Область техники
Данное изобретение относится к композиции, которая вызывает, у хозяина, иммунологическую толерантность к пептидным или белковым действующим агентам, в частности, терапевтическим пептидам, полипептидам или белкам, пептидным или белковым аутоантигенам, пептидам, полипептидам или белкам, вызывающим аллергическую реакцию, или трансплантационным пептидным или белковым антигенам. Изобретение также относится к способу лечения млекопитающего, включая человека.
Известно, что печень содействует вызову иммунной толерантности. Это представлено толеризацией пищевых антигенов в печени и приживляемостью аллотрансплантатов печени. Также была продемонстрирована некоторая антиген-специфическая толерантность к некоторым чужеродным антигенам, попадающим в печень. У E. Breous et al., Hepatology, August 2009, стр. 612-621, описано, что печеночные регулирующие Т-клетки и клетки Купфера являются ключевыми медиаторами толерантности системных Т-клеток к антигенам, поражающим печень мыши. Описано, что в модели связанной с печенью передачи генов, реакции цитотоксических T-лимфоцитов на «не свои» антигены контролируются печеночными регулирующими Т-клетками, которые выделяют иммунодепрессивный цитокиновый интерлейкин IL-10 в ответ на антиген. Кроме того, клетки Купфера являются скорее толерогенными, чем создают иммунную реакцию в этом контексте.
Толерогенная роль клеток Купфера также описана у C. Ju et al., Chem. Res. Toxicol. 2003, 16:1514-1519. См. также A.H. Lau et al., Gut 2003, 52:1075-1078.
Целью данного изобретения является получение композиций, которые могут применяться для вызова иммунной толерантности к множеству пептидных или белковых активных агентов. В частности, его целью является получение специфической иммунной толерантности в отношении одного или нескольких пептидных или белковых активных агентов.
Объектом данного изобретения, поэтому является композиция, которая вызывает, у хозяина, иммунной толерантности к пептидному или белковому активному агенту, где указанная композиция содержит эритроциты, содержащие указанный активный агент. Этим активным агентом может быть терапевтический пептид, полипептид или белок, пептидный или белковый аутоантиген, пептид, полипептид или белок, вызывающий аллергическую реакцию, или трансплантационный пептидный или белковый антиген, и их смеси.
Активный агент может быть природного, синтетического или рекомбинантного происхождения. Термин "содержащая" молекула означает молекулы, которые содержат целевой пептид, полипептид или белок, и другую группу, которая может быть любого происхождения и не вредит действию указанного пептида, полипептида или белка. Например, такая группа включает гаптены.
Не претендуя на теорию, считается, что композиция в соответствии с данным изобретением индуцирует антиген(ы)-специфические Т-клетки (Tregs) и образует иммунодепрессивные цитокины или интерлейкины, в частности, IL-10.
Биологически и/или биотехнологически полученные пептиды, полипептиды или белки все чаще применяют в качестве терапевтических агентов. Однако было обнаружено, что эти агенты могут вызывать гуморальные и/или клеточные иммунные реакции. Последствия иммунной реакции на такой терапевтический агент варьируется от переходного возникновения антител без какой-либо клинической значимости, до тяжелых, угрожающих жизни состояний. Потенциальные клинические последствия включают тяжелые реакции гиперчувствительности, снижение эффективности и стимулирование аутоиммунности, включая антитела к эндогенной форме пептида, полипептида или белка (European Medicines Agency, Committee for Medicinal products for human use (CHMP), Guidelines on immunogenicity assessment of biotechnology-derived therapeutic proteins, Draft, London, 24 January 2007).
Терапевтический пептид, полипептид или белок определяется как молекула, содержащая пептид, полипептид или белок, которая эффективна для лечения патологии, особенно патологии вследствие дефицита, которая может быть скорректирована введением этой молекулы.
В одном варианте, терапевтическим пептидом, полипептидом или белком является антитело. Включен любой его фрагмент.
В другом варианте, терапевтическим пептидом, полипептидом или белком является фактором коагуляции. Включен любой его фрагмент.
В другом варианте, терапевтическим пептидом, полипептидом или белком является фермент. Включен любой его фрагмент.
В другом варианте, терапевтическим пептидом, полипептидом или белком является фактор роста. Включен любой его фрагмент.
Термин «фрагмент» включает любой фрагмент пептида, полипептида или белка, который является эффективным для лечения связанной патологии вместо полной молекулы.
Гликозилированные формы также включены в эти определения. В одном варианте, активным агентом является лизосомальный фермент. Лизосомальным ферментом является такой, который применяют для лечения или корректировки лизосомной болезни накопления через заместительную терапию ферментами (ЗТФ), включая болезнь Помпе (генерализованный гликогеноз II типа), Болезнь Фабри и расстройства мукополисахаридоза МПС I. В качестве примеров можно назвать:
- фермент альфаглюкозидазы, например, Myozyme® для лечения болезни Помпе;
- ларонидазу, например, Aldurazyme® для лечения МПС I;
- альфагалактозидазу A или агальзидазу альфа, например, Fabrazyme® и Replagal® для лечения болезни Фабри.
В другом варианте, активным агентом является фактор коагуляции, применяемый при лечении гемофилии. Фактором коагуляции может быть Фактор Factor VIII, в частности, для лечения гемофилии A. Фактором коагуляции может быть Фактор IX, в частности, для лечения гемофилии B. Фактором коагуляции может быть Фактор VII для лечения обоих типов гемофилии.
Пептидный или белковый аутоантиген определяют как антиген, который является обычной составляющей ткани и у пациента является целью причиняющей вред гуморальной или медиированной клетками иммунной реакции, как в случае аутоиммунной болезни.
В одном варианте, активный агент действует против ревматоидного артрита (РА).
В другом варианте, активный агент действует против рассеянного склероза (РС). Например, активным агентом является миелиновый основной белок.
В другом варианте, активный агент действует против юношеского диабета, такого как диабет 1 типа и ЛАДВ (латентный аутоиммунный диабет взрослых). В качестве примеров могут быть указаны антиген бета-клетки, в частности, декарбоксилаза глутаминовой кислоты (ДГК), про-инсулин и инсулиноподобный фактор роста-2 (ИФР2), и их смеси.
В другом варианте, активный агент действует против увеита. Может быть указан сетчаточный S антиген.
В другом варианте, активный агент действует против воспалительного заболевания толстого кишечника (ВЗТК), такого как болезнь Крона и язвенный колит.
В другом варианте активный агент действует против системной красной волчанки.
В другом варианте, активный агент действует против псориаза.
В другом варианте, активный агент действует против приобретенной миастении гравис. В качестве примера можно указать рецептор ацетилхолина.
Пептид, полипептид или белок, вызывающий аллергическую реакцию определяется как пептид, полипептид или белок, который отвечает за аллергическую реакцию у хозяина, где реакция может включать анафилактический шок.
В одном варианте, таким пептидом, полипептидом или белком, вызывающим аллергическую реакцию, является терапевтический активный пептид, полипептид или белок, указанный выше, где данное изобретение позволяет избежать некоторых или всех аллергических реакций против них и нейтрализовать их.
В другом варианте, пептид, полипептид или белок, вызывающий аллергическую реакцию, имеет пищевое происхождение или может быть любой другой белковой или пептидной молекулой, которая может попадать в кровоток и создавать аллергическую реакцию, например, после перорального проглатывания.
Трансплантационный пептидный или белковый антиген определяется как антиген, который представлен трансплантированной тканью и вовлечен в отторжение трансплантата пациентом, например, болезнь «трансплантат против хозяина» (ТПХ).
В одном варианте, трансплантационный антиген вовлечен в отторжение трансплантата почек.
В одном варианте, трансплантационный антиген вовлечен в отторжение трансплантата сердца.
В одном варианте, трансплантационный антиген вовлечен в отторжение трансплантата печени.
Термин "хозяин" относится, предпочтительно, к человеку, но также к животным, в частности домашним животным (особенно собакам и кошкам) и животным для спорта (особенно лошадям).
В соответствии с данным изобретением, эритроциты содержат, т.е. инкапсулируют, активный агент (АА), что означает, что АА находится практически внутри эритроцитов.
В одном варианте, композиция направлена на антигенпрезентирующие клетки (АПК) ретикулоэндотелиальной системы. Согласно характеристике, эритроциты создают, выбирают или модифицируют таким образом, чтобы нацелить их на антигенпрезентирующие клетки (АПК) ретикулоэндотелиальной системы.
В предпочтительном варианте, композиция нацелена на печень и, особенно, клетки Купфера. Согласно характеристике, эритроциты создают, выбирают или модифицируют таким образом, чтобы нацелить их на печень. Доставка АА в печень стимулирует толерантность печени к АА, особенно АА-специфическую толерантность. Толерогенные АПК печени вовлечены в стимулирование такой толерантности. Эти клетки включают, в основном, клетки Купфера (КК), не зрелые печеночные дендритные клетки и синусоидальные эндотелиальные клетки печени.
В предпочтительном варианте, композицию применяют для подавления провоспалительной реакции АПК. Согласно характеристике, эритроциты предпочтительно создают или модифицируют таким образом, чтобы подавлять провоспалительную реакцию АПК.
В предпочтительном варианте, композиции в соответствии с данным изобретением содержат эритроциты, которые содержат АА и нацелены на печень.
Композиция способствует фагоцитозу таких эритроцитов АПК печени, особенно, КК.
Согласно первому варианту, эритроциты содержат АА и имеют форму иммунного комплекса с иммуноглобулином, который распознает эпитоп на поверхности таких эритроцитов, так, чтобы способствовать фагоцитозу указанных эритроцитов АПК печени, особенно КК.
Композиция также способствует фагоцитозу макрофагами.
Предпочтительно, иммуноглобулином является иммуноглобулин G.
Антителом, которое может применяться для получения адекватной опсонизации, являются анти-Резусные антитела, анти-гликофорин A антитела и анти-РК1 (РК1 - рецептор комплемента 1 типа) антитела. Предпочтительны анти-Резусные антитела.
Согласно другому варианту, доставка в печень и/или ингибирование провоспалительной реакции производится подходящей химической обработкой с применением агентов, которые модифицируют поверхность эритроцитов, и, в частности, мостиковых или поперечно сшивающих агентов, таких как бис(сульфосукцинимидил) суберат (BS3 или BS3), глутаровый альдегид или нейраминидаза.
Согласно другому варианту, доставка в печень и/или ингибирование провоспалительной реакции производится с применением ионофора. Под ионофором понимают, как хорошо известно специалисту в данной области техники, жирорастворимую молекулу, которая позволяет переносить ионы через жировой сэндвич клеточной мембраны. Ионофорами могут быть, в частности, жирорастворимые молекулы, синтезированные микроорганизмами для переноса ионов через жировой сэндвич клеточной мембраны. В общем, ионофор способен образовывать комплекс с ионом и служит в качестве носителя иона.
В одном варианте, ионофор образует комплекс с двухвалентным катионом, таким как кальций. В соответствии с данным изобретением, ионофор может применяться с кальцием, что вызывает увеличение внутриклеточной концентрации кальция и экспозиции фосфатидилсерина, что приводит к раннему старению эритроцитов.
В качестве примера применяют ионофор кальция A23187 (кальцимицин). Полагают, что ионофор, такой как A23187, вызывает повышение внутриклеточных концентраций кальция ЭЦТ, что приводит к старению клеток, и что фагоцитоз состаренных эритроцитов подавляет провоспалительную реакцию. Это согласуется с Romero P.J., Romero E.A., Blood Cells Mol. Dis. 25 (1999) 9-19; и Bratosin D. et al., Cell Death Differ. 8 (2001) 1143-1156.
В конкретном варианте, по крайней мере, объединяют два способа доставки в клетку, и, например, в результате композиция содержат АА-содержащие эритроциты в виде иммунного комплекса, и химически обработана так чтобы обеспечивать их поглощение в печени, и фагоцитоз АПК, в частности, КК.
В одном варианте, эритроциты получают от самого пациента.
В другом варианте, эритроциты получают от донора, совместимого по типу крови.
Композиция в соответствии с данным изобретением может содержать один или более АА в одних и тех же эритроцитах, или каждый в различных эритроцитах.
Методики инкапсулирования активных ингредиентов в эритроциты известны, и основная методика лизис-лигирования, которая является предпочтительной, описана в патентах EP-A-101341 и EP-A-679101, в которых может проконсультироваться специалист в данной области техники. Согласно этой методике, первичное отделение элемента диализа (например, диализная трубка или диализный картридж) непрерывно заполняют суспензией эритроцитов, а вторичное отделение содержит водный раствор, который является гипотоническим по отношению к суспензии эритроцитов, для лизирования эритроцитов; затем, в лигирующем отделении, лигирование эритроцитов индуцируется в присутствии АА повышением осмотического и/или онкотического давления, и затем суспензию эритроцитов, содержащую АА, собирают.
Среди описанных по настоящее время вариантов, предпочтение отдается способу, описанному в WO2006/016247, что позволяет эффективно, воспроизводимо, безопасно и стабильно инкапсулировать АА. Этот способ включает стадии:
1 - суспензия лепешки эритроцитов в изотоническом растворе при уровне гематокрита больше или равном 65%, охлаждение от +1 до +8°C,
2 - измерение осмотической резистентности с применением образца эритроцитов из указанной лепешки эритроцитов,
стадии 1 и 2 могут проводиться в любом порядке (включая параллельный),
3 - лизис и процесс интернационализации АА внутри одной камеры при температуре, постоянно поддерживаемой на уровне от +1 до +8°C, включающий пропускание суспензии эритроцитов при уровне гематокрита более или равном 65%, и гипотонический лизисный раствор охлаждают до +1-8°C, через диализный картридж; и параметры лизиса корректируют согласно измеренной ранее осмотической резистентности; и
4 - процесс лигирования проводят во второй камере, внутри которой температура составляет от +30 до +40°C, и в присутствии гипертонического раствора.
Под "интернализацией" подразумевают проникновение АА внутрь эритроцитов.
В частности, для диализа, лепешку эритроцитов суспендируют в изотоническом растворе при высоком уровне гематокрита, больше или равном 65%, и, предпочтительно, больше или равном 70%, и эту суспензию охлаждают до температуры от около +1 до +8°C, предпочтительно, от +2 до 6°C, обычно в области +4°C. Согласно конкретному варианту, уровень гематокрита составляет от 65% до 80%, предпочтительно от 70% до 80%.
Осмотическую резистентность предпочтительно измеряют в эритроцитах непосредственно перед стадией лизиса. Эритроциты или содержащая их суспензия предпочтительно имеет температуру, близкую или идентичную той, которая выбрана для лизиса. Согласно другой предпочтительной характеристике данного изобретения, измерение осмотической резистентности применяют быстро, т.е. процесс лизиса проводят сразу же после отбора образца. Предпочтительно, период времени между отбором образца и началом лизиса меньше или равен 30 минут, более предпочтительно, меньше или равен 25, и даже меньше или равен 20 минутам.
Подробную методику, по которой проводят лизис, в котором измеряют и принимают во внимание осмотическую резистентность, специалист в данной области техники может посмотреть в WO2006/016247.
Согласно другой характеристике данного изобретения, композиция в соответствии с данным изобретением содержит, на завершающей стадии, суспензию эритроцитов с уровнем гематокрита от около 40% до около 70%, предпочтительно, от около 45% до около 55%, более предпочтительно, около 50%. Предпочтительно ее упаковывают в объеме от около 1 до около 250 мл. Упаковкой предпочтительно является пакет для крови, шприц и подобные, подходящие для переливания или введения крови. Количество инкапсулированного АА, соответствующее медицинскому предписанию, предпочтительно полностью содержатся в пакете для крови, шприце и подобном.
Объектом данного изобретения является также способ вызывания у хозяина иммунной толерантности к пептидному или белковому активному агенту, где указанная композиция содержит эритроциты, содержащие активный агент, выбранный из группы, включающей терапевтический пептид, полипептид или белок, пептидный или белковый аутоантиген, пептид, полипептид или белок, вызывающие аллергическую реакцию, и трансплантационный пептидный или белковый антиген. Этот способ включает введение хозяину эффективного количества композиции в соответствии с данным изобретением, в частности, внутривенно, инъекцией или вливанием, предпочтительно, вливанием.
Согласно одной характеристике изобретения, вводят от около 1 до около 250 мл, предпочтительно, от около 10 до около 250 мл, обычно от около 10 до около 200 мл суспензии эритроцитов. Суспензия имеет подходящий уровень гематокрита, обычно от около 40% до около 70%, предпочтительно, от около 45% до около 55%, наиболее предпочтительно, около 50%. Эритроциты могут оказывать свое собственное толерогенное действие по отношению к активному агенту, который присутствует в то же время (инкапсулированному активному агенту). Высокие количества эритроцитов могут благоприятствовать толерогенному действию. С другой стороны, доставка в печень, как указано выше, позволяет применять низкие дозы эритроцитов. Таким образом, специалист в данной области техники может выбрать оптимальное количество активного агента и эритроцитов, применяемых у пациента, и может принимать во внимание, обработаны или нет эритроциты так, чтобы попадать в печень.
Объектом данного изобретения также является применение композиции в соответствии с данным изобретением для вызова иммунной толерантности, специфической к активному агенту или активным агентам, которые присутствуют во введенных эритроцитах.
Другим объектом изобретения является композиция в соответствии с данным изобретением для применения в качестве лекарственного средства для вызова иммунной толерантности, специфической к активному агенту или активным агентам, которые присутствуют во введенных эритроцитах.
Далее данное изобретение более подробно описано в вариантах, представленных в виде не ограничивающих примеров, которые ссылаются на представленные чертежи:
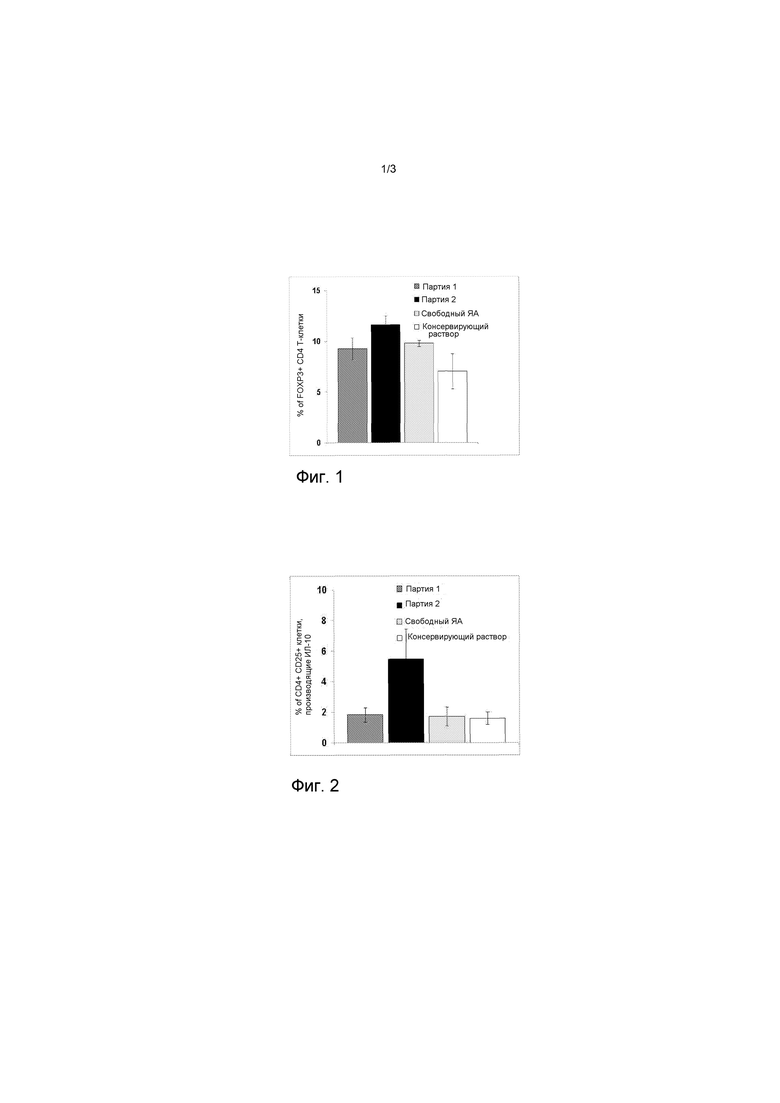
На фигуре 1 представлен график, представляющий процент Т-клеток CD4, экспрессирующих FOXP3.
На фигуре 2 представлен график, представляющий процент регулирующих Т-клеток CD4+ CD25+, производящих IL-10.
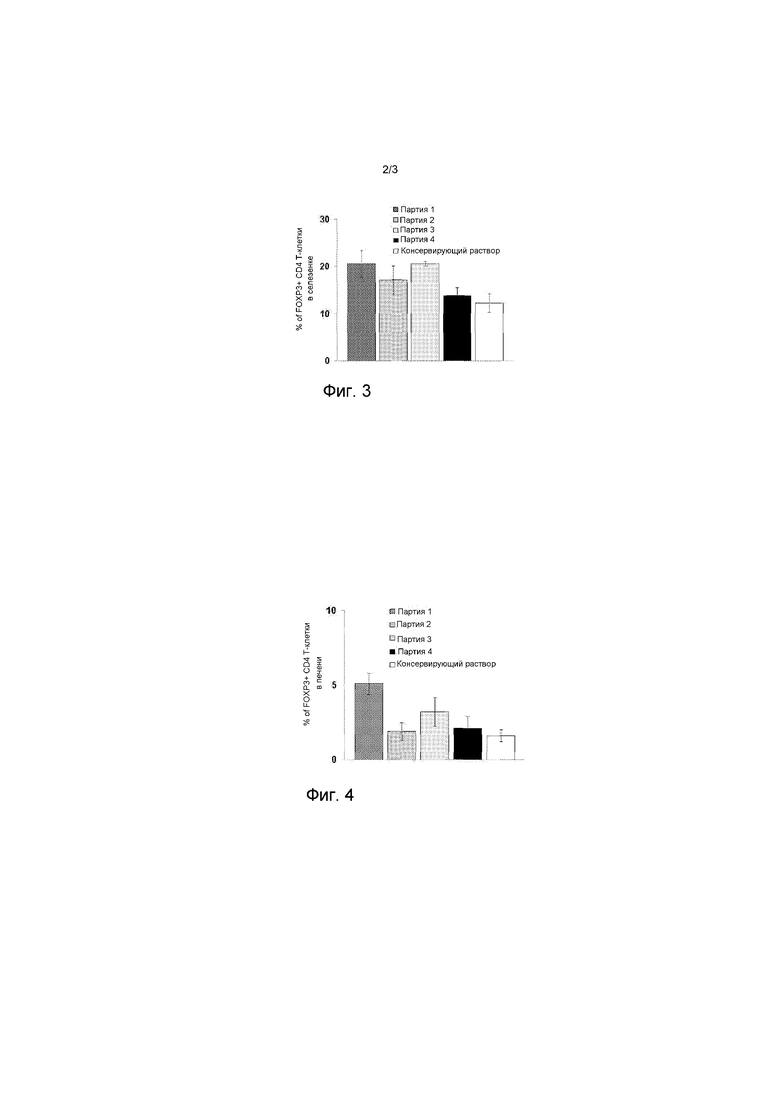
На фигуре 3 представлен график, представляющий процент Т-клеток CD4, экспрессирующих FOXP3 в селезенке.
На фигуре 4 представлен график, представляющий процент Т-клеток CD4, экспрессирующих FOXP3 в печени.
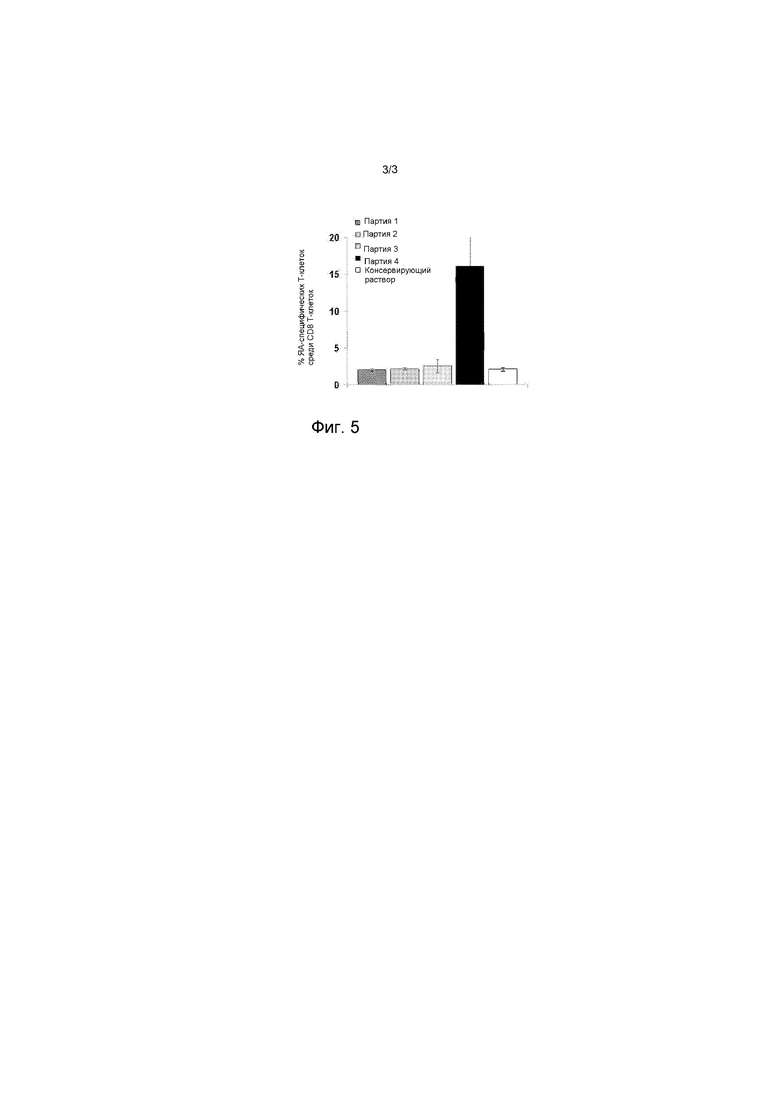
На фигуре 5 представлен график, представляющий процент ЯА-специфических Т-клеток CD8.
Пример 1: Инкапсулирование FITC-декстрана в эритроциты мыши
FITC-декстран фторохром (70 кДа) инкапсулируют в эритроциты, полученные от мыши (мыши OF1) с применением гипотонического диализа на колонке. Кровь центрифугируют и затем промывают 3 раза ФРФБ. Гематокрит доводят до 70% в присутствии FITC-декстрана, добавленного до конечной концентрации 8 мг/мл до диализа. Эритроциты диализируют со скоростью 2 мл/мин вплотную к лизисному буферу, имеющему низкую осмолярность (противоток при 15 мл/мин). Лизированные эритроциты, выходящие из колонки, повторно уплотняют с применением раствора с высокой осмолярностью и инкубируют в течение 30 мин при 37°C. После нескольких промывок ФРФБ, содержащим глюкозу, клетки доводят до гематокрита 50%.
Пример 2. Химическая обработка бис(сульфосукцинимидил) субератом (BS3) 1 мМ эритроцитов, содержащих FITC-декстран
Суспензию эритроцитов с инкапсулированным FITC-декстраном несколько раз промывают перед доведением до 1,7×106 клеток/мкл ФРФБ и смешивают с одним объемом буферного раствора 2 мМ BS3 (раствор BS3 содержит 0,09% глюкозы и фосфатный буфер, pH 7,4), так, чтобы получить конечную концентрацию BS3 1 мМ. Клетки инкубируют в течение 30 минут при комнатной температуре. Реакцию гасят добавлением одного объема 20 мМ Tris-HCl, 140 мМ NaCl. После инкубирования при комнатной температуре в течение 5 минут, смесь центрифугируют при 800 g в течение 5 мин 4°C. Затем клетки дважды промывают ФРФБ, содержащим глюкозу (центрифугирование при 800 g) и один раз SAG-BSA 6% (центрифугирование при 1000 g) в течение 10 мин, перед доведением до гематокрита 50% для составления конечного продукта.
Пример 3. Химическая обработка бис(сульфосукцинимидил) субератом (BS3) 5 мМ эритроцитов, содержащих FITC-декстран
Суспензию эритроцитов с инкапсулированным FITC-декстраном несколько раз промывают перед доведением до 1,7×106 клеток/мкл ФРФБ и смешивают с одним объемом буферного раствора 10 мМ BS3 (раствор BS3 содержит 0,09% глюкозы и фосфатный буфер, pH 7,4), так, чтобы получить конечную концентрацию BS3 5 мМ. Клетки инкубируют в течение 30 минут при комнатной температуре. Реакцию гасят добавлением одного объема 20 мМ Tris-HCl, 140 мМ NaCl. После инкубирования при комнатной температуре в течение 5 минут, смесь центрифугируют при 800 g в течение 5 мин 4°C. Затем клетки дважды промывают ФРФБ, содержащим глюкозу (центрифугирование при 800 g) и один раз SAG-BSA 6% (центрифугирование при 1000 g) в течение 10 мин, перед доведением до гематокрита 50% для составления конечного продукта.
Пример 4. Обработка эритроцитов, содержащих FITC-декстран, A23187 ионофором
Суспензию эритроцитов, содержащих FITC-декстран, один раз промывают буфером A, содержащим Hepes 10 мМ, NaCl 140 мМ, АБС 0,1%, CaCl2 2,5 мМ, и затем суспензию разводят до 1,106 клеток/микролитр с применением буфера A. Ионофор, концентрированный в ДМСО, разводят буфером A и затем добавляют к суспензии клеток для получения финальной концентрации 0,15, 0,2 или 0,3 мкМ. Клетки инкубируют в течение 30 мин при 37°C. Смесь центрифугируют при 800 g в течение 6 мин, 4°C. Затем клетки промывают 2 раза ФРФБ, содержащим глюкозу (центрифугирование при 800 g) и один раз SAG-BSA 6% (центрифугирование при 1000 g), и получают конечные продукты.
Пример 5. Биораспределение FITC-декстрана после инъекции эритроцитов мышам
Получают 5 партий эритроцитов мыши OF1, содержащих FITC-декстран, как описано в примере 1, и обрабатывают или не обрабатывают BS3 или ионофором (согласно примерам 2-4) следующим образом:
Партия 1: без обработки
Партия 2: BS3 1 мМ
Партия 3: BS3 5 мМ
Партия 4: ионофор 0,2 мкМ
Партия 5: ионофор 0,3 мкМ
Каждую партию вводят ВВ при J1 мышам OF1. Мышей умерщвляют через 1 ч 30 мин после инъекции и выделяют кровь, селезенку, печень и костный мозг: аликвоты 50 мкл крови, и для селезенки, печени и костного мозга, аликвоты 50 мкл после измельчения и гомогенизации цельных клеток каждого органа. Аликвоты замораживают т в течение, по крайней мере, 20 минут при -20°C, затем оттаивают медленно при комнатной температуре. Аликвоты, взятые у контрольных мышей, применяют для приготовления стандартного спектра концентраций FITC-декстрана: аликвоты затем лизируют 125 мкл различных концентраций FITC-декстрана для получения стандартного спектра концентраций.
Анализируемые аликвоты образца лизируют с применением 125 мкл дистиллированной воды. Затем к аликвотам добавляют 175 мкл TCA 12%. Эту смесь затем центрифугируют при 15000 g, 10 мин, 4°C. Берут 200 мкл надосадочной кислоты и 500 мкл триэтаноламина 0,4 M добавляют до флуорометрического измерения (возбуждение при 494 нм, эмиссия при 521 нм). Концентрация FITC-декстрана в каждом образце может быть определена с применением стандартного спектра концентраций, и затем может быть определена доля FITC-декстрана, присутствующего в соответствующем органе.
Биораспределение FITC-декстрана через 1 ч 30 мин после инъекции эритроцитов мышам OF1:
Обработка 1 мМ BS3 вызывает попадание эритроцитов в печень и селезенку, в то время как обработка ионофором вызывает попадание эритроцита только в печень. Повышение дозы ионоформа улучшает целевое попадание.
Пример 6. Измерение фагоцитоза в содержащих FITC-декстран эритроцитах мышей
Получают 5 партий эритроцитов мышей OF1, содержащих FITC-декстран, полученных как в Примере 1 и обработанных или не обработанных BS3 или ионофором (в соответствии с примерами 2-4):
Партия 1: без обработки
Партия 2: BS3 1 мМ
Партия 3. BS3 5 мМ
Партия 4: ионофор 0,2 мкМ
Партия 5 ионофор 0,3 мкМ.
Каждую партию вводят ВВ при J1 мышам OF1. Мышей умерщвляют через 1 час 30 мин после инъекции и восстанавливают печень. Флуоресценцию вводят в макрофаги печени, экспрессирующие F4/80 маркер, клетки печени, экспрессирующие маркер CD11b, и дендритные клетки печени, экспрессирующие маркер CD11c, измеряют с применением проточной цитометрии.
Процент клеток печени, в которых в которых фагоцитированы эритроциты, содержащие FITC-декстран, через 1 час 30 мин после введения мышам:
Обработка BS3 и ионофором вызывает эритрофагоцитоз макрофагами (F4/80 и CD11b) и дендритными клетками. Для обработки BS3 процент клеток, которые обработаны фагоцитом, зависит от дозы BS3, применяемой для обработки.
Пример 7. Способ инкапсулирования яичного альбумина в эритроциты мышей и человека
Вариант 1:
Яичный альбумин (белок 45 кДа, альбумин куриного яйца) инкапсулируют в эритроциты мыши (мыши OF1 или мыши C57BL/6) способом гипотонического диализа в диализной трубке. Суспензию эритроцитов промывают несколько раз перед доведением гематокрита до 70% для диализа. Диализ проводят в диализной трубке в лизисном буфере низкой осмолярности в течение от около 1 часа или 30 минут, если диализ происходит после тепловой обработки. Эритроциты повторно уплотняют с применением раствора высокой осмолярности в течение 30 минут. После нескольких промывок конечный продукт помещают в буфер, Sag-маннит, и гематокрит доводят до 50%.
Вариант 2:
Яичный альбумин инкапсулируют в мышиные эритроциты способом гипотонического диализа в диализной колонке. Суспензию эритроцитов промывают несколько раз до доведения гематокрита до 70% для диализа. Диализ проводят в диализной колонке в лизисном буфере низкой осмолярности в течение около 10 мин. После того, как они выйдут из колонки, эритроциты повторно уплотняют с применением раствора высокой осмолярности в течение 30 минут при 37°C. После нескольких промывок конечный продукт помещают в NaCl глюкозный буфер, содержащий глюкозу, SAG-маннит или декомплементарную плазму, и гематокрит доводят до 50%.
Пример 8: Способ инкапсулирования яичного альбумина в эритроциты мышей
Яичный альбумин (Worthington Biochemicai Corporation, Lakewood, NJ) инкапсулируют в эритроциты мышей гипотоническим диализом. Суспензии эритроцитов получают из крови мышей C57BL/6, собранной на литиевый гепарин. Коротко, эритроциты тир раза промывают физиологическим раствором и гематокрит (Hct) крови доводят до 70% перед диализом. В суспензию эритроцитов добавляют ЯА в конечной концентрации 5 или 0,5 мг/мл. Диализ проводят (скорость потока клеток 2 мл/мин) через клеточный лизисный буфер (осмолярность 50 мОсмол/кг), циркулирующий противотоком (15 мл/мин) в 80 половолоконном диализаторе (Gambro, Lyon, France). Эритроциты повторно уплотняют "в текущем режиме" добавлением (конечный объем 10%) гипертонического раствора (1900 мОсмол/кг), содержащего 0,4 г/л аденина (Sigma-Aldrich, Saint-Louts, MI), 15,6 г/л инозина (Sigma-Aldrich), 6,4 г/л пирувата натрия (Sigma-Aldrich), 4,9 г/л дегидрата фосфата мононатрия (Sigma-Aldrich), 10,9 г/л додекагидрата фосфата динатрия (Sigma-Aldrich), 11,5 г/л моногидрата глюкозы (Sigma-Aldrich) и 50 г/л NaCl (Sigma-Aldrich). Эритроциты инкубируют в течение 30 мин при 37°C гипертоническим раствором. После нескольких промывок 0,9% NaCl 0,2% глюкозой (Bioluz, Saint-Jean-de-Luz, France), продукт промывают один раз буфером A, содержащим Hepes 10 мМ, NaCl 140 мМ, АБС 0,1%, CaCl2 2,5 мМ, разбавленным буфером A до 1,106 клеток/мкл, и обрабатывают 0,15 мкМ кальциевым ионофором A23187 (Sigma) в течение 30 мин при 37°C как описано в примере 15. После 3 промывок 0,9% NaCl 0,2% глюкозой конечный продукт повторно суспендируют и его гематокрит доводят до 50% декомплементированной плазмой мышей C57BL/6 mouse plasma (конечный объем 15%). Полученный таким образом продукт хранят при 2-8°C.
Пример 9: Способ инкапсулирования яичного альбумина в эритроциты мышей
Яичный альбумин (Worthington Biochemicai Corporation, Lakewood, NJ) инкапсулируют в эритроциты мышей гипотоническим диализом. Суспензии эритроцитов получают из крови мышей C57BL/6, собранной на литиевый гепарин. Коротко, эритроциты три раза промывают физиологическим раствором и гематокрит (Hct) крови доводят до 70% перед диализом. К суспензии эритроцитов добавляют ЯА в конечной концентрации 5 или 0,5 мг/мл. Диализ проводят (скорость потока клеток 2 мл/мин) через клеточный лизисный буфер (осмолярность 50 мОсмол/кг), циркулирующий противотоком (15 мл/мин) в диализной трубке. После диализа эритроциты повторно уплотняют добавлением (конечный объем 10%) гипертонического раствора (1900 мОсмол/кг), содержащего 0,4 г/л аденина (Sigma-Aldrich, Saint-Louts, MI), 15,6 г/л инозина (Sigma-Aldrich), 6,4 г/л пирувата натрия (Sigma-Aldrich), 4,9 г/л дегидрата фосфата мононатрия (Sigma-Aldrich), 10,9 г/л додекагидрата фосфата динатрия (Sigma-Aldrich), 11,5 г/л моногидрата глюкозы (Sigma-Aldrich) и 50 г/л NaCl (Sigma-Aldrich). Эритроциты инкубируют в течение 30 мин при 37°C гипертоническим раствором и затем химически обрабатывают BS3 как описано в примере 14.
После нескольких промывок 0,9% NaCl 0,2% глюкозой (Bioluz, Saint-Jean-de-Luz, France), конечный продукт повторно суспендируют и его гематокрит доводят до 50% декомплементированной плазмой мышей C57BL/6 mouse plasma (конечный объем 15%). Полученный таким образом продукт хранят при 2-8°C.
Пример 10. Обработка антителами эритроцитов, содержащих яичный альбумин
Суспензию эритроцитов, в которые инкапсулирован яичный альбумин, промывают несколько раз перед доведением 109 клеток/мл для in vivo теста и 108 клеток/мл для in vitro теста. Ее инкубируют с анти-TER119 антителом (10 мкг/мл для in vitro теста и 23 мкг/мл или 5 мкг/мл для in vivo теста) в течение 30 минут при 4°C. После нескольких промывок конечный продукт помещают в буфер с инъецируемым количеством, и гематокрит доводят до 50%.
Пример 11. Измерение фагоцитоза содержащих яичный альбумин эритроцитов дендритными клетками in vitro
Действие обработки антителами на эффективность фагоцитоза эритроцитов, полученных в примере 9, дендритными клетками, измеряют in vitro. Эритроциты метят флуоресцентной меткой, СЭКФ (сукцинимидиловый эфир карбоксифлуоросцеина), в течение 20 мин при 4°C. СЭКФ представляет собой не флуоресцентный краситель, который распространяется через мембрану клетки. Попадая внутрь клетки, молекула становится флуоресцентной после ее расщепления внутриклеточными эстеразами.
Дендритные клетки выделяют из селезенки мышей C57BI/6 с применением магнитных шариков. Эти шарики переносят антитела, которые распознают маркер CD11c, тем самым позволяя выделять фракцию CD11c дендритных клеток.
Меченные или не меченные СЭКФ эритроциты затем инкубируют с дендритными клетками (10×106 клеток/мл) в соотношении 20:1 в конечном объеме 200 мкл/ячейку в круглодонных 96-луночных культуральных планшетах в течение 4 часов при 37°C и 5% CO2. После культивирования в течение 4 часов, эритроциты, не поглощенные дендритными клетками, лизируют с NH4Cl и проводят несколько промывок. Затем измеряют захват СЭКФ флуорохрома дендритными клетками с применением проточной цитометрии (R. Segura et al., J. Immunol, January 2006, 176(1): 441-50).
Тестируют три популяции эритроцитов:
(A) эритроциты с загруженным яичным альбумином и не меченные СЭКФ флуорохромом.
(B) эритроциты с загруженным яичным альбумином и меченные СЭКФ флуорохромом.
(C) эритроциты с загруженным яичным альбумином, обработанные анти-TER119 антителом и меченные СЭКФ.
Результаты
Процент дендритных клеток, имеющих фагоцитозированные флуоресцентные эритроциты
Мышиные эритроциты, загруженные яичным альбумином и обработанные анти-TER119 антителом, были более эффективно фагоцитозированы дендритными клетками, выделенными из селезенки, по сравнению с не обработанными эритроцитами in vitro, через 4 часа совместного культивирования. 36% дендритных клеток фагоцитозировали эритроциты, имеющие антитела, по сравнению с только 27% при отсутствии антител.
Пример 12. Измерение фагоцитоза эритроцитов, содержащих яичный альбумин, макрофагами и дендритными клетками селезенки и печени in vivo у мышей
Это исследование представляет собой аллогенное исследование, так как эритроциты OF1 мышей, содержащие яичный альбумин, впрыскивают не генетически родственным мышам C57BI/6.
Получают три партии 74×107 эритроцитов мышей OF1, загруженных яичным альбумином (пример 9), обработанных анти-TER119 антителом (как описано в примере 10) или не обработанных. Эти партии делят следующим образом:
Партия 1: нет обработки антителом
Партия 2: обработка анти-TER119 антителом.
Каждую партию метят СЭКФ и вводят внутривенно мышам C57BI/6. Через три часа после инъекции берут кровь, селезенку и печень мышей. Процент флуоресцентных эритроцитов, циркулирующих в крови мышей, измеряют проточной цитометрией. Флуоресценцию, введенную в макрофаг селезенки, экспрессирующий F4/80 маркер, в макрофаг печени, экспрессирующий F4/80 маркер, и в дендритные клетки селезенки, экспрессирующие CD11c маркер, измеряют проточной цитометрией.
Результаты
Процент макрофагов или дендритных клеток из селезенки, имеющих фагоцитозированные флуоресцентные эритроциты через 3 часа после инъекции мыши
Через 3 часа после инъекции, мышиные эритроциты, загруженные яичным альбумином и обработанные анти-TER119 антителом, практически не присутствуют в крови мышей (1%), в то время как в крови мышей все еще присутствуют не обработанные, загруженные яичным альбумином эритроциты (4,6%).
Эритроциты, которые были обработаны анти-TER119 антителом, фагоцитозированы F4/80 макрофагами и CD11c дендритными клетками селезенки.
Эритроциты, обработанные анти-TER119 антителом, более эффективно фагоцитозированы F4/80 макрофагами селезенки, по сравнению с не обработанными эритроцитами. 81% макрофагов селезенки фагоцитозировали обработанные антителом эритроциты, и только 28% в необработанной партии (Таблица 4).
Обработанные антителом эритроциты также более эффективно фагоцитозировались CD11c дендритными клетками из селезенки, по сравнению с не обработанными эритроцитами. Соответственно, 22% дендритных клеток фагоцитозировали обработанные антителом эритроциты, по сравнению с только 5% необработанных эритроцитов (Таблица 4).
Процент макрофагов печени, имеющих фагоцитозированные флуоресцентные эритроциты через 3 часа после инъекции мыши
Эритроциты, обработанные анти-TER119 антителом, фагоцитозированы F4/80 макрофагами печени.
Эритроциты, обработанные анти-TER119 антителом, более эффективно фагоцитозированы F4/80 макрофагами печени по сравнению с не обработанными эритроцитами. 50% макрофагов печени фагоцитозируют обработанные антителом эритроциты, по сравнению с только 24% в необработанной партии (таблица 5).
В итоге, связывание антитела с эритроцитами позволяет эффективную доставку эритроцитов в селезенку и печень, и значительно повышает процент дендритных клеток и макрофагов, способных к фагоцитозированию таких эритроцитов.
Пример 13. Измерение процентной доли регулирующих Т-клеток и производство ими противовоспалительного интерлейкина-10 (IL-10) после одной инъекции поли(I:C) и обработанных антителами или не обработанных, содержащих яичный альбумин, эритроцитов у мышей
Целью этого исследования является измерение процентной доли регулирующих Т-клеток у мышей C57BI/6 после инъекции Поли(I:C) и обработанных антителами (анти-TER119) или не обработанных, содержащих яичный альбумин, эритроцитов.
Две партии 30×107 обработанных антителом или не обработанных, содержащих яичный альбумин, эритроцитов от мышей OF1 получают по методике примера 8. В этом исследовании эквивалентное количество уловленного яичного альбумина вводят свободно, и отрицательным контролем является консервирующий раствор эритроцитов (глюкозированный NaCl, содержащий 33% декомплементированной плазмы мышей). Количество свободного или уловленного ЯА, впрыскиваемого мышам, составляет 2 мкг. Количество свободного Поли(I:C), впрыскиваемого мышам, составляет 25 мкг.
Партия 1: содержащие яичный альбумин эритроциты и Поли(I:C)
Партия 2: обработанные антителом, содержащие яичный альбумин, эритроциты и Поли(I:C)
Партии впрыскивают внутривенно мышам C57BI/6 mice (4 мыши в группе). Через семь дней после впрыскивания партии мышей умерщвляют, и их селезенки собирают. Для измерения процента экспрессирующих FOXP3 CD4+ Т-клеток проточной цитометрией (Фигура 1, Таблица 6), используют 2,5×106 клетки селезенки. Коротко, после ЭЦТ лизиса с применением раствора NH4Cl (StemCell Technologies, каталожный номер 7850), клетки селезенки сначала окрашивают PC5-анти-CD4 (Biolegend, номер по каталогу BLE100514) и FITC-анти-CD25 моноклональными антителами (Biolegend, номер по каталогу BLE110569), и затем инкубируют с буферами фиксации и пермеабилизации (Biolegend, номер по каталогу 421303) перед инкубированием с PE-анти-FOXP3 mAb (Biolegend, номер по каталогу 320008) или контролем изотипа.
Для измерения процента регулирующих Т-клеток (CD4+ CD25+), производящих IL-10, проточной цитометрией (Фигура 2). Клетки селезенки (5×106 клеток/мл) культивируют 0,1 мкг/мл пептида ЯА323-339 (Genscript, номер по каталогу 41007-1) в течение 4 часов при 37°C в 5% CO2-воздухе на 24-луночных культуральных планшетах. Через час после начала культивирования, Брефелдин A (Ebioscience, номер по каталогу 420601) добавляют для блокирования секреции цитокина. В конце культивирования, клетки сначала окрашивают PC5-анти-CD4 и FITC-анти-CD25 моноклональными антителами и затем инкубируют буфером фиксации (Biolegend, номер по каталогу 420801) и буфером пермеабилизации (Biolegend, номер по каталогу 421002) перед инкубированием с PE-анти-IL-10 mAb (Biolegend, номер по каталогу 505008) или контролем изотипа.
Процент регулирующих CD4 Т-клеток, экспрессирующих фактор транскрипции FOXP3, значительно повышается после инъекции Поли(I:C) и обработанных антителом, содержащих яичный альбумин, эритроцитов или свободного ЯА по сравнению с контрольными мышами, которым впрыскивали консервирующий раствор (Фигура 1, Таблица 6, тест Стьюдента, p<0,007 и p<0,05, соответственно).
Процент CD4 Т-клеток, экспрессирующих FOXP3
Тем не менее, только регулирующие T-клетки, индуцированные инъекцией обработанных антителом, содержащими ЯА, эритроцитов и Поли(I:C), могут производить противовоспалительный цитокин (IL-10) после повторного стимулирования in vitro с пептидом ЯА (Фигура 2, таблица 7).
Процент регулирующих CD4+ CD25+ Т-клеток, производящих IL-10
В общем, инъекция обработанных антителом, содержащих ЯА, эритроцитов и Поли(I:C) вызывает образование регулирующих Т-клеток, способных вырабатывать IL-10 после повторного стимулирования антигеном.
На Фигуре 1, процент FOXP3+ CD4+ Т-клеток в селезенке определяют проточной цитометрией через 7 дней после внутривенной инъекции мышам C57BL/6 обработанных антителом (черные столбики) или не обработанных (темно-серые столбики), содержащих ЯА, эритроцитов и Поли(I:C) или свободного ЯА и Поли(I:C) (светло-серые столбики) или контрольной среды (белые столбики). Количество свободного или уловленного ЯА, впрыскиваемого мышам, составляло 2 мкг, и количество Поли(I:C) составляло 25 мкг.
На Фигуре 2, образование IL-10 регулирующими CD4+ CD25+ Т-клетками определяют проточной цитометрией после in vitro повторного стимулирования с пептидом ЯА 0,1 мкг/мл клеток селезенки, выделенных у мышей C57BL/6 через 7 дней после внутривенной инъекции обработанных антителом (черные столбики) или не обработанных (темно-серые столбики), содержащих ЯА, эритроцитов и Поли(I:C) или свободного ЯА и Поли(I:C) (светло-серые столбики) или контрольной среды (белые столбики). Количество свободного или уловленного ЯА, впрыскиваемого мышам, составляло 2 мкг, и количество Поли(I:C) составляло 25 мкг.
Пример 14. BS3 обработка эритроцитов, содержащих яичный альбумин
Суспензию эритроцитов, в которые инкапсулирован ЯА (пример 8) несколько раз промывают перед разведением до 1,7×106 клеток/мкл с применением ФРФБ, и смешивают с одним объемом буферного раствора 2 мМ BS3 (раствор BS3 содержит глюкозу, 0,09%, и фосфатный буфер, pH 7,4), с получением конечной концентрации BS3 1 мМ. Клетки инкубируют в течение 30 минут при комнатной температуре. Реакцию гасят добавлением одного объема 20 мМ Tris-HCl, NaCl 140 мМ. После инкубирования при комнатной температуре в течение 5 минут, смесь центрифугируют при 800 g в течение 5 мин, 4°C. Затем клетки промывают три раза NaCl глюкозой (центрифугирование при 800 g) в течение 10 мин, затем доводят до гематокрита 50% декомплементированной плазмой.
Пример 15. Обработка ионофором эритроцитов, содержащих яичный альбумин
Суспензию эритроцитов, в которые инкапсулирован ЯА (пример 8), промывают один раз буфером A, содержащим Hepes 10 мМ, NaCl 140 мМ, АБС 0,1%, CaCl2 2,5 мМ, и затем суспензию разводят до 1,106 клеток/мкл с применением буфера A. Ионофор, концентрированный в ДМСО, разводят буфером A и затем добавляют в суспензию клеток для получения конечной концентрации 0,15 мкМ. Клетки инкубируют в течение 30 мин при 37°C. Смесь центрифугируют при 800 g в течение 6 мин, 4°C. Затем клетки промывают три раза NaCl глюкозой (центрифугирование при 800 g) в течение 10 мин, затем доводят до гематокрита 50% декомплементированной плазмой.
Пример 16. Измерение процента регулирующих Т-клеток в печени и селезенке после одной инъекции содержащих яичный альбумин эритроцитов у мышей, обработанных для доставки в печени и/или для подавления АПК провоспалительной реакции
Целью этого исследования является демонстрация того, что применение ЭЦТ, обработанных для доставки в печень или подавления АПК провоспалительной реакции, в качестве системы доставки антигена, вызывает увеличение процента регулирующих Т-клеток у мышей.
Партии 126×107 обработанных антителом, обработанных BS3 или обработанных ионофором, содержащих яичный альбумин эритроцитов из мышей C57BL/6 получают по методике примера 8. Количество уловленного ЯА, впрыскиваемого мышам, составляет 8 мкг.
Партия 1: Обработанные ионофором, содержащие яичный альбумин эритроциты
Партия 2: обработанные BS3, содержащие яичный альбумин эритроциты
Партия 3: обработанные антителом, содержащие яичный альбумин эритроциты
Партия 4: обработанные антителом, содержащие яичный альбумин эритроциты и Поли(I:С)
Партии впрыскивают внутривенно мышам C57BI/6 (3 мыши в группе). Через семь дней после инъекции партии мышей умерщвляют, и их селезенки и печень собирают. Для измерения процента экспрессирующих FOXP3 CD4+ Т-клеток прочной цитометрией в селезенке (Фигура 3, Таблица 8) и в печени (Фигура 4, Таблица 8), применяют 1×106 и 2,5×106 клеток печени и клеток селезенки. Коротко, после ЭЦТ лизиса с применением раствора NH4Cl (StemCell Technologies, кат. № 7850), клетки сначала окрашивают PC5-анти-CD4 (Biotegend. кат. № BLE100514) и FITC-анти-CD25 моноклональными антителами (Biolegend, кат. № BLE110569), и затем инкубируют с буферами фиксации и пермеабилизации (Biolegend, кат. № 421303), затем инкубируют PE-анти-FOXP3 mAb (Biolegend, кат. № 320008) или контролем изотипа.
На Фигуре 3 процент FOXP3+ CD4+ Т-клеток в селезенке определяют проточной цитометрией через 7 дней после внутривенной инъекции мышам C57BL/6 обработанных ионофором (темно-серный столбик), обработанных BS3 (серый столбик) или обработанных антителом (светло-серый столбик) содержащих ЯА эритроцитов, или обработанных антителом, содержащих ЯА эритроцитов и Поли(I:C) (черный столбик) или контрольной среды (белый столбик). Количество уловленного ЯА, впрыскиваемого мышам составляет 8 мкг.
На Фигуре 4, процент FOXP3+ CD4+ Т-клеток в печени определяют проточной цитометрией через 7 дней после внутривенной инъекции мышам C57BL/6 обработанных ионофором (темно-серный столбик), обработанных BS3 (серый столбик) или обработанных антителом (светло-серый столбик) содержащих ЯА эритроцитов, или обработанных антителом, содержащих ЯА эритроцитов и Поли(I:C) (черный столбик) или контрольной среды (белый столбик). Количество уловленного ЯА, впрыскиваемого мышам составляет 8 мкг.
Для измерения процента ЯА-специфических CD8 Т-клеток проточной цитометрией (Фигура 5, Таблица 9), клетки селезенки окрашивают PC7-анти-CD8 (Biolegend, кат. № BLE100722), FITC-анти-CD3 (Biolegend, кат. № BL E100203) и PC5-анти-C062L моноклональными антителами (Biolegend, кат. № BLE104410) и PE-ЯА-тетрамером (Beckman Coulter, кат. № T20076).
На Фигуре 5, процент ЯА-специфических CD8+ Т-клеток в селезенке определяют проточной цитометрией через 7 дней после внутривенной инъекции мышам C57BL/6 обработанных ионофором (темно-серный столбик), обработанных BS3 (серый столбик) или обработанных антителом (светло-серый столбик) содержащих ЯА эритроцитов, или обработанных антителом, содержащих ЯА эритроцитов и Поли(I:C) (черный столбик) или контрольной среды (белый столбик). Количество уловленного ЯА, впрыскиваемого мышам составляет 8 мкг.
Для измерения процента регулирующих Т-клеток (CD4+ CD25+), производящих IL-10, проточной цитометрией (Таблица 10), клетки селезенки (5×106 клеток/мл) культивируют с 0,1 мкг/мл ЯА323-339 пептидом (Genecript, кат. № 41007-1) или без пептида в течение 4 часов при 37°C в 5% CO2-воздухе в 24-луночных культуральных планшетах. Через один час после начала культивирования, Брефелдин A (Ebiosctence, кат. № 420601) добавляют для блокирования секреции цитокина. В конце культивирования клетки сначала окрашивают PC5-анти-CD4 и FITC-анти-CD25 моноклональными антителами и затем инкубируют буфером фиксации (Biolegend, кат. № 420801) и буфером пермеабилизации (Biolegend, кат. № 421002), затем инкубируют PE-анти-IL-10 mAb (Biolegend, кат. № 505008) или контролем изотипа.
Обработка ионофором является единственной обработкой, которая вызывает значительное увеличение регулирующих CD4 Т-клеток, экспрессирующих FOXP3 и в селезенке, и в печени (Таблица 8 и Фигуры 3 и 4, p<0,05). Обработка антителом вызывает значительное увеличение регулирующих CD4 Т-клеток, экспрессирующих FOXP3, только в селезенке (Таблица 8, Фигура 3, p<0,05).
Процент регулирующих FOXP3 CD4+ Т-клеток в селезенке и печени
(± стандартное отклонение)
Только совместная инъекция Ab-обработанных, содержащих ЯА ЭЦТ и Поли(I:C) вызывает повышение процента ЯА-специфических CD8 Т-клеток (Таблица 9, Фигура 5).
Процент регулирующих ЯА-специфических CD8+ Т-клеток в селезенке
(± стандартное отклонение)
Только совместная инъекция Ab-обработанных, содержащих ЯА ЭЦТ и Поли(I:С) вызывает увеличение процента CD4+ CD25+ T-клеток, производящих IL-10, в селезенке, и это производство не является специфическим к ЯА.
Пример 17. Измерение толерантности к ЯА после трех инъекций содержащих яичный альбумин эритроцитов у мышей, обработанных для доставки в печень и подавления АПК провоспалительной реакции
Целью этого исследования является демонстрация того, что инъекции содержащих ЯА ЭЦТ, обработанных для доставки в печень и подавления АПК провоспалительной реакции, ингибируют реакции ЯА T и B клеток, вызванные ЯА и Поли(I:С).
До обработки образцы крови (200 мкл) у мышей C57BL/6 собирают ретроорбитальной пункцией на трубке сепаратора сыворотка/гель (Becton Dickinson, Microtainer TM SST, ref 365951) с получением пре-иммунной сыворотки. Затем мышам внутривенно вводят три раза в дни -7, -3 и -1 контрольную среду, свободный ЯА или партию обработанных ионофором, содержащих яичный альбумин, эритроцитов, полученных из крови мышей C57BL/6 и согласно методике примера 8 (3-4 мыши в группе). Количество свободного и уловленного ЯА, впрыскиваемого мышам, составляет 120 и 90 мкг, соответственно, на 0 и 21 дни мышам вводят ЯА и Поли(I:С) (100 мкг и 50 мкг/мышь, соответственно). Некоторые мыши получают только контрольную среду. Через шесть дней после последней инъекции образцы крови мышей (200 мкл) собирают с получением пост-иммунной сыворотки, и суспензию СЭКФ-меченных C57BL/6 спленоцитов, представляющих или нет ЯА257-264 пептид в соотношении 1:1 впрыскивают мышам для оценки способности цитотоксических CD8 Т-клеток для лизирования клеток SIINFEKL. Через 16 часов после инъекции клеток ЯА257-264 мышей умерщвляют, и их селезенки собирают.
Для измерения процента активированных и ЯА-специфических CD8 Т-клеток проточной цитометрией (Таблица 11), клетки селезенки окрашивают PC7-анти-CD8 (Biolegend, кат. № BLE100722), FITC-анти-CD3 (Biolegend, кат. № BLE100203) и PC5-анти-CD62L моноклональными антителами (Biolegend, кат. № BLE104410) и PE-ЯА-тетрамером (Beckman Coulter, кат. № T20076).
Для измерения процента ЯА-специфического in vivo лизиса (Таблица 12), процент СЭКФ низких и СЭКФ высоких клеток измеряют проточной цитометрией и определяют по следующей формуле:
% = [1-(доля обработанных мышей/доля не обработанных мышей)]×100, где доля = процент СЭКФ высоких/процент СЭКФ низких
Для измерения анти-ЯА IgG1 и IgG2a титров в сыворотке (Таблица 13), различные разведения пре- и пост-иммунной сыворотки (от 1/50 до 1/36450) инкубируют в 96-луночных планшетах MaxiSorp (Nunc, кат. № 442404), предварительно покрытых ЯА (Serlabo, кат. № WQ-LS003054, 5 мкг/мл). Присутствие анти-ЯА IgG1 и IgG2a обнаруживают инкубированием пероксидазы хрена (HRP), конъюгированной с анти-мышиным IgG1 (Thermo Scientific, кат. № cat PA1-86031, разведение 1/4000) или анти-мышиным IgG2a (Thermo Scientific, кат. № PA1-86039, разведение 1/4000) с последующим инкубированием с тетраметилбензидиновым (ТМБ) субстратом (Biolegend, кат. № 421101). Реакцию останавливают раствором 2N H2SO4 и измеряют оптическую плотность при 450 нм и 630 нм с применением планшетного ридера (Biotek, кат. № ELx808). Данные, полученные при 630 нм, вычитают из данных, полученных при 450 нм, и титр антитела определяют как разведение, для которого оптическая плотность более чем в 3 раза больше О.П., полученной для пре-иммунной сыворотки, разведенной 1/50.
Инъекции обработанных ионофором, содержащих ЯА ЭЦТ способны значительно снижать пролиферацию и активацию ЯА-специфических CD8 Т-клеток, индуцированных ЯА и Поли(I:С) по сравнению с инъекциями ЯА (Таблица 11; p<0,05).
Процент активированных и ЯА-специфических CD8+ Т-клеток в селезенке
Инъекции обработанных ионофором, содержащих ЯА ЭЦТ способны значительно снижать ЯА-специфический клеточный лизис, вызванный ЯА и Поли(I:C) (Таблица 12; p<0,01).
Процент ЯА-специфического in vivo лизиса
Мыши, предварительно обработанные обработанными ионофором, содержащими ЯА ЭЦТ имеют очень низкие и/или не имеют титры анти-ЯА IgG1 и IgG2a антител по сравнению с мышами, предварительно обработанными ЯА (Таблица 13).
Титр анти-ЯА IgG1 и IgG2 антител в сыворотке
Пример 18. Измерение иммунной толерантности к ЯА после инъекций различных количеств содержащих яичный альбумин эритроцитов у мышей
Эксперимент проводят как описано в примере 17, за исключением того, что 2 различных партии обработанных ионофором, содержащих яичный альбумин эритроцитов получают с 2 различными концентрациями ЯА, с получением одной партии, партии 1, с 53250±6800 ЯА молекулами на ЭЦТ, как в примере 17, и другой партии, партии 2, с 8250±840 молекулами на ЭЦТ.
C57BI/8 мышам вводят внутривенно, трижды, в дни -7, -3 и -1 110 мкл или 30 мкл партии 1. 110 мкл партии 2, 110 мкл свободного ЯА или 110 мкл консервирующего раствора. Количество ЯА и ЭЦТ, впрыснутых мышам, представлены в Таблице 14. На день 0 и день 21, мышам вводят ЯА и Поли(I:C) (100 мкг и 50 мкг/мышь, соответственно). Через шесть дней после инъекции мышей умерщвляют, селезенки собирают и количество IgG1, IgG2b и IgG2c измеряют в сыворотке как описано в Примере 17 с применением ПХ, конъюгированной с анти-мышиным IgG2b (Southern Biotech, 1090-05) и анти-мышиным IgG2c (Southern Biotech, 1079-05). Для измерения производства ИФНγ, клетки селезенки (5×106 клеток/мл) сначала культивируют с 0,1 мкг/мл ЯА323-339 пептида (Genscript, кат. № 41007-1) или без пептида в течение 48 часов при 37°C в 5% CO2-воздухе в 24-луночных культуральных планшетах. Затем ИФНγ измеряют в надосадочной жидкости проточной цитометрией с применением Cytometric Bead Array (BO Bioscience, 558296 и 558266). Для измерения процента цитотоксичного T-лимфоцитного антигена 4 (ЦТЛА-4), экспрессирующего CD4+ CD25+ Т-клетки проточной цитометрией, клетки селезенки сначала окрашивают PC5-анти-CD4 (Biolegend, кат. № BLE100514) и FITC-анти-CD25 моноклональными антителами (Biolegend, кат. № BLE110569), и затем фиксируют ФРФБ 1% параформальдегидом (Sigma Aldrich, F1635-25 мл) и пермеабилизуют сапонином 0,3% (Sigma Aldrich, 84510), затем инкубируют PE-анти-ЦТЛА-4 mAb (BO Pharminghen, кат. № 553720) или контролем изотипа.
Количества ЯА и ЭЦТ, впрыскиваемые мышам при каждой инъекции
(30 мкл/мышь)
Мыши, предварительно обработанные Партией 1 высокой дозой, высоким количеством ЯА и ЭЦТ, имели значительно меньшее количество титров анти-ЯА IgG1, IgG2b и IgG2c антител, чем мыши, предварительно обработанные ЯА (Таблица 15, IgG1: p<0,002, IgG2b и IgG2c: p≤0,05). Более того, мыши, обработанные Партией 2, тем же количеством ЭЦТ, но более низкой дозой ЯА (8-11 мкг/мышь), также имели значительно меньше титров анти-ЯА IgG1 и IgG2c антител, чем мыши, обработанные ЯА (Таблица 15, IgG1: p<0,006, и IgG2c: p≤0,05). Однако мыши, обработанные Партией 1 низкой дозой, небольшим количеством ЯА и ЭЦТ, имели значительное количество титров антител. Таким образом, количество ЯА и ЭЦТ, впрыснутое мышам, играет ключевую роль в индуцировании иммунной толерантности.
Титры анти-ЯА IgG1, IgG2b и IgG2c антител в сыворотке
Обработка ЯА и Поли(I:С) вызывает увеличение производства ИФНγ в ответ на стимулирование ЯА. Это увеличение наблюдают у мышей, предварительно обработанных свободным ЯА, но не у мышей, предварительно обработанных обработанными ионофором, содержащими ЯА ЭЦТ Таблица 16, (p<0,005).
Производство ИФНγ спленоцитами, стимулированными ЯА классно 2-ограниченного пептида (среднее ± стандартное отклонение)
+2х ЯА и Поли(I:С)
+2х ЯА и Поли(I:С)
+2х ЯА и Поли(I:С)
+2х ЯА и Поли(I:С)
Так как ЦТЛА-4 является белком, который играет важную регулирующую роль в иммунной системе через передачу ингибирующего сигнала на Т-клетку, его экспрессию измеряют на CD4 CD25 регулирующих Т-клетках проточной цитометрией. Мыши, предварительно обработанные Партией 1 высокой дозой и Партией 2 имеют значительно более высокий процент экспрессии ЦТЛА-4 в CD4 CD25 регулирующих Т-клетках, по сравнению с мышами, предварительно обработанными ЯА и мышами, которые получали только ЯА+Поли(I:C) (Таблица 17, Партия 1: p≤0,03 и p≤0,02, соответственно, и Партия 2: p≤0,05). Более того, мыши, предварительно обработанные Партией 1 высокой дозой, имели значительно более высокую среднюю интенсивность флуоресценции (СИФ) экспрессии ЦТЛА-4 в CD4 CD25 регулирующих Т-клетках (Таблица 17, p≤0,03). Наконец, мыши, предварительно обработанные Партией 1 низкой дозой имели значительно более высокий процент экспрессии ЦТЛА-4 в CD4 CD25 регулирующих Т-клетках, по сравнению с мышами, получавшими только ЯА+Поли(I:C) (Таблица 17, p≤0,04).
Процент и средняя интенсивность флуоресценции (СИФ) ЦТЛА-4 в CD4 CD25 регулирующих Т-клетках (среднее ± стандартное отклонение)
+2х ЯА и Поли(I:С)
+2х ЯА и Поли(I:С)
+2х ЯА и Поли(I:С)
+2х ЯА и Поли(I:С)
В заключение, обработка обработанными ионофором, содержащими антиген ЭЦТ позволяет предотвращать или снижать реакцию антиген-специфических T и B клеток в модели профилактики. Не только количество антигена, но и количество ЭЦТ, вводимое мышам, играет ключевую роль в этой терапии.
| название | год | авторы | номер документа |
|---|---|---|---|
| ИММУНОСТИМУЛИРУЮЩАЯ КОМБИНАЦИЯ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ ГЕПАТИТА С | 2006 |
|
RU2431499C2 |
| КОМПОЗИЦИИ ВАКЦИН И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2600798C2 |
| АНТИГЕННАЯ КОМПОЗИЦИЯ ДЛЯ ИНДУЦИРОВАНИЯ ОТВЕТА ЦИТОТОКСИЧЕСКИХ Т-ЛИМФОЦИТОВ, СПОСОБ ИНДУЦИРОВАНИЯ И СПОСОБ ЛЕЧЕНИЯ БОЛЕЗНИ | 1992 |
|
RU2129439C1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННОГО НОВООБРАЗОВАНИЯ | 2018 |
|
RU2730984C1 |
| МИКРОФЛЮИДИЗИРОВАННАЯ КОМПОЗИЦИЯ ДЛЯ ИНДУКЦИИ СПЕЦИФИЧНОГО ЦИТОТОКСИЧЕСКОГО Т-ЛИМФОЦИТНОГО ИММУННОГО ОТВЕТА И ЕЕ ИСПОЛЬЗОВАНИЕ ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ | 1995 |
|
RU2201253C2 |
| МОДИФИЦИРОВАННЫЕ СУПЕРСПИРАЛЬНЫЕ БЕЛКИ С УЛУЧШЕННЫМИ СВОЙСТВАМИ | 2013 |
|
RU2677799C2 |
| КОМБИНИРОВАННЫЙ ПРЕПАРАТ ДЛЯ УВЕЛИЧЕНИЯ ИММУНОГЕННОСТИ ВАКЦИНЫ (ВАРИАНТЫ) | 2008 |
|
RU2491090C2 |
| Иммуногенная композиция, включающая синтетические пептиды, повторяющие последовательности V3-петли оболочечного белка gp120 ВИЧ1 | 2017 |
|
RU2694576C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ИММУННЫХ ЗАБОЛЕВАНИЙ | 2000 |
|
RU2203682C2 |
| КОМПОЗИЦИЯ ДЛЯ СТИМУЛИРОВАНИЯ СОЗРЕВАНИЯ ДЕНДРИТНЫХ КЛЕТОК, СОДЕРЖАЩАЯ ГИБРИДНЫЙ БЕЛОК Rv2299c/ESAT-6 | 2017 |
|
RU2733886C2 |
Группа изобретений относится к фармацевтической промышленности, а именно к индуцированию, у хозяина, иммунной толерантности к пептидному или белковому активному агенту. Композиция, индуцирующая, у хозяина, иммунную толерантность к пептидному или белковому активному агенту, содержит эритроциты, содержащие пептидный или белковый активный агент. Способ индуцирования иммунной толерантности к пептидному или белковому активному агенту. Вышеописанная композиция позволяет эффективно индуцировать иммунную толерантность. 2 н. и 32 з.п. ф-лы, 5 ил., 17 табл., 18 пр.
1. Композиция, индуцирующая, у хозяина, иммунную толерантность к пептидному или белковому активному агенту, где композиция содержит 1-250 мл эритроцитов, содержащих пептидный или белковый активный агент, где эритроциты содержат модификацию, которая способствует нацеливанию эритроцита на ретикулоэндотелиальную систему печени, причем модификация формируется путем химической обработки эритроцита ионофором и, где пептидный или белковый активный агент содержит (i) терапевтический пептид, полипептид или белок; (ii) пептидный или белковый аутоантиген; (iii) пептид, полипептид или белок, вызывающий аллергическую реакцию; или (iv) трансплантационный пептидный или белковый антиген.
2. Композиция по п.1, где терапевтический пептид, полипептид или белок является антителом или его фрагментом.
3. Композиция по п.1, где терапевтический пептид, полипептид или белок является фактором свертывания крови или его фрагментом.
4. Композиция по п.1, где терапевтический пептид, полипептид или белок является ферментом или его фрагментом.
5. Композиция по п.1, где терапевтический пептид, полипептид или белок является фактором роста или его фрагментом.
6. Композиция по п.4, где ферментом или его фрагментом является лизосомный фермент.
7. Композиция по п.6, где лизосомным ферментом является фермент для заместительной терапии (ЗТФ) при болезни Помпе (генерализованный гликогеноз II типа), болезни Фабри или мукополисахаридозах МПС I.
8. Композиция по п.7, где лизосомный фермент выбирают из группы, состоящей из альфаглюкозидазы, ларонидазы и альфагалактозидазы А и альгазидазы альфа.
9. Композиция по п.3, где фактором свертывания крови является Фактор VII, Фактор VIII или Фактор IX.
10. Композиция по п.1, где пептидным или белковым активным агентом является пептидный или белковый аутоантиген и, где индукция иммунной толерантности предназначена для лечения аутоиммунной болезни.
11. Композиция по п.10, где аутоиммунная болезнь включает ревматоидный артрит (RA).
12. Композиция по п.10, где аутоиммунная болезнь включает рассеянный склероз (MS).
13. Композиция по п.12, где пептидным или белковым аутоантигеном является миелиновый основной белок.
14. Композиция по п.10, где аутоиммунная болезнь включает юношеский диабет.
15. Композиция по п.14, где пептидный или белковый аутоантиген выбирают из группы, состоящей из антигена бета-клеток, про-инсулина, инсулиноподобного фактора роста 2 (IGF2) и их комбинаций.
16. Композиция по п.10, где аутоиммунная болезнь включает увеит.
17. Композиция по п.16, где пептидным или белковым аутоантигеном является сетчаточный S антиген.
18. Композиция по п.10, где аутоиммунная болезнь включает воспалительное заболевание толстого кишечника.
19. Композиция по п.10, где аутоиммунная болезнь включает болезнь Крона.
20. Композиция по п.10, где аутоиммунная болезнь включает язвенный колит.
21. Композиция по п.10. где аутоиммунная болезнь включает системную красную волчанку.
22. Композиция по п.10, где аутоиммунная болезнь включает псориаз.
23. Композиция по п.10, где аутоиммунная болезнь включает приобретенную миастению гравис.
24. Композиция по п.23, где пептидным или белковым аутоантигеном является рецептор ацетилхолина.
25. Композиция по п.1, где пептидным или белковым активным агентом является трансплантационный пептидный или белковый антиген и, где индукция иммунной толерантности предназначена для лечения отторжения трансплантата.
26. Композиция по п.25, где отторжение трансплантата включает отторжение трансплантата почек.
27. Композиция по п.25, где отторжение трансплантата включает отторжение трансплантата сердца.
28. Композиция по п.25, где отторжение трансплантата включает отторжение трансплантата печени.
29. Композиция по п.1, где пептид, полипептид или белок, вызывающий аллергическую реакцию, имеет пищевое происхождение.
30. Композиция по п.1, где ионофор включает кальцимицин.
31. Композиция по п.1, где модификация включает экспозицию фосфатидилсерина.
32. Композиция по п.1, где модификация включает повышенную концентрацию внутриклеточного кальция по сравнению с немодифицированным эритроцитом.
33. Композиция по любому из пп. 1-32, где индукция иммунной толерантности предназначена для лечения провоспалительной реакции антигенпрезентирующих клеток (APCs).
34. Способ индуцирования, у хозяина, иммунной толерантности к пептидному или белковому активному агенту, включающий введение хозяину композиции по любому из предшествующих пунктов.
| WO 9613517 A1, 09.05.1996 | |||
| PRPVATOROV VM, IVANOVA GA The role of erythrocytes in the system of controlled transport of agents //Klinicheskaia Meditsina, 01 Jan 2009, 87(9):4-8 | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| ВОРОБЬЕВ А.А | |||
| и др | |||
| Микробиология | |||
| Москва, "Медицина", 2003, стр.158-159. | |||

