Область изобретения
Настоящее изобретение относится к новому модифицированному полипептиду, обладающему активностью экспортирования О-фосфосерина (OPS), предшественника L-цистеина, полинуклеотиду, кодирующему этот полипептид, О-фосфосерин-продуцирующему микроорганизму, экспрессирующему этот полипептид, способу продуцирования О-фосфосерина с использованием этого микроорганизма и способу продуцирования цистеина или его производного, включающему осуществление взаимодействия между О-фосфосерином, продуцированным вышеупомянутым способом, и сульфидом в присутствии О-фосфосерин-сульфгидрилазы (OPSS) или микроорганизма, экспрессирущего О-фосфосерин-сульфгидрилазу.
Предшествующий уровень техники
L-цистеин, аминокислота, играющая важную роль в метаболизме серы у всех живых организмов, используется не только в синтезе биологических белков, таких как кератин волос, глутатион, биотин, метионин, и других серосодержащих метаболитов, но также в качестве предшественника для биосинтеза кофермента А.
Известные способы продуцирования L-цистеина с использованием микроорганизмов включают: 1) способ биологического превращения D,L-2-аминотиазолин-4-карбоновой кислоты (D,L-ATC) в L-цистеин с использованием микроорганизмов, 2) способ продуцирования L-цистеина путем прямой ферментации с использованием Е. coli (ЕР 0885962 В; Wada М and Takagi Н, Appl. Microbiol. Biochem., 73:48-54, 2006), и 3) способ продуцирования О-фосфосерина путем ферментации с использованием микроорганизмов и превращение О-фосфосерина в L-цистеин путем взаимодействия между О-фосфосерином и сульфидом при каталитическом воздействии О-фосфосерин-сульфгидрилазы (патент Кореи №1381048). В частности, для продуцирования цистеина способом (3) с высоким выходом предшественник, О-фосфосерин, должен продуцироваться в избыточных количествах.
В этом отношении авторы настоящего изобретения приложили значительные усилия для выявления соответствующего фактора экспорта, который может беспрепятственно экспортировать О-фосфосерин, продуцированный в О-фосфосерин-продуцирующем микроорганизме, из клеток. В частности, авторы настоящего изобретения открыли вариант RhtB в качестве белка, обладающего О-фосфосерин-экспортирующей активностью (публикация заявки на патент Кореи №10-2014-0133751), и новый О-фосфосерин-экспортирующий белок (публикация заявки на патент Кореи №10-2014-0133754), и подтвердили, что концентрация О-фосфосерина увеличивалась, когда эти белки были активированы у О-фосфосерин-продуцирующего микроорганизма.
Техническая задача
В этих обстоятельствах авторы настоящего изобретения приложили усилия для выявления О-фосфосерин-экспортирующих белков, которые могут увеличивать продукцию О-фосфосерина с улучшенной О-фосфосерин-экспортирующей активностью, и для разработки их вариантов, и в результате им удалось разработать вариант О-фосфосерин-экспортирующего белка, который может эффективно экспортировать О-фосфосерин из О-фосфосерин-продуцирующего микроорганизма, тем самым выполняя настоящее изобретение.
Техническое решение
Следовательно, задача настоящего изобретения заключается в том, чтобы обеспечить полипептид, обладающий O-фосфосерин(OPS)-экспортирующей активностью.
Задача настоящего изобретения также заключалась в том, чтобы обеспечить полинуклеотид, кодирующий этот полипептид.
Также в задачу настоящего изобретения входило обеспечение О-фосфосерин-продуцирующего микроорганизма рода Escherichia, экспрессирующего этот полипептид.
Также в задачу настоящего изобретения входило обеспечение способа продуцирования О-фосфосерина, включающего культивирование продуцирующего О-фосфосерин микроорганизма.
Также в задачу настоящего изобретения входило обеспечение способа продуцирования цистеина или его производного, включающего осуществление взаимодействия между О-фосфосерином, продуцированным как изложено выше, и сульфидом в присутствии О-фосфосерин-сульфгидрилазы или микроорганизма, экспрессирующего О-фосфосерин-сульфгидрилазу.
Полезные эффекты изобретения
Новый модифицированный полипептид по настоящему изобретению, представленный аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 3 и SEQ ID NO: 5, обладает превосходной OPS-экспортирующей активностью. Соответственно, когда новый полипептид по настоящему изобретению применяют к продуцирующему OPS микроорганизму, это может привести к продукции OPS с высоким выходом, и также к эффективной продукции L-цистеина этим микроорганизмом.
Наилучшее воплощение изобретения
Решение вышеизложенной задачи в настоящем изобретении подробно описано здесь ниже.
В одном аспекте настоящее изобретение относится к полипептиду, обладающему O-фосфосерин(OPS)-экспортирующей активностью, который представлен аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 3 и SEQ ID NO: 5.
В другом аспекте настоящее изобретение относится к применению полипептида, который представлен аминокислотной последовательностью, выбранной группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 3 и SEQ ID NO: 5, для экспортирования О-фосфосерина.
Как его используют здесь, термин "О-фосфосерин" (здесь и далее "OPS") относится к сложному эфиру фосфорной кислоты и серина, который служит в качестве составляющего компонента во многих белках. В частности, OPS является предшественником L-цистеина и может быть превращен в цистеин в результате взаимодействия с сульфидом при каталитическом воздействии сульфгидрилазы OPS (здесь и далее "OPSS").
Как его используют здесь, термин "полипептид, обладающий OPS-экспортирующей активностью", относится к мембранному белку, который обладает активностью экспортирования OPS в клетке наружу из клетки, и в частности, он может относиться к мембранному белку, полученному из Е. coli. Полипептид по настоящему изобретению, обладающий OPS-экспортирующей активностью, может представлять собой транспортер YhhS суперсемейства "major facilitator" (MFS) или его вариант. В частности, полипептид может представлять собой вариант транспортера YhhS MFS, проявляющий улучшенную активность по сравнению с таковой транспортера YhhS MFS дикого типа, который был идентифицирован как белок, обладающий OPS-экспортирующей активностью у Е. coli, где ингибирование роста устранено в условиях, где присутствует избыточное количество OPS.
В частности, полипептид может быть представлен аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 3 и SEQ ID NO: 5, и может включать, без ограничения, мембранные белки, обладающие гомологией последовательности с вышеуказанными последовательностями по меньшей мере 70%, в частности по меньшей мере 80%, более конкретно по меньшей мере 90%, и еще более конкретно по меньшей мере 95%, при условии, что они обладают OPS-экспортирующей активностью, которая по существу является такой же или эквивалентной таковой полипептида. Дополнительно, очевидно, что варианты полипептида, у которых часть последовательности делетирована, модифицирована, заменена или вставлена, должны быть включены в объем настоящего изобретения до тех пор, пока они представляют собой аминокислотные последовательности, имеющие такие гомологии и OPS-экспортирующую активность.
Как его используют здесь, термин "гомология" относится к степени идентичности или эквивалентности с данной полипептидной последовательностью или полинуклеотидной последовательностью, которая может иметь общее эволюционное происхождение или может не иметь общего эволюционного происхождения, и может быть указана в процентах. Как используется здесь, гомологичная последовательность, обладающая такой же или схожей активностью с данной полипептидной последовательностью, может быть указана в терминах "% гомологии". % гомологии может быть подтвержден с использованием стандартного программного обеспечения для расчета таких параметров, как сумма баллов (score), идентичность и сходство, в частности BLAST 2.0, или путем сравнения последовательностей путем экспериментов с гибридизацией по Саузерну в определенных строгих условиях, и определенные строгие условия гибридизации можно определить способом, известным специалисту в данной области техники (например, Sambrook et al., 1989, infra). В примере воплощения по настоящему изобретению, когда две различные аминокислотные последовательности имеют по меньшей мере 21% полипептидной последовательности, совпадающей на протяжении определенной длины аминокислотной последовательности (более конкретно, по меньшей мере примерно 50%, и в частности примерно 75%, 90%, 95%, 96%, 97% или 99%), они "по существу такие же" или "по существу те же самые".
Еще в одном аспекте настоящего изобретения предложен полинуклеотид, кодирующий полипептид, обладающий O-фосфосерин(OPS)-экспортирующей активностью, то есть полинуклеотид, кодирующий вариант полипептида транспортера YhhS MFS. В частности, в настоящем изобретении предложен полинуклеотид, кодирующий полипептид, обладающий О-фосфосерин-экспортирующей активностью, представленный аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 3 и SEQ ID NO: 5.
OPS, YhhS MFS транспортер и так далее являются такими, как описано выше.
Как его используют здесь, термин "полинуклеотид" относится к полимеру нуклеотидов, где нуклеотидные единицы соединены по типу длинной цепи ковалентными связями, и обычно относится к цепи ДНК или РНК, имеющей определенную минимальную длину.
Полинуклеотидная последовательность полипептида, обладающего OPS-экспортирующей активностью, может включать полинуклеотидную последовательность, кодирующую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 3 и SEQ ID NO: 5. В отношении кодонов, предпочтительных для организмов для экспрессии полипептида на основе вырожденности генетического кода, различные модификации полинуклеотида могут быть выполнены в кодирующей области в рамках объема изобретения без изменения аминокислотной последовательности полипептида. Полинуклеотидная последовательность может включать, например, полинуклеотидные последовательности, выбранные из группы, состоящей из SEQ ID NO: 2, SEQ ID NO: 4 и SEQ ID NO: 6. Дополнительно полинуклеотидная последовательность может включать нуклеотидные последовательности, имеющие гомологию последовательности 70% или больше, в частности 80% или больше, более конкретно 90% или больше, еще более конкретно 95% или больше, и наиболее точно 98% или больше, с вышеуказанными последовательностями, которые кодируют полипептиды, по существу обладающие OPS-экспортирующей активностью. Дополнительно, очевидно, что варианты полипептида, в которых часть последовательности делетирована, модифицирована, заменена или вставлена, должны быть включены в объем настоящего изобретения.
Еще в одном аспекте настоящего изобретения предложен микроорганизм, продуцирующий полипептид, обладающий О-фосфосерин-экспортирующей активностью, то есть OPS-продуцирующий микроорганизм экспрессирует варианты полипептида транспортера YhhS MFS, и в частности, микроорганизм рода Escherichia.
Еще в одном аспекте настоящего изобретения предложено применение полипептида, обладающего О-фосфосерин-экспортирующей активностью, то есть продуцирование OPS с использованием микроорганизма, экспрессирующего варианты полипептида транспортера YhhS MFS, и в частности, микроорганизма рода Escherichia.
В настоящем изобретении, OPS, YhhS MFS транспортеры и так далее являются такими же, как описано выше.
Как его используют здесь, термин "OPS-продуцирующий микроорганизм" относится к штамму прокариотического или эукариотического микроорганизма, способного продуцировать OPS in vivo, и в частности, микроорганизма, который может аккумулировать OPS в среде или внутри микроорганизма вследствие генетической манипуляции или природной мутации.
В частности, микроорганизм специально не ограничен, но может представлять собой любой прокариотический или эукариотический микроорганизм, который может экспрессировать полипептид, выбранный из группы, состоящий из SEQ ID NO: 1, SEQ ID NO: 3 и SEQ ID NO: 5, и в частности, может представлять собой прокариотический микроорганизм, например, микробные штаммы, принадлежащие роду Escherichia, роду Erwinia, роду Serratia, роду Providencia, роду Corynebacterium и роду Brevibacterium, и в частности, микроорганизм, принадлежащий роду Escherichia, например Е. coli, но не ограничиваясь ими.
Как здесь используют этот термин, "экспрессия" может быть достигнута путем трансформации с использованием рекомбинантного вектора, который функциональным образом включает полинуклеотид, кодирующий полипептид по настоящему изобретению, или путем вставки полинуклеотида, кодирующего полипептид, в хромосому, но не ограничиваясь этим.
Как его используют здесь, термин "трансформация" относится к процессу введения вектора, включающего полинуклеотид, кодирующий белок-мишень, в клетку-хозяина, тем самым предоставляя возможность экспрессии полинуклеотида, кодирущего белок, в клетке-хозяине. Для трансформированного полинуклеотида не имеет значения, вставлен ли он в хромосому клетки-хозяина и локализован в ней или локализован вне хромосомы при условии, что он может экспрессироваться в клетке-хозяине. Дополнительно полинуклеотид включает ДНК и РНК, которые кодируют белок-мишень. Полинуклеотид может быть вставлен в любой форме с тем, чтобы он мог быть введен в клетку-хозяина и мог бы экспрессироваться в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, которая представляет собой генетическую конструкцию, включающую все существенные элементы, необходимые для самостоятельной экспрессии, но не ограничиваясь этим. Экспрессионная кассета традиционно может включать промотор, функционально связанный с полинуклеотидом, сигнал терминации транскрипции, домен связывания с рибосомой и сигнал терминации трансляции. Экспрессионная кассета может быть в форме вектора экспрессии, способного к саморепликации. Дополнительно полинуклеотид может быть введен в клетку-хозяина как есть и функционально связан с последовательностью, необходимой для его экспрессии в клетке-хозяине.
Дополнительно, как его используют здесь, термин "функционально связанный" относится к функциональной связи между промоторной последовательностью, которая инициирует и опосредует транскрипцию полинуклеотида, кодирующего белок-мишень по настоящему изобретению, и вышеописанной генной последовательностью.
Как его используют здесь, термин "вектор" относится к любому посреднику для клонирования и/или переноса нуклеотидной последовательности в клетку-хозяина. Вектор может представлять собой репликон, с которым может быть связан другой фрагмент ДНК, приводя к репликации объединенного фрагмента. Как его используют здесь, термин "репликон" относится к любой генетической единице, которая функционирует как автономная единица репликации ДНК, то есть к такой, которая может реплицироваться путем саморегуляции (например плазмиды, фаги, космиды, хромосомы и вирусы). Вектор может включать вирусные и невирусные посредники для введения нуклеотида в клетку-хозяина in vivo, ex vivo или in vitro и также может включать миникольцевую ДНК. Например, вектор может включать плазмиды, которые не имеют никакой последовательности бактериальной ДНК. Удаление последовательностей бактериальной ДНК, которые богаты доменами CpG, было проведено для уменьшения экспрессионного "молчания" перенесенного гена, и индуцирования конститутивной экспрессии по сравнению с плазмидным ДНК-вектором (например, Ehrhardt, A. et al. (2003) HumGene Ther 10: 215-25; Yet, N. S. (2002) MoI Ther 5: 731-38; Chen, Z. Y. et al. (2004) Gene Ther 11: 856-64). Дополнительно вектор может включать транспозоны (Annu Rev Genet. 2003; 37: 3-29.) или искусственные хромосомы. В частности, можно использовать векторы pACYC177, pACYC184, pCL1920, pECCG117, pUC19, pBR322 и pMW118 и векторы с их модифицированными промоторами, но не ограничиваясь ими.
Вектор может представлять собой ДНК-конструкцию, включающую полинуклеотидную последовательность полинуклеотида, кодирующего белок-мишень, которая функциональным образом связана с подходящей регуляторной последовательностью так, что белок-мишень может экспрессироваться в подходящем хозяине. Регуляторная последовательность включает промотор, способный инициировать транскрипцию, произвольную операторную последовательность для регуляции транскрипции, последовательность, кодирующую подходящую мРНК домена связывания с рибосомой и последовательность для регуляции транскрипции и трансляции. Вектор, после трансформации им подходящей клетки-хозяина, может реплицироваться или функционировать независимо от генома хозяина или может самостоятельно интегрироваться в геном хозяина.
Вектор, используемый в настоящем изобретении, можно специально не ограничивать при условии, что вектор способен реплицироваться в клетке-хозяине, и можно использовать любой вектор, известный в данной области техники. Примеры вектора могут включать природные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, в качестве фагового вектора или космидного вектора можно использовать pWE15, М13, λMBL3, λMBL4, λIXII, λASHII, λAPII, λt10, λt11, Charon4A, Charon21A и так далее, но не ограничиваясь ими; и в качестве плазмидного вектора можно использовать таковые на основе pBR, pUC, pBluescriptII, pGEM, pTZ, pCL, pET и так далее, но не ограничиваясь ими.
Дополнительно в OPS-продуцирующем микроорганизме активность фосфосерин-фосфатазы (SerB) может быть дополнительно ослаблена по сравнению с ее эндогенной активностью.
SerB обладает активностью превращения OPS в L-серин, и таким образом микроорганизм, модифицированный для уменьшения активности SerB, обладает свойством аккумулировать в себе OPS, таким образом являясь полезным для продукции OPS. SerB может представлять собой белок, имеющий аминокислотную последовательность, представленную в SEQ ID NO: 16, но не ограничиваясь этим. Дополнительно SerB может включать аминокислотную последовательность, имеющую идентичность последовательности 80% или больше, в частности 90% или больше, более конкретно 95% или больше и еще более конкретно 99% или больше, при условии что она проявляет активность SerB, но не ограничиваясь этим. Дополнительно полинуклеотидная последовательность, кодирующая SerB, может иметь полинуклеотидную последовательность, кодирующую аминокислоты, представленные SEQ ID NO: 16.
Что касается кодонов, предпочтительных для организмов для экспрессии полипептида на основе вырожденности генетического кода, то различные модификации полинуклеотида могут быть выполнены в кодирующей области в рамках объема изобретения без изменения аминокислотной последовательности полипептида. Полинуклеотидная последовательность может включать аминокислотную последовательность, например таковую, представленную SEQ ID NO: 17, и может включать нуклеотидные последовательности, имеющие гомологию последовательности 80% с последовательностью и в частности 90% или больше, но не ограничиваясь этим.
Как его используют здесь, термин "ослабление активности" относится к снижению активности белка по сравнению с таковой, которой обладает микроорганизм в его состоянии дикого типа или до модификации, и также он включает случай, когда активность устранена.
"Ослабление" представляет собой концепцию, относящуюся к случаю, когда активность белка уменьшена или устранена по сравнению с эндогенной активностью у микроорганизма вследствие модификации в гене, кодирующем белок, и так далее; к случаю, когда уровень экспрессии белка ниже, чем таковой у штамма дикого типа микроорганизма вследствие ингибирования экспрессии или ингибирования трансляции гена, кодирующего этот же белок, и так далее; к случаю, когда ген вообще не экспрессируется, и к случаю, когда ген экспрессируется, но не проявляет активность.
Ослабление активности белка может быть достигнуто различными способами, хорошо известными в данной области техники. Примеры способов могут включать способ замены в хромосоме кодирующего белок гена на ген, мутированный таким образом, что ферментативная активность может быть снижена, включая случай, когда активность белка устранена; способ модифицирования последовательности регуляции экспрессии гена, кодирующего белок; способ делетирования части или всего гена, кодирующего белок, в хромосоме; способ введения антисмыслового олигонуклеотида (например антисмысловой РНК), что ингибирует трансляцию с мРНК в белок путем комплементарного связывания с транскриптом гена на хромосоме; способ, делающий присоединение рибосомы невозможным вследствие образования вторичной структуры путем искусственного добавления комплементарной последовательности к последовательности Шайна-Дальгарно (SD) на переднем конце последовательности SD гена, кодирующего этот белок; способ конструирования с помощью технологии обратной транскрипции (RTE), при котором промотор добавляют таким образом, чтобы он подвергался обратной транскрипции на 3'-конце открытой рамки считывания (ORF) соответствующей последовательности, и так далее, и также включают их комбинацию, но специально не ограничиваясь этим.
В частности, способ делетирования гена, кодирующего белок, частично или полностью, может быть выполнен путем замены полинуклеотида, кодирующего эндогенный белок-мишень, в хромосоме на полинуклеотид или маркерный ген, имеющий частично делетированную последовательность нуклеиновой кислоты, и используя вектор для вставки хромосом в бактерии. В примере воплощения ген может быть делетирован путем гомологичной рекомбинации. Дополнительно, как его используют здесь, термин "часть", хотя он может варьировать в зависимости от типов полинуклеотида, может в частности относиться к последовательности размером от 1 нуклеотида до 300 нуклеотидов, более конкретно от 1 нуклеотида до 100 нуклеотидов и еще более конкретно от 1 нуклеотида до 50 нуклеотидов, но специально не ограничиваясь этим.
Дополнительно, способ модифицирования последовательности регуляции экспрессии может быть выполнен путем индуцирования модификации в последовательности регуляции экспрессии путем делеции, вставки, консервативной замены, неконсервативной замены или их комбинации, с тем, чтобы дополнительно ослабить активность последовательности регуляции экспрессии; или путем замены последовательности на последовательность нуклеиновой кислоты, обладающей более слабой активностью. Последовательность регуляции экспрессии включает промотор, операторную последовательность, последовательность, кодирующую домен связывания с рибосомой, и последовательность для регуляции транскрипции и трансляции.
Дополнительно способ модифицирования последовательности гена может быть выполнен путем индукции модификации в последовательности гена путем делеции, вставки, консервативной замены, неконсервативной замены или их комбинации с тем, чтобы дополнительно ослабить активность белка; или путем замены последовательности на последовательность гена, улучшенную для обладания более слабой активностью, или на последовательность гена, улучшенную так, чтобы совсем не обладать активностью.
В частности, ослабление активности SerB в настоящем изобретении может быть достигнуто по меньшей мере одним способом, выбранным из группы, состоящей из способа удаления активности SerB; способа замены SerB-кодирующего гена в хромосоме на ген, мутированный таким образом, что активность SerB может быть уменьшена; способа введения модификации в последовательность регуляции экспрессии SerB-кодирующего гена в хромосоме; способа замены последовательности регуляции экспрессии SerB-кодирующего гена на последовательность, обладающую более слабой активностью; способа делетирования SerB-кодирующего гена в хромосоме и способа введения антисмыслового олигонуклеотида, который ингибирует трансляцию с мРНК в белок путем комплементарного связывания с транскриптом SerB-кодирующего гена в хромосоме; способа, делающего присоединение рибосомы невозможным вследствие образования вторичной структуры путем искусственного добавления комплементарной последовательности к последовательности Шайна-Дальгарно (SD) на переднем конце последовательности SD кодирующего белок гена; способа, делающего присоединение рибосомы невозможным вследствие образования вторичной структуры путем искусственного добавления комплементарной последовательности к последовательности Шайна-Дальгарно (SD) на переднем конце последовательности SD SerB-кодирующего гена; и способа конструирования с помощью технологии обратной транскрипции (RTE), который добавляет промотор так, чтобы он подвергался обратной транскрипции на 3'-конце открытой рамки считывания (ORF) соответствующей последовательности.
Дополнительно, OPS-продуцирующий микроорганизм может представлять собой таковой, у которого активности фосфоглицерат-дегидрогеназы (SerA) или фосфосерин-аминотрансферазы (SerC) дополнительно усилены по сравнению с их соответствующими эндогенными активностями.
SerA представляет собой белок, способный превращать 3-фосфоглицерат в 3-фосфогидроксипируват.SerA можно использовать как форму дикого типа или варианта, свободного от ингибирования по типу обратной связи серином. Кроме того, SerC представляет собой белок, способный превращать 3-фосфогидроксипируват в OPS. Соответственно, любой микроорганизм с усиленными активностями SerA и/или SerC можно эффективно использовать в качестве OPS-продуцирующего микроорганизма.
SerA может иметь аминокислотную последовательность, представленную SEQ ID NO: 18 или SEQ ID NO: 19, хотя он не ограничен этим. SEQ ID NO: 18 представляет собой последовательность SerA дикого типа, и SEQ ID NO: 19 представляет собой последовательность варианта, свободного от ингибирования по типу обратной связи серином. Дополнительно, те аминокислотные последовательности, которые имеют по меньшей мере 80% идентичности последовательности с вышеупомянутыми аминокислотами, в частности по меньшей мере 90%, более конкретно по меньшей мере 95%, и еще более конкретно по меньшей мере 99%, могут быть включены при условии, что они проявляют активности SerA дикого типа или вариантов SerA, свободных от ингибирования по типу обратной связи серином, но не ограничиваясь этим. Варианты, свободные от ингибирования по типу обратной связи, представляют собой белки, в которые введена модификация в SerA-кодирующий ген путем вставки, замены и так далее, тем самым делая возможным поддержание активности от ингибирования по типу обратной связи серином или глицином, или имеющие повышенные их активности, и такие варианты, которые свободны от ингибирования по типу обратной связи, уже хорошо известны (Grant GA et al., J. Biol. Chem., 39: 5357-5361, 1999; Grant GA et al., Biochem., 39: 7316-7319, 2000; Grant GA et al., J. Biol. Chem., 276: 17844-17850, 2001; Peters-Wendisch P et al., Appl. Microbiol. Biotechnol., 60: 437-441, 2002; EP Pat. No. EP 0943687 B).
Дополнительно, полинуклеотидная последовательность, кодирующая SerA дикого типа или варианты, свободные от ингибирования по типу обратной связи серином, могут представлять собой полинуклеотидную последовательность, кодирующую любую аминокислотную последовательность, представленную SEQ ID NO: 18 или SEQ ID NO: 19, но не ограничиваясь этим. Вследствие вырожденности генетического кода или учитывая кодоны, предпочтительные для организмов для экспрессии полипептида, различные модификации полинуклеотида могут быть выполнены в кодирующей области в рамках объема изобретения без изменения аминокислотной последовательности полипептида. Полинуклеотидная последовательность может представлять собой, например, любую полинуклеотидную последовательность, представленную SEQ ID NO: 20 или SEQ ID NO: 25, и может иметь нуклеотидную последовательность, имеющую гомологию с этими полинуклеотидными последовательностями по меньшей мере 80%, и в частности по меньшей мере 90%, но не ограничиваясь этим.
SerC может представлять собой белок, имеющий аминокислотную последовательность, которая представлена, например SEQ ID NO: 21, но не ограничиваясь этим. Дополнительно, аминокислотная последовательность, при условии, что она проявляет активность SerC, также может включать аминокислотные последовательности, которые имеют идентичность последовательности с вышеупомянутой аминокислотной последовательностью по меньшей мере 80%, в частности по меньшей мере 90%, более конкретно по меньшей мере 95% и еще более конкретно по меньшей мере 99%, но не ограничиваясь этим.
Дополнительно, полинуклеотидная последовательность, кодирующая SerC, может представлять собой полинуклеотидную последовательность, кодирующую аминокислотную последовательность, представленную SEQ ID NO: 21. Вследствие вырожденности генетического кода или учитывая кодоны, предпочтительные для организмов для экспрессии полипептида, различные модификации полинуклеотида могут быть выполнены в кодирующей области в пределах объема изобретения без изменения аминокислотной последовательности полипептида. Полинуклеотидная последовательность может представлять собой, например, таковую, представленную SEQ ID NO: 22, и может иметь нуклеотидную последовательность, имеющую гомологию с этой полинуклеотидной последовательностью по меньшей мере 80%, и в частности по меньшей мере 90%, но не ограничиваясь этим.
Как его используют здесь, термин "эндогенная активность" относится к активному состоянию полипептида у микроорганизма в состоянии дикого типа или состоянии до модификации. Как его используют здесь, термин "усиление по сравнению с его эндогенной активностью" относится к увеличенной активности полипептида у микроорганизма по сравнению с таковой, которой обладает микроорганизм в состоянии дикого типа или в состоянии до модификации, и представляет собой концепцию, включающую предоставление активности определенного полипептида микроорганизму, который не обладает активностью этого определенного полипептида.
Как его используют здесь, термин "усиление активности" относится, хотя специально не ограничиваясь этим, не только к достижению более сильного действия по сравнению с исходной функцией вследствие увеличения активности самого полипептида, но также к увеличению его активности вследствие увеличения ферментативной активности путем увеличения активности эндогенного гена, амплификации эндогенного гена с помощью внутренних или внешних факторов, замены, модификации или мутации промотора и так далее. В частности, усиление активности может быть выполнено такими способами, как способ увеличения числа копий кодирующего полипептид гена в клетке, способ модификации регуляторной последовательности кодирующего полипептид гена, способ замены кодирующего полипептид гена в хромосоме мутированным геном для увеличения активности полипептида, способ введения модификации в кодирующий полипептид ген в хромосоме для усиления активности полипептида и так далее, но способы не ограничены этим. Эти способы усиления активности можно упомянуть таким же образом для усиления активностей других полипептидов по настоящему изобретению.
В частности, усиление активности по настоящему изобретению может быть достигнуто по меньшей мере одним способом, выбранным из группы, состоящей из способа увеличения числа копий гена, кодирующего SerA или SerC, в клетке; способа введения модификации в регуляторную последовательность гена, кодирующего SerA или SerC, в хромосоме; способа замены регуляторной последовательности гена, кодирующего SerA или SerC, в хромосоме на последовательность, обладающую сильной активностью; способа замены гена, кодирующего SerA или SerC, в хромосоме на мутированный ген для увеличения активности SerA или SerC и способа введения модификации в ген, кодирующий SerA или SerC, в хромосоме для усиления активности SerA или SerC.
В вышесказанном увеличение числа копий гена, хотя специально не ограничиваясь этим, может быть выполнено в форме функциональной связи с вектором или путем вставки в хромосому в клетке-хозяине. В частности, этот способ может быть выполнен путем введения вектора, с которым полинуклеотид, кодирующий белок по настоящему изобретению, связан функциональным образом, и который может реплицироваться и функционировать независимо от хозяина, в клетку хозяина; или введения вектора, с которым полинуклеотид связан функциональным образом, способного вставлять полинуклеотид в хромосому клетки-хозяина, в клетку-хозяина. Вставка полинуклеотида в хромосому может быть выполнена с использованием способа, известного в данной области техники, например путем гомологичной рекомбинации.
Затем модификация последовательности регуляции экспрессии для усиления экспрессии полипептида, хотя специально не ограничиваясь этим, может быть выполнена путем индуцирования модификации в полинуклеотидной последовательности посредством делеции, вставки, консервативной замены, неконсервативной замены или их комбинации с тем, чтобы дополнительно усилить активность последовательности регуляции экспрессии; или путем замены полинуклеотидной последовательности на последовательность нуклеиновой кислоты с более сильной активностью. Последовательность регуляции экспрессии, хотя специально не ограничиваясь этим, может включать промотор, операторную последовательность, последовательность, кодирующую домен связывания с рибосомой, и последовательность для регуляции терминации транскрипции и трансляции и так далее.
Сильный промотор может быть присоединен к верхнему краю экспрессионной единицы полинуклеотида вместо исходного промотора, но не ограничиваясь этим. Примеры известных сильных промоторов могут включать промотор cj1 (патент Кореи №10-0620092), промотор lac, промотор trp, промотор trc, промотор tac, промотор фага лямбда PR, промотор PL и промотор tet, но не ограничиваясь этим.
Дополнительно, модификация полинуклеотидной последовательности в хромосоме, хотя специально не ограничиваясь этим, может быть выполнена путем индуцирования модификации в последовательности регуляции экспрессии полинуклеотидной последовательности путем делеции, вставки, консервативной замены, неконсервативной замены или их комбинации с тем, чтобы дополнительно усилить активность полинуклеотидной последовательности; или путем замены полинуклеотидной последовательности на усиленную полинуклеотидную последовательность, обладающую более сильной активностью.
Обычно введение и усиление активности белка может увеличивать активность или концентрацию соответствующего белка относительно активности или концентрации белка дикого типа или в штамме микроорганизма на начальной стадии на величину по меньшей мере от 1%, 10%, 25%, 50%, 75%, 100%, 150%, 200%, 300%, 400% или 500% максимально до 1000% или 2000%, но не ограничиваясь этим.
Дополнительно, микроорганизм может представлять собой микроорганизм, который дополнительно ослаблен в отношении его способности вводить OPS в клетку или разлагать. В частности, активности PhnC/PhnD/PhnE алкилфосфонат ABC - транспортера (оперон PhnCDE, в частности АТР (аденозинтрифосфат)-связывающий компонент транспортера фосфоната (PhnC; EG 10713)-периплазматический связывающий белковый компонент Pn-транспортера (PhnD; EG 10714)-интегральный мембранный компонент ABC-транспортера алкилфосфоната (PhnE; EG 11283)), щелочной фосфатазы (PhoA) или кислой фосфатазы (AphA) могут быть ослаблены по сравнению с их эндогенными активностями.
Более того, микроорганизм по настоящему изобретению может быть дополнительно усилен активностью нуклеотид-трансгидрогеназы (PntAB; ЕС 1.6.1.1). PntAB, как описано в более раннем источнике (Sauer U Р et al., J Biol Chem.20; 279(8): 6613-9. Epub 2003), вовлечена в метаболизм NADPH и контролирует внутриклеточный окислительно-восстановительный баланс.
В отношении содержания OPS-продуцирующего микроорганизма, описания патента Кореи №1381048 или публикации заявки на патент США №2012-0190081 могут быть использованы в качестве ссылок по настоящему изобретению, в дополнение к таковым, описанным выше.
Еще в одном аспекте настоящего изобретения предложен способ продуцирования OPS, включающий культивирование OPS-продуцирующего микроорганизма для экспрессии полипептида, обладающего OPS-экспортирующей активностью, то есть варианта полипептида транспортера YhhS MFS, в среде.
В настоящем изобретении OPS, полипептид, обладающий OPS-экспортирующей активностью, транспортер YhhS MFS и OPS-продуцирующий микроорганизм являются такими, как описано выше.
В частности, способ может включать культивирование OPS-продуцирующего микроорганизма для экспрессии полипептида, представленного аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 3 и SEQ ID NO: 5, и выделение OPS из OPS-продуцирующего микроорганизма или из среды с вышеизложенной стадии, но способ не ограничен этим.
Как его используют здесь, термин "культивирование" относится к выращиванию микроорганизма в заданных подходящим образом условиях. Процесс культивирования может быть выполнен в соответствии с подходящей средой и условиями для культивирования, известными в данной области техники. Процесс культивирования может быть легко скорректирован для применения специалистом в данной области техники в соответствии с выбранным штаммом. В частности, культивирование может представлять собой периодическое культивирование, непрерывное культивирование и культивирование с подпиткой, но не ограничиваясь этим.
При культивировании рекомбинантного микроорганизма, обладающего пониженной активностью SerB по сравнению с его эндогенной активностью, среда может дополнительно содержать глицин или серии, потому что у рекомбинантного микроорганизма индуцирована потребность в серине. Глицин может быть предоставлен в форме очищенного глицина, глицин-содержащего дрожжевого экстракта или триптона. Концентрация глицина, которая должна быть в среде, обычно составляет от 0,1 г/л до 10 г/л, и в частности от 0,5 г/л до 3 г/л. Дополнительно серии может быть предоставлен в форме очищенного серина, серин-содержащего дрожжевого экстракта или триптона. Концентрация серина, которая должна быть в среде, обычно составляет от 0,1 г/л до 5 г/л, и в частности от 0,1 г/л до 1 г/л.
Примеры источника углерода, которые должны содержаться в среде, могут включать сахариды и углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, мальтоза, крахмал и целлюлоза; масла и жиры, такие как соевое масло, подсолнечное масло, касторовое масло и кокосовое масло; жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота; спирты, такие как глицерол и этанол; и органические кислоты, такие как уксусная кислота. Эти источники углерода можно использовать по отдельности или в комбинации, но не ограничиваясь этим. Примеры источников азота, которые должны содержаться в среде, могут включать органические источники азота, такие как пептон, дрожжевой экстракт, мясной бульон, солодовый экстракт, жидкий кукурузный экстракт и соевую муку; и неорганические источники азота, такие как мочевина, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Эти источники азота можно использовать по отдельности или в комбинации, но не ограничиваясь этим. В качестве источника фосфора культуральная среда может дополнительно содержать дигидрофосфат калия, дикалийгидрофосфат и соответствующие натрийсодержащие соли, но не ограничиваясь этим. Культуральная среда может содержать металлы, такие как сульфат магния и сульфат железа. Дополнительно могут содержаться аминокислоты, витамины и подходящие предшественники. Эти культуральные среды или предшественники можно добавлять в культуру в форме периодической культуры или непрерывной культуры, но не ограничиваясь этим.
Дополнительно рН культуры можно регулировать путем добавления такого соединения как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота и серная кислота, во время культивирования подходящим способом. Дополнительно, образование пузырей можно предотвратить во время культивирования, используя пеногаситель, такой как полигликолевый эфир жирной кислоты. Дополнительно газообразный кислород или газ, содержащий кислород, можно добавлять в культуру для поддержания аэробных условий в культуральной среде; или газообразный азот, газообразный водород или диоксид углерода можно вводить для поддержания анаэробных или микроаэробных условий. Температура в культуре может находиться в диапазоне от 27°С до 37°С, и в частности от 30°С до 35°С. Культивирование можно продолжать до тех пор, пока не будет достигнута продукция желаемого вещества, и в частности от 10 часов до 100 часов, но не ограничиваясь этими иллюстративными примерами.
В настоящем изобретении OPS, продуцированный во время культивирования, может быть далее выделен и очищен. Целевой OPS может быть выделен из культуры с использованием подходящего способа, известного в данной области техники, в соответствии со способом культивирования, например периодического культивирования, непрерывного культивирования и культивирования с подпиткой, но не ограничиваясь этим.
Еще в одном аспекте настоящего изобретения предложен способ продуцирования цистеина или его производного, который включает культивирование OPS-продуцирующего микроорганизма, экспрессирующего полипептид, обладающий OPS-экспортирующий активностью, то есть вариант полипептида транспортера YhhS MFS, в среде для продуцирования О-фосфосерина; и осуществление взаимодействия между продуцированным таким образом OPS и сульфидом в присутствии О-фосфосерин-сульфгидрилазы (OPSS) или микроорганизма, экспрессирующего OPSS.
Дополнительно в настоящем изобретении предложен способ продуцирования цистеина или его производного, включающий культивирование OPS-продуцирующего микроорганизма, экспрессирующего полипептид, обладающий OPS-экспортирующей активностью, то есть вариант полипептида транспортера YhhS MFS, в среде для продуцирования О-фосфосерина; выделение OPS из OPS-продуцирующего микроорганизма или из среды со стадии выше; и осуществление взаимодействия между продуцированным таким образом OPS и сульфидом в присутствии OPSS или микроорганизма, экспрессирующего OPSS.
В настоящем изобретении OPS, полипептид, обладающий OPS-экспортирующей активностью, транспортеры YhhS MFS и OPS-продуцирующий микроорганизм являются такими, как описано выше.
Как его используют здесь, термин "О-фосфосерин-сульфгидрилаза (OPSS)" относится к полипептиду, который катализирует реакцию, в которой тиольная группа (SH) предоставляется для OPS для превращения OPS в цистеин. Впервые этот фермент был обнаружен у Aeropyrum pernix, Mycobacterium tuberculosis, Mycobacterium smegmatics и Trichomonas vaginalis (Mino К and Ishikawa K, FEBS Letters, 551: 133-138, 2003; Burns KE et al., J. Am. Chem. Soc, 127: 11602-11603, 2005). Дополнительно, спектр OPSS включает не только белок OPSS дикого типа, но также варианты, которые включают делению, замену или добавление в части полинуклеотидной последовательности, кодирующей OPSS, которые проявляют активность равную или превышающую биологическую активность белка OPSS дикого типа, и также включает все белки OPSS, раскрытые в патентах Кореи №1381048 и 1208267, и их варианты.
Сульфид для применения по настоящему изобретению может представлять собой любой сульфид, предоставленный не только в твердой форме, обычно используемой в данной области техники, но также в жидкой или газообразной форме вследствие разницы рН, давления и растворимости и, следовательно, может быть превращен в тиольную (SH) группу в форме, например, сульфида (S2-) или тиосульфата (S2O32-). В частности, сульфид для применения по настоящему изобретению может включать Na2S, NaSH, H2S, (NH4)2S и Na2S2O3, которые могут предоставлять тиольную группу для OPS, но не ограничиваясь этим. В этой реакции одна тиольная группа предоставляется для одной реакционноспособной группы OPS с получением одного цистеина или его производного. В этой реакции сульфид добавляют, в частности, в количестве 0,1-3 молярных эквивалентов и, в частности, 1-2 эквивалентов на основе молярной концентрации OPS, но не ограничиваясь этим. Экономически оптимальное превращение может происходить, когда OPS и сульфид, предоставляющий тиольные группы, предоставлены в молярном отношении 1:1 (один к одному).
Кроме того, способ по настоящему изобретению может дополнительно включать выделение и очистку цистеина, продуцированного на вышеописанной реакционной стадии. В частности, целевой цистеин может быть получен путем выделения и очистки из реакционного раствора с использованием подходящей реакции, известной в данной области техники.
Дополнительно, цистеин, продуцированный согласно способу по настоящему изобретению, можно легко синтезировать путем реакции химического синтеза, известной в данной области техники.
Как его используют здесь, термин "производные" относится к похожим соединениям, полученным путем химической модификации части любого соединения. Обычно этот термин относится к соединениям, у которых атом водорода или определенная атомная группа заменены на другой атом водорода или атомную группу.
Как его используют здесь, термин "производные цистеина" относится к соединениям, у которых атом водорода или определенная атомная группа в цистеине заменены на другой атом или атомную группу. Например, производные цистеина могут иметь форму, в которой к атому азота аминогруппы (-NH2) или атому серы тиольной группы (-SH) цистеина присоединен другой атом или атомная группа. Примеры производных цистеина могут включать N-ацетилцистеин (NAC), S-карбоксиметилцистеин (SCMC), Boc-Cys(Me)-OH, (R)-S-(2-амино-2-карбоксиэтил)-L-гомоцистеин, (R)-2-амино-3-сульфопропионовую кислоту, D-2-амино-4-(этилтио)масляную кислоту, 3-сульфино-L-аланин, Fmoc-Cys(Boc-метил)-OH, селено-L-цистеин, S-(2-тиазолил)-L-цистеин, S-(2-тиенил)-L-цистеин, S-(4-толил)-L-цистеин, но не ограничиваясь этим. Из цистеина можно легко синтезировать N-ацетилцистеин (NAC) путем взаимодействия с агентом ацетилирования, и в основных условиях из него можно синтезировать S-карбоксиметилцистеин (SCMC) путем взаимодействия с галогеноуксусной кислотой. Эти производные цистеина применяются главным образом в качестве фармацевтических материалов для средств против кашля, средств, облегчающих кашель, и терапевтических средств против бронхита, бронхиальной астмы, ларингофарингита и так далее.
Еще в одном аспекте настоящего изобретения предложено применение экспортирования OPS полипептидом, который представлен аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 3 и SEQ ID NO: 5.
ПРИМЕРЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Здесь и далее настоящее изобретение будет описано более подробно со ссылкой на следующие Примеры. Однако эти Примеры предназначены исключительно для иллюстративных целей, и их не следует рассматривать как ограничивающие объем настоящего изобретения.
Пример 1: Получение вариантов транспортера YhhS суперсемейства "major facilitator" (MFS)
Для улучшения активности OPS-экспортирующего белка (экспортера) для улучшения OPS-экспортирующей активности OPS-продуцирующего штамма были получены варианты транспортера YhhS суперсемейства "major facilitator" (MFS) (SEQ ID NO: 23), недавно идентифицированного OPS-экспортирующего белка, и yhhS (SEQ ID NO: 24), кодирующего его гена. Подробно процесс описан здесь ниже.
Прежде всего, была сконструирована библиотека вариантов гена yhhS. Для этого была проведена PCR (полимеразная цепная реакция) со случайным мутагенезом (склонная к ошибкам PCR JENA) с использованием пары геноспецифических праймеров (SEQ ID NO: 7 и 8) на основе геномной ДНК Escherichia coli K12_W3110 (АТСС27325) в качестве матрицы. Полученные таким образом в результате мутагенеза генные фрагменты клонировали в векторе pCLPrhtB, где промотор rhtB (SEQ ID NO: 13), который подвергали PCR с использованием пары геноспецифических праймеров (SEQ ID NO: 14 и 15), был вставлен по сайту SacI-EcoRV вектора pCL1920 (GenBank No АВ236930). В частности, вектор pCLPrhtB разрезали EcoRV и PstI, и затем генные фрагменты, полученные в результате мутагенеза, клонировали в него с использованием набора для клонирования In-fusion (Clontech Laboratories, Inc.). Клонирование проводили при 50°С в течение 10 минут, таким образом завершая конструирование плазмидных библиотек вариантов pCL PrhtB yhhS.
Сконструированные таким образом библиотеки рекомбинантных плазмид исследовали путем скрининга с высоким выходом (HTS). В частности, базовый штамм, использованный для скрининга, представлял собой СА07-0012 (KCCM11121P), который представляет собой рекомбинантный микроорганизм, модифицированный для уменьшения активности эндогенной фосфосерин-фосфатазы (SerB) у штамма W3110 Е. coli дикого типа (патент Кореи №10-1381048; публикация заявки на патент США №2012-0190081).
Далее для получения вариантов с улучшенной OPS-экспортирующей активностью сконструированными таким образом плазмидными библиотеками трансформировали базовый штамм СА07-0012 путем электропорации, культивировали в среде, содержащей избыточное количество OPS, и были выбраны три колонии, свободные от ингибирования роста. Затем из этих трех выбранных колоний были получены плазмиды и проанализированы путем технологии секвенирования.
Как таковые были выбраны три варианта yhhS, которые вовлечены в устранение ингибирования роста в условиях добавления OPS, и они получили названия yhhS М2, yhhS М25 и yhhS М45, соответственно.
При анализе нуклеотидных последовательностей yhhS М2, yhhS М25 и yhhS М45 было подтверждено, что yhhS М2 имеет аминокислотную последовательность, представленную SEQ ID NO: 1, yhhS М25 имеет таковую, представленную SEQ ID NO: 3, и yhhS М45 имеет таковую, представленную SEQ ID NO: 5.
Пример 2: Подтверждение OPS-экспортирующей активности вариантов yhhS у OPS-продуцирующего штамма
2-1. Конструирование штамма с усиленным транспортером YhhS MFS с использованием СА07-0012 и оценка OPS-продуиирующей способности
Плазмидами, содержащими три различных варианта, идентифицированные в Примере 1, соответственно, трансформировали СА07-0012, OPS-продуцирующий штамм, путем метода электрического импульса, традиционно используемого в данной области техники. Были сконструированы как таковые OPS-продуцирующие штаммы с введенными вариантами yhhS, то есть CA07-0012/pCL-PrhtB-yhhS М2, CA07-0012/pCL-PrhtB-yhhS М25 и CA07-0012/pCL-PrhtB-yhhS М45, и они получили названия Escherichia coli СА07-0345, Escherichia coli CA07-0344 и Escherichia coli CA07-0346, соответственно. Дополнительно штаммы Escherichia coli CA07-0344, Escherichia coli CA07-0345 и Escherichia coli CA07-0346 были депонированы в Корейском Центре Культур Микроорганизмов (KCCM), признанным как международный орган депонирования согласно Будапештскому договору, 23 июля 2015 года, с номерами доступа KCCM11736P, KCCM11737P и KCCM11738P, соответственно.
Оценивали OPS-продуцирующие способности соответствующих штаммов.
В частности, каждый штамм высевали на твердую среду LB и культивировали в инкубаторе при 33°С в течение ночи. Штаммы, культивированные на твердой среде LB в течение ночи, инокулировали в 25 мл среды для титрования, показанной в Таблице 1 ниже, и культивировали в инкубаторе при 34,5°С на скорости 200 об./мин в течение 48 часов. Результаты представлены в Таблице 2 ниже.
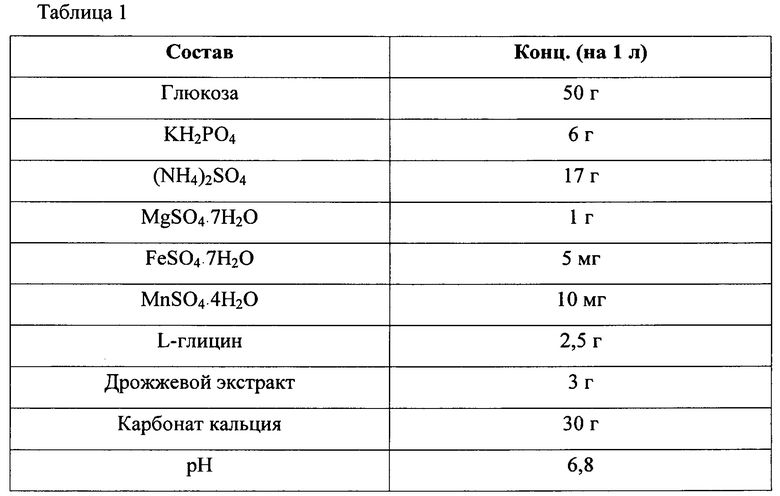
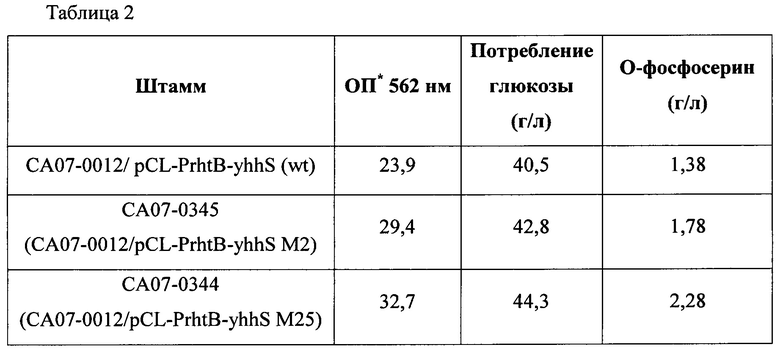

*ОП - оптическая плотность
wt - дикий тип
Как показано в Таблице 2 выше, в случае штаммов с введенными вариантами yhhS по настоящему изобретению, они показывали превосходный результат, демонстрируя увеличение продукции OPS до величины от 128% до 165% по сравнению со штаммом с введенным геном yhhS дикого типа. В частности, вариант yhhS М2 показывал увеличение до 128%, вариант yhhS М25 показывал увеличение до 165%, и вариант yhhS М45 показывал увеличение до 156% по сравнению с диким типом, соответственно.
2-2. Конструирование штамма с усиленным транспортером YhhS MFS с использованием штаммов с усиленными SerA и SerC и оценка OPS-продуцирующей способности
Для повторного подтверждения активности вариантов yhhS по настоящему изобретению использовали CA07-0022/pCL-Prmf-serA*(G336V)-serC (KCCM11103P, патент Кореи №10-1381048), который будучи OPS-продуцирующим штаммом с улучшенной OPS-продуцирующей способностью, имеет повышенную активность D-3-фосфоглицерат-дегидрогеназы (SerA) и 3-фосфосерин-аминотрансферазы (SerC) в пути биосинтеза OPS.
Для конструирования вектора генов pCL-Prmf-serA(G336V)-serC_PrhtB, каждый из вариантов yhhS амплифицировали с использованием пары праймеров (SEQ ID NO: 9 и 10) на основе варианта pCL-PrhtB-yhhS в качестве матрицы, и полученные в результате ампликоны клонировали по сайту рестрикции HindIII вектора pCL-Prmf-serA*(G336V)-serC.
В частности, штаммы, трансформированные каждой из плазмид традиционно применяемым методом электрического импульса, высевали на твердую среду LB и затем культивировали в инкубаторе при 33°С в течение ночи. Штаммы, культивированные на твердой среде LB в течение ночи, инокулировали в 25 мл среды для титрования, показанной в Таблице 1 выше, и культивировали в инкубаторе при 34,5°С на скорости 200 об./мин в течение 48 часов. Результаты показаны в Таблице 3 ниже.
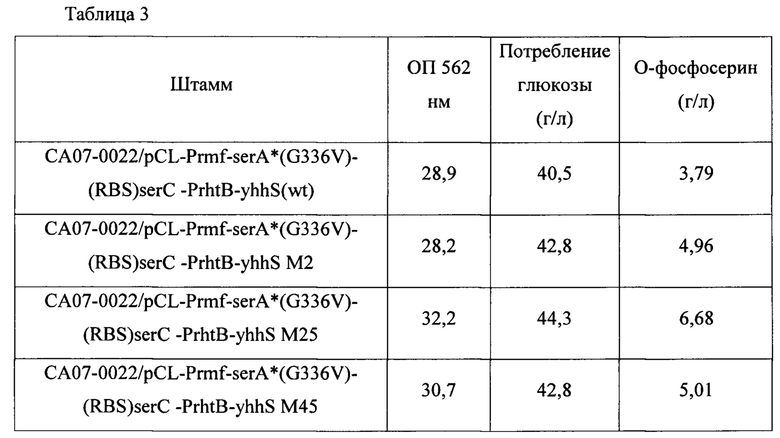
Как показано в Таблице 3 выше, было подтверждено, что когда варианты yhhS по настоящему изобретению были введены в OPS-продуцирующий штамм с усиленным геном биосинтеза OPS, продукция OPS была увеличена до величины от 130% до 176%. Эти результаты показывают, что варианты yhhS по настоящему изобретению можно эффективно использовать для продукции OPS.
2-3. Конструирование штамма с усиленным транспортером YhhS MFS в соответствии с силой промотора на хромосоме и оценка OPS-продуцирующей способности
Дополнительно к вышеизложенным экспериментам для подтверждения того, улучшается ли OPS-экспортирующая активность, когда варианты yhhS введены в хромосому, собственный промотор микроорганизма был заменен на промотор pCJ1 (патент Кореи №10-0620092) и были сконструированы штаммы с введенными вариантами по настоящему изобретению, и была оценена их OPS-продуцирующая способность. Введение промотора pCJ1 и вариантов в хромосому было выполнено с помощью способов, традиционно используемых в данной области техники. Прежде всего, на первой стадии для трансформации рекомбинантный вектор был вставлен в CA07-0022/pCL-Prmf-serA*(G336V)-serC (KCCM11103P, патент Кореи №10-1381048), OPS-продуцирующий штамм, с помощью метода электрического импульса (Appl Microbiol Biotechnol. 1999 Oct; 52(4):541-5). Затем, штаммы, претерпевшие вставку в хромосому путем рекомбинации гомологичных последовательностей, были отобраны в среде, содержащей 25 мг/л канамицина. Отобранные таким образом первичные штаммы подвергали второй стадии -кроссинговеру, и затем были отобраны штаммы, где промотор pCJ1 и варианты были заменены и вектор был удален.
Наконец, наличие замены промотора и варианов трансформированных штаммов было подтверждено путем проведения PCR с использованием пары праймеров (SEQ ID NO: 11 и 12).
Каждый из штаммов высевали на твердую среду LB и затем культивировали в инкубаторе при 33°С в течение ночи. Штаммы, культивированные на твердой среде LB в течение ночи, инокулировали в 25 мл среды для титрования, показанной в Таблице 1 выше, и культивировали в инкубаторе при 34,5°С на скорости 200 об./мин в течение 40 часов. Результаты представлены в Таблице 4 ниже.
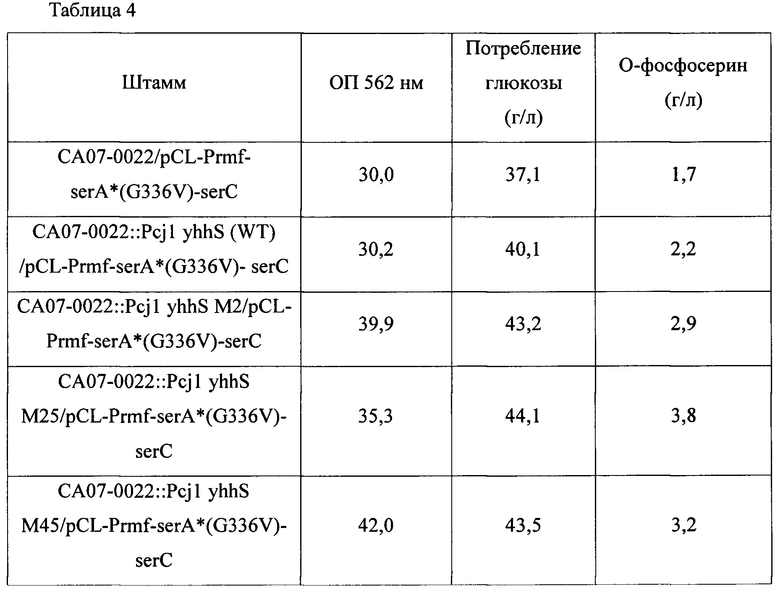
Как показано в Таблице 4 выше, было подтверждено, что когда активность каждого варианта белка в хромосоме была увеличена, уровень продукции OPS вариантом белка демонстрировал максимально 172% по сравнению с таковым штамма с введенным yhhS дикого типа.
На основании вышеизложенного специалист в области техники, к которой принадлежит данное изобретение, понимает, что настоящее изобретение может быть воплощено в других конкретных формах без отступления от технических идей или существенных признаков настоящего изобретения. В этом отношении примеры воплощений, раскрытые здесь, предназначены только для иллюстративных целей и их не следует рассматривать как ограничивающие объем настоящего изобретения. Напротив, настоящее изобретение предназначено охватывать не только примеры воплощений, но также различные альтернативы, модификации, эквиваленты и другие воплощения, которые могут быть включены в концепцию и объем настоящего изобретения как определено в прилагаемой формуле изобретения.
Группа изобретений относится к области биотехнологии. Предложен полипептид, обладающий O-фосфосерин(OPS)-экспортирующей активностью, представленный аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 3 и SEQ ID NO: 5, а также полинуклеотид, кодирующий указанный полипептид. Предложен микроорганизм рода Escherichia, продуцирующий O-фосфосерин, где микроорганизм трансформирован вектором, содержащим полинуклеотид, кодирующий указанный полипептид. Предложены способ продуцирования O-фосфосерина (OPS), способ продуцирования цистеина и способ продуцирования производного цистеина, где указанные способы включают культивирование указанного микроорганизма в среде. Группа изобретений позволяет продуцировать О-фосфосерин с высоким выходом, а также осуществлять эффективную продукцию цистеина и его производного. 6 н. и 5 з.п. ф-лы, 4 табл., 2 пр.
1. Полипептид, обладающий O-фосфосерин(OPS)-экспортирующей активностью, представленный аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 3 и SEQ ID NO: 5.
2. Полинуклеотид, кодирующий полипептид по п. 1.
3. Полинуклеотид по п. 2, где полинуклеотид представлен нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 2, SEQ ID NO: 4 и SEQ ID NO: 6.
4. Микроорганизм рода Escherichia, продуцирующий O-фосфосерин, где микроорганизм трансформирован вектором, содержащим полинуклеотид, кодирующий полипептид по п. 1.
5. Микроорганизм по п. 4, где активность фосфосерин-фосфатазы (SerB) дополнительно ослаблена по сравнению с ее эндогенной активностью.
6. Микроорганизм по п. 4, где активность фосфоглицерат-дегидрогеназы (SerA) или фосфосерин-аминотрансферазы (SerC) дополнительно усилена по сравнению с ее эндогенной активностью.
7. Микроорганизм по п. 4, где микроорганизм рода Escherichia представляет собой Escherichia coli.
8. Способ продуцирования O-фосфосерина (OPS), включающий культивирование микроорганизма по любому из пп. 4-7 в среде.
9. Способ продуцирования цистеина, включающий:
а) культивирование микроорганизма по любому из пп. 4-7 в среде для продуцирования O-фосфосерина (OPS), и
б) осуществление взаимодействия O-фосфосерина, продуцированного на стадии (а), с сульфидом в присутствии O-фосфосерин-сульфгидрилазы (OPSS) или микроорганизма, экспрессирующего O-фосфосерин-сульфгидрилазу.
10. Способ продуцирования производного цистеина, включающий:
а) культивирование микроорганизма по любому из пп. 4-7 в среде для продуцирования O-фосфосерина (OPS),
б) осуществление взаимодействия O-фосфосерина, продуцированного на стадии (а), с сульфидом в присутствии O-фосфосерин-сульфгидрилазы (OPSS) или микроорганизма, экспрессирующего O-фосфосерин-сульфгидрилазу, и
в) превращение цистеина, продуцированного на стадии (б), в производное цистеина.
11. Способ по п. 9 или 10, где сульфид представляет собой по меньшей мере один выбранный из группы, состоящей из Na2S, NaSH, (NH4)2S, H2S и Na2S2O3.
| KR 20140133754 A, 20.11.2014 | |||
| NCBI, GenBank Accession No | |||
| Мешалка для перемешивания двух или более жидкостей | 1946 |
|
SU71170A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| KR 20120041115 A, 30.04.2012 | |||
| KR 20130068135 A, 26.06.2013 | |||
| US 2003008358 A1, 09.01.2003 | |||
| МИКРООРГАНИЗМ, ПРОДУЦИРУЮЩИЙ О-ФОСФОСЕРИН, И СПОСОБ ПОЛУЧЕНИЯ L-ЦИСТЕИНА ИЛИ ЕГО ПРОИЗВОДНЫХ ИЗ О-ФОСФОСЕРИНА С ЕГО ИСПОЛЬЗОВАНИЕМ | 2011 |
|
RU2536250C1 |

