Изобретение относится к биотехнологии и медицине, а именно к онкогематологии, и касается рекомбинантной плазмидной ДНК для использования в качестве калибраторов при проведении ПЦР с обратной транскрипцией (ОТ-ПЦР) и детекцией геноспецифических продуктов в режиме реального времени с целью оценки эффективности проводимой терапии у пациентов с острым миелоидным лейкозом (ОМЛ).
Использование молекулярно-генетических методов исследования, благодаря их специфичности и чувствительности, является ключевым подходом в контроле полной излеченности пациента. Тем не менее, у половины пациентов с ОМЛ не удается идентифицировать информативный генетический маркер. Именно для этой группы пациентов внедрение альтернативных подходов контроля элиминации представляется наиболее актуальным в оценке эффективности терапии.
Гиперэкспрессия гена WT1 была обнаружена при ряде заболеваний крови опухолевой природы, в том числе и при ОМЛ. Высокая экспрессия WT1 встречается у более половины пациентов с ОМЛ и является фактором неблагоприятного прогноза заболевания и высокого риска развития рецидива [Liu-Yin, J. et al. Predictive value of minimal residual disease (MRD) monitoring by RQ-PCR in WT1 positive patients entered in the UK MRC AML-15 Trial. Blood 2008 112, 259.]. Поскольку методом ОТ-ПЦР можно выявить четкие различия в уровне транскрипта WT1 в нормальных и лейкемических клетках, экспрессия WT1 является эффективным маркером в оценке минимальной остаточной болезни у пациентов при отсутствии крупных хромосомных аномалий или других мутаций, характерных для клеток опухолевого клона.
Экспрессия BAALC в зрелых клетках костного мозга и в периферической крови здоровых людей не обнаруживается. Согласно данным литературы, при ОМЛ гиперэкспрессия BAALC встречается у более 50% больных с нормальным кариотипом [Damiani D, Tiribelli М, Franzoni A et al. BAALC overexpression retains its negative prognostic role across all cytogenetic risk groups in acute myeloid leukemia patients. Am J Hematol. 2013 Oct; 88(10): 848-52]. Чрезмерная экспрессия BAALC связана с низкими показателями общей выживаемости и повышенным кумулятивным риском развития рецидива заболевания в ходе терапии.
Гиперэкспрессия гена PRAME была обнаружена у 35-64% пациентов с ОМЛ. На достаточно большой выборке пациентов [Qin YZ, Zhu НН, Liu YR, Wang YZ et al. PRAME and WT1 transcripts constitute a good molecular marker combination for monitoring minimal residual disease in myelodysplastic syndromes. Leuk Lymphoma. 2013 Jul; 54(7): 1442-9] было показано, что уровень транскрипта гена PRAME уменьшается в ходе ремиссии и увеличивается при рецидиве. Интересно, что, в отличие от гена WT1, экспрессия которого ассоциирована с различными генетическими поломками, гиперэкспрессия гена PRAME встречается у пациентов с ОМЛ без крупных транслокаций и с нормальным кариотипом.
Для перечисленных маркеров опухолевого роста подсчет абсолютных количеств копий осуществляется с использованием раздельных наборов калибраторов для анализируемого и референсного генов. Учитывая возможность неравномерной деградации плазмидной ДНК в калибраторах, существует вероятность искажения результатов исследования относительной экспрессии. Вместе с тем, при наличии вероятностных отклонений в оценке копийности плазмидной ДНК методом спетрофотометрии при создании калибраторов на основе последовательных разведений, наличие в последовательности целевого и референсного генов может повысить точность расчетов абсолютных значений относительной экспрессии генов.
Техническим результатом заявленного изобретения является создание плазмидной ДНК, содержащей три последовательности калибраторов, включающих в себя копии целевого и референсного генов, что позволяет нивелировать возникновение возможных погрешностей и оценивать эффективность проводимой терапии в большинстве случаев ОМЛ, в том числе у пациентов без выявленных хромосомных аберраций.
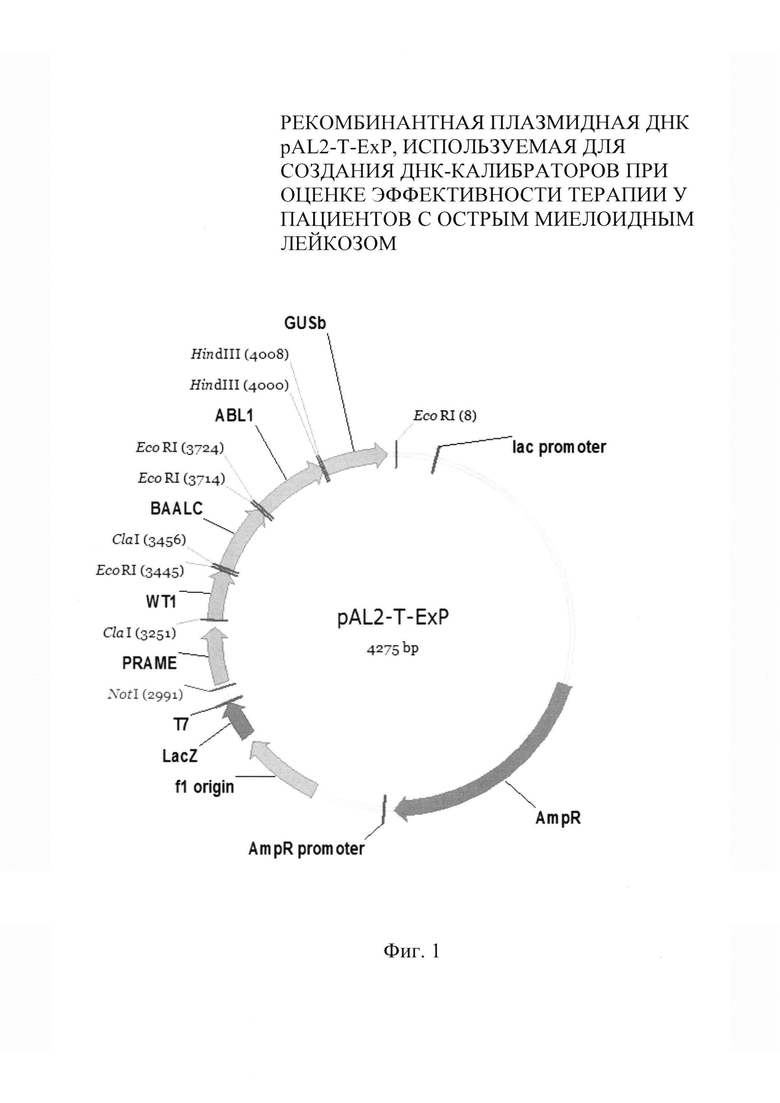
Указанный технический результат обеспечивается тем, что рекомбинантная плазмидная ДНК pAL2-T-ExP, используемая для создания ДНК-калибраторов при оценке эффективности терапии у пациентов с острым миелоидным лейкозом, содержит вставку последовательно сшитых в реакции лигирования участков кДНК генов PRAME, WT1, BAALC, ABL1, GUSB, амплифицируемых в ПЦР-РВ.
Так как совокупная частота выявляемости гиперэкспрессии генов PRAME, WT1, BAALC у пациентов с ОМЛ составляет более 90%, создание унифицированной системы на основе комплексного анализа позволяет оценивать эффективность проводимой терапии практически у всех пациентов с ОМЛ. Вместе с тем, комплексная оценка нескольких опухолевых маркеров представляется целесообразной и в связи с наблюдающимся в ряде случаев изменением профиля экспрессии генов в опухолевой популяции.
Гены PRAME, WT1 и BAALC выступают в качестве маркеров опухолевой прогрессии, а гены ABL1 и GUSB - в качестве нормализаторов экспрессии. Сущность изобретения поясняется фигурами 1-3.
На фиг. 1 представлена физическая карта рекомбинантной плазмиды pAL2-T-ExP со следующими обозначениями: f1 origin - сайт инициации транскрипции одноцепочечного нитевидного фага f1; Т7 - ранний промотор фага Т7 для РНК-полимеразы E. coli; LacZ - ген β-галактозидазы; AmpR - ген устойчивости к антибиотику ампициллину; PRAME - последовательность участка кДНК гена PRAME; WT1 - последовательность участка кДНК гена WT1; BAALC - последовательность участка кДНК гена BAALC; ABL - последовательность участка кДНК гена ABL; GUSB - последовательность участка кДНК GUSb; NotH, ClaI, EcoRI, HindIII - сайты расщепления эндонуклеазами рестрикции;
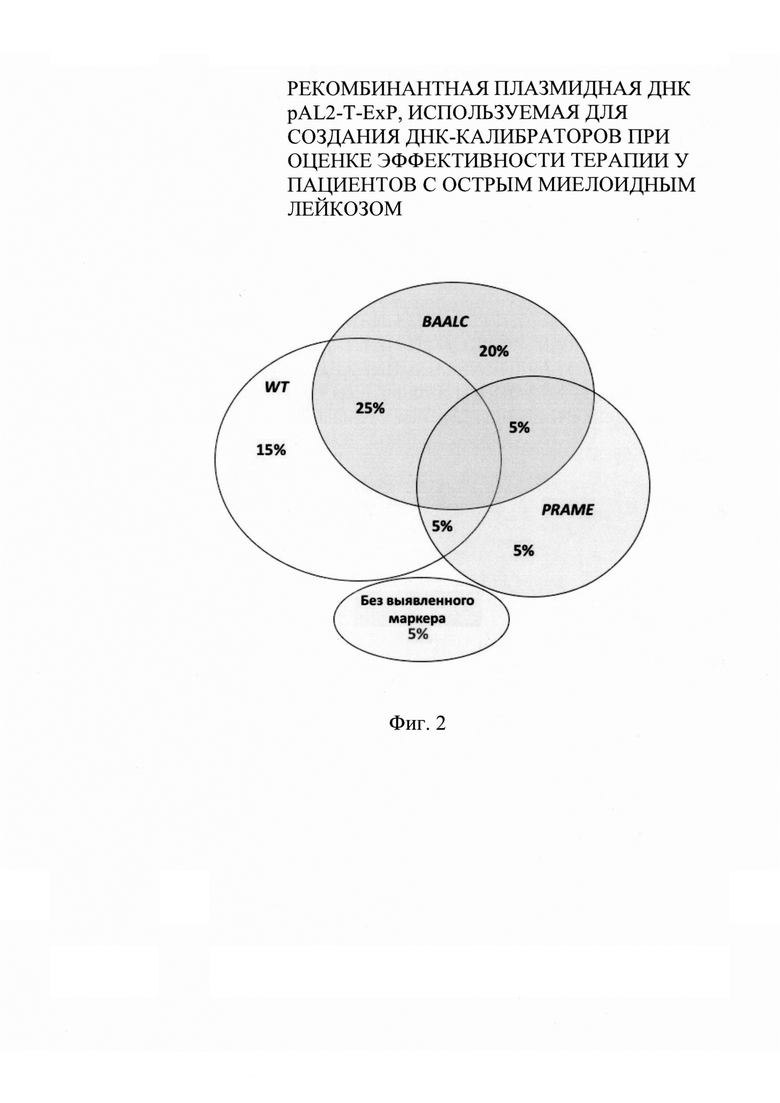
на фиг. 2 - уровни относительной экспрессии гена WT1 у здоровых доноров в костном мозге и периферической крови;
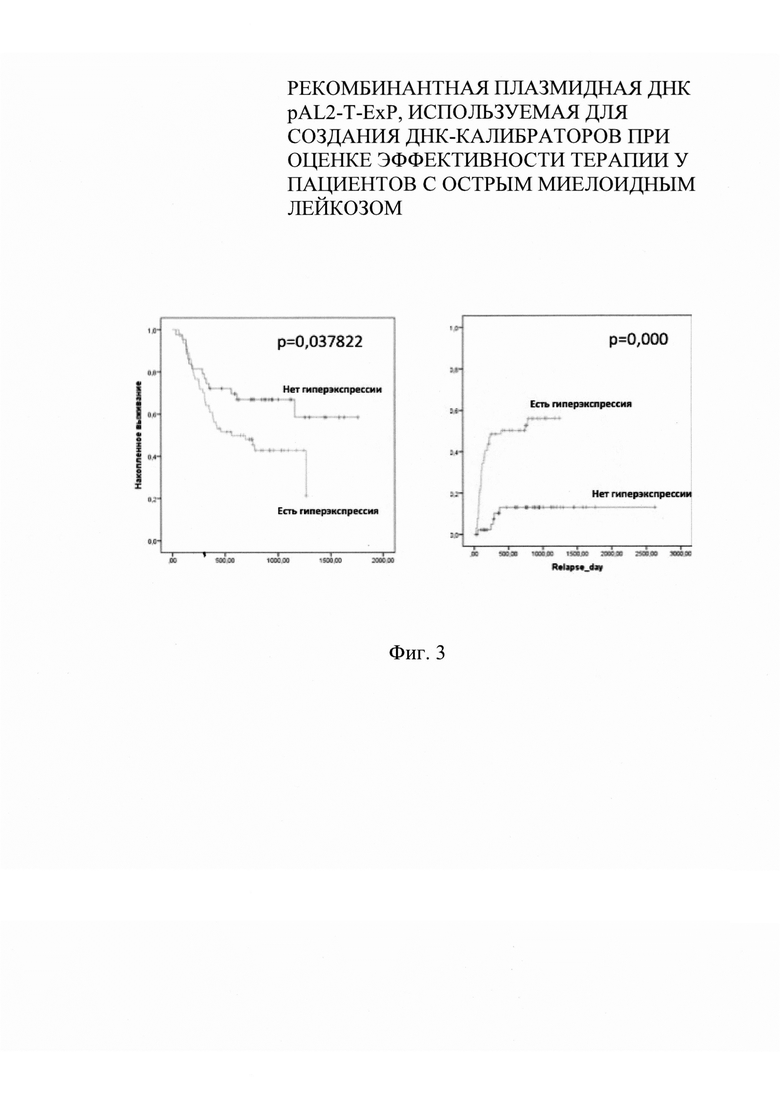
на фиг. 3 - кумулятивный риск развития рецидива в посттрансплантационном периоде на фоне гиперэкспрессии генов WT1, BAALC;
Рекомбинаную плазмидную ДНК pAL2-T-ExP, которая кодирует участки кДНК генов человека PRAME, WT1, BAALC, ABL1, GUSB, получают генно-инженерным методом: путем встраивания ДНК-фрагмента, состоящего из последовательно сшитых в реакции лигирования нуклеотидных последовательностей участков кДНК генов PRAME, WT1, BAALC, ABL1, GUSB, амплифицированных с использованием клинического материала пациентов и здоровых доноров, в вектор pAL2-T.
Исходным генетическим материалом для конструирования плазмиды служат следующие генно-инженерные конструкции:
- плазмидный вектор pAL2-T (Евроген, кат. номер ТА002);
- консервативный участок кДНК гена PRAME, амплифицированного с кДНК, полученной в реакции обратной транскрипции из РНК, выделенной из клинического материала пациентов с ОМЛ;
- консервативный участок кДНК гена WT1, амплифицированного с кДНК, полученной в реакции обратной транскрипции из РНК, выделенной из клинического материала пациентов с ОМЛ;
- консервативный участок кДНК гена BAALC, амплифицированного с кДНК, полученной в реакции обратной транскрипции из РНК, выделенной из клинического материала пациентов с ОМЛ;
- консервативный участок кДНК гена ABL1, амплифицированного с кДНК, полученной в реакции обратной транскрипции из РНК, выделенной из клинического материала здоровых доноров;
- консервативный участок кДНК гена GUSb, амплифицированного с кДНК, полученной в реакции обратной транскрипции из РНК, выделенной из клинического материала здоровых доноров;
- синтетические олигонуклеотиды

Полученная плазмида pAL2-T-ExP, кодирующая участки последовательностей кДНК генов PRAME, WT1, BAALC, ABL1, GUSB, имеет молекулярную массу 1,32 МДа и размер 4275 п.н. и содержит следующие элементы:
- участок плазмиды pAL2-T, размером 3005 п.н., включающий сайт инициации транскрипции одноцепочечного нитевидного фага fl, ранний промотор фага Т7 для РНК-полимеразы Е. coli, запускающий экспрессию рекомбинантного гена, ген устойчивости к ампициллину AmpR для селекции трансформированных плазмидой клеток Е. coli и лактозноый оперон, позволяющий проводить отбор трансформированных бактерий методом сине-белой селекции;
- фрагмент кДНК гена PRAME, амплифицированного с кДНК, полученной в реакции обратной транскрипции из РНК, выделенной из клинического материала пациентов с ОМЛ;
- ClaI/EcoRI фрагмент кДНК гена WT1, амплифицированного с кДНК, полученной в реакции обратной транскрипции из РНК, выделенной из клинического материала пациентов с ОМЛ;
- ClaI/EcoRI фрагмент кДНК гена BAALC, амплифицированного с кДНК, полученной в реакции обратной транскрипции из РНК, выделенной из клинического материала пациентов с ОМЛ;
- EcoRI/HindIII фрагмент кДНК гена ABL1, амплифицированного с кДНК, полученной в реакции обратной транскрипции из РНК, выделенной из клинического материала здоровых доноров;
- EcoRI/HindIII фрагмент кДНК гена GUSb, амплифицированного с кДНК, полученной в реакции обратной транскрипции из РНК, выделенной из клинического материала здоровых доноров;
- уникальный сайт узнавания рестриктазой NotI для линеаризации плазмидной ДНК, имеющий следующие координаты - 2991.
Синтез плазмидной ДНК проводят следующим образом.
Участки кДНК, кодирующие анализируемые методом ОТ-ПЦР последовательности транскриптов, амплифицируют с помощью синтетических олигонуклеотидов, подобранных таким образом, что амплифицируемый в ОТ-ПЦР участок находится внутри клонируемого фрагмента:

ПЦР проводят в объеме 25 мкл с использованием реакционной смеси для ПЦР HF Master Mix (Dialat), содержащей высокоточную Pfu-полимеразу. Концентрация праймеров в реакционной смеси составляет 300 мМ (прямой и обратный), концентрация ионов магния 2,5 мМ, кДНК добавляют в объеме 5 мкл.
Амплифицированные последовательности подвергают электрофоретическому разделению в 1,5% агарозном геле, фрагменты ДНК, размер которых совпадает с теоретическим, вырезают из геля и очищают с помощью набора для очистки фрагментов двухцепочечной ДНК из геля Cleanup Standard (Евроген).
Полученные фрагменты амплифицируют в реакции ПЦР с помощью пар синтетических олигонуклеотидов, подбиранных таким образом, что каждый праймер одним концом комплементарен последовательности одного гена, а другим концом - гену, который будет расположен рядом с ним в генно-инженерной конструкции.

Амплификацию продуктов для клонирования осуществляют методом ПЦР. ПЦР проводят в объеме 25 мкл с использованием реакционной смеси для ПЦР HF Master Mix (Dialat), содержащей высокоточную Pfu-полимеразу. Реакционная смесь содержит праймеры в концентрации 300 мМ (прямой и обратный), ионы магния в концентрации 2,5 мМ. Очищенный из геля фрагмент ДНК добавляют в реакционную смесь в количестве 20-30 нг.
Протокол амплификации:
- первичная денатурация - 2 минуты при 98°С
- амплификационный цикл (×30)
- денатурация - 20 секунд при 98°С
- отжиг праймеров - 30 секунд при 60°С
- элонгация - 20 секунд при 72°С
- финальная элонгация - 2 минуты при 72°С.
Амплифицированные нуклеотидные последовательности подвергают электрофоретическому разделению в 1,5% агарозном геле, очищают с помощью набора для очистки фрагментов двухцепочечной ДНК из геля Cleanup Standard (Евроген) и сшивают в один фрагмент длиной 1505 п.н. с помощью набора для лигирования фрагментов ДНК NEBuilder® HiFi DNA Assembly (NEB). Для этого в одной пробирке смешивают 100 нг целевых фрагменов ДНК и 50 нг вектора pAL2-T, 10 мкл смеси NEB Master Mix и доводят дедионизованной водой до объема 20 мкл. Реакционную смесь инкубируют в течение 60 минут при 50°С и трансформируют при помощи нее химически компетентные клетки и высеивают их на агаризованную ампициллин-содержащую среду LB. Образовавшиеся через сутки колонии тестируют на предмет наличия генно-инженерной вставки с помощью метода скрининговой ПЦР с праймерами WT1_OL_F и ABL-GUSB_OL_R и секвенирования по Сенгеру с помощью универсальных плазмидных праймеров М13. Бактериальные клетки, несущие в себе вектор pAL2-T со вставкой участков кДНК генов ABL1, GUSB, PRAME, WT1, BAALC, инокулируют в 0.5 л жидкой питательной среды LB, содержащей ампициллин в концентрации 100 мкг/мл, и культивируют при 37°С на шейкере в течение 8-12 часов. Затем выделяют плазмидную ДНК из массы бактериальных клеток с помощью набора для выделения плазмидной ДНК Maxiprep Purification Kit (Qiagen). Выделенную плазмидную ДНК pAL2-T-ExP анализируют с помощью рестрикционного анализа по сайтам NotI и HindIII, секвенируют вставки генов ABL1, GUSB, PRAME, WT1, BAALC и хранят при -20°С.
Для использования в качестве ДНК-калибраторов плазмидную ДНК pAL2-T-ExP линеализируют по сайту узнавания рестриктазой NotI, измеряют концентрацию ДНК с помощью спектрофотометра NanoDrop, рассчитывают количество копий плазмидной ДНК по формуле:
N=(С*6.022×1023)/(L*650*1×109),
где N - количество копий ДНК в 1 мкл исходного раствора, С - концентрация плазмидной ДНК в исходном растворе (нг/мкл), L - длина плазмидной ДНК (п.н), и готовят десятичные разведения плазмидной ДНК, содержащие от 10 до 1 ООО 000 копий плазмидной ДНК в 5 микролитрах буфера.
Для амплификации участка гена BAALC используют праймеры:
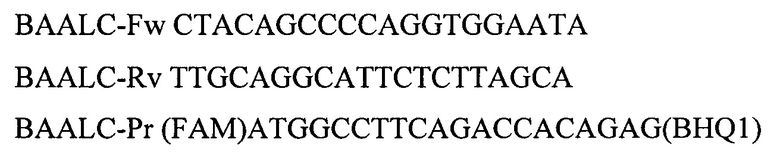
Для амплификации участка гена WT1 используют олигонуклеотидные последовательности:
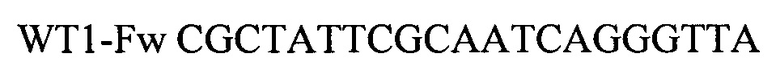
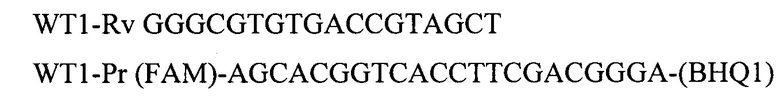
Для амплификации участка гена PRAME используют олигонуклеотидные последовательности:
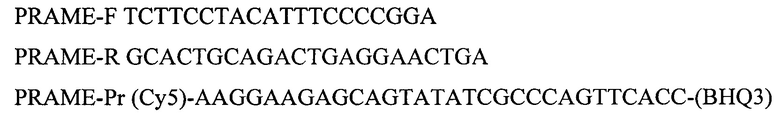
Для амплификации участка гена ABL1 используют олигонуклеотидные последовательности:
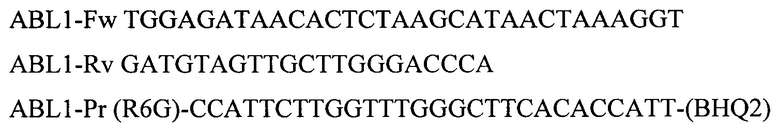
Для амплификации участка гена GUSB используют олигонуклеотидные последовательности:
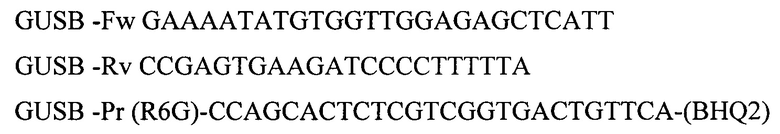
С использованием заявленной рекомбинантной плазмидной ДНК pAL2-T-ExP была проанализирована экспрессия исследуемых генов от 108 пациентов в возрасте от 2 до 60 лет (медиана - 27 лет). Период наблюдения составил в среднем 6 месяцев (от 1 до 47 месяцев). Оценка экспрессии проводилась преимущественно в посттрансплантационном периоде (19 пациентов после родственной аллогенной трансплантациии гемопоэтических стволовых клеток (аллоТГСК), 73 пациента после неродственной аллоТГСК и 16 пациентов после гаплоидентичной аллоТГСК). Помимо этого был обследован материал здоровых доноров (n=10).
Относительную экспрессию генов PRAME, WT1 и BAALC в анализируемых образцах определяли исходя из уравнений калибровочных кривых для соответствующих генов. Для этого в каждую амплификацию включали 5 последовательных убывающих десятичных разведений плазмидной ДНК известной концентрации, содержащей амплифицируемые участки анализируемых генов. Эффективность амплификации определяли как угол наклона калибровочной кривой. Для всех пар праймеров эффективность амплификации составила от 90% до 100%, коэффициент корреляции 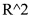 был больше или равен 0,990. Для оценки воспроизводимости значений порогового цикла (Ct) все анализируемые образцы с каждой парой праймеров амплифицировались в двойном повторе. В анализ включались образцы, для которых разница значений Ct между повторами была не более 0,3 цикла.
был больше или равен 0,990. Для оценки воспроизводимости значений порогового цикла (Ct) все анализируемые образцы с каждой парой праймеров амплифицировались в двойном повторе. В анализ включались образцы, для которых разница значений Ct между повторами была не более 0,3 цикла.
Абсолютное количество кДНК каждого гена определяли исходя из уравнения калибровочной кривой при помощи программного обеспечения iQ5 (Bio-Rad). В анализ включали образцы, для которых рассчитанное относительно калибровочной кривой количество кДНК гена ABL1 превышало 10000 копий на реакцию. Уровень относительной экспрессии гена WT1, составляющий более 250 копий гена WT1 на 10000 копий гена ABL рассматривали как гиперэкспрессию маркера. Для гена BAALC под гиперэкспресией понимали выявление более 10 копий транскрипта данного гена на 100 копий гена ABL, для гена PRAME - 500 копий транскрипта на 100 гена ABL.
При оценке уровня относительной экспрессии было выявлено наличие достоверных различий в уровне экспрессии гена WT1 у здоровых доноров в костном мозге и периферической крови, что совпадает с результатами других исследователей [К Inoue, Н Ogawa, Т Yamagami, Т Soma, Y Tani, Т Tatekawa, Y Oji, H Tamaki, T Kyo, H Dohy, A Hiraoka, T Masaoka, T Kishimoto and H Sugiyama. Long-term follow-up of minimal residual disease in leukemia patients by monitoring WT1 (Wilms tumor gene) expression levels. Blood, 1996, 88, 2267-2278].
Исследование информативности предложенной системы анализа указывает на то, что у 95% пациентов обладали хотя бы одним информативным маркером (фиг. 2).
При анализе данных пациентов в посттрансплантационном периоде обращает на себя внимание наличие прямой корреляционной связи между уровнем экспрессии исследуемых генов и выявляемых химерных транскриптов. Для гена WT1 коэффициент корреляции (критерий Спирмена) составил 0,26 (р=0.01), для гена BAALC - 0.27 (р=0.0001), а для гена PRAME - 0.312 (р=0.001). Сопоставимые результаты были получены и при анализе корреляционной зависимости изменения количества клеток опухолевой популяции (бластных клеток). При сопоставлении полученных данных экспрессии с результатами исследования донорского химеризма (приживления клеток донора после аллоТГСК) наблюдалась обратная зависимость, выражающаяся в обратной корреляции - коэффициент корреляции для WT1 составил -0,20 для гена WT1 (р=0.0006), -0.21 для гена BAALC (р=0.00001) и -0,14 для гена PRAME (р-0.002).
При анализе прогностического значения гена PRAME не было выявлено достоверного увеличения развития рецидивов в посттрансплантационном периоде (р=0,14). Вместе с тем нами было показано, что при наличии гиперэкспресии генов WT1 и BAALC достоверно возникает риск развития рецидива у пациентов в посттрансплантационном периоде - р=0,009 и р=0,027 соответственно. При оценке куммулятивного риска развития рецидива после аллоТГСК были получены данные, указывающие на высокую вероятность развития рецидива заболевания на фоне гиперэкспрессии любого из исследуемых генов были получены результаты, указывающие на неблагоприятный прогноз на фоне детекции гиперэкспрессии (фиг. 3).
Таким образом, данное изобретение позволяет повысить точность оценки относительной экспрессии данных генов благодаря конструкции ДНК-калибраторов, совмещающих последовательности всех маркеров. Комплексный анализ генов PRAME, WT1 и BAALC позволяет оценивать эффективность проводимой терапии в большинстве случаев ОМЛ, в том числе у пациентов без выявленных хромосомных аберраций.
Перечень последовательностей



Изобретение относится к биотехнологии и медицине, в частности к рекомбинантной плазмидной ДНК pAL2-T-ExP. Плазмидная ДНК pAL2-T-ExP предназначена для создания ДНК-калибраторов при оценке эффективности терапии у пациентов с острым миелоидным лейкозом. Указанная плазмидная ДНК содержит вставку последовательно сшитых в реакции лигирования участков кДНК генов PRAME, WT1, BAALC, ABL1, GUSB. Изобретение позволяет получать ДНК-калибраторы для оценки эффективности терапии с повышенной точностью у пациентов с острым миелоидным лейкозом. 3 ил.
Рекомбинантная плазмидная ДНК pAL2-T-ExP, используемая для создания ДНК-калибраторов при оценке эффективности терапии у пациентов с острым миелоидным лейкозом, содержащая вставку последовательно сшитых в реакции лигирования участков кДНК генов PRAME, WT1, BAALC, ABL1, GUSB, амплифицируемых в ПЦР-РВ,
| ШАКИРОВА А.И | |||
| и др | |||
| Оценка вероятности развития рецидива острого миелоидного лейкоза в пострансплантационном периоде у пациентов с использованием панели экспрессионных маркеров | |||
| Клеточная терапия и трансплантация, 2016 | |||
| MINETTO P | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Leukemia research, 2015 | |||
| SANTAMARIA C | |||
| et al | |||
| BAALC is an important predictor of refractoriness to chemotherapy and poor survival in intermediate-risk acute myeloid leukemia (AML) | |||
| Annals of hematology, 2010. | |||

