f) по существу не экспрессируют CDX2 или SOX2.
14. Способ по п. 13, в котором клетки со стадии (f) экспрессируют приблизительно менее 10% CDX2 или SOX2.
15. Способ по пп. 11-14, в котором эфриновым лигандом является эфрин A3.
16. Способ по пп. 11-14, в котором эфриновым лигандом является эфрин A4.
17. Способ по пп. 11-14, в котором клетки со стадии (f) экспрессируют инсулин и NKX6.1.
| US 20070154981 A1, 05.07.2007 | |||
| US 20090170198 A1, 02.06.2009 | |||
| HOSOYA M | |||
| et al | |||
| "Induction of differentiation of undifferentiated cells into pancreatic beta cells in vertebrates" | |||
| Int J Dev Bio | |||
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| US 20110014702 A1, 20.01.2011 | |||
| KIM JS | |||
| et al | |||
| "Reprogrammed Piuripotent Stem Cells from Somatic Cells" | |||
| Int J Stem Cells | |||
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| НЕКРАСОВ Е.Д | |||
| и др | |||
| "Индуцированные плюрипотентные стволовые клетки как модель для изучения болезней человека" | |||
| Клеточная трансплантология и тканевая инженерия | |||
| Способ приготовления лака | 1924 |
|
SU2011A1 |

