Область изобретения
Настоящее изобретение относится к микроорганизму, способному продуцировать О-фосфосерин, и к способу получения О-фосфосерина, цистеина или производного цистеина с использованием этого микроорганизма.
Предшествующий уровень техники
L-цистеин, аминокислота, играющая важную роль в метаболизме серы у всех живых организмов, используется не только в синтезе биологических белков, таких как кератин волос, глутатион, биотин, метионин, и других серосодержащих метаболитов, но также в качестве предшественника в биосинтезе кофермента А.
Известные способы получения L-цистеина с использованием микроорганизмов включают: 1) способ биологического превращения D,L-ATC (DL-2-амино-дельта(2)-тиазолин-4-угольная кислота) в L-цистеин с использованием микроорганизмов, 2) способ получения L-цистеина посредством прямой ферментации с использованием Е. coli (ЕР 0885962 В; Wada М and Takagi Н, Appl. Microbiol. Biochem., 73:48-54, 2006) и 3) способ получения О-фосфосерина (ниже "OPS") посредством ферментации с использованием микроорганизмов и превращения OPS в L-цистеин путем взаимодействия OPS с сульфидом при каталитическом действии О-фосфосеринсульфгидрилазы (ниже "OPSS") (патент Кореи 1381048).
В частности, для получения цистеина способом (3) с высоким выходом, предшественник, OPS, должен быть произведен в избыточных количествах. В связи с этим авторы настоящего изобретения выполнили большой объем работ для определения соответствующего фактора экспорта, который позволяет О-фосфосерину, продуцируемому в OPS-продуцирующем микроорганизме, беспрепятственно экспортироваться из клеток.
Описание изобретения
Техническая задача
В этих обстоятельствах авторы настоящего изобретения обнаружили два новых OPS-продуцирующих полипептида, YhhS и MdtD, и подтвердили, что OPS может быть эффективно экспортирован из OPS-продуцирующего микроорганизма путем активации этих двух полипептидов, таким образом осуществляя настоящее изобретение.
Техническое решение
Следовательно, задачей настоящего изобретения является предложение OPS-продуцирующего микроорганизма, у которого активность полипептида, способного экспортировать OPS, увеличена по сравнению с его эндогенной активностью.
Другой задачей настоящего изобретения является предложение способа получения OPS, включающего: культивирование OPS-продуцирующего микроорганизма в среде, и выделение OPS из OPS-продуцирующего микроорганизма или его культуры.
Еще одной задачей настоящего изобретения является предложение применений получения OPS или его экспорта с помощью полипептида.
Еще одной задачей настоящего изобретения является предложение способа получения цистеина или его производного, включающего: а) получение OPS путем культивирования OPS-продуцирующего микроорганизма, у которого активность полипептида, способного экспортировать OPS, увеличена по сравнению с его эндогенной активностью, в среде; и б) взаимодействие OPS, полученного в (а) или в культуре, содержащей OPS с сульфидом, в присутствии OPS-сульфгидрилазы или микроорганизма, способного экспрессировать OPS-сульфгидрилазу.
Преимущества изобретения
Новый полипептид с аминокислотной последовательностью SEQ ID NO: 1 или SEQ ID NO: 2 по настоящему изобретению обладает исключительной OPS-экспортирующей способностью. Соответственно, когда новый полипептид по настоящему изобретению применяют к микроорганизму, способному производить OPS, он может привести к высокому выходу OPS-продукции и также может быть эффективно использован для синтеза L-цистеина и т.д.
Краткое описание графических материалов
На Фиг. 1 показана диаграмма, демонстрирующая результат измерения внутриклеточного уровня OPS посредством высокоэффективной жидкостной хроматографии (ВЭЖХ) после удаления всего OPS, экспортированного из культуры рекомбинантного микроорганизма по настоящему изобретению с усиленной функцией белков YhhS и MdtD.
Лучший вариант изобретения
В одном аспекте в настоящем изобретении предлагается OPS-продуцирующий микроорганизм, у которого активность полипептида, имеющего аминокислотную последовательность SEQ ID NO: 1 или SEQ ID NO: 2 и способного экспортировать О-фосфосерин, увеличена по сравнению с его эндогенной активностью.
Используемый здесь термин "О-фосфосерин" (ниже "OPS") относится к сложному эфиру серина и фосфорной кислоты, который является компонентом многих белков. В частности, OPS является предшественником L-цистеина и может быть превращен в цистеин посредством взаимодействия с сульфидом при каталитическом действии OPS-сульфгидрилазы (ниже называемой "OPSS") (патент Кореи 1381048). Соответственно, он является важным фактором для увеличения производства OPS при получении цистеина и, таким образом, необходимо разработать транспортеры, которые обеспечивают возможность эффективного выделения внутриклеточного OPS из OPS-продуцирующих штаммов.
Используемый здесь термин "полипептид, обладающий активностью экспорта О-фосфосерина" относится к мембранному белку, обладающему активностью экспорта клеточного OPS за пределы клетки и, в частности, может представлять собой мембранный белок, полученный из Е. coli. Два типа мембранных белков были идентифицированы в Е. coli, где ингибирование роста снято в условиях присутствия избыточного количества OPS. В частности, таким образом идентифицированными мембранными белками с OPS-экспортирующей способностью являются транспортер YhhS MFS (суперсемейство мембранных транспортеров), имеющий аминокислотную последовательность SEQ ID NO: 1 и транспортер YegB MFS, имеющий аминокислотную последовательность SEQ ID NO: 2. В настоящем изобретении транспортер YegB MFS может быть использован взаимозаменяемо с MdtD. OPS-экспортирующая способность белка не была известна до тех пор, пока не была впервые подтверждена в настоящем изобретении.
Кроме того, полипептид может иметь аминокислотную последовательность, представленную SEQ ID NO: 1 или SEQ ID NO: 2, и может включать, без ограничения ими, мембранные белки с гомологией последовательности, составляющей по меньшей мере 70%, с вышеуказанными последовательностями, более конкретно по меньшей мере 80%, еще более конкретно по меньшей мере 90%, и еще более конкретно по меньшей мере 95%, при условии что они обладают OPS-экспортирующей способностью, которая является по существу такой же или эквивалентной способности полипептида. Кроме того, очевидно, что варианты полипептидов, в которых часть последовательности делетирована, модифицирована, заменена или вставлена, включены в объем настоящего изобретения, при условии, что они представляют собой аминокислотные последовательности, имеющие такую гомологию и OPS-экспортирующую способность.
Кроме того, полинуклеотидная последовательность полипептида, демонстрирующего OPS-экспортирующую способность, может включать полинуклеотидные последовательности, кодирующие аминокислоты, представленные в SEQ ID NO: 1 или SEQ ID NO: 2. Кроме того, принимая во внимание то, что кодоны, предпочитаемые организмами, экспрессируют полипептиды, исходя из генетической вырожденности кода, в кодирующей области могут присутствовать различные модификации без изменения аминокислотной последовательности полипептида. Полинуклеотидная последовательность может представлять собой последовательность, представленную SEQ ID NO: 3 или SEQ ID NO: 4, и может включать нуклеотидные последовательности, обладающие по меньшей мере 70% гомологией с этими последовательностями, но не ограничена ими.
Используемый здесь термин "гомология" относится к степени идентичности с данной полипептидной последовательностью или полинуклеотидной последовательностью и может быть указана в процентах. При использовании в данном описании изобретения, гомологичная последовательность, обладающая такой же или аналогичной активностью с данной полипептидной последовательностью или полинуклеотидной последовательностью, может быть указана в показателях "% гомологии". % гомологии может быть подтвержден с использованием стандартного программного обеспечения, то есть BLAST 2.0, для расчета таких параметров, как балл, идентичность и сходство, или путем сравнения последовательностей в экспериментах саузерн-гибридизации, и подходящие условия гибридизации, требующие уточнения, могут быть определены способом, известным специалисту в данной области техники (например, Sambrook et al., 1989, ниже).
В типичном воплощении настоящего изобретения было подтверждено, что, когда активности белка YhhS (SEQ ID NO: 1) или белка MdtD (SEQ ID NO: 2) были усилены в микроорганизме, способном продуцировать OPS, этот микроорганизм обладал высокой OPS-экспортирующей способностью в штамм, где белок RhtB был усилен (публикация патентной заявки Кореи 10-2012-0041115) (положительный контроль) или штамм, где активности MFS-транспортеров EmrD или YcaD были усилены (экспериментальная группа). "RhtB" представляет собой мембранный белок, кодируемый геном rhtB, который может экспортировать гомосерин/гомосерин-лактон. Поскольку уже было подтверждено, что усиление активности RhtB в OPS-продуцирующем штамме увеличивает OPS-экспортирующую способность в штамме (патент Кореи 138104), это использовали в качестве положительного контроля. При увеличении активности белка RhtB и белков YhhS и MdtD по настоящему изобретению в OPS-продуцирующем штамме, соответственно, белок RhtB и белки YhhS и MdtD по настоящему изобретению демонстрировали исключительные способности OPS-экспорта относительно белка RhtB. Кроме того, термины "EmrD" и "YcaD" относятся к белкам-транспортерам MFS Е. coli и кодируются геном emrD и геном ycaD, соответственно. EmrD и YcaD, будучи белками, принадлежащими к транспортерам MFS, как и белки YhhS и MdtD, использовали в качестве экспериментальной группы для определения, могут ли другие белки, принадлежащие к транспортерам MFS, также демонстрировать OPS-экспортирующие свойства. В результате было подтверждено, что белки EmrD и YcaD, в отличие от белков YhhS and MdtD, не проявляют OPS-экспортирующих способностей.
В то же время, полипептид по настоящему изобретению обладает OPS-экспортирующей способностью и, таким образом, когда активность этого полипептида усилена по сравнению с его эндогенной активностью в микроорганизме, обладающем OPS-продуцирующей способностью, OPS может эффективно продуцироваться.
Используемый здесь термин "получение OPS" относится не только к продуцированию OPS внутри штамма, но также к экспорту клеточного OPS за пределы клетки, например в среду и, более конкретно, экспорт OPS изнутри клетки наружу.
Используемый здесь термин "эндогенная активность" относится к активному состоянию полипептида в микроорганизме в естественном состоянии, то есть в немодифицированном состоянии.
Используемый здесь термин "усиление по сравнению с его эндогенной активностью" относится к повышенной активности полипептида в микроорганизме по сравнению с активностью, которой он обладает в своем естественном состоянии, и представляет собой понятие, включающее активизацию конкретного полипептида в микроорганизме, который не обладает активностью этого конкретного полипептида.
Используемый здесь термин "усиление активности" относится, хотя и без ограничений ими, не только к получению более значительного эффекта, чем исходная функция, благодаря увеличению активности самого полипептида, но также к увеличению активности белка благодаря увеличению эндогенной активности гена, эндогенной амплификации гена посредством внутренних или внешних факторов, замене, модификации или мутации промотора, и т.д. В частности, усиление активности может быть осуществлено посредством таких методов, как метод увеличения числа копий гена, кодирующего полипептид, в клетке, метод модификации регуляторной последовательности гена, кодирующего полипептид, метод замещения гена, кодирующего полипептид, в хромосоме мутантным геном для увеличения активности полипептида, метод введения модификации в ген, кодирующий полипептид, в хромосоме для усиления активности полипептида, и т.д., но не ограничены ими. Эти методы усиления активности могут относиться также к усилению активностей других полипептидов по настоящему изобретению.
В вышеприведенных методах увеличение числа копий гена, хотя и без ограничения ими, может быть выполнено в состоянии, функционально связанном с вектором, или посредством встраивания в хромосому в клетке-хозяине. В частности, данный метод может быть осуществлен путем введения в клетку хозяина вектора, посредством которого полинуклеотид, кодирующий белок по настоящему изобретению, функционально связан с клеткой-хозяином, и который может воспроизводиться и функционировать независимо от хозяина; или путем введения в клетку-хозяина вектора, с которым полинуклеотид функционально связан, способного встраивать полинуклеотид в хромосому клетки-хозяина. Встраивание полинуклеотида в хромосому может быть выполнено с использованием известного в данной области способа, например посредством гомологичной рекомбинации. Так как вектор по настоящему изобретению может быть встроен в хромосому посредством гомологичной рекомбинации, может быть дополнительно включен селективный маркер для подтверждения такого встраивания. Селективный маркер используют для селекции трансформированной клетки, то есть для того чтобы подтвердить, встроен ли целевой полинуклеотид, и могут быть использованы маркеры, способные обеспечить селективные фенотипы, такие как лекарственная устойчивость, потребность в питательных веществах, устойчивость к цитотоксическим агентам и экспрессия поверхностных белков, без ограничения ими. В условиях воздействия селективными агентами могут выжить только клетки, способные экспрессировать селективные маркеры или экспрессировать другие фенотипические признаки и, таким образом, трансформированные клетки могут быть легко отобраны.
Вектором может представлять собой ДНК-конструкцию, включающую полинуклеотидную последовательность полинуклеотида, кодирующего целевой белок, которая функционально связана с подходящей регуляторной последовательностью, так что целевой белок может экспрессироваться в подходящем хозяине. Регуляторная последовательность включает промотор, способный инициировать транскрипцию, случайную последовательность оператора для регуляции транскрипции, последовательность, кодирующую подходящий мРНК-связывающий домен рибосомы, и последовательность регуляции транскрипции и трансляции. Вектор, после трансформации в подходящую клетку-хозяина, может быть реплицирован, или может функционировать независимо от генома хозяина, или может быть интегрирован в сам геном хозяина.
Вектор, используемый в настоящем изобретении, может не быть конкретно ограничен, при условии, что этот вектор реплицируется в клетке-хозяине, и может быть использован любой вектор, известный в данной области техники. Примеры векторов могут включать природные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, в качестве фагового вектора или космидного вектора могут быть использованы pWE15, М13, λMBL3, λMBL4, λIXII, λASHII, λAPII, λt10, λt11, Charon4A, Charon21A и т.д.; и в качестве плазмидного вектора могут быть использованы векторы на основе pBR, pUC, pBluescriptll, pGEM, pTZ, pCL, pET и т.д. В частности, могут быть использованы векторы pDZ, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118, pCC1BAC и т.д.
Используемый здесь термин "трансформация" относится к процессу введения в клетку-хозяина вектора, включающего полинуклеотид, кодирующий целевой белок, тем самым обеспечивается экспрессия белка, кодируемого полинуклеотидом, в клетке-хозяине. Для трансформированного полинуклеотида не имеет значения встроен он в хромосому клетки-хозяина и расположен в ней или находится вне хромосомы, при условии, что он может экспрессироваться в клетке-хозяине. Кроме того, полинуклеотид включает ДНК и РНК, которые кодируют целевой белок. Полинуклеотид может быть внесен в любой форме, если он может быть введен в клетку-хозяина и экспрессироваться в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, которая представляет собой генетическую конструкцию, включающую все существенные элементы, необходимые для самоэкспрессии, но не ограничивается ею. Экспрессионная кассета может, как правило, включать промотор, функционально связанный с полинуклеотидом, сигнал терминации транскрипции, рибосома-связывающий домен и сигнал терминации трансляции. Экспрессионная кассета может находиться в форме экспрессионного вектора, способного к саморепликации. Кроме того, полинуклеотид может быть введен в клетку-хозяина как есть и функционально связан с последовательностью, существенной для его экспрессии в клетке-хозяине.
Кроме того, используемый здесь термин "функционально связанный" относится к функциональной связи между последовательностью промотора, который инициирует и опосредует транскрипцию полинуклеотида, кодирующего целевой белок по настоящему изобретению, и последовательностью вышеуказанного гена.
Затем, модификация последовательности регуляции экспрессии для увеличения экспрессии полинуклеотида может быть выполнена, без конкретного ограничения ими, путем индукции изменения в полинуклеотидной последовательности посредством делеции, вставки, консервативного замещения, неконсервативного замещения или их комбинации, так, чтобы дополнительно повысить активность последовательности регуляции экспрессии; или путем замены полинуклеотидной последовательности полинуклеотидной последовательностью с более сильной активностью. Последовательность регуляции экспрессии, хотя конкретно не ограничена ими, может включать промотор, последовательность оператора, последовательность, кодирующую рибосома-связывающий домен, и последовательность регулирования терминации транскрипции и трансляции и т.д.
Сильный промотор вместо исходного промотора может быть присоединен к верхнему концу экспрессионной единицы полинуклеотида, без ограничения этим. Примеры известных сильных промоторов могут включать промотор cj1 (патент Кореи 0620092), промотор lac, промотор trp, промотор trc, промотор tac, промотор PR и промотор PL фага лямбда и промотор tet.
Кроме того, модификация полинуклеотидной последовательности в хромосоме, без конкретного ограничения ими, может быть выполнена путем индукции изменения в последовательности регулирования экспрессии полинуклеотидной последовательности посредством делеции, вставки, консервативного замещения, неконсервативного замещения или их комбинации для дополнительного усиления активности полинуклеотидной последовательности; или путем замены полинуклеотидной последовательности улучшенной полинуклеотидной последовательностью с более сильной активностью.
Как правило, введение и усиление активности белка может увеличить активность или концентрацию соответствующего белка относительно активности или концентрации белка дикого типа или в штамме микроорганизма от по меньшей мере 1%, 10%, 25%, 50%, 75%, 100%, 150%, 200%, 300%, 400%, или 500% до максимума 1000% или 2000%, без ограничения этими значениями.
Используемый здесь термин "OPS-продуцирующий микроорганизм" относится прокариотическому или эукариотическому штамму микроорганизма, способного продуцировать в себе OPS, и, в частности, микроорганизма, способного накапливать в себе OPS посредством генной инженерии.
В типичном воплощении настоящего изобретения микроорганизм конкретно не ограничен, но может представлять собой любой прокариотический или эукариотический микроорганизм, который может продуцировать OPS, когда активность полипептида SEQ ID NO: 1 или 2 усилена, и особенно прокариотический микроорганизм. Примеры микроорганизмов могут включать штаммы микроорганизмов, принадлежащие к роду Escherichia, роду Erwinia, роду Serratia, роду Providencia, роду Corynebacterium и роду Brevibacterium. В частности, микроорганизм может представлять собой микроорганизм рода Escherichia. Более конкретно это может быть Е. coli. В частности, микроорганизм Escherichia или рода Corynebacterium может продуцировать OPS и L-серин, так как он содержит белки SerA, SerC и SerB, которые представляют собой ферменты в пути биосинтеза L-серина (Ahmed Zahoor, Computational and Structural Biotechnology Journal, vol. 3, 2012 October; Wendisch VF et al., Curr Opin Microbiol. 2006 Jun; 9(3): 268-74; Peters-Wendisch P et al., Appl Environ Microbiol. 2005 Nov; 71(11): 7139-44).
Кроме того, в OPS-продуцирующем микроорганизме активность фосфосеринфосфатазы (SerB) может быть дополнительно ослаблена по сравнению с ее эндогенной активностью.
SerB обладает активностью превращения OPS в L-серин и, таким образом, микроорганизм, модифицированный с целью снижения активности SerB, обладает свойством накопления в себе OPS, и таким образом является полезным для получения OPS. SerB может представлять собой белок, имеющий аминокислотную последовательность, представленную SEQ ID NO: 17 или SEQ ID NO: 18, но не ограничен ими. Кроме этого, SerB может включать аминокислотную последовательность, имеющую идентичность последовательности 80% или выше, предпочтительно 90% или выше, более предпочтительно 95% или выше, и еще более предпочтительно 99% или выше, при условии, что он демонстрирует активность SerB, но не ограничен ими. Кроме того, полинуклеотидная последовательность, кодирующая SerB, может иметь полинуклеотидную последовательность, кодирующую аминокислоты, представленные в SEQ ID NO: 17 или SEQ ID NO: 18.
Принимая во внимание кодоны, предпочитаемые организмами для экспрессии полипептида, на основе генетической вырожденности кода, различные модификации полинуклеотида могут быть использованы на кодирующем участке в объеме, не изменяющем аминокислотную последовательность полипептида. Полинуклеотидная последовательность может представлять собой аминокислотную последовательность, представленную SEQ ID NO: 19 или SEQ ID NO: 20, и может включать нуклеотидные последовательности, обладающие 80%-ной гомологией с этими последовательностями и, в частности, по меньшей мере 90%-ной гомологией, но без ограничения ими.
Используемый здесь термин "ослабление по сравнению с его эндогенной активностью" относится к уменьшению активности белка по сравнению с его активностью в естественном состоянии и также включает удаление этой активности.
Ослабление представляет собой понятие, относящееся к случаю, когда активность белка снижена по сравнению с исходно присутствующей у микроорганизма из-за модификации гена, кодирующего белок и т.д., к случаю, когда уровень экспрессии общего белка ниже, чем у природного штамма микроорганизма из-за ингибирования экспрессии или ингибирования трансляции гена, кодирующего этот белок, или к случаю, когда ген не экспрессируется совсем, и к случаю, когда ген экспрессируется, но не проявляет активности.
Ослабления или инактивации активности белка можно достигнуть различными методами, хорошо известными в данной области техники. Примеры методов могут включать способ замены гена, кодирующего белок в хромосоме, мутантным геном, с тем чтобы можно было уменьшить ферментативную активность, включая случай удаления активности белка; способ модификации последовательности регуляции экспрессии гена, кодирующего белок; способ удаления части или всего гена, кодирующего белок в хромосоме; способ введения антисмыслового олигонуклеотида (например антисмысловой РНК), который ингибирует трансляцию мРНК в белок посредством комплементарного связывания с транскриптом гена в хромосоме; способ обеспечения невозможности прикрепления рибосомы путем формирования вторичной структуры в результате искусственного добавления последовательности Шайна-Дальгарно (SD) и его комплементарной последовательности на передний конец SD-последовательности гена, кодирующего белок; способ инженерии с обратной транскрипцией (RTE), где промотор добавляют таким образом, чтобы происходила транскрипция в обратном направлении на 3'-конце открытой рамки считывания (ORF) соответствующей последовательности, и т.д., а также включает их комбинацию, но не ограничивается ими.
В частности, способ удаления части или всего гена, кодирующего белок, может быть осуществлен путем замены полинуклеотида, кодирующего эндогенный целевой белок в хромосоме, полинуклеотидом или маркерным геном с частичным удаленной нуклеиновокислотной последовательностью, с использованием вектора для введения хромосом в бактерии. В типичном воплощении ген может быть удален посредством гомологичной рекомбинации. Кроме того, используемый здесь термин "часть", хотя он и может варьироваться в зависимости от типа полинуклеотида, может более конкретно относиться к нуклеотидам в количестве от 1 до 300, более конкретно от 1 нуклеотида до 100 нуклеотидов и еще более конкретно от 1 нуклеотида до 50 нуклеотидов, но не ограничен ими.
Кроме того, способ модификации последовательности регуляции экспрессии может быть осуществлен путем индукции изменений в последовательности регуляции экспрессии с помощью делеции, вставки, консервативного замещения, неконсервативного замещения или их комбинации, так чтобы дополнительно ослабить активность последовательности регуляции экспрессии; или путем замены последовательности нуклеиновокислотной последовательностью с более слабой активностью. Последовательность регуляции экспрессии включает промотор, последовательность оператора, последовательность, кодирующую рибосома-связывающий домен, и последовательность для регуляции транскрипции и трансляции.
Кроме того, способ модификации последовательности гена может быть выполнен посредством индукции изменений в последовательности гена с помощью делеции, вставки, консервативного замещения, неконсервативного замещения или их комбинации, так чтобы дополнительно ослабить активность белка; или посредством замены последовательности последовательностью гена, улучшенной так, чтобы иметь более слабую активность, или последовательностью гена, улучшенной так, чтобы совсем не иметь активности.
Кроме того, OPS-продуцирующий микроорганизм может быть таким, в котором активности фосфоглицератдегидрогеназы (SerA) или фосфосеринаминотрансферазы (SerC) дополнительно усилены по сравнению с их эндогенными активностями.
SerA представляет собой белок, способный превращать 3-фосфоглицерат в 3-фосфогидроксипируват, и для SerA может быть использован белок дикого типа или вариант, где удалена обратная связь на серине. Кроме того, SerC представляет собой белок, способный превращать 3-фосфо-гидроксипируват в OPS. Соответственно, любой микроорганизм с усиленной активностью SerA и/или SerC может быть эффективно использован в качестве OPS-продуцирующего микроорганизма.
SerA может иметь аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 21-26, хотя не ограничен ими. SEQ ID NO: 21 представляет собой последовательность SerA дикого типа, и SEQ ID NO: 22-26 представляют собой последовательности вариантов с удаленной обратной связью на серине. Кроме того, те аминокислотные последовательности, которые обладают по меньшей мере 80% идентичностью последовательности с вышеуказанной аминокислотной последовательностью, предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 95%, и еще более предпочтительно по меньшей мере 99%, могут быть включены при условии, что они проявляют активность SerA дикого типа или вариантов SerA с удаленной обратной связью на серине, но не ограничены ими. Варианты с удаленной обратной связью представляют собой белки, в которых модификация введена в ген, кодирующий SerA, посредством вставки, замены и т.д., и тем самым обеспечивается сохранение активности от ингибирования по типу обратной связи серином или глицином, или имеющие усиленные активности, при этом варианты, у которых удалена обратная связь, уже хорошо известны (Grant GA et al., J. Biol. Chem., 39: 5357-5361, 1999; Grant GA et al., Biochem., 39: 7316-7319, 2000; Grant GA et al., J. Biol. Chem., 276: 17844-17850, 2001; Peters-Wendisch P et al., Appl. Microbiol. Biotechnol., 60: 437-441, 2002; Европейский патент EP 0943687 B).
Кроме того, полинуклеотидная последовательность, кодирующая SerA дикого типа или варианты с удаленной обратной связью на серине, может быть полинуклеотидной последовательностью, кодирующей любую аминокислотную последовательность, представленную SEQ ID NO: 21-26, но не ограничена ими. В связи с вырожденностью генетического кода или принимая во внимание кодоны, предпочитаемыми организмами для экспрессии полипептида, различные модификации полинуклеотида могут быть выполнены в кодирующей области в объеме, не изменяющем аминокислотную последовательность полипептида. Полинуклеотидная последовательность может, например, представлять собой любую из полинуклеотидных последовательностей, представленных SEQ ID NO: 27-32, и может иметь нуклеотидную последовательность, имеющую гомологию по меньшей мере 80% с полинуклеотидными последовательностями и предпочтительно по меньшей мере 90%, но не ограничена ими.
SerC может быть белком, имеющим аминокислотную последовательность, которая, например, представлена SEQ ID NO: 33, но не ограничен им. Кроме того, аминокислотная последовательность, при условии что она проявляет активность SerC, может также включать аминокислотные последовательности, которые имеют идентичность последовательности по меньшей мере 80%, с вышеуказанной аминокислотной последовательностью, предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, и еще более предпочтительно по меньшей мере 99%, но не ограничен ими.
Кроме того, полинуклеотидная последовательность, кодирующая SerC, может представлять собой полинуклеотидную последовательность, кодирующую аминокислотную последовательность, представленную SEQ ID NO: 33. В связи с вырожденностью генетического кода или принимая во внимание кодоны, предпочитаемыми организмами для экспрессии полипептида, могут быть выполнены различные модификации полинуклеотида в кодирующей области в объеме, не изменяющем аминокислотную последовательность полипептида. Полинуклеотидная последовательность может, например, быть представлена SEQ ID NO: 34 и может иметь нуклеотидную последовательность, имеющую гомологию по меньшей мере 80%, с полинуклеотидными последовательностями, и предпочтительно по меньшей мере 90%, но не ограничена ими.
Кроме того, микроорганизм может представлять собой микроорганизм, в котором способность введения OPS в клетку или разложения OPS в клетке дополнительно ослаблена.
Что касается содержания OPS-продуцирующего микроорганизма, то описание в патенте Кореи 1381048 или в публикации патентной заявки США 2012-0190081 могут быть использованы в качестве ссылок для настоящего изобретения в дополнение к описанным выше.
В другом аспекте настоящего изобретения предложен способ получения OPS, включающий культивирование в среде микроорганизма, способного продуцировать О-фосфосерин, в котором усилена активность полипептида, имеющего аминокислотную последовательность SEQ ID NO: 1 или SEQ ID NO: 2 и способного экспортировать О-фосфосерин; и выделение О-фосфосерина из микроорганизма, способного продуцировать О-фосфосерин, или из среды для этого микроорганизма.
Используемый здесь термин "культивирование" относится к выращиванию микроорганизма в должным образом адаптированных условиях. Процесс культивирования может быть выполнен в соответствии с подходящей средой и условиями для культуры, известными в данной области техники. Процесс культивирования может быть легко адаптирован для применения специалистом в данной области техники в соответствии с выбранным штаммом. В частности, культура может представлять собой периодическую культуру, непрерывную культуру и культуру с подпиткой, но не ограничена ими.
При культивировании рекомбинантного микроорганизма, имеющего уменьшенную активность SerB по сравнению с его эндогенной активностью, среда может дополнительно содержать глицин или серии, поскольку индуцируется потребность в серине рекомбинантного микроорганизма. Глицин может быть представлен в виде очищенного глицина, глицин-содержащего дрожжевого экстракта или триптона. Концентрация глицина в среде обычно составляет от 0,1 г/л до 10 г/л и, в частности, от 0,5 г/л до 3 г/л. Кроме того, серии может быть представлен в виде очищенного серина, серин-содержащего дрожжевого экстракта или триптона. Концентрация серина в среде обычно составляет от 0,1 г/л до 5 г/л и, в частности, от 0,1 г/л до 1 г/л.
Примеры источника углерода, содержащегося в среде, могут включать углеводы и сахариды, такие как глюкоза, сахароза, лактоза, фруктоза, мальтоза, крахмал и целлюлоза; масла и жиры, такие как соевое масло, подсолнечное масло, касторовое масло и кокосовое масло; жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота; спирты, такие как глицерин и этанол; и органические кислоты, такие как уксусная кислота. Такие источники углерода могут быть использованы по отдельности или в комбинации, но не ограничены ими. Примеры источников азота, содержащихся в среде, могут включать органические источники азота, такие как пептон, дрожжевой экстракт, подливы, солодовый экстракт, жидкий кукурузный экстракт (CSL) и бобовую муку; и неорганические источники азота, такие как мочевина, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Эти источники азота могут быть использованы по отдельности или в комбинации, но не ограничены ими. В качестве источника фосфора культуральная среда может дополнительно включать однозамещенный фосфорнокислый калий, двузамещенный фосфорнокислый калий и соответствующие натрий-содержащие соли, без ограничения ими. Культуральные среды могут включать металлы, такие как сульфат магния и сульфат железа. Кроме того, могут быть включены аминокислоты, витамины и соответствующие предшественники. Эти культуральные среды или предшественники могут быть добавлены к культуре в виде периодической культуры или непрерывной культуры, без ограничения ими.
Кроме того, рН культуры может быть скорректирован соответствующим образом путем добавления соединения, такого как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота и серная кислота, в процессе культивирования. Кроме того, может быть предотвращено образование пузырьков при культивировании с использованием пеногасителя, такого как полигликолевый эфир жирной кислоты. Кроме того, газообразный кислород или газ, содержащий газообразный кислород, может быть добавлен к культуре для поддержания аэробных условий в культуральной жидкости; воздух не может быть добавлен для поддержания анаэробных условий или микроаэробных условий; или может быть введен газообразный азот, газообразный водород или диоксид углерода. Температура культивирования может составлять от 27°С до 37°С и, в частности, от 30°С до 35°С. Культивирование может быть продолжено вплоть до получения нужного вещества и, в частности, в течение от 10 часов до 100 часов.
В настоящем изобретении OPS, продуцируемый во время культивирования, затем может быть выделен и очищен. Целевой OPS может быть извлечен из культуры подходящим способом, известным в данной области, в соответствии со способом культивирования, например из периодической культуры, непрерывной культуры и культуры с подпиткой, но без ограничения ими.
В другом аспекте настоящего изобретения предложены применения получения OPS и экспорт OPS полипептидом, который имеет аминокислотную последовательность SEQ ID NO: 1 или SEQ ID NO: 2.
В еще одном аспекте настоящего изобретения предложен способ получения цистеина или его производного, включающий а) получение О-фосфосерина (OPS) посредством культивирования в среде микроорганизма, в котором активность полипептида, имеющего аминокислотную последовательность SEQ ID NO: 1 или SEQ ID NO: 2 и способного экспортировать О-фосфосерин, усилена по сравнению с его эндогенной активностью; и б) взаимодействие О-фосфосерина (OPS), полученного в (а) или в культуре, содержащей его, с сульфидом в присутствии О-фосфосеринсульфгидрилазы (OPSS) или микроорганизма, способном ее экспрессировать.
Используемый здесь термин "О-фосфосеринсульфгидрилаза" (ниже "OPSS") относится к полипептиду, катализирующем реакцию, в которой тиольную (SH) группу добавляют к OPS для превращения OPS в цистеин. Фермент был сначала обнаружен в Aeropyrum pernix, Mycobacterium tuberculosis, Mycobacterium smegmatics и Trichomonas vaginalis (Mino К and Ishikawa K, FEBS Letters, 551: 133-138, 2003; Burns KE et al., J. Am. Chem. Soc, 127: 11602-11603, 2005). Кроме того, OPSS включает не только белок OPSS дикого типа, но также варианты, которые включают делецию, замену или добавление в часть полинуклеотидной последовательности, кодирующую OPSS, который демонстрирует активность, которая равна или выше биологической активности белка OPSS дикого типа, например, и включает белки OPSS, раскрытые в патентах Кореи 1381048 и 1208267 и их варианты.
Используемый в настоящем изобретении сульфид может представлять собой любой сульфид, представленный не только в твердой форме, обычно используемой в данной области, но также в жидкой или газообразной форме из-за разницы в рН, давлении и растворимости, и, таким образом, может быть превращен в тиольную (SH) группу в форме, например, сульфида (S2-) или тиосульфата (S2O32-). В частности, сульфид, используемый в настоящем изобретении, может представлять собой Na2S, NaSH, H2S, (NH4)2S или Na2S2O3, который может обеспечить тиольную группу OPS. При взаимодействии предоставляют одну тиольную группу на одну реакционно-способную группу OPS с получением одного цистеина или его производного. При этом взаимодействии сульфид, как правило, добавляют в количестве от 0,1 моль до 3 моль и, в частности, от 1 моль до 2 моль на 1 моль OPS.
Кроме этого способ по настоящему изобретению дополнительно включает выделение и очистку цистеина, полученного в реакции на стадии (б). При этом нужный цистеин может быть извлечен путем выделения и очистки его из реакционного раствора при помощи подходящей реакции, известной в данной области техники.
Кроме того, настоящее изобретение относится к получению OPS с высоким выходом за счет усиления активности полипептида SEQ ID NO: 1 или SEQ ID NO: 2 в OPS-продуцирующем микроорганизме, с последующим взаимодействием полученного таким образом OPS с OPSS, посредством чего эффективно получается цистеин. Полученный таким образом цистеин может быть синтезирован в различные виды производных цистеина посредством реакции химического синтеза, известный в данной области, путем модификации атома водорода или конкретной группы атомов.
Используемый здесь термин "производные" относится к похожим соединениям, полученным посредством химической модификации части любого соединения. Обычно этот термин относится к соединениям, в которых атом водорода или группа атомов заменены другим атомом водорода или группой атомов.
Используемый здесь термин "производные цистеина" относится к соединениям, в которых атом водорода или группа атомов в цистеине заменены другим атомом или группой атомов. Например, производные цистеина могут иметь форму, где к атому азота аминогруппы (-NH2) или к атому серы тиольной группы (-SH) в цистеине присоединен другой атом или атомная группа. Примеры производных цистеина включают N-ацетилцистеин (NAC), S-карбоксиметилцистеин (SCMC), Boc-Cys(Me)-OH, (R)-S-(2-амино-2-карбоксиэтил)-L-гомоцистеин, (R)-2-амино-3-сульфопропионовую кислоту, D-2-амино-4-(этилтио)масляную кислоту, 3-сульфино-L-аланин, Fmoc-Cys(Boc-метил)-ОН, селено-L-цистеин, S-(2-тиазолил)-L-цистеин, S-(2-тиенил)-L-цистеин, 5-(4-толил)-L-цистеин, но не ограничены ими. Цистеин может быть легко синтезирован в N-ацетилцистеин (NAC) посредством реакции с ацетилирующим агентом, и в щелочных условиях он может быть синтезирован в S-карбоксиметилцистеин (SCMC) посредством реакции с галогенуксусной кислотой. Эти производные цистеина используются в основном в качестве фармацевтических веществ для противокашлевых средств; средств, облегчающих кашель, и терапевтических средств при бронхите, бронхиальной астме, ларингофарингите и т.д.
Вариант осуществления изобретения
Ниже в данном документе настоящее изобретение будет описано более подробно со ссылкой на следующие Примеры. Однако, эти Примеры приведены только в целях иллюстрации, и не следует ограничивать данное изобретение этими Примерами.
Пример 1: Идентификация транспортера YhhS MFS и транспортера YegB MFS
Для того чтобы идентифицировать мембранные белки Escherichia coli, вовлеченные в экспорт OPS, была подвергнута скринингу библиотека геномной ДНК Escherichia coli K12_W3110 (АТСС27325).
В частности, для установления условий, в которых OPS ингибирует рост Е. coli, был сконструирован платформенный штамм, продуцирующий OPS. Платформенный штамм для скрининга представлял собой рекомбинантный микроорганизм, модифицированный для снижения активности эндогенной фосфосеринфосфатазы (SerB) в штамме W3110 Е. coli дикого типа и был обозначен как "KCCM11212P" (также называемый как "СА07-0012"; патент Кореи 10-1381048; публикация патентной заявки США 2012-0190081). С использованием OPS-продуцирующего штамма KCCM11212P были установлены оптимальные условия скрининга, показывающие ингибирование роста, путем культивирования KCCM11212P, представляющего собой OPS-продуцирующий штамм, в среде, содержащей OPS.
Затем плазмиды геномной библиотеки W3110 трансформировали в СА07-0012 посредством электропорации (van der Rest et al. 1999) и выбирали колонии, показывающие устранение ингибирования роста в условиях среды, содержащей избыточное количество OPS. Из выбранных колоний были получены плазмиды и их нуклеотидные последовательности были проанализированы посредством секвенирования. В результате были идентифицированы два мембранных белка Е. coli, вовлеченных в устранение ингибирования роста в условиях избыточного количества OPS в среде.
Два мембранных белка Е. coli были идентифицированы как yhhS и mdtD, которые кодируют транспортер YhhS суперсемейства мембранных транспортеров (MFS) (аминокислотная последовательность SEQ ID NO: 1 и нуклеотидная последовательность SEQ ID NO: 3) и транспортер YegB MFS (аминокислотная последовательность SEQ ID NO: 2 и нуклеотидная последовательность SEQ ID NO: 4) соответственно (Pao SS, Paulsen IT, Saier МН (1998). "Major facilitator superfamily." Microbiol Mol Biol Rev 1998; 62 (1); 1-34. PMID: 9529885).
Пример 2: Конструирование yhhS- и mdtD-сверхэкспрессирующих векторов
Для того чтобы исследовать, усиливается ли OPS-экспортирующая способность, когда транспортер YhhS MFS и транспортер YegB MFS, вовлеченные в устранение ингибирования роста посредством OPS, усилены в OPS-продуцирующих штаммах, были сконструированы векторы, которые сверхэкспрессируют каждый из генов. Кроме того, поскольку авторы настоящего изобретения подтвердили, что концентрация OPS увеличивается, когда гомосерин/гомосерин-лактонный транспортер RhtB был усилен в OPS-продуцирующем штамме (патент Кореи 138104), RhtB-усиленный штамм использовали в качестве положительного контроля. Кроме того, также были исследованы мультилекарственные эффлюксные транспортеры EmrD и YcaD MFS, принадлежащие к суперсемейству мембранных транспортеров (MFS), к которому принадлежит МасВ. Таким же образом, как в YhhS и MdtD, были также оценены мультилекарственный эффлюксный транспортер EmrD и YcaD MFS транспортер, которые представляют собой мембранные белки Е. coli, принадлежащие к суперсемейству мембранных транспортеров (MFS). В этом Примере фрагменты гена yhhS (SEQ ID NO: 3, учетный номер: b3473), кодирующего транспортер YhhS MFS, и фрагменты гена mdtD (SEQ ID NO: 4, учетный номер: b2077), кодирующего транспортер YegB MFS, были получены посредством PCR с использованием геномной ДНК из W3770 в качестве матрицы.
Последовательности праймеров, используемых для конструирования сверхэкспрессирующих векторов для каждого из генов мембранных белков, показаны в Таблице 1 ниже.


Более конкретно, реакцию PCR для yhhS выполняли с использованием праймеров SEQ ID NO: 5 и 6, в то время как реакцию PCR для mdtD выполняли с использованием праймеров SEQ ID NO: 7 и 8. Праймеры, используемые в реакциях PCR, были сконструированы на основании информации о гене K12 W3110 (GenBank номер АР 003471), депонированном в NIH GenBank, и окружающих нуклеотидных последовательностях.
Кроме того, фрагменты генов rhtB, emrD и ycaD амплифицировали посредством реакций PCR, используя соответствующие пары праймеров, представленные в Таблице 1 ниже.
Каждый из амплифицированных фрагментов гена обрабатывали рестриктазами EcoRV и HindIII и клонировали в сайты рестрикции EcoRV и HindIII вектора pCL-PrhtB, который включает промотор (PrhtB) гена rhtB Е. coli, встроенный в вектор pCL1920 (GenBank No АВ236930), таким образом конструируя pCL-PrhtB-rhtB, pCL-PrhtB-yhhS, pCL-PrhtB-mdtD, pCL-PrhtB-emrD и pCL-PrhtB-ycaD, соответственно.
Пример 3: Конструирование штамма с усиленными транспортером YhhS MFS и транспортером YegB MFS и оценка OPS-продуцирующей способности
Пример 3-1: Конструирование штамма с усиленными транспортером YhhS MFS и транспортером YegB MFS с использованием СА07-0012 и определение OPS-продуцирующей способности
Каждый из пяти типов плазмид, сконструированных в Примере 2, вводили в OPS-продуцирующий штамм СА07-0012 и затем оценивали OPS-продуцирующую способность полученных штаммов.
Более конкретно, каждый штамм высевали на твердую питательную среду LB и культивировали в течение ночи в инкубаторе при 33°С. Каждый штамм, культивированный в течение ночи на твердой питательной среде LB, инокулировали в 25 мл титровальной среды, показанной в Таблице 2 ниже, и затем инкубировали в инкубаторе при 34,5°С и 200 об/мин в течение 40 часов. Результаты показаны в Таблице 3.


Как показано в Таблице 3 выше, среди случаев введения генов мембранного белка Е. coli в штамм СА07-0012Е. coli, соответственно, штаммы с усиленным rhtB, emrD или ycaD демонстрировали значительные увеличения продукции OPS по сравнению с штаммом СА07-0012 и, в частности, штаммы с усиленными YhhS и MdtD демонстрировали по меньшей мере 150% увеличение концентрации OPS. Напротив, штаммы с усиленным EmrD и YcaD, которые использовались в качестве экспериментальной группы, не показали никакого увеличения концентрации OPS.
Штамм, обозначенный как "C407-0012/pCL-PrhtB-yhhS", был назван "Escherichia coli СА07-0266 (СА07-0266)" и депонирован в Корейском центр культур микроорганизмов, признанном международным органом по депонированию в соответствии с Будапештским договором от 9 декабря 2013 года, под номером KCCM11495P.
Кроме того, штамм, обозначенный как "СА07-0012/pCL-PrhtB-mdtD", был назван "Escherichia coli СА07-0267 (СА07-0267)" и депонирован в Корейском центре культур микроорганизмов, признанном международным органом по депонированию в соответствии с Будапештским договором от 9 декабря 2013 года, под номером KCCM11496P.
Пример 3-2: Конструирование штамма с усиленными транспортером YhhS MFS и транспортером YegB MFS с использованием штамма с усиленным SerA и SerC и определение OPS-продуцирующей способности
Дополнительно, влияние генов мембранного белка Е. coli исследовали с использованием OPS-продуцирующего штамма CA07-0022/pCL-Prmf-SerA*(G336V)-(RBS)SerC (публикация Корейской патентной заявки 10-2012-004111) с усиленными активностями D-3-фосфоглицератдегидрогеназы (SerA) и 3-фосфосеринаминотрансферазы (SerC) в качестве путей биосинтеза OPS. Результаты показаны в Таблице 4 ниже.


Как показано в Таблице 4 выше, было снова подтверждено, что среди штаммов, в которых гены мембранного белка Е. coli были дополнительно введены в штамм CA07-0022/pCL-Prmi-SerA*(G336\/)-(RBS)SerC, штаммы с усиленными YhhS и MdtD по настоящему изобретению и штамм с усиленным RhtB (положительный контроль), показали увеличения в продуцировании OPS по сравнению с происходящим от Е. coli штаммом СА07-0012. В частности, штаммы с усиленным YhhS и MdtD по настоящему изобретению, продемонстрировали по меньшей мере 145%-ное увеличение концентрации OPS, аналогично результатам, показанным в Таблице 3 выше. Напротив, штаммы с усиленными ErmD и YcaD продемонстрировали уменьшение концентрации OPS по сравнению с контрольной группой.
Пример 3-3: Конструирование штамма с усиленными транспортером YhhS MFS и транспортером YegB MFS в соответствии с силой промотора и определение OPS-продуцирующей способности
Дополнительно, для того чтобы исследовать может ли увеличение силы промотора увеличивать экспортирующую способность, мембранные белки YhhS и MdtD с повышенной концентрацией OPS сравнивали с контрольной группой, посредством дополнительного введения генов yhhS и mdtD в CA07-0022/pCL-Prmf-SerA*(G336V)-(RBS)SerC, с использованием промотора trc (Ptrc), который является более сильным промотором, чем промотор rhtB (PrhtB).
Каждый из фрагментов генов yhhS и mdtD обрабатывали рестриктазами EcoRV и HindIII и клонировали в сайты рестрикции EcoRV и HindIII вектора pCL-Ptrc-GFP, который включает промотор trc, встроенный в вектор pCL1920, тем самым конструируя pCL-Ptrc-yhhS и pCL-Ptrc-mdtD соответственно. Затем выполняли реакции PCR, используя каждую плазмиду в качестве матрицы вместе с парами праймеров SEQ ID NO: 15 и SEQ ID NO: 16, и результаты реакции обрабатывали рестриктазой HindIII и затем клонировали в сайт рестрикции HindIII из pCL-Prmf-SerA*(G336V)-(RBS)SerC.


В результате, как показано в Таблице 5 выше, при увеличении экспрессии мембранных белков Е. coli путем усиления промотора, наблюдалось по меньшей мере 150% увеличение выхода по сравнению с контрольной группой и по меньшей мере 120% увеличение по сравнению с использованием промотора rhtB.
Пример 3-4: Конструирование штамма с усиленными транспортером YhhS MFS и транспортером YegB MFS в соответствии с силой промотора в хромосоме и определение OPS-продуцирующей способности
Дополнительно, для того чтобы определить, может ли замена промоторов генов yhhS и mdtD более сильными промоторами в хромосоме усилить экспортирующую способность, оценивали способность продуцировать OPS путем конструирования штамма, где собственный промотор был заменен промотором cj1 (патент Кореи 0620092). Введение промотора cj1 в хромосому Е. coli выполняли обычным методом, как описано ниже. Для замены собственных промоторов yhhS и mdtD в хромосоме, сконструированный рекомбинантный вектор трансформировали в OPS-продуцирующий штамм CA07-0022/pCL-Prmf-Ser4*(G336V)-(RBS)SerC (патент Кореи 138104) и последовательность вышеуказанного промотора в векторе и последовательности собственного промотора были заменены посредством гомологичной рекомбинации, тем самым последовательность промотора cj1 была встроена в хромосому.
Каждый штамм высевали на твердую питательную среду LB и культивировали в течение ночи в инкубаторе при 33°С. Каждый из штаммов, культивированных в течение ночи на твердой питательной среде LB, инокулировали в 25 мл титровальной среды, показанной в Таблице 2 выше, и затем инкубировали в инкубаторе при 34,5°С и 200 об/мин в течение 40 часов. Результаты показаны в Таблице 6 ниже.
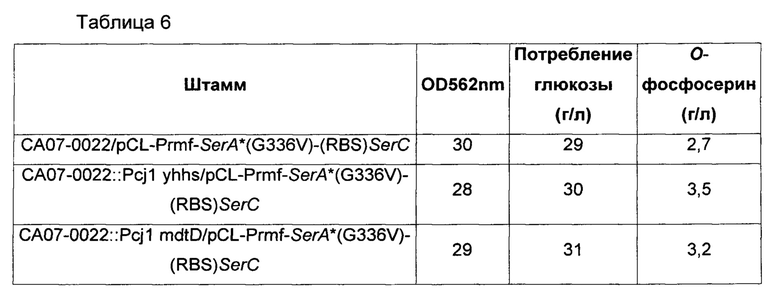
Как показано в Таблице 6 выше, при увеличении экспрессии каждого мембранного белка в хромосоме выход по сравнению с контрольной группой увеличивался вплоть до 130%.
Пример 4: Подтверждение OPS-экспортирующей функции транспортера YhhS MFS и транспортера YegB MFS
Из образцов из колб, в которых продуцирование OPS было подтверждено в Примере 3, после удаления всех OPS, экспортированных в среду, с использованием C0A07-0022/pCL-Prmf-SerA*(G336V)-(RBS)SerC, представляющего собой отрицательный контроль, в котором мембранные белки не были усилены, и образцов CA07-0022/pCL-Prmf-SerA*(G336V)-(RBS)SerC-Ptrc-yhhS и CA07-0022/pCL-Prmf-SerA*(G336V)-(RBS)SerC-Ptrc-mdtD, в которых мембранные белки YhhS и MdtD были усилены, были собраны только клетки и эти клетки были разрушены. Концентрацию OPS в клетке измеряли посредством высокоэффективной жидкостной хроматографии (ВЭЖХ) и результаты показаны на Фиг. 1.
В результате, как показано на Фиг. 1, штаммы с усиленными YhhS и MdtD по настоящему изобретению демонстрировали уменьшение концентрации внутриклеточного OPS на 30%-40% по сравнению с контрольной группой, таким образом подтверждая, что белки Yhhs и MdtD участвуют в экспорте OPS за пределы клетки. Соответственно, было подтверждено, что при усилении белков Yhhs и MdtD клеточный OPS может беспрепятственно экспортироваться наружу клетки, посредством чего повышается OPS-продуцирующую способность.
Из сказанного выше специалисту в данной области техники, к которой относится настоящее изобретение, понятно, что настоящее изобретение может быть воплощено в других конкретных формах без изменения технических идей или существенных характеристик настоящего изобретения. В связи с этим, типичные воплощения, раскрытые здесь, представлены лишь в иллюстративных целях и не должны расцениваться, как ограничивающие объем настоящего изобретения. Напротив, настоящее изобретение предназначено охватывать не только воплощения, представленные в примерах, но и различные альтернативы, модификации, эквиваленты и другие воплощения, которые могут быть включены в пределах сущности и объема настоящего изобретения, как это определено прилагаемой формулой изобретения.



























Изобретение относится к области биохимии и биотехнологии, в частности, к микроорганизму, продуцирующему О-фосфосерин. Указанный микроорганизм отличается тем, что в нем активность полипептида, способного экспортировать О-фосфосерин и имеющего аминокислотную последовательность SEQ ID NO: 1 или SEQ ID NO: 2, усилена по сравнению с его эндогенной активностью. Изобретение также относится к способу получения О-фосфосерина. Настоящий способ включает культивирование указанного микроорганизма в среде и выделение О-фосфосерина из микроорганизма или из среды. Изобретение относится к способу получения цистеина или его производного. Данный способ включает получение О-фосфосерина посредством культивирования указанного микроорганизма в среде и взаимодействие О-фосфосерина с сульфидом в присутствии О-фосфосеринсульфгидрилазы или микроорганизма, способного ее экспрессировать. Настоящее изобретение позволяет получать О-фосфосерин и цистеин с высоким выходом. 3 н.п. и 5 з.п. ф-лы, 1 ил., 6 табл., 4 пр.
1. Микроорганизм, который продуцирует О-фосфосерин (OPS), в котором активность полипептида, имеющего аминокислотную последовательность SEQ ID NO: 1 или SEQ ID NO: 2 и способного экспортировать О-фосфосерин, усилена по сравнению с его эндогенной активностью,
где микроорганизм представляет собой Escherichia sp., Erwinia sp., Serratia sp., Providencia sp., Corynebacterium sp. или Brevibacterium sp.
2. Микроорганизм по п. 1, где активность фосфосеринфосфатазы (SerB) дополнительно ослаблена по сравнению с ее эндогенной активностью.
3. Микроорганизм по п. 1, где активность фосфоглицерат-дегидрогеназы (SerA) или фосфосерин-аминотрансферазы (SerC) дополнительно усилена по сравнению с их эндогенной активностью.
4. Способ получения О-фосфосерина (OPS), включающий:
культивирование в среде микроорганизма, который продуцирует О-фосфосерин, в котором активность полипептида, имеющего аминокислотную последовательность SEQ ID NO: 1 или SEQ ID NO: 2 и способного экспортировать О-фосфосерин, усилена; и
выделение О-фосфосерина из микроорганизма, который продуцирует О-фосфосерин, или из среды для этого микроорганизма,
где микроорганизм представляет собой Escherichia sp., Erwinia sp., Serratia sp., Providencia sp., Corynebacterium sp. или Brevibacterium sp.
5. Способ по п. 4, где у микроорганизма, который продуцирует О-фосфосерин, активность фосфосеринфосфатазы (SerB) дополнительно ослаблена по сравнению с ее эндогенной активностью.
6. Способ по п. 4, где у микроорганизма, который продуцирует О-фосфосерин, активность фосфоглицерат-дегидрогеназы (SerA) или фосфосерин-аминотрансферазы (SerC) дополнительно усилена по сравнению с их эндогенной активностью.
7. Способ получения цистеина или его производного, включающий:
а) получение О-фосфосерина (OPS) посредством культивирования микроорганизма по любому из пп. 1-3 в среде; и
б) взаимодействие О-фосфосерина (OPS), полученного в (а), или культуры, содержащей его, с сульфидом в присутствии О-фосфосеринсульфгидрилазы (OPSS) или микроорганизма, способного ее экспрессировать.
8. Способ по п. 7, где сульфид представляет собой по меньшей мере один сульфид, выбранный из группы, состоящей из Na2S, NaSH, (NH4)2S, H2S и Na2S2O3.
| WO 2012053794 A2, 26.04.2012 | |||
| DATABASE GenBank: WP_001300943.1, 25.05.13 | |||
| Способ для проверки точности хода часов | 1959 |
|
SU130850A1 |
| RU 2010107899 A, 10.09.2011. | |||

