Область техники
[1] Настоящее изобретение относится к новой полифосфат-зависимой глюкокиназе, композиции, содержащей глюкокиназу, и способам получения глюкозо-6-фосфата с использованием глюкокиназы.
[2]
Уровень техники
[3] D-глюкозо-6-фосфат является основным продуктом фосфорилирования в биологическом метаболизме и весьма полезен в промышленности, поскольку его можно превращать в различные полезные метаболиты через путь гликолиза, пентозофосфатный путь и путь биосинтеза гексозаминов. Развитие экономических способов получения глюкозо-6-фосфата имеет большое значение для биологических процессов получения конкретных соединений с высокой добавленной стоимостью из глюкозо-6-фосфата через серию из нескольких ферментативных реакций.
[4] В соответствии с предварительно опубликованными сообщениями, D-глюкозо-6-фосфат получают ферментативным путем с использованием АДФ-зависимой глюкокиназы (EC 2.7.1.147), переносящей β-фосфатную группу аденозиндифосфата (АДФ) на D-глюкозу в качестве сырья, АТФ-зависимой глюкокиназы (EC 2.7.1.2), переносящей γ-фосфатную группу аденозинтрифосфата (АТФ) на D-глюкозу, в качестве сырья, или полифосфат (поли(Pi)n)-зависимой глюкокиназы (EC 2.7.1.63), переносящей фосфатные группы полифосфата (поли(Pi)n) на D-глюкозу, в качестве сырья.
[5] Способ получения глюкозо-6-фосфата с использованием АДФ/АТФ-зависимой глюкокиназы требует дорогостоящих АДФ или АТФ в качестве донора фосфата (см. схему реакции 1) и, таким образом, неблагоприятен в отношении экономической эффективности и стабильности. В попытке преодолеть этот недостаток, АДФ/АТФ-зависимую глюкокиназу используют в комбинации с полифосфат-АМФ/АДФ фосфотрансферазой, способной переносить фосфатные группы с поли(Pi)n на АМФ или АДФ в качестве продукта дефосфорилирования, чтобы извлекать АДФ или АТФ. Однако эта попытка также ограничена в практическом использовании из-за низкой физической и химической (тепло, pH и т. д.) стабильности адениновых нуклеотидов АМФ, АДФ и АТФ.
[6] [Схема реакции 1]
[7] 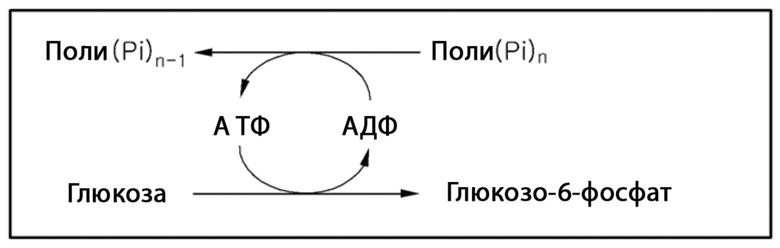
[8] В соответствии со способом с использованием полифосфат-зависимой глюкокиназы, поли(Pi)n используют в качестве донора фосфата, чтобы непосредственно получать глюкозо-6-фосфат (см. схему реакции 2). Использование относительно недорогостоящего и стабильного поли(Pi)n делает этот способ благоприятным с точки зрения экономической и коммерческой эффективности относительно способа с использованием АДФ/АТФ-зависимой глюкокиназы.
[9] [Схема реакции 2]
[10] 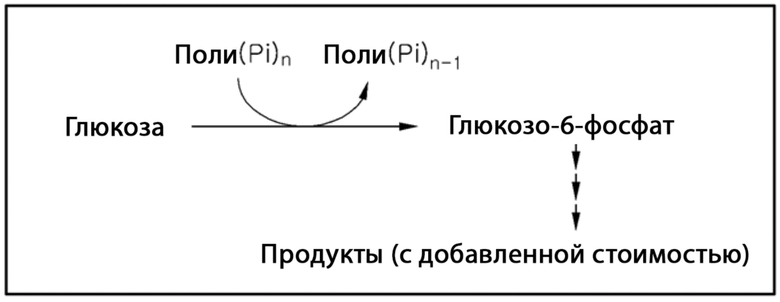
Раскрытие
Техническая проблема
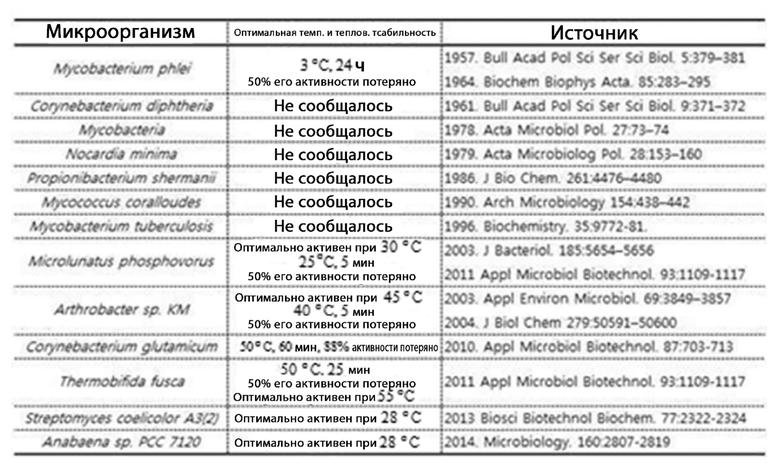
[11] Настоящее изобретение направлено на полифосфат-зависимую глюкокиназу, композицию, содержащую глюкокиназу, и способы получения глюкозо-6-фосфата с использованием глюкокиназы. Стабильность фермента является очень важным требованием в отношении эффективности ферментативного получения конкретного соединения. Однако на сегодняшний день существуют сообщения об ограниченном числе полифосфат-зависимых глюкокиназ, относящихся к настоящему изобретению, у некоторых видов микробов. Большинство выделенных ферментов получали из мезофильных микроорганизмов, что, таким образом, говорит о низкой тепловой стабильности (таблица 1). Настоящее изобретение выполнено с целью решить вышеуказанные проблемы, и одна цель настоящего изобретения состоит в том, чтобы предоставить новую термофильную и терморезистентную полифосфат-зависимую глюкокиназу, получаемую из термофила, композицию, содержащую фермент, и способы получения глюкозо-6-фосфата с использованием фермента.
[12] [Таблица 1]
[13]
[14] Глюкозо-6-фосфат представляет собой основной продукт фосфорилирования в пути гликолиза и пентозофосфатном пути в биологическом метаболизме. Глюкозо-6-фосфат весьма полезен в промышленности, поскольку его можно превращать в различные метаболиты. Более эффективные способы получения глюкозо-6-фосфата необходимы для получения соединений с высокой добавленной стоимостью из глюкозо-6-фосфата через серию из нескольких ферментативных реакций.
[15] АТФ или АДФ обычно используют в качестве донора фосфата для ферментативного превращения глюкозы в глюкозо-6-фосфат в биологическом метаболизме. Однако их высокая стоимость является препятствием для создания эффективных процессов для получения глюкозо-6-фосфата через путь ферментативной реакции. Кроме того, микробная ферментация не подходит для получения глюкозо-6-фосфата, поскольку продуцируемый глюкозо-6-фосфат может легко пересекать клеточные мембраны и подвергаться дефосфорилированию с помощью различных клеточных фосфатаз.
[16] Полифосфат (поли(Pi)n) в качестве донора фосфата в избытке присутствует в природе или может быть экономично получен с помощью химических процессов, и, таким образом, его рассматривают в качестве коммерчески полезного соединения. Следовательно, развитие эффективного способа ферментативного получения глюкозо-6-фосфата из глюкозы с использованием поли(Pi)n имеет большую коммерческую важность.
[17] Однако большинство ранее описанных ферментов для получения глюкозо-6-фосфата с использованием поли(Pi)n вступают в реакцию при относительно низкой температуре и обладают низкой тепловой стабильностью, что ограничивает их применение при получении глюкозо-6-фосфата.
[18]
Техническое решение
[19] Настоящее изобретение направлено на предоставление новой термофильной и терморезистентной полифосфат-зависимой глюкокиназы, получаемой из термофила, и способы получения глюкозо-6-фосфата с использованием фермента.
[20] Подробно описаны несколько аспектов настоящего изобретения.
[21] Один аспект настоящего изобретения предусматривает термофильную и терморезистентную полифосфат-зависимую глюкокиназу, получаемую из рода Anaerolinea.
[22] В частности, полифосфат-зависимая глюкокиназа по настоящему изобретению может иметь аминокислотную последовательность, приведенную в SEQ ID NO. 2. Полифосфат-зависимая глюкокиназа по настоящему изобретению может представлять собой любой белок, который имеет аминокислотную последовательность, обладающую гомологией по меньшей мере 70%, в частности по меньшей мере 80%, более конкретно по меньшей мере 90%, даже более конкретно по меньшей мере 95% с аминокислотной последовательностью, приведенной в SEQ ID NO. 2, и представляет собой белок, по существу идентичный или соответствующий полифосфат-зависимой глюкокиназе. Кроме того, если такая гомологичная последовательность представляет собой аминокислотную последовательность, которая по существу демонстрирует функцию полифосфат-зависимого фосфорилирования глюкозы, даже вариант белка, аминокислотная последовательность которого частично удалена, модифицирована, заменена или дополнена, конечно входит в объем настоящего изобретения.
[23] Как используют в настоящем описании, термин «гомология» относится к степени идентичности или соответствия между заданными полипептидными последовательностями или полинуклеотидными последовательностями, которые могут иметь или не иметь общее эволюционное происхождение, и может быть выражен в виде процентной доли. В настоящем описании, гомологию последовательности, имеющей активность, идентичную или схожую с заданным полипептидом или полинуклеотидной последовательностью, выражают в виде «% гомологии». Например, гомологию можно определять с использованием стандартного программного обеспечения, в частности, BLAST 2.0, для того чтобы вычислять параметры, такие как оценка, идентичность и сходство. Или гомологию можно идентифицировать посредством сравнения последовательностей с помощью эксперимента Саузерн-гибридизации при определенных строгих условиях. Определенные подходящие условия гибридизации можно определять с помощью способов, хорошо известных специалистам в данной области (см. Sambrook et al., 1989, ниже). В одном из вариантов осуществления две аминокислотные последовательности считают «по существу гомологичными» или «по существу схожими», когда по меньшей мере 21% (в частности по меньшей мере приблизительно 50%, в частности приблизительно 75%, 90%, 95%, 96%, 97% или 99%) полипептидов совпадают на протяжении определенной длины аминокислотных последовательностей.
[24] Другой аспект настоящего изобретения предусматривает полинуклеотид, кодирующий терморезистентную полифосфат-зависимую глюкокиназу, получаемую из рода Anaerolinea. В частности, настоящее изобретение может предусматривать полинуклеотидную последовательность, которая кодирует белок, обладающий активностью полифосфат-зависимой глюкокиназы и представленный с помощью SEQ ID NO. 1.
[25] Как используют в настоящем описании, термин «полинуклеотид» относится к полимеру нуклеотидных звеньев, которые связаны ковалентно для того, чтобы формировать длинную цепь. В целом, полинуклеотид обозначает нить ДНК или РНК, длина которой выше предварительно определяемого уровня.
[26] Полинуклеотид, кодирующий белок, имеющий активность полифосфат-зависимой глюкокиназы, может включать полинуклеотидную последовательность, кодирующую аминокислоты, приведенные в SEQ ID NO. 2. Можно создавать различные модификации кодирующей области полинуклеотида до тех пор, пока аминокислотную последовательность полипептида не изменяют из-за вырожденности кодонов или принимая во внимание предпочтительные кодоны в организме, где фермент подлежит экспрессии. Например, полинуклеотид может иметь последовательность, приведенную в SEQ ID NO. 1. Полинуклеотид может иметь нуклеотидную последовательность, обладающую гомологией по меньшей мере 70%, в частности, по меньшей мере 80%, более конкретно по меньшей мере 90%, даже более конкретно по меньшей мере 95%, наиболее конкретно по меньшей мере 98% с последовательностью, приведенной в SEQ ID NO. 1, и может по существу кодировать полипептид, обладающий активностью полифосфат-зависимого фосфорилирования глюкозы. Очевидно, что вариант, аминокислотная последовательность которого частично удалена, модифицирована, заменена или дополнена, также входит в объем настоящего изобретения.
[27] Композиция для получения глюкозо-6-фосфата, содержащая от 1% до 3% по массе глюкозы, от 1% до 10% по массе полифосфата, от 10 Ед/мл до 50 Ед/мл полифосфат-зависимой глюкокиназы и необязательно от 1 мМ до 20 мМ ионов магния (например, MgSO4 или MgCl2) на основании общего объема композиции, позволяет достигать выхода превращения по меньшей мере 70%, более конкретно по меньшей мере 80%, даже более конкретно по меньшей мере 90% в глюкозо-6-фосфат.
[28] В частности, композиция, содержащая 2% по массе глюкозы, 1,5% по массе полифосфата, от 10 Ед/мл до 50 Ед/мл полифосфат-зависимой глюкокиназы и необязательно 10 мМ MgSO4, позволяет достигать выхода превращения по меньшей мере 60%, более конкретно по меньшей мере 70%, даже более конкретно по меньшей мере 80% в глюкозо-6-фосфат.
[29] Композиция для получения глюкозо-6-фосфата, содержащая от 5% до 20% по массе глюкозы, от 5% до 12% по массе полифосфата, от 10 Ед/мл до 50 Ед/мл полифосфат-зависимой глюкокиназы и необязательно от 1 мМ до 20 мМ ионов магния (например, MgSO4 или MgCl2) на основании общего объема композиции, позволяет достигать выхода превращения по меньшей мере 50%, более конкретно по меньшей мере 60%, даже более конкретно по меньшей мере 70% в глюкозо-6-фосфат.
[30] В частности, композиция, содержащая 15% по массе глюкозы, 10% по массе полифосфата, от 10 Ед/мл до 50 Ед/мл полифосфат-зависимой глюкокиназы и необязательно 10 мМ MgSO4 на основании общего объема композиции, позволяет достигать выхода превращения по меньшей мере 50%, более конкретно по меньшей мере 60%, даже более конкретно по меньшей мере 65% в глюкозо-6-фосфат.
[31] Полифосфат-зависимая глюкокиназа может быть активной при температуре от 45°C до 90°C, более конкретно от 55°C до 80°C, наиболее конкретно от 65°C до 70°C.
[32] Полифосфат-зависимая глюкокиназа может быть активной при pH от 4 до 10, наиболее конкретно при pH от 4 до 5.
[33] Активность полифосфат-зависимой глюкокиназы можно усиливать в присутствии ионов магния.
[34] Ионы магния, в частности, могут присутствовать в концентрации от 0,5 мМ до 20 мМ, более конкретно от 0,2 мМ до 10 мМ, даже более конкретно 1 мМ.
[35] Дополнительный аспект настоящего изобретения предусматривает композицию для получения глюкозо-6-фосфата, содержащую полифосфат-зависимую глюкокиназу, описанную в настоящем описании, глюкозу и полифосфат.
[36] Композиция дополнительно может содержать ионы магния. Композицию можно использовать для получения глюкозо-6-фосфата. Ингредиенты, используемые в этом аспекте, и их содержание аналогичны тем, которые описаны в предыдущих и последующих аспектах, и их подробное описание, таким образом, пропущено.
[37] Еще один другой аспект настоящего изобретения предусматривает способ получения глюкозо-6-фосфата из композиции, содержащей полифосфат-зависимую глюкокиназу, описанную в настоящем описании, глюкозу и полифосфат.
[38] Реакцию для получения глюкозо-6-фосфата осуществляют при температуре от 45°C до 90°C и pH от 4 до 10.
[39] Глюкозу можно получать посредством разжижения или осахаривания крахмала или целлюлозы.
[40] Полифосфат служит в качестве донора фосфата, и его примеры включают гексаметафосфат натрия, триполифосфат натрия и гексаметафосфат калия, но не ограничены этим, также они включают те, которые коммерчески доступны.
[41] Глюкозо-6-фосфат можно получать при температуре от 45°C до 90°C, более конкретно от 55°C до 80°C, наиболее конкретно от 65°C до 70°C.
[42] Полифосфат-зависимая глюкокиназа может иметь молекулярную массу от 10 кДа до 100 кДа, в частности от 20 кДа до 50 кДа.
[43] Композиция дополнительно может содержать ионы магния. Например, источником ионов магния может быть MgCl2 или MgSO4. В частности, композиция дополнительно может содержать MgSO4.
[44] Полифосфат-зависимая глюкокиназа может присутствовать в количестве от 10 Ед/мл до 50 Ед/мл.
[45] Глюкоза может присутствовать в количестве от 0,1% до 40% по массе, более конкретно от 1% до 20% по массе, наиболее конкретно от 1% до 10% по массе, на основании общего объема композиции.
[46] Полифосфат может присутствовать в количестве от 0,5% до 25% по массе, более конкретно от 1% до 20% по массе, наиболее конкретно от 1% до 10% по массе, на основании общего объема композиции.
[47] Еще один другой аспект настоящего изобретения предусматривает способ получения глюкозо-6-фосфата из композиции, содержащей полифосфат-зависимую глюкокиназу, описанную в настоящем описании, ферменты для разжижения и осахаривания, крахмал и полифосфат.
[48] Реакцию для получения глюкозо-6-фосфата осуществляют при температуре от 45°C до 90°C и pH от 4 до 10.
[49] Ферменты для разжижения и осахаривания могут представлять собой один или несколько, выбранных из α-амилаз, глюкоамилаз и α-гликозидаз.
[50] Еще один другой аспект настоящего изобретения предусматривает микроорганизм, продуцирующий полифосфат-зависимую глюкокиназу. В частности, микроорганизм по настоящему изобретению относится к роду Escherichia.
[51] Как используют в настоящем описании, термин «микроорганизм, продуцирующий полифосфат-зависимую глюкокиназу» относится к прокариотическому или эукариотическому микробному штамму, который может продуцировать фермент в себе. В частности, микроорганизм, продуцирующий полифосфат-зависимую глюкокиназу, представляет собой микроорганизм, способный накапливать фермент в среде или в себе посредством генетической инженерии или природной мутации.
[52] Микроорганизм конкретно не ограничен и может представлять собой любой, который может экспрессировать полипептид, имеющий последовательность, приведенную в SEQ ID NO. 2. Микроорганизм может представлять собой прокариотический или эукариотический микроорганизм, в частности, прокариотический микроорганизм. Примеры таких прокариотических микроорганизмов включают, но не ограничиваясь этим, микробные штаммы, относящиеся к родам Escherichia, Erwinia, Serratia, Providencia, Corynebacterium и Brevibacterium. В частности, микроорганизм может представлять собой тот, который относится к роду Escherichia. Неограничивающим примером микроорганизма, относящегося к роду Escherichia, является Escherichia coli.
[53] Как используют в настоящем описании, термин «экспрессия» относится к процессу, в котором полинуклеотид, кодирующий полипептид по настоящему изобретению, трансформируют с использованием функционального рекомбинантного вектора или вставляют в хромосому. Экспрессионный процесс конкретно не ограничен.
[54] Как используют в настоящем описании, термин «трансформация» относится ко введению вектора, содержащего полинуклеотид, кодирующий целевой белок, в клетку-хозяина, чтобы экспрессировать белок, кодируемый полинуклеотидом, в клетке-хозяине. Трансфицируемый полинуклеотид или можно вставлять в и располагать в хромосоме клетки-хозяина или он может существовать внехромосомно до тех пор, пока его можно экспрессировать в клетке-хозяине. Полинуклеотид включает ДНК и РНК, кодирующие целевой белок. Полинуклеотид можно вводить в любой форме до тех пор, пока его можно вводить в клетку-хозяина и экспрессировать в ней. Например, полинуклеотид можно вводить в клетку-хозяина в форме экспрессирующей кассеты, которая представляет собой генную конструкцию, содержащую все элементы, необходимые для ее автономной экспрессии, но его форма не ограничена этим. Обычно экспрессирующая кассета содержит промотор, функционально связанный с полинуклеотидом, сигналом терминации транскрипции, доменом связывания рибосомы и сигналом терминации трансляции. Экспрессирующая кассета может быть в форме самореплицирующегося экспрессирующего вектора. Полинуклеотид как он есть можно вводить в клетку-хозяина и функционально связывать с последовательностью, необходимой для экспрессии в клетке-хозяине.
[55] Как используют в настоящем описании, термин «функционально связан» относится к функциональной связи между последовательностью промотора, которая инициирует и опосредует транскрипцию полинуклеотида, кодирующего целевой белок по настоящему изобретению, и последовательностью гена.
[56] Как используют в настоящем описании, термин «вектор» относится к любому носителю для клонирования и/или переноса последовательности оснований в клетку-хозяина. Вектор может представлять собой репликон, к которому можно прикреплять другой сегмент ДНК с тем, чтобы осуществлять репликацию прикрепляемого сегмента. «Репликон» относится к любому генетическому элементу (например, плазмиде, фагу, космиде, хромосоме или вирусу), который выполняет функцию автономной единицы репликации ДНК in vivo, т. е. способен к репликации под своим собственным управлением. Термин «вектор» может включать как вирусные, так и не вирусные носители для введения последовательности оснований в клетку-хозяина in vitro, ex vivo или in vivo. Термин «вектор» также может включать миникольцевую ДНК. Например, вектор может представлять собой плазмиду без бактериальных последовательностей ДНК. Показано, что удаление бактериальных последовательностей ДНК, которые богаты участками CpG, снижает сайленсинг экспрессии трансгена и ведет к более персистирующей экспрессии с плазмидных ДНК векторов (например, Ehrhardt, A. et al. 2003. HumGene Ther 10: 215-25; Yet, et al. 2002. MoI Ther 5: 731-38; Chen et al. 2004, Gene Ther 11: 856-864). Термин «вектор» также может включать транспозоны (Annu Rev Genet. 2003. 37:3-29) или искусственные хромосомы. Конкретные примеры векторов, пригодных для использования в настоящем изобретении, включают, но не ограничиваясь этим, векторы pACYC177, pACYC184, pCL1920, pECCG117, pUC19, pBR322 и pMW118. Варианты этих векторов, например, в которых промоторы мутированы, также можно использовать в настоящем изобретении.
[57] В частности, вектор в настоящем изобретении может представлять собой ДНК конструкцию, содержащую полинуклеотидную последовательность, кодирующую желаемый белок, которая функционально связана с подходящей экспрессионной регуляторной последовательностью, чтобы экспрессировать желаемый белок в подходящей клетке-хозяине. Регуляторная последовательность может включать промотор, который может инициировать транскрипцию, необязательную последовательность оператора для регуляции транскрипции, последовательность, кодирующую подходящий участок мРНК для связывания рибосомы и последовательность, регулирующую терминацию транскрипции и трансляции. После введения вектора в подходящую клетку-хозяина, он может реплицироваться или функционировать независимо от генома хозяина и может сам встраиваться в геном.
[58] Вектор, используемый в настоящем изобретении, конкретно не ограничен до тех пор, пока вектор является реплицируемым в клетке-хозяине. Вектор может представлять собой любой из тех, что известны в данной области. Примеры таких известных векторов включают природные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Фаговые векторы или космидные векторы могут представлять собой, например, pWE15, M13, λE15, λE1515, E15, M13 и Charon21A, но не ограничены этим. Плазмидные векторы могут представлять собой те, что основаны на pBR, pUC, pBluescriptII, pGEM, pTZ, pCL и pET, но не ограничены этим.
[59] Настоящее изобретение также относится к рекомбинантному экспрессирующему вектору, содержащему ген, кодирующий полифосфат-зависимую глюкокиназу.
[60] Настоящее изобретение также относится к Escherichia coli BL21(DE3)/CJ_at_ppgk, трансформированной рекомбинантным экспрессирующим вектором, содержащим последовательность SEQ ID NO. 1. Штамм депонировали в Korean Culture Center of Microorganisms 16 февраля 2016 года под номер депонирования KCCM11814P.
[61] Настоящее изобретение также относится к экономическим способам получения следующих промышленно эффективных соединений из полифосфата и глюкозы или крахмала на основании однореакторного ферментативного превращения с использованием полифосфат-зависимой глюкокиназы и дополнительных функциональных ферментов (например, α-амилаз, глюкоамилаз, α-глюкозидаз, изомераз, альдолаз, синтаз, киназ и фосфатаз).
[62] Примеры таких промышленно эффективных соединений включают, но не ограничиваясь этим, D-глюкозо-1-фосфат, D-фруктозо-6-фосфат, D-фруктозо-1,6-бисфосфат, миоинозитол-3-фосфат, миоинозитол, D-глюкуронат, D-глюкозамин-6-фосфат, D-глюкозамин, N-ацетил-D-глюкозамин-6-фосфат, N-ацетил-D-глюкозамин, N-ацетил-D-маннозамин-6-фосфат, N-ацетил-D-маннозамин, N-ацетилнейраминовую кислоту (сиаловую кислоту), D-маннозо-6-фосфат, D-маннозу, D-тагатозо-6-фосфат, D-тагатозу, D-аллюлозо-6-фосфат, D-аллюлозу, D-глицеральдегид-3-фосфат и дигидроксиацетонфосфат. Промышленно эффективные соединения также могут включать различные соединения, получаемые из глюкозо-6-фосфата.
[63]
Полезные эффекты
[64] Фермент по настоящему изобретению может участвовать в ферментативных реакциях при относительно высоких температурах. Высокая температура реакции увеличивает растворимость D-глюкозы и поли(Pi)n в качестве субстратов, делая возможным использование субстратов в высоких концентрациях. Кроме того, можно увеличивать скорости диффузии материалов и скорость реакции и можно снижать время реакции, достигая увеличенной производительности блока. Кроме того, высокая температура реакции позволяет минимизировать контаминацию, обусловленную чужеродными микроорганизмами во время обработки. Кроме того, тепловую устойчивость фермента, относящегося к настоящему изобретению, можно использовать для того, чтобы легко разрушать рекомбинантные клетки, продуцирующие фермент, используя процессы тепловой обработки. Когда рекомбинантный фермент выделяют и используют, белки, извлекаемые из рекомбинантных экспрессирующих микроорганизмов, также можно избирательно денатурировать и удалять, что делает возможной эффективную очистку фермента.
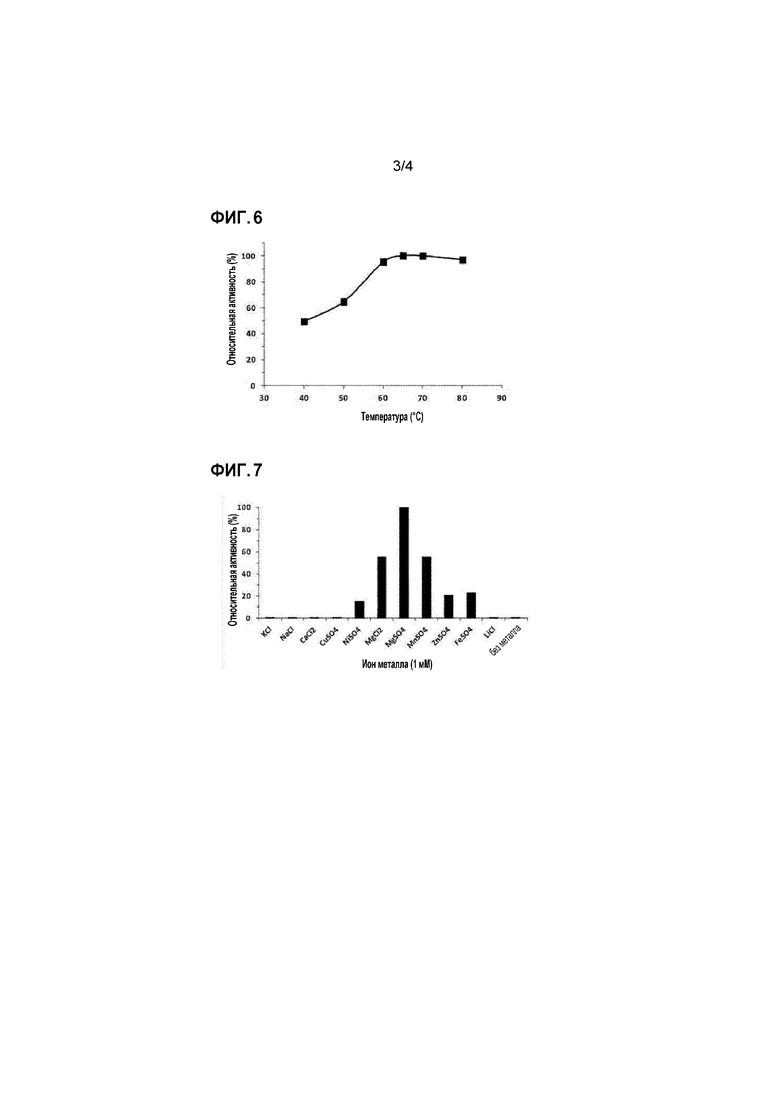
[65]
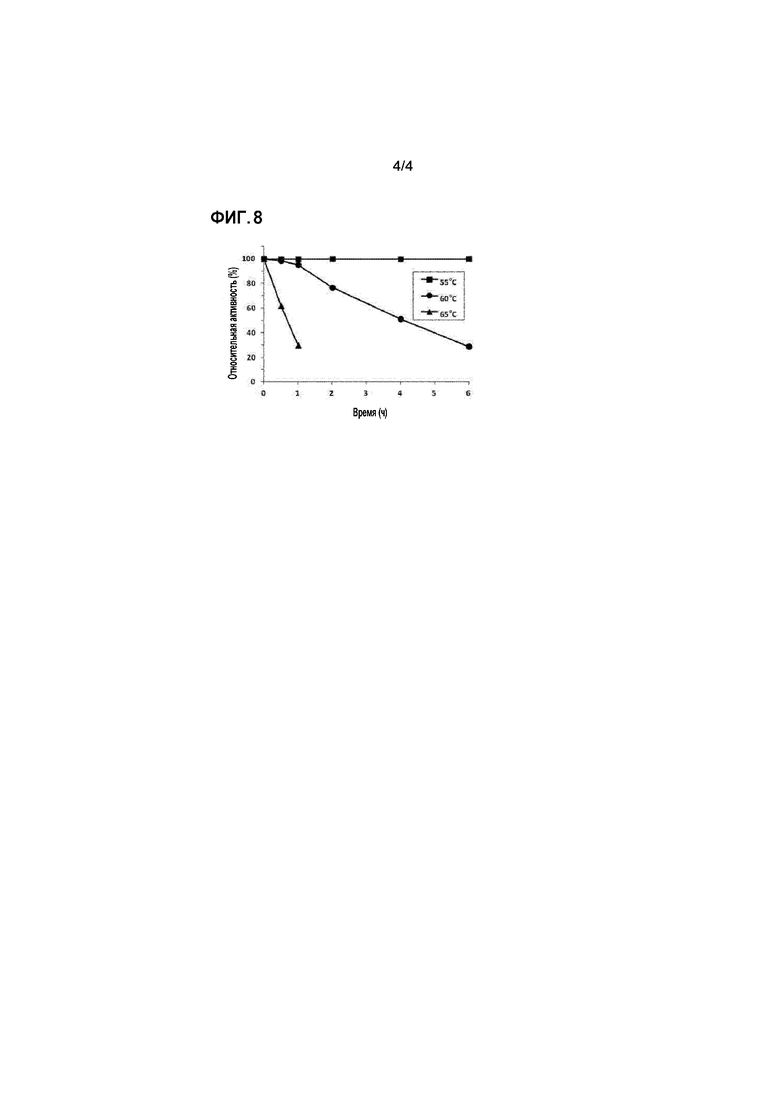
Описание фигур
[66] На фиг. 1 представлена блок-схема, иллюстрирующая способ получения глюкозо-6-фосфата в соответствии с настоящим изобретением.
[67] На фиг. 2 представлена схема реакции для получения глюкозо-6-фосфата из глюкозы и АТФ.
[68] На фиг. 3 представлена схема реакции для получения глюкозо-6-фосфата из глюкозы и полифосфата.
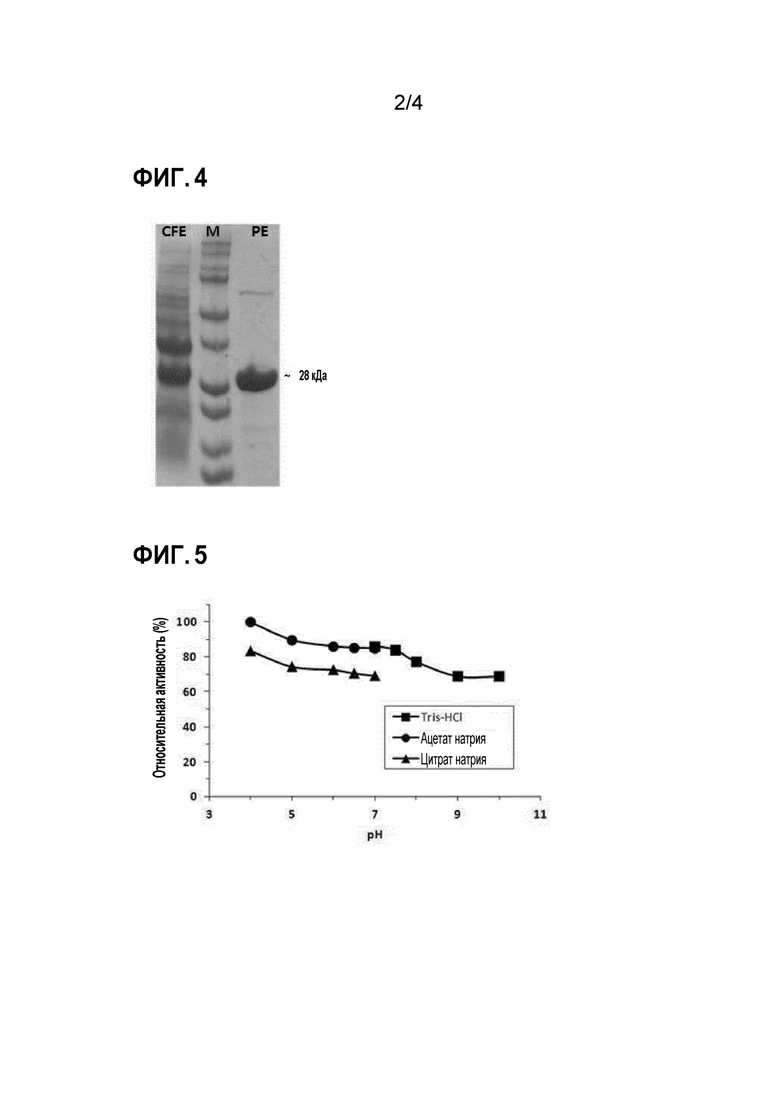
[69] На фиг. 4 представлены изображения геля SDS-PAGE для супернатанта (CFE) после разрушения клеток, маркер размеров (M) и очищенная рекомбинантная полифосфат-зависимая глюкокиназа (PE), которую брали после электрофореза.
[70] На фиг. 5 представлена pH-зависимая активность рекомбинантной полифосфат-зависимой глюкокиназы.
[71] На фиг. 6 представлена зависящая от температуры активность рекомбинантной полифосфат-зависимой глюкокиназы.
[72] На фиг. 7 представлены активности рекомбинантной полифосфат-зависимой глюкокиназы в присутствии ионов металлов различных типов.
[73] На фиг. 8 представлены активности рекомбинантной полифосфат-зависимой глюкокиназы при нагреве до различных температуры.
[74]
Вариант осуществления изобретения
[75] Глюкоза является относительно дешевым источником углерода, и ее можно массово получать из крахмала или целлюлозы. Глюкозу широко используют в качестве базового сырья в процессах химического или биологического превращения для получения различных соединений, которые можно использовать в химической, фармацевтической, косметической и пищевой промышленности.
[76] Однако использование фосфорилированной глюкозы в качестве базового сырья в биологических процессах, в частности, процессах ферментативного превращения, в настоящее время ограничено из-за ее высокой цены.
[77] Глюкозо-6-фосфат является ключевым промышленным метаболитом в метаболизме глюкозы, который можно использовать в качестве базового сырья, которое может индуцировать очень полезные реакции на основании использования различных метаболических ферментов, присутствующих в природе (организмы).
[78] При таких обстоятельствах, настоящее изобретение направлено на предоставление фермента и ферментативных способов для экономического получения глюкозо-6-фосфата, который является сырьем для различных промышленно полезных соединений, из глюкозы и полифосфата.
[79] Использование получаемого глюкозо-6-фосфата и способа получения по настоящему изобретению также может предоставлять функциональные соединения с высокой добавленной стоимостью в фармацевтической, косметической и пищевой промышленности, которые можно получать ферментативными способами.
[80]
[81] [ПРИМЕРЫ]
[82] Пример 1: получение рекомбинантного экспрессирующего вектора, содержащего ген полифосфат-зависимой глюкокиназы, и трансформированного микроорганизма
[83] Чтобы предоставлять новую активную при высоких температурах терморезистентную полифосфат-зависимую глюкокиназу, выделяли ген полифосфат-зависимой глюкокиназы, получаемый из термофильного Anaerolinea thermophila, конструировали рекомбинантный экспрессирующий вектор и получали трансформированный микроорганизм.
[84] В частности, осуществляли скрининг последовательностей генов, относящихся к ферменту по настоящему изобретению, среди последовательностей генов, зарегистрированных в GenBank, и среди них только последовательность гена, полученного из термофильного микроорганизма. На основании зарегистрированной последовательности гена (SEQ ID NO. 1) и аминокислотной последовательности (SEQ ID NO. 2) Anaerolinea thermophila разрабатывали прямой праймер (SEQ ID NO. 3) и обратный праймер (SEQ ID NO. 4). Соответствующий ген амплифицировали из геномной ДНК Anaerolinea thermophila посредством полимеразной цепной реакции (ПЦР) с использованием синтезированных праймеров. Амплифицированный ген полифосфат-зависимой глюкокиназы вставляли в плазмидный вектор pET21a (Novagen) для экспрессии в E. coli с использованием рестрикционных ферментов NdeI и XhoI, чтобы конструировать рекомбинантный экспрессирующий вектор, который называли CJ_at_ppgk. CJ_at_ppgk трансфицировали в штамм E. coli BL21(DE3) с помощью общего способа трансформации (см. Sambrook et al. 1989), чтобы получать трансформированный микроорганизм, который называли E. coli BL21(DE3)/CJ_at_ppgk.
[85]
[86] Пример 2: получение рекомбинантной полифосфат-зависимой глюкокиназы
[87] В этом примере получали рекомбинантную полифосфат-зависимую глюкокиназу. Сначала в культуральную пробирку, содержащую 5 мл жидкой среды LB, инокулировали E. coli BL21(DE3)/CJ_at_ppgk. Инокулят культивировали во встряхиваемом инкубаторе при 37°C до тех пор, пока не достигали поглощения 2,0 при 600 нм. В жидкую среду LB в культуральной колбе добавляли культуральный бульон, после чего следовала основная культура. Когда поглощение культуры при 600 нм достигало 2,0, добавляли 1 мМ IPTG, чтобы индуцировать экспрессию и продуцирование рекомбинантного фермента. Температуру культуры поддерживали на 37°C при перемешивании 200 об./мин. Культуральный бульон центрифугировали на 8000×g при 4°C в течение 20 мин для того, чтобы собирать бактериальные клетки. Собранные бактериальные клетки промывали два раза в 50 мМ буфере Tris-HCl (pH 7,0) и суспендировали в том же буфере. Затем клетки разрушали с использованием ультразвукового гомогенизатора. Клеточный лизат центрифугировали на 13000×g при 4°C в течение 20 мин и брали только супернатант клеточного лизата. Рекомбинантный фермент очищали от супернатанта посредством аффинной хроматографии с гистидиновой меткой. Проводили диализ очищенного рекомбинантного фермента против 50 мМ буфера Tris-HCl (pH 7,0) и затем определяли характеристики.
[88] На фиг. 4 M обозначает маркер размеров, CFE обозначает супернатант после разрушения клеток и PE обозначает очищенный фермент. Обнаруживали, что очищенная рекомбинантная полифосфат-зависимая глюкокиназа имеет молекулярную массу приблизительно 28 кДа, как определяли посредством SDS-PAGE (фиг. 4).
[89]
[90] Пример 3: анализ активности рекомбинантной полифосфат-зависимой глюкокиназы
[91] В этом примере анализировали активность рекомбинантной полифосфат-зависимой глюкокиназы. С этой целью глюкозу (4% (масс./об.)), гексаметафосфат натрия (3% (масс./об.)) и MgCl2 (1 мМ) суспендировали в 50 мМ буфере Tris-HCl (pH 7,0) для того, чтобы получать реакционную композицию для анализа активности. В реакционную композицию добавляли очищенный фермент (0,1 мг/мл). Реакцию оставляли протекать при 60°C в течение 15 мин. Продукт реакции анализировали посредством HPLC при следующих условиях: колонка Aminex HPX-87C (Bio-rad), 80°C, 5 мМ раствор H2SO4 в качестве подвижной фазы и скорость потока 0,6 мл/мин. Глюкозо-6-фосфат обнаруживали и анализировали с использованием детектора показателя преломления.
[92] Результаты анализа выявляли образование глюкозо-6-фосфата из продукта реакции очищенного рекомбинантного фермента.
[93]
[94] Пример 4: анализ pH-зависимой активности рекомбинантной полифосфат-зависимой глюкокиназы
[95] В этом примере изучали влияние pH на активность патентоспособного фермента. С этой целью глюкозу (4% (масс./об.)), гексаметафосфат натрия (3% (масс./об.)) и MgCl2 (1 мМ) суспендировали в 50 мМ буферах с различными уровнями pH (цитрат натрия, pH 4-7; ацетат натрия, pH 4-7; Tris-HCl, pH 7-10) для того, чтобы получать реакционные композиции для анализа эффекта pH. Очищенный фермент (0,1 мг/мл) добавляли в каждую из реакционных композиций. Реакцию оставляли протекать при 60°C в течение 15 мин. После этого образование глюкозо-6-фосфата количественно анализировали посредством HPLC.
[96] Результаты представлены на фиг. 5. Полифосфат-зависимая глюкокиназа, получаемая из Anaerolinea thermophila, по настоящему изобретению демонстрировала максимальную активность около pH 4-5, в отличие от ферментов, о которых сообщалось до сегодняшнего дня. В частности, обнаруживали, что активность фермента выше в натрий-ацетатном буфере, чем в других буферах в соответствующем диапазоне pH. Кроме того, активности фермента в широком диапазоне pH 4-10 составляли≥70% от максимальной активности (фиг. 5).
[97] Новой характеристикой полифосфат-зависимой глюкокиназы по настоящему изобретению является ацидофильность и высокотемпературная активность, что делает возможным эффективное получение глюкозо-6-фосфата из крахмального декстрина, когда патентоспособный фермент используют в комбинации с глюкоамилазой, получаемой из Aspergillus sp. (например, коммерческой глюкоамилазой AMG 300L (Novozymes), получаемой из Aspergillus niger). Коммерческая глюкоамилаза имеет оптимальную активность при pH 4,5 и 60°C. Патентоспособный фермент считают промышленно очень полезным, поскольку его активности в широком диапазоне pH 4-10 составляют≥70% от максимальной активности.
[98] Пример 5: анализ зависящей от температуры активности рекомбинантной полифосфат-зависимой глюкокиназы
[99] В этом примере анализировали зависящую от температуры активность рекомбинантного фермента. С этой целью глюкозу (4% (масс./об.)), гексаметафосфат натрия (3% (масс./об.)) и MgCl2 (1 мМ) суспендировали в 50 мМ буфере Tris-HCl (pH 7,0) для того, чтобы получать реакционную композицию для анализа зависящей от температуры активности рекомбинантного фермента. В реакционную композицию добавляли очищенный фермент (0,1 мг/мл). Реакцию оставляли протекать при 40-80°C в течение 15 мин. После этого образование глюкозо-6-фосфата количественно анализировали посредством HPLC.
[100] Результаты представлены на фиг. 6. Патентоспособный фермент демонстрировал максимальную активность приблизительно при 65-70°C. Кроме того, активности фермента в широком температурном диапазоне 60-80°C составляли≥95% от максимальной активности (фиг. 6).
[101] Известно, что среди полифосфат-зависимых глюкокиназ, о которых сообщалось до сих пор, ферменты, получаемые из Thermobifida fusca, активны и терморезистентны при высокой температуре и, как сообщалось, оптимально активны при температуре 55°C [см. Liao et al. 2012. Appl Microbiol Biotechnol 93:1109-1117].
[102] Следовательно, можно заключить, что получаемая из Anaerolinea thermophila полифосфат-зависимая глюкокиназа по настоящему изобретению более активна при высокой температуре, чем любая полифосфат-зависимая глюкокиназа, о которой сообщали до сих пор, на что указывает ее оптимальная активность при 65-70°C.
[103]
[104] Пример 6: анализ активности рекомбинантной полифосфат-зависимой глюкокиназы в зависимости от типа ионов металла
[105] Известно, то полифосфат-зависимым глюкокиназам, о которых сообщали до сих пор, необходимы ионы металла, такие как Mg2+, Mn2+, Co2+ и Zn2+ для активности. В этом примере изучали влияние ионов металла на активность патентоспособной полифосфат-зависимой глюкокиназы. С этой целью патентоспособный фермент обрабатывали с использованием 10 мМ EDTA, после чего следовал диализ для того, чтобы получать образец фермента. Глюкозу (2% (масс./об.)), гексаметафосфат натрия (1,5% (масс./об.)) и ионы металла (NiSO4, CuSO4, MnSO4, CaCl2, ZnSO4, MgSO4, MgCl2, FeSO4, NaCl, LiCl и KCl, 1 мМ каждое) суспендировали в 50 мМ буфере Tris-HCl (pH 7,0) для того, чтобы получать реакционные композиции. Образец фермента (0,1 мг/мл), не содержащий ионы металла, добавляли в каждую из реакционных композиций. Реакцию оставляли протекать при 60°C в течение 15 мин. После этого образование глюкозо-6-фосфата количественно анализировали посредством HPLC. Активность образца фермента, не обработанного ионами металла, сравнивали с активностями образцов фермента, которые обрабатывали ионами металла.
[106] Как результат, полифосфат-зависимая глюкокиназа, получаемая из Anaerolinea thermophila, демонстрировала потребность в ионах металла, такого как Mg, Mn, Zn, Fe и Ni, для ее активности, как показано на фиг. 7. Ионы магния более эффективны, чем другие ионы металла, что аналогичным образом наблюдали у ферментов, о которых сообщали до сих пор. В частности, обнаруживали, что при добавлении MgSO4 достигали максимальной активности (фиг. 7).
[107] Пример 6: анализ температурной стабильности рекомбинантной полифосфат-зависимой глюкокиназы
[108] Анализировали температурную стабильность патентоспособной полифосфат-зависимой глюкокиназы. С этой целью очищенный рекомбинантный фермент (0,2 мг/мл) нагревали до температуры 55-65°C в течение различных периодов времени, и сравнивали и анализировали остаточные активности.
[109] Глюкозу (4% (масс./об.)), гексаметафосфат натрия (3% (масс./об.)) и MgCl2 (1 мМ) суспендировали в 50 мМ буфере Tris-HCl (pH 7,0) для того, чтобы получать реакционную композицию. Каждый из образцов фермента (0,1 мг/мл), нагретых до различных температур, добавляли в реакционную композицию для анализа остаточной активности. Реакцию оставляли протекать при 60°C в течение 15 мин. После этого образование глюкозо-6-фосфата количественно анализировали посредством HPLC.
[110] Результаты представлены на фиг. 8. Снижение активности фермента не наблюдали при 55°C в течение 6 ч. Фермент терял свою активность приблизительно на 49% после 4 ч при 60°C. Сохранялось приблизительно 62% активности фермента от ее начального значения даже при 65°C в течение 0,5 ч (фиг. 8).
[111] Известно, что ферменты, получаемые из Thermobifida fusca, более терморезистентны, чем любая полифосфат-зависимая глюкокиназа, о которой сообщали до сих пор, и были сообщения о потере их активности (на 50%) после нагрева при 50°C в течение 0,25 ч. Несмотря на то, что ферменты, получаемые из Thermobifida fusca, иммобилизовали для более хорошей термостойкости, их активность снижалась до 50% от их начальной активности после 2 ч [см. Liao et al. 2012. Appl Microbiol Biotechnol 93:1109-1117].
[112] Следовательно, можно заключить, что полифосфат-зависимая глюкокиназа по настоящему изобретению, получаемая из Anaerolinea thermophila, является наиболее термостабильным из ферментов, о которых сообщали до сих пор, поскольку активность патентоспособного фермента сохраняется на уровне приблизительно 51% от ее начального значения даже после нагрева при 60°C в течение 4 ч.
[113] Пример 7: анализ выходов превращения при различных концентрациях субстратов
[114] Анализировали выходы превращения глюкозо-6-фосфата при различных концентрациях глюкозы и гексаметафосфат натрия. С этой целью глюкозу (2-15% (масс./об.)), гексаметафосфат натрия (1,5-11,5% (масс./об.)) и MgSO4 (10 мМ) суспендировали в 50 мМ буфере Tris-HCl (pH 7,0) для того, чтобы получать реакционные композиции. Очищенный фермент (10-50 Ед/мл) добавляли в каждую реакционную композицию. Реакцию оставляли протекать при 55°C в течение 12 ч. После этого образование глюкозо-6-фосфата количественно анализировали посредством HPLC.
[115] Как результат, при использовании 2% (масс./об.) глюкозы и 1,5% (масс./об.) гексаметафосфата натрия достигали выхода превращения 81% после реакции в течение 12 ч. При использовании 5% (масс./об.) глюкозы и 3,5% (масс./об.) гексаметафосфата натрия достигали выхода превращения 78% после реакции в течение 12 ч. При использовании 10% (масс./об.) глюкозы и 7% (масс./об.) гексаметафосфата натрия достигали выхода превращения 77% после реакции в течение 12 ч. При использовании 15% (масс./об.) глюкозы и 10% (масс./об.) гексаметафосфата натрия достигали выхода превращения 65% после реакции в течение 12 ч.
--->
<110> CJ CHEILJEDANG CORPORATION
<120> НОВАЯ ПОЛИФОСФАТ-ЗАВИСИМАЯ ГЛЮКОКИНАЗА И СПОСОБ ПОЛУЧЕНИЯ ГЛЮКОЗО-6-ФОСФАТА С ЕЕ ИСПОЛЬЗОВАНИЕМ
<130> P16-6262
<150> KR 2016/024293
<151> 2016-02-29
<160> 4
<170> KopatentIn 2.0
<210> 1
<211> 762
<212> ДНК
<213> Anaerolinea thermophila
<400> 1
atggggaggc agggcatgga aattttaggg attgatatcg gaggatccgg catcaaaggg 60
gctccggtgg atgtagaaac cggccagtta accgccgagc gataccgctt acccaccccc 120
gaaaatgcct tacctgaaga agtggctctg gtagttgccc aaattgtcga acactttcag 180
tggaaaggtc gtgtaggggc aggatttcct gctgccatca agcacggcgt ggcacagacg 240
gccgcaaaca tccaccctac atggattgga cttcatgctg gcaacctttt cagcgaaaaa 300
tgcggatgtc ctgtctcagt gttgaatgat gcggatgctg ccggactggc ggaaatgatc 360
tttggggcag gaaaaggcca gaaaggggtg gtgctgatga ttaccattgg cactggcatc 420
gggacagccc tgttcaccga tgggatattg gtccctaata ccgagttggg acatattgaa 480
attcggggca aagatgccga acagcgctct tcggaagccg cccgccagcg gaaggattgg 540
acctggcaac aatgggcaaa gcgtctgaat gagcatttgg agcgcctgga agccctgttc 600
tggcccgatt tattcatcct tggtggaggg gcagtaaaaa atcatgaaaa gttcttccct 660
tatctaaaac tgcgtactcc ctttgttgca gcaaaattgg ggaatctggc tgggattgta 720
ggcgcagcgt ggtatgctca cacccaggaa acgcaagcct ga 762
<210> 2
<211> 253
<212> Белок
<213> Anaerolinea thermophila
<400> 2
Met Gly Arg Gln Gly Met Glu Ile Leu Gly Ile Asp Ile Gly Gly Ser
1 5 10 15
Gly Ile Lys Gly Ala Pro Val Asp Val Glu Thr Gly Gln Leu Thr Ala
20 25 30
Glu Arg Tyr Arg Leu Pro Thr Pro Glu Asn Ala Leu Pro Glu Glu Val
35 40 45
Ala Leu Val Val Ala Gln Ile Val Glu His Phe Gln Trp Lys Gly Arg
50 55 60
Val Gly Ala Gly Phe Pro Ala Ala Ile Lys His Gly Val Ala Gln Thr
65 70 75 80
Ala Ala Asn Ile His Pro Thr Trp Ile Gly Leu His Ala Gly Asn Leu
85 90 95
Phe Ser Glu Lys Cys Gly Cys Pro Val Ser Val Leu Asn Asp Ala Asp
100 105 110
Ala Ala Gly Leu Ala Glu Met Ile Phe Gly Ala Gly Lys Gly Gln Lys
115 120 125
Gly Val Val Leu Met Ile Thr Ile Gly Thr Gly Ile Gly Thr Ala Leu
130 135 140
Phe Thr Asp Gly Ile Leu Val Pro Asn Thr Glu Leu Gly His Ile Glu
145 150 155 160
Ile Arg Gly Lys Asp Ala Glu Gln Arg Ser Ser Glu Ala Ala Arg Gln
165 170 175
Arg Lys Asp Trp Thr Trp Gln Gln Trp Ala Lys Arg Leu Asn Glu His
180 185 190
Leu Glu Arg Leu Glu Ala Leu Phe Trp Pro Asp Leu Phe Ile Leu Gly
195 200 205
Gly Gly Ala Val Lys Asn His Glu Lys Phe Phe Pro Tyr Leu Lys Leu
210 215 220
Arg Thr Pro Phe Val Ala Ala Lys Leu Gly Asn Leu Ala Gly Ile Val
225 230 235 240
Gly Ala Ala Trp Tyr Ala His Thr Gln Glu Thr Gln Ala
245 250
<210> 3
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 3
caccatatgg ggaggcaggg catggaaatt ttag 34
<210> 4
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 4
caaactcgag ggcttgcgtt tcctgggtg 29
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВАЯ ПСИКОЗО-6-ФОСФАТ ФОСФАТАЗА, КОМПОЗИЦИЯ ДЛЯ ПОЛУЧЕНИЯ ПСИКОЗЫ, СОДЕРЖАЩАЯ УКАЗАННЫЙ ФЕРМЕНТ, СПОСОБ ПОЛУЧЕНИЯ ПСИКОЗЫ С ИСПОЛЬЗОВАНИЕМ УКАЗАННОГО ФЕРМЕНТА | 2018 |
|
RU2757229C2 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ ГЕКСОЗ | 2018 |
|
RU2766710C2 |
| РЕКОМБИНАНТНОЕ ПОЛУЧЕНИЕ СТЕВИОЛ-ГЛИКОЗИДОВ | 2014 |
|
RU2741103C2 |
| НОВАЯ D-ПСИКОЗО-3-ЭПИМЕРАЗА И СПОСОБ ПОЛУЧЕНИЯ D-ПСИКОЗЫ С ЕЕ ИСПОЛЬЗОВАНИЕМ | 2017 |
|
RU2727903C1 |
| ГЛЮКОАМИЛАЗЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2802790C2 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ D-АЛЛЮЛОЗЫ | 2017 |
|
RU2819709C2 |
| ВАРИАНТЫ ПРОТЕАЗЫ КЛАДЫ APRL И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2733987C2 |
| ГИДРОЛИЗ СТЕВИОЛОВЫХ ГЛИКОЗИДОВ С ПОМОЩЬЮ БЕТА-ГЛЮКОЗИДАЗЫ | 2018 |
|
RU2775697C2 |
| БИОСИНТЕТИЧЕСКОЕ ПОЛУЧЕНИЕ СТЕВИОЛОВОГО ГЛИКОЗИДА РЕБАУДИОЗИДА D4 ИЗ РЕБАУДИОЗИДА E | 2017 |
|
RU2764803C2 |
| ВАРИАНТЫ ПРОТЕАЗ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2790443C2 |
Изобретение относится к биотехнологии и представляет собой композицию для получения глюкозо-6-фосфата, содержащую терморезистентную полифосфат-зависимую глюкокиназу, получаемую из рода Anaerolinea, полифосфат и субстрат i) глюкозу или ii) крахмал, и ферменты для разжижения и осахаривания, где глюкокиназа имеет максимальную активность при 65-70 °C и имеет активность 95% или более относительно максимальной активности во всем диапазоне 60-80 °C. 2 н. и 15 з.п. ф-лы, 8 ил., 1 табл., 7 пр.
1. Композиция для получения глюкозо-6-фосфата, содержащая терморезистентную полифосфат-зависимую глюкокиназу, получаемую из рода Anaerolinea, полифосфат и субстрат i) или ii):
i) глюкозу,
ii) крахмал и ферменты для разжижения и осахаривания,
где глюкокиназа имеет максимальную активность при 65-70 °C и имеет активность 95% или более относительно максимальной активности во всем диапазоне 60-80 °C.
2. Композиция по п. 1, отличающаяся тем, что композиция содержит от 1% до 3% по массе глюкозы, от 1% до 10% по массе полифосфата и от 10 Ед/мл до 50 Ед/мл полифосфат-зависимой глюкокиназы на основании общего объема композиции и достигает выхода превращения по меньшей мере 70% до глюкозо-6-фосфата.
3. Композиция по п. 1, отличающаяся тем, что композиция содержит от 5% до 20% по массе глюкозы, от 5% до 12% по массе полифосфата и от 10 Ед/мл до 50 Ед/мл полифосфат-зависимой глюкокиназы на основании общего объема композиции и достигает выхода превращения по меньшей мере 50% до глюкозо-6-фосфата.
4. Композиция по п. 2 или 3, которая дополнительно содержит ионы магния.
5. Композиция по п. 4, в которой ионы магния присутствуют в концентрации от 0,2 мМ до 20 мМ.
6. Композиция по п. 1, в которой терморезистентная полифосфат-зависимая глюкокиназа получена из рода Anaerolinea, имеет аминокислотную последовательность, приведенную в SEQ ID NO. 2, или экспрессирована с последовательности ДНК, представленной SEQ ID NO. 1.
7. Способ получения глюкозо-6-фосфата, включающий
приведение в контакт терморезистентной полифосфат-зависимой глюкокиназы, полученной из рода Anaerolinea, с глюкозой и полифосфатом,
где глюкокиназа имеет максимальную активность при 65-70 °C и имеет активность 95% или более относительно максимальной активности во всем диапазоне 60-80 °C.
8. Способ по п. 7, в котором глюкозу получают посредством приведения в контакт фермента для разжижения и осахаривания с крахмалом или целлюлозой.
9. Способ по п. 7, в котором полифосфат представляет собой гексаметафосфат натрия.
10. Способ по п. 7, в котором глюкозо-6-фосфат получают при температуре от 45°C до 90°C и/или pH от 4 до 10.
11. Способ по п. 7, в котором активность полифосфат-зависимой глюкокиназы усиливается в присутствии ионов магния.
12. Способ по п. 7, в котором полифосфат-зависимая глюкокиназа присутствует в количестве от 10 Ед/мл до 50 Ед/мл.
13. Способ по п. 7, в котором глюкоза присутствует в количестве от 0,1% до 40% по массе на основании общей массы композиции, которая содержит полифосфат-зависимую глюкокиназу, полифосфат и глюкозу.
14. Способ по п. 7, в котором полифосфат присутствует в количестве от 0,5% до 25% по массе на основании общей массы композиции, которая содержит полифосфат-зависимую глюкокиназу, полифосфат и глюкозу.
15. Способ по п. 7, отличающийся тем, что способ дополнительно включает приведение в контакт, разжижение и осахаривание фермента с крахмалом или целлюлозой до стадии приведения в контакт глюкокиназы с глюкозой и полифосфатом.
16. Способ по п. 8, в котором ферменты для разжижения и осахаривания представляют собой один или несколько, выбранных из α-амилаз, глюкоамилаз и α-гликозидаз.
17. Способ по п. 8, в котором стадия приведения в контакт глюкокиназы с глюкозой и полифосфатом и стадия получения глюкозы представляют собой однореакторное ферментативное превращение.
| Лампа для светолечения | 1927 |
|
SU12029A1 |
| US 20160002672, 07.01.2016 | |||
| РУБЦОВ П.М | |||
| и др | |||
| Глюкокиназа и белок-регулятор глюкокиназы как молекулярные мишени для создания новых противодиабетических препататов, Молекулярная биология, т.49, номер 4, 2015, стр.555. | |||

