Область техники
Настоящее изобретение относится к использованию производного соединения тиохромено[2,3-с]хинолин-12-она для лечения немелкоклеточного рака легких (НМРЛ) с резистентностью к ингибиторам тирозинкиназы рецепторов эпидермального фактора роста.
Уровень техники
Мутации в рецепторе эпидермального фактора роста (РЭФР) считаются перспективными биомаркерами для терапии с использованием ингибиторов тирозинкиназы (ИТК) в качестве лечения немелкоклеточного рака легких (НМРЛ) (См. ссылку 1, 2, 3).
Механизмы первичной резистентности ИТК еще не полностью изучены, но сверхэкспрессия паксиллина (PXN) обеспечивает первичную резистентность к ИТК в НМРЛ посредством модуляции устойчивости белков Мсд-1 и BIM из-за активации внеклеточно регулируемой киназы ERK (См. ссылку 6). Сочетание ИТК с ингибитором ERK селуметинибом улучшает чувствительность ИТК и результаты в клеточных и животных моделях (См. ссылку 7, 8). К сожалению, пока не установлено никаких преимуществ от сочетания ингибитора ERK и ИТК при лечении пациентов с НМРЛ.
Резистентность к ИТК часто встречается у пациентов с РЭФР-мутированным НМРЛ, которые прошли лечение с ИТК, и данная резистентность считается приобретенной (вторичной) (См. ссылку 4, 5). Наиболее распространенная мутация приобретенной резистентности в РЭФР - Т790М в экзоне 20 (См. ссылку 9, 10). Мутация РЭФР-Т790М и амплификация сМЕТ составляют 50-60% и 5-20%, соответственно, наблюдаемой ИТК РЭФР резистентности у пациентов с НМРЛ (См. ссылку 9, 10). Экспрессию белка и фосфорилирование РЭФР-Т790М и сМЕТ связывают с первичной и приобретенной резистентностью к ИТК-целевой терапии у таких пациентов.
Таким образом, разработка нового поколения ингибиторов ИТК РЭФР и сМЕТ является важнейшей стратегией для преодоления ИТК РЭФР резистентности при НМРЛ (См. ссылку 11-19). К сожалению, РЭФР-независимые механизмы приобретенной резистентности к AZD9291, ИТК третьего поколения, уже были обнаружены у РЭФР-Е790М-положительных пациентов с НМРЛ (См. ссылку 20).
У мышиных моделей с раком легких, с выраженными мутациями РЭФР Del19-T790M или L858R-T790M, с одновременной сверхэкспрессией сМЕТ, не было значительной регрессии опухоли в ответ на монотерапию, нацеленную только на РЭФР или сМЕТ (См. ссылку 21). Напротив, комбинированная терапия, которая была одновременно нацелена и на РЭФР, и на сМЕТ, была крайне эффективной против НТК РЭФР-резистентных опухолей, вызванных совместно Del19-T790M или L858R-T790M и сМЕТ. Несмотря на такой многообещающий результат, такой же комбинированный подход с ингибиторами ИТК РЭФР + сМЕТ не имел успеха в клинических испытаниях на людях с НМРЛ с РЭФР-мутациями (См. ссылку 22).
Таким образом, желая разработать направление лечения НМРЛ с РЭФР-мутациями, изобретатель пытается найти двойной ингибитор РЭФР и сМЕТ, чтобы получить лучший эффект от лечения, чем при сочетании ингибитора РЭФР и ингибитора сМЕТ. Это важный вопрос, на решение которого направлено данное изобретение.
Предыдущие патенты TW 1488843, US 8927717 и статьи изобретателя в журналах (См. ссылку 23) показали, что производные соединения тиохромено[2,3-с]хинолин-12-она подавляют разрастание DU-145 и РС-3 НМРЛ. В настоящем изобретении изобретатель пытается исследовать дальше вопрос существования производного соединения тиохромено[2,3-с]хинолин-12-она, которое можно использовать для подавления РЭФР и сМЕТ, чтобы вылечить клетки НМРЛ с резистентностью к ИТК РЭФР.
Краткое изложение сущности изобретения
С одной стороны, настоящее изобретение предлагает способ для лечения немелкоклеточного рака легких с резистентностью к ИТК РЭФР, который заключается во введении терапевтически эффективного количества активного соединения, выбранного из групп, включающих производные соединения тиохромено[2,3-с]хинолин-12-она, их фармацевтически приемлемые соли,
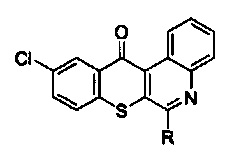
стереоизомеры и энантиомеры, где производное соединение тиохромено[2,3-с]хинолин-12-она имеет следующую структуру:
в которой R группа представляет собой -NH(CH2)n1NH(CH2)n2OH, где n1=3, а n2=2.
Согласно изобретению, резистентность к ИТК включает первичную резистентность к ИТК или приобретенную резистентность к ИТК.
Согласно изобретению, первичная резистентность к ИТК вызвана сверхэкспрессией паксиллина.
Согласно изобретению, приобретенная резистентность к ИТК является мутацией РЭФР Т790М в экзоне 20.
Согласно изобретению, лечение немелкоклеточного рака легких обеспечивается за счет апоптоза.
Согласно изобретению, активное соединение вводится, как минимум, с одной фармацевтически приемлемой основой, разбавителем или наполнителем.
С другой стороны, изобретение предлагает способ подавления HSP90 для лечения немелкоклеточного рака легких с резистентностью к ингибиторам тирозинкиназы рецепторов эпидермального фактора роста, который заключается во введении терапевтически эффективного количества активного соединения, выбранного из групп, включающих производные соединения
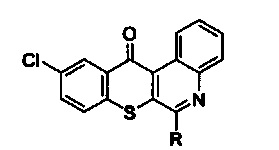
тиохромено[2,3-с]хинолин-12-она, их фармацевтически приемлемые соли, стереоизомеры и энантиомеры, где производное соединение тиохромено[2,3-с]хинолин-12-она имеет следующую структуру:
в которой R группа представляет собой -NH(CH2)n1NH(CH2)n2OH, n1=3, а n2=2.
Согласно изобретению, производное соединение
тиохромено[2,3-с]хинолин-12-она уменьшает экспрессию р-РЭФР, p-Src, pY118-PXN, р-AKT, p-ERK, или р-сМЕТ или увеличивает экспрессию BIM.
Согласно изобретению, где активное соединение вводится, как минимум, с одной фармацевтически приемлемой основой, разбавителем или наполнителем.
С другой стороны, изобретение предлагает способ подавления РЭФР и сМЕТ для лечения немелкоклеточного рака легких с резистентностью к ингибиторам тирозинкиназы рецепторов эпидермального фактора роста, который заключается во введении терапевтически эффективного количества активного соединения, выбранного из групп, включающих производные соединения тиохромено[2,3-с]хинолин-12-она, их фармацевтически приемлемые соли, стереоизомеры и энантиомеры, где производное соединение тиохромено[2,3-с]хинолин-12-она имеет следующую структуру:
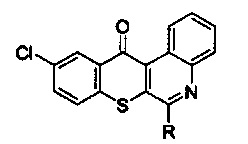
в которой R группа представляет собой -NH(CH2)n1NH(CH2)n2OH, где n1=3, а n2=2.
Согласно изобретению, подавление РЭФР и сМЕТ является уменьшением экспрессии РЭФР и сМЕТ.
Согласно изобретению, подавление РЭФР и сМЕТ является разрушением РЭФР и сМЕТ через убиквитинилирование.
Согласно изобретению, активное соединение вводится, как минимум, с одной фармацевтически приемлемой основой, разбавителем или наполнителем.
Краткое описание графических материалов
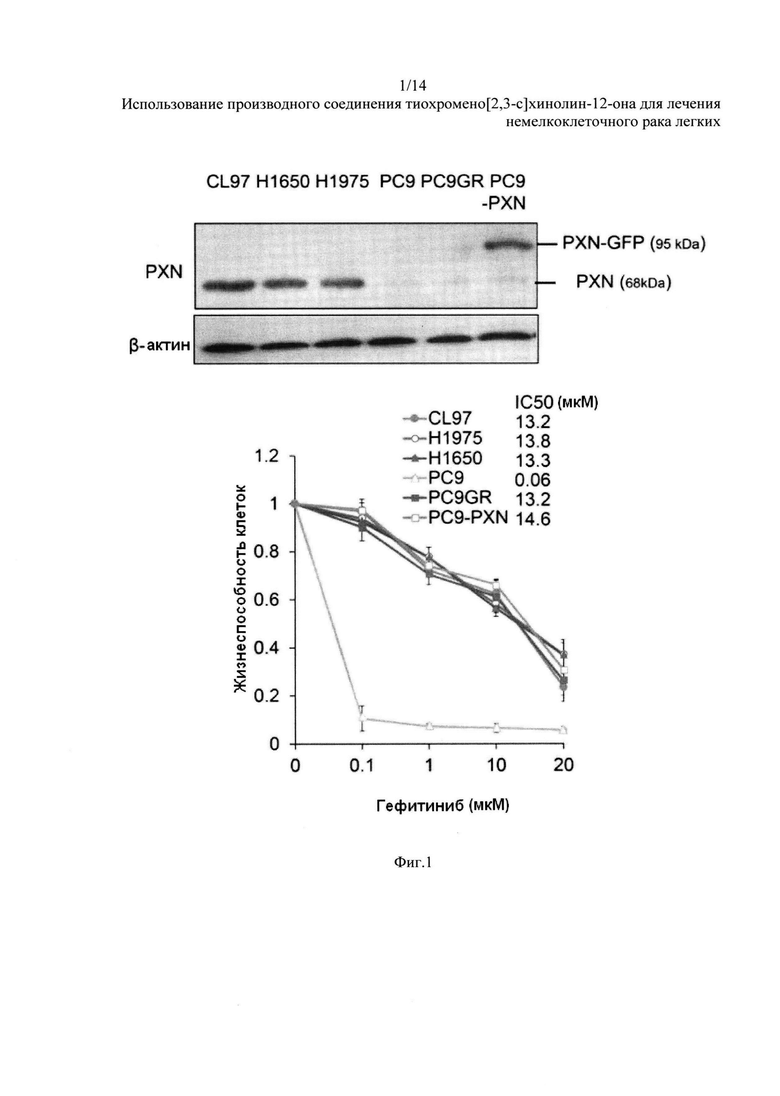
На Фиг. 1 представлено влияние на клетки НМРЛ с мутацией РЭФР посредством гефитиниба. Экспрессия PXN и жизнестойкость клеток с гефитинибом в течение 48 часов среди клеток НМРЛ с мутацией РЭФР.
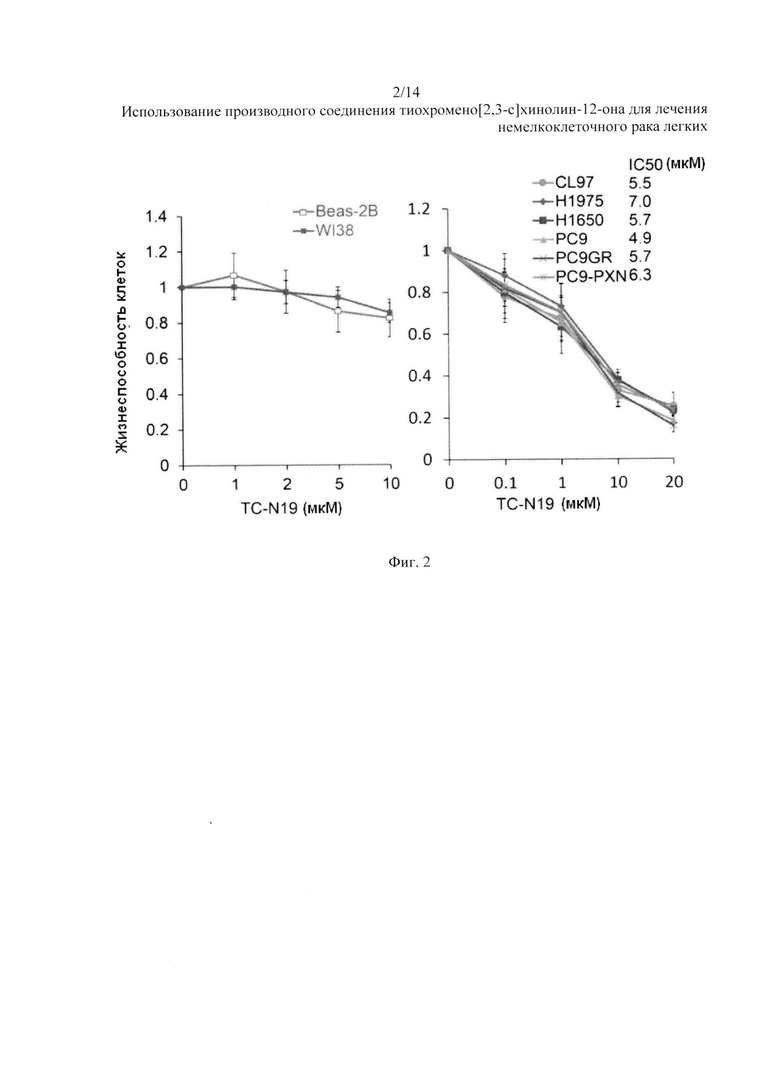
На Фиг. 2 представлено воздействие N19 на клетки НМРЛ с мутацией РЭФР. Жизнестойкость клеток с N19 в течение 48 часов.
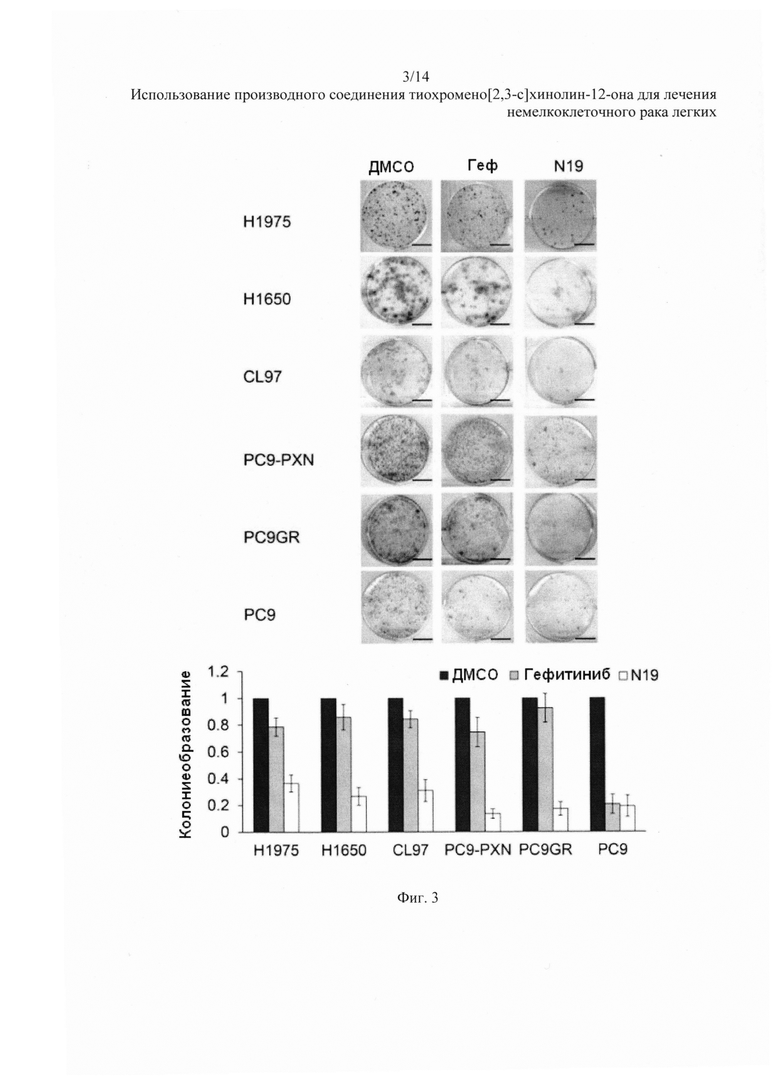
На Фиг. 3 представлено воздействие гефитиниба и N19 на колониеобразование клеток НМРЛ.
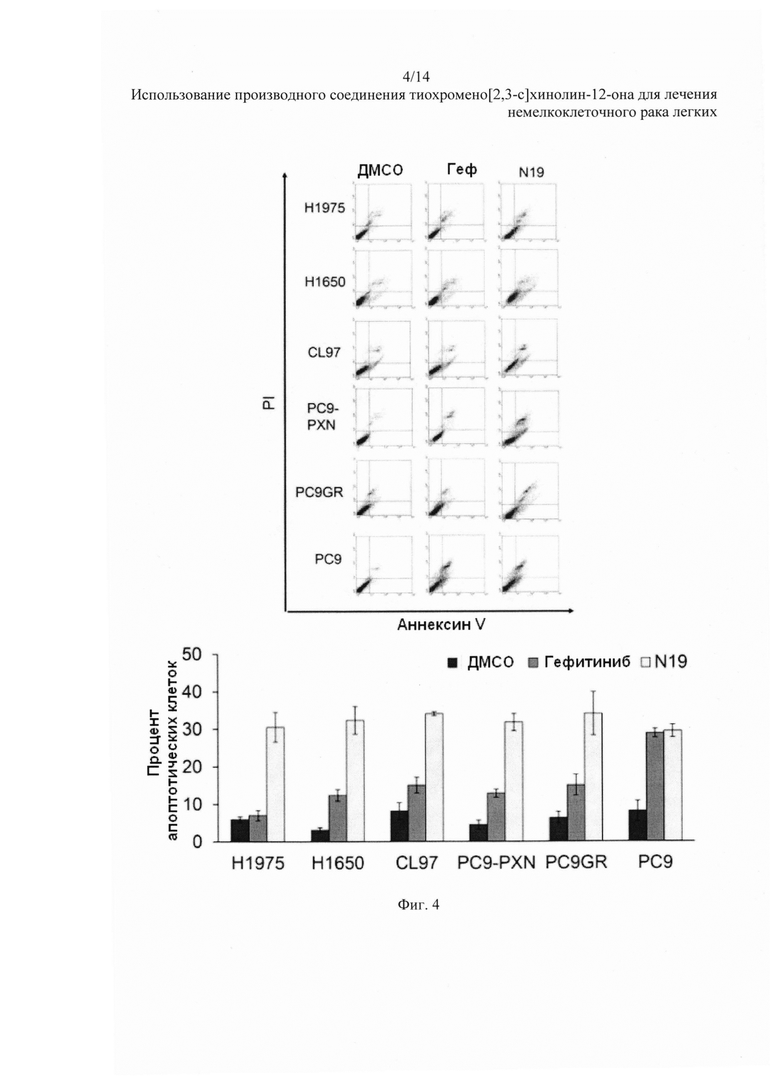
На Фиг. 4 представлено воздействие гефитиниба и N19 на апоптоз клеток НМРЛ.
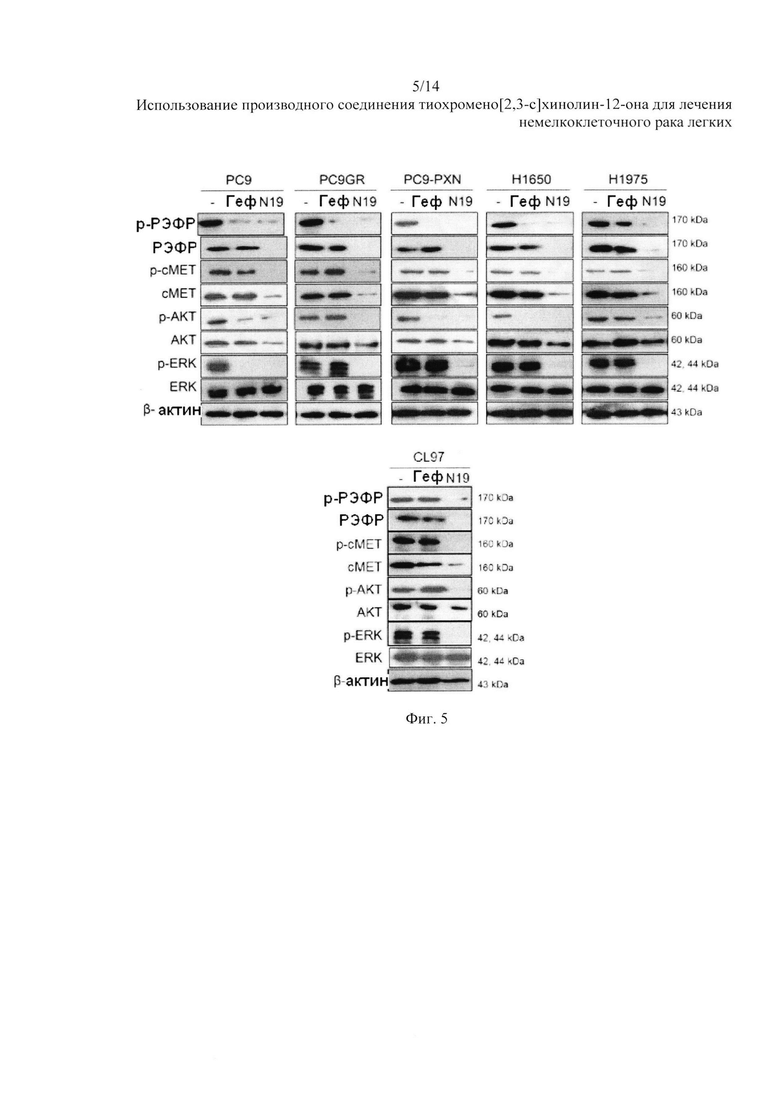
На Фиг. 5 представлено воздействие гефитиниба и N19 на экспрессию белка РЭФР, сМЕТ и их молекул.
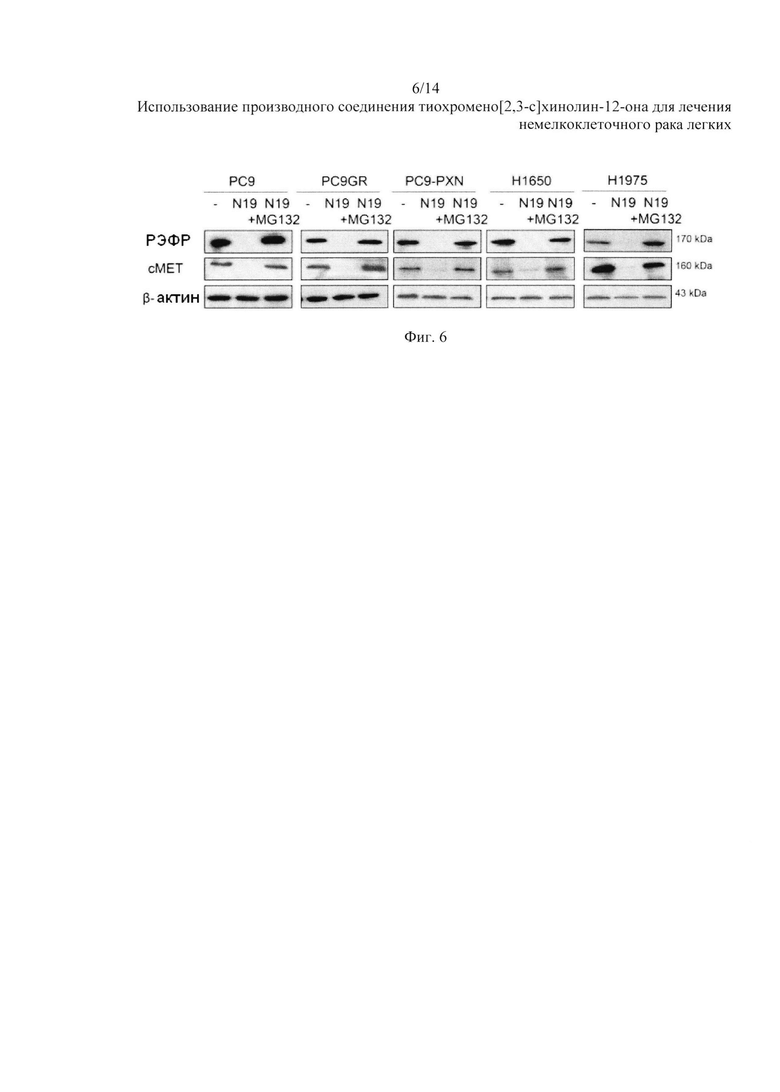
На Фиг. 6 представлено воздействие N19 на РЭФР и сМЕТ через протеасому.
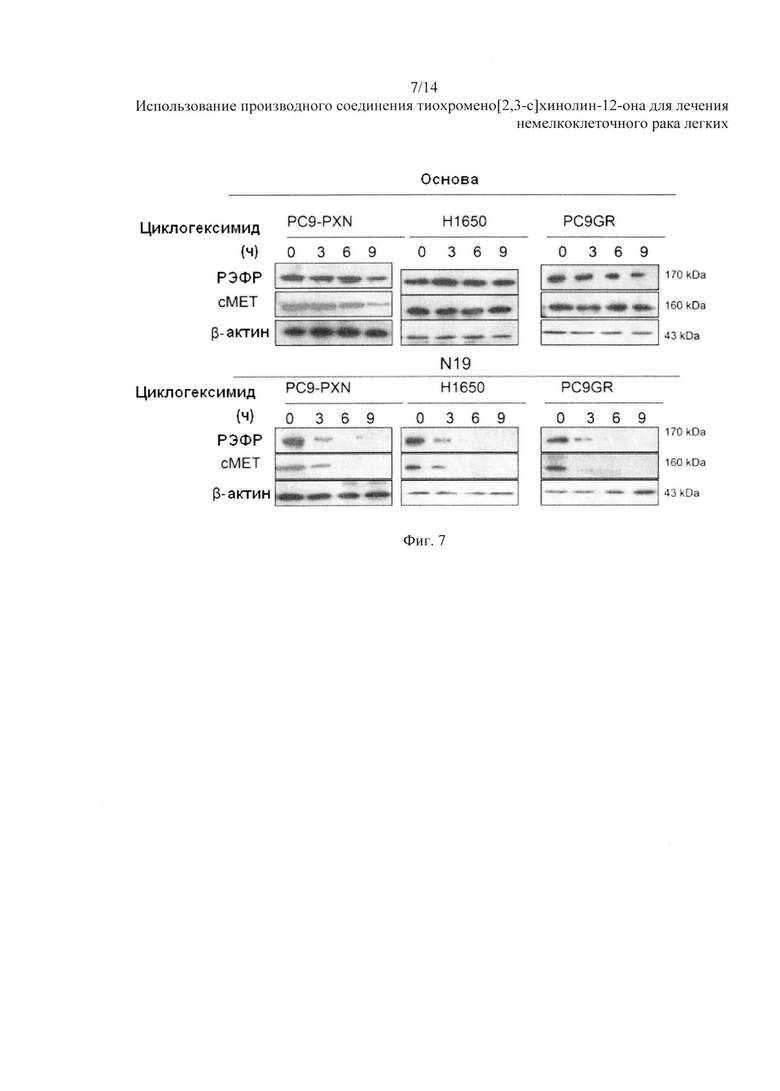
На Фиг. 7 представлено воздействие N19 на эндогенный белок РЭФР и сМЕТ.
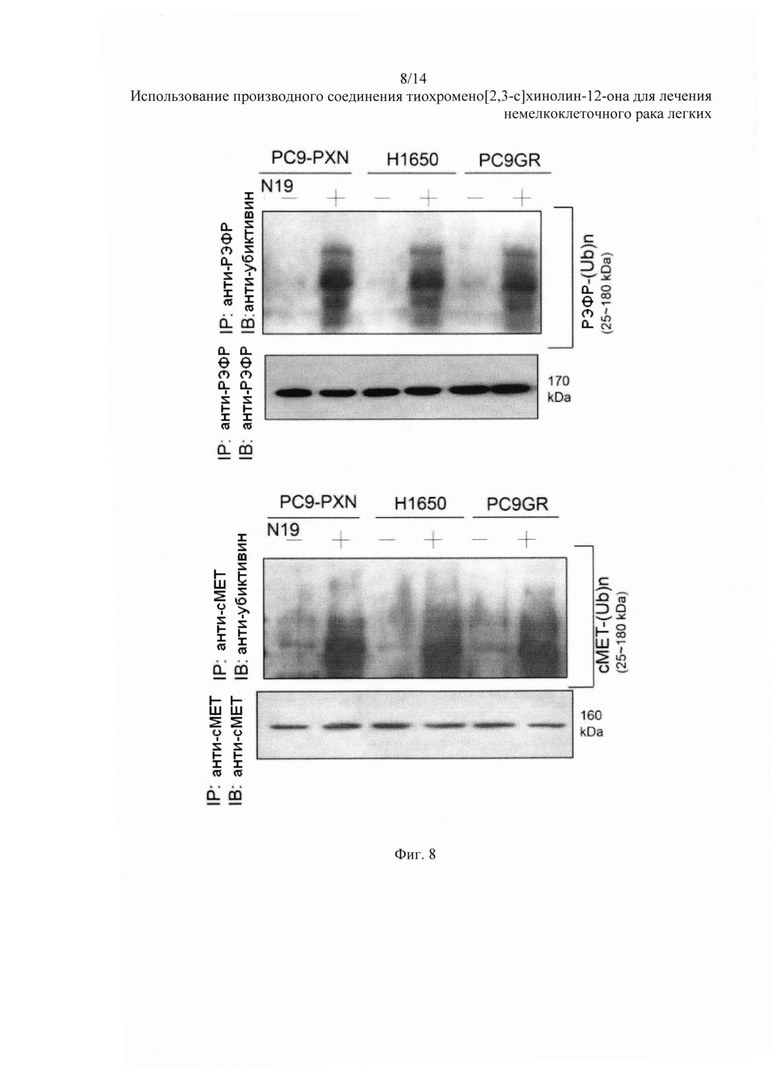
На Фиг. 8 представлено воздействие N19 на полиубиквитинилирование РЭФР и сМЕТ.
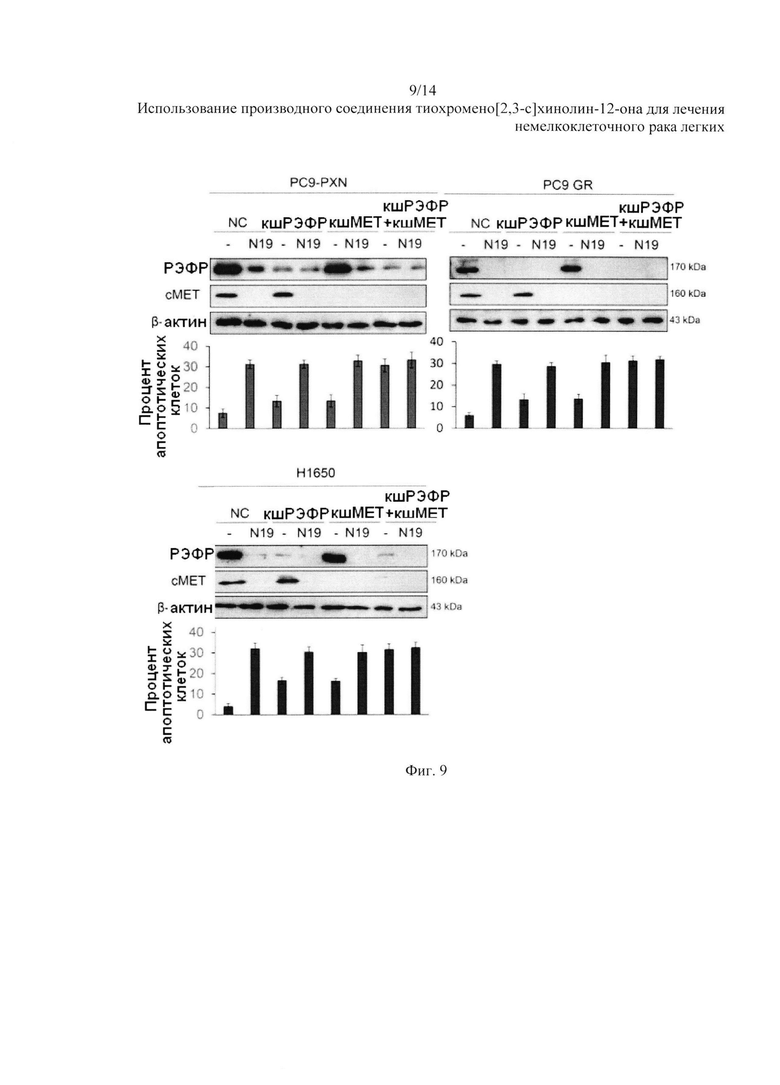
На Фиг. 9 представлено воздействие N19 на РЭФР и сМЕТ и апоптоз.
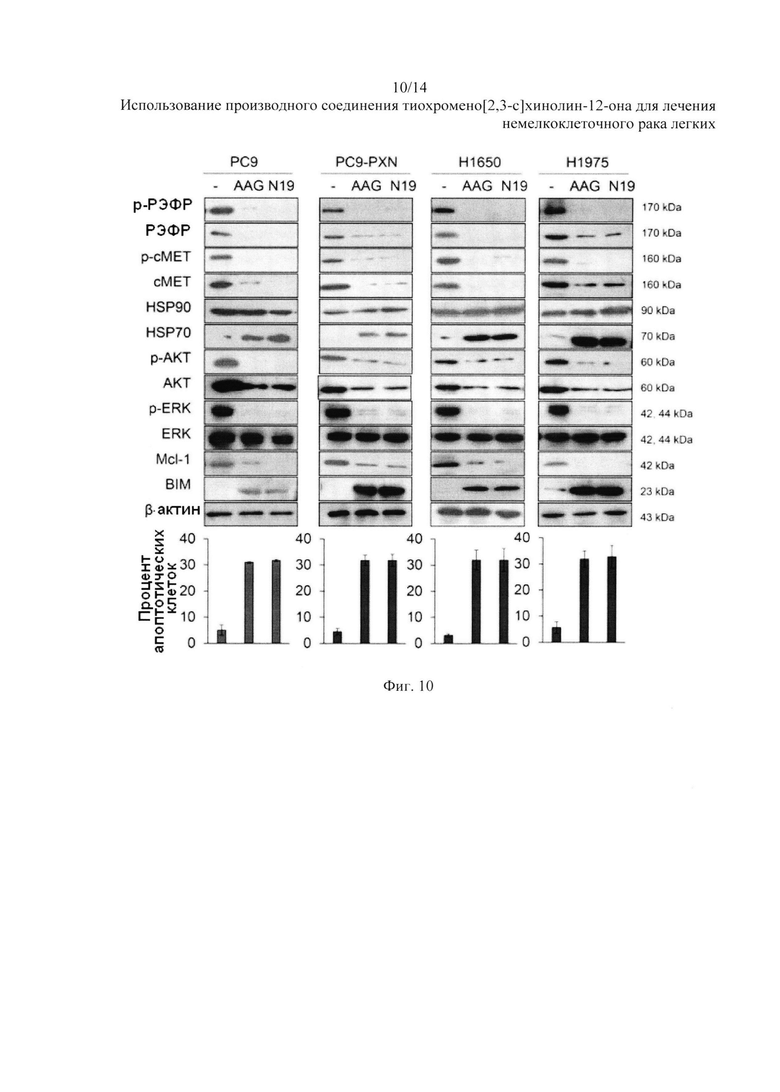
На Фиг. 10 представлено воздействие N19 на разрушение белка РЭФР и сМЕТ.
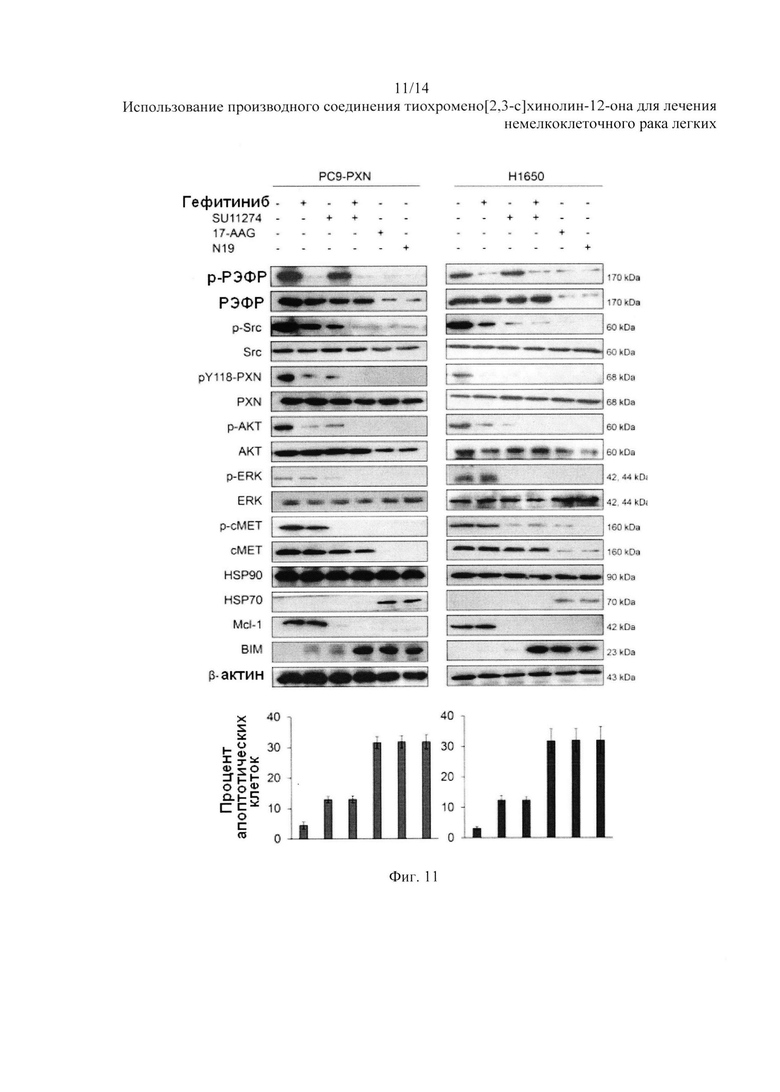
На Фиг. 11 представлено воздействие N19 на разрушение белка РЭФР и сМЕТ.
На Фиг. 12 представлено воздействие N19 на взаимодействие между HSP90 и РЭФР или сМЕТ.
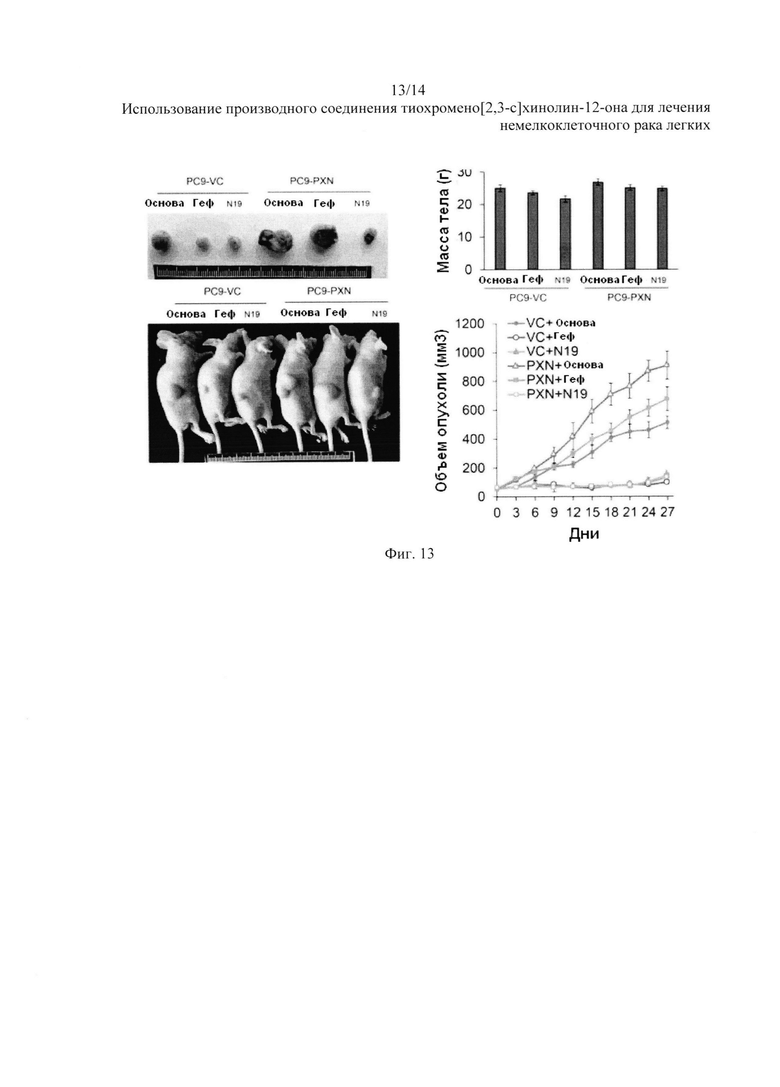
На Фиг. 13 представлено воздействие N19 на рост опухоли.
На Фиг. 14 представлено цитотоксический эффект N19 на клетки сетчатки ARPE-19.
Подробное описание предпочтительного варианта осуществления
В отсутствие иного определения, технические и научные термины в данной инструкции относятся к общим значениям, которые знает обычный технический специалист в этой области.
Термины «лечение», «на лечении» и аналогичные термины относятся к способам, которые могу замедлить, улучшить, уменьшить или обратить заболевание или связанные с ним симптомы, от которых в данный момент страдает пациент, а также к профилактике заболевания или любых возникающих симптомов.
Термин «медицински приемлемый» означает, что материал или соединение должны быть совместимы с другими ингредиентами состава и не должны причинять вред пациенту.
Настоящее изобретение продемонстрировано на следующих примерах, но не ограничивается ими. Лекарства и биоматериалы продаются на рынке и легкодоступны. Ниже представлен просто канал, по которому их можно получить.
Химические вещества и антитела: Гефитиниб, SU11274, 17-DMAG и 17-AAG были получены от компании «Селлеккем» (Selleckchem) (Хьюстон, Техас, США). Все остальные химические вещества были приобретены у компании «Сигма Кемикал» (Sigma Chemical) (Сент-Луис, Миссури, США), если не указано иное. Антитела анти-РЭФР, анти-ERK (общая), анти-фосфорилированная (p)-ERK, анти-AKT (общая), и анти-р-AKT были получены от компании «Селл Сигналинг» (Cell Signaling) (Данверс, Массачусетс, США). Антитела анти-HSP70 и анти-Н8Р90 были получены от компании «Дженетекс» (Genetex) (Ирвайн, Калифорния, США). Все остальные антитела были приобретены у компании «Санта-Круз Байотекнолоджи» (Santa Cruz Biotechnology) (Даллас, Техас, США).
Производное соединение тиохромено[2,3-с]хинолин-12-она, используемое в примерах, имеет следующую структуру:
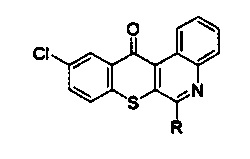
R группа: -NH(CH2)3NH(CH2)2OH, наименование согласно Международному союзу теоретической и прикладной химии: 10-Хлор-6-((3-((2-гидроксиэтил)амино)пропил)амино)-12Н-тиохромено [2,3-с]хинолин-12-он, далее - ТС19 (N19, NSC777201).
Эксперименты на животных моделях: Клетки опухоли были введены подкожно в спину 4-5-недельных самок белых бестимусных мышей. Размер опухоли ксенотрансплантата измерялся каждые 3 дня, и объем опухоли определялся как (длина × ширина2)/2. Когда опухоли выросли до 50 мм, мыши были разделены на следующие группы в случайном порядке: основа (ДМСО), гефитиниб (5 мг/кг) или N19 (5 мг/кг). Лекарства вводили при помощи внутрибрюшинной инъекции каждые 3 дня.
Конструкция плазмидов и трансфекция: Плазмид сверхэкспрессии PXN был любезно предоставлен Доктором Сальгия (Университет Чикаго, Чикаго, Иллинойс, США). РЭФР (TRCN0000121068) и сМЕТ (TRCN0000009850) были куплены у Национального профильного центра РИНА, Академия Синика (Тайбэй, Тайвань). Данные плазмиды были временно трансфицированы в клетки рака легких (1×106) при помощи реагента Turbofect («Форментас» (Formentas), Глен Берни, Мэриленд, США). Через 48 часов клетки были собраны для анализа в последующих экспериментах.
Анализ цитотоксичности МТТ: Перед лечением клетки в экспоненциальной фазе роста были предварительно обработаны плазмидами сверхэкспрессии и нокдауна в течение 24 часов. Цитотоксические эффекты in vitro данного лечения были определены по анализу МТТ
(3-(4,5-диметилтиазол-2-yl)-2,5-дифенилтетразолия бромид) (550 нм), и жизнеспособность клеток была выражена в процентах от контрольных (необработанных) клеток (% контроля).
Анализ с окрашиванием аннексином V-PI: Клетки были собраны посредством трипсинизации и центрифугирования при 1000 × г в течение 5 мин. После ресуспендирования в фиксирующем буферном растворе (10 мМ HEPES-NaOH, 140 мМ NaCl, 2,5 мМ CaCl2) с окончательной плотностью клеток 1-2×106 клеток на мл, 100 мкл суспензии отдельных клеток (1-2×105 клеток) было выдержано вместе с 5 мкл аннексина V-FITC и 5 мкл ингибитора протеазы в течение 15 минут при комнатной температуре в темноте. После добавления 400 мкл фиксирующего буферного раствора образцы были проанализированы при помощи проточного питометра BD FACS Calibur («Би-Ди Байосайенсис» (BD Biosciences), Сан-Хосе, Калифорния, США) в течение 1 часа. Для каждого образца было подсчитано 10000 событий.
Анализ колониеобразования: Посев клеток был произведен в 6-луночных планшетах в среде комплемента в течение ночи. После инкубации культуральные среды были заменены свежей средой, содержащей гефитиниб, N19 и ДМСО в качестве контроля растворителем в течение 48 ч. Обработанные клетки были высеяны в новую среду с добавлением 10% эмбриональной бычьей сыворотки еще на 10 дней. Перед снимками колоний клетки были окрашены 0,01% кристаллвиолета в течение 1 ч при комнатной температуре.
Вестерн-блоттинг и иммунопреципитация: Общие клеточные лизаты были подготовлены в иммунопреципитированном буфере для лизиса (20 мМ Трис, рН 7,5, 100 мМ хлорида натрия, 1% ИГЕПАЛ СА-630, 100 мкМ Na3VO4, 50 мМ NaF, 30 мМ пирофосфата натрия), содержащем полную смесь ингибиторов протеаз с ЭДТА или без ЭДТА («Роше Дайагностикс» (Roche Diagnostics), Базель, Швейцария). Концентрации белка были измерены при помощи анализа белка Bio-Rad («Био-Рад» (Bio-Rad), Ричмонд, Калифорния, США). Белки были подвергнуты иммунопреципитации при помощи 1 мкг первичных антител и гранул белка А («Сигма» (Sigma), Данверс, Массачусетс, США) в течение 4 ч, а затем три раза промыты буфером для лизиса. Образцы белка были разделены посредством 10% геля в присутствии додецилсульфата натрия, перенесены на поливинилидендифторидную мембрану («Миллипор» (Millipore), Билл ерика, Массачусетс, США), и, наконец, был произведен иммуноблоттинг с первичными антителами. Первичные антитела для иммуноблоттинга были использованы в растворе от 1:500 до 1:1000. Вторичные антитела мыши, козла и кролика, конъюгированные с пероксидазой хрена («Санта-Круз Байотекнолоджи» (Santa Cruz Biotechnology), Даллас, Техас, США) были использованы в растворе 1:5000. Белковые сигналы были обнаружены при помощи хемилюминесцентных реагентов («Амершем Фармация» (Amersham Pharmacia), Пискатауэй, Нью-Джерси, США).
Настоящее изобретение продемонстрировано на следующих примерах, но не ограничивается ими.
Пример 1. N19 более эффективна, чем гефитиниб, для апоптозного подавления жизнеспособности клеток и колониеобразования в клетках НМРЛ с мутацией РЭФР
PXN придает первичную резистентность к ИТК в клетках НМРЛ с мутацией РЭФР. Значение IC50 для гефитиниба в шести клеточных линиях НМРЛ с мутацией РЭФР было оценено при помощи анализа МТТ. Значение IC50 для гефитиниба в клетках Н1975, Н1650, CL97 и PC9GR (клетки РС9, резистентные к гефитинибу) варьировалось от 13,2 до 13,8 мкМ. Самое низкое значение IC50 наблюдалось в клетках РС9, но значение IC50 заметно увеличилось (до 14,6 мкМ) за счет эктопической экспрессии PXN в этих клетках (Фиг. 1).
Интересно, что значение IC50 для N19 в данных клеточных линиях варьировалось от 5,5 до 7,0 мкМ, но относительно низкое значение IC50 (4,9 мкМ) для N19 наблюдалось в клетках РС9. Согласно анализу МТТ, N19 не проявила цитотоксичности в нормальных клеточных линиях легких WI38 и Beas-2B (Фиг. 2).
На Фиг. 3 представлен репрезентативный рост колоний на агаровых пластинках. N19 более эффективна, чем гефитиниб, при подавлении колониеобразования во всех типах клеток, кроме клеток РС9 (Фиг. 3, нижняя панель). Репрезентативные апоптозные профили данных клеток после обработки гефитинибом или N19 и анализа с окрашиванием аннексином V-PI показаны на Фиг. 4. Процентное содержание апоптозных клеток было выше после обработки N19, чем после обработки гефитинибом, во всех типах клеток, кроме клеток РС9 (Фиг. 4, нижняя панель).
Данные результаты четко показали, что с N19 получается более сильное подавление жизнеспособности клеток и колониеобразования, чем с гефитинибом, в клетках НМРЛ с мутацией РЭФР, и что такое подавление происходит за счет апоптоза.
Пример 2. N19 убивает клетки НМРЛ с мутацией РЭФР путем апоптоза, вызванного разрушением белков РЭФР и сМЕТ
Мы рассмотрели возможность того, что смерть клеток НМРЛ с мутацией РЭФР в ответ на N19 может происходить за счет разрушения белков РЭФР и сМЕТ. Через сорок восемь часов после обработки гефитинибом (10 мкМ) и N19 (10 мкМ) был собран клеточный лизат и проанализирован с помощью вестерн-блоттинга. Вестерн-блоттинг показал снижение уровней экспрессии р-РЭФР и р-AKT после обработки гефитинибом клеток РС9, РС9 со сверхэкспрессией PXN (PC9-PXN) и HI650, но не после обработки гефитинибом клеток PC9GR и H1975 (Фиг. 5). Уровни экспрессии РЭФР, AKT, ERK, и сМЕТ не изменились после обработки гефитинибом во всех типах клеток. Интересно, что экспрессия р-РЭФР, РЭФР, р-сМЕТ, сМЕТ, р-AKT, AKT и p-ERK была значительно подавлена при обработке N19. Аналогичные изменения в экспрессии белка наблюдались в ответ на обработку гефитинибом или N19 клеток CL97 с высокой экспрессией PXN.
Тем не менее, снижение экспрессии РЭФР и сМЕТ от N19 в данных видах клеток было обращено в присутствии MG132, по сравнению с клетками без обработки N19 (Фиг. 6).
Клетки PC9-PXN, H1650 и PC9GR были обработаны 100 мкг/мл циклогексимида в течение указанного количества часов с или без 5 мкМ N19. Уровни белков РЭФР и сМЕТ были оценены с помощью вестерн-блоттинга. Опыт «пульс-чейз» с клетками PC9-PXN, H1650 и PC9GR показал, что экспрессия белков РЭФР и сМЕТ постепенно снижалась от N19 в течение указанных временных интервалов; тем не менее, уровни обоих белков остались относительно без изменений после обработки контрольным растворителем (Фиг. 7).
Клетки PC9-PXN, H1650 и PC9GR были обработаны 5 мкМ N19 в течение 5 часов. Полиубиквитинирование РЭФР и сМЕТ было оценено с помощью вестерн-блоттинга в иммунопреципитации. Убиквитиновый лэддер белков РЭФР и сМЕТ был очевиден в данных видах клеток после обработки N19, но отсутствовал в контрольном растворителе (Фиг. 8). Таким образом, разрушение белков РЭФР и сМЕТ от N19 возникает за счет посттрансляционного механизма, опосредованного убиквитин-протеасомой, а не за счет модуляции аппарата для трансляции.
Затем мы проверили, действительно ли апоптоз клеток, вызванный N19, возникал за счет разрушения белков РЭФР и сМЕТ. Три вида клеток (PC9-PXN, PC9GR и HI650) были трансфицированы при помощи короткой шпилечной РНК (кшРНК) РЭФР (кшРЭФР) и кш-с-МЕТ, по отдельности или в сочетании, а затем обработаны 5 мкМ N19 в течение 24 часов. Вестерн-блоттинг показал, что уровни экспрессии РЭФР и сМЕТ были значительно снижены после обработки N19 в данных видах клеток, трансфицированных неспецифической кшРНК (NC, Фиг. 9). Затем экспрессия белка РЭФР и сМЕТ была снижена в клетках, трансфицированных кш-РЭФР, кш-с-МЕТ, или кш-РЭФР+ кш-с-МЕТ и подвергнутых обработке N19 (Фиг. 9). Апоптоз клеток был измерен при помощи анализа с окрашиванием аннексином V-PI с использованием проточной цитометрии. Процент апоптозных клеток варьировался при различной обработке и зависел от уровней экспрессии РЭФР и сМЕТ в данных видах клеток. Результаты показали, что N19 убивает клетки НМРЛ с мутацией РЭФР посредством апоптоза, преимущественно за счет разрушения белков РЭФР и сМЕТ убиквитин-протеасомами.
Пример 3. Вероятно, N19 действует в качестве ингибитора HSP90 и убивает клетки НМРЛ с мутацией РЭФР за счет одновременного разрушения белков РЭФР и сМЕТ убиквитин-протеасомами.
РЭФР и сМЕТ - это клиентские белки HSP90. Поэтому мы предположили, что N19 может действовать в качестве ингибитора HSP90 и убивать клетки НМРЛ с мутацией РЭФР за счет разрушения белков РЭФР и сМЕТ убиквитин-протеасомами. Через сорок восемь часов после обработки 17-AAG (5 мкМ) или N19 (5 мкМ), клеточные лизаты были собраны и проанализированы на предмет изменения сигналов с помощью вестерн-блоттинга с указанными антителами. Вестерн-блоттинг показал, что экспрессия РЭФР и сМЕТ в клетках РС9, PC9-PXN, H1650 и H1975 почти полностью подавлена ингибитором HSP90 17-AAG или N19. Уровни экспрессии р-AKT и p-ERK в данных видах клеток были также сильно снижены после обработки 17-AAG или N19 (Фиг. 10), в то время как такая же обработка значительно повысила уровень экспрессии HSP70 в данных видах клеток. Рост Мс1-1 и снижение BIM в данных видах клеток был обращен за счет обработки 17-AAG или N19. Изменения в уровнях белков Мс1-1 и BIM соответствовали проценту апоптозных клеток, наблюдающихся после обработки 17-AAGmraN19.
Клетки PC9-PXN и HI 650 были обработаны гефитинибом, ингибитором сМЕТ SU11274, сочетанием гефитиниба + SU11274, 17-AAG или N19, чтобы определить, можно ли преодолеть резистентность к ИТК, опосредованную PXN, при помощи гефитиниба + SU11274, а также 17-AAG или N19. Клетки были обработаны гефитинибом, SU11274, гефитинибом + SU11274, 17-AAG или N19 в течение 48 часов. Апоптоз клеток был измерен при помощи анализа с окрашиванием аннексином V-PI с использованием проточной цитометрии. Клеточные лизаты были собраны и проанализированы на предмет изменения сигналов с помощью вестерн-блоттинга с указанными антителами. Вестерн-блоттинг показал, что уровни экспрессии р-РЭФР, p-Src, pY118-PXN, р-АКТ, p-ERK и р-сМЕТ были значительно снижены от гефитиниба + SU11274, 17-AAG или N19 в обоих видах клеток, но лишь частично снижены от одного гефитиниба или SU11274 (Фиг. 11). Появление экспрессии HSP70 наблюдалось в обоих видах клеток, но только после обработки 17-AAG или N19, а не других видов обработки (Фиг. 11). Экспрессия Мс1-1 и BIM была обращена за счет гефитиниба + SU 11274, и такое обращение также наблюдалось после обработки 17-AAG или N19. Аналогично высокий процент апоптозных клеток наблюдался в обоих видах клеток после обработки гефитинибом + SU11274, 17-AAG или N19, но не после обработки одним только гефитинибом или SU11274 (Фиг. 11). Клетки Н1650 были обработаны N19 (10 мкМ) в течение 43 часов, а затем MG132 в течение еще 5 часов. Клеточные лизаты были подвергнуты иммунопреципитации с анти-HSP90-коньюгированными гранулами. Иммунопреципитаты были оценены для определения экспрессии РЭФР и сМЕТ с помощью вестерн-блоттинга. β-актин использовался в качестве контроля для нанесения. Анализ с иммунопреципитацией показал, что взаимодействие HSP90 с РЭФР или сМЕТ в клетках HI 650 снизилось при обработке N19 в присутствии или в отсутствии MG132 (Фиг. 12). Данные результаты показывают, что N19, вероятно, действует в качестве ингибитора HSP90 и убивает клетки НМРЛ с мутацией РЭФР посредством апоптоз за счет разрушения белков РЭФР и сМЕТ.
Затем мы сравнили подавление роста подкожных опухолей у бестимусных мышей, вызванных РС9-РХ№стабильным клоном, за счет лечения с N19 и гефитинибом. Репрезентативная опухолевая масса и мыши с подкожными опухолями показаны на Фиг. 13. На фотографиях изображены репрезентативные опухолевые массы у мышей после введения препарата в течение 27 дней через внутрибрюшинную инъекцию. Репрезентативные образцы опухоли были иссечены на 27-й день. Воздействие N19 на массу тела мышей. Среднее значение ± стандартное отклонение были рассчитаны по объему опухоли и массе тела пяти бестимусных мышей в каждой группе. Опухолевая масса у бестимусных мышей, вызванная PC9-VC- или РС9-PXN-стабильным клоном, была почти полностью подавлена N19. Подавляющее действие гефитиниба также наблюдалось в опухолевой массе бестимусных мышей, вызванной клетками векторного контроля. Тем не менее, опухолевая масса у бестимусных мышей, вызванная РС9-PXN-стабильным клоном или PC9-VC клетками, постепенно уменьшилась за 27 дней, пока мыши были подвергнуты лечению гефитинибом или контрольным растворителем. Масса тела мышей не изменилась во время лечения N19 или гефитинибом в течение 27-дневного исследования. Аналогичное подавляющее действие наблюдалось у N19 для опухолевой массы, вызванной PC9GR, в клеточных и животных моделях. Данные результаты четко показали, что N19 полностью подавляет рост опухоли, вызванной РС9-PXN-стабильным клоном и клетками PC9GR. Поэтому мы полагаем, что N19 эффективно подавляет образование ксенотрансплантата опухоли с первичной и приобретенной резистентностью к ИТК.
Пример 4. Анализ ретинальной цитотоксичности
Известно, что все существующие ингибиторы HSP90 обладают ретинальной цитотоксичностью, и провести клинические опыты невозможно, поэтому мы используем анализ МТТ, чтобы узнать, обладает ли N19 цитотоксичностью для клетки сетчатки ARPE-19. Анализ МТТ и анализ с окрашиванием аннексином V-PI были выполнены на клетках ARPE, подвергнутых обработке N19, 17-AAG или 17-DMAG. Все данные были собраны по трем независимым экспериментам. Среднее значение и стандартное отклонение были указаны в виде столбца с планками погрешностей. Как показано на Фиг. 14, N19 в концентрации 10 мкМ (данная концентрация может эффективно подавлять HSP90) не обладает цитотоксичностью для клеток ARPE-19, но другие ингибиторы HSP90 17-AAG или 17-DMAG проявляют цитотоксичность для клеток ARPE-19.
Данное изобретение решает проблему прошлого лечения НМРЛ, ингибитор РЭФР и ингибитор сМЕТ пока не эффективны для пациентов с НМРЛ с мутацией ИТК РЭФР, и все ингибиторы HSP90 (в обратном направлении от РЭФР, сМЕТ) имеют недостаток в виде цитотоксичности для клеток сетчатки. Изобретение оказывает следующее воздействие : 1) N19 обладает терапевтическим эффектом при НМРЛ с первичной резистентностью к ИТК из-за сверхэкспрессии PXN, 2) N19 обладает терапевтическим эффектом при НМРЛ с приобретенной резистентностью к ИТК из-за мутации Т790М, 3) N19 разрушает РЭФР и сМЕТ за счет убиквитинилирования и ведет к апоптозу НМРЛ с мутацией РЭФР, 4) N19 является двойным ингибитором EGF и сМЕТ, и 5) N19 является ингибитором HSP90 и не обладает цитотоксичностью для клеток сетчатки. В качестве ингибитора HSP90, N19 может преодолевать резистентность РЭФР к ИТК и убивать НМРЛ с мутацией РЭФР за счет разрушения РЭФР и сМЕТ убиквитин-протеасомами. По сравнению с другими ингибиторами HSP90, N19 не обладает цитотоксичностью для клеток сетчатки.
Вышеуказанные данные индивидуальны для практических случаев использования данного изобретения. Практические случаи ограничивают объем запатентованного изобретения. Все равные эффективные применения и изменения, соответствующие духу изобретения, должны быть включены в объем запатентованного изобретения.
Множественный эффект, описанный выше, полностью соответствует требованиям новизны и прогрессивной законной монополии. Таким образом, в соответствии с законом мы просим одобрить настоящее изобретение в целях поощрения.
Список литературы
1. Мок TS. Personalized medicine in lung cancer: what we need to know. Nat Rev Clin Oncol 2011; 8: 661-668.
2. Hang Z, Stiegler AL, Boggon TJ, Kobayashi S, Halmos B. EGFR-mutated lung cancer: a paradigm of molecular oncology. Oncotarget 2010; 1: 497-514.
3. Rosell R, Moran T, Queralt C, Porta R, Cardenal F, Camps С et al. Screening for epidermal growth factor receptor mutations in lung cancer. N Engl J Med 2009; 361: 958-967.
4. Gazdar AF. Activating and resistance mutations of EGFR in non-small-cell lung cancer: role in clinical response to EGFR tyrosine kinase inhibitors. Oncogene 2009; 28(Suppl 1): S24-S31.
5. Tan CS, Gilligan D, Pacey S. Treatment approaches for EGFR-inhibitor-resistant patients with non-small-cell lung cancer. Lancet Oncol 2015; 16:e447-e459.
6. Wu DW, Chen CY, Chu CL, Lee H. Paxillin confers resistance to tyrosine kinase inhibitors in EGFR-mutant lung cancers via modulating BIM and Mcl-1 protein stability. Oncogene 2015; 35: 621-630.
7. Huang MH, Lee Ж, Chang YJ, Tsai HH, Lin YL, Lin AM et al. MEK inhibitors reverse resistance in epidermal growth factor receptor mutation lung cancer cells with acquired resistance to gefitinib. Mol Oncol 2013; 7: 112-120.
8. Eberlein CA, Stetson D, Markovets AA, Al-Kadhimi KJ, Lai Z, Fisher PR et al. Acquired resistance to the mutant-selective EGFR inhibitor AZD9291 is associated with increased dependence on RAS signaling in preclinical models. Cancer Res 2015; 75: 2489-2500.
9. Engelman JA, Zejnullahu K, Mitsudomi T, Song Y, Hyland C, Park JO et al. MET amplification leads to gefitinib resistance in lung cancer by activating ERBB3 signaling. Science 2007; 316: 1039-1043.
10. Bean J, Brennan C, Shih JY, Riely G, Viale A, Wang L et al. MET amplification occurs with or without T790M mutations in EGFR mutant lung tumors with acquired resistance to gefitinib or erlotinib. Proc Natl Acad Sci USA 2007; 104: 20932-20937.
11. Cross DA, Ashton SE, Ghiorghiu S, Eberlein C, Nebhan CA, Spitzler PJ et al. AZD9291, an irreversible EGFR TKI, overcomes T790M-mediated resistance to EGFR inhibitors in lung cancer. Cancer Discov 2014; 4: 1046-1061.
12. Nanjo S, Yamada T, Nishihara H, Takeuchi S, Sano T, Nakagawa T et al. Ability of the Met kinase inhibitor crizotinib and new generation EGFR inhibitors to overcome resistance to EGFR inhibitors. PLoS One 2013; 8: e84700.
13. Janne PA, Yang JC, Kim DW, Planchard D, Ohe Y, Ramalingam SS et al. AZD9291 in EGFR inhibitor-resistant non-small-cell lung cancer. N Engl J Med 2015; 372: 1689-1699.
14. Szokol B,  P,
P,  I, Baska F,
I, Baska F,  C, Greff Z et al. Discovery and biological evaluation of novel dual EGFR/c-Met inhibitors. ACS Med Chem Lett 2014; 5: 298-303.
C, Greff Z et al. Discovery and biological evaluation of novel dual EGFR/c-Met inhibitors. ACS Med Chem Lett 2014; 5: 298-303.
15. Takeuchi S, Wang W, Li Q, Yamada T, Kita K, Donev IS et al. Dual inhibition of Met kinase and angiogenesis to overcome HGF-induced EGFR-TKI resistance in EGFR mutant lung cancer. Am J Pathol 2012; 181: 1034-1043.
16. Rho JK, Choi YJ, Kim SY, Kim TW, Choi EK, Yoon SJ et al. MET and AXL inhibitor NPS-1034 exerts efficacy against lung cancer cells resistant to EGFR kinase inhibitors because of MET or AXL activation. Cancer Res 2014; 74: 253-262.
17. Nakade J, Takeuchi S, Nakagawa T, Ishikawa D, Sano T, Nanjo S et al. Triple inhibition of EGFR, Met, and VEGF suppresses regrowth of HGF-triggered, erlotinib-resistant lung cancer harboring an EGFR mutation. J Thorac Oncol 2014; 9: 775-783.
18. Sano Y, Hashimoto E, Nakatani N, Abe M, Satoh Y, Sakata К et al. Combining onartuzumab with erlotinib inhibits growth of non-small cell lung cancer with activating EGFR mutations and HGF overexpression. Mol Cancer Ther 2015; 14: 533-541.
19. Sequist LV, Rolfe L, Allen AR. Rociletinib in EGFR-mutated non-small-cell lung cancer. N Engl J Med 2015; 373: 578-579.
20. Planchard D, Loriot Y,  F, Gobert A, Auger N, Lacroix L et al. EGFR-independent mechanisms of acquired resistance to AZD9291 in EGFR T790M-positive NSCLC patients. Ann Oncol 2015; 26: 2073-2078.
F, Gobert A, Auger N, Lacroix L et al. EGFR-independent mechanisms of acquired resistance to AZD9291 in EGFR T790M-positive NSCLC patients. Ann Oncol 2015; 26: 2073-2078.
21. Xu L, Kikuchi E, Xu C, Ebi H, Ercan D, Cheng KA et al. Combined EGFR/MET or EGFR/HSP90 inhibition is effective in the treatment of lung cancers codriven by mutant EGFR containing T790M and MET. Cancer Res 2012; 72: 3302-3311.
22. Sequist LV, von Pawel J, Garmey EG, Akerley WL, Brugger W, Ferrari D et al. Randomized phase II study of erlotinib plus tivantinib versus erlotinib plus placebo in previously treated non-small-cell lung cancer. J Clin Oncol 2011; 29: 3307-3315.
23. Chen TC, Wu CL, Lee CC, Chen CL, Yu DS, Huang HS. Structure-based hybridization, synthesis and biological evaluation of novel tetracyclic heterocyclic azathioxanthone analogues as potential antitumor agents. Eur J Med Chem 2015; 103: 615-627.


Группа изобретений относится к медицине, а именно к онкологии, и может быть использована для лечения немелкоклеточного рака легких (НМРЛ) с резистентностью к ингибиторам тирозинкиназы рецепторов эпидермального фактора роста. Способы по изобретению включают введение производного соединения тиохромено[2,3-с]хинолин-12-она - N19. Использование изобретений позволяет лечить резистентный НМРЛ за счет подавления HSP90, рецепторов эпидермального фактора роста и сМЕТ без ретинальной цитотоксичности. 3 н. и 10 з.п. ф-лы, 4 пр., 14 ил.
1. Способ для лечения немелкоклеточного рака легких с резистентностью к ингибиторам тирозинкиназы рецепторов эпидермального фактора роста, который заключается во введении терапевтически эффективного количества активного соединения, выбранного из групп, включающих производные соединения тиохромено[2,3-с]хинолин-12-она, их фармацевтически приемлемые соли, стереоизомеры и энантиомеры, где производное соединение тиохромено[2,3-с]хинолин-12-она имеет следующую структуру:
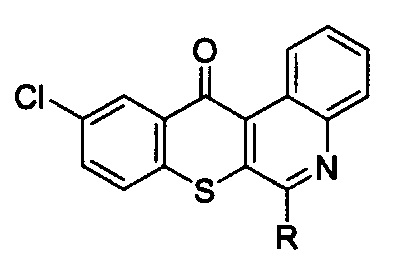
в которой R группа представляет собой -NH(CH2)n1NH(CH2)n2OH, где n1 = 3 и n2 = 2.
2. Способ по п. 1, где резистентность к ингибиторам тирозинкиназы включает первичную резистентность к ингибиторам тирозинкиназы или приобретенную резистентность к ингибиторам тирозинкиназы.
3. Способ по п. 2, где первичная резистентность к ингибиторам тирозинкиназы вызвана сверхэкспрессией паксиллина.
4. Способ по п. 2, где приобретенная резистентность к ингибиторам тирозинкиназы является мутацией рецепторов эпидермального фактора роста Т790М в экзоне 20.
5. Способ по п. 1, где лечение немелкоклеточного рака легких обеспечивается за счет апоптоза.
6. Способ по п. 1, где активное соединение вводится, как минимум, с одной фармацевтически приемлемой основой, разбавителем или наполнителем.
7. Способ подавления HSP90 для лечения немелкоклеточного рака легких с резистентностью к ингибиторам тирозинкиназы рецепторов эпидермального фактора роста, который заключается во введении терапевтически эффективного количества активного соединения, выбранного из групп, включающих производные соединения тиохромено[2,3-с]хинолин-12-она, их фармацевтически приемлемые соли, стереоизомеры и энантиомеры, где производное соединение тиохромено[2,3-с]хинолин-12-она имеет следующую структуру:
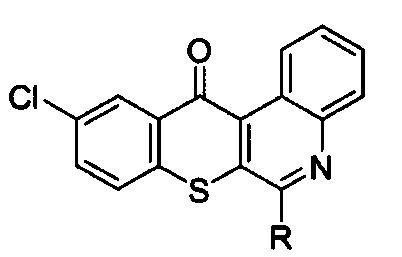
в которой R группа представляет собой -NH(CH2)n1NH(CH2)n2OH, где n1 = 3 и n2 = 2.
8. Способ по п. 7, где данное производное соединение тиохромено[2,3-с]хинолин-12-она уменьшает экспрессию р-РЭФР, p-Src, pY118-PXN, р-AKT, p-ERK, или р-сМЕТ или увеличивает экспрессию BIM.
9. Способ по п. 7, где активное соединение вводится, как минимум, с одной фармацевтически приемлемой основой, разбавителем или наполнителем.
10. Способ подавления рецепторов эпидермального фактора роста и сМЕТ для лечения немелкоклеточного рака легких с резистентностью к ингибиторам тирозинкиназы рецепторов эпидермального фактора роста, который заключается во введении терапевтически эффективного количества активного соединения, выбранного из групп, включающих производные соединения тиохромено[2,3-с]хинолин-12-она, их фармацевтически приемлемые соли, стереоизомеры и энантиомеры, где производное соединение тиохромено[2,3-с]хинолин-12-она имеет следующую структуру:
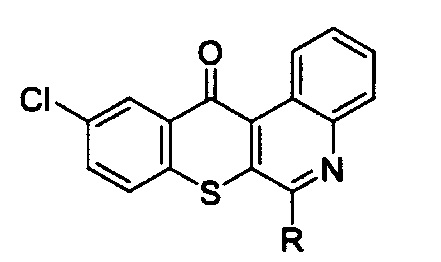
в которой R группа представляет собой -NH(CH2)n1NH(CH2)n2OH, где n1 = 3 и n2 = 2.
11. Способ по п. 10, где подавление рецепторов эпидермального фактора роста и сМЕТ является уменьшением экспрессии рецепторов эпидермального фактора роста и сМЕТ.
12. Способ по п. 10, где подавление рецепторов эпидермального фактора роста и сМЕТ является разрушением рецепторов эпидермального фактора роста и сМЕТ через убиквитинилирование.
13. Способ по п. 10, где активное соединение вводится, как минимум, с одной фармацевтически приемлемой основой, разбавителем или наполнителем.
| US 8927717 B1, 06.01.2015 | |||
| СПОСОБ ЛЕЧЕНИЯ РАКА, НЕСУЩЕГО МУТАЦИИ EGFR | 2007 |
|
RU2492864C2 |
| НОВОЕ ПРОИЗВОДНОЕ ИНДОЛА, СОДЕРЖАЩЕЕ КАРБАМОИЛЬНУЮ ГРУППУ, УРЕИДНУЮ ГРУППУ И ЗАМЕЩЕННУЮ ОКСИГРУППУ | 2009 |
|
RU2503661C2 |
| WU D.-W | |||
| et al | |||
| Способ изготовления электрических сопротивлений посредством осаждения слоя проводника на поверхности изолятора | 1921 |
|
SU19A1 |
| Токарный резец | 1924 |
|
SU2016A1 |
| Токарный резец | 1924 |
|
SU2016A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |

