Область техники, к которой относится изобретение
Настоящее изобретение относится к варианту L-арабинозоизомеразы из Thermotoga neapolitana DSM 5068, которая получена с помощью белковой инженерии и обладает повышенной активностью превращения D-галактозы в D-тагатозу, к микроорганизму, который экспрессирует вариант L-арабинозоизомеразы, и к способу получения D-тагатозы из D-галактозы с применением такого микроорганизма.
Уровень техники
D-тагатоза является моносахаридом, которая имеет сладость, равную приблизительно 90% сладости сахара, и обладает такими свойствами, как низкая калорийность и неспособность вызывать кариес. Она может применяться в качестве диетического подсластителя, который в отличие от обычных подсластителей не вызывает различные заболевания у взрослых.
Благодаря таким свойствам D-тагатоза рассматривается в качестве заменителя сахара и, как известно, обладает большим потенциалом на продовольственном рынке. Однако тагатоза является редким сахаром, который мало распространен в природе, но содержится в молочных продуктах или некоторых растениях в очень малых количествах. Поэтому для применения тагатозы в качестве низкокалорийного функционального подсластителя необходимо разработать технологию, позволяющую получать тагатозу.
D-тагатоза была получена способом химической изомеризации из D-галактозы при использовании Ca(OH)2 в качестве катализатора компанией Arla Foods Ingredients Inc. в 2003 году и выпущена под товарным знаком "Gaio-тагатоза". Однако известно, что процесс химический изомеризации превосходен с точки зрения выхода изомеризационного превращения, но имеет недостатки, связанные с трудностью выделения и очистки, а также сложностью процесса, и, таким образом, общий выход процесса ниже, чем в случае процесса ферментативной изомеризации.
L-арабинозоизомераза (EC 5.3.1.5) является ферментом, который катализирует реакцию изомеризации превращения L-арабинозы в L-рибулозу. Кроме того, известно, что L-арабинозоизомераза не только превращает L-арабинозу (которая является ее естественным субстратом) в L-рибулозу, но также и D-галактозу (которая является субстратом, структурно подобным L-арабинозе) в D-тагатозу.
Наиболее важный фактор, способный обеспечить повышение производительности процесса производства D-тагатозы из D-галактозы с применением L-арабинозоизомеразы, состоит в разработке фермента, который обладает хорошой реакционной способностью и может быть успешно применен в процессе получения путем модификации изомеразы. Поскольку повышение производительности играет важную роль в максимизации прибыли при уменьшении издержек производства и успешности бизнеса, сохранялась потребность в модификации арабинозоизомеразы.
Арабинозоизомераза из Thermotoga neapolitana DSM 5068, который является термофильным микроорганизмом, обладает очень высокой термостабильностью, но требуется ее дополнительное улучшение, чтобы гарантировать экономическую производительность арабинозоизомеразы, которая сопоставима с производительностью глюкозоизомеразы.
Как правило, способы получения вариантов ферментов с целью повышения активности ферментов или создания ферментов, активных в отношении новых субстратов, в основном подразделяются на способ случайного мутагенеза и способ рационального дизайна. Способ случайного мутагенеза широко применяется, поскольку его можно использовать без необходимости в специальной информации в отношении целевого фермента. Впрочем, для этого требуется система скрининга, способная обрабатывать очень большое количество различных ферментов. С другой стороны, модификация ферментов с помощью рационального дизайна не требует никакой специальной системы скрининга, поскольку она дает лишь ограниченное число различных ферментов. Однако в случае рационального дизайна факторы, которые определяют каталитический механизм, способность связывания субстрата или субстратную специфичность целевого фермента, требуется тщательно исследовать.
Техническая задача
Таким образом, заявитель попытался увеличить субстратную специфичность L-арабинозоизомеразы из Thermotoga neapolitana DSM 5068 в отношении D-галактозы путем изменения трехмерной структуры L-арабинозоизомеразы на основе белковой инженерии, молекулярного моделирования и анализа механизма ферментативной реакции таким образом, чтобы арабинозоизомераза, обладающая потенциалом получения тагатозы, могла обеспечивать промышленное получение тагатозы.
Цель настоящего изобретения состоит в предоставлении варианта арабинозоизомеразы из Thermotoga neapolitana DSM 5068, который обладает повышенной превращающей активностью, и нуклеотидной последовательности гена, кодирующей указанный вариант.
Другая цель настоящего изобретения состоит в предоставлении рекомбинантного вектора, включающего нуклеотидную последовательность гена, и микроорганизма рода Corynebacterium, трансформированного указанным рекомбинантным вектором.
Еще одна цель настоящего изобретения состоит в предоставлении способа получения D-тагатозы из D-галактозы с применением варианта арабинозоизомеразы или трансформированного микроорганизма, или культуры трансформированного микроорганизма.
Техническое решение
Для достижения вышеуказанных целей в настоящем изобретении предложен вариант арабинозоизомеразы, обладающий повышенной активностью превращения D-галактозы в D-тагатозу, при этом вариант арабинозоизомеразы содержит замену лейцина в положении 469 на пролин и замену на аминокислоту кроме фенилаланина аминокислоты в положении 275 арабинозоизомеразы из Thermotoga neapolitana DSM 5068, и нуклеотидная последовательность гена, кодирующая вариант арабинозоизомеразы.
В настоящем изобретении также предложен рекомбинантный вектор, включающий нуклеотидную последовательность гена, и микроорганизм рода Corynebacterium, трансформированный рекомбинантным вектором.
В настоящем изобретении также предложен способ получения D-тагатозы из D-галактозы с применением варианта арабинозоизомеразы или трансформированного микроорганизма, или культуры трансформированного микроорганизма.
Полезные эффекты
Согласно настоящему изобретению продукция D-тагатозы может быть увеличена с применением микроорганизма рода Corynebacterium, трансформированного новым вариантом арабинозоизомеразы или нуклеотидной последовательностью гена, кодирующей вариант арабинозоизомеразы, что обеспечивает уменьшение издержек производства и инвестиций в инфраструктуру.
Описание фигур
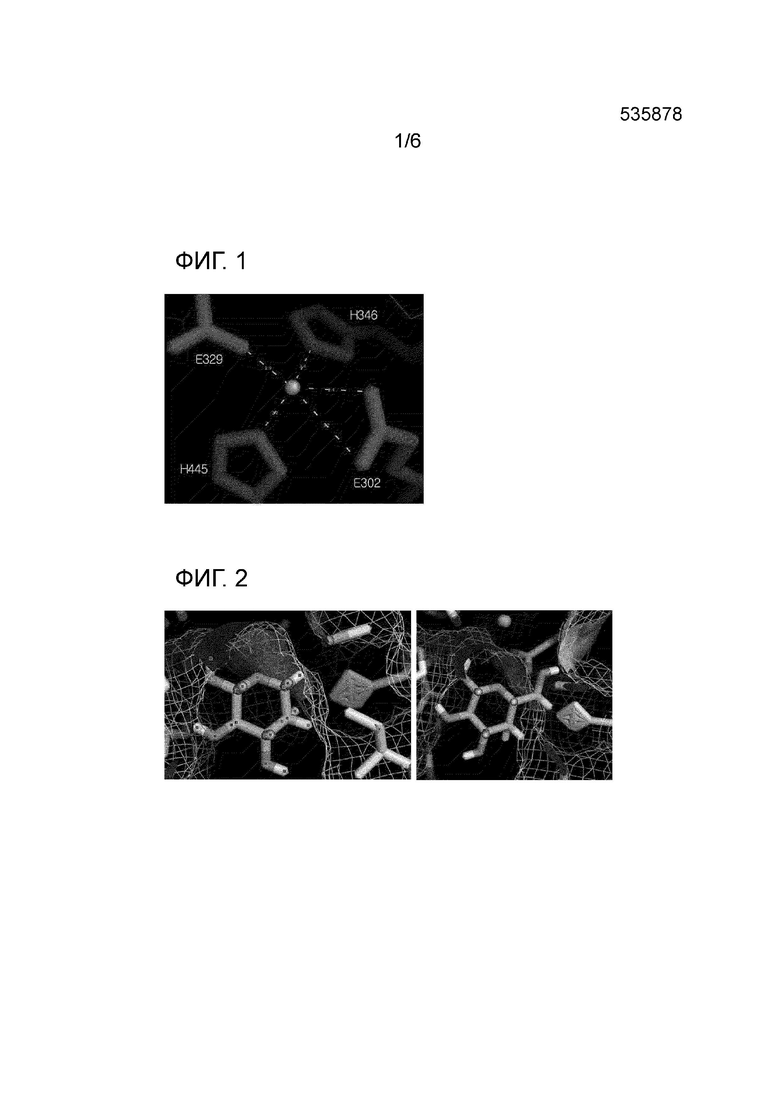
На Фиг. 1 показана структура, состоящая из активного центра и иона марганца в качестве кофактора L-арабинозоизомеразы из Thermotoga neapolitana.
На Фиг. 2 показаны участки L-арабинозы и D-галактозы, которые связываются с активным центром L-арабинозоизомеразы, и функциональные группы L-арабинозы и D-галактозы, которые взаимодействуют с активным центром L-арабинозоизомеразы. Показано, что углерод 6 D-галактозы вызывает стерическое затруднение с остатком фенилаланина в положении 275 L-арабинозоизомеразы.
На Фиг. 3 показаны результаты, полученные при насыщающем мутагенезе остатка 275 L-арабинозоизомеразы и сравнении относительной активности вариантов, отобранных с помощью цистеин-карбазольного метода, посредством реакции с развитием цвета. Было обнаружено значительное количество вариантов, показывающих более высокую активность, чем у контроля.
На Фиг. 4 показаны структуры отобранных вариантов (F275V/L469P, F275M/L469P и F275I/L469P), предсказанных с помощью методики молекулярного моделирования.
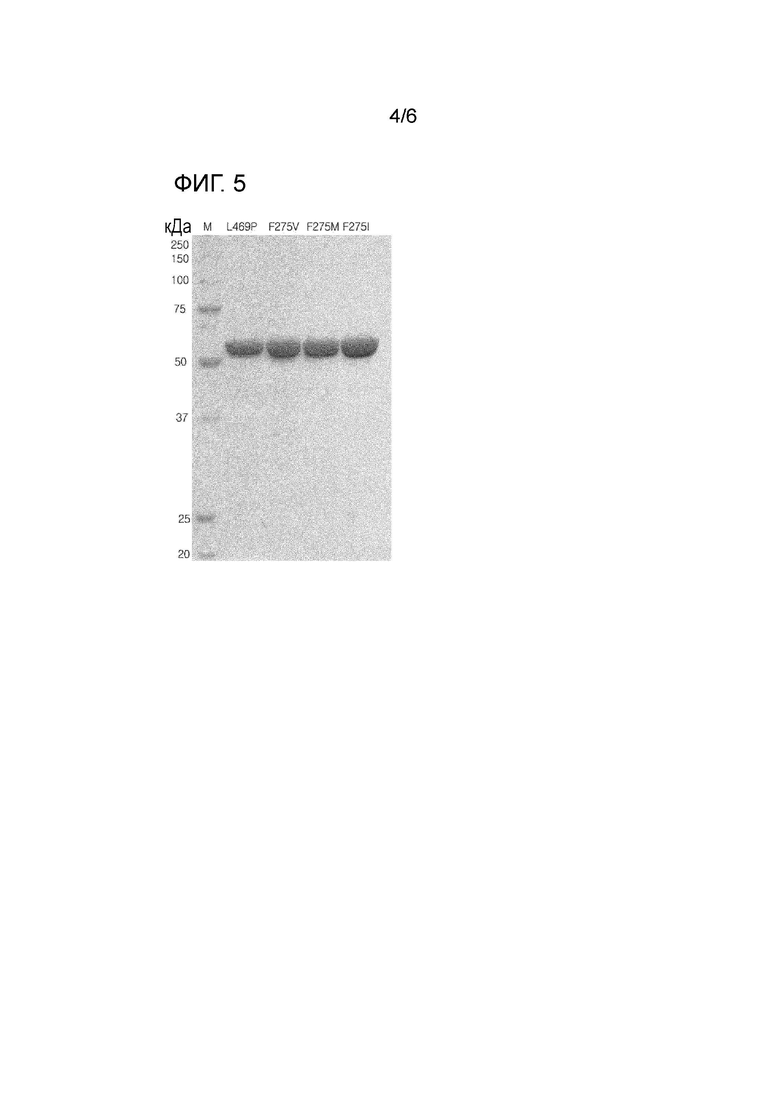
На Фиг. 5 показаны результаты анализа с помощью электрофореза в ДСН-ПААГ выделенных и очищенных вариантов L-арабинозоизомеразы Thermotoga neapolitana.
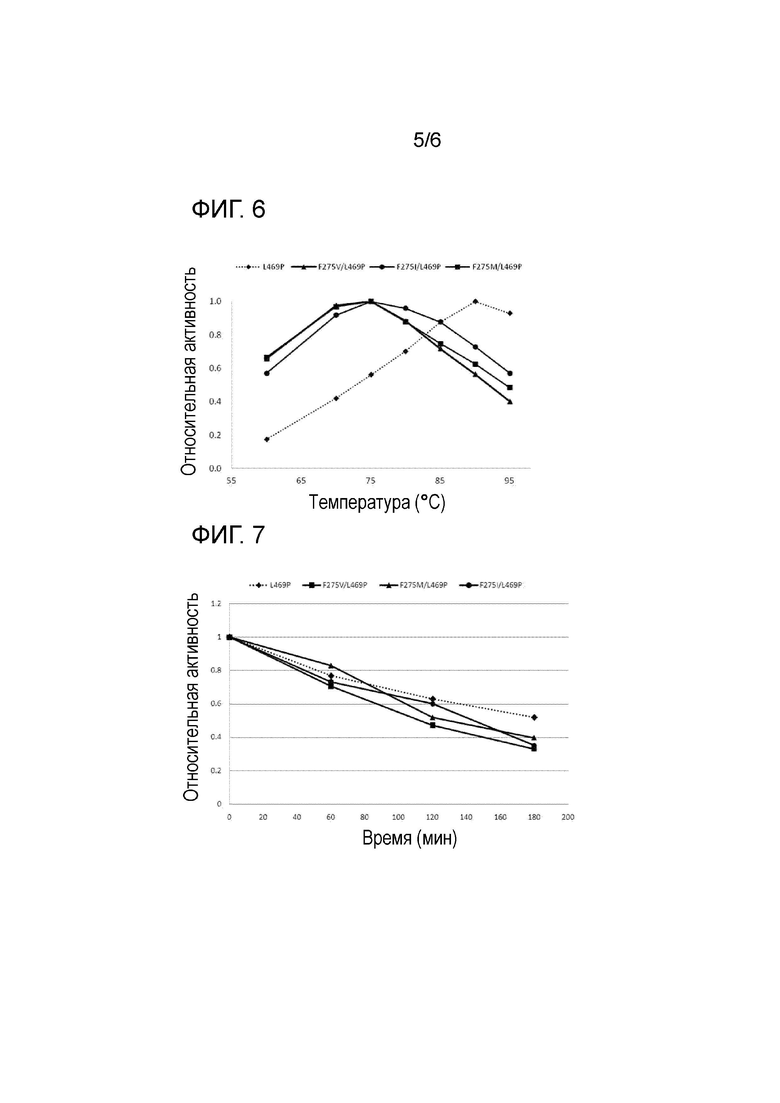
На Фиг. 6 показаны результаты оценки относительной активности отобранных вариантов при различных температурах для определения оптимальных температур вариантов.
На Фиг. 7 показаны результаты измерения термостабильности отобранных вариантов при 95°C в зависимости от времени.
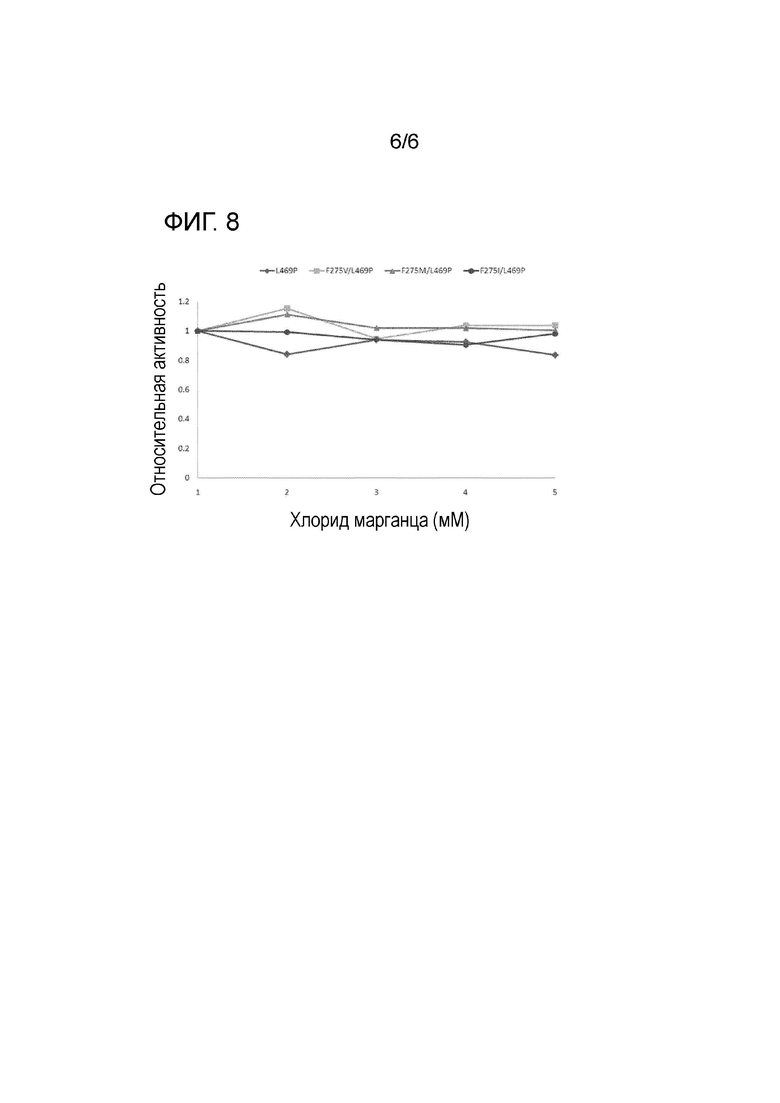
На Фиг. 8 показаны результаты оценки относительной активности отобранных вариантов для определения зависимости вариантов от марганца.
Вариант осуществления изобретения
В варианте осуществления настоящего изобретения предложен вариант арабинозоизомеразы из Thermotoga neapolitana DSM 5068, который обладает повышенной превращающей активностью, и нуклеотидная последовательность гена, кодирующая указанный вариант.
В предпочтительном варианте осуществления настоящего изобретения предложен вариант арабинозоизомеразы, обладающий повышенной активностью превращения D-галактозы в D-тагатозу, при этом вариант арабинозоизомеразы содержит замену лейцина в положении 469 на пролин и замену на аминокислоту кроме фенилаланина аминокислоты в положении 275 арабинозоизомеразы из Thermotoga neapolitana DSM 5068, и нуклеотидная последовательность гена, кодирующая указанный вариант арабинозоизомеразы.
При использовании в настоящем описании, выражение "арабинозоизомераза, которая превращает D-галактозу в D-тагатозу" означает фермент, который катализирует реакцию изомеризации с использованием D-галактозы в качестве субстрата для продукции D-тагатозы.
Вариант арабинозоизомеразы согласно настоящему изобретению предпочтительно содержит замену аминокислоты в положении 275 арабинозоизомеразы на аминокислоту, имеющую неполярную алифатическую боковую цепь.
При использовании в настоящем описании, выражение "аминокислота, имеющая неполярную алифатическую боковую цепь", означает аланин, валин, изолейцин, лейцин, метионин или пролин.
Предпочтительно аминокислота в положении 275 арабинозоизомеразы заменена любой аминокислотой, выбранной из группы, состоящей из валина, метионина и изолейцина.
Предпочтительно вариант арабинозоизомеразы согласно настоящему изобретению дополнительно содержит замену лейцина в положении 469 арабинозоизомеразы на пролин.
При использовании в настоящем описании, термин "замена" означает замену аминокислоты в определенном положении другой аминокислотой с получением мутации. Подходящий метод мутагенеза может быть любым методом, который может использоваться специалистами для этой цели. В частности, метод мутагенеза может быть методом насыщающего мутагенеза, методом случайного мутагенеза или методом сайт-направленного мутагенеза (Evolutionary molecular 15 engineering based on RNA replication, Pure Appl. Chem. 1984, 56:967-978; Promoters selected from random DNA-sequences, Proc. Natl. Acad. Sci. USA, 1986, 83:7405-7409; Mutants generated by the insertion of random oligonucleotides into the active-site of the beta-lactamase gene, Biochemistry 1989, 28:5703-5707).
Предпочтительно в варианте арабинозоизомеразы согласно настоящему изобретению аминокислота в положении 275 заменена с использованием метода насыщающего мутагенеза, и лейцин в положении 469 заменен с использованием метода сайт-направленного мутагенеза.
В варианте осуществления настоящего изобретения случайный мутагенез был выполнен с использованием арабинозоизомеразы дикого типа (имеющей аминокислотную последовательность SEQ ID NO: 1 и нуклеотидную последовательность SEQ ID NO: 6) из Thermotoga neapolitana DSM 5068 в качестве матрицы, в результате чего были получены варианты, обладающие улучшенными ферментативными характеристиками, и генетическая информация о вариантах. Варианты были проанализированы совместно, и в результате было обнаружено, что вариация аминокислотной последовательности C-концевой области арабинозоизомеразы влияла на увеличение ферментативной активности.
Кроме того, аминокислоты в C-концевой области, составляющей активный центр арабинозизомеразы дикого типа и варианта арабинозоизомеразы, проанализировали с помощью молекулярного моделирования. В результате было обнаружено, что вариант содержал замену лейцина в положении 469 арабинозоизомеразы дикого типа на пролин, и таким образом бета-лист в положении 18 белка исчезал, при этом угол основной цепи наклонялся, тогда как трехмерная структура альфа-спирали в положении 17 смещалась к телу белка, что указывает на то, что структура белка была изменена.
Исходя из вышеописанных результатов, лейцин в положении 469 арабинозоизомеразы дикого типа был заменен пролином при использовании сайт-направленного мутагенеза, с получением варианта (L469P) (имеющего аминокислотную последовательность SEQ ID NO: 2 и нуклеотидную последовательность SEQ ID NO: 7). Вариант инкубировали, затем измеряли его активность, и в результате было обнаружено, что данный вариант показал более высокую активность в отношении субстрата галактозы по сравнению с арабинозоизомеразой дикого типа.
В варианте осуществления настоящего изобретения, с целью получения фермента, обладающего повышенной превращающей активностью по сравнению с вариантом арабинозоизомеразы (L469P), были выбраны главные остатки субстрат-связывающей области и активной области фермента, и был оценен механизм реакции, в результате чего была выбрана аминокислота в положении 275. Мутации были введены в положение 275 при использовании метода насыщающего мутагенеза, после чего варианты подвергли скринингу, отбрав таким образом варианты, обладающие повышенной превращающей активностью.
Отобранные варианты секвенировали, и в результате было обнаружено, что варианты содержали замены аминокислоты в положении 275 на валин (L469P/F275V), метионин (L469P/F275M) и изолейцин (L469P/F275I), соответственно. Каждый из этих трех вариантов трансформировали в микроорганизмы рода Corynebacterium, и реакцию изомеризации проводили при культивировании указанных микроорганизмов. В результате обнаружено, что все три варианта показали увеличенную активность по сравнению с вариантом L469P, содержащим замену лейцина в положении 469 на пролин.
Для исследования свойств этих трех вариантов, экспрессированные арабинозоизомеразы выделяли из микроорганизмов рода Corynebacterium, культивированных при вышеописанных условиях. Было обнаружено, что очищенные белки показали арабинозоизомеразную активность в отношении D-галактозы. Кроме того, с помощью электрофореза в ДСН-ПААГ было обнаружено, что очищенные белки имели молекулярные массы, соответствующие молекулярной массе арабинозоизомеразы.
Используя очищенные варианты ферментов, измеряли оптимальную температуру, термостабильность, использование ионов металлов и ферментативную активность различных ферментов. В результате было обнаружено, что эти три варианта показали наиболее высокую активность при 75°C, но при этом несколько уменьшилась термостабильность, использование ионов металлов существенно не отличалось, и удельные активности вариантов арабинозоизомеразы приблизительно в 5,5 раз (F275V/L469P), в 5 раз (F275M/L469P) и в 3,9 раза (F275I/L469P), соответственно, превышали активность варианта L469P.
В варианте осуществления настоящего изобретения предложена нуклеотидная последовательность гена, кодирующая вариант арабинозоизомеразы.
Нуклеотидная последовательность гена может быть любой последовательностью, выбранной из группы, состоящей из SEQ ID NO: 8, SEQ ID NO: 9 и SEQ ID NO: 10.
Варианты арабинозоизомеразы согласно настоящему изобретению могут иметь нуклеотидные последовательности генов, кодирующие белки, обладающие гомологией по меньшей мере 80%, предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 95% и наиболее предпочтительно по меньшей мере 97%, с аминокислотными последовательностями SEQ ID NO: 3-5, при условии, что арабинозоизомеразная активность вариантов арабинозоизомеразы согласно настоящему изобретению может быть сохранена или увеличена. Наиболее предпочтительно варианты арабинозоизомеразы согласно настоящему изобретению имеют нуклеотидные последовательности генов, представленные в SEQ ID NO: 8-10.
При использовании в настоящем описании, термин "гомология" относится к идентичности между двумя аминокислотными последовательностями. Гомология может быть определена при использовании методов, известных специалистам в данной области техники, например, BLAST 2.0, который вычисляет такие параметры, как оценку, идентичность или подобие.
Кроме того, полинуклеотиды согласно настоящему изобретению могут быть вариантами, кодирующими варианты арабинозоизомеразы, которые могут гибридизоваться с полинуклеотидами, представленными в SEQ ID NO: 8-10, или зондами из таких полинуклеотидов при строгих условиях, и которые нормально функционируют.
При использовании в настоящем описании термин "строгие условия" означает условия, которые обеспечивают специфичную гибридизацию между полинуклеотидами. Например, гибридизацию проводят в гибридизационном буфере (3,5> SSC, 0,02% фиколла, 0,02% поливинилпирролидона, 0,02% бычьего сывороточного альбумина, 2,5 мМ NaH2PO4 (pH 7), 0,5% ДСН, 2 мМ ЭДТА) при 65°C ("Molecular Cloning", A Laboratory Manual, J. Sambrook et al., Editors, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989 или Current Protocols in Molecular Biology, F.M. Ausubel et al., Editors, John Wiley & Sons, Inc., New York). В данном случае, SSC обозначает 0,15 М хлорида натрия/0,15 М цитрата натрия (pH 7). После гибридизации мембрану, на которую перенесена ДНК, промывают 2> SSC при комнатной температуре, и затем промывают 0,1-0,5> SSC/0,1×ДСН при температуре 68°C.
В варианте осуществления настоящего изобретения предложен вариант изомеразы, имеющий любую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 3, SEQ ID NO: 4 и SEQ ID NO: 5. В частности, вариант является мутантом арабинозоизомеразы, который содержит замену аминокислоты в положении 275 на любую аминокислоту, выбранную из группы, состоящей из валина, метионина и изолейцина, а также содержит замену аминокислоты в положении 469 на пролин.
Впрочем, вариант изомеразы не ограничивается этим, поскольку аминокислотная последовательность фермента, демонстрирующего активность полипептида, может отличаться в зависимости от видов или штаммов микроорганизмов. В частности, вариант настоящего изобретения может быть мутантом или искусственным мутантом, кодирующим полипептид, имеющий аминокислотную последовательность, включающую замену, делецию, вставку или добавление одной или нескольких аминокислот в одном или более положениях кроме положений 275 и 469 аминокислотной последовательности любой из SEQ ID NO: 3-5, при условии, что активность арабинозоизомеразы может быть сохранена или увеличена.
При использовании в настоящем описании термин "несколько аминокислот" означает 2-20 аминокислот, предпочтительно 2-10 аминокислот и более предпочтительно 2-5 аминокислот, в зависимости от типа или положений остатков аминокислот в трехмерной структуре белка.
Кроме того, замены, делеции, вставки, добавления или инверсии аминокислот могут включать природные мутанты или искусственные варианты, основанные на индивидуальных различиях и/или видовых различиях микроорганизма, экспрессирующего арабинозоизомеразу.
В варианте осуществления настоящего изобретения также предложен рекомбинантный вектор, включающий полинуклеотид, функционально связанный с ним.
При использовании в настоящем описании термин "вектор" относится к конструкции ДНК, содержащей нуклеотидную последовательность гена, кодирующего целевой белок, функционально связанную с подходящей регуляторной последовательностью, что позволяет экспрессировать целевой ген в подходящей клетке-хозяине. Регуляторная последовательность включает промотор, способный инициировать транскрипцию, какой-либо оператор для регуляции этой транскрипции, последовательность, кодирующую подходящий мРНК участок связывания рибосомы, и последовательность для регуляции терминации транскрипции и трансляции. После трансформации подходящего организма-хозяина вектор может реплицироваться или функционировать независимо от генома организма-хозяина, или может интегрироваться в геном непосредственно.
Вектор, который применяется в настоящем изобретении, специально не ограничен и может быть любым вектором, известным в уровне техники, при условии, что он может реплицироваться в организме-хозяине. Примеры обычно используемых векторов могут включать природные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги.
Кроме того, вектор, который применяется в настоящем изобретении, является вектором, способным к трансформации клеток-хозяев, которая приводит к вставке полинуклеотида, кодирующего целевой белок, в хромосому клетки-хозяина. Определенные примеры вектора включают, без ограничения, шаттл-вектор pECCG112, который может самореплицироваться в обоих направлениях в E. coli и бактериях корине-типа (Kap-Soo, Noh, Kor. Jour. Microb iol. July 1991, p 149-154).
Кроме того, полинуклеотид, кодирующий эндогенный целевой белок в хромосоме, может быть заменен новым полинуклеотидом с помощью вектора для вставки в бактериальную хромосому. Вставка полинуклеотида в хромосому может быть выполнена любым методом, известным в уровне техники, например, гомологичной рекомбинацией.
Поскольку вектор настоящего изобретения может быть вставлен в хромосому посредством гомологичной рекомбинации, он может также включать селективный маркер для подтверждения его вставки в хромосому. Селективный маркер используется для отбора клетки, трансформированной вектором, то есть подтверждения вставки целевого полинуклеотида. Селективный маркер, который применяется в настоящем изобретении, может быть выбран из маркеров, которые обеспечивают селектируемые фенотипы, такие как устойчивость к лекарственному средству, ауксотрофия, устойчивость к цитотоксическим средствам или поверхностная экспрессия белков. Только клетки, экспрессирующие селективный маркер, могут выживать или демонстрировать различные фенотипы в среде, обработанной селективным агентом, и таким образом могут быть отобраны трансформированные клетки.
В варианте осуществления настоящего изобретения также предложен микроорганизм рода Corynebacterium, трансформированный рекомбинантным вектором.
При использовании в настоящем описании термин "трансформация" означает введение вектора, включающего полинуклеотид, кодирующий целевой белок, в клетку-хозяина, с тем чтобы экспрессировать белок, кодируемый полинуклеотидом, в клетке-хозяине. Трансформированные полинуклеотиды включают все гены, вставленные в хромосому клетки-хозяина или расположенные вне хромосомы, при условии, что они могут экспрессироваться в клетке-хозяине. Кроме того, полинуклеотиды включают ДНК и РНК, которые кодируют целевой белок. При условии, что полинуклеотид может быть введен в клетку-хозяина и экспрессироваться в ней, ген может быть введен в любой форме. Например, полинуклеотид может быть введен в клетку-хозяина в форме кассеты экспрессии, которая является полинуклеотидной конструкцией, включающей все элементы для экспрессии гена. Кассета экспрессии включает промотор, который функционально связан с геном, сигнал терминации транскрипции, участок связывания рибосомы и сигнал терминации трансляции. Кассета экспрессии может быть в форме вектора экспрессии, способного к саморепликации. Полинуклеотид может быть также введен в клетку-хозяина сам по себе и может быть функционально связан с последовательностью, необходимой для экспрессии в клетке-хозяине.
Микроорганизм настоящего изобретения включает любой из прокариотических микроорганизмов и эукариотических микроорганизмов, при условии, что он может экспрессировать вариант изомеразы. Например, он может включать микроорганизм, относящийся к роду Escherichia, роду Erwinia, роду Serratia, роду Providencia, роду Corynebacterium или роду Brevibacterium. Предпочтительно микроорганизм настоящего изобретения является микроорганизмом, относящимся к роду Corynebacterium. Более предпочтительно он является Corynebacterium glutamicum.
В примере настоящего изобретения Corynebacterium glutamicum, обладающую способностью продуцировать L-аминокислоту, трансформировали вектором, имеющим нуклеотидную последовательность каждого SEQ ID NO: 8, 9 и 10, и сконструированные штаммы были названы Corynebacterium glutamicum pFIS-1-TNAI-2, pFIS-1-TNAI-3 и pFIS-1-TNAI-4, соответственно, и были депонированы в Корейском центре культур микроорганизмов (Korean Culture Center of Microorganisms, 361-221, Honje 1-dong, Seodaemun-gu, Seoul, South Korea), международном депонирующем органе, 14 февраля 2013 года, под регистрационными номерами KCCM11378P, KCCM11379P и KCCM11380P, соответственно. В варианте осуществления настоящего изобретения также предложена культура микроорганизма рода Corynebacterium.
Культура может быть неразведенной культурой, включающей клетки микроорганизма, или может быть микробной клеткой, полученной при удалении супернатанта культуры или концентрировании культуры. Состав среды для культивирования микроорганизма может включать не только обычные компоненты, требуемые для культуры микроорганизмов рода Corynebacterium, но также компоненты, оказывающие синергическое воздействие на рост микроорганизмов рода Corynebacterium, и может быть легко подобран специалистами в данной области техники. Кроме того, культура может находиться в жидком или сухом состоянии, и способы сушки культуры включают, без ограничения, воздушную сушку, естественную сушку, сушку распылением и лиофильную сушку.
Микроорганизм рода Corynebacterium согласно настоящему изобретению можно культивировать любым стандартным способом. В частности, микроорганизм можно культивировать путем его инокулирования в среду, которая полностью или частично содержит сахарозу или глюкозу в качестве источника углерода. Процесс культивирования можно вести в подходящей среде и условиях культивирования, известных в уровне техники. Этот процесс культивирования может быть легко изменен любым специалистом в данной области в зависимости от типа выбранного штамма. Примеры процесса культивирования включают, без ограничения, периодическое культивирование, непрерывное культивирование и культивирование с подпиткой. Среда, которая используется при культивировании микроорганизма настоящего изобретения, должна надлежащим образом соответствовать потребностям микроорганизма настоящего изобретения.
В частности, среда, которая используется в настоящем изобретении, содержит сахарозу или глюкозу в качестве основного источника углерода. Кроме того, патока, содержащая высокую концентрацию сахарозы, также может использоваться в качестве источника углерода. Кроме того, может использоваться подходящее количество различных источников углерода. Предпочтительно используется очищенная глюкоза. Примеры источников азота, которые могут использоваться в настоящем изобретении, включают органические источники азота, такие как пептон, дрожжевой экстракт, мясной экстракт, солодовый экстракт, кукурузный сироп и соевую муку, и неорганические источники азота, такие как мочевину, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Предпочтительно используется пептон. Указанные источники азота могут использоваться отдельно или в комбинации. Среда может содержать дигидрофосфат калия, гидрофосфат калия и соответствующие натрийсодержащие соли в качестве источников фосфора. Кроме того, среда может содержать соль металла, такую как сульфат магния или сульфат железа. Кроме того, питательная среда может содержать аминокислоты, витамины и подходящие предшественники. Указанные среды или предшественники можно добавлять в среду в периодическом или непрерывном режиме.
Среду обычно поддерживают при температуре в пределах от 27°C до 37°C и предпочтительно от 30°C до 37°C. Культивирование микроорганизма можно продолжать до тех пор, пока не будет получен требуемый уровень белка. Предпочтительно период культивирования составляет от 10 до 100 часов.
В настоящем изобретении предложен способ получения D-тагатозы, включающий реакцию раствора, содержащего D-галактозу, с источником ионов металла, выбранных из группы, состоящей из ионов марганца, ионов магния и ионов цинка, в присутствии варианта арабинозоизомеразы, обладающего активностью превращения D-галактозы в D-тагатозу, трансформированного микроорганизма рода Corynebacterium или культуры трансформированного микроорганизма рода Corynebacterium, с получением в результате D-тагатозы.
Чтобы обеспечить введение субстрата в трансформированный микроорганизм рода Corynebacterium или культуру трансформированного микроорганизма рода Corynebacterium, микробные клетки, получаемые при центрифугировании трансформированного микроорганизма или культуры, можно обработать поверхностно-активным веществом, лизоцимом или ксилолом. Предпочтительно микробные клетки можно обработать 0,1% POESA.
Раствор, содержащий D-галактозу, которая используется в настоящем изобретении, может быть выбран из группы, состоящей из очищенной D-галактозы, полученной из биомассы D-галактозы и D-галактозы, полученной при гидролизе лактозы, но не ограничивается этим.
Арабинозоизомераза является металлоферментом, который использует ион металла в качестве кофактора. Ион металла может быть выбран из группы, состоящей из ионов марганца, ионов магния и ионов цинка, но не ограничен, и может быть любым ионом металла, который может связываться с изомеразой для выполнения реакции изомеризации. В частности, источник иона марганца включает хлорид марганца; источник иона магния включает хлорид магния; и источник иона цинка включает хлорид цинка; впрочем, объем настоящего изобретения не ограничивается этими источниками ионов металлов.
Реакционный раствор для получения D-тагатозы содержит буферную систему для поддержания pH, такую как Трис-буфер или фосфатный буфер. Предпочтительно он содержит Трис-буфер (pH 6,5-7,5). Хлорид марганца, хлорид магния или хлорид цинка содержатся в концентрации от 0,1 мМ до 10 мМ, и предпочтительно от 1 мМ до 5 мМ. Субстрат D-галактозу добавляют в количестве 1-300 г/л и предпочтительно 18-300 г/л, и реакцию изомеризации инициируют при температуре от 60°C до 95°C, предпочтительно от 70°C до 80°C и более предпочтительно от 72°C до 78°C, с получением в результате D-тагатозы.
В дальнейшем настоящее изобретение будет описано более подробно со ссылкой на примеры. Специалистам в данной области будет очевидно, что эти примеры предназначены исключительно в иллюстративных целях и не должны ограничивать объем настоящего изобретения.
Примеры
Пример 1: Моделирование белка арабинозоизомеразы Thermotoga neapolitana для конструирования фермента, обладающего увеличенной превращающей активностью, и отбор основных аминокислотных мутаций
Поскольку трехмерная структура арабинозоизомеразы дикого типа из Thermotoga neapolitana DSM 5068 пока не была установлена, предсказание трехмерной структуры выполняли с помощью методики молекулярного моделирования при использовании в качестве матрицы арабинозоизомеразы Escherichia coli, трехмерная структура которой уже была установлена и которая обладает высокой гомологией последовательности. Для предсказания трехмерной структуры использовали методику сравнительного моделирования, и структурная модель была получена при использовании модуля APM (Tripos, USA) в пакете молекулярного моделирования.
Методика сравнительного моделирования является способом, который наиболее часто используют для предсказания трехмерных структур белков. Если аминокислотная последовательность требуемого белка обладает высоким подобием с последовательностью другого белка, трехмерная структура которого была известна, трехмерная структура требуемого белка может быть легко предсказана при использовании методики сравнительного моделирования, и в этом случае точность предсказания очень высока.
Структура арабинозоизомеразы E. coli, используемой в данном Примере, являлась структурой тримерного типа, зарегистрированной как 2HXG.pdb в Protein Data Bank (PDB).
На основе результатов моделирования было предсказано, что арабинозоизомераза дикого типа (имеющая аминокислотную последовательность SEQ ID NO: 1 и нуклеотидную последовательность SEQ ID NO: 6) из Thermotoga neapolitana DSM 5068 обладала бы очень высоким подобием с арабинозоизомеразой E. coli не только в отношении нуклеотидной последовательности, но также и дву- и трехмерных структур. В нескольких исследованиях арабинозоизомеразы E. coli сообщали информацию о субстрат-связывающем участке, кофакторе ионе марганца (Mn2+) и главных аминокислотах ((Manjasetty & Chance, J. Mol. Biol., 2006. 360:297-309). На основе такой информации были отобраны главные остатки изомеразы из Thermotoga neapolitana DSM 5068.
Для анализа главных аминокислотных остатков проводили выравнивание последовательностей, моделирование молекулярного докинга и анализ механизма реакции. Выравнивание последовательностей выполняли при использовании алгоритма clustalW (//www.ebi.ac.uk/Tools/msa/clustalw2/) на основе информации о последовательностях 10 различных L-арабинозоизомераз и арабинозоизомеразы Thermotoga neapolitana.
Результаты выравнивания последовательностей указывают, что последовательности металлсвязывающего участка и активного центра арабинозоизомеразы из Thermotoga neapolitana DSM 5068 являлись высококонсервативными, как и у других изомераз. Изомераза из Thermotoga neapolitana DSM 5068 имела активный центр, включающий аминокислотные остатки E302 (глутаминовая кислота в положении 302), E329 (глутаминовая кислота в положении 329), H346 (гистидин в положении 346) и H445 (гистидин в положении 445) и ионы марганца (Фиг. 1), при этом аминокислотные остатки E302, E329, H346 и H445 влияли на связывание иона марганца. В частности, было предсказано, что остатки E302 и E329 являются наиболее важными факторами, которые способствуют протеканию реакции изомеризации (Manjasetty & Chance, J. Mol. Biol., 2006. 360:297-309).
Предполагали, что субстратная специфичность арабинозоизомеразы Thermotoga neapolitana будет определяться в зависимости от размера и морфологии активного центра, а также свойств аминокислотных остатков активного центра. С помощью моделирования молекулярного докинга (surflexDock; Tripos, USA), выполненного с использованием исходного субстрата L-арабинозы, были отобраны остатки, важные при распознавании субстрата. Кроме того, с помощью анализа механизма реакции (Adrian J. Mulholland, Drug Discov. Today. 2005. 10(20):1393-402) было выбрано наиболее подходящее положение связывания D-галактозы. На основе такого выбора был выбран аминокислотный остаток, имеющий высокую возможность препятствовать связыванию между активным центром арабинозоизомеразы и D-галактозой.
Аминокислота в положении 275 состояла из фенилаланина, который имеет ароматическую боковую цепь, относительно большой размер и является менее гибким, при этом было предсказано, что аминокислота в положении 275 вызовет стерическое затруднение с углеродом 6 D-галактозы (Фиг. 2). Поскольку L-арабиноза, пентоза, демонстрирует по существу такую же структуру, что и D-галактоза, за исключением того, что количество атомов углерода меньше на один, чем в D-галактозе, предполагали, что реакционная способность арабинозоизомеразы с D-галактозой могла быть значительно увеличена посредством только замены аминокислоты в положении 275 другой аминокислотой.
В качестве аминокислот, способных заменить фенилаланин, были выбраны валин, метионин и изолейцин, которые имеют полярность по сравнению с фенилаланином, имеют относительно малый размер и очень гибкие, и, таким образом, как ожидают, будут оказывать меньшее воздействие на полную структуру арабинозоизомеразы, при минимальном отталкивании с углеродом 6 D-галактозы.
Поскольку эти выбранные аминокислоты состоят из неполярной алифатической цепи в отличие от фенилаланина, включающего ароматическую цепь, предполагали, что эти аминокислоты могли заменить фенилаланин, и при этом они будут структурно свободными.
Структуры вариантов (имеющих аминокислотные последовательности SEQ ID NO: 3-5 и нуклеотидные последовательности SEQ ID NO: 8-10), содержащих точечные мутации, были предсказаны с помощью методики молекулярного моделирования. В результате было предсказано, что влияние мутаций на полную структуру арабинозоизомеразы будет незначительным.
Кроме того, с помощью случайного мутагенеза, выполненного при использовании арабинозоизомеразы дикого типа из Thermotoga neapolitana DSM 5068 в качестве матрицы, были получены варианты, демонстрирующие улучшенные ферментативные характеристики, а также генетическая информация о вариантах. Варианты были проанализированы совместно, и в результате было обнаружено, что изменение аминокислотной последовательности C-концевой области арабинозоизомеразы привело к увеличению ферментативной активности.
Явление, вызвавшее значительное изменение структуры цепи C-концевой области арабинозоизомеразы, проанализировали с помощью методики молекулярного предсказания. В результате предположили, что реакционная способность арабинозоизомеразы в отношении D-галактозы может быть значительно увеличена даже посредством только замены лейцина на пролин в положении 469 арабинозоизомеразы дикого типа.
Пример 2: Получение сконструированных вариантов арабинозоизомеразы
(1) Замена лейцина на пролин в положении 469
Лейцин в положении 469 арабинозоизомеразы дикого типа из Thermotoga neapolitana DSM 5068 заменяли пролином с помощью метода сайт-направленного мутагенеза при использовании специфичных праймеров.
В качестве праймеров использовали N-концевой праймер (SEQ ID NO: 13) и C-концевой праймер (SEQ ID NO: 14), которые являются олигонуклеотидами, содержащими комплементарные нуклеотидные последовательности с мутацией. При использовании плазмидной ДНК в качестве матрицы, плазмиду, содержащую новую мутацию, амплифицировали и синтезировали в пробирке, а затем ДНК дикого типа удаляли путем расщепления рестриктазой Dpn I. Другими словами, ДНК дикого типа, используемая в качестве матрицы, представляла собой ДНК, выделенную из E. coli, и была расщеплена Dpn I, которая распознает и расщепляет Gm6ATC, но при этом ДНК, синтезированная в пробирке, не была расщеплена.
ДНК трансформировали в E. coli DH5 альфа с получением варианта гена, и затем нуклеотидную последовательность варианта гена проанализировали с целью подтверждения, что мутация прошла надлежащим образом. Вариант гена трансформировали в Corynebacterium glutamicum ATCC 13032 с получением рекомбинантного штамма, который затем назвали L469P. Рекомбинантный штамм использовали в качестве контроля.
(2) Замена аминокислоты в положении 259 аминокислотой кроме фенилаланина
Чтобы вызвать дополнительную мутацию в сконструированном варианте L469P, вектор, клонированный с изомеразой, подвергали насыщающему мутагенезу с использованием пары праймеров, содержащих мутацию.
Подобранная пара праймеров была подобрана таким образом, чтобы кодон аминокислоты в положении 275 был заменен NNS (N: A, T, G или C; и S: G или C). Варианты, полученные таким способом, могут включать 20 видов аминокислот (SEQ ID NO: 11 и SEQ ID NO: 12). Такие варианты дали одиночные колонии при трансформации, и изменения активности при 20 аминокислотных мутациях в соответствующих положениях могли быть надежно подтверждены с помощью скрининга одиночных колоний и анализа их активности.
В частности, для получения библиотеки, включающей 20 видов аминокислот, проводили полимеразную цепную реакцию (ПЦР) при использовании 100 мкл/мл плазмиды pECCG117-CJ1-TNAI_L469P (выложенная корейская патентная публикация 10-2010-0016948) в качестве матрицы и прямого и обратного праймеров. Реакцию ПЦР проводили при условиях, показанных в Таблицах 1 и 2 ниже. Библиотеку, полученную с помощью реакции ПЦР, трансформировали в штамм E. coli K12 DH5 α, получив колонии. Используемую плазмиду экспрессировали в штамме E. coli и тестировали ее активность в E. coli, поскольку она могла реплицироваться и экспрессироваться и в E. coli, и в бактериях корине-типа.
Активности изомераз, экспрессированных из 110 колоний, полученных, как описано, анализировали с помощью цистеин-карбазольного метода (Dische, Z., and E. Borenfreund., A New Spectrophotometric Method for the Detection and Determination of Keto Sugars and Trioses, J. Biol. Chem., 192:583-587, 1951), и в результате мог быть обнаружен ряд клонов, обладающих более высокой активностью, чем контроль (L469P) (Фиг. 3).
Из них отобрали и секвенировали 10 колоний, которые по результатам измерения имели наиболее высокую активность. В результате, как ожидалось, можно было наблюдать, что активность повышалась в порядке: валин, метионин и изолейцин (аминокислотные последовательности SEQ ID NO: 3-5 и нуклеотидные последовательности SEQ ID NO: 8-10). Кроме того, мутации в аминокислотах помимо аминокислоты в положении 275 не были обнаружены, что указывает на то, что остаток фенилаланина в положении 275 вызывает ингибирование реакционной способности D-галактозы.
Для исследования, оказывает ли мутация какое-либо воздействие на изомеразу, структуры вариантов предсказывали с помощью методики молекулярного моделирования (Фиг. 4). Результаты предсказания структур показали, что все три аминокислоты заменяли фенилаланин в положении 275, не вызывая существенных изменений в дву- и трехмерных структурах. Таким образом, можно было ожидать, что реакционная способность изомеразы в отношении D-галактозы увеличится без значительных увеличений термостабильности и других производственных показателей.
Варианты трансформировали в Corynebacterium glutamicum ATCC 13032, получив рекомбинантные штаммы. Рекомбинантные штаммы назвали "Corynebacterium glutamicum pFIS-1-TNAI-2, pFIS-1-TNAI-3 и pFIS-1-TNAI-4", соответственно, и депонировали в Корейском центре культур микроорганизмов (361-221, Honje 1-dong, Seodaemun-gu, Seoul, South Korea), международном депонирующем органе, 14 февраля 2013 года под регистрационными номерами KCCM11378P, KCCM11379P и KCCM11380P, соответственно.
Таблица 1: ПЦР с насыщающим мутагенезом
Таблица 2: условия реакции ПЦР
Пример 3: Экспрессия вариантов арабинозоизомеразы в микроорганизмах рода Corynebacterium
Для оценки степени увеличения активности отобранных вариантов и применимости вариантов для фактического производства D-тагатозы три варианта экспрессировали в микроорганизмах рода Corynebacterium и проводили исследования производительности и связанных с производством показателей.
Три варианта, отобранные в Примере 2, трансформировали в Corynebacterium glutamicum ATCC 13032, получив рекомбинантные штаммы. Эти рекомбинантные штаммы культивировали в среде (20 г/л глюкозы, 10 г/л полипептона, 10 г/л дрожжевого экстракта, 10 г/л сульфата аммония, 5,2 г/л KH2PO4, 10,7 г/л K2HPO4, 0,5 г/л MgSO4, 1,5 г/л мочевины, 1,8 мг/л D-биотина, 9 мг/л тиамина, 9 мг/л пантотената Ca, 60 мг/л ниацинамида), содержащей 50 мкг/мл канамицина, при 30°C в течение 20 часов для индукции экспрессии мутантов рекомбинантной арабинозоизомеразы.
Для измерения активности экспрессированных арабинозоизомераз, культуры центрифугировали при 8000 g в течение 10 минут для сбора бактериальных клеток, которые затем ресуспендировали в 50 мМ Трис-HCl (pH 7,5) буфере. Суспендированные клетки обрабатывали 0,1% POESA при комнатной температуре в течение 1 часа для ослабления клеточной стенки. Затем центрифугирование выполняли снова при вышеописанных условиях для сбора клеток, которые затем ресуспендировали в смешанном растворе 300 г/л D-галактозы, 5 мМ хлорида марганца и 50 мМ Трис-HCl (pH 7,5) при концентрации 4% (в/об). Суспензии позволяли реагировать при 75°C в течение 1 часа и проводили ВЭЖХ анализ (WATERS HPLC, EMPOWER system, колонка WATERS SugarPak ID 6.5-L 300 мм, рефрактометрический детектор 2414) для определения количества D-галактозы и D-тагатозы.
Результаты одного часа реакции показали, что контроль L469P давал 35 г/л D-тагатозы, тогда как вариант F275V/L469P показал производительность 121 г/л⋅ч, вариант F275M/L469P показал производительность 117 г/л⋅ч и вариант F275I/L469P показал 101 г/л⋅ч.
Пример 4: Выделение вариантов арабинозоизомеразы, экспрессированных в коринебактериях
200 мл рекомбинантного штамма, включающего каждый из трех вариантов (F275V/L469P, F275M/L469P и TNAI-F275I/L469P), активности которых были подтверждены, и контроль L469P культивировали после инокулирования в 2 л колбе при таких же условиях, как описано в Примере 3. Для исследования, экспрессировались ли арабинозоизомеразы, активности изомераз измеряли при использовании части каждой культуры при таких же условиях, описанных в Примере 3.
Бактериальные клетки, полученные из каждой культуры, ресуспендировали в 20 мМ Трис-HCl буфере (pH 7,5), содержащем 0,1 мМ хлорида марганца, и разрушали с использованием клеточного гомогенизатора высокого давления серии T (4,0 кВт: Constant systems, UK). Для удаления эндогенных белков кроме изомераз, клетки подвергали термической обработке при 75°C в течение 20 минут. Термически обработанный клеточный дебрис удаляли с помощью центрифугирования при 8000 g в течение 10 минут, а затем клеточный дебрис и липиды дополнительно удаляли с помощью ультрацентрифугирования (ультрацентрифуга BECKMAN COULTER Optima L-80 XP) при 60000 g.
Полученный в результате клеточный экстракт очищали с помощью анионообменной хроматографии (Mono QTM 10/100GL, GE Healthcare). Очищенный клеточный экстракт предварительно уравновешивали связывающим раствором (50 мМ NaCl, 0,1 мМ хлорида марганца, 20 мМ Трис-HCl (pH 7,5), и затем избыточное количество клеточного экстракта связывали и фракционировали с повышением соотношения элюирующего раствора [1 М NaCl, 0,1 мМ хлорида марганца, 20 мМ Трис-HCl (pH 7,5). Фракции, показывающие активность в отношении субстрата (D-галактозы), отбирали с помощью цистеин-карбазол-сернокислотного метода, и затем анализировали с помощью ДСН-ПААГЭ.
В результате можно было заметить, что очищенный белок имел молекулярную массу приблизительно 56 кДа, что соответствует известной молекулярной массе арабинозоизомеразы Thermotoga neapolitana. Фракция, имеющая самую высокую степень очистки, была выбрана с помощью ДСН-ПААГЭ, и высокая концентрация NaCl была удалена из нее при использовании обессоливающей колонки PD-10 (GE Healthcare). Полученный в результате очищенный фермент анализировали с помощью ДСН-ПААГЭ (Фиг. 5). Выделенный и очищенный белок определяли количественно при помощи анализа Брэдфорд, при этом BSA (бычий сывороточный альбумин) использовали в качестве стандартного белка.
Пример 5: Работы по исследованию вариантов арабинозоизомеразы
Было обнаружено, что три варианта арабинозоизомеразы, полученные в Примере выше, показали значительно более высокую активность по сравнению с вариантом, имеющим замену лейцина на пролин в положении 469. На основе данного результата были проведены эксперименты по параметрам, связанным с условиями реакции, влияющими на фактическую продукцию D-тагатозы.
5-1: Исследование оптимальной температуры
Арабинозоизомераза Thermotoga neapolitana, термофильный фермент, имеет относительно высокую термостабильность и оптимальную температуру. Температура, подходящая для продукции D-тагатозы из D-галактозы, составляет от 55°C до 75°C. При температуре ниже 55°C появляются проблемы, вызванные контаминацией гетерологичными штаммами, а при температуре выше 75°C возникают проблемы в отношении стабильности получаемой D-тагатозы. Арабинозоизомераза дикого типа или контроль L469P, оптимальная температура реакции для которых составляет 85°C, имеет недостаток, связанный с тем, что она показывает относительно низкую активность при температуре, при которой может применяться способ получения.
Для исследования оптимальных температур для вариантов арабинозоизомеразы, очищенных в Примере 4, каждый из очищенных ферментов добавляли к 100 мМ субстрата D-галактозы, и их активность измеряли в 50 мМ Трис-HCl буфере (pH 7,5), содержащем 1 мМ хлорида марганца (MnCl2), при температуре от 60°C до 90°C с интервалами 5°C.
Измерение активности выполняли с помощью цистеин-карбазольного метода. Как показано на Фиг. 6, результаты измерения ферментативной активности в зависимости от температуры показали, что оптимальная температура реакции трех вариантов арабинозоизомераз составила 75°C, что было на 10°C ниже, чем у L469P. Таким образом, было обнаружено, что активность изомераз в диапазоне температур, в котором может применяться способ получения, может быть в целом высокой, и что диапазон температур применения способа получения может быть более широким.
Кроме того, эти три варианта показали сходные температурные профили, что указывает на то, что свойства остатка фенилаланина в положении 275 влияли на оптимальную температуру реакции. Таким образом, можно заметить, что необходимо обеспечить достаточную гибкость, чтобы стерическое затруднение фенилаланина в положении 275 с D-галактозой в ходе реакции изомеразы с D-галактозой могло быть сведено к минимуму. При повышении температуры молекулярное движение фенильного остатка фенилаланина и окружающих остатков может быть более активным, и таким образом стерическое затруднение с углеродом 6 D-галактозы может быть уменьшено. В то же время можно ожидать, что оптимальная температура может быть сформирована в пределах диапазона, который не оказывает значительного влияния на трех- и четырехмерные структуры белка. Таким образом, может присутствовать изменение оптимальной температуры вариантов, стерическое затруднение которых было существенно снижено в результате мутации в положении 275. Впрочем, необходим дополнительный анализ и эксперименты для научного подтверждения этого факта.
5-2: Исследование термостабильности
Для исследования термостабильности вариантов арабинозоизомеразы каждый из очищенных ферментов добавляли к раствору 50 мМ Трис-HCl (pH 7,5) и 1 мМ хлорида марганца в концентрации 20 мкг/мл и инкубировали в термостатируемой водяной бане при 95°C в течение 180 минут. Ферментативную реакцию проводили при использовании ферментного раствора, с отбором проб в различные точки времени для измерения остаточной активности фермента.
Как показано на Фиг. 7, результаты исследования термостабильности показали, что варианты арабинозоизомеразы демонстрировали уменьшение остаточной активности с течением времени при 95°C, однако различие в термостабильности между вариантами не было значимым. Для получения дополнительных количественных данных измеряли полупериод существования активности каждого варианта. В результате было обнаружено, что L469P имел полупериод существования приблизительно 3 часа, а варианты, имеющие мутацию в положении 275, имели полупериод существования приблизительно 2 часа (Таблица 3). Термостабильность вариантов, имеющих мутацию в положении 275, как было измерено, являлась относительно низкой, однако считается, что различие в термостабильности не является значимым и может быть в достаточной мере скомпенсировано увеличением активности и снижением температуры процесса.
Таблица 3: Полупериод существования арабинозоизомераз, измеренный при 95°C
5-3: Исследование изменения активности, вызванного влиянием ионов металлов
Многие ферменты требуют присутствия ионов металлов для катализа. Поэтому для исследования зависимости термостабильности вариантов арабинозоизомеразы настоящего изобретения от ионов металлов исследовали изменение активности вариантов в присутствии ионов металлов при использовании каждого из очищенных ферментов.
Для исследования изменения активности фермента в зависимости от концентрации фермента, каждый из очищенных ферментов добавляли к 100 мМ субстрата D-галактозы и измеряли активность каждого фермента в 50 мМ Трис-HCl (pH 7,5) буферном растворе, содержащем 1-5 мМ хлорида марганца (MnCl2) при 75°C в течение 10 минут (Фиг. 8). В результате можно было заметить, что все варианты показали аналогичные активности в диапазоне концентраций хлорида марганца, используемом в эксперименте, и что влияние ионов марганца на активность изомераз при концентрации хлорида марганца, необходимой для активности фермента, не было значимым.
5-4: Исследование активности ферментов
Для исследования скорости реакции ферментов удельные активности вариантов арабинозоизомеразы измеряли при температуре реакции 75°C. В частности, каждый из ферментов добавляли к 1 мМ хлорида марганца и 100 мМ D-галактозы и измеряли реакционную способность 1 мг фермента при pH 7,5 и 75°C. Удельную активность каждого фермента во время реакции измеряли при указанных выше условиях в течение 10 минут, и в результате было показано, что удельные активности вариантов, имеющих мутацию в положении 275, приблизительно в 5,5 раз (F275V), 5 раз (F275M) и 3,9 раза (F275I), соответственно, превышали удельную активность L469P (Таблица 4).
Таблица 4: Удельные активности вариантов
Регистрационные номера
Депонирующий орган: Корейский центр культур микроорганизмов;
Регистрационный номер: KCCM11378P;
Дата депонирования: 14 февраля 2013 года.
Депонирующий орган: Корейский центр культур микроорганизмов;
Регистрационный номер: KCCM11379P;
Дата депонирования: 14 февраля 2013 года.
Депонирующий орган: Корейский центр культур микроорганизмов;
Регистрационный номер: KCCM11380P;
Дата депонирования: 14 февраля 2013 года.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИЯ ДЛЯ ПОЛУЧЕНИЯ ТАГАТОЗЫ И СПОСОБ ПОЛУЧЕНИЯ ТАГАТОЗЫ ИЗ ФРУКТОЗЫ | 2015 |
|
RU2701669C2 |
| Новая термостабильная тагатоза-6-фосфатфосфатаза и способ получения тагатозы с ее использованием | 2017 |
|
RU2722208C1 |
| Новая термостабильная фруктозо-6-фосфат-3-эпимераза и способ получения аллюлозы с ее использованием | 2017 |
|
RU2733427C2 |
| Новый промотор и его применение | 2017 |
|
RU2733425C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТАГАТОЗЫ С ИСПОЛЬЗОВАНИЕМ ОЛИГОСАХАРИДА СОИ | 2009 |
|
RU2451688C2 |
| Вариант белка внутренней мембраны и способ получения целевого продукта с его использованием | 2020 |
|
RU2804941C1 |
| НОВАЯ ПОЛИФОСФАТ-ЗАВИСИМАЯ ГЛЮКОКИНАЗА И СПОСОБ ПОЛУЧЕНИЯ ГЛЮКОЗО-6-ФОСФАТА С ЕЕ ИСПОЛЬЗОВАНИЕМ | 2017 |
|
RU2730602C2 |
| Новый вариант цитратсинтазы и способ получения L-аминокислот c его применением | 2022 |
|
RU2836733C1 |
| Новый полипептид и способ продуцирования IMP с его использованием | 2018 |
|
RU2725193C1 |
| НОВЫЙ ВАРИАНТ САХАРОФОСФАТ-ИЗОМЕРАЗЫ/ЭПИМЕРАЗЫ И СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА С ЕГО ПРИМЕНЕНИЕМ | 2021 |
|
RU2793436C1 |





























Изобретение относится к области биотехнологии и биохимии, в частности к L-арабинозоизомеразе, обладающей увеличенной активностью превращения D-галактозы в D-тагатозу по сравнению с арабинозоизомеразой, содержащей замещение лейцина в положении 469 пролином, или арабинозоизомеразой дикого типа. Указанная L-арабинозоизомераза содержит замену аминокислоты в положении 275 аминокислотой, имеющей неполярную алифатическую боковую цепь, кроме фенилаланина, и замену аминокислоты в положении 469 пролином в аминокислотной последовательности, представленной в SEQ ID NO: 1. Изобретение также относится к микроорганизму рода Corynebacterium, продуцирующему указанную L-арабинозоизомеразу, и способу получения D-тагатозы с использованием указанного микроорганизма. Изобретение позволяет получать D-тагатозу с высокой эффективностью. 5 н. и 7 з.п. ф-лы, 4 табл., 8 ил., 5 пр.
1. L-арабинозоизомераза, обладающая увеличенной активностью превращения D-галактозы в D-тагатозу по сравнению с арабинозоизомеразой, содержащей замещение лейцина в положении 469 пролином, или арабинозоизомеразой дикого типа, где L-арабинозоизомераза содержит замену аминокислоты в положении 275 аминокислотой, имеющей неполярную алифатическую боковую цепь, кроме фенилаланина, и замену аминокислоты в положении 469 пролином в аминокислотной последовательности, представленной в SEQ ID NO: 1.
2. L-арабинозоизомераза по п.1, где аминокислоту, имеющую неполярную алифатическую боковую цепь, выбирают из группы, состоящей из валина, метионина и изолейцина.
3. Полинуклеотид, кодирующий L-арабинозоизомеразу по п. 1 или 2.
4. Полинуклеотид по п.3, который имеет нуклеотидную последовательность SEQ ID NO: 8, SEQ ID NO: 9 или SEQ ID NO: 10.
5. Микроорганизм рода Corynebacterium, продуцирующий L-арабинозоизомеразу по п.1, трансформированный рекомбинантным вектором, включающим полинуклеотид по п. 3.
6. Микроорганизм по п.5, который представляет собой Corynebacterium glutamicum.
7. Микроорганизм по п.5, который представляет собой Corynebacterium glutamicum pFIS-1-TNAI-2 (KCCM 11378P), pFIS-1-TNAI-3 (KCCM 11379P) или pFIS-1-TNAI-4 (KCCM 11380P).
8. Способ получения D-тагатозы, включающий превращение D-галактозы в D-тагатозу с использованием L-арабинозоизомеразы по п.1, с реагированием раствора, содержащего D-галактозу и источник ионов металлов, в присутствии L-арабинозоизомеразы.
9. Способ по п.8, где источник ионов металла выбирают из группы, состоящей из иона марганца, иона магния и иона цинка в присутствии микроорганизма.
10. Способ получения D-тагатозы, включающий культивирование микроорганизма по любому из пп.5-7, с реагированием раствора, содержащего D-галактозу, с источником ионов металла в присутствии микроорганизма.
11. Способ по п.10, где источник ионов металла, выбирают из группы, состоящей из хлорида марганца, хлорида магния и хлорида цинка, в присутствии микроорганизма.
12. Способ по п.11, где хлорид марганца добавляют в концентрации 0,1-10 мМ.
| KR 20100016948 A, 16.02.2010 | |||
| OH H.J | |||
| et al | |||
| Increase in D-tagatose production rate by site-directed mutagenesis of L-arabinose isomerase from Geobacillus thermodenitrificans | |||
| Biotechnology Letters, 2006 | |||
| RHIMI M | |||
| et al | |||
| Облицовка комнатных печей | 1918 |
|
SU100A1 |
| Journal of Bacteriology, 2007 | |||
| WO 2004007738 A1, 22.01.2004 | |||
| СПОСОБ ПОЛУЧЕНИЯ ТАГАТОЗЫ С ИСПОЛЬЗОВАНИЕМ ОЛИГОСАХАРИДА СОИ | 2009 |
|
RU2451688C2 |

