Область техники, к которой относится изобретение
Настоящее изобретение относится к варианту белка YjeH, который представляет собой белок внутренней мембраны, и к способу получения целевого продукта с его использованием.
Предшествующий уровень техники
О-Ацетилгомосерин действует как предшественник метионина, который является одной из незаменимых аминокислот в организме человека. Известно, что гомосерин, который является предшественником и метионина, и треонина, преобразуется в треонин через фосфогомосерин и преобразуется в метионин через ацетилгомосерин.
Метионин используют не только как кормовую и пищевую добавку, но также в качестве сырьевого ингредиента для приготовления растворов для инфузии и фармацевтических препаратов и получают путем химического и биологического синтеза. Был опубликован способ получения L-метионина в две стадии из предшественника L-метионина, полученного путем ферментации, посредством ферментативного превращения (Международная публикация WO 2008/013432).
В этом процессе, осуществляемом в две стадии, О-сукцинилгомосерин и О-ацетилгомосерин используют в качестве предшественников метионина, и очень важно получить О-ацетилгомосерин с высоким выходом в целях экономичного массового производства.
Раскрытие изобретения
Техническая задача
В результате интенсивных усилий по повышению продукции О-ацетилгомосерина или гомосерина авторы настоящего изобретения обнаружили вариант белка внутренней мембраны, обладающий усиленной активностью экспорта О-ацетилгомосерина или гомосерина, в результате чего было осуществлено настоящее изобретение.
Техническое решение
В настоящем изобретении предложен вариант белка YjeH, который представляет собой белок внутренней мембраны, включающий i) замену аминокислоты, соответствующей 92-му положению, аспарагином, ii) замену аминокислоты, соответствующей 351-му положению, лейцином, или iii) замену в обоих положениях i) и ii), в аминокислотной последовательности SEQ ID NO: 1, и обладающий гомологией 95% или более, но менее 100% с SEQ ID NO: 1.
В настоящем изобретении предложен полинуклеотид, кодирующий этот вариант.
В настоящем изобретении предложен вектор, включающий этот полинуклеотид.
В настоящем изобретении предложен микроорганизм, включающий по меньшей мере одно из: варианта белка внутренней мембраны YjeH, включающего i) замену аминокислоты, соответствующей 92-му положению, аспарагином, ii) замену аминокислоты, соответствующей 351-му положению, лейцином, или iii) замену в обоих положениях i) и ii), в аминокислотной последовательности SEQ ID NO: 1, и обладающего гомологией 95% или более, но менее 100% с SEQ ID NO: 1; полинуклеотида, кодирующего этот вариант; и вектора, кодирующего этот полинуклеотид.
В настоящем изобретении предложен способ получения целевого продукта, включающий культивирование микроорганизма в культуральной среде, где микроорганизм включает по меньшей мере одно из: варианта белка внутренней мембраны YjeH, включающего i) замену аминокислоты, соответствующей 92-му положению, аспарагином, ii) замену аминокислоты, соответствующей 351-му положению, лейцином, или iii) замену в обоих положениях i) и ii), в аминокислотной последовательности SEQ ID NO: 1, и обладающего гомологией 95% или более, но менее 100% с SEQ ID NO: 1; полинуклеотида, кодирующего этот вариант; и вектора, кодирующего этот полинуклеотид.
В настоящем изобретении предложен способ получения метионина, включающий получение О-ацетилгомосерина путем культивирования микроорганизма в культуральной среде и превращения О-ацетилгомосерина в метионин путем взаимодействия О-ацетилгомосерина с сульфидом, где микроорганизм включает по меньшей мере одно из: варианта белка внутренней мембраны YjeH, включающего i) замену аминокислоты, соответствующей 92-му положению, аспарагином, ii) замену аминокислоты, соответствующей 351-му положению, лейцином, или iii) замену в обоих положениях i) и ii), в аминокислотной последовательности SEQ ID NO: 1, и обладающего гомологией 95% или более, но менее 100% с SEQ ID NO: 1; полинуклеотида, кодирующего этот вариант; и вектора, кодирующего этот полинуклеотид.
В настоящем изобретении предложен способ получения глюфосината, включающий получение О-ацетилгомосерина или гомосерина путем культивирования микроорганизма в культуральной среде и превращения О-ацетилгомосерина или гомосерина в глюфосинат, где микроорганизм включает по меньшей мере одно из: варианта белка внутренней мембраны YjeH, включающего i) замену аминокислоты, соответствующей 92-му положению, аспарагином, ii) замену аминокислоты, соответствующей 351-му положению, лейцином, или iii) замену в обоих положениях i) и ii), в аминокислотной последовательности SEQ ID NO: 1, и обладающего гомологией 95% или более, но менее 100% с SEQ ID NO: 1; полинуклеотида, кодирующего этот вариант; и вектора, кодирующего этот полинуклеотид.
В настоящем изобретении предложена композиция для получения гомосерина, включающая микроорганизм или культуру микроорганизма.
В настоящем изобретении предложена композиция для получения О-ацетилгомосерина, включающая микроорганизм или культуру микроорганизма.
В настоящем изобретении предложено применение варианта белка YjeH внутренней мембраны для получения О-ацетилцистеина или гомосерина.
В настоящем изобретении предложено применение микроорганизма для продукции О-ацетилцистеина или гомосерина.
Полезные эффекты изобретения
О-Ацетилгомосерин или гомосерин можно получать с высокими выходами путем культивирования микроорганизма, обладающего способностью продуцировать О-ацетилгомосерин или гомосерин, модифицированного с использованием варианта YjeH, представляющего собой белок внутренней мембраны, в соответствии с описанием настоящего изобретения по сравнению с немодифицированными белками.
Краткое описание графических материалов
ФИГ. 1 представляет собой схематическое изображение плазмиды pDCM2.
Лучший способ осуществления изобретения
Настоящее изобретение будет раскрыто подробно. В то же время каждое описание и воплощение изобретения, раскрытое в описании настоящего изобретения, можно применять к различным представленным в нем описаниям и воплощениям. Иными словами, в объем описания настоящего изобретения включены все комбинации различных компонентов, раскрытых в описании настоящего изобретения. Кроме того, объем описания настоящего изобретения не должен ограничиваться приведенными ниже описаниями.
В одном аспекте описания настоящего изобретения для достижения вышеописанных целей предложен вариант белка YjeH внутренней мембраны, включающий i) замену аминокислоты, соответствующей 92-му положению, аспарагином, ii) замену аминокислоты, соответствующей 351-му положению, лейцином, или iii) замену в обоих положениях i) и ii), в аминокислотной последовательности SEQ ID NO: 1, и обладающий гомологией 95% или более, но менее 100% с SEQ ID NO: 1.
Вариант белка YjeH внутренней мембраны может обладать способностью к экспорту О-ацетилгомосерина или гомосерина, т.е. активностью экспорта О-ацетилгомосерина или гомосерина.
Используемый в настоящем документе термин «гомосерин» представляет собой α-аминокислоту, имеющую гидроксильную группу в боковой цепи. Известно, что гомосерин как промежуточное соединение биосинтеза треонина и метионина в микроорганизмах и растениях образуется из аспарагиновой кислоты 4-полуальдегида и имеет химическую формулу C4H9NO3. Гомосерин может быть преобразован в О-ацетилцистеин гомосеринацетилтрансферазой в присутствии ацетилкоэнзима А (ацетил-КоА).
Используемый в настоящем документе термин «О-ацетилгомосерин» относится к ацетильному производному L-гомосерина, который является специфичным промежуточным материалом в пути биосинтеза метионина в микроорганизмах. Известно, что О-ацетилгомосерин образуется путем взаимодействия между гомосерином и ацетил-КоА, катализируемого гомосеринацетилтрансферазой. Он имеет химическую формулу С6Н11NO4. О-Ацетилгомосерин может также называться О-ацетил-L-гомосерином.
В частности, в варианте белка в соответствии с описанием настоящего изобретения i) аминокислота, соответствующая 92-му положению, может быть заменена другой аминокислотой, ii) аминокислота, соответствующая 351-му положению, может быть заменена другой аминокислотой, или iii) обе аминокислоты, соответствующие 92-му положению и 351-му положению, могут быть заменены другими аминокислотами в аминокислотной последовательности SEQ ID NO: 1, но без ограничений. Замена аминокислот может представлять собой i) замену аминокислоты, соответствующую 92-му положению, аспарагином, или ii) замену аминокислоты, соответствующую 351-му положению, лейцином, и более конкретно вариант белка может представлять собой вариант белка YjeH внутренней мембраны, в котором i) аминокислота, соответствующая 92-му положению, заменена аспарагином, ii) аминокислота, соответствующая 351-му положению, заменена лейцином, или iii) аминокислота, соответствующая 92-му положению, заменена аспарагином, а аминокислота, соответствующая 351-му положению, заменена лейцином, в аминокислотной последовательности SEQ ID NO: 1, но без ограничений.
Используемый в настоящем документе термин «белок внутренней мембраны» относится к белку, который является членом семейства белков эффлюкса аминокислот (Amino Acid Efflux, ААЕ), входящего в суперсемейство белков-переносчиков аминокислот-полиаминов-органокатионов (АРС), являющихся посредниками экспорта О-ацетилгомосерина и/или гомосерина. Предположительно этот белок содержит 12 транс мембранных α-спиралей, 10 из которых образуют складку инвертированных повторов, которая характерна для суперсемейства АРС. Белок внутренней мембраны может представлять собой, например, белок, включающий аминокислотную последовательность SEQ ID NO: 1. Белок, включающий аминокислотную последовательность SEQ ID NO: 1, может использоваться взаимозаменяемо с белком, имеющим аминокислотную последовательность SEQ ID NO: 1, или белок, состоящий из аминокислотной последовательности SEQ ID NO: 1. В описании настоящего изобретения белок внутренней мембраны может использоваться взаимозаменяемо с О-ацетилгомосерин- или гомосерин-экспортирующим белком, белком, обладающим способностью к экспорту О-ацетилгомосерина или гомосерина, белком, обладающим О-ацетилгомосерин- или гомосерин-экспортирующей активностью, белком YjeH или YjeH.
В описании настоящего изобретения SEQ ID NO: 1 может относиться к аминокислотной последовательности белка YjeH, который представляет собой белок внутренней мембраны. В частности, SEQ ID NO: 1 может представлять собой аминокислотную последовательность белка внутренней мембраны, кодируемого геном yjeH и обладающего О-ацетилгомосерин- или гомосерин-экспортирующей активностью. Аминокислотная последовательность SEQ ID NO: 1 может быть получена из известной базы данных GenBank Национального центра биотехнологической информации (NCBI). Например, аминокислотная последовательность может иметь происхождение из Escherichia coli (Е. coli), но без ограничений, и любая последовательность, обладающая активностью, идентичной активности этой аминокислотной последовательности, включена без ограничений. В дополнение к этому, хотя белок внутренней мембраны в описании настоящего изобретения может быть определен как белок, включающий аминокислотную последовательность SEQ ID NO: 1, этот белок внутренней мембраны не исключает мутацию, которая может произойти естественным путем или путем добавления незначащей последовательности выше или ниже аминокислотной последовательности SEQ ID NO: 1, или ее молчащую мутацию. Если белок обладает активностью, идентичной или эквивалентной активности белка, содержащего аминокислотную последовательность SEQ ID NO: 1, он явным образом принадлежит к белку внутренней мембраны по описанию настоящего изобретения. Например, белок внутренней мембраны по описанию настоящего изобретения может представлять собой белок, состоящий из аминокислотной последовательности SEQ ID NO: 1 или из аминокислотной последовательности, обладающей по меньшей мере 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5% или 99,7% гомологией или идентичностью с ней. Также будет очевидно, что любой белок, имеющий аминокислотную последовательность, включающую делецию, модификацию, замену или добавление в части аминокислотной последовательности, входит в объем описания настоящего изобретения, если аминокислотная последовательность сохраняет вышеописанную гомологию или идентичность и эффекты, эквивалентные эффектам этого белка.
Таким образом, хотя в описании настоящего изобретения используются выражения «белок или полипептид, имеющий аминокислотную последовательность, представленную заданной последовательностью SEQ ID NO» и «белок или полипептид, включающий аминокислотную последовательность, представленную заданной последовательностью SEQ ID NO», очевидно, что любой белок, имеющий аминокислотную последовательность, включающую делецию, модификацию, замену или добавление в части аминокислотной последовательности, может также использоваться в описании настоящего изобретения, если этот белок обладает активностью, идентичной или эквивалентной активности полипептида, состоящей из заданной аминокислотной последовательности. Например, очевидно, что «белок, включающий аминокислотную последовательность SEQ ID NO: 1» или «полипептид, включающий аминокислотную последовательность SEQ ID NO: 1», относится к «белку, состоящему из аминокислотной последовательности SEQ ID NO: 1», или «полипептиду, состоящему из аминокислотной последовательности SEQ ID NO: 1», если первый обладает такой же или эквивалентной активностью, как последний.
Используемый в настоящем документе термин «вариант» относится к белку, полученному путем консервативной замены и/или модификации по меньшей мере одной аминокислоты, отличающейся от аминокислоты указанной последовательности при сохранении ее функций или свойств. Вариант может иметь аминокислотную последовательность, отличающуюся от идентифицированной последовательности вследствие замены, делеции или добавления нескольких аминокислот. Такие варианты, как правило, могут быть идентифицированы путем модификации по меньшей мере одной аминокислоты вышеуказанной аминокислотной последовательности белка и оценки свойств модифицированного белка. Таким образом, активность варианта может быть усилена, может быть не изменена или снижена относительно нативного белка. В дополнение к этому некоторые варианты могут включать варианты, из которых удален по меньшей мере один участок, такой как N-концевая лидерная последовательность или трансмембранный домен. Другие варианты могут включать варианты, в которых участок удален с N- и/или С-конца зрелого белка. Термин «вариант» может быть также использован взаимозаменяемо с другими терминами, такими как модификация, модифицированный белок, модифицированный полипептид, мутант, мутеин и дивергент, а также любыми терминами, используемыми для указания вариации, которые могут быть также использованы без ограничений. В свете целей описания настоящего изобретения способность варианта может быть усилена относительно нативного или немодифицированного белка, но без ограничений.
Используемый в настоящем документе термин «консервативная замена» относится к замене одной аминокислоты другой аминокислотой, обладающей подобным структурным и/или химическим свойством. Например, вариант может включать по меньшей мере одну консервативную замену при сохранении по меньшей мере одной биологической активности. Такая замена аминокислоты может, как правило, происходить на основе сходства полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфипатического характера остатка. Например, аминокислоты с электрическим зарядом и с боковыми цепями включают положительно заряженные (основные) аминокислоты, такие как аргинин, лизин и гистидин, и отрицательно заряженные (кислые) аминокислоты, такие как глутаминовая кислота и аспарагиновая кислота; а незаряженные аминокислоты с боковыми цепями включают неполярные аминокислоты, такие как глицин, аланин, валин, лейцин, изолейцин, метионин, фенилаланин, триптофан и пролин, и полярные, или гидрофильные аминокислоты, такие как серии, треонин, цистеин, тирозин, аспарагин и глутамин. Среди неполярных аминокислот ароматические аминокислоты включают фенилаланин и триптофан.
Варианты могут также включать делецию или добавление аминокислот, которые оказывают минимальное влияние на свойства и вторичную структуру полипептида. Например, полипептид может быть конъюгирован с сигнальной (или лидерной) последовательностью на N-конце белка, которая котрансляционно или посттрансляционно направляет перенос белка. Полипептид может быть также конъюгирован с другой последовательностью или линкером для идентификации, очистки или синтеза полипептида.
В описании настоящего изобретения «заменена другой аминокислотой» конкретно не ограничено, если аминокислота после замены отличается от аминокислоты до замены.
В описании настоящего изобретения «заменена другой аминокислотой» конкретно не ограничено, если аминокислота после замены отличается от аминокислоты до замены. В частности, аминокислота может быть замещена любой из лизина, гистидина, глутаминовой кислоты, аспарагиновой кислоты, глицина, аланина, валина, лейцина, изолейцина, метионина, фенилаланина, триптофана, пролина, серина, треонина, цистеина, тирозина, аспарагина и глутамина, в отличие от предыдущей аминокислотной последовательности.
В частности, в аминокислотной последовательности SEQ ID NO: 1 фенилаланин, который представляет собой аминокислоту, соответствующую 92-му положению, может быть заменен аспарагином, либо фенилаланин, который представляет собой аминокислоту, соответствующую 351-му положению, может быть заменен лейцином. В то же время в описании настоящего изобретения очевидно, что, если не используется выражение «заменен другой аминокислотой», выражение «заменен определенной аминокислотой» означает, что аминокислота после замены отличается от аминокислоты до замены.
Замещенный аминокислотный остаток может включать не только природную аминокислоту, но также и неприродную аминокислоту. Неприродная аминокислота может представлять собой, например, D-аминокислоту, (гомо)-аминокислоту, (бета-гомо)-аминокислоту, N-метил-аминокислоту, α-метил-аминокислоту и редкую аминокислоту (например, цитруллин или нафтилаланин), но без ограничений. В то же время в описании настоящего изобретения очевидно, что, если не используется выражение «заменена другой аминокислотой», выражение «заменена определенной аминокислотой» означает, что аминокислота после замены отличается от аминокислоты до замены.
Вариант может представлять собой такой, в котором по меньшей мере одна из аминокислот, соответствующая 92-му положению и 351-му положению, заменена аминокислотой, отличающейся от аминокислоты до замены в аминокислотной последовательности SEQ ID NO: 1, либо заменена незаряженной кислотой с боковой цепью, отличающейся от той, которая была до замены, но без ограничений.
В частности, вариант может представлять собой вариант белка, в котором i) аминокислота, соответствующая 92-му положению, может быть заменена другой аминокислотой, ii) аминокислота, соответствующая 351-му положению, может быть заменена другой аминокислотой, или iii) обе аминокислоты, соответствующие 92-му положению и 351-му положению, могут быть заменены другими аминокислотами в аминокислотной последовательности SEQ ID NO: 1, но без ограничений. Замена аминокислот может представлять собой i) замену 92-й аминокислоты аспарагином, или ii) замену 351-й аминокислоты лейцином. Более конкретно вариант может представлять собой вариант белка YjeH внутренней мембраны, в котором i) 92-я аминокислота заменена аспарагином, ii) 351-я аминокислота заменена лейцином, или iii) 92-я аминокислота заменена аспарагином, и 351-я аминокислота заменена лейцином в аминокислотной последовательности SEQ ID NO: 1, но без ограничений.
Используемый в настоящем документе термин «соответствует (чему-то)» относится к аминокислотному остатку в указанном положении в белке или полипептиде или к аминокислотному остатку, подобному, идентичному или гомологичному остатку, указанному в белке или полипептиде. Используемый в настоящем документе термин «соответствующая область» в целом относится к области, подобной ей в родственном белке или в белке сравнения.
В описании настоящего изобретения для положения аминокислотного остатка белка, используемого в описании настоящего изобретения, может использоваться конкретная нумерация. Например, положение, соответствующее аминокислотному остатку белка по описанию настоящего изобретения, может быть перенумеровано в результате приведения аминокислотной последовательности сравниваемого белка в соответствие с аминокислотной последовательностью белка по описанию настоящего изобретения.
Вариант белка YjeH внутренней мембраны, предложенный в описании настоящего изобретения, может относиться к варианту, обладающему усиленной способностью к экспорту О-ацетилгомосерина или гомосерина по сравнению с немодифицированным белком в результате замены аминокислоты в определенном положении в вышеописанном белке внутренней мембраны, обладающем способностью к экспорту О-ацетилгомосерина или гомосерина.
Вариант белка YjeH внутренней мембраны, включающий i) замену аминокислоты, соответствующей 92-му положению, аспарагином, ii) замену аминокислоты, соответствующей 351-му положению, лейцином, или замену в обоих положениях i) и ii) в аминокислотной последовательности SEQ ID NO: 1 может включать любую из аминокислотных последовательностей SEQ ID NO: 35 и 36, в частности состоящих из любой из аминокислотных последовательностей SEQ ID NO: 35 и 36, более конкретно состоящих из любой из аминокислотных последовательностей SEQ ID NO: 35 и 36, но без ограничений.
В дополнение к этому вариант может включать аминокислотные последовательности SEQ ID NO: 35 и 36 или аминокислотную последовательность, в которой по меньшей мере любая одна аминокислота из 92-й и 351-й аминокислот фиксирована в аминокислотных последовательностях SEQ ID NO: 35 и 36, и имеющие по меньшей мере 80% гомологию с любой из аминокислотных последовательностей SEQ ID NO: 35 и 36, но без ограничений. В частности, вариант по описанию настоящего изобретения может включать полипептид, имеющий 80%, 90%, 95%, 96%, 97%, 98% или 99% гомологию или идентичность с любой из аминокислотных последовательностей SEQ ID NO: 35 и 36. Также очевидно, что любой белок, имеющий аминокислотную последовательность, включающую делецию, модификацию, замену или добавление в другой части аминокислотной последовательности, кроме 92-го положения и 351-го положения, входит в объем описания настоящего изобретения, если аминокислотная последовательность сохраняет вышеописанную гомологию или идентичность и эффекты, эквивалентные эффектам этого белка.
Используемый в настоящем документе термин «гомология» или «идентичность» относится к степени соответствия между двумя данными аминокислотными последовательностями, или последовательностями оснований, и может быть выражен в процентах. Эти термины «гомология» и «идентичность» часто могут использоваться взаимозаменяемо.
Гомологию или идентичность консервативных полинуклеотидов или полипептидов можно определить с помощью стандартного алгоритма приведения последовательностей в соответствие, вместе с которым можно использовать штрафы за гэпы по умолчанию, установленные программой. По существу гомологичные или идентичные последовательности могут гибридизоваться друг с другом на по меньшей мере около 50%, 60%, 70%, 80% или 90% полноразмерной последовательности или по всей полноразмерной последовательности в в условиях умеренной или высокой жесткости. В гибрид изо ванных полинуклеотидах можно также рассматривать полинуклеотиды, включающие вырожденный кодон вместо кодона.
Гомологию, подобие или идентичность последовательности между двумя определенными полипептидами или полинуклеотидами можно определить, используя любой известный компьютерный алгоритм, такой как программа FASTA, используя параметры по умолчанию, которые введены, например, Pearson et al. (1988) Рrос. Natl. Acad. Sci. USA 85: 2444. Альтернативно можно использовать алгоритм Нидлмана-Вунша (1970, J. Mol. Biol. 48: 443 453), выполняемый в программе Needleman из пакета программ The European Molecular Biology Open Software Suite (EMBOSS) (Rice et al., 2000, Trends Genet. 16: 276-277) (версия 5.0.0 или более новая) для этого определения (включая пакет программ GCG (Devereux, J. et al., Nucleic Acids Research 12: 387 (1984)), BLASTP, BLASTN, FASTA (Atschul, S.F. et al., J MOLEC BIOL 215: 403 (1990); Guide to Huge Computers, Martin J. Bishop, ed., Academic Press, San Diego, 1994, и CARILLO et al. (1988) SIAM J Applied Math 48: 1073). Например, гомологию, подобие или идентичность можно определить, используя программу BLAST из базы данных Национального центра биотехнологической информации, или ClustalW.
Гомологию, подобие или идентичность между полинуклеотидами или полипептида можно определить, например, путем сравнения информации о последовательности, используя компьютерную программу GAP, например программу, введенную Needleman et al. (1970), J Mol Biol. 48: 443, например, как описано в Smith and Waterman, Adv. Appl. Math (1981) 2: 482. Кратко, программа GAP определяет подобие как число приведенных в соответствие символов (т.е. нуклеотидов или аминокислот), которые являются подобными, деленное на общее число символов в более короткой из двух последовательностей. Параметры по умолчанию для программы GAP могут включать: (1) матрица двоичных сравнений (содержащая значения 1 для идентичности и 0 для неидентичности), матрица взвешенных сравнений Gribskov et al. (1986) Nucl. Acids Res. 14: 6745, описанная в Schwartz and Dayhoff, eds., Atlas Of Protein Sequence And Structure, National Biomedical Research Foundation, pp. 353-358 (1979) (или матрица замен EDNAFULL (версия EMBOSS NCBI NUC 4.4)); (2) штраф 3.0 за каждый гэп и дополнительный штраф 0.10 за каждый символ в каждом гэпе (или штраф за открытие гэпа 10 и штраф за удлинение гэпа 0,5); и (3) отсутствие штрафа за концевые гэпы.
Гомологию, подобие или идентичность между двумя определенными полинуклеотидами или полипептидами также можно идентифицировать путем сравнения их последовательностей методом Саузерн-гибридизации в заданных условиях жесткости, и эти заданные условия жесткости входят в объем технологии и могут быть заданы методом, хорошо известным обычному специалисту в данной области техники (например, J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989; F.M. Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York).
Используемый в настоящем документе термин «вариант белка внутренней мембраны» или «вариант белка YjeH внутренней мембраны» может обладать способностью к экспорту О-ацетилгомосерина или гомосерина и может использоваться взаимозаменяемо с модифицированным полипептидом, обладающим способностью к продукции О-ацетилгомосерина или гомосерина, О-ацетилгомосерин- или гомосерин-продуцирующим модифицированным полипептидом, модифицированным полипептидом, обладающим О-ацетилгомосерин- или гомосерин-продуцирующей способностью, модифицированным полипептидом, обладающим активностью экспорта О-ацетилгомосерина или гомосерина, вариантом с активностью экспорта О-ацетилгомосерина или гомосерина, О-ацетилгомосерин- или гомосерин-экспортирующим вариантом, модифицированным гомосерин-экспортирующим белком, модифицированным О-ацетилгомосерин-экспортирующим белком, модифицированным YjeH, вариантом YjeH, YjeH-вариантом, YjeH-мутантом и т.п.
В дополнение к этому белок может иметь происхождение из штаммов, относящихся к роду Escherichia, роду Erwinia, роду Serratia, роду Providencia, роду Corynebacterium, роду Pseudomonas, роду Leptospira, роду Salmonella, роду Brevibacterium, роду Hypomononas, роду Chromobacterium и роду Norcardia, или из дрожжей и грибов, в частности, иметь происхождение из штаммов, относящихся к роду Escherichia, роду Corynebacterium и роду Leptospira, и из дрожжей, и более конкретно иметь происхождение из штаммов, относящихся к роду Escherichia, например Escherichia coli (Е. coli), но без ограничений.
Вариант белка YjeH внутренней мембраны может включать мутацию в 92-м положении и/или 351-м положении в аминокислотной последовательности SEQ ID NO: 1, и любой вариант, включающий добавление аминокислоты в аминокислотную последовательность SEQ ID NO: 1 или делецию из нее может входить в объем описания настоящего изобретения, если аминокислота, соответствующая 92-му положению и/или 351-му положению от N-конца SEQ ID NO: 1 заменена. Вариант белка YjeH внутренней мембраны может включать аминокислотную последовательность, включающую замену 92-й аминокислоты и/или 351-й аминокислоты другой аминокислотой в аминокислотной последовательности SEQ ID NO: 1 и может обладать усиленной активностью по сравнению с белком внутренней мембраны, включающим аминокислотную последовательность SEQ ID NO: 1 или не включающую мутацию. Вариант белка YjeH внутренней мембраны может иметь гомологию или идентичность на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99%, 99,5%, 99,7%или более и менее чем 100% с SEQ ID NO: 1.
Мутация 92-й или 351-й аминокислоты может представлять собой замену 92-й аминокислоты аспарагином или замену 351-й аминокислоты лейцином.
В частности, вариант белка YjeH внутренней мембраны может включать i) замену 92-й аминокислоты аспарагином, ii) замену 351-й аминокислоты лейцином, или замену обеих аминокислот i) и ii) в аминокислотной последовательности SEQ ID NO: 1 и может обладать усиленной активностью по сравнению с белком, включающим аминокислотную последовательность SEQ ID NO: 1 или с белком внутренней мембраны, имеющим происхождение из микроорганизма дикого типа без модификации.
В другом аспекте описания настоящего изобретения предложен полинуклеотид, кодирующий вариант белка YjeH внутренней мембраны.
Белок YjeH внутренней мембраны и его вариант являются такими, как описано выше.
Используемый в настоящем документе термин «полинуклеотид» относится к нити ДНК или РНК, имеющий определенную минимальную или большую длину, в виде полимера нуклеотидов, в котором нуклеотидные мономеры связаны друг с другом ковалентными связями, образуя длинную цепь; более конкретно полинуклеотид может представлять собой полинуклеотидный фрагмент, кодирующий вариант.
Полинуклеотид может включать любую нуклеотидную последовательность, кодирующую вариант белка YjeH внутренней мембраны по описанию настоящего изобретения, без ограничений. В описании настоящего изобретения ген, кодирующий аминокислотную последовательность белка YjeH внутренней мембраны, представляет собой ген yjeH, который может иметь происхождение из Escherichia coli (Е. coli) без ограничений.
В частности, полинуклеотид по описанию настоящего изобретения может включать различные модификации, выполненные в кодирующей области, при условии, что аминокислотная последовательность, экспрессирующаяся с кодирующей области в результате вырожденности кодонов или с учетом кодонов, предпочтительных в живом организме, в котором экспрессируется белок, не изменяется. В частности, может быть включена любая полинуклеотидная последовательность, кодирующая вариант белка YjeH внутренней мембраны, в котором 92я или 351я аминокислота заменена другой аминокислотой в аминокислотной последовательности SEQ ID NO: 1, без ограничений.
В дополнение к этому может быть включена любая последовательность, кодирующая белок внутренней мембраны, в котором 92-я или 351-я аминокислота аминокислотной последовательности SEQ ID NO: 1 заменена другой аминокислотой путем гибридизации с зондом, который может быть получен из последовательностей известных генов, например последовательностей, комплементарных полноразмерной нуклеотидной последовательности или ее части, в жестких условиях, без ограничений. Термин «жесткие условия» относится к условиям, обеспечивающим возможность специфичной гибридизации между полинуклеотидами. Такие условия раскрыты подробно в известных документах (например, J. Sambrook et al.). Например, эти условия могут включать условия выполнения гибридизации между генами, обладающими высокой гомологией или идентичностью, например, гомологией или идентичностью 40% или более, в частности 90% или более, более конкретно 95% или более, еще более конкретно 97% или более и наиболее конкретно 99% или более, без выполнения гибридизации между генами, обладающие более низкой гомологией, чем упомянутые выше значения гомологии или идентичности, или с выполнением гибридизации один раз, в частности два или три раза, в общепринятых условиях отмывки для Саузерн-гибридизации при концентрации соли и температуре 60°С, 1×раствор цитрата и хлорида натрия (SSC) и 0,1% додецилсульфат натрия (ДСН), в частности 60°С, 0,1×SSC, 0,1% ДСН, и более конкретно 68°С, 0,1×SSC и 0,1% ДСН.
Для гибридизации требуется, чтобы два полинуклеотид а имели комплементарные последовательности, хотя основания могут не совпадать, в зависимости от степени жесткости гибридизации. Термин «комплементарный» используется для описания соотношения между основаниями нуклеотидов, способных гибридизоваться друг с другом. Например, что касается ДНК, аденин комплементарен тимину, а цитозин комплементарен гуанину. Таким образом, в описание настоящего изобретения может быть включена не только по существу подобная нуклеиновая кислота, но также изолированный фрагмент нуклеиновой кислоты, но комплементарный полноразмерной последовательности.
В частности, полинуклеотиды, обладающие гомологией или идентичностью, могут быть выявлены с использованием вышеописанных условий гибридизации и с использованием процесса гибридизации при значении Тm 55°С. Значение Тm может также составлять, но без ограничений, 60°С, 63°С или 65°С, и может быть соответствующим образом скорректировано специалистами в данной области техники в соответствии с предусмотренными целями.
Соответствующая степень жесткости для гибридизации полинуклеотидов может зависеть от длин и степени комплементарности полинуклеотидов, а также от показателей, хорошо известных специалистам в данной области техники (Sambrook et al., см. выше, 9.50-9.51, 11.7-11.8).
В другом аспекте описания настоящего изобретения предложен вектор, включающий полинуклеотид, кодирующий вариант белка YjeH внутренней мембраны.
Белок YjeH внутренней мембраны, его вариант и полинуклеотид являются такими, как описано выше.
Вектор по описанию настоящего изобретения может представлять собой конструкцию ДНК, содержащую полинуклеотид, кодирующий целевой полипептид и функционально связанный с подходящей регуляторной областью экспрессии (регуляторной последовательностью экспрессии) таким образом, чтобы создать возможность экспрессии целевого полипептида в подходящей клетке-хозяине. Регуляторная последовательность экспрессии может включать промотор, способный инициировать транскрипцию, любую последовательность оператора для регуляции транскрипции, последовательность, кодирующую подходящий сайт связывания рибосомы на мРНК, и последовательность для регуляции терминации транскрипции и трансляции. При трансформации подходящей клетки-хозяина вектором этот вектор может реплицироваться или функционировать независимо от генома клетки-хозяина или может интегрировать в геном человека.
При использовании в настоящем документе термин «экспрессия» включает любой процесс, связанный с продукцией полипептида, такой как транскрипция, посттранскрипционная модификация, трансляция, посттрансляционная модификация и секреция и т.д., без ограничений.
При использовании в настоящем документе термин «экспрессионный вектор» относится к линейной или кольцевой молекуле нуклеиновой кислоты, включающей кодирующую последовательность и функционально связанную регуляторную последовательность для ее экспрессии.
При использовании в настоящем документе термин «функционально связанный» относится к расположению регуляторной последовательности в соответствующем положении для регуляции экспрессии кодирующей последовательности. Таким образом, «функционально связанный» включает связывание между регуляторной областью функционального домена, обладающей известной или требуемой активностью, такой как промотор, стоп-кодон, сигнальная последовательность, либо энхансер и целевая последовательность (ген или полипептид) регуляции экспрессии, секреции или функционирования целевой последовательности в соответствии с известной или требуемой активностью.
Вектор, используемый в описании настоящего изобретения, конкретно не ограничен, и можно использовать любой известный вектор. Примеры известных векторов могут включать природные или рекомбинантные плазмиды, космиды, вирусы и бактериофаги. Например, в качестве фагового или космидного вектора можно использовать pWE15, М13, MBL3, MBL4, IXII, ASHII, АРII, t10, t11, Charon4A и Charon21A и т.д., а в качестве плазмидного вектора можно использовать векторы на основе pBR, на основе pUC, на основе pBluescriptII, на основе pGEM, на основе pTZ, на основе pCL и на основе рЕТ. В частности, можно использовать векторы pDCM2, pACYC177, pACYC184, pCL, pECCG117, pUC19, pBR322, pMW118 и pCC1BAC. Более конкретно вектор, используемый в описании настоящего изобретения, может представлять собой pDCM2 (ФИГ. 1, SEQ ID NO: 37), сконструированный для вставки и замены гена на хромосоме Corynebacterium, но без ограничений, и можно использовать любой экспрессионный вектор.
Например, полинуклеотид, кодирующий целевой полипептид, может быть вставлен в хромосому с использованием вектора для вставки в хромосому клетки. Вставка полинуклеотида в хромосому может быть выполнена любым известным в данной области техники способом, например гомологичной рекомбинацией, без ограничений. Полинуклеотид может дополнительно включать селективный маркер для подтверждения вставки в хромосому. Селективный маркер используют для отбора клеток, трансформированных вектором, то есть для подтверждения вставки требуемой молекулы нуклеиновой кислоты, и примеры селективного маркера могут включать маркеры, обеспечивающие селективные фенотипы, такие как лекарственная устойчивость, потребность в питательном веществе, резистентность к цитотоксическим агентам или экспрессия поверхностного полипептида. В окружающей среде, обработанной селективным агентом, только клетки, экспрессирующие селективный маркер, способны выживать или проявлять различные фенотипы, и, таким образом, можно отобрать трансформированные клетки.
В другом аспекте описания настоящего изобретения предложен микроорганизм, включающий по меньшей мере одно из: варианта белка внутренней мембраны YjeH, включающего i) замену аминокислоты, соответствующей 92-му положению, аспарагином, ii) замену аминокислоты, соответствующей 351-му положению, лейцином, или iii) замену в обоих положениях i) и ii), в аминокислотной последовательности SEQ ID NO: 1, и обладающего гомологией 95% или более, но менее 100% с SEQ ID NO: 1; полинуклеотида, кодирующего этот вариант; и вектора, кодирующего этот полинуклеотид.
SEQ ID NO: 1, белок YjeH внутренней мембраны, его вариант, полинуклеотид, кодирующий этот вариант, и вектор, включающий этот полинуклеотид, являются такими, как описано выше.
Используемый в настоящем документе термин «микроорганизм, включающий вариант белка YjeH внутренней мембраны» относится к микроорганизму с усиленной способностью экспортировать О-ацетилгомосерин или гомосерин, полученному путем введения варианта белка YjeH внутренней мембраны в микроорганизм, обладающий слабой способностью экспортировать О-ацетилгомосерин или гомосерин. В частности, микроорганизм представляет собой микроорганизм, экспрессирующий вариант белка YjeH внутренней мембраны, включающий модификацию по меньшей мере одной аминокислоты в аминокислотной последовательности SEQ ID NO: 1, и модификация аминокислоты может включать замену 92й или 351й аминокислоты с N-конца другой аминокислотой.
В свете целей описания настоящего изобретения внеклеточный экспорт О-ацетилгомосерина или гомосерина микроорганизма, включающего по меньшей мере любое одно из варианта белка YjeH внутренней мембраны; полинуклеотида, кодирующего этот вариант; и вектора, включающего полинуклеотид, увеличивается, и, таким образом, этот микроорганизм отличается тем, что его способность продуцировать О-ацетилгомосерин или гомосерин усиливается. Хотя микроорганизмы дикого типа или немодифицированные микроорганизмы не могут экспортировать или могут продуцировать следовые количества О-ацетилгомосерина или гомосерина, способность продуцировать О-ацетилгомосерин или гомосерин может быть усилена в соответствии с описанием настоящего изобретения за счет увеличения экспортируемых количеств О-ацетилгомосерина или гомосерина путем введения по меньшей мере любого одного из варианта белка YjeH внутренней мембраны; полинуклеотида, кодирующего этот вариант; и вектора, включающего этот полинуклеотид.
Используемый в настоящем изобретении термин «немодифицированный микроорганизм» не исключает штаммы, включающие мутацию, которая может возникать в микроорганизмах естественных путем, и относится к штамму или микроорганизму дикого типа, не включающему вариант белка YjeH внутренней мембраны. «Немодифицированный микроорганизм» может использоваться взаимозаменяемо со «штаммом до модификации», «микроорганизмом до модификации», «немутированным штаммом», «немутированным микроорганизмом», «немодифицированный штаммом» или «микроорганизмом сравнения».
Используемый в настоящем документе термин «белок, который должен экспрессироваться/экспрессируется» означает состояние, в котором целевой белок введен в микроорганизм или модифицирован для экспрессии в микроорганизме. В том случае, если целевой белок присутствует в микроорганизме, активность этого белка можно усилить по сравнению с активностью эндогенного белка или белка до модификации. В свете объектов описания настоящего изобретения «целевой белок» может представлять собой вышеописанный вариант белка YjeH внутренней мембраны.
В частности, термин «введение белка» относится к обеспечению активности конкретного белка в микроорганизме, который не имеет этого белка, или к усилению активности белка по сравнению с собственной активностью белка или с его активностью до модификации. Например, введение белка может относиться к введению в хромосому полинуклеотида, кодирующего конкретный белок, или к введению в микроорганизм вектора, включающего полинуклеотид, кодирующий этот конкретный белок, в результате чего экспрессируется активность этого белка. В дополнение к этому «усиление активности» может означать, что активность конкретного белка микроорганизма усиливается по сравнению с собственной активностью или активностью до модификации. Термин «собственная активность» может относиться к активности конкретного белка, которой обладает родительский штамм до трансформации, когда микроорганизм трансформируют естественным или искусственным природным вариантом.
В частности, усиление активности по описанию настоящего изобретения может быть достигнуто по меньшей мере одним способом, выбранным из группы, состоящей из: способа увеличения числа копий гена, кодирующего вариант белка; способа введения мутации в регуляторную последовательность экспрессии гена, кодирующего вариант белка; способа замещения регуляторной последовательности экспрессии гена последовательностью, кодирующей вариант белка и обладающей более сильной активностью; способа замещения гена, кодирующего белок внутренней мембраны дикого типа на хромосоме, геном, кодирующим вариант белка; способа дополнительного введения мутации в ген, кодирующий вариант белка, для усиления активности варианта белка; и способа введения варианта белка, без ограничений.
В приведенном выше описании увеличение числа копий гена, хотя конкретных ограничений нет, можно выполнять в состоянии функциональной связи с вектором или в состоянии, встроенном в хромосому клетки-хозяина. В частности, увеличение числа копий гена можно выполнять путем введения вектора, с которым функционально связан полинуклеотид, кодирующий вариант белка по описанию настоящего изобретения, и который может реплицироваться и функционировать в клетке-хозяине независимо от клетки-хозяина. Альтернативно в клетку вводят вектор, с которым функционально связан полинуклеотид, способен встраивать полинуклеотид в хромосому клетки-хозяина. Встраивание полинуклеотида в хромосому может быть выполнено любым известным в данной области техники способом, например гомологичной рекомбинации.
Модификация экспрессионной регуляторной последовательности для повышения уровня экспрессии полинуклеотида может быть выполнена посредством, но без конкретных ограничений, индукции мутации на последовательности путем делеции, инсерции, неконсервативной замены, консервативной замены или любой их комбинации, чтобы дополнительно усилить активность регуляторной последовательности экспрессии или замены последовательностью, обладающей более сильной активностью. Регуляторная последовательность экспрессии может включать промотор, последовательность оператора, последовательность, кодирующую подходящий сайт связывания рибосомы, и последовательность для регуляции терминации транскрипции и трансляции.
Более сильный промотор, чем собственный промотор, может быть связан выше экспрессионной единицы полинуклеотида без ограничений. Примеры более сильного промотора, известные в данной области техники, могут включать промоторы CJ1-CJ7 (US 7662943 В2), промотор lac, промотор trp, промотор trc, промотор tac, промотор PR фага лямбда, промотор PL, промотор tet, промотор gapA, промотор SPL7, промотор SPL13 (sm3) (US 10584338 В2), промотор O2 (US 10273491 В2), промотор tkt и промотор уссА, без ограничений.
В дополнение к этому модификация полинуклеотидной последовательности на хромосоме может быть выполнена путем индукции мутации в регуляторной последовательности экспрессии путем делеции, инсерции, неконсервативной замены, консервативной замены или любой их комбинации, чтобы дополнительно усилить активность полинуклеотидной последовательности, или путем замещения нуклеотидной последовательности модифицированной нуклеотидной последовательностью, обладающей более сильной активностью, но без ограничений.
Введение белка и усиление активности может повысить активность или концентрацию белка по сравнению с активностью или концентрацией штамма дикого типа или немодифицированного штамма микроорганизма, без ограничений.
В описании настоящего изобретения микроорганизм, включающий вариант белка YjeH внутренней мембраны, может представлять собой рекомбинантный микроорганизм, полученный путем трансформации с использованием вектора, включающего полинуклеотид, кодирующий вариант YjeH белка внутренней мембраны, но без ограничений.
Используемый в настоящем документе термин «трансформация» относится к процессу введения в клетку-хозяина вектора, включающего полинуклеотид, кодирующий целевой белок, таким образом, чтобы кодируемый этим полинуклеотидом белок экспрессировался в клетке-хозяине. Трансформированный полинуклеотид может находиться либо в форме, встроенной в хромосому клетки-хозяина, либо в форме, локализованной вне хромосомы, и в любом случае белок экспрессируется в клетке-хозяине. В дополнение к этому полинуклеотид включает ДНК и/или РНК, кодирующую целевой белок. Полинуклеотид может быть введен в клетку-хозяина при условии, что полинуклеотид введен в клетку-хозяина и белок экспрессируется в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, которая представляет собой генную конструкцию, включающую все незаменимые элементы, требующиеся для самостоятельной экспрессии. Экспрессионная кассета может, как правило, включать промотор, сигнал терминации транскрипции, сайт связывания рибосомы и сигнал терминации трансляции, которые функционально связаны с полинуклеотидом. Экспрессионная кассета может находиться в форме самостоятельно реплицирующегося экспрессионного вектора. Полинуклеотид может быть также введен в клетку-хозяина в его первоначальной форме и функционально связан с последовательностью, требующейся для экспрессии в клетке-хозяине, без ограничений.
Типы микроорганизма, обладающего способностью к продукции гомосерина, конкретно не ограничены, если этот микроорганизм способен продуцировать О-ацетилгомосерин или гомосерин, и в частности микроорганизм может представлять собой любой микроорганизм, относящийся к роду Corynebacterium, роду Escherichia, роду Enterobacter, роду Erwinia, роду Serratia, роду Providencia и роду Brevibacterium, более конкретно микроорганизм, относящийся к роду Corynebacterium.
Более конкретно микроорганизм, относящийся к роду Corynebacterium, может представлять собой Corynebacterium glutamicum, Corynebacterium ammoniagenes, Corynebacterium crudilactis, Corynebacterium deserti, Corynebacterium efficiens, Corynebacterium callunae, Corynebacterium stationis, Corynebacterium singulare, Corynebacterium halotolerans, Corynebacterium striatum, Corynebacterium pollutisoli, Corynebacterium imitans, Corynebacterium testudinoris или Corynebacterium flavescens, a также можно использовать без ограничений любые микроорганизмы, относящиеся к роду Corynebacterium.
В описании настоящего изобретения родительский штамм микроорганизма может представлять собой микроорганизм, в котором ген, ослабляющий путь биосинтеза О-ацетилгомосерина или гомосерина, дополнительно инактивирован, или путь биосинтеза О-ацетилгомосерина или гомосерина усилен, чтобы повысить продукцию О-ацетилгомосерина или гомосерина, без ограничений.
В частности, чтобы инактивировать ген, ослабляющий путь биосинтеза О-ацетилгомосерина или гомосерина, может быть, например, ослаблена или инактивирована экспрессия гена metB (SEQ ID NO: 17), кодирующего цистатионин-гшилш-синтазу, вовлеченную в путь деградации О-ацетилгомосерина, или гена metY (SEQ ID NO: 22), кодирующего О-ацетилгомосерин(тиол)-лиазу, вовлеченную в путь деградации О-ацетилгомосерина.
Чтобы усилить путь биосинтеза О-ацетилгомосерина или гомосерина, например, можно ввести генетическую мутацию в ген lysC (SEQ ID NO: 27), кодирующий аспартокиназу, либо можно амплифицировать экспрессию гена metX (SEQ ID NO: 32), кодирующего О-ацетилгомосеринтрансферазу.
Тем не менее данное воплощение не ограничено этим, и способность к продукции О-ацетилгомосерина или гомосерина можно усилить любыми известными способами регуляции экспрессии гена.
Используемый в настоящем документе термин «усиление/повышение» представляет собой понятие, включающее повышение активности по сравнению с собственной активностью.
Усиление или повышение активности гена может быть достигнуто путем применения различных способов, хорошо известных в данной области техники. Примеры этих способов могут включать по меньшей мере один, выбранный из группы, состоящей из: способа увеличения числа копий гена; способа введения мутации в регуляторную последовательность экспрессии гена; способа замещения регуляторной последовательности экспрессии гена последовательностью, обладающей более сильной активностью; способа дополнительного введения мутации в ген с целью усиления активности гена; способа введения в микроорганизм чужеродного гена; и любой их комбинации, без ограничений.
Используемый в настоящем документе термин «инактивация» белка означает, что активность белка ослаблена по сравнению с собственной активностью, либо что белок не обладает активностью.
Инактивация белка может быть достигнута путем применения различных способов, хорошо известных в данной области техники. Примеры способов могут включать: способ делетирования части или полноразмерного гена, кодирующего белок на хромосоме, включающий удаление активности белка; способ замещения гена, кодирующего белок на хромосоме, мутантным геном для снижения ферментативной активности; способ индукции мутации в регуляторной последовательности экспрессии гена, кодирующего белок на хромосоме; способ замещения регуляторной последовательности экспрессии гена, кодирующего белок, последовательностью, обладающей более слабой активностью или не обладающей активностью (например, способ замещения промотора гена более слабым промотором, чем эндогенный промотор); способ делетирования части или полноразмерного гена, кодирующего белок на хромосоме; способ введения антисмыслового нуклеотида (например, антисенс-РНК), который ингибирует трансляцию с мРНК в белок посредством комплементарного связывания с транскриптом гена на хромосоме; способ создания невозможности присоединения рибосомы за счет формирования вторичной структуры путем искусственного присоединения комплементарной последовательности к последовательности Шайна - Дальгарно (ШД) выше ШД последовательности гена, кодирующего белок; и способ конструирования путем обратной транскрипции (RTE), который присоединяет промотор обратной транскрипции к 3' концу открытой рамки считывания (ОРС) соответствующей последовательности, и их комбинацию, но без конкретных ограничений.
В другом аспекте описания настоящего изобретения предложен способ получения целевого продукта, включающий культивирование микроорганизма в культуральной среде, где микроорганизм включает по меньшей мере одно из: варианта белка внутренней мембраны YjeH, включающего i) замену аминокислоты, соответствующей 92-му положению, аспарагином, ii) замену аминокислоты, соответствующей 351-му положению, лейцином, или iii) замену в обоих положениях i) и ii), в аминокислотной последовательности SEQ ID NO: 1, и обладающего гомологией 95% или более, но менее 100% с SEQ ID NO: 1; полинуклеотида, кодирующего этот вариант; и вектора, кодирующего этот полинуклеотид.
SEQ ID NO: 1, белок YjeH внутренней мембраны, его вариант, полинуклеотид, кодирующий этот вариант, вектор, включающий этот полинуклеотид, и включающий их микроорганизм являются такими, как описано выше.
Целевой продукт может представлять собой О-ацетилгомосерин или гомосерин, но без ограничений.
Используемый в настоящем документе термин «культивирование» относится к выращиванию микроорганизма в отрегулированной соответствующим образом окружающей среде. Процесс культивирования по описанию настоящего изобретения можно выполнять в соответствии с подходящей средой и условиями культивирования, известными в данной области техники. Процесс культивирования может быть легко отрегулирован для использования специалистом в данной области техники в соответствии с отбираемым штаммом. Культивирование микроорганизма может быть выполнено в периодическом процессе, непрерывном процессе, периодическом процессе с подпиткой и т.д. без ограничений.
Используемый в настоящем документе термин «культуральная среда» относится к субстанции, в которой смешаны питательные компоненты, требующиеся для культивирования микроорганизма, в качестве основных ингредиентов и дополнены питательными компонентами и факторами роста, а также водой, которые необходимы для выживания и роста. В частности, хотя культуральная среда и другие условия культивирования для выращивания микроорганизма по настоящему изобретению конкретно не ограничены, если культуральная среда широко используется при культивировании микроорганизмов; микроорганизм по описанию настоящего изобретения можно культивировать в обычной среде, содержащей подходящие источники углерода, источники азота, источники фосфора, неорганические соединения, аминокислоты и/или витамины, в аэробных условиях при регуляции температуры, рН и т.д.
В описании настоящего изобретения в качестве источников углерода можно использовать углеводы, такие как глюкоза, фруктоза, сахароза и мальтоза; сахарные спирты, такие как маннит и сорбит, органические кислоты, такие как пировиноградная кислота, молочная кислота и лимонная кислота; и аминокислоты, такие как глутаминовая кислота, метионин и лизин. В дополнение к этому можно использовать натуральные органические питательные компоненты, такие как гидролизаты крахмала, мелассы, сырые мелассы, рисовые отруби, маниок съедобный, тростниково-сахарная багасса и кукурузный сироп, и в частности углеводы, такие как глюкоза и предварительно стерильно обработанные мелассы (т.е. мелассы, преобразованные для снижения содержания сахаров), а также можно использовать подходящие количества любых других источников углерода без ограничений. Эти источники углерода можно использовать по отдельности или в виде комбинации по меньшей мере двух из них.
В качестве источников азота можно использовать неорганические источники азота, такие как аммиак, сульфат аммония, хлорид аммония, ацетат аммония, фосфат аммония, карбонат аммония и нитрат аммония; и органические источники азота, такие как аминокислоты, например глутаминовая кислота, метионин и глутамин, пептон, NZ-амин, мясной экстракт, дрожжевой экстракт, солодовый экстракт, кукурузный сироп, гидролизат казеина, рыба или продукты ее деградации, а также обезжиренный соевый жмых или продукты его деградации. Эти источники азота можно использовать по отдельности или в виде комбинации по меньшей мере двух из них.
В качестве источников фосфора можно использовать однозамещенный фосфат калия, двузамещенный фосфат калия или соответствующие им натрийсодержащие соли. В качестве неорганических соединений можно использовать хлорид натрия, хлорид кальция, хлорид железа, сульфат магния, сульфат железа, сульфат марганца, карбонат кальция и т.п. Дополнительно можно также включать аминокислоты, витамины и/или подходящий предшественник. Питательную среду и предшественники можно добавлять в культуральную среду в периодическом или непрерывном процессе без ограничений.
В описании настоящего изобретения в процессе культивирования микроорганизма к культуре можно добавлять соединения, такие как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота и серная кислота, для доведения рН питательной среды. В процессе культивирования для подавления образования пены можно также добавлять пеногаситель, такой как полигликоле вый эфир жирной кислоты. В дополнение к этому в культуральную среду можно вводить кислород или кислородсодержащий газ для поддержания культуральной среды в аэробном состоянии, а в культуры можно вводить газы азот, водород или диоксид углерода для поддержания анаэробных и микроаэробных условий без введения каких-либо других газов, но воплощение не ограничено этим.
Температуру культуральной среды можно поддерживать при 25°С - 50°С, более конкретно при 30°С - 37°С без ограничений. Культивирование можно продолжать до получения требуемого количества продукта, в частности в течение от 10 часов до 100 часов, без ограничений.
Целевой продукт, полученный в результате культивирования, может экспортироваться в культуральную среду или оставаться в клетках.
Способ может дополнительно включать выделение целевого продукта из культуральной среды или микроорганизма.
Выделение целевого продукта, полученного на стадии культивирования описания настоящего изобретения, может быть выполнено путем сбора целевого продукта из культуры с использованием любого известного способа, выбранного в соответствии со способом культивирования. Например, можно использовать центрифугирование, фильтрование, анионообменную хроматографию, кристаллизацию и высокоэффективную жидкостную хроматографию (ВЭЖХ), и целевая аминокислота может быть выделена из питательной среды или микроорганизма с использованием любого подходящего способа в данной области техники без ограничений. Культура может также называться культуральной жидкостью.
Стадия выделения может также включать процесс очистки, который может быть выполнен с использованием подходящего способа, хорошо известного в данной области техники. Таким образом, выделенный целевой продукт может быть очищен из ферментационного бульона микроорганизма, включающего целевой продукт (Introduction to Biotechnology and Genetic Engineering, A.J. Nair., 2008).
В другом аспекте описания настоящего изобретения предложен способ получения метионина, включающий: получение О-ацетилгомосерина путем культивирования микроорганизма в культуральной среде; и превращения О-ацетилгомосерина в метионин путем взаимодействия О-ацетилгомосерина с сульфидом, где микроорганизм включает по меньшей мере одно из: варианта белка внутренней мембраны YjeH, включающего i) замену аминокислоты, соответствующей 92-му положению, аспарагином, ii) замену аминокислоты, соответствующей 351-му положению, лейцином, или iii) замену в обоих положениях i) и ii), в аминокислотной последовательности SEQ ID NO: 1, и обладающего гомологией 95% или более, но менее 100% с SEQ ID NO: 1; полинуклеотида, кодирующего этот вариант; и вектора, кодирующего этот полинуклеотид.
SEQ ID NO: 1, белок YjeH внутренней мембраны, его вариант, полинуклеотид, кодирующий этот вариант, вектор, включающий этот полинуклеотид, включающий их микроорганизм и О-ацетилгомосерин являются такими, как описано выше.
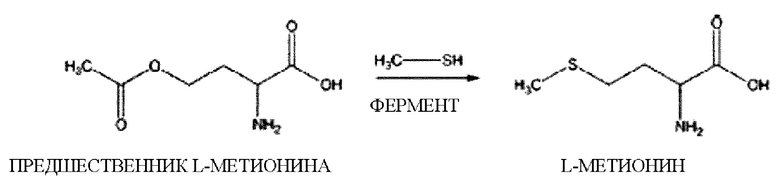
В описании настоящего изобретения О-ацетилгомосерин в качестве предшественника метионина может быть превращен в метионин посредством взаимодействия с сульфидом.
Взаимодействие с сульфидом относится к процессу образования L-метионина из О-сукцинилгомосерина с использованием любого известного способа. Например, L-метионин может быть получен путем взаимодействия метил меркаптана в качестве сульфида с О-ацетилгомосерином. В дополнение к этому можно добавлять катализатор или фермент или проводить реакцию в микроорганизме, включающем фермент, для повышения скорости реакции и повышения выхода.
«Сульфид» может представлять собой, например, метилмеркаптан, и метилмеркаптан может представлять собой любые производные метилмеркаптана, способные обеспечивать атомы серы, такие как диметилсульфид (ДМС), раскрытые в международной публикации WO 2010/098629), а также натрия метилмеркаптан (CH3S-Na) в жидкой фазе и метилмеркаптан (CH3SH) в газообразном или жидком состоянии.
В качестве фермента, используемого в реакции превращения метионина, можно использовать цистатионин-γ-синтазу, О-ацетилгомосерин-сульфгидрилазу или О-сукцинилгомосерин-сульфгидрилазу, которые известны в данной области техники.
Процесс реакции превращения метионина может быть выражен следующей схемой реакции:
О-ацетил-L-гомосерин+CH3SH↔L-метионин+ацетат
В этой реакции остаток CH3S метилмеркаптана замещается остатком ацетата О-ацетилгомосерина с получением L-метионина.
Эту реакцию можно проводить при температуре от 20°С до 45°С, при этом поддерживая рН реакционной смеси в диапазоне от 6 до 7, и можно добавлять пиридоксаль-5'-фосфат в качестве кофермента.
Превращение предшественника L-метионина в L-метионин в присутствии метилмеркаптана является ферментативной реакцией и раскрыто в международной публикации № WO 2008/013432. В международной публикации № WO 2008/013432 представлено подробное описание свойств и получения метионинпревращающих ферментов, обладающих соответствующей активностью для превращения предшественника L-метионина в L-метионин в присутствии метилмеркаптана. Такой подходящий метионинпревращающий фермент можно получить биотехнологическими методами, используя экспрессию гена.
Последовательность гена, кодирующего фермент, обладающий метионинпревращающей активностью, может быть получен, но без ограничений, из базы данных NCBI (США) и банка данных ДНК (KEGG, Япония). Для биологического превращения ген можно клонировать из полученной последовательности гена, а затем вводить в экспрессионный вектор. В частности, ген может экспрессироваться в активной форме с вектора pCL-CJI (CJ, Корея), который представляет собой экспрессионный вектор Е. coli в рекомбинантных штаммах, а экспрессированный белок может быть получен из раствора фермента, в котором лизируют клетки, обработанные ультразвуком. И штамм, экспрессирующий метионинпревращающий фермент, и экспрессированный метионинпревращающий фермент можно непосредственно использовать в реакции превращения метионина.
Для инициации реакции метионинпревращающий фермент, экспрессируемый с гена или из экспрессирующего его штамма микроорганизма, можно непосредственно, частично или полностью смешивать с супернатантом ферментации или с ферментационным бульоном, в котором накапливается L-метионин. Таким образом, реакцию можно инициировать путем добавления метионинпревращающего фермента или экспрессирующего его штамма микроорганизма в ферментационный раствор, в котором накапливается О-ацетилгомосерин, и добавления к нему метилмеркаптана.
В частности, О-ацетилгомосерин, который накапливается в ферментационном растворе, может превращаться в L-метионин под действием цистатионин-γ-синтазы, О-ацетилгомосеринсульфгидрилазы или О-сукцинилгомосеринсульфгидрилазы.
Реакция превращения в метионин может быть подтверждена с использованием DTNB (5,5-дитиобис(2-нитробензойная кислота), Sigma, США), и продукт реакции можно анализировать методом ВЭЖХ. В процессе реакции превращения в метионин можно дополнительно получить побочные продукты, такие как уксусная кислота, без отдельного процесса производства, посредством взаимодействия между метилмеркаптаном и О-ацетилгомосерином.
Ферментативная реакция между предшественником L-метионина и метилмеркаптаном может быть представлена следующей схемой реакции:
Способ получения метионина может легко определить обычный специалист в данной области техники при условии известных оптимальных условий культивирования и условий активации фермента. Конкретные условия и среды для культивирования являются такими, как определено выше.
В другом аспекте описания настоящего изобретения предложен способ получения глюфосината, включающий получение О-ацетилгомосерина или гомосерина путем культивирования микроорганизма в культуральной среде и превращения О-ацетилгомосерина или гомосерина в глюфосинат, где микроорганизм включает по меньшей мере одно из: варианта белка внутренней мембраны YjeH, включающего i) замену аминокислоты, соответствующей 92-му положению, аспарагином, ii) замену аминокислоты, соответствующей 351-му положению, лейцином, или iii) замену в обоих положениях i) и ii), в аминокислотной последовательности SEQ ID NO: 1, и обладающего гомологией 95% или более, но менее 100% с SEQ ID NO: 1; полинуклеотида, кодирующего этот вариант; и вектора, кодирующего этот полинуклеотид.
SEQ ID NO: 1, белок YjeH внутренней мембраны, его вариант, полинуклеотид, кодирующий этот вариант, вектор, включающий этот полинуклеотид, включающий их микроорганизм и гомосерин являются такими, как описано выше.
Используемый в настоящем документе термин «глюфосинат» относится к широкому ряду натуральных гербицидов, продуцируемых несколькими видами почвенных бактерий Streptomyces, и также известен под названием «фосфинотрицин».
В описании настоящего изобретения О-ацетилгомосерин или гомосерин можно превращать в глюфосинат посредством множества стадий, включая галогенирование.
Реакция превращения в глюфосинат раскрыта, например, в CN 108516991 А. В частности, реакция превращения в глюкосинат может включать: (1) стадию выполнения азеотропной дегидратации гомосерина (формула II) с использованием органического растворителя в присутствии кислотного катализатора; и стадию добавления галогенирующего агента к азеотропно дегидратированному гомосерину и выполнения галогенирования с получением L-3,6-бис(2-галогенэтил)-2,5-дионпиперазина (формула III); (2) стадию добавления диэфира или метилфосфитного диэфира метилфосфиновой кислоты к L-3,6-бис(2-галогеноэтил)-2,5-дионпиперазину и выполнения реакции Аббова (Abbov) с использованием растворителя с высокой температурой кипения в присутствии катализатора с получением соединения формулы IV; и (3) стадию выполнения реакции гидролиза путем растворения соединения формулы IV в кислоте и нагревания раствора, удаления растворителя после завершения реакции, добавления к нему спирта и выполнения реакции превращения путем добавления к ней алкиленоксида с получением глюфосината (формула I).
Реакция превращения в глюфосинат с использованием гомосерина в качестве предшественника может быть представлена с помощью следующей схемы:
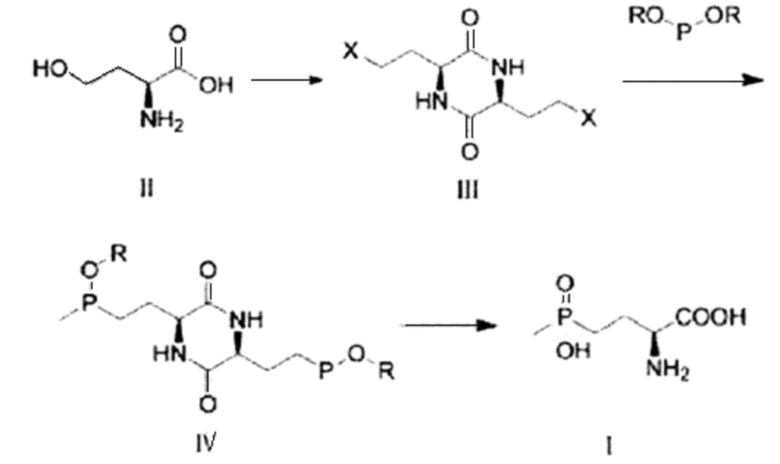
Способ получения глюфосината может быть выполнен с использованием гомосерина, полученного путем биологической ферментации, в качестве предшественника согласно описанию настоящего изобретения. Глюфосинат может быть получен посредством получения L-3,6-бис(2-галогенэтил)-2,5-дионпиперазина взаимодействием с метилфосфонием путем азеотропной дегидратации и галогенирования, выполнения реакции Аббова, а затем выполнения реакции гидролиза, как описано выше.
В другом аспекте описания настоящего изобретения предложена композиция для получения гомосерина, включающая микроорганизм или культуру микроорганизма в культуральной среде, где микроорганизм включает по меньшей мере одно из: варианта белка внутренней мембраны YjeH, включающего i) замену аминокислоты, соответствующей 92-му положению, аспарагином, ii) замену аминокислоты, соответствующей 351-му положению, лейцином, или iii) замену в обоих положениях i) и ii), в аминокислотной последовательности SEQ ID NO: 1, и обладающего гомологией 95% или более, но менее 100% с SEQ ID NO: 1; полинуклеотида, кодирующего этот вариант; и вектора, кодирующего этот полинуклеотид.
В другом аспекте описания настоящего изобретения предложена композиция для получения О-ацетилгомосерина, включающая микроорганизм или культуру микроорганизма в культуральной среде, где микроорганизм включает по меньшей мере одно из: варианта белка внутренней мембраны YjeH, включающего i) замену аминокислоты, соответствующей 92-му положению, аспарагином, ii) замену аминокислоты, соответствующей 351-му положению, лейцином, или iii) замену в обоих положениях i) и ii), в аминокислотной последовательности SEQ ID NO: 1, и обладающий гомологией 95% или более, но менее 100% с SEQ ID NO: 1; полинуклеотида, кодирующего этот вариант; и вектора, кодирующего этот полинуклеотид.
SEQ ID NO: 1, белок YjeH внутренней мембраны, его вариант, полинуклеотид, кодирующий этот вариант, вектор, включающий этот полинуклеотид, включающий их микроорганизм, гомосерин и О-ацетилгомосерин являются такими, как описано выше.
Композиция для получения О-ацетилгомосерина или гомосерина может относиться к композиции, используемой для продукции О-ацетилгомосерина или гомосерина вариантом белка YjeH внутренней мембраны по описанию настоящего изобретения. Эта композиция может включать вариант белка YjeH внутренней мембраны или компонент, способный выполнять функцию белка YjeH внутренней мембраны без ограничений. Вариант белка YjeH внутренней мембраны может находиться в форме вставки в вектор, способный к экспрессии функционально связанного с ним гена в клетке-хозяине.
Композиция может дополнительно включать криопротектор или эксципиент. Криопротектор или эксципиент может представлять собой природное или не встречающееся в природе вещество, но без ограничений. В другом примере криопротектор или эксципиент может представлять собой вещество, с которым микроорганизм не вступает в контакт в природе и которое в природе не включено в этот микроорганизм, но без ограничений.
В другом аспекте описания настоящего изобретения предложено применение варианта белка YjeH внутренней мембраны для продукции О-ацетилгомосерина или гомосерина.
В другом аспекте описания настоящего изобретения предложено применение микроорганизма для получения О-ацетилгомосерина или гомосерина, где микроорганизм включает по меньшей мере одно из: варианта белка внутренней мембраны YjeH, включающего i) замену аминокислоты, соответствующей 92-му положению, аспарагином, ii) замену аминокислоты, соответствующей 351-му положению, лейцином, или iii) замену в обоих положениях i) и ii), в аминокислотной последовательности SEQ ID NO: 1, и обладающего гомологией 95% или более, но менее 100% с SEQ ID NO: 1; полинуклеотида, кодирующего этот вариант; и вектора, кодирующего этот полинуклеотид.
Гомосерин, О-ацетилгомосерин, белок YjeH внутренней мембраны, его вариант, полинуклеотид, кодирующий этот вариант, вектор, включающий этот полинуклеотид, и включающий их микроорганизм являются такими, как описано выше.
Способ осуществления изобретения
Далее описание настоящего изобретения будет приведено более подробно со ссылкой на следующие примеры. Тем не менее следующие примеры представлены исключительно для иллюстрации описания настоящего изобретения, и объем описания настоящего изобретения не ограничен ими.
Пример 1: Конструирование плазмиды
Плазмида (pDCM2, ФИГ. 1, SEQ ID NO: 37) была разработана для встраивания в хромосому Corynebacterium и замещения гена на ней и синтезирована при использовании службы синтеза генов компании Bionics Co., Ltd. Плазмида была разработана таким образом, что содержала фермент рестрикции, эффективный для клонирования, со ссылкой на статью, относящуюся к системе sacB, общеизвестной в данной области техники (Gene, 145 (1994) 69-73). Синтезированная плазмида pDCM2 обладает следующими характеристиками.
1) В связи с тем, что точка начала репликации активна только в Е. coli, плазмида самостоятельно реплицируется только в Е. coli и не может самостоятельно реплицироваться в Corynebacterium.
2) Плазмида содержит ген устойчивости к канамицину в качестве селективного маркера.
3) Плазмида содержит ген левансукразы (sacB) в качестве вторичного положительного селективного маркера.
4) В окончательно полученном штамме не сохраняется генетической информации, имеющей происхождение из плазмиды pDCM2.
Пример 2: Конструирование штаммов с введенным вариантом экзогенного мембранного белка (YjeH) и оценка способности к продукции О-ацетилгомосерина и гомосерина
2-1. Конструирование штаммов с введенным экзогенным мембранным белком (YjeH) и вариантом YjeH
В целях оценки эффективности YjeH, который введен в Corynebacterium glutamicum АТСС13032 в качестве экзогенного мембранного белка и О-ацетилгомосерин-экспортирующего белка был получен вектор для встраивания в хромосому, включающий ген yjeH (SEQ ID NO: 2) кодирующий YjeH, имеющий происхождение из E. coli.
В частности, в целях конструирования вектора с делецией транспозазы были разработаны пара праймеров (SEQ ID NO: 4 и 5) для амплификации 5' вышележащей области и пара праймеров (SEQ ID NO: 6 и 7) для амплификации 3' нижележащей области по отношению к гену, кодирующему транспозазу (SEQ ID NO: 3, № гена NCg12335). Праймеры SEQ ID NO: 5 и 6 были разработаны так, что они пересекались друг с другом. В этой области была локализована последовательность фермента рестрикции SmaI. Последовательности праймеров представлены в таблице 1 ниже.

ПЦР выполняли с использованием хромосомы дикого типа (WT) АТСС13032 в качестве матрицы и праймеров SEQ ID NO: 4 и 5 и SEQ ID NO: 6 и 7. ПЦР выполняли в следующих условиях. После денатурации при 95°С в течение 5 минут повторяли 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и полимеризации при 72°С в течение 30 секунд, а затем выполняли полимеризацию при 72°С в течение 7 минут. В результате получили фрагменты ДНК: 851 п. о. 5' вышележащей области и 847 п. о. 3' нижележащей области по отношению к области делеции гена NCg12335.
ПЦР выполняли с использованием двух амплифицированных фрагментов ДНК в качестве матриц и праймеров SEQ ID NO: 4 и 7. ПЦР выполняли в следующих условиях. После денатурации при 95°С в течение 5 минут повторяли 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и полимеризации при 72°С в течение 90 секунд, а затем выполняли полимеризацию при 72°С в течение 7 минут. В результате были амплифицированы фрагмент ДНК 1648 п. о., включающий область, способную делетировать ген, кодирующий транспозазу (SEQ ID NO: 3, № ген NCg12335).
Полученные продукты ПЦР клонировали путем слияния, используя вектор pDCM2, обработанный ферментом рестрикции SmaI, и набор реактивов для клонирования In-Fusion® HD Cloning Kit (Clontech). E. coli DH5α трансформировали клонированным вектором, и трансформированные Е. coli высевали методом мазка на твердую среду Лурия-Бертани (LB), содержащую канамицин (25 мг/л). Колонии, трансформированные плазмидой со вставкой целевого гена, отбирали с помощью ПЦР и получили плазмиду с использованием метода экстракции плазмиды. Наконец, был сконструирован рекомбинантный вектор pDCM2-ΔNCg12335, включающий клонированную кассету делеции NCg12335.
В целях оценки эффективности О-ацетилхолин-экспортирующего белка был разработан вектор для вставки в хромосому, включающий ген (SEQ ID NO: 2), кодирующий YjeH, имеющий происхождение из Е. coli. С этой целью был сконструирован вектор, экспрессирующий ген yjeH, с использованием промотора CJ7 (SEQ ID NO: 8, US 7662943 В2). Были разработаны пара праймеров (SEQ ID NO: 9 и 10) для амплификации промоторной области CJ7 и пара праймеров (SEQ ID NO: 11 и 12) для амплицикации области yjeH Е. coli. Последовательности праймеров представлены в таблице 2 ниже.

ПЦР выполняли с использованием pECCG117-PCJ7-gfp (US 7662943 В2) в качестве матрицы и праймеров SEQ ID NO: 9 и 10, а также выполняли ПЦР с использованием хромосомы дикого типа Е. coli в качестве матрицы и праймеров SEQ ID NO: 11 и 12 соответственно. ПЦР выполняли в следующих условиях. После денатурации при 95°С в течение 5 минут повторяли 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и полимеризации при 72°С в течение 90 секунд, а затем выполняли полимеризации при 72°С в течение 7 минут. В результате получили фрагмент ДНК 360 п. о. промоторной области CJ7 и фрагмент ДНК 1297 п. о. гена yjeН E. coli.
ПЦР выполняли с использованием двух амплифицированных фрагментов ДНК в качестве матриц и праймеров SEQ ID NO: 9 и 12. ПЦР выполняли в следующих условиях. После денатурации при 95°С в течение 5 минут повторяли 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и полимеризации при 72°С в течение 90 секунд, а затем выполняли полимеризацию при 72°С в течение 7 минут. В результате были амплицифированы фрагмент ДНК 1614 п.о., включающий область, в которой находились вставки промотора CJ7 и гена. yjeH.
Фрагмент ДНК с делецией гена, полученный с помощью ПЦР, клонировали в векторе pDCM2-ΔNCg12335, обработанном ферментом рестрикции SmaI, с использованием набора реактивов In-Fusion® HD Cloning Kit (Clontech). Был окончательно сконструирован рекомбинантный вектор pDCM2-ΔNCg12335::PCJ7-yjeH (eco, WT).
В частности, фенилаланин (Phe), который представляет собой 92-ю аминокислоту аминокислотной последовательности YjeH, был заменен аспарагином (Asn) (F92N), используя плазмиду pDCM2-ΔNCg12335::PCJ7-yjeH (eco, WT) в качестве матрицы и праймеры SEQ ID NO: 13 и 14. Сконструированная плазмида, включающая ген, кодирующий вариант YjeH (F92N), была названа pDCM2-ΔNCg12335::PCJ7-yjeH (eco, F92N).
Фенилаланин (Phe), который представляет собой 351-ю аминокислоту аминокислотной последовательности YjeH, был заменен лейцином (F351L) с использованием плазмиды pDCM2-ΔNCg12335::PCJ7-yjeH (eco, WT) в качестве матрицы и праймеров SEQ ID NO: 15 и 16. Сконструированная плазмида, включающая ген, кодирующий вариант YjeH (F351L), была названа pDCM2-ΔNCg12335::PCJ7-yjeH (eco, F351L). Последовательности праймеров представлены в таблице 3 ниже.
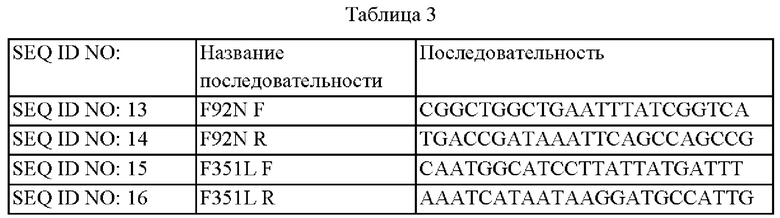
Штамм АТСС13032 трансформировали каждой из конструкций pDCM2-ΔNCg12335, pDCM2-ΔNCg12335::PCJ7-yjeH (eco, WT), pDCM2-ΔNCg12335::PCJ7-yjeH (eco, F92N) и pDCM2-ΔNCg12335::PCJ7-yjeH (eco, F351L) с использованием электроимпульсного метода и подвергали процессу вторичного кроссинговера с получением АТСС13032 ΔNCg12335, АТСС13032 ΔNCg12335::PCJ7-yjeH (eco, WT), ATCC13032 ΔNCg12335::PCJ7-yjeH (eco, F92N) и ATCC13032 ΔNCg12335::PCJ7-yjeH (eco, F351L), в которых ген NCg12335 был делетирован из хромосомы. Вставку инактивированного гена NCg12335 и вновь введенного гена Е. coli yjeH идентифицировали путем выполнения ПЦР с использованием праймеров SEQ ID NO: 4 и 7 и сравнения результатов с результатами для АТСС13032, в котором ген NCg12335 не был инактивирован.
2-2. Оценка способности к продукции О-ацетилгомосерина
В целях сравнения О-ацетилгомосерин (О-АН)-продуцирующей способности АТСС13032 ΔNCg12335, АТСС13032 ΔNCg12335::PCJ7-yjeH (eco, WT), ATCC13032 ΔNCg12335::PCJ7-yjeH (eco, F92N) и ATCC13032 ΔNCg12335::PCJ7-yjeH (eco, F351L), которые были сконструированы в примере 2-1, и штамма дикого типа АТСС13032 анализировали О-ацетилгомосерин, содержащийся в культурах, следующим методом.
Одну микробиологическую петлю для инокуляции каждого из штаммов инокулировали в колбу с угловыми перегородками вместимостью 250 мл, содержащую 25 мл описанной ниже среды, а затем культивировали при встряхивании при 33°С в течение 20 часов при 200 об/мин. Концентрации О-ацетилгомосерина анализировали с использованием ВЭЖХ, и анализируемые концентрации показаны в таблице 4.
Среда для продукции О-ацетилгомосерина (рН 7.2)
30 г глюкозы, 2 г KН2РО4, 3 г мочевины, 40 г (NH4)2SO4, 2,5 г пептона, 5 г (10 мл) кукурузного сиропа (CSL, Sigma), 0,5 г MgSO4⋅7H2O и 20 г СаСО3 (в пересчете на 1 л дистиллированной воды)
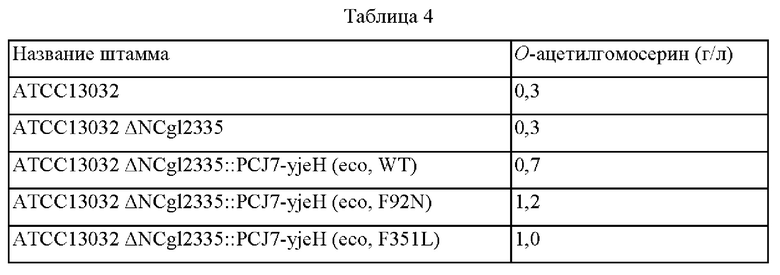
В результате было подтверждено, что при культивировании АТСС13032 в качестве контроля О-ацетил-L-гомосерин накапливался в концентрации 0,3 г/л, и, как показано в таблице 4, делеция гена NCg12335 (транспозаза) не влияла на продукцию О-ацетил-L-гомосерина. В частности, было подтверждено, что при экспрессии гена yjeH дикого типа О-ацетил-L-гомосерин накапливался в концентрации 0,7 г/л, а при экспрессии вариантов гена yjeH О-ацетил-L-гомосерин накапливался в концентрациях 1,2 г/ли 1,0 г/л.
Таким образом, на основании представленных выше результатов было подтверждено, что гены дикого типа и вариант yjeH по описанию настоящего изобретения приводят к значимому повышению продукции представляющей интерес аминокислоты в результате экспорта О-ацетилгомосерина из АТСС13032.
Штаммы АТСС13032 ΔNCg12335::PCJ7-yjeH (eco, F92N) и АТСС13032 ΔNCg12335::PCJ7-yjeH (eco, F351L) были названы Corynebacterium glutamicum СМ04-0651 и Corynebacterium glutamicum CM04-0652 соответственно и депонированы в Корейском центре культур микроорганизмов согласно Будапештскому договору 4 декабря 2019 г. под номерами доступа KССМ12633Р и KССМ12634Р соответственно.
2-3. Оценка способности к продукции гомосерина
В целях сравнения гомосерин-продуцирующей способности АТСС13032 ΔNCg12335, АТСС13032 ΔNCg12335::PCJ7-yjeH (eco, WT), АТСС13032 ΔNCg12335::PCJ7-yjeH (eco, F92N) и ATCC13032 ΔNCg12335::PCJ7-yjeH (eco, F351L), которые были сконструированы в примере 2-1, и штамма дикого типа АТСС13032 анализировали гомосерин, содержащийся в культурах, следующим методом.
Одну микробиологическую петлю для инокуляции каждого из штаммов инокулировали в колбу с угловыми перегородками вместимостью 250 мл, содержащую 25 мл описанной ниже среды, а затем культивировали при встряхивании при 33°С в течение 20 часов при 200 об/мин. Концентрации гомосерина анализировали с использованием ВЭЖХ, и анализируемые концентрации показаны в таблице 5.
Среда для продукции гомосерина (рН 7.2)
30 г глюкозы, 2 г KН2РО4, 3 г мочевины, 40 г (NH4)2SO4, 2,5 г пептона, 5 г (10 мл) кукурузного сиропа (CSL, Sigma), 0,5 г MgSO4⋅7H2O и 20 г СаСО3 (в пересчете на 1 л дистиллированной воды)
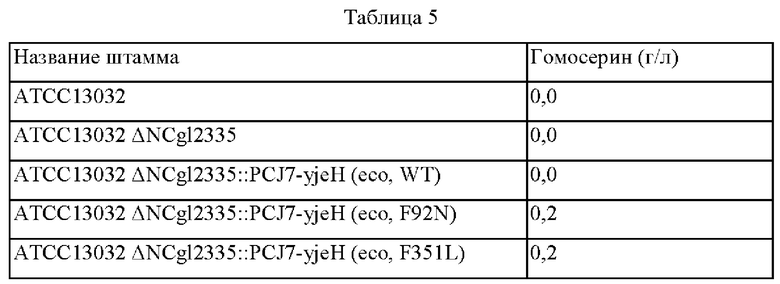
В результате было подтверждено, что при культивировании АТСС13032 в качестве контроля гомосерин не накапливался (концентрация 0,0 г/л), и, как показано в таблице 5, делеция гена NCg12335 (транспозаза) не влияла на продукцию гомосерина. В частности, было подтверждено, что при экспрессии гена yjеН дикого типа гомосерин не накапливался (концентрация 0,0 г/л), а при экспрессии вариантов гена yjeH гомосерин накапливался в концентрациях 0,2 г/л и 0,2 г/л.
Таким образом, на основании представленных выше результатов было подтверждено, что вариант тепа. yjeH по описанию настоящего изобретения приводит к значимому повышению продукции представляющей интерес аминокислоты в результате экспорта гомосерина из АТСС13032.
Пример 3: Конструирование штамма с усиленной способностью к продукции О-ацетилгомосерина и оценка способности к продукции О-ацетилгомосерина и гомосерина
3-1. Инактивация цистатионин-гамма (γ)-синтазы (MetB)
С помощью ПЦР с использованием хромосомной ДНК Corynebacterium glutamicum АТСС13032 в качестве матрицы был получен ген metB, кодирующий цистатионин-γ-синтазу, вовлеченную в путь деградации О-ацетилгомосерина. Информация о нуклеотидной последовательности гена metB (NCBI № Ncg12360, SEQ ID NO: 17) была получена из GenBank Национальных институтов здравоохранения (США), и на ее основе были синтезированы праймеры (SEQ ID NO: 18 и 19), включающие N-конец гена metB и линкер, и праймеры (SEQ ID NO: 20 и 21), включающие его С-конец и линкер. Последовательности праймеров представлены в таблице 6 ниже.

ПЦР выполняли с использованием хромосомной ДНК АТСС13032 в качестве матрицы и праймеров SEQ ID NO: 18 и 19 и SEQ ID NO: 20 и 21. В качестве полимеразы использовали ДНК полимеразу высокой надежности PfuUltra™ (Stratagene) и выполняли ПЦР путем повторения 30 циклов денатурации при 96°С в течение 30 секунд, отжига при 53°С в течение 30 секунд и полимеризации при 72°С в течение 1 минуты. В результате получили: амплифицированный ген 558 п. о., содержащий N-конец гена metB и линкер, и амплифицированный ген 527 п. о., содержащий С-конец гена metB и линкер, соответственно.
ПЦР выполняли с использованием двух амплифицированных генов, полученных, как описано выше, в качестве матриц путем повторения 10 циклов денатурации при 96°С в течение 60 секунд, отжига при 50°С в течение 60 секунд и полимеризации при 72°С в течение 1 минуты, а затем добавления праймеров SEQ ID NO: 18 и 21 с последующей полимеризацией 20 раз. В результате была получена кассета инактивации 1064 п. о., включающая N-конец-линкер-С-конец гена metB. Полученные продукты ПЦР обрабатывали ферментами рестрикции XbaI и SalI и лигировали с вектором pDCM2, обработанным ферментами рестрикции XbaI и SaiI, для клонирования с получением конечной конструкции рекомбинантного вектора pDCM2-ΔmetB с клонированной кассетой инактивации metB.
АТСС13032 трансформировали сконструированным вектором pDCM2-ΔmetB с использованием электроимпульсного метода и подвергали процессу вторичного кроссинговера с получением АТСС13032 ΔmetB, в котором ген metB был инактивирован в хромосоме. Инактивированный ген metB был, в конечном счете, подтвержден путем сравнения с АТСС13032, в котором ген metB не был инактивирован, после выполнения ПЦР с использованием праймеров SEQ ID NO: 18 и 21.
3-2. Инактивация O-ацетилгомосерин(тнол)-лиазы (MetY)
С помощью ПЦР с использованием хромосомной ДНК Corynebacterium glutamicum АТСС13032 в качестве матрицы был получен ген metY, кодирующий О-ацетилгомосерин(тиол)-лиазу, вовлеченную в путь деградации О-ацетилгомосерина. Информация о нуклеотидной последовательности гена metY (NCBI № Ncg10625, SEQ ID NO: 22) была получена из GenBank Национальных институтов здравоохранения (США), и на ее основе были синтезированы праймеры (SEQ ID NO: 23 и 24), включающие N-конец гена metY и линкер, и праймеры (SEQ ID NO: 25 и 26), включающие его С-конец и линкер. Последовательности праймеров представлены в таблице 7 ниже.

ПЦР выполняли с использованием хромосомной ДНК АТСС13032 в качестве матрицы и праймеров SEQ ID NO: 23 и 24 и SEQ ID NO: 25 и 26. В качестве полимеразы использовали ДНК полимеразу высокой надежности PfuUltra™ (Stratagene) и выполняли ПЦР путем повторения 30 циклов денатурации при 96°С в течение 30 секунд, отжига при 53°С в течение 30 секунд и полимеризации при 72°С в течение 1 минуты. В результате получили: амплифицированный ген 548 п.о., содержащий N-конец гена metY и линкер, и амплифицированный ген 550 п.о., содержащий С-конец гена metY и линкер, соответственно. ПЦР выполняли с использованием двух амплифицированных генов, полученных, как описано выше, в качестве матриц путем повторения 10 циклов денатурации при 96°С в течение 60 секунд, отжига при 50°С в течение 60 секунд и полимеризации при 72°С в течение 1 минуты, а затем добавления праймеров SEQ ID NO: 23 и 26 с последующей полимеризацией 20 раз. В результате была получена кассета инактивации 1077 п.о., включающая N-конец линкер С-конец гена metY. Полученные продукты ПЦР обрабатывали ферментами рестрикции XbaI и SalI и лигировали с вектором pDCM2, обработанным ферментами рестрикции XbaI и SalI, для клонирования с получением конечной конструкции рекомбинантного вектора pDCM2-ΔmetY с клонированной кассетой инактивации metY.
Штамм АТСС13032 ΔmetB трансформировали сконструированным вектором pDCM2-ΔmetY с использованием электроимпульсного метода и подвергали процессу вторичного кроссинговера с получением АТСС13032 ΔmetB ΔmetY, в котором ген metY инактивирован в хромосоме. Инактивированный ген metY был, в конечном счете, подтвержден путем сравнения с АТСС13032, в котором ген metY не был инактивирован, после выполнения ПЦР с использованием праймеров SEQ ID NO: 23 и 26.
3-3. Введение варианта аспартокиназы (LysC)
В целях максимального повышения продукции О-ацетилгомосерина в полученный из Corynebacterium glutamicum АТСС13032 ген lysC (SEQ ID NO: 27), кодирующий аспартокиназу, вводили мутацию (L377K) для усиления экспрессии гена lysC и ослабления ингибирования L-лизином и L-треонином по механизму обратной связи (US 10662450 В2). В целях получения вектора, включающего вариант гена lysC, была разработана пара праймеров (SEQ ID NO: 28 и 29) для амплификации 5' вышележащей области и пара праймеров (SEQ ID NO: 30 и 31) для амплификации 3' нижележащей области по отношению к положению мутации. Вставку областей ферментов рестрикции XbaI и SalI вводили с обоих концов праймеров SEQ ID NO: 28 и 31, а праймеры SEQ ID NO: 29 и 30 были разработаны так, что они пересекались друг с другом, и нуклеотидная замена была локализована в этой области. Последовательности праймеров являются такими, как представлено в таблице 8 ниже.
ПЦР выполняли с использованием Corynebacterium glutamicum АТСС13032 в качестве матрицы и праймеров SEQ ID NO: 28 и 29 и SEQ ID NO: 30 и 31. ПЦР выполняли в следующих условиях. После денатурации при 95°С в течение 5 минут повторяли 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и полимеризации при 72°С в течение 30 секунд, а затем выполняли полимеризацию при 72°С в течение 7 минут. В результате получили фрагмент ДНК 512 п.о. 5' вышележащей области и фрагмент ДНК 522 п.о. 3' нижележащей области по отношению к мутации гена lysC.
ПЦР выполняли с использованием двух амплифицированных фрагментов ДНК в качестве матриц и праймеров SEQ ID NO: 28 и 31. ПЦР выполняли в следующих условиях. После денатурации при 95°С в течение 5 минут повторяли 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и полимеризации при 72°С в течение 60 секунд, а затем выполняли полимеризацию при 72°С в течение 7 минут. В результате амплифицировали фрагмент ДНК 1011 п.о., включающий вариант гена lysC (L377K), кодирующий вариант аспартокиназы, в котором лейцин в 377-м положении был заменен лизином.
Фрагмент lysC (L377K), полученный с помощью ПЦР, обрабатывали ферментами рестрикции XbaI и SalI на обоих концах и лигировали с вектором pDCM2, обработанным ферментами рестрикции XbaI и SalI, после чего его клонировали и окончательно получили рекомбинантный вектор pDCM2-lysC (L377K) с клонированной кассетой замены lysC (L377K).
Штамм АТСС13032 ΔmetB ΔmetY трансформировали полученным вектором pDCM2-lysC (L377K) электроимпульсным методом и подвергали процессу вторичного кроссинговера с получением Corynebacterium glutamicum АТСС13032 ΔmetB ΔmetY lysC (L377K) с введенным вариантом гена lysC на хромосоме. Вариант гена был окончательно идентифицирован с помощью ПЦР, выполненной с использованием праймеров SEQ ID NO: 28 и 31, а затем сравнивали последовательность с последовательностью гена lysC дикого типа методом секвенирования.
3-4. Конструирование плазмиды с введением О-ацетилгомосеринтрансферазы (MetX)
В целях амплификации гена, кодирующего О-ацетилгомосеринтрансферазу (MetX), была получена информация о нуклеотидной последовательности гена metX (NCBI № NCg10624, SEQ ID NO: 32) из GenBank Национальных институтов здравоохранения (США), фермент рестрикции BamHI был вставлен в оба конца каждого из праймеров (SEQ ID NO: 33 и 34)), основанных на этой последовательности, для амплификации области от промотора (локализованного примерно на 300 п.о. выше стартового кодона) до терминатора (локализованного примерно на 100 п.о. ниже стоп-кодона. ПЦР выполняли в следующих условиях. После денатурации при 95°С в течение 5 минут повторяли 30 циклов денатурации при 95°С в течение 30 секунд, отжига при 55°С в течение 30 секунд и полимеризации при 72°С в течение 90 секунд, а затем выполняли полимеризацию при 72°С в течение 7 минут. В результате получили фрагмент ДНК 1546 п. о. кодирующей области гена metX. Вектор pECCG117 (KR 10-0057684 В1) и фрагмент ДНК metX обрабатывали ферментом рестрикции BamHI, лигировали с помощью ДНК лигазы и клонировали с получением плазмиды, названной pECCG117-metX WT. Последовательности праймеров являются такими, как представлено в таблице 9 ниже.
3-5. Конструирование штамма-продуцента О-ацетилгомосерина и оценка способности к продукции О-ацетилгомосерина
После получения вектора pECCG117-metX WT в примере 3-4 его вводили в Corynebacterium glutamicum АТСС13032 ΔmetB ΔmetY lysC (L377K), используя электроимпульсный метод, штамм высевали методом мазка на селективной среде, содержащей канамицин (25 мг/л), и культивировали до получения трансформантов.
Для сравнения О-ацетилгомосерин-продуцирующей способности между штаммами, эти штаммы культивировали описанным ниже способом и анализировали О-ацетилгомосерин, содержащийся в культурах.
Одну микробиологическую петлю для инокуляции каждого из штаммов инокулировали в колбу с угловыми перегородками вместимостью 250 мл, содержащую 25 мл описанной ниже среды, а затем культивировали при встряхивании при 37°С в течение 20 часов при 200 об/мин. Концентрации О-ацетилгомосерина анализировали с использованием ВЭЖХ, и анализируемые концентрации показаны в таблице 10.
Среда для продукции О-ацетилгомосерина (рН 7.2)
30 г глюкозы, 2 г KН2РО4, 3 г мочевины, 40 г (NH4)2SO4, 2,5 г пептона, 5 г (10 мл) кукурузного сиропа (CSL, Sigma), 0,5 г MgSO4⋅7H2O, 400 мг метионина и 20 г СаСО3 (в пересчете на 1 л дистиллированной воды)

В результате было подтверждено, что при культивировании АТСС13032 ΔmetB ΔmetY lysC (L377K) в качестве контроля О-ацетил-L-гомосерин накапливался в концентрации 0,7 г/л, а при культивировании штамма АТСС13032 ΔmetB ΔmetY lysC (L377K)/pECCG117-metX WT О-ацетил-L-гомосерин накапливался в концентрации 1,3 г/л, как показано в таблице 10.
Таким образом, повышение продукции О-ацетил-L-гомосерина в штамме АТСС13032 ΔmetB ΔmetY lysC (L377K)/pECCG117-metX WT было подтверждено на основании представленных выше результатов.
3-6. Оценка способности к продукции гомосерина
В Corynebacterium glutamicum АТСС13032 ΔmetB ΔmetY lysC (L377K), сконструированный в примере 3-3, вводили вектор pECCG117-metX WT, сконструированный в примере 3-4, используя электроимпульсный метод, высевали методом мазка на селективную среду, содержащую (25 мг/л) канамицина и культивировали до получения трансформантов.
Для сравнения гомосерин-продуцирующей способности между штаммами эти штаммы культивировали описанным ниже способом и анализировали гомосерин, содержащийся в культурах.
Одну микробиологическую петлю для инокуляции каждого из штаммов инокулировали в колбу с угловыми перегородками вместимостью 250 мл, содержащую 25 мл описанной ниже среды, а затем культивировали при встряхивании при 37°С в течение 20 часов при 200 об/мин. Концентрации гомосерина анализировали с использованием ВЭЖХ, и анализируемые концентрации показаны в таблице 11.
Среда для продукции гомосерина (рН 7.2)
30 г глюкозы, 2 г KН2РО4, 3 г мочевины, 40 г (NH4)2SO4, 2,5 г пептона, 5 г (10 мл) кукурузного сиропа (CSL, Sigma), 0,5 г MgSO4⋅7H2O, 400 мг метионина и 20 г СаСО3 (в пересчете на 1 л дистиллированной воды)

В результате было подтверждено, что при культивировании АТСС13032 ΔmetB ΔmetY lysC (L377K) в качестве контроля гомосерин накапливался в концентрации 0,2 г/л, а при культивировании штамма АТСС13032 ΔmetB ΔmetY lysC (L377K)/pECCG117-metX WT гомосерин накапливался в концентрации 0,2 г/л, как показано в таблице 11.
Таким образом, на основании этих результатов было подтверждено, что штамм АТСС13032 ΔmetB ΔmetY lysC (L377K)/pECCG117-metX WT характеризовался таким же уровнем продукции гомосерина.
Пример 4: Конструирование штаммов с введенным вариантом экзогенного мембранного белка (YjeH), обладающего усиленной способностью к продукции О-ацетилгомосерина, и оценка способности к продукции О-ацетилгомосерина
4-1. Введение варианта экзогенного белка (YjeH)
Штамм Corynebacterium glutamicum АТСС13032 ΔmetB ΔmetY lysC (L377K), сконструированный в примере 3-3, трансформировали векторами pDCM2-ΔNCg12335, pDCM2-ΔNCg12335::PCJ7-yjeH (eco, WT), pDCM2-ΔNCg12335::PCJ7-yjeH (eco, F92N) и pDCM2-ΔNCg12335::PCJ7-yjeH (eco, F351L), ранее сконструированными в примере 2-1, и подвергали процессу вторичного кроссинговера с получением АТСС13032 ΔmetB ΔmetY lysC (L377K) ΔNCg12335, ATCC13032 ΔmetB ΔmetY lysC (L377K) ΔNCg12335::PCJ7-yjeH (eco, WT), ATCC13032 ΔmetB ΔmetY lysC (L377K) ΔNCg12335::PCJ7-yjeH (eco, F92N) и АТСС13032 ΔmetB ΔmetY lysC (L377K) ΔNCg12335::PCJ7-yjeH (eco, F351L), из хромосомы которых был удален ген NCg12335 (транспозаза). Встраивание инактивированного гена NCg12335 и вновь введенного гена yjeH Е. coli идентифицировали путем выполнения ПЦР с использованием праймеров SEQ ID NO: 4 и 7 и сравнения результатов с результатами для АТСС13032, в котором ген NCg12335 не был инактивирован.
4-2. Конструирование штамма-продуцента О-ацетилгомосерина и оценка способности к продукции О-ацетилгомосерина
После получения вектора pECCG117-metX WT в примере 3-4 его вводили в АТСС13032 ΔmetB ΔmetY lysC (L377K) ΔNCg12335, ATCC13032 ΔmetB ΔmetY lysC (L377K) ΔNCg12335::PCJ7-yjeH (eco, WT), ATCC13032 ΔmetB ΔmetY lysC (L377K) ΔNCg12335::PCJ7-yjeH (eco, F92N) и АТСС13032 ΔmetB ΔmetY lysC (L377K) ΔNCg12335::PCJ7-yjeH (eco, F351L), полученные в примере 4-1, используя электроимпульсный метод, штаммы высевали методом мазка на селективную среду, содержащую (25 мг/л) канамицина и культивировали до получения трансформантов.
Для сравнения продуцирующей способности О-ацетилгомосерина между штаммами эти штаммы культивировали описанным ниже способом и анализировали О-ацетилгомосерин, содержащийся в культурах.
Одну микробиологическую петлю для инокуляции каждого из штаммов инокулировали в колбу с угловыми перегородками вместимостью 250 мл, содержащую 25 мл среды для продуцирования О-ацетилгомосерина (рН 7,2) примера 3-5, а затем культивировали при встряхивании при 33°С в течение 20 часов при 200 об/мин. Концентрации О-ацетилгомосерина анализировали с использованием ВЭЖХ, и анализируемые концентрации показаны в таблице 12.


В результате было подтверждено, что при культивировании Corynebacterium glutamicum АТСС13032 ΔmetB ΔmetY lysC (L377K) в качестве контроля О-ацетил-L-гомосерин накапливался в концентрации 1,3 г/л, и, как показано в таблице 12, делеция гена NCg12335 (транспозаза) не влияла на продукцию О-ацетил-L-гомосерина. В частности, было подтверждено, что при экспрессии гена yjeH дикого типа О-ацетил-L-гомосерин накапливался в концентрации 2,1 г/л, а при экспрессии вариантов тепа. yjeH О-ацетил-L-гомосерин накапливался в концентрациях 2,7 г/л и 2,3 г/л.
Таким образом, на основании представленных выше результатов было подтверждено, что гены дикого типа и вариант yjeH по описанию настоящего изобретения приводят к значимому повышению продукции представляющей интерес аминокислоты в результате экспорта О-ацетил-L-гомосерина из Corynebacterium glutamicum АТСС13032.
4-3. Оценка способности к продукции гомосерина
После получения вектора pECCG117-metX WT в примере 3-4 его вводили в АТСС13032 ΔmetB ΔmetY lysC (L377K) ΔNCg12335, ATCC13032 ΔmetB ΔmetY lysC (L377K) ΔNCg12335::PCJ7-yjeH (eco, WT), ATCC13032 ΔmetB ΔmetY lysC (L377K) ΔNCg12335::PCJ7-yjeH (eco, F92N) и АТСС13032 ΔmetB ΔmetY lysC (L377K) ΔNCg12335::PCJ7-yjeH (eco, F351L), полученные в примере 4-1, используя электроимпульсный метод, штаммы высевали методом мазка на селективную среду, содержащую (25 мг/л) канамицина и культивировали до получения трансформантов.
Для сравнения гомосерин-продуцирующей способности между штаммами эти штаммы культивировали описанным ниже способом и анализировали гомосерин, содержащийся в культурах.
Одну микробиологическую петлю для инокуляции каждого из штаммов инокулировали в колбу с угловыми перегородками вместимостью 250 мл, содержащую 25 мл описанной ниже среды, а затем культивировали при встряхивании при 33°С в течение 20 часов при 200 об/мин. Концентрации гомосерина анализировали с использованием ВЭЖХ, и анализируемые концентрации показаны в таблице 13.
Среда для продукции гомосерина (рН 7.2)
30 г глюкозы, 2 г KН2РО4, 3 г мочевины, 40 г (NH4)2SO4, 2,5 г пептона, 5 г (10 мл) кукурузного сиропа (CSL, Sigma), 0,5 г MgSO4⋅7H2O, 400 мг метионина и 20 г СаСО3 (в пересчете на 1 л дистиллированной воды)
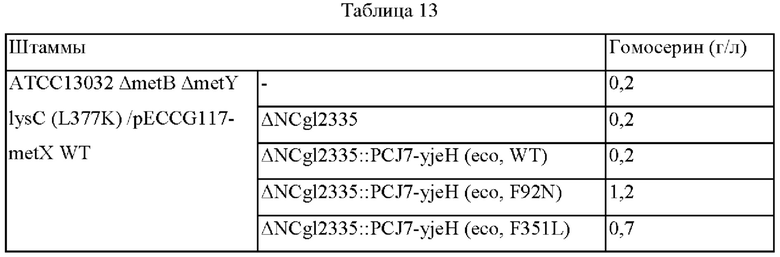
В результате было подтверждено, что при культивировании Corynebacterium glutamicum АТСС13032 ΔmetB ΔmetY lysC (L377K) в качестве контроля гомосерин накапливался в концентрации 0,2 г/л, и, как показано в таблице 13, делеция гена NCg12335 (транспозаза) не влияла на продукцию гомосерина. В частности, было подтверждено, что при экспрессии гена yjeH дикого типа гомосерин накапливался в концентрации 0,2 г/л, а при экспрессии вариантов гена уjеН гомосерин накапливался в концентрациях 1,2 г/л и 0,7 г/л.
Таким образом, на основании представленных выше результатов было подтверждено, что гены дикого типа и вариант yjeH по описанию настоящего изобретения приводят к значимому повышению продукции представляющей интерес аминокислоты в результате экспорта гомосерина из Corynebacterium glutamicum АТСС13032.
Пример 5: Получение метионина (МЕТ) с использованием реакции превращения О-ацетилгомосерина, продуцируемого штаммом
После получения вектора pECCG117-metX WT в примере 3-4 его вводили в АТСС13032 ΔmetB ΔmetY lysC (L377K) ΔNCg12335, ATCC13032 ΔmetB ΔmetY lysC (L377K) ΔNCg12335::PCJ7-yjeH (eco, WT), ATCC13032 ΔmetB ΔmetY lysC (L377K) ΔNCg12335::PCJ7-yjeH (eco, F92N) и АТСС13032 ΔmetB ΔmetY lysC (L377K) ΔNCg12335::PCJ7-yjeH (eco, F351L), полученные в примере 4-1, используя электроимпульсный метод, штаммы высевали методом мазка на селективную среду, содержащую (25 мг/л) канамицина и культивировали до получения транс фор мантов.
В соответствии со способом, раскрытом в поданном ранее патентном документе (WO 2008/013432), был также клонирован ген metZ, кодирующий О-сукцинилгомосеринсульфгидрилазу, имеющий происхождение из Pseudomonas aeruginosa и Pseudomonas putida, который должен использоваться для превращения О-ацетилгомосерина в метионин. ПЦР выполняли с использованием хромосомы каждого штамма в качестве матрицы и праймеров для получения гена metZ из Pseudomonas aeruginosa и Pseudomonas putida, как описано ниже: 30 циклов денатурации при 94°С в течение 30 секунд, отжига при 55°С в течение 30 секунд, удлинения при 72°С в течение 2 минут с получением фрагментов ДНК. Полученные фрагменты ДНК расщепляли NdeI/PacI и клонировали в векторе pCL-CJ1 (CJ, Корея), расщепленном теми же ферментами. Клетки Е. coli W3110 трансформировали клонированным вектором и культивировали на чашке Петри со средой LB, содержащей 50 мкг/л спектиномицина, а затем отбирали колонии. Отобранными колониями инокулировали 3 мл среды LB, содержащей 50 мкг/л спектиномицина, и культивировали в течение ночи при 37°С. Клетки выделяли из культуры, промывали 0,1 М буферным раствором фосфата калия (рН 7,5), суспендировали в 200 мкл буферного раствора фосфата калия и лизировали путем обработки ультразвуком 5 раз через 30-секундные интервалы. Клеточный лизат центрифугировали при 12 000 об/мин в течение 10 мин, и в полученном супернатанте определяли количественно общий уровень белка с использованием раствора Bio-Rad для количественного определения белков (Bio-Rad, США). Экспрессию белка идентифицировали методом электрофореза в полиакриламидном геле с додецилсульфатом натрия (ДСН-ПААГ). Супернатант, полученный из клеточного экстракта, использовали для реакции ферментативного превращения.
Также был клонирован ген metZ, кодирующий О-сукцинилгомосеринсульфгидрилазу, имеющий происхождение от Hyphomonas neptunium. ПЦР выполняли, используя хромосому Hyphomonas neptunium в качестве матрицы и праймер таким же образом, как описано выше для получения фрагментов ДНК. Полученные фрагменты ДНК расщепляли NdeI/AvrII и клонировали в векторе pCL-CJ1 (СJ, Корея), расщепленном теми же ферментами. Супернатант, полученный с использованием клонированного вектора, использовали таким же образом, как описано выше, и использовали в реакции ферментативного превращения.
Каждым из штаммов W3110, трансформированных экспрессионным вектором metZ, имеющего происхождение из Pseudomonas, и трансформированных экспрессионным вектором metZ, имеющего происхождение из Hyphomonas, инокулировали чашку с питательной средой LB, содержащей спектиномицин в качестве антибиотика, и культивировали в течение ночи при температуре от 30°С до 40°С. Затем отдельной колонией инокулировали 40 мл среды LB, содержащей спектиномицин, и культивировали в течение 5 часов при температуре от 30°С до 40°С. Штаммы, экспрессирующие metZ происхождения из Pseudomonas и metZ происхождения из Hypomononas, культивировали в емкости для ферментации вместимостью 1 л при 600-900 об/мин и температуре от 30°С до 40°С в течение 15-30 часов с получением ферментационных бульонов. Питательная среда, используемая при культивировании, описана ниже. Клетки полученного ферментационного бульона лизировали путем обработки ультразвуком для получения растворов ферментов.
Среда 2XYT
10 г дрожжевого экстракта, 16 г триптофана, 40 г глюкозы и 50 г спектиномицина (в пересчете на 1 л дистиллированной воды)
Реакцию превращения в метионин выполняли путем добавления раствора фермента О-сукцинилгомосеринсульфгидрилазы происхождения из Pseudomonas и Hyphomonas к культуре трансформанта, продуцирующего О-ацетилгомосерин. После добавления 0,1 л раствора фермента к 2,0 л культуры О-ацетилгомосерина, из которой не были удалены клетки, добавляли 0,3 л 15% раствора натрия метилмеркаптана для инициации реакции. Через 2 часа ферментационный бульон извлекали, удаляли из него клетки и идентифицировали концентрации полученного метионина методом ВЭЖХ. Анализируемые концентрации представлены в таблице 14 ниже.

Приведенное выше описание настоящего изобретения представлено с целью иллюстрации, и специалистам в данной области техники должно быть понятно, что могут быть выполнены различные изменения и модификации без изменения технической концепции и существенных признаков настоящего изобретения. Таким образом, ясно, что описанные выше воплощения являются иллюстративными во всех аспектах и не ограничивают настоящее изобретение. Таким образом, объем данного описания определяется не подробным описанием, а формулой изобретения и его эквивалентами, а также всеми вариантами в рамках объема формулы изобретения, и его эквиваленты следует трактовать как включенные в объем описания.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CJ CheilJedang Corporation
<120> Модифицированный белок внутренней мембраны и способы получения
целевого продукта с его использованием
<130> OPA20205
<150> KR 10-2019-0172055
<151> 2019-12-20
<160> 37
<170> KoPatentIn 3.0
<210> 1
<211> 418
<212> PRT
<213> Неизвестно
<220>
<223> Escherichia coli YjeH
<400> 1
Met Ser Gly Leu Lys Gln Glu Leu Gly Leu Ala Gln Gly Ile Gly Leu
1 5 10 15
Leu Ser Thr Ser Leu Leu Gly Thr Gly Val Phe Ala Val Pro Ala Leu
20 25 30
Ala Ala Leu Val Ala Gly Asn Asn Ser Leu Trp Ala Trp Pro Val Leu
35 40 45
Ile Ile Leu Val Phe Pro Ile Ala Ile Val Phe Ala Ile Leu Gly Arg
50 55 60
His Tyr Pro Ser Ala Gly Gly Val Ala His Phe Val Gly Met Ala Phe
65 70 75 80
Gly Ser Arg Leu Glu Arg Val Thr Gly Trp Leu Phe Leu Ser Val Ile
85 90 95
Pro Val Gly Leu Pro Ala Ala Leu Gln Ile Ala Ala Gly Phe Gly Gln
100 105 110
Ala Met Phe Gly Trp His Ser Trp Gln Leu Leu Leu Ala Glu Leu Gly
115 120 125
Thr Leu Ala Leu Val Trp Tyr Ile Gly Thr Arg Gly Ala Ser Ser Ser
130 135 140
Ala Asn Leu Gln Thr Val Ile Ala Gly Leu Ile Val Ala Leu Ile Val
145 150 155 160
Ala Ile Trp Trp Ala Gly Asp Ile Lys Pro Ala Asn Ile Pro Phe Pro
165 170 175
Ala Pro Gly Asn Ile Glu Leu Thr Gly Leu Phe Ala Ala Leu Ser Val
180 185 190
Met Phe Trp Cys Phe Val Gly Leu Glu Ala Phe Ala His Leu Ala Ser
195 200 205
Glu Phe Lys Asn Pro Glu Arg Asp Phe Pro Arg Ala Leu Met Ile Gly
210 215 220
Leu Leu Leu Ala Gly Leu Val Tyr Trp Gly Cys Thr Val Val Val Leu
225 230 235 240
His Phe Asp Ala Tyr Gly Glu Lys Met Ala Ala Ala Ala Ser Leu Pro
245 250 255
Lys Ile Val Val Gln Leu Phe Gly Val Gly Ala Leu Trp Ile Ala Cys
260 265 270
Val Ile Gly Tyr Leu Ala Cys Phe Ala Ser Leu Asn Ile Tyr Ile Gln
275 280 285
Ser Phe Ala Arg Leu Val Trp Ser Gln Ala Gln His Asn Pro Asp His
290 295 300
Tyr Leu Ala Arg Leu Ser Ser Arg His Ile Pro Asn Asn Ala Leu Asn
305 310 315 320
Ala Val Leu Gly Cys Cys Val Val Ser Thr Leu Val Ile His Ala Leu
325 330 335
Glu Ile Asn Leu Asp Ala Leu Ile Ile Tyr Ala Asn Gly Ile Phe Ile
340 345 350
Met Ile Tyr Leu Leu Cys Met Leu Ala Gly Cys Lys Leu Leu Gln Gly
355 360 365
Arg Tyr Arg Leu Leu Ala Val Val Gly Gly Leu Leu Cys Val Leu Leu
370 375 380
Leu Ala Met Val Gly Trp Lys Ser Leu Tyr Ala Leu Ile Met Leu Ala
385 390 395 400
Gly Leu Trp Leu Leu Leu Pro Lys Arg Lys Thr Pro Glu Asn Gly Ile
405 410 415
Thr Thr
<210> 2
<211> 1257
<212> ДНК
<213> Неизвестно
<220>
<223> Escherichia coli yjeH
<400> 2
atgagtggac tcaaacaaga actggggctg gcccagggca ttggcctgct atcgacgtca 60
ttattaggca ctggcgtgtt tgccgttcct gcgttagctg cgctggtagc gggcaataac 120
agcctgtggg cgtggcccgt tttgattatc ttagtgttcc cgattgcgat tgtgtttgcg 180
attctgggtc gccactatcc cagcgcaggc ggcgtcgcgc acttcgtcgg tatggcgttt 240
ggttcgcggc ttgagcgagt caccggctgg ctgtttttat cggtcattcc cgtgggtttg 300
cctgccgcac tacaaattgc cgccgggttc ggccaggcga tgtttggctg gcatagctgg 360
caactgttgt tggcagaact cggtacgctg gcgctggtgt ggtatatcgg tactcgcggt 420
gccagttcca gtgctaatct acaaaccgtt attgccggac ttatcgtcgc gctgattgtc 480
gctatctggt gggcgggcga tatcaaacct gcgaatatcc cctttccggc acctggtaat 540
atcgaactta ccgggttatt tgctgcgtta tcagtgatgt tctggtgttt tgtcggtctg 600
gaggcatttg cccatctcgc ctcggaattt aaaaatccag agcgtgattt tcctcgtgct 660
ttgatgattg gtctgctgct ggcaggatta gtctactggg gctgtacggt agtcgtctta 720
cacttcgacg cctatggtga aaaaatggcg gcggcagcat cgcttccaaa aattgtagtg 780
cagttgttcg gtgtaggagc gttatggatt gcctgcgtga ttggctatct ggcctgcttt 840
gccagtctca acatttatat acagagcttc gcccgcctgg tctggtcgca ggcgcaacat 900
aatcctgacc actacctggc acgcctctct tctcgccata tcccgaataa tgccctcaat 960
gcggtgctcg gctgctgtgt ggtgagcact ttggtgattc atgctttaga gatcaatctg 1020
gacgctctta ttatttatgc caatggcatc tttattatga tttatctgtt atgcatgctg 1080
gcaggctgta aattattgca aggacgttat cgactactgg cggtggttgg cgggctgtta 1140
tgcgttctgt tactggcaat ggtcggctgg aaaagtctct atgcgctgat catgctggcg 1200
gggttatggc tgttgctgcc aaaacgaaaa acgccggaaa atggcataac cacataa 1257
<210> 3
<211> 267
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Tn2335
<400> 3
gtggcctaca ccttcgacca cgtcgtcgcc tggcgctggt gcaccaaaga agacgcctac 60
aactacaccc acctcttcga tcaactccaa ccacccttaa tcgtgaccac cgacggacaa 120
aaaaggcgca ctcaaagcca tcaccacgac ctggccgaca acgaaaatcc aacgctgcct 180
cgtccacgtc aaacgcaacg tccaaaaaca cgtcacccta agacccgtgc tgagctcgcc 240
gaaaagcact ccggggtctc tccttga 267
<210> 4
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Tn_5 F
<400> 4
tgaattcgag ctcggtaccc caccgacgcg catctgcct 39
<210> 5
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Tn_5 R
<400> 5
ggtgtggtga ctttcagcag ttcccggggg ggaggaggca tgtggtgttg 50
<210> 6
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Tn_3 F
<400> 6
caacaccaca tgcctcctcc cccccgggaa ctgctgaaag tcaccacacc 50
<210> 7
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Tn_3 R
<400> 7
gtcgactcta gaggatcccc ctcccaaacc attgaggaat gg 42
<210> 8
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CJ7
<400> 8
agaaacatcc cagcgctact aatagggagc gttgaccttc cttccacgga ccggtaatcg 60
gagtgcctaa aaccgcatgc ggcttaggct ccaagatagg ttctgcgcgg ccgggtaatg 120
catcttcttt agcaacaagt tgaggggtag gtgcaaataa gaacgacata gaaatcgtct 180
cctttctgtt tttaatcaac atacaccacc acctaaaaat tccccgacca gcaagttcac 240
agtattcggg cacaatatcg ttgccaaaat attgtttcgg aatatcatgg gatacgtacc 300
caacgaaagg aaacactc 318
<210> 9
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CJ7_yjeH F
<400> 9
acaccacatg cctcctcccc agaaacatcc cagcgctac 39
<210> 10
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CJ7_yjeH R
<400> 10
agttcttgtt tgagtccact catagtgttt cctttcgttg ggt 43
<210> 11
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> yjeH F
<400> 11
acccaacgaa aggaaacact atgagtggac tcaaacaaga actg 44
<210> 12
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> yjeH R
<400> 12
gactttcagc agttcccggg ttatgtggtt atgccatttt ccgg 44
<210> 13
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> F92N F
<400> 13
cggctggctg aatttatcgg tca 23
<210> 14
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> F92N R
<400> 14
tgaccgataa attcagccag ccg 23
<210> 15
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> F351L F
<400> 15
caatggcatc cttattatga ttt 23
<210> 16
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> F351L R
<400> 16
aaatcataat aaggatgcca ttg 23
<210> 17
<211> 1161
<212> ДНК
<213> Неизвестно
<220>
<223> Corynebacterium glutamicum metB
<400> 17
ttgtcttttg acccaaacac ccagggtttc tccactgcat cgattcacgc tgggtatgag 60
ccagacgact actacggttc gattaacacc ccaatctatg cctccaccac cttcgcgcag 120
aacgctccaa acgaactgcg caaaggctac gagtacaccc gtgtgggcaa ccccaccatc 180
gtggcattag agcagaccgt cgcagcactc gaaggcgcaa agtatggccg cgcattctcc 240
tccggcatgg ctgcaaccga catcctgttc cgcatcatcc tcaagccggg cgatcacatc 300
gtcctcggca acgatgctta cggcggaacc taccgcctga tcgacaccgt attcaccgca 360
tggggcgtcg aatacaccgt tgttgatacc tccgtcgtgg aagaggtcaa ggcagcgatc 420
aaggacaaca ccaagctgat ctgggtggaa accccaacca acccagcact tggcatcacc 480
gacatcgaag cagtagcaaa gctcaccgaa ggcaccaacg ccaagctggt tgttgacaac 540
accttcgcat ccccatacct gcagcagcca ctaaaactcg gcgcacacgc agtcctgcac 600
tccaccacca agtacatcgg aggacactcc gacgttgttg gcggccttgt ggttaccaac 660
gaccaggaaa tggacgaaga actgctgttc atgcagggcg gcatcggacc gatcccatca 720
gttttcgatg catacctgac cgcccgtggc ctcaagaccc ttgcagtgcg catggatcgc 780
cactgcgaca acgcagaaaa gatcgcggaa ttcctggact cccgcccaga ggtctccacc 840
gtgctctacc caggtctgaa gaaccaccca ggccacgaag tcgcagcgaa gcagatgaag 900
cgcttcggcg gcatgatctc cgtccgtttc gcaggcggcg aagaagcagc taagaagttc 960
tgtacctcca ccaaactgat ctgtctggcc gagtccctcg gtggcgtgga atccctcctg 1020
gagcacccag caaccatgac ccaccagtca gctgccggct ctcagctcga ggttccccgc 1080
gacctcgtgc gcatctccat tggtattgaa gacattgaag acctgctcgc agatgtcgag 1140
caggccctca ataaccttta g 1161
<210> 18
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> metB_N_del F
<400> 18
tctagacgcc cgcatactgg cttc 24
<210> 19
<211> 37
<212> ДНК
<213> Искусственная последовательность
<220>
<223> metB_N_del R
<400> 19
cccatccact aaacttaaac agatgtgatc gcccggc 37
<210> 20
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> metB_C_del F
<400> 20
tgtttaagtt tagtggatgg ggaagaacca cccaggcc 38
<210> 21
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> metB_C_del R
<400> 21
gtcgaccaat cgtccagagg gcg 23
<210> 22
<211> 1314
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Corynebacterium glutamicum metY
<400> 22
atgccaaagt acgacaattc caatgctgac cagtggggct ttgaaacccg ctccattcac 60
gcaggccagt cagtagacgc acagaccagc gcacgaaacc ttccgatcta ccaatccacc 120
gctttcgtgt tcgactccgc tgagcacgcc aagcagcgtt tcgcacttga ggatctaggc 180
cctgtttact cccgcctcac caacccaacc gttgaggctt tggaaaaccg catcgcttcc 240
ctcgaaggtg gcgtccacgc tgtagcgttc tcctccggac aggccgcaac caccaacgcc 300
attttgaacc tggcaggagc gggcgaccac atcgtcacct ccccacgcct ctacggtggc 360
accgagactc tattccttat cactcttaac cgcctgggta tcgatgtttc cttcgtggaa 420
aaccccgacg accctgagtc ctggcaggca gccgttcagc caaacaccaa agcattcttc 480
ggcgagactt tcgccaaccc acaggcagac gtcctggata ttcctgcggt ggctgaagtt 540
gcgcaccgca acagcgttcc actgatcatc gacaacacca tcgctaccgc agcgctcgtg 600
cgcccgctcg agctcggcgc agacgttgtc gtcgcttccc tcaccaagtt ctacaccggc 660
aacggctccg gactgggcgg cgtgcttatc gacggcggaa agttcgattg gactgtcgaa 720
aaggatggaa agccagtatt cccctacttc gtcactccag atgctgctta ccacggattg 780
aagtacgcag accttggtgc accagccttc ggcctcaagg ttcgcgttgg ccttctacgc 840
gacaccggct ccaccctctc cgcattcaac gcatgggctg cagtccaggg catcgacacc 900
ctttccctgc gcctggagcg ccacaacgaa aacgccatca aggttgcaga attcctcaac 960
aaccacgaga aggtggaaaa ggttaacttc gcaggcctga aggattcccc ttggtacgca 1020
accaaggaaa agcttggcct gaagtacacc ggctccgttc tcaccttcga gatcaagggc 1080
ggcaaggatg aggcttgggc atttatcgac gccctgaagc tacactccaa ccttgcaaac 1140
atcggcgatg ttcgctccct cgttgttcac ccagcaacca ccacccattc acagtccgac 1200
gaagctggcc tggcacgcgc gggcgttacc cagtccaccg tccgcctgtc cgttggcatc 1260
gagaccattg atgatatcat cgctgacctc gaaggcggct ttgctgcaat ctag 1314
<210> 23
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> metY_N_del F
<400> 23
tctagaccat cctgcaccat ttag 24
<210> 24
<211> 37
<212> ДНК
<213> Искусственная последовательность
<220>
<223> metY_N_del R
<400> 24
cccatccact aaacttaaac acgctcctgc caggttc 37
<210> 25
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> metY_C_del F
<400> 25
tgtttaagtt tagtggatgg gcttggtacg caaccaagg 39
<210> 26
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> metY_C_del R
<400> 26
gtcgacgatt gctccggctt cgg 23
<210> 27
<211> 1266
<212> ДНК
<213> Неизвестно
<220>
<223> Corynebacterium glutamicum lysC
<400> 27
gtggccctgg tcgtacagaa atatggcggt tcctcgcttg agagtgcgga acgcattaga 60
aacgtcgctg aacggatcgt tgccaccaag aaggctggaa atgatgtcgt ggttgtctgc 120
tccgcaatgg gagacaccac ggatgaactt ctagaacttg cagcggcagt gaatcccgtt 180
ccgccagctc gtgaaatgga tatgctcctg actgctggtg agcgtatttc taacgctctc 240
gtcgccatgg ctattgagtc ccttggcgca gaagcccaat ctttcacggg ctctcaggct 300
ggtgtgctca ccaccgagcg ccacggaaac gcacgcattg ttgatgtcac tccaggtcgt 360
gtgcgtgaag cactcgatga gggcaagatc tgcattgttg ctggtttcca gggtgttaat 420
aaagaaaccc gcgatgtcac cacgttgggt cgtggtggtt ctgacaccac tgcagttgcg 480
ttggcagctg ctttgaacgc tgatgtgtgt gagatttact cggacgttga cggtgtgtat 540
accgctgacc cgcgcatcgt tcctaatgca cagaagctgg aaaagctcag cttcgaagaa 600
atgctggaac ttgctgctgt tggctccaag attttggtgc tgcgcagtgt tgaatacgct 660
cgtgcattca atgtgccact tcgcgtacgc tcgtcttata gtaatgatcc cggcactttg 720
attgccggct ctatggagga tattcctgtg gaagaagcag tccttaccgg tgtcgcaacc 780
gacaagtccg aagccaaagt aaccgttctg ggtatttccg ataagccagg cgaggctgcg 840
aaggttttcc gtgcgttggc tgatgcagaa atcaacattg acatggttct gcagaacgtc 900
tcttctgtag aagacggcac caccgacatc accttcacct gccctcgttc cgacggccgc 960
cgcgcgatgg agatcttgaa gaagcttcag gttcagggca actggaccaa tgtgctttac 1020
gacgaccagg tcggcaaagt ctccctcgtg ggtgctggca tgaagtctca cccaggtgtt 1080
accgcagagt tcatggaagc tctgcgcgat gtcaacgtga acatcgaatt gatttccacc 1140
tctgagattc gtatttccgt gctgatccgt gaagatgatc tggatgctgc tgcacgtgca 1200
ttgcatgagc agttccagct gggcggcgaa gacgaagccg tcgtttatgc aggcaccgga 1260
cgctaa 1266
<210> 28
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> lysC_L377K_5 F
<400> 28
tcctctagag ctgcgcagtg ttgaatacg 29
<210> 29
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> lysC_L377K_5 R
<400> 29
aggtggaaat cttttcgatg ttc 23
<210> 30
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> lysC_L377K_3 F
<400> 30
gaacatcgaa aagatttcca cct 23
<210> 31
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> lysC_L377K_3 R
<400> 31
gactctagag ttcacctcag agacgatta 29
<210> 32
<211> 1134
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Corynebacterium glutamicum metX
<400> 32
atgcccaccc tcgcgccttc aggtcaactt gaaatccaag cgatcggtga tgtctccacc 60
gaagccggag caatcattac aaacgctgaa atcgcctatc accgctgggg tgaataccgc 120
gtagataaag aaggacgcag caatgtcgtt ctcatcgaac acgccctcac tggagattcc 180
aacgcagccg attggtgggc tgacttgctc ggtcccggca aagccatcaa cactgatatt 240
tactgcgtga tctgtaccaa cgtcatcggt ggttgcaacg gttccaccgg acctggctcc 300
atgcatccag atggaaattt ctggggtaat cgcttccccg ccacgtccat tcgtgatcag 360
gtaaacgccg aaaaacaatt cctcgacgca ctcggcatca ccacggtcgc cgcagtactt 420
ggtggttcca tgggtggtgc ccgcacccta gagtgggccg caatgtaccc agaaactgtt 480
ggcgcagctg ctgttcttgc agtttctgca cgcgccagcg cctggcaaat cggcattcaa 540
tccgcccaaa ttaaggcgat tgaaaacgac caccactggc acgaaggcaa ctactacgaa 600
tccggctgca acccagccac cggactcggc gccgcccgac gcatcgccca cctcacctac 660
cgtggcgaac tagaaatcga cgaacgcttc ggcaccaaag cccaaaagaa cgaaaaccca 720
ctcggtccct accgcaagcc cgaccagcgc ttcgccgtgg aatcctactt ggactaccaa 780
gcagacaagc tagtacagcg tttcgacgcc ggctcctacg tcttgctcac cgacgccctc 840
aaccgccacg acattggtcg cgaccgcgga ggcctcaaca aggcactcga atccatcaaa 900
gttccagtcc ttgtcgcagg cgtagatacc gatattttgt acccctacca ccagcaagaa 960
cacctctcca gaaacctggg aaatctactg gcaatggcaa aaatcgtatc ccctgtcggc 1020
cacgatgctt tcctcaccga aagccgccaa atggatcgca tcgtgaggaa cttcttcagc 1080
ctcatctccc cagacgaaga caacccttcg acctacatcg agttctacat ctaa 1134
<210> 33
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> metX F
<400> 33
ggatcccctc gttgttcacc cagcaacc 28
<210> 34
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> metX R
<400> 34
ggatcccaaa gtcacaacta cttatgttag 30
<210> 35
<211> 418
<212> PRT
<213> Искусственная последовательность
<220>
<223> YjeH F92N
<400> 35
Met Ser Gly Leu Lys Gln Glu Leu Gly Leu Ala Gln Gly Ile Gly Leu
1 5 10 15
Leu Ser Thr Ser Leu Leu Gly Thr Gly Val Phe Ala Val Pro Ala Leu
20 25 30
Ala Ala Leu Val Ala Gly Asn Asn Ser Leu Trp Ala Trp Pro Val Leu
35 40 45
Ile Ile Leu Val Phe Pro Ile Ala Ile Val Phe Ala Ile Leu Gly Arg
50 55 60
His Tyr Pro Ser Ala Gly Gly Val Ala His Phe Val Gly Met Ala Phe
65 70 75 80
Gly Ser Arg Leu Glu Arg Val Thr Gly Trp Leu Asn Leu Ser Val Ile
85 90 95
Pro Val Gly Leu Pro Ala Ala Leu Gln Ile Ala Ala Gly Phe Gly Gln
100 105 110
Ala Met Phe Gly Trp His Ser Trp Gln Leu Leu Leu Ala Glu Leu Gly
115 120 125
Thr Leu Ala Leu Val Trp Tyr Ile Gly Thr Arg Gly Ala Ser Ser Ser
130 135 140
Ala Asn Leu Gln Thr Val Ile Ala Gly Leu Ile Val Ala Leu Ile Val
145 150 155 160
Ala Ile Trp Trp Ala Gly Asp Ile Lys Pro Ala Asn Ile Pro Phe Pro
165 170 175
Ala Pro Gly Asn Ile Glu Leu Thr Gly Leu Phe Ala Ala Leu Ser Val
180 185 190
Met Phe Trp Cys Phe Val Gly Leu Glu Ala Phe Ala His Leu Ala Ser
195 200 205
Glu Phe Lys Asn Pro Glu Arg Asp Phe Pro Arg Ala Leu Met Ile Gly
210 215 220
Leu Leu Leu Ala Gly Leu Val Tyr Trp Gly Cys Thr Val Val Val Leu
225 230 235 240
His Phe Asp Ala Tyr Gly Glu Lys Met Ala Ala Ala Ala Ser Leu Pro
245 250 255
Lys Ile Val Val Gln Leu Phe Gly Val Gly Ala Leu Trp Ile Ala Cys
260 265 270
Val Ile Gly Tyr Leu Ala Cys Phe Ala Ser Leu Asn Ile Tyr Ile Gln
275 280 285
Ser Phe Ala Arg Leu Val Trp Ser Gln Ala Gln His Asn Pro Asp His
290 295 300
Tyr Leu Ala Arg Leu Ser Ser Arg His Ile Pro Asn Asn Ala Leu Asn
305 310 315 320
Ala Val Leu Gly Cys Cys Val Val Ser Thr Leu Val Ile His Ala Leu
325 330 335
Glu Ile Asn Leu Asp Ala Leu Ile Ile Tyr Ala Asn Gly Ile Phe Ile
340 345 350
Met Ile Tyr Leu Leu Cys Met Leu Ala Gly Cys Lys Leu Leu Gln Gly
355 360 365
Arg Tyr Arg Leu Leu Ala Val Val Gly Gly Leu Leu Cys Val Leu Leu
370 375 380
Leu Ala Met Val Gly Trp Lys Ser Leu Tyr Ala Leu Ile Met Leu Ala
385 390 395 400
Gly Leu Trp Leu Leu Leu Pro Lys Arg Lys Thr Pro Glu Asn Gly Ile
405 410 415
Thr Thr
<210> 36
<211> 418
<212> PRT
<213> Искусственная последовательность
<220>
<223> YjeH F351L
<400> 36
Met Ser Gly Leu Lys Gln Glu Leu Gly Leu Ala Gln Gly Ile Gly Leu
1 5 10 15
Leu Ser Thr Ser Leu Leu Gly Thr Gly Val Phe Ala Val Pro Ala Leu
20 25 30
Ala Ala Leu Val Ala Gly Asn Asn Ser Leu Trp Ala Trp Pro Val Leu
35 40 45
Ile Ile Leu Val Phe Pro Ile Ala Ile Val Phe Ala Ile Leu Gly Arg
50 55 60
His Tyr Pro Ser Ala Gly Gly Val Ala His Phe Val Gly Met Ala Phe
65 70 75 80
Gly Ser Arg Leu Glu Arg Val Thr Gly Trp Leu Phe Leu Ser Val Ile
85 90 95
Pro Val Gly Leu Pro Ala Ala Leu Gln Ile Ala Ala Gly Phe Gly Gln
100 105 110
Ala Met Phe Gly Trp His Ser Trp Gln Leu Leu Leu Ala Glu Leu Gly
115 120 125
Thr Leu Ala Leu Val Trp Tyr Ile Gly Thr Arg Gly Ala Ser Ser Ser
130 135 140
Ala Asn Leu Gln Thr Val Ile Ala Gly Leu Ile Val Ala Leu Ile Val
145 150 155 160
Ala Ile Trp Trp Ala Gly Asp Ile Lys Pro Ala Asn Ile Pro Phe Pro
165 170 175
Ala Pro Gly Asn Ile Glu Leu Thr Gly Leu Phe Ala Ala Leu Ser Val
180 185 190
Met Phe Trp Cys Phe Val Gly Leu Glu Ala Phe Ala His Leu Ala Ser
195 200 205
Glu Phe Lys Asn Pro Glu Arg Asp Phe Pro Arg Ala Leu Met Ile Gly
210 215 220
Leu Leu Leu Ala Gly Leu Val Tyr Trp Gly Cys Thr Val Val Val Leu
225 230 235 240
His Phe Asp Ala Tyr Gly Glu Lys Met Ala Ala Ala Ala Ser Leu Pro
245 250 255
Lys Ile Val Val Gln Leu Phe Gly Val Gly Ala Leu Trp Ile Ala Cys
260 265 270
Val Ile Gly Tyr Leu Ala Cys Phe Ala Ser Leu Asn Ile Tyr Ile Gln
275 280 285
Ser Phe Ala Arg Leu Val Trp Ser Gln Ala Gln His Asn Pro Asp His
290 295 300
Tyr Leu Ala Arg Leu Ser Ser Arg His Ile Pro Asn Asn Ala Leu Asn
305 310 315 320
Ala Val Leu Gly Cys Cys Val Val Ser Thr Leu Val Ile His Ala Leu
325 330 335
Glu Ile Asn Leu Asp Ala Leu Ile Ile Tyr Ala Asn Gly Ile Leu Ile
340 345 350
Met Ile Tyr Leu Leu Cys Met Leu Ala Gly Cys Lys Leu Leu Gln Gly
355 360 365
Arg Tyr Arg Leu Leu Ala Val Val Gly Gly Leu Leu Cys Val Leu Leu
370 375 380
Leu Ala Met Val Gly Trp Lys Ser Leu Tyr Ala Leu Ile Met Leu Ala
385 390 395 400
Gly Leu Trp Leu Leu Leu Pro Lys Arg Lys Thr Pro Glu Asn Gly Ile
405 410 415
Thr Thr
<210> 37
<211> 5803
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pDCM2
<400> 37
gttcgcttgc tgtccataaa accgcccagt ctagctatcg ccatgtaagc ccactgcaag 60
ctacctgctt tctctttgcg cttgcgtttt cccttgtcca gatagcccag tagctgacat 120
tcatccgggg tcagcaccgt ttctgcggac tggctttcta cgtgttccgc ttcctttagc 180
agcccttgcg ccctgagtgc ttgcggcagc gtgaagctag cttttatcgc cattcgccat 240
tcaggctgcg caactgttgg gaagggcgat cggtgcgggc ctcttcgcta ttacgccagc 300
tggcgaaagg gggatgtgct gcaaggcgat taagttgggt aacgccaggg ttttcccagt 360
cacgacgttg taaaacgacg gccagtgaat tcgagctcgg tacccgggga tcctctagag 420
tcgacctgca ggcatgcaag cttggcgtaa tcatggtcat agctgtttcc tgtgtgaaat 480
tgttatccgc tcacaattcc acacaacata cgagccggaa gcataaagtg taaagcctgg 540
ggtgcctaat gagtgagcta actcacatta attgcgttgc gctcactgcc cgctttccag 600
tcgggaaacc tgtcgtgcca gctgcattaa tgaatcggcc aacgcgcggg gagaggcggt 660
ttgcgtattg ggcgctcttc cgcttcctcg ctcactgact cgctgcgctc ggtcgttcgg 720
ctgcggcgag cggtatcagc tcactcaaag gcggtaatac ggttatccac agaatcaggg 780
gataacgcag gaaagaacat gtgagcaaaa ggccagcaaa aggccaggaa ccgtaaaaag 840
gccgcgttgc tggcgttttt ccataggctc cgcccccctg acgagcatca caaaaatcga 900
cgctcaagtc agaggtggcg aaacccgaca ggactataaa gataccaggc gtttccccct 960
ggaagctccc tcgtgcgctc tcctgttccg accctgccgc ttaccggata cctgtccgcc 1020
tttctccctt cgggaagcgt ggcgctttct caatgctcac gctgtaggta tctcagttcg 1080
gtgtaggtcg ttcgctccaa gctgggctgt gtgcacgaac cccccgttca gcccgaccgc 1140
tgcgccttat ccggtaacta tcgtcttgag tccaacccgg taagacacga cttatcgcca 1200
ctggcagcag ccactggtaa caggattagc agagcgaggt atgtaggcgg tgctacagag 1260
ttcttgaagt ggtggcctaa ctacggctac actagaagga cagtatttgg tatctgcgct 1320
ctgctgaagc cagttacctt cggaaaaaga gttggtagct cttgatccgg caaacaaacc 1380
accgctggta gcggtggttt ttttgtttgc aagcagcaga ttacgcgcag aaaaaaagga 1440
tctcaagaag atcctttgat cttttctacg gggtctgacg ctcagtggaa cgaaaactca 1500
cgttaaggga ttttggtcat gagattatca aaaaggatct tcacctagat ccttttgggg 1560
tgggcgaaga actccagcat gagatccccg cgctggagga tcatccagcc ctgatagaaa 1620
cagaagccac tggagcacct caaaaacacc atcatacact aaatcagtaa gttggcagca 1680
tcacccgacg cactttgcgc cgaataaata cctgtgacgg aagatcactt cgcagaataa 1740
ataaatcctg gtgtccctgt tgataccggg aagccctggg ccaacttttg gcgaaaatga 1800
gacgttgatc ggcacgtaag aggttccaac tttcaccata atgaaataag atcactaccg 1860
ggcgtatttt ttgagttatc gagattttca ggagctgata gaaacagaag ccactggagc 1920
acctcaaaaa caccatcata cactaaatca gtaagttggc agcatcaccc gacgcacttt 1980
gcgccgaata aatacctgtg acggaagatc acttcgcaga ataaataaat cctggtgtcc 2040
ctgttgatac cgggaagccc tgggccaact tttggcgaaa atgagacgtt gatcggcacg 2100
taagaggttc caactttcac cataatgaaa taagatcact accgggcgta ttttttgagt 2160
tatcgagatt ttcaggagct ctttggcatc gtctctcgcc tgtcccctca gttcagtaat 2220
ttcctgcatt tgcctgtttc cagtcggtag atattccaca aaacagcagg gaagcagcgc 2280
ttttccgctg cataaccctg cttcggggtc attatagcga ttttttcggt atatccatcc 2340
tttttcgcac gatatacagg attttgccaa agggttcgtg tagactttcc ttggtgtatc 2400
caacggcgtc agccgggcag gataggtgaa gtaggcccac ccgcgagcgg gtgttccttc 2460
ttcactgtcc cttattcgca cctggcggtg ctcaacggga atcctgctct gcgaggctgg 2520
ccggctaccg ccggcgtaac agatgagggc aagcggatgg ctgatgaaac caagccaacc 2580
aggaagggca gcccacctat caaggtgtac tgccttccag acgaacgaag agcgattgag 2640
gaaaaggcgg cggcggccgg catgagcctg tcggcctacc tgctggccgt cggccagggc 2700
tacaaaatca cgggcgtcgt ggactatgag cacgtccgcg agggcgtccc ggaaaacgat 2760
tccgaagccc aacctttcat agaaggcggc ggtggaatcg aaatctcgtg atggcaggtt 2820
gggcgtcgct tggtcggtca tttcgaaaaa ggttaggaat acggttagcc atttgcctgc 2880
ttttatatag ttcantatgg gattcacctt tatgttgata agaaataaaa gaaaatgcca 2940
ataggatatc ggcattttct tttgcgtttt tatttgttaa ctgttaattg tccttgttca 3000
aggatgctgt ctttgacaac agatgttttc ttgcctttga tgttcagcag gaagctcggc 3060
gcaaacgttg attgtttgtc tgcgtagaat cctctgtttg tcatatagct tgtaatcacg 3120
acattgtttc ctttcgcttg aggtacagcg aagtgtgagt aagtaaaggt tacatcgtta 3180
ggcggatcaa gatccatttt taacacaagg ccagttttgt tcagcggctt gtatgggcca 3240
gttaaagaat tagaaacata accaagcatg taaatatcgt tagacgtaat gccgtcaatc 3300
gtcatttttg atccgcggga gtcagtgaac aggtaccatt tgccgttcat tttaaagacg 3360
ttcgcgcgtt caatttcatc tgttactgtg ttagatgcaa tcagcggttt catcactttt 3420
ttcagtgtgt aatcatcgtt tagctcaatc ataccgagag cgccgtttgc taactcagcc 3480
gtgcgttttt tatcgctttg cagaagtttt tgactttctt gacggaagaa tgatgtgctt 3540
ttgccatagt atgctttgtt aaataaagat tcttcgcctt ggtagccatc ttcagttcca 3600
gtgtttgctt caaatactaa gtatttgtgg cctttatctt ctacgtagtg aggatctctc 3660
agcgtatggt tgtcgcctga gctgtagttg ccttcatcga tgaactgctg tacattttga 3720
tacgtttttc cgtcaccgtc aaagattgat ttataatcct ctacaccgtt gatgttcaaa 3780
gagctgtctg atgctgatac gttaacttgt gcagttgtca gtgtttgttt gccgtaatgt 3840
ttaccggaga aatcagtgta gaataaacgg atttttccgt cagatgtaaa tgtggctgaa 3900
cctgaccatt cttgtgtttg gtcttttagg atagaatcat ttgcatcgaa tttgtcgctg 3960
tctttaaaga cgcggccagc gtttttccag ctgtcaatag aagtttcgcc gactttttga 4020
tagaacatgt aaatcgatgt gtcatccgca tttttaggat ctccggctaa tgcaaagacg 4080
atgtggtagc cgtgatagtt tgcgacagtg ccgtcagcgt tttgtaatgg ccagctgtcc 4140
caaacgtcca ggccttttgc agaagagata tttttaattg tggacgaatc aaattcagaa 4200
acttgatatt tttcattttt ttgctgttca gggatttgca gcatatcatg gcgtgtaata 4260
tgggaaatgc cgtatgtttc cttatatggc ttttggttcg tttctttcgc aaacgcttga 4320
gttgcgcctc ctgccagcag tgcggtagta aaggttaata ctgttgcttg ttttgcaaac 4380
tttttgatgt tcatcgttca tgtctccttt tttatgtact gtgttagcgg tctgcttctt 4440
ccagccctcc tgtttgaaga tggcaagtta gttacgcaca ataaaaaaag acctaaaata 4500
tgtaaggggt gacgccaaag tatacacttt gccctttaca cattttaggt cttgcctgct 4560
ttatcagtaa caaacccgcg cgatttactt ttcgacctca ttctattaga ctctcgtttg 4620
gattgcaact ggtctatttt cctcttttgt ttgatagaaa atcataaaag gatttgcaga 4680
ctacgggcct aaagaactaa aaaatctatc tgtttctttt cattctctgt attttttata 4740
gtttctgttg catgggcata aagttgcctt tttaatcaca attcagaaaa tatcataata 4800
tctcatttca ctaaataata gtgaacggca ggtatatgtg atgggttaaa aaggatcacc 4860
ccagagtccc gctcagaaga actcgtcaag aaggcgatag aaggcgatgc gctgcgaatc 4920
gggagcggcg ataccgtaaa gcacgaggaa gcggtcagcc cattcgccgc caagctcttc 4980
agcaatatca cgggtagcca acgctatgtc ctgatagcgg tccgccacac ccagccggcc 5040
acagtcgatg aatccagaaa agcggccatt ttccaccatg atattcggca agcaggcatc 5100
gccatgggtc acgacgagat cctcgccgtc gggcatccgc gccttgagcc tggcgaacag 5160
ttcggctggc gcgagcccct gatgctcttc gtccagatca tcctgatcga caagaccggc 5220
ttccatccga gtacgtgctc gctcgatgcg atgtttcgct tggtggtcga atgggcaggt 5280
agccggatca agcgtatgca gccgccgcat tgcatcagcc atgatggata ctttctcggc 5340
aggagcaagg tgagatgaca ggagatcctg ccccggcact tcgcccaata gcagccagtc 5400
ccttcccgct tcagtgacaa cgtcgagaca gctgcgcaag gaacgcccgt cgtggccagc 5460
cacgatagcc gcgctgcctc gtcttggagt tcattcaggg caccggacag gtcggtcttg 5520
acaaaaagaa ccgggcgccc ctgcgctgac agccggaaca cggcggcatc agagcagccg 5580
attgtctgtt gtgcccagtc atagccgaat agcctctcca cccaagcggc cggagaacct 5640
gcgtgcaatc catcttgttc aatcatgcga aacgatcctc atcctgtctc ttgatcagat 5700
cttgatcccc tgcgccatca gatccttggc ggcaagaaag ccatccagtt tactttgcag 5760
ggcttcccaa ccttaccaga gggcgcccca gctggcaatt ccg 5803
<---
Изобретение относится к биотехнологии, в частности к варианту белка YjeH, который представляет собой белок внутренней мембраны, к микроорганизму, включающему его, и к способу получения целевого продукта с его использованием. Микроорганизм можно использовать для получения O-ацетилгомосерина, гомосерина, метионина или глюфосината. Изобретение позволяет получать O-ацетилгомосерин, гомосерин, метионин или глюфосинат с высокой степенью эффективности. 11 н. и 4 з.п. ф-лы, 1 ил., 14 табл., 5 пр.
1. Белок, обладающий активностью белка YjeH внутренней мембраны, включающий i) замену аминокислоты, соответствующей 92му положению, аспарагином, ii) замену аминокислоты, соответствующей 351му положению, лейцином, или iii) замену в обоих положениях i) и ii) в аминокислотной последовательности SEQ ID NO: 1, и обладающий гомологией 95% или более, но менее 100% с SEQ ID NO: 1.
2. Белок по п. 1, обладающий способностью экспортировать O-ацетилгомосерин или гомосерин.
3. Полинуклеотид, кодирующий белок по п. 1.
4. Экспрессионный вектор, содержащий полинуклеотид по п. 3.
5. Микроорганизм для получения O-ацетилгомосерина, гомосерина, метионина или глюфосината, содержащий по меньшей мере любое одно из: белка, включающего i) замену аминокислоты, соответствующей 92му положению, аспарагином, ii) замену аминокислоты, соответствующей 351му положению, лейцином, или iii) замену в обоих положениях i) и ii), в аминокислотной последовательности SEQ ID NO: 1, и обладающего гомологией 95% или более, но менее 100% с SEQ ID NO: 1; и полинуклеотида, кодирующего этот белок.
6. Микроорганизм по п. 5, обладающий усиленной способностью продуцировать O-ацетилгомосерин или гомосерин по сравнению с немодифицированным микроорганизмом.
7. Микроорганизм по п. 5, который относится к роду Corynebacterium.
8. Способ получения целевого продукта, включающий культивирование микроорганизма в культуральной среде, где микроорганизм содержит по меньшей мере любое одно из: белка, включающего i) замену аминокислоты, соответствующей 92му положению, аспарагином, ii) замену аминокислоты, соответствующей 351му положению, лейцином, или iii) замену в обоих положениях i) и ii), в аминокислотной последовательности SEQ ID NO: 1, и обладающего гомологией 95% или более, но менее 100% с SEQ ID NO: 1; и полинуклеотида, кодирующего этот белок, где целевой продукт представляет собой O-ацетилгомосерин или гомосерин.
9. Способ по п. 8, дополнительно включающий выделение целевого продукта из культуральной среды или микроорганизма.
10. Способ получения метионина, включающий:
получение O-ацетилгомосерина путем культивирования микроорганизма в культуральной среде, где микроорганизм содержит по меньшей мере любое одно из: белка, включающего i) замену аминокислоты, соответствующей 92му положению, аспарагином, ii) замену аминокислоты, соответствующей 351му положению, лейцином, или iii) замену в обоих положениях i) и ii), в аминокислотной последовательности SEQ ID NO: 1, и обладающего гомологией 95% или более, но менее 100% с SEQ ID NO: 1; и полинуклеотида, кодирующего этот вариант; и превращение O-ацетилгомосерина в метионин путем взаимодействия O-ацетилгомосерина с сульфидом.
11. Способ получения глюфосината, включающий:
получение O-ацетилгомосерина или гомосерина путем культивирования микроорганизма в культуральной среде, где микроорганизм содержит по меньшей мере любое одно из: белка, включающего i) замену аминокислоты, соответствующей 92му положению, аспарагином, ii) замену аминокислоты, соответствующей 351му положению, лейцином, или iii) замену в обоих положениях i) и ii), в аминокислотной последовательности SEQ ID NO: 1, и обладающего гомологией 95% или более, но менее 100% с SEQ ID NO: 1; и полинуклеотида, кодирующего этот белок; и превращение O-ацетилгомосерина или гомосерина в глюфосинат.
12. Композиция для получения гомосерина, содержащая микроорганизм по п. 5 или культуру микроорганизма и эксципиент.
13. Композиция для получения O-ацетилгомосерина, содержащая микроорганизм по п. 5 или культуру микроорганизма и эксципиент.
14. Применение белка по п. 1 для получения O-ацетилгомосерина или гомосерина.
15. Применение микроорганизма по п. 5 для получения O-ацетилгомосерина или гомосерина.
| ВАРИАНТ O-АЦЕТИЛГОМОСЕРИН-СУЛЬФГИДРИЛАЗЫ И СПОСОБ ПОЛУЧЕНИЯ L-МЕТИОНИНА С ИСПОЛЬЗОВАНИЕМ ЭТОГО ВАРИАНТА | 2016 |
|
RU2694041C1 |
| МИКРООРГАНИЗМ, ПРОДУЦИРУЮЩИЙ О-АЦЕТИЛГОМОСЕРИН, И СПОСОБ ПОЛУЧЕНИЯ О-АЦЕТИЛГОМОСЕРИНА С ИСПОЛЬЗОВАНИЕМ ЭТОГО МИКРООРГАНИЗМА | 2015 |
|
RU2614253C1 |
| Микроорганизм, продуцирующий O-ацетилгомосерин, и способ получения O-ацетилгомосерина с использованием этого микроорганизма | 2016 |
|
RU2706535C2 |
| WO 2012087039 A2, 28.06.2012 | |||
| CA 3091741 A1, 05.12.2019 | |||
| US 20130236934 A1, 12.09.2013 | |||
| AU 2015264911 B2, 23.11.2017. | |||
