Область, к которой относится изобретение
Настоящее изобретение относится к способам и композициям, связанным с модуляцией устойчивости клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к композициям и способам для применения одного или нескольких генов или белков cas для модулирования устойчивости клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к способам и композициям, которые находят применение при разработке и использовании комбинаций штаммов и ротаций заквасочных культур. В дополнительных вариантах осуществления, настоящее изобретение относится к способам мечения и/или идентификации бактерий. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к способам применения локусов CRISPR для определения потенциальной вирулентности фага против клетки и применения CRISPR-cas для модулирования генетической последовательности фага для повышенного уровня вирулентности. В еще одних вариантах осуществления, настоящее изобретение относится к средствам и композициям для разработки и применения фагов в качестве агентов биологического контроля.
Уровень техники
Культуры и, в частности, заквасочные культуры широко используются в пищевой промышленности при изготовлении подвергнутых брожению продуктов, включая молочные продукты (например, йогурт, пахта и сыр), мясные продукты, хлебобулочные продукты, вино и растительные продукты. Получение культур трудоемко, занимает большое пространство и оборудование, и при нем существует значительный риск загрязнения вызывающими порчу бактериями и/или фагами во время стадий размножения. Несостоятельность бактериальных культур вследствие инфекции и размножения бактериофагов (фагов) представляет собой большую проблему при промышленном использовании бактериальных культур. Существует много различных типов фагов, и продолжают появляться новые штаммы. Кроме того, имеется потребность в способах и композициях для прослеживания бактерий, используемых в таких культурах. Действительно, несмотря на достижения в разработке культур, сохраняется потребность в усовершенствовании культур для применения в промышленности.
Краткое описание сущности изобретения
Настоящее изобретение относится к способам и композициям, связанным с модуляцией устойчивости клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к композициям и способам для применения одного или нескольких генов или белков cas для модулирования устойчивости клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции. В некоторых вариантах осуществления, настоящее изобретение относится к способам и композициям, которые находят применение при разработке и использовании комбинаций штаммов и ротаций заквасочных культур. В дополнительных вариантах осуществления, настоящее изобретение относится к способам мечения и/или идентификации бактерий. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к способам применения локусов CRISPR для определения потенциальной вирулентности фага против клетки и применению CRISPR-cas для модулирования генетической последовательности фага для повышенного уровня вирулентности. В еще одних вариантах осуществления, настоящее изобретение относится к средствам и композициям для разработки и применения фагов в качестве агентов биологического контроля.
Настоящее изобретение относится к способам для генерирования, по меньшей мере, одного устойчивого к бактериофагам вариантного штамма, содержащим стадии: (а) воздействие на материнский бактериальный штамм, содержащий, по меньшей мере, от части локуса CRISPR, по меньшей мере, до одной последовательности нуклеиновой кислоты для получения смеси бактерий, содержащей, по меньшей мере, один устойчивый к бактериофагам вариантный штамм, содержащий модифицированный локус CRISPR; (b) выбора устойчивого к бактериофагам вариантного штамма из смеси бактерий; (с) выбора устойчивых к бактериофагам вариантных штаммов, содержащих дополнительный фрагмент нуклеиновой кислоты в модифицированном локусе CRISPR из устойчивых к бактериофагам штаммов, выбранных на стадии (b); и (d) выделения, по меньшей мере, одного устойчивого к бактериофагам вариантного штамма, где штамм содержит дополнительный фрагмент нуклеиновой кислоты в модифицированном локусе CRISPR. В некоторых предпочтительных вариантах осуществления, способы, кроме того, включают стадию сравнения локуса CRISPR или его части материнского бактериального штамма и модифицированного локуса CRISPR устойчивого к бактериофагам вариантного штамма для идентификации устойчивых к бактериофагам вариантных штаммов, содержащих, по меньшей мере, один дополнительный фрагмент нуклеиновой кислоты в модифицированном локусе CRISPR, который отсутствует в локусе CRISPR материнского бактериального штамма. В некоторых особенно предпочтительных вариантах осуществления, способы, кроме того, включают стадию выбора устойчивых к бактериофагам вариантных штаммов, содержащих дополнительный фрагмент нуклеиновой кислоты в модифицированном локусе CRISPR. В некоторых вариантах осуществления, материнский бактериальный штамм подвергается воздействию двух или более последовательностей нуклеиновых кислот. В некоторых вариантах осуществления, материнский бактериальный штамм подвергается одновременному воздействию двух или более последовательностей нуклеиновых кислот, тогда как в некоторых альтернативных вариантах осуществления, материнский бактериальный штамм последовательно подвергается воздействию двух или более последовательностей нуклеиновых кислот. В некоторых особенно предпочтительных вариантах осуществления, материнский бактериальный штамм подвергается воздействию последовательности нуклеиновых кислот посредством инфекции, по меньшей мере, одним бактериофагом, содержащим последовательность нуклеиновой кислоты. В некоторых других предпочтительных вариантах осуществления, по меньшей мере, один бактериофаг выбран из группы семейств вирусов, состоящей из: Corticoviridae, Cystoviridae, Inoviridae, Leviviridae, Microviridae, Myoviridae, Podoviridae, Siphoviridae и Tectiviridae. В некоторых дополнительных предпочтительных вариантах осуществления, по меньшей мере, один бактериофаг представляет собой естественно встречающийся бактериофаг, тогда как в некоторых других предпочтительных вариантах осуществления, по меньшей мере, один бактериофаг представляет собой мутированный бактериофаг, полученный посредством селективного давления с использованием устойчивого к бактериофагам бактериального штамма. В еще одних предпочтительных вариантах осуществления, материнский бактериальный штамм подвергается воздействию нуклеиновой кислоты посредством естественного механизма захвата нуклеиновых кислот. В некоторых вариантах осуществления, естественный механизм захвата нуклеиновых кислот включает естественную компетентность. В некоторых дополнительных вариантах осуществления, естественный механизм захвата нуклеиновых кислот материнского бактериального штамма осуществляется конъюгацией или трансформацией. В еще одних вариантах осуществления, устойчивый к бактериофагам штамм представляет собой нечувствительный к бактериофагам мутант. В еще одних дополнительных вариантах осуществления, материнский бактериальный штамм представляет собой нечувствительный к бактериофагам мутант. В некоторых других вариантах осуществления, конец 5' и/или конец 3' локуса CRISPR материнского бактериального штамма сравнивается с модифицированным локусом CRISPR устойчивого к бактериофагам вариантного штамма. В некоторых других вариантах осуществления, конец 5' и/или конец 3', по меньшей мере, первого повтора CRISPR или, по меньшей мере, первого спейсера CRISPR локуса CRISPR материнского бактериального штамма сравнивается с модифицированным локусом CRISPR устойчивого к бактериофагам вариантного штамма. В еще одних вариантах осуществления, устойчивый к бактериофагам вариантный штамм содержит, по меньшей мере, один дополнительный фрагмент нуклеиновой кислоты в модифицированном локусе CRISPR. В некоторых дополнительных вариантах осуществления, по меньшей мере, часть локуса CRISPR материнского бактериального штамма и, по меньшей мере, часть модифицированного локуса CRISPR устойчивого к бактериофагам вариантного штамма сравниваются амплификацией, по меньшей мере, части локуса CRISPR и, по меньшей мере, части модифицированного локуса CRISPR, для получения амплифицированной последовательности локуса CRISPR и амплифицированной последовательности модифицированного локуса CRISPR. В еще одних вариантах осуществления, амплификация проводится с использованием PCR (полимеразной цепьевой реакции). В некоторых предпочтительных вариантах осуществления, по меньшей мере, часть локуса CRISPR материнского бактериального штамма и, по меньшей мере, часть модифицированного локуса CRISPR устойчивого к бактериофагам вариантного штамма сравниваются секвенированием, по меньшей мере, части локуса CRISPR и, по меньшей мере, части модифицированного локуса CRISPR. В некоторых особенно предпочтительных вариантах осуществления, способы, кроме того, включают стадию секвенирования амплифицированной последовательности локуса CRISPR и амплифицированной последовательности модифицированного локуса CRISPR. В некоторых дополнительных вариантах осуществления, дополнительный фрагмент нуклеиновой кислоты в модифицированном локусе CRISPR представляет собой дополнительный элемент повтора-спейсера. В некоторых предпочтительных вариантах осуществления, дополнительный элемент повтора-спейсера содержит, по меньшей мере, 44 нуклеотида. В некоторых альтернативных предпочтительных вариантах осуществления, дополнительный элемент повтора-спейсера содержит от примерно 44 до примерно 119 нуклеотидов. Однако не предполагается, что настоящее изобретение должно ограничиваться этими определенными диапазонами размера, поскольку другие размеры могут использоваться в настоящем изобретении, как описано в настоящей заявке. В некоторых вариантах осуществления, дополнительный элемент повтора-спейсера содержит, по меньшей мере, одну нуклеотидную последовательность, которая имеет, по меньшей мере, 95% идентичность с повтором CRISPR в локусе CRISPR материнского бактериального штамма. В некоторых других вариантах осуществления, дополнительный элемент повтора-спейсера содержит, по меньшей мере, одну нуклеотидную последовательность, которая имеет, по меньшей мере, 95% идентичность с нуклеотидной последовательностью в геноме, по меньшей мере, одного бактериофага. В некоторых особенно предпочтительных вариантах осуществления, материнский бактериальный штамм представляет собой промышленно применимый штамм. В некоторых дополнительных вариантах осуществления, материнский бактериальный штамм восприимчив к инфекции, по меньшей мере, одним бактериофагом. В некоторых других предпочтительных вариантах осуществления, материнский бактериальный штамм содержит культуру, выбранную из заквасочных культур, пробиотических культур и культур пищевых добавок. В некоторых предпочтительных вариантах осуществления, материнский бактериальный штамм содержит штамм, полученный из культуры. В некоторых особенно предпочтительных вариантах осуществления, культура представляет собой заквасочную культуру, пробиотическую культуру и/или культуру пищевой добавки. В еще одних дополнительных вариантах осуществления, материнский бактериальный штамм выбран из Escherichia, Shigella, Salmonella, Erwinia, Yersinia, Bacillus, Vibrio, Legionella, Pseudomonas, Neisseria, Bordetella, Helicobacter, Listeria, Agrobacterium, Staphylococcus, Streptococcus, Enterococcus, Clostridium, Corynebacterium, Mycobacterium, Treponema, Borrelia, Francisella, Brucella, Campylobacter, Klebsiella, Frankia, Bartonella, Rickettsia, Shewanella, Serratia, Enterobacter, Proteus, Providencia, Brochothrix, Bifidobacterium, Brevibacterium, Propionibacterium, Lactococcus, Lactobacillus, Pediococcus, Leuconostoc и Oenococcus
Настоящее изобретение также относится, по меньшей мере, к одному устойчивому к бактериофагам вариантному штамму, полученному с использованием представленных в настоящем описании способов. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к устойчивым к бактериофагам вариантным штаммам, где устойчивый к бактериофагам вариантный штамм представляет собой промышленно применимый штамм, который представляет собой, по меньшей мере, один компонент заквасочной культуры, пробиотической культуры, культуры пищевой добавки и/других полезных культур.
Настоящее изобретение также относится к композициям, содержащим устойчивый к бактериофагам вариантный штамм, полученный с использованием представленных в настоящем описании способов. В некоторых вариантах осуществления, настоящее изобретение относится к композициям, содержащим, по меньшей мере, два устойчивых к бактериофагам вариантных штамма, полученных с использованием представленных в настоящем описании способов. Настоящее изобретение также относится к пищевым продуктам и/или кормам, содержащим, по меньшей мере, одну из этих композиций. Настоящее изобретение также относится к способам получения пищевых продуктов и/или кормов, включающим добавление, по меньшей мере, одной из этих композиций к пище или корму. Настоящее изобретение также относится к заквасочным культурам, пробиотическим культурам, культурам пищевых добавок и другим полезным культурам, которые содержат, по меньшей мере, одну из этих композиций. Настоящее изобретение также относится к способам ферментации, включающим добавление, по меньшей мере, одной из этих композиций к заквасочной культуре. В некоторых вариантах осуществления, настоящее изобретение также относится к способам ферментации, включающим добавление, по меньшей мере, одной из этих композиций к среде ферментации в условиях, при которых происходит брожение компонентов среды ферментации. В некоторых вариантах осуществления, на брожение не воздействует присутствие бактериофагов. В некоторых вариантах осуществления, среда ферментации представляет собой пищевой продукт. В некоторых предпочтительных вариантах осуществления, пищевой продукт представляет собой диетический продукт. В некоторых особенно предпочтительных вариантах осуществления, диетический продукт представляет собой молоко. В некоторых других вариантах осуществления, по меньшей мере, две различные композиции, содержащие два или более устойчивых к бактериофагам вариантных штамма, последовательно подвергаются воздействию среды ферментации.
Настоящее изобретение также относится к способам уменьшения вредной популяции бактериофагов в среде ферментации, включающим воздействие на среду ферментации, по меньшей мере, одного устойчивого к бактериофагам вариантного штамма, полученного с использованием представленных в настоящем описании способов в условиях, при которых уменьшается популяция бактериофагов.
Настоящее изобретение также относится к способам генерирования, по меньшей мере, одного устойчивого к бактериофагам вариантного штамма, включающим стадии: (а) воздействие на материнский бактериальный штамм, содержащий, по меньшей мере, от части локуса CRISPR, по меньшей мере, до одной последовательности нуклеиновой кислоты для получения смеси бактерий, содержащей, по меньшей мере, один устойчивый к бактериофагам вариантный штамм, содержащий модифицированный локус CRISPR; (b) выбора устойчивого к бактериофагам вариантного штамма из смеси бактерий; (с) сравнения локуса CRISPR или его части материнского бактериального штамма и модифицированного локуса CRISPR устойчивого к бактериофагам вариантного штамма, содержащего, по меньшей мере, один дополнительный фрагмент нуклеиновой кислоты в модифицированном локусе CRISPR, который отсутствует в локусе CRISPR материнского бактериального штамма; (d) выбора устойчивых к бактериофагам вариантных штаммов, содержащих дополнительный фрагмент нуклеиновой кислоты в модифицированном локусе CRISPR; (е) анализа, по меньшей мере, одного дополнительного фрагмента нуклеиновой кислоты в модифицированном локусе CRISPR для идентификации, по меньшей мере, одного устойчивого к бактериофагам вариантного штамма; и (f) выделения, по меньшей мере, одного устойчивого к бактериофагам вариантного штамма.
Настоящее изобретение также относится к способам генерирования мутантов фагов, избегающих CRISPR, включающим: (а) получение, по меньшей мере, одного материнского фага и устойчивого к фагам бактериального штамма, содержащего, по меньшей мере, один локус CRISPR, где локус CRISPR содержит последовательность нуклеиновой кислоты, которая, по меньшей мере, примерно на 95% идентична, по меньшей мере, одной протоспейсерной последовательности в геноме, по меньшей мере, одного материнского фага; (b) воздействие, по меньшей мере, на один материнский фаг устойчивого к фагам бактериального штамма в таких условиях, чтобы был получен, по меньшей мере, один вариант фага; и (с) отбор, по меньшей мере, одного варианта фага, где, по меньшей мере, один вариант фага проявляет способность инфицировать устойчивый к фагам бактериальный штамм и представляет собой мутант фага, избегающий CRISPR. В некоторых вариантах осуществления, устойчивый к фагам бактериальный штамм представляет собой устойчивый к бактериофагам вариантный штамм, полученный с использованием представленных в настоящем описании способов. В некоторых вариантах осуществления, способы, кроме того, включают стадию сравнения, по меньшей мере, части, по меньшей мере, одной протоспейсерной последовательности и мотива CRISPR, расположенного около, по меньшей мере, одной протоспейсерной последовательности в варианте фага, по меньшей мере, с одной протоспейсерной последовательностью и мотивом CRISPR материнского фага. В еще одних дополнительных вариантах осуществления, способы, кроме того, включают стадию выбора вариантных фагов, которые инфицируют устойчивый к фагам бактериальный штамм, где вариантные фаги содержат мутанты фагов, избегающих CRISPR, и где фаги, избегающие CRISPR, содержат, по меньшей мере, одну мутацию, по меньшей мере, в одной протоспейсерной последовательности и/или в мотиве CRISPR мутантов фагов, избегающих CRISPR. В еще одних дополнительных вариантах осуществления, способы постоянно повторяются один или несколько раз с использованием мутантов фагов, избегающих CRISPR и другого устойчивого к фагам CRISPR бактериального штамма, содержащего, по меньшей мере, один локус CRISPR, где локус CRISPR содержит последовательность нуклеиновой кислоты, которая, по меньшей мере, на 95% идентична, по меньшей мере, одной протоспейсерной последовательности мутантов фагов, избегающих CRISPR. В еще одних дополнительных вариантах осуществления, по меньшей мере, один бактериофаг выбран из группы семейств вирусов, состоящей из Corticoviridae, Cystoviridae, Inoviridae, Leviviridae, Microviridae, Myoviridae, Podoviridae, Siphoviridae и Tectiviridae. В некоторых предпочтительных вариантах осуществления, устойчивый к фагам бактериальный штамм выбран из Escherichia, Shigella, Salmonella, Erwinia, Yersinia, Bacillus, Vibrio, Legionella, Pseudomonas, Neisseria, Bordetella, Helicobacter, Listeria, Agrobacterium, Staphylococcus, Enterococcus, Clostridium, Cаmplyobacter, Corynebacterium, Mycobacterium, Treponema, Borrelia, Francisella, Brucella, Klebsiella, Frankia, Bartonella, Rickettsia, Shewanella, Serratia, Enterobacter, Proteus, Providencia, Brochothrix, Bifidobacterium, Brevibacterium, Propionibacterium, Lactococcus, Lactobacillus, Pediococcus, Leuconostoc, Streptococcus и Oenococcus.
Настоящее изобретение также относится к мутантам фагов, избегающих CRISPR, полученным с использованием представленных в настоящем описании способов. В некоторых вариантах осуществления, мутанты фагов, избегающих CRISPR, содержат две или более мутации, присутствующие, по меньшей мере, в двух протоспейсерных последовательностях и/или в мотиве CRISPR.
Настоящее изобретение также относится к мутантам фагов, избегающих CRISPR, где геном мутантов фагов, избегающих CRISPR, в целом создан методом генной инженерии для включения мутаций, по меньшей мере, в одном протоспейсере и/или мотиве CRISPR. В некоторых вариантах осуществления, по меньшей мере, один мотив CRISPR мутирован в мутантах фагов, избегающих CRISPR, тогда как в некоторых альтернативных вариантах осуществления, по меньшей мере, один мотив CRISPR подвергнут делеции в мутантах фагов, избегающих CRISPR. Настоящее изобретение также относится к композициям, содержащим, по меньшей мере, один мутант фага, избегающий CRISPR.
Настоящее изобретение также относится к способам регулирования бактериальных популяций в продукте, включающим воздействие на композиции, содержащие, по меньшей мере, один мутант фага, избегающий CRISPR, среды ферментации, где среда ферментации содержит, по меньшей мере, одну популяцию нежелательных бактерий, в таких условиях, что популяция нежелательных бактерий уменьшается, и среда ферментации используется для генерирования продукта. В некоторых вариантах осуществления, продукт выбран из пищевых продуктов, кормов, косметических изделий, продуктов для личного ухода, гигиенических продуктов, ветеринарных продуктов и пищевых добавок. В еще одних некоторых вариантах осуществления, способы повторяются, по меньшей мере, однократно, и другие композиции и/или композиции, содержащие другие мутанты фагов, избегающих CRISPR, используются при ротации.
В некоторых вариантах осуществления, настоящее изобретение относится к способам и композициям для применения одного или нескольких генов или белков cas для модулирования устойчивости в клетке против нуклеиновой кислоты-мишени или продукта ее транскрипции. В некоторых дополнительных вариантах осуществления, настоящее изобретение относится к композициям и способам применения последовательности рекомбинантной нуклеиновой кислоты, содержащей, по меньшей мере, один ген cas, и, по меньшей мере, два повтора CRISPR вместе, по меньшей мере, с одним спейсером CRISPR, где, по меньшей мере, один спейсер CRISPR является гетерологичным, по меньшей мере, к одному гену cas, и/или, по меньшей мере, к двум повторам CRISPR, для модулирования устойчивости к нуклеиновой кислоте-мишени или продукты ее транскрипции. В еще одних дополнительных вариантах осуществления, настоящее изобретение относится, по меньшей мере, к одной последовательности нуклеиновой кислоты, содержащей, по меньшей мере, один ген cas.
В еще одних вариантах осуществления, настоящее изобретение относится, по меньшей мере, к одной последовательности нуклеиновой кислоты, содержащей, по меньшей мере, один ген cas и, по меньшей мере, два повтора CRISPR. В некоторых вариантах осуществления, настоящее изобретение относится к последовательности нуклеиновой кислоты, содержащей, по меньшей мере, один ген cas и, по меньшей мере, один спейсер CRISPR. В еще одних вариантах осуществления, настоящее изобретение относится к последовательности нуклеиновой кислоты, содержащей, по меньшей мере, один ген cas и, по меньшей мере, один спейсер CRISPR и, по меньшей мере, два повтора CRISPR. В еще одних вариантах осуществления, настоящее изобретение относится к последовательности рекомбинантной нуклеиновой кислоты, содержащей, по меньшей мере, один ген cas и, по меньшей мере, два повтора CRISPR вместе, по меньшей мере, с одним спейсером CRISPR, где спейсер CRISPR является гетерологичным, по меньшей мере, к одному гену cas, и/или, по меньшей мере, к двум повторам CRISPR.
Настоящее изобретение также относится к конструктам, содержащим одну или несколько последовательностей нуклеиновых кислот, описанных в настоящей заявке. В еще одних дополнительных вариантах осуществления, настоящее изобретение относится к векторам, содержащим одну или несколько последовательностей нуклеиновых кислот или один или несколько конструктов, описанных в настоящей заявке. В еще одних вариантах осуществления, настоящее изобретение относится к клеткам, содержащим последовательность нуклеиновой кислоты или конструкт, или вектор, описанные в настоящей заявке.
Настоящее изобретение также относится к способам модулирования (например, придания или увеличения) устойчивости клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции, включающим стадии: (i) идентификации последовательности (например, консервативной последовательности) в организме (в некоторых вариантах осуществления, это последовательность, которая существенна для функции или выживания организма); (ii) получения спейсера CRISPR, который гомологичен идентифицированной последовательности; (iii) получения нуклеиновой кислоты (например, рекомбинантной нуклеиновой кислоты), содержащей, по меньшей мере, один ген cas, и, по меньшей мере, два повтора CRISPR вместе со спейсером; CRISPR; и (iv) введения нуклеиновой кислоты в клетку, таким образом, для придания клетке устойчивости к нуклеиновой кислоте-мишени или к продукту ее транскрипции.
Настоящее изобретение также относится к способам модулирования (например, придания или увеличения) устойчивости клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции, включающим стадии: (i) идентификации одного или нескольких спейсеров CRISPR или псевдо-спейсеров CRISPR в организме, устойчивом к нуклеиновой кислоте-мишени или продукту ее транскрипции; (ii) получения рекомбинантной нуклеиновой кислоты, содержащей, по меньшей мере, один ген или белок cas, и, по меньшей мере, два повтора CRISPR вместе с идентифицированными одним или несколькими спейсерами; и (iii) введения рекомбинантной нуклеиновой кислоты в клетку, таким образом, для придания клетке устойчивости к нуклеиновой кислоте-мишени или к продукту ее транскрипции.
Настоящее изобретение также относится к способам модулирования (например, придания или увеличения) устойчивости клетки, содержащей, по меньшей мере, один или несколько генов или белков cas и два или более повтора CRISPR к нуклеиновой кислоте-мишени или продукту ее транскрипции, включающим стадии: (i) идентификации одного или нескольких спейсеров CRISPR в организме, устойчивом к нуклеиновой кислоте-мишени или продукту ее транскрипции; и (ii) модификации последовательности одного или нескольких спейсера(ов) CRISPR в клетке так, чтобы спейсер(ы) CRISPR имел гомологию спейсеру(ам) CRISPR в организме.
Настоящее изобретение также относится к способам модулирования (например, придания или увеличения) устойчивости клетки, содержащей, по меньшей мере, один или несколько генов или белков cas и два или более повтора CRISPR к нуклеиновой кислоте-мишени или продукту ее транскрипции, включающим стадии: (i) идентификации одного или нескольких спейсеров CRISPR в организме, который по существу устойчив к нуклеиновой кислоте-мишени или продукту ее транскрипции; и (ii) модификации последовательности, по меньшей мере, одного или нескольких спейсера(ов) CRISPR в клетке так, чтобы спейсер(ы) CRISPR имел сниженную степень гомологии спейсеру(ам) CRISPR в организме.
Настоящее изобретение также относится к способам модулирования (например, придания или увеличения) устойчивости клетки, содержащей, по меньшей мере, один или несколько генов или белков cas и два или более повтора CRISPR к нуклеиновой кислоте-мишени или продукту ее транскрипции, включающим модификацию одного или нескольких генов или белков cas и/или двух или более повторов CRISPR в клетке.
Настоящее изобретение также относится к способам идентификации спейсера CRISPR или псевдо-спейсера CRISPR для применения при модулировании устойчивости клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции, включающим стадии: (i) получения клетки, содержащей, по меньшей мере, два повтора CRISPR, и, по меньшей мере, один ген или белок cas; (ii) идентификации спейсера CRISPR или псевдо-спейсера CRISPR в организме, который по существу устойчив к нуклеиновой кислоте-мишени или продукту ее транскрипции; (iii) идентификации последовательности спейсера CRISPR в клетке с тем, чтобы спейсер имел гомологию со спейсером CRISPR организма; и (iv) определения того, модулирует ли клетка устойчивость к нуклеиновой кислоте-мишени или продукту ее транскрипции, где модулирование устойчивости клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции указывает на то, что спейсер CRISPR модулирует устойчивость клетки.
Настоящее изобретение также относится к способам идентификации гена cas для применения при модулировании устойчивости клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции, включающим стадии: (i) получения клетки, содержащей, по меньшей мере, один спейсер CRISPR и, по меньшей мере, два повтора CRISPR, (ii) генной инженерии клетки с тем, чтобы она содержала, по меньшей мере, один ген cas; и (iii) определения того, модулирует ли клетка устойчивость к нуклеиновой кислоте-мишени или продукту ее транскрипции, где модулирование устойчивости клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции указывает на то, что ген cas может применяться для модулирования устойчивости клетки.
Настоящее изобретение также относится к способам идентификации повтора CRISPR для применения при модулировании устойчивости клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции, включающим стадии: (i) получения клетки, содержащей, по меньшей мере, один спейсер CRISPR и, по меньшей один ген cas;(ii) генной инженерии клетки с тем, чтобы она содержала повтор CRISPR; и (iii) определения того, модулирует ли клетка устойчивость к нуклеиновой кислоте-мишени или продукту ее транскрипции, где модулирование устойчивости клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции указывает на то, что повтор CRISPR может применяться для модулирования устойчивости клетки.
Настоящее изобретение также относится к способам идентификации функциональной комбинации гена cas и повтора CRISPR, включающим стадии: (а) определения последовательностей гена cas и повтора CRISPR; (b) идентификации одного или нескольких кластеров генов cas, по данным определения анализом сравнения последовательностей; (с) идентификации одного или нескольких кластеров повторов CRISPR; и (d) комбинирования тех генов cas и повторов CRISPR, которые укладываются в пределы одного и того же кластера, где комбинация последовательностей гена cas и повтора CRISPR, в пределы одного и того же кластера указывает на то, что комбинация представляет собой функциональную комбинацию.
Настоящее изобретение также относится к способам модулирования фаготипа бактериальной клетки, содержащего один или несколько генов или белков cas и два или более повторов CRISPR, включающим стадии: (i) идентификации одного или нескольких псевдо-спейсеров CRISPR в геномной последовательности бактериофага, к которой должна модулироваться устойчивость; и (ii) модификации последовательности одного или нескольких спейсеров CRISPR бактериальной клетки с тем, чтобы спейсер(ы) CRISPR бактериальной клетки имел гомологию с псевдо-спейсером (спейсерами) CRISPR бактериофага, устойчивость к которому должна модулироваться.
Настоящее изобретение также относится к способам модулирования (например, придания или увеличения) устойчивости бактериальной клетки к бактериофагу, включающим стадии: (i) идентификации последовательности (например, консервативной последовательности) в бактериофаге (предпочтительно, последовательности, существенной для функции или выживания бактериофага); (ii) получения спейсера CRISPR, который гомологичен идентифицированной последовательности; (iii) получения нуклеиновой кислоты, содержащей, по меньшей мере, один ген cas и, по меньшей мере, два повтора CRISPR вместе со спейсером CRISPR; и (iv) введение нуклеиновой кислоты в бактериальную клетку, таким образом, делая бактериальную клетку устойчивой к нуклеиновой кислоте-мишени или продукту ее транскрипции.
Настоящее изобретение также относится к способам модулирования (например, придания или увеличения) устойчивости бактериальной клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции в бактериофаге, включающим стадии: (i) идентификации одного или нескольких псевдо-спейсеров CRISPR в геноме бактериофага, который способен обеспечить устойчивость к нуклеиновой кислоте-мишени или продукту ее транскрипции; (ii) получения рекомбинантной нуклеиновой кислоты, содержащей, по меньшей мере, один ген cas и, по меньшей мере, два повтора CRISPR вместе с идентифицированными одним или несколькими псевдо-спейсерами CRISPR; и (iii) введения рекомбинантной нуклеиновой кислоты в бактериальную клетку, таким образом, делая бактериальную клетку устойчивой к нуклеиновой кислоте-мишени или продукту ее транскрипции.
Настоящее изобретение также относится к способам модулирования устойчивости бактериальной клетки, содержащей один или более генов или белков cas и два или более повторов CRISPR к нуклеиновой кислоте-мишени или продукту ее транскрипции в бактериофаге, включающим стадии: (i) идентификации одного или нескольких псевдо-спейсеров CRISPR в бактериофаге, который способен обеспечить устойчивость к нуклеиновой кислоте-мишени или продукту ее транскрипции; (ii) идентификации одного или нескольких спейсеров CRISPR в бактериальной клетке, в которой должна модулироваться устойчивость; и (iii) модификации последовательности спейсера(ов) CRISPR в бактериальной клетке, в которой должна модулироваться устойчивость, с тем, чтобы спейсер(ы) CRISPR имел более высокую степень гомологии с псевдо-спейсером (спейсерами) CRISPR бактериофага, к которому должна модулироваться устойчивость.
Настоящее изобретение также относится к способам определения устойчивости клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции, включающим идентификацию одной или нескольких функциональных комбинаций повтора CRISPR-cas и одного или нескольких спейсеров CRISPR в клетке.
Настоящее изобретение также относится к клеткам, полученным или получаемым с использованием представленных в настоящем описании способов. В некоторых вариантах осуществления. настоящее изобретение относится к спейсерам CRISPR или псевдо-спейсерам CRISPR, полученным или получаемым способами, описанными в настоящей заявке.
В некоторых вариантах осуществления, настоящее изобретение относится к генам cas, полученным или получаемым способами, описанными в настоящей заявке. В некоторых других вариантах осуществления, настоящее изобретение относится к повторам CRISPR, полученным или получаемым способами, описанными в настоящей заявке. В еще одних вариантах осуществления, настоящее изобретение относится к функциональным комбинациям, полученным или получаемым способами, описанными в настоящей заявке. В еще одних вариантах осуществления, настоящее изобретение относится к рекомбинантным локусам CRISPR, содержащим, по меньшей мере, один спейсер CRISPR или псевдо-спейсер CRISPR, и/или, по меньшей мере, один ген cas, и/или, по меньшей мере, один повтор CRISPR и/или функциональную комбинацию.
В некоторых вариантах осуществления, настоящее изобретение относится к способам применения клеток, по меньшей мере, одного спейсера CRISPR или псевдо-спейсера CRISPR, по меньшей мере, одного гена cas, по меньшей мере, одного повтора CRISPR или их функциональной комбинации для модулирования устойчивости клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции.
В некоторых других вариантах осуществления, настоящее изобретение относится к клеточным культурам, содержащим, по меньшей мере, одну клетку, по меньшей мере, один спейсер CRISPR или псевдо-спейсер CRISPR, по меньшей мере, один ген cas, по меньшей мере, один повтор CRISPR или функциональную комбинацию для модулирования устойчивости клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции.
В некоторых других вариантах осуществления, настоящее изобретение относится к пищевым продуктам и/или корму, содержащих представленные в настоящем описании культуры. В некоторых других вариантах осуществления, настоящее изобретение относится к способам получения пищевых продуктов и/или корма, содержащих представленные в настоящем описании культуры. В других дополнительных вариантах осуществления, настоящее изобретение относится к пищевым продуктам и/или корму, полученным или получаемым представленным в настоящем описании способами. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к способам применения культур, представленных в настоящем описании, для получения пищевых продуктов и/или корма.
Настоящее изобретение, кроме того, относится к нуклеотидным последовательностям, содержащим или состоящим из последовательностей, представленных в любой из SEQ ID NOS:7-10 и SEQ ID NOS:359-405, а также их вариантов, фрагментов, гомологов и производных. Настоящее изобретение также относится к аминокислотам, кодирующимся представленными в настоящем описании нуклеотидными последовательностями. В еще одних вариантах осуществления, настоящее изобретение относится к конструктам и/или векторам, содержащим одну или несколько представленных в настоящем описании нуклеотидных последовательностей. Настоящее изобретение также относится к клеткам-хозяевам, содержащим, по меньшей мере, один из представленных в настоящем описании конструктов и/или нуклеотидных последовательностей.
В некоторых вариантах осуществления, один или несколько генов или белков cas используются в комбинации с двумя или более повторов CRISPR. В некоторых других вариантах осуществления, один или несколько генов или белков cas и/или два или более повторов CRISPR получены из одной и той же клетки. В некоторых дополнительных вариантах осуществления, один или несколько генов или белков cas и два или более повторов CRISPR естественно совместно встречаются в одной и той же клетке. В еще некоторых других вариантах осуществления, один или несколько генов или белков используются в комбинации с одни или несколькими спейсерами CRISPR.
В некоторых вариантах осуществления, спейсер(ы) CRISPR получен из другого организма, чем клетка, из которой получены один или несколько генов или белков cas и два или более повторов CRISPR.
В некоторых вариантах осуществления, спейсер получен из клетки, которая устойчива к нуклеиновой кислоте-мишени. В некоторых вариантах осуществления, спейсер CRISPR представляет собой синтетическую последовательность нуклеиновой кислоты. В некоторых других вариантах осуществления, спейсер(ы) CRISPR имеют 100% идентичность с нуклеиновой кислотой-мишенью, по меньшей мере, по длине корового спейсера.
В некоторых вариантах осуществления, один или несколько генов или белков cas применяются в комбинации, по меньшей мере, с одним или несколькими спейсерами CRISPR и, по меньшей мере, с двумя или более повторами CRISPR. В некоторых вариантах осуществления, нуклеиновая кислота-мишень или продукт ее транскрипции получены из ДНК бактериофага. В еще одних вариантах осуществления, нуклеиновая кислота-мишень или продукт ее транскрипции получены, по меньшей мере, из одной плазмиды. В некоторых других вариантах осуществления, нуклеиновая кислота-мишень или продукт ее транскрипции получены, по меньшей мере, из одной ДНК подвижного генетического элемента. В некоторых вариантах осуществления, нуклеиновая кислота-мишень или продукт ее транскрипции получены из перемещаемого элемента и/или вставочной последовательности. В некоторых альтернативных вариантах осуществления, нуклеиновая кислота-мишень или продукт ее транскрипции получены из гена устойчивости к антибиотикам/противомикробным препаратам. В некоторых альтернативных вариантах осуществления, нуклеиновая кислота-мишень или продукт ее транскрипции получены из нуклеиновой кислоты, кодирующей, по меньшей мере, один фактор вирулентности. В некоторых предпочтительных вариантах осуществления, фактор вирулентности включает токсины, интерналины, гемолизины и/или другие факторы вирулентности.
В некоторых вариантах осуществления настоящего изобретения, один или несколько генов или белков cas и два или более повторов CRISPR получены из одной и той же клетки. В некоторых альтернативных вариантах осуществления, один или несколько генов или белков cas и два или более повторов CRISPR естественно встречаются в одной и той же клетке. В еще одних вариантах осуществления, спейсеры CRISPR получены из организма, отличного от клетки, из которой получены один или несколько генов cas и/или два или более повторов CRISPR. В некоторых вариантах осуществления, клетка представляет собой клетку-реципиента или клетку-хозяина.
В некоторых вариантах осуществления, один или несколько генов или белков cas и/или два или более повторов CRISPR получены из одной и той же клетки. В некоторых вариантах осуществления, спейсеры получены из организма, отличного от клетки, содержащей один или несколько генов или белков cas и/или два или более повторов CRISPR.
В некоторых вариантах осуществления, один или несколько генов или белков cas и/или два или более повторов CRISPR естественно совместно встречаются в одной и той же клетке.
В некоторых вариантах осуществления, модификация включает вставку одного или нескольких спейсеров CRISPR в клетку. В некоторых вариантах осуществления, модификация включает генетическую инженерию спейсера CRISPR в клетке. В некоторых вариантах осуществления, спейсер клетки имеет 100% гомологию со спейсером CRISPR или псевдо-спейсером CRISPR организма. В некоторых вариантах осуществления, весь или часть спейсера в клетке модифицирован. В некоторых вариантах осуществления, модификация включает модификацию рекомбинантного спейсера. В некоторых вариантах осуществления, модификация происходит посредством спонтанной мутации или мутагенеза. В некоторых вариантах осуществления, по меньшей мере, один или несколько спейсеров CRISPR в клетке подвергнуты делеции. В некоторых вариантах осуществления, один или несколько генов cas подвергнуты делеции. В некоторых вариантах осуществления, CRISPR и/или один или несколько генов cas подвергнуты делеции. В некоторых вариантах осуществления, один или несколько генов или белков cas и/или два или более повторов CRISPR подвергнуты делеции. В некоторых вариантах осуществления, нуклеотидные последовательности гена cas и повтора CRISPR получены из одного и того же или разных штаммов. В некоторых вариантах осуществления, нуклеотидные последовательности гена cas и повтора CRISPR получены из одного и того же или разных видов.
В некоторых вариантах осуществления, нуклеотидные последовательности гена cas и повтора CRISPR получены из одного и того же или разных родов. В некоторых вариантах осуществления, нуклеотидные последовательности гена cas и повтора CRISPR получены из одного и того же или разных организмов.
В некоторых вариантах осуществления настоящего изобретения, нуклеиновая кислота-мишень в бактериофаге представляет собой высоко консервативную последовательность нуклеиновой кислоты. В некоторых вариантах осуществления, нуклеиновая кислота-мишень в бактериофаге кодирует белок специфичности хозяина. В некоторых других вариантах осуществления, нуклеиновая кислота-мишень в бактериофаге кодирует белок, который является существенным для выживания, репликации или роста бактериофага. В некоторых других вариантах осуществления, нуклеиновая кислота-мишень в бактериофаге кодирует геликазу, примазу, головной или хвостовой структурный белок, белок с консервативным доменом (например, голин, лизин и т.д.) или, по меньшей мере, одну консервативную последовательность среди важных генов фага.
В некоторых вариантах осуществления, способ определения устойчивости клетки к нуклеиновой кислоте-мишени или ее продуктов транскрипции включает дополнительную стадию сравнения последовательности одного или нескольких спейсеров CRISPR в клетке с последовательностью нуклеиновой кислоты-мишени. В некоторых альтернативных вариантах осуществления, способ определения устойчивости клетки к нуклеиновой кислоте-мишени или ее продуктов транскрипции включает дополнительную стадию определения профиля устойчивости клетки.
В некоторых вариантах осуществления, культура представляет собой заквасочную культуру или пробиотическую культуру.
Настоящее изобретение также относится к «меченым бактериям», которые устойчивы к фагу (т.е. «нечувствительные к бактериофагу мутанты», “BIM”). В некоторых вариантах осуществления, настоящее изобретение относится к бактериям, содержащим одну или более последовательностей, происходящих, по меньшей мере, из одного генома бактериофага, который интегрирован в локус CRISPR бактерий. Полученная из фага последовательность обеспечивает метку, которая может идентифицироваться локализацией и/или последовательностью, и/или соседней последовательностью.
В некоторых вариантах осуществления, настоящее изобретение относится к дуплицированным последовательностям (например, дуплицированным повторам CRISPR), которые происходят из материнской бактерии и также интегрированы повторно, последовательно, одновременно или по существу одновременно наряду с последовательностью, происходящей из генома бактериофага.
Кроме того, настоящее изобретение относится к способам, которые содействуют интеграции одной или нескольких других последовательностей бактериофага в локус CRISPR бактериального штамма. В некоторых вариантах осуществления, интеграция других последовательностей бактериофага в локус CRISPR бактериального штамма представляет собой случайное явление. Таким образом, это не всегда тот же локус генома бактериофага, который интегрирован в локус CRISPR бактерии. Однако после его интеграции, он сохраняется и, таким образом, становится отчетливой меткой для мечения и/или прослеживания бактерии. Соответственно, одна или несколько последовательностей, происходящие из генома бактериофага, являются не только новыми для локуса CRISPR материнской бактерии, но также меткой, которая является уникальной для каждой бактерии. Поэтому настоящее изобретение относится к способам мечения (например, маркировки) и/или идентификации бактерий.
В некоторых вариантах осуществления, способы по настоящему изобретению являются «натуральными» и не приводят к получению генетически модифицированных организмов. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к способам мечения бактерий, включающим стадии: (а) воздействия бактериофага на материнскую бактерию; (b) выбора мутанта, нечувствительного к бактериофагу; (с) сравнения локуса CRISPR или его части из материнской бактерии и мутанта, нечувствительного к бактериофагу; и (d) отбора меченой бактерии, содержащей дополнительный фрагмент ДНК в локусе CRISPR, который не присутствует в материнской бактерии.
Настоящее изобретение также относится к меченым бактериям, полученным с использованием способов по настоящему изобретению. В некоторых вариантах осуществления, настоящее изобретение относится к клеточным культурам, содержащим, по меньшей мере, один меченый бактериальный штамм. В еще одних вариантах осуществления, настоящее изобретение относится к пищевому продукту и/или корму, содержащему меченые бактерии, включая без ограничения клеточные культуры, содержащие такие меченые бактерии. Настоящее изобретение также относится к способам получения пищевого продукта и/или корма, содержащего, по меньшей мере, один меченый бактериальный штамм. В некоторых вариантах осуществления, способы включают добавление, по меньшей мере, одного меченого бактериального штамма или клеточную культуру к пищевому продукту и/или корму.
Настоящее изобретение также относится к способам генерирования вариантов CRISPR, содержащим стадии: (а) воздействия бактериофага на материнскую бактерию; (b) выбора бактерии, устойчивой к бактериофагу (т.е. «мутанта, нечувствительного к бактериофагу»); (с) сравнения локуса CRISPR или его части из материнской бактерии и мутанта, нечувствительного к бактериофагу; (d) отбора меченой бактерии, содержащей дополнительный фрагмент ДНК в локусе CRISPR, который не присутствует в материнской бактерии; и (е) выделения и/или клонирования, и/или секвенирования дополнительного фрагмента ДНК. Настоящее изобретение также относится к вариантам CRISPR, полученным с использованием описанных в настоящей заявке способов. В некоторых особенно предпочтительных вариантах осуществления, варианты CRISPR представляют собой устойчивые к фагам мутантные штаммы, которые имеют модифицированный локус CRISPR с дополнительным спейсером.
В некоторых дополнительных вариантах осуществления, настоящее изобретение также относится к способам применения, по меньшей мере, одной нуклеотидной последовательности, полученной или получаемой из бактериофага, для маркировки и/или идентификации бактерий, где нуклеотидная последовательность интегрирована в пределах локуса CRISPR материнской бактерии.
В еще одних вариантах осуществления, настоящее изобретение относится к способам применения, по меньшей мере, одной нуклеотидной последовательности мечения и/или идентификации бактерии, где нуклеотидная последовательность получена или может быть получена: (а) воздействием бактериофага на материнскую бактерию; (b) выбором мутанта, нечувствительного к бактериофагу; (с) сравнением локуса CRISPR или его части из материнской бактерии и мутанта, нечувствительного к бактериофагу; и (d) отбором меченой бактерии, содержащей дополнительный фрагмент ДНК в локусе CRISPR, который не присутствует в материнской бактерии. В еще одних дополнительных вариантах осуществления, настоящее изобретение относится к способам идентификации меченой бактерии, включающим стадию скрининга бактерии для выявления дополнительного фрагмента ДНК в пределах локуса CRISPR бактерии, в качестве еще одного аспекта настоящего изобретения.
В еще одних вариантах осуществления, настоящее изобретение относится к способам идентификации меченых бактерий, включающим стадии: (а) скрининга бактерий для выявления дополнительного фрагмента ДНК в локусе CRISPR; (b) определения нуклеотидной последовательности дополнительного фрагмента ДНК; (с) сравнения нуклеотидной последовательности дополнительного фрагмента ДНК с базой данных меченых бактерий, полученных или получаемых способом по настоящему изобретению; и (d) идентификации в базе данных меченых бактерий нуклеотидной последовательности, которая соответствует дополнительному фрагменту ДНК.
В некоторых предпочтительных вариантах осуществления, конец 5' и/или конец 3' локуса CRISPR материнской бактерии сравнивается с мечеными бактериями. В некоторых альтернативных, предпочтительных вариантах осуществления, сравнивается, по меньшей мере, первый повтор CRISPR или первый спейсер CRISPR (например, кор первого спейсера CRISPR) на конце 5' локуса CRISPR. В еще одних вариантах осуществления, сравнивается, по меньшей мере, последний повтор CRISPR последнего спейсера CRISPR (например, кор последнего спейсера CRISPR) на конце 3' локуса CRISPR.
В некоторых предпочтительных вариантах осуществления, способы по настоящему изобретению включают стадию выбора меченой бактерии, содержащей дополнительный фрагмент ДНК на конце 5' и/или на конце 3' локуса CRISPR, который не присутствует в материнской бактерии. В некоторых альтернативных, предпочтительных вариантах осуществления, способы, кроме того, включают воздействие на материнскую бактерию двух или более бактериофагов или одновременно, или последовательно. В еще одних вариантах осуществления, локус CRISPR или его часть из материнской бактерии и нечувствительный к бактериофагу мутант сравниваются амплификацией локуса CRISPR или его части из материнской бактерии и/или нечувствительного к бактериофагу мутанта. В некоторых особенно предпочтительных вариантах осуществления, амплификация выполняется с использованием PCR. В некоторых альтернативных вариантах осуществления, локус CRISPR или его часть из материнской бактерии и нечувствительный к бактериофагу мутант сравниваются секвенированием локуса CRISPR или его части из материнской бактерии и/или нечувствительного к бактериофагу мутанта. В некоторых предпочтительных вариантах осуществления, локус CRISPR или его часть из материнской бактерии и нечувствительный к бактериофагу мутант сравниваются амплификацией и затем секвенированием локуса CRISPR или его части из материнской бактерии и/или нечувствительного к бактериофагу мутанта. В некоторых альтернативных предпочтительных вариантах осуществления, дополнительный фрагмент ДНК имеет длину, по меньшей мере, 44 нуклеотида. В некоторых дополнительных предпочтительных вариантах осуществления, выбирается меченая бактерия, содержащая два или более дополнительных фрагмента ДНК. В еще одних дополнительных вариантах осуществления, дополнительный фрагмент ДНК содержит, по меньшей мере, одну нуклеотидную последовательность, которая имеет, по меньшей мере, примерно 95% идентичность, или, предпочтительно, 100% идентичность с повтором CRISPR в локусе CRISPR материнской бактерии. В еще одних вариантах осуществления, дополнительный фрагмент ДНК содержит, по меньшей мере, одну нуклеотидную последовательность, которая имеет, по меньшей мере, примерно 95% идентичность, и, в некоторых вариантах осуществления, предпочтительно, примерно 100% идентичность с нуклеотидной последовательностью в геноме бактериофаге, используемом для выбора меченой бактерии. В некоторых вариантах осуществления, настоящее изобретение также относится, по меньшей мере, к одному дополнительному фрагменту ДНК, который содержит первую нуклеотидную последовательность и вторую нуклеотидную последовательность, где, по меньшей мере, одна из нуклеотидных последовательностей имеет примерно 95% идентичность, и, в некоторых вариантах осуществления, предпочтительно, примерно 100% идентичность с нуклеотидной последовательностью в геноме бактериофаге, используемом для выбора меченой бактерии.
В некоторых вариантах осуществления, настоящее изобретение относится к материнским бактериям, которые подходят для применения в качестве культур затравки, пробиотических культур и/или пищевых добавок. В некоторых вариантах осуществления, материнские бактерии выбраны из подходящего рода, включая без ограничения Escherichia, Shigella, Salmonella, Erwinia, Yersinia, Bacillus, Vibrio, Legionella, Pseudomonas, Neisseria, Bordetella, Helicobacter, Listeria, Agrobacterium, Staphylococcus, Streptococcus, Enterococcus, Clostridium, Corynebacterium, Mycobacterium, Treponema, Borrelia, Francisella, Brucella, Bifidobacterium, Brevibacterium, Propionibacterium, Lactococcus, Lactobacillus, Pediococcus, Leuconostoc и Oenococcus. В некоторых вариантах осуществления, бактериофаг выбран из подходящего семейства вирусов, включая без ограничения Corticoviridae, Cystoviridae, Inoviridae, Leviviridae, Microviridae, Myoviridae, Podoviridae, Siphoviridae и Tectiviridae. В некоторых вариантах осуществления, настоящее изобретение относится к клеточным культурам, которые выбраны из заквасочных культур, пробиотических культур и/или пищевых добавок. В некоторых особенно предпочтительных вариантах осуществления, настоящее изобретение относится к способам идентификации меченых бактерий, включающим стадию сравнения, по меньшей мере, одного дополнительного фрагмента ДНК с базой данных последовательностей бактериофагов и/или базой данных бактериальных последовательностей.
Настоящее изобретение также относится к штаммам S. thermophilus, содержащим последовательность, полученную или получаемую из бактериофага, где последовательность содержит спейсер CRISPR из штамма DGCC7778 Streptococcus thermophilus, именуемого в настоящем описании SEQ ID NO:680 (caacacattcaacagattaatgaagaatac; SEQ ID NO:680).
В еще одних дополнительных вариантах осуществления, настоящее изобретение относится к штаммам S. thermophilus, содержащим последовательность, полученную или получаемую из бактериофага, где последовательность содержит SEQ ID NO:680 ниже по ходу транскрипции (например, непосредственно ниже по ходу транскрипции) повтора CRISPR, по меньшей мере, в одном локусе CRISPR.
В других вариантах осуществления, настоящее изобретение относится к штаммам S. thermophilus, содержащим последовательность, полученную или получаемую из бактериофага, где последовательность содержит SEQ ID NO:680 ниже по ходу транскрипции (например, непосредственно ниже по ходу транскрипции) первого повтора CRISPR, по меньшей мере, в одном локусе CRISPR.
В других вариантах осуществления, настоящее изобретение относится к штаммам S. thermophilus, содержащим последовательность, полученную или получаемую из бактериофага, где последовательность содержит спейсер CRISPR (5'-3') из штамма DGCC7778 S. thermophilus (tccactcacgtacaaatagtgagtgtactc; SEQ ID NO:681). В еще одних дополнительных вариантах осуществления, настоящее изобретение относится к штаммам S. thermophilus, содержащим последовательность, полученную или получаемую из бактериофага, где последовательность содержит SEQ ID NO:681 ниже по ходу транскрипции (например, непосредственно ниже по ходу транскрипции) первого повтора CRISPR, по меньшей мере, в одном локусе CRISPR.
В еще одних вариантах осуществления, настоящее изобретение к штаммам S. thermophilus, содержащим последовательность, полученную или получаемую из бактериофага, где последовательность содержит SEQ ID NO:683:
Последовательность CRISPR (5'-3') штамма DGCC7710-RH1 Streptococcus thermophilus


В еще одних вариантах осуществления, настоящее изобретение относится к штаммам S. thermophilus, содержащими последовательность, полученную или получаемую из бактериофага, где последовательность содержит SEQ ID NO:683 ниже по ходу транскрипции (например, непосредственно ниже по ходу транскрипции) первого повтора CRISPR, по меньшей мере, в одном локусе CRISPR. Штаммы S. thermophilus, содержащие последовательность, полученную или получаемую из бактериофага, где последовательность содержит SEQ ID NO:685 (т.е. 5'-TACGTTTGAAAAGAATATCAAATCAATGA-3').
В еще одних вариантах осуществления, настоящее изобретение относится к штаммам S. thermophilus, содержащими последовательность, полученную или получаемую из бактериофага, где последовательность содержит SEQ ID NO:685 ниже по ходу транскрипции (например, непосредственно ниже по ходу транскрипции) первого повтора CRISPR, по меньшей мере, в одном локусе CRISPR.
В некоторых вариантах осуществления, настоящее изобретение относится к способам и композициям, которые находят применение при разработке и использовании комбинаций штаммов и ротаций заквасочных культур. В некоторых дополнительных вариантах осуществления, изобретение относится к применению одновременно одной или нескольких CRISPR BIMs) в заквасочных культурах. В некоторых других вариантах осуществления, изобретение относится к применению одной или нескольких CRISPR BIMs в ротационной схеме. В некоторых других вариантах осуществления, изобретение относится к применению одной или нескольких комбинаций CRISP BIMs в ротационной схеме.
Настоящее изобретение также относится к средству для анализа CRISPR организма-мишени для обеспечения возможности сравнения спейсерных последовательностей с геномом фага биологического контроля. В некоторых вариантах осуществления, настоящее изобретение относится к средству для прогнозирования вирулентности фага и выбора, по меньшей мере, одного фага биологического контроля, по меньшей мере, против одного микроорганизма-мишени.
Настоящее изобретение также относится к способам и композициям для использования CRISPR-cas (т.е. натурального мутагенеза, в некоторых предпочтительных вариантах осуществления) для конструкции, по меньшей мере, одного варианта устойчивого к фагам CRISPR, по меньшей мере, одного микроорганизма-мишени, который затем применяется для генерирования мутантного фага, который передает устойчивость CRISPR-cas посредством мутации в пределах фига, соответствующей последовательности, выбранной из спейсерных последовательностей, псевдоспейсерных последовательностей, проксимальной последовательности, мотивов распознавания и т.д., для усиления вирулентности фага. В некоторых предпочтительных вариантах осуществления, фаг с усиленной вирулентностью находит применение в качестве агентов биологического контроля.
В некоторых вариантах осуществления, настоящее изобретение относится к композициям и способам, подходящим для получения фага, имеющего усиленную вирулентность, по сравнению с материнским фагом. В некоторых вариантах осуществления, по меньшей мере, один клонированный спейсер вводится в активный локус CRISPR-cas, для получения устойчивого к фагу варианта клетки для использования при генерировании мутантного фага. В некоторых предпочтительных вариантах осуществления, способы включают введение последовательности, которая служит в качестве специфической мишени последовательности генома фага (например, области, которая высоко восприимчива в качестве мишени спейсера или последовательности распознавания для включения хозяина спейсера). В некоторых дополнительных вариантах осуществления, настоящее изобретение относится к способам и композициям для прямой инженерии фага с тем, чтобы последовательность генома, соответствующая спейсеру, соответственно мутировалась.
Настоящее изобретение также относится к способам для направления эволюции данного фага с использованием приобретенной устойчивости CRISPR к фагу соответствующего штамма-хозяина для создания более вирулентного и поэтому эффективного агента биологического контроля.
Описание фигур
На фиг. 1 схематически показано, что интеграция спейсера CRISPR в локус CRISPR S. thermophilus обеспечивает сопротивляемость против бактериофага, с которым спейсер CRISPR проявляет идентичность. Материнский DGCC7710 чувствителен к фагу, а BIM DGCC7710RH1 устойчив к фагу. BIM DGCC7710RH1 имеет новый спейсер (Sn) в локусе CRISPR, который проявляет 100% идентичность с последовательностью фага. Как показано на стадии (В), штамм стимулируется фагом 858, и выбирается устойчивый к фагу мутант. Как показано на стадии (С), локус CRISPR 1 мутанта имеет дополнительный спейсер, который имеет 100% идентичность с областью 31.921-31.950 пар оснований фага.
На фиг. 2 схематически показано, что интеграция спейсера CRISPR в локус CRISPR S. thermophilus обеспечивает сопротивляемость против бактериофага, с которым спейсер CRISPR проявляет идентичность. Материнский DGCC7710 чувствителен к фагу, а BIM DGCC7710RH2 устойчив к фагу. BIM DGCC7710RH2 имеет новый спейсер (Sn) в локусе CRISPR, который проявляет 100% идентичность с последовательностью фага. Как показано на стадии (В), штамм стимулируется фагом 858, и выбирается устойчивый к фагу мутант. Как показано на стадии (С), эксперимент независимо повторялся и выбирался устойчивый мутант. Локус CRISPR 1 мутанта имеет дополнительный спейсер (отличный от такового в RH1), который имеет 100% идентичность с областью 17.125-17.224 пар оснований фага.
На фиг. 3 графически представлена иллюстрация получения конструкта CASIKO, в которой ген cas1 разорван гомологичной рекомбинацией.
На фиг. 4 графически представлено получение конструкта с использованием рестрикционного фермента для генерирования конструкта RT из конструкта S1S2. Имеются рестрикционные сайты BglI в пределах повтора, которые позволяют вырезать «среднюю» часть. После ферментативного переваривания, лигазу использовали для соединения вместе двух концевых кусочков, таким образом, генерируя новый конструкт, который имеет RT, но не имеет спейсеров.
На фиг. 5 графически представлена интеграция конструкта RT.
На фиг. 6 графически представлена иллюстрация созданного генной инженерией конструкта S1S2 с использованием специфических праймеров и повторных реакций PCR. Первая панель иллюстрирует все использованные праймеры и протокол для первых двух реакций PCR (реакции №1 с затравками Р1 и Р2 и реакции №2 с праймерами Р2 и Р3). На второй панели показаны продукты PCR, полученные в результате первых двух реакций PCR, с продуктом реакции №1 слева и продуктом реакции №2 справа. На третьей панели показана третья реакция PCR с использованием комбинации продуктов первых двух реакций PCR в качестве матрицы для третьей реакции PCR, и праймера Р1 первой реакции наряду с праймером Р4 второй реакции. На четвертой панели показан продукт PCR №3, который технически генерирует конструкт S1S2.
На фиг. 7 графически представлены детали структуры праймера для праймеров 2 и 3, которые содержат ключевые последовательности для эксперимента, полученные из спейсеров, идентичных последовательностям фага (продукты PCR, полученные из этих праймеров PCR, генерируют спейсеры, которые в конечном счете обеспечат устойчивость к фагам).
На фиг. 8 графически представлена интеграция конструкта S1S2.
На фиг. 9 графически представлен общий вид локуса CRISPR S. thermophilus, вновь приобретенного спейсера в устойчивых к фагам мутантах, и соответствующая чувствительность к фагам. Наверху находится локус CRISPR1 DGCC7710 (WT, дикого типа). Область повтора/спейсера WT представлена посередине: повторы (черные ромбики), спейсеры (нумерованные серые квадраты), лидерный (L, белый квадрат) и концевой повтор (Т, черный ромб). В нижней части слева детально показано содержание спейсера на лидерной стороне локуса у устойчивых к фагам мутантов с вновь приобретенными спейсерами (белые квадраты, S1-S14). Справа представлена чувствительность каждого штамма к фагам 858 и 2972 в виде гистограммы эффективности бляшкообразования (ЕОР), которая представляет отношение количества бляшек мутантного штамма к количеству бляшек штаммов дикого типа.
На фиг. 10 представлена инженерия спейсера CRISPR, инактивация гена cas и соответствующая чувствительность к фагу. I, мутант WTФ858+S1S2, II, мутант WTФ858+S1S2ΔCRISPR1, где CRISPR1 был подвергнут делеции; III, мутант WTФ858+S1S2::pR, где CRISPR1 был вытеснен и замещен уникальным повтором; IV, WTФ2972+S4::pS1S2, мутант штамма WTФ2972+S4, где CRISPR был дуплицирован и замещен вариантом, содержащим S1 и S2; WTФ858+S152::pcas5- с инактивированным cas5; VI, WTФ858+S152::pcas7- с инактивированным cas7. pOR1 указывает инактивированную плазмиду (12). Чувствительность к фагам каждого фага в отношении фагов 858 и 2972 представлена в нижней части в виде гистограммы эффективности бляшкообразования (ЕОР).
На фиг. 11 схематически представлено построение конструкта S1S2.
На фиг. 12 схематически представлена конструкция WTФ858+S1S2ΔCRISPR1.
На фиг. 13 представлено совмещение спейсера CRISPR S1 с соответствующей геномной областью фага 858 и двумя мутантными фагами, которые избежали устойчивости к CRISPR штамма WTФ858+S1S2.
На фиг. 14 схематически представлена конструкция устойчивого к фагам варианта первого уровня. Каждый вариант имеет один дополнительный спейсер в пределах его CRISPR. Дополнительные спейсеры не связаны друг с каждым из других (например, каждый имеет другую последовательность). Все спейсеры происходят из фага Р.
На фиг. 15 схематически представлен второй уровень устойчивых к фагам вариантов, проявляющих увеличенную устойчивость к фагам. Конечные варианты (A1.n и A2.n) происходят от штамма А и имеют последовательную интеграцию дополнительных спейсеров в пределах его CRISPR, причем все спейсеры отличаются от каждого из других и происходящих от фага Р.
На фиг. 16 схематически представлен второй уровень устойчивых к фагам вариантов, проявляющих увеличенную устойчивость к фагам. Конечный вариант (A1pqr) происходит от штамма А и имеет последовательную интеграцию дополнительных спейсеров в пределах его CRISPR, происходящих от 3 различных фагов (т.е. от фага P, Q и R).
На фиг. 17 схематически представлен локус CRISPR1 (панель А) и локус CRISPR3 (панель В) штаммов S. thermophilus, описанных в примерах с 7 по пример 16. Названия штаммов представлены на левой стороне чертежа. Черные стрелки представляют повторы CRISPR, “R” обозначает Повтор и “RT” обозначает Концевой Повтор. Серые стрелки, пронумерованные с 1 до 32 в части А и с 1 до 12 в части В представляют соответственно спейсеры CRISPR1 и спейсеры CRISPR3, как они находятся в DGCC7710. Белые стрелки, пронумерованные с S4 по S35, представляют дополнительные спейсеры CRISPR, специфичные для описанных штаммов.
На фиг. 18 схематически показан вариант осуществления настоящего изобретения, при котором маркирующая последовательность и повтор CRISPR интегрированы на одном конце локуса CRISPR. На панели А указаны локус CRISPR и элементы, включающие повторы (R), спейсеры (S), лидер выше по ходу транскрипции и трейлер ниже по ходу транскрипции с концевым повтором (RT), примыкающим к трейлеру, и гены cas, находящиеся поблизости (здесь, гены 4 cas, называемые с cas1 по cas4, нарисованы не в масштабе). Гены cas могут находиться на любом конце или расщепляться и присутствовать на обоих концах. Кроме того, гены cas могут локализоваться на любой из двух нитей ДНК. На панели В показана последовательность фага при фрагментах последовательности (Sn), используемых в качестве дополнительного спейсера (т.е. маркирующей последовательности). На панели С показана вставка нового спейсера (Sn) (т.е. маркирующей последовательности) на одном конце локуса CRISPR (близко к лидеру в данном примере на конце 5' локуса CRISPR), между двумя повторами. На панели D представлено сравнение содержания локуса CRISPR между материнской и мутантной бактерией (т.е. меченой бактерией) с новым спейсером (Sn) (т.е. маркирующей последовательностью), интегрированным на одном конце локуса CRISPR (близко к лидеру в данном примере), между повторами. Новый спейсер (Sn) составляет маркирующую последовательность, которая специфична для мутантной бактерии (т.е. меченой бактерии). В некоторых вариантах осуществления, применение этого способа приводит к добавлению одного или нескольких спейсеров из последовательности фага.
На фиг. 19 представлено схематическое изображение варианта осуществления, в котором две маркирующие последовательности и два повтора CRISPR интегрированы на одном конце локуса CRISPR. На панели А, (А) указаны локус CRISPR и элементы, включающие повторы (R), спейсеры (S), лидер выше по ходу транскрипции и трейлер ниже по ходу транскрипции с концевым повтором (RT), примыкающим к трейлеру, и гены cas, находящиеся поблизости (здесь, гены 4 cas, называемые с cas1 по cas4, нарисованы не в масштабе). Гены cas могут находиться на любом конце или расщепляться и присутствовать на обоих концах. Гены cas могут локализоваться на любой из двух нитей ДНК. На панели В последовательность фага показана черным цветом при двух фрагментах последовательности (Sn и Sn'), используемых в качестве дополнительных спейсеров (т.е. маркирующих последовательностей). На панели С показана вставка новых спейсеров (т.е. маркирующих последовательностей) (Sn и Sn') на одном и том же конце локуса CRISPR (близко к лидеру в данном примере на конце 5'), каждый из которых находится между двумя повторами. На панели D представлено сравнение содержания локуса CRISPR между материнской и мутантной бактерией (т.е. меченой бактерией) с двумя новыми спейсерами (Sn и Sn'), интегрированными на одном и том же конце локуса CRISPR (близко к лидеру в данном примере на конце 5'), при каждом, локализующимся между повторами. Новые спейсеры Sn и Sn' составляют маркирующую последовательность, которая специфична для мутанта. В некоторых вариантах осуществления, этот способ приводит к добавлению одного или нескольких спейсеров из последовательности фага.
На фиг. 20 представлен график, показывающий эволюцию количества фагов в молоке, содержащем 107 кое/мл D2972 во время ферментации с DGCC7710 (черные ромбы) или с DGCC9836 (пустые квадраты). Молоко представляло собой 10% (масс/об) молочный порошок в воде. Температура инкубации составляла 42°С.
На фиг. 21 представлен график, показывающий эволюцию кумулированного количества фага на WTphi2972+S20 и WTphi2972+S21 в молоке, содержащем 107 кое/мл D2972 во время ферментации, инокулированного WTphi2972+S20 (пунктир) или WTphi2972+S21 (светло-серый цвет), или с WTphi2972+S20 (пунктир), и с WTphi2972+S21 (темно-серый цвет). Молоко представляло собой 10% (масс/об) молочный порошок в воде. Температура инкубации составляла 42°С.
На фиг. 22 представлен Web Logo для мотива CRISPR1 NNAGAAW (SEQ ID NO:696).
На фиг. 23 представлено совмещение выбранных протоспейсеров CRISPR3 и фланкирующих областей и web lgo для мотива CRISPR3 NGGNG (SEQ ID NO:723). На этом чертеже, представлены

Описание изобретения
Настоящее изобретение относится к способам и композициям, связанным с модулированием устойчивости клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к композициям и способам для применения одного или нескольких генов или белков cas для модулирования устойчивости клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к способам и композициям, которые находят применение при разработке и применении комбинаций штаммов и ротаций заквасочных культур. В дополнительных вариантах осуществления, настоящее изобретение относится к способам мечения и/или идентификации бактерий. В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к применению локусов CRISPR для определения потенциальной вирулентности фага против клетки и применению CRISPR-cas для модулирования генетической последовательности фага с целью увеличения уровня вирулентности. В еще одних вариантах осуществления, настоящее изобретение относится к средствам и композициям для разработки и применения фагов в качестве агентов биологического контроля.
Streptococcus thermophilus представляет собой грамотрицательный бактериальный вид с низким G+C, который является ключевым видом, используемым для сбраживания систем молочных культур для получения йогурта и сыра. Сравнительные анализы геномики близко родственных штаммов S. thermophilus ранее выявили, что генетический полиморфизм в первую очередь возникает в гипервариабельных локусах, таких как опероны eps и rps, а также в двух локусах кластерных, вставленных с равномерными промежутками коротких палиндромных повторов (CRISPR) (см., например, Jansen et al., Mol. Microbiol., 43:1565 [2002]; Bolotin et al., Microbiol., 151:2551 [2005] и Bolotin et al., Nat. Biotechnol., 22:1554 [2004]). Как более детально описано в настоящей заявке, локусы CRISPR обычно состоят из нескольких не смежных прямых повторов, разделенных отрезками вариабельных последовательностей, называемыми спейсерами, и часто соседствуют с генами cas (ассоциированными с CRISPR). Хотя функция локусов CRISPR биологически не была установлена, анализы in silico спейсеров выявили гомологию последовательности с инородными элементами, включая последовательности бактериофагов и плазмид (см., например, приведенные ссылки на работы Bolotin et al., Microbiol; Mojica et al. и Pourcel et al.). На основании исключительно анализов in silico, было выдвинуто несколько гипотез, предлагающих роли генов CRISPR и cas, которые включают обеспечение иммунитета против инородных генетических элементов, посредством механизма, основанного на интерференции РНК (см. Makarova et al., Biol. Direct., 1:7 [2006]). Однако не предполагается, что настоящее изобретение ограничивается каким-либо конкретным механизмом и/или средством действия.
Современные стратегии, используемые в промышленности для минимизации инфекции бактериофагом и итоговой несостоятельности бактериальных культур, включают применение: (i) смешанных заквасочных культур; и (ii) применение чередующихся штаммов, имеющих различные профили восприимчивости к фагам (т.е. ротацию штаммов). Традиционно, заквасочные культуры, используемые в молочной промышленности, представляют собой смеси штаммов молочнокислых бактерий. Сложная композиция смешанных заквасочных культур гарантирует то, что обеспечивается определенный уровень устойчивости. Однако повторное субкультивирование культур смешанных штаммов ведет к непрогнозируемым изменениям распределения отдельных штаммов и, в конечном счете, часто к доминированию нежелательного штамма. Это в свою очередь может привести к увеличенной восприимчивости к атаке фагов и риску несостоятельности ферментации.
Ротация выбранных бактериальных штаммов, которые чувствительны к различным фагам, представляет собой другой подход, используемый в настоящее время для ограничения развития фагов. Однако трудно и обременительно идентифицировать и выбрать достаточное число штаммов, имеющих различные профили фаготипа для обеспечения эффективной и надежной программы ротации. Кроме того, непрерывное использование штаммов требует тщательного контроля для выявления новых инфекционных фагов и требуют быстрого замещения инфицированного штамма устойчивым бактериальным штаммом. На промышленных предприятиях, где большие количества основной массы заквасочных культур получают задолго до использования, такая быстрая реакция обычно невозможна. Так, предпринималось несколько попыток для повышения устойчивости культур для использования в промышленности.
Кроме того, хотя было бы полезно иметь заквасочные культуры, которые являются мечеными с тем, чтобы можно было определить их происхождение, это не было сделано. Действительно, хотя возможна вставка синтетического олигонуклеотида в штамм для его маркировки или мечения с использованием технологий рекомбинантной ДНК, меченый штамм считался бы генетически модифицированным организмом, и мог бы вследствие этого столкнуться в законодательными проблемами при промышленных масштабах применения. Таким образом, в данной области существует потребность в натуральных способах и композициях, подходящих для введения в бактерии необычной последовательности, которая могла бы использоваться для идентификации и/или прослеживания бактерий.
Бактериофаги представляют собой самую обширную биологическую сущность (что может оспариваться) на планете (см. Breitbart and Rohwer, Trends Microbiol., 13:278 [2005]). Их повсеместное распространение и многочисленность оказывают важное воздействие на микробную экологию и эволюцию бактериальных геномов (см., Chibani-Chennoufi et al., J. Bacteriol., 186:3677 [2004]). Следовательно, у бактерий развилось разнообразие механизмов естественной защиты, которые нацелены на различные стадии жизненного цикла фагов, заметно блокируя адсорбцию, предотвращая инъекцию ДНК, ограничивая вход ДНК и абортивных инфекционных систем. Эти противовирусные барьеры также могут подвергаться генной инженерии и манипуляциям для улучшения контроля популяций фагов (см., например, Chibani-Chennoufi et al., см. выше; и Sturino and Klaenhammer, Nat. Rev. Microbiol., 4:395 [2006]).
Многочисленные бактерии были выбраны людьми и обширно использовались для ферментации и биотехнологических процессов. К сожалению, одомашненные бактерии, используемые в промышленности, часто восприимчивы к атаке фагов, включая те роды и виды, которые широко используются в качестве молочных культур (см. Brussiw, Ann. Rev. Microbiol., 55:283 [2001]). Соответственно, в промышленности были разработаны различные стратегии борьбы с фагами на основе разнообразия штаммов, нечувствительных к бактериофагам мутантов и плазмид, несущих механизмы устойчивости к фагам.
Определения
Пока нет других определений, в настоящем описании все технические и научные термины, используемые в настоящей заявке, имеют такое же значение, как общепонятно среднему специалисту в данной области, для которого предназначено настоящее изобретение. Хотя в осуществлении описанного в настоящей заявке изобретения находят применение любые способы и материалы, аналогичные или эквивалентные тем, которые описаны в настоящей заявке, в настоящей заявке описаны иллюстративные способы и материалы. Пока контекст ясно не указывает на иное, используемые в настоящем описании термины в единственном числе включают соответствующую форму множественного числа. Пока нет других указаний, нуклеиновые кислоты написаны слева направо при ориентации от 5' к 3'; аминокислотные последовательности написаны слева направо в ориентации от амино к карбокси, соответственно. Следует понимать, что данное изобретение не ограничивается описанными конкретными методологией, протоколами и реагентами, поскольку они могут варьировать, в зависимости от контекста, в котором они используются специалистами в данной области.
Предполагается, что любое максимальное числовое ограничение, приведенное по всему данному описанию, включает каждое более низкое числовое ограничение, как если бы такое более низкое числовое ограничение было ясно написано в настоящем описании. Каждое минимальное числовое ограничение, приведенное по всему данному описанию, должно включать каждое более высокое числовое ограничение, как если бы такие более высокие числовые ограничения были ясно описаны в настоящем описании. Каждый числовой диапазон, представленный по всему настоящему описанию, должен включать каждый более узкий диапазон, который подпадает в пределы такого более широкого числового диапазона, как если бы такие более узкие числовые диапазоны были ясно описаны в настоящем описании.
Используемый в настоящем описании термин «естественно встречающийся» относится к элементам и/или процессу, который происходит в природе.
Используемый в настоящем описании термин «конструкт», «конъюгат, «кассета» и «гибрид» включает нуклеотидную последовательность, прямо или опосредованно присоединенную к другой последовательности (например, регуляторной последовательности, такой как промотер). В некоторых вариантах осуществления, настоящее изобретение относится к конструктам, содержащим нуклеотидную последовательность, функционально связанную с такой регуляторной последовательностью. Термин «функционально связанная» относится к положению прямой мутации и супрессора, где описанные компоненты находятся в связи, позволяющей им функционировать их предназначенным образом. Регуляторная последовательность, «функционально связанная» с кодирующей последовательностью, лигируется таким образом, чтобы была достигнута экспрессия кодирующей экспрессии в условиях, совместимым с контрольной последовательностью. Используемый в настоящем описании термин «регуляторные последовательности» включает промотеры и энхансеры, и другие сигналы регуляции экспрессии. Используемый в настоящем описании термин «промотер» используется в обычном смысле, понимаемом в данной области, например, сайт связывания РНК полимеразы. В некоторых вариантах осуществления, конструкты содержат или экспрессируют маркер, который обеспечивает возможность выбора конструкта нуклеотидной последовательности, например, в бактерии. Существуют различные маркеры. Которые могут использоваться, например, те маркеры, которые обеспечивают устойчивость к антибиотикам/противомикробным средствам.
В некоторых вариантах осуществления, конструкт содержит вектор (например, плазмиду). В некоторых других вариантах осуществления, настоящее изобретение относится к вектору, содержащему один или несколько конструктов или последовательностей, описанных в настоящей заявке. Используемый в настоящем описании термин «вектор» включает векторы экспрессии, векторы трансформации и челночные векторы. Термин «вектор трансформации» означает конструкт, способный переноситься от одного организма к другому организму, который может быть того же вида или может представлять собой другой вид. Конструкты, которые способны переноситься от одного вида к другому, иногда именуются «челночными векторами». В некоторых вариантах осуществления, векторы трансформируются в подходящую клетку-хозяин, как описано в настоящей заявке. В некоторых вариантах осуществления, векторы являются плазмидными или фаговыми векторами, обеспеченными происхождением репликации, необязательно, промотером для экспрессии полинуклеотида, и, необязательно, регулятором промотера. В некоторых вариантах осуществления, векторы содержат одну или несколько отбираемых маркерных нуклеотидных последовательностей. Наиболее подходящие системы отбора для промышленных микроорганизмов представляют собой те, которые образуются группой маркеров отбора, которые не требуют мутации в организме хозяина. В некоторых вариантах осуществления, векторы используются in vitro (например, для получения РНК или для трансфекции или трансформации клетки-хозяина). В некоторых вариантах осуществления, полинуклеотиды включены в рекомбинантный вектор (обычно, реплицируемый вектор), такой как вектор клонирования или экспрессии. Вектор находит применение при репликации нуклеиновой кислоты в совместимой клетке-хозяине.
Введение нуклеиновой кислоты (например, фага, конструкта или вектора () в клетку может быть осуществлено различными способами. Например, в некоторых вариантах осуществления, могут найти применение трансдукция, трансформация, трансфекция фосфатом кальция, трансфекция, опосредованная DEAE-декстраном, трансфекция, опосредованная катионным липидом, электропорообразование, трансдукция или инфекция. Действительно, любой подходящий способ, известный в данной области, находит применение в настоящем изобретении. В некоторых вариантах осуществления, клетки, содержащие экзогенную нуклеиновую кислоту (введенную посредством фага, конструкта или вектора) выбираются для использования любого подходящего способа, известного в данной области.
Сведения о трансформации клеток хорошо документированы в данной области, например, см. (Molecular Cloning: A Laboratory Manual, 2nd edition, 1989, Cold Spring Harbor Laboratory Press) и Ausubel et al., Current Protocols in Molecular Biology (1995), John Wiley & Sons, Inc. В контексте введения нуклеиновой кислоты в клетку, в некоторых вариантах осуществления, предпочтительно, чтобы термин «введение» означал одно или несколько из трансформации, трансфекции, конъюгации или трансдукции. В некоторых особенно предпочтительных вариантах осуществления, на бактериальные штаммы (например, материнские бактериальные штаммы, вариантные бактериальные штаммы и т.д.) «воздействует», по меньшей мере, один фаг, так, чтобы нуклеиновая кислота фага была введена в клетки бактериального штамма.
Используемые в настоящем описании термины «последовательность нуклеиновой кислоты», «нуклеотидная последовательность» и «нуклеиновая кислота» относятся к любой последовательности нуклеиновой кислоты, включая ДНК, РНК, геномную, синтетическую, рекомбинантную (например, кДНК). Предполагается, что эти термины охватывают двухнитевые и/или однонитевые последовательности, либо представляющие смысловую или антисмысловую нить или их комбинацию. Последовательности рекомбинантной нуклеиновой кислоты получают применением любых подходящих методик рекомбинантной ДНК. В некоторых вариантах осуществления, как описано в настоящей заявке, предоставленные последовательности нуклеиновой кислоты включают генные последовательности, которые кодируют CRISPR, Cas и другие последовательности. Действительно, как используется в контексте, настоящее изобретение охватывает последовательности нуклеиновых кислот, которые кодируют различные последовательности CRISPR, включая без ограничения спейсеры, псевдоспейсеры, лидеры и т.д., а также последовательности cas и другие бактериальные и фаговые («бактериофаговые») последовательности нуклеиновых кислот.
Термины «молекула нуклеиновой кислоты, кодирующая», «последовательность нуклеиновой кислоты, кодирующая», «последовательность ДНК, кодирующая» относятся к порядку или последовательности дезоксирибонуклеотидов вдоль нити дезоксирибонуклеиновой кислоты. Порядок этих дезоксирибонуклеотидов определяет порядок аминокислот вдоль полипептидной (белковой) цепи. Таким образом, последовательность ДНК кодирует аминокислотную последовательность.
Используемый в настоящем описании в контексте введения аминокислотной последовательности в клетку термин «введенная» относится к любому способу, подходящему для переноса последовательности нуклеиновой кислоты в клетку. Такие способы введения включают без ограничения слияние протопласта, трансфекцию, трансформацию, сопряженное связывание и трансдукцию. В некоторых особенно предпочтительных вариантах осуществления, нуклеиновая кислота вводится в клетки-реципиенты после инфекции клеток бактериофагом (бактериофагами).
В некоторых вариантах осуществления, последовательности нуклеиновых кислот и нуклеиновые кислоты, предоставленные в настоящем изобретении, являются изолированными или по существу очищенными. Под «изолированной» или «по существу очищенной» подразумевается, что молекулы нуклеиновой кислоты или их биологически активные фрагменты или варианты, гомологи или производные по существу лишены компонентов, обычно обнаруживаемых в ассоциации с нуклеиновой кислотой в натуральном состоянии. Такие компоненты включают без ограничении другой клеточный материал, культуральные среды, материалы, полученные в результате рекомбинантной продукции, и различные химические соединения, используемые при химическом синтезе нуклеиновых кислот.
В некоторых вариантах осуществления, «изолированная» последовательность нуклеиновой кислоты или нуклеиновая кислота или обычно лишена последовательностей нуклеиновой кислоты, которые фланкируют представляющую интерес нуклеиновую кислоту ы геномной ДНК организма, из которого была получена нуклеиновая кислота (например, кодирующие последовательности, присутствующие на концах 5' или 3'). Однако молекула может включать некоторые дополнительные основания или фрагменты, которые не оказывают повреждающего воздействия на основные характеристики композиции.
Используемый в настоящем описании термин «модификация» относится к изменениям, сделанным в пределах последовательностей нуклеиновой кислоты и/или аминокислоты. В некоторых вариантах осуществления, модификации осуществляются с использованием методов генетической инженерии (например, рекомбинантных), тогда как в других вариантах осуществления, модификации производятся с использованием естественно встречающихся генетических механизмов. Предполагается, что вся или часть последовательности будет модифицирована с использованием способов по настоящему изобретению. В некоторых предпочтительных вариантах осуществления, модифицированные нуклеиновые кислоты включают один или несколько естественно встречающихся или рекомбинантно полученных спейсеров CRISPR, генов или белков cas, повторов CRISPR, локусы CRISPR, а также нуклеиновых кислот бактериофага. Любой подходящий способ, известный в данной области, находит применение в настоящем изобретении, включая без ограничения применение PCR, клонирования, направленного на сайт мутагенеза и т.д. Действительно, выпускаемые наборы находят применение в настоящем изобретении. В некоторых вариантах осуществления, используются синтетические олигонуклеотиды. В некоторых вариантах осуществления, используются способы, такие как гомологическая рекомбинация (например, для вставки или делеции спейсеров CRISPR). В некоторых вариантах осуществления, генетическая инженерия включает активацию одной или нескольких последовательностей нуклеиновых кислот (например, локусов CRISPR, повторов CRISPR, спейсеров CRISPR, генов или белков cas, функциональных комбинаций генов cas и повторов CRISPR или их комбинаций).
В некоторых вариантах осуществления, один или несколько спейсеров CRISPR или псевдо-спейсеров CRISPR вставляются, по меньшей мере, в один локус CRISPR. В еще одних вариантах осуществления, модификация не прерывает один или несколько генов cas, по меньшей мере, одного локуса CRISPR. В других вариантах осуществления, один или несколько генов cas остаются интактными. В еще одних дополнительных вариантах осуществления, модификация не прерывает один или несколько повторов CRISPR, по меньшей мере, одного локуса CRISPR. В некоторых вариантах осуществления, один или несколько повторов CRISPR остаются интактными. В некоторых других вариантах осуществления, один или несколько спейсеров CRISPR вставляются в локус CRISPR или в пределы, по меньшей мере, одного локуса CRISPR. В некоторых других вариантах осуществления, один или несколько спейсеров или псевдо-спейсеров CRISPR вставляются на конце 5', по меньшей мере, одного локуса CRISPR.
В некоторых вариантах осуществления, модификация включает введение, по меньшей мере, одного спейсера CRISPR или псевдо-спейсера CRISPR в клетку (например, клетку-реципиент). В некоторых других вариантах осуществления, модификация включает введение, по меньшей мере, одного или нескольких спейсеров CRISPR или псевдо-спейсеров CRISPR в (например, для модификации или замещения) один или несколько спейсеров CRISPR или клетку-реципиент. В некоторых вариантах осуществления, спейсеры CRISPR клетки являются одинаковыми, тогда как в других вариантах осуществления, они являются различными. В некоторых вариантах осуществления, модификация включает вставку, по меньшей мере, одного спейсера CRISPR или псевдо-спейсера CRISPR из организма донора в клетку-реципиент. В некоторых вариантах других осуществления, модификация включает вставку одного или нескольких спейсеров CRISPR или псевдо-спейсеров CRISPR из организма донора в клетку-реципиент в условиях, подходящих для модификации или замещения одного или нескольких спейсеров CRISPR или псевдо-спейсеров CRISPR клетки-реципиента. В некоторых вариантах осуществления, один или несколько спейсеров CRISPR или псевдо-спейсеров CRISPR из организма донора вставляются в один или более, предпочтительно, два или более повтора CRISPR клетки. В некоторых предпочтительных вариантах осуществления, по меньшей мере, одна функциональная комбинация повтора CRISPR-cas остается интактной в клетке.
В некоторых других вариантах осуществления, происходит вставка, примыкающая к одному или нескольким (предпочтительно, двум или более) спейсерам или псевдо-спейсерам CRISPR. Используемый в настоящем описании термин «примыкающая» означает «рядом» в его самом широком смысле и включает «непосредственно примыкающая» к одному или нескольким спейсерам или псевдо-спейсерам CRISPR клетки-реципиента. (т.е. спейсер(ы) CRISPR или псевдо-спейсер(ы) CRISPR вставляются так, чтобы между спейсерами не было промежуточных нуклеотидов).
В других дополнительных вариантах осуществления, спейсер(ы) CRISPR или псевдо-спейсер(ы) CRISPR вставляется так, чтобы было, по меньшей мере, примерно 2, примерно 3, примерно 4, примерно 5, примерно 6, примерно 7, примерно 8, примерно 9, примерно 10, примерно 15, примерно 20, примерно 25, примерно 30, примерно 35, примерно 40, примерно 45, примерно 50 примерно 55, примерно 60, примерно 65, примерно 70, примерно 75, примерно 80, примерно 85, примерно 90, примерно 95, примерно 100, примерно 200, примерно 300, примерно 400, примерно 500, примерно 600, примерно 700, примерно 800, примерно 900, примерно 1000 примерно 10000, примерно 100000 или примерно 1000000, или более промежуточных нуклеотидов между спейсерами.
В некоторых других вариантах осуществления, промежуточный нуклеотид именуется «лидерной последовательностью». В настоящем описании эти термины используются взаимозаменяемо. Лидерная последовательность может иметь разную длину у разных бактерий. В некоторых вариантах осуществления, лидерная последовательность имеет длину, по меньшей мере, из примерно 20, примерно 25, примерно 30, примерно 35, примерно 40, примерно 45, примерно 50 примерно 55, примерно 60, примерно 65, примерно 70, примерно 75, примерно 80, примерно 85, примерно 90, примерно 95, примерно 100, примерно 200, примерно 300, примерно 400 или примерно 500, или более нуклеотидов. В некоторых предпочтительных вариантах осуществления, лидерная последовательность находится между геном cas (на конце 3') и первым повтором CRISPR (на конце 5') локуса CRISPR. В некоторых вариантах осуществления, лидерная последовательность имеет длину примерно 20-500 нуклеотидов.
В некоторых вариантах осуществления, один или несколько спейсеров CRISPR или псевдо-спейсеров CRISPR из донорского организма вставляются, примыкая к одному или нескольким генам cas клетки-реципиента, где гены cas являются одинаковыми или различными. В некоторых дополнительных вариантах осуществления, один или несколько спейсеров CRISPR или псевдо-спейсеров CRISPR из донорского организма вставляются, примыкая к одним и тем же или различным спейсерам клетки-реципиента.
В другом варианте осуществления, каждый один или несколько спейсеров CRISPR или псевдо-спейсеров CRISPR - таких как один или несколько спейсеров CRISPR или псевдо-спейсеров CRISPR из донорского организма - вставляется, примыкая к одним и тем же или другим повторам CRISPR клетки. В другом варианте осуществления, каждый один или несколько спейсеров CRISPR или псевдо-спейсеров CRISPR - таких как один или несколько спейсеров CRISPR или псевдо-спейсеров CRISPR из донорского организма - вставляется, примыкая к одним и тем же или другим генам cas клетки-реципиента.
В некоторых других вариантах осуществления, последовательность одного или нескольких спейсеров CRISPR из донорского организма предоставляются в условиях, при которых клетка-реципиент модифицируется таким образом, что спейсер CRISPR имеет гомологию со спейсером CRISPR или псевдо-спейсером CRISPR донорского организма. В некоторых вариантах осуществления, спейсер CRISPR имеет 100% гомологию со спейсером CRISPR донорского организма.
В некоторых вариантах осуществления, спейсер(ы) CRISPR или псевдо-спейсеры CRISPR содержат ДНК или РНК геномного, синтетического или рекомбинантного происхождения. В некоторых вариантах осуществления, спейсер(ы) CRISPR или псевдо-спейсеры CRISPR являются двухнитевыми, тогда как в других вариантах осуществления, они являются однонитевыми, представляющими или смысловую, или антисмысловую нить, или их комбинации. Предусматривается, что спейсер(ы) CRISPR или псевдо-спейсеры CRISPR должны получаться применением методик ДНК (например, рекомбинантной ДНК), как описано в настоящей заявке.
В некоторых вариантах осуществления, модификация включает вставку одного или нескольких спейсеров CRISPR или псевдо-спейсеров CRISPR из организма донора, который (которые) по существу устойчивы к нуклеиновой кислоте-мишени или продуктам ее транскрипции, в один или несколько локусов CRISPR по существу чувствительной клетки. В некоторых вариантах осуществления, вставка происходит в области или между функциональной комбинацией, по меньшей мере, двух повторов CRISPR и, по меньшей мере, одного гена cas по существу в чувствительной клетке. В некоторых вариантах осуществления, модификация включает модифицирование (например, мутацию) клетки-реципиента (например, плазмидной ДНК или геномной ДНК), так что один или несколько генов cas создаются в ДНК клетки. В некоторых вариантах осуществления, гены cas клонируются в конструкт, плазмиду или вектор и т.д., который затем трансформируется в клетку с использованием любого подходящего способа.
В некоторых вариантах осуществления, модификация включает модифицирование (например, мутацию) ДНК клетки-реципиента (например, такой как ДНК плазмиды или геномной ДНК), так что один или несколько, предпочтительно, два или более повторов CRISPR создаются в ДНК клетки. В некоторых вариантах осуществления, повторы CRISPR клонируются в конструкт, плазмиду или вектор и т.д., который затем трансформируется в клетку с использованием подходящего способа.
В некоторых вариантах осуществления, модификация включает модифицирование (например, мутацию) ДНК клетки-реципиента (например, ДНК плазмиды или геномной ДНК), так что один или несколько функциональных комбинаций cas-повторов CRISPR могут быть клонированы в конструкт, плазмиду или вектор, который затем трансформируется в клетку с использованием подходящего способа.
В некоторых вариантах осуществления, модификация включает модифицирование (например, мутацию) ДНК клетки-реципиента (например, ДНК плазмиды или геномной ДНК), так что один или несколько спейсеров CRISPR создаются в ДНК клетки. В некоторых вариантах осуществления, спейсеры CRISPR могут клонироваться в конструкт, плазмиду или вектор, который затем трансформируется в клетку с использованием подходящего способа. В некоторых вариантах осуществления, спейсер CRISPR фланкируется двумя повторами CRISPR (т.е. спейсер CRISPR имеет, по меньшей мере, один повтор CRISPR с каждой стороны).
В некоторых вариантах осуществления, модификация включает вставку одного или нескольких спейсеров CRISPR (например, гетерологичных спейсеров CRISPR) вблизи (например, примыкая или непосредственно примыкая) к одному или нескольким генам cas и/или лидерной последовательности. Таким образом, в некоторых вариантах осуществления, организация естественно встречающегося локуса CRISPR поддерживается после вставки одного или нескольких спейсеров CRISPR.
Используемый в настоящем описании термин «нуклеиновая кислота-мишень» относится к любой последовательности нуклеиновой кислоты или продукту ее транскрипции, устойчивость к которой в клетке (например, клетке-реципиенте) модулируется. В некоторых вариантах осуществления, устойчивость направлена против самой последовательности нуклеиновой кислоты-мишени. Преимущественно, это придает устойчивость клетке к организму донора, из которого может быть получена нуклеиновая кислота-мишень (нуклеиновые кислоты-мишени). Таким образом, в некоторых вариантах осуществления, вставка псевдо-спейсера CRISPR, полученного из бактериофага, или спейсера (спейсеров) CRISPR, который (которые) является (являются) комплементарными или гомологичными одному или нескольким псевдо-спейсеру (спейсерам) CRISPR, в клетку-реципиент, придает устойчивость к бактериофагу. Таким образом, в некоторых предпочтительных вариантах осуществления, вставка между двумя повторами CRISPR псевдо-спейсера CRISPR, полученного из бактериофага, или спейсера (спейсеров) CRISPR, который (которые) является (являются) комплементарными или гомологичными одному или нескольким псевдо-спейсеру (спейсерам) CRISPR, в клетку-реципиент, придает устойчивость к бактериофагу. В еще одном аспекте, предоставляется способ модулирования устойчивости клетки-реципиента к нуклеиновой кислоте или продукту ее транскрипции.
Настоящее изобретение также относится к способам определения профиля устойчивости клетки к нуклеиновой кислоте-мишени. Используемый в настоящем описании термин «профиль устойчивости» означает одну или несколько сущностей, к которым устойчива или чувствительна клетка. Соответственно, в некоторых вариантах осуществления, профиль устойчивости клетки отражает, что клетка устойчива к первому бактериофагу, чувствительна ко второму бактериофагу, устойчива к первому мобильному генетическому элементу и чувствительна к первому гену устойчивости к антибиотикам и т.д.
В некоторых вариантах осуществления, выявляются и/или секвенируются один или несколько генов или белков cas CRISPR, один или несколько повторов CRISPR, один или несколько генов cas, одна или несколько функциональных комбинаций cas-повторов CRISPR, один или несколько спейсеров CRISPR, и/или один или несколько спейсеров CRISPR и т.д. внутри клетки с тем, чтобы прогнозировать/определить вероятный профиль устойчивости конкретной клетки. В некоторых вариантах осуществления, выявляются и/или секвенируются один или несколько спейсеров CRISPR внутри клетки с тем, чтобы прогнозировать/определить вероятный профиль устойчивости конкретной клетки. Подходящие способы выявления включают без ограничения PCR, гибридизацию ДНК-ДНК, гибридизацию ДНК-РНК, микрочипы ДНК и т.д. Действительно, предполагается, что любой подходящий способ найдет применение в настоящем изобретении. В дополнительных вариантах осуществления, вероятный профиль устойчивости конкретной бактериальной клетки к одному или нескольким бактериофагам используется в качестве фактора прогноза лизотипа для микробного отбора. В некоторых дополнительных вариантах осуществления, один или сколько генов Cas и/или один или несколько повторов CRISPR секвенируются в дополнение к одному или нескольким спейсерам CRISPR, для верификации совместимости комбинации гена cas-повтора CRISPR или для идентификации новых пар совместимых комбинаций cas/повтора.
Используемый в настоящем описании термин «модулирование устойчивости» относится к подавлению, снижению, уменьшению, индукции, приданию, восстановлению, подъему, увеличению или другому воздействию на устойчивость клетки к нуклеиновой кислоте-мишени, как взято в контексте.
Используемый в настоящем описании термин «устойчивость», не предназначен для подразумевания, что клетка на 100% устойчива к нуклеиновой кислоте-мишени или продукту ее транскрипции.
Используемый в настоящем описании термин «устойчивость к нуклеиновой кислоте-мишени или продукту ее транскрипции» означает, что устойчивость придается клетке или организму (например, фагу), которая включает или продуцирует нуклеиновую кислоту-мишень или продукт ее транскрипции. В некоторых вариантах осуществления, минимальный компонент, требуемый для придания иммунитета или устойчивости к нуклеиновой кислоте-мишени или продукту ее транскрипции, представляет собой, по меньшей мере, один ген cas (или один белок Cas) и, по меньшей мере, два повтора CRISPR, фланкирующие спейсер.
В некоторых вариантах осуществления, настоящее изобретение относится к способам для модулирования (например, придания или увеличения) устойчивости клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции, включающим стадии: идентификации последовательности (например, консервативной последовательности) в организме (предпочтительно, последовательность, которая существенна для функции или выживания организма); получения спейсера CRISPR, который содержит последовательность, гомологичную (например, на 100% идентичную) идентифицированной последовательности; получения нуклеиновой кислоты, содержащей, по меньшей мере, один ген cas, и, по меньшей мере, два повтора CRISPR вместе со спейсером CRISPR; и (iv) трансформирования клетки нуклеиновой кислотой, таким образом, для придания клетке устойчивости к нуклеиновой кислоте-мишени или к продукту ее транскрипции.
Используемый в настоящем описании термин «консервативная последовательность» в контексте идентификации консервативной последовательности в организме необязательно должна быть консервирована в прямом смысле, поскольку достаточно знание одной последовательности из данного организма. Кроме того, последовательность необязательно должна быть частью важной сущности. Однако в некоторых вариантах осуществления, консервативная последовательность представляет собой последовательность, которая существенна для функции и/или выживания, и/или репликации, и/или инфекционности и тому подобных организма или клетки. В некоторых вариантах осуществления, консервативная последовательность содержит геликазу, примазу. Головной или хвостовой структурный белок, белок с консервативным доменом (например, холинг, лизин и другие), или консервативные последовательности среди важных генов фага.
В некоторых других вариантах осуществления, настоящее изобретение относится к способам для модулирования (например, придания или увеличения) устойчивости клетки к нуклеиновой кислоте-мишени или продукту ее транскрипции, включающим стадии: идентификации одного или нескольких спейсеров CRISPR в организме, устойчивом к нуклеиновой кислоте-мишени или продукту ее транскрипции, получения рекомбинантной нуклеиновой кислоты, содержащей, по меньшей мере, один ген cas, и, по меньшей мере, два повтора CRISPR вместе с идентифицированным одним или несколькими спейсерами; и трансформации клетки рекомбинантной нуклеиновой кислотой, таким образом, для придания клетке-реципиенту устойчивости к нуклеиновой кислоте-мишени или к продукту ее транскрипции.
В некоторых вариантах осуществления, настоящее изобретение относится к способам для модулирования (например, придания или увеличения) устойчивости клетки, содержащей, по меньшей мере, один или несколько генов или белков cas и один или несколько, предпочтительно, два или более повторов CRISPR к нуклеиновой кислоте-мишени или продукту ее транскрипции, включающим стадии: идентификации одного или нескольких спейсеров CRISPR в организме, устойчивом к нуклеиновой кислоте-мишени или продукту ее транскрипции; и модификации последовательности одного или нескольких спейсера (спейсеров) CRISPR в клетке так, чтобы спейсер(ы) CRISPR имел гомологию спейсеру (спейсерам) CRISPR в организме. В некоторых вариантах осуществления, один или несколько спейсеров CRISPR в клетке-реципиенте модифицируются (например, методами генной инженерии) так, чтобы спейсер(ы) CRISPR имели гомологию с одним или несколькими спейсером (спейсерами) CRISPR в организме донора, который по существу устойчив к нуклеиновой кислоте-мишени или продукту ее транскрипции, для придания клетке устойчивости к нуклеиновой кислоте-мишени. В некоторых предпочтительных вариантах осуществления, один или несколько генов или белков cas и один или несколько, предпочтительно, два или более повторов CRISPR в клетке представляют собой функциональную комбинацию, как описано в настоящее заявке.
Методы генной инженерии включают подходящие методы, известные в данной области, включая без ограничения добавление (например, вставку), делецию (например, удаление) или модификацию (например, мутацию) последовательности одного или нескольких спейсеров CRISPR и/или одного или нескольких псевдо-спейсеров CRISPR в клетке так, чтобы спейсер CRISPR имел гомологию (например, увеличенную гомологию после генетической инженерии) с одним или несколькими спейсерами CRISPR организма донора. Этот этап инженерии приводит к получению клетки, которая является по существу чувствительной к нуклеиновой кислоте-мишени или продукту ее транскрипции, являясь по существу устойчивой к нуклеиновой кислоте-мишени или продукту ее транскрипции.
В некоторых дополнительных вариантах осуществления, настоящее изобретение относится к способам уменьшения или снижения устойчивости клетки-реципиента, содержащей один или несколько генов или белков cas и один или несколько, предпочтительно, два или более повторов CRISPR к нуклеиновой кислоте-мишени или продукту ее транскрипции.
В некоторых вариантах осуществления, способы включают стадии: идентификации одного или нескольких спейсеров CRISPR в организме, который по существу устойчив к нуклеиновой кислоте-мишени или продукту ее транскрипции; и модификации последовательности, по меньшей мере, одного или нескольких спейсера(ов) CRISPR в клетке так, чтобы спейсер(ы) CRISPR имел сниженную степень гомологии со спейсером (спейсерами) CRISPR в организме.
В других вариантах осуществления, способы модулирования (например, уменьшения) устойчивости клетки, содержащей один или несколько генов или белков cas и один или несколько, предпочтительно, два или более повторов CRISPR к нуклеиновой кислоте-мишени или продукту ее транскрипции включают стадии: идентификации спейсера CRISPR или псевдо-спейсера CRISPR в организме, содержащем нуклеиновую кислоту-мишень или продукт ее транскрипции, устойчивость к которой должна модулироваться; и идентификации спейсера CRISPR в организме, в котором должна модулироваться устойчивость; и (iii) адаптации последовательности спейсера CRISPR в организме, у которого должна модулироваться устойчивость, с тем, чтобы спейсер CRISPR имел более низкую степень гомологии спейсера CRISPR или псевдо-спейсера CRISPR организма, содержащего нуклеиновую кислоту-мишень или продукт ее транскрипции, устойчивость к которой должна модулироваться.
Один или несколько спейсеров CRISPR по существу в устойчивой клетке подвергаются генной инженерии для придания клетке чувствительности к нуклеиновой кислоте-мишени. Методы генной инженерии. Которые находят применение, включают без ограничения добавление (например, вставку), делецию (например, удаление) или модификацию одной или нескольких функциональных комбинаций повтора CRISPR-cas или их частей или фрагментов в по существу устойчивой клетке и/или добавление (например, вставку), делецию (например, удаление) или модификацию одного или нескольких спейсеров CRISPR или их частей, или фрагментов в по существу устойчивой клетке. Эта стадия инженерии приводит к получению клетки, которая по существу устойчива к нуклеиновой кислоте-мишени или продукту ее транскрипции, становясь по существу чувствительной к нуклеиновой кислоте-мишени или продукту ее транскрипции.
В некоторых вариантах осуществления, для придания чувствительности клетке, предусматривается, что один или несколько спейсеров CRISPR, один или несколько генов cas, один или несколько, предпочтительно, два или более повторов CRISPR и/или одна или несколько функциональных комбинаций повтора CRISPR-cas из по существу устойчивой клетки будет удален, подвергнут делеции или модифицирован с тем, чтобы больше не была придана устойчивость. В некоторых вариантах осуществления, клетки, которые чувствительны к нуклеиновой кислоте или продукту ее транскрипции, получают так, чтобы их уровни внутри данной культуры (например, заквасочной культуры) могли при желании модулироваться (например, снижаться). Таким образом, в некоторых вариантах осуществления, разрабатываются заквасочные культуры, содержащие два или более бактериальных штамма, с тем, чтобы все члены культуры были чувствительны к одному и тому же агенту (например, одному и тому же бактериофагу). Таким образом, когда наступает время, когда больше нежелательно, чтобы культура была живой, то культура контактирует с тем же одним агентом для уничтожения всех членов культуры. В некоторых вариантах осуществления, чувствительность клеток модулируется к одному или нескольким агентам (например, фагам) с тем, чтобы агент уничтожал только определенную долю клеток в данной культуре (например, примерно 10, примерно 20, примерно 30, примерно 40, примерно 50, примерно 60, примерно 70, примерно 80, примерно 90 или примерно 95% клеток в культуре).
В некоторых вариантах осуществления, клетка-реципиент подвергается генной инженерии с тем, чтобы она содержала спейсер CRISPR или последовательность, соответствующую псевдо-спейсеру CRISPR, посредством этого придавая клетке устойчивость к нуклеиновой кислоте-мишени или продукту ее транскрипции. Клетка подходящим образом подвергается генной инженерии с тем, чтобы спейсер CRISPR или последовательность, соответствующая псевдо-спейсеру CRISPR, использовалась вместе с функциональной комбинацией повтора CRISPR-cas, как описано в настоящей заявки.
В некоторых вариантах осуществления, клетка, устойчивая к нуклеиновой кислоте-мишени или продукту ее транскрипции, подвергается генной инженерии с тем, чтобы спейсер CRISPR, придающий иммунитет против нуклеиновой кислоты-мишени или продукта ее транскрипции, был вставлен в клетку, которая содержит функциональную комбинацию повтора CRISPR-cas, посредством этого придавая клетке устойчивость к нуклеиновой кислоте-мишени или продукту ее транскрипции.
В некоторых других вариантах осуществления, определяется последовательность одного или нескольких спейсеров CRISPR или псевдо-спейсеров CRISPR клетки, которая устойчива к нуклеиновой кислоте-мишени или продукту ее транскрипции. Затем клетка-реципиент подвергается генной инженерии с тем, чтобы она содержала последовательность спейсера CRISPR и функциональную комбинацию повтора CRISPR-cas, посредством этого придавая клетке устойчивость к нуклеиновой кислоте-мишени или продукту ее транскрипции.
В некоторых дополнительных вариантах осуществления, получают спейсер CRISPR из клетки-реципиента и функциональную комбинацию повтора CRISPR-cas из той же или другой клетки (например, той же или другой клетки-реципиента). Затем еще одна клетка-реципиент подвергается генной инженерии с тем, чтобы она содержала последовательность спейсера CRISPR и функциональную комбинацию повтора CRISPR-cas, посредством этого придавая клетке устойчивость к нуклеиновой кислоте-мишени или продукту ее транскрипции.
В некоторых вариантах осуществления, устойчивость направлена против продукта транскрипции последовательности нуклеиновой кислоты-мишени (например, транскрипта последовательности нуклеиновой кислоты-мишени, в частности, РНК или мРНК), транскрипта (например, транскрипта смысловой и антисмысловой РНК) или продукта транскрипции полипептида. В некоторых вариантах осуществления, это придает клетке сопротивляемость против донорского организма, из которого получен продукт транскрипции.
В некоторых вариантах осуществления, нуклеотидная последовательность-мишень содержит ДНК или РНК геномного, синтетического или рекомбинантного происхождения. В некоторых других вариантах осуществления, нуклеотидная последовательность является двухнитевой, тогда как в другом варианте осуществления, она является однонитевой, представляя или смысловую, или антисмысловую нить, или их комбинации. В еще одних вариантах осуществления, нуклеотидная последовательность является такой же, как естественно встречающаяся форма, тогда как в других вариантах осуществления, она получена из нее. В еще одних вариантах осуществления, последовательность-мишень нуклеиновой кислоты получена из гена. В некоторых других вариантах осуществления, последовательность-мишень нуклеиновой кислоты получена из варианта, гомолога, фрагмента или производного гена. В некоторых предпочтительных вариантах осуществления, нуклеиновая последовательность-мишень получена из бактериофага. В некоторых предпочтительных вариантах осуществления, нуклеиновая последовательность-мишень получена из ДНК плазмиды. В некоторых предпочтительных вариантах осуществления, нуклеиновая последовательность-мишень получена из мобильного генетического элемента. В некоторых дополнительных вариантах осуществления, нуклеиновая последовательность-мишень получена из перемещаемого элемента или последовательности вставки. В других дополнительных вариантах осуществления, нуклеиновая последовательность-мишень получена из гена, который придает устойчивость. В некоторых других вариантах осуществления, нуклеиновая последовательность-мишень получена из гена, который придает устойчивость к антибиотику или противомикробному средству. В некоторых предпочтительных вариантах осуществления, нуклеиновая последовательность-мишень получена из фактора вирулентности. В некоторых дополнительных вариантах осуществления, нуклеиновая последовательность-мишень получена из токсина, интерналина или гемолизина.
В некоторых предпочтительных вариантах осуществления, нуклеиновая последовательность-мишень или продукт ее транскрипции получен из одной или нескольких бактерий. Таким образом, в некоторых предпочтительных вариантах осуществления, устойчивость бактериальных клеток модулируется с использованием способов и композиций по настоящему изобретению. В некоторых предпочтительных вариантах осуществления, нуклеиновая последовательность-мишень получена из гена, ассоциированного с устойчивостью к переносу плазмид у бактерий. В некоторых вариантах осуществления, один или несколько спейсеров CRISPR в клетке модифицируются так, чтобы спейсер CRISPR клетки имел гомологию к спейсеру CRISPR и/или псевдо-спейсеру CRISPR, содержащемуся в ДНК плазмиды бактериальной клетки, посредством этого обеспечивая сопротивляемость против конкретной плазмиды (плазмид). Таким образом, предотвращается перенос инородной ДНК в клетку. В некоторых предпочтительных вариантах осуществления, мишенью являются определенные области внутри ДНК плазмиды с тем, чтобы обеспечить иммунитет против ДНК плазмиды. Например, в некоторых вариантах осуществления, мишенью являются последовательности внутри происхождения репликации плазмиды или последовательности внутри генов, кодирующих репликационные белки.
В некоторых вариантах осуществления, настоящее изобретение относится к способам, включающим стадии: идентификации спейсера CRISPR или псевдо-спейсера CRISPR, полученного из ДНК плазмиды бактериальной клетки, против которых должна модулироваться устойчивость; и модификации последовательности спейсера CRISPR в клетке, в которой должна модулироваться устойчивость, с тем чтобы спейсер CRISPR имел гомологию к спейсеру CRISPR и/или псевдо-спейсеру CRISPR, содержащемуся в ДНК плазмиды бактериальной клетки.
В еще одних вариантах осуществления, настоящее изобретение относится к способам придания устойчивости клетке против переноса плазмиды, включающим стадии: идентификации спейсера CRISPR и/или псевдо-спейсера CRISPR, полученного из ДНК плазмиды; идентификации одной или нескольких функциональных комбинаций повтора CRISPR-гена cas в клетке, которая по существу чувствительна к плазмиде; и генной инженерии одного или нескольких локусов CRISPR в по существу чувствительной клетке с тем, чтобы они содержали один или несколько спейсеров CRISPR и/или псевдо-спейсеров CRISPR из плазмиды, посредством этого делая клетку устойчивой.
В некоторых вариантах осуществления, нуклеотидная последовательность-мишень получена из гена, ассоциированного с устойчивостью к одному или нескольким мобильным генетическим элементам. В некоторых вариантах осуществления, конкретные спейсеры CRISPR и/или псевдо-спейсеры CRISPR, полученные из одного или нескольких мобильных генетических элементов, добавляются в пределы локуса CRISPR клетки с тем, чтобы обеспечить сопротивляемость против мобильных генетических элементов (например, перемещаемых элементов и последовательности вставок), таким образом, предотвращая перенос инородной ДНК и генетического дрейфа. В некоторых вариантах осуществления, мишенями являются конкретные области в пределах транспозонов и последовательностей вставок с тем, чтобы обеспечить иммунитет против мобильных генетических элементов. Например, в некоторых вариантах осуществления, мишени включают без ограничения конъюгативные транспозоны (Tn916), транспозоны II класса (Tn501), последовательности вставок (IS26) и гены треспосазы.
В некоторых вариантах осуществления, настоящее изобретение относится к способам, включающим стадии: идентификации спейсера CRISPR и/или псевдо-спейсера CRISPR, полученного из одного или нескольких мобильных генетических элементов клетки, против которых должна модулироваться устойчивость, и модификации спейсера CRISPR в клетке, в которой должна модулироваться устойчивость, с тем, чтобы спейсер CRISPR и/или псевдо-спейсер CRISPR клетки имел гомологию к спейсеру CRISPR, содержащемуся в мобильном генетическом элементе (элементах) клетки.
В еще одних вариантах осуществления, настоящее изобретение относится к способам придания устойчивости клетке против одного или нескольких мобильных генетических элементов, включающим стадии: идентификации спейсера CRISPR и/или псевдо-спейсера CRISPR, полученного из одного или нескольких мобильных генетических элементов; идентификации одной или нескольких функциональных комбинаций повтора CRISPR-гена cas в клетке, которая по существу чувствительна к одному или нескольким мобильным генетическим элементам; и генной инженерии одного или нескольких локусов CRISPR в по существу чувствительной клетке с тем, чтобы они содержали или имели гомологию к одному или нескольким спейсерам CRISPR и/или псевдо-спейсерам CRISPR из одного или нескольких мобильных генетических элементов, для придания клетке устойчивости.
В некоторых вариантах осуществления, нуклеотидная последовательность-мишень получена из гена, ассоциированного с устойчивостью к антибиотикам и/или противомикробным средствам. Используемый в настоящем описании термин «противомикробная» относится к любой композиции, которая уничтожает или ингибирует рост или репродукцию микроорганизмов. Предполагается, что термин охватывает антибиотики (т.е. композиции, продуцируемые другими микроорганизмами), а также синтетически продуцируемые композиции. Гены устойчивости к противомикробным средствам включают без ограничения blatem, blarob, blashv, aadB, aacCl, aacC2, аасСЗ, aacA4, mecA, vanA, vanH, vanX, satA, aacA-aphH, vat, vga, msrA sul и/или int. Гены устойчивости к противомикробным средствам включают без ограничения к родам Escherichia, Klebsiella, Pseudomonas, Proteus, Streptococcus, Staphylococcus, Enterococcus, Haemophilus и Moraxella мобильным генетическим элементам. Гены устойчивости к противомикробным средствам также включают гены, которые получены из бактериальных видов, включающих без ограничения Escherichia coli, Klebsiella pneumoniae, Pseudomonas aeruginosa, Proteus mirabilis, Streptococcus pneumoniae, Staphylococcus aureus, Staphylococcus epidermidis, Enterococcus faecalis, Staphylococcus saprophyticus, Streptococcus pyogenes, Haemophilus influenzae и Moraxella catarrhalis. В некоторых вариантах осуществления, конкретные спейсеры CRISPR и/или псевдо-спейсеры CRISPR, полученные из генов, кодирующих устойчивость к противомикробным средствам, добавляются в пределы локуса CRISPR клетки-реципиента в условиях, в которых предотвращается перенос генов устойчивости к противомикробным средствам (т.е. маркеров). В некоторых вариантах осуществления, мишени также включают vanR, (т.е. устойчивости к ванкомицину), tetR (т.е. устойчивости к тетрациклину) и/или факторы устойчивости, которые обеспечивают устойчивость к бета-лактамазе.
В некоторых вариантах осуществления, настоящее изобретение относится к способам, включающим стадии: идентификации одного или нескольких спейсеров CRISPR и/или псевдо-спейсеров CRISPR, полученных из клетки, которая содержит один или несколько генов устойчивости к противомикробным средствам или маркеров; и модификации последовательности спейсера CRISPR в клетке, которая не содержит или не экспрессирует гены устойчивости к противомикробным средствам или маркеры, с тем, чтобы спейсер CRISPR клетки имел гомологию к одному или нескольким спейсерам CRISPR и/или псевдо-спейсерам CRISPR, содержащимся в клетке, которая содержит один или несколько генов устойчивости к противомикробным средствам.
В еще одних вариантах осуществления, настоящее изобретение относится к способам модулирования приобретения маркеров устойчивости к противомикробным средствам у клетки, включающим стадии: идентификации одного или нескольких спейсеров CRISPR и/или псевдо-спейсеров CRISPR, полученных из клетки, которая содержит один или несколько генов устойчивости к противомикробным средствам или маркеров; идентификации одного или нескольких локусов CRISPR в клетке, которая не содержит или не экспрессирует гены устойчивости к противомикробным средствам или маркеры; и модификации последовательности спейсера CRISPR в клетке, которая не содержит или не экспрессирует гены устойчивости к противомикробным средствам или маркеры, с тем, чтобы спейсер CRISPR и/или псевдо-спейсер CRISPR имел гомологию к спейсеру CRISPR, содержащемуся в клетке, устойчивой к переносу генов, придающих устойчивость к одному или нескольким противомикробным средствам.
В некоторых вариантах осуществления, нуклеотидная последовательность-мишень получена, по меньшей мере, из одного гена, ассоциированного с фактором (факторами) вирулентности. В некоторых вариантах осуществления, конкретные спейсеры CRISPR и/или псевдо-спейсеры CRISPR, полученные из генов, кодирующих факторы вирулентности, добавляются в пределы бактериального локуса CRISPR для обеспечения сопротивляемости против переноса генов, придающих вирулентность бактериям. В некоторых вариантах осуществления, мишенями являются факторы, которые обычно вносят вклад в микробную вирулентность (например, у патогенов), такие как токсины, интерналины, гемолизины и другие факторы вирулентности.
Настоящее изобретение также относится к способам, включающим стадии: идентификации одного или нескольких спейсеров CRISPR и/или псевдо-спейсеров CRISPR, полученных из клетки, которая содержит один или несколько факторов вирулентности; и модификации последовательности спейсера CRISPR в клетке, которая не содержит или не экспрессирует фактор(ы) вирулентности, с тем, чтобы спейсер CRISPR клетки имел гомологию к одному или нескольким спейсерам CRISPR и/или псевдо-спейсерам CRISPR, содержащимся в клетке, которая содержит один или несколько факторов вирулентности.
В еще одних вариантах осуществления, настоящее изобретение относится к способам придания клетке сопротивляемости против одного или нескольких фактора (факторов) вирулентности или маркера (маркеров), включающим стадии: идентификации одного или нескольких спейсеров CRISPR и/или псевдо-спейсеров CRISPR, полученных из одного или нескольких фактора (факторов) вирулентности или маркера (маркеров); идентификации одной или нескольких функциональных комбинаций повтора CRISPR-cas в клетке, которая по существу чувствительна к одному или нескольким фактору (факторам) вирулентности или маркеру (маркерам); и генной инженерии одного или нескольких локусов CRISPR в по существу чувствительной клетке с тем, чтобы они включали один или несколько спейсеров CRISPR и/или псевдо-спейсеров CRISPR из одного или нескольких фактора (факторов) вирулентности или маркера (маркеров) для придания клетке устойчивости.
Настоящее изобретение охватывает применение их вариантов, гомологов, производных и фрагментов, включая варианты, гомологи, производные и фрагменты локусов CRISPR, спейсеров CRISPR, псевдо-спейсеров CRISPR, генов или белков cas, повторов CRISPR, функциональных комбинаций повтора CRISPR-гена cas, последовательностей-мишеней нуклеиновых кислот или продуктов их транскрипции.
Используемый термин «вариант» используется для обозначения естественно встречающегося полипептида или нуклеотидной последовательности, которая отличается от последовательности дикого типа.
Термин «фрагмент» указывает на то, что полипептид или нуклеотидная последовательность содержит фракцию последовательности дикого типа. Он может содержать один или несколько больших примыкающих отрезков последовательности или множество маленьких отрезков. Последовательность может также содержать элементы последовательности, например, она может представлять собой гибридный белок с другим белком. Предпочтительно, последовательность содержит, по меньшей мере, 50%, предпочтительнее, по меньшей мере, 65%, предпочтительнее, по меньшей мере, 80%, предпочтительнее, по меньшей мере, 85%, предпочтительнее, по меньшей мере, 90%, предпочтительнее, по меньшей мере, 95%, предпочтительнее, по меньшей мере, 96%, предпочтительнее, по меньшей мере, 97%, предпочтительнее, по меньшей мере, 98%, наиболее предпочтительно, по меньшей мере, 99% последовательности дикого типа.
Предпочтительно, фрагмент сохраняет 50%, предпочтительнее, 60%, предпочтительнее, 70%, предпочтительнее, 80%, предпочтительнее, 85%, предпочтительнее, 90%, предпочтительнее, 95%, предпочтительнее, 96%, предпочтительнее, 97%, предпочтительнее, 98% или, наиболее предпочтительно, 99% активности полипептида или нуклеотидной последовательности дикого типа.
Предпочтительно, спейсер CRISPR или псевдо-спейсер CRISPR содержит, по меньшей мере, 50%, предпочтительнее, по меньшей мере, 65%, предпочтительнее, по меньшей мере, 80%, предпочтительнее, по меньшей мере, 85%, предпочтительнее, по меньшей мере, 90%, предпочтительнее, по меньшей мере, 95%, предпочтительнее, по меньшей мере, 96%, предпочтительнее, по меньшей мере, 97%, предпочтительнее, по меньшей мере, 98%, наиболее предпочтительно, по меньшей мере, 99% последовательности дикого типа. Предпочтительно, спейсер CRISPR сохраняет 50%, предпочтительнее, 60%, предпочтительнее, 70%, предпочтительнее, 80%, предпочтительнее, 85%, предпочтительнее, 90%, предпочтительнее, 95%, предпочтительнее, 96%, предпочтительнее, 97%, предпочтительнее, 98% или, наиболее предпочтительно, 99% активности полипептида или нуклеотидной последовательности дикого типа.
Предпочтительно, ген cas сохраняет, по меньшей мере, 50%, предпочтительнее, по меньшей мере, 65%, предпочтительнее, по меньшей мере, 80%, предпочтительнее, по меньшей мере, 85%, предпочтительнее, по меньшей мере, 90%, предпочтительнее, по меньшей мере, 95%, предпочтительнее, по меньшей мере, 96%, предпочтительнее, по меньшей мере, 97%, предпочтительнее, по меньшей мере, 98%, наиболее предпочтительно, по меньшей мере, 99% последовательности дикого типа. Предпочтительно, спейсер CRISPR сохраняет 50%, предпочтительнее, 60%, предпочтительнее, 70%, предпочтительнее, 80%, предпочтительнее, 85%, предпочтительнее, 90%, предпочтительнее, 95%, предпочтительнее, 96%, предпочтительнее, 97%, предпочтительнее, 98% или, наиболее предпочтительно, 99% активности полипептида или нуклеотидной последовательности дикого типа.
Предпочтительно, белок cas сохраняет, по меньшей мере, 50%, предпочтительнее, по меньшей мере, 65%, предпочтительнее, по меньшей мере, 80%, предпочтительнее, по меньшей мере, 85%, предпочтительнее, по меньшей мере, 90%, предпочтительнее, по меньшей мере, 95%, предпочтительнее, по меньшей мере, 96%, предпочтительнее, по меньшей мере, 97%, предпочтительнее, по меньшей мере, 98%, наиболее предпочтительно, по меньшей мере, 99% последовательности дикого типа. Предпочтительно, белок cas сохраняет 50%, предпочтительнее, 60%, предпочтительнее, 70%, предпочтительнее, 80%, предпочтительнее, 85%, предпочтительнее, 90%, предпочтительнее, 95%, предпочтительнее, 96%, предпочтительнее, 97%, предпочтительнее, 98% или, наиболее предпочтительно, 99% активности полипептида или нуклеотидной последовательности дикого типа.
Предпочтительно, повтор CRISPR сохраняет, по меньшей мере, 50%, предпочтительнее, по меньшей мере, 65%, предпочтительнее, по меньшей мере, 80%, предпочтительнее, по меньшей мере, 85%, предпочтительнее, по меньшей мере, 90%, предпочтительнее, по меньшей мере, 95%, предпочтительнее, по меньшей мере, 96%, предпочтительнее, по меньшей мере, 97%, предпочтительнее, по меньшей мере, 98%, наиболее предпочтительно, по меньшей мере, 99% последовательности дикого типа. Предпочтительно, спейсер CRISPR сохраняет 50%, предпочтительнее, 60%, предпочтительнее, 70%, предпочтительнее, 80%, предпочтительнее, 85%, предпочтительнее, 90%, предпочтительнее, 95%, предпочтительнее, 96%, предпочтительнее, 97%, предпочтительнее, 98% или, наиболее предпочтительно, 99% активности полипептида или нуклеотидной последовательности дикого типа.
Предпочтительно, функциональная комбинация повтора CRISPR-cas сохраняет, по меньшей мере, 50%, предпочтительнее, по меньшей мере, 65%, предпочтительнее, по меньшей мере, 80%, предпочтительнее, по меньшей мере, 85%, предпочтительнее, по меньшей мере, 90%, предпочтительнее, по меньшей мере, 95%, предпочтительнее, по меньшей мере, 96%, предпочтительнее, по меньшей мере, 97%, предпочтительнее, по меньшей мере, 98%, наиболее предпочтительно, по меньшей мере, 99% последовательности дикого типа. Предпочтительно, функциональная комбинация повтора CRISPR-cas сохраняет 50%, предпочтительнее, 60%, предпочтительнее, 70%, предпочтительнее, 80%, предпочтительнее, 85%, предпочтительнее, 90%, предпочтительнее, 95%, предпочтительнее, 96%, предпочтительнее, 97%, предпочтительнее, 98% или, наиболее предпочтительно, 99% активности полипептида или нуклеотидной последовательности дикого типа.
Предпочтительно, последовательность-мишень нуклеиновой кислоты сохраняет, по меньшей мере, 50%, предпочтительнее, по меньшей мере, 65%, предпочтительнее, по меньшей мере, 80%, предпочтительнее, по меньшей мере, 85%, предпочтительнее, по меньшей мере, 90%, предпочтительнее, по меньшей мере, 95%, предпочтительнее, по меньшей мере, 96%, предпочтительнее, по меньшей мере, 97%, предпочтительнее, по меньшей мере, 98%, наиболее предпочтительно, по меньшей мере, 99% последовательности дикого типа. Предпочтительно, последовательность-мишень нуклеиновой кислоты сохраняет 50%, предпочтительнее, 60%, предпочтительнее, 70%, предпочтительнее, 80%, предпочтительнее, 85%, предпочтительнее, 90%, предпочтительнее, 95%, предпочтительнее, 96%, предпочтительнее, 97%, предпочтительнее, 98% или, наиболее предпочтительно, 99% активности полипептида или нуклеотидной последовательности дикого типа.
В некоторых вариантах осуществления, фрагмент представляет собой функциональный фрагмент. Под «функциональным фрагментом» молекулы подразумевается фрагмент, сохраняющий или обладающий по существу такой же биологической активностью как интактная молекула, Во всех случаях, фрагмент молекулы сохраняет, по меньшей мере, 10%, и, по меньшей мере, примерно 25%, примерно 50%, примерно 75%, примерно 80%, примерно 85%, примерно 90%, примерно 95%, примерно 96%, примерно 97%, примерно 98%, или примерно 99% биологической активности интактной молекулы.
Термин «гомолог» означает сущность, имеющую определенную гомологию с обсуждаемыми аминокислотными последовательностями и обсуждаемыми нуклеотидными последовательностями. В настоящем описании, термин «гомология» может приравниваться к «идентичности».
В настоящем контексте, гомологичная последовательность воспринимается с включением аминокислотной последовательности, которая может быть, по меньшей мере, на 75, 85 или 90% идентична, предпочтительно, по меньшей мере, на 95%, 96%, 97%, 98% или 99% идентична обсуждаемой последовательности. Хотя гомология может также рассматриваться с точки зрения аналогии (т.е. аминокислотных остатков, имеющих аналогичные химические свойства/функции), в контексте настоящего изобретения предпочтительно выражать гомологию с точки зрения идентичности последовательностей.
В настоящем контексте, гомологичная последовательность воспринимается с включением нуклеотидной последовательности, которая может быть, по меньшей мере, на 75, 85 или 90% идентична, предпочтительно, по меньшей мере, на 95%, 96%, 97%, 98% или 99% идентична обсуждаемой последовательности. Хотя гомология может также рассматриваться с точки зрения аналогии (т.е. аминокислотных остатков, имеющих аналогичные химические свойства/функции), в контексте настоящего изобретения, предпочтительно выражать гомологию с точки зрения идентичности последовательностей.
Сравнения гомологии могут проводиться визуально, или обычнее, с помощью общедоступных программ сравнения последовательностей. Эти имеющиеся в продаже компьютерные программы могут рассчитывать % гомологии между двумя или более последовательностями.
Процент (%) гомологии может быть рассчитан по смежным последовательностям (т.е. одна последовательность совмещается с другой последовательностью, и каждая аминокислота в последовательности непосредственно сравнивается с соответствующей аминокислотой в другой последовательности, каждый раз в одном остатке). Это называется совмещением «без зазоров». Обычно, такие совмещения без зазоров выполняются только по относительно небольшому числу остатков.
Хотя это очень простой и согласованный способ, он не учитывает, что, например, в идентичной по другим параметрам паре, одна вставка или делеция вызовет исключение из совмещения, таким образом, возможно приводя к значительному снижению % гомологии, когда выполняется полное совмещение. Следовательно, большинство способов сравнения предназначены для получения оптимальных совмещений, которые учитывают возможные вставки и делеции без ненужного снижения общей балльной оценки гомологии. Это достигается вставкой «гэпов» в совмещение последовательностей с попыткой максимизации местной гомологии.
Однако эти более сложные способы предназначают «наказания за гэпы» каждому гэпу, который встречается при совмещении, с тем, чтобы для одного и того же числа идентичных аминокислот, совмещение последовательностей с насколько возможно малым числом гэпов, отражающим более высокую связь между двумя сравниваемыми последовательностями, достигнет более высокой балльной оценки, чем последовательности с множеством гэпов. «Стоимость аффинных гэпов» обычно используется для осуществления относительно высокого наказания за существование гэпа и меньшего наказания за каждый последующий остаток в гэпе. Это наиболее часто используемая система балльной оценки гэпов. Высокие наказания за гэпы, конечно, дадут оптимизированные совмещения с меньшим количеством гэпов. Большинство программ совмещения обеспечивают возможность модификации наказаний за гэпы. Однако предпочтительно использование величин по умолчанию при использовании такого программного обеспечения для сравнений последовательностей. Например, при использовании пакета программного обеспечения GCG Wisconsin Bestfit, наказание за гэпы по умолчанию для аминокислотных последовательностей составляет -12 за гэп и -4 за каждый выступающий конец.
Поэтому расчет максимального % гомологии, во-первых, требует получения оптимального совмещения с учетом наказаний за гэпы. Подходящей компьютерной программой для проведения такого совмещения является пакет программного обеспечения GCG Wisconsin Bestfit (Висконсинский университет, США; Devereux et al., 1984, Nucleic Acids Research 12:387). Примеры другого программного обеспечения, которое может выполнить сравнение последовательностей, включают без ограничения пакеты программного обеспечения BLAST (см. Ausubel et al., 1999, там же - глава 18), FASTA (Atschul et al., 1990, J. Mol. Biol., 403-410), набор инструментов сравнения GENEWORKS и CLUSTAL. И BLAST, и FASTA имеются для автономного поиска и поиска в реальном масштабе времени (см. Ausubel et al., 1999, там же, стр. 7-58 до 7-60). Однако для некоторых видов применения, предпочтительно использование программы GCG Bestfit. Инструмент, называемый BLAST 2 Sequences, имеется для сравнения белков и нуклеотидных последовательностей (см. FEMS Microbiol Lett 1999 174(2): 247-50; FEMS Microbiol Lett 1999 177(1): 187-8).
Хотя конечный % гомологии может быть измерен с точки зрения идентичности, сам процесс совмещения обычно не основан на сравнении пар по принципу «все или ничего». Вместо этого, в целом используется шкальная матрица балльной оценки подобия, которая определяет балльные оценки для каждого попарного сравнения на основании химического подобия или эволюционной дистанции. Примером такой обычно используемой матрицы является матрица BLOSUM62 - матрица по умолчанию для набора программ BLAST. В программах GCG Wisconsin в целом используются или публичные величины по умолчанию, или, при поставке, заказная таблица символьных сравнений (дополнительные детали представлены в руководстве по применению). Для некоторых видов применения, предпочтительно использование публичных величин по умолчанию для пакета GCG. Или в случае использования другого программного обеспечения, - матрицы по умолчанию - такой как BLOSUM62.
После того как программное обеспечение дало оптимальное совмещение, можно рассчитать % гомологии, предпочтительно % идентичности последовательностей. Программное обеспечение обычно делает это как часть сравнения последовательностей и генерирует числовой результат.
Если используются наказания за зазоры при определении идентичности последовательностей, то возможно применение следующих параметров:
Для сравнения полипептидных последовательностей могут использоваться следующие структуры: наказание за создание гэпа 3,0 и наказание за продолжение гэпа 0,1. Степень идентичности в отношении аминокислотной последовательности может определяться, по меньшей мере, по 5 смежным аминокислотам, определяться, по меньшей мере, по 10 смежным аминокислотам, по меньшей мере, по 15 смежным аминокислотам, по меньшей мере, по 20 смежным аминокислотам, по меньшей мере, по 30 смежным аминокислотам, по меньшей мере, по 40 смежным аминокислотам, по меньшей мере, по 50 смежным аминокислотам, или, по меньшей мере, по 60 смежным аминокислотам. Последовательности могут также иметь делеции, вставки или замещения аминокислотных остатков, которые продуцируют молчащее изменение и приводят к функционально эквивалентному веществу. Преднамеренные аминокислотные замещения могут производиться на основании подобия полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфифатической природы остатков, пока сохраняется активность вторичного связывания вещества. Например, отрицательно заряженные аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту; положительно заряженные аминокислоты включают лизин и аргинин; и аминокислоты с незаряженными полярными головными группами, имеющие одинаковые величины гидрофильности, включают лейцин, изолейцин, валин, глицин, аланин, аспарагин, глутамин, серин, треонин, фенилаланин и тирозин.
Консервативные замещения могут производиться, например, в соответствии с представленной ниже таблицей. Аминокислоты в одном и том же блоке во втором столбце, а предпочтительно, в одной и той же строке в третьем столбце могут замещаться друг другом:
Настоящее изобретение также охватывает гомологичное замещение (и замещение, и замена используются в настоящем описании для обозначения взаимной замены существующего аминокислотного остатка с альтернативным остатком), которое может происходить, т.е. замещение похожего на похожий остаток, например, основного на основный, кислотный на кислотный, полярный на полярный и т.д. Может также происходить не гомологичное замещение, т.е. от одного класса остатка на другой, или альтернативно, вовлекающее включение не натуральных аминокислот - таких как орнитин (далее именуемый Z), орнитин диаминомасляной кислоты (далее именуемый B), норлейцин орнитин (далее именуемый О), пирииламин, тиенилаланин, нафтилаланин и фенилглицин.
Замещения, которые могут также производиться не натуральными аминокислотами, включают: альфа* и альфа-дизамещенные* аминокислоты, N-алкиламинокислоты*, молочную кислоту*, галоидные производные натуральных аминокислот - такие как трифтортирозин*, п-Cl-фенилаланин*, п-Br-фенилаланин*, п-1-фенилаланин*, L-аллил-глицин*, β-аланин*, L-α-аминомасляная кислота*, L-γ-аминомасляная кислота*, L-α-аминоизомасляная кислота*, L-ε-аминокапроновая кислота#, 7-аминогептаноевая кислота*, L-метионинсульфон#*, L-норлейцин*, L-норвалин*, п-нитро-L-фенилаланин*, L-гидроксипролин#, L-тиопролин*, метильные производные фенилаланина(Phe) - такие как 4-метил-Phe*, пентаметил-Phe*, L-Phe(4-амино)#, L-Tyr(метил)*, L-Phe(4-изопропил)*, L-Tic(1,2,3,4-тетрагидроизохинолин-3-карбоксильная кислота)*, L-диаминопропионовая кислота # и L-Phe(4-бензил)*. Обозначение * использовалось с целью приведенного выше обсуждения (относительно гомологичного или не гомологичного замещения) для указания гидрофобной природы производного, тогда как # для указания гидрофильной природы производного, #* указывает на амфипатические характеристики.
Вариантные аминокислотные последовательности включают подходящие группы спейсеров, которые подходят для вставки между любыми двумя аминокислотными остатками последовательности, включая алкильные группы - такие как метильная, этильная или пропильная группы - в дополнение к аминокислотным спейсерам - таким как остатки глицина или β-аланина. Другая форма вариации включает присутствие одного или нескольких аминокислотных остатков в пептоидной форме будет хорошо понятна специалистам в данной области. Во избежание сомнения, «пептоидная форма» используется для обозначения вариантных аминокислотных остатков, где на атоме азота остатка находится α-карбоновая заместительная группа, а не α-углерод. Протеазы для получения пептидов в пептоидной форме хорошо известны в данной области.
Нуклеотидные последовательности для применения в настоящем изобретении могут включать в себя синтетические или модифицированные нуклеотиды. В данной области известен ряд различных типов модификации олигонуклеотидов. Они включают основные цепи метилфосфоната и фосфортиоата и/или добавление цепей акридина или полилизина на концах 3' и/или 5' молекулы. В целях настоящего изобретения, следует понимать, что нуклеотидные последовательности могут быть модифицированы любым способом, имеющимся в данной области. Такие модификации могут проводиться для усиления активности in vivo или продолжительности жизни нуклеотидных последовательностей, используемых в настоящем изобретении.
CRISPRs
CRISPRs (кластерные, вставленные с равномерными промежутками короткие палиндромные повторы), также известные как SPIDRs (Прямые повторы с промежуточными вставленными спейсерами) составляют семейство недавно описанных локусов ДНК, которые обычно специфичны для определенного вида бактерий. Локус CRISPR представляет собой отдельный класс вставленных в промежутки повторов коротких последовательностей (Ishino et al, J. Bacteriol., 169:5429-5433 [1987] и Nakata et al., J. Bacteriol., 171:3553-3556 [1989]). Аналогичные вставленные в промежутки SSRs были идентифицированы в Haloferax mediterranei, Streptococcus pyogenes, Anabaena и Mycobacterium tuberculosis (см., Groenen et al., Mol. Microbiol., 10:1057-1065 [1993]; Hoe et al., Emerg. Infect. Dis., 5:254-263 [1999]; Masepohl et al., Biochim. Biophys. Acta 1307:26-30 [1996]; и Mojica et al., Mol. Microbiol., 17:85-93 [1995]). Локусы CRISPR отличаются от SSRs структурой повторов, которые были названы короткими, расположенными через равномерные промежутки (Janssen et al., OMICS J. Integ. Biol., 6:23-33 [2002]; и Mojica et al., Mol. Microbiol., 36:244-246 [2000]). Повторы представляют собой короткие элементы, которые встречаются кластерами, которые всегда расположены с равномерными промежутками, образуемыми необычными вставленными последовательностями с постоянной длиной (Mojica et al., [2000], см. выше). Хотя последовательности повторов высоко консервативны между штаммами, число вставленных в промежутки повторов и последовательности областей спейсеров отличаются от штамма к штамму (van Embden et al., J. Bacteriol., 182:2393-2401 [2001]).
Локусы CRISPR состоят из коротких и высоко консервативных частично палиндромных повторов ДНК, обычно от 24 до 40 пар оснований, содержащих внутренние и концевые инвертированные повторы длиной до 11 пар оснований. Хотя были выявлены изолированные элементы, они в целом располагаются кластерами (примерно до 20 или более на геном) повторных единиц, расположенных с промежутками, образованными необычными вставленными последовательностями из 20-58 пар оснований. К настоящему времени, до 20 отдельных локусов CRISPR были обнаружены в пределах одной хромосомы.
CRISPRs в целом однородны в пределах данного генома, причем большинство из них являются идентичными. Однако имеются примеры гетерогенности, например, Archea (Mojica et al., [2000], см. выше).
Используемый в настоящем описании термин «локус CRISPR» относится к сегменту ДНК, который включает все повторы CRISPR, начиная с первого нуклеотида первого повтора CRISPR и заканчивая последним нуклеотидом последнего (концевого) повтора CRISPR.
Хотя биологическая функция локусов CRISPR неизвестна, были предложены некоторые гипотезы. Например, было высказано предположение, что они участвуют в присоединении хромосомы к клеточной структуре, или в репликации хромосом и разделении репликона (Jansen et al., OMICS 6:23-33 [2002]; Jansen et al., Mol. Microbiol., 43:1565-1575 [2002]; и Pourcel et al., Microbiol., 151:653-663 [2005]). Mojica et al. (Mojica et al., J. Mol. Evol., 60:174-182 [2005]) выдвинули гипотезу, что CRISPR может участвовать в обеспечении специфического иммунитета против инородной ДНК, а Piurcel et al. (см. выше) выдвинули гипотезу, что CRISPRs представляют собой структуры, которые способны захватывать кусочки инородной ДНК, как часть механизма защиты. Bolotin et al. (см. выше) свидетельствуют о том, что элементы спейсера CRISPR представляют собой следы прошлых внедрений внехромосомных элементов, и высказывают гипотезу, что они обеспечивают клетку иммунитетом против инфекции фагов, а в более общем смысле, против экспрессии инородной ДНК, путем кодирования антисмысловой РНК. Bolotin et al. (см. выше) также свидетельствуют о том, что гены cas необходимы для образования CRISPR. Однако не предполагается, что настоящее изобретение ограничивается каким-либо конкретным механизмом, функцией, теорией средством действия.
Геном Streptococcus thermophilus LMG18311 содержит 3 локуса CRISPR (34 повтора), состоящие из 36 пар оснований повторяющиеся последовательности различны у CRISPR1 (34 повтора), CRISPR2 (5 повторов) и CRISPR3 (одна последовательность). Тем не менее они отлично сохранены в пределах каждого локуса. Повторы CRISPR1 и CRISPR2 соответственно разделены промежуточными 33 и 4 последовательностями длиной 30 пар оснований. Все эти промежуточные последовательности отличаются друг от друга. Они также отличаются от последовательностей, обнаруживаемых у штамма CNRZ1066 (41 промежуточных последовательностей в пределах CRISPR1) и у штамма LMD-9 (16 в пределах CRISPR1 и 8 в пределах CRISPR3), которые обе представляют собой S. thermophilus.
В данной области известны различные способы идентификации локусов CRISPR. Например, Jensen et al. (Jensen et al., [2002]. См. выше) описывают основанный на компьютерном анализе подход, при котором происходит поиск рамок CRISPR в нуклеотидых последовательностях с использованием программы PATSCAN Отдела математики и Компьютерной Науки в Аргонской Национальной Лаборатории, Argonne, IL., USA. Алгоритм, который использовали для идентификации мотивов CRISPR, представлял собой p1 = a…bc…dp1c…dplc…dp1, где a и b нижний и верхний предел размера повтора и p1 и c, и d представляли собой нижний и верхний предел размера последовательностей спейсера. Величины a, b, c и d могут варьировать от примерно 15 до и 70 пар оснований с приростами 5 оснований. В некоторых предпочтительных вариантах осуществления, локусы CRISPR идентифицируются с использованием нанесения точек на график (например, путем использования компьютерной программы Dotter).
Любой подходящий способ, известный в данной области, находит применение при анализе аналогии последовательности. Например, анализ может выполняться с использованием программы NCBI BLAST с базой данных микробных геномов и GenBank, как известно в данной области. Кроме того, нуклеотидные последовательности, включая те, которые представлены в настоящем описании, включены в базы данных (например, сайт интернета GenBank или JGI genome). Используемый в настоящем описании термин «выше по ходу транскрипции» означает в направлении 5', а «ниже по ходу транскрипции» означает в направлении 3'.
В дополнительных вариантах осуществления, в способах по настоящему изобретению используются процедуры амплификации (см., например, Mojica et al., [2005], см. выше; и Pourcel et al., [2005], см. выше). Амплификация желаемой ДНК может достигаться любым способом, известным в данной области, включая полимеразную реакцию синтеза цепи (PCR). «Амплификация» относится к получению дополнительных копий последовательности нуклеиновой кислоты. Это в целом проводится с использованием технологий PCR, хорошо известных в данной области. «Полимеразная реакция синтеза цепи» (или полимеразная цепная реакция) («PCR») хорошо известна специалистам в данной области. В настоящем изобретении, олигонуклеотидные праймеры предназначены для использования в реакциях PCR для амплификации всего или части локуса CRISPR.
Термин «праймер» относится к олигонуклеотиду, встречающемуся или естественно как в очищенном рестрикционном переваре, или полученному синтетически, который способен действовать в качестве точки инициации синтеза при помещении в условиях, в которых индуцируется синтез продукта выступающего конца праймера, который является комплементарным к нити нуклеиновой кислоты (т.е. в присутствии нуклеотидов и индуцирующего агента, такого как ДНК-полимераза, и при подходящей температуре и рН). В некоторых вариантах осуществления, праймер является однонитевым для максимальной эффективности при амплификации, хотя в других вариантах осуществления, праймер является двухнитевым. В некоторых вариантах осуществления, праймер представляет собой олигодезоксирибонуклеотид. Праймер должен быть достаточно длинным для обеспечения возможности синтеза продуктов выступающего конца в присутствии индуцирующего агента. Точная длина праймера зависит от многих факторов, включая температуру, источник праймера и применяемый способ. Праймеры для PCR обычно имеют длину, по меньшей мере, примерно 10 нуклеотидов, а наиболее типично, имеют длину, по меньшей мере, примерно 20 нуклеотидов. Способы и проведения PCR хорошо известны в данной области и включают без ограничения способы с использованием спаренных праймеров, интрагенных праймеров, одиночных специфических праймеров, дегенеративных праймеров, ген-специфических праймеров, вектор-специфических праймеров, частично ошибочно спаренных праймеров и т.д.
В некоторых предпочтительных вариантах осуществления настоящего изобретения, локус CRISPR или его часть из материнской бактерии и меченая бактерия сравниваются с использованием любого подходящего способа, известного в данной области. В некоторых предпочтительных вариантах осуществления настоящего изобретения, локус CRISPR или его часть из материнской бактерии и меченая бактерия сравниваются амплификацией локуса CRISPR или его части. В дополнение к хорошо известным циклическим способам амплификации (например, PCR, реакция цепи лигазы и т.д.), другие способы, включая без ограничения способы изотермической амплификации, находят применение в настоящем изобретении. Хорошо известные способы изотермической амплификации, которые находят применение в настоящем изобретении, включают без ограничения амплификацию со смещением нити (SDA), Q-бета-репликазу, амплификацию последовательности на основе нуклеиновой кислоты (NASBA) и аутоустойчивую реплификацию последовательности.
В некоторых других предпочтительных вариантах осуществления настоящего изобретения, локус CRISPR или его часть из материнской бактерии и меченая бактерия сравниваются амплификацией и затем секвенированием локусов CRISPR или их части. В некоторых вариантах осуществления, сравниваются одни концы локусов CRISPR, тогда как в других вариантах осуществления, сравниваются и концы 3', и концы 5' локусов. В некоторых других вариантах осуществления, сравниваются одни концы (например, концы 5') локусов CRISPR. В еще одних вариантах осуществления, сравниваются, по меньшей мере, последний повтор CRISPR на конце 3' локуса CRISPR и/или, по меньшей мере, последний спейсер CRISPR (например, кор последнего спейсера CRISPR) на конце 3' локуса CRISPR и/или, по меньшей мере, первый повтор CRISPR на конце 5' локуса CRISPR и/или, по меньшей мере, первый спейсер CRISPR (например, кор первого спейсера CRISPR) и конец 5' локуса CRISPR. В некоторых предпочтительных вариантах осуществления, сравниваются, по меньшей мере, первый повтор CRISPR на конце 5' локуса CRISPR, и/или, по меньшей мере, первый спейсер CRISPR (например, кор первого спейсера CRISPR) на конце 5' локуса CRISPR. В некоторых дополнительных предпочтительных вариантах осуществления, сравниваются, по меньшей мере, последний спейсер (например, по меньшей мере, кор последнего спейсера CRISPR) на конце 3' локуса CRISPR и/или, по меньшей мере, первый спейсер CRISPR (например, по меньшей мере, кор первого спейсера CRISPR) на конце 5' локуса CRISPR. В некоторых других предпочтительных вариантах осуществления, сравниваются, по меньшей мере, первый спейсер CRISPR (например, кор первого спейсера CRISPR) на концах 5' локусов CRISPR.
В некоторых вариантах осуществления, локусы CRISPR содержат ДНК, тогда как в других вариантах осуществления, локусы CRISPR содержат РНК. В некоторых вариантах осуществления, нуклеиновая кислота имеет геномное происхождение, тогда как в других вариантах осуществления, она имеет синтетическое или рекомбинантное происхождение. В некоторых вариантах осуществления, локусы CRISPR являются двухнитевыми, тогда как в других вариантах осуществления, они являются однонитевыми, представляющими или смысловую, или антисмысловую нить, или их комбинацию. В некоторых вариантах осуществления, локусы CRISPR получают использованием методик рекомбинантной ДНК, как раскрыто в настоящем описании.
Настоящее изобретение также относится к способам генерирования вариантов CRISPR. Эти варианты экспрессируются, выделяются, клонируются и/или секвенируются с использованием любого подходящего способа, известного в данной области. В некоторых особенно предпочтительных вариантах осуществления, варианты CRISPR представляют собой устойчивые к фагам мутантные штаммы, которые имеют модифицированный локус CRISPR с дополнительным спейсером. В некоторых особенно дополнительных вариантах осуществления, эти варианты находят применение в качестве мишеней для целей выявления/идентификации, или для создания методами инженерии сопротивляемости против молекул нуклеиновой кислоты. В еще одних вариантах осуществления, эти варианты находят применение в разработке агентов биологического контроля.
В контексте настоящего изобретения, локус CRISPR ориентирован, как описано ниже. Лидерный CRISPR представляет собой сегмент консервативной ДНК определенного размера. Ориентация локуса CRISPR устанавливается с использованием следующих характеристик:
положение CRISPR относительно соседних генов cas (последовательностей, ассоциированных с CRISPR); CRISPR1 локализуется ниже по ходу транскрипции от 4 генов cas (генов str0657, str0658, str0659 и str0660 в пределах последовательности хромосомы CNRZ1066);
эта последовательность повтора может образовывать вторичную структуру «шпильки», хотя она не является полностью палиндромной, и обратную комплементарную последовательность;
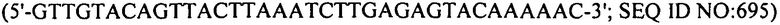 (отличается от прямой последовательности
(отличается от прямой последовательности 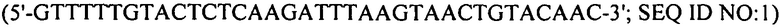 . В целом, конец 5' прямой последовательности богаче нуклеотидами G и T, чем конец 5' обратной комплементарной последовательности. Кроме того, поскольку спаривание оснований G-T лучше, чем спаривание оснований А-С, то структура «шпильки» в целом прочнее на прямой нити; и
. В целом, конец 5' прямой последовательности богаче нуклеотидами G и T, чем конец 5' обратной комплементарной последовательности. Кроме того, поскольку спаривание оснований G-T лучше, чем спаривание оснований А-С, то структура «шпильки» в целом прочнее на прямой нити; и
используемое в настоящем описании положение концевого повтора представляет собой концевой повтор, который проявляет вариацию последовательности на ее конце 3', в целом является концевым повтором.
Лидерная последовательность CRISPR представляет собой консервативный сегмент ДНК определенного размера, который локализуется непосредственно выше по ходу транскрипции от первого повтора. Например, лидерная последовательность CRISPR1 S. thermophilus представляет собой сегмент ДНК, начинающийся непосредственно после терминирующего кодона гена str0660, и заканчивающийся непосредственно перед первым повтором. Лидерная последовательность CRISPR локализуется на конце 5' локуса CRISPR. Лидерная последовательность CRISPR локализуется непосредственно выше по ходу транскрипции первого повтора CRISPR локуса CRISPR.
Трейлерная последовательность CRISPR представляет собой консервативный сегмент ДНК определенного размера, который локализуется непосредственно ниже по ходу транскрипции от концевого повтора. Например, трейлерная последовательность CRISPR1 S. thermophilus представляет собой сегмент ДНК, начинающийся непосредственно после концевого повтора, и заканчивающийся непосредственно перед терминирующим кодоном гена str0661 (локализующемся на противоположной нити ДНК). Трейлерная последовательность CRISPR локализуется непосредственно ниже по ходу транскрипции концевого повтора.
Например, лидерная последовательность CRISPR и трейлерная последовательность CRISPR в локусе CRISPR1 штамма CNRZ1066 Streptococcus thermophilus представляют собой:
Лидерная последовательность CRISPR:
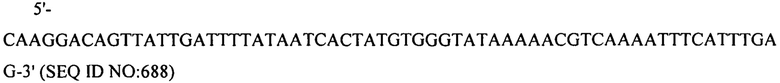
Трейлерная последовательность CRISPR:
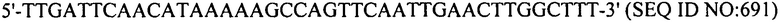
Лидерная последовательность CRISPR соответствует положениям с 625038 по 625100, а трейлерная последовательность CRISPR соответствует положениям с 627845 по 627885 в полном геноме (СР000024) S. Thermophilus.
Используемый в настоящем описании термин «выше по ходу транскрипции» означает в направлении 5', а «ниже по ходу транскрипции» означает в направлении 3'. Используемый в настоящем описании термин «его часть» в контексте локуса CRISPR означает, по меньшей мере, примерно 10 нуклеотидов, примерно 20 нуклеотидов, примерно 24 нуклеотида, примерно 30 нуклеотидов, примерно 40 нуклеотидов, примерно 44 нуклеотида, примерно 50 нуклеотидов, примерно 60 нуклеотидов, примерно 70 нуклеотидов, примерно 80 нуклеотидов, примерно 90 нуклеотидов, примерно 98 нуклеотидов или даже примерно 100, или более нуклеотидов (например, по меньшей мере, примерно 44-98 нуклеотидов) локуса CRISPR. В некоторых предпочтительных вариантах осуществления, термин «его положение» означает, по меньшей мере, примерно 10 нуклеотидов, примерно 20 нуклеотидов, примерно 24 нуклеотида, примерно 30 нуклеотидов, примерно 40 нуклеотидов, примерно 44 нуклеотида, примерно 50 нуклеотидов, примерно 60 нуклеотидов, примерно 70 нуклеотидов, примерно 80 нуклеотидов, примерно 90 нуклеотидов, примерно 98 нуклеотидов или даже примерно 100, или более нуклеотидов (например, по меньшей мере, примерно 44-98 нуклеотидов) с одного или обоих концов (т.е. концов 5' и/или 3') локуса CRISPR. В некоторых предпочтительных вариантах осуществления, термин «его положение» относится, по меньшей мере, примерно к первым 44 нуклеотидам конца 5' локуса CRISPR или примерно к последним 44 нуклеотидам конца 3' локуса CRISPR.
В некоторых других вариантах осуществления, термин «его положение» в контексте локуса CRISPR означает, по меньшей мере, примерно 10 нуклеотидов, примерно 20 нуклеотидов, примерно 24 нуклеотида, примерно 30 нуклеотидов, примерно 40 нуклеотидов, примерно 44 нуклеотида, примерно 50 нуклеотидов, примерно 60 нуклеотидов, примерно 70 нуклеотидов, примерно 80 нуклеотидов, примерно 90 нуклеотидов, примерно 98 нуклеотидов или примерно 100, или более нуклеотидов (например, по меньшей мере, примерно 44-98 нуклеотидов) ниже по ходу транскрипции от первого нуклеотида первого повтора CRISPR на конце 5' локуса CRISPR или выше по ходу транскрипции от первого нуклеотида первого повтора CRISPR на конце 3' локуса CRISPR. В некоторых предпочтительных вариантах осуществления, термин «его положение» относится, по меньшей мере, примерно к первым 44 нуклеотидам ниже по ходу транскрипции от первого нуклеотида первого повтора CRISPR на конце 5' локуса CRISPR или, по меньшей мере, примерно к 44 нуклеотидам выше по ходу транскрипции от последнего нуклеотида последнего повтора CRISPR на конце 3' локуса CRISPR.
В некоторых вариантах осуществления, минимальный размер дуплицированной последовательности составляет примерно 24 нуклеотида, а минимальный размер маркировочной последовательности составляет примерно 20 нуклеотидов. Таким образом, в некоторых предпочтительных вариантах осуществления, термин «его положение» в контексте локуса CRISPR означает, по меньшей мере, 44 нуклеотида.
В некоторых вариантах осуществления, максимальный размер дуплицированной последовательности составляет примерно 40 нуклеотидов, а максимальный размер маркировочной последовательности составляет примерно 58 нуклеотидов. Таким образом, в некоторых вариантах осуществления, термин «его положение» при использовании в контексте локуса CRISPR означает, по меньшей мере, 98 нуклеотидов. В некоторых предпочтительных вариантах осуществления, термин «его положение» в контексте локуса CRISPR означает, по меньшей мере, примерно 44-98 нуклеотидов.
При сравнении локуса CRISPR или его части из материнской бактерии и меченой бактерии, сравниваются, по меньшей мере, примерно 10 нуклеотидов, примерно 20 нуклеотидов, примерно 24 нуклеотида, примерно 30 нуклеотидов, примерно 40 нуклеотидов, примерно 44 нуклеотида, примерно 50 нуклеотидов, примерно 60 нуклеотидов, примерно 70 нуклеотидов, примерно 80 нуклеотидов, примерно 90 нуклеотидов, примерно 98 нуклеотидов или примерно 100 (например, по меньшей мере, примерно 44-98 нуклеотидов) локуса CRISPR.
В некоторых предпочтительных вариантах осуществления, сравниваются, по меньшей мере, примерно 10 нуклеотидов, примерно 20 нуклеотидов, примерно 24 нуклеотида, примерно 30 нуклеотидов, примерно 40 нуклеотидов, примерно 44 нуклеотида, примерно 50 нуклеотидов, примерно 60 нуклеотидов, примерно 70 нуклеотидов, примерно 80 нуклеотидов, примерно 90 нуклеотидов, примерно 98 нуклеотидов или примерно 100 (например, по меньшей мере, примерно 44-98 нуклеотидов) на конце 5' локуса CRISPR или на конце 3' локуса CRISPR. В некоторых предпочтительных вариантах осуществления, сравниваются, по меньшей мере, первые примерно 44 нуклеотида на конце 5' локуса CRISPR или, по меньшей мере, первые примерно 44 нуклеотида на конце 3' локуса CRISPR.
В некоторых вариантах осуществления, сравниваются, по меньшей мере, примерно 10 нуклеотидов, примерно 20 нуклеотидов, примерно 24 нуклеотида, примерно 30 нуклеотидов, примерно 40 нуклеотидов, примерно 44 нуклеотида, примерно 50 нуклеотидов, примерно 60 нуклеотидов, примерно 70 нуклеотидов, примерно 80 нуклеотидов, примерно 90 нуклеотидов, примерно 98 нуклеотидов или примерно 100, или более нуклеотидов (например, по меньшей мере, примерно 44-98 нуклеотидов) ниже по ходу транскрипции от первого нуклеотида первого повтора CRISPR на конце 5' локуса CRISPR или выше по ходу транскрипции от последнего нуклеотида последнего повтора CRISPR на конце 3' локуса CRISPR. В некоторых предпочтительных вариантах осуществления, сравниваются, по меньшей мере, первые примерно 44 нуклеотида ниже по ходу транскрипции от первого нуклеотида первого повтора CRISPR на конце 5' локуса CRISPR или примерно 44 нуклеотида выше по ходу транскрипции от последнего нуклеотида последнего повтора CRISPR на конце 3' локуса CRISPR.
В некоторых вариантах осуществления, минимальный размер дуплицированной последовательности составляет примерно 24 нуклеотида, а минимальный размер маркировочной последовательности составляет примерно 20 нуклеотидов. В некоторых предпочтительных вариантах осуществления, мере, сравниваются, 44 нуклеотида. В некоторых альтернативных вариантах осуществления, минимальный размер дуплицированной последовательности составляет примерно 40 нуклеотидов, а максимальный размер маркировочной последовательности составляет примерно 58 нуклеотидов. В некоторых предпочтительных вариантах осуществления, сравниваются, по меньшей мере, 98 нуклеотидов. В некоторых альтернативных предпочтительных вариантах осуществления, сравниваются, по меньшей мере, примерно 44-98 нуклеотидов.
Используемый в настоящем описании термин «повтор CRISPR» имеет обычное значение, используемое в данной области (т.е. множественные короткие прямые повторы, которые не проявляют или проявляют очень небольшую изменчивость последовательности в пределах данного локуса CRISPR). Используемый в настоящем описании в контексте термин «повтор CRISPR» синономичен термину «CRISPR».
Локус CRISPR содержит один или несколько повторов CRISPR, и имеются спейсеры CRISPR. Таким образом, повтор CRISPR соответствует повторяемым последовательностям в пределах локуса CRISPR. Например, за исключением концевого повтора, типичная последовательность повтора последовательности CRISPR1 S. thermophilus представляет собой:
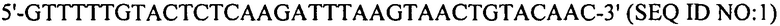
Наблюдались точечные изменения последовательности этого повтора, но они были очень редкими. По сравнению с этой типичной последовательностью повтора, концевая последовательность повтора всегда проявляет одинаковое изменение на конце 3'. Наблюдались также точечные изменения последовательности этого концевого повтора, но они также были очень редкими. Повторы CRISPR могут естественно возникать в материнской бактерии. Номера доступа в генном банке последовательностей CRISPR1 включают: 
Как дополнительно детально описано в настоящей заявке, дуплицированная последовательность производится, может производиться, получается или может получаться из материнской бактерии. В некоторых предпочтительных вариантах осуществления, последовательность содержит геномную ДНК материнской бактерии. В некоторых особенно предпочтительных вариантах осуществления, дуплицированная последовательность CRISPR (например, в одном и том же локусе CRISPR) интегрируется повторно, последовательно, одновременно или по существу одновременно наряду с маркирующей последовательностью в материнскую бактерию для получения меченой бактерии.
Число нуклеотидов в повторе составляет в целом от примерно 20 до 40 пар оснований (например, 36 пар оснований), но в других вариантах осуществления, составляет от примерно 20 до примерно 39 пар оснований, от примерно 20 до примерно 37 пар оснований, от примерно 20 до примерно 35 пар оснований, от примерно 20 до примерно 33 пар оснований, от примерно 20 до примерно 30 пар оснований, от примерно 21 до примерно 40 пар оснований, от примерно 21 до примерно 39 пар оснований, от примерно 21 до примерно 37 пар оснований, от примерно 23 до примерно 40 пар оснований, от примерно 23 до примерно 39 пар оснований, от примерно 23 до примерно 37 пар оснований, от примерно 25 до примерно 40 пар оснований, от примерно 25 до примерно 39 пар оснований, от примерно 25 до примерно 37 пар оснований, от примерно 25 до примерно 35 пар оснований или от примерно 28 или 29 пар оснований.
Число нуклеотидов в повторе составляет в целом от примерно 20 до примерно 40 пар оснований, но может составлять от примерно 20 до примерно 39 пар оснований, от примерно 20 до примерно 37 пар оснований, от примерно 20 до примерно 35 пар оснований, от примерно 20 до примерно 33 пар оснований, от примерно 20 до примерно 30 пар оснований, от примерно 21 до примерно 40 пар оснований, от примерно 21 до примерно 39 пар оснований, от примерно 21 до примерно 37 пар оснований, от примерно 23 до примерно 40 пар оснований, от примерно 23 до примерно 39 пар оснований, от примерно 23 до примерно 37 пар оснований, от примерно 25 до примерно 40 пар оснований, от примерно 25 до примерно 39 пар оснований, от примерно 25 до примерно 37 пар оснований, от примерно 25 до примерно 35 пар оснований или от примерно 28 или 29 пар оснований. Число повторов может находиться в диапазоне от примерно 1 до примерно 140, от примерно 1 до примерно 100, от примерно 2 до примерно 100, от примерно 5 до примерно 100, от примерно 10 до примерно 100, от примерно 15 до примерно 100, от примерно 20 до примерно 100, от примерно 25 до примерно 100, от примерно 30 до примерно 100, от примерно 35 до примерно 100, от примерно 40 до примерно 100, от примерно 45 до примерно 100, от примерно 50 до примерно 100, от примерно 1 до примерно 135, от примерно 1 до примерно 130, от примерно 1 до примерно 125, от примерно 1 до примерно 120, от примерно 1 до примерно 115, от примерно 1 до примерно 110, от примерно 1 до примерно 105, от примерно 1 до примерно 100, от примерно 1 до примерно 95, от примерно 1 до примерно 90, от примерно 1 до примерно 80, от примерно 1 до примерно 70, от примерно 1 до примерно 60, от примерно 1 до примерно 50, от примерно 10 до примерно 140, от примерно 10 до примерно 130, от примерно 10 до примерно 120, от примерно 10 до примерно 110, от примерно 10 до примерно 95, от примерно 10 до примерно 90, от примерно 20 до примерно 80, от примерно 30 до примерно 70, от примерно 30 до примерно 60, от примерно 30 до примерно 50, от примерно 30 до примерно 40 или примерно 32.
В некоторых других вариантах осуществления, число нуклеотидов в повторе составляет от примерно 20 до примерно 39 пар оснований, от примерно 20 до примерно 37 пар оснований, от примерно 20 до примерно 35 пар оснований, от примерно 20 до примерно 33 пар оснований, от примерно 20 до примерно 30 пар оснований, от примерно 21 до примерно 40 пар оснований, от примерно 21 до примерно 39 пар оснований, от примерно 21 до примерно 37 пар оснований, от примерно 23 до примерно 40 пар оснований, от примерно 23 до примерно 39 пар оснований, от примерно 23 до примерно 37 пар оснований, от примерно 25 до примерно 40 пар оснований, от примерно 25 до примерно 39 пар оснований, от примерно 25 до примерно 37 пар оснований, от примерно 25 до примерно 35 пар оснований или от примерно 28 или 29 пар оснований.
В некоторых вариантах осуществления, число повторов находится в диапазоне от примерно 1 до примерно 144, от примерно 1 до примерно 100, от примерно 2 до примерно 100, от примерно 5 до примерно 100, от примерно 10 до примерно 100, от примерно 15 до примерно 100, от примерно 20 до примерно 100, от примерно 25 до примерно 100, от примерно 30 до примерно 100, от примерно 35 до примерно 100, от примерно 40 до примерно 100, от примерно 45 до примерно 100, от примерно 50 до примерно 100, от примерно 1 до примерно 135, от примерно 1 до примерно 130, от примерно 1 до примерно 125, от примерно 1 до примерно 120, от примерно 1 до примерно 115, от примерно 1 до примерно 110, от примерно 1 до примерно 105, от примерно 1 до примерно 100, от примерно 1 до примерно 95, от примерно 1 до примерно 90, от примерно 1 до примерно 80, от примерно 1 до примерно 70, от примерно 1 до примерно 60, от примерно 1 до примерно 50, от примерно 10 до примерно 140, от примерно 10 до примерно 130, от примерно 10 до примерно 120, от примерно 10 до примерно 110, от примерно 10 до примерно 95, от примерно 10 до примерно 90, от примерно 20 до примерно 80, от примерно 30 до примерно 70, от примерно 30 до примерно 60, от примерно 30 до примерно 50, от примерно 30 до примерно 40 или примерно 30, 31, 32, 33, 34 или 35 повторов.
В некоторых вариантах осуществления, число повторов находится в диапазоне от примерно 2 до примерно 140, от примерно 2 до примерно 100, от примерно 5 до примерно 100, от примерно 10 до примерно 100, от примерно 15 до примерно 100, от примерно 20 до примерно 100, от примерно 25 до примерно 100, от примерно 30 до примерно 100, от примерно 35 до примерно 100, от примерно 40 до примерно 100, от примерно 45 до примерно 100, от примерно 50 до примерно 100.
В некоторых других вариантах осуществления, число повторов находится в диапазоне от примерно 2 до примерно 135, от примерно 2 до примерно 130, от примерно 2 до примерно 125, от примерно 2 до примерно 120, от примерно 2 до примерно 115, от примерно 2 до примерно 110, от примерно 2 до примерно 105, от примерно 2 до примерно 100, от примерно 2 до примерно 95, от примерно 2 до примерно 90, от примерно 2 до примерно 80, от примерно 2 до примерно 70, от примерно 2 до примерно 60, от примерно 2 до примерно 50, от примерно 2 до примерно 40, от примерно 2 до примерно 30, от примерно 2 до примерно 20, от примерно 2 до примерно 10, от примерно 2 до примерно 9, от примерно 2 до примерно 8, от примерно 2 до примерно 7, от примерно 2 до примерно 6, от примерно 2 до примерно 5, от примерно 2 до примерно 4, от примерно 2 до примерно 3.
В некоторых вариантах осуществления, повторы CRISPR содержат ДНК, тогда как в других вариантах осуществления, повторы CRISPR содержат РНК. В некоторых вариантах осуществления, нуклеиновая кислота имеет геномное происхождение, тогда как в других вариантах осуществления, она имеет синтетическое или рекомбинантное происхождение. В некоторых вариантах осуществления, гены повторов CRISPR являются двухнитевыми или однонитевыми, представляющими смысловую или антисмысловую нить или их комбинацию. В некоторых вариантах осуществления, гены повторов CRISPR получаются путем применения методик рекомбинантной ДНК (например, рекомбинантной ДНК), как описано в настоящей заявке.
В некоторых вариантах осуществления, один или несколько повторов CRISPR используются для генной инженерии клетки (например, клетки-реципиента). В некоторых предпочтительных вариантах осуществления, один или несколько, предпочтительно, два или более повторов CRISPR, используются для генной инженерии клетки (например, клетки-реципиента), что в комбинации с одним или несколькими генам или белками cas и одним или несколькими спейсерами CRISPR модулирует устойчивость клетки против нуклеиновой кислоты-мишени или продукта ее транскрипции. Например, в некоторых вариантах осуществления, повтор(ы) CRISPR вставляются в ДНК клетки (например, плазмиду и/или геномную ДНК клетки-реципиента), с использованием любого подходящего способа, известного в данной области. В дополнительных вариантах осуществления, повтор(ы) CRISPR находят применение в качестве матрицы для модификации (например, мутации) ДНК клетки (например, плазмиды и/или геномной ДНК клетки-реципиента) с тем, чтобы создать или подвернуть генной инженерии повтор(ы) CRISPR в ДНК клетки. В дополнительных вариантах осуществления, повтор(ы) CRISPR присутствуют, по меньшей мере, в одном конструкте, по меньшей мере, в одной плазмиде и/или, по меньшей мере, в одном векторе и т.д. В других вариантах осуществления, повторы CRISPR вводятся в клетку с использованием любого подходящего способа, известного в данной области.
В дополнительных вариантах осуществления, настоящее изобретение относится к способам идентификации повтора CRISPR для использования при модулировании устойчивости клетки против нуклеиновой кислоты-мишени или продукта ее транскрипции, включающим стадии: (i) получения клетки, содержащей, по меньшей мере, один спейсер CRISPR и, по меньшей мере, один ген cas; (ii) генной инженерии клетки с тем, чтобы она содержала повтор CRISPR; и (iii) определения того, модулирует ли клетка сопротивляемость против нуклеиновой кислоты-мишени или продукта ее транскрипции, где модулирование устойчивости клетки против нуклеиновой кислоты-мишени или продукта ее транскрипции указывает на то, что повтор CRISPR может применяться для модулирования устойчивости.
В некоторых других вариантах осуществления, один или несколько генов или белков cas используются вместе или в комбинации с одним или несколькими, предпочтительно, двумя или более повторов CRISPR и, необязательно, одного или нескольких спейсеров CRISPR. В некоторых особенно предпочтительных вариантах осуществления, один или несколько ген(ов) или белок (белки) cas и повтор(ы) CRISPR образуют функциональную комбинацию, как описано ниже. В некоторых вариантах осуществления, повторы CRISPR содержат любой из нуклеотидов, представленных в SEQ ID NOS:1-22. SEQ ID NOS:1-12 получены из S.thermophilus, SEQ ID NOS:13-16 получены из Streptococcus agalactiae, SEQ NO:17 получена из S.mutans и SEQ ID NOS:18-22 получены из S. pyogenes.


Спейсер CRISPR
Используемый в настоящем описании термин «спейсер CRISPR» охватывает не повторяющиеся спейсерные последовательности, которые обнаруживаются между множественными короткими прямыми повторами (т.е. повторами CRISPR) или локусами CRISPR. В некоторых вариантах осуществления настоящего изобретения, «спейсер CRISPR» относится к сегменту нуклеиновой кислоты, который фланкируется двумя повторами CRISPR. Было обнаружено, что последовательности спейсеров CRISPR часто имеют значимые сходства с разнообразными подвижными молекулами ДНК (например, бактериофагами и плазмидами). В некоторых предпочтительных вариантах осуществления, спейсеры CRISPR локализуются между двумя идентичными повторами CRISPR. В некоторых вариантах осуществления, спейсеры CRISPR идентифицируются анализом последовательности во фрагментах секвенирования, локализующихся между двумя повторами CRISPR. В некоторых предпочтительных вариантах осуществления, спейсер CRISPR естественно присутствует между двумя идентичными множественными короткими прямыми повторами, которые являются палиндромными.
Представляет интерес, что клетки, несущие эти спейсеры CRISPR, неспособны инфицироваться молекулами ДНК, содержащими последовательности, гомологичные спейсерам (Mojica et al., 2005). В некоторых предпочтительных вариантах осуществления, спейсер CRISPR гомологичен нуклеиновой кислоте-мишени или продукту ее транскрипции, или идентифицированной последовательности. Хотя гомологию можно также рассматривать с точки зрения подобия, в контексте настоящего изобретения предпочтительно выражать гомологию с точки зрения идентичности последовательностей. Считается, что гомологичная последовательность включает спейсер CRISPR, который может быть, по меньшей мере, примерно на 70, примерно на 75, примерно на 85 или примерно на 90% идентичен, или, по меньшей мере, примерно на 91, примерно на 92, примерно на 93, примерно на 94, примерно на 95, примерно на 96, примерно на 97, примерно на 98, примерно на 99% идентичен последовательности нуклеиновой кислоты-мишени или продукта ее транскрипции или идентифицированной последовательности. В некоторых предпочтительных вариантах осуществления, спейсер CRISPR примерно на 100% идентичен последовательности нуклеиновой кислоты-мишени. Отмечается также, что число спейсеров CRISPR в данных локусах или локусе CRISPR может варьироваться между видами. Кроме того, число спейсеров находится в диапазоне от примерно 1 до примерно 140, от примерно 1 до примерно 100, от примерно 2 до примерно 100, от примерно 5 до примерно 100, от примерно 10 до примерно 100, от примерно 15 до примерно 100, от примерно 20 до примерно 100, от примерно 25 до примерно 100, от примерно 30 до примерно 100, от примерно 35 до примерно 100, от примерно 40 до примерно 100, от примерно 45 до примерно 100 или от примерно 50 до примерно 100. В некоторых предпочтительных вариантах осуществления, число спейсеров находится в диапазоне от примерно 1 до примерно 135, от примерно 1 до примерно 130, от примерно 1 до примерно 120, от примерно 1 до примерно 115, от примерно 1 до примерно 110, от примерно 1 до примерно 105, от примерно 1 до примерно 100, от примерно 1 до примерно 95, от примерно 1 до примерно 90, от примерно 1 до примерно 80, от примерно 1 до примерно 70, от примерно 1 до примерно 60, от примерно 1 до примерно 50, от примерно 1 до примерно 40, от примерно 1 до примерно 30, от примерно 1 до примерно 20, от примерно 1 до примерно 10, от примерно 1 до примерно 9, от примерно 1 до примерно 8, от примерно 1 до примерно 7, от примерно 1 до примерно 6, от примерно 1 до примерно 5, от примерно 1 до примерно 4, от примерно 1 до примерно 3 или от примерно 1 до примерно 2. В некоторых предпочтительных вариантах осуществления, спейсеры CRISPR идентифицируются анализом последовательностей в виде фрагментов секвенирования ДНК, локализующихся между двумя повторами.
Как описано в настоящей заявке, настоящее изобретение относится к способам и композициям, которые содействуют применению одного или нескольких генов или белков cas в комбинации с одним или несколькими, предпочтительно, двумя или более, повторами CRISPR, подходящими для придания специфичности иммунитета, по меньшей мере, одному спейсеру CRISPR в клетке-реципиенте. В некоторых предпочтительных вариантах осуществления, по меньшей мере, один из генов или белков cas и, по меньшей мере, один повтор CRISPR используются в функциональных комбинациях для придания специфичности иммунитета, по меньшей мере, к одному спейсеру CRISPR в клетке.
Используемый в настоящем описании термин «специфичность иммунитета» означает, что иммунитет придан против определенной последовательности нуклеиновой кислоты или продукта ее транскрипции с использованием специфической последовательности спейсера CRISPR или псевдо-спейсера CRISPR. Как указано в настоящем описании, данный спейсер CRISPR не придает сопротивляемость против любой последовательности нуклеиновой кислоты или продукта ее транскрипции, но только тем последовательностям, против которых спейсер CRISPR или псевдо-спейсер CRISPR является гомологичным (например, тем, которые идентичны примерно на 100%).
В некоторых вариантах осуществления, спейсер(ы) CRISPR получаются из донорского организма, который отличается от клетки-реципиента. В некоторых вариантах осуществления, клетки-доноры и реципиенты представляют собой различные бактериальные штаммы, виды и/или роды. В некоторых предпочтительных вариантах осуществления, по меньшей мере, один из генов или белков cas и/или, по меньшей мере, один из повторов CRISPR получены из организмов, отличных от организма реципиента. В некоторых предпочтительных вариантах осуществления, переносятся, по меньшей мере, два из повтора CRISPR. В некоторых дополнительных предпочтительных вариантах осуществления, спейсеры CRISPR получаются из организма, который гетерологичен для клетки-реципиента или другой донорской клетки, из которой получены, по меньшей мере, один из генов и/или белков cas и/или, по меньшей мере, один из повторов CRISPR. В некоторых альтернативных предпочтительных вариантах осуществления, спейсеры CRISPR получены из организма, который является гомологичным для клетки-реципиента или другой донорской клетки, из которой получены, по меньшей мере, один из генов и/или белков cas и/или, по меньшей мере, один из повторов CRISPR. В некоторых предпочтительных вариантах осуществления, спейсер(ы) CRISPR сконструирован(ы) и получен(ы) с использованием рекомбинантных способов, известных в данной области. Действительно, предполагается, что спейсеры CRISPR будут получены с использованием любого подходящего способа, известного в данной области.
В некоторых вариантах осуществления, спейсеры CRISPR гетерологичны к клетке-реципиенту, из которой получены, по меньшей мере, один из генов и/или белков cas и/или, по меньшей мере, один, а в некоторых вариантах осуществления, два или более повтора CRISPR. В некоторых альтернативных вариантах осуществления, спейсеры CRISPR гомологичны к клетке-реципиенту, из которой получены, по меньшей мере, один из генов и/или белков cas и/или, по меньшей мере, один, а в некоторых вариантах осуществления, два или более повтора CRISPR. Действительно, предполагается, что любой из элементов, используемых в способах, будет гетерологичным или гомологичным. В некоторых вариантах осуществления, где используются множественные элементы (например, любые комбинации спейсера (спейсеров) CRISPR, повтора (повторов) CRISPR, гена (генов) cas и белка (белков) Cas), некоторые элементы являются гомологичными друг другу, а некоторые элементы являются гетерологичными друг другу (например, в некоторых вариантах осуществления, спейсер(ы) CRISPR и гены cas являются гомологичными, но повтор(ы) CRISPR является/являются гетерологичными). Таким образом, в некоторых вариантах осуществления, спейсер CRISPR не является естественно ассоциированным с повтором CRISPR и/или генами cas, и/или функциональной комбинацией повтора CRISPR-гена cas. Действительно, предполагается, что любая комбинация гетерологичных и гомологичных элементов находят применение в настоящем изобретении. В других дополнительных вариантах осуществления, клетки-доноры и клетки-реципиенты являются гетерологичными, тогда как в других вариантах осуществления, они являются гомологичными. Предполагается также, что элементы, содержащиеся внутри клеток-доноров и клеток-реципиентов, будут гомологичными и/или гетерологичными. Элементы (например, спейсеры CRISPR) вводятся в плазмиду и/или геномную ДНК клетки-реципиента с использованием любого подходящего способа, известного в данной области.
В некоторых предпочтительных вариантах осуществления, по меньшей мере, один спейсер CRISPR используется для генной инженерии клетки (например, клетки-реципиента). В некоторых дополнительных вариантах осуществления, один или несколько спейсеров CRISPR используются в комбинации с одним или несколькими генами cas и/или одного или нескольких, предпочтительно, двух или более повторов CRISPR (в некоторых предпочтительных вариантах осуществления, используются одна или несколько их функциональных комбинаций) для модулирования устойчивости клетки против нуклеиновой кислоты-мишени или продукта ее транскрипции, для получения клетки, подвергнутой генной инженерии. В некоторых других вариантах осуществления, спейсеры CRISPR используются в качестве матрицы, для модификации (например, мутации) на ней плазмиды и/или геномной ДНК клетки (например, клетки-реципиента), с тем, чтобы были созданы спейсеры CRISPR в ДНК клетки. В некоторых вариантах осуществления, спейсер(ы) CRISPR клонируется (клонируются), по меньшей мере, в один конструкт, плазмиду или вектор, которыми затем клетка-реципиент трансформируется с использованием любого подходящего способа, известного в данной области.
В некоторых других вариантах осуществления, настоящее изобретение относится к способам идентификации спейсера CRISPR для использования при модулировании устойчивости клетки против нуклеиновой кислоты-мишени или продукта ее транскрипции, включающим стадии: получения клетки, содержащей, по меньшей мере, два повтора CRISPR, и, по меньшей мере, один ген или белок cas; идентификации, по меньшей мере, одного спейсера CRISPR в организме (например, организме донора); модификации последовательности спейсера CRISPR клетки с тем, чтобы он имел гомологию со спейсером CRISPR организма донора, содержащего нуклеиновую кислоту-мишень; и определения того, модулирует ли клетка сопротивляемость против нуклеиновой кислоты-мишени, где модулирование устойчивости клетки против нуклеиновой кислоты-мишени или продукта ее транскрипции указывает на то, что спейсер CRISPR модулирует устойчивость клетки против нуклеиновой кислоты-мишени.
В некоторых предпочтительных вариантах осуществления, спейсеры CRISPR содержат или состоят из нуклеотидной последовательности, в любой одной или нескольких SEQ ID NOS:23-460 и/или SEQ ID NOS:522-665. SEQ ID NOS:23-339, 359-408, 522-665 получены из S.thermophilus, тогда как SEQ ID NOS:340-358 получены из S.vestibularis, SEQ ID NOS:409-446 получены из S.agalactiae, SEQ ID NOS:447-452 получены из S.mutans и SEQ ID NOS:453-460 получены из S.pyogenes.











В особенно предпочтительных вариантах осуществления, спейсеры CRISPR фланкируются двумя повторами CRISPR (т.е. спейсер CRISPR имеет, по меньшей мере, один повтор CRISPR с каждой стороны). Хотя не предполагается, что настоящее изобретение ограничивается каким-либо конкретным механизмом, теорией или гипотезой, предусматривается, что далее данный спейсер CRISPR получен с конца 5' локуса CRISPR, содержащего ген(ы) cas и/или лидерную последовательность, чем ниже устойчивость, приданная этим спейсером CRISPR. Таким образом, в некоторых вариантах осуществления настоящего изобретения, модифицируются один или несколько из первых 100 спейсеров CRISPR с конца 5' локуса CRISPR, тогда как в других вариантах осуществления, модифицируются один или несколько из первых 50 спейсеров CRISPR с конца 5' локуса CRISPR. В некоторых дополнительных вариантах осуществления, модифицируются один или несколько из первых 40 спейсеров CRISPR с конца 5' локуса CRISPR, тогда как в некоторых других вариантах осуществления, модифицируются один или несколько из первых 30 спейсеров CRISPR с конца 5' локуса CRISPR, а в других дополнительных вариантах осуществления, модифицируются один или несколько из первых 20 спейсеров CRISPR с конца 5' локуса CRISPR, и в еще одних вариантах осуществления, модифицируются один или несколько из первых 15 спейсеров CRISPR с конца 5' локуса CRISPR. В некоторых предпочтительных вариантах осуществления, модифицируются один или несколько из первых 10 спейсеров CRISPR с конца 5' локуса CRISPR. Как указано в настоящем описании, различные бактерии имеют различные количества спейсеров CRISPR, таким образом, в некоторых вариантах осуществления, модифицируются различные спейсеры.
Кор спейсера CRISPR
Для определенного типа CRISPR внутри микробного вида, спейсер CRISPR обычно представлен определенной преобладающей длиной, хотя размер может варьировать. Было обнаружено, что типы CRISPR, описанные к настоящему времени, содержат спейсер определенной преобладающей длины от примерно 20 пар оснований до примерно 58 пар оснований.
Используемый в настоящем описании термин «кор спейсера CRISPR» относится к длине самого короткого наблюдаемого спейсера в пределах типа CRISPR. Таким образом, например, в пределах типа I CRISPR S.thermophilus (CRISPR1), преобладающая длина спейсера составляет 30 пар оснований при меньшинстве спейсеров размером от 28 пар оснований до 32 пар оснований. Так, у типа I CRISPR S.thermophilus кор спейсера CRISPR определяется как непрерывный фрагмент секвенирования длиной 28 пар оснований.
В некоторых предпочтительных вариантах осуществления настоящего изобретения, кор спейсера CRISPR гомологичен нуклеиновой кислоте-мишени, продукту ее транскрипции или идентифицированной последовательности по длине коровой последовательности. Как указано выше, хотя гомология может также рассматриваться с точки зрения сходства, в некоторых предпочтительных вариантах осуществления настоящего изобретения, гомология выражается с точки зрения идентичности последовательности. Таким образом, в некоторых вариантах осуществления, гомологичная последовательность охватывает кор спейсера CRISPR, который может быть идентичным, по меньшей мере, примерно на 90%, или идентичным, по меньшей мере, примерно на 91, примерно на 92, примерно на 93, примерно на 94, примерно на 95, примерно на 96, примерно на 97, примерно на 98 или примерно на 99% нуклеиновой кислоте-мишени, продукту ее транскрипции или идентифицированной последовательности по длине коровой последовательности. В некоторых особенно предпочтительных вариантах осуществления, кор спейсера CRISPR примерно на 100%, идентичен нуклеиновой кислоте-мишени, продукту ее транскрипции или идентифицированной последовательности по длине коровой последовательности.
Во время создания настоящего изобретения, анализировали последовательности CRISPR различных штаммов S.thermophilus, включая близко родственные промышленные штаммы и устойчивые к фагам варианты. Различия числа и типа спейсеров наблюдались главным образом в локусе CRISPR1. Примечательно, что, как оказалось, чувствительность к фагам коррелирует с содержанием спейсера CRISPR1. В частности, содержание спейсера было почти идентичным между материнскими штаммами и устойчивыми к фагам производными, за исключением дополнительных спейсеров, присутствующих в последнем. Эти данные свидетельствуют о потенциальной связи между присутствием дополнительных спейсеров и различиями, наблюдаемыми в чувствительности данного штамма к фагам. Это наблюдение стимулировало исследование происхождения и функции дополнительных спейсеров, присутствующих в устойчивых к фагам мутантах.
Используемый в настоящем описании термин «псевдо-спейсер CRISPR» относится к последовательности нуклеиновой кислоты, присутствующей в организме (например, организме донора, включая без ограничения бактериофаг), которая является предпочтительно существенной для функции и/или выживания, и/или репликации, и/или инфекционности и т.д., и которая содержит последовательность спейсера CRISPR. В некоторых вариантах осуществления, псевдо-спейсеры CRISPR находят применение при получении последовательностей спейсеров CRISPR, которые комплементарны или гомологичны псевдо-спейсеру CRISPR. В некоторых особенно предпочтительных вариантах осуществления, эти последовательности находят применение при модулировании устойчивости.
В некоторых вариантах осуществления, по меньшей мере, один псевдо-спейсер CRISPR и спейсер(ы) CRISPR, который (которые) комплементарны или гомологичны, по меньшей мере, одному псевдо-спейсеру (спейсерам) CRISPR, используются для генной инженерии клетки-реципиента. В некоторых предпочтительных вариантах осуществления, по меньшей мере, один псевдо-спейсер CRISPR или спейсер(ы) CRISPR, который (которые) комплементарны или гомологичны, по меньшей мере, одному псевдо-спейсеру (спейсерам) CRISPR, используются в комбинации с одним или несколькими генами или белками cas и/или с одним или несколькими повторами CRISPR (например, одной или несколькими их комбинациями) для генной инженерии клетки-реципиента с тем, чтобы устойчивость клетки-реципиента была модулирована против нуклеиновой кислоты-мишени или продукта ее транскрипции.
В некоторых вариантах осуществления, псевдо-спейсер CRISPR или спейсер(ы) CRISPR, который (которые) комплементарны или гомологичны, по меньшей мере, одному псевдо-спейсеру (спейсерам) CRISPR, вставляются в плазмиду и/или геномную ДНК клетки-реципиента с использованием любого подходящего способа, известного в данной области.
В некоторых дополнительных вариантах осуществления, псевдо-спейсеры CRISPR используются в качестве матрицы, для модификации (например, мутации) на ней плазмиды и/или геномной ДНК клетки-реципиента, с тем, чтобы были созданы спейсеры CRISPR в плазмиде и/или геномной ДНК клетки. В некоторых других вариантах осуществления, псевдо-спейсеры CRISPR или спейсер(ы) CRISPR, который (которые) комплементарен (комплементарны) или гомологичен (гомологичны) одному или нескольким псевдо-спейсеру (спейсерам) CRISPR клонируются в конструкт, плазмиду и/или вектор, и т.д., вводится/вводятся в клетку-хозяин с использованием любого подходящего способа, известного в данной области.
CAS и гены cas
Используемый в настоящем описании термин «ген cas» имеет обычное значение, используемое в данной области, и относится к одному или нескольким генам cas, которые в целом соединены, ассоциированы или расположены рядом или поблизости от фланкирующих локусов CRISPR. Сравнительный обзор семейства белков Cas представлен в документе Haft et al. (Haft et al., PLoS. Comput. Biol., 1(6): e60 [2005]), в котором описаны 41 вновь распознанное семейство генов, ассоциированных с CRISPR (cas), в дополнение к четырем ранее известным семействам генов. Как указано в настоящем описании, системы CRISPR относятся к различным классам с различными типами повторов, наборами генов и диапазонами видов. Как указано в настоящем описании, число генов cas в данном локусе CRISPR может варьировать между видами.
В некоторых вариантах осуществления, настоящее изобретение относится к способам и композициям для использования одного или нескольких генов или белков cas, отдельно или в любой комбинации с одним или несколькими спейсерами CRISPR для модулирования устойчивости клетки (например, клетки-реципиенте) против нуклеиновой кислоты-мишени или продукта ее транскрипции.
В некоторых вариантах осуществления, один или несколько генов и/или белков cas естественно встречаются в клетке-реципиенте и один или несколько гетерологичных спейсеров интегрируется/интегрируются или вставляются, примыкая к одному или нескольким генам или белкам cas. В некоторых вариантах осуществления, один или несколько генов и/или белков cas является/являются гетерологичными клетке-реципиенту, и один или несколько спейсеров является/являются гомологичными или гетерологичными. В некоторых предпочтительных вариантах осуществления, спейсеры интегрируются или вставляются, примыкая к одному или нескольким генам или белкам cas.
В некоторых других вариантах осуществления, настоящее изобретение относится к способам и композициям для применения одного или нескольких генов или белков cas и, по меньшей мере, двух повторов CRISPR для модулирования устойчивости в клетке (например, клетке-реципиенте) против нуклеиновой кислоты-мишени или продукта ее транскрипции.
В других дополнительных вариантах осуществления, настоящее изобретение относится к способам и композициям для применения одного или нескольких генов или белков cas и, по меньшей мере, двух повторов CRISPR и, по меньшей мере, одного спейсера CRISPR для модулирования устойчивости в клетке (например, клетке-реципиенте) против нуклеиновой кислоты-мишени или продукта ее транскрипции.
Структуры CRISPR обычно обнаруживаются вблизи четырех генов, называемых с cas1 по cas4. Самое обычное расположение генов представляет собой cas3-cas4-cas1-cas2. Представляется, что белок Cas3 является геликазой, тогда как напоминает семейство RecB экзонуклеаз и содержит обогащенный цистеином мотив, свидетельствующий о связывании ДНК. Cas1 в целом является высоко основным и представляет собой единственный белок Cas, постоянно обнаруживаемый у всех видов, которые содержат локусы CRISPR. Cas2 предстоит еще охарактеризовать, cas1-4 обычно характеризуются их близким расположением к локусам CRISPR и их широким распределением по бактериальным и археальным видам. Хотя не все гены cas1-4 ассоциированы со всем локусами CRISPR, все они обнаруживаются во множественных подтипах.
Кроме того, у многих бактериальных видов имеется другой кластер из ассоциированных со структурами CRISPR трех генов, именуемых в настоящем описании cas1B, cas5 и сas6 (см. Bolotin et al., [2005], выше). Отмечается, что номенклатура генов cas еще составляется. Так, представленный в настоящем описании текст должен восприниматься в контексте. В некоторых вариантах осуществления, ген cas выбран из cas1, cas2, сas3 cas4, cas1B, сas5 и/или сas6. В некоторых предпочтительных вариантах осуществления, ген cas представляет собой cas1. В еще одних вариантах осуществления, ген cas выбран из фрагментов, вариантов, гомологов и/или их производных cas1, cas2, сas3 cas4, cas1B, сas5 и/или сas6. В некоторых дополнительных вариантах осуществления, комбинация двух или более генов cas находят применение, включая любые подходящие комбинации, включая те, которые представлены в документе WO 07/025097, включенном в настоящее описание путем ссылки. В некоторых вариантах осуществления, предоставляется множество генов cas. В некоторых вариантах осуществления, имеется множество различных и/или одинаковых генов cas. В некоторых вариантах осуществления, имеется множество различных и/или одинаковых генов cas или любая их комбинация, как представлено в документе WO 07/025097.
В некоторых вариантах осуществления, гены cas содержат ДНК, тогда как в других вариантах осуществления, гены cas содержат РНК. В некоторых вариантах осуществления, нуклеиновая кислота имеет геномное происхождение, тогда как в других вариантах осуществления, она имеет синтетическое или рекомбинантное происхождение. В некоторых вариантах осуществления, гены cas являются двухнитевыми или однонитевыми, представляющими или смысловую, или антисмысловую нить, или их комбинацию. В некоторых вариантах осуществления, гены cas получены использованием методик рекомбинантной ДНК (например, рекомбинантной ДНК), как описано в настоящей заявке.
Как описано в настоящей заявке, в некоторых вариантах осуществления, ген cas содержит фрагмент гена cas (т.е. этот фрагмент гена cas содержит часть последовательности дикого типа). В некоторых вариантах осуществления, последовательность содержит, по меньшей мере, примерно 30%, по меньшей мере, примерно 40%, по меньшей мере, примерно 50%, по меньшей мере, примерно 60%, по меньшей мере, примерно 65%, по меньшей мере, примерно 70%, по меньшей мере, примерно 75%, по меньшей мере, примерно 80%, по меньшей мере, примерно 85%, по меньшей мере, примерно 90%, по меньшей мере, примерно 95%, по меньшей мере, примерно 96%, по меньшей мере, примерно 97%, по меньшей мере, примерно 98% или, по меньшей мере, примерно 99% последовательности дикого типа.
В некоторых вариантах осуществления, предпочтительно, чтобы ген cas представлял собой ген cas, который является самым близким к лидерной последовательности или первому повтору CRISPR на конце 5' локуса CRISPR - такого как cas4 или cas6.
В некоторых вариантах осуществления, белок Cas выбран из Cas1, Cas2, Cas3, Cas4, Cas1B, Cas5 и/или Cas6, а также их фрагменты, варианты, гомологи и/или производные. В некоторых других вариантах осуществления, белок Cas выбран из Cas1, Cas2, Cas3, Cas4, Cas1B, Cas5 и/или Cas6, и их комбинаций, как описано в документе WO 07/025097. В еще одних вариантах осуществления, белок Cas выбран из одного или нескольких из Cas1, Cas2, Cas3, Cas4, Cas1B, Cas5 и/или Cas6, или множества одинаковых или различных белков Cas, в любом подходящем количестве и/или комбинации.
Термин «белок Cas» также охватывает множество белков Cas (например, от примерно 2 до примерно 12 белков Cas, предпочтительнее, от примерно 3 до примерно 11 белков Cas, предпочтительнее, от примерно 4 до примерно 10 белков Cas, предпочтительнее, от примерно 4 до примерно 9 белков Cas, предпочтительнее, от примерно 4 до примерно 8 белков Cas, а предпочтительнее, от примерно 4 до примерно 7 белков Cas; например, 4, 5, 6 или 7 белков Cas).
В некоторых вариантах осуществления, белки Cas кодируются генами cas, содержащими ДНК, тогда как в других вариантах осуществления, cas содержат РНК. В некоторых вариантах осуществления, нуклеиновая кислота имеет геномное происхождение, тогда как в других вариантах осуществления, она имеет синтетическое или рекомбинантное происхождение. В некоторых вариантах осуществления, гены cas являются двухнитевыми или однонитевыми, представляющими смысловую или антисмысловую нить или их комбинации. В некоторых вариантах осуществления, гены cas получаются использованием методик рекомбинантной ДНК (например, рекомбинантной ДНК), как описано в настоящей заявке.
Настоящее изобретение также относится к способам идентификации гена cas для применения при модулировании устойчивости клетки против нуклеиновой кислоты-мишени или продукта ее транскрипции, включающим стадии: получения клетки, содержащей, по меньшей мере, один спейсер CRISPR и, по меньшей мере, два повтора CRISPR; генной инженерии клетки с тем, чтобы она содержала, по меньшей мере, один ген cas; и определения того, модулирует ли клетка сопротивляемость против нуклеиновой кислоты-мишени или продукта ее транскрипции, где модулирование устойчивости клетки против нуклеиновой кислоты-мишени или продукта ее транскрипции указывает на то, что ген cas может применяться для модулирования устойчивости клетки.
В некоторых других вариантах осуществления, настоящее изобретение относится к способам и одному или нескольким генам cas, которые могут применяться при генной инженерии клеток (например, клетки-реципиента). В некоторых предпочтительных вариантах осуществления, один или несколько генов cas используются для генной инженерии клетки (например, клетки-реципиента), которая в комбинации с одним или несколькими, предпочтительно, двумя или более повторами CRISPR и один или несколько спейсеров CRISPR находит применение при модулировании устойчивости клетки против нуклеиновой кислоты-мишени или продукта ее транскрипции. Например, в некоторых вариантах осуществления, ген(ы) cas вставляется/вставляются в ДНК клетки (например, плазмиду и/или геномную ДНК клетки-реципиента) с использованием любого подходящего способа, известного в данной области. В некоторых дополнительных вариантах осуществления, гены cas используются в качестве матрицы для модификации (например, мутации) ДНК клетки (например, плазмиды и/или геномной ДНК клетки-реципиента) с тем, чтобы гены cas создавались или формировались в ДНК клетки. В некоторых вариантах осуществления, гены cas присутствуют, по меньшей мере, в одном конструкте, по меньшей мере, в одной плазмиде, и/или, по меньшей мере, в одном векторе, который (которые) затем вводится (вводятся) в клетку с использованием любого подходящего способа, известного в данной области.
В некоторых вариантах осуществления, гены cas содержат, по меньшей мере, один кластер cas, выбранный из любой одной или нескольких из SEQ ID NOS:461, 466, 473, 478, 488, 493, 498, 504, 509 и 517. В других вариантах осуществления, гены cas содержат, любую одну или несколько из SEQ ID NOS: 462-465, 467-472, 474-477, 479-487, 489-492, 494-497, 499-503, 505-508, 510-517, используемых отдельно или вместе с любой подходящей комбинацией. В некоторых предпочтительных вариантах осуществления, кластер(ы) используется/используются в комбинации с одним или несколькими, предпочтительно, двумя или более повторами CRISPR и, необязательно, одним или несколькими спейсерами CRISPR. В некоторых дополнительных вариантах осуществления, один или несколько генов или белков cas используется/используются в подходящих комбинациях.
Как указано в настоящем описании, данный набор генов или белков cas всегда связан с данной повторной последовательностью в пределах определенного локуса CRISPR. Таким образом, представляется, что гены или белки cas специфичны для данного повтора ДНК (т.е. гены или белки cas и повторные последовательности из функциональной пары).
Соответственно, определенные комбинации одного или нескольких генов или белков cas и одного или нескольких, предпочтительно, двух или более повторов CRISPR используются в порядке для того, чтобы спейсер CRISPR придал сопротивляемость против нуклеиновой кислоты-мишени или продукта ее транскрипции в клетке (например, клетке-реципиенте). Соответственно, к удивлению, было обнаружено, что невозможно просто использовать любые гены или белки cas, или любые повторы CRISPR. Вместо этого, признаком настоящего изобретения является то, что комбинация является функциональной.
В контексте описанной в настоящей заявке комбинации повтора CRISPR-гена cas, термин «функциональная» означает, что комбинация способна придать устойчивость нуклеиновой кислоте-мишени или гомологичному продукту ее транскрипции при использовании вместе со спейсером CRISPR, который совмещается с нуклеиновой кислотой-мишенью или продуктом ее транскрипции или гомологичен им. Используемые в настоящем описании термины «функциональная комбинация повтора CRISPR-cas» и «функциональная комбинация повтора CRISPR-гена cas» включает функциональную комбинацию, в которой cas представляет собой ген cas или белок Cas.
Подходящим образом, один или несколько генов или белков cas и/или один или несколько, предпочтительно, два или более повторов CRISPR получены из одной и той же клетки (например, одной и той же клетки-реципиента). В некоторых вариантах осуществления, при использовании в контексте, термин «производимая» синонимичен термину «получаемая». В некоторых предпочтительных вариантах осуществления, при использовании в контексте, термин «происходящая» также синонимичен «полученная», так как не предполагается, что настоящее изобретение определенно ограничивается элементами, которые являются «происходящими». В некоторых вариантах осуществления, при использовании в контексте, термин «происходящие» синонимичен термину «получаемые».
В некоторых вариантах осуществления, один или несколько генов или белков cas и/или один или несколько, предпочтительно, два или более повторов CRISPR получены из одного и того же локуса CRISPR в пределах генома или плазмиды, предпочтительно, генома или плазмиды одного и того же штамма, вида или рода. В некоторых других вариантах осуществления, один или несколько генов или белков cas и/или один или несколько, предпочтительно, два или более повторов CRISPR получены из одного и того же локуса CRISPR в пределах одного генома или плазмиды, предпочтительно, одного генома или плазмиды одного и того же штамма, вида или рода. В некоторых вариантах осуществления, один или несколько генов или белков cas и/или один или несколько, предпочтительно, два или более повторов CRISPR совместно встречаются естественно. В других дополнительных вариантах осуществления, один или несколько генов или белков cas и/или один или несколько, предпочтительно, два или более повторов CRISPR совместно встречаются естественно в одной и той же клетке (например, клетке-реципиенте). В других вариантах осуществления, один или несколько генов или белков cas и/или один или несколько, предпочтительно, два или более повторов CRISPR совместно встречаются естественно в одном и том же геноме клетки (например, клетки-реципиента). В других дополнительных вариантах осуществления, один или несколько генов или белков cas и/или один или несколько, предпочтительно, два или более повторов CRISPR совместно встречаются естественно в одном и том же геноме штамма, вида или рода. В некоторых других предпочтительных вариантах осуществления, настоящее изобретение относится к любой подходящей комбинации нуклеиновой кислоты, состоящей по существу, по меньшей мере, из двух повторов CRISPR и, по меньшей мере, одного гена или белка cas.
В некоторых вариантах осуществления, термин «состоит по существу из» относится к комбинации, по меньшей мере, двух повторов CRISPR и, по меньшей мере, одного гена или белка cas, и исключая, по меньшей мере, один дополнительный компонент локуса CRISPR (например, в отсутствие одного или нескольких спейсера (спейсеров) CRISPR и/или в отсутствие одной или нескольких обычной (обычных) лидерной последовательности (последовательностей) локуса CRISPR. В некоторых альтернативных вариантах осуществления, термин «состоит по существу из» относится к комбинации, по меньшей мере, двух повторов CRISPR и, по меньшей мере, только одного гена или белка cas, и исключая, все другие компоненты локуса CRISPR (например, естественно встречающегося локуса CRISPR). В некоторых других вариантах осуществления, термин «состоит по существу из» относится к комбинации, по меньшей мере, только одного гена или белка cas, и исключая все другие компоненты локуса CRISPR (например, естественно встречающегося локуса CRISPR). В некоторых других вариантах осуществления, термин «состоит по существу из» относится к комбинации, по меньшей мере, двух повторов CRISPR и, по меньшей мере, только одного гена или белка cas, и исключая, по меньшей мере, один другой компонент локуса CRISPR, предпочтительно, исключая, по меньшей мере, один другой компонент естественно встречающегося локуса CRISPR. В некоторых еще одних вариантах осуществления, термин «состоит по существу из» относится к комбинации, по меньшей мере, двух повторов CRISPR и, по меньшей мере, одного гена или белка cas, при условии, что отсутствует, по меньшей мере, один другой компонент естественного локуса CRISPR (например, по существу отсутствует). Таким образом, предполагается, что термин находит использование в контексте. В некоторых вариантах осуществления, настоящее изобретение относится к любой подходящей комбинации, по меньшей мере, двух повторов CRISPR и, по меньшей мере, одного гена или белка cas, при условии, что отсутствуют все другие компоненты локуса CRISPR (например, по существу отсутствуют), предпочтительно, что отсутствуют все другие компоненты локуса CRISPR естественной комбинации повтора (повторов) CRISPR и одного или нескольких генов или белков cas, используются в комбинации или вместе с одним или несколькими спейсерами CRISPR. В некоторых дополнительных вариантах осуществления, один или несколько генов или белков cas используются в комбинации или вместе, по меньшей мере, с одним или несколькими спейсерами CRISPR и, по меньшей мере, с одним или несколькими, предпочтительно, двумя или более повторами CRISPR. В некоторых вариантах осуществления, спейсер(ы) CRISPR представлены или получены из организма (например, организма донора), который отличен от клетки (например, клетки-реципиента), из которой получены один или несколько генов или белков cas, и/или один или несколько, предпочтительно, два или более повторов CRISPR.
Предоставлены различные расположения повтора (повторов) CRISPR и гена (генов) или белка (белков) cas, особенно, функциональных комбинаций повтора CRISPR-cas. В некоторых вариантах осуществления, комбинация содержит, состоит или по существу состоит, по меньшей мере, из любого количества из примерно 9, примерно 10, примерно 11, примерно 12, примерно 13, примерно 14, примерно 15, примерно 16, примерно 17, примерно 18, примерно 19 или примерно 20 повторов CRISPR, в комбинации с любым количеством из примерно 1, примерно 2, примерно 3, примерно 4, примерно 5, примерно 6, примерно 7, примерно 8, примерно 9, примерно 10, примерно 11, примерно 12, примерно 13, примерно 14, примерно 15, примерно 16, примерно 17, примерно 18, примерно 19 или примерно 20 генов или белков cas, (например, 16 повторов CRISPR и 12 генов или белков cas, или 18 повторов CRISPR и 20 генов или белков cas, или любых их комбинаций). Настоящее изобретение относится к повтору (повторам) CRISPR и гену (генам) cas, расположенным различным образом, как представлено в документе WO 07/025097. В некоторых вариантах осуществления, в которых комбинация гена cas и повтора CRISPR содержит более одного гена cas, следует понимать, что повтор CRISPR вставляется на конце 3' генов cas, на конце 5' генов cas или между генами cas, при условии, что, по меньшей мере, один из генов cas остается функциональным.
В некоторых вариантах осуществления, первая комбинация повтора CRISPR-гена или белка cas (содержащая, по меньшей мере, один ген или белок cas, и по меньшей мере, два повтора CRISPR, где оба получены из одного и того же локуса CRISPR в пределах генома) используется в комбинации со второй комбинацией повтора CRISPR-гена или белка cas (содержащей, по меньшей мере, один ген или белок cas, и по меньшей мере, два повтора CRISPR, где оба получены из одного и того же или другого локуса CRISPR в пределах генома). Соответственно, в этих вариантах осуществления изобретения, первая и вторая комбинации получены из одних и тех же или различных локусов CRISPR в пределах генома. Таким образом, в некоторых вариантах осуществления, первая и вторая комбинации повтора CRISPR-гена или белка cas получены из различных геномов (например, из различных геномов в пределах одного и того же кластера), как более детально описано далее в настоящей заявке.
В еще одних вариантах осуществления настоящего изобретения, первая и/или вторая комбинация повтора CRISPR-гена или белка cas (содержащая, по меньшей мере, один ген cas и, по меньшей мере, два повтора CRISPR, полученных из одного и того же локуса CRISPR в пределах генома) используются в комбинации примерно с 3, примерно с 4, примерно с 5, примерно с 6, примерно с 7, примерно с 8, примерно с 9 или примерно с 10, или более комбинаций повтора CRISPR-гена или белка cas (каждой содержащей, по меньшей мере, один ген или белок cas и, по меньшей мере, два повтора CRISPR, полученные из одних и тех же или других локусов CRISPR в пределах генома). Соответственно, в этих вариантах осуществления изобретения, комбинации получены из одних и тех же или других локусов CRISPR в пределах генома. В некоторых других вариантах осуществления изобретения, комбинации получены из различных геномов (например, различных геномов в пределах одного и того же кластера), как более детально описано далее в настоящей заявке.
Таким образом, в некоторых вариантах осуществления, для того, чтобы комбинация повтора CRISPR-гена или белка cas придавала устойчивость, повтор(ы) CRISPR и ген(ы) или белок (белки) cas совместно естественно встречаются в пределах данного локуса CRISPR генома. В некоторых вариантах осуществления, повтор(ы) CRISPR и ген(ы) или белок (белки) cas совместно естественно встречаются в пределах данного локуса CRISPR генома. В некоторых вариантах осуществления, эти функциональные комбинации, взятые вместе, придают сопротивляемость против нуклеиновой кислоты-мишени или продукта ее транскрипции.
В некоторых других вариантах осуществления, настоящее изобретение относится к способам идентификации функциональной комбинации гена или белка cas и повтора CRISPR, включающим стадии: анализа последовательностей (например, последовательностей нуклеиновой кислоты или белка) гена или белка cas и повтора CRISPR; идентификации одного или нескольких кластеров генов или белков cas; идентификации одного или нескольких кластеров повторов CRISPR; и комбинирования тех генов или белков cas и последовательностей повторов CRISPR, которые укладываются в пределы одного и того же кластера.
В некоторых дополнительных вариантах осуществления, настоящее изобретение относится к способам идентификации функциональной комбинации гена или белка cas и повтора CRISPR для использования при модулировании устойчивости клетки против нуклеиновой кислоты-мишени или продукта ее транскрипции, включающим стадии: получения клетки, держащей комбинацию одного или нескольких генов или белков cas и одного или нескольких, предпочтительно, двух или более повторов CRISPR; генной инженерии клетки с тем, чтобы она содержала один или несколько спейсеров CRISPR; и определения того, модулирует ли клетка сопротивляемость против нуклеиновой кислоты-мишени, где модулирование устойчивости клетки против нуклеиновой кислоты-мишени или продукта ее транскрипции указывает на то, что комбинация может применяться для модулирования устойчивости клетки против нуклеиновой кислоты-мишени.
В некоторых вариантах осуществления, последовательности гена и/или белка cas и/или повтора CRISPR получены из одних и тех же или других штаммов, видов, родов и/или организмов. В некоторых вариантах осуществления, комбинация содержит ДНК и/или РНК геномного, рекомбинантного и/или синтетического происхождения. В некоторых вариантах осуществления, повтор(ы) CRISPR содержит (содержат) ДНК и/или РНК геномного, рекомбинантного и/или синтетического происхождения. В некоторых вариантах осуществления, ген(ы) cas содержит (содержат) ДНК и/или РНК геномного, рекомбинантного и/или синтетического происхождения. Действительно, предполагается, что настоящее изобретение охватывает любую комбинацию ДНК и/или РНК для каждого из элементов (например, ген cas и/или повтор CRISPR. В некоторых вариантах осуществления, элементы анализируются с использованием любых подходящих способов, известных в данной области. В некоторых вариантах осуществления, повтор CRISPR и/или ген cas являются двухнитевыми, тогда как в других вариантах осуществления, каждый из них является однонитевым, представляющим или смысловую, или антисмысловую нить, или их комбинации.
В некоторых вариантах осуществления, одна или несколько описанных в настоящей заявке функциональных комбинаций используются для генной инженерии клетки (например, клетки-реципиента). В некоторых предпочтительных вариантах осуществления, одна или несколько функциональных комбинаций используются для генной инженерии клетки (например, клетки-реципиента), что в комбинации с одним или несколькими спейсерами CRISPR находит применение при модулировании устойчивости клетки против нуклеиновой кислоты-мишени или продукта ее транскрипции. В некоторых вариантах осуществления, функциональные комбинации вставляются в ДНК клетки-реципиента (например, плазмиду или геномную ДНК клетки) с использованием любых подходящих способов, известных в данной области. В некоторых дополнительных вариантах осуществления, функциональные комбинации используются в качестве матрицы для модификации (например, мутации) ДНК клетки-реципиента (например, ДНК плазмиды и/или геномной ДНК) с тем, чтобы создать функциональные комбинации в ДНК клетки. В еще одних дополнительных вариантах осуществления, функциональные комбинации клонируются в конструкт, плазмиду или вектор и т.д., который затем трансформируется в клетку с использованием способов, таких как описанные в настоящей заявке и известные в данной области.
В некоторых вариантах осуществления, функциональная комбинация получается или может быть получена способом, включающим стадии: анализа последовательностей гена cas и повтора CRISPR; идентификации одного или нескольких кластеров генов cas; идентификации одного или нескольких кластеров повторов CRISPR; и комбинирования последовательностей тех генов cas и повторов CRISPR, которые укладываются в пределы одного и того же кластера, где комбинация последовательностей гена cas и повтора CRISPR в пределах одного и того же кластера указывает на то, что комбинация представляет собой функциональную комбинацию.
Как указано выше, к удивлению, было обнаружено, что невозможно просто обменивать комбинации повтора CRISPR-cas между любыми клетками (например, любыми штаммами, видами или родами), так как это необязательно приводит к получению функциональных комбинаций повтора CRISPR-cas. Действительно, для того, чтобы комбинация (комбинации) повтора CRISPR-cas были функциональными, они должны быть совместимыми. Таким образом, предусматривается, что невозможно включить гены cas или повторы CRISPR между различными локусами CRISPR пока они из одного и того же кластера. Даже более удивительно, что кластеры не соблюдают филогению «организма». В частности, в пределах одного организма может быть несколько CRISPR. В результате, считается, что функциональная комбинация повтора CRISPR-cas требует, чтобы комбинация была включена в пределы кластера, в отличие от включения внутрь организма.
Во избежание сомнения, термин «кластер», используемый в настоящем описании, не относится к кластеру генов, локализующихся в одном и том же локусе (обычно образующем оперон), но к выходу из анализа сравнения последовательностей (например, сравнительного анализа множества последовательностей и/или совмещений множественных последовательностей и/или анализа нанесением точек на график). Соответственно, в некоторых вариантах осуществления, кластерный анализ локусов CRISPR выполняется с использованием различных способов, которые известны в данной области (например, такого как описанный в настоящей заявке анализ нанесением точек на график или множественное совмещение) с последующим расчетом дендрограммы. В некоторых вариантах осуществления, кластер представляет собой класс, семейство или группу последовательностей.
Преимущественно, использование естественно совместно встречающихся комбинации (комбинаций) повтора CRISPR-cas обеспечивает взаимообмен комбинации и в пределах, и между данным видом, посредством этого, обеспечивая возможность получения методами генной инженерии устойчивости одного штамма, используя комбинацию из другого штамма.
Бактериофаг
Используемый в настоящее описании термин «бактериофаг» (или «фаг») имеет обычное значение, понятное в данной области (т.е. вирус, который селективно инфицирует один или несколько бактериальных видов). Многие бактериофаги специфичны к определенному роду или виду, или штамму бактерий. В некоторых предпочтительных вариантах осуществления, фаги способны инфицировать материнские бактерии и/или клетки-хозяева. В некоторых вариантах осуществления, бактериофаги являются вирулентными для материнской бактерии. В некоторых вариантах осуществления, фаги являются литическими, тогда как в других вариантах осуществления, фаги являются лизогенными.
Литический бактериофаг представляет собой тот, который следует по литическому пути посредством завершения литического цикла, а не вступления в лизогенный путь. Литический бактериофаг подвергается вирусной репликации, ведущей к лизису клеточной мембраны, разрушению клетки и высвобождению потомства частиц бактериофага, способных инфицировать другие клетки.
Лизогенный бактериофаг представляет собой тот, который способен к вступлению в лизогенный путь, при котором бактериофаг становится покоящейся, пассивной частью генома клетки посредством предшествующего завершения его литического цикла.
Бактериофаги, которые находят применение в настоящем изобретении, включают без ограничения бактериофаги, которые относятся к любому из следующих семейств вирусов: Corticoviridae, Cystoviridae, Inoviridae, Leviviridae, Microviridae, Myoviridae, Podoviridae, Siphoviridae или Tectiviridae. В некоторых вариантах осуществления, особое применение находит бактериофаг, который инфицирует бактерии, являющиеся патогенными для растений и/или животных (включая людей). В некоторых особенно предпочтительных вариантах осуществления, модулируется устойчивость клетки против бактериофага.
В некоторых особенно предпочтительных вариантах осуществления, бактериофаги по настоящему изобретению включают без ограничения бактериофаги, способные инфицировать бактерию, которая естественно содержит один или несколько локусов CRISPR. Локусы CRISPR были идентифицированы в более чем 40 прокариотах (см., например, Jansen et al., Mol. Microbiol., 43:1565-1575 [2002]; и Mojica et al., [2005]), включая без ограничения Aeropyrum, Pyrobaculum, Sulfolobus, Archaeoglobus, Halocarcula, Methanobacterium, Methanococcus, Methanosarcina, Methanopyrus, Pyrococcus, Picrophilus, Thermoplasma, Corynebacterium, Mycobacterium, Streptomyces, Aquifex, Porphyromonas, Chlorobium, Thermus, Bacillus, Listeria, Staphylococcus, Clostridium, Thermoanaerobacter, Mycoplasma, Fusobacterium, Azarcus, Chromobacterium, Neisseria, Nitrosomonas, Desulfovibrio, Geobacter, Myxococcus, Campylobacter, Wolinella, Acinetobacter, Erwinia, Escherichia, Legionella, Methylococcus, Pasteurella, Photobacterium, Salmonella, Xanthamonas, Yersinia, Treponema и Thermotoga.
В некоторых вариантах осуществления, бактериофаги включают без ограничения те бактериофаги, которые способны инфицировать бактерии, относящиеся к следующим родам: Escherichia, Shigella, Salmonella, Erwinia, Yersinia, Bacillus, Vibrio, Legionella, Pseudomonas, Neisseria, Bordetella, Helicobacter, Listeria, Agrobacterium, Staphylococcus, Streptococcus, Enterococcus, Clostridium, Corynebacterium, Mycobacterium, Treponema, Borrelia, Francisella, Brucella и Xanthomonas.
В других дополнительных вариантах осуществления, бактериофаги включают без ограничения те бактериофаги, которые способны инфицировать (или трансдуцировать) молочнокислые бактерии, Bifidobacterium, Brevibacterium, Propionibacterium, Lactococcus, Streptococcus, Lactobacillus (например, L. acidophilus), Enterococcus, Pediococcus, Leuconostoc и Oenococcus.
В еще одних вариантах осуществления, бактериофаги включают без ограничения те бактериофаги, которые способны инфицировать Lactococcus lacti (например, L. lactis подвид lactis и L. lactis подвид cremoris, и L. lactis подвид lactis biovar diacetylactis), Streptococcus thermophilus, Lactobacillus delbrueckii подвид bulgaricus, Lactobacillus helveticus, Bifidobacterium lactis, Lactobacillus acidophilus, Lactobacillus casei, Bifidobacterium infantis, Lactobacillus paracasei, Lactobacillus salivarius, Lactobacillus plantarum, Lactobacillus reuteri, Lactobacillus gasseri, Lactobacillus johnsonii или Bifidobacterium longum.
В еще одних вариантах осуществления, бактериофаги включают без ограничения те бактериофаги, которые способны инфицировать любые ферментативные бактерии, восприимчивые к разрушению инфекцией бактериофагом, включая без ограничения способы получения антибиотиков, аминокислот и растворителей. Продукты, полученные ферментацией, которые, как известно, испытывали инфекцию бактериофагом, и соответствующие инфицированные ферментационные бактерии включают домашний сыр (Lactococcus lactis подвид lactis, Lactococcus lactis подвид cremoris), йогурт (Lactobacillus delbrueckii подвид bulgaricus, Streptococcus thermophilus), швейцарский сыр (S. thermophilus, Lactobacillus lactis, Lactobacillus helveticus), голубой сыр (Leuconostoc cremoris), итальянский сыр (L. Bulgaricus, S. thermophilus), viili (Lactococcus lactis подвид cremoris, Lactococcus lactis подвид lactis biovar diacetylactis, Leuconostoc cremoris), якулт (Lactobacillus casei), казеин (Lactococcus lactis подвид cremoris), натто ((Bacillus subtilis var. natto), вино (Leuconostoc oenos), саке (Leuconostoc mesenteroides), полимиксин (Bacillus polymyxa), колистин (Bacillus colistrium), бацитрацин (Bacillus licheniformis), L-глутаминовую кислоту (Brevibacterium lactofermentum, Microbacterium ammoniaphilum) и ацетон и бутанол (Clostridium acetobutylicum, Clostridium saccharoperbutylacetonicum).
В некоторых предпочтительных вариантах осуществления, бактерии, которые находят применение в настоящем изобретении, включают без ограничения S. thermophilus, L. delbrueckii, подвид bulgaricus и/или L. acidophilus.
В некоторых особенно предпочтительных вариантах осуществления, бактериофаги включают без ограничения те бактериофаги, которые способны инфицировать бактерии, которые содержат один или несколько гетерологичных локусов CRISPR. В некоторых вариантах осуществления, бактериофаги включают один или несколько гетерологичных локусов CRISPR и/или один или несколько гетерологичных генов cas. и/или один или несколько гетерологичных повторов CRISPR и/или один или несколько гетерологичных спейсеров CRISPR.
Инфекция бактерий фагом возникает в результате инъекции или переноса фага ДНК в клетки. В некоторых вариантах осуществления, инфекция ведет к экспрессии (т.е. транскрипции и трансляции) нуклеиновой кислоты бактериофага внутри клетки и продолжению жизненного цикла бактериофага. В некоторых вариантах осуществления, включающих рекомбинантный бактериофаг, также экспрессируются рекомбинантные последовательности внутри генома фага (например, нуклеиновые кислоты-репортеры).
Было обнаружено, что последовательности спейсеров CRISPR в прокариотах часто имеют значимые сходства с разнообразными молекулами ДНК, включая такие генетические элементы как хромосомы, бактериофаги и конъюгативные плазмиды. Сообщалось, что клетки, несущие эти спейсеры CRISPR, неспособны инфицироваться молекулами ДНК, содержащими последовательности, гомологичные спейсерам (см. Mojica et al., [2005]).
В некоторых вариантах осуществления настоящего изобретения, один или несколько псевдо-спейсеров, полученных из ДНК бактериофага, или спейсер(ы) CRISPR, который (которые) является (являются) комплементарными или гомологичными одному или нескольким псевдо-спейсеру (спейсерам) CRISPR, добавляются в пределы локуса CRISPR клетки (например, клетки-реципиента), для модулирования (например, обеспечения) сопротивляемости против определенного бактериофага, таким образом, по существу предотвращая атаку фага.
В некоторых предпочтительных вариантах осуществления, мишенью являются определенные области внутри генома фага для получения псевдо-спейсеров, включая без ограничения гены, кодирующие белки специфичности хозяина, включая те, которые обеспечивают распознавание хозяина определенного фага, такие как геликаза, примаза, головные или хвостовые структурные белки, белки с консервативным доменом (например, холинг, лизин и другие) или консервативные последовательности среди важных генов фага.
Любая нуклеиновая кислота, происходящая из генома фага, может придать иммунитет против фага при вставке, например, между двумя повторами в активном локусе CRISPR. В некоторых вариантах осуществления, иммунитет более «эффективный», когда спейсер CRISPR соответствует внутренней последовательности гена фага. В некоторых особенно предпочтительных вариантах осуществления, иммунитет получается еще более «эффективным», когда ген кодирует «основной белок (например, антирецептор).
В некоторых предпочтительных вариантах осуществления, настоящее изобретение относится к способам придания клетке (например, бактериальной клетке) сопротивляемости против бактериофага, включающим стадии: (а) предоставления одного или нескольких псевдо-спейсеров CRISPR, по меньшей мере, из одного бактериофага; (b) идентификации одной или нескольких функциональных комбинаций повтора CRISPR-cas в клетке, по меньшей мере, в одной клетке, которая по существу чувствительна к бактериофагу; и (с) генной инженерии одного или нескольких локусов CRISPR в по существу чувствительной клетке с тем, чтобы они содержали один или несколько псевдо-спейсеров CRISPR из бактериофага или один или несколько спейсер(ов) CRISPR, который (которые) является (являются) комплементарными или гомологичными одному или нескольким псевдо-спейсеру (спейсерам) CRISPR для придания клетке устойчивости.
В еще одних дополнительных вариантах осуществления, настоящее изобретение относится к способам придания клетке (например, бактериальной клетке) сопротивляемости против бактериофага, включающим стадии: (а) предоставления одного или нескольких псевдо-спейсеров CRISPR, по меньшей мере, из одного бактериофага; (b) идентификации одной или нескольких функциональных комбинаций повтора CRISPR-cas, по меньшей мере, в одной клетке, которая по существу чувствительна к бактериофагу; и (с) вставки одного или нескольких псевдо-спейсеров CRISPR из бактериофага или одного или нескольких спейсера (спейсеров) CRISPR, который (которые) является (являются) комплементарными или гомологичными одному или нескольким псевдо-спейсеру (спейсерам) CRISPR по существу в чувствительную клетку с тем, чтобы придать клетке по существу устойчивости к бактериофагу.
В еще одних дополнительных вариантах осуществления, настоящее изобретение относится к способам модулирования изотипа бактериальной клетки, включающим стадии: (а) предоставления одного или нескольких псевдо-спейсеров CRISPR, по меньшей мере, из одного бактериофага; (b) идентификации одной или нескольких функциональных комбинаций повтора CRISPR-cas в клетке, по меньшей мере, в одной клетке, которая по существу чувствительна к бактериофагу; и (с) генной инженерии одного или нескольких локусов CRISPR в по существу чувствительной клетке с тем, чтобы они содержали один или несколько псевдо-спейсеров CRISPR из бактериофага или один или несколько спейсер(ов) CRISPR, который (которые) является (являются) комплементарными или гомологичными одному или нескольким псевдо-спейсеру (спейсерам) CRISPR.
В еще одних других вариантах осуществления, настоящее изобретение относится к способам модулирования изотипа бактериальной клетки, включающим стадии: (а) предоставления одного или нескольких псевдо-спейсеров CRISPR, по меньшей мере, из одного бактериофага; (b) идентификации одной или нескольких функциональных комбинаций повтора CRISPR-cas, по меньшей мере, в одной клетке, которая по существу чувствительна к бактериофагу; и (с) вставки одного или нескольких псевдо-спейсеров CRISPR из бактериофага или одного или нескольких спейсера (спейсеров) CRISPR, который (которые) является (являются) комплементарными или гомологичными одному или нескольким псевдо-спейсеру (спейсерам) CRISPR по существу в чувствительную клетку.
В еще одних дополнительных вариантах осуществления, настоящее изобретение относится к способам придания клетке (например, бактериальной клетке) сопротивляемости против бактериофага, включающим стадии: (i) идентификации псевдо-спейсера CRISPR в бактериофаге, содержащем нуклеиновую кислоту-мишень или продукт ее транскрипции, против которого предстоит модулировать устойчивость; и (ii) модификации последовательности спейсера CRISPR клетки с тем, чтобы спейсер CRISPR клетки имел гомологию с псевдо-спейсером CRISPR бактериофага, содержащего нуклеиновую кислоту-мишень.
В еще одних вариантах осуществления, настоящее изобретение относится к способам придания клетке (например, бактериальной клетке) сопротивляемости против бактериофага, включающим стадии: (i) идентификации псевдо-спейсера CRISPR в бактериофаге, содержащем нуклеиновую кислоту-мишень или продукт ее экспрессии, против которого предстоит модулировать устойчивость; и (ii) модификации последовательности спейсера CRISPR клетки с тем, чтобы спейсер CRISPR клетки имел 100% гомологию или идентичность с псевдо-спейсером CRISPR бактериофага, содержащего нуклеиновую кислоту-мишень.
В еще одних дополнительных вариантах осуществления, настоящее изобретение относится к способам модулирования изотипа бактериальной клетки, включающим стадии: (i) идентификации псевдо-спейсера CRISPR в бактериофаге, содержащем нуклеиновую кислоту-мишень или продукт ее экспрессии, против которого предстоит модулировать устойчивость; и (ii) модификации последовательности спейсера CRISPR клетки с тем, чтобы спейсер CRISPR клетки имел гомологию с псевдо-спейсером CRISPR бактериофага, содержащего нуклеиновую кислоту-мишень.
В еще одних вариантах осуществления, настоящее изобретение относится к способам модулирования изотипа бактериальной клетки, включающим стадии: (i) идентификации псевдо-спейсера CRISPR в бактериофаге, содержащем нуклеиновую кислоту-мишень или продукт ее экспрессии, против которого предстоит модулировать устойчивость; и (ii) модификации последовательности спейсера CRISPR клетки с тем, чтобы спейсер CRISPR клетки имел 100% гомологию или идентичность с псевдо-спейсером CRISPR бактериофага, содержащего нуклеиновую кислоту-мишень.
В некоторых вариантах осуществления, спейсер CRISPR бактериальной клетки имеет 100% гомологию или идентичность с последовательностью (например, такой как псевдо-спейсера CRISPR) в бактериофаге, содержащем нуклеиновую кислоту-мишень.
В некоторых альтернативных вариантах осуществления, спейсер CRISPR бактериальной клетки образует составную часть локуса CRISPR, содержащего функциональную комбинацию повтора CRISPR-cas, как описано в настоящей заявке.
В некоторых особенно предпочтительных вариантах осуществления, нуклеиновая кислота-мишень или продукт ее транскрипции в бактериофаге представляет собой высоко консервативную последовательность нуклеиновой кислоты. В некоторых других предпочтительных вариантах осуществления, нуклеиновая кислота-мишень или продукт ее транскрипции в бактериофаге представляет собой ген, кодирующий белок специфичности хозяина. В некоторых дополнительных вариантах осуществления, нуклеиновая кислота-мишень или продукт ее транскрипции в бактериофаге кодирует фермент, который существенен для выживания, репликации и/или роста бактериофага. В других дополнительных вариантах осуществления, нуклеиновая кислота-мишень или продукт ее транскрипции в бактериофаге кодирует геликазу, примазу, головной или хвостовой структурный белок или белок с консервативным доменом (например, холинг, лизин и т.д.).
В некоторых предпочтительных вариантах осуществления, получаются бактериальные клетки, которые имеют «сниженную восприимчивость к размножению бактериофага или инфекции им». Этот термин, используемый в настоящем описании, относится к бактериям как имеющим низкую или отсутствующую восприимчивость к размножению бактериофага или инфекции им, по сравнению с бактериями дикого типа при культивировании (например, в молочной среде).
В некоторых вариантах осуществления, бактериальные клетки проявляют «низкую восприимчивость к размножению бактериофага». Этот термин относится к уровню размножения бактериофага в бактерии, который ниже уровня, который вызвал бы вредное воздействие на культуру в данный период времени. Такие вредные воздействия на культуру включают без ограничения отсутствие сворачивания молока во время производства ферментированных молочных продуктов (например, йогурта или сыра), неадекватное или медленное снижение рН во время производства ферментированных молочных продуктов (например, йогурта или сыра), медленное созревание сыра и/или порча текстуры пищевого продукта до такой степени, что он становится неаппетитным или неподходящим для употребления.
Для эквивалентного набора условий культивирования, бактериальная восприимчивость к бактериофагу по настоящему изобретению в целом выражается в сравнении с бактериями дикого типа. В некоторых вариантах осуществления, бактерии имеют примерно в 100 раз более низкую (Эффективность бляшкообразования [EOP]=10-2), предпочтительно, примерно в 1000 раз более низкую (EOP=10-3), предпочтительнее, примерно в 10000 раз более низкую (EOP=10-4), а наиболее предпочтительно, примерно в 100000 раз более низкую (EOP=10-5). В некоторых предпочтительных вариантах осуществления, уровень размножения бактериофагов в культуре измеряется примерно после 14 часов инкубации культуры, предпочтительнее, примерно после 12 часов, а еще предпочтительнее, примерно после 7 часов, еще предпочтительнее, примерно после 6 часов, еще предпочтительнее, примерно после 5 часов, а наиболее предпочтительно, примерно после 4 часов.
В дополнительных вариантах осуществления, настоящее изобретение относится к способам придания чувствительности клетки (например, бактериальной клетки) против бактериофага, включающим стадии: (а) предоставления псевдо-спейсера CRISPR, по меньшей мере, из одного бактериофага; (b) идентификации одной или нескольких функциональных комбинаций повтора CRISPR-cas в клетке, которая по существу устойчива к бактериофагу; и (с) генной инженерии одного или нескольких локусов CRISPR по существу в чувствительной клетке с тем, чтобы они содержали один или несколько псевдо-спейсеров или один или несколько спейсер (спейсеров) CRISPR, который (которые) является (являются) комплементарными или гомологичными одному или нескольким псевдо-спейсеру (спейсерам) CRISPR, которые имеют сниженную степень гомологии, по сравнению с одним или несколькими локусами CRISPR по существу в устойчивой клетке.
В других вариантах осуществления, настоящее изобретение относится к способам модулирования (например, снижения) лизотипа клетки (например, бактериальной клетки), содержащей один или несколько генов или белков cas и один или несколько, предпочтительно, два или более повтора CRISPR, включающим стадии: (i) идентификации псевдо-спейсера CRISPR в бактериофаге, против которого предстоит модулировать устойчивость; и (ii) модификации последовательности спейсера CRISPR клетки с тем, чтобы спейсер CRISPR клетки имел сниженную степень гомологии с псевдо-спейсером CRISPR бактериофага, содержащего нуклеиновую кислоту-мишень.
В еще одних вариантах осуществления, настоящее изобретение относится к способам модулирования (например, снижения или уменьшения) устойчивости бактериальной клетки, содержащей один или несколько генов или белков cas и один или несколько, предпочтительно, два или более повтора CRISPR, против бактериофага, включающим стадии: (i) идентификации одного или нескольких псевдо-спейсеров CRISPR в бактериофаге, против которого предстоит модулировать устойчивость; и (ii) модификации спейсера CRISPR в бактериальной клетке, у которой предстоит модулировать устойчивость, с тем, чтобы спейсер CRISPR имел более низкую степень гомологии с псевдо-спейсером (спейсерами) CRISPR бактериофага, против которого предстоит модулировать устойчивость.
В некоторых вариантах осуществления, спейсер CRISPR клетки имеет сниженную степень гомологии (например, уменьшение гомологии примерно, на 1, примерно на 2, примерно на 3, примерно на 4, примерно на 5, примерно на 6, примерно на 7, примерно на 8, примерно на 9, примерно на 10, примерно на 15, примерно на 20, примерно на 25, примерно на 30, примерно на 35, примерно на 40, примерно на 45, примерно на 50, примерно на 55, примерно на 60, примерно на 65, примерно на 70, примерно на 75, примерно на 80, примерно на 90 или примерно на 95%), по сравнению с псевдо-спейсером (спейсерами) CRISPR бактериофага, против которого предстоит модулировать устойчивость.
В некоторых вариантах осуществления, бактериальные клетки получаются с использованием способов по настоящему изобретению, так, чтобы клетки имели «повышенную восприимчивость к размножению бактериофага». Используемый в настоящем описании, этот термин относится к бактериям, которые имеют увеличенную или более высокую восприимчивость к размножению бактериофага, по сравнению с бактериями дикого типа при культивировании (например, в молочной среде).
В некоторых вариантах осуществления, термин «высокая восприимчивость к размножению бактериофага» относится к уровню размножения бактериофага в бактерии, который выше уровня, который вызвал бы вредное воздействие на культуру в данный период времени. Такие вредные воздействия на культуру включают без ограничения отсутствие сворачивания молока во время производства ферментированных молочных продуктов (например, йогурта или сыра), неадекватное или медленное снижение рН во время производства ферментированных молочных продуктов (например, йогурта или сыра), медленное созревание сыра и/или порча текстуры пищевого продукта до такой степени, что он становится неаппетитным или неподходящим для употребления.
Для эквивалентного набора условий культивирования, бактериальная восприимчивость к бактериофагу по настоящему изобретению в целом выражается в сравнении с бактериями дикого типа. В некоторых вариантах осуществления, бактерии имеют примерно в 100 раз более низкую (Эффективность бляшкообразования [EOP]=10-2), предпочтительно, примерно в 1000 раз более низкую (EOP=10-3), предпочтительнее, примерно в 10000 раз более низкую (EOP=10-4), а наиболее предпочтительно, примерно в 100000 раз более низкую (EOP=10-5). В некоторых предпочтительных вариантах осуществления, уровень размножения бактериофагов в культуре измеряется примерно после 14 часов инкубации культуры, предпочтительнее, примерно после 12 часов, а еще предпочтительнее, примерно после 7 часов, еще предпочтительнее, примерно после 6 часов, еще предпочтительнее, примерно после 5 часов, а наиболее предпочтительно, примерно после 4 часов.
В некоторых предпочтительных вариантах осуществления, спейсер CRISPR фланкируется двумя повторами CRISPR (т.е. спейсер CRISPR имеет, по меньшей мере, один повтор CRISPR с каждой стороны).
В некоторых вариантах осуществления настоящего изобретения, материнская бактерия (например, «материнский бактериальный штамм») подвергается воздействию (например, повторно, последовательно, одновременно или по существу одновременно) более чем одного бактериофага (например, смеси одного или нескольких фагов). В некоторых вариантах осуществления, материнский бактериальный штамм чувствителен к каждому из бактериофагов, который подвергается воздействию в смеси, тогда как в других вариантах осуществления, бактериальный штамм чувствителен к некоторым из бактериофагов, но устойчив к другим.
Используемый в настоящем описании термин «маркированная последовательность» относится к части дополнительного фрагмента ДНК, который получен из генома одного или нескольких бактериофагов (например, «плюс-цепи» генома одного или нескольких бактериофагов), которые воздействуют на материнскую бактерию в соответствии со способами по настоящему изобретению, и используются в качестве метки или маркировки (например, обеспечивающей необычную метку или необычную маркировку).
Маркировочная последовательность представляет собой обычно последовательность, которая является естественно встречающейся в бактериофаге. Предпочтительно, маркировочная последовательность имеет идентичность с естественно встречающейся в бактериофаге последовательностью, по меньшей мере, примерно 90%, примерно 95%, примерно 96%, примерно 97%, примерно 98%, примерно 99% (например, геноме бактериофага, из которого он получен). В наиболее предпочтительных вариантах осуществления, маркировочная последовательность имеет идентичность примерно 100% (например, геноме бактериофага, из которого он получен).
В некоторых вариантах осуществления, маркировочная последовательность имеет идентичность менее чем примерно 40%, примерно 30%, примерно 20%, примерно 10%, примерно 5%, примерно 4%, примерно 3%, примерно 2%, примерно 1% или примерно 0% с любыми другими спейсерами CRISPR или корами спейсеров CRISPR в одном или нескольких локусах CRISPR меченой бактерии.
В некоторых вариантах осуществления, маркировочная последовательность имеет идентичность менее чем примерно 40%, примерно 30%, примерно 20%, примерно 10%, примерно 5%, примерно 4%, примерно 3%, примерно 2%, примерно 1% или примерно 0% с любой другой последовательностью в одном или нескольких локусах CRISPR меченой бактерии.
В некоторых дополнительных вариантах осуществления, маркировочная последовательность, которая идентична последовательности (например, такой как спейсер CRISPR) в локусе CRISPR бактерии. В некоторых других вариантах осуществления, маркировочная последовательность, которая идентична имеет последовательность, которая идентична последовательности (например, такой как спейсер CRISPR) в локусе CRISPR бактерии, в сторону от полиморфизма одного или нескольких одиночных нуклеотидов (например, полиморфизма одного или двух одиночных нуклеотидов).
В некоторых предпочтительных вариантах осуществления, маркировочная последовательность имеет длину, по меньшей мере, примерно 20 нуклеотидов, тогда как в некоторых особенно предпочтительных вариантах осуществления, она имеет длину от примерно 20 до примерно 58 нуклеотидов.
В некоторых особенно предпочтительных вариантах осуществления, по меньшей мере, одна маркировочная последовательность интегрирована в материнскую бактерию. В некоторых дополнительных вариантах осуществления, также интегрируется, по меньшей мере, одна дуплицированная последовательность (например, дуплицированная последовательность повтора CRISPR), полученная из генома материнской бактерии или одной или нескольких из плазмид материнской бактерии (например, мегаплазмид). Не предполагается, что настоящее изобретение ограничивается любым конкретным механизмом или теорией. Однако считается, что копируется или замещается, по меньшей мере, одна дуплицированная последовательность из генома материнской бактерии. В частности, считается, что обычно дуплицирована последовательность повтора CRISPR в локусе CRISPR, и маркировочная последовательность интегрирована в геном бактерии непосредственно за (т.е. ниже по ходу транскрипции) новым дуплицированным повтором CRISPR.
В некоторых особенно предпочтительных вариантах осуществления, по меньшей мере, одна дуплицированная последовательность представляет собой последовательность повтора CRISPR, которая имеет идентичность, по меньшей мере, примерно 90%, примерно 95%, примерно 96%, примерно 97%, примерно 98% или примерно 99% с повторами CRISPR в одном или нескольких локусах CRISPR материнской бактерии и/или меченой бактерии. Наиболее предпочтительно, по меньшей мере, одна дуплицированная последовательность представляет собой последовательность повтора CRISPR, которая имеет идентичность, по меньшей мере, примерно 100% с повторами CRISPR в одном или нескольких локусах CRISPR материнской бактерии и/или меченой бактерии. В некоторых предпочтительных вариантах осуществления, дуплицированная последовательность имеет длину, по меньшей мере, 24 нуклеотида, тогда как в некоторых особенно предпочтительных вариантах осуществления, дуплицированная, она имеет длину от примерно 24 до примерно 40 нуклеотидов.
В некоторых предпочтительных вариантах осуществления, по меньшей мере, одна маркировочная последовательность и, по меньшей мере, одна дуплицированная последовательность интегрирована в материнскую бактерию. Не предполагается, что настоящее изобретение ограничивается любым конкретным механизмом или теорией. Однако считается, что каждый раз, когда маркировочная последовательность интегрируется в геном материнской бактерии, это сопровождается повторной, последовательной, одновременной или по существу одновременной интеграцией, по меньшей мере, одной дуплицированной последовательности. Соответственно, по меньшей мере, одна пара последовательностей, содержащих маркировочную последовательность и дуплицированную последовательность, интегрирована в материнскую бактерию, посредством этого, приводя к получению меченой бактерии.
В некоторых предпочтительных вариантах осуществления, по меньшей мере, одна маркировочная последовательность и, по меньшей мере, одна дуплицированная последовательность интегрируются по соседству друг с другом. Предпочтительнее, по меньшей мере, одна маркировочная последовательность и, по меньшей мере, одна дуплицированная последовательность интегрированы, непосредственно примыкая друг с другом так, что между последовательностями нет промежуточных нуклеотидов.
В некоторых вариантах осуществления, дуплицированная последовательность прикреплена, связана или слита с одним концом (например, концом 5' или концом 3') маркировочной последовательности. Предпочтительно, дуплицированная последовательность связана, присоединена или слита с концом 5' маркировочной последовательности. Соответственно, после интеграции одной пары последовательностей, дуплицированная последовательность представляет собой первую последовательность на конце 5' локуса CRISPR, а маркировочная последовательность будет второй (например, следующей) последовательностью в локусе CRISPR, ниже по ходу транскрипции дуплицированной последовательности. В некоторых предпочтительных вариантах осуществления, последовательности непосредственно прикреплены, связаны или непосредственно слиты так, что между дуплицированной последовательностью и маркировочной последовательностью нет промежуточных нуклеотидов.
Таким образом, в некоторых вариантах осуществления, пара дуплицированной последовательности и маркировочной последовательности интегрируется в геном материнской бактерии для получения меченой бактерии. Дуплицированная последовательность происходит, может происходить, получена или может быть получена из генома материнской бактерии, а маркировочная последовательность происходит, может происходить, получена или может быть получена из генома бактериофага, который используется для инфицирования материнской бактерии.
К удивлению, было даже обнаружено, что в некоторых вариантах осуществления, множественные пары последовательностей интегрированы в геном материнской бактерии. В соответствии с этими вариантами осуществления, множественные пары содержат первую пару, содержащую дуплицированную последовательность и маркировочную последовательность, и вторую пару, содержащую вторую дуплицированную последовательность и вторую маркировочную последовательность. Вторая дуплицированная последовательность обычно содержит такую же последовательность (например, с идентичностью более чем примерно 95%, примерно 96%, примерно 97%, примерно 98%, примерно 99% или примерно 100%) как первая дуплицированная последовательность. Маркировочная последовательность обычно содержит последовательность, отличную (например, с идентичностью менее чем примерно 40%, примерно 30%, примерно 20%, примерно 10%, примерно 5%, примерно 4%, примерно 3%, примерно 2%, примерно 1% или примерно 0%) от первой маркировочной последовательности. Это также имеет место в вариантах осуществления, в которые интегрированы дополнительные пары последовательностей.
Соответственно, конфигурация множественных пар обычно содержит:
[дуплицированную последовательность - маркировочную последовательность]n,
где n=2, 3, 4, 5, 6 или более.
Предпочтительно, конфигурация множественных пар обычно содержит:
[повтор CRISPR - маркировочную последовательность]n,
где n=2, 3, 4, 5, 6 или более.
В некоторых вариантах осуществления, конфигурация множественных пар обычно содержит:
5'-[дуплицированную последовательность - маркировочную последовательность]n-3'
где n=2, 3, 4, 5, 6 или более.
Предпочтительно, конфигурация множественных пар обычно содержит:
5'-[ повтор CRISPR - маркировочную последовательность]n-3'
где n=2, 3, 4, 5, 6 или более.
Соответственно, в некоторых вариантах осуществления, множественные пары интегрированы в материнскую бактерию. В некоторых вариантах осуществления, маркировочная последовательность интегрируется, примыкая к: (i) дуплицированной последовательности, которая является гомологичной (например, идентичной) естественно встречающейся последовательности в материнской бактерии; (ii) дуплицированной последовательности, которая является гомологичной (например, идентичной) естественно встречающейся последовательности в локусе CRISPR материнской бактерии; или (iii) предпочтительнее, дуплицированной последовательности, которая является гомологичной (например, идентичной) естественно встречающемуся повтору CRISPR в локусе CRISPR материнской бактерии.
После каждого воздействия на материнскую бактерию данного бактериофага в независимых экспериментах, маркировочная последовательность в каждой из меченой бактерии представляет другую нуклеотидную последовательность, посредством этого, создавая последовательность, которая является необычной для каждой бактерии. Таким образом, без связи с какой-либо конкретной теорией, считается, что после воздействия данного бактериофага на материнскую бактерию, маркировочная последовательность, которая интегрирована в материнскую бактерию, очевидно, случайно выбрана из генома бактериофага. Однако не предполагается, что настоящее изобретение ограничивается случайными явлениями интеграции.
Преимущественно, эти удивительные данные используются в контексте настоящего изобретения в силу того, что случайно выбранная маркировочная последовательность обеспечивает необычный ярлык или метку в меченой бактерии. К удивлению, было также обнаружено, что когда на одну и ту же материнскую бактерию воздействует один и тот же бактериофаг, то маркировочная последовательность, которая интегрируется в независимых/отдельных экспериментах, имеет другую последовательность, посредством этого, приводя к получению необычной метки в меченой бактерии после каждого воздействия.
В некоторых вариантах осуществления, случайно выбранная маркировочная последовательность идентифицируется в меченой бактерии на основании одного или нескольких из следующих свойств маркировочной последовательности:
(1) Локализация маркировочной последовательности в одном или нескольких локусах CRISPR нечувствительного к бактериофагу мутанта. Как описано в настоящей заявке, маркировочная последовательность обычно локализуется на одном и/или обоих концах (например, конце 5' и/или конце 3', предпочтительнее, конце 5') локуса CRISPR меченой бактерии;
(2) Маркировочная последовательность имеет высокую степень гомологии или идентичности (например, идентичность 100%) с последовательностью генома бактериофага, который воздействовал на материнскую бактерию; и/или
(3) Маркировочная последовательность слита, связана или прикреплена (например, непосредственно слита, связана или прикреплена), по меньшей мере, с одной последовательностью (например, повтором CRISPR), которая дуплицирована из генома материнской бактерии. Обычно, как описано в настоящей заявке, эта дополнительная пара последовательностей локализуется на одном или обоих концах (например, конце 5' и/или конце 3', предпочтительнее, конце 5') локуса CRISPR меченой бактерии.
В некоторых описанных в настоящей заявке вариантах осуществления маркировки/метки, одна или несколько дуплицированных последовательностей (например, дуплицированный повтор CRISPR из материнской бактерии) интегрируются в локус CRISPR материнской бактерии. В некоторых предпочтительных вариантах осуществления, одна или несколько описанных в настоящей заявке пар дуплицированной последовательности - маркировочной последовательности интегрируются в локус CRISPR материнской бактерии. В некоторых вариантах осуществления, маркировочная последовательность (последовательности) и/или дуплицированная последовательность (последовательности) интегрированы в пределы локуса CRISPR материнской бактерии. В некоторых других вариантах осуществления, маркировочная последовательность (последовательности) и/или дуплицированная последовательность (последовательности) интегрированы на одном или обоих концах локуса CRISPR материнской бактерии. В еще одних вариантах осуществления, маркировочная последовательность (последовательности) и/или дуплицированная последовательность (последовательности) интегрированы на обоих концах локуса CRISPR материнской бактерии так, что последовательности находятся на конце 5' и конце 3' локуса CRISPR. Одна из дуплицированных последовательностей будет обычно представлять собой первую последовательность на конце 5' локуса CRISPR, а маркировочная последовательность будет находиться непосредственно ниже по ходу транскрипции от дуплицированной последовательности. Другая дуплицированная последовательность будет представлять собой последнюю последовательность на конце 3' локуса CRISPR, а маркировочная последовательность будет находиться непосредственно выше по ходу транскрипции от дуплицированной последовательности.
В некоторых вариантах осуществления, маркировочная последовательность (последовательности) и/или дуплицированная последовательность (последовательности) интегрированы в один или несколько локусов CRISPR. В еще одних вариантах осуществления, маркировочная последовательность (последовательности) и/или дуплицированная последовательность (последовательности) интегрированы на одном конце локуса CRISPR материнской бактерии так, что последовательность (последовательности) находятся на конце 3' локуса CRISPR. Дуплицированная последовательность будет представлять собой последнюю последовательность на конце 3' локуса CRISPR, а маркировочная последовательность будет находиться непосредственно выше по ходу транскрипции от дуплицированной последовательности. Предпочтительно, маркировочная последовательность (последовательности) и/или дуплицированная последовательность (последовательности) интегрированы на одном конце локуса CRISPR материнской бактерии так, что последовательности находятся на конце 5' локуса CRISPR. Дуплицированная последовательность представляет собой первую последовательность на конце 5' локуса CRISPR, а маркировочная последовательность находится непосредственно ниже по ходу транскрипции от дуплицированной последовательности.
Как описано в настоящей заявке, маркировочная последовательность (последовательности) представляет собой специфичную для штамма метку в том смысле, что маркировочная последовательность, которая интегрирована или вставлена из бактериофага в материнскую бактерию, отличается каждый раз, когда материнская бактерия (например, та же материнская бактерия) подвергается воздействию бактериофага (например, того же бактериофага). Следовательно, маркировочная последовательность находит применение в качестве необычной метки для данного бактериального штамма.
Маркировочная последовательность (последовательности) и/или дуплицированная последовательность (последовательности) интегрированы в один или несколько различных локусов CRISPR, тогда как в других вариантах осуществления, две или более различные маркировочные последовательности и/или дуплицированные последовательности интегрированы в один локус CRISPR, а в еще одних вариантах осуществления, каждая из двух или более различных маркировочных последовательностей и/или дуплицированных последовательностей интегрированы в два или более различных локусов CRISPR. Каждая из маркировочных последовательностей из каждого бактериофага и/или каждая из дуплицированных последовательностей (дуплицированного повтора CRISPR) из материнской бактерии может интегрироваться в один и тот же локус CRISPR.
В некоторых вариантах осуществления, каждая из маркировочных последовательностей и/или каждая из дуплицированных последовательностей интегрируются на одном или обоих концах одного и того же локуса CRISPR. В некоторых других вариантах осуществления, каждая из маркировочных последовательностей и/или каждая из дуплицированных последовательностей интегрируются на конце 5' и/или 3' одного и того же локуса CRISPR. Предпочтительно, каждая из маркировочных последовательностей и/или каждая из дуплицированных последовательностей интегрируются на конце 5' одного и того же локуса CRISPR. В некоторых вариантах осуществления, каждая из маркировочных последовательностей и/или каждая из дуплицированных последовательностей из материнской бактерии интегрируются повторно, одновременно или по существу одновременно. В некоторых вариантах осуществления, каждая из маркировочных последовательностей и/или каждая из дуплицированных последовательностей из материнской бактерии интегрируются последовательно, посредством чего первая маркировочная последовательность и/или первая дуплицированная последовательность интегрируется в материнскую бактерию. Вторая маркировочная последовательность из второго бактериофага и/или другая дуплицированная последовательность затем интегрируется в материнскую бактерию. Подходящим образом, маркировочная последовательность и/или дуплицированная последовательность интегрируется в хромосомную ДНК материнской бактерии.
В некоторых вариантах осуществления, каждая из маркировочных последовательностей и/или каждая из дуплицированных последовательностей интегрированы в один конец (например, конец 5') одного и того же локуса CRISPR, примыкая (например, рядом) друг к другу. Таким образом, в некоторых вариантах осуществления, каждая из маркировочных последовательностей и/или каждая из дуплицированных последовательностей интегрированы последовательно, посредством чего первые последовательности интегрированы в материнскую бактерию на одном конце (например, в пределах или на конце 5' и/или конце 3') локуса CRISPR. Затем вторая маркировочная последовательность и/или дуплицированная последовательность может интегрироваться в материнскую бактерию, примыкая (например, непосредственно примыкая) к первой паре последовательностей. В некоторых вариантах осуществления, вторые последовательности интегрируются в материнскую бактерию, примыкая (например, непосредственно примыкая) к концу 5' или концу 3' первых последовательностей. Предпочтительно, вторые последовательности интегрируются в материнскую бактерию, примыкая (например, непосредственно примыкая) к концу 3' первых последовательностей и т.д. В некоторых вариантах осуществления, каждая из последовательностей интегрируются, примыкая (например, рядом) друг к другу в пределах конца 3' и/или конца 5' одного и того же локуса CRISPR материнской бактерии. В некоторых предпочтительных вариантах осуществления, каждая из последовательностей интегрируются, примыкая (например, рядом) друг к другу на конце 5' одного и того же локуса CRISPR материнской бактерии. Предпочтительнее, каждая из последовательностей интегрируется, примыкая (например, рядом) друг к другу выше по ходу транскрипции конца 5' локуса CRISPR материнской бактерии. Предпочтительнее, каждая из последовательностей интегрируется, примыкая (например, рядом) друг к другу выше по ходу транскрипции 5' повтора CRISPR локуса CRISPR материнской бактерии. Наиболее предпочтительнее, каждая из последовательностей интегрируется, примыкая (например, рядом) друг к другу выше по ходу транскрипции первого 5' повтора CRISPR локуса CRISPR материнской бактерии.
Меченые бактерии
Используемый в настоящем описании термин «меченые бактерии» и «меченая бактерия» относится к материнским бактериям или материнскому бактериальному штамму, в котором один или несколько локусов CRISPR или их часть были модифицированы (например, мутированы) таким образом, чтобы они были нечувствительны к одному или нескольким бактериофагам, который мог на них воздействовать. Как в дополнительных деталях описано в настоящей заявке, в некоторых вариантах осуществления на меченую бактерию воздействует более чем один бактериофаг (например, или повторно, последовательно или одновременно), так что она накапливает одну или несколько геномных модификаций в пределах одного или нескольких локусов CRISPR таким образом, что они становятся нечувствительными к каждому из бактериофагов, которые на них воздействовали.
Для инфицирования клеток, бактериофаг инжектирует или переносит его нуклеиновую кислоту в клетку при нуклеиновой кислоте фага существующей независимо от генома клетки. В некоторых вариантах осуществления, инфекция приводит к экспрессии (т.е. транскрипции и трансляции) нуклеиновой кислоты бактериофага внутри клетки и продолжению жизненного цикла бактериофага.
В некоторых вариантах осуществления, после воздействия бактериофага, меченая бактерия имеет сниженную или отсутствующую восприимчивость к инфекции бактериофагом и/или его размножению, по сравнению с материнской бактерией. Используемый в настоящем описании термин «сниженная или отсутствующая восприимчивость к инфекции бактериофагом и/или его размножению» означает, что уровень инфекции бактериофагом и/или его размножения в меченой бактерии не вызывает вредное воздействие на меченую бактерию.
Таким образом, в некоторых вариантах осуществления настоящего изобретения, материнская бактерия не уничтожается после воздействия бактериофага вследствие мутации материнской бактерии таки образом, что она становится чувствительной к бактериофагу.
В некоторых вариантах осуществления, меченая бактерия нечувствительна или по существу нечувствительна к дальнейшей инфекции бактериофагом и/или к его размножению. В дополнительных вариантах осуществления, меченая бактерия нечувствительна или по существу нечувствительна к одному или нескольким механизмам, которые бактериофаг использует для инфекции и/или размножения в бактерии. В еще одних вариантах осуществления, меченая бактерия нечувствительна или по существу нечувствительна ко всем механизмам, которые бактериофаг использует для инфекции и/или размножения в бактерии. В других дополнительных вариантах осуществления, у меченой бактерии развивается один или несколько механизмов, которые ослабляют, инактивируют или разрушают бактериофаг во время цикла инфекции. В некоторых других вариантах осуществления, настоящее изобретение относится к меченым штаммам, выбранным стандартными процедурами скрининга, которые известны в данной области, для выделения нечувствительных к бактериофагу мутантов.
В некоторых вариантах осуществления настоящего изобретения, меченая бактерия, содержащая маркировочную последовательность в локусе CRISPR, которая не присутствует в материнской бактерии, выбирается после сравнения локуса CRISPR или его части из материнской бактерии и меченой бактерии.
В некоторых предпочтительных вариантах осуществления, выбирается меченая бактерия, содержащая в пределах или на конце 5' и/или конце 3' локуса CRISPR дополнительный фрагмент ДНК, который не присутствует в материнской бактерии. Предпочтительнее, выбирается меченая бактерия, содержащая маркировочную последовательность, примыкающую (например, непосредственно примыкающую) к концу 3' вновь дуплицированной последовательности в локусе CRISPR меченой бактерии, которая не присутствует в материнской бактерии. Наиболее предпочтительно, выбирается меченая бактерия, содержащая маркировочную последовательность, примыкающую (например, непосредственно примыкающую) к концу 3' первого повтора CRISPR локуса CRISPR в меченой бактерии, которая не присутствует в материнской бактерии.
В некоторых вариантах осуществления, выделяется и/или клонируется маркировочная последовательность (например, одна или несколько маркировочных последовательностей). В некоторых других вариантах осуществления, секвенируется маркировочная последовательность (например, одна или несколько маркировочных последовательностей). Эти варианты осуществления обеспечивают преимущества, поскольку они не только предоставляют информацию о локализации маркировочной последовательности в пределах локуса CRISPR, но также его специфической последовательности. В некоторых вариантах осуществления, эта информация хранится в базе данных, посредством этого обеспечивая необычную метку для данной бактерии, а также средство для последующего прослеживания и/или идентификации бактерии.
Когда известна последовательность маркировочной последовательности в меченой бактерии, одна маркировочная последовательность находит применение при идентификации бактерий. С использованием различных способов, которые известны в данной области и описаны в настоящей заявке, определяются последовательность и/или локализация маркировочной последовательности. Эта последовательность затем сопоставляется, например, с базой данных бактериальных последовательностей и/или базой данных последовательностей бактериофагов, и/или базой данных меток/маркировок для идентификации бактерии.
Донорский организм
Используемый в некоторых вариантах осуществления термин «донорский организм» относится к организму или клетке, из которых получены повтор CRISPR и/или ген cas, и/или их комбинация (комбинации) и/или спейсеры CRISPR. В некоторых вариантах осуществления, термин «донорский организм» относится к организму или клетке, из которых получены один или несколько, предпочтительно, два или более повтора CRISPR и/или один или несколько генов cas и/или их комбинации (комбинация) и/или спейсеров CRISPR. Они могут быть одинаковыми или различными. В некоторых вариантах осуществления, спейсер CRISPR и/или псевдоспейсер CRISPR получены синтетически. В еще одних вариантах осуществления, донорский организм или клетка содержит один или несколько спейсеров CRISPR, которые придают специфический иммунитет против нуклеиновой кислоты-мишени и продукту ее транскрипции. В дополнительных вариантах осуществления, донорский организм или клетка, из которых получены ген cas и/или повтор CRISPR, и/или их комбинация, представляет собой также клетку/организм реципиента для рекомбинантного локуса CRISPR. Они могут быть одинаковыми или различными. В других вариантах осуществления, донорский организм или клетка, из которых получен спейсер CRISPR, представляет собой также клетку/организм реципиента для рекомбинантного локуса CRISPR. Они могут быть одинаковыми или различными. В вариантах осуществления, в которых донорский организм представляет собой бактериальную клетку, то донорский организм обычно содержит спейсер CRISPR, который придает специфический иммунитет против нуклеиновой кислоты-мишени или продукта ее транскрипции. В некоторых вариантах осуществления, организм представляет собой бактериальную клетку, тогда как в других вариантах осуществления, он представляет собой бактериофаг.
Клетка-хозяин
Используемый в настоящем описании термин «клетка-хозяин» относится к любой клетке, которая содержит комбинацию, конструкт или вектор и им подобный, в соответствии с настоящим изобретением. В некоторых вариантах осуществления, клетки-хозяева трансформированы или трансфецированы нуклеотидной последовательностью, содержащейся в векторе (например, векторе клонирования). В некоторых вариантах осуществления, нуклеотидная последовательность может переноситься в векторе для репликации и/или экспрессии нуклеотидной последовательности. Клетки выбираются для того, чтобы быть совместимыми с вектором и в некоторых вариантах осуществления, прокариотическими (например, бактериальными) клетками.
Клетка-реципиент
Используемый в настоящем описании термин «клетка-реципиент» относится к любой клетке, в которой модулируется или должна модулироваться сопротивляемость против нуклеиновой кислоты-мишени или продукта ее транскрипции. В некоторых вариантах осуществления, клетка-реципиент относится к любой клетке, содержащей рекомбинантную нуклеиновую кислоту в соответствии с настоящим изобретением. В некоторых вариантах осуществления, клетка-реципиент содержит один или несколько, предпочтительно, два или более повторов CRISPR и один или несколько генов или белков cas. Подходящим образом, повторы CRISPR и генов или белков cas образуют функциональную комбинацию в клетке-реципиенте, как описано в настоящей заявке. В некоторых других вариантах осуществления, клетка-реципиент содержит один или несколько модифицированных повторов CRISPR и/или один или несколько модифицированных генов или белков cas. Подходящим образом, модифицированные повторы CRISPR и/или модифицированные гены или белки cas образуют функциональную комбинацию в клетке-реципиенте, как описано в настоящей заявке. В некоторых вариантах осуществления, клетка-реципиент содержит один или несколько полученных методами генной инженерии повторов CRISPR и/или один или несколько полученных методами генной инженерии генов или белков cas. Подходящим образом, полученные методами генной инженерии повторы CRISPR и/или полученные методами генной инженерии гены или белки cas образуют функциональную комбинацию в клетке-реципиенте, как описано в настоящей заявке. В некоторых альтернативных вариантах осуществления, клетка-реципиент содержит один или несколько рекомбинантных повторов CRISPR и/или один или несколько рекомбинантных генов или белков cas. Подходящим образом, рекомбинантные повторы CRISPR и/или рекомбинантные гены или белки cas образуют функциональную комбинацию в клетке-реципиенте, как описано в настоящей заявке. В еще одних дополнительных вариантах осуществления, клетка-реципиент содержит один или несколько естественно встречающихся повторов CRISPR и один или несколько естественно встречающихся генов или белков cas. Подходящим образом, повтор(ы) CRISPR и ген(ы) или белки cas образуют функциональную комбинацию.
В некоторых вариантах осуществления, клетка-реципиент содержит комбинации одного или нескольких модифицированных, полученных методами генной инженерии, рекомбинантных или естественно встречающихся повторов CRISPR и одного или нескольких модифицированных, полученных методами генной инженерии, рекомбинантных или естественно встречающихся генов или белков cas. Подходящим образом, один или несколько модифицированных, полученных методами генной инженерии, рекомбинантных или естественно встречающихся повтор(ов) CRISPR или одного или нескольких модифицированных, полученных методами генной инженерии, рекомбинантных или естественно встречающихся генов или белков cas образуют функциональную комбинацию.
В некоторых вариантах осуществления, клетка-реципиент представляет собой прокариотическую клетку. В некоторых предпочтительных вариантах осуществления, клетка-реципиент представляет собой бактериальную клетку. Подходящие бактериальные клетки описаны в настоящей заявке. В некоторых вариантах осуществления, бактериальная клетка выбрана из вида молочнокислых бактерий, вида a Brevibacterium, вида a Propionibacterium, вида a Lactococcus, вида a Streptococcus, вида a Lactobacillus, включая вида Enterococcus, вида Pediococcus, вида a Leuconostoc и вида Oenococcus. Подходящие виды включают без ограничения Lactococcus lactis, включая Lactococcus lactis подвид lactis и Lactococcus lactis подвид cremoris, Lactococcus lactis подвид cremoris, Leuconostoc sp., Lactococcus lactis подвид lactis biovar, Streptococcus thermophilus, Lactobacillus delbrueckii подвид bulgaricus и Lactobacillus helveticus, Bifidobacterium lactis, Lactobacillus acidophilus, Lactobacillus casei.
В некоторых вариантах осуществления, в которых предстоит модулировать устойчивость клетки, бактериальная клетка используется для ферментации мяса (включая говядину, свинину и мясо птицы), включая без ограничения молочнокислые бактерии, Pediococcus cerevisiae, Lactobacillus plantarum, Lactobacillus brevis, Micrococcus species, Lactobacillus sakei, Lactobacillus curvatus, Pediococcus pentosaceus, Staphylococcus xylosus и Staphylococcus vitulinus и их смеси, как известно в данной области. В некоторых альтернативных вариантах осуществления, для ферментации овощей (например, моркови, огурцов, помидоров, перца и капусты) используется бактериальная клетка, включая без ограничения Lactobacillus plantatum, Lactobacillus brevis, Leuconostoc mesenteroides, Pediococcus pentosaceus и их смеси, как известно в данной области. В некоторых альтернативных вариантах осуществления, бактериальная клетка используется для ферментации теста, полученного из злаков (например, пшеницы, ржи, риса, овса, ячменя и кукурузы). В некоторых других вариантах осуществления, бактериальная клетка используется для продукции вина. Обычно, это достигается брожением фруктового сока, обычно виноградного сока. В еще одних вариантах осуществления, бактериальная клетка используется для сбраживания молока для получения сыра (например, Lactobacillus delbrueckii подвид bulgaricus, Lactobacillus helveticus, Streptococcus thermophilus, Lactococcus lactis подвид lactis, Lactococcus lactis подвид cremoris, Lactococcus lactis подвид lactis biovar diacetylactis, Lactococcus, Bifidobacterium и Enterococcus и их смеси), как известно в данной области. В некоторых других вариантах осуществления, бактериальная клетка используется для ферментации яиц (например, Pediococcus pentosaceus, Lactobacillus plantarum и их смеси), как известно в данной области. В некоторых других вариантах осуществления, бактериальная клетка используется в косметических или фармацевтических композициях.
В некоторых вариантах осуществления, клетка, у которой предстоит модулировать устойчивость, представляет собой бактерию, которая естественно содержит один или несколько локусов CRISPR. Локусы CRISPR были идентифицированы в более 40 прокариотах (см. Haft et al., [2005], выше), включая без ограничения Aeropyrum, Pyrobaculum, Sulfolobus, Archaeoglobus, Halocarcula, Methanobacterium, Methanococcus, Methanosarcina, Methanopyrus, Pyrococcus, Picrophilus, Thermoplasma, Corynebacterium, Mycobacterium, Streptomyces, Aquifex, Porphyromonas, Chlorobium, Thermus, Bacillus, Listeria, Staphylococcus, Clostridium, Thermoanaerobacter, Mycoplasma, Fusobacteriutn, Azarcus, Chromobacterium, Neisseria, Nitrosomonas, Desulfovibrio, Geobacter, Myxococcus, Campylobacter, Wolinella, Acinetobacter, Erwinia, Escherichia, Legionella, Methylococcus, Pasteurella, Photobacterium, Salmonella, Xanthamonas, Yersinia, Treponema и Thermotoga.
Штамм материнской бактерии
Используемые в настоящем описании термины «материнская бактерия» и «материнские бактерии» и «материнский штамм» относятся к любой бактерии/бактериям/штаммам, на который (которые) воздействует один или несколько бактериофагов. В некоторых вариантах осуществления, бактериофаги являются вирулентными для материнского бактериального штамма, тогда как в других вариантах осуществления, они не являются вирулентными. В некоторых особенно предпочтительных вариантах осуществления, материнские бактерии чувствительны к вирулентному фагу. В некоторых предпочтительных вариантах осуществления, материнский штамм инфицирован бактериофагом. В некоторых особенно предпочтительных вариантах осуществления, инфекция фагом придает материнской бактерии/штамму или его субпопуляции нечувствительность к дальнейшей инфекции бактериофагом. В некоторых предпочтительных вариантах осуществления, инфекция «материнской бактерии» одним или несколькими бактериофагами приводит к созданию меченого штамма, который может быть выбран на основании его нечувствительности к бактериофагу. В некоторых предпочтительных вариантах осуществления, «устойчивые к бактериофагу мутанты» представляют собой бактерии, которые маркированы или мечены в соответствии со способами по настоящему изобретению. В некоторых вариантах осуществления, материнские бактерии представляют собой бактериальные штаммы дикого типа. В некоторых предпочтительных вариантах осуществления, материнские бактерии представляют собой бактериальные штаммы дикого типа бактерий, которые ранее не были инфицированы каким-либо бактериофагом. В некоторых предпочтительных вариантах осуществления, материнские бактерии представляют собой бактериальные штаммы дикого типа бактерий, которые ранее не были маркированы или мечены, тогда как в некоторых альтернативных вариантах осуществления, материнские бактерии представляют собой устойчивые к бактериофагу мутанты, которые были ранее маркированы или мечены.
В некоторых особенно предпочтительных вариантах осуществления, материнская бактерия выбрана из любой бактерии, которая естественно содержит один или несколько локусов CRISPR. Как указано выше, локусы CRISPR были идентифицированы более чем в 40 прокариотах (см. Haft et al., [2005], выше), включая без ограничения Aeropyrum, Pyrobaculum, Sulfolobus, Archaeoglobus, Halocarcula, Methanobacterium, Methanococcus, Methanosarcina, Methanopyrus, Pyrococcus, Picrophilus, Thermoplasma, Corynebacterium, Mycobacterium, Streptomyces, Aquifex, Porphyromonas, Chlorobium, Thermus, Bacillus, Listeria, Staphylococcus, Clostridium, Thermoanaerobacter, Mycoplasma, Fusobacterium, Azarcus, Chromobacterium, Neisseria, Nitrosomonas, Desulfovibrio, Geobacter, Myxococcus, Campylobacter, Wolinella, Acinetobacter, Erwinia, Escherichia, Legionella, Methylococcus, Pasteurella, Photobacterium, Salmonella, Xanthamonas, Yersinia, Treponema и Thermotoga.
В некоторых вариантах осуществления, материнская бактерия содержит один или несколько гетерологичных спейсеров CRISPR, один или несколько повторов CRISPR и/или один или несколько гетерологичных генов cas. В некоторых альтернативных вариантах осуществления, материнская бактерия содержит один или несколько гетерологичных локусов CRISPR, предпочтительно, один или несколько полных локусов CRISPR. В некоторых других вариантах осуществления, материнская бактерия естественно содержит один или несколько локусов CRISPR, а также содержит один или несколько гетерологичных спейсеров CRISPR, один или несколько гетерологичных повторов CRISPR, и/или один или несколько гетерологичных генов cas. В некоторых дополнительных вариантах осуществления, материнская бактерия естественно содержит один или несколько локусов CRISPR, а также содержит один или несколько гетерологичных локусов CRISPR, предпочтительно, один или несколько полных локусов CRISPR.
В некоторых предпочтительных вариантах осуществления, устойчивая к фагу субпопуляция, созданная воздействием на материнские бактерии, по меньшей мере, одного фага, представляет собой чистую культуру. Однако не предполагается, что настоящее изобретение ограничивается чистыми культурами бактериальных штаммов, вариантов и фага. Действительно, предполагается, что настоящее изобретение охватывает смешанные культуры клеток и фага. В некоторых вариантах осуществления, смешанная культура представляет собой смесь различных мутантов, соответствующих различным явлениям интеграции в одних и тех же и/или в различных локусах CRISPR.
Хотя не предполагается, что настоящее изобретение так ограничивается, предпочтительные материнские бактериальные рода представляют собой Streptococcus и Lactobacillus. Действительно, предполагается, что любые бактериальные виды найдут применение в настоящем изобретении, включая без ограничения Escherichia, Shigella, Salmonella, Erwinia, Yersinia, Bacillus, Vibrio, Legionella, Pseudomonas, Neisseria, Bordetella, Helicobacter, Listeria, Agrobacterium, Staphylococcus, Streptococcus, Enterococcus, Clostridium, Corynebacterium, Mycobacterium, Treponema, Borrelia, Francisella, Brucella, Bifidobacterium, Brevibacterium, Propionibacterium, Lactococcus, Lactobacillus, Enterococcus, Pediococcus, Leuconostoc, Oenococcus и/или Xanthomonas. В некоторых вариантах осуществления, материнские бактерии содержатся или получены из молочнокислых бактерий, включая без ограничения Bifidobacterium, Brevibacterium, Propionibacterium, Lactococcus, Streptococcus, Lactobacillus (например, L. acidophilus), Enterococcus, Pediococcus, Leuconostoc и/или Oenococcus. В других вариантах осуществления, материнские бактерии содержатся или получены из молочнокислых бактерий, включая без ограничения Lactococcus lactis {например, L. lactis подвид lactis и L. lactis подвид cremoris, и L. lactis подвид lactis biovar diacetylactis), L. delbrueckii подвид bulgaricus, L. helveticus, L. acidophilus, L. casei, L. paracasei, L. salivarius, L. plantarum, L. reuteri, L. gasseri, L. johnsonii, Bifidobacterium lactis, B. infantis, B. longum и/или Streptococcus thermophilus.
В некоторых вариантах осуществления настоящего изобретения, материнская бактерия представляет собой «бактерию пищевого сорта» (т.е. бактерию, которая используется и в целом рассматривается как безопасная для применения при получении и/или производстве пищевых продуктов и/или корма). В некоторых предпочтительных вариантах осуществления, материнская бактерия подходит для использования в качестве заквасочной культуры, пробиотической культуры и/или пищевой добавки. В дополнительных вариантах осуществления, материнская бактерия находит применение для ферментации мяса (включая говядину, свинину, баранину и мясо птицы), включая без ограничения молочнокислые бактерии, Pediococcus cerevisiae, Lactobacillus plantarum, L. brevis, L. sakei, L. curvatus, Micrococcus species, Pediococcus pentosaceus, Staphylococcus xylosus, S. vitulinus и их смеси (см., например, Knorr (ed.), Food Biotechnology, at 538-39 [1987]; и Pederson, Microbiology of Fermented Foods, at 210-34, 2d ed., [1979]; и патент США № 2225783, полностью включенные в настоящее описание путем ссылки). В других дополнительных вариантах осуществления, материнская бактерия находит применение для ферментации овощей (например, моркови, огурцов, помидоров, перца и капусты), включая без ограничения L. plantatum, L. brevis, Leuconostoc mesenteroides, Pediococcus pentosaceus и их смеси (см., например, Knorr, см. выше; Pederson, см. выше; и патенты США №№ 3024116, 3403032, 3932674 и 3897307). В еще одних вариантах осуществления, материнская бактерия находит применение для ферментации теста, полученного из злаков (например, пшеницы, ржи, риса, овса, ячменя и кукурузы). В еще одних вариантах осуществления, материнская бактерия находит применение для продукции вина посредством брожения фруктового сока (например, виноградного сока). В некоторых дополнительных вариантах осуществления, материнская бактерия находит применение для сбраживания молока (например, L. delbrueckii подвид bulgaricus, L. acidophilus, S. thermophilus и их смеси (см. Knorr, см. выше; Pederson, см. выше, на стр. 105-35). В некоторых предпочтительных вариантах осуществления, материнская бактерия находит применение для производства сыра, включая без ограничения L. delbrueckii подвид bulgaricus, L. helveticus, L. lactis подвид lactis, L. lactis подвид cremoris, L. lactis подвид lactis biovar diacetylactis, S. thermophilus, Bifidobacterium Enterococcus и их смеси (см. Knorr, см. выше; Pederson, см. выше, на стр. 135-51). В еще одних вариантах осуществления, материнская бактерия находит применение для ферментации яиц, включая без ограничения Pedicoccus pentosaceus, Lactobacillus plantarum и их смеси (см. Knorr выше). В некоторых вариантах осуществления, материнская бактерия находит применение при ферментации для производства различных продуктов, включая без ограничения чеддер и домашний сыр (например. L. lactis, подвид lactis, L. lactis, подвид cremoris), йогурт (L. delbrueckii, подвид bulgaricus и S. thermophilus), швейцарский сыр (например, S. thermophilus, L. lactis и L. helveticus), сыр с добавлением культур Penicillum (Leuconostoc cremoris), итальянский сыр (L. bulgaris и S. thermophilus), виили {L. lactis подвид cremoris, L. lactis подвид lactis biovar diacetylactis, Leuconostoc cremoris), якулт (L. casei), казеин (L. lactis подвид cremoris), натто ((Bacillus subtilis вариант натто), вино (Leuconostoc oenos), саке (Leuconostoc mesenteroides), полимиксин (Bacillus polymyxa), колистин (Bacillus colistrium), бацитрацин (Bacillus licheniformis), L-глутаминовую кислоту (Brevibacterium lactofermentum и Micobacteriukm ammoniaphilum) и ацетон. и бутанол (Clostridium acetobutyricum и Clostridium saccharoperbutylacetonicum). В некоторых предпочтительных вариантах осуществления, материнские бактериальные виды выбраны из S. thermophilus, L. delbrueckii, подвид bulgaricus и/или L. acidophilus.
В еще одних вариантах осуществления, материнские бактерии находят применение в способах, включая без ограничения производство антибиотиков, производство аминокислот, производство растворителей и производство других экономически полезных материалов. В еще одних вариантах осуществления, материнские бактерии находят применение в косметических, терапевтических и/или фармацевтических композициях. В некоторых вариантах осуществления, композиции имеют определенные виды активности, включая без ограничения регенерацию кожи, включая без ограничения свойства против морщин, удаления старых рубцов, восстановление тканей, пораженных ожогом, содействие заживлению кожи, удаление пигментных пятен и т.д. В некоторых вариантах осуществления, композиции или стимулируют, или ингибируют рост ногтей и волос. В некоторых дополнительных вариантах осуществления, композиции содержат, по меньшей мере, одну микробную культуру и/или меченую бактерию, и/или клеточную культуру, полученную с использованием способов и композиций по настоящему изобретению.
В других вариантах осуществления, материнские бактерии представляют собой мутанты, нечувствительные к бактериофагу. Таким образом, в некоторых вариантах осуществления, материнские бактерии нечувствительны к одному или нескольким бактериофагам. В некоторых предпочтительных вариантах осуществления, материнская бактерия не представляет собой мутант, нечувствительный к бактериофагу, для бактериофага, воздействие которого предстоит во время использования настоящего изобретения.
Заквасочные культуры
Заквасочные культуры широко используются в пищевой промышленности при изготовлении ферментированных продуктов, включая молочные продукты (например, йогурт и сыр), а также мясных продуктов, хлебобулочных продуктов, вина и растительных продуктов.
Заквасочные культуры, используемые при изготовлении многих ферментированных молочных, сырных и сливочно-масляных продуктов, включают культуры бактерий, в целом классифицируемых как молочнокислые бактерии. Такие бактериальные заквасочные культуры придают специфические признаки различным молочным продуктам путем выполнения ряда функций.
Выпускаемые не концентрированные культуры бактерий именуются в промышленности «материнскими культурами» и размножаются на производственном участке, например, молочном заводе, перед добавлением в пищевой исходный материал, такой как молоко, для сбраживания. Заквасочная культура, размножающаяся на производственном участке для инокуляции в пищевой исходный материал, именуется «основной массой закваски».
Подходящие заквасочные культуры для использования в настоящем изобретении включают любой организм, который используется в пищевой, косметической или фармацевтической промышленности (т.е. «промышленно используемые культуры» или «промышленно используемые штаммы»).
Заквасочные культуры получают методиками, хорошо известными в данной области (см., например, патент США № 4621058, включенный в настоящее описание путем ссылки). В некоторых вариантах осуществления, заквасочные культуры получают введением инокулята, например, бактерии, в ростовую среду (например, среду или продукт ферментации) для получения инокулированной среды и инкубации инокулированной среды для получения заквасочной культуры.
Высушенные заквасочные культуры получают методиками, хорошо известными в данной области (см., например, патенты США №№ 4423079 и 4140800). Любая подходящая форма высушенных заквасочных культур находит применение в настоящем изобретении, включая твердые препараты (например, таблетки, пилюли, капсулы, присыпки, гранулы и порошки), которые являются смачиваемыми, высушенными распылением, высушенными замораживанием или лиофилизированными. В некоторых вариантах осуществления, высушенные заквасочные культуры для использования в настоящем изобретении представлены или в форме глубоко замороженных пилюль или в форме высушенного замораживанием порошка. Высушенные заквасочные культуры в глубоко замороженной пилюле или высушенном замораживанием порошке получают в соответствии с любым подходящим способом, известным в данной области.
В некоторых вариантах осуществления, заквасочные культуры, используемые в настоящем изобретении, представлены в форме концентратов, которые содержат по существу высокую концентрацию одного или нескольких бактериальных штаммов. В некоторых вариантах осуществления, концентрации растворяются в воде или ресуспендируются в воде или других подходящих растворителях (например, соответствующей ростовой среде, минеральном масле или растительном масле). Высушенные заквасочные культуры по настоящему изобретению в форме концентратов получают в соответствии со способами, известными в данной области (например, центрифугированием, фильтрацией или комбинацией таких методик).
В некоторых вариантах осуществления, заквасочная культура подходит для использования в молочной промышленности. При использовании в молочной промышленности, заквасочная культура часто выбирается из вида молочнокислых бактерий, вида Bifidobacterium, вида Brevibacterium, вида Propionibaqcterium. Подходящие заквасочные культуры группы молочнокислых бактерий включают обычно используемые штаммы вида Lactococcus, вида Streptococcus, вида Lactobacillus, включая Lactobacillus acidophilus, вид Enterococcus, вид Pediococcus, вид Leuconostoc и вид Oenococcus.
Культуры молочнокислых бактерий обычно используются при изготовлении ферментированных молочных продуктов (например, пахты, йогурта или сметаны) и при изготовлении масла и сыра (например, бри (мягкого сыра) или хаварти (полумягкого сыра из коровьего молока)). Виды Lactococcus включают широко используемую Lactococcus lactis, включая Lactococcus lactis подвид, lactis и Lactococcus lactis, подвид cremoris.
Другие виды молочнокислых бактерий включают Leuconostoc sp., Streptococcus thermophilus, Lactobacillus delbrueckii подвид bulgaricus и Lactobacillus helveticus. Кроме того, пробиотические штаммы (например, вид Lactococcus) включают широко используемые Lactococcus lactis, включая Lactococcus lactis, подвид lactis и Lactococcus lactis, подвид cremoris.
Мезофильные культуры молочнокислых бактерий, обычно используемых при изготовлении ферментированных молочных продуктов, таких как пахта, йогурт или сметана, и при изготовлении масла и сыра (например, бри или хаварти). Другие виды Lactococcus включают Lactococcus lactis подвид cremoris, Lactococcus lactis, Leuconostoc sp., Lactococcus lactis подвид lactis biovar, Streptococcus thermophilus, Lactobacillus delbrueckii подвид bulgaricus и Lactobacillus helveticus. Кроме того, в некоторых вариантах осуществления, пробиотические штаммы, такие как Bifidobacterium lactis, Lactobacillus acidophilus, Lactobacillus casei, добавляются во время изготовления для усиления вкуса и запаха или укрепления здоровья.
Культуры молочнокислых бактерий, обычно используемые при изготовлении сыров чеддер и «Монтерей джек», включают Streptococcus thermophilus, Lactococcus lactis, подвид lactis и Lactococcus lactis, подвид cremoris, или их комбинации.
Термофильные культуры, обычно используемые при изготовлении итальянских сыров, таких как «паста филата» или пармезан, включают Streptococcus thermophilus и Lactobacillus delbrueckii, подвид bulgaricus. Другие виды Lactobacillus (например, Lactobacillus helveticus) добавляются во время изготовления для получения желательного вкуса и запаха.
В некоторых предпочтительных вариантах осуществления, организм заквасочной культуры содержит или состоит из генетически модифицированного штамма, полученного в соответствии с представленными в настоящем описании способами, из одной из указанных выше молочнокислых бактерий или любого другого штамма заквасочной культуры.
Выбор организмов для заквасочной культуры по изобретению будет зависеть от конкретного типа продуктов, которые предстоит получить и обработать. Так, например, для изготовления сыра и масла, широко используются мезофильные культуры вида Lactococcus, вида Leuconоstoc и вида Lactobacillus, в то время как для йогурта и других ферментированных молочных продуктов, обычно используются термофильные штаммы вид Streptococcus и вида Lactobacillus.
В некоторых вариантах осуществления, заквасочная культура представляет собой сушеную заквасочную культуру, дегидратированную заквасочную культуру, замороженную заквасочную культуру или концентрированную заквасочную культуру. В некоторых вариантах осуществления, заквасочная культура используется при прямой инокуляции среды или продукта ферментации.
В некоторых вариантах осуществления, заквасочная культура содержит чистую культуру (т.е. содержащую только один бактериальный штамм). В некоторых альтернативных вариантах осуществления, заквасочная культура представляет собой смешанную культуру (т.е. содержащую, по меньшей мере, два различных бактериальных штамма).
Молочнокислые бактерии
Особенно подходящие заквасочные культуры, в частности, сушеные заквасочные культуры для использования в настоящем изобретении, содержат молочнокислые бактерии.
Используемый в настоящем описании термин «молочнокислые бактерии» относится к грамположительным, микроаэрофильным или анаэробными бактериями, которые ферментируют сахар с получением кислот, включая молочную кислоту в качестве преимущественно продуцируемой кислоты, уксусную кислоту, муравьиную кислоту и пропионовую кислоту. Наиболее промышленно используемые молочнокислые бактерии обнаруживаются среди видов Lactococcus, таких как вид Lactococcus lactis, вид Lactobacillus, вид Bifidobacterium, вид Streptococcus, вид Leuconostoc, вид Pediococcus и вид Propionibacterium.
Заквасочные культуры по настоящему изобретению могут содержать один или несколько видов молочнокислых бактерий, таких как Lactococcus lactis, Lactobacillus delbrueckii, подвид bulgaricus, и Streptococcus thermophilus, или их комбинации.
Заквасочные культуры молочнокислых бактерий обычно используются в пищевой промышленности в виде культур смешанных штаммов, содержащих один или несколько видов. Для ряда культур смешанных штаммов, таких как заквасочные культуры йогурта, содержащие штаммы Lactobacillus delbrueckii, подвид bulgaricus, и Streptococcus thermophilus, существует симбиотическая связь между видами, где продукция молочной кислоты больше, по сравнению с культурами молочнокислых бактерий одного штамма (см., например, Rajagopal et al., J. Dairy Sci., 73:894-899 [1990]).
Продукты
Подходящие продукты для использования в настоящем изобретении включают без ограничения пищевые продукты, косметические продукты или фармацевтические продукты. Любой продукт, который получен из культуры или содержит ее, предусмотрен в соответствии с настоящим изобретением. Они включают без исключения фрукты, бобы, кормовые культуры и растения, включая полученные продукты, зерно и полученные из зерна продукты, молочные пищевые продукты и продукты, полученные из молочных пищевых продуктов, мясо, птица, морские пищевые продукты, косметические и фармацевтические продукты.
Термин «пищевой продукт» используется в широком смысле и включает корма, пищевые продукты, пищевые ингредиенты, пищевые добавки и функциональные пищевые продукты.
Используемый в настоящем описании термин «пищевой ингредиент» включает состав, который добавляется или может добавляться к пищевым продуктам, и включает составы, которые могут использоваться на низких уровнях в широком разнообразии продуктов, которые требуют, например, подкисления или эмульгирования.
Используемый в настоящем описании термин «функциональный пищевой продукт» означает пищевой продукт, который способен обеспечить не только питательный эффект и/или вкусовое удовлетворение, но также способен обеспечить потребителю дополнительный благоприятный эффект. Хотя нет законодательного определения функционального пищевого продукта, большинство сторон, которые имеют интерес в данной области, согласны, что выпускаемые пищевые продукты оказывают специфические воздействия на здоровье.
Термин «пищевой продукт» охватывает пищевой продукт для людей, а также пищевой продукт для животных (т.е. корм). В предпочтительном аспекте, пищевой продукт предназначен для употребления людьми.
В некоторых вариантах осуществления, клетки, описанные в настоящей заявке, содержат или добавляются к ингредиенту пищевого продукта, пищевой добавке или функциональному пищевому продукту. В некоторых вариантах осуществления, пищевой продукт представлен в жидкой форме (например, растворе), в форме геля, эмульсии или твердого вещества, что определяется способом применения и/или введения.
Клетки, описанные в настоящей заявке, находят применение при получении пищевых продуктов, таких как один или несколько из: кондитерских продуктов, молочных продуктов, мясных продуктов, продуктов из мяса птицы, рыбных продуктов и хлебобулочных продуктов. В некоторых вариантах осуществления, бактерии находят применение в качестве ингредиентов слабоалкогольных напитков, фруктовых соков, безалкогольных напитков, содержащих сывороточный белок, оздоровительных чаев, напитков какао, молочных напитков, напитков с молочнокислыми бактериями, йогуртов, питьевых йогуртов, вина и т.д.
Предоставляется также способ получения пищевого продукта, причем способ включает смешивание клеток в соответствии с настоящим изобретением с ингредиентом пищевого продукта (таким как исходный материал для пищевого продукта). Способ для получения пищевого продукта также является другим аспектом настоящего изобретения.
Подходящим образом, пищевой продукт, описанный в настоящей заявке, представляет собой молочный продукт. В некоторых предпочтительных вариантах осуществления, молочный продукт представляет собой йогурт, сыр (например, кислый мягкий сыр, твердый сыр, полутвердый сыр, домашний сыр и т.д.), пахту, свежий восточно-европейский сыр, сметану, кефир, густые сливки, ферментированный напиток на основе сыворотки, кумыс, молочный напиток или йогуртовый напиток.
Используемый в настоящем описании термин «пища» имеет очень широкий смысл, поскольку он предназначен для охвата пищи для людей, а также для животных, кроме человека (т.е. корма). В некоторых предпочтительных вариантах осуществления, пища предназначена для употребления людьми. Используемый в настоящем описании термин «корм» включает необработанный и переработанный растительный материал и не растительный материал. Этот термин охватывает любой корм, подходящий для потребления животным, включая без ограничения скотом, домашней птицей, рыбой, ракообразными и/или домашними животными.
Создание устойчивых к фагам штаммов и заквасочных культур
Во время разработки настоящего изобретения, были выяснены: устойчивость к фагу с участием CRISPR-генов cas, а также их роль в устойчивости к входящей чужеродной ДНК и влияние спейсеров, вставленных в пределы CRISPR, на специфичность этой устойчивости. Важно, что настоящее изобретение относится к способам и композициям для создания устойчивых к фагам штаммов и заквасочных культур. В некоторых вариантах осуществления, на материнский штамм «А» воздействует фаг «Р», и выбирается устойчивый к фагу вариант (Вариант «А1.0»). Производится анализ Варианта А1.0 (например, с использованием PCR и/или секвенирования ДНК) для подтверждения присутствия дополнительного вставленного спейсера в пределах локуса CRISPR. Затем определяется нуклеотидная последовательность дополнительного спейсера (Спейсера Sp1.0). Обычно, спейсер Sp1.0 представляет собой фрагмент размером приблизительно из 30 нуклеотидов из фага Р, и он обеспечивает устойчивость к фагу Р и связанным фагам («связанные фаги» представляют собой те, которые содержат последовательность спейсера в своих геномах и определяют семейство фагов).
Независимо от первого воздействия фага, тот же штамм А подвергается воздействию того же фага Р, и выбирается второй устойчивый к фагу вариант (Вариант А2.0). Вариант А2.0 выбран для того, чтобы также иметь дополнительный спейсер (Спейсер Sp2.0), вставленный в пределах локуса CRISPR, но с последовательностью спейсера Sp2.0, отличающейся от последовательности спейсера Sp1.0. Обычно, спейсер Sp2.0 представляет собой фрагмент размером приблизительно из 30 нуклеотидов из фага Р, и он обеспечивает устойчивость к фагу Р и связанным фагам. Аналогичным образом, в некоторых вариантах осуществления, с варианта А3.0 до варианта Ах.0 генерируются посредством воздействия на один и тот же штамм А одного и того же фага Р. Все варианты «А» выбраны для того, чтобы был дополнительный спейсер, вставленный (Спейсер Sp3.0 до Spx.0) в пределах локуса CRISPR, но с последовательностью всех спейсеров «Sp», отличающейся от каждой из других. Обычно спейсеры «Sp» представляют собой фрагменты размером приблизительно 30 нуклеотидов из фага Р, и все обеспечивают устойчивость к фагу Р и связанным фагам.
Хотя эти варианты могут использоваться, они ограничены с точки зрения объема их устойчивости. Таким образом, в некоторых вариантах осуществления, имела бы преимущество разработка устойчивых к фагам штаммам второго уровня. Действительно, имела бы преимущество дальнейшая разработка этих устойчивых к фагам вариантов путем увеличения и расширения их устойчивости к фагам. Обычно, можно оценить, что уровень устойчивости будет составлять приблизительно уровень устойчивости одной мутации, происходящей внутри генома фага, в пределах последовательности, соответствующей спейсеру (т.е. от примерно 10-4 до 10-6). Следовательно, устойчивые к фагам штаммы, которые аккумулируют различные спейсеры в пределах локуса CRISPR, имеют повышенный уровень устойчивости к фагу, содержащему последовательность этих спейсеров внутри их генома (т.е. поскольку должны произойти множественные одиночные мутации внутри генома фага).
В некоторых вариантах осуществления, варианты второго уровня продуцируются выделением мутированного фага посредством воздействия фага Р на вариант А1.0. Обычно, этот мутированный фаг (фаг P1.0) имеет мутацию (делецию, точечную мутацию и т.д.) в геноме в пределах области, содержащей последовательность спейсера Sp1.0. Вариант А1.0 чувствителен к фагу Р1.0. Затем, вариант А1.0 подвергается воздействию фага Р1.0 и выбирается устойчивый к фагу вариант (Вариант А1.1). Вариант А1.1 также выбирается так, чтобы он имел дополнительный спейсер, вставленный (Спейсер Sp1.1) в пределах локуса CRISPR, но с последовательностью спейсера Sp1.1, отличающейся от последовательностей спейсеров Sp1.0, от Sp2.0 до Spx.0. Обычно, спейсер Sp1.1 представляет собой фрагмент размером приблизительно 30 нуклеотидов из фага Р1.0, и он должен обеспечить устойчивость к фагу Р1.0 и связанным фагам. Вариант А1.1 устойчив к фагу Р1.0 и предпочтительно, имеет увеличенную устойчивость к фагу Р ввиду аккумуляции спейсера Sp1.0 и Sp1.1.
В дополнительных вариантах осуществления, вновь мутированный фаг (фаг Р1.1) генерируется посредством воздействия фага Р1.0 на вариант А1.1. Затем, после воздействия фага Р1.1 на вариант А1.1, получается новый вариант А1.2, который содержит один новый дополнительный спейсер (Sp1.2). Спейсер обеспечивает устойчивость к фагу Р1.1 и, предпочтительно, увеличивает устойчивость к фагу Р1.0 и Р (т.е. вследствие аккумуляции спейсеров Sp1.0, Sp1.1, Sp1.2). В других дополнительных вариантах осуществления, различные спейсеры (например, 2, 3 или 4) повторно аккумулируются внутри штамма А посредством варианта А1, затем варианта А1.1, затем варианта А1.2 и т.д. для получения варианта, высоко устойчивого к фагам (вариант А1.n). В еще одних вариантах осуществления, дополнительные различные спейсеры могут аккумулироваться в одном и том же штамме посредством варианта А2, затем варианта А2.1, затем варианта А2.2 и т.д. для параллельного генерирования другого варианта штамма А, высоко устойчивого к фагам (вариант А2.n). Такая же стратегия находит применение с вариантами от А3 до Ах.0.
В некоторых вариантах осуществления, предоставляются штаммы, которые устойчивы к более чем одному семейству фагов. Поскольку данный штамм может быть чувствителен к более чем одному семейству фагов, в некоторых вариантах осуществления, желательно увеличить устойчивость штаммов к множеству семейств фагов введением дополнительного спейсера (спейсеров) в пределы локуса CRISPR, происходящего из других семейств фагов (см. фиг. 16). Например, фаги P, Q и R являются репрезентативными фагами из трех семейств фагов, способных инфицировать штамм А. С использованием описанного выше и в данном разделе способа, получают вариант, устойчивые ко всем трем семействам фагов. В некоторых вариантах осуществления, фаг Р используется для генерирования варианта А1р (содержащего спейсер Sp1), который устойчив к фагу Р. Затем, на вариант А1р воздействует фаг Q, и выбирается фаг, устойчивый к фагу (Вариант A1pq). Вариант A1pq имеет один дополнительный спейсер (Sq1), вставленный внутрь локуса CRISPR. Обычно, спейсер Sq1 представляет собой фрагмент размером приблизительно 30 нуклеотидов из фага Q, и он обеспечивает устойчивость к фагу Q и связанным фагам. Вариант A1pq устойчив и к фагу Р, и к фагу Q. Затем, вариант A1pq подвергается воздействию фага R, и выбирается вариант, устойчивый к фагу (Вариант A1pqr). Вариант A1pqr имеет третий дополнительный спейсер (Sr1), вставленный внутрь локуса CRISPR. Обычно, спейсер Sr1 представляет собой фрагмент размером приблизительно 30 нуклеотидов из фага R, и он также обеспечивает устойчивость к фагу R и связанным фагам. Вариант A1pqr устойчив ко всем трем фагам.
В дополнительных вариантах осуществления, описанные выше способы используются в комбинации для получения увеличенной или расширенной устойчивости к фагам. В некоторых особенно предпочтительных вариантах осуществления, эти варианты имеют высокие уровни устойчивости к множественным семействам фагов. В еще одних вариантах осуществления, получаются штаммы, которые устойчивы к определенным фагам или семействам фагов, которые проблематичны в определенных фабриках и/или ферментерах.
Опосредованный CRISPR иммунитет и виды применения для устойчивых к фагам штаммов
В отличие от положений предшествующего уровня техники, которые высказывают гипотезу, что CRISPR или спейсеры CRISPR могут участвовать в придании специфического иммунитета, настоящее изобретение частично основано на удивительных данных, что гены или белки cas требуются для иммунитета против нуклеиновой кислоты-мишени или продукта ее транскрипции. Однако не предполагается, что настоящее изобретение ограничивается каким-либо определенным механизмом, функцией или средством действия.
Еще более удивительно, что во время создания настоящего изобретения, было обнаружено, что один или несколько генов или белков cas связаны с двумя или более повторами CRISPR в пределах локусов CRISPR. Другими словами, представляется, что гены или белки cas специфичны для данного повтора CRISPR ДНК, означая, что гены или белки cas и повторная последовательность образуют функциональную пару. Соответственно, один или несколько спейсеров CRISPR находят применение с одной или несколькими из этих функциональных пар (т.е. спейсеров CRISPR и генов cas) для модулирования устойчивости клетки против нуклеиновой кислоты-мишени или продукта ее транскрипции.
В одном варианте осуществления, для того, чтобы один или несколько спейсеров CRISPR придали клетке иммунитет, повтор(ы) CRISPR и ген(ы) или белки cas образуют функциональную комбинацию (т.е. повтор(ы) CRISPR и ген(ы) или белки cas являются совместимыми).
В дополнительных предпочтительных вариантах осуществления, настоящее изобретение относится к генам/белкам cas, которые влияют на устойчивость бактерий к фагам. В дополнительных других предпочтительных вариантах осуществления, настоящее изобретение относится, по меньшей мере, к двум повторам CRISPR и, по меньшей мере, к одному гену/белку cas, которые могут применяться при прогнозе, определении и/или модификации бактериальной устойчивости к фагам. Действительно, настоящее изобретение относится к способам модификации лизотипа (т.е. устойчивость/чувствительность к различным фагам) бактерий. Следовательно, идентификация и выявление локусов CRISPR в клетках и фагах обеспечивает средство для определения, прогноза и модификации профиля устойчивости клеток, а также взаимодействий между фагом и хозяином.
Преимущественно, применение одного или нескольких локусов CRISPR, двух или более повторов CRISPR, одного или нескольких генов или белков cas и/или одного или нескольких спейсеров CRISPR при генетической инженерии, обеспечивает средство для получения устойчивых или чувствительных вариантов клеток для использования в пределах широкого разнообразия видов применения в биотехнологической промышленности.
Как более детально обсуждается ниже, фаги представляют собой естественные паразиты бактерий, которые могут развиться во время ферментации. После инфекции фагами, бактерии уничтожаются, что нарушает процесс ферментации. При молочной ферментации, эти инфекции фагами часто оказывают существенные экономические воздействия в диапазоне от сниженного качества ферментированного продукта до полной потери продукта.
Для преодоления проблем, связанных с фагами, компании, производящие заквасочные культуры, разработали разнообразные стратегии. Традиционные программы получения заквасочных культур зависели от ротационных стратегий защиты от фагов (PDRS) для минимизации неудач вследствие атаки фагов (см., например, Klaenhammer, Adv. Appl. Microbiol., 30:1 [1984]; Lawrence et al., J. Dairy Res. 43:141 [1976); и Whitehead и Hunter, J. Dairy Res., 15:112 [1947]). Эти стратегии основаны на множественных, генетически не связанных штаммах, которые, вероятно, проявляют другой спектр чувствительности к фагам (т.е. другие лизотипы). Когда фаг появляется во время процесса ферментации с использованием определенного штамма, то вместо этого для ферментации используется штамм, который в идеале имеет другой лизотип (т.е. с другим типом чувствительности к фагам). Однако исторически доказано, что трудно идентифицировать достаточные количества различных лизотипов для успешного использования этих стратегий. Действительно, многие штаммы, представляющие промышленный интерес, проявляют редкие функциональные признаки (например, быстро окисляющие текстурирующие S. thermophilus). Кроме того, не все штаммы проявляют соответствующие признаки для получения в качестве заквасочных культур. Кроме того, ввиду их редкости и увеличения размера молочных фабрик, эти штаммы интенсивно используются.
Существуют дополнительные проблемы, связанные с традиционными стратегиями ротации заквасочных культур. Хотя некоторые штаммы при введении не атакуются существующими фагами, фаги часто в конечном счете появляются вследствие мутации, модификации и конструирования фагов, которые атакуют вновь введенный штамм (см., например, Heap и Lawrence, N.Z.J. Dairy Sci. Technol., 11:16 [1976]; Limsowtin и Terzaghi, N.Z.J. Dairy Sci. Technol., 11:251 [1976]; Pearce, N.Z.J. Dairy Sci. Technol., 13:166 [1978]; и Sanders и Klaenhammer, Appl. Environ. Microbiol., 40:500 [1980]). Кроме того, во многих случаях, длительность существования и заквасочная активность ротаций сложных штаммов является непрогнозируемой и часто ведет к ранней несостоятельности (см., например, Limsowtin et al., N.Z.J. Dairy Sci. Technol.,13:1 [1977]; и Thunell et al., J. Dairy Sci., 64, 2270 [1981]). Кроме того, длительные ротации, вовлекающие многочисленные штаммы, увеличивают уровень и разнообразие фага, заражающего растение (см., например, Heap и Lawrence, N.Z.J. Dairy Sci. Technol., 12:213 [1981]; Lawrence et al., J. Dairy Sci., 61:1181 [1978]; и Thunell et al., J. Dairy Sci. 64, 2270 [1981]).
Для борьбы с пролиферацией фагов, традиционные программы заквасочных культур зависели от применения штаммов, проявляющих такие же или подобные технологические функциональные свойства, но различную чувствительность к фагам. Эти штаммы используются при ротации для выполнения успешной ферментации. Эти программы традиционно основаны на множественных генетически не связанных штаммах, которые в последующем проявляют другой спектр чувствительности к фагу (лизотип). В альтернативных подходах (см., например, патент США № 5593885) используются программы заквасочных культур на основе использования наборов изогенных штаммов, которые проявляют другую чувствительность к фагам, вместо генетически не связанных штаммов, проявляющих другие лизотипы. Используемый в настоящем описании термин «набор изогенных штаммов» определяет штаммы, которые идентичны с хромосомной точки зрения, но что каждый отличается присутствием одного или нескольких механизмов устойчивости к фагам, которые являются исходящими из плазмиды. В такой программе ротации заквасочных культур, когда фаг появляется во время процесса ферментации с использованием определенного штамма, штамм, который в идеале относится к другому лизотипу (т.е. с другим спектром чувствительности к фагам), который используется в качестве замещения для ферментации. Благодаря этому другому лизотипу, на второй штамм не воздействуют фаги, которые остаются покоящимися в окружающей среде. Затем большая часть популяции покоящихся фагов смываются последующей ферментацией и санацией, и искореняется ко времени, когда первый штамм снова используется для ферментации, если система работает в предназначенном режиме.
Настоящее изобретение относится к усовершенствованным способам и композициям, подходящим для решения этих проблем в ферментационной промышленности. Действительно, настоящее изобретение относится к усовершенствованным способам и композициям для ферментационной промышленности и, в частности, для молочной промышленности с выбором штаммов, подходящих для удовлетворения потребностей стратегий ротации для защиты от фагов. Кроме того, настоящее изобретение относится к способам и композициям, подходящим для использования штаммов, имеющих лизотипы, которые адаптированы к определенной среде фага. В частности, настоящее изобретение относится к способам и композициям, подходящим для направления эволюции данного штамма в различные лизотипы для получения штаммов, которые отличаются друг от друга только спектром их чувствительности к фагам (лизотипа). Это различие лизотипа является функцией системы CRISPR-cas, как описано в настоящей заявке. В некоторых предпочтительных вариантах осуществления, различные лизотипы получаются посредством «модулирования» устойчивости к фагу. В некоторых особенно предпочтительных вариантах осуществления, хотя лизотипы являются различными, штаммы этого типа имеют идентичный метаболизм (например, углерода, азота и т.д.) и, таким образом, идентичные функции (например, подкисление, запах и вкус, текстуру и т.д.). Это обеспечивает средство для амплификации конструкции ротации закваски. Кроме того, возможности промышленной переработки устойчивого к фагу штамма являются идентичными (например, питательные потребности, устойчивость к операции переработки и т.д.), таким образом, снижая потребность в разработке определенных производственных процессов. Действительно, настоящее изобретение относится к способам и композициям, подходящим для минимизации отказов ферментации вследствие атаки фагов. В некоторых вариантах осуществления, способы и композиции представлены для получения заквасочных культур с высокой устойчивостью к фагам путем ассоциации множественных устойчивых к фагам штаммов, отличающихся их лизотипом. В некоторых альтернативных вариантах осуществления, способы и композиции представлены для получения заквасочных культур со строго идентичными промышленными функциями для использования при ротационной молочной ферментации. В других вариантах осуществления, представлены способы и композиции, которые походят для замещения существующих заквасок путем предотвращения частых атак фагов на молочных заводах, внедрением нового бактериального штамма, который устойчив к фагам, участвующим в этой атаке фага. В некоторых вариантах осуществления, эти способы и композиции используются повторно для борьбы с последовательными атаками фагов.
В некоторых дополнительных вариантах осуществления, заквасочная культура представляет собой смешанную бактериальную культуру. В некоторых особенно предпочтительных вариантах осуществления, закваска содержит равные количества множественных (т.е. по меньшей мере, 2) устойчивых к фагам вариантов, которые отличаются только по их CRISPR и их чувствительности к фагам. В некоторых вариантах осуществления, эти варианты относятся к первому уровню устойчивых к фагам вариантов (например, к вариантам А1.0 плюс А2.0, как описано выше). В некоторых предпочтительных вариантах осуществления, варианты выбраны из вариантов на втором уровне устойчивых к фагам вариантов (например, вариантов А1.4 плюс А2.4, как описано выше). В некоторых особенно предпочтительных вариантах осуществления, варианты выбраны среди третьего уровня устойчивых к фагам вариантов. В таких смешанных бактериальных культурах, когда один из вариантов атакуется данным фагом, другие варианты не должны атаковаться фагом ввиду их отличных чувствительностей к фагам, и не осуществляются неблагоприятные воздействия на ферментацию.
В некоторых других вариантах осуществления, используется основная закваска и поддерживающая закваска. Основная закваска составлена из одного штамма. В некоторых вариантах осуществления, этот штамм относится к первому уровню устойчивых к фагам вариантов, тогда как в других предпочтительных вариантах осуществления, штамм относится ко второму уровню, а в еще одних более предпочтительных вариантах осуществления, штамм относится к третьему уровню. В некоторых предпочтительных вариантах осуществления, поддерживающая закваска основана на устойчивом к фагам варианте, полученном независимо из одного и того же материнского штамма. Этот второй устойчивый к фагам вариант отличается от других вариантов его CRISPR и относится к первому уровню устойчивых к фагам вариантов, хотя в других предпочтительных вариантах осуществления, штамм относится ко второму уровню, а в еще одних других более предпочтительных вариантах осуществления, штамм относится к третьему уровню. Например, в некоторых вариантах осуществления, основная закваска изготовлена из варианта А1.4, а поддерживающая закваска изготовлена из штамма А2.4. После первого появления фага во время ферментации основной закваской, закваска удаляется и замещается поддерживающей закваской. В некоторых предпочтительных вариантах осуществления, третья закваска также получается в качестве поддерживающей закваски, которая должна служить поддержкой для поддержки. В некоторых предпочтительных вариантах осуществления, каждая из заквасок изготовлена из множественно устойчивых к фагам вариантов.
В еще одних вариантах осуществления, настоящее изобретение относится к способам и композициям, подходящим для стратегий ротации. В некоторых вариантах осуществления, вместо удаления закваски, часто атакованной фагами, закваски используются циклическим образом, даже если наблюдается атака фага. Эта стратегия ограничивает количество подлежащих разработке заквасок. В некоторых особенно предпочтительных вариантах осуществления, каждая закваска изготовлена из штаммов, устойчивых к множеству фагов, вместо одного. Это обеспечивает увеличенное противодействие появляющемуся фагу. В еще одних вариантах осуществления, предоставляются закваски, изготовленные по заказу. В некоторых предпочтительных вариантах осуществления, продуцируются устойчивые к фагам варианты для специфической борьбы с фагами, которые присутствуют на данном ферментативном заводе или установке.
Типирование
В еще одном аспекте настоящее изобретение относится к способу идентификации (например, типирования) меченой бактерии.
В одном варианте осуществления, стадия идентификации выполняется амплификацией (например, амплификации PCR) локуса CRISPR или его части.
Первый праймер может быть предназначен для гибридизации с последовательностью, которая локализуется выше по ходу транскрипции от первого повтора CRISPR локуса CRISPR. В качестве примера, первый праймер может гибридизироваться с соседним геном, который локализуется выше по ходу транскрипции от локуса CRISPR.
Второй праймер может гибридизироваться ниже по ходу транскрипции, по меньшей мере, от первого спейсера CRISPR или, по меньшей мере, от кора первого спейсера CRISPR. Второй праймер может гибридизироваться так далеко как с трейлером или даже в расположенном ниже по ходу транскрипции соседним геном. Предпочтительно, второй праймер гибридизируется в пределах локуса CRISPR. Предпочтительно, второй праймер гибридизируется, по меньшей мере, частично с расположенным ниже по ходу транскрипции спейсером CRISPR или кором спейсера CRISPR.
После амплификации маркировочная последовательность может быть идентифицирована с использованием различных способов, которые известны в данной области.
В качестве примера, маркировочная последовательность может быть идентифицирована определением рестрикционного типа продукта амплификации. Соответственно, после амплификации ДНК, содержащей локус CRISPR или ее часть, она может быть переварена (например, разрезана) одним или несколькими рестрикционными ферментами.
Используемый в настоящем описании термин «рестрикционные ферменты» относится к ферментам (например, бактериальным ферментам), каждый из которых разрезает двухнитевую ДНК в специфической нуклеотидной последовательности или около нее. Рестрикционные ферменты хорошо известны в данной области и могут быть легко получены, например, из разнообразных промышленных источников (например, New England Biolabs, Inc., Beverly, Massachusetts). Аналогичным образом, способы использования рестрикционных ферментов также в целом хорошо известны и общеприняты в данной области. Могут использоваться рестрикционные ферменты, которые продуцируются между 10 и 24 фрагментами ДНК при разрезании локуса CRISPR или его части. Примеры таких ферментов включают без ограничения, например, в виде полос при гель-электрофорезе. Рестрикционные ферменты могут использоваться для создания Полиморфизма Длин Рестрикционного Фрагмента (RFLP).
RFLP генерируются разрезанием («ограничением») молекулы ДНК рестрикционной эндонуклеазой. Были изолированы многие сотни таких ферментов, которые естественно вырабатываются бактериями. По существу, бактерии используют такие ферменты в качестве защитной системы для распознавания, а затем расщепления (рестрикции) любых инородных молекул ДНК, которые могут поступить в бактериальную клетку (например, вирусной инфекции). Было обнаружено, что каждый из многих сотен различных рестрикционных ферментов разрезает (т.е. «расщепляет» или «ограничивает») ДНК в другой последовательности 4 основных нуклеотидов (A, T, G, C), которые составляют молекулы ДНК, например, один фермент может специфически и только распознавать последовательность A-A-T-G-A-C, тогда как другой может специфически и только распознавать последовательность G-T-A-C-T-A и т.д. В зависимости от необычного участвующего фермента, такие распознаваемые последовательности могут варьировать по длине от столь небольшого количества как 4 нуклеотида до количества, достигающего 21 нуклеотида. Чем больше распознаваемых последовательностей, тем к меньшему числу рестрикционных фрагментов это приведет, поскольку чем больше сайт распознавания, тем ниже вероятность того, что он будет повторно обнаружен по всей ДНК.
В качестве еще одного примера, маркирующая последовательность может быть идентифицирована определением или также определением различия размера продукта амплификации.
Разделение может быть достигнуто любым способом, подходящим для выделения ДНК, включая без ограничения гель-электрофорез, высокоэффективную жидкостную хроматографию (ВЭЖХ), масс спектроскопию и использование микрожидкостного устройства. В одном варианте осуществления, продукты амплификации или фрагменты ДНК отделяются электрофорезом на агарозном геле. Гель-электрофорез отделяет заряженные молекулы различного размера скоростью их движения через стационарный гель под влиянием электрического тока. Эти выделенные продукты амплификации или фрагменты ДНК могут быть легко визуализированы, например, окрашиванием этидием бромидом и визуализацией геля под УФ освещением. Тип образования полос отражает размеры рестрикционной переваренной ДНК или продуктов амплификации.
В качестве еще одного примера, маркировочная последовательность может быть идентифицирована секвенированием продуктов амплификации.
Последовательность амплифицированных продуктов может быть получена любым способом, известным в данной области, включая способы автоматического и ручного секвенирования. См., например, Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Plainview, New York; Roe et al. (1996) DNA Isolation и Sequencing (Essential Techniques Series, John Wiley & Sons).
Способы гибридизации также охватываются объемом настоящего изобретения, или использующие молекулу нуклеиновой кислоты в качестве зонда, или молекулу нуклеиновой кислоты, способную гибридизироваться с определенной нуклеотидной последовательностью. См., например, Sambrook et al. (1989) Molecular Cloning: Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Plainview, New York).
В методиках гибридизации, зонд(ы) гибридизации могут представлять собой фрагменты геномной ДНК, продукты, амплифицированные PCR, или другие олигонуклеотиды, и могут содержать всю или часть известной нуклеотидной последовательности. Кроме того, он может быть мечен выявляемой группой, такой как 32Р, или любым другим выявляемым маркером, таким как другие радиоизотопы, флюоресцентное соединение, фермент или ко-фактор фермента. Термин «меченый» в отношении зонда предназначен для охвата прямого мечения зонда соединением (т.е. физического связывания) выявляемого вещества с зондом, а также непрямого мечения зонда взаимодействием с другим реагентом, который подвергнут прямому мечению. Примеры непрямого мечения включают концевое мечение зонда ДНК биотином с тем, чтобы его можно было выявить флюоресцентно меченым стрептавидином.
Также охватываются способы, включающие методики гибридизации для выявления или дифференцировки бактериальных штаммов. Они включают без ограничения саузерн блоттинг (см., например, Vaqn Embden et al. (1993) J. Clin. Microbiol. 31:406-409), анализ подвижности сдвига (см., например, опубликованную заявку на патент США № 20030219778), анализы секвенирования с использованием олигонуклеотидных чипов (см., например, Pease et al. (1994) Proc. Natl. Acad. Sci. USA 91:5022-5026), сполиготипирование (см., например, Kamerbeek et al. (1997) J. Clin. Microbiol. 35:907-914), Флюоресцентную Гибридизацию In Situ (FISH) (см., например, Amann et al. N(1990) J. Bacteriol. 172:762-770) анализы гетеродуплексного слежения или анализ гетеродуплексной подвижности (см., например, White et al. (2000) J. Clin. Microbiol. 38:477-482).
Маркировочная последовательность, которая идентифицируется, может сравниваться с базой данных последовательностей фагов и/или базой данных бактериальных последовательностей. Обычно, маркировочная последовательность будет сопоставима с одной или несколькими последовательностями в базе данных последовательностей фагов, но не в базе данных бактериальных последовательностей.
Поскольку новые меченые бактерии получают с использованием способов, описанных в настоящей заявке, может быть создана база данных меток, позволяющая проводить специфическую идентификацию бактерий, которые были мечены.
В некотором аспекте изобретение относится к применению последовательности, полученной или получаемой из бактериофага (например, при получении меченой бактерии) для мечения и/или идентификации бактерии, где указанная последовательность интегрирована на одном конце локуса CRISPR материнской бактерии.
В еще одном аспекте изобретение относится к применению последовательности, полученной или получаемой из бактериофага (например, при получении меченой бактерии) для мечения и/или идентификации бактерии, где указанная последовательность содержит: (i), по меньшей мере, одну последовательность, которая гомологична (например, идентична) повтору CRISPR в локусе CRISPR указанной бактерии; и (ii) маркировочную последовательность.
В еще одном аспекте изобретение относится к применению последовательности для мечения и/или идентификации бактерии (например, при получении меченой бактерии), где указанная последовательность получается или может быть получена: (а) воздействием бактериофага на материнскую бактерию; (b) выбором нечувствительного к бактериофагу мутанта; (с) сравнением локуса CRISPR или его части из материнской бактерии и нечувствительного к бактериофагу мутанта; и (d) выбора последовательности в локусе CRISPR или его части нечувствительного к бактериофагу мутанта, которая не присутствует в материнской бактерии.
CRISPR и эукариоты
Как описано в настоящей заявке, было показано, что CRISPR обеспечивает сопротивляемость против нуклеиновой кислоты, входящей в прокариоты. В частности, было показано, что спейсеры CRISPR, проявляющие гомологию к вирусной ДНК (например, нуклеиновой кислоте бактериофага), обеспечивают сопротивляемость против вируса, разделяющего идентичность последовательности, по меньшей мере, с одной спейсерной последовательностью. Однако также предусматривается, что система CRISPR, включающая гены и/или белки cas наряду со спейсерами, повторами, лидером и трейлером к клеткам, в настоящее время не содержащим локусы CRISPR, найдет применение при обеспечении сопротивляемости против нуклеиновой кислоты de novo. Действительно, в некоторых вариантах осуществления, такие манипуляции находят применение с различными эукариотами, включая без ограничения других животных, грибы и т.д. Предусматривается, что система CRISPR переносится в эукариотические клетки с использованием любого подходящего способа, известного в данной области. Включая без ограничения трансформацию через плазмиды. В этих вариантах осуществления, локусы CRISPR, а также при необходимости сигналы транскрипции/трансляции, включены в ДНК плазмиды, все для экспрессии и функции последовательностей в эукариотических клетках.
В некоторых дополнительных вариантах осуществления, спейсерные последовательности подвергаются генной инженерии с тем, чтобы они имели идентичность с вирусными последовательностями, представляющими интерес инфицирования вовлеченного хозяина. В некоторых предпочтительных вариантах осуществления, эти способы и композиции обеспечивают сопротивляемость клетки-хозяина против вирусов, которые разделяют идентичность последовательности со спейсером CRISPR, введенным в клетку. В некоторых особенно предпочтительных вариантах осуществления, вирусы включают без ограничения ВИЧ, ортомиксовирусы, парамиксовирусы, псевдомиксовирусы, RSV (вирус саркомы Рауша), гриппа, кори, ветряной оспы, краснухи, коронавирусы, вирусы гепатита, калицивирусы, поксвирусы, герпесвирусы, аденовирусы, паповавирусы, папилломавирусы, энтеровирусы, арбовирусы, рабдовирусы, аренавирусы, арбовирусы, риновирусы, реовирусы, коронавирусы, реовирусы, ротавирусы, ретровирусы и т.д. В еще одних вариантах осуществления, специфическое нацеливание высоко консервативных последовательностей нуклеиновых кислот в спейсерах CRISPR обеспечивает повышенную сопротивляемость против таких вирусов у эукариотических клеток. В некоторых особенно предпочтительных вариантах осуществления, эукариотические клетки представляют собой клетки человека.
CRISPR и генерирование устойчивых к фагам мутантов
Во время разработки настоящего изобретения, были проведены эксперименты для определения того, изменены ли локусы CRISPR во время естественного генерирования устойчивых к фагам мутантов. Была выбрана модельная система фага-хозяина, состоящая из чувствительного к фагам штамма S. thermophilus дикого типа, широко используемого в молочной промышленности, DGCC7710 (WT) и двух различных, но близко родственных вирулентных бактериофагов, выделенных из промышленных образцов йогурта, а именно фага 858 и фага 2972 (Levesque et al., Appl. Environ. Microbiol., 71:4057 [2005]). Девять устойчивых к фагам мутантов были независимо генерированы стимуляцией штамма WT фагом 858, фагом 2972 или одновременно обоими, и анализировались их локусы CRISPR. Различия согласованно наблюдались в локусе CRISPR1, где от 1 до 4 дополнительных спейсеров были вставлены рядом с 32 спейсерами, присутствующими в штамме WT (см. фиг. 9). Представляется, что добавление новых спейсеров в ответ на инфекцию фагом поляризуется в направлении к концу локуса CRISPR. Это согласуется с предыдущими наблюдениями гипервариабельности спейсеров на лидерном конце локуса CRISPR у различных штаммов (см., например, Pourcel et al., Microbiol., 151:653 [2005]; и Lillestol et al, Archaea 2:59 [2006]). Анализ последовательности дополнительных спейсеров, вставленных в локус CRISPR1 различных устойчивых к фагам мутантов, выявил подобие последовательностям, обнаруженным внутри геномов фагов, используемых при стимуляции. Подобия наблюдались по всем геномам фага, в большинстве функциональных модулях и на кодирующей, и на не кодирующих нитях. Представлялось, что определенной мишенью не была конкретная последовательность, ген или функциональная группа. Эти результаты указывают на то, что после того как локус CRISPR1 становился устойчивым к бактериофагам, он был модифицирован интеграцией новых спейсеров, очевидно полученных из ДНК фага. Однако не предполагается, что настоящее изобретение ограничивается каким-либо определенным механизмом.
К удивлению, наблюдалось, что некоторые штаммы были устойчивы к обоим фагам, тогда как другие были устойчивы только к фагу, используемому при провокационной пробе (см. фиг. 9). Представлялось, что профиль устойчивости к фагам коррелирует с содержанием спейсеров, посредством чего штаммы со спейсерами, такими как спейсеры S3, S6 и S7, проявляющими 100% идентичность с консервативными последовательностями в обоих фагах, были устойчивы к обоим фагам. Напротив, когда полиморфизм нуклеотидов наблюдался между последовательностью спейсера и фага (от 1 до 15 SNP (полиморфизма одиночных нуклеотидов) по 29 или 30 нуклеотидам), представлялось, что спейсер, такой как спейсеры S1, S2, S4, S5 и S8, не обеспечивал устойчивость (см. фиг. 9).
Кроме того, когда вставлялись несколько спейсеров (S9-S14), уровни устойчивости к фагам были выше. Эти данные указывают на то, что локус CRISPR1 подвержен динамичным и быстрым эволюционным изменениям, запускаемым воздействием фага. Эти результаты указывают на то, что локусы CRISPR действительно могут изменяться во время генерирования устойчивых к фагам мутантов и устанавливают связь между содержанием CRISPR и чувствительностью к фагам. Таким образом, предусматривается, что присутствие спейсера CRISPR, идентичного последовательности фага, обеспечивает сопротивляемость против фагов, содержащих эту конкретную последовательность.
Для определения того, определяет ли содержание локуса CRISPR устойчивость к фагам, локус CRISPR1 был изменен добавлением и делецией спейсеров, и тестировалась чувствительность штамма к фагам. Все конструкты были генерированы и интегрированы в хромосому S. thermophilus с использованием способов, известных в данной области (см., например, Russell and Klaenhammer, Appl. Environ. Microbiol., 67:4361 [2001]). Спейсеры и повторы в локусе CRISPR1 штамма WTФ858+S1S2 удаляли и замещали одним повтором без какого-либо спейсера. Полученный штамм WTФ858+S1S2 ΔCRISPR1 был чувствителен к фагу 858, указывая на то, что устойчивость к фагам первоначального устойчивого к фагам мутанта (WTФ858+S1S2) была, вероятно, связана с присутствием S1 и S2 (см. фиг. 10).
Кроме того, для определения того, обеспечивает ли добавление спейсеров устойчивость к фагам, локус CRISPR1 штамма WTФ858+S4 замещали только вариантом, содержащим спейсеры S1 и S2. Затем тестировали чувствительность полученного конструкта к фагам. Полученный штамм WTФ858+S4::pS1S2 приобретал повышенную устойчивость к фагу 858, свидетельствуя о том, что эти два спейсера способны обеспечить устойчивость к фагам de novo (см. фиг. 10). Эти наблюдавшиеся модификации устанавливают связь между содержанием спейсера CRISPR и устойчивостью к фагам.
В процессе генерирования штамма WTФ858+S1S2 ΔCRISPR1, WTФ858+S1S2::pR, был создан вариант, который содержит вектор интеграции с одним повтором, вставленным между генами cas и нативным локусом CRISPR1 (см. фиг. 10). Неожиданно, штамм WTФ858+S1S2::pR был чувствителен к фагу 858, хотя спейсеры S1 и S2 продолжали присутствовать на хромосоме (см. фиг. 10). Аналогичным образом, конструкт WTФ2972+S4::pS1S2 утратил чувствительность к фагу 2972, хотя спейсер S4 присутствует в хромосоме (см. фиг. 10). Эти результаты указывали на то, что одни спейсеры не обеспечивали устойчивость и, вероятно, для того, чтобы быть эффективными, они должны быть в определенном генетическом контексте.
Хотя ранние эксперименты свидетельствовали об участии в репарации ДНК (Makarova et al., Nucl. Acids Res., 30:482 [2002]), современная гипотеза состоит в том, что гены cas (Jansen et al., Mol. Microbiol., 43:1565 [2002]; и Haft et al., PloS Comput. Biol., l:e60 [2005]) участвуют в опосредованном CRISPR иммунитете (Makarova et al., Biol. Direct. 1:7 [2006]). В дополнительных экспериментах, два гена cas в штамме WTФ858+S1S2 были инактивированы, а именно, cas5 (COG3513) и cas7, которые эквивалентны соответственно str0657/stu0657 и str0660/stu0660 (Bolotin et al., Nat. Biotechnol., 22:1554 [2004]; и Bolotin et al., Microbiol., 151:2551 [2005]). Инактивация cas5 приводила к утрате устойчивости к фагу (см. фиг. 10). Кроме того, возможно, что cas5 действует в качестве нуклеазы, поскольку он содержит мотив нуклеазы типа HNH. Напротив, инактивация cas7 не изменяла устойчивость к фагу 858 (см. фиг. 10). Однако не предполагается, что настоящее изобретение ограничивается каким-либо конкретным механизмом. Кроме того, эксперименты для генерирования мутантов CRISPR1, устойчивых к фагу, из нокаута cas7, не имели эффекта. Хотя нет намерения, чтобы настоящее изобретение ограничивалось каким-либо конкретным механизмом, это может быть связано с тем, что Cas7 участвует в синтезе и/или вставке новых спейсеров и дополнительных повторов.
После тестирования чувствительности устойчивых к фагам мутантов, было обнаружено, что образование бляшек было резко снижено, но что относительно небольшая популяции бактериофагов сохраняла способность инфицировать мутанты. Далее анализировались варианты фага, полученные из фага 858, которые сохраняли способность инфицировать WTФ858+S1S2. В частности, были исследованы последовательности области генома, соответствующие дополнительным спейсерам S1 и S2, в двух вирулентных вариантах фага. В обоих случаях, последовательность генома варианта фага была мутирована, и в последовательности, соответствующей спейсеру S1, были идентифицированы два различных вида полиморфизма одиночных нуклеотидов (см. фиг. 13).
В целом, представляется, что у прокариотов развилась система «иммунитета» на основе нуклеиновой кислоты, посредством чего специфичность определяется содержанием спейсера CRISPR, тогда как устойчивость обеспечивается ферментным механизмом Cas. Кроме того, были высказаны суждения, что некоторые из генов cas, которые непосредственно не обеспечивают устойчивость, действительно участвуют во вставке дополнительных спейсеров и повторов CRISPR в качестве части адаптивного «иммунного» ответа. Эта система на основе нуклеиновой кислоты контрастирует с основанными на аминокислотах параллельными системами в эукариотах, посредством чего адаптивный иммунитет не является наследуемым. Наследуемая природа спейсеров CRISPR обосновывает применение локусов CRISPR в качестве мишеней для эволюционных, типирующих и сравнительных геномных исследований (см. Pourcel et al., supra; Groenen et al., Mol. Microbiol., 10:1057 [1993]; Mongodin et al., J. Bacteriol., 187:4935 [2005]; и DeBoy et al., J. Bacteriol., 188:2364 [2006]). Ввиду того, что эта система реагирует на окружающую среду фага, она, вероятно, играет существенную роль в прокариотической эволюции и экологии и обеспечивает историческую перспективу воздействия фагов, а также в качестве прогностического инструмента для определения чувствительности к фагам. Однако нет намерения, чтобы настоящее изобретение ограничивалось каким-либо конкретным механизмом. Тем не менее настоящее изобретение относится к способам и композициям для использования системы CRISPR/cas в качестве средства защиты от вирусов, а также для потенциального уменьшения диссеминации подвижных генетических элементов и приобретения нежелательных признаков, таких как гены противомикробной сопротивляемости и маркеры вирулентности. В некоторых вариантах осуществления, из перспективы эволюции фага, кроме того, предусматривается, что интегрированные последовательности фага внутрь локусов CRISPR также обеспечат дополнительные точки прикрепления для содействия рекомбинации во время последующих инфекций фагами, таким образом, увеличивая генный пул, к которому имеют доступ фаги (см. Hendrix et al., Proc. Natl. Acad. Sci. USA 96:2192 [1999]). Поскольку локусы CRISPR обнаруживаются в большинстве бактериальных родов и являются вездесущими у Archea (см. Jansen et al., supra; Lillestol et al., supra; и Goode и Bickerton J. Mol. Evol., 62:718 [2006]), они обеспечивают новое понимание взаимоотношения и совместно направленной эволюции между прокариотами и их пожирателями.
Фаги биологического контроля
Настоящее изобретение также относится к способам и композициям для разработки фагов в качестве средств биологического контроля. Как указано в настоящем описании, бактерии могут стать устойчивыми к атаке фагов путем включения последовательностей (спейсеров), полученных из фагов, в активные локусы CRISPR. Фаг может избежать этой устойчивости мутацией внутри последовательности генома, соответствующей спейсеру или последовательности распознавания мотива CRISPR, для получения системы Cas-CRISPR. Посредством повторных циклов стимуляции фагами для создания устойчивых к фагам производных, опосредованных CRISPR штамма-хозяина, и выделения мутантов избегания фагов, настоящее изобретение относится к фагам, которые были подвергнуты изменению в пределах последовательностей-мишеней CRISPR и/или предполагаемых сайтов распознавания CRISPR, которые направляют вставку спейсера. Кроме того, настоящее изобретение относится к фагам, которые были синтетически сконструированы, с тем, чтобы была устранена последовательность мотива CRISPR для данной системы Cas-CRISPR. Эти «измененные» фаги, применяемые в виде смеси или в «последовательной ротационной схеме», снижают способность бактерий-мишеней адаптировать устойчивость через систему CRISPR. Действительно, настоящее изобретение относится к разнообразному набору вирулентных фагов для использования в качестве средств биологического контроля. В особенно предпочтительных вариантах осуществления, мишенью в этом разнообразии является направляемый CRISPR механизм устойчивости к фагам с тем, чтобы способность организма хозяина быстро развивать сопротивляемость против атаки фагов (через CRISPR) резко снижалась или устранялась. Введение разнообразных фагов или в виде смеси, или при последовательной ротации, дополнительно снижает возможность организма хозяина адаптироваться или развить направляемую CRISPR устойчивость к фагам.
Фаги представляют собой естественные противомикробные агенты, которые интенсивно исследовались в качестве терапевтического средства, альтернативного антибиотикам. Этот интерес был недавно возобновлен ввиду пролиферации патогенов, устойчивым к многим антибиотикам. Как и с антибиотиками, бактерии развили множественные механизмы преодоления атаки фагов. Настоящее изобретение относится к способам и композициям, включающим использование Cas-CRISPR в опосредовании устойчивости к фагам для генерирования популяции разнообразных фагов, для создания синтетических фагов, лишенных последовательностей мотива CRISPR, а также к способам для введения такого фага, которые снизят способность организма-мишени развивать сопротивляемость против фага.
Как детально описано в настоящей заявке, системы Cas-CRISPR раскрыты в широком диапазоне организмов, которые включают примеры патогенных родов. После инфекции фагом, может быть обнаружено, что бактерии, избегающие лизиса, содержат новую спейсерную последовательность (последовательности) в пределах локуса CRISPR. Новый спейсер имеет обычно определенную длину, которая характерна для данного локуса CRISPR, и получена из генома атакующего фага, к которому он придает устойчивость. Поскольку уровень устойчивости, придаваемый одним спейсером, часто неполный, то фаг может избежать этого механизма. Анализ «избегающих фагов» указывает на то, что геномы были мутированы в соответствующей спейсерной последовательности, обнаруживаемой в устойчивом варианте хозяина, или вблизи ее. Кроме того, «избегающие фаги» являются полностью вирулентными для опосредованного CRISPR варианта хозяина, из которого они были получены.
Один необычный аспект терапевтического фага, отличающий его от традиционных антибиотиков, состоит в способности экспоненциально размножаться в сочетании с инфицированными бактериями. Хотя он может иметь преимущество с точки зрения фармакологической перспективы, он также обеспечивает уникальные возможности для фага развиться в направлении адаптивной реакции бактерий-мишеней на атаку фагов.
У бактерий развились несколько механизмов защиты против вирулентного фага. После инфекции фагом, анализ выживших бактерий обнаружил, что некоторые изоляты имели вставленный новый спейсерный элемент в пределах их резидентного локуса CRISPR, последовательность которого была идентична последовательности, обнаруженной в соответствующем геноме фага. При стимуляции фагом, эти варианты первого поколения с опосредованной CRISPR устойчивостью к фагам образуют бляшки; фаги которых, как было обнаружено, полностью инфективны и для материнской бактерии, и для производного. Анализ этих фагов «избегания CRISPR» указал на то, что их геномы были мутированы в последовательности, соответствующей спейсеру CRISPR, содержащемуся в устойчивом к фагу варианте, или в проксимальной последовательности, которая, как считается, направляет вставку спейсера и идентифицирована как мотив CRISPR, специфичный для данной системы Cas-CRISPR. Поэтому, фаг «избегания CRISPR» потенциально более вирулентный, чем материнские бактерии или вариант первого поколения.
Как указано выше, локусы CRISPR были идентифицированы в нескольких родах/видах, которые включают примеры известных патогенов и вредных микроорганизмов. Также, как описано в настоящей заявке, настоящее изобретение относится к способам и композициям для использования локусов CRISPR в комбинации с белками Cas для придания «иммунитета» к внедряющейся инородной ДНК, в частности, бактериофагам. Также, как описано в настоящей заявке, бактериальные штаммы, включающие «активные» локусы CRISPR-cas, содержащие спейсер, который идентичен соответствующей последовательности внутри генома фага (т.е. «протоспейсер»), придают этому бактериальному штамму устойчивость к фагу. В некоторых особенно предпочтительных вариантах осуществления, последовательности генома фага биологического контроля известны. В некоторых особенно предпочтительных вариантах осуществления, выделенный микроорганизм-мишень исследуется на присутствие локусов CRISPR. В некоторых предпочтительных вариантах осуществления, находит применение PCR с использованием специфических праймеров для консервативных последовательностей, которые фланкируют локусы CRISPR микроорганизма-мишени. В некоторых предпочтительных вариантах осуществления, продукт(ы) амплификации секвенируются в сравнении с последовательностью генома фага биологического контроля. В некоторых предпочтительных вариантах осуществления, генерирование вариантов, устойчивых к CRISPR фагу, и анализ спейсеров/протоспейсеров предоставляет средство для идентификации специфического мотива CRISPR. После идентификации, информация о последовательности используется для конструирования и синтеза фага, лишенного мотива CRISPR. Таким образом, полученный в результате фаг чувствителен к опосредованной CRISPR-cas устойчивости. При этих оценках, отсутствие спейсеров с подобием геному фага, указывает на восприимчивость микроорганизма-мишени к фагу биологического контроля. Таким образом, фаг биологического контроля имеет большую степень вирулентности и эффективности в качестве агента биологического контроля.
Настоящее изобретение относится к способам и композициям, подходящим для применения в пищевой, кормовой, медицинской и ветеринарной промышленности для генерирования фагов с более широким диапазоном хозяев и способа их применения для более эффективного биологического контроля бактерий. Настоящее изобретение относится к средству для получения достаточного количества измененных фагов (в ответ на CRISPR) для значимого снижения способности нативных бактерий развивать эффективную устойчивость, опосредованную CRISPR. Настоящее изобретение также относится к способам применения/введения, структурированным так, чтобы скорость развития нативных бактерий была значительно снижена.
Экспериментальный раздел
Следующие примеры представлены для демонстрации и дополнительной иллюстрации определенных предпочтительных вариантов осуществления и аспектов настоящего изобретения, и их не следует рассматривать как ограничивающие его объем.
В следующем экспериментальном разделе, следующие аббревиатуры обозначают: °С (градусы Цельсия); об/мин (обороты в минуту); Н2О (вода); HCl (хлористоводородная кислота); аа (аминокислота); bp (пар оснований); kb (тысяч пар оснований); кД (килодальтон); мкг и ug (микрограмм); мг (миллиграмм); нг (нанограмм); мкл и ul (миллилитры); мм (миллиметры); нм (нанометры); мкм и um (микрометры); М (моли); мМ (миллимоли); ЕД (единицы); ВВ (вольты); ММ (молекулярная масса); сек (секунды); мин (минута/минуты); ч (час/часы); MOI (множественность инфекции); ЕОР (эффективность бляшкообразования); MgCl2 (хлорид магния); NaCl (хлорид натрия); OD420 (оптическая плотность при 420 нм); PAGE (электрофорез на полиакриламидном геле); EtOH (этанол); PBS (солевой раствор с фосфатным буфером [159 мМ NaCl, 10 мМ буфера фосфата натрия, рН 7,2]); SDS (додецилсульфат натрия); Tris (трис(гидроксиметил)аминометан); масс/об (масса к объему); об/об (объем к объему); Amicon (Amicon, Inc., Beverly, MA); АТСС (Американская коллекция типовых культур, Manassas, VA); Amersham (Amersham Biosciences, Inc., Piscataway, NJ); NEB (New England Biolabs, Beverly, MA); Becton Dickinson (Becton Dickinson Labware, Lincoln Park, NJ); BioRad (BioRad, Richmond, CA); Clontech (CLONTECH Laboratories, Palo Alto, CA); Difco (Difco Laboratories, Detroit, MI); GIBCO BRL or Gibco BRL (Life Technologies, Inc., Gaithersburg, MD); Sigma (Sigma Chemical Co., St. Louis, MO); и Sorvall (Sorvall Instruments, a subsidiary of DuPont Co., Biotechnology Systems, Wilmington, DE).
Пока нет других указаний, в настоящем изобретении используются обычные методики химии, молекулярной биологии, микробиологии, рекомбинантной ДНК и иммунологии, которые может осуществить средний специалист в данной области. Такие методики хорошо известны специалистам в данной области.
Используемое в настоящем описании обозначение DGCC7710 также именуется “WT” (диким типом); DGCC7710RH1 также обозначается как “DGCC7710-RH1” и “RH1”; DGCC7710RH2 также обозначается как “DGCC7710-RH2” и “RH-2”; DGCC7778cas1 также обозначается как “DGCC7778cas1KO”, “CASIKO” и “cas1 KO”; DGCC7778cas4 также обозначается как “DGCC7778cas4KO”; DGCC7778 также обозначается как “WTФ858+S1S2”; DGCC7778RT также обозначается как “WTФ858+S1S2::pR”; DGCC7778RT' также обозначается как “WTФ858+S1S2ΔCRISPR1”; DGCC7710-R2 также обозначается как “WTФ2972+S4”; и DGCC7710-R2S1S2 также обозначается как “WTФ2972+S4::pS1S2.”
Пример 1
Манипуляция со специфичными для фага спейсерами
В этом примере, описаны эксперименты, проведенные для манипулирования специфичными для фага спейсерами. В некоторых экспериментах, специфичный для фага спейсер вставляли в существующий, функциональный CRISPR для обеспечения устойчивости к соответствующему фагу. Использованный бактериальный штамм представлял собой Streptococcus thermophilus ST0089, а фаг представлял собой фаг 2972. S. thermophilus ST0089 является промышленно важным штаммом, используемым при изготовлении йогурта. Он в целом поддается манипулированию и восприимчив к хорошо известному вирулентному фагу 2972.
Локусы CRISPR были определены в штамме ST0089. Это было преимущественно определено секвенированием всего генома ST0089. Альтернативно, локусы CRISPR идентифицируются посредством PCR с использованием наборов затравок с последовательностями, идентичными ранее идентифицированным элементам CRISPR S. thermophilus.
После идентификации, определяли последовательности локусов CRISPR, а также проксимальных областей, содержащих релевантные гены cas.
Для дальнейшего манипулирования выбирали, по меньшей мере, один определенный локус CRISPR-cas. Функциональность этого локуса определяли посредством анализа in silico спейсерных областей и их гомологий с последовательностями ДНК фага (т.е. отсутствия и/или присутствия спейсерных последовательностей и корреляции с инфективностью фага штаммом ST0089). При отсутствии этой корреляции, функциональность предполагали на основании присутствия всех документированных элементов (т.е. повторов, спейсеров, лидерных последовательностей и генов cas, которые предположительно кодируют белки полной длины).
Подходящую спейсерную последовательность (последовательности) выбирали из генома фага 2972. Критерии, использованные для выбора спейсера, в целом основывались на длине спейсеров в пределах выбранного локуса CRISPR и идентичности (предпочтительно, приблизительно 100%) последовательности фага. Действительно, любая подходящая последовательность фага находит применение в различных вариантах осуществления настоящего изобретения.
В некоторых вариантах осуществления, химически синтезировали единицу CRISPR, состоящую из спейсерной последовательности фага 2972, фланкированной двумя повторяющимися элементами (идентичными выбранному локусу CRISPR). По определению, эта синтетическая «единица CRISPR» имеет длину приблизительно 100 bp и является слишком короткой для обеспечения интеграции в локус CRISPR.
Поэтому, дополнительную фланкирующую ДНК конструировали наряду с единицей CRISPR. Для облегчения интеграции, получали минимум 500 bp идентичной локусу CRISPR-мишени, гомологичной ДНК, которая фланкирует синтетическую единицу CRISPR.
В дополнительных вариантах осуществления, имеются множественные подходы. В одном варианте осуществления, конкурирует с добавлением нового спейсера в существующий CRISPR. В некоторых альтернативных вариантах осуществления, весь локус CRISPR замещается синтетической единицей CRISPR.
Полученный в результате интегрант CRISPR верифицировали, используя секвенирование ДНК локуса CRISPR перед биологическим тестированием. Кроме того, тестировали типы чувствительности к фагам интегранта CRISPR против фага 2972 и сравнивали с материнским штаммом.
Сконструированный интегрант CRISPR успешно продемонстрировал прямую корреляцию между присутствием специфического спейсера в пределах соответствующего контекста CRISPR-cas.
В дополнительных экспериментах, выполняли вставку спейсера, гомологичного ДНК фага, в клетку-реципиент. В этих экспериментах, новый спейсер CRISPR был сконструирован из ДНК фага (со 100% идентичностью ДНК фага) в пределах анти-рецепторного гена и вставлен в клетку в локус CRISPR. Анти-рецепторный ген являлся мишенью, потому было обнаружено, что спейсеры CRISPR из других штаммов проявляют подобие анти-рецепторным генам фага. 4 штамма, несущих спейсеры, проявляющие идентичность анти-рецепторным генам фага, были устойчивыми к определенному фагу. Мутант подвергали воздействию фага и обнаружили, что он является устойчивым.
В дополнительных экспериментах, спейсер вставляли в первоначального хозяина, но не в локус CRISPR. Полученный мутант сохранял чувствительность к фагу. Таким образом, эти эксперименты показали, что спейсер должен находиться в определенной среде в пределах контекста CRISPR и генов cas.
В других экспериментах, определенный спейсер CRISPR был подвергнут делеции из естественно встречающегося локуса CRISPR. Эта делеция устраняла иммунитет против данного фага, и хозяин становился чувствительным (т.е. терял устойчивость) к фагу, к которому был гомологичен спейсер. Результаты этих экспериментов представлены на фиг. 10.
В еще одних дополнительных экспериментах, цельные комбинации повтора CRISPR-cas были вставлены в клетку-реципиент для обеспечения иммунитета против входящей нуклеиновой кислоты.
В дополнительных экспериментах, плазмиду, содержащую спейсер CRISPR, получали, используя способы, изложенные в настоящем описании. Попытки переноса этой плазмиды в клетки, которые содержат тот же спейсер, были безуспешными. Однако плазмиды, которые не содержат спейсер, могут быть трансформированы в клетки. На фиг. 11 и 12 иллюстрируются эти результаты.
В других экспериментах, производили обмен комбинаций CRISPR-cas, присутствующих в двух различных штаммах. Было показано, что этот обмен спейсеров модифицировал их фенотипы (чувствительность/устойчивость к фагу). Как указано в настоящем описании, когда S1S2 вводится в штамм с S4, включается чувствительность к фагу (см. фиг. 10).
В дополнительных экспериментах, получали различные комбинации cas-повтора CRISPR. Для функциональности требуются не только гены или белки cas, но и специфические пары cas-повтора CRISPR. Когда гены или белки cas предоставлены из другого локуса CRISPR, то штамм остается чувствительным к фагу.
В еще одних дополнительных экспериментах, подвергали делеции один или несколько генов cas (из функциональной единицы CRISPR-cas). Гены cas необходимы для обеспечения иммунитета. Мутанты cas еще чувствительны к фагу, несмотря на присутствие спейсера, идентичного ДНК фага. В этих экспериментах, делеции подвергались cas5 (ранее известный как cas1) cas7 (ранее известный как cas4). Было показано, что для устойчивости требуется cas5. Кроме того, было показано, что для интеграции новых спейсеров требуется cas7.
В дополнительных экспериментах, гены cas представлены хозяину в транс форме. Когда осуществляется нокаут гена cas, иммунитет восстанавливается.
Пример 2
Интеграция спейсера (спейсеров) CRISPR
В этих экспериментах, было показано, что интеграция спейсера CRISPR в локус CRISPR обеспечивает сопротивляемость против бактериофага, к которому спейсер CRISPR проявляет идентичность. В этих экспериментах, получали штамм DGCC7710RH1 S. thermophilus.
Штамм DGCC7710-RH1 Streptococcus thermophilus (депонированный во французской «Национальной коллекции культур микроорганизмов» под номером CNCM 1-2423), имеет, по меньшей мере, 3 локуса CRISPR: CRISPR1, CRISPR2 и CRISPR3. В штаммах CNRZ1066 и LMG18311 S. thermophilus, для которых известна полная последовательность генома (Bolotin et al., Microbiol., 151:2551-2561 [2005]), CRISPR1 локализуется в том же хромосомном локусе: между str0660 (или stu0660) и str0661 (или stu0661).
У штамма DGCC7710, CRISPR1 также локализуется в том же хромосомном локусе, между высоко подобными генами. CRISPR1 штамма DGCC7710 содержит 33 повтора (включая концевой повтор) и таким образом, 32 спейсера.
Все эти спейсеры отличаются друг от друга. Большинство из этих спейсеров являются новыми (т.е. ранее не описанными в пределах локусов CRISPR), но четыре спейсера около трейлера CRISPR1, идентичны уже известным спейсерам CRISPR1:
- 28-й спейсер DGCC7710 на 100% идентичен 31-му повтору спейсера CRISPR1 штамма CNRZ1575 (номер доступа в генном банке DQ072991);
- 30-й спейсер DGCC7710 на 100% идентичен 27-му спейсеру CRISPR1 штамма CNRZ703 (номер доступа в генном банке DQ072990);
- 31-й спейсер DGCC7710 на 100% идентичен 28-му спейсеру CRISPR1 штамма CNRZ703 (номер доступа в генном банке DQ072990);
- 32-й спейсер DGCC7710 на 100% идентичен 30-му спейсеру CRISPR1 штамма CNRZ703 (номер доступа в генном банке DQ072990).
Во время разработки настоящего изобретения, штамм DGCC7710RH1 S. thermophilus был выделен в виде естественного устойчивого к фагу мутанта с использованием DGCC7710 в качестве материнского штамма, а фага D858 в качестве вирулентного фага. Использовали D858, бактериофаг, относящийся к семейству вирусов Siphoviridae.
CRISPR1 штамма DGCC7710-RH1 содержит 34 повтора (включая концевой повтор), и, таким образом, 33 спейсера. При сравнении с последовательностью CRISPR1 штамма DGCC7710 S. thermophilus, последовательность CRISPR1 штамма DGCC7710-RH1 S. thermophilus имеет один дополнительный новый спейсер (и один дополнительный повтор, который фланкирует новый спейсер) на одном конце локуса CRISPR (т.е. близко к лидеру, на конце 5' локуса CRISPR). Все другие спейсеры локуса CRISPR1 оставались неизменными.
Последовательность CRISPR1 (5'-3') штамма DGCC7710-RH1 представлена ниже:

В указанной выше последовательности, лидер имеет последовательность:
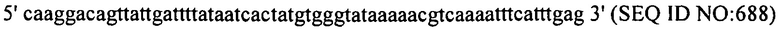
Интегрированная последовательность
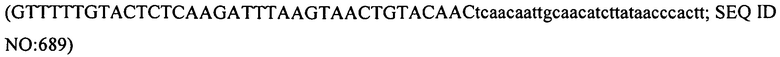 ,
,
содержащая повтор CRISPR, показана в выделении с прописными буквами, а спейсер CRISPR (т.е. маркировочная последовательность) – в выделении со строчными буквами; обе показаны выше серым цветом. Последовательности концевого повтора и трейлера повтора CRISPR показаны ниже:
Концевой повтор: 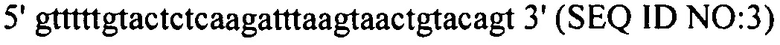
Последовательность трейлера: 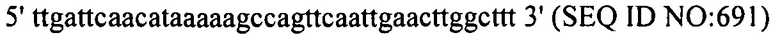
Последовательность нового спейсера
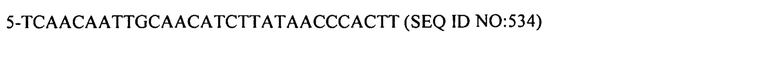
существует в пределах генома фага D858.
Последовательность спейсера обнаруживается между положениями 31921 и 31950 bp (т.е. на «плюс-цепи») генома D858 (и имеет 100% идентичность с геномной последовательностью по 30 нуклеотидам):

Новый спейсер, который интегрирован в локус CRISPR1 штамма DGCC7710-RH1 S. thermophilus, придает этому штамму устойчивость к фагу D858, как показано на фиг. 1 и таблице 2-1.






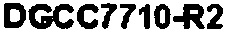
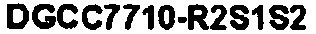
2 Чувствительность штамма к фагу: S = чувствительный, R = устойчивый, по данным анализов пятен и бляшек
3 Гомология между новым спейсером мутанта и последовательностью ДНК фага, использованного для генерирования мутанта.
Фаги сохраняли способность адсорбироваться к мутантам.
Кроме того, во время разработки настоящего изобретения, штамм DGCC7710-RH2 S. thermophilus был выделен в виде естественного устойчивого к фагу мутанта с использованием штамма DGCC7710 S. thermophilus в качестве материнского штамма, и фага D858 в качестве вирулентного фага.
CRISPR1 штамма DGCC7710-RH2 S. thermophilus содержит 34 повтора (включая концевой повтор), и, таким образом, 33 спейсера. При сравнении с последовательностью CRISPR1 штамма DGCC7710 S. thermophilus, последовательность CRISPR1 штамма DGCC7710-RH2 S. thermophilus имеет один дополнительный новый спейсер (и один дополнительный повтор, который фланкирует новый спейсер) на одном конце локуса CRISPR (т.е. близко к лидеру, на конце 5' локуса CRISPR). Все другие спейсеры локуса CRISPR1 оставались неизменными.
Последовательность CRISPR1 (5'-3') штамма DGCC7710-RH2 представлена ниже:


В указанной выше последовательности, лидерная последовательность представляет собой:
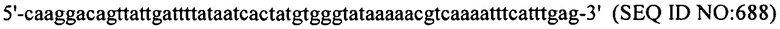
Интегрированная последовательность, содержащая повтор CRISPR, показана в выделении с прописными буквами, а спейсер CRISPR (т.е. маркировочная последовательность) – в выделении со строчными буквами; обе показаны выше серым цветом 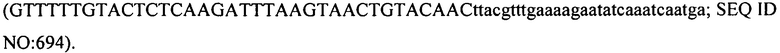 . Последовательности концевого повтора и трейлера повтора CRISPR показаны ниже:
. Последовательности концевого повтора и трейлера повтора CRISPR показаны ниже:
Концевой повтор: 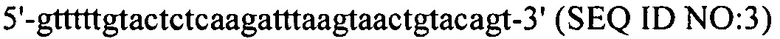
Последовательность трейлера: 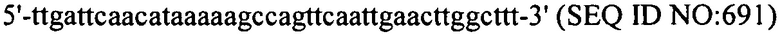 .
.
Было показано, что последовательность нового спейсера существует в пределах генома фага D858.
Последовательность спейсера (SEQ ID NO:535) обнаруживается между положениями 17215 и 17244 bp (т.е. на «плюс цепи») генома D858 (и имеет 100% идентичность D858 с геномной последовательностью по 30 нуклеотидам):

Новый спейсер, который интегрирован в локус CRISPR1 штамма DGCC7710-RH2 S. thermophilus, придает штамму DGCC7710-RH2 S. thermophilus устойчивость к фагу D858, как показано на фиг. 2 и таблице 2-1 (см. также фиг. 10).
Пример 3
Интеграция и нокаут конструкта
В этом примере описаны способы, использованные для интеграции и нокаута конструкта.
Штаммы, использованные в этих экспериментах представляли собой:
материнский штамм DGCC7710 S. thermophilus, чувствительный к фагам 858 и 2972
мутант CRISPR DGCC7778 S. thermophilus, устойчивый к 858
DGCC7778cas1KO S. thermophilus
DGCC7778cas4KO S. thermophilus
DGCC7778RT S. thermophilus
DGCC7778RT' S. thermophilus
мутант CRISPR DGCC7710R2 S. thermophilus, устойчивый к 2972
DGCC7710R2S1S2 S. thermophilus
E. coli EC1,000, обеспеченный pOR128 (см. Russell and Klaenhammer, Appl. Environ. Microbiol., 67:4361-4364 [2001]).
Escherichia coli pCR2.1 TOPO, обеспеченный pTOPO (см. Каталог Invitrogen #K4500-01).
Следующие плазмиды использовались в этих экспериментах:
pTOPO, плазмида, использованная для субклонирования различных конструктов
pTOPOcas1KO содержит интегральный фрагмент cas1
pTOPOcas4KO содержит интегральный фрагмент cas4
pTOPOS1S2 содержит конструкт спейсера S1S2
pTOPO RT содержит конструкт концевого повтора RT
pOR128 представляет собой плазмиду, использованную для интеграции различных конструктов в хромосому штаммов S. thermophilus.
pORcas1KO содержит интегральный фрагмент cas1
pORcas4KO содержит интегральный фрагмент cas4
pOR1S1S2 содержит конструкт спейсера S1S2
purist содержит конструкт концевого повтора RT
Следующие затравки использовались в этих экспериментах:
Cas1
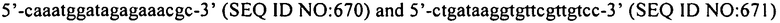
Cas4
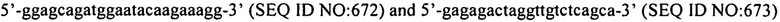
S1S2 и RT
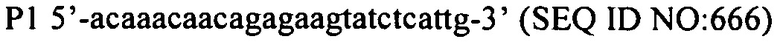

Штаммы и фаги были получены из коллекции культур Danisco Culture Collection, или из указанных в ссылках источников (Russell и Klaenhammer, Appl. Environ. Microbiol., 67:43691-4364 [2001]; и Levesque et al., Appl. Environ. Microbiol., 71:4057-4068 [2005]).
Получение, очистку и тесты фагов проводили с использованием способов, известных в данной области (см., например, Duplessis et al., Virol., 340:192-208 [2005]; и Levesque et al., Appl. Environ. Microbiol., 71:4057-4068 [2005]).
Штаммы S. thermophilus выращивали при 37°С или 42°С в среде M17 (Difco) с добавкой 0,5% лактозы или сахарозы. Для инфекции фагом, к среде перед инфекцией фагом добавляли 10 мм CaCl2, как известно в данной области (см., например, приведенные выше ссылки на работы Duplessis et al. и Levesque et al.).
Ферменты, использованные для проведения рестрикционных перевариваний и PCR, закупали у компании Invitrogen и использовали в соответствии с инструкциями производителя. PCR проводили на термоциклическом устройстве Eppendorf Mastercycler Gradient, как известно в данной области (см., например, Barrangou et al., Appl. Environ. Microbiol., 68:2877-2884 [2002]).
Инактивацию генов и сайт-специфическую вставку в хромосому S. thermophilus плазмиды посредством гомологичной рекомбинации проводили путем субклонирования в систему pCR2.1 TOPO Invitrogen, последующего клонирования в системе pOR1 с использованием E. Coli в качестве хозяина, и конструкты в конечном счете очищали и трансформировали в S. thermophilus, как описано ранее (Russell and Klaenhammer, см. выше).
Интеграция конструкта RT
Используя генную инженерию конструкта RT, как показано на фиг. 4, конструкт вставляли непосредственно после Cas4, как показано на фиг. 5. Материнский DGCC7778 устойчив к фагу 858. Материнский штамм имеет два спейсера (S1 и D2), которые идентичны ДНК фага 858. Полученный штамм (RT) утратил устойчивость к фагу 858. Этот результат указывает на то, что гены cas должны находиться в непосредственной близости к спейсеру (спейсерам) для придания устойчивости. Как показано на фиг. 3, материнский DGCC7778 был подвергнут генной инженерии с тем, чтобы разрушить ген cas1, что приводит к потере устойчивости, означая, что cas1 необходим для придания устойчивости. Как показано на фиг. 3, материнский DGCC7778 был подвергнут генной инженерии с тем, чтобы разрушить ген cas4. Кроме того, конструкт S1S2 был интегрирован в материнский DGCC7710, как показано на фиг. 6-8.
Пример 4
Выделение устойчивых к фагам мутантов и подтверждение последовательностей CRISPR
В этом примере описаны способы, использованные при выделении устойчивых к фагам мутантов и подтверждении последовательностей CRISPR. Устойчивые к фагам мутанты S. thermophilus были получены стимуляцией штамма-хозяина DGCC7710 дикого типа (также именуемого “RD534”) фагом 2972 и/или фагом 858 (Levesque et al., Appl. Environ. Microbiol., 71:4057 [2005]). Штамм-хозяин выращивали при 42°С в 10 мл бульона М17 с добавкой 0,5% лактозы (LM17). Когда оптическая плотность (600 нм) достигала 0,3, добавляли фаги и 10 мМ хлорида кальция при конечной концентрации соответственно 107 бляшкообразующих единиц/мл и 50 мМ. Содержащую фаги культуру инкубировали при 42°С в течение 24 часов и контролировали на наличие лизиса. Затем, 100 мкл лизата инокулировали в 10 мл свежей LM17. Оставшийся лизат центрифугировали, и осадок после центрифугирования инокулировали в другую пробирку, содержащую 10 мл свежей LM17. Эти две культуры инкубировали при 42°С в течение 16 часов. Наконец, эти культуры разбавляли и высевали на LM17. Изолированные колонии тестировали на чувствительность к фагам, как известной в данной области (см. Moineau et al., Can. J. Microbiol., 38:875 [1992]). Локусы CRISPR устойчивых изолятов верифицировали секвенированием продуктов PCR, и используя релевантную информацию о геноме фага, известную в данной области (см. Levesque et al., Appl. Environ. Microbiol., 71:4057 [2005]).
Пример 5
Генная инженерия спейсеров CRISPR
В этом примере описаны способы, использованные для генной инженерии спейсеров CRISPR. Ферменты, использованные для проведения рестрикционных перевариваний и PCR, закупали у компании Invitrogen и использовали в соответствии с инструкциями производителя. PCR проводили на термоциклическом устройстве Eppendorf Mastercycler Gradient, как известно в данной области.
Инактивацию генов и сайт-специфическую вставку плазмид посредством гомологичной рекомбинации в хромосоме S. thermophilus проводили субклонированием в систему pCR2.1 TOPO (Invitrogen) путем последующего клонирования в системе pOR1 с использованием E. Coli в качестве хозяина, и конструкты в конечном счете очищали и трансформировали в S. thermophilus, как описано ранее (Russell and Klaenhammer, Appl. Environ. Microbiol.., 67:4261 [2001]).
ДНК из мутантного WTФ858+S1S2 использовали в качестве матрицы для амплификации двух различных фрагментов PCR с использованием Р1 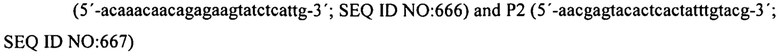
в одной реакции и Р3 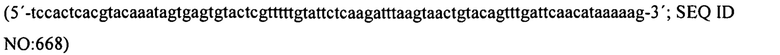
и Р4 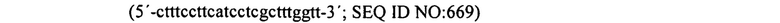
в другой реакции. Оба продукта PCR в последующем использовали в качестве матриц в другой реакции PCR с использованием праймеров Р1 и Р4 для генерирования конструкта S1S2, в соответствии с фиг. 11.
Конструкт S1S2 субклонировали в систему pCR2.1 TOPO. Этот конструкт переваривали NotI и HindIII и в последующем клонировали в pOR1 в сайты NotI и HindIII, получая конструкт pS1S2. Интеграция pS1S2 в локус CRISPR1 WTФ2972+S4 происходила посредством гомологичной рекомбинации на конце посредством гомологичной рекомбинации на конце 3' cas7 для генерирования WTФ2972+S4:: pS1S2.
Конструкт pR генерировали, используя конструкт pS1S2 в качестве матрицы. В частности, конструкт S1S2, субклонированный в pCR2.1 TOPO, переваривали, используя BsrGI, который вырезает в пределах повтора CRISPR. Затем перевар повторно лигировали, и плазмиду, содержащую один повтор и не содержащую спейсер, использовали в последующем для клонирования в pORI, используя NotI и HindIII, генерируя pR. Интеграция pR в хромосому WTФ858+S1S2 на конце 3' cas7 посредством гомологичной рекомбинации генерировала WTФ858+S1S2::pR, мутант, где локус CRISPR1 замещен, и, вместо него, вставлен необычный повтор.
Мутант WTФ858+S1S2::pR в последующем выращивали в отсутствие эритромицина, и чувствительные к антибиотикам варианты анализировали для выявления мутанта, который имел полную делецию локуса CRISPR1. Делецию получали в результате гомологичной рекомбинации, происходящей на конце 3' ORF (в отличие от явления рекомбинации, происходящего на конце 3' cas7, которая бы привела к восстановлению штамма WTФ858+S1S2), генерируя WTФ858+S1S2ΔCRISPR1 (см. фиг. 12), мутант, где локус CRISPR1 не подвергнут делеции (см. также фиг. 10).
Пример 6
Инактивация генов cas
Для инактивации cas5, внутренний отрезок cas5 длиной 801 bp амплифицировали PCR, используя праймеры 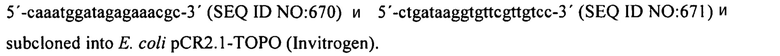
Этот конструкт переваривали EcoRV и HindIII и в последующем клонировали в pORI в сайтах EcoRV и HindIII. Интеграция этого конструкта в ген cas5 WTФ858+S1S2 происходила посредством гомологичной рекомбинации внутреннего отрезка гена, приводя к получению WTФ858+S1S2::pcas5-.
Аналогичным образом, внутренний отрезок cas7 длиной 572 bp амплифицировали PCR, используя праймеры 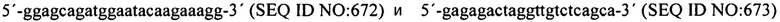
и субклонировали в pCR2.1-TOPO E. Coli (Invitrogen). Этот конструкт переваривали EcoRV и HindIII и в последующем клонировали в pORI в сайтах EcoRV и HindIII. Интеграция этого конструкта в ген cas7 WTФ858+S1S2 происходила посредством гомологичной рекомбинации внутреннего отрезка гена, приводя к получению WTФ858+S1S2::pcas7- (см. фиг. 10-12).
Пример 7
Естественные способы вставки дополнительной последовательности в локус CRISPR
В этом примере описаны способы естественного вызова вставки дополнительной последовательности в пределы локуса CRISPR бактериального штамма. Используемый в настоящем описании термин «дополнительная последовательность» определяется как спейсерная последовательность, ассоциированная с последовательностью повтора CRISPR. Конкретнее, «дополнительная последовательность» частично происходит из донорского фага, способного инфицировать бактерию-мишень, и, частично, из дупликации последовательности повтора CRISPR. Введение ДНК донорского фага в бактериальную клетку происходит в результате инфекции клетки донорским фагом. Выбор клеток, которые содержат дополнительную последовательность, осуществляется посредством селекционного давления донорским фагом с тем, чтобы выбранные модифицированные клетки были устойчивыми к фагу.
В этих экспериментах материнский штамм был подвергнут воздействию донорского фага, и был выбран устойчивый к фагу материнский штамм (т.е. вариантный штамм). Вариантный штамм анализировали (например, PCR и/или секвенированием ДНК) для подтверждения присутствия дополнительной последовательности в пределах локуса CRISPR. Определяли нуклеотидную последовательность дополнительной последовательности. Обычно, дополнительная последовательность представляет собой фрагмент размером приблизительно 30 нуклеотидов из донорского фага, ассоциированный (слитый) с последовательностью повтора CRISPR, и он придает устойчивость к донорскому фагу.
В некоторых экспериментах, материнский штамм предварительно культивировали в течение ночи в среде на молочной основе при 42°С. Затем среду на молочной основе инокулировали при 0,1% (об./об.) предварительной культурой материнского штамма и суспензией донорского фага при MOI 10. После 6 ч инкубации при 42°С, разведения культуры высевали на питательную среду для получения изолированных колоний. Затем изоляты тестировали на их устойчивость к донорскому фагу (любой подходящий способ, известный в данной области, находит применение в этих экспериментах). Затем вариантные штаммы анализировали на присутствие дополнительной последовательности в пределах одного из их локусов CRISPR.
Локусы CRISPR амплифицировали PCR, и нуклеотидные последовательности полученных продуктов PCR определяли секвенированием ДНК, используя стандартные способы PCR и секвенирования, известные в данной области. Затем эти последовательности сравнивали с последовательностями материнского штамма с использованием способов, известных в данной области.
В некоторых экспериментах, DGCC7710 использовали в качестве материнского штамма, и D2792 использовали в качестве донорского фага. Штамм DGCC7710 материнской S. thermophilus подвергали воздействию донорского фага D2792, как описано выше. Получали вариантный штамм, называемый WTphi2972+S6 (см. таблицу 7-1). Таблица 7-1 также включает результаты для вариантных штаммов, описанных в других примерах. В таблице 7-1, ЕОР выражена относительно фага D2792. Пока нет иного определения, расположение дополнительной последовательности в геноме фага дано относительно фага D2792.
Таблица 7.1
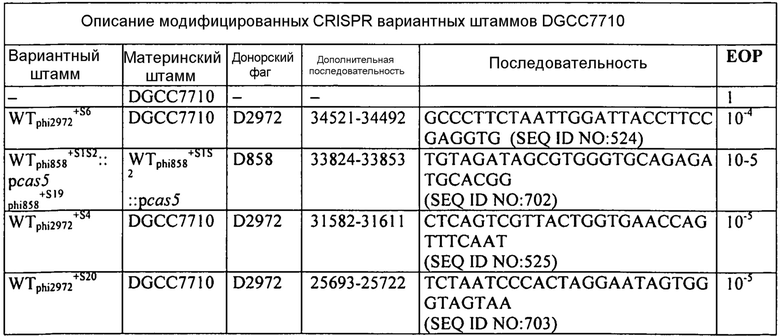

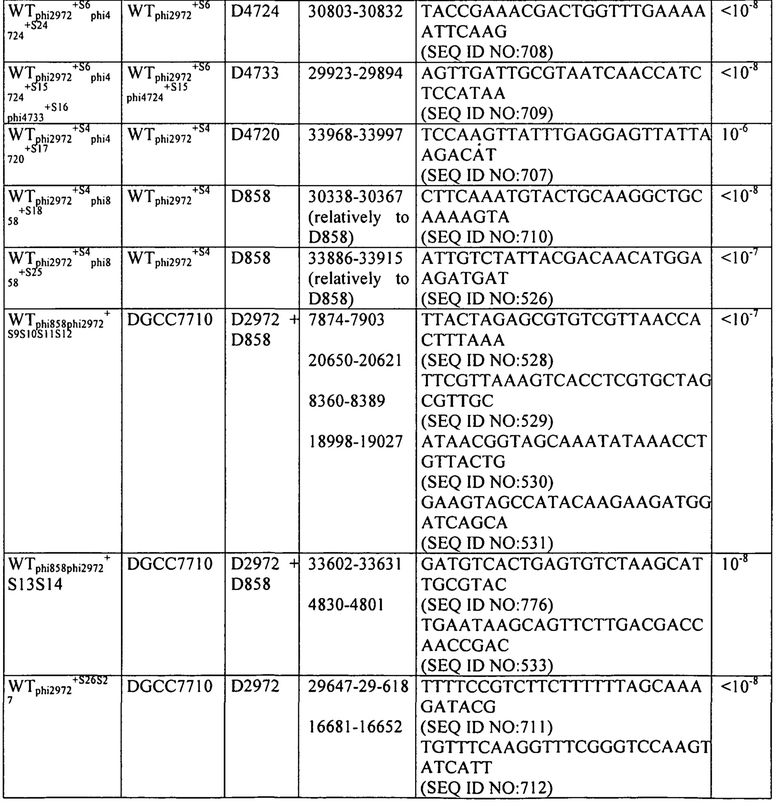


Этот вариант проявил устойчивость к D2792, так как эффективность бляшкообрзования (ЕОР) D2792 на WTphi2972+S6 была снижена на 4 логарифма. ДНК экстрагировали из WTphi2972+S6, и его локус CRISPR1 анализировали PCR, как известно в данной области (см., например, Bolotin et al. [2005] выше), с использованием комбинации одного прямого праймера (или YC70 и/или 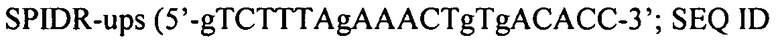
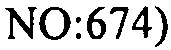
и одного обратного праймера (или YC31 и/или


Определяли последовательность продукта PCR и сравнивали с последовательностью локуса CRISPR1 DGCC7710. По сравнению с DGCC7710, было обнаружено, что WTphi2972+S6 отличается добавлением одной спейсерной последовательности размером 30 bp на конце 5' области его CRISPR1 и дупликацией последовательности повтора, как показано на фиг. 14. Сравнение дополнительной последовательности с последовательностью генома D2972 показывает, что новая спейсерная последовательность на 100% идентична последовательности генома D2972 от нуклеотида 34521 до нуклеотида 34492.
В некоторых дополнительных экспериментах, WTphi858+S1S2::pcas5 использовали в качестве материнского штамма, а D858 использовали в качестве донорского фага. Полученный вариантный штамм, называемый WTphi858+S1S2::pcas5phi858+S19 (см. таблицу 7-1) был устойчив к D858 при ЕОР, сниженной на 5 логарифмов. ДНК экстрагировали из WTphi858+S1S2::pcas5phi858+S19, и его локус CRISPR3 анализировали PCR, используя один прямой праймер 
и один обратный праймер 
Определяли последовательность продукта PCR и сравнивали с последовательностью локуса CRISPR3 WTphi858+S1S2::pcas5. По сравнению с WTphi858+S1S2::pcas5, WTphi858+S1S2::pcas5phi858+S19 отличается добавлением одной спейсерной последовательности размером 30 bp на конце 5' области его CRISPR3 и дупликацией последовательности повтора. Сравнение дополнительной последовательности с последовательностью генома D858 показало, что новая спейсерная последовательность на 100% идентична последовательности генома D858 от нуклеотида 33824 до нуклеотида 33853.
В других дополнительных экспериментах, DGCC7809 использовали в качестве материнского штамма, а D3743 использовали в качестве донорского фага. Полученный вариантный штамм, называемый DGCC7809phiD3743+S28 (см. таблицу 7-2) был устойчив к D3743 при ЕОР, сниженной на 8 логарифмов. ДНК экстрагировали из DGCC7809phiD3743+S28, и его локус CRISPR3 анализировали PCR, используя один прямой праймер
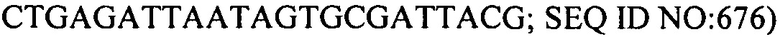
и один обратный праймер 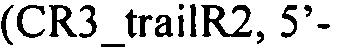
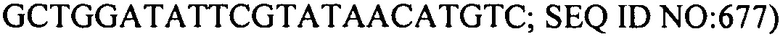 .
.
Определяли последовательность продукта PCR и сравнивали с последовательностью локуса CRISPR3 ST0189. По сравнению с DGCC7809, DGCC7809phiD3743+S28 отличается добавлением одной спейсерной последовательности размером 29 bp на конце 5' области его CRISPR3 и дупликацией последовательности повтора. Последовательность фага D3743 неизвестна, однако сравнение дополнительной последовательности с последовательностью генома других стрептококковых фагов показывает, что новая спейсерная последовательность на 100% идентична последовательности генома фага DТ1 от нуклеотида 6967 до нуклеотида 6996.
В других дополнительных экспериментах, DGCC3198 использовали в качестве материнского штамма, а D4241 использовали в качестве донорского фага. Полученный вариантный штамм, называемый DGCC3198phi4241+S29 (см. таблицу 7-2) был устойчив к D4241 при ЕОР, сниженной на 8 логарифмов. ДНК экстрагировали из DGCC3198phi4241+S1, и его локус CRISPR1 анализировали PCR, используя один прямой праймер (или YC70 и/или 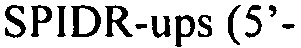
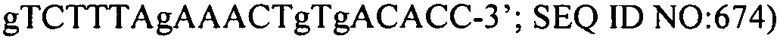
и один обратный праймер (или YC31 и/или 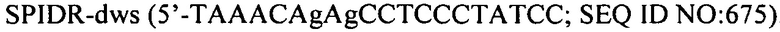
Определяли последовательность продукта PCR и сравнивали с последовательностью локуса CRISPR1 DGCC3198. По сравнению с DGCC3198, DGCC3198phi4241+S29 отличается добавлением одной спейсерной последовательности размером 30 bp на конце 5' области его CRISPR1 и дупликацией последовательности повтора. Последовательность фага D4241 неизвестна, однако сравнение дополнительной последовательности с последовательностью генома других стрептококковых фагов показывает, что новая спейсерная последовательность на 100% идентична последовательности генома фага DТ1 от нуклеотида 3484 до нуклеотида 3455.
Ниже в таблице 7-2 представлено описание модифицированных CRISPR вариантных штаммов из DGCC7809 и из DGCC3198. В этой таблице, ЕОР выражена относительно донорских фагов. Расположение дополнительной последовательности в фаге дано относительно фага DT1.
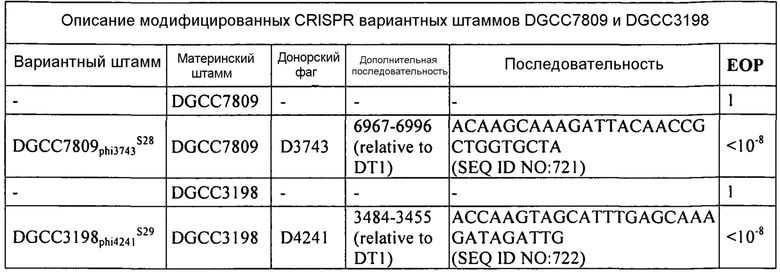
Пример 8
Выбор набора модифицированных CRISPR вариантных штаммов из одного и того же материнского штамма
В этом примере, описаны способы, использованные для выбора набора вариантных штаммов из одного и того же материнского штамма, отличающихся их дополнительной последовательностью, происходящей из одного и того же фага. Поскольку различные части донорского фага находят применение в качестве источников для дополнительных последовательностей, из данного донорского фага могут быть генерированы множественные различные вариантные штаммы. Кроме того, каждый вариантный штамм имеет другую дополнительную последовательность. Следовательно, в дополнение к вариантному штамму, описанному в примере 7, из одного и того же материнского штамма были разработаны множественные штаммы. В некоторых экспериментах, эти дополнительные штаммы были генерированы воздействием на штамм-реципиент одного и того же донорского фага. Предусматривалось, что различные полученные вариантные штаммы представляли различный спектр чувствительности к фагам.
В независимых культурах, материнский штамм был подвергнут воздействию одного и того же донорского фага. Для каждой культуры, был выделен и проанализирован один устойчивый к фагу вариант, как описано в примере 7. Дополнительные последовательности в каждом из вариантных штаммов сравнивали друг с другом. Спектр чувствительности вариантных штаммов к донорскому фагу и другим фагам определяли, используя классические микробиологические способы, известные в данной области. Затем сравнивали спектры чувствительности различных штаммов. Выбранными вариантными штаммами были те, которые проявляли дополнительную последовательность (последовательности) и различные спектры чувствительности к фагам.
В некоторых экспериментах, проводили отбор различных вариантных штаммов DGCC7710, используя D2972 в качестве одного донорского фага. Материнский штамм DGCC7710 подвергали воздействию донорского фага D2972 в четырех независимых культурах, как описано в примере 7. Из каждой культуры был выделен вариантный штамм и соответственно назван WTphi2972+S4, WTphi2972+S20, WTphi2972+S21 и WTphi2972+S22 (см. таблицу 7-1).
Эти вариантные штаммы проявили устойчивость к D2972, поскольку эффективность бляшкообразования (ЕОР) D2972 на указанных четырех устойчивых к фагам вариантах была снижена на 3-5 логарифмов. ДНК экстрагировали из WTphi2972+S4, WTphi2972+S20, WTphi2972+S21 и WTphi2972+S22 и анализировали PCR, используя способы, известные в данной области (см., например, Bolotin et al. [2005] выше), с использованием комбинации одного прямого праймера (или YC70 и/или 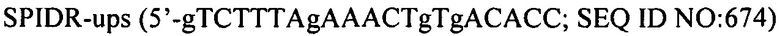
и одного обратного праймера (или YC31 и/или 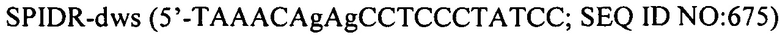
Определяли последовательность продуктов PCR и сравнивали с последовательностью локуса CRISPR1 DGCC7710. По сравнению с DGCC7710, WTphi2972+S4, WTphi2972+S20, WTphi2972+S21 и WTphi2972+S22 отличаются добавлением спейсерной последовательности размером 30 bp на конце 5' области его CRISPR1 и дупликацией последовательности повтора, как показано на фиг. 17. Сравнение этих новых спейсерных последовательностей с последовательностью генома D2972 показывает, что новые спейсерные последовательности на 100% идентичны последовательности генома D2972 соответственно от нуклеотида 31582 до нуклеотида 31611, от нуклеотида 25693 до нуклеотида 25722, от нуклеотида 27560 до нуклеотида 27589 и от нуклеотида 24624 до нуклеотида 24653. Было обнаружено, что все четыре дополнительных спейсера отличались друг от друга и отличались от спейсеров, описанных в примере 7.
Пример 9
Естественные способы, использованные для вставки второй дополнительной последовательности в локус CRISPR
В этом примере, описаны естественные способы, использованные для вызова вставки второй дополнительной последовательности в локус CRISPR. После вставки второй дополнительной последовательности из данного донорского фага в бактериальный локус CRISPR, вариантный штамм становится устойчивым или, по меньшей мере, менее чувствительным к этому фагу. Поэтому, способ, описанный в примере 7, больше не эффективен для вставки дополнительных последовательностей в локус CRISPR вариантного штамма. Например, этот способ нельзя применять к вариантному штамму WTphi2972+S6 (в качестве материнского штамма) с использованием D2972 в качестве донорского фага, потому что WTphi2972+S6 имеет значимо сниженную чувствительность к D2972 (см. пример 7).
В некоторых экспериментах, эта проблема была преодолена использованием мутированного донорского фага, полученного из D2972, который включает, по меньшей мере, одну специфическую модификацию в пределах его генома (т.е. «мутированного фага»). Этот мутированный фаг был выбран воздействием на донорский фаг вариантного штамма с тем, чтобы модификация (т.е. мутация) материнского фага придавала ему вирулентность для вариантного штамма.
В некоторых экспериментах, мутированный фаг имел мутацию в его геноме в пределах области, содержащей последовательность дополнительного спейсера, которая является частью дополнительной последовательности в вариантном штамме. Вариантный штамм был чувствителен к этому мутированному фагу. Вариантный штамм был подвергнут воздействию мутированного фага, и был выбран новый устойчивый к фагу вариант (вариант 2-го поколения) вариантного штамма. Вариант 2-го поколения анализировали, используя подходящие способы, известные в данной области (например, PCR и секвенирование), для подтверждения присутствия дополнительной последовательности в пределах локуса CRISPR. Определяли нуклеотидную последовательность дополнительной последовательности. В некоторых экспериментах, было обнаружено, что дополнительная последовательность содержит фрагмент размером приблизительно 30 нуклеотидов из мутированного фага, который обеспечивает устойчивость к мутированному фагу.
В некоторых экспериментах, вариантный штамм предварительно культивировали в течение ночи в соответствующей среде на основе молока при 42°С. Затем подходящую среду на основе молока инокулировали предварительной культурой вариантного штамма в концентрации примерно 106 бляшкообразующих единиц/мл и суспензией донорского фага при MOI более чем 100. Культуру инкубировали в течение ночи при 42°С и затем центрифугировали. Надосадочную жидкость собирали и фильтровали с использованием фильтра с размером пор 0,45 мкм. Разведения отфильтрованного супернатанта использовали для инокуляции питательных агаровых сред, засеянных вариантным штаммом, для получения изолированных бляшек фага с использованием любого подходящего способа, известного в данной области. Изолированные бляшки культивировали на вариантном штамме в жидких питательных средах с использованием любого подходящего способа, известного в данной области. Суспензию мутированного фага получали фильтрацией культуры через фильтр с размером пор 0,45 мкм. Затем мутированный фаг использовали, как описано выше (см. пример 7), для вызова вставки второй дополнительной спейсерной последовательности в локусе CRISPR вариантного штамма.
В некоторых экспериментах, WTphi2972+S6 (см. пример 7 и таблицу 7-1) использовали в качестве материнского штамма и D4724 использовали в качестве донорского фага. Вариантный штамм WTphi2972+S6 культивировали в присутствии высокой концентрации фага D2972. Мутированный фаг D4724 выделяли бляшкообразованием из супернатанта из этой культуры на штамме WTphi2972+S6 с использованием описанных выше способов. Верифицировали вирулентность мутированного фага D4724 на WTphi2972+S6. Вариантный штамм WTphi2972+S6 подвергали воздействию фага D4724 в культуре, как описано в примере 7. Получали устойчивый к фагу вариантный штамм, названный WTphi2972+S6phi4724+S15 (см. таблицу 7-1).
По сравнению с WTphi2972+S6, этот вариантный штамм проявил повышенную устойчивость к D2972, поскольку эффективность бляшкообразования (ЕОР) D2972 на WTphi2972+S6phi4724+S15 была снижена более чем на 8 логарифмов (вместо 4 логарифмов); кроме того, его устойчивость была также увеличена, по сравнению с WTphi2972+S6, поскольку он проявляет некоторую устойчивость к D4724 (см. таблицу 9-1). ДНК экстрагировали из WTphi2972+S6phi4724+S15, и его локус CRISPR1 анализировали PCR, как описано выше, с использованием тех же комбинаций праймеров, как описано выше. Определяли последовательность продукта PCR и сравнивали с последовательностью локуса CRISPR1 WTphi2972+S6. По сравнению с WTphi2972+S6, WTphi2972+S6phi4724+S15 отличается добавлением спейсерной последовательности размером 30 bp на конце 5' области его CRISPR1 и дупликацией последовательности повтора, как показано на фиг. 17. Сравнение этой дополнительной спейсерной последовательности с последовательностью генома D2972 показало, что вторая дополнительная спейсерная последовательность на 100% идентична последовательности генома D2972 от нуклеотида 1113 до нуклеотида 1142.
Из независимых культур, с использованием идентичных экспериментальных условий были выделены и анализировали вариантные штаммы WTphi2972+S6phi4724+S17 и WTphi2972+S6phi4724+S24 (см. таблица 7-1). По сравнению с WTphi2972+S6, эти вариантные штаммы проявляли повышенную устойчивость к D2972, поскольку эффективность бляшкообразования (ЕОР) D2972 на WTphi2972+S6phi4724+S17 и WTphi2972+S6phi4724+S24 была снижена более чем на 8 логарифмов для обоих вариантных штаммов; и их устойчивость также была увеличена, по сравнению с WTphi2972+S6, поскольку они проявляли некоторую устойчивость к D4724 (см. таблицу 9-1). Кроме того, эти вариантные штаммы проявляют дополнительные спейсерные последовательности в CRISPR1, которые на 100% идентичны последовательности генома D2972 соответственно от нуклеотида 33968 до нуклеотида 33997 и от нуклеотида 30803 до нуклеотида 30832.
В дополнительных экспериментах, WTphi2972+S6phi4724+S15 использовали в качестве материнского штамма и D4733 использовали в качестве донорского фага. Описанные выше способы использовали для генерирования мутированного фага D4733 из фага D4724. Затем фаг D4733 использовали для получения устойчивого к фагу вариантного штамма из WTphi2972+S6phi4724+S15. Полученный вариантный штамм был назван WTphi2972+S6phi4724+S15phi4733+S16 (см. таблицу 7-1). Этот вариантный штамм содержит одну дополнительную последовательность, включающую спейсерную последовательность, которая на 100% идентична последовательности их генома D2972, от нуклеотида 29923 до нуклеотида 29894. Он проявлял повышенную устойчивость к D2972, поскольку эффективность бляшкообразования (ЕОР) D2972 на WTphi2972+S6phi4724+S15phi4733+S16 была снижена более чем на 8 логарифмов, и его устойчивость распространялась на фаг D4733 (см. таблицу 9-1). В таблице 9-1 представлено описание устойчивости к фагу модифицированных CRISPR вариантных штаммов из DGCC7710. В этой таблице, «nd» указывает на то, что результаты не определялись.
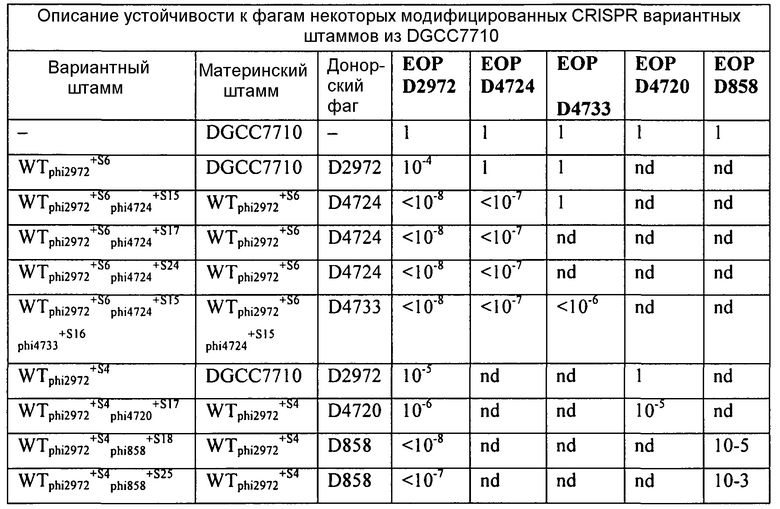
В еще одних экспериментах, WTphi2972+S4 использовали в качестве материнского штамма и D4720 использовали в качестве донорского фага. Используя те же способы, которые описаны выше, генерировали мутированный фаг D4720 из фага D2972. Фаг D4720 использовали для получения устойчивого к фагу варианта из WTphi2972+S4. Полученный вариантный штамм был назван WTphi2972+S4phi4720+S17 (см. таблицу 7-1). Этот вариантный штамм содержит одну дополнительную последовательность, включающую спейсерную последовательность, которая на 100% идентична последовательности из генома D2972, от нуклеотида 33968 до 33997. Он проявлял повышенную устойчивость к D2972, поскольку эффективность бляшкообразования (ЕОР) D2972 на WTphi2972+S4phi4724+S17 была снижена на 6 логарифмов (по сравнению с 5 логарифмами), и его устойчивость была распространена на фаг D4720 (см. таблицу 9-1).
Пример 10
Альтернативные естественные способы для вставки второй дополнительной последовательности (последовательностей) в локусы CRISPR
В этом примере, описаны альтернативные естественные способы, которые могут использоваться для вставки второй дополнительной последовательности в локус CRISPR. Известно, что данный материнский штамм может быть чувствителен более чем к одному семейству фагов. Разнообразие чувствительности преимущественно использовалось для вставки дополнительных последовательностей в локус CRISPR вариантного штамма, как описано в настоящей заявке. В этих экспериментах, второй донорский фаг был выбран тестированием вирулентности выбора фагов на материнском штамме и на вариантном штамме (штаммах). Представляющие интерес вторые донорские фаги представляли собой те, которые были вирулентными для обоих штаммов. Предусматривалось, что эти фаги, вероятно, представляли бы семейство фагов, отличное от семейств, представленных первоначальным донорским фагом. После его выбора, второй донорский фаг использовали для инфицирования вариантного штамма. Как описано в способах выше, устойчивый к фагу вариантный штамм второго поколения был выделен и тестирован на наличие дополнительной последовательности в локусе CRISPR.
В этих экспериментах, коллекцию фагов (или содержащих фаги образцов) тестировали против материнского штамма с использованием классических микробиологических способов, известных в данной области. Затем фаги (или образцы), которые являются вирулентными для материнского штамма, тестировали против вариантного штамма с использованием тех же способов. Один фаг (или образец), который был вирулентным для вариантного штамма, выбирали в качестве второго донорского фага. В случае образцов, содержащих фаги, один вирулентный фаг очищали до однородности на вариантном штамме с использованием классических микробиологических способов, известных в данной области. В некоторых экспериментах, определяли последовательность второго донорского фага. В некоторых экспериментах, затем использовали второй донорский фаг, как описано выше (см. пример 7) для вызова вставки второй дополнительной последовательности в локус CRISPR вариантного штамма.
В некоторых экспериментах, WTphi2972+S4 (см. пример 8 и таблицу 7-1) использовали в качестве материнского штамма и D858 использовали в качестве донорского фага. После тестирования различных фагов, было обнаружено, что штамм DGCC7710 чувствителен и к фагу D2972, и к фагу D858. Кроме того, было обнаружено, что D858 является вирулентным против вариантного штамма WTphi2972+S4. Полученный вариантный штамм был назван WTphi2972+S4. Поэтому, в некоторых экспериментах фаг D858 был выбран в качестве второго фага.
Вариантный штамм WTphi2972+S4 был подвергнут воздействию второго донорского фага D858, как описано в примере 7. Получали устойчивый к фагу вариантный штамм, названный WTphi2972+S4phi858+S18 (см. таблицу 7-1), который устойчив к D858 (см. таблицу 9-1). Этот штамм проявляет повышенную устойчивость к D2972, поскольку эффективность бляшкообразования D2792 на WTphi2972+S4phi858+S18 была снижена более чем на 8 логарифмов (по сравнению с 5 логарифмами для WTphi2972+S4, см. таблицу 9-1). ДНК экстрагировали из WTphi2972+S4phi858+S18, и его локус CRISPR1 анализировали PCR, используя такие же способы и праймеры, как описано ранее. Определяли последовательность продукта PCR и сравнивали с последовательностью локуса CRISPR WTphi2972+S4. По сравнению с WTphi2972+S4, WTphi2972+S4phi858+S18 отличается добавлением спейсерной последовательности размером 30 bp на конце 5' области его CRISPR1 и дупликацией последовательности повтора, как показано на фиг. 17. Сравнение этой дополнительной последовательности с последовательностью генома D858 показало, что вторая дополнительная спейсерная последовательность на 100% идентична последовательности генома D858 от нуклеотида 30338 до нуклеотида 30367.
В независимой экспериментальной работе также получали другой вариантный штамм, названный WTphi2972+S4phi4720+S25 (см. таблицу 7-1), используя этот способ. Этот вариантный штамм содержит одну дополнительную последовательность, включающую спейсерную последовательность, которая на 100% идентична последовательности из генома D858 от нуклеотида 33886 до 33915. Он проявляет повышенную устойчивость к D2972, поскольку эффективность бляшкообразования D2792 на WTphi2972+S4phi4724+S25 была снижена более чем на 7 логарифмов (см. таблицу 9-1).
Пример 11
Генез модифицированного CRISPR вариантного штамма, устойчивого к множественным фагам, множественными вставками дополнительных последовательностей в локусы CRISPR
В этом примере описана разработка штамма, устойчивого к множественным фагам, посредством повторного добавления последовательностей фага в локусы CRISPR, поскольку добавление 2 последовательностей фага в локусы CRISPR недостаточно для придания данному штамму устойчивости ко всем фагам. Например, было обнаружено, что штамм WTphi2972+S4phi858+S18 (описанные в примере 10) чувствителен к множеству других фагов. В процессе разработки устойчивого ко многим фагам штамма, материнский штамм был подвергнут воздействию второго фага для выбора вариантного штамма второго поколения, который устойчив к обоим фагам. Затем, последний вариантный штамм был подвергнут повторному воздействию фагов, к которым он был еще чувствителен, до тех пор, пока не был получен конечный вариантный штамм, который был устойчив ко всем доступным фагам.
С использованием способов, известных в данной области, был идентифицирован набор из 10 эталонных фагов, которые являются репрезентативными разнообразия фагов, которые способны развиться на штамме DGCC7710, а именно, фагов D858, D1126, D2766, D2972, D3288, D3821, D4083, D4752, D4753 и N1495. Как описано в примере 7, DGCC7710 был подвергнут воздействию фага D2972 для генерирования вариантного штамма DGCC9705. Было обнаружено, что DGCC9705 устойчив к фагу D2766 и D4752. В дополнение к фагу D2972, но был еще чувствителен к другим фагам, как показано в таблице 11-1. DGCC9705 описан в таблице 11-1 и на фиг. 17. DGCC9705 представляет 1 дополнительную последовательность в CRISPR1 и 1 дополнительную последовательность в CRISPR3. Анализ последовательности локуса CRISPR1 и локуса CRISPR3 проводили в соответствии со способами, описанными в примере 7. Определяли последовательности продуктов PCR и сравнивали с последовательностями локусов CRISPR1 и 3 DGCC7710. DGCC9705 представляет 1 дополнительный спейсер в его локусе CRISPR1 и один дополнительный спейсер в его локусе CRISPR3. Спейсерные последовательности идентичным последовательностям из фага D2972. С использованием тех же способов, DGCC9705 был затем подвергнут воздействию фага D3821, и затем был выделен вариантный штамм DGCC9726. В дополнение к устойчивости к D2972, DGCC9726 обладает устойчивостью к фагам D858, D3821, D4083 и N1495 (см. таблицу 11-1). DGCC9726 имеет 1 дополнительную последовательность в его CRISPR1, по сравнению с DGCC9705 (см. таблицу 7-1 и фиг. 17). Дополнительная спейсерная последовательность идентична последовательности из D2972. Посредством воздействия фага D3288 на штамм DGCC9726, был выделен DGCC9733. Штамм DGCC9733 дополнительно устойчив к фагу D3288 и D1126 (см. таблицу 11-1). DGCC9733 имеет 1 дополнительную последовательность в его CRISPR1, по сравнению с DGCC9726 (см. таблицу 7-1 и фиг. 17). Эта спейсерная последовательность имеет такую же идентичность (идентичность 25 из 30 пар оснований) как последовательность стрептококкового фага 7201. Наконец, путем последнего повторного воздействия фага D4753, был выделен DGCC9836, который устойчив ко всем фагам (см. таблицу 11-1). DGCC9836 имеет 2 дополнительных спейсерных последовательности в его локусе CRISPR1 и 2 дополнительных спейсерных последовательности в его локусе CRISPR3 (см. таблицу 7-1 и фиг. 17). Одна спейсерная последовательность идентична последовательности в фаге D2972, а 3 другие спейсерные последовательности идентичны последовательностям в фаге D858.
В таблице 11-1 представлены данные относительно чувствительности к фагам модифицированного CRISPR вариантного штамма DGCC9836 и промежуточных модифицированных CRISPR вариантных штаммов. В этой таблице, “S” указывает чувствительность и “R” указывает устойчивость.
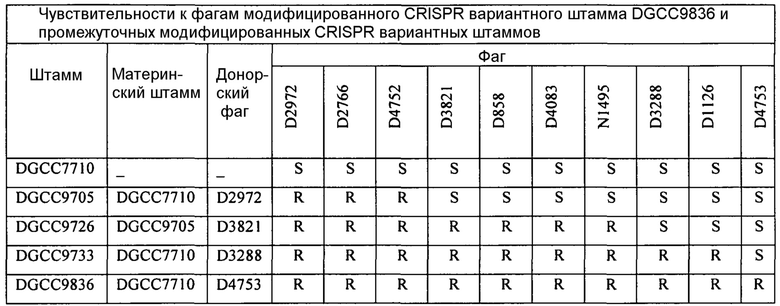
Пример 12
Естественный способ вставки множественных дополнительных последовательностей в локус CRISPR
В этом примере описаны способы вставки множественных дополнительных последовательностей в локусы CRISPR. В этих способах, вместо повторного использования различных фагов, материнский штамм подвергали воздействию смеси, содержащей множественные фаги. Коллекцию фагов тестировали против множественных штаммов с использованием классических микробиологических способов для определения спектра их хозяев. Отбирали фаги, которые были вирулентными для материнского штамма, но которые имели другой спектр хозяев. Отобранные фаги смешивали и использовали в представленных выше способах (см. пример 7) для вызова вставки дополнительных последовательностей в локусы CRISPR вариантного штамма.
В некоторых экспериментах, DGCC7710 использовали в качестве материнского штамма, а D858 и D2972 использовали в качестве донорских фагов. После тестирования различных фагов, было обнаружено, что штамм DGCC7710 чувствителен и к фагу D2972, и к фагу D858. Однако D2972 и D858 проявляли различные спектры хозяев при тестировании на штамме DGCC7778, свидетельствуя о том, что эти два фага были различными.
Материнский штамм DGCC7710 подвергали воздействию смеси фага D858 и D2972, как описано в примере 7. Получали устойчивый к фагу вариантный штамм, названный WTphi858phi2972+S9S10S11S12 (см. таблицу 7-1). Он проявляет устойчивость к D858, поскольку эффективность бляшкообразования D858 на WTphi858phi2972+S9S10S11S12 была снижена более чем на 7 логарифмов, а также устойчивость к D2972, поскольку эффективность бляшкообразования D2972 на WTphi858phi2972+S9S10S11S12 была снижена более чем на 7 логарифмов. ДНК экстрагировали из WTphi858phi2972+S9S10S11S12, и его локусы CRISPR1 анализировали PCR, используя такие же способы и праймеры, как описано выше. Определяли последовательность продуктов PCR и сравнивали с последовательностью локуса CRISPR1 и локуса CRISPR3 DGCC7710. По сравнению с DGCC7710, WTphi858phi2972+S9S10S11S12 отличается добавлением 4 спейсерных последовательностей размером 30 bp на конце 5' области его CRISPR1 и дупликацией последовательностей повторов, как показано на фиг. 17. Сравнение дополнительных спейсерных последовательностей с последовательностью генома D2972 показало, что дополнительные спейсерные последовательности на 100% идентичны последовательности D2972 от нуклеотида 7874 до нуклеотида 7903, от нуклеотида 20650 до нуклеотида 20621, от нуклеотида 8360 до нуклеотида 8389 и от нуклеотида 18998 до нуклеотида 19027.
В других экспериментах штамм WTphi858phi2972+S13S14 (см. таблицу 7-1) также получали в соответствии с этими способами. Он проявляет устойчивость к D858, поскольку эффективность бляшкообразования D858 на WTphi858phi2972+S13S14 была снижена на 7 логарифмов, и устойчивость к D2972, поскольку эффективность бляшкообразования D2972 на WTphi858phi2972+S13S14 была снижена на 8 логарифмов. Сравнение дополнительных спейсерных последовательностей с последовательностью генома D2972 показало, что дополнительные спейсерные последовательности на 100% идентичны последовательности D2972 от нуклеотида 33602 до нуклеотида 33361 и от нуклеотида 4830 до нуклеотида 4801.
Пример 13
Естественный способ вставки множественных дополнительных последовательностей в локус CRISPR
В этом примере описаны способы борьбы с фагами при ферментации путем использования вариантного штамма, а не материнского (т.е. дикого типа, реципиентного) штамма. Таким образом, этот пример представляет еще одно описание преимуществ, обеспечиваемых вариантными штаммами.
В некоторых экспериментах, выполняли сравнение штамма DGCC7710 со штаммом WTphi2972+S20 и штаммом WTphi2972+S26S27 при ферментации молока в присутствии фага D2972. DGCC7710 представляет собой промышленный штамм, используемый при ферментации молока. Штамм WTphi2972+S20 описан в таблице 7-1 и в примере 8 и проявляет в своем локусе CRISPR1 дополнительный спейсер, по сравнению со штаммом DGCC7710. Штамм WTphi2972+S20 проявляет повышенную устойчивость к D2972, по сравнению с DGCC7710. WTphi2972+S26S27 представляет собой другой вариант, проявляющий некоторую устойчивость к D2972 (описанную в таблице 7-1) и имеет 2 дополнительных спейсера в своем локусе CRISPR1.
Серийные ферментации выполняли с каждым штаммом. Сначала, среду 10% молочного порошка (масс/об) засевали 1% (об/об) предварительной культуры испытанного штамма и 104 бляшкообразующих единиц/мл фага D2972. Культуру инкубировали при 42°С в течение 6 ч. После первой ферментации, проводили вторую ферментацию. Использовали точно такие же условия ферментации, за исключением того, что добавляли 0,1% объема ферментата предыдущей ферментации (перед добавлением, ферментат фильтровали с использованием фильтра с размером пор 0,45 мкм). Затем выполняли последующие ферментации при таких же экспериментальных условиях как условия, использованные для второй ферментации. Все ферментации регистрировали импедансометрией. В конце каждой ферментации, тестировали сворачиваемость молока и выполняли титрование фагов с использованием способов, известных в данной области.
В случае ферментации молока в отсутствие фага, изменение импеданса при DGCC7710 было выше 2500 мкс в пределах 6 часов. В присутствии D2972 (при DGCC7710 очень чувствительном к фагам), фаги D2972 достигали высокого уровня популяции в течение первой культуры, и ферментация не смогла вызвать сворачивание молока. Изменение импеданса через 6 часов было всегда ниже 500 мкс. Напротив, ферментация молока WTphi2972+S20 в присутствии D2972 обеспечила возможность сворачивания молока, по меньшей мере, до 3-ей субкультуры и отмеченного медленного развития уровня фага. Изменение импеданса увеличилось более чем до 2500 мкс до 3-ей субкультуры. Это демонстрирует, что вариантный штамм WTphi2972+S20 более целесообразен, чем материнский штамм DGCC7710, для подкисления молока в присутствии фагов. Кроме того, ферментация молока WTphi2972+S26S27 обеспечила возможность сворачивания молока, по меньшей мере, до последней субкультуры без развития фага. Кроме того, увеличение импеданса увеличилось более чем до 2500 мкс также до последней субкультуры. Это демонстрирует, что вариантный штамм WTphi2972+S26S27 более целесообразен, чем материнский штамм DGCC7710, и даже более целесообразен, чем WTphi2972+S20, для подкисления молока в присутствии фагов. Эксперименты выполняли в двух повторностях, и их результаты представлены в таблице 13-1.
 первое испытание второе испытание
первое испытание второе испытание первое испытание
первое испытание
второе испытание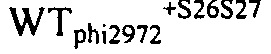 первое испытание второе испытание
первое испытание второе испытание
Во второй серии экспериментов, было воспроизведено сравнение штамма DGCC7710 со штаммом WTphi2972+S20 и штаммом WTphi2972+S26S27 при ферментации молока в присутствии фага D2972. Кроме того, был исследован штамм DGCC9836. Штамм DGCC9836 представляет собой даже развитый вариантный штамм DGCC7710, что является результатом множественной стимуляции фагами. Этот штамм проявляет 5 дополнительных спейсеров в его локусе CRISPR1 и 3 дополнительных спейсера в его локусе CRISPR3 (см. пример 11 и фиг. 17). DGCC9836 устойчив ко всем тестированным фагам.
Эксперименты проводили как описано выше. Результаты представлены в таблице 13-2. Что касается первой серии экспериментов, то ферментация молока WTphi2972+S20 в присутствии D2972 обеспечила возможность сворачивания молока до 5-ой субкультуры, и измеряли медленное развитие уровня фагов. Изменение импеданса увеличилось более чем до 2500 мкс в течение 5 первых субкультур, демонстрируя, что фаги не воздействуют на подкисление молока. При 6-ой субкультуре, уровень фагов значимо увеличился, и было нарушено брожение молока. Для двух других вариантных штаммов, воздействия на брожение молока не было в течение всех 6 субкультур, фаги никогда не развивались, и регистрируемое изменение импеданса было всегда выше 2500 мкс.
Эти эксперименты демонстрируют, что штаммы, содержащие, по меньшей мере, одну дополнительную спейсерную последовательность в локусе CRISPR1, обеспечивают возможность ферментации молока даже в присутствии фагов. Ферментация молока обеспечивалась в еще большей степени, когда штаммы имеют более одной дополнительной спейсерной последовательности в локусах CRISPR.
Пример 14
Борьба с фагами при ферментации с использованием комбинации модифицированных CRISPR вариантных штаммов
В этом примере описаны способы борьбы с фагами при ферментации путем использования комбинации вариантных штаммов, вместо использования одного штамма. Таким образом, этот пример иллюстрирует одновременное использование одного или нескольких вариантных штаммов (т.е. комбинации вариантных штаммов). Действительно, смеси штаммов проявляют такие же функциональные возможности, но при таких видах применения могут использоваться другие типы чувствительности к фагам. Например, 2 или 3 или даже более описанных в настоящей заявке вариантных штаммов могут использоваться при таких видах применения. Применение комбинации вариантных штаммов с различными добавленными спейсерными последовательностями в их локусах CRISPR обеспечивает возможность ферментации для облегчения сопротивляемости любым появляющимся мутантным фагам.
В некоторых экспериментах, сравнения проводили между одним штаммом WTphi2972+S21 и комбинацией 3 штаммов (а именно, WTphi2972+S20, WTphi2972+S21 и WTphi2972+S22), использованными при ферментации молока в присутствии фага D2972. Штаммы WTphi2972+S20, WTphi2972+S21 и WTphi2972+S22 описаны в таблице 7-1 и примере 8. Они представляют собой независимые вариантные штаммы DGCC7710. Каждый вариантный штамм проявляет в своем локусе CRISPR1 отчетливую дополнительную спейсерную последовательность (которая произошла из фага D2972), по сравнению со штаммом DGCC7710.
Серийные ферментации выполняли или одним штаммом WTphi2972+S21, или комбинацией из трех штаммов. Сначала, среду 10% молочного порошка (масс/об) засевали 1% (об/об) предварительной культуры одного штамма или комбинации штаммов и с 104 бляшкообразующих единиц/мл фага D2972. Культуру инкубировали при 42°С в течение 6 ч. После первой ферментации, проводили вторую ферментацию. Использовали точно такие же условия ферментации, за исключением того, что добавляли 0,1% объема ферментата предыдущей ферментации (перед добавлением, ферментат фильтровали с использованием фильтра с размером пор 0,45 мкм). Затем выполняли последующие ферментации, используя такие же экспериментальные условия как условия, использованные для второй ферментации. Все ферментации регистрировали импедиметрией. В конце каждой ферментации, тестировали сворачиваемость молока и выполняли титрование фагов с использованием способов, известных в данной области. Эксперименты проводили в двух повторностях, и их результаты представлены в таблице 14-1.
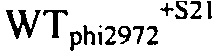 второе испытание
второе испытание
первое испытание второе испытание
Ферментация молока, проведенная WTphi2972+S21 в присутствии фагов, оказалась несостоятельной при третьей субкультуре в обоих испытаниях. Это было показано отсутствием сворачивания молока и высоким снижением изменения импеданса после 6 часов ферментации. Напротив, несмотря на некоторое развитие фага D2972, ферментации успешно проводились до пятой субкультуры, когда использовалась смесь трех штаммов. Сворачивание молока регистрировали во всех культурах, и изменение импеданса в пределах 6 часов инкубации никогда не было ниже 3000 мкс.
Эти эксперименты демонстрируют, что применение комбинации вариантных штаммов, имеющих, по меньшей мере, одну дополнительную спейсерную последовательность в ее локусе CRISPR1, обеспечивает возможность ферментации молока в присутствии фагов, по сравнению с использованием одного вариантного штамма.
Пример 15
Борьба с фагами при ферментации с использованием ротации модифицированных CRISPR вариантных штаммов
В дополнительных экспериментах, вариантные штаммы имели такие же функциональные возможности, но другие типы чувствительности к фагам. Таким образом, в этом примере эксперименты, проведенные при повторном/последовательном применении нескольких различных штаммов (т.е. модифицированных CRISPR вариантных штаммов) последовательно в схеме ротации.
В некоторых экспериментах, выполняли сравнение одного штамма WTphi2972+S21 и штаммов WTphi2972+S20 WTphi2972+S21 и WTphi2972+S22, применяемых последовательно (при ротации) при ферментации молока в присутствии фага D2972. Первую ферментацию молока проводили штаммом WTphi2972+S20. Затем для второй ферментации использовали штамм WTphi2972+S22, и штамм WTphi2972+S21 использовали для третьей ферментации. Затем проводили четвертую ферментацию, снова используя штамм WTphi2972+S20, с последующей ферментацией штаммом WTphi2972+S22, затем штаммом WTphi2972+S21 и так далее. Штаммы WTphi2972+S20, WTphi2972+S21 и WTphi2972+S22 описаны выше в таблице 7-1. Они представляют собой независимые вариантные штаммы DGCC7710. Каждый вариантный штамм проявляет отчетливую дополнительную спейсерную последовательность в локусе CRISPR1, по сравнению со штаммом DGCC7710, который был получен из фага D2972.
Серийные ферментации выполняли с использованием тех же экспериментальных способов, которые описаны в примере 14. Эксперименты проводили в трех повторностях. И результаты представлены в таблице 15-1. Серийные ферментации, инокулированные одним WTphi2972+S20, были успешными до 3-й субкультуры, как показано величинами изменения импеданса выше 3000 мкс и сворачиванием молока. Следующие субкультуры не смогли свернуть молоко, и были зарегистрированы высокие величины фагов. Напротив, серийные ферментации, выполненные инокуляцией молока при ротации 3 различными вариантными штаммами (WTphi2972+S20, WTphi2972+S21 и WTphi2972+S22), были успешными до десятой субкультуры. В этих экспериментальных условиях, фаги не могли размножаться и оставались на низких уровнях. Эти результаты указывают на то, что использование ротации вариантных штаммов обеспечивает повышенную устойчивость к фагам во время ферментации, по сравнению с применением одного вариантного штамма.
Таблица 15-1
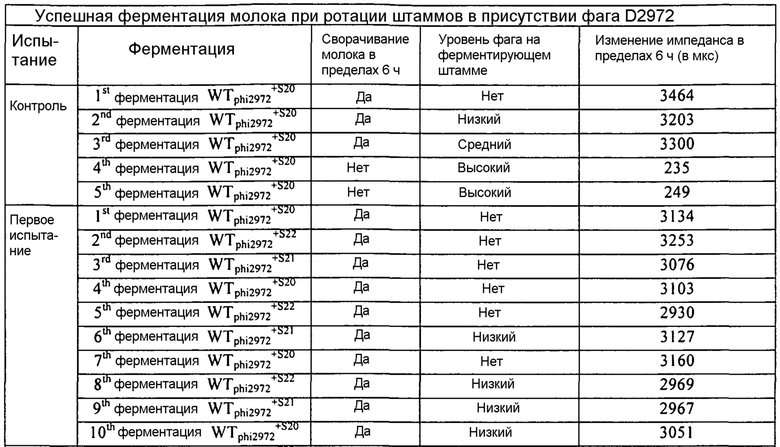
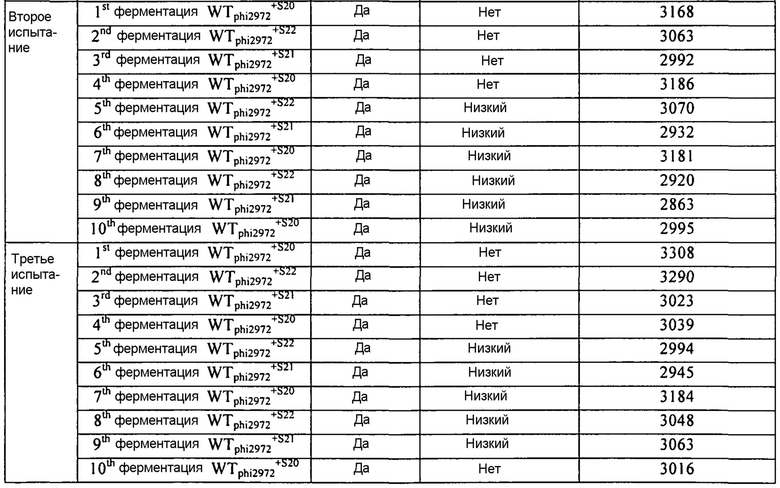
Пример 16
Уменьшение и регуляция популяции фагов с использованием модифицированных CRISPR вариантных штаммов
В этом примере описаны эксперименты, проведенные для определения способности модифицированного CRISPR штамма разрушать фаги, устойчивость к которым была у него получена. В частности, эксперименты были спланированы для определения того, уменьшится ли популяция фагов до не выявляемого уровня во время ферментации модифицированным CRISPR штаммом.
В некоторых экспериментах, DGCC9836 (описанный в примере 11 и на фиг. 17) использовали для выполнения ферментации молока в присутствии фага D2972, по сравнению с ферментацией, проведенной его материнским штаммом DGCC7710 в присутствии D2972. Среду 10% молочного порошка (масс/об) засевали приблизительно 106 бляшкообразующих единиц/мл предварительной культуры тестированного штамма и 107 бляшкообразующих единиц/мл фага D2972. Культуру инкубировали при 42°С в течение 24 ч. В различные точки времени, брали аликвотное количество и популяцию фага измеряли, используя двухслойную агаровую пластину, засеянную DGCC7710 с использованием стандартных способов, известных в данной области. Результаты представлены на фиг. 20. При ферментации молока DGCC7710, фаг D2972 размножался, достигая популяции более 108 бляшкообразующих единиц/мл. Напротив, во время ферментации DGCC9836, популяция фага D2972 постепенно уменьшалась до очень низкого уровня (120 бляшкообразующих единиц/мл) после 6 часов инкубации, и была почти не выявляемой после 24 часов инкубации. Этот последний результат свидетельствует о том, что фаги были разрушены во время процесса ферментации вариантным штаммом DGCC9836.
Способность вариантного штамма разрушать фаги, а также отсутствие чувствительности к фагам представляет дополнительное преимущество, по сравнению с традиционной программой ротации заквасочной культуры, к которой штаммы нечувствительны, но безвредны для фагов. Действительно, путем использования вариантных штаммов, искоренение покоящихся фагов произойдет посредством комбинации вымывания фагов (как при ротации с использованием традиционной заквасочной культуры) и разрушения фагов.
В других экспериментах, вариантные штаммы, проявляющие некоторую, но неполную устойчивость к фагу D2972, были ассоциированы при ферментации молока в присутствии D2972. Отобранные вариантные штаммы включали WTphi2972+S20 и WTphi2972+S21, как описано в примере 8 и в таблице 7-1. Эти штаммы проявляют снижение ЕОР для фага D2972 примерно 5 логарифмов. Ферментации молока выполняли, как описано выше (скорость бактериальной инокуляции 106 бляшкообразующих единиц/мл; скорость инокуляции фага 107 бляшкообразующих единиц/мл). Ферментации молока проводили или WTphi2972+S20, или WTphi2972+S21, или смесью двух штаммов. В различные точки времени, регистрировали популяцию фагов. С этой целью, брали аликвотное количество, и популяцию фага измеряли, используя двухслойную агаровую пластину, засеянную или WTphi2972+S20, или WTphi2972+S21, с использованием стандартных способов, известных в данной области. Результаты представлены на фиг. 21, которая указывает сумму фагов, выявленных на WTphi2972+S20 и на WTphi2972+S21, для каждой из ферментаций молока. При использовании для ферментации одного штамма (WTphi2972+S20 или WTphi2972+S21), количество выявленных фагов во время инокуляции составляло примерно 100 бляшкообразующих единиц/мл (вследствие 5 логарифмов снижения ЕОР). После культивирования, это количество фагов возросло до 106 или 107 (соответственно), соответствуя размножению фагов на 4-5 логарифмов. Фактор размножения фагов был гораздо ниже (2 логарифма) для ферментации молока, инокулированного 2 штаммами. Действительно, количество фагов возросло от 100 бляшкообразующих единиц/мл до максимума примерно 104 бляшкообразующих единиц/мл. Эти результаты заключительно показывают, что во время совместного культивирования 2 вариантных штаммов, скорость размножения фагов значимо снижается, по сравнению со скоростью размножения фагов в культуре, достигаемой одиночными вариантными штаммами.
Пример 17
Вставка спейсеров
В этом примере описаны способы и композиции, используемые для вставки двух спейсеров в DGCC7710 S. thermophilus. Штамм DGCC7710 S. thermophilus (депонированный во французской «Национальной коллекции культур микроорганизмов» под номером CNCM 1-2423), имеет, по меньшей мере, 3 локуса CRISPR: CRISPR1, CRISPR2 и CRISPR3. В CNRZ1066 и LMG18311, для которых известна полная последовательность генома (см. Bolotin et al., [2004] выше), CRISPR1 локализуется в том же хромосомном локусе: между str0660 (или stu0660) и str0661 (или stu0661). В штамме DGCC7710, CRISPR1 также локализуется в том же хромосомном локусе, между аналогичными генами. CRISPR1 штамма DGCC7710 содержит 33 повтора (включая концевой повтор), и, таким образом, 32 спейсера (см. фиг. 19). Все эти спейсеры отличаются друг от друга. Большинство из этих спейсеров ранее не были описаны как находящиеся в локусах CRISPR, но четыре спейсера, близких к трейлеру CRISPR1, идентичны известным спейсерам CRISPR1. Например, 28-й спейсер DGCC7710 на 100% идентичен 31-му спейсеру DGCC7710 штамма CNRZ1575 (номер доступа в генном банке DQ072991); 30-й спейсер DGCC7710 на 100% идентичен 27-му спейсеру CRISPR1 CNRZ703 (номер доступа в генном банке DQ072990); 31-й спейсер DGCC7710 на 100% идентичен 28-му спейсеру CRISPR1 штамма CNRZ703 (номер доступа в генном банке DQ072990); и 32-й спейсер DGCC7710 на 100% идентичен 30-му спейсеру CRISPR1 штамма CNRZ703 (номер доступа в генном банке DQ072990). Последовательность CRISPR1 (5'-3') штамма DGCC7710 показана ниже в SEQ ID NO:678:


D858, фаг, использованный в этих экспериментах, представляет собой бактериофаг, относящийся к семейству Siphoviridae вирусов. Хотя его геномная последовательность была полностью определена, ее, очевидно, еще предстоит опубликовать. Этот фаг является вирулентным для штамма DGCC7710 S. thermophilus. Штамм DGCC7778 S. thermophilus был выделен в виде естественного, устойчивого к фагу мутанта, с использованием DGCC7710 в качестве материнского штамма, и фага D858 в качестве вирулентного фага. CRISPR1 штамма DGCC7778 содержит 35 повторов (включая концевой повтор) и, таким образом, 34 спейсера. По сравнению с последовательностью CRISPR1 DGCC7710, последовательность CRISPR1 DGCC7778 имеет два дополнительных, прилегающих новых спейсера (и, конечно, два дополнительных повтора, которые фланкируют новые спейсеры) на одном конце локуса CRISPR (т.е. близко к лидеру). Все другие спейсеры локуса CRISPR1 являются неизменными. Последовательность CRISPR1 (5'-3') штамма DGCC7778 показана ниже в SEQ ID NO:678:

В случае DGCC7778, первый спейсер 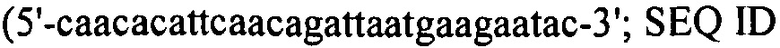
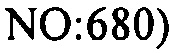 и второй спейсер
и второй спейсер  составляют специфическую для штамма метку, которая идентифицирует этот меченый штамм. Определено, что последовательность обоих новых спейсеров существует внутри генома фага D858. Последовательность второго нового спейсера обнаруживается между положениями 25471 и 25442 bp (т.е. на «минус-цепи») генома D858, с одним ошибочным спариванием (96,7% идентичных нуклеотидов по 30 нуклеотидам).
составляют специфическую для штамма метку, которая идентифицирует этот меченый штамм. Определено, что последовательность обоих новых спейсеров существует внутри генома фага D858. Последовательность второго нового спейсера обнаруживается между положениями 25471 и 25442 bp (т.е. на «минус-цепи») генома D858, с одним ошибочным спариванием (96,7% идентичных нуклеотидов по 30 нуклеотидам).
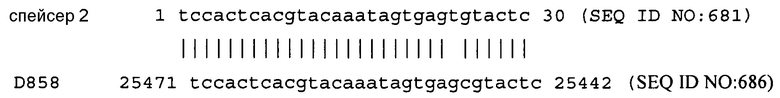
Последовательность первого спейсера обнаруживается между положениями 31481 и 31410 bp (т.е. на «плюс-цепь») генома D858 (100% идентичных нуклеотидов по 30 нуклеотидам).

Хотя нет намерений, что настоящее изобретение ограничивается каким-либо конкретным механизмом или теорией, предусматривается, что два новых спейсера, присутствующих в локусе CRISPR1 DGCC7778, требуются для придания штамму DGCC7778 новой устойчивости к фагу D858). Спейсер «2» (обнаруживаемый в DGCC7778) сначала вставлялся в локус CRISPR1 DGCC7710 (33 повтора и 32 спейсера) на одном конце этого локуса CRISPR, вместе с одним повтором. Эта вставка дала мутант, нечувствительный к бактериофагу (промежуточный штамм), меченый этим дополнительным новым спейсером (таким образом, несущий 34 повтора и 33 спейсера). Этот спейсер был получен из генома D858, но во время процесса вставки, вероятно, произошла ошибка репликации или ошибка обратной транскрипции, приводящая к точечной мутации. Ввиду неправильного спаривания (т.е. 1 ошибочного спаривания) между этим вновь приобретенным спейсером и меченой последовательностью фага, эффективность устойчивости этого промежуточного штамма к фагу D858 была низкой. Второе явление вставки спейсера происходило в этом промежуточном штамме (более устойчивом к фагу D858, чем материнский штамм DGCC7710, но не «полностью» устойчивый ввиду ошибочно спаривания), приводя к вставке второго нового спейсера (т.е. спейсера «1», как обнаруживается в DGCC7778) на том же конце локуса CRISPR1, вместе с одним повтором. Эта вторая вставка дала новый устойчивый к бактериофагу мутант, который был выделен и назван DGCC7778. DGCC7778 более устойчив к D858, чем промежуточный штамм, и, конечно, гораздо более устойчив, чем материнский штамм DGCC7710, ввиду присутствия спейсера «1», и который на 100% идентичен последовательности меченого фага.
Пример 18
Способ мечения DGCC7710 и отбор меченого штамма DGCC7778
В этом примере, описываются способы, использованные для мечения DGCC7710 и отбора меченого штамма DGCC7778. Штамм DGCC7710 инфицировали/подвергали стимуляции фагом D858 инокуляцией пастеризованного молока штаммом DGCC7710 в количестве примерно 2×106 колониеобразующих единиц/мл и фагом D858 в количестве примерно 1×105 колониеобразующих единиц/мл. Инокулированное молоко культивировали в течение 12 часов при 35°С. После инкубации, жизнеспособные бактерии (т.е. те, которые, вероятно, представляют собой нечувствительные к бактериофагу мутанты) были выделены на не селективной среде (молочно-агаровые пластины) при 35°С с использованием соответствующего разведения инфицированной культуры. Один изолят, называемый DGCC7778, размножался в жидкой среде М17-глюкозы при 35°С, и его ДНК экстрагировали, используя классический протокол экстракции ДНК, как известно в данной области.
Экстракт ДНК амплифицировали, используя PCR, как известно в данной области (см., например, Bolotin et al. [2005], см. выше), с применением комбинации одного прямого праймера ((или yc70 и/или 
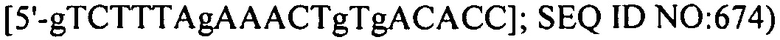
и одного обратного праймера (или yc31 и/или 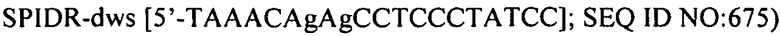
Определяли последовательность продуктов PCR и сравнивали с последовательностью локуса CRISPR DGCC7710.
Пример 19
Получение второго меченого штамма
В этом примере, описываются способы, использованные для получения второго меченого штамма. Штамм DGCC7710-RH1 S. thermophilus выделяли в качестве естественного устойчивого к фагу мутанта с использованием штамма DGCC7710 в качестве материнского штамма и фага D858 в качестве вирулентного фага.
CRISPR1 штамма DGCC7710-RH1 содержит 34 повтора (включая концевой повтор) и, таким образом 33 спейсера. При сравнении с последовательностью CRISPR1 штамма DGCC7710 S. thermophilus, последовательность CRISPR1 штамма DGCC7710-RH1 S. thermophilus имеет один дополнительный новый спейсер (т.е. маркировочную последовательность) (и, конечно, один дополнительный повтор, который фланкирует новый спейсер) на одном конце локуса CRISPR (т.е. близко к лидеру, на конце 5' локуса CRISPR). Все другие спейсеры локуса CRISPR1 являются неизменными. Последовательность CRISPR1 (5'-3') штамма DGCC7710-RH1 представляет собой:

Лидерная последовательность представляет собой 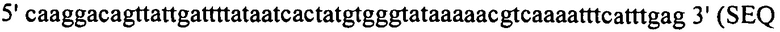
 .
.
Интегрированная последовательность 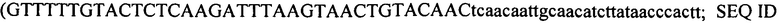
 , показанная серым цветом, включающая повтор CRISPR (выделение с прописными буквами), а спейсер CRISPR (т.е. маркировочную последовательность), который показан в выделении со строчными буквами. Показаны концевой повтор
, показанная серым цветом, включающая повтор CRISPR (выделение с прописными буквами), а спейсер CRISPR (т.е. маркировочную последовательность), который показан в выделении со строчными буквами. Показаны концевой повтор 
 и трейлерная последовательность
и трейлерная последовательность 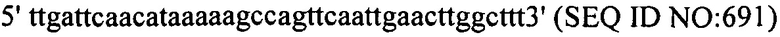 .
.
Соответственно, в случае штамма DGCC7710-RH1 S. thermophilus, спейсер 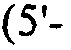
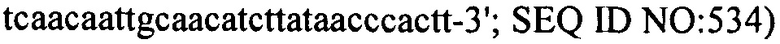 составляет специфическую для штамма маркировочную последовательность, которая идентифицирует этот мутантный штамм (т.е. меченую бактерию). Последовательность спейсера обнаруживается между положениями 31921 и 31950 bp (т.е. на «плюс-цепи») генома D858 (и имеет 100% идентичность с последовательностью генома D858 по 30 нуклеотидам):
составляет специфическую для штамма маркировочную последовательность, которая идентифицирует этот мутантный штамм (т.е. меченую бактерию). Последовательность спейсера обнаруживается между положениями 31921 и 31950 bp (т.е. на «плюс-цепи») генома D858 (и имеет 100% идентичность с последовательностью генома D858 по 30 нуклеотидам):

Пример 20
Получение третьего меченого штамма
В этом примере, описываются способы, использованные для получения третьего меченого штамма. Штамм DGCC7710-RH2 S. thermophilus выделяли в качестве естественного устойчивого к фагу мутанта с использованием штамма DGCC7710 в качестве материнского штамма и фага D858 в качестве вирулентного фага. CRISPR1 штамма DGCC7710-RH2 S. thermophilus содержит 34 повтора (включая концевой повтор) и, таким образом 33 спейсера. При сравнении с последовательностью CRISPR1 штамма DGCC7710 S. thermophilus, последовательность CRISPR1 штамма DGCC7710-RH2 S. thermophilus имеет один дополнительный новый спейсер (т.е. маркировочную последовательность) (и, конечно, один дополнительный повтор, который фланкирует новый спейсер) на одном конце локуса CRISPR (т.е. близко к лидеру, на конце 5' локуса CRISPR). Все другие спейсеры локуса CRISPR1 являются неизменными.
Последовательность CRISPR1 (5'-3') штамма DGCC7710-RH2 представляет собой:

Лидерная последовательность представляет собой 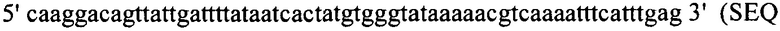
 .
.
Интегрированная последовательность 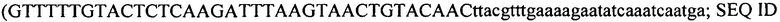
 , показанная серым цветом, включающая повтор CRISPR (выделение с прописными буквами), а спейсер CRISPR (т.е. маркировочную последовательность), который показан в отделении со строчными буквами. Показаны концевой повтор
, показанная серым цветом, включающая повтор CRISPR (выделение с прописными буквами), а спейсер CRISPR (т.е. маркировочную последовательность), который показан в отделении со строчными буквами. Показаны концевой повтор 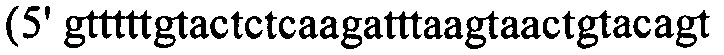
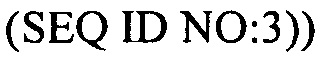 и трейлерная последовательность
и трейлерная последовательность 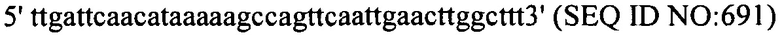 .
.
Таким образом, в случае штамма DGCC7710-RH2 Streptococcus thermophilus, спейсер 
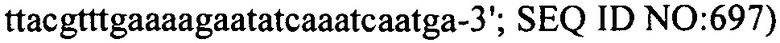 составляет специфическую для штамма метку, которая идентифицирует этот мутантный штамм (т.е. меченую бактерию). Было показано, что последовательность нового спейсера существует в пределах генома фага D858. Последовательность спейсера обнаруживается между положениями 17215 и 17244 bp (т.е. на «плюс-цепи») генома фага D858 (и имеет 100% идентичность с последовательностью генома D858 по 30 нуклеотидам):
составляет специфическую для штамма метку, которая идентифицирует этот мутантный штамм (т.е. меченую бактерию). Было показано, что последовательность нового спейсера существует в пределах генома фага D858. Последовательность спейсера обнаруживается между положениями 17215 и 17244 bp (т.е. на «плюс-цепи») генома фага D858 (и имеет 100% идентичность с последовательностью генома D858 по 30 нуклеотидам):

Новый спейсер, интегрированный в локус CRISPR1 штамма DGCC7710-RH2 S. thermophilus придает штамму DGCC7710-RH2 S. thermophilus новую устойчивость к фагу D858.
Пример 21
Построение фага «избегания CRISPR» из устойчивых к фагам бактериальных вариантов
В этом примере описаны способы построения фагов избегания CRISPR. Сначала конструируются устойчивые к фагам варианты хозяев, как описано в примерах выше. В этих экспериментах, на материнский штамм «А» воздействует фаг «Р» и отбирается устойчивый к фагу вариант (вариант «А1.0»). Вариант А1.0 анализируется (пример, PCFR и/или секвенированием ДНК) для подтверждения присутствия дополнительного вставленного спейсера в пределах локуса CRISPR. Затем определяется нуклеотидная последовательность дополнительного спейсера (спейсера Sp1.0). Обычно, спейсер Sp1.0 представляет собой фрагмент размером приблизительно 30 нуклеотидов из фага Р, и он придает устойчивость к фагу Р и родственным фагам («родственные фаги» представляют собой те, которые содержат в своем геноме последовательность спейсера, и определяют семейство фагов).
Независимо от воздействия первого фага, один и тот же штамм А подвергается воздействию одного и того же фага Р, и выбирается второй устойчивый к фагу вариант (вариант А2.0). Вариант А2.0 выбран для наличия также вставленного дополнительного спейсера (спейсера Sp2.0) в пределах локуса CRISPR, но при последовательности спейсера Sp2.0, отличающегося от последовательности спейсера Sp1.0. Обычно, спейсер Sp2.0 представляет собой фрагмент размером приблизительно 30 нуклеотидов из фага Р, и он придает устойчивость к фагу Р и родственным фагам. Аналогичным образом, в некоторых вариантах осуществления, генерируются варианты от А3.0 до Ах.0 посредством воздействия одного и того же фага Р на один и тот же штамм А. Все варианты «А» также выбраны для наличия вставленного дополнительного спейсера (спейсера от Sp3.0 до Spх.0) в пределах локуса CRISPR, но при последовательности всех спейсеров «Sp», отличающихся друг от друга. Обычно спейсеры «Sp» представляют собой фрагменты размером приблизительно 30 нуклеотидов из фага Р, и они все придают устойчивость к фагу Р и родственным фагам.
Обычно, можно оценить, что уровень устойчивости будет приблизительно составлять уровень устойчивости одной мутации, происходящей в пределах генома фага в пределах последовательности, соответствующей спейсеру (т.е. грубо от 10-4 до 10-6). Таким образом, фаги, которые избегают опосредованной CRISPR устойчивости, легко выделить. Мутированные фаги генерируются посредством воздействия фага Р на вариант А1.0. Обычно, мутированный фаг «избегания CRISPR» (Р1.0) содержит, по меньшей мере, одну мутацию в пределах своего генома, соответствующую последовательности спейсера Sp1.0 (например, делецию (делеции), точечную мутацию (мутации) и т.д.), или в некоторых предпочтительных вариантах осуществления, в области, фланкирующей Sp1.0, плюс или минус 200 bp, соответствующей мотиву CRISPR. Вариант А1.0 должен быть чувствительным к фагу Р1.0. Аналогичным образом, независимо генерированные устойчивые к фагу Р варианты (вариант А2.0, от А3.0 до Ах.0), которые содержат необычные спейсеры (соответственно, Sp2.0, от Sp3.0 до Spx.0) аналогичным образом стимулируются фагом Р для генерирования соответствующих мутантных фагов (соответственно, Р2.0, от Р3.0 до Рх.0). В последующем, может генерироваться пул мутантных вирулентных фагов, чьи геномы были подвергнуты специфической мутации в последовательность, предполагаемой как спейсер CRISPR.
Действительно, фаг D2792 представляет полностью вирулентный фаг биологического контроля против штамма DGCC7710 S. thermophilus (WT). Напротив, анализ локуса CRISPR родственных штаммов WTphi2972+S6, WTphi2972+S4, WTphi2972+S20, WTphi2972+S21 и WTphi2972+S22, указывает на присутствие спейсерной последовательности, которая аналогична последовательностям, обнаруживаемым в фаге D2792, что указывает на то, что фаг D2792 имеет сниженную вирулентность в отношении этих штаммов. Данные бляшкообразования (см. таблицу 7.1) подтверждают сниженную вирулентность фага D2792 в отношении этих штаммов. В отношении штамма WTphi2972+S6, который характеризовался как устойчивый к фагу D2792 вследствие присутствия соответствующего спейсера CRISPR, скрининг связанных с D2792 фагов на полную увеличенную вирулентность идентифицировал фаги D4724 и D4733 как перспективные агенты в качестве агентов биологического контроля (см. таблицу 7-1).
В дополнительных экспериментах, штамм DGCC7710 подвергался воздействию фага D2792 для генерирования устойчивого варианта WTphi2972+S6. Когда штамм WTphi2972+S6 подвергался воздействию фага D2792, можно было выделить мутантный фаг, такой как D4724. Было обнаружено, что этот фаг D4724 полностью вирулентный для DGCC7710 и WTphi2972+S6. При втором повторении, на WTphi2972+S6 воздействовали фагом D4724 для генерирования устойчивого варианта WTphi2972+S6phi4724+S15. После воздействия D4724 на этот штамм, были идентифицированы мутантные фаги, такие как D4733, которые являются полностью вирулентными в отношении DGCC7710 и WTphi2972+S6. В некоторых вариантах осуществления, последовательные повторения используются для генерирования фага с желательным уровнем вирулентности.
Дополнительные примеры мутантов фага представлены на фиг. 13. На этом чертеже, показаны мутантные фаги 858-А и 858-В, полученные из материнского фага D858. Мутации соответствуют спейсеру S1 из WTФ858+S1S2, стимулированному D858.
В еще одних примерах, полностью вирулентные мутанты фага, где мутация идентифицируется в мотиве CRISPR, показаны в таблице 20-1. В этой таблице показаны нуклеотидные последовательности в фагах дикого типа и мутантных фагах, которые соответствуют спейсерам, вновь приобретенным мутантами S. thermophilus. Мотив AGAAW выделен серым цветом. Каждая мутация выделена жирным шрифтом и подчеркнута. * указывает на делецию. В этой таблице представлены последовательности для пар устойчивого к фагу CRISPR варианта и вирулентного мутанта фага: DGCC7710Ф858+S1/фаг 2972.S3C, DGCC7710Ф2972+S4/фаг 2972.S4А или фаг 2972.S4C, DGCC7710Ф2972+S6/фаг 2972.S6А и DGCC7710Ф2972+S4 Ф858+S32/фаг 858.S32А или фаг 858.S32D. В этой таблице новый спейсер соответствует SEQ ID NO:535 (DGCC7710Ф858+S3).
Таблица 20-1


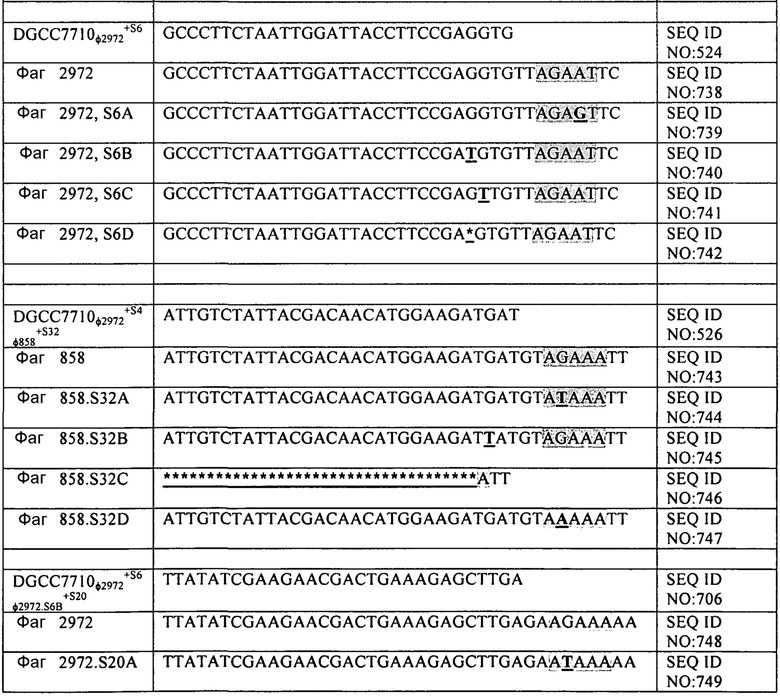
Пример 22
Фаг «избегания CRISPR» второго уровня
В этом примере описаны эксперименты по построению Фагов «избегания CRISPR» второго уровня (т.е. с множественными мутациями, направленными на множественные спейсеры).
Посредством повторного процесса создания вариантов с устойчивостью к фагам, опосредованной CRISPR, с последующим выделением мутированного («избегания CRISPR») фага, способного преодолеть механизм cas-CRISPR, возможно создание фага, который был «предварительно адаптирован» множественными мутациями против потенциальной устойчивости, опосредованной CRISPR.
В некоторых вариантах осуществления, варианты второго уровня получают выделением мутированного фага посредством воздействия фага Р на вариант А1.0. Обычно, этот мутированный фаг (фаг Р1.0) имеет мутацию (делецию, точечную мутацию и т.д.) в его геноме в пределах области, содержащей последовательность спейсера Sp1.0 или в пределах области, фланкирующей Sp1.0, плюс или минус фрагмент из 20 bp, соответствующий мотиву CRISPR. Вариант А1.0 чувствителен к фагу Р1.0. Затем, вариант А1.0 подвергается воздействию фага Р1.0, и выбирается устойчивый к фагу вариант (вариант А1.1) (см. фиг. 15). Вариант А1.1 также выбирается так, чтобы он имел дополнительный спейсер (спейсер Sp1.1) в пределах локуса CRISPR, но при последовательности спейсера Sp1.1 отличающейся от последовательности спейсеров Sp1.0, от Sp2.0 до Spх.0. Обычно, спейсер Sp1.1 представляет собой фрагмент размером приблизительно 30 нуклеотидов из фага Р1.0 и придает устойчивость к фагу Р1.0 и родственным фагам. Вариант А1.1 устойчив к фагу Р1.0 и предпочтительно имеет повышенную устойчивость к фагу Р ввиду накопления спейсеров Sp1.0 и Sp1.1.
В дополнительных вариантах осуществления, вновь мутированный фаг (фаг Р1.1) генерируется посредством воздействия фага Р1.0 на вариант А1.1. Затем, после воздействия фага Р1.0 на вариант А1.1, получают новый вариант А1.2, который содержит один новый дополнительный спейсер (Sp1.2). Этот спейсер придает устойчивость к фагу Р1.1 и, предпочтительно, увеличивает устойчивость к фагу Р1.0 и Р (т.е. ввиду накопления спейсеров Sp1.0, Sp1.1, Sp1.2). Фаг Р1.1 является полностью инфективным в отношении материнского штамма А, а также варианты А1.0 и А1.1.
В еще одних дополнительных вариантах осуществления, другие спейсеры (например, 2, 3 или 4) повторно накапливаются внутри штаммов с А по вариант А1, затем вариант А1.1, затем вариант А1.2 и т.д. для получения варианта, высоко устойчивого к фагам (вариант А1.n). В еще одних вариантах осуществления, дополнительные другие спейсеры могут накапливаться в одном и том же штамме до варианта А2, затем варианта А2.1, затем варианта А2.2 и т.д. для генерирования другого варианта штамма А, параллельно высоко устойчивого к фагам (вариант А2.n). Такая же стратегия находит применение с вариантами от А3.0 до Ах.0.
После повторного процесса создания устойчивых к фагам вариантов CRISPR и выделения мутантного фага «избегания CRISPR» (например, воздействия фага Р1.1 на вариант А1.1, создающего новый вариант А1.2, который содержит один новый дополнительный спейсер (Sp1.2), из которого выделяется мутантный фаг (Р1.2), который является полностью вирулентным для варианта А1.2, А1.1, А1.0 и материнского штамма А.
В некоторых вариантах осуществления, комбинаторные мутации накапливаются повторной конструкцией бактериальных вариантов, комбинирующих различные спейсеры (например, Sp2.0, Sp3.0-Spx.0), воздействием соответствующего мутантного фага первого уровня (Р2.0, Р3.0-Рх.0) и выделением мутантных фагов второго уровня.
Примеры повторных комбинаторных мутаций, создающих варианты, устойчивые к CRISPR фагу, и мутантный фаг «избегания CRISPR» показан в таблице 22-1. В этой таблице представлен список новых спейсеров, обнаруженных в CRISPR1, и соответствующая область в фагах 2972, 858 и DT1. В этой таблице, «а» указывает на области ДНК, которые на 100% идентичны между фагами 858 и 2972. «Положение 5'» относится к положению 5' протоспейсера в геноме фага. Подчеркнутые и заштрихованные нуклеотиды в последовательности фотоспейсера указывают на ошибочные спаривания между фагом и спейсером. Звездочка (*) указывает делецию. «Фланкирующая 3' область» указывает последовательность, фланкирующую 3', в геноме фага. Ошибочные спаривания в мотиве AGAAW подчеркнуты и заштрихованы в серый цвет. В колонке, обозначенной «Нить/Модуль», модулями транскрипции являются «Е (ранние экспрессированные гены); «М» (средние экспрессированные гены) и “L” (поздние экспрессированные гены).

DGCC7710 подвергали воздействию фага 2972 для создания устойчивого к фагу варианта CRISPR DGCC7710Ф2972+S6, из которого был генерирован мутант избегания CRISPR фага 2972.S6В. Воздействие фага 2972.S6В на DGCC7710Ф2972+S6 создало устойчивый к фагу вариант CRISPR DGCC7710Ф2972+S6a2972S56В+S20, из которого был выделен мутант избегания CRISPR фага 2972.S20A.
В некоторых вариантах осуществления, предоставляются штаммы, которые устойчивы более чем к одному семейству фагов. Поскольку данный штамм может быть чувствительным более чем к одному семейству фагов, то в некоторых вариантах осуществления, желательно увеличить устойчивость штамма к множествам семейств фагов введением дополнительного спейсера (спейсеров) в пределы локуса CRISPR, происходящего из других семейств фагов (см. фиг. 16). Например, фаги P, Q и R являются репрезентативными фагами из трех семейств фагов, способных инфицировать штамм А. Используя описанный в настоящей заявке способ, получают варианты, устойчивые ко всем трем семействам фагов. В некоторых вариантах осуществления, фаг Р используется для генерирования варианта А1р (содержащего спейсер Sp1), который устойчив к фагу Р. Затем, вариант А1р подвергается воздействию фага Q, и выбирается устойчивый к фагу вариант (вариант А1pq). Вариант А1pq один дополнительный спейсер (Sq1), вставленный в пределы локуса CRISPR. Обычно, спейсер Sq1 представляет собой фрагмент размером приблизительно 30 нуклеотидов из фага Q, и он придает устойчивость к фагу Q и родственным фагам. Вариант А1pq устойчив и к фагу P, и к фагу Q. Затем вариант А1pq подвергается воздействию фага R, и выбирается устойчивый к фагу вариант (вариант А1pqr). Вариант А1pqr имеет третий дополнительный спейсер (Sr1), вставленный в пределы локуса CRISPR. Обычно, спейсер Sr1 представляет собой фрагмент размером приблизительно 30 нуклеотидов из фага R, и он также придает устойчивость к фагу R и родственным фагам. Вариант А1pqr устойчив ко всем трем фагам. В некоторых особенно предпочтительных вариантах осуществления, вариант также устойчив к родственным фагам.
Эти фаги избегания CRISPR находят применение в качестве фагов биологического контроля и терапевтических фагов. Как описано выше, посредством процесса создания опосредованных CRISPR устойчивых к фагам вариантов, воздействия фага и выделения вирулентного фага «избегания CRISPR», генерируется смесь видов фагов, которые содержат одну и/или множественные мутации, нацеленные против одной и/или множественных последовательностей генома фага, которые являются потенциальными спейсерами CRISPR-мишенями. Поскольку мишени в виде бактерий-хозяев могут стать устойчивыми к фагам посредством включения одного или множественных спейсеров, и учитывая, что механизм Cas-CRISPR может быть преодолен посредством мутации в пределах генома фага, соответствующей таким спейсерам, то использование смеси фагов, содержащих различные мутации, снижает частоту приобретения отдельной бактерией успешного приобретения новых спейсеров и пролиферации.
В дополнительном варианте осуществления, анализ протоспейсера и фланкирующих областей, по данным определения по спейсерам у соответствующих устойчивых к фагам вариантов CRISPR, облегчает идентификацию мотива CRISPR для определенного CRISPR. В примере CRISPR1 DGCC7710, устойчивые к фагу варианты, содержащие спейсеры S1-S33, были генерированы после стимуляции фагом 2972 или 858. Совмещение протоспейсера и фланкирующих областей из генома фагов 2972 или 858, которые соответствуют спейсерам S1-S33, с использованием программного обеспечения Clustal X, идентифицировало мотив CRISPR1 в виде NNAGAAW (SEQ ID NO:696), и визуализировалось с использованием WebLogo (фиг. 22).
В еще одном примере, устойчивые к фагу варианты CRISPR3 были получены из DGCC7710 после стимуляции фагами 858 и 3821, и LMD-9 после стимуляции фагом 4241. Совмещение протоспейсера и фланкирующих областей из геномов соответствующих фагов с соответствующими спейсерами соответствующих устойчивых к фагам вариантов CRISPR 3, идентифицировало мотив CRISPR 3 в виде NGGNG (SEQ ID NO:723) (фиг. 23).
Анализ для выявления присутствия специфического мотива CRISPR обеспечивает средство для идентификации локализации предполагаемых протоспейсеров в пределах генома или другой определенной последовательности (например, плазмиды или другого мобильного генетического элемента). В примере секвенированных фагов 858, 2972 и DT1, анализ распределения мотива CRISPR 1 AGAAW, идентифицировал локализацию потенциальных протоспейсеров в пределах соответствующих геномов. Используя дегенерацию генетического кода и/или применение консервативных аминокислотных замещений, каждый мотив AGAAW был устранен в процессе химического синтеза генома, как описано для фага ØХ174, как известно в данной области. Таким образом, фаг становился нечувствительным к системе устойчивости Cas-CRISPR. Таким образом, молекула ДНК, лишенная специфических мотивов CRISPR, нечувствительна к соответствующей системе Cas-CRISPR.
Эти фаги и «коктейли» множественных типов фагов находят применение в стратегиях ротации (например, определенном последовательном введении фага). В качестве расширения применения одного коктейля, составленного из фага, содержащего различные спейсерные мутации, в некоторых вариантах осуществления определенным последовательным образом используются множественные вирулентные фаги, каждый из которых содержит другую спейсерную мутацию. Например, используя набор фагов «избегания CRISPR» (Р1.0, Р2.0 и Р3.0 или Р1.0, Р1.1, Р1.2 или некоторые их комбинации), каждый фаг применяется отдельно и в определенной последовательности и ротации (P.1.0>P.2.0>P.3.0>P.1.0,P2.0> и т.д.) с тем, чтобы минимизировать вероятность развития у бактерий-мишеней опосредованной CRISPR устойчивости к фагу. Аналогичным образом, набор из коктейля фагов (т.е. каждый фаг внутри коктейля, а также каждый коктейль имеет необычную комбинацию мутаций) находит применение последовательно и путем ротации. В некоторых вариантах осуществления, фаг и/или коктейль содержит одно семейство фагов, тогда как в других вариантах осуществления, фаг и/или коктейль содержит множество семейств фагов.
Пример 23
Функциональные комбинации
В этом примере представлены различные функциональные комбинации, которые находят применение в настоящем изобретении. Только в качестве примера, следующие функциональные комбинации могут применяться в соответствии с настоящим изобретением.
Функциональная комбинация №1
Последовательности cas: с SEQ ID NO:461 по SEQ ID NO:465 и с SEQ ID NO:473 по SEQ ID NO:477 (все из которых являются последовательностями S. thermophilus), как представлено ниже:








с последовательностями повтора: с SEQ ID NO:1 по SEQ ID NO:10
Функциональная комбинация №2
Последовательности cas: с SEQ ID NO:466 по SEQ ID NO:472 и с SEQ ID NO:478 по SEQ ID NO:487 (все из которых являются последовательностями S. thermophilus), как представлено ниже:









с последовательностями повтора: SEQ ID NO:11 и/или SEQ ID NO:12.
Функциональная комбинация №3
Последовательности cas: с SEQ ID NO:488 по SEQ ID NO:508 и с SEQ ID NO:517 по SEQ ID NO:521, как показано ниже. SEQ ID NOS:488-497 из S. agalactiae, тогда как SEQ ID NOS:498-503 из S. mutans, и SEQ ID NOS:504-508, 517-521 из S. pyogenes.




















с последовательностями повтора: с SEQ ID NO:13 по SEQ ID NO:19
Функциональная комбинация №4
Последовательности cas: с SEQ ID NO:509 по SEQ ID NO:516 (все из которых из S. pyogenes), как показано ниже
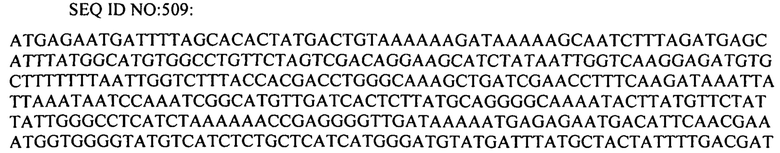





с последовательностями повтора: SEQ ID NO:20 и SEQ ID NO:22.
Все патенты и публикации, приведенные в описании, указывают на уровни осведомленности специалистов в данной области, к которым относится это изобретение. Все патенты и публикации включены в настоящее описание путем ссылки в такой же степени, как если бы каждая отдельная публикация была специально и отдельно указана как включенная путем ссылки.
Специалисты в данной области легко поймут, что настоящее изобретение достаточно адаптировано для достижения целей, а также получения указанных присущих ему преимуществ. Описанные в настоящей заявке способы и композиции представляют собой репрезентативные варианты осуществления. Иллюстрирующие, а не ограничивающие объем изобретения. Для специалиста в данной области очевидно, в описанное в настоящей заявке изобретение могут быть внесены различные замещения и модификации без отхода от объема и сущности изобретения.
Иллюстративно раскрытое в настоящем описании изобретение может быть подходящим образом осуществлено в отсутствие любого элемента или элементов, ограничения или ограничений, которые конкретно не раскрыты в настоящем описании. Использованные термины и выражения применяются как термины описания, а не ограничения, и нет намерения, что использование таких терминов и выражений исключает любые эквиваленты показанных и описанных признаков или их частей, но признается, что в пределах объема изобретений возможны различные модификации. Таким образом, следует понимать, что хотя настоящее изобретение было конкретно раскрыто предпочтительными вариантами осуществления и необязательными признаками, специалисты в данной области могут прибегнуть к модификации и изменению раскрытых в настоящем описании концепций, и считается, что такие модификации и изменения входят в объем настоящего изобретения.
В настоящем описании изобретение представлено в широком и родовом аспекте. Каждая из более узких видовых и подродовых группировок в пределах родового описания также составляет часть изобретения. Это также включает родовое описание изобретения при условии ограничения или отрицательном ограничении, устраняющем любой предмет из рода, независимо от того, указан ли конкретно в настоящем описании или нет подвергнутый ограничению материал.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ГЕНЕРИРОВАНИЯ ЗАКВАСОЧНОЙ КУЛЬТУРЫ, ЗАКВАСОЧНАЯ КУЛЬТУРА И СПОСОБ ФЕРМЕНТАЦИИ С ЕЕ ИСПОЛЬЗОВАНИЕМ | 2008 |
|
RU2531343C2 |
| ПОСЛЕДОВАТЕЛЬНОСТИ CRISPR БИФИДОБАКТЕРИЙ | 2009 |
|
RU2570562C2 |
| КОМПОНЕНТЫ СИСТЕМЫ CRISPR-CAS, СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ МАНИПУЛЯЦИИ С ПОСЛЕДОВАТЕЛЬНОСТЯМИ | 2013 |
|
RU2701662C2 |
| КОНСТРУИРОВАНИЕ СИСТЕМ, СПОСОБЫ И ОПТИМИЗИРОВАННЫЕ НАПРАВЛЯЮЩИЕ КОМПОЗИЦИИ ДЛЯ МАНИПУЛЯЦИИ С ПОСЛЕДОВАТЕЛЬНОСТЯМИ | 2013 |
|
RU2701850C2 |
| СИСТЕМЫ CRISPR-CAS И СПОСОБЫ ИЗМЕНЕНИЯ ЭКСПРЕССИИ ПРОДУКТОВ ГЕНОВ | 2013 |
|
RU2687451C1 |
| НУКЛЕАЗА CPF1 ИЗ БАКТЕРИИ Ruminococcus bromii, МОЛЕКУЛА ДНК ИЛИ РНК, КОДИРУЮЩАЯ НУКЛЕАЗУ, ВЕКТОР, СОДЕРЖАЩИЙ УКАЗАННУЮ МОЛЕКУЛУ ДНК, СИСТЕМА CRISPR/CPF1, СОДЕРЖАЩАЯ УКАЗАННУЮ НУКЛЕАЗУ И НАПРАВЛЯЮЩУЮ РНК, КЛЕТКА-ХОЗЯИН ДЛЯ ПОЛУЧЕНИЯ НУКЛЕАЗЫ CPF1, СПОСОБ ПОЛУЧЕНИЯ НУКЛЕАЗЫ CPF1 И ЕЕ ПРИМЕНЕНИЕ | 2022 |
|
RU2816876C1 |
| СИСТЕМЫ CRISPR-CAS И СПОСОБЫ ИЗМЕНЕНИЯ ЭКСПРЕССИИ ПРОДУКТОВ ГЕНОВ | 2013 |
|
RU2796273C2 |
| НОВЫЕ ФЕРМЕНТЫ И СИСТЕМЫ CRISPR | 2016 |
|
RU2777988C2 |
| КОМПОЗИЦИИ И СПОСОБЫ С УЧАСТИЕМ НУКЛЕИНОВЫХ КИСЛОТ, НАЦЕЛЕННЫХ НА НУКЛЕИНОВЫЕ КИСЛОТЫ | 2014 |
|
RU2662932C2 |
| МУТАЦИИ ФЕРМЕНТА CRISPR, УМЕНЬШАЮЩИЕ НЕЦЕЛЕВЫЕ ЭФФЕКТЫ | 2016 |
|
RU2752834C2 |
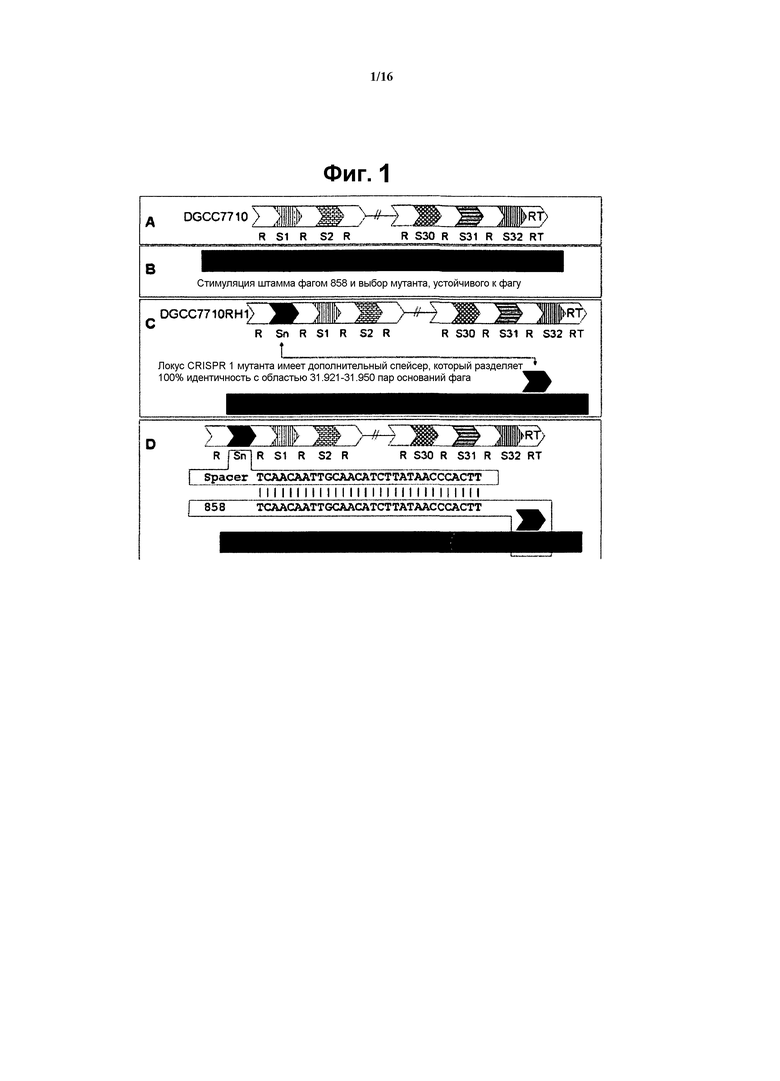

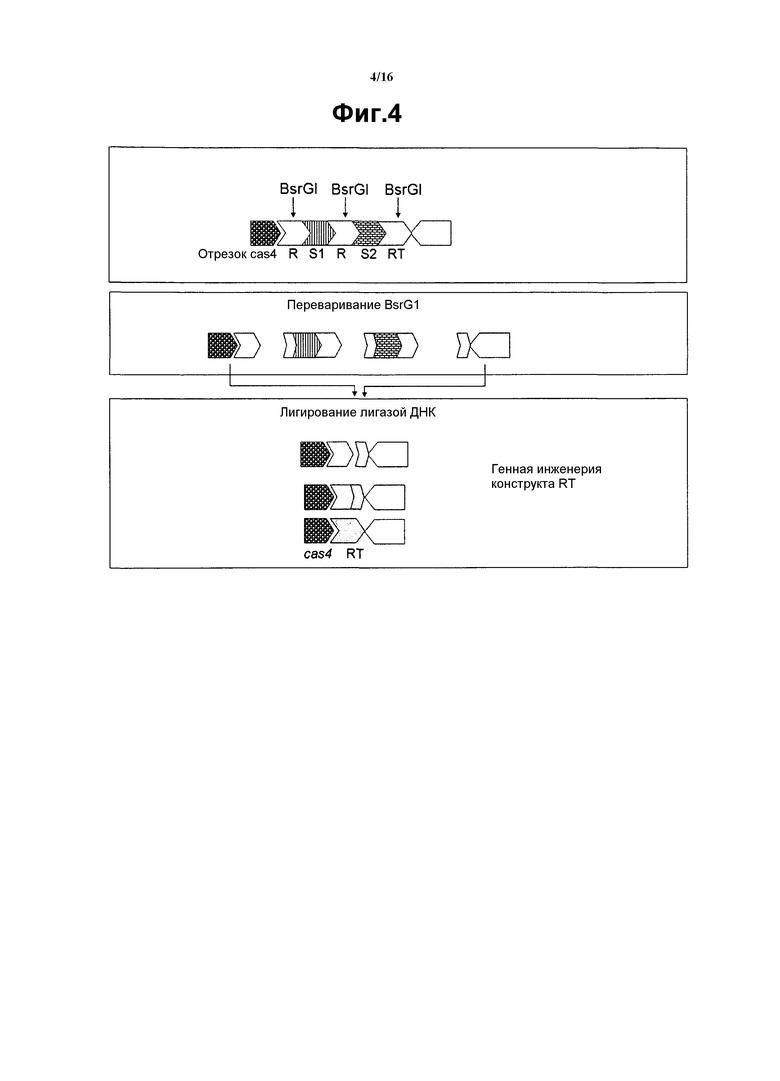
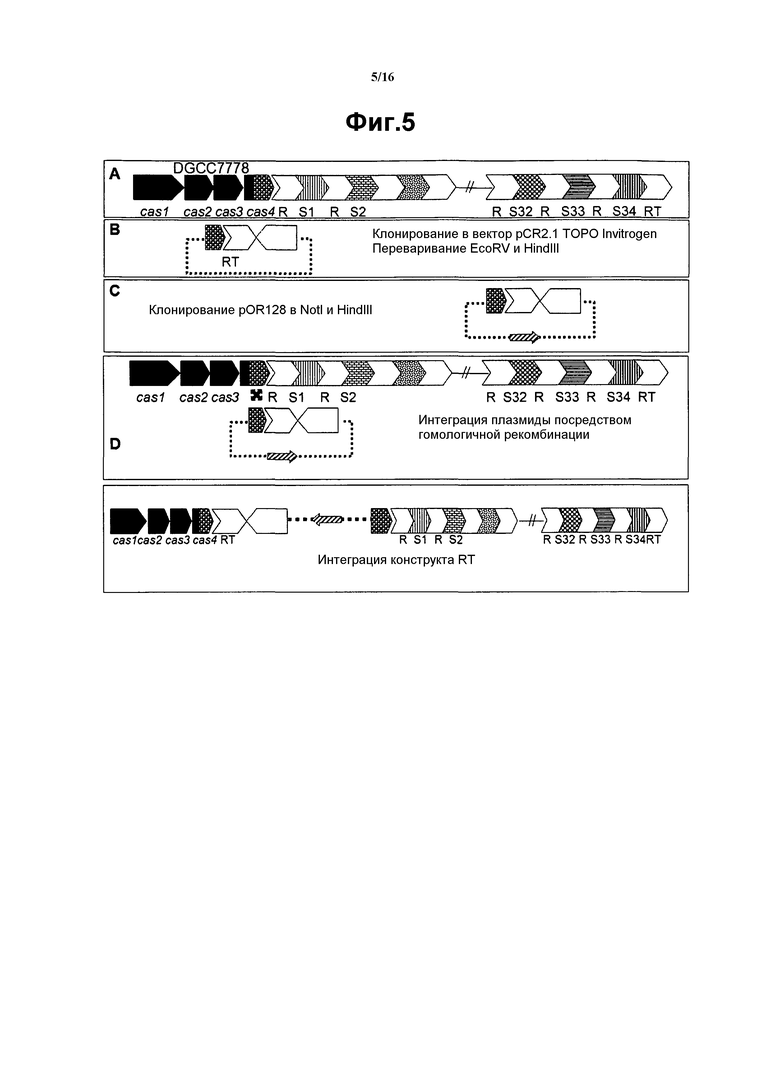
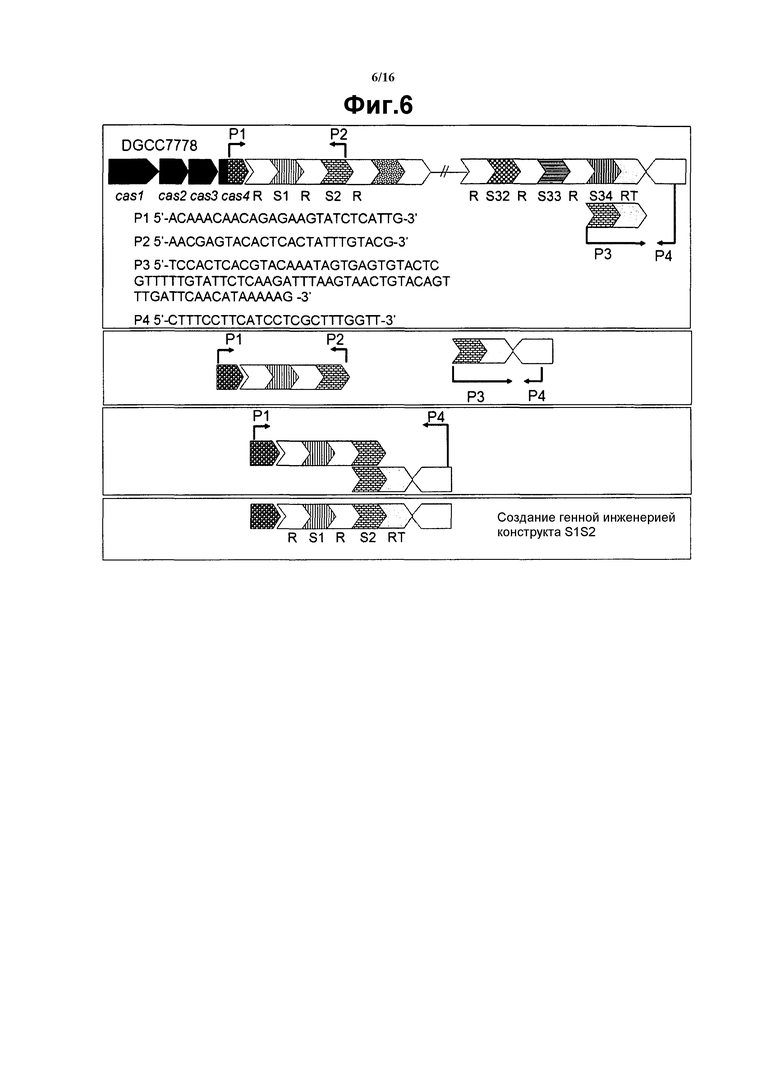

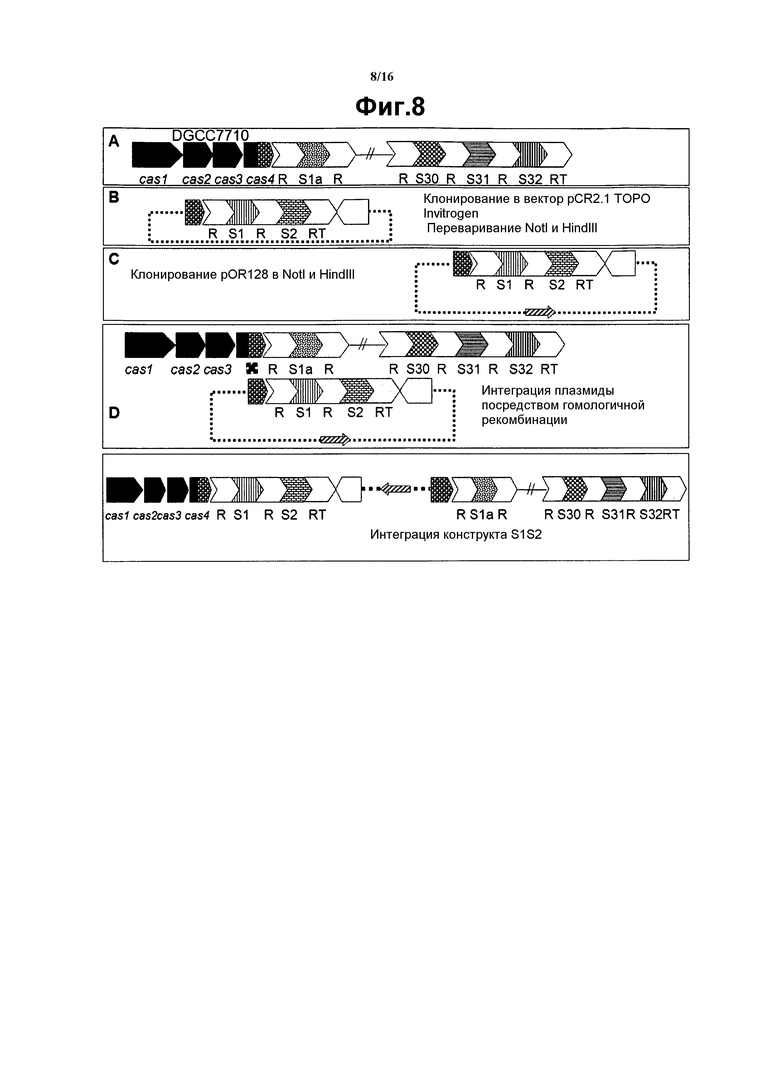

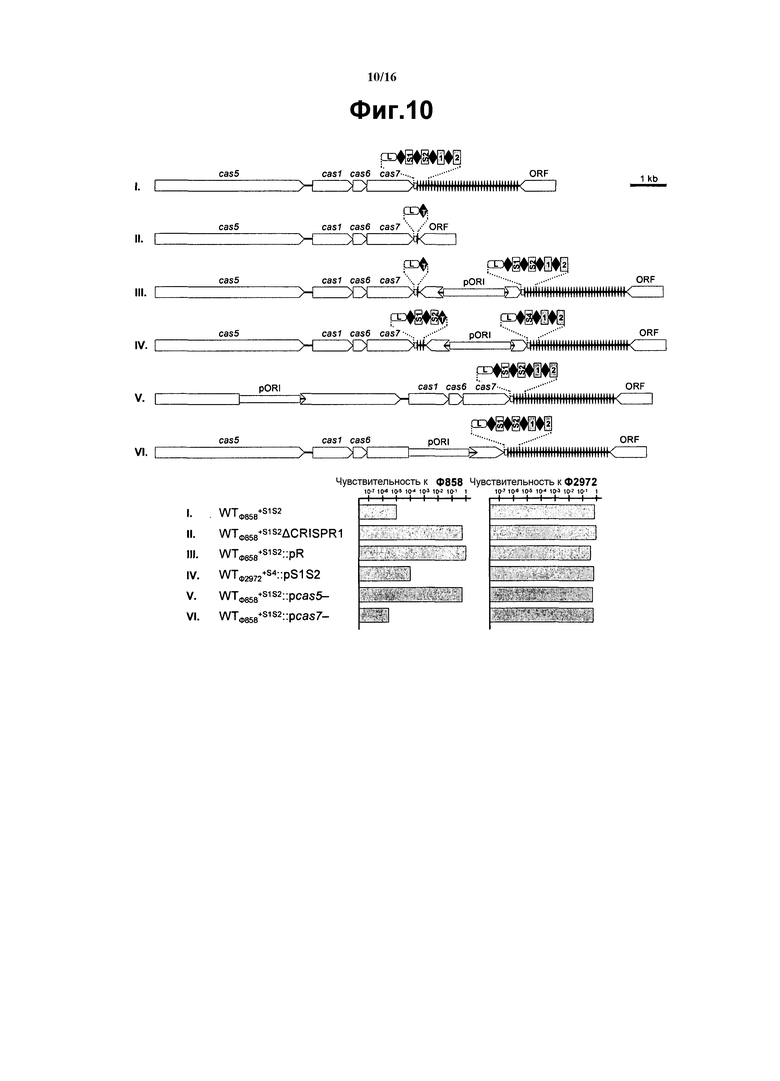
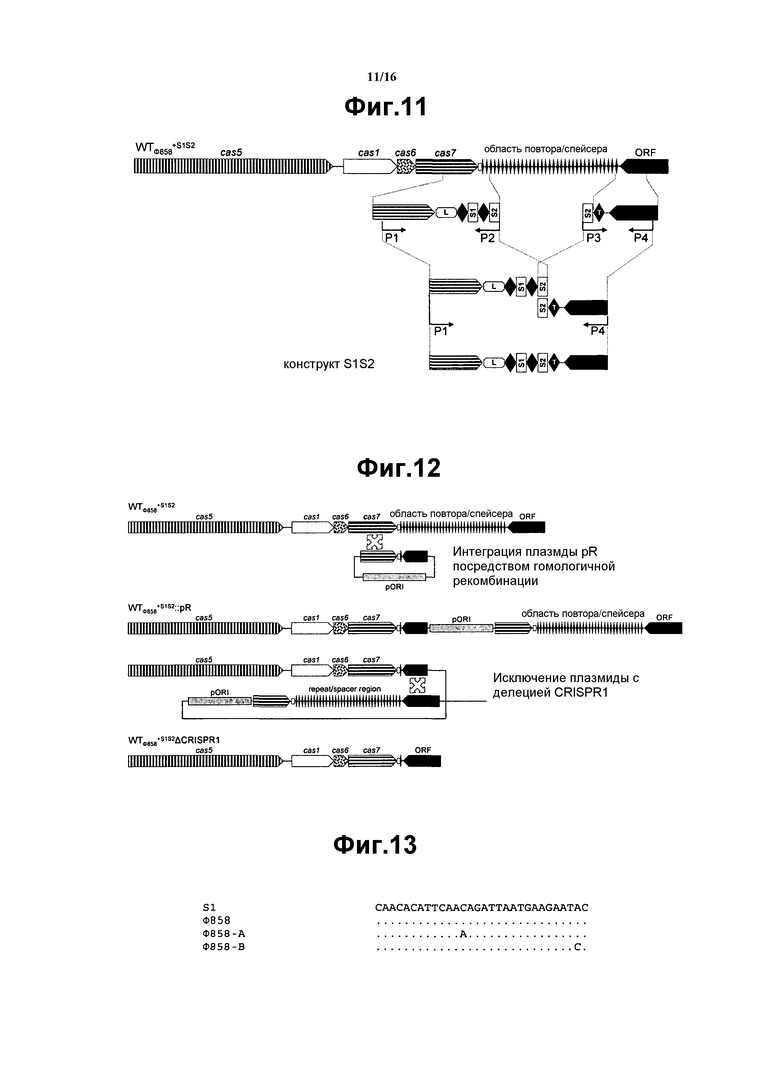
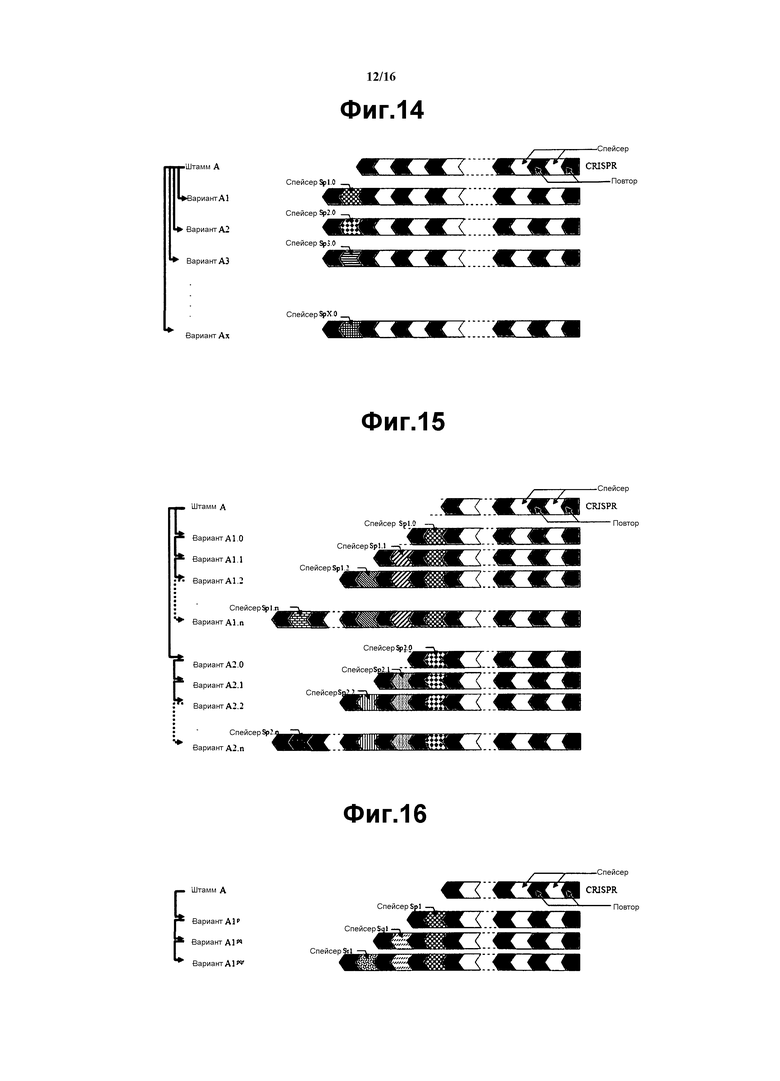

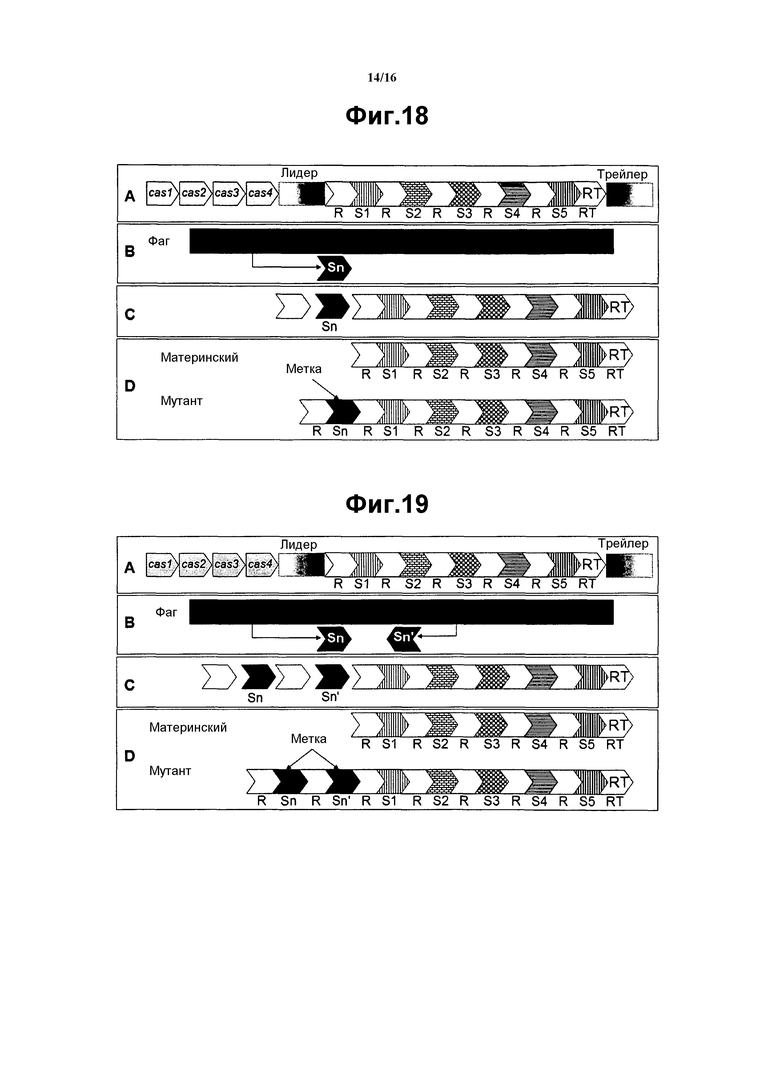
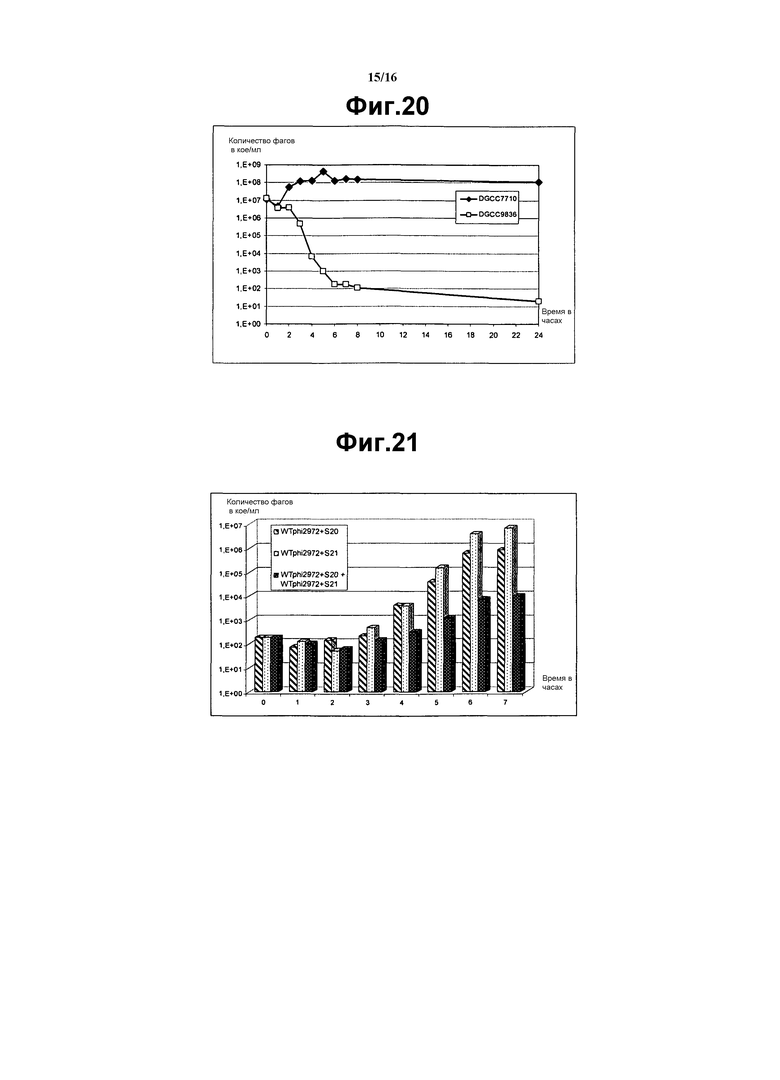
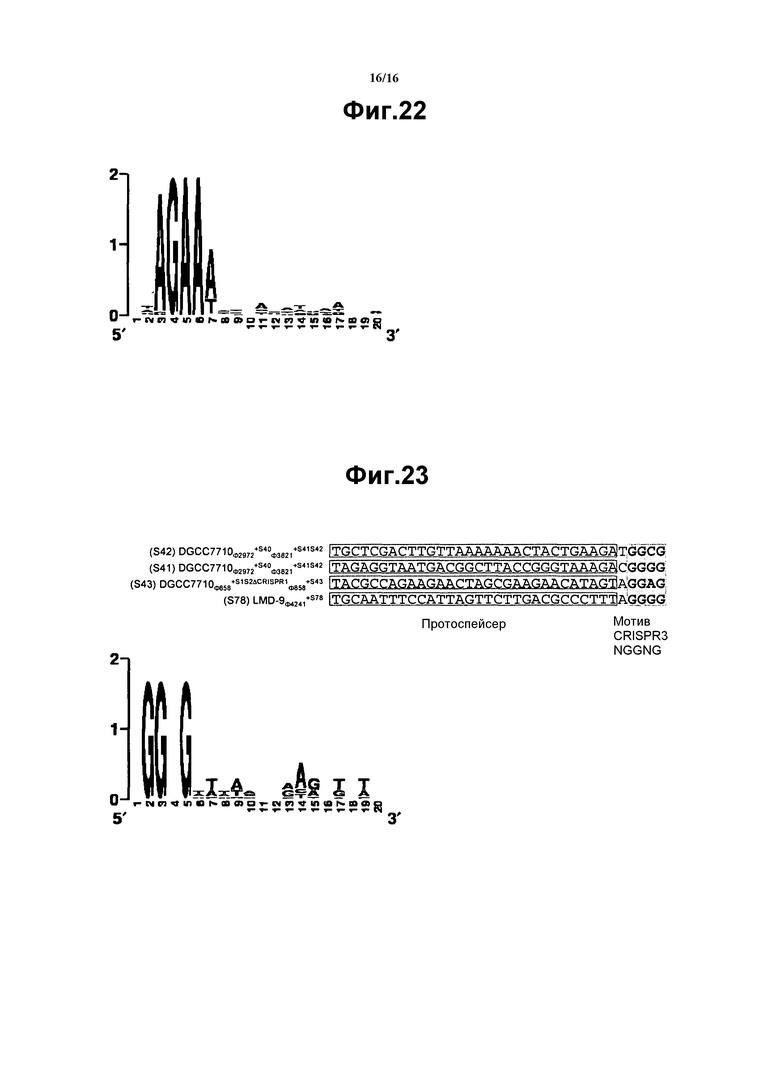
Изобретение относится к области пищевой промышленности. Описан способ получения заквасочной культуры, включающей по крайней мере два различных устойчивых к бактериофагам вариантных штамма Streptococcus thermophiles. В одном из вариантов способ включает стадию воздействия на материнский бактериальный штамм Streptococcus thermophiles, содержащий локус CRISPR, по меньшей мере одним бактериофагом семейства Siphoviridae, способным инфицировать указанный материнский штамм, для получения смеси бактерий, содержащей устойчивые к бактериофагам вариантные штаммы. Изобретение расширяет арсенал средств для получения заквасочных культур. 23 з.п. ф-лы, 23 ил., 10 табл., 23 пр.
1. Способ получения заквасочной культуры, включающей по крайней мере два различных устойчивых к бактериофагам вариантных штамма Streptococcus thermophilus, где способ включает стадии:
(a) воздействия на материнский бактериальный штамм Streptococcus thermophilus, содержащий локус CRISPR, по меньшей мере одним бактериофагом семейства Siphoviridae, способным инфицировать указанный материнский штамм, для получения смеси бактерий, содержащей устойчивые к бактериофагам вариантные штаммы;
(а1) отбора устойчивых к бактериофагам вариантных штаммов из смеси бактерий по (а);
(а2) отбора из устойчивых к бактериофагам вариантных штаммов по (а1) по меньшей мере одного устойчивого к бактериофагам вариантного штамма, который содержит локус CRISPR, модифицированный добавлением по меньшей мере одного спейсера относительно локуса CRISPR материнского штамма, где отбор осуществляют путем сравнения локуса CRISPR или его части материнского бактериального штамма с локусом CRISPR устойчивого к бактериофагам вариантного штамма;
(а3) выделения указанного по меньшей мере одного устойчивого к бактериофагам вариантного штамма по (а2);
(b) воздействия на материнский бактериальный штамм Streptococcus thermophilus, содержащий локус CRISPR, по меньшей мере одним бактериофагом семейства Siphoviridae, способным инфицировать указанный материнский штамм, для получения смеси бактерий, содержащей другие устойчивые к бактериофагам вариантные штаммы;
(b1) отбора устойчивых к бактериофагам вариантных штаммов из смеси бактерий по (b);
(b2) отбора из устойчивых к бактериофагам вариантных штаммов по (b1) по меньшей мере одного устойчивого к бактериофагам вариантного штамма, который содержит локус CRISPR, модифицированный добавлением по меньшей мере одного спейсера относительно локуса CRISPR материнского штамма, где отбор осуществляют путем сравнения локуса CRISPR или его части материнского бактериального штамма с локусом CRISPR устойчивого к бактериофагам вариантного штамма, и где по меньшей мере один спейсер отличается от по меньшей мере одного спейсера по (а2);
(b3) выделения по меньшей мере одного устойчивого к бактериофагам вариантного штамма по (b2);
(с) объединения по меньшей мере одного устойчивого к бактериофагам вариантного штамма по (а2) с по меньшей мере одним устойчивым к бактериофагам вариантным штаммом по (b2) с получением заквасочной культуры.
2. Способ по п. 1, где на указанный материнский бактериальный штамм воздействуют двумя или более бактериофагами.
3. Способ по п. 1, где на указанный материнский бактериальный штамм одновременно воздействуют двумя или более бактериофагами.
4. Способ по п. 1, где на указанный материнский бактериальный штамм последовательно воздействуют двумя или более бактериофагами.
5. Способ по п. 1, где указанный материнский бактериальный штамм инфицирован бактериофагом.
6. Способ по п. 5, где указанный по меньшей мере один бактериофаг является бактериофагом из семейства Siphoviridae.
7. Способ по п. 1, где стадии (а2) и (b2) включают сравнение конца 5’ и/или конца 3’ локуса CRISPR материнского бактериального штамма с локусом CRISPR устойчивого к бактериофагам вариантного штамма.
8. Способ по п. 1, где стадии (а2) и (b2) включают сравнение конца 5’ и/или конца 3’ первого спейсера CRISPR локуса CRISPR материнского бактериального штамма с локусом CRISPR устойчивого к бактериофагам вариантного штамма.
9. Способ по п. 1, где стадии (а2) и (b2) включают амплификацию, по меньшей мере, части локуса CRISPR материнского бактериального штамма и, по меньшей мере, части локуса CRISPR устойчивого к бактериофагам вариантного штамма.
10. Способ по п. 9, где указанная амплификация проводится с использованием полимеразной цепной реакции.
11. Способ по п. 1, где стадии (а2) и (b2) включают секвенирование, по меньшей мере, части локуса CRISPR материнского бактериального штамма и, по меньшей мере, части локуса CRISPR устойчивого к бактериофагам вариантного штамма.
12. Способ по п. 9, дополнительно включающий стадию секвенирования по меньшей мере части локуса CRISPR, амплифицированного из материнского бактериального штамма, и по меньшей мере части локуса CRISPR, амплифицированного из устойчивого к бактериофагам вариантного штамма.
13. Способ по п. 1, где дополнительный спейсер в локусе CRISPR устойчивого к бактериофагам вариантного штамма образует часть элемента повтора-спейсера.
14. Способ по п. 13, где элемент повтора-спейсера содержит по меньшей мере примерно 44 нуклеотидов.
15. Способ по п. 13, где элемент повтора-спейсера содержит по меньшей мере примерно от 44 до примерно 119 нуклеотидов.
16. Способ по п. 13, где последовательность повтора элемента повтора-спейсера имеет по меньшей мере примерно 95% идентичность повтору CRISPR локуса CRISPR материнского бактериального штамма.
17. Способ по п. 13, где последовательность спейсера элемента повтора-спейсера имеет по меньшей мере примерно 95% идентичность нуклеотидной последовательности в геноме по меньшей мере одного бактериофага, используемого на стадии (а) или (b).
18. Способ по п. 1, где материнский бактериальный штамм представляет собой промышленно применимый штамм.
19. Способ по п. 18, где материнский бактериальный штамм восприимчив к инфекции, по меньшей мере, одним бактериофагом.
20. Способ по п. 18, где материнский бактериальный штамм подходит для использования в качестве заквасочной культуры, пробиотической культуры и/или культуры пищевых добавок.
21. Способ по п. 1, где материнский бактериальный штамм является чувствительным к каждому из бактериофагов, которым воздействуют на смесь.
22. Способ по п. 1, где материнский бактериальный штамм является чувствительным к некоторым из бактериофагов, которым воздействуют на смесь.
23. Способ по п. 1, где устойчивый к бактериофагам вариантный штамм отличается от материнского бактериального штамма двумя или более различными спейсерами, где каждый интегрирован в два или более отдельных локуса CRISPR.
24. Способ по п. 1, где устойчивый к бактериофагам вариантный штамм отличается от материнского бактериального штамма двумя или более различными спейсерами, где каждый интегрирован в один локус CRISPR.
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| US 5593885, 14.01.1997 | |||
| Штамм 122-АНТАгОНиСТ пОСТОРОННЕй МиКРОфлОРыСыРА | 1979 |
|
SU823423A1 |

