Область изобретения
Настоящее изобретение относится к рекомбинантному микроорганизму с контролируемой экспрессией NADH:хинон-оксидоредуктазы и к способу получения О-фосфосерина, цистеина и производных цистеина с использованием этого микроорганизма.
Предшествующий уровень техники
L-цистеин представляет собой аминокислоту, которая играет важную роль в метаболизме серы у всех живых организмов. Его используют в синтезе белков, таких как кератин волос и тому подобные, глутатиона, биотина, метионина и других серосодержащих метаболитов, и также он служит в качестве предшественника для биосинтеза коэнзима А. Известные способы получения L-цистеина с использованием микроорганизмов включают: 1) способ биологического превращения D,L-2-аминотиазолин-4-карбоновой кислоты в L-цистеин с использованием микроорганизмов, 2) способ получения L-цистеина путем прямой ферментации с использованием Е. coli (ЕР 0885962 В; Wada, М. and Takagi, Н., Appl. Microbiol. Biochem., 73:48-54, 2006) и 3) способ получения О-фосфосерина путем ферментации с использованием микроорганизмов, и превращения О-фосфосерина в L-цистеин путем проведения реакции между O-фосфосерином и сульфидом при каталитическом действии O-фосфосерин-сульфгидрилазы (US 2012-0190081 А1).
В частности, для продукции цистеина способом 3) с высоким выходом предшественник, О-фосфосерин, необходимо получить в избыточном количестве.
Краткое изложение сущности изобретения
Техническая задача
Техническая задача настоящего изобретения заключается в том, чтобы обеспечить рекомбинантный микроорганизм с регулируемой экспрессией NADH:хинон-оксидоредуктазы и способ получения O-фосфосерина, цистеина и производных цистеина с использованием этого микроорганизма.
Техническое решение
Одна задача настоящего изобретения заключается в предоставлении рекомбинантного микроорганизма рода Escherichia, обладающего повышенной активностью NADH:хинон-оксидоредуктазы и O-фосфосерин-продуцирующей способностью.
Другая задача настоящего изобретения заключается в предоставлении способа получения O-фосфосерина, включающего культивирование O-фосфосерин-продуцирующего микроорганизма по настоящему изобретению в среде.
Еще одна задача настоящего изобретения заключается в предоставлении способа получения цистеина или его производного, включающего:
а) культивирование O-фосфосерин-продуцирующего микроорганизма с повышенной активностью NADH:хинон-оксидоредуктазы в среде с получением О-фосфосерина или среды, содержащей О-фосфосерин; и
б) взаимодействие О-фосфосерина, полученного на стадии (а), или среды, содержащей O-фосфосерин, с сульфидом в присутствии O-фосфосерин-сульфгидрилазы (OPSS) или микроорганизма, экспрессирующего указанную O-фосфосерин-сульфгидрилазу.
Полезные эффекты изобретения
OPS(O-фосфосерин)-продуцирующий микроорганизм, обладающий повышенной активностью NADH:хинон-оксидоредуктазы, по настоящему изобретению может продуцировать OPS с высокой эффективностью.
Подробное изложение предпочтительных воплощений
Далее настоящее изобретение описано более подробно. Между тем, каждое описание и воплощение, раскрытое здесь, можно применять к каждому другому описанию и воплощению. То есть, объем настоящего изобретения охватывает все комбинации различных элементов, раскрытых здесь. Далее, объем настоящего изобретения не ограничен определенным описанием, раскрытым ниже. Дополнительно, несколько статей и патентных документов процитировано в настоящем описании. Содержание процитированных статей и патентных документов включено сюда посредством ссылки во всей своей полноте, и уровень техники, к которой принадлежит настоящее изобретение, и содержание настоящей заявки далее описано более подробно.
В одном аспекте настоящего изобретения предложен O-фосфосерин-продуцирующий микроорганизм, обладающий повышенной активностью NADH:хинон-оксидоредуктазы.
Как его используют здесь, термин "О-фосфосерин" (здесь и далее, "OPS") относится к сложному эфиру фосфорной кислоты и серина, который служит составным компонентом для многих белков. В частности, OPS представляет собой предшественник L-цистеина и может быть превращен в цистеин путем проведения реакции с сульфидом при каталитическом действии OPS-сульфгидрилазы (здесь и далее, "OPSS"), но не ограничиваясь этим (US 2012-0190081 А1).
Как его используют здесь, термин "NADH:хинон-оксидоредуктаза" (здесь и далее, "Nuo") относится к ферменту, который уменьшает количество хинона в мембране клетки путем окисления NADH в электронно-транспортной цепи микроорганизмов. Белок-фермент также может быть обозначен как NADH-дегидрогеназа-1 (NDH-1). Ген, кодирующий этот белок, может представлять собой, например, кластер генов nuoABCEFGHIJKLMN, но не ограничиваясь этим. Кластер генов nuoABCEFGHIJKLMN составляет оперон nuo, и его экспрессию может регулировать промотор перед опероном и полинуклеотид в сайте связывания с рибосомой. В настоящем изобретении термин "ген nuoABCEFGHIJKLMN" можно использовать взаимозаменяемо с выражением "ген, кодирующий NADH:хинон-оксидоредуктазу", "ген nuoABCEFGHIJKLMN", "оперон nuo" и "ген nuo".
В настоящем изобретении термин "оперон" относится к функциональной единице ДНК, включающей группу генов, экспрессия которых регулируется одной последовательностью регуляции экспрессии, в частности, одним промотором. мРНК, транскрибируемая с оперона, может представлять собой полицистроиную мРНК, в которой одна молекула мРНК кодирует один или более чем один белок, или моноцистронную мРНК, в которой одна молекула мРНК кодирует один белок.
Nuo представляет собой комплекс из 13 белковых субъединиц (NuoA, NuoB, NuoC, NuoE, NuoF, NuoG, NuoH, Nuol, NuoJ, NuoK, NuoL, NuoM, NuoN). Два типа оперонов, nuoABCEFGHIJKL и nuoMN используют в качестве матрицы при трансляции каждой белковой субъединицы. Структуру оперона nuo можно найти в ЕсоСус (www.biocyc.org) (No. доступа EG12082). Известно, что оперон nuo включает структурный ген и область регуляции экспрессии. Термин "область регуляции экспрессии" в отношении оперона nuo относится к области, расположенной выше по ходу транскрипции от структурного гена, составляющего оперон nuo, и, следовательно, способной регулировать экспрессию этого структурного гена. Область регуляции экспрессии оперона nuo может включать промотор (промотор nuoA и/или промотор nuoM), исключая структурный ген и оператор, и, в частности, может включать промотор.
Оперон nuo может включать нуклеотидную последовательность, кодирующую аминокислотную последовательность, имеющую 70%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, или 99%, или более чем 99% гомологии или идентичности с аминокислотными последовательностями SEQ ID NO: 26 - SEQ ID NO: 38. В частности, оперон nuo может включать структурный ген, кодирующий аминокислотную последовательность SEQ ID NO: 26 - 38 или последовательность, гомологичную или идентичную этой последовательности и проявляющую соответствующую функцию; область регуляции экспрессии, которая регулирует экспрессию последовательности структурного гена. Последовательности SEQ ID NO: 26 - 38 можно подтвердить по известной базе данных NCBI Genbank.
В частности, оперон nuo может представлять собой нуклеотид с последовательностью SEQ ID NO: 1 и/или имеющей 70%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, или 99%, или более чем 99% гомологии или идентичности с SEQ ID NO: 1. Кроме того, очевидно, что любая нуклеотидная последовательность, часть которой делетирована, модифицирована, заменена или добавлена, также может попадать в объем настоящего изобретения при условии, что нуклеотидная последовательность обладает указанной степенью гомологии или идентичности и проявляет функцию, соответствующую функции оперона nuo.
Как его используют здесь, термин "гомология или идентичность" относится к степени соответствия между двумя данными аминокислотными последовательностями или нуклеотидными последовательностями, и она может быть выражена в процентах. Термины "гомология" и "идентичность" часто можно использовать взаимозаменяемо друг с другом.
Гомологию или идентичность последовательностей консервативных полинуклеотидов или полипептидов можно определить с помощью стандартных алгоритмов выравнивания и можно использовать штраф за пропуск, установленный используемой программой по умолчанию. По существу, обычно ожидают, что гомологичные или идентичные последовательности гибридизуются по всей длине или по меньшей мере примерно на 50%, 60%, 70%, 80% или 90% всей длины последовательностей в умеренно строгих или очень строгих условиях. Также учитывают полинуклеотиды, которые содержат вырожденные кодоны вместо кодонов в гибридизующихся полинуклеотидах.
Степень гомологии или идентичности полипептидных или полинуклеотидных последовательностей можно определить, например, с помощью алгоритма BLAST согласно описанию в литературе (смотрите Karlin and Altschul, Pro. Natl. Acad. Sci. USA, 90, 5873 (1993)), или FASTA от Pearson (смотрите Methods Enzymol, 183, 63, 1990). Ha основании алгоритма BLAST разработана программа, известная как BLASTN или BLASTX (смотрите: http://www.ncbi.nlm.nih.gov). Далее, обладают ли какие-либо аминокислотные или полинуклеотидные последовательности гомологией, сходством или идентичностью друг с другом, можно определить путем сравнения этих последовательностей в эксперименте с гибридизацией по Саузерну в заданных строгих условиях, и заданные подходящие условия гибридизации находятся в пределах компетенции специалиста в данной области техники, и их можно легко определить способом, хорошо известным специалисту в данной области техники (например, J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F. M. Ausubel et al., Current Protocols in Molecular Biology).
Микроорганизм по настоящему изобретению не ограничен по типу при условии, что он может продуцировать OPS, и может представлять собой любой прокариотический или эукариотический микроорганизм, в частности прокариотический микроорганизм. Прокариотический микроорганизм может включать штаммы микроорганизмов, принадлежащих роду Escherichia, роду Erwinia, роду Serratia, роду Providencia, роду Cory neb acterium и роду Brevibacterium, в частности штаммы микроорганизма, принадлежащего роду Escherichia, и более конкретно Escherichia coli, но не ограничиваясь ими. В частности, в случае, когда микроорганизм принадлежит роду Escherichia, OPS и L-серин могут продуцироваться с помощью SerA, SerC и SerB, представляющих собой ферменты пути биосинтеза L-серина (Ahmed Zahoor, Computational and structural biotechnology journal, Vol.3, 2012 October; Wendisch V. F. et al., Curr Opin Microbiol. 2006 Jun;9(3):268-74; Peters-Wendisch P. et al, Appl Environ Microbiol. 2005 Nov;7 1(11):7 139-44).
Термин "О-фосфосерин-продуцирующий микроорганизм" по настоящему изобретению относится к микроорганизму, обладающему естественной способностью продуцировать О-фосфосерин, или микроорганизму, где способность продуцировать О-фосфосерин придана родительскому штамму, не обладающему способностью продуцировать О-фосфосерин. В частности, микроорганизм может представлять собой О-фосфосерин-продуцирующий микроорганизм с повышенной активностью Nuo, вызванной природной или искусственной генетической модификацией. Для задач настоящего изобретения О-фосфосерин-продуцирующий микроорганизм может представлять собой любой микроорганизм, способный продуцировать О-фосфосерин путем повышения активности Nuo способом, раскрытым в настоящем изобретении. В настоящем изобретении термин "О-фосфосерин-продуцирующий микроорганизм" можно использовать взаимозаменяемо с терминами "микроорганизм, продуцирующий О-фосфосерин" или "микроорганизм, обладающий способностью продуцировать О-фосфосерин".
В одном воплощении О-фосфосерин-продуцирующий микроорганизм по настоящему изобретению может представлять собой генетически модифицированный микроорганизм или рекомбинантный микроорганизм, у которого активность Nuo повышена, вследствие чего повышается желаемая О-фосфосерин-продуцирующая способность, но не ограничиваясь этим.
Как его используют здесь, термин "повышение активности" белка означает, что активность белка увеличена по сравнению с его эндогенной активностью. Термин "эндогенная активность" относится к активности определенного белка, изначально присутствующего у родительского штамма до трансформации или у немодифицированного микроорганизма, когда признак изменен путем генетической модификации, вызванной природными или искусственными факторами, и его можно использовать взаимозаменяемо с термином "активность до модификации". Термин "увеличение" при сравнении активности белка с его эндогенной активностью обозначает, что активность белка повышена по сравнению с активностью определенного белка, изначально присутствующего у родительского штамма до трансформации или у немодифицированного микроорганизма. Например, родительский штамм может представлять собой Escherichia coli АТСС27325. В другом примере родительский штамм может представлять собой штамм, включающий модификацию для увеличения OPS-продуцирующей способности, например, СА07-0012 (КССМ 11212Р; US 2012-0190081 А) или СА07-4821, но не ограничиваясь этим.
"Повышение активности" может быть достигнуто путем введения чужеродного белка или путем повышения активности эндогенного белка и может, в частности, быть достигнуто путем повышения активности эндогенного белка. Повышение активности белка можно подтвердить по повышению уровня активности белка, уровня экспрессии или количества продукта, продуцированного с участием этого белка.
Повышение активности может быть осуществлено различными способами, хорошо известными в данной области техники, и эти способы не ограничены при условии, что они обеспечивают повышение активности белка-мишени по сравнению с этой активностью у микроорганизма до модификации. В частности, можно использовать хорошо известные специалисту в данной области техники методы генетической инженерии и/или белковой инженерии, представляющие собой общепринятые методы молекулярной биологии, но способ не ограничен ими (например, Sitnicka et al. Functional Analysis of Genes. Advances in Cell Biology. 2010, Vol.2. 1-16, Sambrook et al. Molecular Cloning 2012 и так далее).
В настоящем изобретении белок, направленный на повышение активности, то есть, целевой белок, может представлять собой Nuo, но белок не ограничен этим при условии, что он представляет собой "белок, образующий протон-движущую силу путем потребления NADH" или "белок, потребляющий NADH", и оба белка закодированы опероном nuo.
Повышение активности Nuo по настоящему изобретению может включать повышение активности одной или более чем одной из белковых субъединиц, составляющих белковый комплекс Nuo.
В частности, повышение активности белка по настоящему изобретению может быть достигнуто путем:
1) увеличения числа внутриклеточных копий гена, кодирующего белок;
2) модификации области регуляции экспрессии гена, кодирующего белок на хромосоме;
3) модификации нуклеотидной последовательности, кодирующей инициирующий кодон или 5'-UTR транскрипта гена, кодирующего белок;
4) модификации аминокислотной последовательности, приводящей к повышению активности белка;
5) модификации полинуклеотидной последовательности, кодирующей белок, приводящей к повышению активности белка (например, модификации последовательности гена, кодирующего белок, приводящей к кодированию белка, модифицированного с повышением активности);
6) введения чужеродного полинуклеотида, проявляющего активность белка, или кодон-оптимизированного варианта полинуклеотида этого полинуклеотида;
7) кодон-оптимизации полинуклеотида, кодирующего белок;
8) анализа третичной структуры белка и на основе этого анализа выбора и модификации сайта экспозиции или химической модификации этого сайта; или
9) комбинации двух или более чем двух методик, выбранных из вышеизложенных пунктов 1 - 8), но не ограничиваясь этим.
В частности, 1) способ увеличения числа внутриклеточных копий гена, кодирующего белок, может быть выполнен любым способом, известным в данной области техники, например, путем введения вектора, функционально связанного с геном, кодирующим белок, и способного реплицироваться и функционировать независимо от клетки-хозяина, в клетку-хозяина. Дополнительно, способ можно осуществить путем введения вектора, функционально связанного с геном и способным вставлять этот ген в хромосому клетки-хозяина, в клетку-хозяина, но не ограничиваясь этим.
Как его используют здесь, термин "вектор" относится к ДНК-конструкции, содержащей полинуклеотидную последовательность, кодирующую целевой белок, функционально связанный с подходящей регуляторной последовательностью, так чтобы обеспечить возможность экспрессии целевого белка в подходящей клетке-хозяине. Последовательность регуляции экспрессии может включать промотор, способный инициировать транскрипцию, любую операторную последовательность для регуляции транскрипции, последовательность, кодирующую подходящую мРНК сайта связывания с рибосомой, и последовательность регуляции терминации транскрипции и трансляции. После трансформации подходящей клетки-хозяина вектор может реплицироваться или функционировать независимо от генома хозяина или может интегрировать в геном хозяина.
Вектор, используемый в настоящем изобретении, специально не ограничен при условии, что он может реплицироваться, и можно использовать любой вектор, известный в данной области техники. Примеры обычно используемых векторов могут включать природные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, в качестве фагового вектора или космидного вектора можно использовать pWE15, М13, MBL3, MBL4, IXII, ASHII, APII, t10, t11, Charon4A и Charon21A и так далее; и в качестве плазмидного вектора можно использовать векторы на основе pBR, pUC, pBluescriptII, pGEM, pTZ, pCL, pSK, pSKH и pET и так далее. В частности можно использовать векторы pCL, pSK, pSKH130, pDZ, pACYC177, pACYC184, pECCG117, pUC19, pBR322, pMW118, pCC1BAC.
Вставку полинуклеотида в хромосому можно выполнять любым способом, известным в данной области техники, например, гомологичной рекомбинацией, но не ограничиваясь этим.
Как его используют здесь, термин "трансформация" относится к введению рекомбинантного вектора, содержащего полинуклеотид, кодирующий целевой белок, в клетку-хозяина так, чтобы белок, кодируемый этим полинуклеотидом, мог экспрессироваться в клетке-хозяине. При условии, что полинуклеотид, которым трансформировали, может экспрессироваться в клетке-хозяине, не имеет значения, интегрирован ли полинуклеотид, которым трансформировали, в хромосому клетки-хозяина и расположен на ней, или он расположен экстрахромосомно, и оба случая могут быть включены. Способ трансформации вектором включает любой способ введения нуклеиновой кислоты в клетку и может быть выбран из подходящих стандартных методов, известных в данной области техники, в зависимости от клетки-хозяина. Например, трансформацию можно проводить путем электропорации, осаждения фосфатом кальция (CaPO4), осаждения хлоридом кальция (CaCl2), микроинъекции, полиэтиленгликолевым методом (PEG), DEAE-декстрановым методом, катионно-липосомным методом, методом с ацетатом лития-DMSO и так далее, но способ не ограничен этими.
Далее, как его используют здесь, термин "функционально связанный" обозначает, что полинуклеотидная последовательность функционально связана с промоторной последовательностью или областью регуляции экспрессии, которая инициирует и опосредует транскрипцию полинуклеотида, кодирующего целевой белок по настоящему изобретению. Функциональную связь можно выполнять с использованием технологии рекомбинантной ДНК, хорошо известной в данной области техники, и сайт-специфическое разрезание и сшивание ДНК можно выполнять с использованием ферментов для разрезания и сшивания и так далее, известных в данной области техники, но не ограничиваясь этим.
2) Способ модификации последовательности регуляции экспрессии гена, кодирующего белок на хромосоме, может быть выполнен любым способом, известным в данной области техники, например, путем индуцирования модификации в этой последовательности путем делеции, вставки, неконсервативной или консервативной замены в этой последовательности нуклеиновой кислоты или их комбинации для дополнительного повышения активности последовательности регуляции экспрессии, или путем замены этой последовательности на последовательность нуклеиновой кислоты, обладающую более сильной активностью, или путем вставки такой последовательности. Последовательность регуляции экспрессии может включать промотор, операторную последовательность, последовательность, кодирующую сайт связывания с рибосомой, и последовательность, регулирующую терминацию транскрипции и трансляции и так далее, но специально не ограничиваясь этим. В частности, способ может включать вставку сильного гетерологичного промотора ниже по ходу транскрипции от исходного промотора, но не ограничиваясь этим.
Примеры известного сильного промотора могут включать промоторы CJ1 - CJ7 (US 7662943 B2), промотор lac, промотор trp, промотор trc, промотор tac, промотор фага лямбда PR, промотор PL, промотор tet и промотор rmf и так далее, но не ограничиваясь ими, и может включать все замены на более сильный промотор по сравнению эндогенным промотором.
3) Способ модификации нуклеотидной последовательности инициирующего кодона или 5'-UTR транскрипта гена, кодирующего белок, можно выполнять любым способом, известным в данной области техники, например, путем замены эндогенного инициирующего кодона белка на другой инициирующий кодон, обеспечивающий более высокий уровень экспрессии этого белка по сравнению с эндогенным инициирующим кодоном, но не ограничиваясь этим.
Способы модификации аминокислотной последовательности или полинуклеотидной последовательности 4) и 5) можно осуществить любым способом, известным в данной области техники, например, путем индуцирования модификации в этой последовательности путем делеции, вставки, неконсервативной или консервативной замены в этой полинуклеотидной последовательности или комбинации вышеперечисленного для дополнительного повышения активности полинуклеотидной последовательности, или путем замены этой последовательности на полинуклеотидную последовательность, модифицированную так, чтобы иметь более сильную активность. Эту замену можно осуществить, в частности, путем вставки гена в хромосому путем гомологичной рекомбинации, но не ограничиваясь этим.
Используемый здесь вектор может дополнительно включать селективный маркер для подтверждения вставки в хромосому. Селективный маркер предназначен для отбора клеток, трансформированных вектором, то есть, для подтверждения вставки гена, предназначенного для введения, и можно использовать маркеры, обеспечивающие селектируемые фенотипы, такие как устойчивость к лекарственным средствам, ауксотрофность, устойчивость к цитотоксическим агентам или экспрессия поверхностных белков, но селектируемый маркер не ограничен этими. Только клетки, экспрессирующие селективный маркер, способны выживать или демонстрировать отличающиеся фенотипы в условиях окружающей среды, обработанной агентом селекции, что позволяет отбирать трансформированные клетки.
6) Способ введения чужеродного полинуклеотида, обеспечивающего активность белка, можно выполнять любым способом, известным в данной области техники, например, путем введения в клетку-хозяина чужеродного полинуклеотида, кодирующего белок, проявляющий такую же активность как указанный белок или близкую активность, или кодон-оптимизированного варианта этого полинулеотида. Чужеродный белок можно использовать без ограничения независимо от его происхождения или последовательности при условии, что он проявляет такую же активность как этот белок или близкую активность. Дополнительно, введенный в клетку-хозяина чужеродный полинуклеотид может быть кодон-оптимизирован, так чтобы обеспечить оптимальную транскрипцию и трансляцию в клетке-хозяине. Введение может выполнять специалист средней квалификации в данной области техники путем выбора соответствующего способа трансформации, известного в данной области техники, и экспрессия введенного полинуклеотида в клетке-хозяине обеспечивает возможность продукции белка, тем самым увеличивая его активность.
7) Способ кодон-оптимизации полинуклеотида, кодирующего белок, может быть выполнен путем кодон-оптимизации эндогенного полинуклеотида для повышения уровня транскрипции или трансляции в клетке-хозяине или путем оптимизации его кодонов таким образом, чтобы обеспечить возможность оптимизированной транскрипции и трансляции чужеродного полинуклеотида в клетке-хозяине.
8) Способ анализа третичной структуры белка и на основе этого анализа выбора и модификации сайта экспозиции или химической модификации сайта экспозиции, может быть выполнен, например, путем сравнения информации о последовательности анализируемого полипептида с базой данных, в которой хранится информация об известных белках, для определения образцов белков-кандидатов в соответствии со степенью сходства последовательностей, и, соответственно, подтверждения структуры на основе этой информации, для выбора и трансформации или модификации сайта экспозиции, подлежащего модификации, или его химической модификации.
В одном воплощении повышение активности белка можно выполнить любым одним или более чем одним способом из вышеизложенных пунктов 1) - 2).
В любом из вышеизложенных воплощений повышение активности NADH:хинон-оксидоредуктазы по настоящему изобретению может представлять собой увеличение экспрессии оперона nuo. В любом из вышеизложенных воплощений повышение активности NADH:хинон-оксидоредуктазы может включать последовательность регуляции экспрессии гена с повышенной активностью, находящуюся выше по ходу транскрипции от гена, кодирующего эту NADH:хинон-оксидоредуктазу. В частности, положение "выше по ходу транскрипции от гена, кодирующего NADH:хинон-оксидоредуктазу" может быть выше по ходу транскрипции от гена nuoA. В одном воплощении повышение активности NADH:хинон-оксидоредуктазы может представлять собой повышение уровня экспрессии одного или более чем одного структурного гена, присутствующего в опероне, например 2 или более чем 2, 3 или более чем 3 или всей последовательности структурных генов вследствие модификации одной последовательности регуляции экспрессии, присутствующей в опероне nuo, кодирующем белковый комплекс Nuo. В частности, модификация последовательности регуляции экспрессии может представлять собой дополнительную вставку последовательности регуляции экспрессии гена с повышенной активностью между эндогенным промотором оперона nuo и геном nuoA; например, последовательность регуляции экспрессии гена может представлять собой промотор, но не ограничиваясь этим.
Такое повышение активности белка может означать, что активность белка или его концентрация увеличена по сравнению с активностью или концентрацией белка, экспрессируемого у микроорганизма дикого типа или у микроорганизма до модификации, или что количество продукта, продуцированного с участием этого белка, увеличено, но не ограничиваясь этим.
Как его используют здесь, термин "штамм до модификации" или "микроорганизм до модификации" не исключает штамм, содержащий мутацию, которая может произойти естественным путем у микроорганизма, и может относиться к самому штамму природного типа или штамму до изменения признака вследствие генетической модификации, вызванной природными или искусственными факторами. В настоящем изобретении модификация признаков может представлять собой повышение активности Nuo. Термины "штамм до модификации" или "микроорганизм до модификации" можно использовать взаимозаменяемо с терминами "немутированный штамм", "немодифицированный штамм", "немутированный микроорганизм", "немодифицированный микроорганизм" или "референтный микроорганизм".
Микроорганизм по настоящему изобретению обеспечивает дополнительное повышение способности продуцировать OPS и/или способности выводить OPS из клетки; или может включать модификации, повышающие способность расщеплять и/или потреблять OPS.
Примеры модификаций, повышающих способность продуцировать OPS и/или способность экспортировать OPS из клеток или повышать способность расщеплять и/или потреблять OPS, могут включать понижение активности фосфосерин фосфатазы (SerB), усиление активности фосфосерин-экспортирующего белка (YhhS) или комбинацию этих модификаций, но не ограничиваясь этим.
Как его используют здесь, термин "понижение" в отношении активности полипептида представляет собой комплексное понятие, включающее как активность, сниженную относительно его эндогенной активности, так и отсутствие активности. Термин "понижение активности" можно использовать взаимозаменяемо с такими терминами как инактивация, недостаточность, понижающая регуляция, понижение, снижение, уменьшение активности и так далее.
"Понижение активности" также может включать случай, когда собственная активность полипептида уменьшена по сравнению с активностью полипептида, исходно присутствующего у микроорганизма, или устранена вследствие мутации полинуклеотида, кодирующего этот полипептид; случай, когда общий уровень активности и/или концентрация (уровень экспрессии) внутриклеточного полипептида понижен(а) по сравнению с природным штаммом вследствие ингибирования экспрессии гена полинуклеотида, кодирующего этот полипептид, или ингибирования его трансляции в полипептид и так далее; случай, когда полинуклеотид совсем не экспрессируется; и/или случай, когда активность полипептида не наблюдается даже при экспрессии полинуклеотида. Как его используют здесь, термин "эндогенная активность" относится к активности определенного полипептида, исходно присутствующего у родительского штамма до трансформации, микроорганизма дикого типа или немодифицированного микроорганизма, когда признак изменен вследствие генетической модификации, вызванной природными или искусственными факторами, и его можно использовать взаимозаменяемо с выражением "активность до модификации". Выражение "активность полипептида "подавлена недостаточна, уменьшена, подвержена понижающей регуляции, снижена или ослаблена" при сравнении с эндогенной активностью этого полипептида обозначает, что активность полипептида уменьшена относительно активности определенного полипептида, исходно присутствующего у родительского штамма до трансформации или у немодифицированного микроорганизма.
Понижение активности полипептида можно получить любым способом, известным в данной области техники, но способ не ограничен этим, и его можно достичь путем применения различных способов, хорошо известных в данной области техники (например, Nakashima N et al., Bacterial cellular engineering by genome editing and gene silencing. Int J Mol Sci. 2014;15(2):2773-2793; Sambrook et al. Molecular Cloning 2012; и так далее).
В частности, понижение активности полипептида по настоящему изобретению можно получить путем:
1) делетирования части гена или всего гена, кодирующего этот полипептид;
2) модификации области регуляции экспрессии (последовательности регуляции экспрессии), с тем чтобы понизить уровень экспрессии гена, кодирующего этот полипептид;
3) модификации аминокислотной последовательности, составляющей полипептид, с тем чтобы устранить или ослабить активность этого полипептида (например, введения делеции/замены/добавления одной или более чем одной аминокислоты в аминокислотную последовательность);
4) модификации последовательности гена, кодирующего полипептид, с тем чтобы устранить или ослабить активность этого полипептида (например, введения делеции/замены/добавления одного или более чем одного нуклеотида в нуклеотидную последовательность гена полипептида для кодирования полипептида, модифицированного с устранением или понижением активности полипептида);
5) модификации нуклеотидной последовательности, кодирующей инициирующий кодон или 5'-UTR транскрипта гена, кодирующего полипептид;
6) введения антисмыслового олигонуклеотида (например, антисмысловой РНК), который комплементарно связывается с транскриптом гена, кодирующего полипептид;
7) добавления последовательности, комплементарной последовательности Шайна-Дальгарно (SD), на переднем конце последовательности SD гена, кодирующего полипептид, для образования вторичной структуры, приводящей к ингибированию присоединения рибосомы;
8) конструирования с помощью обратной транскрипции, посредством которого добавляют промотор, предназначенный для обратной транскрипции, на 3'-конце открытой рамки считывания (ORF) последовательности гена, кодирующего полипептид, или
9) комбинации двух или более чем двух способов, выбранных из 1) - 8) выше, но специально не ограничиваясь этим.
Например:
1) Способ делетирования части гена или всего гена, кодирующего полипептид, может быть выполнен путем делетирования всего полинуклеотида, кодирующего эндогенный целевой полипептид, на хромосоме или путем замены этого полинуклеотида на полинуклеотид с делецией части нуклеотидов или на ген-маркер.
2) Способ модификации области регуляции экспрессии (последовательности регуляции экспрессии) может быть выполнен путем введения модификации в область регуляции экспрессии (последовательность регуляции экспрессии) путем введения делеции, инсерции, неконсервативной замены или консервативной замены или их комбинации; или путем замены последовательности на последовательность, обладающую более слабой активностью. Область регуляции экспрессии может включать промотор; последовательность оператора; последовательность, кодирующую сайт связывания с рибосомой, и последовательность для регуляции транскрипции и трансляции, но не ограничиваясь ими.
Далее, 5) способ модификации нуклеотидной последовательности, кодирующей инициирующий кодон или 5'-UTR транскрипта гена, кодирующего полипептид, может быть выполнен, например, путем замены этой нуклеотидной последовательности на нуклеотидную последовательность, кодирующую другой инициирующий кодон, обеспечивающий более низкий уровень экспрессии полипептида, чем эндогенный инициирующий кодон, но не ограничиваясь этим.
3) и 4) Способы модификации аминокислотной последовательности или полинуклеотидной последовательности могут быть выполнены путем введения модификации в последовательность путем делеции, инсерции, неконсервативной или консервативной замены в аминокислотной последовательности полипептида или полинуклеотидной последовательности, кодирующей этот полипептид, или их комбинации для понижения активности полипептида, или путем замены последовательности на аминокислотную последовательность или полинуклеотидную последовательность, модифицированную для понижения активности, или аминокислотную последовательность или полинуклеотидную последовательность, модифицированную для устранения активности, но не ограничиваясь этим. Например, экспрессия гена может быть ингибирована или понижена путем введения мутации в полинуклеотидную последовательность для образования стоп-кодона, но не ограничиваясь этим.
6) Способ введения антисмыслового олигонуклеотида (например, антисмысловой РНК), который комплементарно связывается с транскриптом гена, кодирующим полипептид, можно найти в литературе (Weintraub, Н. et at, Antisense-RNA as a molecular tool for genetic analysis, Reviews - Trends in Genetics, Vol.1(1) 1986).
7) Способ добавления последовательности, комплементарной последовательности Шайна-Дальгарно (SD) на переднем конце последовательности SD гена, кодирующего полипептид, для образования вторичной структуры, таким образом обеспечивающий ингибирование присоединения рибосомы путем ингибирования трансляции мРНК или уменьшения его скорости.
8) Конструирование с помощью обратной транскрипции (RTE), при котором добавляют промотор, предназначенный для обратной транскрипции, на 3'-конце открытой рамки считывания (ORF) последовательности гена, кодирующего полипептид, можно выполнить путем создания антисмыслового нуклеотида, комплементарного транскрипту гена, кодирующему этот полипептид, для понижения его активности.
SerB по настоящему изобретению обладает активностью по превращению OPS в L-серин, и следовательно микроорганизм, модифицированный для ослабления активности SerB, обладает свойством накапливать в себе OPS, таким образом, являясь полезным для продукции OPS. SerB по настоящему изобретению может представлять собой белок, имеющий аминокислотную последовательность, представленную SEQ ID NO: 2, или включающий такую последовательность, или может представлять собой белок, состоящий из аминокислотной последовательности, представленной SEQ ID NO: 2, или по существу состоящий из этой последовательности, но не ограничиваясь этим. Дополнительно SerB по настоящему изобретению может иметь или включать аминокислотную последовательность, имеющую 70%, 80%, 90%, 95% или 99% или более чем 99% гомологии или идентичности аминокислотной последовательности, представленной SEQ ID NO: 2, при условии, что он проявляет активность SerB. Более того, SerB по настоящему изобретению может состоять или по существу состоять из аминокислотной последовательности, имеющей 70%, 80%, 90%, 95% или 99% или более чем 99% гомологии или идентичности аминокислотной последовательности, представленной SEQ ID NO: 2, но не ограничиваясь этим. Дополнительно полинуклеотид, кодирующий SerB, может иметь нуклеотидную последовательность SEQ ID NO: 3, кодирующую аминокислотную последовательность, представленную SEQ ID NO: 2, или включать такую последовательность. Далее, полинуклеотид, кодирующий SerB, может состоять или по существу состоять из нуклеотидной последовательности, кодирующей аминокислотную последовательность, представленную SEQ ID NO: 2. Полинуклеотид, кодирующий SerB, по настоящему изобретению может подвергаться различным модификациям в кодирующей области в объеме, при котором не изменяется аминокислотная последовательность белка SerB вследствие вырожденности генетического кода или с учетом кодонов, предпочтительных для организма, предназначенного для экспрессии белка SerB. Полинуклеотид, кодирующий SerB, по настоящему изобретению может иметь нуклеотидную последовательность, имеющую 70%, 80%, 90%, 95% или 99% или более чем 99% и менее чем 100% гомологии или идентичности нуклеотидной последовательности SEQ ID NO: 3 или включать такую последовательность. Дополнительно, полинуклеотид, кодирующий SerB, по настоящему изобретению может состоять или по существу состоять из нуклеотидной последовательности, имеющей 70%, 80%, 90%, 95%, или 99%, или более чем 99% и менее чем 100% гомологии или идентичности нуклеотидной последовательности SEQ ID NO: 3, но не ограничиваясь этим.
YhhS по настоящему изобретению обладает активностью по экспортированию OPS, и следовательно микроорганизм, модифицированный для усиления активности YhhS, обладает свойством экспортировать OPS, таким образом, являясь полезным для продукции OPS. YhhS по настоящему изобретению может представлять собой белок, имеющий аминокислотную последовательность, представленную SEQ ID NO: 4, или включающий такую последовательность, или может представлять собой белок, состоящий или по существу состоящий из аминокислотной последовательности, представленной SEQ ID NO: 4, но не ограничиваясь этим. Дополнительно YhhS по настоящему изобретению может иметь аминокислотную последовательность, имеющую 70%, 80%, 90%, 95%, или 99%, или более чем 99% гомологии или идентичности аминокислотной последовательности, представленной SEQ ID NO: 4, или включать такую последовательность при условии, что он проявляет активность YhhS. Более того, YhhS по настоящему изобретению может состоять или по существу состоять из аминокислотной последовательности, имеющей 70%, 80%, 90%, 95%, или 99%, или более чем 99% гомологии или идентичности аминокислотной последовательности, представленной SEQ ID NO: 4, но не ограничиваясь этим. Дополнительно, полинуклеотид, кодирующий YhhS, может иметь нуклеотидную последовательность, кодирующую аминокислотную последовательность, представленную SEQ ID NO: 4, или включать такую последовательность. Далее, полинуклеотид, кодирующий YhhS, может состоять или по существу состоять из нуклеотидной последовательности, кодирующей аминокислотную последовательность, представленную SEQ ID NO: 4. Полинуклеотид, кодирующий YhhS, по настоящему изобретению может претерпевать различные модификации в кодирующей области в объеме, при котором не изменяется аминокислотная последовательность белка YhhS вследствие вырожденности генетического кода или с учетом кодонов, предпочтительных для организма, предназначенного для экспрессии белка YhhS. Полинуклеотид, кодирующий YhhS, по настоящему изобретению может иметь нуклеотидную последовательность, имеющую 70%, 80%, 90%, 95%, или 99%, или более чем 99% и менее чем 100% гомологии или идентичности нуклеотидной последовательности SEQ ID NO: 5 или включать такую последовательность. Дополнительно полинуклеотид, кодирующий YhhS, по настоящему изобретению может состоять или по существу состоять из нуклеотидной последовательности, имеющей 70%, 80%, 90%, 95%, или 99%, или более чем 99% и менее чем 100% гомологии или идентичности нуклеотидной последовательности SEQ ID NO: 5, но не ограничиваясь этим.
В одном воплощении микроорганизм, включающий модификации для усиления способности продуцировать OPS и/или способности экспортировать OPS из клетки; или усиливать способность расщеплять и/или потреблять OPS, может представлять собой СА07-0012 (КССМ11212Р; US 2012-0190081 А) или СА07-4821, но не ограничиваясь этим. Что касается состава OPS-продуцирующего микроорганизма дополнительно к вышеизложенному в настоящем изобретении можно использовать описание патента KR No. 1381048 или заявки US No. 2012-0190081 в качестве ссылки.
В другом аспекте настоящего изобретения предложен способ получения OPS, включающий культивирование OPS-продуцирующего микроорганизма с повышенной активностью NADH:хинон-оксидоредуктазы в среде.
Микроорганизм является таким, как описано выше.
Как его используют здесь, термин "культивирование" обозначает выращивание микроорганизма по настоящему изобретению в подходящим образом контролируемых условиях окружающей среды. Процесс культивирования по настоящему изобретению можно проводить в подходящей культуральной среде и в условиях культивирования, известных в данной области техники. Специалист в данной области техники легко может наладить такой процесс культивирования в соответствии со штаммом, который нужно выбрать. В частности, культивирование может представлять собой периодическое культивирование, непрерывное культивирование и/или периодическое культивирование с подпиткой, но не ограничиваясь этим.
Среда для культивирования микроорганизма может дополнительно содержать глицин и серии. Глицин может быть представлен в форме очищенного глицина, глицинсодержащего дрожжевого экстракта или триптона. Концентрация глицина, содержащегося в среде, обычно составляет от 0,1 г/л до 10 г/л и, в частности, от 0,5 г/л до 3 г/л. Дополнительно, серии может быть предоставлен в форме очищенного серина, серинсодержащего дрожжевого экстракта или триптона. Концентрация серина, содержащегося в среде, обычно составляет от 0,1 г/л до 5 г/л, и в частности от 0,1 г/л до 1 г/л.
Примеры источника углерода, содержащегося в среде, могут включать сахариды и углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, мальтоза, крахмал и целлюлоза; масла и жиры, такие как соевое масло, подсолнечное масло, касторовое масло и кокосовое масло; жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота; спирты, такие как глицерин и этанол, и органические кислоты, такие как уксусная кислота. Эти источники углерода можно использовать по отдельности или в комбинации, но не ограничиваясь ими.
Примеры источника азота, содержащегося в среде, может включать органические источники азота, такие как пептон, дрожжевой экстракт, мясной бульон, солодовый экстракт, жидкий кукурузный экстракт и бобовая мука; и неорганические источники азота, такие как мочевина, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Эти источники азота можно использовать по отдельности или в комбинации, но не ограничиваясь ими.
Примеры источников фосфора, содержащихся в среде, могут включать дигидрофосфат калия, гидрофосфат калия или соответствующие натрийсодержащие соли, но не ограничиваясь ими.
Дополнительно культуральная среда может включать соли металлов, такие как сульфат магния или сульфат железа, и может дополнительно включать аминокислоты, витамины и подходящие предшественники. Эти культуральные среды или предшественники можно добавлять в культуру при периодическом культивировании или при непрерывном культивировании, но не ограничиваясь этим.
рН культуры можно регулировать путем добавления такого соединения как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота и серная кислота при культивировании подходящим образом. Дополнительно, образование пены во время культивирования можно предупреждать с помощью пеногасителя, такого как полигликолевый сложный эфир жирных кислот. Далее, газ кислород или кислородсодержащий газ можно вводить в культуру для поддержания аэробных условий в культуре; или газ азот, газ водород или углекислый газ можно вводить или можно не вводить никакой газ для поддержания анаэробных или микроаэробных условий. Температура в культуре может находиться в диапазоне от 25 до 40°С, в частности, от 30 до 35°С. Культивирование можно продолжать до получения нужного количества полезного вещества, и в частности от 10 до 100 часов, но не ограничиваясь этими иллюстративными примерами.
Настоящее изобретение может дополнительно включать стадию получения среды перед стадией культивирования в способе по настоящему изобретению, но не ограничиваясь этим.
Способ может дополнительно включать стадию выделения OPS из культуральной среды или из микроорганизма. Стадию выделения можно дополнительно включать после стадии культивирования.
В способе выделения OPS по настоящему изобретению желаемый OPS можно выделять из культурального раствора с использованием подходящих методов, известных в данной области техники, в зависимости от способа культивирования. Например, можно использовать такие методы как центрифугирование, фильтрование, ионообменная хроматография, кристаллизация и HPLC (высокоэффективная жидкостная хроматография), и желаемый OPS можно выделять из среды или из микроорганизма с использованием подходящего способа, известного в данной области техники.
Далее, стадия выделения может дополнительно включать процесс очистки, который можно выполнять с помощью подходящего способа, известного в данной области техники. Таким образом, выделенный OPS может находиться в очищенном состоянии или в среде ферментации микроорганизмов, содержащей OPS. Дополнительно выделение OPS можно эффективно выполнять путем добавления подходящего способа, известного в данной области техники, до и после стадии культивирования и до и после стадии выделения.
Еще в одном аспекте настоящего изобретения предложен способ получения цистеина или его производного, включающий:
а) культивирование О-фосфосерин-продуцирующего микроорганизма с повышенной активностью NADH:хинон-оксидоредуктазы в среде с получением О-фосфосерина или среды, содержащей О-фосфосерин; и
б) взаимодействие О-фосфосерина, полученного на стадии (а) или среды, содержащей этот О-фосфосерин, с сульфидом в присутствии О-фосфосерин-сульфгидрилазы (OPSS) или микроорганизма, экспрессирующего О-фосфосерин-сульфгидрилазу.
Стадии (а) и (б) не обязательно ограничены по порядку, то есть их можно выполнять непрерывно или последовательно, и может отсутствовать промежуток времени между этими стадиями, и эти стадии можно выполнять одновременно или с интервалом в несколько секунд, несколько минут, несколько часов, несколько суток.
В частности, способ может представлять собой способ получения цистеина или его производного: включая культивирование OPS-продуцирующего микроорганизма с повышенной активностью Nuo в среде для получения OPS или в среде, содержащей OPS; и взаимодействие О-фосфосерина, полученного на стадии (а), или среды, содержащей О-фосфосерин, с сульфидом в присутствии О-фосфосерин-сульфгидрилазы (OPSS) или микроорганизма, экспрессирующего эту О-фосфосерин-сульфгидрилазу. Повышение активности NADH:хинон-оксидоредуктазы и О-фосфосерин-продуцирующий микроорганизм являются такими, как описано выше.
Как его используют здесь, термин "производное" относится к близкородственным соединениям, полученным путем химической модификации части любого соединения. Обычно этот термин относится к соединениям, в которых атом водорода или определенная группа атомов заменен(а) на другой атом или группу атомов.
Как его используют здесь, термин "производное цистеина" относится к соединениям, в которых атом водорода или определенная группа атомов в молекуле цистеина замещена другим атомом или группой атомов. Например, производные цистеина могут находиться в форме, в которой к атому азота аминогруппы (-NH2) или атому серы тиольной группы (-SH) в молекуле цистеина присоединен(а) другой атом или группа атомов, и примеры производных цистеина могут включать NAC (N-ацетилцистеин), SCMC (S-карбоксиметилцистеин), Boc-Cys(Me)-OH, (R)-S-(2-амино-2-карбоксиэтил)-L-гомоцистеин, (R)-2-амино-3-сульфопропионовую кислоту, D-2-амино-4-(этилтио)масляную кислоту, 3-сульфино-L-аланин, Fmoc-Cys(Boc-метил)-OH, селено-L-цистеин, S-(2-тиазолил)-L-цистеин, S-(2-тиенил)-L-цистеин, S-(4-толил)-L-цистеин и так далее, но не ограничиваясь ими.
При условии, что цистеин получен в соответствии со способом по настоящему изобретению, превращение в производные цистеина можно легко осуществить путем преобразования в различные производные цистеина способом, хорошо известным в данной области техники.
В частности, способ получения производных цистеина может дополнительно включать превращение цистеина, полученного на стадии (б), в производное цистеина. Например, из цистеина можно синтезировать N-ацтилцистеин (NAC) путем проведения реакции с ацетилирующим агентом или из него можно синтезировать S-карбоксиметилцистеин (SCMC) путем проведения реакции с галогенуксусной кислотой в основных условиях, но не ограничиваясь этим.
Эти производные цистеина используют главным образом в качестве фармацевтических веществ для противокашлевых препаратов; препаратов, облегчающих кашель, и терапевтических агентов для лечения бронхита, бронхиальной астмы, ларингофарингита и так далее, но не ограничиваясь этим.
Как его используют здесь, термин "О-фосфосерин-сульфгидрилаза (OPSS)" относится к ферменту, который катализирует реакцию, посредством которой OPS при участии тиольной группы (группа SH) превращается в цистеин. Возможно, этот фермент был впервые обнаружен у Aeropyrum pernix, Mycobacterium tuberculosis, Mycobacterium smegmatics и Trichomonas vaginalis (Mino K. and Ishikawa K., FEBS Letters, 551:133-138, 2003; Bums К. E. et al, J. Am. Chem. Soc, 127:11602-11603, 2005). Дополнительно, OPSS может включать не только белки OPSS дикого типа, но также варианты белков, включающие делецию, замену или добавление в части полинуклеотидной последовательности, кодирующей OPSS, демонстрирующие активность, равную или более высокую, чем биологическая активность белков OPSS дикого типа, и также может включать все белки OPSS, раскрытые в US 2012-0190081 A1 и US 9127324 В2, и их варианты.
Сульфид для использования по настоящему изобретению может представлять собой любой сульфид, предоставленный не только в твердой форме, обычно используемой в данной области техники, но также в жидкой или газообразной форме вследствие различия в рН, давлении и растворимости, и следовательно может быть превращен в тиольную (SH) группу, в форме, например, сульфида (S2-) или тиосульфата (S2O32-) без ограничения. В частности, сульфид может включать Na2S, NaSH, H2S, (NH4)2S, NaSH и Na2S2O3, которые могут предоставлять тиольную группу для OPS, но не ограничиваясь этим. В этой реакции необходима одна тиольная группа на одну активную группу OPS для получения одной молекулы цистеина или его производного. В этой реакции сульфид добавляют, в частности, в количестве от 0,1 до 3 молярных эквивалентов, и в частности от 1 до 2 молярных эквивалентов на основе молярной концентрации OPS, но не ограничиваясь этим.
Еще в одном аспекте настоящего изобретения предложен способ получения OPS-продуцирующего микроорганизма, включающий модификацию микроорганизма рода Escherichia для повышения активности NADH:хинон-оксидоредуктазы.
Еще в одном аспекте настоящего изобретения предложено применение микроорганизма, обладающего повышенной активностью NADH:хинон-оксидоредуктазы, для получения О-фосфосерина, цистеина или их производных.
NADH:хинон-оксидоредуктаза, повышение ее активности, микроорганизмы и так далее являются такими, как описано выше.
Примеры
Настоящее изобретение будет описано более подробно посредством Примеров. Однако, специалисту в области техники, к который принадлежит настоящее изобретение, очевидно, что эти Примеры предназначены исключительно для иллюстративных целей, и не предназначены каким-либо образом ограничивать объем настоящего изобретения.
Пример 1: Оценка OPS-продуцирующей способности штамма с мутированной областью регуляции экспрессии Nuo
В результате более раннего исследования было обнаружено, что средние уровни транскрипции генов nuoA, rmf и idi у OPS-продуцирующего штамма-хозяина составляют 8986, 32205 и 631, соответственно, и конкретные значения для каждого типа культуры представлены в Таблице 1.

Было подтверждено, что среднее значение уровня транскрипции rmf было в 3,6 раза больше этого значения для nuoA, и среднее значение уровня транскрипции idi составляло 0,07-ую часть этого значения для nuoA. Эти результаты показывают, что промотор гена rmf представляет собой сильный промотор, и промотор гена idi представляет собой слабый промотор по сравнению с промотором nuo А.
Следовательно, сравнивали OPS-продуцирующую способность микроорганизмов при регуляции экспрессии NADH:хинон-оксидоредуктазы путем вставки промотора rmf и промотора idi с различной активностью в оперон nuo (SEQ ID NO: 1) у OPS-продуцирующего микроорганизма.
1-1: Конструирование плазмиды для усиления области регуляции экспрессии Nuo
Фрагменты гена в области выше по ходу транскрипции от промотора дикого типа гена nuo получали с помощью пары праймеров, имеющих нуклеотидные последовательности SEQ ID NO: 14 и 15, и фрагменты гена в области ниже по ходу транскрипции от этого промотора получали с помощью пары праймеров, имеющих нуклеотидные последовательности SEQ ID NO: 18 и 19, на основе хромосомной ДНК Е. coli АТСС27325 в качестве матрицы. Дополнительно, промоторную область гена rmf получали с помощью пары праймеров, имеющих нуклеотидные последовательности SEQ ID NO: 16 и 17, на основе хромосомной ДНК.Е. coli АТСС27325 в качестве матрицы.
Для получения этих фрагментов проводили ПЦР с использованием ДНК-полимеразы Pfu-X Solg™, и ПЦР проводили в следующих условиях ПЦР-амлификации: денатурация при 95°С в течение 2 минут с последующими 30 циклами денатурации при 95°С в течение 30 секунд, отжиг при 60°С в течение 30 секунд и полимеризация при 72°С в течение 60 секунд и затем полимеризация при 72°С в течение 5 минут.
Фрагменты выше по ходу транскрипции и фрагменты ниже по ходу транскрипции от промотора nuo и фрагменты промотора rmf, полученные в вышеизложенном процессе, клонировали в вектор pSKH130 (SEQ ID NO: 39, Публикация США No. 2020-0048619) для хромосомной трансформации, расщепленный ферментом рестрикции EcoRV, используя набор для клонирования "in-fusion" (Clontech Laboratories, Inc.) с получением рекомбинантной плазмиды, названной SKH130 Prmf-nuoA.
1-2: Конструирование плазмиды для ослабления области регуляции экспрессии Nuo
Фрагменты гена в области выше по ходу транскрипции промотора дикого типа гена nuo получали с помощью пары праймеров, имеющих нуклеотидные последовательности SEQ ID NO: 14 и 20, и фрагменты гена в области ниже по ходу транскрипции от этого промотора получали с помощью пары праймеров, имеющих нуклеотидные последовательности SEQ ID NO: 23 и 19, на основе хромосомной ДНК Е. coli АТСС27325 в качестве матрицы. Дополнительно, область промотора гена idi получали с помощью пары праймеров, имеющих нуклеотидные последовательности SEQ ID NO: 21 и 22, на основе хромосомной ДНК£". coli АТСС27325 в качестве матрицы.
ПЦР проводили в таких же условиях как в Примере 1-1 для получения фрагментов, и клонирование выполняли таким же образом как в Примере 1-1 с использованием этих фрагментов с получением рекомбинантной плазмиды. Полученная конструкция получила название pSKH130_Pidi-nuoA.
Последовательности праймеров, используемых в Примерах 1-1 и 1-2, показаны в Таблице 2.


1-3: Конструирование штамма, мутантного по области регуляции экспрессии Nuo
pSKH130 Prmf-nuoA, полученной в Примере 1-1, трансформировали СА07-0012 (КССМ 11212Р, Публикация США No. 2012-0190081), обладающий OPS-продуцирующей способностью, методом электропорации (Appl. Microbiol. Biotechnol. (1999) 52:541-545) и проводили вторичный кроссинговер, таким образом получая штамм СА07-4826, в который вставлена нуклеотидная последовательность промотора гена rmf на конце нуклеотидной последовательности промотора дикого типа гена nuo.
В частности, вектор pSKH130 содержал репликон R6K, зависимый от белка PI (ген pir), ген SacB (Levansucrase) и ген устойчивости к канамицину. После получения желаемого штамма с использованием R6K и канамицина при первом кроссинговере антибиотики удаляли из среды, содержащей сахарозу, для получения штамма.
Вставку нуклеотидной последовательности промотора rmf у СА07-4826 подтверждали методом ПЦР и геномного секвенирования с использованием пары праймеров SEQ ID NO: 24 и 25.
Таким же образом, pSKH130 Pidi-nuoA, полученной в Примере 1-2, трансформировали методом электропорации (Appl. Microbiol. Biotechnol. (1999) 52:541-545) и проводили вторичный кроссинговер, таким образом получая штамм СА07-4827, у которого нуклеотидная последовательность промотора гена idi вставлена на конце нуклеотидной последовательности промотора дикого типа гена nuo. Вставку нуклеотидной последовательности промотора idi у СА07-4827 подтверждали путем ПЦР и геномного секвенирования с использованием пары праймеров SEQ ID NO: 24 и 25. Последовательности используемых праймеров показаны в Таблице 3.
1-4: Сравнение OPS-продгцирующей способности штаммов с усиленным и ослабленным опероном nuo с использованием среды для титрования
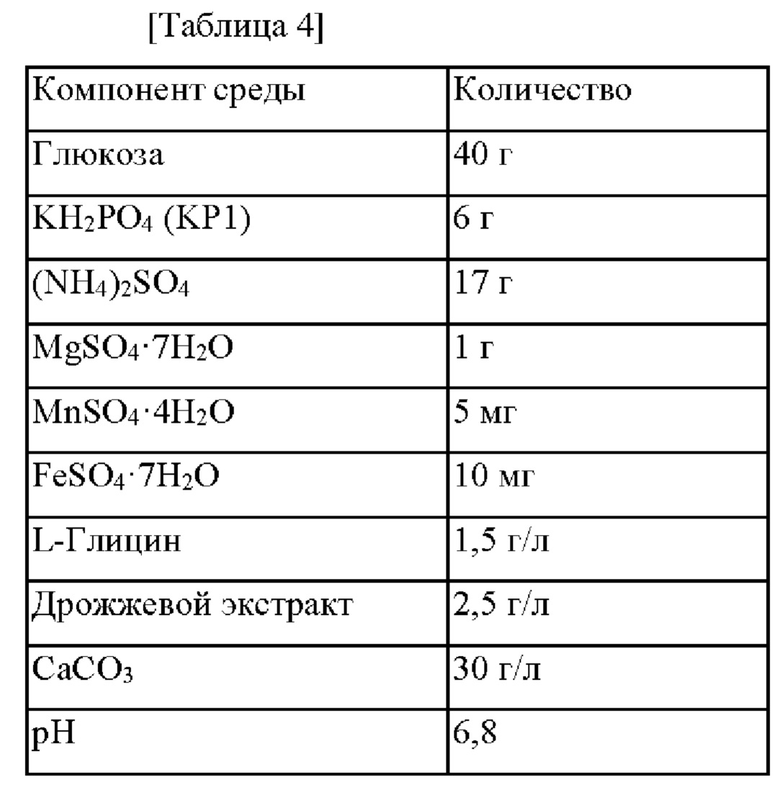
Для оценки OPS-продуцирующей способности двух штаммов, СА07-4826 и СА07-4827, полученных в Примере 1-3, и контрольного штамма СА07-0012 использовали следующую среду (Таблица 4).
В частности, каждый из штаммов высевали на твердую среду LB и культивировали в инкубаторе при 33°С в течение ночи. Штаммы, культивированные на твердой среде LB в течение ночи, инокулировали в 25 мл среды для титрования, показанной в Таблице 3 ниже, и затем культивировали в инкубаторе при скорости 200 об/мин в течение 48 часов при 33°С. Концентрация полученного OPS представлена в Таблице 5.
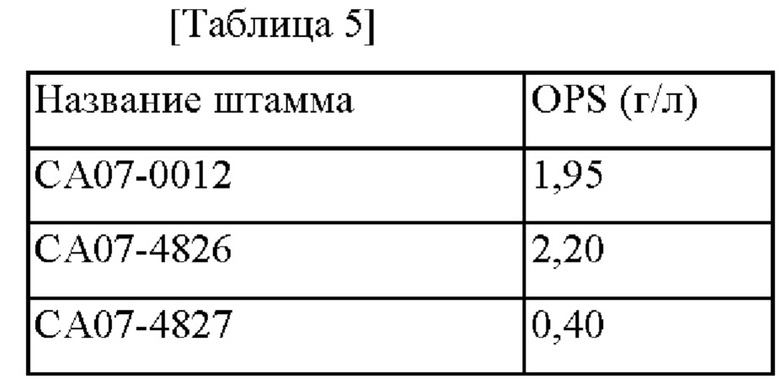
СА07-4826, у которого оперон nuo был усилен промотором rmf, демонстрировал OPS-продуцирующую способность, улучшенную примерно на 12,8% по сравнению с родительским штаммом, и СА07-4827, у которого оперон nuo был ослаблен промотором idi, демонстрировал OPS-продуцирующую способность, пониженную примерно на 79,5% по сравнению с родительским штаммом.
Пример 2: Оценка OPS-продуцирующей способности в соответствии с усилением оперона nuo у штамма с повышенной OPS-продуцирующей способностью
Было проведено исследование того, происходит ли дальнейшее повышение OPS-продуцирующей способности, когда оперон Nuo усилен у штамма с повышенной OPS-экспортирующей способностью. Для этого OPS-продуцирующую способность оценивали при дополнительном усилении оперона nuo у штамма, усиленного белком YhhS (SEQ ID NO: 4, Патент США No. 10323262 В2), обладающего OPS-экспортирующей способностью.
2-1: Конструирование плазмиды для усиления YhhS у OPS-продуцирующего штамма
Фрагменты гена в области выше по ходу транскрипции от промотора дикого типа гена yhhS получали с помощью пары праймеров, имеющих нуклеотидные последовательности SEQ ID NO: 6 и 7, и фрагменты гена в области ниже по ходу транскрипции от промотора дикого типа гена yhhS получали с помощью пары праймеров, имеющих нуклеотидные последовательности SEQ ID NO: 8 и 9, на основе хромосомной ДНК Е. coli АТСС27325 в качестве матрицы. Дополнительно промотор trc (Ptrc) получали с помощью пары праймеров SEQ ID NO: 8 и 9 на основе pCL_Ptrc-gfp(US 2017-0247727 Al) в качестве матрицы. Последовательности используемых праймеров представлены в Таблице 6 ниже.

Для получения этих фрагментов проводили ПЦР с использованием ДНК полимеразы Pfu-X Solg™, и ПЦР проводили в следующих условиях ПЦР-амлификации: денатурация при 95°С в течение 2 минут с последующими 30 циклами денатурации при 95°С в течение 30 секунд, отжига при 60°С в течение 30 секунд и полимеризации при 72°С в течение 60 секунд и затем полимеризация при 72°С в течение 5 минут.
Фрагменты выше по ходу транскрипции и ниже по ходу транскрипции от промотора yhhS и фрагмент промотора trc, полученные в вышеизложенном процессе, клонировали в вектор для хромосомной трансформации, расщепленный ферментом рестрикции EcoRV, с использованием набора для клонирования "in-fusion" (Clontech Laboratories, Inc.) для получения рекомбинантной плазмиды, и она получила название PSKH130_Ptrc-yhhS.
2-2: Конструирование штамма, усиленного по YhhS
pSKH130_Ptrc-yhhS, полученной в Примере 2-1, трансформировали СА07-0012 методом электропорации (Appl. Microbiol. Biotechnol. (1999) 52:541-545) и проводили вторичный кроссинговер, таким образом получая штамм СА07-4821, у которого нуклеотидная последовательность промотора trc вставлена на конце нуклеотидной последовательности промотора дикого типа гена yhhS. Вставку нуклеотидной последовательности промотора trc у штамма СА07-4821 подтверждали методом ПЦР и геномного секвенирования с использованием пары праймеров SEQ ID NO: 12 и 13 (Таблица 7).

2-3: Оценка OPS-продуцирующей способности штамма, усиленного по YhhS, с использованием среды для титрования
Для оценки OPS-продуцирующей способности штамма СА07-4821, полученного в Примере 2-2, и его родительского штамма, СА07-0012, оценку проводили таким же образом как в Примере 1-4 с использованием среды (Таблица 4).
В результате было подтверждено, что СА07-4821 демонстрирует OPS-продуцирующую способность, улучшенную примерно на 26,7% по сравнению с СА07-0012, и результаты представлены в Таблице 8.

2-4: Конструирование штамма с усиленными YhhS и опероном nuo
pSKH130_Prmf-nuoA, полученным в Примере 1-1, трансформировали СА07-4821, полученный в Примере 2-3, методом электропорации (Appl. Microbiol. Biotechnol. (1999) 52:541 545) и проводили вторичный кроссинговер, таким образом получая штамм СА07-4828, у которого нуклеотидная последовательность промотора гена rmf вставлена на конце нуклеотидной последовательности промотора дикого типа гена nuo.
Вставку нуклеотидной последовательности промотора rmf у СА07-4828 подтверждали с помощью ПЦР и геномного секвенирования с использованием пары праймеров SEQ ID NO: 24 и 25.
2-5: Оценка OPS-продуцирующей способности штамма с усиленными Vhhs и опероном nuo с использованием среды для титрования
Для оценки OPS-продуцирующей способности СА07-4828 оценку проводили таким же образом как в Примере 1-4 с использованием СА07-4821 в качестве контроля, и результаты показаны в Таблице 9.
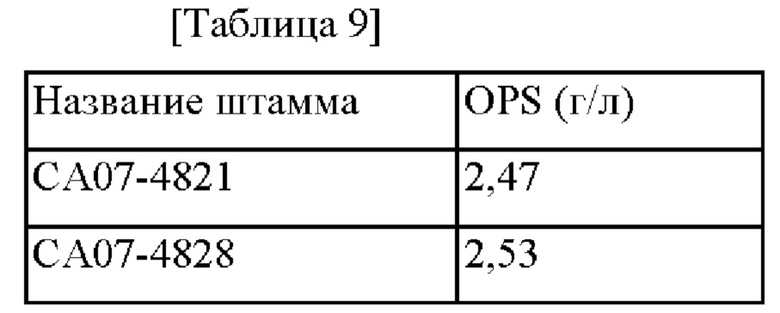
В случае СА07-4828, когда YhhS и оперон nuo одновременно усилены, OPS-продуцирующая способность была усилена примерно на 2,4% по сравнению с СА07-4821, у которого усилен только YhhS. Следовательно, было подтверждено, что усиление оперона nuo дополнительно увеличивает OPS-продуцирующую способность даже у штамма с повышенной OPS-продуцирующей способностью.
На основании вышеизложенного специалист в области техники, к которой принадлежит настоящее изобретение, понимает, что настоящее изобретение может быть воплощено в других определенных формах без изменения технических идей или существенных характеристик настоящего изобретения. В этом отношении раскрытые здесь примеры воплощений предназначены исключительно для иллюстративных целей, и их не следует рассматривать как ограничивающие объем настоящего изобретения. Напротив, объем настоящего изобретения охватывает не только примеры воплощений, но также различные альтернативы, модификации, эквиваленты и другие воплощения, которые могут быть включены в рамках основной идеи и объема настоящего изобретения, как определено прилагаемой формулой изобретения.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
--->
<110> CJ CheilJedang Corporation
<120> РЕКОМБИНАНТНЫЙ МИКРООРГАНИЗМ С КОНТРОЛИРУЕМОЙ ЭКСПРЕССИЕЙ
NADH:ХИНОН-ОКСИДОРЕДУКТАЗЫ И СПОСОБ ПОЛУЧЕНИЯ O-ФОСФОСЕРИНА, ЦИСТЕИНА
И ЕГО ПРОИЗВОДНОГО С ИСПОЛЬЗОВАНИЕМ ЭТОГО МИКРООРГАНИЗМА
<130> OPA22034
<150> KR 10-2021-0081785
<151> 2021-06-23
<160> 39
<170> KoPatentIn 3.0
<210> 1
<211> 15655
<212> DNA
<213> Artificial Sequence
<220>
<223> оперон nuo
<220>
<221> misc_feature
<222> (1)..(630)
<223> промотор nuoA
<220>
<221> misc_feature
<222> (631)..(633)
<223> инициирующий кодон nuoA
<220>
<221> misc_feature
<222> (1090)..(1092)
<223> инициирующий кодон nuoB
<220>
<221> misc_feature
<222> (1846)..(1848)
<223> инициирующий кодон nuoC
<220>
<221> misc_feature
<222> (3651)..(3653)
<223> инициирующий кодон nuoE
<220>
<221> misc_feature
<222> (4148)..(4150)
<223> инициирующий кодон nuoF
<220>
<221> misc_feature
<222> (5532)..(5534)
<223> инициирующий кодон nuoG
<220>
<221> misc_feature
<222> (8261)..(8263)
<223> инициирующий кодон nuoH
<220>
<221> misc_feature
<222> (9253)..(9255)
<223> nuoI инициирующий кодон
<220>
<221> misc_feature
<222> (9807)..(9809)
<223> nuoJ инициирующий кодон
<220>
<221> misc_feature
<222> (10358)..(10360)
<223> инициирующий кодон nuoK
<220>
<221> misc_feature
<222> (10657)..(10659)
<223> инициирующий кодон nuoL
<220>
<221> misc_feature
<222> (12662)..(12664)
<223> инициирующий кодон nuoM
<220>
<221> misc_feature
<222> (14378)..(14380)
<223> инициирующий кодон nuoN
<400> 1
tcgacggacg atagataatt cctgagacaa tagtgtaaaa aacgagccgc tgggggtgtt 60
ttaaacccca gcggcttttt tttagtaaaa atccttcatc gcatagtgca gaacgtacaa 120
aactgttcat ttttcaacca ccagagattc acgtcccgaa cgcacaaata atcgcctgaa 180
aaaaatcacc tgctacaatg atgttaaaaa atacgtaaat ttgttgctgc gtttttgtta 240
tggaatgaca aaagcgtgtc acagatcaag aaaatactct cttttggggg gaggaatcga 300
cacaaattcc tgtcaaaata gacccgtatt ttttccattg cttcacaacg gacacgattc 360
aacaacatct aattatcctg gagtcgtcaa ggatctgggg tgaaagggca ttaaatgcta 420
atggtgttga tattatgtaa actaatgtga agaaactttt gttaaagttg acaaaaggtt 480
atagaaagga gtaaaaaacc acatcaatta gctgtataaa agaatttcta cagtgattgt 540
aaggtttttt ttattcctcc ccatgaatcg atgtggcgtc catctgccgt gaagagcagt 600
gaatctggcg ctacttttga tgagtaagca atgagtatgt caacatccac tgaagtcatc 660
gctcatcact gggcattcgc tatctttctt atcgttgcca ttggcctgtg ttgcctgatg 720
ctggtaggcg gttggttttt aggcggtcgc gcacgcgcga ggtcgaaaaa cgtgccgttt 780
gaatccggta tcgactcggt cggctccgcc cgcttacgcc tgtccgccaa gttttatctg 840
gtggccatgt tcttcgttat cttcgacgtt gaagcgctgt atctgttcgc atggtcaacc 900
tctatccgcg aaagcggctg ggtaggcttt gtggaagctg caatttttat ttttgtgtta 960
ctggcaggtc tggtttatct ggtgcgtatt ggcgcgctgg actggacgcc cgcgcgttca 1020
cgccgcgagc gtatgaaccc ggaaacgaac agtatcgcta atcgtcaacg ctaaccgcga 1080
ggcattaaga tggattatac gctcacccgc atagatccca acggtgagaa cgaccgttac 1140
cccctgcaaa agcaggagat cgtaaccgac cctctggagc aagaagttaa caaaaacgtg 1200
tttatgggca agctcaatga catggttaac tggggtcgta aaaactcaat ttggccgtat 1260
aacttcggtc tttcctgctg ttacgttgag atggtgactt cgtttaccgc ggtgcatgac 1320
gtggcgcgtt ttggcgcaga agtattgcgt gcttcgccgc gtcaggctga cctgatggtg 1380
gttgcaggaa cctgctttac caaaatggca ccggttattc agcgtctgta tgaccagatg 1440
ctggaaccaa aatgggttat ctcaatgggt gcctgtgcca actctggtgg tatgtacgat 1500
atttattccg ttgtgcaggg cgtcgataaa ttcatcccgg ttgatgtgta tatcccgggc 1560
tgcccgccgc gtcctgaagc gtacatgcag gcactgatgc tgttgcagga atctatcggc 1620
aaagaacgtc gtccgctctc ctgggtggtt ggcgatcagg gcgtttatcg cgccaatatg 1680
caatcagagc gcgaacgcaa gcgcggtgaa cgcattgccg taactaacct gcgtacacct 1740
gacgagattt aatttgcgcc tgtcggcaaa gggatttttc ttcgcttatt cctaaatcta 1800
tttcgcgaag cttactgcgc cgacagtcac cacggaccat ttgcaatggt gaacaatatg 1860
accgacttaa ccgcgcaaga acccgcctgg cagacccgcg atcatcttga tgatccggtg 1920
attggcgaac tgcgcaaccg ttttgggccg gatgccttta ctgttcaggc gactcgcacc 1980
ggggttcccg ttgtgtggat caagcgtgaa caattactgg aagttggcga tttcttaaag 2040
aaactgccga aaccttacgt catgctgttt gacttacacg gcatggacga acgtctgcgc 2100
acacaccgcg aagggttacc tgccgcggat ttttccgttt tctaccatct gatttctatc 2160
gatcgtaacc gcgacatcat gctgaaggtg gcgctggcag aaaacgacct gcacgtaccg 2220
accttcacca aactgttccc gaacgctaac tggtatgagc gtgaaacctg ggatctgttt 2280
ggcattactt tcgacggtca cccgaacctg cgacgcatca tgatgccgca aacctggaaa 2340
ggtcacccgc tgcgtaaaga ttatccggcg cgcgctaccg aattctcgcc gtttgagctg 2400
accaaagcca aacaggatct ggagatggaa gccctgacct tcaaaccgga agagtggggg 2460
atgaagcgcg gcaccgaaaa cgaggacttc atgttcctca acctcggtcc gaaccacccg 2520
tcggcgcacg gggctttccg tatcgttttg caactcgatg gcgaagagat tgtcgactgc 2580
gtaccagaca tcggttacca ccaccgtggt gcggagaaaa tgggcgaacg ccagtcctgg 2640
cacagctaca ttccgtatac tgaccgtatc gaatacctcg gcggctgcgt taacgaaatg 2700
ccttacgtgc tggcggtaga gaaactggcc gggatcaccg tgccggatcg cgttaacgtc 2760
attcgcgtta tgctctccga actgttccgc atcaacagtc acctgctgta tatctcgacc 2820
tttattcagg acgtcggcgc aatgacgcca gtgttcttcg cctttaccga tcgtcagaaa 2880
atttacgatc tggtggaagc aatcactggt ttccgtatgc acccggcgtg gttccgtatt 2940
ggcggcgtag cgcacgacct gccgcgcggc tgggatcgcc tgctgcgtga gttcctcgac 3000
tggatgccga aacgtctggc gtcttacgag aaagcggcgc tgcaaaacac cattctgaaa 3060
ggtcgttccc agggcgttgc cgcctatggc gcgaaagagg cgctggagtg gggcaccact 3120
ggcgcgggcc tgcgtgctac cgggatcgac ttcgacgtgc gtaaggcgcg tccttattct 3180
ggctatgaaa acttcgactt tgaaatcccg gtgggtggtg gcgtttctga ctgctacacc 3240
cgcgtaatgc ttaaagtgga agagctgcgc cagagtctgc gcattcttga gcagtgcctc 3300
aacaacatgc cggaaggccc gttcaaagcg gatcacccgc tgaccacgcc gccgccgaaa 3360
gagcgcacgc tgcaacatat cgaaaccctg atcacccact tcctgcaagt gtcgtggggt 3420
ccggtgatgc ctgccaatga atctttccag atgattgagg cgaccaaagg gatcaacagt 3480
tactacctga ccagcgacgg cagcaccatg agttaccgca cccgtgttcg taccccgagc 3540
tttgcgcatt tgcagcaaat tccggcggcg atccgcggca gcctggtgtc tgacctgatt 3600
gtttatctgg gcagtatcga ttttgttatg tcagatgtgg accgctaatt atgcacgaga 3660
atcaacaacc acaaaccgag gcttttgagc tgagtgcggc agagcgtgaa gcgatcgagc 3720
acgagatgca ccactacgaa gacccgcgtg cggcgtccat tgaagcgctg aaaatcgttc 3780
agaagcagcg tggctgggtg ccggatggtg cgatccacgc gatcgccgat gtgctgggta 3840
ttccggcaag cgacgtcgaa ggtgtggcaa cgttctacag tcagatcttc cgccagccgg 3900
ttggtcgcca tgtgatccgt tattgtgaca gcgtggtctg tcatatcaac ggttatcagg 3960
gtattcaggc ggcgctcgag aaaaagctga acatcaaacc agggcaaacg acatttgatg 4020
gccgctttac gctgctgcca acttgctgcc tggggaactg tgataaaggg ccaaacatga 4080
tgatcgatga ggacactcac gcgcatctga ccccggaagc gatccctgaa ctgctggagc 4140
ggtataaatg aaaaacatta tccgtactcc cgaaacgcat ccgctgacct ggcgtctgcg 4200
cgatgacaaa cagccagtgt ggctggacga ataccgcagc aaaaacggtt acgaaggcgc 4260
gcgtaaggcg ctgaccgggc tgtctccgga cgaaatcgtt aatcaggtaa aagacgctgg 4320
tctgaaaggg cgcggcggcg cgggcttctc gactggcctg aaatggagcc tgatgccgaa 4380
agacgaatcc atgaacatcc gttacctgct gtgtaatgcc gatgaaatgg agccgggcac 4440
ctataaagac cgcctgttga tggagcaact gccgcacctg ctggtggaag gtatgctcat 4500
ctccgcgttt gcgctgaaag cttaccgtgg ctacatcttc ctgcgtggcg aatatatcga 4560
agcggcagtt aatctgcgcc gtgccattgc cgaagccacc gaagcgggtc tgcttggcaa 4620
aaacattatg ggaacaggtt tcgatttcga actgttcgtc cataccgggg cagggcgcta 4680
catctgcggg gaagaaacag cgttaatcaa ctccctggaa ggacgtcgtg ctaacccacg 4740
ctcgaagcca cccttcccgg caacctccgg cgcatggggt aaaccgacct gtgtcaacaa 4800
cgtcgaaacc ctgtgtaacg ttccggcgat cctcgctaac ggcgtggagt ggtatcagaa 4860
catctcgaaa agtaaagatg ctggcaccaa gctgatgggc ttctccggtc gggtgaaaaa 4920
tccgggactg tgggaactgc cgttcggcac caccgcacgc gagatcctcg aagattacgc 4980
cggtggtatg cgtgatggtc tgaaatttaa agcctggcag ccaggcggcg cggggactga 5040
cttcctgacc gaagcgcacc ttgatctgcc gatggaattc gaaagtatcg gtaaagcggg 5100
cagccgtctg ggtacggcgc tggcgatggc ggttgaccat gagatcaaca tggtgtcgct 5160
ggtgcgtaac ctggaagagt ttttcgcccg tgagtcctgc ggctggtgta cgccgtgccg 5220
cgacggtctg ccgtggagcg tgaaaattct gcgtgcgctg gagcgtggtg aaggtcagcc 5280
gggcgatatc gaaacacttg agcaactgtg tcgattctta ggcccgggta aaactttctg 5340
tgcccacgca cctggtgcag tggagccgtt acagagcgcc atcaaatatt tccgcgaaga 5400
atttgaggcg ggaatcaaac agccgttcag caatacccat ttgattaatg ggattcagcc 5460
gaacctgctg aaagagcgct ggtaaccgaa tttcgattaa cgctcagtct ctgactgaga 5520
aaactggaag catgctaatg gctacaattc atgtagacgg caaagaatac gaggtcaacg 5580
gagcggacaa cctgctggaa gcttgtctgt ctctgggcct tgatattcct tacttttgct 5640
ggcatccggc gctgggaagt gtcggtgctt gccgccagtg tgcggtgaag caataccaaa 5700
acgcggaaga cacgcgtggt cgcctggtga tgtcctgtat gacaccggct tccgatggca 5760
cctttatttc cattgacgac gaagaagcga aacagttccg tgaaagcgtg gtcgagtggt 5820
tgatgaccaa ccacccgcac gactgtccgg tatgtgaaga gggcggtaac tgccatcttc 5880
aggatatgac tgtgatgacc ggacacagct tccgtcgcta ccgtttcacc aaacgtaccc 5940
accgtaatca ggatttgggg ccattcatct ctcacgaaat gaaccgctgc atcgcctgct 6000
accgctgtgt gcgttactac aaagattacg ctgacggtac agatctgggc gtttacggtg 6060
cgcacgacaa cgtctacttc ggtcgcccgg aagacggcac gctggaaagc gaattttccg 6120
gtaacctggt cgaaatttgc ccgaccggcg tatttaccga caaaacgcac tccgagcgtt 6180
acaaccgtaa atgggatatg cagtttgcgc cgagcatctg ccagcaatgt tccatcggct 6240
gtaacatcag ccccggtgaa cgttacggcg aactgcgtcg tatcgaaaac cgttacaacg 6300
gtacggtaaa ccactacttc ctctgcgacc gtggtcgttt cggttacggt tacgtcaacc 6360
tgaaggatcg tccgcgtcag ccagtacagc gtcgtggcga tgatttcatt accctcaacg 6420
ccgaacaggc aatgcagggc gcggcagata ttctgcgtca gtcgaagaaa gtgatcggta 6480
ttggttctcc gcgtgccagc gtggaaagca actttgcgct gcgtgaactg gtgggcgaag 6540
aaaacttcta caccggtatc gctcacggtg agcaggaacg tctgcaactg gcgctgaaag 6600
tgctgcgtga aggcggcatt tatactccgg ctctgcgcga aatcgaatct tacgatgcgg 6660
tactggtgct gggcgaagac gttacccaga ccggcgcgcg cgtcgcgctg gcagtgcgtc 6720
aggctgtgaa aggtaaagcg cgcgaaatgg cggcagcaca gaaagtggct gactggcaga 6780
ttgcggcaat cctcaacatc ggtcaacgtg cgaagcatcc gctgtttgtt accaacgttg 6840
atgacacccg tctggatgat atcgcggcgt ggacttaccg cgcaccggtt gaagatcagg 6900
cgcgtttagg ttttgccatc gcccatgcgc tggataactc tgcaccagcg gttgacggta 6960
tcgaacctga gctgcaaagc aaaatcgacg tcatcgtgca ggcactggca ggtgcgaaga 7020
aaccgttgat tatctccggg acgaacgccg gtagcttaga ggtgattcag gcggcggcta 7080
acgtcgcgaa agccctgaaa ggtcgcggcg ctgacgtcgg tatcaccatg attgcccgtt 7140
ccgtcaacag catggggctg ggcattatgg gtggcggttc gcttgaagaa gcgttaaccg 7200
aactggaaac cggacgcgcc gacgcggtgg tggtgttgga aaacgatctg catcgtcacg 7260
cttctgctat ccgcgtgaat gctgcgctgg ctaaagcacc gctggtgatg gtggttgatc 7320
atcaacgcac agcgattatg gaaaacgccc atctggtact ttctgctgcc agctttgctg 7380
aaagcgacgg tacggtgatc aacaacgaag gccgcgccca acgtttcttc caggtttacg 7440
atcctgctta ttacgacagc aaaactgtca tgctggaaag ctggcgctgg ttacactcgc 7500
tgcacagcac cctgctgagc cgtgaagtgg actggacgca gctcgaccat gtgattgacg 7560
ctgttgtggc gaaaatcccg gaactggcag gtatcaaaga tgctgcgccg gatgcgacat 7620
tccgtattcg tgggcagaaa ctggcccgtg aaccgcaccg ttacagcggt cgtaccgcca 7680
tgcgcgccaa tatcagcgtt catgagccgc gtcagccgca ggatattgac accatgttca 7740
ccttctcgat ggaaggtaac aaccagccga ctgcgcaccg ttcgcaagtg ccgtttgcct 7800
gggcgccggg ctggaactcc ccgcaggcgt ggaacaaatt ccaggacgaa gtgggcggca 7860
aactgcgctt tggcgatccg ggcgtgcgtc tgtttgaaac cagcgaaaat ggtctggatt 7920
acttcaccag cgtaccggca cgcttccagc cgcaggacgg gaaatggcgt atcgcgccgt 7980
attaccacct gtttggcagc gatgaattgt cacagcgtgc tccggtcttc cagagccgta 8040
tgccgcagcc gtacatcaaa ctcaacccag cggatgccgc gaagttgggt gtgaacgcag 8100
gtacacgcgt ctcctttagt tacgatggca acacggtcac gctgccggtt gaaatcgccg 8160
aaggactgac ggcagggcag gtgggcttgc cgatgggtat gtccggcatt gctccggtgc 8220
tggctggcgc gcatcttgag gatctcaagg aggcacaaca atgagttgga tatcaccgga 8280
actgattgag atcctgctga ccatcctcaa agcggtggtg atcctgctgg tggttgtcac 8340
ctgcggggca ttcatgagct ttggcgaacg tcgcctgctg ggtctgttcc agaaccgtta 8400
cggacctaac cgtgttggct ggggcggttc gctccagctg gttgcggaca tgatcaaaat 8460
gttctttaaa gaagactgga tcccgaaatt ctcggatcgc gtcatcttta ccctggcacc 8520
gatgattgcc tttacctcgc tgctgctggc ctttgcgatt gtgccagtca gtccgggttg 8580
ggtggttgcc gacctgaaca tcgggatttt gttcttcctg atgatggcag gtctggcggt 8640
ttacgcggtg ctgtttgcgg gctggtcaag taacaacaaa tactcgttgc tgggtgcgat 8700
gcgtgcttct gcgcagaccc tgagctacga agtgttcctc gggctttcct tgatgggcgt 8760
ggtggcgcag gccggttcat tcaacatgac cgacatcgtc aacagccagg cgcatgtgtg 8820
gaacgttatc ccgcaattct ttggttttat tacctttgcc atcgcgggcg tggcggtatg 8880
tcaccgtcac ccgtttgacc agccggaagc cgagcaggaa ctggcggatg gttaccacat 8940
tgaatattcc ggtatgaagt tcggtctgtt cttcgtgggt gagtacatcg ggattgtgac 9000
catctctgca ttgatggtga cgctgttctt cggtggctgg caaggcccgt tgttaccgcc 9060
attcatctgg ttcgcgctga aaaccgcgtt ctttatgatg atgttcattt tgattcgtgc 9120
gtcgttaccg cgtccgcgtt atgaccaggt aatgtccttc ggctggaaaa tctgcctgcc 9180
gctgacgctg atcaacttgc tggtaacggc ggctgtcatt ctctggcagg cgcaataagg 9240
ggcaataaga ccatgacctt aaaagaattg ttagtaggtt tcggcaccca ggttcgtagt 9300
atctggatga tcggcctgca cgcgttcgcc aaacgcgaaa cgcgaatgta cccggaagag 9360
ccggtctatc tgccgccccg ttatcgtggt cgtatcgttc tgacccgcga cccggacggc 9420
gaagagcgtt gcgtagcctg taacctctgc gcggtagcct gcccggtcgg ctgtatctcg 9480
ctgcaaaaag cagaaaccaa agacggtcgc tggtacccgg aatttttccg catcaacttc 9540
tcacgctgca ttttctgtgg tctgtgcgaa gaagcctgtc cgaccacggc gattcagtta 9600
accccggatt tcgaaatggg ggaatacaag cgccaggatc tggtttacga gaaagaggat 9660
ctgctgatct ccggtccggg caaatacccg gaatataact tctaccggat ggcaggtatg 9720
gcaatcgacg gcaaagataa gggcgaagca gagaacgaag ccaagcctat cgacgtcaag 9780
agcctgttac cgtaaggaga ggggcaatgg agttcgcttt ttatatctgt ggcctgatag 9840
ccatacttgc gaccttgcga gtgatcaccc ataccaatcc ggtacacgca ctgctgtacc 9900
tgattatttc gctgctggcg atttccgggg tgttcttctc actgggcgct tacttcgccg 9960
gtgcgctgga aattatcgtc tacgcgggtg ccattatggt gctgttcgtg ttcgtggtga 10020
tgatgctcaa cctgggcggt tcagaaatcg aacaggaacg ccagtggctg aaaccgcagg 10080
tgtggattgg tccggcaatt ttgtcggcca tcatgctggt ggtgattgtt tacgccatcc 10140
tcggtgttaa cgatcagggt atcgacggta cgccaatcag tgctaaagca gtgggtatta 10200
cgctgttcgg gccttacgta ctggcggtgg aactggcttc tatgctgctg ctcgcaggtc 10260
tggttgtggc cttccacgtc ggtcgtgaag agcgtgcggg tgaagtgctg agcaatcgta 10320
aagacgacag cgcgaaaaga aaaacggagg agcacgcatg atccccttac aacatggact 10380
gatcctcgcg gcaatcttat tcgttcttgg cttaaccggt ctggttatcc gtcgcaatct 10440
gctgtttatg ttgattggtc tggaaatcat gattaacgcc tccgcgctgg ccttcgtggt 10500
cgccggaagc tactggggcc agaccgacgg tcaggtgatg tacattctcg ccatcagcct 10560
cgcggcggca gaagcgagta tcggccttgc gctgctgctg caacttcacc gtcgtcgcca 10620
gaacctgaac atcgattcag taagtgagat gcgcggatga acatgcttgc cttaaccatt 10680
attttgccat tgattggctt cgtcctgctg gcattctccc gtgggcgctg gtctgaaaac 10740
gtctcggcga tcgtcggcgt aggctctgtg ggcctggcgg cgctggtaac cgcctttatc 10800
ggcgttgatt tcttcgctaa cggcgagcag acatacagcc agccgctgtg gacgtggatg 10860
tcggtaggcg actttaacat cggttttaac ctggtgctgg acggcctgtc gctgaccatg 10920
ctctcggtgg tcactggtgt gggtttcctt attcacatgt acgcctcctg gtatatgcgc 10980
ggtgaagagg gctactctcg cttcttcgct tacaccaacc tgttcatcgc cagcatggtg 11040
gttctggtgc ttgccgacaa cctgctgctg atgtacctcg gctgggaagg cgtgggcctg 11100
tgctcctatc tgctgatcgg gttctattac accgatccga agaatggcgc agcggcaatg 11160
aaagcgttcg tcgtgacccg tgtgggtgac gtgttcctcg ctttcgcact gttcattctt 11220
tacaacgaac tgggcaccct gaacttccgc gaaatggtgg aactggcacc agcgcacttt 11280
gctgacggca ataacatgct gatgtgggcg acgctgatgc tgctgggcgg tgcggtcggt 11340
aaatctgcgc agttgccgtt gcagacatgg cttgccgacg cgatggcggg cccgacgcct 11400
gtctccgcgc tgatccacgc cgcaaccatg gtaaccgcgg gtgtctacct gatcgcccgt 11460
acccacggcc tgttcctgat gacgccggaa gttctgcatc tggtgggtat tgtcggggcg 11520
gttacgctgc tgctggccgg ttttgccgcg ctggtacaga ccgacatcaa acgtgttctc 11580
gcttactcta ccatgagcca gattggctac atgttcctcg cgcttggcgt gcaggcatgg 11640
gatgcggcga ttttccactt gatgacccac gcgttcttta aagcgctgct gttcctggca 11700
tccggttccg tcattctggc ctgccatcac gaacagaaca tcttcaagat gggcggtctg 11760
cgtaaatcta ttccgctggt ttatctctgc ttcctggtgg gcggcgcagc actgtcggca 11820
ctaccgctgg tcactgcggg cttcttcagt aaggatgaga tcctcgcggg tgcgatggcg 11880
aatggtcata tcaatctgat ggtggcaggt ctggtcggtg cgtttatgac ctcgctctac 11940
accttccgta tgattttcat cgtcttccac ggaaaagaac aaattcacgc tcacgccgtg 12000
aaaggggtaa ctcacagcct gccgctgatt gtgctgctga tcctttccac cttcgttggc 12060
gcactgattg taccgccgct gcagggcgtg cttccgcaaa cgacggaact ggcgcacggc 12120
agcatgttga ccctggaaat tacctctggc gtggtcgcgg tggtcggcat tctgctggca 12180
gcctggctgt ggctgggtaa acgtactctg gtgacctcca tcgccaacag tgcgccgggc 12240
cgtctgctgg gcacctggtg gtacaacgcc tggggatttg actggctgta tgacaaagtg 12300
ttcgtcaagc cgttcctggg tattgcctgg ttgctgaaac gcgatccgct gaactcaatg 12360
atgaacatcc cggctgtcct ttcccgcttt gcaggtaaag gtctgctgtt aagtgagaac 12420
ggctatctgc gctggtatgt ggcatccatg agcatcggtg cggtcgtggt gctggcactg 12480
ttgatggtac tgcgttgagt taaggattgt gggattgccc ctgggcatat aagaacgtag 12540
gtcggataag ccgccttcgc ggtgcatccg acactacccc cagtccgaag agaattttct 12600
gtcgtgagaa ttcgttgaaa atccggtcct gacgggactt ttacaaggaa taaagatcgc 12660
catgttacta ccctggctaa tattaattcc ctttattggc ggcttcctgt gctggcagac 12720
cgaacgcttt ggcgtcaagg tgccacgctg gatcgcgctg atcaccatgg gattgacgct 12780
ggcgctgtcg ctgcaactgt ggttgcaggg cggttattca ctgacgcaat ccgccggaat 12840
tccgcagtgg cagtctgaat tcgacatgcc gtggatcccg cgttttggta tctctattca 12900
tctcgccatt gacgggctgt cgctgctgat ggtcgtgctg accggtctgc tcggtgtgct 12960
ggcggtacta tgttcgtgga aagagatcga aaaatatcag ggcttcttcc acctcaacct 13020
gatgtggatc ctgggcggcg ttatcggcgt gttccttgcc atcgacatgt tcctgttctt 13080
cttcttctgg gaaatgatgc tggtgccgat gtacttcctg atcgcactgt gggggcataa 13140
agcctctgac ggtaaaacgc gtatcacggc ggcaaccaag ttcttcattt acacccaggc 13200
gagtggcctg gtgatgttga ttgccatcct ggcgctggtt tttgttcact acaatgcgac 13260
cggcgtctgg accttcaact atgaagagct gctgaatacg ccaatgtcca gtggtgtgga 13320
atacctgtta atgctgggct tcttcatcgc cttcgcagtc aaaatgccgg tggttccgct 13380
gcatggctgg ctgccggatg cgcactccca ggctccgacc gccggttccg ttgacctcgc 13440
ggggatcttg ctgaaaactg ccgcttacgg tttgctgcgt ttctccctgc cgctgttccc 13500
gaacgcgtcg gcagagttcg cgccaattgc tatgtggctg ggtgttatcg gcatcttcta 13560
cggtgcgtgg atggccttcg cccagaccga tatcaaacgt ctgatcgcct acacctcggt 13620
ttcccacatg ggcttcgtgc tgattgctat ctacaccggc agccagttgg cctaccaggg 13680
cgcggtaatc cagatgattg cgcacggttt gtcggcggcg ggtctgttta ttctttgtgg 13740
tcagctttat gaacgtatcc atacccgcga catgcgcatg atgggcggtc tgtggagcaa 13800
gatgaaatgg ctgccagcac tgtcgctgtt ctttgcggtg gcaacgcttg ggatgcctgg 13860
caccggtaac ttcgtcggcg aatttatgat tctgttcggc agcttccagg ttgtaccggt 13920
gattaccgtt atctctacct ttgggctggt ctttgcatct gtttattcgc tggcgatgtt 13980
acatcgcgct tacttcggta aagcgaaaag ccagattgcc agccaggaac tgccagggat 14040
gtcgctgcgt gagctgttta tgatcctgtt gctggtggtg ctgctggtac tgctgggctt 14100
ctatccgcag ccgattctgg atacctcgca ctccgcgatt ggcaatatcc agcagtggtt 14160
tgttaattcc gttactacta caaggccgta aatcgccatg acaataactc cacaaaacct 14220
gatcgcactg ctaccgttgc tgatcgtcgg cttgacggtg gtggttgtga tgctctccat 14280
tgcgtggcga cgcaatcatt tcctcaacgc tacgctctcg gttattgggc ttaacgcggc 14340
gctggtttcg ctctggtttg ttggccaggc gggcgctatg gacgttacgc cgctgatgcg 14400
cgttgatggt ttcgccatgc tttacaccgg gctggtattg ttggcgagcc tcgccacctg 14460
tactttcgcc tacccgtggc ttgaaggcta taacgacaac aaggatgagt tctacctgtt 14520
ggtgttaatt gccgcgctgg gcgggatcct gctggcgaat gccaaccatc tggcgtctct 14580
gttcctcggt atcgaactga tctctttgcc gctgtttggc ctggtcggtt acgctttccg 14640
ccagaaacgt tcactggaag ccagtatcaa atacaccatc ctttctgccg cagcgtcttc 14700
tttcctgctg tttggtatgg cgctggtgta tgcgcagtct ggcgacctgt cgtttgtcgc 14760
gttgggtaaa aaccttggcg acggtatgct caacgagccg ctgttgctgg caggtttcgg 14820
cctgatgatt gttggcctcg gcttcaaact ctctctggtg ccgttccacc tgtggacgcc 14880
agacgtatac cagggcgcgc ctgcgccggt ttccactttc ctggcgacgg cgagcaaaat 14940
cgctatcttc ggtgtggtga tgcgtctgtt cctctacgca ccggtgggtg acagcgaagc 15000
gattcgcgtg gtgctggcga ttatcgcctt tgcctccatc atcttcggta acctgatggc 15060
gctgagccag accaatatca aacgtctgct cggttactca tctatctctc acctcggcta 15120
tctgctggta gcgctgattg cgctgcaaac cggcgagatg tcgatggaag cggtaggggt 15180
ttacctggcc ggttatctgt tcagcagcct cggcgcgttc ggcgtggtca gcctgatgtc 15240
cagcccgtat cgtggcccgg atgctgattc cctgttctct taccgcggtc tgttctggca 15300
tcgtccgatc ctcgcggcag tgatgacggt gatgatgctg tctctggccg gtatcccgat 15360
gacgctgggc tttatcggta agttctacgt gctggcggtc ggtgtccagg cacacttgtg 15420
gtggctggtg ggtgccgtgg ttgtcggttc ggcaatcggc ctctactact acctgcgcgt 15480
ggcggtgagc ctgtatcttc acgccccgga acaaccgggt cgcgatgcac catcaaactg 15540
gcagtacagc gcgggcggta tcgttgtgct gatctctgca ctgttggtac tggtgctggg 15600
tgtatggcca caaccgctga ttagcattgt gcgtttggca atgccgctga tgtaa 15655
<210> 2
<211> 322
<212> PRT
<213> Artificial Sequence
<220>
<223> SerB
<400> 2
Met Pro Asn Ile Thr Trp Cys Asp Leu Pro Glu Asp Val Ser Leu Trp
1 5 10 15
Pro Gly Leu Pro Leu Ser Leu Ser Gly Asp Glu Val Met Pro Leu Asp
20 25 30
Tyr His Ala Gly Arg Ser Gly Trp Leu Leu Tyr Gly Arg Gly Leu Asp
35 40 45
Lys Gln Arg Leu Thr Gln Tyr Gln Ser Lys Leu Gly Ala Ala Met Val
50 55 60
Ile Val Ala Ala Trp Cys Val Glu Asp Tyr Gln Val Ile Arg Leu Ala
65 70 75 80
Gly Ser Leu Thr Ala Arg Ala Thr Arg Leu Ala His Glu Ala Gln Leu
85 90 95
Asp Val Ala Pro Leu Gly Lys Ile Pro His Leu Arg Thr Pro Gly Leu
100 105 110
Leu Val Met Asp Met Asp Ser Thr Ala Ile Gln Ile Glu Cys Ile Asp
115 120 125
Glu Ile Ala Lys Leu Ala Gly Thr Gly Glu Met Val Ala Glu Val Thr
130 135 140
Glu Arg Ala Met Arg Gly Glu Leu Asp Phe Thr Ala Ser Leu Arg Ser
145 150 155 160
Arg Val Ala Thr Leu Lys Gly Ala Asp Ala Asn Ile Leu Gln Gln Val
165 170 175
Arg Glu Asn Leu Pro Leu Met Pro Gly Leu Thr Gln Leu Val Leu Lys
180 185 190
Leu Glu Thr Leu Gly Trp Lys Val Ala Ile Ala Ser Gly Gly Phe Thr
195 200 205
Phe Phe Ala Glu Tyr Leu Arg Asp Lys Leu Arg Leu Thr Ala Val Val
210 215 220
Ala Asn Glu Leu Glu Ile Met Asp Gly Lys Phe Thr Gly Asn Val Ile
225 230 235 240
Gly Asp Ile Val Asp Ala Gln Tyr Lys Ala Lys Thr Leu Thr Arg Leu
245 250 255
Ala Gln Glu Tyr Glu Ile Pro Leu Ala Gln Thr Val Ala Ile Gly Asp
260 265 270
Gly Ala Asn Asp Leu Pro Met Ile Lys Ala Ala Gly Leu Gly Ile Ala
275 280 285
Tyr His Ala Lys Pro Lys Val Asn Glu Lys Ala Glu Val Thr Ile Arg
290 295 300
His Ala Asp Leu Met Gly Val Phe Cys Ile Leu Ser Gly Ser Leu Asn
305 310 315 320
Gln Lys
<210> 3
<211> 969
<212> DNA
<213> Artificial Sequence
<220>
<223> serB
<400> 3
atgcctaaca ttacctggtg cgacctgcct gaagatgtct ctttatggcc gggtctgcct 60
ctttcattaa gtggtgatga agtgatgcca ctggattacc acgcaggtcg tagcggctgg 120
ctgctgtatg gtcgtgggct ggataaacaa cgtctgaccc aataccagag caaactgggt 180
gcggcgatgg tgattgttgc cgcctggtgc gtggaagatt atcaggtgat tcgtctggca 240
ggttcactca ccgcacgggc tacacgcctg gcccacgaag cgcagctgga tgtcgccccg 300
ctggggaaaa tcccgcacct gcgcacgccg ggtttgctgg tgatggatat ggactccacc 360
gccatccaga ttgaatgtat tgatgaaatt gccaaactgg ccggaacggg cgagatggtg 420
gcggaagtaa ccgaacgggc gatgcgcggc gaactcgatt ttaccgccag cctgcgcagc 480
cgtgtggcga cgctgaaagg cgctgacgcc aatattctgc aacaggtgcg tgaaaatctg 540
ccgctgatgc caggcttaac gcaactggtg ctcaagctgg aaacgctggg ctggaaagtg 600
gcgattgcct ccggcggctt tactttcttt gctgaatacc tgcgcgacaa gctgcgcctg 660
accgccgtgg tagccaatga actggagatc atggacggta aatttaccgg caatgtgatc 720
ggcgacatcg tagacgcgca gtacaaagcg aaaactctga ctcgcctcgc gcaggagtat 780
gaaatcccgc tggcgcagac cgtggcgatt ggcgatggag ccaatgacct gccgatgatc 840
aaagcggcag ggctggggat tgcctaccat gccaagccaa aagtgaatga aaaggcggaa 900
gtcaccatcc gtcacgctga cctgatgggg gtattctgca tcctctcagg cagcctgaat 960
cagaagtaa 969
<210> 4
<211> 405
<212> PRT
<213> Artificial Sequence
<220>
<223> YhhS
<400> 4
Met Pro Glu Pro Val Ala Glu Pro Ala Leu Asn Gly Leu Arg Leu Asn
1 5 10 15
Leu Arg Ile Val Ser Ile Val Met Phe Asn Phe Ala Ser Tyr Leu Thr
20 25 30
Ile Gly Leu Pro Leu Ala Val Leu Pro Gly Tyr Val His Asp Val Met
35 40 45
Gly Phe Ser Ala Phe Trp Ala Gly Leu Val Ile Ser Leu Gln Tyr Phe
50 55 60
Ala Thr Leu Leu Ser Arg Pro His Ala Gly Arg Tyr Ala Asp Ser Leu
65 70 75 80
Gly Pro Lys Lys Ile Val Val Phe Gly Leu Cys Gly Cys Phe Leu Ser
85 90 95
Gly Leu Gly Tyr Leu Thr Ala Gly Leu Thr Ala Ser Leu Pro Val Ile
100 105 110
Ser Leu Leu Leu Leu Cys Leu Gly Arg Val Ile Leu Gly Ile Gly Gln
115 120 125
Ser Phe Ala Gly Thr Gly Ser Thr Leu Trp Gly Val Gly Val Val Gly
130 135 140
Ser Leu His Ile Gly Arg Val Ile Ser Trp Asn Gly Ile Val Thr Tyr
145 150 155 160
Gly Ala Met Ala Met Gly Ala Pro Leu Gly Val Val Phe Tyr His Trp
165 170 175
Gly Gly Leu Gln Ala Leu Ala Leu Ile Ile Met Gly Val Ala Leu Val
180 185 190
Ala Ile Leu Leu Ala Ile Pro Arg Pro Thr Val Lys Ala Ser Lys Gly
195 200 205
Lys Pro Leu Pro Phe Arg Ala Val Leu Gly Arg Val Trp Leu Tyr Gly
210 215 220
Met Ala Leu Ala Leu Ala Ser Ala Gly Phe Gly Val Ile Ala Thr Phe
225 230 235 240
Ile Thr Leu Phe Tyr Asp Ala Lys Gly Trp Asp Gly Ala Ala Phe Ala
245 250 255
Leu Thr Leu Phe Ser Cys Ala Phe Val Gly Thr Arg Leu Leu Phe Pro
260 265 270
Asn Gly Ile Asn Arg Ile Gly Gly Leu Asn Val Ala Met Ile Cys Phe
275 280 285
Ser Val Glu Ile Ile Gly Leu Leu Leu Val Gly Val Ala Thr Met Pro
290 295 300
Trp Met Ala Lys Ile Gly Val Leu Leu Ala Gly Ala Gly Phe Ser Leu
305 310 315 320
Val Phe Pro Ala Leu Gly Val Val Ala Val Lys Ala Val Pro Gln Gln
325 330 335
Asn Gln Gly Ala Ala Leu Ala Thr Tyr Thr Val Phe Met Asp Leu Ser
340 345 350
Leu Gly Val Thr Gly Pro Leu Ala Gly Leu Val Met Ser Trp Ala Gly
355 360 365
Val Pro Val Ile Tyr Leu Ala Ala Ala Gly Leu Val Ala Ile Ala Leu
370 375 380
Leu Leu Thr Trp Arg Leu Lys Lys Arg Pro Pro Glu His Val Pro Glu
385 390 395 400
Ala Ala Ser Ser Ser
405
<210> 5
<211> 1218
<212> DNA
<213> Artificial Sequence
<220>
<223> yhhS
<400> 5
atgcccgaac ccgtagccga acccgcgcta aacggattgc gcctgaattt gcgcattgtc 60
tctatagtca tgtttaactt cgccagctac ctcaccatcg ggttgccgct cgctgtatta 120
ccgggctatg tccatgatgt gatgggcttt agcgccttct gggcaggatt ggttatcagc 180
ctgcaatatt tcgccacctt gctgagccgc cctcatgccg gacgttacgc cgattcgctg 240
ggacccaaaa agattgtcgt cttcggttta tgcggctgct ttttgagcgg tctggggtat 300
ctgacggcag gattaaccgc cagtctgcct gtcatcagcc tgttattact ttgcctgggg 360
cgcgtcatcc ttgggattgg gcaaagtttt gccggaacgg gatcgaccct atggggcgtt 420
ggcgtggttg gctcgctgca tatcgggcgg gtgatttcgt ggaacggcat tgtcacttac 480
ggggcgatgg cgatgggtgc gccgttaggc gtcgtgtttt atcactgggg cggcttgcag 540
gcgttagcgt taatcattat gggcgtggcg ctggtggcca ttttgttggc gatcccgcgt 600
ccgacggtaa aagccagtaa aggcaaaccg ctgccgtttc gcgcggtgct tgggcgcgtc 660
tggctgtacg gtatggcgct ggcactggct tccgccggat ttggcgtcat cgccaccttt 720
atcacgctgt tttatgacgc taaaggttgg gacggtgcgg ctttcgcgct gacgctgttt 780
agctgtgcgt ttgtcggtac gcgtttgtta ttccctaacg gcattaaccg tatcggtggc 840
ttaaacgtag cgatgatttg ctttagcgtt gagataatcg gcctgctact ggttggcgtg 900
gcgactatgc cgtggatggc gaaaatcggc gtcttactgg cgggggccgg gttttcgctg 960
gtgttcccgg cattgggtgt agtggcggta aaagcggttc cgcagcaaaa tcagggggcg 1020
gcgctggcaa cttacaccgt atttatggat ttatcgcttg gcgtgactgg accactggct 1080
gggctggtga tgagctgggc gggcgtaccg gtgatttatc tggcggcggc gggactggtc 1140
gcaatcgcgt tattactgac gtggcgatta aaaaaacggc ctccggaaca cgtccctgag 1200
gccgcctcat catcttaa 1218
<210> 6
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> yhhS UP F
<400> 6
ctgcaggaat tcgatgaagg taaccacttt ttccg 35
<210> 7
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> yhhS UP R
<400> 7
gagttgcagc aagcggagga tcaccacatt tttac 35
<210> 8
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Ptrc F
<400> 8
aatgtggtga tcctccgctt gctgcaactc tctca 35
<210> 9
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Ptrc R
<400> 9
tacgggttcg ggcatgatag ctctcctgtg tgaaa 35
<210> 10
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> yhhS Down F
<400> 10
cacaggagag ctatcatgcc cgaacccgta gccga 35
<210> 11
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> yhhS Down R
<400> 11
gtcgactagc gtgatagcgg tttgccttta ctggc 35
<210> 12
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> yhhSP Conf F
<400> 12
agttgagcaa aaacgccagt 20
<210> 13
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> yhhSP Conf R
<400> 13
gccataccgt acagccagac 20
<210> 14
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> UP F1
<400> 14
ctgcaggaat tcgatcaacc accagagatt cacgt 35
<210> 15
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> UP R1
<400> 15
gcttcctgac tccagtgctt actcatcaaa agtag 35
<210> 16
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Prmf F
<400> 16
tttgatgagt aagcactgga gtcaggaagc cgctt 35
<210> 17
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Prmf R
<400> 17
tgttgacata ctcatgcctc gtttccctca tactg 35
<210> 18
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Down F1
<400> 18
tgagggaaac gaggcatgag tatgtcaaca tccac 35
<210> 19
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Down R1
<400> 19
acggtgagaa cgaccgttac atcacgctag tcgac 35
<210> 20
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> UP R2
<400> 20
tcatttttct tcagctgctt actcatcaaa agtag 35
<210> 21
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Pidi F
<400> 21
tttgatgagt aagcagctga agaaaaatga gcatg 35
<210> 22
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> Pidi R
<400> 22
tgttgacata ctcataattt ctcacatgta attctgatc 39
<210> 23
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<223> Down F2
<400> 23
tacatgtgag aaattatgag tatgtcaaca tccac 35
<210> 24
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> NuoP Conf F
<400> 24
gagagctacc ataatccgtg 20
<210> 25
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> NuoP Conf R
<400> 25
catctcaacg taacagcagg 20
<210> 26
<211> 147
<212> PRT
<213> Artificial Sequence
<220>
<223> NuoA
<400> 26
Met Ser Met Ser Thr Ser Thr Glu Val Ile Ala His His Trp Ala Phe
1 5 10 15
Ala Ile Phe Leu Ile Val Ala Ile Gly Leu Cys Cys Leu Met Leu Val
20 25 30
Gly Gly Trp Phe Leu Gly Gly Arg Ala Arg Ala Arg Ser Lys Asn Val
35 40 45
Pro Phe Glu Ser Gly Ile Asp Ser Val Gly Ser Ala Arg Leu Arg Leu
50 55 60
Ser Ala Lys Phe Tyr Leu Val Ala Met Phe Phe Val Ile Phe Asp Val
65 70 75 80
Glu Ala Leu Tyr Leu Phe Ala Trp Ser Thr Ser Ile Arg Glu Ser Gly
85 90 95
Trp Val Gly Phe Val Glu Ala Ala Ile Phe Ile Phe Val Leu Leu Ala
100 105 110
Gly Leu Val Tyr Leu Val Arg Ile Gly Ala Leu Asp Trp Thr Pro Ala
115 120 125
Arg Ser Arg Arg Glu Arg Met Asn Pro Glu Thr Asn Ser Ile Ala Asn
130 135 140
Arg Gln Arg
145
<210> 27
<211> 220
<212> PRT
<213> Artificial Sequence
<220>
<223> NuoB
<400> 27
Met Asp Tyr Thr Leu Thr Arg Ile Asp Pro Asn Gly Glu Asn Asp Arg
1 5 10 15
Tyr Pro Leu Gln Lys Gln Glu Ile Val Thr Asp Pro Leu Glu Gln Glu
20 25 30
Val Asn Lys Asn Val Phe Met Gly Lys Leu Asn Asp Met Val Asn Trp
35 40 45
Gly Arg Lys Asn Ser Ile Trp Pro Tyr Asn Phe Gly Leu Ser Cys Cys
50 55 60
Tyr Val Glu Met Val Thr Ser Phe Thr Ala Val His Asp Val Ala Arg
65 70 75 80
Phe Gly Ala Glu Val Leu Arg Ala Ser Pro Arg Gln Ala Asp Leu Met
85 90 95
Val Val Ala Gly Thr Cys Phe Thr Lys Met Ala Pro Val Ile Gln Arg
100 105 110
Leu Tyr Asp Gln Met Leu Glu Pro Lys Trp Val Ile Ser Met Gly Ala
115 120 125
Cys Ala Asn Ser Gly Gly Met Tyr Asp Ile Tyr Ser Val Val Gln Gly
130 135 140
Val Asp Lys Phe Ile Pro Val Asp Val Tyr Ile Pro Gly Cys Pro Pro
145 150 155 160
Arg Pro Glu Ala Tyr Met Gln Ala Leu Met Leu Leu Gln Glu Ser Ile
165 170 175
Gly Lys Glu Arg Arg Pro Leu Ser Trp Val Val Gly Asp Gln Gly Val
180 185 190
Tyr Arg Ala Asn Met Gln Ser Glu Arg Glu Arg Lys Arg Gly Glu Arg
195 200 205
Ile Ala Val Thr Asn Leu Arg Thr Pro Asp Glu Ile
210 215 220
<210> 28
<211> 600
<212> PRT
<213> Artificial Sequence
<220>
<223> NuoC
<400> 28
Met Val Asn Asn Met Thr Asp Leu Thr Ala Gln Glu Pro Ala Trp Gln
1 5 10 15
Thr Arg Asp His Leu Asp Asp Pro Val Ile Gly Glu Leu Arg Asn Arg
20 25 30
Phe Gly Pro Asp Ala Phe Thr Val Gln Ala Thr Arg Thr Gly Val Pro
35 40 45
Val Val Trp Ile Lys Arg Glu Gln Leu Leu Glu Val Gly Asp Phe Leu
50 55 60
Lys Lys Leu Pro Lys Pro Tyr Val Met Leu Phe Asp Leu His Gly Met
65 70 75 80
Asp Glu Arg Leu Arg Thr His Arg Glu Gly Leu Pro Ala Ala Asp Phe
85 90 95
Ser Val Phe Tyr His Leu Ile Ser Ile Asp Arg Asn Arg Asp Ile Met
100 105 110
Leu Lys Val Ala Leu Ala Glu Asn Asp Leu His Val Pro Thr Phe Thr
115 120 125
Lys Leu Phe Pro Asn Ala Asn Trp Tyr Glu Arg Glu Thr Trp Asp Leu
130 135 140
Phe Gly Ile Thr Phe Asp Gly His Pro Asn Leu Arg Arg Ile Met Met
145 150 155 160
Pro Gln Thr Trp Lys Gly His Pro Leu Arg Lys Asp Tyr Pro Ala Arg
165 170 175
Ala Thr Glu Phe Ser Pro Phe Glu Leu Thr Lys Ala Lys Gln Asp Leu
180 185 190
Glu Met Glu Ala Leu Thr Phe Lys Pro Glu Glu Trp Gly Met Lys Arg
195 200 205
Gly Thr Glu Asn Glu Asp Phe Met Phe Leu Asn Leu Gly Pro Asn His
210 215 220
Pro Ser Ala His Gly Ala Phe Arg Ile Val Leu Gln Leu Asp Gly Glu
225 230 235 240
Glu Ile Val Asp Cys Val Pro Asp Ile Gly Tyr His His Arg Gly Ala
245 250 255
Glu Lys Met Gly Glu Arg Gln Ser Trp His Ser Tyr Ile Pro Tyr Thr
260 265 270
Asp Arg Ile Glu Tyr Leu Gly Gly Cys Val Asn Glu Met Pro Tyr Val
275 280 285
Leu Ala Val Glu Lys Leu Ala Gly Ile Thr Val Pro Asp Arg Val Asn
290 295 300
Val Ile Arg Val Met Leu Ser Glu Leu Phe Arg Ile Asn Ser His Leu
305 310 315 320
Leu Tyr Ile Ser Thr Phe Ile Gln Asp Val Gly Ala Met Thr Pro Val
325 330 335
Phe Phe Ala Phe Thr Asp Arg Gln Lys Ile Tyr Asp Leu Val Glu Ala
340 345 350
Ile Thr Gly Phe Arg Met His Pro Ala Trp Phe Arg Ile Gly Gly Val
355 360 365
Ala His Asp Leu Pro Arg Gly Trp Asp Arg Leu Leu Arg Glu Phe Leu
370 375 380
Asp Trp Met Pro Lys Arg Leu Ala Ser Tyr Glu Lys Ala Ala Leu Gln
385 390 395 400
Asn Thr Ile Leu Lys Gly Arg Ser Gln Gly Val Ala Ala Tyr Gly Ala
405 410 415
Lys Glu Ala Leu Glu Trp Gly Thr Thr Gly Ala Gly Leu Arg Ala Thr
420 425 430
Gly Ile Asp Phe Asp Val Arg Lys Ala Arg Pro Tyr Ser Gly Tyr Glu
435 440 445
Asn Phe Asp Phe Glu Ile Pro Val Gly Gly Gly Val Ser Asp Cys Tyr
450 455 460
Thr Arg Val Met Leu Lys Val Glu Glu Leu Arg Gln Ser Leu Arg Ile
465 470 475 480
Leu Glu Gln Cys Leu Asn Asn Met Pro Glu Gly Pro Phe Lys Ala Asp
485 490 495
His Pro Leu Thr Thr Pro Pro Pro Lys Glu Arg Thr Leu Gln His Ile
500 505 510
Glu Thr Leu Ile Thr His Phe Leu Gln Val Ser Trp Gly Pro Val Met
515 520 525
Pro Ala Asn Glu Ser Phe Gln Met Ile Glu Ala Thr Lys Gly Ile Asn
530 535 540
Ser Tyr Tyr Leu Thr Ser Asp Gly Ser Thr Met Ser Tyr Arg Thr Arg
545 550 555 560
Val Arg Thr Pro Ser Phe Ala His Leu Gln Gln Ile Pro Ala Ala Ile
565 570 575
Arg Gly Ser Leu Val Ser Asp Leu Ile Val Tyr Leu Gly Ser Ile Asp
580 585 590
Phe Val Met Ser Asp Val Asp Arg
595 600
<210> 29
<211> 166
<212> PRT
<213> Artificial Sequence
<220>
<223> NuoE
<400> 29
Met His Glu Asn Gln Gln Pro Gln Thr Glu Ala Phe Glu Leu Ser Ala
1 5 10 15
Ala Glu Arg Glu Ala Ile Glu His Glu Met His His Tyr Glu Asp Pro
20 25 30
Arg Ala Ala Ser Ile Glu Ala Leu Lys Ile Val Gln Lys Gln Arg Gly
35 40 45
Trp Val Pro Asp Gly Ala Ile His Ala Ile Ala Asp Val Leu Gly Ile
50 55 60
Pro Ala Ser Asp Val Glu Gly Val Ala Thr Phe Tyr Ser Gln Ile Phe
65 70 75 80
Arg Gln Pro Val Gly Arg His Val Ile Arg Tyr Cys Asp Ser Val Val
85 90 95
Cys His Ile Asn Gly Tyr Gln Gly Ile Gln Ala Ala Leu Glu Lys Lys
100 105 110
Leu Asn Ile Lys Pro Gly Gln Thr Thr Phe Asp Gly Arg Phe Thr Leu
115 120 125
Leu Pro Thr Cys Cys Leu Gly Asn Cys Asp Lys Gly Pro Asn Met Met
130 135 140
Ile Asp Glu Asp Thr His Ala His Leu Thr Pro Glu Ala Ile Pro Glu
145 150 155 160
Leu Leu Glu Arg Tyr Lys
165
<210> 30
<211> 445
<212> PRT
<213> Artificial Sequence
<220>
<223> NuoF
<400> 30
Met Lys Asn Ile Ile Arg Thr Pro Glu Thr His Pro Leu Thr Trp Arg
1 5 10 15
Leu Arg Asp Asp Lys Gln Pro Val Trp Leu Asp Glu Tyr Arg Ser Lys
20 25 30
Asn Gly Tyr Glu Gly Ala Arg Lys Ala Leu Thr Gly Leu Ser Pro Asp
35 40 45
Glu Ile Val Asn Gln Val Lys Asp Ala Gly Leu Lys Gly Arg Gly Gly
50 55 60
Ala Gly Phe Ser Thr Gly Leu Lys Trp Ser Leu Met Pro Lys Asp Glu
65 70 75 80
Ser Met Asn Ile Arg Tyr Leu Leu Cys Asn Ala Asp Glu Met Glu Pro
85 90 95
Gly Thr Tyr Lys Asp Arg Leu Leu Met Glu Gln Leu Pro His Leu Leu
100 105 110
Val Glu Gly Met Leu Ile Ser Ala Phe Ala Leu Lys Ala Tyr Arg Gly
115 120 125
Tyr Ile Phe Leu Arg Gly Glu Tyr Ile Glu Ala Ala Val Asn Leu Arg
130 135 140
Arg Ala Ile Ala Glu Ala Thr Glu Ala Gly Leu Leu Gly Lys Asn Ile
145 150 155 160
Met Gly Thr Gly Phe Asp Phe Glu Leu Phe Val His Thr Gly Ala Gly
165 170 175
Arg Tyr Ile Cys Gly Glu Glu Thr Ala Leu Ile Asn Ser Leu Glu Gly
180 185 190
Arg Arg Ala Asn Pro Arg Ser Lys Pro Pro Phe Pro Ala Thr Ser Gly
195 200 205
Ala Trp Gly Lys Pro Thr Cys Val Asn Asn Val Glu Thr Leu Cys Asn
210 215 220
Val Pro Ala Ile Leu Ala Asn Gly Val Glu Trp Tyr Gln Asn Ile Ser
225 230 235 240
Lys Ser Lys Asp Ala Gly Thr Lys Leu Met Gly Phe Ser Gly Arg Val
245 250 255
Lys Asn Pro Gly Leu Trp Glu Leu Pro Phe Gly Thr Thr Ala Arg Glu
260 265 270
Ile Leu Glu Asp Tyr Ala Gly Gly Met Arg Asp Gly Leu Lys Phe Lys
275 280 285
Ala Trp Gln Pro Gly Gly Ala Gly Thr Asp Phe Leu Thr Glu Ala His
290 295 300
Leu Asp Leu Pro Met Glu Phe Glu Ser Ile Gly Lys Ala Gly Ser Arg
305 310 315 320
Leu Gly Thr Ala Leu Ala Met Ala Val Asp His Glu Ile Asn Met Val
325 330 335
Ser Leu Val Arg Asn Leu Glu Glu Phe Phe Ala Arg Glu Ser Cys Gly
340 345 350
Trp Cys Thr Pro Cys Arg Asp Gly Leu Pro Trp Ser Val Lys Ile Leu
355 360 365
Arg Ala Leu Glu Arg Gly Glu Gly Gln Pro Gly Asp Ile Glu Thr Leu
370 375 380
Glu Gln Leu Cys Arg Phe Leu Gly Pro Gly Lys Thr Phe Cys Ala His
385 390 395 400
Ala Pro Gly Ala Val Glu Pro Leu Gln Ser Ala Ile Lys Tyr Phe Arg
405 410 415
Glu Glu Phe Glu Ala Gly Ile Lys Gln Pro Phe Ser Asn Thr His Leu
420 425 430
Ile Asn Gly Ile Gln Pro Asn Leu Leu Lys Glu Arg Trp
435 440 445
<210> 31
<211> 910
<212> PRT
<213> Artificial Sequence
<220>
<223> NuoG
<400> 31
Met Leu Met Ala Thr Ile His Val Asp Gly Lys Glu Tyr Glu Val Asn
1 5 10 15
Gly Ala Asp Asn Leu Leu Glu Ala Cys Leu Ser Leu Gly Leu Asp Ile
20 25 30
Pro Tyr Phe Cys Trp His Pro Ala Leu Gly Ser Val Gly Ala Cys Arg
35 40 45
Gln Cys Ala Val Lys Gln Tyr Gln Asn Ala Glu Asp Thr Arg Gly Arg
50 55 60
Leu Val Met Ser Cys Met Thr Pro Ala Ser Asp Gly Thr Phe Ile Ser
65 70 75 80
Ile Asp Asp Glu Glu Ala Lys Gln Phe Arg Glu Ser Val Val Glu Trp
85 90 95
Leu Met Thr Asn His Pro His Asp Cys Pro Val Cys Glu Glu Gly Gly
100 105 110
Asn Cys His Leu Gln Asp Met Thr Val Met Thr Gly His Ser Phe Arg
115 120 125
Arg Tyr Arg Phe Thr Lys Arg Thr His Arg Asn Gln Asp Leu Gly Pro
130 135 140
Phe Ile Ser His Glu Met Asn Arg Cys Ile Ala Cys Tyr Arg Cys Val
145 150 155 160
Arg Tyr Tyr Lys Asp Tyr Ala Asp Gly Thr Asp Leu Gly Val Tyr Gly
165 170 175
Ala His Asp Asn Val Tyr Phe Gly Arg Pro Glu Asp Gly Thr Leu Glu
180 185 190
Ser Glu Phe Ser Gly Asn Leu Val Glu Ile Cys Pro Thr Gly Val Phe
195 200 205
Thr Asp Lys Thr His Ser Glu Arg Tyr Asn Arg Lys Trp Asp Met Gln
210 215 220
Phe Ala Pro Ser Ile Cys Gln Gln Cys Ser Ile Gly Cys Asn Ile Ser
225 230 235 240
Pro Gly Glu Arg Tyr Gly Glu Leu Arg Arg Ile Glu Asn Arg Tyr Asn
245 250 255
Gly Thr Val Asn His Tyr Phe Leu Cys Asp Arg Gly Arg Phe Gly Tyr
260 265 270
Gly Tyr Val Asn Leu Lys Asp Arg Pro Arg Gln Pro Val Gln Arg Arg
275 280 285
Gly Asp Asp Phe Ile Thr Leu Asn Ala Glu Gln Ala Met Gln Gly Ala
290 295 300
Ala Asp Ile Leu Arg Gln Ser Lys Lys Val Ile Gly Ile Gly Ser Pro
305 310 315 320
Arg Ala Ser Val Glu Ser Asn Phe Ala Leu Arg Glu Leu Val Gly Glu
325 330 335
Glu Asn Phe Tyr Thr Gly Ile Ala His Gly Glu Gln Glu Arg Leu Gln
340 345 350
Leu Ala Leu Lys Val Leu Arg Glu Gly Gly Ile Tyr Thr Pro Ala Leu
355 360 365
Arg Glu Ile Glu Ser Tyr Asp Ala Val Leu Val Leu Gly Glu Asp Val
370 375 380
Thr Gln Thr Gly Ala Arg Val Ala Leu Ala Val Arg Gln Ala Val Lys
385 390 395 400
Gly Lys Ala Arg Glu Met Ala Ala Ala Gln Lys Val Ala Asp Trp Gln
405 410 415
Ile Ala Ala Ile Leu Asn Ile Gly Gln Arg Ala Lys His Pro Leu Phe
420 425 430
Val Thr Asn Val Asp Asp Thr Arg Leu Asp Asp Ile Ala Ala Trp Thr
435 440 445
Tyr Arg Ala Pro Val Glu Asp Gln Ala Arg Leu Gly Phe Ala Ile Ala
450 455 460
His Ala Leu Asp Asn Ser Ala Pro Ala Val Asp Gly Ile Glu Pro Glu
465 470 475 480
Leu Gln Ser Lys Ile Asp Val Ile Val Gln Ala Leu Ala Gly Ala Lys
485 490 495
Lys Pro Leu Ile Ile Ser Gly Thr Asn Ala Gly Ser Leu Glu Val Ile
500 505 510
Gln Ala Ala Ala Asn Val Ala Lys Ala Leu Lys Gly Arg Gly Ala Asp
515 520 525
Val Gly Ile Thr Met Ile Ala Arg Ser Val Asn Ser Met Gly Leu Gly
530 535 540
Ile Met Gly Gly Gly Ser Leu Glu Glu Ala Leu Thr Glu Leu Glu Thr
545 550 555 560
Gly Arg Ala Asp Ala Val Val Val Leu Glu Asn Asp Leu His Arg His
565 570 575
Ala Ser Ala Ile Arg Val Asn Ala Ala Leu Ala Lys Ala Pro Leu Val
580 585 590
Met Val Val Asp His Gln Arg Thr Ala Ile Met Glu Asn Ala His Leu
595 600 605
Val Leu Ser Ala Ala Ser Phe Ala Glu Ser Asp Gly Thr Val Ile Asn
610 615 620
Asn Glu Gly Arg Ala Gln Arg Phe Phe Gln Val Tyr Asp Pro Ala Tyr
625 630 635 640
Tyr Asp Ser Lys Thr Val Met Leu Glu Ser Trp Arg Trp Leu His Ser
645 650 655
Leu His Ser Thr Leu Leu Ser Arg Glu Val Asp Trp Thr Gln Leu Asp
660 665 670
His Val Ile Asp Ala Val Val Ala Lys Ile Pro Glu Leu Ala Gly Ile
675 680 685
Lys Asp Ala Ala Pro Asp Ala Thr Phe Arg Ile Arg Gly Gln Lys Leu
690 695 700
Ala Arg Glu Pro His Arg Tyr Ser Gly Arg Thr Ala Met Arg Ala Asn
705 710 715 720
Ile Ser Val His Glu Pro Arg Gln Pro Gln Asp Ile Asp Thr Met Phe
725 730 735
Thr Phe Ser Met Glu Gly Asn Asn Gln Pro Thr Ala His Arg Ser Gln
740 745 750
Val Pro Phe Ala Trp Ala Pro Gly Trp Asn Ser Pro Gln Ala Trp Asn
755 760 765
Lys Phe Gln Asp Glu Val Gly Gly Lys Leu Arg Phe Gly Asp Pro Gly
770 775 780
Val Arg Leu Phe Glu Thr Ser Glu Asn Gly Leu Asp Tyr Phe Thr Ser
785 790 795 800
Val Pro Ala Arg Phe Gln Pro Gln Asp Gly Lys Trp Arg Ile Ala Pro
805 810 815
Tyr Tyr His Leu Phe Gly Ser Asp Glu Leu Ser Gln Arg Ala Pro Val
820 825 830
Phe Gln Ser Arg Met Pro Gln Pro Tyr Ile Lys Leu Asn Pro Ala Asp
835 840 845
Ala Ala Lys Leu Gly Val Asn Ala Gly Thr Arg Val Ser Phe Ser Tyr
850 855 860
Asp Gly Asn Thr Val Thr Leu Pro Val Glu Ile Ala Glu Gly Leu Thr
865 870 875 880
Ala Gly Gln Val Gly Leu Pro Met Gly Met Ser Gly Ile Ala Pro Val
885 890 895
Leu Ala Gly Ala His Leu Glu Asp Leu Lys Glu Ala Gln Gln
900 905 910
<210> 32
<211> 325
<212> PRT
<213> Artificial Sequence
<220>
<223> NuoH
<400> 32
Met Ser Trp Ile Ser Pro Glu Leu Ile Glu Ile Leu Leu Thr Ile Leu
1 5 10 15
Lys Ala Val Val Ile Leu Leu Val Val Val Thr Cys Gly Ala Phe Met
20 25 30
Ser Phe Gly Glu Arg Arg Leu Leu Gly Leu Phe Gln Asn Arg Tyr Gly
35 40 45
Pro Asn Arg Val Gly Trp Gly Gly Ser Leu Gln Leu Val Ala Asp Met
50 55 60
Ile Lys Met Phe Phe Lys Glu Asp Trp Ile Pro Lys Phe Ser Asp Arg
65 70 75 80
Val Ile Phe Thr Leu Ala Pro Met Ile Ala Phe Thr Ser Leu Leu Leu
85 90 95
Ala Phe Ala Ile Val Pro Val Ser Pro Gly Trp Val Val Ala Asp Leu
100 105 110
Asn Ile Gly Ile Leu Phe Phe Leu Met Met Ala Gly Leu Ala Val Tyr
115 120 125
Ala Val Leu Phe Ala Gly Trp Ser Ser Asn Asn Lys Tyr Ser Leu Leu
130 135 140
Gly Ala Met Arg Ala Ser Ala Gln Thr Leu Ser Tyr Glu Val Phe Leu
145 150 155 160
Gly Leu Ser Leu Met Gly Val Val Ala Gln Ala Gly Ser Phe Asn Met
165 170 175
Thr Asp Ile Val Asn Ser Gln Ala His Val Trp Asn Val Ile Pro Gln
180 185 190
Phe Phe Gly Phe Ile Thr Phe Ala Ile Ala Gly Val Ala Val Cys His
195 200 205
Arg His Pro Phe Asp Gln Pro Glu Ala Glu Gln Glu Leu Ala Asp Gly
210 215 220
Tyr His Ile Glu Tyr Ser Gly Met Lys Phe Gly Leu Phe Phe Val Gly
225 230 235 240
Glu Tyr Ile Gly Ile Val Thr Ile Ser Ala Leu Met Val Thr Leu Phe
245 250 255
Phe Gly Gly Trp Gln Gly Pro Leu Leu Pro Pro Phe Ile Trp Phe Ala
260 265 270
Leu Lys Thr Ala Phe Phe Met Met Met Phe Ile Leu Ile Arg Ala Ser
275 280 285
Leu Pro Arg Pro Arg Tyr Asp Gln Val Met Ser Phe Gly Trp Lys Ile
290 295 300
Cys Leu Pro Leu Thr Leu Ile Asn Leu Leu Val Thr Ala Ala Val Ile
305 310 315 320
Leu Trp Gln Ala Gln
325
<210> 33
<211> 180
<212> PRT
<213> Artificial Sequence
<220>
<223> NuoI
<400> 33
Met Thr Leu Lys Glu Leu Leu Val Gly Phe Gly Thr Gln Val Arg Ser
1 5 10 15
Ile Trp Met Ile Gly Leu His Ala Phe Ala Lys Arg Glu Thr Arg Met
20 25 30
Tyr Pro Glu Glu Pro Val Tyr Leu Pro Pro Arg Tyr Arg Gly Arg Ile
35 40 45
Val Leu Thr Arg Asp Pro Asp Gly Glu Glu Arg Cys Val Ala Cys Asn
50 55 60
Leu Cys Ala Val Ala Cys Pro Val Gly Cys Ile Ser Leu Gln Lys Ala
65 70 75 80
Glu Thr Lys Asp Gly Arg Trp Tyr Pro Glu Phe Phe Arg Ile Asn Phe
85 90 95
Ser Arg Cys Ile Phe Cys Gly Leu Cys Glu Glu Ala Cys Pro Thr Thr
100 105 110
Ala Ile Gln Leu Thr Pro Asp Phe Glu Met Gly Glu Tyr Lys Arg Gln
115 120 125
Asp Leu Val Tyr Glu Lys Glu Asp Leu Leu Ile Ser Gly Pro Gly Lys
130 135 140
Tyr Pro Glu Tyr Asn Phe Tyr Arg Met Ala Gly Met Ala Ile Asp Gly
145 150 155 160
Lys Asp Lys Gly Glu Ala Glu Asn Glu Ala Lys Pro Ile Asp Val Lys
165 170 175
Ser Leu Leu Pro
180
<210> 34
<211> 184
<212> PRT
<213> Artificial Sequence
<220>
<223> NuoJ
<400> 34
Met Glu Phe Ala Phe Tyr Ile Cys Gly Leu Ile Ala Ile Leu Ala Thr
1 5 10 15
Leu Arg Val Ile Thr His Thr Asn Pro Val His Ala Leu Leu Tyr Leu
20 25 30
Ile Ile Ser Leu Leu Ala Ile Ser Gly Val Phe Phe Ser Leu Gly Ala
35 40 45
Tyr Phe Ala Gly Ala Leu Glu Ile Ile Val Tyr Ala Gly Ala Ile Met
50 55 60
Val Leu Phe Val Phe Val Val Met Met Leu Asn Leu Gly Gly Ser Glu
65 70 75 80
Ile Glu Gln Glu Arg Gln Trp Leu Lys Pro Gln Val Trp Ile Gly Pro
85 90 95
Ala Ile Leu Ser Ala Ile Met Leu Val Val Ile Val Tyr Ala Ile Leu
100 105 110
Gly Val Asn Asp Gln Gly Ile Asp Gly Thr Pro Ile Ser Ala Lys Ala
115 120 125
Val Gly Ile Thr Leu Phe Gly Pro Tyr Val Leu Ala Val Glu Leu Ala
130 135 140
Ser Met Leu Leu Leu Ala Gly Leu Val Val Ala Phe His Val Gly Arg
145 150 155 160
Glu Glu Arg Ala Gly Glu Val Leu Ser Asn Arg Lys Asp Asp Ser Ala
165 170 175
Lys Arg Lys Thr Glu Glu His Ala
180
<210> 35
<211> 100
<212> PRT
<213> Artificial Sequence
<220>
<223> NuoK
<400> 35
Met Ile Pro Leu Gln His Gly Leu Ile Leu Ala Ala Ile Leu Phe Val
1 5 10 15
Leu Gly Leu Thr Gly Leu Val Ile Arg Arg Asn Leu Leu Phe Met Leu
20 25 30
Ile Gly Leu Glu Ile Met Ile Asn Ala Ser Ala Leu Ala Phe Val Val
35 40 45
Ala Gly Ser Tyr Trp Gly Gln Thr Asp Gly Gln Val Met Tyr Ile Leu
50 55 60
Ala Ile Ser Leu Ala Ala Ala Glu Ala Ser Ile Gly Leu Ala Leu Leu
65 70 75 80
Leu Gln Leu His Arg Arg Arg Gln Asn Leu Asn Ile Asp Ser Val Ser
85 90 95
Glu Met Arg Gly
100
<210> 36
<211> 613
<212> PRT
<213> Artificial Sequence
<220>
<223> NuoL
<400> 36
Met Asn Met Leu Ala Leu Thr Ile Ile Leu Pro Leu Ile Gly Phe Val
1 5 10 15
Leu Leu Ala Phe Ser Arg Gly Arg Trp Ser Glu Asn Val Ser Ala Ile
20 25 30
Val Gly Val Gly Ser Val Gly Leu Ala Ala Leu Val Thr Ala Phe Ile
35 40 45
Gly Val Asp Phe Phe Ala Asn Gly Glu Gln Thr Tyr Ser Gln Pro Leu
50 55 60
Trp Thr Trp Met Ser Val Gly Asp Phe Asn Ile Gly Phe Asn Leu Val
65 70 75 80
Leu Asp Gly Leu Ser Leu Thr Met Leu Ser Val Val Thr Gly Val Gly
85 90 95
Phe Leu Ile His Met Tyr Ala Ser Trp Tyr Met Arg Gly Glu Glu Gly
100 105 110
Tyr Ser Arg Phe Phe Ala Tyr Thr Asn Leu Phe Ile Ala Ser Met Val
115 120 125
Val Leu Val Leu Ala Asp Asn Leu Leu Leu Met Tyr Leu Gly Trp Glu
130 135 140
Gly Val Gly Leu Cys Ser Tyr Leu Leu Ile Gly Phe Tyr Tyr Thr Asp
145 150 155 160
Pro Lys Asn Gly Ala Ala Ala Met Lys Ala Phe Val Val Thr Arg Val
165 170 175
Gly Asp Val Phe Leu Ala Phe Ala Leu Phe Ile Leu Tyr Asn Glu Leu
180 185 190
Gly Thr Leu Asn Phe Arg Glu Met Val Glu Leu Ala Pro Ala His Phe
195 200 205
Ala Asp Gly Asn Asn Met Leu Met Trp Ala Thr Leu Met Leu Leu Gly
210 215 220
Gly Ala Val Gly Lys Ser Ala Gln Leu Pro Leu Gln Thr Trp Leu Ala
225 230 235 240
Asp Ala Met Ala Gly Pro Thr Pro Val Ser Ala Leu Ile His Ala Ala
245 250 255
Thr Met Val Thr Ala Gly Val Tyr Leu Ile Ala Arg Thr His Gly Leu
260 265 270
Phe Leu Met Thr Pro Glu Val Leu His Leu Val Gly Ile Val Gly Ala
275 280 285
Val Thr Leu Leu Leu Ala Gly Phe Ala Ala Leu Val Gln Thr Asp Ile
290 295 300
Lys Arg Val Leu Ala Tyr Ser Thr Met Ser Gln Ile Gly Tyr Met Phe
305 310 315 320
Leu Ala Leu Gly Val Gln Ala Trp Asp Ala Ala Ile Phe His Leu Met
325 330 335
Thr His Ala Phe Phe Lys Ala Leu Leu Phe Leu Ala Ser Gly Ser Val
340 345 350
Ile Leu Ala Cys His His Glu Gln Asn Ile Phe Lys Met Gly Gly Leu
355 360 365
Arg Lys Ser Ile Pro Leu Val Tyr Leu Cys Phe Leu Val Gly Gly Ala
370 375 380
Ala Leu Ser Ala Leu Pro Leu Val Thr Ala Gly Phe Phe Ser Lys Asp
385 390 395 400
Glu Ile Leu Ala Gly Ala Met Ala Asn Gly His Ile Asn Leu Met Val
405 410 415
Ala Gly Leu Val Gly Ala Phe Met Thr Ser Leu Tyr Thr Phe Arg Met
420 425 430
Ile Phe Ile Val Phe His Gly Lys Glu Gln Ile His Ala His Ala Val
435 440 445
Lys Gly Val Thr His Ser Leu Pro Leu Ile Val Leu Leu Ile Leu Ser
450 455 460
Thr Phe Val Gly Ala Leu Ile Val Pro Pro Leu Gln Gly Val Leu Pro
465 470 475 480
Gln Thr Thr Glu Leu Ala His Gly Ser Met Leu Thr Leu Glu Ile Thr
485 490 495
Ser Gly Val Val Ala Val Val Gly Ile Leu Leu Ala Ala Trp Leu Trp
500 505 510
Leu Gly Lys Arg Thr Leu Val Thr Ser Ile Ala Asn Ser Ala Pro Gly
515 520 525
Arg Leu Leu Gly Thr Trp Trp Tyr Asn Ala Trp Gly Phe Asp Trp Leu
530 535 540
Tyr Asp Lys Val Phe Val Lys Pro Phe Leu Gly Ile Ala Trp Leu Leu
545 550 555 560
Lys Arg Asp Pro Leu Asn Ser Met Met Asn Ile Pro Ala Val Leu Ser
565 570 575
Arg Phe Ala Gly Lys Gly Leu Leu Leu Ser Glu Asn Gly Tyr Leu Arg
580 585 590
Trp Tyr Val Ala Ser Met Ser Ile Gly Ala Val Val Val Leu Ala Leu
595 600 605
Leu Met Val Leu Arg
610
<210> 37
<211> 509
<212> PRT
<213> Artificial Sequence
<220>
<223> NuoM
<400> 37
Met Leu Leu Pro Trp Leu Ile Leu Ile Pro Phe Ile Gly Gly Phe Leu
1 5 10 15
Cys Trp Gln Thr Glu Arg Phe Gly Val Lys Val Pro Arg Trp Ile Ala
20 25 30
Leu Ile Thr Met Gly Leu Thr Leu Ala Leu Ser Leu Gln Leu Trp Leu
35 40 45
Gln Gly Gly Tyr Ser Leu Thr Gln Ser Ala Gly Ile Pro Gln Trp Gln
50 55 60
Ser Glu Phe Asp Met Pro Trp Ile Pro Arg Phe Gly Ile Ser Ile His
65 70 75 80
Leu Ala Ile Asp Gly Leu Ser Leu Leu Met Val Val Leu Thr Gly Leu
85 90 95
Leu Gly Val Leu Ala Val Leu Cys Ser Trp Lys Glu Ile Glu Lys Tyr
100 105 110
Gln Gly Phe Phe His Leu Asn Leu Met Trp Ile Leu Gly Gly Val Ile
115 120 125
Gly Val Phe Leu Ala Ile Asp Met Phe Leu Phe Phe Phe Phe Trp Glu
130 135 140
Met Met Leu Val Pro Met Tyr Phe Leu Ile Ala Leu Trp Gly His Lys
145 150 155 160
Ala Ser Asp Gly Lys Thr Arg Ile Thr Ala Ala Thr Lys Phe Phe Ile
165 170 175
Tyr Thr Gln Ala Ser Gly Leu Val Met Leu Ile Ala Ile Leu Ala Leu
180 185 190
Val Phe Val His Tyr Asn Ala Thr Gly Val Trp Thr Phe Asn Tyr Glu
195 200 205
Glu Leu Leu Asn Thr Pro Met Ser Ser Gly Val Glu Tyr Leu Leu Met
210 215 220
Leu Gly Phe Phe Ile Ala Phe Ala Val Lys Met Pro Val Val Pro Leu
225 230 235 240
His Gly Trp Leu Pro Asp Ala His Ser Gln Ala Pro Thr Ala Gly Ser
245 250 255
Val Asp Leu Ala Gly Ile Leu Leu Lys Thr Ala Ala Tyr Gly Leu Leu
260 265 270
Arg Phe Ser Leu Pro Leu Phe Pro Asn Ala Ser Ala Glu Phe Ala Pro
275 280 285
Ile Ala Met Trp Leu Gly Val Ile Gly Ile Phe Tyr Gly Ala Trp Met
290 295 300
Ala Phe Ala Gln Thr Asp Ile Lys Arg Leu Ile Ala Tyr Thr Ser Val
305 310 315 320
Ser His Met Gly Phe Val Leu Ile Ala Ile Tyr Thr Gly Ser Gln Leu
325 330 335
Ala Tyr Gln Gly Ala Val Ile Gln Met Ile Ala His Gly Leu Ser Ala
340 345 350
Ala Gly Leu Phe Ile Leu Cys Gly Gln Leu Tyr Glu Arg Ile His Thr
355 360 365
Arg Asp Met Arg Met Met Gly Gly Leu Trp Ser Lys Met Lys Trp Leu
370 375 380
Pro Ala Leu Ser Leu Phe Phe Ala Val Ala Thr Leu Gly Met Pro Gly
385 390 395 400
Thr Gly Asn Phe Val Gly Glu Phe Met Ile Leu Phe Gly Ser Phe Gln
405 410 415
Val Val Pro Val Ile Thr Val Ile Ser Thr Phe Gly Leu Val Phe Ala
420 425 430
Ser Val Tyr Ser Leu Ala Met Leu His Arg Ala Tyr Phe Gly Lys Ala
435 440 445
Lys Ser Gln Ile Ala Ser Gln Glu Leu Pro Gly Met Ser Leu Arg Glu
450 455 460
Leu Phe Met Ile Leu Leu Leu Val Val Leu Leu Val Leu Leu Gly Phe
465 470 475 480
Tyr Pro Gln Pro Ile Leu Asp Thr Ser His Ser Ala Ile Gly Asn Ile
485 490 495
Gln Gln Trp Phe Val Asn Ser Val Thr Thr Thr Arg Pro
500 505
<210> 38
<211> 425
<212> PRT
<213> Artificial Sequence
<220>
<223> NuoN
<400> 38
Met Asp Val Thr Pro Leu Met Arg Val Asp Gly Phe Ala Met Leu Tyr
1 5 10 15
Thr Gly Leu Val Leu Leu Ala Ser Leu Ala Thr Cys Thr Phe Ala Tyr
20 25 30
Pro Trp Leu Glu Gly Tyr Asn Asp Asn Lys Asp Glu Phe Tyr Leu Leu
35 40 45
Val Leu Ile Ala Ala Leu Gly Gly Ile Leu Leu Ala Asn Ala Asn His
50 55 60
Leu Ala Ser Leu Phe Leu Gly Ile Glu Leu Ile Ser Leu Pro Leu Phe
65 70 75 80
Gly Leu Val Gly Tyr Ala Phe Arg Gln Lys Arg Ser Leu Glu Ala Ser
85 90 95
Ile Lys Tyr Thr Ile Leu Ser Ala Ala Ala Ser Ser Phe Leu Leu Phe
100 105 110
Gly Met Ala Leu Val Tyr Ala Gln Ser Gly Asp Leu Ser Phe Val Ala
115 120 125
Leu Gly Lys Asn Leu Gly Asp Gly Met Leu Asn Glu Pro Leu Leu Leu
130 135 140
Ala Gly Phe Gly Leu Met Ile Val Gly Leu Gly Phe Lys Leu Ser Leu
145 150 155 160
Val Pro Phe His Leu Trp Thr Pro Asp Val Tyr Gln Gly Ala Pro Ala
165 170 175
Pro Val Ser Thr Phe Leu Ala Thr Ala Ser Lys Ile Ala Ile Phe Gly
180 185 190
Val Val Met Arg Leu Phe Leu Tyr Ala Pro Val Gly Asp Ser Glu Ala
195 200 205
Ile Arg Val Val Leu Ala Ile Ile Ala Phe Ala Ser Ile Ile Phe Gly
210 215 220
Asn Leu Met Ala Leu Ser Gln Thr Asn Ile Lys Arg Leu Leu Gly Tyr
225 230 235 240
Ser Ser Ile Ser His Leu Gly Tyr Leu Leu Val Ala Leu Ile Ala Leu
245 250 255
Gln Thr Gly Glu Met Ser Met Glu Ala Val Gly Val Tyr Leu Ala Gly
260 265 270
Tyr Leu Phe Ser Ser Leu Gly Ala Phe Gly Val Val Ser Leu Met Ser
275 280 285
Ser Pro Tyr Arg Gly Pro Asp Ala Asp Ser Leu Phe Ser Tyr Arg Gly
290 295 300
Leu Phe Trp His Arg Pro Ile Leu Ala Ala Val Met Thr Val Met Met
305 310 315 320
Leu Ser Leu Ala Gly Ile Pro Met Thr Leu Gly Phe Ile Gly Lys Phe
325 330 335
Tyr Val Leu Ala Val Gly Val Gln Ala His Leu Trp Trp Leu Val Gly
340 345 350
Ala Val Val Val Gly Ser Ala Ile Gly Leu Tyr Tyr Tyr Leu Arg Val
355 360 365
Ala Val Ser Leu Tyr Leu His Ala Pro Glu Gln Pro Gly Arg Asp Ala
370 375 380
Pro Ser Asn Trp Gln Tyr Ser Ala Gly Gly Ile Val Val Leu Ile Ser
385 390 395 400
Ala Leu Leu Val Leu Val Leu Gly Val Trp Pro Gln Pro Leu Ile Ser
405 410 415
Ile Val Arg Leu Ala Met Pro Leu Met
420 425
<210> 39
<211> 4695
<212> DNA
<213> Artificial Sequence
<220>
<223> pSKH130
<400> 39
tcgaggccgc gattaaattc caacatggat gctgatttat atgggtataa atgggctcgc 60
gataatgtcg ggcaatcagg tgcgacaatc tatcgattgt atgggaagcc cgatgcgcca 120
gagttgtttc tgaaacatgg caaaggtagc gttgccaatg atgttacaga tgagatggtc 180
agactaaact ggctgacgga atttatgcct cttccgacca tcaagcattt tatccgtact 240
cctgatgatg catggttact caccactgcg atccccggga aaacagcatt ccaggtatta 300
gaagaatatc ctgattcagg tgaaaatatt gttgatgcgc tggcagtgtt cctgcgccgg 360
ttgcattcga ttcctgtttg taattgtcct tttaacagcg atcgcgtatt tcgtctcgct 420
caggcgcaat cacgaatgaa taacggtttg gttgatgcga gtgattttga tgacgagcgt 480
aatggctggc ctgttgaaca agtctggaaa gaaatgcata agcttttgcc attctcaccg 540
gattcagtcg tcactcatgg tgatttctca cttgataacc ttatttttga cgaggggaaa 600
ttaataggtt gtattgatgt tggacgagtc ggaatcgcag accgatacca ggatcttgcc 660
atcctatgga actgcctcgg tgagttttct ccttcattac agaaacggct ttttcaaaaa 720
tatggtattg ataatcctga tatgaataaa ttgcagtttc atttgatgct cgatgagttt 780
ttctaatcag aattggttaa ttggttgtaa cactggcaga gcattacgct gacttgacgg 840
gacggcggct ttgttgaata aatcgaactt ttgctgagtt gaaggatcag atcacgcatc 900
ttcccgacaa cgcagaccgt tccgtggcaa agcaaaagtt caaaatcacc aactggtcca 960
cctacaacaa agctctcatc aaccgtggct ccctcacttt ctggctggat gatggggcga 1020
ttcaggcctg gtatgagtca gcaacacctt cttcacgagg cagacctcag cgctcaaaga 1080
tgcaggggta aaagctaacc gcatctttac cgacaaggca tccggcagtt caacagatcg 1140
ggaagggctg gatttgctga ggatgaaggt ggaggaaggt gatgtcattc tggtgaagaa 1200
gctcgaccgt cttggccgcg acaccgccga catgatccaa ctgataaaag agtttgatgc 1260
tcagggtgta gcggttcggt ttattgacga cgggatcagt accgacggtg atatggggca 1320
aatggtggtc accgcgcgta atacgactca ctatagggcg aattggagct ccaccgcggt 1380
ggcggccgct ctagacttta cggtatcgcc gctcccgatt cgcagcgcat cgccttctat 1440
cgccttcttg acgagttctt ctgagcggga ctctggggtt cgctagagga tcgatccttt 1500
ttaacccatc acatatacct gccgttcact attatttagt gaaatgagat attatgatat 1560
tttctgaatt gtgattaaaa aggcaacttt atgcccatgc aacagaaact ataaaaaata 1620
cagagaatga aaagaaacag atagattttt tagttcttta ggcccgtagt ctgcaaatcc 1680
ttttatgatt ttctatcaaa caaaagagga aaatagacca gttgcaatcc aaacgagagt 1740
ctaatagaat gaggtcgaaa agtaaatcgc gcgggtttgt tactgataaa gcaggcaaga 1800
cctaaaatgt gtaaagggca aagtgtatac tttggcgtca ccccttacat attttaggtc 1860
tttttttatt gtgcgtaact aacttgccat cttcaaacag gagggctgga agaagcagac 1920
cgctaacaca gtacataaaa aaggagacat gaacgatgaa catcaaaaag tttgcaaaac 1980
aagcaacagt attaaccttt actaccgcac tgctggcagg aggcgcaact caagcgtttg 2040
cgaaagaaac gaaccaaaag ccatataagg aaacatacgg catttcccat attacacgcc 2100
atgatatgct gcaaatccct gaacagcaaa aaaatgaaaa atatcaagtt cctgaattcg 2160
attcgtccac aattaaaaat atctcttctg caaaaggcct ggacgtttgg gacagctggc 2220
cattacaaaa cgctgacggc actgtcgcaa actatcacgg ctaccacatc gtctttgcat 2280
tagccggaga tcctaaaaat gcggatgaca catcgattta catgttctat caaaaagtcg 2340
gcgaaacttc tattgacagc tggaaaaacg ctggccgcgt ctttaaagac agcgacaaat 2400
tcgatgcaaa tgattctatc ctaaaagacc aaacacaaga atggtcaggt tcagccacat 2460
ttacatctga cggaaaaatc cgtttattct acactgattt ctccggtaaa cattacggca 2520
aacaaacact gacaactgca caagttaacg tatcagcatc agacagctct ttgaacatca 2580
acggtgtaga ggattataaa tcaatctttg acggtgacgg aaaaacgtat caaaatgtac 2640
agcagttcat cgatgaaggc aactacagct caggcgacaa ccatacgctg agagatcctc 2700
actacgtaga agataaaggc cacaaatact tagtatttga agcaaacact ggaactgaag 2760
atggctacca aggcgaagaa tctttattta acaaagcata ctatggcaaa agcacatcat 2820
tcttccgtca agaaagtcaa aaacttctgc aaagcgataa aaaacgcacg gctgagttag 2880
caaacggcgc tctcggtatg attgagctaa acgatgatta cacactgaaa aaagtgatga 2940
aaccgctgat tgcatctaac acagtaacag atgaaattga acgcgcgaac gtctttaaaa 3000
tgaacggcaa atggtacctg ttcactgact cccgcggatc aaaaatgacg attgacggca 3060
ttacgtctaa cgatatttac atgcttggtt atgtttctaa ttctttaact ggcccataca 3120
agccgctgaa caaaactggc cttgtgttaa aaatggatct tgatcctaac gatgtaacct 3180
ttacttactc acacttcgct gtacctcaag cgaaaggaaa caatgtcgtg attacaagct 3240
atatgacaaa cagaggattc tacgcagaca aacaatcaac gtttgcgcca agcttcctgc 3300
tgaacatcaa aggcaagaaa acatctgttg tcaaagacag catccttgaa caaggacaat 3360
taacagttaa caaataaaaa cgcaaaagaa aatgccgatg ggtaccgagc gaaatgaccg 3420
accaagcgac gcccaacctg ccatcggatc ccccgggctg caggaattcg atatcacgct 3480
agtcgaccta gctagcatat ggggagatct actagtaaag catgccaatt ggtattctat 3540
agtgtcacct aaatcgtatg tgtatgatac ataaggttat gtattaattg tagccgcgtt 3600
ctaacgacaa tatgtacaag cctaattgtg tagcatctgg cttactgaag cagaccctat 3660
catctctctc gtaaactgcc gtcagagtcg gtttggttgg acgaaccttc tgagtttctg 3720
gtaacgccgt cccgcacccg gaaatggtca gcgaaccaat cagcagggtc atcgctagcc 3780
catggctaat tcccatgtca gccgttaagt gttcctgtgt cactcaaaat tgctttgaga 3840
ggctctaagg gcttctcagt gcgttacatc cctggcttgt tgtccacaac cgttaaacct 3900
taaaagcttt aaaagcctta tatattcttt tttttcttat aaaacttaaa accttagagg 3960
ctatttaagt tgctgattta tattaatttt attgttcaaa catgagagct tagtacgtga 4020
aacatgagag cttagtacgt tagccatgag agcttagtac gttagccatg agggtttagt 4080
tcgttaaaca tgagagctta gtacgttaaa catgagagct tagtacgtga aacatgagag 4140
cttagtacgt actatcaaca ggttgaactg ctgatcttca gatcctctac gccggacgca 4200
tcgtggccgg atcttgcggc cgcaaaaatt aaaaatgaag ttttaaatca atctaaagta 4260
tatatgagta aacttggtct gacagttacc aatgcttaat cagtgaggca ccaataactg 4320
ccttaaaaaa actagcgctg aggtctgcct cgtgaagaag gtgttgctga ctcataccag 4380
gcctgaatcg ccccatcatc cagccagaaa gtgagggagc cacggttgat gagagctttg 4440
ttgtaggtgg accagttggt gattttgaac ttttgctttg ccacggaacg gtctgcgttg 4500
tcgggaagat gcgtgatctg atccttcaac tcagcaaaag ttcgatttat tcaacaaagc 4560
cacgttgtgt ctcaaaatct ctgatgttac attgcacaag ataaaaatat atcatcatga 4620
acaataaaac tgtctgctta cataaacagt aatacaaggg gtgttatgag ccatattcaa 4680
cgggaaacgt cttgc 4695
<---
Изобретение относится к биотехнологии. Предложено применение рекомбинантного микроорганизма рода Escherichia для получения O-фосфосерина, где указанный микроорганизм обладает повышенной активностью NADH:хинон-оксидоредуктазы по сравнению с ее эндогенной активностью, и где повышение активности NADH:хинон-оксидоредуктазы по сравнению с ее эндогенной активностью достигнуто путем увеличения экспрессии оперона nuo. Также предложены способы получения О-фосфосерина, цистеина и производного цистеина с использованием указанного микроорганизма. Также предложены рекомбинантные микроорганизмы рода Escherichia, обладающие повышенной активностью NADH:хинон-оксидоредуктазы по сравнению с ее эндогенной активностью и обладающие O-фосфосерин-продуцирующей способностью, где повышение активности NADH:хинон-оксидоредуктазы по сравнению с ее эндогенной активностью достигнуто путем увеличения экспрессии оперона nuo, в которых дополнительно понижена активность фосфосеринфосфатазы (SerB) по сравнению с ее эндогенной активностью или повышена активность O-фосфосерин-экспортирующего белка (YhhS) по сравнению с его эндогенной активностью. Изобретение обеспечивает высокопродуктивное продуцирование О-фосфосерина. 6 н. и 6 з.п. ф-лы, 9 табл., 2 пр.
1. Применение рекомбинантного микроорганизма рода Escherichia для получения O-фосфосерина, где указанный микроорганизм обладает повышенной активностью NADH:хинон-оксидоредуктазы по сравнению с ее эндогенной активностью, и где повышение активности NADH:хинон-оксидоредуктазы по сравнению с ее эндогенной активностью достигнуто путем увеличения экспрессии оперона nuo.
2. Применение по п. 1, где повышение активности NADH:хинон-оксидоредуктазы по сравнению с ее эндогенной активностью достигнуто путем включения последовательности регуляции экспрессии гена с повышенной активностью по сравнению с ее эндогенной активностью, находящейся выше по ходу транскрипции от гена, кодирующего NADH:хинон-оксидоредуктазу.
3. Применение по п. 1, где повышение активности NADH:хинон-оксидоредуктазы по сравнению с ее эндогенной активностью достигнуто путем включения промотора гена с повышенной активностью по сравнению с его эндогенной активностью, находящейся выше по ходу транскрипции от гена, кодирующего NADH:хинон-оксидоредуктазу.
4. Применение по п. 2, где выше по ходу транскрипции от гена, кодирующего NADH:хинон-оксидоредуктазу, представляет собой выше по ходу транскрипции от гена nuoA.
5. Рекомбинантный микроорганизм рода Escherichia, обладающий повышенной активностью NADH:хинон-оксидоредуктазы по сравнению с ее эндогенной активностью и обладающий O-фосфосерин-продуцирующей способностью,
где повышение активности NADH:хинон-оксидоредуктазы по сравнению с ее эндогенной активностью достигнуто путем увеличения экспрессии оперона nuo,
где в указанном микроорганизме дополнительно понижена активность фосфосеринфосфатазы (SerB) по сравнению с ее эндогенной активностью.
6. Рекомбинантный микроорганизм рода Escherichia, обладающий повышенной активностью NADH:хинон-оксидоредуктазы по сравнению с ее эндогенной активностью и обладающий O-фосфосерин-продуцирующей способностью,
где повышение активности NADH:хинон-оксидоредуктазы по сравнению с ее эндогенной активностью достигнуто путем увеличения экспрессии оперона nuo,
где в указанном микроорганизме дополнительно повышена активность O-фосфосерин-экспортирующего белка (YhhS) по сравнению с его эндогенной активностью.
7. Применение по п. 1, где микроорганизм представляет собой Escherichia coli.
8. Способ получения O-фосфосерина, включающий культивирование рекомбинантного микроорганизма рода Escherichia в среде,
где указанный микроорганизм обладает повышенной активностью NADH:хинон-оксидоредуктазы по сравнению с ее эндогенной активностью и обладает O-фосфосерин-продуцирующей способностью, и
где повышение активности NADH:хинон-оксидоредуктазы по сравнению с ее эндогенной активностью достигнуто путем увеличения экспрессии оперона nuo.
9. Способ по п. 8, дополнительно включающий выделение O-фосфосерина из культуральной среды или из микроорганизма.
10. Способ получения цистеина, включающий:
а) культивирование O-фосфосерин-продуцирующего микроорганизма с повышенной активностью NADH:хинон-оксидоредуктазы по сравнению с ее эндогенной активностью в среде с получением O-фосфосерина или среды, содержащей O-фосфосерин,
где повышение активности NADH:хинон-оксидоредуктазы по сравнению с ее эндогенной активностью достигнуто путем увеличения экспрессии оперона nuo; и
б) взаимодействие O-фосфосерина, полученного на стадии (а), или среды, содержащей O-фосфосерин, с сульфидом в присутствии O-фосфосерин-сульфгидрилазы (OPSS) или микроорганизма, экспрессирующего O-фосфосерин-сульфгидрилазу.
11. Способ получения производного цистеина, включающий:
а) культивирование O-фосфосерин-продуцирующего микроорганизма с повышенной активностью NADH:хинон-оксидоредуктазы по сравнению с ее эндогенной активностью в среде с получением O-фосфосерина или среды, содержащей O-фосфосерин,
где повышение активности NADH:хинон-оксидоредуктазы по сравнению с ее эндогенной активностью достигнуто путем увеличения экспрессии оперона nuo;
б) взаимодействие O-фосфосерина, полученного на стадии (а), или среды, содержащей O-фосфосерин, с сульфидом в присутствии O-фосфосерин-сульфгидрилазы (OPSS) или микроорганизма, экспрессирующего O-фосфосерин-сульфгидрилазу; и
в) превращение цистеина, полученного на стадии (б), в производное цистеина.
12. Способ по п. 10 или 11, где сульфид представляет собой по меньшей мере один, выбранный из группы, состоящей из Na2S, NaSH, (NH4)2S, H2S и Na2S2O3.

