РОДСТВЕННЫЕ ЗАЯВКИ И ВКЛЮЧЕНИЕ В КАЧЕСТВЕ ССЫЛКИ
[001] По настоящей заявке испрашивается приоритет временной заявке на патент США № 62/181663, поданной 18 июня 2015 года и 62/245264, поданной 22 октября 2015 года.
[002] Все документы, цитируемые или упоминаемые в процитированных в настоящей заявке документах, вместе с любыми инструкциями производителей, описаниями, характеристиками и технологической картой продукта, для любого из продуктов, упомянутых в настоящем описании или в любом документе, включенном в настоящее описание в качестве ссылки, включены в настоящее описание в качестве ссылки и могут быть использованы при осуществлении изобретения. Более конкретно, все упоминаемые документы включены в качестве ссылки в той же степени, как если бы каждый отдельный документ был конкретно и индивидуально указан как включенный в качестве ссылки.
ЗАЯВЛЕНИЕ О ФЕДЕРАЛЬНОМ СПОНСИРОВАНИИ ИССЛЕДОВАНИЯ
[003] Настоящее изобретение было сделано при поддержке правительственных грантов под номерами MH100706, MH110049, DK097768, GM10407, присужденными Национальным Институтом Здравоохранения. Правительство обладает определенными правами на изобретение.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
[004] Настоящее изобретение относится к системам, способам и композициям, применяющимся для контроля экспрессии генов, включая нацеливание на последовательности, такое как изменение транскрипции генов или редактирование нуклеиновых кислот, в котором могут использоваться векторные системы, относящиеся к коротким палиндромным повторам, регулярно расположенным кластерами (CRISPR), и их компонентам.
УРОВЕНЬ ТЕХНИКИ, К КОТОРОМУ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[005] Последние достижения в способах секвенирования генома и анализа получаемых последовательностей существенно увеличили скорость каталогизации и картирования генетических факторов, ассоциированных с широким спектром биологических функций и заболеваний. Технологии направленного внесения изменений в геном необходимы для систематического поиска генетических вариаций, являющихся причинами заболеваний, а также для развития синтетической биологии, применений в биотехнологии и медицине. Несмотря на существование различных техник редактирования генома, таких как сконструированные цинковые пальцы, подобные активатору транскрипции эффекторы (TALE), хоминг-мегануклеазы, остается потребность в новых технологиях инженерии генома, в которых используются новейшие стратегии и молекулярные механизмы, и которые являются доступными, легко устанавливаемыми, масштабируемыми и способными к нацеливанию на множественно положений в пределах эукариотического генома. Это может обеспечить основной ресурс для новых подходов в генной инженерии и биотехнологии.
[006] Системы адаптивного иммунитета бактерий и архей CRISPR-Cas демонстрируют огромное разнообразие белкового состава и архитектуры геномных локусов. Локусы системы CRISPR-Cas включают более 50 семейств генов, и не существует строго универсальных генов, что указывает на быструю эволюцию и чрезвычайное разнообразие архитектуры локусов. На данный момент с использованием комплексного подхода примерно 395 профилей 93 Cas-белков были исчерпывающе идентифицированы как гены cas. Для классификации систем CRISPR-Cas используют профили генов и характерные особенности архитектуры локусов. В последней предложенной классификации системы CRISPR-Cas в самом общем виде разделены на два класса: системы 1 класса состоят из многосубъединичных эффекторных комплексов, а в системах 2 класса эффекторный модуль состоит из одного белка, например, белка Cas9 (фиг.1A и 1B). Новые эффекторные белки, ассоциированные с системами CRISPR-Cas 2 класса, могут служить мощными инструментами для геномной инженерии и прогнозирование предполагаемых новых эффекторных белков и их конструирование и оптимизация представляются важными.
[007] Цитирование или указание на какой-либо документ в настоящей заявке не является признанием того, что такой документ доступен в качестве уровня техники для настоящего изобретения.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[008] Существует острая необходимость разработки альтернативных и надежных систем и техник для нацеливания на нуклеиновые кислоты и полинуклеотиды (например, ДНК или РНК, а также любые их гибриды и производные) с широким спектром практических применений. Настоящее изобретение направлено на решение этой задачи и имеет ряд связанных с этим преимуществ. Обогащение существующего репертуара технологий нацеливания на представляющую интерес последовательность ДНК или РНК (таргетинга) в геноме и эпигеноме новыми системами, представленными в настоящем описании, может привести к преобразованиям не только в изучении, но и в изменении или редактировании определенных участков-мишеней путем прямого обнаружения, анализа и модификации. Чтобы эффективно использовать рассматриваемые в данной заявке системы геномного или эпигеномного нацеливания на ДНК или РНК без неблагоприятных последствий, важно понимать основные принципы инженерии и оптимизации этих инструментов нацеливния на ДНК или РНК.
[009] Настоящее изобретение относится к способу модификации последовательностей, ассоциированных с локусом-мишенью или находящихся непосредственно в нем, причем способ включает доставку в указанный локус не встречающейся в природе или сконструированной композиции, содержащей эффекторный белок локусов системы CRISPR-Cas типа V и один или более компонентов-нуклеиновых кислот, где эффекторный белок формирует комплекс с одним или более компонентами-нуклеиновыми кислотами и при связывании указанного комплекса с представляющим интерес локусом эффекторный белок индуцирует модификацию последовательностей, ассоциированных с представляющим интерес локусом или находящихся непосредственно в нем. В предпочтительном варианте осуществления модификация представляет собой внесение разрыва цепи. В предпочтительном варианте осуществления последовательности в локусе-мишени, ассоциированные с ним или находящиеся непосредственно в нем, включают ДНК и эффекторный белок кодируется локусами CRISPR-Cas подтипа V-A, или локусами CRISPR-Cas подтипа V-B, или локусами CRISPR-Cas подтипа V-C.
[0010] Будет понятно, что термины "фермент Cas", "фермент CRISPR", "белок CRISPR", "белок Cas" и "CRISPR Cas" используются обычно взаимозаменяемо и в каждом случае, когда они упоминаются в настоящем описании, обозначают новые эффекторные белки CRISPR, описанные далее в настоящем описании, если иное не следует из контекста, в частности при конкретном упоминании Cas9. Эффекторные белки CRISPR, описанные в настоящем описании, предпочтительно являются эффекторными белками C2c1 или C2c3.
[0011] Изобретение относится к способу модификации последовательностей, ассоциированных с или находящихся в представляющем интерес локусе-мишени, причем способ включает доставку в указанные последовательности, ассоциированные с или находящиеся в локусе не встречающейся в природе или сконструированной способами инженерии композиции, содержащей эффекторный белок локусов C2c1 или C2c3 и один или более компонентов, являющихся нуклеиновыми кислотами, где эффекторный белок C2c1 или C2c3 образует комплекс с одним или более компонентами, являющимися нуклеиновыми кислотами, и при связывании указанного комплекса с представляющим интерес локусом эффекторный белок индуцирует модификацию последовательностей, ассоциированных с или находящихся в представляющем интерес локусе-мишени. В предпочтительном варианте осуществления модификация представляет собой внесение разрыва цепи. В предпочтительном варианте осуществления эффекторный белок C2c1 или C2c3 образует комплекс с одним компонентом, являющимся -нуклеиновой кислотой, преимущественно сконструированным способами инженерии или не встречающимся в природе компонентом, являющимся нуклеиновой кислотой. Индукция модификации последовательностей, ассоциированных с или находящихся в представляющем интерес локусе-мишени, может направляться комплексом эффекторный белок C2c1 или C2c3-нуклеиновая кислота. В предпочтительном варианте осуществления изобретения указанным компонентом, являющимся нуклеиновой кислотой, является РНК CRISPR (cr-RNA). В предпочтительном варианте осуществления компонентом, являющимся нуклеиновой кислотой, является зрелая cr-РНК или направляющая РНК, где зрелая cr-РНК или направляющая РНК содержит спейсерную последовательность (или направляющую последовательность) и содержащую прямые повторы последовательность либо ее производные. В предпочтительном варианте осуществления спейсерная последовательность или ее производное включает последовательность-затравку, причем последовательность-затравка имеет критическое значение для распознавания и/или гибридизации с последовательностью локуса-мишени. В предпочтительном варианте осуществления изобретения последовательности, ассоциированные с или находящиеся в целевом представляющем интерес локусе, содержат линейную или суперспиральную ДНК.
[0012] Аспекты изобретения связаны с образованными эффекторным белком C2c1 или C2c3 комплексами, включающими один или более компонентов, являющихся не встречающимися в природе, или сконструированными способами инженерии, или модифицированными, или оптимизированными нуклеиновыми кислотами. В предпочтительном варианте осуществления изобретения компонент, являющийся нуклеиновой кислотой, комплекса может содержать направляющую последовательность, связанную с содержащей прямые повторы последовательностью, где содержащая прямые повторы последовательность содержит одну или более шпилечных структур или оптимизированные вторичные структуры. В определенных вариантах осуществления изобретения прямые повторы имеют длину минимум 16 нуклеотидов, а также одну шпилечную структуру. В следующих вариантах осуществления изобретения прямые повторы могут иметь длину более 16 нуклеотидов, предпочтительно 17 нуклеотидов, а также одну или более шпилечных структур или оптимизированных вторичных структур. В предпочтительном варианте осуществления изобретения прямой повтор может быть модифицирован таким образом, чтобы он содержал один или более связывающих белки РНК-аптамеров. В предпочтительном варианте осуществления изобретения один или более аптамеров могут быть включены в качестве части оптимизированной вторичной структуры. Такие аптамеры могут быть способны к связыванию белка оболочки бактериофага. Белок оболочки бактериофага может быть выбран из группы, включающей Qβ, F2, GA, fr, JP501, MS2, M12, R17, BZ13, JP34, JP500, KU1, M11, MX1, TW18, VK, SP, FI, ID2, NL95, TW19, AP205, ϕСb5, ϕСb8r, ϕСb12r, ϕСb23r, 7s и PRR1. В предпочтительном варианте осуществления изобретения используется белок оболочки бактериофага MS2. Изобретение также относится к компонентам вышеназванных комплексов, являющимся нуклеиновыми кислотами длиной 30 или более, 40 или более, а также 50 или более нуклеотидов.
[0013] Изобретение относится к способам редактирования генома, причем способ включает два или более этапа нацеливания на эффекторный белок C2c1 или C2c3 и расщепления. В определенных вариантах осуществления изобретения первый этап включает расщепление эффекторным белком C2c1 или C2c3 последовательностей, ассоциированных с локусом-мишенью, находящимся на большом расстоянии от последовательности-затравки, в то время как второй этап включает расщепление эффекторным белком C2c1 или C2c3 последовательностей в локусе-мишени. В предпочтительных вариантах осуществления изобретения первый этап нацеливания эффекторного белка C2c1 или C2c3 приводит к инсерции-делеции, в то время как второй этап нацеливания эффекторного белка C2c1 или C2c3 может реаприроваться посредством направляемой гомологией репарацией (HDR). В наиболее предпочтительном варианте осуществления изобретения один или более этапов нацеливания эффекторного белка C2c1 или C2c3 приводит к образованию разрыва с липкими концами, который может быть репарирован вставкой матрицы репарации.
[0014] Изобретение относится к способам редактирования или модификации генома, ассоциированных с представляющим интерес локусом-мишенью или проводимых непосредственно в нем, где способ подразумевает введение комплекса эффекторного белка C2c1 или C2c3 в любой желательный тип клеток, прокариотических или эукариотических клеток, в результате чего комплекс эффекторного белка C2c1 или C2c3 эффективно функционирует, встраивая ДНК-вставку в геном эукариотической или прокариотической клетки. В предпочтительных вариантах осуществления клетка представляет собой эукариотическую клетку, а геном представляет собой геном млекопитающих. В предпочтительных вариантах осуществления встраивание ДНК-вставки облегчается с помощью механизмов вставки гена, основанных на негомологичном соединении концов (НСК- NHEJ). В предпочтительных вариантах осуществления ДНК-вставка представляет собой экзогенно введенную ДНК-матрицу или матрицу репарации. В одном предпочтительном варианте осуществления экзогенно введенную ДНК-матрицу или матрицу репарации доставляют с комплексом эффекторного белка C2c1 или C2c3 или одним компонентом или полинуклеотидным вектором для экспрессии любого компонента комплекса. В еще более предпочтительном варианте эукариотическая клетка представляет собой не делящуюся клетку (например, не делящуюся клетку, в которой редактирование генома посредством гомологичной репарации (HDR) является особенно сложным). В предпочтительных спочсобах редактирования генома в клетках человека эффекторные белки C2c1 или C2c3 могут включать, но не ограничиваются ими, конкретные типы эффекторных белков C2c1 или C2c3, описанных в настоящем описании.
[0015] Изобретение также относится к способу модификации представляющего интерес локуса-мишени, причем способ включает доставку в указанный локус не встречающейся в природе или сконструированной способами инженерии композиции, содержащей эффекторный белок локусов С2с1 и один или более компонентов, являющихся нуклеиновыми кислотами, где эффекторный белок С2с1 образует комплекс с одним или более компонентами, являющимися нуклеиновой кислотой, и при связывании указанного комплекса с представляющим интерес локусом эффекторный белок индуцирует модификацию представляющего интерес локуса-мишени. В предпочитаемом варианте осуществления изобретения такой модификацией является разрыв цепи.
[0016] Изобретение также относится к способу модификации представляющего интерес локуса-мишени, причем способ включает доставку в указанный локус не встречающейся в природе или сконструированной способами инженерии композиции, содержащей эффекторный белок локусов С2с3 и один или более компонентов, являющихся нуклеиновыми кислотами, где эффекторный белок С2с3 образует комплекс с одним или более компонентами, являющимися нуклеиновой кислотой, и при связывании указанного комплекса с представляющим интерес локусом эффекторный белок индуцирует модификацию представляющего интерес локуса-мишени. В предпочитаемом варианте осуществления изобретения такой модификацией является разрыв цепи.
[0017] В таких способах представляющий интерес локус-мишень может являться частью молекулы ДНК. В таких способах представляющий интерес локус-мишень может быть частью молекулы ДНК in vitro.
[0018] В таких способах представляющий интерес локус-мишень может быть частью молекулы ДНК в клетке. Такая клетка может быть как прокариотической, так и эукариотической. Такая клетка может быть клеткой млекопитающего. Такая клетка млекопитающего может быть клеткой отличного от человека примата, клеткой коровы, свиньи, грызуна или мыши. Такая клетка может быть клеткой домашней птицы, рыбы или креветки. Такая клетка может быть растительной клеткой, полученной из таких растений как маниок, кукуруза, сорго, пшеница или рис. Такая клетка растения может также быть клеткой водоросли, дерева или овоща. Модификации, внесенные в такую клетку в соответствии с настоящим изобретением, могут быть выполнены таким образом, что такая клетка и ее потомки будут изменены с целью улучшить производство биологических продуктов, таких как антитела, крахмал, спирт или другой желаемый продукт клетки. Такие внесенные в клетку модификации в соответствии с настоящим изобретением могут быть способны приводить к тому, что клетка или ее дочерние клетки будут иметь свойство производить измененный продуцируемый биологический продукт.
[0019] Клетка млекопитающего может быть клеткой отличного от человека млекопитающего, например, клеткой примата, коровы, овцы, свиньи, собаки, грызуна, представителя семейства Leporidae, клеткой такого млекопитающего как обезьяна, клеткой коровы, овцы, свиньи, собаки, кролика, крысы или мыши. Клетка может быть эукариотической клеткой не являющегося млекопитающим животного, такого как домашняя птица (например, курицы), позвоночной рыбы (например, лосося) или беспозвоночного (например, устрицы, двустворчатого моллюска, лобстера, креветки). Такая клетка может быть также клеткой растения. Такая растительная клетка может быть получена из однодольного растения или двудольного растения, а также сельскохозяйственного растения или злака, такого как маниок, кукуруза, сорго, соя, пшеница, овес или рис. Такая растительная клетка может быть также быть клеткой водоросли, дерева или растения-продуцента, фрукта или овоща (например, дерева такого как цитрусовые деревья, например, апельсина, грейпфрута или лимона; персика или нектарина; яблони и груши; орехоносного дерева, такого как миндаль, грецкий орех или фисташка; пасленового растения; растения рода Brassica; растения рода Lactuca; растения рода Spinacia; растения рода Capsicum; хлопка, табака, спаржи, моркови, капусты, брокколи, цветной капусты, томата, баклажана, перца, латука, шпината, клубники, черники, малины, ежевики, винограда, кофе, какао и т.д.).
[0020] Изобретение также относится к сконструированному способами инженерии белку CRISPR, комплексу, композиции, системе, вектору, клетке или клеточной линии по изобретению для использования в модификации представляющего интерес локуса в клетке. Указанная модификация предпочтительно включает приведение клетки в контакт с любой из вышеописанных композиций или любой из вышеописанных систем. Также изобретение относится к применению сконструированного способами инженерии белка CRISPR, комплекса, композиции, системы, вектора, клетки или клеточной линии по изобретению для изготовления лекарственного средства для модификации представляющего интерес локуса в клетке.
[0021] Также изобретение относится к сконструированному способами инженерии белку CRISPR, комплексу, композиции, системе, вектору, клетке или клеточной линии по изобретению для применения в модификации представляющего интерес локуса в клетке. Также изобретение относится к применению сконструированного способами инженерии белка CRISPR, комплекса, композиции, системы, вектора, клетки или клеточной линии по изобретению при получении лекарственного средства для модификации представляющего интерес локуса в клетке. Указанная модификация предпочтительно включает приведение клетки в контакт с любой из вышеописанных композиций или любой из вышеописанных систем.
[0022] Изобретение относится к способу модификации представляющего интерес локуса-мишени, причем способ включает доставку в указанный локус не встречающейся в природе или сконструированной способами инженерии композиции, содержащей эффекторный белок локусов CRISPR-Cas типа IV и один или более компонентов, являющихся нуклеиновыми кислотами, где эффекторный белок образует комплекс с одним или более компонентами, являющимися нуклеиновыми кислотами, причем после связывания указанного комплекса с представляющим интерес локусом эффекторный белок индуцирует модификацию представляющего интерес локуса-мишени. В предпочитаемом варианте осуществления изобретения такой модификацией является разрыв цепи.
[0023] В предпочтительном варианте осуществления представляющий интерес локус-мишень представляет собой ДНК.
[0024] В таких способах представляющий интерес локус-мишень может являться частью молекулы ДНК или молекулы РНК. В предпочитаемом варианте осуществления изобретения представляющий интерес локус-мишень представляет собой РНК.
[0025] В таких методах представляющий интерес локус-мишень, может находиться в молекуле ДНК внутри клетки. Клетка может быть прокариотической или эукариотической клеткой. Клетка может быть клеткой млекопитающего. Такая клетка млекопитающего может быть клеткой отличного от человека млекопитающего, например, клеткой примата, коровы, овцы, свиньи, собаки, грызуна, представителя семейства Leporidae, клеткой такого млекопитающего как обезьяна, клеткой коровы, овцы, свиньи, собаки, кролика, крысы или мыши. Такая клетка может быть эукариотической клеткой не являющегося млекопитающим животного, такого как домашняя птица (например, курица), позвоночная рыба (например, лосось) или беспозвоночное (например, устрица, двустворчатый моллюск, лобстер, креветка). Такая клетка может быть также клеткой растения. Такая растительная клетка может быть получена из однодольного растения или двудольного растения, а также сельскохозяйственного растения или злака, такого как маниок, кукуруза, сорго, соя, пшеница, овес или рис. Такая растительная клетка может быть также клеткой водоросли, дерева или растения-продуцента, фрукта или овоща (например, дерева, такого как цитрусовые деревья, например, апельсин, грейпфрут или лимон; персик или нектарин; яблоня или груша; орехоносное дерево, такое как миндаль, грецкий орех или фисташка; пасленового растения; растения рода Brassica; растения рода Lactuca; растения рода Spinacia; растения рода Capsicum; хлопка, табака, спаржи, моркови, капусты, брокколи, цветной капусты, томата, баклажана, перца, латука, шпината, клубники, черники, малины, ежевики, винограда, кофе, какао и т.д.)
[0026] В любом из описанных способов представляющий интерес локус-мишень может представлять собой геномный или эпигеномный локус. В любом из описанных способов указанный комплекс может доставляться несколькими направляющими молекулами для мультиплексного применения. В любом из описанных способов может быть использовано более одного белка(ов).
[0027] В предпочтительных вариантах осуществления изобретения биохимическое или in vitro или in vivo расщепление последовательностей, связанных с или непосредственно находящихся в представляющем интерес локусе-мишени, происходит без предполагаемой трансактивирующей последовательности cr-РНК (tracr РНК), например, посредством расщепления при помощи эффекторного белка C2c1 или C2c3. В других вариантах осуществления изобретения расщепление может происходить при помощи предполагаемой трансактивирующей последовательности cr-РНК (tracr РНК), например, посредством расщепления другими эффекторными белками семейства CRISPR.
[0028] В любом из описанных способов эффекторный белок (например, C2c1 или C2c3) и компоненты, являющиеся нуклеиновыми кислотами, могут быть предоставлены посредством одной или более полинуклеотидных молекул, кодирующих белок и/или компонент(ы), являющийся нуклеиновой кислотой, причем одна или более полинуклеотидных молекул функционально приспособлены для экспрессии белка и/или компонента(ов), являющегося нуклеиновой кислотой. Одна или более полинуклеотидных молекул могут содержать один или более регуляторных элементов, функционально приспособленных для экспрессии белка и/или компонента(ов), являющегося нуклеиновой кислотой. Одна или более полинуклеотидных молекул может находиться в одном или более векторах. Изобретение охватывает такие полинуклеотидные молекулы, например, полинуклеотидные молекулы, функционально приспособленные для экспрессии белка и/или компонента(ов), являющегося нуклеиновой кислотой, а также указанного вектора (ов).
[0029] Изобретение также относится к сконструированному способами инженерии белку CRISPR, комплексу, композиции, системе, вектору, клетке или клеточной линии по изобретению для применения для модификации представляющего интерес локуса-мишени в клетке. Указанная модификация предпочтительно включает приведение клетки в контакт с любой из вышеописанных композиций или любой из вышеописанных систем. Изобретение также относится к применению сконструированного белка CRISPR, комплекса, композиции, системы, вектора, клетки или клеточной линии по изобретению для получения лекарственного средства для модификации локуса-мишени в клетке.
[0030] Также изобретение относится к сконструированному способами инженерии белку CRISPR, комплексу, композиции, системе, вектору, клетке или клеточной линии по изобретению для применения для модификации представляющего интерес локуса-мишени в клетке. Изобретение также относится к применению сконструированного белка CRISPR, комплекса, композиции, системы, вектора, клетки или клеточной линии по изобретению для получения лекарственного средства для модификации представляющего интерес локуса-мишени. Указанная модификация предпочтительно включает приведение клетки в контакт с любой из вышеописанных композиций или любой из вышеописанных систем.
[0031] Также изобретение относится к способу модификации представляющего интерес локуса-мишени, причем указанный способ включает доставку в указанный локус не встречающейся в природе или сконструированной способами инженерии композиции, содержащей эффекторный белок C2c2 локуса и один или более компонентов, являющихся нуклеиновыми кислотами, где эффекторный белок C2c2 образует комплекс с одним или более компонентами, являющимися нуклеиновыми кислотами, и при связывании указанного комплекса с представляющим интерес локусом эффекторный белок осуществляет модификацию представляющего интерес локуса-мишени. В предпочтительном варианте осуществления модификация представляет собой внесение одноцепочечного разрыва.
[0032] В таких способах интересующий целевой локус может содержаться в молекуле ДНК in vitro. В таких методах интересующий целевой локус может содержаться в молекуле ДНК внутри клетки. Предпочтительно, в таких способах представляющий интерес локус-мишень может быть включен в молекулу РНК in vitro. Также предпочтительно, чтобы в таких способах представляющий интерес локус-мишень мог быть включен в молекулу РНК внутри клетки. Клетка может быть прокариотической или эукариотической клеткой. Клетка может быть клеткой млекопитающего. Клетка может быть клеткой грызуна. клетка может быть клеткой мыши.
[0033] Изобретение также относится к сконструированному белку CRISPR, комплексу, композиции, системе, вектору, клетке или клеточной линии по изобретению для применения для модификации представляющего интерес локуса в клетке. Указанная модификация предпочтительно включает приведение клетки в контакт с любой из вышеописанных композиций или любой из вышеописанных систем. Изобретение также обеспечивает использование сконструированного способами инженерии белка CRISPR, комплекса, композиции, системы, вектора, клетки или клеточной линии в соответствии с изобретением при получении лекарственного средства для модификации представляющего интерес локуса в клетке.
[0034] Также изобретение относится к сконструированному белку CRISPR, комплексу, композиции, системе, вектору, клетке или клеточной линии для применения для модификации представляющего интерес локуса в клетке согласно изобретению. Изобретение также относится к применению сконструированного способами инженерии белка CRISPR, комплекса, композиции, системы, вектора, клетки или клеточной линии при получении лекарственного средства по изобретению для модификации интересующего локуса в клетке. Указанная модификация предпочтительно включает приведение клетки в контакт с любой из вышеописанных композиций или любой из вышеописанных систем.
[0035] В любом из описанных способов представляющий интерес локус-мишень может быть геномным или эпигеномным представляющим интерес локусом. В любом из описанных способов комплекс может быть доставлен с использованием множественных направляющих молекул для мультиплексного применения. В любом из описываемых способов может быть использовано более одного белка(ов).
[0036] В других аспектах изобретения компоненты, являющиеся нуклеиновыми кислотами, могут содержать предполагаемую последовательность РНК CRISPR (crRNA) и/или предполагаемую трансактивирующую последовательность cr-РНК (tracr-РНК). В некоторых вариантах осуществления расщепление, такое как биохимическое или in vitro расщепление, или расщепление в клетках, может происходить без предполагаемой трансактивирующей последовательности cr-РНК (tracr РНК). В других вариантах осуществления расщепление, такое как биохимическое или in vitro расщепление, или расщепление в клетках, может происходить с предполагаемой трансактивирующей последовательностью cr-РНК (tracr-РНК);
[0037] В некоторых вариантах осуществления, где эффекторный белок представляет собой эффекторный белок локусов CRISPR-Cas типа V, такой как эффекторный белок C2c1 локусов CRISPR-Cas или эффекторный белок C2c3 локусов CRISPR-Cas, предпочтительно эффекторный белок C2c1 локусов CRISPR-Cas, компоненты, являющиеся нуклеиновыми кислотами, могут содержать предполагаемую последовательность РНК CRISPR (cr-РНК) и предполагаемую транс-активирующую последовательность cr-РНК (tracr РНК).
[0038] В дальнейших аспектах изобретения компоненты, являющиеся нуклеиновыми кислотами, могут содержать предполагаемую последовательность РНК CRISPR (cr-РНК) и не включать какую-либо предполагаемую транс-активирующую последовательность cr-РНК (tracr-РНК). В качестве неограничивающего примера, заявители предполагают, что в таких случаях пре-cr-РНК может содержать вторичную структуру, достаточную для процессинга, чтобы получить зрелую cr-РНК, а также для нагрузки эффекторного белка посредством cr-РНК. В качестве неограничивающего примера, такая вторичная структура может содержать, по существу состоять из или состоять из шпильки внутри пре-cr-РНК, более конкретно, в пределах прямого повтора.
[0039] В некоторых вариантах осуществления, где эффекторный белок представляет собой эффекторный белок локусов CRISPR-Cas типа VI, такой как эффекторный белок C2c2, компоненты, являющиеся нуклеиновыми кислотами, могут содержать предполагаемую последовательность типа CRISPR (cr-РНК) и не включать какую-либо предполагаемую транс-активирующую последовательность cr-РНК (tracr РНК).
[0040] В любом из описанных способов эффекторный белок и компоненты, являющиеся нуклеиновыми кислотами, могут быть доставлены посредством одной или более полинуклеотидных молекул, кодирующих белок и/или компоненты, являющиеся нуклеиновыми кислотами, и где одна или более полинуклеотидных молекул функционально организованы для экспрессии белка и/или компонента(ов), являющегося нуклеиновой кислотой. Одна или более полинуклеотидных молекул могут содержать один или более регуляторных элементов, функционально организованных для экспрессии белка и/или компонента, являющегося нуклеиновой кислотой. Одна или более полинуклеотидных молекул могут быть включены в один или более векторов. В любом из описанных способов представляющий интерес локус-мишень может являться геномным или эпигеномным локусом. В любом из описанных способов комплекс может доставляться посредством нескольких направляющих молекул для мультиплексного применения. В любом из описанных способов может быть использовано более одного белка(ов).
[0041] В любом из описанных способов разрыв цепи может быть одноцепочечным или двухцепочечным разрывом.
[0042] Регуляторные элементы могут включать индуцибельные промоторы. Полинуклеотиды и/или векторные системы могут включать индуцибельные системы.
[0043] В любом из описанных способов одна или более полинуклеотидных молекул могут быть включены в систему доставки, или один или более векторов могут быть включены в систему доставки.
[0044] В любом из описанных способов такую не встречающуюся в природе или сконструированную способами инженерии композицию можно доставлять при помощи липосом и частиц, включая наночастицы, экзосом, микровезикул, генных пушек или одного или более вирусных векторов, например, при помощи молекулы нуклеиновой кислоты или вирусного вектора.
[0045] Также изобретение относится к не встречающейся в природе или сконструированной способами инженерии композиции, которая представляет собой композицию, имеющую характеристики, описанные в настоящем описании или определенные в любом из описанных в настоящем описании способов.
[0046] Также изобретение относится к сконструированному способами инженерии белку CRISPR, комплексу, композиции, системе, вектору, клетке или клеточной линии по изобретению для использования в терапии.
[0047] В определенных вариантах осуществления изобретение, таким образом, относится к не встречающейся в природе или сконструированной способами инженерии композиции, такой как, в частности, композиция, способная к или приспособленная для модификации представляющего интерес локуса-мишени, причем указанная композиция содержит эффекторный белок локуса CRISPR-Cas типа V и один или более компонентов, являющихся нуклеиновыми кислотами, где эффекторный белок образует комплекс с одним или более компонентами, являющимися нуклеиновыми кислотами, и при связывании указанного комплекса с представляющим интерес локусом эффекторный белок вызывает модификацию представляющего интерес локуса-мишени. В определенных вариантах осуществления эффекторный белок может кодироваться подтипом V-A локусов CRISPR-Cas или подтипом V-B локусов CRISPR-Cas или подтипом V-C локусов CRISPR-Cas. В определенных вариантах осуществления эффекторный белок может представлять собой эффекторный белок C2c1 или эффекторный белок C2c3.
[0048] Таким образом, в некоторых вариантах осуществления изобретение относится к встречающейся в природе или сконструированной способами инженерии композиции, в частности, композиции, способной или приспособленной для модификации представляющего интерес локуса-мишени, причем указанная композиция включает эффекторный белок локусов CRISPR-Cas типа VI и один или более компонентов, являющихся нуклеиновыми кислотами, причем такой эффекторный белок формирует комплекс с одним или более компонентами, являющимися нуклеиновыми кислотами, и при связывании указанного комплекса с представляющим интерес локусом такой эффекторный белок индуцирует модификацию представляющего интерес локуса-мишени. В определенных вариантах осуществления изобретения такой эффекторный белок может быть эффекторным белком локусов С2с2.
[0049] Также в следующем аспекте настоящее изобретение относится к не встречающейся в природе или сконструированной способами инженерии композиции, в частности, композиции, способной или приспособленной для модификации представляющего интерес локуса-мишени, причем указанная композиция включает: (a) молекулу направляющей РНК (или комбинацию молекул направляющих РНК, например, первую молекулу направляющей РНК-гида или вторую молекулу направляющей РНК) или нуклеиновую кислоту, кодирующую молекулу направляющей РНК (или одну или более нуклеиновых кислот, кодирующих комбинацию молекул направляющих РНК); (b) эффекторный белок локусов CRISPR-Cas типа V или нуклеиновую кислоту, кодирующую эффекторный белок локусов CRISPR-Cas типа V. В определенных вариантах осуществления эффекторный белок может кодироваться подтипом V-A локусов CRISPR-Cas или подтипом V-B локусов CRISPR-Cas или подтипом V-C локусов CRISPR-Cas. В определенных вариантах осуществления эффекторный белок может представлять собой эффекторный белок C2c1 локусов CRISPR Cas или эффекторный белок C2c3 локусов CRISPR Cas.
[0050] Еще один аспект изобретения также относится к не встречающейся в природе или сконструированной способами инженерии композиции, такой как, в частности, композиция, способная к или приспособленная для модификации представляющнего интерес локуса-мишени, причем указанная композиция содержит: (a) молекулу направляющей РНК (или комбинацию молекул направляющей РНК, например, первую молекулу направляющей РНК и вторую молекулу направляющей РНК) или нуклеиновую кислоту, кодирующую комбинацию молекул направляющих РНК; (b) эффекторный белок локусов CRISPR-Cas типа VI или нуклеиновую кислоту, кодирующую эффекторный белок локусов CRISPR Cas типа VI. В некоторых вариантах осуществления эффекторный белок может представлять собой эффекторный белок C2c2.
[0051] В следующем аспекте изобретение также относится к не встречающейся в природе или сконструированной способами инженерии композиции, содержащей: (I) одну или более полинуклеотидных последовательностей системы CRISPR-Cas, содержащих (а) направляющую последовательность, способную к гибридизации с последовательностью-мишенью в полинуклеотидном локусе, (b) последовательность-партнер tracr, (c) последовательность tracr-РНК и (II) вторую полинуклеотидную последовательность, кодирующую эффекторный белок локусов CRISPR-Cas типа V, где при транскрипции последовательность-партнер tracr гибридизуется с последовательностью tracr-РНК и направляющая последовательность направляет специфическое для последовательности связывание комплекса CRISPR с последовательностью-мишенью и где комплекс CRISPR содержит эффекторный белок локусов CRISPR-Cas типа V в комплексе с (1) направляющей последовательностью, которая гибридизируется с последовательностью-мишенью, и (2) последовательностью-партнером tracr, которая гибридизируется с последовательностью tracr-РНК. В определенных вариантах осуществления эффекторный белок может кодироваться подтипом V-A локусов CRISPR-Cas или подтипом V-B локусов CRISPR-Cas или подтипом V-C локусов CRISPR-Cas. В определенных вариантах осуществления эффекторный белок может представлять собой эффекторный белок C2c1 локусов CRISPR-Cas или эффекторный белок C2c3 локусов CRISPR-Cas.
[0052] В следующем аспекте изобретение также относится к не встречающейся в природе или сконструированной способами инженерии композиции, содержащей: (I) одну или более полинуклеотидных последовательностей системы CRISPR-Cas, содержащих (а) направляющую последовательность, способную к гибридизации с последовательностью-мишенью в полинуклеотидном локусе, (b) последовательность-партнер tracr, (c) последовательность tracr-РНК и (ii) вторую полинуклеотидную последовательность, кодирующую эффекторный белок локусов CRISPR Cas типа VI, где при транскрипции последовательность-спутник tracr гибридизуется c последовательностью tracr-РНК и направляющая последовательность направляет специфическое для последовательности связывание комплекса CRISPR с последовательностью-мишенью и где комплекс CRISPR содержит эффекторный белок локусов CRISPR Cas типа VI в комплексе с (1) направляющей последовательностью, которая гибридизируется с последовательностью-мишенью, и (2) последовательностью-партнером tracr, которая гибридизируется с последовательностью tracr-РНК. В определенных вариантах осуществления эффекторный белок может кодироваться подтипом V-A локусов CRISPR-Cas или подтипом V-B локусов CRISPR-Cas или подтипом V-C локусов CRISPR-Cas. В некоторых вариантах осуществления эффекторный белок может представлять собой эффекторный белок C2c2 локусов CRISPR-Cas.
[0053] В определенных вариантах осуществления tracr-РНК может не требоваться. Следовательно, в некоторых вариантах осуществления изобретение также относится к не встречающейся в природе или сконструированной способами инженерии композиции, содержащей: (I) одну или более полинуклеотидных последовательностей системы CRISPR-Cas, содержащих (а) направляющую последовательность, способную к гибридизации с последовательностью-мишенью в полинуклеотидном локусе, и (b) последовательность прямого повтора, и (II) вторую полинуклеотидную последовательность, кодирующую эффекторный белок локусов CRISPR-Cas типа V или типа VI, где при транскрипции направляющая последовательность управляет специфическим для последовательности связыванием комплекса CRISPR с последовательностью-мишенью, и где комплекс CRISPR включает эффекторный белок локусов CRISPR-Cas типа V или типа VI в комплексе с (I) направляющей последовательностью, которая гибридизована с последовательностью-мишенью, и (2) последовательностью прямого повтора. В определенных вариантах осуществления изобретения эффекторный белок типа V может кодироваться подтипами V-A локусов CRISPR-Cas, или подтипами V-B локусов CRISPR-Cas, или подтипами V-C локусов CRISPR-Cas. В определенных вариантах осуществления эффекторный белок может представлять собой эффекторный белок C2c1 локусов CRISPR-Cas или эффекторный белок C2c3 локусов CRISPR-Cas. Предпочтительно, чтобы эффекторный белок представлял собой эффекторный белок локусов CRISPR-Cas типа VI. Более предпочтительно, чтобы эффекторный белок был эффекторным белком C2c2 локусов CRISPR-Cas. Без наложения ограничений, заявители предполагают, что в таких случаях последовательность прямого повтора может содержать вторичную структуру, достаточную для загрузки эффекторного белка посредством cr-РНК. В качестве неограничивающего примера, такая вторичная структура может включать, по существу состоять или состоять из шпильки внутри прямого повтора.
[0054] Также изобретение относится к векторной системе, включающей один или более векторов, причем один или более векторов содержат одну или более полинуклеотидных молекул, кодирующих компоненты не встречающейся в природе или сконструированной способами инженерии композиции, которая представляет собой композицию, обладающую характеристиками, которые определены в любом из описанных в настоящем описании способов.
[0055] Также изобретение относится к системе доставки, содержащей один или более векторов или одну или более полинуклеотидных молекул, причем один или более векторов или полинуклеотидных молекул включают одну или более полинуклеотидных молекул, кодирующих компоненты не встречающейся в природе или сконструированной способами инженерии композиции, которая представляет собой композицию, имеющую характеристики, описанные в настоящем описании или определенными в любом из описанных способов.
[0056] Также изобретение относится к не встречающейся в природе или сконструированной способами инженерии композиции, или одному или более полинуклеотидам, кодирующим компоненты указанной композиции, или к вектору либо системе доставки, содержащим один или более полинуклеотидов, кодирующих компоненты указанной композиции, для использования в терапевтическом способе лечения. Такой терапевтический способ лечения может включать генное или геномное редактирование или генную терапию.
[0057] Изобретение также относится к сконструированному белку CRISPR, комплексу, композиции, системе, вектору, клетке или клеточной линии по изобретению для использования в терапии заболеваний, нарушений или инфекционных заболеваний у индивидуума, нуждающегося в этом. Заболевание, расстройство или инфекция могут включать вирусную инфекцию. Вирусная инфекция может представлять собой гепатит В (HBV). Изобретение также относится к использованию сконструированного белка CRISPR, комплекса, композиции, системы, вектора, клетки или клеточной линии по изобретению для получения лекарственного средства для лечения заболевания, расстройства или инфекции у индивидуума, нуждающегося в этом. Болезнь, расстройство или инфекция могут включать вирусную инфекцию.
[0058] Также изобретение относится к вычислительным (биоинформатическим) способам и алгоритмам прогнозирования новых систем CRISPR-Cas 2 класса и идентификации их компонентов.
[0059] Также изобретение относится к способам и композициям, в которых один и несколько аминокислотных остатков эффекторного белка могут быть модифицированы, например, в сконструированном способами инженерии или не встречающемся в природе эффекторном белке, или C2c1 или C2c3. В одном из вариантов осуществления изобретения модификация может включать мутацию одного или более аминокислотных остатков эффекторного белка. Одна или более мутаций могут быть локализованы в одном или более каталитических доменах эффекторного белка. Нуклеазная активность такого эффекторного белка может быть снижена или отсутствовать по сравнению с эффекторным белком, не имеющим указанную одну или более мутаций. Такой эффекторный белок может быть не способен направлять расщепление цепи ДНК или РНК в представляющем интерес локусе-мишени. Предпочтительный вариант осуществления включает две таких мутаций. В предпочтительном варианте осуществления изобретения такие один или более аминокислотных остатков модифицированы в эффекторном белке C2c1 или C2c3, например, в сконструированном способами инженерии или не встречающемся в природе эффекторном белке, или C2c1 или C2c3.
[0060] Также изобретение относится к одной или более мутациям или к двум или более мутациям в каталитически активном домене эффекторного белка, содержащего домен RuvC. В некоторых вариантах осуществления изобретения такой домен RuvC может являться одним из доменов RuvCI, RuvCII или RuvCIII, либо каталитически активным доменом, гомологичным доменам RuvCI, RuvCII и RuvCIII и т. д. или любому соответствующему домену, как описано в любом из описанных в настоящем описании способов. В некоторых вариантах осуществления такие одна или более мутаций могут быть локализованы в каталитически активном домене эффекторного белка, являющемся доменом HEPN, или каталитически активном домене, гомологичном домену HEPN. Такой эффекторный белок может иметь один или более доменов, являющихся сигналом ядерной локализации (NLS). Такие один или более гетерологичных функциональных доменов могут содержать по меньшей мере два или более доменов NLS. Такие один или более доменов NLS могут быть расположены в конце или вблизи конца последовательности эффекторного белка (например, C2c1 или C2c3), а в случае двух и более NLS каждый из этих двух может быть расположен вблизи конца эффекторного белка (например, C2c1 или C2c3). Такие один или более гетерологичных функциональных доменов могут являться одним или более доменами активации транскрипции. В предпочтительном варианте осуществления домен активации транскрипции может включать VP64. Такие один или более функциональных доменов могут являться одним или более доменами репрессии транскрипции. В предпочтительном варианте осуществления такой домен репрессии транскрипции является доменом KRAB или доменом SID (например, SID4X). Такие один или более гетерологичных функциональных доменов могут являться одним или более нуклеазными доменами. В предпочтительном варианте осуществления нуклеазный домен является доменом Fok1.
[0061] Также изобретение относится к одному или более гетерологичным функциональным доменам следующих типов активности: метилазная активность, деметилазная активность, активация транскрипции, репрессия транскрипции, активность фактора терминации транскрипции, модификация гистонов, нуклеазная активность, расщепление одноцепочечной РНК, расщепление двухцепочечной РНК, расщепление одноцепочечной ДНК, расщепление двухцепочечной ДНК и связывание нуклеиновых кислот. По меньшей мере один или более гетерологичных функциональных доменов могут находиться в N-конце эффекторного белка или вблизи него, при этом также по меньшей мере один или более гетерологичных функциональных доменов могут находиться в C-конце эффекторного белка или вблизи него. Такой один или более гетерологичных функциональных доменов могут быть слиты с таким эффекторным белком. Такие один или более гетерологичных функциональных доменов могут быть присоединены к такому эффекторному белку. Такие один или более гетерологичных функциональных доменов могут быть соединены с таким эффекторным белком посредством связующей группы.
[0062] Также изобретение относится к эффекторному белку, включающему эффекторный белок из организма, относящегося к одному из следующих родов: Streptococcus, Campylobacter, Nitratifractor, Staphylococcus, Parvibaculum, Roseburia, Neisseria, Gluconacetobacter, Azospirillum, Sphaerochaeta, Lactobacillus, Eubacterium, Corynebacter, Carnobacterium, Rhodobacter, Listeria, Paludibacter, Clostridium, Lachnospiraceae, Clostridiaridium, Leptotrichia, Francisella, Legionella, Alicyclobacillus, Methanomethyophilus, Porphyromonas, Prevotella, Bacteroidetes, Helcococcus, Letospira, Desulfovibrio, Desulfonatronum, Opitutaceae, Tuberibacillus, Bacillus, Brevibacilus, Methylobacterium или Acidaminococcus. Такой эффекторный белок может представлять собой химерный эффекторный белок, первый фрагмент которого получен из первого ортолога эффекторного белка, а второй фрагмент - из второго ортолога эффекторного белка, причем первый и второй ортологи эффекторного белка различаются. По меньшей мере один из первого и второго ортологов эффекторного белка может представлять собой эффекторный белок одного из следующих организмов: Streptococcus, Campylobacter, Nitratifractor, Staphylococcus, Parvibaculum, Roseburia, Neisseria, Gluconacetobacter, Azospirillum, Sphaerochaeta, Lactobacillus, Eubacterium, Corynebacter, Carnobacterium, Rhodobacter, Listeria, Paludibacter, Clostridium, Lachnospiraceae, Clostridiaridium,Leptotrichia, Francisella, Legionella, Alicyclobacillus, Methanomethyophilus, Porphyromonas, Prevotella, Bacteroidetes, Helcococcus, Letospira, Desulfovibrio, Desulfonatronum, Opitutaceae, Tuberibacillus, Bacillus, Brevibacillus, Methylobacterium или Acidaminococcus.
[0063] В некоторых вариантах осуществления изобретения такой эффекторный белок, в частности, эффекторный белок локусов типа V, а более конкретно эффекторный белок локусов типа V-B, в частности C2c1p, может происходить из, быть получен из или являться производным метаболизма бактерий, относящегося к таким таксонам как Bacilli, Verrucomicrobia, альфа-протеобактерии или дельта-протеобактерии. В некоторых вариантах осуществления такой эффекторный белок, в частности, эффекторный белок локусов типа V, более конкретно эффекторный белок локусов V-B типа, в частности C2c1p, может происходить из, быть выделен из или являться производным метаболизма бактерий, относящихся к одному из следующих родов: Alicyclobacillus, Desulfovibrio, Desulfonatronum, Opitutaceae, Tuberibacillus, Bacillus, Brevibacillus, Desulfatirhabdium, Citrobacter, и Methylobacterium. В некоторых вариантах осуществления такой эффекторный белок, в частности, эффекторный белок локусов типа V, более конкретно эффекторный белок локусов типа V-B, в частности C2c1p, может происходить из, быть выделен из или являться производным метаболизма следующих бактерий: Alicyclobacillus acidoterrestris (например, ATCC 49025), Alicyclobacillu scontaminans (например, DSM 17975), Desulfovibr ioinopinatus (например, DSM 10711), Desulfonatronum thiodismutans (например, штамм MLF-1), бактерии Opitutaceae TAV5, Tuberibacillus calidus (например, DSM 17572), Bacillus thermoamylovorans (например, штамм B4166), Brevibacillus sp. CF112, Bacillus sp. NSP2.1, Desulfatirhabdium butyrativorans (например, DSM 18734), Alicyclobacillus herbarius (например, DSM 13609), Citrobacter freundii (например, ATCC 8090), Brevibacillus agri (например, BAB-2500), Methylobacterium nodulans (например, ORS 2060). В некоторых вариантах осуществления такой эффекторный белок, в частности, эффекторный белок локусов типа V, более конкретно эффекторный белок локусов типа V-B, в частности C2c1p, может происходить из, быть выделен из являться производным метаболизма бактерий из перечня, приведенного в таблице на фиг.41A-B.
[0064] В некоторых вариантах осуществления такой эффекторный белок, а именно белок локусов типа V, более конкретно эффекторный белок локусов типа V-B, в частности C2c1p, может содержать, по существу состоять или состоять из аминокислотной последовательности, выбранной из перечня, включающего аминокислотные последовательности, представленные на множественном выравнивании последовательностей на фиг.13D-H.
[0065] В некоторых вариантах осуществления локус типа V-B, как подразумевается в настоящем описании, может кодировать слитую конструкцию Cas1-Cas4, Cas2 и эффекторный белок С2с1p. В некоторых вариантах осуществления локус типа V-B, как подразумевается в настоящем описании, может примыкать к последовательности CRISPR. Характерная организация локусов типа V-B проиллюстрирована на фиг.9 и фиг.41A-B.
[0066] В некоторых вариантах осуществления белок Cas1, кодируемый локусом типа V-B, как подразумевается в настоящем описании, может группироваться с системой типа I-U. На фиг.10A и 10B и 10C-V проиллюстрировано дерево Cas1, включая Cas1, кодируемый репрезентативными локусами типа V-B.
[0067] В определенных вариантах осуществления изобретения эффекторный белок, конкретно эффекторный белок локусов типа V, более конкретно эффекторный белок локусов типа V-B, а частности - C2c1p, например, природный C2c1p, может иметь длину от приблизительно 1100 до приблизительно 1500 аминокислот, например, от приблизительно 1100 до приблизительно 1200, или от приблизительно 1200 до приблизительно 1300 аминокислот, или от приблизительно 1300 до приблизительно 1400 аминокислот, или от приблизительно 1400 до приблизительно 1500 аминокислот, например, приблизительно 1100, приблизительно 1200, приблизительно 1300, приблизительно 1400, или приблизительно 1500 аминокислот.
[0068] В некоторых вариантах осуществления изобретения эффекторный белок, конкретно эффекторный белок локусов типа V, более конкретно эффекторный белок локусов типа V-B, в частности - C2c1p, и предпочтительно C-концевая часть указанного эффекторного белка, содержит три каталитических мотива RuvC-подобной нуклеазы (т.е. RuvCI, RuvCII и RuvCIII). В некоторых вариантах осуществления изобретения указанный эффекторный белок и предпочтительно С-концевая часть указанного эффекторного белка может далее содержать область, соответствующую мостиковой спирали (также известной как богатый аргинином кластер), которая в белке Cas9 участвует в связывании cr-РНК. В некоторых вариантах осуществления изобретения указанный эффекторный белок и предпочтительно C-концевая часть указанного белка может далее включать домен, содержащий цинковый палец, который может быть неактивным (т.е. который не связывает цинк, например, в котором Zn-связывающие остатки цистеина отсутствуют). В некоторых вариантах осуществления изобретения указанный эффекторный белок и предпочтительно C-концевая часть указанного эффекторного белка может содержать три каталитических мотива RuvC-подобной нуклеазы (т.е. RuvCI, RuvCII и RuvCIII), область, соответствующую мостиковой спирали, домен, содержащий цинковый палец, предпочтительно в следующем порядке, от N- к C-концу: RuvCI-мостиковая спираль-RuvCII-цинковый палец-RuvCIII См. Фиг.11, фиг.12 и фиг.13A и 13C для иллюстрации доменной архитектуры представителей эффекторных белков типа V-B.
[0069] В некоторых вариантах осуществления изобретения локусы типа V-B, как описано в настоящем описании, могут содержать CRISPR-повторы длиной от 30 до 40 п.н., более типично от 34 до 38 п.н., еще более типично - от 36 до 37 п.н., например, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39 или 40 п.н.
[0070] В некоторых вариантах осуществления изобретения эффекторный белок, конкретно эффекторный белок локусов типа V, более конкретно - эффекторный белок локусов типа V-C, в частности - C2c3p может происходить, быть изолирован или получен из бактериального метагенома, выбранного из бактериальных метагеномов, перечисленных в таблице на фиг.43 A-B.
[0071] В некоторых вариантах осуществления изобретения эффекторный белок, конкретно эффекторный белок локуcов типа V, более конкретно эффекторный белок локусов типа V-C, в частности - C2c3p, может включать, состоять в основном из или состоять только из аминокислотной последовательности, выбранной из группы аминокислотных последовательностей, показанных на множественном выравнивании последовательностей на фиг.131.
[0072] В некоторых вариантах осуществления изобретения локус типа V-C, как описано в настоящем описнаии, может кодировать эффекторный белок Cas1 и C2c3p. См. Фиг.14 и фиг.43A-B для иллюстрации организации характерных локусов типа V-С.
[0073] В некоторых вариантах осуществления изобретения белок Cas1, кодируемый локусом типа V-С, как описано в настоящем описании, может образовывать кластеры с системой типа I-B. См. Фиг.10A и 10B и фиг.10C-W, иллюстрирующие дерево Cas-белков, включающее белок Cas1, кодируемый характерными локусами типа V-С.
[0074] В некоторых вариантах осуществления изобретения эффекторный белок, а именно белок локусов типа V, более конкретно эффекторный белок локусов типа V-С, еще более конкретно - C2c3p, в частности C2c3p, может иметь длину от приблизительно 1100 до приблизительно 1500 аминокислот, например, от приблизительно 1100 до приблизительно 1200 аминокислот, или от приблизительно 1200 до приблизительно 1300 аминокислот, или от приблизительно 1300 до приблизительно 1400 аминокислот, или от приблизительно 1400 до приблизительно 1500 аминокислот, например, приблизительно 1100, приблизительно 1200, приблизительно 1300, приблизительно 1400 или приблизительно 1500 аминокислот, или как минимум приблизительно 1100, как минимум приблизительно 1200, как минимум приблизительно 1300, как минимум приблизительно 1400 или как минимум приблизительно 1500 аминокислот.
[0075] В некоторых вариантах осуществления изобретения эффекторный белок, а именно эффекторный белок локусов типа V, более конкретно эффекторный белок локусов типа V-С, в частности - C2c3p, предпочтительно C-концевая часть указанного эффекторного белка, включает три каталитических мотива RuvC-подобной нуклеазы (т.е., RuvCI, RuvCII и RuvCIII). В некоторых вариантах осуществления изобретения указанный эффекторный белок, предпочтительно C-концевая часть указанного эффекторного белка, может содержать область, соответствующую мостиковой спирали (также известной как богатый аргинином кластер), которая в белке Cas9 участвует в связывании cr-РНК. В некоторых вариантах осуществления изобретения указанный эффекторный белок, предпочтительно C-концевая часть указанного эффекторного белка, может далее включать домен цинкового пальца. Предпочтительно сохранение Zn-связывающих остатков цистеина в C2c3p. В некоторых вариантах осуществления изобретения указанный эффекторный белок, предпочтительно C-концевая часть указанного эффекторного белка, может включать три каталитических мотива RuvC-подобной нуклеазы (т.е., RuvCI, RuvCII и RuvCIII), область, соответствующую мостиковой спирали, и домен цинкового пальца, предпочтительно в следующем порядке с N к C концу: RuvCI-мостиковая спираль-RuvCII-цинковый палец-RuvCIII. См. Фиг.13A и 13C для иллюстрации характерной доменной архитектуры эффекторных белков типа V-С.
[0076] В некоторых вариантах осуществления изобретения локусы типа V-С, как предполагается в настоящем описании, могут содержать CRISPR-повторы длиной от 20 до 30 п.н., более типично от 22 до 27 п.н. длиной, и еще более типично 25 п.н. длиной, например, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, или 30 п.н.
[0077] В некоторых вариантах осуществления изобретения эффекторный белок, а именно эффекторный белок локусов типа VI, в частности C2c2p, может происходить, быть выделенными или полученным из бактерий, принадлежащих к таксонам альфа-протеобактерии, Bacilli, Clostridia, Fusobacteria и Bacteroidetes. В некоторых вариантах осуществления изобретения эффекторный белок, конкретнее эффекторный белок локусов типа VI, в частности - C2c2p, может происходить, быть выделенным или полученным из бактерий, принадлежащих к одному из следующих родов: Lachnospiraceae, Clostridium, Carnobacterium, Paludibacter, Listeria, Leptotrichia и Rhodobacter. В некоторых вариантах осуществления изобретения эффекторный белок, а именно эффекторный белок локусов типа VI, в частности C2c2p, может происходить, быть выделенным или полученным из следующих видов бактерий: Lachnospiraceaebacterium MA2020, Lachnospiraceaebacterium NK4A179, Clostridiumaminophilum (например, DSM 10710), Lachnospiraceaebacterium NK4A144, Carnobacteriumgallinarum (например, DSM 4847 штамм MT44), Paludibacterpropionicigenes (например, WB4), Listeriaseeligeri (например, серовар ½b штамм SLCC3954), Listeriaweihenstephanensis (например, FSL R9-0317 c4), Listerianewyorkensis (например, штамм FSL M6-0635), Leptotrichiawadei (например, F0279), Leptotrichiabuccalis (например, DSM 1135), Leptotrichiasp. Oraltaxon 225 (например, str. F0581), Leptotrichiasp. Oraltaxon 879 (например, штамм F0557), Leptotrichiashahii (например, DSM.19757), Rhodobactercapsulatus (например, SB 1003, R121, или DE442). В некоторых вариантах осуществления эффекторный белок, более конкретно эффекторный белок локусов типа V, еще более конкретно - C2c2p может происходить, быть выделенным или полученным из видов бактерий, перечисленных в таблице в фиг.42А-В.
[0078] В некоторых вариантах осуществления изобретения эффекторный белок, а именно, эффекторный белок локусов типа VI, в частности - C2c2p, может содержать, состоять только из или включать аминокислотную последовательность, выбранную из последовательностей, показанных на множественном выравнивании на фиг.13J-N.
[0079] В некоторых вариантах осуществления изобретения локус типа VI, как предполагается в рамках настоящего изобретения, может кодировать эффекторные белки Cas1, Cas2 и C2c2p. В некоторых вариантах осуществления изобретения локус типа V-С, как предполагается в рамках настоящего изобретения, может содержать последовательность CRISPR. В некоторых вариантах осуществления изобретения локус типа V-С, как предполагается в рамках настоящего изобретения, может содержать ген c2c2 и последовательность CRISPR, и не содержать гены cas1 и cas2. См. Фиг.15 и фиг.42A-В для иллюстрации характерной организации локусов типа VI.
[0080] В некоторых вариантах осуществления изобретения белок Cas1, кодируемый локусом типа VI, как предполагается в рамках настоящего изобретения, может образовывать кластеры с поддеревом типа II, включая небольшую ветвь типа III-A, или внутри системы типа III-A. См. Фиг.10A и 10B и фиг.10C-W, на которых изображено дерево Cas1, включающее белок Cas1, кодируемый типичными локусами типа VI.
[0081] В некоторых вариантах осуществления изобретения эффекторный белок, конкретно эффекторный белок локусов типа VI, в частности C2c2p, например природный C2c2p, может быть от приблизительно 1000 до приблизительно 1500 аминокислотных остатков длиной, таким как от приблизительно 1100 до приблизительно 1400 аминокислот длиной, например, от приблизительно 1000 до приблизительно 1100 аминокислот длиной, от приблизительно 1100 до приблизительно 1200 аминокислот длиной, или от приблизительно 1200 до приблизительно 1300 аминокислот длиной, или от приблизительно 1300 до приблизительно 1400 аминокислот длиной, или от приблизительно 1400 до приблизительно 1500 аминокислот длиной, например, приблизительно 1000, приблизительно 1100, приблизительно 1200, приблизительно 1300, приблизительно 1400 или приблизительно 1500 аминокислот длиной.
[0082] В некоторых вариантах осуществления изобретения эффекторный белок, а именно эффекторный белок локусов типа VI, а частности C2c2p, содержит как минимум один или предпочтительно как минимум два, наиболее предпочтительно ровно два, консервативных мотива RxxxxH. Каталитические RxxxxH-мотивы характерны для HEPN-доменов (домен, связывающийся с ДНК, присутствующий у эукариот и прокариот). Следовательно, в некоторых вариантах осуществления изобретения эффекторный белок, а именно эффекторный белок локусов типа VI, в частности - C2c2p, содержит как минимум один или предпочтительно как минимум два, наиболее предпочтительно ровно два, HEPN-домена. См. Фиг.11 и фиг.13B для иллюстрации характерной доменной архитектуры эффекторных белков типа VI. В некоторых вариантах осуществления изобретения HEPN-домены могут обладать РНК-азной активностью. В других вариантах осуществления изобретения HEPN-домены могут обладать ДНК-азной активностью.
[0083] В некоторых вариантах осуществления изобретения локусы типа VI, как предполагается в рамках настоящего изобретения, могут содержать CRISPR-повторы длиной от 30 до 40 п.н., более типично длиной от 35 до 39 п.н., например, длиной 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, или 40 п.н.
[0084] В некоторых вариантах осуществления изобретения последовательность, прилегающая к протоспейсеру (PAM), или PAM-подобный мотив управляет связыванием эффекторного белкового комплекса с представляющим интерес локусом-мишенью, как описано настоящем описании. В некоторых вариантах осуществления изобретения, PAM может представлять собой 5'-PAM (т.е. быть расположенным выше 5'-конца протоспейсера). В других вариантах осуществления изобретения PAM может представлять собой 3'-PAM (т.е. быть расположенным ниже 5'-конца протоспейсера).
[0085] В предпочтительном варианте осуществления изобретения эффекторный белок, конкретно эффекторный белок локусов типа V, более конкретно эффекторный белок локусов типa V-B, еще более конкретно - C2c1p, может распознавать 5'-PAM. В некоторых вариантах осуществления изобретения эффекторный белок, конкретно эффекторный белок локусов типа V, более конкретно - эффекторный белок локусов типа V-B, еще более конкретно - C2c1p, может распознавать 5'-PAM, который является 5'-TTN- или 5'-ATTN-последовательностью, где N - это A, C, G или T. В некоторых предпочтительных вариантах осуществления изобретения эффекторный белок может быть белком C2c1p бактерии Alicyclobacillus acidoterrestris, более предпочтительно - белком ATCC 49025 C2c1p бактерии Alicyclobacillus acidoterrestris, в котором 5'-PAM представлен 5'-последовательностью TTN, где N - это A, C, G или T, более предпочтительно где N - это A, G или T. В других наилучших вариантах осуществления изобретения эффекторный белок является белком C2c1p бактерии Bacillus thermoamylovorans, более предпочтительно - белком C2c1p штамма B4166 бактерии Bacillusthermo amylovorans, в котором 5'-PAM представлен 5'-последовательностью ATTN, где N - это A, C, G или T.
[0086] В некоторых вариантах осуществления изобретения фермент CRISPR сконструирован способами инженерии и может содержать одну и несколько мутаций, которые уменьшают или полностью устраняют нуклеазную активность.
[0087] Мутации также могут быть внесены в соседние остатки, например, в аминокислоты, близкие к указанным выше, которые участвуют в нуклеазной активности. В некоторых вариантах осуществления инактивируется только домен RuvC, а в других вариантах осуществления инактивируется другой предполагаемый нуклеазный домен, при этом эффекторный белковый комплекс функционирует как никаза и расщепляет только одну цепь ДНК. В некоторых вариантах осуществления два варианта C2c1 или C2c3 (каждый из которых представляет собой другую никазу) используются для увеличения специфичности, два варианта никазы используется для расщепления ДНК на мишени (где обе никазы расщепляют цепочку ДНК, минимизируя или полностью устраняя нецелевые модификации, где только одна цепь ДНК расщепляется и затем подвергается репарации). В предпочтительных вариантах осуществления эффекторный белок C2c1 или C2c3 расщепляет последовательности, ассоциированные с или непосредственно находящиеся в представляющем интерес локусе-мишени, в качестве гомодимера, содержащего две молекулы эффекторного белка C2c1 или C2c3. В предпочтительном варианте осуществления гомодимер может содержать две молекулы эффекторного белка C2c1 или две молекулы эффекторного белка C2c3 или смесь C2c1 и C2c3, содержащую различные мутации в соответствующих доменах RuvC.
[0088] Изобретение относится к способам использования двух или более никаз, в частности, к способу, предполагающему использование двойной или сдвоенной никазы. В некоторых аспектах и вариантах осуществления может быть доставлен один тип никазы C2c1 или C2c3, например, модифицированная C2c1 или C2c3 или модифицированная никаза C2c1 или C2c3, как описано в настоящем описании. Это приводит к тому, что ДНК-мишень связывается двумя C2c1 или двумя C2c3-никазами или смесью C2c1 и C2c3-никаз. Кроме того, также предусматривается, что могут быть применены различные ортологи, например, никаза C2c1 или C2c3 на одной цепи (например, кодирующей цепи) ДНК и ортолог на некодирующей или противоположной цепи ДНК. Ортолог может быть, но не ограничивается, никазой Cas9, такой как никаза SaCas9 или никаза SpCas9. Может быть предпочтительно использовать два разных ортолога, которые требуют разных PAM, а также могут иметь разные требования к направляющим последовательностям, что позволит обеспечить более высокую степень контроля при использовании изобретения. В некоторых вариантах осуществления расщепление ДНК будет включать по меньшей мере четыре типа никаз, причем каждый тип будет направляться в иную нуклеотидною последовательность ДНК-мишени, причем каждая пара никаз вносит первый разрыв цепи в одну цепь ДНК, а вторая вносит разрыв цепи во вторую цепь ДНК. При таких способах по меньшей мере две пары одноцепочечных разрывов вносятся в ДНК-мишень, где при внесении первой и второй пары одноцепочечных разрывов в спирали вырезаются последовательности-мишнени между первой и второй парой однонитевых разрывов. В некоторых вариантах осуществления один или оба ортолога являются контролируемыми, т.е. индуцируемыми.
[0089] В некоторых вариантах осуществления изобретения направляющая РНК или зрелая cr-РНК содержит, состоит в основном из или состоит только из прямой повторяющейся последовательности и направляющей последовательности или спейсера. В некоторых вариантах осуществления изобретения направляющая РНК или зрелая cr-РНК состоит из частичного прямого повтора размером 19 нуклеотидов с последующей направляющей последовательностью или спейсером размером 23-25 нуклеотидов. В некоторых вариантах осуществления эффекторный белок представлен эффекторным белком C2c1 или C2c3 и требует направляющей последовательности длиной как минимум 16 нуклеотидов для достижения поддающегося обнаружению расщепления ДНК и направляющей последовательности размером минимум 17 нуклеотидов для достижения эффективного расщепления ДНК in vitro. В некоторых вариантах осуществления прямая повторяющаяся последовательность расположена выше (т.е. на 5'-конце) направляющей последовательности или спейсера. В предпочтительном варианте осуществления последовательность затравки (т.е. последовательность, необходимая для распознавания и/или гибридизации с последовательностью локуса-мишени) направляющей РНК C2c1 или C2c3 находится примерно в пределах первых 5 нуклеотидов на 5'-конце направляющей последовательности или спейсера.
[0090] В предпочтительных вариантах осуществления изобретения зрелая cr-РНК содержит шпилечную структуру или оптимизированную шпилечную структуру или любую оптимизированную вторичную структуру. В предпочтительных вариантах осуществления изобретения зрелая cr-РНК содержит шпилечную структуру или оптимизированную шпилечную структуру в прямой повторяющейся последовательности, где шпилечная структура или оптимизированная шпилечная структура важна для расщепления. В некоторых вариантах осуществления изобретения зрелая cr-РНК предпочтительно включает единичную шпилечную структуру. В некоторых вариантах осуществления изобретения прямая повторяющаяся последовательность предпочтительно включает единичную шпилечную структуру. В некоторых вариантах осуществления изобретения активность расщепления комплекса эффекторного белка может быть модифицирована путем внесения мутаций, которые влияют на структуру дуплекса РНК шпилечной структуры. В предпочтительных вариантах осуществления изобретения могут быть внесены мутации, поддерживающие структуру дуплекса РНК шпилечной структуры, благодаря которым будет сохранятся ферментативная активность, осуществляемая комплексом эффекторного белка. В других предпочтительных вариантах осуществления изобретения мутации, нарушающие структуру дуплекса РНК шпилечной структуры, могут быть внесены для полного удаления ферментативной активности, осуществляемой комплексом эффекторного белка.
[0091] Также изобретение относится к нуклеотидной последовательности, кодирующей эффекторный белок, являющейся кодон-оптимизированной для экспрессии в эукариотическом организме или эукариотической клетке посредством любого из описанных в настоящем описании способов или композиций. В одном варианте осуществления изобретения кодон-оптимизированный эффекторный белок представляет собой любой C2c1 или C2c3, обсуждаемый в настоящем описании, и подвергнута оптимизации кодонов для удобства использования в эукариотической клетке или организме, например, такой клетке или организме, как описано в настоящем описании в иных местах, например, но не ограничиваясь этим, в клетке дрожжей, или клетке или организме млекопитающего, включая клетку мыши, клетку крысы, и клетку человека или не являющегося человеком эукариотического организма, например, растения.
[0092] В некоторых вариантах осуществления изобретения по меньшей мере один сигнал ядерной локализации (NLS) присоединен к последовательностям нуклеиновых кислот, кодирующим эффекторные белки C2c1 или C2c3. В предпочтительных вариантах осуществления изобретения по меньшей мере одна или более C-концевых или N-концевых последовательностей NLS присоединены (следовательно, молекулы нуклеиновых кислот, кодирующие эффекторный белок C2c1 или C2c3, могут включать код для последовательностей NLS, чтобы экспрессируемый продукт имел присоединенную или связанную последовательность NLS). В предпочтительном варианте осуществления изобретения C-концевые NLS присоединены для оптимальной экспрессии и нацеливания в ядро эукариотических клеток, предпочтительно клеток человека. В предпочтительном варианте осуществления изобретения кодон-оптимизированным эффекторным белком является C2c1 или C2c3 и длина спейсера направляющей РНК составляет от 15 до 35 нуклеотидов. В некоторых вариантах осуществления изобретения длина спейсера направляющей РНК - по меньшей мере 16 нуклеотидов, такая как по меньшей мере 17 нуклеотидов. В некоторых вариантах осуществления изобретения длина спейсера составляет от 15 до 17 нуклеотидов, от 17 до 20 нуклеотидов, от 20 до 24 нуклеотидов, например, 20, 21, 22, 23 или 24 нуклеотидов, от 23 до 25 нуклеотидов, например, 23, 24 или 25 нуклеотидов, от 24 до 27 нуклеотидов, 27-30 нуклеотидов, 30-35 нуклеотидов или 35 нуклеотидов или длиннее. В некоторых вариантах осуществления изобретения кодон-оптимизированным эффекторным белком является C2c1 или C2c3, и длина прямого повтора направляющей РНК составляет по меньшей мере 16 нуклеотидов. В некоторых вариантах осуществления изобретения кодон-оптимизированным эффекторным белком является C2c1 или C2c3, и длина прямого повтора направляющей РНК составляет от 16 до 20 нуклеотидов, например, 16, 17, 18, 19 или 20 нуклеотидов. В определенных предпочтительных вариантах осуществления изобретения длина прямого повтора направляющей РНК равна 19 нуклеотидам.
[0093] Настоящее изобретение также охватывает способы для доставки множественных компонентов, являющихся нуклеиновыми кислотами, где каждый компонент, являющийся нуклеиновой кислотой, специфичен к различным представляющим интерес локусам-мишеням для изменения множественных представляющих интерес локусов-мишеней. Компонент, являющийся нуклеиновой кислотой, комплекса может включать один или более связывающих белок РНК-аптамеров. Один или более аптамеров могут быть способны к связыванию с белком оболочки бактериофага. Белок оболочки бактериофага может быть выбран из группы, включающей: Qβ, F2, GA, fr, JP501, MS2, M12, R17, BZ13, JP34, JP500, KU1, М11, MX1, TW18, VK, SP, FI, ID2, NL95, TW19, AP205, ϕСb5, ϕСb8r, ϕCb12r, ϕСb23r, 7s и PRR1. В предпочтительном варианте осуществления изобретения белок оболочки бактериофага представлен белком MS2. Изобретение также предусматривает, что компонент, являющийся нуклеиновой кислотой, комплекса имеет длину 30 или больше, 40 или больше или 50 или больше нуклеотидов.
[0094] Изобретение также относится к клеткам, компонентам и/или системам по настоящему изобретению, содержащим незначительное количество катионов, присутствующих в клетках, компонентах и/или системах. В основном таким катионом является магний, такой как Mg2+. Катион может присутствовать в следовом количестве. Предпочтительный диапазон концентрации для катиона Mg2+ составляет приблизительно от 1 мМ до 15 мМ. Предпочтительная концентрация может составлять около 1 мМ для клеток, компонентов и/или систем на основе человека и примерно от 10 мМ до 15 мМ для клеток, компонентов и/или систем на основе бактерий. См., например, статью Gasiunas et al., PNAS, опубликованную через интернет 4 сентября 2012 года, www.pnas.org/cgi/doi/10.1073/pnas. 1208507109.
[0095] Соответственно, задачей изобретения не является охватить в своих рамках какой-либо ранее известный продукт, процесс получения продукта или способ применения, таким образом, заявители сохраняют за собой право и настоящим подтверждают отказ от какого-либо ранее известного продукта, процесса или способа. Далее отмечено, что изобретение не намеревается охватить в своих рамках какой-либо продукт, процесс получения или способ применения продукта, которые не соответствует письменному описанию и разрешающим условиям USPTO (35 Свод Законов США 112, параграф первый) или EPO (Статья 83 EPC), так что Заявители сохраняют за собой право и настоящим подтверждают отказ от каого-либо ранее описанного продукта, процесса получения или способа применения продукта. В практике изобретения следует руководствоваться Статьей 53 (c) EPC и Правилом 28 (b) и (c) EPC. Ничто в настоящем описании не может быть истолковано как обещание.
[0096] Отмечено, что в настоящей заявке и особенно в формуле изобретения и/или параграфах термины, такие как "содержать", "содержавшийся", "содержащий" и т.п., могут иметь значение, приписанное им в американском Патентном Законе; например, они могут означать "включает", "включенный", "включающий", и т.п.; термины, такие как "по существу состоящий из" и "по существу состоит из" имеют значение, приписанное им в американском Патентном праве, например, они включают элементы, не указанные явно, но исключают элементы, которые учтены в предшествующих заявках или которые влияют на основные или новые характеристики изобретения.
[0097] В следующем аспекте изобретение относится к эукариотической клетке, включающей модифицированный представляющий интерес локус-мишень, где представляющий интерес локус-мишень модифицирован согласно любому из способов, описанных в настоящем описании. Еще один аспект изобретения относится к созданию клеточной линии указанной клетки. Следующий аспект изобретения предусматривает создание многоклеточного организма, включающего одну или более указанных клеток.
[0098] В некоторых вариантах осуществления изобретения модификация представляющего интерес локуса-мишени может приводить к появлению эукариотической клетки с измененной экспрессией продукта по меньшей мере одного гена; эукариотической клетки с измененной экспрессией продукта по меньшей мере одного гена, причем экспрессия этого продукта увеличена; эукариотической клетки с измененной экспрессией продукта по меньшей мере одного гена, причем экспрессия этого продукта уменьшена; или эукариотической клетки, содержащей отредактированный геном.
[0099] В некоторых вариантах осуществления изобретения эукариотическая клетка может быть клеткой млекопитающего или человека.
[00100] В дальнейших вариантах осуществления изобретения не встречающиеся в природе или сконструированные способами инженерии композиции, векторные системы или системы доставки, как описано в настоящем описании, могут использоваться для сайт-специфического нокаута генов; сайт-специфического редактирования генома; специфической интерференции последовательности ДНК; или мультиплексной инженерии генома.
[00101] Также изобретение относится к получению генного продукта из клетки, клеточной линии или организма, как описано в настоящем описании. В некоторых вариантах осуществления изобретения количество экспрессированного генного продукта может быть больше или меньше, чем количество генного продукта в клетке без измененной экспрессии или отредактированного генома. В некоторых вариантах осуществления изобретения генный продукт может быть изменен по сравнению с генным продуктом из клетки без измененной экспрессии или отредактированного генома.
[00102] Эти и другие варианты осуществления изобретения раскрыты в, очевидны из или охвачены следующим Подробным Описанием.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[00103] Новые признаки изобретения подробно сформулированы в приложенной формуле изобретения. Лучшее понимание признаков и преимуществ данного изобретения будет достигнуто при ознакомлении со следующим подробным описанием, которое включает материалы, иллюстрирующие принципы изобретения, и сопроводительные Фиг.к ним:
[00104] На фиг.1A и 1B изображена новая классификация систем CRISPR-Cas. Класс 1 включает многосубъединичные эффекторные комплексы cr-РНК (Cascade), и Класс 2 включает односубъединичные эффекторные комплексы cr-РНК (Cas9-подобные). На фиг.1B приведено другое описание новой классификации систем CRISPR-Cas.
[00105] На фиг.2 изображена молекулярная организация систем CRISPR-Cas.
[00106] На фиг.3 изображены структуры эффекторных комплексов типов I и III: общая архитектура/общее происхождение прослеживается несмотря на обширную дивергенцию последовательностей.
[00107] На фиг.4 изображена CRISPR-Cas как система, основанная на мотивах распознавания РНК (RRM).
[00108] На фиг.5 изображена филогению Cas1, где показан главный аспект эволюции системы CRISPR-Cas - рекомбинация эффекторных модулей cr-РНК в целях адаптации.
[00109] На фиг.6 представлен перечень систем CRISPR-Cas, а именно распространение типов/подтипов систем CRISPR-Cas среди архей и бактерий.
[00110] На фиг.7 изображен алгоритм для идентификации Cas-кандидатов.
[00111] На фиг.8A и 8B изображена организация полных локусов систем CRISPR-Cas 2 класса. Обозначены три подтипа типа II и подтипы V-A, V-B и V-C и тип VI. Подсемейства на основе филогении Cas1 также обозначены. Схема включает только общие гены, представленные в каждом подтипе; дополнительные гены, присутствующие в некоторых вариантах, опущены. Красным прямоугольником обозначен вырожденный повтор. Серые стрелки показывают направление транскрипции последовательности CRISPR. PreFran, Prevotella-Francisella. На фиг.8B приведено другое описание организации полных локусов нескольких систем CRISPR-Cas класса 2.
[00112] На фиг.9 изображены окрестности C2c1, т.е. геномная архитектура локусов CRISPR-Cas C2c1. Обозначено количество повторов в последовательностях CRISPR. Для каждого геномного контига обозначен числовой ID Genebank и координаты локуса.
[00113] На Фиг.10A и 10B изображены представления о дереве последовательностей белка Cas1. Дерево на фиг.10B было построено на основе множественного выравнивания 1498 последовательностей Cas1, которые содержали 304 филогенетически информативных позиции. Ветви, соответствующие системам класса 2 подчеркнуты: голубым - тип II; оранжевым - подтип V-A; красным - подтип V-B; коричневым - подтип V-C; фиолетовым - тип VI. Вставки показывают расширенные ветви новых (под)типов. Значения поддержки бутстрэпа приведены в процентах и показаны только для некоторых соответствующих ветвей.
[00114] На рисунках 10C-10W показано полное дерево Cas1, которое схематично показано на фиг.10B в формате Newick с видовыми названиями и значениями поддержки бутстрэпа. Дерево было построено с помощью программы FastTree (опции "-gamma -wag"). Множественное выравнивание последовательностей Cas1 было отфильтровано с порогом однородности 0,1 и порогом возникновения промежутка 0,5 до реконструкции дерева.
[00115] На фиг.11 изображена доменная организация семейств класса 2.
[00116] На фиг.12 изображены участки гомологии TnpB белков класса 2.
[00117] На Фиг.13A-13N представлено другое изображение доменной архитектуры и консервативных мотивов эффекторных белков класса 2. Фиг.13A иллюстрирует типы II и V: нуклеазы-производные TnpB. Верхний ряд на рисунке демонстрирует нуклеазы RuvC из Thermos thermophilus (PDB ID: 4EP5) с обозначением каталитических аминокислотных остатков. Ниже показана архитектура каждого домена, приведено выравнивание консервативных мотивов для отдельных представителей соответствующих семейств белков (единственная последовательность для RuvC). Каталитические остатки показаны белыми буквами на черном фоне, консервативные гидрофобные остатки выделены желтым; консервативные остатки малых аминокислот выделены зеленым; в выравнивании спирального мостика положительно заряженные остатки выделены красным. Предсказанная вторичная структура показана ниже выровненных последовательностей: H обозначает α-спирали, Е обозначает выпрямленную конформацию (β-структура). Малоконсервативные последовательности спейсеров между блоками выравнивания обозначены числами. Фиг.13B иллюстрирует тип VI: белки содержат 2 HEPN-домена, которые могут демонстрировать РНК-азную активность. Верхние блоки выравнивания содержат отдельные описанные ранее домены HEPN, нижние блоки содержат каталитические мотивы из эффекторных белков типа VI. Обозначения как на фиг.13 A. Фиг.13C иллюстрирует ближайшие гомологи новых эффекторных белков типа V из числа кодируемых транспозонами белков: непересекающиеся наборы гомологов. Фиг.13D-H иллюстрирует множественное выравнивание семейства белка C2c1. Данное выравнивание выполнено с использованием программы MUSCLE и изменено вручную на основе локальных попарных выравниваний, полученных с помощью PSI-BLAST. Для каждой последовательности приведен номер-идентификатор GenBank (GI) и систематическое название организма. Вторичная структура предсказана с помощью Jpred и показана ниже последовательности, использованной в качестве последовательности-запроса (обозначения: H - альфа-спираль, E - бета-структура). CONSENSUS рассчитан для каждого столбца выравнивания путем сравнения суммы парных весов каждого столбца для гомогенных столбцов (совпадающий остаток во всех выровненных последовательностях) и случайного столбца с порогом гомогенности 0,8. Мотивы активного сайта RuvC-подобного домена показаны ниже выравнивания. На фиг.13I представлено множественное выравнивание для семейства белка C2c3. Данное выравнивание выполнено с использованием программы MUSCLE. Для каждой последовательности обозначены внутренний приписанный ей номер и номер-идентификатор GenBank (GI) метагеномного контига, кодирующего соответствующий белок С2с3. Вторичная структура предсказана с помощью Jpred и показана ниже выравнивания (обозначения: H - альфа-спираль, E - бета-структура). CONSENSUS рассчитан для каждого столбца выравнивания путем сравнения суммы парных весов каждого столбца для гомогенных столбцов (совпадающие остатки во всех выровненных последовательностях) и случайного столбца с порогом гомогенности 0,8. Мотивы активного сайта RuvC-подобного домена показаны ниже выравнивания С-концевого домена. Фиг.13J-N иллюстрирует множественное выравнивание семейства белка C2c2. Выравнивание выполнено с использованием программы MUSCLE и изменено вручную на основе локальных выравниваний с помощью PSI-BLAST. Для каждой последовательности обозначен номер-идентификатор GenBank (GI) и систематическое название организма. Вторичная структура предсказана с помощью Jpred и показана ниже последовательности, использованной в качестве последовательности-запроса (обозначения: H - альфа-спираль, E - бета-структура). CONSENSUS рассчитан для каждого столбца выравнивания путем сравнения суммы парных весов каждого столбца для гомогенных столбцов (совпадающие остатки во всех выровненных последовательностях) и случайного столбца с порогом гомогенности 0,8. Мотивы активных сайтов домена HEPN показаны ниже выравнивания.
[00118] На фиг.14 изображены окрестности C2c3, т.е. геномная архитектура C2c3-локусов системы CRISPR-Cas. Указано количество повторов в последовательностях CRISPR. Для каждого геномного контига обозначены номер-идентификатор GenBank и координаты локуса.
[00119] На фиг.15 изображены окрестности C2c2, т.е. геномная архитектура C2c2-локусов системы CRISPR-Cas. Указано количество повторов в последовательностях CRISPR. Для каждого геномного контига обозначены номер-идентификатор GenBank и координаты локуса.
[00120] На фиг.16 изображен мотив RxxxxH домена HEPN семейства C2c2.
[00121] На фиг.17 изображен C2C1: 1. Alicyclobacillus acidoterrestris ATCC 49025.
[00122] На фиг.18 изображен C2C1: 4. Desulfonatronum thiodismutans штамма MLF-1.
[00123] На фиг.19 изображен C2C1: 5. бактерии Opitutaceae TAV5.
[00124] На фиг.20 изображен C2C1: 7. Bacillus thermoamylovorans штамма B4166.
[00125] На фиг.21 изображен C2C1: 9. Bacillussp. NSP2.1.
[00126] На фиг.22 изображен C2C2: 1. бактерии Lachnospiraceae MA2020.
[00127] На фиг.23 изображен C2C2: бактерии 2. Lachnospiraceae NK4A179.
[00128] На фиг.24 изображен C2C2: 3. [Clostridium] aminophilum DSM 10710.
[00129] На фиг.25 изображен C2C2: 4. бактерии Lachnospiraceae NK4A144.
[00130] На фиг.26 изображен C2C2: 5. Carnobacterium gallinarum DSM 4847.
[00131] На фиг.27 изображен C2C2: 6. Carnobacterium gallinarum DSM 4847
[00132] На фиг.28 изображен C2C2: 7. Paludibacter propionicigenes WB4.
[00133] На фиг.29 изображен C2C2: 8. Listeria seeligeri серовара 1/2b.
[00134] На фиг.30 изображен C2C2: 9. Listeria weihenstephanensis FSL R9-0317.
[00135] На фиг.31 изображен C2C2: 10. Listeria bacterium FSL M6-0635.
[00136] На фиг.32 изображен C2C2: 11. Leptotrichia wadei F0279.
[00137] На фиг.33 изображен C2C2: 12. Leptotrichia wadei F0279.
[00138] На фиг.34 изображен C2C2: 14. Leptotrichia shahii DSM 19757.
[00139] На фиг.35 изображен C2C2: 15. Rhodobacter capsulatus SB 1003.
[00140] На фиг.36 изображен C2C2: 16. Rhodobacter capsulatus R121.
[00141] На фиг.37 изображен C2C2: 17. Rhodobacter capsulatus DE442.
[00142] На фиг.38 изображено дерево DR-последовательностей.
[00143] На фиг.39 изображено дерево белков C2c2.
[00144] Фиг.40A-40D содержит таблицу, включающую описания 63 больших кодирующих белки генов вблизи генов cas1, идентифицированных с помощью описанного в настоящем описании вычислительного способа. Описанные в настоящем описании представители новых подтипов (V-B, V-C, VI) выделены цветом. Последовательности белков для представителей типа V-B и типа IV, кодирующих AUXO014641567.1, AUXO011689375.1, AUXO011689375.1, AIJXO011277409.1, AUXO014986615.1 не проанализированы, т.к. для этих последовательностей не может быть установлена видовая принадлежность.
[00145] Фиг.41A-41M содержат таблицу, демонстрирующую результаты анализа локусов V- B типа (кодирующих белок C2c1). * cas1cas4 - ген, содержащий домены cas4 и cas1; CRISPR - CRISPR-повторы; SOS - гены SOS-ответа; unk - гипотетический белок; > - направление кодирующей последовательности гена; [D] - вырожденный повтор (определены, где возможно); [T] - tracr-РНК. Фиг.41C-J иллюстрирует анализ CRISPR-последовательностей для локусов V-B типа (кодирующих белки C2c1), как описано в настоящем описании (раздел CRISPR является частью базового результата работы pilercr (см. описание результата работы на сайте pilercr: http://www.drive5.com/pHercr/); фолдинг повторов выполнен с помощью mfold (см. описание результата работы на сайте mfold: http://mfold.ma.albany.edu/?q=mfQld/DNA-Folding-FGrm); результаты фолдинга повторов и анализа CRISPR-последовательностей помещены после детального описания каждого случая; расположение CRISPR см. по ссылке в таблице на фиг.41 A-В. Фиг.41K иллюстрирует классификацию CRISPRmap CRISPR-повторов локусов типа V-B (кодирующих белок C2c1) как описано в настоящем описании с помощью CRISPRmap (детали см. http://rna.informatik.uni-freiburg.de/CRISPRmap/Input.jsp). Фиг.41L иллюстрирует вырожденные повторы локусов V-B типа (кодирующие белок C2c1) обнаруженные, как описано в настоящем описании, с помощью алгоритма поиска CRISPR-последовательностей (http://crispr.u-psud.fr/Server/). Столбец нормальных повторов содержит нормальный повтор, столбец спейсеров - последний спейсер, столбец downstream - область низлежащих последовательностей, начиная с вырожденного повтора (250 п.н.); номер последовательности соответствует номеру последовательности CRISPR в соответствующем локусе (см. Таблицу на фиг.41 A-В); выделенная желтым область имеет полное совпадение между нормальным повтором и вырожденным повтором (при несовпадении с другой частью вырожденного повтора). Фиг.41M иллюстрирует предсказанные структуры tracr-РНК, образующие пары оснований с этими повторами. Tracr-РНК для Alicyclobacillus acidoterrestric был идентифицирован с помощью секвенирования РНК. Для остальных локусов предполагаемые tracr-РНК идентифицированы на основе присутствия последовательности антипрямого повтора (DR) (Anti-DR). Последовательности антипрямых повторов были идентифицированы с помощью Geneious (www.geneious.com) путем поиска последовательностей внутри каждого соответствующего локуса CRISPR с высокой гомологией к DR. 5'- и 3'-концы каждой предполагаемой tracr-РНК были определены с помощью вычислительного прогнозирования сайтов старта бактериальной транскрипции и терминации с помощью соответственно BPROM (www.softberry.com) и ARNOLD (rna.ig-mors.u-psud.fr/toolbox/amold/). Предсказания кофолдинга получены с помощью Geneious, 5'-концы выделены синим, 3'-концы выделены оранжевым.
[00146] Фиг.42A-42N содержит таблицу, демонстрирующую результаты анализа локусов типа VI (кодирующих белок C2c2). *CRISPR - CRISPR повторы; unk - гипотетический белок; > - направление кодирующей последовательности гена; [D] - вырожденный повтор (определены, где возможно); [T] - tracr-РНК, фиг.42C-I демонстрируют результаты анализа CRISPR-последовательностей локусов типа VI (кодирующих белок C2c2) как описано в настоящем описании (раздел CRISPR является частью базового результата работы pilercr (см. описание результата работы на сайте pilercr: http://www.drive5.com/pilercr/); фолдинг повторов выполнен с помощью mfold (см. описание результата работы на сайте mfold: http://mfold.rna.albany.edu/?q=mfQld/DNA-Folding-Form); результаты фолдинга повторов и анализа CRISPR-последовательностей помещены после детального описания каждого случая; см. расположение CRISPR по ссылке в таблице на фиг.42 A-В. Фиг.42J иллюстрирует классификацию CRISPRmap CRISPR повторов локусов типа VI (кодирующих белок C2c2) с помощью, как описано в настоящем описании CRISPRmap (детали см. http://rna.informatik.uni-freiburg.de/CRISPRmap/Input.jsp). Фиг.42K-L иллюстрирует вырожденные повторы локусов типа VI (кодирующие белок C2c2) как описано в настоящем описании, обнаруженные с помощью алгоритма по поиску CRISPR-последовательностей (http://crispr.u-psud.fr/Server/). Столбец нормальных повторов содержит нормальный повтор, столбец спейсеров - последний спейсер, столбец downstream - область низлежащих последовательностей, начиная с вырожденного повтора (250 п.н.); номер последовательности соответствует числовому значению CRISPR-последовательности в соответствующем локусе (см. таблицу на фиг.42 A-В); выделенная желтым область имеет полное совпадение между нормальным повтором и вырожденным повтором (при несовпадении с другой частью вырожденного повтора). Фиг.42M-N иллюстрирует предсказанные структуры tracr-РНК, образующие пары оснований с этими повторами. Предполагаемые tracr-РНК были идентифицированы на основе присутствия последовательности антипрямого повтора (DR) (Anti-DR). Последовательности антипрямых повторов были идентифицированы с помощью Geneious (www.geneious.com) путем поиска последовательностей внутри каждого соответствующего локуса CRISPR с высокой гомологией с DR. 5'- и 3'-концы каждой предполагаемой tracr-РНК были определены с помощью вычислительного прогнозирования сайтов старта бактериальной транскрипции и терминации с помощью BPROM (www.softberry.com) и ARNOLD (rna.ig-mors.u-psud.fr/toolbox/amold/) соответственно. Предсказания кофолдинга получены с помощью Geneious, 5'-концы выделены синим, 3'-концы выделены оранжевым.
[00147] Фиг.43A-43F содержит таблицу, демонстрирующую результаты анализа локусов типа V-С (кодирующих белок C2c3). *CRISPR - CRISPR-повторы; unk - гипотетический белок; > - направление кодирующей последовательности гена; [D] - вырожденный повтор (определены, где возможно). Фиг.43C - демонстрируют результаты анализа CRISPR-последовательностей локусов типа V-С (кодирующих белок C2c3) как описано в настоящем описании (раздел CRISPR является частью базового результата работы pilercr (см. описание результата работы на сайте pilercr: http://www.drive5.com/pilercr/); фолдинг повторов выполнен с помощью mfold (см. описание результата работы на сайте mfold: http://mfold.ma.albany.edu/?q=mfQld/DNA-Folding-Form); результаты фолдинга повторов и анализа CRISPR-последовательностей помещены после детального описания каждого случая; см. расположение CRISPR по ссылке в таблице на фиг.43 A-В. Показаны статистически значимые совпадения BLAST для спейсера с прокариотами и их вирусами. Фиг.43E иллюстрирует классификацию CRISPRmap CRISPR повторов локусов типа V-С (кодирующих белок C2c3) как описано в настоящем описании с помощью CRISPRmap (детали см. http://rna.informatik.uni-freiburg.de/CRISPRmap/Input.jsp). Фиг.43F иллюстрирует вырожденные повторы локусов типа V-С (кодирующие белок C2c3) как описано в настоящем описании, обнаруженные с помощью алгоритма по поиску CRISPR-последовательностей (http://crispr.u-psud.fr/Server/). Столбец нормальных повторов содержит нормальный повтор, столбец спейсера - последний спейсер, столбец downstream - область низлежащих последовательностей, начиная с вырожденного повтора (250 п.н.); номер последовательности соответствует номеру CRISPR-последовательности в соответствующем локусе (см. таблицу на фиг.43 A-В); выделенная желтым область имеет полное совпадение между нормальным повтором и вырожденным повтором (при несовпадении с другой частью вырожденного повтора).
[00148] На фиг.44A-44E приведен полный список локусов CRISPR-Cas в геномах, где были обнаружены белки C2с1 или C2c2. Гены белков C2c1 и C2c2 выделены желтым.
[00149] На фиг.45A-45C проиллюстрировано выравнивание локусов Listeria, кодирующих предполагаемую систему CRISPR-Cas типа VI. Выровненный синтетический участок, соответствующий контигу AODJ01000004.1 Listeria weihenstephanensis FSL R9-0317 с координатами 42281-46274 и контигу JNFB01000012.1 Listeria newyorkensis, штамм FSL M6-0635, с координатами 169489-173541. Обозначения: ген C2c2 выделен голубым, повторы CRISPR - пурпурным, вырожденные повторы - пурпурным, спейсеры - жирным.
[00150] На фиг.46A-46D представлено функциональное подтверждение локуса C2c1 Alicyclobacillus acideoterrestris. Фиг.46A: секвенирование РНК демонстрирует высокую экспрессию локуса C2c1 Alicyclobacillus acideoterrestris при помощи процессированной cr-РНК, включающей прямой повтор длиной 14 нуклеотидов на 5'-конце и спейсер длиной 20 нуклеотидов. Предполагаемая tracr-РНК длиной 79 нуклеотидов стабильно экспрессируется в том же направлении, что и кластер генов cas. Фиг.46B: Анализ с использованием нозерн-блоттинга РНК, экспрессируемой с эндогенного локуса (М), и минимальной последовательности первого спейсера (S) показывают процессированные cr-РНК с прямым повтором на 5'-конце и наличие небольшой предполагаемой tracr-РНК. Стрелки указывают положения зондов и их направление. Фиг.46C: Совместный фолдинг прямого повтора cr-РНК и предполагаемой tracr-РНК in silico показывает стабильную вторичную структуру и комплементарность между двумя РНК. 5'-основания окрашены в синий цвет, а 3'-основания окрашены в оранжевый цвет. Фиг.46D: Гетерологичная экспрессия локуса Alicyclobacillus acideoterrestris C2c1 в pACYC-184, трансформированном в E. coli, дает идентичные результаты с экспрессией, наблюдаемой в эндогенном штамме (фиг.46А). Процессированные cr-РНК имеют прямой повтор DR длиной 14 нуклеотидов на 5'-конце и спейсер длиной 20 нуклеотидов, и предполагаемая tracr-РНК длиной 79 нуклеотидов экспрессируется стабильно.
[00151] На фиг.47A-47C представлена идентификация соседнего с протоспейсером мотива (PAM) для фермента C2c1 из Alicyclobacillus acideoterrestris. Фиг.47A: Схема скрининга для определения PAM. Фиг.47B: Истощение на левом 5'-конце библиотеки PAM показывает наличие TTM PAM на 5'-конце. Истощение определяется как логарифм по основанию 2, причем количество PAM выше порога 3,5 используются для расчета оценки энтропии для каждой позиции. Фиг.47C: Подтверждение PAM для C2c1 из Alicycclobacillus acideoterrestris путем измерения интерференции с восемью различными PAM. PAM, соответствующие мотиву TTN, демонстрируют наличие истощения, измеренное при помощи метода колониеобразующих единиц (КОЭ).
[00152] На фиг.48A-F представлена охарактеризация условия, в которых C2c1 из Alicyclobacillus acideoterrestris производит расщепление in vitro. Фиг.48А: расщепление in vitro мишени EMX1 лизатом клеток человека, экспрессирующим C2c1 из Alicyclobacillus acideoterrestris, показывает, что нацеливание C2c1 Alicyclobacillus acideoterrestris in vitro надежно и зависит от tracr-РНК. Ненацеливающая cr-РНК. (cr-РНК 2) не может расщеплять мишень EMX1, тогда как cr-РНК 1, нацеливающая на EMX1, обеспечивает сильное расщепление в присутствии Mg++ и слабое расщепление в отсутствие Mg++. Фиг.48B: расщепление мишени EMX1 in vitro в присутствии tracr-РНК различной длины идентифицирует типы длиной 78 нуклеотидов в качестве минимальной длины tracr-РНК, также отмечается повышение эффективности расщепления для формы длиной 91 нуклеотидов. Фиг.48C: Анализ температурной зависимости расщепления мишени EMX1 in vitro показывает, что оптимальный температурный диапазон расщепления AacC2c1 составляет от 40 до 55°C. Фиг.48D: Подтверждение in vitro требований к PAM белка AacC2c1 с четырьмя различными PAM. Те PAM, которые были комплементарны мотиву TTN, расщеплялись эффективно. Фиг.48E: При фолдинге in silico химерной sg-РНК AacC2c1 наблюдается формирование стабильной структуры с прямым повтором: обратное соединение сегментов, происходящих из tracr-РНК (отмечено красным) и cr-РНК (отмечено черным). Фиг.48F: Сравнение расщепления мишеней in vitro в присутствии cr-РНК-tracr-РНК AacC2c1 и sg-РНК показывает сходную эффективность расщепления.
[00153] На фиг.49A-49C представлено функциональное подтверждение локуса C2c1 Bacillus thermoamylovorans. Фиг.49A: Гетерологичная экспрессия локуса C2c1 Bacillus thermoamylovorans в E. coli. Предполагаемая tracr-РНК экспрессируется в значительной степени и процессируется до 91 нуклеотидов. Процессированные cr-РНК также присутствуют и имеют прямой повтор длиной 14 нуклеотидов и спейсер длиной 19 нуклеотидов на 5'-конце. Фиг. 49B: при кофолдинге in silico прямого повтора cr-РНК и предполагаемой tracr-РНК показано образование стабильной вторичной структуры и наличие комплементарности между двумя РНК. 5'-концы окрашены в синий цвет, а 3'-концы окрашены в оранжевый цвет. Фиг.49C: тстощение на 5' конце левой PAM библиотеки показывает наличие ATTN PAM на 5'-конце. Истощение измеряется как логарифм по основанию 2, и количество PAM выше порога 3,5 используется для расчета оценки энтропии для каждой позиции.
[00154] На фиг.50 представлены все результаты считывания для локуса C2C1Bacillus sp.
[00155] На фиг.51 представлены отфильтрованные результаты считывания от 0 до 55 пар оснований в локусе C2C1 Bacillus sp.
[00156] На фиг.52 представлен кофолдинг последовательности прямого повтора и tracr-РНК, соответствующих локусу C2C1 B. sp.
[00157] На фиг.53 проиллюстрирован эволюционный сценарий для систем CRISPR-Cas. Предполагается, что белок Cas8 возник путем инактивации Cas10 (обозначен белым X), что сопровождалось значительным ускорением эволюции. Сокращения: TR, концевые повторы; TS, концевые последовательности, HD, семейство эндонуклеаз HD; HNH, семейство эндонуклеаз HNH, RuvC, семейство эндонуклеаз RuvC; HEPN, предполагаемая эндорибонуклеаза суперсемейства HEPN. Показанные серым области генов и белков обозначают последовательности, которые были закодированы в соответствующих мобильных элементах, но были утрачены в ходе эволюции систем CRISPR-Cas.
[00158] Чертежи приведены в настоящем описании исключительно в иллюстративных целях и не обязательно выполнены с соблюдением масштаба.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[00159] Прежде чем будут описаны способы по настоящему изобретению, следует понимать, что настоящее изобретение не ограничивается отдельными описанными в настоящем описании способами, компонентами, продуктами или их комбинациями, в силу того, что такие способы, продукты и комбинации, безусловно, могут, бесспорно, варьироваться. Необходимо понимать и что используемая в настоящем описании терминология не ставит своей целью наложение ограничений, поскольку рамки настоящего изобретения ограничиваются только прилагаемой формулой изобретения.
[00160] В целом, понятие "система CRISPR-Cas или CRISPR", используемое также в предшествующих документах, таких как W0 2014/093622 (PCT/US2013/074667), используется для совместного обозначения транскриптов и других компонентов, участвующих в экспрессии или управлении активностью CRISPR-ассоциированных ("Cas") генов, включая последовательности, кодирующие ген Cas, последовательность tracr (трансактивирующая CRISPR) (например, tracr-РНК или активная частичная tracr-РНК), последовательность tracr-помощника (включащая "прямой повтор" и процессируемый tracr-РНК частичный прямой повтор в составе эндогенной системы CRISPR), направляющую последовательность (также называемую "спейсером" в составе эндогенной системы CRISPR) или "молекулу(ы) РНК", как этот термин использован в настоящем описании (например, молекулу(ы) РНК, направляющую Cas, такие как Cas9, например, CRISPR-РНК и трансактивирующую (tracr) РНК или одиночную направляющую РНК (sgRNA) (химерная РНК)) или другие последовательности и транскрипты локуса CRISPR. В целом, система CRISPR характеризуется компонентами, которые способствуют формированию комплекса CRISPR на участке последовательности-мишени (в составе эндогенной системы CRISPR также называемый протоспейсером). В контексте формирования комплекса CRISPR, "последовательность-мишень" означает последовательность, для которой целенаправленно разработана комплементарная направляющая последовательность, причем гибридизация между последовательностью-мишенью и направляющей последовательностью способствует образованию комплекса CRISPR. Последовательность-мишень может включать любой полинуклеотид, в том числе полинуклеотиды РНК или ДНК. В некоторых вариантах осуществления изобретения последовательность-мишень расположена в ядре или цитоплазме клетки. В некоторых вариантах осуществления изобретения прямые повторения могут быть идентифицированы in silico путем поиска повторяющихся мотивов, которые удовлетворяют любому или всем из следующих критериев: 1. обнаружены в пределах области геномной последовательности размером 2 т.п.н., фланкирующей локусы CRISPR II типа; 2. длина от 20 до 50 п.н.; и 3. разделяющие их промежутки длиной 20-50 п.н. В некоторых вариантах осуществления изобретения могут быть использованы любые 2 из этих критериев, например, 1 и 2, 2 и 3 или 1 и 3. В некоторых вариантах осуществления изобретения могут быть использованы все 3 критерия.
[00161] В вариантах осуществления изобретения такие понятия как зрелая cr-РНК, единственная направляющая РНК, направляющая последовательность и направляющая РНК, т.е. РНК, способная направлять Cas к целевому локуса генома, используются взаимозаменяемо в предшествующих цитируемых документах, таких как W0 2014/093622 (PCT/US2013/074667). В целом, направляющей последовательностью является любая полинуклеотидная последовательность, имеющая комплементарность к полинуклеотиду-мишени, достаточную для гибридизации с последовательностью-мишенью и направления последовательности на прямое специфическое связывание комплекса CRISPR с последовательностью-мишенью. В некоторых вариантах осуществления изобретения уровень комплементарности между направляющей последовательностью и соответствующей ей последовательностью-мишенью при условии оптимального выравнивания с использованием надлежащего алгоритма выравнивания близок или превосходит значения 50%, 60%, 75%, 80%, 85%, 90%, 95%, 97,5%, 99% или выше. Оптимальное выравнивание может быть определено с использованием любого надлежащего алгоритма для выравнивания последовательностей включая, но не ограничиваясь следующими: алгоритм Смита-Ватермана, алгоритм Нидлмана-Вунша, алгоритмы, основанные на преобразовании Барроуза-Уилера (например, Burrows Wheeler Aligner), ClustalW, Clustal X, BLAST, Novoalign (Novocraft Technologies; доступен на www.novocraft.com), ELAND (Illumina, Сан-Диего, Калифорния, США), SOAP (доступен на soap.genomics.org.cn) и Maq (доступен на maq.sourceforge.net). В некоторых вариантах осуществления направляющая последовательность имеет длину, примерно равную или превышающую 5, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 35, 40, 45, 50, 75 или более нуклеотидов. В некоторых вариантах осуществления направляющая последовательность имеет длину, меньшую 75, 50, 45, 40, 35, 30, 25, 20, 15, 12 или менее нуклеотидов. Предпочтительная длина направляющей последовательности составляет 10-30 нуклеотидов. Способность направляющей последовательности направлять специфически определяемое последовательностью связывание комплекса CRISPR с последовательностью-мишенью может быть оценена любой подходящей методикой. Например, компоненты системы CRISPR, достаточные для формирования комплекса CRISPR, включая проверяемую направляющую последовательность, могут быть доставлены в клетку-хозяина, содержащую соответствующую последовательность-мишень, в том числе путем трансфекции с использованием векторов, кодирующих компоненты последовательности CRISPR с последующей количественной оценкой локализации предпочтительного расщепления последовательности-мишени, в частности, с использование анализа, как описано в настоящем описании. Сходным образом расщепление последовательности нуклеотида-мишени может быть оценено in vitro при использовании последовательности-мишени, компонентов комплекса CRISPR, включая проверяемую направляющую последовательность и направляющую последовательность-контроль, отличную от тестируемой направляющей последовательности при сравнении уровня или скорости расщепления последовательности-мишени между реакциями с участием тестируемой и контрольной направляющих последовательностей. Другие способы анализа также допустимы и могут быть выбраны специалистом в данной области.
[00162] В классических системах CRISPR-Cas уровень комплементарности между последовательностью направляющей молекулы и соответствующей ей последовательностью-мишенью может быть примерно равен или превышать 50%, 60%, 75%, 80%, 85%, 90%, 95%, 97,5%, 99% или 100%; направляющая молекула или РНК или sg-РНК могут быть иметь длину, примерно равную или превышающую 5, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 35, 40, 45, 50, 75 или более нуклеотидов; либо направляющая молекула или РНК или sg-РНК могут иметь длину, меньшую чем приблизительно 75, 50, 45, 40, 35, 30, 25, 20, 15, 12 или менее нуклеотидов; и преимущественно tracr-РНК имеет длину 30 или 50 нуклеотидов. Однако в одном из своих аспектов изобретение призвано уменьшить нецелевые взаимодействия, например, уменьшить взаимодействие направляющей молекулы с молекулой-мишенью с низкой комплементраностью. Действительно, в примерах показано, что в изобретение вовлекает мутации, делающие систему CRISPR-Cas способной различать последовательности-мишени и последовательности, не являющиеся мишенями, с комплементарностью более 80% до примерно 95%, например, с комплементарностью 83%-84%, 88-89% или 94-95% (к примеру, различая молекулу-мишень, имеющую 18 нуклеотидов молекулы, не являющейся мишенью, длиной в 18 нуклеотидов с 1, 2 или 3 нарушениями комплементарности). В соответствии с этим, в контексте настоящего изобретения уровень комплементарности между направляющей последовательностью и соответствующей ей молекулой-мишенью превышает 94,5%, 95%, 95,5%, 96%, 96,5%, 97%, 97,5%, 98%, 98,5%, 99%, 99,5%, 99,9% или 100%. Для молекулы, не являющейся мишенью, он составляет менее 100%, 99,9%, 99,5%, 99%, 99%, 98,5%, 98%, 97,5%, 97%, 96,5%, 96%, 95,5%, 95%, 94,5%, 94%, 93%, 92%, 91%, 90%, 89%, 88%, 87%, 86%, 85%, 84%, 83%, 82%, 81% или 80% комплементарности между ее последовательностью и направляющей молекулой, причем предпочтительно, чтобы не являющаяся мишенью молекула имела комплементарность последовательности 100%, 99,9%, 99,5%, 99%, 99%, 98,5%, 98%, 97,5%, 97%, 96,5%, 96%, 95,5%, 95% или 94,5% с направляющей молекулой.
[00163] В особенно предпочтительных вариантах осуществления настоящего изобретения направляющая РНК (способная направлять Cas к локусу-мишени) может включать (1) направляющую последовательность, способную к гибридизации с геномным локусом-мишенью в эукариотической клетке; (2) последовательность tracr; и (3) последовательность-партнер tracr. Компоненты (1)-(3) могут находится на одной РНК, т.е. sg-РНК (расположенной в направлении от 5' к 3'-концу), либо tracr-РНК может являться РНК, отличной от РНК, содержащей направляющую последовательность и последовательность tracr. Tracr гибридизуется с последовательностью-партнером tracr и направляет комплекс CRISPR/Cas к последовательности-мишени.
[00164] Аспекты изобретения касаются идентификации и конструирования способами инженерии новых эффекторных белков, связанных с системами CRISPR-Cas класса 2. В предпочтительном варианте осуществления эффекторный белок включает односубъединичный эффекторный модуль. В будущем варианте осуществления изобретения эффекторный белок является функциональным в прокариотических или эукариотических клетках для применения in vitro, in vivo или ex vivo. Один аспект изобретения охватывает вычислительные способы и алгоритмы для прогнозирования новых систем CRISPR-Cas класса 2 и определения составляющих их компонентов.
[00165] В одном из вариантов осуществления вычислительный способ идентификации новых локусов системы CRISPR-Cas класса 2 включает следующие шаги: обнаружение всех контигов, кодирующих белок Cas1; идентификация всех предсказанных генов, кодирующих белки, в пределах 20 т.п.н. от гена cas1; сравнение идентифицированных генов со специфическими профилями белков Cas и предсказание последовательностей CRISPR путем отбора неклассифицированных возможных локусов CRISPR-Cas, содержащих белки с последовательностями длиной более 500 аминокислот (>500 а.к.); анализ отобранных кандидатов с использованием анализов PSI-BLAST и HHPred для выделения и идентификации новых локусов CRISPR-Cas класса 2. В дополнение к вышеупомянутым шагам возможные кандидаты могут быть дополнительно проанализированы путем поиска гомологов по метагеномным базам данных.
[00166] Одним из аспектов обнаружения всех контигов, кодирующих белок Cas1, является использование программы GenemarkS для предсказания генов, как более подробно описано в статье "GeneMarkS: a self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions," John Besemer, Alexandre Lomsadze and Mark Borodovsky, Nucleic Acids Research (2001) 29, pp 2607-2618, включенной в настоящее описание в качестве ссылки.
[00167] В одном аспекте проводится идентификация всех предсказанных генов, кодирующих белки, с помощью сравнения идентифицированных генов со специфическими профилями белков Cas и их аннотирования согласно Базе Данных Консервативных Доменов NCBI (CDD), которая состоит из коллекции хорошо аннотированных многочисленных моделей выравнивания последовательностей для древних доменов и полноразмерных белков. Они доступны в виде позиционных весовых матриц (PSSM) для быстрой идентификации консервативных доменов в последовательностях белков посредством анализа RPS-BLAST. CDD включает домены, курируемые NCBI, которые используют информацию о 3D-структуре, чтобы точно определить границы доменов и обеспечить понимание взаимоотношений между последовательностью/структурой/функцией, а также модели домена, импортированные из многих внешних баз данных (Pfam, SMART, COG, PRK, TIGRFAM). В следующем аспекте для предсказания последовательностей CRISPR использовалась программы PILER-CR, которая является общедоступным программным обеспечением для поиска CRISPR-повторов, как описано в статье "PILER-CR: fast and accurate identification of CRISPR repeats", Edgar, R.C., BMC Bioinformatics, Jan 20:8:18(2007), включенной в настоящее описание в качестве ссылки.
[00168] В следующем аспекте проводят индивидуальный анализ каждого случая с использованием PSI-BLAST (Средство поиска основного локального выравнивания с учетом позиции). Анализ PSI-BLAST использует позиционную весовую матрицу (PSSM) или профиль множественного выравнивания последовательностей, обнаруженных выше заданного весового порога при белок-белковом анализе BLAST. Данная матрица PSSM используется, чтобы далее искать новые совпадения в базе данных и обновляется для последующих итераций с этими недавно обнаруженными последовательностями. Таким образом, PSI-BLAST является способом обнаружения отдаленных отношений между белками.
[00169] В другом аспекте индивидуальный анализ каждого случая предполагает использование HHpred, способа поиска по базам данных последовательностей и предсказания структуры, столь же простого в использовании, как BLAST или PSI-BLAST, и в то же время намного более чувствительного при нахождении отдаленных гомологов. Фактически, чувствительность способа HHpred конкурентоспособна по отношению к самым мощным серверам для предсказания структуры, доступным в настоящее время. HHpred является первым сервером, основанным на попарном сравнении профилей скрытых марковских моделей (СММ). В то время как большинство обычных способов поиска последовательностей выполняют поиск по базам данных последовательностей, таким как UniProt или NR, HHpred выполняет поиск по базам данных выравнивания, таким как Pfam или SMART. Это значительно упрощает получение списка совпадений со многим семействами последовательностей вместо разрозненных единичных последовательностей. Все основные общедоступные базы данных профилей и выравниваний доступны через HHpred. HHpred использует в качестве единичного поискового запроса последовательность или множественное выравнивание. В течение всего нескольких минут HHpred представляет результаты поиска в легком для чтения формате, подобном формату PSI-BLAST. Параметры поиска включают локальное или глобальное выравнивание и весовое сходство вторичной структуры. HHpred может произвести попарные выравнивания последовательностей поискового запроса и шаблона, объединенное множественное выравнивание поискового запроса и шаблона (например, для транзитивного поиска), а также модели 3D-структуры, вычисленные программным обеспечением MODELLER на основе выравниваний HHpred.
[00170] Термин "система нацеливания на нуклеиновые кислоты", где нуклеиновая кислота представляет собой ДНК или РНК, и в некоторых аспектах может также относится к гибридам РНК-ДНК или их производным, относится собирательно к транскриптам и другим элементам, участвующим в экспрессии или направлении активности нацеливания на ДНК или РНК генов, ассоциированных с системой CRISPR ("Cas"), которые могут содержать последовательности, кодирующие белки Cas, нацеленные ДНК или РНК, и направляющую РНК, нацеленную ДНК или РНК, включая последовательность cr-РНК (cr-РНК) и (в системе CRISPR-Cas9, но не всех системах) последовательность РНК, транс-активирующей систему CRISPR/Cas (tracr-РНК), или другие последовательности и транскрипты локуса CRISPR, нацеленные на ДНК или РНК. В целом система нацеливания на РНК характеризуется элементами, способствующими формированию нацеленного на РНК комплекса на участке последовательности РНК, являющейся мишенью. В контексте образования нацеленного на ДНК или РНК комплекса, термин "последовательность-мишень" относится к последовательности ДНК или РНК, которой комплементарна направляющая РНК, нацеливающая на ДНК или РНК, где гибридизация между последовательностью-мишенью и направляющей РНК, нацеливающей на РНК, способствует образованию нацеливающего на РНК комплекса. В некоторых вариантах осуществления изобретения последовательность-мишень расположена в ядре или цитоплазме клетки.
[00171] В одном из вариантов осуществления изобретения новые нацеленные на ДНК системы, также называемые нацеленными на ДНК CRISPR-Cas или CRISPR-Cas-нацеленными на ДНК системами в контексте настоящей заявки, основаны на идентифицированных белках Cas типа V (например, подтип V-A и подтип V-B), не требующих получения специализированных белков для специфических последовательностей РНК, а содержащих всего один эффекторный белок или фермент, который может быть запрограммирован молекулой РНК для распознавания определенной ДНК-мишени; иными словами, белок может быть привлечен к определенной ДНК-мишени с помощью указанной молекулы РНК. Аспекты изобретения относятся, в частности, к нацеленным на ДНК РНК-направляемым системам CRISPR C2c1 или C2c3.
[00172] В одном аспекте изобретения новые системы нацеливания на РНК, также называемые РНК- или РНК-нацеленные CRISPR/Cas или система нацеливания на РНК с помощью системы CRISPR-Cas, описанные в настоящем описании, основаны на идентифицированных белках Cas типа VI, не требующих создания индивидуальных белков для нацеливания на определенные последовательности РНК, кроме единственного фермента, который может быть запрограммирован молекулой РНК для узнавания конкретной ДНК-мишени, иными словами, фермент может быть предназначен для работы с конкретной ДНК-мишенью с помощью указанной молекулы РНК.
[00173] Как используют в настоящем описании, белок Cas или фермент CRISPR относится к любым белкам, представленным в новой классификации систем CRISPR-Cas. В предпочтительном варианте осуществления настоящее изобретение относится к эффекторным белкам, идентифицированным в локусах CRISPR-Cas типа V, например, к кодирующим Cpf1 локусам, обозначаемым как подтип V-A. В настоящее время локусы подтипа V-A включают cas1, cas2, отдельный ген, обозначенный как cpf1 и кассету CRISPR. Cpf1 (ассоциированный с CRISPR белок Cpf1, подтип PREFRAN) - крупный белок (примерно 1300 аминокислот), содержащий подобный RuvC нуклеазный домен, гомологичный соответствующему домену Cas9, наряду с соответствующим богатому аргинином кластеру Cas9. Однако Cpf1 не имеет нуклеазного домена HNH, присутствующего во всех белках Cas9, а подобный RuvC домен является непрерывным в последовательности Cpf1, в противоположность Cas9, где он содержит длинные вставки, включая домен HNH. В соответствии с этим, в отдельных вариантах осуществления изобретения фермент CRISPR-Cas включает только подобный RuvC нуклеазный домен.
[00174] В предпочтительном варианте осуществления настоящее изобретение относится к композициям и системам, содержащим эффекторные белки, идентифицированные в локусах C2c1, обозначаемые как подтип V-B. В настоящем описании C2c1 относится к 1 кандидату 2 класса. Все локусы C2c1 кодируют слитую конструкцию Cas1-Cas4, Cas2, и крупного белка, который заявители обозначили как C2c1p, и обычно она прилегает к кассете CRISPR.
[00175] В предпочтительном варианте осуществления настоящее изобретение относится к эффекторному белку, идентифицированному в локусах CRISPR-Cas типа VI, например, локусах C2c2. В настоящем описании C2c2 означает 2 кандидата 2 класса. Локусы C2c2 включают гены cas1 и cas2, наряду с крупным белком, обозначенным Заявителями как C2c2p и кассету CRISPR; однако, в отличие от C2c1p, C2c2p часто кодирует следующую за кассетой CRISPR последовательность, но не cas1-cas2.
[00176] В предпочтительном варианте осуществления настоящее изобретение относится к композициям и системам, содержащим эффекторные белки, идентифицированные в локусах С2с3, обозначенные как подтип V-C. Все локусы C2c3 кодируют слитую конструкцию Cas1-Cas4, Cas2 и крупного белка, обозначенного Заявителями как C2c3p.
[00177] Аспекты настоящего изобретения также включают спосбы и применение композиций и систем, описанных в настоящем описании, для геномной инженерии, например, для изменения или управления экспрессией одного или более генов или одного или более генных продуктов в прокариотических или эукариотических клетках in vitro, in vivo или ex vivo.
[00178] Системы нацеливания на нуклеиновые кислоты, векторные системы, векторы и композиции, описанные в настоящем описании, могут использоваться в практике нацеливания на нуклеиновые кислоты, изменения или модификация синтеза генных продуктов, таких как белки, расщепления нуклеиновых кислот, редактирования нуклеиновых кислот, сплайсинга нуклеиновых кислот, переноса нуклеиновых кислот-мишеней, отслеживания нуклеиновых кислот-мишеней, выделения нуклеиновых кислот-мишеней, визуализации нуклеиновых кислот-мишеней и т.д.
[00179] Аспекты изобретения также охватывают способы и применения композиций и систем, описанных в настоящем описании, в генной инженерии, например, для изменения или манипулирования экспрессией одного или более генов или одного или более генных продуктов, в прокариотических или эукариотических клетках, in vitro, in vivo или ex vivo.
[00180] Способы по настоящему изобретению, описанные в настоящем описании, подразумевают внесение одной или более мутаций в эукариотическую клетку (in vitro, т.е. в выделенную эукариотическую клетку), как описано в настоящем описании, включая доставку в клетку вектора, как описано в настоящем описании. Мутации могут включать инсерцию, делецию или замену одного или более нуклеотидов в каждой последовательности-мишени клетки/клеток с помощью направляющей/их РНК или молекулы/молекул sg-РНК. Мутации могут включать инсерцию, делецию или замену 1-75 нуклеотидов в каждой/ых указанной/ых клетке/ах с помощью направляющей/их РНК или молекулы/молекул sg-РНК. Мутации могут включать инсерцию, делецию или замену 1, 5, 10, 11, 12, 13, 14, 15, 16, 1/, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 35, 40, 45, 50 или 75 нуклеотидов в каждой/ых указанной/ых клетке/ах с помощью направляющей/их РНК или молекулы/молекул sg-РНК. Мутации могут включать инсерцию, делецию или замену 10, 11, 12, 13, 14, 15, 16, 17, Ж 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 35, 40, 45, 50 или 75 нуклеотидов в каждой/ых указанной/ых клетке/ах с помощью направляющей/их РНК или молекулы/молекул sg-РНК. Мутации могут включать инсерцию, делецию или замену 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 35, 40, 45, 50 или 75 нуклеотидов в каждой/ых указанной/ых клетке/ах с помощью направляющей/их РНК или молекулы/молекул sg-РНК. Мутации могут включать инсерцию, делецию или замену 40, 45, 50, 75, 100, 200, 300, 400 или 500 нуклеотидов в каждой/ых указанной/ых клетке/ах с помощью направляющей/их РНК или молекулы/молекул sg-РНК.
[00181] Для минимизации токсичности и снижения возможности проявления нецелевых эффектов является важным контроль концентрации доставляемой мРНК Cas и направляющей РНК. Оптимальные концентрации мРНК Cas и направляющей РНК могут быть определены путем тестирования различных концентраций в клеточных или отличных от человеческих эукариотических моделях на животных и с применением способов глубокого секвенирования для анализа степени модификации в потенциальных нецелевых геномных локусах. Другим альтернативным способом минимизации уровня токсичности и нецелевых эффектов, возможна доставка мРНК никазы Cas (например, Cas9 S. pyogenes с мутацией D10A) с двумя направляющими РНК, нацеленными на участок-мишень. Направляющие последовательности и стратегии минимизации токсичности и нецелевых эффектов могут использоваться, как описано в WO 2014/093622 (PCT/US 2013/074667) или при помощи мутации, как описано в настоящем описании.
[00182] Обычно в контексте эндогенной системы CRISPR образование комплекса CRISPR (включающего направляющую последовательность, гибридизованную с последовательностью-мишенью, находящейся в комплексе с одним или более белками Cas) приводит к расщеплению одной или обеих цепей вблизи последовательности-мишени (например, на расстоянии 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50 или более пар оснований от нее) или в ней самой. Без связи с теорией, последовательность tracr, которая может содержать или состоять из части последовательности tracr дикого типа (например, примерно или более чем 20, 26, 32, 45, 48, 54, 63, 67, 85 или более нуклеотидов последовательности tracr дикого типа), может также образовывать часть комплекса CRISPR, в частности, путем гибридизации по меньшей мере части последовательности tracr с полной последовательностью-партнером tracr или ее частью, функционально связанной с направляющей последовательностью.
[00183] Предпочтительно, чтобы молекула нуклеиновой кислоты, кодирующая Cas, представляла сбой кодон-оптимизированную последовательность кодонам Cas. Примером оптимизированной по кодонам последовательности в данном случае является последовательность, оптимизированная для экспрессии в эукариоте, например, человеке (т.е. являющаяся оптимизированной для экспрессии в человеке) или другом эукариоте, животном или млекопитающем, как обсуждается в настоящем описании; см, например, кодон-оптимизированную для человека последовательность SaCas9 в WO 2014/093622 (PCT/US2013/074667). Хотя это является предпочтительным, понятно, что возможны другие примеры и известная оптимизация кодонов для хозяина вида, отличного от человека, или оптимизация кодонов для конкретных органов. В некоторых вариантах осуществления, кодирующая фермент последовательность кодирует Cas и кодон-оптимизирована для экспрессии в конкретных клетках, например, эукариотических клетках. Такие эукариотические клетки могут принадлежать или быть выделены из конкретного организма, в частности млекопитающего, включая, но не ограничиваясь этим: человека, отличного от человека эукариота, животного или млекопитающего, как описано в настоящем описании, например, мыши, крысы, кролика, собаки, домашнего скота или отличного от человека млекопитающего или примата. В некоторых вариантах осуществления, способы модификации генетической принадлежности зародышевой линии человека и/или способы модификации генетической принадлежности животных, которые с большой долей вероятности могут причинить им страдания без получения существенной медицинской пользы для человека или животных, а также получаемые в результате таких способов животные, могут быть исключены. В целом под оптимизацией кодонов понимается процесс изменения последовательности с целью усиления экспрессии в клетках заданного организма путем замены по меньшей мере одного кодона (например, примерно или более 1, 2, 3, 4, 5, 10, 15, 20, 25, 50 или большего количества кодонов) нативной последовательности на кодоны, которые более часто или наиболее часто используются в генах таковой клеток целевого организма при сохранении исходной аминокислотной последовательности. Различные виды демонстрируют конкретное смещение в частоте встречаемости отдельных кодонов для отдельных аминокислот. Смещение частоты кодонов (различие в использовании кодонов между различными организмами) зачастую коррелирует с эффективностью трансляции матричной РНК (мРНК), которая в свою очередь считается зависящей, в числе прочего, от свойств транслируемых кодонов и доступности отдельных молекул транспортной РНК (тРНК). Преобладание конкретных тРНК в клетке обычно соответствует тем кодонам, которые используются наиболее часто при синтезе пептидов. В соответствии с этим, гены могут быть разработаны с учетом оптимизации кодонов, делающей экспрессию генов оптимальной в некотором организме. Таблицы используемых кодонов доступны, к примеру, в "Codon Usage Database", доступной по ссылке www.kazusa.orjp/codon/, причем такие таблицы могут быть адаптированы рядом способов. См. Nakamura, Y., et al. "Codon usage tabulated from the international DNA sequence databases: status for the year 2000" Nucl. Acids Res. 28:292 (2000). Также доступны компьютерные алгоритмы оптимизации кодонов для конкретной последовательности и экспрессии в конкретной клетке-хозяине, такие как Gene Forge (Aptagen; Джакобус, Пенсильвания, США). В некоторых вариантах осуществления один или более кодонов (например, 1, 2, 3, 4, 5, 10, 15, 20, 25, 50 или более, либо все кодоны) в последовательности, кодирующей Cas, соответствуют наиболее часто используемым кодонам для определенной аминокислоты.
[00184] В определенных вариантах осуществления изобретения способы, как описано в настоящем описании, могут включать предоставление трансгенной по Cas клетки, в которой одна или более нуклеиновых кислот, кодирующих одну или более молекул направляющих РНК, предоставлены или введены, функционально связанными в клетке с регуляторным элементом, содержащим промотор одного или более представляющих интерес генов. Термин "трансгенная по Cas клетка", как используют в настоящем описании, относится к клетке, в частности, эукариотической клетке, в геном которой встроен ген Cas. Природа, тип или происхождение такой клетки не ограничивается особым образом, согласно данному изобретению. Также способ введения трансгена Cas в клетку может различаться и являться любым из способов, доступных в данной области. В некоторых вариантах осуществления изобретения трансгенная клетка Cas получена путем введения трансгена Cas в выделенную клетку. В некоторых других вариантах осуществления изобретения трансгенная клетка Cas может быть получена путем выделения клеток из трансгенного организма Cas. В качестве не ограничивающего примера, трансгенная клетка Cas, как используют в настоящем описании, может быть получена из трансгенного по Cas эукариотического организма, такого как эукориотический организм с встроенным Cas. См. WO 2014/093622 (PCT/US13/74667), включенную в настоящее описание в качестве ссылки. Способы согласно патенту США № 8771985, выданному Sangamo BioSciences, Inc. и Sigma-Aldrich Co. LLC, и публикации патента США № 20110265198, закрепленной за Sangamo BioSciences, Inc., направленные на нацеливание на локус Rosa, могут быть изменены для использования системы CRISPR Cas данного изобретения. Способы публикации патента США № 20130236946, закрепленных за Cellectis, направленные на нацеливание на локус Rosa, могут также быть изменены для использования системы CRISPR-Cas по настоящему изобретению. В качестве следующего примера приводится ссылка на статью Platt et. al. (Cell; 159(2):440-455 (2014)) с описанием мыши со вставкой Cas9 мыши, включенную в настоящее описание в качестве ссылки. Трансген Cas, кроме того, может включать кассету Lox-Stop-polyA-Lox(LSL), что делает экспрессию Cas управляемой с помощью рекомбиназы Cre. Альтернативно, клетка с трансгенным Cas может быть получена путем введения трансгена Cas в выделенную клетку. Системы доставки трансгенов хорошо известны в данной области. Примером может служить доставка трансгена Cas, например, в эукариотическую клетку посредством вектора (например, AAV, аденовируса, лентивируса) и/или частицы и/или наночастицы, как также описано в другой части настоящего описания.
[00185] Для квалифицированного специалиста будет понятно, что клетка, такая как трансгенная по Cas клетка в используемом в настоящем описании значении, может содержать дальнейшие изменения генома помимо наличия встроенного гена Cas или мутаций, являющихся результатом последовательность-специфического действия Cas, в комплексе с РНК, способной направлять Cas к локусу-мишени, например, такие как одна или более онкогенных мутаций, как в качестве неограничивающего примера описано в Platt et al. (2014), Chen et al., (2014), или Kumar et al. (2009).
[00186] В некоторых вариантах осуществления последовательность Cas является слитой с одной или более последовательностями сигнала ядерной локализации (NLS), в частности примерно с или более чем примерно с 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более NLS. В некоторых вариантах осуществления Cas содержит примерно или более 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более NLS на N-конце или вблизи него, примерно или более 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более NLS на C-конце или их комбинацию (например, ноль или по меньшей мере один или более NLS на N-конце и ноль или по меньшей мере один или более NLS на С-конце). В случае присутствия более одного NLS каждый из них может быть выбран независимо от других, в частности, отдельный NLS может быть представлен более чем одной копией и/или в комбинации с другим одним или более NLS, представленного одной или более копиями. В предпочитаемом варианте осуществления Cas содержит не более 6 NLS. В некоторых вариантах осуществления NLS считается N- или C-концевым, если ближайшая к концу аминокислота NLS находится на расстоянии 1, 2, 3, 4, 5, 10, 15, 20, 25, 30, 40, 50 или более аминокислот полипептидной цепи от N- или C-конца. Неограничивающие примеры NLS включают последовательности NLS, полученные на основе: NLS большого T-антигена вируса SV40, имеющего аминокислотную последовательность PKKKRKV (SEQ ID NO: X); NLS из нуклеоплазмина (например, состоящая из двух частей NLS нуклеоплазмана с последовательностью KRPAATKKAGQAKKKK) (SEQ ID NO: X); NLS c-myc, имеющего аминокислотную последовательность PAAKRVKLD (SEQ ID NO: X) или RQRRNELKRSP(SEQ ID NO: X); NLS hRNPA1 M9, имеющего последовательность NQ S SNFGPMKGGNFGGRS SGPYGGGGQYFAKPRNQGGY (SEQ ID NO: X); последовательности RMRIZFKNKGKDTAELRRRRVEVSVELRKAKKDEQILKRRNV (SEQ ID NO: X) из домена импортина-альфа; последовательности VSRKRPRP (SEQ ID NO: X) и PPKKARED (SEQ ID NO: X) T-белка миомы; последовательности POPKKKPL (SEQ ID NO: X) p53 человека; последовательности SALIKKKKKMAP (SEQ ID NO: X) c-abi IV мыши, последовательности DRLRR (SEQ ID NO: X) и PKQKKRK (SEQ ID NO: X) вируса гриппа NS1; последовательности RKLKKKIKKL (SEQ ID NO: X) антигена дельта вируса гепатита; последовательности REKKKFLKRR (SEQ ID NO: X) белка Mx1 мыши; последовательности KRKGDEVDGVDEVAKKKSKK (SEQ ID NO: X) поли(АДФ-рибоза) полимеразы человека и последовательности RKCLQAGMNLEARKTKK (SEQ ID NO: X) рецепторов глюкокортикоидных стероидных гормонов (человека). В целом, такая одна или более NLS имеют достаточную силу для того, чтобы вызвать накопление Cas в заметных количествах в ядре эукариотической клетки. В целом сила активности ядерной локализации может определяться рядом NLS в белках Cas, конкретным(и) использованным(и) NLS или комбинацией этих факторов. Например, поддающаяся обнаружению метка может быть связана с Cas таким образом, чтобы визуализировать локализацию в клетке, в частности, за счет сочетания способов определения локализации ядра (например, специфичный ядерный краситель, такой как DAPI). Клеточные ядра могут быть также выделены из клеток, содержимое которых может затем быть проанализировано с помощью любого подходящего способы обнаружения белков, такого как иммуногистохимия, вестерн-блоттинг или анализ активности ферментов. Накопление в ядре может быть определено косвенно, в частности, с помощью анализа эффекта образования комплексов CRISPR (например, анализ расщепления ДНК или мутаций в последовательности-мишени, или анализ изменения экспрессии генов вследствие образования комплексов CRISPR и/или активности ферментов Cas), в сравнении с контролем, не подвергнутым воздействию Cas или комплекса или подвергнутым действию Cas, лишенного одной или более NLS.
[00187] В некоторых аспектах изобретение включает такие векторы, например, для доставки или введения в клетку Cas и/или РНК, способной направлять Cas к локусу-мишени (т.е. направляющей РНК), а также для воспроизведения этих компонентов (например, в прокариотических клетках). Как используют в настоящем описании, "вектор" представляет собой инструмент, позволяющий или облегчающий перенос некоторого объекта из одного окружения в другое. Он представляет собой репликон, такой как плазмида, фаг или космида, в который может быть вставлен другой участок ДНК таким образом, чтобы добиться репликации вставленного участка. В целом вектор способен к репликации при условии ассоциации с надлежащими элементами контроля. В целом понятие "вектор" обозначает молекулу нуклеиновой кислоты, способную транспортировать другую нуклеиновую кислоту, с которой она связана. Неограничивающими примерами векторов являются молекулы нуклеиновых кислот, являющихся одноцепочечными, двухцепочечными или частично двухцепочечными; молекулы нуклеиновых кислот, имеющие один или более свободных концов, не имеющие свободных концов (например, кольцевые); молекулы нуклеиновых кислот, включающие ДНК, РНК или одновременно и ДНК и РНК и другие известные в данной области разновидности полинуклеотиды. Одним из типов векторов является "плазмида", представляющая собой кольцевую двухцепочечную петлю ДНК, в которую могут быть вставлены дополнительные участки ДНК, в том числе с использованием стандартных способов клонирования. Другим типом вектора является вирусный вектор, содержащий полученную из вирусной ДНК или РНК последовательность, необходимую для упаковки в вирус (например, ретровирусы, дефективные в отношении репликации ретровирусы, аденовирусы, аденовирусы, ретровирусы и аденоасоциированные вирусы (AAV, AAV) с нарушенной репликацией). Вирусные векторы также включают полинуклеотиду, помещенную в вирус с целью трансфекции в клетку-хозяина. Некоторые векторы способны к автономной репликации в клетке хозяина, в которую они введены (например, бактериальные векторы, имеющие бактериальный ориджин репликации и эписомные векторы млекопитающих). Другие векторы (например, векторы млекопитающих, отличные от эписомных) интегрируются в геном клетки хозяина при введении в клетку хозяина и вследствие этого реплицируются вместе с геномом хозяина. Более того, некоторые векторы способны управлять экспрессией генов, с которыми они непосредственно связаны. Такие векторы называются в настоящем описании "экспрессирующими векторами". Векторы, которые приводят к экспрессии в эукариотической клетке, могут быть называться в настоящем описании "эукариотическми экспрессирующими векторами". Распространенной формой экспрессирующих векторов в способах рекомбинантных ДНК часто являются плазмиды.
[00188] Рекомбинантные экспрессирующие векторы могут включать нуклеиновую кислоту по изобретению в форме, подходящей для экспрессии нуклеиновой кислоты в клетке-хозяине, что означает, что рекомбинантные экспрессирующие векторы включают один или более регуляторных элементов, которые могут быть подобраны на основе клеток-хозяев, которые будут использоваться для экспрессии, т.е. функционально связаны с последовательностью нуклеиновых кислот, которая будет экспрессирована. В рекомбинантном векторе экспрессии "функционально связанный" должно означать, что представляющая интерес нуклеотидная последовательность связана с регуляторным элементом(элементами) таким образом, что обеспечивает экспрессию нуклеотидной последовательности (например, в системе транскрипции/трансляции in vitro или в клетке-хозяине, когда вектор введен в клетку-хозяина). Относительно рекомбинации и способов клонирования, упоминается заявка на патент США № 10/815730, опубликованная 2 сентября 2004 года как US 2004-0171156 A1, содержание которой включено в настоящее описание в качестве ссылки в полном объеме.
[00189] Вектор(ы) может включать регуляторный элемент(ы), например, промотор(ы). Вектор(ы) может включать последовательности, кодирующие белки Cas и/или единственную направляющую последовательность РНК, однако возможно также по меньшей мере 3, или 8, или 16, или 32, или 48, или 50 последовательностей, кодирующих направляющие последовательности РНК (например, sg-РНК), например 1-2, 1-3, 1-4 1-5, 3-6, 3-7, 3-8, 3-9, 3-10, 3-8, 3-16, 3-30, 3-32, 3-48, 3-50 РНК (например, sg-РНК). В отдельном векторе может содержаться промотор для каждой РНК (например, sg-РНК), предпочтительно до 16 молекул РНК (sg-РНК) и в случае если отдельный вектор обеспечивает более чем 16 РНК (например, sg-РНК), один или более промоторов могут обеспечить экспрессию более одной РНК (например, sg-РНК), например, в присутствии 32 РНК (например, sg-РНК), каждый промотор может обеспечить экспрессию двух РНК (например, sg-РНК) и в присутствии 48 РНК (например, sg-РНК), каждый промотор может обеспечить экспрессию трех РНК (например, sg-РНК). На основе простой арифметики и хорошо утвердившихся протоколов клонирования, а также концепций данного изобретения квалифицированный в данной области специалист может с легкостью применить данное изобретение к молекуле(ам) РНК (например, sg-РНК) для подходящего часто используемого вектора, такого как AAV, и подходящего промотора, в частности, промотора U6, например, U6 - sg-РНК. Например, максимальный возможный объем упаковки в AAV составляет ~4,7 т.п.н. Длина отдельно U6-sg-РНК (включая сайты рестрикции для клонирования) составляет 361 п.н. Следовательно, квалифицированный специалист может легко поместить примерно 12-16, например, 13 кассет U6-sg-РНК в один вектор. Он может быть собран с помощью любых подходящих методов, в частности, "стратегии золотых ворот" для сборки TALE (http://www.genome-engineering.org/taleffectors/). Квалифицированный специалист может также использовать стратегию тандемного гида для увеличения количества U6-sg-РНК примерно в 1,5 раза, например, для увеличения с 12-16, например 13, до 18-24, например, примерно 19 U6-sg-РНК. Следовательно, квалифицированный в данной области специалист может легко получить примерно 18-24, например, примерно 19 конструкций промотор-РНК, например, U6-sg-РНК в одном векторе, например, вектора AAV. Другие средства увеличения количества промоторов и РНК приведены в WO 2016/205749 PCT/US2016/038238, например, молекула(ы) sg-РНК в векторе для использования одного промотора (например, U6) для экспрессии последовательностей РНК, например, sg-РНК, разделенных способными к расщеплению последовательностей. Дальнейшие способы увеличения количества конструкций промотор-РНК, например sg-РНК в векторе, состоят в экспрессии последовательности промотор-РНК, например, sg-РНК в векторе, разделенных способными к расщеплению последовательностями в составе интрона кодирующей последовательности или гена; в этом случае предпочтительно использование промотора полимеразы II, который может усилить экспрессию и сделать возможной транскрипцию длинной РНК специфично к типу ткани (см., например, http://nar.oxfordjournals.org/content/34/7/e53.short, http://www.nature.com/mt/journal/vl6/n 9/abs/mt20081 44a. html). В предпочтительном варианте осуществления изобретения в вектор AAV может быть упакован тандем U6-sg-РНК, нацеленный приблизительно на вплоть до 50 генов. Соответственно, исходя из знаний в данной области и руководства в настоящей заявке, квалифицированный специалист может создавать и использовать вектор(ы), например, единичный вектор, экспрессирующий множественные РНК или другие направляющие молекулы под контролем, оперативно или функционально связанный с одним или более промоторами, особенно для числа РНК или других направляющих молекул, обсуждаемого в настоящем описании, без какого-либо излишнего экспериментирования.
[00190] Последовательности, кодирующие направляющие РНК, например sg-РНК, и/или последовательности, кодирующие белки Cas, могут быть функционально или непосредственно связаны с регуляторным элементом(элементами) и, следовательно, регуляторный элемент(элементы) управляет их экспрессией. Промотор(промоторы) может быть конститутивным промотором (промоторами) и/или временным промотором(промоторами), и/или индуцируемым промотором(промоторами), и/или тканеспецифическим промотором(промоторами). Промотор может быть выбран из группы, состоящей из РНК-полимераз: pol I, pol II, pol III, T7, U6, H1, ретровирусного LTR-промотора (LTR) вируса саркомы Рауса (RSV), промотора цитомегаловируса (CMV), промотора вируса SV40, промотора дигидрофолатредуктазы, промотора β-актина, промотора фосфоглицераткиназы (PGK) и промотора EF1α. Предпочтительным промотором является промотор U6.
[00191] В целом система CRISPR-Cas9 является такой, как используется в вышеприведенных документах, таких как WO 2014/093622(PCT/US2013/074667) и относится в совокупности к транскриптам и других элементов, участвующих в экспрессии или направляющих активность ассоциированного с CRISPR ("Cas") фермента, например, Cas9, включая последовательности, кодирующие фермент Cas (нацеленный на ДНК и/или РНК) или участвующие в его доставке, последовательность tracr (трансактивирующая CRISPR) (например, tracr РНК или активную частичную tracr РНК), последовательность-партнер tracr (включающую "прямой повтор" и tracr-РНК-процессированный частичный прямой повтор в контексте эндогенной системы CRISPR), направляющую последовательность (также называемую "спейсером" в контексте эндогенной системы CRISPR), или "молекулу(ы) РНК" в используемом в настоящем описании значении (например, молекула(ы) РНК, направляющая Cas9, как, например, CRISPR РНК (cr-РНК) и трансактивирующая cr-РНК (tracr РНК) или единая направляющая РНК (sg-РНК) (химерная РНК) или другие последовательности и транскрипты локуса CRISPR. В целом, система CRISPR характеризуется наличием элементов, стимулирующих образование комплекса CRISPR в участке последовательности-мишени (также называемых протоспейсерами в контексте эндогенной системы CRISPR). В контексте образования комплекса CRISPR, "последовательность-мишень" означает последовательность, для нацеливания на которую разработана направляющая последовательность, в частности, имеющая комплементарность, причем гибридизация между последовательностью-мишенью и направляющей последовательностью стимулирует образование комплекса CRISPR. Определенный участок направляющей последовательности, опосредующий значимость комплементарности к последовательности-мишени для активности по расщеплению, обозначен в настоящем описании как последовательность-затравка (seed-sequence). Последовательность-мишень может включать любой полинуклеотид, такой как полинуклеотиды ДНК или РНК, и находиться в составе представляющего интерес локуса-мишени. В некоторых вариантах осуществления изобретения последовательность-мишень локализована в ядре или цитоплазме клетки. Описанное в настоящем описании изобретение охватывает новые эффекторные белки 2 класса систем CRISPR-Cas, среди которых Cas 9 является иллюстративным эффекторным белком и, следовательно, понятия, используемые в настоящей заявке при описании новых эффекторных белков, могут коррелировать с понятиями, используемыми для описания систем CRISPR-Cas9.
[00192] Локусы системы CRISPR-Cas насчитывают более 50 семейств генов и не имеют строго универсальных генов. Следовательно, для локусов системы CRISPR-Cas ни одно эволюционное дерево не будет единственно верным, и для идентификации новых семейств генов необходим многоплановый подход. На данный момент для 395 профилей 93 белков Cas исчерпывающе идентифицированы гены cas. Для классификации систем CRISPR-Cas используют профили генов и характерные особенности архитектуры локусов. Новая классификация систем CRISPR-Cas предложена на фиг.1A и 1B. 1 класс включает многосубъединичные эффекторные комплексы cr-РНК (Cascade), и 2 класс включает односубъединичные эффекторные комплексы cr-РНК (Cas9-подобные). На фиг.2 изображена молекулярная организация систем CRISPR-Cas. На фиг.3 изображена структура эффекторных комплексов типов I и III: общая архитектура/общее происхождение прослеживается несмотря на обширную дивергенцию последовательностей. На фиг.4 CRISPR-Cas изображена как систему, основанную на мотивах распознавания РНК (RRM). На фиг.5 изображена филогения белка Cas1, показан главный аспект эволюции системы CRISPR-Cas - рекомбинация эффекторных модулей cr-РНК в целях адаптации. На фиг.6 представлен перечень систем CRISPR-Cas, а именно распространение типов/подтипов систем CRISPR-Cas среди архей и бактерий.
[00193] Действие системы CRISPR-Cas обычно делится на три этапа: (1) адаптация или интеграция спейсера, (2) процессинг первичного транскрипта локуса CRISPR (пре-cr-РНК) и созревание cr-РНК, которая включает спейсер и вариабельные регионы, соответствующие фрагментам CRISPR-повторов на 5'- и 3'-концах, и (3) РНК- или ДНК-интерференция. Два белка, Cas1 и Cas2, которые присутствуют в подавляющем большинстве известных систем CRISPR-Cas, достаточны для вставки спейсеров в кассеты CRISPR. Эти два белка формируют комплекс, который необходим для процесса адаптации; деятельность эндонуклеазы Cas1 необходима для интеграции спейсера, тогда как Cas2 выполняет неферментативную функцию. Комплекс Cas1-Cas2 представляет собой высококонсервативный модуль "обработки информации" системой CRISPR-Cas, который квазиавтономен от остальной системы. (См. Annotation and Classification of CRISPR-Cas Systems. Makarova KS, Koonin EV. Methods Mol Biol. 2015;1311:47-75).
[00194] Ранее описанные системы 2 класса, а именно, типа II и предполагаемого типа V, состояли только из трех или четырех генов в Cas-опероне, а именно, генов cas1 и cas2, составляющих модуль адаптации (пара генов cas1-cas2 не участвует в интерференции), а также единственного многодоменного эффекторного белка, ответственного за интерференцию, но также способствующего процессингу пре-cr-РНК и адаптации, и часто четвертого гена с неохарактеризованными функциями, который необязателен, по меньшей мере, в некоторых системах типа II (и в некоторых случаях четвертый ген - cas4 (данные биохимических или in silico экспериментов показывают, что cas4 принадлежит к суперсемейству нуклеаз PD-(DE)xK с кластером из трех остатков цистеина на C-конце; обладает 5'-экзонуклеазной активностью для оцДНК), или csn2, который кодирует инактивированную АТФ-азу). В большинстве случаев последовательность CRISPR и ген определенной разновидности РНК, известной как tracr-РНК, транс-закодированная небольшая cr-РНК, прилегают к Cas-оперонам класса 2. tracr-РНК частично гомологична повторам в соответствующей последовательности CRISPR и необходима для процессинга пре-cr-РНК, который катализируется РНК-азой III, широко распространенным бактериальным ферментом, не связанным с локусами системы CRISPR-Cas.
[00195] Cas1 является самым консервативным белком, который присутствует в большинстве систем CRISPR-Cas и эволюционирует медленнее, чем другие белки Cas. Соответственно, филогения Cas1 использовалась для классификации систем CRISPR-Cas. Результаты биохимических экспериментов или экспериментов in silico показывают, что Cas1 представляет собой металл-зависимую дезоксирибонуклеазу. Делеции в гене Cas1 в геноме E. coli приводят к увеличению чувствительности к повреждению ДНК и ослаблению расхождения хромосом, как описано в "A dual function of the CRISPR-Cassystem in bacterial antivirus immunity and DNA repair," Babu M et al. Mol Microbiol 79:484-502 (2011). Результаты биохимических экспериментов или in silico экспериментов показывают, что Cas2 является РНК-азой, характерной для регионов, насыщенных U, и является двухцепочечной ДНК-азой.
[00196] Аспекты изобретения касаются идентификации и конструирования способами инженерии новых эффекторных белков, связанных с системами CRISPR-Cas класса 2. В предпочтительном варианте осуществления эффекторный белок включает односубъединичный эффекторный модуль. В будущем варианте осуществления изобретения эффекторный белок является функциональным в прокариотических или эукариотических клетках для применения in vitro, in vivo или ex vivo. Один аспект изобретения охватывает вычислительные способы и алгоритмы для прогнозирования новых систем CRISPR-Cas класса 2 и определения составляющих их компонентов.
[00197] В одном из вариантов осуществления вычислительный способ идентификации новых локусов системы CRISPR-Cas класса 2 включает следующие шаги: обнаружение всех контигов, кодирующих белок Cas1; идентификация всех предсказанных генов, кодирующих белки, в пределах 20 т.п.н. от гена cas1, более конкретно, в пределах 20 т.п.н. от начала гена cas1 и 20 т.п.н. от конца гена cas1; сравнение идентифицированных генов со специфическими профилями белков Cas и предсказание последовательностей CRISPR путем отбора частичных и/или неклассифицированных возможных локусов CRISPR-Cas, содержащих белки с последовательностями длиной более 500 аминокислот (>500 а.к.); анализ отобранных кандидатов с использованием анализов PSI-BLAST и HHPred для выделения и идентификации новых локусов CRISPR-Cas класса 2. В дополнение к вышеупомянутым шагам возможные кандидаты могут быть дополнительно проанализированы путем поиска гомологов по метагеномным базам данных.
[00198] Одним из аспектов обнаружения всех контигов, кодирующих белок Cas1, является использование программы GenemarkS для предсказания генов, как более подробно описано в статье "GeneMarkS: a self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions," John Besemer, Alexandre Lomsadze and Mark Borodovsky, Nucleic Acids Research (2001) 29, pp 2607-2618, включенной в настоящее описание в качестве ссылки.
[00199] В одном аспекте проводится идентификация всех предсказанных генов, кодирующих белки, с помощью сравнения идентифицированных генов со специфическими профилями белков Cas и их аннотирования согласно Базе Данных Консервативных Доменов NCBI (CDD), которая состоит из коллекции хорошо аннотированных многочисленных моделей выравнивания последовательностей для древних доменов и полноразмерных белков. Они доступны в виде позиционных весовых матриц (PSSM) для быстрой идентификации консервативных доменов в последовательностях белков посредством анализа RPS-BLAST. CDD включает домены, курируемые NCBI, которые используют информацию о 3D-структуре, чтобы точно определить границы доменов и обеспечить понимание взаимоотношений между последовательностью/структурой/функцией, а также модели домена, импортированные из многих внешних баз данных (Pfam, SMART, COG, PRK, TIGRFAM). В следующем аспекте для предсказания последовательностей CRISPR использовалась программы PILER-CR, которая является общедоступным программным обеспечением для поиска CRISPR-повторов, как описано в статье "PILER-CR: fast and accurate identification of CRISPR repeats", Edgar, R.C., BMC Bioinformatics, Jan 20:8:18(2007), включенной в настоящее описание в качестве ссылки.
[00200] В следующем аспекте проводят индивидуальный анализ каждого случая с использованием PSI-BLAST (Средство поиска основного локального выравнивания с учетом позиции). Анализ PSI-BLAST использует позиционную весовую матрицу (PSSM) или профиль множественного выравнивания последовательностей, обнаруженных выше заданного весового порога при белок-белковом анализе BLAST. Данная матрица PSSM используется, чтобы далее искать новые совпадения в базе данных и обновляется для последующих итераций с этими недавно обнаруженными последовательностями. Таким образом, PSI-BLAST является способом обнаружения отдаленных отношений между белками.
[00201] В другом аспекте индивидуальный анализ каждого случая предполагает использование HHpred, способа поиска по базам данных последовательностей и предсказания структуры, столь же простого в использовании, как BLAST или PSI-BLAST, и в то же время намного более чувствительного при нахождении отдаленных гомологов. Фактически, чувствительность способа HHpred конкурентоспособна по отношению к самым мощным серверам для предсказания структуры, доступным в настоящее время. HHpred является первым сервером, основанным на попарном сравнении профилей скрытых марковских моделей (СММ). В то время как большинство обычных способов поиска последовательностей выполняют поиск по базам данных последовательностей, таким как UniProt или NR, HHpred выполняет поиск по базам данных выравнивания, таким как Pfam или SMART. Это значительно упрощает получение списка совпадений со многим семействами последовательностей вместо разрозненных единичных последовательностей. Все основные общедоступные базы данных профилей и выравниваний доступны через HHpred. HHpred использует в качестве единичного поискового запроса последовательность или множественное выравнивание. В течение всего нескольких минут HHpred представляет результаты поиска в легком для чтения формате, подобном формату PSI-BLAST. Параметры поиска включают локальное или глобальное выравнивание и весовое сходство вторичной структуры. HHpred может произвести попарные выравнивания последовательностей поискового запроса и шаблона, объединенное множественное выравнивание поискового запроса и шаблона (например, для транзитивного поиска), а также модели 3D-структуры, вычисленные программным обеспечением MODELLER на основе выравниваний HHpred.
[00202] Термин "система нацеливания на нуклеиновые кислоты", где нуклеиновая кислота представляет собой ДНК или РНК, и в некоторых аспектах может также относится к гибридам РНК-ДНК или их производным, относится собирательно к транскриптам и другим элементам, участвующим в экспрессии или направлении активности нацеливания на ДНК или РНК генов, ассоциированных с системой CRISPR ("Cas"), которые могут содержать последовательности, кодирующие белки Cas, нацеленные ДНК или РНК, и направляющую РНК, нацеленную ДНК или РНК, включая последовательность cr-РНК (cr-РНК) и (в некоторых, но не всех системах) последовательность РНК, транс-активирующей систему CRISPR/Cas (tracr-РНК), или другие последовательности и транскрипты локуса CRISPR, нацеленные на ДНК или РНК. В целом система нацеливания на РНК характеризуется элементами, способствующими формированию нацеленного на ДНК или РНК комплекса на участке последовательности ДНК или РНК, являющейся мишенью. В контексте образования нацеленного на ДНК или РНК комплекса, термин "последовательность-мишень" относится к последовательности ДНК или РНК, которой комплементарна направляющая РНК, нацеливающая на ДНК или РНК, где гибридизация между последовательностью-мишенью и направляющей РНК, нацеливающей на РНК, способствует образованию нацеливающего на РНК комплекса. В некоторых вариантах осуществления изобретения последовательность-мишень расположена в ядре или цитоплазме клетки.
[00203] В одном аспекте изобретения новые системы нацеливания на ДНК, также называемые ДНК- или ДНК-нацеленными CRISPR/Cas или системой нацеливания на ДНК с помощью системы CRISPR-Cas, описанные в настоящем описании, основаны на идентифицированных белках Cas типа V (например, подтипа V-A, подтипа V-B и подтипа V-C), не требующих создания индивидуальных белков для нацеливания на конкретные последовательности ДНК, а вместо этого единственный эффекторный белок или фермент может быть запрограммирован молекулой РНК для узнавания конкретной ДНК-мишени, иными словами, фермент может быть привлечен к конкретной ДНК-мишени с использованием указанной молекулы РНК. Аспекты изобретения, в частности, относятся к нацеленным на ДНК направляемым РНК системам CRISPR C2c1 или C2c3.
[00204] В одном аспекте изобретения новые системы нацеливания на РНК, также называемые РНК- или РНК-нацеленные CRISPR/Cas или система нацеливания на РНК с помощью системы CRISPR-Cas, описанные в настоящем описании, основаны на идентифицированных белках Cas типа VI, не требующих создания индивидуальных белков для нацеливания на определенные последовательности РНК, кроме единственного фермента, который может быть запрограммирован молекулой РНК для узнавания конкретной ДНК-мишени, иными словами, фермент может быть предназначен для работы с конкретной ДНК-мишенью с помощью указанной молекулы РНК.
[00205] Системы нацеливания на нуклеиновые кислоты, векторные системы, векторы и композиции, описанные в настоящем описании, могут использоваться в практике нацеливания на нуклеиновые кислоты, изменения или модификация синтеза генных продуктов, таких как белки, расщепления нуклеиновых кислот, редактирования нуклеиновых кислот, сплайсинга нуклеиновых кислот, переноса нуклеиновых кислот-мишеней, отслеживания нуклеиновых кислот-мишеней, выделения нуклеиновых кислот-мишеней, визуализации нуклеиновых кислот-мишеней и т.д.
[00206] Как используют в настоящем описании, белок Cas или фермент CRISPR обозначают любые белки, представленные в новой классификации систем CRISPR-Cas. В предпочтительном варианте осуществления настоящее изобретение охватывает эффекторные белки, идентифицированные в локусах CRISPR-Cas типа V, при этом следует отметить, что Cpf1 кодирует локусы, обозначаемые как подтип V-A. В настоящее время локусы подтипа V-A включают cas1, cas2, отдельный ген, обозначенный как cpf1 и кассету CRISPR. Cpf1 (ассоциированный с CRISPR белок Cpf1, подтип PREFRAN) представляет собой крупный белок (примерно 1300 аминокислот), содержащий подобный RuvC нуклеазный домен, гомологичный соответствующему домену Cas9, наряду с аналогом характерного богатого аргинином кластера Cas9. При этом Cpf1 лишен нуклеазного домена HNH, имеющегося у всех белков Cas9, и подобный RuvC домен является непрерывным, в противоположность Cas9, где он содержит длинные вставки, содержащие домен HNH. C2c1 и C2c3 сходны с Cpf1 в том отношении, что также кодируют локусы CRIPSR типа II класса V. В соответствии с этим, в некоторых вариантах осуществления фермент CRISPR-Cas содержит только подобный RuvC нуклеазный домен. Ген Cpf1 обнаружен в нескольких различающихся бактериальных геномах, в основном в одном и том же локусе, содержащем гены cas1, cas2, cas4 и кассету CRISPR (например, FNFX1_1431-FNFX1_1428 из Francisella cf. novicida Fx1). Таким образом, структурная организация Cpf1 представляется схожей с типом II-B. Более того, сходным с Cas9 образом, белок Cpf1 содержит легко идентифицируемый C-концевой участок, гомологичный транспозону ORF-B и включающий активную подобную RuvC нуклеазу, богатый аргинином участок и цинковый палец (отсутствующий у Cas9). Однако, в отличие от Cas9, Cpf1 представлен также в нескольких геномах, не содержащих CRISPR-Cas и его сравнительно высокое сходство с ORF-B предполагает, что он может являться компонентом транспозона. Было предложено, что если эта система CRISPR-Cas является истинной и Cpf1 является функциональным аналогом Cas9, то таким образом ее можно отнести к новому типу CRISPR-Cas, а именно, типу V (см. Annotation and Classification ofCRISPR-Cas Systems. Makarova KS, Koonin EV. Methods Mol Biol. 2015;1311:47-75). Однако, как описано в настоящем описании, Cpf1 относят к подтипу V-A для того, чтобы отличать его от C2c1p и C2c3p, не имеющих идентичную доменную структуру и, следовательно, относящихся соответственно к подтипам V-B и V-C.
[00207] В предпочтительном варианте осуществления настоящее изобретение относится к композициям и системам, включающим эффекторные белки, идентифицированные в локусах C2c1, выделенных в подтип V-B. В данном случае C2c1 относится к 1 кандидату из 2 класса. Все локусы C2c1 кодируют слитый белок Cas1-Cas4, Cas2 и крупный белок, обозначенный заявителями как C2c1p, обычно располагающийся вблизи кассеты CRISPR.
[00208] В предпочтительном варианте осуществления настоящее изобретение относится к композициям и системам, включающим эффекторные белки, идентифицированные в локусах C2c3, выделенных в подтип V-C. В данном случае C2c3 относится к 3 кандидату из 2 класса. Локусы C2c3 кодируют Cas1 и крупный белок, обозначенный как C2c3p.
[00209] В предпочтительном варианте осуществления настоящее изобретение охватывает эффекторные белки, относящиеся к типу VI локусов системы CRISPR-Cas, например, локусам C2c2. В настоящем описании C2c2 относится к кандидату 2 класса 2. Локусы C2c2 включают гены cas1 и cas2 наряду с крупным белком, который заявители обозначают как C2c2p, и последовательности CRISPR; однако, в отличие от C2c1p, C2c2p часто кодируется вблизи последовательностей CRISPR, но не генами cas1-cas2 (для сравнения фиг.9 и фиг.15).
[00210] Аспекты изобретения также охватывают способы и применение описанных в настоящем описании композиций и систем в геномной инженерии, например, для изменения или управления экспрессией одного или более генов или одного или более продуктов генов в прокариотических или эукариотических клетках in vitro, in vivo или ex vivo.
[00211] В вариантах осуществления изобретения понятия "направляющая последовательность" и "направляющая РНК" используются взаимозаменяемо, как и в упоминаемых выше документах, таких как WO 2014/093622 (PCT/US2013/074667). В широком смысле, направляющая последовательность представляет собой любую нуклеотидную последовательность, имеющую достаточную комплементарность с полинуклеотидной последовательностью-мишенью для гибридизации с последовательностью-мишенью и обеспечения специфичного к последовательности связывания комплекса CRISPR с последовательностью-мишенью. В некоторым вариантах осуществления, уровень комплементаности между направляющей последовательностью и соответствующей последовательностью-мишенью при условии оптимального выравнивания с использованием надлежащего алгоритма выравнивания составляет примерно или превышает 50%, 60%, 75%, 80%, 85%, 90%, 95%, 97,5%, 99% или более. Оптимальное выравнивание может быть определено с использованием любого подходящего алгоритма для выравнивания последовательностей, не ограничивающими примерами которого являются алгоритм Смита-Ватермана, алгоритм Нидлмана-Вунша, алгоритмы, основанные на преобразовании Барроуза-Виллера (например, Burrows Wheeler Aligner), ClustalW, Clustal X, BLAT, Novoalign (Novocraft Technologies; доступен на www.novocraft.com), ELAND (Illumina, San Diego, CA), SOAP (доступен на soap.genomics.org.cn) и Maq (доступен на maq.sourceforge.net). В некоторых вариантах осуществления направляющая последовательность имеет длину примерно или более 5, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 35, 40, 45, 50, 75 или более нуклеотидов. В некоторых вариантах осуществления, направляющая последовательность имеет длину менее чем примерно 75, 50, 45, 40, 35, 30, 25, 20, 15, 12 или менее нуклеотидов. Предпочтительно, чтобы направляющая последовательность имела длину 10-30 нуклеотидов. Способность направляющей последовательности направлять специфичное к последовательности связывание комплекса CRISPR с последовательностью-мишенью может быть количественно оценена любым подходящим способом анализа. Например, компоненты системы CRISPR, достаточные для образования комплекса CRISPR, включая тестируемую направляющую последовательность, могут быть доставлены в клетку, содержащую соответствующую последовательность-мишень, например, путем трансфекции векторами, кодирующими компоненты последовательности CRISPR, с последующей количественной оценкой сайтов преимущественного расщепления последовательности-мишени, в частности, анализом Surveyor, как описано в настоящем описании. Сходным образом, расщепление полинуклеотидной последовательности-мишени может быть оценено в пробирке, содержащей последовательность-мишень, компоненты комплекса CRISPR, включая тестируемую направляющую последовательность и контрольную направляющую последовательность, при сравнении связывания или скорости расщепления последовательности-мишени для реакций с тестовой и контрольной направляющими последовательностями. Другие способы анализа также возможны и будут понятны квалифицированному специалисту в данной области. В некоторых вариантах осуществления последовательность-мишень является последовательностью в геноме клетки. Иллюстративные последовательности-мишени включают те, которые являются уникальными в геноме-мишени.
[00212] В целом и в описании настоящего изобретения понятие "вектор" обозначает молекулу нуклеиновой кислоты, способную транспортировать другую нуклеиновую кислоту, с которой она связана. Векторы включают, но не ограничиваются ими, молекулы нуклеиновых кислот, являющиеся одноцепочечными, двухцепочечными или частично двухцепочечными; нуклеиновые кислоты, которые включают один или более свободных концов, не имеют свободных концов (например, кольцевые); нуклеиновые кислоты, которые представляют собой ДНК, РНК или ту и другую и другие разновидности полинуклеотидов, известных в данной области. Один из типов векторов называется "плазмидой", что означает кольцевую двухцепочечную ДНК-петлю, в которую могут быть внесены дополнительные участки ДНК, в том числе стандартными способами молекулярного клонирования. Другим типом векторов является вирусный вектор, в котором находятся последовательности, полученные из вирусных ДНК или РНК в векторе для упаковки в вирус (например, ретровирусы, дефектные по репликации ретровирусы, аденовирусы, дефектные по репликации аденовирусы и аденоассоциированные вирусы). Вирусные векторы также включают полинуклеотиды, переносимые вирусом для трансфекции клетки хозяина. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальный ориджин репликации и векторы эписом млекопитающих). Другие векторы (например, неэписомные векторы млекопитающих) встраиваются в геном клетки-хозяина при введении в клетку хозяина и благодаря этому реплицируются вместе с геномом хозяина. Более того, некоторые векторы способны направлять экспрессию генов, с которыми они непосредственно связаны. Такие векторы называются в настоящем описании "экспрессирующими векторами". Векторы для экспрессии и обеспечивающие экспрессию в эукариотической клетке могут называться в настоящем описании "эукариотическими экспрессирующими векторами". Часто используемыми векторами в способах рекомбинантных ДНК являются плазмиды.
[00213] Рекомбинантные экспрессирующие векторы могут включать нуклеиновую кислоту по изобретению в форме, подходящей для экспрессии такой нуклеиновой кислоты в клетке-хозяине, что означает, что такие рекомбинантные экспрессирующие векторы содержат один или более регуляторных элементов, которые могут быть выбраны в соответствии с используемыми для экспрессии клетками хозяина и которые функционально связаны с последовательностью нуклеиновой кислоты, которая должна быть экспрессирована. В рекомбинантном векторе экспрессии "функционально связанный" должно означать, что нуклеотидная последовательность-мишень связана с регуляторным(и) элементом(ами) таким образом, что экспрессия такой нуклеотидной последовательности является возможной (например, в системе транскрипции/трансляции in vitro или в клетке хозяина при введении вектора в клетку хозяина).
[00214] Термин "регуляторный элемент" используется для обозначения промоторов, энхансеров, участков внутренней посадки рибосомы (IRES) и других элементов контроля экспрессии (например, сигналы терминации транскрипции, такие как сигналы полиаденилирования и поли(U)-последовательности). Такие регуляторные элементы описаны, например, в Goeddel, GENE EXPRESSION TECHNOLOGY: METHODS IN ENZYMOLOGY 185, Academic Press, San Diego, Calif. (1990). Регуляторные элементы включают те, что обеспечивают конститутивную экспрессию нуклеотидной последовательности в различных типах клеток, и те, что обеспечивают экспрессию нуклеотидной последовательности только в определенных клетках-хозяевах (например, тканеспецифические регуляторные последовательности). Тканеспецифический промотор может обеспечивать экспрессию преимущественно в желаемой целевой ткани, такой как мышца, нейрон, кость, кожа, кровь, конкретных органах (например, печень, поджелудочная железа) или конкретных типах клеток (например, лимфоцитах). Регуляторные элементы могут также управлять экспрессией в зависимости от времени, например в зависимости от стадии клеточного цикла или стадии развития, и при этом могут быть также специфичными к тканям или типу клеток. В некоторых вариантах осуществления изобретения вектор включает один или более промоторов РНК-полимеразы III (например, 1, 2, 3, 4, 5 или более промоторов РНК-полимеразы III), один или более промоторов РНК-полимеразы II (например, 1, 2, 3, 4, 5 или более промоторов РНК-полимеразы II), один или более промоторов РНК-полимеразы I (например, 1, 2, 3, 4, 5 или более промоторов РНК-полимеразы I) или их комбинацию. Примеры промоторов РНК-полимеразы III включают, но не ограничиваются ими, промоторы U6 и H1. Примеры промоторов РНК-полимеразы II включают, но не ограничиваются ими, промоторы вируса саркомы Рауса (RSV) (необязательно с энхансером RSV), промотор цитомегаловируса (CMV), (необязательно с энхансером CMV) [см., например, Boshart et al., Cell, 41:521-530 (1985)], промотор SV40, промотор дигидрофолатредуктазы, промотор p-актина, промотор фосфоглицеролкиназы (PGK) и промотор EF1a. Термин "регуляторный элемент" относится и к элементам-энхансерам, таким как WPRE; энхансеры CMV; R-U5'-сегмент в LTR HTLV-I (Mol. Cell. Biol., Vol. 8(1), p. 466-472, 1988); энхансер SV40 и последовательность интрона между 2 и 3 экзонами P-глобина кролика (Proc. Natl, Acad. Sci. USA., Vol. 78(3), p. 1527-31, 1981). Квалифицированный специалист в данной области оценит, что конструкция вектора экспрессии может зависеть от таких факторов как выбранная клетка-хозяин, трансфекция которой будет производиться, желаемый уровень экспрессии и т.д. Вектор может быть введен в клетки таким образом, чтобы получались транскрипты, белки или пептиды, включая слитые белки или пептиды, кодируемые нуклеиновыми кислотами, как описано в настоящем описании (например, транскрипты системы коротких палиндромных повторов, регулярно расположенных кластерами (CRISPR), их белки, ферменты, мутантные формы, слитые белки и т.д.)
[00215] Преимущественные векторы включают лентивирусы и аденоассоциированные вирусы, и типы таких векторов могут также быть выбраны для нацеливания на конкретные типы клеток.
[00216] Как используют в настоящем описании, термин "cr-РНК" или "направляющая РНК" или "единственная направляющая РНК" или "sg-РНК" или "один или более компонентов нуклеиновых кислот" эффекторного белка локуса типа V или типа VI системы CRISPR Cas включает любую полинуклеотидную последовательность, имеющую достаточную комплементарность с последовательностью нуклеиновой кислоты-мишени для гибридизации с последовательностью нуклеиновой кислоты-мишени и прямого, специфичного к последовательности связывания комплекса, нацеленного на нуклеиновую кислоту, с последовательностью нуклеиновой кислоты-мишени. В некоторых вариантах осуществления изобретения степень комплементарности при оптимальном выравнивании с использованием подходящего алгоритма выравнивания приблизительно равна 50%, 60%, 75%, 80%, 85%, 90%, 95%, 97,5%, 99% или более. Оптимальное выравнивание может быть проведено с использованием любого подходящего алгоритма выравнивания последовательностей, примеры которого включают, но не ограничиваются ими алгоритм Смита-Ватермана, алгоритм Нидлмана-Вунша, алгоритмы, основанные на преобразовании Барроуза - Уилера (например, Burrows Wheeler Aligner), ClustalW, Clustal X, BLAST, Novoalign (Novocraft Technologies; доступен на www.novocraft.com), ELAND (Illumina, Сан-Диего, Калифорния, США), SOAP (доступен на soap.genomics.org.cn) и Maq (доступен на maq.sourceforge.net). Способность направляющей последовательности (в пределах направляющей РНК, нацеливающей на нуклеиновые кислоты) управлять прямым специфичным к последовательности связыванием комплекса, нацеленного на нуклеиновые кислоты, может быть оценена с помощью любого подходящего способа анализа. Например, компоненты системы CRISPR, нацеленной на нуклеиновые кислоты, достаточные для формирования комплекса, нацеленного на нуклеиновую кислоту, включая направляющую последовательность, подлежащую тестированию, могут быть предоставлены клетке-хозяину, имеющей соответствующую последовательность нуклеиновой кислоты-мишени, например, путем трансфекции векторами, кодирующими компоненты комплекса, нацеленного на нуклеиновую кислоту, с последующей оценкой предпочтительного нацеливания (например, расщепления) в последовательности нуклеиновой кислоты-мишени, например, в анализе Survey, описанном в настоящем описании. Аналогично расщепление последовательности нуклеиновой кислоты-мишени может быть оценено в пробирке путем представления клетке последовательности нуклеиновой кислоты-мишени, компонентов комплекса, нацеленного на нуклеиновые кислоты, включая направляющую последовательность, подлежащую тестированию, и контрольную направляющую последовательность, отличающаяся от тестируемой направляющей последовательности, и сравнения связывания или уровня расщепления последовательности-мишени между реакциями тестовой и контрольной направляющими последовательностями. Другие способы анализа возможны и понятны специалисту в данной области. Направляющая последовательность, и, следовательно, направляющая РНК, нацеливающая на нуклеиновые кислоты, могут быть выбраны для нацеливания на любую последовательность нуклеиновой кислоты-мишени. Последовательность-мишень может быть представлена ДНК. Последовательность-мишень может быть представлена любой последовательностью РНК. В некоторых вариантах осуществления изобретения последовательность-мишень может представлять собой последовательность в молекуле РНК, выбранной из группы, состоящей из матричной РНК (мРНК), пре-мРНК, рибосомной РНК (рРНК), транспортной РНК (тРНК), микроРНК, малой интерферирующей РНК (миРНК), малой ядерной РНК (мяРНК), малой ядрышковой РНК (мякРНК), двухцепочечной РНК (дцРНК), некодирующей РНК (нкРНК), длинной некодирующей РНК (днкРНК) и малой цитоплазматической РНК (мцРНК). В некоторых предпочтительных вариантах осуществления изобретения последовательность-мишень может представлять собой последовательность в молекуле РНК, выбранную из группы, состоящей из мРНК, пре-мРНК и рРНК. В некоторых предпочтительных вариантах осуществления изобретения последовательность-мишень может представлять собой последовательность в молекуле РНК, выбранную из группы, состоящей из нкРНК и днкРНК. В некоторых более предпочтительных вариантах осуществления изобретения последовательность-мишень может представлять собой последовательность в молекуле мРНК или молекуле пре-мРНК.
[00217] В некоторых вариантах осуществления изобретения направляющая РНК, нацеленная на нуклеиновые кислоты, выбрана таким образом, чтобы вторичная структура направляющей РНК, нацеленной на РНК, была более компактной. В некоторых вариантах осуществления изобретения приблизительно или меньше чем приблизительно 75%, 50%, 40%, 30%, 25%, 20%, 15%, 10%, 5%, 1%, или менее нуклеотидов направляющей РНК, нацеленной на нуклеиновую кислоту, участвует в самокомплементарном спаривании оснований при оптимальном фолдинге. Оптимальный фолдинг может быть определен любым подходящим алгоритмом выявления фолдинга полинуклеотида. Некоторые программы основаны на вычислении минимальной свободной энергии Гиббса. Примером одного такого алгоритма является mFold, описанный Zuker и Stiegler (Nucleic Acids Res. 9 (1981), 133-148). В качестве примера другого алгоритма фолдинга можно привести онлайн вебсервер RNAfold, разработанный в Институте Теоретической Химии в Венском университете, который используют центроидный алгоритм предсказания структуры (см., например, A.R. Gruber et al, 2008, Cell 106(1): 23-24; и PA Carr and GM Church, 2009, Nature Biotechnology 27(12): 1151-62).
[00218] В определенных вариантах осуществления изобретения направляющая РНК или cr-РНК может включать, по существу состоять из или состоять из последовательности прямого повтора (DR) и последовательности направляющей молекулы или последовательности спейсера. В некоторых вариантах осуществления изобретения направляющая РНК или cr-РНК может включать, по существу состоять из или состоять из последовательности прямого повтора (DR), слитой или связанной с последовательностью направляющей молекулы или последовательностью спейсера. В некоторых вариантах осуществления изобретения последовательность прямого повтора (DR) может быть расположена в восходящем направлении (т.е. 5') от последовательности направляющей молекулы или последовательности спейсера. В других вариантах осуществления изобретения последовательность прямого повтора может быть расположена в нисходящем направлении (т.е. 3') от последовательности направляющей молекулы или последовательности спейсера.
[00219] В определенных вариантах осуществления изобретения cr-РНК включает шпилечную структуру, предпочтительно единственную шпилечную структуру. В некоторых вариантах осуществления изобретения шпилечная структура, предпочтительно единственная шпилечная структура, формирует последовательность прямого повтора.
[00220] В определенных вариантах осуществления изобретения длина спейсера направляющей РНК составляет от 15 до 35 нуклеотидов. В определенных вариантах осуществления изобретения длина спейсера направляющей РНК составляет по меньшей мере 15 нуклеотидов. В некоторых вариантах осуществления изобретения длина спейсера составляет от 15 до 17 нуклеотидов, например, 15, 16 или 17 нуклеотидов, от 17 до 20 нуклеотидов, например, 17, 18, 19 или 20 нуклеотидов, от 20 до 24 нуклеотидов, например, 20, 21, 22, 23 или 24 нуклеотидов, от 23 до 25 нуклеотидов, например, 23, 24 или 25 нуклеотидов, от 24 до 27 нуклеотидов, например, 24, 25, 26 или 27 нуклеотидов, от 27-30 нуклеотидов, например, 27, 28, 29 или 30 нуклеотидов, 30-35 нуклеотидов, например, 30, 31, 32, 33, 34 или 35 нуклеотидов или 35 нуклеотидов или более.
[00221] Термин "последовательность tracr-РНК" или аналогичный термин включает любой полинуклеотид, который имеет достаточную комплементарность к cr-РНК для последующей гибридизации. В целом, степень комплементарности относится к оптимальному выравниванию последовательности sca и последовательности tracr по длине наиболее короткой из двух последовательностей. Оптимальное выравнивание может быть определено любым подходящим алгоритмом выравнивания и, кроме того, оно может учитывать вторичную структуру, например, самокомплементарность в пределах последовательности sca или последовательности trascr. В некоторых вариантах осуществления степень комплементарности между последовательностью tracr и последовательностью sca по длине более короткой из двух при оптимальном выравнивании составляет около или около 25%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 97,5%, 99% или выше. В некоторых вариантах осуществления tracr-РНК может не потребоваться.
[00222] Заявители также провели пробный эксперимент для проверки способности белков типа V, таких как C2c1 или C2c3, к нацеливанию и расщеплению ДНК. Этот эксперимент во многом повторяет подобную работу для гетерологичной экспрессии StCas9 в E. coli (Sapranauskas, R. et al. Nucleic Acids Res 39, 9275-9282 (2011)). Заявители ввели плазмиду, содержащую одновременно PAM и ген устойчивости к антибиотикам, в гетерологичный штамм E. coli, и затем поместили на среду, содержащую соответствующий антибиотик. В случае расщепления ДНК плазмиды заявители не наблюдали жизнеспособных колоний.
[00223] Более подробно анализ ДНК-мишени может быть проведен следующим образом. В анализе используются два штамма E.coli. Один из штаммов имеет плазмиду, которая кодирует локус эндогенного эффекторного белка бактериального штамма. Другой штамм имеет пустую плазмиду (например, pACYC184, контрольный штамм). Все возможные PAM длиной 7 или 8 п.н. представлены на плазмиде устойчивости к антибиотикам (pUC19 с геном устойчивости к ампициллину). PAM расположена рядом с последовательностью протоспейсера 1 (ДНК-мишень для первого спейсера в локусе эндогенного эффекторного белка). Были клонированы две библиотеки PAM. Одна библиотека содержит последовательности 5'-конца протоспейсера, состоящие из 8 случайных п.н. (например, всего 65 536 различных PAM=комплексность). Другая библиотека содержит последовательности 3'-конца протоспейсера, состоящие из 7 случайных п.н. (например, полная комплексность - 16 384 различных PAM). Обе библиотеки были клонированы, чтобы иметь в среднем 500 плазмид, содержащих каждый возможный PAM. Тестовый и контрольный штаммы были трансформированы библиотеками 5'-PAM и 3'-PAM в ходе отдельных трансформаций, трансформированные клетки были помещены в культуру на среду, содержавшую ампициллин. Распознавание и последующее разрезание/интерференция посредством плазмиды делают клетку неустойчивой к ампициллину и прекращают ее рост. Приблизительно через 12 ч после трансформации все колонии, образованных тестовым и контрольным штаммами, были собраны и из них была выделена плазмидная ДНК. Плазмидная ДНК использовалась в качестве матрицы для ПЦР-амплификации и последующего глубокого секвенирования. Представленность всех PAM в нетрансформированных библиотеках показала ожидаемую представленность PAM в трансформированных клетках. Представленность всех PAM, найденных в штаммах контроля, показала фактическую представленность. Представленность всех PAM в тестовом штамме показала, какие PAM не распознаются ферментом. Сравнение с контрольным штаммом позволяет извлекать последовательности с обедненным PAM.
[00224] В некоторых вариантах осуществления изобретения CRISPR-Cas9, уровень комплементарности между последовательностями tracr-РНК и cr-РНК присутствует по длине наиболее короткой из них при наиболее оптимальном выравнивании. Как описано в настоящем описании, в некоторых вариантах осуществления настоящего изобретения tracr-РНК не требуется. В некоторых вариантах ранее описанных систем CRISPR-Cas (например, систем CRISPR-Cas9) схемы сконструированных химерных направляющих РНК (sgRNA) могут включать по меньшей мере 12 п.н. дуплексной структуры между cr-РНК и tracr-РНК, однако в описанных в настоящем описании системах CRISPR Cpf1 такие химерные РНК (chi-РНК) невозможны, поскольку система не использует tracrRNA.
[00225] В определенных вариантах осуществления изобретения зрелые cr-РНК включают элемент последовательности, происходящий из последовательности повтора a (5'-метка) локуса CRISP, которая является важной для фукнции. См. Marraffini et al., 28-Jan-2010, Nature Letters 463 (568-572), которая включена в качестве ссылки.
[00226] В некоторых вариантах осуществления степень комплементарности между последовательностью tracr-РНК и последовательностью cr-РНК по длине более короткой из них при оптимальном выравнивании составляет приблизительно или больше около 25%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 97,5%, 99% или выше. В некоторых вариантах осуществления последовательность tracr составляет приблизительно или больше, чем приблизительно 5, 6, 7, 8, 9, 10, II, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 40, 50 или более нуклеотидов в длину. В некоторых вариантах осуществления последовательность tracr и последовательность cr-РНК содержатся в одном транскрипте, так что гибридизация между ними создает транскрипт, имеющий вторичную структуру, такую как шпилька. В одном из вариантов осуществления изобретения транскрипт или транскрибируемая полинуклеотидная последовательность имеет по меньшей мере две или более шпилек. В предпочтительных вариантах осуществления транскрипт имеет две, три, четыре или пять шпилек. В следующем варианте осуществления изобретения транскрипт имеет не более пяти шпилек. В структуре шпильки часть последовательности с 5'-стороны от конечного "N" и выше петли соответствует последовательности tracr, а часть последовательности с 3'-стороны петли соответствует последовательности tracr. В некоторых вариантах осуществления tracr-РНК может не потребоваться.
[00227] В некоторых вариантах осуществления ранее описанных систем CRISPR-Cas (например, CRISPR-Cas9) конструкции сконструированных химерных направляющих РНК (sgRNA) могут включать по меньшей мере 12 п.н. дуплексной структуры crRNA и tracrRNA, однако в системах CRISPR Cpf1 такие химерные РНК (chi-РНК) невозможны, поскольку система не использует tracr-РНК.
[00228] Для минимизации токсичности и неспецифических эффектов важно управлять концентрацией доставленной направляющей РНК. Оптимальные концентрации направляющей РНК, нацеленной на нуклеиновую кислоту, могут быть определены путем тестирования различных концентраций в клеточных моделях или эукариотических моделях животных, не являющихся человеком, глубокое секвенирование может быть использовано для анализа степени модификации в потенциальных геномных локусах, не являющихся мишенями. Концентрация для доставки in vivo должна быть подобрана так, чтобы обеспечивать наибольший уровень целевой модификации мишени при минимальном уровне нецелевой модификации. Предпочтительным является получение системы нацеливания на нуклеиновые кислоты из системы CRISPR типа V/типа VI. В некоторых вариантах осуществления изобретения один или более элементов системы нацеливания на нуклеиновые кислоты получены из конкретного организма, включающего эндогенную систему нацеливания на РНК. В предпочтительных вариантах осуществления изобретения система нацеливания на РНК представляет собой систему CRISPR типа V. В конкретных вариантах осуществления изобретения фермент Cas типа V, нацеленный на РНК, представляет собой C2c1 или C2c3. Примеры белков Cas включают, но не ограничиваются ими, Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas6, Cas7, Cas8, Cas9 (также известный как Csn1 и Csx12), Cas10, Csy1, Csy2, Csy3, Cse1, Cse2, Csc1, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx15, Csf1, Csf2, Csf3, Csf4, их гомологи или модифицированные версии. В вариантах осуществления изобретения белок типа V, такой как C2c1 или C2c3, упомянутый в настоящем описании, также включает гомологи или ортологи белков типа V, таких как C2c1 или C2c3. Термины "ортолог" (также обозначаемый в настоящем описании как "ortologue") и "гомолог" (также обозначаемый в настоящем описании как "homologue") хорошо известны в данной области. Посредством дальнейшего руководства "гомолог" белка как используют в настоящем описании, представляет собой белок того же самого вида, который выполняет ту же самую или подобную функцию, что и белок, гомологом которого он является. Гомологичные белки могут иметь сходную или частично сходную структуру, но это не обязательно. "Ортолог" белка, как используют в настоящем описании, является белком другого вида, который выполняет ту же самую или подобную функцию, что и белок, ортологом которого он является. Ортологичные белки могут иметь сходную или частично сходную структуру, но это не обязательно. В конкретных вариантах осуществления изобретения гомологи или ортологи белков типа V, таких как C2c1 или C2c3, упомянутый в настоящем описании, имеют гомологию или идентичность последовательностей по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, например, по меньшей мере 95% с белками типа V, такими как C2c1 или C2c3. В следующих вариантах осуществления изобретения гомологи или ортологи белков типа V, таких как C2c1 или C2c3, упомянутый в настоящем описании, имеют идентичность последовательности по меньшей мере 80%, более предпочтительно по меньшей мере 85%, еще более предпочтительно по меньшей мере 90%, например, по меньшей мере 95% с белками типа V дикого типа, такими как C2c1 или C2c3.
[00229] В варианте осуществления изобретения белки Cas типа V, нацеленные на РНК, могут быть ортологами C2c1 или C2c3 организмов из родов, которые включают, но не ограничены следующими: Corynebacter, Sutterella, Legionella, Treponema, Filifactor, Eubacterium, Streptococcus, Lactobacillus, Mycoplasma, Bacteroides, Flaviivola, Flavobacterium, Sphaerochaeta, Azospirillum, Gluconacetobacter, Neisseria, Roseburia, Parvibaculum, Staphylococcus, Nitratifractor, Mycoplasma и Campylobacter. Вид организма, принадлежащего к данным родам, может быть обсужден в данной заявке иным образом.
[00230] Некоторые способы идентификации ортологов ферментов системы CRISPR-Cas могут включать идентификацию tracr-последовательности в геномах-мишенях. Идентификация tracr-последовательностей может быть связана со следующими шагами: поиск прямых повторов или последовательности tracr-помощника в базе данных, чтобы идентифицировать область CRISPR, включающую фермент CRISPR; поиск гомологичных последовательностей в области CRISPR, фланкирующем фермент CRISPR в смысловом и антисмысловом направлениях; поиск терминаторов транскрипции и вторичных структур; идентификация любой последовательности, которая не является прямым повтором или последовательностью tracr-помощника, но имеет больше чем 50% идентичность с прямым повтором или последовательностью tracr-помощника как потенциальная tracr-последовательность; поиск последовательностей терминатора транскрипции, ассоциированных с потенциальной tracr-последовательностью.
[00231] Следует принимать во внимание, что любой из функциональных видов активности, описанных в настоящем описании, может быть встроена способами инженерии в ферменты CRISPR из других ортологов, включая химерные ферменты, включающие фрагменты многих ортологов. Примеры таких ортологов описаны в настоящем описании. Таким образом, химерные ферменты могут включать фрагменты ортологов фермента CRISPR организма, который включает, но не ограничен следующими: Corynebacter, Sutterella, Legionella, Treponema, Filifactor, Eubacterium, Streptococcus, Lactobacillus, Mycoplasma, Bacteroides, Flaviivola, Flavobacterium, Sphaerochaeta, Azospirillum, Gluconacetobacter, Neisseria, Roseburia, Parvibaculum, Staphylococcus, Nitratifractor, Mycoplasma и Campylobacter. Химерный фермент может включать первый фрагмент и второй фрагмент, фрагменты могут происходить из ортологов фермента CRISPR организмов, принадлежащих к родам, упомянутым в настоящем описании, или видам, упомянутым в настоящем описании; предпочтительно фрагменты происходят из ортологов фермента CRISPR различных видов.
[00232] В вариантах осуществления изобретения, эффекторный белок типа V/типа VI, нацеленный на РНК, в частности, белок C2c2 C2c1/C2c3, упоминаемый в настоящем описании, также охватывает функциональный вариант C2c1/C2c3 или его гомолог, или ортолог. "Функциональный вариант" белка, как используют в настоящем описании, относится к варианту такого белка, который сохраняет, по меньшей мере частично, активность этого белка. Функциональные варианты могут включать мутанты (которые могут иметь инсерции, делеции или замены), включая полиморфы, и т.д. Также в функциональные варианты включены продукты слияния такого белка с другим, обычно неродственным, белком, нуклеиновой кислотой, полипептидом или пептидом. Функциональные варианты могут быть естественными или искусственными. Предпочтительные варианты осуществления изобретения могут включать не встречающийся в природе или сконструированный способами инженерии эффекторный белок типа V/типа VI, нацеленный на РНК, например, C2c1/C2c3, или его ортолог или гомолог.
[00233] В варианте осуществления изобретения кодон молекулы (молекул) нуклеиновой кислоты, кодирующей эффекторный белок типа V/типа VI, нацеленный на РНК, например, C2c1/C2c3, или его ортолог или гомолог, может быть оптимизирован для экспрессии в эукариотической клетке. Эукариотический организм может быть таким, как описано в настоящем описании. Молекула (молекулы) нуклеиновой кислоты может быть сконструирована способами инженерии или не встречаться в природе.
[00234] В варианте осуществления изобретения эффекторный белок типа V/типа VI, нацеленный на РНК, например, C2c1/C2c3 (или его ортолог или гомолог), может включать одну или более мутаций (следовательно, молекула (молекулы) нуклеиновой кислоты, кодирующая этот белок, имеет ту же самую мутацию (мутации)). Мутации могут быть внесены искусственно и могут включать, но не ограничиваться ими, одну или более мутациий в каталитическом домене. Примеры каталитических доменов в отношении фермента Cas9 могут включать, но не ограничиваться ими, домены RuvC I, RuvC II, RuvC III и HNH.
[00235] В варианте осуществления изобретения белок типа V/типа VI, например, C2c1 или C2c3, или его ортолог или гомолог, может включать одну или более мутаций. Мутации могут быть внсены искусственно и могут включать, но не ограничиваться ими, одну или более мутаций в каталитическом домене. Примеры каталитических доменов в отношении фермента Cas могут включать, но не ограничиваться ими, домены RuvC I, RuvC II, RuvC III, HNH и домены HEPN.
[00236] В одном варианте осуществления изобретения белок типа V/типа VI, например, C2c1 или C2c3, или его ортолог или гомолог, может использоваться в качестве универсального белка, связывающего нуклеиновые кислоты, в слиянии или будучи функционально связанным с функциональным доменом. Иллюстративные функциональные домены могут включать, но не ограничиваются ими, инициатор трансляции, активатор трансляции, репрессор трансляции, нуклеазы, в частности, рибонуклеазы, сплайсосомы, магнитные гранулы, индуцируемый/контролируемый светом домен или химически индуцируемый/контролируемый домен.
[00237] В некоторых вариантах осуществления изобретения ферментативная активность неизмененного эффекторного белка, нацеленного на нуклеиновые кислоты, заключается в расщеплении. В некоторых вариантах осуществления изобретения эффекторный белок нацеливания на РНК может направлять расщепление одной или обеих нуклеиновых кислот (ДНК или РНК) цепи в местоположении около последовательности-мишени, таком как в пределах последовательности-мишени и/или в пределах последовательности, комплементарной последовательности-мишени, или в последовательностях, ассоциированных с последовательностью-мишенью. В некоторых вариантах осуществления изобретения белок Cas, нацеленный на нуклеиновую кислоту, может направлять расщепление одной или обеих цепей ДНК или РНК в пределах приблизительно 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 50, 100, 200, 500 или более пар оснований от первого или последнего нуклеотида последовательности-мишени. В некоторых вариантах осуществления изобретения расщепление может быть тупым, т.е. производить тупые концы. В некоторых вариантах осуществления изобретения расщепление может быть ступенчатым, т.е. производить липкие концы. В некоторых вариантах осуществления изобретения расщепление может быть ступенчатым с 5'-выступающими концами, например, 5'-выступающими концами длиной 1-5 нуклеотидов. В некоторых вариантах осуществления изобретения расщепление может быть ступенчатым с 3'-выступающими концами, например, 3'-выступающими концами длиной 1-5 нуклеотидов. В некоторых вариантах осуществления изобретения вектор кодирует белок Cas, нацеленный на нуклеиновые кислоты, который может быть мутантным по сравнению с соответствующим ферментом дикого типа, таким образом, что мутантный белок Cas, нацеленный на нуклеиновую кислоту, имеет пониженную способность расщеплять одну или обе цепи ДНК или РНК полинуклеотида-мишени, содержащего последовательность-мишень. В качестве еще одного примера, мутации могут быть внесены в два или более каталитических домена белка Cas (RuvC I, RuvC II и RuvC III или домен HNH или домен HEPN), чтобы мутантный белок Cas практически утратил активность расщепления РНК. Как описано в настоящем описании, мутации могут быть внесены в соответствующие каталитические домены эффекторного белка C2c1 или C2c3, чтобы активность расщепления ДНК мутантного эффекторного белка C2c1 или C2c3 была недостаточной или существенно уменьшенной. В некоторых вариантах осуществления изобретения эффекторный белок, нацеленный на нуклеиновую кислотe, считается имеющий существенный недостаток активности расщепления РНК, когда активность расщепления РНК мутантного фермента составляет не больше чем 25%, 10%, 5%, 1%, 0,1%, 0,01% или менее активности расщепления нуклеиновой кислоты немутантной формы фермента; например, когда активность расщепления нуклеиновой кислоты мутантной формы равна нулю или незначительна по сравнению с немутантной формой. Эффекторный белок может быть идентифицирован в отношении общего класса ферментов, которые являются гомологичными самой большой нуклеазе со множественными нуклеазными доменами системы CRISPR типа V/типа VI. Наиболее предпочтительно использование эффекторного белка типа V/типа VI, такого как C2c1/C2c3. Под "происходящим" заявители подразумевают, что происходящий фермент в значительной степени основан, в смысле наличия высокой степени гомологии последовательности, на ферменте дикого типа, но что он некоторым образом мутирован (модифицирован), как известно в данной области или как описано в настоящем описании.
[00238] Снова, следует принимать во внимание, что термины "Cas и фермент CRISPR", "белок CRISPR" и "белок Cas" обычно используются взаимозаменяемо и во всех случаях упоминания в настоящем описании относится относятся по аналогии к новым эффекторным белкам CRISPR, далее описанными в настоящем описании, если иное не очевидно, например, при конкретной отсылке к Cas9. Как упомянуто выше, многие нумерации остатков, используемые в настоящем описании, относятся к эффекторному белку локуса CRISPR типа V/типа VI. Однако следует принимать во внимание, что это изобретение включает еще множество эффекторных белков из других видов микроорганизмов. В некоторых вариантах осуществления изобретения, Cas может конститутивно присутствовать, или индуцируемо присутствовать, или кондиционально присутствовать, или быть введенным, или быть доставленным. Оптимизация Cas может использоваться для усиления функций или разработки новых функций, могут быть получены химерные белки Cas. Cas может использоваться в качестве универсального белка, связывающего нуклеиновые кислоты.
[00239] Как правило, в контексте эндогенной системы, нацеленной на нуклеиновую кислоту, образование комплекса, нацеленного на нуклеиновую кислоту (включающего направляющую РНК, гибридизованную с последовательностью-мишенью, и образующую комплекс с одним или более нацеленными на нуклеиновую кислоту эффекторными белками) приводит к расщеплению одной или обеих цепей ДНК или РНК в пределах или рядом (например, в 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50, или более пар оснований от) с последовательностью-мишенью. Как используют в настоящем описании, термин "последовательность (последовательности), ассоциированная с локусом-мишень", относится к последовательностям вблизи последовательности-мишени (например, в пределах 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50 или более пар оснований от последовательности-мишени, где последовательность-мишень находится в локусе-мишени).
[00240] Примером кодон-оптимизированной последовательности в данном случае является последовательность, оптимизированная для экспрессии в эукариотическом организме, например, в человеке (т.е. являющаяся оптимизированной для экспрессии в человеке) или другом эукариотическом организме, животном или млекопитающем, как описано в настоящем описании; см., например, кодон-оптимизированную для человека последовательность SaCas9 в WO 2014/093622 (PCT/US2G13/074667) в качестве примера кодон-оптимизированной последовательности (как это известно из уровня техники и настоящего описания, кодон оптимизированная кодирующая молекула(ы) нуклеиновой(ых) кислот(ы), особенно в случае эффекторного белка (например, C2c2), входит в компетенцию квалифицированного специалиста). В то время как именно представленное выше описание является предпочтительным, будет также понятно, что другие примеры являются возможными и известна оптимизация кодонов для видов хозяев, отличных от человека, или оптимизация кодонов для определенных органов. В некоторых вариантах осуществления кодирующая фермент последовательность, в которой закодирован нацеленный на ДНК/РНК белок Cas, является кодон-оптимизированной для экспрессии в определенных клетках, в частности, эукариотических клетках. Такие эукариотические клетки конкретного организма, или могут происходить из него, который может быть млекопитающим, включая, но не ограничиваясь этим человека, отличного от человека эукариотического организма, животного или млекопитающего, как описано в настоящем описании, например, мышь, крысу, кролика, собаку, домашний скот или отличного от человека млекопитающего или примата. В некоторых вариантах осуществления процессы модификации генетической принадлежности эмбриональной линии клеток человеческих индивидов и/или процессы модификации генетической принадлежности эмбриональной линии клеток животных, которые с большой долей вероятности могут принести им страдания без получения существенной пользы для медицинских приложений для человека или животных, а также получаемые при этих процессах животные, могут быть исключены. В целом, под оптимизацией кодонов понимается процесс изменения последовательности нуклеиновой кислоты для усиления экспрессии в клетках-мишенях хозяина путем замены по меньшей мере одного кодона (например, примерно или более чем 1, 2, 3, 4, 5, 10, 15, 20, 25, 50 или более кодонов) нативной последовательности кодонами, которые более часто или наиболее часто используются в генах этой клетки-хозяина при сохранении нативной аминокислотной последовательности. Различные виды демонстрируют определенное предпочтение для определенных кодонов конкретных аминокислот. Предпочтение кодонов (различия в использовании кодонов между организмами) часто коррелирует с эффективностью трансляции матричных РНК (мРНК), что в свою очередь считается зависящим от, в числе прочего, от свойств транслируемых кодонов или доступности молекул определенной транспортной РНК (тРНК). Преобладание определенных тРНК обычно отражает наиболее часто используемые при синтезе пептидов кодоны. В соответствии с этим, гены могут быть адаптированы для оптимальной экспрессии генов в конкретном организме на основе оптимизации кодонов. Таблицы частот использования кодонов свободно доступны, например, в базе "Codon Usage Database", доступной на www.kazusa.oijp/codon/, причем такие таблицы могут быть адаптированы различными способами. См. Nakamura, Y. et al. "Codon usage tabulated from the international DNA sequence databases: status for the year 2000" Nucl. Acids Res, 28:292 (2000). Также доступны компьютерные алгоритмы для оптимизации кодонов для конкретной последовательности для экспрессии в определенной клетке-хозяине, в частности, такие как Gene Forge (Aptagen; Джакобус, Пенсильвания, США). В некоторых вариантах осуществления один или более кодонов (например, 1, 2, 3, 4, 5, 10, 15, 20, 25, 50, более или все кодоны) в последовательности, кодирующей нацеленный на ДНК/РНК белок Cas соответствуют наиболее часто используемым кодонам для конкретной аминокислоты. Что касается использования кодонов в дрожжах, приводится ссылка на онлайн-базу данных Yeast Genome, доступную на http:.Avww.yaastyeoome.org/eommuniyvcodon_rsage.shtml, или Codon selection in yeast, Bennetzen and Hall, J. Biol Chem. 1982 Mar 25;257(6):3026-31. Что касается использования кодонов в растениях, включая водоросли, прилагается ссылка на Codon usage in higher plants, green algae, and cyanobacteria, Campbell and Gowri, Plant Physiol. 1990 Jan; 92(1): 1-11. а также на Codon usage in plant genes, Murray et al, Nucleic Acids Res. 1989 Jan 25;17(2):477-98 или Selection on the codon bias of chloroplast and cyanelle genes in different plant and algal lineages, Morton BR, J. Mol. Evol. 1998 Apr;46(4):449-59.
[00241] В некоторых вариантах осуществления вектор кодирует нацеленный на нуклеиновую кислоту эффекторный белок, такой как нацеленный на РНК эффекторный белок типа V, в частности C2c2, либо ортолог или гомолог такового, содержащий одну или более последовательностей сигнала ядерной локализации (NLS), к примеру примерно или более 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более NLS. В некоторых вариантах осуществления такой нацеленный на РНК эффекторный белок содержит примерно или более чем 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более NLS на N-конце или вблизи него, примерно или более чем 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более NLS на C-конце или вблизи него или их сочетание (например, ноль или по меньшей мере один или более NLS на N-конце или более NLS на C-конце). В случае наличия более одной NLS, каждая из них может быть отобрана независимо от других, в частности, отдельная NLS может присутствовать в виде более чем одной копии и/или в сочетании с одной или более NLS, представленной одной или более копией. В некоторых вариантах осуществления NLS считается находящейся вблизи N- или C- конца, когда ближайшая аминокислота NLS находится на расстоянии менее примерно 1, 2, 3, 4, 5, 10, 15, 20, 25, 30, 40, 50 или более аминокислот полипептидной цепи от N-конца или C-конца. Неограничивающие примеры NLS включают последовательности NLS, полученные из: NLS большого T-антигена вируса SV40, имеющий аминкоислотную последовательность PKKKRKV; NLS нуклеоплазмина (например, двухкомпонентная NLS, имеющий последовательностью KRPAATKKAGQAKKKK); NLS c-myc, имеющий аминокислотную последовательность PAAKRVKLD или RQRRNELКRSP; NLS hRNPA1 M9, имеющий аминокислотную последовательность NQSSNFGPMKGGNFGGRSSGPYGGGGQYFAKPRNQGGY; последовательность RMRIZFKNKGKDTAELRRRRVEVSVELRKAKKDEQILKRRNV домена IBB импортина-альфа; последовательности VSRKRPRP и PPKKARED T-белка миомы; последовательность РОРKKKPL p53 человека; последовательность SALIKKKKKMAP c-ab1 IV мыши; последовательности DRLRR и PKQKKRK вируса гриппа NS1; последовательность RKLKKKIKKL дельта-антигена вируса гепатита; последовательность REKKKFLKRR белка Mx1 мыши; последовательность KRKGDEVDGVDEVAKKKSKK поли(АДФ-рибоза)-полимеразы и последовательность RKCLQAGMNLEARKTKK глюкокортикоида рецепторов стероидных гормонов. В целом, одна или более NLS являются достаточными, чтобы вызвать накопление нацеленного на ДНК/РНК белка Cas в поддающихся обнаружению количествах в ядре эукариотической клетки. В целом, выраженность активности ядерной локализации может быть результатом деятельности ряда NLS в нацеленном на нуклеиновые кислоты эффекторном белке, использования определенных NLS или сочетания этих факторов. Отслеживание накопления в ядре может быть выполнено с помощью любого подходящего способа. Например, отслеживаемый маркер может быть слит с нацеленным на нуклеиновую кислоту белком, так что его расположение в клетке может быть визуализировано, в частности, в комбинации со способами определения расположения ядра (например, с помощью специфичного для ядра красителя, такого как DAPI). Ядра клеток могут также быть выделены из клеток, содержимое которых может затем быть проанализировано с использованием любого подходящего способа отслеживания белка, такого как иммунногистохимия, вестерн-блоттинг или анализ активности белка. Накопление в ядре может быть определено и косвенными способами, например, путем анализа эффекта образования нацеленного на нуклеиновые кислоты комплекса (например, изменения активности экспрессии при образовании нацеленного на ДНК или РНК комплекса и/или активности белка Cas по нацеливанию на РНК), при сравнении с контролем, не подвергнутым воздействию нацеленного на нуклеиновую кислоту белка Cas или нацеленного на нуклеиновую кислоту комплекса, или подвергнутым действию нацеленного на нуклеиновую кислоту белка Cas, лишенного одного или более NLS. В предпочитаемом варианте описываемые в настоящем описании комплексы и системы эффекторного белка C2c1 или C2c3 и комплексы и системы эффекторного белка C2c1 или C2c3 включают NLS, присоединенный к C-концу белка.
[00242] В некоторых вариантах осуществления один или более векторов, управляющих экспрессией одного или более систем нацеливания на нуклеиновые кислоты вводят в клетку таким образом, что экспрессия элементов системы нацеливания на нуклеиновую кислоту управляет образованием нацеленного на нуклеиновую кислоту комплекса на одном или более участках-мишенях. Например, нацеленный на нуклеиновую кислоту эффекторный белок и нацеливающая на нуклеиновую кислоту направляющая РНК могут быть функционально связаны с отдельными регуляторными элементами в отдельных векторах. Молекула(ы) системы, нацеленной на нуклеиновую кислоту, может быть доставлена к трансгенныму эффекторному белку, нацеленному на нуклеиновую кислоту, являющемуся белком животного или млекопитающего, конститутивно, индуцибельно или кондиционально экспрессирующего эффекторный белок, нацеленный на нуклеиновую кислоту, или животного или млекопитающего, экспрессирующего эффекторный белок, нацеленный на нуклеиновую кислоту иным образом, или имеющего клетки, содержащие эффекторный белок, нацеленный на нуклеиновую кислоту, находящийся там вследствие предшествующего введения вектора или векторов, кодирующих и экспрессирующих in vivo эффекторный белок, нацеленный на нуклеиновую кислоту. Альтернативно этому, два или более элементов, экспрессированных при участии одного или различных регуляторных элементов, могут быть введены в единый вектор, с одним или более дополнительных векторов, обеспечивающих какие-либо компоненты системы, нацеленной на нуклеиновую кислоту, не включенных в первый вектор. Компоненты системы, нацеленной на нуклеиновую кислоту, объединенные в единый вектор, могут быть собраны в любой подходящей ориентации, например, в которой один элемент расположен в восходящем направлении относительно 5'-конца или в нисходящем направлении относительно 3'-конца второго элемента. Кодирующая последовательность одного элемента может быть расположена на той же или противоположной цепи кодирующей последовательности второго элемента и иметь одну и ту же или противоположную ориентацию. В некоторых вариантах осуществления один и тот же промотор контролирует экспрессию транскриптов, кодирующих эффекторный белок, нацеленный на нуклеиновую кислоту, и направляющую РНК, нацеленную на нуклеиновую кислоту, встроенных в одну или более последовательностей интронов (например, каждая в отдельном интроне, две или более по меньшей мере в одном интроне или все в одном и том же интроне). В некоторых вариантах осуществления эффекторный белок, нацеленный на нуклеиновую кислоту, и нацеливающая на нуклеиновую кислоту направляющая РНК могут быть функционально связаны с или экспрессироваться с одного и того же промотора. Средства доставки, векторы, частицы, наночастицы, составы и их компоненты для экспрессии одного или более элементов системы, нацеленной на нуклеиновую кислоту, используются в вышеупомянутых документах, в частности, в WO 2014/093622 (PCT/US2013/074667). В некоторых вариантах осуществления вектор содержит одну или более участков вставок, таких как последовательности узнавания эндонуклеаз рестрикции (также называемые "сайтом клонирования"). В некоторых вариантах осуществления один или более участков вставок (например, примерно или более чем 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более участков вставок) расположены в восходящем и/или нисходящем направлении относительно одной или более элементов последовательностей одного или более векторов. В некоторых вариантах осуществления вектор, содержащий участок-вставку в восходящем направлении последовательности-партнера tracr, и необязательно в нисходящем направлении от регуляторного элемента, функционально связанного с последовательностью-партнером tracr, таким образом, что после внесения направляющей (гидовой) последовательности в участок-вставку при экспрессии направляющей последовательности, направляет специфичное к последовательности связывание нацеленного на нуклеиновую кислоту комплекса с последовательностью-мишенью в эукариотической клетке. В некоторых вариантах осуществления вектор включает два или более участков вставок, что делает возможным введение в качестве вставок направляющих последовательностей в каждый участок. При такой организации две или более направляющих последовательностей могут включать две или более копий единичной направляющей последовательности, две или более различных направляющих последовательностей или их комбинацию. При использовании множества различных направляющих последовательностей единственная конструкция экспрессии может быть использована для нацеливания активности связывания нуклеиновых кислот на множественно различных соответствующих последовательностей-мишеней в клетке. К примеру, единственный вектор может включать примерно или более 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20 или более направляющих последовательностей. В некоторых вариантах осуществления, может быть предоставлено примерно или более 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более таких содержащих направляющую последовательность векторов, и необязательно они также могут быть доставлены в клетку. В некоторых вариантах осуществления вектор содержит регуляторный элемент, функционально связанный с кодирующей фермент последовательностью. Эффекторный белок, нацеленный на нуклеиновую кислоту или направляющая РНК или направляющие РНК, нацеливающие на нуклеиновую кислоту, могут быть доставлены независимо; предпочтительно, чтобы по меньшей мере один(одна) из них был(а) доставлен(а) посредством комплекса частиц или наночастиц. мРНК эффекторного белка, нацеленного на нуклеиновую кислоту, может быть доставлена прежде направляющей РНК, нацеливающей на нуклеиновую кислоту, для того, чтобы обеспечить время для экспрессии эффекторного белка, нацеленного на нуклеиновую кислоту. мРНК эффекторного белка, нацеленного на нуклеиновую кислоту, может быть введена на 1-12 часов (предпочтительно примерно 2-6 часов) раньше введения направляющей РНК, нацеливающей на нуклеиновую кислоту. Альтернативно этому, мРНК нацеленного на нуклеиновую кислоту эффекторного белка и направляющая РНК, нацеливающая на нуклеиновую кислоту, могут быть введены одновременно. Предпочтительно, когда вторая усиливающая доза направляющей РНК вводится на протяжении 1-12 часов (предпочтительно примерно 2-6 часов) после начального введения мРНК нацеленного нуклеиновую кислоту эффекторного белка+направляющей РНК. Дополнительные введения нацеленного на нуклеиновую кислоту эффекторного белка могут оказаться полезными, чтобы достичь наиболее эффективных уровней модификации генома.
[00243] В одном аспекте изобретение относится к способам использования одного или более компонентов системы нацеливания на нуклеиновую кислоту. Такой нацеленный на нуклеиновую кислоту комплекс по изобретению обеспечивает эффективный инструмент модификации ДНК- или РНК-мишени, являющейся одно- или двухцепочечной, линейной или суперспирализованной. Такой нацеленный на нуклеиновую кислоту комплекс по изобретению имеет разнообразные преимущества, включая модификацию (например, удаление, вставка, перемещение, инактивация, активация) ДНК- или РНК-мишени в множестве типов клеток. Как таковой нацеленный на нуклеиновую кислоту комплекс по изобретению имеет широкий спектр применений, например, в генной терапии, фармакологическом скрининге, диагностике заболеваний и их прогнозировании. Иллюстративный нацеленный на нуклеиновую кислоту комплекс включает нацеленный на ДНК или РНК эффекторный белок в комплексе с направляющей РНК, гибридизованной с последовательностью-мишенью в представляющем интерес локусе-мишени.
[00244] В одном из вариантов осуществления настоящее изобретение относится к способу расщепления РНК-мишени. Данный способ может включать модификацию РНК-мишени с использованием нацеленного на нуклеиновую кислоту комплекса, который связывает РНК-мишень и осуществляет расщепление указанной РНК-мишени. В одном из вариантов осуществления нацеленный на нуклеиновую кислоту комплекс по изобретению при введении в клетку может вносить разрыв (например, одноцепочечный или двухцепочечный разрыв) в последовательность РНК. Например, такой способ может быть использован для расщепления связанной с заболеванием РНК в клетке. Например, экзогенная РНК-матрица, включающая последовательность, которую необходимо встроить, фланкированная вышележащей последовательностью и нижележащей последовательностью, может быть введена в клетку. Такие вышележащие и нижележащие последовательности характеризуются сходством последовательности с обоими концами участка встраивания в РНК. При желании, донорная РНК может представлять собой мРНК. Экзогенная РНК-матрица содержит последовательность, которую необходимо встроить (например, мутированную РНК). Последовательность для встраивания может представлять собой последовательность, эндогенную или экзогенную для клетки. Примерами последовательностей для встраивания являются кодирующая белок РНК или некодирующая РНК (например, микроРНК). Таким образом, последовательность для встраивания может быть функционально связана с надлежащей последовательностью или последовательностями контроля. Альтернативно, встраиваемая последовательность может выполнять регуляторную функцию. Вышележащие и нижележащие последовательности в экзогенной РНК выбирают так, чтобы стимулировать рекомбинацию между РНК-мишенью и донорной РНК. Вышележащая последовательность является последовательностью РНК, имеющей сходство последовательности с последовательностью РНК в восходящем направлении от целевого участка интеграции. Сходным образом, нижележащая последовательность является последовательностью РНК, имеющей сходство последовательности с последовательностью РНК в нисходящем направлении от целевого участка интеграции. Вышележащая и нижележащая последовательности в экзогенной РНК-матрице могут иметь 75%, 80%, 85%, 90%, 95% или 100% идентичность последовательности с последовательностью РНК-мишени. Предпочтительно, чтобы вышележащая и нижележащая последовательности в экзогенной РНК-матрице имели примерно 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с последовательностью РНК-мишени. В некоторых способах вышележащая и нижележащая последовательности экзогенной РНК-матрицы имеют 99% или 100% идентичность последовательности с последовательности РНК-мишени. Вышележащие или нижележащие последовательности могут содержать от примерно 20 п.н. до примерно 2500 п.н., например, примерно 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1100, 1200, 1300, 1400, 1500, 1600, 1700, 1800, 1900, 2000, 2100, 2200, 2300, 2400 или 2500 п.н. В некоторых способах иллюстративная вышележащая или нижележащая последовательность имеет от примерно 200 п.н. до примерно 2000 п.н., от примерно 600 п.н. до примерно 1000 п.н. или более конкретно от примерно 700 п.н. до примерно 1000 п.н. В некоторых способах экзогенная РНК-матрица может дополнительно содержать маркер. Такой маркер может облегчить скрининг целевых встраиваний. Примеры подходящих маркеров включают участки рестрикции, флуоресцентные белки или метки, по которым возможен отбор. Экзогенная РНК-матрица по изобретению может быть получена с помощью рекомбинантных способов (см., например, Sambrook et al., 2001 и Ausubel et al., 1996). Согласно способу модификации РНК-мишени путем встраивания экзогенной РНК-мишени разрыв (например, двухцепочечный или одноцепочечный разрыва в одноцепочечной или двухцепочечной ДНК или РНК) вносится в последовательность ДНК или РНК нацеленным на нуклеиновую кислоту комплексом, причем такой разрыв репарируется за счет гомологичной рекомбинации с экзогенной РНК-матрицы таким образом, что матрица оказывается встроенной в РНК-мишень. Присутствие двухцепочечного разрыва облегчает встраивание матрицы. В других вариантах осуществления настоящее изобретение относится к способу модификации экспрессии РНК в эукариотической клетке. Такой способ включает увеличение или уменьшение экспрессии полинуклеотида-мишени путем использования нацеленного на нуклеиновую кислоту комплекса, связывающего ДНК или РНК (например, мРНК или пре-мРНК). В некоторых способах РНК-мишень может быть инактивирована для того, чтобы влиять на модификацию экспрессии в клетке. Например, при связывании нацеленного на РНК комплекса с последовательностью-мишенью в клетке РНК-мишень инактивируется так, что такая последовательность не транслируется, кодируемый ей белок не образуется или такая последовательность не функционирует так, как последовательность дикого типа. Например, белок, или транскрипты микроРНК или пре-микроРНК не образуются. РНК-мишень нацеленного на РНК комплекса может представлять собой любую РНК, эндогенную или экзогенную для данной эукариотической клетки. Например, РНК-мишень может представлять собой РНК, находящуюся в ядре эукариотической клетки. РНК-мишень может представлять собой последовательность (например, мРНК или пре-мРНК), кодирующую генный продукт (например, белок) или некодирующую последовательность (например, нкРНК, lnc-РНК, тРНК или рРНК). Примеры РНК-мишени включают последовательность, ассоциированную с сигнальными биохимическими путями, например, ассоциированную с сигнальными биохимическими путями РНК. Примеры РНК-мишени включают РНК, ассоциированные с заболеваниями. "Ассоциированные с заболеваниями" РНК обозначает любые РНК, используемые для синтеза продуктов трансляции на аномальном уровне или в аномальной форме в клетках, полученных из затронутых заболеванием тканей в сравнении с тканями или клетками не затронутого заболеванием контроля. Это может быть РНК, транскрибированная с гена, которая экспрессируется на аномально высоком уровне; это может быть РНК, транскрибированная с гена, который экспрессируется на аномально низком уровне, причем измененная экспрессия коррелирует с появлением и/или прогрессированием заболевания. Ассоциированные с заболеванием РНК также упоминают как РНК, транскрибированные с гена, несущего мутацию(и) или генетическую вариацию, которые напрямую ответственны или связаны с неравновесностью сцепления с геном(ами), который(ые) вовлечен в этиологию заболевания. Продукт трансляции может быть известен или не известен и его количество может иметь нормальный или аномальный уровень. РНК-мишень нацеленного на РНК комплекса может представлять собой любую РНК, эндогенную или экзогенную для такой эукариотической клетки. Например, РНК-мишень нацеленного на РНК комплекса может представлять собой РНК, находящуюся в ядре эукариотической клетки. Такая РНК-мишень может быть последовательностью (например, мРНК или пре-мРНК), кодирующей продукт гена (например, белок) или некодирующей последовательностью (например, нкРНК, lnc-РНК, тРНК или рРНК).
[00245] В некоторых вариантах осуществления изобретения способ может позволять нацеленному на нуклеиновую кислоту комплексу связываться с ДНК- или РНК-мишенью, чтобы произвести расщепление указанной ДНК- или РНК-мишени, таким образом, изменяя ДНК- или РНК-мишень, где нацеленный на нуклеиновые кислоты комплекс включает нацеленный на нуклеиновые кислоты эффекторный белок в комплексе с направляющей РНК, гибридизованной с последовательностью-мишенью в указанной ДНК- или РНК-мишени. В одном аспекте изобретение относится к способу модификации экспрессии ДНК или РНК в эукариотической клетке. В некоторых вариантах осуществления изобретения способ позволяет нацеленному на нуклеиновые кислоты комплексу связываться с ДНК или РНК, таким образом, что указанное связывание приводит к увеличению или уменьшению экспрессии указанной ДНК или РНК; где нацеленный на нуклеиновые кислоты комплекс включает нацеленный на нуклеиновые кислоты эффекторный белок в комплексе с направляющей РНК. Применимы соображения и условия, сходные с теми, что указаны выше для способов модификации ДНК- или РНК-мишени. В действительности, варианты взятия образцов, культивирования и повторного введения применяются во многих аспектах настоящего изобретения. В одном аспекте изобретение относится к способам модификации ДНК- или РНК-мишени в эукариотической клетке, которая может происходить in vivo, ex vivo или in vitro. В некоторых вариантах осуществления изобретения способ включает взятие клетки или популяции клеток у человека или животного, не являющегося человеком, и модификацию этой клетки или клеток. Культивирование ex vivo может происходить на любой стадии. Клетка или клетки могут быть повторно введены в животное, не являющееся человеком, или растение. Для повторно введенных клеток особо предпочтительно, чтобы эти клетки являлись стволовыми клетками.
[00246] Действительно, в любом аспекте изобретения нацеленный на нуклеиновую кислоту комплекс может включать нацеленный на нуклеиновую кислоту эффекторный белок в комплексе с направляющей РНК, которая гибридизованной с последовательностью-мишенью.
[00247] Изобретение относится к инженерии и оптимизации систем, способов и композиций, используемых для контроля экспрессии генов, включающим нацеливание на последовательности ДНК и РНК, которые связаны с нацеленной на нуклеиновые кислоты системой и ее компонентами. В предпочтительных вариантах осуществления изобретения эффекторным белком (ферментом) является белок типа V, такой как C2c1 или C2c3. Преимущество способов по настоящему изобретению состоит в том, что система CRISPR минимизирует или избегает неспецифического связывания и его побочных эффектов. Эта система достигается с использованием систем, организованных так, что они имеют высокую степень специфичности к ДНК- или РНК-мишени.
[00248] Что касается нацеленного на нуклеиновую кислоту комплекса или системы, предпочтительно, чтобы tracr-последовательность имела одну или более шпилек и составляла 30 или более нуклеотидов в длину, 40 или более нуклеотидов в длину или 50 или более нуклеотидов в длину; длина последовательности cr-РНК варьировалась между 10 и 30 нуклеотидами, а нацеленный на нуклеиновую кислоту эффекторный белок принадлежал к эффекторным белкам типа V.
[00249] В некоторых модификациях изобретения эффекторный белок может представлять собой C2c1p из Alicyclobacillus sp., предпочтительно, C2c1p из Alicyclobacillus acidoterrestris, более предпочтительно C2c1p Alicyclobacillus acidoterrestris ATCC 49025, и последовательность cr-РНК может иметь длину 34 нуклеотида и содержать 5'-концевой прямой повтор (DR) длиной 14 нуклеотидов и спейсер длиной 20 нуклеотидов.
[00250] В некоторых модификациях изобретения эффекторный белок может представлять собой C2c1p из Bacillus sp., предпочтительно, C2c1p из Bacillus thermoamylovorans, более предпочтительно C2c1p Bacillus thermoamylovorans strain B4166, и последовательность cr-РНК может иметь длину 33 нуклеотида и содержать 5'-концевой прямой повтор (DR) длиной 14 нуклеотидов и спейсер длиной 29 нуклеотидов.
[00251] В некоторых модификациях эффекторный белок может быть эффекторным белком локусов типа V-B, более конкретно C2c1p, и последовательность cr-РНК может иметь длину от 27 до 40 нуклеотидов, предпочтительно от 29 до 38 нуклеотидов или от 30 до 37 нуклеотидов, более предпочтительно от 31 до 36 нуклеотидов или от 32 до 35 нуклеотидов, наиболее предпочтительно 33 или 34 нуклеотидов. например, cr-РНК может включать, состоять по большей части или состоять только из прямого повтора (DR), предпочтительно 5'-концевого DR длиной от 12 до 16 нуклеотидов, предпочтительно от 13 до 15 нуклеотидов, еще более предпочтительно 14 нуклеотидов, а также спейсер длиной от 15 до 24 нуклеотидов, предпочтительно от 16 до 23 нуклеотидов, более предпочтительно от 17 до 22 нуклеотидов, еще более предпочтительно от 18 до 21 нуклеотидов и наиболее предпочтительно 19 или 20 нуклеотидов.
[00252] В некоторых вариантах осуществления изобретения эффекторный белок может представлять собой C2c2p Listeria sp., предпочтительно C2c2p Listeria seeligeria, более предпочтительно C2c2p Listeria seeligeria серовара 1/2b штамма SLCC3954, и последовательность cr-РНК может составлять 44-47 нуклеотидов в длину с прямыми повторами (DR) на 5'-конце длиной 29 нуклеотидов и спейсером длиной от 15 нуклеотидов до 18 нуклеотидов.
[00253] В некоторых вариантах осуществления изобретения эффекторный белок может представлять собой C2c2p Leptotrichia sp., предпочтительно C2c2p Leptotrichia shahii, более предпочтительно C2c2p Leptotrichia shahii DSM 19757, и последовательность cr-РНК может составлять 42-58 нуклеотидов в длину с прямым повтором на 5'-конце длиной 28 нуклеотидов и спейсер длиной от 14 нуклеотидов до 28 нуклеотидов.
[00254] В некоторых вариантах осуществления изобретения эффекторный белок может относиться к эффекторным белкам локусов типа VI, более конкретно C2c2p, последовательность cr-РНК может иметь длину 36-63 нуклеотидов, предпочтительно от 37 нуклеотидов до 62 нуклеотидов или от 38 нуклеотидов до 61 нуклеотидов или от 39 нуклеотидов до 60 нуклеотидов, более предпочтительно от 40 нуклеотидов до 59 нуклеотидов или от 41 нуклеотидов до 58 нуклеотидов, наиболее предпочтительно от 42 нуклеотидов до 57 нуклеотидов. Например, cr-РНК может включать, по существу состоять из или состоять из прямого повтора (DR), предпочтительно на 5'-конце длиной от 26 нуклеотидов до 31 нуклеотидов, предпочтительно от 27 нуклеотидов до 30 нуклеотидов, еще более предпочтительно 28 нуклеотидов или 29 нуклеотидов в длину, и спейсера длиной от 10 нуклеотидов до 32 нуклеотидов, предпочтительно от 11 нуклеотидов до 31 нуклеотидов, более предпочтительно от 12 нуклеотидов до 30 нуклеотидов, еще более предпочтительно от 13 нуклеотидов до 29 нуклеотидов, и наиболее предпочтительно от 14 нуклеотидов до 28 нуклеотидов.
[00255] В некоторых вариантах осуществления эффекторный белок может представлять собой белок C2c1p из Alicyclobacillus sp., предпочтительно C2c1p из Alicyclobacillus acidoterrestris, более предпочтительно C2c1p из Alicyclobacillus acidoterrestris ATCC 49025 и последовательность cr-РНК может иметь длину по меньшей мере 78, например, может иметь длину 79 нуклеотидов или длину более 79 нуклеотидов, например, может иметь длину по меньшей мере 80 нуклеотидов, по меньшей мере 90 нуклеотидов, по меньшей мере 100 нуклеотидов, по меньшей мере 110 нуклеотидов, по меньшей мере 120 нуклеотидов, по меньшей мере 130 нуклеотидов, по меньшей мере 140 нуклеотидов, по меньшей мере 1500 нуклеотидов или более.
[00256] В некоторых вариантах осуществления эффекторный белок может представлять собой белок C2c1p из Bacillus sp., предпочтительно C2c1p из Bacillus thermoamylovorans, более предпочтительно C2c1p из Bacillus thermoamylovorans, штамм strain B4166 и последовательность tracr-РНК может иметь длину примерно 91 нуклеотид, например, 91 нуклеотид в длину.
[00257] В некоторых вариантах эффекторный белок может являться эффекторным белком локусов типа V-B, более конкретно C2c1p, и последовательность tracr-РНК может иметь длину по меньшей мере 60 нуклеотидов, в частности, по меньшей мере 65 нуклеотидов в длину, или по меньшей мере 70 нуклеотидов в длину, в частности от 60 до 70 нуклеотидов в длину, или от 60 до 70 нуклеотидов в длину, от 70 до 80 нуклеотидов в длину, от 80 до 90 нуклеотидов в длину, от 90 до 100 нуклеотидов в длину, от 100 до 110 нуклеотидов в длину, от 110 до 120 нуклеотидов в длину, от 120 до 130 нуклеотидов в длину, от 130 до 140 нуклеотидов в длину, от 140 до 150 нуклеотидов в длину или более 150 нуклеотидов в длину. См. иллюстрирующие примеры на фиг.17-21.
[00258] В некоторых вариантах осуществления изобретения эффекторный белок может относиться к эффекторным белкам локусов типа VI, более конкретно C2c2p, и последовательность tracr-РНК может иметь длину по меньшей мере 60 нуклеотидов, такую как по меньшей мере 65 нуклеотидов или по меньшей мере 70 нуклеотидов, такую как от 60 нуклеотидов до 70 нуклеотидов, или от 60 нуклеотидов до 70 нуклеотидов, или от 70 нуклеотидов до 80 нуклеотидов, или от 80 нуклеотидов до 90 нуклеотидов, или от 90 нуклеотидов до 100 нуклеотидов, или от 100 нуклеотидов до 110 нуклеотидов, или от 110 нуклеотидов до 120 нуклеотидов, или от 120 нуклеотидов до 130 нуклеотидов, или от 130 нуклеотидов до 140 нуклеотидов, или от 140 нуклеотидов до 150 нуклеотидов, или больше, чем 150 нуклеотидов. См. иллюстративные примеры на фиг.22-37.
[00259] В определенных вариантах осуществления изобретения эффекторный белок может относиться к эффекторным белкам локусов типа VI, более конкретно C2c2p, поэтому tracr-РНК для расщепления может не требоваться.
[00260] Использование двух различных аптамеров (каждый ассоциирован с отдельной нацеливающей на нуклеиновую кислоту направляющей РНК) позволяет использование слитых слитой конструкции активатор-адаптерный белок и слитых конструкций репрессор-адаптерный белок с различными нацеливающими на нуклеиновые кислоты направляющими РНК, для активации экспрессии только одной ДНК или РНК, подавляя другую. Их, наряду с их различными направляющими РНК можно вводить совместно, или по существу совместно, в мультиплексном подходе. Большое количество таких модифицированных нацеленных на нуклеиновые кислоты направляющих РНК может использоваться одновременно, например 10 или 20 или 30 и т.д., в то время как только одна (или по меньшей мере минимальное число) молекула эффекторного белка должна быть доставлена, так как сравнительно небольшое количество молекул эффекторного белка может использоваться с большим количеством модифицированных направляющих молекул. Адаптерный белок может быть ассоциирован (предпочтительно связан или слит с) одним или более активаторами или одним или более репрессорами. Например, адаптерный белок может быть ассоциирован с первым и вторым активаторами. Первый и второй активаторы могут быть одинаковыми, но предпочтительно они являются различными активаторами. Можно использовать три и более или даже четыре и более активатора (или репрессора), однако допустимый размер упаковки ограничивает их число 5 различными функциональными доменами. Использование линкеров является предпочтительным над прямым слиянием с адаптерным белком, где два или больше функциональных домена ассоциированы с адаптерным белком. Подходящие линкеры могут включать линкер GlySer.
[00261] Также предусматривается, что нацеленный на нуклеиновые кислоты комплекс эффекторного белка и направляющей РНК в целом может быть ассоциирован с двумя или более функциональными доменами. Например, может быть два или более функциональных домена, связанных с нацеленным на нуклеиновые кислоты эффекторным белком, или может быть два или более функциональных домена, связанных с направляющей РНК (через один или более адаптерных белков), или может быть один или более функциональных доменов, связанных с нацеленным на нуклеиновые кислоты эффекторным белком, и один или более функциональных доменов, связанных с направляющей РНК (посредством одного или более адаптерных белков).
[00262] Слитая конструкция адаптерного белка и активатора или репрессора может включать линкер. Например, могут использоваться линкеры GlySer GGGS. Они могут использоваться в повторах по 3 ((GGGGS)3) или 6, 9 или даже 12 или более, чтобы обеспечить подходящие длину, как требуется. Линкеры могут использоваться между направляющими РНК и функциональным доменом (активатор или репрессор), или между нацеленным на нуклеиновые кислоты эффекторным белком и функциональным доменом (активатор или репрессор). Линкеры используют для придания белку необходимой "механической гибкости". Изобретение охватывает нацеленный на нуклеиновую кислоту комплекс, включающий нацеленный на нуклеиновую кислоту эффекторный белок и направляющую РНК, где нацеленный на нуклеиновую кислоту эффекторный белок включает по меньшей мере одну мутацию, такую, что у нацеленного на нуклеиновую кислоту белка Cas есть не более 5% активности нацеленного на нуклеиновую кислоту белка Cas, не имеющего по меньшей мере одной мутации и необязательно по меньшей мере одну или более последовательностей ядерной локализации; направляющая РНК включает последовательность направляющей молекулы, способную к гибридизации с последовательностью-мишенью в представляющей интерес РНК в клетке; и где нацеленный на нуклеиновую кислоту эффекторный белок ассоциирован с двумя или более функциональными доменами, или по меньшей мере одна шпилечная структура направляющей РНК изменена вставкой отдельной последовательности (последовательностей) РНК, которая связана с одним или более адаптерными белками, и где адаптерный белок связан с двумя или более функциональными доменами; или нацеленный на нуклеиновую кислоту эффекторный белок ассоциирован с одним или более функциональными доменами, и по меньшей мере одна шпилечная структура направляющей РНК изменена вставкой отдельной последовательности (последовательностей) РНК, которая связана с одним или более адаптерными белками, и где адаптерный белок связан с одним или более функциональными доменами.
Мутации ферментов, уменьшающие нецелевые эффекты
[00263] В одном аспекте изобретение относится к не встречающемуся в природе или сконструированному способами инженерии ферменту CRISPR, предпочтительно ферменту CRISPR 2 класса, предпочтительно ферменту CRISPR типа V или VI, как описано в настоящем описании, в частности, предпочтительно, но не ограничиваясь этим, к C2c1 или C2c3, как описано в настоящем описании, имеющим одну или более мутаций, приводящих к уменьшению нецелевых эффектов, т.е. к улучшенным ферментам CRISPR для использования с целью вызвать модификации локусов-мишеней, но снижающих или устраняющих нецелевую активность, в частности, в комплексе с направляющими РНК, к усовершенствованным ферментам CRISPR для повышения активности ферментов CRISPR, в частности, в комплексе с направляющими РНК. Следует понимать, что мутантные ферменты, описанные ниже, могут быть использованы в любом из способов по изобретению, как описано в настоящем описании. Любые из этих способов, продуктов, композиций и способов применения, описанных в настоящем описании, в равной степени применимы совместно с мутантными ферментами CRISPR, как это более подробно описано ниже. Следует понимать, что в аспектах и вариантах осуществления изобретения, описанных в настоящем описании, при упоминании или описании C2c1 или C2c3 в качестве фермента CRISPR предпочтительно, чтобы восстановление функциональной системы CRISPR-Cas не требовало или не было зависимо от последовательности tracr и/или прямой повтор находился в 5'(восходящем)-направлении от направляющей последовательности (мишени или спейсера).
[00264] В качестве дальнейшего руководства приводятся следующие конкретные аспекты и варианты осуществления.
[00265] Авторы изобретения неожиданно установили, что возможны модификации ферментов CRISPR, сообщающие им сниженную нецелевую активность по сравнению с немодифицированными ферментами и/или увеличенную целевую активность по сравнению с немодифицированными ферментами CRISPR. Таким образом, в определенных аспектах изобретения в рамках настоящего изобретения предусматриваются улучшенные ферменты CRISPR, которые могут использоваться для широкого круга применений по модификации генов. Также в рамках настоящего изобретения предусмтриваются комплексы CRISPR, композиции и системы, а также способы и способы применения, все из которых включают описанные в настоящем описании модифицированные ферменты CRISPR.
[00266] В настоящем описании термин "Cas" может означать "C2c1", "C2c3" или фермент CRISPR. Термины "C2c1p" и "C2c1" используются взаимозаменяемо и термины "C2c3p" и "C2c3" используются взаимозаменяемо. Буква p в C2c1p и C2c3p служит обозначением белка. В контексте этого аспекта изобретения C2c1, C2c3 или фермент CRISPR является мутантным или модифицированным, "согласно чему фермент комплекса CRISPR имеет сниженную способность модифицировать один или более не являющийся мишенью локус по сравнению с немодифицированным ферментом" (или сходные утверждения); и при прочтении настоящего описания термины "C2c1", "C2c3","Cas" или "фермент CRISPR" и т.п. призваны включать мутантные или модифицированные C2c1, C2c3, Cas или фермент CRISPR по изобретению, т.е. "в связи с чем фермент комплекса CRISPR имеет сниженную способность модифицировать один или более нецелевых локусов по сравнению с немодифицированным ферментом" (или сходные утверждения).
[00267] В одном аспекте предусматриваются сконструированные способами инженерии белки C2c1 или C2c3 согласно приведенному в настоящем описании определению, в частности, C2c1 или C2c3, где белки в комплексе с молекулой нуклеиновой кислоты, включающей РНК, для образования комплекса CRISPR, причем в случае пребывания в составе комплекса CRISPR молекула нуклеиновой кислот нацеливает на один или более полинуклеотидных локусов-мишеней и белок содержит по меньшей мере одну модификацию по сравнению с немодифицированным белком C2c1 или немодифицированным белком C2c3, причем комплекс CRISPR, содержащий модифицированный белок, имеет измененную активность по сравнению с таковой комплекса, содержащего немодифицированный белок C2c1 или немодифицированный белок C2c3. Следует понимать, что при упоминании в настоящем описании "белка" CRISPR предпочтительно, чтобы белок C2c1 или белок C2c3 являлись модифицированным ферментом CRISPR (например, имели увеличенную или сниженную (или отсутствующую) ферментативную активность) в частности включая, но не ограничиваясь ими, C2c1 или C2c3. Термин "белок CRISPR" может использоваться взаимозаменяемо с термином "фермент CRISPR" независимо от наличия в белке внесенных изменений, в частности, увеличенной или сниженной (или отсутствующей) ферментативной активности по сравнению с белком CRISPR дикого типа.
[00268] В одном аспекте измененная активность сконструированного белка CRISPR включает измененные свойства связывания в отношении молекулы нуклеиновой кислоты, включающей РНК или локусы полинуклеотида-мишени, измененную кинетику связывания в отношении молекулы нуклеиновой кислоты, включающей РНК или локусы полинуклеотида-мишени или измененную специфичность связывания в отношении молекулы нуклеиновой кислоты, включающей РНК или локусы полинуклеотида-мишени по сравнению локусами полинуклеотида, не являющегося мишенью.
[00269] В некоторых вариантах осуществления немодифицированный Cas имеет активность расщепления ДНК, такую как активность C2c1 или C2c3. В некоторый вариантах осуществления Cas обеспечивает расщепление одной или обеих цепей в участке последовательности-мишени, в частности, внутри последовательности-мишени и/или в участке, комплементарном последовательности-мишени. В некоторых модификациях изобретения Cas обеспечивает расщепление одной или обеих цепей на расстоянии до 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 50, 100, 200, 500 или более нуклеотидов от первого или последнего нуклеотида последовательности-мишени. В некоторых вариантах осуществления вектор кодирует Cas, являющийся мутантным по сравнению с соответствующим ферментом дикого типа таким образом, что мутированный Cas лишен способности расщеплять одну или обе цепи полинуклеотида-мишени, содержащего последовательность-мишень. В некоторых вариантах осуществления Cas считается по существу лишенным какой-либо активности расщепления ДНК в случае, если активность мутантного фермента в отношении расщепления составляет не более чем приблизительно 25%, 10%, 5%, 1%, 0,1%, 0,01% или менее относительно активности расщепления ДНК немутантной формы фермента; к примеру, активность расщепления ДНК у мутантной формы отсутствует или пренебрежимо мала в сравнении с немутантной формой. Таким образом, Cas может включать одну или более мутаций и может быть использован как общий ДНК-связывающий белок со слиянием с функциональным доменом или без него. Такие мутации могут быть внесены искусственно или являться мутациями с приобретением или потерей функции. В одном из аспектов изобретения фермент Cas может быть слит с другим белком, например, TAG и/или индуцибельным/контролируемым доменом, в частности, химически индуцибельным/контролируемым доменом. Cas по настоящему изобретению могут представлять собой химерные белки Cas; например, Cas, имеющий усиленную функцию за счет того, что является химерным. Химерные белки Cas могут быть новыми Cas, содержащими фрагменты из более чем одного встречающегося в природе Cas. Они могут включать химерные белки, содержащие N-концевой фрагмент(ы) одного из гомологов Cas9 с C-концевым фрагментом(ами) другого гомолога Cas. Такой Cas может быть доставлен в клетку в форме мРНК. Экспрессия Cas может происходить под контролем индуцибельного промотора. Безусловной целью настоящего изобретения является избежание использование известных мутаций. В действительности, утверждение "согласно которому фермент комплекса CRISPR имеет сниженную способность модифицировать один или более локусов, не являющихся мишенями, в сравнении с немодифицированным ферментом и/или в связи с чем фермент комплекса CRISPR имеет увеличенную способность модифицировать один или более локусов-мишеней в сравнении с немодифицированным ферментом" (или подобные утверждения) не имеет целью описание мутаций, единственным результатом которых являются никаза, "мертвый" Cas или известные мутации Cas9. Однако не подразумевается, что модификация(и) или мутация(и) по настоящему изобретению, "в результате чего фермент комплекса CRISPR имеет сниженную способность модифицировать один или более локусов, не являющихся мишенями, по сравнению с немодифицированным ферментом и/или в результате чего фермент комплекса CRISPR имеет увеличенную способность модифицировать один или более локусов-мишеней по сравнению с немодифицированным ферментом" (или подобные утверждения) не могут сочетаться с мутациями, приводящими к тому, что фермент является никазой или нефункциональным ("убитым") ферментом. Такой "мертвый" фермент может иметь увеличенную способность к связыванию нуклеиновой кислот. Такая никаза может являться улучшенной никазой. Например, замена нейтральной аминокислот(ы) в и/или рядом с бороздой и/или других заряженных остатков в других участках Cas, расположенных в непосредственной близости к нуклеиновой кислоте (например, ДНК, кДНК, РНК, gРНК) на положительно заряженные аминокислоты может привести к тому, что "в связи с этим фермент комплекса CRISPR будет иметь сниженную способность к модификации одного или более локусов, не являющихся мишенями, по сравнению с немодифицированным ферментом", например, более активному расщеплению. Поскольку в связи с этим могут усилиться как целевое, так и нецелевое расщепление (сверхрасщепление C2c1 или C2c3), возможно использование этой методики вместе с тем, что в данной области называется tru-направляющей или tru-sg-РНК (см., например, Fu et at., "Improving CRISPR - Cas nuclease specificity using truncated guide RNAs," Nature Biotechnology 32, (2014) doi: 10,1038/nbt.2808, Received 17 November 2013, Accepted 06 January 7 2014, Published online 26 January 2014, Corrected online 29 January 2014) для усиления целевой активности без усиления нецелевого расщепления для получения сверхрасщепляющих никаз или в сочетании с мутацией, делающей Cas "убитым" для молекулы со сверхсвязыванием.
[00270] В определенных вариантах осуществления измененная активность сконструированных белков C2c1 или C2c3 включает увеличенную эффективность нацеливания или уменьшенное нецелевое связывание. В определенных вариантах осуществления измененная активность сконструированных белков C2c1 или C2c3 включает модифицированную активность расщепления.
[00271] В определенных вариантах осуществления измененная активность включает измененные свойства связывания в отношении молекулы нуклеиновой кислоты, содержащей РНК или локусы полинуклеотида-мишени, измененную кинетику связывания в отношении молекулы нуклеиновой кислоты, содержащей РНК или локусы полинуклеотида-мишени или измененную специфичность связывания в отношении молекулы нуклеиновой кислоты, включающей РНК или локусы полинуклеотида-мишени или измененную специфичность связывания в отношении молекулы нуклеиновой кислоты, содержащей РНК или локусы полинуклеотида-мишени в сравнении с локусами полинуклеотида, не являющегося мишенью.
[00272] В определенных вариантах осуществления измененная активность включает увеличение эффективности нацеливания или уменьшение нецелевого связывания. В некоторых вариантах осуществления измененная активность включает усиление активности расщепления в отношении локусов полинуклеотида-мишени. В некоторых модификациях изменение активности включает уменьшение активности расщеплениz в отношении локусов полинуклеотида-мишени. В некоторых вариантах осуществления измененная активность включает усиление активности расщепления локусов полинуклеотида, не являющегося мишенью.
[00273] В соответствии с этим в определенных вариантах осуществления существует увеличенная специфичность в отношении локусов полинуклеотида-мишени по сравнению локусами полинуклеотида, не являющегося мишенью. В других вариантах осуществления специфичность в отношении локусов полинуклеотида-мишени в сравнении с локусами полинуклеотида, не являющегося мишенью, уменьшена.
[00274] В одном из аспектов изобретения измененная активность сконструированного способами инженерии белка C2c1 или C2c3 включает измененную кинетику хеликазной активности.
[00275] В одном из аспектов изобретения сконструированные способами инженерии белки C2c1 или C2c3 включают модификацию, которая изменяет связывание белка с нуклеиновой кислотой, включающей РНК, или цепью локусов полинуклеотида-мишени или цепью локусов полинуклеотида, не являющегося мишенью. В одном из аспектов изобретения сконструированные способами инженерии белки C2c1 или C2c3 включают модификацию, изменяющую образование комплекса CRISPR.
[00276] В некоторых вариантах осуществления модифицированные белки C2c1 или C2c3 включают модификацию, изменяющую нацеливание молекулы нуклеиновой кислоты на локусы полинуклеотида. В определенных вариантах изобретения модификация включает мутацию в участке белка, который задействован в связывании с молекулой нуклеиновой кислоты. В некоторых вариантах осуществления такая модификация включает мутацию в участке белка, который задействован в связывании с цепью локусов полинуклеотида-мишени. В некоторых вариантах осуществления такая модификация включает мутацию в участке белка, который задействован в связывании с цепью локусов полинуклеотида, не являющегося мишенью. В некоторых вариантах осуществления модификация или мутация включает уменьшение отрицательного заряда в участке белка, который участвует в связывании молекулы нуклеиновой кислоты, включающей РНК, цепи локусов полинуклеотида-мишени или цепи локусов полинуклеотида, не являющегося мишенью. В некоторых вариантах осуществления модификация или мутация включает уменьшение отрицательного заряда в участке белка, который задействован с связывании молекулы нуклеиновой кислоты, включающей РНК, цепи локусов полинуклеотида-мишени или цепи локусов полинуклеотида, не являющегося мишенью. В некоторых вариантах осуществления модификация или мутация включает увеличенный положительный заряд в участке белка, который задействован в связывании с молекулой нуклеиновой кислоты, включающей РНК, цепи локусов полинуклеотида-мишени или цепи локусов полинуклеотида, не являющегося мишенью. В некоторых вариантах осуществления модификация или мутация включает увеличенный отрицательный заряд в участке белка, который задействован в связывании молекулы нуклеиновой кислоты, включающей РНК, цепи локусов полинуклеотида-мишени или цепи локусов полинуклеотида, не являющегося мишенью. В некоторых вариантах осуществления модификация или мутация увеличивает пространственные затруднения взаимодействия между белком и молекулой нуклеиновой кислоты, содержащей РНК, цепью локусов целевого полинуклеотида или цепью локусов нецелевого полинуклеотида. В некоторых вариантах осуществления модификация или мутация включает замену Lys, His, Arg, Glu, Asp, Ser, Gly или Thr. В некоторых вариантах осуществления модификация или мутация включает замену на Gly, Ala, He, Glu или Asp. В некоторых случаях модификация или мутация заключается в замене аминокислоты в борозде связывания.
[00277] В одном аспекте настоящее изобретение относится к не встречающемуся в природе ферменту CRISPR, как определено в настоящем описании, такому как C2c1 или C2c3, где фермент образует комплекс с направляющей РНК с образованием комплекса CRISPR, направляющая РНК, когда она в составе комплекса CRISPR, нацеливает на одного или более локусов полинуклеотида-мишени и фермент вносит изменения в локусы этого полинуклеотида и фермент имеет по меньшей мере одну модификацию, в соответствии с чем фермент в составе комплекса CRISPR имеет сниженную способность модифицировать один или более локусов, не являющихся мишенью, по сравнению с немодифицированным белком, и в соответствии с чем фермент в составе комплекса CRISPR имеет увеличенную способность модифицировать один или более локусов-мишеней в сравнении с немодифицированным ферментом.
[00278] В любом из таких не встречающихся в природе ферментов CRISPR модификация может включать модификацию одного или более аминокислотных остатков фермента.
[00279] В любом из таких не встречающихся в природе ферментов CRISPR модификация может включать модификацию одного или более аминокислотных остатков, локализованных в участке, содержащем остатки, являющиеся положительно заряженными в немодифицированном ферменте.
[00280] В любом из таких не встречающихся в природе ферментов CRISPR модификация может включать модификацию одного или более аминокислотных остатков, являющихся положительно заряженными в немодифицированном ферменте.
[00281] В любом из таких не встречающихся в природе ферментов CRISPR модификация может включать модификацию одного или более остатков аминокислот, не являющихся положительно заряженными в немодифицированном ферменте.
[00282] Модификация может включать модификацию одной или более аминокислот, не имеющих заряда в немодифицированном белке.
[00283] Модификация может включать модификацию одной или более аминокислот, имеющих отрицательный заряд в немодифицированном белке.
[00284] Модификация может включать модификацию одной или более аминокислот, являющихся гидрофобными в немодифицированном белке.
[00285] Модификация может включать модификацию одной или более аминокислот, являющихся полярными в немодифицированном белке.
[00286] В некоторых описанных выше не встречающихся в природе ферментах CRISPR модификация может включать модификацию одного или более остатков, расположенных в борозде связывания.
[00287] В некоторых описанных выше не встречающихся в природе ферментах CRISPR модификация может включать модификацию одного или более остатков, расположенных вне борозды связывания.
[00288] В некоторых описанных выше не встречающихся в природе ферментах CRISPR модификация может включать модификацию одного или более остатков, где такие один или более остатков могут являться аргинином, гистидином или лизином.
[00289] В любом из описанных выше не встречающихся в природе ферментов CRISPR такой фермент может быть модифицирован мутацией указанных одного или более остатков.
[00290] В некоторых из описанных выше не встречающихся в природе ферментов CRISPR фермент модифицирован мутацией одного или более указанных остатков, где мутация представляет собой замену остатка немодифицированного фермента остатком аланина.
[00291] В некоторых из описанных выше не встречающихся в природе ферментов CRISPR фермент модифицирован мутацией одного или более указанных остатков, где мутация представляет собой замену остатка немодифицированного фермента остатком глутаминовой кислоты.
[00292] В некоторых из описанных выше не встречающихся в природе ферментов CRISPR фермент модифицирован мутацией одного или более указанных остатков, где мутация представляет собой замену остатка немодифицированного фермента остатком серина, треонина, аспарагина или глутамина.
[00293] В некоторых из описанных выше не встречающихся в природе ферментов CRISPR фермент модифицирован мутацией одного или более указанных остатков, где мутация представляет собой замену остатка немодифицированного фермента остатком аланина, глицина, изолейцина, лейцина, метионина, фенилаланина, триптофана, тирозина или валина.
[00294] В некоторых из описанных выше не встречающихся в природе ферментов CRISPR фермент модифицирован мутацией одного или более указанных остатков, где мутация представляет собой замену остатка немодифицированного фермента остатком полярной аминокислоты.
[00295] В некоторых из описанных выше не встречающихся в природе ферментов CRISPR фермент модифицирован мутацией одного или более указанных остатков, где мутация представляет собой замену остатка немодифицированного фермента остатком, не являющимся остатком полярной аминокислоты.
[00296] В некоторых из описанных выше не встречающихся в природе ферментов CRISPR фермент модифицирован мутацией одного или более указанных остатков, где мутация представляет собой замену остатка немодифицированного фермента остатком отрицательно заряженной аминокислоты.
[00297] В некоторых из описанных выше не встречающихся в природе ферментов CRISPR фермент модифицирован мутацией одного или более указанных остатков, где мутация представляет собой замену остатка немодифицированного фермента остатком, не являющимся остатком отрицательно заряженной аминокислоты.
[00298] В некоторых из описанных выше не встречающихся в природе ферментов CRISPR фермент модифицирован мутацией одного или более указанных остатков, где мутация представляет собой замену остатка немодифицированного фермента остатком не несущей заряд аминокислоты.
[00299] В некоторых из описанных выше не встречающихся в природе ферментов CRISPR фермент модифицирован мутацией одного или более указанных остатков, где мутация представляет собой замену остатка немодифицированного фермента остатком, не являющимся остатком не несущей заряд аминокислоты.
[00300] В некоторых из описанных выше не встречающихся в природе ферментов CRISPR фермент модифицирован мутацией одного или более указанных остатков, где мутация включает замену остатка немодифицированного фермента остатком гидрофобной аминокислоты.
[00301] В случае некоторых из вышеописанных не встречающихся в природе ферментов CRISPR фермент модифицирован мутацией одного или более упомянутых остатков, где мутация может являться заменой остатка немодифицированного фермента аминокислотным остатком, который не является гидрофобным аминокислотным остатком.
[00302] В определенных вариантах осуществления эффекторный белок может являться белком C2c1p Alicyclobacillus sp., или, более предпочтительно, C2c1p Alicyclobacillus acidoterrestris, или, еще более предпочтительно, C2c1p Alicyclobacillus acidoterrestris, ATCC 49025. В определенных вариантах осуществления эффекторный белок может быть белком C2с1p Bacillus sp., или, более предпочтительно, C2e1p Bacillus thermoamylovorans, или, еще более предпочтительно, C2c1p Bacillus thermoamylovorans штамма B4I66. В некоторых вариантах осуществления эффекторный белок может быть эффекторным белком локусов типа V-B, в частности, C2c1p.
[00303] В некоторых вариантах осуществления белок C2c1 или C2c3 содержат один или более доменов с сигналом ядерной локализации (NLS). В некоторых вариантах осуществления белок C2c1 или C2c3 содержит не менее двух NLS.
[00304] В некоторых вариантах осуществления изобретения белок C2c1 или C2c3 представляет собой химерный белок CRISPR, состоящий из первого фрагмента из первого ортолога CRISPR, и второго фрагмента из второго ортолога CRISPR, причем первый и второй ортологи CRISPR различаются.
[00305] В некоторых вариантах осуществления изобретения фермент модифицирован или содержит модификации, например, содержит, по существу состоит или состоит из модификаций, полученных посредством мутаций любого из перечисленных в настоящем описании остатков или соответствующего остатка в соответствующем ортологе: либо фермент содержит или по существу состоит или состоит из модификаций любого (одиночная модификация), двух (двойная), трех (тройная), четырех (четырехкратная) положений или большего числа положений, в соответствии с описанием настоящего изобретения в настоящей заявке, или соответствующих остатков или положений в ортологе фермента CRISPR; например, фермент содержит или по существу состоит или состоит из модификации любых перечисленных в настоящем описании остатков C2c1 или C2c3 или соответствующего остатка или позиции в ортологе фермента CRISPR. В таком ферменте каждый остаток может быть модифицирован заменой на остаток аланина.
[00306] Заявители недавно описали способ получения ортологов Cas9 с повышенной специфичностью (Slaymaker и др. 2015 "Rationally engineered Cas9 nucleases with improved specificity"). Эта стратегия может быть использована для повышения специфичности ортологов C2c1 или C2c3. Приоритетными остатками для мутагенеза предпочтительно являются все положительно заряженные остатки в пределах RuvC-домена. Дополнительными остатками являются положительно заряженные остатки, являющиеся консервативными в различных ортологах.
[00307] В некоторых вариантах осуществления специфичность C2c1 или C2c3 может быть повышена за счет мутаций остатков, которые стабилизируют цепь ДНК, не являющейся мишенью.
[00308] В любом из не встречающихся в природе ферментов CRISPR:
(a) может существовать одно неспаренное основание между последовательностью-мишенью и соответствующей последовательностью в одном или более не являющихся мишенью локусах; и/или
(b) могут существовать два, три, четыре или более неспаренных оснований между последовательностью-мишенью и соответствующей последовательностью в одном или более не являющихся мишенью локусах; и/или
(c) указанные в (b) два, три, четыре или более неспаренных оснований являются смежными.
[00309] В случае любого из встречающихся в природе ферментов CRISPR фермент в комплексе CRISPR может обладать сниженной способностью к модификации одного или более не являющихся мишенью локусов по сравнению с немодифицированным ферментом, при этом фермент в комплексе CRISPR обладает повышенной способностью к модификации указанных локусов-мишеней по сравнению с немодифицированным ферментом.
[00310] В случае любого из встречающихся в природе ферментов CRISPR относительная разница между способностью фермента в комплексе CRISPR к модификации локуса-мишени и способностью к модификации по меньшей мере одного не являющегося мишенью локуса может быть увеличена по сравнению с немодифицированным ферментом.
[00311] В случае любого из не встречающихся в природе ферментов CRISPR CRISPR-фермент может содержать одну или более дополнительных мутаций, при этом одна или более дополнительных мутаций находятся в одном или более каталитически активных доменах.
[00312] В случае таких не встречающихся в природе ферментов CRISPR фермент CRISPR может обладать сниженной нуклеазной активностью, по сравнению с ферментом, в котором отсутствуют указанные одна или более дополнительные мутации, или может не обладать ей.
[00313] В случае некоторых таких не встречающихся в природе ферментов CRISPR фермент CRISPR не осуществляет расщепление одной или другой цепи ДНК в участке последовательности-мишени.
[00314] В случае, когда фермент CRISPR содержит одну или более дополнительных мутаций в одном или более каталитически активных доменах одна или более дополнительных мутаций могут находиться в каталитически активном домене фермента CRISPR, включая домены RuvCI, RuvCII и RuvCIII.
[00315] Не ограничиваясь теорией, в одном аспекте изобретения описанные способы и мутации предусматривают увеличение конформационных перестроек доменов ферментов CRISPR (например, доменов C2c1 и C2c3) в конформационные состояния, которые приводят к расщеплению в участках-мишенях и снижение вероятности этих конформационных состояний в участках, не являющихся мишенями. Расщепление ДНК-мишени ферментом CRISPR происходит в виде серии последовательных скоординированных стадий. Вначале PAM-связывающий домен узнает последовательность PAM с 5'-стороны от ДНК-мишени. После связывания с PAM первые 10-12 нуклеотидов последовательности-мишени (последовательность-затравка) проверяются на комплементарность комплекса направляющей РНК (гРНК) с ДНК, и этот процесс зависит от разделения цепей ДНК. Если нуклеотиды затравочной последовательности комплементарны гРНК, оставшаяся ДНК оказывается неспаренной и гРНК гибридизуется с цепью ДНК-мишени по всей длине. Бороздки, не являющиеся мишенью, могут стабилизировать цепь ДНК-мишени и способствовать расплетанию за счет неспецифических взаимодействий с положительно заряженным фосфатным остовом ДНК. Взаимодействия РНК с комплементарной ДНК и фермента CRISPR с некомплементарной ДНК способствуют тому, что ДНК расплетается, конкурируя с обратной гибридизацией комплементарной и некомплементарной цепей ДНК. Другие домены ферментов CRISPR также могут влиять на конформацию нуклеазных доменов, например, линкеры, соединяющие различные домены. Соответственно, приведенные методы и мутации затрагивают без ограничений RuvCI, RuvCIll, RuvCIII, а также линкеры. Конформационные изменения, например, C2c1 или C2c3, обусловленные связыванием с ДНК-мишенью, включая связывание с последовательностью-затравкой и взаимодействия с цепями ДНК, являющимися и не являющимися мишенью, определяют, будут ли домены расположены так, чтобы инициировать нуклеазную активность. Таким образом, приведенные в настоящем описании способы и мутации демонстрируют и обеспечивают возможность модификаций, выходящих за пределы узнавания PAM и спаривания оснований РНК и ДНК.
[00316] В одном из аспектов изобретение относится к нуклеазам CRISPR, как определено в настоящем описании, например, C2c1 или C2c3, обладающим улучшенным равновесием, смещенным в сторону образования конформаций, стимулирующих расщепление при целевых взаимодействиях, и/или равновесием, смещенным в сторону снижения вероятности образования конформаций, связанных с расщеплением при нецелевых взаимодействиях. В одном аспекте изобретение относится к нуклеазе Cas (например, C2c1 или C2c3) с улучшенной редактирующей активностью, т.е. нуклеазе Cas (например, C2c1 и C2c3), которая принимают конформацию, способствующую нуклеазной активности при нахождении на участке-мишени и снижающую ее вероятность при нахождении на участке, не являющемся мишенью. Авторы статьи (Sternberg et al., Nature 327(7576): 110-3, doi: 10.1038/naturel3344, опубликованной через интернет 28 октября 2013 года. Epub 2013 Oct 28) использовали способ FRET (флуоресцентный резонансный перенос энергии), чтобы определить относительную ориентацию каталитических доменов Cas9 при ассоциации с ДНК, являющейся и не являющейся мишенью, и эти эксперименты могут быть экстраполированы на ферменты CRISPR по настоящему изобретению (например, C2c1 или C2c3).
[00317] Настоящее изобретение относится к способам и мутациям для модулирования нуклеазной активности и/или специфичности с использованием модифицированных направляющих РНК. Как описано, нецелевая нуклеазная активность может быть повышена или снижена. Кроме того, можно повышать или снижать специфичность - соотношение целевой активности к нецелевой. Модифицированные направляющие РНК включают, но не ограничиваются ими, укороченные направляющие РНК, нефункциональные направляющие РНК, химически модифицированные направляющие РНК, соединенные с функциональными доменами направляющие РНК, модифицированные направляющие РНК, содержащие функциональные домены, содержащие аптамеры направляющие РНК, содержащие адаптерные белки направляющие РНК, направляющие РНК, содержащие добавленные или модифицированные петли. В некоторых вариантах осуществления один или более функциональных доменов ассоциирован с мертвыми направляющими РНК (dРНК). В некоторых вариантах осуществления комплекс dРНК с ферментом CRISPR осуществляет регуляцию генов с помощью функционального домена в пределах генного локуса, в то время как направляющая РНК направляет расщепление ДНК ферментом CRISPR в другом локусе. В некоторых вариантах осуществления dРНК выбирают для максимизации селективности регуляции представляющего интерес генного локуса по сравнению с нецелевой регуляцией. В некоторых вариантах осуществления dРНК выбираются так, чтобы максимально повысить уровень регуляции генов и минимизировать расщепление ДНК-мишени.
[00318] С целью дальнейшего обсуждения, указание на функциональный домен означает в настоящем описании функциональный домен, связанный с ферментом CRISPR, или функциональный домен, связанный с адаптерным белком.
[00319] При применении настоящего изобретения на практике, петли гРНК могут быть удлинены без взаимодействия с белком Cas9 (например, C2c1 или C2c3) за счет вставки отдельной петли(петель) РНК или отдельной последовательности(ей), которые могут привлекать адаптерные белки, способные связываться с отдельной петлей(петлями) РНК или отдельной последовательностю(ями). Адаптерные белки могут включать, но не ограничиваются ими, комбинации ортогональные РНК-связывающие белки/аптамеры, которые существуют в многообразии белков оболочки бактериофагов. Список таких белков оболочки бактериофагов включает, но не ограничивается ими, следующие: Qβ, F2, GA, fr, JP501, M12, R17, BZ13, JP34, JP500, KU1, М11, MX1, TW18, VK, SP, F1, ID2, NL95, TW19, AP205, ϕСb5, ϕСb8г, ϕСb12r, ϕСb2Зr, 7s и PRR1. Адаптерные белки или ортогональные РНК-связывающие белки могут дальше привлекать эффекторные белки или слитые белки, которые состоят из одного или более функциональных доменов. В некоторых вариантах осуществления функциональный домен может быть выбран из группы, включающий: транспозазный домен, интегразный домен, рекомбиназный домен, резольвазный домен, инвертазный домен, протеазный домен, ДНК-метилтрансферазный домен, домен ДНК-гидроксиметилазы, домен ДНК-деметилазы, гистонацетилазный домен, домен гистоновой деацетилазы, нуклеазный домена, репрессорный домен, активационный домен, домен, содержащий сигнал ядерной локализации, домен белков, осуществляющих транскрипционную регуляцию или рекрутирующих транскрипционный комплекс, домен, ассоциированный с поглощением веществ клеткой, домен, связывающий нуклеиновые кислоты, антигенпрезентирующий домен, гистон-модифицирующий домен, привлекающий модифицирующие гистоны белки: ингибитор гистонмодифицирующих ферментов, гистоновую метилтрансферазу, гистоновую деметилазу, гистоновую киназу, гистоновую фосфатазу, гистоновую рибозилазу, гистоновую дерибозилазу, гистоновую убиквитиназу, гистоновую деубиквитиназу, гистоновую биотиназу, протеазу гистоновых хвостов. В некоторых предпочтительных вариантах осуществления функциональный домен может являться трансактивационным доменом, таким как, но не ограничиваясь ими, VP64, p65, MyoD1, HSF1, RTA, SET7/9 или гистонацетилтрансферазный домен. В некоторых вариантах осуществления функциональный домен может представлять собой транскрипционно репрессорным доменом, предпочтительно KRAB. В некоторых вариантах осуществления транскрипционно репрессорным доменом могут являться SID или конкатемеры SID (например SID4X). В некоторых вариантах осуществления функциональным доменом может являться домен, осуществляющий эпигенетические модификации, так что предусматривается фермент, осуществляющий эпигенетические модификации. В некоторых вариантах осуществления функциональный домен может быть активационным доменом, который может являться активационным доменом P65. В некоторых вариантах осуществления функциональный домен может быть деаминазой, такой как цитидиндеаминаза. Цитидиндеаминаза может быть направлена на нуклеиновую кислотe-мишень, где она осуществляет преобразование цитидина в уридин, что приводит к замене C (цитозина) на Т (тимин) (и G (гуанина) на А (аденин) в комплементарной цепи). В таком варианте осуществления нуклеотидная замена может быть осуществлена без расщепления ДНК.
[00320] В одном аспекте настоящее изобретение также относится к способам и мутациям для модулирования связывающей активности (например, C2c1 или C2c3) и/или специфичности связывания Cas. В некоторых вариантах осуществления используются белки Cas (например, C2c1 или C2c3), у которых отсутствует нуклеазная активность и/или специфичность связывания. В некоторых вариантах осуществления используются модифицированные направляющие РНК, которые способствуют связыванию, но не стимулируют нуклеазную активность нуклеазы Cas (например, C2c1 или C2c3). В некоторых вариантах осуществления уровень целевого связывания может быть повышен или снижен. Также в некоторых вариантах осуществления уровень нецелевого связывания может быть повышен или снижен. Более того, может быть повышенная или сниженная специфичность, определяемая отношением целевого связывания к нецелевому.
[00321] В некоторых вариантах осуществления снижение уровня нецелевого расщепления обеспечивается дестабилизацией разделения цепей, в частности, внесением мутаций в ферменты C2c1 или C2c3, снижающих положительный заряд в областях, взаимодействущих с ДНК (как описано здесь и далее на примере Cas9 в статье Slaymaker et al. 2016 (Science, 1;351(6268):84-8). В дальнейших вариантах осуществления снижение нецелевого расщепления обеспечивается внесением мутаций в ферменты C2c1 или C2c3, которые влияют на взаимодействие между цепью-мишенью и последовательностью направляющей РНК, в частности, разрушая взаимодействие между C2c1 или C2c3 с фосфатным остовом цепи ДНК-мишени таким образом, чтобы оставалась целевая специфичная активность, но снижалась нецелевая активность (как описано для Cas9 в статье Kleinstiver et al. 2016, Nature, 28;529(7587):490-5). В некоторых вариантах осуществления изобретения нецелевая активность снижена за счет модификации C2c1 (или модифицикации C2c3), при этом изменяется взаимодействие как цепи-мишени, так и цепи, не являющейся мишенью, по сравнению с C2c1 дикого типа (или C2c3 дикого типа).
[00322] Способы и мутации, которые могут быть использованы в различных комбинациях, чтобы повысить или снизить активность и/или специфичность целевой активности по отношению к нецелевой, или повысить или снизить связывание и/или специфичность целевого связывания по отношению к нецелевому, могут быть использованы для компенсации или усиления эффекта мутаций или модификаций, внесенных для усиления других эффектов. Такие мутации или модификации, внесенные для усиления других эффектов, включают мутации и модификации Cas (например, C2c1 или C2c3) и/или мутации или модификации, внесенные в направляющие РНК. В определенных вариантах осуществления способы и мутации используются в отношении химически модифицированной направляющей РНК. Примеры химических модификаций направляющей РНК включают, но не ограничиваются ими, включение 2'-O-метила (M), 2'-O-метил-3'-фосфоротиоата (MIS) или 2'-O-метил-3'-тио-PACE (MSP) в один или более концевых нуклеотидов. Такие химически модифицированные направляющие РНК могут быть более стабильными и активными по сравнению с немодифицированными направляющими РНК, хотя соотношение целевой и нецелевой специфичности предсказать нельзя. (См. Hendel, 2015, Nat Biotechnol. 33(9):985-9, doi: 10.1038/nfot.3290., опубликованную через интернет 29 июня 2015 года). Химически модифицированные направляющие РНК включают, но не ограничиваются ими, РНК с фосфоротиоатными связями и нуклеотиды закрытых нуклеиновых кислот (LNA), содержащие метиленовый мостик между 2'-4'-атомами углерода рибозного кольца. Способы и мутации согласно изобретению используются для модуляции активности нуклеазы (например, C2c1 или C2c3) Cas (или C2c3) и или связывания с химически модифицированными направляющими РНК.
[00323] В одном аспекте изобретение относится к способам и мутациям для модулирования связывания и/или специфичности связывания белков Cas (например, C2c1 или C2c3) по изобретению, как определено в настоящем описании, включая функциональные домены, такие как нуклеазы, активаторы транскрипции, репрессоры транскрипции и т.п. Например, белок Cas (например, C2c1 или C2c3) может быть создан так, чтобы у него отсутствовала нуклеазная активность или нуклеазная активность была изменена или снижена за счет внесения мутаций, таких как мутации C2c1 или C2c3. Белки Cas (например, C2c1 или C2c3) без нуклеазной активности могут быть полезны для доставки функциональных доменов, направляемой РНК и специфической для последовательности ДНК. Изобретение относится к способам и мутациям для модулирования связывания белков Cas (например, C2c1 или C2c3). В одном из вариантов осуществления функциональный домен содержит VP64, обеспечивая направляемый РНК фактор транскрипции. В другом варианте осуществления функциональный домен содержит FokI, что обеспечивает опосредованную РНК нуклеазную активность. Это упомянуто в публикации патента США 2014/0356959, публикации патента США 2014/0342456, публикации патента США 2015/0031132 и Mali, P. et al., 2013, Science 339(6121):823-6, doi: 10.1126/science. 1.232033, опубликованной через интернет 3 января 2013 года, и за счет изложенных в настоящем описании идей изобретение охватывает способы и материалы этих документов, применяемых в привязке к изложенным в настоящем описании идеям. В некоторых вариантах осуществления целевое связывание увеличено. В некоторых вариантах осуществления нецелевое связывание уменьшено. В некоторых вариантах осуществления уровень целевого связывания снижен. В некоторых вариантах осуществления уровень нецелевого связывания повышен. Соответственно, изобретение также обеспечивает увеличение или снижение специфичности целевого связывания по отношению к нецелевому у функционализованных Cas-связывающих белков (например, C2c1 или C2c3).
[00324] Использование Cas (например, C2c1 или C2c3) в качестве направляемого РНК связывающего белка не ограничивается использованием Cas (например, C2c1 или C2c3) без нуклеазной активности. Ферменты Cas (например, C2c1 или C2c3) с присутствующей нуклеазной активностью могут также функционировать как направляемые РНК связывающие белки при использовании с конкретными направляющими РНК. Например, короткие направляющие РНК и направляющие РНК, содержащие нуклеотиды, не соответствующие мишени, могут стимулировать направляемое РНК связывание Cas (например, C2c1 или C2c3) с последовательностью-мишенью при низком уровне расщепления или в его отсутствие (см., например, Dahlman, 2015, Nat Biotechnol. 33(11):1159-1161, doi: 10,1038/nbt.3390, опубликовано через интернет 05 октября 2015). В некоторых вариантах осуществления изобретение относится к способам и мутациям для модулирования связывания белков Cas (например, C2c1 или C2c3), обладающих нуклеазной активностью. В некоторых вариантах осуществления уровень целевого связывания повышен. В некоторых вариантах осуществления уровень нецелевого связывания снижен. В некоторых вариантах осуществления уровень целевого связывания снижен. В некоторых вариантах осуществления уровень нецелевого связывания повышен. В некоторых вариантах осуществления увеличена или уменьшена специфичность целевого связывания по отношению к нецелевому связыванию. В некоторых вариантах осуществления нуклеазная активность фермента, включающего направляющую РНК и Cas (например, C2c1 или C2c3), также модулируется.
[00325] Образование гетеродуплекса РНК-ДНК важно для наличия активности и специфичности расщепления по всей области-мишени, а не только области затравочной последовательности, наиболее близкой по расположению к PAM. Таким образом, у укороченных направляющих РНК снижен уровень активности и уровень специфичности расщепления. В одном аспекте изобретение относится к способу и мутациям для увеличения активности и специфичности расщепления с использованием измененных направляющих РНК.
[00326] Изобретение также демонстрирует, что модификации нуклеазной специфичности Cas (например, C2c1 или C2c3) могут быть осуществлены одновременно с изменениями области-мишени. Мутанты Cas (например, C2c1 или C2c3) могут быть созданы так, чтобы они обладали как увеличенной целевой специфичностью, так и адаптивными изменениями в узнавании PAM, например, при выборе мутаций, которые изменяют специфичность PAM и комбинировании этих мутаций с мутациями бороздки, не являющей мишенью, которые могут увеличивать (или, при желании, снижать) специфичность к последовательностям-мишеням по отношению к не являющимся мишенями последовательностям. В одном таком варианте осуществления остаток PI-домена мутирован так, чтобы улучшить узнавание представляющей интерес последовательности PAM, в то время как одна или более аминокислот бороздки, не являющейся мишенью, мутированы так, чтобы изменить целевую специфичность. Способы и модификации Cas (например, C2c1 или C2c3), описанные в настоящем описании, могут быть использованы для того, чтобы предотвратить потерю специфичности, возникающую при изменении узнавания PAM, повысить увеличенный уровень специфичности, возникающий при изменении узнавания PAM, предотвратить увеличение уровня специфичности, возникающее при изменении узнавания PAM, или способствовать потере специфичности, возникающей при изменении узнавания PAM.
[00327] Эти способы и мутации могут быть использованы для любых ферментов Cas (например, C2c1 или C2c3) с измененным узнаванием PAM. Неограничивающие примеры включенных PAM являются такими, как описано в настоящем описании.
[00328] В следующих вариантах осуществления изобретения в способах и мутациях используются модифицированные белки.
[00329] В случае любого из не встречающихся в природе ферментов CRISPR фермент CRISPR может содержать один или более гетерологичных функциональных доменов.
[00330] Один или более гетерологичных функциональных доменов могут включать один или более доменов с сигналом ядерной локализации (NLS). Один или более гетерологичных функциональных доменов могут включать по меньшей мере два или более NLS.
[00331] Один или более гетерологичных функциональных доменов могут включать один или более трансактивационных доменов. Трансактивационным доменом может являться VP64.
[00332] Один или более гетерологичных функциональных доменов могут содержать один или доменов репрессии транскрипции. Домен репрессии транскрипции может включать домен KRAB или домен SID.
[00333] Один или более гетерологичных функциональных доменов могут включать один или более нуклеазных доменов. Одним или более нуклеазными доменами могут являться FokI.
[00334] Один или более гетерологичных функциональных доменов могут обладать только одной или более из следующих функций: метилазная активность, деметилазная активность, трансактивационная активность, активность репрессии транскрипции, активность в отношении высвобождения транскрипционных факторов, активностью модификации гистонов, нуклеазной активностью, активностью расщепления одноцепочечной РНК, активностью расщепления двуцепочечной РНК, активностью расщепления одноцепочечной ДНК, активностью расщепления двуцепочечной ДНК и активностью связывания нуклеиновых кислот.
[00335] По меньшей мере один или более гетерологичных функциональных доменов могут находиться рядом или на N-конце фермента и/или рядом или на С-конце фермента.
[00336] Один или более гетерологичных функциональных доменов могут быть слиты с ферментом CRISPR или связаны с ферментом CRISPR или соединены с ферментом CRISPR через линкерную часть.
[00337] В любом из не встречающихся в природе ферментов CRISPR фермент CRISPR может включать фермент CRISPR из организмов рода, такого как: Alicyclobacillus, Desulfovibrio, Desulfonatronum, Opitutaceae, Tuberibacillus, Bacillus, Brevibacillus, Desulfatirhabdium, Citrobacter и Methylobacterium.
[00338] В случае любого из не встречающихся в природе ферментов CRISPR фермент CRISPR может включать химерный фермент Cas (например, C2c1 или C2c3), содержащий первый фрагмент первого ортолога Cas (например, C2c1 или C2c3) и второй фрагмент второго ортолога Cas (например, C2c1 или C2c3), причем первый и второй ортологи Cas (например, C2c1 или C2c3) различаются. По меньшей мере один из ортологов (первый и/или второй) Cas (например, C2c1 или C2c3) могут представлять собой Cas (например, C2c1 или C2c3) таких видов как: Alicyclobacillus acidoterrestris (например, ATCC 49025), Alicyclobacillus contaminans (например, DSM 17975), Desulfovihrio inopinatus (например, DSM 10711), Desulfonatronum thiodismutans (например, штамм MLF-1), бактерии Opitutaceae TAV5, Tuberibacillus calidus (например, DSM 17572), Bacillus thermoamylovorans (например, штамм B4166), Brevibacillus sp. CF112, Bacillus sp. NSP2.1, Desulfatirhabdium butyrativorans (например, DSM 18734), Alicyclobacillus herbarius (например, DSM 13609), Citrobacter freundii (например, ATCC 8090), Brevibacillus agri (например, BAB-2500) и Methylobacterium nodulans (например, ORS 2060).
[00339] В любом из не встречающихся в природе ферментов CRISPR нуклеотидная последовательность, кодирующая фермент CRISPR, может быть кодон-оптимизированной для экспрессии в эукариотах.
[00340] В любом из не встречающихся в природе ферментов CRISPR клетка может быть эукариотической клеткой или прокариотической клеткой, причем комплекс CRISPR функционален в клетке, и при этом фермент комплекса CRISPR обладает сниженной способностью к модификации одного или более локусов клетки, не являющихся мишенью, по сравнению с немодифицированным ферментом, и/или фермент в комплексе CRISPR обладает повышенной способностью к модификации одного или более локусов клетки, не являющихся мишенью, по сравнению с немодифицированным ферментом.
[00341] Соответственно, в одном аспекте изобретение относится к эукариотической клетке, содержащей искусственный белок или систему CRISPR, как определено в настоящем описании.
[00342] В определенных вариантах осуществления изобретения способы, как описано в настоящем описании, могут включать доставку Cas (например, C2c1 или C2c3) в трансгенную клетку, в которой одна или более нуклеиновых кислот, кодирующих одну или более молекул направляющих РНК, доставленных или непосредственно введенных, связаны в клетке с регуляторным элементом, содержащим промотор одного или более генов-мишеней. Термин "трансгенная по Cas клетка", как используют в настоящем описании, относится к клетке, в частности, эукариотической клетке, в геном которой встроен ген Cas. Природа, тип или происхождение такой клетки не ограничивается особым образом, согласно данному изобретению. Также способ введения трансгена Cas в клетку может различаться и являться любым из способов, доступных в данной области. В некоторых вариантах осуществления изобретения трансгенная клетка Cas получена путем введения трансгена Cas в выделенную клетку. В некоторых других вариантах осуществления изобретения трансгенная клетка Cas может быть получена путем выделения клеток из трансгенного организма Cas. В качестве не ограничивающего примера, трансгенная клетка Cas, как используют в настоящем описании, может быть получена из трансгенного по Cas эукариотического организма, такого как эукориотический организм с встроенным Cas. См. WO 2014/093622 (PCT/US13/74667), включенную в настоящее описание в качестве ссылки. Способы публикации патентов США № 20120017290 и 20110265198, закрепленных за Sangamo BioSciences, Inc., направленные на нацеливание на локус Rosa, могут быть изменены для использования системы CRISPR Cas данного изобретения. Способы публикации патента США № 20130236946, закрепленных за Cellectis, направленные на нацеливание на локус Rosa, могут также быть изменены для использования системы CRISPR-Cas по настоящему изобретению. В качестве следующего примера приводится ссылка на статью Platt et. al. (Cell; 159(2):440-455 (2014)) с описанием мыши со вставкой Cas9 мыши, которая включена в настоящее описание в качестве ссылки и которая может быть экстраполирована на охарактеризованные в настоящем описании ферменты CRISPR по настоящему изобретению. Трансген Cas, кроме того, может включать кассету Lox-Stop-polyA-Lox(LSL), что делает экспрессию Cas управляемой с помощью рекомбиназы Cre. Альтернативно, клетка с трансгенным Cas может быть получена путем введения трансгена Cas в выделенную клетку. Системы доставки трансгенов хорошо известны в данной области. Примером может служить доставка трансгена Cas, напримр, в эукариотическую клетку посредством вектора (например, AAV, аденовируса, лентивируса) и/или частицы и/или наночастицы, как также описано в другой части настоящего описания.
[00343] Для квалифицированного специалиста будет понятно, что клетка, такая как трансгенная по Cas клетка в используемом в настоящем описании значении, может содержать дальнейшие изменения генома помимо наличия встроенного гена Cas или мутаций, являющихся результатом последовательность-специфического действия Cas, в комплексе с РНК, способной направлять Cas к локусу-мишени, например, такие как одна или более онкогенных мутаций, как в качестве неограничивающего примера описано в Platt et al. (2014), Chen et al., (2014), или Kumar et al. (2009).
[00344] Также изобретение относится к композиции, содержащей сконструированный способами инженерии белок CRISPR, описанный в настоящем описании, как описано в данном разделе.
[00345] Изобретение также относится к не встречающейся в природе сконструированной способами инженерии композиции, содержащей комплекс CRISPR-Cas, содержащий любой из не встречающихся в природе ферментов CRISPR, описанных выше.
[00346] В одном из аспектов изобретение относится к векторной системе, включающей один или более векторов, причем один или более векторов содержат:
а) первый регуляторный элемент, функционально связанный с одной или более нуклеотидными последовательностями, кодирующими сконструированный способам инженерии белок CRISPR, как определено в настоящем описании, и необязательно
b) второй регуляторный элемент, функционально связанный с одной или более нуклеотидными последовательностями, кодирующими одну или более нуклеиновых кислот, включая направляющую РНК, содержащую направляющую последовательность, последовательность прямого повтора, и необязательно где компоненты (а) и (b) расположены в одном векторе или разных векторах.
[00347] Изобретение также относится к не встречающейся в природе композиции, включающей:
систему доставки, функционально оптимизированную для доставки в клетку компонентов комплекса CRISPR-Cas или одной или более полинуклеотидных последовательности, содержащих или кодирующих указанные компоненты, при этом указанный комплекс CRISPR-Cas должен быть функционален внутри клетки.
компоненты комплекса CRISPR-Cas или одна или более полинуклеотидных последовательностей, кодирующих для транскрипции и/или трансляции в клетке компоненты комплекса CRISPR-Cas, включающие:
не встречающиеся в природе ферменты CRISPR (например, сконструированные способами инженерии C2c1 или C2c3), описанные в настоящем описании;
направляющую РНК CRISPR-Cas, содержащую направляющую последовательность, последовательность прямого повтора, при этом фермент в комплексе CRISPR обладает сниженной способностью модифицировать один или более локусов, не являющихся мишенью, по сравнению с немодифицированным ферментом, и/или фермент в комплексе CRISPR обладает увеличенной способностью модифицировать один или более локусов-мишеней, по сравнению с немодифицированным ферментом.
[00348] В одном аспекте изобретение также относится к системе, содержащей указанный сконструированный способами инженерии белок CRISPR, такой как описан в этом разделе.
[00349] В любой из таких композиций система доставки может включать: дрожжевую систему, систему липофекции, систему микроинъекций, генную пушку, виросомы, липосомы, иммунолипосомы, поликатионы, конъюгаты нуклеиновых кислот с липидами или искусственные вирионы, охарактеризованные в настоящем описании и в других источниках.
[00350] В любой из таких композиций система доставки может включать векторную систему, содержащую один или более векторов, причем компонент (II) содержит первый регуляторный элемент, функционально связанный с полинуклеотидной последовательностью, которая содержит направляющую последовательность и последовательность прямого повтора, и необязательно компонент (I) содержит второй регуляторный элемент, функционально связанный с полинуклеотидной последовательностью, кодирующий фермент CRISPR.
[00351] В любой из таких композиций система доставки может включать векторную систему, содержащую один или более векторов, причем компонент (II) содержит первый регуляторный элемент, функционально связанный с последовательностью и последовательностью прямого повтора, а компонент (I) содержит второй регуляторный элемент, функционально связанный с полинуклеотидной последовательностью, кодирующий фермент CRISPR.
[00352] В любой из таких композиций, композиция может включать более одной направляющей РНК, и каждая направляющая РНК имеет отличающуюся мишень, обеспечивая мультиплексный подход.
[00353] В любой из таких композиций полинуклеотидная последовательность(и) может находиться на одном векторе.
[00354] Изобретение также относится к не встречающейся в природе векторной системе коротких палиндромных повторов, регулярно расположенных кластерами CRISPR-Cas ((CRISPR-CRISPR-ассоциированный белок (Cas) (CRISPR-Cas)), включающей один или более векторов, которые содержат:
а) первый регуляторный элемент, функционально связанный с полинуклеотидной последовательностью, кодирующей не встречающийся в природе фермент CRISPR любой из конструкций по изобретению, описанных в настоящем описании;
b) второй регуляторный элемент, функционально связанный с полинуклеотидной последовательностью, кодирующей одну или более направляющих РНК, и эта направляющая РНК содержит направляющую последовательность, последовательность прямого повтора, где: компоненты (а) и (b) расположены в одном или разных векторах, образуется комплекс CRISPR, направляющие РНК нацелены на полинуклеотидные локусы-мишени, и фермент действует в отношении полинуклеотидных локусов-мишеней, и фермент в комплексе CRISPR обладает сниженной способностью модифицировать один или более локусов, не являющихся мишенью, по сравнению с немодифицированным ферментом, и/или этот фермент в комплексе CRISPR обладает повышенной способностью модифицировать один или более локусов-мишеней по сравнению с немодифицированным ферментом.
[00355] В такой системе компонент (II) может содержать первый регуляторный элемент, функционально связанный с полинуклеотидной последовательностью, которая содержит направляющую последовательность, последовательность прямого повтора, и где компонент (II) может содержать второй регуляторный элемент, функционально связанный с полинуклеотидной последовательностью, кодирующей фермент CRISPR. В такой системе, когда это возможно, направляющая РНК может включать химерную РНК.
[00356] В такой системе компонент (I) может содержать первый регуляторный элемент, функционально связанный с направляющей последовательностью и последовательностью прямого повтора, тогда как компонент (II) может содержать второй регуляторный элемент, функционально связанный с полинуклеотидной последовательностью, кодирующей не встречающийся в природе фермент CRISPR. Такая система может включать более одной направляющей РНК и каждая направляющая РНК может нацеливать на различные мишени, таким образом обеспечивая мультиплексный подход. Компоненты (a) и (b) могут быть расположены на одном и том же векторе.
[00357] В любой из таких систем, включающих векторы, один или более векторов могут содержать один или более вирусных векторов, таких как ретровирусный, лентивирусный, аденовирусный, аденоассоциированный вирус или вирус простого герпеса.
[00358] В любой из таких систем, содержащих регуляторные элементы, по меньшей мере один из указанных регуляторных элементов может содержать тканеспецифический промотор. Тканеспецифический промотор может направлять экспрессию в клетке крови млекопитающего, клетке печени млекопитающего или глаза млекопитающего.
[00359] В любой из описанных выше смесей или систем последовательность прямого повтора может содержать один или более связывающих белки РНК-аптамеров. Один или более аптамеров могут находиться в тетрапетле. Один или более аптамеров могут обладать способностью связывать белок оболочки бактериофага MS2.
[00360] В любых из описанных выше составов или систем клетка может являться эукариотической клеткой или прокариотической клеткой: где комплекс CRISPR функционален в клетке и фермент комплекса CRISPR обладает сниженной способностью модифицировать один или более локусов, не являющихся мишенями, по сравнению с немодифицированным ферментом, и/или этот фермент в комплексе CRISPR обладает увеличенной способностью модифицировать один или более локусов-мишеней по сравнению с немодифицированным ферментом.
[00361] Изобретение также относится к комплексу CRISPR любой из описанных выше композиций или любой из описанных выше систем.
[00362] Изобретение также относится к способу модификации представляющего интерес локуса в клетке, включающему приведение в контакт клетки с любым из описанных в настоящем описании сконструированных способами инженерии ферментов CRISPR (например, сконструированные способами инженерии C2c1 или C2c3), композициями или любыми описанными в настоящем описании системами или векторными системами, либо клетка содержит любой из описанных в настоящем описании комплексов CRISPR. В случае таких методов клетка может являться эукариотической клеткой или прокариотической клеткой. В случае таких способов клетка может представлять собой прокариотическую клетку или эукариотическую клетку, предпочтительно эукариотическую клетку. В таких способах организм может содержать клетку. В таких способах организм может не быть человеком или другим животным.
[00363] Любой из таких способов может быть реализован ex vivo или in vitro.
[00364] В некоторых вариантах осуществления изобретения нуклеотидная последовательность, кодирующая по меньшей мере одно из упомянутой направляющей РНК или белка Cas, функционально связана в клетке с регуляторным элементом, содержащим промотор представляющего интерес гена, причем экспрессия по меньшей мере одного компонента системы CRISPR-Cas индуцируется промотором представляющего интерес гена. "Функционально связанный" означает, что нуклеотидная последовательность, кодирующая направляющую РНК и/или белок Cas, связана с регуляторным элементом(ами) таким образом, что происходит экспрессия нуклеотидной последовательности, как также упоминается в настоящем описании. Термин "регуляторный элемент" также описан в настоящем описании. Согласно изобретению, регуляторный элемент содержит промотор представляющего интерес гена, предпочтительно такой как промотор эндогенного представляющего интерес гена. В некоторых вариантах осуществления промотор находится в его эндогенном геномном положении. В таких вариантах осуществления нуклеиновая кислота, кодирующая CRISPR и/или Cas, находится в условиях транскрипционной регуляции промотором представляющего интерес гена в его природном геномном положении. В других конкретных вариантах осуществления промотор предоставляется на (отдельной) молекуле нуклеиновой кислоты, такой как вектор или плазмида, или другой внехромосомной нуклеиновой кислоте, т.е. промотор не присутствует в его природном геномном положении. В некоторых вариантах осуществления промотор интегрирован в геном не в его природном геномном положении.
[00365] В любом из подобных способов, указанные модификации включают модулирование экспрессии генов. Упомянутое модулирование экспрессии генов может включать активацию экспрессии генов и/или репрессию экспрессии генов. Соответственно, в одном аспекте изобретение относится к способу модулирования экспрессии генов, где способ включает введение в клетку сконструированного способами инженерии белка или системы CRISPR, как описано в настоящем описании.
[00366] Также изобретение относится к способу лечения заболевания, расстройства или инфекции в индивидуума, нуждающемуся в этом, включающему введение эффективного количества любого из сконструированных способами инженерии ферментов CRISPR (например, C2c1 или C2c3), композиций, систем или комплексов CRISPR, описываемых в настоящем описании. Заболевания, расстройства или инфекции могут включать вирусную инфекцию. Вирусная инфекция может являться вирусом гепатита В (HBV).
[00367] Изобретение также относится к использованию одного из сконструированных способами инженерии ферментов CRISPR (например, C2c1 или C2c3), композиций, систем или комплексов CRISPR, описанных выше, для генной или геномной инженерии.
[00368] Изобретение также относится к способу изменения экспрессии представляющего интерес геномного локуса в клетке млекопитающего, включая взаимодействие клетки со сконструированными способами инженерии ферментами CRISPR (например, C2c1 или C2c3), смесями, системами или комплексами CRISPR, описанными в настоящем описании, и тем самым доставку CRISPR-Cas (вектор) и обеспечения возможности образоваться комплексу CRISPR-Cas и связаться с мишенью, и определение, изменяется ли экспрессия изменяемого геномного локуса, например, происходит ли увеличение или снижение экспрессии или модификация продукта гена.
[00369] Изобретение также относится к любому сконструированному способами инженерии ферменту CRISPR (например, C2c1 или C2c3), композиции, системе или комплексу CRISPR, описанным выше, для терапевтического применения. Терапевтическое применение может быть предназначено для редактирования гена или генома или для генной терапии.
[00370] В некоторых вариантах осуществления активность искусственных ферментов CRISPR (например, C2c1 или C2c3), описанных в настоящем описании, включает расщепление геномной ДНК, необязательно приводящее к снижению транскрипции гена.
[00371] В одном аспекте изобретение относится к выделенной клетке с измененной экспрессией генного локуса способом, описанным в настоящем описании, где экспрессия изменяется по сравнению с клеткой, не подвергнутой изменению экспрессии генного локуса этим способом. В родственном аспекте изобретение относится к клеточной линии, полученной из такой клетки.
[00372] В одном аспекте изобретение относится к способу модификации организма или не являющегося человеком организма путем манипуляции с последовательностью-мишенью в представляющем интерес геномном локусе, например, гемопоэтической стволовой клетки (HSC), в частности, в которой представляющий интерес геномный локус ассоциирован с мутацией, связанной с аберрантной экспрессией белка или с заболеванием или состоянием, включающему:
доставку в HSC, например, за счет приведения HSC в контакт с частицей, содержащей не встречающуюся в природе композицию, включающую в себя:
полинуклеотидную последовательность направляющей РНК системы CRISPR-Cas, содержащую:
а) направляющую последовательность, способную гибридизоваться с последовательностью-мишенью в HSC;
б) прямой повтор; и
фермент CRISPR, необязательно содержащий по меньшей мере одну или более последовательностей ядерной локализации;
где направляющая последовательность направляет специфическое для последовательности связывание комплекса CRISPR с последовательностью-мишенью, и
где комплекс CRISPR содержит фермент CRISPR в комплексе с (1) направляющей последовательностью, которая гибридизуется с последовательностью-мишенью, и
указанный способ необязательно может включать доставку матрицы для HDR, например, посредством частицы, контактирующей с HSC, содержащей матрицу для HDR, или посредством контакта HSC с другой частицей, содержащей матрицу HDR, причем матрица для HDR обеспечивает экспрессию нормальной или менее аберрантной формы белка; где "нормальная форма" представляет собой белок дикого типа и "аберрантная форма" представляет собой белок, вызывающий развитие заболевания или состояния, и
необязательно способ может включать выделение или получение HSC из организма или не являющегося человеком организма, необязательно увеличение популяции HSC, приведения частиц(ы) в контакт с HSC для получения популяции модифицированных HSC, необязательно увеличение популяции модифицированных HSC и необязательно введение модифицированных HSC в организм или не являющийся человеком организм.
[00373] Одним из аспектов изобретения является способ модификации организма или не являющегося человеком организма путем манипуляции с последовательностью-мишенью в представляющем интерес геномном локусе и, в частности, в HSC, например, в случае, когда представляющий интерес геномный локус ассоциирован с мутацией, связанной с аберрантной экспрессией белка или заболеванием или состоянием, включая доставку к HSC, в частности, путем связывания с HSC частиц, содержащих не встречающуюся в природе или сконструированную способами инженерии композицию, которая содержит:
I.(а) направляющую последовательность, способную гибридизоваться с последовательностью-мишенью в HSC, и
(b) по меньшей мере один или более прямых повторов, и
II. фермент CRISPR, необязательно содержащий один или более NLS, причем направляющая последовательность направляет специфическое для последовательности связывание комплекса CRISPR, включающего фермент CRISPR в комплексе с направляющей последовательностью, которая гибридизуется с последовательностью-мишенью, и
указанный способ необязательно может включать доставку матрицы для HDR, например, посредством частицы, контактирующей с HSC, содержащей матрицу для HDR, или посредством контакта HSC с другой частицей, содержащей матрицу HDR, причем матрица для HDR обеспечивает экспрессию нормальной или менее аберрантной формы белка; где "нормальная форма" представляет собой белок дикого типа и "аберрантная форма" представляет собой белок, вызывающий развитие заболевания или состояния, и
необязательно способ может включать выделение или получение HSC из организма или не являющегося человеком организма, необязательно увеличение популяции HSC, приведения частиц(ы) в контакт с HSC для получения популяции модифицированных HSC, необязательно увеличение популяции модифицированных HSC и необязательно введение модифицированных HSC в организм или не являющийся человеком организм.
[00374] Доставляться может один или более полинуклеотидов, кодирующих любой один или более или все из комплексов CRISPR, предпочтительно связанных с одним или более регуляторными элементами для экспрессии in vivo, например, посредством частиц, содержащих вектор, содержащий полинуклеотид(ы), функционально связанный с регуляторным элементом(и). Любая из этих полинуклеотидных последовательностей, кодирующая фермент CRISPR, направляющую последовательность, прямой повтор, может представлять собой РНК. Понятно, что если упомянут полинуклеотид, являющийся РНК, про который указано, что он включает признак, такой как прямой повтор, то РНК также включает этот признак. Если полинуклеотид представляет собой ДНК и про него указано, что он включает признак, такой как последовательность прямого повтора, то последовательность ДНК транскрибируется или может быть транскрибирована в РНК, включающую этот признак, о которой идет речь. Когда признак представляет собой белок, такой как фермент CRISPR, упоминаемая последовательность ДНК или РНК является или может быть транслированной (и в случае ДНК - сначала транскрибированной)
[00375] В некоторых вариантах осуществления изобретение относится к способу модификации организма, в частности, млекопитающего, включая млекопитающее или организм, являющиеся или не являющиеся человеком, с помощью манипуляции с последовательностью-мишенью в представляющем интерес геномном локусе и, в частности, в HSC, например, в случае, когда представляющий интерес геномный локус ассоциирован с мутацией, связанной с аберрантной экспрессией белка или заболеванием или состоянием, включая доставку в HSC, в частности, путем приведения HSC в контакт с не встречающейся в природе или сконструированной способами инженерии композиции, которая содержит одну или более частиц, содержащих вирусный вектор (вирусные векторы), плазмидный вектор (плазмидные векторы) или вектор(ы) с молекулой нуклеиновой кислоты (например, РНК), функционально кодирующие указанную композицию, для ее экспрессии, при этом композиция включает:
(А) I. первый регуляторный элемент, функционально связанный с полинуклеотидной последовательностью РНК системы CRISPR-Cas, где полинуклеотидная последовательность содержит
(а) направляющую последовательность, способную гибридизоваться с последовательностью-мишенью в эукариотической клетке,
(b) последовательность прямого повтора, и
II. второй регуляторный элемент, функционально связанный с кодирующей последовательностью, кодирующей фермент CRISPR, содержащий по меньшей мере один или более сигналов ядерной локализации (или необязательно по меньшей мере один или более сигналов ядерной локализации (NLS), поскольку некоторые варианты осуществления могут не включать NLS)
причем (а), (b) и (в) расположены в ориентации от 5' к 3'-концу, причем компоненты I и II расположены на одной или разных векторах системах, при этом транскрибированная направляющая последовательность направляет специфическое для последовательности связывание комплекса CRISPR с последовательностью-мишенью, причем комплекс CRISPR содержит фермент CRISPR в комплексе с направляющей РНК, гибридизованной с последовательностью-мишенью, или
(B) не встречающуюся в природе или сконструированную способами инженерии композицию, содержащую векторную систему, включающую один или более векторов, содержащих:
I. первый регуляторный элемент, функционально связанный с
(а) направляющей последовательностью, способной гибридизоваться с целевой последовательностью в эукариотической клетке,
(b) по меньшей мере одну или более последовательностей прямого повтора, и
II. второй регуляторный элемент, функционально связанный с кодирующей фермент последовательностью, кодирующей CRISPR-фермент, и необязательно, если применимо,
где компоненты I и II расположены на одном или разных векторах системы, при этом транскрибированная направляющая последовательность направляет специфическое для последовательности связывание комплекса CRISPR с последовательностью-мишенью, причем комплекс CRISPR содержит фермент CRISPR в комплексе с направляющей последовательностью, которая гибридизуется с последовательностью-мишенью; способ необязательно может включать доставку матрицы для HDR, например, посредством частицы, контактирующей с HSC, содержащей матрицу для HDR, или посредством контакта HSC с другой частицей, содержащей матрицу HDR, причем матрица для HDR обеспечивает экспрессию нормальной или менее аберрантной формы белка; где "нормальная форма" представляет собой белок дикого типа и "аберрантная форма" представляет собой белок, вызывающий развитие заболевания или состояния, и необязательно способ может включать выделение или получение HSC из организма или не являющегося человеком организма, необязательно увеличение популяции HSC, приведения частиц(ы) в контакт с HSC для получения популяции модифицированных HSC, необязательно увеличение популяции модифицированных HSC и необязательно введение модифицированных HSC в организм или не являющийся человеком организм. В некоторых вариантах осуществления компоненты I, II и III расположены на одном векторе. В некоторых вариантах осуществления компоненты I и II расположены на одном векторе, в то время как компонент III расположен на другом векторе. В некоторых вариантах осуществления компоненты I и III расположены на одном векторе, в то время как компонент II расположен на другом векторе. В некоторых вариантах осуществления компоненты II и III расположены на одном векторе, в то время как компонент I расположен на другом векторе. В некоторых вариантах осуществления компоненты I, II и III расположены на разных векторах. Изобретение также относится к вирусной или плазмидной векторной системе, описанную в настоящем описании.
[00376] Под манипуляциями с последовательностью-мишень заявители также подразумевают эпигенетические манипуляции с последовательностью-мишенью. Это может быть манипуляция с состоянием хроматина последовательности-мишени, такая как модификация состояния метилирования последовательности-мишени (т.е. внесение или устранение метилирования или паттерна метилирования или CpG-островков), модификация гистонов, повышение или снижение доступности последовательности-мишени или облегчение сворачивания в трехмерную структуру. Понятно, что если упоминается способ модификации организма или млекопитающего, включая организм человека или организм млекопитающего, не являющегося человеком, с помощью манипуляций с последовательностью-мишенью в представляющем интерес геномном локусе, подразумевается, что это может относиться к организму (или млекопитающему) в целом или только к одной клетке или популяции клеток из этого организма (если организм многоклеточный). В случае человека, к примеру, заявители предусматривают, среди прочего, единственную клетку или популяцию клеток, и они могут быть предпочтительно модифицированы ex vivo и затем введены обратно. В этом случае может быть необходима биопсия или другая ткань или биологическая жидкость. Стволовые клетки также предпочтительны в этом отношении. Но, разумеется, также предусматриваются варианты осуществления in vivo. Это изобретение в особенности обладает преимуществами в случае HSC.
[00377] В некоторых вариантах осуществления изобретение относится к способу модификации организма или не являющегося человеческим организма с помощью манипуляции с первой и второй последовательностями-мишенями на противоположных цепях дуплекса ДНК в представляющем интерес геномном локусе HSC, в частности, в частности, где представляющий интерес геномный локус ассоциирован с мутацией, связанной с аберрантной белковой экспрессией или с заболеванием или синдромом, включающему доставку, например, посредством приведения HSC в контакт с частицей(ами), содержащей не встречающуюся в природе или сконструированную способами инженерии композицию, содержащую:
I. полинуклеотидную последовательность направляющей РНК первой системы CRISPR-Cas (например, C2c1 или C2c3), причем первая полинуклеотидная последовательность содержит:
(a) первую направляющую последовательность, способную гибридизоваться с первой последовательностью-мишеью,
(b) первую последовательность прямого повтора, и
II. полинуклеотидную последовательность направляющей РНК второй системы CRISPR-Cas (например, C2c1 или C2c3), причем вторая полинуклеотидная последовательность содержит:
(a) вторую направляющую последовательность, способную гибридизоваться со второй последовательностью-мишенью,
(b) вторую последовательность прямого повтора, и
III. полинуклеотидную последовательность, кодирующую фермент CRISPR, содержащий по меньшей мере один или более сигналов ядерной локализации и содержащий одну или более мутаций, причем (а), (b) и (c) расположены в порядке от 5' к 3'-концу; или
IV. продукт(ы) экспрессии одного или более компонентов от I до III, например, первую и вторую последовательность прямого повтора, фермент CRISPR;
при этом при транскрипции первая и вторая направляющая последовательности направляют специфическое для последовательности связывание первого и второго комплекса CRISPR с первой и второй последовательностью-мишенью, соответственно, причем первый комплекс CRISPR включает фермент CRISPR в комплексе с (1) первой направляющей последовательностью, которая гибридизуется с первой последовательностью-мишенью, и второй комплекс CRISPR включает фермент CRISPR в комплексе со (1) второй направляющей последовательностью, которая гибидизуется со второй последовательностью-мишенью, причем полинуклеотидная последовательность, кодирующая CRISPR-фермент, может являться ДНК или РНК, при этом первая направляющая последовательность направляет расщепление одной цепи дуплекса ДНК вблизи первой целевой последовательности, а вторая направляющая последовательность направляет расщепление другой цепи дуплекса ДНК вблизи второй целевой последовательности, вызывая образование двуцепочечного разрыва, таким образом, модифицируя организм или организм, не принадлежащий человеку; метод может необязательно включать доставку матрицы для репарации путем гомологичной рекомбинации, в частности, посредством частиц, связывающихся с HSC или связывающихся с HSC посредством другой частицы и содержащих матрицу для репарации путем гомологичной рекомбинации, причем матрица для репарации путем гомологичной рекомбинации обеспечивает экспрессию нормальной или менее аберрантной формы белка; при этом «нормальная форма» - это белок дикого типа, и «аберрантная форма» - это белок, вызывающий развитие болезни или синдрома; также необязательно метод может включать выделение или получение HSC из организма или организма, не являющегося человеком, необязательно - размножение популяции HSC, проведение связывания частиц(ы) с HSC для получения популяции модифицированных HSC, необязательно - размножение популяции модифицированных HSC, необязательно - введение модифицированных HSC в организм или организм, не являющийся человеком. В некоторых методах изобретения любой или все полинуклеотидные последовательности кодируют CRISPR-фермент, первую и вторую направляющая последовательности, первый и второй прямые повторы. В дальнейших реализациях изобретения полинуклеотиды, кодирующие последовательность, кодирующую CRISPR-фермент, первую и вторую направляющая последовательности, первый и второй прямые повторы, являются РНК и будут доставляться посредством липосом, наночастиц, экзосом, микровезикул или генной пушки, но обладает наибольшими преимуществами доставка посредством частиц. В конкретных реализациях изобретения последовательности первого и второго повторов обладают 100% уровнем идентичности. В некоторых реализациях изобретения полинуклеотиды могут содержаться в векторной системе, включающей один или более векторов. В предпочтительных вариантах реализации первый CRISPR-фермент содержит одну или более мутации, такие что фермент является никазой комплементарной цепи, и второй CRISPR-фермент содержит одну или более мутации, такие что фермент является никазой некомплементарной цепи. Альтернативно, первый CRISPR-фермент содержит одну или более мутации, такие что фермент является никазой некомплементарной цепи, и второй CRISPR-фермент содержит одну или более мутации, такие что фермент является никазой комплементарной цепи. В предпочтительных методах изобретения первая направляющая последовательность, направляющая расщепление одной цепи дуплекса ДНК вблизи первой целевой последовательности, и вторая направляющая последовательность, направляющая расщепление одной цепи дуплекса ДНК вблизи второй целевой последовательности, приводят к образованию липких концов с 5'- выступающими нуклеотидами (оверхэнгов). В реализациях изобретения липкий конец с 5'- выступающими нуклеотидами составляет почти 200 пар оснований, предпочтительно, не более 100 пар оснований или, еще более предпочтительно, не более 50 пар оснований. В реализациях изобретения липкий конец с 5'- выступающими нуклеотидами (оверхэнг) составляет не менее 26 пар оснований, предпочтительно, не менее 30 пар оснований и, наиболее предпочтительно, 34-50 пар оснований.
[00378] Данное изобретение в некоторых вариантах осуществления предусматривает способ модификации организма или организма, не являющегося человеческим, с помощью манипуляции с первой и второй целевыми последовательностями на противоположных цепях дуплекса ДНК в представляющем интерес геномном локусе HSC, в частности, в частности, в которой интересующий геномный локус ассоциирован с мутацией, связанной с аберрантной белковой экспрессией или с заболеванием или синдромом, включая достаку, например, за счет связывания HSC с частицей (частицами), содержащими не встречающуюся в природе или искусственную композицию, включающей:
I. первый регуляторный элемент, функционально связанный с
(а) первой направляющей последовательностью, способной гибридизоваться с первой целевой последовательностью в эукариотической клетке,
(b) по меньшей мере одну или более последовательность прямого повтора, и
II. второй регуляторный элемент, функционально связанный со
(а) второй направляющей последовательностью, способной гибридизоваться со второй целевой последовательностью в эукариотической клетке,
(b) по меньшей мере одну или более последовательность прямого повтора, и
III. третий регуляторный элемент, функционально связанный с кодирующей фермент последовательностью, кодирующей CRISPR-фермент (например, C2el или C2c3),
IV. продукт экспрессии одного или более компонентов от I до IV, в частности, первый и второй прямые повторы, CRISPR-фермент;
при этом компоненты I, II, III и IV расположены на одном или разных векторах системы, где при транскрипции первая и вторая направляющая последовательности направляют специифичное для последовательности связывание первого и второго CRISPR-комплексов к первой и второй целевым последовательностям, соответственно, причем первый CRISPR-комплекс включает CRISPR-фермент в комплексе с первой направляющей последовательностью, гибридизованной с первой целевой последовательностью, и второй CRISPR-комплекс включает CRISPR-фермент в комплексе со второй направляющей последовательностью, гибридизованной со второй целевой последовательностью, причем полинуклеотидная последовательность, кодирующая CRISPR-фермент, может являться ДНК или РНК, при этом первая направляющая последовательность направляет расщепление одной цепи дуплекса ДНК вблизи первой целевой последовательности, а вторая направляющая последовательность направляет расщепление другой цепи дуплекса ДНК вблизи второй целевой последовательности, вызывая образование двуцепочечного разрыва, таким образом, модифицируя организм или организм, не принадлежащий человеку; метод может необязательно включать доставку матрицы для репарации путем гомологичной рекомбинации, в частности, посредством частиц, связывающихся с HSC или связывающихся с HSC посредством другой частицы и содержащих матрицу для репарации путем гомологичной рекомбинации, причем матрица для репарации путем гомологичной рекомбинации обеспечивает экспрессию нормальной или менее аберрантной формы белка; при этом «нормальная форма» - это белок дикого типа, и «аберрантная форма» - это белок, вызывающий развитие болезни или синдрома; также необязательно метод может включать выделение или получение HSC из организма или организма, не являющегося человеком, необязательно - размножение популяции HSC, проведение связывания частиц(ы) с HSC для получения популяции модифицированных HSC, необязательно - размножение популяции модифицированных HSC, необязательно - введение модифицированных HSC в организм или организм, не являющийся человеком.
[00379] Изобретение также предоставляет векторную систему, описанную здесь. Система может включать один, два, три или четыре различных вектора, со всеми комбинациями локализации предусмотренных компонентов, например, компоненты I, II, III и IV могут находиться в одном векторе; компоненты I, II, III и IV могут находиться на разных векторах; компоненты I, II, III и IV могут находиться на двух или трех разных векторах, в любых возможных комбинациях. В некоторых методах изобретения любая или все полинуклеотидные последовательности, кодирующие CRISPR-фермент, первая и вторая направляющая последовательности, первый и второй прямые повторы - это РНК. В дальнейших реализациях изобретения последовательности первого и второго повторов обладают 100% уровнем идентичности. В предпочтительных вариантах реализации, первый CRISPR-фермент содержит одну или более мутации, при которых (которой) фермент является никазой комплементарной цепи, и второй CRISPR-фермент содержит одну или более мутации, при которых (которой) фермент является никазой некомплементарной цепи. Альтернативно, первый фермент может быть никазой некомплементарной цепи, и второй фермент может быть никазой некомплементарной цепи. В дальнейших реализациях изобретения, один или более вирусных векторов будут доставляться посредством липосом, наночастиц, экзосом, микровезикул или генной пушки; но доставка частицами обладает наибольшими преимуществами.
[00380] В предпочтительных способах данного изобретения первая направляющая последовательность, направляющая расщепление одной цепи дуплекса ДНК вблизи первой целевой последовательности, и вторая направляющая последовательность, направляющая расщепление одной цепи дуплекса ДНК вблизи второй целевой последовательности, приводят к образованию липкого конца с 5'- выступающими нуклеотидами (оверхэнга). В реализациях изобретения липкий конец с 5'- выступающими нуклеотидами (оверхэнг) составляет почти 200 пар оснований, предпочтительно, не более 100 пар оснований или, еще более предпочтительно, не более 50 пар оснований. В реализациях изобретения липкий конец с 5'- выступающими нуклеотидами (овехэнг) составляет не менее 26 пар оснований, предпочтительно, не менее 30 пар оснований и, наиболее предпочтительно, 34-50 пар оснований.
[00381] Изобретение в некоторых вариантах осуществления предусматривает способ модификации интересующего геномного локуса, в частности, HSC, например, тогда, когда интересующий геномный локус ассоциирован с мутацией, связанной с аберрантной экспрессией белка или заболеванием или синдромом, включая доставку к HSC частицей (частицами), содержащей белок Cas, содержащий одну или более мутации и две гРНК, мишенями которых являются первая и вторая цепи молекулы ДНК в HSC, соответственно, причем гРНК нацеливает на молекулу ДНК, и белок Cas вносит ник-разрывы в каждую (первую и вторую) цепь молекулы ДНК, причем мишень в HSC изменяется; и при этом белок Cas и две направляющие РНК не встречаются в природе вместе, и метод может необязательно включать доставку матрицы для репарации путем гомологичной рекомбинации, например, посредством частиц, связывающихся с HSC или связывающихся с HSC посредством другой частицы и содержащих матрицу для репарации путем гомологичной рекомбинации, причем матрица для репарации путем гомологичной рекомбинации обеспечивает экспрессию нормальной или менее аберрантной формы белка; при этом «нормальная форма» - это белок дикого типа, и «аберрантная форма» - это белок, вызывающий развитие болезни или синдрома, и необязательно: способ может включать выделение или получение HSC из организма или организма, не являющегося человеком, необязательно - размножение популяции HSC, проведение связывания частиц(ы) с HSC для получения популяции модифицированных HSC, необязательно - размножение популяции модифицированных HSC, необязательно - введение модифицированных HSC в организм или организм, не являющийся человеком. В предпочтительных методах изобретения никазная активность белка Cas в отношении каждой (первой и второй) цепей молекулы ДНК приводит к образованию липкого конца с 5'- выступающими нуклеотидами (оверхэнга). В реализациях изобретения липкий конец с 5'- выступающими нуклеотидами (оверхэнг) составляет не менее 26 пар оснований, предпочтительно, не менее 30 пар оснований и, наиболее предпочтительно, 34-50 пар оснований. В аспекте изобретения кодонный состав белка Cas оптимизирован для экспрессии в эукариотической клетке, предпочтительно в клетке млекопитающего или человеческой клетке. Аспекты изобретения связаны со снижением экспрессии продукта гена или дальнейшим введением матричного полинуклеотида в молекулу ДНК, кодирующую продукт гена, или точным вырезанием прерывающей последовательности, позволяющим отжечься друг на друга двум 5'-оверхенгам с последующим лигированием, или изменением активности или функции продукта гена или увеличением экспрессии продукта гена. При реализации изобретения продукт гена является белком.
[00382] В некоторых вариантах осуществления изобретение предусматривает метод модификации интересующего геномного локуса, например, в HSC, в частности, в которой интересующий геномный локус ассоциирован с мутацией, связанной с аберрантной белковой экспрессией или с заболеванием или синдромом, включая доставку, например, за счет связывания HSC с частицей (частицами), включающей:
а) первый регуляторный элемент, функционально связанный с каждой из двух направляющих РНК CRISPR-Cas систем которые нацелены, соответственно, на первую и вторую цепи двуцепочечной молекулы ДНК в HSC, и
b) второй регуляторный элемент, функционально связанный с белком Cas (например, C2c1 или C2c3), или
в) продукт(ы) экспрессии a) или b),
причем компоненты (а) и (b) расположены на одном или разных векторах системы, мишенью гРНК является молекула ДНК HSC, и белок Cas вносит ник-разрывы в каждую (первую и вторую) цепь молекулы ДНК HSC; белок Cas и две гРНК не встречаются в природе вместе; способ может дополнительно включать доставку матрицы для репарации путем гомологичной рекомбинации, например, посредством частиц, связывающихся с HSC или связывающихся с HSC посредством другой частицы и содержащих матрицу для репарации путем гомологичной рекомбинации, причем матрица для репарации путем гомологичной рекомбинации обеспечивает экспрессию нормальной или менее аберрантной формы белка; при этом «нормальная форма» - это белок дикого типа, и «аберрантная форма» - это белок, вызывающий развитие болезни или синдрома, и необязательно: способ может включать выделение или получение HSC из организма или организма, не являющегося человеком, необязательно - размножение популяции HSC, проведение связывания частиц(ы) с HSC для получения популяции модифицированных HSC, необязательно - размножение популяции модифицированных HSC, необязательно - введение модифицированных HSC в организм или организм, не являющийся человеком. В аспекте изобретения направляющие РНК могут содержать направляющие последовательности, слитые с последовательностями прямых повторов. Аспекты изобретения связаны со снижением экспрессии продукта гена или дальнейшим введением матричного полинуклеотида в молекулу ДНК, кодирующую продукт гена, или точным вырезанием прерывающей последовательности, позволяющим отжечься друг на друга двум 5'-оверхенгам с последующим лигированием, или изменением активности или функции продукта гена или увеличением экспрессии продукта гена. В реализации изобретения продукт гена является белком. В предпочтительных вариантах осуществления вектора системы являются вирусными векторами. В дальнейших реализациях изобретения полинуклеотиды, кодирующие последовательность, кодирующую CRISPR-фермент, первую и вторую направляющая последовательности, первый и второй прямые повторы, являются РНК и будут доставляться посредством липосом, наночастиц, экзосом, микровезикул или генной пушки, но обладает преимуществами доставка посредством частиц. В одном аспекте относится к способу модификации целевого полинуклеотида в HSC. В некоторых вариантах осуществления метод позволяет CRISPR-комплексу связываться с целевым полинуклеотидом, чтобы обеспечить расщепление целевого полинуклеотида, таким образом, модифицируя целевый полинуклеотид, причем CRISPR-комплекс содержит CRISPR-фермент в комплексе с направляющей последовательностью, способной гибридизоваться с целевой последовательностью в составе указанного целевого полинуклеотида; при этом указанная направляющая последовательность соединена с последовательностью прямого повтора. В некоторых вариантах осуществления упомянутое расщепление включает расщепление одной или двух цепей в месте локализации целевой последовательности упомянутым CRISPR-ферментом. В некоторых вариантах осуществления упомянутое расщепление приводит к снижению уровня транскрипции целевого гена. В некоторых вариантах осуществления способ далее включает репарацию упомянутого расщепленного целевого полинуклеотида за счет гомологичной рекомбинации с экзогенным матричным полинуклеотидом, причем упомянутая репарация приводит к мутации, включающей вставку, делецию или замену одного или более нуклеотидов в упомянутом целевом полинуклеотиде. В некоторых вариантах осуществления указанная мутация приводит к одной или более аминокислотным заменам в образующемся белке при экспрессии гена, содержащего целевую последовательность. В некоторых вариантах осуществления метод включает введение одного или более векторов или последующая экспрессия продукта (продуктов) в HSC, в частности, посредством частиц(ы), причем один или более векторов индуцируют экспрессию одного или более компонентов из списка: CRISPR-фермента, направляющую последовательность, соединенную с последовательностью прямого повтора. В некоторых вариантах осуществления упомянутые векторы доставляются, в частности, к HSC субъекта. В некоторых вариантах осуществления реализуются указанные модификации HSC в клеточной культуре. В некоторых вариантах осуществления метод далее включает выделение упомянутых HSC из субъекта перед упомянутыми модификациями. В некоторых вариантах осуществления метод далее включает введение упомянутых HSC и/или клеток, полученных из них, обратно упомянутому субъекту.
[00383] В одном из вариантов относится к способу получения, в частности, HSC, содержащих мутантный ген, ассоциированный с заболеванием. В некоторых вариантах осуществления такой ген, ассоциированный с заболеванием, может быть ассоциирован с риском возникновения или развития заболевания. В некоторых вариантах осуществления метод включает (а) введение одного или более векторов или последующая экспрессия продукта (продуктов), в частности, посредством частиц, в HSC, причем один или более векторов индуцируют экспрессию одного или более компонентов из списка: CRISPR-фермента, направляющую последовательность, соединенную с последовательностью прямого повтора; и (b) обеспечение связывания CRISPR-комплекса с целевым полинуклеотидом, способствующим расщепление целевого полинуклеотида внутри упомянутого гена, асоциированного с заболеванием, причем CRISPR-комплекс включает CRISPR-фермент в комплексе с направляющей последовательностью, способной гибридизоваться с целевой последовательностью в составе указанного целевого полинуклеотида, и, необязательно, если это применимо, таким образом, приводя к получению HSC, содержащих мутантный ген, ассоциированный с заболеванием. В некоторых вариантах осуществления упомянутое расщепление включает расщепление одной или двух цепей в месте локализации целевой последовательности упомянутым CRISPR-ферментом. В некоторых вариантах осуществления упомянутое расщепление приводит к снижению уровня транскрипции целевого гена. В некоторых вариантах осуществления способ далее включает репарацию упомянутого расщепленного целевого полинуклеотида за счет гомологичной рекомбинации с экзогенным матричным полинуклеотидом, причем упомянутая репарация приводит к мутации, включающей вставку, делецию или замену одного или более нуклеотидов в упомянутом целевом полинуклеотиде. В некоторых вариантах осуществления упомянутая мутация приводит к одной или более аминокислотным заменам в образующемся белке при экспрессии гена, содержащего целевую последовательность. В некоторых вариантах осуществления модифицированные HSC вводятся в животное, что, таким образом, приводит к получению модельного организма.
[00384] В одном из аспектов относится к способу модифицикации целевого полинуклеотида в, к примеру, HSC. В некоторых вариантах осуществления метод позволяет CRISPR-комплексу связываться с целевым полинуклеотидом, чтобы обеспечить расщепление целевого полинуклеотида, таким образом, модифицируя целевый полинуклеотид, причем CRISPR-комплекс содержит CRISPR-фермент в комплексе с направляющей последовательностью, способной гибридизоваться с целевой последовательностью в составе указанного целевого полинуклеотида, при этом указанная направляющая последовательность соединена с последовательностью прямого повтора. В других вариантах реализации изобретение предоставляет модификацию экспрессии полинуклеотида в эукариотической клетке, котопая происходит, например, от HSC. Способ изобретения включает увеличение или снижение экспрессии целевого полинуклеотида с использованием CRISPR-комплекса, который связывается с полинуклеотидом в HSC; обладает наибольшими преимуществами доставка CRISPR-комплекса частицей (частицами).
[00385] В некоторых способах целевый полинуклеотид может быть инактивирован, чтобы действовать на экспрессию в, например, гемапоэтических стволовых клетках. Например, при связывании CRISPR-комплекса с целевой последовательностью в клетке целевый полинуклеотид инактивируется, так что последовательность не траскрибируется, кодируемый белок не вырабатывается или последовательность не функционирует таким образом, как функционирует последовательность дикого типа.
[00386] В некоторых вариантах осуществления изобретения, указанная РНК CRISPR-Cas-системы, например, гРНК может быть модифицирована; например, включать аптамеры и функциональный домен. Аптамер - это синтетический нуклеотид, который связывается со специфической молекулой-мишенью; например, молекула нуклеиновой кислоты, которая была создана путем повторных раундов селекции in vitro, или методом SELEX (systematic evolution of ligands by exponential enrichment, систематическая эволюция лигандов экспоненциальным обогащением), чтобы связывать различные молекулярные мишени, такие как малые молекулы, белки, нуклеиновые кислоты и даже клетки, ткани и организмы. Аптамеры удобны для использования, поскольку характеристики узнавания ими молекул сравнимы с таковыми у антител. В дополнение к их способности к тонкому распознаванию, преимущества аптамеров перед антителами включают низкую или отсутствующую иммуногенность при терапевтическом применении. Соответственно, при применении изобретения, фермент и/или РНК могут включать функциональный домен.
[00387] В некоторых вариантах осуществления функциональный домен представляет собой трансактивационный домен, предпочтительно VP64. В некоторых вариантах осуществления изобретения функциональный домен представляет собой домен репрессии транскрипции, предпочтительно KRAB. В некоторых вариантах осуществления изобретения домен репрессии транскрипции представляет собой SID или конкатемеры SID (например, SID4X). В некоторых вариантах осуществления изобретения функциональный домен является эпигенетическим доменом модификации, обеспечивающим работу эпигенетического фермента модификации. В некоторых вариантах осуществления изобретения функциональный домен представляет собой доме активации, который может быть доменом активации P65. В некоторых вариантах осуществления функциональный домен может обладать нуклеазной активностью. В одном из вариантов осуществления функциональный домен включает Fok1.
[00388] Изобретение также относится к клетке in vitro или ex vivo, содержащей любой из модифицированных ферментов CRISPR, композиций или комплексов, описанных выше, полученные любыми из вышеописанных способов. Клетка может быть как эукариотической, так и прокариотической. Изобретение также относится к потомству таких клеток. Изобретение также относится к продукту любой клетки или любого такого потомства, где продукт является продуктом указанного одного или более локусов-мишеней, модифицированных ферментом CRISPR или комплексом CRISPR. Продукт может представлять собой пептид, полипептид или белок. Некоторые из этих продуктов могут быть модифицированы модифицированным ферментом CRISPR или комплексом CRISPR. В случае некоторых таких модифицированных продуктов продукт локуса-мишени физически отличается от продукта указанного локуса-мишени, который не был модифицирован модифицированным ферментом CRISPR или комплексом CRISPR.
[00389] Изобретение также относится к полинуклеотидной молекуле, содержащей полинуклеотидную последовательность, кодирующую любой из не встречающихся в природе ферментов CRISPR, описанных выше.
[00390] Любой такой полинуклеотид может далее содержать один или более регуляторных элементов, которые функционально связаны с полинуклеотидной последовательностью, кодирующей ферменты CRISPR, не встречающиеся в природе.
[00391] В любом таком полинуклеотиде, который содержит один или более регуляторных элементов, один или более регуляторных элементов могут быть функционально организован для экспрессии не встречающегося в природе фермента CRISPR в эукариотической клетке. Эукариотическая клетка может быть клеткой человека. Эукариотическая клетка может клеткой грызуна, возможно, клеткой мыши. Эукариотическая клетка может представлять собой дрожжевую клетку. Эукариотическая клетка может быть клеткой яичника китайского хомячка (CHO). Эукариотическая клетка может быть клеткой насекомого.
[00392] В любом такой полинуклеотиде, который содержит один или более регуляторных элементов, один или более регуляторных элементов могут быть функционально оптимизированы для экспрессии не встречающихся в природе ферментов CRISPR в прокариотической клетке.
[00393] В любом таком полинуклеотиде, который содержит один или более регуляторных элементов, один или более регуляторных элементов могут быть функционально оптимизированы для экспрессии в системе in vitro.
[00394] Изобретение также относится к экспрессирующему вектору, содержащему одну из описанных выше полинуклеотидных молекул. Изобретение также относится к такой полинуклеотидной молекуле(молекулам), например, полинуклеотидным молекулам, функционально организованным для экспрессии компонента(ов), являющегося белом и/или нуклеиновой кислотой, а также к такому вектору(векторам).
[00395] Кроме того, изобретение относится к способу внесения мутаций в Cas (например, C2c1 или C2c3) или мутантный или модифицированный Cas (например, C2c1 или C2c3), который является ортологом ферментов CRISPR, по изобретению, описанному в настоящем описании, включающему определение аминокислоты (аминокислот) в этом ортологе, которая может находиться вблизи или соприкасаться с молекулой нуклеиновой кислоты, например, ДНК, РНК, гРНК и т.д., и/или аминокислоты(аминокислот), аналогичных или соответствующих охарактеризованной в настоящем описании аминокислоте(аминокислотам) в ферментах CRISPR по изобретению, описанным в настоящем описании, для мутаций и/или модификаций, или синтез, или получение, или экспрессию ортолога(ортологов), состоящие или по существу состоящие из модификации(модификаций) и/или мутации(й), или внесение мутации, как описано в настоящем описании, например, модификацию, например, замену или мутацию нейтральной аминокислоты на заряженную, в частности, положительно заряженную аминокислоту, например, аланин. Модифицированный таким образом ортолог может быть использован в системе CRISPR-Cas; и молекула(ы) нуклеиновой кислоты, экспрессирующая его, может быть использована в векторе или других системах доставки, которые доставляют молекулы или кодируют компоненты систем CRISPR-Cas, описанных в настоящем описании.
[00396] В одном из аспектов изобретение обеспечивает эффективную целевую активность и минимизированную нецелевую активность. В одном аспекте изобретение обеспечивает эффективное целевое расщепление белком CRISPR и минимизированное нецелевое расщепление белком CRISPR. В одном аспекте изобретение обеспечивает специфическое для направляющей РНК связывание белка CRISPR в локусе гена без расщепления ДНК. В одном аспекте изобретение обеспечивает эффективное направляемое направляющей РНК целевое связывание белка CRISPR в локусе гена и минимизированное нецелевое связывание белка CRISPR. Соответственно, в этом аспекте изобретение обеспечивает специфическую регуляцию гена-мишени. В одном аспекте изобретение относится к специфическому для направляющей РНК связыванию фермента CRISPR в генном локусе в отсутствие расщепления ДНК. Соответственно, в одном аспекте изобретение обеспечивает расщепление в одном генном локусе и генную регуляцию в другом генном локусе с использованием единственного фермента CRISPR. В одном аспекте изобретение обеспечивает ортогональную активацию и/или ингибирование и/или расщепление множественных мишеней с использованием одного или более белков и/или ферментов CRISPR.
[00397] В другом варианте настоящее изобретение относится к способу функционального скрининга генома в совокупности клеток ex vivo или in vivo, включая введение или экспрессию библиотеки, содержащей множество направляющих РНК (гРНК) системы CRISPR-Cas, и при этом скрининг далее включает использование фермента CRISPR, причем комплекс CRISPR модифицирован так, чтобы содержать гетерологичный функциональный домен. В одном аспекте изобретение относится к способу скринига генома, включающему введение в хозяина или экспрессию в организме-хозяине библиотеки in vivo. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором активатор вносится в организм хозяина или экспрессируется в организме хозяина. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором активатор связан с белком CRISPR. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором активатор связан с N-концом или C-концом белка CRISPR. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором активатор связан с петлей в направляющей РНК. В одном аспекте изобретение относится к способу, описанному в настоящем описании, включающий репрессор, который вносится в организм хозяина или экспрессируется в организме хозяина. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором скрининг включает изменение и обнаружение активации гена, ингибирования гена или расщепления в локусе.
[00398] В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором организм-хозяин представляет собой эукариотическую клетку. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором организм-хозяин представляет собой клетку млекопитающего. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором организм-хозяин представляет собой эукариотическую клетку, не являющуюся клеткой человека. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором эукариотическая клетка, не являющаяся человеческой, представляет собой клетку млекопитающего, не являющегося человеком. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором клетка млекопитающего, не являющегося человеком, может включать, но не ограничиваться ими, клетки приматов, быков, птиц, свиней, собак, грызунов, зайцевых, таких как клетки обезьян, коров, овец, свиньи, собаки, кролика, крысы или мыши. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором клетка может быть клеткой эукариотического организма, не являющегося млекопитающим, такого как домашняя птица (например, цыпленок), позвоночная рыба (например, лосось) или моллюск (например, устрица, лобстер, креветка). В одном аспекте изобретение относится к способу, описанному в настоящем описании, где эукариотическая клетка, не являющаяся человеческой, может представлять собой растительную клетку. Растительная клетка может принадлежать однодольному или двудольному растению или культурному или зерновому растению, такому как маниока, кукуруза, сорго, соя, пшеница, овес или рис. Растительная клетка также может принадлежать водоросли, дереву или продовольственному растению, фрукту или овощу (например, деревья, такие как цитрусовые деревья, в частности, апельсиновые, грейпфрутовые или лимонные; персиковые или нектариновые деревья, яблони или груши; орехи, такие как миндаль, фундук или фисташка; паслен; растения рода Brassiccr, растения рода Lactuca, растения рода Spinada, растения рода Capsicum, хлопок, табак, спаржа, морковь, капуста, брокколи, цветная капуста, томат, баклажан, перец, салат, шпинат, малина, клубника, голубика, черника, виноград, кофе, какао и т.д.).
[00399] В одном из аспектов изобретение относится к способу, описанному в настоящем описании, включающему доставку комплексов CRISPR-Cas или их компонента(компонентов) или молекул(ы) нуклеиновой кислоты, кодирующей их, где указанная нуклеиновая кислота функционально связана с регуляторными последовательностями и экспрессируется in vivo. В этом аспекте изобретение относится к способу, описанному в настоящем описании, в котором экспрессия in vivo происходит посредством лентивируса, аденовируса или аденоассоциированного вируса. В этом варианте изобретение относится к способу, как описано в настоящем описании, в котором доставка происходит через частицу, наночастицу, липид или проникающий в клетку пептид (CPP).
[00400] В конкретных вариантах осуществления может быть интересно направить комплекс CRISPR-Cas в хлоропласт. Во многих случаях это нацеливание может быть осуществлено в присутствии N-концевого удлинения, называемого транзитным пептидом хлоропластов (CTP) или транзитным пептидом пластид. Хромосомные трансгены из бактериальных источников должны иметь последовательность, кодирующую последовательность CTP, слитую с последовательностью, кодирующей экспрессируемый полипептид, если экспрессируемый полипептид должен быть направлен в растительную пластиду (т.е. хлоропласт). Соответственно, локализация экзогенного полипептида в хлоропласте часто достигается за счет функционального связывания полинуклеотидной последовательности, кодирующей последовательность CTP, с 5'-областью полинуклеотида, кодирующего экзогенный пептид. Однако аминокислотная последовательность CTP и близлежащих к N-концу пептида последовательностей может влиять на эффективность процессинга. Другие варианты нацеливания в хлоропласт, которые описаны, представляют собой сигнальную последовательность cab-m7 кукурузы (патент США 7022896, WO 97/41228), сигнальную последовательность глутатион-редуктазы гороха (WO 97/41228) и CTP, описанный в US2009029861.
[00401] В одном аспекте изобретение относится к паре комплексов CRISPR-Cas, каждый из которых содержит направляющую РНК (гРНК), включающую направляющую последовательность, способную к гибридизации с последовательностью-мишенью в представляющем интерес геномном локусе клетки, где по меньшей мере одна петля каждой одиночной направляющей последовательности (sg-РНК) модифицирована вставками отдельной последовательности(последовательностей) РНК, которые связываются с одним или более адаптерными белками, причем адаптерный белок ассоциирован с одним или более функциональными доменами, при этом каждая гРНК каждого комплекса CRISPR-Cas содержит функциональный домен, обладающей активностью расщепления ДНК. В одном аспекте изобретение относится к парным комплексам CRISPR-Cas, таким как описано в настоящем описании, где активность расщепления ДНК обеспечивается нуклеазой Fok1.
[00402] В этом аспекте изобретение относится к способу расщепления последовательности-мишени в представляющем интерес геномном локусе, включающему доставку комплексов CRISPR-Cas или их компонентов(компонента) или молекул(ы) нуклеиновой кислоты, кодирующей их, причем указанная нуклеиновая кислота функционально связана с регуляторной последовательностью и экспрессируется in vivo. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором доставка происходит за счет лентивируса, аденовируса или аденоассоциированного вируса. В этом аспекте изобретение относится к способу, описанному в настоящем описании, или к парным комплексам CRISPR-Cas, описанным в настоящем описании, в которых последовательность-мишень для первого комплекса пары находится на первой цепи двуцепочечной ДНК, и последовательность-мишень для второго комплекса пары находится на второй цепи двуцепочечной ДНК. В этом аспекте изобретение относится к способу, описанному в настоящем описании, или парным комплексам CRISPR-Cas, описанным в настоящем описании, в котором последовательности-мишени для первого и второго комплекса находятся вблизи друг друга таким образом, что ДНК разрезается так, чтобы обеспечивать гомологичную репарацию. В одном аспекте этот способ или эти парные комплексы CRISPR-Cas могут включать такие комплексы CRISPR-Cas, каждый из которого содержит фермент CRISPR, в который внесены мутации таким образом, что он обладает не более чем 5% нуклеазной активности немутантного фермента CRISPR.
[00403] В одном аспекте изобретение относится к библиотеке, способу или комплексу, как описано в настоящем описании, где гРНК модифицирована так, что она имеет по меньшей мере одну некодирующую функциональную петлю, например, где по меньшей мере одна некодирующая функциональная петля является репрессирующей; например, где по меньшей мере одна некодирующая функциональная петля содержит Alu.
[00404] В одном аспекте изобретение относится к способу изменения или модификации экспрессии продуктов генов. Упомянутый способ может включать внесение в клетку, содержащую или экспрессирующую молекулу ДНК, кодирующий продукт гена, не встречающейся в природе системы CRISPR-Cas, содержащей белок Cas и направляющую РНК, мишенью которой является молекула ДНК, где направляющая РНК нацелена на молекулу ДНК, кодирующую продукт гена, и белок Cas расщепляет молекулу ДНК, кодирующую продукт гена, причем экспрессия продукта гена изменяется; и при этом белок Cas и направляющая РНК вместе не встречаются в природе. Изобретение далее включает белок Cas, являющийся кодон-оптимизированным для экспрессии в эукариотической клетке. В предпочтительном варианте осуществления эукариотическая клетка представляет собой клетку млекопитающего, и в еще более предпочитаемом варианте осуществления клетка млекопитающего представляет собой клетку человека. В следующем варианте осуществления изобретения экспрессия генного продукта снижена.
[00405] В одном аспекте изобретение относится к измененным клеткам и потомству этих клеток, а также к продуктам этих клеток. Белки и системы CRISPR-Cas (например, C2c1) и по настоящему изобретению могут быть использованы для получения клеток, содержащих модифицированный локус-мишень. В некоторых вариантах осуществления способ обеспечивает связывание комплекса, мишенью которого является нуклеиновая кислота, с ДНК-мишенью или РНК-мишенью, для расщепления упомянутой ДНК-мишени или РНК-мишени, таким образом, модифицируя упомянутую ДНК-мишень или РНК-мишень, где комплекс, мишенью которого является нуклеиновая кислота, включает эффекторный белок, мишенью которого является нуклеиновая кислота, в комплексе с направляющей РНК, гибридизованной с последовательностью-мишенью, в рамках упомянутой ДНК-мишени или РНК-мишени. В одном аспекте изобретение относится к способу репарации генного локуса в клетке. В другом аспекте относится к способу изменения экспрессии ДНК или РНК в генном локусе клетки. В некоторых вариантах осуществления способ обеспечивает связывание комплекса, мишенью которого является нуклеиновая кислота, с ДНК-мишенью или РНК-мишенью, так что упомянутое связывание приводит к увеличению или снижению уровня экспрессии указанной ДНК или РНК, причем комплекс, мишенью которого является нуклеиновая кислота, включает эффекторный белок, мишенью которого является нуклеиновая кислота, в комплексе с направляющей РНК. Для модификации ДНК-мишени или РНК-мишени применимы соображения и условия, сходные с теми, что описаны выше. Фактически, эти условия отбора, культивирования и обратного введения применимы в разных аспектах настоящего изобретения. В одном аспекте изобретение относится к способу модификации ДНК или РНК в эукариотической клетке, который может быть осуществлен in vivo, ex vivo или in vitro. В некоторых вариантах осуществления способ включает отбор клетки или популяции клеток из организма человека или организма, не являющегося человеческим, и модификацию клетки или клеток. Культивирование может происходить на любой стадии ex vivo. Такие клетки могут быть, без любых ограничений, растительной клеткой, животной клеткой, конкретным типом клетки любого организма, включая стволовые клетки, иммунные клетки, Т-клетки, В-клетки, дендритные клетки, клетки сердечно-сосудистой системы, эпителиальные клетки и т.п. Клетки могут быть модифицированы согласно изобретению для получения продуктов генов, например, в контролируемых количествах, которые могут быь увеличены или снижены в зависимости от цели использования, или мутированы. В определенных вариантах осуществления генетический локус клетки репарируется. Клетка или клетки даже могут быть повторно введены в организм, не являющийся человеческим, или растение. В случае повторно введенных клеток может быть предпочтительным использование стволовых клеток.
[00406] В однои из аспектов изобретение относится к клеткам, которые временно содержат системы CRISPR или их компоненты. Например, белки или ферменты CRISPR и нуклеиновые кислоты временно вводят в клетки, и генетический локус изменяется, после чего следует снижение количества одного из компонентов системы CRISPR. Затем, клетки, потомство клеток и организмы, содержащие клетки, получившие генетические изменения с помошью CRISPR, содержат меньшее количество одного или более компонентов системы CRISPR или более не содержат ни одного из компонентов системы CRISPR. Одним неограничивающим примером является самоинактивирующася cистема CRISPR-Cas, как описано в настоящем описании. Таким образом, изобретение относится к клеткам, предшественникам клеток и организмам, содержащим клетки, которые содержат один или более генетических локусов, измененных посредством системы CRISPR, но в которых по существу отсутствует один или более компонентов системы CRISPR. В некоторых вариантах осуществления компоненты системы CRISPR отсутствуют. Такие клетки, ткани и организмы преимущественно содержат желаемое или выбранное генетическое изменение, но утратили компоненты CRISPR-Cas или их остатки, которые потенциально могут неспецифически действовать и вести к возникновению проблем безопасноти или препятствовать одобрению регулирующим учреждением. Также изобретение относится к продуктам деятельности клеток, организмов, предшественников клеток и организмов.
Индуцибельные системы CRISPR-Cas C2c1 или C2c3 ("разделенный C2c1" или "разделенный C2c3")
[00407] В одном из аспектов изобретение относится к не встречающейся в природе или сконструированной способами инженерии индуцибельной системе CRISPR-Cas C2c1 или C2c3, включающей:
первую слитую конструкцию C2c1 или C2c3, связанную с первой половиной индуцибельного димера, и вторую слитую конструкцию C2c1 или C2c3, связанную со второй половиной индуцибельного димера;
при этом первая слитая конструкция C2c1 или C2c3 функционально связана с одним или более сигналами ядерной локализации;
при этом вторая слитая конструкция C2c1 или C2c3 функционально связана с одним или более сигналами ядерной локализации;
при этом контакт с индуцирующим источником энергии сводит первую и вторую половины индуцибельного димера вместе;
при этом сведение первой и второй половин индуцибельного димера вместе позволяет первой и второй слитым конструкциям C2c1 составлять функциональную систему CRISPR-Cas C2c1, или первой и второй слитым конструкциям C2c3 составлять функциональную систему CRISPR-Cas C2c3;
при этом система CRISPR-Cas C2c1 или система CRISPR-Cas C2c3 включают направляющую РНК (гРНК), включающую направляющую последовательность, способную к гибридизации с последовательностью-мишенью в представляющем интерес геномном локусе клетки; и
при этом функциональная система CRISPR-Cas C2c1 или система CRISPR-Cas C2c3 связывается с последовательностью-мишенью и необязательно редактирует геномный локус для изменения экспрессии гена.
[00408] В одном из возможных аспектов изобретения в индуцибельной системе CRISPR-Cas C2c1 индуцибельный димер является, или содержит, или по существу состоит или состоит из индуцибельного гетеродимера. В одном из возможных аспектов в индуцибельной системе CRISPR-Cas C2c1 первая половина или первая часть или первый фрагмент индуцибельного гетеродимера является, содержит, по существу состоит или состоит из белка FKBP, необязательно FKBP12. В одном из возможных аспектов изобретения в индуцибельной системе CRISPR-Cas C2c1 вторая половина или вторая часть или второй фрагмент индуцибельного гетеродимера является, содержит, по существу состоит или состоит из белка FRB. В одном из возможных аспектов изобретения в индуцибельной системе CRISPR-Cas C2c1 структура первой слитой конструкции C2c1 является, содержит, по существу состоит или состоит из NES-N-концевой области C2c1-FRB-NES. В одном из возможных аспектов изобретения в индуцибельной системе CRISPR-Cas C2c1 структура второй слитой конструкции C2c1 является, содержит, по существу состоит или состоит из NLS-C-концевой области C2c1-FRB-NLS. В одном из возможных аспектов в индуцибельной системе CRISPR-Cas C2c1 может присутствовать линкер, отделяющий часть C2c1 от половины, части или фрагмента индуцибельного димера. В одном из возможных аспектов в индуцибельной системе CRISPR-Cas C2c1 источник индукции является, содержит, по существу состоит или состоит из рапамицина. В одном из возможных аспектов в индуцибельной системе CRISPR-Cas C2c1 индуцибельный димер является индуцибельным гомодимером. В одном из возможных аспектов в C2c1 индуцибельной системе CRISPR-Cas C2c1 представляет собой AacC2c1. В одном из возможных аспектов в индуцибельной системе CRISPR-Cas C2c1 один или более функциональных доменов ассоциированы с одной или обеими частями C2c1, в частности, функциональные домены могут необязательно включать транскрипционный активатор, транскрипционный репрессор или нуклеазу, такую как нуклеаза Fok1. В одном из возможных аспектов в индуцибельной системе CRISPR-Cas C2c1 функциональная система CRISPR-Cas C2c1 связывается с последовательностью-мишенью, и фермент является мертвым C2c1, необязательно имеющим нуклеазную активность, сниженную по меньшей мере на 97% или на 100% (соответственно, имеющим не более 3% нуклеазной активности или преимущественно 0% нуклеазной активности), по сравнению с C2c1, не содержащим ни одну мутацию. Кроме того, изобретение относится и один из возможных аспектов изобретения предоставляет собой полинуклеотид, кодирующий индуцибельную систему CRISPR-Cas C2c1, описанную в настоящем описании.
[00409] В одном из возможных аспектов изобретения в индуцибельной системе CRISPR-Cas C2c3 индуцибельный димер является, или содержит, или по существу состоит или состоит из индуцибельного гетеродимера. В одном из возможных аспектов в индуцибельной системе CRISPR-Cas C2c3 первая половина или первая часть или первый фрагмент индуцибельного гетеродимера является, или содержит, или по существу состоит или состоит из белка FKBP, необязательно FKBP12. В одном из возможных аспектов изобретения в индуцибельной системе CRISPR-Cas C2c3 вторая половина или вторая часть или второй фрагмент индуцибельного гетеродимера является, или содержит, или по существу состоит или состоит из белка FRB. В одном из возможных аспектов изобретения в индуцибельной системе CRISPR-Cas C2c3 структура первой слитой конструкции C2c3 является, или содержит, или по существу состоит или состоит из NES-N-концевой области C2c3-FRB-NES. В одном из возможных аспектов изобретения в индуцибельной системе CRISPR-Cas C2c3 структура второго слитой конструкции C2c3 является, или содержит, или по существу состоит или состоит из NLS-C-концевой области C2c3-FRB-NLS. В одном из возможных аспектов в индуцибельной системе CRISPR-Cas C2c1 может присутствовать линкер, отделяющий часть C2c3 от половины, части или фрагмента индуцибельного димера. В одном из возможных аспектов в индуцибельной системе CRISPR-Cas C2c1 источник индукции является, или содержит, или по существу состоит или состоит из рапамицина. В одном из возможных аспектов в индуцибельной системе CRISPR-Cas C2c3 индуцибельный димер является индуцибельным гомодимером. В одном из возможных аспектов в индуцибельной системе CRISPR-Cas C2c3 один или более функциональных доменов ассоциированы с одной или обеими частями C2c3, в частности, функциональные домены могут необязательно включать транскрипционный активатор, транскрипционный репрессор или нуклеазу, такую как нуклеаза Fok1. В одном из возможных аспектов в индуцибельной системе CRISPR-Cas C2c3 функциональная система CRISPR-Cas C2c3 связывается с последовательностью-мишенью и фермент является мертвым C2c3, необязательно имеющим нуклеазную активность, сниженную по меньшей мере на 97% или на 100% (соответственно, имеющим не более 3% нуклеазной активности или преимущественно 0% нуклеазной активности), по сравнению с C2c3, не содержащим ни одну мутацию. Кроме того, изобретение относится и один из возможных аспектов изобретения предоставляет собой полинуклеотид, кодирующий индуцибельную систему CRISPR-Cas C2c3, описанную в настоящем описании.
[00410] В одном из возможных аспектов изобретения изобретение относится к вектору для доставки первой слитой конструкции C2c1, связанной с первой половиной или частью или фрагментом индуцибельного димера и функционально связанной с одним или более сигналами ядерной локализации, как описано в настоящем описании. В одном из возможных аспектов изобретение относится к вектору для доставки второй слитой конструкции C2c1, связанной со второй половиной или частью или фрагментом индуцибельного димера и функционально связанной с одним или более сигналами ядерного экспорта.
[00411] В одном из возможных аспектов изобретения изобретение относится к вектору для доставки первой слитой конструкции C2c3, связанной с первой половиной или частью или фрагментом индуцибельного димера и функционально связанной с одним или более сигналами ядерной локализации, как описано в настоящем описании. В одном из возможных аспектов изобретение относится к вектору для доставки второй слитой конструкции C2c3, связанной со второй половиной или частью или фрагментом индуцибельного димера и функционально связанной с одним или более сигналами ядерного экспорта
[00412] В одном из возможных аспектов изобретения изобретение относится к вектору для доставки как первой слитой конструкции C2c1, слитой с первой половиной или частью или фрагментом индуцибельного димера и функционально связанной с одним или более сигналами ядерной локализации, так и второй слитой конструкции C2c1, связанной со второй половиной или частью или фрагментом индуцибельного димера и функционально связанного с одним или более сигналами ядерного экспорта, как описано в настоящем описании.
[00413] В одном из возможных аспектов изобретения изобретение относится к вектору для доставки как первой слитой конструкции C2c3, слитой с первой половиной или частью или фрагментом индуцибельного димера и функционально связанной с одним или более сигналами ядерной локализации, так и второй слитой конструкции C2c3, связанной со второй половиной или частью или фрагментом индуцибельного димера и функционально связанного с одним или более сигналами ядерного экспорта, как описано в настоящем описании.
[00414] В одном из возможных аспектов вектор может быть единственной плазмидой или экспрессирующей кассетой.
[00415] В одном из возможных аспектов изобретение относится к эукариотической клетке-хозяину или клеточной линии, трансформированным любым из векторов, описанных в настоящем описании, или экспрессирующим индуцибельную систему CRISPR-Cas C2c1 или C2c3, описанную в настоящем описании.
[00416] В одном из возможных аспектов изобретение относится к трансгенному организму, трансформированному одним из векторов, описанных в настоящем описании, или экспрессирующему индуцибельную систему CRISPR-Cas C2c1 или C2c3, описанную в настоящем описании, или к потомкам этого организма. В одном из возможных аспектов изобретение относится к модельному организму, который конститутивно экспрессирует индуцибельную систему CRISPR-Cas C2c1 или C2c3, описанную в настоящем описании.
[00417] В одном из возможных аспектов изобретение относится к не встречающейся в природе или сконструированной способами инженерии системе CRISPR-Cas C2c1, включающей: первую слитую конструкцию C2c1 конструкт, связанную с первой половиной индуцибельного гетеродимера, и вторую слитую конструкцию C2c1, связанную со второй половиной индуцибельного гетеродимера, причем первая слитая конструкция C2c1 функционально связана с одним или более сигналами ядерной локализации, причем вторая слитая конструкция C2c1 функционально связана с одним или более сигналами ядерной локализации NES, причем при взаимодействии с источником индукции первая и вторая половины индуцибельного гетеродимера объединяются, причем объединение первой и второй половин индуцибельного гетеродимера позволяет первой и второй слитым конструкциям C2c1 составить функциональную систему CRISPR-Cas C2c1, причем система CRISPR-Cas C2c1 включает направляющую РНК (гРНК), содержащую направляющую последовательность, способную гибридизоваться с последовательностью-мишенью в представляющем интерес геномном локусе клетки, и причем функциональная система CRISPR-Cas C2c1 редактирует геномный локус, изменяя экспрессию гена.
[00418] В одном из возможных аспектов изобретение относится к не встречающейся в природе или сконструированной способами инженерии системе CRISPR-Cas C2c3, включающей: первую слитую конструкцию C2c3 конструкт, связанную с первой половиной индуцибельного гетеродимера, и вторую слитую конструкцию C2c3, связанную со второй половиной индуцибельного гетеродимера, причем первая слитая конструкция C2c3 функционально связана с одним или более сигналами ядерной локализации, причем вторая слитая конструкция C2c3 функционально связана с одним или более сигналами ядерной локализации NES, причем при взаимодействии с источником индукции первая и вторая половины индуцибельного гетеродимера объединяются, причем объединение первой и второй половин индуцибельного гетеродимера позволяет первой и второй слитым конструкциям C2c3 составить функциональную систему CRISPR-Cas C2c3, причем система CRISPR-Cas C2c3 включает направляющую РНК (гРНК), содержащую направляющую последовательность, способную гибридизоваться с последовательностью-мишенью в представляющем интерес геномном локусе клетки, и причем функциональная система CRISPR-Cas C2c3 редактирует геномный локус, изменяя экспрессию гена.
[00419] В одном из возможных аспектов изобретение относится к способу лечения индивидуума, нуждающегося в этом, включающему индукцию редактирования гена путем трансформации индивидуума полинуклеотидом, описанным в настоящем описании, или любым из векторов, описанных в настоящем описании, и введение индивидууму источника индукции. Изобретение относится к использованию такого полинуклеотида или вектора в производстве лекарственных средств, например, такого как лекарственное средство для лечения индивидуума или для такого способа лечения индивидуума. Изобретение относится к полинуклеотиду, описанному в настоящем описании, или любому из векторов, описанных в настоящем описании, для использования в способе лечения индивидуума, нуждающегося в этом, включающем индукцию редактирования гена путем трансформации индивидуума полинуклеотидом, описанным в настоящем описании или любым векторов, описанных в настоящем описании, и введение индивидууму источника индукции. В одном из возможных аспектов способа предоставляется матрица репарации, например, доставляемая вектором, содержащим указанную матрицу репарации.
[00420] В одном из возможных аспектов изобретение относится к способу лечения индивидуума, нуждающегося в этом, включающему индукцию активации или репрессии транскрипции гена путем трансформации индивидуума полинуклеотидом, описанным в настоящем описании, или любым из векторов, описанных в настоящем описании, где полинуклеотид или вектор кодирует или содержит каталитически неактивный белок C2c1 или неактивный белок C2c3 и один или более ассоциированных с ним функциональных доменов, описанных в настоящем описании; кроме того, способ включает введение индивидууму источника индукции. Изобретение относится к использованию такого полинуклеотида или вектора в производстве лекарственного средства, например, такого как лекарственное средство для лечения индивидуума или для такого способа лечения индивидуума. Изобретение относится к полинуклеотиду, описанному в настоящем описании, или любому из векторов, описанных в настоящем описании, для использования в способе лечения индивидуума, нуждающегося в этом, включающем индукцию активации или репрессии транскрипции гена путем трансформации индивидуума полинуклеотидом, описанным в настоящем описании, или любым из векторов, описанных в настоящем описании, и введение индивидууму источника индукции.
[00421] Соответственно, изобретение, среди прочего, охватывает гомодимеры, а также также гетеродимеры, мертвый C2c1 или C2c1, по существу не обладающий нуклеазной активностью, в частности, за счет мутаций, системы или комплексы, в которых присутствует один или более NLS и/или один или более NES; функциональный(-ые) домен(ы), соединенные с разделенным C2c1; способы, включая способы лечения, и применение.
[00422] Соответственно, изобретение, среди прочего, охватывает гомодимеры, а также также гетеродимеры, мертвый C2c3 или C2c3, по существу не обладающий нуклеазной активностью, в частности, за счет мутаций, системы или комплексы, в которых присутствует один или более NLS и/или один или более NES; функциональный(-ые) домен(ы), соединенные с разделенным C2c3; способы, включая способы лечения, и применение.
[00423] Понятно, что, когда в настоящем описании упоминается C2c1, белок C2c1 или фермент C2c1, это включает разделенный C2c1 по настоящему изобретению. В одном из аспектов изобретение относится к способу изменения или модификации экспрессии продукта гена. Указанный способ может включать введение в клетку, содержащую или экспрессирующую молекулу ДНК, кодирующую продукт гена, сконструированной не встречающейся в природе системы CRISPR-Cas C2c1, содержащей белок C2c1 и направляющую РНК, мишенью которой является молекула ДНК, где направляющая РНК нацелена на молекулу ДНК, кодирующую продукт гена, и белок C2c1 расщепляет молекулу ДНК, кодирующую продукт гена, за счет чего изменяется экспрессия продукта гена; причем белок C2c1 и направляющая РНК не встречаются вместе в природе. Изобретение относится к направляющей РНК, содержащей направляющую последовательность, соединенную с последовательностью DR. Кроме того, изобретение предусматривает оптимизацию кодонного состава белка C2c1 для экспрессии в эукариотической клетке. В предпочтительном варианте осуществления эукариотическая клетка является клеткой млекопитающего, и в более предпочтительном варианте осуществления клетка млекопитающего является клеткой человека. В следующем варианте осуществления изобретения экспрессия продукта гена снижается.
[00424] Понятно, что, когда в настоящем описании упоминается C2c3, белок C2c3 или фермент C2c3, это включает разделенный C2c3 по настоящему изобретению. В одном из аспектов изобретение относится к способу изменения или модификации экспрессии продукта гена. Указанный способ может включать введение в клетку, содержащую или экспрессирующую молекулу ДНК, кодирующую продукт гена, сконструированной не встречающейся в природе системы CRISPR-Cas C2c3, содержащей белок C2c3 и направляющую РНК, мишенью которой является молекула ДНК, где направляющая РНК нацелена на молекулу ДНК, кодирующую продукт гена, и белок C2c3 расщепляет молекулу ДНК, кодирующую продукт гена, за счет чего изменяется экспрессия продукта гена; причем белок C2c3 и направляющая РНК не встречаются вместе в природе. Изобретение относится к направляющей РНК, содержащей направляющую последовательность, соединенную с последовательностью DR. Кроме того, изобретение предусматривает оптимизацию кодонного состава белка C2c3 для экспрессии в эукариотической клетке. В предпочтительном варианте осуществления эукариотическая клетка является клеткой млекопитающего, и в более предпочтительном варианте осуществления клетка млекопитающего является клеткой человека. В следующем варианте осуществления изобретения экспрессия продукта гена снижается.
[00425] В одном аспекте изобретение относится к сконструированной способами инженерии не встречающейся в природе системе CRISPR-Cas C2c1, содержащей белок C2c1 и направляющую РНК, мишенью которой является молекула ДНК, кодирующая продукт гена в клетке, причем направляющая РНК нацелена на молекулу ДНКа, кодирующую продукт гена, и белок C2c1 расщепляет молекулу ДНК, кодирующую продукт гена, за счет чего изменяется экспрессия продукта гена; причем белок C2c1 и направляющая РНК не встречаются вместе в природе; и причем система включает разделенный C2c1 по настоящему изобретению. Изобретение относится к направляющей РНК, содержащей направляющую последовательность, соединенную с последовательностью DR. Кроме того, изобретение предусматривает оптимизацию кодонного состава белка C2c1 для экспрессии в эукариотической клетке. В предпочтительном варианте осуществления эукариотическая клетка является клеткой млекопитающего, и в более предпочтительном варианте осуществления клетка млекопитающего является клеткой человека. В следующем варианте осуществления изобретения экспрессия продукта гена снижается.
[00426] В одном аспекте изобретение относится к сконструированной способами инженерии не встречающейся в природе системе CRISPR-Cas C2c3, содержащей белок C2c3 и направляющую РНК, мишенью которой является молекула ДНК, кодирующая продукт гена в клетке, причем направляющая РНК нацелена на молекулу ДНКа, кодирующую продукт гена, и белок C2c3 расщепляет молекулу ДНК, кодирующую продукт гена, за счет чего изменяется экспрессия продукта гена; причем белок C2c3 и направляющая РНК не встречаются вместе в природе; и причем система включает разделенный C2c3 по настоящему изобретению. Изобретение относится к направляющей РНК, содержащей направляющую последовательность, соединенную с последовательностью DR. Кроме того, изобретение предусматривает оптимизацию кодонного состава белка C2c3 для экспрессии в эукариотической клетке. В предпочтительном варианте осуществления эукариотическая клетка является клеткой млекопитающего, и в более предпочтительном варианте осуществления клетка млекопитающего является клеткой человека. В следующем варианте осуществления изобретения экспрессия продукта гена снижается.
[00427] В другом аспекте изобретение относится к сконструированной не встречающейся в природе системе, содержащей один или более векторов, содержащих первый регуляторный элемент, функционально связанный с направляющей РНК системы CRISPR-Cas C2c1 или C2c3, мишенью которой является молекула ДНК, кодирующая продукт гена, и второй регуляторный элемент, функционально связанный с белком C2c1 или белком C2c3; включая разделенный C2c1 или C2c3 по настоящему изобретению. Компоненты (а) и (b) могут быть находиться в одном или разных векторах системы. Направляющая РНК осуществляет нацеливание на молекулу ДНК, кодирующую продукт гена в клетке, и белок C2c1 или белок C2c3 расщепляет молекулу ДНК, кодирующую продукт гена, за счет чего экспрессия продукта гена изменяется; и причем белок C2c3 или белок C2c1 и направляющая РНК не встречаются вместе в природе. Изобретение относится к направляющей РНК, содержащей направляющую последовательность, соединенную с последовательностью DR. Кроме того, изобретение относится к белку C2c1 или белку C2c3, кодон-оптимизированным для экспрессии в эукариотической клетке. В предпочтительном варианте осуществления эукариотическая клетка является клеткой млекопитающего, и в более предпочтительном варианте осуществления клетка млекопитающего является клеткой человека. В следующем варианте осуществления изобретения экспрессия продукта гена снижается.
[00428] В одном аспекте изобретение относится к системе векторов, содержащей один или более векторов. В некоторых вариантах осуществления система включает: а) первый регуляторный элемент, функционально связанный с последовательностью DR и одним или более участками вставки для вставки одной или более направляющих последовательностей ниже последовательности DR, причем при экспрессии направляющая последовательность обеспечивает специфическое для последовательности связывание комплекса CRISPR-Cas C2c1 или комплекса CRISPR-Cas C2c3 с последовательностью-мишенью в эукариотической клетке, причем комплекс CRISPR-Cas C2c1 или CRISPR-Cas C2c3 содержит C2c1 или C2c3 в комплексе с (1) направляющей последовательностью, которая гибридизована с последовательностью-мишенью, и (2) последовательностью DR; и (b) второй регуляторный элемент, функционально связанный с кодирующей фермент последовательностью, кодирующей упомянутый фермент C2c1 или C2c3, содержащий последовательность ядерной локализации; причем компоненты (а) и (b) расположены в одном и нескольких векторах системы; включая разделенные C2c1 или C2c3. В некоторых вариантах осуществления компонент (а) далее включает две или более направляющих последовательностей, функционально связанных с первым регуляторным элементом, причем при экспрессии две или более направляющих последовательности обеспечивают специфическое для последовательности связывание C2c1 или C2c3 с различными последовательностями-мишенями в эукариотической клетке.
[00429] В некоторых вариантах осуществления комплекс CRISPR-Cas C2c1 или CRISPR-Cas C2c3 содержит одну или более последовательностей ядерной локализации достаточной силы для индукции накопления упомянутого комплекса CRISPR-Cas C2c1 или CRISPR-Cas C2c3 в количествах, позволяющих его обнаружить в ядре эукариотической клетки. Без связи с теорией полагают, что последовательность ядерной локализации не является необходимой для активности комплекса CRISPR-Cas C2c1 или CRISPR-Cas C2c3 в эукариотических организмах, но что включение таких последовательностей повышает активность системы, особенно в отношении направления молекул нуклеиновых кислот в ядро.
[00430] В некоторых вариантах осуществления фермент C2c1 представляет собой C2c1 видов бактерий, выбранных из группы, состоящей из: Alicyclobacillus acidoterrestris (например, ATCC 49025), Alicyclobacillus contaminans (например, DSM I7975), Desulfovibrio inopinatus (например, DSM I07II), Desulfonatronum thiodismutans (например, штамм MLF-1), бактерия Opitutaceae TAV5, Tuberibacillus calidus (например, DSM I7572), Bacillus thermoamylovorans (например, штамм B4I66), Brevibacillus sp. CFII2, Bacillus sp. NSP2.I, Desulfatirhabdium butyrativorans (например, DSM I8734), Alicyclobacillus herbarius (например, DSM I3609), Citrobacter freundii (например, ATCC 8090), Brevibacillus agri (например, BAB-2500) и Methylobacterium nodulans (например, ORS 2060). Фермент может быть гомологом или ортологом C2c1 или C2c3. В некоторых вариантах осуществления белки C2c1 или C2c3 являются кодон-оптимизированными для экспрессии в эукариотической клетке. В некоторых вариантах осуществления C2c1 или C2c3 направляет расщепление одной или двух цепей в участке последовательности-мишени. В предпочтительном варианте осуществления разрыв цепи представляет собой ступенчатое разрезание с 5'-выступающими нуклеотидами. В некоторых вариантах осуществления первый регуляторный элемент представляет собой промотор полимеразы III. В некоторых вариантах осуществления второй регуляторный элемент представляет собой промотор полимеразы II. В некоторых вариантах осуществления минимальная длина прямого повтора составляет 16 нуклеотидов, и он содержит единственную шпильку. В дальнейших вариантах осуществления длина прямого повтора превышает 16 нуклеотидов, предпочтительно превышает 17 нуклеотидов, и он содержит более одной шпильки или обладает оптимизированной вторичной структурой.
[00431] В одном аспекте изобретение относится к эукариотической клетке-хозяину, содержащей а) первый регуляторный элемент, функционально связанный с последовательностью прямого повтора и одним или более участками вставки для вставки одной или более направляющих последовательностей ниже последовательности DR, причем при экспрессии направляющая последовательность направляет специфическое для последовательности связывание комплекса CRISPR-Cas C2c1 или комплекса CRISPR-Cas C2c3 с последовательностью-мишенью в эукариотической клетке, причем комплекс CRISPR-Cas C2c1 или CRISPR-Cas C2c3 содержит C2c1 или C2c3 в комплексе с (1) направляющей последовательностью, которая гибридизована с последовательностью-мишенью, и (2) последовательностью DR; и (b) второй регуляторный элемент, функционально связанный с кодирующей фермент последовательностью, кодирующей упомянутый фермент C2c1 или C2c3, содержащий последовательность ядерной локализации. В некоторых вариантах осуществления клетка-хозяин содержит компоненты (а) и (b); включая разделенные C2c1 или C2c3. В некоторых вариантах осуществления компонент (а) или (b) или компоненты (а) и (b) стабильно встраиваются в геном эукариотической клетки-хозяина. В некоторых вариантах осуществления компонент (а) далее содержит две или более направляющих последовательностей, функционально связанных с первым регуляторным элементом, причем при экспрессии две или более направляющие последовательности направляют специфическое для последовательности связывание C2c1 или C2c3 с различными последовательностями-мишенями в эукариотической клетке. В некоторых вариантах осуществления белки C2c1 или C2c3 являются кодон-оптимизированными для экспрессии в эукариотической клетке. В некоторых вариантах осуществления C2c1 или C2c3 направляет расщепление одной или двух цепей в участке последовательности-мишени. В предпочтительном варианте осуществления разрыв цепи представляет собой ступенчатое разрезание с 5'-выступающими нуклеотидами. В некоторых вариантах осуществления первый регуляторный элемент представляет собой промотор полимеразы III. В некоторых вариантах осуществления второй регуляторный элемент представляет собой промотор полимеразы II. В некоторых вариантах осуществления минимальная длина прямого повтора составляет 16 нуклеотидов, и он содержит единственную шпильку. В дальнейших вариантах осуществления длина прямого повтора превышает 16 нуклеотидов, предпочтительно превышает 17 нуклеотидов, и он содержит более одной шпильки или обладает оптимизированной вторичной структурой. В одном аспекте изобретение относится к эукариотическому организму, не являющемуся человеческим, предпочтительно многоклеточному эукариотическому организму, включающему эукариотическую клетку-хозяина согласно описанным вариантам осуществления. В других аспектах изобретение относится к эукариотическому организму, предпочтительно многоклеточному эукариотическому организму, включающему эукариотическую клетку-хозяина, согласно описанным вариантам осуществления. В некоторых вариантах осуществления этих аспектов организм может быть животным, например, млекопитающим. Также организм может быть членистоногим, в частности, насекомым. Организм также может быть растением. Кроме того, организм может представлять собой гриб.
[00432] В одном аспекте изобретение относится к набору, содержащему один или более компонентов, описанных в настоящем описании. В некоторых вариантах осуществления набор включает систему векторов и инструкцию по применению набора. В некоторых вариантах осуществления система включает а) первый регуляторный элемент, функционально связанный с последовательностью прямого повтора и одним или более участками вставки для вставки одной или более направляющих последовательностей ниже последовательности DR, причем при экспрессии направляющая последовательность направляет специфическое для последовательности связывание комплекса CRISPR-Cas C2c1 или комплекса CRISPR-Cas C2c3 с последовательностью-мишенью в эукариотической клетке, причем комплекс CRISPR-Cas C2c1 или CRISPR-Cas C2c3 содержит C2c1 или C2c3 в комплексе с (1) направляющей последовательностью, которая гибридизована с последовательностью-мишенью, и (2) последовательностью DR; и (b) второй регуляторный элемент, функционально связанный с кодирующей фермент последовательностью, кодирующей упомянутый фермент C2c1 или C2c3, содержащий последовательность ядерной локализации; преимущественно включая разделенный C2c1 или C2c3. В некоторых вариантах осуществления набор содержит компоненты (а) и (b), расположенные на одном или более векторах системы. В некоторых вариантах осуществления компонент (а) далее содержит две или более направляющих последовательностей, функционально связанных с первым регуляторным элементом, причем при экспрессии две или более направляющих последовательностей направляют специфическое для последовательности связывание C2c1 или C2c3 с различными последовательностями-мишенями в эукариотической клетке. В некоторых вариантах осуществления C2c1 или C2c3 содержит одну или более последовательностей ядерной локализаци достаточной силы, чтобы индуцировать накопление упомянутого C2c1 или C2c3 в поддающихся обнаружению количествах в ядре эукариотической клетки. В некоторых вариантах осуществления фермент C2c1 представляет собой C2c1 видов бактерий, выбранных из группы, состоящей из: Alicyclobacillus acidoterrestris (например, ATCC 49025), Alicyclobacillus contaminans (например, DSM I7975), Desulfovibrio inopinatus (например, DSM I07II), Desulfonatronum thiodismutans (например, штамм MLF-1), бактерия Opitutaceae TAV5, Tuberibacillus calidus (например, DSM I7572), Bacillus thermoamylovorans (например, штамм B4I66), Brevibacillus sp. CFII2, Bacillus sp. NSP2.I, Desulfatirhabdium butyrativorans (например, DSM I8734), Alicyclobacillus herbarius (например, DSM I3609), Citrobacter freundii (например, ATCC 8090), Brevibacillus agri (например, BAB-2500) и Methylobacterium nodulans (например, ORS 2060). Фермент может быть гомологом или ортологом C2c1 или C2c3. В некоторых вариантах осуществления белки C2c1 или C2c3 являются кодон-оптимизированными для экспрессии в эукариотической клетке. В некоторых вариантах осуществления C2c1 или C2c3 направляет расщепление одной или двух цепей в участке последовательности-мишени. В предпочтительном варианте осуществления разрыв цепи представляет собой ступенчатое разрезание с 5'-выступающими нуклеотидами. В некоторых вариантах осуществления первый регуляторный элемент представляет собой промотор полимеразы III. В некоторых вариантах осуществления второй регуляторный элемент представляет собой промотор полимеразы II. В некоторых вариантах осуществления минимальная длина прямого повтора составляет 16 нуклеотидов, и он содержит единственную шпильку. В дальнейших вариантах осуществления длина прямого повтора превышает 16 нуклеотидов, предпочтительно превышает 17 нуклеотидов, и он содержит более одной шпильки или обладает оптимизированной вторичной структурой.
[00433] В одном из возможных аспектов изобретение относится к модификациям полинуклеотида-мишени в эукариотической клетке. В некоторых вариантах осуществления способ включает позволение комплексу CRISPR-Cas C2c1 или C2c3 связаться с полинуклеотидом-мишенью, чтобы обеспечить расщепление упомянутого полинуклеотида-мишени, тем самым модифицируя полинуклеотид-мишень, при этом комплекс CRISPR-Cas C2c1 или C2c3 содержит C2c1 или C2c3 в комплексе с направляющей последовательностью, гибридизованной с последовательностью-мишенью в пределах упомянутого полиуклеотида-мишени, причем указанный полиуклеотид-мишень соединен с последовательностью прямого повтора. В некоторых вариантах осуществления указанное расщепление включает расщепление одной или двух цепей в участке указанной последовательности-мишени указанным C2c1 или C2c3; включая разделенный C2c1 или разделенный C2c3 по настоящему изобретению. В некоторых вариантах осуществления упомянутое расщепление приводит к снижению уровня транскрипции гена-мишени. В некоторых вариантах осуществления способ далее включает репарацию упомянутого расщепленного полинуклеотида-мишени за счет гомологичной рекомбинации с экзогенным матричным полинуклеотидом, причем упомянутая репарация приводит к мутации, включающей вставку, делецию или замену одного или более нуклеотидов в упомянутом полинуклеотиде-мишени. В некоторых вариантах осуществления указанная мутация приводит к одной или более аминокислотным заменам в белке, образующемся при экспрессии гена, содержащего последовательность-мишень. В некоторых вариантах осуществления способ далее включает доставку одного или более векторов в упомянутую эукариотическую клетку, причем один или более векторов индуцируют экспрессию одного или более из C2c1 или C2c3 и направляющей последовательности, связанной с последовательностью DR. В некоторых вариантах осуществления упомянутые векторы доставляются в эукариотическую клетку, находящуюся в индивидууме. В некоторых вариантах осуществления упомянутая модификация происходит в указанной эукариотической клетке в клеточной культуре. В некоторых вариантах осуществления способ далее включает выделение указанной эукариотической клетки из индивидуума перед упомянутой модификацией. В некоторых вариантах осуществления способ далее включает обратное введение указанной эукариотической клетки и/или клеток, полученных из нее впоследствии, упомянутому субъекту.
[00434] В одном аспекте изобретение относится к способу модификации экспрессии полинуклеотида в эукариотической клетке. В некоторых вариантах осуществления способ включает позволение комплексу CRISPR-Cas C2c1 или C2c3 связаться с полинуклеотидом, так чтобы указанное связывание приводило к увеличению или снижению экспрессии упомянутого полинуклеотида, причем комплекс CRISPR-Cas C2c1 или C2c3 содержит C2c1 или C2c3 в комплексе с направляющей последовательностью, способной гибридизоваться с последовательностью-мишенью в пределах упомянутого полиуклеотида-мишени, причем указанный полинуклеотид-мишень соединен с последовательностью прямого повтора; это включает разделенный C2c1 или разделенный C2c3 по настоящему изобретению. В некоторых вариантах осуществления способ далее включает доставку с помощью одного или более векторов в упомянутую эукариотическую клетку, причем один или более векторов индуцируют экспрессию одного или более из C2c1 или C2c3 и направляющей последовательности, соединенной с последовательностью DR.
[00435] В одном аспекте изобретение относится к способу получения модельной эукариотической клетки, содержащей мутантный ген, ассоциированный с заболеванием. В некоторых вариантах осуществления такой ген, ассоциированный с заболеванием, может быть ассоциирован с риском возникновения или развития заболевания. В некоторых вариантах осуществления способ включает (а) введение одного или более векторов в эукариотическую клетку, где один или более векторов запускают экспрессию одного или более из C2c1 или C2c3 и направляющей последовательности, соединенной с последовательностью прямого повтора; и (b) позволение комплексу CRISPR-Cas C2c1 или C2c3 связаться с полинуклеотидом-мишенью, обеспечивая расщепления полинуклеотида-мишени внутри упомянутого гена, ассоциированного с заболеванием, причем CRISPR-комплекс C2c1 или C2c3 включает C2c1 или C2c3 в комплексе с (1) направляющей последовательностью, гибридизованной с последовательностью-мишенью в пределах указанного полинуклеотида-мишени, и (2) последовательностью DR, таким образом, приводя к получению модельной эукариотической клетки, содержащей мутантный ген, ассоциированный с заболеванием, это включает разделенный C2c1 или C2c3 по настоящему изобретению. В некоторых вариантах осуществления C2c1 или C2c3 обеспечивает расщепление одной или двух цепей в участке последовательности-мишени. В предпочтительном варианте осуществления разрыв цепи представляет собой ступенчатое разрезание с 5'-выступающими нуклеотидами. В некоторых вариантах осуществления упомянутое расщепление приводит к снижению транскрипции гена-мишени. В некоторых вариантах осуществления способ далее включает репарацию упомянутого расщепленного целевого полинуклеотида за счет гомологичной рекомбинации с экзогенным матричным полинуклеотидом, причем упомянутая репарация приводит к мутации, включающей вставку, делецию или замену одного или более нуклеотидов в упомянутом полинуклеотиде-мишени. В некоторых вариантах осуществления упомянутая мутация приводит к одной или более аминокислотным заменам в образующемся белке при экспрессии гена, содержащего последовательность-мишень.
[00436] В одном из возможных аспектов изобретение относится к способу разработки биологически активного агента, который модулирует события передачи сигнала в клетке, ассоциированные с геном, связанным с заболеванием. В некоторых вариантах осуществления такой ген, связанный с заболеванием, может быть ассоциирован с риском возникновения или развития заболевания. В некоторых вариантах осуществления способ включает а) приведение исследуемого соединения в контакт с модельной клеткой согласно любому из описанных вариантов осуществления; и b) определение изменения данных, указывающего на снижение или увеличение числа событий передачи сигнала в клетке, связанное с упомянутой мутацией в упомянутом гене, ассоциированном с заболеванием, тем самым разрабатывая биологически активный агент, который модулирует события передачи сигнала в клетке, ассоциированные с геном, связанным с заболеванием.
[00437] В одном из возможных аспектов изобретение относится к рекомбинантному полинуклеотиду, содержащему направляющую последовательность ниже последовательности прямого повтора, причем при экспрессии направляющая последовательность направляет специфическое для последовательности связывание комплекса CRISPR-Cas C2c1 или комплекса CRISPR-Cas C2c3 с соответствующей последовательностью-мишенью в эукариотической клетке. В некоторых вариантах осуществления последовательность-мишень является протоонкогеном или онкогеном.
[00438] В одном из возможных аспектов изобретение относится к способу селекции одной или более клетки(ок), причем способ включает: введение одного или более векторов в клетку(и), причем один или более векторов индуцируют экспрессию одного или более из C2c1 или C2c3, направляющей последовательности, связанной с последовательностью прямого повтора, и редактирующей матрицы; причем редактирующая матрица содержит одну или более мутаций, которые предотвращают расщепление белком C2c1 или C2c3, что позволяет осуществить гомологичную рекомбинацию редактирующей матрицы с помощью полинуклеотида-мишени в клетке, подлежащей селекции; это позволяет комплексу CRISPR-Cas C2c1 или C2c3 связываться с полинуклеотидом-мишенью так, чтобы осуществить расщепление упомянутого полинуклеотида-мишени, при этом комплекс CRISPR-Cas C2c1 или C2c3 содержит C2c1 или C2c3 в комплексе с (1) направляющей последовательностью, гибридизованной с последовательностью-мишенью в пределах упомянутого полиуклеотида-мишени, и (2) последовательностью прямого повтора, причем связывание комплекса CRISPR-Cas C2c1 или C2c3 с полинуклеотидом-мишенью индуцирует клеточную гибель, таким образом, обеспечивая селекцию одной или более клеток, в которые внесена мутация; это включает разделенный C2c1 или разделенный C2c3 по настоящему изобретению. В другом предпочтительном варианте осуществления клетка, которая подвергается селекции, является эукариотической клеткой. Аспекты изобретения позволяют выбрать конкретные клетки без применения селективного маркера или двухступенчатого процесса, который может включать систему контрселекции.
[00439] В настоящей заявке встречается фраза "этот включает разделенный C2c1 или разделенный C2c3 по настоящему изобретению" или сходный текст для того, чтобы указать на то, что C2c1 в вариантах осуществления настоящего изобретения может представлять собой разделенный C2c1 и C2c3 может представлять собой разделенный C2c3, как описано в настоящем описании.
[00440] В одном из возможных аспектов изобретение относится к не встречающейся в природе или сконструированной способами инженерии системе CRISPR-Cas C2c1, включающей первую слитую конструкцию C2c1, связанную с первой половиной индуцибельного гетеродимера, и вторую слитую конструкцию C2c1, связанную со второй половиной индуцибельного гетеродимера, причем первая слитая конструкция C2c1 функционально связана с одним или более сигналами ядерной локализации, причем вторая слитая конструкция C2c1 функционально связана с одним или более сигналами ядерного экспорта, причем при взаимодействии с источником индукции первая и вторая половины индуцибельного гетеродимера объединяются, причем объединение первой и второй половин индуцибельного гетеродимера позволяет первой и второй слитым конструкциям C2c1 составить функциональную систему CRISPR-Cas C2c1, причем система CRISPR-Cas C2c1 включает направляющую РНК (гРНК), содержащую направляющую последовательность, способную гибридизоваться с последовательностью-мишенью в представляющем интерес геномном локусе клетки, и причем функциональная система CRISPR-Cas C2c1 редактирует геномный локус, изменяя экспрессию гена. В одном из возможных аспектов изобретения в индуцибельной системе CRISPR-Cas C2c1 первая половина индуцибельного гетеродимера является FKBP12 и вторая половина индуцибельного гетеродимера является FRB. В другом варианте осуществления источником энергии индукции является рапамицин.
[00441] В одном из возможных аспектов изобретение относится к не встречающейся в природе или сконструированной способами инженерии системе CRISPR-Cas C2c3, включающей первую слитую конструкцию C2c3, связанную с первой половиной индуцибельного гетеродимера, и вторую слитую конструкцию C2c3, связанную со второй половиной индуцибельного гетеродимера, причем первая слитая конструкция C2c3 функционально связана с одним или более сигналами ядерной локализации, причем вторая слитая конструкция C2c3 функционально связана с одним или более сигналами ядерного экспорта, причем при взаимодействии с источником индукции первая и вторая половины индуцибельного гетеродимера объединяются, причем объединение первой и второй половин индуцибельного гетеродимера позволяет первой и второй слитым конструкциям C2c3 составить функциональную систему CRISPR-Cas C2c3, причем система CRISPR-Cas C2c3 включает направляющую РНК (гРНК), содержащую направляющую последовательность, способную гибридизоваться с последовательностью-мишенью в представляющем интерес геномном локусе клетки, и причем функциональная система CRISPR-Cas C2c3 редактирует геномный локус, изменяя экспрессию гена. В одном из возможных аспектов изобретения в индуцибельной системе CRISPR-Cas C2c3 первая половина индуцибельного гетеродимера является FKBP12 и вторая половина индуцибельного гетеродимера является FRB. В другом варианте осуществления источником энергии индукции является рапамицин.
[00442] Источником энергии индукции может считаться простой индуктор или димеризующий агент. Термин "источник энергии индукции" для согласованности используется на протяжении всего настоящего описания. Источник энергии индукции (или индуктор) восстанавливает C2c1 или C2c3. В некоторых вариантах осуществления источник энергии индукции объединяет две части C2c1 или C2c3 за счет действия двух половин индуцибельного димера. Следовательно, две половины индуцибельного димера объединяются в присутствии источника энергии индукции. Две половины димера не димеризуются (не образуют димер) без источника энергии индукции.
[00443] Таким образом, две половины индуцибельного димера кооперируют с источником энергии индукции для димеризации димера. Это, в свою очередь, восстанавливает C2c1 или C2c3 объединением первой и второй частей C2c1 или C2c3.
[00444] Каждая слитая конструкция фермента CRISPR, включает часть разделенного C2c1 или разделенного C2c3. Они являются слитыми, предпочтительно через линкер, такой как Gly-Ser, описанный в настоящем описании, с одной из двух половин димера. Две половины димера могут быть по существу одинаковыми мономерами, которые вместе образуют гомодимер, или могут быть разными мономерами, которые вместе образуют гетеродимер. В таком случае два мономера можно считать половиной полного димера.
[00445] C2c1 или C2c3 разделены том смысле, что две части фермента C2c1 или C2c3 по существу составляют функциональный C2c1 или C2c3. Этот C2c1 или C2c3 может функционировать как фермент, модифицирующий геном (при формировании комплекса с ДНК-мишенью и направляющей нуклеиновой кислотой), такой как никаза или нуклеаза (разрезающая обе цепи ДНК), или это может быть мертвый C2c1 или мертвый C2c3, который по существу является ДНК-связывающим белком с очень низкой или отсутствующей каталитической активностью, обычно за счет мутации(й) в каталитическом домене.
[00446] Две части разделенного C2c1 или разделенного C2c3 могут считаться N-концевой частью и С-концевой частью разделенного C2c1 или разделенного C2c3. Слияние обычно происходит в точке разделения C2c1 или C2c3. Другими словами, и С-конец N-концевой части разделенного C2c1 или разделенного C2c3 слит с одной из половин димера, в то время как N-конец С-концевой части слит с другой половиной димера.
[00447] C2c1 или C2c3 не должны быть разделены в том смысле, что разрыв создается вновь. Точка разделения обычно конструируется in silico и клонируется в конструкты. Вместе две части конструкции разделенного C2c1 или разделенного C2c3, N-концевая часть и С-концевая часть, образуются целый C2c1 или C2c3, включающий предпочтительно по меньшей мере 70% или более аминокислот дикого типа (или нуклеотидов, кодирующих их), предпочтительно по меньшей мере 80% или более, предпочтительно по меньшей мере 90% или более, предпочтительно по меньшей мере 95% или более, предпочтительно по меньшей мере 99% или более аминокислот дикого типа (или нуклеотидов, кодирующих их). Являются возможными укорочения и предусматриваются мутанты. Нефункциональные домены могут быть полностью удалены. Важным является то, что две части могут быть объединены и что желаемая функция C2c1 или C2c3 восстановлена.
[00448] Димер может быть гомодимером или гетеродимером.
[00449] Один или более, предпочтительно два, NLS можно использовать в качестве функционально связанных с первой конструкцией C2c1. Один или более, предпочтительно два, NES можно использовать в качестве функционально связанных с первой конструкцией C2c1. NLS и/или NES предпочтительно фланкируют слитую конструкцию разделенный C2c1-димер (т.е. половина димера), т.е. NLS может располагаться на N-конце первой конструкции C2c1, и NLS может располагаться на С-конце первой конструкции C2c1. Аналогично, NES может располагаться на N-конце второй конструкции C2c1, и NES может располагаться на С-конце второй конструкции C2c1. Будет понятно, что, когда упоминается N'- или C'-конец, это соответствует 5'- и 3'-концам соответствующей нуклеотидной последовательности.
[00450] Один или более, предпочтительно два, NLS можно использовать в качестве функционально связанных с первой конструкцией C2c3. Один или более, предпочтительно два, NES можно использовать в качестве функционально связанных с первой конструкцией C2c3. NLS и/или NES предпочтительно фланкируют слитую конструкцию разделенный C2c3-димер (т.е. половина димера), т.е. NLS может располагаться на N-конце первой конструкции C2c3, и NLS может располагаться на С-конце первой конструкции C2c3. Аналогично, NES может располагаться на N-конце второй конструкции C2c3, и NES может располагаться на С-конце второй конструкции C2c3. Будет понятно, что, когда упоминается N'- или C''-конец, это соответствует 5'- и 3'-концам соответствующей нуклеотидной последовательности.
[00451] Предпочтительной является следующая организация первой конструкции C2c1: 5'-NLS-(N-концевая часть C2c1)-линкер-(первая половина димера)-NLS-3' или следующая организация конструкции C2c3: 5'-NLS-(N-концевая часть C2c3)-линкер-(первая половина димера)-NLS-3'. Предпочтительной является следующая организация второй конструкции C2c1: 5'-NES-(С-концевая часть C2c1)-линкер-(вторая половина димера)-NES-3' или следующая организация второй конструкции C2c3: 5'-NES-(С-концевая часть C2c3)-линкер-(вторая половина димера)-NES-3'.
[00452] В некоторых вариантах осуществления один или более NES, функциональной связанный со второй конструкцией C2c1, могут быть заменены на NLS. Однако обычно это не является предпочтительным, и в других вариантах осуществления сигнал локализации, функционально связанный со вторым C2c1, представляет собой NES. В некоторых вариантах осуществления один или более NES, функционально связанных со второй конструкцией C2c3, может быть заменен на NLS. Однако обычно это не является предпочтительным, и в других вариантах осуществления сигнал локализации, функционально связанный со вторым C2c3, представляет собой NES.
[00453] Было бы желательно, чтобы NES был функционально связан с N-концевым фрагментом разделенного C2c1 или разделенного C2c3 и чтобы NLS был функционально связан с C-концевым фрагментом разделенного C2c1 или разделенного C2c3. Однако может быть предпочтительной организация, когда NLS функционально связан с N-концевым фрагментом разделенного C2c1 или разделенного C2c3 и NES функционально связан с C-концевым фрагментом разделенного C2c1 или разделенного C2c3.
[00454] NES обеспечивает локализацию второй слитой конструкции C2c1 или C2c3 вне ядра по меньшей мере до тех пор, пока присутствует источник энергии индукции (например, по меньшей мере до тех пор, пока энергетический источник предоставлен индуктору для выполнения его функции). Присутствие индуктора стимулирует димеризацию двух слитых конструкций C2c1 или C2c3 в цитоплазме и делает термодинамически выгодной ядерную локализацию димеризованных первого и второго слитых белков C2c1 или C2c3. Без связи с теорией, заявители полагают, что NES ограничивает вторую слитую конструкцию C2c1 или C2c3 цитоплазмой (т.е. выводит из ядра). NLS на первой слитой конструкции C2c1 или C2c3 способствует его ядерной локализации. В обоих случаях заявители используют NES или NLS, чтобы сдвинуть равновение (равновесие ядерного транспорта) в желаемую сторону. Димеризация обычно происходит вне ядра (очень малая доля ее может произойти в ядре), и NLS на димеризованном комплексе сдвигает равновесие ядерного транспорта в сторону ядерной локализации, так что димеризованный и, таким образом, восстановленный белок C2c1 или C2c3 входит в ядро.
[00455] Способность заявителей восстановить функцию разделенного C2c1 или разделенного C2c3 является преимущественной. Для доказательства концепции используется временная трансфекция и в присутствии источника энергии индукции димеризация происходит на фоновом уровне. Активность не обнаруживается при раздельных фрагментах C2c1 или C2c3. Затем используется стабильная экспрессия посредством лентивирусной доставки используется для разработки и демонстрации того, что подход разделенного C2c1 или C2c3 может быть использован.
[00456] Этот предоставленный подход разделенного C2c1 или разделенного C2c3 обладает преимуществами, поскольку он обеспечивает индуцибельный характер активности C2c1 или C2c3, таким образом, обеспечивая контроль во времени. Более того, могут быть использованы различные последовательности локализации (т.е. предпочтительные NES и NLS), чтобы снизить уровень фоновой активности самособирающихся комплексов. Также могут быть использованы тканеспецифические промоторы, к примеру, один промотор для каждой из слитых конструкций C2c1 или C2c3, для тканеспецифического нацеливания, таким образом, обеспечивая пространственный контроль. Два различных тканеспецифических промотора могут быть использованы, если требуется более тонкая регуляция. Такой же подход может быть использован в отношении промоторов, специфичных к определенной стадии развития, или может быть использована комбинация тканеспецифических промоторов и промоторов, специфичных к определенной стадии развития. Причем одна пара слитых конструкций C2c1 или C2c3 (первая и вторая) находится под контролем (т.е. функционально связана или включает) тканеспецифического промотора, в то время как другие первая и вторая конструкции C2c1 или C2c3 находятся под контролем (т.е. функционально связана или включает) промотора, специфичного к определенной стадии развития.
[00457] Индуцибельная система CRISPR-Cas C2c1 или C2c3 включает одну или более последовательностей ядерной локализации (NLS), как описано в настоящем описании, например, функционально связанных с первой слитой конструкцией C2c1 или C2c3. Эти последовательности ядерной локализации в идеальном случае обладают достаточной силой, чтобы индуцировать накопление указанной слитой конструкции C2c1 или C2c3 в ядре эукариотической клетки в поддающихся обнаружению количествах. Без связи с теорией, полагают, что последовательность ядерной локализации не является необходимой для активности комплекса CRISPR-Cas C2c1 или CRISPR-Cas C2c3 в эукариотах, но также что включение таких последовательностей повышает активность системы, особенно посредством направления молекул нуклеиновой кислоты в ядро, и способствует функционированию представленной двухкомпонентной системы.
[00458] Аналогично, вторая слитая конструкция C2c1 или C2c3 функционально связана с последовательностью ядерного экспорта. Действительно, она может быть связана с одной или более NES. Другими словами, число используемых NES в случае второй слитой конструкции C2c1 или C2c3 предпочтительно составляет 1, 2 или 3. Обычно являются предпочтительными 2 NES, но в некоторых вариантах осуществления достаточно и, таким образом, является предпочтительным 1. Подходящие примеры NLS и NES известны в данной области. Например, предпочтительный сигнал ядерного экспорта (NES) принадлежит белку тирозинкиназе 2 человека. Предпочтительные сигналы являются видоспецифическими.
[00459] В системе, в которой используются FRB и FKBP, предпочтительно, чтобы FKBP был фланкирован последовательностями ядерной локализации (NLS). В системе, в которой используются FRB и FKBP, предпочтительным расположением является: N-концевой C2c1-FRB-NES:C-концевой C2c1-FKBP-NLS или N-концевой C2c3-FRB-NES:C-концевой C2c3-FKBP-NLS. Таким образом, первая слитая конструкция C2c1 или C2c3 содержит С-концевую часть C2c1 или C2c3 и вторая слитая конструкция C2c1 или C2c3 содержит N-концевую часть C2c1 или C2c3.
[00460] Другим преимущественным аспектом настоящего изобретения является то, что оно может быть быстро запущено, т.е. обладает быстрым ответом. Без связи с теорией полагают, что активность C2c1 или C2c3 может быть индуцирована путем димеризации существующей (уже присутствующей) слитой конструкции (за счет контакта с источником энергии индукции) быстрее, чем путем экспрессии (особенно трансляции) новой слитой конструкции. Как таковые, первая и вторая слитые конструкции C2c1 или C2c3 конструкты могут экспрессироваться в клетке-мишени заранее, т.е. до того, как активность C2c1 или C2c3 необходима. Активность C2c1 или C2c3 может контролироваться во времени и затем запускаться добавлением источника энергии индукции, который в идеальном случае действует быстрее (димеризуя гетеродимер и, таким образом, обеспечения активность C2c1 или C2c3), чем посредством экспрессии (включая индукцию транскрипции) C2c1 или C2c3, доставляемого, например, с помощью вектора.
[00461] Термины "C2c1", или "фермент C2c1", или "фермент CRISPR" используются в настоящем описании взаимозаменяемо, если не очевидно обратное. Термины "C2c3", или "фермент C2c3", или "фермент CRISPR" используются в настоящем описании взаимозаменяемо, если не очевидно обратное.
[00462] Заявители демонстрируют, что C2c1 или C2c3 могут быть разделены на два компонента, которые восстанавливают функциональную нуклеазу, будучи объединенными. Используя чувствительные к рапамицину димеризационные домены, заявители создают химически индуцируемый C2c1 или C2c3 для контроля во времени C2c1-зависимого редактирования генома или C2c3-зависимого редактирования генома или модулирования транскрипции. Другими словами, заявители демонстрируют, что C2c1 или C2c3 могут становиться химически индуцируемыми, будучи разделенными на два фрагмента, причем для контроля сборки C2c1 или C2c3 могут быть использованы чувствительные к рапамицину димеризационные домены. Заявители показывают, что объединенные C2c1 или C2c3 могут быть использованы для того, чтобы опосредовать редактирование генома (за счет нуклеазной/никазной активности), как и для модулирования транскрипции (за счет ДНК-связывающего домена - так называемого "мертвого C2c1" или "мертвого C2c3").
[00463] Как таковое, использование димеризационных доменов, чувствительных к рапамицину, является предпочтительным. Обратная сборка может быть определена с помощью восстановления связывающей активности. Когда C2c1 или C2c3 функционирует как никаза или образует двуцепочечный разрыв, в настоящем описании представлены подходящие проценты для сравнения с диким типом.
[00464] Обработка рапамицином может длиться 12 суток. Доза может составлять 200 нМ. Эта временная и/или молярная дозировка может являться примером подходящей дозы для линии эмбриональных клеток почки человека 293FT (HEK293FT) и может быть использована для других клеточных линий. Это число может быть изменено для терапевтического использования in vivo, будучи выраженным, например, в мг/кг. Однако, также предусматривается, что может быть использована также стандартная доза приема рапамицина индивидуумом. Под стандартной дозировкой подразумевается нормальная терапевтическая доза рапамицина или доза, используемая при основном назначении (т.е. доза, используемая, когда рапамицин принимается для предотвращения отторжения органов).
[00465] Следует заметить, что предпочтительная организация частей C2c1-FRB/FKBP или C2c3-FRB/FKBP такова, что части находятся по отдельности и неактивны до тех пор, пока индуцированная рапамицином димеризации FRB и FKBP не приводит к сборке функциональной полноразмерной нуклеазы C2c1 или C2c3. Таким образом, предпочтительно, чтобы слитая конструкция C2c1 или C2c3 была связана с первой половиной индуцибельного гетеродимера.
[00466] Чтобы ограничить фрагмент C2c1(N)-FRB или C2c3(N)-FRB цитоплазмой, где меньше вероятность димеризации с локализованным в ядре фрагментом C2c1(C)-FKBP или C2c3(C)-FKBP, предпочтительно использовать для C2c1(N)-FRB один NES из белка тирозинкиназы 2 человека (C2c1(N)-FRB-NES) или для C2c3(N)-FRB один NES из белка тирозинкиназы 2 человека (C2c3(N)-FRB-NES). В присутствии рапамицина C2c1(N)-FRB-NES димеризуется с C2c1(C)-FKBP-2xNLS или C2c3(N)-FRB-NES димеризуется с C2c3(C)-FKBP-2xNLS, восстанавливая полноразмерный белок C2c1 или C2c3, который смещает баланс ядерного транспорта в сторону импорта в ядро и позволяет нацеливание на ДНК.
[00467] Высокая дозировка C2c1 или C2c3 может увеличить частоту инсерций-делеций в последовательностях, не являющихся мишенями, которые имеют немного несоответствующих направляющей цепи оснований. Такие последовательности являются особенно чувствительными, если несоответствующие основания не последовательны и/или находятся снаружи от последовательности-затравки направляющей нуклеиновой кислоты. Соответственно, временной контроль активности C2c1 или C2c3 может быть использован для снижения дозировки в длительных экспериментах по экспрессии и, следовательно, может обеспечить уменьшение нецелевых инсерций-делеций по сравнению с конститутивно активным C2c1 или C2c3.
[00468] Предпочтительной является вирусная доставка. В частности, предусмотрена доставка с помощью лентивирусного вектора или вектора аденоассоциированного вируса. Заявители создают разделенный C2c1 или разделенный C2c3 лентивирусный конструкт, подобный плазмиде lentiCRISPR. Эти разделенные части должны быть достаточно малы, чтобы соответствовать ограничениям в размере, накладываемым аденоассоциированным вектором длиной приблизительно 4.7 т.п.о.
[00469] Заявители показывают, что стабильная низкокопийная экспрессия разделенного C2c1 или разделенного C2c3 может быть использована для индукции значительных инсерций-делеций в локусе-мишени без значительных мутаций в участках, не являющихся мишенями. Заявители клонируют фрагменты C2c1 (2 части на основе разделенной системы 5, описанной в настоящем описании) или фрагменты C2c3.
[00470] Также можно использовать мертвый C2c1 или C2c3, включающий трансактивационный домен VP64, например, добавляемый к C2c1(C)-FKBP-2xNLS (убитый-C2c1(C)-FKBP-2xNLS-VP64) или C2c3(C)-FKBP-2xNLS (убитый-C2c3(C)-FKBP-2xNLS-VP64). Активация транскрипции индуцируется посредством VP64 в присутствии рапамицина, индуцируя димеризацию слитой конструкции C2c1(C)-FKBP и слитой конструкции C2c1(N)-FRB или слитой конструкции C2c3(C)-FKBP и слитой конструкции C2c3(N)-FRB. Другими словами, заявители проверяют возможность индукции разделенного белка мертвый C2c1-VP64 или разделенного белка мертвый C2c3-VP64 и демонстрируют, что активация транскрипции индуцируется разделенным белком мертвый C2c1-VP64 или разеделнным белком мертвый C2c3-VP64 в присутствии рапамицина. Как таковой, предоставленный индуцибельный C2c1 или C2c3 может быть связан с одним или более функциональными доменами, таким как транскрипционный активатор или репрессор, или нуклеаза (такая как FokI). Функциональный домен может быть связан или слит с одной частью разделенного C2c1 или разделенного C2c3.
[00471] Предпочтительная организация является следующей: первая конструкция организована следующим образом: 5'-1-й сигнал локализации-(N-концевая часть C2c1)-линкер-(1-я часть димера)- 1-й сигнал локализации-3' или первая конструкция C2c3 организована следующим образом: 5'-1-й сигнал локализации-(N-концевая часть C2c3)-линкер-(1-я часть димера)-1-й сигнал локализации-3', и вторая конструкция C2c1 организована следующим образом: 5'-2-й сигнал локализации-(2-я половина димера)-линкер-(С-концевая часть C2c1)-2-й сигнал локализации-функциональный домен-3' или вторая конструкция C2c3 организована следующим образом: 5'-2-й сигнал локализации-(2-я половина димера)-линкер-(С-концевая часть C2c3)-2-й сигнал локализации-функциональный домен-3'. Здесь функциональный домен размещен на 3'-конце второй конструкции C2c1 или C2c3. В качестве альтернативы функциональный домен может быть размещен на 5'-конце первой конструкции C2c1 или C2c3. Один или более функциональных доменов могут быть размещены на 3'-конце или 5'-конце, или на обоих концах. Подходящий промотор предпочтительно находится в конструкциях выше этих последовательностей. Сигналы локализации могут представлять собой NLS или NES, но разные сигналы не могут находиться на одной конструкции.
[00472] В одном из возможных аспектов изобретение относится к индуцибельной системе CRISPR-Cas C2c1 или C2c3, описанной в настоящем описании, в которой C2c1 или C2c3 обладает сниженной по меньшей мере на 97% или 100% нуклеазной активностью, по сравнению с C2c1 или C2c3, не содержащим по меньшей мере одну мутацию.
[00473] Соответственно, также предпочтительно, чтобы C2c1 или C2c3 был мертвым C2c1 или мертвым C2c3. В идеальном случае, разделение должно быть таким, чтобы не затрагивался каталитический домен(ы). В случае мертвого C2c1 или мертвого C2c3 предполагается, что связывание с ДНК происходит, но расщепление или никазная активность отсутствует.
[00474] В одном из возможных аспектов изобретение относится к индуцибельной системе CRISPR-Cas C2c1 или C2c3, описанной в настоящем описании, в которой один или более функциональных доменов связаны с C2c1 или C2c3. Этот функциональный домен может быть связан (например, связан или слит с) одной частью разделенного C2c1 или обеими частями, или одной частью разделенного C2c3 или обеими частями. Один из них может быть связана с каждой из двух частей разделенного C2c1 или разделенного C2c3. Следовательно, они могут быть обычно предоставлены как части первой и/или второй слитой конструкции C2c1 или C2c3, будучи слитыми с этой конструкцией. Функциональные домены обычно слиты через линкер, такой как линкер GlySer, описанный в настоящем описании. Один или более функциональных доменов могут представлять собой трансактивационный домен или транскрипционный репрессорный домен. Хотя функциональные домены могут различаться, предпочтительно, чтобы все функциональные домены были либо трансактивационными, либо репрессорными доменами, и чтобы они не использовались в одной смеси.
[00475] Трансактивационный домен может представлять собой: VP64, p65, MyoDl, HSF1, RTA или SET7/9.
[00476] В одном из возможных аспектов изобретение относится к индуцибельной системе CRISPR-Cas C2c1 или C2c3, описанной в настоящем описании, в которой один или более функциональных доменов, связанных с C2c1 или C2c3, являются транскрипционным репрессорным доменом.
[00477] В одном из возможных аспектов изобретение относится к индуцибельной системе CRISPR-Cas C2c1 или C2c3, описанной в настоящем описании, в которой репрессорным доменом является домен KRAB.
[00478] В одном из возможных аспектов изобретение относится к индуцибельной системе CRISPR-Cas C2c1 или C2c3, описанной в настоящем описании, в которой транскрипционным репрессорным доменом является домен NuE, домен NcoR, домен SID или домен SID4X.
[00479] В одном из возможных аспектов изобретение относится к индуцибельной системе CRISPR-Cas C2c1 или C2c3, описанной в настоящем описании, в которой один или более функциональных доменов связаны с адаптерным белком, обладающим одним или более видами активности, включая метилазную активность, деметилазную активность, трансактивационную активность, транскрипционную репрессорную активность, активность в отношении высвобождения транскрипционных факторов, гистонмодифицирующую активность, РНК-расщепляющую активность, ДНК-расщепляющую активность, ДНК-интегразную активность или активность в отношении связывания нуклеиновых кислот.
[00480] Гистонмодифицирующие домены также являются предпочтительными в некоторых вариантах осуществления. Типовые гистонмодифицирующие домены обсуждаются далее. Транспозазные домены, домены аппарата, осуществляющего гомологичную рекомбинацию, рекомбиназные домены и/или интегразные домены также являются предпочтительными в качестве функциональных доменов в рамках настоящего изобретения. В некоторых вариантах осуществления активность в отношении интеграции в ДНК предусматривает домены аппарата, осуществляющего гомологичную рекомбинацию, интегразные домены, рекомбиназные домены и/или транспозазные домены.
[00481] В одном из возможных аспектов изобретение относится к индуцибельной системе CRISPR-Cas C2c1 или C2c3, описанной в настоящем описании, расщепляющая ДНК активность которой происходит за счет наличия нуклеазы.
[00482] В одном из возможных аспектов изобретение относится к индуцибельной системе CRISPR-Cas C2c1 или C2c3, описанной в настоящем описании, в которой нуклеаза представляет собой нуклеазу FokI.
[00483] Использование таких функциональных доменов, которые являются предпочтительными для системе с разделенным C2c1 или разделенным C2c3 по настоящему изобретению, также подробно обсуждается в статье Konermaim et al. ("Genome-scale transcriptional activation with an engineered CRISPR-Cas9 complex" Nature, опубликованная 11 декабря 2014 года).
[00484] Система по настоящему изобретению может использоваться с любой направляющей последовательностью.
[00485] В некоторых вариантах осуществления могут использоваться модифицированные направляющие последовательности. Особенно предпочтительны направляющие последовательности, являющиеся реализацией концепции из упомянутой статьи Konermann Nature 11 Dec 2014. Эти направляющие последовательности модифицированы так, что в них добавлены связывающиеся с белком части РНК (такие как аптамеры). Соответствующие РНК-связывающие белковые домены могут использоваться для узнавания РНК и привлечения к направляющей последовательности функциональных доменов, таких как описанные в настоящем описании. Это может быть использовано, в первую очередь, вместе в случае мертвого C2c1 или мертвого C2c3, приводя к транскрипционной активации или репрессии или расщеплению ДНК нуклеазами, такими как FokI. Использование таких направляющих последовательностей в комбинации с мертвым C2c1 или мертвым C2c3 обладает высокой эффективностью, особенно в случае, когда C2c1 или C2c3 сам связан с собственным функциональным доменом, как описано в настоящем описании. Когда индуцируется восстановление мертвого C2c1 или мертвого C2c3 (с или без собственного связанного функционального домена) в соответствии с настоящим изобретением, т.е. разделенного C2c1 или разделенного C2c3, этот инструмент особенно полезен.
[00486] Направляющая РНК (гРНК), которая предпочтительно используется в рамках настоящего изобретения, может содержать направляющую последовательность, способную гибридизоваться с последовательностью-мишенью в представляющем интерес геномном локусе клетки, причем гРНК модифицирована путем вставки отдельной последовательности(ей) РНК, связывающейся с одним или более адаптерными белками, при этом адаптерный белок связан с одним или более функциональными доменами. C2c1 или C2c3 может содержать по меньшей мере одну мутацию, такую что фермент C2c1 или C2c3 обладает не более чем 5% нуклеазной активности фермента C2c1 или C2c3, не содержащего мутацию; и/или по меньшей мере один или более NLS. Также предусматривается не встречающаяся в природе или сконструированная способами инженерии композиция, включающая: одну или более направляющих РНК (гРНК), содержащих направляющую последовательность, способную гибридизоваться с последовательностью-мишенью в представляющем интерес геномном локусе клетки, фермент C2c1 или C2c3, содержащий по меньшей мере один или более NLS, причем фермент C2c1 или C2c3 может содержать по меньшей мере одну мутацию, такую что фермент C2c1 или C2c3 обладает не более чем 5% нуклеазной активности фермента C2c1 или C2c3, не содержащего по меньшей мере одну мутацию, при этом по меньшей мере гРНК модифицирована путем вставки отдельной последовательности(ей) РНК, связывающейся с одним или более адаптерными белками, при этом адаптерный белок связан с одним или более функциональными доменами.
[00487] гРНК предпочтительно модифицированы путем вставки отдельной последовательности(ей) РНК, которая связывается с одним или более адаптерными белками. Вставка отдельной последовательности(ей) РНК, которая связывается с одним или более адаптерными белками, является предпочтительно последовательностями аптамеров, специфичных к одному или более адаптерным белкам. Адаптерный белок может являться: MS2, PP7, Qβ, F2, GA, fr, JP501, M12, R17, BZ13, JP34, JP50G, KU1, МU, MX1, TW18, VK, SP, FI, ID2, NL95, TW19, AP205, ϕСb5, ϕСb8г, ϕСb12г, ϕСb2Зr, 7s и PRR1. Могут быть полезны клеточные линии, стабильно экспрессирующие, среди прочего, разделенный мертвый C2c1 или разделенный мертвый C2c3.
[00488] Заявители показывают, что белок C2c1 или C2c3 может быть разделен на два различных фрагмента, который составляет функциональную полноразмерную нуклеазу C2c1 или C2c3 при объединении с помощью химической индукции. Архитектура разделенного C2c1 или разделенного C2c3 будет полезна при различных способах применения. Например, разделенный C2c1 может позволить генетические стратегии ограничения активности C2c1 определенными клеточными популяциями, путем помещения каждого фрагмента под контроль тканеспецифического промотора или разделенный C2c3 может позволить генетические стратегии ограничения активности C2c1 определенными клеточными популяциями, путем помещения каждого фрагмента под контроль другого тканеспецифического промотора. Кроме того, могут быть использованы различные химически индуцируемые димеризационные домены, такие как APA и гиббереллин.
[00489] Источником энергии индукции предпочтительно является химическая индукция.
[00490] Положение или место разделение представляет собой точку, в которой первая часть фермента C2c1 или C2c3 отделена от второй части. В некоторых вариантах осуществления первая часть включает или кодирует аминокислоты с 1 по Х, в то время как вторая часть включает аминокислоты с Х+1 по последнюю. В этом примере нумерация непрерывна, но это может быть не всегда необходимо, поскольку аминокислоты (или кодирующие их нуклеотиды) могут быть отрезаны с каждого из разделенных концов, так чтобы присутствовала достаточная ДНК-связывающая активность и, необязательно, ДНК-никазная или расщепляющая активность, например, по меньшей мере 40%, 50%, 60%, 70%, 80%, 90% или 95% активность, по сравнению с C2c1 дикого типа или C2c3 дикого типа.
[00491] Иллюстративная нумерация, описанная в настоящем описании, может относиться к белку дикого типа, предпочтительно C2c1 дикого типа или C2c3 дикого типа. Однако предусматривается, что могут использоваться мутанты C2c1 дикого типа, такого как белок C2c1 Bacillus, или мутанты C2c3 дикого типа. Нумерация может не соответствовать точно нумерации C2c1 или C2c3, в частности, могут использоваться некоторые N- и C-концевые укорочения или делеции, но это можно учесть с помощью стандартных инструментов для выравнивания последовательностей. Ортологи также являются предпочтительными инструментами выравнивания последовательностей.
[00492] Таким образом, положение разделения может быть выбрано с использованием стандартных способов, например, на основе данных кристаллической структуры и/или предсказания структуры на основе компьютерного моделирования.
[00493] В идеальном случае положение разделения должно находиться внутри петли. Предпочтительно, чтобы положение разделения находилось там, где прерывание аминокислотной последовательности не приводит к частичному или полному разрушению структурного элемента (например, альфа-спирали или бета-листа). Часто лучшими вариантами являются неструктурированные участки (участки, которые не видны в кристаллических структурах, поскольку эти участки не структурированы в достаточной степени, чтобы быть "замороженными" в кристалле). Заявители могут для примера делать разделения в неструктурированных участках, которые экспонированы на поверхность C2c1 или C2c3.
[00494] Заявители могут следовать следующей методике, которая предоставлена в качестве предпочтительного примера и руководства. Поскольку неструктурированные участки не видны на кристаллических структурах, заявители соотносят окружающую аминокислотную последовательность кристалла с первичной аминокислотной последовательностью C2c1 или C2c3. Каждый неструктурированный участок может состоять из, к примеру, 3-10 аминокислот, которые не видны в кристаллической структуре. В таком случае заявители делают разделение в этом участке аминокислотной последовательности. Чтобы включить больше потенциальных участков разделения, заявители включают разделения, расположенные на петлях снаружи от C2c1 или C2c3, используя те же критерии, что и в случае неструктурированных участков.
[00495] В некоторых вариантах осуществления положение разделения находится на наружной петле C2c1 или C2c3. В других предпочтительных вариантах осуществления положение разделения находится в неструктурированном участке C2c1 или C2c3. Неструктурированный участок обычно представляет собой высокоподвижный участок снаружи от петли, структура которого не может быть определена на основе кристаллической решетки.
[00496] Как только положение разделения идентифицировано, могут быть спроектированы подходящие композиции.
[00497] Обычно NES на находится на N-конце первой части разделенной аминокислотной последовательности (или на 5'-концевом нуклеотиде, кодирующем ее). В таком случае NLS расположен на С-конце второй части разделенной аминокислотной последовательности (или на 3'-концевом нуклеотиде, кодирующем ее), таким образом, первая слитая конструкция C2c1 или C2c3 может быть функционально связана с одним или более NES, а вторая слитая конструкция C2c1 или C2c3 может быть функционально связана с одним или более NLS.
[00498] Конечно, может предусматриваться обратный порядок, при котором NLS находится на N-конце первой части разделенной аминокислотной последовательности (или на 5'-концевом нуклеотиде, кодирующем ее). В таком случае NES расположен на С-конце второй части разделенной аминокислотной последовательности (или на 3'-концевом нуклеотиде, кодирующем ее), таким образом, первая слитая конструкция C2c1 или C2c3 может быть функционально связана с одним или более NLS, и вторая слитая конструкция C2c1 или C2c3 может быть функционально связана с одним или более NES.
[00499] Разделенные системы, в которых две части (каждая из сторон разделенной системы) имеют приблизительно одинаковую длину, могут обладать преимуществами в отношении упаковки. Например, считается, что поддерживать стехиометрию между двумя частями проще, если транскрипты имеют приблизительно одинаковый размер.
[00500] В конкретных примерах N- и С-концевые части кодон-оптимизированного C2c1 человека, такого как C2c1, слиты с димеризационными доменами FRB и FKBP, соответственно. Такая организация может быть предпочтительной. Их можно поменять местами (т.е. N-конец слит с FKBP и C-конец слит с FRB). В конкретных примерах N- и С-концевые части кодон-оптимизированного C2c3 человека, такого как C2c3, слиты с димеризационными доменами FRB и FKBP, соответственно. Такая организация может быть предпочтительной. Их можно поменять местами (т.е. N-конец слит с FKBP и C-конец слит с FRB).
[00501] Использование линкеров, таких как (GGGGS)3, является предпочтительным для отделения фрагмента C2c1 или фрагмента C2c3 от димеризационного домена. Остатки глицина являются наиболее гибкими, и остатки серина увеличивают вероятность того, что линкер будет находиться на внешней поверхности белка. В качестве альтернативы могут быть использованы линкеры (GGGGS)6, (GGGGS)9 или (GGGGS)12. Другими предпочтительными альтернативами являются GGGGS)1, (GGGGS)2, (GGGGS)4, (GGGGS)5, (GGGGS)7, (GGGGS)8, (GGGGS)10 или (GGGGS)11.
[00502] Например, (GGGGS)3 может быть включен между N-концом фрагмента C2c1 или фрагмента C2c3 и FRB. Например, (GGGGS)3 может быть включен между FKBP и C-концом фрагмента C2c1 или фрагмента C2c3.
[00503] Возможны также и альтернативные линкеры, но считается, что высокоподвижные линкеры функционируют лучше всего, обеспечивая максимальную вероятность объединения C2c1 или C2c3 и, таким образом, восстановления активности C2c1 или C2c3. Одной из альтернатив является использование в качестве линкера NLS нуклеоплазмина.
[00504] Линкер также может использоваться для соединения C2c1 или C2c3 и любого функционального домена. Опять же, между C2c1 или C2c3 или функциональным доменом может быть использован линкер (GGGGS)3 (или 6-, 9- или 12-кратные повторы) или NLS нуклеоплазмина.
[00505] Предусматриваются альтернативы системе FRB/FKBP, например, система АВА и гиббереллин.
[00506] Соответственно, предпочтительными примерами семейства FKBP являются любые из следующих индуцибельных систем: FKBP, который димеризуется с кальциневрином А (CNA) в присутствии FK506, FKBP, который димеризуется с CyP-Fas в присутствии FKCsA, FKBP, который димеризуется с FRB в присутствии рапамицина, GyrB, который димеризуется с GryB в присутствии кумермицина, GAI, который димеризуется с GID1 в присутствии гибберреллина, Snap-tag, который димеризуется с HaloTag в присутствии HaXS.
[00507] Альтернативы внутри семейства FKBP также являются предпочтительными. Например, FKBP, который гомодимеризуется (т.е. один FKBP димеризуется с другим FKBP) в присутствии FK1012. Таким образом, также предусматривается не встречающаяся в природе или сконструированная способами инженерии система C2c1 или CRISPR-Cas C2c3, содержащая:
первую слитую конструкцию C2c1 или первую слитую конструкцию C2c3, связанную с первой половиной индуцибельного гомодимера, и вторую слитую конструкцию C2c1 или вторую слитую конструкцию C2c3, связанную со второй половиной индуцибельного гомодимера,
причем первая слитая конструкция C2c1 или первая слитая конструкция C2c3 функционально связана с одним или более NLS, причем первая слитая конструкция C2c1 или первая слитая конструкция C2c3 функционально связана с (необязательно одним или более) сигналом(ами) ядерного экспорта,
причем взаимодействие с источником энергии индукции объединяет первую и вторую половины индуцибельного гомодимера,
при этом объединение первой и второй половин индуцибельного гомодимера позволяет первой и второй слитым конструкциям C2c1 восстановить функциональную систему CRISPR-Cas C2c1 или первой и второй слитым конструкциям C2c3 восстановить функциональную систему CRISPR-Cas C2c3,
причем система C2c1 или CRISPR-Cas C2c3 содержит направляющую РНК (гРНК), содержащую направляющую последовательность, способную гибридизоваться с последовательностью-мишенью в геномном локусе-мишени клетки, и функциональная система C2c1 или CRISPR-Cas C2c3 связывается с последовательностью-мишенью и необязательно редактирует геномный локус, изменяя экспрессию гена.
[00508] В одном варианте осуществления гомодимер предпочтительно представляет собой FKBP, и источником энергии индукции предпочтительно является FK1012. В другом варианте осуществления гомодимер предпочтительно представляет собой GryB, и источником энергии индукции предпочтительно является кумермицин. В другом варианте осуществления гомодимер предпочтительно представляет собой АВА, и источником энергии индукции предпочтительно является гиббереллин.
[00509] В других вариантах осуществления димер является гетеродимером. Предпочтительными вариантами гетеродимеров являются любые из следующих индуцибельных систем: FKBP, который димеризуется с кальциневрином А (CNA) в присутствии FK506, FKBP, который димеризуется с CyP-Fas в присутствии FKCsA, FKBP, который димеризуется с FRB в присутствии рапамицина, GyrB, который димеризуется с GryB в присутствии кумермицина, GAI, который димеризуется с GID1 в присутствии гибберреллина, Snap-tag, который димеризуется с HaloTag в присутствии HaXS.
[00510] Заявители использовали FKBP/FRB, поскольку эта система хорошо охарактеризована и оба домена достаточно малы (менее 100 аминокислот), что облегчает упаковку. Более того, рапамицин используется в течение длительного времени, и его побочные эффекты хорошо известны. Большие димеризационные домены (более 300 а.к.) также должны работать, но этом случае могут требоваться более длинные линкеры, чтобы сделать возможным восстановление C2c1 или C2c3.
[00511] В статье Paulmurugan and Gambhir (Cancer Res, August 15, 2005 65; 7413) обсуждается история системы FRB/FKBP/рапамицин. Другой полезной статьей является статья Crabtree et al. (Chemistry & Biology 13, 99-107, Jan 2006).
[00512] Например, конструируют один вектор, одну экспрессирующую кассету (плазмиду). гРНК находится под контролем промотора U6. Используются два различных разделенных C2c1 или разделенных C2c3. Конструкция разделенного C2c1 или разделенного C2c3 основана на первой слитой конструкции C2c1 или C2c3, фланкируемой NLS, в которой FKBP присоединен к С-концевой части разделенного C2c1 или разделенного C2c3 через глицин-сериновый линкер; и на второй слитой конструкции C2c1 или C2c3, фланкируемой NES, в которой FRB присоединен к N-концевой части разделенного C2c1 или разделенного C2c3 через глицин-сериновый линкер. Разделение первой и второй слитых конструкций C2c1 происходит во время транскрипции с помощью Р2А. Уровень образования инсерций-делеций в случае разделенного C2c1 или разделенного C2c3 в присутствии рапамицина является приблизительно таким же, как и в случае дикого типа, но значительно ниже, чем в случае дикого типа в отсутствие рапамицина.
[00513] Соответственно, предусматривается единственный вектор. Вектор содержит:
первую слитую конструкцию C2c1 или первую слитую конструкцию C2c3, соединенную с первой половиной индуцибельного димера, и вторую слитую конструкцию C2c1 или вторую слитую конструкцию C2c3, соединенную со второй половиной индуцибельного димера,
причем первая слитая конструкция C2c1 или первая слитая конструкция C2c3 функционально связана с одним или более сигналами ядерной локализации, причем первая слитая конструкция C2c1 или первая слитая конструкция C2c3 функционально связана с одним или более сигналами ядерного экспорта,
причем взаимодействие с источником энергии индукции объединяет первую и вторую половины индуцибельного гомодимера,
при этом объединение первой и второй половин индуцибельного гомодимера позволяет первой и второй слитым конструкциям C2c1 восстановить функциональную систему CRISPR-Cas C2c1 или первой и второй слитым конструкциям C2c3 восстановить функциональную систему CRISPR-Cas C2c3,
причем система C2c1 или CRISPR-Cas C2c3 содержит направляющую РНК (гРНК), содержащую направляющую последовательность, способную гибридизоваться с последовательностью-мишенью в геномном локусе-мишени клетки, и функциональная система C2c1 или CRISPR-Cas C2c3 связывается с последовательностью-мишенью и необязательно редактирует геномный локус, изменяя экспрессию гена.
Эти элементы предпочтительно предоставляются на единственной конструкции, например, в составе экспрессирующей кассеты.
[00514] Первая слитая конструкция C2c1 или первая слитая конструкция C2c3 предпочтительно фланкирована по меньшей мере одним сигналом ядерной локализации на каждом конце. Вторая слитая конструкция C2c1 или вторая слитая конструкция C2c3 предпочтительно фланкирована по меньшей мере одним сигналом ядерного экспорта на каждом конце.
[00515] Также предусматривается способ лечения индивидуума, нуждающегося в этом, включающий индукцию редактирования гена путем трансформации индивидуума полинуклеотидом, кодирующим систему, или любым векторов, описанных в настоящем описании, и введение индивидууму источника энергии индукции. В одном из возможных аспектов применения указанного способа также может быть предоставлена матрица репарации, например, доставляемая вектором, содержащим указанную матрицу репарации.
[00516] Также предусматривается способ лечения индивидуума, нуждающегося в этом, включающий индукцию редактирования гена путем трансформации индивидуума полинуклеотидом, кодирующим систему, или любым векторов, описанных в настоящем описании, причем упомянутый полинуклеотид кодирует или включает каталитически неактивный C2c1 или C2c3 и один или более ассоциированных функциональныъ доменов; кроме того, способ включает введение индивидууму источника энергии индукции.
[00517] Также предусматриваются композиции, содержащие систему по настоящему изобретению для применения в упомянутом способе лечения. Также предусматривается способ использования системы по настоящему изобретению в производстве лекарственных средств для таких способов лечения.
[00518] Примеры синдромов, которые можно лечить с помощью системы по настоящему изобретению, описаны в настоящем описании или в документах, цитированных в настоящем описании.
[00519] Единственный вектор может включать агент, разделяющий траснкрипт, например, Р2А. Р2А разделяет транскрипт на два, разделяя первую и вторую слитые конструкции C2c1 или первую и вторую слитые конструкции C2c3. Разделение происходит за счет пропускания рибосомы. По сути, рибосома пропускает аминокислоту во время трансляции, что вызывает обрыв аминокислотной цепи и приводит к образованию двух различных полипептидов/белков. Использование единственного вектора также полезно, когда низкая фоновая активность не является необходимой, но желательна высокая активность индукции.
[00520] Одним из примеров может служить поколение линий клональных эмбриональных стволовых клеток. Нормальная процедура состоит во временной трансфекции плазмидами, кодирующими C2c1 дикого типа или C2c1-никазы, или C2c3 дикого типа или C2c3-никазы. Эти плазмиды продуцируют молекулы C2c1 или C2c3, которые остаются активными в течение нескольких дней и имеют более высокую вероятность нецелевой активности. Использование единственного экспрессинного вектора с разделенным C2c1 или разделенным C2c3 позволяет ограничить "высокий" уровень активности C2c1 или C2c3 до более короткого периода времени (например, одна доза индуктора, такого как рапамицин). Без непрерывного (ежедневного) введения индуктора (например, рапамицина) активность единственного экспрессионного вектора с разделенным C2c1 или разделенным C2c3 является низкой, таким образом, снижая вероятность возникновения нежелательных нецелевых эффетов.
[00521] Пик индуцированной активности C2c1 или C2c3 обладает преимуществами в некоторых вариантах осуществления и может быть легко достигнут с использованием единственного доставляющего вектора, но это также возможно при использовании двойной векторной системы (каждый вектор доставляет одну половину разделенного C2c1 или разделенного C2c3). Пик может представлять собой высокую активность и короткий период времени, обычно совпадающий со временем полужизни индуктора.
[00522] Соответственно, предусматривается способ получения линий клональных стволовых клеток, включающий трансфекцию одной или более эмбриональных стволовых клеток полинуклеотидом, кодирующим систему по настоящему изобретению, или одним из векторов по настоящему изобретению для экспрессии разделенного C2c1 или разделенного C2c3 и введение или взаимодействие одной или более стволовых клеток с источником энергии индукции по настоящему изобретению для индукции восстановления C2c1 или C2c3. Может быть предоставлена матрица репарации.
[00523] Так же как и в случае всех способов, описанных в настоящем описании, желательно, чтобы подходящая гРНК или направляющие молекулы требовались.
[00524] В случае, когда функциональные домены и т.п. "ассоциированы" с одной и второй частью фермента, они обычно представляют собой слитые конструкции. Термин "ассоциирован с" используется в настоящем описании в отношении того, как одна молекула ассоциирует с другой, например, между частями C2c1 и функциональным доменом или между частями C2c3 и функциональным доменом. В случае белок-белковых взаимодействий эта ассоциация может рассматриваться с точки зрения узнавания, подобно тому, как антитело распознает эпитоп. Альтернативно, один белок может быть ассоциирован с другим белком слиянием обоих белков, например, когда одна субъединица слита с другой субъединицей. Слияние обычно осуществляется добавлением аминокислотной последовательности одной субъединицы к аминокислотной последовательности другой, к примеру, за счет сплайсинга обеих нуклеотидных последовательностей, кодирующих каждый белок или субъединицу. Альтернативно, это может, по существу, рассматриваться как связывание двух молекул или прямое соединение, как например, слитый белок; в любом случае в состав слитого белка может входить линкер между двумя представляющими интерес субъединицами (т.е. между ферментом и функциональным доменом или между адаптерным белком и функциональным доменом). Таким образом, в некоторых вариантах осуществления части C2c1 или C2c3 связаны с функциональными доменами, поскольку они слиты друг с другом необязательно через промежуточный линкер. Примеры линкеров включают упомянутые линкеры GlySer.
[00525] Другие примеры индукторов включают свет и гормоны. В случае света, индуцибельные димеры могут являться гетеродимерами и содержать первую индуцируемую светом половину димера и вторую (или комплементарную) индуцируемую светом половину димера. Предпочтительным примером первой и второй индуцируемых светом половин димера является система CIBI и CRY2. Домен CIBI представляет собой гетеродимерный белок, связывающийся с светочувствительным криптохромом 2 (CRY2).
[00526] В другом примере синяя отвечающая на свет магнитная димеризационная система (pMag и nMag) может быть слита с двумя частями разделенного белка C2c1 или разделенного белка C2c3. В ответ на стимуляцию светом pMag и nMag димеризуются и происходит сборка C2c1 или C2c3. Например, такая система описана в отношении Cas9 в статье Nihongaki et al. (Nat. Bjofechnol. 33, 755-790, 2015).
[00527] Изобретение предусматривает, что источником энергии индукции может быть тепловая энергия, ультразвук, энергия электромагнитного излучения или химическая энергия. В предпочтительном варианте осуществления изобретения источник энергии индукции может представлять собой абсцизовую кислоту (АВА), доксициклин (DOX), кумат, рапамицин, 4-гидрокситамоксифен (4-ОНТ), эстроген или экдизон. Изобретение предусматривает по меньшей мере один переключатель, который может быть выбран из группы, включающей системы, индуцируемые антибиотиками, системы, индуцируемые электромагнитным излучением, индуцибельные системы на основе ядерных рецепторов и индуцибельные системы на базе гормонов. В более предпочтительном варианте осуществления может быть выбран по меньшей мере один переключатель из группы, включающей тетрациклин/доксициклиновую (Tet/DOX) индуцибельную систему, индуцируемую светом систему, индуцируемую АВА систему, куматовую репрессор/операторную индуцибельную систему, 4-ОНТ/экстрогеновую индуцибельную систему, индуцибельную систему на основе экдизона и индуцируемую FKBP12/FRAP (комплексом FKBP12-рапамицин) систему. Такие индукторы также обсуждаются в настоящем описании и в PCT/US20I3/051418, включенной в настоящее описание в качестве ссылки.
[00528] В целом, C2c1 или C2c3, либо дикого типа, либо никазы, либо мертвые C2c1/C2c3 (связанные или не связанные с функциональными доменами) могут применяться с использованием подхода разделенного C2c1 или разделенного C2c3 по настоящему изобретению.
[00529] В качестве следующего примера, могут быть созданы слитые конструкции разделенного C2el или C2c3 с флуоресцентным белком, таким как GFP. Это могло бы позволить визуализировать геномные локусы (см. "Imaging of Genomic Loci in Living Human Cells by an Optimized CRISPR/Cas System" Chen В et al. Cell 2013), но индуцируемым способом. По существу, в некоторых вариантах осуществления одна или более частей C2сl или C2c3 могут быть связаны (в частности, слиты) с флуоресцентным белком, например, GFP.
[00530] Следующие эксперименты направлены на то, чтобы выяснить, есть ли различие в нецелевом расщеплении между C2сl дикого типа и разделенным C2c1 или C2с3 дикого типа и разделенным C2c3. Для этого заявители используют временную трансфекцию плазмид с C2c1 wt и разделенным C2c1 или с C2c3 wt или разделенным C2c3 и осуществляют сбор в различные моменты времени. Заявители проводят поиск нецелевой активации после нахождения набора образцов, в которых целевое разрезание находится в пределах 5%. Заявители создают клеточные линии со стабильной экспрессией фермента дикого типа или разделенного фермента (C2c1 или C2с3) без направляющих нуклеиновых кислот (с помощью лентивируса). После селекции посредством антибиотика доставляются направляющие нуклеиновые кислоты с помощью отдельного лентивируса, и проводят сбор клеток в различные моменты времени для определения целевого и нецелевого разрезания.
[00531] Заявители вводят дестабилизирующую последовательность (PEST, см. "Use of mRNA- and protein- destabilizing elements to develop a highly responsive reporter system" Voon DC et al. Nucleic Acids Research 2005) в фрагмент FRB(N)C2c1-NES или фрагмент FRB(N)C2c3-NES, что способствует более быстрой деградации и последующему снижению стабильности комплекса разделенный мертвый C2c1-VP64 или комплекса разделенный мертвый C2c3-VP64.
[00532] Такие дестабилизирующие последовательности, описанные в других местах настоящей заявки (включая PEST), могут обладать преимуществами для использования в системах разделенного C2c1 или разделенного C2c3.
[00533] Получены клеточные линии, стабильно экспрессирующие разделенный мертвый C2c1-VP64 и MS2-p65-HSFl+направляющие последовательности. Скрининг по устойчивости к PLZ может продемонстирировать, что необратимая, своевременная транскрипционная активация может быть эффективна для скрининга низкомолекулярных веществ. Этот подход может обладать преимуществами в случае, когда разделенный мертвый C2c1-VP64 является необратимым.
[00534] Получены клеточные линии, стабильно экспрессирующие разделенный мертвый C2c3-VP64 и MS2-p65-HSFl+направляющие последовательности. Скрининг по устойчивости к PLZ может продемонстирировать, что необратимая, своевременная транскрипционная активация может быть эффективна для скрининга низкомолекулярных веществ. Этот подход может обладать преимуществами в случае, когда разделенный мертвый C2c3-VP64 является необратимым.
[00535] В одном аспекте изобретение относится к не встречающейся в природе или сконструированной способами инженерии системе CRISPR-Cas C2c1 или C2c3, которая может включать по меньшей мере один переключатель, при этом активность упомянутой системы CRISPR-Cas C2c1 или C2c3 регулируется контактом по меньшей мере с одним источником энергии индукции в качестве переключателя. В одном варианте осуществления изобретения регуляция в отношении по меньшей мере одного переключателя или активность упомянутой системы CRISPR-Cas C2c1 или C2c3 может быть активирована, усилена, прекращена или репрессирована. Первым эффектом может являться одно или более событий импорта в ядро, экспорта из ядра, привлечения вторичного компонента (такого как эффекторная молекула), конформационных изменений (белка, ДНК или РНК), расщепления, высвобождения груза (такого как присоединенная молекула или кофактор), ассоциации или диссоциации. Второй эффект может представлять собой одно или более событий активации, усиления, терминации или репрессии регуляции по меньшей мере одного переключателя или активности упомянутой системы CRISPR-Cas C2c1 или C2c3.
[00536] В другом аспекте изобретения система CRISPR-Cas C2c1 или C2c3 может далее включать по меньшей мере один сигнал ядерной локализации (NLS), сигнал ядерного экспорта (NES), функциональный домен, подвижный линкер, мутацию, делецию, изменение или укорачивание. Один или более NLS, NES или функциональных доменов могут быть активированы или инактивированы в определенных условиях. В другом варианте осуществления мутация может представлять собой одну или более мутаций в области гомологии фактору транскрипции, мутаций в ДНК-связывающем домене (таких как мутация основных остатков основного мотива спираль-петля-спираль), мутаций в эндогенном NLS или мутаций в эндогенном NES. Изобретение предусматривает, что источник энергии индукции может представлять собой тепловую энергию, ультразвук, энергию электромагнитного излучения или химическую энергию. В предпочтительном варианте осуществления изобретения источник энергии индукции может представлять собой антибиотик, низкомолекулярное соединение, гормон, производное гормона, стероид или стероидное производное, или в более предпочтительном варианте осуществления источник энергии индукции может представлять собой абсцизовую кислоту (АВА), доксициклин (DOX), кумат, рапамицин, 4-гидрокситамоксифен (4-ОНТ), эстроген или экдизон. Изобретение предусматривает, что по меньшей мере один переключатель может быть выбран из группы, состоящей из индуцибельных систем на основе антибиотиков, индуцибельных систем на основе электромагнитного излучения, индуцибельных систем на основе низкомолекулярных соединений, индуцибельных систем на основе ядерных рецепторов, индуцибельных систем на основе гормонов. В более предпочтительных вариантах осуществления по меньшей мере один переключатель может быть выбран из группы, состоящей из тетрациклин/доксициклиновой (Tet/DOX) индуцибельной системы, индуцируемой светом системы, индуцируемой АВА системы, куматовой индуцибельной системы репрессор/оператор, 4-ОНТ/экстрогеновой индуцибельной системы, индуцибельной системы на базе экдизона и индуцируемуой FKBP12/FRAP (комплексом FKBP12-рапамицин) системы.
[00537] Аспекты регуляции, подробно изложенные в настоящей заявке, относятся по меньшей мере к одному или более переключателю(ям). Как используют в настоящем описании, термин "переключатель" относится к системе или набору компонентов, которые действуют согласованно, вызывая изменение, охватывающее все аспекты биологической функции, такой как активация, репрессия, усиление или терминация этой функции. В одном аспекте термин "переключатель" охватывает генетические переключатели, которые включают основные компоненты белков, участвующих в генной регуляции, и конкретные последовательности ДНК, узнаваемые этими белками. В одном аспекте переключатели связаны с индуцибельными и репрессорными системами, используемыми в генной регуляции. В целом, индуцибельная система может быть выключена до тех пор, пока не будет присутствовать некоторая молекула (называемая индуктором), которая обеспечивает генную экспрессию. Эту молекулу называют "индуцирующей экспрессию". Способ, которым это явление опосредовано, зависит от регуляторных механизмов, так и различий типов клеток. Репрессорная система включена, за исключением случаев, когда присутствует некоторая молекула (называемая корепрессором), которая супрессирует генную экспрессию. Эту молекулу называют "репрессирующей экспрессию". То, как это происходит, зависит от регуляторных механизмов, а также различий в типах клеток. Как используют в настоящем описании, термин "индуцибельный" может заключать в себе все аспекты переключения независимо от вовлеченного в это молекулярного механизма. Соответственно, переключатель, предусматриваемый изобретением, может включать, но не ограничиваться ими, индуцибельные системы на основе антибиотиков, индуцибельные системы на основе электромагнитного излучения, индуцибельные системы на основе низкомолекулярных соединений, индуцибельные системы на основе ядерных рецепторов, индуцибельные системы на основе гормонов. В более предпочтительных вариантах осуществления переключатель может быть выбран из группы, состоящей из тетрациклин/доксициклиновой (Tet/DOX) индуцибельной системы, индуцируемой светом системы, индуцируемой АВА системы, куматовой индуцибельной системы репрессор/оператор, 4-ОНТ/экстрогеновой индуцибельной системы, индуцибельной системы на основе экдизона и индуцируемой FKBP12/FRAP (комплексом FKBP12-рапамицин) системы.
[00538] Система CRISPR-Cas C2c1 или C2c3 по настоящему изобретению может быть сконструирована для модулирования или изменения экспрессии индивидуальных эндогенных генов в точно определенные временные и пространственные промежутки. Система CRISPR-Cas C2c1 или C2c3 может быть разработана, чтобы связываться с промоторными последовательностями гена-мишени и изменять экспрессию гена. C2c1 или C2c3 может быть разделен на две части, причем одна половина слита с одной половиной гетеродимера криптохрома-2 (CIB1), в то время как оставшийся партнер криптохрома слит с другой половиной C2c1 или C2c3. В некоторых аспектах также в систему CRISPR-Cas C2c1 или C2c3 может быть включен транскрипционный репрессорный домен. Эффекторные домены могут представлять собой как активаторы, такие как VP16, VP64 или p65, так и репрессоры, такие как KRAB, EnR и SID. При отсутствии стимуляции одна половина белка C2c1-криптохром-2 или белка C2c3-криптохром-2 локализуется в промоторной области гена-мишени, но не связана с CIB1-эффекторным белком. При стимуляции светом синей области спектра криптохром-2 активируется и подвергается конформационным изменениям и становится доступным его связывающий домен. CIB1, в свою очередь, связывается с криптохромом-2, что приводит к локализации второй половины C2c1 или второй половины C2c3 в промоторной области гена-мишени и инициации редактирования генома, которое может приводить к суперэкспресии или сайленсингу гена. Аспекты LITE далее описаны в статье Liu, H et al., Science, 2008 and Kennedy M et al. Nature Methods 2010, содержание которой включен в настоящее описание в качестве ссылки в полном объеме.
[00539] Активаторные и репрессорные домены, которые могут далее модулировать функции, могут быть выбраны, исходя из вида, силы, механизма, длительности, размера или любого числа других параметров. Предпочтительные эффекторные домены включают, но не ограничиваются ими, транспозазный домен, интегразный домен, рекомбиназный домен, резольвазный домен, инвертазный домен, протеазный домен, ДНК-метилтрансферазный домен, домен ДНК-деметилазы, гистонацетилазный домен, домен гистоновой деацетилазы, нуклеазный домен, репрессорный домен, активационный домен, домен, содержащий сигнал ядерной локализации, домен белков, рекрутирующих транскрипционные белки, домен, ассоциированный с поглощением веществ клеткой, домен, связывающий нуклеиновые кислоты, антигенпрезентирующий домен.
[00540] Существуют также несколько способов получения химически индуцибельных систем:
1. Система на основе ABI-PYL, индуцируемая абсцизовой кислотой (ABA) (см., например, вебсайт stke.sciencemag.org/cgi/content/abstract/sigtrans;4/1.64/rs2),
2. Система на основе FKBP-FRB, индуцируемая рапамицином (или аналогами на основе рапамицина) (см., например, вебсайт nature.com/nmeth/joumal/v2/n6/full/nmeth763.html),
3. Система на основе GID1-GAI, индуцируемая гиббереллином (GA) (см., например, вебсайт nature.com/nchembio/journal/v8/n5/full/nchembio.922.html).
[00541] Другая система, предусматриваемая настоящим изобретением, представляет собой химически индуцируемую систему на основе изменения внутриклеточной локализации. Заявители также предлагают систему CR1SPR-Cas C2c1 или C2c3, разработанную для нацеливания на геномный локус-мишень, причем фермент CR1SPR-Cas C2c1 или C2c3 разделен на две слитых конструкции, которые далее связаны с различными частями химически или энергетически чувствительного белка. Химически или энергетически чувствительный белок индуцирует изменения во внутриклеточной локализации любой половины фермента C2c1 или C2c3 (т.е. транспорт любой половины фермента C2c1 или C2c3 из цитоплазмы в ядро клетки) при связывании химического соединения или переносе энергии химически или энергетически чувствительному белку. Этот транспорт слитых конструкций из одного внутриклеточного компартмента или органеллы, в которых активность секвестрируется в силу отсутствия субстрата для восстановленной системы CRISPR-Cas C2c1 или C2c3, в другой, в котором присутствует субстрат, позволяет компонентам объединиться и восстановить ее функциональную активность и затем провзаимодействовать с субстратом-мишенью (т.е. геномной ДНК в ядре млекопитающего) и привести к активации или репрессии экспрессии гена-мишени.
[00542] Предусматриваются другие индуцибельные системы, такие как, но не ограничиваются ими, регуляция тяжелыми металлами [Mayo KE et al., Cell AC 29:99-108; Searle PF et al., Mol Cell Biol. 1985, 5:1480-1489 and Brinster RL et al., Nature (London) 1982, 296:39-42], стероидными гормонами [Hynes NE et al., Proc Natl Acad Sci USA 181, 78:2038-2042; Klock G et al., Nature. (London) 1987, 329:734-736 и Lee F et al.. Nature (London) 1981, 294:228-232.], тепловым шоком [Nouer L: Heat Shock Response. Boca Raton, FL: CRC; 1991] и другими разработанными реагентами [Mullick A, Massie B: Transcription, translation and the control of gene expression. In Encyclopedia of Cell Technology Edited by: Speir RE. Wiley; 2000:1140-1164 and Fussenegger M,. Biotechnol Prog 2001, 17:1-51]. Однако в случае этих индуцибельных промоторов млекопитающих есть ограничение, заключающееся в «утечках» промоторов в «выключенном» состоянии и плейотропный эффект индукторов (теплового шока, тяжелых металлов, глюкокортикоидов и др.). Использование гормонов насекомых (экдизона) предложено в попытке снизить влияние на внутриклеточные процессы в клетках млекопитающих [No D et al., Proc Natl Acad Sci USA 1996, 93:3346-3351]. В другой предпочтительной и хорошо продуманной системе в качестве индуктора используется рапамицин [Rivera VM et al., Nat Med 1996. 2:1028-1032], но иммуносупрессорное действие рапамицина является основным ограничением для его использования in vivo, и поэтому было необходимо найти биологически инертное соединение [Saez E et al., Proc Natl Acad Sci USA 2000, 97:14512-14517] для регуляции генной экспрессии.
[00543] В конкретных вариантах осуществления системы редактирования генов, описанные в настоящем описании, находятся под контролем закодированного переключателя-убийцы, который является механизмом, эффективно обеспечивающим гибель клетки-хозяина, когда условия в клетке изменяются. Это обеспечивается введением гибридных транскрипционных факторов семейства LacI-GalR, для включения которых требуется присутствие IPTG (Chan et al. 2015 Nature Nature Chemical Biology doi:10.1038/nchembio.1979), что может быть использовано для индукции гена, кодирующего фермент, необходимый для выживания клетки. Комбинируя различные транскрипционные факторы, чувствительные к различным химическим веществам, можно создавать "код". Эта система может быть использована для временного и пространственного контроля распространения генетических модификаций, индуцируемых CRISPR, что может представлять интерес, когда необходимо избежать "выхода" GMO из предполагаемой ниши.
Доставка в целом
Редактирование гена или изменение локуса-мишени с помощью C2c1 или C2c3
[00544] Двуцепочечный разрыв или одноцепочечный разрыв в одной из цепей должен находиться достаточно близко к целевой позиции, таким образом, чтобы происходила репарация. В одном из возможных вариантов осуществления расстояние не должно превышать 50, 100, 200, 300, 350 или 400 нуклеотидов. Без связи с теорией, полагают, что разрыв должен находиться достаточно близко к целевой позиции, чтобы разрыв находился внутри области, которая предназначена для удаления посредством экзонуклеаз при удалении концов. Если расстояние между целевой позицией и разрывом слишком велико, мутация может не быть включена в удаление концов и, следовательно, не может быть репарирована, поскольку последовательность матричной нуклеиновой кислоты может быть использована для репарации последовательности только в пределах области удаления концов.
[00545] В одном из возможных вариантов осуществления, в котором направляющая РНК и молекула типа V/типа VI, в частности, C2c1/C2c3, или их ортолог или гомолог, предпочтительно нуклеаза C2c1 или C2c3, индуцирует двуцепочечный разрыв для индукции гомологичной репарации, участок расщепления находится в пределах 0-200 нуклеотидов (например, 0-175, 0-150, 0-125, 0-100, 0-75, 0-50, 0-25, 25-200, 25-175, 25-150, 25-125, 25-100, 25-75, 25-50, 50-200, 50-175, 50-150, 50-125, 50-100, 50-75, 75-200, 75-175, 75-150, 75-1 25, 75-100 нуклеотидов) от целевой позиции. В одном из возможных вариантов осуществления сайт расщепления находится в пределах 0-100 нуклеотидов (например, 0-75, 0-50, 0-25, 25-100, 25-75, 25-50, 50-100, 50-75 или 75-100 нуклеотидов) от целевой позиции. В следующем варианте осуществления две или более направляющих РНК в комплексе с C2c1 или C2c3, или их ортологом или гомологом, могут быть использованы для внесения множественных разрывов для индукции гомологичной репарации.
[00546] Плечо гомологии должно охватывать по меньшей мере тот участок, в котором может произойти удаление концов, например, чтобы обеспечить поиск удаленными одноцепочечными выступающими нуклеотидами комплементарного участка в пределах донорской матрицы. Полная длина может быть ограничена такими параметрами как размер плазмиды или пределы упаковывания с помощью вируса. В одном из возможных вариантов осуществления плечо гомологии может не достигать повторяющихся элементов. Типичная длина плеча гомологии включает по меньшей мере 50, 100, 250, 500, 750 или 1000 нуклеотидов.
[00547] Упомянутая здесь целевая позиция относится к участку в нуклеиновой кислоте-мишени или гене-мишени (например, хромосоме), который модифицируется молекулой типа V/типа VI, в частности, C2c1/C2c3 или их ортологом или гомологом, предпочтительно в ходе процесса, зависящего от молекулы C2c1 или C2c3. Например, в целевой позиции может происходить расщепление нуклеиновой кислоты-мишени молекулой модифицированного C2c1 или C2c3, и модификация, направляемая матричной нуклеиновой кислотой. Модификация может представлять собой, например, коррекциую (репарациую) целевой позиции. В одном из возможных вариантов осуществления целевая позиция может являться сайтом между двумя нуклеотидами, например, прилегающими нуклеотидами, на нуклеиновой кислоте, к которой добавляются один или более нуклеотидов. Целевая позиция может содержать один или более нуклеотидов, которые подвергаются изменениям, например, репарации с помощью матричной нуклеиновой кислоты. В одном из возможных вариантов осуществления целевая позиция может находиться в пределах последовательности-мишени (например, последовательности, с которой связывается направляющая). В одном из возможных вариантов осуществления целевая позиция находится выше или ниже последовательности-мишени (например, последовательности, с которой связывается направляющая РНК).
[00548] Матричная нуклеиновая кислота, как этот термин используют в настоящем описании, относится к последовательности нуклеиновой кислоты, которая может быть использована в сочетании с молекулой типа V/типа VI, предпочтительно молекулой C2c1/C2c3 и молекулой направляющей РНК, чтобы изменить структуру целевой позиции. В одном из возможных вариантов осуществления нуклеиновая кислота-мишень модифицирована так, чтобы она содержала часть или всю последовательности матричной нуклеиновой кислоты, обычно рядом или в самом сайте (самих сайтах) расщепления. В одном из возможных вариантов осуществления матричная нуклеиновая кислота является двуцепочечной. В одном из возможных вариантов осуществления матричная нуклеиновая кислота представляет собой ДНК. В другом варианте осуществления матричная нуклеиновая кислота является одноцепочечной ДНК.
[00549] В одном из возможных вариантов осуществления матричная нуклеиновая кислота изменяет структуру целевой позиции, участвуя в гомологичной рекомбинации. В одном из возможных вариантов осуществления матричная нуклеиновая кислота изменяет структуру целевой позиции. В одном из возможных вариантов осуществления матричная нуклеиновая кислота приводит к включению модифицированного или не встречающегося в природе основания в нуклеиновую кислоту-мишень.
[00550] Матричная последовательность может подвергаться разрывам, опосредованным или катализируемым рекомбинацией с последовательностью-мишенью. В одном из возможных вариантов осуществления матричная нуклеиновая кислота может содержать последовательность, соответствующую участку на последовательности-мишени, который подвергается опосредованному C2c1 или C2c3 расщеплению. В одном из возможных вариантов осуществления матричная нуклеиновая кислота может содержать последовательность, соответствующую которая соответствует как первому участку в последовательности-мишени, расщепляемому в рамках первого опосредованного C2c1 или C2c3 события, так и второму участку в последовательности-мишени, расщепляемому в рамках второго опосредованного C2c1 или C2c3 события.
[00551] В одном из возможных вариантов осуществления матричная нуклеиновая кислота может содержать последовательность, которая приводит к изменению в кодирующей последовательности транслируемой последовательности, например, последовательность, которая приводит к замене одной аминокислоты на другую в белковом продукте, например, трансформируя мутантный аллель в аллель дикого типа, трансформируя аллель дикого типа в мутантный аллель и/или внося стоп-кодон, вставку аминокислотного остатка или делецию аминокислотного остатка или нонсенс-мутацию. В одном из возможных вариантов осуществления матричная нуклеиновая кислота может содержать последовательность, которая приводит к изменению в некодирующей последовательности, например, изменению в экзоне или 5'- или 3'-нетранслируемой области (НТО) или нетранскрибируемой области. Такие изменения могут включать изменение в регуляторном элементе, например, промоторе, энхансере, или изменение в цис- или транс-регуляторном элементе.
[00552] Матричная нуклеиновая кислота, обладающая гомологией с целевой позицией в гене-мишени, может быть использована для изменения структуры последовательности-мишени. Матричная нуклеиновая кислота может быть использована для изменения нежелательной структуры, например, нежелательного или мутантного нуклеотида. Матричная нуклеиновая кислота может содержать последовательность, которая при интеграции приводит к: снижению активности позитивного регуляторного элемента; увеличению активности негативного регуляторного элемента; снижению экспрессии гена; увеличению экспрессии гена; увеличению устойчивости к синдрому или заболеванию; увеличению устойчивости к вирусному заражению; корректировке мутации или изменению нежелательного аминокислотного остатка, предоставляющего, увеличивающего, предотвращающего или снижающего биологическое свойство продукта гена, например, увеличение ферментативной активности фермента или увеличение способности продукта гена взаимодействовать с другой молекулой.
[00553] Матричная нуклеиновая кислота может содержать последовательность, которая приводит к изменениям в последовательности 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или более нуклеотидов последовательности-мишени. В одном из возможных вариантов осуществления матричная нуклеиновая кислота может иметь длину 20+/-10, 30+/-10, 40+/-10, 50+/-10, 60+/-10, 70+/-10, 80+/-10, 90+/-10, 100-+/-10, 110+/-10, 120+/-10, 130+/-10, 140+/-10, 150+/-10, 160+/-10, 170+/-10, 1 80+/-10, 190+/-10, 200+/-10, 210+/-10 или 220+/-10 нуклеотидов. В одном из возможных вариантов осуществления матричная нуклеиновая кислота может иметь длину 30+/-20, 40+/-20, 50+/-20, 60+/-20, 70+/- 20, 80+/-20, 90+/-20, 100+/-20, 1 10+/-20, 120+/-20, 130+Л20, 140+/-20, I 50+/-20, 160+/-20, 170+/-20, 180+/-20, 190+/-20, 200+/-20, 210+/-20, 220+/-20 нуклеотидов. В одном из возможных вариантов осуществления матричная нуклеиновая кислота может иметь длину 10-1000, 20-900, 30-800, 40-700, 50-600, 50-500, 50-400, 50-300, 50-200 или 50-100 нуклеотидов.
[00554] Матричная нуклеиновая кислота включает следующие компоненты: [5'-плечо гомологии]-[последовательность замены]-[3'-плечо гомологии]. Плечо гомологии обеспечивают рекомбинацию в хромосоме, заменяя, таким образом, нежелательный элемент, например мутацию или мотив, с помощью последовательности замены. В одном варианте осуществления гомологические плечи фланкируют самые отдаленные участки расщепления. В одном из вариантов осуществления 3'-конец 5'-плеча гомологии представляет собой положение рядом с 5'-концом последовательности замены. В варианте осуществления 5'-плечо гомологии может удлиняться по меньшей мере 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов с 5'-стороны от 5'-конца последовательности замены. В варианте осуществления 5'-конец 3'-плеча гомологии представляет собой положение рядом с 3'-концом последовательности замены. В варианте осуществления 3'-плечо гомологии может включать по меньшей мере на 10, 20, 30, 40, 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500 или 2000 нуклеотидов с 3'-стороны от 3'-конца последовательности замены.
[00555] В определенных вариантах осуществления одно или оба плеча гомологии могут быть укорочены, чтобы избежать включения определенных повторяющихся последовательностей. Например, 5'-плечо гомологии может быть укорочено, чтобы избежать включения повторяющейся последовательности. В других вариантах осуществления 3'-плечо гомологии может быть укорочено, чтобы избежать включения повторяющейся последовательности. В некоторых вариантах осуществления как 5'-, так и 3'-плечо гомологии может быть укорочено, чтобы избежать включения определенных повторяющихся последовательностей.
[00556] В определенных вариантах осуществления матричная нуклеиновая кислота для коррекции мутаций может быть сконструирована для использования в качестве одноцепочечных олигонуклеотидов. При использовании одноцепочечных олигонуклеотидов 5'- и 3'-плечо гомологии может иметь длину в диапазоне вплоть до 200 нуклеотидов, например, по меньшей мере 25, 50, 75, 100, 125, 150, 175 или 200 нуклеотидов.
Опосредованное системой комплекса эффекторного белка C2c1 или C2c3 негомологичное соединение концов
[00557] В некоторых вариантах осуществления индуцированное нуклеазами негомологичное соединение концов (NHEJ) может быть использовано для обеспечения специфических нокаутов генов. Индуцированное нуклеазами NHEJ также может быть использовано для удаления последовательности в гене-мишени. В целом, NHEJ репарирует двуцепочечный разрыв в ДНК путем соединения двух концов; однако, в целом, исходная последовательность восстанавливается только в случае, если два совместимых конца, точно в том виде, в котором они были образованы при двуцепочечном разрыве, точно лигированы. Концы двуцепочечного разрыва ДНК часто являются объектом ферментативных процессов, что приводит к добавлению или удалению нуклеотидов в случае одной или обеих цепей перед воссоединением концов. Это приводит к присутствию мутаций с вставками и/или делециями (инсерций-делеций) в последовательности ДНК в участке NHEJ-зависимой репарации. Две трети этих мутаций обычно изменяют рамку считывания и, следовательно, приводят к продукции нефункционального белка. Кроме того, мутации, которые сохраняют рамку считывания, но которые заключаются во вставке или делеции значительной части последовательности, могут сделать белок нефункциональным. Это зависит от локуса, поскольку вероятно, что мутации в критических функциональных доменах будут приводить к более значительным последствиям, что в некритических участках белков. Мутации в виде инсерций-делеций, внесенные посредством NHEJ, непредсказуемы по своей природе; однако в случае конкретного разрыва конкретные последовательности инсерций-делеций обладают большими преимуществами и более представлены в популяции, наиболее вероятно, за счет коротких участков микрогомологии. Длина делеций может значительно варьировать; наиболее часто она находится в диапазоне 1-50 нуклеотидов, но она может и превышать 50 нуклеотидов, например, достигая длины, превышающей 100-200 нуклеотидов. Вставки обычно являются более короткими и часто включают короткие дупликации последовательности, непосредственно окружающие сайт разрыва. Однако возможно обеспечить длинные вставки, и в этих случаях вставленная последовательность часто может отслеживаться в других участках генома или на плазмидной ДНК, присутствующей в клетках.
[00558] Поскольку NHEJ-зависимая репарация является мутагенным процессом, она может использоваться для удаления мотивов последовательностей, если образование специфической финальной последовательности не требуется. Если мишень двуцепочечного разрыва должна быть рядом с короткой последовательностью-мишенью, делеционные мутации, вызываемые NHEJ-зависимой репарацией, часто охватывают и, следовательно, удаляют нежелательные нуклеотиды. Для делеции более длинных сегментов ДНК внесение двух двуцепочечных разрывов, одного с каждой стороны последовательности, может приводить к негомологичному соединению концов (NHEJ) с последующим полным удаление лежащей между ними последовательности. Оба этих подхода могут быть использованы для удаления определенных последовательностей ДНК; однако природа негомологичного соединения концов, ведущая к высокой вероятности ошибок, может вызывать образование инсерций-делеций в участке репарации.
[00559] Обе индуцирующие двуцепочечный разрыв типа V/типа VI, молекулы в частности, C2c1/C2c3, или их ортологи или гомологи, предпочтительно молекулы C2c1 или C2c3, или индуцирующие одноцепочечный разрыв (никазы) молекулы типа V/типа VI, в частности, C2c1/C2c3, или их ортологи или гомологи, предпочтительно молекулы C2c1 или C2c3 могут быть использованы в описанных в настоящем описании способах и композициях для образования инсерций-делеций, опосредованных NHEJ-репарацией. NHEJ-опосредованные инсерции-делеции, нацеленные на ген, например кодирующую область, например, раннюю кодирующую область гена-мишени, могут быть использованы для нокаута (то есть устранения экспрессии) гена-мишени. Например, ранняя кодирующая область гена-мишени включает последовательность сразу после сайта начала транскрипции в пределах первого экзона кодирующей последовательности или в пределах 500 пар оснований от точки начала транскрипции (например, менее 500, 450, 400, 350, 300, 250, 200, 150, 100 или 50 нуклеотидов).
[00560] В одном из возможных вариантов осуществления, в которых направляющая РНК и молекула типа V/типа VI, в частности, C2c1/C2c3, или их ортолог или гомолог, предпочтительно нуклеаза C2c1 или C2c3, индуцирует двуцепочечный разрыв для индукции инсерций-делеций, опосредованных репарацией посредством негомологичного соединения концов (NHEJ), направляющие РНК могут быть сконструированы так, чтобы позиция двуцепочечного находилась вблизи нуклеотида целевой позиции. В одном из возможных вариантов осуществления сайт расщепления может находиться в пределах 0-500 нуклеотидов от целевой позиции (например, меньше 500, 400, 300, 200, 100, 50, 40, 30, 25, 20, 15, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 нуклеотидов от целевой позиции).
[00561] В одном из возможных вариантов осуществления, в которых две направляющих РНК в комплексе с молекулами типа V/типа VI, в частности, C2c1/C2c3, или их ортологами или гомологами, предпочтительно никазами C2c1 или C2c3, индуцируют два одноцепочечных разрыва для индукции инсерций-делеций, опосредованных репарацией посредством негомологичного соединения концов (NHEJ), две направляющих РНК могут быть разработаны так, чтобы позиции двух одноцепочечных разрывов были таковы, чтобы обеспечить репарацию нуклеотида в целевой позиции посредством NHEJ.
Комплексы эффекторного белка C2c1 или C2c3 могут доставлять функциональные эффекторные белки
[00562] В отличие от нокаута гена, опосредованного CRISPR-Cas, который перманентно элиминирует экспрессию мутантного гена на уровне ДНК, нокдаун с помощью CRISPR-Cas позволяет временно снизить экспрессию гена за счет использования искусственных транскрипционных факторов. Мутации ключевых остатков в обоих расщепляющих ДНК доменах белка C2c1 или C2c3 приводят к образованию каталитически неактивного C2c1 или C2c3. Каталитически неактивный C2c1 или C2c3 образует комплекс с направляющей РНК и локализуется на последовательности ДНК, определяемой доменом-мишенью направляющей РНК, однако не расщепляет ДНК-мишень. Слияние неактивного белка C2c1 или C2c3 с эффекторным доменом, например, транскрипционным репрессорным доменом, позволяет привлекать эффектор к любому участку ДНК, определяемому направляющей РНК. В определенных вариантах осуществления C2c1 или C2c3 могут быть слиты с транскрипционным репрессорным доменом и привлечены к промоторному участку гена. В частности, что касается генной репрессии, предполагается, что блокирование участка связывания эндогенного транскрипционного фактора может способствовать снижению уровня экспрессии гена. В другом варианте осуществления неактивный C2c1 или C2c3 может быть слит с белком, модицифицирующим хроматин. Изменение состояния хроматина может приводить к снижению экспрессии гена-мишени.
[00563] В одном из возможных вариантов осуществления молекула направляющей РНК может быть нацелена на известные транскрипционные респонсивные элементы (например, промоторы, энхансеры и т.п.), известные активирующие последовательности, находящиеся выше точки начала транскрипции, и/или последовательности известной или неизвестной функции, про которые предполагают, что они могут контролировать экспрессию целевой ДНК.
[00564] В некоторых способах полинуклеотид-мишень может быть инактивирован, чтобы осуществить модификацию экспрессии в клетке. Например, при связывании комплекса CRISPR с последовательностью-мишенью в клетке, полинуклеотид-мишень инактивируется так, что последовательность не транскрибируется, кодируемый белок не продуцируется или последовательность не функционирует так, как функционирует последовательность дикого типа. Например, кодирующая последовательность белка или микроРНК может быть инактивирована так, чтобы белок не продуцировался.
[00565] В некоторых вариантах осуществления фермент CRISPR содержит одну или более мутаций, выбранных из перечня, состоящего из замен D917A, E1006A и D1225A, и/или одну или более мутаций в домене RuvC фермента CRISPR, или представляет собой другую мутацию, описанную в настоящем описании. В некоторых вариантах осуществления фермент CRISPR содержит одну или более мутаций в каталитическом домене, в котором во время транскрипции последовательность прямого повтора формирует одиночную шпильку и направляющая последовательность направляет специфическое для последовательности связывание с последовательностью-мишенью комплекса CRISPR, причем фермент далее включает функциональный домен. В некоторых вариантах осуществления функциональный домен представляет собой транскрипционный репрессорный домен, предпочтительно KRAB. В некоторых вариантах осуществления транскрипционный репрессорный домен представляет собой SID и конкатемеры SID (SID4X). В некоторых вариантах осуществления функциональный домен представляет собой домен, вносящий эпигенетические модификации, если предоставлен белок, осуществляющий эпигенетические модификации. В некоторых вариантах осуществления функциональный домен представляет собой активационный домен, который может являться активационным доменом Р65.
Доставка комплекса эффекторного белка C2c1 или C2c3 или его компонентов
[00566] В настоящем описании, а также из уровня техники, предусматривается, что TALE-нуклеазы, системы CRISPR-Cas или их компоненты, или их молекулы нуклеиновых кислот (в том числе, например, матрицу HDR), или молекулы нуклеиновых кислот, кодирующие или доставляющие их компоненты, могут быть доставлены системой доставки, описанной в настоящем описании как в общем виде, так и более подробно.
[00567] Векторная доставка, например, плазмидная, вирусная доставка: фермент CRISPR, например белок типа V, такой как C2c1 или C2c3, и/или любая из РНК по настоящему изобретению, например направляющая РНК, могут быть доставлены с использованием любого подходящего вектора, например, плазмидных или вирусных векторов, таких как аденоассоциированный вирус (AAV), лентивирус, аденовирус или другие типы вирусных векторов или их комбинации. Эффекторные белки и одна или более направляющих РНК могут быть упакованы в один или более векторов, например, плазмидные или вирусные векторы. В некоторых вариантах осуществления изобретения вектор, например, плазмидный или вирусный вектор, доставляется в целевую ткань, например, путем внутримышечной инъекции, в то время как в других случаях доставка осуществляется внутривенно, трансдермально, интраназально, перорально, через слизистые оболочки или другими способами доставки. Такая доставка может быть в виде единичной дозы либо в виде нескольких доз. Специалисту в данной области понятно, что фактическая дозировка, которая должна быть доставлена, может значительно варьироваться в зависимости от ряда факторов, таких как выбор вектора, тип клетки-мишени, самого организма или ткани, общее состояние пациента, желаемая степень трансформации/модификации, способ введения, режим введения, тип желаемой трансформации/модификации и т.д.
[00568] Такой препарат может в дальнейшем включать, например, носитель (в составе которого вода, водный раствор хлорида натрия, этанол, глицерин, лактоза, сахароза, фосфат кальция, желатин, декстран, агар, пектин, арахисовое масло, кунжутное масло и т.д.), растворитель, применяемый в фармакологии носитель (например, фосфатный солевой буфер), применяемый в фармакологии наполнитель и/или другие соединения, известные в данной области. Препарат, кроме того, может содержать одну или более применяемых в фармакологии солей, например, таких как соль неорганической кислоты, такая как гидрохлорид, гидробромид, фосфат, сульфат и т.д., и соли органических кислот, такие как ацетаты, пропионаты, малонаты, бензоаты и т.д. Кроме того, также использоваться могут вспомогательные вещества, такие как смачивающие или эмульгирующие, буферные растворы, гели или гелеобразователи, вкусовые добавки, красители, микросферы, полимеры, вещества, образующие суспензии и т.д. Кроме того, один или более других стандартно применяемых фармацевтических добавок, таких как консерванты, влагоудерживающие добавки, суспенизирующие добавки, поверхностно-активные добавки, антиоксиданты, противослеживающие добавки, наполнители, хелатирующие добавки, вещества оболочки, химические стабилизаторы и т.д. могут также быть использованы, особенно если дозировка имеет разбавленную форму. Подходящие иллюстративные компоненты включают микрокристаллическую целлюлозу, натрий карбокисметилцеллюлозу, полисорбат 80, фенилэтиловый спирт, хлорбутанол, сорбат калия, сорбиновую кислоту, двуокись серы, пропилгаллат, парабены, этилванилин, глицерин, фенол, парахлорофенол, желатин, альбумин и их комбинацию. Полное обсуждение применяемых в фармакологии наполнителей доступно в REMINGTON'S PHARMACEUTICAL SCIENCES (Mack Pub, Co., N.J. 1991), который включен в настоящее описание в качестве ссылки.
[00569] В одном варианте осуществления изобретения, описанном в настоящем описании, доставка осуществляется с помощью аденовируса, который может быть доставлен в виде единичной вспомогательной дозы, содержащей по меньшей мере 1×105 частиц (также называемых единицами частиц, еч) аденовирусного вектора. В одном варианте осуществления изобретения настоящего изобетения, предпочтительная доза составляет по меньшей мере приблизительно 1×106 частиц (например, приблизительно 1×106-1×1012 частиц), более предпочтительно по меньшей мере приблизительно 1×107 частиц, более предпочтительно по меньшей мере приблизительно 1×108 частиц (например, приблизительно 1×108-1×1011 частиц или приблизительно 1×108-1×1012 частиц), и наиболее предпочтительно по меньшей мере приблизительно 1×109 частиц (например, приблизительно 1×109-1×1010 частиц или приблизительно 1×109-1×1012 частиц), или даже по меньшей мере приблизительно 1×1010 частиц (например, приблизительно 1×1010-1×1012 частиц) аденовирусного вектора. Альтернативно, доза включает не больше чем приблизительно 1×1014 частиц, предпочтительно не больше чем приблизительно 1×1013 частиц, еще более предпочтительно не больше чем приблизительно 1×1012 частиц, еще более предпочтительно не больше чем приблизительно 1×1011 частиц, и наиболее предпочтительно не больше чем приблизительно 1×1010 частиц (например, не больше чем приблизительно 1×109 частиц). Таким образом, доза может содержать единственную дозу аденовирусного вектора, например, с приблизительно 1×106 единиц частиц (еч), приблизительно 2×106 еч, приблизительно 4×106 еч, 1×107 еч, приблизительно 2×107 еч, приблизительно 4×107 еч, приблизительно 1×108 еч, приблизительно 2×108 еч, приблизительно 4×108 еч, приблизительно 1×109 еч, приблизительно 2×109 еч, приблизительно 4×109 еч, приблизительно 1×1010 еч, приблизительно 2×1010 еч, приблизительно 4×1010 еч, приблизительно 1×1011 еч, приблизительно 2×1011 еч, приблизительно 4×1011 еч, приблизительно 1×1012 еч, приблизительно 2×1012 еч или приблизительно 4×1012 еч аденовирусного вектора. См., например, аденовирусные векторы в заявке на патент США № 8454972, B2 Nabel, et al., поданной 4 июня 2013 года, включенной в настоящее описание в качестве ссылки, и в ее колонке 29, строки 36-58. В одном варианте осуществления изобретения, описанном в настоящем описании, аденовирус доставляется посредством нескольких доз.
[00570] В описанном в настоящем описании варианте осуществления изобретения доставка осуществляется через AAV. Терапевтически эффективная дозировка для in vivo доставки AAV человеку, как полагают, находится в диапазоне от приблизительно 20 приблизительно до 50 мл солевого раствора, содержащего от приблизительно 1×1010 до приблизительно 1×1010 раствора функционального AAV/мл. Дозировка может быть скорректирована для уравновешивания терапевтического эффекта и каких-либо побочных эффектов. В одном варианте осуществления изобретения, описанном в настоящем описнии, доза AAV обычно находится в следующем диапазоне концентраций: от приблизительно 1×105 до 1×1050 геномов AAV, от приблизительно 1×108 до 1×1020 геномов AAV, от приблизительно 1×1010 до приблизительно 1×1016 геномов или от приблизительно 1×1011 до приблизительно 1×1016 геномов AAV. Дозировка для человека может составлять приблизительно 1×1016 геномов AAV. В таких концентрациях можно доставить от приблизительно 0,001 мл до приблизительно 100 мл, от приблизительно 0,05 до приблизительно 50 мл или от приблизительно 10 до приблизительно 25 мл раствора-носителя. Другие эффективные дозировки могут быть легко определены квалифицированным специалистом в данной области посредством стандартных испытаний, на основе которых строится кривая зависимости ответа от дозы. См., например, заявку на патент США № 8404658 B2 Hajjar, et al., поданную 26 марта 2013 года, колонка 27, строки 45-60.
[00571] В одном варианте осуществления изобретения, описанном в настоящем описании, доставка производится через плазмиду. В случае таких плазмидных конструкций их дозировка должна быть достаточной для того, чтобы вызвать ответ. Например, подходящее количество ДНК-плазмиды в конструкциях плазмиды может составлять от приблизительно 0,1 до приблизительно 2 мг, или от приблизительно 1 пг до приблизительно 10 пг на человека массой 70 кг. Плазмиды по изобретению обычно включают (i) промотор: (ii) последовательность, кодирующую нацеленный на нуклеиновую кислоту фермент CRISPR, функционально связанный с указанным промотором; (iii) селективный маркер; (iv) ориджин репликации; и (v) терминатор транскрипции, находящийся ниже и функционально связанный с (ii). Плазмида может также кодировать РНК-компоненты комплекса CRISPR, но один или более из них может кодироваться другим вектором.
[00572] Дозы, описанные в настоящем описании, основаны на средней массе человека, равной 70 кг. Определение частоты введения входит в компетенции практикующего медицинского или ветеринарного специалиста (например, врач, ветеринар) или ученого-специалиста в данной области. Также отмечается, что мыши, используемые в экспериментах, как правило, имеют массу приблизительно 20 г, и результаты экспериментов на мышах могут быть экстраполированы на индивида массой 70 кг.
[00573] В некоторых вариантах осуществления изобретения молекулы РНК по изобретению доставляют в составе липосом или в составе, содержащим липофектин и т.п. и могут быть получены с помощью способов, известных специалисту в данной области. Такие способы описаны, например, в патентах США № 5593972, 5589466 и 5580859, которые включены в настоящее описание в качестве ссылки в полном объеме. Системы доставки для на усиленной и улучшенной доставки миРНК в клетки млекопитающих были разработаны (см., например, Shen et al. FEBS Let. 2003, 539:111-114; Xia et al., Nat. Biotech. 2002, 20:1006-1010; Reich et al., Mol. Vision. 2003, 9: 210-216; Sorensen et al., J. Mol. Biol. 2003, 327: 761-766; Lewis et al., Nat. Gen. 2002, 32: 107-108 и Simeoni et al., NAR 2003, 31, 11: 2717-2724) и могут быть использованы в рамках настоящего изобретения. Недавно миРНК была успешно использована для ингибирования экспрессии генов у приматов (см., например, Tolentino et al., Retina 24(4):660) и может также быть применена в рамках настоящего изобретения.
[00574] Действительно, РНК-доставка является эффективным способом для доставки in vivo. Возможно доставить нацеленный на нуклеиновую кислоту Cas, белок Cas9 и направляющую РНК - гРНК (и, например, матрицу для репарации HR) в клетки с использованием липосом или частиц. Таким образом, доставка нацеленного на нуклеиновую кислоту белка Cas/ фермента CRISPR, такого как Cas-белок Cas9 и/или доставка направляющих РНК по изобретения может производится в форме РНК с использованием микровезикул, липосом или частиц. Например, мРНК Cas и направляющая РНК могут быть упакованы в липосомальные частицы для доставки in vivo. Липосомальные реактивы для трансфекции, такие как липофектамин производства Life Technologies и другие реактивы, доступные на рынке, могут эффективно доставить молекулы РНК в печень.
[00575] Предпочтительно, чтобы средства доставки РНК также включали доставку РНК через наночастицы (Cho, S, Goldberg, M, Son, S, Xu, Q, Yang, F, Mei, Y, Bogatyrev, S, Langer, R, and Anderson, D, Lipid-like nanoparticles for small interfering RNA delivery to endothelial cells, Advanced Functional Materials, 19; 3112-3118, 2010) или экзосомы (Schroeder, A, Levins, C, Cortez, C, Langer, R, and Anderson, D, Lipid-based nanotherapeutics for siRNA delivery, Journal of Internal Medicine, 267: 9-21, 2010, PMID: 20059641). Действительно, экзосомы, как показано, особенно полезны при доставке миРНК, системы, обладающей некоторым сходством с системой нацеливания на РНК. Например, El-Andaloussi S et al. ("Exosome-mediated delivery of siRNA in vitro and in vivo Nat Protoc. 2012 Dec;7(12):2112-26. doi: 10.1038/nprot.2012.131. Epub 2012 Nov 15.) описали, что экзосомы являются многообещающим инструментом доставки лекарственных средств через различные биологические барьеры и могут использоваться для доставки миРНК in vitro и in vivo. Данный подход состоит в производстве экзосом-мишеней посредством трансфекции вектора экспрессии, включающего экзосомный белок, слитый с лигандом-пептидом. Экзосомы далее проходят очистку и для отделения от клеточного супернатанта, далее РНК загружают в экзосомы. Доставка или введение согласно изобретению могут быть произведены с применением экзосом, к примеру, но не ограничиваясь этим, в мозг. Витамин Е (α-токоферол) может быть связан с нацеленным на нуклеиновую кислоту белком Cas и доставлен в мозг вместе с липопротеином высокой плотности (HDL), например, аналогично тому, как это было сделано Uno et al. (HUMAN GENE THERAPY 22:711-719 (June 2011)) для доставки милой интерферирующей РНК (миРНК)) в мозг. Мышам проводились вливание с применением осмотического микронасоса (модель 1007D; Alzet, Cupertino, Калифорния, США) фосфатного солевого буфера (PBS) или свободных ToсsiBACE или Toc-siBACE/HDL и подсоединяли к набору для вливаний в мозг (Brain Infusion Kit 3, Alzet). Используемая для вливаний в мозг канюля помещалась примерно на 0,5 мм позади брегмы по центральной линии для вливания в третий дорзальный желудочек. Uno et al. установили, что не более 3 нмоль Toc-siRNA с HDL могут вызывать целевое уменьшение в степени, сравнимой с таковой при применении интракраниального способа введения (ICV). Сходная дозировка нацеленного на нуклеиновые кислоты эффекторного белка, конъюгированного с α-токоферолом и введенного совместно с нацеленным на мозг HDL, предполагается для человека в рамках настоящего изобретения, например, может предполагаться от примерно 3 нмоль до примерно 3 мкмоль нацеленного на нуклеиновую кислоту эффекторного белка, нацеленного в мозг. Zou et al. (HUMAN GENE THERAPY 22:465-475 (April 2011)) описывают способ опосредованной лентивирусом доставки кшРНК, нацеленной на PKCγ для подавления экспрессии генов in vivo в спинном мозге крыс. Zou et all. вводили примерно 10 мкл рекомбинантного лентивируса, имеющего титр 1×109 трансдуцирующих частиц (тч)/мл, через интратекальный катетер. Сходная дозировка нацеленного на нуклеиновую кислоту эффекторного белка, экспрессированного с лентивирусного вектора, нацеленного в мозг, также может предполагаться в рамках настоящего изобретения, например, также предполагается примерно 10-50 мл нацеленного на нуклеиновую кислоту эффекторного белка, направленного в мозг, с титром лентивируса 1×109 трансдуцирующих частиц (тч)/мл.
[00576] В отношении локальной доставки в мозг, возможны различные ее пути. Например, материал может быть доставлен в стриатум, например, путем инъекции. Инъекция может быть произведена стереотактически при трепанации черепа.
[00577] Повышение эффективности NHEJ или HR также является полезным для такой доставки. Предпочтительно, чтобы эффективность NHEJ была усилена одновременно экспрессируемыми ферментами процессинга концов, такими как Trex2 (Dumitrache et al. Genetics. 2011 August: 188(4): 787-797). Предпочтительно, чтобы эффективность HR была увеличена временным ингибированием машинерией NHEJ, в частности, Ku70 и Ku86. Эффективность HR может также быть увеличена путем одновременной экспрессии прокариотических или эукариотических ферментов рекомбинации, таких как RecBCD, RecA.
Общая характеристика Упаковки и Промоторов
[00578] Способы упаковки молекул нуклеиновых кислот, кодирующих нацеленный на нуклеиновую кислоту эффекторный белок (такой как белок типа V, например C2c1 или C2c3), например, ДНК, в векторы, например, вирусные векторы, для опосредования модификации генома in vivo включают:
Для достижения опосредованного NHEJ нокаута гена:
Вектор на основе одного вируса:
Вектор, содержащий две или более кассет экспрессии:
Промотор-направляющая РНК-терминатор
Промотор-направляющая РНК (N-конец)-терминатор (до предела размера вектора)
Двойной вирусный вектор:
Вектор 1, содержащий одну экпрессирующую кассету для управления экспрессией нацеленного на нуклеиновую кислоту эффекторного белка (такого как белок типа V, например, C2c1 или C2c3)
Промотор-молекула нуклеиновой кислоты, кодирующая нацеленный на нуклеиновую кислоту эффекторный белок-терминатор
Вектор 2, содержащий одну или более экспрессирующих кассет для управления экспрессией одной или более направляющей(их) РНК
Промотор-направляющая РНК 1-терминатор
Промотор-направляющая РНК 1 (N-конец)-терминатор (до предела длины вектора)
Для опосредования гомологичной репарации.
В дополнение к подходам с одиночным и двойными векторами, дополнительный вектор используется для доставки матрицы репарации на основе гомологии.
[00579] Промотор, используемый для управления экспрессией нуклеиновой кислоты, нацеленной на нуклеиновую кислоту эффекторным белком (таким как белок типа V, такой как C2c1 или C2c3) может включать:
ITR AAV может выступать в качестве промотора: это предпочтительно для того, чтобы избавиться от дополнительного промоторного элемента (который занимает место в векторе). Освобожденное в результате этого дополнительное пространство может быть использовано, чтобы управлять экспрессией дополнительных элементов (направляющая РНК и т.д.). Кроме того, активность ITR относительно слабее, так что он может использоваться для уменьшения потенциальной токсичности из-за сверхэкспрессии нацеленного на нуклеиновые кислоты эффекторного белка (такого как белок типа V, такой как C2c1 или C2c3).
Для повсеместной экспрессии можно использовать промоторы: CMV, CAG, CBh, PGK, SV40, тяжелых или легких цепей ферритина, и т.д.
Для экспрессии в мозге или другой экспрессии в ЦНС можно использовать промоторы: промотор синапсина I для всех нейронов, CaMKIIalpha для возбуждающих нейронов, GAD67, GAD65 или VGAT для GABA-ергических нейронов, и т.д.
Для экспрессии в печени можно использовать промотор альбумина.
Для экспрессии в легких можно использовать SP-B.
Для эндотелиальных клеток можно использовать ICAM.
Для гемопоэтических клеток можно использовать IFN-бета или CD45.
Для остеобластов можно использовать OG-2.
[00580] Промотор, используемый для нацеливания направляющей РНК, может включать:
промотор РНК-полимеразы III, такой как U6 или H1,
использование промотора РНК-полимеразы II и интронных кассет экспрессии направляющей РНК.
Аденоассоциированный вирус (AAV)
[00581] Нацеленный на нуклеиновую кислоту эффекторный белок (такой как белок типа V, такой как C2c1 или C2c3) и одна или более направляющих РНК могут быть доставлены с использованием аденоассоциированного вируса (AAV), лентивируса, аденовируса или других плазмидных или вирусных типов векторов, в частности, с использованием лекарственных форм и доз, приведенных, например, в заявках на патент США №№ 8454972 (лекарственные формы, дозы для аденовируса), 8404658 (лекарственные формы, дозы для AAV) и 5846946 (лекарственные формы, дозы для ДНК-плазмид) и клинических испытаниях и публикациях на тему клинических испытаний, включающих лентивирус, AAV и аденовирус. Для примера, для AAV способ введения, лекарственная форма и доза приведены в заявке на патент США № 8454972 и в клинических испытаниях, включающих AAV. Для аденовируса способ введения, лекарственная форма и доза приведены в заявке на патент США № 8404658 и в клинических испытаниях, включающих аденовирус. Для доставки плазмиды способ введения, лекарственная форма и доза приведены в заявке на патент США № 5846946 и в клинических испытаниях, включающих плазмиды. Дозы могут быть разработаны на основе среднестатистического человека весом 70 кг (например, взрослого мужчину), и могут быть адаптированы для пациентов, индивидов, млекопитающих различного веса и вида. Частота введения находится в пределах компетенции медицинского или ветеринарного практика (например, врача, ветеринара), в зависимости от обычных факторов включая возраст, пол, общее состояние здоровья, другие состояния пациента или индивида, и конкретное представляющее интерес состояние и симптомы. Вирусные векторы могут быть введены в ткань-мишень. Для специфической для типа клетки модификации генома экспрессия нацеленного на нуклеиновую кислоту эффекторного белка (такого как белок типа V, такой как C2c1 или C2c3) может находиться под контролем специфического для типа клетки промотора. Например, специфическая для печени экспрессия может происходить под контролем промотора альбумина, и специфическая для нейронов экспрессия (например, для нацеливания на расстройства центральной нервной системы) может происходить под контролем промотора синапсина I.
[00582] С точки зрения доставки in vivo AAV предпочтительнее других вирусных векторов по двум причинам:
-низкая токсичность (возможно, благодаря способу очистки, не требующему ультрацентрифугирования частей клетки, которые могут вызывать иммунную реакцию), и
-низкая вероятность возникновения инсерционного мутагенеза, потому что AAV не интегрируется в геном хозяина.
[00583] Предел упаковывания AAV составляет 4,5 или 4,75 т.п.н. Это означает, что нацеленный на нуклеиновую кислоту эффекторный белок (такой как белок типа V, такой как C2c1 или C2c3), а также промотор и терминатор транскрипции должны быть помещены один и тот же вирусный вектор. Следовательно, варианты осуществления изобретения включают использование гомологов нацеленного на нуклеиновую кислоту эффекторного белка (такого как белок типа V, такого как C2c1 или C2c3), которые короче.
[00584] Что касается AAV, то AAV может представлять собой AAV1, AAV2, AAV5 или любую комбинацию. Можно выбрать ААV, исходя из клеток, на которые проводят нацеливание; например, можно выбрать серотипы AAV 1, 2, 5 или AAV1, AAV2, AAV5 с гибридным капсидом или любую их комбинацию для нацеливания в клетки головного мозга или нейроны; можно выбрать AAV4 для нацеливания в сердечную ткань, AAV8 пригоден для доставки в печень. В рамках настоящего изобретения предпочтительными являются индивидуальные промоторы и векторы. Соответствие определенных серотипов AAV и типов клеток (см. Grimm, D. et al, J. Virol. 82: 5887- 5911 (2008)) является следующим:
jLd jLu jh Au
Лентивирус
[00585] Лентивирусы представляют собой сложные ретровирусы, способные заражать и экспрессировать свои гены как в митотических, так и в постмитотических клетках. Наиболее известным лентивирусом является вирус иммунодефицита человека (ВИЧ), который использует гликопротеины вирусной оболочки других вирусов для нацеливания в широкий диапазон типов клеток.
[00586] Лентивирусы могут быть получены следующим образом. После клонирования pCasES10 (который содержит основу трансферной плазмиды лентивируса), HEK293FT при небольшом числе пересевов (p=5) были посеяны в колбе T-75 до 50% смыкания монослоя за день до трансфекции в DMEM с 10% эмбриональной бычьей сывороткой и без антибиотиков. Через 20 часов питательные среды заменяли на среду OptiMEM (без сыворотки) и трансфекция проводили через 4 часа. Клетки трансфицировали 10 мкг лентивирусной плазмиды для переноса (pCasES10) и следующими упаковочными плазмидами: 5 мкг pMD2.G (псевдотип VSV-g) и 7,5 мкг psPAX2 (gag/pol/rev/tat). Трансфекцию проводили в 4 мл OptiMEM со средством доставки катионных липидов (50 мкл Lipofectamine 2000 и 100 мкл реагента Plus). Через 6 часов питательную среду заменяли на DMEM без антибиотика с 10% эмбриональной бычьей сывороткой. В этих способах используется сыворотку во время культивирования, однако предпочтительными являются способы без сыворотки.
[00587] Лентивирус может быть очищен следующим образом. Вирусные супернатанты собирали через 48 часов. Супернатанты сначала очищали от дебриса и пропускали через 0,45-мкм фильтр с низким связыванием белков (PVDF). Затем их ультрацентрифугировали в течение 2 часов при скорости 24000 об/мин. Затем осадки с вирусом ресуспендировали в 50 мкл DMEM в течение ночи при 4°C. Потом их были разделяли на аликвоты и сразу же замораживали при -80°C.
[00588] В другом варианте осуществления изобретения также предусматривается минимальный вектор на основе лентивируса животного, отличного от примата, основанный на вирусе инфекционной анемии лошадей (EIAV), особенно для генотерапии глаза (см., например, Balagaan, J Gene Med 2006; 8; 275-285). В другом варианте осуществления изобретения также предусматривается RetinoStat®, лентивирусный вектор для генной терапии на основе вируса инфекционной анемии лошадей, экспрессирующий ангиостатические белки эндостатин и ангиостатин, доставляемый через субретинальную инъекцию для лечения возрастной дегенерации желтого пятна (см., например, Binley et al., HUMAN GENE THERAPY 23:980-991 (September 2012)) и этот вектор может быть изменен для использования в систему нацеливания на нуклеиновую кислоту по настоящему изобретению.
[00589] В другом варианте осуществления изобретения самоинактивирующиеся лентивирусные векторы с миРНК, нацеленной на экзон, общий для генов tat/rev ВИЧ, локализованной в ядрышке TAR-ловушки и анти-CCR5-специфического рибозима типа "hammerhead" (см, например, DiGiusto et al. (2010) Sci Transl Med 2:36ra43), может использоваться и/ или быть адаптирован для использования в нацеленной на нуклеиновую кислоту системе по настоящему изобретению. Минимум 2,5×106 CD34+ клеток на килограмм веса пациента могут быть получены предварительным стимулированием в течение 16-20 часов в питательной среде X-VIVO 15 (Lonza), содержащей 2 мкМ L-глютамина, фактор стволовых клеток (100 нг/мл), лиганд Flt-3 (Flt-3L) (100 нг/мл) и тромбопоэтин (10 нг/мл) (CellGenix) в плотности 2×106 клеток/мл. Предварительно стимулированные клетки могут быть трансдуцированы лентивирусным вектором при множественности инфекции, равной 5, в течение 16-24 часов в 75-см2 колбах для культивирования тканей, покрытых фибронектином (25 мг/см2) (RetroNectin, Takara Bio Inc.).
[00590] Лентивирусые векторы описаны для лечения для болезни Паркинсона, см., например, публикацию патентных заявок США № 20120295960, № 7303910 и № 7351585. Лентивирусые векторы описаны как способ лечения заболеваний глаз, см., например, публикации патентных заявок США № 20060281180, 20090007284, US20110117189; US20090017543; US20070054961, US20100317109. Лентивирусные векторы были также описаны как средство доставки в мозг, см., например, публикации патентных заявок США № US20110293571; US20110293571, US20040013648, US20070025970, US20090111106 и патент США № US7259015.
Доставка РНК
[00591] Доставка РНК: нацеленный на нуклеиновую кислоту белок Cas, например, белок типа V, такой как C2c1 или C2c3, и/или направляющая РНК, могут также быть доставлены в форме РНК. мРНК нацеленного на нуклеиновую кислоту белка Cas (например, белка типа V, такого как C2c1 или C2c3) может быть получена с использованием транскрипции in vitro. мРНК нацеленного на нуклеиновую кислоту белка Cas (например, белка типа V, такого как C2c1 или C2c3) может быть синтезирована с использованием ПЦР-кассеты, содержащей следующие элементы: промотор T7-последовательность Козака (GCCACC)-эффекторный белок-3'-нетранслируемая область (UTR) полиА-последовательности бета-глобина (последовательность из 120 или более остатков аденина). Такая кассета может быть использована для транскрипции с участием Т7-полимеразы. Направляющие РНК могут также быть транскрибированы путем транскрипции in vitro с кассеты, содержащей промотор T7-GG-последовательность направляющей РНК.
[00592] Для усиления экспрессии и уменьшения возможной токсичности последовательность, кодирующая нацеленный на нуклеиновую кислоту эффекторный белок, и/или направляющая РНК могут быть модифицированы так, чтобы содержать один или более модифицированных нуклеозидов, например, с использованием псевдо-U или 5-метил-C.
[00593] Способы доставки мРНК в настоящее время представляются особенно перспективными для доставки в печень.
[00594] Значительные клинические усилия по доставке РНК были сфокусированы на РНК-i или антисмысловых молекулах, однако такие системы могут быть адаптированы для доставки РНК с целью осуществления настоящего изобретения. В соответствии с этим следует прочесть ссылки на РНК-I и т.д., приведенные ниже.
Системы доставки частиц и/или составов:
[00595] Несколько типов систем доставки частиц и/или составов нашли применение в широком спектре биомедицинских применений. В целом, частица определяется как малый объект, ведущий себя как целая единица в отношении ее перемещения и свойств. Далее частицы подразделяются в соответствии с диаметром. Крупные частицы соответствуют диапазону от 2500 до 10000 нанометров. Тонкие частицы имеют размер от 100 до 2500 нанометров. Ультратонкие частицы или наночастицы в целом от 1 до 100 нанометров в размере. Основой для выбора предела в 100 нм является проявление новых свойств, отличающих частицы от насыпного материала, которые обычно развиваются при переходном масштабе длины менее 100 нм.
[00596] Как используют в настоящем описании, система доставки частиц/лекарственная форма определяется как любая биологическая система доставки/лекарственная форма, которая включает частицу в соответствии с настоящим изобретением. Частица в соответствии с настоящем изобретением представляет собой любой объект, имеющий самое большое измерение (например, диаметр) менее 100 микронов (мкм). В некоторых вариантах осуществления изобретения частицы по изобретению имеют самое большое измерение менее 10 мкм. В некоторых вариантах осуществления частицы по изобретению имеют самое большое измерение меньше, чем 2000 нанометров (нм). В некоторых вариантах осуществления частицы по изобретени имеют самое большое измерение меньше, чем 1000 нанометров (нм). В некоторых вариантах осуществления частицы по изобретению имеют самое большое измерение меньше, чем 900 нм, 800 нм, 700 нм, 600 нм, 500 нм, 400 нм, 300 нм, 200 нм или 100 нм. Как правило, частицы по изобретению имеют самое большое измерение (например, диаметр) 500 нм или меньше. В некоторых вариантах осуществления частицы по изобретению имеют самое большое измерение (например, диаметр) 250 нм или меньше. В некоторых вариантах осуществления частицы по изобретению имеют самое большое измерение (например, диаметр) 200 нм или меньше. В некоторых вариантах осуществления частицы по изобретению имеют самое большое измерение (например, диаметр) 150 нм или меньше. В некоторых вариантах осуществления изобретения разработанные частицы имеют самое большое измерение (например, диаметр) 100 нм или меньше. Меньшие частицы, например, имеющие самое большое измерение 50 нм или меньше используется в некоторых вариантах осуществления изобретения. В некоторых вариантах осуществления изобретения частицы по изобретению имеют самое большое измерение в диапазоне от 25 нм до 200 нм.
[00597] Охарактеризацию частиц (включая, например, морфологию, измерение, и т.д.) проводят с использованием ряд различных способов. Общие способы представляют собой электронную микроскопию (ТЕМ, SEM), атомно-силовую микроскопию (AFM), динамическое рассеяние света (DLS), рентгеновскую фотоэлектронную спектроскопию (XPS), порошковую рентгеновскую дифракцию (XRD), инфракрасную спектроскопию с преобразованием Фурье (FTIR), времяпролетную матрично-активированную лазерную десорбцию/ионизацию (MALDI-TOF), оптическую спектроскопию, интерферометрию двойной поляризации и ядерный магнитный резонанс (NMR). Охарактеризация (измерение размеров) может быть проведена относительно нативных частиц (т.е. перед загрузкой) или после загрузки груза (в настоящем описании груз относится, например, к одному или более компонентам системы CRISPR-Cas, например, ферменту CRISPR, или мРНК, или направляющей РНК, или любой их комбинации, и может включать дополнительные носители и/или эксципиенты), чтобы обеспечить частицы оптимального размера для доставки в любом способе применения настоящего изобретения in vitro, ex vivo и/или in vivo. В определенных предпочтительных вариантах осуществления изобретения охарактеризация размеров частиц (например, диаметра) основана на измерении с использованием динамического рассеяния света (DLS). Упоминаются заявка на патент США № 8709843; заявка на патент США № 6007845, патент США № 5855913; патент США № 5985309; заявка на патент США № 5543158; и публикация James E. Dahlman and Carmen Barnes et al. Nature Nanotechnology (2014) опубликованная через интернет 11 мая 2014 года, doi:10.1038/nnano.2014.84, касающаяся частицы, способов их получения и применения и измерения.
[00598] Системы доставки частиц в рамках настоящего изобретения могут быть представлены в любой форме, включая, но не ограничиваясь ими, твердую, полутвердую, эмульсию или коллоидные частицы. По существу любые такие системы доставки, описанные в настоящем описании, включая, но не ограничиваясь ими, например, системы, основанные на липидах, липосомах, мицеллах, микровезикулах, экзосомах или генных пушках, могут быть предоставлены для доставки частиц в рамках настоящего изобретения.
Частицы
[00599] мРНК фермента CRISPR и направляющая РНК могут быть доставлены одновременно с использованием частиц или липидных вирусных оболочек; например, фермент CRISPR и РНК по изобретению, например, в виде комплекса, могут быть доставлены с помощью частиц согласно Dahlman et al., WO2015089419 A2 и документам, цитируемых в них, таких как 7C1 (см., например, работу James E. Dahlman and Carmen Barnes et al. Nature Nanotechnology (2014) опубликованную через интернет 11 мая 2014 года, doi:10.1038/nnano.2014.84), например, частица доставки, включающая липид или липидоид и гидрофильный полимер, например, катионный липид, и гидрофильный полимер, например, такой, где катионный липид включает 1,2-диолеоил-3-триметиламмоний-пропан (DOTAP) или 1,2-дитетрадеканоил-sn-глицеро-3-фосфохолин (DMPC) и/или где гидрофильный полимер включает этиленгликоль или полиэтиленгликоль (PEG); и/или где частица далее включает холестерин (например, частица состава 1=DOTAP 100, DMPC 0, PEG 0, холестерин 0; состава номер 2=DOTAP 90, DMPC 0, PEG 10, холестерин 0; состава номер 3=DOTAP 90, DMPC 0, PEG 5, холестерин 5), где частицы сформированы c использованием эффективного, многоступенчатого процесса, где сначала, эффекторный белок и РНК смешивают вместе, например, в молярном отношении 1:1, например, при комнатной температуре, например, в течение 30 минут, например, в стерильном, свободном от нуклеаз 1X PBS; и отдельно, DOTAP, DMPC, PEG и холестерин, как применимо для состава, растворяют в спирте, например, 100% этаноле; и, эти два раствора смешивают вместе, чтобы сформировать частицы, содержащие комплексы).
[00600] мРНК нацеленных на нуклеиновые кислоты эффекторных белков (таких как белки типа V, таких как C2c1 или C2c3) и направляющая РНК могут быть доставлены одновременно с использованием частиц или липидов вирусных оболочек.
[00601] Например, Su X, Fricke J, Kavanagh DG, Irvine DJ ("In vitro and in vivo mRNA delivery using lipid-enveloped pH-responsive polymer nanoparticles" Mol Pharm. 2011 Jun 6;8(3):774-87. doi: 10.1021/mpl00390w. Epub 2011 Apr 1) описывают биоразлагаемые структурированные частицы c ядром из поли(β-аминоэфира) (PBAE), окруженным фосфолипидным бислоем. Они были разработаны для доставки мРНК in vivo. pH-зависимый компонент PBAE был выбран, чтобы способствовать эндосомному разрушению, в то время как поверхностный липидный слой был выбран, чтобы минимизировать токсичность ядра поликатиона. Следовательно, такой способ предпочтителен для доставки РНК по настоящему изобретению.
[00602] В одном варианте осуществления изобретения рассмотрены частицы на основе самосборки биоадгезивных полимеров, которые могут быть применены для доставки пептидов в головной мозг различными способами доставки: пероральным, внутривенным, назальным. Также предусматриваются другие варианты осуществления, такие как пероральная и окулярная доставка гидрофобных лекарств. Технология молекулярной оболочки включает спроектированную полимерную оболочку, которая защищена и доставлена к области заболевания (см., например, Mazza, M. et al. ACSNano, 2013. 7(2): 1016-1026; Siew, A., et al. Mol Pharm, 2012. 9(1): 14-28; Lalatsa, A., et al. J Contr Rel, 2012. 161(2):523-36; Lalatsa, A., et al., Mol Pharm, 2012. 9(6): 1665-80; Lalatsa, A., et al. Mol Pharm, 2012. 9(6): 1764-74, Garrett, N.L., et al. J Biophotonics, 2012. 5(5-6):458-68; Garrett, N.L., et al. J Raman Spect, 2012. 43(5):681-688, Ahmad, S., et al. J Royal Soc Interface 2010. 7:S423-33; Uchegbu, I.F. Expert Opin Drug Deliv, 2006. 3(5):629-40; Qu, X. et al. Biomacromolecules, 2006. 7(12):3452-9 и Uchegbu, IF., et al. Int J Pharm, 2001. 224:185-199). Предусматриваются дозы приблизительно 5 мг/кг с единичными или множественными дозами, в зависимости от ткани-мишени.
[00603] В одном варианте осуществления частицы, которые могут доставить РНК в злокачественную клетку, чтобы остановить рост опухоли, разработанные лабораторией Dan Anderson в MIT, могут использоваться/и или адаптированы для нацеленной на нуклеиновую кислоту системы по настоящему изобретению. В частности, лаборатория Anderson разработала полностью автоматизированные, комбинаторные системы для синтеза, очистки, охарактеризации и составляения новых биоматериалов и нанолекарственных форм. См., например, Alabi et al., Proc Natl Acad Sci U S A. 2013 Aug 6; 110(32): 12881-6; Zhang et al., Adv Mater. 2013 Sep 6;25(33):4641-5; Jiang et al., Nano Lett. 2013 Mar 13;13(3): 1059-64; Karagiannis et al., ACS Nano. 2012 Oct 23;6(10):8484-7; Whitehead et al., ACS Nano. 2012 Aug 28;6(8):6922-9 and Lee et al., Nat Nanotechnol. 2012 Jun 3,7(6):389-93.
[00604] Заявка на патент США 20110293703 касается липидоидных комплексов, особенно полезных при введении полинуклеотидов, которые могут быть применены для доставки нацеленной на нуклеиновую кислоту системы по настоящему изобретению. В одном аспекте аминоспиртовые липидоидные соединения комбинируют с агентом для доставки в клетку или индивидууму для формирования микрочастиц, наночастиц, липосом или мицелл. Агент, доставляемый частицами, липосомами или мицеллами, может быть в форме газа, жидкости или твердого тела, и агент может быть полинуклеотидом, белком, пептидом или низкомолекулярным соединением. Аминоспиртовые липидоидные соединения могут быть объединены с другими аминоспиртовыми липидоидными соединениями, полимерами (синтетическими или естественными), поверхностно-активными веществами, холестерином, углеводами, белками, липидами, и т.д. для формирования частиц. Затем эти частицы могут быть произвольно объединены с фармацевтическим наполнителем для получения фармацевтической композиции.
[00605] В заявке на патент США № 20110293703 также описаны способы получения аминоспиртовое липидоидное соединение. Одному или более эквивалентам амина позволяют реагировать с одним или более эквивалентами соединений с эпоксидной концевой группой, что при подходящих условиях позволяет сформировать аминоспиртовое липидоидное соединение по настоящему изобретению. В некоторых вариантах осуществления изобретения все аминогруппы амина полностью реагируют с эпоксидными концевыми группами с образованием третичных аминов. В других вариантах осуществления изобретения все аминогруппы амина не полностью реагируют с эпоксидными концевыми группами, таким образом, приводя к образованию первичных или вторичных аминов в аминоспиртовом липидоидном соединении. Эти первичные или вторичные амины сохраняют способность реагировать с другими электрофилами, такими как различные соединения с эпоксидной концевой группой. Как может оценить специалист в данной области, реакция амина с недостатком соединения с эпоксидными группами приводит ко множеству различных аминоспиртовых липидоидных соединений с различным числом хвостовых частей. Определенные амины могут быть полностью функционализированы двумя соединениями с эпоксидной концевой группой с образованием двух хвостовых частей, в то время как другие молекулы не являются полностью функционализированными соединениями с эпоксидной концевой группой с образованием хвостовых частей. Например, диамин или полиамин могут включать один, два, три, или четыре полученных из соединений с эпоксидной концевой группой хвостовых частей, присоединенных к разным аминогруппам молекулы, что приводит к образованию первичных, вторичных, и третичных аминов. В некоторых вариантах осуществления изобретения все аминогруппы не полностью функционализируются. В некоторых вариантах осуществления изобретения используются два соединения с эпоксидной концевой группой одного типа. В других вариантах осуществления изобретения используются два или более различных соединений с эпоксидными концевыми группами. Синтез аминоспиртовых липидоидных соединений может происходить с растворителем или без него, и синтез может быть выполнен при более высоких температурах в пределах 30-100°C, предпочтительно приблизительно при 50-90°C. Подготовленные аминоспиртовые липидоидные соединения необязательно могут быть очищены. Например, смесь аминоспиртовых липидоидных соединений может быть очищена, чтобы обеспечить аминоспиртовое липидоидное соединение с конкретным числом полученных из эпоксида составных хвостовых частей. Или смесь может быть очищена, чтобы привести к определенному стерео- или региоизомеру. Аминоспиртовые липидоидные соединения также могут быть алкилированы с использованием алкилгалогенида (например, метилиодида) или другого алкилирующего агента, и/или они могут быть ацилированы.
[00606] В заявке на патент США № 20110293703 также описаны библиотеки аминоспиртовых липидоидных соединений, полученных способами по изобретению. Эти аминоспиртовые липидоидные соединения могут быть получены и/или исследованы с использованием высокопроизводительных способов, включающих жидкостные манипуляторы, роботы, микропланшеты для титрования, компьютеры, и т.д. В некоторых вариантах осуществления изобретения аминоспиртовые липидоидные соединения подвергают скринингу в отношении их способности трансфицировать полинуклеотиды или другие агенты (например, белки, пептиды, маленькие молекулы) в клетку.
[00607] Заявка на патент США № 20130302401 касается класса поли(бета-аминоспиртов) (PBAA), которые получают с помощью комбинаторной полимеризации. Разработанные PBAA могут использоваться в биотехнологии и биомедицинских способах применения как покрытия (такие как покрытия пленок или многослойных пленок для медицинских устройств или имплантатов), добавки, материалы, наполнители, агенты, не подверженные биологическому обрастанию, агенты для микроструктурирования и агенты клеточной инкапсуляции. Использование PBAA в качестве поверхностных покрытий позволило индуцировать разные уровни воспаления, как in vitro, так in vivo, в зависимости от химической структуры. Большое химическое разнообразие этого класса материалов позволило идентифицировать полимерные покрытия, которые ингибируют активацию макрофагов in vitro. Кроме того, эти покрытия уменьшают привлечение клеток воспаления и уменьшают фиброз после подкожной имплантации карбоксилированных микрочастиц полистирола. Эти полимеры могут использоваться для формирования капсулы комплекса полиэлектролита для инкапсуляции клетки. У изобретения может также быть много других биологических способов применения, таких как антибактериальные покрытия, доставка ДНК или миРНК, и тканевая инженерия на основе стволовых клеток. Рекомендации заявки на патент США № 20130302401 могут быть применены к нацеленной на нуклеиновые кислоты системе по настоящему изобретению.
[00608] В другом варианте осуществления изобретения предусматриваются липидные наночастицы (LNP). Малая интерферирующая РНК против антитранстиретина была заключена в капсулу липидных наночастиц и доставлена в человеческие клетки (например, см. Coelho et al., N Engl J Med 2013;369:819-29), и такая система может быть а.даптирована и применена для нацеленной на нуклеиновую кислоту системы по настоящему изобретению. Рассмотренные дозы составляют от приблизительно 0,01 до приблизительно 1 мг на кг массы тела при доставке внутривенно. Для снижения риска инфузионных реакций предусматриваются такие лекарственные препараты как дексаметазон, ацетаминофен, дифенгидрамин, цетиризин и ранитидин. Рассмотрено применение в виде множественных доз, составляющих приблизительно 0,3 мг на килограмм массы тела каждые 4 недели до пяти доз.
[00609] Показана высокая эффективность LNP для доставки миРНК в печень (см, например, Tabemero et al, Cancer Discovery, April 2013, Vol. 3, No. 4, pages 363-470), следовательно, LNP могут использоваться для доставки РНК, кодирующей нацеленный на нуклеиновую кислоту эффекторный белок к печени. Дозировка может составлять приблизительно четыре дозы по 6 мг/кг LNP каждые две недели. Tabemero et al. продемонстрировали, что регресс опухоли наблюдался после первых 2 циклов LNP при дозировке на уровне 0,7 мг/кг, к концу 6 циклов пациент достиг частичной ремиссии с полной регрессией метастазов лимфатических узлов и существенным уменьшением опухолей печени, полная ремиссия была достигнута после 40 доз, потом ремиссия сохранилась, лечение было закончено после 26 месяцев. У двух пациентов с RCC и внепеченочными участками заболевания, включая почку, легкое и лимфатические узлы, которые прогрессировали после предшествующей терапии с ингибиторами каскада VEGF, было стабильное заболевание во всех областях в течение приблизительно 8-12 месяцев, и пациент с PNET и метастазами печени продолжил лечение в дополнительном исследовании в течение 18 месяцев (36 доз) со стабильным заболеванием.
[00610] Однако заряд LNP также должен быть принят во внимание. Катионные липиды комбинируются с отрицательно заряженными липидами для индукции небислойных структур для облегчения внутриклеточной доставки. Поскольку заряженные LNP быстро покидают кровоток после внутривенной инъекции, были разработаны ионизируемые катионные липиды со значениями pKa ниже 7 (см., например, Rosin et al, Molecular Therapy, vol. 19, no. 12, pages 1286-2200, Dec. 2011). Отрицательно заряженные полимеры, такие как РНК могут быть загружены в LNP при низких значениях pH (например, pH 4), когда ионизируемые липиды имеют положительный заряд. Однако при физиологических значениях pH LNP имеют низкий поверхностный заряд, совместимый с более длительным временем циркуляции. Исследование было сосредоточено на четырех видах ионизируемых катионных липидов, а именно, 1,2-дилинеоил-3-диметиламмоний-пропан (DLinDAP), 1,2-дилинолеилокси-3-N, N-диметиламинопропан (DLinDMA), 1,2-дилинолеилокси-кето-N,N-диметил-3-аминопропан (DLinKDMA) и 1,2-дилинолеил-4-(2-диметиламиноэтил)-[1,3]-диоксолан (DLinKC2-DMA). Было показано, что системы миРНК LNP, содержащие эти липиды, показывают значительно отличающиеся свойства подавления экспрессии генов в гепатоцитах in vivo с эффективностью, варьирующей согласно ряду DLinKC2-DMA>DLinKDMA>DLinDMA>>DLinDAP при использовании моделей подавления экспрессии гена фактора VII (см., например, Rosin et al, Molecular Therapy, vol, 19, no. 12, pages 1286-2200, Dec, 2011). Может быть рассмотрена дозировка 1 мкг/мл LNP или РНК CRISP-Cas внутри или в ассоциации с LNP, особенно для лекарственного препарата, содержащего DLinKC2-DMA.
[00611] Подготовка LNP и инкапсуляция CRISPR-Cas может быть использована или адаптирована из Rosin et al, Molecular Therapy, vol. 19, no. 12, pages 1286-2200, Dec, 2011. Катионные липиды 1,2-дилинеоил-3-диметиламмоний-пропан (DLinDAP), 1,2-дилинолеилокси-3-N, N-диметиламинопропан (DLinDMA), 1,2-дилинолеилокси-кето-N,N-диметил-3-аминопропан (DLinKDMA), 1,2-дилинолеил-4-(2-диметиламиноэтил)-[1,3]-диоксолан (DLinKC2-DMA), (3-o-[2"-(метоксиполиэтиленгликоль 2000)сукциноил]-1,2-димиристоил-sn-гликоль (PEG-S-DMG) и R-3-[(ко-метокси-поли(этиленгликоль)2000) карбамоил]-1,2-димиристилоксипропил-3-амин (PEG-C-DOMG) могут быть приобретены у Tekmira Pharmaceuticals (Ванкувер, Канада) или синтезированы. Холестерин может быть приобретен у Sigma (Сент-Луис, Миссури). РНк конкретного нацеленного на нуклеиновую кислоту комплекса (CRISPR-Cas) может быть заключена в капсулу из LNP, содержащую DLinDAP, DLinDMA, DLinK-DMA и DLinKC2-DMA (катионный липид:DSPC:CHOL:PEGS-DMG или PEG-C-DOMG в молярных отношениях 40:10:40:10). Необязательно 0,2% SP-DiOC18 (Invitrogen, Берлингтон, Канада) может быть включен в состав, чтобы оценить клеточный захват, внутриклеточную доставку и биораспределение. Инкапсуляция может быть выполнена путем растворения липидной смеси, состоящей из катионного липида:DSPC:холестерола:PEG-c-DOMG (молярное отношение 40:10:40:10), в этаноле до конечной концентрации липида 10 ммоль/л. Этот раствор липида в этаноле может быть добавлен по каплям к цитрату с концентрацией 50 ммоль/л, pH 4,0, для формирования многослойных везикул в целях достижения конечной концентрации этанола 30% по объему. Большие однослойные везикулы могут быть сформированы после пропускания многослойных везикул через два сложенных поликарбонатных фильтра Nuclepore на 80 нм с использованием экструдера (Northern Lipids, Ванкувер, Канада). Инкапсуляция может быть достигнута добавлением РНК, растворенной до концентрации 2 мг/мл, в 50 ммоль/л цитрата, pH 4,0, содержащего 30% этанола по объему, по каплям к предварительно сформированным большим однослойным пузырькам и инкубации при температуре 31°C в течение 30 минут с постоянным перемешиванием до конечного отношения РНК/липид по весу, равного 0,06/1. Удаление этанола и нейтрализация полученного состава буфером могут быть выполнены с помощью диализа против натрий-фосфатного буфера (PBS), pH 7,4, в течение 16 часов с использованием восстановленных целлюлозных мембран для диализа Spectra/Por 2. Гранулометрический состав может быть определен динамическим рассеянием света с использованием гранулометра NICOMP 370, режима везикул/интенсивности и Гауссовской аппроксимации (Nicomp Particle Sizing, Санта-Барбара, Калифорния). Размер частиц для всех трех систем LNP может составлять ~70 нм в диаметре. Эффективность инкапсуляции РНК может быть определена удалением свободной РНК с использованием колонки VivaPureD Mini (Sartorius Stedim Biotech) из образцов, собранных до и после диализа. Инкапсулированная РНК может быть извлечена из элюированных частиц и определена количественно на уровне 260 нм. Отношение РНК к липидам может быть определено измерением содержания холестерина в везикулах с использованием ферментативного анализа с холестерином E от Wako Chemicals USA (Ричмонд, Вирджиния). Вместе с обсуждаемыми в настоящем описании LNP и липидами PEG, пегилированные липосомы или LNP так же подходят для доставки нацеленной на нуклеиновую кислоту системы или ее компонентов.
[00612] Получение липидных наночастиц (LNP) возможно производить согласно и/или адаптировано из Rosin et al, Molecular Therapy, vol. 19, no. 12, pages 1286-2200, Dec. 2011. Предварительно подготовленный раствор липидной смеси (с общей концентрацией липидов 20,4 мг/мл) может быть получен в этаноле, содержащем DLinKC2-DMA, DSPC и холестерин в молярных соотношениях 50:10:38,5. Ацетат натрия может быть добавлен в предварительно подготовленный раствор липидной смеси в молярном соотношении 0,75:1 (ацетат натрия: DLinKC2-DMA). Липиды могут быть далее гидратированы с использованием смеси 1,85 объема цитратного буфера (10 ммоль/л, pH 3,0) при активном перемешивании, приводящем к спонтанному образования липосом в водном буфере, содержащем 35% этанол. Раствор липосом может быть инкубирован при 37°C, чтобы сделать возможным зависимое от времени увеличение размеров частиц. Аликвоты могут быть удалены в разные моменты времени в ходе инкубации для исследования изменений размера липосом с помощью способа динамического светорассеяния (Zetasizer Nano ZS, Malvern Instruments, Worcestershire, Великобритания). При достижении желаемого размера частиц водный раствор PEG и липидов (исходный раствор=10 мг/мл PEG-DMG в 35% (объем/объем) этанола) может быть добавлен к смеси липосом для достижения конечной концентрации PEG 3,5% от общего липида. При добавлении PEG-липидов липосомы должны быть примерно их размера и эффективно подавляя дальнейший рост. Далее РНК может быть добавлена к пустым липосомам при соотношении РНК к общим липидам примерно 1:10 (вес:вес) с последующей инкубацией на протяжении примерно 30 мин при 37°C для образования загруженных липидных наночастиц (LNP). Такая смесь может быть в дальнейшем подвергнута диализу на протяжении ночи в PBS) и фильтрации через 0,45-пм фильтр-шприц.
[00613] Конструкции сферических нуклеиновых кислот (Spherical Nucleic Acid, SNA™) и другие частицы (в частности, частицы золота) также предусматриваются в качестве средства доставки нацеленных на нуклеиновую кислоту систем к предполагаемым мишеням. Значительный объем данных демонстрирует полезность конструкций терапевтических сферических нуклеиновых кислот от AuraSense (SNA™), основанных на функционализированных нуклеиновой кислотой золотых частицах.
[00614] Литература, которая может быть использована в совокупности с изложенными в настоящем описании идеями, включает: Cutler et al., J. Am. (Chem. Soc. 2011 133:9254-9257, Hao et al., Small. 2011 7:3158-3162, Zhang et al., ACS Nano. 2011 5:6962-6970, Cutler et al., J. Am. Chem. Soc. 2012 134:1376-1391, Young et al., Nano Lett. 2012 12:3867-71, Zheng et al., Proc. Natl. Acad. Sci. USA. 2012 109:11975- 80, Mirkin, Nanomedicine 2012 7:635-638 Zhang et al., J. Am. Chem. Soc. 2012 134:16488- 1691, Weintraub, Nature 2013 495:S14-S16, Choi et al., Proc. Natl. Acad. Sci. USA. 2013 110(19):7625-7630, Jensen et al., Sci. Transl. Med. 5, 209ral52 (2013) и Mirkin, et al., Small, 10:186-192.
[00615] Самособирающиеся частицы с РНК могут быть произведены из полиэтиленимина (PEI), связанного с PEG через пептидный лиганд Arg-Gly-Asp (RGB), присоединенный к дальнему концу PEG. Такая система была использована, например, в качестве средства для нацеливания на сосуды опухоли, экспрессирующей интегрины, и доставки миРНК, ингибирующей экспрессию рецептора факторов роста эндотелия сосудов 2 (VEGFR2), и достижения, таким образом, ангиогенеза в опухоли (см., например, Sehiffelers et al, Nucleic Acids Research, 2004, Vol. 32, No. 19). Наноплексы могут быть приготовлены смешиванием равных объемов водных растворов катионного полимера и нуклеиновой кислоты для получения чистого молярного избытка ионизируемого азота (полимер) относительно фосфата (нуклеиновая кислота) в диапазоне от 2 до 6. Электростатические взаимодействия между катионными полимерами и нуклеиновой кислотой приводят к образованию полиплексов со средним значением распределения размеров примерно 100 нм, в связи с чем в настоящем описании они названы наноплексами. Предусматривается дозировка примерно от 100 до 200 мг РНК комплекса, нацеленного на нуклеиновую кислоту, для доставки в самособирающихся частицах согласно Sehiffelers et al.
[616] Наноплексы, описанные Bartlett et al. (PNAS, September 25, 2007,vol. 104, no. 39) также могут быть применены в рамках настоящего изобретения. Наноплексы, описанные Bartlett et al., получают смешиванием равных объемов водных растворов катионного полимера и нуклеиновой кислоты для получения чистого молярного избытка ионизируемого азота (полимера) по отношению к фосфату (нуклеиновая кислота) в диапазоне от 2 до 6. Электростатические взаимодействия между катионными полимерами и нуклеиновой кислотой приводят к образованию полиплексов со средним значением распределения размеров примерно 100 нм, в связи с чем здесь они названы наноплексами. DOTA-миРНК, описанная Bartlett et al., была синтезирована следующим образом: 1,4,7,10-тетразациклододекан-1,4,7,10-тетрауксусной кислоты моно (N-гидроксисукцинимид эфир) (DOTA-эфир NHS) был заказан у Macrocyclics (Даллас, Техас, США). Кодирующая цепь аминомодифицированной РНК со 100-кратным молярным избытком эфира DOTA-NHS в карбонатном буфере (pH 9) была помещена в пробирку для микроцентрифугирования. Реакция содержимого была проведена при перемешивании на протяжении 4 ч при комнатной температуре. Связанные DOTA-кодирующая РНК были осаждены этанолом, ресуспенидрованы в воде и прошли отжиг на немодифицированную некодирующую цепь для получения DOTA-миРНК. Все жидкости были предварительно обработаны Chelex-100 (Bio-Rad, Hercules, Калифорния, США) для удаления следов загрязнения металлом. Нацеленные на Tf и ненацеленные частицы миРНК могут быть образованы с использованием содержащих циклодекстрин поликатионов. Обычно частицы образуются в воде при соотношении зарядов (+/-), равном 3, и концентрации миРНК 0,5 г/л. Один процент молекул адамантан-PEG на поверхности нацеленных частиц был модифицирован (адамантан-PEG-Tf). Частицы были суспендированы в 5% (масса/объем) полученного раствора переносчика глюкозы для инъекций.
[00617] Davis et al. (Nature, Vol 464, 15 April 2010) провели клинические испытания РНК с использованием системы доставки нацеленных частиц (регистрационный номер клинического испытания NCT00689065). Пациенты с солидным типом злокачественной опухли, нечувствительной к стандартным способам лечения, получали дозировки направленных частиц в дни 1, 3, 8 и 10 21-дневного курса путем 30-минутного внутривенного вливания. Такие частицы включают, по существу состоят или состоят из синтетической системы доставки, содержащей: (1) линейный полимер на основе циклодекстрина (CDP), (2) лиганд, нацеленный на белок трансферрин человека (TF), находящийся снаружи наночастицы для активации рецепторов TF (TFR) на поверхности злокачественных клеток, (3) гидрофильный полимер (полиэтиленгликоль, PEG) используется для увеличения стабильности наночастиц в биологических жидкостях) и (4) миРНК, разработанная для уменьшения экспрессии RRM2 (используемая в клинической практике последовательность ранее была обозначена как siR2B+5). На протяжении долгого времени было известно, что TFR активнее экспрессируется в клетках злокачественных опухолей, поэтому RRM2 является доказанная мишень терапии против злокачественной опухоли. Такие частицы (используемая в клинической практики версия обозначена как CALAA-01) продемонстрировали хорошую переносимость в исследованиях с различными дозировками на отличных от человека приматах. Хотя ранее один пациент с хроническим миелоидным лейкозом получал миРНК путем доставки липосомами, клинические испытания Davis et al. являются первичными для систематической доставки миРНК с нацеленной системой доставки и с целью лечения пациентов с солидными злокачественными опухолями. Для проверки того, может ли система направленной доставки обеспечить эффективную доставку функциональной миРНК в опухоли у человека, Davis et al. исследовали материалы биопсии трех пациентов из трех групп по дозировке: пациенты A, В и C, каждый из которых имел метастазирующую меланому и получал CALAA-01 в дозах 18, 24 и 30 мг/м2 миРНК соответственно. Сходные дозы могут также быть предположены для нацеленной на нуклеиновую кислоту системы по настоящему изобретению. Доставка согласно изобретению может быть достигнута с использованием частиц, содержащих линейный полимер на основе циклодекстрина (CDP), лиганд, нацеленный на белок трансферрин человека (TF), находящийся снаружи наночастицы для активации рецепторов TF (TFR) на поверхности злокачественных клеток и/или гидрофильный полимер (например, полиэтиленгликоль (PEG), используемый для повышения стабильности в биологических жидкостях).
[00618] В отношении настоящего изобретения, предпочтительно наличие одного или более компонентов нацеленного на нуклеиновую кислоту комплекса, например, нацеленного на нуклеиновую кислоту эффекторного белка или мРНК, либо направляющая РНК, доставленных при помощи частиц или липидной упаковки. Другие системы доставки или векторы могут быть использованы в совокупности с частицами в соответствующих вариантах изобретения.
[00619] В целом под "наночастицей" понимают любую частицу с диаметром менее 1000 нм. В определенных предпочитаемых вариантах осуществления наночастицы по настоящему изобретению имеют размер вдоль длинной оси (например, в диаметре) 500 нм или менее. В других предпочитаемых вариантах, описываемых в данном изобретении, наночастицы имеют наибольший размер в диапазоне между 25 нм и 200 нм. В других предпочитаемых вариантах описываемые в данном изобретении наночастицы имеют вдоль длинной оси размер 100 нм или менее. В других предпочитаемых вариантах описываемые в данном изобретении наночастицы имеют размер от 35 нм до 60 нм.
[00620] Частицы, относящиеся к настоящему изобретению, могут быть доставлены в различных формах, например, твердые частицы (например, металлы как серебро, золото, железо, титан), неметаллы, твердые частицы на основе липидов, полимеры, суспензии частиц или их комбинации. Могут быть получены частицы из металлов, диэлектриков или полупроводников, как и гибридные структуры (например, с ядром и оболочкой). Частицы из полупроводниковых материалов могут быть также маркированы квантовыми точками, если имеют размер, достаточно малый (обычно менее 10 нм) для возможности квантования электронных уровней. Такие наночастицы используются в биомедицинских применениях в качестве носителей для лекарств или агентов для визуализации и могут быть адаптированы для сходных целей в рамках настоящего изобретения.
[00621] Были произведены полутвердые и мягкие частицы, входящие в объем настоящего изобретения. Частицей-прототипом полутвердой природы является липосома. Различные типы липосомных частиц в настоящее время используются в клинической практике в качестве систем доставки для противораковых лекарств и вакцин. Наполовину гидрофильные и наполовину гидрофобные частицы называются янус-частицами и они особенно эффективны для стабилизации эмульсий. Они способны к самостоятельной сборке на поверхностях раздела вода/масло и действуют как твердые поверхностно-активные вещества.
[00622] В патенте США № 8709843, включенном в настоящее описание в качестве ссылки, предложена система доставки лекарств для нацеленной доставки содержащих терапевтический агент частиц к тканям, клеткам и внутриклеточным компартментам. Настоящее изобретение относится к нацеленным частицам, содержащим полимер, связанный с поверхностно-активным веществом, гидрофильный полимер или липид.
[00623] В патенте США № 6007845, включенном в настоящее описание в качестве ссылки, предложены частицы, имеющие ядро из мультиблочного сополимера, образованное путем ковалентного связывания мультифункционального соединения с одним или более гидрофильными полимерами, и содержащие биологически активный материал.
[00624] В патенте США № 5855913, включенном в настоящее описание в качестве ссылки, предложена композиция частиц, включающая аэродинамически легкие частицы с плотностью менее 0,4 г/см3 и средним диаметром от 5 пм до 30 пм, имеющих на поверхности поверхностно-активное вещество для доставки лекарств в систему легких.
[00625] В патенте США № 5985309, включенном в настоящее описание в качестве ссылки, предложены частицы, содержащие поверхностно-активное вещество и/или гидрофильный или гидрофобный комплекс с положительно или отрицательно заряженным лекарственным веществом или агентом для диагностики и заряженную частицу с противоположным зарядом для доставки в легкие.
[00626] В патенте США № 5543158, включенном в настоящее описание в качестве ссылки, предложены биоразлагаемые инъецируемые частицы с биоразлагаемым твердым ядром, содержащим биологически активный материал и остатки поли(алкиленгликоля) на поверхности.
[00627] В WO2012135025 (также опубликованной как US20120251560), включенной в настоящем описании в качестве ссылки, описаны полимеры конъюгированного полиэтиленимина (PEI) и конъюгированных азамакроциклов (вместе называемых "конъюгированный липомер" или "липомеры"). В некоторых вариантах осуществления может быть предусмотрено, что такие способы и материалы из цитируемых в настоящем описании документов, например, конъюгированные липомеры, могут быть использованы в контексте нацеленной на нуклеиновую кислоту системы для достижения in vitro, ex vivo и in vivo изменений генома для модификации экспрессии генов, а также экспрессии белков.
[00628] В одном варианте осуществления частица может быть модифицированным эпоксидом липидным полимером, предпочтительно 7C1 (см., например, James E. Dahlman and Carmen Barnes et al. Nature Nanotechnology (2014) опубликованная через интернет 11 мая 2014 года, doi:10.1038/nnano.2014.84). C71 был синтезирован в реакции Cl5 липидов с концевыми эпоксидными группами с PEI600 при молекулярном соотношении 14:1 и составлен с C14PEG2000 для получения частиц (диаметр между 35 и 60 нм), стабильных в растворе PBS (фосфатно-солевой буфер, PBS) на протяжении не менее 40 дней.
[00629] Модифицированный эпоксидными группами липид-полимер может быть использован для доставки нацеленной на нуклеиновую кислоту системы по настоящему изобретению в клетки легких, сердечно-сосудистой системы или почек, однако, квалифицированный специалист в данной области может адаптировать систему для доставки к другим органам-мишеням. Предполагаются дозировки в диапазоне от приблизительно 0,05 до приблизительно 0,6 мг/кг. Также предусматриваются дозировки, вводимые на протяжении нескольких суток или недель с общей дозой приблизительно 2 мг/кг.
Экзосомы
[00630] Экзосомы являются эндогенными нановезикулами, транспортирующими РНК или белки, которые могут доставлять РНК в головной мозг и другие органы-мишени. Для уменьшения иммуногенности Alvarez-Erviti et al. (2011, Nat Biotechnol 29: 341) использовали полученные ими собственные дендритные клетки мышей для получения экзосом. Нацеливание в головной мозг было достигнуто инженерией дендритных клеток, приводящей к экспрессии Lamp2b, экзосомального мембранного белка, слитого со специфическим пептидом нейронов RVG. Очищенные экзосомы были загружены экзогенной РКН с помощью электропорации. миРНК GAPDH, доставленная введенными внутривенно нацеленными на RVG экзосомами в нейроны, микроглию, олигодендроциты в головном мозге вызывает нокдаун конкретного гена. Предварительная обработка экзосомами RVG не ослабляла нокдаун, неспецифический захват в других тканях не наблюдался. Терапевтический потенциал опосредованной экзосомами доставки миРНК был продемонстрирован сильным нокдауном мРНК (60%) и белка (62%) ВACE1, терапевтической мишени болезни Альцгеймера.
[00631] Для получения совокупности иммунологически инертных экзосом Alvarez-Erviti et al. взяли пробы костного мозга мышей инбредной линии C57BL/6 с однородным гаплотипом по главному комплексу гистосовместимости (MHC). Поскольку незрелые дендритные клетки производят большие объемы экзосом, лишенных активаторов Т-клеток, таких как MHC-II и CD86, Alvarez-Erviti et al. отбирали дендритные клетки с использованием гранулоцитарно-макрофагального колониестимулирующего фактора (GM-CSF) на протяжении 7 дней. Экзосомы были очищены от культурального супернатанта на следующий день с использованием стандартных протоколов ультрацентрифугирования. Полученные экзосомы были физически однородны с распределением размеров, имеющим пик, соответствующий диаметру 80 нм, как установлено путем анализа траекторий частиц (NTA) и электронной микроскопией. Alvarez-Erviti et al. получили 6-12 мкг экзосом (измерено на основе концентрации белка) на 106 клеток.
[00632] Далее Alvarez-Erviti et al. исследовали возможность загрузки модифицированных экзосом экзогенным грузом при помощи протоколов электропорации, адаптированных для применения в наномасштабе. Поскольку электропорация мембранных частиц в наноразмерном масштабе недостаточно охарактеризована, неспецифическая маркированная Cy5 РНК была использована для оптимизации протокола электропорации эмпирически. Количество упакованной РНК было оценено после ультрацентрифугирования и лизиса экзосом. Электропорация при 400 В и 125 пФ привела к максимальному удержанию РНК и использовалась во всех последующих экспериментах.
[00633] Alvarez-Erviti et al. вводили 150 мкг каждой миРНК BACE1, упакованной в 150 пг экзосом RVG, нормальным мышам C57BL/6 и сравнивали эффективность нокдауна с четырьмя контролями: интактные мыши, мыши, которым вводились только экзосомы RVG, мыши, которым вводились миРНК BACE1 в комплексе с in vivo реагентом катионных липосом и мыши, которым вводились миРНК BACE1 в комплексе с RVG-9R, пептидом RVG, связанным с 9 остатками D-аргинина, электростатически связанными с миРНК. Образцы кортикальной ткани были проанализированы после 3 дней введения, был обнаружен существенный нокдаун белка (45%, P<0,05, против 62%, P<0,01) у получавших экзосомы как с миРНК-RVG-9R, так и с миРНК-RVG, что было следствием существенного уменьшения уровней мРНК BACE1 (66% [+ или -] 15%, P<0,001 и 61% [+ или -] 13% соответственно, P<0,01). Более того, заявители продемонстрировали существенное уменьшение (55%, P<0,05) общих уровней бета-амилоида 1-42, главного компонента амилоидных бляшек при патологии болезни Альцгеймера в случае животных, которым вводили экзосомы RVG. Наблюдаемое уменьшение превосходило уменьшение для бета-амилоида 1-40 у нормальных мышей после внутрижелудочкового введения ингибиторов BACE1. Alvarez-Erviti et al. осуществили быструю амплификацию 5'-концов кДНК (RACE) для продукта расщепления BACE1, что предоставило доказательства опосредованного РНК-интерференцией нокдауна за счет миРНК.
[00634] Наконец, Alvarez-Erviti et al. исследовали, вызывают ли экзосомы РНК-RVG иммунный ответ in vivo путем измерения концентраций L-6, IP-10, TNFα и IFN-α в сыворотке. После введения экзосомам для всех цитокинов были зарегистрированы несущественные изменения, сходные с возникающими при введении реагента для трансфекции миРНК, и противоположные возникающим при обработке миРНК-RVG-9R, которая стимулирует высокую секрецию IL-6, что подтверждает иммунологическую инертность при введении экзосом. Учитывая, что экзосомы содержат только 20% миРНК, доставка RVG-экзосомами представляется более эффективной, чем доставка RVG-9R при достижении сравнимого уровня нокдауна мРНК с использованием меньшего в 5 раз количества миРНК без соответствующего уровня иммунной стимуляции. Данный эксперимент продемонстрировал терапевтический потенциал технологии RVG-экзосом, которая может подходить для долговременного сайленсинга генов, связанных с нейродегенеративными заболеваниями. Система доставки с использованием экзосом по Alvarez-Erviti et al. может быть применена для доставки нацеленной на нуклеиновую кислоту системы по настоящему изобретению к мишеням терапии, особенно в случае нейродегенеративных заболеваний. Доза в диапазоне примерно от 100 до 1000 мг нацеленной на нуклеиновую кислоту системы, упакованной в от ~100 до ~1000 мг RVG-экзосом, может быть предусмотрена в рамках настоящего изобретения.
[00635] El-Andaloussi et al. (Nature Protocols 7,2112-2126(2012)) установили, каким образом полученные из культивируемых клеток экзосомы могут быть использованы для доставки РНК in vitro и in vivo. Данный протокол вначале описывает получение нацеленных экзосом путем трансфекции экспрессирующего вектора, содержащего экзосомальный белок, слитый с пептидным лигандом. Далее El-Andaloussi et al. описывают очистку и охарактеризацию экзосом из супернатанта трансфицированных клеток. Далее El-Andaloussi et al. детализируют ключевые шаги загрузки РНК в экзосомы. В конце El-Andaloussi et al. кратко описывают использование экзосом для эффективной in vitro и in vivo доставки РНК в головной мозг мыши. Примеры ожидаемых результатов, для которых количественно оценена опосредованная экзосомами доставка путем функционального анализа и визуализации, также приведены. Полный протокол занимает ~3 недели. Доставка или введение согласно изобретению могут быть выполнены с использованием экзосом, полученных из дендритных клеток пациента. В соответствии с описанными в настоящем описании идеями, это может быть использовано в практике использования изобретения.
[00636] В другом варианте осуществления рассматриваются экзосомы плазмы из Wahlgren et al. (Nucleic Acids Research, 2012, Vol. 40, No. 17 el30). Экзосомы являются наноразмерными везикулами (размером 30-90 нм), производимыми многими типами клеток, включая дендритные клетки (DC), В-клетки, T-клетки, тучные клетки, эпителиальные клетки и клетки опухоли. Такие везикулы образуются путем впячивания и отделения поздних эндосом и далее освобождаются во внеклеточную среду при слиянии с плазматической мембраной. Поскольку экзосомы в естественном состоянии переносят РНК между клетками, такое их свойство может быть полезно для генной терапии, и исходя из данного описания экзосомы могут быть использованы при применении настоящего изобретения.
[00637] Экзосомы из плазмы могут быть получены путем центрифугирования фракции тромбоцитов и белых клеток крови при 900g на протяжении 20 мин для выделения плазмы с последующим отбором супернатанта, центрифугирования при 300g на протяжении 10 мин для удаления клеток и при 16500g на протяжении 30 мин с последующей фильтрацией через 0,22-мм фильтр. Экзосомы осаждаются при ультрацентрифугировании при 120000g на протяжении 70 мин. Химическая трансфекция миРНК в экзосомы выполнена согласно рекомендациям производителя для начального набора для РНК-i человек/мышь (RNAi Human/Mouse Starter Kit, Quiagen, Хильден, Германия). миРНК добавляется к 100 мл PBS в конечной концентрации 2 ммоль/мл. После добавления реагента для трансфекции HiPerFect смесь инкубируется на протяжении 10 мин при нормальных условиях. Для того, чтобы удалить избыток мицелл, экзосомы вновь выделяют с использованием альдегид/сульфатных латексных шариков. Химическая трансфекция нацеленной на нуклеиновую кислоту системы в экзосомы может быть проведена сходным образом с таковой для миРНК. Такие экзосомы могут быть сокультивированы с моноцитами и лимфоцитами, выделенными из периферической крови здоровых доноров. Следовательно, можно предположить, что экзосомы, содержащие нацеленную на нуклеиновые кислоты систему, могут быть введены в моноциты и лимфоциты и аутологично вновь введены в человека. В соответствии с этим, доставка или введение в соответствии с изобретением могут быть выполнены с применением экзосом плазмы.
Липосомы
[00638] Доставка или введение по изобретению могут быть выполнены с помощью липосом. Липосомы представляют собой сферические везикулярные структуры, состоящие из одно- или многослойного липидного бислоя, окружающего внутренние водные компартменты, и относительно непроницаемого внешнего липофильного фосфолипидного бислоя. Липосомы привлекают значительное внимание как переносчики для доставки лекарственных средств, потому что они биологически совместимы, нетоксичны, могут доставлять и гидрофильные и липофильные молекулы препарата, защитить свой груз от деградации ферментами плазмы и переносить свой груз через биологические мембраны и гемато-энцефалический барьер (ГЭБ) (см., например, Spuch and Navarro, Journal of Drug Delivery, vol. 2011, Article ID 469679, 12 pages, 2011. doi: 10.1155/2011/469679 для обзора).
[00639] Липосомы могут состоять из нескольких различных типов липидов; однако, для производства липосом в качестве переносчиков препаратов обычно используются фосфолипиды. Липосомы образуются самопроизвольно при смешивании липидной пленки с водным раствором, но этот процесс может быть ускорен с помощью механического воздействия в виде встряхивания при помощи гомогенизатора, генератора ультразвука или экструдера (см.. например, Spuch and Navarro, Journal of Drug Delivery, vol. 2011, Article ID 469679, 12 pages, 2011. doi: 10.1155/2011/469679 для обзора).
[00640] Несколько других добавок могут быть добавлены к липосомам, чтобы изменить их структуру и свойства. Например, холестерин или сфингомиелин могут быть добавлены к липосомальной смеси для стабилизации структуры липосом и предотвращения потери внутреннего груза. Далее, липосомы могут быть получены из гидрированного фосфатидилхолина яйца или фосфатидилхолина яйца, холестерина и диацетилфосфата, и средние размеры везикул могут быть доведены до приблизительно 50 и 100 нм. (см., например, Spuch and Navarro, Journal of Drug Delivery, vol. 2011, Article ID 469679, 12 pages, 2011. doi: 10.1155/2011/469679 для подробного обзора).
[00641] Лекарственная форма липосомы может состоять, главным образом, из естественных фосфолипидов и липидов, таких как 1,2-дистеароил-sn-глицеро-3-фосфатидилхолин (DSPC), сфингомиелин, фосфатидилхолины яйца и монозиалоганглиозид. Так как эта лекарственная форма составлена только из фосфолипидов, при создании липосом возникло много проблем, в том числе их нестабильность в плазме. Было предпринято несколько попыток преодолеть эти проблемы, в основном путем изменения состава липидной мембраны. Одна из этих попыток заключалась в добавлении холестерина. Добавление холестерина к обычным лекарственным формам замедляет высвобождение инкапсулированного биологически активного вещества в плазму, или 1,2-диолеоил-sn-глицеро-3-фосфоэтаноламин (DOPE) увеличивает стабильность (см., например, Spuch and Navarro, Journal of Drug Delivery, vol. 2011, Article ID 469679, 12 pages, 2011. doi: 10.1155/2011/469679 для обзора).
[00642] В особенно предпочтительном варианте осуществления изобретения желательно использование липосом, действующих по принципу "троянского коня" (также известный как "молекулярные троянские кони"), и протоколы можно найти по ссылке http://cshprotocols.cshlp.org/content/2010/4/pdb.prot5407.long. Эти частицы обеспечивают доставку трансгена ко всему головному мозгу после внутрисосудистой инъекции. Без ограничения, считается, что нейтральные липидные частицы со специфичными антителами, связанными с их поверхностью, способны пересекать гематоэнцефалический барьера путем эндоцитоза. Заявитель утверждает, что липосомы, действующие по принципу "троянского коня", могут быть использованы для доставки семейства нуклеаз CRISPR к головному мозгу путем внутрисосудистой инъекции, что позволило бы создавать животных с трансгенным головным мозгом без потребности в манипуляции с эмбрионами. Приблизительно 1-5 г ДНК или РНК могут быть использованы для введения in vivo в липосомах.
[00643] В другом варианте осуществления изобретения нацеленную на нуклеиновую кислоту систему или ее компоненты можно доставить при помощи липосом, например, стабильной нуклео-липидной частицы (SNALP) (см., например, Morissey et al., Nature Biotechnology, Vol. 23, No. 8, August 2005). Предусматриваются ежедневные внутривенные инъекции приблизительно 1, 3 или 5 мг/кг/сутки конкретной нацеленной на нуклеиновую кислоту системы, нацеленной в SNALP. Ежедневное лечение может быть проведено приблизительно за три дня и затем еженедельно в течение приблизительно пяти недель. В другом варианте осуществления изобретения также предусматривается конкретная нацеленная на нуклеиновую кислоту система, заключенная в SNALP, введенная внутривенной инъекцией в дозах приблизительно 1 или 2,5 мг/кг (см., например, Zimmerman et al., Nature Letters, Vol. 441, 4 May 2006). Состав SNALP может содержать липиды 3-N-[(вметоксиполи(этиленгликоль)2000)карбамоил]-1,2-димиристилокси-пропиламин (PEG-C-DMA), 1,2-дилинолеилокси-N,N-диметил-3-аминопропан (DLinDMA), 1,2-дистеароил-sn-глицеро-3-фосфохолин (DSPC) и холестерин, в соотношении 2:40:10:48 молярных процентов (см., например, Zimmerman et al., Nature Letters, Vol. 441, 4 May 2006).
[00644] В другом варианте осуществления было доказано, что стабильные нуклео-липидные частицы (SNALP) являются эффективными молекулами для доставки в сильно васкуляризированные опухоли печени, полученные из HepG2, но не в слабо васкуляризированные опухоли печени, образованные из НСТ-116 (см., например, Li, Gene Therapy (2012) 19, 775-780). Липосомы SNALP могут быть получены смешением D-Lin-DMA и PEG-C-DMA с дистероилфосфатидилхолином (DSPC), холестерином и миРНК, использованных в соотношении 25:1 липид/миРНК и молярным соотношением 48/40/10/2 для холестерина/D-Lin-DMA/DSPC/PEG-C-DMA. Полученные липосомы SNALP имеют размер примерно 80-100 нм.
[00645] В еще одном варианте осуществления стабильные частицы нуклеиновая кислота-липид (SNALP) могут включать синтетический холестерин (Sigma-Aldrich, Сент-Луис, Миссури, США), дипальмитоилфосфатидилхолин (Avanti Polar Lipids, Alabaster, Алабама, США), 3-N-[(w-метокси-поли(этиленгликоль)2000)карбамоил]-1,2-димиристоилоксипропиламин и катионный 1,2-дилиеноилокси-3-N,N-диметиламинопропанан (см., например, Geisbert et al., Lancet 2010; 375: 1896-905). Может предусматриваться дозировка примерно 2 мг/кг нацеленной на нуклеиновую кислоту системы, введенная, например, внутривенной болюсной инфузий.
[00646] В еще одном варианте осуществления изобретения SNALP может включать синтетический холестерин (Sigma-Aldrich), 1,2-дистеароил-sn-глицеро-3-фосфохолин (DSPC; Avanti Polar Lipids Inc.), PEG-cDMA, и 1,2-дилинолеилокси-3-(N;N-диметил)аминопропан (DLinDMA) (см, например, Judge, J. Clin. Invest. 119:661-673 (2009)). Составы, используемые для исследований in vivo, могут включать конечное соотношение липид/РНК по массе приблизительно 9:1.
[00647] Безопасность нанолекарств на основе РНК-i была рассмотрена Barros and Gollob из Alnylam Pharmaceuticals (см., например, Advanced Drug Delivery Reviews 64 (2012) 1730-1737). Стабильная нуклео-липидная частица кислоты (SNALP) состоит из четырех различных липидов: ионизируемого липида (DLinDMA), который является катионным при низких значениях pH, нейтрального липида-помощника, холестерина и способного к диффузии липида на основе полиэтиленгликоля (PEG). Частица составляет приблизительно 80 нм в диаметре и имеет нейтральный заряд при физиологическом pH. Ионизируемый липид служит для связывания липида с анионной РНК во время формирования частицы. При увеличении кислотности внутри эндосомы, ионизируемый липид становится положительно заряженным и опосредует слияние SNALP с мембраной эндосомы, высвобождая РНК в цитоплазму. PEG-содержащий липид стабилизирует частицу и уменьшает агрегирование во время сборки, впоследствии обеспечивая нейтральную гидрофильную поверхность, которая улучшает фармакокинетические свойства.
[00648] На данный момент две клинические программы были начаты с использованием смесей SNALP с РНК. Tekmira Pharmaceuticals недавно закончили фазу I клинического испытания с применением единственной дозы SNALP-ApoB на взрослых добровольцах с повышенным уровнем холестерина LDL. ApoB преимущественно экспрессируется в печени и тощей кишке и необходим для сборки и секреции VLDL и LDL. Семнадцать участников исследования получили единственную дозу SNALP-ApoB (постепенное увеличение дозы на протяжении 7 уровней). Не было никаких доказательств токсичности для печени (ожидались как потенциальная ограничивающая дозу токсичность на основе доклинических исследований). Один (из двух) испытуемых при получении самой высокой дозы испытал гриппоподобные симптомы, согласующиеся со стимуляцией иммунной системы, и было принято решение завершить испытание.
[00649] Alnylam Pharmaceutcals так же занимались продвижением ALN-TTR01, который использует технологию SNALP, описанную выше, и нацелен на продукцию гепатоцитами мутанта и TTR дикого типа для лечения амилоидоза TTR (ATTR). Были описаны три синдрома ATTR: семейная амилоидная полинейропатия (FAP) и семейная амилоидная кардиомиопатия (FAC) - оба вызываемые аутосомными доминирующими мутациями в TTR; и старческий системный амилоидоз (SSA), вызываемый TTR дикого типа. Недавно была закончена фаза I плацебо-контролируемых испытаний на пациентах с ATTR единственный дозы ALN-TTR01 c ее постепенным увеличением. ALN-TTR01 был введен в виде 15-минутного внутривенного вливания 31 пациенту (23 с препаратом исследования и 8 с плацебо) в диапазоне дозы от 0,01 до 1,0 мг/кг (на основе миРНК). Испытуемые хорошо переносили лечение без значительных увеличений функции печени в тестах. Инфузионные реакции были отмечены в 3 из 23 пациентов при дозе более 0,4 мг/кг; все реагировали на замедление скорости вливания и все продолжили участие в исследовании. Минимальное и временное повышение уровня цитокинов IL-6, IP-10 и IL-1ra в сыворотке крови было отмечено у двух пациентов при самой высокой дозе 1 мг/кг (как предполагалось исходя из доклинических испытаний и исследований на приматах). Понижая уровень TTR в сыворотке, ожидаемый фармакодинамический эффект от введения ALN-TTR01, наблюдался при дозе 1 мг/кг.
[00650] В еще одном варианте осуществления изобретения SNALP может быть собран с помощью растворения катионного липида, DSPC, холестерина и PEG-липида, например, в этаноле, например, в молярном отношении 40:10:40:10, соответственно (см., например, Semple et al., Nature Biotechnology, Volume 28 Number 2 February 2010, pp. 172-177). Липидная смесь была добавлена к водному буферу (50-мМ цитрата, pH 4) до достижения окончательной концентрации этанола и липидов 30% (по объему) и 6,1 мг/мл, соответственно, и была уравновешена при 22°C в течение 2 минут перед экструзией. Гидратированные липиды были экструдированы через два сложенных фильтра с размером пор 80 нм (Nuclepore) при 22°C с использованием Lipex Extruder (Northern Lipids) до достижения диаметра везикулы 70-90 нм, как определено динамическим анализом рассеяния света. Это обычно требовало 1-3 проходов. миРНК (растворенная в 50 мМ цитрата, водном растворе с pH 4, содержащем 30% этанола), была добавлена к предварительно уравновешенным (35°C) везикулам со скоростью 5 мл/минуту при смешивании. После достижения окончательного целевого отношения миРНК/липид, равного 0,06 (вес/вес), смесь была инкубирована в течение 30 минут при 35°C, чтобы обеспечить реорганизацию везикул и инкапсуляцию миРНК. Потом этанол был удален и внешний буфер заменен на PBS (155 мМ NaCl, 3 мМ Na2HP04, 1 мМ KH2PO4, pH 7,5) либо диализом, либо фильтрованием из тангенциального потока. миРНК была заключена в капсулу из SNALP с использованием контролируемого пошагового процесса растворения. Липидными компонентами KC2-SNALP были DLin-KC2-DMA (катионный липид), дипальмитоилфосфатидилхолин (DPPC; Avanti Polar Lipids), синтетический холестерин (Sigma) и PEG-C-DMA в молярном отношении 57,1:7,1:34,3:1,4. После формирования нагруженных частиц SNALP были диализированы против PBS, фильтрат был простерилизован через 0,2 мкм-фильтр перед использованием. Средние размеры частиц составляли 75-85 нм, и 90-95% миРНК были инкапсулированы в липидных частицах. Окончательное отношение миРНК/липид в составах, использовавшихся для in vivo тестирования, составляло ~0,15 (вес/вес). Системы LNP-миРНК, содержавшие миРНК против фактора VII, были сразу растворены до соответствующих концентраций в стерильном PBS перед использованием, и составы были введены внутривенно через боковую хвостовую вену в суммарном объеме 10 мл/кг. Этот способ и эти системы доставки могут быть экстраполированы на нацеленную на нуклеиновую кислоту систему по настоящему изобретению.
[00651] Другие катионные липиды, такие как аминолипид 2,2-дилинолеил-4-диметиламиноэтил-[1,3]-диоксолан (DLin-KC2-DMA) могут быть использованы, чтобы инкапсулировать нацеленную на нуклеиновую кислоту систему, компоненты этой системы или молекулу (молекулы) нуклеиновой кислоты, кодирующие компоненты этой системы, например, подобные миРНК (см., например, Jayaraman, Angew. Chem. Int. Ed. 2012, 51, 8529-8533) и, следовательно, могут использоваться в практике изобретения. Может быть рассмотрен предварительно сформированная везикула со следующим липидным составом: аминолипид, дистеароилфосфатидилхолин (DSPC), холестерин и (R)-2,3-бис(октадецилокси)пропил-1-(метоксиполи(этиленгликоль)2000)пропилкарбамат (PEG-липид) в молярном отношении 40/10/40/10, соответственно, и отношении мРНК фактора свертывания крови VII (FVII)/общее количество липидов, приблизительно равном 0,05 (в весовом отношении). Чтобы гарантировать узкое распределение размеров частиц в диапазоне 70-90 нм и низкий индекс полидисперсности 0,11+0,04 (n=56), частицы могут быть экструдированы до трех раз через 80-нм мембраны до добавления направляющей РНК. Могут быть использованы частицы, содержащие сильнодействующий аминолипид 16, в котором молярное отношение четырех липидных компонентов равно 16, DSPC, холестерин и PEG-липид (50/10/38,5/1,5), которое может быть далее оптимизировано, чтобы увеличить активность in vivo.
[00652] Michael S D Kormann et al. ("Expression of therapeutic proteins after delivery of chemically modified mRNA in mice: Nature Biotechnology, Volume:29, Pages: 154-157 (2011)) описывают использование липидных оболочек для доставки РНК. Использование липидных оболочек также предпочтительно в данном изобретении.
[00653] В другом варианте осуществления изобретения липиды могут быть смешаны с нацеленной на нуклеиновую кислоту системой по настоящему изобретению, ее компонентом(ами) или молекулой(ами) нуклеиновой(ых) кислот(ы), кодирующей таковой(ые), для образования липидных наночастиц (LNP). Липиды включают, без наложения ограничений, DLin-KC2- DMA4, C12-200 и колипиды дистероилфосфатидилхолина, холестерин и PEG-DMG могут быть смешаны с нацеленной на РНК системой вместо миРНК (см., например, Novobrantseva, Molecular Therapy-Nucleic Acids (2012) I, e4; doi:10.1038/mtna.2011.3) с использованием процедуры спонтанного образования везикул. Молярное отношение компонентов может быть примерно 50/10/38,5/1,5 (DLin-KC2-DMA или C12-200/дистероилфосфатидилхолин/холестерин/PEG-DMG). Конечное весовое соотношение липид:РНК может составлять 12:1 и 9:1 в случае липидных частиц (LNP) с DLin-KC2-DMA и C12-200, соответственно. Такие составы могут содержать частицы со средним диаметром ~80 нм при эффективности захвата >90%. Может предусматриваться доза 3 мг/кг.
[00654] Tekmira располагает портфолио, содержащим примерно 95 семейств патентов в США и в других странах, относящихся к различным аспектам липидных наночастиц (LNP) и их смесей (см., например, патенты США № 7982027; 7799565; 8058069; 8283333; 7901708; 7745651; 7803397; 8101741; 8188263; 7915399; 8236943 и 7838658 и европейские патенты № 1766035; 1519714; 1781593 и 1664316), все могут быть использованы и/или адаптированы для использования в настоящем изобретении.
[00655] Нацеленная на нуклеиновую кислоту система или ее компоненты или молекула(ы), кодирующая(ие) таковые, могут быть доставлены в упакованном в микросферы PLGA виде таким образом, как описано в опубликованных патентных публикациях США 20130252281, 20130245107 и 20130244279 (приписываются Modema Therapeutics), относящихся к аспектам составления композиций, включающих молекулы модифицированной нуклеиновой кислоты, которые могут кодировать белок, предшественник белка или частично или полностью процессированную форму белка или предшественника белка. Смесь может иметь молярное соотношение 50:10:38,5:1,5:3,0 (катионный липид: фузогенный липид:холестерин:PEG-липид). PEG-липид может быть выбран, но не ограничиваясь ими, из PEG-c-DOMG, PEG-DMG. Фузогенным липидом может являться DSPC. См. также Schrum et aJ., Delivery and Formulation of Engineered Nucleic Acids, опубликованное приложение US 20120251618.
[00656] Технологии Nanomerics призваны преодолеть трудности, связанные с биодоступностью для широкого круга терапевтических применений, включая низкомолекулярные гидрофобные лекарства, пептиды и основанные на нуклеиновых кислотах препараты (плазмиды, миРНК, микроРНК). Конкретные пути введения, для которых показаны значительные преимущества данной технологии, включают пероральное введение, транспортировка через гематоэнцефалический барьер, доставка в солидные опухоли, а также глаз. См., например, Mazza et al., 2013, ACS Nano. 2013 Feb 26;7(2): 1016-26; Uchegbu and Slew, 2013, J Pharm Sci. 102(2):305-10 и Lalatsa et al., 2012, J Control Release, 2012 Jul 20; 161 (2):523-36.
[00657] В публикации патентной заявки США № 20050019923 описываны катионные дендримеры для доставки биоактивных молекул, таких как молекулы полинуклеотидов, пептидов или полипептидов, и/или фармакологических агентов, в тело млекопитающего. Такие дендримеры подходят для направления доставки биоактивных молекул, например, в печень, селезенку, легкие, почку или сердце (или даже в головной мозг). Дендримеры представляют собой трехмерные макромолекулы, полученные за несколько этапов из простых разветвленных единиц-мономеров, природа и функциональность которых может легко контролироваться и изменяться. Дендримеры синтезируют пyтем повторяющегося добавления структурных единиц к мультифункциональному ядру (дивергентный способ синтеза) или их наращивания в направлении мультифункционального ядра (конвергентный способ синтеза), и каждое добавление к трехмерной оболочке структурных единиц приводит к образованию следующего поколения дендримеров. Полипропилениминовые дендимеры образуются, начиная с диаминобутанового ядра, к которому присоединяются вдвое больше аминогруппп в реакции Михаэля с присоединением акрилонитрила к первичным аминам и последующей гидрогенизацией нитрилов. Это приводит к удвоению количества аминогрупп. Полипропилениминовые дендримеры содержат 100% протонируемых азотов и до 64 терминальных аминогрупп (поколение 5, DAB 64). Протонируемыми группами обычно являются аминогруппы, которые способны присоединять протоны при нейтральном pH. Использование дендримеров в качестве агентов доставки генов было в значительной степени сфокусировано на использовании полиамидоамина, и фосфорсодержащие соединения со смесью аминов/амидов или N-P(02)S как конъюгирующих единиц, соответственно, при этом не было сообщений об использовании предшествующих поколений полипропилениминовых дендримеров для доставки генов. Полипропилениминовые дендримеры также исследовались в качестве чувствительных к pH систем контролируемого высвобождения для доставки лекарств и для их упаковки в качестве гостевых молекул при их химической модификации периферическими аминокислотными группами. Цитотоксичность и взаимодействие полипропилениминовых дендримеров с ДНК, а также эффективность трансфекции DAB 64 также были изучены.
[00658] Патентная публикация США № 20050019923 основана на наблюдении о том, что, в противоречие с более ранними сообщениями, катионные дендримеры, такие как полипропилениминовые дендримеры, демонстрируют подходящие свойства, такие как специфическое нацеливание и низкая токсичность, для использования для направленной доставки биоактивных молекул, таких как молекулы генетического материала. В дополнение к этому, производные катионных дендримеров также демонстрируют подходящие свойства для направленной доставки биоактивных молекул. См. также Bioactive Polymers, опубликованная заявка США 20080267903, в которой описано: "Различные полимеры, включая катионные полиаминные полимеры и дендримерные полимеры, как было показано, обладают антипролиферативной активностью, и следовательно могут быть полезны для лечения заболеваний, характеризуемых нежелательной пролиферацией клеток, таких как новообразования и опухоли, воспалительные заболевания (включая аутоиммунные заболевания), псориаз и атеросклероз. Такие полимеры могут быть использованы как самостоятельно в качестве активных агентов или в качестве "контейнера" для доставки других терапевтических агентов, таких как молекулы лекарств или нуклеиновые кислоты для генной терапии. В таких случаях собственная присущая полимеру противоопухолевая активность может дополнять активность доставляемого агента. Утверждения из данной патентной публикации могут быть использованы в совокупности с приведенными в настоящем описании идеями для доставки нацеленной на нуклеиновую кислоту систем(ы) или ее компонента(ов), или кодирующую таковые молекулу(ы) нуклеиновой кислоты.
Сверхзаряженные белки
[00659] Сверхзаряженные белки представляют собой класс полученных способами инженерии или природных белков с исключительно высоким положительным или отрицательным суммарным теоретическим зарядом, которые могут использоваться для доставки нацеленной на нуклеиновую кислоту системы(систем), или ее компонента(ов), или молекулы(молекул) нуклеиновой кислоты, кодирующей их. Несущие как сверхположительный, так и сверхотрицательный заряд белки демонстрируют выдающуюся устойчивость к вызванному термически или химически агрегированию. Сверхположительно заряженные белки также способны проникать в клетки млекопитающих. Связывание с этими белками груза, такого как ДНК плазмиды, РНК или другие белки, может сделать возможной функциональную доставку этих макромолекул в клетки млекопитающих in vitro и in vivo. Лаборатория под руководством David Liu сообщила о создании и описании сверхзаряженных белков в 2007 года (Lawrence et. al., 2007, Journal of the American Chemical Society 129, 10110-10112).
[00660] Отличная от вирусной доставка РНК и ДНК плазмиды в клетки млекопитающих представляет ценность как для исследований, так и для терапевтических применений (Akinc et al., 2010, Nat. Biotech. 26, 561-569). Очищенный белок +36 GFP (или другой сверхположительно заряженный белок) смешивали с РНК на соответствующих питательных средах без сыворотки и оставляли для образования комплексов перед добавлением к клеткам. Добавление сыворотки на этой стадии ингибирует формирование сверхзаряженных комплексов белок-РНК и снижает эффективность лечения. Приведенный ниже протокол продемонстрировал эффективность для ряда клеточных линий (McNaughton et al., 2009, Proc. Natl. Acad. Sci, USA 106, 6111-6116). Однако экспериментальные эксперименты, изменяющие дозу белка и РНК, должны быть выполнены, чтобы оптимизировать процедуру специфичных клеточных линий. Однако следует провести пробные эксперименты с изменениями дозы белка и РНК для оптимизации использования процедуры применительно к конкретным клеточным линиям.
(1) За один день до обработки высевают 1×105 клеток на лунку в 48-луночный планшет.
(2) В день обработки растворяют очищенный белок +36 GFP в питательных средах без сыворотки до конечной концентрации 200 нМ. Добавляют РНК до конечной концентрации 50 нМ. Перемешивают при помощи устройства для встряхивания и инкубируют при комнатной температуре в течение 10 минут.
(3) Во время инкубации удаляют питательную среду клеток и промывают PBS один раз.
(4) После инкубации +36 GFP с РНК к клеткам добавляют комплексы белок-РНК.
(5) Инкубируют клетки с комплексами при 37°C в течение 4 часов.
(6) После инкубации отбирают питательные среды и промывают три раза раствором гепарина в PBS (20 Е/мл). Инкубируют клетки с содержащими сыворотку питательными средами на протяжении 48 ч или более в зависимости от анализа активности.
(7) Анализируют клетки при помощи иммуноблоттинга, к-ПЦР, анализа фенотипа или другого соответствующего способа.
[00661] Лаборатория под руководством David Liu также установила, что белок +36 GFP является эффективным агентом доставки плазмид в различные клетки. Поскольку ДНК плазмиды является большим грузом, чем миРНК, для эффективного образования комплексов с плазмидами требуется пропорционально большее количество белков +36 GFP. Для эффективной доставки плазмиды заявители доставки разработали вариант +36 GFP, несущий C-концевую метку HA2, пептида, для которого известна способность разрушать эндосомы, полученного из белка гемагглютинина вируса гриппа. Следующий протокол продемонстрировал эффективность для ряда клеток, однако как и рекомендовано выше, необходимо, чтобы дозы ДНК плазмиды и сверхзаряженного белка были оптимизированы для конкретных клеточных линий и различных применений способа доставки.
(1) За день до обработки высевают по 1×105 клеток на лунку в 48-луночный планшет.
(2) В день обработки растворяют очищенный белок+36 GFP в питательных средах без сыворотки до конечной концентрации 2 нМ. Добавляют 1 мг ДНК плазмиды. Перемешивают с помощью устройства для встряхивания и инкубируют при комнатной температуре на протяжении 10 мин.
(3) Во время инкубации отбирают питательные среды клеток и промывают PBS один раз.
(4) После инкубации +36 GFP и ДНК плазмиды осторожно добавляют комплексы белок-ДНК к клеткам.
(5) Инкубируют клетки с комплексами при 37°C на протяжении 4 ч.
(6) После инкубации удаляют питательные среды, и промывают клетки PBS. Инкубируют клетки в содержащих сыворотку питательных средах и инкубируют следующие 24-48 часов.
(7) Анализируют доставку плазмиды (например, по управляемой плазмидой экспрессии генов) в зависимости от ситуации.
[00662] См. также, например, McNaughton et al., Proc. Natl. Acad. Sci. USA 106, 6111-6116 (2009); Cronican et al., ACS Chemical Biology 5, 747-752 (2010); Cronican et al., Chemistry & Biology 18, 833-838 (2011); Thompson et al., Methods in Enzymology 503, 293-319 (2012); Thompson, D.B., et al., Chemistry & Biology 19 (7), 831-843 (2012). Способы с применением сверхзаряженных белков могут использоваться и/или адаптированы для доставки нацеленной на нуклеиновую кислоту системы по настоящему изобретению. Такие системы Dr. Lui и приведенные в настоящей заявке документы в совокупности с изложенными в настоящем описании идеями могут использоваться при доставке нацеленной на нуклеиновую кислоту систем(ы) или ее компонента(ов) или кодирующей молекул(ы) нуклеиновой кислот.
Пептиды, проникающие в клетки (CPP)
[00663] В еще одном варианте осуществления изобретения рассмотрено использование пептидов, проникающих в клетки (CPP), для доставки системы CRISPR-Cas. CPP представляют собой короткие пептиды, которые облегчают захват клетками различных молекулярных грузов (от наноразмерных частиц до маленьких химических молекул и больших фрагментов ДНК). Термин "груз" как используют в настоящем описании, включает, но не ограничивается ими, состоящую из терапевтических агентов, диагностических зондов, пептидов, нуклеиновых кислот, антисмысловых олигонуклеотидов, плазмид, белков, частиц, включая наночастицы, липосом, хромофоров, низкомолекулярных соединений и радиоактивных материалов. Аспекты изобретения подразумевают также, что груз может также включать любой компонент системы CRISPR-Cas или целую функциональную систему CRISPR-Cas. В одном аспекте изобретение относится к следующим способам доставки желаемого груза индивиду, включающим: (a) получение комплекса, включающего пептид, который способен проникать в клетки, и желаемый груз, и (b) пероральное, внутрисуставное, внутрибрюшинное, интратекальное, внутриартериальное, интраназальное, внутрипаренхимное, подкожное, внутримышечное, внутривенное, дермальное, ректальное или местное введение комплекса индивиду. Груз связывают с пептидами посредством химической связи (ковалентной) или посредством нековалентных взаимодействий.
[00664] Функцией проникающих в клетки пептидов (CPP) является доставка груза в клетки - процесс, который обычно осуществляется посредством эндоцитоза груза, доставленного к эндосомам живых клеток млекопитающих. Проникающие в клетку пептиды (CPP) имеют различные размеры, последовательность аминокислот и заряд, но все CPP объединяет одна отличительная особенность, которая состоит в способности перемещаться через плазменную мембрану и облегчать доставку различных молекулярных грузов в цитоплазму или органеллы. Такое перемещение CPP может быть разделено на три главные группы в соответствии с механизмами входа: прямое проникновение через мембрану, опосредованный эндоцитозом вход и перемещение с формированием переходной структуры. CPP нашли многочисленные способы применения в медицине в качестве агентов доставки лекарственных средств при лечении различных заболеваний, включая противораковые средства и средства для лечения вирусных заболеваний, а также контрастирующих агентов для маркировки клетки. Примеры последних включают использование в качестве переносчика GFP, контрастирующих агентов для МРТ или квантовых точек. CPP имеют большой потенциал в качестве векторов как in vitro, так и in vivo доставки для исследований и использования в медицине. CPP, как правило, либо имеют аминокислотный состав с высоким относительным содержанием положительно заряженных аминокислот, таких как лизин или аргинин, либо содержат последовательности, имеющие чередующийся паттерн полярных/заряженных аминокислот и неполярных, гидрофобных аминокислот. Эти два типа структур обозначают как поликатионные или амфифильные соответственно. Третий класс CPP представляет собой гидрофобные пептиды, содержащие только неполярные остатки с низким суммарным зарядом или имеющие гидрофобные аминокислотные группы, которые крайне важны для клеточного захвата. Один из первых обнаруженных CPP - трансактивирующий активатор транскрипции (Tat) вируса иммунодефицита человека 1 (ВИЧ-1), который эффективно захватывается из питательных сред многочисленными типами культивируемых клеток. Позднее количество известных CPP значительно увеличилось, и были получены небольшие молекулярные синтетические аналоги, более эффективные при трансдукции белка. CPP включают, но не ограничиваются ими: пенетратин, Tat (48-60), транспортан и (R-AhX-R4) (Ahx=аминогексаноил).
[00665] В патенте США 8372951 описан CPP, полученный из эозинофильного катионного белка (ECP), который демонстрирует высокую эффективность проникания в клетку и низкую токсичность. Также описаны особенности доставки CPP с грузом в представителях позвоночных. Будущие аспекты CPP и их доставки описаны в патентах США 8575305; 8614194 и 8044019. CPP может использоваться для доставки системы CRISPR-Cas или ее компонентов. Использование CPP для доставки системы CRISPR-Cas или ее компонентов также описано в рукописи "Gene disruption by cell-penetrating peptide-mediated delivery of Cas9 protein and guide RNA", by Suresh Ramakrishna, Abu-Bonsrah Kwaku Dad, Jagadish Beloor, et al. Genome Res. 2014 Apr 2, [Epub в печати], включенной в настоящее описание в качестве ссылки в полном оъеме, где продемонстрировано, что лечение рекомбинантным белком Cas9, конъюгированным с CPP, и направляющими РНК в комплексе с CPP приводит к генным нарушениям эндогенного характера в клеточных линиях человека. В статье белок Cas9 был связан с CPP через тиоэфирную связь, тогда как направляющая РНК была в комплексе с CPP, формируя уплотненные, положительно заряженные частицы. Было показано, что одновременная и последовательная обработка клеток человека, включая эмбриональные стволовые клетки, фибробласты кожи, клетки HEK293T, клетки HeLa и эмбриональные клетки карциномы, модифицированным Cas9 и направляющей РНК, ведет к эффективным генным изменениям с уменьшением количества нецелевых мутаций по сравнению с трансфекцией плазмидами.
Имплантируемые устройства
[00666] В другом варианте осуществления изобретения имплантируемые устройства также предусматриваются для доставки нацеленной на нуклеиновую кислоту системы, ее компонента (компонентов) или молекулы (молекул) нуклеиновых кислот, кодирующих их. Например, в публикации патентной заявки США 20110195123 описано имплантируемое медицинское устройство, которое местно элюирует лекарственное средство и находится в человеке длительное время, включая несколько типов такого устройства, способы лечения и способы имплантации. Устройство, включающее полимерную подложку, такую как матрица, например, которая используется в качестве корпуса устройства, и лекарственное средство, и в некоторых случаях дополнительные каркасные материалы, такие как металлы или дополнительные полимеры и материалы, чтобы увеличить видимость и визуализацию. Имплантируемое устройство может быть предпочтительно для местной доставки лекарственного средства в течение длительного периода, когда препарат доставляется непосредственно во внеклеточный матрикс (ECM) пораженной заболеванием области, такой как опухоль, область воспаления, дегенерации, или для симптоматического лечения, или к травмированным клеткам гладкой мускулатуры, или для профилактики. Одним типом лекарственного средства является РНК, как описано выше, и эта система может быть использована/и или адаптирована для нацеленной на нуклеиновую кислоту системы по настоящему изобретению. Способы имплантации в некоторых вариантах осуществления изобретения представляют собой существующие способы имплантации, которые разработаны и используются в настоящее время для других способов лечения, включая брахитерапию и биопсию при помощи иглы. В таких случаях размеры нового импланта, описанного в этом изобретении, подобны оригинальному импланту. Как правило, несколько устройств могут быть имплантированы во время одной лечебной процедуры.
[00667] В публикации патента США 20110195123 описано лекарственное средство, которое обеспечивает имплантируемую или вставную систему, включая системы, применимые для полостей, таких как брюшная полость, и/или любого другого типа введения, в которых система доставки лекарственного средства не закреплена или прикреплена, содержащую биостабильную и/или разлагаемую и/или биоабсорбируемую полимерную подложку, которая в некоторых случаях может представлять собой, например, матрицу. Следует отметить, что термин "установка" также включает имплантацию. Система доставки лекарственного средства предпочтительно реализуется как "Loder", как описано в публикации патента США 20110195123.
[00668] Полимер или множество полимеров являются биосовместимыми, включающими агент и/или множество агентов, что позволяет высвобождать агент с контролируемой скоростью, причем общий объем полимерного субстрата, такого как матрица, например, в некоторых вариантах осуществления необязательно и предпочтительно не превышают максимального объема, который позволяет достичь терапевтического уровня агента. В качестве неограничивающего примера такой объем предпочтительно находится в диапазоне от 0,1 м3 до 1000 мм3, как того требует объем для загрузки агента. "Loder" в некоторых случаях может быть более крупным, например, когда он включен в устройство, размер которого определяется функциональностью, например, но без ограничения, коленный сустав, кольцо шейки матки и тому подобное.
[00669] Система доставки лекарственного средства (для доставки состава) сконструирована в некоторых вариантах осуществления, чтобы предпочтительно использовать разлагаемые полимеры, где основным механизмом высвобождения является объемная эрозия; или в некоторых вариантах осуществления используются нерастворимые или медленно деградирующие полимеры, в которых основным механизмом высвобождения является диффузия, а не объемная эрозия, так что наружная часть функционирует как мембрана, а ее внутренняя часть функционирует как резервуар для лекарственных средств, который практически не испытывает влияния окружения в течение длительного периода времени (например, от недели до нескольких месяцев). Могут также использоваться комбинации различных полимеров с различными механизмами высвобождения. Градиент концентрации на поверхности предпочтительно эффективно поддерживается постоянным в течение значительной части полного периода высвобождения лекарственного средства, и поэтому скорость диффузии является фактически постоянной (называемой диффузией "нулевого режима"). Под термином "постоянная" подразумевается скорость диффузии, которая предпочтительно поддерживается выше нижнего порога терапевтической эффективности, но которая может по-прежнему в некоторых случаях иметь начальный всплеск и/или может колебаться, например увеличиваясь и уменьшаясь в определенной степени. Скорость диффузии предпочтительно поддерживают в течение длительного периода времени, и ее можно считать постоянной до определенного уровня для оптимизации терапевтически эффективного периода, например эффективного периода подавления экспрессии генов.
[00670] Система доставки лекарственного средства необязательно и предпочтительно предназначена для защиты терапевтического агента на основе нуклеотидов от деградации, будь то химическая по своей природе или из-за действия ферментов и других факторов в организме индивида.
[00671] Система доставки лекарственного средства, описанная в публикации патента США 20110195123, в некоторых случаях связана с чувствительными и/или активирующими устройствами, которые работают во время и/или после имплантации устройства, посредством неинвазивных и/или минимально инвазивных способов активации и/или ускорения/замедления, например, необязательно включая, но не ограничиваясь ими, нагревание и охлаждение, лазерные лучи и ультразвук, включая фокусированные ультразвуковые и/или радиочастотные (RF) способы или устройства.
[00672] Согласно некоторым вариантам публикации патента США 010195123, область для местной доставки может необязательно включать целевые области, характеризующиеся высокой аномальной пролиферацией клеток и подавленным апоптозом, включая опухоли, активные и хронические воспаления и инфекции, включая аутоиммунные заболевания, дегенерацию тканей, включая мышцы и нервную ткань, хронические боли, участки дегенерации и места переломов костей и других ран для улучшения регенерации ткани и поврежденной сердечной, гладкой и поперечно-полосатой мускулатуры.
[00673] Область для имплантации композиции или целевая область, предпочтительно представляет собой радиус, область и/или объем, который достаточно мал, для целенаправленной локальной доставки. Например, целевой участок произвольно имеет диаметр в диапазоне приблизительно от 0,1 мм до приблизительно 5 см.
[00674] Местоположение целевой области должно быть выбрано предпочтительно для максимальной терапевтической эффективности. Например, композицию системы доставки лекарственных средств (необязательно с устройством для имплантации, как описано выше) необязательно и предпочтительно имплантируют непосредственно в саму опухоль или ее ближайшее окружение или связанные с ней кровеносные сосуды.
[00675] Например, композицию (необязательно вместе с устройством) необязательно имплантируют непосредственно в или вблизи поджелудочной железы, предстательной железы, молочных желез, печени, через сосок, в сосудистую систему и т.д.
[00676] Положение целевой области может быть выбрано из группы, включающей, по существу состоящей или состоящей из следующих примеров (не налагающих ограничений, поскольку любой участок тела может подходить для имплантации Loder): 1. дегенеративные участки мозга в базальных ганглиях, белом и сером веществе при заболеваниях Паркинсона или Альцгеймера, 2. позвоночник, например, в случае бокового амиотрофического склероза (ALS); 3. шейка матки для предотвращения инфекции вирусом папилломы человека (HPV); 4. суставы с острым и хроническим воспалением; 5. кожа, например, в случае псориаза; 6. симпатические и сенсорные нервы для получения обезболивающего эффекта; 7. внутрикостная имплантация; 8. области острой и хронической инфекции; 9. внутривлагалищная имплантация; 10. внутреннее ухо - слуховая система, лабиринт внутреннего уха, вестибулярная система; 11. внутритрахеальная имплантация; 12. внутрисердечная, коронарная, эпикардиальная имплантация; 13. имплантация в мочевой пузырь; 14. имплантация в желчную систему; 15. паренхимная ткань, включающая, не ограничиваясь, почкой, печенью, селезенкой; 16. лимфатические узлы; 17. слюнные железы; 18. десны; 19. внутрисуставная имплантация (в суставы); 20. внутриглазная имплантация; 21. ткани головного мозга; 22. желудочки мозга; 23. полости, включая брюшную полость (включая, не ограничиваясь, рак яичника); 24. внутрь пищевода и 25. ректально.
[00677] В некоторых случаях введение системы (например, устройства, содержащего состав), связано с инъекцией материала во внеклеточный матрикс в целевой области и вблизи этой области для изменения локального значения pH, и/или температуры, и/или других биологических факторов, влияющих на диффузию препарата и/или фармакокинетику во внеклеточном матриксе целевой области и вблизи нее.
[00678] В некоторых случаях, согласно некоторым вариантам осуществления изобретения, высвобождение указанного агента может быть связано с сенсорами и/или активирующими устройствами, управление которыми производится до и/или во время и/или после имплантации, неинвазивными и/или минимально инвазивными и/или другими способами активации и/или способами ускорения/замедления, включая использование лазерных лучей, радиации, нагревания и охлаждения, и ультразвуковое воздействие, включая фокусированный ультразвук и/или радиочастотные способы или устройства, а также химические активаторы.
[00679] Согласно другим вариантам осуществления изобретения, описанным в публикации патента США 20110195123, используемый препарат предпочтительно включает РНК, например для локализованных случаев рака молочной железы, поджелудочной железы, мозга, почки, мочевого пузыря, легкого и простаты, как описано ниже. Несмотря на то, что в качестве примера приводится РНК-интерференция, многие лекарственные препараты применимы при инкапсулирования в "Loder", и могут использоваться совместно с данным изобретением при условии, что такие лекарственные препараты сами по себе могут быть инкапсулированы в субстрат "Loder", например, такой как матрица, и эта система может быть использована и/или адаптирована для доставки нацеленной на нуклеиновую кислоту системы по настоящему изобретению.
[00680] В качестве другого примера конкретного применения можно привести нервные и мышечные дегенеративные заболевания, развивающиеся из-за аномальной экспрессии генов. Локальная доставка РНК может проявлять терапевтическое действие свойства для изменения аномальной экспрессии генов. Локальная доставка антиапоптотических, противовоспалительных и антидегенеративных лекарственных препаратов, включая небольшие по размеру молекулы препаратов и макромолекулы может также быть терапевтической. В таких случаях "Loder "применяется для длительного высвобождения с постоянной скоростью и/или через специализированное устройство, которое имплантируется отдельно. Все это может быть использовано и/или адаптировано для нацеленной на нуклеиновую кислоту системы по настоящему изобретению.
[00681] В качестве еще одного примера конкретного применения могут быть приведены психиатрические и когнитивные заболевания, которые можно лечить с помощью модификаторов активности генов. Генный нокдаун является вариантом лечения этих заболеваний. "Loder", местно доставляющие терапевтические агенты к участкам центральной нервной системы, являются терапевтическими вариантами лечения психиатрических и когнитивных заболеваний включая, но не ограничиваясь ими, психоз, биполярные расстройства, невротические заболевания и поведенческие расстройства. "Loder" также могут местно доставлять лекарственные препараты, включая небольшие молекулы лекарственных препаратов и макромолекулы после имплантации в конкретные области головного мозга. Все это может быть использовано и/или адаптировано для нацеленной на нуклеиновую кислоту системы по настоящему изобретению.
[00682] Другим примером конкретного применения является подавление экспрессии врожденных и/или адаптивных медиаторов иммунитета в локальных участках, что позволяет предотвратить отторжение трансплантатов. Локальная доставка РНК и иммуномодулирующих реактивов с использованием Loder, имплантированного в пересаженный орган и/или область имплантации, способствует локальному иммунному подавлению, отражая иммуноциты, такие как CD, активированные в отношении пересаженного органа. Все это может быть использовано/и или адаптировано для нацеленной на нуклеиновую кислоту системы по настоящему изобретению.
[00683] В качестве другого примера конкретного применения можно привести факторы роста сосудов, включая VEGF, ангиогенин и другие, необходимые для неоваскуляризации. Локальная доставка факторов, пептидов, пептидомиметиков или подавление их репрессоров является важным терапевтическим способом; подавление репрессоров и локальная доставка факторов, пептидов, макромолекул и небольших молекул лекарств, стимулирующих ангиогенез с помощью "Loder", являются терапевтическими для периферических, системных и сердечно-сосудистых заболеваний.
[00684] Способ введения, такой как имплантация, может уже использоваться для других типов имплантации тканей и/или для введения и/или для отбора проб тканей, в некоторых случаях без модификаций, или, альтернативно, в некоторых случаях только с незначительными модификациями в таких способах. Такие способы включают, но не ограничиваются ими, способы брахитерапии, биопсии, эндоскопии вместе с/или без применения ультразвука, такие как ретроградная холангиопанкреатография (ERCP,) стереотаксические способы для тканей головного мозга, лапароскопию, включая имплантацию лапароскопом в суставы, органы брюшной полости, стенку мочевого пузыря и полости тела.
[00685] Обсуждаемая в настоящем описании технология имплантируемых устройств может быть использована в рамках настоящего изобретения и, следовательно, благодаря этой информации и знаниям в данной области, система CRISPR-Cas, ее компоненты, их молекулы нуклеиновых кислот, или кодирующие или обеспечивающие их компоненты могут быть доставлены через имплантируемое устройство.
Пациент-специфические способы скрининга
[00686] Нацеленная на нуклеиновую кислоту система, которая нацелена на РНК, например, тринуклеотидные повторы, может быть использована для скрининга пациентов или образцов клеток/тканей пациентов на предмет наличия таких повторов. Повторы могут быть мишенью РНК системы нацеливания на нуклеиновую кислоту, и при наличии связывания с ней с помощью системы нацеливания на нуклеиновую кислоту, это связывание может быть обнаружено, тем самым указывая на то, что такой повтор присутствует. Таким образом, систему нацеливания на нуклеиновую кислоту можно использовать для скрининга пациентов или образцов клеток/тканей пациентов на наличие повторов. Затем пациенту можно назначить подходящее соединение (соединения) для лечения этого состояния; или может быть введена система нацеливания на нуклеиновую кислоту для связывания, которое приводит к инсерции, делеции или мутации и облегчению состояния.
[00687] Изобретение относится к нуклеиновым кислотам для связывания последовательностей РНК-мишеней.
мРНК эффекторного белка CRISPR и направляющая РНК
[00688] мРНК эффекторного белка CRISPR и направляющая РНК могут также доставляться отдельно. мРНК эффекторного белка системы CRISPR можно доставить до направляющей РНК, чтобы дать время для экспрессии белка CRISPR. мРНК эффекторного белка системы CRISPR можно вводить за 1-12 часов (предпочтительно приблизительно 2-6 часов) до введения направляющей РНК.
[00689] Альтернативно, мРНК эффекторного белка CRISPR и направляющую РНК можно вводить вместе. Предпочтительно вторую усиливающую дозу направляющей РНК можно вводить через 1-12 часов (предпочтительно приблизительно 2-6 часов) после первоначального введения мРНК эффекторного белка CRISPR+направляющей РНК.
[00690] Эффекторный белок CRISPR согласно настоящему изобретению, а именно эффекторный белок C2c1 или C2c3, иногда упоминается в настоящем описании как фермент CRISPR. Понятно, что эффекторный белок основан или получен из фермента, поэтому термин "эффекторный белок", безусловно, включает "фермент" в некоторых вариантах осуществления изобретения. Однако также следует понимать, что эффекторный белок может, как требуется в некоторых вариантах осуществления, связывать ДНК или РНК, но не обязательно разрезать или вносить одноцепочечный разрыв, что является функцией "мертвого" эффекторного белка Cas.
[00691] Дополнительное введение мРНК эффекторного белка CRISPR и/или направляющей РНК может быть полезным для достижения наиболее эффективных уровней модификации генома. В некоторых вариантах осуществления фенотипическое изменение предпочтительно является результатом модификации генома, когда мишенью является генетическое заболевание, особенно в способах терапии, и предпочтительно, когда обеспечивается матрица репарации для коррекции или изменения фенотипа.
[00692] В некоторых вариантах осуществления заболевания, которые могут быть мишенью, включают заболевания, которые связаны с дефектами сплайсинга.
[00693] В некоторых вариантах осуществления клеточные мишени включают гемопоэтические стволовые клетки/клетки-предшественники (CD34+); Т-клетки человека; и клетки сетчатки - например, клетки-предшественники фоторецепторов.
[00694] В некоторых вариантах осуществления гены-мишени включают: ген бета-глобина человека - HBB (для лечения серповидноклеточной анемии, включая стимуляцию генной конверсии (с использованием близкородственного гена HBD в качестве эндогенной матрицы)); CD3 (Т-клетки) и CEP920 - сетчатка (глаза).
[00695] В некоторых вариантах осуществления болезни-мишени также включают: злокачественную опхуоль; серповидноклеточную анемию (основанную на точечной мутации); ВИЧ; бета-талассемию; и офтальмологические или глазные заболевания - например, амавроз Лебера (LCA), вызываемый дефектом сплайсинга.
[00696] В некоторых вариантах осуществления способы доставки включают: опосредованную катионными липидами "прямую" доставку комплекса фермент-гид (рибонуклеопротеин) и электропорацию плазмидной ДНК.
[00697] Способы по изобретению могут дополнительно включать доставку матриц, таких как матрицы репарации, которые могут представлять собой дцОДН или оцОДН, см. ниже. Доставка матриц может быть произведена одновременно или отдельно от доставки любого или всех эффекторных белков CRISPR или направляющих молекул, и через один и тот же механизм доставки или любой другой. В некоторых вариантах осуществления предпочтительно, чтобы матрица доставлялась вместе с направляющей молекулой и также предпочтительно эффекторным белком CRISPR. Примером может служить вектор AAV.
[00698] Способы по изобретению могут дополнительно включать: (а) доставку в клетку двухцепочечного олигодезоксинуклеотида (дцОДН), включающего выступающие концы, комплементарные выступающим концам, созданным упомянутым разрывом двойной цепи, причем указанный дцОДН встраивается в локус-мишень, или - (б) доставку в клетку одноцепочечного олигодезоксинуклеотида (оцОДН), где указанный оцОДН действует как матрица для гомологичной направленной репарации разрыва двойной цепи. Способы по изобретению могут быть предназначены для профилактики или лечения заболевания у индивида, когда указанное заболевание вызвано дефектом в указанном локусе-мишени. Способы по изобретению могут осуществляться in vivo в индивидууме или ex vivo на клетке, взятой у индивида, где в некоторых случаях указанную клетку возвращают индивиду.
[00699] Для минимизации токсичности и неспецифических эффектов важно контролировать концентрацию доставляемых мРНК эффекторного белка CRISPR и направляющей РНК. Оптимальные концентрации мРНК эффекторного белка CRISPR и направляющей РНК могут быть определены путем тестирования различных концентраций на клеточной или животной модели и с использованием глубокого секвенирования для анализа степени модификации в потенциально нецелевых геномных локусах. Например, для направляющей последовательности, нацеленной на 5'-GAGTCCGAGCAGAAGAAGAA-3' в гене EMX1 генома человека, глубокое секвенирование может быть использовано для оценки уровня модификации в следующих двух нецелевых локусах: 1: 5'-GAGTCCTAGC AGGAGAAG AA-3' и 2: 5'-GAGTCTAAGCAGAAGAAGAA-3'. Концентрация, которая дает наивысший уровень модификации мишени, минимизируя уровень внецелевой модификации, должна быть выбрана для доставки in vivo.
Индуцируемые системы
[00700] В некоторых вариантах осуществления изобретения эффекторный белок CRISPR может составлять компонент индуцируемой системы. Индуцируемый характер системы позволяет пространственно-временный контроль редактирования генов или экспрессии генов с использованием энергии. Форма энергии может включать, но не ограничивается ими, электромагнитное излучение, звуковую энергию, химическую энергию и тепловую энергию. Примеры индуцируемой системы включают стимулирующие тетрациклин промоторы (Tet-On или Tet-Off), системы двугибридной активации транскрипции с малой молекулой (FKBP, ABA и т.д.) или светоиндуцируемые системы (фитохром, LOV-домены или криптохром). В одном варианте осуществления эффекторный белок CRISPR может быть частью светоиндуцируемого эффектора транскрипции (LITE) для прямого изменения транскрипционной активности последовательность-специфическим образом. Светоиндуцируемые компоненты могут включать эффекторный белок CRISPR, светочувствительный гетеродимер цитохрома (например, Arabidopsis thaliana) и домен активации/репрессии транскрипции. Другие примеры индуцибельных связывающих ДНК белков и способы их применения приведены в US 61/736465 и US 61/721283 и WO 2014018423 A2, которые включены в настоящее описание в качестве ссылки в полном объеме.
Самоинактивирующиеся cистемы
[00701] Как только все копии гена в геноме клетки отредактированы, продолжение экспрессии эффекторного белка CRISPR/C2c1 или C2c3 в такой клетке больше не требуется. Действительно, длительная экспрессия является нежелательной с точки зрения нецелевых эффектов на не являющихся мишенью участках генома и т.д. Таким образом, является полезной ограниченная по времени экспрессия. Одним из подходов является индуцируемая экспрессия, но, кроме того, заявители сконструировали cамоинактивирующуюся систему CRISPR, которая основана на использовании некодирующей направляющей последовательности-мишени в самом векторе CRISPR. Таким образом, после начала экспрессии система CRISPR произведет собственное разрушение, но прежде чем разрушение завершится, у системы будет время, чтобы отредактировать геномные копии гена-мишени (который в случае боычной точечной мутации в диплоидной клетке требует не более двух редактирований). Просто самоинактивирующаяся система CRISPR-Cas включает дополнительную РНК (т.е. направляющую РНК), которая нацелена на кодирующую последовательность самого эффекторного белка CRISPR или нацелена на одну или более некодирующих направляющих последовательностей-мишеней, комплементарных уникальным последовательностям, находящимся в одном или более из следующих полжоений:
(a) в промоторе, управляющем экспрессией некодирующих элементов РНК,
(b) в промоторе, управляющем экспрессией гена эффекторного белка C2c1 или C2c3,
(c) в 100 п.о. от кодона начала трансляции ATG в последовательности, кодирующей эффекторный белок C2c1 или C2c3,
(d) в инвертированном концевом повторе (iTR) вирусного вектора доставки, например, в геноме AAV.
[00702] Кроме того, такая РНК может быть доставлена посредством вектора, например, отдельного вектора или того же самого вектора, который кодирует комплекс CRISPR. При доставке посредством отдельного вектора РНК CRISPR, нацеленная на экспрессию Cas, может быть введена последовательно или одновременно. При последовательном введении, РНК CRISPR, нацеленная на экспрессию Cas, должна быть доставлена после РНК CRISPR, которая предназначена, например, для генного редактирования или генной инженерии. Этот период может продолжаться минуты (например, 5 минут, 10 минут, 20 минут, 30 минут, 45 минут, 60 минут). Этот период может продолжаться часы (например, 2 часа, 4 часа, 6 часов, 8 часов, 12 часов, 24 часа). Этот период может продолжаться дни (например, 2 дня, 3 дня, 4 дня, 7 дней). Этот период может продолжаться недели (например, 2 недели, 3 недели, 4 недели). Этот период может продолжаться месяцы (например, 2 месяца, 4 месяца, 8 месяцев, 12 месяцев). Этот период может продолжаться годы (2 года, 3 года, 4 года). Там, где направляющая РНК нацеливает на последовательности, кодирующие экспрессию белка Cas, эффекторный белок становится препятствием, и система становится самоинактивирующейся. Таким же образом РНК CRISPR, нацеленная на экспрессию Cas, применяемую посредством, например липосом, липофекции, частиц, микровезикул, как описано в настоящемописании, может быть введена последовательно или одновременно. Точно так же самоинактивация может использоваться для инактивации одной или более направляющих РНК, использовавшихся для нацеливания на одну или более мишеней.
[00703] В некоторых аспектах предусматривается единственная гРНК, способная к гибридизации с низлежащей последовательностью по отношению к старт-кодону эффекторного белка CRISPR, таким образом, после промежутка времени наблюдается потеря экспрессии эффекторного белка CRISPR. В некоторых вариантах предлагаются одна или более гРНК, способных к гибридизации с одним или более кодирующими или некодирующими участками полинуклеотида, кодирующего систему CRISPR-Cas, в результате чего после промежутка времени происходит инактивация одного или более, или в некоторых случаях всех, систем CRISPR-Cas. В некоторых аспектах системы, и не ограничиваясь теорией, клетка может включать множество комплексов CRISPR-Cas, где первая подгруппа комплексов CRISPR включает первую направляющую РНК, способную к нацеливанию на геномный локус или локусов для редактирования, и вторая подгруппа комплексов CRISPR включает по меньшей мере одну вторую направляющую РНК, способную к нацеливанию на полинуклеотид, кодирующий систему CRISPR-Cas, где первая подгруппа комплексов CRISPR-Cas опосредует редактирование геномного локуса или локусов, являющихся мишенью, и вторая подгруппа комплексов CRISPR в итоге инактивирует систему CRISPR-Cas, таким образом инактивируя дальнейшую экспрессию CRISPR-Cas в клетке.
[00704] Таким образом изобретение отностся к системе CRISPR-Cas, включающей один или более векторов для доставки в эукариотическую клетку, где вектор(ы) кодирует: (i) эффекторный белок CRISPR; (ii) первую направляющую РНК, способную к гибридизации с последовательностью-мишенью в клетке; (iii) вторую направляющую РНК, способную к гибридизации с одной или более последовательностями-мишенями в векторе, кодирующем эффекторный белок CRISPR, таким образом отличаясь только направляющей последовательностью, где при экспрессии в клетке первая направляющая РНК направляет специфическое для последовательности связывание первого комплекса CRISPR с последовательностью-мишенью в клетке, вторая направляющая РНК направляет специфическое для последовательности связывание второго комплекса CRISPR к последовательности-мишенью в векторе, который кодирует эффекторный белок CRISPR; комплексы CRISPR включают эффекторный белок CRISPR, связанный с направляющей РНК, так что направляющая РНК может гибридизироваться с ее последовательностью-мишенью; и второй комплекс CRISPR инактивирует систему CRISPR-Cas для предотвращения продолжительной экспрессии эффекторного белка CRISPR клеткой.
[00705] Следующие характеристики вектора(ов), кодируемого фермента, направляющих последовательностей, и т.д. раскрыты в других разделах здесь. Система может кодировать (i) фермент CRISPR, более конкретно C2c1 или C2c3; (ii) первую направляющую РНК, включающую последовательность, способную к гибридизации с первой последовательностью-мишенью в клетке, (ii) вторую направляющую РНК, способную к гибридизации с вектором, который кодирует фермент CRISPR. Точно так же фермент может включать одну или более последовательностей NLS и т.д.
[00706] Различные кодирующие последовательности (эффекторный белок CRISPR и направляющие РНК) могут быть включены в единственный вектор или в несколько векторов. Например, возможно закодировать эффекторный белок в одном векторе и различные последовательности РНК в другом векторе, или закодировать эффекторный белок и одну направляющую РНК в одном векторе и оставшуюся направляющую РНК в другом векторе, или возможна любая другая перестановка. В целом предпочтительна система с использованием всего одного или двух различных векторов.
[00707] При использовании нескольких векторов, возможно доставлять их в неравных количествах, и идеально с избытком вектора, кодирующего первую направляющую РНК относительно второй направляющей РНК, таким образом помогая задерживать конечную инактивацию системы CRISPR, чтобы произошло редактирование генома.
[00708] Первая направляющая РНК может быть нацелена на любую представляющую интерес последовательность-мишень в геноме, как описано в других разделах здесь. Вторая направляющая РНК нацелена на последовательность в векторе, который кодирует эффекторный белок C2c1 или C2c3 системы CRISPR, и таким образом инактивирует экспрессию эффекторного белка c этого вектора. Таким образом, последовательность-мишень в векторе должна быть способна инактивировать экспрессию. Подходящие последовательности-мишени могут находиться, например, вблизи или в старт-кодоне трансляции кодирующей последовательности эффекторного белка C2c1 или C2c3, в некодирующей последовательности в промоторе, управляющем экспрессией некодирующих элементов РНК, в промоторе, управляющем экспрессией гена эффекторного белка C2c1 или C2c3, в 100 п.о. от старт-кодона трансляции ATG в кодирующей последовательности Cas, и/или в инвертированном концевом повторе (iTR) вирусного вектора доставки, например, в геноме AAВ. Двухцепочечный разрыв около этого участка может вызывать изменение структуры в кодирующей последовательности Cas, вызывая потерю экспрессии белка. Альтернативная последовательность-мишень для "самоинактивирующейся" направляющей РНК иметь целью редактирование/инактивацию регуляторных областей/последовательностей, необходимых для экспрессии системы CRISPR с эффекторным белком C2c1 или C2c3 или для стабильности вектора. Например, если промотор для кодирующей Cas последовательности разрушен, тогда транскрипция может ингибироваться или прекращаться. Точно так же, если вектор включает последовательности для репликации, поддержания или стабильности, тогда можно осуществлять нацеливание на них. Например, в векторе AAВ полезная последовательность-мишень находится в инвертированных концевых повторах (iTR). Другие полезные последовательности для нацеливания могут представлять собой последовательности промотора, участки полиаденилирования и т.д.
[00709] Кроме того, если направляющие РНК будут экспрессироваться в формате матрицы, то "самоинактивирующиеся" направляющие РНК, которые нацелены на оба промотора одновременно, приведут к вырезанию находящихся между ними нуклеотидов из конструкции, экспрессирующей CRISPR-Cas, эффективно приводя к ее полной инактивации. Точно так же вырезание промежуточных нуклеотидов будет приводить к результату в случаях, где направляющие РНК нацелены на оба инвертированных концевых повтора (ITR), или нацелены на два или более других компонентов CRISPR-Cas одновременно. Самоинактивация, как объяснено в настоящем описании, в целом применима для регулирования работы систем CRISPR-Cas. Например, самоинактивация, как объяснено в настоящем описании, может быть применена для репарации мутаций системой CRISPR, например, нарушений, вызванных экспансией повторов. В результате самоинактивации репарация при помощи системы CRISPR обладает временной активностью.
[00710] Добавление ненацеливающих нуклеотидов к 5'-концу (например, 1-10 нуклеотидов, предпочтительно 1-5 нуклеотидов) "самоинактивирующейся" направляющей РНК может использоваться для задержки ее процессинга и/или изменения ее эффективности в качестве средства обеспечения редактирования в геномном локусе-мишени до прекращения работы системы CRISPR-Cas.
[00711] В одном аспекте самоинактивирующейся системы AAV-CRISPR-Cas плазмиды, совместно экспрессирующие одну или более направляющих РНК, нацеленных на представляющие интерес геномные последовательности (например, 1-12, 1-5, 1-10, 1-15, 1-20, 1-30), могут быть сконструированы с "самоинактивирующимися" направляющими РНК, которые нацелены на последовательность SpCas9 непосредственно в или вблизи сконструированного старт-кодона ATG (например, в пределах 5 нуклеотидов, в пределах 15 нуклеотидов, в пределах 30 нуклеотидов, в пределах 50 нуклеотидов, в пределах 100 нуклеотидов). Также направляющая РНК может осуществлять нацеливание на регуляторную последовательность в области промотора U6. Направляющие РНК, управляемые U6, могут быть сконструированы в формате матрицы, таким образом, что одновременно может высвобождаться несколько последовательностей направляющей РНК. Псле первоначальной доставки в ткань/клетки, являющиеся мишенью, (покинувшие клетку) направляющие РНК начинают накапливаться, в то время как уровни Cas повышаются в ядре. Комплексы Cas со всеми направляющими РНК опосредуют геномное редактирование и самоинактивацию плазмид CRISPR-Cas.
[00712] Одним из аспектов самоинактивирующейся системы CRISPR-Cas является экспрессия отдельно или в формате тандемной матрице от 1 до 4 или более различных направляющих последовательностей; например, до приблизительно 20 или приблизительно 30 направляющих последовательностей. Каждая индивидуальная самоинактивирующаяся направляющая последовательность может осуществлять нацеливание на отдельную мишень. Таковая может быть процессирована из, например, одного химерного транскрипта pol3. Могут использоваться промоторы Pol3, такие как промоторы U6 или Hl. Также могут использоваться промоторы Pol2, такие как описанные в настоящем описании. Последовательности инвертированных концевых повторов (iTR) могут фланкировать промотор Pol3-направляющую (направляющие) РНК-промотор Pol2-Cas.
[00713] Одним из аспектов транскрипта с тандемной матрицей является то, что одна или более направляющих РНК редактируют одну или более мишень(ей), в то время как одно или более самоинактивирующихся направляющих РНК инактивирует систему CRISPR-Cas. Таким образом, например, описанная система CRISPR-Cas для репарации экспансии повторов может быть непосредственно объединена с самоинактивирующейся системой CRISPR-Cas, описанной в настоящем описании. У такой системы может быть, например, две направляющих РНК, нацеливающих на учасок-мишень для редактирования, а также по меньшей мере третья направляющая РНК, нацеливающая на самоинактивацию CRISPR-Cas. См. заявку с серийным номером № PCT/US2014/069897 под названием "Compositions And Methods Of Use Of Crispr-Cas Systems In Nucleotide Repeat Disorders", опубликованную 12 декабря 2014 года в качестве WO/2015/089351.
[00714] Направляющая РНК может быть контрольной направляющей РНК. Например, она может быть сконструирована для нацеливания на последовательность нуклеиновой кислоты, кодирующую сам фермент CRISPR, как описано в US2015232881Al, описание которого включено в настоящее описание в качестве ссылки. В некоторых вариантах осуществления изобретения система или композиция может содержать только направляющую РНК, сконструированную для нацеливания на последовательность нуклеиновой кислоты, кодирующую фермент CRISPR. Кроме того, система или композиция может содержать направляющую РНК, сконструированную для нацеливания на последовательность нуклеиновой кислоты, кодирующую фермент CRISPR, а также последовательность нуклеиновой кислоты, кодирующую фермент CRISPR и, необязательно, вторую направляющую РНК и, далее необязательно, матрицу репарации. Вторая направляющая РНК может быть основной мишенью системы или композиции CRISPR (такой как терапевтическая, диагностическая, предназначенная для нокаута и т.д., как определено в настоящем описании). Таким образом система или композиция являются самоинактивирующимися. Это проиллюстрировано в отношении Cas9 в US2015232881A1 (также изданном как WO2015070083(Al), упоминаемой в настоящем описании, и может быть экстраполировано на C2c1 или C2c3.
Ферменты согласно изобретению используемые в мультиплексном (тандемном) подходе нацеливания
[00715] Авторы изобретения показали, что ферменты CRISPR, как определено в настоящем описании, могут использовать более одной направляющей РНК, не теряя активности. Это позволяет использовать ферменты CRISPR, системы или комплексы, как определено в настоящем описании, для нацеливания на несколько ДНК, генов или локусов, являющихся мишенями, посредством одного фермента, системы или комплекса, как определено в настоящем описании. Направляющие РНК могут быть расположены тандемно, и необязательно разделены последовательностью нуклеотидов, такой как прямой повтор, как определено здесь. Положение различных направляющих РНК в тандеме не влияет на активность. Отмечено, что термины "система CRISPR-Cas", "Комплекс CRISPR Cas" "комплекс CRISPR" и "система CRISPR" используются взаимозаменяемо. Также термины "фермент CRISPR", "фермент Cas", или "фермент CRISPR-Cas", могут использоваться взаимозаменяемо. В предпочтительных вариантах осуществления изобретения указанный фермент CRISPR, фермент CRISPR-Cas или фермент Cas является C2c1 или C2c3 или любым из модифицированных или мутированных их вариантов, описанных в настоящем описании в других разделах.
[00716] В одном из аспектов изобретение относится к не встречающемуся в природе или сконструированному способами инженерии ферменту CRISPR, предпочтительно ферменту CRISPR 2 класса, предпочтительно ферменту CRISPR типа V или VI, как описано в настоящем описании, такому как, но не ограничиваясь ими, C2c1 или C2c3, описанные в настоящем описании, для применения для тандемного или мультиплексного нацеливания. Нужно понимать, что в данном подходе может использоваться любой из ферментов CRISPR (или CRISPR-Cas или Cas), комплексов или систем согласно изобретению, как описано здесь в других разделах. Любой из способов, продуктов, композиций и применения, как описано в настоящем описании, в равной степени применим в мультиплексном или тандемном подходе к нацеливанию, как подробно описано ниже. В качестве дальнейшего руководства предложены следующие конкретные аспекты и варианты осуществления изобретения.
[00717] В одном из аспектов изобретение относится к применению фермента C2c1 или C2c3, комплекса или системы, как определено в настоящем описании, для нацеливания на несколько локусов генов. В одном из вариантов осуществления изобретения это может быть осуществлено при помощи нескольких (тандемных или мультиплексных) последовательностей направляющей РНК (гРНК).
[00718] В одном из аспектов изобретение относится к способам применения одного или более элементов фермента C2c1 или C2c3, комплекса или системы, как определено в настоящем описании, для тандемного или мультиплексного нацеливания, где указанная система CRISPR включает несколько последовательностей направляющей РНК. Предпочтительно, чтобы указанные последовательности гРНК были разделены последовательностями нуклеотидов, такими как прямой повтор, как определено в настоящем описании.
[00719] Фермент C2c1 или C2c3, система или комплекс, как определено в настоящем описании, обеспечивают эффективное средство для модификации нескольких полинуклеотидов-мишеней. Фермент C2c1 или C2c3, система или комплекс, как определено в настоящем описании, имеет большое разнообразие применений, включая модификацию (например, делецию, инсерцию, транслокацию, инактивацию, активацию) одного или более полинуклеотидов-мишеней в разнообразных типах клеток. Как таковой фермент C2c1 или C2c3, система или комплекс по изобретению, как определено в настоящем описании, имеет широкий спектр применений, например, в генной терапии, скрининге лекарственных средств, диагностике заболеваний и прогнозировании, включая нацеливание на несколько локусов генов посредством единственной системы CRISPR.
[00720] В одном из аспектов изобретение относится к ферменту C2c1 или C2c3, системе или комплексу, как определено в настоящем описании, т.е. комплексу CRISPR-Cas C2c1 или C2c3, имеющему белок C2c1 или C2c3, к тому же имеющему по меньшей мере один домен дестабилизации, ассоциированный с ним, и несколько направляющих РНК, которые нацелены на несколько молекул нуклеиновых кислот, такие как молекулы ДНК, посредством чего каждая из указанных нескольких направляющих РНК специфически нацелена на соответствующую молекулу нуклеиновой кислоты, например, молекулу ДНК. Каждая молекула нуклеиновой кислоты, являющаяся мишенью, например, молекула ДНК, может кодировать продукт гена или содержать локус. Следовательно, использование нескольких направляющих РНК позволяет нацеливание на несколько локусов или более генов. В некоторых вариантах осуществления изобретения фермент C2c1 или C2c3 может расщеплять молекулу ДНК, кодирующую продукт гена. В некоторых вариантах осуществления изобретения экспрессия продукта гена изменена. Белок C2c1 или C2c3 и направляющие РНК в природе не встречаются вместе. Изобретение охватывает направляющие РНК, включающие тандемно расположенные направляющие последовательности. Изобретение далее охватывает кодирующие последовательности для белка C2c1 или C2c3, являющиеся кодон-оптимизированными для экспрессии в эукариотической клетке. В предпочтительном варианте осуществления изобретения эукариотическая клетка является клеткой млекопитающего, клеткой растения или клеткой дрожжей, и в более предпочтительном варианте осуществления изобретения клетка млекопитающего является клеткой человека. Экспрессия продукта гена может быть уменьшена. Фермент C2c1 или C2c3 может являться частью системы CRISPR. или комплекса, который далее включает, тандемно расположенные направляющие РНК (гРНК), включающие серию из 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 25, 25, 30 или более 30 направляющих последовательностей, каждая из которых способна к специфичной гибридизации к последовательностью-мишенью в геномном локусе-мишени в клетке. В некоторых вариантах осуществления изобретения функциональная система или комплекс CRISPR C2c1 или C2c3 связывается с несколькими последовательностями-мишенями. В некоторых вариантах осуществления изобретения функциональная система CRISPR или комплекс могут редактировать несколько последовательностей-мишеней, например, последовательности-мишени могут включать геномный локус, и в некоторых вариантах осуществления изобретения может происходить изменение экспрессии гена. В некоторых вариантах осуществления изобретения функциональная система CRISPR или комплекс могут включать дополнительные функциональные домены. В некоторых вариантах осуществления изобретение относится к способу изменения или модификации экспрессии нескольких продуктов генов. Способ может включать введение в клетку, содержащую указанные нуклеиновые кислоты, являющиеся мишенью, например, молекулы ДНК, или содержащую и экспрессирующую нуклеиновые кислоты, являющиеся мишенью, например, молекулы ДНК; например, нуклеиновые кислоты, являющиеся мишенями, могут кодировать продукты генов или обеспечивать экспрессию продуктов генов (например, регуляторные последовательности).
[00721] В предпочтительных вариантах осуществления изобретения фермент CRISPR, используемый для мультиплексного нацеливания, является C2c1 или C2c3, или системой или комплексом CRISPR, которые включают C2c1 или C2c3. В некоторых вариантах осуществления изобретения фермент CRISPR, используемый для мультиплексного нацеливания, представляет собой AacC2c1, или система или комплекс CRISPR, используемые для мультиплексного нацеливания, включают AacC2c1. В некоторых вариантах осуществления изобретения фермент CRISPR является AacC2c1, или система или комплекс CRISPR включают AacC2c1. В некоторых вариантах осуществления изобретения фермент С2с1, используемый для мультиплексного нацеливания, расщепляет обе цепи ДНК, внося двухцепочечный разрыв (DSB). В некоторых вариантах осуществления изобретения фермент CRISPR, используемый для мультиплексного нацеливнаия, является никазой. В некоторых вариантах осуществления изобретения фермент C2c1 или C2c3, используемый для мультиплексного нацеливания, является двойной никазой. В некоторых вариантах осуществления изобретения фермент C2c1 или C2c3, используемый для мультиплексного нацеливания, является ферментом C2c1 или C2c3, таким как фермент C2c1 или C2c3, как определено в настоящем описании.
[00722] В некоторых основных вариантах осуществления изобретения фермент C2c1 или C2c3, используемый для мультиплексного нацеливания, связан с одним или более функциональными доменами. В некоторых более конкретных вариантах осуществления изобретения фермент CRISPR, используемый для мультиплексного нацеливания, является мертвым C2c1 или мертвым C2c3, как определено в настоящем описании.
[00723] В одном из аспектов настоящее изобретение относится к средству для доставки фермента C2c1 или C2c3, системы или комплекса для использования во множественном нацеливании, как определено в настоящем описании, или полинуклеотидов, определенных в настоящем описании. Неограничивающие примеры таких средств доставки включают, например, частицы(ы), доставляющие компонент(ы) комплекса, вектор(ы), включающие полинуклеотид(ы), обсуждаемые в настоящем описании (например, кодирующие ферменты CRISPR, обеспечивающие нуклеотиды, кодирующие комплекс CRISPR). В некоторых вариантах осуществления изобретения вектор может быть плазмидой или вирусным вектором, таким как ААV или лентивирус. Временная трансфекция с плазмидами, например, в клетки HEK может быть преимущественной, особенно учитывая ограничения ААV в отношении размера и что, в то время как C2c1 или C2c3 помещаются в ААV, с дополнительными направляющими РНК может быть достигнут верхний предел.
[00724] Также предусматривается модель, которая конститутивно экспрессирует фермент C2c1 или C2c3, комплекс или систему, используемые в рамках настоящего изобретения для мультиплексного нацеливания. Организм может быть трансгенным и может быть трансфицирован векторами по настоящему изобретению или может быть потомком организма, трансфицированного таким образом. В следующем аспекте настоящее изобретение относится к композициям, включающим фермент CRISPR, систему и комплекс, как определено в настоящем описании, или полинуклеотиды или векторы, описанные в настоящем описании. Также предусматриваются системы CRISPR C2c1 или C2c3 или комплексы, включающие несколько направляющих РНК, предпочтительно организованные в формате тандема. Указанные различные направляющие РНК могут быть разделены последовательностями нуклеотидов, такими как прямые повторы.
[00725] Также предусматривается способ лечения индивидуума, например, индивидуума, нуждающегося в этом, включающий индукцию генного редактирования посредством трансформации индивидуума полинуклеотидами, кодирующими систему CRISPR C2c1 или C2c3 или комплекс, или любым из полинуклеотидов или векторов, описанных в настоящем описании и вводимых субъекту. Также предусматривается подходящая матрица репарации, например доставлена вектором, включающим указанную матрицу репарации. Также предусматривается способ лечения индивидуума, например, индивидуума, нуждающегося в этом, включающий индукцию активации или репрессии транскрипции нескольких локусов-мишеней посредством трансформации индивидуума полинуклеотидами или векторами, описанными в настоящем описании, где указанные полинуклеотиды или векторы кодируют или включают фермент C2c1 или C2c3, комплекс или систему, включающие несколько направляющих РНК, предпочтительно расположенных тандемно. Если какой-либо лечение происходит ex vivo, например в клеточной культуре, тогда понятно, что термин "индивидуум" может быть заменен фразой "клетка или клеточная культура".
[00726] Также предлагаются композиции, включающие фермент C2c1 или C2c3, комплекс или систему, включающие множественные направляющие РНК, предпочтительно тандемно расположенные, или полинуклеотид или вектор, кодирующий или включающий указанный фермент C2c1 или C2c3, комплекс или систему, включающие множественные направляющие РНК, предпочтительно тандемно расположенные, для применения в способах лечения, как определено в настоящем описании. Может предусматриваться набор, включающий такие композиции. Также предусматривается использование указанной композиции для изготовления лекарственных средств для таких способов лечения. Также изобретение предусматривает использование системы CRISPR C2c1 или C2c3 в скрининге, например, скрининге приобретения функции. Клетки, которые искусственным образом вынудили сверхэкспрессировать ген, способны со временем снизить экспрессию гена (обратное восстановление равновесия), например, посредством отрицательной обратной связи. Ко времени начала скрининга экспрессия нерегулируемого гена снова может снизиться. Использование индуцибельного активатора C2c1 или C2c3 позволяет вызывать транскрипцию прямо перед скринингом и поэтому минимизирует вероятность ложноотрицательных результатов. Таким образом, при использовании настоящего изобретения в скрининге, например, скрининге приобретения функции, вероятность ложных отрицательных результатов может быть сведена к минимуму.
[00727] В одном из аспектов изобретение относится к сконструированной способами инженерии не встречающейся в природе системе CRISPR, включающей белок C2c1 или C2c3 и несколько направляющих РНК, каждая из которых специфически нацелена на молекулу ДНК, кодирующую продукт гена в клетке, посредством чего каждая из нескольких направляющих РНК нацелена на конкретную молекулу ДНК, кодирующую продукт гена, и белок C2c1 или C2c3 расщепляет молекулу ДНК-мишень, кодирующую продукт гена, посредством чего экспрессия продукта гена изменяется, и где белок CRISPR и направляющие РНК в природе не встречаются вместе. Изобретение охватывает несколько направляющих РНК, включая несколько направляющих последовательностей, предпочтительно отделенные нуклеотидной последовательностью, такой как прямой повтор. В одном варианте осуществления изобретения белок CRISPR является белком CRISPR-Cas типа V или VI, и в более предпочтительном варианте осуществления изобретения белок CRIPSR является белком C2c1 или C2c3. Кроме того, изобретение включает белок C2c1 или C2c3, являющийся кодон-оптимизированным для экспрессии в эукариотической клетке. В предпочтительном варианте осуществления изобретения эукариотическая клетка является клеткой млекопитающего, и в более предпочтительном варианте осуществления изобретения клетка млекопитающего является клеткой человека. В дальнейшем варианте осуществления изобретения уменьшена экспрессия продукта гена.
[00728] В другом аспекте изобретение относится к сконструированной способами инженерии не встречающейся в природе векторной системе, включающей один или более векторов, включающих первый регуляторный элемент, функционально связанный со несколькими направляющими РНК системы CRISPR C2c1 или C2c3, каждая из которых специфически нацелена на молекулу ДНК, кодирующую продукт гена, и второй регуляторный элемент, функционально связанный с последовательностью, кодирующей белок CRISPR. Оба регуляторных элемента могут быть расположены на том же самом векторе или на различных векторах системы. Несколько направляющих РНК нацелено на несколько молекул ДНК, кодирующих несколько продуктов генов в клетке, и белок CRISPR может расщеплять несколько молекул ДНК, кодирующих продукты генов (он может расщеплять одну или обе цепи или по существу не иметь никакой нуклеазной активности), посредством чего экспрессия нескольких продуктов генов может быть изменена; и где белок CRISPR и несколько направляющих РНК в природе не встречаются вместе. В предпочтительном варианте осуществления изобретения белок CRISPR является белком C2c1 или C2c3, необязательно кодон-оптимизированным для экспрессии в эукариотической клетке. В предпочтительном варианте осуществления изобретения эукариотическая клетка является клеткой млекопитающего, клеткой растения или клеткой дрожжей, и в более предпочтительном варианте осуществления изобретения клетка млекопитающего является клеткой человека. В следующем варианте осуществления изобретения экспрессия каждого из нескольких продуктов генов изменена, предпочтительно уменьшена.
[00729] В одном из аспектов изобретение относится к векторной системе, включающей один или более векторов. В некоторых вариантах осуществления изобретения система включает: (a) первый регуляторный элемент, функционально связанный с последовательностью прямого повтора и одним или более участками вставки для вставки одной или более направляющих последовательностей выше или ниже последовательностей прямого повтора, где, когда экспрессируются, одна или более направляющих последовательностей управляют специфическим для последовательности связыванием комплекса CRISPR с одной или более нацеливающими последовательностями в эукариотической клетке, где комплекс CRISPR включает фермент C2c1 или C2c3 в комплексе с одной или более направляющими последовательностями, гибридизованными с одной или более последовательностями-мишенями, и (b) второй регуляторный элемент, функционально связанный с кодирующей фермент последовательностью, кодирующей указанный фермент C2c1 или C2c3, предпочтительно включая по меньшей мере одну последовательность ядерной локализации и/или по меньшей мере одну последовательность NES; где компоненты (a) и (b) расположены на одной и той же или на различных векторах системы. В некоторых вариантах осуществления изобретения компонент (a) далее включает две или более направляющих последовательностей, функционально связанных с первым регуляторным элементом, где при экспрессии каждая из двух или более направляющих последовательностей управляет специфическим для последовательности связыванием комплекса CRISPR C2c1 или C2c3 c различными последовательностями-мишенями в эукариотической клетке. В некоторых вариантах осуществления изобретения комплекс CRISPR включает одну или более последовательностей ядерной локализации и/или один или более NES достаточной силы, чтобы стимулировать накопление указанного комплекса CRISPR C2c1 или C2c3 в поддающемся обнаружению количестве внутри или снаружи ядра эукариотической клетки. В некоторых вариантах осуществления изобретения первый регуляторный элемент является промотором полимеразы III. В некоторых вариантах осуществления изобретения второй регуляторный элемент является промотором полимеразы II. В некоторых вариантах осуществления изобретения каждая из направляющих последовательностей имеет длину по меньшей мере 16, 17, 18, 19, 20, 25 нуклеотидов, или 16-30, или 16-25, или 16-20 нуклеотидов.
[00730] Рекомбинантные экспрессирующие векторы могут включать полинуклеотиды, кодирующие фермент C2c1 или C2c3, систему или комплекс для применения в множественном нацеливании, как определено в настоящем описании, в форме, подходящей для экспрессии такой нуклеиновой кислоты в клетке-хозяине, что означает, что такие рекомбинантные экспрессирующие векторы содержат один или более регуляторных элементов, которые могут быть выбраны в соответствии с используемыми для экспрессии клетками хозяина и которые функционально связаны с последовательностью нуклеиновой кислоты, которая должна быть экспрессирована. В рекомбинантном векторе экспрессии "функционально связанный" должно означать, что нуклеотидная последовательность-мишень связана с регуляторным(и) элементом(ами) таким образом, что экспрессия такой нуклеотидной последовательности является возможной (например, в системе транскрипции/трансляции in vitro или в клетке хозяина при введении вектора в клетку хозяина).
[00731] В некоторых вариантах осуществления изобретения клетка-хозяин временно или постоянно трансфицирована одним или более векторами, включающими полинуклеотиды, кодирующие фермент C2c1 или C2c3, систему или комплекс для использования во множественном нацеливании, как определено в настоящем описании. В некоторых вариантах осуществления изобретения трансфицирована клетка, естественным образом встречающаяся у индивидуума. В некоторых вариантах осуществления изобретения клетка, которая трансфицирована, получена от индивидуума. В некоторых вариантах осуществления изобретения клетка происходит от клеток, полученных от индивидуума, таких как клеточная линия. Большое разнообразие клеточных линий для культуры тканей известно в данной области и иллюстрируются в настоящем описании. Доступны клеточные линии из множества источников, известных квалифицированным специалистам в данной области (см., например, American Type Culture Collection (ATCC) (Manassus, Вирджиния)). В некоторых вариантах осуществления изобретения клетка, трансфицированная одним или более векторами, включающими полинуклеотиды, кодирующие фермент C2c1 или C2c3, систему или комплекс для использования во множественном нацеливании, как определено в настоящем описании, используется для создания новой клеточной линии, включающей одну или более происходящих из вектора последовательностей. В некоторых вариантах осуществления изобретения клетка, временно трансфицированная компонентами системы CRISPR C2c1 или C2c3 или комплекса для использования во множественном нацеливании, как описано в настоящем описании (например, посредством временной трансфекции одного или более векторов или трансфекции РНК), и модифицированная посредством активности системы CRISPR C2c1 или C2c3 или комплекса, используется для получения новой клеточной линии, включающей клетки, содержащие модификацию, но лишенные какой-либо другой экзогенной последовательности. В некоторых вариантах осуществления изобретения клетки, временно или постоянно трансфицированные одним или более векторами, включающими полинуклеотиды, кодирующие фермент C2c1 или C2c3, систему или комплекс для использования во множественном нацеливании, как определено в настоящем описании, или клеточные линии, полученные из таких клеток, используются в оценке одного или более исследуемых соединений.
[00732] Термин "регуляторный элемент" определен в настоящем описании в других разделах.
[00733] Преимущественные векторы включают лентивирусы и адено-ассоциированные вирусы, и такие типы векторов также могут быть отобраны для нацеливания на конкретные типы клеток.
[00734] В одном из аспектов изобретение относится к эукариотической клетке-хозяину, включающей (a) первый регуляторный элемент, функционально связанный с последовательностью прямого повтора и одним или более участками вставки для вставки одной или более последовательностей направляющей РНК выше или ниже (в зависимости от того, что применимо) относительно последовательности прямого повтора, где, когда экспрессируется, направляющая последовательность(и) управляет последовательность-специфическим связыванием комплекса CRISPR C2c1 или C2c3 c соответствующей последовательностью(и)-мишенью в эукариотической клетке, где комплекс CRISPR C2c1 или C2c3 включает фермент C2c1 или C2c3 в комплексе с одной или более направляющими последовательностями, гибридизованными с соответствующей последовательностью(и)-мишенью; и/или (b) второй регуляторный элемент, функционально связанный с кодирующей фермент последовательностью, кодирующей указанный фермент C2c1 или C2c3, включающий предпочтительно по меньшей мере одну последовательность ядерной локализации и/или NES. В некоторых вариантах осуществления изобретения клетка-хозяин включает компоненты (a) и (b). В некоторых вариантах осуществления изобретения компонент (a), компонент (b) или компоненты (a) и (b) стабильно встраиваются в геном эукариотической клетки-хозяина. В некоторых вариантах осуществления изобретения компонент (a) далее включает две или более направляющих последовательностей, функционально связанных с первым регуляторным элементом, и необязательно разделенных прямым повтором, где при экспрессии каждая из двух или более направляющих последовательностей управляет специфическим для последовательности связыванием комплекса CRISPR C2c1 или C2c3 с различными последовательностями-мишенями в эукариотической клетке. В некоторых вариантах осуществления изобретения фермент C2c1 или C2c3 включает одну или более последовательностей ядерной локализации и/или последовательностей ядерного экспорта (NES) достаточной силы, чтобы стимулировать накопление указанного фермента CRISPR в поддающемся обнаружению количестве внутри и/или снаружи ядра эукариотической клетки.
[00735] В некоторых вариантах осуществления изобретения фермент C2c1 или C2c3 является ферментом системы CRISPR типа V или VI. В некоторых вариантах осуществления изобретения фермент C2c1 является ферментом AacC2c1. В некоторых вариантах осуществления изобретения фермент C2c1 получен из Alicyclobacillus acidoterrestris (например, ATCC 49025), Alicyclobacillus contaminans (например, DSM 17975), Desulfovibrio inopinatus (например, DSM 10711), Desulfonatronum thiodismutans (например, штамм MLF-1), бактерий Opitutaceae TAV5, Tuberibacillus calidus (например, DSM 17572), Bacillus thermoamylovorans (например, штамм B4166), Brevibacillus sp. CF112, Bacillus sp. NSP2.1, Desulfatirhabdium butyrativorans (например, DSM 18734), Alicyclobacillus herbarius (например, DSM 13609), Citrobacter freundii (например, ATCC 8090), Brevibacillus agri (например, BAB-2500), Methylobacterium nodulans (например, ORS 2060), и может включать дальнейшие изменения или мутации C2c1, как определено в настоящем описании, и может быть химерным C2c1. В некоторых вариантах осуществления изобретения фермент C2c1 или C2c3 кодон-оптимизирован для экспрессии в эукариотической клетке. В некоторых вариантах осуществления изобретения фермент CRISPR управляет расщеплением одной или двух цепей в определенном положении последовательности-мишени. В некоторых вариантах осуществления изобретения первый регуляторный элемент является промотором полимеразы III. В некоторых вариантах осуществления изобретения второй регуляторный элемент является промотором полимеразы II. В некоторых вариантах осуществления изобретения одна или более направляющих последовательностей (каждая) составляют по меньшей мере 16, 17, 18, 19, 20, 25 нуклеотидов, или 16-30, или 16-25, или 16-20 нуклеотидов в длину. Когда используется несколько направляющих РНК, они предпочтительно разделены последовательностью прямого повтора. В одном из аспектов изобретение относится к эукариотическому организму, не являющемуся человеком; предпочтительно многоклеточному эукариотическому организму, включающему эукариотическую клетку-хозяина согласно любому из описанных вариантов осуществления изобретения. В других аспектах изобретение относится к эукариотическому организму, предпочтительно многоклеточному эукариотическому организму, включая эукариотическую клетку-хозяина согласно любому из описанных вариантов осуществления изобретения. В некоторых вариантах осуществления организм согласно этим аспектам изобретения может быть животным, например млекопитающим. Кроме того, организм может быть членистоногим, таким как насекомое. Организм также может быть растением. Кроме того, организм может быть грибом.
[00736] В одном из аспектов изобретение относится к набору, включающему один или более компонентов, описанных в настоящем описании. В некоторых вариантах осуществления изобретения набор включает векторную систему и инструкции по применению набора. В некоторых вариантах осуществления изобретения векторная система включает (a) первый регуляторный элемент, функционально связанный с последовательностью прямого повтора и одним или более участками вставки для вставки одной или более направляющих последовательностей выше или ниже (в зависимости от того, что применимо) относительно последовательности прямого повтора, где при экспрессии направляющая последовательность управляет специфическим для последовательности связыванием комплекса CRISPR C2c1 или C2c3 c последовательностью-мишенью в эукариотической клетке, где комплекс CRISPR C2c1 включает фермент C2c1 или C2c3 в комплексе с направляющей последовательностью, гибридизованной с последовательностью-мишенью; и/или (b) второй регуляторный элемент, функционально связанный с кодирующей фермент последовательностью, кодирующей указанный фермент C2c1 или C2c3, включающий последовательность ядерной локализации. В некоторых вариантах осуществления изобретения набор включает компоненты (a) и (b), расположенные на одном и том же или на различных векторах системы. В некоторых вариантах осуществления изобретения набор (a) далее включает две или более направляющих последовательности, функционально связанных с первым регуляторным элементом, где при экспрессии каждая из этих двух или более направляющих последовательностей управляет специфическим для последовательности связыванием комплекса CRISPR с различными последовательностями-мишенями в эукариотической клетке. В некоторых вариантах осуществления изобретения фермент C2c1 или C2c3 включает одну или более последовательностей ядерной локализации достаточной силы, чтобы стимулировать накопление указанного фермента CRISPR в обнаружимом количестве в ядре эукариотической клетки. В некоторых вариантах осуществления изобретения фермент CRISPR является ферментом системы CRISPR типа V или VI. В некоторых вариантах осуществления изобретения фермент CRISPR является ферментом C2c1 или C2c3. В некоторых вариантах осуществления изобретения фермент C2c1 получен из Alicyclobacillus acidoterrestris (например, ATCC 49025), Alicyclobacillus contaminans (например, DSM 17975), Desulfovibrio inopinatus (например, DSM 10711), Desulfonatronum thiodismutans (например, штамм MLF-1), бактерии Opitutaceae TAV5, Tuberibacillus calidus (например, DSM 17572), Bacillus thermoamylovorans (например, штамм B4166), Brevibacillus sp. CF112, Bacillus sp. NSP2.1, Desulfatirhabdium. butyrativorans (например, DSM 18734), Alicyclobacillus herbarius (например, DSM 13609), Citrobacter freundti (например, ATCC 8090), Brevibacilhis agri (например, BAB-2500), Methylobacterium nodulans (например, ORS 2060) (например, модифицированный, чтобы иметь или быть связанными по меньшей мере с одним доменом дестабилизации («DD»), и может включать дальнейшее изменение или мутацию C2c1, и может быть химерным C2c1. В некоторых вариантах осуществления изобретения фермент DD-CRISPR кодон-оптимизирован для экспрессии в эукариотической клетке. В некоторых вариантах осуществления изобретения фермент DD-CRISPR управляет расщеплением одной или двух цепей в определенном положении последовательности-мишени. В некоторых вариантах осуществления изобретения фермент DD-CRISPR лишен ферментативной активности расшепления цепи ДНК (например, имеет не более, чем 5% нуклеазной активности по сравнению с ферментом дикого типа или ферментом, не имеющим мутации или изменения, которое уменьшает нуклеазную активность). В некоторых вариантах осуществления изобретения первый регуляторный элемент является промотором полимеразы III. В некоторых вариантах осуществления изобретения второй регуляторный элемент является промотором полимеразы II. В некоторых вариантах осуществления изобретения направляющая последовательность составляет по меньшей мере 16, 17, 18, 19, 20, 25 нуклеотидов, или 16-30 или 16-25 или 16-20 нуклеотидов в длину.
[00737] В одном из аспектов изобретение относится к способу модификации нескольких полинуклеотидов-мишеней в клетке-хозяине, такой как эукариотическая клетка. В некоторых вариантах осуществления изобретения способ позволяет комплексу CRISPR C2c1 или C2c3 связываться с несколькими полинуклеотидами-аишенями, например, производя расщепление указанных нескольких полинуклеотидов-мишеней, таким образом модифицируя несколько полинуклеотидов-мишеней, где комплекс CRISPR C2c1 или C2c3 включает фермент C2c1 или C2c3 в комплексе со несколькими направляющими последовательностями, каждая из которых гибридизована с определенной последовательностью-мишенью в указанном полинуклеотиде-мишени, где указанные несколько направляющих последовательностей связаны с последовательностью прямого повтора. В некоторых вариантах осуществления изобретения указанное расщепление включает расщепление одной или двух цепей в определенном положении каждой последовательности-мишени указанным ферментом C2c1 или C2c3. В некоторых вариантах осуществления изобретения указанное расщепление приводит к уменьшению транскрипции нескольких генов-мишеней. В некоторых вариантах осуществления изобретения способ далее включает репарацию одного или более указанных расщепленных полинуклеотидов-мишеней посредством гомологичной рекомбинации с экзогенным матричным полинуклеотидом, где указанная репарация приводит к мутации, включающей инсерцию, делецию или замену одного или более нуклеотидов в одном или более указанных полинуклеотидах-мишенях. В некоторых вариантах осуществления изобретения указанные мутации приводят к одному или более изменениям в аминокислотной последовательности белка, экспрессируемого с гена, включающего одну или более последовательностей-мишеней. В некоторых вариантах осуществления изобретения способ далее включает доставку одного или более векторов в указанную эукариотическую клетку, где один или более векторов осуществляют экспрессию одного или более из фермента C2c1 или C2c3 и нескольких последовательностей направляющих РНК, связанных с последовательностью прямого повтора. В некоторых вариантах осуществления изобретения указанные векторы доставляются в эукариотическую клетку в индивидууме. В некоторых вариантах осуществления изобретения указанные модификация происходят в указанной эукариотической клетке в клеточной культуре. В некоторых вариантах осуществления изобретения способ далее включает извлечение указанной эукариотической клетки из индивидуума до произведения указанной модификации. В некоторых вариантах осуществления изобретение способ далее включает обратное введение указанной эукариотической клетки и/или клеток, полученных из нее, указанному индивидууму.
[00738] В одном из аспектов изобретение относится к способу модификации экспрессии нескольких полинуклеотидов в эукариотической клетке. В некоторых вариантах осуществления изобретения способ позволяет комплексу CRISPR C2c1 или C2c3 связываться со несколькими полинуклеотидами, таким образом, таким образом, что указанное связывание приводит к увеличению или уменьшению экспрессии указанных полинуклеотидов; где комплекс CRISPR C2c1 или C2c3 включает фермент C2c1 или C2c3 в комплексе со несколькими направляющими последовательностями, каждая из которых специфически гибридизирована со своей собственной последовательностью-мишенью в указанном полинуклеотиде, где указанные направляющие последовательности связаны с последовательностью прямого повтора. В некоторых вариантах осуществления изобретения способ далее включает доставку одного или более векторов в указанные эукариотические клетки, где один или более векторов осуществляют экспрессию одного или более из фермента C2c1 или C2c3 и нескольких направляющих последовательностей, связанных с последовательностями прямого повтора.
[00739] В одном из аспектов изобретение относится к рекомбинантному полинуклеотиду, включающему несколько последовательностей направляющей РНК выше или ниже (в зависимости от того, что применимо) относительно последовательности прямого повтора, где каждая из направляющих последовательностей, когда экспрессируется, управляет специфическим для последовательности связыванием комплекса CRISPR C2c1 или C2c3 с соответствующей ему последовательностью-мишенью, присутствующей в эукариотической клетке. В некоторых вариантах осуществления изобретения последовательность-мишень является вирусной последовательностью, присутствующей в эукариотической клетке. В некоторых вариантах осуществления изобретения последовательность-мишень является протоонкогеном или онкогеном.
[00740] Аспекты изобретения охватывают не встречающуюся в природе или сконструированную способами инженерии композицию, которая может включать направляющую РНК (гРНК), включающую направляющую последовательность, способную к гибридизации с последовательностью-мишенью в представляющем интерес геномном локусе в клетке и фермент C2c1 или C2c3, как определено в настоящем описании, который может включать по меньшей мере одну или более последовательностей ядерной локализации.
[00741] Один из аспектов изобретения охватывает способы модификации представляющего интерес геномного локуса для изменения экспрессии гена в клетке посредством введения в клетку любой из композиций, описанных в настоящем описании.
[00742] Одним из аспектов изобретения является то, что вышеупомянутые элементы содержатся в единственной композиции или содержатся в индивидуальных композициях. Эти композиции могут быть успешно применены к хозяину, чтобы выявить функциональный эффект на геномном уровне.
[00743] Как используется в настоящем описании, термин «направляющая РНК» или «гРНК» имеет такое же значение, как в других разделах, и включает любую полинуклеотидную последовательность, имеющую достаточную комплементарность с последовательностью нуклеиновой кислоты, являющейся мишенью, чтобы гибридизироваться с последовательностью нуклеиновой кислоты, являющейся мишенью, и управлять специфическим для последовательности связыванием нацеленного на нуклеиновые кислоты комплекса с последовательностью нуклеиновой кислоты, являющейся мишенью. Каждая гРНК может быть сконструирована, чтобы включать множественные сайты узнавания для связывания (например, аптамеры) специфичные к одному или различным адаптерным белкам. Каждая гРНК может быть сконструирована для связывания с промоторной областью -1000-+1 нуклеотид выше сайта начала транскрипции (т.е. TSS), предпочтительно в участке -200 нуклеотид. Это расположение служит для улучшения функциональных доменов, которые влияют на активацию генов (например, активаторы транскрипции) или ингибирование генов (например, репрессоры транскрипции). Модифицированная гРНК может содержать одну или более модифицированных гРНК, нацеливающих на один или более локусов-мишеней (например, по меньшей мере 1 гРНК, по меньшей мере 2 гРНК, по меньшей мере 5 гРНК, по меньшей мере 10 гРНК, по меньшей мере 20 гРНК, по меньшей мере 30 гРНК, по меньшей мере 50 гРНК), находящихся в композиции. Указанные множественные последовательности гРНК могут быть организованы в формате тандема и предпочтительно разделены прямым повтором.
[00744] Таким образом, каждый из гРНК, фермента CRISPR, как определено в настоящем описании, может индивидуально находиться в композиции и вводиться хозяину индивидуально или совместно. Альтернативно, эти компоненты могут содержаться в единой композиции для введения хозяину. Введение хозяину может быть выполнено посредством вирусных векторов, известных квалифицированному специалисту или описанных в настоящем описании, для доставки хозяину (например, лентивирусный вектор, аденовирусный вектор, вектор ААV). Как объяснено в настоящем описании, использование различных селективных маркеров (например, для селекции лентивирусной гРНК) и концентрация гРНК (например, в зависимости от того, используется ли несколько гРНК) может быть преимущественным для выявления улучшенного эффекта. На основании этой концепции существует несколько вариантов выявления событий в геномном локусе, включая расщепление ДНК, активацию и инактивацию генов. Используя предложенные композиции, квалифицированный специалист в данной области может преимущественно и специфически осуществлять нацеливание на один или более локусов с использованием одинаковых или различных функциональных доменов для выявления одного или более событий в геномном локусе. Композиции могут быть применены в большом разнообразии способов скрининга библиотек в клетках и функциональном моделировании in vivo (например, активация генов длинными межгенными некодирующими РНК (lincРНК) и идентификация функций; моделирование приобретения функции; моделирование потери функции; использование композиций изобретения для создания клеточных линий и трансгенных животных в целях оптимизации и скрининга).
[00745] Настоящее изобретение охватывает использование композиций по настоящему изобретению для создания и использования трансгенных по зависимой от условий или индуцибельной системе CRISPR клеток/животных; см., например, Platt et al., Cell (2014), 159(2): 440-455 или патентную публикацию PCT, цитируемую в настоящем описании, такую как WO 2014/093622 (PCT/US2013/074667). Например, клетки или животные, такие как животные, не являющиеся человеком, например, позвоночные животные или млекопитающие, такие как грызуны, например, мыши, крысы или другие лабораторные или не лабораторные животные, например, кошки, собаки, овцы, и т.д., могут быть "нокин" (knock-in), в результате чего животное в зависимости от условий или индуцируемо экспрессирует C2c1 или C2c3, как в статье Platt et al. Таким образом, клетка-мишень или животное-мишень включают фермент CRISRP (например, C2c1 или C2c3) в зависимости от условий или индуцируемо (например, в форме Cre-зависимой композиции), после экспрессии вектора, введенного в клетку-мишень, экспрессия вектора индуцирует или создает условия для экспрессии фермента CRISRP (например, C2c1 или C2c3) в клетке-мишени. Посредством применения руководства и композиции, как определено в настоящем описании, с известным способом создания комплекса CRISPR, индуцируемые события в геноме также являются аспектом настоящего изобретения. Примеры таких индуцируемых событий описаны в настоящем описании в других разделах.
[00746] В некоторых вариантах осуществления фенотипическое изменение предпочтительно является результатом модификации генома, когда мишенью является генетическое заболевание, особенно в способах терапии, и предпочтительно, когда обеспечивается матрица репарации для коррекции или изменения фенотипа.
[00747] В некоторых вариантах осуществления заболевания, которые могут быть мишенью, включают заболевания, которые связаны с дефектами сплайсинга.
[00748] В некоторых вариантах осуществления клеточные мишени включают гемопоэтические стволовые клетки/клетки-предшественники (CD34+); Т-клетки человека; и клетки сетчатки - например, клетки-предшественники фоторецепторов.
[00749] В некоторых вариантах осуществления гены-мишени включают: ген бета-глобина человека - HBB (для лечения серповидноклеточной анемии, включая стимуляцию генной конверсии (с использованием близкородственного гена HBD в качестве эндогенной матрицы)); CD3 (Т-клетки) и CEP920 - сетчатка (глаза).
[00750] В некоторых вариантах осуществления болезни-мишени также включают: злокачественную опхуоль; серповидноклеточную анемию (основанную на точечной мутации); ВИЧ; бета-талассемию; и офтальмологические или глазные заболевания - например, амавроз Лебера (LCA), вызываемый дефектом сплайсинга.
[00751] В некоторых вариантах осуществления способы доставки включают: опосредованную катионными липидами "прямую" доставку комплекса фермент-гид (рибонуклеопротеин) и электропорацию плазмидной ДНК.
[00752] Способы, продукты и применения, описанные в настоящем описании, могут использоваться в нетерапевтических целях. Кроме того, любой из способов, описанных в настоящем описании, может быть применен in vitro и ex vivo.
[00753] В одном из аспектов предусматривается не встречающаяся в природе или сконструированная способами инженерии композиция, включающая:
I. две или более полинуклеотидных последовательностей системы CRISPR-Cas, содержащих
(a) первую направляющую последовательность, способную к гибридизации с первой последовательностью-мишенью в локусе полинуклеотида,
(b) вторую направляющую последовательность, способную к гибридизации со второй последовательностью-мишенью в локусе полинуклеотида,
(c) последовательность прямого повтора, и
II. фермент C2c1 или вторую полинуклеотидную последовательность, кодирующую его, или фермент C2c3 или вторую полинуклеотидную последовательность, кодирующую его,
где при транскрипции первая и вторая направляющие последовательности управляют специфическим для последовательности связыванием первого и второго комплекса CRISPR C2c1 или C2c3 c первыми и вторыми последовательностями-мишенями, соответственно,
где первый комплекс CRISPR включает фермент C2c1 или фермент C2c3 в комплексе с первой направляющей последовательностью, которая способна к гибридизации с первой последовательностью-мишенью,
где второй комплекс CRISPR включает фермент C2c1 или фермент C2c3 в комплексе со второй направляющей последовательностью, которая способна к гибридизации со второй последовательностью-мишенью, и
где первая направляющая последовательность управляет расщеплением одной цепи дуплекса ДНК вблизи первой последовательности-мишени, и вторая направляющая последовательность управляет расщеплением другой цепи вблизи второй последовательности-мишени, вызывая двойной разрыв цепи, таким образом модифицируя организм или организм, не являющийся животным или не являющийся человеком. Точно так же могут предусматриваться композиции, включающие более двух направляющие РНК, например, каждая РНК специфична к одной мишени и все РНК организованы тандемно в композиции, или системе CRISPR, или комплексе, как описано в настоящем описании.
[00754] В другом варианте осуществления изобретения C2c1 или C2c3 доставляется в клетку в виде белка. В другом и особенно предпочтительном варианте осуществления изобретения C2c1 или C2c3 доставляется в клетку в виде белка или в виде нуклеотидной последовательности, кодирующей его. Доставка к клетке в виде белка может включать доставку комплекса рибонуклеопротеина (RNP), где белок образует комплекс с несколькими направляющими последовательностями.
[00755] В одном из аспектов предусматриваются клетки-хозяева и клеточные линии, модифицированные или включающие композиции, системы или модифицированные ферменты по настоящему изобретению, включая стволовые клетки и их потомков.
[00756] В одном из аспектов предусматриваются способы клеточной терапии, где, например, одну клетку или популяцию клеток отбирают или культивируют, эту клетку или клетки модифицируют ex vivo, как описано в настоящем описании, и потом повторно вводят (извлеченные клетки) или вводят (культивируемые клетки) в организм. Стволовые клетки, эмбриональные или индуцированные плюрипотентные или тотипотентные стволовые клетки, также особенно предпочтительны в этом отношении. Но, конечно, также предусматриваются варианты осуществления изобретения in vivo.
[00654] Способы по изобретению могут дополнительно включать доставку матриц, таких как матрицы репарации, которые могут представлять собой дцОДН или оцОДН, см. ниже. Доставка матриц может быть произведена одновременно или отдельно от доставки любого или всех эффекторных белков CRISPR или направляющих молекул, и через один и тот же механизм доставки или любой другой. В некоторых вариантах осуществления предпочтительно, чтобы матрица доставлялась вместе с направляющей молекулой и также предпочтительно эффекторным белком CRISPR. Примером может служить вектор AAV.
[00655] Способы по изобретению могут дополнительно включать: (а) доставку в клетку двухцепочечного олигодезоксинуклеотида (дцОДН), включающего выступающие концы, комплементарные выступающим концам, созданным упомянутым разрывом двойной цепи, причем указанный дцОДН встраивается в локус-мишень, или - (б) доставку в клетку одноцепочечного олигодезоксинуклеотида (оцОДН), где указанный оцОДН действует как матрица для гомологичной направленной репарации разрыва двойной цепи. Способы по изобретению могут быть предназначены для профилактики или лечения заболевания у индивида, когда указанное заболевание вызвано дефектом в указанном локусе-мишени. Способы по изобретению могут осуществляться in vivo в индивидууме или ex vivo на клетке, взятой у индивида, где в некоторых случаях указанную клетку возвращают индивиду.
[00759] Изобретение также охватывает продукты, полученные с использованием фермента CRISPR, или фермента Cas, или фермента C2c1, или фермента C2c3, или фермента CRISPR-CRISPR, или системы CRISPR-Cas, или системы CRISPR-C2c1, или системы CRISPR-C2c3 для использования в тандемном или множественном нацеливании, как определено в настоящем описании.
Наборы
[00760] В одном из аспектов изобретение предлагает наборы, содержащие любой один или более элементов, описанных для вышеупомянутых способов и композиций. В некоторых вариантах осуществления набор включает векторную систему, как описано в настоящем описании и в инструкциях по применению наборов. Элементы могут быть предоставлены индивидуально или в комбинациях и могут быть предоставлены в любом подходящем контейнере, таком как флакон, бутылка или тюбик, в некоторых вариантах осуществления изобретения набор включает инструкции на одном или более языках, например на более чем одном языке. Инструкции могут быть специальными для приложений и способов, описанных в настоящем описании.
[00761] В некоторых вариантах осуществления изобретения набор включает один или более реактивов для использования в способе, в котором используется один или более элементов, описанных в настоящем описании. Реактивы могут быть предоставлены в любом подходящем контейнере. Например, в наборе может быть предоставлен один или более буферов для хранения или проведения реакций. Реактивы могут быть предложены в форме, которая применима в конкретном анализе, или в форме, которая требует добавления одного или более других компонентов перед использованием (например, в концентрате или лиофилизированной форме). Буфер может быть любым буфером, включая, но не ограничиваясь ими, натрий-карбонатный буфер, натрий-бикарбонатный буфер, боратный буфер, Tris-буфер, буфер MOPS, буфер HEPES и их комбинациями. В некоторых вариантах осуществления изобретения буфер является щелочным. В некоторых вариантах осуществления изобретения буфер имеет pH от приблизительно 7 до приблизительно 10. В некоторых вариантах осуществления изобретения набор включает один или более олигонуклеотидов, соответствующих направляющей последовательности, для вставки в вектор для функционального связывания направляющей последовательности и регуляторного элемента. В некоторых вариантах осуществления изобретения набор включает гомологичный полинуклеотид матрицы рекомбинации. В некоторых вариантах осуществления изобретения набор включает один или более векторов и/или один или более полинуклеотидов, описанных в настощоящем описании. Набор преимущественно позволяет предоставить все элементы систем изобретения по изобретению.
[00762] В одном из аспектов изобретение относится к способам применения одного или более элементов системы CRISPR. Комплекс CRISPR по изобретению обеспечивает эффективное средство модификации полинуклеотида-мишени. Комплекс CRISPR по изобретению имеет большое разнообразие полезных применений, включая модификацию (например, удаление, вставку, перемещение, инактивацию, активацию) полинуклеотида-мишени в разнообразных типах клеток. Как таковой, комплекс CRISPR по изобретению имеет широкий спектр применений, например, в генотерапии, скрининге лекарств, диагностике заболеваний и прогнозировании. Типичный комплекс CRISPR включает эффекторный белок CRISPR в комплексе с направляющей последовательностью, гибридизованной с последовательностью-мишенью в полинуклеотиде-мишени. В определенных вариантах осуществления изобретения последовательность прямого повтора связана с направляющей последовательностью.
[00763] В одном из вариантов осуществления изобретения настоящее изобретение относится к способу расщепления полинуклеотида-мишени. Способ включает модификацию полинуклеотида-мишени с использованием комплекса CRISPR, который связывается с полинуклеотидом-мишенью и производит расщепление указанного полинуклеотида-мишени. Как правило, комплекс CRISPR по изобретению при введении в клетку создает разрыв (например, одноцепочечный или двухцепочечный разрыв) в последовательности генома. Например, способ может использоваться для расщепления гена заболевания в клетке.
[00764] Разрыв, внесенный комплексом CRISPR, может быть восстановлен процессами репарации, такими как негомологичное соединение концов (NHEJ), подверженное ошибкам, или гомологичная репарация (HDR) с высокой точностью. Во время этих процессов репарации экзогенная полинуклеотидная матрица может быть встроена в последовательность генома. В некоторых способах для модификации последовательности генома используется процесс HDR. Например, экзогенная полинуклеотидная матрица, содержащая последовательность, подлежащую встраиванию, фланкированную нижележащей последовательностью и вышележащей последовательностью, вводится в клетку. Вышележащая и нижележащая последовательности имеют сходство с каждой стороной сайта интеграции в хромосоме.
[00765] Где желаемо, донорный полинуклеотид может представлять собой ДНК, например, плазмидную ДНК, бактериальную искусственную хромосому (ВАС), дрожжевую искусственную хромосому (YAC), вирусный вектор, линейный фрагмент ДНК, фрагмент ПЦР, "голую" нуклеиновую кислоту или нуклеиновую кислоту в комплексе с носителем для доставки, таким как липосома или полоксамер.
[00766] Экзогенная полинуклеотидная матрица включает последовательность, которая будет встроена (например, мутированный ген). Последовательность для интеграции может быть последовательностью, эндогенной или экзогенной по отношению к клетке. Примеры последовательностей для интеграции включают полинуклеотиды, кодирующие белок или некодирующую РНК (например, микроРНК). Таким образом, последовательность для интеграции может быть функционально связана с соответствующей контрольной последовательностью или последовательностями. Альтернативно, последовательность для интеграции может иметь регуляторную функцию.
[00767] Вышележащие и нижележащие последовательности в экзогенной полинуклеотидной матрице выбирают, чтобы обеспечить рекомбинацию между интересующей хромосомной последовательностью и донорным полинуклеотидом. Вышележащая последовательность является последовательностью нуклеиновых кислот, имеющей сходство с последовательностью генома, вышележащей относительно целевого участка интеграции. Точно так же нисходящая последовательность является последовательностью нуклеиновых кислот, имеющей сходство с хромосомной последовательностью, нижележащей относительно целевого участка интеграции. У вышележащих и нижележащих последовательностей в экзогенной полинуклеотидной матрице может быть 75%, 80%, 85%, 90%, 95%, или 100% сходство последовательности с последовательностью генома, являющейся мишенью. Предпочтительно, когда вышележащие и нижележащие последовательности в экзогенной полинуклеотидной матрице имеют приблизительно 95%, 96%, 97%, 98%, 99%, или 100% сходство последовательности с последовательностью генома, являющейся мишенью. В некоторых методах вышележащие и нижележащие последовательности в экзогенной полинуклеотидной матрице имеют приблизительно 99% или 100% сходства последовательности с последовательностью генома, являющейся мишенью.
[00768] Вышележащая или нижележащая последовательность может включать от приблизительно 20 п.о. до приблизительно 2500 пн, например, приблизительно 50, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1100, 1200, 1300, 1400, 1500, 1600, 1700, 1800, 1900, 2000, 2100, 2200, 2300, 2400, или 2500 п.о. В некоторых способах типичная вышележащая или нижележащая последовательность имеет от приблизительно 200 п.о. до приблизительно 2000 п.о., от приблизительно 600 п.о. до приблизительно 1000 п.о., или более конкретно от приблизительно 700 п.о. до приблизительно 1000 п.о.
[00769] В некоторых методах экзогенная полинуклеотидная матрица может дополнительно включать маркер. Такой маркер может облегчить скрининг на заданную интеграцию. Примеры подходящих маркеров включают участки рестрикции, флуоресцентные белки или выбираемые маркеры. Экзогенная полинуклеотидная матрица изобретения может быть сконструирована с использованием рекомбинантных методов (см., например, Sambrook et al, 2001 and Ausubel et al., 1996).
[00770] В типичном способе модификации полинуклеотида-мишени посредством интеграции экзогенной полинуклеотидной матрицы, двухцепочечный разрыв вносится в последовательность генома комплексом CRISPR, разрыв восстанавливается путем гомологичной рекомбинации с экзогенной полинуклеотидной матрицей, таким образом, что матрица интегрируется в геном. Присутствие двухцепочечного разрыва облегчает интеграцию матрицы.
[00771] В других вариантах осуществления настоящее изобретение относится к способу модификации экспрессии полинуклеотида в эукариотической клетке. Способ включает увеличение или уменьшение экспрессии полинуклеотида-мишени при помощи комплекса CRISPR, который связывается с полинуклеотидом.
[00772] В некоторых методах полинуклеотид-мишень может быть инактивирован, чтобы произвести модификацию экспрессии в клетке. Например, после связывания комплекса CRISPR с последовательностью-мишенью в клетке, полинуклеотид-мишень инактивируется таким образом, что последовательность не транскрибируется, кодируемый белок не синтезируется, или последовательность не функционирует как последовательность дикого типа. Например, кодирующая последовательность белка или микроРНК могут быть инактивированы таким образом, что синтез белка прекращается.
[00773] В некоторых способах контрольная последовательность может быть инактивирована таким образом, что она больше не функционирует как контрольная последовательность. Как используют в настоящем описании, "контрольная последовательность" относится к любой последовательности нуклеиновой кислоты, которая производит транскрипцию, трансляцию или обеспечивает доступность последовательности нуклеиновой кислоты. Примеры контрольной последовательности включают промотор, терминатор транскрипции, и энхансер. Инактивированная последовательность-мишень может включать делеционную мутацию (т.е. делецию одного или более нуклеотидов), инсерционную мутацию (т.е. инсерцию одного или более нуклеотидов) или нонсенс-мутацию (т.е. замена единственного нуклеотида на другой нуклеотид, таким образом, что получается стоп-кодон). В некоторых методах инактивация последовательности-мишени приводит к "нокауту" последовательности-мишени.
Иллюстративные способы применения системы CRISPR Cas
[00774] Изобретение относится к не встречающейся в природе или сконструированной способами инженерии композиции, или одному или более полинуклеотидам, кодирующим компоненты указанной композиции, или векторам или системам доставки, содержащим один или более полинуклеотидов, кодирующих компоненты указанной композиции, для применения для модификации клетки-мишени in vivo, ex vivo или in vitro, и это может быть осуществлено способом, который изменяет клетку таким образом, что после модификации потомки или клеточная линия модифицированной клетки CRISPR сохраняет измененный фенотип. Модифицированные клетки и их потомки могут быть частью многоклеточного организма, такого как растение или животное, с применением системы CRISPR ex vivo к желаемым типам клеток. Изобретение CRISPR может представлять собой терапевтический способ лечения. Терапевтический способ лечения может включать редактирование гена или генома, или генную терапию.
Использование инактивированного фермента CRISPR C2c1 или С2с3 для способов обнаружения, таких как FISH
[00775] В одном из аспектов изобретение относится к сконструированной способами инженерии не встречающейся в природе системе CRISPR-Cas, включающую каталитически инактивированный белок Cas, описанный в настоящем описании, предпочтительно инактивированный C2c1 (dC2c1) или инактивированный C2c3 (dC2c3), и к применению этой системы в способах обнаружения, таких как флюоресцентная гибридизация in situ (FISH). dC2c1 или dC2c3, лишенный способности производить двухцепочечные разрывы двойной цепи ДНК, может быть связан с маркером, таким как флуоресцентный белок, такой как усиленный зеленый флуоресцентный белок (eEGFP) и совместно экспрессирован с небольшими направляющими РНК для нацеливания на перицентрические, центрические и теломерные повторы in vivo. Системы с dC2c1 или dC2c3 могут использоваться для визуализации как повторяющихся последовательностей, так и индивидуальных генов в геноме человека. Такие новые применения маркированных систем CRISPR-Cas могут быть важны для визуализации клеток и изучения функциональной ядерной архитектуры, особенно в случае маленького объема ядра или сложных 3D-структур. (Chen B, Gilbert LA, Cimini BA, Schnitzbauer J, Zhang W, Li GW, Park J, Blackburn EH, Weissman JS, Qi LS, Huang B. 2013. Dynamic imaging of genomic loci in living human cells by an optimized CRISPR/Cas Cell 155(7): 1479-91, doi: 10.1016/j.cel1.2013.12.001)
Модификация Мишени посредством системы или комплекса CRISPR Cas (например, комплекса C2c1/C2c3-РНК)
[00776] В одном аспекте изобретение относится к способам модификации полинуклеотида-мишени в эукариотической клетке, которая может быть in vivo, ex vivo или in vitro. В некоторых вариантах осуществления способ включает отбор образца клетки или популяции клеток у человека или животного, не являющегося человеком, и модификацию клетки или клеток. Культивирование может происходить на любом этапе ex vivo. Клетка или клетки могут быть повторно введены в животное или растение, отличное от человека. Для вновь введенных клеток особенно предпочтительно, чтобы клетки были стволовыми клетками.
[00777] В некоторых вариантах осуществления способ включает предоставление возможности CRISPR-комплексу связываться с полинуклеотидом-мишенью для осуществления расщепления указанного полинуклеотида-мишени, в результате чего происходит модификация полинуклеотида-мишени, где комплекс CRISPR содержит эффекторный белок CRISPR и направляющую последовательность, гибридизованную или способную к гибридизации с последовательностью-мишенью в указанном полинуклеотиде-мишени.
[00778] В одном из вариантов изобретение относится к способу модификации экспрессии полинуклеотида в эукариотической клетке. В некоторых вариантах осуществления способ включает предоставление комплексу CRISPR возможности связываться с полинуклеотидом, так что указанное связывание приводит к увеличению или уменьшению экспрессии указанного полинуклеотида; где комплекс CRISPR содержит эффекторный белок CRISPR, объединенный с направляющей последовательностью, гибридизованной или способной к гибридизации с последовательностью-мишенью в указанном полинуклеотиде. Аналогичные соображения и условия применяются, как указано выше, для способов модификации полинуклеотида-мишени. Фактически, эти варианты отбора проб, культивирования и повторного введения применяются во всех аспектах настоящего изобретения.
[00779] Действительно, в любом аспекте изобретения комплекс CRISPR может содержать эффекторный белок CRISPR в комплексе с направляющей последовательностью, гибридизованной или способной к гибридизации с последовательностью-мишенью. Аналогичные соображения и условия применяются, как указано выше, для способов модификации полинуклеотида-мишени.
[00780] Таким образом, любой из не встречающихся в природе эффекторных белков CRISPR, описанных в настоящем описании, имеет по меньшей мере одну модификацию, и, тем самым, эффекторный белок обладает определенными улучшенными возможностями. В частности, любой из эффекторных белков способен образовывать комплекс CRISPR с направляющей РНК. Когда такой комплекс образуется, направляющая РНК способна связываться с полинуклеотидной последовательностью-мишенью, и эффекторный белок способен модифицировать локус-мишень. Кроме того, эффекторный белок в комплексе CRISPR уменьшает способность модифицировать один или более локусов-мишеней по сравнению с немодифицированным ферментом/эффекторным белком.
[00781] Кроме того, модифицированные ферменты CRISPR, описанные в настоящем описании, охватывают ферменты которые обеспечивают повышение способности эффекторного белка комплекса CRISPR модифицировать один или более локусов-мишеней по сравнению с немодифицированным ферментом/эффекторным белком. Такая функция может быть предоставлена отдельно или предоставлена в сочетании с вышеописанной уменьшенной способностью модифицировать один или более локусов, не являющихся мишенями. Любые такие эффекторные белки могут быть снабжены любой из дополнительных модификаций в эффекторном белке CRISPR, как описано в настоящем описании, например, в сочетании с любой активностью, обеспечиваемой одним или более связанными гетерологичными функциональными доменами, любыми другими мутациями для снижения активности нуклеазы и т.п.
[00782] В предпочтительных вариантах осуществления изобретения модифицированный эффекторный белок CRISPR обладает сниженной способностью модифицировать один или более локусов, не являющихся мишенями, по сравнению с немодифицированным ферментом/эффекторным белком и повышает способность модифицировать один или более локусов-мишеней по сравнению с немодифицированным ферментом/эффекторным белком. В сочетании с дополнительными модификациями эффекторного белка может быть достигнута значительно увеличенная специфичность. Например, комбинация таких предпочтительных вариантов осуществления с одной или более дополнительными мутациями предусматривается в том случае, если одна или более дополнительных мутаций находятся в одном или более активных каталитически активных доменах. Такие дополнительные мутации каталитического домена могут сообщать никазную функциональность, как подробно описано в настоящем описании. В таких эффекторных белках повышенная специфичность может быть достигнута благодаря улучшенной специфичности в отношении активности эффекторного белка.
[00783] Модификации для уменьшения нецелевых эффектов и/или усиления целевых эффектов, как описано выше, могут быть сделаны для аминокислотных остатков, расположенных в положительно заряженной области/желобке, расположенных между доменами RuvC-III и HNH. Понятно, что любой из описанных выше функциональных эффектов может быть достигнут путем модификации аминокислот в вышеупомянутом желобке, а также путем модификации аминокислот, смежных с этим желобком или вне него.
[00784] Дополнительные функциональные возможности, которые могут быть внесены способами инженерии в модифицированные эффекторные белки CRISPR, как описано в настоящем описании, включают следующие. 1. модифицированные эффекторные белки CRISPR, которые нарушают взаимодействия ДНК:белок, не влияя на третичную или вторичную структуру белка. Это включает остатки, которые контактируют с любой частью дуплекса РНК:ДНК. 2. модифицированные эффекторные белки CRISPR, которые ослабляют внутрибелковые взаимодействия путем поддержания C2c1 или C2c3 в конформации, необходимой для нуклеазного разрезания в ответ на связывание ДНК (целевое или нецелевое). Например, модификация, которая мягко ингибирует, но все равно допускает конформацию нуклеазы домена HNH (расположена на расщепляющемся фосфате), 3. модифицированные эффекторные белки CRISPR, которые усиливают внутрибелковые взаимодействия путем поддержания C2c1 или C2c3 в конформации, ингибирующей активность нуклеазы в реакции связывания ДНК (целевой или нецелевой). Например: модификация, которая стабилизирует HNH-домен в конформации расщепляющегося фосфата. Любое такое дополнительное функциональное усиление может быть обеспечено в сочетании с любой другой модификацией эффекторного белка CRISPR, как подробно описано в данной заявке в другом пункте.
[00785] Любая из описанных в настоящем описании улучшенных функциональных возможностей может быть внесена способами инженерии в любой эффекторный белок CRISPR, такой как эффекторный белок C2c1 или C2c3. Однако будет понятно, что любая из описанных в настоящем описании функциональных возможностей может быть внесена способами инженерии в эффекторные белки C2c1 или C2c3 из других ортологов, включая химерные эффекторные белки, содержащие фрагменты из множества ортологов.
Нуклеиновые кислоты, аминокислоты и белки, регуляторные последовательности, векторы и т.д.
[00786] В рамках изобретения используются нуклеиновые кислоты для связывания последовательностей ДНК-мишеней. Это перспективно, так как нуклеиновые кислоты намного легче и дешевле производить, чем белки, а также специфичность может варьироваться в зависимости от длины участка, гомологии которого требуется. Например, не требуется сложное трехмерное позиционирование нескольких пальцев. Термины "полинуклеотид", "нуклеотид", "нуклеотидная последовательность", "нуклеиновая кислота" и "олигонуклеотид" используются взаимозаменяемо. Они относятся к полимерной форме нуклеотидов любой длины, либо дезоксирибонуклеотидов, либо рибонуклеотидов, или их аналогов. Полинуклеотиды могут иметь любую трехмерную структуру и могут выполнять любую из возможных известных или неизвестных функций. Ниже приведены неограничивающие примеры полинуклеотидов: кодирующие или некодирующие области гена или фрагмента гена, локусы (локус), определенные из анализа сцепления, экзоны, интроны, информационные РНК (мРНК), транспортные РНК (тРНК), рибосомные РНК (рРНК), малые интерферирующие РНК (миРНК), короткие шпилечные РНК (кшРНК), микроРНК, рибозимы, кДНК, рекомбинантные полинуклеотиды, разветвленные полинуклеотиды, плазмиды, векторы, выделенная ДНК любой последовательности, выделенная РНК любой последовательности, зонды нуклеиновых кислот и праймеры. Этот термин также охватывает структуры, подобные нуклеиновой кислоте, с синтетическими скелетами, см., например, Eckstein, 1991; Baserga et al., 1992; Milligan, 1993; WO 97/03211; WO 96/39154; Mata, 1997; Strauss-Soukup, 1997; and Samstag, 1996. Полинуклеотид может содержать один или более модифицированных нуклеотидов, таких как метилированные нуклеотиды и аналоги нуклеотидов. При их наличии, модификации нуклеотидной структуры могут происходить до или после сборки полимера. Последовательность нуклеотидов может быть прервана ненуклеотидными компонентами. Полинуклеотид может быть дополнительно модифицирован после полимеризации, например, путем конъюгации с меченым компонентом. Используемый в настоящем описании термин "дикий тип" является термином, понятным специалистам, и означает типичную форму организма, штамма, гена или характеристик, которая существует в природе, в отличие от мутантных или вариантных форм. "Дикий тип" может быть базовой линией. Как используют в настоящем описании, термин "вариант" следует понимать как проявление качеств, отличающихся от существующих в природе. Термины "не встречающиеся в природе" или "сконструированные способами инженерии" используются взаимозаменяемо и указывают на участие руки человека. Термины, относящиеся к молекулам нуклеиновой кислоты или полипептидам, означают, что молекула нуклеиновой кислоты или полипептид по меньшей мере по существу свободна от по меньшей мере одного другого компонента, с которым они естественно связаны в природе и существуют в природе. "Комплементарность" относится к способности нуклеиновой кислоты образовывать водородную связь (связи) с другой последовательностью нуклеиновой кислоты либо традиционным спариванием оснований Уотсона-Крика, либо другими нетрадиционными типами. Процентная комплементарность обозначает процентное содержание остатков в молекуле нуклеиновой кислоты, которые могут образовывать водородные связи (например, уотсон-криковское спаривание оснований) со второй последовательностью нуклеиновой кислоты (например, 5, 6, 7, 8, 9, 10 из 10 50%, 60%, 70%, 80%, 90% и 100% комплементарные). "Полностью комплементарные" означает, что все смежные остатки последовательности нуклеиновой кислоты связаны водородной связью с таким же количеством смежных остатков во второй последовательности нуклеиновой кислоты. "По существу комплементарные", как используется в настоящем описании, относится к степени комплементарности, которая составляет по меньшей мере 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99% или 100% в области 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 30, 35, 40, 45, 50 или более нуклеотидов или относится к двум нуклеиновым кислотам, которые гибридизуются в жестких условиях. Как используют в настоящем описании, термин "жесткий условия" для гибридизации относится к условиям, при которых нуклеиновая кислота, имеющая комплементарность с последовательностью-мишенью, преимущественно гибридизуется с последовательностью-мишенью и по существу не гибридизуется с последовательностями, не являющимися мишенями. Жесткие условия, как правило, зависят от последовательности и меняются в зависимости от ряда факторов. В общем, чем длиннее последовательность, тем выше температура, при которой последовательность специфически гибридизуется с ее целевой последовательностью. Неограничивающие примеры жестких условий подробно описаны в Tijssen (1993), Laboratory Techniques In Biochemistry And Molecular Biology-Hybridization With Nucleic Acid Probes Part I, Second Chapter "Overview of principles of hybridization and the strategy of nucleic acid probe assay", Elsevier, N.Y. Когда приводится полинуклеотидная последовательность, то также предусматриваются комплементарные или частично комплементарные последовательности. Они предпочтительно способны гибридизироваться с упоминаемой последовательностью в очень жестких условиях. Как правило, для максимизации скорости гибридизации выбирают условия относительно низкой гибридизации: примерно на 20-25° C ниже, чем термическая температура плавления (Tm). Tm представляет собой температуру, при которой 50% конкретной последовательности-мишени гибридизуется с идеально комплементарным зондом в растворе при определенной ионной силе и рН. Как правило, для того, чтобы требовать по меньшей мере ~85% нуклеотидной комплементарности гибридизованных последовательностей, выбирают очень жесткие условия промывки примерно на 5-15°C ниже, чем Tm. Чтобы требовать по меньшей мере приблизительно 70% нуклеотидной комплементарности гибридизованных последовательностей, выбирают умеренно жесткие условия промывки примерно на 15-30°C ниже, чем Tm. Условия высокой разрешимости (очень низкая жесткость) могут быть на 50°C ниже Tm, что обеспечивает высокий уровень ошибочного соответствия между гибридизованными последовательностями. Специалисты в данной области понимают, что другие физические и химические параметры на этапах гибридизации и промывки также могут быть изменены для влияния на результат детектируемого сигнала гибридизации с определенного уровня гомологии между последовательностями-мишенями и зондами. Предпочтительные очень жесткие условия включают инкубацию в 50% формамиде, 5 x SSC и 1% SDS при 42°C или инкубацию в 5×SSC и 1% SDS при 65°C с промывкой 0,2×SSC и 0,1% SDS при 65°C. "Гибридизация" относится к реакции, в которой один или более полинуклеотидов реагируют с образованием комплекса, который стабилизируется посредством водородной связи между основаниями нуклеотидных остатков. Водородное связывание может происходить по принципу спаривания оснований Уотсона-Крика, хугстиновского связывания или любым другим способом, специфичным для последовательности. Комплекс может содержать две цепи, образующие дуплексную структуру, три или более цепей, образующих многоцепочечный комплекс, одну цепь, которая гибридизируется сама с собой, или любую их комбинацию. Реакция гибридизации может представлять собой стадию в более широком процессе, таком как инициирование ПЦР или расщепление полинуклеотида ферментом. Последовательность, способная к гибридизации с данной последовательностью, называется "комплементарной" данной последовательности. Используемый в настоящем описании термин "геномный локус" или "локус" (множественное число: локусы) представляет собой специфическое местоположение гена или последовательности ДНК на хромосоме. "Ген" относится к участкам ДНК или РНК, которые кодируют полипептид или цепь РНК, которая играет функциональную роль в организме и, следовательно, является молекулярной единицей наследственности в живых организмах. Для целей настоящего изобретения можно считать, что гены включают области, которые регулируют продукцию гена, независимо от того, являются ли такие регулятивные последовательности "смежными" с кодирующими и/или транскрибируемыми последовательностями. Соответственно, ген включает, но не обязательно ограничивается ими, последовательности промоторов, терминаторы, последовательносты регуляторов трансляции, такими как сайты связывания рибосом и внутренние сайты входа рибосомы, энхансеры, сайленсеры, изоляторы, пограничные элементы, точки начала репликации, сайты прикрепления матрикса и области контроля локуса. Как используют в настоящем описании, термин "экспрессия геномного локуса" или "экспрессия гена" представляет собой процесс, посредством которого информация, заключенная в гене, используется для синтеза функционального генного продукта. Продукты экспрессии генов часто являются белками, но в генах, не содержащих белок, таких как гены рРНК или гены тРНК, продукт представляет собой функциональную РНК. Процесс экспрессии генов используется всеми известными живыми организмами - эукариотами (включая многоклеточные организмы), прокариотами (бактериями и археями) и вирусами для выработки функциональных продуктов для выживания. Как используют в настоящем описании, термин "экспрессия" гена или нуклеиновой кислоты охватывает не только экспрессию клеточного гена, но также транскрипцию и трансляцию нуклеиновой кислоты (кислот) в системах клонирования и в любом другом контексте. Как используют в настоящем описании, термин "экспрессия" также относится к способу, посредством которого полинуклеотид транскрибируется из ДНК-матрицы (например, в мРНК или другой транскрипт РНК), и/или к процессу, посредством которого транскрибированная мРНК затем транслируется в пептиды, полипептиды или белки. Транскрипты и кодированные полипептиды могут в совокупности обозначаться как "генный продукт". Если полинуклеотид получен из геномной ДНК, экспрессия может включать сплайсинг мРНК в эукариотической клетке. Термины "полипептид", "пептид" и "белок" используют в настоящем описании взаимозаменяемо для обозначения полимеров аминокислот любой длины. Полимер может быть линейным или разветвленным, он может содержать модифицированные аминокислоты и может быть прерван неаминокислотами. Термины также включают модифицированный аминокислотный полимер; например, образование дисульфидной связи, гликозилирование, липидацию, ацетилирование, фосфорилирование или любые другие манипуляции, такие как конъюгация с меченым компонентом. Как используют в настоящем описании, термин "аминокислота" включает природные и/или неприродные или синтетические аминокислоты, включая глицин и оба оптических изомера D или L, а также аминокислотные аналоги и пептидомиметики. Как используют в настоящем описании, термин "домен" или "белковый домен" относится к части белковой последовательности, которая может существовать и функционировать независимо от остальной белковой цепи. Как описано для аспектов данного изобретения, идентичность последовательности связана с гомологией последовательности. Сравнения гомологии могут проводиться на глаз или, чаще всего, с помощью легкодоступных программ сравнения последовательностей. Эти коммерчески доступные компьютерные программы могут вычислять процентную (%) гомологию между двумя или более последовательностями и также могут вычислять идентичность последовательностей, которой обладают две или более последовательностей аминокислот или нуклеиновых кислот.
[00787] В определенных вариантах изобретения термин "направляющая РНК" относится к полинуклеотидной последовательности, содержащей одну или более предполагаемых или идентифицированных tracr-последовательностей и предполагаемых или идентифицированных последовательностей cr-РНК или направляющей последовательности. В конкретных вариантах осуществления "направляющая РНК" включает предполагаемую или идентифицированную последовательность cr-РНК или направляющую последовательность. В других вариантах осуществления направляющая РНК не включает предполагаемую или идентифицированную tracr-последовательность.
[00788] Как используют в настоящем описании, термин "дикий тип" является термином, понятным квалифицированным специалистам, и означает типичную форму организма, штамма, гена или характеристики в том виде, как они встречаются в природе, в отличие от мутантных или вариантных форм. "Дикий тип" может быть базовой линией.
[00789] Как используют в настоящем описании, термин "вариант" следует понимать как проявление свойств, которые имеют характерные черты, по которым эти свойства отличаются от природных свойств.
[00790] Термины "неприродный" или "сконструированный способами инженерии" используются взаимозаменяемо и указывают на вмешательство человека. Эти термины, относящиеся к молекулам нуклеиновой кислоты или полипептидам, означают, что молекула нуклеиновой кислоты или полипептид по меньшей мере в некоторой степени лишены одного другого компонента, с которым они естественным образом ассоциированы в своем природном виде. Во всех аспектах и вариантах осуществления, независимо от того, включают ли они эти термины или нет, следует понимать, что использование этих терминов необязательно и, следовательно, не существует предпочтения по их включению. Кроме того, термины "неприродный" или "полученный способами инженерии" могут использоваться взаимозаменяемо и поэтому могут применяться по отдельности или вместе, и то или другое может заменить упоминание обоих вместе. В частности, "полученный способами инженерии" предпочтительнее вместо "неприродный" или "неприродный и/или полученный способами инженерии".
[00791] Гомология последовательностей может быть определена любой из ряда компьютерных программ, известных в данной области, например BLAST или FASTA et al. Подходящей для выполнения такого выравнивания компьютерной программой является пакет GCG Wisconsin Bestfit (Университет Висконсина, США, University of Wisconsin, U.S.A; Devereux et al., 1984, Nucleic Acids Research 12:387). Примеры другого программного обеспечения, способного выполнять сравнения последовательностей, включают, без наложения ограничений, пакет BLAST (см. Ausubel et al., 1999 ibid - Chapter 18), FASTA (Atschul et al., 1990, J. Mol. Biol., 403-410) и набор инструментов сравнения GENEWORKS. Как BLAST, так и FASTA доступны для поиска оффлайн и онлайн (см. Ausubel et al., 1999 ibid, pages 7-58 to 7-60). При этом предпочтительно использование программы GCG Bestfit. Процент (%) гомологии последовательно может быть рассчитан для перекрывающихся последовательностей, т.е. одна последовательность выровнена против другой и аминокислота или нуклеотид одной последовательности напрямую сравнивается с соответствующей другой аминокислотой или нуклеотидом другой последовательности по одной(му) за раз. В этом случае его называют "выравнивание без пропусков". Обычно такое выравнивание без пропусков выполнятся только для сравнительно небольшого числа остатков. Хотя это очень простой и логичный способ, он не способен рассматривать, к примеру, случаи идентичных в других отношениях пары последовательностей, где одна инсерция или делеция может привести к тому, что последующий аминокислотный остаток окажется вне рассмотрения выравнивания, тем самым приводя к возможному большому уменьшению % гомологии при выполнении глобального выравнивания. В результате большинство способов сравнения последовательностей разработаны для проведения оптимального выравнивания с учетом возможных инсерций и делеций без необоснованного наложения штрафа общей гомологии или балла идентичности. Это достигается путем вставления "пропусков" в выравнивании последовательности с целью максимизации локальной гомологии или идентичности. Однако такие более сложные способы налагают "штраф за внесение пропуска" для каждого пропуска, возникающего в выравнивании, так что для того же числа идентичных аминокислот возможно добавление нескольких пропусков - что отражает более близкое родство между двумя сравниваемыми последовательностями - и позволяет достичь более высокого балла, чем в случае одной со многими пропусками. "Цена сродства пропусков" обычно используется для наложения сравнительно большого штрафа за присутствие пропуска и меньшее - за каждый последующий остаток в пропуске. Это наиболее часто используемая система оценки пропусков. Высокие штрафы за пропуски могут, разумеется, давать в результате оптимизированные выравнивания с меньшим числом пропусков. Большинство программ выравнивания допускают внесение изменений в штрафы за пропуски. Однако предпочтительно использование исходных значений при использовании такого программного обеспечения для сравнения последовательностей. Например, при использовании пакета GCG Wisconsin Bestfit исходный штраф за пропуск в аминокислотных последовательностях равен -12 за каждый пропуск и -4 за каждую вставку. Расчет максимального % гомологии, таким образом, сначала требует проведения оптимального выравнивания с рассмотрением штрафов за пропуски. Подходящая компьютерная программа для проведения такого выравнивая - пакет GCG Wisconsin Bestfit (Devereux et al., 1984 Niic. Acids Research 12 p387). Примерами другого программного обеспечения, способного выполнять сравнение последовательностей являются, но не ограничиваются ими, пакеты BLAST (см. Ausubel et al., 1999 Short Protocols in Molecular Biology, 4to Ed. - Chapter 18), FASTA (Altschul et al., 1990 J. Mol Biol. 403-410) и набор инструментов сравнения GENEWORKS. Как BLAST, так и FASTA доступны для поиска оффлайн и онлайн (см. Ausubel et al., 1999 ibid, pages 7-58 to 7-60). Однако для решения некоторых задач предпочтительно использование программы GCG Bestfit. Новый инструмент, называющийся BLAST 2 Sequences, также используется для сравнения белковых и нуклеотидных последовательностей (см. FEMS Microbiol Lett. 1999 174(2): 247-50; FEMS Microbiol Lett. 1999 177(1): 187-8 и веб-сайт Национального Центра Биотехнологии на веб-сайте Национального Института Здравоохранения США). Хотя конечный % гомологии может быть измерен в смысле идентичности, процесс проведения выравнивания сам по себе не основан на сравнении "все-или-ничего" пар. Вместо этого обычно используется масштабированная матрица оценки сходства для расчета баллов для каждого попарного сравнения на основе химического сходства или эволюционного расстояния. Примером такой часто используемой матрицы является матрица BLOSUM62 - матрица, используемая по умолчанию для набора программ BLAST. Программы GCG Wisconsin обычно используют либо общедоступные значения по умолчанию, либо пользовательскую таблицу сравнения символов при ее наличии (см. руководство пользователя для дальнейших деталей). Для решения некоторых задач предпочтительно использование общедоступных значений по умолчанию пакета GCG или, в случае другого программного обеспечения, матрицы по умолчанию, такой как BLOSUM62. Альтернативно этому, процент гомологии может быть рассчитан с использованием свойства множественного выравнивания из DNASIS™ (Hitachi Software), основанного на алгоритме, аналогичном CLUSTAL (Higgins DG & Sharp PIVI (1988), Gene 73(1), 237-244). После того как программное обеспечение нашло оптимальное выравнивание, становится возможным рассчитать % гомологии, предпочтительно % идентичности последовательности. Данное программное обеспечение обычно производит это в ходе сравнения последовательностей и приводит численный результат. Последовательности могут также иметь делеции, инсерции или замены аминокислотных остатков, что приводит к изменению без уведомления и получению в результате функционально эквивалентных состояний. Преднамеренные замены аминокислот могут быть сделаны на основе сходства в свойствах аминокислот (таких как полярность, заряд, растворимость, гидрофобность, гидрофильность и/или амфипатическая природа остатков) и, как следствие, полезны для объединения аминокислот в функциональные группы. Аминокислоты могут быть сгруппированы на основе свойств только их боковых цепей. Однако более целесообразно также включать данные о мутациях. Наборы аминокислот, полученные таким образом, с большой вероятностью являются консервативными в связи с их структурой. Такие наборы могут быть описаны в форме диаграмм Венна (Livingstone C.D. and Barton G.J. (1993) "Protein sequence alignments: a strategy for the hierarchical analysis of residue conservation" Comput. Appl. Biosci. 9: 745-756) (Taylor W.R. (1986) "The classification of amino acid conservation" J. Theor. Biol 119, 205-218). Могут быть произведены консервативные замены, например в соответствии с таблицей ниже, которая описывает общепринятую группировку аминокислот в виде диаграммы Венна.
[792] Термины "субъект", "индивид" и "пациент" используются в настоящем описании взаимозаменяемо для обозначения позвоночного, предпочтительно млекопитающего, более предпочтительно человека. Млекопитающие включают, но не ограничиваются ими, мышей, обезьян, людей, сельскохозяйственных животных, спортивных животных и домашних животных. Также охватываются ткани, клетки и их потомки, полученные от биологического объекта in vivo или культивируемые in vitro.
[00793] Термины "терапевтический агент", "терапевтически способный агент" или "лечебный агент" используются взаимозаменяемо и относятся к молекуле или соединению, которые придают какой-либо полезный эффект при введении индивиду. Полезный эффект включает обеспечение диагностических определений; облегчение протекания заболевания, симптома, расстройства или патологического состояния; уменьшение или предотвращение возникновения заболевания, симптома, расстройства или состояния; и в целом противодействие заболеванию, симптому, расстройству или патологическому состоянию.
[00794] Как используют в настоящем описании, термины "лечение" или "облегчение" или "улучшение" используются взаимозаменяемо. Эти термины относятся к подходу для получения полезных или желаемых результатов, включая, но не ограничиваясь ими, терапевтический эффект и/или профилактическую пользу. Под терапевтическим эффектом понимается любое терапевтически значимое улучшение или воздействие на одно или более заболеваний, состояний или симптомов, подлежащих лечению. Для профилактической пользы конструкции могут вводиться индивиду, подверженному риску развития конкретного заболевания, состояния или симптома, или индивиду, демонстрирующему один или более физиологических симптомов заболевания, хотя болезнь, состояние или симптом, возможно, еще не проявились.
[00795] Термин "эффективное количество" или "терапевтически эффективное количество" относится к количеству агента, достаточному для получения полезных или желаемых результатов. Терапевтически эффективное количество может варьироваться в зависимости от одного или более факторов: индивид и состояние заболевания, подвергаемого лечению, вес и возраст индивида, тяжесть заболевания, способ введения и т.п., которые могут быть легко определены специалистом в данной области. Этот термин также относится к дозе, которая обеспечит визуализацию для обнаружения любым из способов визуализации, описанных в настоящем описании. Конкретная доза может варьироваться в зависимости от одного или более факторов: выбранный конкретный агент, режим дозирования, который следует соблюдать, введение комбинации с другими соединениями, время введения, подлежащая визуализации ткань и физическая система доставки.
[00796] В практике настоящего изобретения используются, если не указано иначе, общепринятые способы иммунологии, биохимии, химии, молекулярной биологии, микробиологии, клеточной биологии, геномики и рекомбинантных ДНК, которые входят в компетенции специалистов. См. Sambrook, Fritsch and Maniatis, MOLECULAR CLONING: A LABORATORY MANUAL, 2nd edition (1989); CURRENT PROTOCOLS IN MOLECULAR BIOLOGY (F, M. Ausubel, et al. eds., (1987)); the series METHODS IN ENZYMOLOGY (Academic Press, Inc.): PCR 2: A PRACTICAL APPROACH (M.J. MacPherson, B.D. Flames and G.R. Taylor eds. (1995)), Harlow and Lane, eds. (1988) ANTIBODIES, A LABORATORY MANUAL, and ANIMAL CELL CULTURE (R.I. Freshney, ed. (1987)).
[00797] Несколько аспектов изобретения относятся к векторным системам, содержащим один или более векторов, или к векторам как таковым. Векторы могут быть разработаны для экспрессии транскриптов CRISPR (например, транскриптов нуклеиновых кислот, белков или ферментов) в прокариотических или эукариотических клетках. Например, транскрипты CRISPR могут быть экспрессированы в бактериальных клетках, таких как Escherichia coli, клетках насекомых (с использованием экспрессирующих векторов бакуловируса), дрожжевых клетках или клетках млекопитающих. Подходящие клетки-хозяева обсуждаются далее в Goeddel, GENE EXPRESSION TECHNOLOGY: METHODS IN ENZYMOLOGY 185, Academic Press, San Diego, Calif. (1990). Альтернативно, рекомбинантный экспрессирующий вектор можно транскрибировать и транслировать in vitro, например, с использованием регуляторных последовательностей промотора T7 и полимеразы T7.
[00798] Варианты осуществления изобретения включают последовательности (как полинуклеотидные, так и полипептидные), которые могут содержать гомологичную замену ("замещение" используются в настоящем описании для обозначения обмена существующего аминокислотного остатка или нуклеотида с альтернативным остатком или нуклеотидом), которая может происходить, например, для подобной замены в случае аминокислот, таких как основные для основных, кислотные для кислотных, полярные для полярных и т. д. Негомологичное замещение может также происходить, то есть из одного класса остатков в другой или, в качестве альтернативы, включение неестественных аминокислот, таких как орнитин (далее обозначаемый как Z), орнитин диаминомасляной кислоты (далее называемый B), норлейцин орнитин (далее называемый O), пироариланин, тиенилаланин, нафтилаланин и фенилглицин. Вариантные аминокислотные последовательности могут включать подходящие спейсерные группы, которые могут быть вставлены между любыми двумя аминокислотными остатками последовательности, включая алкильные группы, такие как метил-, этил- или пропильные группы, в дополнение к аминокислотным спейсерам, таким как остатки глицина или β-аланина. Специалисты в данной области могут хорошо понимать дальнейшую форму вариации, которая включает присутствие одного или более аминокислотных остатков в пептоидной форме. Во избежание сомнений, "пептоидная форма" используется для обозначения вариантов аминокислотных остатков, в которых α-углеродная заместительная группа находится на атоме азота остатка, а не на α-углероде. Способы получения пептидов в пептоидной форме известны в данной области, например, Simon RJ et al., PNAS (1992) 89 (20), 9367-9371 и Horwell DC, Trends Biotechnol. (1995) 13 (4), 132-134.
[00799] Моделирование гомологии: соответствующие остатки в других ортологах C2c2 могут быть идентифицированы с помощью способов Zhang et al., 2012 (Nature, 490 (7421): 556-60) и Chen et al., 2015 (PLoS Comput Biol; 11 (5): e1004248) - способ вычисления белок-белкового взаимодействия (PP1) для прогнозирования взаимодействий, опосредованных поверхностями контакта домен-мотив. PrePPI (Predicting PPI), основанный на структуре способ прогнозирования PPI, объединяет структурные доказательства с неструктурными данными с использованием байесовской статистической модели. Способ включает взятие пары белков запроса и использование структурного выравнивания для идентификации структурных представителей, соответствующим либо их экспериментально определенным структурам, либо гомологичным моделям. Структурное выравнивание далее используется для идентификации как близких, так и отдаленных структурных соседей путем рассмотрения глобальных и локальных геометрических отношений. Всякий раз, когда два соседа структурных представителей образуют комплекс, указанный в Protein Data Bank, это определяет шаблон для моделирования взаимодействия между двумя белками запросов. Модели комплекса создаются путем наложения репрезентативных структур на соответствующего структурного соседа в шаблоне. Этот подход далее описан в Dey et al, 2013 (Prot Sci, 22: 359-66).
[00800] Для целей настоящего изобретения амплификация означает любой способ, использующий праймер и полимеразу, способную реплицировать последовательность-мишень с разумной точностью. Амплификацию можно проводить с помощью природных или рекомбинантных ДНК-полимераз, таких как ДНК-полимераза TaqGold™, T7, фрагмент Кленова ДНК-полимеразы E.coli и обратная транскриптаза. Предпочтительным способом амплификации является ПЦР.
[00801] В некоторых вариантах изобретение включает векторы. Как используют в настоящем описании, термин "вектор" представляет собой инструмент, который позволяет или облегчает передачу объекта из одной среды в другую. Это репликон, такой как плазмида, фаг или космида, в которые может быть вставлен другой сегмент ДНК, чтобы вызвать репликацию вставленного сегмента. Как правило, вектор способен к репликации, когда он связан с соответствующими элементами управления. В общем, термин "вектор" относится к молекуле нуклеиновой кислоты, способной транспортировать другую нуклеиновую кислоту, с которой она была связана. Векторы включают, но не ограничиваются, молекулами нуклеиновых кислот, которые являются однонитевыми, двухцепочечными или частично двухцепочечными; молекулами нуклеиновых кислот, которые содержат один или более свободных концов, или не содержат свободных концов (например, круглые); молекулы нуклеиновых кислот, которые содержат ДНК, РНК или и то, и другое; и другие разновидности полинуклеотидов, известных в данной области. Один тип вектора представляет собой "плазмиду", которая относится к круглой двухцепочечной петле ДНК, в которую могут быть добавлены дополнительные сегменты ДНК стандартными способами молекулярного клонирования. Другим типом вектора является вирусный вектор, в котором в векторе для упаковки в вирус присутствуют вирусные ДНК или РНК (например, ретровирусы, ретровирусы с дефектами репликации, аденовирусы, аденовирусы с дефектами репликации и аденоассоциированные вирусы (AAV)). Вирусные векторы также включают полинуклеотиды, переносимые вирусом для трансфекции в клетку-хозяина. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую они вводятся (например, бактериальные векторы, имеющие бактериальную точку начала репликации и эписомальные векторы млекопитающих). Другие векторы (например, неэписомальные векторы млекопитающих) интегрируются в геном клетки-хозяина, после введения в клетку-хозяина и, таким образом, реплицируются вместе с геномом хозяина. Более того, некоторые векторы способны направлять экспрессию генов, с которыми они функционально связаны. Такие векторы упоминаются в настоящем описании как "экспрессирующие векторы". Обычные экспрессирующие векторы, использующиеся в способах рекомбинантной ДНК, часто имеют форму плазмид.
[00802] Рекомбинантные экспрессирующие векторы могут содержать нуклеиновые кислоты по изобретению в форме, подходящей для экспрессии нуклеиновых кислот в клетке-хозяине, что означает, что рекомбинантные экспрессирующие векторы включают один или более регуляторных элементов, которые могут быть выбраны на основе клеток-хозяев, используемых для экспрессии, и которые функционально связаны с последовательностью нуклеиновых кислот, которая должна быть экспрессирована. Внутри рекомбинантного экспрессирующего вектора "функционально связанный" означает, что целевая нуклеотидная последовательность связана с регуляторным элементом(ами) таким образом, который позволяет экспрессию нуклеотидной последовательности (например, в системе in vitro транскрипции/трансляции или в клетке-хозяине, когда вектор вводится в клетку-хозяина). Что касается способов рекомбинации и клонирования, может быть упомянута заявка на патент США 10/815730, опубликованная 2 сентября 2004 года в качестве US 2004-0171156 A1, содержание которой включено в настоящее описание в качестве ссылки в полом объеме.
[00803] Аспекты изобретения относятся к бицистронным векторам для направляющей РНК и РНК дикого типа, модифицированных или мутированных эффекторных белков/ферментов CRISPR (например, C2c2). Бицистронные экспрессирующие векторы направляющей РНК и РНК дикого типа, модифицированных или мутированных эффекторных белков/ферментов CRISPR (например, C2c1 или C2c3). В целом и, в частности, в этом варианте осуществления и диком типе, модифицированные или мутированные эффекторные белки/ферменты CRISPR (например, C2c1 или C2c3) предпочтительно находятся под контролем промотора CBh. РНК может предпочтительно находиться под контролем промотора РНК-полимеразы III, такого как промотор U6. В идеале эти два промотора сочетаются.
[00804] В некоторых вариантах осуществления предусмотрена петля в направляющей РНК. Это может быть шпилька или тетрапетля. Петля предпочтительно представляет собой GAAA, но не ограничивается этой последовательностью или даже длиной 4 п.о. Действительно, предпочтительные последовательности формирования петли для использования в структурах шпильки имеют длину четыре нуклеотида и наиболее предпочтительно имеют последовательность GAAA. Однако могут использоваться более длинные или более короткие последовательности петель, а также альтернативные последовательности. Последовательности предпочтительно включают нуклеотидный триплет (например, AAA) и дополнительный нуклеотид (например, C или G). Примеры последовательностей, формирующих петли, включают CAAA и AAAG.
[00805] При осуществления любого из способов, описанных в настоящем описании, подходящий вектор может быть введен в клетку или эмбрион посредством одного или более способов, известных в данной области, включая, но не ограничиваясь ими, микроинъекцию, электропорацию, сонопорацию, биолистику, опосредованную фосфатом кальция трансфекцию, катионную трансфекцию, трансфекцию липосом, трансфекцию дендримера, трансфекцию теплового шока, нуклеофекцию, магнитотрансфекцию, липофекцию, импалефекцию, оптическую трансфекцию, запатентованное усиленное агентом поглощение нуклеиновых кислот, доставку посредством липосом, иммунолипосом, виросом или искусственных вирионов. В некоторых способах вектор вводится в эмбрион путем микроинъекции. Вектор или векторы могут быть введены посредством микроинъекции в ядро или цитоплазму эмбриона. В некоторых способах вектор или векторы могут быть введены в клетку путем нуклеофекции.
[00806] Термин "регуляторный элемент" используется для обозначения промоторов, энхансеров, участков внутренней посадки рибосомы (IRES) и других элементов управления экспрессией (например, сигналов терминации транскрипции, таких как сигналы полиаденилирования и последовательности поли-U). Такие регуляторные элементы описаны, например, в Goeddel, GENE EXPRESSION TECHNOLOGY: METHODS IN ENZYMOLOGY 185, Academic Press, San Diego, Calif. (1990). Регуляторы включают как те, которые напрямую определяют конститутивную экспрессию нуклеотидной последовательности во многих типах клеток-хозяев, так и те, которые управляют экспрессией нуклеотидной последовательности только в определенных клетках-хозяевах (например, тканеспецифические регуляторные последовательности). Тканеспецифический промотор может управлять экспрессией в основном в желаемой интересующей ткани, такой как мышца, нейрон, кость, кожа, кровь, специфические органы (например, печень, поджелудочная железа) или конкретные типы клеток (например, лимфоциты). Регуляторные элементы также могут управлять экспрессией в зависимости от времени, например, в зависимости от стадии клеточного цикла или стадии развития, и могут быть или не быть специфичным для тканей или клеток. В некоторых вариантах осуществления вектор содержит один или более промоторов ДНК-полимеразы III (например, 1, 2, 3, 4, 5 или более промоторов ДНК-полимеразы III), один или более промоторов ДНК-полимеразы II (например, 1, 2, 3, 4, 5 или более промоторов ДНК-полимеразы II), один или более промоторов ДНК-полимеразы I (например, 1, 2, 3, 4, 5 или более промоторов ДНК-полимеразы I) или их комбинации. Примеры промоторов ДНК-полимеразы III включают, но не ограничиваются ими, промоторы U6 и HI. Примеры промоторов ДНК-полимеразы II включают, но не ограничиваются ими, ретровирусный промотор LTR RAR-саркомы Рауса (RSV) (необязательно с энхансером RSV), промотор цитомегаловируса (CMV) (необязательно с энхансером CMV) [см., например, Boshart et al., Ceil, 41: 521-530 (1985)], промотор SV40, промотор дигидрофолатредуктазы, промотор 3-актина, промотор фосфоглицерин-киназы (PGK) и промотор EF1a. Кроме того, термин "регуляторный элемент" охватывает энхансерные элементы, такие как WPRE; энхансеры CMV; сегмент R-U5' в LTR HTLV-I (Mol, Cell. Biol, том 8 (1), стр. 466-472, 1988); энхансер SV40; и интронная последовательность между экзонами 2 и 3 белка кролика P-глобина (Proc. Natl. Acad. Sci. USA., т. 78 (3), p, 1527-31, 1981). Специалистам в данной области будет понятно, что конструкция вектора экспрессии может зависеть от таких факторов, как выбор клетки-хозяина, которая должна быть трансфицирована, а также желаемый уровень экспрессии и т.д. Вектор может быть введен в клетки таким образом, чтобы они продуцировали транскрипты, белки или пептиды, включая слитые белки или пептиды, кодируемые нуклеиновыми кислотами, как описано в настоящем описании (например, транскрипты системы коротких палиндромных повторов, регулярно расположенных кластерами (CRISPR), их белки, ферменты, их мутантные формы, их слитые белки и т.д.). Что касается регуляторных последовательностей, то может быть упомянута заявка на патент США 10/491026, содержание которой включено в настоящее описание в качестве ссылки в полном объеме. Что касается промоторов, упоминается публикация РСТ WO 2011/028929 и Приложение США 12/511940, содержание которых включено в настоящее в качестве ссылки в полном объеме.
[00807] Векторы могут быть сконструированы для экспрессии транскриптов CRISPR (например, транскриптов нуклеиновых кислот, белков или ферментов) в прокариотических или эукариотических клетках. Например, транскрипты CRISPR могут быть экспрессированы в бактериальных клетках, таких как Escherichia coli, клетках насекомых (с использованием экспрессирующих векторов бакуловируса), дрожжевых клетках или клетках млекопитающих. Подходящие клетки обсуждаются далее в Goeddel, GENE EXPRESSION TECHNOLOGY: METHODS IN ENZYMOLOGY185, Academic Press, San Diego, Calif. (1990). Альтернативно, рекомбинантный экспрессирующий вектор можно транскрибировать и транслировать in vitro, например, с использованием регуляторных последовательностей промотора T7 и полимеразы T7.
[00808] Векторы могут введены и увеличены в количестве в прокариотических организмах или прокариотических клетках. В некоторых вариантах осуществления прокариотический организм используют для амплификации копий вектора, который должен быть введен в эукариотическую клетку или в качестве промежуточного вектора при образовании вектора, который должен быть введен в эукариотическую клетку (например, амплификация плазмиды как части вирусной векторная упаковочная система). В некоторых вариантах осуществления прокариотический организм используется для амплификации копий вектора и экспрессии одной или более нуклеиновых кислот, например, для обеспечения источника одного или более белков для доставки в клетку-хозяина или организм-хозяин. Экспрессия белков в прокариотах чаще всего происходит в Escherichia coli с векторами, содержащими конститутивные или индуцируемые промоторы, которые направляют экспрессию либо слитых, либо неслитых белков. Слитые векторы встраивают ряд аминокислот в кодируемый им белок, например, на N-конце рекомбинантного белка. Такие слитые векторы могут служить одной или более целям, таким как: (i) увеличение экспрессии рекомбинантного белка; (ii) увеличение растворимости рекомбинантного белка; и (iii) содействие очистке рекомбинантного белка, действуя как лиганд в аффинной очистке. Часто в слитых экспрессирующих векторах вводится сайт протеолитического расщепления на стыке области слияния и рекомбинантного белка, чтобы обеспечить отделение рекомбинантного белка от слитого фрагмента после очистки слитого белка. Такие ферменты и их родственные последовательности распознавания включают фактор Ха, тромбин и энтерокиназу. Примеры экспрессирующих векторов слияния включают pGEX (Pharmacia Biotech Inc, Smith and Johnson, 1988. Gene 67: 31-40), pMAL (New England Biolabs, Beverly, Mass.) И pRIT5 (Pharmacia, Piscataway, NJ), которые производят слияние глютатион S-трансферазы (GST), связывающего мальтозу белка E или белка A, соответственно, с целевым рекомбинантным белком.
[00809] Примеры подходящих индуцибельных неслитых экспрессирующих векторов E.coli включают pTrc (Amrann et al., (1988) Gene 69: 301-315) и pET lid (Studier et al., GENE EXPRESSION TECHNOLOGY: METHODS IN ENZYMOLOGY 185, Academic Press, Сан-Диего, Калифорния (1990) 60-89).
[00810] В некоторых вариантах осуществления вектор представляет собой экспрессирующий вектор дрожжей. Примеры экспрессирующих векторов в дрожжах Saccharomyces cerivisae включают pYepSeel (Baldari, et al., 1987. EMBO J. 6: 229-234), pMFa (Kuijan and Herskowitz, 1982. Cell 30: 933-943), pJRY88 (Schultz et al., 1987. Gene 54: 113-123), pYES2 (Invitrogen Corporation, San Diego, CA.) и picZ (InVitrogen Corp, Сан-Диего, Калифорния).
[00811] В некоторых вариантах осуществления вектор управляет экспрессией белка в клетках насекомых с использованием экспрессирующих векторов бакуловируса. Бакуловирусные векторы, доступные для экспрессии белков в культивируемых клетках насекомых (например, клетки SF9), включают серию pAc (Smith, et al., 1983. Mol. Cell. Biol., 3: 2156-2165) и серию pVL (Lucklow and Summers, 1989. Virology 170: 31-39).
[00812] В некоторых вариантах осуществления вектор способен приводить к экспрессии одной или более последовательностей в клетках млекопитающих с использованием вектора экспрессии в млекопитающих. Примеры экспрессирующих векторов млекопитающих включают pCDM8 (Seed, 1987. Nature 329: 840) и pMT2PC (Kaufman, et al., 1987. EMBO J. 6: 187-195). При использовании в клетках млекопитающих функции управления вектором экспрессии обычно выполняются одним или более регуляторными элементами. Например, обычно используемые промоторы получают из полиомы, аденовируса 2, цитомегаловируса, вируса SV40 и других, описанных в настоящем описании и известных в данной области. Другие подходящие системы экспрессии как для прокариотических, так и для эукариотических клеток можно изучить по: см., например, главы 16 и 17 Sambrook et al. MOLECULAR CLONING: A LABORATORY MANUAL, 2nd ed., Laboratory Cold Spring Harbor, Laboratory Cold Spring Harbour Press, Cold Spring Harbour, NY, 1989.
[00813] В некоторых вариантах осуществления рекомбинантный экспрессирующий вектор млекопитающих способен направлять экспрессию нуклеиновых кислот предпочтительно в конкретном типе клеток (например, тканеспецифические регуляторные элементы используются для экспрессии нуклеиновой кислоты). Специфические для ткани регуляторные элементы известны в данной области. Не ограничивающие примеры подходящих тканеспецифических промоторов включают промотор альбумина (специфический для печени: Pinkert, et al., 1987. Genes Dev. 1: 268-277), лимфоидспецифические промоторы (Calarne and Eaton, 1988. Adv. Immunol 43: 235-275), в частности, промоторы рецепторов Т-клеток (Winoto and Baltimore, 1989. EMBO J. 8: 729-733) и иммуноглобулины (Baneiji, et al., 1983. Cell 33: 729-740; Queen and Baltimore, 1983. Cell 33: 741-748), нейроспецифические промоторы (например, промотор нейрофиламентов, Byrne and Ruddle, 1989. Proc. Natl. Acad. Sci. USA, 86: 5473-5477), специфические для поджелудочной железы промоторы (Edlund, et al., 1985. Science 230: 912-916) и промоторы молочной железы, специфические для молочной железы (например, промотор молочной сыворотки, патент США № 4873,316 и публикация Европейской заявки № 264,166). Регулируемые развитием организма промоторы также включают, например, hox-промоторы мыши (Kessel and Grass, 1990. Science 249: 374-379) и промотор α-фетопротеина (Campes and Tilghman, 1989. Гены Dev. 3: 537-546). Что касается этих прокариотических и эукариотических векторов, то может быть упомянут патент США 6750509, содержание которого включено в настоящее описание в качестве ссылки в полном объеме полноте. Другие варианты осуществления изобретения могут относиться к использованию вирусных векторов, в отношении которых упоминается заявка на патент США 13/1002085, содержание которой включено в настоящее описание в качестве ссылки в полном объеме. Специфичные для ткани регуляторные элементы известны в данной области, и в этой связи упоминается патент США 7776321, содержание которого включено в настоящее описание в качестве ссылки в полном объеме.
[00814] В некоторых вариантах осуществления регуляторный элемент функционально связан с одним или более элементами системы CRISPR, чтобы управлять экспрессией одного или более элементов системы CRISPR. В целом, CRISPR (короткие палиндромные повторы, регулярно расположенные кластерами), также известный как SPIDR (рассеянные между спейсерами прямые повторы), представляют собой семейство локусов ДНК, которые обычно специфичны для конкретных видов бактерий. Локус CRISPR содержит отчетливый класс чередующихся коротких повторов последовательности (SSR), которые были описаны в E. coli (Ishino et al., J. Bacteriol. 169: 5429-5433 [1987] и Nakata et al., J. Bacteriol., 171: 3553- 3556 [1989]) и ассоциированных генов. Аналогичные участки SSR были идентифицированы в Haloferax mediterranei, Streptococcus pyogenes, Anabaena и Mycobacterium tuberculosis (см. Groenen et al., Mol. Microbiol., 10: 1057-1065 [1993], Hoe et al., Emerg. Infect. Dis., 5: 254-263 [1999], Masepohl et al., Biochim Biophys Acta 1307: 26-30 [1996], Mojica et al., Mol. Microbiol., 17: 85-93 [1995]). Локусы CRISPR, как правило, отличаются от других SSR структурой повторов, которые называются короткими регулярно прерывающимися повторами (SRSR) (Janssen et al., OMICS J. Integ. Biol., 6: 23-33 [2002], и Mojica et al., Mol. Microbiol., 36: 244-246 [2000]). В общем случае, повторы представляют собой короткие элементы, которые образуют кластеры, которые регулярно распределены по уникальным перемежаемым последовательностям с примерно постоянной длиной (Mojica et al., [2000], см. выше). Хотя повторяющиеся последовательности очень консервативны для разных штаммов, количество прерывающихся повторов и последовательностей спейсерных областей обычно отличаются для разных штаммов (van Embden et al., J. Bacteriol, 182: 2393-2401 [2000]). Локусы CRISPR были идентифицированы более чем у 40 прокариотов (см., Например, Jansen et al., Mol. Microbiol., 43: 1565-1575 [2002] и Mojica et al., [2005]), включая, но не ограничиваясь, Aeropyrum, Pyrobacidum, Sulfolobus, Archaeoglobus, Halocarcula, Methanobacterium, Methanococcus, Methanosarcina, Methanopyrus, Pyrococcus, Picrophilus, Thermoplasma, Corynebacterium, Mycobacterium, Streptomyces, Aquifex, Porphyromonas, Chlorobium, Thermus, Bacillus, Listeria, Staphylococcus, Clostridium, Thermobacterium, Mycoplasma, Fusobacterium, Azarcus, Chromobacterium, Neisseria, Nitrosomonas, Desidfovibrio, Geobacter, Myxococcus, Campylobacter, Wolinella, Acinetobacter, Erwinia, Escherichia, Legionella, Methylococcus, Pasteurella, Photobacterium, Salmonella, Xanthomonas, Yersinia, Treponema и Thermotoga.
[00815] В целом понятие "система нацеливания на нуклеиновую кислоту", используемая в настоящем изобретении, относится в совокупности к транскриптам и другим элементам, участвующим в экспрессии или направляющим активность нацеливания на нуклеиновых кислоты CRISPR-ассоциированных ("Cas") генов (также называемых в настоящем описании эффекторными белками), включая последовательности, кодирующие белок (эффекторный), нацеленный на нуклеиновую кислоту, и направляющую РНК (содержащую последовательность cr-РНК и трансактивирующую последовательность РНК (tracr-РНК) системы CRISPR/Cas) или другие последовательности и транскрипты из локуса CRISPR, нацеленного на нуклеиновые кислоты. В некоторых вариантах осуществления один или более элементов системы нацеливания на нуклеиновую кислоту получены из системы CRISPR, нацеленной на нуклеиновые кислоты, типов V/VI. В некоторых вариантах осуществления один или более элементов системы нацеливания на нуклеиновую кислоту получают из конкретного организма, включающего эндогенную систему CRISPR, нацеленную на нуклеиновые кислоты. В целом система нацеливания на нуклеиновую кислоту характеризуется элементами, которые способствуют образованию комплекса, нацеленного на нуклеиновую кислоту, на участке последовательности-мишени. В контексте формирования комплекса, нацеленного на нуклеиновую кислоту, "последовательность-мишень" относится к последовательности, для которой предназначена направляющая последовательность, и которая имеет комплементарность, необходимую для гибридизации между последовательностью-мишенью и направляющей РНК, что способствует образованию комплекса с ДНК или РНК. Полная комплементарность не обязательна, если имеется достаточная комплементарность, вызывающая гибридизацию и способствующая образованию комплекса, нацеленного на нуклеиновую кислоту. Последовательность-мишень может содержать полинуклеотиды РНК. В некоторых вариантах осуществления последовательность-мишень находится в ядре или цитоплазме клетки. В некоторых вариантах осуществления последовательность-мишень может находиться внутри органелл эукариотической клетки, например митохондрий или хлоропластов. Последовательность или матрица, которая может быть использована для рекомбинации в локусе-мишени, содержащем последовательность-мишень, упоминается как "редактирующая матрица или "редактирующая РНК" или "редактирующая последовательность". В аспектах изобретения экзогенная РНК-матрица может упоминаться как матрица редактирования. В любом аспекте изобретения рекомбинация представляет собой гомологичную рекомбинацию.
[00816] Как правило, в контексте эндогенной системы нацеливания на нуклеиновую кислоту образование комплекса, нацеленного на нуклеиновую кислоту (содержащего направляющую РНК, гибридизованную с последовательностью-мишенью, в комплексе с одним или более эффекторными белками, нацеленными на нуклеиновую кислоту) приводит к расщеплению одной или обеих цепей РНК (примерно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50 или более пар оснований) целевой последовательности. В некоторых вариантах осуществления один или более векторов, управляющих экспрессией одного или более элементов системы нацеливания на нуклеиновую кислоту, вводят в клетку, так что экспрессия элементов системы нацеливания на нуклеиновую кислоту руководит образованием комплекса, нацеленного на нуклеиновую кислоту, в одном или более участках-мишенях. Например, эффекторный белок, нацеленный на нуклеиновую кислотe, и направляющая РНК могут быть функционально связаны с отдельными регуляторными элементами на отдельных векторах. Альтернативно, два или более элемента, экспрессируемые с участием одних и тех же или разных регуляторных элементов, могут быть объединены в одном векторе с одним или более дополнительными векторами, обеспечивающими другие компоненты системы нацеливания на нуклеиновую кислоту, не включенные в первый вектор. Компоненты системы нацеливания на нуклеиновую кислоту, объединенные в одном векторе, могут быть расположены в любой подходящей ориентации, такой как один элемент, расположенный в направлении 5'-конца относительно (в восходящем направлении) или 3'-конца относительно (в нисходящем направлении) второго элемента. Кодирующая последовательность одного элемента может быть расположена на той же или противоположной цепи кодирующей последовательности второго элемента и быть ориентирована в том же или противоположном направлении. В некоторых вариантах осуществления один промотор руководит экспрессией транскрипта, кодирующего эффекторный белок, нацеленный нуклеиновую кислоту, и направляющей РНК, встроенной в одну или более интронных последовательностей (например, каждая в отдельном интроне, два или более по меньшей мере в одном интроне или все в одном интроне). В некоторых вариантах осуществления изобретения эффекторный белок, нацеленный на нуклеиновую кислоту, и направляющая РНК функционально связаны и экспрессируются с одного и того же промотора.
[00817] В общем, направляющая последовательность представляет собой любую полинуклеотидную последовательность, имеющую достаточную комплементарность с полинуклеотидной последовательностью-мишенью для гибридизации с последовательностью-мишенью и прямого специфического для последовательности связывания комплекса, нацеленного на нуклеиновую кислоту, с последовательностью-мишенью. В некоторых вариантах осуществления степень комплементарности между направляющей последовательностью и соответствующей ей последовательностью-мишенью при оптимальном выравнивании с использованием подходящего алгоритма выравнивания составляет приблизительно или более чем 50%, 60%, 75%, 80%, 85%, 90%, 95%, 97,5%, 99% и более. Оптимальное выравнивание может быть определено с использованием любого подходящего алгоритма для выравнивания последовательностей, неограничивающие примеры которого, включают алгоритм Смита-Уотермана, алгоритм Нидлмана-Вунча, алгоритмы, основанные на преобразовании Берроуза-Уилера (например, Burrows Wheeler Aligner), ClustalW, Clustal X, BLAST, Novoalign (Novocraft Technologies, ELAND (iIllumina, Сан-Диего, Калифорния), SOAP (доступно на soap.genornics.org.cn) и Maq (доступно на maq.sourceforge.net). В некоторых вариантах осуществления, направляющая последовательность составляет приблизительно или больше, чем приблизительно 5, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 35, 40, 45, 50, 75 или более нуклеотидов в длину. В некоторых вариантах осуществления изобретения направляющая последовательность меньше чем приблизительно 75, 50, 45, 40, 35, 30, 25, 20, 15, 12 или менее нуклеотидов в длину. Способность направляющей последовательности управлять специфическим для последовательности связыванием нацеленного на нуклеиновую кислоту комплекса с последовательностью-мишенью может быть оценена любым подходящим способом. Например, компоненты нацеленной на нуклеиновую кислоту системы, достаточные для образования данного комплекса, включая направляющую последовательность, подлежащую тестированию, могут быть доставлены в клетку, имеющую соответствующую последовательность-мишень, например, с помощью трансфекции векторами, кодирующими компоненты нацеленной на нуклеиновые кислоты последовательности CRISPR с последующей оценкой предпочтительного расщепления в пределах или вблизи последовательности-мишени, такой как анализ Surveyor, описанный в настоящем описании. Аналогично, расщепление полинуклеотидной последовательности-мишени (или последовательности вблизи нее) может быть оценено в пробирке путем предоставления последовательности-мишени, компонентов нацеленного на нуклеиновую кислоту комплекса, включая направляющую последовательность, подлежащую тестированию, и контрольной направляющей последовательности, отличной от тестируемой направляющей последовательности, и сравнения связывания или скорости расщепления происходящего непосредственно в или вблизи последовательности-мишени между реакциями тестовой и контрольной направляющих последовательностей. Другие способы анализа также возможны и могут быть разработаны специалистами в данной области.
[00818] Направляющая последовательность может быть выбрана так, чтобы нацеливать на любую последовательность-мишень. В некоторых вариантах осуществления последовательность-мишень представляет собой последовательность внутри транскрипта гена или мРНК.
[00819] В некоторых вариантах осуществления последовательность-мишень представляет собой последовательность в геноме клетки.
[00820] В некоторых вариантах осуществления выбирается направляющая последовательность для компактизации вторичной структуры внутри направляющей последовательности. Вторичная структура может быть определена любым подходящим алгоритмом фолдинга полинуклеотидов. Некоторые программы основаны на вычислении минимальной свободной энергии Гиббса. Примером такого алгоритма является mFold, как описано Zuker и Stiegler (Nucleic Acids Res., 9 (1981), 133-148). Другим примером алгоритма выявления фолдинга является вебсервер RNAfold, разработанный в Институте теоретической химии Венского университета, с использованием алгоритма прогнозирования центроида (см., например, AR Gruber era, 2008, Cell 106 (1): 23-24; PA Carr and GM Church, 2009, Nature Biotechnology 27 (12): 1151-62). Другие алгоритмы могут быть найдены в заявке США серийного номера TBA (номер патентного реестра 44790.11.2022, широкая ссылка В1-2013/004A); включенной в настоящее описание в качестве ссылки.
[00821] В некоторых вариантах осуществления предусматривается матрица рекомбинации. Матрица рекомбинации может быть компонентом другого вектора, описанного в настоящем описании, может содержаться в отдельном векторе или предоставляться в виде отдельного полинуклеотида. В некоторых вариантах осуществления матрица рекомбинации разработана так, чтобы служить матрицей гомологичной рекомбинации рядом с последовательностью-мишенью или в пределах последовательности-мишени, которая расщепляется или в которую вносится ник-разрыв нацеленным на нуклеиновую кислоту эффекторным белком, будучи частью нацеленного на нуклеиновую кислоту комплекса. Матричный полинуклеотид может обладать любой подходящей длиной, составляющей приблизительно или превышающей 10, 15, 20, 25, 50, 75, 100, 150, 200, 500, 1000 или более нуклеотидов. В некоторых вариантах осуществления матричный полинуклеотид комплементарен части полинуклеотида, содержащего последовательность-мишень. При оптимальном выравнивании матричный полинуклеотид может перекрываться с одним или более нуклеотидами последовательности-мишени (например, приблизительно или более 1, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100 или более нуклеотидов). В некоторых вариантах осуществления при оптимальном выравнивании матричного полинуклеотида с полинуклеотидом, содержащим последовательность-мишень, ближайший нуклеотид матричной последовательности находится в пределах 1, 5, 10, 15, 20, 25, 50, 75, 100, 200, 300, 400, 500, 1000, 5000, 10000 или более нуклеотидов от последовательности-мишени.
[00822] В некоторых вариантах осуществления эффекторный белок, нацеленный на нуклеиновую кислоту, является частью слитого белка, содержащего один или более гетерологичных белковых доменов (например, приблизительно или более 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более доменов в дополнение к эффекторному белку, нацеленному на нуклеиновую кислоту). В некоторых вариантах осуществления эффекторный белок/фермент CRISPR является частью слитого белка, содержащего один или более гетерологичных белковых доменов (например, приблизительно или более 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, или более доменов в дополнение к ферменту CRISPR). Эффекторный белок/фермента CRISPR может содержать любую дополнительную белковую последовательность и необязательно линкерную последовательность между любыми двумя доменами. Примеры белковых доменов, которые могут быть слиты с эффекторным белком, включают, но не ограничиваются ими, метки эпитопов, последовательности репортерного гена и белковые домены, имеющие одну или более из следующих активностей: метилазная, деметилазная активность, активность активации транскрипции, активность подавления транскрипции, активность фактора терминации транскрипции, активность модификации гистонов, активность расщепления РНК и активность связывания нуклеиновых кислот. Неограничивающие примеры меток эпитопов включают метки гистидина (His), метки V5, метки FLAG, метки гемагглютинина (HA) гриппа, метки Мус, метки VSV-G и теги тиоредоксина (Trx). Примеры репортерных генов включают, но не ограничиваются следующими: глутатион-S-трансфераза (GST), пероксидаза хрена (HRP), бета-галактозидаза ацетитрансферазы хлорамфеникола (CAT), бета-глюкуронидаза, люцифераза, зеленый флуоресцентный белок (GFP), HcRed, DsRed, голубой флуоресцентный белок (CFP), желтый флуоресцентный белок (YFP) и аутофлуоресцентные белки, включая синий флуоресцентный белок (BFP). Эффекторный белок, нацеленный на нуклеиновую кислоту, может быть слит с последовательностью гена, кодирующей белок или фрагмент белка, который связывает молекулы ДНК или связывает другие клеточные молекулы, включая, но не ограничиваясь, следующими: мальтоза-связывающий белок (MBP), S-tag, слитые конструкции ДНК-связывающего домена (DBD) Lex A, слитые конструкции ДНК-связывающего домена GAL4 и слитые конструкции белка BP16 вируса простого герпеса (HSV). Дополнительные домены, которые могут быть частью слитого белка, включающего эффекторный белок, нацеленный на нуклеиновую кислоту, описаны в патентной публикации США 20110059502, включенной в настоящее описание в качестве ссылки. В некоторых вариантах осуществления меченый эффекторный белок, нацеленный на нуклеиновую кислоту, используется для идентификации местоположения последовательности-мишени.
[00823] В некоторых вариантах осуществления фермент CRISPR может являться компонентом индуцируемой системы. Индуцируемый характер системы обеспечивает пространственно-временный контроль над редактированием генов или экспрессией генов с использованием энергии. Форма энергии может включать, но не ограничивается ими, электромагнитное излучение, звуковую энергию, химическую энергию и тепловую энергию. Примеры индуцируемой системы включают индуцируемые тетрациклином промоторы (Tet-On или Tet-Off), двугибридные системы активации транскрипции с малой молекулой (FKBP, ABA и т. д.), светоиндуцируемые системы (фитохром, LOV-домены или криптохром). В варианте одном осуществления изобретения фермент CRISPR может быть частью светоиндуцируемого эффектора транскрипции (LITE) для непосредственного изменения транскрипционной активности, специфичного к последовательности. Светоиндуцируемые компоненты могут включать фермент CRISPR, светочувствительный гетеродимер цитохрома (например, Arabidopsis thaliana) и домен активации/подавления транскрипции. Другие примеры индуцируемых связывающих ДНК белков и способы их использования приведены в US 61/736465 и US 61/721283 и WO 2014/018423 и US8889418, US8895308, US20140186919, US20140242700, US20140273234, US20140335620, WO2014093635, которые включены в настоящее описание в качестве ссылок в полном объеме.
Доставка
[00824] В некоторых вариантах осуществления настоящее изобретение относится к способам, включающим доставку одного или более полинуклеотидов, таких как или один или более векторов, как описано в настоящем описании, один или более его транскриптов и/или один или более белков, транскрибируемых с них, в клетку. В некоторых аспектах изобретение, кроме того, относится к клеткам, полученным такими способами, и организмам (таким как животные, растения или грибы), содержащим или полученным из таких клеток. В некоторых вариантах осуществления эффекторный белок, нацеленный на нуклеиновую кислоту, в сочетании с (и, возможно, в составе комплекса) с направляющей РНК доставляется в клетку. Обычные вирусные и невирусные способы переноса генов могут быть использованы для введения нуклеиновых кислот в клетки млекопитающих или в целевые ткани. Такие способы могут быть использованы для введения нуклеиновых кислот, кодирующих компоненты системы нацеливания на нуклеиновую кислоту, в клетки культуры или организма. Системы доставки невирусных векторов включают ДНК-плазмиды, РНК (например, транскрипт вектора, описанного в настоящем описании), "голую" нуклеиновую кислоту и нуклеиновую кислоту в комплексе со средством доставки, таким как липосома. Системы вирусной доставки векторов включают ДНК и РНК-вирусы, которые имеют либо эписомальные, либо интегрированные геномы после доставки в клетку. Обзор способов генной терапии см. в Anderson, Science 256: 808-813 (1992); Nab el & Feigner, TIBTECH 11: 211-217 (1993), Mitani & Caskey, TIBTECH 11: 162-166 (1993); Dillon, TIBTЕСН 11: 167-175 (1993); Miller, Nature 357: 455-460 (1992); Van Brunt, Biotechnology 6 (10): 1149-1154 (1988); Vigne, Restorative Neurology and Neuroscience 8: 35-36 (1995), Kremer & Perricaudet, British Medical Bulletin 51 (1): 31-44 (1995); Haddada et al., В Current Topics in Microbiology and Immunology, Doerfler and Bomm (eds) (1995) и Yu et al., Gene Therapy 1: 13-26 (1994).
[00825] Способы невирусной доставки нуклеиновых кислот включают липофекцию, нуклеофекцию, микроинъекцию, генные пушки, виросомы, липосомы, иммунолипосомы, поликатионы или конъюгаты липидов с нуклеиновыми кислотами, "голую" ДНК, искусственные вирионы и усиленное агентом поглощение ДНК. Липофекция описана, например, в патентах США № 5049386, 4946787; и 4897355), и липофекционные реагенты продаются на коммерческой основе (например, Transfectam™ и Lipofectin™). Катионные и нейтральные липиды, которые пригодны для эффективного распознавания рецепторов липофекции полинуклеотидов, включают таковые Feigner, WO 91/17424; WO 91/16024. Доставка может осуществляться в клетки (например, in vitro или ex vivo) или ткани-мишени (например, in vivo).
[00826] Получение комплексов липидов с нуклеиновыми кислотами, включая целевые липосомы, такие как иммунолипидные комплексы, хорошо известно специалисту в данной области (см., например, Crystal, Science 270: 404-410 (1995), Blaese et al., Cancer Gene Ther. 2: 291-297 (1995), Behr et at., Bioconjugate Chem., 5: 382-389 (1994), Remy et al., Bioconjugate Chem., 5: 647-654 (1994), Gao et al., Gene Therapy 2: 710-722 (1995), Ahmad et al., Cancer Res. 52: 4817-4820 (1992), патенты США №№ 4186183, 4217344, 4235871, 4261975, 4485054, 4501728, 4774085, 4837028 и 4946787).
[00827] В применении РНК или ДНК-вирусных систем для доставки нуклеиновых кислот используются современные способы нацеливания вируса на конкретные клетки в организме и передачи вирусного генома в ядро. Вирусные векторы могут вводиться непосредственно пациентам (in vivo) или их можно использовать для обработки клеток in vitro, а модифицированные клетки могут быть введены пациентам (ex vivo). Обычные вирусные системы могут включать ретровирусные, лентивирусные, аденовирусные, аденоассоциированные и вирусные вирусы простого герпеса для переноса генов. Встраивание в геном хозяина возможно с помощью способов переноса генов ретровирусов, лентивирусов и аденоассоциированных вирусов, часто приводящих к длительной экспрессии введенного трансгена. Кроме того, высокая эффективность трансдукции наблюдалась во многих других типах клеток- и тканей-мишеней.
[00828] Тропизм ретровируса может быть изменен путем включения чужеродных белков оболочки, что позволяет расширить потенциальную популяцию клеток, которые могут стать мишенью генного воздействия. Лентивирусные векторы представляют собой ретровирусные векторы, которые способны трансформировать или инфицировать не делящиеся клетки и, как правило, имеют высокие вирусные титры. Поэтому выбор ретровирусной системы переноса генов будет зависеть от ткани-мишени. Ретровирусные векторы состоят из цис-действующих длинных концевых повторов с упаковочной емкостью до 6-10 т.п.н. чужеродной последовательности. Минимальные цис-действующие LTR достаточны для репликации и упаковки векторов, которые затем используются для интеграции терапевтического гена в клетку-мишень для обеспечения постоянной трансгенной экспрессии. Широко используемые ретровирусные векторы включают векторы, основанные на вирусе лейкемии мышей (MuLV), вирусе лейкемии гиббонов (GaLV), вирусе иммунодефицита обезьян (SIV), вирусе иммунодефицита человека (ВИЧ) и их комбинациях (см., например, Buchscher et al., J. Virol, 66: 2731-2739 (1992), Johann et al., J. Virol. 66: 1635-1640 (1992), Sommnerfelt et al., Virol. 176: 58-59 (1990), Wilson et al., J. Virol, 63: 2374-2378 (1989), Miller et al., J. Virol. 65: 2220-2224 (1991), PCT/US94/05700). В тех вариантах, где предпочтительна временная экспрессия, могут быть использованы аденовирусные системы. Аденовирусные векторы способны к очень высокой эффективности трансдукции во многих типах клеток и не требуют деления клеток. С такими векторами был получен высокий титр и уровни экспрессии. Этот вектор может быть получен в больших количествах в относительно простой системе. Векторы на основе аденоассоциированных вирусов ("AAV") также могут быть использованы для трансдукции клеток целевыми нуклеиновыми кислотами, например, при продуцировании in vitro нуклеиновых кислот и пептидов, а также для процедур генной терапии in vivo и ex vivo (см., например, West et al., Virology 160: 38-47 (1987), US 4797368, WO 93/24641, Kotin, Human Gene Therapy 5: 793-801 (1994), Muzyczka, J. Clin. Invest.: 1351 (1994). Конструирование рекомбинантных векторов AAV описано в ряде публикаций, включая патент США № 5173414, Tratschin et al. Mol. Cell, Biol., 5: 3251-3260 (1985), Tratschin, et al., Mol. Cell, Biol., 4: 2072-2081 (1984), Hermonat & Muzyczka, PNAS 81: 6466-66470 (1984) и Samulski et al., J. Virol. 63: 03822-3828 (1989).
Варианты для ДНК/РНК, или ДНК/ДНК, или РНК/РНК, или белка/РНК
[00829] В некоторых вариантах осуществления компоненты CRISPR-систем могут доставляться в различных формах, таких как комбинации ДНК/РНК, или РНК/РНК, или белок/РНК. Например, C2c1 или C2c3 может быть доставлен как ДНК-кодирующий полинуклеотид или как РНК-кодирующий полинуклеотид или как белок. Направляющая молекула может быть доставлена как ДНК-кодирующий полинуклеотид или как РНК. Предусмотрены все возможные комбинации, включая смешанные формы доставки.
[00830] В некоторых вариантах осуществления предусматриваются все такие комбинации (ДНК/РНК, или ДНК/ДНК, или РНК/РНК, или белок/РНК).
[00831] В некоторых вариантах осуществления, когда C2c1 или C2c3 доставляют в форме белка, возможна предварительная сборка с одной или более направляющими молекулами.
Наноклубки
[00832] Кроме того, система CRISPR может доставляться с использованием наноклубков, например, описанных в Sun W et al, Cocoon-like self-degradable DNA nanoclew for anticancer drag delivery., J Am Chem Soc. 2014 Oct 22; 136(42): 14722-5. doi: 10.1021/ja5088024. Epub 2014 Oct 13; или в Sun W et al, Self- Assembled DNA Nanoclews for the Efficient Delivery of CRISPR-Cas9 for Genome Editing., Angew Chem Int Ed En), 2015 Oct 5;54(41): 12029-33. doi: 10.1002/anie.201506030. Epub 2015 Aug 27.
[00833] В практическом применении настоящего изобретения используются, если не указано обратное, общепринятые способы иммунологии, биохимии, химии, молекулярной биологии, микробиологии, клеточной биологии, геномики и технологии рекомбинантной ДНК, которые являются стандартными для специалистов в данной области. См. See Sambrook, Fritsch and Maniatis, MOLECULAR CLONING: A LABORATORY MANUAL, 2nd edition (1989); CURRENT PROTOCOLS IN MOLECULAR BIOLOGY (F. M. Ausubel, et al. eds., (1987)); серия METHODS IN ENZYMOLOGY (Academic Press, Inc.): PCR 2: A PRACTICAL APPROACH (M.J. MacPherson, B.D. Hanies and G.R. Taylor eds. (1995)), Harlow and Lane, eds. (1988) ANTIBODIES, A LABORATORY MANUAL, AND ANIMAL CELL CULTURE (R.I. Freshney, ed. (1987)).
Модели генетических и эпигенетических условий
[00834] Способ по изобретению может быть использован для создания растения, животного или клетки, которые могут быть использованы для моделирования и/или изучения генетических или эпигенетических условий, представляющих интерес, например, посредством модели представляющих интерес мутаций или заболеваний. Как используют в настоящем описании, термин "заболевание" относится к заболеванию, расстройству или симптому у индивида. Например, способ по изобретению может быть использован для создания животного или клетки, которые включают модификацию в одной или более последовательностях нуклеиновых кислот, связанных с заболеванием, или растение, животное или клетку, в которых экспрессия одной или более последовательностей нуклеиновых кислот, связанных с заболеванием, изменена. Такая последовательность нуклеиновых кислот может кодировать связанную с заболеванием последовательность белка или может быть связанной с заболеванием последовательностью контроля. Соответственно, понятно, что в вариантах осуществления изобретения растение, индивид, пациент, организм или клетка могут быть не являющимися человеком пациентом, организмом или не происходящей от человека клеткой. Таким образом, изобретение относится к растению, животному или клетке, полученным способами по настоящему изобретению, или к их потомству. Потомство может быть клоном произведенного растения или животного или может быть результатом полового размножения путем скрещивания с другими особями одного и того же вида для внедрения других желательных признаков в их потомство. Клетка может находиться in vivo или ex vivo в случае многоклеточных организмов, особенно животных или растений. В случае, когда клетка культивируется, клеточная линия может быть создана, если соответствующие условия культивирования удовлетворяются, и предпочтительно, если клетка подходяще приспособлена для этой цели (например, стволовая клетка). Также предусматриваются бактериальные клеточные линии, полученные с соответствии с изобретением. Таким образом, клеточные линии также предусматриваются.
[00835] В некоторых способах модель болезни может быть использована для изучения эффектов мутаций на животное или клетку, развития и/или прогрессирования заболевания с использованием мер, обычно используемых при изучении этого заболевания. Альтернативно, такая модель болезни полезна для изучения влияния фармацевтически активных соединений на заболевание.
[00836] В некоторых способах модель болезни может быть использована для оценки эффективности потенциальной стратегии генной терапии. То есть связанный с болезнью ген или полинуклеотид может быть модифицирован таким образом, что развитие заболевания и/или его прогрессирование ингибируются или уменьшаются. В частности, способ включает модификацию связанного с заболеванием гена или полинуклеотида таким образом, что образуется измененный белок и, как результат, животное или клетка имеют измененный ответ. Соответственно, в некоторых способах генетически модифицированное животное можно сравнить с животным, предрасположенным к развитию заболевания, что даст возможность оценить влияние генной терапии.
[00837] В другом варианте осуществления настоящее изобретение относится к способам разработки биологически активного агента, который изменяет сигнал клеток, связанных с геном болезни. Способ включает приведение в контакт испытуемого соединения с клеткой, содержащей один или более векторов, управляющих экспрессией одного или более ферментов CRISPR, и последовательность прямого повтора, связанную с направляющей последовательностью; и обнаружение изменения в показаниях, которое указывает на уменьшение или увеличение сигнала клетки, связанного, например, с мутацией в гене болезни, содержащемся в клетке.
[00838] Модель клетки или модель на животных может быть создана в сочетании со способом изобретению для диагностики изменения клеточной функции. Такая модель может быть использована для изучения влияния последовательности генома, модифицированной комплексом CRISPR по изобретению, на интересующую клеточную функцию. Например, модель клеточной функции может использоваться для изучения влияния модифицированной последовательности генома на внутриклеточную передачу сигнала или внеклеточную передачу сигнала. Альтернативно, модель клеточной функции может использоваться для изучения эффектов модифицированной последовательности генома на сенсорное восприятие. В некоторых таких моделях модифицирована одна или более геномных последовательностей, связанных с сигнальным биохимическим путем в модели.
[00839] Было исследовано несколько моделей заболеваний. К ним относятся исследование de novo генов риска аутизма CHD8, KATNAL2 и SCN2A; и гена синдромного аутизма (синдром Ангельмана) UBE3A. Конечно, эти гены и результирующие модели аутизма предпочтительнее, но служат для того, чтобы показать широкую применимость изобретения для генов и соответствующих моделей.
[00840] Измененная экспрессия одной или более геномных последовательностей, связанных с сигнальным биохимическим путем, может быть определена путем анализа разности уровней мРНК соответствующих генов между тестовой модельной клеткой и контрольной клеткой, когда они контактируют с кандидатом. Альтернативно, дифференциальная экспрессия последовательностей, связанных с сигнальным биохимическим путем, определяется путем обнаружения различий в количестве кодируемого полипептида или генного продукта.
[00841] Для анализа индуцированного агентом изменения уровня транскриптов мРНК или соответствующих полинуклеотидов нуклеиновую кислоту, содержащуюся в образце, сначала экстрагируют в соответствии со стандартными способами в данной области. Например, мРНК может быть выделена с использованием различных литических ферментов или химических растворов в соответствии с методиками, изложенными в Sambrook et al. (1989) или экстрагирована путем связывания нуклеиновой кислоты со смолами в соответствии с прилагаемыми инструкциями производителей. Затем мРНК, содержащуюся в образце экстрагированной нуклеиновой кислоты, выявляют с помощью способов амплификации или обычных анализов гибридизации (например, нозерн-блоттинга) в соответствии со способами, широко известными в данной области или на основе способов, описанных в настоящем описании.
[00842] Для целей настоящего изобретения амплификация означает любой способ, использующий праймер и полимеразу, способную реплицировать последовательность-мишень с достаточной точностью. Амплификацию можно проводить с помощью природных или рекомбинантных ДНК-полимераз, таких как ДНК-полимераза TaqGold™, T7, фрагмент Кленова ДНК-полимеразы E.coli и обратная транскриптаза. Предпочтительным способом амплификации является ПЦР. В частности, выделенная РНК может быть подвергнута анализу с использованием обратной транскрипции, которая сопряжена с количественной полимеразной цепной реакцией (ОТ-РЦР), чтобы количественно определить уровень экспрессии последовательности, связанной с сигнальным биохимическим путем.
[00843] Определение уровня экспрессии гена может проводиться в реальном времени в анализе с использованием амплификации. В одном из аспектов амплифицированные продукты могут быть непосредственно визуализированы с помощью флуоресцентных связывающих ДНК агентов, включая, но не ограничиваясь ими: интеркаляторы ДНК и связывающиеся с желобками ДНК агенты. Поскольку количество интеркаляторов, включенных в молекулы двухцепочечной ДНК, как правило, пропорционально количеству амплифицированных продуктов ДНК, удобно определять количество амплифицированных продуктов путем количественной оценки флуоресценции интеркалировавшего красителя с использованием стандартно применяемых оптических приборов, используемых в данной области. ДНК-связывающие красители, подходящие для применения с этой целью, включает SYBR green, SYBR blue, DAPI, пропидия йодид, hoechst, SYBR gold, бромид этидия, акридиновые красители, дауномицин, хлорохин, дистамицин D, хромомицин, митрамицин, полипиридины рутения, антрамицин и т.п.
[00844] В другом аспекте в реакции амплификации могут быть использованы другие флуоресцентные метки, такие как специфические для последовательности зонды для облегчения обнаружения и количественного определения амплифицированных продуктов. Количественная амплификация на основе использования зонда зависит от специфического для последовательности определения желаемого продукта амплификации. Она использует флуоресцентные целеспецифичные зонды (например, зонды TaqMan®), что приводит к повышению специфичности и чувствительности. Способы осуществления количественной амплификации на основе использования зонда хорошо известны в данной области и описаны в патенте США № 5210015.
[00845] В еще одном варианте аспекте могут быть выполнены обычные анализы гибридизации с использованием зондов гибридизации, гомологичных последовательностям, связанным с сигнальным биохимическим путем. Обычно допускается формирование зондами стабильных комплексов с последовательностями, связанными с сигнальным биохимическим путем, содержащимися в биологическом образце, полученном от исследуемого индивидуума, в ходе реакции гибридизации. Квалифицированному специалисту в данной области должно быть понятно, что, когда антисмысловая последовательность используется в нуклеиновой кислоте-зонде, полинуклеотиды-мишени, представленные в образце, выбирают так, чтобы они были комплементарными антисмысловым последовательностям нуклеиновых кислот. Напротив, когда нуклеотидный зонд является смысловой последовательностью нуклеиновой кислоты, в качестве полинуклеотида-мишени выбирают комплементарную к последовательностям смысловую последовательность нуклеиновой кислоты.
[00846] Гибридизация может выполняться в условиях, различающихся по уровню жесткости. Подходящие условия гибридизации для практики настоящего изобретения таковы, что взаимодействие при распознавании зондом последовательностей, ассоциированных с сигнальным биохимическим путем, является достаточно специфичным и достаточно стабильным. Условия, которые повышают жесткость условий реакции гибридизации, широко известны и опубликованы в данной области. См., например, (Sambrook, et al., (1989), Nonradioactive In Situ Hybridization Application Manual, Boehringer Mannheim, second edition). Анализ гибридизации может быть проведен с использованием зондов, иммобилизованных на любом твердом носителе, включая, но не ограничиваясь, нитроцеллюлозу, стекло, силикон и множество генных матриц. Является предпочтительным проведение анализа гибридизации на генных чипах высокой плотности, как описано в патенте США № 5445934.
[00847] Для удобного обнаружения комплексов зонд-мишень, образующихся во время анализа гибридизации, нуклеотидные зонды могут быть конъюгированы с поддающейся обнаружению меткой. Поддающиеся обнаружению метки, подходящие для использования в рамках настоящего изобретения, включают любую конструкцию, детектируемую фотохимическими, биохимическими, спектроскопическими, иммунохимическими, электрическими, оптическими или химическими способами. В данной области известно большое количество подходящих поддающихся обнаружению меток, которые включают флуоресцентные или хемилюминесцентные метки, метки радиоактивного изотопа, ферментативные или другие лиганды. В предпочтительных вариантах осуществления, вероятно, может быть желательным использовать флуоресцентную метку или метку фермента, такую как дигоксигенин, β-галактозидаза, уреаза, щелочная фосфатаза или пероксидаза, комплекс авидин/биотин.
[00848] Способы обнаружения, используемые для обнаружения или количественной оценки гибридизации, обычно зависят от выбора метки. Например, радиоактивные метки могут быть обнаружены с использованием фотопленки или прибора Phosphoimager. Флуоресцентные маркеры могут быть обнаружены и количественно определены с помощью фотоприемника для обнаружения испускаемого света. Ферментативные метки обычно обнаруживают путем обеспечения фермента субстратом и измерения количества полученного продукта реакции, создаваемого воздействием фермента на субстрат; и, наконец, колориметрические метки обнаруживаются путем простой визуализации цветной метки.
[00849] Индуцированное агентом изменение экспрессии последовательностей, связанных с сигнальным биохимическим путем, также может быть определено путем изучения соответствующих продуктов гена. Определение уровня белка обычно включает: а) проведение реакции белка, содержащегося в биологическом образце, с агентом, который специфически связывается с белком, ассоциированным с сигнальным биохимическим путем; и (b) идентификацию любого образовавшегося комплекса агент-белок. В одном аспекте данного варианта осуществления агент, который специфически связывает белок, связанный с сигнальным биохимическим путем, представляет собой антитело, предпочтительно моноклональное антитело.
[00850] Реакцию проводят путем связывания агента с образцом белков, ассоциированных с сигнальным биохимическим путем, полученным из пробных образцов, в условиях, которые позволяют образование комплекса агентом и белками, ассоциированными с сигнальным биохимическим путем. Образование комплекса может быть обнаружено прямо или косвенно в соответствии со стандартными методиками в данной области. При использовании способа прямого обнаружения агенты снабжаются поддающейся обнаружению меткой, и непрореагировавшие агенты могут быть удалены из комплекса; количество неиспользованной метки тем самым позволяет определить количество образовавшегося комплекса. Для такого способа предпочтительно выбирать метки, которые остаются связанными с агентом даже при жестких условиях промывания. Предпочтительно, чтобы метка не мешала реакции связывания. В альтернативном варианте процедура косвенного обнаружения может использовать агент, который содержит метку, введенную либо химическим, либо ферментативным путем. Желательная метка обычно не препятствует связыванию или стабильности полученного комплекса полипептида и агента. Однако метка обычно предназначена для того, чтобы быть доступной для эффективного связывания антителом и, следовательно, генерировать распознаваемый сигнал.
[00851] В данной области известно большое количество меток, подходящих для определения уровней белка. Неограничивающие примеры включают радиоизотопы, ферменты, коллоидные металлы, флуоресцентные соединения, биолюминесцентные соединения и хемилюминесцентные соединения
[00852] Количество комплексов агент-полипептид, образующихся во время реакции связывания, может быть количественно определено стандартными количественными способами анализа. Как проиллюстрировано выше, образование комплекса агент-полипептид может быть непосредственно измерено по количеству метки, оставшейся в участке связывания. В альтернативном варианте белок, связанный с сигнальным биохимическим путем, тестируют на его способность конкурировать с меченым аналогом за сайты связывания специфического агента. В этом конкурентном анализе количество захваченной метки обратно пропорционально количеству белковых последовательностей, ассоциированных с сигнальным биохимическим путем, присутствующих в тестируемом образце.
[00853] Ряд способов анализа белка, основанных на общих принципах, изложенных выше, доступен в данной области. Они включают, но не ограничиваются ими, радиоиммунные способы анализа, ELISA (иммунорадиометрические анализ с ферментным связыванием), "сэндвич-иммуноанализ", иммунорадиометрический анализ, иммунологический анализ in situ (с использованием, например, коллоидного золота, фермента или радиоизотопной метки), анализ с использованием вестерн-блоттинга, анализ иммунопреципитации, иммунофлуоресцентный анализ и SDS-PAGE.
[00854] Антитела, которые специфически распознают или связывают с белки, связанные с сигнальным биохимическим путем, являются предпочтительными для проведения вышеупомянутых анализов белка. При желании можно использовать антитела, которые распознают конкретный тип посттрансляционных модификаций (например, индуцирбельные модификации сигнального биохимического пути). Посттрансляционные модификации включают, но не ограничиваются ими: гликозилирование, липидирование, ацетилирование и фосфорилирование. Эти антитела могут быть приобретены у коммерческих поставщиков. Например, доступны антитела к антифосфотирозину, которые специфически распознают тирозин-фосфорилированные белки, ряда поставщиков, включая Invitrogen и Perkin Elmer. Антитела к антифосфотирозину особенно полезны для обнаружения белков, которые по-разному фосфорилируются по остаткам тирозина в ответ на стресс ER. Такие белки включают, но не ограничиваются этим, эукариотический фактор инициации трансляции 2 альфа (eIF-2a). Альтернативно, эти антитела могут быть получены с использованием традиционных технологий поликлональных или моноклональных антител путем иммунизации животного-хозяина или клетки, продуцирующей антитела, целевым белком, который демонстрирует желаемую посттрансляционную модификацию.
[00855] При практическом осуществлении способа введения в индивида может быть желательным выявить паттерн экспрессии белка, связанного с сигнальным биохимическим путем в различных тканях организма, в разных типах клеток и/или в разных субклеточных структурах. Эти исследования могут быть выполнены с использованием специфичных для ткани, специфичных для клеток или субклеточных структур антител, способных связываться с маркерами белка, которые предпочтительно экспрессируются в определенных тканях, типах клеток или субклеточных структурах.
[00856] Измененная экспрессия гена, связанного с сигнальным биохимическим путем, также может быть определена путем изучения изменения активности генных продуктов в сравнении с контрольной клеткой. Анализ вызванного агентом изменения активности белка, связанного с сигнальным биохимическим путем, будет зависеть от биологической активности и/или пути передачи сигнала, который исследуется. Например, когда белок является киназой, изменение его способности фосфорилировать субстрат (субстраты) ниже в том же пути можно определить с помощью различных анализов, известных в данной области. Типичные способы анализа включают, однако не ограничиваются такими как: иммуноблоттинг и иммунопреципитацию антителами, такими как антитела к антифосфотирозину, которые распознают фосфорилированные белки. Кроме того, активность киназы может быть обнаружена с помощью высокопроизводительных хемилюминесцентных способов анализа, таких как AlphaScreen™ (доступный от Perkin Elmer) и eTag™ (Chan-Hui, et al., (2003) Clinical Immunology 111: 162-174).
[00857] Если белок, связанный с сигнальным биохимическим путем, является частью сигнального каскада, приводящего к колебаниям внутриклеточного рН, в качестве репортерных молекул можно использовать чувствительные к рН молекулы, такие как флуоресцентные рН-индикаторы. В другом примере, где белок, связанный с сигнальным биохимическим путем, является ионным каналом, можно контролировать флуктуации мембранного потенциала и/или концентрации внутриклеточных ионов. Ряд коммерческих наборов и высокопроизводительных устройств особенно подходит для быстрого и надежного скрининга модуляторов ионных каналов. Типичные устройства включают FLIPR™ (Molecular Devices, Inc.) и VIPR (Aurora Biosciences). Эти приборы способны одновременно обнаруживать реакции в более чем 1000 лунках образцов микропланшета и обеспечивать измерение и функциональные данные в режиме реального времени в течение секунд или даже миллисекунд.
[00858] При использовании любого из описанных в настоящем описании способов подходящий вектор может быть введен в клетку или эмбрион с использованием одного или более способов, известных в данной области, включая, но не ограничиваясь этим, микроинъекции, электропорацию, образование пор с использованием ультразвука, биолистику, опосредованную фосфатом кальция трансфекцию, катионную трансфекцию, липосомную трансфекцию, трансфекцию с использованием дендримеров, трансфекцию с использованием теплового шока, трансфекцию-нуклеофекцию, магнитофекцию, липофекцию, импалефекцию, оптическую трансфекцию, захват нуклеиновых кислот патентованным агентом и доставку с использованием липосом, иммунолипосом, вирусом и искусственных вирионов. В некоторых способах вектор вводится в эмбрион путем микроинъекции. Вектор или векторы могут быть введены посредством микроинъекции в ядро или цитоплазму эмбриона. В некоторых способах вектор или векторы могут быть введены в клетку путем нуклеофекции.
[00859] Полинуклеотид-мишень комплекса CRISPR может быть любым полинуклеотидом, эндогенным или экзогенным для эукариотической клетки. Например, полинуклеотид-мишень может представлять собой полинуклеотид, который находится в ядре эукариотической клетки. Полинуклеотид-мишень может представлять собой последовательность, кодирующую генный продукт (например, белок) или некодирующую последовательность (например, регуляторный полинуклеотид или мусорную ДНК).
[00860] Примеры полинуклеотидов-мишеней включают последовательность, связанную с сигнальным биохимическим путем, например, сигнальным биохимическим путем, ассоциированным с геном или полинуклеотидом. Примеры полинуклеотидов-мишеней включают связанный с заболеванием ген или полинуклеотид. Понятие ген или полинуклеотид, "ассоциированный с заболеванием" относится к любому гену или полинуклеотиду, который производит продукты транскрипции или трансляции на аномальном уровне или в аномальной форме в клетках, полученных из тканей, пораженных болезнью, по сравнению с контрольными тканями или клетками, не пораженными заболеванием. Это может быть ген, который экспрессируется на аномально высоком уровне; это может быть ген, который экспрессируется на аномально низком уровне, причем изменения уровня экспрессии коррелируют с возникновением и/или прогрессированием заболевания. Ассоциированный с заболеванием ген также относится к гену, имеющему мутацию(и) или генетическую вариацию, которая непосредственно ответственна или находится в неравновесной взаимосвязи с геном (генами), который ответственен за этиологию заболевания. Продукты транскрипции или трансляции могут быть известны или неизвестны и могут быть на нормальном или аномальном уровне.
[00861] Полинуклеотид-мишень комплекса CRISPR может быть любым полинуклеотидом, эндогенным или экзогенным, в эукариотической клетке. Например, полинуклеотид-мишень может быть полинуклеотидом, находящимся в ядре эукариотической клетки. Полинуклеотид-мишень может представлять собой последовательность, кодирующую генный продукт (например, белок) или некодирующую последовательность (например, регуляторный полинуклеотид или мусорную ДНК). Не ограничиваясь теорией, считается, что последовательность-мишень должна быть связана с PAM (мотив, находящийся вблизи протоспесера); то есть короткой последовательностью, распознаваемой комплексом CRISPR. Точные требования к последовательности и длине для последовательности PAM различаются в зависимости от используемого фермента CRISPR, но последовательность PAM обычно находится в 2-5 парах оснований от протоспейсера (то есть последовательности-мишени). Примеры последовательностей PAM приведены в разделе примеров ниже, и квалифицированный специалист в данной области сможет идентифицировать дальнейшие последовательности PAM для использования с данным ферментом CRISPR.
[00862] Полинуклеотид-мишень комплекса CRISPR может включать ряд ассоциированных с заболеванием генов и полинуклеотидов, а также генов и полинуклеотидов, ассоциированных с сигнальными биохимическими путями, как указано в предварительных патентных заявках США 61/736527 и 61/748427, обе под названием "SYSTEMS METHODS AND COMPOSITIONS FOR SEQUENCE MANIPULATION", поданных 12 декабря 2012 года и 2 января 2013 года, соответственно, и заявке РСТ PCT/US2013/074667, озаглавленной "DELIVERY, ENGINEERING AND OPTIMIZATION OF SYSTEMS, METHODS AND COMPOSITIONS FOR SEQUENCE MANIPULATION AND THERAPEUTIC APPLICATIONS", поданной 12 декабря 2013 года, содержание каждой из которых включено в настоящее описание в качестве ссылки в полном объеме.
[00863] Примеры полинуклеотидов-мишеней включают последовательность, ассоциированную с сигнальным биохимическим путем, например, ген или нуклеотид, ассоциированный с сигнальным биохимическим путем. Примеры полинуклеотидов-мишеней включают ассоциированный с заболеванием ген или полинуклеотид. Ген или полинуклеотид, "ассоциированный с заболеванием", относится к любому гену или полинуклеотиду, который дает продукты транскрипции или трансляции на аномальном уровне или в аномальной форме в клетках, полученных из тканей, пораженных заболеванием, по сравнению с контрольными тканями или клетками, не пораженными заболеванием. Это может быть ген, который экспрессируется на аномально высоком уровне; это может быть ген, который экспрессируется на аномально низком уровне, когда измененная экспрессия коррелирует с возникновением и/или прогрессированием заболевания. "Ассоциированный с заболеванием ген" также относится к гену, обладающему мутацией (мутациями) или генетической вариацией, которая непосредственно ответственна или находится в неравновесии по сцеплению с геном(ами), который отвечает за этиологию заболевания. Транскрибированные или транслированные продукты могут быть известны или неизвестны и могут быть на нормальном или ненормальном уровне.
Полногеномный скрининг посредством нокаута
[00864] Экспериментальные белковые комплексы CRISPR, описанные в настоящем описании, могут быть использованы для проведения эффективного и экономически эффективного функционального геномного скрининга. При таком скрининге можно использовать геномные библиотеки, основанные на эффекторном белке CRISPR. Такой скрининг и библиотеки могут обеспечить определение функций генов, клеточных путей, в которые вовлечены гены, и то, как какое-либо изменение экспрессии гена может привести к определенному биологическому процессу. Преимущество настоящего изобретения состоит в том, что система CRISPR позволяет избежать нецелевого связывания и возникающих в результате неспецифических эффектов. Это достигается с использованием систем, организованных с высокой степенью специфичности последовательности к ДНК-мишени. В предпочтительных вариантах осуществления изобретения комплексы эффекторных белков CRISPR представляют собой комплексы эффекторных белков C2c1 или C2c3.
[00865] В вариантах осуществления изобретения полногеномная библиотека может содержать множество направляющих РНК для C2c1 или C2c3, как описано в настоящем описании, включая направляющие последовательности, которые способны нацеливать на множество последовательностей-мишеней во множестве геномных локусов в популяции эукариотических клеток. Популяция клеток может представлять собой популяцию эмбриональных стволовых (ES) клеток. Последовательность-мишень в геномном локусе может быть некодирующей последовательностью. Некодирующая последовательность может быть интронной, регуляторной последовательностью, сайтом сплайсинга, 3-'UTR, 5'-UTR или сигналом полиаденилирования. Функция одного или более генных продуктов может быть изменена путем нацеливания. Нацеливание может привести к нокауту функции гена. Нацеливание на генный продукт может включать более одной направляющей РНК. Нацеливание на генный продукт может быть осуществлено посредством 2, 3, 4, 5, 6, 7, 8, 9 или 10 направляющих РНК, предпочтительно от 3 до 4 на ген. Нецелевые модификации могут быть сведены к минимуму за счет использования ступенчатых разрывов двойной цепи, образуемых эффекторными белковыми комплексами C2c1 или C2c3, или с использованием способов, аналогичных тем, которые используются в системах CRISPR-Cas9 (см., например, DNA targeting specificity of RNA-guided Cas9 nucleases. Hsu, P., Scott, D., Weinstein, J., Ran, FA., Konermann, S., Agarwala, V., Li, Y., Fine, E., Wu, X., Shalem, O., Cradick, TX, Marraffmi, LA., Bao, G., & Zhang, F. Nat Biotechnol doi: 10.1038/nbt.2647 (2013)), включенной в настоящее описание в качестве ссылки. Нацеливание может охватывать приблизительно 100 или более последовательностей. Нацеливание может охватывать приблизительно 1000 или более последовательностей. Нацеливание может составлять приблизительно 20000 или более последовательностей. Нацеливание может охватывать весь геном. Нацеливание может охватывать ряд последовательностей-мишеней, ориентированных на соответствующий или желательный процесс. Процесс может быть иммунным процессом. Процесс может быть процессом клеточного деления.
[00866] Один из вариантов изобретения охватывает геномную библиотеку, которая может содержать множество направляющих РНК для C2c1 или C2c3, которые могут содержать направляющие последовательности, которые могут нацеливать на множество последовательностей-мишеней во множестве (геномных) локусов, приводя к нокауту функции гена. Эта библиотека может потенциально содержать направляющие РНК, которые нацелены на каждый ген в геноме организма.
[00867] В некоторых вариантах осуществления изобретения организм или индивид является эукариотическим организмом (включая млекопитающее, включая человека), или эукариотическим организмом, не являющимся человеком, или животным, не являющимся человеком, или млекопитающим, не являющимся человеком. В некоторых вариантах осуществления организм или индивид является животным, не являющимся человеком, и может быть членистоногим, например насекомым, или может быть нематодой. В некоторых способах изобретения организм или индивид является растением. В некоторых способах по изобретению организм или субъект представляет собой млекопитающее или млекопитающее, не являющееся человеком. Млекопитающее, не являющееся человеком, может быть, например, грызуном (предпочтительно мышью или крысой), копытным или приматом. В некоторых способах по изобретению организм или субъект является водорослью, включая микроводоросли, или грибом.
[00868] Нокаут функции гена может включать введение в каждую клетку в популяции клеток векторной системы из одного или более векторов, содержащих сконструированную, не встречающуюся в природе систему эффекторного белка C2c1 или C2c3, содержащую I. эффекторный белок C2c1 или C2c3, и II. одну или более направляющих РНК, где компоненты I и II могут находиться на одном или разных векторах системы, встраивающих компоненты I и II в каждую клетку, причем направляющая последовательность нацелена на уникальный ген в каждой клетке, где эффекторный белок C2c1 или C2c3 является функционально связанным с регуляторным элементом, где при транскрипции направляющая РНК, содержащая направляющую последовательность, обеспечивает последовательность-специфическое связывание системы эффекторного белка C2c1 или C2c3 с последовательностью-мишенью в геномных локусах уникального гена, вызывая расщепление геномных локусов эффекторным белком C2c1 или C2c3, и подтверждение различных случаев нокаута во множестве уникальных генов в каждой клетке популяции, тем самым, получая библиотеку клеток с нокаутом гена. Изобретение подразумевает, что популяция клеток представляет собой популяцию эукариотических клеток, и в предпочтительном варианте осуществления популяция клеток представляет собой популяцию эмбриональных стволовых клеток (ES).
[00869] Один или более векторов могут быть плазмидными векторами. Вектор может представлять собой один вектор, содержащий эффекторный белок C2c1 или C2c3, одноцепочечную направляющую РНК и необязательно селективный маркер для клеток-мишеней. Без связи с теорией, одновременная доставка эффекторного белка C2c1 или C2c3и одноцепочечной направляющей РНК с помощью одного вектора позволяет применять этот способ к любому типу клеток-мишеней без необходимости сначала получать клеточные линии, которые экспрессируют эффекторный белок C2c1 или C2c3. Регулирующим элементом может быть индуцибельный промотор. Индуцируемый промотор может быть промотором, индуцируемым доксициклином. В некоторых способах по изобретению экспрессия направляющей последовательности находится под контролем промотора Т7 и направляется экспрессией Т7-полимеразы. Подтверждение различных вызывающих нокаут мутаций может быть осуществлено полноэкзомным секвенированием. Вызывающая нокаут мутация может быть достигнута в 100 или более уникальных генах. Вызывающая нокаут мутация может быть достигнута в 1000 или более уникальных генах. Вызывающая нокаут мутация может быть достигнута в 20000 или более уникальных генов. Вызывающая нокаут мутация может быть достигнута и во всем геноме. Нокаут функции гена может быть достигнут во множестве уникальных генов, которые функционируют в определенном физиологическом процессе или состоянии. Процесс или состояние может быть иммунным процессом или состоянием. Процесс или состояние может быть процессом или состоянием деления клеток.
[00870] Изобретение также относится к наборам, включающим обширные библиотеки геномов, упомянутые в настоящем описании. Набор может содержать один контейнер, содержащий векторы или плазмиды, содержащие библиотеку по изобретению. Набор может также содержать панель, содержащую отобранный комплект уникальных направляющих РНК системы эффекторных белков C2c1 или C2c3, содержащих направляющие последовательности из библиотеки по изобретению, причем отобранный комплект соответствует конкретному физиологическому состоянию. В рамках изобретения подразумевается, что нацеливанию подвергается приблизительно 100 или более последовательностей, приблизительно 1000 или более последовательностей или приблизительно 20000 или более последовательностей или весь геном. Кроме того, панель последовательностей-мишеней может быть сфокусирована на соответствующем или желательном процессе, таком как иммунный процесс или деление клеток.
[00871] В дополнительном аспекте изобретения эффекторный белок C2c1 или C2c3 может содержать одну или более мутаций и может быть использован в качестве общего ДНК-связывающего белка с или без слияния с функциональным доменом. Мутации могут быть искусственно введенными мутациями или мутациями приобретения или потери функции. Мутации были охарактеризованы, как описано в настоящем описании. В одном аспекте изобретения функциональный домен может быть доменом активации транскрипции, который может быть VP64. В других аспектах изобретения функциональный домен может быть доменом подавления транскрипции, который может быть KRAB или SID4X. Другие аспекты изобретения относятся к мутантному эффекторному белку C2c1 или C2c3, слитому с доменами, которые включают, но не ограничиваются, ими: активатор транскрипции, репрессор, рекомбиназу, транспозазу, ремоделирующий гистон, деметилазу, ДНК-метилтрансферазу, криптохром, светоиндуцируемый/контролируемый домен или химически индуцируемый/контролируемый домен. Некоторые способы по изобретению могут включать индукцию экспрессии генов-мишеней. В одном варианте осуществления индукция экспрессии путем нацеливания на множество последовательностей-мишеней во множестве геномных локусов в популяции эукариотических клеток осуществляется с использованием функционального домена.
[00872] В практике настоящего изобретения, использующего комплексы эффекторного белка C2c1 или C2c3, могут использоваться способы, в которых используются системы CRISPR-Cas9, и может быть приведена ссылка на:
[00873] Genome-Scale CRISPR-Cas9 Knockout Screening in Human Cells. Shalem, O., Sanjana, NE., Hartenian, E., Shi, X., Scott, DA., Mikkelson, T., Heckl, D., Ebert, BE., Root, DE., Doench, JG., Zhang, F. Science Dec 12. (2013). Опубликовано в окончательной редакции как: Science. 2014 Jan 3; 343(6166): 84-87
[00874] В Shalem et al. описан новый способ исследования функции гена в масштабе всего генома. Их исследования показали, что доставка полногеномной библиотеки с нокаутом CRISPR-Cas9 (GeCKO), нацеленной на 18080 генов с 64751 уникальными направляющими последовательностями, позволила скрининг как с отрицательной, так и с положительной селекцией в клетках человека. Во-первых, авторы показали использование библиотеки GeCKO для идентификации генов, необходимых для жизнеспособности клеток в злокачественных и плюрипотентных стволовых клетках. Затем, с использованием модели меланомы, авторы провели скрининг генов, утрата которых связана с резистентностью к вемурафенибу, препарату, который ингибирует мутантную протеинкиназу BRAF. Их исследования показали, что кандидаты с самым высоким рейтингом включают ранее подтвержденные гены NF1 и MED 12, также как и новые высокие результаты: NF2, CUL3, TADA2B и TADA1. Авторы наблюдали высокий уровень согласованности между независимыми направляющими РНК, нацеленными на один и тот же ген, и высокий показатель подтверждения высоких результатов, и, таким образом, продемонстрировали перспективность скрининга генома с помощью Cas9.
[00875] См. также патент США №20140357530; и публикацию патентной заявки РСТ W02014093701, включенные в настоящее описание в качестве ссылки. Также см. пресс-релиз NIH от 22 октября 2015 года под названием "Researchers identify potential alternative to CRISPR-Cas genome editing tools: New Cas enzymes shed light on evolution of CRISPR-Cas systems", включенный в настоящее описание в качестве ссылки.
Функциональные изменения и скрининг
[00 876] В другом аспекте осуществления настоящее изобретение относится к способу функциональной оценки и скрининга генов. Использование системы CRISPR согласно настоящему изобретению для точной доставки функциональных доменов, для активации или подавления генов или для изменения эпигенетического состояния путем точного изменения сайта метилирования в конкретном локусе-мишени может быть выполнено с использованием одной или более направляющих РНК для одной клетки, или популяции клеток, или библиотеки, примененной к геному в совокупности клеток ex vivo или in vivo, включая введение или экспрессию библиотеки, содержащей множество направляющих молекул РНК (sg-РНК), и где скрининг дополнительно включает использование эффекторного белка C2c1 или C2c3, причем комплекс CRISPR, содержащий эффекторный белок C2c1 или C2c3, модифицирован таким образом, чтобы содержать гетерологичный функциональный домен. В одном аспекте изобретение обеспечивает способ скрининга генома, включающий введение хозяину или экспрессию в хозяине библиотеки in vivo. В одном аспекте изобретение относится к способу, описанному в настоящем описании, дополнительно включающему активатор, вводимый хозяину или экспрессируемый в хозяине. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором активатор присоединен к эффекторному белку C2c1 или C2c3. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором активатор присоединен к N-концу или C-концу эффекторного белка C2c1 или C2c3. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором активатор присоединен к петле sg-РНК. В одном аспекте изобретение относится к способу, описанному в настоящем описании, дополнительно включающему репрессор, вводимый хозяину или экспрессируемый в хозяине. В одном аспекте изобретение относится к способу, описанному в настоящем описании, где скрининг включает изменение и обнаружение активации гена, ингибирования гена или расщепления в конкретном локусе.
[00877] В одном аспекте изобретение обеспечивает эффективную целевую активность и минимизирует нецелевую активность. В одном аспекте изобретение обеспечивает эффективное целевое расщепление эффекторным белком C2c1 или C2c3 и сводит к минимуму нецелевое расщепление эффекторным белком C2c1 или C2c3. В одном аспекте изобретение обеспечивает направляющее специфическое связывание эффекторного белка C2c1 или C2c3 в локусе гена без расщепления ДНК. Соответственно, в одном аспекте изобретение обеспечивает мишень-специфическую генную регуляцию. В одном аспекте изобретение обеспечивает направляющее специфическое связывание эффекторного белка C2c1 или C2c3 в локусе гена без расщепления ДНК. Соответственно, в одном варианте изобретение предусматривает расщепление в одном локусе гена и регуляцию гена в другом локусе гена с использованием одного эффекторного белка C2c1 или C2c3. В одном аспекте изобретение обеспечивает ортогональную активацию, и/или ингибирование, и/или расщепление множества мишеней с использованием одного или более эффекторных белков C2c1 или C2c3 и/или ферментов.
[00878] В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором клетка-хозяин является эукариотической клеткой. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором клетка-хозяин представляет собой клетку млекопитающего. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором хозяин представляет собой эукариотический организм, отличный от человека. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором эукариотический организм, отличный от человека, является млекопитающим, отличным от человека. В одном аспекте изобретение относится к способу, описанному в настоящем описании, где млекопитающее, отличное от человека, представляет собой мышь. Одним аспектом изобретения является способ, описанный в настоящем описании, включающий доставку комплексов эффекторных белков C2c1 или C2c3 или их компонента(ов) или молекулы(молекул) нуклеиновой кислоты, кодирующей их, где указанная молекула(ы) нуклеиновой кислоты функционально связана(ы) с регуляторной последовательностью(ми) и экспрессируется in vivo. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором экспрессия in vivo осуществляется с использованием лентивируса, аденовируса или AAV. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором доставка осуществляется через частицу, наночастицу, липид или проникающий в клетку пептид (CPP).
[00879] В одном аспекте изобретение относится к паре комплексов CRISPR, включающих эффекторный белок C2c1 или C2c3, каждый из которых содержит направляющую молекулу РНК (sg-РНК), содержащую направляющую последовательность, способную к гибридизации с последовательностью-мишенью в геномном локусе-мишени в клетке, где по меньшей мере одна петля каждой sg-РНК модифицирована вставкой отдельной последовательности(ей) РНК, которая связана с одним или более адаптерными белками, и где адаптерный белок связан с одним или более функциональными доменами, где каждая sg-РНК каждого комплекса эффекторного белка C2c1 или C2c3 включает функциональный домен, обладающий активностью расщепления ДНК. В одном аспекте изобретение относится к парным комплексам эффекторного белка C2с1 или C2c3, как описано в настоящем описании, где активность расщепления ДНК обусловлена нуклеазой Fok1.
[00880] В одном аспекте изобретение относится к способу разрезания последовательности-мишени в представляющем интерес геномном локусе, включающему доставку к клетке комплексов эффекторного белка C2c1 или C2c3, или его компонента или молекулы(молекул) нуклеиновой кислоты, кодирующей их, причем молекула(ы) нуклеиновой кислоты функционально связана с регуляторной последовательностью (последовательностями) и экспрессируется in vivo. В одном аспекте изобретение относится к способу, описанному в настоящем описании, в котором доставка производится с использованием лентивируса, аденовируса или AAV. В одном аспекте изобретение относится к способу, описанному в настоящем описании, или парному комплексу эффекторного белка C2c1 или C2c3, как описано в настоящем описании, в котором последовательность-мишень для первого комплекса пары находится на первой цепи двухцепочечной ДНК и последовательность-мишень для второго комплекса пары находится на второй цепи двухцепочечной ДНК. В одном аспекте изобретение относится к способу, описанному в настоящем описании, или парному комплексу эффекторного белка C2c1 или C2c3, как описано в настоящем описании, в котором последовательности-мишени первого и второго комплексов находятся вблизи друг друга, так что ДНК разрезается таким образом, чтобы облегчить направляемую гомологией репарацию. В одном аспекте способ настоящего изобретения может дополнительно включать введение в клетки матрицы ДНК. В одном аспекте описанный в настоящем описании способ или описанные в настоящем описании парные комплексы эффекторного белка C2c1 или C2c3 могут вовлекать случаи, где каждый комплекс эффекторного белка C2c2 имеет эффекторный фермент C2c1 или C2c3, в который внесена мутация таким образом, что он имеет не более чем приблизительно 5% активности нуклеазы эффекторного фермента C2c1 или C2c3, который не имеет мутации.
[00881] В одном аспекте изобретение относится к библиотеке, способу или комплексу, как описано в настоящем описании, в которых одноцепочечная направляющая sg-РНК модифицирована, чтобы иметь по меньшей мере одну некодирующую функциональную шпильку, например, где по меньшей мере одна некодирующая функциональная шпилька является репрессивнной; например, в которой, по меньшей мере одна некодирующая функциональная петля содержит Alu.
[00882] В одном аспекте изобретение относится к способу изменения или модификации экспрессии продукта гена. Указанный способ может включать введение в клетку, содержащую и экспрессирующую молекулу ДНК, кодирующую продукт гена, сконструированной, не встречающейся в природе системы CRISPR, содержащей эффекторный белок C2c1 или C2c3 и направляющую РНК, которая нацелена на молекулу ДНК, в результате чего направляющая РНК осуществляет нацеливание на молекулу ДНК, кодирующую продукт гена, и эффекторный белок C2c1 или C2c3 расщепляет молекулу ДНК, кодирующую продукт гена, в результате чего экспрессия продукта гена изменяется, и где эффекторный белок C2c1 или C2c3 и направляющая РНК не встречаются вместе в естественных условиях. Изобретение охватывает направляющую РНК, содержащую направляющую последовательность, связанную с последовательностью прямого повтора. Кроме того, изобретение подразумевает, что эффекторный белок C2c1 или C2c3 является кодон-оптимизированным для экспрессии в эукариотической клетке. В предпочтительном варианте осуществления эукариотическая клетка представляет собой клетку млекопитающего, а в более предпочтительном варианте осуществления клетка млекопитающего представляет собой клетку человека. В дальнейшем варианте осуществления изобретения экспрессия продукта гена уменьшается.
[00883] В некоторых вариантах осуществления один или более функциональных доменов связаны с эффекторным белком C2c1 или C2c3. В некоторых вариантах осуществления один или более функциональных доменов связаны с адаптерным белком, например, с использованием модифицированных направляющих молекул Konnerman et al. (Nature 517, 583-588, 29 января 2015 года). В некоторых вариантах осуществления один или более функциональных доменов связаны с "мертвой" одноцепочечной направляющей (sg-РНК, d-РНК). В некоторых вариантах осуществления комплекс d-РНК с активным эффекторным белком C2c1 или C2c3 обеспечивает регуляцию гена посредством функционального домена в локусе гена, тогда как sg-РНК обеспечивает расщепление ДНК активным эффекторным белком C2c1 или C2c3 в другом локусе, например, как описано аналогично для систем CRISPR-Cas9 в Dahlman et al., "Orthogonal gene control with a catalytically active Cas9 nuclease" (в печати). В некоторых вариантах осуществления d-РНК выбирают так, чтобы максимизировать селективность регуляции представляющего интерес локуса гена по сравнению с неспецифической регуляцией. В некоторых вариантах осуществления d-РНК выбирают так, чтобы максимизировать регулирование гена-мишени и минимизировать расщепление мишени.
[00884] Для целей последующего обсуждения "функциональный домен" может представлять собой функциональный домен, связанный с эффекторным белком C2c1 или C2c3, или функциональный домен, связанный с адаптерным белком.
[00885] В некоторых вариантах осуществления один или более функциональных доменов представляют собой NLS (последовательность ядерной локализации) или NES (ядерный экспортный сигнал). В некоторых вариантах осуществления один или более функциональных доменов являются доменом активации транскрипции, включающим VP64, p65, MyoD1, HSF1, RTA, SET7/9 и гистонацетилтрансферу. Другие случаи упоминания в настоящем описании доменов активации (или доменов-активаторов) в отношении доменов, которые связаны с ферментом CRISPR, включают любой известный домен активации транскрипции и, в частности, VP64, p65, MyoD1, HSF1, RTA, SET7/9 или гистонацетилтрансферазу.
[00886] В некоторых вариантах осуществления один или более функциональных доменов являются доменами подавления транскрипции. В некоторых вариантах осуществления доменами подавления транскрипции является KRAB-домен. В некоторых вариантах осуществления домен подавления транскрипции является доменом NuE, доменом NcoR, доменом SID или доменом SID4X.
[00887] В некоторых вариантах осуществления один или более функциональных доменов имеют одну или более функциональных активностей, включающих метилазную активность, деметилазную активность, активность активации транскрипции, активность подавления транскрипции, активность фактора терминации транскрипции, активность модификации гистонов, активность расщепления РНК, активность расщепления ДНК, активность интеграции ДНК или активность связывания нуклеиновых кислот.
[00888] Домены модификации гистонов также предпочтительны в некоторых вариантах осуществления. Типичные домены модификации гистонов обсуждаются ниже. В качестве функциональных доменов в рамках настоящего изобретения также предпочтительными являются домены транспозазы, домены HR (гомологичной рекомбинации), домены рекомбиназы и/или интегразы. В некоторых вариантах осуществления активность интеграции ДНК включает домены HR, домены интегразы, домены рекомбиназы и/или домены транспозазы. В некоторых вариантах предпочтительны домены гистонацетилтрансферазы.
[00889] В некоторых вариантах осуществления активность расщепления ДНК обусловлена нуклеазой. В некоторых вариантах осуществления нуклеаза содержит нуклеазу Fok1. См. "Dimeric CRISPR RNA-guided Fok1 nucleases for highly specific genome editing", Shengdar Q. Tsai, Nicolas Wyvekens, Cyd Khayter, Jennifer A. Foden, Vishal Thapar, Deepak Reyon, Mathew J. Goodwin, Martin J. Aryee, J. Keith Joung Nature Biotechnology 32(6): 569-77 (2014), которая относится к димерным Fok1-нуклеазам, направляемым РНК, которые распознают расширенные последовательности и могут редактировать эндогенные гены с высокой эффективностью в клетках человека.
[00890] В некоторых вариантах осуществления один или более функциональных доменов присоединены к эффекторному белку C2c1 или C2c3, так что при связывании с одноцепочечной направляющей РНК (sg-РНК) и мишенью функциональный домен находится в пространственной ориентации, позволяющей функциональному домену выполнять свойственную ему функцию.
[00891] В некоторых вариантах осуществления один или более функциональных доменов присоединены к адаптерному белку, так что после связывания эффекторного белка C2c1 или C2c3 с одноцепочечной направляющей РНК (sg-РНК) и мишенью функциональный домен находится в пространственной ориентации, позволяющей функциональному домену выполнять свойственную ему функцию.
[00892] В одном аспекте изобретение относится к композиции, описанной в настоящем описании, в которой один или более функциональных доменов присоединены к эффекторному белку C2c1 или C2c3 или адаптерному белку посредством линкера, необязательно линкера GlySer, как описано в настоящем описании.
[00893] Эндогенное подавление транскрипции часто опосредуется ферментами, модифицирующими хроматин, такими как гистон-метилтрансферазы (ГМТ) и деацетилазы (HDAC). Репрессивные эффекторные домены гистонов известны, и их иллюстративный список приведен ниже. В таблице предпочтение отдавалось белкам и функциональным укороченным частям небольшого размера для облегчения эффективной упаковки вирусов (например, посредством AAV). В общем, однако, домены могут включать домены деацетилазы (HDAC), гистон-метилтрансферазы (HMT) и ингибиторы гистонацетилтрансферазы (HAT), а также белки, рекрутирующие HDAC и HMT. Функциональный домен может быть или включать в некоторых вариантах осуществления эффекторные домены HDAC, рекрутирующие эффекторные домены HDAC, эффекторные домены гистонацетилтрансферазы (HMT), рекрутирующие эффекторные домены гистон-метилтрансферазы (HMT), рекрутирующие эффекторные домены гистонацетилтрансферазы.
Эффекторные домены HDAC
(Gregoretti)
H4K16Ac
H3K56Ac
(Hnisz)
(Landry)
(Wilson)
H4K16Ac
(Gertz)
(Zhu)
H3K56Ac
(Tennen)
[894] Соответственно, репрессорные домены по настоящему изобретению могут быть выбраны из гистон-метилтрансферазы (ГМТ), гистондезацетилаз (HDAC), ингибиторов гистонацетилтрансферазы (HAT), а также привлекающих HDAC и HMT белков.
[00895] Домен HDAC может быть любым из приведенных в таблице выше: HDAC8, RPD3, MesoLo4, HDAC11, HDT1, SIRT3, HST2, CobB, HST2, SIRT5, Sir2A или SIRT6.
[00896] В некотором варианте осуществления функциональный домен может быть рекрутирующим эффекторным доменом HDCP. Предпочтительные примеры включают те, которые приведены в таблице ниже, а именно MeCP2, MBD2b, Sin3a, NcoR, SALL1, RCOR1. NcoR проиллюстрирован в примерах настоящего описания и, хотя использование этого белка предпочтительно, предполагается, что другие представители класса также будут полезны.
Таблица HDAC рекрутирующие эффекторные домены
комплекс
(Nan)
(Boeke)
(Laherty)
взаимодействие с HDAC1
(Zhang)
(Lauberth)
(Gu, Ouyang)
[897] В некотором варианте осуществления функциональный домен может быть эффекторным доменом метилтрансферазы (HMT). Предпочтительные примеры включают те, которые приведены в таблице ниже, а именно NUE, vSET, EHMT2/G9A, SUV39H1, dim-5, KYP, SUVR4, SET4, SET1, SETD8 и TgSET8. NUE проиллюстрированы в примерах настоящего описания и, хотя использование этого белка предпочтительно, предполагается, что другие представители класса также будут полезны.
Таблица эффекторных доменов гистонметилтрансферазы (HMT)
H3, H4
(Pennini)
SET2
G9A
H3K9,
H3K27
H1K25me1
(Tachibana)
Пре-SET
SET,
Пост-SET
(Snowden)
Пре-SET
SET,
Пост-SET
9
(Rathert)
Пре-SET
SET,
Пост-SET
(подсемейство SUVH)
(Jackson)
(подсемейство SUVR)
(Thorstensen)
Пре-SET
SET,
Пост-SET
(Vielle)
(Vielle)
(Couture)
SET
(Sautel)
SET
[898] В некоторых вариантах осуществления функциональный домен может представлять собой рекрутирующий эффекторный домен гистон-метилтрансферазы (HMT). Предпочтительные примеры включают примеры, приведенные в таблице ниже, а именно Hp1a, PHF19 и NIPP1.
[899] Таблица эффективности рекрутирующих эффекторных доменов гистонметилтрансферазы (HMT).
комплекс
1
(Hathawa)
chromoshad
0
Линкер GGSG+(500-580)
(Ballaré)
PHD2
1
(Jin)
EED
[900] В некоторых вариантах осуществления функциональный домен может быть ингибиторным эффекторным доменом гистонацетилтрансферазы. Предпочтительные примеры включают SET/TAF-1β, перечисленные в таблице ниже.
[901] Таблица ингибиторных эффекторных доменов гистонацетилтрансферазы.
комплекс
musculus
(Cervoni)
[902] Также является предпочтительным нацеливание на эндогенные (регуляторные) элементы контроля (такие как энхансеры и сайленсеры), дополнительно к промотору или элементам, расположенным проксимально относительно промотора. Таким образом, изобретение может также использоваться для нацеливания на эндогенные элементы контроля (включая энхансеры и сайленсеры) в дополнение к нацеливанию на промотор. Эти элементы контроля могут быть расположены в восходящем- и нисходящем-направлении от точки начала транскрипции (TSS), начиная с 200 п.н. от TSS до 100 т.п.н от нее. Нацеливание на известные элементы контроля может быть использовано для активации или подавления гена-мишени. В некоторых случаях один элемент управления может влиять на транскрипцию нескольких генов-мишеней. Таким образом, нацеливания на один элемент контроля может использоваться для управления транскрипцией нескольких генов одновременно.
[00903] Нацеливание на предполагаемые элементы контроля с другой стороны, (например, путем охвата области предполагаемого элемента контроля, а также от 200 п.н. до 100 т.п.н. вокруг элемента) может использоваться как средство подтверждения таких элементов (путем измерения транскрипции представляющего интерес гена) или для обнаружения новых элементов контроля (например, путем охвата области, находящейся на расстоянии 100 т.п.н. в восходящем и нисходящем направлении от TSS представляющего интерес гена). Кроме того, нацеливание на предполагаемые элементы контроля может быть полезно в контексте понимания генетических причин заболевания. Многие мутации и распространенные SNP-варианты, связанные с фенотипами заболевания, расположены за пределами кодирующих областей. После нацеливания на такие области с помощью систем активации или подавления, описанных в настоящем описании, можно провести определение данных транскрипции для либо a) набора предполагаемых мишеней (например, набора генов, расположенных в непосредственной близости от элемента контроля), либо b) для всего транскриптома, например, с помощью секвенирования РНК или микрочипа. Это позволило бы идентифицировать вероятные гены-кандидаты, вовлеченные в фенотип болезни. Такие гены-кандидаты могут быть полезны в качестве новых мишеней лекарственных препаратов.
[00904] В настоящем описании упоминаются ингибиторы гистонацетилтрансферазы (HAT). Однако в некоторых вариантах осуществления альтернатива заключается в том, что один или более функциональных доменов содержат ацетилтрансферазу, предпочтительно гистонацетилтрансферазу. Они полезны в области эпигеномики, например, в способах исследования эпигенома. Способы исследования эпигенома могут включать, например, нацеливание на эпигеномные последовательности. Нацеливание на эпигемомные последовательностей может включать направляющую молекулу, направленную на эпигеномную последовательность-мишень. В некоторых вариантах осуществления эпигеномная последовательность-мишень может включать последовательность промотора, сайленсера или энхансера.
[00905] Использование функционального домена, связанного с эффекторным белком C2c1 или C2c3, как описано в настоящем описании, предпочтительно мертвым эффекторным белком C2c1 или C2c3, более предпочтительно мертвым эффекторным белком AacC2c1 или C2c3, для нацеливания на эпигеномные последовательности возможно для активации или подавления промоторов, сайленсеров или энхансеров.
[00906] Примеры ацетилтрансфераз известны, но могут включать, в некоторых вариантах осуществления, гистонацетилтрансферазы. В некоторых вариантах осуществления гистонацетилтрансфераза может содержать каталитическое ядро ацетилтрансферазы р300 человека (Gerbasch & Reddy, Nature Biotech, 6th April 2015).
[00907] В некоторых предпочтительных вариантах осуществления функциональный домен связан с мертвым эффекторным белком C2c1 или C2c3 для нацеливания на и активации эпигеномных последовательностей, таких как промоторы или энхансеры. Одна или более направляющих молекул, нацеленных на такие промоторы или энхансеры, также могут быть предоставлены для управления связыванием фермента CRISPR с такими промоторами или энхансерами.
[00908] Термин "ассоциированный с" используется в настоящем описании в отношении ассоциации функционального домена с эффекторным белком C2c1 или C2c3 или адаптерным белком. Он используется в отношении того, как одна молекула "ассоциирует" (связывается) с другой, например, между адаптерным белком и функциональным доменом, или между эффекторным белком C2c1 или C2c3 и функциональным доменом. В случае таких белок-белковых взаимодействий эту ассоциацию можно рассматривать с точки зрения распознавания, подобно тому, как антитело распознает эпитоп. Альтернативно, один белок может быть связан с другим белком посредством слияния двух субъединиц, например, одна субъединица является слитой с другой субъединицей. Слияние обычно происходит путем добавления одной аминокислотной последовательности к другой, например, путем совместного сплайсинга нуклеотидных последовательностей, которые кодируют каждый белок или субъединицу. Альтернативно, это может по существу рассматриваться как связывание двух молекул или прямое образование связи, такое как слитый белок. В любом случае слитый белок может включать линкер между представляющими интерес субъединицами (то есть между ферментом и функциональным доменом или между адаптерным белком и функциональным доменом). Таким образом, в некоторых вариантах осуществления эффекторный белок C2c1 или C2c3 или адаптерный белок ассоциирован с функциональным доменом путем связывания с ним. В других вариантах осуществления эффекторный белок C2c1 или C2c3 или адаптерный белок ассоциирован с функциональным доменом, потому что оба слиты вместе, необязательно посредством промежуточного линкера.
[00909] Присоединение функционального домена или слитого белка может производиться посредством линкера, например, гибкого глицин-серинового (GlyGlyGlySer) или (GGGS)3, либо жесткого альфа-спирального линкера, в частности, (Ala(GluAlaAlaAlaLys)Ala). Предпочтительно для разделения доменов белка или пептида в рамках настоящего изобретения используются линкеры, подобные (GGGGS)3. (GGGGS)3 является предпочтительным поскольку он является сравнительно длинным линкером (15 аминокислот). Остатки глицина являются наиболее гибкими, остатки серина усиливают вероятность пребывания линкера снаружи белка. В качестве предпочтительных альтернатив могут использоваться (GGGGS)6 (GGGGS) или (GGGGS)12. Другими предпочтительными вариантами являются (GGGGS)1, (GGGGS)2, (GGGGS)4, (GGGGS)5, (GGGGS)7, (GGGGS)8, (GGGGS)10 или (GGGGS)n. Альтернативные линкеры также доступны, однако линкеры с высокой гибкостью считаются функционирующими наилучшим образом для обеспечения возможности для 2 частей C2c1 или C2c3 сблизиться и, таким образом, восстановить активность C2c1 или C2c3. Альтернативным является, в частности, использование NLS в качестве линкера. Например, линкер может также быть находиться между C2c1 и любым функциональным доменом или между C2c3 и любым функциональным доменом. Вновь, линкер (GGGGS)3 (или его разновидности, содержащие 6, 9 или 12 повторов) или NLS нуклеоплазмина могут быть использованы здесь в качестве линкера между C2c1 и функциональным доменом или между C2c3 и функциональным доменом.
Насыщающий мутагенез
[00910] Система(ы) эффекторного белка C2c1 или C2c3, описанная в настоящем описании, может быть использована для выполнения насыщающего или глубоко сканирующего мутагенеза геномных локусов в сочетании с клеточным фенотипом, например, для определения критических минимальных признаков и дискретных уязвимостей функциональных элементов, необходимых для экспрессии генов, лекарственной устойчивости и купирования заболевания. Под "насыщающим" или "глубоко сканирующим мутагенезом" подразумевается, что каждое или по существу каждое основание ДНК разрезается внутри геномных локусов. В популяцию клеток можно ввести библиотеку направляющих РНК эффекторных белков C2c1 или C2c3. Библиотека может быть введена таким образом, что каждая клетка получает одну направляющую молекулу РНК (sg-РНК). В случае, когда библиотека вводится путем трансдукции вирусного вектора, как описано в настоящем описании, используется низкая множественность инфекции. Библиотека может включать sg-РНК, нацеленные на каждую последовательность в восходящем направлении от последовательности, прилегающей к протоспейсеру (PAM) в геномном локусе. Библиотека может включать по меньшей мере 100 непересекающихся геномных последовательностей в восходящем направлении от PAM для каждых 1000 пар оснований в геномном локусе. Библиотека может включать sg-РНК, нацеливающие на последовательности в восходящем направлении по меньшей мере от одной отличающейся PAM. Системы эффекторного белка C2c1 или C2c3 могут включать более одного белка C2c1 или C2c3. Можно использовать любой эффекторный белок C2c1 или C2c3, как описано в настоящем описании, включая ортологи или сконструированные эффекторные белки C2c1 или C2c3, которые распознают различные последовательности PAM. Частота не являющихся мишенями сайтов для sg-РНК может составлять менее 500. Оценка нецелевого связывания sg-РНК необходима для отбора sg-РНК с наименьшим уровнем возможного связывания с не являющимися мишенями участками. Любой фенотип, который, как установлено, связан с разрезанием участке-мишени sg-РНК, может быть подтвержден с использованием sg-РНК, нацеленных на один и тот же сайт в одном эксперименте. Валидация участка-мишени также может быть выполнена с использованием модифицированного эффекторного белка C2c1 или C2c3, как описано в настоящем описании, и двух sg-РНК, нацеливающих на участок-мишень в геноме. Без связи с теорией, можно утверждать, что участок-мишень действительно установлен, если изменение фенотипа наблюдается в экспериментах по проверке достоверности.
[00911] Геномные локусы могут включать по меньшей мере одну непрерывную геномную область. По меньшей мере одна непрерывная геномная область может включать и весь геном. По меньшей мере одна непрерывная геномная область может содержать функциональный элемент генома. Функциональный элемент может находиться в некодирующей области, кодирующем гене, интронной области, промоторе или энхансере. По меньшей мере одна непрерывная геномная область может содержать по меньшей мере 1 т.п.н., предпочтительно по меньшей мере 50 т.п.н. геномной ДНК. По меньшей мере одна непрерывная геномная область может содержать сайт связывания фактора транскрипции. По меньшей мере одна непрерывная геномная область может содержать область, гиперчувствительную к расщеплению ДНК-азой I. По меньшей мере одна непрерывная геномная область может содержать усиливающий транскрипцию энхансер или подавляющий транскрипцию репрессорный элемент. По меньшей мере одна непрерывная геномная область может содержать сайт, обогащенный эпигенетической сигнатурой. По меньшей мере одна непрерывная область геномной ДНК может содержать эпигенетический инсулятор. По меньшей мере, одна непрерывная геномная область может содержать две или более непрерывных геномных областей, которые физически взаимодействуют. Геномные области, которые взаимодействуют, могут определяться "технологией 4C". 4C-технология позволяет провести скрининг всего генома непредвзятым образом для сегментов ДНК, которые физически взаимодействуют с выбранным фрагментом ДНК, как описано в Zhao et al. ((2006) Nat Genet 38, 1341-7) и в патенте США 8642295, которые включены в настоящее описание в качестве ссылок в полном объеме. Эпигенетическая сигнатура может представлять собой ацетилирование, метилирование, убиквитинилирование, фосфорилирование гистонов, метилирование ДНК или отсутствие всего вышеперечисленного.
[00912] Система(ы) эффекторного белка C2c1 или C2c3 для насыщающего или глубоко сканирующего мутагенеза может быть использована в популяции клеток. Система эффекторного белка C2c1 или C2c3 может использоваться в эукариотических клетках, включая, но не ограничиваясь ими, клетки млекопитающих и растений. Популяция клеток может состоять из прокариотических клеток. Популяция эукариотических клеток может быть популяцией эмбриональных стволовых (ES) клеток, нейронов, эпителиальных клеток, иммунных клеток, эндокринных клеток, мышечных клеток, эритроцитов, лимфоцитов, растительных клеток или дрожжевых клеток.
[00913] В одном аспекте настоящее изобретение относится к способу скрининга функциональных элементов, связанных с изменением фенотипа. Библиотека может быть введена в популяцию клеток, которые адаптированы для содержания эффекторного белка C2c1 или C2c3. Клетки могут быть рассортированы по меньшей мере на две основные группы на основе фенотипа. Фенотип может включать экспрессию гена, рост или жизнеспособность клеток. Может быть определена относительная представленность направляющих РНК, присутствующих в каждой группе, причем геномные сайты, связанные с изменением фенотипа, определяются по представленности направляющих РНК, присутствующих в каждой группе. Изменение фенотипа может быть изменением экспрессии гена-мишени. Экспрессия гена-мишени может быть повышена, понижена, или полностью выключена (нокаут гена). Клетки могут быть отсортированы в группы с высокой и низкой экспрессией. Популяция клеток может включать репортерную конструкцию, которая определяет фенотип. Конструкция репортера может включать поддающийся обнаружению маркер. Клетки могут быть отсортированы с помощью поддающегося обнаружению маркера.
[00914] В другом аспекте настоящее изобретение относится к способу скрининга геномных сайтов, связанных с устойчивостью к химическому соединению. Химическое соединение может быть лекарственным средством или пестицидом. Библиотека может быть введена в популяцию клеток, адаптированных к содержанию эффекторного белка C2c1 или C2c3, где каждая клетка популяции содержит не более одной направляющей РНК; популяцию клеток обрабатывают химическим соединением; и представленность направляющих РНК определяют после обработки химическим соединением в более поздний момент времени по сравнению с ранним моментом времени, при котором геномные участки, связанные с устойчивостью к химическому соединению, определяются по обогащению направляющими РНК. Представленность sg-РНК может быть определена способами глубокого секвенирования.
[00915] Полезными в практике настоящего изобретения с использованием комплексов эффекторных белков C2c1 или C2c3 являются способы, в которых используются системы CRISPR-Cas9, и может быть приведена ссылка на статью под названием "BCL11A enhancer dissection by Cas9-mediated in situ saturating mutagenesis". Canver, M.C., Smith,E.C., Sher, F., Pinello, L., Sanjana, N.E., Shalem, O., Chen, D.D., Schupp, P.G., Vinjamur, D.S., Garcia, S.P., Luc, S., Kurita, R., Nakamura, Y., Fujiwara, Y., Maeda, T., Yuan, G., Zhang, F., Grkin, S.H., & Bauer, D.E. DOI:10.1038/naturel552I, published online September 16, 2015, которая включена в настоящее описание в качестве ссылки и кратно обсуждается ниже.
[00916] Canver et al. использует новые объединенные библиотеки направляющих cr-РНК-Cas9, чтобы выполнять насыщающий мутагенез in situ человеческих и мышиных эритроидных энхансеров BCL11A, ранее идентифицированных как энхансер, ассоциированный с уровнем фетального гемоглобина (HbF), и мышиный ортолог которого необходим для экспрессии эритроида BCL11A. Этот подход выявил критические минимальные признаки и дискретную уязвимость этих энхансеров. Благодаря редактированию первичных человеческих предшественников и трансгенеза мышей, авторы подтвердили, что эритроидный энхансер BCL11A может быть использован в качестве мишени для реиндукции HbF. Авторы создали подробную карту энхансера, которая может быть использована при терапевтическом редактировании генома.
Способы использования систем C2c1 или C2c3 для модификации клетки или организма
[00917] В некоторых вариантах осуществления изобретение относится к способу модификации клетки или организма. Клетка может быть прокариотической или эукариотической. Клетка может быть клеткой млекопитающего. Клетка может быть не принадлежащей человеку клеткой млекопитающего, клеткой примата, коровы, свиньи, грызуна или мыши. Клетка может быть эукариотической клеткой не млекопитающего, такой как клетка домашней птицы, рыбы или креветки. Клетка также может быть растительной клеткой. Растительная клетка может быть клеткой зерновой культуры, такой как маниока, кукуруза, сорго, пшеница или рис. Растительная клетка может быть клеткой водоросли, дерева или овоща. Модификация, введенная в клетку согласно настоящему изобретению, может быть такой, что клетка и потомки клетки изменяются для улучшения производства биологических продуктов, таких как антитела, крахмал, спирт или другой желаемый клеточной продукции. Модификация, введенная в клетку согласно настоящему изобретению, может быть такой, что клетка и ее потомки включают изменение, которое изменяет продуцируемый ими биологический продукт.
[00918] Описываемая система может содержать один или более различных векторов. В одном аспекте изобретения эффекторный белок является кодон-оптимизированным для экспрессии в желаемом типе клеток, предпочтительно эукариотических клеток, предпочтительно клеток млекопитающего или клеток человека.
[00919] Упаковочные клетки обычно используются для получения вирусных частиц, которые способны инфицировать клетку-хозяина. Такие клетки включают клетки 293, которые упаковывают аденовирусы и клетки ψ2 или клетки РА317, которые упаковывают ретровирусы. Вирусные векторы, используемые в генной терапии, обычно получают путем создания клеточной линии, которая производит упаковку векторов нуклеиновой кислоты в вирусные частицы. Векторы обычно содержат минимальные вирусные последовательности, необходимые для упаковки и последующей интеграции в клетку-хозяина, при этом другие вирусные последовательности заменяются кассетой экспрессии для полинуклеотида(ов), подлежащего(их) экспрессии. Отсутствующие функции вируса обычно поставляются в процессе доставки упаковочными клетками линии. Например, векторы AAV, используемые в генной терапии, обычно имеют только последовательности ITR из генома AAV, которые необходимы для упаковки и интеграции в геном хозяина. Вирусная ДНК упаковывается с помощью клеточной линии, которая содержит вспомогательную плазмиду, кодирующую другие гены AAV, а именно rep и cap, но не содержащую последовательность ITR. Линия клеток также может быть инфицирована аденовирусом в качестве вспомогательной частицы. Вспомогательный вирус способствует репликации вектора AAV и экспрессии генов AAV из вспомогательной плазмиды. Вспомогательная плазмида не может быть упакована в больших количествах из-за отсутствия последовательности ITR. Контаминация аденовирусом может быть уменьшена, например, с помощью термообработки, к которой аденовирус более чувствителен, чем AAV. Квалифицированным специалистам в данной области известны дополнительные способы доставки нуклеиновых кислот в клетки. См., например, US20030087817, включенную в настоящее описание в качестве ссылки.
Доставка
[00920] Изобретение охватывает по меньшей мере один компонент комплекса CRISPR, например РНК, доставляемый посредством по меньшей мере одного комплекса наночастиц. В некоторых аспектах изобретение относится к способам, включающим доставку одного или более полинуклеотидов, в частности, одного или более векторов, как описано в настоящем описании, одного или более их транскриптов и/или одного или более белков, транскрибированных с них, в клетку-хозяина. В некоторых аспектах изобретение также относится к клеткам, полученным с помощью таких способов, и животным, содержащим указанные клетки или полученные из указанных клеток. В некоторых вариантах осуществления изобретения фермент CRISPR в сочетании (необязательно в составе комплекса) с направляющей последовательностью доставляется в клетку. Традиционные способы переноса генов, как основанные, так и не основанные на использовании вирусов, могут использоваться для введения нуклеиновых кислот в клетки млекопитающих или ткани-мишени. Такие способы могут быть использованы для введения нуклеиновых кислот, кодирующих компоненты системы CRISPR, в клетки в культуре или организме хозяина. Отличные от вирусных системы доставки включают ДНК-плазмиды, РНК (например, транскрипт описываемого в настоящей спецификации вектора), "голую" нуклеиновую кислоту и нуклеиновую кислоту в комплексе со средством доставки, таким как липосома. Системы доставки на основе вирусных векторов включают ДНК- или РНК-вирусы, которые имеют эписомальные или интегрированные геномы после доставки в клетку. Обзор методик генной терапии см. в Anderson, Science 256:808-813 (1992); Nabel & Feigner, TIBTECH 1.1:21.1-217 (1993); Mitam & Caskey, TIBTECH 11:162-166 (1993); Dillon, TIBTECH 11:167-175 (1993); Miller, Nature 357:455-460 (1992); Van Brunt, Biotechnology 6(10): 1149-1154 (1988); Vigne, Restorati ve Neurology and Neuroscience 8:35-36 (1995); Kremer & Perricaudet, British Medical Bulletin 51(1):31-44 (1995); Haddada et al., in Current Topics in Microbiology and Immunology Doerfler and Bohm (eds) (1995); и Yu et al., Gene Therapy 1:13-26 (1994).
[00921] Способы невирусной доставки нуклеиновых кислот включают липофекцию, нуклеофекцию, микроинъекцию, генные пушки, виросомы, липосомы, иммунолипосомы, поликатионы или конъюгаты липидов с нуклеиновыми кислотами, "голую" ДНК, искусственные вирионы и усиленное агентом поглощение ДНК. Липофекция описана, например, в патентах США № 5049386, 4946787; и 4897355), и липофекционные реагенты продаются на коммерческой основе (например, Transfectam™ и Lipofectin™). Катионные и нейтральные липиды, которые пригодны для эффективного распознавания рецепторов липофекции полинуклеотидов, включают таковые Feigner, WO 91/17424; WO 91/16024. Доставка может осуществляться в клетки (например, in vitro или ex vivo) или ткани-мишени (например, in vivo).
[00922] Получение комплексов липидов с нуклеиновыми кислотами, включая нацеленные липосомы, такие как иммунолипидные комплексы, хорошо известно специалисту в данной области (см., например, Crystal, Science 270: 404-410 (1995), Blaese et al., Cancer Gene Ther. 2: 291-297 (1995), Behr et at., Bioconjugate Chem., 5: 382-389 (1994), Remy et al., Bioconjugate Chem., 5: 647-654 (1994), Gao et al., Gene Therapy 2: 710-722 (1995), Ahmad et al., Cancer Res. 52: 4817-4820 (1992), патенты США №№ 4186183, 4217344, 4235871, 4261975, 4485054, 4501728, 4774085, 4837028 и 4946787).
[00923] В применении РНК или ДНК-вирусных систем для доставки нуклеиновых кислот используются современные способы нацеливания вируса на конкретные клетки в организме и передачи вирусного груза в ядро. Вирусные векторы могут вводиться непосредственно пациентам (in vivo) или их можно использовать для обработки клеток in vitro, а модифицированные клетки могут быть введены пациентам (ex vivo). Обычные вирусные системы могут включать ретровирусные, лентивирусные, аденовирусные, аденоассоциированные и вирусные вирусы простого герпеса для переноса генов. Встраивание в геном хозяина возможно с помощью способов переноса генов ретровирусов, лентивирусов и аденоассоциированных вирусов, часто приводящих к длительной экспрессии введенного трансгена. Кроме того, высокая эффективность трансдукции наблюдалась во многих других типах клеток- и тканей-мишеней.
[00924] Тропизм ретровируса может быть изменен путем включения чужеродных белков оболочки, что позволяет расширить потенциальную популяцию клеток-мишеней. Лентивирусные векторы представляют собой ретровирусные векторы, которые способны трансформировать или инфицировать не делящиеся клетки и, как правило, имеют высокие вирусные титры. Поэтому выбор ретровирусной системы переноса генов будет зависеть от ткани-мишени. Ретровирусные векторы состоят из цис-действующих длинных концевых повторов с упаковочной емкостью до 6-10 т.п.н. чужеродной последовательности. Минимальные цис-действующие LTR достаточны для репликации и упаковки векторов, которые затем используются для интеграции терапевтического гена в клетку-мишень для обеспечения постоянной трансгенной экспрессии. Широко используемые ретровирусные векторы включают векторы, основанные на вирусе лейкемии мышей (MuLV), вирусе лейкемии гиббонов (GaLV), вирусе иммунодефицита обезьян (SIV), вирусе иммунодефицита человека (ВИЧ) и их комбинациях (см., например, Buchscher et al., J. Virol, 66: 2731-2739 (1992), Johann et al., J. Virol. 66: 1635-1640 (1992), Sommnerfelt et al., Virol. 176: 58-59 (1990), Wilson et al., J. Virol, 63: 2374-2378 (1989), Miller et al., J. Virol. 65: 2220-2224 (1991), PCT/US94/05700).
[00925] В другом варианте осуществления предусматриваются псевдотипированные оболочкой везикуловируса Cocal ретровирусные векторные частицы (см., например, публикацию патента США № 20120164118, зарегистрированную Fred Hutchinson Cancer Research Center). Вирус Cocal принадлежит к роду Vesiculovirus и является возбудителем везикулярного стоматита у млекопитающих. Исходно вирус Cocal был выделен из клещей в Тринидаде, Honkers et al. Am. J. Vet. Res. 25:236-242 (1964)), и случаи инфекции были установлены в Тринидаде, Бразилии и Аргентине у насекомых, крупного рогатого скота и лошадей. Многие везикуловирусы, заражающие млекопитающих, были выделены из зараженных естественным образом членистоногих, что предполагает их трансмиссивность. Антитела к везикуловирусам распространены у людей, живущих в промышленных регионах, где данные вирусы эндемичны и получены в лабораториях; заражение людей обычно приводит к симптомам, сходных с гриппом. Гликопротеин оболочки вируса Cocal на аминокислотном уровне имеет 71,5% идентичности с VSV-G Indiana, и филогенетическое сравнение гена оболочки везикуловирусов демонстрирует, что вирус Cocal является серологически отличным от штаммов VSV-G Indiana, но имеет наибольшее родство с ним (Jonkers et al. Am. J. Vet. Res. 25:236-242 (1964) and Travassos da Rosa et al. Am. J. Tropical Med. & Hygiene 33:999-1006 (1984)). Псевдотипированные оболочкой везикуловируса Cocal ретровирусные векторные частицы могут включать, например, лентивирусные, альфаретровирусные, бетаретровирусные, гаммаретровирусные и эпсилон ретровирусные векторные частицы, которые могут содержать ретровирусные Gag, Pol и/или один или более вспомогательный/ых белок/белков, а также белок оболочки везикуловируса Cocal. В некоторых аспектах изобретения Gag, Pol и вспомогательные белки являются лентивирусными и/или гаммаретровирусными. Изобретение охватывает AVV, содержащие или состоящие по большей части из молекул экзогенных нуклеиновых кислот, кодирующих систему CRISPR, например, многообразие кассет, включающее или состоящее из первой кассеты, включающей или по существу состоящей из промотора, нуклеиновой кислоты, молекулы, кодирующей ассоциированный с CRISPR (Cas) белок (предполагаемые белки-нуклеазы или хеликазы), например, C2c1 или C2c3 и терминатор, и 2 или более, предпочтительно до предела объема емкости упаковки, например, в общей сложности пять (включая первую кассету) кассет, содержащих или по существу состоящих из промотора, молекулы нуклеиновой кислоты, кодирующей направляющую РНК (гРНК) и терминатор (например, каждая кассета может быть схематически представлена как промотор-гРНК1-терминатор, промотор-гРНК2-терминатор … промотор-гРНК(N)-терминатор (где N - количество, которое может быть вставлено, представляет собой верхний предел упаковки вектора), или два или более отдельных rAAV, каждый из которых содержит одну или более кассет системы CRISPR, например, первый rAAV, содержащий первую кассету, включающую или по существу состоящую из промотора, молекулы нуклеиновой кислоты, кодирующей Cas, например, Cas (C2c1 или C2c3) и терминатор, и второй rAAV, содержащий многообразие из четырех кассет, содержащих или по существу состоящих из промотора, молекулы нуклеиновой кислоты, кодирующей направляющую РНК (гРНК) и терминатор (например, каждая кассета может быть схематически представлена как промотор-gРНК1-терминатор, промотор-gРНК2-терминатор … промотор-gРНК(N)-терминатор (где N - количество, которое может быть вставлено, представляет собой верхний предел упаковки вектора). Поскольку rAAV является ДНК-содержащим вирусом, молекулы нуклеиновой кислоты, описанные в настоящем описании в отношении AAV или rAAV предпочтительно представляет собой ДНК. В некоторых модификациях промотор предпочтительно является промотором cинапсина I (hSyn). Дополнительные способы доставки нуклеиновых кислот в клетку известны специалисту в данной области. См., например, US20030087817, включенный в настоящее описание в качестве ссылки.
[00926] В некоторых вариантах осуществления клетку-хозяина временно или постоянно трансфицируют одним или более векторами, описанными в настоящем описании. В некоторых вариантах осуществления клетку трансфицируют в том виде, в каком она присутствует у индивидуума естественным образом. В некоторых вариантах осуществления изобретения клетку, которую трансфицируют, извлекают из субъекта, такого как клеточная линия. В данной области известно большое разнообразие клеточных линий для тканевой культуры. Примеры клеточных линий включают, но не ограничиваются ими: C8161, CCRF-CEM, MOLT, mIMCD-3, NHDF, HeLa-S3, Huh1, Huh4, Huh7, HUVEC, HASMC, HEKn, HEKa, MiaPaCell, Panc1, PC-3, TF1, CTLL-2, C1R, Rat6, CV1, RPTE, A10, T24, J82, A375, ARH-77, Calu1, SW480, SW620, SKOV3, SK-UT, CaCo2, P388D1, SEM-K2, WEHI-231, HB56, TIB55, Jurkat, J45.01, LRMB, Bc1-1, ВС-3, IC21, DLD2, Raw264.7, NRK, NRK-52E, MRC5, MEF, Hep G2, HeLa B, HeLa T4, COS, COS-1, COS-6, COS-M6A, эпителий почки обезьяны BS-C-1, фибробласты эмбрионов мыши BALB/3T3, 3T3 Swiss, 3T3-L1, 132-d5 фетальные фибробласты человека, фибробласты мыши 10.1, 293-Т, 3T3, 721, 9L, A2780, A2780ADR, A2780cis, A172, A20, A253, A431, A-549, ALC, B16, B35, BCP-1, BEAS-2B, bEnd.3, BHK-21, BR 293, ВхРСЗ, C3H-10T1/2, C6/36, Cal-27, CHO, CHO-7, CHO-IR, CHO-K1, CHO-K2, CHO-T, CHO Dhfr -/-, COR-L23, COR-L23/CPR, COR-L23/5010, COR-L23/R23, COS-7, COV-434, CML T1, CMT, CT26, D17, DH82, DU145, DuCaP, EL4, EM2, EM3, EMT6/AR1, EMT6/AR10.0, FM3, H1299, H69, HB54, HB55, HCA2, HEK-293, HeLa, Hepa1c1c7, HL-60, HMEC, HT-29, Jurkat, JY клетки, клетки K562, Ku812, KCL22, KG1, KYO1, LNCap, Ma-Mel 1-48, MC-38, MCF-7, MCF-10A, MDA-MB-231, MDA-MB-468, MDA-MB-435, MDCK II, MDCK II, MOR/0.2R. MONO-MAC 6, MTD-1A, MyEnd, NCI-H69/CPR, NCI-H69/LX10, NCI-H69/LX20, NCI-H69/LX4, NIH-3T3, NALM-1, NW-145, OPCN /OPCT клеточные линии, Peer, PNT-1A/PNT 2, RenCa, RIN-5F, RMA/RMAS, клетки Saos-2, Sf-9, SkBr3, T2, T-47D, T84, THP1, U373, U87, U937, VCaP, клетки Vero, WM39, WT-49, X63, YAC-1, YAR и их трансгенные разновидности. Клеточные линии доступны из множества источников, известных квалифицированным специалистам в данной области (см., например, Американскую коллекцию типовых культур (ATCC) (Manassus, Va.)). В некоторых вариантах осуществления клетка, трансфицированная одним или более векторами, описанными в настоящем описании, используется для получения новой клеточной линии, содержащей одну или более последовательностей, полученных из вектора. В некоторых вариантах осуществления клетка, временная трансфицированная компонентами системы, нацеленной на нуклеиновую кислоту, как описано в настоящем описании (например, путем временной трансфекции одного или более векторов или трансфекции РНК) и модифицированная посредством активности комплекса нацеливания на нуклеиновую кислоту, используется для создания новой клеточной линии, содержащей клетки, имеющие модификацию, но не имеющие какой-либо другой экзогенной последовательности. В некоторых вариантах осуществления изобретения клетки, временно или постоянно трансфицированные одним или более векторами, описанными в настоящем описании, или клеточные линии, полученные из таких клеток, используются для оценки одного или более тестируемых соединений.
[00927] В некоторых вариантах осуществления изобретения один или более векторов, описанных в настоящем описании, используются для получения трансгенного животного, отличного от человека, или трансгенного растения. В некоторых вариантах осуществления трансгенное животное представляет собой млекопитающее, такое как мышь, крыса или кролик. В некоторых вариантах осуществления организм или субъект является растением. В некоторых вариантах осуществления организм, субъект или растение являются водорослями. Способы получения трансгенных растений и животных известны в данной области и обычно начинаются с использования способа трансфекции клеток, как описано в настоящем описании. В другом варианте осуществления устройство для доставки жидкости игольчатой матрицей (см., например, публикацию патента США № 20110230839 за авторством Fred Hutchinson Cancer Research Center) может рассматриваться как средство для доставки CRISPR Cas в твердые ткани. Устройство согласно публикации патента США № 20110230839 для доставки жидкости в твердую ткань может состоять из множества игл, объединенных в матрицу; ряда резервуаров, каждый из которых связан потоком жидкости с соответствующей иглой из матрицы игл, а также множества приводов, функционально сопряженных с соответствующими им резервуарами и настроенных таким образом, чтобы контролировать давление жидкости в резервуаре. В некоторых вариантах осуществления каждый из множества приводов может включать один из множества поршней, один конец каждого из множества поршней соединен с соответствующим ему одному из множества резервуаров, и в некоторых дальнейших вариантах осуществления поршни из множества поршней функционально соединены вместе соответствующим вторым концом таким образом, что возможно одновременное надавливание на них. Некоторые дальнейшие варианты осуществления могут включать пусковой механизм поршня, настроенный таким образом, чтобы надавливать на множество поршней с избираемой варьирующейся скоростью. В других модификациях каждый из множества приводов может включать один из множества передающих жидкость линий, имеющих первый и второй конец, первый конец каждой из множества передающих жидкость линий сопряжен с соответствующим ему из множества резервуаров. В других вариантах осуществления такое устройство может включать источник давления жидкости и каждый из множества приводов включает гидравлическую передачу между источником давления и соответствующему ему из множества резервуаров. В следующих вариантах осуществления источник давления жидкости может содержать по меньшей мере один из следующих элементов: компрессор, вакуумный накопитель, перистальтический насос, главный цилиндр, микроструйный насос и клапан. В другом варианте осуществления каждая из множества игл может содержать множество отверстий, распределенных по ее длине.
[00928] В одном аспекте изобретение относится к способам модификации полинуклеотида-мишени в эукариотической клетке. В некоторых вариантах осуществления такой способ включает позволение комплекса, нацеленного на нуклеиновую кислоту, связываться с полинуклеотидом-мишенью для расщепления указанного полинуклеотида-мишени, тем самым модифицируя полинуклеотид-мишень, причем комплекс, нацеленный на нуклеиновую кислоту, включает эффекторный белок, нацеленный на нуклеиновую кислоту, и направляющую РНК, гибридизованную с последовательностью-мишенью в указанном полинуклеотиде-мишени.
[00929] В одном аспекте изобретение относится к способу модификации экспрессии полинуклеотида в эукариотической клетке. В некоторых вариантах осуществления изобретения способ включает позволение комплексу, нацеленному на нуклеиновую кислоту, связываться с полинуклеотидом, так что указанное связывание приводит к увеличению или уменьшению экспрессии указанного полинуклеотида; где комплекс, нацеленный на нуклеиновую кислоту, содержит эффекторный белок, нацеленный на нуклеиновую кислоту, связанный с направляющей РНК, которая гибридизована с последовательностью-мишенью в указанном полинуклеотиде.
[00930] Компоненты комплекса CRISPR могут быть доставлены в сочетании с транспортирующими частями (адаптированными, например, из подходов, описанных в патентах США № 8106022, 8313772). Стратегии доставки нуклеиновой кислоты могут, например, быть использованы для улучшения доставки направляющей РНК, матричных РНК или кодирующих ДНК, в которых закодированы компоненты комплекса CRISPR. Например, молекулы РНК могут включать в себя модифицированные нуклеотиды РНК для повышения стабильности, снижения иммуностимуляции и/или улучшения специфичности (см. Deleavey, Glen F. et al., 2012, Chemistry & Biology, Volume 19, Issue 8, 937-954; Zalipsky, 1995, Advanced Drug Delivery Reviews 16. 157-182; Caliceti and Veronese, 2003, Advanced Drug Delivery Reviews 55: 1261-1277). Описаны различные структуры, которые могут быть использованы для модификации нуклеиновых кислот, таких как гРНК, с целью более эффективной доставки, в частности, обратимо нейтрализующие заряд модификации остова фосфотриэфира, которые могут быть адаптированы для модификации гРНК, обеспечивающей ей более высокую гидрофобность и меньшую анионность, облегчая вход в клетку (Meade BR et al., 2014, Nature Biotechnology 32,1256-1261). В дальнейших альтернативных вариантах осуществления отдельные мотивы РНК могут быть полезны для опосредования трансфекции клеток (Magalhaes M, et al., Molecular Therapy (2012); 20 3, 616-624). Сходным образом, аптамеры могут быть адаптированы для доставки компонентов комплекса CRISPR, например, путем присоединения аптамеров к молекулам гРНК (Tan W. et al., 2011, Trends in Biotechnology, December 2011, Vol. 29, No. 12).
[00931] В некоторых вариантах осуществления конъюгат трехантенного N-ацетилгалактозамина (GalNAc) с олигонуклеотидными компонентами может быть использован для улучшения доставки, например, доставки в заданные типы клеток, например, гепатоциты (см. WO2014118272, включенные в настоящее описание в качестве ссылки; Nair, JK et al., 2014, Journal of the American Chemical Society 136 (49), 16958-16961). Он может рассматриваться как основанная на молекуле сахара частица, и дальнейшие подробности, затрагивающие другие основанные на использование частиц системы доставки и/или составы приведены в настоящем описании, таким образом, GalNAc может рассматриваться как частица согласно приведенному в настоящем описании определению, так что главные способы использования и другие вопросы, например, затрагивающие доставку названных частиц, распространяются также на частицы GalNAc. Стратегия получения конъюгатов в растворе может быть, например, использована для присоединения трехантенных кластеров (с молекулярным весом ~2000), активированных в качестве эфиров PFP (пентафлуорофенил) c 5'-гексамино-модифицированным олигонуклеотидам (5'-HA ASO, молекулярная масса ~8000 Да; Ostergaard et al., Bioconjugate Chem., 2015, 26 (8), pp 1451-1455). Сходным образом, описано применение поли(акрилатных полимеров для доставки нуклеиновых кислот in vivo (см. WO2013158141, включенную в настоящее описание в качестве ссылки). В следующих альтернативных вариантах осуществления предварительное добавление наночастиц CRISPR (или белковых комплексов) к естественно присутствующим в плазме белкам плазмы может использоваться для улучшения способов доставки (Akinc A et al, 2010, Molecular Therapy vol. 18 no. 7, 1357-1364).
[00932] Доступны способы скрининга для применения с целью идентификации средств усиления доставки, например, скрининг химических библиотек (Gilleron J. et al., 2015, Nucl. Acids Res. 43 (16): 7984-8001). Также описаны подходы для количественной оценки эффективности средств доставки, в частности, липидные наночастицы, которые могут быть использованы для идентификации эффективных средств доставки компонентов CRISPR (см. Sahay G. et al., 2013, Nature Biotechnology 31, 653-658).
[00933] В некоторых вариантах осуществления доставка белковых компонентов CRISPR может быть облегчена за счет добавления функциональных пептидов к белку, в частности, пептидов, изменяющих гидрофобность, например, для того, чтобы улучшить функциональность in vivo. Белки, являющиеся компонентами CRISPR, сходным образом могут быть модифицированы для облегчения протекания химических реакций. Например, возможно добавление аминокислот к белку, имеющему группы, подвергнутые модификации способом клик-химии (Nikie I. et al., 2015, Nature Protocols 10,780-791). В таких вариантах осуществления участвующая в клик-реакции группа может далее быть использована для добавления широкого круга альтернативных структур, в частности, поли(этиленгликоля) с целью повышения стабильности, проникающих в клетку пептидов, РНК-аптамеров, липидов и углеводов, в частности, GalNAc. В дальнейших альтернативных вариантах белковый компонент CRISPR может быть адаптирован для проникновения в клетку (см. Svensen et al., 2012, Trends in Pharmacological Sciences, Vol. 33, No. 4), например, путем присоединения к белку проникающих в клетку пептидов (см. Kauffman, W. Berkeley et al., 2015, Trends in Biochemical Sciences, Volume 40, Issue 12, 749-764; Koren and Torchilin, 2012, Trends in Molecular Medicine, Vol. 18, No. 7). В следующих альтернативных вариантах осуществления пациентам или индивидуумам могут предварительно быть введены соединения или составы, способствующие последующей доставке компонентов CRISPR.
Комплексы эффекторного белка C2c1 и C2c3 могут использоваться в растениях
[00934] Система(ы) эффекторного белка C2c1 или C2c3 (например, одиночная или мультиплексированная) может использоваться в сочетании с новейшими достижениями в геномике культур. Описанную в настоящем описании систему можно использовать для выполнения результативного и экономически эффективного исследования или манипуляции с геном или геномом растений, например, для быстрого исследования, и/или отбора, и/или выбора, и/или сравнения, и/или манипуляций, и/или трансформации растительных генов или геномов, например, для создания, идентификации, разработки, оптимизации или присвоения определенной характеристики(характеристик) или признака (набора признаков) растениям или для трансформации генома растения. Таким образом, может быть улучшено производство растений, а именно новых растений с новыми комбинациями признаков или характеристик или новых растений с улучшенными признаками. Система или системы из эффекторного белка C2c1 или C2c3 может использоваться на растениях при использовании следующих способов: сайт-направленной интеграции (SDI) или редактирования генов (GE), близкого обратного скрещивания (NRB) или обратного скрещивание (RB). Аспекты использования описанных в настоящем описании эффекторных систем C2c1 или C2c3 могут быть аналогичны использованию систем CRISPR-Cas (например, CRISPR-Cas9) в растениях. В этой связи заслуживает упоминания веб-сайт Университета Аризоны "CRISPR-PLANT" (http: /www.genome.arizona.edu/crispr/) (поддерживается PennState и AGI). Варианты осуществления изобретения могут быть использованы при редактировании генома в растениях или в тех случаях, где ранее применялись способы с использованием иРНК или аналогичных способов редактирования генома; см., например, Nekrasov, "Plant genome editing made easy: targeted mutagenesis in model and crop plants using the CRISPR-Cas system," Plant Methods 2013, 9:39 (doi: 10.1186/1746-4811-9-39); Brooks, "Efficient gene editing in tomato in the first generation using the CRISPR-Cas9 system," Plant Physiology September 2014 pp 114.247577; Shan, "Targeted genome modification of crop plants using a CRISPR-Cas system," Nature Biotechnology 31, 686-688 (2013); Feng, "Efficient genome editing in plants using a CRISPR/Cas system," Cell Research (2013) 23:1229-1232 doi:10.1038/cr.2013.114; published online 20 August 2013; Xie, "RNA-guided genome editing in plants using a CRISPR-Cas system," Mol Plant. 2013 Nov;6(6):1975- 83. doi: 10.1093/mp/sstl 19. Epub 2013 Aug 17; Xu, "Gene targeting using the Agrobacterium tumefaciens-mediated CRISPR-Cas system in rice," Rice 2014, 7:5 (2014), Zhou et af., "Exploiting SNPs for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-coumarate: CoA ligase specificity and Redundancy," New Phytologist (2015) (Forum) 1-4 (доступен через интернет по ссылке wmv.newphytologist.com); Caliando et al, "Targeted DNA degradation using a CRISPR device stably carried in the host genome, NATURE COMMUNICATIONS 6:6989,DOI: 10.1038/ncomms7989, vvwvv.nature.com/naturecommunications DOI: 10.1038/ncomms7989; патент США № 6603061 - Agrobacterium-Mediated Plant Transformation Method; патент США № 7868149 - Plant Genome Sequences and Uses Thereof, и US 2009/0100536 - Transgenic Plants with Enhanced Agronomic Traits, содержание каждого из которых включено в настоящее описание в качестве ссылки в полном объеме. В практическом воплощении изобретения, содержание и раскрытие Morrell et al. "Crop genomics: advances and applications", NatRevGenet. 2011 Dec 29; 13(21:85-96; включено в настоящее описание в качестве ссылки, в том числе в отношении того, как данное изобретение может быть применено к растениям. Соответственно, данная ссылка может также применяться и к клеткам животных, с учетом возможных различий, к растительным клеткам, если не указано иное; и в этом случае ферменты, обладающие сниженными неспецифическими эффектами и системы, использующие такие ферменты, могут быть использованы применительно к растениям, в том числе к тем, которые упомянуты в настоящей заявке.
Применение систем CRISPR C2c1 или C2c3 в растениях и дрожжах
Определение:
[00935] В целом термин "растение" касается любых разнообразных фотосинтетических, эукариотических, одноклеточных или многоклеточных организмов царства Растения (Plantae), для которых характерен рост путем клеточного деления, наличие хлоропластов, и клеточных стенок, состоящих из целлюлозы. Термин "растение" включает однодольные и двудольные растения. Более конкретно, предполагается, что растения включают, но не ограничиваются ими, покрытосеменные и голосеменные растения, такие как акация, люцерна, амарант, яблоко, абрикос, артишок, ясень, спаржа, авокадо, банан, ячмень, бобы, свекла, береза, бук, ежевика, черника, брокколи, брюссельская капуста, капуста, канола, мускусная дыня, морковь, маниока, цветная капуста, кедр, хлебные злаки, сельдерей, каштан, вишня, китайская капуста, цитрусовые, клементин, клевер, кофе, кукуруза, хлопок, вигна (коровий горох), огурец, кипарис, баклажан, вяз, эндивий, эвкалипт, фенхель, инжир, ель, герань, виноград, грейпфрут, земляной орех, физалис, тсуга, гикори, кудрявая капуста (кале), киви, кольраби, лиственница, салат-латук, лук-порей, лимон, лайм, робиния, сосна, адиантум, маис, манго, клен, дыня, просо, гриб, горчица, орехи, дуб, овес, масличная пальма, бамия, лук, апельсин, декоративное растение или цветок или дерево, папайя, пальма, петрушка, пастернак, горох, персик, арахис, груша, сфагнум, перец, хурма, голубиный горох (каян), сосна, ананас, подорожник, слива, гранат, картофель, тыква, радиккио, редька, рапс, малина, рис, рожь, сорго, сафлор, ива, соя, шпинат, ель, патиссон, земляника, сахарная свекла, сахарный тростник, подсолнечник, батат, сладкая кукуруза, мандарин, чай, табак, помидор, деревья, тритикале, газонные травы, репа, виноградная лоза, грецкий орех, водяной кресс, арбуз, пшеница, ямс, тис и цукини. Термин растение также охватывает водоросли (Algae), которые являются, главным образом, фотоавтотрофами, объединенными, прежде всего, отсутствием корней, листьев и других органов, которые характеризуют высшие растения.
[00936] Способы редактирования генома с использованием системы C2c1 или C2c3, как описано в настоящем описании, могут использоваться для придания желаемых свойств по существу любому растению. Большое разнообразие растений и систем растительных клеток с желаемыми физиологическими и агрономическими характеристиками, описанными в настоящем описании, может быть разработано с использованием конструкции из нуклеиновых кислот, описанных в настоящем описании, и различных упомянутых выше способов преобразования. В предпочтительных вариантах осуществления изобретения целевые растения и растительные клетки для инженерии включают, но не ограничиваются ими, однодольные и двудольные растения, такие как зерновые культуры, включая хлебные культуры (например, пшеница, кукуруза, рис, просо, ячмень), фруктовые культуры (например, помидор, яблоко, груша, клубника, апельсин), кормовые культуры (например, люцерна), овощные культуры (например, морковь, картофель, сахарная свекла, ямс), листовые овощные культуры (например, салат, шпинат); цветущие растения (например, петуния, роза, хризантема), хвойные деревья и (например, соснa, пихта, ель), растения, используемые в фиторемедиации (например, растения, накапливающие тяжелые металлы); масличные культуры (например, подсолнечник, рапс) и растения, используемые в экспериментальных целях (например, Arabidopsis). Таким образом, способы и системы CRISPR-Cas могут использоваться в широком спектре растений, например, в двудольных растениях, принадлежащих к следующим отрядам: Magnoliales, llliciales, Laurales, Piperales, Aristochiales, Nymphaeales, Ranunculales, Papaverales, Sarraceniaceae, Trochodendrales, Hamamelidales, Eucomiales, Leitneriales, Myricales, Fagales, Casuarinales, Caryophyllales, Batales, Polygonales, Plumbaginales, Dilleniales, Theales, Malvales, Urticales, Lecythidales, Violales, Salicales, Capparales, Ericales, Diapensales, Ebenales, Primulales, Rosales, Fabales, Podostemales, Haloragales, Myrtales, Cornales, Proteales, Santales, Rafflesiales, Celastrales, Euphorbiales, Rhamnales, Sapindales, Juglandales, Geraniales, Polygalales, Umbellales, Gentianales, Polemoniales, Lamiales, Plantaginales, Scrophulariales, Campanulales, Rubiales, Dipsacales и Asterales; способы и системы CRISPR-Cas могут использоваться в однодольных растениях, таких как растения, принадлежащие к следующим отрядам: Alismatales, Hydrocharitales, Najadales, Triuridales, Commelinales, Eriocaulales, Restionales, Poales, Juncales, Cyperales, Typhales, Bromeliales, Zingiberales, Arecales, Cyclanthales, Pandanales, Arales, Lilliales и Orchidales, или в растениях, принадлежащих к голосеменным, например, принадлежащих к отрядам: Pinales, Ginkgoales, Cycadales, Araucariales, Cupressales и Gnetales.
[00937] Системы CRISPR C2c1 или C2c3 и способы их применения, описанные в настоящем описании, могут использоваться в широком диапазоне видов растений, включенных в неограниченный список двудольных, однодольных или голосеменных растений, принадлежащих к следующим родам: Atropa, Alseodaphne, Anacardium, Arachis, Beilschmiedia, Brassica, Carthamus, Cocculus, Croton, Cucumis, Citrus, Citrullus, Capsicum, Catharanthus, Cocos, Coffea, Cucurbita, Daucus, Duguetia, Eschscholzia, Ficus, Fragaria, Glaucium, Glycine, Gossypium, Helianthus, Hevea, Hyoscyamus, Lactuca, Landolphia, Linum, Litsea, Lycopersicon, Lupinus, Manihot, Majorana, Malus, Medicago, Nicotiana, Olea, Parthenium, Papaver, Persea, Phaseolus, Pistacia, Pisum, Pyrus, Prunus, Raphanus, Ricinus, Senecio, Sinomenium, Stephania, Sinapis, Solanum, Theobroma, Trifolium, Trigonella, Vicia, Vinca, Vitis, and Vigna; and the genera Allium, Andropogon, Eragrostis, Asparagus, Avena, Cynodon, Elaeis, Festuca, Festulolium, Heterocallis, Hordeum, Lemna, Lolium, Musa, Oryza, Panicum, Pannesetum, Phleum, Poa, Secale, Sorghum, Triticum, Zea, Abies, Cunninghamia, Ephedra, Picea, Pirns и Pseudotsuga.
[00938] Системы CRISPR C2c1 или C2c3 и способы их применения могут также использоваться в широком спектре "водорослей" или "клеток водорослей"; включая, например, водоросли, отобранные из нескольких эукариотических филюмов, включая следующие: Rhodophyta (красные водоросли), Chlorophyta (зеленые водоросли), Phaeophyta (коричневые водоросли), Bacillariophyta (диатомовые водоросли), Eustigmatophyta и динофлагелляты, а также прокариотический филюм Cyanobacteria (сине-зеленые водоросли). Термин "водоросли" включает, например, водоросли, отобранные из следующих родов: Amphora, Anabaena, Ankistrodesmus, Botryococcus, Chaetoceros, Chlamydomonas, Chlorella, Chlorococcum, Cyclotella, Cylindrotheca, Dunaliella, Emiliana, Euglena, Hematococcus, Isochrysis, Monochrysis, Monoraphidium, Nannochloris, Nannochloropsis, Navicula, Nephrochloris, Nephroselmis, Nitzschia, Nodularia, Nostoc, Oochromonas, Oocystis, Oscillatoria, Pavlova, Phaeodactylum, Playtmonas, Pleurochrysis, Porphyra, Pseudoanabaena, Pyramimonas, Stichococcus, Synechococcus, Synechocystis, Tetraselmis, Thalassiosira и Trichodesmium.
[00939] Часть растения, т.е. "растительная ткань" может быть обработана согласно способам по настоящему изобретению, чтобы получить растение c улучшенными свойствами. Понятие "растительная ткань" также включает растительные клетки. Термин "растительная клетка", как используют в настоящем описании, относится к отдельным единицам живого растения, как в интактном целом растении, так и в изолированной форме, выращенной в культуре тканей in vitro, на питательных средах или агаре, в суспензии на питательной среде или буфере или в качестве части более высокоорганизованных единиц, например, таких как растительная ткань, орган растения или целое растение.
[00940] Термин "протопласт" относится к растительной клетке, защитная клеточная стенка которой была полностью или частично удалена, например, механическим или ферментативным способом, что привело к появлению интактной биохимически компетентной единице живого растения, которая способна заново сформировать клеточную стенку, пролиферировать, регенерировать и вырасти в целое растение при надлежащих условиях выращивания.
[00941] Термин "трансформация" в широком смысле относится к процессу генетической модификации растения-хозяина с помощью введения ДНК посредством Agrobacteria или одного из ряда химических или физических способов. Как используют в настоящем описании, термин "растение-хозяин" относится к растениям, включая любые клетки, ткани, органы или потомство растений. Много подходящих растительных тканей или растительных клеток могут быть преобразованы и включают, но не ограничены, следующими: протопласты, соматические эмбрионы, пыльца, листья, саженцы, стебли, каллусы, столоны, микроклубни и побеги. Понятие "растительная ткань" также относится к любому клону такого растения, семени, потомству, черенкам, возникших в результате полового или вегетативного размножения, и потомкам любого из них, такие как черенки или семена.
[00942] Термин "трансформированный", как используют в настоящем описании, относится к клетке, ткани, органу или организму, в которые была введена чужеродная конструкция, такая как молекула ДНК. Введенная молекула ДНК может быть интегрирована в ДНК клетки-реципиента, ткани, органа или организма, таким образом, что введенная молекула ДНК передается последующему потомству. В этих вариантах осуществления изобретения "трансформированная" или "трансгенная" клетка или растение также могут включать потомство клетки или растения и потомство, полученное с помощью искусственного разведения, использующего такое преобразованное растение в качестве родителя в скрещивании, и демонстрирующее измененный фенотип в результате присутствия введенной молекулы ДНК. Предпочтительно, трансгенное растение фертильно и способно к передаче введенной ДНК потомству посредством полового размножения.
[00943] Термин "потомство", такое как потомство трансгенного растения, относится к тому, что порождается, происходит от или получено из растения или трансгенного растения. Введенная молекула ДНК также может быть временно введена в клетку реципиента, таким образом, что последующее потомство не наследует введенную молекулу ДНК и, таким образом, не считается "трансгенным". Соответственно, как используют в настоящем описании, "нетрансгенное" растение или растительная клетка является растением, не содержащим чужеродную ДНК, устойчиво интегрированную в его геном.
[00944] Термин "промотор растения", как используют в настоящем описании, является промотором, способным к инициированию транскрипции в растительных клетках, независимо от того, происходит ли он из растительной клетки. Иллюстративные подходящие промоторы растений включают, но не ограничиваются ими, промоторы, полученные из растений, вирусов растения и бактерий, таких как Agrobacterium или Rhizobium, которые включают гены, экспрессируемые в растительных клетках.
[00945] Как используют в настоящем описании, "клетка гриба" относится к любым организмам из царства грибов, включая Ascomycota, Basidiomycota, Blastocladiomycota, Chytridiomycota, Glomeromycota, Microsporidia и Neocallimastigomycota. Клетки грибов могут включать дрожжи, плесени и филаментные грибы. В некоторых модификациях изобретения клеткой гриба является дрожжевая клетка.
[00946] Как используют в настоящем описании, термин "дрожжевая клетка" относится к любой клетке гриба в родов Ascomycota и Basidiomycota. Дрожжевые клетки могут включать почкующиеся дрожжевые клетки, делящиеся дрожжевые клетки и клетки плесени. Не будучи ограниченным этими организмами, многие типы дрожжей, используемых в лабораторных и промышленных условиях, являются частью рода Ascomycota. В некоторых вариантах осуществления изобретения дрожжевая клетка является клеткой S. cerevisiae, Kluyveromyces marxianus или Issatchenkia orientalis cell. Другие дрожжевые клетки могут включать, но не ограничиваться ими, Candida spp. (например, Candida albicans), Yarrowia spp. (например, Yarrowia lipolytica), Pichia spp. (например, Pichia pastoris), Kluyveromyces spp. (например, Kluyveromyces laсtis и Kluyveromyces marxianus), Neurospora spp. (например, Neurospora crassa), Fusarium spp. (например, Fusarium oxysporum), и Issatchenkia spp. (например, Issatchenkia orientalis, также именуемые Pichia kudriavzevii и Candida acidothermophilum). В некоторых вариантах осуществления изобретения клетка гриба является нитевидной клеткой гриба. Как используют в настоящем описании, термин "нитевидная клетка гриба" относится к любому типу клетки гриба, которая растет в виде нитей, т.е. гиф или мицелия. Примеры волокнистых клеток гриба могут включать, но не ограничиваться ими, Aspergillus spp. (например, Aspergillus niger), Trichoderma spp. (например, Trichoderma reesei), Rhizopus spp., (например, Rhizopus oryzae) и Mortierella spp. (например, Mortierella isabellina).
[00947] В некоторых вариантах осуществления изобретения клетка гриба является промышленным штаммом. Как используют в настоящем описании, "промышленный штамм" относится к любому штамму клетки гриба, используемому в или выделенному из производственного процесса, например, производства продукта в коммерческих или промышленных масштабах. Понятие "промышленный штамм" может относиться к виду гриба, который, как правило, используется в производственном процессе, или оно может относиться к изоляту вида гриба, который также может использоваться в непромышленных целях (например, лабораторных исследованиях). Примеры производственных процессов могут включать брожение (например, в производстве продуктов питания или напитков), дистилляцию, производство биотоплива, производство различных соединений и полипептидов. Примеры промышленных штаммов могут включать, но не ограничиваться ими, JAY270 и АТСС4124.
[00948] В некоторых вариантах осуществления изобретения клетка гриба является полиплоидной клеткой. Как используют в настоящем описании, понятие "полиплоидная" клетка может относиться к любой клетке, геном в которой присутствует больше, чем в одной копии. Понятие "полиплоидная клетка" может относиться к клетке, которая естественным образом существует в полиплоидном состоянии, или к клетке, полиплоидное состояние генома которой было вызвано искусственно (например, посредством специфического регулирования, изменения, инактивации, активации, или модификации мейоза, цитокинеза, или репликации ДНК). "Полиплоидная клетка" может относиться к клетке, весь геном которой является полиплоидным, или к клетке, которая является полиплоидом в конкретном геномном локусе-мишени. Чтобы не привязываться к теории, считается, что изобилие направляющих РНК может ограничивать скорость генной инженерии полиплоидных клеток в большей степени, чем гаплоидных клеток, и, таким образом, в способах использования системы CRISPR с белком С2с1 или С2с2, описанных в настоящем описании, могут использоваться преимущества определенного типа клетки гриба.
[00949] В некоторых вариантах осуществления изобретения клетка гриба является диплоидной клеткой. Как используют в настоящем описании, понятие "диплоидная" клетка может относиться к любой клетке, геном которой присутствует в двух копиях. Диплоидная клетка может относиться к типу клетки, которая естественным образом существует в диплоидном состоянии, к клетке, диплоидное состояние генома которой было вызвано искусственно (например, посредством специфического регулирования, изменения, инактивации, активации или модификации мейоза, цитокинеза или репликации ДНК). Например, штамм S. cerevisiae, S228C, может находиться в гаплоидном или диплоидном состоянии. "Диплоидная" клетка может относиться к клетке, весь геном которой является диплоидным, или к клетке, которая является диплоидной в конкретном геномном локусе-мишени. В некоторых вариантах осуществления изобретения клетка гриба является гаплоидной клеткой. Как используют в настоящем описании, "гаплоидная" клетка может относиться к любой клетке, геном которой присутствует в одной копии. Гаплоидная клетка может относиться к типу клетки, которая естественно находится в гаплоидном состоянии, или к клетке, гаплоидное состояние генома которой было вызвано искусственно (например, посредством специфичного регулирования, изменения, инактивации, активации или модификации мейоза, цитокинеза или репликации ДНК). Например, штамм S. cerevisiae, S228C, может находиться в гаплоидном или диплоидном состоянии. "Гаплоидная" клетка может относиться к клетке, весь геном которой является гаплоидным, или к клетке, которая является гаплоидной в конкретном геномном локусе-мишени.
[00950] Как используют в настоящем описании, "экспрессирующий вектор дрожжей" относится к нуклеиновой кислоте, которая содержит одну или более последовательностей, кодирующих РНК и/или полипептид, и может далее содержать любые желаемые элементы, которые управляют экспрессией нуклеиновой кислоты (кислот), а также любыми элементами, которые обеспечивают репликацию и сохранение вектора экспрессии в дрожжевой клетке. В данной области известно множество подходящих экспрессирующих векторов дрожжей и их особенностей; например, различные векторы и способы иллюстрированы в Yeast Protocols, 2nd edition, Xiao, W., ed. (Humana Press, New York, 2007) and Buckholz, R.G. and Gleeson, M.A. (1991) Biotechnology (NY) 9(11): 1067-72. Векторы дрожжей могут содержать, без ограничения, центромерную (CEN) последовательность, автономно реплицирующуюся последовательность (ARS), промотор, такой как промотор РНК-полимеразы III, функционально связанный с целевой последовательностью или геном, терминатор, такой как терминатор РНК-полимеразы III, ориджин репликации и маркерный ген (например, ауксотрофии, антибиотика или другие выбранные маркеры). Примеры экспрессирующих векторов для использования в дрожжах могут включать плазмиды, дрожжевые искусственные хромосомы, плазмиды 2μ, дрожжевые интегративные плазмиды, дрожжевые репликативные плазмиды, челночные векторы и эписомы.
Стабильная интеграция компонентов системы CRISPR С2с1 или С2с3 в геном растений и растительных клеток
[000001] В конкретных вариантах осуществления изобретения предусматривается, что полинуклеотиды, кодирующие компоненты системы CRISPR С2с1 или С2с3 вводятся для стабильной интеграции в геном растительной клетки. В этих вариантах осуществления изобретения конструкция вектора трансформации или системы экспрессии может быть отрегулирована в зависимости от того, когда, где и при каких условиях направляющая РНК и/или ген С2с1 или С2с3 экспрессируются.
[000002] В конкретных вариантах осуществления изобретения предусматривается введение компонентов системы CRISPR С2с1 или С2с3 стабильно в геномную ДНК растительной клетки. Дополнительно или альтернативно, предусматривается введение компонентов системы CRISPR С2с1 или С2с3 для стабильной интеграции в ДНК органелл растений, таких как, но не ограниченных ими, пластиды, митохондрии или хлоропласты.
[000003] Система экспрессии для стабильной интеграции в геном растительной клетки может содержать один или более следующих элементов: элемент промотора, который может использоваться для экспрессии РНК и/или фермента С2с1 или С2с3 в растительной клетке; 5'-нетранслируемые области для увеличения экспрессии; элемент интрона для дальнейшего увеличения экспрессии в определенных клетках, таких как клетки однодольного растения, сайт множественного клонирования, чтобы обеспечить удобные сайты рестрикции для вставки направляющей РНК и/или последовательности гена С2с1 или С2с3 и других желаемых элементов; и 3'-нетранслируемые области для обеспечения эффективной терминации экспрессируемого транскрипта.
[000004] Элементы системы экспрессии могут быть на одной или более конструкциях экспрессии, которые являются либо кольцевыми, такими как плазмида или вектор трансформации, либо линейными, такими как двойная спираль ДНК.
[000005] В конкретном варианте система экспрессии CRISPR С2с1 или С2с3 включает, по меньшей мере:
(a) нуклеотидную последовательность, кодирующую направляющую РНК (гРНК), которая гибридизируется с последовательностью-мишенью в растении, и где направляющая РНК включает направляющую последовательность и последовательность прямого повтора;
(b) нуклеотидную последовательность, кодирующую белок С2с1 или С2с3,
[00951] где компоненты (a) или (b) расположены на одной и той же или на различных конструкциях, и посредством чего различные нуклеотидные последовательности могут находиться под контролем одного и того же или различных регуляторных элементов, действующих в растительной клетке.
[00952] Конструкция (конструкции) ДНК, содержащая компоненты системы CRISPR С2с1 или С2с3 и, когда применимо, последовательность матрицы, может быть введена в геном растения, часть растения или растительную клетку рядом обычных способов. Процесс обычно включает этапы отбора подходящей клетки или ткани, введение конструкции (конструкций) в клетку или ткань и регенерацию растительных клеток или растений из клеток или тканей.
[00953] В конкретных вариантах осуществления изобретения конструкция ДНК может быть введена в растительную клетку с использованием способов, таких как, но не ограничиваясь этим, электропорация, микроинъекция, аэрозольная инъекция протопластов растительной клетки, или конструкции ДНК могут быть введены непосредственно в растительную ткань с использованием биолистических способов, таких как бомбардировка частицами ДНК (см. также Fu et al., Transgenic Res, 2000 Feb;9(1):11 -9). Бомбардировка клеток частицами основана на ускорении частиц, покрытых геном(генами)-мишенью, приводящем к проникновению частиц в протоплазму и типично стабильной интеграции в геном, (см., например, Klein et al., Nature (1987), Klein et al., Bio/Technology (1992), Casas et al., Proc. Natl. Acad. Sci. USA (1993)).
[00954] В конкретных вариантах осуществления изобретения конструкции ДНК, содержащие компоненты системы CRISPR С2с1 или С2с3, могут быть введены в растение с помощью агробактериальной трансформации. Конструкции ДНК могут быть объединены с подходящими регионами фланкирования T-ДНК и введены в обычный вектор Agrobacterium tumefaciens. Чужеродная ДНК может быть включена в геном растений путем заражения растений или инкубирования протопластов растения с бактериями Agrobacterium, содержащими одну или более Ti-плазмид (индуцирующая опухоль плазмида) (см., например, Fraley et al., (1985), Rogers et al., (1987) и патент США № 5563055).
Промоторы растений
[00955] Для обеспечения подходящей экспрессии в растительной клетке, компоненты системы CRISPR С2с1 или С2с3, описанной в настоящем описании, как правило, помещены под контроль промотора растения, т.е. промотора, действующего в растительных клетках. Предусматривается использование различных типов промоторов.
[00956] Конститутивный промотор растения - промотор, который способен экспрессировать открытую рамку считывания (ORF), которую он контролирует во всех или почти всех тканях растения во время всех или почти всех стадий развития растения (называется "конститутивной экспрессией"). Одним не ограничивающим примером конститутивного промотора является промотор 35S вируса мозаики цветной капусты. Понятие "регулируемый промотор" относится к промоторам, которые управляют экспрессией генов не конститутивно, а регулируемым во времени и/или пространстве образом, и включает тканеспецифичные, тканепредпочтительные и индуцируемые промоторы. Различные промоторы могут управлять экспрессией гена в различных тканях или типах клеток, или на различных стадиях развития, или в ответ на различные условия окружающей среды. В конкретных вариантах осуществления изобретения один или более компонентов системы CRISPR С2с1 или С2с3 экспрессируется под контролем конститутивного промотора, такого как промотор 35S вируса мозаики цветной капусты. Тканепредпочтительные промоторы могут быть использованы для нацеливания на увеличенную экспрессию в определенных типах клеток в определенных тканях растения, например клеток сосудов в листьях или корнях или в определенных клетках семени. Примеры конкретных промоторов для использования в системе CRISPR С2с1 или С2с3 можно найти в Kawamata et al., (1997) Plant Cell Physiol 38:792-803; Yamamoto et al., (1997) Plant J 12:255- 65; Hire et al., (1992) Plant Mol Biol 20:207-18,Kuster et al., (1995) Plant Mol Biol 29:759-72, и Capana et al., (1994) Plant Mol Biol 25:681-91.
[00957] Иллюстратвные индуцибельные промоторы, обеспечивающие пространственно-временной контроль редактирования или экспрессии генов, могут использовать энергию. Форма энергии может включать, но не ограничиваться этим, звуковую энергию, электромагнитную радиацию, химическую энергию и/или тепловую энергию. Примеры индуцируемых систем включают тетрациклин-индуцируемые промоторы (Tet-On или Tet-Off), двухгибридные системы активации транскрипции, использующие небольшие молекулы (FKBP, ABA, и т.д.), или системы, индуцируемые светом (фитохромы, домены LOV или криптохромы), такие как эффектор транскрипции, индуцируемый светом (LITE), которые управляют изменениями в транскрипционной активности последовательность-специфическим образом. Компоненты индуцируемой светом системы могут включать фермент CRISPR С2с1 или С2с3 светочувствительный гетеродимер цитохрома (например, из Arabidopsis thaliana), и транскрипционный домен активации/репрессии. Другие примеры индуцируемых ДНК-связывающих белков и способов их применения приведены в US 61/736465 и US 61/721283, которые включены в настоящее описание в качестве ссылки в полном объеме.
[00958] В определенных вариантах осуществления изобретения временная или индуцируемая экспрессия может быть достигнута при помощи, например, регулируемых химическими стимулами промоторов, т.е. таких, для которых применение экзогенного химического агента вызывает экспрессию генов. Модуляция экспрессии генов может также быть достигнута при использовании репрессируемых химическими стимулами промоторов, т.е. таких, для которых применение химического агента подавляет экспрессию генов. Такие индуцируемые химическими стимулами промоторы включают, не ограничиваются ими, промотор кукурузы ln2-2, активируемый антидотами бензолсульфонамидных гербицидов (De Veylder et al., (1997) Plant Cell Physiol 38:568-77), промотор кукурузы GST (GST-lI-27, W093/01294), активированный гидрофобными электрофильными соединениями, используемыми в общих гербицидах, наносимых до всхода урожая, и промотор табака PR 1 (Ono et al., (2004) Biosci Biotechnol Biochem 68:803-7), активируемый салициловой кислотой. Промоторы, активность которых регулируется антибиотиками, такие как индуцируемые тетрациклином и репрессируемые тетрациклином промоторы (Gatz et al, (1991) Mol Gen Genet 227:229-37; патенты США № 5814618 и 5789156) также могут быть использованы в рамках насоящего изобретения.
Перемещение к определенным органеллам и/или экспрессия в определенных органеллах растения
[00959] Система экспрессии может включать элементы для транслокации в и/или экспрессии в определенной органелле растения.
Нацеливание в хлоропласты
[00960] В конкретных вариантах осуществления изобретения предусматривается использование системы CRISPR С2с1 или С2с3 для того, чтобы специфически изменить гены хлоропласта или обеспечить экспрессию в хлоропласте. С этой целью используются способы трансформации хлоропласта или компартментализации компонентов системы CRISPR С2с1 или С2с3 в хлоропласте. Например, введение генетических модификаций в геноме пластида может снизить проблемы биологической безопасности, такие как перенос генов через пыльцу.
[00961] Способы трансформации хлоропласта известны в данной области и включают баллистическую трансформацию с бомбардировкой частицами, обработку PEG и микроинъекцию. Кроме того, можно использовать способы, включающие перемещение кассет трансформации от ядерного генома к пластиде, согласно описанию в WO2010061186.
[00962] Альтернативно этому, предусматривается нацеливание одного или более компонентов CRISPR С2с1 или С2с3 в хлоропласт растения. Это достигается за счет встраивания в конструкцию экспрессии последовательности, кодирующей транзитный пептида хлопласта (CTP) или транзитный пептид пластида, функционально связанный с 5'-концевой областью последовательности, кодирующей белок С2с1 или С2с3. CTP удаляется на этапе процессинга во время перемещения в хлоропласт. Квалифицированному специалисту известно о нацеливания экспрессируемых белков в хлоропласт (см., например, Protein Transport into Chloroplasts, 2010, Annual Review of Plant Biology, Vol. 61: 157-180). В таких вариантах осуществления изобретения также желательно нацеливание направляющей РНК в хлоропласт растения. Способы и конструкции, которые могут использоваться для транслокации направляющей РНК в хлоропласт благодаря последовательности локализации в хлоропласте, описаны, например, в US 20040142476 и включены в настоящее описание в качестве ссылки. Такие изменения конструкций могут быть введены в системы экспрессии по настоящему изобретению с целью эффективного перемещения направляющей(их) РНК, нацеливающей С2с1 или С2с3.
Введение полинуклеотидов, кодирующих систему CRISPR С2с1 или С2с3 в водорослевые клетки
[00963] Трансгенные водоросли (или другие растения, такие как рапс) могут быть особенно полезными для производства растительных масел или биотоплива, такого как спирты (особенно метанол и этанол) или другие продукты. Они могут быть модифицированы способами инженерии таким образом, чтобы экспрессировать или сверхэкспрессировать большие объемы масла или спиртов для использования в отраслях промышленности, использующих нефть или биотопливо.
[00964] В US 8945839 описан способ модификации способами инженерии вида микроводорослей (клетки Chlamydomonas reinhardtii) с использованием Cas9. Используя подобную технику, способы с использованием описанной в настоящем описании системы CRISPR С2с1 или С2с3 могут быть применены для видов рода Chlamydomonas и других водорослей. В определенных вариантах осуществления изобретения С2с1 или С2с3 и направляющая РНК введены в водоросли, в которых они экспрессируются, с использованием вектора экспрессии С2с1 или С2с3 под контролем конститутивного промотора, такого как Hsp70A-Rbc S2 или бета-2-тубулина. Направляющая РНК необязательно может быть доставлена с использованием вектора, содержащего промотор T7. Альтернативно этому, в клетки водоросли может быть доставлена РНК Cas9 и транскрибированная in vitro направляющая РНК. Квалифицированному специалисту доступны протоколы электропорации, в том числе стандартный рекомендуемый протокол из набора GeneArt Chlamydomonas Engineering kit.
[00965] В конкретных вариантах осуществления используемая в рамках настоящего изобретения эндонуклеаза представляет собой разделенный фермент C2c1 или разделенный фермент C2c3. Разделенные ферменты C2c1 или C2c3 предпочтительно используется в водорослях для направленной модификации генома, как описано для Cas9 в WO 2015086795. Использование разделенных систем C2c1 или C2c3 особенно подходит для случая индуцируемого метода нацеливания на геном и позволяет избежать потенциального токсического эффекта сверхэкспрессии C2c1 или C2c3 в клетках водорослей. В определенных вариантах осуществления названные разделенные домены C2c1 или C2c3 (домены RuvC и HNH) могут быть одновременно или последовательно введены в клетку таким образом, что указанный разделенный домен(ы) C2c1 или C2c3 производят процессинг последовательности нуклеиновой кислоты-мишени в клетке водоросли. Уменьшенный размер разделенного C2c1 или C2c3 в сравнении с C2c1 или C2c3 дикого типа делает возможным применение других способов доставки системы CRISPR в клетки, в частности, использование проникающих в клетку пептидов, как описано в настоящем описании. Этот способ представляет особый интерес для получения генетически модифицированных водорослей.
Введение полинуклеотидов, кодирующих компоненты C2c1 или C2c3 в дрожжевые клетки
[00966] В определенных вариантах осуществления изобретения изобретение относится к применению системы CRISPR C2c1 или C2c3 для редактирования генома дрожжевых клеток. Способы трансформации дрожжевых клеток, которые могут быть использованы для введения полинуклеотидов, кодирующих компоненты системы CRISPR C2c1 или C2c3, известны квалифицированному специалисту и рассмотрены в Kawai et al., 2010, Bioeng Bugs. 2010 Nov-Dec, 1(6): 395-403). Неограничивающие примеры включают трансформацию дрожжевых клеток обработкой ацетатом лития (с возможностью дальнейшей обработки ДНК-переносчиком и PEG), баллистического способа или электропорации.
Временная экспрессия компонентов системы CRISPR C2c1 или C2c3 в растениях и растительной клетке
[00967] В определенных вариантах осуществления изобретения предусматривается, что направляющая РНК и/или ген C2c1 или C2c3 экспрессируется в растительной клетке. В таких вариантах осуществления изобретения система CRISPR C2c1 или C2c3 способна обеспечить модификацию гена-мишени только тогда, когда и направляющая РНК, и белок C2c1 или C2c3 присутствуют в клетке, причем в дальнейшем возможен контроль модификации генома. Поскольку экспрессия фермента C2c1 или C2c3 является временной, растения, регенерирующие из таких растительных клеток, как правило, не содержат чужеродной ДНК. В конкретных вариантах осуществления изобретения фермент C2c1 или C2c3 стабильно экспрессируется растительной клеткой и направляющая последовательность экспрессируется временно.
[00968] В конкретных вариантах осуществления изобретения компоненты системы CRISPR C2c1 или C2c3 могут быть введены в растительные клетки с использованием вирусного вектора растений (Scholthof et al. 1996, Annu Rev Pbytopathol. 1996;34:299-323). В следующих конкретных вариантах осуществления указанный вирусный вектор является вектором ДНК-вируса. Например, геминивируса (geminivirus) (например, вирус листовой курчавости капусты, вирус желтой карликовости фасоли, вирус карликовости пшеницы, вирус листовой курчавости томата, вирус полосатости кукурузы, вирус листовой курчавости табака или вирус золотой мозаики томата), или нановируса (nanovirus) (например, вирус желтого некроза бобовых). В других конкретных вариантах указанный вирусный вектор является вектором РНК-вируса, например, тобравируса (tobravirus) (например, вирус погремковости табака, вирус табачной мозаики), потексвируса (potexvirus) (например, X-вирус картофеля) или гордеивируса (hordeivirus) (например, вирус полосатой мозаики ячменя). Геномы репликации вирусов растений являются неинтегральными векторами.
[00969] В некоторых вариантах осуществления вектор, используемый для временной экспрессии конструкций CRISPR C2c1 или C2c3, может быть, в частности, вектором pEAQ, сконструированным специально для опосредованной Agrobacterium временной экспрессии в протопласте (Sainsbury F. et al., Plant Biotechnol J. 2009 Sep:7(7):682-93). Точное нацеливание на локализацию в геноме было продемонстрировано с использованием вектора на основе модифицированного вируса листовой курчавости капусты (CaLCuV) для экспрессии направляющих РНК в стабильно траснгенных растениях, экспрессирующих фермент CRISPR (Scientific Reports 5, Article number: 14926 (2015), doi: 10.1038/srep14926).
[00970] В конкретных вариантах осуществления изобретения фрагменты двухцепочечной ДНК, кодирующие направляющую РНК и/или ген C2c1 или C2c3, могут быть временно введены в клетку растения. В таких вариантах осуществления внесенные двухцепочечные фрагменты ДНК присутствуют в количествах, достаточных для модификации клетки, но не остаются после окончания рассматриваемого периода времени или после одного или более клеточных делений. Способы прямого переноса ДНК известны квалифицированному специалисту (см., например, Davev et al. Plant Mol Biol 1989 Sep;13(3):273-85.)
[00971] В других вариантах осуществления изобретения полинуклеотид, кодирующий белок C2c1 или C2c3, вводят в растительную клетку с его последующей трансляцией и процессированием в клетке, синтезирующей этот белок в количестве, достаточном для изменения клетки (в присутствии по меньшей мере одной молекулы направляющей РНК), но который не остается после окончания рассматриваемого периода времени или после одного или более клеточных делений. Способы введения мРНК в протопласт растения известны квалифицированному специалисту ((см., например, Gallie, Plant Cell Reports (1993), 13; 119-122).
[00972] Tакже предусматриваются комбинации различных описанных выше способов.
Доставка компонентов CRISPR C2c1 или C2c3 в растительную клетку
[00973] В конкретных вариантах осуществления изобретения требуется доставить один или более компонентов системы CRISPR C2c1 или C2c3 напрямую в растительную клетку. Это представляет интерес, среди прочего, для получения нетрансгенных растений (см. ниже). В некоторых вариантах осуществления изобретения, один или более компонентов C2c1 или C2c3 получен(ы) вне растения или растительной клетки и доставлен(ы) в клетку. Например, в некоторых вариантах осуществления изобретения белок C2c1 или C2c3 получен in vitro до введения в растительную клетку. Белок C2c1 или C2c3 может быть получен различными известными квалифицированному в данной области специалисту способами, в том числе рекомбинантным получением. После экспрессии белок C2c1 или C2c3 выделяют, подвергают рефолдингу в случае необходимости, очищают и необязательно из него удаляют меток для очистки, такой как His-метка. Когда неочищенный, частично очищенный или более полно очищенный белок C2c1 или C2c3 получен, его можно вводить в растительную клетку.
[00974] В определенных вариантах осуществления изобретения белок C2c1 или C2c3 объединен с направляющей молекулой РНК, нацеливающей на ген-мишень, для образования предварительно собранного рибонуклеопротеина.
[00975] Отдельные компоненты или сам предварительно собранный рибонуклеопротеин могут быть введены в растительную клетку путем электропорации, бомбардировки частицами, покрытыми продуктом гена, ассоциированным с C2c1 или C2c3, химической трансфекции или некоторых другими средств транспорта через клеточную мембрану. Например, была продемонстрирована трансфекция протопласта растения с предварительно собранным рибонуклеопротеином CRISPR с целью подтвердить модификацию генома растения (как описано в Woo et at. Nature Biotechnology, 2015; DQI: 1.0.1038/nbt.3389).
[00976] В конкретных вариантах осуществления изобретения система CRISPR C2c1 или C2c3 может быть введена в растительные клетки с использованием наночастиц. Компоненты, такие как белок, нуклеиновая кислота или их сочетание, могут быть загружены или упакованы в наночастицы и применены к растениям (как описано, например, в WO 2008042156 и US 20130185823). В частности, варианты осуществления изобретения включают наночастицы, в которые загружена или в которых содержится молекула(ы) ДНК, кодирующая белок C2c1 или C2c3, молекулы ДНК, кодирующие направляющую РНК, и/или выделенная направляющая РНК, как описано в WO2015089419.
[00977] Следующим средством введения одного или более компонентов систем CRISPR C2c1 или C2c3 в растительную клетку является использование проникающих в клетку пептидов (CPP). В соответствии с этим отдельные варианты осуществления изобретения включают композиции, содержащие проникающие в клетку пептиды, связанные с белком C2c1 или C2c3. В конкретных вариантах осуществления настоящего изобретения белок C2c1 или C2c3 и/или направляющая РНК сопряжены с одним или более CPP для их эффективной доставки в протопласт растения (также см. Ramakrishna, Genome Res. 2014 Jun;24(6):1020-7, для Cas9 в клетках человека). В других вариантах осуществления ген нацеленного на РНК белка и/или направляющая(ие) РНК кодируются одной или более кольцевыми или некольцевыми молекулой(ами), сопряженными с одним или более CPP для доставки в протопласт растения. Протопласты растения далее регенерируют в растительные клетки, а далее в растения. CPP в целом представляют собой короткие пептиды длиной менее 35 аминокислот полученные либо из белка, либо из химерных последовательностей, способных транспортировать биомолекулы через клеточную мембрану вне зависимости от рецепторов. CPP могут представлять собой катионные пептиды, пептиды, имеющие гидрофобные последовательности, амфипатические пептиды, пептиды, богатые пролином, и антимикробные последовательности, а также химерные или двухкомпонентные пептиды (Pooga and Langel, 2005). CPP способны проникать через биологические мембраны и при этом запускать движение различных биомолекул через мембраны клеток в цитоплазму и улучшать их внутриклеточное перемещение, тем самым облегчая взаимодействие биомолекулы с мишенью. Примеры CPP в числе прочих включают: Tat, ядерный белок-активатор транскрипции, необходимый для репликации вируса ВИЧ типа I, пенетратин, сигнальная пептидная последовательность фактора роста фибробластов (FGF) Капоши, сигнальная пептидная последовательность интегрина β3; последовательность пептида полиаргинина Args, богатые аргинином молекулярные транспортеры, пептид "sweet arrow".
Использование систем CRISPR C2c1 или C2c3 для получения генетически модифицированных нетрансгенных растений
[00978] В определенных вариантах осуществления способы, описанные в настоящем описании, используются для модификации эндегенных генов или модификации их экспрессии без введения на постоянной основе в геном растения каких-либо чужеродных генов, в том числе кодирующих компоненты CRISPR, чтобы избежать присутствия чужеродной ДНК в геноме растения. Это может представлять интерес в связи с тем, что нормативные требования к нетрансгенным растениям являются менее строгими.
[00979] В конкретных вариантах осуществления изобретения это обеспечивается временной экспрессией компонентов CRISPR C2c1 или C2c3. В некоторых вариантах осуществления один или более компонентов CRISPR экспрессируются с одного или более вирусных векторов, которые производят достаточное количество белка C2c1 или C2c3 и направляющей РНК для того, чтобы стабильно и непрерывно обеспечивать модификацию представляющего интерес гена в соответствии со способом, описанным в настоящем описании.
[00980] В конкретных вариантах осуществления временная экспрессия конструкций CRISPR C2c1 или C2c3 осуществляется в протопластах растения и, таким образом, не происходит их встраивание в геном. Экспрессия с ограниченной продолжительностью может быть достаточной для того, чтобы система CRISPR C2c1 или C2c3 была способна модифицировать ген-мишени, как описано в настоящем описании.
[00981] В некоторых вариантах осуществления изобретения различные компоненты системы CRISPR C2c1 или C2c3 введены в растительную клетку, протопласт или растительную ткань по отдельности или в виде смеси с помощью молекул для доставки в форме частиц, таких как наночастицы или молекулы CPP, как описано выше.
[00982] Экспрессия компонентов CRISPR C2c1 или C2c3 может вызывать направленную модификацию генома за счет либо прямого действия нуклеаз C2c1 или C2c3 и необязательно введения ДНК-матрицы, либо за счет модификации генов-мишеней при помощи системы CRISPR C2c1 или C2c3, как это описано в настоящей заявке. Различные стратегии, описанные выше, делают возможным опосредованное C2c1 или C2c3 редактирование генома, не требующее введение компонентов CRISPR C2c1 или C2c3 в геном растения. Компоненты, которые временно введены в растительную клетку, обычно не сохраняются после кроссинговера.
Обнаружение модификаций в геноме растений - селективные маркеры
[00983] В некоторых вариантах осуществления, в которых способ включает модификацию эндогенного гена-мишени в геноме растения, любой подходящий способ может быть использован для того, чтобы после того, как растение, часть растения или растительная клетка инфицированы или трансфицированы системой CRISPR C2c1 или C2c3, определить, имело ли место нацеливание на ген или направленный мутагенез по сайту-мишени. В случае если способ включает введение трансгена, трансформированная растительная клетка, каллус или растение могут быть идентифицированы и выделена за счет отбора или скрининга модифицированного растительного материала на предмет наличия трансгена или признаков, кодируемых трансгеном. Для идентификации растений или растительных клеток-трансформантов, содержащих встроенные генные конструкции или эндогенные модификации ДНК, могут быть использованы физические или биохимические способы. Такие способы включают, но не ограничиваются ими: 1) саузерн=анализ или ПЦР-амплификация для обнаружения и определения структуры рекомбинантной ДНК-вставки или модифицированных эндогенных генов; 2) нозерн-блоттинг, защита от РНК-азы S1, удлинение праймеров или амплификация ПЦР с обратной транскриптазой для обнаружения и изучения транскриптов РНК генных конструкций; 3) ферментный анализ для определения активности ферментов или рибозимов, где такие генные продукты кодируются генной конструкцией или на экспрессию влияет генетическая модификация; 4) гель-электрофорез белков, методики вестерн-блота, иммунопреципитация, иммуноанализ, связанный с активностью белков, когда генные конструкции или эндогенные продукты гена являются белками. Дополнительные методики, такие как гибридизация in situ, ферментное окрашивание, иммуноокрашивание могут также использоваться для детектирования присутствия или экспрессии рекомбинантных конструкций или обнаружения модификации эндогенного гена в конкретных органах и тканях растения. Способы проведения всех данных способов анализа хорошо известны квалифицированному в данной области специалисту.
[00984] Дополнительно (или альтернативно) экспрессирующая система, кодирующая компоненты CRISPR C2c1 или C2c3, обычно сконструирована таким образом, чтобы включать один или более селективных маркеров, которые обеспечивают инструмент для выделения или эффективного отбора клеток, содержащих и/или модифицированных системой CRISPR C2c1 или C2c3 на ранних стадиях и в больших масштабах.
[00985] В случае опосредованной Agrobacterium трансформации кассета-маркер может быть расположенной вблизи или между границ фланкирующей Т-ДНК и может находиться внутри бинарного вектора. В другом варианте осуществления такая кассета-маркер может находиться вне Т-ДНК. Селективная кассета-маркер также может находиться внутри или вблизи границ Т-ДНК, в то время как экспрессирующая кассета может находиться в другом месте внутри второй Т-ДНК на бинарном векторе (например, система 2 Т-ДНК).
[00986] Для бомбардировки частицами или трансформации протопластов система экспрессии может включать один или более выделенных линейных фрагментов или может являться частью большей конструкции, которая может содержать бактериальные элементы репликации, бактериальные селективные маркеры или другие поддающиеся обнаружению элементы. Такая экспрессирующая кассета(ы), содержащая полинуклеотиды, кодирующие направляющую молекулу и/или C2c1 или C2c3, может быть физически соединена с кассетой-маркером или может находиться в смеси с второй молекулой нуклеиновой кислоты, кодирующей кассету-маркер. Кассета-маркер состоит из необходимых для экспрессии поддающегося обнаружению или селективного маркера элементов, которые делают возможным эффективную селекцию трансформированных клеток.
[00987] Процедура отбора клеток, основанная на селективном маркере, зависит от природы гена-маркера. В определенных вариантах осуществления используется селективный маркер, т.е. маркер, делающий возможным прямой отбор клеток на основе экспрессии маркера. Селективный маркер может обеспечивать положительный или отрицательный отбор и быть зависимым или не зависимым от присутствия внешних субстратов (Miki et al. 2004, 107(3): 193-232). Более распространен случай, когда гены устойчивости к антибиотику или гербициду используются в качестве маркера, тем самым отбор осуществляется путем выращивания растительного материала на средах, содержащих ингибирующие концентрации антибиотика или гербицида, устойчивость к которым придает ген-маркер. Примерами таких генов являются гены, которые придают устойчивость к антибиотикам, таким как гигромицин (hpt) и канамицин (nptII), и гены, которые придают устойчивость к гербицидам, таким как фосфинотрицин (bar) и хлоросульфорон (als).
[00988] Трансформированные растения и растительные клетки могут также быть идентифицированы с помощью скрининга активности видимого маркера, обычно фермента, способного к процессингу окрашенного субстрата (например, β-глюкуронидаза, люцифераза, гены В или Cl). Такая методология селекции и скрининга хорошо известна специалисту в данной области.
Культуры и регенерация растений
[00989] В определенных вариантах осуществления растительная клетка, которая имеет модифицированный геном, произведенный или полученный любым из описанных в настоящей спецификации способов, может быть культивирована для регенерации целого растения, имеющего модифицированный генотип и, как следствие, желаемый фенотип. Традиционно применяемые методики регенерации основаны на манипулировании определенными фитогормонами в среде для роста культуры ткани и обычно основаны на биоцидном и/или гербицидном маркере, который вводится вместе с желаемой нуклеотидной последовательностью. В определенных дальнейших модификациях изобретения регенерация растения достигается на основе культивируемых протопластов, каллусов растения, эксплантов, органов, пыльцы, эмбрионов или их частей (см., например, Evans et. al. (1983), Handbook of Plant Cell Culture, Klee et al (1987) Ann. Rev. of Plant Phys.).
[00990] В определенных вариантах осуществления трансформированные или улучшенные растения, как описано в настоящей спецификации, могут быть самоопыляющимися для обеспечения семян для гомозиготных улучшенных растений по настоящему изобретению (гомозиготные для модификации ДНК) или могут быть скрещены с нетрансгенными растениями или иными улучшенными растениями для того, чтобы обеспечить семена для гетерозиготных растений. В случае если рекомбинантная ДНК была введена в растительную клетку, получаемое в результате такого скрещивания растение является гетерозиготным в отношении рекомбинантной молекулы ДНК. Такие растения, как гомозиготные, так и гетерозиготные, полученные путем скрещивания улучшенных растений и содержащие генетические модификации (которые могут представлять собой рекомбинантную ДНК) в настоящем описании называются "потомством". Растения-потомство представляют собой растения, происходящие от исходного трансгенного растений и содержащие модификации генома или введенную с помощью способов, описанных в настоящем описании, рекомбинантную молекулу ДНК. Альтернативно этому генетически модифицированные растения могут быть получены с помощью одного из описанных выше способов с использованием фермента C2c1 или C2c3, в соответствии с чем чужеродная ДНК не встраивается в геном. Потомство таких растений, полученных путем последующей селекционной работы, может также содержать генетические модификации. Селекционная работа выполняется с помощью любых широкого применяемых способов селекции для различных сельскохозяйственных культур (например, Allard, Principles of Plant Breeding, John Wiley & Sons, NY, U. of CA, Davis, CA, 50-98 (1960).
Получение растений с улучшенными агрономическими характеристиками
[00991] Системы CRISPR, основанные на C2c1 или C2c3, описанные в настоящем описании, могут быть использованы для внесения направленных двухцепочечных или одноцепочечных разрывов и/ил введения систем активаторов или репрессоров гена и, не ограничиваясь этим, могут быть использованы для нацеливания на гены, замены генов, направленного мутагенеза, направленных делеций или инсерций, направленных инверсий и/или транслокаций. За счет совместных экспрессии нескольких нацеливающих РНК, направленных на несколько модификаций в одной клетке, возможно обеспечить мультиплексированные модификации генома. Данная технология может быть использована для высокоточного конструирования растений с улучшенными свойствами, включая улучшенные пищевые характеристики, увеличенную устойчивость к заболеваниям или устойчивость к стрессу биологической или абиотической природы и увеличенную продукцию имеющих коммерческое значения продуктов или гетерологических соединений.
[00992] В определенных вариантах осуществления системы CRISPR C2c1 или C2c3, как описано в настоящем описании, используются для внесения напралвенных двухцепочечных разрывов (DSB) в эндогенную последовательность ДНК. DSB активируют пути репарации ДНК в клетке, что может быть использовано для достижения желаемых модификаций последовательности ДНК вблизи положения разрыва. Это представляет интерес в случае, если инактивация эндогенных генов может обеспечить или внести вклад в желаемую характеристику. В определенных вариантах осуществления гомологичная рекомбинация с последовательностью-матрицей может быть осуществлена в участке DSB для того, чтобы внести представляющий интерес ген.
[00993] В определенных вариантах осуществления системы CRISPR C2c1 или C2c3 могут быть использованы в качестве связывающего нуклеиновую кислоту белка в общем понимании с его слиянием или функциональным связыванием с функциональным доменом для активации и/или репрессии эндогенных генов растения. Примеры традиционно применяемых функциональных доменов могут включать, не ограничиваться ими, инициатор трансляции, активатор трансляции, репрессор трансляции, нуклеазы, в частности, рибонуклеазы, сплайсосому, микрогранулы, активируемый/контролируемый светом домен или активируемый/контролируемый химическими стимулами домен. В случае таких модификаций изобретения белок C2c1 или C2c3 обычно содержит по меньшей мере одну мутацию, в частности такую, что он имеет не более 5% активности белка C2c1 или C2c3, не имеющих такой по меньшей мере одной мутации; направляющая РНК включает направляющую последовательность, способную к гибридизации с последовательностью-мишенью.
[00994] Описанные в настоящем описании способы в целом обеспечивают получение улучшенных растений, имеющих одну или более желательных характеристик по сравнению с растениями дикого типа. В определенных вариантах осуществления полученные растения, растительные клетки или части растений являются трансгенными растениями, содержащими экзогенную последовательность ДНК, встроенную в геном всех или части клеток растения. В конкретных вариантах осуществления получают нетрансгенные генетически модифицированные растения, части растений или клетки в том смысле, что в геном любых из клеток растения не включена экзогенная последовательность ДНК. В таких вариантах осуществления изобретения улучшенные растения является нетрансгенными. В случае если обеспечена только модификация некоторого эндогенного гена и чужеродные гены не внесены и не поддерживаются в геноме растения, полученные в результате генетически модифицированные культуры не содержат чужеродных генов и, таким образом, по существу должны рассматриваться как нетрансгенные. Различные применения систем CRISPR C2c1 или C2c3 для редактирования растительных геномов более детально описаны ниже:
Введение одного или более чужеродных генов для сообщения представляющей интерес агрономической характеристики
[00995] Настоящее изобретение относится к способам редактирования генома или модификации последовательностей, ассоциированных с или расположенных в представляющих интерес локусах-мишенях, где такой способ включает введение комплекса эффекторного белка C2c1 или C2c3 в растительную клетку, в связи с чем комплекс эффекторного белка C2c1 или C2c3 эффективно встраивает ДНК-вставку, например, кодирующую интересующий чужеродный ген в геном растительной клетки. В предпочтительных вариантах осуществления такой интеграции ДНК-вставки способствует HR с экзогенно введенной матрицей ДНК или матрицей репарации. Обычно экзогенно введенная матрица ДНК или матрица репарации доставляются совместно с комплексом эффекторного белка C2c1 или C2c3, или одним компонентом или полинуклеотидным вектором для экспрессии компонента комплекса.
[00996] Системы CRISPR C2c1 или C2c3, предусматриваемые в рамках настоящего изобретения, делают возможной направленную доставку генов. Становится все более очевидным, что эффективность экспрессии представляющего интерес гена в большой степени определяется локализацией встраивания в геноме. Способы по настоящему изобретению делают возможной направленную интеграцию чужеродного гена в желаемое положение в геноме. Локализация может быть выбрана на основе информации ранее полученных вставок или может быть выбрана с помощью способов, описанных в настоящем описании.
[00997] В определенных вариантах осуществления способы, описанные в настоящем описании, включают (a) введение в клетку комплекса CRISPR C2c1 или C2c3, содержащего направляющую РНК, включающую прямой повтор и направляющую последовательность, где направляющая последовательность гибридизуется c последовательностью-мишенью, являющейся эндогенной для растительной клетки; (b) введение в растительную клетку эффекторной молекулы C2c1 или C2c3, которая образует комплекс с направляющей РНК, где направляющая последовательность гибридизуется с последовательностью-мишенью и вызывает двухцепочечный разрыв в или вблизи последовательности, на которую нацеливает направляющая; (c) введение в клетку нуклеотидной последовательности, кодирующей матрицу репарации HDR, которая кодирует представляющий интерес ген и которая вводится в место локализации двухцепоченого разрыва в результате HDR. В определенных вариантах осуществления изобретения этап введения может включать доставку в растительную клетку одного или более полинуклеотидов, кодирующих эффекторной белок C2c1 или C2c3, направляющую РНК и матрицу репарации. В определенных вариантах осуществления эти полинуклеотиды доставляются в клетку ДНК-содержащим вирусом (например, джеминивирусом) или РНК-содержащим вирусом (например, тобравирусом). В определенных вариантах осуществления этапы введения включают доставку в растительную клетку Т-ДНК, содержащей одну или более полинуклеотидных последовательностей, кодирующих эффекторный белок C2c1 или C2c3, направляющую РНК и матрицу репарации, где доставка опосредована Agrobacterium. Последовательность нуклеиновой кислоты, кодирующей эффекторный белок C2c1 или C2c3 может быть функционально соединена с промотором, в частности, конститутивным промотором (например, промотором 35S вируса мозаики цветной капусты) или специфическим для клетки либо индуцибельным промотором. В определенных вариантах осуществления полинуклеотид вводится с помощью бомбардировки микрочастицами. В определенных вариантах осуществления способ также включает скрининг растительной клетки после этапов введения с целью определить, была ли матрица репарации, т.е. представляющий интерес ген, успешно введена. В определенным вариантах осуществления данные способы включают этап регенерации растения из растительной клетки. В последующих вариантах осуществления такие способы включают скрещивание растения для получения потомства с желаемыми генетическими свойствами. Примеры чужеродных генов, кодирующих представляющую интерес характеристику, перечислены ниже.
b) Редактирование эндогенных генов для сообщения представляющего интерес агрономического признака
[00998] Настоящее изобретение относится к способам редактирования или модификации последовательностей, ассоциированных с или расположенных в представляющем интерес локусе-мишени, где такой способ включает введение комплекса эффекторного белка C2c1 или C2c3 в растительную клетку, в связи с чем комплекс C2c1 или C2c3 модифицирует экспрессию эндогенного гена растения. Этого можно достичь различными путями. В определенных вариантах осуществления желательно прекращение экспрессии эндогенного гена и комплекс CRISPR C2c1 или C2c3 используется для нацеливания и расщепления эндогенного гена для того, чтобы модифицировать экспрессию гена. В таких вариантах осуществления способы, описанные в настоящем описании, включают (a) введение в растительную клетку комплекса C2c1 или C2c3, включающего направляющую РНК, содержащую прямой повтор и направляющую последовательность, где направляющая последовательность гибридизуется с последовательностью-мишенью внутри представляющего интерес гена в геноме растительной клетки; (b) введение в клетку эффекторного белка C2c1 или C2c3, который при связывании с направляющей РНК включает направляющую последовательность, которая гибридизована с последовательностью-мишенью, и вносит двухцепочечный разрыв в или вблизи последовательности, на которую нацеливает направляющая последовательность. В определенных вариантах осуществления этап введения может включать доставку в растительную клетку одного или более полинуклеотидов, кодирующих эффекторный белок C2c1 или C2c3 и направляющую РНК.
[00999] В определенных вариантах осуществления полинуклеотиды доставляются в клетку ДНК-содержащим вирусом (например, джеминивирусом) или РНК-содержащим вирусом (например, тобравирусом). В определенных вариантах осуществления этапы введения включают доставку в растительную клетку Т-ДНК, содержащей одну или более полинуклеотидных последовательностей, кодирующих эффекторный белок C2c1 или C2c3 и направляющую РНК, где доставка опосредована Agrobacterium. Последовательность нуклеиновой кислоты, кодирующей компоненты системы эффекторного белка C2c1 или C2c3, может быть функционально соединена с промотором, в частности, конститутивным промотором (например, промотором 35S вируса мозаики цветной капусты) или специфическим для клетки либо индуцибельным промотором. В определенных вариантах осуществления полинуклеотид вводится с помощью бомбардировки микрочастицами. В определенных вариантах осуществления способ также включает скрининг растительной клетки после этапов введения с целью определить, была ли матрица репарации, т.е. представляющий интерес ген, успешно введена. В определенным вариантах осуществления данные способы включают этап регенерации растения из растительной клетки. В последующих вариантах осуществления такие способы включают скрещивание растения для получения потомства с желаемыми генетическими свойствами.
[001000] В конкретных вариантах осуществления способов, описанных выше, устойчивые к заболеваниям зерновые культуры, получают посредством целенаправленной мутации генов восприимчивости к заболеванию или генов, кодирующих отрицательные регуляторы (например, ген Mio) генов защиты растений. В конкретном варианте устойчивые к гербицидам зерновые культуры получают путем целенаправленной замены конкретных нуклеотидов в генах растений, таких как те, которые кодируют ацетолактатсинтазу (ALS) и протопорфириногеноксидазу (PPO). В конкретных вариантах осуществления устойчивые к засухе и засолению зерновые культуры получают путем целенаправленной мутации в генах, кодирующих отрицательные регуляторы устойчивости к абиотическому стрессу, зерновые с низким содержанием амилозы - путем целенаправленной мутации гена Waxy, рис или другие зерновые с уменьшенной прогорклостью -путем целенаправленной мутации основных генов липазы в алейроновом слое, и т.д. Более обширный список эндогенных генов, кодирующих интересующие признаки, приведен ниже.
c) модулирование эндогенных генов системой CRISPR C2c1 или CRISPR C2c3 для получения представляющих интерес сельскохозяйственных характеристик
[001001] Также в рамках настоящего изобретения предусматриваются способы модулирования (т.е. активации или подавления), экспрессии эндогенных генов с использованием белка C2c1 или C2c3, описанного в настоящем описании. В таких способах используется определенная последовательность(и) РНК, которую встраивают в геном растения посредством комплекса C2c1 или C2c3. Более подробно, определенная последовательность(и) РНК связывается с двумя или более адаптерными белками (например, аптамерами), посредством чего каждый адаптерный белок связан с одним или более функциональными доменами и где по меньшей мере один из одного или более функциональных доменов, связанных с адаптерным белком, имеет один или более типов активности, включающих метилазную активность, деметилазную активность, активность активации транскрипции, активность репрессии транскрипции, активность фактора терминации транскрипции, активность модификации гистонов, активность интеграции ДНК, активность расщепления РНК, активность расщепления ДНК или активность связывания нуклеиновых кислот. Функциональные домены используются для модуляции экспрессии эндогенных генов растений для получения желаемых характеристик. Как правило, в этих вариантах осуществления изобретения эффекторный белок C2c1 или C2c3 имеет одну или более мутаций, таким образом, чтобы он имел не более 5% нуклеазной активности эффекторного белка C2c1 или C2c3, не имеющего ни одной мутации.
[001002] В конкретных вариантах осуществления способы, описанные в настоящем описании, включают следующие стадии: (a), введение в клетку комплекса CRISPR C2c1 или CRISPR C2c3, включающего направляющую РНК, содержащую прямой повтор и направляющую последовательность, где направляющая последовательность гибридизируется с последовательностью-мишенью, которая является эндогенной по отношению к растительной клетке; (b) введение в растительную клетку эффекторной молекулы C2c1 или C2c3, которая образует комплекс с направляющей РНК, когда направляющая последовательность гибридизируется с последовательностью-мишенью; и где или направляющая РНК модифицирована, чтобы включать определенную последовательность РНК (аптамер), связывающуюся с функциональным доменом, и/или эффекторный белок C2c1 или C2c3 модифицирован для связывания с функциональным доменом. В конкретных вариантах осуществления изобретения, стадия введения может включать доставку в растительную клетку одного или более полинуклеотидов, кодирующих (модифицированный) эффекторный белок C2c1 или C2c3 и (модифицированную) направляющую РНК. Детали компонентов системы CRISPR C2c1 или C2c3 для использования в этих методах описаны в других разделах настоящего изобретения.
[001003] В конкретных вариантах осуществления полинуклеотиды доставляются в клетку посредством ДНК-вируса (например, джеминивирусом) или РНК-вирусом (например, тобравирусом). В конкретных вариантах осуществления стадия введения включают доставку в растительную клетку T-ДНК, содержащей одну или более полинуклеотидных последовательностей, кодирующих эффекторный белок C2c1 или C2c3 и направляющую РНК, где доставка осуществляется посредством Agrobacterium. Последовательность нуклеиновых кислот, кодирующая один или более компонентов системы CRISPR C2c1 или C2c3 CRISPR, может быть функционально связанной c промотором, таким как конститутивный промотор (например, промотор 35S вируса мозаики цветной капусты), или специфический для клетки или индуцибельный промотор. В конкретных вариантах осуществления полинуклеотид вводится путем бомбардировки микрочастицами. В конкретных вариантах осуществления метод далее включает скрининг растительной клетки после введения, чтобы определить, была ли модифицирована экспрессия интересующего гена. В конкретных вариантах осуществления методы включают стадию регенерации растения из растительной клетки. В дальнейших вариантах осуществления методы включают скрещивание растений для получения, генетически желаемой линии растений. Более обширный список эндогенных генов, кодирующих представляющие интерес характеристики, упомянут ниже.
Использование С2с1 или С2с3 для модификации полиплоидных растений
[001004] Многие растения являются полиплоидами, что означает, что они содержат дуплицированные копии своих геномов - иногда целых шесть, как в пшенице. Согласно настоящему изобретению, методы, использующие эффекторный белок C2c1 или C2c3 системы CRISPR, могут быть "мультиплексированы" для воздействия на все копии гена или нацеливания на десятки генов сразу. Например, в конкретных вариантах осуществления способы по настоящему изобретению используют, чтобы одновременно вносить мутацию потери функции в различные гены, ответственные за подавление защиты от заболевания. В конкретных вариантах осуществления методы данного изобретения используются для одновременного подавления экспрессии последовательностей нуклеиновых кислот TaMLO-A1, TaMLO-B1 и TaMLO-D1 в растительной клетке пшеницы и регенерации растения пшеницы из нее, чтобы обеспечить устойчивость пшеницы к мучнистой росе (см. также WO2015109752).
Иллюстративные гены, ответственные за агрономические характеристики
[000006] Как описано в настоящем описании выше, в конкретных вариантах осуществления изобретение охватывает использование системы C2c1 или C2c3 CRISPR, как описано в настоящем описании, для инсерции представляющей интерес ДНК, включая один или более способных к экспрессии генов. В дальнейших конкретных вариантах изобретение охватывает способы и инструменты, использующие систему C2c1 или C2c3, как описано в настоящем описании, для частичной или полной делеции одного или более экспрессируемых растениями генов. В других дальнейших конкретных вариантах изобретение охватывает способы и инструменты, использующие систему C2c1 или C2c3, как описано в настоящем описании, для обеспечения модификации одного или более экспрессируемых растением генов мутацией, заменой, инсерцией, одного или более нуклеотидов. В других конкретных вариантах изобретение охватывает использование системы CRISPRC2c1 или C2c3, как описано в настоящем описании, для обеспечения модификации экспрессии одного или более экспрессируемых растением генов путем специфической модификации одного или более регуляторных элементов, управляющих экспрессией указанных генов.
[000007] В конкретных вариантах осуществления изобретение охватывает способы, которые включают введение экзогенных генов и/или нацеливание на эндогенные гены и их регуляторные элементы, такие как упомянуты ниже:
[000008] 1. Гены, придающие устойчивость к вредителям или заболеваниям:
- Гены устойчивости к заболеваниям растений. Растение может быть трансформировано клонированными генами устойчивости для создания способами инженерии растений, устойчивых к специфичным штаммам патогенов. См., например, Jones et al., Science 266:789 (1994) (клонирование гена томата Cf-9 для устойчивости к Cladosporium fulvum); Martin et al., Science 262:1432 (1993) (ген томата Pto для устойчивости к Pseudomonas syringae pv, томат кодирует протеинкиназу); Mindrinos et al, Cell 78:1089 (1994) (ген RSP2 Arabidopsmay be для устойчивости к Pseudomonassyringae).
- Гены, сообщающие устойчивость к вредителю, такому как нематода кисты сои. См., например, заявку PCT WO 96/30517, заявку PCT WO 93/19181.
- Белки Bacillus thuringiensis см., например, Geiser et al. Gene 48:109 (1986).
- Лектины, см., например, VanDamme et al. Plant Molec. Biol. 24:25 (1994).
- Белки, связывающие витамины, такие как авидин, см. заявку PCTUS93/06487, в которой описано использование авидина и гомологов авидина в качестве ларвицида против насекомых-вредителей.
- Ингибиторы ферментов, такие как ингибиторы протеазы или протеиназы, или ингибиторы амилазы. См., например, Abe et al., J. Biol. Chem. 262:16793 (1987), Huub et al. Plant Molec. Biol. 21:985 (1993), Sumitani et al., Biosci. Biotech. Biochem. 57:1243 (1993) и патент США № 5494813.
- Специфические для насекомых гормоны или феромоны, такие как экдистероид или ювенильный гормон, их варианты, миметики на их основе или их антагонисты или агонисты. См., например, Nature 344:458 (1990).
- Специфические для насекомых пептиды или нейропептиды, которые в результате экспрессии нарушают физиологию затронутого вредителя. Например, Regan, J. Biol. Chem. 269:9 (1994) and Pratt et al., Biochem. Biophys. Res. Comm. 163:1243 (1989). См. также патент США № 5266317.
- Специфические для насекомых яды, производимые в природе змеями, осами или любыми другими организмами. Например, см. Pang et al., Gene 116: 165 (1992).
- Ферменты, ответственные за гипераккумуляцию монотерпена, сесквитерпена, стероида, гидроксамовой кислоты, производную фенилпропаноида или другой небелковой молекулы с инсектицидной активностью.
- Ферменты, вовлеченные в модификацию, включая пост-трансляционную модификацию, биологически активной молекулы, например, гликолитического фермента, протеолитического фермента, липолитического фермента, нуклеазы, циклазы, трансаминазы, эстеразы, гидролазы, фосфатазы, киназы, фосфорилазы, полимеразы, эластазы, хитиназы и глюканазы, естественной или синтетической. См. заявку PCT WO93/02197, Kramer et al., Insect Biochem. Molec. Biol. 23:691 (1993) и Kawalleck et al., Plant Molec. Biol 21:673 (1993).
- Молекулы, стимулирующие трансдукцию сигнала. Например, см. Botella et al., Plant Molec. Biol. 24:757 (1994), and Griess et al., Plant Physiol. 104:1467 (1994).
- Инвазивные вирусные белки или комплексный токсин на их основе. См. Beachy et al., Ann. rev. Phytopathol. 28:451 (1990).
- Белки, задерживающие развитие, продуцируемые в природе патогеном или паразитом. См. Lamb et al., Вio/Technology 10:1436 (1992) и Toubart. et al., Plant J. 2:3(57 (1992).
- Белки, задерживающие развитие, продуцируемые в природе растением. Например, Logemann et al., Bio/Technology 10:305 (1992).
- В растениях патогены часто являются специфическими для хозяина. Например, некоторые виды Fusarium вызывают увядание томатов, но поражает только томаты, а другие разновидности Fusarium поражают только пшеницу. Растения имеют врожденный (конститутивный) и приобретенный (индуцированный) фитоиммунитет для защиты от большинства патогенов. Мутации и события рекомбинации в поколениях растений приводят к генетической изменчивости, которая приводит к восприимчивости, тем более что патогены воспроизводятся с большей частотой, чем растения. Растения могут иметь устойчивость, не связанную с хозяином, например, хозяин и патоген несовместимы, или частичную устойчивость против всех рас патогена, которой, как правило, управляют много генов, и/или также полную устойчивость к некоторым расам патогенов, но не к другим расам. Такой устойчивостью, как правило, управляют несколько генов. Используя способы и компоненты системы CRISPR-C2c1 или CRISPR-C2c3, теперь существует новый инструмент для индуцирования специфичных мутаций замедленного действия и превентивного характера. Соответственно, можно проанализировать геном источников генов устойчивости, и в растениях, имеющих желаемые характеристики или свойства, использовать способ и компоненты системы CRISPRC2c1 или C2c3, чтобы вызвать повышение экспрессии генов устойчивости. Настоящие системы могут сделать это с большей точностью, чем предыдущие мутагенные агенты и, следовательно, ускорить и улучшить программы селекции растения.
[000009] 2. Гены, вовлеченные в болезни растений, такие как перечисленные в WO 2013046247:
- Болезни риса: Magnaporthe grisea, Cochliobolus miyabeanus, Rhizoctonia solani, Gibberella fujikuroi; Болезни пшеницы: Erysiphe graminis, Fusarium graminearum, F. avenaceum, F. culmorum, Microdochium nivale, Puccinia striiformis, P. graminis, P. recondita, Micronectriella nivale, Typhula sp., Ustilago tritici, Tilletia caries, Pseudocercosporella herpotrichoides, Mycosphaerella graminicola, Stagonospora nodorum, Pyrenophora tritici-repentis; Болезни ячменя: Erysiphe graminis, Fusarium graminearum, F. avenaceum, F. culmorum, Microdochium nivale, Puccinia striiformis, P. graminis, P. hordei, Ustilago nuda, Rhynchosporium secalis, Pyrenophora teres, Cochliobolus sativus, Pyrenophora graminea, Rhizoctonia solani; Болезни кукурузы: Ustilago maydis, Cochliobolus heterostrophus, Gloeocercospora sorghi, Puccinia polysora, Cercospora zeae-maydis, Rhizoctonia solani.
- Болезни цитрусов: Diaporthe citri, Elsinoe fawcetti, Penicillium digitatum, P. italicum, Phytophthora parasitica, Phytophthora citrophthora; Болезни яблока: Monilinia mali, Valsa ceratosperraa, Podosphaera leucotricha, Alternaria alternata яблочный патотип, Venturia inaequalis, Colletotrichum acutatum, Phytophtora cactorum;
- Болезни груши: Venturia nashicola, V. pirina, Alternaria alternata. Японский грушевый патотип, Gymnosporangium haraeanum, Phytophtora cactorum;
- Болезни персика: Monilinia fructicola, Cladosporium carpophilum, Phomopsis sp.;
- Болезни винограда: Elsinoe ampelina, Glomerella cingulata, Uninula necator, Phakopsora ampelopsidis, Guignardia bidwellii, Plasmopara viticola;
- Болезни хурмы: Gloesporium kaki, Cercospora kaki, Mycosphaerela nawae;
- Болезни тыквы: Colletotrichum lagenarium, Sphaerotheca fuliginea, Mycosphaerella melonis, Fusarium oxysporum, Pseudoperonospora cubensis, Phytophthora sp., Pythium sp.;
- Болезни томата: Alternaria solani, Cladosporium fulvum, Phytophthora infestans;
- Болезни баклажана: Phomopsis vexans, Erysiphe cichoracearum;
- Болезни овощей семейства Крестоцветные: Alternaria japonica, Cercosporella brassicae, Plasmodiophora brassicae, Peronospora parasitica;
- Болезни лука-батуна: Puccinia allii, Peronospora destructor;
- Болезни сои: Cercospora kikuchii, Elsinoe glycines, Diaporthe phaseolorum var. sojae, Septoria glycines, Cercospora sojina, Phakopsora pachyrhizi, Phytophthora sojae, Rhizoctonia solani, Corynespora casiicola, Sclerotinia sclerotiorum;
- Болезни фасоли: Colletrichum lindemthianum;
- Болезни арахиса: Cercospora personata, Cercospora arachidicola, Sclerotium rolfsii;
- Болезни гороха: Erysiphe pisi;
- Болезни картофеля: Alternaria solani, Phytophthora infestans, Phytophthora erythroseptica, Spongospora subterranea, f. sp. Subterranea;
- Болезни клубники: Sphaerotheca humuli, Glomerella cingulata;
- Болезни чая: Exobasidium reticulatum, Elsinoe leucospila, Pestalotiopsis sp., Colletotriсhum theae-sinensis;
- Болезни табака: Alternaria longipes, Erysiphe cichoracearum, Colletotrichum tabacum, Peronospora. tabacina, Phytophthora nicotianae;
- Болезни рапса: Sclerotinia sclerotiorum, Rhizoctonia solani;
- Болезни хлопка: Rhizoctonia solani;
- Болезни свеклы: Cercospora beticola, Thanatephorus cucumeris, Aphanomyces cochlioides;
- Болезни розы: Diplocarpon rosae, Sphaerotheca pannosa, Peronospora sparsa;
- Болезни хризантем и семейства Сложноцветные: Bremia lactuca, Septoria chrysanthemi-indici, Puccinia horiana;
- Болезни различных растений: Pythium. aphanidermatum, Pythium debarianum, Pythium graminicola, Pythium irregulare, Pythium ultimum, Botrytis cinerea, Sclerotinia sclerotiorum;
- Болезни редиса: Alternaria brassicicola;
- Болезни зойсии: Sclerotinia homeocarpa, Rhizoctonia solani;
- Болезни бананов: Mycosphaerella fijiensis, Mycosphaerella musicola;
- Болезни подсолнечника: Plasmopara halstedii;
- Болезни семян или болезни наначальной стадии роста различных растений, вызываемых Aspergillus spp., Penicillium spp., Fusarium spp., Gibberella spp., Tricoderma spp., Thielaviopsis spp., Rhizopus spp., Mucor spp., Corticium spp., Rhoma spp., Rhizoctonia spp., Diplodia. spp., и т.п.;
- Вирусные заболевания различных растений, опосредованных Polymixa spp., Olpidium spp., и т.п.
[000010] 3. Примеры генов, придающих устойчивость к гербицидам;
- Устойчивость к гербицидам, которые ингибируют точки роста или меристему, такую как имидазолинон или сульфонилмочевина, например, Lee et al., EMBOJ. 7:1241 (1988), и Miki et al., Theor. Appl. Genet. 80:449 (1990), соответственно.
- Устойчивость к глифосату (устойчивость передается, например, мутантными генами 5-енолпирувилшикимат-3-фосфатсинтазы (EPSP), генами aroA и генамиглифосатацетилтрансферазы (GAT), соответственно), или устойчивость к другим соединениям с фосфоногруппой, таким как глюфосинат (гены фосфинотрицинацетилтрансферазы (PAT) из видов Streptomyces, включая Streptomyces hygroscopicus и Streptomyces viridichromogenes), и к пиридинокси-илифенокси-пропионовым кислотам и циклогексенам посредством генов, кодирующих ингибитор ACC-азы. См., например, патент США № 4940835 и патент США № 6248876, патент США № 4769061, EP№ 0 333 033 и патент США № 4975374. См. также № 0242246, DeGreef et al., Bio/Technology 7:61 (1989), Marshall et al., Theor. Appl. Genet. 83:435 (1992), WO 2005012515 Castle et. al. и WO 2005107437.
- Устойчивость к гербицидам, ингибирующим фотосинтез, такие как триазин (гены psbA и. gs+) или бензонитрил (геннитрилазы), иглутатион S-трансфераза в Przibila et al., Plant Cell 3:169 (1991), патент США № 4810648 и Hayes et al., Biochem. J. 285: 173 (1992).
- Гены, кодирующие Ферменты, детоксифицирующие гербицид, или мутантный фермент глутаминсинтетазы, устойчивый к ингибированию, например, в патентной заявке США с серийным номером № 11/760602. В ином случае детоксифицирующий фермент является ферментом, кодирующим фосфинотрицин ацетилтрансферазу (такую как белок bar или pat из видов Streptomyces). Фосфинотрицин ацетилтрансферазы описаны, например, в патентах США № 5561236; 5648477, 5646024; 5273894; 5637489; 5276268; 5739082; 5908810 и 7112665.
- Ингибиторы гидроксифенилпируват-диоксигеназы (HPPD), то есть, встречающиеся в природе ферменты, устойчивые к HPPD, или гены, кодирующие мутированный или химерный фермент HPPD, как описано в WO 96/38567, WO 99/24585, и WO 99/24586, WO 2009/144079, WO 2002/046387 или в патенте США № 6768044.
[000011] 4. Примеры генов, вовлеченных в устойчивость к абиотическому стрессу:
- Трансген, способный к уменьшению экспрессии и/или активности гена поли(АДФ-рибоза)-полимеразы (PARP) в растительных клетках или растениях, как описано в WO00/04173 или, WO/2006/045633.
- Трансгены, способные к уменьшению экспрессии и/или активности кодирующих PARG генов растений или клеток растений, как описано, например; в WO2004/090140.
- Трансгены, кодирующие функциональный в растениях фермент "спасательного" пути синтеза никотинамидадениндинуклеотида, включая никотинамидазу, никотинат фосфорибозилтрансферазу, мононуклеотид-аденилтрансферазу никотиновой кислоты, никотинамидадениндинуклеотид-синтетазу или никотинамидфосфорибозил-трансферазу, как описано, например, в EP 04077624.7, WO2006/133827, PCT/EP07/002433, EP 1999263, или WO2007/107326.
- Ферменты, участвующие в биосинтезе углеводов, включают ферменты, описанные, например, в EP 0571427, WO 95/04826, EP 0719338, WO 96/15248, WO 96/19581, WO 96/27674, WO 97/11188, WO 97/26362, WO 97/32985, WO 97/42328, WO 97/44472, WO 97/45545, WO 98/27212, WO 98/40503, WO99/58688, WO 99/58690, WO 99/58654, WO 00/08184, WO 00/08185, WO 00/08175, WO 00/28052, WO 00/77229, WO 01/12782, WO 01/12826, WO 02/101059, WO 03/071860, WO 2004/056999, WO 2005/030942, WO 2005/030941, WO 2005/095632, WO 2005/095617, WO 2005/095619, WO 2005/095618, WO 2005/123927, WO 2006/018319, WO 2006/103107, WO 2006/108702, WO 2007/009823, WO 00/22140, WO 2006/063862, WO 2006/072603, WO 02/034923, EP 06090134.5, EP 06090228.5, EP 06090227.7, EP 07090007.1, EP 07090009.7, WO 01/14569, WO 02/79410, WO 03/33540, WO 2004/078983, WO 01/19975, WO 95/26407, WO 96/34968, WO 98/20145, WO 99/12950, WO 99/66050, WO 99/53072, патенте США № 6734341, WO 00/11192, WO 98/22604, WO 98/32326, WO 01/98509, WO 01/98509, WO 2005/002359, патенте США № 5824790, патенте США № 6013861, WO94/04693, WO94/09144, WO94/11520, WO95/35026 или WO97/20936 или ферменты, участвующие в синтезе полифруктанов, особенно типа инулина и левана, как описано в EP 0663956, WO 96/01904, WO 96/21023, WO 98/39460, иWO 99/24593, синтезе альфа-1,4-глюканов, как описано вWO 95/31553, US 2002031826, патенте США № 6284479, патенте США № 5712107, WO 97/47806, WO 97/47807, WO 97/47808 и WO 00/14249, синтезе альфа-1,6-разветвленных альфа-1,4-глюканов, как описано в WO 00/73422, синтезе альтернана, как описано, например, в WO 00/47727, WO 00/73422, EP 06077301.7, патенте США № 5908975 и EP 0728213, синтезе гиалуронана, описанного в качестве примера в WO 2006/032538, WO 2007/039314, WO 2007/039315, WO 2007/039316, JP 2006304779 и WO 2005/012529.
- Гены, повышающие устойчивость к засухе. Например, в WO 2013122472 описано, что отсутствие или сниженный уровень функционального белка убиквитинлигазы (UPL), более конкретно UPL3, приводят к уменьшению потребности в воде или улучшению устойчивости к засухе указанного растения. Другие примеры трансгенных растений с увеличенной устойчивостью к засухе описаны, например, в US2009/0144850, US2007/0266453, и WO2002/083911. В US2009/0144850 описано растение, имеющее фенотип устойчивости к засухе благодаря измененной экспрессии нуклеиновой кислоты DR02. В US2007/0266453 описано растение, имеющее фенотип устойчивости к засухе благодаря измененной экспрессии нуклеиновой кислоты DR03, и WO 2002/083911 описано растение, имеющее увеличенную устойчивость к засухе благодаря уменьшению активности переносчика ABC, который экспрессируется в замыкающих клетках устьиц. Другим примером является работа Kasuga с соавторами (1999), которая описывает сверхэкспрессию комплементарной ДНК (кДНК), кодирующей DREB1A в трансгенных растениях, что активировало экспрессию многих генов устойчивости к стрессу при нормальных условиях выращивания и привело к улучшению устойчивости к засухе, засолению и замораживанию. Однако экспрессия DREB1A также привела к серьезному замедлению роста при нормальных условиях выращивания (Kasuga (1999) NatBiotechnol 17(3) 287-291).
[000012] В следующих конкретных вариантах хлебные злаки могут быть улучшены путем влияния на конкретные характеристики растений. Это может быть осуществлено, например, путем разработки устойчивых к пестицидам растений, улучшения устойчивости растений к болезням, улучшения устойчивости растений к насекомым и нематодам, улучшения устойчивости растений к паразитарным сорнякам, улучшения устойчивости растений к засухе, улучшения пищевой ценности растений, улучшения устойчивости растений к стрессу, избегания самоопыления, увеличения усвояемости биомассы растительных кормов, увеличения урожайности зерна и т.д. Несколько конкретных неограничивающих примеров предложены в настоящем описании ниже.
[001005] В дополнение к целенаправленному внесению мутаций в одиночные гены, комплексы CRISPR с C2c1 или C2c3 могут быть разработаны для целенаправленного внесения мутаций в множественные гены, делеции хромосомных фрагментов, сайт-специфичной интеграции трансгенов, сайт-направленного мутагенеза in vivo, и точной замены гена или аллеля в растениях. Поэтому способы, описанные в настоящем описании, имеют широкое применение для обнаружения и проверки генов, мутационной и цисгенной селекции, и гибридной селекции. Эти заявки облегчают производство нового поколения генетически модифицированных зерновых культур с различными улучшенными агрономическими характеристиками, такими как устойчивость к гербицидам, устойчивость к болезням, устойчивость к абиотическому стрессу, высокая урожайность и высшее качество.
Использование гена C2c1 или C2c3 для создания мужских стерильных растений
[001006] Гибридные растения, как правило, имеют выгодные агрономические характеристики по сравнению с инбредными растениями. Однако для самоопыляемых растений создание гибридов может быть затруднено. В различных типах растений были определены гены, которые важны для фертильности растений, более конкретно мужской фертильности. Например, для кукурузы было определено, что по меньшей мере два гена важны для фертильности (Amitabh Mohanty International Conference on New Plant Breeding Molecular Technologies Technology Development And Regulation, Oct 9-10, 2014, Jaipur, India; Svitashev et al. Plant Physiol. 2015 Oct; 1.69(2):93 N45; Djukanovic et al. PlantJ. 2013 Dee;76(5):888-99). Способы, описанные в настоящем описании, могут использоваться для нацеливания на гены, необходимые для мужской фертильности, чтобы производить мужские стерильные растения, которые легко скрещиваются с образованием гибридов. В конкретных вариантах осуществления система CRISPRc C2c1 или C2c3, описанная в настоящем описании, используется для целенаправленного мутагенеза цитохром P450-подобного гена (MS26) или гена мегануклеазы (MS45), таким образом, обеспечивая мужскую стерильность растений кукурузы. Растения кукурузы, генетически измененные таким образом, могут использоваться в программах гибридной селекции.
Увеличение фертильной стадии в растениях
[001007] В конкретных вариантах осуществления способы, описанные в настоящем описании, используются для продления фертильной стадии растения, такого как рис. Например, на ген фертильной стадии риса, такой как Ehd3, можно осуществлять нацеливание для внесения мутации в ген, и всходы могут быть отобраны для длительной регенерации фертильной стадии растения (как описано в CN 104004782).
Использование C2c1 или C2c3 для создания наследственной изменчивости в представляющей интерес культуре
[001008] Доступность дикой гермоплазмы и наследственной изменчивости в хлебных злаках является ключом к программам их улучшения, но доступное разнообразие гермоплазм хлебных злаков ограничено. Настоящее изобретение относится к способам создания разнообразия наследственной изменчивости в представляющей интерес гермоплазме. В этом способе применения систем CRISPR C2c1 или C2c3 предполагается введение в растительные клетки библиотеки направляющих РНК, нацеленных на различные места в геноме растения, вместе с эффекторным белком C2c1 или C2c3. Таким образом, может быть создана коллекция точечных мутаций и генных нокаутов в масштабах размера генома. В конкретных вариантах осуществления способы включают создание части растения или растения из клеток, полученных таким образом, и скрининг клеток в отношении представляющего интерес признака. Гены-мишени могут включать как кодирующие, так и некодирующие участки. В конкретных вариантах осуществления признаком является устойчивость к стрессу, и способ представляет собой способ создания устойчивых к стрессу разновидностей культур.
Использование C2c1 или C2c3 для влияния на созревание фруктов
[001009] Дозревание - нормальная фаза в процессе созревания фруктов и овощей. Спустя только несколько дней после начала дозревания, фрукт или овощ становится несъедобным. Этот процесс приносит значительные убытки для фермеров и потребителей. В конкретных вариантах осуществления способы по настоящему изобретению используются для уменьшения синтеза этилена. Это обеспечивается одним или более из следующих способов: a. подавление экспрессии гена АЦК-синтазы. АЦК (1-аминоциклопропан -l-карбоксильная кислота)-синтаза является ферментом, ответственным за преобразование S-аденозилметионина (SAM) в АЦК; предпоследнему шагу в биосинтезе этилена. Экспрессии фермента затрудняется, когда антисмысловая ("зеркальное отображение") или усеченная копия синтезированного гена вставляется в геном растения, b. Инсерция гена АЦК-дезаминазы. Ген, кодирующий фермент, получен из Pseudomonas chlororaphis, обыкновенной непатогенной почвенной бактерии. Этот фермент преобразовывает AЦК в другое соединение, таким образом, уменьшая количество AЦК, доступного для синтеза этилена; c. Инсерция гена SAM-гидролазы. Этот подход подобен таковому с АЦК-дезаминазой, где синтез этилена прекращается, когда количество предшествующего ему метаболита уменьшается; в этом случае SAM преобразуется в гомосерин. Ген, кодирующий фермент, получен из бактериофага T3 E. coli, и d. Подавление экспрессии гена АЦК-оксидазы. АЦК-оксидаза является ферментом, катализирующим окисление AЦК до этилена, последний шаг в биосинтетическом пути этилена. Используя способы, описанные в настоящем описании, регуляция по типу отрицательной обратной связи гена оксидазы AЦК приводит к подавлению синтеза этилена, таким образом задерживая дозревание фруктов. В конкретных вариантах осуществления, дополнительно или альтернативно к модификациям, описанным выше, способы, описанные в настоящем описании, используются для модификации этиленовых рецепторов, для вмешательства в этиленовые сигналы, получаемые фруктами. В конкретных вариантах осуществления экспрессия гена ETR1, кодирующего связывающий этилен белок, модифицирована, более конкретно - подавлена. В конкретных вариантах осуществления, дополнительно или альтернативно модификациям, описанным выше, способы, описанные в настоящем описании, используются для модификации экспрессии гена, кодирующего полигалактуроназу (PG), которая является ферментом, ответственным за разрушение пектина, вещества, поддерживающего целостность клеточной стенки растительной клетки. Разрушение пектина происходит в начале процесса дозревания, что приводит к размягчению фруктов. Соответственно, в конкретных вариантах осуществления, описанные в настоящем описании способы используются для внесения мутации в ген PG или подавления активации гена PG, чтобы уменьшить количество синтезированного фермента PG, таким образом, замедляя деградацию пектина.
[001010] Таким образом, в конкретных вариантах осуществления способы включают использование системы CRISPR c C2c1 или C2c3 для осуществления одной или более модификаций генома растительной клетки, таких как описано выше, и регенерации растения из этих клеток. В конкретных вариантах осуществления растение является томатом.
Увеличение жизни хранения растений
[001011] В конкретных вариантах осуществления способы по изобретению используют для модификации генов, участвующих в производстве соединений, которые затрагивают срок хранения части растения или растения. Более подробно, модификация вносится в ген, который предотвращает накопление восстанавливающихся сахаров в картофельных клубнях. После высокотемпературной обработки эти восстанавливающиеся сахара реагируют со свободными аминокислотами, что приводит к образованию коричневых продуктов с горьким вкусом и повышенным содержанием акриламида, который является потенциальным канцерогеном. В конкретных вариантах осуществления изобретения способы, описанные в настоящем описании, используют, чтобы уменьшить или препятствовать экспрессии гена вакуолярной инвертазы (VInv), кодирующей белок, который расщепляет сахарозу до глюкозы и фруктозы (Clasen et al. DOI: 10 1111/pbi. 12370).
Использование системы CRISPR C2c1 или C2c3 для обеспечения увеличения ценности
[000013] В конкретных вариантах осуществления система C2c1 или C2c3 CRISPR используется для производства сельскохозяйственных зерновых культур с улучшенными питательными свойствами. В конкретных вариантах осуществления, способы, описанные в настоящем описании, адаптированы для производства "функциональных продуктов", т.е. модифицированных продуктов питания или их ингредиентов, которые могут быть более полезны для здоровья. Кроме традиционных питательных веществ они содержат "нутрицевтик", т.е. вещество, которые можно считать продуктом питания или частью продукта питания, приносящим пользу для здоровья, включая профилактику и лечение заболеваний. В конкретных вариантах осуществления нутрицевтики полезны для предотвращения и/или лечения одного или более типов рака, диабета, сердечно-сосудистых заболеваний и гипертонии.
[000014] Примеры зерновых культур с улучшенными питательными свойствами включают (Newell-McGloughlin, Plant Physiology, July 2008, Vol. 147, pp. 939-953):
- модифицированное качество белка, содержание и/или аминокислотный состав, которые были описаны для Байя (Luciani et al. 2005, Florida Genetics Conference Poster), канолы (Roesler et al., 1997, Plant Physiol 113 75-81), Кукурузы (Cromwell et al, 1967, 1969 JAnim Sci 26 1325-1331, O'Quin et al. 2000 JAnim Sci 78 2144-2149, Yang et al. 2002, Transgenic Res 11 11-20, Young et al. 2004, Plant J 38 910-922), картофеля (Yu J and Ao, 1997 Acta Bot Sin 39 329-334; Chakraborty et al. 2000, Proc Natl Acad Sci USA 97 3724-3729; Li et al., 2001 Chin Sci Bull 46 482-484), риса (Katsube et al. 1999, Plant Physiol 120 1063-4074), сои (Dinkins et al. 2001, Rapp 2002, In Vitro Cell Dev Biol Plant 37 742-747), Батата (Egnin. and Prakash 1997, In Vitro Cell Dev Biol 33 52A).
- содержание незаменимых аминокислот, что было описано для канолы (Falco et al. 1995, Bio/Technology 13 577-582), люпина (White et al. 2001, J Sci Food Agric 81 147-154), кукурузы (Lai and Messing, 2002, Agbios 2008 GM crop database (March 11, 2008)), картофеля (Zeh et al. 2001, Plant Physiol 127 792-802), сорго (Zhao et al. 2003, Kluwer Academic Publishers, Dordrecht, The Netherlands, pp 413-416), Сои (Falco et al. 1995 Bio/Technology 13 577-582; Galdi et al. 2002 Crit Rev Plant Sci 21:167-204).
- Масла и жирные кислоты, такие как для канолы (Dehesh et al. (1996) Plant J 9 167-172 [PubMed]; Del Vecchio (1996) INFORM International News on Fats, Oils and Related Materials 7 230-243; Roesler et al. (1997) Plant Physiol 113 75 81 [PMC free article] [PubMed]; Froman and Ursin (2002, 2003) Abstracts of Papers of The American Chemical Society 223 U35; James et al. (2003) Am J Clin Nutr 77 1140-1145 [PubMed]; Agbios (2008, выше), хлопка (Chapman et al. (2001). J Am. OH Chem. Soc 78 941-947; Liu et al. (2002) J Am Coll Nutr 21 205S-21 IS [PubMed]; O'Neill (2007) Australian Life Scientist http://www.biotechnews.com.au/index.php/id;866694817;fp;4;fpid;2 (June 17, 2008), льна (Abbadi et al., 2004, Plant Cell 16: 2734-2748), кукурузы (Young et al., 2004, Plant J 38 910-922), масличной пальмы (Jalani et al. 1997, J Am Oil Chem Soc 74 1451 1455; Parveez, 2003, AgBiotechNet 113 1-8), Риса (Anai et al., 2003, Plant Cell Rep 21 988-992), Сои (Reddy and Thomas, 1996, Nat Biotechnol 14 639-642; Kinney and Kwolton, T9Q8. Blackie Academic and Professional, London, pp 193-213), подсолнечника (Arcadia, Biosciences 2008).
- Углеводы, такие как фруктаны, описанные для цикория (Smeekens (1997) Trends Plant Sci 2 286-287, Sprenger et al. (1997) FEBS Lett 400 355-358, Sévenier et al. (1998) Nat Biotechnol 16 843-846), кукурузы (Caimi et al. (1996) Plant Physiol 110 355-363), картофеля (Hellwege et al.,1997 Plant J 12 1057-1065), сахарной свеклы (Smeekens et al. 1997, above); инулин, как описано для картофеля (Hellewege et al. 2000, Proc Natl Acad Sci USA 97 8699-8704), крахмал, как описано для риса (Schwall et al. (2000) Nat Biotechnol 18 551-554, Chiang et al. (2005) Mol Breed 15 125-143).
- Витамины и каратиноиды, как описано для канолы (Shintani and DellaPenna (1998) Science 282 2098-2100), кукурузы (Rocheford et al. (2002) J Am Coll Nutr 21 191S-198S, Cahoon et al. (2003) Nat Biotechnol 21 1082-1087, Chen et al. (2003) Proc Natl Acad Sci USA 100 3525-3530), горчичного семени (Shewmaker et al. (1999) Plant J 20 401- 412), картофеля (Ducreux et al., 2005, J Exp Bot 56 81-89), Риса (Ye et al. (2000) Science 287 303-305), клубники (Agius et al. (2003), Nat Biotechnol 21 177-181), томата (Rosati et al. (2000) Plant J 24 413-419, Fraser et al. (2001) J Sci Food Agric 81 822-827, Mehta et al. (2002) Nat Biotechnol 20 613-618, Diaz de la Garza et al. (2004) Proc Natl Acad Sci USA 101 13720-13725, Enfissi et al. (2005) Plant Biotechnol J 3 17-27, DellaPenna (2007) Proc Natl Acad Sci USA 104 3675-3676).
- Функциональные вторичные метаболиты, такие как описаны для яблока (стильбены, Szankowski et al. (2003) Plant Cell Rep 22: 141-149), люцерны (ресвератрол, Hipskind and Paiva (2000) Mol Plant Microbe Interact 13 551-562), киви (ресвератрол, Kobayashi et al. (2000) Plant Cell Rep 19 904-910), кукурузы и сои (флавоноиды, Yu et al. (2000) Plant Physiol 124 781-794), картофеля (антоцианы, алкалоиды и гликоиды, Lukaszewicz et al. (2004) J Agric Food Chem. 52 1526-1533), риса (флавоноиды и ресвератрол, Stark-Lorenzen et al. (1997) Plant Cell Rep 16 668-673, Shin et al. (2006) Plant Biotechnol J 4 303-315), томата (ресвератрол, хлорогеновая кислота, флавоноиды, стильбены, Rosati. et al. (2000) above, Muir et al. (2001) Nature 19 470-474, Niggeweg et al. (2004) Nat Biotechnol 22 746-754, Giovinazzo et al. (2005) Plant Biotechnol J 3 57-69), пшеницы (кофейная и феруловая кислоты, ресвератрол, United Press International (2002)); и
- Минеральные вещества, такие как описаны для люцерны (фитаза, Austin-Phillips et al. (1999) http://www.molecularfarming.com/nonmedical.html), салата-латука (железо, Goto et al. (2000) TheorApplGenet 100 658-664), риса (железо, Lucca et al. (2002) JAmCollNutr 21 184S-190S), кукурузы, сои и пшеницы (фитаза, Drakakaki et al. (2005) Plant Mol Biol 59 869-880, Denbow et al. (1998) Poult Sci 77 878-881, Brinch-Pedersen et al. (2000) Mol Breed 6 195-206).
[000015] В конкретных вариантах осуществления признак повышения ценности связан с предусматриваемой пользой для здоровья соединений, присутствующих в растении. Например, в конкретных вариантах осуществления, культура с добавленной стоимостью получена с применением способов по изобретению для модификации или индукции/увеличения синтеза одного или более следующих соединений:
- Каротиноиды, такие как α-каротин, содержащийся в моркови, который нейтрализует свободные радикалы, которые могут наносить ущерб клеткам, или β-каротин, содержащийся в различных фруктах и овощах, который нейтрализует свободные радикалы
- Лютеин, содержащийся в зеленых овощах, который способствует поддержанию здорового зрения
- Ликопин, содержащийся в томатах и продуктах из томатов, который, как считается, снижает риск рака предстательной железы
- Зеаксантин, содержащийся в цитрусовых и кукурузе, который способствует поддержанию здорового зрения
- Диетические волокна, такие как нерастворимые волокна, содержащиеся в пшеничных отрубях, которые могут снижать риск рака молочной железы и/или рака толстой кишки и β-глюканы, содержащиеся в овсяном зерне, растворимые волокна, содержащиеся в Psylium и цельных зерновых культурах, которые могут снижать риск сердечно-сосудистых заболеваний (CVD)
- Жирные кислоты, такие как ω-3-жирные кислоты, которые могут снижать риск сердечно-сосудистых заболеваний и улучшать умственные и зрительные функции, Конъюгированная линолевая кислота, которая может улучшать форму тела, снижать риск развития некоторых видов рака и GLA, который может снижать риск воспаления при злокачественной опухоли и сердечно-сосудистых заболеваниях (CVD), может улучшать форму тела
- Флавоноиды, такие как гидроксициннаматы, содержащиеся в пшенице, могут иметь Антиокcидантную активность, снижать риск дегенеративных заболеваний, флавонолы, катехины и танины, содержащиеся во фруктах и овощах, нейтрализуют свободные радикалы и могут снижать риск рака
- Глюкозинолаты, индолы, изотиоцианаты, такие как сульфорафан, содержащийся в Крестоцветных овощах (брокколи, листовой капусте), хрене, которые нейтрализуют свободные радикалы, могут снижать риск рака
- Фенолы, такие как стильбены, содержащиеся в винограде, могут снижать риск дегенеративных заболеваний, заболеваний сердца, а также рака, могут положительно влиять на продолжительность жизни, кофейная кислота и ферульная кислота, содержащиеся в овощах и цитрусовых, имеют подобную антиоксидантную активность, могут снижать риск дегенеративных заболеваний, заболеваний сердца, и заболеваний глаз и эпикатехин, содержащийся в какао, имеет подобную антиоксидантную активность, может снижать риск дегенеративных заболеваний и заболеваний сердца
- Растительные станолы/стерины, содержащиеся в кукурузе, сое, пшенице и деревянных маслах, которые могут снижать риск ишемической болезни сердца, понижая уровень холестерина в крови
- Фруктаны, инулины, фруктоолигосахариды, содержащиеся в топинамбуре, луке-шалоте, луковом порошке, который может улучшать здоровье желудочно-кишечного тракта
- Сапонины, содержащиеся в сое, могут понижать уровень холестерина ЛПНП
- Белок сои, содержащийся в сое, может снижать риск заболеваний сердца
- Фитоэстрогены, такие как изофлавоны, содержащиеся в сое, могут уменьшать симптомы менопаузы, такие как приливы, могут снижать риск развития остеопороза и сердечно-сосудистых заболеваний (CVD) и лигнаны, содержащиеся во льне, ржи и овощах, могут защищать от заболеваний сердца и некоторых случаев рака, могут понижать холестерин ЛПНП, общий холестерин.
- Сульфиды и тиолы, такие как диаллилсульфид, содержащийся в луке, чесноке, маслинах, луке-порее, и сквален и аллилметилтрисульфид, дитиолтионы, содержащиеся в крестоцветных овощах, которые могут понижать уровень холестерина ЛПНП, помогают поддерживать здоровье иммунной системы
- Танины, такие как проантоцианидины, содержащиеся в клюкве, какао, могут улучшать здоровье мочевых путей, снижать риск развития сердечно-сосудистых заболеваний (CVD) и высокого кровяного давления
- и т.д.
[0016] Кроме того, способы по изобретению также предусматривают модификацию функциональности белка/крахмала, срока годности, вкуса/эстетических характеристик, качества волокон и аллергенов, антинутриентов и признаки снижения содержания токсинов.
[000017] Соответственно, изобретение охватывает способы производства растений с увеличенной питательной ценностью, причем указанные способы включают введение в растительную клетку гена, кодирующего фермент, вовлеченный в синтез компонента увеличенной питательной ценности, с использованием системы CRISPR C2c1 или C2c3, как описано в настоящем описании, и регенерации растения из указанной растительной клетки, где указанное растение характеризуется увеличением экспрессии указанного компонента увеличенной питательной ценности. В конкретных вариантах система CRISPRC2c1 или C2c3 используется для модификации эндогенного синтеза этих соединений косвенно, например, модификации одного или более транскрипционных факторов, управляющих метаболизмом этого соединения. Способы введения интересующего гена в растительную клетку и/или модификации эндогенного гена с использованием системы CRISPRC2c1 или C2c3 описаны в настоящем описании выше.
[000018] Некоторые конкретные примеры модификаций в растениях, которые были модифицированы для приобретения признака увеличенной ценности включают: растения с модифицированным метаболизмом жирных кислот, например, путем трансформации растения с антисмысловым геном стеарил-ACP-десатуразы для увеличения содержания стеариновой кислоты в растении. См. Knultzon et al., Proc. Natl. Acad. Sci. U.S.A. 89:2624 (1992). Другой пример включает уменьшение содержания фитата, например путем клонирования и затем повторного ввода ДНК, ассоциированной с единственным аллелем, которая может быть ответственным за мутантов кукурузы, характеризуемых низкими уровнями фитиновой кислоты. См. Raboy et al, Maydica 35:383 (1990).
[001012] Аналогично экспрессия генов кукурузы (Zea mays) Tfs C1 и R, которые регулируют производство флавоноидов в алейроновых слоях кукурузы под контролем сильного промотора, привела к высокой скорости накопления антоцианинов в арабидопсисе (Arabidopsisthaliana), предположительно посредством активации всего пути (Bruce et al., 2000, PlantCell 12:65-80). DellaPenna (Welsch et al., 2007 AnnaRevPlantBiol 57: 711-738) обнаружил, что Tf RAP2.2 и взаимодействующий с ним SINAT2, увеличивали каротиногенез в листьях Арабидопсиса. Экспрессия Tf Dof1 вызвало-регулирование генов, кодирующих для синтеза углеродного скелета, заметный рост содержания аминокислот и сокращение уровня Glc в трансгенном Арабидопсисе (Yanagisawa, 2004 PlansCellPhysiol 45: 386-391), и DOF Tf AtDof1.1 (OBP2) интенсифицировал все шаги глюкозинолатного биосинтетического пути в Арабидопсисе (Skirycz et al., 2006 Plant J 47: 10-24).
Уменьшение содержания аллергенов в растениях
[001013] В конкретных вариантах осуществления способы, описанные в настоящем описании, используют, чтобы произвести растения со сниженным уровнем аллергенов, делая их более безопасными для потребителя. В конкретных вариантах осуществления способы включают модификацию экспрессии одного или более генов, ответственных за производство аллергенов растения. Например, в конкретных вариантах осуществления способы включают понижающую регуляцию экспрессии гена Lolp5 в растительной клетке, такой как растительная клетка райграсса и регенерирующее из нее растение, чтобы уменьшить аллергенность пыльцы указанного растения (Bhalla et al. 1999, Proc. Natl. Acad. Sci. USA Vol. 96: 1.1.676-11680).
[001014] Аллергия на арахис и аллергия на бобовые в общем являются реальной и серьезной медицинской проблемой. Система эффекторного белка C2c1 или C2c3 по настоящему изобретению может использоваться для идентификации, а затем редактирования или подавления генов, кодирующих аллергенные белки таких бобовых. Не ограничиваясь такими генами и белками, Nicolaou et al. идентифицировали аллергенные белки в арахисе, сое, чечевице, горохе, люпине, зеленой фасоли и бобах мунг. См., Nicolaou et al., Current Opinion in Allergy and Clinical Immunology 2011; 11 (3):222).
Способы скрининга представляющих интерес эндогенных генов
[001015] Способы, описанные в настоящем описании, далее позволяют проводить идентификацию ценных генов, кодирующих ферменты, вовлеченные в производство компонентов увеличенной питательной ценности или в целом генов, затрагивающих представляющие интерес агрономические признаки, среди видов, типов и всего царства растений. Посредством выборочного нацеливания, например, на гены, кодирующие ферменты метаболических путей в растениях с использованием системы CRISPRC2c1 или C2c3, как описано в настоящем описании, могут быть определены гены, ответственные за определенные пищевые аспекты растения. Точно так же, посредством выборочного нацеливания на гены, которые могут затрагивать желательную агрономическую характеристику, могут быть определены соответствующие гены. Соответственно, настоящее изобретение охватывает способы скрининга генов, кодирующих ферменты, вовлеченные в синтез соединений с определенной питательной ценностью и/или агрономическими признаками.
Следующие применения системы CRISPR C2c1 или C2c3 в растениях и дрожжах
Использование системы CRISPR C2c1 или C2c3 при производстве биотоплива
[001016] Под термином "биотопливо" в настоящем описании понимается альтернативное топливо, полученное из растений и других растительных ресурсов. Возобновляемое биотопливо может быть извлечено из органического вещества, энергия которого была запасена в ходе процесса фиксации углерода или получена посредством использования или конверсии биомассы. Эта биомасса может быть использована в качестве биотоплива или может быть переведена в форму удобных содержащих энергию веществ за счет термической, химической или биохимической переработки. Такая переработка позволит получить топливо в твердой, жидкой или газообразной форме. Существует два типа биотоплива: биоэтанол и биодизель. Биоэтанол получают в основном путем сбраживания углеводов - целлюлозы (крахмала), получаемых главным образом из кукурузы и сахарного тростника. Биодизель, с другой стороны, производится главным образом из масличных культур, таких как рапс, масличная пальма и соя. Биотопливо используется, главным образом, в качестве топлива для транспортных средств.
Улучшение свойств растения для производства биотоплива
[001017] В конкретных вариантах осуществления способы с использованием системы CRISPR C2c1 или C2c3, как описано в настоящем описании, используют для изменения свойств клеточной стенки с целью облегчения доступа ключевых агентов гидролиза и более эффективного высвобождения сахаров, используемых для брожения. В некоторых вариантах осуществления изобретения может быть модифицирован биосинтез целлюлозы и/или лигнина. Целлюлоза является главным компонентом клеточной стенки. Биосинтез целлюлозы и лигнина регулируются согласованно. Уменьшая долю лигнина в растении можно увеличить долю целлюлозы. В некоторых вариантах осуществления изобретения описанные в настоящем описании способы используются для снижения уровня биосинтеза лигнина в растении с целью увеличения доли сбраживаемых углеводов. Конкретнее, способы, описанные в настоящем описании, используются для снижения активности по меньшей мере первого гена биосинтеза лигнина, выбранного из группы, состоящей из 4-кумарат-3-гидроксилазы (C3H), фенилаланинаммонийлиазы (PAL), циннамат-4-гидроксилазы (C4H), гидрокслициннамоилттрансферазы (HCT), O-метилтрансферазы кофейной кислоты (COMT), коффеоил-КоA-3-O-метилтрансферазы (CCoAOMT), ферулат-5-гидроксилазы (F5H), дегидрогеназы циннамилового спирта (CAD), циннамоил-КоA-редуктазы (CCR), 4-кумароат-КоA-лигазы (4CL), монолигнол-лигнин-специфической гликозилтрансферазы и альдегиддегидрогеназы (ALDH), как описано в WO 2008064289 A2.
[001018] В некоторых вариантах осуществления изобретения описанные в настоящем описании способы используются для производства биомассы растения, образующей более низкие уровни уксусной кислоты при сбраживании (см. также WO 2010096488). Более конкретно, способы, описанные в настоящем описании, используют для внесения мутаций в гомологи CasIL для снижения ацетилирования полисахаридов.
Модификация дрожжей для производства биотоплива
[001019] В конкретных вариантах осуществления изобретения предлагаемый фермент C2c1 или C2c3 используется для производства биоэтанола в рекомбинантных микроорганизмах. Например, ферменты C2c1 или C2c3 могут быть использованы для получения микроорганизмов, таких как дрожжи, для производства биотоплива или биополимеров из сбраживаемых сахаров и, возможно, для разложения лигноцеллюлозы, полученной из растений, извлеченных из отходов сельского хозяйства в качестве источника сбраживаемых сахаров. Конкретнее, изобретение относится к способам, в которых комплекс CRISPR C2c1 или C2c3 используется для внесения в микроорганизмы чужеродных генов, необходимых для синтеза биотоплива, и/или изменения эндогенных генов, которые могут препятствовать синтезу биотоплива. Более конкретно, такие способы подразумевают введение в микроорганизм, такой как дрожжи, одной или более нуклеотидных последовательностей, кодирующих ферменты, участвующие в преобразовании пирувата в этанол или иной целевой продукт. В определенных вариантах осуществления такие способы обеспечивают стимуляцию экспрессии одного или более ферментов, позволяющих микроорганизмам разлагать целлюлозу, таких как целлюлаза. В других последующих вариантах осуществления нацеленный на РНК комплекс CRISPR, используется, чтобы подавить эндогенные метаболические процессы, конкурирующие с процессом производства биотоплива.
[001020] Соответственно, в более конкретных вариантах осуществления способы, описанные в настоящем описании, используются, чтобы модифицировать микроорганизм следующим образом:
[001021] введение по меньшей мере одной гетерологичной нуклеиновой кислоты или увеличение экспрессии по меньшей мере одной эндогенной нуклеиновой кислоты, кодирующей фермент, разрушающий клеточную стенку растений, чтобы указанный микроорганизм был способен к экспрессии указанной нуклеиновой кислоты и к синтезу и секреции указанного фермента, разрушающего клеточную стенку растений;
[001022] введение по меньшей мере одной гетерологичной нуклеиновой кислотой или увеличение экспрессии по меньшей мере одной эндогенной нуклеиновой кислоты, кодирующей фермент, который преобразует пируват в ацетальдегид, необязательно объединенный по меньшей мере с одной гетерологичной нуклеиновой кислотой, кодирующей фермент, который преобразует ацетальдегид в этанол так, чтобы указанная клетка-хозяин была способна к экспрессии указанной нуклеиновой кислоты; и/или
[001023] модификация по меньшей мере одной нуклеиновой кислоты, кодирующей фермент в метаболическом пути в указанной клетке-хозяине, где указанный путь производит метаболит кроме ацетальдегида из пирувата или этанола из ацетальдегида, и где указанная модификация приводит к уменьшению производства указанного метаболита, или введение по меньшей мере одной нуклеиновой кислоты, кодирующей ингибитор указанного фермента.
Модификация водорослей и растений для производства растительных масел или биотоплива
[1024] Трансгенные водоросли или другие растения, такие как рапс, могут быть особенно востребованы при производстве растительных масел или биотоплива, например, такого как спирты (особенно метанол и этанол). Они могут быть целенаправленно разработаны для экспрессии или сверхэкспрессии больших объемов масел или спиртов для использования в отраслях промышленности, связанных с нефтью или биотопливом.
[001025] Согласно конкретным вариантам осуществления изобретения, система CRISPR C2c1 или C2c3 используется для производства богатых липидами диатомовых водорослей, полезных в производстве биотоплива.
[001026] В конкретных вариантах осуществления предусматривается специфическая модификация генов, которые вовлечены в модификацию количества липидов и/или качества липидов, произведенных клеткой водоросли. Примеры генов, кодирующих ферменты, вовлеченные в пути синтеза жирных кислот, могут кодировать белки, имеющие, например, ацетил-КoA-карбоксилазу, синтазу жирных кислот, 3-кетоацил-ацил-переносящий белок-синтазу III, глицерол-3-фосфат дегидрогеназу (G3PDH), еноил-ацил-переносящий белок-редуктазу, глицерол-3-фосфат-ацилтрансферазу, лизофосфатид-ацилтрансферазу или диацилглицерол-ацилтрансферазу, фосфолипид:диацилглицерол-ацилтрансферазу, фосфатидат-фосфатазу, тиоэстеразу жирных кислот, такие тиоэстераза пальмитоил-белка или ферментативная активность в отношении яблочной кислоты. В дальнейших вариантах осуществления изобретения предусматривается создать диатомовые водоросли, в которых увеличено накопление липидов. Это может быть достигнуто посредством нацеливания на гены, уменьшающие катаболизм липидов. Особенный интерес для использования в способах по настоящему изобретению представляют гены, вовлеченные в активацию как триацилглицерина, так и свободных жирных кислот, а также гены, непосредственно вовлеченные в β-окисление жирных кислот, такие как ацил-КоА-синтетаза, 3-кетоацил-КоА-тиолаза, активность ацил-КоА-оксидазы и фосфоглюкомутаза. Система CRISPR C2c1 или C2c3 и способы, описанные в настоящем описании, могут использоваться для специфической активации таких генов в диатомовых водорослях для повышения содержания в них липидов.
[001027] Организмы, такие как дрожжи и микроводоросли, широко используются в синтетической биологии. Stovicek et al. (Metab.Eng. Comm., 2015; 2:13) описывают редактирование генома промышленных дрожжей, например Saccharomyces cerevisae, для эффективного создания устойчивых штаммов в целях промышленного производства. Stovicek использовал кодон-оптимизированную систему CRISPR-Cas9 для дрожжей, чтобы одновременно производить как разрыв аллели эндогенного гена, так и нокин гетерологичного гена. Cas9 и гРНК экспрессировали в геномных или эписомальных положениях на векторе. Авторы также показали, что эффективность разрушения гена может быть улучшена путем оптимизации уровней экспрессии Cas9 и гРНК. Hlavova et al. (Biotechnol, Adv., 2015) описали развитие видов или штаммов микроводорослей с использованием таких способов, как CRISPR, для нацеливания на ядерные и хлоропластные гены в целях инсерционного мутагенеза и скрининга. Те же самые плазмиды и векторы могут быть применены к системам C2c1 или C2c3 по настоящему изобретению.
[001028] В US 8945839 описан способ модификации способами инженерии вида микроводорослей (клетки Chlamydomonas reinhardtii) с использованием Cas9. Используя подобную технику, способы с использованием описанной в настоящем описании системы CRISPR C2c1 или C2c3 могут быть применены для видов рода Chlamydomonas и других водорослей. В определенных вариантах осуществления изобретения белок C2c1 или C2c3 и направляющая(ие) РНК введены в водоросли, в которых они экспрессируются, с использованием вектора экспрессии белка C2c1 или C2c3 под контролем конститутивного промотора, такого как Hsp70A-Rbc S2 или бета-2-тубулина. Направляющая РНК необязательно может быть доставлена с использованием вектора, содержащего промотор T7. Альтернативно этому, в клетки водоросли может быть досталвена мРНК C2c1 или C2c3 и транскрибированная in vitro направляющая РНК. Квалифицированному специалисту доступны протоколы электропорации, в том числе стандартный рекомендуемый протокол из набора GeneArt Chlamydomonas Engineering kit.
Использование C2c1 или C2c3 для создания микроорганизмов, способных производить жирные кислоты
[001029] В конкретных вариантах осуществления способы по изобретению используются для создания генетически модифицированных микроорганизмов, способных к производству сложных эфиров жирных кислот, таких как сложные метиловые эфиры жирных кислот ("FAME") и этиловые эфиры жирных кислот ("FAEE").
[001030] Как правило, клетки-хозяева могут быть сконструированы для производства сложных эфиров жирных кислот из источника углерода, такого как спирт, находящегося в среде, посредством экспрессии или сверхэкспрессии гена, кодирующего тиоэстеразу, гена, кодирующего ацил-КоА-синтазу, и гена, кодирующего синтазу сложного эфира. Соответственно, способы, описанные в настоящем описании, используются для модификации микроорганизмов, чтобы сверхэкспрессировать или ввести ген тиоэстеразы, ген, кодирующий ацил-КоА-синтазу, и ген, кодирующий синтазу сложного эфира. В конкретных вариантах осуществления ген тиоэстеразы отобран из tesA, 'tesA, tesB, fatB, fatB2, fatB3, fatA1 или fatA. В конкретных вариантах осуществления ген, кодирующий афил-КоА-синтазу, отобран из fadDJadK, BH3103, pf1-4354, EAV15023, fadD1, fadD2, RPC_4074, fadDD35, fadDD22, faa39, или идентифицированного гена, кодирующего фермент, имеющий те же самые свойства. В конкретных вариантах осуществления ген, кодирующий синтазу сложного эфира, является геном, кодирующим синтазу/ацил-КоА:диацилглицерил-ацилтрансферазу из Simmondsiachinensis, Acinetobacter sp. ADP, Alcanivoraxborkumensis, Pseudomonasaeruginosa, Fundibacterjadensis, Arabidopsisthaliana, или Alkaligeneseutrophus или их вариант. Дополнительно или альтернативно, способы, описанные в настоящем описании, используются для уменьшения экспрессии в указанном микроорганизме по меньшей мере одного из гена, кодирующего ацил-КоА-дегидрогеназу, гена, кодирующего белковый рецептор внешней мембраны, и гена, кодирующего транскрипционный регулятор биосинтеза жирных кислот. В конкретных вариантах осуществления один или более из этих генов инактивированы, например, внесением мутации. В конкретных вариантах осуществления ген, кодирующий ацил-КоА-дегидрогеназу, представляет собой fadE. В конкретных вариантах осуществления ген, кодирующий транскрипционный регулятор биосинтеза жирных кислот, кодирует репрессор транскрипции ДНК, например, fabR.
[001031] Дополнительно или альтернативно, указанный микроорганизм модифицирован, чтобы уменьшить экспрессию по меньшей мере одного гена, кодирующего, пируват-формиат-лиазу, гена, кодирующего лактат-дегидрогеназу или обоих этих генов. В конкретных вариантах осуществления ген, кодирующий пируват-формиат-лиазу, представляет собой pflB. В конкретных вариантах осуществления ген, кодирующий лактат-дегидрогеназу, является IdhA. В конкретных вариантах осуществления один или более из этих генов инактивированы, например, введением в них мутации.
[001032] В конкретных вариантах осуществления микроорганизм выбирается из следующих родов: Escherichia, Bacillus, Lactobacillus, Rhodococcus, Synechococcus, Synechoystis, Pseudomonas, Aspergillus, Trichoderma, Neurospora, Fusarium, Humicola, Rhizomucor, Kluyveromyces, Pichia, Mucor, Myceliophtora, Penicillium, Phanerochaete, Pleurotus, Trameles, Chrysosporium, Saccharomyces, Stenotrophamonas, Schizosaccharomyces, Yarrowia или Streptomyces.
Использование C2c1 или C2c3 для создания микроорганизмов, способных производить органические кислоты
[001033] Способы, описанные в настоящем описании, далее используются для конструирования способами инженерии микроорганизмов, способных производить органические кислоты, более конкретно, из пентозных или гексозных сахаров. В конкретных вариантах осуществления способы включают введение в микроорганизм экзогенного гена LDH. В конкретных вариантах осуществления производство органических кислот в указанных микроорганизмах дополнительно или альтернативно увеличено путем инактивации эндогенных генов, кодирующих белки, вовлеченные в эндогенный метаболический путь, который производит метаболит кроме представляющей интерес органической кислоты, и/или где эндогенный метаболический путь использует органическую кислоту. В конкретных вариантах осуществления модификация гарантирует, что производство метаболита кроме представляющей интерес органической кислоты уменьшено. Согласно конкретным вариантам, способы используются для внесения по меньшей мере одной внесенной способами инженерии делеции в ген и/или инактивации эндогенного пути, в котором органическая кислота используется или ген кодирует продукт, вовлеченный в эндогенный путь, производящий метаболит кроме интересующей органической кислоты. В конкретных вариантах осуществления эта по меньшей мере одна искусственно созданная делеция в гене или инактивация находятся в одном или более генах, кодирующих фермент, отобранный из группы, состоящей из пируват-декарбоксилазы (pdc), фумарат-редуктазы, алкогольдегидрогеназы(adh), ацетальдегиддегидрогеназы, фосфоенолпируват-карбоксилазы (ppc), D-лактат-дегидрогеназы (d-ldh), L-лактат-дегидрогеназы (l-ldh), лактат-2-монооксигеназы. В дальнейших вариантах осуществления изобретения по меньшей мере одна внесенная способами инженерии делеция и/или инактивация гена находятся в эндогенном гене, кодирующем пируват-декарбоксилазу (pdc).
[001034] В дальнейших вариантах осуществления изобретения микроорганизм сконструирован для производства молочной кислоты, и по меньшей мере одна внесенная способами инженерии генная делеция и/или инактивация находится в эндогенном гене, кодирующем лактат-дегидрогеназу. Дополнительно или альтернативно, микроорганизм включает по меньшей мере одну внесенную способами инженерии делецию или инактивацию гена в эндогенном гене, кодирующем цитохром-зависимую лактат-дегидрогеназу, такую как цитохром B2-зависимая L-лактат-дегидрогеназа.
Использование C2c1 или C2c3 для создания улучшенных использующих ксилозу или целлобиозу штаммов дрожжей
[001035] В конкретных вариантах осуществления система CRISPR C2c1 или C2c3 может быть применена для отбора улучшенных использующих ксилозу или целлобиозу штаммов дрожжей. ПЦР пониженной точности может использоваться для амплификации одного (или более) генов, участвующих в путях утилизации ксилозы или путях утилизации целлобиозы. Примеры генов, вовлеченных в пути утилизации ксилозы или пути утилизации целлобиозы, могут включать, но не ограничиваться ими, пути, описанные в Ha, S.J. et al. (2011) Proc. Natl Acad. Sci. USA 108(2):504-9 and Galazka, J.M., et al. (2010) Science 330(6000):84-6. Полученные библиотеки молекул двухцепочечной ДНК, каждая из которых включает случайную мутацию в таком отобранном гене, может быть ко-трансформирована с компонентами системы CRISPR C2c1 или C2c3 в штамм дрожжей (например, S288C), и могут быть отобраны штаммы с повышенной способностью утилизировать ксилозу или целлобиозу, как описано в WO2015138855.
Использование C2c1 или C2c3 в создании улучшенных штаммов дрожжей для биосинтеза изопреноидов
[001036] Tadas Jakociunas et al. описал успешное применение мультиплексной системы CRISPR/Cas9 для конструирования способами геномной инженерии до 5 различных геномных локусов за один шаг трансформации в пекарских дрожжах Saccharomyces cerevisiae (Metabolic Engineering Volume 28, March 2015, Pages 213 -222), что привело к созданию штаммов с высоким производством мевалоната, ключевого промежуточного соединения промышленно важного пути биосинтеза изопреноидов. В конкретных вариантах осуществления система CRISPR C2c1 или C2c3 может быть применена в мультиплексном способе геномной инженерии, как описано в настоящем описании, для идентификации дополнительных высокопроизводительных штаммов дрожжей для применения в синтезе изопреноидов.
Использование C2c1 или C2c3 для создания производящих молочную кислоту штаммов дрожжей
[001037] Другой вариант осуществления изобретения охватывает успешное применение мультиплексной системы C2c1 или C2c3 CRISPR. По аналогии с Vratislav Stovicek et al. (Metabolic Engineering Communications, Volume 2, December 2015, Pages 13-22), улучшенные производящие молочную кислоту штаммы могут быть разработаны и получены путем единственного трансформационного события. В конкретном варианте система C2c1 или C2c3 CRISPR используется для одновременной инсерции гетерологичного гена лактат-дегидрогеназы и разрушение двух эндогенных генов PDC1 и PDC5.
Следующие применения системы CRISPR C2c1 или C2c3 в растениях
[001038] В конкретных вариантах осуществления система CRISPR, и предпочтительно система CRISPR C2c1 или C2c3, описанная в настоящем описании, может использоваться для визуализации динамики генетических элементов. Например, с помощью CRISPR можно визуализировать как повторяющиеся, так и неповторяющиеся геномные последовательности, изменение длины теломеры и движения теломеры и отслеживать динамику генных локусов на протяжении клеточного цикла (Chen et al., Cell, 2013). Эти способы также могут быть применены к растениям.
[001039] Другие применения системы CRISPR, и предпочтительно системы CRISPR C2c1 или C2c3, описанной в настоящем описании, заключаются в скрининге для положительного отбора целевого разрушения гена in vitro и in vivo (Malina et al.., Genes and Development, 2013). Эти способы также могут быть применены к растениям.
[001040] В конкретных вариантах осуществления слияние инактивированных эндонуклеаз C2c1 или C2c3 с ферментами модификации гистонов может вносить направленные изменения в сложный эпигеном (Rusk et al., Nature Methods, 2014). Эти способы также могут быть применены к растениям.
[001041] В конкретных вариантах осуществления система CRISPR, и предпочтительно система CRISPR C2c1 или C2c3, описанная в настоящем описании, может использоваться для очищения конкретной части хроматина и определения связанных белков, чтобы выяснить их регуляторные роли в транскрипции (Waldrip et al., Epigenetics, 2014). Эти способы также могут быть применены к растениям.
[001042] В конкретных вариантах осуществления изобретения предполагается его использование в качестве терапии для удаления вирусов из растительных систем, поскольку оно позволяет расщеплять как вирусную ДНК, так и вирусную РНК. Предыдущие исследования, проведенные для систем человека, продемонстрировали успешное использование CRISPR для нацеливания на вирус гепатита С, содержащий одноцепочечную РНК (A. Price, et al., Proc. Natl. Acad. Sci, 2015), а также на вирус гепатита B, содержащий двухцепочечную ДНК (V. Ramanan, et al., Sci. Rep, 2015). Эти способы могут также быть адаптированы для использования нацеленной на РНК системы CRISPR в растениях.
[001043] В конкретных вариантах осуществления настоящее изобретение может использоваться для изменения сложности генома. В следующем конкретном варианте система CRISPR, и предпочтительно система CRISPR C2c1 или C2c3, описанная в настоящем описании, может использоваться, для разрушения или изменения числа хромосом и создания гаплоидных растений, которые содержат хромосомы только от одного родителя. В таких растениях можно индуцировать дупликацию хромосом с преобразованием в диплоидные растения, содержащие только гомозиготные аллели (Karimi-Ashtiyani et al., PNAS, 2015; Anton et al. Nucleus, 2014). Эти способы могут также быть применены к растениям.
[001044] В конкретных вариантах осуществления система CRISPR C2c1 или C2c3, описанная в настоящем описании, может использоваться для саморасщепления. В этих вариантах осуществления изобретения промотор фермента C2c1 или C2c3 и гРНК может быть конститутивным промотором, и вторая гРНК вводится в составе той же самой кассеты трансформации, но управляется индуцибельным промотором. Эта вторая гРНК может быть сконструирована, чтобы вызывать специфическое расщепление в гене C2c1 или C2c3 с образованием нефункционального C2c1 или C2c3. В следующем конкретном варианте вторая гРНК вызывает расщепление на обоих концах кассеты трансформации, что приводит к удалению кассеты из генома хозяина. Эта система предлагает контролируемую продолжительность воздействия фермента Cas на клетку и далее минимизирует нецелевое редактирование. Кроме того, расщепление обоих концов кассеты CRISPR/Cas может использоваться для производства свободных от трансгена растений T0 с биаллельными мутациями (как описано для Cas9, например, Moore et al., NucleicAcidsResearch, 2014; Schaeffer et al., PlantScience, 2015). Способы Moore et al. могут быть применены к системам CRISPR C2c1 или C2c3, описанным в настоящем описании.
[001045] Sugano et al. (Plant Cell Physiol. 2014 Mar;55(3):475-81. doi: 10.1093/pcp/pcu014. Epub 2014 Jan 18) сообщает о применении CRISPR-Cas9 для направленного мутагенеза печеночника Marchantiapolymorpha L., который стал использоваться как модель для экспериментов для изучения эволюции высших растений. Промотор U6 M. polymorpha идентифицировали и клонировали для экспрессии гРНК. Последовательность, являющаяся мишенью гРНК, была разработана для нарушения работы гена, кодирующего фактор ауксина 1 (ARF1) у M. polymorpha. Используя трансформацию при помощи Agrobacterium, Sugano et al. получили устойчивые мутанты в фазе гаметофита для M. polymorpha. При использовании CRISPR-Cas9 сайт-направленный мутагенез in vivo был получен с помощью промотора мозаики цветной капусты 35S или промотора M. polymorphaEF1α для экспрессии Cas9. Изолированные мутантные индивидуумы, демонстрирующие ауксин-устойчивый фенотип, не были химерами. Более того, стабильные мутанты были получены путем бесполого размножения растений T1. Многочисленные аллели arf1 легко получали с использованием направленного мутагенеза на основе системы CRIPSR/Cas9. Способы Sugano et al. могут быть использованы для системы эффекторных белков C2c1 или C2c3 по настоящему изобретению.
[001046] Kabadi et al. (Nucleic Acids Res. 2014 Oct 29;42(19):e147. doi: 10.1093/nar/gku749. Epub 2014 Aug 13) разработали единую лентивирусную систему, чтобы экспрессировать вариант Cas9, ген-репортер и до четырех sg-РНК с независимых промоторов РНК-полимеразы III, которые включены в вектор с помощью достаточно удобного способа клонирования GoldenGate. Каждая sg-РНК была эффективно экспрессирована и может опосредовать редактирование мультиплексного гена и устойчивую активацию транскрипции в иммортализованных и первичных клетках человека. Способы Kabadi et al. могут быть использованы для системы эффекторных белков C2c1 или C2c3 по настоящему изобретению.
[001047] Ling et al. (BMC Plant Biology 2014, 14: 327) разработали бинарный векторный набор CRISPR-Cas9 на основе каркаса pGreen или pCAMBIA, а также гРНК. Этот набор инструментов не требует каких-либо дополнительных ферментов рестрикции помимо ВsaI для создания конечных конструкций с высокой эффективностью всего за одну стадию клонирования. Этот инструментарий был подтвержден с использованием протопластов кукурузы, трансгенных линий кукурузы и трансгенных линий Arabidopsis и, как было показано, демонстрирует высокую эффективность и специфичность воздействия. Однако, что видится еще более важным, с использованием этого инструментария были обнаружены направленные мутации трех генов Arabidopsis в трансгенных ростках поколения T1. Более того, мутации с множественными генами могут быть унаследованы следующим поколением. Набор векторных модулей (направляющая РНК) использовался в качестве инструментария для мультиплексного редактирования генома в растениях. Инструментарий Ling et al. может быть использован для системы эффекторных белков C2c1 или C2c3 по настоящему изобретению.
[001048] Протоколы для направленного редактирования генома растений c помощью CRISPR-Cas9 также доступны в томе 1284 серии Methods in Molecular Biology стр. 239-255, 10 февраля 2015 года. Описана подробная методика проектирования, конструирования и оценки двойных гРНК для геномного редактирования, опосредованного Cas9, кодон-оптимизированным для экспрессии в растениях (pcoCas9), с использованием протопластов Arabidopsis thaliana и Nicotiana benthamiana в качестве модельных клеточных систем. Стратегии применения системы CRISPR-Cas9 для внесения направленных модификаций в геном целых растений также обсуждены. Протоколы, описанные в этой главе, могут быть применены к системе эффекторных белков C2c1 или C2c3 по настоящему изобретению.
[001049] Ma et al. (Mol Plant, 2015 Aug 3, 8 (8): 1274-84. Doi: 10.1016./j.molp.2015.04.007) сообщают о надежной векторной системе CRISPR-Cas9, использующей кодон-оптимизированный для растений ген Cas9 для удобного и высокоэффективного мультиплексного редактирования генома в однодольных и двудольных растениях. Ma et al. разработали специальные процедуры на основе ПЦР, для быстрого получения множественных экспрессирующих кассет для sg-РНК, которые могут быть собраны в бинарные векторы CRISPR-Cas9 за один раунд клонирования путем лигирования по типу GoldenGate или сборки Гиббса (GibsonAssembly). В этой системе Ma et al. отредактировали 46 целевых сайтов риса со средней скоростью мутации 85,4%, в основном в биаллельном и гомозиготном состоянии. Ma et al. приводят примеры моделирования генной мутации - потери функции в растениях риса T0 и Arabidopsis T1 путем одновременного нацеливания на несколько (до восьми) членов семейства генов, множественно генов в биосинтетическом пути или множество участков в одном гене. Способы Ma et al. могут быть использованы для системы эффекторных белков C2c1 или C2c3 по настоящему изобретению.
[001050] Lowder et al. (PlantPhysiol. 2015 Aug 21. doi: pp.00636.2015) также разработали набор инструментов CRISPR-Cas9, позволяющий редактировать мультиплексный геном и транскрипционную регуляцию экспрессированных генов, генов с подавленной экспрессией или некодирующих генов в растениях. Этот набор инструментов предоставляет исследователям протокол и реагенты для быстрой и эффективной сборки функциональных конструкций T-ДНК CRISPR-Cas9 для однодольных и двудольных растений с использованием способов клонирования GoldenGate и Gateway. Он предоставляется с полным набором возможных инструментов, включая мультиплексированное редактирование генов и активацию транскрипции или репрессирование эндогенных генов растений. Технология трансформации на основе Т-ДНК имеет фундаментальное значение для современной биотехнологии растений, генетики, молекулярной биологии и физиологии. Поэтому, авторы изобретения разработали способ сборки Cas9 (WT, никазы или dCas9) и одной или более гРНК в представляющий интерес конечный Т-ДНК-вектор. Способ сборки основан как на типе сборки GoldenGate, так и на рекомбинации MultiSite Gateway. Для сборки требуются три модуля. Первый модуль представляет собой исходный вектор Cas9, который содержит гены Cas9 без промоторов или производные от него гены, фланкированные сайтами attL1 и attR5. Второй модуль представляет собой вектор записи гРНК, который содержит входную гРНК. Экспрессирующие кассеты фланкированы сайтами attL5 и attL2. Третий модуль включает attR1-attR2-содержащие конечные Т-ДНК-векторы, которые обеспечивают промоторы выбора для экспрессии Cas9. Инструментарий Lowder et al. могут быть применен к системе эффекторных белков C2c1 или C2c3 согласно настоящему изобретению.
[001051] В предпочтительном варианте осуществления изобретения растение может быть деревом. Настоящее изобретение также может использовать описанную в настоящем описании систему CRISPR-Cas для трав (см., например, Belhaj et al., Plant Methods 9: 39 и Harrison et al., Genes&Development, 28: 1859-1872). В особенно предпочтительном варианте осуществления система CRISPR-Cas по настоящему изобретению может быть нацелена на однонуклеотидный полиморфизм (SNP) в деревьях (см., например, Zhou et al., NewPhytologist, Volume 208, Issue 2, pages 298-301, October 2015). Zhou et al. применили систему CRISPR-Cas на древесном многолетнем Populus с использованием семейства генов 4-кумарата:КоA-лигазы (4CL) в качестве примера и достигли 100% мутационной эффективности для двух преобразованных таким образом генов 4CL, при этом каждый трансформированный организм был исследован на предмет переноса двуаллельных модификаций. В исследовании Zhou et al. система CRISPR-Cas9 была очень чувствительна к однонуклеотидным полиморфизмам (SNP), поскольку расщепление третьего гена 4CL устранялось из-за SNP в последовательности-мишени. Эти способы могут быть применены к системе эффекторных белков C2c1 или C2c3 согласно настоящему изобретению.
[001052] Способы Zhou et al. (New Phytologist, Volume 208, Issue 2, pages 298-301, October 2015) можно применить к настоящему изобретению следующим образом. Два гена 4CL, 4CL1 и 4CL2, связанные с биосинтезом лигнина и флавоноидов, соответственно, являются мишенью для редактирования CRISPR-Cas9. Populustremula x alba клон 717-1B4, обычно используемый для трансформации, отличается от генотипа Populus trichocarpa. Таким образом, гРНК 4CL1 и 4CL2, сконструированные на основе эталонного генома, могут быть проанализированы при помощи собственно полученных данных 717 секвенирований РНК, чтобы убедиться в отсутствии SNP, которые могут ограничивать эффективность Cas. Также предусматривается третья гРНК, предназначенная для 4CL5, дубликата 4CL1. Соответствующие 717 последовательностей содержат один SNP в каждом аллеле вблизи/внутри PAM, оба из которых, как ожидается, устраняют нацеливание посредством 4CL5-гРНК. Все три сайта-мишени гРНК расположены в пределах первого экзона. Для 717 трансформаций гРНК экспрессируется с промотора Medicago U6.6 вместе с кодон-оптимизированным Cas человека под контролем промотора CaMV 35S в бинарном векторе. Трансформация только с помощью вектора Cas может служить в качестве контроля. Случайно выбранные линии 4CL1 и 4CL2 подвергаются секвенированию ампликонов. Затем данные обрабатываются, а биаллельные мутации подтверждаются во всех случаях. Эти способы могут быть применены к системе эффекторных белков C2c1 или C2c3 согласно настоящему изобретению..
[001053] В растениях патогены часто специфичны по отношению к хозяину. Например, Fusarium oxysporum f. sp. lycopersici поражает только томат, а F. oxysporum f. dianthii Pucciniagraminis f. sp. tritici только пшеницу. Растения имеют естественную и индуцируемую защиту, чтобы противостоять большинству патогенов. Мутации и явления рекомбинации между поколениями растений приводят к генетической изменчивости, которая приводит к восприимчивости к болезням, особенно в том случае, когда патогены воспроизводятся с большей частотой, чем растения. В растениях может наблюдаться "нехозяйская" устойчивость, например, когда хозяин и патоген не совместимы. Также может наблюдаться горизонтальная устойчивость, например, частичная устойчивость к аллелям расы патогенных организмов, обычно контролируемая многими генами, и вертикальная устойчивость, например, полная устойчивость к некоторым расам патогена, но не к другим расам, обычно контролируемым несколькими генами. На уровне отношений "ген против гена" растения и патогены развиваются вместе, происходит извечное нарушение и восстановление баланса: генетические изменения в одном приводят к нарушениям в другом. Соответственно, используя природную вариативность генома, селекционеры объединяют в одном растении наиболее полезные гены для урожайности, качества, однородности, выносливости, сопротивления различным болезням. Источниками генов устойчивости являются местные или иностранные сорта, старые сорта негибридного происхождения, родственники диких растений и сорта с индуцированной мутацией, например, после обработки растительного материала мутагенными агентами. Так, в соответствии с настоящим изобретением селекционерам предлагается новый инструмент стимуляции мутаций. Соответственно, специалист в данной области сможет проанализировать геномы разных сортов - возможных источников резистентных генов, имеющих желаемые характеристики или признаки, и сможет использовать настоящее изобретение, чтобы индуцировать появление генов устойчивости со значительно большей точностью, чем предыдущие мутагенные агенты, что, следовательно, приведет к ускорению и улучшению селекции растений.
Улучшение свойств растений и дрожжей
[001054] Настоящее изобретение также относится к растениям и дрожжевым клеткам, которые могут быть получены с помощью способов, описанных в настоящем описании. Растения с улучшенными свойствами, полученные описанными в настоящем описании способами, могут быть полезны для производства продуктов питания или кормов через экспрессию генов, обеспечивающих, в частности, устойчивость к вредителям, гербицидам, засухе, низким или высоким температурам, чрезмерному увлажнению и т.д.
[001055] Растения с улучшенными свойствами, полученные описанными в настоящем описании способами, в особенности зерновые культуры и водоросли, могут быть произведены в качестве продуктов питания и кормов для обеспечения, например, более высокого содержания белков, углеводов, питательных веществ или витаминов по сравнения с таковыми для дикого типа. В этом отношении предпочтительны растения с улучшенными свойствами, в особенности бобовые и клубненосные.
[001056] Улучшенные водоросли или другие растения, такие как рапс, могут быть особенно полезными для производства растительных масел или биотоплива, например такого, как спирты (особенно метанол и этанол). Они могут быть модифицированы способам инженерии для экспрессии или сверхэкспрессии высоких уровней масел или спиртов с целью использования в отраслях промышленности, связанных с нефтью или биотопливом.
[1057] Также изобретение относится к улучшению составляющих частей растений. Такие части растения включают, но не ограничиваются ими, листья, стебли, корни, клубни, семена, эндосперм, яйцеклетки и пыльцу. Такие части растения, как предусматривается в рамках настоящего изобретения, могут быть жизнеспособными, нежизнеспособными, регенерируемыми и/или нерегенерируемыми.
[001058] Также настоящее изобретение охватывает получение растительных клеток и растений, полученных согласно способам по изобретению. Гаметы, семена, зародыши, либо зиготические либо соматические, потомство или гибриды растений, несущих генетическую модификацию, которые получены способами традиционного скрещивания, также входят в рамки настоящего изобретения. Такие растения могут содержать гетерологическую или чужеродную последовательность ДНК, встроенную в последовательность-мишень или ее заменяющую. Альтернативно, такие растения могут содержать только одну модификацию (мутация, делеция, инсерция, замена) в одном или более нуклеотидах. По существу такие растения могут отличаться от исходных растений только наличием конкретной модификации.
[001059] Таким образом, изобретение относится к растению, животному или клетке, полученным способами по настоящему изобретению, или к их потомству. Потомство может быть клоном полученного растения или животного или может быть результатом полового размножения путем скрещивания с другими особями того же вида для дальнейшей интрогрессии желаемых признаков потомству. Клетка может существовать in vivo или ex vivo в случае многоклеточных организмов, в частности, животных и растений.
[001060] Настоящее изобретение также может быть применено и в других сельскохозяйственных областях, таких как, например, фермерство и животноводство. Например, у свиней есть много особенностей, которые делают их привлекательными биомедицинскими моделями, особенно в регенеративной медицине. В частности, свиньи с тяжелым комбинированным иммунодефицитом (SCID) могут служить подходящими моделями для регенеративной медицины, ксенотрансплантации и развития опухолей и помогут в разработке методов лечения пациентов с SCID человека. Lee et al., (Proc Natl Acad Sci USA. 2014 May 20, 11 ((20): 7260-5) использовали нуклеазы с доменами аналогов активаторов транскрипции (TALEN) для получения целевых модификаций гена активации рекомбинации (RAG) 2, включая те, которые воздействовали на оба аллеля, в соматических клетках с высокой эффективностью. В подобной системе можно использовать эффекторный белок C2c1 или C2c3.
[001061] Способы Lee et al. (Proc Natl Acad Set USA. 2014 May 20, 111 (20): 7260-5) могут быть применены в настоящем изобретении следующим образом. Свиньи с мутацией получают путем направленной модификации RAG2 в клетках фибробластов плода, после чего проводят SCNT и перенос эмбрионов. Конструкции, кодирующие CRISPR-Cas, и репортер вводят путем электропорации в клетки фибробластов плода. Через 48 ч трансфицированные клетки, экспрессирующие зеленый флуоресцентный белок, сортируют в отдельные лунки 96-луночного планшета в предполагаемой концентрации одна клетка на лунку. Скрининг целевой модификации RAG2 проводят путем амплификации фрагмента геномной ДНК, фланкирующего любые участки разрезания CRISPR с последующим секвенированием продуктов ПЦР. После скрининга и проверки на отсутствие нецелевых мутаций клетки с целевой модификацией RAG2 используются для SCNT. Полярное тело вместе с частью соседней цитоплазмы ооцита, предположительно содержащее пластинку метафазы II, удаляют и донорскую клетку помещают в перивителлин. Восстановленные эмбрионы затем подвергают электропорации, чтобы соединить донорскую клетку с ооцитом, а затем химически активируют. Активированные эмбрионы инкубируют в среде Porcine Zygote Medium 3 (PZM3) с 0,5 Scriptaid (S7817, Sigma-Aldrich) в течение 14-16 часов. Затем эмбрионы промывают, чтобы удалить Scriptaid и культивируют в PZM3 до переноса в яйцеводы суррогатных свиней.
[001062] Настоящее изобретение также применимо для модификации посредством SNP других животных, таких как коровы. Tan et al, (Proc Natl Acad Sci USA, 2013 Oct 8, 110 (41): 16526-16531) расширили набор инструментов для редактирования генов в области животноводства путем включения нуклеаз с доменами аналогов активаторов транскрипции и кластерных коротких палиндромных повторов, регулярно расположенных группами (CRISPR)/Cas9-стимулированную гомологичную репарацию (HDR) с использованием плазмид, rAAV и олигонуклеотидных матриц. Ген-специфические последовательности гРНК клонировали в вектор Church lab gRNA vector (Addgene ID: 41824) в соответствии с их способами (Mali P, et al., (2013) RNA-Guided Human Genome Engineering via Cas9. Science 339(6121):823-826). Нуклеазу Cas9 получали либо путем совместной трансфекцией с плазмидой hCas9 (Addenene ID: 41815), либо мРНК, синтезированной с RCIScript-hCas9. Эту RCIScript-hCas9 конструировали путем субклонирования фрагмента XbaI-AgeI из плазмиды hCas9 (охватывающей сДНК hCas9) в плазмиду RCIScript.
[001063] Heo et al. (Stem Cells Dev. 2015 Feb 1;24(3):393-402. doi: 10.1089/scd.2014.0278. Epub 2014 Nov 3) сообщили о высокоэффективном нацеливании на геном быка с использованием бычьих плюрипотентных клеток и кластерных коротких палиндромных повторов, регулярно расположенных группами. Сначала Heo et al. получили индуцированные плюрипотентные стволовые клетки ((iPSC) из бычьих соматических фибробластов посредством эктопической экспрессии факторов яманаки и обработки ингибитором GSK3p и MEK (2i). Heo et al увидели, что бычьи iPSC похожи на наивные плюрипотентные стволовые клетки в отношении экспрессии генов и потенциала развития в тератомах. Более того, нуклеаза CRISPR-Cas9, специфичная для локуса NANOG у быков, показала высокую эффективность при редактировании бычьего генома в бычьих iPSC и эмбрионов.
[001064] Igenity® проводит профильный анализ животных, например, коров, с целью достижения и передачи признаков, имеющих промысловую важность, таких как состав туши, качество туши, материнские и репродуктивные характеристики, и средний привес в день. Комплексный профильный анализ Igenity® начинается с определения ДНК-маркеров (чаще всего однонуклеотидных полиморфизмов или SNP). Все ДНК маркеры в профиле Igenity® определялись независимыми ученым в научно-исследовательских институтах, включая университеты, исследовательские организации и правительственные организации, например, министерство сельского хозяйства США. Затем проводилось подтверждение маркеров на выборке Igenity®. Igenity® использует множество источников выборки, которые представляют различные условия производства и биологические типы, часто сотрудничая с партнерами по промышленности, например, коровье-телячьим, кормовым, упаковочным сегментом индустрии по производству говядины, что позволяет собрать редкие фенотипы, обычно недоступные. Широко доступны базы данных генома крупного рогатого скота, см., например, Программу координации генома крупного рогатого скота NAGRP (http://www.anirnaigenorne.org/cattle/rnaps/db.htnai). Таким образом, настоящее изобретение может быть применено к целевым бычьим SNP. Специалист в данной области техники может использовать вышеуказанные протоколы для направленного внесения SNP и применять их к бычьим SNP, как описано, например, Tan et al. или Heo et al.
[001065] Qingjian Zou et al. (Journal of Molecular Cell Biology, Advance Access published October 12, 2015) продемонстрировали увеличение мышечной массы собак путем нацеливания на первый экзон собачьего гена миостатина (MSTN) -негативного регулятора массы мускулов и скелета. Сначала подтверждали эффективность sg-РНК с помощью сотрансфекции sg-РНК, направленной на MSTN, и вектора Cas9 в эмбриональные фибробласты собаки. Затем получили MSTN KO собак путем микроинъекции мРНК Cas9 и sg-РНК MSTN в эмбрионы с нормальной морфологией и аутотрансплантацией зигот в яйцевод той же самки собаки. Нокаутированные щенки показали явный мускульный фенотип в области бедер по сравнению с однопометными животными дикого типа. Подобный результат может быть получен также с помощью CRISPR C2c1 или C2c3, описанных в настоящем описании.
Скот - свиньи
[001066] Мишени для вируса в домашнем скоте в некоторых вариантах осуществления могут включать CD163 свиней; например, в макрофагов свиней CD163 ассоциирован с инфекцией (которая, как считается, связана с проникновением вирусных клеток) с помощью PRRSv (свиной репродуктивный и респираторный вирус, артеривирус). Инфекция PRRSv, особенно свиных альвеолярных макрофагов (обнаруженная в легких), приводит к ранее неизлечимому свиному синдрому ("Загадочная болезнь свиней" или "болезнь голубого уха"), которая вызывает симптомы, включающие репродуктивную недостаточность, потерю веса и высокие показатели смертности у домашних свиней. Инфекции, вызываемые условно-патогенными организмами, а также благоприятными условиями, например, энзоотическая пневмония, менингит и отек уха, считаются вызванными иммунодефицитом на фоне потери активности макрофагов. Они также приводят к значительными финансовым и экологическим последствиям из-за повышенного приема антибиотиков и финансовых потерь ($660 млн. в год).
[001067] Как сообщили Kristin M Whitworth и Dr Randall Prather et al. (Nature Biotech 3434 published online 07 December 2015) в университете Миссури и в сотрудничестве с Genus Pic, на CD163 было осуществлено нацеливание с использованием CRISPR-Cas9, и потомки свиней с отредактированным геномом были устойчивы к PRRSv. Один самец и одна самка с мутациями экзона 7 были скрещены с целью получения потомства. У самца-основателя была делеция 11 п.о. в экзоне 7 на одном аллеле, что приводит к мутации рамки и пропусканию аминокислоты 45 в области 5 и, следовательно, преждевременному стоп-кодону на аминокислоте 64. Другой аллель имел 2 п.о. в экзоне 7 и делеции 377 п.о. в предшествующем интроне, которые, как предсказывалось, приводят к экспрессии первых 49 аминокислот домена 5 с последующим преждевременном стоп-кодоном на аминокислоте 85. У свиноматки было 7 п.о. на одном аллеле, что, как было предсказано, при транскрипции должно было привести к экспрессии первых 48 аминокислот домена 5, за которыми следует преждевременный стоп-кодон на аминокислоте 70. Другой аллель свиньи был неамплифицируемым. Было спрогнозировано, что после селекции потомство будет представлять собой нулевое животное (CD163 -/-), то будет иметь есть нокаут CD163.
[001068] Соответственно, в некоторых вариантах осуществления белки CRISPR могут быть нацелены на альвеолярные макрофаги свиней. В некоторых вариантах осуществления белки CRISPR могут быть нацелены на CD163 свиньи. В некоторых вариантах осуществления CD163 свиньи может быть нокаутирован посредством введения DSNB или вставок и делеций, например, направленной делеции или модификации экзона 7, включая одну или более из описанных выше, или в других областях гена, например, делеции или модификации экзона 5.
[001069] Предусматриваются отредактированные свиньи и их потомство, например, свинья с нокаутом CD163. Это может быть осуществлено для целей животноводства, разведения или модулирования (например, модель на свиньях). Также предусматривается сперма с нокаутрованным геном.
[001070] CD163 является представителем суперсемейства фагоцитарных рецепторов, богатых цистеином (SRCR). Как показывают исследования in vitro, домен 5 белка SRCR представляет собой домен, отвечающий за распаковку и высвобождение вирусного генома. Следовательно, на другие члены суперсемейства SRCR также может быть осуществлено нацеливание с целью повышения резистентности к другим вирусам. PRRSV также является членом группы артеривирусов млекопитающих, которая также включает вирус роста мышиной лактатдегидрогеназы, вирус геморрагической лихорадки обезьяны и вирус конского артерита. Артеривирусы обладают важными свойствами патогенеза, включая тропизм макрофагов и способность вызывать как тяжелое заболевание, так и персистирующую инфекцию. Соответственно, может быть осуществлено нацеливание на артеривирусы и, в частности, вирус повышения уровня лактатдегидрогеназы мышей, вирус геморрагической лихорадки обезьян и вирус лошадиного артерита, например, через CD163 свиней или его гомологи у других видов, а также в моделях на мышах, обезьянах и лошадях, и также предусматривается нокаут.
[001071] Действительно, этот подход может быть расширен на вирусы или бактерии, которые вызывают другие болезни скота, которые могут передаваться людям, такие как штаммы вируса свиного гриппа (SIV), которые включают грипп C и подтипы гриппа A, известные как H1N1, H1N2, H2N1, H3N1, H3N2 и H2N3, а также пневмония, менингит и отек, упомянутые выше.
Терапевтическое нацеливание посредством направляемого РНК комплекса эффекторных белков C2c1 или C2c3
[001072] Как будет очевидно, предусматривается, что систему по настоящему изобретению можно использовать для нацеливания на любую представляющую интерес полинуклеотидную последовательность. Настоящее изобретение относится к не встречающейся в природе или сконструированной способами инженерии композиции, или одному или более полинуклеотидам, кодирующих компоненты указанной композиции, или к векторным системам или системам доставки, содержащим один или более полинуклеотидов, кодирующих компоненты указанной композиции, для применения в модификации клетки-мишени in vivo, ex vivo или in vitro, и она может быть осуществлена способом, который изменяет клетку таким образом, что после модификации потомства или клеточной линии модифицированная посредством CRISPR клетка сохраняет измененный фенотип. Модифицированные клетки и потомство могут быть частью многоклеточного организма, такого как растение или животное с применением системы CRISPR ex vivo или in vivo в отношении желаемого типа клеток. CRISPR по изобретению может быть терапевтическим способом лечения. Терапевтический способ лечения может включать редактирования гена или генома, или генную терапию.
Лечение от патогенов, таких как бактериальные, грибковые и паразитарные патогены
[001073] Настоящее изобретение также может применяться для лечения от бактериальных, грибковых и паразитарных патогенов. Большинство усилий исследователей сосредоточено на разработке новых антибиотиков, которые после разработки, тем не менее, будут подвержены тем же проблемам лекарственной резистентности. Изобретение относится к новым альтернативам на основе CRISPR, которые преодолевают эти трудности. Кроме того, в отличие от существующих антибиотиков, лечение на основе CRISPR может быть специфическим для патогена, индуцируя гибель бактериальных клеток патогена-мишени, избегая при этом гибели полезных бактерий.
[001074] Jiang et al. ("RNA-guided editing of bacterial genomes using CRISPR-Cas systems," Nature Biotechnology vol. 31, p. 233-9, March 2013) использовали систему CRISPR-Cas9, чтобы мутировать или вызывать гибель S. pneumoniae и E. coli. Эта работа, которая представляет точные мутации в геномах, основывается на двойном расщеплении участка генома, являющегося мишенью, направляемого РНК и Cas9, с целью обеспечения гибели немутантных клеток и обхода необходимости выбора маркеров или систем против отбора. Cистемы CRISPR могут использоваться для способствования регрессии лекарственной резистентности и исключить перенос резистентности между штаммами. Bickard et al. показали, что Cas9, перепрограммированный на нацеливание на гены вирулентности, убивает вирулентный, но не авирулентный, S. aureus. Перепрограммируя нуклеазу для генов устойчивости к антибиотикам, являющихся мишенью, осуществляли разрушение стафилококковых плазмид, которые содержали гены устойчивости к антибиотикам и имммунизировали против распространения генов резистентности плазмиды (см. Bikard et al, "Exploiting CRISPR-Cas nucleases to produce sequence-specific antimicrobials," Nature Biotechnology vol. 32, 1146-1150, doi: 1Q.1038/nbt.3043, published online 05 October 2014). Bikard показал, что противомикробные средства на основе CRISPR-Cas9 функционируют, уничтожая S. aureus в модели колонизации кожи мыши. Аналогично, Yosef et al. использовали CRISPR для нацеливания на гены, которые кодируют ферменты), придающие резистентность к β-лактамным антибиотикам (см. Yousef et al., "Temperate and lytic bacteriophages programmed to sensitize and kill antibiotic-resistant bacteria," Proc. Nail Acad. Sci. USA, vol. 112, p. 72.67-72.72, doi: 10.1073/pnas. 1500107112, опубликованную через интернет 18 мая 2015 года).
[001075] Системы CRISPR можно использовать для редактирования геномов паразитов, устойчивых к другим генетическим подходам. Например, было показано, что система CRISPR-Cas9 вносит двухцепочечный разрыв в геном Plasmodium yoelii (см., Zhang et al., "Efficient Editing of Malaria Parasite Genome Using the CRISPR/Cas9 System," mBio. vol. 5, e01414-14, Jul-Aug 2014). Ghorbal et al. ("Genome editing in the human malaria parasite Plasmodium falciparum using the CRISPR-Cas9 system," Nature Biotechnology, vol. 32., p. 819-821, doi: 10.1038/nbt.2925, опубликованную через интернет 1 июня 2014 года) модифицировали последовательности двух генов: orc1 и kelch13, которые предположительно участвуют в подавлении гена и возникающей устойчивости к артемизинину, соответственно. Паразиты, которые были изменены в соответствующих участках, восстанавливались с очень высокой эффективностью, несмотря на отсутствие прямой селекции по модификации, что указывает на то, что с помощью этой системы можно вносить нейтральные или даже вредные мутации. CRISPR-Cas9 также используется для модификации геномов других патогенных паразитов, включая Toxoplasma gondii (см. Shen et al., "Efficient gene disruption in diverse strains of Toxoplasma gondii, using CRISPR/CAS9," mBio vol. 5:e01114-14, 2014; and SidiJk et al., "Efficient Genome Engineering of Toxoplasma, gondii Using CRISPR/Cas9," PLoS One vol. 9, el00450, doi: 10.1371/joumal.pone.0100450, опубликованную через интернет 27 июня 2014 года).
[001076] Vyas et al ("A Candida albicans CRISPR system permits genetic engineering of essential genes and gene families," Science Advances, vol. 1, el500248, DOI: 10.1126/seiadv.1500248, 3 апреля 2015 года) использовали систему CRISPR для преодоления давних препятствий в генной инженерии C. albicans и эффективно внесли мутацию в одном эксперименте в обе копии нескольких разных генов. В организме, где несколько механизмов способствуют лекарственной резистентности, Vyas получил гомозиготные двойные мутанты, которые больше не проявляли гиперрезистентность к флуконазолу или циклогексимиду, демонстрируемую родительским клиническим изолятом Can90. Vyas также получил гомозиготные мутации с потерей функциональности в основных генах C. albicans путем создания условных аллелей. Нуль-аллели DСR1, который необходим для обработки рибосомной РНК, являются летальными при низкой температуре, но жизнеспособны при высокой температуре. Vyas использовал шаблон восстановления, в котором вводилась нонсенс-мутация и изолировались мутанты dcr1/dcr1, которые не росли при 16°C.
[001077] Систему CRISPR по настоящему изобретению использовали в P. falciparum путем разрушения хромосомного локуса. Ghorbal et al. ("Genome editing in the human malaria parasite Plasmodium falciparum using the CRISPR.-Cas9 system.". Nature Biotechnology, 32, 81.9-821 (2014), DOI: 10.1038/nbt.2925, June 1, 2014) использовали систему CRISPR для внесения определенных генных нокаутов и однонуклеотидных замен в геноме малярии. Чтобы адаптировать систему CRISPR-Cas9 к P. falciparum, Ghorbal et al. сконструировали экспрессирующий вектор для контроля регуляторных элементов плазмодия в эписоме pUFl-Cas9, которая также несет селективный маркер ydhodh, который обеспечивает резистентность к DSM1, ингибитору дигидрогенатдегидрогеназы P. falciparum (PfDHODH), и для транскрипции sg-РНК использовали регуляторные элементы на основе малой ядерной (sn)РНК U6 P. facciparum, разместив направляющую РНК и донорную ДНК-матрицу для репарации на основе гомологичной рекомбинации на одной и той же плазмиде, pL7. См. также Zhang С. V al. ("Efficient editing of malaria parasite genome using the CRISPR/Cas9 system", MBio, 2014 Jul 1, 5(4):E01414-I4, doi: 10. EI.28/MbKO.O1.414-M) and Wagner et al. ("Efficient CRISPR-Cas9-mediated genome editing in Plasmodium falciparum, Nature Methods 11, 915-918 (2014), DOI: 10.1038/nmeth.3063).
Лечение от патогенов, таких как вирусные патогены, такие как ВИЧ
[001078] Cas-опосредованное редактирование генома может быть использовано для внесения защитных мутаций в соматические ткани для борьбы с негенетическими или комплексными заболеваниями. Например, NHEJ-опосредованная инактивация рецептора CCR5 в лимфоцитах (Lombardo et al., Nat Biotechnol. 2007 Nov; 25(11): 1298-306) может стать плодотворной стратегией в борьбе с ВИЧ-инфекцией, а делеция PC5K9 (Cohen et al., Nat Genet. 2005 Feb; 37(2): 1(51-5) или ангиопоэтина (Musunuru et al., N Engl J Med. 2010 Dec 2; 363(231:2220-7) может оказать терапевтический эффект при статин-резистентнй гиперхолестеролемии или гиперлипидемии. Хотя эти мишени могут быть удалены путем sg-РНК-опосредованного нокаута белка, уникальным преимуществом NHEJ-опосредованной инактивации является способность достичь постоянной терапевтической пользы без необходимости продолжения лечения. Как и при всех способах генной терапии, безусловно, важно удостовериться, что каждое предлагаемое терапевтическое использование имеет благоприятное соотношение пользы и риска.
[001079] Было показано, что гидродинамическая доставка плазмидной ДНК, кодирующей Cas9, и направлящей РНК вместе с матрицей репарации в печень в модели тирозинемии на взрослых мышах, скорректировала мутацию Fah гена и восстановила экспрессию белка Fah дикого типа в ~1 из 250 клеток(Nat Biotechnol. 2014 Jun; 32(6):551-3). Кроме того, в клинических испытаниях были успешно использованы нуклеазы с цинковыми пальцами для борьбы с ВИЧ-инфекцией путем нокаута рецептора CCR5 ex vivo. У всех пациентов снизился уровень ДНК ВИЧ, и у одного из четырех пациентов РНК ВИЧ стала неопределяемой (Tebas et al., N Engl J Med. 2014 Mar 6; 370(10):901-10). Оба результата показывают перспективы использования программируемых нуклеаз как новой терапевтической платформы.
[001080] В другом варианте осуществления изобретения самоинактивирующиеся лентивирусные векторы с миРНК, нацеленный экзон, общий для генов tat/rev ВИЧ, локализующейся в ядрышке TAR-ловушкой и анти-CCR5-специфичным рибозимом типа "hammerhead" (см, например, DiGiusto et al. (2010) Sci Transl Med 2:36ra43), может использоваться и/или быть адаптирован для использования в нацеленной на нуклеиновые кислоты системе по настоящему изобретению. Минимум 2,5×106 CD34+ клеток на килограмм массы тела пациента может быть собрано и подвергнуто предварительному стимулированию в течение 16-20 часов в питательной среде X-VIVO 15 (Lonza), содержащей 2 мкМ/L глютамина, фактор стволовой клетки (100 нг/мл), лиганд Flt-3 (Flt-3L) (100 нг/мл) и тромбопоэтин (10 нг/мл) (CellGenix) при плотности 2×106 клеток/мл. Предварительно стимулированные клетки можно трансдуцировать лентивирусным вектором с множественностью инфекции, равной 5, в течение 16-24 часов в 75-см2 флаконах для культивирования тканей, покрытых фибронектином (25 мг/см2) (RetroNectin, Takara Bio Inc.).
[01081] Специалисты в данной области, используя имеющуюся в данной области информацию и идеи в соответствии с настоящим изобретением, могут исправить HSC в отношении иммунодефицитного состояния, такого как ВИЧ/СПИД, путем приведения HSC в контакт с системой CRISPR-Cas9, которая нацелена на и нокаутирует CCR5. Направляющая РНК (и, предпочтительно, подход с двумя направляющими молекулами, например, парой различных направляющих РНК, например, направляющими РНК, нацеленными на два клинически значимых гена B2M и CCR5 в первичные CD4+ T-клетки и CD34+ гемопоэтические стволовые клетки и клетки-предшественники (HSPC) человека), которая нацелена на и нокаутирует ССR5, и содержащие белки С2с1 или С2с3 частицы, приводят в контакт с HSC. Такие вступившие в контакт клетки можно вводить и, если необходимо, обрабатывать/увеличивать в количестве (см. Cartier., также см. Kiem, "Hematopoietic stem cell-based gene therapy for HIV disease", Cell Stem Cell Feb 3, 2012; 10(2): 137-147, включенные в настоящее описание в качестве ссылки вместе цитируемыми в них документами. Mandal et al., "Efficient Ablation of Genes in Human Hematopoietic Stem and Effector Cells using CRISPR/Cas9," Cell Stem Cell, Volume 15, Issue 5, p643--652, 6 November 2014, включенную в настоящее описание в качестве ссылки вместе с цитируемыми в ней документами. Также может быть упомянута Ebina, "CRlSPR7Cas9 system, to suppress HIV-1 expression by editing HIV-1 integrated proviral DNA" SCIENTIFIC REPORTS | 3: 2510 | DOI: 10.1038/srep02510, включенная в настоящее описание в качестве ссылки вместе с цитируемыми в ней документами, в качестве другого средства борьбы с ВИЧ/СПИДом с использованием системы CRISPR C2c1 или С2с3.
[001082] Основанием для редактирования генома для лечения ВИЧ является наблюдение, что гомозиготные индивидуумы по мутациям в CCR5 с потерей функции, клеточным корецепторе для вируса, обладают высокой устойчивостью к инфекции и в остальном являются здоровыми, что указывает на то, что попытка воссоздать данную мутацию может быть безопасным и эффективным способом лечения [Liu, R., et al. Cell 86, 367-377 (1996)]. Эта идея была подтверждена клиническими испытаниями, когда ВИЧ-инфицированному пациенту была проведена трансплантация аллогенного костного мозга от донора, гомозиготного по мутации CCR5 с потерей функции, что привело к не поддающимся обнаружению уровням ВИЧ и восстановлению нормальных показателей количества CD4 Т-клеток [Flutter, G., et al. The New England Journal Of Medicine 360, 692-698 (2009)]. Хотя трансплантация костного мозга не является реалистичным методом лечения для большинства пациентов с ВИЧ из-за стоимости и потенциальной реакции на трансплант, терапия ВИЧ, которая преобразует собственные Т-клетки пациента в мутантные по CCR5, является желательной.
[001083] Предшествующие исследования с использованием ZFN и HNEI для нокаута CCR5 в гуманизированной модели ВИЧ на мышах показали, что трансплантация CD4 T-клеток с отредактированным CCR5, снижала вирусную нагрузку и количество CD4 T-клеток [Perez, E.E., et al. Nature Biotechnology 26, 808-816 (2008)]. Важно отметить, что эти модели также показали, что ВИЧ-инфекция привела к селекции нуль-клеток по CCR5, указывая на то, что редактирование дает преимущество в выносливости и потенциально позволяет небольшому количеству отредактированных клеток обеспечить терапевтический эффект.
[001084] В результате этого и других перспективных доклинических исследований терапия редактирования генома, которая нокаутирует CCR5 в Т-клетках пациента, далее была протестирована на людях [Holt, N., et al. Nature Biotechnology 28, 839-847 (2010); Li, L., et al. Molecular therapy: the journal of the American Society of Gene Therapy 21, 1259-1269 (2013)]. В недавних клинических испытаниях фазы I, CD4+ Т-клетки у пациентов с ВИЧ были выделены, отредактированы посредством ZFN, предназначенными для нокаутирования гена CCR5, и аутологично пересажены обратно пациентам [Tebas, P., et al. The New England Journal Of Medicine 370, 901-910 (2014)].
[001085] В другом исследовании Mandal et al., Cell. Stem. Cell, Volume 15, Issue 5, p643-652, 6 November 2014), CRISPR-Cas9 нацеливали на два клинически значимых гена, B2M и CCR5, в CD4+ Т-клетках и CD34+ гемопоэтических стволовых клетках и клетках-предшественниках (HSPC) человека. Использование одиночных направляющих РНК привело к высокоэффективному мутагенезу в HSPC, но не затронуло Т-клетки. Подход с двумя направляющими РНК повысил эффективность делеции генов в обоих типах клеток. HSPC, подвергшиеся редактированию генома посредством CRISPR-Cas9, сохранили потенциал в отношении множества ростков. Прогнозируемые мутации на и вне мишени, были исследованы с помощью секвенированияе с захватом мишени в HSPC, и низкие уровни мутагенеза вне мишени наблюдались только в одном участке. Эти результаты демонстрируют, что CRISPR-Cas9 может эффективно устранять гены в HSPC с минимальным мутагенезом вне мишени, что имеет широкую применимость в терапии на основе гемопоэтических клеток.
[001086] Wang et al. ((PLoS One, 2014 Dec 26;9(12):el 15987. doi: 10.1371/joumal.pone.Ol 15987) обеспечили сайленсинг CCR5 через ассоциированный с CRISPR белок 9 (Cas9) и единственную направляющую РНК (направляющие РНК) с использованием лентивирующих векторов, экспрессирующих Cas9 и направляющие РНК против CCR5. Wang et al. показали, что единичная циклическая трансдукция лентивирусных векторов, экспрессирующих Cas 9 и направляющие РНК против CCR5 в восприимчивые к ВИЧ-1 CD4+ клетки человека, приводит к высоким частотам разрушения гена CCR5. Клетки с нарушенным геном CCR5 не только устойчивы к R5-тропическому ВИЧ-1, в том числе передаваемые/основополагающие изоляты (T/F) ВИЧ-1, но также обладают селективным преимуществом по сравнению с клетками с неразрушенным CCR5 во время R5-тропической инфекции ВИЧ-1. Геномные мутации в потенциальных участках вне мишени, которые высоко гомологичны этим направляющим РНК против CCR5 в стабильно трансдуцированных клетках даже спустя 84 дня после трансдукции, не были обнаруживались с помощью анализа с эндонуклеазой I T7.
[001087] Fine et al. (Sci Rep. 2015 Jui 1;5:10777. doi: 10.1038/'srep10777) идентифицировали двухкассетную систему, экспрессирующую фрагменты белка Cas9 S. pyogenes (SpCas9), которые соединяются вместе в клетке с образованием функционального белка, способного к сайт-специфическому расщеплению ДНК. С помощью специфических направляющих цепей CRISPR, Fine et al. продемонстрировали эффективность этой системы при расщеплении генов HBB и CCR5 в клетках человека HEK-293T в виде единственного Cas9 и в виде пары никаз Cas9. Транс-сплайсированный SpCas9 (tsSpCas9) продемонстрировал ~35% активность нуклеазы по сравнению с SpCas9 дикого типа (wtSpCas9) при стандартных дозах трансфекции, но имел значительно сниженную активность при более низких уровнях дозирования. Существенно уменьшенная длина открытой рамки считывания tsSpCas9 по отношению к wtSpCas9 потенциально позволяет упаковывать более сложные и более длинные генетические элементы в вектор AAV, включая тканеспецифические промоторы, мультиплексную экспрессию направляющей РНК и слияние эффекторных доменов с SpCas9.
[001088] Li et al. (J Gen Virol. 2015 Aug;96(8):2381-93. doi: 10.1099/vir.0.000139. Epub 2015 Apr 8) продемонстрировали, что CRISPR-Cas9 может эффективно опосредовать редактирование локуса CCR5 в клеточных линиях, в результате чего индуцируется экспрессия CCR5 на поверхности клетки. Секвенирования нового поколения показало, что различные мутации были внесены вокруг спрогнозированного участка расщепления CCR5. Для каждой из трех наиболее эффективных направляющих РНК, которые были проанализированы, никаких значительных эффектов вне цели не было обнаружено в 15 наиболее перспективных участков. Конструируя химерные аденовирусы Ad5F35, несущие компоненты CRISPR-Cas9, Li et al. эффективно трансдуцировали первичные CD4+ Т-лимфоциты и нарушили экспрессию CCR5, и положительно трансдуцированные клетки получали резистентность к ВИЧ-1.
[001089] Специалист в данной области может применять любой из способов, изложенных выше, например, Holt, N, et al. Nature biotechnology 28, 839-847 (2010), Li, L., et al. Molecular therapy: the journal of the American Society of Gene Therapy 21, 1259-1269 (2013), Mandal et al., Cell Stem Cell, Volume 15, Issue 5, p643 65g, 6 November 2014, Wang et al. (PLoS One. 2014 Dec 26;9(12):e115987. doi: 10.1371/joumal.pone.0115987), Fine et al. (Sci Rep. 2015 Jul 1,5:10777. doi: 10.1038/srep 10777) and Li et al. (J Gen Virol. 2015 Aug;96(8): 23 81-93. doi: 10.1099/vir.0.000139. Epub 2015 Apr 8) для нацеливания на CCR5 при помощи системы CRISPR cas по настоящему изобретению.
Нацеливание на патогенs, например, вирусные патогены, такие как HBV
[001090] Настоящее изобретение также может применяться для лечения вируса гепатита B (HBV). Тем не менее, система CRISPR Cas должна быть адаптирована, чтобы избежать недостатков применения РНК-i, таких как риск перенасыщения эндогенных путей малой РНК, например, путем оптимизации дозы и последовательности (см., Grimm et al., Nature vol. 441, 26 May 2006). Например, предполагаются низкие дозы, такие как приблизительно 1-10×1014 частиц на человека. В другом варианте осуществления система CRISPR Cas, направленная на HBV, может вводиться в липосомах, таких как стабильная частица нуклеиновая кислота-липид (SNALP) (см., например, Morrissey et al., Nature Biotechnology, Vol. 23, No, 8, August 2005). Предусматриваются ежедневные внутривенные инъекции приблизительно в дозе 1, 3 или 5 мг/кг/сутки CRISPR Cas, нацеленного на РНК HBV в SNALP. Ежедневное лечение может длиться более трех дней, а затем продолжаться еженедельно в течение примерно пяти недель. В другом варианте осуществления система Chen et al. (Gene Therapy (2007) 14, 11-19) могут быть использованы и/или адаптированы для системы CRISPR Cas согласно настоящему изобретению. Chen et al. использовал двухцепочечный псевдотипированный вектор на основе аденоассоциированного вируса 8 fdsAAV2/8) для доставки кшРНК. Единственное введение вектора dsAAV2/8 (1×1012 векторных геномов на мышь), несущего HBV-специфическую кшРНК, эффективно подавляло устойчивый уровень белка HBV, мРНК и репликативной ДНК в печени трансгенных мышей HBV, что приводило к снижению нагрузки HBV вплоть до 2-3 Log10 в кровотоке. Значительное подавление HBV поддерживалось в течение по меньшей мере 120 дней после введения вектора. Терапевтический эффект кшРНК был зависимым от последовательности мишени и не включал активацию интерферона. Для настоящего изобретения система CRISPR Cas, направленная на HBV, может быть клонирована в вектор AAV, такой как вектор dsAAV2/8, и может быть введена человеку, например, в дозе приблизительно 1×1015 векторных геномов до примерно 1×106 векторных геномов на человека. В другом варианте осуществления способ Wooddell et al. (Molecular Therapy, vol. 21, no 5, 973-985, May 2013) могут быть использованы и/или адаптированы к системе CRISPR Cas по настоящему изобретению, Woodell et al. показывают, что простая совместная инъекция направленного на гепатоцит связанного с N-ацетилгалактозамином мелиттин-подобного пептида (NAG-MLP) с тропной к печени конъюгированной с холестерином миРНК (chol-siРНК), нацеленной на фактор VII свертывания (F7), приводит к эффективному нокдауну F7 у мышей и не являющихся человеком приматов без изменений в клинической химии или индукции цитокинов. Используя переходные и трансгенные модели на мышах для инфекции HBV, Wooddell et al. показали, что одна совместная инъекция NAG-MLP с мощными chol-миРНК, направленными на консервативные последовательности HBV, приводила к репрессии, составляющей несколько порядков, вирусной РНК, белков и вирусной ДНК с большой продолжительностью действия. В настоящем изобретении могут быть предусмотрены внутримозговые совместные инъекции, например, около 6 мг/кг NAG-MLP и 6 мг/кг HBV-специфического CRISPR Cas. В альтернативном варианте около 3 мг/кг NAG-MLP и 3 мг/кг HBV-специфического CRISPR Cas может быть доставлено в первый день с последующим введением примерно 2-3 мг/кг NAG-MLP и 2-3 мг/кг HBV-специфического CRISPR Cas через две недели.
[001091] Lin et al. (Mol Ther Nucleic Acids. 2014 Aug 19;3:el86. doi: 10.1Q38/mtna.2014.38) разработали восемь гРНК против HBV генотипа A. С помощью специфичных к HBV гРНК система CRISPR-Cas9 значительно уменьшала производство коровых и поверхностных белков HBV в клетках Huh-7, трансфицированных вектором экспрессии HBV. Среди восьми подвергнутых скринингу гРНК были идентифицированы две эффективные. Одна гРНК, нацеливающая на консервативную последовательность HBV, действовала против разных генотипов. Используя модель гидродинамической персистенции HBV на мышах, Lin et al. дополнительно продемонстрировали, что эта система может расщеплять внутрипеченочную содержащую геном HBV плазмиду и способствовать ее очистке in vivo, что приводит к снижению уровней поверхностного антигена сыворотки. Эти данные свидетельствуют о том, что система CRISPR-Cas9 может нарушить HBV-экспрессирующие матрицы как in vitro, так и in vivo, что указывает на ее потенциал к полному излечению стойкой инфекции HBV.
[001092] Dong et al. (Antiviral Res., 2015, 118: 110-7. Doi: 10.1016/j.antiviral.2015.03.015, Epub 2015 Apr 3) использовали систему CRISPR-Cas9 для нацеливания на геном HBV и эффективного ингибирования инфекции HBV. Dong et al. синтезировали четыре единичных направляющих РНК, направленных на консервативные области HBV. Экспрессия этих направляющих РНК с Cas9 уменьшила продукцию вируса в клетках Huh7, а также в HBV-репликационной клетке HepG2.2.15. Dong et al. также продемонстрировали, что прямое расщепление посредством CRISPR-Cas9 и опосредованный расщеплением мутагенез произошли в ccc-ДНК HBV трансфицированных клеток. В модели на мышах, несущих ccc-ДНК HBV, инъекция направляющих плазмид RNA-Cas9 через хвостовую вену приводила к низкому уровню ccc-ДНК и белка HBV.
[001093] Liu et al. (J Gen Virol, 2015, август, 96 (8): 2252-61. Doi: 10.1099/vir.0.000159. Epub 2015 Apr 22) разработали восемь направляющих РНК (гРНК), которые осуществляли нацеливание на консервативные области различных генотипов HBV, что могло значительно ингибировать репликацию HBV как in vitro, так и in vivo, чтобы исследовать возможность использования системы CRISPR-Cas9 для разрушения ДНК-матриц HBV. HBV-специфическая система гРНК/C2c1 или /C2c3 могла ингибировать репликацию HBV разных генотипов в клетках и уровень вирусной ДНК значительно снижался посредством одной системы гРНК/C2c1 или /C2c3 и устранялся посредством комбинации различных систем гРНК/C2c1 или /C2c3.
[001094] Wang et al. (World J Gastroenterol, 2015, август 28, 21 (32): 9554-65. Doi: 10.3748/wjg.v21.i32.9554) разработали 15 гРНК против HBV генотипов A-D. Были выбраны 11 комбинаций двух вышеуказанных гРНК (двойных гРНК), охватывающих регуляторную область HBV. Эффективность каждой гРНК и 11 двойных гРНК в отношении подавлении репликации HBV (генотипов A-D) исследовали путем количественного определения поверхностного антигена HBV (HBsAg) или антигена (HBeAg) в супернатанте культуры. Разрушение вектора, экспрессирующего HBV, исследовали в клетках HuH7, совместно трансфицированных двойными гРНК и вектором, экспрессирующим HBV, с использованием полимеразной цепной реакции (ПЦР) и способом секвенирования, а разрушение ccc-ДНК исследовали в клетках HepAD38 с использованием осаждения с KCl, безопасное для плазмиды АТФ-зависимое расщеление ДНК-азой (PSAD), амплификации по типу катящегося кольца и комбинированного способа количественной ПЦР. Цитотоксичность этих гРНК оценивали посредством анализа митохондриального тетразолия. Все гРНК могли значительно снизить синтез HBsAg или HBeAg в супернатанте культуры, который зависит от области, на которую была нацелена гРНК. Все двойные гРНК могли эффективно подавлять продукцию HBsAg и/или HBeAg HBV генотипов A-D, а эффективность двойных гРНК в подавлении продукции HBsAg и/или HBeAg была значительно увеличена по сравнению с единственной гРНК. Кроме того, путем прямого секвенирования ПЦР авторы изобретения подтвердили, что эти двойные гРНК могут специфически разрушать HBV-экспрессирующую матрицу, удаляя фрагмент между сайтами расщепления двух используемых гРНК. Самое главное, количественного гРНК-5 и гРНК-12 могла не только эффективно подавлять продукцию HBsAg и/или HBeAg, но также разрушать резервуары ccc-ДНК в клетках HepAD38.
[001095] Karimova et al. (Sci Rep. 2015 Sep 3;5:13734. doi: 10.1038/srepl3734) идентифицировали перекрестно-генотипные консервативные последовательности HBV в S и X-области генома HBV, которые были специфически и эффективно нацелены на расщепление никазой Cas9. Этот подход нарушал не только эписомальную ccc-ДНК и интегрированные в хромосому участки-мишени HBV в линиях репортерных клеток, но также и репликацию HBV в хронически и de novo инфицированных клеточных линиях гепатомы.
[001096] Специалист в данной области может использовать любое из вышепредставленных исследований, например, Lin et al. (Mol Ther Nucleic Acids, 2014 Aug 19;3:e186, doi: 10.1038/mtna.2014.38), Dong et al. (Antiviral Res. 2015 Jun;l 18:110-7. doi: 10.1016/j.antiviral.2015.03.015. Epub 2015 Apr 3), Liu et al. (J Gen Virol. 2015 Aug;96(8):2252-61. doi: 10.1099/vir.0.000159. Epub 2015 Apr 22), Wang et al. (World J Gastroenterol. 2015 Aug 28;21(32):9554-65. doi: 10.3748/wig.v21.132.9554) и Karimova et al. (Sci Rep. 2015 Sep 3;5:13734. doi: 10.1038/srepl3734) для нацеливания на HBV при помощи системы CRISPR cas по настоящему изобретению.
[001097] Хроническая инфекция вирусом гепатита (HBV) распространена, смертельна и редко излечивается из-за стойкости вирусной эписомальной ДНК (ccc-ДНК) в инфицированных клетках. Ramanan et al. (Ramanan V, Shiomai A, Cox DB, Schwartz RE, Michailidis E, Bhatta A, Scott IDA, Zhang F, Rice CM, Bhatia SN,.Sci Rep. 2 015 2 июня, 5: 10833. doi: 10.1038/srep10833, опубликованная через интернет 2 июня 2015 года) показали, что система CRISPR/Cas9 может быть специфически нацелена на и расщеплять консервативные области в геноме HBV, что приводит к устойчивому подавлению экспрессии и репликации вирусных генов. При устойчивой экспрессии Cas9 и соответственно выбранных направляющих РНК они продемонстрировали расщепление ccc-ДНК посредством Cas9 и резкое снижение как уровня ccc-ДНК, так и других параметров экспрессии и репликации вирусных генов. Таким образом, они показали, что непосредственное нацеливание на вирусную эписомальную ДНК является новым терапевтическим подходом для контроля над вирусом и, возможно, для излечения пациентов. Это также описано в WO2015089465 Al, из Tire Broad Institute и др., содержание которых включено в настоящее описание в качестве ссылки.
[001098] Таким образом, нацеливание на вирусную ДНК при HBV является предпочтительным в некоторых вариантах осуществления изобретения.
[001099] Настоящее изобретение также может быть применено для лечения патогенов, например, бактериальных, грибковых и паразитарных патогенов. Большинство усилий исследователей были сосредоточены на разработке новых антибиотиков, которые после разработки, тем не менее, подвержены тем же проблемам развития устойчивости к ним. Изобретение предусматривает новые альтернативы на основе CRISPR, которые преодолевают эти трудности. Кроме того, в отличие от существующих антибиотиков, лечение на основе CRISPR может быть смоделировано таким образом, чтобы оно было специфическим для патогена и индуцировало гибель бактериальных клеток патогена-мишени, но никак не влияло на полезные бактерии.
[001100] Настоящее изобретение также может применяться для лечения вируса гепатита С (HCV). К системе CRISPR-Cas могут применяться способы Roelvinki et al. (Molecular Therapy vol. 20 no. 9, 1737-1749 Sep 2012). Например, вектор AAV, такой как AAV8, может представлять собой предполагаемый вектор, и может быть предусмотрена дозировка примерно 1,25×1011-1,25×1013 векторных геномов на килограмм массы тела. Настоящее изобретение также может быть применено для лечения патогенных микроорганизмов, например, бактерий, грибов и паразитов. Большинство усилий исследователей были сосредоточены на разработке новых антибиотиков, которые после разработки, тем не менее, подвержены тем же проблемам развития устойчивости к ним. Изобретение предусматривает новые альтернативы на основе CRISPR, которые преодолевают эти трудности. Кроме того, в отличие от существующих антибиотиков, лечение на основе CRISPR может быть смоделировано таким образом, чтобы оно было специфическим для патогена и индуцировало гибель бактериальных клеток патогена-мишени, но никак не влияло на полезные бактерии.
Jiang et al. ("RNA-guided editing of bacterial genomes using CRISPR-Cas systems," Nature Biotechnology vol. 31, p. 233-9, March 2013) использовали систему CRISPR-Cas9, чтобы мутировать или вызывать гибель S. pneumoniae и E. coli. Эта работа, которая представляет точные мутации в геномах, основывается на двойном расщеплении участка генома, являющегося мишенью, направляемого РНК и Cas9, с целью обеспечения гибели немутантных клеток и обхода необходимости выбора маркеров или систем против отбора. Cистемы CRISPR могут использоваться для способствования регрессии лекарственной резистентности и исключить перенос резистентности между штаммами. Bickard et al. показали, что Cas9, перепрограммированный на нацеливание на гены вирулентности, убивает вирулентный, но не авирулентный, S. aureus. Перепрограммируя нуклеазу для генов устойчивости к антибиотикам, являющихся мишенью, осуществляли разрушение стафилококковых плазмид, которые содержали гены устойчивости к антибиотикам и имммунизировали против распространения генов резистентности плазмиды (см. Bikard et al, "Exploiting CRISPR-Cas nucleases to produce sequence-specific antimicrobials," Nature Biotechnology vol. 32, 1146-1150, doi: 1Q.1038/nbt.3043, published online 05 October 2014). Bikard показал, что противомикробные средства на основе CRISPR-Cas9 функционируют, уничтожая S. aureus в модели колонизации кожи мыши. Аналогично, Yosef et al. использовали CRISPR для нацеливания на гены, которые кодируют ферменты), придающие резистентность к β-лактамным антибиотикам (см. Yousef et al., "Temperate and lytic bacteriophages programmed to sensitize and kill antibiotic-resistant bacteria," Proc. Nail Acad. Sci. USA, vol. 112, p. 72.67-72.72, doi: 10.1073/pnas. 1500107112, опубликованную через интернет 18 мая 2015 года).
[001075] Системы CRISPR можно использовать для редактирования геномов паразитов, устойчивых к другим генетическим подходам. Например, было показано, что система CRISPR-Cas9 вносит двухцепочечный разрыв в геном Plasmodium yoelii (см., Zhang et al., "Efficient Editing of Malaria Parasite Genome Using the CRISPR/Cas9 System," mBio. vol. 5, e01414-14, Jul-Aug 2014). Ghorbal et al. ("Genome editing in the human malaria parasite Plasmodium falciparum using the CRISPR-Cas9 system," Nature Biotechnology, vol. 32., p. 819-821, doi: 10.1038/nbt.2925, опубликованную через интернет 1 июня 2014 года) модифицировали последовательности двух генов: orc1 и kelch13, которые предположительно участвуют в подавлении гена и возникающей устойчивости к артемизинину, соответственно. Паразиты, которые были изменены в соответствующих участках, восстанавливались с очень высокой эффективностью, несмотря на отсутствие прямой селекции по модификации, что указывает на то, что с помощью этой системы можно вносить нейтральные или даже вредные мутации. CRISPR-Cas9 также используется для модификации геномов других патогенных паразитов, включая Toxoplasma gondii (см. Shen et al., "Efficient gene disruption in diverse strains of Toxoplasma gondii, using CRISPR/CAS9," mBio vol. 5:e01114-14, 2014; and SidiJk et al., "Efficient Genome Engineering of Toxoplasma, gondii Using CRISPR/Cas9," PLoS One vol. 9, el00450, doi: 10.1371/joumal.pone.0100450, опубликованную через интернет 27 июня 2014 года).
[001076] Vyas et al ("A Candida albicans CRISPR system permits genetic engineering of essential genes and gene families," Science Advances, vol. 1, el500248, DOI: 10.1126/seiadv.1500248, 3 апреля 2015 года) использовали систему CRISPR для преодоления давних препятствий в генной инженерии C. albicans и эффективно внесли мутацию в одном эксперименте в обе копии нескольких разных генов. В организме, где несколько механизмов способствуют лекарственной резистентности, Vyas получил гомозиготные двойные мутанты, которые больше не проявляли гиперрезистентность к флуконазолу или циклогексимиду, демонстрируемую родительским клиническим изолятом Can90. Vyas также получил гомозиготные мутации с потерей функциональности в основных генах C. albicans путем создания условных аллелей. Нуль-аллели DOR1, который необходим для обработки рибосомной РНК, являются летальными при низкой температуре, но жизнеспособны при высокой температуре. Vyas использовал шаблон восстановления, в котором вводилась нонсенс-мутация и изолировались мутанты dcr1/dcr1, которые не росли при 16°C.
Лечение заболеваний с генетическими или эпигенетическими аспектами
[001104] Системы CRISPR-Cas по настоящему изобретению могут быть использованы для коррекции генетических мутаций, которые ранее предпринимались с ограниченным успехом с использованием TALEN и ZFN и были идентифицированы как потенциальные мишени для систем Cas9, в том числе, как и в опубликованных заявках EditasMedicine, описывающих способы использования Cas9 для нацеливания на локусы, чтобы с помощью генной терапии вылечивать болезни, в том числе, в WO 2015/048577 CRISPR-RELATED METHODS AND COMPOSITIONS Gluckmannet al, WO 2015/070083 CRISPR-RELATED METHODS AND COMPOSITIONS WITH GOVERNING gRNAS, Glucksmann et al. В некоторых вариантах осуществления предусматривается лечение, профилактика или диагностика первичной открытой глаукомы (POAG). Мишень предпочтительно представляет собой ген MYOC. Это описано в WO2015153780, содержание которой включено в настоящее описание в качестве ссылки.
[001105] Упоминается WO2015/134812 CRISPR/CAS-RELATED METHODS AND COMPOSITIONS FOR TREATING USHER SYNDROME AND RETINITIS PIGMENTOSA Maeder et al. В соответствии с идеями, описанными в настоящем описании, изобретение охватывает способы и материалы этих документов, применяемые в сочетании с идеями, поисанными в настоящем описании. В аспекте глазной и слуховой генной терапии способы и конструкции для лечения синдрома Ашера и пигментного ретинита могут быть адаптированы к системе CRISPR-Cas по настоящему изобретению (см., например, WO02015/134812). В одном варианте осуществления изобретение WO 2015/134812 включает лечение или задержку начала или прогрессирования синдрома Ашера типа IIА (USH2A, USHIIA) и пигментного ретинита 39 (RP39) путем редактирования генов, например, с использованием CRISPR-Cas9-опосредованных спсобов для исправления делеции гуанина в положении 2299 в гене USH2A (например, замена удаленного остатка гуанина в положении 2299 гена USH2A). Аналогичный эффект может быть достигнут с помощью C2c1 или C2c3. В родственном аспекте нацеливание на мутацию осуществляется путем расщепления одной или более нуклеазами, одной или более никазами или их комбинацией, например, для индуцирования HDR с помощью донорной матрицы, которая корректирует точечную мутацию (например, один нуклеотид, например, делеция гуанина). Изменение или коррекция мутантного гена USH2A может быть опосредована любым механизмом. Типичные механизмы, которые могут быть связаны с изменением (например, коррекцией) мутантного гена USH2A, включают, но не ограничиваются ими, негомологичное концевое соединение, опосредованное микрогомологией концевое соединение (MMEJ), восстановление, связанное с гомологией (например, опосредованное эндогенной донорной матрицей), SDSA (отжиг цепи, зависящий от синтеза), одноцепочечный отжиг или одноцепочечная инвазия. В одном варианте осуществления способ, используемый для лечения синдрома Ашера и пигментного ретинита, может включать сбор данных о мутации, имеющейся у индивидуума, например, путем секвенирования соответствующей части гена USH2A.
[001106] Также упоминается WO 2015/138510 и в соответствии идеями, описанными в настоящем описании, изобретение (с использованием системы CRISPR-Cas9) предусматривает лечение или задержку начала или прогрессирования врожденного амароза Лебера 10 (LCA 10). LCA 10 вызывается мутацией в гене CEP290, например, c.2991+1655, заменой аденина на гуанин в гене CEP290, что приводит к внесению скрытого сайта сплайсинга в интроне 26. Она представляет собой мутацию на нуклеотиде 1655 интрона 26 CEP290, например, замену А на G. CEP290 также известен как: CT87; MKS4; POC3; rd16, BBS14; JBTS5; LCAJO; NPHP6; SLSN6 и 3H11Ag (см., например, WO 2015/138510). В аспекте генной терапии изобретение включает внесение одного или более разрывов вблизи места расположения мишени LCA (например, c.2991+1655, замена А на G) по меньшей мере в одном аллеле гена CEP290. Изменение заденного положения LCA10 означает (1) индуцированное разрывом внесение инсерции-делеции (также упоминаемое в настоящем описании как опосредованное NHEJ внесение инсерции-делеции) в непосредственной близости или в заданном положении LCA10 (например, C.2991+1655A-G) или (2) делецию, вызванную разрывом (также упоминаемая в настоящем описании как NHEJ-опосредованная делеция) геномной последовательности, включающую мутацию в заданном положении LCA10 (например, C.2991+1655A-G). Оба подхода приводят к потере или разрушению скрытого участка сплайсинга в результате мутации в заданном положении LCA 10.
[001107] Исследователи рассматривают возможность использования генной терапии для лечения широкого спектра заболеваний. Системы CRISPR по настоящему изобретению на основе эффекторного белка C2c1 или C2c3 предусматриваются для таких терапевтических целей, включая, но не ограничиваясь ими, дополнительные иллюстративные области-мишени и способы доставки, как показано ниже. Некоторые примеры состояний или заболеваний, которые могут быть эффективно вылечены с использованием настоящей системы, включены в примеры генов, и ткже представлены ссылки, включенные в настоящее описание и в настоящее время ассоциируемые с этими состояниями. Представленные гены и состояния не являются исчерпывающими.
Лечение заболеваний системы кровообращения
[001108] Настоящее изобретение также предусматривает доставку системы CRISPR-Cas, в частности, новых белковых систем CRISPR, описанных в настоящем описании, в клетки крови или гемопоэтические стволовые клетки. Плазматические экзосомы согласно Wahlgren et al. (Nucleic Acids Research, 2012, Vol. 40, No. 17 e130) были описаны ранее и могут быть использованы для доставки системы CRISPR Cas в кровь. Система нацеливания на нуклеиновую кислоту по настоящему изобретению также предусматривается для лечения гемоглобинопатий, таких как талассемия и серповидноклеточная анемия. См., например, международную патентную публикацию № WO 2013/126794 для потенциальных мишеней, на которые может быть осуществлено нацеливание системы CRISPR Cas по настоящему изобретению.
[001109] В Drakopoulou, "Review Article, The Ongoing Challenge of Hematopoietic Stem Cell-Based Gene Therapy for β-Thalassemia", Stem Cells International, Volume 2011, Article ID 987980, 10 pages, doi: 10.4061/2011/987980, включенной в настоящее описание в качестве ссылыки вместе с цитированными в ней документами, как если бы они были изложены полностью, описано изменение HSC с использованием лентивируса, который доставляет ген для β-глобина или γ-глобина. В отличие от использования лентивируса, c использованием знаний в данной области и идей, описанных в настоящем описании, квалифицированный специалист может скорректировать HSC в отношении β-талассемии с использованием системы CRISPR-Cas, которая нацелена на и исправляет мутацию (например, с подходящей матрицей HDR, которая доставляет кодирующую последовательность для β-глобина или γ-глобина, преимущественно не-серповидного β-глобина или γ-глобина); в частности, направляющая РНК может нацеливать на мутацию, которая приводит к β-талассемии, а HDR может обеспечивать кодирование для правильной экспрессии β-глобина или γ-глобина. Направляющую РНК, которая нацелена на мутацию, и содержащую белок Cas частицу, приводят в контакт с HSC, несущими мутацию. Частица также может содержать подходящую матрицу HDR, чтобы корректировать мутации для правильной экспрессии β-глобина или γ-глобина; или HSC можно приводить в контакт со второй частицей или вектором, которые содержат или доставляет матрицу HDR. Клетки, приведенные в контакт таким образом, можно вводить и необязательно обрабатывать/увеличивать в количестве, см. Cartier. В этой связи упоминаются Cavazzana, "Outcomes of Gene Therapy for β-Thalassemia Major via Transplantation of Autologous Hematopoietic Stem Cells Transduced Ex Vivo with a Lentiviral βA-T87Q-Globin Vector." tif2014.org/abstractFiles/Jean%20Antoine%20Ribeil. Abstract.pdf; Cavazzana-Calvo, "Transfusion independence and HMGA2 activation after gene therapy of human β-thalassaemia", Nature 467, 318-322 (16 September 2010) doi:10.1038/n.ature09328; Nienhuis, "Development of Gene Therapy for Thalassemia, Cold Spring Harbor Perpsectives in Medicine, doi: 10.n01/cshperspect.a011833 (2012), LentiGlobin BB305, a lentiviral vector containing an engineered β-globin gene (PA-T87Q); и Xie et al., "Seamless gene correction of β-thalassaemia mutations in patient-specific iPSCs using CRISPR/Cas9 and piggyback" Genome Research gr.1713427.114 (2014) http://www.genome.org/cgi/doi/10.1101/gr.173427.114 (Cold Spring Harbor Laboratory Press); которые являются предметом работы Cavazzana, связанной с β-талассемией человека, и предметом работы Xie, и они включены в настоящее описание в качестве ссылки со всеми документами, цитированными в них или связанными с ним. В рамках настоящего изобретения матрица HDR может быть прдеоставлена HSC для экспрессии сконструированного гена β-глобина (например, βA-T87Q) или β-глобина согласно Xie.
[001110] Xu et al. (SciRep. 2015 Jul9;5:12065. doi: 10.1038/srepl2065) разработали TALEN и CRISPR-Cas9, для непосредственного нацеливания на сайт мутации интрона 2 IVS2-654 в гене глобина. Xu et al. наблюдали различные частоты двунитевых разрывов (DSB) в локусах IVS2-654 с использованием TALEN и CRISPR-Cas9, а TALEN опосредовали более высокую эффективность нацеливания на гомологичные гены по сравнению с CRISPR-Cas9 в сочетании с донором транспозона piggyBac. Кроме того, для CRISPR-Cas9 наблюдалось большее количество нецелевых событий по сравнению с TALEN. Наконец, iPSC-клоны, скорректированные TALEN, были выбраны для дифференцировки эритробластов с использованием системы совместного культивирования с OP9 и обнаружили относительно более высокую транскрипцию HBB, чем в нескорректированных клетках.
[001111] Song et al. (Stem Cells Dev. 2015 May 1;24(9): 1053-65. doi: 10, 1089/scd.2014.0347. Epub 2015 Feb 5) использовали CRISPR/Cas9 для исправления β-Thal iPSC; клетки со скорректированными генами демонстрировали нормальные кариотипы и полную плюрипотентность, поскольку эмбриональные стволовые клетки человека (hESC) не продемонстрировали нецелевых эффектов. Затем Song et al. оценили эффективность дифференцирования генетически скорректированных P-Thal iPSC. Song et al. обнаружили, что при гемопоэтической дифференцировке генетически скорректированные P-Thal iPSC продемонстрировали повышенную долю эмбриоидных телец и различные процентные доли гемопоэтических предшественников. Что еще более важно, генетически скорректированные линии β-Thal iPSC восстанавливали экспрессию HBB и уменьшали производство активных форм кислорода по сравнению с нескорректированной группой. В исследовании Song et al. было показано, что эффективность гемопоэтической дифференцировки β-Thal iPSC значительно улучшилась после корректировки системой CRISPR-Cas9. Аналогичные способы могут быть применены с использованием описанных в настоящем описании систем CRISPR-Cas, например, систем, содержащих эффекторные белки C2c1 или C2c3.
[001112] Серповидноклеточная анемия является аутосомно-рецессивным генетическим заболеванием, в котором эритроциты принимают форму серпа. Это вызвано заменой одного основания в гене β-глобина, который расположен на коротком плече хромосомы 11. В результате вместо глутаминовой кислоты образуется валин, вызывая образование серповидного гемоглобина (HbS). Это приводит к образованию искаженной формы эритроцитов. Из-за этой аномальной формы мелкие кровеносные сосуды могут закупориваться, нанося серьезные повреждения тканям кости, селезенки и кожи. Это может привести к болевым эпизодам, частым инфекциям, синдрому "руки-стопы" или даже множественной органной недостаточности. Искаженные эритроциты также более восприимчивы к гемолизу, что приводит к серьезной анемии. Как и в случае β-талассемии, серповидноклеточная анемия может быть скорректирована путем изменения HSC с помощью системы CRISPR-Cas. Система позволяет специфически редактировать геном клетки, разрезая ее ДНК, а затем позволяя ей восстанавливать себя. Белок Cas вводится и направляется РНК к точке мутации, а затем разрезает ДНК в этой точке. Одновременно вводится нормальная версия последовательности. Эта последовательность используется собственной системой репарации для зашивания индуцированного разреза. Таким образом, CRISPR-Cas позволяет корректировать мутацию в ранее полученных стволовых клетках. Обладая знаниями данной области и идеями, описанными в настоящем описании, квалифицированный специалист может скорректировать HSC в отношении серповидноклеточной анемии с использованием системы CRISPR-Cas, которая нацелена на и корректирует мутацию (например, с подходящей матрицей HDR, которая обеспечивает кодирующую последовательность для β-глобина, преимущественно не-серповидного β-глобина); в частности, направляющая РНК может нацеливать на мутацию, которая вызывает серповидноклеточную анемию, и HDR может обеспечивать кодирование для правильной экспрессии β-глобина. Направляющую РНК, которая нацелена на мутацию, и частицу, содержащую белок Cas, приводят в контакт с HSC, несущими мутацию. Частица также может содержать подходящую матрицу HDR, чтобы корректировать мутации для правильной экспрессии β-глобина; или HSC можно приводить в контакт со второй частицей или вектором, которые содержат или доставляют матрицу HDR. Таким образом приведенные в контакт клетки могут быть введены или необязательно обработаны/увеличены в количестве. Матрица HDR может обеспечивать экспрессию в HSC сконструированного гена β-глобина (например, βA-T87Q) или β-глобин, как в Xie.
[001113] В Williams, "Broadening the Indications for Hematopoietic Stem Cell Genetic Therapies," Cell Stem Cell 13:263-264 (2013), включенной в настоящее описание в качестве ссылыки вместе с цитированными в ней документами, как если бы они были изложены полностью, описан опосредованный лентивирусом перенос гена в клетки HSC/P у пациентов с метахроматической лейкодистрофией (MLD), генетическим заболеванием, вызываемым дефицитом арилсульфатазы A (ARSA), что приводит к демиелинизации нервов; и перенос генов, опосредованных лентивирусом, в HSC у пациентов с синдромом Вискотта-Олдрича (WAS) (пациенты с дефектным белком WAS, являющимся эффектором малой ГТФ-азы CDC42, который регулирует функцию цитоскелета в линиях клеток крови и, таким образом, они страдают иммунодефицитом с рецидивирующими инфекциями, аутоиммунными симптомами и тромбоцитопенией с аномально малыми и дисфункциональными тромбоцитами, приводящими к чрезмерному кровотечению и повышенному риску лейкемии и лимфомы). В отличие от использования лентивируса, используя знания в данной области и идеи, описанные в настоящем описании, квалифицированный специалист может скорректировать HSC в отношении MLD (дефицит арилсульфатазы А (ARSA)) с использованием системы CRISPR-Cas, которая нацелена на и исправляет мутацию (дефицит арилсульфатазы А (ARSA)) (например, с подходящей матрицей HDR, которая обеспечивает кодирующую последовательность для ARSA); в частности, направляющая РНК может быть нацелена на мутацию, которая приводит к MLD, а HDR может обеспечивать кодирование для правильной экспрессии ARSA. Направляющую РНК, которая нацелена на мутацию, и частицу, содержащую белок Cas, приводят в контакт с HSC, несущими мутацию. Частица также может содержать подходящую матрицу HDR, чтобы корректировать мутации для правильной экспрессии ARSA; или HSC можно приводить в контакт со второй частицей или вектором, которые содержат или доставляют матрицу HDR. Таким образом приведенные в контакт клетки могут быть введены или необязательно обработаны/увеличены в количестве. В отличие от использования лентивируса, используя знания в данной области и идеи, описанные в настоящем описании, квалифицированный специалист может скорректировать HSC в отношении WAS с использованием системы CRISPR-Cas, которая нацелена на и исправляет мутацию (дефицит белка WAS) (например, с подходящей матрицей HDR, которая обеспечивает кодирующую последовательность для WAS); в частности, направляющая РНК может нацеливать на мутацию, которая приводит к дефициту WAS, а HDR может обеспечивать кодирование для правильной экспрессии WAS. Направляющую РНК, которая нацелена на мутацию, и частицу, содержащую белок Cas, приводят в контакт с HSC, несущими мутацию. Частица также может содержать подходящую матрицу HDR, чтобы корректировать мутации для правильной экспрессии WAS; или HSC можно приводить в контакт со второй частицей или вектором, которые содержат или доставляют матрицу HDR. Таким образом приведенные в контакт клетки могут быть введены или необязательно обработаны/увеличены в количестве.
[001114] В Watts, "Hematopoietic Stem Cell Expansion and Gene Therapy" Cytotherapy 13(10): 1164-1171. doi: 10.3109/14653249.2011.620748 (2011), включенной в настоящее описание в качестве ссылыки вместе с цитированными в ней документами, как если бы они были изложены полностью, описана генная терапия гемопоэтических стволовых клеткок (HSC), например вирус-опосредованная генная терапия HSC, в качестве в высокой степени привлекательного варианта лечения многих расстройств, включая гематологические состояния, иммунодефициты, такие как ВИЧ/СПИД, и другие генетические нарушения, такие как болезни лизосомного хранения, включая SCID-X1, ADA-SCID, β-талассемию, X-связанный CGD, синдром Вискотта-Олдрича, анемия Фанкони, адренолейкодистрофия (ALD) и метахроматическая лейкодистрофия (MLD).
[001115] Патентные публикации США №№ 20110225664, 20110091441, 20100229252, 20090271881 и 20090222937, закрепленные за Cellectis, относятся к вариантам CREI, где по меньшей мере один из двух мономеров I-CreI имеет по меньшей мере два заместителя, по одному в каждом из двух функциональных поддоменов центрального домена LAGLIDADG (SEQ ID NO: 26), расположенного соответственно в положениях 26-40 и 44-77 I-CreI, причем указанный вариант способен расщеплять последовательность ДНК-мишени из гена гамма-цепи рецептора человека (IL2RG) человека, также называемого общим геном гамма-цепи рецептора цитокинов или гамма C. Последовательности-мишени, идентифицированные в публикациях US 2011225664, 20110091441, 20100229252, 20090271881 и 20090222937, могут быть использованы для системы нацеливания на нуклеиновые кислоты по настоящему изобретению.
[001116] Тяжелый комбинированный иммуннодефицит (SCID) является следствием дефекта созревания лимфоцитов T, всегда связанного с функциональным дефектом в лимфоцитах В (Cavazzana-Calvo et al., Annu. Rev. Med., 2005, 56, 585-602; Fischer et al., Immunol. Rev., 2005, 203, 98-109). Общая встречаемость заболевания оценивается как 1 на 75000 рождений. Пациенты с не подвергнутым лечению SCID имеют множественные оппортунистические инфекции микроорганизмами и, как правило, не живут более одного года, SCID можно лечить с помощью аллогенной передачи гемопоэтических стволовых клеток от семейного донора. Гистосовместимость с донором может варьироваться широко. В случае дефицита аденозиндеаминазы (ADA), одной из форм SCID, пациентов можно лечить путем инъекции рекомбинантного фермента аденозиндезаминазы.
[001117] С тех пор как было показано, что ген ADA является мутантным у пациентов с SCID (Giblettetah, Lancet, 1972, 2, 1067-1069), было идентифицировано несколько других генов, участвующих в SCID (Cavazzana-Calvo et al., Annu. Rev. Med., 2005, 56, 585-602; Fischer et al., ImmunolRev., 2005, 203, 98-109). Для SCID существует четыре основные причины: (i) наиболее частая форма SCID, SCID-X1 (X-связанный SCID или X-SCID), вызвана мутацией в гене IL2RG, что приводит к отсутствию зрелых Т-лимфоцитов и NK-клеток. IL2RG кодирует белок γ-C (Noguchi, et al., Cell, 1.993, 73, 147-157), общий компонент по меньшей мере пяти рецепторных комплексов интерлейкина. Эти рецепторы активируют несколько мишеней через киназу JAK3 (Macchietah, Nature, 1995, 377, 65-68), инактивация которой приводит к тому же синдрому, что и инактивация гамма-C; (ii) мутация в гене ADA приводит к дефекту в метаболизме пуринов, который является летальным для предшественников лимфоцитов, что, в свою очередь, приводит практически к отсутствию В, Т и NK-клеток; (iii) V (D) J рекомбинация является существенной стадией созревания иммуноглобулинов и рецепторов Т-лимфоцитов (TCR). Мутации в активирующем рекомбинацию гене 1 и 2 (RAG1 и RAG2) и Artemis - трех генов, участвующих в этом процессе, приводят к отсутствию зрелых Т-лимфоцитов; и (iv) сообщалось также о мутациях в других генах, таких как CD45, участвующих в Т-клеточных каскадов, хотя они представляют меньшую часть случаев (Cavazzana-Calvo et al., Ann. Rev. Med., 2005, 56, 585- 602, Fischer et al., Immunol, Rev., 2005, 203, 98-109). С тех пор, как были идентифицированы их генетические основы, различные формы SCID стали парадигмой для подходов к генной терапии (Fischer et al., Immunol. Rev., 2005, 203, 98-109) по двум основным причинам. Во-первых, как и во всех заболеваниях крови, может предусматриваться лечение ex vivo. Гемопоэтические стволовые клетки (HSC) могут быть выделены из костного мозга и могут сохранить свои плюрипотентные свойства на протяжении нескольких клеточных делений. Поэтому их можно обрабатывать in vitro, а затем повторно вводить в организм пациента, где они заселяют костный мозг. Во-вторых, поскольку созревание лимфоцитов нарушается у пациентов с SCID, исправленные клетки имеют селективное преимущество. Поэтому небольшое количество исправленных клеток может восстановить функциональную иммунную систему. Эта гипотеза несколько раз подтверждалась: (i) частичным восстановлением иммунных функций, связанных с реверсированием мутаций у пациентов с SCID (Hirschhom et al., Nat. Genet., 1996, 13, 290-295; Stephan et al., N. Engl J. Med., 1996, 335, 1563-1567, Bousso et al., Proc. Natl., Acad. Set. USA, 2000, 97, 274-278, Wada et al., Proc. Natl. Acad. Set., 2001, 98, 8697-8702, Nishikomori et al., Blood, 2004, 103, 4565-4572), (ii) коррекцией дефицита SCTD-X1 in vitro в гемопоэтических клетках (Candotti et. Ah, Blood, 1996, 87, 3097-3102, Cavazzana-Calvo et al., Blood, 1996, Blood, 88, 3901-3909; Taylor et al., Blood, 1996, 87, 3103-3107; Hacein-Bey et al., Blood, 1998, 92, 4090-4097), (iii) коррекцией дефицита SCID-X1 (Soudais et al., Blood, 2000, 95, 3071-3077; Tsai et al., Blood, 2002, 100, 72-79), JAK-3 (Bunting et ah, Nat. Med., 1998, 4, 58-64, Bunting et al., Hum. Gene Ther., 2000, 11, 2353-2364) и RAG2 (Yates et al., Blood, 2002, 100, 3942-3949) in vivo в моделях на животных, и (iv) результатом клинических испытаний генной терапии (Cavazzana-Calvo et al., Science, 2000, 288, 669-672; Aiuti et al., Nat. Med., 2002; 8, 423-425, Gaspar et al., Lancet. 2004, 364, 2181-2187).
[001118]Патентная публикация США № 20110182867, присвоенная Корпорации детского медицинского центра, а также президенту и стипендиатам Гарвардского колледжа, относится к методам и использованиям модуляции фетального гемоглобина, экспрессии (HbF) в гемопоэтических клетках-предшественниках с помощью ингибиторов экспрессии или активности BCL11A, таких как RNAi и антитела. Мишени, раскрытые в опубликованной патентной публикации США № 20110182867, такой как BCL11A, могут быть подвергнуты нацеливанию системой CRISPR Cas по настоящему изобретению для модуляции экспрессии фетального гемоглобина. См. Также Bauer et al. (Science 11 October 2013: том 342, № 6155, pp. 253-257) и Xu et al. (Science 18 ноября 2011: том 334, № 6058 стр. 993-996) для дополнительных мишеней BCL11A.
[001119] Используя знания в данной области и идеи, описанные в настоящем описании, квалифицированный специалист может скорректировать HSC в отношении гематологических расстройств, таких как β-талассемия, гемофилия или генетическое лизосомальное заболевание.
Доставка HSC и редактирование гемопоэтических стволовых клеток; и конкретные состояния
[001120] Термин «гемопоэтическая стволовая клетка» или "HSC" включает в широком значении те клетки, которые считаются HSC, например, клетки крови, которые приводят к появлению всех других клеток крови и происходят из мезодермы; расположенные в красном костном мозге, который содержится в сердцевине большинства костей, HSC по изобретению включают клетки, имеющие фенотип гемопоэтических стволовых клеток, идентифицированных по небольшим размером, отсутствию маркеров линии (lin) и маркеров, которые принадлежат к кластерам дифференциации, например: CD34, CD38, CD90, CD133, CD105, CD45, а также e-kit - рецептор фактора стволовых клеток. Гемопоэтические стволовые клетки отрицательны по маркерам, которые используются для выявления приверженности линии, и, таким образом, они называются Lin- и во время их очистки посредством FACS содержат до 14 различных зрелых маркеров крови, например CD13 и CD33 для миелоидных клеток, CD71 для эритроидных клеток, CD19 для В-клеток, CD61 для мегакариоцитов и т. д. для человека, и B220 (CD45 мыши) для В-клеток, Mac-1 (CD11b/CD18) для моноцитов, Gr-1 для гранулоцитов, Ter119 для эритроидов, IL7Ra, CD3, CD4, CD5, CD8 для Т-клеток и т. д. Маркеры HSC мыши: CD34lo/-, SCA-1+, Thy1.1+/lo, CD38+, C-kit+, lin- и маркеры HSC человека: CD34+, CD59+, Thyl/CD90+, CD38lo/-, C-kit/CD117+ и lin-. HSC идентифицируются маркерами. Следовательно, в вариантах осуществления, обсуждаемых в настоящем описании, HSC могут быть CD34+ клетками. HSC также могут представлять собой гемопоэтические стволовые клетки, которые являются CD34-/CD38-. Стволовые клетки, которые могут быть лишены c-kit на поверхности клетки, которые рассматриваются в данной области как HSC, входят в объем изобретения, также как и клетки CD133+, которые аналогично рассматриваются как HSC в данной области.
[001121] Система CRISPR-Cas (например, C2c1 или C2c3) может быть сконструирована для нацеливания на генетический локус или локусы в HSC. Можно получить белок Cas (например, C2c1 или C2c3), преимущественно кодон-оптимизированный для эукариотической клетки и особенно клетки млекопитающего, например клетки человека, HSC, и sg-РНК, нацеливающую на локус или локусы в HSC, например на ген EMX1. Они могут быть доставлены через частицы. Частицы могут быть образованы белком Cas (например, C2c1 или C2c3), смешанным с гРНК. Смесь гРНК и белка Cas можно смешивать, например, со смесью, содержащей или по существу состоящей из поверхностно-активного вещества, фосфолипида, биодеградируемого полимера, липопротеина и спирта, посредством чего могут формироваться частицы, содержащие гРНК и Cas (например, C2c1 или C2c3). Изобретение предусматривает, получение частиц таким образом и частицы, полученные таким способом, а также их применение.
[001122] В более общем случае частицы могут быть сформированы с использованием эффективного процесса. Во-первых, белок Cas (например, C2c1 или C2c3) и гРНК, нацеленная на ген EMX1 или контрольный ген LacZ, могут быть смешаны вместе в подходящем молярном соотношении, например, от 3:1 до 1:3 или от 2:1 до 1:2 или 1:1, при подходящей температуре, например, 15-30°С, 20-25°С, комнатной температуре, в течение подходящего периода времени, например 15-45, 30 минут, преимущественно в стерильном, свободном от нуклеаз буфере, например, 1X PBS. Отдельно компоненты частиц, такие как или содержащие: поверхностно-активное вещество, например катионный липид, 1,2-диолеоил-3-триметиламмонийпропан (DOTAP); фосфолипид, например димиристоилфосфатидилхолин (DMPC); биодеградируемый полимер, такой как полимер этиленгликоля или PEG, и липопротеин, такой как липопротеин низкой плотности, например холестерин, могут быть растворены в спирте, преимущественно С1-6-алкиловом спирте, таком как метанол, этанол, изопропанол, например 100% этанол. Эти два раствора могут быть смешаны вместе с образованием частиц, содержащих комплексы Cas (например, C2c1 или C2c3)-гРНК. В некоторых вариантах осуществления частица может содержать матрицу HDR. Это может быть частица, совместно вводимая с белоксодержащей частицей гРНК+Cas (например, C2c1 или C2c3) или помимо контактирования HSC с белоксодержащей частицей гРНК+Cas (например, C2c1 или C2c3) HSC контактирует с частицей, содержащей матрицу HDR; или HSC приводят в контакт с частицей, содержащей все из гРНК, Cas (например, C2c1 или C2c3) и матрицы HDR. Матрицу HDR можно вводить отдельным вектором, в результате чего в первом случае частица проникает в клетку HSC, а отдельный вектор также проникает в клетку, где геном HSC модифицируется гРНК+Cas (например, C2c1 или C2c3) и также присутствует матрицы HDR, в котором, например, геномные локусы модифицируются HDR, что может привести к исправлению мутации.
[001123] После образования частиц HSC в 96-луночных планшетах могут быть трансфицированы посредством 15 мкг белка Cas (например, C2c1 или C2c3) на лунку. HSC могут быть собраны через три дня после трансфекции, и может быть определено количество инсерций и делеций (инсерций-делеций) в локусе EMX1.
[001124] Это иллюстрирует, как HSC могут быть модифицированы с использованием CRlSPR-Cas (например, C2c1 или C2c3), нацеленным на геномный локус или локусы, представляющие интерес в HSC. HSC, которые должны быть модифицированы, могут быть in vivo, то есть в организме, например, человека или других эукариот, например, животных, таких как рыбы (данио-рерио), млекопитающих, в частности, приматов (шимпанзе, макак), грызунов (мышей, кроликов, крыс) собак, домашнего скота, птиц. HSC, которые должны быть модифицированы, могут быть in vitro, то есть вне такого организма. Далее, модифицированные HSC могут использоваться ex vivo, то есть одна или более HSC такого организма могут быть получены или выделены из организма, возможно, HSC могут быть увеличены в количестве, HSC модифицированы конструкцией, включающей CRISPR-Cas (например, C2c1 или C2c3), которые нацелены на генетический локус или локусы в HSC, например, путем контактирования HSC с конструкцией, например, где последняя содержит частицу, содержащую фермент CRISPR, и одну или более гРНК, которая нацелена на генетический локус или локусы в HSC, такие как частица, полученная или получаемая при смешивании смеси гРНК и белка Cas (например, C2c1 или C2C3) со смесью, содержащей или по существу состоящей из поверхностно-активного вещества, фосфолипида, биодеградируемого полимера, липопротеина и спирта (где одна или более гРНК нацелены на генетический локус или локусы в HSC), необязательно увеличивая в количестве полученные модифицированные HSC и вводя в организм результирующие модифицированные HSC. В некоторых случаях выделенные или полученные HSC могут быть из первого организма, такого как организм того же вида, что и второй организм, а второй организм может представлять собой организм, которому вводят полученные модифицированные HSC, например, первый организм может быть донором (таким как родственник (родитель или родной брат)) для второго организма. Модифицированные HSC могут иметь генетические модификации, чтобы устранить или облегчить, или уменьшить симптомы заболевания или состояния индивидуума, субъекта или пациента. Модифицированные HSC, например, в случае донорства первого организма второму, могут иметь генетические модификации, чтобы HSC имели один или более белков, таких как поверхностные маркеры или белки, больше похожие на маркеры второго организма. Модифицированные HSC могут иметь генетические модификации для имитации болезни или состояния индивидуума или субъекта или пациента и могут повторно вводиться в организм, не являющийся человеком, для получения моделей на животных. Увеличение HSC в количестве входит в компетенцию квалифицированного специалиста, на основании настоящего описания и знания в данной области, см., например, Lee, "Improved ex vivo expansion of adult hematopoietic stem cells by overcoming CUL4-mediated degradation of HOXB4." Blood. 2013 May 16;121(20):4082-9. doi: 10.1182/blood-2012-09- 455204. Epub 2013 Mar 21.
[001125] Как указано, для улучшения активности могут быть предварительно созданы комплексы гРНК с белком Cas (например, C2c1 или C2c3), перед тем как они окажутся в составе полного комплекса в частице. Составы могут созданы, исходя из различных молярных соотношений различных компонентов, про которые известно, что они могут способствовать доставке нуклеиновых кислот в клетки (например, 1,2-диолеил-3-триметиламмоний-пропан (DOTAP) или 1,2-дитетрадеканоил-sn-3-фосфохолин (DMPC), и/или в котором гидрофильный полимер содержит этиленгликоль или полиэтиленгликоль (PEG) и/или в котором частица далее содержит холестерин). Например, частица состава DOTAP 100, DMPC 0, PEG 0, холестерол 0; или DOTAP 90, DMPC 0, PEG 10, холестерин 0; или DOTAP 90, DMPC 0, PEG 5, холестерин 5, или DOTAP 100, DMPC 0, PEG 0, холестерин 0. Соответственно, изобретение предусматривает добавление гРНК, белка Cas (например, C2c1 или C2c3) и компонентов, формирующих частицу, а также частицы, получаемые при таком добавлении.
[001126] В предпочтительном варианте осуществления частицы, содержащие комплексы Cas (например, C2c1 или C2c3) с гРНК, могут быть образованы путем смешивания белка Cas (например, C2c1 или C2c3) и одной или более гРНК, предпочтительно в молярном соотношении фермента и направляющей РНК, равном 1:1. Отдельно различные компоненты, известные способностью способствовать доставке нуклеиновых кислот (например, DOTAP, DMPC, PEG и холестерин), растворяют, предпочтительно в этаноле. Два раствора смешивают вместе, чтобы сформировать частицы, содержащие комплексы Cas (например, C2c1 или C2c3) с гРНК. После образования частиц комплексы Cas (например, C2c1 или C2c3) с гРНК могут быть трансфицированы в клетки (например, HSC). Может применяться баркодирование. Частицы, Cas- и/или гРНК могут быть баркодированы.
[001127] Изобретение в одном из вариантов осуществления предусматривает способ получения частицы, содержащей белок Cas (например, C2c1 или C2c3) и гРНК, включающий смешение смеси гРНК и белка Cas (например, C2c1 или C2c3) со смесью, содержащей или по существу состоящей из поверхностно активных веществ, фосфолипидов, биодеградируемых липопротеина и спирта. Изобретение относится к частице, содержащей белок Cas (например, C2c1 или C2c3) и гРНК, согласно этому способу. Изобретение в одном из вариантов осуществления предусматривает использование частицы в способе модификации геномного локуса-мишени, или организма, или организма, не являющегося человеческим, путем манипуляции с последовательностью-мишенью в геномном локусе-мишени, включая приведение клетки, содержащей геномный локус-мишень, в контакт с частицей, где гРНК нацеливает на геномный локус-мишень; или способ модификации геномного локуса-мишени, или организма, или организма, не являющегося человеческим, путем манипуляции с последовательностью-мишенью в геномном локусе-мишени, включая приведение клетки, содержащей геномный локус-мишень, в контакт с частицей, где гРНК нацелена на геномный локус-мишень. В этих вариантах осуществления геномный локус-мишень преимущественно является геномным локусом в HSC.
[001128] Соображения для терапевтического применения: фактором, учитываемым в геном-редактирующей терапии является выбор нуклеазы, специфичной к последовательности, такой как вариант нуклеазы C2c1 или C2c3. Каждый вариант нуклеазы может обладать своим уникальным набором преимуществ и недостатков, многие из которых должны быть сбалансированы в контексте лечения, чтобы максимизировать терапевтическую эффективность. До сих пор два терапевтических подхода к редактированию нуклеазами продемонстрировали значительные перспективы: разрушение гена и коррекция гена. Разрушение гена включает стимуляцию репарации путем объединения негомологичных концов (NHEJ) для создания направленных инсерций-делеций в генетических элементах, что часто приводит к вызывающим потерю функции мутациям, которые полезны для пациентов. Напротив, для коррекции гена используется направляемая гомологией репарация (HDR), чтобы напрямую обратить мутацию, вызывающую заболевание, восстановить функцию, при этом сохраняя физиологическую регуляцию скорректированного элемента. Направляемая гомологией репарация также может быть использована для вставки терапевтического трансгена в определенный безопасный локус генома ("безопасную гавань"), чтобы восстановить потерянную функцию гена. Для того чтобы специфичная редактирующая терапия была эффективной, в популяции клеток-мишеней должен быть достигнут достаточно высокий уровень модификаций, чтобы обратить вспять симптомы заболевания. Этот "порог" терапевтической модификации определяется выживаемостью отредактированных клеток после лечения и количеством продукта гена, необходимого для обращения вспять симптомов. С учетом выживаемости клеток, редактирование создает три потенциальных результата для обработанных клеток относительно их неотредактированных копий: увеличение выживаемости, снижение выживаемости и отсутствие влияния на выживаемость. В случае увеличения выживаемости, например, при лечении X-сцепленного тяжелого иммунодефицита, модифицированные гемопоэтические клетки-предшественники селективно увеличиваются в количестве по сравнению с их неотредактированными копиями. X-сцепленный тяжелый иммунодефицит является заболеванием, вызванным мутациями в гене IL2RG, функционирование которого необходимо для правильного развития гемопоэтической лимфоцитарной линии [Leonard, W.J., et al. Immunological reviews 138, 61-86 (1994); Kaushansky, K. & Williams, W.J. Williams hematology, (McGraw-Hill Medical, New York, 2010)]. Во время клинических испытаний пациенты, получавшие вирусную генную терапию для лечения X-сцепленного тяжелого иммунодефицита при редком примере спонтанной коррекции мутации, связанной с X-сцепленным тяжелым иммунодефицитом, скорректированные гемопоэтические прогениторные клетки могут преодолевать этот блок развития и размножаться, по сравнению с их неотредактированными копиями, способствуя лечению [Bousso, P., et al. Proceedings of the National Academy of Sciences of the United States of America 97, 274-278 (2000); Hacein-Bey-Abina, S, et al. The New England journal of medicine 346, 1185-1193 (2002); Gaspar, H.B., et al. Lancet 364, 2181-2187 (2004)]. В этом случае, когда отредактированная клетка обладает селективным преимуществом, даже низкое число отредактированных клеток могут быть увеличено в количестве, обеспечивая терапевтическую пользу для пациента. Напротив, редактирование других гемопоэтических заболеваний наподобие гранулематозного расстройства, может не вызывать изменений выживаемости отредактированных гемопоэтических клеток-предшественников, увеличивая порог терапевтической модификации. Гранулематозное расстройство вызвано мутациями в генах, кодирующих белки фагоцитарной оксидазы, которые обычно используются нейтрофилами для образования активных форм кислорода, чтобы убивать патогены [Mukherjee, S. & Thrasher, A.J. Gene 525, 17-4-181 (2013)]. Поскольку дисфункция этих генов не влияет на выживаемость или развитие гемопоэтических клеток-предшественников, а только на способность зрелых гемопоэтических клеток бороться с инфекцией, маловероятно, чтобы происходила предпочтительная экспансия отредактированных клеток при этом заболевании. Действительно, при гранулематозном расстройстве во время испытаний генной терапии в отредактированных гемопоэтических клетках-предшественниках не наблюдалось селективного преимущества, что приводило к трудностям при длительной имплантации клеток [Malech, H.L., et al. Proceedings of the National Academy of of the United States of America 94, 12133-12138 (1997); Kang, H.J., et al. Molecular therapy: the journal of the American Society of Gene Therapy 19, 2092-2101 (2011)]. По существу, может требоваться значительно более высокий уровень редактирования для лечения заболеваний наподобие гранулематозного расстройства, при котором редактирование не создает преимущества для выживаемости, относительно заболеваний, при которых редактирование создает преимущества для выживаемости целевых клеток. Если редактирование лишает клетки преимущества для их выживаемости, как могло бы быть в случае восстановления функции гена опухолевого супрессора в опухолевой клетке, модифицированные клетки будут вытесняться неотредактированными клетками, из-за чего терапевтические преимущества оказались бы ниже относительно уровня редактирования. Этот последний класс заболеваний мог бы быть особенно трудным для лечения с помощью геномной редактирующей терапии.
[001129] В дополнение к выживаемости клеток, на минимальный уровень терапевтического редактирования генома, которое должно быть осуществлено для обращения вспять симптомов, также влияет количество продукта гена, необходимое для лечения заболевания. Гемофилия В является одним из заболеваний, при которых небольшое изменение количества продукта гена может приводить к значительным изменениям в клиническом результате. Это заболевание вызывается мутациями в гене, кодирующем фактор IX, белок, который в норме секретируется из печени в кровь, где он функционирует как компонент каскада реакций свертывания крови. Клиническая тяжесть гемофилии В связана с уровнем активности фактора IX. В то время как тяжесть заболевания связана с активностью ниже 1% от нормальной активности, более мягкие формы заболевания связаны активностью, превышающей 1% от нормальной активности фактора IX [Kaushansky, K. & Williams, W.J. Williams hematology, (McGraw-Hill Medical, New York, 2010), Lofqvist, T,, et al. Journal Of Internal Medicine 241, 395-400 (1997)]. Это позволяет предположить, что редактирующая терапия, которая может восстановить экспрессию фактора IX даже в небольшом проценте клеток печени, может оказать значительное влияние на клинический результат. Исследование, в котором используются ZFN для коррекции модели гемофилии В на мышах сразу после рождения, показало, что 3-7% достаточно для обращения вспять симптомов заболевания, обеспечивая доклинические доказательства для этой гипотезы [Li, H., et al. Nature Alb, 217-221 (2011)].
[001130] Расстройства, при которых малое изменение уровня продукта гена может влиять на клинический результат, и заболевания, при которых у отредактированных клеток появляются преимущества, связанные с выживанием, являются идеальными мишенями для терапии, связанной с редактированием генома, поскольку порог для терапевтической модификации достаточно низкий, чтобы обеспечить высокую вероятность успеха в условиях современных технологий. Использование в качестве мишеней этих заболеваний в настоящее время привело к успеху в редактирующей терапии на уровне доклинических исследований и фазе I клинических испытаний. Манипуляции для улучшения репарации двухцепочечных разрывов и доставки нуклеазы необходимы, чтобы экстраполировать эти многообещающие результаты на заболевания, при которых у отредактированных клеток нет преимуществ, связанных с выживанием, или при которых для лечения необходим больший уровень продукта гена. Таблица внизу показывает некоторые примеры применения редактирования генома для терапевтических моделей, и ссылки в таблице ниже и документы, цитированные в этих ссылках, включены в настоящем описании, как если бы они были изложены полностью.
510, 235-240 (2014)
тирозинемия
[001131] Обращение к терапии каждого из состояний из таблицы с использованием системы CRISPR-Cas (например, C2c1 или C2c3) с помощью коррекции, опосредованной HDR, или вставки корректной последовательности гена, опосредованной HDR, преимущественно за счет системы доставки из настоящего изобретения, например, системы доставки частиц, входит в компетенцию квалифицированного специалиста на основании настоящего описания и знаний в данной области. Таким образом, один из возможных вариантов осуществления предусматривает взаимодействие HSC, несущих мутации, связанные с гемофилией В, тяжелым комбинированным иммунодефицитом или наследственной тирозинемией, с частицей, несущей гРНК и белок Cas (например, C2c1 или C2c3), направленными на геномный локус-мишень в случае гемофилии В, тяжелого комбинированного иммунодефицита (например, Х-сцепленного тяжелого комбинированного иммунодефицита, тяжелого комбинированного иммунодефицита, связанного с недостаточностью аденозиндеаминазы) или наследственной тирозинемии (например, как в случае Li, Genovese or Yin). Частица может также содержать подходящую матрицу HDR для коррекции мутации; или HSC могут взаимодействовать со второй частицей или вектором, содержащим или доставляющим матрицу направляемой гомологией репарации. В этом отношении упоминается, что гемофилия В является сцепленным с Х-хромосомой рецессивным заболеванием, вызванным связанными с потерей функции мутациями в гене, кодирующим фактор IX, ключевой компонент каскада системы свертывания крови. Восстановление активности фактора IX выше 1% от его уровня у тяжелобольных может трансформировать заболевание в его значительно более мягкую форму, поскольку профилактическое введение рекомбинантного фактора IX в организм таких пациентов с раннего возраста для достижения такого уровня активности в значительной степени облегчает клинические осложнения. Обладая знаниями в данной области и идеями, описанными в настоящей заявке, квалифицированный специалист может корректировать HSC как в случае гемофилии В с использованием системы CRISPR-Cas (например, C2c1 или C2c3), которая направленно корректирует мутацию (Х-сцепленное рецессивное заболевание, вызванное вызывающей потерю функции мутацией в гене, кодирующем фактор IX) (например, с помощью подходящей матрицы HDR, которая доставляет кодирующую последовательность фактора IX), в частности, гРНК может быть направлена на мутацию, вызывающую развитие гемофилии В, и HDR может обеспечивать кодирование правильной экспрессии фактора IX. Частица, содержащая гРНК, направленную на мутацию, и белок Cas (например, C2c1 или C2c3), взаимодействует с несущими мутацию HSC. Частица может также содержать подходящую матрицу HDR, чтобы скорректировать мутацию для правильной экспрессии фактора IX, или HSC может взаимодействовать со второй частицей, которая содержит или доставляет матрицу HDR. Провзаимодействовавшие таким образом клетки могут быть введены; и необязательно обработаны/увеличены в количестве (см. статью Cartier, обсуждаемую в настоящем описании).
[001132] В статье Cartier, "MINI-SYMPOSIUM: X-Linked Adrenoleukodystrophypa, Hematopoietic Stem. Cell Transplantation and Hematopoietic Stem Cell Gene Therapy in X-Linked Adrenoleukodystrophy," Brain Pathology 20 (2010) 857-862, включенной в настоящее описание в качестве ссылки в полном объеме с документами, которые в ней цитируются, утверждается, что аллогенная трансплантация гемопоэтических стволовых клеток (HSC) используется для доставки нормального лизосомального фермента в мозг пациента с болезнью Гурлер, и обсуждается генная терапия с помощью HSC для лечения адренолейкодистрофии. У двух пациентов периферические CD34+ клетки были собраны после мобилизации гранулоцитарно-макрофагальным колониестимулирующим фактором (G-CSF) и трансдуцированы энхансером вируса миелопролиферативной саркомы, участок негативной регуляции был удален, участок связывания праймера dl587rev был заменен в (MND)-ALD лентивирусном векторе. CD34+ клетки пациентов были трансдуцированы (MND)-ALD вектором в течение 16 ч в присутствии низкой концентрации цитокинов. Трансдуцированные CD34+ клетки были заморожены после трансдукции, чтобы провести на 5% клеток различные тесты на безопасность, которые включают, в частности, три анализа на репликационную компетентность лентивируса. Эффективность трансдукции CD34+ клеток варьировалась от 35% до 50% со средним значением числа интегрированных копий лентивируса между 0,65 и 0,7. После разморозки трансдуцированных CD34+ клеток пациентам было введено более 4.106 трансдуцированных CD34+ клеток/кг, после чего следовала миелоаблация с использованием бисульфана или циклофосфамида. HSC пациента были удалены, чтобы улучшить приживление HSC с корректированными генами. Гематологическое восстановление у двух пациентов происходило между 13 и 15 сутками. Почти полное иммунологическое восстановление происходило спустя 12 месяцев в случае первого пациента и спустя 9 месяцев в случае второго пациента. В отличие от использования лентивируса, квалифицированный специалист на основании знаний в данной области и идей, описанных в настоящем описании, может корректировать HSC при адренолейкодистрофии, используя систему CRISPR-Cas (например, C2c1 или C2c3), которая направленно корректирует мутацию (например, с помощью подходящей матрицы направляемой гомологией репарации); в частности, гРНК может быть направлена на мутации в гене ABCD1, расположенном на Х-хромосоме, которая кодирует ALD, лизосомальный мембранный белок-транспортер, и матрица HDR может обеспечить кодирование правильной экспрессии белка. Частица, содержащая гРНК, направленную на мутацию, и белок Cas (например, C2c1 или C2c3), взаимодействует с несущими мутацию HSC, например, с CD34+ клетками, несущими мутацию, согласно Cartier. Частица может также содержать подходящую матрицу HDR, чтобы скорректировать мутацию для правильной экспрессии лизосомального мембранного белка-транспортера, или HSC может взаимодействовать со второй частицей или вектором, который или которая содержит или доставляет матрицу HDR. Провзаимодействовавшие таким образом клетки могут быть обработаны, согласно Cartier. Провзаимодействовавшие таким образом клетки могут быть введены, согласно Cartier.
[001133] Упомянут патент WO 2015/148860, и за счет идей, описанных в настоящем описании, изобретение относится к способам и материалам этого документа, применимым в сочетании с идеями, описанными в настоящем описании. В аспекте генной терапии заболевания крови, методы и композиции для лечения β-талассемии могут быть адаптированы для CRISPR-Cas систем по настоящему изобретению (см, например, WO 2015/148860). В одном из возможных вариантов осуществления WO 2015/148860 включает лечение или профилактику β-талассемии или ее симптомов, например, посредством изменения гена В-клеточного CLL/лимфомы 11А (BCL11A). Ген BCL11A также известен как ген В-клеточного CLL/лимфомы 11А, BCLI1A-L, BCLI1A-S, BCL11AXL, CTIP1, HBFQTL5 и ZNF. BCLI1A кодирует белок с мотивом "цинковые пальцы", который вовлечен в регуляцию экспрессии генов глобинов. Изменяя ген BCL11A (например, один или оба аллеля гена BCL11A), можно увеличить уровень γ-глобина. γ-глобин может заменить β-глобин в комплексе гемоглобина и эффективно доставлять кислород в ткани таким образом, улучшая фенотип, характерный для β-талассемии.
[001134] Также упоминается патент WO 2015/148863, и за счет идей, описанных в настоящем описании, предусматриваются способы и материалы этого документа, которые могут быть адаптированы для систем CRISPR-Cas по настоящему изобретению. В одном из возможных аспектов лечения и предотвращения серповидноклеточной анемии, которая является наследственным заболеванием крови, WO 2015/148863 предусматривает изменение гена BCL11A. Путем изменения гена BCL11A (например, одного или обоих аллелей гена BCLI 1A), можно увеличить уровень γ-глобина. γ-глобин может заменить β-глобин в комплексе гемоглобина и эффективно доставлять кислород в ткани, таким образом, улучшая фенотип, характерный для серповидноклеточной анемии.
[001135] В одном из возможных аспектов изобретения предусматриваются способы и композиции, включающие редактирование последовательности нуклеиновой кислоты-мишени или модулирование экспрессии последовательности нуклеиновой кислоты-мишени, и их применение в связи с иммунотерапией онкологических заболеваний путем адаптации системы CRISPR-Cas по настоящему изобретению. Приводится ссылка на применение генной терапии в WO 2015/161276, которая включает способы и композиции, которые могут быть использованы для воздействия на Т-клеточную пролиферацию, выживаемость и/или функции путем изменения одного или более генов, экспрессируемых в Т-клетках, например, генов CBLB и/или PTPN6, генов FAS и/или BID, CTLA4 и/или PDCDI и/или TRAC и/или TRBC.
[001136] Т-клетки с химерным антигенным рецептором 19 (CAR19) демонстрируют противолейкозные эффекты у пациентов со злокачественными опухолями. Однако пациенты с лейкозами часто не имеют достаточно T-клеток для сбора, что означает, что лечение должно вовлекать модифицированные T-клетки доноров. Соответственно, представляет интерес создание банка донорских Т-клеток. В статье Qasim et al. ("First Clinical Application of Talen Engineered Universal CAR19 T Cells in B-ALL" ASH 57th Annual Meeting and Exposition, Dec. 5-8, 2015, Abstract 2046 (https://ash.confex.com/ash/2015/webprogram/Paper81653.html published online November 2015) обсуждается модификация CAR19 Т-клеток для предотвращения риска реакции "трансплантат против хозяина" посредством нарушения экспрессии Т-клеточного рецептора и нацеливания на CD52. Более того, было осуществлено нацеливание на клетки CD52 так, что они становились нечувствительными к алемтузумабу, и это позволяло алемтузумабу предотвращать реакцию отторжения хозяйским организмом CAR19 клеток с несовпадением человеческого лейкоцитарного антигена (HLA). Исследователи использовали самоинактивирующийся лентивирусный вектор 3-го поколения, кодирующий 4g7 CAR19 (CD19 scFv-4-1BB-CD3ζ), связанный с RQR8, затем вводили клетки посредством электропорации две пары мРНК TALEN для множественного нацеливания на оба локуса: локус α-константной цепи Т-клеточных рецепторов (TCR) и локус гена CD52. Совокупность клеток, которые продолжали экспрессировать TCR после увеличения в количестве ex vivo, истощали с помощью методики истощения α/β TCR CliniMacs, и получали продукт, состоящий из Т-клеток (UCART19) с экспрессией TCR <1% TCR, 85% которых экспрессировали CAR19 и 64% становились CD52-негативными. Модифицированные CAR19 Т-клетки вводили для лечения пациента с рецидивом острого лимфобластного лейкоза. Идеи, описанные в настоящем описании, обеспечивают эффективные способы предоставления модифицированных HSC и их потомков, включая, но не ограничиваясь ими, клетки миелоидной и лимфоидной линий клеток крови, включая Т-клетки, В-клетки, моноциты, макрофаги, нейтрофилы, базофилы, эозинофилы, эритроциты, дендритные клетки и мегакариоциты или тромбоциты, или натуральные киллеры и их предшественники и производные клетки. Такие клетки могут быть модифицированы путем нокина, нокаута или других модулирующих агентов, например, для удаления или модулирования CD52, описанного выше, или других мишеней, таких как, но не ограничиваясь ими, CXCR4 и PD-1. Таким образом, композиции, клетки и способы по изобретению могут быть использованы для модулирования иммунных ответов и для лечения, но не ограничиваясь этим, злокачественных заболеваний, вирусных инфекций и иммунных заболеваний, в сочетании с модификацией введения Т-клеток и других клеток пациенту.
[001137] Упоминается патент WO 2015/148670, и за счет идей, описанных в настоящем описании, изобретение предусматривает способы и материалы этого документа, применимые в сочетании с идеями, изложенными в настоящем описании. В аспекте генной терапии предусматриваются способы и композиции для редактирования последовательности-мишени, родственной или связанной с вирусом иммунодефицита человека (ВИЧ) и синдромом приобретенного иммунодефицита (СПИД). В связанном аспекте описываемое в настоящем описании изобретение предусматривает предотвращение и лечение ВИЧ-инфекции и СПИДа путем внесения одной или более мутаций в ген С-С хемокинового рецептора типа 5 (CCR5). CCR5 также известен как CKR5, CCR-5, CD195, CKR-5, CCCKR5, CMKBR5, 1DDM22 и CC-CKR-5. В следующем аспекте описываемое в настоящем описании изобретение предусматривает предотвращение или снижение ВИЧ-инфекции или снижение способности ВИЧ попадать в клетку-хозяина, например, у уже инфицированных индивидуумов. Приводимые в качестве примера клетки-хозяева ВИЧ включают, но не ограничиваются ими, CD4 клетки, T клетки, клетки лимфоидной ткани, ассоциированной с кишечником (GALT), макрофаги, дендритные клетки, клетки миелоидных предшественников и микроглию. Попадание вируса в клетку требует взаимодействия вирусных гликопротеинов gp41 и gpl20 одновременно с CD4 рецептором и корецептором, например, CCR5. Если корецептор, например CCR5, не присутствует на поверхности клеток-хозяев, вирус не может связываться и попадать в хозяйские клетки. Развитие заболевания таким образом затормаживается. Путем нокаута или нокдауна CCR5 в клетках-хозяевах, например, путем введения защитной мутации (такой как Δ32 мутация CCR5), предотвращается попадание ВИЧ в хозяйские клетки.
[001138] Х-сцепленная хроническая гранулематозная болезнь представляет собой наследственное заболевание защитной системы хозяина за счет отсутствия или снижения активности фагоцитарной NADPH-оксидазы. Предусматривается использование системы CR1SPR-Cas (C2c1 или C2c3), которая направленно корректирует мутацию (отсутствующую или сниженную активность фагоцитарной NADPH-оксидазы) (например, с помощью HDR-матрицы, которая доставляет кодирующую последовательность для фагоцитарной NADPH-оксидазы); в частности, гРНК может быть направлена на мутацию, которая дает начало Х-сцепленной гранулематозной болезни (недостаточность фагоцитарной NADPH-оксидазы), и HDR-репарация может обеспечить кодирование для правильной экспрессии фагоцитарной NADPH-оксидазы. Частицу, содержащая гРНК, направленную на мутацию, и белок Cas (например, C2c1 или C2c3), приводят в контакт с несущими мутацию HSC. Частица может также содержать подходящую HDR-матрицу, чтобы скорректировать мутацию для правильной экспрессии фагоцитарной NADPH-оксидазы, или HSC может взаимодействовать со второй частицей или вектором, которые содержат или доставляет HDR-матрицу. Провзаимодействовавшие таким образом клетки могут быть введены; и необязательно обработаны/увеличены в количестве (см. Cartier).
[001139] Анемия Фанкони: мутации по меньшей мере 15 генов (FANCA, FANCB, FANCC, FANCD1/BRCA2, FANCD2, FANCE, FANCF, FANCG, FANCI, FANCJ/BACTI1/BRIP1, FANCL/PHF9/POG, FANCM, FANCN/PALB2, FANCO/RadSlC и FANCP/SLX4/BTBD12) могут вызывать анемию Фанкони. Белки, образуемые этими генами, вовлечены в клеточный процесс, известный как сигнальный путь анемии Фанкони. Сигнальный путь анемии Фанкони включается (активируется), когда процесс создания новых копий ДНК, называемый репликацией ДНК, блокируется за счет повреждений ДНК. Сигнальный путь анемии Фанкони отправляет определенные белки к поврежденному участку, запуская репарацию ДНК, так что репликация ДНК может продолжаться. Сигнальный путь анемии Фанкони особенно чувствителен к конкретным типам повреждений ДНК, известным как межцепочечные сшивки. Межцепочечные сшивки происходят, когда два строительных блока ДНК (нуклеотида) на противоположных цепях ДНК аномально прикреплены друг к другу или соединены вместе, что останавливает процесс репликации ДНК. Межцепочечные сшивки могут быть вызваны встраиванием токсичных соединений, образуемых в организме, или при лечении некоторыми противоопухолевыми соединениями. Восемь белков, ассоциированных с анемией Фанкони, группируются вместе, образуя комплекс, известный как коровый FA-комплекс. Коровый FA-комплекс активирует два белка, называемых FANCD2 и FANCF. Активация этих двух белков привлекает белки репарации ДНК к участку межцепочечной сшивки, так что сшивка может быть удалена и репликация ДНК сможет продолжаться. В частности, коровый FA-комплекс является ядерным мультипротеиновым комплексом, состоящим из FANCA, FANCB, FANCC, FANCE, FANCF, FANCG, FANCL и FANCM, функционирующим как убиквитинлигаза и опосредующим активацию ID-комплекса, который является гетеродимером, состоящим из FANCD2 и FANCI. Будучи моноубиквитинилированным, он взаимодействует с классическими опухолевыми супрессорами ниже по сигнальному пути анемии Фанкони, включая взаимодействия FANCD1/BRCA2, FANCN/PALB2, FANCJ/BRIP1 и FANCO/Rad51C, и таким образом, вносит вклад в репарацию ДНК за счет гомологичной рекомбинации. 80-90% случаев анемии Фанкони происходят за счет мутации от одного до трех генов: FANCA, FANCC и FANCG. Эти гены предоставляют инструкции для образования компонентов корового FA-комплекса. Мутации в таких генах, ассоциированных с коровым FA-комплексом, сделают комплекс нефункциональным и нарушат весь сигнальный путь анемии Фанкони. В результате повреждение ДНК не будет репарироваться эффективно, и межцепочечные сшивки будут со временем накапливаться. В статье Geiselhart, "Review Article, Disrupted Signaling through the Fanconi Anemia Pathway Leads to Dysfunctional Hematopoietic Stem Cell Biology: Underlying Mechanisms and Potential Therapeutic Strategies," Anemia Volume 2012 (2012), Article ID 265790, http://dx.doi.org/10. 1155/2012/265790 обсуждается анемия Фанкони и эксперименты на животных, включающие интрафеморальную инъекцию лентивируса, кодирующего ген FANCC, что приводило к коррекции HSC in vivo. Предусматривается использование системы CRISPR-Cas (C2c1 или C2c3), которая направлена на одну или более мутаций, ассоциированных с анемией Фанкони, например, системы CRISPR-Cas (C2c1 или C2c3), содержащей гРНК или HDR-матрицу(ы), которые соответственно направлены на одну или более мутаций FANCA, FANCC или FANCG, которые вызывают анемию Фанкони, и обеспечивает корректную экспрессию одного или более FANCA, FANCC или FANCG; например, гРНК может быть направлена на мутацию в FANCC, и HDR-репарация (направляемая гомологией репарация) может обеспечивать кодирование правильной экспрессии FANCC. Частицу, содержащую гРНК, направленную на мутацию(и) (например, одну или более, вовлеченные в развитие анемии Фанкони, такие как мутации(я) одного или более из FANCA, FANCC или FANCG) и белок Cas (C2c1 или C2c3) приводят в контакт с HSC, несущими мутацию(и). Частица может также содержать подходящую HDR-матрицу, чтобы скорректировать мутацию для правильной экспрессии одного или более белков, вовлеченных в развитие анемии Фанкони, или HSC может взаимодействовать со второй частицей или вектором, которые содержат или доставляют HDR-матрицу. Провзаимодействовавшие таким образом клетки могут быть введены и необязательно обработаны/увеличены в количестве (см. Cartier).
[001140] Упоминаемая в настоящем описании частица (например, содержащая гРНК и Cas (например, C2c1 или C2c3), необязательно HDR-матрицу(ы); например для заболеваний, таких как гемофилия В, тяжелый комбинированный иммунодефицит (SCID), Х-сцепленный тяжелый комбинированный иммунодефицит, тяжелый комбинированный иммунодефицит, связанный с недостаточностью аденозиндеаминазы, наследственная тирозинемия, β-талассемия, Х-сцепленная гранулематозная болезнь, синдром Вискотта-Олдрича, анемия Фанкони, адренолейкодистрофия, метахроматическая лейкодистрофия, ВИЧ-инфекция/СПИД, синдром иммунодефицита, гематологические заболевания или генетическая лизосомная болезнь накопления) преимущественно получают или могут получить при добавлении смеси гРНК и белка Cas (C2c1 или C2c3) (необязательно содержащей HDR-матрицу(ы) или такую смесь, содержащую только HDR-матрицу(ы), если желательна отдельная частица для матриц(ы)) к смеси, содержащей или по существу состоящей или состоящей из поверхностно активных веществ, фосфолипидов, биодеградируемых липопротеина и спирта (причем одна или более гРНК нацелены на геномный(е) локус(ы) в нейрональных стволовых клетках).
[001141] Действительно, изобретение особенно подходит для лечения генетических гемопоэтических заболеваний путем геномного редактирования и синдромов иммунодефицита, таких как генетические синдромы иммунодефицита, особенно за счет использования технологий частиц, описанных в настоящем описании. Генетические иммунодефициты являются заболеваниями, при которых могут быть успешными вмешательства путем редактирования генома, согласно настоящему изобретению. Причины включают: гемопоэтические клетки, подмножеством которых являются иммунные клетки, являются терапевтически доступными. Они могут быть удалены из организма и трансплантированы аутологично или аллогенно. Далее, конкретные генетические иммунодефициты, например, тяжелый комбинированный иммунодефицит, пролиферативно невыгодны для иммунных клеток. Коррекция генетических повреждений, вызывающих тяжелый комбинированный иммунодефицит, путем редких спонтанных "обратных" мутаций указывает на то, что коррекция даже одного предшественника лимфоцита может быть достаточной для восстановления функций иммунитета у пациентов (см. Bousso, P., et al. Diversity, functionality, and stability of the T cell repertoire derived in vivo from a single human T cell precursor. Proceedings of the- National Academy of Sciences of the United States of America 97, 274-278 (2000)). Селективное преимущество отредактированных клеток обеспечивает то, что даже низкий уровень редактирования приводит к терапевтическому эффекту. Этот эффект настоящего изобретения может быть виден при тяжелом комбинированном иммунодефиците, синдроме Вискотта-Олдрича и других упомянутых здесь состояниях, включая другие генетические болезни гемопоэтической системы, такие как α- и β-талассемия, при которых недостаточность гемоглобина отрицательно влияет на выживаемость предшественников эритроцитов.
[001142] Активность репарации NHEJ и HDR значительно варьируется в зависимости от типа клетки и состояния клетки. NHEJ-репарация не регулируется в значительной степени в зависимости от клеточного цикла и эффективна в различных типах клеток, что обеспечивает высокий уровень нарушения генов в доступных популяциях клеток-мишеней. Напротив, HDR-репарация происходит в основном во время S/G2-фазы и, следовательно, ограничивается клетками, которые активно делятся, что ограничивает мишени лечения, требующие точных геномных модификаций, митотическими клетками [Ciccia, A. & Elledge, S.J. Molecular cell 40, 179-204 (2010); Chapman, J.R., et al. Molecular cell 47, 497-510 (2012)].
[001143] Эффективность коррекции при HDR-репарации может регулироваться эпигенетическим состоянием или последовательностью локуса-мишени или конфигурацией используемой специфичной матрицы репарации (одно- или двухцепочечной, с длинными или короткими гомологичными плечами). [Hacein-Bey-Abina, S., et al. The New England journal of medicine 346, 1185-1193 (2002); Gaspar, H.B., et al. Lancet 364, 2181-2187 (2004); Beumer, K.J., et al. G3 (2013)]. Относительная активность машинерии NHEJ-репарации и HDR-репарации в клетках-мишенях может также влиять на на эффективность коррекции гена, поскольку эти пути могут конкурировать за репарацию двухцепочечных разрывов Beumer, K.J., et al. Proceedings of the National Academy of Sciences of the United Slates of America 105, 19821-19826 (2008)], HDR-репарация также налагает ограничения на доставку, которых нет в случае стратегий NHEJ-репарации, поскольку она требует одновременной доставки нуклеаз и матриц репарации. При практическом применении эти ограничения до сих пор приводили к низкому уровню HDR-репарации в терапевтически значимых типах клеток. Клиническое применение поэтому во многом сфокусировано на стратегиях NHEJ-репарации для лечения заболеваний, хотя доклинические исследования по проверке концепции HDR-репарации сейчас описаны для модели гемофилии В на мышах и наследственной тирозинемии [Li, H., et al. Nature 475, 217-221 (2011); Yin, H, et al. Nature biotechnology 32, 551-553 (2014)].
[001144] Любое данное применение геномного редактирования может включать комбинации белков, молекул малых РНК и/или матриц репарации, что делает доставку множества этих частей значительно более сложной, чем терапия низкомолекулярными соединениями. Разработаны две основные стратегии доставки инструментов редактирования генома ex vivo и in vivo. При обработках ex vivo нарушенные клетки, ассоциированные с заболеванием, удаляются из организма, редактируются и трансплантируются обратно пациенту. Редактирование ex vivo обладает тем преимуществом, что позволяет четко определить популяцию клеток-мишеней и определить конкретную дозировку терапевтических молекул, доставляемых в клетки. Последнее соображение может быть особенно важным, когда есть угроза нецелевых модификаций, поскольку титрование количества нуклеазы может снизить уровень таких мутаций (Hsu et al., 2013). Другим преимуществом подходов ex vivo является обычно высокий уровень редактирования, который может быть достигнут за счет разработки эффективных систем доставки белков и нуклеиновых кислот в клетки в культуре в рамках исследований и применения в генной терапии.
[001145] У подходов ex vivo могут быть недостатки, которые ограничивают применение до малого числа заболеваний. Например, клетки-мишени должны быть способны выжить во время манипуляций вне организма. В случае многих тканей, таких как ткани мозга, культивирование клеток вне организма является главной проблемой, поскольку клетки или не выживают, или теряют свойства, необходимые для их функций in vivo. Таким образом, в свете настоящего изобретения и знаний в данной области возможна терапия ex vivo в случае тканей с популяциями стволовых клеток взрослых, таких как гемопоэтическая система, поддающихся культивированию и манипуляциям ex vivo посредством системы CRISPR-Cas (C2c1 или C2c3). [Bunn, H.F. & Aster, J. Pathophysiology Of Blood Disorders, (McGraw-Hill, New York, 2011)].
[001146] Геномное редактирование in vivo включает прямую доставку редактирующих систем в типы клеток в их ткани. Редактирование in vivo позволяет лечить заболевания, при которых популяция пораженных клеток не поддается манипуляциям ex vivo. Более того, доставка in situ нуклеаз в клетки позволяет лечить сразу множество тканей и клетки множества типов. Эти свойства, вероятно, позволяют применять обработки in vivo к более широкому спектру заболеваний, чем подходы к терапии ex vivo.
[001147] На сегодняшний день, редактирование in vivo было во многом достигнуто благодаря использованию вирусных векторов с определенным тканеспецифическим тропизмом. Такие векторы в настоящий момент ограничены с точки зрения размера переносимого груза и тропизма, ограничивая этот тип терапии системами органов, в которых трансдукция клинически эффективными векторами является эффективной, такими как печень, мышцы и глаза [Kotterman, M.A. & Schaffer, D.V. Nature reviews. Genetics 15, 445-451 (2014); Nguyen, T.H. & Ferry, N. Gene therapy 11 Suppl 1, S76-84 (2004), Boye, S.E., et al. Molecular therapy: the journal of the American Society of Gene Therapy 21, 509-519 (2013)].
[001148] Потенциальным барьером для доставки in vivo является иммунный ответ, который может спровоцироваться в ответ на высокое количество вируса, необходимо для лечения, но этот феномен не является уникальным для геномного редактирования и часто наблюдается при других способах генной терапии на основе вирусов [Bessis, N., et al. Gene therapy 11 Suppl 1, S10-17 (2004)]. Также возможно, что пептиды из редактирующих нуклеаз сами презентируются на молекулах MHC класса I, стимулируя иммунный ответ, хотя в доклинических исследованиях мало подтверждений этому. Другой значительной проблемой в связи с этим способом терапии является контроль распространения и впоследствии дозировки редактирующих геном нуклеаз in vivo, что приводит к нецелевым профилям мутаций, которые может быть сложно предсказать. Однако с учетом настоящего изобретения и знаний в данной области, использование способов терапии на базе вирусов и частиц для использования в лечении онкологических заболеваний, модифицикации HSC in vivo, например, с помощью доставки частицей или вирусом, входит в компетенцию квалифицированного специалиста.
[001149] Редактирующая терапия ex vivo: долгосрочные клинические исследования по очистке, культивированию и трасплантации гематопоээтических клеток позволили заболеваниям крови, таким как СПИД, анемия Фанкони, синдром Вискотта-Олдрича и серповидноклеточной анемии, оказаться в фокусе редактирующей терапии ex vivo. Другая причина сфокусироваться на HSC - это существование относительно высокоэффективных систем доставки, благодаря предшествовавшим попыткам разработать генную терапию заболеваний крови. С учетом этих преимуществ, данный способ терапии может применяться для заболеваний, при которых отредактированные клетки обладают преимуществом, связанным с выживанием, поэтому малое число привитых отредактированных клеток может размножаться и лечить заболевание. Одно такое заболевание представляет собой ВИЧ, при котором инфекция приводит к невыгодности выживания CD4+ T-клеток.
[001150] Область использования основанной на ex vivo редактировании терапии недавно расширилась и включила в себя стратегии коррекции генов. Препятствующие HDR ex vivo факторы были преодолены в недавней статье Genovese и соавторов, которым удалось выполнить генную коррекцию мутированного гена IL2RG в гемопоэтических стволовых клетках (HSC), полученных от пациента, страдающего X-сцепленным тяжелым комбинированным иммунодефицитом (SCID-X1) [Genovese, P., et al. Nature 510, 235240 (2014)]. Genovese et. al успешно осуществили генную коррекцию в HSC с использованием мультимодальной стратегии. Сначала проводилась трансдукция HSC с применением лентивируса со сниженной интеграцией, содержащего матрицу HDR, кодирующую терапевтическую кДНК для IL2RG. После трансдукции проводилась электропорация клеток с мРНК, кодирующей ZFN, нацеленную на "горячую точку" мутаций в IL2RG для того, чтобы стимулировать основанную на HDR генную коррекцию. С целью увеличения скорости HDR используются оптимизированные условия культивирования, стимулирующие деление HSC. При оптимизированных условиях культивирования нуклеазы и матрицы HDR, несущие корректированные гены HSC от пациента с SCID-X1, были получены в культуре со скоростью, имеющей терапевтическую значимость. HSC от имеющих заболевание индивидов, к которым была применена та же самая процедура генной коррекции, могут поддерживать долговременный гемопоэз в мышах, являющихся золотым стандартом функций HSC, HSC способны производить гемопоэтические клетки всех типов и могут быть аутологично трансплантированы, что делает их исключительно ценной популяцией клеток для использования при всех гемопоэтических генетических заболеваний [Weissman, I.L. & Shizuru, J.A, Blood 112, 3543-3553 (2008)]. Прошедшие генную коррекцию в HSC могут, в принципе, быть использованы для лечения широкого круга генетических заболеваний крови, что делает настоящее исследование воодушевляющим прорывом в терапевтическом редактировании генома.
[001151] Терапия на основе редактирования in vivo: редактирование in vivo может быть успешно применено, как следует из настоящей Заявки и представлений данной области. В случае систем органов, для которых доставка осуществляется успешно, к настоящему времени достигнут ряд многообещающих успехов доклинических испытаний. Первый пример успешной терапии на основе редактирования in vivo был показан для модели гемофилии В на мышах [Li, H., et al. Nature 475, 217-221 (2011)]. Как отмечалось раннее, гемофилия В является X-сцепленным рецессивным заболеванием, вызванным мутацией типа "потеря функции" в гене, кодирующем фактор IX, являющийся ключевым компонентом каскада свертывания крови. Возвращение уровня активности Фактора IX до значений свыше 1% от уровня такового у индивидов с тяжелой патологией может перевести болезнь в значительно более мягкую форму, введение рекомбинантного Фактора IX таким пациентам в качестве профилактики начиная с раннего возраста для того, чтобы достигнуть таких уровней в значительной степени снижает выраженность клинических осложнений [Lofqvist, T. et al. Journal of internal medicine 241, 395-400 (1997)]. Таким образом, только низкие уровни коррекции гена HDR необходимы для изменения клинических исходов пациентов. Помимо этого, фактор IX синтезируется и секретируется печенью, органом, который может быть эффективно трансдуцирован посредством вирусных векторов, кодирующих системы редактирования.
[001152] С использованием гепатотропных аденоассоциированных вирусных (AAV) серотипов, кодирующих ZFN и корректирующую матрицу HDR, возможно добиться генной коррекции мутированного гуманизированного гена Факторв IX в печени мыши достигающей 7% [Li, H., et al. Nature 475, 217-221 (2011)]. Результатом этого стало улучшение кинетики тромбообразования, являющейся мерой функции каскада свертывания крови, впервые продемонстрировавшей, что терапия на основе редактирования in vivo не только возможна, но также эффективна. Как обсуждается в настоящем описании, квалифицированный специалист должен руководствоваться изложенными в настоящей спецификации идеями и знаниями в данной области, например, работами Li, для решения лечения гемофилии В с помощью содержащей частицу матрицы HDR и системы CRISPR-Cas (C2c1 или C2c3), которая нацелена на мутацию X-сцепленного рецессивного заболевания для того, чтобы обратить вспять мутацию с потерей функции.
[001153] Опираясь на данное исследование, другие группы ученых недавно использовали редактирование генома in vivo в печени с помощью CRlSPR-Cas для успешного лечения наследственной тирозинемии в модели на мышах и создания мутаций, обеспечивающих защиту против сердечно-сосудистых заболеваний. Эти два различных применения показывают универсальность данного подхода в случае заболеваний, включающих дисфункцию печени. [Yin, H., et al. Nature biotechnology, 32, 551-553 (2014); Ding, Q., et al. Circulation research 115, 488-492 (2014)]. Применение редактирования in vivo к другим системам органов необходимо для обоснования того, что данная стратегия является широко применимой. В настоящее время предпринимаются усилия по оптимизации как вирусных и невирусных векторов для того, чтобы расширить круг заболеваний, терапию которых возможно проводить с помощью такого подхода [Kotterman, M.A. & Schaffer, D.V. Nature reviews. Genetics 15, 445-451 (2014); Yin, H., et al. Nature reviews. Genetics 15, 541-555 (2014)]. Согласно настоящему описанию, квалифицированный специалист должен использовать приведенные здесь идеи и знания в данной области, например, работы Yin, для того чтобы применять при лечении наследственной тирозинемии содержащую частицы матрицу HDR и систему CRISPR-Cas (C2c1 или C2c3), нацеленную на эту мутацию.
[001154] Направленная делеция, терапевтические применения: может быть предпочтительной направленная делеция генов. Следовательно, предпочтительными являются гены, вовлеченные в иммунодефицитные состояния, гематологические расстройства или генетические лизосомные болезни накопления, например, гемофилию B, тяжелый комбинированный иммунодефицит (SCID), X-сцепленный тяжелый комбинированный иммунодефицит (SCID-X1), тяжелый комбинированный иммунодефицит с дефицитом аденозиндезаминазы (ADA-SCID), наследственную тирозинемию, β-талассемия, X-сцепленный хронический гранулематоз (CGD), синдром Вискотта-Олдрича, анемию Фанкони, адренолейкодистрофия (ALD), метахроматическую лейкодистрофию (MLD), ВИЧ/СПИД, другие метаболические заболевания, гены, кодирующие неправильно свернутые белки, вовлеченные в заболевания, гены, приводящие к потере функции, вовлеченные в заболевания; в целом, мутации, на которые может быть осуществлено нацеливание в HSC с применением любой описанной в настоящем описании системы доставки в системой на основе частиц, рассматриваются как предпочтительные.
[001155] В настоящем изобретении иммуногенность фермента CRISPR может быть снижена, в частности, с использованием подхода, впервые описанного Tangri et al. в отношении эритропоэтина и получившего дальнейшее развитие. В соответствии с этим направленная эволюция или рациональное конструирование могут быть использованы для снижения иммуногенности фермента CRISPR (например, C2c1 или C2c3) в организме хозяина (человека или других видов).
[001156] Редактирование генома: системы CRISPR/Cas (C2c1 или C2c3) по настоящему изобретению могут быть использованы для коррекции генетических мутаций, предыдущие попытки которой были преприняты ранее с ограниченным успехом при помощи TALEN, ZFN и лентивирусов, включая рассматриваемые в настоящем описании; см. также WO2013163628.
Лечение заболеваний головного мозга, центральной нервной и иммунной систем
[001157] Настоящее изобретение также предусматривает доставку систем CRISPR-Cas в головной мозг или нейроны. Например, РНК-интерференция (РНК-i) предоставляет возможности для терапии этих расстройств путем снижения экспрессии HTT, гена, вызывающего болезнь Хантингтона (см., например, McBride et al. Molecular Therapy vol. 19 no. 12 Dec. 2011, pp. 2152-2162), следовательно, заявители утверждают, что ее можно использовать вместе с и/или адаптировать к системе CRISPR-Cas. Система CRISPR-Cas может быть получена при помощи алгоритма снижения потенциала нецелевого действия антисмысловых последовательностей. Последовательности CRISPR-Cas могут быть нацелены на последовательность экзона 512 мыши, резус или хантингин человека и быть экспрессированы с помощью вирусного вектора, в частности, AAV. Животным, включая человека, могут быть произведены примерно три микроинъекции на одно полушарие (всего шесть инъекций): первая на 1 мм ростральнее передней комиссуры (12 мкл) и две остальные инъекции (12 и 10 мкл, соответственно), локализованные на 3 и 6 мм каудальнее первой инъекции, содержащие 1e12 мг/мл AAV со скоростью примерно 1 мкл/мин, причем игла остается на месте еще на 5 мин, чтобы позволить введенному раствору распространиться от кончика иглы.
[001158] DiFiglia et al. (PNAS, October 23, 2007, vol. 104, no. 43, 17204-17209) сделали наблюдение, что единичное введение в стриатум взрослого миРНК, нацеленных на HTT, может вызывать сайленсинг мутантного HTT, ослаблять патологию нейронов и отсрочить проявление патологического фенотипа поведения, наблюдаемого при быстро развивающейся форме полученной с помощью вируса трансгенной модели HD. В работе DiFiglia производилось введение мышам интрастриально 2 мкл Су3-меченной cc-миРНК-HTT или неконъюгированной миРНК-HTT в концентрации 10 мкМ. Сходная дозировка CRISPR Cas, нацеленного на HTT, может предусматриваться в рамках настоящего изобретения для человека, например, приблизительно 5-10 мл 10 мкМ CRISPR Cas, нацеленного на HTT, можно вводить интрастриально.
[001159] В другом случае Boudreau et al. (Molecular Therapy vol. 17 no. 6 June 2009) вводили 5 мкл рекомбинантного векторов AAV серотипа 2/1, экспрессирующих вирус с HTT-специфичной iРНК (в концентрации 4×1012 геномов вируса/мл) в стриатум. Сходная дозировка CRISPR Cas, нацеленного на HTT, может предусматриваться в рамках настоящего изобретения для человека, например, приблизительно 10-20 мл раствора CRISPR Cas, нацеленного на HTT, с концентрацией 4×1012 вирусных геномов на мл, могут вводиться интрастриально.
[001160] Другим примером является CRISPR Cas, который нацелен на HTT и вводится непрерывно (см., например, Yu et al., Cell 150, 895-908, August 31, 2012). Yu et al. использовали осмотические насосы, вливающие 0,25 мкл/ч (Model 2004) для доставки оц-миРНК или фосфатно-солевой буфер со скоростью 300 мг/сутки (PBS) (Sigma Aldrich) на протяжении 28 суток, и насосы, сконструированные для доставки 0,5 мкл/час (Model 2002), использовались для вливания 75 мг/день положительного контроля MOE ASO на протяжении 14 суток. Насосы (Durect Corporation) заполняли оц-миРНК или MOE, растворенными в стерильном PBS, и далее инкубировали при 37°C на протяжении 24 или 48 часов (Model 2004) перед имплантацией. В качестве анестезии мышам вводился 2,5% изофлуоран, срединный разрез производился в основании черепа. Стереотаксически направляемая имплантация канюли служила для ее введения в правый латеральный желудочек, далее она герметично закреплялась клеем Loctite. Катетер, присоединенный к осмотической мини-помпе, соединялся с канюлей; помпа помещалась подкожно в области средней лопатки. На место разреза накладывали нейлоновые швы 5,0. Сходная дозировка CRISPR Cas, нацеленного на HTT, может предусматриваться в рамках настоящего изобретения для человека, например, возможно введение приблизительно от 500 до 1000 г CRISPR Cas, нацеленного на HTT, ежедневно.
[001161] В другом примере непрерывного введения, Stiles et al. (Experimental Neurology 233 (2012) 463-471) имплантировали внутрь паренхимы катетер с титановым кончиком иглы в правый путамен. Катетер соединялся с насосом SynchroMed® II Pump (Medtronic Neurological, Миннеаполис, Миннесота), имплантированным подкожно в брюшную полость. После 7 суток вливания фосфатного солевого буфера со скоростью 6 мкл/суток, насосы повторно наполняли тестируемым образцом и программировали на непрерывное вливание на протяжении 7 дней. Примерно от 2,3 до 11,52 мг миРНК в день вливались с изменяющейся скоростью введения, равной примерно от 0,1 до 0,5 мкл/мин. Сходная дозировка CRISPR Cas, нацеленного на HTT, может предусматриваться в рамках настоящего изобретения для человека, например, возможно введение приблизительно от 20 до 200 мг CRISPR Cas, нацеленного на HTT. В другом примере способы патентной публикации США № 20130253040, зарегистрированной Sangamo, могут также быть адаптированы на основе TALES для нацеленной на нуклеиновые кислоты системы по настоящему изобретению с целью лечения болезни Хантингтона.
[001162] Другим примером служат способы патентной публикации США № 20130253040 (WO2013130824), зарегистрированной Sangamo, которые также могут быть адаптированы с TALES к системе CRISPR Cas по настоящему изобретению с целью лечения болезни Хантингтона.
[001163] В WO2015089354 A1, зарегистрированной Broad Institute et al., включенной в настоящее описание в качестве ссылки, описаны мишени для болезни Хантингтона (HD). Возможные гены-мишени комплекса CRISPR применительно к болезни Хантингтона: PRKCE; IGF1; EP300; RCOR1; PRKCZ; HDAC4 и TGM2. В соответствии с этим в некоторых вариантах осуществления настоящего изобретения один или более из списка PRKCE; IGF1; EP300; RCOR1; PRKCZ; HDAC4 и TGM2 могут быть выбраны в качестве мишеней для болезни Хантингтона.
[001164] Другие заболевания связанны с тринуклеотидными повторами. Таковые могут включать любые из следующих: категория I включает болезнь Хантингтона (HD) и спиномозжечковые атаксии; экспансии категории II отличаются фенотипически от гетерогенных экспансий, которые обычно имеют малую протяженность, но также находятся в экзонах генов; категория III включает синдром хрупкой X-хромосомы, миотоническую дистрофию, две спиномозжечковые атаксии, ювенильную миоклоническую эпилепсию и атаксию Фридрейха.
[001165] Следующий аспект изобретения относится к использованию системы CRISPR-Cas для коррекции дефектов в генах EMP2A и EMP2B, которые были идентифицированы как ассоциированные с болезнью Лафоры. Болезнь Лафоры является аутосомно-рецессивным заболеванием, которое характеризуется прогрессивным миоклональной эпилепсией, которая может начинаться с эпилептических припадков в подростковом возрасте. Редкие случаи данного заболевания могут быть вызваны мутациями в не идентифицированных к настоящему времени генах. Данное заболевание вызывает припадки, спазмы мышц, нарушения ходьбы, деменцию и, в конечном итоге, смерть. В настоящее время нет способов лечения, продемонстрировавших эффективность в отношении прогрессирования заболевания. Cистема CRISPR-Cas также может быть нацелена на другие генетические аномалии, ассоциированные с эпилепсией, и лежащие в основе генетические механизмы описаны в Genetics of Epilepsy and Genetic Epilepsies, edited by Giuliano Avanzini, Jeffrey L. Noebels, Mariani Foundation Paediatric Neurology: 20; 2009).
[001166] Способы патентной публикации США № 20110158957, зарегистрированной Sangamo BioSciences, Inc., вовлеченные в инактивации генов Т-клеточных рецепторов (TCR), могут также быть модифицированы для системы CRISPR-Cas по настоящему изобретению. В другом примере способы патентной публикации США № 20100311124, зарегистрированной Sangamo BioSciences, Inc., и способы патентной публикации США № 20110225664, зарегистрированной Cellectis, которые в обоих случаях вовлечены в инактивацию экспрессии гена глутаминсинтетазы, могут также быть модифицированы для системы CRISPR-Cas по настоящему изобретению.
[001167] Выбор способов доставки в головной мозг включает инкапсуляцию фермента CRISPR и направляющей РНК в форме ДНК или РНК в липосомы и их конъюгацию с молекулярными "троянскими конями" для доставки через гемато-энцефалический барьер (ГЭБ). Молекулярные "троянские кони" продемонстрировали эффективность для доставки векторов экспрессии B-gal в головной мозг приматов, отличных от человека. Тот же подход может быть использован для доставки при помощи векторов, содержащих фермент CRISPR и направляющую РНК. Например, Xia CF and Boado RJ, Pardridge WM ("Antibody-mediated targeting of siRNA via the human insulin receptor using avidin-biotin technology." Mol Pharm. 2009 May- Jun;6(3):747-51. doi: 10.1021/mp800194) описывает, каким образом доставка малых интерферирующих РНК (миРНК) в клетки культуры и in vivo может быть возможна при комбинированном применении рецептор-специфичных моноклональных антител (mAb) и технологии авидина и биотина. Авторы также сообщили о том, что в силу того, что связь между нацеливающим mAb и миРНК является стабильной при использовании технологии авидина-биотина, эффекты РНК-i в отдаленных участках, в частности, в головном мозге, наблюдаются in vivo после внутривенного введения нацеленной миРНК.
[001168] Zhang et al. (Mol Ther. 2003 Jan;7(l):11-8.)) описывают плазмиды экспрессии, кодирующие репортерные конструкции, такие как люцифераза, инкапсулированные внутри вируса, содержащего пегилированные иммунолипосомы размером 85 нм, которые нацелены на головной мозг макака-резус in vivo, посредством моноклональных антител (MAb) к рецептору инсулина человека (HIR). HIRMAb делает возможным перенос липосомой экзогенного гена для его последующего трансцитоза через гемато-энцефалический барьер и эндоцитоза через плазматическую мембрану нейрона после внутривенного введения. Уровень экспрессии гена люциферазы в головном мозге был в 50 раз выше в случае макака-резус в сравнении с мышью. Протекание нейронной экспрессии гена бета-галактозидазы в значительной части головного мозга было продемонстрировано при помощи гистохимии и конфокальной микроскопии. Авторы указывают, что данный подход делает возможным обратимый трансгенез у взрослых на протяжении 24 часов. В соответствии с этим использование иммунолипосом является предпочтительным. Они могут быть использованы в сочетании с антителами для нацеливания на конкретные ткани или белки клеточной поверхности.
[001169] В патентной публикации США № 20110023153 описывано использование нуклеаз типа цинковых пальцев для генетической модификации клеток, животных и белков, ассоциированных с болезнью Альцгеймера (AD). После модификации клетки и животные могут в дальнейшем быть протестированы с применением известных способов для исследования эффектов направленных мутаций на развитие и/или прогрессирование AD с применением широко используемых при изучении AD показателей, таких как, но не ограничиваясь ими, обучение и память, тревожность, депрессия, зависимость, чувствительные и моторные функции, а также способы анализа, оценивающие поведенческие, функциональные, патологические, метаболические и биохимические функции.
[001170] Настоящая заявка охватывает редактирование любых хромосомных последовательностей, кодирующих белки, ассоциированные с AD. Связанные с AD белки обычно выбирают на основе показанной в экспериментах ассоциации связанных с AD белков с заболеванием AD. Например, скорость образования или циркулирующая концентрация связанного с AD белка может быть повышена или понижена в популяции, имеющей расстройства, связанные с AD, в сравнении с популяцией без AD. Различия в уровнях белков могут быть оценены с использованием методик протеомики, включая, но не ограничиваясь ими, вестерн-блоттинг, иммуногистохимическое окрашивание, иммуноферментный анализа (ELISA) и масс-спектрометрию. Альтернативно этому связанные с AD белки могут быть идентифицированы путем получения профилей генной экспрессии для генов, кодирующих белки, с применением методик геномики, включающих, не ограничиваясь таковыми, анализ на микрочипах ДНК, серийный анализ экспрессии генов (SAGE) и количественную полимеразную цепную реакцию в реальном времени (кПЦР).
[001171] Примерами ассоциированных с болезнью Альцгеймера белков являются рецептор липопротеина очень низкой плотности (VLDLR), кодируемый геном VLDLR, подобный убиквитину модифицирующий активирующий белок 1 (UBA1), кодируемый геном UBA1, или белок, NEDD8-активирующий каталитическую субъединицу E1 фермента (UBE1C), кодируемый, например, геном UBA3.
[001172] В качестве неограничивающего примера, ассоциированные с AD белки могут включать, но не ограничиваться ими, следующие: кодируемый хромосомной последовательностью белок ALAS2, дельта-аминолевуленатсинтаза 2 (ALAS2), ABCA1 ATP-связывающий кассетный транспортер (ABCA1), ангиотензин-превращающий фермент I ACE (ACE), предшественник аполипопротеина E APOE (APOE), белок-предшественник амилоида APP (APP), белок аквапорин I AQP1 (AQP1), зависимый от Myc-бокса взаимодействующий белок I BIN1 или соединяющий интегрирующий белок 1 (BIN1), нейротрофический фактор головного мозга BDNF (BDNF), подобный бутирофилину белок 8 BTNL8 (BTNL8), открытая рамка считывания 49 хромосомы 1 C10RF49, кадгерин-4 CDH4, бета-2 субъединица рецептора ацетилхолина CHRNB2, подобный CKLF белок 2 CKLFSF2, содержащий трансмембранный домен MARVEL (CKLFSF2), лектиновый домен типа C, представитель семейства 4 CLEC4E (CLEC4E), белок кластерин CLU (также называемый J), рецептор комплемента эритроцитов 1 CR1 (CR1, также называемый CD35, рецептор C3b/C4b и рецептор иммунной адгезии), рецептор комплемента эритроцитов CR1L (CR1L), рецептор гранулоцитного колониестимулирующего фактора 3 CSF3R (CSF3R), цистатин C или цистанин 3 CST3, цитохром P450 2C CYP2C, ассоциированная со смертью протеинкиназа 1 DAPK1 (DAPK1), рецептор эстрогена 1 ESR1, фрагмент Fc рецептора IgA FCAR (FCAR, также называемый CD89), фрагмент Fc IgG, рецептор низкой плотности IIIb FCGR3B (FCGR3B или CD16b), рецептор свободных жирных кислот 2 FFA2 (FFA2), фибриноген FGA (фактор I), ассоциированный с GRB2 белок связывания 2 GAB2 (GAB2), ассоциированный с GRB2 белок связывания 2 GAB2 (GRB2), подобный галанину пептид GALP, сперматогенная глицеральдегид-3-фосфат дегидрогеназа GAPDHS (GAPDHS), GMPB - GMBP, гаптоглобин HP (HP), рецептор 5-гидрокситриптофана (серотонина) 7 (зависимый от аденилатциклазы) FITR7, фермент деградации инсулина IDE, IF127 - IF127, интерферон, альфа-индуцируемый белок 6 IFI6 (IFI6), индуцируемый интерфероном белок с повторами тетратрикопептида 1 IFIT2 (IFIT2), IL1RN - антагонист рецептора интерлейкина-1 (IL-IRA), рецептор интерлейкина 8 ILSRA (ILSRA), рецептор альфа (IL8RA или CD181), рецептор интерлейкина 8 IL8RB (IL8RB), бета (IL8RB), Jagged I JAG1 (JAG1) калиевый канал прямого выпрямления, подсемейство J, представитель 15 KCNJ15 (KCNJ15), белок, связанный с рецептором липопротеина низкой плотности 6 LRP6 (LRP6), ассоциированный с микротрубочками белок тау МАРТ (МАРТ), регулирующая сродство MAP/микротрубочки киназа 4 MARK4 (MARK4) фосфопротеин 1 фазы M MPHOSPH1, 5,10-метилентетрагидрофолатредуктаза MTHFR, индуцированный интерферонов связываюзих GTP белок MX2, нибрин NBN, также называемый NBN, никастрин NCSTN, рецептор ниацина 2 NIACR2 (NIACR2, также называемый GPR109B), никотинамиднуклеотидаденилилтрансфераза 3 NMNAT3, нейротримин NTM (HINT), ороскумоид 1 ORM1 (ORM1) или гликопротеин 1 альфа-1-кислоты, пуринорецептор P2Y 13 P2RY13 (P2RY13), никотинамидфосфорибозилтрансфераза PBEF1 (NAmPRTase или Nampt), также называемая усиливающим образование колоний предшественников B-клеток фактором 1 (PBEF1) или висфатином, фосфоенолпируваткарбоксилаза PCK1, связывающий фосфатидилинозитол клатрин, участвующий в ассоциации (PICALM), активатор плазминогена типа урокиназы PLAU (PLAU), плексин PLXNC1 (PLXNC1), прионный белок PRNP, белок пресенилин 1 PSEN1 (PSEN1), белок пресенилин 2 PSEN2 (PSEN2), белок рецептора белковой тирозинфосфатазы типа A PTPRA (PTPRA), Ral GEF с доменом PH и связывающим SH3 мотивом 2 RALGPS2 (RALGPS2), белок, подобный регулятору сигналинга с участием G-белков 2, регулятор связанного с сигналингом с участием G-белков 2 RGSL2 (RGSL2), связывающий селен белок 1 SELENBP1 (SELNBP1), митоферрин 1 SLC25A37, связанный с сортилином рецептор L (класс DLR), содержащий повторы A белок SORL1 (SORL1), трансферрин TF, митохондриальный фактор транскрипции A TFAM, фактор некроза опухолей TNF, суперсемейство рецептора фактора некроза опухолей, представитель 10C TNFRSF10C (TNFRSF10C), суперсемейство рецептора фактора некроза опухолей (TRAIL), представитель 10a TNFSF10 (TNFSF10), подобный убиквитину активирующий белок-модификатор 1 UBA1 (UBA1), UBA3, NEDD8-активирующий каталитическую субъединицу фермента E1 (UBE1C), убиквитин В UBB (UBB), убиквитин-1 UBQLN1, убиквитин карбоксилтерминальная эстераза белка L1 UCHL1 (UCHL1), убиквитин карбоксилтерминальная гидролиаза изозима белка L3 UCHL3 (UCHL3), белок рецептора липопротеина очень низкой плотности VLDLR (VLDLR).
[001173] В иллюстративных вариантах осуществления белки, ассоциированные с AD, хромосомную последовательность которых подвергают редактированию, могут принадлежать белку рецептора липопротеина очень низкой плотности (VLDLR), кодируемому геном, подобному убиквитину активирующего фермента-модификатора 1 (UBA1), кодируемому геном UBA1 NEDD8-активирующему каталитическую субъединицу фермента E1 белку, кодируемому геном UBA3, белку аквапорину 1 (AQP1), кодируемому геном AQP1, белку убиквитин-карбоксилтерминальной эстеразе L1 (UCHL1), кодируемому геном UCHL1, белку убиквитин-карбоксилтерминальной гидролазе изозима L3 (UCHL3), кодируемому геном UCHL3, белку убиквитину В (UBB), кодируемому геном UBB, ассоциированному с микротрубочками белку тау (МАРТ), кодируемому геном МАРТ, белку тирозинфосфатазного рецептора типа A (PTPRA), кодируемому геном PTPRA, связывающему фосфатидилинозитол белку, участвующему в сборке клатрина (P1CALM), кодируемому геном P1CALM, белку кластерину (называемому также аполипопротеин J), кодируемому геном CLU, белку пресенелину 1, кодируемому геном PSEN1, белку пресенелину 2, кодируемому геном PSEN2, связанному с сортилином рецептору L (класс DLR), содержащему повторы A белку (SORL1), кодируемому геном SORL1, белку-предшественнику амилоида (APP), кодируемому геном APP, предшественнику аполипопротеина E (APOE), кодируемому геном APOE или мозговому нейротрофическому фактору (BDNF), кодируемому геном BDNF. В иллюстративных вариантах осуществления генетически модифицированными животными являются крысы, и подвергшаяся редактирования хромосомная последовательность, кодирующая белок, ассоциированный AD, может являться следующей: APP - белок-предшественник амилоида NM_019288, AQP1 - белок аквапорин 1 (AQP1) NM_012778, BDNF - мозговой нейротрофический фактор NM_012513, CLU - белок кластерин (так называемый NM_053021 - аполипопротеин J), МАРТ - ассоциированный с микротрубочками белок тау NM_017212 (МАРТ), PICALM - связывающий фосфатидилинозитол белок, участвующий в сборке клатрина NM_053554 (P1CALM), PSEN1 - белок пресенелин 1 (PSEN1) NM_019163, PSEN2 - белок пресенелин 2 (PSEN2) NM_031087, PTPRA - белок рецептора тирозинфосфатазы типа A NM_012763 (PTPRA), SORL1 - связанный с сортилином рецептор L (класс DLR, NM_053519), содержащий повторы A белок SORL1 XM_001065506, белок XM_217115, UBA1 - подобный убиквитину активирующий белок-модификатор 1 (UBA1) NM_001014080, UBA3 - белок, NEDD8-активирующий каталитическую субъединицу фермента E1 (NEDD8- activating enzyme E1 catalytic subunit protein, UBE1C, NM_057205), UBB - белок убиквитин В (UBB, NM_138895), UCHL1 - убиквитин карбоксилтерминальная эстераза белка L1 (UCHL1), UCHL3 убиквитин карбоксилтерминальная гидролиаза изозима белка L3 (UCHL3) NM_001110165, VLDLR - белок рецептора липопротеина очень низкой плотности (VLDLR) NM_013155.
[001174] Животное или клетка может содержать 1, 2, 3, 4, 5, 6, 7, 8, 9,10, 11, 12, 13, 14, 15 или более прерванных хромосомных последовательностей, кодирующих белок, ассоциированный с AD, и ноль, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 или более хромосомно интегрированных последовательностей, кодирующий белок, ассоциированный с AD.
[001175] Отредактированная или интегрированная хромосомная последовательность может быть модифицирована таким образом, чтобы кодировать измененный белок, ассоциированный с AD. Для ряда мутаций в связанных с AD хромосомных последовательностей установлена ассоциация с AD. Например, миссенс-мутация V717I (т.е. замена валина в положении 717 на изолейцин) в APP вызывает семейную форму AD. Несколько мутаций в белке пресенилин-1, в частности, H163R (т.е. замена гистидина в положении 163 на аргинин), A246E (т.е. замена аланина в положении 246 на глутамат), L286V (т.е. замена лейцина в положении 286 на валин) и C410Y (т.е. замена цистеина в положении 410 на тирозин) вызывают семейную форму болезни Альцгеймера 3 типа. Мутации в белке пресенелине-2, в частности, N141I (т.е. замена аспарагина в положении 141 на изолейцин), M239V (т.е. замена метионина 239 на валин) и D439A (т.е. замена аспартата 439 на аланин) вызывают семейную форму болезни Альцгеймера 4 типа. Другие ассоциации генетических вариантов связанных с AD генов и заболеванием известны в настоящей области. См., например, Waring et al. (2008) Arch. Neurol. 65:329-334, содержание которых включено в настоящее описание в качестве ссылки.
Связанные с секретазами нарушения
[001176] В публикации патента США № 20110023146 описано применение нуклеаз с цинковыми пальцами для генетической модификации клеток, животных или белков, ассоциированных с заболеваниями, связанными с секретазами. Секретазы необходимы для процессинга предшественников белков до их биологически активных форм. Дефекты в различных компонентах метаболических путей секретазы вносят вклад в различные заболевания, в частности, тех, которые имеют отличительные признаки - амилоидогенез и амилоидные бляшки, в частности, болезнь Альцгеймера (AD).
[001177] Связанные с секретазой нарушения и белки, ассоциированные с такими заболеваниями, представляют собой разнообразный набор белков, влияющих на предрасположенность к многочисленным заболеваниям, наличие таких заболеваний, их выраженность или их комбинацию. Настоящее изобретение включает редактирование любых хромосомных последовательностей, кодирующих белки, ассоциированные с расстройствами, связанными с секретазами. Связанные с нарушениями функционирования секретаз белки, выбраны на основе показанной в экспериментах ассоциации белков, ассоциированных с расстройствами, связанными с секретазами, с заболеванием, связанным с секретазами. Например, скорость образования или циркулирующая концентрация белка, ассоциированного с расстройством, связанным с секретазами, может быть повышена или понижена в популяции, имеющей расстройства, связанные с секретазами, в сравнении с популяцией без связанных с секретазами расстройств. Различия в уровнях белков могут быть оценены с использованием методик протеомики, включая, без наложения ограничений, вестерн-блоттинг, иммуногистохимическое окрашивание, твердофазный иммуноферментный анализ (ELISA) и масс-спектрометрию. Альтернативно этому связанные с ассоциированными с секретазами нарушениями белки могут быть идентифицированы путем получения профилей генной экспрессии для генов, кодирующих белки, с применением методик геномики, включающих, не ограничиваясь таковыми, анализ на микрочипах ДНК, серийный анализ экспрессии генов (SAGE) и количественную полимеразную цепную реакцию в реальном времени (кПЦР).
[001178] В качестве неограничивающего примера, белки, ассоциированные со связанными с секретазами нарушениями, включают PSENEN (гомолог энхансера пресенелина 2 (C. elegans)), CTSB (катепсин B), PSEN1 (пресенелин 1), APP (белок-предшественник бета-амилоида (A4)), APH1B (гомолог B дефекта передней глотки 1 (anterior pharynx defective 1 homolog В, C. elegans)), PSEN2 (пресенелин 2 (болезнь Альцгеймера 4)), BACE1 (расщепляющий APP по бета-сайту фермент 1), ITM2B (интегральный мембранный белок 2B), CTSD (катепсин D), NOTCH1 (гомолог Notch 1, ассоциированный с транслокацией (Drosophila)), TNF (фактор некроза опухолей (суперсемейство TNF, представитель 2)), INS (инсулин), DYT10 (дистония 10), ADAM 17 (металлопептидазный домен ADAM 17), APOE (аполипопротеин E), ACE (конвертирующий ангиотензин I фермент (пептидил-дипептидаза A) 1), STN (статин), TP53 (белок опухоли p53), IL6 (интерлейкин 6 (интерферон бета 2)), NGFR (рецептор фактора роста нервов (суперсемейство TNFR, представитель 16)), IL IB (интерлейкин I, бета), ACHE (ацетилхолинэстераза (Yt группа крови)), CTNNB1 (катенин (ассоциированный с кадерином белок), бета 1, 88 кДа), IGF1 (инсулиноподобный фактор роста 1 (соматомедин C)), IFNG (интерферон, гамма), NRG1 (нейрегулин 1), CASP3 (каспаза 3, связанная с апоптозом цистеинпептидаза), МАРК 1 (митогенактивирюущеая протеинкиназа 11), CDH1 (кадерин 1, тип 1, E- кодерин (эпителиалный)), APBB1 (связывающий белок-предшественник бета-амилоида (A4), семейство B, представитель 1 (Fe65)), HMGCR (3-гидрокси-3-метилглутарил-кофермент A редуктаза), CREB1 (белок 1, связывающийся с цАМФ-зависимым элементом), PTGS2 (простагландин эндопероксидсинтаза 2 (простагландин G/H синтаза и циклооксигеназа)), HES1 (ген hairy и энхансер splt 1, (Drosophila)), CAT (каталаза), TGFB1 (трансформирующий фактор роста бета 1), ENO2 (енолаза 2 (гамма, нейрональная)), ERBB4 (v-erb-a гомолог 4 вирусного онкогена эритробластной лейкемии (птичий)), TRAPPC10 (комплекс 10 частицы транспорта белка), MAOB (моноаминоксидаза B), NGF (фактор роста нервов (бета-полимпептид)), MMP12 (матриксная металлопептидаза 12 (эластаза макрофагов)), JAG1 (jagged 1 (синдром Алажиля)), CD40LG (лиганд CD40), PPARG (активированный пролифератором пероксисомы рецептор гамма), FGF2 (фактор роста фибробластов 2 (основной)), IL3 (интерлейкин 3 (колониестимулирующий фактор, несколько), LRP1 (белок 1, связанный с рецептором липопротеина низкой плотности), NOTCH4 (гомолог 4 Notch (Drosophila)), MAPK8 (митогенактивируемая протеинкиназа 8), PREP (пролилэндопептидаза), NOTCH3 (гомолог 3 Notch 3 (Drosophila)), PRNP (прионный белок), CTSG (катепсин G), EGF (эпидермальный фактор роста (бета-урогастрон)), REN (ренин), CD44 (молекула CD44 (по индийской классификации групп крови)), SEEP (селектин P (140-кДа белок мембранных гранул, антиген CD62)), GHR (рецептор гормона роста), ADCYAP1 (активирующий аденилатциклазу полипептид 1 (гипофизарный)), INSR (рецептор инсулина), GFAP (глиальный фибриллярный кислый белок), MMP3 (матриксная металлопептидаза 3 (стромелизин 1, прожелатиназа)), MAPK10 (митогенактивируемая протеинканаза 10), SP1 (транскрипционный фактор Sp1), MYC (гомологичный y-myc миелоцитоматозный вирусный онкоген (птичий)), CTSE (катепсин E), PPARA (активируемый профилератором пероксисом рецептор альфа), JUN (онкоген jun), TEMPI (ингибитор металлопептидазы TEMP 1), ILS (интерлейкин 5 (колониестимулирующий фактор, эозинофил)), ILIA (интерлейкин 1, альфа), MMP9 (матриксная металлопептидаза 9 (желатиназа B, 92-кДа желатиназа, 92-кДа коллагеназа IV типа)), HTR4 (рецептор 5-гирокситриптамина (серотонина) 4), HSPG2 (гепарансультфат протеогликан 2), KRAS (v-Ki-ras2 гомолог вирусного онкогена крысиной саркомы Кирстена), CYCS (цитохром c, соматический), SMG1 (SMG1 гомолог связанной с фосфатидилинозитол 3- киназой киназа (C. elegans)), IL1R1 (рецептор интерлейкина 1, тип I), PROK1 (прокинетицин 1), MAPK3 (митогенактивируемая протеинканаза 3), NTRK1 (рецептор нейроторфической тирозинкиназы типа 1), IL13 (интерлейкин 13), MME (мембранная металлоэндопептидаза), TKT (транскетолаза), CXCR2 (рецептор хемокина 2 (мотив C-X-C)), IGF1R (рецептор инсулиноподобного фактора роста 1), KARA (рецептор ретиноевой кислоты, альфа), CREBBP (связывающий CREB белок), PTGS1 (простагландинэндопероксид синтаза 1 (простагландин G/H синтаза и циклооксигеназа)), GALT (галактозо-1-фосфат уридилтрансфераза), CHRM1 (холинергический рецептор, мускариновый 1), ATXN1 (аткасин 1), PAWR (PRKC, апоптоз, WT1, регулятор), NOTCH2 (гомолог 2 Notch (Drosophila)), M6PR (рецептор маннозо-6-фосфата (катионзависимый)), CYP46A1 (цитохром P450, семейство 46, подсемейство A, полипептид 1), CSNK1 D (казеинкиназа 1, дельта), MAPK14 (митогенактивируемая протеинканаза 14), PRG2 (протеогликан 2, костный мозг (активатор естественных киллеров, главный основный белок)), PRKCA (протеинкиназа C, альфа), LI CAM (молекула клеточной адгезии LI), CD40 (молекулы CD40, рецептор суперсемейства TNF, представитель 5), NRH2 (ядерный рецептор подсемейства 1, группа I, представитель 2), JAG2 (jagged 2), CTNND1 (катенин (кадеринассоциированный белок), дельта 1), CDH2 (кадерин 2, тип 1, N-кадерин (нейрональный)), CMA1 (химаза 1, тучные клетки), SORT (сортилин 1), DLK1 (дельтаподобны гомолог 1 (Drosophila)), THEM4 (суперсемейства тиоэстерезы, представитель 4), JUP (плакоглобин участка соединения), CD46 (молекула D46, регуляторный белок комплемента), CCL11 (лиганд хемокина (мотив C-C)11), CAV3 (кавеолин 3), RNASES (рибонуклеаза, семейство РНКаз A, 3 (катионный белок эозинофилов)), HSPA8 (белок теплового шока белок 8 массой 70 кДа), CASP9 (каспаза 9, связанный с апоптозом цистеиновая пептидазаза), CYP3A4 (цитохром P450, семейство 3, подсемейство A, полипептид 4), CCR3 (рецептор хемокина (мотив C-C) 3), TFAP2A (транскрипционный фактор AP-2 альфа (активирующий связывающий энхансер белок 2 альфа)), SCP2 (белок-переносчик стирола 2), CDK4 (циклинзависимая киназа 4), HIF1A (альфа-субъединица фактора (щелочной транскрипционный фактор спираль-петля-спираль), индуцируемого гипоксией 1), TCF7L2 (7-подобный транскрипионный фактор 2 (специфичный для T-клеток, бокс HMG)), IL1R2 (рецептор интерлейкина 1, тип II), B3GALTL (подобный 1,3-галактозилтрансферазе), MDM2 (Mdm2 гомолог связывающего p53 белок (мыши)), RELA (v-rel гомолог A онкогена вируса ретикулоэндотелиоза (птичий)), CASP7 (каспаза 7, связанная с апоптозом цистеиновая пептидаза), IDE (инсулиндеградирующий фермент), FABP4 (связывающий жирные кислоты белок 4, адипоцит), CASK (кальций/кальмодулин-зависимая сериновая протеинкиназа (семейство MAGUK)), ADCYAP1R1 (рецептор активирующего аденилатиклазу полипептида 1 (гипофиз) типа type I), ATF4 (активирующий транскрипционный фактор 4 (отвечающий на tax (tax-responsive) энхансерный элемент B67)), PDGFA (полипептид альфа фактора роста тромбоцитов), C21 или 03 (открытая рамка считывания 33 хромосомы 21), SCG5 (секретогранин V (белок 7B2)), RNF123 (белок c цинковыми пальцами 123), NFKB1 (ядерный фактор энхансера гена легкого полипептида каппа в B-клетках 1), ERBB2 (v-erb-b2 гомолог 2 вирусного онкогена эритробластной лейкемии, полученные из нейро/глиобластомы гомолог онкогена (птичий)), CAV1 (кавеолин 1, белок ямки, 22 кДа), MMP7 (матриксная металлопептидаза 7 (матрилизин, утерин)), TGFA (трансформирующий фактор роста, альфа), RXRA (ретиноидный рецептор X, альфа), STX1A (синтаксин 1A (мозг)), PSMC4 (субъединиа 26S АТФ-азы 4 протеосомы (просома, мультикаталитическая протеаза), P2RY2 (пуринергический рецептор P2Y, сопряженный с G-белком, 2), TNFRSF21 (семейства фактора некроза опухоли, представитель 21), DLG1 (диски, большой гомолог 1 (Drosophila)), NUMBL (гомолог, подобный гену numb (Drosophila)), SPN (сиалофорин), PLSCR1 (фосфолипидскрамблаза 1), UBQLN2 (убиквитин 2), UBQLN1 (убиквитин 1), PCSK7 (предшественник конвертазы субтилизин/кексин типа 7), SPON1 (spondin 1, extracellular matrix protein), SILV (гомолог silver (мышь)), QPCT (глутамилпептид циклотрансфераза), HESS (hairy и энхансер split 5 (Drosophila)), GCC1 (GRIP и содержащий coiled-coil домен 1) или любую их комбинацию.
[001179] Генетически модифицированное животное или клетка могут содержать 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более прерывающихся хромосомных последовательностей, кодирующих белок, ассоциированный с заболеваниями, связанными с секретазами, и ноль, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более интегрированных в хромосому последовательностей, кодирующих прерывающийся белок, ассоциированный со связанными с секретазой заболеваниями.
ALS
[001180] В патентной публикации США № 20110023144 описано применение нуклеаз с цинковыми пальцами к генетически модифицированным клеткам, животным и белкам, ассоциированным с амиотрофическим латеральным склерозом (ALS). ALS характеризуется постепенной стабильной дегенерацией некоторых нервных клеток в коре головного мозга, стволе головного мозга и спинном мозге, которые вовлечены в произвольные движения.
[001181] Заболевания, связанные с моторными нейронами и белки, ассоциированные с такими заболеваниями, представляют собой разнообразный набор белков, которые вызывают подверженность развитию заболеваний моторных нейронов, выраженность заболевания моторных нейронов или их любую комбинацию. Настоящее изобретение охватывает редактирование любых хромосомных последовательностей, кодирующих белки, ассоциированные с ALS, специфическим заболеванием моторных нейронов. Ассоциированные с ALS белки обычно выявляют на основе экспериментальной ассоциации связанных с ALS белков с ALS. Например, скорость образования или циркулирующая концентрация связанного с ALS белка может быть повышена или понижена в популяции с ALS в сравнении с популяцией без ALS. Различия в уровнях белков могут быть оценены с использованием методик протеомики, включая, без наложения ограничений, вестерн-блоттинг, иммуногистохимическое окрашивание, твердофазный иммуноферментный анализ (ELISA) и масс-спектрометрию. Альтернативно этому связанные с ALS белки могут быть идентифицированы путем получения профилей генной экспрессии для генов, кодирующих белки, с применением методик геномики, включающих, не ограничивающихся ими, анализ на микрочипах ДНК, серийный анализ экспрессии генов (SAGE) и количественную полимеразную цепную реакцию в реальном времени (кПЦР).
[001182] В качестве неограничивающего примера, ассоциированные с ALS белки включают, но не ограничиваются ими, следующие: SOD1 супероксиддисмутаза 1, ALS3 растворимый сенатаксин амиотрофического латерального склероза 3 SETX, ALS5 5 FUS амиотрофического латерального склероза, в виде химеры в саркоме, ALS7 - амиотрофического латерального склероза 7, ALS2 амиотрофического латерального склероза, DPP6 дипептидилпептидаза 6 склероза 2, простагландинполипептид эндопероксидсинтаза 1 тяжелого PTGS1 нейрофиламента NEFH, SLC1A2 растворимый переносчик семейства 1, TNFRSF10B фактор некроза опухолей (подсемейство глиальных рецепторов с высокой афинностью, глутаматный транспортер), представитель 10b, представитель 2, PRPH периферин, HSP90AA1 90-кДА белок теплового шока альфа (цитозольный), класс A, представитель 1, GRIA2 глутаматный рецептор, IFNG интерферон, гамма ионотропный, AMPA 2 S100B S100 кальцийсвязывающий, FGF2 белок B фактора роста фибробластов 2, A0X1 альдегидоксидаза 1, CS цитратсинтаза, TARDBP TAR ДНК-связыващий белок, TXN тиоредоксин, RAPH1 Ras ассоциация, MAP3K5 митогенактивирущий белок (RaIGDS/AF-6) и домены гомологии 1 киназы 5 плекстрина, NBEAL1 нейробичин-подобный 1, GPX1 глутатионпероксидаза 1, ICA1L аутоантиген клеток островка, RAC1 субстрат связанного с ras C3 подобного 1,69-кДа ботулинотоксину токсина 1, МАРТ ассоиированный с микротрубочками, ITPR2 трифосфатный рецептор инозитол-1,4,5-белка тау, тип 2, ALS2CR4 амиотрофического латерального склероза, GLS глутаминаза склероза 2 (ювенильная) хромосомного региона, кандидат 4, ALS2CR8 амиотрофического латерального, CNTFR - рецептор хромосомного региона цилиарного нейротрофического фактора склероза (ювенильный), кандидат 8, ALS2CR11 амиотрофического латерального склероза, FOLH1 хромосоный регион фолатгидролазы 1 склероза 2 (ювенильный), кандидат 11, FAM117B семейство с последовательностью P4HB пролил-4-гидроксилазы, сходство 117, представитель В бета полипептид, CNTF цилиарный, нейротрофический фактор, SQSTM1 секвестосома 1, STRADB связанная с STE20 киназа, NAIP семейство NLR, ингибиторный белок адаптора бета апоптоза, YWHAQ тирозин 3-SLC33A1 растворимый переносчик семейства 33 монооксигеназа/триптофан (транспортер ацетил-CoA), представитель 1 активирующего 5-монооксигеназу белка, тета-полипептид, TRAK2 белок транспорта, FIG. 4 FIG. 4 гомолог, SACI липидфосфатазный домен, связывающий кинезин 2, содержащий NIF3L1, NIF3, NGGI, взаимодействующий INA, интемексин, нейрональный фактор 3 типа 1, белок промежуточного филамента, альфа PARD3B par-3 (разделяющий) COX8A дефективный гомолог B цитохромоксидазы c, субъединица VIHA, CDK15 циклинзависимая киназа, HECW1, HECT, C2 и WW 15 содержащая домен убиквитин E3 протеинлигаза 1, NOS1 синтаза оксида азота 1, MET протоонкогне met, SOD2 супероксиддисмутаза 2, HSPB1 митохондриальный белок теплового шока 1, 27-кДа, NEFL нейрофиламент, легкий, CTSB полипептид катепсина В, ANG ангиогенин, HSPA8 рибонуклеаза теплового шока, 70-кДа, РНКаза A белка 8 семейства 5, 5 VAPB, VAMP (ассоциированный с везикулярным-ESR1 рецептором эстрогена 1 мембранный белок)-ассоциированный белок В и C, SNCA синуклеин, альфа, HGF фактор роста гепатоцитов, CAT каталаза, ACTB актин, бета, NEFM нейрофиламент, средний, TH полипептид тирозингидроксилазы, BCL2 В-клеток,l CLL/лимфома 2, FAS Fas (рецептор суперсемейства TNF, представитель 6), CASP3 каспаза 3, связанная с апоптозом CLU-кластерином цистеиновая пептидаза, SMN1 выживания моторных нейронов, G6PD глюкозо-6-фосфат 1, теломерная дегидрогеназа, BAX, ассоциированный с BCL2 X, HSF1 транскрипционный фактор теплового шока 1, RNF19A белок цинковых пальцев AA, JUN онкоген jun, ALS2CR12 амиотрифического латериального склероза, HSPA5 70-кДа белок теплового шока склероза 2 (ювенильный), белок 5 хромосомного региона, кандидат 12, MAPK14 митогенактивируемый белок, IL10 интерлейкин 10 киназа 14, APEX1 нуклеаза APEX, TXNRD1 тиоредоксинредуктаза 1 (мультифункциональный домен репарации ДНК) 1, NOS2 синтаза оксида азота 2, TIMP1 TIMP индуцибельный ингибитор металлопептидазы 1, CASP9 каспаза 9, апоптоз- XIAP X-связанный ингибитор связанной цистеиновой пептидазы апоптоза, GLG1 гликопротеин комплекса Гольджи 1, EPO эритропоэтин, VEGFA фактор роста эндотелия сосудов фактор A, эластин ELN factor A, GDNF ядерный фактор NFE2L2, полученный из глиальных клеток, группа (производный эритроидный нейротрофический фактор 2), 2 SLC6A3 растворимый переносчик, семейство 6, HSPA4 70-кДа белок теплового шока (белок-переносчик нейромедиаторов 4, допамин), представитель 3, APOE аполипопротеин E, PSMB8 субъединица бета протеосомы (просома, мультикаталитическая протеаза), тип 8, DCTN1 динактин 1, TIMP A3 TIMP металлопептидазный ингибитор 3, KIFAP3 кинезинассоциированный SLC1A1 растворимый переносчик семейства 1, белок 3 (нейрональный/эпителиальный глутаматный транспортер высокой афинности, система Xag), представитель 1, SMN2 циклин C2 выживания моторных нейронов CCNC, мембранный белок центромер MPP4, STUB1, STIP1 гомологии и U-пальмитолеированный содержащий бокс 4 белок 1, ALS2 амилоид бета (A4), PRDX6 белок-предшественник пероксиредоксина 6, SYP синаптофизин, CABIN1 связывающий кальцийнейрин белок 1, CASP1 каспаза 1, апоптоз-GART фосфорибозилглицинамин-связанная цистеиндеформилтрансфераза, пептидаза фосфорибозилглицинаминдесинтетаза, фосфорибозиламиноимидазолсинтетаза, CDK5 циклинзависимая киназа 5, ATXN3 атаксин 3, RTN4 ретикулон 4, C1QB компонент комплемента 1, субкомпонент q, В цепь фактора роста нервов VEGFC, HTT рецептор хантингина, PARK7 болезни Паркинсона 7, XDH ксантиндегидрогеназа, GFAP глиальный фибриллярный кислотный белок, MAP2 ассоциированный с микротрубочками белок 2, CYCS цитохром c, соматический, FCGR3B Fc фрагмент IgG, низкой афинности IIIb, CCS медный шаперон убиквитиноподобной супероксиддисмутазы 5 UBL5, MMP9 матриксная металлопептидаза 9, SLC18A3 растворимый переносчик семейства 18 ((везикулярный аетилхолин), представитель 3, TRPM7 транзиентный рецептор, HSPB2 катионный канал теплового шока 27-кДа, белок 2 подсемейства M, представитель 7, AKT1 v-akt мышиной тимомы, DERL1 семейство подобных Der1 доменов, гомолог вирусного онкогена 1, предстаувитель 1, CCL2 хемокин(мотив C--C), NGRN нейгрин, нейритный лиганд 2, связанная с почкованием GSR гутатион редуктаза, TPPP3 семейство стимулирующего полимеризацию тубулина белка, представитель 3, APAF1 пептидаза апоптоза, BTBD10 активирующи домен BTB (POZ) фактор 1, содержащий 10 GLUD1 глутамат, CXCR4 хемокин (мотив C--X--C), рецептор 4 дегирогеназы 1, SLC1A3 растворимый переносчик семейства 1, FLT1 связанный с fms тирозиновый (глиальный глутаматный транспортер высокой афинности), представитель 3, PON1 параоксоназа 1, рецепотр андрогена AR, L1P ингибиторный фактор лейкемии, ERBB3 v-erb-b2 гомолог 3 вирусного онкогена эритроблатсный лейкемии, LGALS1 лектин, галактозид, CD44 связывающий CD44, растворимый белок 1, TP53 белок опухоли p53, TLR3 толл-подобный рецептор 3, GR1A1 глутаматный рецептор, GAPDH глицеральдегид-3-ионотропный, АМРА 1 фосфатдегидрогеназа, GR1K1 глутаматный рецептор, DES десмин, ионотропный, каинат 1, CHAT холинацетилтрансфераза, FLT4 связанный с fms триозинкиназа 4, CHMP2B модифицирующий хроматин, BAG1 ассоциированный с BCL2 белок, 2B атаноген, MT3 металлотионенин 3, CHRNA4 холинергически рецептор, никотиновый, альфа 4, GSS - глутатионсинтетаза BAK1, антагонист BCL2/киллер 1, KDR домен вставки киназы, GSTP1 рецептор глутатион S-трансферазы (рецептор тирозинкиназы типа III pi 1), OGG1 8-оксогуанин ДНК, IL6 интерлейкин 6 (интерферон, гликозилаза бета 2).
[001183] Животное или клетка могут включать 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более прерывающихся хромосомных последовательностей, кодирующих белок, ассоциированный с ALS и нуль, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более хромосомно интегрированных последовательностей, кодирующих прерывающийся белок, ассоциированный с ALS. Предпочтительные белки, ассоциированные с ALS, включают SOD1 (супероксиддисмутаза 1), ALS2 (белок амиотрофического латерального склероза 2), FUS (слитый в саркоме), TARDBP (TAR ДНК связывающий белок), VAGFA (фактор роста эндотелия сосудов A), VAGFB (фактор роста эндотелия сосудов B) и VAGFC (фактор роста эндотелия сосудов C) и любую их комбинацию.
Аутизм
[001184] В публикации патента США № 20110023145 описано применение нуклеаз с цинковыми пальцами к генетически модифицированным клеткам, животным или белкам, ассоциированным с расстройствами аутистического спектра (ASD). Расстройства аутистического спектра (ASD) представляют собой группу заболеваний, характеризующихся количественным ухудшением социальных взаимодействий и коммуникации, и фиксированными повторяющимися и стереотипными паттернами поведения, интересов и активности. Три заболевания: аутизм, синдром Аспергера (AS) и первазивное расстройство развития без дополнительных уточнений (PDD-NOS) представляют собой спектр одного расстройства с варьирующей степенью тяжести, связанного интеллектуального функционирования и состояние здоровья. ASD являются преимущественно генетическими заболеваниями с наследственностью около 90%.
[001185] В публикации патента США № 20110023145 описано редактирование любых хромосомных последовательностей, кодирующих белки, ассоциированные с ASD, к которым могут применяться системы CRISPR-Cas по настоящему изобретению. Связанные с ASD белки обычно устанавливаются на основе показанной в экспериментах ассоциации связанных с ASD белками с встречаемостью или проявлением ASD. Например, скорость образования или циркулирующая концентрация связанного с ASD белка может быть повышена или понижена в популяции, имеющей ASD в сравнении с популяцией без ASD. Различия в уровнях белков могут быть оценены с использованием методик протеомики, включая, но не ограничиваясь ими, вестерн-блоттинг, иммуногистохимическое окрашивание, иммуноферментный анализ (ELISA) и масс-спектрометрию. Альтернативно этому связанные с ASD белки могут быть идентифицированы путем получения профилей генной экспрессии для генов, кодирующих белки, с применением методик геномики, включающих, не ограничиваясь таковыми, анализ на микрочипах ДНК, серийный анализ экспрессии генов (SAGE) и количественную полимеразную цепную реакцию в реальном времени (кПЦР).
[001186] Не ограничивающие примеры патологических состояний или расстройств, которые могут быть ассоциированы с белками, связанными с ASD, включают аутизм, синдром Аспергера (AS), первазивное расстройство развития без дополнительных уточнений (PDD-NOS), синдром Ретта, туберозный склероз, фенилкетонурия, синдром Смита-Лемли-Опица и синдром хрупкой X-хромосомы. В качестве не ограничивающего примера, белки, ассоциированные с ASD включают, не ограничиваясь, следующие: ATP10C аминофосфолипид, MET рецептор MET, транспортирующий АТФазу тирозинкиназу (ATP10C), BZRAP1 MGLUR5 (GRM5) метаботропный глутаматный рецептор 5, (MGLUR5) CDH10 кадерин-10, MGLUR6 (GRM6) метаботропный глутаматный рецептор 6 (MGLUR6), CDH9 кадерин-9 NLGN1, нейролигин-1, CNTN4 контактин-4, NLGN2 нейролигин-2, CNTNAP2 контакин-ассоциированный, SEMA5A тип нейролигина-3 белок 2 (CNTNAP2), DHCR7 7-дегилрохолестерин, NLGN4X нейролигин-4, связанный с X-редуктазой (DHCR7) DOC2A, двойной подобный C2-домен, NLGN4Y нейролигин-4, связанный с Y-содержащим белком альфа, DPP6 дипепидил, NLGN5 нейролигин-5, подобный аминопептидазе белок 6, EN2 engrailed 2 (EN2), NRCAM молекула адгезии нейрональных клеток (NRCAM), MDGA2 умственной отсталости при хрупкой X-хромосоме, NRXN1 - нейрексин-1 1 (MDGA2), FMR2 (AFF2), семейство AF4/FMR2 представитель 2 OR4M2 обонятельный рецептор (AFF2), 4M2 FOXP2 белок с боксом Forkhead P2, OR4N4 обонятельный рецептор (FOXP2), 4N4 FXR1 умственной отсталости при хрупкости X-хромосомы, OXTR рецептор окситоцина, аутосомальный (OXTR) гомолог 1, (FXR1) FXR2 умственной отсталости при хрупкой X -хромосоме, PAH фенилаланин гидроксилаза (PAH), гомолог 2, (FXR2) GABRA1 гамма-аминомасляная кислота, PTEN фосфатаза и рецептор, гомолог тензина, субъединица альфа-1 (GABRA1) (PTEN), GABRA5 GABAA (рецептор аминомасляной кислоты PTPRZ1) рецептор альфа 5 тирозин-белковая субъединица, (GABRA5) фосфатаза зета (PTPRZ1), GABRBI рилиновый рецептор аминомасляной кислоты RELN, субъединиа бета-1 (GABRB1), GABRB3 GABAA рецептор бета 3 субъединицы белка L10 (гамма-аминомасляной кислоты RPL10 рибосомы 60S), L10 (GABRB3), GABRG1 гамма-аминомасляная кислота, SEMA5A рецептор семафорина-5A, субъединица гамма-1, (SEMA5A) (GABRG1) HIRIP3 HIRA-взаимодействующий белок 3, SEZ6L2 группа связанного с припадками гомолога 6 (мышь), 2 HOXA1 белок гомеобокса Hox-A1, SHANKS SH3 и (HOXA1) домены множественных анкирин-повторов 3 (SHANKS), 1L6 интерлейкин- 6, SHBZRAP1 SH3 и домены множественных анкириновых повторов 3 (SHBZRAP1), LAMB1 ламинин, субъединица бета-1, SLC6A4 транспортер серотонина (LAMB1) (SERT), MAPK3 митогенактивируемый белок, TAS2R1 киназа вкусового рецептора 3, тип 2, представитель 1, TAS2R1 MAZ ассоциированный с Myc цинковый палец, TSC1 белок туберозного склероза 1, MDGA2 домен MAM, содержащий TSC2, белок гликозилфосфатидилинозитол 2, якорь 2 (MDGA2), MECP2 связывающий метил CpG, UBE3A белок убиквитин, 2, (MECP2) лигаза E3A (UBE3A), MECP2 связывающий метил CpG WNT2, белок типа Wingless 2 (MECP2), MMTV семейства сайта интеграции, представитель 2 (WNT2).
[001187] Тип белка, ассоциированного с ASD, хромосомная последовательность которого подвергается редактирования, может и будет варьировать. В предпочтительных вариантах осуществления, белки, ассоциированные с ASD, хромосомные последовательности которых могут редактироваться, могут быть следующими: связанный с бензодиазепиновым рецептором (периферическим) белок 1 (BZRAP1), кодируемый геном BZRAP1, белок из семейства AF4/FMR2, представитель 2 (AFF2), кодируемый геном AMA (также называемым MFR2), аутосомальный белок-гомолог 1 умственной отсталости при хрупкой X-хромосоме (FXR1), кодируемый геном FXR1, аутосомальный белок-гомолог 2 умственной отсталости при хрупкой X-хромосоме (FXR2), кодируемый геном FXR2, домен MAXI, содержащий гликозилфосфатидилинозитол якорный белок 2 (MDGA2), кодируемый геном MDGA2, связывающий метил-CpG белок 2 (XIECP2), кодируемый геном MECP2, метаботропный рецептор глутамата 5 (MGLUR5), кодируемый геном MGLUR5-1 (также называемый GRM5), белок нейрексин 1, кодируемый геном NRXN1 или белок семафорин-5A (SEMA5A), кодируемый геном SEMA5A. В служащем примером варианте осуществления генетически модифицированным животным является крыса, редактируемая хромосомная последовательность, кодирующей белок, ассоциированный с ASD, входит в следующий список: BZRAP1 бензодиазепиновый рецептор XM_002727789, (периферический) ассоциированный XM_001081125, AFF2 (FMR2), представитель 2 семейства AF4/FMR2 XM_219832, (AFF2) XM__001054673, FXR1 умствеенной отсталости при хрупкой X-хромосоме NM_001012179, аутосомальный гомолог 1 (FXR1) FXR2 умственной отсталости при хрупкой X-хромосоме NX_001100647, аутосомальный гомолог 2 (FXR2) MDGA2, домен MAXI, содержащий якорь 2 гликозилфосфатидилинозитола NM_199269, (MDGA2) MECP2 связывающий метил-CpG белок 2 NM_022673, (MECP2) MGLUR5 метаботропный глутаматный рецептор 5 NM_017012 (GRM5) (MGLUR5), NRXN1 нейрексин-1 NM_021767 SEMA5A, семафорин-5A (SEMA5A) NM_001107659.
Нарушения, связанные с экспансией тринуклеотидных повторов
[001188] В публикации патента США № 20110016540 описано использование нуклеаз цинковых пальцев для генетической модификации клеток, животных и белков, ассоциированных с заболеваниями экспансии тринуклеотидных повторов. Заболевания экспансии тринуклеотидных повторов являются сложными, прогрессирующими заболеваниями, которые затрагивают нейробиологию развития и часто влияют на когнитивную сферу и сенсорно-моторные функции.
[001189] Белки экспансии тринуклеотидных повторов являются разнообразным набором белков, ассоциированных с подверженностью развитию заболевания экспансии тринуклеотидных повторов, наличием заболевания экспансии тринуклеотидных повторов или их комбинации. Заболевания экспансии тринуклеотидных повторов разделяют на 2 категории в зависимости от типа повторов. Наиболее распространенным повтором является триплет CAG, который, в случае присутствия в кодирующей области гена, кодирует аминокислоту глутамин (Q). Следовательно, такие заболевания называют полиглуматиновыми (polyQ) заболеваниями и включают следующие заболевания: болезнь Хантингтона (HD); спинобульбарная мышечная атрофия (SBMA), спиномозжечковые атаксии (SCA, типы 1, 2, 3, 6, 7 и 17), дентаторубро-паллидолюисова атрофия (DRPLA). Остальные заболевания тринуклеотидных повторов либо не подразумевают присутствия триплета CAG, либо триплет CAG не расположен кодирующей области гена и, как следствие, называются неполиглутаминовыми заболеваниями. Неполиглутаминовые заболевания включают синдром хрупкой X-хромосому (FRAXA); умственную отсталость при хрупкой X-хромосоме (FRAXE); атаксию Фридрейха (FRDA); миотоническую дистрофию (DM) и спиномозжечковую атаксию (SCA, типы 8 и 12).
[001190] Связанные с заболеваниями экспансии тринуклеотидных повторов белки, обычно устанавливают на основе показанной в экспериментах ассоциации связанных с заболеваниями экспансии тринуклеотидных повторов белков с заболеванием экспансии тринуклеотидных повторов. Например, скорость образования или циркулирующая концентрация связанного заболеваниями экспансии тринуклеотидных повторов белка может быть повышена или понижена в популяции, имеющей заболевание экспансии тринуклеотидных повторов в сравнении с популяцией без заболеваний экспансии тринуклеотидных повторов. Различия в уровнях белков могут быть оценены с использованием методик протеомики, включая, но не ограничиваясь ими, вестерн-блоттинг, иммуногистохимическое окрашивание, твердофазный иммуноферментный анализ (ELISA) и масс-спектрометрию. Альтернативно этому связанные с заболеваниями экспансии тринуклеотидных повторов белки могут быть идентифицированы путем получения профилей генной экспрессии для генов, кодирующих белки, с применением методик геномики, включающих, не ограничиваясь таковыми, анализ на микрочипах ДНК, серийный анализ экспрессии генов (SAGE) и количественная полимеразная цепная реакция в реальном времени (кПЦР).
[001191] Не ограничивающими примерами белков, ассоциированных с заболеваниями экспансии тринуклеотидных повторов, являются: AR (рецептор андрогена), FMR1 (умственной отсталости при хрупкой X-хромосоме1), HTT (хантингин), DMPK (протеинкиназа миотонической дистрофии), FXN (фратаксин), ATXN2 (атаксин 2), ATN1 (атрофин 1), FENl (структуроспеифичная эндонуклеаза flap 1), TNRC6A (содержащий тринуклеотидный повтор 6A), PABPN1 (связывающий поли(A) ядерный белок 1), JPH3 (юнктофилин 3), MED15 (субъединица 15 медиаторного комплекса), ATXN1 (атаксин 1), ATXN3 (атаксин 3), TBP (связывающий TATA-бокс белок), CACNA1A (кальциевый канал, потенциалозависимый, тип P/Q, субъединица альфа 1A), ATXN80S (ATXN8 обратной цепи (некодирующей)), PPP2R2B (протеинфосфатаза 2, регуляторная субъединица B, бета), ATXN7 (атаксин 7), TNRC6B (содержащий тринуклеотидный повтор 6B), TNRC6C (содержащий тринуклеотидный повтор 6C), CELF3 (CUGBP, Elav-подобное семейство, представитель 3), MAB21L1 (mab-21-подобный I (C. elegans)), MSH2 (гомолог mutS 2, рак прямой кишки, неполипозного типа 1 (E. coli)), TMEM185A (трансмемранный белок 185A), SIX5 (SIX гомеобокс 5), CNPY3 (гомолог canopy 3 (zebrafish)), FRAXE (хрупкий сайт, тип фолиевой кислоты, редкий, fra(X)(q28) E), GNB2 (связывающий нуклоетид гуанин белок (G-белок), бета-полипептид 2), RPL14 (рибосомальный белок L14), ATXN8 (атаксин 8), INSR (рецептор инсулина), TTR (транстиретин), EP400 (связывающий E1A белок p400), GIGYF2 (взаимодействующий с GRB10 GYT белок 2), OGGI (8-оксогуанин ДНК гликозилаза), STC1 (станниокальцин 1), CNDPI (карнозин дипептидаза 1 (семейство металлопептидазы M20)), C10orf2 (открытая рамка считывания 2 хромосомы 10), MAML3 подобный mastermind 3 (Drosophila), DKC1 (дискерин врожденного дискератоза 1), PAXIP1 PAX взаимодействующий (с доменом активации транскрипции домен) белок 1, CASK (кальций/кальмодулин-зависимая протеинкиназа (семейство MAGUK)), МАРТ (ассоциированный с микротрубочками белок тау), SP1 (транскрипионный фактор Sp1), POLG (ДНК-зависимая РНК-полимераза гамма), AFF2 (семейство AF4/FMR2, представитель 2), THBS1 (тромбоспондин 1), TP53 (белок опухолей p53), ESR1 (рецептор эстрогена 1), CGGBP1 (связывающий повторы триплета CGG белок 1), ABT1 (активатор базальной транскрипии 1), KLK3 (связанная с каликреином пептидаза 3), PRNP (прионный белок), JUN (онкоген jun), KCNN3 (калиевый средней/малой проводимости активируемый кальцием канал, подсемейство N, представитель 3), BAX (ассоциированный с BCL2 белок X), FRAXA (хрупкий сайт, тип фолиевой кислоты, редкий, fra(X)(q27.3), A (макроорхидизм при умственной отсталости)), KBTBD10 (повтор kelch и содержащий домен BTB (POZ) 10), MBNL1 (подобный muscleblind (Drosophila)), RAD51 (гомолог RAD51 (гомолог RecA, E. coli) (S. cerevisiae)), NCOA3 (коактиватор ядерного рецептора 3), ERDA1 (домен экспансированных повторов, CAG/CTG 1), TSC1 (туберозного склероза 1), COMIP (белок олигомерно матрикса хряща), GCLC (глутаматцистеиновая лигаза, каталитическая RRAD (связанная с Ras, ассоциированная с диабетом), MSH3 (гомолог mutS 3 (E. coli)), DRD2 (дофаминовый рецептор D2), CD44 (молекула CD44 (по индийской системе групп крови)), CTCF (связывающий CCCTC фактор (белок цинковый палец)), CCND1 (циклин DI), CLSPN (гомолог класпина (Xenopus laevis)), MEF2A (энхансерный фактор миоцитов 2A), PTPRU (протеиновая тирозинфосфатаза, тип рецептора U), GAPDH (глицеральдегид-3-фосфат дегидрогеназа), TRIM22 (трехчастный содержащий мотив 22), WT1 (опухоли Вилмса 1), AHR (рецептор арилуглеводорода), GPX1 (глутатионпероксидаза 1), TPMT (тиопурин-S- метилтрансфераза), NDP (болезнь Норри (псевдоглиома)), ARX (связанный с aristaless гомеобокс), MUS81 (MUS81 гомолог эндонуклеазы (S. cerevisiae)), TYR (тирозиназа (окулокутанный альбинизм LA)), EGR1 (раннего ростового ответа 1), UNG (урацил-ДНК гликозилаза), NUMBL (подобный гомологу numb (Drosophila)), FABP2 (связывающий жирные кислоты белок 2, кишечный), EN2 (engrailed, ген, содержащий гомеобокс 2), CRYGC (кристалин гамма C), SRP14 (14-кДа сигнальная частица узнавания (гомологичный Alu РНК-связывающий белок)), CRYGB (кристаллин, гамма B), PDCD1 (программируемой клеточной смерти 1), HOXA1 (гомеобокс Al), ATXN2L (подобный атаксину 2), PMS2 (PMS2 постмейотического усиления сегрегации 2 (S. cerevisiae)), GLA (галактозидаза, альфа), CBL (Cas-Br-M (мышиный) экотропическая ретровирусная трансформирующая последовательность), FTH1 (ферритин, тяжелый полипептид 1), IL12RB2 (рецептор интерлейкина 12, бета 2), OTX2 (гомеобокс orthodenticle 2), HOXA5 (гомеобокс A5), POLG2 (ДНК-зависимая РНК-полимераза, гамма 2, дополнительная субъединица), DLX2 (гомеобокс distal-less 2), SIRPA. (сигнальный регуляторный белок альфа), OTX1 (orthodenticle-гомеобокс 1), AHRR (репрессор арилуглеводородного рецептора), MANF (мезенцефальный происходящий из астроцитов нейротрофический фактор), TMEM158 (трансмембранный белок 158 (ген/псевдоген)) и ENSG00000078687.
[001192] Предпочтительные белки, ассоциированные с заболеваниями, связанными с экспансией тринуклеотидных повторов, включают HTT (хантингин), AR (рецептор андрогена), FXN (фратаксин), Atxn3 (атаксин), Atxn1 (атаксин), Atxn2 (атаксин), Atxn7 (атаксин), Atxn10 (атаксин), DMPK (протеинкиназа миотонической дистрофии), Atn1 (атрофин 1), СВР (связывающий creb белок), VLDLR (рецептор липопротеинов очень низкой плотности) и их комбинацию.
Лечение заболеваний слуха
[001193] Настоящее изобретение также предусматривает доставку системы CRISPR-Cas в одно или оба уха.
[001194] Внимание исследователей уделяется вопросу о том, может ли генная терапия быть использована для того, чтобы улучшить существующие способы лечения глухоты, а именно кохлеарные импланты. Частой причиной глухоты является утрата или повреждение волосковых клеток, что вызывает нарушение передачи сигнала к слуховым нейронам. В таких случаях кохлеарные импланты могут использоваться для того, чтобы обеспечить реакцию на звук и передачу электрических сигналов к нервным клеткам. Однако такие нейроны часто дегенерируют и втягиваются из улитки при уменьшении количества факторов роста, выделяемых поврежденными волосковыми клетками.
[001195] В патентной заявке США 20120328580 описана инъекцию фармакологической смеси в ухо (например, введение в ухо), как, например, введение в просвет улитки (например, в среднюю лестницу (Scala media), лестницу преддверия (Scala vestibulae) и барабаннуюю лестницу (Scala tympani)), например, с использование шприца, например, одноразового шприца. К примеру, одно или более соединений, описанных в настоящей спецификации, могут быть введены с помощью интратимпанальной инъекции (в частности, в среднее ухо), и/или инъекций во наружное, среднее и/или внутреннее ухо. Такие методы широко применяются в данной области, например, для введения стероидов и антибиотиков в ухо человека. Инъекция может производиться, в частности, через круглое окно в ухе или капсулу улитки. Другие способы введения во внутреннее ухо также известны в данной (см., например, Salt and Plontke, Drug Discovery Today, 10:1299-1306, 2005).
[001196] При использовании другого способа введения фармакологическая смесь может быть введена in situ с помощью катетера или насоса. Катетер или насос могут, например, направлять фармакологическую смесь в кохлеарную полость или круглое окно в ухе и/или просвет прямой кишки. Широко применяемыми для доставки лекарств аппаратами и методами, которые подходят для введения одного или более описанных в настоящей спецификации соединений в ухо, например, ухо человека, как это описано в McKenna et al., (публикация США № 2006/0030837) и Jacobsen et al. (патент США № 7206639). В некоторых вариантах осуществления катетер или насос могут быть помещены, например, в ухе (например, в наружном, среднем и/или внутреннем ухе) пациента в ходе хирургического вмешательства. В некоторых модификациях изобретения катетер или насос могут быть помещены, например, в ухо (например, в наружном, среднем и/или внутреннем ухе) пациента без необходимости в хирургическом вмешательстве.
[001197] Альтернативно или в дополнительно к этому один или более компонентов, описанных в настоящей спецификации, может быть введен в сочетании с механическим устройством, таким как кохлеарный имплант и слуховой аппарат, которые помещают в наружное ухо. Широко используемый кохлеарный имплант, подходящий для использования вместе с настоящим изобретением, описан Edge et al. (публикация США № 2007/0093878).
[001198] В некоторых вариантах осуществления описанные выше способы введения могут быть использованы в произвольном порядке, одновременно или попеременно.
[001199] Альтернативно или в дополнение к этому настоящее изобретение может быть осуществлено в соответствии с любыми одобренными Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов (Food and Drug Administration) методами, например, описанными в Руководстве по стандартам данных (Data Standards Manual) CDER версии номер 004 (которая доступна на fda.give/cd.er/dsmZDRG/drg003 01. htm).
[001200] В целом, способы клеточной терапии, описанные в заявке на патент США 20120328580, могут быть применены для того, чтобы привести к полной или частичной дифференцировке клетки или применены по отношению к зрелым типам клеток, например, внутреннего уха (например, волосковых клеток) in vitro. Клетки, полученные в результате применения таких способов, затем могут быть трансплантированы или имплантированы пациенту, нуждающемуся в таком лечении. Способы культивирования клеток, необходимые для осуществления этих методов, включая способы идентификации и выбора подходящих типов клеток, способы содействия полной или частичной дифференцировке выбранных клеток, способы идентификации полных или частично дифференцированных типов клеток, а также способы имплантации полных или частично дифференцированных клеток описаны ниже.
[001201] Клетки, подходящие для использования в настоящем изобретении, включают, но не ограничиваются, клетками, способными полностью или частично дифференцироваться в зрелую клетку внутреннего уха, например, волосковую клетку (например, внутреннюю и/или внешнюю волосковую клетку) при контакте, например, in vitro, с одним или более соединениями, описанными в настоящей заявке. Примеры клеток, способных дифференцироваться в волосковые клетки, включают, но не ограничиваются, стволовыми клетками (например, стволовыми клетками внутреннего уха, взрослыми стволовыми клетками, стволовыми клетками костного мозга, эмбриональными стволовыми клетками, мезенхимальными стволовыми клетками, стволовыми клетками кожи, индуцированными стволовыми клетками iPS и стволовыми клетками, полученные из жировой ткани), клетками-предшественниками (например, клетками-предшественниками внутреннего уха), вспомогательными клетками (например, клетками Дейтерса, столбчатыми клетками, внутренними фаланговыми клетками, клетками тектума и клетками Хенсена) и/или зародышевыми клетками. Использование стволовых клеток для замены сенсорных клеток внутреннего уха описано в Li et al. (публикация США № 2005/0287127) и Li et al. (патент США № 11/953,797). Использование стволовых клеток, полученных из костного мозга, для замены сенсорных клеток внутреннего уха описано в Edge et al., PCT/US2007/084654. Использование индуцируемых стволовых клеток (iPS) клеток описано, например, в Takahashi et al. Cell, Volume 131, Issue 5, Pages 861-872 (2007); Takahashi and Yamanaka, Cell 126, 663-76 (2006); Okita et al., Nature 448, 260-262 (2007); Yu, J. et al., Science 318(5858):1917-1920 (2007), Nakagawa et al., Nat. Biotechnol. 26:101-106 (2008); and Zaehres and Scholer, Cell 131(5):834- 835 (2007). Такие подходящие клетки можно идентифицировать, анализируя (например, качественно или количественно) наличие одного или более тканеспецифических генов. Например, экспрессия гена может быть обнаружена путем обнаружения белкового продукта одного или более тканеспецифических генов. Способы обнаружения белка включают окрашивающие белки (например, с использованием экстрактов клеток или целых клеток) с использованием антител против соответствующего антигена. В этом случае подходящим антигеном является белковый продукт тканеспецифической экспрессии гена. Хотя, в целом, первичное антитело (то есть антитело, которое связывает антиген) может быть меченым, более частым является (и улучшает визуализацию) использование второго антитела к первому (например, анти-IgG). Это второе антитело конъюгировано либо с флуорохромами, либо с соответствующими ферментами для колориметрических реакций, либо с золотыми шариками (для электронной микроскопии), либо с системой биотина-авидина, так что можно определить местоположение первичного антитела и, следовательно, антигена.
[001202] Молекулы CRISPR Cas по настоящему изобретению могут быть доставлены в ухо путем прямого внесения фармацевтической композиции в наружное ухо с композициями, модифицированными из опубликованной заявки США 20110142917. В некоторых вариантах осуществления фармацевтическая композиция доставляется в ушной канал. Доставка в ухо может также называться ушной или проводимой в ухо доставкой.
[001203] В некоторых вариантах осуществления изобретения молекулы РНК по изобретению поставляются в композициях липосомы или липофектина и тому подобных и могут быть получены способами, хорошо известными специалистам в данной области. Такие способы описаны, например, в патентах США № 5599772, 5589466 и 5580859, которые включены в настоящее описание в качестве отсылки.
[001204] Системы доставки, нацеленные специфично на расширенную и улучшенную доставку миРНК в клетки млекопитающих, были разработаны (см., например, Shen et al. FEBS Let. 2003, 539:111-114; Xia et al., Nat. Biotech. 2002, 20:1006-1010; Reich et al., Mol. Vision. 2003, 9: 210-216; Sorensen et al., J. Mol. Biol. 2003, 327: 761-766; Lewis et al., Nat. Gen. 2002, 32: 107- 108 и Simeoni et al., NAR 2003, 31, 11: 2717-2724) и могут быть применены к данному изобретению. Недавно миРНК была успешно использована для ингибирования экспрессии генов у приматов (см., например, Tolentino et al., Retina 24(4):660) и может также быть применена к данному изобретению.
[001205] Qi et al. описывает способы эффективной трансфекции миРНК во внутреннее ухо через интактное круглое отверстие согласно новой протеоидной технологии доставки, которая может быть применена к системе нацеливания на нуклеиновые кислоты согласно настоящему изобретению (см., например, Qi et al., Gene Therapy (12013), 1-9). В частности, доставка двухцепочечных РНК-связывающих доменов ТАТ (TAT-DRBD), которые могут трансфицировать меченную Cy3 миРНК в клетки внутреннего уха, включая внутренние и внешние волосковые клетки, полукружные каналы (crista ampullaris), слуховые пятна: овального мешочка (macula utriculi) и пятно сферического мешочка (macula sacculi), посредством интактного проникновения в круглое отверстие была успешной для двухцепочечных миРНК in vivo для лечения различных заболеваний внутреннего уха и сохранения слуховой функции. Приблизительно 40 мкл 10 мМ РНК можно рассматривать как оптимальную дозировку для введения в ухо.
[001206] Согласно Rejali et al. (Hear Res, 2007 Jun, 228 (l-2): 180-7), функционирование кохлеарного имплантата может быть улучшена путем хорошей сохранности спиральных ганглиозных нейронов, которые являются объектом электрической стимуляции имплантатом. Ранее было показано, что нейротропный фактор мозга (BDNF) повышает выживаемость спирального ганглия в экспериментально оглушенных ушах. Rejali et al. протестировали модифицированную конструкцию кохлеарного имплантационного электрода, которая включала покрытие из клеток фибробластов, трансдуцированных вирусным вектором с вставкой гена BDNF. Чтобы выполнить этот тип передачи гена ex vivo, Rejali et al. трансдуцировали фибробласты морской свинки с аденовирусом с вставкой из кассеты BDNF и удостоверились, что эти клетки секретируют BDNF, а затем прикрепили BDNF-секретирующие клетки к кохлеарному имплантату с помощью агарозного геля и имплантировали электрод в барабанную лестницу (scala tympani). Rejali et al.. определили, что экспрессирующие BDNF электроды смогли сохранить значительно больше спиральных ганглиозных нейронов в базальных витках улитки после 48 дней имплантации по сравнению с контрольными электродами и продемонстрировали возможность объединения лечения при помощи кохлеарной имплантации с переносом гена ex vivo для усиления выживаемости нейронов спирального ганглия. Такая система может быть применена к системе нацеливания на нуклеиновые кислоты, представленной в изобретении, для доставки к уху.
[001207] Mukherjea et al. (Antioxidants & Redox Signaling Volume 13, Number 5, 2010), продемонстрировали, что нокдаун N0X3 с использованием короткой интерферирующей (si) РНК блокировал ототоксичность цисплатина, о чем свидетельствует защита внешних волосковых клеток (OHC) от повреждения и снижение пороговых сдвигов в слуховой реакции ствола головного мозга (ABR). Различные дозы siNOX3 (0,3, 0,6 и 0,9 мкг) вводили крысам, а экспрессию NOX3 оценивали с помощью ОТ-ПЦР в реальном времени. Самая низкая доза используемой NOX3 миРНК (0,3 мкг) не показала никакого ингибирования мРНК N0X3 по сравнению с транстимпаническим введением скремблированной миРНК или необработанными слуховыми улитками. Однако введение более высоких доз миРНК N0X3 (0,6 и 0,9 мкг) уменьшало экспрессию N0X3 по сравнению с контрольной скремблированной миРНК. Такая система может быть применена к системе CRISPR Cas согласно настоящему изобретению для транстимпанического введения при дозе от примерно 2 мг до примерно 4 мг CRISPR Cas для введения человеку.
[001208] Jung et al. (Molecular Therapy, vol. 21 no. 4, 834-841 apr. 2013) демонстрируют, что уровень Hes5 в эллиптическом мешочке уменьшался после применения миРНК и что количество волосковых клеток в указанном эллиптическом мешочке было значительно больше, чем в контроле. Данные показывают, что технология с применением миРНК может быть полезна для индукции восстановления и регенерации во внутреннем ухе и что сигнальный путь Notch является потенциально полезной мишенью для специфического ингибирования экспрессии генов. Jung et al. вводили 8 мкг миРНК Hes5 в объеме 2 мкл, полученном путем добавления стерильного нормального физиологического раствора к лиофилизированной миРНК к вестибулярному эпителию уха. Такая система может быть применена к системе нацеливания нуклеиновых кислот согласно настоящему изобретению для введения в вестибулярный эпителий уха с дозой от примерно 1 до примерно 30 мг CRISPR Cas для введения человеку.
Нацеливание на гены в неделящихся клетках (нейроны и мышцы)
[001209] Неделящиеся (особенно неделящиеся, полностью дифференцированные) типы клеток представляют проблемы для нацеливания на гены или генной инженерии, например, поскольку гомологичная рекомбинация (HR) обычно подавляется в фазе клеточного цикла G1. Однако, изучая механизмы, с помощью которых клетки контролируют нормальные системы репарации ДНК, Durocher обнаружил ранее неизвестный переключатель, который удерживает HR в "выключеном" состоянии в неделящихся клетках и разработал стратегию для переключения этого переключателя обратно. Orthwein et al. (лаборатория Daniel Durocher в больнице Mount Sinai в Оттаве, Канада) недавно опубликовали (Nature 16142, опубликовано через интернет 9 декабря 2015 года) что подавление HR может быть отменено, и, таким образом, нацеливание на гены как в клетках почек (293T), так и в клетках остеосаркомы (U2OS) проведено успешно. Известно, что опухолевые супрессоры BRCA1, PALB2 и BRAC2 способствуют восстановлению двухцепочечных разрывов ДНК (DSB) путем гомологичной рекомбинации (HR). Они обнаружили, что образование комплекса BRCA1 с PALB2-BRAC2 регулируется сайтом убиквитина на PALB2, так, что такое действие регулируется на указанном сайте с помощью убиквитин-лигазы E3. Эта убиквитин-лигаза E3 состоит из KEAP1 (белка, взаимодействующего с PALB2) в комплексе с куллином-3 (CUL3)-RBX1. Убиквитиляция PALB2 подавляет его взаимодействие с BRCA1 и противодействует деубиквитиназе USP11, которая сама по себе контролируется клеточным циклом. Восстановление взаимодействия BRCA1-PALB2 в сочетании с активацией резекции (удаления) концов ДНК являются достаточными для индукции гомологичной рекомбинации в G1, что можно измерить рядом способов, включая анализ нацеливания на гены на основе CRISPR-Cas9, направленный на USP11 или KEAP1 (экспрессируемый из вектора pX459). Однако, когда взаимодействие BRCA1-PALB2 восстанавливалось в грамотрицательных G1-клетках с использованием либо истощения KEAP1, либо экспрессии мутанта PALB2-KR, было обнаружено достоверное увеличение случаев нацеливания на гены.
[001210] Таким образом, в некоторых вариантах предпочтительна реактивация HR в особенности в неделящихся клетках полностью дифференцированных типов клеток. В некоторых вариантах осуществления обеспечение взаимодействия BRCA1-PALB2 является предпочтительным. В некоторых вариантах осуществления целевая клетка является неделящейся. В некоторых вариантах осуществления клетка-мишень представляет собой нейрон или мышечную клетку. В некоторых вариантах осуществления нацеливание на клетку-мишень происходит in vivo. В некоторых вариантах осуществления клетка находится в G1 и HR подавлена. В некоторых вариантах осуществления предполагается использование KEAP1-истощения, например, ингибирования экспрессии активности KEAP1, что является предпочтительным. KEAP1-истощение может быть достигнуто посредством миРНК, например, как показано в Orthwein et al. Альтернативно, экспрессия мутанта PALB2-KR (без всех восьми остатков Lys в домене взаимодействия BRCA1) является предпочтительной и используется либо отдельно, либо в сочетании с истощением KEAP1. PALB2-KR взаимодействует с BRCA1 независимо от периода клеточного цикла, таким образом, продвижение или восстановление взаимодействия BRCA1-PALB2, особенно в клетках G1, является предпочтительным в некоторых вариантах осуществления, особенно там, где клетки-мишени не делятся, или где удаление и возврат (нацеливание на ген ex vivo) проблематично, например, в нейронах и мышечных клетках. миРНК KEAP1 можно получить от ThermoFischer. В некоторых вариантах реализации в клетку G1 может быть доставлен комплекс BRCA1-PALB2. В некоторых вариантах осуществления изобретения может быть стимулирована деубиквитинирование PALB2, например, путем увеличения экспрессии деубиквитиназы USP11, поэтому предполагается, что конструкция может быть предоставлены для стимуляции или повышения регуляции экспрессии или активности деубиквитиназы USP11.
Лечение заболеваний глаз
[001211] Настоящее изобретение также предполагает доставку системы CRISPR-Cas в один или оба глаза.
[001212] В еще одном варианте изобретения система CRISPR-Cas может использоваться для коррекции глазных дефектов, которые возникают из нескольких генетических мутаций, описанных подробно далее в Genetic Diseases of the Eye, Second Edition, edited by Elias I. Traboulsi, Oxford University Press, 2012.
[001213] Для введения в глаза особенно предпочтительны лентивирусные векторы, в частности вирусы инфекционной анемии лошадей (EIAV).
[001214] В одном из вариантов осуществления предусматриваются также минимальные лентивирусные векторы, на основе вируса инфекционной анемии лошадей (EIAV), особенно для глазной генной терапии (см., например, Balagaan, J Gene Med 2006; 8: 275-285, Published online 21 November 2005 in Wiley InterScience (www.interscience.wiley.com). DOF 10.1002/jgm.845). Предполагается, что векторы имеют промотор цитомегаловируса (CMV), индуцирующий экспрессию гена-мишени. Интракамеральные, субретинальные, интраокулярные и интравитреальные инъекции (см., например, Balagaan, J Gene Med 2006; 8: 275-285, Published online 21 November 2005 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/jgm.845). Интраокулярные инъекции могут выполняться с помощью операционного микроскопа. Для субретинальных и интравитреальных инъекций глаза могут быть пролабированы под небольшим давлением и под наблюдением и при осмотре, которые визуализируются с использованием системы контактных линз, состоящей из капли раствора соединительной среды, на роговице, покрытой покровным стеклом микроскопа. Для субретинальных инъекций кончик 10-мм 34-калибральной иглы, установленный на шприце с галоидом 5 мкл, может быть продвинут под непосредственной визуализацией через высшую экваториальную склеру тангенциально к заднему полюсу до тех пор, пока отверстие иглы не будет видно в субретинальное пространство. Затем может быть введено 2 мкл векторной суспензии для получения выступающей буллезной отслойки сетчатки, что подтверждает введение субретинального вектора. Этот подход создает самогерметичную склеротомию, позволяющую удерживать векторную суспензию в субретинальном пространстве до тех пор, пока она не будет поглощена пигментным эпителием сетчатки (RPE), обычно в течение 48 часов после процедуры. Эту процедуру можно повторить в нижнем полушарии, чтобы произвести там отслоение сетчатки. Этот способ приводит к экспонированию примерно 70% нейросенсорной сетчатки и пигментного эпителия сетчатки (RPE) к суспензии вектора. Для интравитреальных инъекций наконечник иглы можно продвигать через склеру 1 мм сзади к корнеосклеральному лимбу и вводить 2 мкл векторной суспензии в стекловидную полость. Для внутрикамеральных инъекций наконечник иглы может быть продвинут через корнеосклеральный лимбальный парацентез, направленный к центральной роговице, и может быть введено 2 мкл векторной суспензии. Эти векторы могут быть инъецированы в титрах 1,0-1,4×1010 или 1,0-1,4×109 трансдуцирующих единиц (TU)/мл.
[001215] В другом варианте осуществления изобретения, RetinoStat®, основанная на вирусе инфекционной анемии лошадей лентивирусная генотерапия использует вектор, экспрессирующий ангиостатические белки эндостатин и ангиостатин, доставляемые посредством субретинальную инъекцию для лечения сетчатой формы возрастной макулодегенерации (см, например, Binley et al., HUMAN GENE THERAPY 23:980-991 (September 2012)). Такой вектор может быть изменен для использования в нацеленной на нуклеиновые кислоты системе по настоящему изобретению. Каждый глаз можно лечить RetinoStat® в дозировке 1,1×105трансдуцирующих единиц на глаз (TU/глаз) в общем объеме 100 мкл.
[001216] В другом варианте осуществления E1-, частичный E3-, E4-удаленный аденовирусный вектор может быть предусмотрен для доставки в глаз. Двадцать восемь пациентов с развитой неоваскулярной возрастной макулодистрофией (AMD) получили одну интравитреальную инъекцию E1-, частичного E3-, E4-удаленного аденовирусного вектора, экспрессирующего пигментный фактор эпителиального происхождения (AdPEDF II) (см. например, Campochiaro et al., Human Gene Therapy 17:167-176 (February 2006). Были исследованы дозы от 106 до 109,5 единиц (PU) и не было показано никаких серьезных побочных эффектов, связанных с AdPEDF II, и отсутствовала токсичность, накладывающая ограничения на дозу препарата (см., например, Campochiaro et al., Human Gene Therapy 17:167-176 (February 2006)). Аденовирусный перенос окулярного гена при помощи вектора, по-видимому, является продуктивным способом для лечения глазных заболеваний и может быть применен к системе CRISPR Cas.
[001217] В другом варианте осуществления система sd-rxRNA® RXi Pharmaceuticals может быть использована и/или адаптирована для доставки CRISPR-Cas в глаз. Данная система подразумевает одно интравитреальное введение 3 мкг sd-rxRNA, которое приводит к последовательному снижению уровня мРНК PPIB в течение 14 дней. Система sd-rxRNA® может быть применена к системе нацеливания на нуклеиновые кислоты согласно настоящему изобретению, предполагая дозу приблизительно от 3 до 20 мг CRISPR, вводимую человеку.
[001218] Millington-Ward et al. (Molecular Therapy, vol. 19 no. 4, 642-649 apr. 2011) описывают использование аденоассоциированных вирусов (AAV) для доставки супрессора родопсина, основанного на РНК-интерференции (РНК-i), и кодон-модифицированного гена родопсина, устойчивого к супрессии вследствие нуклеотидных изменений в вырожденных позициях в сайте-мишени РНК интерференции. Инъекция в глаз дозы 6,0×108 вирусных частиц (вч) или 1,8×1010 вч AAV была произведена субретинентально Millington-Ward et al. ААV-векторы Millington-Ward et al. могут быть применены к системе CRISPR Cas согласно настоящему изобретению, с предполагаемой дозой от примерно 2×1011 до примерно 6×1013 вирусных частиц (вч), вводимую человеку.
[001219] Dalkara et al. (Sci Transl Med 5, 189ra76 (2013)) также обращается к направленной эволюции in vivo для создания AAV-вектора, который доставляет версии дефектных генов дикого типа по всей сетчатке после неинвазивной инъекции в стекловидное тело глаза. Dalkara описывает 7-мерную пептидную библиотеку и AAV-библиотеку, сконструированную путем ДНК-шаффлинга генов cap из AAV1, 2, 4, 5, 6, 8 и 9. Библиотеки rcAAV и векторы rAAV, экспрессирующие GFP под воздействием промотора CAG или Rho, были упакованы и, были получены устойчивые к воздействию дезоксирибонуклеаз геномные титры при помощи количественной ПЦР. Библиотеки были объединены, и были проведены два раунда эволюции, каждый из которых состоял из первоначальной диверсификации библиотек, за которой следовали три этапа отбора in vivo. На каждом таком этапе мышам P30 rho-GFP интравитреально вводили 2 мл очищенной иодиксанолом фосфатно-буферной солевой (PBS)-диализованной библиотеки с геномным титром около 1×1012 вирусных геномов (вг)/мл. AAV-векторы Dalkara et al. могут быть применены к системе нацеливания нуклеиновой кислоты по настоящему изобретению, предусматривающей дозу приблизительно от 1×1015 до приблизительно 1×1016 вирусных геномов (вг)/мл, вводимую человеку.
[001220] В другом варианте может быть осуществлено нацеливание на ген родопсина для лечения пигментного ретинита (RP), в этом варианте система согласно патентной публикации США № 20120204282 закрепленная за Sangamo BioSciences Inc. может быть модифицирована в соответствии с системой CRISPR-Cas по настоящему изобретению.
[001221] В одном из вариантов осуществления способы патентной публикации США № 20130183282, закрепленные за Cellectis, которые заключаются в методах расщепления последовательности-мишени из гена родопсина человека, также могут быть модифицированы для системы нацеливания на нуклеиновые кислоты согласно настоящему изобретению.
[001222] Патентная публикация США № 20130202678, закрепленная за Academia Sinica, относится к способам лечения ретинопатий и угрожающих зрению офтальмологических расстройств, связанных с доставкой гена Puf-A (который экспрессируется в ганглии сетчатки и пигментированных клетках тканей глаза и показывает уникальную антиапоптозную активность) в субретинальное или интравитреальное пространство глаза. В частности, желательными мишенями являются zgc: 193933, prdm1a, spata2, tex10, rbb4, ddx3, zp2.2, Blimp-1 и HtrA2, на все из которых может быть нацелена система нацеливания нуклеиновых кислот согласно настоящему изобретению.
[001223] Wu (Cell Stem Cell, 13: 659-62, 2013) разработали направляющую РНК, которая направляла Cas9 к мутации в единственной паре оснований, вызывавшей катаракту у мышей, где она индуцировала расщепление ДНК. Затем, используя либо другой аллель дикого типа, либо олигонуклеотиды, вводимые в зиготы, механизмы репарации корректировали последовательность сломанного аллеля и исправляли генетический дефект, вызывающий катаракту у мутантной мыши.
[001224] В патентной публикации США № 20120159653 описано использование нуклеаз с цинковыми пальцами для генетической модификации клеток, животных и белков, связанных с макулярной дегенерацией (МD). Макулярная дегенерация (MD) является основной причиной нарушения зрения у пожилых людей, но также является определяющим симптомом детских заболеваний, таких как болезнь Штаргардта, дистрофия глазного дна Сорсби, и фатальные нейродегенеративные болезни у детей, с проявлением первых признаков заболевания в младенчестве. Дегенерация макулы (желтого пятна) приводит к потере зрения в центре поля зрения (макулы), поскольку повреждается сетчатка. В настоящее время существующие животные модели не повторяют основные признаки болезни таким образом, наблюдающиеся у людей. Доступные животные модели, содержащие мутантные гены, кодирующие белки, связанные с MD, также приводят к проявлению в значительной степени разнообразных фенотипов, что усложняет переход к изучению заболевания у человека и создает проблемы для разработки терапии.
[001225] В одном аспекте патентная публикация США № 20120159653 относится к редактированию любых хромосомных последовательностей, которые кодируют белки, связанные с развитием MD, которые могут быть применены к системе нацеливания нуклеиновых кислот согласно настоящему изобретению. Белки, связанные с MD, обычно выбираются экспериментальным путем, связывая белок, ассоциированный с MD, с расстройством МD. Например, скорость синтеза или циркулирующая концентрация белка, связанного с МD, может быть повышена или понижена в популяции, имеющей расстройство MD, относительно популяции, у которой отсутствует расстройство MD. Различия в уровнях белка могут быть оценены с использованием методов протеомики, включая, но не ограничиваясь ими: вестерн-блоттинг, иммуногистохимическое окрашивание, иммуноферментный анализ (ELISA) и масс-спектрометрию. Альтернативно, белки, связанные с МD, могут быть идентифицированы путем получения профилей генной экспрессии генов, кодирующих белки, при помощи геномных методов, включая, но не ограничиваясь ими, анализ на микрочипах ДНК, последовательный анализ экспрессии генов (SAGE) и количественную полимеразную цепную реакцию в реальном времени (кПЦР).
[001226] В качестве неограничивающего примера белки, связанные с МD, включают, но не ограничиваются ими: ATP-связывающую кассету, член 4 (ABCA4) подсемейства A (ABC1), член 4 ACHM1-ахроматопсии (палочковая монохромазия), 1 ApoE аполипопротеин E (ApoE), C1QTNF5 (CTRP5) Clq и связанный с фактором некроза опухоли белок 5 (C1QTNF5), C2 компонент комплемента 2 (C2), компоненты комплемента C3 (C3), CCL2 лиганд хемокинов (CC-мотив) 2 (CCL2), CCR2 рецептор 2 хемокинов (CC мотив) (CCR2), CD36 кластер дифференциации 36, CFB фактор комплемента В, CFH фактор комплемента CFH, фактор комплемента 1, связанный с Н CFHR1, фактор комплемента 3, связанный с Н CFHR3, циклический нуклеотид закрытого канала бета 3 CNGB3, CP-целлюлоплазмин (CP), CRP C-реактивный белок (CRP), CST3 цистатин C или цистатин 3 (CST3), CTSD катепсин D (CTSD), CX3CR1 рецептор 1 хемокинов (C-X3-C-мотив), ELOVL4 белок удлинение очень длинноцепочечных жирных кислот 4, эксцизионной репарации ДНК ERCC6, белок кросс-дополняющего дефицита грызунов, группа комплемента 6, фибулин-5 FBLN5, фибулин 6 FBLN6, FSCN2 фасцин (FSCN2), HMCN1 гемицентрин 1, HTRA1 HTRA сериновая пептидаза 1 (HTRA1), IL-6 интерлейкин 6, IL-8 интерлейкин 8, LOC387715, гипотетический белок PLEKHA1, член 1 семейства А, содержащего домен гомологии плекстрина (PLEKHA1), PROM1 проминин 1 (PROM1 или CD133), PRPH2 периферин-2, RPGR регулятор GTP-азы пигментного ретинита, SERPING1 ингибитор серпиновой пептидазы, член 1 клады G (Cl-ингибитор), TCOF1, Treacle TIMP3 ингибитор металлопротеиназы 3 (TIMP3), Toll-подобный рецептор 3 TLR3.
[001227] Тип белка, связанного с MD, хромосомная последовательность которого отредактирована, может и будет изменяться. В предпочтительных вариантах осуществления белки, связанные с MD, хромосомная последовательность которыхподвергается редактированию, могут представлять АТФ-связывающую кассету, член 4 (ABCA4) подсемейства A (ABC1), кодируемый геном ABCR, аполипопротеин E (APOE), кодируемый ген APOE, лиганд хемокинов (C-C мотив) 2 (CCL2), кодируемый геном CCL2, рецептор хемокина 2 (C-C мотив) (CCR2), кодируемый геном CCR2, церулоплазмин (CP), кодируемый геном CP, катепсин D (CTSD), кодируемый геном CTSD, или ингибитор металлопротеиназы 3 (TIMP3), кодируемый геном TIMP3. В иллюстративном варианте осуществления генетически модифицированное животноеявляется крысой, а отредактированная хромосомная последовательность, кодирующая белок, связанный с MD, может быть: (ABCA4) ATP-связывающая кассета, подгруппа NM_000350 подсемейства A (ABC1), член 4 APOE аполипопротеин E NM138828 (APOE), CCL2 лиганд 2 хемокинов (C-C NM_031530 мотив) (CCL2), CCR2 хемокин (C-C NM_021866 мотив) рецептор 2 (CCR2), CP церулоплазмин (CP) NM_012532, CTSD катепсин D (CTSD) NM_134334, ингибитор металлопротеиназы TIMP3 NM_012886 3 (TIMP3). Животное или клетка могут содержать 1, 2, 3, 4, 5, 6, 7 или более нарушенных хромосомных последовательностей, кодирующих белок, ассоциированный с MD, а также ноль, 1, 2, 3, 4, 5, 6, 7 или более хромосомно-интегрированные последовательностей, кодирующих разрушенный белок, связанный с MD.
[001228] Измененная или интегрированная хромосомная последовательность может быть модифицирована для кодирования измененного белка, связанного с MD. Несколько мутаций в хромосомных последовательностях были связаны с MD. Неограничивающие примеры мутаций в хромосомных последовательностях, связанных с MD, включают те, которые могут вызывать MD, в том числе в белке ABCR, E471K (т.е. глутамат в положении 471 заменен на лизин), R1129L (то есть аргинин в положении 11129 заменен на лейцин), T1428M (т.е. треонин в положении 1428 заменен на метионин), R1517S (т.е. аргинин в положении 1517 заменен на серин), I1562T (т.е. изолейцин в положении 1562 заменен на треонин), а G1578R (т.е. глицин в положении 1578 изменен на аргинин); в белке CCR2 V64I (то есть валин в положении 192 заменен на изолейцин); в белке CP G969B (т.е. глицин в положении 969 заменен на аспарагин или аспартат); в белке TIMP3 S156C (т.е. серин в положении 156 заменен на цистеин), G166C (т.е. глицин в положении 166 заменен на цистеин), G167C (т.е. глицин в положении 167 заменен на цистеин), Y168C (т.е. тирозин в положении 168 изменен на цистеин), S17C (т.е. серин в положении 170 заменен на цистеин), Y172C (т.е. тирозин в положении 172 заменен на цистеин) и S181C (т.е. серин в положении 181 заменен на цистеин). Также и другие ассоциации генетических вариантов в MD-ассоциированных генах и заболеваниях известны в данной области.
Лечение заболеваний кровеносной и мышечной систем
[001229] Настоящее изобретение также предусматривает доставку системы CRISPR-Cas, описанной здесь, например, систем эффекторных белков C2c1 или C2c3 в сердце. Для доставки в сердце предпочтительным является аденоассоциированный вирус миокарда (AAVM), в частности AAVM41, который показал преимущественный перенос генов в сердце (см., например, Lin-Yanga et al., PNAS, March 10, 2009, vol, 106, no. 10). Доставка может быть системной или локальной. Для системного введения предусматривается дозировка около 1-10×1014 векторных геномов. Смотрите, например, Eulalio et al. (2012) Nature 492: 376 and Somasuntharam et al. (20013) Biomaterials 34: 7790.
[001230] Например, в патентной публикации США № 20110023139 описано использование нуклеаз цинковых пальцев для генетической модификации клеток, животных и белков, связанных с сердечно-сосудистыми заболеваниями. Сердечно-сосудистые заболевания обычно включают высокое артериальное давление, сердечные приступы, сердечную недостаточность и инсульт и TIA. Любая хромосомная последовательность, вовлеченная в сердечно-сосудистое заболевание или белок, кодируемый любой хромосомной последовательностью, участвующий в сердечно-сосудистых заболеваниях, может быть использованы в способах, описанных в настоящем описании. Белки, связанные с сердечно-сосудистыми заболеваниями, обычно выбираются на основе экспериментальной ассоциации связанного с сердечно-сосудистыми заболеваниями белка с развитием сердечно-сосудистых заболеваний. Например, скорость синтеза или циркулирующая концентрация связанного с сердечно-сосудистым заболеванием белка может быть повышена или понижена в популяции, имеющей сердечно-сосудистое расстройство, относительно популяции, не имеющей сердечно-сосудистого расстройства. Различия в уровнях белка могут быть оценены с использованием методов протеомики, включая, но не ограничиваясь ими: вестерн-блоттинг, иммуногистохимическое окрашивание, иммуноферментный анализ (ELISA) и масс-спектрометрию. Альтернативно, белки, связанные с сердечно-сосудистыми заболеваниями, могут быть идентифицированы путем получения профилей экспрессии генов, кодирующих белки, с использованием геномных методов, включая, но не ограничиваясь ими, анализ на микрочипах ДНК, последовательный анализ экспрессии генов (SAGE) и количественную полимеразную цепную реакцию в реальном времени (кПЦР).
[001231] В качестве примера хромосомная последовательность может включать, но не ограничивается, следующими: IL1B (интерлейкин 1, бета), XDH (ксантиндегидрогеназа), TP53 (белок опухолей p53), PTGIS (простагландин 12 (простациклин) синтаза), MB (миоглобин), IL4 (интерлейкин 4), ANGPT1 (ангиопоэтин 1), ABCG8 (АТФ-связывающая кассета, подсемейство G (WHITE), член 8), CTSK (катепсин K), PTGIR (простагландин 12 (простациклин) рецептор (IP)), KCNJ11 (калиевый канал внутреннего выпрямления, подсемейство J, член 11), INS (инсулин), CRP (C-реактивный белок, связанный с пентраксином), PDGFRB (рецептор фактора роста тромбоцитов, бета-полипептид), CCNA2 (циклин A2), PDGFB (бета-полипептид фактора роста тромбоцитов (вирусный онкогенный гомолог саркомы обезьян (v-sis)), KCNJ5 (калиевый канал внутреннего выпрямления, подсемейство J, член 5), KCNN3 (кальций-активируемые калиевые каналы средней/малой проводимости, подсемейство N, член 3), CAPN10 (кальпаин 10), PTGES (простагландин-E-синтаза), ADRA2B (адренергический, альфа-2B-, рецептор), ABCG5 (ATФ-связывающая кассета, подгруппа G (WHITE), элемент 5), PRDX2 (пероксиредоксин 2), CAPN5 (кальпаин 5), PARP14 (семейство поли (ADФ-рибоза), полимераз, член 14), MEX3C (мекс-3 гомолог С (С. elegans)), ACE фермент превращения ангиотензина I (пептидилдипептидаза А) 1, ФНО (фактор некроза опухолей (надсемейство TNF, член 2)), IL6 (интерлейкин 6 (интерферон, бета 2)), STN (статин), SERPINE1 (ингибитор серпиновой пептидазы, клада E (нексин, ингибитор активатора плазминогена типа 1), член 1), ALB (альбумин), ADIPOQ (адипонектин, C1Q и содержащий домен коллагена), APOB (аполипопротеин В (включая Ag (x) антиген)), APOE (аполипопротеин E) LEP (лептин), MTHFR (5,10-метилентетрагидрофолатредуктаза (NADPH)), APOA1 (аполипопротеин A-I), EDN1 (эндотелин I), NPPB (предшественник натрийуретического пептида B), NOS3 (синтаза оксида азота 3 (эндотелиальная клетка)), PPARG (гамма-рецептор, активируемый пролифератором пероксисом), PLAT (активатор плазминогена, ткань), PTGS2 (простагландин-эндопероксид-синтаза 2 (простагландин G/H-синтаза и циклооксигеназа)), CETP (белок переноса холестеринового эфира, плазма), AGTR1 (рецептор ангиотензина II, тип 1), HMGCR (3-гидрокси-3-4-метилглютарил-кофермент A редуктаза), IG F1 (инсулиноподобный фактор роста 1 (соматомедин C)), SELE селектин Е E), REN (ренин), PPARA (рецептор альфа,активируемый пролифератором пероксисом), PON1 (параоксоназа 1), KNG1 (кининоген 1), CCL2 (хемокин (CC мотив) лиганд 2), LPL (липопротеинлипаза), VWF (фактор фон Виллебранда), F2 (фактор коагуляции II (тромбин)), ICAM1 (молекула межклеточной адгезии 1), TGFB1 (трансформирующий фактор роста, бета 1), NPPA (предшественник натрийуретического пептида A), IL10 (интерлейкин 10), EPO (эритропоэтин), SOD1 (супероксиддисмутаза 1, растворимый), VCAM1 (васкулярная молекула клеточной адгезии 1), IFNG (интерферон, гамма), LPA (липопротеин, Lp (a)), MPO (миелопероксидаза), ESR1 (рецептор эстрогена 1), MAPK1 (митоген-активированная протеинкиназа 1), HP (гаптоглобин), F3 (фактор коагуляции III (тромбопластин, тканевой фактор)), CST3 (цистатин C), COG2 (компонент олигомерного комплекса Гольджи 2), MMP9 (матриксная металлопептидаза 9 (желатиназа B, 92-кДа желатиназа, коллагеназа IV типа), SERPINC1 (ингибитор серпиновой пептидазы, клада C (антитромбин), член 1), F8 (фактор коагуляции VIII, прокоагулянтный компонент), HMOX1 (гемоксигеназа (дециклирующая) 1), APOC3 (аполипопротеин C-III), IL8 (интерлейкин 8), PROK1 (прокинетицин 1), CBS (цистатионин-бета-синтаза), NOS2 (синтаза оксида азота 2, индуцибельная), TLR4 (толл-подобный рецептор 4), SELF (селектин P (гранульный мембранный белок 140 кДа, антиген CD62)), ABCA1 (АТФ-связывающая кассета, подсемейство А (АВС 1), член 1), AGT (ангиотензиноген (ингибитор серпиновой пептидазы, клада A, член 8)), LDLR (рецептор липопротеинов низкой плотности), GPT (глутамат-пируват трансаминаза (аланинаминотрансфераза)), VEGFA (фактор роста эндотелия сосудов A), NR3C2 (подсемейство ядерных рцепторов 3, группа C, член 2), IL18 (интерлейкин 18 (интерферон-гамма-индуцирующий фактор)), NOS1 (синтаза оксида азота 1 (нейронная)), NR3C1 (подсемейство ядерных рецепторов 3, группа C, член 1 (глюкокортикоидный рецептор)), FGB (бета-цепь фибриногена), HGF (фактор роста гепатоцитов (гепапоэтин A, фактор рассеяния)), IL1A (интерлейкин 1, альфа), RETN (резистин), AKT1 (мышиный тимома вирусный онкогенный гомолог (v-akt) 1), LIPC (липаза, печеночная), HSPD1 (60 кДа белок теплового шока 1 (шаперонин)), МАРК14 (митоген-активируемая протеинкиназа 14), SPP1 (секретируемый фосфопротеин 1), ITGB3 (интегрин, бета 3 (тромбоцитарный гликопротеин 111a, антиген CD61)), CAT (каталаза), UTS2 (уротензин 2), THBD (тромбомодулин), F10 (фактор коагуляции X), CP (церулоплазмин (ферроксидаза)), TNFRSF11B (надсемейство рецепторов фактора некроза опухолей, член 11b), EDNRA (рецептор эндотелина типа A), EGFR (рецептор эпидермального фактора роста (вирусный онкогенный гомолог эритробластного лейкоза (v-erb-b), птичий)), MMP2 (матриксная металлопептидаза 2 (желатиназа A, 72 кДа желатиназа, 12 кДа коллагеназа IV типа), PLG (плазминоген), NPY (нейропептид Y), RHOD (семейство генов ras, член D), MAPK8 (митоген-активируемая протеинкиназа 8), MYC (v-myc myelocytomatosis вирусный онкогенный гомолог миелоцитоматоза (v-myc) (птичий)), FN1 (фибронектин 1), CMA1 (химаза 1, тучная клетка), PLAU (активатор плазминогена, урокиназа), GNB3 (гуанин-нуклеотид связывающий белок (G-белок), бета-полипептид 3), ADRB2 бета-2-адренорецептор, поверхностный), APOA5 (аполипопротеин A-V), SOD2 (супероксиддисмутаза 2, митохондриальная), F5 (фактор коагуляции V (проакцелерин, лабильный фактор)), VDR (рецептор витамина D (1,25-дигидроксивитамин D3)), ALOX5 (арахидонатная 5-липоксигеназа), HLA-DRB1 (главный комплекс гистосовместимости, класс II, DR бета I), PARP1 (поли (AДФ-рибоза) полимераза 1), CD40LG (лиганд CD40), PON2 (параоксоназа 2), AGER (рецептор конечных продуктов гликозилирования), IRS1 (субстрат рецептора инсулина 1), PTGS1 (простагландин-эндопероксид-синтаза 1 (простагландин G/H-синтаза и циклооксигеназа)), ECE1 (эндотелин-конвертирующий фермент 1), F7 (фактор свертывания крови VII (ускоритель конверсии протромбина сыворотки)), URN (антагонист рецептора интерлейкина 1), EPHX2 (эпоксидгидролаза 2, цитоплазматическая), IGFBP1 (инсулиноподобный белок, связывающий фактор роста 1), МАРК 10 (митоген-активируемая протеинкиназа 10), FAS (Fas (суперсемейство рецепторов TNF, член 6)), ABCB1 (ATФ-связывающая кассета, подсемейство (MDR/TAP), член 1), JUN (jun-онкоген), IGFBP3 (инсулиноподобный белок, связывающий фактор роста 3), CD14 (молекула CD14), PDE5A (фосфодиэстераза 5A, cGMP-специфичная), AGTR2 (рецептор ангиотензина II, тип 2), CD40 (молекула CD40, член суперсемейства TNF-рецепторов 5), LCAT (лецитинхолестеринацилтрансфераза), CCR5 (рецептор хемокина (C-C мотив) 5), MMP1 (матриксная металлопептидаза 1 (интерстициальная коллагеназа)), TIMP1 (ингибитор металлопептидазы TIMP 1), ADM (адреномедуллин), DYT10 (дистония 10), STAT3 (сигнальный белок и активатор транскрипции 3 (фактор ответа острой фазы), MMP3 (матриксная металлопептидаза 3 (стромелизин 1, прожелатиназа)), ELN (эластин), USF1 (фактор транскрипции upstream 1), CFH (фактор комплемента H), HSPA4 (70-кДа белок теплового шока 4), MMP12 (матриксная металлопептидаза 12 (макрофагальная эластаза)), MME (мембранная металлоэндопептидаза), F2R (рецептор фактора свертывания крови II (тромбина)), SELL (селектин L), ANXA5 (аннексин A5), ADRB1 (бета-1-адренорецептор), CYBA (цитохром b-245, альфа-полипептид), FGA (альфа-цепь фибриногена), GGT1 (гамма-глутамилтрансфераза 1), LIPG (липаза, эндотелиальная), HIF1A (фактор, индуцируемый гипоксией 1, альфа-субъединица (основной фактор транскрипции с доменом "спираль-петля-спираль")), CXCR4 (рецептор хемокина (мотив C-X-C)), PROC (белок C (инактиватор факторов свертывания крови Va и VIIIa)), SCARB1 (скавенджер-рецептор класса B, член 1), CD79A (молекула CD79a, ассоциированная с иммуноглобулином альфа), PLTP (белок переноса фосфолипидов), ADD1 (аддуктин 1 (альфа)), FGG (гамма-цепь фибриногена), SAA1 (сывороточный амилоид Al), KCNH2 (потенциал-зависимый калиевый канал, подсемейство H (связанные с eag), член 2), DPP4 (дипептидилпептидаза 4), G6PD (глюкозо-6-фосфатдегидрогеназа), NPR1 (натрийуретический пептидный рецептор A/гуанилатциклаза A (рецептор атрионатриуретического пептида A)), VTN (витронектин), KIAA0101 (KIAA0101), FOS (гомолог вирусного онкогена мышиной остеосаркомы FBJ), TLR2 (толл-подобный рецептор 2), РРIG (пептидил-пролил-изомераза G (циклофилин G)), IL1R1 (рецептор интерлейкина 1, тип I), AR (андрогеновый рецептор), CYP1A1 (цитохром P450, семейство 1, подсемейство A, полипептид 1), SERPINA1 (ингибитор серпиновой пептидазы, клада A (альфа-1 антипротеиназа, антитрипсин), член 1), MTR (5-метилтетрагидрофолат-гомоцистеин метилтрансфераза), RBP4 (ретинол-связывающий белок 4, плазма), APOA4 (аполипопротеин A-IV), CDKN2A (ингибитор циклинзависимой киназы 2A (меланома, p16, ингибирует CDK4)), FGF2 (фактор роста фибробластов 2 (основной)), EDNRB (рецептор эндотелина типа B), ITGA2 (интегрин, альфа 2 (CD49B, альфа-2 субъединица рецептора VLA-2)), CABIN1 (связывающий кальциневрин белок 1), SHBG (глобулин, связывающий половые гормоны), HMGB1 (белок из группы с высокой подвижностью 1), HSP90B2P (90 кДа белок теплового шока бета (Grp94), член 2 (псевдоген)), CYP3A4 (цитохром P450, семейство 3, подсемейство A, полипептид 4), GJA1 (белок щелевого соединения, альфа 1, 43 кДа), CAV1 (кавеолин 1, белок кавеолы, 22 кДа), ESR2 (рецептор эстрогена 2 (ER beta)), LTA (лимфотоксин альфа (надсемейство TNF, член 1)), GDF15 (фактор дифференцировки роста 15), BDNF (нейротрофический фактор мозга), CYP2D6 (цитохром P450, семейство 2, подсемейство D, полипептид 6), NGF (фактор роста нервов (бета-полипептид)), SP1 (фактор транскрипции Spl), TGIF1 (фактор гомеобокс 1, индуцированный TGFB), SRC (гомолог вирусного онкогена саркомы (Шмидт-Руппин А-2) v-src (птичья)), EGF (эпидермальный фактор роста (бета-урогастрон)), PIK3CG (фосфоинозитид-3-киназа, каталитический, гамма-полипептид), HLA-A (главный комплекс гистосовместимости, класс I, A), KCNQ1 (потенциал-зависимый калиевый канал, KQT-подобное подсемейство, член 1), CNR1 (каннабиноидный рецептор 1 (мозг)), FBN1 (фибриллин 1), CHKA (холинкиназа альфа), BEST1 (бестрофин 1), APP (белок предшественника бета-амилоида (A4)), CTNNB1 (катенин (связанный с кадгерином белок), бета 1, 88 кДа), IL2 (интерлейкин 2), CD36 (молекула CD36 (рецептор тромбоспондина)), PRKAB1 (протеинкиназа, AMФ-активируемая, некаталитическая субъединица бета 1), TPO (пероксидаза щитовидной железы), ALDH7A1 (семейство альдегиддегидрогеназы 7, член A1), CX3CR1 (рецептор хемокина (C-X3-C мотив) 1), TH (тирозингидроксилаза), F9 (фактор свертывания крови IX), GH1 (гормон роста 1), TF (трансферрин), HFE (гемохроматоз), IL17A (интерлейкин 17A), PTEN (гомолог фосфатазы и тензина), GSTM1 (глутатион S-трансфераза mu 1), DMD (дистрофин), GATA4 (GATA-связывающий белок 4), F13A1 (фактор свертывания крови XIII, полипептид А1), TTR (транстиретин), FABP4 (связывающий жирные кислоты белок 4, адипоцит), PON3 (параоксоназа 3), APOC1 (аполипопротеин C-I), INSR (инсулиновый рецептор), TNFRSF1B (семейство рецепторов фактора некроза опухолей, член 1B), HTR2A (рецептор 5-гидрокситриптамина (серотонина) 2A), CSF3 (колониестимулирующий фактор 3 (гранулоцит)), CYP2C9 (цитохром P450, семейство 2, подсемейство C, полипептид 9), TXN (тиоредоксин), CYP11B2 (цитохром P450, семейство 11, подсемейство B, полипептид 2), PTH (паратиреоидный гормон), CSF2 (колониестимулирующий фактор 2 (гранулоцит-макрофаг)), KDR (рецептор киназного вставочного домена(рецептор тирозинкиназы типа III), PLA2G2A (фосфолипаза A2, группа IIА (тромбоциты, синовиальная жидкость)), B2M (бета-2-микроглобулин), THBS1 (тромбоспондин 1), GCG (глюкагон), RHOA (семейство генов гомологии ras, член A), ALDH2 (семейство альдегиддегидрогеназы 2 (митохондриальный)), TCF7L2 (транскрипционный фактор 7-подобный 2 (специфичный для Т-клеток, группа генов с высокой мобильностью - HMG-box)), BDKRB2 (рецептор брадикинина B2), NFE2L2 (ядерный фактор 2, подобный эритроидному деривату), NOTCH1 (гомолог Notch 1, связанный с транслокацией (Drosophila)), UGT1A1 (семейство УДФ-глюкуронозилтрансферазы 1, полипептид Al), IFNA1 (интерферон, альфа I), PPARD (дельта-рецептор, активируемый пероксисомным пролифератором), SIRT1 (сиртуин (гомолог silent mating type information regulation 2) 1 (S. cerevisiae)), GNRH1 (гонадотропин-рилизинг-гормон 1 (лютеинизирующий гормон)), PAPPA (ассоциированный с беременностью протеин-A плазмы, паппализин 1), ARR3 (аррестин 3, сетчатка (X-аррестин)), NPPC (предшественник натрийуретического пептида C), AHSP (стабилизирующий белок альфа-гемоглобина), PTK2 (белок PTK2 тирозинкиназа 2), IL13 (интерлейкин 13), MTOR (механистическая мишень рапамицина (серин/треонинкиназа)), ITGB2 (интегрин, бета 2 (комплементарный компонент 3 рецептора 3 и субъединицы 4)), GSTT1 (глутатион S-трансфераза тета 1), IL6ST (сигнальный белок интерлейкина 6 (gpl30, рецептор онкостатина M)), CPB2 (карбоксипептидаза B2 (плазма)), CYP1A2 (цитохром P450, семейство 1, подсемейство A, полипептид 2), HNF4A (ядерный фактор гепатоцитов 4, альфа), SLC6A4 (семейство транспортеров растворенных веществ 6 (транспортер нейротрансмиттеров, серотонин), член 4), PLA2G6 (фосфолипаза A2, группа VI (цитозольная, не зависящая от кальция)), TNFSF11 (надсемейство фактора некроза опухолей (лиганд), член 11), SLC8A1 (семейство транпортеров растворенных веществ 8 (натрий/кальциевый обменник), член 1), F2RL1 (фактор свертывания крови II (тромбин), подобный рецептору 1), AKR1A1 (альдокеторедуктаза семейства 1, член Al (альдегидредуктаза), ALDH9A1 (альдегиддегидрогеназа семейства 9, член Al), BGLAP (белок костного гамма-карбоксиглутамата (gla)), MTTP (микросомальный белок переноса триглицеридов), MTRR (редуктаза 5-метилтетрагидрофолат-гомоцистеинметилтрансферазы), SULT1A3 (семейство сульфотрансфераз, цитозольный, 1A, фенол-предпочтительный, член 3), RAGE (почечный опухолевый антиген), C4B (компонент комплемента 4B (группа крови Chido), P2RY12 (пуринергический рецептор P2Y, G-белок связанный, 112.), RNLS (реналаза, FAD-зависимая аминоксидаза), CREB1 (белок, связывающий цАМФ-отзывчивый элемент 1), POMC (проопиомеланокортин), RAC1 (связанный с ras субстрат ботулотоксина C3 1 (семейство rho, небольшой ГТФ-связывающий белок Racl)), LMNA (ламин NC), CD59 (молекула CD59, регуляторный белок комплемента), SCN5A (потенциал-зависимый натриевый канал, тип V, альфа-субъединица), CYP1B1 (цитохром P450, семейство 1, подсемейство B, полипептид 1), MIF (ингибитор миграции макрофагов) (фактор ингибирования гликозилирования)), MMP13 (матриксная металлопептидаза 13 (коллагеназа 3)), TIMP2 (ингибитор металлопептидазы ингибитор TIMP 2), CYP19A1 (цитохром P450, семейство 19, подсемейство A, полипептид 1), CYP21A2 (цитохром P450, семейство 21, подсемейство A, полипептид 2), PTPN22 (протеиновая тирозинфосфатаза, не рецепторный тип 22 (лимфоидный)), MYH14 (миозин, тяжелая цепь 14, немышечный), MBL2 (маннозосвязывающий лектин (белок C) 2, растворимый (опсонический дефект)), SELPLG (лиганд P-селектина), AOC3 (аминоксидаза, содержащая медь 3 (сосудистый адгезионный белок 1)), CTSL1 (катепсин L1), PCNA (ядерный антиген пролиферирующих клеток), IGF2 (инсулиноподобный фактор роста 2 (соматомедин A)), ITGB1 (интегрин, бета 1 (рецептор фибронектина, бета-полипептид, антиген CD29 включает MDF2, MSK12)), CAST (кальпастатин), CXCL12 (хемокин (мотив C-X-C) лиганд 12 (фактор стромальных клеток 1)), IGHE (тяжелая константная цепь эпсилон иммуноглобулина), KCNE1 (потенциал-зависимый калиевый канал, семейство, связанное с Isk, член 1), TFRC (рецептор трансферрина (p90, CD71)), COL1A1 (коллаген, тип I, альфа 1), COL1A2 (коллаген, тип I, альфа 2), IL2RB (рецептор интерлейкина 2, бета), PLA2G10 (фосфолипаза A2, группа X), ANGPT2 (ангиопоэтин 2), PROCR (рецептор белка C, эндотелиальный (EPCR)), NOX4 (-НАДФH-оксидаза 4), HAMP (антимикробный пептид гепсидин), PTPN11 (протеиновая тирозинфосфатаза, не рецепторный тип 11), SLC2A1 (семейство транспортеров растворенных веществ 2 (облегченный переносчик глюкозы), член 1), IL2RA (рецептор интерлейкина 2, альфа), CCL5 (хемокин (C-C мотив) лиганд 5), IRF1 (интерферон регуляторный фактор 1), CFLAR (CASP8- и FADD-подобные регуляторы апоптоза), CALCA (связанный с кальцитонином полипептид альфа), EIF4E (фактор инициации трансляции эукариот 4E), GSTP1 (глутатион S-трансфераза pi 1), JAK2 (Янус-киназа2), CYP3A5 (цитохром P450, семейство 3, подсемейство A, полипептид 5), HSPG2 (гепарансульфат-протеогликан 2), CCL3 (хемокин (C-C мотив) лиганд 3), MYD88 (ген миелоидной дифференцировки первичного ответа (88)), VIP (вазоактивный интестинальный пептид), SOAT1 (стерол-O-ацилтрансфераза 1), ADRBK1 (киназа бета-адренорецептора 1), NR4A2 (подсемейство 4 ядерных рецепторов, группа A, член 2), MMP8 (матричная металлопептидаза 8 (нейтрофильная коллагеназа)), NPR2 (натрийуретический пептидный рецептор B/гуанилатциклаза В (атрионатриуретический пептидный рецептор B)), GCH1 (ГТФ-циклогидролаза 1), EPRS (глутамил-пролил-тРНК-синтетаза), PPARGC1A (гамма-активатор, активируемый пероксисомным пролифератором, коактиватор 1 альфа), F12 (фактор свертывания крови XII (фактор Хагемана)), PECAM1 (молекула адгезии тромбоцитов/эндотелиальных клеток), CCL4 (хемокин (C-C мотив) лиганд 4), SERPINA3 (ингибитор серпиновой пептидазы, клада A (альфа-1 антипротеиназа, антитрипсин), член 3), CASR (кальций-чувствительный рецептор), GJA5 (белок щелевого контакта, альфа 5, 40 кДа), FABP2 (белок, связывающий жирные кислоты 2, кишечный), TTF2 (фактор терминации транскрипции, РНК-полимераза II), PROS1 (белок S (альфа)), CTF1 (кардиотрофин 1), SGCB (саркогликан, бета (43 кДа, дистрофин-ассоциированный гликопротеин)), YME1L1 (YME1-подобный 1 (S. cerevisiae)), CAMP (антимикробный пептид кателицидин), ZC3H12A (цинковый палец CCCH-типа, содержащий 12А), AKR1B1 (альдокеторедуктаза 1 семейства, член B1 (альдозоредуктаза)), DES (десмин), MMP7 (матричная металлопептидаза 7 (матрилизин, маточный), AHR (арильный углеводородный рецептор), CSF1 (колониестимулирующий фактор 1 (макрофагальный)), HDAC9 (гистондеацетилаза 9), CTGF (фактор роста соединительной ткани), KCNMA1 (кальций-активируемый калиевый канал высокой проводимости, подсемейство M, член альфа 1), UGT1A (семейство UDP УДФ-глюкуронозилтрансфераза 1 семейства, комплексный локус полипептида A), PRKCA (протеинкиназа C, альфа), COMT (катехол-бета-метилтрансфераза), S100B (кальций-связывающий белок S100 B), EGR1 (early growth factor 1), PRL (пролактин), IL15 (интерлейкин 15), DRD4 (дофаминовый рецептор D4), CAMK2G (кальций/кальмодулинзависимая протеинкиназа II гамма), SLC22A2 (семейство транспортеров растворенных веществ 22 (транспортер органических катионов) член 2), CCL11 (хемокин (C-C мотив) лиганд 11), PGF (плацентарный фактор роста B321), THPO (тромбопоэтин), GP6 (гликопротеин VI (тромбоцитарный)), TACR1 (рецептор тахикинина 1), NTS (нейротензин), HNF1A (HNF1 гомеобокс A), SST (соматостатин), KCND1 (потенциал-зависимый калиевый канал Shal-связанного подсемейства, член 1), LOC646627 (ингибитор фосфолипазы), TBXAS1 (тромбоксан-А-синтаза 1 (тромбоцитарный)), CYP2J2 (цитохром P450, семейство 2, подсемейство J, полипептид 2), TBXA2R (рецептор тромбоксана A2), ADH1C (алкогольдегидрогеназа 1C(класс I), гамма-полипептид), ALOX12 (арахидонат-12-липоксигеназа), AHSG (альфа-2-HS-гликопротеин), BHMT (бетаин-гомоцистеин-метилтрансфераза), GJA4 (белок щелевого контакта, альфа 4, 37 кДа), SLC25A4 (семейство транспортеров растворенных веществ 25 (митохондриальный переносчик; аденин-нуклеотидный транслокатор), член 4), ACLY (АТФ-цитратлиаза), ALOX5AP (белок, активирующий арахидонат-5-липоксигеназа-), NUMA1 (белок митотического аппарата ядра 1), CYP27B1 (цитохром P450, семейство 27, подсемейство B, полипептид 1), CYSLTR2 (рецептор цистеинил- лейкотриена 2), SOD3 (супероксиддисмутаза 3, внеклеточная), LTC4S (лейкотриенC4-синтаза), UCN (урокортин), GHRL (препропептид грелина/обестатина), APOC2 (аполипопротеин C-II), CLEC4A (семейство лектиновых доменов C-типа 4, член A), KBTBD10 (белок, содержащий kelch-повтор и домен BTB (POZ) 10), TNC (тенасцин C), TYMS (тимидилатсинтетаза), SHC1 (SHC (содержащий домен гомологии src)-трансформирующий белок 1), LRP1 (белок 1, связанный с рецептором липопротеинов низкой плотности), SOCS3 (супрессор цитокинового сигнала 3), ADH1B (алкогольдегидрогеназа 1B (класс I), бета-полипептид), KLK3 (калликреин-связанная пептидаза 3), HSD11B1 (11 бета-гидроксистероид--дегидрогеназа 1), VKORC1 (комплекс эпоксид-редуктазы витамина К, субъединица 1), SERPINB2 (ингибитор серпиновой пептидазы, клада В (овальбумин), член 2), TNS1 (тензин 1), RNF19A (ring-белок 19A), EPOR (рецептор эритропоэтина), ITGAM (интегрин, альфа M (компонент комплемента рецептор 3 субъединица 3)), РITХ2 (парноподобный гомеодомен 2), MAPK7 (митоген-активируемая протеинкиназа 7), FCGR3A (Fc фрагмент IgG, низкое сродство 111a, рецептор (CD16a)), LEPR (рецептор лептина), ENG (эндоглин), GPX1 (глутатионпероксидаза 1), GOT2 (глутамат-оксалоацетат-трансаминаза 2, митохондриальная (аспартатаминотрансфераза 2)), HRH1 (рецептор гистамина H1), NR112 (подсемейство ядерного рецептора 1, группа I, член 2), CRH (кортикотропин-рилизинг-гормон), HTR1A (рецептор 5- гидрокситриптамина (серотонина) lA), VDAC1 (потенциал-зависимый анионный канал 1), HPSE (гепараназа), SFTPD (белок D), TAP2 (транспортер 2, АТФ-связывающая кассета, подгруппа В (MDR/TAP)), RNF123 (ring-белок 123), PTK2B (белок PTK2B-тирозинкиназа 2-бета), NTRK2 (нейротрофическая тирозинкиназа, рецептор, тип 2), IL6R (рецептор интерлейкина 6), ACHE (ацетилхолинэстераза (группа крови Yt)), GLP1R (рецептор глюкагоноподобного пептида 1), GHR (рецептор гормона роста), GSR (глутатионредуктаза), NQO1 (НАД (Ф) H дегидрогеназа, хинон 1), NR5A1 (подсемейство 5 ядерных рецепторов, группа A, член 1), GJB2 (белок щелевого контакта, бета 2, 26 кДа), SLC9A1 (семейство транспортеров растворенных веществ 9 (натрий/водородный обменник), член 1), MAOA (моноаминоксидаза A), PCSK9 (пропротеиновая конвертаза субтилизин-кексинового типа 9), FCGR2A (Fc-фрагмент IgG, низкое сродство IIa, рецептор (CD32)), SERPINF1 (ингибитор серпиновой пептидазы, клада F (альфа-2-антиплазмин, пигментный эпителий-происходящий фактор), член 1), EDN3 (эндотелин 3), DHFR (дигидрофолатредуктаза), GAS6 (growth arrest specific 6), SMPD1 (сфингомиелин-фосфодиэстераза 1, кислая лизосомальная), UCP2 (разобщающий белок 2 (митохондриальный, переносчик протонов)), TFAP2A (транскрипционный фактор AP-2 альфа (активирующий энхансер-связывающий белок 2 альфа), C4BPA (белок, связывающий компонент комплемента 4, альфа), SERPINF2 (ингибитор серпиновой пептидазы, клада F (альфа-2-антиплазмин, пигментный эпителий-происходящий фактор), член 2), TYMP (тимидинфосфорилаза), ALPP (щелочная фосфатаза, плацентарная (изофермент Реган)), CXCR2 (хемокиновый (C-X-C мотив) рецептор 2), SLC39A3 (семейство транспортеров растворенных веществ 39 (переносчик цинка), член 3), ABCG2 (АТФ-связывающая кассета, подсемейство G (WHITE), член 2), ADA (аденозиндезаминаза), JAK3 (Янус-киназа 3), HSPA1A (70 кДа белок теплового шока 1A), FASN (синтаза жирных кислот), FGF1 (фактор роста фибробластов 1 (кислотный)), F11 (фактор свертывания крови XI), ATP7A (АТФаза, перенос Cu++, альфа-полипептид), CRI (рецептор компонента комплемента (3b/4b) 1 (группа крови Knops)), GFAP (глиальный фибриллярный кислый белок), ROCK1 (Rho-ассоциированная содержащая домен coiled-coilпротеинкиназа 1), MECP2 (метил- CpG-связывающий белок 2 (синдром Ретта)), MYLK (киназа легких цепей миозина), BCHE (бутирилхолинэстераза), LIPE (липаза, гормон- чувствительная), PRDX5 (пероксиредоксин 5), ADORA1 (рецептор аденозина A1), WRN (синдром Вернера, RecQ хеликаза-подобный), CXCR3 (хемокиновый (C-X-C-мотив) рецептор 3), CD81 (молекула CD81), SMAD7 (член семейства SMAD 7), LAMC2 (ламинин, гамма 2), MAP3K5 (митоген-активируемая протеинкиназа к 5), CHGA (хромогранин A (паратиреоидный секреторный белок 1)), IAPP (островковый амилоидный полипептид), RHO (родопсин), ENPP1 (эктонуклеотид пирофосфатаза/фосфодиэстераза 1), PTHLH (паратиреоидный гормон-родственный белок, NRG1 (нейрегулин 1), VEGFC (фактор роста эндотелия сосудов C), ENPEP (глутамил-аминопептидаза (аминопептидаза A)), CEBPB (CCAAT/энхансер-связывающий белок (C/EBP), бета), NAGLU (N-ацетилглюкозаминидаза, альфа-), F2RL3 (фактор свертывания крови II (тромбин) рецептороподобный 3), CX3CL1 (хемокин (монотив C-X3-C) 1), BDKRB1 (рецептор брадикинина B1), ADAMTS13 (ADAM-металлопептидаза с мотивом тромбоспондина типа 1, 13), ELANE (эластаза, экспрессируемая нейтрофилами), ENPP2 (эктонуклеотид пирофосфатаза/фосфодиэстераза 2), CISH (индуцируемый цитокинами SH2-содержащий белок), GAST (гастрин), MYOC (миоцилин, индуцируемый трабекулярной сетью глюкокортикоидный ответ), ATP1A2 (АТФаза, перенос Na +/K +, альфа2-полипептид), NF1 (нейрофибромин 1) GJB1 (белок щелевых контактов, бета 1, 32 кДа), MEF2A (фактор энхансера миоцитов 2A), VCL (винкулин), BMPR2 (рецептор костного морфогенетического белка, тип II (серин/треонинкиназа)), TUBB (тубулин, бета) CDC42 (белок клеточного цикла 42 (ГТФ-связывающий белок, 25 кДа)), KRT18 (кератин 18), HSF1 (фактор транскрипции белков теплового шока 1), MYB (v-myb гомолог вирусного онкогена миелобластоза v-myb (птичий)), PRKAA2 (протеинкиназа, AMФ-активируемая, альфа-2-каталитическая субъединица), ROCK2 (Rho-ассоциированная содержащая домен coiled-coil протеинкиназа 2), TFPI (ингибитор пути тканевого фактора (липопротеин-ассоциированный коагуляционный ингибитор)), PRKG1 (протеинкиназа, цГМФ-зависимая, тип I), BMP2 (костный морфогенетический белок 2), CTNND1 (катенин (ассоциированный с кадгерином белок), дельта 1), CTH (цистатионаза (цистатионин-гамма-лиаза)), CTSS (катепсин S), VAV2 (фактор обмена гуаниновых нуклеотидов vav 2), NPY2R (нейропептидный Y-рецептор Y2), IGFBP2 (белок, связывающий инсулиноподобный фактор роста 2, 36 кДа), CD28 (молекула CD28), GSTA1 (глутатион-S-трансфераза альфа 1), PP1A (пептидилпролилизомераза A (циклофилин A)), APOH (аполипопротеин H (бета-2-гликопротеин I)), S100A8 (S100 кальций-связывающий белок A8), IL11 (интерлейкин 11), ALOX15 (арахидонат-15-липоксигеназа), FBLN1 (фибулин 1), NR1H3 (подсемейство ядерных рецепторов 1, группа H, член 3), SCD (стеароил-CoA-десатураза (дельта-9-десатураза)), GIP (желудочный ингибиторный полипептид), CHGB (хромогранин В (секретогранин 1)), PRKCB (протеинкиназа C, бета), SRD5A1 (стероид-5-альфа-редуктаза, альфа-полипептид 1 (3-оксо-5-альфа-стероид- дельта-4-дегидрогеназа альфа 1)), HSD11B2 (гидроксистероид-(11-бета)-дегидроге аза 2), CALCRL (кальцитонин-рецептор-подобный), GALNT2 (UDP-N-ацетил-альфа-D-галактозамин: полипептид N-ацетилгалактозаминилтрансфераза 2 (GalNAc-T2)), ANGPTL4 (ангиопоэтин-подобный 4), KCNN4 (кальций-активируемые калиевые каналы средней/малой проводимости, подсемейство N, член 4), PIK3C2A (фосфоинозитид-3-киназа, класс 2, альфа-полипептид), HBEGF (гепарин-связывающий эпидермальный фактор роста-подобный фактор роста), CYP7A1 (цитохром P450, семейство 7, подсемейство A, полипептид 1), HLA-DRB5 (главный комплекс гистосовместимости, класс II, DR бета 5), BNIP3 (BCL2/аденовирус E1B 19 кДа-взаимодействующий белок 3), GCKR (регулятор глюкокиназы (гексокиназы 4)), S100A12 (S100 кальций-связывающий белок A12), PADI4 (пептидиларгининдеиминаза, тип IV), HSPA14 (70 кДа белок теплового шока 14), CXCR1 (хемокиновый (C-X-C-мотив) рецептор 1), H19 (H19, импринтируемый транскрипт, выраженный по материнской линии (небелковое кодирование)), KRTAP19-3 (связанный с кератином белок 19-3), IDDM2 (инсулинозависимый сахарный диабет 2), RAC2 (связанный с ras субстрат ботулотоксина C3 2 (семейство rho, небольшой ГТФ-связывающий белок Rac2)), RYR1 (рианодиновый рецептор 1 (скелетный)), CLOCK (clock-гомолог (мышь)), NGFR (рецептор фактора роста нервов (суперсемейство TNFR, член 16)), DBH (дофамин бета-гидроксилаза (дофамин бета-монооксигеназа)), CHRNA4 (холинергический рецептор, никотиновый, альфа 4), CACNA1C (потенциал-зависимый кальциевый канал, L-тип, альфа-1C-субъединица), PRKAG2 (протеинкиназа, AMФ-активируемая, некаталитическая субъединица гамма-2), CHAT (холин-ацетилтрансфераза), PTGDS (простагландин D2-синтаза 21 кДа (мозг)), NR1H2 (подсемейство ядерных рецепторов 1, группа H, член 2), ТЕК (ТЕК-тирозинкиназа, эндотелиальная), VEGFB (фактор роста эндотелия сосудов B), MEF2C (фактор-энхансера миоцитов 2C), MAPKAPK2 (митоген-активируемая протеинкиназа-активируемая протеинкиназа 2), TNFRSF11A (надсемейство рецепторов фактора некроза опухолей, член 11a, активатор NFKB), HSPA9 (70 кДа белок теплового шока 9 (морталин)), CYSLTR1 (рецептор цистеинил-лейкотриена 1), MAT1A (метионин-аденозилтрансфераза I, альфа), OPRL1 (опиатный рецептор-подобный 1), IMPA1 (инозитол (мио)-1 (или 4)-монофосфатаза 1), CLCN2 (хлоридный канал 2), DLD (дигидролипамид-дегидрогеназа), PSMA6 (субъединица протеасомы (просома, макропоин), альфа-тип, 6), PSMB8 (субъединица протеасомы (просома, макропоин), бета-тип, 8 (большая многофункциональная пептидаза 7)), CHI3L1 (хитиназа 3-подобный белок 1 (хрящевой гликопротеин-39)), ALDH1B1 (семейство альдегиддегидрогеназ 1, член B1), PARP2 (поли (ADФ-рибоза) полимераза 2), STAR (стероидогенный острый регуляторный белок) LBP (липополисахарид-связывающий белок), ABCC6 (АТФ-связывающая кассета, подгруппа C (CFTR/MRP), член 6), RGS2 (регулятор G-белковой сигнализации 2, 24 кДа), EFNB2 (эфрин-B2), GJB6 (белок щелевых контактов, бета 6, 30 кДа), APOA2 (аполипопротеин A-II), AMPD1 (аденозинмонофосфатдезаминаза 1), DYSF (дисферлин, мышечная дистрофия поясов конечностей 2B (аутосомно-рецессивная)), FDFT1 (фарнезилдифосфат-фарнезилтрансфераза 1), EDN2 (эндотелин 2), CCR6 (хемокиновый (C-C мотив) рецептор 6), GJB3 (белок щелевого контакта, бета 3, 31 кДа), IL1RL1 (интерлейкин 1-рецептор-подобный белок 1), ENTPD1 (эктонуклеозидтрифосфат дифосфогидролазы 1), BBS4 (синдром Бардета-Бидля 4), CELSR2 (кадгерин, EGF LAG семипроходный рецептор G-типа 2 (гомолог фламинго, дрозофила)), F11R (F11-рецептор), RAPGEF3 (фактор обмена гуанинового нуклеотида (GEF) 3), HYAL1 (гиалуроноглюкозаминидаза 1), ZNF259 (белок цинкового пальца 259), ATOX1 (гомолог ATX1 антиоксидантного белка 1 (дрожжи)), ATF6 (фактор активации транскрипции 6) KHK (кетогексокиназа (фруктокиназа)), SAT1 (спермидин/спермин N1-ацетилтрансфераза 1), GGH (гамма-глутамилгидролаза (конъюгаза, фолилполигаммаглутамил-гидролаза)), TIMP4 (ингибитор металлопептидазы TIMP 4), SLC4A4 (семейство транспортеров растворенных веществ 4, натрий, бикарбонатный котранспортер, член 4), PDE2A (фосфодиэстераза 2A, стимулируема цГМФ), PDE3B (фосфодиэстераза 3B, ингибируемая цГМФ), FADS1 (десатураза жирных кислот 1), FADS2 (десатураза жирных кислот 2), TMSB4X (тимозин бета 4, X-связанный), TXNIP (тиоредоксин-взаимодействующий белок), LIMS1 (LIМ и антиген-подобные домены стареющих клеток 1), RHOB (семейство генов гомологии ras, член B), LY96 (лимфоцитарный антиген 96), FOXO1 (белок forkhead box O1), PNPLA2 (белок, содержащий пататин-подобный фосфолипазный домен 2), TRH (тиреотропин-рилизинг- гормон), GJC1 (белок щелевых контактов, гамма 1 45 кДа), SLC17A5 (семейство транспортеров растворенных вещест 17 (переносчик анионов/сахаров), член 5), FTO (ассоциированный с жировой массой и ожирением), GJD2 (белок щелевых контактов, дельта 2, 36 кДа), PSRC1 (пролин/серин-богатый домен coiled-coil 1), CASP12 (каспаза 12 (ген/псевдоген)), GPBAR1 (рецептор желчной кислоты, связанный с белками G 1), PXK (домен PX, содержащий серин/треонинкиназу), IL33 (интерлейкин 33), TRIB1 (гомолог tribbles 1 (дрозофила)), PBX4 (гомеобокс пре-B-клеток лейкемии 4), NUPRI (ядерный белок, регулятор транскрипции, 1), 15-Sep (15 кДа селенопротеин), CILP2 (белок промежуточного слоя хряща 2), TERC (компонент теломеразы РНК), GGT2 (гамма-глутамилтрансфераза 2), MT-CO1 (митохондриально кодируемая цитохромоксидаза с I) и UOX (уратоксидаза, псевдоген). Любая из этих последовательностей может быть мишенью для системы CRISPR-Cas, например, для устранения мутации.
[001232] В еще одном варианте осуществления хромосомная последовательность может дополнительно быть выбрана из Pon1 (параоксоназа 1), LDLR (рецептор LDL), ApoE (аполипопротеин E), Apo B-100 (аполипопротеин B-100), ApoA (аполипопротеин (а)), ApoA1 (аполипопротеин A1), CBS (цистатионB-синтаза), гликопротеина IIb/IIb, MTHRF (5,10-метилентетрагидрофолатредуктазы (НАДФH) и их комбинации. В одной итерации хромосомные последовательности и белки, кодируемые хромосомными последовательностями, участвующие в развитии сердечно-сосудистых заболеваний могут быть выбраны из CacnalC, Sodl, Pten, Ppar (alpha), Apo E, лептина и их комбинаций в качестве мишени или мишеней для системы CRISPR-Cas.
Лечение заболеваний печени и почек
[001233] Настоящее изобретение также предусматривает доставку системы CRISPR-Cas, описанной в настоящем описании, например, систем эффекторных белков C2c1 или C2c3, в печень и/или почку. Стратегии доставки, чтобы вызвать клеточное поглощение терапевтической нуклеиновой кислоты, включают физическую силу или векторные системы, такие как доставка на основе вирусов, липидов или комплексов или наноносители. Из изначальных приложений, обладающих меньшей клинической значимостью, когда нуклеиновые кислоты были доставлены в клетки почек при помощи систематического впрыскивания под высоким давлением (безыгольные инъекции), широкий диапазон генных терапевтических вирусных и невирусных носителей уже применяется для нацеливания на посттранскрипционные события в разных моделях болезней почек на животных in vivo (Csaba Revesz and Péter Hamar (2011). Delivery Methods to Target RNAs in the Kidney, Gene Therapy Applications, Prof. Chunsheng Kang (Ed,), ISBN:978-953-307-541-9, InTech, Available from: http://www.intechopen.com/books/gene-therapy-a.pplications/delivery-methods-to-target-rnas-inthe-kidney) Способы доставки в почку могут включать способы, описанные в Yuan et al. (Am J Physiol Renal Physiol 295: F605-F617, 2008), которые исследовали, может ли до ставка in vivo малых интерферирующих РНК (миРНК), нацеленных на 12/15-липоксигеназный (12/15-LO) путь метаболизма арахидоновой кислоты, улучшить состояние при повреждениях почек и диабетической нефропатии (DN) на модели сахарного диабета типа I у мышей, которым инъецировали стрептозоцин. Для достижения большего доступа in vivo и экспрессии миРНК в почках Yuan et al. использовали двухцепочечны еолигонуклеотидые РНК 12/15-LO, конъюгированные с холестерином. Около 400 мкг миРНК вводили мышам подкожно. Способы Yuang et al. могут быть применены к системе CRISPR Cas согласно настоящему изобретению, предусматривается 1-2 г подкожной инъекции CRISPR Cas, конъюгированного с холестерином, человеку для доставки в почки.
[001234] Molitoris et al. (J AmSocNephrol 20: 1754-1764, 2009) использовали проксимальные канальцевые клетки (PTC) в качестве области реабсорбции олигонуклеотидов в почках, чтобы проверить эффективность миРНК, нацеленной на p53 - ключевой белок в апоптотическом пути, для предотвращения повреждения почек. Голая синтетическая миРНК к p53 вводится внутривенно через 4 часа после ишемического повреждения, и наиболее успешно защищает как PTC, так и функции почек. Данные Molitoris et al. показывают, что быстрая доставка миРНК в клетки проксимальных канальцев следует за внутривенным введением. Для получения кривой доза-ответ крысам вводили следующие дозы siP53, 0,33; 1, 3 или 5 мг/кг, в указанные промежутки времени, что приводило к кумулятивным дозам 1,32; 4, 12 и 20 мг/кг соответственно. Все испытуемые дозы миРНК дали эффект снижения SCr на первый день, при этом более высокие дозы оказались эффективны в течение примерно пяти дней по сравнению с контрольными крысами с ишемией, получавшими фосфатно-солевой буфер (PBS). Совокупные дозы 12 и 20 мг/кг обеспечивали лучший защитный эффект. Способы Molitoris et al. могут быть применены к системе нацеливания нуклеиновых кислот согласно настоящему изобретению, предусматривающей совокупные дозы 12 и 20 мг/кг человеку для доставки в почки.
[001235] Thompson et al. (Nucleic Acid Therapeutics, Volume 22, Number 4, 2012) раскрывают токсикологические и фармакокинетические свойства синтетической малой интерферирующей РНК 15NP после внутривенного введения грызунам и нечеловеческим приматам. 15NP предназначен для воздействия на путь РНК-интерференции (РНК-i), чтобы временно ингибировать экспрессию проапоптотического белка р53 и разрабатывается для защиты клеток от острых ишемических/реперфузионных травм, таких как острая почечная травма, которые могут возникать во время базовой кардиохирургии и отсроченной функции трансплантата, которая может происходить после трансплантации почек. Дозы 800 мг/кг 15NP у грызунов и 1000 мг/кг 15NP у не являющихся человеком приматов должны были вызывать побочные эффекты, которые у обезьян привели к различным типам эффектов на кровь: субклиническая активация комплемента и несколько увеличенное время свертывания крови. У крыс никаких побочных эффектов не наблюдалось при введении крысиного аналога 15NP, что указывает на то, что эти эффекты, вероятно, представляют собой эффекты класса синтетических РНК-дуплексов, а не токсичность, связанную с предполагаемой фармакологической активностью 15NP. В совокупности эти данные подтверждают возможность прохождения клинических испытаний внутривенного введения 15NP для сохранения функции почек после острой ишемии/реперфузионного повреждения. Доза, не вызывающая наблюдаемых побочных эффектов (NOAEL), у обезьян составляла 500 мг/кг. Никаких эффектов на сердечно-сосудистые, респираторные и неврологические параметры не наблюдалось у обезьян после внутривенного введения при дозах до 25 мг/кг. Таким образом, подобная доза может быть предусмотрена для внутривенного введения CRISPR Cas в почки человека.
[001236] Shimizu et al. (JAmSocNephrol 21: 622-633, 2010) разработали систему для нацеливания доставки малых интерферирующих РНК (миРНК) в клубочки (гломерулы) с использованием поли (этилен-гликоль)-поли (L-лизин). Комплекс миРНК/наноноситель был приблизительно от 10 до 20 нм в диаметре, что который позволило бы ему перемещаться через фенестрированный эндотелий для доступа к мезангию после внутрибрюшинной инъекции флуоресцентных комплексов миРНК/наноноситель. Shimizu et al., наблюдали циркуляцию миРНК в кровотоке в течение длительного времени. Повторное внутрибрюшинное введение митоген-активированной протеинкиназы 1 (MAPK1) комплекса миРНК/наноноситель подавляло гломерулярную мРНК MAPK1 и экспрессию белка в мышиной модели гломерулонефрита. Для исследования накопления миРНК, меченых Cy5 миРНК в комплексе с PIC-наноносителями (0,5 мл, 5 нмоль содержания миРНК), голых Cy5-меченых миРНК (0,5 мл, 5 нмоль) или Cy5-меченых миРНК, инкапсулированных в HVJ-E (0,5 мл, 5 нмоль содержимого миРНК) вводили мышам BALBc. Способ Shimizu et al. может быть применен к системе нацеливания нуклеиновых кислот согласно настоящему изобретению, предусматривающей дозу около 10-20 микромоль CRISPRCas в комплексе с нанопереносчиками в количестве примерно 1-2 литра для внутрибрюшинного введения и доставки в почки человека.
[001237] Способы доставки к почкам суммированы в следующей таблице:
pp. (14883-14888)
J Pathol, (Oct 2008), Voi. 173, No. 4, pp. (973-980)
Soc Nephrol, (Dec 2006), Voi. 17, No. 12, pp. (3336-3346)
1 Controlled Release, (Jul
2005), Voi. 105, No. 3, pp. (318-331)
and Experimental Therapeutics, (Feb 2004), Vol. 308, No. 2, pp. (688-693)
Nature
Biotechnology,
(Sep 2007), Vol. 25, No. 10, pp. (1149-1157)
20, No. 8, pp. (1754-1764)
DOTAP/DO
PE/DOPE-
PEG2000
2011), Vol. 16, No.
3, pp. (217-226))
2008), Vol. 295, pp. (F605-F617)
Cancer Res, (May 2009), Vol. 69, No. 10, pp.
(4244-4251)
Pharmacology,
(Apr 2009), Vol. 236, No. 1, pp. (97-108)
Nephrology, (Apr 2010), Vol. 21, No. 4, pp. (622-633)
(May-Jun 2009), Vol. 6, No. 3, pp. (727-737)
mNOX-E36
[001238] Предусматривается нацеливание в клетки печени. Это может быть in vitro или in vivo. Предпочтительными являются гепатоциты. Доставка белка CRISPR, такого как C2c1 или C2c3, здесь может быть осуществлена посредством вирусных векторов, особенно векторов AAV (и, в частности, AAV2/6). Они могут вводиться внутривенно.
[001239] Предпочтительной мишенью для печени, будь то in vitro или in vivo, является ген альбумина. Это так называемая «безопасная гавань», поскольку альбумин экспрессируется на очень высоких уровнях, и поэтому допускается некоторое снижение синтеза альбумина после успешного редактирования гена. Также предпочтительно то, что высокие уровни экспрессии, наблюдаемые с помощью промотора/энхансера альбумина, позволяют получать подходящие уровни нормального или трансгенного синтеза (со вставленной матрицы донора), даже если отредактирована только небольшая часть гепатоцитов.
[001240] Wechsler et al. было показано (сообщение 57th Annual Meeting and Exposition of the American Society of Hematology - аннотация доступна в интернете по адресу https:.//ash.confex.com./ash/2015/webprogram/Paper86495.html и представлена 6 декабря 2015 года), что интрон 1 альбумина является подходящим участком-мишенью. В их работе были использованы цинковые пальцы для разрезания ДНК в данном участке-мишени и подходящие направляющие последовательности могут быть получены, чтобы направлять расщепление в том же месте с помощью белка CRISPR.
[001241] Использование мишеней в высоко экспрессируемых генах (гены с высокоактивными энхансерами/промоторами), таких как альбумин, также может позволить использование донорной матрицы без промотора, как было показано Wechsler et al., и это также широко применимо для других органов, кроме печени. Известны и другие примеры высоко экспрессируемых генов.
Ассоциированные с печенью заболевания крови, особенно гемофилия и в частности гемофилия В
[000019] Успешное редактирование генов гепатоцитов у мышей (как in vitro, так и in vivo) и у не являющихся человеком приматов (in vivo), показало, что лечение заболеваний крови посредством редактирования генов/генной инженерии в гепатоцитах возможно. В частности, показана экспрессия гена F9 (hF9) человека в гепатоцитах нечеловеческих приматов, что указывает и на возможность лечения гемофилии В у людей.
[000020] Wechsler et al. сообщили на 57th Annual Meeting and Exposition of the American Society of Hematology (аннотация представлена 6 декабря 2015 года и доступна в интернете по адресу https://ash.confex.com/ash/2015/webprogrammpaper86495.html), что они успешно экспрессировали человеческий F9 (hF9) из гепатоцитов у нечеловеческих приматов с помощью редактирования гена in vivo. Это было достигнуто с использованием 1) двух нуклеаз с цинковыми пальцами, нацеленных на интрон 1 локуса альбумина, и 2) конструкции донорной матрицы человеческого F9. Цинковые пальцы и донорная матрица были закодированы в отдельных гепатотропных серотипах ы 2/6 (AAV2/6) аденоассоциированного вируса, вводимых внутривенно, что приводило к целенаправленной вставке скорректированной копии гена hF9 в локус альбумина в части печеночных гепатоцитов.
[000021] Локус альбумина был выбран в качестве «безопасной гавани», поскольку производство этого наиболее распространенного белка плазмы превышает 10 г/день, а умеренное снижение этого уровня хорошо переносится. Гепатоциты с отредактированным геномом продуцировали в терапевтических количествах нормальный hFIX (hF9) в отличие от альбумина, который управляется высокоактивным энхансером/промотором альбумина. Показана целевая интеграция трансгена hF9 в локусе альбумина и сплайсинге этого гена в транскрипт альбумина.
[000022] Исследования на мышах: мышам C57BL/6 вводили средство доставки(n=20) или векторы AAV2/6 векторы (n=25), кодирующие мышиные суррогатные реагенты в количестве 1,0×1013 векторных геномов (вг)/кг путем инъекции в хвостовую вену. Анализ ELISA плазмы hFIX у обработанных мышей показал пиковые уровни в 50-1053 нг/мл, которые поддерживались на протяжении 6-месячного исследования. Анализ активности FIX на основе мышиной плазмы подтвердил биологическую активность, соизмеримую с уровнями экспрессии.
[000023] Исследования на нечеловеческих приматах: однократная внутривенная совместная инфузия векторов AAV2/6, кодирующих целевые альбумин специфические нуклеазы цинковых пальцев, нацеленные на NHP, и донорного человеческого F9 в количестве 1×1013вг/кг (n=5/группа) привела к > 50 нг/мл (> 1% от нормы) в этой большой животной модели. Использование более высоких доз AAV2/6 (до 1,5×1014 вг/кг) приводило к количеству hFIX в плазме до 1000 нг/мл (или 20% от нормы) у нескольких животных и до 2000 нг/мл (или 50% от нормального) у одного животного на протяжении всего исследования (3 месяца).
[000024] Лечение хорошо переносилось у мышей и нечеловеческих приматов без значительных токсикологических результатов, связанных с лечением AAV2/6 ZFN +донор у обоих видов в терапевтических дозах. Sangamo (Калифорния, США) с тех пор обратился к FDA и получил разрешение на проведение первого в мире клинического испытания на людях для применения в области редактирования генома in vivo. Это основывается на одобрении EMEA в отношении генной терапии Glybera при лечении дефицита липопротеиновой липазы.
[000025] Соответственно, в некоторых вариантах осуществления предпочтительно использовать любое или все из следующего:
* AAV (особенно AAV2/6) векторы, предпочтительно вводимые внутривенно;
* Альбумин в качестве мишени для редактирования генов/введения трансгена/матрицы - особенно в интроне 1 альбумина;
* донорная матрица человеческого F9; и/или «беспромоторная донорная матрица».
Гемофилия В.
[000026] Соответственно, в некоторых вариантах осуществления предпочтительно, чтобы настоящее изобретение использовалось для лечения гемофилии B. Как таковое, предпочтительно, чтобы была предусмотрена матрица и чтобы она являлась человеческим геном F9. Понятно, что шаблон hF9 содержит правильную или "верную" версию hF9, так чтобы лечение было эффективным.
[000027] В альтернативном варианте осуществления версия F9 гемофилии В может быть доставлена таким образом, чтобы создать модельный организм, клетку или клеточную линию (например, клетку, клеточную линию или модельный организм на основе мыши или нечеловеческого примата), модельный организм, клетку или клеточную линию, имеющие или несущие фенотип гемофилии В, т.е. неспособность продуцировать F9 дикого типа.
Гемофилия A
[000028] В некоторых вариантах осуществления ген F9 (фактор IX) может быть заменен геном F8 (фактор VIII), описанным выше, что приводит к лечению гемофилии A (посредством обеспечения правильного гена F8) и/или созданию модельного организма, клетки или клеточной линии с гемофилией А (путем предоставления неправильной версии гена F8 гемофилии A).
Гемофилия C
[001242] В некоторых вариантах осуществления ген F9 (фактор IX) может быть заменен геном F11 (фактор XI), описанным выше, что приводит к лечению гемофилии C (посредством обеспечения правильного гена F11) и/или созданию модельного организма, клетки или клеточной линии с гемофилией C(путем предоставления неправильной версии гена F11 гемофилии С).
Лечение заболеваний эпителия и легких
[001243] Настоящее изобретение предусматривает доставку систем CRISPR-Cas, описанных в настоящем описании, например, систем эффекторных белков C2c1 или C2c3, в одно или оба легких.
[001244] Хотя векторы на основе AAV-2 были первоначально предложены для доставки CFTR в воздухоносные пути CF, другие серотипы, такие как AAV-1, AAV-5, AAV-6 и AAV-9, демонстрируют улучшенную эффективность переноса генов во множестве моделей эпителия легкого (см., например, Li et al, Molecular Therapy, vol. 17 no. 12, 2067-2077 Dec 2009). Было продемонстрировано, что AAV-1 в 100 раз эффективнее, чем AAV-2 и AAV-5 при трансдуцировании эпителиальных клеток дыхательных путей человека in vitro, хотя AAV-1 трансдуцировал воздухоносный эпителий трахеи мыши in vivo с эффективностью, равной эффективности AAV -5. Другие исследования показали, что AAV-5 в 50 раз эффективнее AAV-2 при доставке генов в эпителий дыхательных путей человека (HAE) in vitro и значительно более эффективен в эпителии дыхательных путей мышей in vivo. Было также показано, что AAV-6 более эффективен, чем AAV-2 в эпителиальных клетках дыхательных путей человека in vitro и мышиных дыхательных путях in vivo. Более поздний изолят AAV-9 показал большую эффективность переноса генов, чем AAV-5 в носовом и альвеолярном эпителии мыши in vivo с экспрессией гена, обнаруженной по прошествии более 9 месяцев, позволяет предположить, что AAV может обеспечить долгосрочную экспрессию гена in vivo. Именно это является желательным свойством для вектора доставки гена CFTR. Кроме того, было продемонстрировано, что AAV-9 может быть введен в мышиное легкое без потери экспрессии CFTR и минимальных иммунных последствиях. CF и не-CF HAE-культуры могут быть инокулированы на апикальной поверхности с помощью 100 мл векторов AAV в течение нескольких часов (см., например, Liet al., Molecular Therapy, vol. 17 no, 12, 2067-2077 Dec 2009). MOI может варьироваться от 1×103 до 4×105 векторных геномов/клетка, в зависимости от концентрации вируса и целей экспериментов. Вышеупомянутые векторы предназначены для доставки и/или введения настоящего изобретения.
[001245] Zamora et al. (Am JRespirCritCareMed Vol. 183. pp. 531-538, 2011) описали пример применения РНК-интерференционной терапии для лечения инфекционного заболевания человека, а также рандомизированное исследование противовирусного препаратана инфицированных респираторно-синцитиальным вирусом (RSV) реципиентов трансплантированного легкого. Zamora et al. провели рандомизированное двойное слепое исследование с использованием в качестве контроля эффекта плацебо у реципиентов LTX с инфекцией респираторных путей RSV. Пациентам было разрешено получать стандартное лечение при RSV. Аэрозольный ALN-RSV01 (0,6 мг/кг) или плацебо вводили ежедневно в течение 3 дней. Это исследование демонстрирует, что лечение на основе РНК-интерференции, нацеленное на RSV, можно безопасно проводить реципиентам LTX с инфекцией RSV. Три ежедневных дозы ALN-RSV01 не приводили к обострению симптомов респираторного тракта, ухудшению функции легких и проявлению системных воспалительных эффектов, таких как индукция цитокинов или CRP. Фармакокинетика показала только низкое временное системное воздействие после ингаляции, что согласуется с данными доклинических испытаний на животных, показывающими, что ALN-RSV01, вводимый внутривенно или путем ингаляции, быстро выводится из круга кровообращения посредством обработки экзонуклеазами и почечной экскреции. Способы Zamora et al. могут быть применены к системе нацеливания нуклеиновых кислот согласно настоящему изобретению, и для настоящего изобретения может быть рассмотрен аэрозольный CRISPR Cas, например, с дозировкой 0,6 мг/кг.
[001246] Индивидуумы, получающие терапию при заболеваниях легких, могут, например, получать фармацевтически эффективное количество аэрозольной системы AAV-вектора на легкое, доставляемое эндобронхиально при спонтанном дыхании. Таким образом, аэрозольная доставка предпочтительна для доставки AAV в целом. Для доставки можно использовать аденовирус или AAV-частицу. Подходящие генные конструкции, каждая из которых функционально связана с одной или более регуляторными последовательностями, могут быть клонированы в вектор доставки. В этом случае в качестве примеров приводятся следующие конструкции: промотор Cbh или EF1a для Cas (C2c1 или C2c3), промотор U6 или H1 для направляющей РНК): Предпочтительно использование направляющей последовательности CFTRdelta508, матрицы восстановления для мутации deltaF508 и кодон-оптимизированного фермента C2c1 или C2c3, необязательно с одним или более сигналами или последовательностью ядерной локализации (NLS, например, двумя (2) NLS. Также предусматриваются конструкции без NLS.
Лечение заболеваний мышечной системы
[001247] Настоящее изобретение предполагает введение системы CRISPR-Cas, например, систем из эффекторных белков C2c1 or C2c3 в мышцы.
[001248] Bortolanza et al. (MolecularTherapyvol. 19 no. 11, 2055-2064 Nov. 2011) показали, что системная доставка кассет экспрессии для РНК-интерференции у мыши FRG1 после начала фасциально-мышечной дистрофии (FSHD) привела к дозозависимому длительному нокдауну FRG1 без признаков токсичности. Bortolanza et al. обнаружили, что однократная внутривенная инъекция 5×1012 вг rAAV6-sh1FRG1 сохраняет мышечную гистопатологию и мышечную функцию FRG1у мышей. Аналогично, 200 мл, содержащих 2×1012 или 5×1012 вг вектора в физиологическом растворе, вводили в хвостовую вену с использованием шприца Terumo-25. Способ Bortolanza et al. может быть применен к AAV, экспрессирующему CRISPR Cas, и человеку может быть введена доза приблизительно 2×1015 или 2×1016 вг вектора.
[001249] Dumonceaux et al. (Molecular Therapy vol. 18 no. 5, 881-887 May 2010) ингибировали путь миостатина, используя способ РНК-интерференции, направленной против мРНК рецепторов остатина (sh-AcvRIIb) AcvRIIb. Восстановление квазидистрофина было опосредовано векторизованной методикой пропуска U7 (U7-DYS). Аденоассоциированные векторы, несущие либо только конструкцию sh-AcvrIIb, либо только конструкцию U7-DYS, либо комбинацию обеих конструкций, были введены в переднюю большеберцовую мышцу мышей mdx с дистрофией. Инъекции выполняли, используя 1011 вирусных геномов AAV. Способ Dumonceaux et al. может быть применен к AAV, экспрессирующему CRISPR Cas, и введение человеку может быть осуществлено в дозе, например, от приблизительно 1014 до приблизительно 1015 вг вектора.
[001250] Kinouchi et al. (GeneTherapy (2008) 15, 1126-1130) сообщают об эффективности доставки малой интерферирующей РНК in vivo в скелетную мышечную ткань нормальных или больных мышей посредством образования наночастиц химически немодифицированных миРНК с ателоколлагеном (ATCOL). ATCOL-опосредованное местное применение миРНК, нацеленное на миостатин, отрицательный регулятор роста скелетных мышц, в мышиных скелетных мышцах или внутривенно, вызвало заметное увеличение мышечной массы в течение нескольких недель после применения. Эти результаты подразумевают, что ATCOL-опосредованное применение миРНК является мощным инструментом для будущего терапевтического использования при таких заболеваниях, как мышечная атрофия. Msts-миРНК (конечная концентрация, 10 мМ) смешивали с ATCOL (конечная концентрация для местного применения, 0,5%) (AteloGene, Kohken, Токио, Япония) в соответствии с инструкциями производителя. После анестезии мышей (20-недельный самец C57BL/6) с применением Nembutal (25 мг/кг, внутрибрюшинно), комплекс Mst-миРНК/ATCOL вводили в жевательную мышцу и двуглавую мышцу бедра. Метод Kinouchi et al. может быть применен к CRISPR Casс введением в организм человека, например, дозы от 500 до 1000 мл 40 мкМ раствора в мышцу. Hagstrom et al. (MolecularTherapy, т. 10, No 2, August 2004) описывают внутрисосудистую, невирусную методологию, которая обеспечивает эффективную и повторяемую доставку нуклеиновых кислот в мышечные клетки (миофибриллы) во всех мышцах конечностей млекопитающих. Процедура включает инъекцию голой плазмидной ДНК или миРНК в дистальную вену конечности, которая временно изолирована жгутом или манжетами для измерения кровяного давления. Доставка нуклеиновой кислоты в миофибриллы облегчается быстрой инъекцией в достаточном объеме, чтобы обеспечить экстравазацию раствора нуклеиновой кислоты в мышечную ткань. Высокие уровни экспрессии трансгена в скелетных мышцах достигались как у мелких, так и у крупных животных с минимальной токсичностью. Также было получено доказательство доставки миРНК в мышцы конечностей. Для внутривенной инъекции плазмидной ДНК у макак-резус, зажим из трех составляющих был соединен с двумя шприцевыми насосами (модель PHD 2000, HarvardInstruments), каждый из которых имелоднн шприц. Через пять минут после инъекции папаверина вводили пДНК (от 15,5 до 25,7 мг в 40-100 мл изотонического раствора) со скоростью 1,7 или 2,0 мл/с. Доза может быть увеличена для плазмидной ДНК, экспрессирующей CRISPR Cas согласно настоящему изобретению, с инъекцией от 300 до 500 мг в 800-2000 мл физиологического раствора для человека. Для инъекций аденовирусного вектора крысе 2×109 инфекционных частиц были инъецированы в 3 мл нормального солевого раствора (NSS). Доза может быть увеличена для аденовирусного вектора, экспрессирующего CRISPR Cas согласно настоящему изобретению, с инъекцией около 1×1013 инфекционных частиц, инъецированных в 10 литрах NSS для человека. Для миРНК крысу инъецировали в большую подкожную вену 12,5 мкг миРНК, а приматов инъецировали в большую подкожную вену 750 мкг миРНК. Доза может быть увеличена для CRISPR Cas согласно настоящему изобретению, например, при инъекции от примерно 15 до 50 мг в большую подкожную вену человека.
[001251] См. также, например, WO2013163628 A2, "Genetic Correction of Mutated Genes", опубликованную заявку Duke University, описывающую попытки по исправлению, например, мутаций сдвига рамки считывания, которая вызывает преждевременное появление стоп-кодона и усеченный генный продукт, который может быть скорректирован с помощью опосредуемого нуклеазой негомологичного соединения концов, таких, как те, которые отвечают за мышечную дистрофию Дюшенна ("DMD"), рецессивное, смертельное, связанное с X-хромосомой заболевание, которое приводит к дегенерации мышц из-за мутаций в гене дистрофина. Большинство мутаций дистрофина, которые вызывают DMD, представляют делеции экзонов, которые нарушают рамку считывания и вызывают преждевременную терминацию трансляции в гене дистрофина. Дистрофин представляет собой цитоплазматический белок, который обеспечивает структурную стабильность дистрогликанового комплекса клеточной мембраны, который отвечает за регулирование целостности и функции мышечных клеток. Ген дистрофина или "ген DMD", используются в настоящем описании взаимозаменяемо, и представляет собой 2,2 млн п.н. в локусе Xp21. Первичная транскрипция составляет около 2400 т.п.н., а зрелая мРНК составляет около 14 т.п.н. 79 экзонов кодируют белок, который содержит более 3500 аминокислот. Эксон 51 часто примыкает к делециям, сдвигающим рамку считывания у пациентов с DMD и является целью клинических испытаний для исключения экзонов на основе олигонуклеотидов. Клиническое исследование исключающего экзон 51 соединения недавно сообщило о значительной функциональной пользе через 48 недель, причем дистрофин-положительных волокон было в среднем на 47% больше по сравнению с исходным уровнем. Мутации в экзоне 51 идеально подходят для постоянной коррекции путем редактирования генома на основе негомологичного соединения концов (NHEJ).
[001252] Способы, описанные в патентной публикации США № 20130145487, закрепленной за Cellectis, относятся к вариантам использования мегануклеазы для расщепления последовательности-мишени из гена дистрофина (DMD) человека, и также могут быть модифицированы для системы нацеливания на нуклеиновые кислоты согласно настоящему изобретению.
[001253] Настоящее изобретение также предусматривает доставку системы CRISPR-Cas, описанной в настоящем описании, например, систем эффекторных белков, таких как C2c1 или C2c3, в кожу.
[001254] Hickerson et al. (MolecularTherapy-NucleicAcids (2013) 2, el29) описали моторизированное устройство с микроиголками для доставки самодоставляющейся (sd) миРНК в кожу человека или мыши. Основной проблемой перевода миРНК терапии кожи в клинику является разработка эффективных систем доставки. Значительные усилия были вложены в различные технологии доставки в кожу, но показали ограниченный успех. В клиническом исследовании, в котором кожа была обработана миРНК, боль, связанная с использованием мельчайших игл для подкожных инъекций, не позволила зарегистрировать дополнительных пациентов для исследования, что подчеркивает необходимость улучшения способов доставки, которые были бы ориентированы на пациента и вызывали небольшие болевые ощущения или вообще были безболезненными для пациента. Микроиглы представляют собой эффективный способ доставки больших заряженных соединений, включая миРНК, через первичный барьер, роговой слой, и обычно считаются менее болезненными, чем обычные иглы для подкожных инъекций. Было показано, что моторизованные устройства "штампующего" типа, в том числе устройство моторизированной микроиглы (MMNA), используемое Hickerson et al, безопасны. На безволосых мышах было показано, что использование таких устройств вызывает небольшую боль или отсутствие боли, о чем свидетельствует (i) широкое распространение использование в косметической промышленности и (ii) ограниченное тестирование, в котором почти все добровольцы обнаружили, что использование устройства было гораздо менее болезненным, чем прививка от гриппа, предполагая, что доставка миРНК с использованием этого устройства приведет к гораздо меньшей боли, чем это было в предыдущем клиническом исследовании при использовании игл для подкожных инъекций. Устройство MMNA (продаваемое как Triple-M или Tri-M от Bomtech Electronic Co, Сеул, Южная Корея) было адаптировано для доставки миРНК в кожу мыши и человека. Раствор самодоставляющихся миРНК (до 300 мкл 0,1 мг/мл РНК) вводили в камеру одноразовой кассеты Tri-M (Bomtech), которую устанавливали на глубину 0,1 мм. Для лечения кожи человека деидентифицированную кожу: (полученную сразу же после хирургических процедур) вручную растягивали и прикрепляли к пробковой платформе перед обработкой. Все внутрикожные инъекции проводились с использованием шприца для инсулина с иглой диаметром 0,5 дюйма 28 калибра. Устройство MMNA и способ Hickerson et al. могут быть использованы и/или адаптированы для доставки CRISPR Cas согласно настоящему изобретению, например, в дозе до 300 мкл 0,1 мг/мл CRISPR Cas в кожу.
[001255] Статья Leachman et al. (MolecularTherapy, vol. 18 no. 2, 442-446 Feb. 2010) относится к клиническому испытанию фазы Ib для лечения редкого расстройства кожи пахионии врожденной (pachyonychia congenita) (ПВ), аутосомно-доминантного синдрома, включающего пальмоплантарную кератодермию, приводящую к нетрудоспособности, при помощи малой интерферерующей РНК (миРНК). Эта миРНК, называемая TD101, специфически и эффективно нацелена на мутантную мРНК кератина 6a (K6a) N171K, не влияя на мРНК K6a дикого типа.
[001256] Zheng et al. (PNAS, 24 июля 2012 г., т. 109, № 30, 11975-11980) показывают, что сферические конъюгаты наночастиц нуклеиновой кислоты (SNA-NC), золотые сердцевины, окруженные плотной оболочкой в высокой степени ориентированной ковалентно иммобилизованной миРНК, свободно проникают почти в 100% кератиноцитов in vitro, в кожу мыши и эпидермиса человека в течение нескольких часов после применения. Zheng et al. продемонстрировали, что однократное применение 25 нМ SNA-NC для рецепторов эпидермального фактора роста (EGFR) в течение 60 ч демонстрирует эффективное разрушение этого гена в коже человека. Подобную дозу можно предусмотреть для CRISPR Cas, иммобилизованного в SNA-NC для введения в кожу.
Злокачественная опухоль
[001257] В некоторых вариантах осуществления изобретения предусматривается лечение, профилактика или диагностика злокачественной опухоли. Мишенью предпочтительно является один или более из генов FAS, BID, CTLA4, PDCD1, CBLB, PTPN6, TRAC или TRBC. Злокачественная опухоль может представлять собой одну или более из лимфом, хронического лимфоцитарного лейкоза (CLL), В-клеточного лимфобластного лейкоза (B-ALL), острого лимфобластного лейкоза, острого миелоидного лейкоза, неходжкинской лимфомы (NHL), диффузной крупноклеточной лимфомы (DLCL), множественной миеломы, карциномы почек (RCC), нейробластомы, колоректального рака, рака молочной железы, рака яичников, меланомы, саркомы, рака предстательной железы, легких, пищевода, гепатоцеллюлярной карциномы, рака поджелудочной железы, астроцитомы, мезотелиомы, рака головы и шеи и медуллобластомы. Это может быть реализовано с помощью сконструированного химерного антигена Т-клеточного рецептора (СAR). Это описано в WO2015161276, содержание которого включено в настоящее описание в качестве ссылки и описано в настоящем описании ниже.
[001258] Гены-мишени, подходящие для лечения или профилактики злокачественной опухоли, могут включать, в некоторых вариантах, те, которые описаны в WO 02015048577, содержание которых включено в настоящее описание в качестве ссылки.
Синдром Ашера или пигментный ретинит-39
[001259] В некоторых вариантах осуществления предусматривается лечение, профилактика или диагностика синдрома Ашера или пигментного ретинита-39 (retinitis pigmentosa-39). Мишень предпочтительно представляет собой ген USH2A. В некоторых вариантах осуществления предусматривается коррекция G-делеции в позиции 2299 (2299delG). Это описано в WO2015134812A1, содержание которой включено в настоящее описание в качестве ссылки.
Муковисцидоз (CF)
[001260] В некоторых вариантах осуществления предусматривается лечение, профилактика или диагностика муковисцидоза. Мишенью предпочтительно является SCNN1A или ген CFTR. Это описано в W02015157070, содержание которой включено в настоящее описание в качестве ссылки.
[001261] Schwank et al. (CellS:.emCell, 13: 653-58, 2013) использовали CRISPR-Cas9 для коррекции дефекта, связанного с муковисцидозом, в стволовых клетках человека. Мишенью группы был ген ионного канала, рецептор трансмембранного проводника муковисцидоза (CFTR). Делеция в CFTR приводит к неправильному сворачиванию белка у пациентов с муковисцидозом. Используя культивируемые кишечные стволовые клетки, полученные из образцов клеток у двух детей с муковисцидозом, Schwank et al. смогли исправить дефект, используя CRISPR вместе с донорской плазмидой, содержащей последовательность для репарации инсерции. Затем исследователи вырастили клетки в кишечные "органоиды" или миниатюрные кишки и показали, что они функционируют нормально. В этом случае примерно половина клоновых органоидов подвергалась надлежащей генетической коррекции.
ВИЧ и СПИД
[001262] В некоторых вариантах осуществления предусматривается лечение, профилактика или диагностика ВИЧ и СПИДа. Мишенью при ВИЧ предпочтительно является ген CCR5. Это описано в WO2015148670A1, содержание которой включено в настоящее описание в качестве ссылки.
Бета-Талассемия
[001263] В некоторых вариантах осуществления предусматривается лечение, профилактика или диагностика бета-талассемии. Мишень предпочтительно представляет собой ген BCL11A. Это описано в WO2015148860, описание которой включено в настоящее описание в качестве ссылки.
Серповидноклеточная анемия
[001264] В некоторых вариантах осуществления предусматривается лечение, профилактика или диагностика серповидноклеточной анемии (SCD). Мишень предпочтительно представляет собой ген HBB или BCL11A. Это описано в WO2015148863, описание которого включено в настоящее описание в качестве ссылки.
Вирус простого герпеса 1 и 2
[001265] В некоторых вариантах осуществления предоставляется лечение, профилактика или диагностика HSV-1 (вирус простого герпеса 1). Мишень предпочтительно представляет собой ген UL19, UL30, UL48 или UL50 в HSV-1. Это описано в WO2015153789, описание которой включено в настоящее описание в качестве ссылки.
[001266] В других вариантах осуществления предусматривается лечение, профилактика или диагностика HSV-2 (вирус простого герпеса 2). Мишень предпочтительно представляет собой ген UL19, UL30, UL48 или UL50 в HSV-2. Это описано в изобретении WO2015153791, содержание которой включено в настоящее описание в качестве ссылки.
[001267] В некоторых вариантах осуществления предусматривается лечение, профилактика или диагностика первичной открытой угловой глаукомы (ПОАГ). Мишень предпочтительно представляет собой ген MYOC. Это описано в WO2015153780, содержание которой включено в настоящее описание в качестве ссылки.
Адоптивная клеточная терапия
[001268] Настоящее изобретение также предполагает использование системы CRISPR-Cas, описанной в настоящем описании, например. C2c1 или C2c3, чтобы модифицировать клетки для адоптивной терапии. Соответственно, аспекты настоящего изобретения включают адоптивную передачу клеток иммунной системы, таких как T-клетки, специфичных для выбранных антигенов, таких как связанные с опухолью антигены (см. Maus et al., 2014, Adoptive Immunotherapy for Cancer or Viruses, Annual Review of Immunology, Vol. 32: 189-225; Rosenberg and Restifo, 2015, Adoptive cell transfer as personalized immunotherapy for human cancel, Science Vol, 348 no. 6230 pp. 62-68; and, Restifo et al., 2015, Adoptive immunotherapy for cancer: harnessing the T cell response. Nat. Rev. Immunol. 12(4): 269-281; and Jenson and Riddell, 2014, Design and implementation of adoptive therapy with chimeric antigen receptor-modified T cells. Immunol Rev. 257(1): 127-144). Например, можно использовать различные стратегии для генетической модификации Т-клеток путем изменения специфичности Т-клеточного рецептора (TCR), например, путем введения новых TCRα и β-цепей с выбранной пептидной специфичностью (см. патент США № 8697854; публикации патентных заявок РСТ: W02003020763, W02004033685, WQ2004044004, W02005114215, W02006000830, W02008038002, W02008074322, W02005113595, WO2006125962, WO2013166321, WO2013039889, WO2014018863, WO2014083173, патент США 8088379).
[001269] В качестве альтернативы или дополнения к модификации TCR, химерные рецепторы антигенов (CAR) могут быть использованы для получения иммуноположительных клеток, таких как Т-клетки, специфичных для выбранных мишеней, таких как злокачественные клетки, с широким спектром рецепторных химерных конструкций, описанных (см. 5843728, 5851828, 5912170, 6004811, 6284240, 6392013, 6410014, 6753162, 8211422 и публикацию PCT WO9215322). Альтернативные конструкции CAR могут быть охарактеризованы как принадлежащие к последующим поколениям. CAR первого поколения обычно состоят из одноцепочечного вариабельного фрагмента антитела, специфичного к антигену, например, содержащего VL, связанного с VH специфического антитела, связанного гибким линкером, например, с помощью домена-петли CD8α и трансмембранного домена CD8α, с трансмембранным и внутриклеточным доменам сигнализации, такими как CD3cξ, так и FcRγ (sсFν-CD3ξ или scFv-FcRγ, см. патент США 7741465, патент США № 5912172, патент США № 5906936). CAR второго поколения включают внутриклеточные домены одной или более костимулирующих молекул, таких как CD28, OX40 (CD134) или 4-1BB (CD137) в эндодомене (например, scFv-CD28/OX4O/4-lBB-CD3Q, см. в патентах США №№ 8911993, 8916381, 8975071, 9017584, 9102760, 9102761), CAR третьего поколения включают комбинацию костимулирующих эндодоменов, таких как CD3ξ-цепь, CD97, CD11a-CD18, CD2, ICOS, CD27, CD154, CDS, OX40, 4-1BB, или CD28 (например, scFv-CD28-4-1BB-CD3ξ или scFv-CD28-OX40-CD3ξ, см. патент США 8906682, патент США № 8399645, патент США № 5686281; публикацию РСТ № WO2014134165; публикацию PCT № W02012079000). Альтернативно, костимуляция может быть организована путем экспрессии CAR в антигенспецифических Т-клетках, выбранных таким образом, чтобы их можно было активировать и расширять после взаимодействия их нативного αβTCR, например, с антигеном на профессиональных антиген-представляющих клетках, с сопутствующей костимуляцией. Кроме того, могут быть предусмотрены дополнительные модифицированные рецепторы на иммуночувствительных клетках, например, для улучшения нацеливания Т-клеточной атаки и/или минимизации побочных эффектов.
[001270] Для трансформации иммуноположительных клеток-мишеней могут быть использованы альтернативные способы, такие как слияние протопластов, липофекция, трансфекция или электропорация. Можно использовать широкий спектр векторов, таких как ретровирусные векторы, лентивирусные векторы, аденовирусные векторы, аденоассоциированные вирусные векторы, плазмиды или транспозоны, такие как транспозон Sleeping Beauty (см. патенты США №№ 6489458, 7148203, 7166682, 7985739; 8227432), для введения CAR, например, с использованием антиген-специфических CAR 2-го поколения, сигнализирующих посредством CD3ξ, и либо CD28, либо CD137. Вирусные векторы могут, например, включать векторы на основе ВИЧ, SV40, EBV, HSV или BPV.
[001271] Клетки, которые являются мишенями для последующей трансформации, могут, например, включать Т-клетки, естественные киллеры (NK), цитотоксические Т-лимфоциты (CTL), регуляторные Т-клетки, эмбриональные стволовые клетки человека, опухолевые инфильтрационные лимфоциты (TIL) или плюрипотентную стволовую клетку, из которых могут быть дифференцированы лимфоидные клетки. Т-клетки, экспрессирующие желаемый CAR, могут быть отобраны, например, посредством cокультивирования с γ-облученными активируемыми и размножающимися клетками (AaPC), которые совместно экспрессируют антиген злокачественной опухоли и костимулирующие молекулы. Может быть осуществлено увеличение в количестве сконструированных CARТ-клеток, например, путем совместного культивирования на AaPC в присутствии растворимых факторов, таких как IL-2 и IL-21. Такое увеличение в количестве может быть выполнено, например, таким образом, чтобы обеспечить память CAR+ T-клеток (которые могут, например, анализироваться при помощи неферментативной цифровой матрицы и/или мультипанельной проточной цитометрии). Таким образом, могут быть предоставлены CARТ-клетки, которые обладают специфической цитотоксической активностью против антигенсвязывающих опухолей (необязательно в сочетании с получением желаемых хемокинов, таких как интерферон-γ). CAR Т-клетки такого типа могут, например, использоваться на моделях животных, например, для того, чтобы уничтожать опухолевые ксенотрансплантаты.
[001272] Следующие подходы могут быть адаптированы для обеспечения способов лечения и/или увеличения выживаемости индивидуума, имеющего заболевание, такого как новообразование, например, путем введения эффективного количества иммуноположительной клетки, содержащей рецептор, распознающий антиген, который связывает выбранный антиген, где связывание активирует иммунореактивную клетку, тем самым вылечивая или предотвращая болезнь (такую как новообразование, патогенная инфекция, аутоиммунное расстройство или реакция на аллогенный трансплантат). Дозировки в терапии с использованием Т-клеток могут, например, включать введение от 106 до 109 клеток/кг, с или без курса лимфодеплеции, например, с циклофосфамидом.
[001273] В одном варианте осуществления лечение можно проводить пациентам, проходящим иммуносупрессивное лечение. Клетки или популяция клеток могут быть устойчивыми по меньшей мере к одному иммунодепрессанту из-за инактивации гена, кодирующего рецептор для такого иммунодепрессанта. Не ограниченное только теорией иммунодепрессивное лечение должно способствовать выбору и расширению отвечающих за иммунный ответ Т-клеток в соответствии с изобретением внутри самого пациента.
[001274] Введение клеток или популяций клеток в соответствии с настоящим изобретением может быть осуществлено любым удобным способом, в том числе путем аэрозольной ингаляции, инъекции, приема внутрь, трансфузии, имплантации или трансплантации. Клетки или популяция клеток могут быть введены пациенту подкожно, внутрикожно, внутриопухольно, интраназально, интрамедуллярно, внутримышечно, путем внутривенной или интралимфатической инъекции или внутрибрюшинно. В одном варианте осуществления композиции клеток по настоящему изобретению. Предпочтительно вводят путем внутривенной инъекции.
[001275] Введение клеток или популяции клеток может состоять из введения 104-109 клеток на кг массы тела, предпочтительно от 105 до 106 клеток/кг массы тела, включая все целочисленные значения количества клеток в этих диапазонах. Дозирование в Т-клеточной CAR терапии может, например, включать введение от 106 до 109 клеток/кг, с или без курса деплеции клеток, например, при помощи циклофосфамида. Клетки или популяцию клеток можно вводить в виде одной или более доз. В другом варианте осуществления эффективное количество клеток вводят в виде разовой дозы. В другом варианте осуществления эффективное количество клеток вводят в виде более чем одной дозы в течение определенного периода времени. Сроки введения находятся в пределах компетенции лечащего врача и зависят от клинического состояния пациента. Клетки или популяция клеток могут быть получены из любого источника, такого как банк крови или донор. В то время как индивидуальные потребности различаются, определение оптимальных диапазонов эффективного количества данного типа клеток для конкретного заболевания или условий находится в пределах компетенции специалиста в данной области. Эффективное количество означает количество, которое обеспечивает терапевтическое или профилактическое преимущество. Назначенная дозировка будет зависеть от возраста, здоровья и веса получателя, вида одновременно получаемого лечения, если таковое имеется, частоты лечения и характера желаемого эффекта.
[001276] В другом варианте осуществления эффективное количество клеток или композиции, содержащей эти клетки, вводят парентерально. Введение может быть внутривенным. Введение может быть непосредственно произведено путем инъекции внутрь опухоли.
[001277] Для защиты от возможных побочных реакций искусственные иммуноположительные клетки могут быть оснащены трансгенным предохранительным переключателем в форме трансгена, который делает клетки уязвимыми для воздействия на конкретный сигнал. Например, можно использовать ген тимидинкиназы вируса простого герпеса (TK), например, путем введения в аллогенные Т-лимфоциты, используемые в качестве инфузий донорных лимфоцитов после трансплантации стволовых клеток (Greco et al., Improving the safety of cell therapy with the TK-suicide gene. Front. Pharmacol. 2015: 6: 95). В таких клетках введение нуклеозидного пролекарства, такого как ганцикловир или ацикловир, вызывает гибель клеток. Альтернативные конструкции предохранительного выключателя включают индуцибельную каспазу, например, инициированную введением мелкомолекулярного димеризатора, который объединяет две нефункциональные молекулы icasp9 для образования активного фермента. Был описан широкий спектр альтернативных подходов к осуществлению контроля клеточной пролиферации (см. публикацию патента США № 20130071414, публикацию патентной заявки PCTWO2011146862, публикацию патентной заявки PCTWO2014011987, публикацию патентной заявки PCTWO2013040371, Zhou et al. BLOOD, 2014, 123/25: 3895-3905; Di Stasi et al, The New England Journal of Medicine 2011; 365: 1673-1683; Sadelain M, New England Journal of Medicine, 2011; 365: 1735-173; Ramos et al., Stem. Cells 28 (6): 1107-15 (2010)).
[001278] В дальнейшем усовершенствовании адоптивной терапии возможно редактирование генома с помощью системы CRISPR-Cas. Как описано в настоящем описании, для адаптации иммуноположительных клеток к альтернативным реализациям, например, для предоставления отредактированных CARТ-клеток (см. Poirot et al., 2015, Multiplex genome edited T-cell manufacturing platform. for "off-the-shelf" adoptive T-cell immunotherapies, CancerRes 75 (18); 3853). Например, иммуноположительные клетки могут быть удалены, для удаления экспрессии какого-либо или всего класса молекул HLA типа II и/или типа I или нокаута выбранных генов, которые могут ингибировать желаемый иммунный ответ, такой как ген PD1.
[001279] Клетки могут быть изменены с помощью любой системы CRISPR и способа ее применения, описанного в настоящей заявке. Системы CRISPR могут быть доставлены в иммунную клетку любым способом, описанным в настоящем описании. В предпочтительных вариантах осуществления клетки редактируются ex vivo и далее доставляются индивидууму, нуждающемуся в этом. Клетки иммунной системы, CARТ-клетки или любые клетки, используемые для переноса клеток для данного вида терапии, могут быть отредактированы. Редактирование может быть выполнено для устранения потенциальных аллореактивных Т-клеточных рецепторов (TCR), разрушения мишени химиотерапевтического агента, блокирования иммунной точки контроля, активации Т-клеток и/или усиления дифференцировки и/или пролиферации функционально истощенных или дисфункциональных CD8+ Т-клеток (см. публикации патентов PCT: WO2013176915, WO2014059173, WO2014172606, WO2014184744 и WO2014191128). Редактирование может привести к инактивации гена.
[001280] При инактивации гена, предполагается, что интересующий ген не экспрессируется в функциональной белковой форме. В конкретном варианте осуществления система CRISPR специфически катализирует расщепление в одном гене-мишени, тем самым инактивируя указанный ген. Вызываемые разрывы цепи нуклеиновой кислоты обычно восстанавливаются через различные механизмы гомологичной рекомбинации или не гомологичного соединения концов (NHEJ). Однако NHEJ является несовершенным процессом восстановления, который часто приводит к изменениям последовательности ДНК на месте расщепления. Восстановление через не гомологичное соединение концов (NHEJ) часто приводит к небольшим инсерциям или делециям (Indel) и может быть использовано для создания специфических нокаутов гена. Клетки, в которых произошел мутагенез, индуцированный расщеплением, могут быть идентифицированы и/или выбраны известными способами в данной области.
[001281] Т-клеточные рецепторы (TCR) являются рецепторами на поверхности клетки, которые участвуют в активации Т-клеток в ответ на появление антигена. TCR обычно состоит из двух цепей: α и β, которые собираются с последующим образованием гетеродимера и ассоциируется с CD3-трансдуцирующими субъединицами, с образованием комплекса рецептора Т-клеток, присутствующего на поверхности клетки. Каждая α и β цепочка TCR состоит из иммуноглобулиноподобной N-концевой переменной (V) и константной (C) области, гидрофобного трансмембранного домена и короткой цитоплазматической области. Что касается молекул иммуноглобулина, переменная область цепочек α и β генерируется V (D) J, создавая большое разнообразие антигенспецифических особенностей в популяции Т-клеток. Однако, в отличие от иммуноглобулинов, которые распознают интактный антиген, Т-клетки активируются обработанными пептидными фрагментами в сочетании с молекулой МНС, вводя дополнительное изменение в распознавание антигена Т-клетками, известное как МНС-рестрикция. Распознавание различий МНС между донором и реципиентом через рецептор Т-клеток приводит к пролиферации Т-клеток и потенциальному развитию болезни "трансплантат против хозяина" (GVHD). Инактивация TCR может приводить к элиминации TCR с поверхности Т-клеток, предотвращающих распознавание аллоантигена и, таким образом, к развитию отторжения (GVHD). Однако нарушение TCR обычно приводит к элиминации составляющей сигнального компонента CD3 и изменяет путь дальнейшей экспансии Т-клеток.
[001282] Было показано, что аллогенные клетки быстро отвергаются иммунной системой хозяина. Было продемонстрировано, что аллогенные лейкоциты, присутствующие в необлученной крови, будут сохраняться в течение не более 5-6 дней (Boni, Muranski et al, 2008 Blood 1; H2 (12): 4746-54). Таким образом, чтобы предотвратить отторжение аллогенных клеток, иммунная система хозяина обычно должна быть в некоторой степени подавлена. Тем не менее, несмотря на простоту адоптивной передачи клеток, использование иммунодепрессантов также оказывает пагубное влияние и на вводимые терапевтические Т-клетки. Поэтому для эффективного использования подхода адоптивной иммунотерапии в этих условиях вводимые клетки должны быть устойчивыми к иммунодепрессивному лечению. Таким образом, в конкретном варианте осуществления настоящее изобретение дополнительно включает стадию модификации Т-клеток, чтобы сделать их устойчивыми к иммунодепрессанту, предпочтительно путем инактивации по меньшей мере одного гена, кодирующего мишень иммунодепрессанта. Иммунодепрессивный агент является агентом, который подавляет иммунную функцию одним из нескольких механизмов действия. Иммунодепрессивный агент может быть, но не ограничен, ингибитором кальциневрина, мишенью рапамицина, блокатором α-цепи рецептора интерлейкина-2, ингибитором инозин-монофосфатдегидрогеназы, ингибитором дигидрофолиевой кислоты редуктазы, кортикостероидом или иммуносупрессивным антиметаболитом. Настоящее изобретение позволяет придавать иммунодепрессивную резистентность Т-клеткам для иммунотерапии путем инактивации мишени иммунодепрессанта в Т-клетках. В качестве неограничивающих примеров мишеней для иммуносупрессивного агента могут быть использованы рецепторы для иммуносупрессивного агента, такие как: CD52, глюкокортикоидный рецептор (GR), член семейства семейств FKBP и член семейства семейств циклофилина.
[001283] Иммунные точки контроля представляют собой ингибирующие пути, которые замедляют или останавливают иммунные реакции и предотвращают чрезмерное повреждение тканей при неконтролируемой активности иммунных клеток. В определенных вариантах осуществления целевым объектом иммунной контрольной точки является ген запрограммированной смерти 1 (PD-l или CD279) (PDCD1). В других вариантах осуществления иммунной точкой контроля является цитотоксический Т-лимфоцитарный антиген (CTLA-4). В дополнительных вариантах осуществления являющейся мишенью иммунной точкой контроля является другой член суперсемейства CD28 и CTLA4 Ig, такой как BTLA, LAGS, ICOS, PDL1 или KIR. В дополнительных вариантах осуществления настоящего изобретения являющаяся мишенью иммунная точка контроля является членом надсемейства TNFR, таким как CD40, OX40, CD137, GITR, CD27 или TIM-3.
[001284] Дополнительные иммунные точки контроля включают содержащую домен Src-гомологии 2 тирозинфосфатазу 1 (SHP-1) (Watson HA, et al., SHP-1: the next checkpoint target for cancer immunotherapy? Biochem Soc Trans. 2016 Apr 15;44(2):356-62). SHP-1 является широко экспрессируемой ингибирующей белковой тирозинфосфатазой (PTP). В Т-клетках она является отрицательным регулятором антигензависимой активации и пролиферации. Она представляет собой цитозольный белок и, следовательно, не поддается способам, опосредованным антителами, но ее роль в активации и пролиферации делает его привлекательной мишенью для генетической манипуляции в стратегиях адаптивного переноса, таких как Т-клетки с химерным рецептором антигена (CAR). Иммунные контрольные точки могут также включать Т-клеточный иммунорецептор с доменами Ig и ITEM (TIGIT/Vstm3/WUCAM/VSIG9) и VISTA (LeMercierI, et al., (2015) Beyond CTLA-4 and PD-1, the generation of negative checkpoint regulators. Front. Immunol. 6:418).
[001285] WO2014172606 относится к применению MT1 и/или ингибиторов MT1 для увеличения пролиферации и/или активности израсходованных CD8+ Т-клеток и для уменьшения выработки CD8+ Т-клеток (например, снижения функционально истощенных или невосприимчивых иммунных клеток CD8+). В некоторых вариантах осуществления металлотионеины нацелены на редактирование генов в адоптивно перенесенных Т-клетках.
[001286] В некоторых вариантах осуществления мишени для редактирования гена могут быть по меньшей мере одним локусом-мишенью, участвующим в экспрессии иммунного контрольного белка. Такие мишени могут включать, но не ограничиваются следующмими:CTLA4, PPP2CA, PPP2CB, PTPN6, PTPN22, PDCD1, ICOS (CD278), PDL1, KIR, LAG3, HAVCR2, BTLA, CD 160, TIGIT, CD96, CRTAM, LAIRI, SIGLEC7, SIGLEC9, CD244 (2B4), TNFRSF10B, TNFRSF10A, CASP8, CASP10, CASP3, CASP6, CASP7, FADD, FAS, TGFBRI1, TGFRBRI, SMAD2, SMAD3, SMAD4, SMAD10, SKI, SKIL, TG1F1, IL10BA, IL10RB, HM0X2, IL6R, IL6ST, EIF2AK4, CSK, PAG1, SIT1, FOXP3, PRDM1, BATF, VISTA, GUCY1A2, GUCY1A3, GUCY1B2, GUCYIB3, MT1, MT2, CD40, OX40, CD137, GITR, CD27, SHP-l или TIM-3.В предпочтительных вариантах осуществления мишенью является локус гена, участвующий в экспрессии генов PD-1 или CTLA-4,. В других предпочтительных вариантах осуществления мишенью являются следущие комбинации генов, но ограничиваясь ими: PD-1 и TIGIT.
[001287] В других вариантах редактируются по меньшей мере два гена. Пары генов могут включать, но не ограничиваются ими: PD1 и TCRα, PD1 и TCRβ, CTLA-4 и TCRα, CTLA-4 и TCRβ, LAG3 и TCRα, LAG3 и TCRβ, Tim3 и TCRα, Tim3 и TCRβ, BTLA и TCRα, BTLA и TCRβ, BY55 и TCRα, BY55 и TCRβ, TIGIT и TCRα, TIGIT и TCRβ, B7H5 и TCRα, B7H5 и TCRβ, LAIRI и TCRα, LAIRI и TCRβ, SIGLEC10 и TCRα, SIGLEC10 и TCRβ, 2B4 и TCRα, 2B4 и TCRβ.
[001288] Независимо от того, до или после генетической модификации Т-клеток, Т-клетки могут быть активированы и проведена экспансия, как правило, с использованием способов, описанных, например в патентах США 6352694; 6534055; 6905680; 5858358; 6887466; 6905681; 7144575; 7232566; 7175843; 5883223; 6905874; 6797514; 6867041; и 7572631. Экспансия Т-клеток может быть проведена как in vitro так и in vivo.
[001289] Практика настоящего изобретения использует, если не указано иначе, общепринятые способы иммунологии, биохимии, химии, молекулярной биологии, микробиологии, клеточной биологии, геномики и рекомбинантных ДНК, которые входят в компетенции специалистов. См. Sambrook, Fritsch and Maniatis, MOLECULAR CLONING: A LABORATORY MANUAL, 2nd edition (1989); CURRENT PROTOCOLS IN MOLECULAR BIOLOGY (F, M. Ausubel, et al. eds., (1987)); the series METHODS IN ENZYMOLOGY (Academic Press, Inc.): PCR 2: A PRACTICAL APPROACH (M.J. MacPherson, B.D. Flames and G.R. Taylor eds. (1995)), Harlow and Lane, eds. (1988) ANTIBODIES, A LABORATORY MANUAL, and ANIMAL CELL CULTURE (R.I. Freshney, ed. (1987)).
[001290] Практика настоящего изобретения использует, если не указано иначе, общепринятые методы создания поколения генетически модифицированных мышей. Sec Marten H. Hofker and Jan van Deursen, TRANSGENIC MOUSE METHODS AND PROTOCOLS, 2nd edition (2011).
Пусковые факторы генов
[001291] Настоящее изобретение также предполагает использование системы CRISPR-Cas, описанной в настоящем описании, например, эффекторного белка C2c1 или C2c3, системы для обеспечения пусковых факторов генов, управляемых РНК, например, в системах, аналогичных описанным элементам, запускающим экспрессию генов, например описанных в публикации патента PCT WO 2015/105928. Системы такого типа могут, например, обеспечивать способы изменения эукариотических клеток зародышевой линии путем введения в клетку последовательности нуклеиновой кислоты, кодирующей ДНК-нуклеазу, нацеленную на РНК, и одну или более направляющих РНК. Направляющие РНК могут быть сконструированы так, чтобы быть комплементарными одному или более местам мишеней на геномной ДНК клетки зародышевой линии. Последовательность нуклеиновой кислоты, кодирующая ДНК-нуклеазу, ориентированную на РНК, и последовательность нуклеиновой кислоты, кодирующую направляющие РНК, могут быть предоставлены на конструкциях между фланкирующими последовательностями с промоторами, расположенными так, что клетка зародышевой линии может экспрессировать ДНК-нуклеазу, нацеленную на РНК, и направляющие РНК вместе с любыми желаемыми последовательности кодирования необходимого продукта, которые также расположены между фланкирующими последовательностями. Фланкирующие последовательности, как правило, включают последовательность, которая идентична соответствующей последовательности на выбранной хромосоме-мишени, так что фланкирующие последовательности работают с компонентами, кодируемыми конструкцией, чтобы облегчить введение последовательностей конструкции посторонней нуклеиновой кислоты в геномную ДНК, чтобы разрезать представляющий интерес сайт с помощью таких механизмов, как гомологичная рекомбинация, чтобы сделать зародышевую клетку гомозиготной по отношению к чужой нуклеиновой кислоте. Таким образом, системы запуска генов способны интрогрессировать желаемые гены-грузы во всей размножающейся популяции (Gantz et al., 2015, Highly efficient Cas9-mediated gene drive for population modification of the malaria vector mosquito Anopheles PNAS 2015, опубликованная до печати 23 ноября 2015 года, doi: 10.1073/pnas. 1521077112; Esvelt et al., 2014). В некоторых вариантах осуществления могут быть выбраны последовательности-мишени, которые имеют мало потенциальных участков нецелевого назначения в геноме. Ориентация нескольких сайтов в локусе-мишени с использованием нескольких направляющих РНК может увеличить частоту разрезания и затруднить эволюцию устойчивых к пусковым факторам аллелей. Укороченные направляющие РНК могут снижать нецелевое разрезание. Вместо одиночной нуклеазы можно использовать парные никазы для дальнейшего увеличения специфичности. Конструкции запуска генов могут включать последовательности грузов, кодирующие регуляторы транскрипции, например, для активации генов гомологичной рекомбинации и/или подавления негомологичного концевого соединения. Участки-мишени могут быть выбраны в пределах важного гена, так что события негомологичного соединения концов могут вызывать летальность, а не создавать резистентный к пусковому фактору аллель. Конструкции пусковых факторов генов могут быть спроектированы для работы в диапазоне температур Cho et al. 2013, Rapid and Tunable Control of Protein Stability in Caenorhabditis elegans Using a Small Molecule, PLoS ONE 8(8): e72393. doi: 10.1371 /journal.pone.0072393).
Ксенотрансплантация
[001292] Настоящее изобретение также предусматривает использование системы CRISPR-Cas, описанной в настоящем описании, например, C2c1 или C2c3, чтобы обеспечить РНК-направленные ДНК-нуклеазы, адаптированные для использования в модифицированных тканях для трансплантации. Например, нуклеазы ДНК, основанные на РНК, могут быть использованы для нокаута, нокдауна или разрушения выбранных генов у животного, такого как трансгенная свинья (например, линия свиней, трансгенных по гену гемоксигеназы-1 человека), например, нарушая экспрессию генов, которые кодируют эпитопы, распознаваемые иммунной системой человека, т.е. гены ксеноантигена. Гены-кандидаты свиньи для нарушения могут, например, включать гены α(1,3)-галактозилтрансферазы и гидроксилазы цитидин-монофосфат--N-ацетилнейраминовой кислоты (см. публикацию PCT WO 2014/066505). Кроме того, могут быть разрушены гены, кодирующие эндогенные ретровирусы, например гены, кодирующие все эндогенные ретровирусы свиней (см. Yang et al., 2015, Genome-wide inactivation of porcine endogenous retroviruses (PERVs), Science 27 November 2015: Vol. 350 no. 6264 pp. 1101-1104). Кроме того, РНК-направленные ДНК-нуклеазы могут использоваться для нацеливания на сайт для интеграции дополнительных генов донорских животных ксенотрансплантата, таких как ген CD55 человека, для улучшения защиты от особенно острого отторжения.
Общие вопросы генной терапии
[001293] Примеры генов и полинуклеотидов, связанных с болезнями, а также конкретную информацию о болезнях можно получить в McKusick-Nathans Institute of Genetic Medicine, Johns Hopkins University (Baltimor, Md.) и в National Center for Biotechnology Information, National Library of Medicine, (Bethesda Md.)), которые доступны через интернет.
[001294] Мутации в этих генах и путях могут приводить к продукции неправильных белков или белков в неподходящих количествах, что влияет на функцию. Другие примеры генов, заболеваний представлены в предварительной заявке на патент США 61/736527, поданной 12 декабря 2012 года, включенной в качестве ссылки. Подобные гены, белки и пути могут быть подвергнуты нацеливанию полинуклеотидом комплекса CRISPR по настоящему изобретению. Примеры связанных с болезнями генов и полинуклеотидов перечислены в таблицах 1 и 2. Примеры генов и полинуклеотидов, связанных с сигнальными биохимическими каскадами, перечислены в таблице 3.
Таблица 1
Таблица 2
Таблица 3
[001295] Варианты осуществления изобретения также относятся к способам и конструкциям, связанным с нокаутом генов, амплификацией генов и исправлением конкретных мутаций, ассоциированных с нестабильностью повторов ДНК и неврологическими заболеваниям (Robert D. Wells, Tetsuo Ashizawa, Genetic Instabilities and Neurological Diseases, Second Edition, Academic Press, Oct 13, 2011-Medical). Было обнаружено, что конкретные аспекты последовательностей тандемных повторов ответственны более чем за двадцать заболеваний человека (New insights into repeat instability: role of RNA-DNA hybrids. Mclvor El, Polak U, Napierala M. RNA. Biol. 2010 Sep-Oct; 7(5):551-8). Указанные системы эффекторных белков могут быть использованы, чтобы корректировать эти дефекты геномной нестабильности.
[001296] Некоторые дальнейшие аспекты изобретения относятся к корректировке дефектов, связанных с широким диапазоном генетических заболеваний, которые далее описаны на сайте Национального института здоровья в тематическим подразделе "Генетические заболевания". Генетические заболевания головного мозга могут включать (но этим не ограничиваются) адренолейкодистрофию, агенез мозолистого тела, синдром Аикарди, болезнь Альперса, болезнь Альцгеймера, синдром Барта, болезнь Баттена, CADASIL, мозжечковую дегенерацию, болезнь Фабри, болезнь Герстмана-Штраусслера-Шейнкера, болезнь Хантингтона и другие болезни триплетных повторов, болезнь Лея, синдром Леша-Нихена, болезнь Менкеса, митохондриальные миопатии, кольпоцефалия NINDS. Эти заболевания описаны далее на сайте Национального института здоровья в подразделе "Генетические заболевания".
Разработка и применение Cas9
[001297] Настоящее изобретение может быть дополнительно проиллюстрировано и расширено при помощи описания разработки и использования CRISPR-Cas9, и, в частности, с точки зрения доставки белкового комплекса CRISPR и использования эндонуклеазы, направляемой РНК, в клетках и организмах, как изложено в следующих статьях, включенных посредством ссылок:
> Multiplex genome engineering using CRISPR/Cas systems. Cong, L., Ran, F.A., Cox, D., Lin, S., Barretto, R., Habib, N., Hsu, P.D., Wu, X., Jiang, W., Marraffini, L.A., & Zhang, F. Science Feb 15;339(6121):819-23 (2013);
> RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Jiang W., Bikard D., Cox D., Zhang F, Marraffini LA. Nat Biotechnol Mar;31(3):233-9 (2013);
> One-Step Generation of Mice Carrying Mutations in Multiple Genes by CRISPR/Cas- Mediated Genome Engineering. Wang H., Yang H., Shivalila CS., Dawlaty MM., Cheng AW., Zhang F., Jaenisch R. Cell May 9;153(4):910-8 (2013);
> Optical control of mammalian endogenous transcription and epigenetic states. Konermann S, Brigham MD, Trevino AE, Hsu PD, Heidenreich M, Cong L, Platt RT Scott DA, Church GM, Zhang F. Nature. Aug 22;500 (7463):472-6. doi: 10.1G38/Naturel2466. Epub 2013 Aug 23 (2013);
> Double Nicking by RNA-Guided CRISPR Cas9 for Enhanced Genome Editing Specificity, Ran, FA., Hsu, PD., Lin, CY., Gootenberg, JS., Konermann, S., Trevino, AE., Scott, DA., Inoue, A., Matoba, S., Zhang, Y., & Zhang, F. Cell Aug 28. pii: S0092- 8674(13)01015-5 (2013-A);
> DNA targeting specificity of RNA-guided Cas9 nucleases. Hsu, P., Scott, D., Weinstein, J., Ran, FA., Konermann, S., Agarwala, V., Li, Y., Fine, E., Wu, X., Shalem, O., Cradick, TJ., Marraffini, LA., Bao, G., & Zhang, F. Nat Biotechnol doi: 10.1038/nbt.2647 (2013);
> Genome engineering using the CRISPR-Cas9 system. Ran, FA., Hsu, PD., Wright, J., Agarwala, V., Scott, DA., Zhang, F. Nature Protocols Nov;8(l l):2281-308 (2013-B);
> Genome-Scale CRISPR-Cas9 Knockout Screening in Human Cells. Shalem, O., Sanjana, NE., Hartenian, E., Shi, X., Scott, DA., Mikkelson, T., Heckl, D., Ebert, BL., Root, DE., Doench, JG., Zhang, F. Science Dec 12. (2013). [Epub ahead of print];
- Crystal staicture of cas9 in complex with guide RNA and target DNA. Nishimasu, H., Ran, FA., Hsu, PD., Konermann, S., Shehata, SI, Dohmae, N., Ishitani, R., Zhang, F., Nureki, (), Cell Feb 27, 156(5):935-49 (2014);
- Genome-wide binding of the CRISPR endonuclease Cas9 in mammalian cells. Wu X., Scott DA., Kriz AJ., Chiu AC., Hsu PD., Dadon DB, Cheng AW., Trevino AE, Konermann S., Chen S., Jaenisch R., Zhang F., Sharp PA. Nat Bioteehnol. Apr 20. doi: 10. lQ38/nbt.2889 (2014);
- CRISPR-Cas9 Knockin Mice for Genome Editing and Cancer Modeling. Platt RJ, Chen S, Zhou Y, Yim MJ, Swiech L, Kempton HR, Dahlman JE, Parnas G, Eisenhaure TM, Jovanovic M, Graham DB, Jhunjhunwala S, Heidenreich M, Xavier RJ, Danger R, Anderson DG, Hacohen N, Regev A, Feng G, Sharp PA, Zhang F. Cell 159(2): 440-455 DOI: 10.1016/j. cell.2014.09.014(2014);
- Development and Applications of CRISPR-Cas9 for Genome Engineering, Hsu PD, Lander ES, Zhang F., Cell. Jun 5,157(6): 1262-78 (2014).
- Genetic screens in human cells using the CRISPR/Cas9 system, Wang T, Wei JJ, Sabatini DM, Lander ES., Science. January 3; 343(6166): 80-84. doi: 10.1126/science. 1246981 (2014);
- Rational design of highly active sgRNAs for CRISPR-Cas9-mediated gene inactivation, Doench JG, Hartenian E, Graham DB, Tothova Z, Hegde M, Smith I, Sullender M, Ebert BL, Xavier RJ, Root DE., (published online 3 September 2014) Nat Bioteehnol. Dec;32(12): 1262-7 (2014);
- In vivo interrogation of gene function in the mammalian brain using CRISPR-Cas9, Swiech L, Heidenreich M, Banerjee A, Habib N, Li Y, Trombetta J, Sur M, Zhang F., (published online 19 October 2014) Nat Bioteehnol, Jan,33(1): 102-6 (2015);
- Genome-scale transcriptional activation by an engineered CRISPR-Cas9 complex, Konermann S, Brigham MD, Trevino AE, Joung J, Abudayyeh OO, Barcena C, Hsu PD, Habib N, Gootenberg JS, Nishimasu H, Nureki O, Zhang F., Nature. Jan 29;517(7536):583-8 (2015).
- A split-Cas9 architecture for inducible genome editing and transcription modulation, Zetsche B, Volz SE, Zhang F., (published online 02 February 2015) Nat Bioteehnol. Feb;33(2): 139-42 (2015);
- Genome-wide CRISPR Screen in a Mouse Model of Tumor Growth and Metastasis, Chen S, Sanjana NE, Zheng K, Shalem O, Lee K, Shi X, Scott DA, Song J, Pan JQ, Weissleder R, Lee H, Zhang F, Sharp PA. Cell 160, 1246-1260, March 12, 2015 (multiplex screen in mouse), and
- In vivo genome editing using Staphylococcus aureus Cas9, Ran FA, Cong L, Yan WX, Scott DA, Gootenberg JS, Kriz AJ, Zetsche B, Shalem Q, Wu X, Makarova KS, Koonin EV, Sharp PA, Zhang F., (published online 01 April 2015), Nature. Apr 9;520(7546): 186-91 (2015).
- Shalem et. al, "High-throughput functional genomics using CRISPR-Cas9," Nature Reviews Genetics 16, 299-311 (May 2015).
- Xu et al., "Sequence determinants of improved CRISPR sgRNA design," Genome Research 25, 1147-1157 (August 2015).
- Parnas et al., "A Genome-wide CRISPR Screen in Primary Immune Cells to Dissect Regulatory Networks," Cell 162, 675-686 (July 30, 2015).
- Ramanan et al., CRISPR/Cas9 cleavage of viral DNA efficiently suppresses hepatitis В virus," Scientific Reports 5:10833, doi: 10.1038/srepl0833 (June 2, 2015),
- Nishimasu et al., Crystal Structure of Staphylococcus aureus Cas9," Cell 162, 1113-1126 (Aug, 27, 2015),
- BCL11A enhancer dissection by Cas9-mediated in situ saturating mutagenesis, Canver et al., Nature 527(7577): 192-7 (Nov. 12, 2015) doi: 10.1038/naturel5521. Epub 2015 Sep 16.
- Cpf1 Is a Single RNA-Guided Endonuclease of a Class 2 CRISPR-Cas System, Zetsche et al..Cell 163, 759-71 (Sep 25, 2015),
- Discovery and Functional Characterization of Diverse Class 2 CRISPR-Cas Systems.Shmakov et al, Molecular Cell, 60(3), 385-397 doi: 10.1016/j.molcel.2015,10.008 Epub October 22, 2015.
- Rationally engineered Cash nucleases with improved specificity, Slaymaker et al., Science 2015 Dec 1. pii: aad5227. [Epub ahead of print],
каждый из которых включен в настоящее описание в качестве ссылки, может рассматриваться в практике настоящего изобретения и кратко обсуждается ниже:
- Cong et al. разработали системы CRISPR-Cas типа II для использования в эукариотических клетках на основе как Streptococcus thermophilus Cas9, так и Streptococcus pyogenes Cas9 и показали, что нуклеазы Cas9 могут быть направлены короткой РНК, чтобы индуцировать точное расщепление ДНК в клетках человека и мыши. Их исследование также показало, что Cas9, превращенный в создающий однонитевые разрывы фермент, может быть использован для облегчения гомологической репарации в эукариотических клетках с минимальной мутагенной активностью. Кроме того, их исследование показало, что несколько направляющих последовательностей могут быть закодированы в единую последовательность CRISPR, чтобы обеспечить одновременное редактирование нескольких участков на эндогенных геномных локусах в геноме млекопитающих, демонстрируя легкость в программировании и широкую применимость технологии нуклеазы, основанной на РНК. Эта возможность использовать РНК для программирования последовательность-специфичного расщепления ДНК в клетке определила новый класс инструментов для геномной инженерии. Эти исследования также показали, что другие локусы CRISPR, вероятно, могут быть трансплантированы в клетки млекопитающих для опосредования расщепления в целевых участках генома млекопитающих. Важно отметить, что в будущем некоторые аспекты системы CRISPR-Cas могут быть дополнительно улучшены для повышения ее эффективности и универсальности.
- Jiang et al. использовали короткие палиндромные повторы, регулярно расположенные кластерами (CRISPR), связанные с эндонуклеазой Cas9, объединенные с двойными РНК, для введения точных мутаций в геномы Streptococcus pneumoniae и Escherichia coli. Этот подход основан на двойной РНК: расщепление, направляемое Cas9, на целевом геномном участке уничтожает немутированные клетки и позволяет обойти необходимость выбора маркеров или систем отрицательного отбора. В исследовании сообщается о перепрограммировании специфики комплекса двойной РНК: Cas9 путем изменения последовательности короткой CRISPRРНК (cr-РНК) для внесения одно- и мультинуклеотидные изменений в матрицы редактирования. Исследование показало, что одновременное использование двух cr-РНК обеспечивало мультиплексный мутагенез. Кроме того, когда подход использовался в сочетании с рекомбинацией, у S. pneumoniae почти 100% клеток, которые были восстановлены с использованием описанного подхода, содержали желаемую мутацию, а в E.coli 65% клеток, которые были восстановлены, содержали мутацию.
- Wang et al., 2013 использовали систему CRISPR/Cas для одношаговой генерации мышей, несущих мутации в нескольких генах, которые традиционно генерировались на нескольких стадиях путем последовательной рекомбинации в эмбриональных стволовых клетках и/или трудоемким скрещиванием мышей с единственная мутация. Система CRISPR/Cas значительно ускорит исследование in vivo функционально избыточных генов и эпистатических генных взаимодействий.
- Konermann et al. (2013) рассмотрели необходимость в универсальных и надежных технологий для оптической и химической модуляции ДНК-связывающих доменов на основе фермента CRISPR Cas9, а также подобных эффекторам активаторов транскрипции (TALE).
- Ran et al. (2013-А) описали подход, в котором объединены никазный мутант Cas9 с парными направляющими РНК для введения целенаправленных двунитевых разрывов. Это касается нуклеазыCas9 из микробной системы CRISPR-Cas, которая нацелена на конкретные геномные локусы с помощью направляющей последовательности, которая может переносить определенные несоответствия с ДНК-мишенью и тем самым способствовать нежелательному нецелевому мутагенезу. Поскольку отдельные однонитевые разрывы в геноме восстанавливаются с высокой точностью, одновременное внесение однонитевых разрывов с помощью надлежащим образом смещенных направляющих РНК требуется для двухцепочечных разрывов и увеличивает количество специфично распознаваемых оснований для целевого расщепления. Авторы продемонстрировали, что использование парного внесения одноцепочечных разрывов может снизить нецелевую активность в 50-100 раз в клеточных линиях и облегчить нокаут генов у зигот мышей без ущерба для эффективности расщепления мишеней. Эта универсальная стратегия позволяет использовать широкий спектр приложений для редактирования генома, которые требуют высокой специфичности.
- Hsu et al. (2013) охарактеризовали специфичность нацеливания, осуществляемого SpCas9 в клетках человека, чтобы получить данные о выборе целевых сайтов и избегать нецелевого воздействия. В исследовании оценивали> 700 вариантов направляющей РНК и индуцированные SpCas9 уровни инсерции/делеции (indel-мутации)на > 100 предсказанных геномных нецелевых локусов в клетках 293T и 293FT. Авторы показали, что SpCas9 допускают несоответствия между направляющей РНК и ДНК-мишенью в разных положениях зависимым от последовательности образом, чувствительны к числу, положению и распределению несоответствий. Авторы также показали, что SpAas9-опосредованное расщепление не подвержено влиянию метилирования ДНК и что доза SpCas9 и одноцепочечной направляющей РНК (sgRNA) может быть титрована для минимизации нецелевой модификации. Кроме того, для облегчения применения приложений для инженерии генома млекопитающих авторы разработали программного обеспечения для выбора и проверки целевых последовательностей, а также нецелевого анализа.
- Ran et al. (2013-B) описали набор инструментов для Cas9-опосредованного редактирования генома путем негомологичного соединения концов (NHEJ) или гомологичной репарации (HDR) в клетках млекопитающих, а также создания модифицированных клеточных линий для последующих функциональных исследований. Для минимизации нецелевого расщепления, авторы описали стратегию внесения двойных одноцепочечных разрывов, использующую никазный мутант Cas9 с парными направляющими РНК. Протокол, предоставленный авторами, экспериментально выработал руководящие принципы для выбора целевых участков, оценки эффективности расщепления и анализа нецелевой активности. Исследования показали, что, начиная с разработки мишени, модификации генов могут быть достигнуты всего за 1-2 недели, а модифицированные клональные клеточные линии могут быть получены в течение 2-3 недель.
- Shalem et al. описали новый способ исследования функции гена в масштабе всего генома. Их исследования показали, что доставка полногеномной библиотеки нокаутов CRISPR-Cas9 (GeCKO), нацеленной на 18080 генов с 64751 уникальными направляющими последовательностями, позволила провести скрининг c помощью как отрицательного, так и положительного отбора в клетках человека. Во-первых, авторы показали использование библиотеки GeCKO для идентификации генов, необходимых для жизнеспособности клеток при раке и плюрипотентных стволовых клеток. Далее, в модели меланомы авторы провели скрининг для поиска генов, потеря которых связана с резистентностью к вемурафенибу, терапевтическому препарату, который ингибирует мутантную протеинкиназу ВRAF. Их исследования показали, что кандидаты высшего ранга включали ранее проверенные гены NF1 и MED12, а также новые: NF2, CUL3, TADA2B и TADA1. Авторы наблюдали высокий уровень согласованности между независимыми направляющими РНК, нацеленными на один и тот же ген, и высоким показателем подтверждения полученного результата, и, таким образом, продемонстрировали возможность скрининга целого генома с помощью Cas9.
- Nishimasu et al. исследовали кристаллической структуре Cas9 Streptococcus pyogenes в комплексе с одноцепочечной направляющей РНК (sg-РНК) и его ДНК-мишени при разрешении 2,5 A°. Обнаружена "двулопастная" архитектура структуры, состоящая из субъединицы для распознавания мишени и нуклеазной субъединицы, вмещающих гетеродуплекс sg-РНК:ДНК в положительно заряженном желобке на их поверхности. В то время как субъединица ("доля") для распознавания необходима для связывания sg-РНК и ДНК, нуклеазная субъединица ("доля") содержит домены HNH и RuvC-нуклеазы, которые должным образом расположены для расщепления комплементарных и не комплементарных цепей ДНК-мишени соответственно. Нуклеазная субъединица также содержит карбоксильный концевой домен, ответственный за взаимодействие с последовательностью, прилегающей к протоспейсеру (PAM). Эта структура с высоким разрешением и сопутствующие функциональные анализы выявили молекулярный механизм РНК-направленного нацеливания на ДНК, осуществляемого Cas9, таким образом прокладывая путь для рационального проектирования новых, универсальных технологий редактирования генома.
- Wu et al. картировали во всем геноме сайты связывания каталитически неактивного Cas9 (dCas9) из Streptococcus pyogenes, нагруженных одноцепочечными направляющими РНК (sg-РНК) в эмбриональных стволовых клетках мыши (mESC). Авторы показали, что каждая из четырех sg-РНК тестировала мишени dCas9 между десятками и тысячами геномных участков, часто характеризующихся 5-нуклеотидной областью-затравкой в sg-РНК и NGG-соседним мотивом, прилегающим к протоспейсеру (PAM). Недоступность хроматина уменьшает связывание dCas9 с другими сайтами с соответствующими "seed"-последовательностями; таким образом, 70% нецелевых сайтов связаны с генами. Авторы показали, что целенаправленное секвенирование 295 сайтов связывания dCas9 в mESC, трансфицированных каталитически активным Cas9, выявило только один сайт, мутированный выше фоновых уровней. Авторы предложили модель с двумя состояниями для связывания и расщепления Cas9, в которой совпадение "seed"-последовательности вызывает связывание, но для расщепления требуется обширное спаривание оснований с ДНК-мишенью.
- Platt et al. создали Cre-зависимую мышь с нокином Cas9. Авторы продемонстрировали как in vivo, так и ex vivo редактирование генома с использованием аденоассоциированного вируса (AAV), лентивируса или опосредованной частицами доставки направляющей РНК в нейроны, иммунные клетки и эндотелиальные клетки.
- Hsu et al. (2014) - обзорная статья, в которой обсуждается история CRISPR-Cas9 от йогурта до редактирования генома, включая генетический скрининг клеток.
- Wang et al. (2014) относится к объединенному способу генетического скрининга, основанному на потере функциональности, подходящему как для положительного, так и для отрицательного отбора, который использует полногеномную лентивирусную библиотеку одноцепочечной направляющей РНК (sg-РНК).
- Doench et al. создали пул одноцепочечных направляющих РНК (sg-РНК), чередуя все возможные целевые сайты группы из шести эндогенных мышиных и трех эндогенных генов человека и количественно оценили их способность продуцировать нулевые аллели их целевого гена путем окрашивания антителом и проточной цитометрии. Авторы показали, что оптимизация PAM улучшила активность, а также предоставила интерактивный инструмент для разработки одноцепочечных направляющих sg-РНК.
- Swiech et al. продемонстрировали, что AAV-опосредованное редактирование генома с помощью SpCas9 может позволить провести ретроспективные генетические исследования функции гена в головном мозге.
- Konermann et al. (2015) обсуждают возможность присоединения нескольких эффекторных доменов, например активаторов транскрипции, функциональных и эпигеномных регуляторов в соответствующих положениях на направляющей молекуле, таких как шпилька или тетрапетля с линкерами и без них.
- Zetsche et al. демонстрируют, что фермент Cas9 можно разделить на два и, следовательно, можно контролировать сборку Cas9 для активации.
- Chen et al. c помощью мультиплексного полногеномного скрининга in vivo посредством CRISPR-Cas9 обнаружили у мышей гены, регулирующие метастазы в легкие.
- Ran et al. (2015) исследовали SaCas9 и его способность редактировать геномы и продемонстрировали, что нельзя экстраполировать биохимические анализы. Shalem et al. (2015) описаны способы использования каталитически неактивные слияния Cas9 (dCas9) для синтетического подавления (CRISPRi) или активации (CRISPRa) экспрессии и продемонстрированы преимущества использованияCas9 для полногеномного скрининга, включая упорядоченный и объединенные скрининг, нокаутные подходы, инактивирующие геномные локусы, и стратегии, модулирующие активность транскрипции.
Окончательные изменения
- Shalem et al. (2015) описали способы использования каталитически неактивные слияния Cas9 (dCas9) для синтетического подавления (CRISPRi) или активации (CRISPRa) экспрессии и продемонстрированы преимущества использования Cas9 для полногеномного скрининга, включая упорядоченный и объединенные скрининг, нокаутные подходы, инактивирующие геномные локусы, и стратегии, модулирующие активность транскрипции.
- Xu et al. (2015) оценили характеристики последовательности ДНК, которые способствуют эффективности одноцепочечной направляющей РНК (sg-РНК) в скрининге на основе CRISPR. Авторы исследовали эффективность нокаута CRISPR/Cas9 и предпочтение нуклеотидов на участке расщепления. Авторы также обнаружили, что предпочтительные последовательности для CRISPR-интерференции/CRISPR-активации (CRISPRi/a) существенно отличаются от таковых для нокаута CRISPR/Cas9.
- Paraas et al. (2015) ввели полногеномные объединенные библиотеки CRISPR-Cas9 в дендритные клетки для идентификации генов, контролирующих индукцию фактора некроза опухолей (Tnf) бактериальным липополисахаридом (LPS). Известные регуляторы сигналинга Tlr4 и ранее неизвестные кандидаты были идентифицированы и классифицированы на три функциональных модуля с явным влиянием на канонические ответы на LPS.
- Ramananetal (2015) продемонстрировали расщепление вирусной эписомальной ДНК (cccDNA) в инфицированных клетках. Геном HBV существует в ядрах инфицированных гепатоцитов в виде двухцепочечных эписомальных ДНК длиной 3,2 т.п.н., которые называются ковалентно замкнутой кольцевой ДНК (cccDNA), которая является ключевым компонентом жизненного цикла HBV, репликация которого не ингибируется современной терапией. Авторы показали, что одноцепочечные направляющие молекулы РНК (sg-РНК), специально предназначенные для высококонсервативных областей HBV, эффективно подавляют репликацию вируса и обедненную cccDNA.
- Nishimasu et al. (2015) сообщили о кристаллических структурах SaCas9 в комплексе с одной направляющей РНК (sg-РНК) и ее двухцепочечными ДНК-мишенями, содержащими 5'-TTGAAT-3' PAM-последовательность и 5'-TTGGGT-3' PAM-последовательность. Структурное сравнение SaCas9 с SpCas9 выявило как консервативность структуры, так и расхождение, объяснив отчетливую специфичность их PAM и ортологическое распознавание sg-РНК.
- Canver et al. (2015) продемонстрировали функциональное исследование не кодирующих геномных элементов на основе CRISPR-Cas9. Авторы разработали объединенные библиотеки направляющей РНК CRISPR-Cas9 для насыщающего мутагенеза insitu энхансеров BCL11A человека и мыши, которые выявили важные особенности этих энхансеров.
- Zetsche et al. (2015) сообщили о характеристике Cpf1, нуклеазы класса 2 CRISPR из Francisellanovicida U112, имеющей особенности, отличные от Cas9. Cpf1 представляет собой единственную РНК-направляемую эндонуклеазу, не имеющую tracr-РНК, использующую T-богатый мотив, прилегающий к протоспейсеру, и расщепляющую ДНК путем ступенчатого двухцепочечного разрыва.
- Shmakov et al. (2015) сообщили о трех разных классах систем CRISPR-Cas. Два фермента системы CRISPR (C2c1 и C2c3) содержат RuvC-подобные эндонуклеазные домены, отдаленно связанные с Cpf1. В отличие от Cpf1, C2c1 зависит как от cr-РНК, так и от tracr-РНК для расщепления ДНК. Третий фермент (C2c2) содержит два предсказанных РНК-азных домена HEPN и является независимым от tracr-РНК.
- Slaymaker et al. (2016) сообщили об использовании структурной белковой инженерии для улучшения специфичностиCas9 Streptococcus pyogenes (SpCas9). Авторы разработали варианты "расширенной специфичности" SpCas9 (eSpCas9), которые поддерживали устойчивое целевое расщепление с уменьшением нецелевых эффектов.
[001298] Кроме того, "Dimeric CRISPR RNA-guided Fok1 nucleases for highly specific genome editing", Shengdar Q. Tsai, Nicolas Wyvekens, CydKhayter, Jennifer A. Foden, Vishal Thapar, Deepak Reyon, Mathew J, Goodwin, Martin J, Aryee, J. Keith Joung Nature Biotechnology 32(6): 569-77 (2014), относится к димерным РНК-направленным Fok1-нуклеазам, которые распознают расширенные последовательности и могут редактировать эндогенные гены с высокой эффективностью в клетках человека.
[001299] Что касается общей информации о системах CRISPR-Cas, их компонентах и доставке таких компонентов, включаяс пособы, материалы, транспортные средства доставки, векторы, частицы, AAV, а также их изготовление и использование, в том числе что касается количества и формул, все полезное на практике для настоящего изобретения, приводится в настоящем описании в качестве ссылки на следующие номера патентов США: № 8999641, 8993233, 8945839, 8932814, 8906616, 8895308, 8889418, 8889356, 8871445, 8865406, 8795965, 8771945 и 8697359; публикация заявки на патент США US 2014-0310830 (US APP.Ser. No. 14/105031), US 2014-0287938 A1 (US App. Ser. No. 14/213991), US 2014- 0273234 A1 (U.S. App, Ser. No. 14/293674), US2014-0273232 A1 (U.S. App. Ser. No. 14/290575), US 2014-0273231 (U.S. App. Ser. No. 14/259420), US 2014-0256046 A1 (U.S. App. Ser. No. 14/226274), US 2014-0248702 A1 (U.S. App. Ser. No. 14/258458), US 2014-0242700 A1 (U.S. App. Ser. No. 14/222930), US 2014-0242699 A1 (U.S. App. Ser. No, 14/183512), US 2014-0242664 A1 (U.S. App. Ser. No. 14/104990), US 2014-0234972 A1 (U.S, App. Ser. No. 14/183471), US 2014-0227787 A1 (U.S. App. Ser. No. 14/256912), US 2014-0189896 A1 (U.S. App. Ser. No. 14/105035), US 2014-0186958 (U.S. App. Ser. No. 14/105017), US 2014-0186919 Al (U.S. App. Ser. No. 14/104977), US 2014-0186843 A1 (U.S. App. Ser. No. 14/104900), US 2014-0179770 A1 (US. App. Ser. No. 14/104837) и US 2014-0179006 A1 (US. App. Ser. No. 14/183,486), US 2014-0170753 (US App. Ser. No. 14/183429), US 2015-0184139 (US. App. Ser. No. 14/324960), 14/054414; Европейские патенты: EP 2 764 103 (EP13824232.6), EP 2 784 162 (EP14170383.5) и EP 2 771 468 (EP13818570.7); и публикации патентных заявок PCT: WO 2014/093661 (PCT/US2013/074743), WO 2014/093694 (PCT/US2013/074790), WO 2014/093595 (PCT/US2013/074611), WO 2014/093718 (PCTAJS2013/074825), WO 2014/093709 (PCT/US2013/074812), WO 2014/093622 (PCT/US2013/074667), WO 2014/093635 (PCT/US2013/074691), WO 2014/093655 (PCT/US2013/07473 6), WO 2014/093712 (PCT/US2013/074819), WO 2014/093701 (PCT/US2013/074800), WO 2014/018423 (PCT/US2013/051418), WO 2014/204723 (PCT/US2014/041790), WO 2014/204724 (PCT/US2014/041800), WO 2014/204725 (PCTAJS2014/041803), WO 2014/204726 (PCT.AJS2014/041804), WO 2014/204727 (PCT/US2014/041806), WO 2014/204728 (PCT/US2014/041808), WO 2014/204729 (PCT/US2014/041809), WO 2015/089351 (PCT/US2014/069897), WO 2015/089354 (PCT/US2014/069902), WO 2015/089364 (PCT/US2014/069925), WO 2015/089427 (PCT7US2014/070068), WO 2015/089462 (PCT/US2014/070127), WO 2015/089419 (PCT/US2014/070057), WO 2015/089465 (PCT/US2014/070135), WO 2015/089486 (PCT/US2014/070175), PCT/US2015/051691, PCT/US2015/051830. Также упоминается предварительная патентная заявка США 61/758468; 61/802174; 61/806375; 61/814263; 61/819803 и 61/828130, поданная 30 января 2013 года; 15 марта 2013 года; 28 марта 2013 года; 20 апреля 2013 года; 6 мая 2013 года и 28 мая 2013 года соответственно. Также упоминается предварительная патентная заявка США 61/836123, поданная 17 июня 2013 года. См. Также дополнительные заявки на патент США 61/835931, 61/835936, 61/836127, 61/836101, 61/836080 и 61/835973, поданные 17 июня 2013 года. Дальнейшая ссылка делается на временные заявки на патент США 61/862468 и 61/862355, поданные 5 августа 2013 года, 61/871301, поданная 28 августа 2013 года, 61/960777, поданную 25 сентября 2013 года и 61/961980, поданную 28 октября 2013 года. Далее делается ссылка на: PCX/US2014/041803, PCT/US2014/041809, PCT/US2014/041804 и PCT/US2014/041806, каждый из которых подан 10 июня 2014 года; PCT/US2014/041808, поданную 11 июня 2014 года; и PCT/US2014/62558, поданную 28 октября 2014 года, и в патенты США №: 61/915148, 61/915150, 61/915153, 61/915203, 61/915251, 61/915301, 61/915267, 61/915260 и 61/915397, поданных 12 декабря 2013 года; 61/757972 и 61/768959, поданные 29 января 2013 года и 25 февраля 2013 года; 61/835936, 61/836127, 61/836101, 61/836080, 61/835973 и 61/835931, поданные 17 июня 2013 года; 62/010888 и 62/010879, поданные 11 июня 2014 года; 62/010329 и 62/010441, поданные 10 июня 2014 года; 61/939228 и 61/939242, каждая из которых была опубликована 12 февраля 2014 года, 61/980012, поданную 15 апреля 2010 года; 62/038358, поданную 17 августа 2014 года; 62/054490, 62/055484, 62/055460 и 62/055487, поданные 25 сентября 2014 года, и 62/069243, поданную 27 октября 2014 года. Ссылка делается на временную заявку США 61/930214, поданную 22 января 2014 года
[001300] Упоминается также заявка США 62/180 709, поданная 17 июня 2015 года, PROTECTED GUIDE RNAS (PGRNAS); заявка США 62/091455, поданная 12 декабря 2014 года, PROTECTED GUIDE RNAS (PGRNAS); заявка США 62/096708, поданная 24 декабря 2014 года, PROTECTED GUIDE RNAS (PGRNAS); заявка США 62/091462, поданная 12 декабря 2014 года, DEAD GUIDES FOR CRISPR TRANSCRIPTION FACTORS; заявка США 62/096324, поданная 23 декабря 2014 года, 62/180681, 17 июня 2015 года, и 62/237496, 5 октября 2015 года, DEAD GUIDES FOR CRISPR TRANSCRIPTION FACTORS; заявка США 62/091456, 12 декабря 2014 года и 62/180692, 17 июля 2015 года, ESCORTED AND FUNCTIONALIZED GUIDES FOR CRISPR-CAS SYSTEMS, заявка США 62/091461, 12 декабря 2014 года, DELIVERY, USE AND THERAPEUTIC APPLICATIONS OF THE CRISPR-CAS SYSTEMS AND COMPOSITIONS FOR GENOME EDITING AS TO HEMATOPOETIC STEM CELLS (I ISC's); заявка США 62/094903, 19 декабря 2014 года, UNBIASED IDENTIFICATION OF DOUBLE-STRAND BREAKS AND GENOMIC REARRANGEMENT BY GENOME-WISE INSERT CAPTURE SEQUENCING; заявка США 62/096761, 24 декабря 2014 года, ENGINEERING OF SYSTEMS, METHODS AND OPTIMIZED ENZYME AND GUIDE FUNCTIONAL-CRISPR COMPLEXES; заявка США 62/087475, 4 декабря 2014 года, FUNCTIONAL SCREENING WITH OPTIMIZED FUNCTIONAL CRISPR-CAS SYSTEMS; заявка США 62/055487, 25 сентября 2014 года, FUNCTIONAL SCREENING WITH OPTIMIZED FUNCTIONAL CRISPR-CAS SYSTEMS; заявка США 62/087546, 4 декабря 2014 года, и 62/181687, MULTIFUNCTIONAL CRISPR COMPLEXES AND/OR OPTIMIZED ENZYIFUNCTIONAL-CRISPR COMPLEXES; и заявка США 62/098285, 30 декабря 2014 года, CRISPR MEDIATED IN VIVO MODELING AND GENETIC SCREENING OF TUMOR GROWTH AND METASTASIS.
[001301] Упоминаются следующие заявки США 62/181659, 18 июня 2015 года, и 62/207318, 19 августа 2015 года, ENGINEERING AND OPTIMIZATION OF SYSTEMS, METHODS, ENZYME AND GUIDE SCAFFOLDS OF CAS9 ORTHOLOGS AND VARIANTS FOR SEQUENCE MANIPULATION. Упоминаются заявки США 62/181675, 18 июня 2015 года и номер патентного реестра - 46783.01.2128, поданные 22 октября 2015 года, NOVEL CRISPR ENZYMES AND SYSTEMS, заявка США 62/232067, 24 сентября 2015 года, заявка США 62/205733, 16 августа 2015 года, заявка США 62/201542, 5 августа 2015 года, заявка США 62/193507, 16 июля 2015 года, и заявка США 62/181739, 18 июня 2015 года, каждая из которых озаглавлена NOVEL CRISPR ENZYMES AND SYSTEMS, а также в заявке США 62/245270, 22 октября 2015 года, NOVEL CRISPR ENZYMES AND SYSTEMS. Также упоминается заявка США 61/939256, 12 февраля 2014 года, и WO 2015/089473 (PCT/US2014/070152), 12 декабря 2014 года, каждая из которых озаглавлена ENGINEERING OF SYSTEMS, METHODS AND OPTIMIZED GUIDE COMPOSITIONS WITH NEW ARCHITECTURES FOR SEQUENCE MANIPULATION. Также упоминаются PCT.AJS2015/045504, 15 августа 2015 года, заявка США 62/180699, 17 июня 2015 года, и заявка США 62/038358, 17 августа 2014, каждая из которых озаглавлена GENOME EDITING USING CAS9 NICKASES.
[001302] Каждый из этих патентов, патентных публикаций и заявлений и всех документов, цитируемых в нем или во время их делопроизводства ("приложенные к делу документы"), и всех документов, цитируемых или упоминаемых в приведенных в данном приложении документах, вместе с любыми инструкциями, описаниями, спецификациями и технологическими картами продуктов для любых продуктов, упомянутых в нем или в любом документе и включенных в настоящее описание посредством ссылки, включены в настоящее описание в качестве ссылки и могут быть использованы в практике изобретения. Все документы (например, эти патенты, патентные публикации и заявки и приведенные в приложении документы) включены сюда путем отсылки так же как если бы каждый отдельный документ был специально и индивидуально указан для включения в качестве отсылки.
[001303] Кроме того, упоминается заявка PCT PCT/US14/70057, номер патентного реестра 47627.99.2060 и BI-2013/107, озаглавленная "DELIVERY, USE AND THERAPEUTIC APPLICATIONS OF THE CRISPR-CAS SYSTEMS AND COMPOSITIONS FOR TARGETING DISORDERS AND DISEASES USING PARTICLE DELIVERY COMPONENTS" (притязающая на приоритет по одной или более или всех предварительных заявок на патент США: 62/054490, поданная 24 сентября 2014 года, 62/010441, поданная 10 июня 2014 года, и 61/915118, 61/915215 и 61/915148, каждая из которых подана 12 декабря 2013 года) ("PCT для доставки частиц"), включенной в настоящее описание в качестве отсылки, в отношении способа получения sg-РНК и,tkrf Cas9, включающего смешивание смеси, содержащей sg-РНК и белок Cas9 (и необязательно матрицу HDR) со смесью, содержащей или состоящей только из или состоящей из поверхностно-активного вещества, фосфолипида, биоразлагаемого полимера, липопротеина и спирта; и частицы из такого процесса. Например, когда белок Cas9 и sg-РНК смешивали вместе при подходящем молярном соотношении, например, от 3:1 до 1:3 или от 2:1 до 1:2 или 1:1 при подходящей температуре, например 15-30°С, например, 20-25°C, например, комнатной температуре, в течение подходящего времени, например 15-45, например 30 минут, преимущественно в стерильном, свободном от нуклеазы буфере, например IX PBS. Отдельно компоненты частиц, такие как или содержащие: поверхностно-активное вещество, например, катионный липид, например, 1,2-диолеоил-3-триметиламмонийпропан (DOTAP); фосфолипид, например димиристоилфосфатидилхолин (DMPC); биодеградируемый полимер, такой как полимер этиленгликоля или PEG, или липопротеин, такой как липопротеин низкой плотности, например холестерин, были растворены в спирте, преимущественно C1-6-алкиловом спирте, таком как метанол, этанол, изопропанол, например 100% этанол. Оба раствора смешивали вместе до образования частиц, содержащих комплексы Cas9-sg-РНК. Соответственно, sg-РНК может быть предварительно скомбинирована с белком Cas9 перед формированием всего комплекса в частице. Препараты могут быть получены с другим молярным соотношением различных компонентов, которые, как известно, способствуют доставке нуклеиновых кислот в клетки (например, 1,2-диолеоил-3-триметиламмоний-пропан (DOTAP), 1,2-дитретраканоил-s-глицеро-3-фосфохолин (DMPC), полиэтиленгликоль (PEG) и холестерин). Например, vолярные соотношения DOTAP: DMPC: PEG: холестерина могут быть DOTAP 100, DMPC 0, PEG 0, холестерин 0; или DOTAP 90, DMPC 0, PEG 10, холестерин 0; или DOTAP 90, DMPC 0, PEG 5, холестерин 5. DOTAP 100, DMPC 0, PEG 0, холестерин 0. Настоящее описание соответственно охватывает смешение sg-РНК, белка Cas9 и компонентов, которые образуют частицу; а также частиц от такого смешивания. Варианты настоящего изобретения могут включать частицы; например, частицы, используемые в способе, аналогичном способу PCT доставки частиц, например, путем смешивания смеси, содержащей sg-РНК и/или Cas9, как в настоящем изобретении, и компонентов, которые образуют частицу, например, как в для PCT доставки частиц, образуют частицу и частицы из такой смеси (или, конечно, другие частицы, содержащие sg-РНК и/или Cas9, как в настоящем изобретении).
[001304] Настоящее изобретение дополнительно проиллюстрировано в следующих примерах, которые приведены только для иллюстративных целей и не предназначены для ограничения изобретения каким-либо образом.
Пример 1: Происхождение и эволюция систем адаптивного иммунитета
[001305] Классификация и аннотация систем CRISPR-Cas в геномах архей и бактерий. Локусы CRISPR-Cas имеют более чем 50 семейств генов, при этом не существует строго универсальных генов, происходит быстрая эволюция и наблюдается значительное разнообразие архитектуры локусов. Поэтому не существует возможного древа и комплексный подход особенно необходим. Сейчас существует подробная идентификация cas-генов в рамках 395 профилей для 93 cas белков. Классификация включает профили экспрессии генов и профили архитектуры локусов
[001306] Последняя классификация систем CRISPR-Cas представлена на фиг.1. Класс l включает мультисубъединичные эффекторные комплексы cr-РНК (Cascade), а класс 2 включает односубъединичные эффекторные комплексы cr-РНК (Cas-9 типа). Фиг.2 представляет молекулярную организацию CRISPR-Cas. Фиг.3 демонстрирует структуры эффекторных комплексов I и III типов: они имеют общую структуру/ общую природу несмотря на значительные различия в последовательностях. Фиг.4 показывает CRISPR-Cas как систему, основанную на мотивах узнавания РНК (RRM). На фиг.5 представлена филогения cas1, где рекомбинация адаптационного и cr-РНК эффекторого модулей показывают основной аспект эволюции систем CRISPR-Cas. На фиг.6 представлен перечень систем CRISPR-Cas, в особенности распределение типов и подтипов CRISPR-Cas среди архей и бактерий.
[001307] Белок Сas1 не всегда связан только с системами CRISPR-Cas, поэтому возможно, что существуют две ветви отдельных "соло" Сas 1, что говорит о том, что возможны различия как в функционировании, так и в происхождении, а также возможно обнаружение новых мобильных элементов (см. Makarova, Krupovic, Koonin, Frontiers Genet 2014). Геномная организация трех семейств казпозонов может предоставить возможные ответы. Вдобавок к cas 1 и PolB, казпозоны имеют различные гены включая нуклеазы (Krupovic et al. BMC Biology 2014). Одно семейство имеет праймированную белком полимеразу, другое семейство имеет РНК-праймированную полимеразу. В добавок к представителям типов Euryarchaeota и Thaumarchaeota, казпозоны, обнаруженные в нескольких бактериях предполагают горизонтальный перенос генов. Казпозон Cas1 (транспозаза/интеграза) предполагает наличие общей базальной филогенетической группы в развитии Cas1.
[001308] Бактерии и археи используют CRISPR - систему для адаптивной иммунной защиты в клетках про- и эукариот посредством геномной манипуляции. Белок cas 1 представляет собой уже существующий способ геномной манипуляции. Существуют похожие механизмы интеграции в каcпозоны и CRISPR, в особенности репликационно-заваисимое приобретение посредством процессов копирования и вставки, а не вырезания и вставки (Krupovic et al. BMC Biology 2014). Cas1 - есть эталонный образец интегразы (Nunez JK, Lee AS, Engelman A, Doudna JA. Integrase-mediated spacer acquisition during CRISPR-Cas adaptive immunity. Nature. 2015 Feb 18). Есть определенное сходство между терминальными обратными повторами каcпозонов и врожденным локусом иммунитета (Koonin, Kmpovic, Nature Rev Genet, 2015). Эволюция систем адаптивного иммунитета в прокариотах и животных возможно шла параллельными путями при помощи интеграции транспозонов в локусы врожденного иммунитета (Koonin, Krupovic, Nature Rev Genet, 2015). Транспозаза RAG1 (ключевой энзим V(D)J-рекомбинации позвоночных) возможно произошел из транспозонов класса Transib (Kapitonov VV, Jurka J.. RAG1 core and V(D)J recombination signal sequences were derived from Transib transposons.. PLoS Biol. 2005 Jun;3(6):el81), однако ни один из транспозонов класса Transibs не кодирует белок RAG2. RAG1 и RAG2 кодирующие транспозоны описаны в Kapitonov, Koonin, Biol Direct 2015 и филогения транспозазы класса Transib представлена в Kapitonov, Koonin, Biol Direct 2015. Защитное уничтожение ДНК в инфузориях эволюционировало из транспозона PiggyMAc, что вместе с РНК-i образуют врожденный иммунитет (Swart EC, Nowacki M. The eukaryotic way to defend and edit genomes by sRNA-targeted DNA deletion. Ann N Y Acad Sci. 2015).
[001309] Относительная стабильность классификации означает, что наиболее часто встречающиеся варианты CRISPR-Cas уже известны. Однако существование редких и до сих пор не включенных в классификацию вариантов также говорит о том, что дополнительные типы и подтипы все ждут своего описания (Makarova et al, 2015. Evolutionary classification of CRISPR-Cas systems and cas genes).
[001310] Транспозоны играют ключевую роль в эволюции адаптивного иммунитета и других систем, участвующих в изменении ДНК. Класс 1 CRISPR-Cas возник из транспозонов, однако это касается только адаптационного функционального модуля системы. Класс 2 CRISPR-Cas имеет и адаптационную и эффекторную функции, модули которых могут происходить из разных транспозонов.
Пример 2: Новые предсказанные системы CRISPR-Cas класса 2 и доказательства их независимого происхождения из транспозонов
[001311] Системы CRISPR-Cas адаптивного иммунитета бактерий и архей демонстрируют огромное разнообразие состава Cas белков, а также разновидной структурой геномных локусов. Однако эти системы разделяются в целом на два класса: класс 1, имеющий мульти-протеиновый (многоблоковый) эффекторный комплекс и Класс 2, где эффекторный комплекс состоит из одного большого белка (одноблоковый комплекс), например белка cas-9 (фиг.1А и 1В). Заявители разработали простую биоинформатическую методику (фиг.7) для обработки все время пополняемых геномных и метагеномных баз данных, сопровождающихся углублением понимания систем CRISPR-cas для поиска возможных кандидатов для системы CRISPR-Cas класса 2. Анализ базы данных полных (полностью отсеквенированных) геномов при помощи разработанной нами вычислительной методики привело к индентефикации трех новых вариантов, каждый из которых представлен в различных бактериях и имеет в своем составе гены cas1 и cas2, а также третий ген, кодирующий большой белок, который как предполагается функционирует как эффекторный модуль. В первом из этих локусов, возможный эффекторный белок (C2c1p) имеет похожий на RuvC науклеазный домен и напоминает ранее описанный белок Cpf1, предсказанный эффектор для типа V CRISPR -cas систем, соответственно новая предсказанная система классифицируется как подтип V-B. Всестороннее сравнение белковых последовательностей показало, что эффекторные белки Cas9 Cpf1 и C2Cip, содержащие домен RuvC, возникли из различных групп кодируемых транспозонами TnpB-белков. Вторая группа новых кандидатов для локусов CRISPR-Cas включает большой белок, содержащий два HEPN домена с предсказанной РНК-азной активностью. Учитывая новизну предсказанного эффекторного белка, эти локусы были причислены к новому VI типу CRISPR-Cas, который наиболее вероятно способен к нацеливанию на РНК. В целом, результаты анализа показывают, что системы CRISPR-Cas класса 2 возникли различными, часто не зависимыми друг от друга путями, путем соединения различных кодирующих Cas1-Cas2 адаптационных модулей с эффекторными белками, образованными от различных мобильных генетических элементов. Путь эволюции с наибольшей вероятностью привел к появлению различных вариантов систем класса 2, некоторые из которых еще не открыты.
[001312] Системы адаптивного иммунитета CRISPR-Cas присутствуют в геномах 45% бактерий и 90% архей и они демонстрируют значительное разнообразие в составе cas белков, их последовательности, а также архитектуры геномных локусов. Согласно архитектуре эффекторных комплексов cr-РНК, эти системы подразделяются на два класса: Класс 1 имеет мультисубъединичный эффекторный комплекс, а Класс 2 - единственный эффекторный белок (фиг.1A и 1B). Системы класса 1 встречаются намного чаще и более разнообразны, чем системы класса 2. В настоящей классификации Класс 1 представлен 12-ю отдельными подтипами, кодируемыми многочисленным геномами бактерий и архей, а то время как класс 2 включает 3 подтипа система 2 типа и возможный V тип, который в целом можно обнаружить в 10% отсеквенированных бактериальных геномах (вместе с единственным геномом архей включающим возможную систему данного типа). Системы класса 2 обычно содержат только три или четыре гена в опероне cas, а именно пару генов cas1 ~cas2, которые участвуют в адаптации, но не в интерференции, один многодоменный эффекторный белок, который отвечает за интерференцию, но также способствует процессингу и адаптации пре-cr-РНК. Также часто имеется и четвертый ген с еще неизвестными функциями, которые могут встречаться, по меньшей мере, в некоторых системах II типа. В большинстве случаев CRISPR кассета и ген для определенного типа РНК, известного как tracr-РНК (трансактивирующая небольшая cr-РНК), находятся рядом с оперонами класса 2 (Chylinski, 2014). Tracr-РНК частично гомологична повторам в соответствующей области CRISPR кассеты и имеет важное значение для процессинга пре-cr-РНК, которая катализируется RNAse III, распространенным ферментом бактерий, не связанным с локусами CRISPR-cas (Deltcheva, 2011)(Chylinski, 20I4;Chylinski, 2013).
[001313] Эффекторный белок второго типа систем CRISPR-Cas - Cas9 был изучен в мельчайших подробностях в отношении его функций и структуры. В различных бактериях, белки Cas9 включают от ~950 до более чем 1600 аминокислот, а размер варьируются от 950 до 1400 аминокислот. Он содержит два типа нуклеазных доменов, а именно RuvC подобный домен (РНК-аза H консервативный домен) и HNH (MrcA подобный консервативный домен) домен (Makarova, 2011). Кристаллическая структура cas9 показывает "двулопастную" организацию белка с явно выраженным участком узнавания и нуклеазной активности, при этом последний связывается и с RuvC и с HNH доменами (Nishimasu, 2014)(Jinek, 2014). Каждый из нуклеазных доменов Cas9 необходим для разрезания одной из цепочек ДНК-мишени (Jinek, 2012; Sapranauskas, 2011). Недавно было показано что Cas9 участвует во всех трех стадиях CRISPR иммунного ответа, то есть не только приводит к расщеплению ДНК-мишени (стадия интерференции) но также участвует в стадиях адаптации и процессинга пре-cr-РНК (Jinek, 2012). Более конкретно было показано, что определенный домен в участке, обладающем науклеазной активностью узнает и связывается с последовательностью, прилегающей к протоспейсеру (PAM) в вирусной ДНК на стадии адаптации (Nishimasu, 2014)(Jinek, 2014)(Heler, 2015; Wei, 2015). На стадии иммунного ответа CRISPR, Cas9 формирует комплекс с белками Cas l и Cas2, которые непосредственно участвуют в встраивании новых спейсеров во всех системах CRISPR-Cas (Heier, 2015; Wei, 2015).
[001314] Белок Cas9, соединенный с tracr-РНК недавно стал ключевым инструментом в создании нового поколения способов редактирования геномов и генной инженерии (Gasiunas, 2013; Mali, 2013; Sampson, 2014; Cong, 2015). Такая ценность Cas9 в генной инженерии основывается на том, что во втором типе CRISPR-Cas в отличие от других систем CRISPR-Cas все операции необходимые для узнавания ДНК-мишени и ее последующего расщепления сосредоточены в рамках одного, но большого мультидоменного белка. Эта особенность систем II типа значительно упрощает разработку эффективных инструментов манипуляции генома. Важно и то, что не все варианты cas9 похожи. На сегодняшний момент большая часть работы по искомому белку была выполнена на cas9 из Streptococcus pyogenes но и другие виды могут предложить существенные преимущества. Что касается рассматриваемого случая последние эксперименты с cas9, который на 300 аминокислот короче чем S. pyogenes позволили упаковать в адено-ассоциированный вирусный вектор, что привело к существенному улучшению возможностей использования системы CRISPR- Cas для редактирования генома in vivo (Ran, 2015).
[001315] Cистемы CRISPR-Cas второго типа в настоящее время подразделяются на три подтипа ((II-A, II-B и II-C) (Makarova, 2011) (Fonfara, 2014; Chylinski, 2013; Chylinski, 2014). Помимо генов cas1, cas2 и cas9, которые характерны для всех локусов II типа, подтип II-A характеризуется дополнительным геном, csn2, который кодирует инактивированную АТФазу (Nam, 2011, Koo, 2012, Lee, 2012), роль которой в приобретении новых спейсеров еще изучена слабо (Barrangou, 2007; Arslan, 2013) (Heler, 2015). В системах подтипа II-B отсутствует csn2, но вместо этого содержится ген cas4, который наоборот более типичен для систем типа I и кодирует 5'-3' экзонуклеазу семейства recB, которая способствует накоплению спейсеров путем генерации рекомбиногенных концов ДНК (Zhang, 2012) (Lemak, 2013, Lemak, 2014). Гены cas1 и cas2 подтипа II-B наиболее тесно связаны с соответствующими белками систем I типа CRISPR-Cas, что подразумевает рекомбинантное происхождение этого подтипа (Chylinski, 2014).
[001316] Подтип II-C систем CRISPR-Cas демонстрирует минимальное разнообразие и состоит только из генов Cas1, cas2 и cas9 (Chylinski, 2013; Koonin, 2013; Chylinski, 2014). Примечательно, однако, что в приобретении спейсера Campylobacter jejuni системами типа II-C требуется участие Cas4, кодируемого бактериофагом (Hooton, 2014). Другой отличительной особенностью подтипа II-C является образование некоторых cr-РНК посредством транскрипции, включая транскрипцию из внутренних альтернативных промоторов, что отличается от процессинга, наблюдаемого во всех других описанных экспериментально системах CRISPR-Cas (Zhang, 2013).
[001317] Cуществование типа V систем CRISPR-Cas было недавно предсказано путем сравнительного анализа бактериальных геномов. Эти предполагаемые новые системы CRISPR-Cas представлены в нескольких бактериальных геномах, в частности, из рода Francisella и одной археи Methanomethylophilus alvus (Vestergaard, 2014). Все предполагаемые локусы типа V включают Cas1, cas2, отдельный ген, обозначенный как cpf1, и CRISPR кассету (Schunder, 20I3) (Макарова, 2015). Cpf1 представляет собой большой белок (приблизительно 1300 аминокислот), который содержит RuvC-подобный нуклеазный домен, гомологичный соответствующему домену Cas9, а также аналог типичного богатого аргинином кластера Cas9. Однако Cpf1 не обладает нуклеазным доменом HNH, который присутствует во всех белках Cas9, и RuvC-подобный домен является прилегает к последовательности Cpf1, в отличие от Cas9, где он содержит длинные вставки, включая домен HNH (Chylinski, 2014; Makarova, 2015). Эти основные отличия в доменных архитектурах Cas9 и Cpf1 предполагают, что системы содержащие Cpf1, следует классифицировать как новый тип. Состав предполагаемых систем типа V подразумевает, что Cpf1 представляет собой одноблоковый эффекторный комплекс, и, соответственно, эти системы относятся ко второму классу систем CRISPR-Cas. Некоторые из предполагаемых локусов типа V кодируют Cas4 и соответственно напоминают подтипы П-В локусов, тогда как другие не имеют Cas4 и, следовательно, аналогичны подтипу II-C.
[001318] Недавно было показано, что самыми близкими гомологами белков Cas9 и Cpf1 являются белки TnpB, которые кодируются транспозонами семейства IS605 и содержат RuvC-подобный нуклеазный домен, а также домен цинкового пальца, который имеет аналог в Cpf1. Кроме того, были идентифицированы гомологи TnpB, которые содержат домен HNH, вставленный в RuvC-подобный домен и показывающий высокое сходство последовательностей с Cas9. Роль TnpB в транспозонах остается неопределенной, поскольку было показано, что этот белок не требуется для транспозиции.
[001319] Учитывая гомологию Cas9 и Cpf1 и белков, кодируемых транспозонами, заявители предположили, что системы CRISPR-Cas класса 2 могут возникать многократно в результате рекомбинации между транспозоном и локусом Cas1-cas2. Соответственно, заявители разработали простую вычислительную стратегию для идентификации геномных локусов, которые могут быть кандидатами для вариантов второго класса. Настоящей заявкой Заявители описывают первое применение этого подхода, в результате которого идентифицируются три группы таких кандидатов, два из которых кажутся принадлежащими к различным подтипам типа V, тогда как третий, по-видимому, подходит к VI типу. Новые варианты систем CRISPR-Cas класса 2 представляют очевидный интерес так как могут быть использованы в качестве инструментов для редактирования и регуляции экспрессии.
[001320]Стратегия поиска по базам данных для обнаружения новых кандидатов для CRISPR-Cas локуса класса 2. Заявители использовали простой биоинформатический подход для обнаружения новых претендентов для систем CRISPR-Cas класса 2 (фиг.7. схема). Поскольку подавляющее большинство локусов CRISPR-Cas включает ген cas1 (Makarova, 2011, Makarova, 2015) а последовательность Cas1 является наиболее консервативной для всех белков Cas (Takeuchi, 2012), Заявители предположили, что cas1 является лучшей из возможных зацепок для определения и поиска возможных новых локусов при помощи PSI-BLAST. После обнаружения всех контигов, кодирующих Cas1, путем поиска следующих баз данных WGS (содержащий весь геном) и NT (нуклеотидных) в NCBI, кодирующие белок гены были предсказаны с использованием GenemarkS в районе 20 т.п.н. вышележащих и нижележащих последовательностей от гена Cas1. Эти предсказанные гены были аннотированы с использованием профилей по базе данных NCBI Conserved Domain Database (CDD) и специфических Cas-белковых профилей (Makarova et al., 2015, Nat Rev Microbiol. 2015, doi: 10.1038/nrmiero3569), а CRISPR кассеты были предсказаны с использованием программы PILER- CR. Эта процедура дала возможность определения обнаруженных локусов CRISPR-Cas в рамках уже известных подтипов. Частичные и/или не поддающиеся классификации возможные локусы CRISPR-Cas, содержащие большие (> 500 ак) белки, были выбраны в качестве возможных кандидатов для новых систем класса 2 основываясь на характерном присутствии таких больших одноблоковых эффекторных белков в системах типов II и V (Cas9 и Cpf1, соответственно). Все 63 возможных локуса, обнаруженные с использованием этих критериев (перечислены в таблице, представленной на фиг.40A-D), анализировались далее каждый в отдельности с использованием программ PSI-BLAST и HHpred. Последовательности белка, кодированные в локусах-кандидатах, далее использовались в качестве запроса для поиска метагеномных баз данных для дополнительных гомологов, при этом обнаруженные длинные контиги анализировались, как указано выше. В этом вычислительном конвейере было получено в общей сложности 53 новых локуса (некоторые из первоначально идентифицированных 63 локусов-кандидатов были отброшены как ложные, в то время как несколько неполных локусов с отсутствующим cas1 были добавлены) с характерными особенностями CRISPR-Cas, систем класса 2 которые, основываясь на природе предсказанных эффекторных белков, могли бы быть классифицированы на три группы локусов (см. Фиг.8А и 8В, фиг.9, 14 и 15 и фиг.41А-В, 42А-В и 43А-В) Хотя и бактериофаги, заражающие бактерии, которые могли бы иметь недавно открытые системы CRISPR-Cas класса 2, на практике неизвестны, для каждой из этих систем авторы изобретения обнаружили спейсеры, которые соответствуют фагам или предсказанным профагам.
[001321] Используя вышеописанную вычислительную стратегию, Заявители обнаружили три новые системы CRISPR-Cas класса 2, а именно C2e1 и C2c3, которые классифицируются как подтипы ранее описанного предполагаемого типа V и C2c2, который Заявители приписывают новому предполагаемому VI типу. Заявители представляют несколько стратегий доказательства того, что эти локусы кодируют функциональные системы CRISPR-Cas. Основываясь на сравнении геномов, авторы изобретения определили фаго-специфические спейсеры для каждой из трех предполагаемых новых систем, а также показали, что наборы спейсеров полностью различаются даже в близкородственных бактериальных геномах, что указывает на активный, функционирующий иммунитет. Многие из этих новых систем встречаются в бактериальных геномах, которые не содержат других локусов CRISPR-Cas, что говорит о том, что системы типа V и типа VI могут функционировать автономно. Более того, даже когда другие системы CRISPR-Cas были идентифицированы в одних и тех же геномах, ассоциированные повторяющиеся структуры были явно отличны от других типов в типах V и VI, что свидетельствует об их независимой функциональности.
[001322] Предполагаемая система типа V-B. Первая группа возможных локусов, условно обозначенная как C2c1 (класс 2, кандидат 1), представлена в бактериальных геномах четырех основных таксонов, включая Bacilli, Verrucomicrobia, альфа-протеобактерии и дельта-протеобактерии (фиг.8A-B "Организация полных локусов систем класса 2" фиг.41А-В). Все локусы C2c1 кодируют слитый белок Cas1-Cas4, Cas2 и большой белок, обозначаемый Заявителями как C2c1p и, как правило, находятся рядом с CRISPR кассетой (фиг.9, окрестности C2c1, фиг.41A-B). В филогенетическом дереве Cas1, соответствующий белковый кластер соединяется с системой типа I-U (фиг.10A и 10B, фиг.10C-W, дерево Cas1) и является единственным, в котором обнаружено слияние Cas1-Cas4. Длина белков C2c1p, идентифицированных здесь, варьируется 1100 до 1500 аминокислот, например, может состоять примерно из 1200 аминокислот. HHpred поиск обнаруживает значительное сходство между С-концевой частью белков C2c1p и подмножеством белков TnpB закодированных в транспозонах семейства IS6G5 (фиг.13А и 13С). Напротив, не было обнаружено никакого существенного сходства между C2c1p и Cas9 или Cpf1, которые аналогичны другим группам белков TnpB (Chylinski, 2014) (Makarova, 2015; Makarova, 2015). Таким образом, доменная архитектура C2c1p аналогична домену Cpf1 и отличается от архитектуры Cas9 (фиг.13A), хотя все три белка Cas, по-видимому, эволюционировали из семейства TnpB (фиг.11 "Организация домена семейств 2-го класса"; фиг.13А). N-концевая область C2c1p не имеет существенного сходства с другими белками. Прогнозирование вторичной структуры указывает на то, что этот регион принимает преимущественно альфа-спиральную конформацию. Два сегмента сходства с TnpB охватывают три каталитических мотива типа нуклеазы, подобной RuvC, с диагностической сигнатурой D..E..D каталитических аминокислотных остатков (Aravind et al, 2000, Nucleic Acids Res, том 28, 3417-3432) (фиг.12, "Области гомологии TnpB в белках класса 2"); область, соответствующая мостиковой спирали (также известная как богатый аргинином кластер), который в белке Cas9 участвует в связывании cr-РНК; и небольшую область, которая, по-видимому, является аналогом цинкового пальца TnpB (однако в большинстве белков C2c1 отсутствуют Zn-связывающие цистеиновые остатки, что указывает на то, что такие белки не связывают цинк, более того, C2c1 содержит множественные вставки и делеции в этой области, что указывает на функциональную дивергенцию (фиг.13А,фиг.13D-H,фиг.131). Сохранение каталитических остатков (фиг.13А) явно указывает на то, что гомологичные домены RuvC всех этих белков являются активными нуклеазами. N-концевые области C2c1 не показывают значительного сходства с любыми известными белками. Предсказание вторичной структуры показывают, что N-концевые области белков C2c1 принимают смешанную α/β-конформацию (фиг.13D-H,фиг.1 131). Сходство доменных архитектур C2c1p и Cpf1 предполагает, что локусы C2c1 лучше всего классифицировать как подтип VB, при этом локусы кодирующие Cpf1 - как подтип VA.
[001323] Несмотря на то, что гены cas1, связанные с этой системой, очень похожи, повторы CRISPR в соответствующих кассетах являются сильно гетерогенными, хотя все они имеют длину 36-37 п. н. и могут быть классифицированы как неструктурированные (энергия фолдинга AG составляет -0,5-4,5 ккал/моль, тогда как высоко палиндромная CRISPR имеют АГ ниже -7 ккал/моль). Согласно схеме CRISPR (Lange, 2013) несколько повторов V-B подтипа имеют некоторую последовательность или структурное сходство с повторами II типа (фиг.41А-Т). Однако большинство повторов нельзя было отнести к известным семействам последовательностей или структур они могут быть отнесены к четырем из 6 суперклассов.
[001324] Рассматривая возможность того, что предположительные CRISPR-Cas подтипа V-B структурно аналогичны системам II типа, Заявители предприняли попытку идентифицировать tracr-РНК в соответствующих геномных локусах.
[001325] Сравнение спейсеров CRISPR кассет типа V-B с небезызбыточной базой данных нуклеотидных последовательностей привело к обнаружению нескольких совпадений с различными бактериальными геномами. В частности, один из спейсеров из Alicyclobacillus acidoterrestris и один из спейсеров от Brevibacillus agri соответствовали неопределенным генам в пределах прогнозируемых профагов, интегрированных в соответствующие бактериальные геномы (фиг.41А-L).
[001326] Предполагаемые системы типа VI. Вторая группа потенциальных локусов CRISPR-Cas, обозначенная как C2c2 (класса 2, кандидат 2), была обнаружена в геномах 5 основных бактериальных таксонов, включая альфа-протеобактерии, бациллы, клостридии, фузобактерии и бактероиды (фиг.8А-В. "Организация полных локусов систем 2 класса" на фиг.42А-В). Определенное число локусов C2c2 включают cas1 и cas2 гены, а также большой белок (C2c2p), который не демонстрирует сходства последовательностей с C2c1, Cpf1 или Cas9 и CRISPR кассетой; однако, в отличие от C2c1, C2c2p часто кодируется рядом с CRISPR кассетой, но не Cas1-cas2 (фиг.15, окрестности C2c2, фиг.42A-B). Хотя в вычислительной стратегии авторов изобретения изначально идентифицированные локусы C2c2 охватывали гены cas1 и cas2, последующие поиски показали, что большинство таких локусов может состоять только из гена c2c2 и CRISPR-кассеты. Такие, по-видимому, неполные локусы могут либо кодировать дефектные системы CRISPR-Cas, либо могут функционировать в купе с адаптационным модулем, закодированным в ином месте генома, как это было, к примеру обнаружено для некоторых систем типа III (Majumdar et al., 2015, RNA, vol. 21, 1147- 1158). В филогенетическом дереве Cas1 белки Cas1 из локусов C2c2 распределены среди двух клад. Первая клада включает Cas1 от Clostridia и расположена в поддереве II типа вместе с небольшой ветвью типа III-А (фиг.10А и 10B, фиг.10C-V, дерево Cas1). Второй клад состоит из белков Cas1 из локусов C2c2 Leptotrichia и расположен внутри смешанной ветви, которая в основном содержит белки Cas1 из систем CRISPR-Cas типа III-A. Поиск по базе данных с использованием HHpred и PSI-BLAST не обнаружил сходства последовательностей между C2c2p и другими белками. Тем не менее, проверка множественных выравниваний белковых последовательностей C2c2p привела к идентификации двух строго консервативных мотивов RxxxxH, которые характерны для доменов HEPN (нуклеотид-связывающий участок высших эукариотических и прокариотических организмов) (Anantharaman et al., 2013, Biol Direct, том 8, 15, Grynberg et al., 2003, Trends in biochemical sciences, vol 28, 224-226) (фиг.11 и фиг.13В, фиг.13J-N). Предсказание вторичной структуры белка указывает на то, что эти мотивы находятся в структурных контекстах, совместимых с доменной структурой HEPN, также как и в целом предсказание вторичной структуры для соответствующих части C2c2p. Домены HEPN представляют собой небольшие (-150 а/к) альфа-спиральные домены с различными последовательностями, но с крайне консервативными каталитическими мотивами, для которых была показана или предсказана РНК-азная активность, а также, что они часто связаны с различными системами защиты (Anantharaman, 2013) (фиг.13B и 16, Мотив HEPN RxxxxH в семействе C2c2). Последовательности доменов HEPN мало консервативны, за исключением каталитического мотива RxxxxH. Хотя последовательности двух предполагаемых HEPN-доменов C2c2 мало похожи на другие HEPN-домены, за исключением каталитических мотивов RxxxxH, тип домена в значительной степени подкрепляется предсказанием вторичной структуры, которые указывают на то, что каждый мотив находится в рамках совместимых структурных контекстов (фиг.13B, фиг.13J-N). Кроме того, предсказанная вторичная структура всей последовательности для каждого предполагаемого домена также согласуется с укладкой домена HEPN (фиг.13J-N). Таким образом, представляется вероятным, что C2c2p содержит два активных домена HEPN. Домен HEPN не является новым для систем CRISPR-Cas, поскольку он часто ассоциирован с доменом CARF (CRISPR-Associated Rossmann Fold) в белках Csm6 и Csx1, которые присутствуют во многих системах CRISPR-Cas типа III (Makarova, 2014). Эти белки не относятся ни к адаптационным модулям, ни к эффекторным комплексам, но предполагается, что они способны выполнять некоторые вспомогательные, но еще не охарактеризованные функции в родственных системах CRISPR, а именно, они могут быть компонентами связанного модуля иммунитета, который присутствует в большинстве систем CRISPR-Cas, и работает при запрограммированной гибели клеток (апоптозе), а также выполняет регулятивные функции во время CRISPR ответа (Koonin, 2013; Makarova, 2012; Makarova, 2013). Однако C2c2p отличается от Csmo и Csx1 тем, что этот больший белок является единственным общим белком, кодируемым в локусах C2c2, за исключением Cas1 и Cas2. Таким образом, представляется вероятным, что C2c2p является эффектором в предполагаемых новых системах CRISPR-Cas, а HEPN-домены соответственно являются их каталитическими остатками. Вне предсказанных доменов HEPN последовательность C2c2p не обнаруживала сходства с другими белками и, согласно прогнозам, принимала смешанную альфа/бета вторичную структуру без заметного сходства с любыми известными способами фолдинга белка (фиг.13J-N).
[001327] Кассеты CRISPR в локусах C2c2 весьма неоднородны, с длиной от 35 до 39 п.н. и неструктурированны (энергия фолдинга от -0,9 до 4,7 ккал/моль). Согласно картированию CRISPR (Lange, 2013), эти CRISPR не принадлежат ни к одному из установленных структурных классов и относятся к 3 из 6 суперклассов. Только CRISPR от Listeria seeligeri можно было приписать семейству последовательностей номер 24, которое обычно ассоциировано с системами типа II-C (фиг.42A-L).
[001328] Aнализ спейсеров локусов C2c2 идентифицировал одну 30 нуклеотидную область, идентичную геномной последовательности Listeria weihenstephanensis, и два неполных совпадения с геномами бактериофагов, в частности, спейсер из Listeria weihenstephanensis совпадал с хвостовым геном бактериофага Listeria (фиг.42A-L).
[001329] Учитывая уникальность предсказанного эффекторного комплекса C2c2, эти системы могут быть классифицированы как предположительный тип VI систем CRISPR-Cas. Более того, поскольку все экспериментально охарактеризованные и ферментативно активные HEPN-домены являются РНК-азами, системы типа VI вероятно, действуют на уровне РНК, таких как мРНК,
[001330] Предполагаемые системы типа V-С. Третья группа локусов-кандидатов включает исключительно метагеномные последовательности и, следовательно, не может быть отнесена к конкретным таксонам. Эти локусы включают только два кодирующих белок гена, которые кодируют Cas1 и большой белок, обозначенный C2c3 (класса 3, кандидат 3) (фиг.8A "Организация полных локусов систем 2 класса"; фиг.14, окрестности C2c3, фиг.44A-Б). Белки C2c3 находятся в том же диапазоне размеров, что и Cpf1 и C2c1, и аналогично содержат TnpB-гомологичный домен на своих C-концах, который, в отличие от соответствующего домена C2c1, продемонстрировал ограниченное, но значительное сходство с Cpf1 (фиг.13А и 13С). Области гомологичности TnpB C2c3 содержат три каталитических мотива типа RuvC-подобной нуклеазы, с диагностической триадой D..E..D каталитических аминокислотных остатков (Aravind et al., 2000, выше), область, соответствующую мостиковой спирали (также известную как богатый аргинином кластер), который участвует в cr-РНК-связывании в Cas9, и небольшую область, которая представляется соответствующей "цинковому пальцу" TnpB (Zn-связывающие остатки цистеина сохраняются в C2c3). Сохранение каталитических остатков явно свидетельствует о том, что гомологичные домены RuvC всех этих белков являются активными нуклеазами. N-концевые области C2c1 и C2c3 не демонстрируют существенного сходства друг с другом или с любыми известными белками. Предсказание вторичной структуры показывает, что N-концевые области белков C2c3 принимают смешанную конформацию cx/j3. Таким образом, общие архитектуры доменов C2c1 и C2c3 и, в частности, организация домена RuvC аналогичны областям Cpf1, но отличаются от архитектуры Cas9. Это говорит о том, что локусы C2c1 и C2c3 правильнее классифицировать как подтипы V-B (см. выше) и V-C, соответственно, с локусами кодирующими Cpf1 на настоящий момент обозначенными подтипом V-A,
[001331] Среди c2c3 локусов только один содержит CRISPR кассету с нехарактерно короткими спейсерами длиной 17-18 нуклеотидов. Повторы в кассетах имеют длину 25 п.н. и оказывается, что они неструктурированные и имеют энергию 1,6 ккал/моль (фиг.43A-F).
[001332] Спейсеры из единственного континга C2c3, содержащего CRISPR кассету, слишком коротки, чтобы иметь статистически значимые совпадения. Тем не менее, было обнаружено несколько совпадений с последовательностями из предсказанных профагов (Фиг.43 A-F).
[001333] Подтипы белков TnpB имеющие значительное сходство с известным (Cas9) и тремя описанными здесь предполагаемыми эффекторами класса 2 (Cpf1, C2c1 и C2c3) не совпадали (фиг.13А и 13С). Хотя расхождение в последовательности TnpB-подобных доменов слишком велико для обеспечения надежного филогенетического анализа, эти данные свидетельствуют о том, что четыре известных в настоящее время больших эффекторных белка класса 2, Cas9, Cpf1, C2c1 и C2c3 развивались независимо от генов различных мобильных генетических элементов.
[001334] Хотя большинство спейсеров в новых локусах CRISPR-Cas, описанных в настоящем описании, не были в значительной степени похожи на любые доступные последовательности, существование спейсеров, совпадающих с геномом фага, подразумевает, что эти локусы могут кодировать активные функциональные адаптивные системы иммунитета. Малая доля фаго-специфических спейсеров типична для систем CRISPR-Cas и, скорее всего, указывает на их динамическую эволюцию и малую долю вирусного разнообразия, которая представлена в современных базах данных последовательностей. Эта интерпретация соотносится с наблюдением, что близкородственные бактериальные штаммы, кодирующие гомологичные локусы CRISPR-Cas, обычно содержат несвязанные коллекции спейсеров, примером которых являются локусы C2c2 из Listeria weihemtephanensis и Listeria newyorkensis (фиг.45А-С).
[001335] Заявители применили простую и понятную вычислительную стратегию для прогнозирования новых CRISPR-cas систем класса 2. Ранее описанные системы класса 2, а именно тип II и предполагаемый тип V, состояли из генов cas1 и cas2 (а в некоторых случаях и cas4), содержащих адаптационный модуль и один большой белок, который составляет эффекторный модуль. Поэтому заявители предположили, что любой геномный локус, содержащий cas1 и большой белок, может быть потенциальным кандидатом на новую систему класса 2 и заслуживает тщательного исследования. Такой анализ с использованием чувствительных способов сравнения белковых последовательностей привел к идентификации трех наиболее вероятных кандидатов, два из которых являются отдельными подтипами ранее описанного предполагаемого типа V (подтипы VB и VC), тогда как третий может быть классифицирован как новый предполагаемый тип VI, в силу наличия нового предсказанного эффекторного белка. Многие из этих новых систем встречаются в бактериальных геномах, которые не содержат других локусов CRISPR-Cas (фиг.44А-E), что указывает на возможность автономного функционирования систем типа V и типа VI. Рассматриваемые в настоящем описании локусы-кандидаты были проверены с помощью функциональных способов, которые выявили экспрессию и процессинг соответствующих CRISPR-кассет, производящих зрелые cr-РНК, определили предполагаемую tracr-РНК (где возможно), продемонстрировали интерференцию при экспрессии в E.coli, определили PAM и выявили минимальные компонентов, необходимые для расщепления лизата.
[001336] Системы типа V кодируют предсказанные эффекторные белки, которые по общей доменной архитектуре напоминают Cas9, но, в отличие от Cas9, RuvC-подобные домены Cpf1, C2c1 и C2c3 являются соседними и не имеют вставок, характерных для Cas9, в частности домена нуклеазы HNH. Наличие одного вместо двух доменов нуклеазы указывает на то, что эффекторные белки типа V механически отличаются от Cas9, в которых домены HNH и RuvC отвечают за расщепление комплементарных и не комплементарных цепей ДНК-мишени соответственно (Chen et al., 2014, The Journal of biological chemistry7, vol. 289, 13284- 13294, Gasiunas et al., 2012, Proceedings of the National Academy of Sciences of the United States of America, vol. 109, E2579-2586; Jinek et al., 2012, Science, vol. 337, 816-821). Предсказанные белки-эффекторы типа V могут образовывать димеры, в которых два домена, подобные RuvC, могут расщеплять противоположные цепи молекулы-мишени.
[001337] В основе предполагаемых систем CRISPR-Cas типа VI по-видимому лежит новый эффекторный белок, который содержит два предсказанных домена HEPN, которые, подобно ранее характеризованным доменам HEPN, могут обладать РНК-азной активностью, что системы типа VI могут быть нацелены на и расщеплять мРНК. Ранее сообщалось о нацеливании на мРНК для некоторых систем типа CRISPR-Cas типа III (Hale et al., 2014, Genes Dev, том 28, 2432-2443, Hale et al, 2009, Cell, vol. 139, 945-956, Peng et al., 2015, Nucleic acid research, vol. 43, 406-417). Альтернативная возможность заключается в том, что C2c2 является первой ДНК-азой в суперсемействе HEPN, возможно, с двумя доменами HEPN, каждый из которых расщепляет одну цепь ДНК. Таким образом, кажется возможным преобразование C2c1 и C2c2 в инструменты редактирования генома с различными классами целей.
[001338] Чтобы проверить функциональность этих систем CRISPR-Cas класса 2, заявители показали, что две C2c1 CRISPR-кассеты экспрессируются, процессируются в зрелые cr-РНК и способны к интерференции в случае экспрессии в E. coli. Эти эксперименты выявили более характеристик локуса C2c'l, включая: (i) процессируемые прямые повторы (DR) на 5'-конце cr-РНК, (ii) 5' PAM и (iii) наличие короткой РНК с гомологией повтор-антиповтор по отношению к процессируемому 5' DR, то есть предполагаемая tracr-РНК. Открытие процессируемых DR на 5'-конце cr-РНК и 5' PAM поддерживает такое развитие событий при котором C2c1 происходит из систем класса 1, потому что эти системы демонстрируют процессинг 5'-концов (I типа и III типа) и 5'-концевой последовательности PAM (I типа) (Mojica et al., 2009, Microbiology, vol. 155, 733-740; Makarova et al., 2011, Nat Rev Microbioln, т. 9, 467-477). В данном случае для С2с1 идентифицирована AT-богатая последовательность PAM, в противоположность ГЦ-богатым последовательностям PAM хорошо охарактеризованных систем II класса. Для локусов C2c1, охарактеризованных в настоящем описании экспериментально, Заявители определили cr-РНК, которые обрабатываются до длины, при которой сохраняется возможность к связыванию и кофолдингу с предполагаемыми tracr-РНКs, что указывает на то, что tracr-РНКs могут быть вовлечены и, возможно, даже необходимы для образования комплексов. Затем авторы изобретения использовали экспрессию C2c1 в культуре клеток человека для экспериментальной верификации того, что в этих условиях участвовала tracr-РНК и была необходима для in vitro расщепления ДНК-мишени конкретной исследуемой нуклеазой C2c1.
[001339] Заявители также показали, что, когда C2c2 локус из L.seeligeri экспрессируется в E.coli, он процессируется в cr-РНК с прямым повтором (DR) на 5'-конце 29 нуклеотидов длиной, похожие результаты были получены для локуса C2c2 из L.shahii. В этом случае, вырожденный повтор находится в начале последовательности, а не в конце, как это типично для большинства CRISPR-последовательностей, и последовательность и гены cas транскрибируются сонаправленно. Заявители не обнаружили предполагаемую tracr-РНК в данных секвенирования РНК C2c2 РНК. Однако прогнозируемая вторичная структура прямого повтора (DR) 29 нуклеотидов длиной показывает стабильную "шпильку", которая может быть потенциально важной для образования комплекса с эффекторным белком C2c2.
[001340] В сочетании с результатами предыдущих анализов, (Chylinski, 2014; Makarova, 2011), идентификация новых систем CRISPR-Cas класса 2 обнаруживает главную тенденцию в развитии систем CRISPR-Cas класса 2. Эффекторные белки двух или трех типов в этом классе, по-видимому, эволюционировали из пула мобильные генетических элементов, которые кодируют белки TnpB, содержащие RuvC-подобный домен. Последовательности RuvC-подобных доменов TnpB и гомологичных доменов эффекторных белков класса 2 слишком сильно дивергировали для надежного филогенетического анализа. Тем не менее, для Cas9, эффекторного белка системы II Типа, специфичная группа предков, по-видимому, легко идентифицируется, а именно семейство TnpB-подобных белков, особенно обильных в цианобактериях, которые показывают относительно высокое сходство с Cas9 и разделяют с Cas9 всю доменную архитектуру, а именно RuvC-подобные домены, нуклеазные домены HNH и богатую аргинином мостиковую спираль (Chylinski, 2014) (Рисунок 11, Фиг.13A и 13B, "Доменная организация семейств 2 класса"; Рисунок 12, Фиг.13A и 13B, "области гомологии TnpB в белках 2 класса"). В отличии от Cas9, невозможно отнести Cpf1, C2c1, и C2c3 к определенному семейству, несмотря на сохранение всех мотивов, сконцентрированных в каталитических остатках RuvC-подобных нуклеаз, эти белки показывают только ограниченное сходство с родовыми профилями TnpB. Однако, учитывая, что C2сlp не показывает обнаружимого сходства последовательностей с Cpf1, что Cpf1, C2c1, и C2c3 содержат различные вставки между мотивами RuvC и явно неродственными N-концевыми областями, наиболее вероятно, что Cpf1, C2c1, и C2c3 возникли независимо из различных семейств в пуле кодирующих элементов TnpB.
[001341] Любопытно, что белки TnpB, во-видимому, "предопределены" для использования в эффекторных комплексах систем CRISPR-Cas класса 2, так что они, по-видимому, были рекрутированы во множестве разных случаев. По-видимому, такая полезность белков TnpB связана с их предсказанной способностью разрезать одноцепочечную ДНК, будучи связанными с молекулой РНК через R-богатую мостиковую спираль, которая, как это было показано, в Cas9 связывает cr-РНК (Jinek, 2014; Nishimasu, 2014; Anders et al., 2014, Nature, vol. 513, 569-573). Функции TnpB в жизненных циклах соответствующих транспозонов плохо изучены. Эти белки не нужны для транспозиции, и в одном случае было показано, что белок TnpB снижает транспозицию (Pasternak, 2013), но их механизм действия остается неизвестным. Экспериментальное исследование TnpB, вероятно, прольет свет на механические аспекты систем CRISPR-Cas класса 2. Стоит отметить, что механизмы Cpf1 и C2c1 могут быть похожими друг на друга, но они существенно отличаются от Cas9, так как в первых двух белках отсутствует домен HNH, который в Cas9 отвечает за разрыв одной из цепей ДНК-мишени (Gasiunas, 2012)(Jinek, 2012)(Chen, 2014). Соответственно, использование Cpf1 и C2c1 может привести к дополнительным возможностям редактирования генома.
[001342] С точки зрения эволюции поразительно, что системы CRISPR-Cas класса 2, по-видимому, полностью произошли из различных мобильных генетических элементов, учитывая недавние данные о вероятном возникновении генов cas1 из отдельного семейства транспозонов (Koonin, 2015, Krupovic, 2014). Более того, вероятное независимое происхождение эффекторных белков из разных семейств TnpB, наряду с различной филогенетической близостью соответствующих cas1 белков, настоятельно свидетельствует о том, что системы класса 2 развивались неоднократно посредством сочетания различных адаптационных модулей и полученных из транспозонов нуклеаз, что привело к появлению эффекторных белков. Этот способ эволюции, по-видимому, является окончательным проявлением модульности, характерной для эволюции системы CRISPR-Cas (Makarova, 2015), что подразумевает, что в природе могут существовать дополнительные комбинации адаптационного и эффекторного модулей.
[001343] Предсказанные системы CRISPR-Cas типа VI содержат предсказанный эффекторный белок, в составе которого присутствуют два предсказанных домена HEPN, которые, вероятно, обладают РНК-азной активностью. Эти домены HEPN не являются частями эффекторных комплексов в других системах CRISPR-Cas, но задействованы в осуществления различных защитных функций, включая прогнозируемую вспомогательную роль (Anantharaman, 2013) (Makarova, 2015) в различных системах CRISPR Cas. Присутствие HEPN-доменов в качестве каталитических единиц предсказанного эффекторного модуля подразумевает, что системы типа VI нацеливаются на и расщепляют мРНК. Ранее для систем CRISPR-Cas III Типа уже сообщалось о нацеливании на мРНК (Hale, 2014; Hale, 2009) (Peng, 2015). Несмотря на то, что домены HEPN до сих пор не были обнаружены в настоящих транспозонах, они характеризуются высокой горизонтальной подвижностью и являются неотъемлемой частью мобильных элементов, таких как системы токсин-антитоксин (Anantharaman, 2013). Таким образом, предполагаемые системы типа VI, похоже, соответствуют общей парадигме модульной эволюции CRISPR-Cas класса 2 от мобильных элементов, и ожидается, что новые варианты и новые типы систем будут обнаружены путем анализа данных геномики и метагеномики. Учитывая, что белок C2c2 не связан с другими эффекторами класса 2 (все они содержат RuvC-подобные домены, даже при условии, отдаленного родства), открытие типа VI можно считать подтверждением независимого происхождения других вариантов класса 2.
[001344] Учитывая вырисовывающийся сценарий эволюции систем класса 2 из мобильных элементов, представляется полезным изучить общую эволюцию локусов CRISPR-Cas и, в частности, вклад мобильных элементов в этот процесс (фиг.53). Предшественники адаптивной иммунной системы, скорее всего, возникли из-за вставки каспозона (транспозона, кодирующего Cas), примыкающего к локусу, который кодировал примитивную систему врожденного иммунитета (iKoonin и Krupovic, 2015, Nature reviews Genetics, vol. 16, 184-192; Krupovic et al., 2014, BMC Biology, vol. 12, 36). Важным результатом было также включение системы токсин-антитоксин, которая передавала ген cas2 и могла появиться либо в предковом каспазоне, либо в изменяющемся локусе адаптивного иммунитета (фиг.51).
[001345] Учитывая чрезвычайно широкое распространение систем класса 1 в археях и бактериях и распространение в них древних доменов RRM (мотив распознавания РНК, RNA Recognition Motif - RRM), не возникает сомнений, что предковая система принадлежала к классу 1 (фиг.51). Скорее всего, предковая архитектура напоминала существующий тип III и включала ферментативно активный белок Cas10 (Makarova et al, 2011, Biol Direct, том 6, 38; Makarova et al, 2013, Biochem Soc Trans, том 41, 1392-1400). Белок Cas10 представляет собой гомолог ДНК-полимераз семейства В и нуклеотидных циклаз семейства GGDEF, который показывает значительное сходство последовательностей с этими ферментами и сохраняет все каталитические аминокислотные остатки (Makarova et al., 2011, Biol Direct, том 6, 38, Makarova et al., 2006, Biol Direct, том 1, 7). Структурный анализ подтвердил наличие в Cas10 полимеразоциклазоподобного домена и дополнительно выявил второй, вырожденный и, по-видимому, неактивный домен этого семейства (Khachatryan et al., 2015, Phys Rev Lett, vol. 114, 051801; Shao et al., 2013, Structure, vol. 21, 376-384, Zhu and Ye, 2012, FEBS Lett, том 586, 939-945). Точный характер каталитической активности Cas10 остается неясным, но было показано, что каталитические остатки полимеразоциклазоподобного домена являются существенными для расщепления ДНК-мишени (Saniai et al., 2015, Cell, vol. 161, 1164-1174). Белки Cas8, присутствующие в системах типа CRISPR-Cas I Типа, сходны по размерам с Cas10 и занимают эквивалентные положения в эффекторных комплексах (Jackson et al., 2014, Science, vol. 345, 1473-1479; Jackson and Wiedenheft, 2015, Mol Cell, vol. 58, 722-728, Staals et al., 2014, Molecular cell, vol. 56, 518-530), что наводящий на размышления об эволюционном родстве между большими субъединицами эффекторных комплексов III Типа и I Типа. Более конкретно, белки Cas8, которые сильно расходятся в последовательности между подтипами I Типа, могут быть каталитически неактивными производными Cas10 (Makarova et al., 2011, Biol Direct, том 6, 38, Makarova et al., 2015). Этот сценарий предполагает правдоподобную направленность эволюции, от подобного типу III предкового системы класса 1 до систем типа I. Дивергенция систем типа III и типа I могла быть ускорена за счет получения гелиазы Cas3 во время возникновения типа I (фиг.53). Различные типы и подтипы класса 2 затем эволюционировали посредством множественных замещений блока гена, кодирующего эффекторные комплексы класса 1, путем введения транспозируемых элементов, кодирующих различные нуклеазы (фиг.53). Эта конкретная направленность эволюции вытекает из наблюдения того, что адаптационные модули разных вариантов класса 2 происходят из разных типов 1-го класса (фиг.10А и 10В).
[001346] Системы класса 2 CRISPR-Cas, по-видимому, полностью получены из разных мобильных элементов. В частности, по-видимому, было, по меньшей мере, два (в подтипе V-C), но обычно три или, в случае II Типа, даже четыре представителя мобильных элементов: (i) предковый каспозон, (ii) модуль токсин-антитоксин, который дал начало Cas 2 (iii) транспозируемый элемент, во многих случаях кодирующий TnpB, который был предком эффекторного комплекса класса 2, и (iv) в случае II Типа нуклеаза HNH могла быть пожертвована предковому транспозону с помощью самосплайсирующегося интрона группы I или группы II (Stoddard, 2005, Q Rev Biophys, vol. 38, 49-95) (фиг.53). Локусы предполагаемого типа V-C, описанные в настоящем описании, кодируют конечную минимальную систему CRISPR-Cas, единственную в настоящее время идентифицированную, которой не хватает Cas2; по-видимому, сильно расходящиеся подтипы V-C Cas1-белков способны самостоятельно формировать адаптационный комплекс без вспомогательной субъединицы Cas2. Многократное возникновение систем класса 2 из мобильных элементов представляет собой окончательное проявление модульности, характерное для эволюции CRISPR-Cas (Makarova et al., 2015).
[001347] Демонстрация того, что различные разновидности систем класса 2 CRISPR-Cas, независимо от того, что они эволюционировали из разных транспозонов, подразумевает, что дополнительные варианты и новые типы еще предстоит идентифицировать. Хотя большинство, если не все новые системы CRISPR-Cas, как ожидается, будут редкими, они могут использовать новые стратегии и молекулярные механизмы и могут стать важным ресурсом для новых, разнообразных приложений в области инженерии генома и биотехнологии.
[001348] Модульная эволюция является ключевой особенностью систем CRISPR-Cas. Этот способ эволюции, по-видимому, наиболее выражен в системах класса 2, которые развиваются благодаря сочетанию адаптационных модулей от различных других систем CRISPR-Cas с эффекторными белками, которые, как представляется, рекрутируются из мобильных элементов в нескольких независимых случаях. Учитывая экстремальное разнообразие мобильных элементов в бактериях, представляется вероятным, что эффекторные модули систем класса 2 CRISPR-Cas также очень разнообразны. Здесь заявители использовали простой вычислительный метод для определения трех новых вариантов систем CRISPR-Cas, но многие другие, вероятно, будут содержаться в бактериальных геномах, которые еще не были секвенированы. Хотя большинство, если не все из этих новых систем CRISPR-Cas, как ожидается, будут редкими, они могут использовать новые стратегии и молекулярные механизмы и станут важным ресурсом для новых приложений в области инженерии генома и биотехнологии.
[001349] Программа TBLASTN со значением отсечки 0,01 и отключенной фильтрацией низкой сложности была использована для поиска для профиля Cas1 (Makarova et al., 2015) в качестве запроса к базе данных NCSI WGS. Последовательности контигов или полных геномов, в которых был обнаружено попадание (результат с высокими баллами), были извлечены из той же базы данных. Область вокруг гена Cas1 (область 20 т.п.н. от начала гена Cas1 и 20 т.п.н. от конца гена Cas1) была извлечена и транслирована с использованием GeneMarkS (Besemer et al, 2001, выше). Для предсказанных белков каждой Cas1-кодирующей области производился поиск с использованием набора профилей из базы данных CDD (Marchler-Bauer, 2009) и конкретным профилям белков Cas (Makarova et al., 2015) с использованием программы RPS-BLAST (Marchler-Bauer et al, 2002, Nucleic Acids Res, том 30, 281-283). Процедура определения полноты локусов CRISPR и классификации систем CRISPR-Cas в существующие типы и подтипы (Makarova et al., 2015), разработанные ранее, были применены к каждому локусу.
[001350] CRISPRmap (Lange, 2013) использовался для повторной классификации.
[001351] Частичные и/или неклассифицированные локусы, которые охватывали белки размером более 500 аминокислот, анализировали на индивидуальной основе. В частности, для каждого предсказанного белка, кодируемый этими локусами, был произведен поиск с использованием итеративных поисков профилей с PSI-BLAST (Altschul, 1997), и композиционная статистика с отключенной фильтрацией низкой сложности для поиска удаленных аналогичных последовательностей против базы данных "безызбыточных" (NR) белков NCBF. Для каждого идентифицированного "безызбыточного" белка производился поиск в базе данных WGS с использованием программы TBLAST (Altschul, 1997). Программа HHpred использовалась с параметрами по умолчанию для идентификации сходства отдаленной последовательности (Soding, 2005), используя в качестве запросов все белки, идентифицированные в поисках BLAST. Множественные выравнивания последовательностей были построены с использованием MUSCLE (Edgar, 2004) и MAFFT (Katoh and Standiey, 2013, Mol Biol Evol, vol, 30, 772-780). Филогенетический анализ проводился с использованием программы FastTree с эволюционной моделью WAG и дискретной гамма-моделью с 20 категориями скоростей (Price et al., 2010, PLoS One, том 5, e9490). Вторичная структура белка была предсказана с использованием Jpred 4 (Drozdetskiy, 2015).
[001352] CRISPR-повторы были обнаружены при помощи PILER-CR (Edgar, 2007, выше) или для вырожденных повторов - CRISPRfinder (Grissa et al., 2007, Nucleic Acids Res, vol. 35, W52-57). Программа Mfold (Zukler, 2003, Nucleic Acids Res, vol 31, 3406-3415) были использованы для обнаружения наиболее стабильных структур для повторяющихся последовательностей.
[001353] Был выполнен поиск последовательностей спейсеров был выполнен по базам данных NCBI: NR and WGS database при помощи MEGABLAST (Morgulis et al., 2008, Bioinformatics, vol. 24, 1757-1764) с заданными параметрами по умолчанию за исключением того, что параметр wordsize был задан на 20
[001354] Выбранные кандидаты генов
[001355] ГенID: A; Тип гена:C2C1; Организм: 5. бактерия Opitutaceae TAV5; Длина спейсера - мода (диапазон): 34 (33-37);
DR1:
DR2: нет;
tracr-РНК1:
tracr-РНК2: нет;
Последовательность белка:
[001356] ID гена: B; Тип гена: C2C1; Организм: 7. Bacillus thermoamylovorans штамма B4166; Длина спейсера - мода (диапазон): 37 (35-38);
DR1:
DR2: нет; tracr-РНК1;
tracr-РНК2: нет;
Последовательность белка:
[001357] ID гена: C; Тип гена: C2C1; Организм: 9. Bacillus sp. NSP2.1; Длина спейсера - мода (диапазон): 36 (35-42);
DR1:
DR2: нет; tracr-РНК1:
tracr-РНК2: нет;
Последовательность белка:
[001358] ID гена: D; Тип гена: C2C2; Организм: 4. Бактерия Lachnospiraceae NK4A144 G619; Длина спейсера - мода (диапазон): 35;
DR1:
DR2:
tracr-РНК1: нет; tracr-РНК2: нет; Последовательность белка:
[001359] ID гена: E; Тип гена: C2C2; Организм: 8. Listeria seeligeri серовар 1/2b штамм SLCC3954; Длина спейсера - мода (диапазон): 30;
DR1:
DR2: нет; tracr-РНК1:
tracr-РНК2: нет; Последовательность белка:
[001360] ID гена: F; Тип гена: C2C2;
Организм: 12. Leptotrichiawadei F0279;
Длина спейсера - мода(диапазон): 31;
DR1:
DR2: нет;
tracr-РНК1:
tracr-РНК2:
Последовательность белка:
[001361] ID гена: G; Тип гена: C2C2;
Организм: l4. Leptotrichiashahii DSM 19757 B031;
Длина спейсера - мода (диапазон): 30 (30-32);
DR1:
DR2: нет;
tracr-РНК1:
tracr-РНК2: нет;
Последовательность белка:
[001362] ID гена: H;
Тип гена: Cpf1; Организм: Francisellaularensis subsp. novicida U112, Длина спейсера - мода (диапазон): 31;
DR1:
DR2: нет;
tracr-РНК1:
tracr-РНК2: нет;
Последовательность белка:
[001363] Гены для синтеза
[001364] Для генов от А-Н заявители оптимизируют гены для экспрессии в человеке и добавляют следующую последовательность ДНК в конец каждого гена. Обратите внимание, что эта последовательность ДНК содержит стоп-кодон (подчеркнутый), поэтому стоп-кодон не добавляется к последовательности, оптимизированной кодоном гена:
[001365] Для оптимизации избегайте следующих сайтов рестрикции: BamHI, EcoRI, HindIII, BsmBI, BsaI, BbsI, AgeI, XhoI, Ndel, NotI, Kpnl, BSrGI, SpeI, XbaI, NheI
[001366] Эти гены клонированы в простой экспрессирующий вектор млекопитающих:
[001367] A
[001368]
[001371] C
[001372]
[001373] D
[001374]
[001377] F
[001378]
[001379] G
[001380]
[001381] H
[001382]
[001383] Локусы от A до G клонированы и вставлены в плазмиду с низким количеством копий. Используется вектор, не содержащий имеет резистентности к Amp.
[01384] A-локус
[01385]
Пример 3: Дальнейшая оценка C2c2p и ассоциированных компонентов
Пример 3. Дальнейшая оценка C2c1 и связанных с ней компонентов
[001398] Заявители продемонстрировали в описанных экспериментах, что локусы 2 класса CRISPR-Cas экспрессируются с образованием зрелой cr-РНК и кодированием предполагаемых tracr-РНК и функциональных направленных на РНК нуклеаз.
[001399] Заявители проанализировали репрезентативный локус C2c1, то есть локус C2c1 Alticclobacillis acidoterrestris (AacC2c1) CRISPR, и провели РНК-секвенирование (RNA-seq) и нозерн-блоттинг, чтобы охарактеризовать его транскрипционную активность.
[001400] Для секвенирования РНК Alicyclobacillus acidoterrestris (ATCC 49025) культивировали с использованием суспензии ATCC Medium 1.655 при 50°C и 300 об/мин. E. coli, содержащие гетерологичные конструкции, культивировали в суспензии в среде Лурия с добавлением соответствующих антибиотиков при 37°C и 300 об/мин. Оба штамма выращивали в аэробных условиях и собирали в стационарной фазе роста.
[001401] Чтобы получить локус AacC2c1 для гетерологичной экспрессии, геномную ДНК выделили из Alicyclobacillus acidoterrestris с использованием Qiagen DNeasy Blood and Tissue kit, а локус C2c1 амплифицировали ПЦР для клонирования в pACYC-184.
[001402] РНК была выделена из бактерий в стационарной фазе, сначала бактерии были ресуспендированы в TRIzol, а затем гомогенизированы с шариками циркония/кремния (BioSpec Products) в BeadBeater (BioSpec Products) в течение 7 одноминутных циклов. Общая РНК была очищена из гомогенизированных образцов с помощью протокола Miniprep Direct-Zol RNA (Zymo), обработаны ДНК-азой TURBO DNAase (Life Technologies), и 3'-дефосфорилирована T4-полинуклеотид-киназой (New England Biolabs). рРНК удаляли с помощью набора для удаления бактериальной рРНК Ribo-Zero (Illumina). Библиотеки секвенирования РНК были получены из рРНК-обедненной РНК с использованием модификации ранее описанного способа секвенирования cr-РНК (Heidriсh et al., 2015, Methods Mol Biol, vol. 1311, 1-21). Кратко, транскрипты получили поли-А-хвост с помощью поли-(A)-полимеразы E. coli (New England Biolabs), были лигированы с 5'-РНК-адаптерами с использованием T4 РНК-лигазы 1 (оцРНК-лигаза), высокой концентрации (High Concentration, New England Biolabs) и обратно транскрибированы многотемпературной обратной транскриптазой AffmityScript (Agilent Technologies). кДНК была амплифицирована ПЦР с помощью штрих-кодированных праймеров с использованием полимеразы Herculase II (Agilent Technologies).
[001403] Готовые библиотеки кДНК секвенировали на MiSeq (Alumina). Считывания из каждого образца были идентифицированы на основе ассоциированных с ними штрих-кодов и выровнены в соответствии с эталонным геномом RefSeq с использованием BWA (Li and Durbin, 2009, Bioinformatics, vol. 25, 1754-1760). Попарные выравнивания были использованы для выделения последовательностей целых транскриптов с помощью инструментов Picard (http://broadinstitute.github.io/picard) и эти последовательности были проанализированы с использованием Geneious 8.1.5.
[001404] Полнотранскриптомный анализ RNAseq Aac показал, что транскрипция массива CRISPR происходит в той же ориентации, что и у генов Cas (фиг.17). Избыточность отображения транскриптов, картируемых на массив CRISPR, снижалось в направлении от 5' к 3' (фиг.46A). Это согласуется с результатами нозерн-блоттинга заявителей. Массив CRISPR продемонститровал надежность процессинга cr-РНК длиной 34 нуклеотидов с 5'-прямым повтором из 14 нуклеотидов (DR) и спейсером из 20 нуклеотидов (фиг.46A).
[001405] Заявители идентифицировали избыточную малую РНК размером 79 нуклеотидов, закодированную между геном cas2 и массивом CRISPR, и транскрибируемую в той же ориентации, что и массив CRISPR (фиг.46А и В). Внутренняя область этой малой РНК содержит последовательность, которая является высококомплементарной процессированной повторяющейся последовательности CRISPR (антиповтору), позволяя предположить, что этот малый транскрипт является tracr-РНК. При со-сворачивании in silico процессированного повтора CRISPR из 14 нуклеотидов с этой предполагаемой tracr-РНК образуется стабильная предсказанная вторичная структура (фиг.46C).
[001406] Заявители также экспрессировали локус C2c1 Alicyclobacillus acidoterrestris (ATCC 49025) в E. coli и анализировали его экспрессию с использованием Нозерн-блоттинга. Процедура проводилась, по существу, как описано в Pougach and Severinov, 2012 (Methods Mol Biol, vol. 905, 73-86). Клетки E. coli BL21 AI трансформировали плазмидой pACYCduet-1, содержащей контролируемый T7-промотором оперон cas Alicyclobacillus acidoterrestris, и плазмидой pCDF-1b, содержащей минимальную кассету CRISPR с одним спейсером. Общая РНК была экстрагирована из 5 мл клеток E.coli, индуцированных 1 мМ арабинозы/0,2 мМ IPTG и выращенных до OD600 0,8-1,0. Клетки лизировали с помощью 5-минутной обработки с использованием Max Bacterial Enhancement Reagent с последующей очисткой РНК реагентом TRIzol (Thermo Fisher Scientific). 15 пг общей РНК разделяли в денатурирующей 8 М мочевине - 12% полиакриламидном геле и электрофоретически переносили на мембрану Hybond-XL (GE Healthcare) с использованием Mini Trans-Blot Electrophoretic Transfer Cell (Bio-Rad). Мембрану высушивали, а затем подвергали кросс-линкингу ультрафиолетовым излучением. Раствор для гибридизации ExpHyb (Clontech) использовали для гибридизации в соответствии с инструкциями производителя в течение 1 часа при 40°C с олигонуклеотидными зондами с 32P-мечеными концами.
[001407] Основываясь на наблюдении, что предполагаемая tracr-РНК в A. acidoterrestris содержит характерную последовательность антиповтора, заявители стремились предсказать потенциальные tracr-РНК для остальной части идентифицированных локусов C2c1, а также C2c2 и C2c3 путем поиска последовательностей анти-повторов в каждом локусе. Во многих локусах CRISPR-Cas повтор, расположенный на дальнем от промотора конце последовательности CRISPR, является вырожденным и имеет последовательность, которая явно отличается от остальных повторов (Biswas et al., 2014, Bioinformatics, vol. 30, 1805 -1813). Такие вырожденные повторы были обнаружены в нескольких системах C2c1 и C2c2 (фиг.9, 14 и 15), что позволило заявителям предсказать направление транскрипции последовательности. Интегрируя эту информацию, были идентифицированы предполагаемые tracr-РНК для 4 из 13 локусов C2c1 и 4 из 17 локусов C2c2 (фиг.9, 14 и 15, фиг.41A-M, фиг.42A-N). В некоторых подтипах II-В и II-С последовательности CRISPR транскрибируется в противоположном направлении, начиная с вырожденного повтора (Sampson et al., 2013, Nature, vol. 497, 254-257; Zhang et al, 2013, Mol Cell, vol. 50, 488-503). Соответственно, авторы изобретения попытались предсказать tracr-РНК в разных положениях по отношению к последовательности CRISPR, но не смогли идентифицировать дополнительные последовательности tracr-РНК-кандидатов. Разумеется, предсказание tracr-РНК для других локусов было затруднено из-за сочетания таких факторов, как неполная комплементарность повторов, отсутствие связанной последовательности CRISPR и/или потенциальная неполнота локусов. Кроме того, существует вероятность того, что не все системы CRISPR класса 2 требуют tracr-РНК.
[001408] Учитывая надежную экспрессию транскриптов в локусе Aac и идентификацию процессированной tracr-РНК и cr-РНК, заявители стремились проверить функциональность фермента AacC2c1. С этой целью был разработан интерференционный скрининг, чтобы определить, являются ли эти локусы активными, и идентифицировать необходимый соседний с протоспейсером мотив (PAM), который определяет, где эффекторный белок будет расщепляться в известных системах типа II (фиг.47А). Была сконструирована библиотека плазмид, дающих устойчивость к ампициллину и несущих протоспейсеры, фланкированный 7 случайными нуклеотидами нуклеотидов. Хотя наличие фрагмента DR на 5'-концевой стороне cr-РНК предполагает наличие 5'-PAM, были проанализированы отдельные библиотеки со случайными последовательностями длиной 7 нуклеотидов, расположенными вверху или внизу от протоспейсера. Более 10 колоний были объединены для представления всех 16384 возможных последовательностей PAM. Клетки E. coli, несущие либо плазмиду локуса AacC2c1 (фиг.46D), либо контрольную плазмиду (pACYC184), трансформировали 5'- или 3'-PAM-библиотеками.
[001409] В частности, рандомизированные библиотеки плазмид PAM были сконструированы с использованием синтезированных олигонуклеотидов (IDT), состоящих из 7 рандомизированных нуклеотидов либо выше, либо ниже от мишени спейсера 1. (22) Рандомизированные олигонуклеотиды одноцепочечной ДНК были сделаны двухцепочечными путем отжига со коротким праймером и с использованием большого Фрагмента Кленова для синтеза второй цепи. Продукт dsDNA собирали в линеаризованный PUC19 с использованием Gibson-клонирования. Stabl3 E. coli клетки трансформировали клонированными продуктами и собирали и собрали более 107 клеток. Плазмидную ДНК выделяли с использованием набора Qiagen maxi. 360 нг объединенной библиотеки трансформировали в клетки E. coli, трансформированные локусом AacC2c1, и pACYC-184. После трансформации клетки высевали на чашки с LB-агаром, дополненные ампициллином и хлорамфениколом, и выращивали при 37°C. После 16 часов роста > 4×106 клеток и ДНК плазмиды выделяли с использованием набора Qiagen maxi-prep. Целевая область PAM была амплифицирована и секвенирована с помощью Illumina. MiSeq с одноконцевыми 150 циклами.
[001410] В рамках этой экспериментальной установки протоспейсеры с PAM, которые распознаются комплексом AacC2c1, будут уничтожены, что делает клетки чувствительными к ампициллину и, как ожидается, будет демонстрировать обеднение на экране. После ночного роста бактерий целевые плазмиды выделили и подвергли глубокому секвенированию. Эффективность каждого PAM определяли путем вычисления его относительного уровня в образце локуса AacC2c1 по сравнению с контрольным образцом.
[001411] В то время как скрининг PAM длиной 3 нуклеотида не показал значительного истощения PAM, скрининг библиотеки 5'-PAM привел к идентификации 364 PAM, уровень которые был значительно снижен; все эти PAM имели последовательность NNNNTTN (фиг.47B). Все же были несколько более предпочтительны основания, отличные от С, в 5'-положении, непосредственно прилегающем к протоспейсеру, эти результаты показывают, что 5-TTN-мотив распознается комплексом AacC2c1.
[001412] Предложенный PAM был проверен с использованием первого спейсера локуса AacC2c1 и всех четырех TTN PAM (РИС. 47C). Последовательности, соответствующие как PAM, так и не-PAM, клонировали в расщепленный pUC19 и лигировали с T4-лигазой (ферментами). Компетентную E. coli с плазмидой локуса AacC2c1 или плазмидой pACYCl84 трансформировали 20 г PAM-плазмиды и высевали на чашки с агаром LB, дополненным ампициллином и хлорамфениколом. Через 18 часов колонии подсчитывали с помощью OpenCFU (Geissmann 2013, PLoS ONE, vol 8, e54072). Результаты этого эксперимента подтверждают, что для интерференции требуется 5'TTN PAM, и эта интерференция несколько снижается в случае 5'TTC PAM (фиг. 47C).
[001413] Заявители затем попытались выяснить, можно ли экспрессировать C2c1-нуклеазу с использованием клеток человека и одновременно исследовать ее требования к tracr-РНК. С этой целью Заявители получили ДНК-конструкцию с кодон-оптимизированным AacC2c1 человека и собрали белковый лизат клеток HEK293, экспрессирующих белок. В частности, был разработан кодон-оптимизированный для экспрессии в клетке человека белок C2c1, несущий N-терминальный NLS. Последовательность белка была синтезирована и клонирована в экспрессионную плазмиду pcDNA3.1 Genscript. Используя реагент Lipofectamine 2000 (Life Technologies), 2000 нг экспрессионных плазмид трансфицировали в клетки HEK293FT (6-луночные планшеты при 90% конфлюэнтности). Через 48 часов клетки промывали DPBS (Life Technologies), лизировали с использованием буфера для лизиса (20 мМ Hepes pH 7,5, 100 мМ KC1, 5 мМ MgCL, 1 мМ DTT, 5% глицерина, 0,1% Triton X-100, IX cOmplete Protease Inhibitor Cocktail (Roche)]. После обработки ультразвуком в течение 10 минут в ультразвуковых аппаратах Biorupter (Diagenod) (50%-ный рабочий цикл на высоком уровне) лизат центрифугировали и супернатант замораживали при -80°С для последующего использования в анализах на расщепление.
[001414] Основываясь на самом длинном транскрипте cr-РНК, наблюдаемом при секвенировании РНК, заявители разработали cr-РНК с прямым повторным (DR) длиной 20 нуклеотидов и спейсером длиной 22 нуклеотида, нацеленным на область локуса EMX1 размером 639-п.о., который был амплифицирован ПЦР из геномной ДНК человека. Так как A. acidoterrestris оптимально растет при 50°C (Chang and Kang, 2004, Crit Rev Microbiol, vol. 30, 55-7-4), заявители предположили, что белок AacC2c1 может лучше всего работать при этой температуре. Следовательно, расщепление проводили с использованием лизата млекопитающих из клеток, экспрессирующих белок C2c1, при 50°C, если не указано иное, в буфере для расщепления (NEBuffer 3, 5 мМ DTT) в течение 1 часа. В каждой реакции расщепления использовали 200 мкг ДНК-мишени и эквимолярное соотношение cr-РНК tracr-РНК (500 мкг кРНК). РНК предварительно отжигали нагреванием при 95°C и медленно охлаждали до 4°C. ДНК-мишень состояла либо из геномных ПЦР-ампликонов из гена EMX1, либо из первого протоспейсерного локуса AacC2c1, клонированного в pUC19. Реакционную смесь очищали с помощью ПЦР. (Qiagen) и прогоняли на 2% агарозных E-гелях (Life Technologies).
[001415] При 50°C заявители показали, что самая длинная экспрессируемая tracr-РНК, наблюдаемая в экспериментах RNAseq (151 нуклеотидов), необходима для расщепления ДНК при использовании лизата клеток C2c1 и что расщепление является специфичным к комплементарной области между кРНК и ДНК-мишенью (фиг.48A). Поскольку эксперименты с РНК-seq дали предполагаемые транскрипты tracr-РНК переменной длины (фиг.46A), авторы настоящего изобретения протестировали серию 3'-укороченных транскриптов tracr-РНК и обнаружили, что самая короткая tracr-РНК, обеспечивающая расщепление при использовании лизата C2c1 клеток, представляет собой последовательность из 78 нуклеотидов (фиг.48B). Используя эту минимальную tracr-РНК, авторы изобретения показали, что 50°C - действительно оптимальная температура расщепления, и показали отсутствие наблюдаемого расщепления при температуре ниже 40°C с использованием лизата клеток C2c1 (фиг.48C).
[001416] Чтобы дополнительно подтвердить требования к PAM для C2c1, Заявители разработали кРНК для конструкциями для валидации протоспейсера-1 РАМ, используемых в описанных выше экспериментах (фиг.47C) и продемонстрировали расщепление для TTT, TTA и TTC PAM, но не GGA с использованием tracr-РНК из 78-нуклеотидов (фиг.4DD).
[001417] Учитывая обнаружение специфичной для последовательности, перенаправляемой активности расщепления ДНК для нуклеазы AacC2c1, заявители предположили, что, подобно Cas9, дуплекс cr-РНК tracr-РНК C2RRRRRTRRANA может быть переработан в однонаправленную РНК (sg-РНК) путем присоединения 3' tracr-РНК из 78 нуклеотидов к 5'-концу cr-РНК (фиг.48E). Активность в отношении расщепления мишени, аналогичная полученной в случае дуплекса cr-РНК tracr-РНК, наблюдалась в случае sg-РНК как с плазмидными мишенями EMX1, так и с протоспейсером 1 (фиг.48F). Эти эксперименты показали, что лизат из клеток человека, экспрессирующих C2c1, может расщеплять ДНК-мишень, идентифицировали температурный оптимум фермента и продемонстрировали необходимость в дуплексе cr-РНК tracr-РНК или sg-РНК и 5'-PAM для активности нуклеазы AacC2c1.
[001418] В дальнейших экспериментах заявители изучают доменную структуру и организацию, а также экспрессию матрицы CRISPR в разных бактериальных штаммах, содержащих локусы CRISPR, обозначенные как подтип V-B, как, например, показано на фиг.8А и 8В. Заявители обладают оптимизированными методами для получения локусов C2c1 из других бактериальных штаммов.
[001419] В дальнейших экспериментах заявители разрабатывают клонирование pACYC из изолированных геномных бактериальных штаммов и выращенных бактериальных штаммов для проведения экспериментов по расщеплению ДНК/РНК в E. coli с эффекторными белками, кодируемыми локусами CRISPR.
[001420] В дальнейших экспериментах заявители разрабатывают библиотеки PAM приманки для оценки способности разрезания ДНК описанным здесь эффекторным белком C2c1p. Заявители разрабатывают эксперименты по разрезанию ДНК, основанные на разрезе транскрипта гена устойчивости.
[001421] В дальнейших экспериментах заявители тестируют функцию в клетках млекопитающих с использованием U6 ПЦР продуктов: спейсер (DR-спейсер-DR) (в некоторых аспектах спейсеры могут упоминаться как cr-РНК или направляющая РНК или как аналогичный термин, как описано в этой заявке) и tracr для указанных штаммов, указанных как подтип V-B на фиг.8А и 8В.
[001422] Пример 4: Дальнейшая оценка ортологоа локуса C2c1
Заявители также скринировали локус C2c1 из Bacillus thermoamylovorans (Bth): заявители экспрессировали локус B. Thermoamylovorans C2c1 (BthC2c1.) в E. coli и собирали данные РНК-Seq, демонстрирующие образование транскрипта. Локус BthC2c1 был синтезирован с помощью Genscript и клонирован в вектор pET-28. Клетки, содержащие плазмиды, были сделаны компетентными с использованием Z-компетентного набора (Zymo). E. coli, содержащие гетерологичные конструкции, культивировали в суспензии в среде Лурия с добавлением соответствующих антибиотиков при 37°C и 300 об/мин. Бактерии выращивали в аэробных условиях и собирали в стационарной фазе роста.
[001423] РНК была изолирована, и проведено РНК-секвенирование с помощью методов, изложенных в примере 3.
[001424] Полнотранскриптомное секвенирование синтезированного BthC2c1. локуса, клонированного в pET-28 в E. coli, выявило значительный процессинг обоих как спейсеров, а также экспрессию РНК длиной 91 нуклеотид (фиг.49А), которая обладает вторичной структурой и мотивом повтор-антиповтор-повтор, схожим с Aac tracr-РНК (фиг.49B). Предполагаемая последовательность tracr может быть tracr длиной ~91 нуклеотидов, имеющим следующую последовательность:
CGAGGUUCUGUCUUUUGGUCAGGACAACCGUCUAGCUAUAAGUGCUGCAGGGGU
GUGAGAAACUCCUAUUGCUGGACGAUGUCUCUUUUAU
[001425] В варианте исполнения изобретения последовательность прямого повтора (DR) может быть процессирована до полинуклеотида длиной 15 п.о., имеющего последовательность UACGAGGCAUUAGCAC.
[001426] Заявители определили консенсусный PAM для BthC2c1 из скрининга для обнаружения PAM. Заявители трансформировали библиотеку PAM с соответствующим спейсером в клетки E. coli, содержащие локус BthC2c1, и сравнили истощение с pET-28. Скрининг PAM осуществляли так же, как указано для скрининга AACC2c1 PAM, за исключением того, что после трансформации клетки высевали на чашки с LB-агаром, дополненным ампициллином и канамицином. Этот скрининг показал, что BthC2c1 использует 5' T-богатый PAM с консенсусной последовательностью ATTN (фиг.49C).
[001427] Заявители также экспрессировали локус C2c1 B. sp. в E. coli и получили данные RNAseq, демонстрирующие образование транскриптов. Стрелка на фиг.50 указывает предполагаемую tracr-РНК, связанную с локусом, в то время как показаны все риды. Предполагаемая последовательность tracr может быть tracr длиной 101 нуклеотид, имеющей следующую последовательность:
UCAGUCGCACGCUAUAGGCCAUAAGUCGACUUACAUAUCCGUGCGUGUGCAUUAU
GGGCCCAUCCACAGGUCUAUUCCCACGGAUAAUCACGACUUUCCAC
[001428] В одном из возможных вариантов осуществления изобретения последовательность прямого повтора (DR) может быть процессирована до 20 нуклеотидов, имеющих последовательность GAAAGCUUCGUGGUUAGCAC (фиг.51). Заявители также показали со-сворачивание DR и tracr-РНК, связанное с локусом C2c1 B. sp. (фиг.52)
Пример 5: Модули адаптации новых систем класса 2, вероятно, эволюционировали независимо от разных подразделений класса 1
[001429] Cas1, специализированная ДНК-аза и интеграза, которая играет центральную роль в адаптации CRISPR (Nunez et al., 2014, Nature, Nature structural & molecular biology, vol. 21, 528-534; Nunez et al., 2015, vol. 519, 193-8), является наиболее консервативным белком Cas (Takeuchi et al., 2012, J Bacteriol, vol. 194, 1216-1225) и единственным, для которого возможен комплексный филогенетический анализ (Makarova et al., 2011b, Nat; Rev Microbiol, vol.9, 467-477, Makarova et al., 2015). В филогенетическом дереве Cas1 предполагаемый подтип VB (C2c1) является в значительной степени монофилетической группой и кластеры с типом IU (фиг.10А и 10В, фиг.10C-V). Среди всех (предполагаемых) локусов CRISPR-Cas только типы I-U и C2c1 кодируют слияние белков Cas1 и Cas4. Вместе этот общую характеристику и филогенетическое сродство Cas1 настоятельно указывают на то, что модуль адаптации подтипа V-B происходит от адаптивного типа I-U. Тип V-C Cas1 является наиболее отличным вариантом последовательностей Cas1, обнаруженным на сегодняшний день, как показано длинной ветвью в филогенетическом дереве (фиг.10А и 10В). В дереве Cas1 ветвь V-C типа находится в подтипе I-B (фиг. 10A и 10B), хотя позиционирование такой быстро эволюционирующей группы должно приниматься с осторожностью. Белки типа VI Cas1 распределены между двумя кладами. Первая группа включает Cas1 из Leptotrichia и расположена в поддереве типа II вместе с небольшой ветвью типа III-A. Вторая группа включает белки Cas1 из локусов C2c2 Clostridia и относится к смешанной ветви, которая в основном содержит белки Cas1 из систем типа CRISPR-Cas типа III-A (фиг.10A и 10B).
[001430] Хотя Cas2 является небольшим и относительно малоконсервативным белком, для которого трудно получить надежную филогению, все имеющиеся данные указывают на эволюционную когерентность адаптационного модуля, то есть на коэволюцию генов cas1 и cas2 (Chylinsky et al., 2014), Nucleic Acids Res., Vol.42, 6091-105, Norais et al., 2013, RNA Biol, том 10, 659-670). Собрав все вместе и не ограничиваясь какой-либо гипотезой, эти данные показывают, что адаптационные модули новых систем класса 2 CRISPR-Cas исходят из разных вариантов класса 1.
[001431] Хотя предпочтительные варианты осуществления настоящего изобретения были продемонстрированы и описаны в настоящем описании, специалистам в данной области будет очевидно, что такие варианты осуществления предоставляются только в качестве примера. Многочисленные вариации, изменения и замены теперь могут быть разработаны специалистами в данной области без отступления от изобретения. Следует понимать, что различные альтернативы вариантам осуществления изобретения, описанным в настоящем описании, могут быть использованы при осуществлении изобретения. Предполагается, что нижеследующая формула изобретения определяет объем изобретения и что способы и структуры, входящие в объем этих притязаний и их эквиваленты, будут охвачены таким образом.
--->
SEQUENCE LISTING
<110> THE BROAD INSTITUTE INC.
MASSACHUSETTS INSTITUTE OF TECHNOLOGY
PRESIDENT AND FELLOWS OF HARVARD COLLEGE
RUTGERS, THE STATE UNIVERSITY OF NEW JERSEY
SKOLKOVO INNOVATION OF SCIENCE AND TECHNOLOGY
<120> NOVEL CRISPR ENZYMES AND SYSTEMS
<130> 47627.99.2136
<140> US 15/842,073
<141> 2017-12-14
<150> PCT/US2016/038238
<151> 2016-06-17
<150> 62/245,264
<151> 2015-10-22
<150> 62/181,663
<151> 2015-06-18
<160> 1786
<170> PatentIn version 3.5
<210> 1
<211> 7
<212> PRT
<213> Simian virus 40
<400> 1
Pro Lys Lys Lys Arg Lys Val
1 5
<210> 2
<211> 16
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Nucleoplasmin bipartite NLS sequence"
<400> 2
Lys Arg Pro Ala Ala Thr Lys Lys Ala Gly Gln Ala Lys Lys Lys Lys
1 5 10 15
<210> 3
<211> 9
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
C-myc NLS sequence"
<400> 3
Pro Ala Ala Lys Arg Val Lys Leu Asp
1 5
<210> 4
<211> 11
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
C-myc NLS sequence"
<400> 4
Arg Gln Arg Arg Asn Glu Leu Lys Arg Ser Pro
1 5 10
<210> 5
<211> 38
<212> PRT
<213> Homo sapiens
<400> 5
Asn Gln Ser Ser Asn Phe Gly Pro Met Lys Gly Gly Asn Phe Gly Gly
1 5 10 15
Arg Ser Ser Gly Pro Tyr Gly Gly Gly Gly Gln Tyr Phe Ala Lys Pro
20 25 30
Arg Asn Gln Gly Gly Tyr
35
<210> 6
<211> 42
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
IBB domain from importin-alpha sequence"
<400> 6
Arg Met Arg Ile Glx Phe Lys Asn Lys Gly Lys Asp Thr Ala Glu Leu
1 5 10 15
Arg Arg Arg Arg Val Glu Val Ser Val Glu Leu Arg Lys Ala Lys Lys
20 25 30
Asp Glu Gln Ile Leu Lys Arg Arg Asn Val
35 40
<210> 7
<211> 8
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Myoma T protein sequence"
<400> 7
Val Ser Arg Lys Arg Pro Arg Pro
1 5
<210> 8
<211> 8
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Myoma T protein sequence"
<400> 8
Pro Pro Lys Lys Ala Arg Glu Asp
1 5
<210> 9
<211> 8
<212> PRT
<213> Homo sapiens
<400> 9
Pro Gln Pro Lys Lys Lys Pro Leu
1 5
<210> 10
<211> 12
<212> PRT
<213> Mus musculus
<400> 10
Ser Ala Leu Ile Lys Lys Lys Lys Lys Met Ala Pro
1 5 10
<210> 11
<211> 5
<212> PRT
<213> Influenza virus
<400> 11
Asp Arg Leu Arg Arg
1 5
<210> 12
<211> 7
<212> PRT
<213> Influenza virus
<400> 12
Pro Lys Gln Lys Lys Arg Lys
1 5
<210> 13
<211> 10
<212> PRT
<213> Hepatitis delta virus
<400> 13
Arg Lys Leu Lys Lys Lys Ile Lys Lys Leu
1 5 10
<210> 14
<211> 10
<212> PRT
<213> Mus musculus
<400> 14
Arg Glu Lys Lys Lys Phe Leu Lys Arg Arg
1 5 10
<210> 15
<211> 20
<212> PRT
<213> Homo sapiens
<400> 15
Lys Arg Lys Gly Asp Glu Val Asp Gly Val Asp Glu Val Ala Lys Lys
1 5 10 15
Lys Ser Lys Lys
20
<210> 16
<211> 17
<212> PRT
<213> Homo sapiens
<400> 16
Arg Lys Cys Leu Gln Ala Gly Met Asn Leu Glu Ala Arg Lys Thr Lys
1 5 10 15
Lys
<210> 17
<211> 4
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 17
Gly Gly Gly Ser
1
<210> 18
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 18
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 19
<211> 30
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 19
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25 30
<210> 20
<211> 45
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 20
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
20 25 30
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
35 40 45
<210> 21
<211> 60
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 21
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
20 25 30
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
35 40 45
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
50 55 60
<210> 22
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 22
Gly Gly Gly Gly Ser
1 5
<210> 23
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 23
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 24
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 24
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 25
<211> 25
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 25
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25
<210> 26
<211> 35
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 26
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
20 25 30
Gly Gly Ser
35
<210> 27
<211> 40
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 27
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
20 25 30
Gly Gly Ser Gly Gly Gly Gly Ser
35 40
<210> 28
<211> 50
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 28
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
20 25 30
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
35 40 45
Gly Ser
50
<210> 29
<211> 55
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 29
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
20 25 30
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
35 40 45
Gly Ser Gly Gly Gly Gly Ser
50 55
<210> 30
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<220>
<221> MOD_RES
<222> (2)..(2)
<223> Aminohexanoyl
<220>
<221> MOD_RES
<222> (5)..(5)
<223> Aminohexanoyl
<220>
<221> MOD_RES
<222> (8)..(8)
<223> Aminohexanoyl
<220>
<221> MOD_RES
<222> (11)..(11)
<223> Aminohexanoyl
<400> 30
Arg Xaa Arg Arg Xaa Arg Arg Xaa Arg Arg Xaa Arg
1 5 10
<210> 31
<211> 20
<212> DNA
<213> Homo sapiens
<400> 31
gagtccgagc agaagaagaa 20
<210> 32
<211> 20
<212> DNA
<213> Homo sapiens
<400> 32
gagtcctagc aggagaagaa 20
<210> 33
<211> 20
<212> DNA
<213> Homo sapiens
<400> 33
gagtctaagc agaagaagaa 20
<210> 34
<211> 12
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 34
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser
1 5 10
<210> 35
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
peptide"
<400> 35
Ala Glu Ala Ala Ala Lys Ala
1 5
<210> 36
<211> 9
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
'LAGLIDADG' family motif peptide"
<400> 36
Leu Ala Gly Leu Ile Asp Ala Asp Gly
1 5
<210> 37
<211> 5
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
'GGDEF' family motif peptide"
<400> 37
Gly Gly Asp Glu Phe
1 5
<210> 38
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 38
gccgcagcga augccguuuc acgaaucguc aggcgg 36
<210> 39
<211> 75
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 39
gcuggagacg uuuuuugaaa cggcgagugc ugcggauagc gaguuucucu uggggaggcg 60
cucgcggcca cuuuu 75
<210> 40
<211> 1388
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 40
Met Ser Leu Asn Arg Ile Tyr Gln Gly Arg Val Ala Ala Val Glu Thr
1 5 10 15
Gly Thr Ala Leu Ala Lys Gly Asn Val Glu Trp Met Pro Ala Ala Gly
20 25 30
Gly Asp Glu Val Leu Trp Gln His His Glu Leu Phe Gln Ala Ala Ile
35 40 45
Asn Tyr Tyr Leu Val Ala Leu Leu Ala Leu Ala Asp Lys Asn Asn Pro
50 55 60
Val Leu Gly Pro Leu Ile Ser Gln Met Asp Asn Pro Gln Ser Pro Tyr
65 70 75 80
His Val Trp Gly Ser Phe Arg Arg Gln Gly Arg Gln Arg Thr Gly Leu
85 90 95
Ser Gln Ala Val Ala Pro Tyr Ile Thr Pro Gly Asn Asn Ala Pro Thr
100 105 110
Leu Asp Glu Val Phe Arg Ser Ile Leu Ala Gly Asn Pro Thr Asp Arg
115 120 125
Ala Thr Leu Asp Ala Ala Leu Met Gln Leu Leu Lys Ala Cys Asp Gly
130 135 140
Ala Gly Ala Ile Gln Gln Glu Gly Arg Ser Tyr Trp Pro Lys Phe Cys
145 150 155 160
Asp Pro Asp Ser Thr Ala Asn Phe Ala Gly Asp Pro Ala Met Leu Arg
165 170 175
Arg Glu Gln His Arg Leu Leu Leu Pro Gln Val Leu His Asp Pro Ala
180 185 190
Ile Thr His Asp Ser Pro Ala Leu Gly Ser Phe Asp Thr Tyr Ser Ile
195 200 205
Ala Thr Pro Asp Thr Arg Thr Pro Gln Leu Thr Gly Pro Lys Ala Arg
210 215 220
Ala Arg Leu Glu Gln Ala Ile Thr Leu Trp Arg Val Arg Leu Pro Glu
225 230 235 240
Ser Ala Ala Asp Phe Asp Arg Leu Ala Ser Ser Leu Lys Lys Ile Pro
245 250 255
Asp Asp Asp Ser Arg Leu Asn Leu Gln Gly Tyr Val Gly Ser Ser Ala
260 265 270
Lys Gly Glu Val Gln Ala Arg Leu Phe Ala Leu Leu Leu Phe Arg His
275 280 285
Leu Glu Arg Ser Ser Phe Thr Leu Gly Leu Leu Arg Ser Ala Thr Pro
290 295 300
Pro Pro Lys Asn Ala Glu Thr Pro Pro Pro Ala Gly Val Pro Leu Pro
305 310 315 320
Ala Ala Ser Ala Ala Asp Pro Val Arg Ile Ala Arg Gly Lys Arg Ser
325 330 335
Phe Val Phe Arg Ala Phe Thr Ser Leu Pro Cys Trp His Gly Gly Asp
340 345 350
Asn Ile His Pro Thr Trp Lys Ser Phe Asp Ile Ala Ala Phe Lys Tyr
355 360 365
Ala Leu Thr Val Ile Asn Gln Ile Glu Glu Lys Thr Lys Glu Arg Gln
370 375 380
Lys Glu Cys Ala Glu Leu Glu Thr Asp Phe Asp Tyr Met His Gly Arg
385 390 395 400
Leu Ala Lys Ile Pro Val Lys Tyr Thr Thr Gly Glu Ala Glu Pro Pro
405 410 415
Pro Ile Leu Ala Asn Asp Leu Arg Ile Pro Leu Leu Arg Glu Leu Leu
420 425 430
Gln Asn Ile Lys Val Asp Thr Ala Leu Thr Asp Gly Glu Ala Val Ser
435 440 445
Tyr Gly Leu Gln Arg Arg Thr Ile Arg Gly Phe Arg Glu Leu Arg Arg
450 455 460
Ile Trp Arg Gly His Ala Pro Ala Gly Thr Val Phe Ser Ser Glu Leu
465 470 475 480
Lys Glu Lys Leu Ala Gly Glu Leu Arg Gln Phe Gln Thr Asp Asn Ser
485 490 495
Thr Thr Ile Gly Ser Val Gln Leu Phe Asn Glu Leu Ile Gln Asn Pro
500 505 510
Lys Tyr Trp Pro Ile Trp Gln Ala Pro Asp Val Glu Thr Ala Arg Gln
515 520 525
Trp Ala Asp Ala Gly Phe Ala Asp Asp Pro Leu Ala Ala Leu Val Gln
530 535 540
Glu Ala Glu Leu Gln Glu Asp Ile Asp Ala Leu Lys Ala Pro Val Lys
545 550 555 560
Leu Thr Pro Ala Asp Pro Glu Tyr Ser Arg Arg Gln Tyr Asp Phe Asn
565 570 575
Ala Val Ser Lys Phe Gly Ala Gly Ser Arg Ser Ala Asn Arg His Glu
580 585 590
Pro Gly Gln Thr Glu Arg Gly His Asn Thr Phe Thr Thr Glu Ile Ala
595 600 605
Ala Arg Asn Ala Ala Asp Gly Asn Arg Trp Arg Ala Thr His Val Arg
610 615 620
Ile His Tyr Ser Ala Pro Arg Leu Leu Arg Asp Gly Leu Arg Arg Pro
625 630 635 640
Asp Thr Asp Gly Asn Glu Ala Leu Glu Ala Val Pro Trp Leu Gln Pro
645 650 655
Met Met Glu Ala Leu Ala Pro Leu Pro Thr Leu Pro Gln Asp Leu Thr
660 665 670
Gly Met Pro Val Phe Leu Met Pro Asp Val Thr Leu Ser Gly Glu Arg
675 680 685
Arg Ile Leu Leu Asn Leu Pro Val Thr Leu Glu Pro Ala Ala Leu Val
690 695 700
Glu Gln Leu Gly Asn Ala Gly Arg Trp Gln Asn Gln Phe Phe Gly Ser
705 710 715 720
Arg Glu Asp Pro Phe Ala Leu Arg Trp Pro Ala Asp Gly Ala Val Lys
725 730 735
Thr Ala Lys Gly Lys Thr His Ile Pro Trp His Gln Asp Arg Asp His
740 745 750
Phe Thr Val Leu Gly Val Asp Leu Gly Thr Arg Asp Ala Gly Ala Leu
755 760 765
Ala Leu Leu Asn Val Thr Ala Gln Lys Pro Ala Lys Pro Val His Arg
770 775 780
Ile Ile Gly Glu Ala Asp Gly Arg Thr Trp Tyr Ala Ser Leu Ala Asp
785 790 795 800
Ala Arg Met Ile Arg Leu Pro Gly Glu Asp Ala Arg Leu Phe Val Arg
805 810 815
Gly Lys Leu Val Gln Glu Pro Tyr Gly Glu Arg Gly Arg Asn Ala Ser
820 825 830
Leu Leu Glu Trp Glu Asp Ala Arg Asn Ile Ile Leu Arg Leu Gly Gln
835 840 845
Asn Pro Asp Glu Leu Leu Gly Ala Asp Pro Arg Arg His Ser Tyr Pro
850 855 860
Glu Ile Asn Asp Lys Leu Leu Val Ala Leu Arg Arg Ala Gln Ala Arg
865 870 875 880
Leu Ala Arg Leu Gln Asn Arg Ser Trp Arg Leu Arg Asp Leu Ala Glu
885 890 895
Ser Asp Lys Ala Leu Asp Glu Ile His Ala Glu Arg Ala Gly Glu Lys
900 905 910
Pro Ser Pro Leu Pro Pro Leu Ala Arg Asp Asp Ala Ile Lys Ser Thr
915 920 925
Asp Glu Ala Leu Leu Ser Gln Arg Asp Ile Ile Arg Arg Ser Phe Val
930 935 940
Gln Ile Ala Asn Leu Ile Leu Pro Leu Arg Gly Arg Arg Trp Glu Trp
945 950 955 960
Arg Pro His Val Glu Val Pro Asp Cys His Ile Leu Ala Gln Ser Asp
965 970 975
Pro Gly Thr Asp Asp Thr Lys Arg Leu Val Ala Gly Gln Arg Gly Ile
980 985 990
Ser His Glu Arg Ile Glu Gln Ile Glu Glu Leu Arg Arg Arg Cys Gln
995 1000 1005
Ser Leu Asn Arg Ala Leu Arg His Lys Pro Gly Glu Arg Pro Val
1010 1015 1020
Leu Gly Arg Pro Ala Lys Gly Glu Glu Ile Ala Asp Pro Cys Pro
1025 1030 1035
Ala Leu Leu Glu Lys Ile Asn Arg Leu Arg Asp Gln Arg Val Asp
1040 1045 1050
Gln Thr Ala His Ala Ile Leu Ala Ala Ala Leu Gly Val Arg Leu
1055 1060 1065
Arg Ala Pro Ser Lys Asp Arg Ala Glu Arg Arg His Arg Asp Ile
1070 1075 1080
His Gly Glu Tyr Glu Arg Phe Arg Ala Pro Ala Asp Phe Val Val
1085 1090 1095
Ile Glu Asn Leu Ser Arg Tyr Leu Ser Ser Gln Asp Arg Ala Arg
1100 1105 1110
Ser Glu Asn Thr Arg Leu Met Gln Trp Cys His Arg Gln Ile Val
1115 1120 1125
Gln Lys Leu Arg Gln Leu Cys Glu Thr Tyr Gly Ile Pro Val Leu
1130 1135 1140
Ala Val Pro Ala Ala Tyr Ser Ser Arg Phe Ser Ser Arg Asp Gly
1145 1150 1155
Ser Ala Gly Phe Arg Ala Val His Leu Thr Pro Asp His Arg His
1160 1165 1170
Arg Met Pro Trp Ser Arg Ile Leu Ala Arg Leu Lys Ala His Glu
1175 1180 1185
Glu Asp Gly Lys Arg Leu Glu Lys Thr Val Leu Asp Glu Ala Arg
1190 1195 1200
Ala Val Arg Gly Leu Phe Asp Arg Leu Asp Arg Phe Asn Ala Gly
1205 1210 1215
His Val Pro Gly Lys Pro Trp Arg Thr Leu Leu Ala Pro Leu Pro
1220 1225 1230
Gly Gly Pro Val Phe Val Pro Leu Gly Asp Ala Thr Pro Met Gln
1235 1240 1245
Ala Asp Leu Asn Ala Ala Ile Asn Ile Ala Leu Arg Gly Ile Ala
1250 1255 1260
Ala Pro Asp Arg His Asp Ile His His Arg Leu Arg Ala Glu Asn
1265 1270 1275
Lys Lys Arg Ile Leu Ser Leu Arg Leu Gly Thr Gln Arg Glu Lys
1280 1285 1290
Ala Arg Trp Pro Gly Gly Ala Pro Ala Val Thr Leu Ser Thr Pro
1295 1300 1305
Asn Asn Gly Ala Ser Pro Glu Asp Ser Asp Ala Leu Pro Glu Arg
1310 1315 1320
Val Ser Asn Leu Phe Val Asp Ile Ala Gly Val Ala Asn Phe Glu
1325 1330 1335
Arg Val Thr Ile Glu Gly Val Ser Gln Lys Phe Ala Thr Gly Arg
1340 1345 1350
Gly Leu Trp Ala Ser Val Lys Gln Arg Ala Trp Asn Arg Val Ala
1355 1360 1365
Arg Leu Asn Glu Thr Val Thr Asp Asn Asn Arg Asn Glu Glu Glu
1370 1375 1380
Asp Asp Ile Pro Met
1385
<210> 41
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 41
guccaagaaa aaagaaauga uacgaggcau uagcac 36
<210> 42
<211> 107
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 42
cuggacgaug ucucuuuuau uucuuuuuuc uuggaucuga guacgagcac ccacauugga 60
cauuucgcau ggugggugcu cguacuauag guaaaacaaa ccuuuuu 107
<210> 43
<211> 1108
<212> PRT
<213> Bacillus thermoamylovorans
<400> 43
Met Ala Thr Arg Ser Phe Ile Leu Lys Ile Glu Pro Asn Glu Glu Val
1 5 10 15
Lys Lys Gly Leu Trp Lys Thr His Glu Val Leu Asn His Gly Ile Ala
20 25 30
Tyr Tyr Met Asn Ile Leu Lys Leu Ile Arg Gln Glu Ala Ile Tyr Glu
35 40 45
His His Glu Gln Asp Pro Lys Asn Pro Lys Lys Val Ser Lys Ala Glu
50 55 60
Ile Gln Ala Glu Leu Trp Asp Phe Val Leu Lys Met Gln Lys Cys Asn
65 70 75 80
Ser Phe Thr His Glu Val Asp Lys Asp Val Val Phe Asn Ile Leu Arg
85 90 95
Glu Leu Tyr Glu Glu Leu Val Pro Ser Ser Val Glu Lys Lys Gly Glu
100 105 110
Ala Asn Gln Leu Ser Asn Lys Phe Leu Tyr Pro Leu Val Asp Pro Asn
115 120 125
Ser Gln Ser Gly Lys Gly Thr Ala Ser Ser Gly Arg Lys Pro Arg Trp
130 135 140
Tyr Asn Leu Lys Ile Ala Gly Asp Pro Ser Trp Glu Glu Glu Lys Lys
145 150 155 160
Lys Trp Glu Glu Asp Lys Lys Lys Asp Pro Leu Ala Lys Ile Leu Gly
165 170 175
Lys Leu Ala Glu Tyr Gly Leu Ile Pro Leu Phe Ile Pro Phe Thr Asp
180 185 190
Ser Asn Glu Pro Ile Val Lys Glu Ile Lys Trp Met Glu Lys Ser Arg
195 200 205
Asn Gln Ser Val Arg Arg Leu Asp Lys Asp Met Phe Ile Gln Ala Leu
210 215 220
Glu Arg Phe Leu Ser Trp Glu Ser Trp Asn Leu Lys Val Lys Glu Glu
225 230 235 240
Tyr Glu Lys Val Glu Lys Glu His Lys Thr Leu Glu Glu Arg Ile Lys
245 250 255
Glu Asp Ile Gln Ala Phe Lys Ser Leu Glu Gln Tyr Glu Lys Glu Arg
260 265 270
Gln Glu Gln Leu Leu Arg Asp Thr Leu Asn Thr Asn Glu Tyr Arg Leu
275 280 285
Ser Lys Arg Gly Leu Arg Gly Trp Arg Glu Ile Ile Gln Lys Trp Leu
290 295 300
Lys Met Asp Glu Asn Glu Pro Ser Glu Lys Tyr Leu Glu Val Phe Lys
305 310 315 320
Asp Tyr Gln Arg Lys His Pro Arg Glu Ala Gly Asp Tyr Ser Val Tyr
325 330 335
Glu Phe Leu Ser Lys Lys Glu Asn His Phe Ile Trp Arg Asn His Pro
340 345 350
Glu Tyr Pro Tyr Leu Tyr Ala Thr Phe Cys Glu Ile Asp Lys Lys Lys
355 360 365
Lys Asp Ala Lys Gln Gln Ala Thr Phe Thr Leu Ala Asp Pro Ile Asn
370 375 380
His Pro Leu Trp Val Arg Phe Glu Glu Arg Ser Gly Ser Asn Leu Asn
385 390 395 400
Lys Tyr Arg Ile Leu Thr Glu Gln Leu His Thr Glu Lys Leu Lys Lys
405 410 415
Lys Leu Thr Val Gln Leu Asp Arg Leu Ile Tyr Pro Thr Glu Ser Gly
420 425 430
Gly Trp Glu Glu Lys Gly Lys Val Asp Ile Val Leu Leu Pro Ser Arg
435 440 445
Gln Phe Tyr Asn Gln Ile Phe Leu Asp Ile Glu Glu Lys Gly Lys His
450 455 460
Ala Phe Thr Tyr Lys Asp Glu Ser Ile Lys Phe Pro Leu Lys Gly Thr
465 470 475 480
Leu Gly Gly Ala Arg Val Gln Phe Asp Arg Asp His Leu Arg Arg Tyr
485 490 495
Pro His Lys Val Glu Ser Gly Asn Val Gly Arg Ile Tyr Phe Asn Met
500 505 510
Thr Val Asn Ile Glu Pro Thr Glu Ser Pro Val Ser Lys Ser Leu Lys
515 520 525
Ile His Arg Asp Asp Phe Pro Lys Phe Val Asn Phe Lys Pro Lys Glu
530 535 540
Leu Thr Glu Trp Ile Lys Asp Ser Lys Gly Lys Lys Leu Lys Ser Gly
545 550 555 560
Ile Glu Ser Leu Glu Ile Gly Leu Arg Val Met Ser Ile Asp Leu Gly
565 570 575
Gln Arg Gln Ala Ala Ala Ala Ser Ile Phe Glu Val Val Asp Gln Lys
580 585 590
Pro Asp Ile Glu Gly Lys Leu Phe Phe Pro Ile Lys Gly Thr Glu Leu
595 600 605
Tyr Ala Val His Arg Ala Ser Phe Asn Ile Lys Leu Pro Gly Glu Thr
610 615 620
Leu Val Lys Ser Arg Glu Val Leu Arg Lys Ala Arg Glu Asp Asn Leu
625 630 635 640
Lys Leu Met Asn Gln Lys Leu Asn Phe Leu Arg Asn Val Leu His Phe
645 650 655
Gln Gln Phe Glu Asp Ile Thr Glu Arg Glu Lys Arg Val Thr Lys Trp
660 665 670
Ile Ser Arg Gln Glu Asn Ser Asp Val Pro Leu Val Tyr Gln Asp Glu
675 680 685
Leu Ile Gln Ile Arg Glu Leu Met Tyr Lys Pro Tyr Lys Asp Trp Val
690 695 700
Ala Phe Leu Lys Gln Leu His Lys Arg Leu Glu Val Glu Ile Gly Lys
705 710 715 720
Glu Val Lys His Trp Arg Lys Ser Leu Ser Asp Gly Arg Lys Gly Leu
725 730 735
Tyr Gly Ile Ser Leu Lys Asn Ile Asp Glu Ile Asp Arg Thr Arg Lys
740 745 750
Phe Leu Leu Arg Trp Ser Leu Arg Pro Thr Glu Pro Gly Glu Val Arg
755 760 765
Arg Leu Glu Pro Gly Gln Arg Phe Ala Ile Asp Gln Leu Asn His Leu
770 775 780
Asn Ala Leu Lys Glu Asp Arg Leu Lys Lys Met Ala Asn Thr Ile Ile
785 790 795 800
Met His Ala Leu Gly Tyr Cys Tyr Asp Val Arg Lys Lys Lys Trp Gln
805 810 815
Ala Lys Asn Pro Ala Cys Gln Ile Ile Leu Phe Glu Asp Leu Ser Asn
820 825 830
Tyr Asn Pro Tyr Glu Glu Arg Ser Arg Phe Glu Asn Ser Lys Leu Met
835 840 845
Lys Trp Ser Arg Arg Glu Ile Pro Arg Gln Val Ala Leu Gln Gly Glu
850 855 860
Ile Tyr Gly Leu Gln Val Gly Glu Val Gly Ala Gln Phe Ser Ser Arg
865 870 875 880
Phe His Ala Lys Thr Gly Ser Pro Gly Ile Arg Cys Ser Val Val Thr
885 890 895
Lys Glu Lys Leu Gln Asp Asn Arg Phe Phe Lys Asn Leu Gln Arg Glu
900 905 910
Gly Arg Leu Thr Leu Asp Lys Ile Ala Val Leu Lys Glu Gly Asp Leu
915 920 925
Tyr Pro Asp Lys Gly Gly Glu Lys Phe Ile Ser Leu Ser Lys Asp Arg
930 935 940
Lys Leu Val Thr Thr His Ala Asp Ile Asn Ala Ala Gln Asn Leu Gln
945 950 955 960
Lys Arg Phe Trp Thr Arg Thr His Gly Phe Tyr Lys Val Tyr Cys Lys
965 970 975
Ala Tyr Gln Val Asp Gly Gln Thr Val Tyr Ile Pro Glu Ser Lys Asp
980 985 990
Gln Lys Gln Lys Ile Ile Glu Glu Phe Gly Glu Gly Tyr Phe Ile Leu
995 1000 1005
Lys Asp Gly Val Tyr Glu Trp Gly Asn Ala Gly Lys Leu Lys Ile
1010 1015 1020
Lys Lys Gly Ser Ser Lys Gln Ser Ser Ser Glu Leu Val Asp Ser
1025 1030 1035
Asp Ile Leu Lys Asp Ser Phe Asp Leu Ala Ser Glu Leu Lys Gly
1040 1045 1050
Glu Lys Leu Met Leu Tyr Arg Asp Pro Ser Gly Asn Val Phe Pro
1055 1060 1065
Ser Asp Lys Trp Met Ala Ala Gly Val Phe Phe Gly Lys Leu Glu
1070 1075 1080
Arg Ile Leu Ile Ser Lys Leu Thr Asn Gln Tyr Ser Ile Ser Thr
1085 1090 1095
Ile Glu Asp Asp Ser Ser Lys Gln Ser Met
1100 1105
<210> 44
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 44
guucgaaagc uuaguggaaa gcuucguggu uagcac 36
<210> 45
<211> 69
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 45
cacggauaau cacgacuuuc cacuaagcuu ucgaauuuua ugaugcgagc auccucucag 60
gucaaaaaa 69
<210> 46
<211> 1108
<212> PRT
<213> Bacillus sp.
<400> 46
Met Ala Ile Arg Ser Ile Lys Leu Lys Leu Lys Thr His Thr Gly Pro
1 5 10 15
Glu Ala Gln Asn Leu Arg Lys Gly Ile Trp Arg Thr His Arg Leu Leu
20 25 30
Asn Glu Gly Val Ala Tyr Tyr Met Lys Met Leu Leu Leu Phe Arg Gln
35 40 45
Glu Ser Thr Gly Glu Arg Pro Lys Glu Glu Leu Gln Glu Glu Leu Ile
50 55 60
Cys His Ile Arg Glu Gln Gln Gln Arg Asn Gln Ala Asp Lys Asn Thr
65 70 75 80
Gln Ala Leu Pro Leu Asp Lys Ala Leu Glu Ala Leu Arg Gln Leu Tyr
85 90 95
Glu Leu Leu Val Pro Ser Ser Val Gly Gln Ser Gly Asp Ala Gln Ile
100 105 110
Ile Ser Arg Lys Phe Leu Ser Pro Leu Val Asp Pro Asn Ser Glu Gly
115 120 125
Gly Lys Gly Thr Ser Lys Ala Gly Ala Lys Pro Thr Trp Gln Lys Lys
130 135 140
Lys Glu Ala Asn Asp Pro Thr Trp Glu Gln Asp Tyr Glu Lys Trp Lys
145 150 155 160
Lys Arg Arg Glu Glu Asp Pro Thr Ala Ser Val Ile Thr Thr Leu Glu
165 170 175
Glu Tyr Gly Ile Arg Pro Ile Phe Pro Leu Tyr Thr Asn Thr Val Thr
180 185 190
Asp Ile Ala Trp Leu Pro Leu Gln Ser Asn Gln Phe Val Arg Thr Trp
195 200 205
Asp Arg Asp Met Leu Gln Gln Ala Ile Glu Arg Leu Leu Ser Trp Glu
210 215 220
Ser Trp Asn Lys Arg Val Gln Glu Glu Tyr Ala Lys Leu Lys Glu Lys
225 230 235 240
Met Ala Gln Leu Asn Glu Gln Leu Glu Gly Gly Gln Glu Trp Ile Ser
245 250 255
Leu Leu Glu Gln Tyr Glu Glu Asn Arg Glu Arg Glu Leu Arg Glu Asn
260 265 270
Met Thr Ala Ala Asn Asp Lys Tyr Arg Ile Thr Lys Arg Gln Met Lys
275 280 285
Gly Trp Asn Glu Leu Tyr Glu Leu Trp Ser Thr Phe Pro Ala Ser Ala
290 295 300
Ser His Glu Gln Tyr Lys Glu Ala Leu Lys Arg Val Gln Gln Arg Leu
305 310 315 320
Arg Gly Arg Phe Gly Asp Ala His Phe Phe Gln Tyr Leu Met Glu Glu
325 330 335
Lys Asn Arg Leu Ile Trp Lys Gly Asn Pro Gln Arg Ile His Tyr Phe
340 345 350
Val Ala Arg Asn Glu Leu Thr Lys Arg Leu Glu Glu Ala Lys Gln Ser
355 360 365
Ala Thr Met Thr Leu Pro Asn Ala Arg Lys His Pro Leu Trp Val Arg
370 375 380
Phe Asp Ala Arg Gly Gly Asn Leu Gln Asp Tyr Tyr Leu Thr Ala Glu
385 390 395 400
Ala Asp Lys Pro Arg Ser Arg Arg Phe Val Thr Phe Ser Gln Leu Ile
405 410 415
Trp Pro Ser Glu Ser Gly Trp Met Glu Lys Lys Asp Val Glu Val Glu
420 425 430
Leu Ala Leu Ser Arg Gln Phe Tyr Gln Gln Val Lys Leu Leu Lys Asn
435 440 445
Asp Lys Gly Lys Gln Lys Ile Glu Phe Lys Asp Lys Gly Ser Gly Ser
450 455 460
Thr Phe Asn Gly His Leu Gly Gly Ala Lys Leu Gln Leu Glu Arg Gly
465 470 475 480
Asp Leu Glu Lys Glu Glu Lys Asn Phe Glu Asp Gly Glu Ile Gly Ser
485 490 495
Val Tyr Leu Asn Val Val Ile Asp Phe Glu Pro Leu Gln Glu Val Lys
500 505 510
Asn Gly Arg Val Gln Ala Pro Tyr Gly Gln Val Leu Gln Leu Ile Arg
515 520 525
Arg Pro Asn Glu Phe Pro Lys Val Thr Thr Tyr Lys Ser Glu Gln Leu
530 535 540
Val Glu Trp Ile Lys Ala Ser Pro Gln His Ser Ala Gly Val Glu Ser
545 550 555 560
Leu Ala Ser Gly Phe Arg Val Met Ser Ile Asp Leu Gly Leu Arg Ala
565 570 575
Ala Ala Ala Thr Ser Ile Phe Ser Val Glu Glu Ser Ser Asp Lys Asn
580 585 590
Ala Ala Asp Phe Ser Tyr Trp Ile Glu Gly Thr Pro Leu Val Ala Val
595 600 605
His Gln Arg Ser Tyr Met Leu Arg Leu Pro Gly Glu Gln Val Glu Lys
610 615 620
Gln Val Met Glu Lys Arg Asp Glu Arg Phe Gln Leu His Gln Arg Val
625 630 635 640
Lys Phe Gln Ile Arg Val Leu Ala Gln Ile Met Arg Met Ala Asn Lys
645 650 655
Gln Tyr Gly Asp Arg Trp Asp Glu Leu Asp Ser Leu Lys Gln Ala Val
660 665 670
Glu Gln Lys Lys Ser Pro Leu Asp Gln Thr Asp Arg Thr Phe Trp Glu
675 680 685
Gly Ile Val Cys Asp Leu Thr Lys Val Leu Pro Arg Asn Glu Ala Asp
690 695 700
Trp Glu Gln Ala Val Val Gln Ile His Arg Lys Ala Glu Glu Tyr Val
705 710 715 720
Gly Lys Ala Val Gln Ala Trp Arg Lys Arg Phe Ala Ala Asp Glu Arg
725 730 735
Lys Gly Ile Ala Gly Leu Ser Met Trp Asn Ile Glu Glu Leu Glu Gly
740 745 750
Leu Arg Lys Leu Leu Ile Ser Trp Ser Arg Arg Thr Arg Asn Pro Gln
755 760 765
Glu Val Asn Arg Phe Glu Arg Gly His Thr Ser His Gln Arg Leu Leu
770 775 780
Thr His Ile Gln Asn Val Lys Glu Asp Arg Leu Lys Gln Leu Ser His
785 790 795 800
Ala Ile Val Met Thr Ala Leu Gly Tyr Val Tyr Asp Glu Arg Lys Gln
805 810 815
Glu Trp Cys Ala Glu Tyr Pro Ala Cys Gln Val Ile Leu Phe Glu Asn
820 825 830
Leu Ser Gln Tyr Arg Ser Asn Leu Asp Arg Ser Thr Lys Glu Asn Ser
835 840 845
Thr Leu Met Lys Trp Ala His Arg Ser Ile Pro Lys Tyr Val His Met
850 855 860
Gln Ala Glu Pro Tyr Gly Ile Gln Ile Gly Asp Val Arg Ala Glu Tyr
865 870 875 880
Ser Ser Arg Phe Tyr Ala Lys Thr Gly Thr Pro Gly Ile Arg Cys Lys
885 890 895
Lys Val Arg Gly Gln Asp Leu Gln Gly Arg Arg Phe Glu Asn Leu Gln
900 905 910
Lys Arg Leu Val Asn Glu Gln Phe Leu Thr Glu Glu Gln Val Lys Gln
915 920 925
Leu Arg Pro Gly Asp Ile Val Pro Asp Asp Ser Gly Glu Leu Phe Met
930 935 940
Thr Leu Thr Asp Gly Ser Gly Ser Lys Glu Val Val Phe Leu Gln Ala
945 950 955 960
Asp Ile Asn Ala Ala His Asn Leu Gln Lys Arg Phe Trp Gln Arg Tyr
965 970 975
Asn Glu Leu Phe Lys Val Ser Cys Arg Val Ile Val Arg Asp Glu Glu
980 985 990
Glu Tyr Leu Val Pro Lys Thr Lys Ser Val Gln Ala Lys Leu Gly Lys
995 1000 1005
Gly Leu Phe Val Lys Lys Ser Asp Thr Ala Trp Lys Asp Val Tyr
1010 1015 1020
Val Trp Asp Ser Gln Ala Lys Leu Lys Gly Lys Thr Thr Phe Thr
1025 1030 1035
Glu Glu Ser Glu Ser Pro Glu Gln Leu Glu Asp Phe Gln Glu Ile
1040 1045 1050
Ile Glu Glu Ala Glu Glu Ala Lys Gly Thr Tyr Arg Thr Leu Phe
1055 1060 1065
Arg Asp Pro Ser Gly Val Phe Phe Pro Glu Ser Val Trp Tyr Pro
1070 1075 1080
Gln Lys Asp Phe Trp Gly Glu Val Lys Arg Lys Leu Tyr Gly Lys
1085 1090 1095
Leu Arg Glu Arg Phe Leu Thr Lys Ala Arg
1100 1105
<210> 47
<211> 35
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 47
guuuugagaa uagcccgaca uagagggcaa uagac 35
<210> 48
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 48
guuaugaaaa cagcccgaca uagagggcaa uagaca 36
<210> 49
<211> 1334
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 49
Met Lys Ile Ser Lys Val Asp His Thr Arg Met Ala Val Ala Lys Gly
1 5 10 15
Asn Gln His Arg Arg Asp Glu Ile Ser Gly Ile Leu Tyr Lys Asp Pro
20 25 30
Thr Lys Thr Gly Ser Ile Asp Phe Asp Glu Arg Phe Lys Lys Leu Asn
35 40 45
Cys Ser Ala Lys Ile Leu Tyr His Val Phe Asn Gly Ile Ala Glu Gly
50 55 60
Ser Asn Lys Tyr Lys Asn Ile Val Asp Lys Val Asn Asn Asn Leu Asp
65 70 75 80
Arg Val Leu Phe Thr Gly Lys Ser Tyr Asp Arg Lys Ser Ile Ile Asp
85 90 95
Ile Asp Thr Val Leu Arg Asn Val Glu Lys Ile Asn Ala Phe Asp Arg
100 105 110
Ile Ser Thr Glu Glu Arg Glu Gln Ile Ile Asp Asp Leu Leu Glu Ile
115 120 125
Gln Leu Arg Lys Gly Leu Arg Lys Gly Lys Ala Gly Leu Arg Glu Val
130 135 140
Leu Leu Ile Gly Ala Gly Val Ile Val Arg Thr Asp Lys Lys Gln Glu
145 150 155 160
Ile Ala Asp Phe Leu Glu Ile Leu Asp Glu Asp Phe Asn Lys Thr Asn
165 170 175
Gln Ala Lys Asn Ile Lys Leu Ser Ile Glu Asn Gln Gly Leu Val Val
180 185 190
Ser Pro Val Ser Arg Gly Glu Glu Arg Ile Phe Asp Val Ser Gly Ala
195 200 205
Gln Lys Gly Lys Ser Ser Lys Lys Ala Gln Glu Lys Glu Ala Leu Ser
210 215 220
Ala Phe Leu Leu Asp Tyr Ala Asp Leu Asp Lys Asn Val Arg Phe Glu
225 230 235 240
Tyr Leu Arg Lys Ile Arg Arg Leu Ile Asn Leu Tyr Phe Tyr Val Lys
245 250 255
Asn Asp Asp Val Met Ser Leu Thr Glu Ile Pro Ala Glu Val Asn Leu
260 265 270
Glu Lys Asp Phe Asp Ile Trp Arg Asp His Glu Gln Arg Lys Glu Glu
275 280 285
Asn Gly Asp Phe Val Gly Cys Pro Asp Ile Leu Leu Ala Asp Arg Asp
290 295 300
Val Lys Lys Ser Asn Ser Lys Gln Val Lys Ile Ala Glu Arg Gln Leu
305 310 315 320
Arg Glu Ser Ile Arg Glu Lys Asn Ile Lys Arg Tyr Arg Phe Ser Ile
325 330 335
Lys Thr Ile Glu Lys Asp Asp Gly Thr Tyr Phe Phe Ala Asn Lys Gln
340 345 350
Ile Ser Val Phe Trp Ile His Arg Ile Glu Asn Ala Val Glu Arg Ile
355 360 365
Leu Gly Ser Ile Asn Asp Lys Lys Leu Tyr Arg Leu Arg Leu Gly Tyr
370 375 380
Leu Gly Glu Lys Val Trp Lys Asp Ile Leu Asn Phe Leu Ser Ile Lys
385 390 395 400
Tyr Ile Ala Val Gly Lys Ala Val Phe Asn Phe Ala Met Asp Asp Leu
405 410 415
Gln Glu Lys Asp Arg Asp Ile Glu Pro Gly Lys Ile Ser Glu Asn Ala
420 425 430
Val Asn Gly Leu Thr Ser Phe Asp Tyr Glu Gln Ile Lys Ala Asp Glu
435 440 445
Met Leu Gln Arg Glu Val Ala Val Asn Val Ala Phe Ala Ala Asn Asn
450 455 460
Leu Ala Arg Val Thr Val Asp Ile Pro Gln Asn Gly Glu Lys Glu Asp
465 470 475 480
Ile Leu Leu Trp Asn Lys Ser Asp Ile Lys Lys Tyr Lys Lys Asn Ser
485 490 495
Lys Lys Gly Ile Leu Lys Ser Ile Leu Gln Phe Phe Gly Gly Ala Ser
500 505 510
Thr Trp Asn Met Lys Met Phe Glu Ile Ala Tyr His Asp Gln Pro Gly
515 520 525
Asp Tyr Glu Glu Asn Tyr Leu Tyr Asp Ile Ile Gln Ile Ile Tyr Ser
530 535 540
Leu Arg Asn Lys Ser Phe His Phe Lys Thr Tyr Asp His Gly Asp Lys
545 550 555 560
Asn Trp Asn Arg Glu Leu Ile Gly Lys Met Ile Glu His Asp Ala Glu
565 570 575
Arg Val Ile Ser Val Glu Arg Glu Lys Phe His Ser Asn Asn Leu Pro
580 585 590
Met Phe Tyr Lys Asp Ala Asp Leu Lys Lys Ile Leu Asp Leu Leu Tyr
595 600 605
Ser Asp Tyr Ala Gly Arg Ala Ser Gln Val Pro Ala Phe Asn Thr Val
610 615 620
Leu Val Arg Lys Asn Phe Pro Glu Phe Leu Arg Lys Asp Met Gly Tyr
625 630 635 640
Lys Val His Phe Asn Asn Pro Glu Val Glu Asn Gln Trp His Ser Ala
645 650 655
Val Tyr Tyr Leu Tyr Lys Glu Ile Tyr Tyr Asn Leu Phe Leu Arg Asp
660 665 670
Lys Glu Val Lys Asn Leu Phe Tyr Thr Ser Leu Lys Asn Ile Arg Ser
675 680 685
Glu Val Ser Asp Lys Lys Gln Lys Leu Ala Ser Asp Asp Phe Ala Ser
690 695 700
Arg Cys Glu Glu Ile Glu Asp Arg Ser Leu Pro Glu Ile Cys Gln Ile
705 710 715 720
Ile Met Thr Glu Tyr Asn Ala Gln Asn Phe Gly Asn Arg Lys Val Lys
725 730 735
Ser Gln Arg Val Ile Glu Lys Asn Lys Asp Ile Phe Arg His Tyr Lys
740 745 750
Met Leu Leu Ile Lys Thr Leu Ala Gly Ala Phe Ser Leu Tyr Leu Lys
755 760 765
Gln Glu Arg Phe Ala Phe Ile Gly Lys Ala Thr Pro Ile Pro Tyr Glu
770 775 780
Thr Thr Asp Val Lys Asn Phe Leu Pro Glu Trp Lys Ser Gly Met Tyr
785 790 795 800
Ala Ser Phe Val Glu Glu Ile Lys Asn Asn Leu Asp Leu Gln Glu Trp
805 810 815
Tyr Ile Val Gly Arg Phe Leu Asn Gly Arg Met Leu Asn Gln Leu Ala
820 825 830
Gly Ser Leu Arg Ser Tyr Ile Gln Tyr Ala Glu Asp Ile Glu Arg Arg
835 840 845
Ala Ala Glu Asn Arg Asn Lys Leu Phe Ser Lys Pro Asp Glu Lys Ile
850 855 860
Glu Ala Cys Lys Lys Ala Val Arg Val Leu Asp Leu Cys Ile Lys Ile
865 870 875 880
Ser Thr Arg Ile Ser Ala Glu Phe Thr Asp Tyr Phe Asp Ser Glu Asp
885 890 895
Asp Tyr Ala Asp Tyr Leu Glu Lys Tyr Leu Lys Tyr Gln Asp Asp Ala
900 905 910
Ile Lys Glu Leu Ser Gly Ser Ser Tyr Ala Ala Leu Asp His Phe Cys
915 920 925
Asn Lys Asp Asp Leu Lys Phe Asp Ile Tyr Val Asn Ala Gly Gln Lys
930 935 940
Pro Ile Leu Gln Arg Asn Ile Val Met Ala Lys Leu Phe Gly Pro Asp
945 950 955 960
Asn Ile Leu Ser Glu Val Met Glu Lys Val Thr Glu Ser Ala Ile Arg
965 970 975
Glu Tyr Tyr Asp Tyr Leu Lys Lys Val Ser Gly Tyr Arg Val Arg Gly
980 985 990
Lys Cys Ser Thr Glu Lys Glu Gln Glu Asp Leu Leu Lys Phe Gln Arg
995 1000 1005
Leu Lys Asn Ala Val Glu Phe Arg Asp Val Thr Glu Tyr Ala Glu
1010 1015 1020
Val Ile Asn Glu Leu Leu Gly Gln Leu Ile Ser Trp Ser Tyr Leu
1025 1030 1035
Arg Glu Arg Asp Leu Leu Tyr Phe Gln Leu Gly Phe His Tyr Met
1040 1045 1050
Cys Leu Lys Asn Lys Ser Phe Lys Pro Ala Glu Tyr Val Asp Ile
1055 1060 1065
Arg Arg Asn Asn Gly Thr Ile Ile His Asn Ala Ile Leu Tyr Gln
1070 1075 1080
Ile Val Ser Met Tyr Ile Asn Gly Leu Asp Phe Tyr Ser Cys Asp
1085 1090 1095
Lys Glu Gly Lys Thr Leu Lys Pro Ile Glu Thr Gly Lys Gly Val
1100 1105 1110
Gly Ser Lys Ile Gly Gln Phe Ile Lys Tyr Ser Gln Tyr Leu Tyr
1115 1120 1125
Asn Asp Pro Ser Tyr Lys Leu Glu Ile Tyr Asn Ala Gly Leu Glu
1130 1135 1140
Val Phe Glu Asn Ile Asp Glu His Asp Asn Ile Thr Asp Leu Arg
1145 1150 1155
Lys Tyr Val Asp His Phe Lys Tyr Tyr Ala Tyr Gly Asn Lys Met
1160 1165 1170
Ser Leu Leu Asp Leu Tyr Ser Glu Phe Phe Asp Arg Phe Phe Thr
1175 1180 1185
Tyr Asp Met Lys Tyr Gln Lys Asn Val Val Asn Val Leu Glu Asn
1190 1195 1200
Ile Leu Leu Arg His Phe Val Ile Phe Tyr Pro Lys Phe Gly Ser
1205 1210 1215
Gly Lys Lys Asp Val Gly Ile Arg Asp Cys Lys Lys Glu Arg Ala
1220 1225 1230
Gln Ile Glu Ile Ser Glu Gln Ser Leu Thr Ser Glu Asp Phe Met
1235 1240 1245
Phe Lys Leu Asp Asp Lys Ala Gly Glu Glu Ala Lys Lys Phe Pro
1250 1255 1260
Ala Arg Asp Glu Arg Tyr Leu Gln Thr Ile Ala Lys Leu Leu Tyr
1265 1270 1275
Tyr Pro Asn Glu Ile Glu Asp Met Asn Arg Phe Met Lys Lys Gly
1280 1285 1290
Glu Thr Ile Asn Lys Lys Val Gln Phe Asn Arg Lys Lys Lys Ile
1295 1300 1305
Thr Arg Lys Gln Lys Asn Asn Ser Ser Asn Glu Val Leu Ser Ser
1310 1315 1320
Thr Met Gly Tyr Leu Phe Lys Asn Ile Lys Leu
1325 1330
<210> 50
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 50
guuuuagucc ucuuucauau agagguaguc ucuuac 36
<210> 51
<211> 99
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 51
augaaaagag gacuaaaacu gaaagaggac uaaaacacca gauguggaua acuauauuag 60
uggcuauuaa aaauucgucg auauuagaga ggaaacuuu 99
<210> 52
<211> 1120
<212> PRT
<213> Listeria seeligeri
<400> 52
Met Trp Ile Ser Ile Lys Thr Leu Ile His His Leu Gly Val Leu Phe
1 5 10 15
Phe Cys Asp Tyr Met Tyr Asn Arg Arg Glu Lys Lys Ile Ile Glu Val
20 25 30
Lys Thr Met Arg Ile Thr Lys Val Glu Val Asp Arg Lys Lys Val Leu
35 40 45
Ile Ser Arg Asp Lys Asn Gly Gly Lys Leu Val Tyr Glu Asn Glu Met
50 55 60
Gln Asp Asn Thr Glu Gln Ile Met His His Lys Lys Ser Ser Phe Tyr
65 70 75 80
Lys Ser Val Val Asn Lys Thr Ile Cys Arg Pro Glu Gln Lys Gln Met
85 90 95
Lys Lys Leu Val His Gly Leu Leu Gln Glu Asn Ser Gln Glu Lys Ile
100 105 110
Lys Val Ser Asp Val Thr Lys Leu Asn Ile Ser Asn Phe Leu Asn His
115 120 125
Arg Phe Lys Lys Ser Leu Tyr Tyr Phe Pro Glu Asn Ser Pro Asp Lys
130 135 140
Ser Glu Glu Tyr Arg Ile Glu Ile Asn Leu Ser Gln Leu Leu Glu Asp
145 150 155 160
Ser Leu Lys Lys Gln Gln Gly Thr Phe Ile Cys Trp Glu Ser Phe Ser
165 170 175
Lys Asp Met Glu Leu Tyr Ile Asn Trp Ala Glu Asn Tyr Ile Ser Ser
180 185 190
Lys Thr Lys Leu Ile Lys Lys Ser Ile Arg Asn Asn Arg Ile Gln Ser
195 200 205
Thr Glu Ser Arg Ser Gly Gln Leu Met Asp Arg Tyr Met Lys Asp Ile
210 215 220
Leu Asn Lys Asn Lys Pro Phe Asp Ile Gln Ser Val Ser Glu Lys Tyr
225 230 235 240
Gln Leu Glu Lys Leu Thr Ser Ala Leu Lys Ala Thr Phe Lys Glu Ala
245 250 255
Lys Lys Asn Asp Lys Glu Ile Asn Tyr Lys Leu Lys Ser Thr Leu Gln
260 265 270
Asn His Glu Arg Gln Ile Ile Glu Glu Leu Lys Glu Asn Ser Glu Leu
275 280 285
Asn Gln Phe Asn Ile Glu Ile Arg Lys His Leu Glu Thr Tyr Phe Pro
290 295 300
Ile Lys Lys Thr Asn Arg Lys Val Gly Asp Ile Arg Asn Leu Glu Ile
305 310 315 320
Gly Glu Ile Gln Lys Ile Val Asn His Arg Leu Lys Asn Lys Ile Val
325 330 335
Gln Arg Ile Leu Gln Glu Gly Lys Leu Ala Ser Tyr Glu Ile Glu Ser
340 345 350
Thr Val Asn Ser Asn Ser Leu Gln Lys Ile Lys Ile Glu Glu Ala Phe
355 360 365
Ala Leu Lys Phe Ile Asn Ala Cys Leu Phe Ala Ser Asn Asn Leu Arg
370 375 380
Asn Met Val Tyr Pro Val Cys Lys Lys Asp Ile Leu Met Ile Gly Glu
385 390 395 400
Phe Lys Asn Ser Phe Lys Glu Ile Lys His Lys Lys Phe Ile Arg Gln
405 410 415
Trp Ser Gln Phe Phe Ser Gln Glu Ile Thr Val Asp Asp Ile Glu Leu
420 425 430
Ala Ser Trp Gly Leu Arg Gly Ala Ile Ala Pro Ile Arg Asn Glu Ile
435 440 445
Ile His Leu Lys Lys His Ser Trp Lys Lys Phe Phe Asn Asn Pro Thr
450 455 460
Phe Lys Val Lys Lys Ser Lys Ile Ile Asn Gly Lys Thr Lys Asp Val
465 470 475 480
Thr Ser Glu Phe Leu Tyr Lys Glu Thr Leu Phe Lys Asp Tyr Phe Tyr
485 490 495
Ser Glu Leu Asp Ser Val Pro Glu Leu Ile Ile Asn Lys Met Glu Ser
500 505 510
Ser Lys Ile Leu Asp Tyr Tyr Ser Ser Asp Gln Leu Asn Gln Val Phe
515 520 525
Thr Ile Pro Asn Phe Glu Leu Ser Leu Leu Thr Ser Ala Val Pro Phe
530 535 540
Ala Pro Ser Phe Lys Arg Val Tyr Leu Lys Gly Phe Asp Tyr Gln Asn
545 550 555 560
Gln Asp Glu Ala Gln Pro Asp Tyr Asn Leu Lys Leu Asn Ile Tyr Asn
565 570 575
Glu Lys Ala Phe Asn Ser Glu Ala Phe Gln Ala Gln Tyr Ser Leu Phe
580 585 590
Lys Met Val Tyr Tyr Gln Val Phe Leu Pro Gln Phe Thr Thr Asn Asn
595 600 605
Asp Leu Phe Lys Ser Ser Val Asp Phe Ile Leu Thr Leu Asn Lys Glu
610 615 620
Arg Lys Gly Tyr Ala Lys Ala Phe Gln Asp Ile Arg Lys Met Asn Lys
625 630 635 640
Asp Glu Lys Pro Ser Glu Tyr Met Ser Tyr Ile Gln Ser Gln Leu Met
645 650 655
Leu Tyr Gln Lys Lys Gln Glu Glu Lys Glu Lys Ile Asn His Phe Glu
660 665 670
Lys Phe Ile Asn Gln Val Phe Ile Lys Gly Phe Asn Ser Phe Ile Glu
675 680 685
Lys Asn Arg Leu Thr Tyr Ile Cys His Pro Thr Lys Asn Thr Val Pro
690 695 700
Glu Asn Asp Asn Ile Glu Ile Pro Phe His Thr Asp Met Asp Asp Ser
705 710 715 720
Asn Ile Ala Phe Trp Leu Met Cys Lys Leu Leu Asp Ala Lys Gln Leu
725 730 735
Ser Glu Leu Arg Asn Glu Met Ile Lys Phe Ser Cys Ser Leu Gln Ser
740 745 750
Thr Glu Glu Ile Ser Thr Phe Thr Lys Ala Arg Glu Val Ile Gly Leu
755 760 765
Ala Leu Leu Asn Gly Glu Lys Gly Cys Asn Asp Trp Lys Glu Leu Phe
770 775 780
Asp Asp Lys Glu Ala Trp Lys Lys Asn Met Ser Leu Tyr Val Ser Glu
785 790 795 800
Glu Leu Leu Gln Ser Leu Pro Tyr Thr Gln Glu Asp Gly Gln Thr Pro
805 810 815
Val Ile Asn Arg Ser Ile Asp Leu Val Lys Lys Tyr Gly Thr Glu Thr
820 825 830
Ile Leu Glu Lys Leu Phe Ser Ser Ser Asp Asp Tyr Lys Val Ser Ala
835 840 845
Lys Asp Ile Ala Lys Leu His Glu Tyr Asp Val Thr Glu Lys Ile Ala
850 855 860
Gln Gln Glu Ser Leu His Lys Gln Trp Ile Glu Lys Pro Gly Leu Ala
865 870 875 880
Arg Asp Ser Ala Trp Thr Lys Lys Tyr Gln Asn Val Ile Asn Asp Ile
885 890 895
Ser Asn Tyr Gln Trp Ala Lys Thr Lys Val Glu Leu Thr Gln Val Arg
900 905 910
His Leu His Gln Leu Thr Ile Asp Leu Leu Ser Arg Leu Ala Gly Tyr
915 920 925
Met Ser Ile Ala Asp Arg Asp Phe Gln Phe Ser Ser Asn Tyr Ile Leu
930 935 940
Glu Arg Glu Asn Ser Glu Tyr Arg Val Thr Ser Trp Ile Leu Leu Ser
945 950 955 960
Glu Asn Lys Asn Lys Asn Lys Tyr Asn Asp Tyr Glu Leu Tyr Asn Leu
965 970 975
Lys Asn Ala Ser Ile Lys Val Ser Ser Lys Asn Asp Pro Gln Leu Lys
980 985 990
Val Asp Leu Lys Gln Leu Arg Leu Thr Leu Glu Tyr Leu Glu Leu Phe
995 1000 1005
Asp Asn Arg Leu Lys Glu Lys Arg Asn Asn Ile Ser His Phe Asn
1010 1015 1020
Tyr Leu Asn Gly Gln Leu Gly Asn Ser Ile Leu Glu Leu Phe Asp
1025 1030 1035
Asp Ala Arg Asp Val Leu Ser Tyr Asp Arg Lys Leu Lys Asn Ala
1040 1045 1050
Val Ser Lys Ser Leu Lys Glu Ile Leu Ser Ser His Gly Met Glu
1055 1060 1065
Val Thr Phe Lys Pro Leu Tyr Gln Thr Asn His His Leu Lys Ile
1070 1075 1080
Asp Lys Leu Gln Pro Lys Lys Ile His His Leu Gly Glu Lys Ser
1085 1090 1095
Thr Val Ser Ser Asn Gln Val Ser Asn Glu Tyr Cys Gln Leu Val
1100 1105 1110
Arg Thr Leu Leu Thr Met Lys
1115 1120
<210> 53
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 53
guuuuagucc ccuucguuuu ugggguaguc uaaauc 36
<210> 54
<211> 113
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 54
gauuuagagc accccaaaag uaaugaaaau uugcaauuaa auaaggaaua uuaaaaaaau 60
gugauuuuaa aaaaauugaa gaaauuaaau gaaaaauugu ccaaguaaaa aaa 113
<210> 55
<211> 70
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 55
auuuagauua ccccuuuaau uuauuuuacc auauuuuucu cauaaugcaa acuaauauuc 60
caaaauuuuu 70
<210> 56
<211> 1389
<212> PRT
<213> Leptotrichia wadei
<400> 56
Met Gly Asn Leu Phe Gly His Lys Arg Trp Tyr Glu Val Arg Asp Lys
1 5 10 15
Lys Asp Phe Lys Ile Lys Arg Lys Val Lys Val Lys Arg Asn Tyr Asp
20 25 30
Gly Asn Lys Tyr Ile Leu Asn Ile Asn Glu Asn Asn Asn Lys Glu Lys
35 40 45
Ile Asp Asn Asn Lys Phe Ile Arg Lys Tyr Ile Asn Tyr Lys Lys Asn
50 55 60
Asp Asn Ile Leu Lys Glu Phe Thr Arg Lys Phe His Ala Gly Asn Ile
65 70 75 80
Leu Phe Lys Leu Lys Gly Lys Glu Gly Ile Ile Arg Ile Glu Asn Asn
85 90 95
Asp Asp Phe Leu Glu Thr Glu Glu Val Val Leu Tyr Ile Glu Ala Tyr
100 105 110
Gly Lys Ser Glu Lys Leu Lys Ala Leu Gly Ile Thr Lys Lys Lys Ile
115 120 125
Ile Asp Glu Ala Ile Arg Gln Gly Ile Thr Lys Asp Asp Lys Lys Ile
130 135 140
Glu Ile Lys Arg Gln Glu Asn Glu Glu Glu Ile Glu Ile Asp Ile Arg
145 150 155 160
Asp Glu Tyr Thr Asn Lys Thr Leu Asn Asp Cys Ser Ile Ile Leu Arg
165 170 175
Ile Ile Glu Asn Asp Glu Leu Glu Thr Lys Lys Ser Ile Tyr Glu Ile
180 185 190
Phe Lys Asn Ile Asn Met Ser Leu Tyr Lys Ile Ile Glu Lys Ile Ile
195 200 205
Glu Asn Glu Thr Glu Lys Val Phe Glu Asn Arg Tyr Tyr Glu Glu His
210 215 220
Leu Arg Glu Lys Leu Leu Lys Asp Asp Lys Ile Asp Val Ile Leu Thr
225 230 235 240
Asn Phe Met Glu Ile Arg Glu Lys Ile Lys Ser Asn Leu Glu Ile Leu
245 250 255
Gly Phe Val Lys Phe Tyr Leu Asn Val Gly Gly Asp Lys Lys Lys Ser
260 265 270
Lys Asn Lys Lys Met Leu Val Glu Lys Ile Leu Asn Ile Asn Val Asp
275 280 285
Leu Thr Val Glu Asp Ile Ala Asp Phe Val Ile Lys Glu Leu Glu Phe
290 295 300
Trp Asn Ile Thr Lys Arg Ile Glu Lys Val Lys Lys Val Asn Asn Glu
305 310 315 320
Phe Leu Glu Lys Arg Arg Asn Arg Thr Tyr Ile Lys Ser Tyr Val Leu
325 330 335
Leu Asp Lys His Glu Lys Phe Lys Ile Glu Arg Glu Asn Lys Lys Asp
340 345 350
Lys Ile Val Lys Phe Phe Val Glu Asn Ile Lys Asn Asn Ser Ile Lys
355 360 365
Glu Lys Ile Glu Lys Ile Leu Ala Glu Phe Lys Ile Asp Glu Leu Ile
370 375 380
Lys Lys Leu Glu Lys Glu Leu Lys Lys Gly Asn Cys Asp Thr Glu Ile
385 390 395 400
Phe Gly Ile Phe Lys Lys His Tyr Lys Val Asn Phe Asp Ser Lys Lys
405 410 415
Phe Ser Lys Lys Ser Asp Glu Glu Lys Glu Leu Tyr Lys Ile Ile Tyr
420 425 430
Arg Tyr Leu Lys Gly Arg Ile Glu Lys Ile Leu Val Asn Glu Gln Lys
435 440 445
Val Arg Leu Lys Lys Met Glu Lys Ile Glu Ile Glu Lys Ile Leu Asn
450 455 460
Glu Ser Ile Leu Ser Glu Lys Ile Leu Lys Arg Val Lys Gln Tyr Thr
465 470 475 480
Leu Glu His Ile Met Tyr Leu Gly Lys Leu Arg His Asn Asp Ile Asp
485 490 495
Met Thr Thr Val Asn Thr Asp Asp Phe Ser Arg Leu His Ala Lys Glu
500 505 510
Glu Leu Asp Leu Glu Leu Ile Thr Phe Phe Ala Ser Thr Asn Met Glu
515 520 525
Leu Asn Lys Ile Phe Ser Arg Glu Asn Ile Asn Asn Asp Glu Asn Ile
530 535 540
Asp Phe Phe Gly Gly Asp Arg Glu Lys Asn Tyr Val Leu Asp Lys Lys
545 550 555 560
Ile Leu Asn Ser Lys Ile Lys Ile Ile Arg Asp Leu Asp Phe Ile Asp
565 570 575
Asn Lys Asn Asn Ile Thr Asn Asn Phe Ile Arg Lys Phe Thr Lys Ile
580 585 590
Gly Thr Asn Glu Arg Asn Arg Ile Leu His Ala Ile Ser Lys Glu Arg
595 600 605
Asp Leu Gln Gly Thr Gln Asp Asp Tyr Asn Lys Val Ile Asn Ile Ile
610 615 620
Gln Asn Leu Lys Ile Ser Asp Glu Glu Val Ser Lys Ala Leu Asn Leu
625 630 635 640
Asp Val Val Phe Lys Asp Lys Lys Asn Ile Ile Thr Lys Ile Asn Asp
645 650 655
Ile Lys Ile Ser Glu Glu Asn Asn Asn Asp Ile Lys Tyr Leu Pro Ser
660 665 670
Phe Ser Lys Val Leu Pro Glu Ile Leu Asn Leu Tyr Arg Asn Asn Pro
675 680 685
Lys Asn Glu Pro Phe Asp Thr Ile Glu Thr Glu Lys Ile Val Leu Asn
690 695 700
Ala Leu Ile Tyr Val Asn Lys Glu Leu Tyr Lys Lys Leu Ile Leu Glu
705 710 715 720
Asp Asp Leu Glu Glu Asn Glu Ser Lys Asn Ile Phe Leu Gln Glu Leu
725 730 735
Lys Lys Thr Leu Gly Asn Ile Asp Glu Ile Asp Glu Asn Ile Ile Glu
740 745 750
Asn Tyr Tyr Lys Asn Ala Gln Ile Ser Ala Ser Lys Gly Asn Asn Lys
755 760 765
Ala Ile Lys Lys Tyr Gln Lys Lys Val Ile Glu Cys Tyr Ile Gly Tyr
770 775 780
Leu Arg Lys Asn Tyr Glu Glu Leu Phe Asp Phe Ser Asp Phe Lys Met
785 790 795 800
Asn Ile Gln Glu Ile Lys Lys Gln Ile Lys Asp Ile Asn Asp Asn Lys
805 810 815
Thr Tyr Glu Arg Ile Thr Val Lys Thr Ser Asp Lys Thr Ile Val Ile
820 825 830
Asn Asp Asp Phe Glu Tyr Ile Ile Ser Ile Phe Ala Leu Leu Asn Ser
835 840 845
Asn Ala Val Ile Asn Lys Ile Arg Asn Arg Phe Phe Ala Thr Ser Val
850 855 860
Trp Leu Asn Thr Ser Glu Tyr Gln Asn Ile Ile Asp Ile Leu Asp Glu
865 870 875 880
Ile Met Gln Leu Asn Thr Leu Arg Asn Glu Cys Ile Thr Glu Asn Trp
885 890 895
Asn Leu Asn Leu Glu Glu Phe Ile Gln Lys Met Lys Glu Ile Glu Lys
900 905 910
Asp Phe Asp Asp Phe Lys Ile Gln Thr Lys Lys Glu Ile Phe Asn Asn
915 920 925
Tyr Tyr Glu Asp Ile Lys Asn Asn Ile Leu Thr Glu Phe Lys Asp Asp
930 935 940
Ile Asn Gly Cys Asp Val Leu Glu Lys Lys Leu Glu Lys Ile Val Ile
945 950 955 960
Phe Asp Asp Glu Thr Lys Phe Glu Ile Asp Lys Lys Ser Asn Ile Leu
965 970 975
Gln Asp Glu Gln Arg Lys Leu Ser Asn Ile Asn Lys Lys Asp Leu Lys
980 985 990
Lys Lys Val Asp Gln Tyr Ile Lys Asp Lys Asp Gln Glu Ile Lys Ser
995 1000 1005
Lys Ile Leu Cys Arg Ile Ile Phe Asn Ser Asp Phe Leu Lys Lys
1010 1015 1020
Tyr Lys Lys Glu Ile Asp Asn Leu Ile Glu Asp Met Glu Ser Glu
1025 1030 1035
Asn Glu Asn Lys Phe Gln Glu Ile Tyr Tyr Pro Lys Glu Arg Lys
1040 1045 1050
Asn Glu Leu Tyr Ile Tyr Lys Lys Asn Leu Phe Leu Asn Ile Gly
1055 1060 1065
Asn Pro Asn Phe Asp Lys Ile Tyr Gly Leu Ile Ser Asn Asp Ile
1070 1075 1080
Lys Met Ala Asp Ala Lys Phe Leu Phe Asn Ile Asp Gly Lys Asn
1085 1090 1095
Ile Arg Lys Asn Lys Ile Ser Glu Ile Asp Ala Ile Leu Lys Asn
1100 1105 1110
Leu Asn Asp Lys Leu Asn Gly Tyr Ser Lys Glu Tyr Lys Glu Lys
1115 1120 1125
Tyr Ile Lys Lys Leu Lys Glu Asn Asp Asp Phe Phe Ala Lys Asn
1130 1135 1140
Ile Gln Asn Lys Asn Tyr Lys Ser Phe Glu Lys Asp Tyr Asn Arg
1145 1150 1155
Val Ser Glu Tyr Lys Lys Ile Arg Asp Leu Val Glu Phe Asn Tyr
1160 1165 1170
Leu Asn Lys Ile Glu Ser Tyr Leu Ile Asp Ile Asn Trp Lys Leu
1175 1180 1185
Ala Ile Gln Met Ala Arg Phe Glu Arg Asp Met His Tyr Ile Val
1190 1195 1200
Asn Gly Leu Arg Glu Leu Gly Ile Ile Lys Leu Ser Gly Tyr Asn
1205 1210 1215
Thr Gly Ile Ser Arg Ala Tyr Pro Lys Arg Asn Gly Ser Asp Gly
1220 1225 1230
Phe Tyr Thr Thr Thr Ala Tyr Tyr Lys Phe Phe Asp Glu Glu Ser
1235 1240 1245
Tyr Lys Lys Phe Glu Lys Ile Cys Tyr Gly Phe Gly Ile Asp Leu
1250 1255 1260
Ser Glu Asn Ser Glu Ile Asn Lys Pro Glu Asn Glu Ser Ile Arg
1265 1270 1275
Asn Tyr Ile Ser His Phe Tyr Ile Val Arg Asn Pro Phe Ala Asp
1280 1285 1290
Tyr Ser Ile Ala Glu Gln Ile Asp Arg Val Ser Asn Leu Leu Ser
1295 1300 1305
Tyr Ser Thr Arg Tyr Asn Asn Ser Thr Tyr Ala Ser Val Phe Glu
1310 1315 1320
Val Phe Lys Lys Asp Val Asn Leu Asp Tyr Asp Glu Leu Lys Lys
1325 1330 1335
Lys Phe Lys Leu Ile Gly Asn Asn Asp Ile Leu Glu Arg Leu Met
1340 1345 1350
Lys Pro Lys Lys Val Ser Val Leu Glu Leu Glu Ser Tyr Asn Ser
1355 1360 1365
Asp Tyr Ile Lys Asn Leu Ile Ile Glu Leu Leu Thr Lys Ile Glu
1370 1375 1380
Asn Thr Asn Asp Thr Leu
1385
<210> 57
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 57
guuuuagucc ccuucgauau uggggugguc uauauc 36
<210> 58
<211> 95
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 58
auugaugugg uauacuaaaa auggaaaauu guauuuuuga uuagaaagau guaaaauuga 60
uuuaauuuaa aaauauuuua uuagauuaaa guaga 95
<210> 59
<211> 1300
<212> PRT
<213> Leptotrichia shahii
<400> 59
Met Ser Ile Tyr Gln Glu Phe Val Asn Lys Tyr Ser Leu Ser Lys Thr
1 5 10 15
Leu Arg Phe Glu Leu Ile Pro Gln Gly Lys Thr Leu Glu Asn Ile Lys
20 25 30
Ala Arg Gly Leu Ile Leu Asp Asp Glu Lys Arg Ala Lys Asp Tyr Lys
35 40 45
Lys Ala Lys Gln Ile Ile Asp Lys Tyr His Gln Phe Phe Ile Glu Glu
50 55 60
Ile Leu Ser Ser Val Cys Ile Ser Glu Asp Leu Leu Gln Asn Tyr Ser
65 70 75 80
Asp Val Tyr Phe Lys Leu Lys Lys Ser Asp Asp Asp Asn Leu Gln Lys
85 90 95
Asp Phe Lys Ser Ala Lys Asp Thr Ile Lys Lys Gln Ile Ser Glu Tyr
100 105 110
Ile Lys Asp Ser Glu Lys Phe Lys Asn Leu Phe Asn Gln Asn Leu Ile
115 120 125
Asp Ala Lys Lys Gly Gln Glu Ser Asp Leu Ile Leu Trp Leu Lys Gln
130 135 140
Ser Lys Asp Asn Gly Ile Glu Leu Phe Lys Ala Asn Ser Asp Ile Thr
145 150 155 160
Asp Ile Asp Glu Ala Leu Glu Ile Ile Lys Ser Phe Lys Gly Trp Thr
165 170 175
Thr Tyr Phe Lys Gly Phe His Glu Asn Arg Lys Asn Val Tyr Ser Ser
180 185 190
Asn Asp Ile Pro Thr Ser Ile Ile Tyr Arg Ile Val Asp Asp Asn Leu
195 200 205
Pro Lys Phe Leu Glu Asn Lys Ala Lys Tyr Glu Ser Leu Lys Asp Lys
210 215 220
Ala Pro Glu Ala Ile Asn Tyr Glu Gln Ile Lys Lys Asp Leu Ala Glu
225 230 235 240
Glu Leu Thr Phe Asp Ile Asp Tyr Lys Thr Ser Glu Val Asn Gln Arg
245 250 255
Val Phe Ser Leu Asp Glu Val Phe Glu Ile Ala Asn Phe Asn Asn Tyr
260 265 270
Leu Asn Gln Ser Gly Ile Thr Lys Phe Asn Thr Ile Ile Gly Gly Lys
275 280 285
Phe Val Asn Gly Glu Asn Thr Lys Arg Lys Gly Ile Asn Glu Tyr Ile
290 295 300
Asn Leu Tyr Ser Gln Gln Ile Asn Asp Lys Thr Leu Lys Lys Tyr Lys
305 310 315 320
Met Ser Val Leu Phe Lys Gln Ile Leu Ser Asp Thr Glu Ser Lys Ser
325 330 335
Phe Val Ile Asp Lys Leu Glu Asp Asp Ser Asp Val Val Thr Thr Met
340 345 350
Gln Ser Phe Tyr Glu Gln Ile Ala Ala Phe Lys Thr Val Glu Glu Lys
355 360 365
Ser Ile Lys Glu Thr Leu Ser Leu Leu Phe Asp Asp Leu Lys Ala Gln
370 375 380
Lys Leu Asp Leu Ser Lys Ile Tyr Phe Lys Asn Asp Lys Ser Leu Thr
385 390 395 400
Asp Leu Ser Gln Gln Val Phe Asp Asp Tyr Ser Val Ile Gly Thr Ala
405 410 415
Val Leu Glu Tyr Ile Thr Gln Gln Ile Ala Pro Lys Asn Leu Asp Asn
420 425 430
Pro Ser Lys Lys Glu Gln Glu Leu Ile Ala Lys Lys Thr Glu Lys Ala
435 440 445
Lys Tyr Leu Ser Leu Glu Thr Ile Lys Leu Ala Leu Glu Glu Phe Asn
450 455 460
Lys His Arg Asp Ile Asp Lys Gln Cys Arg Phe Glu Glu Ile Leu Ala
465 470 475 480
Asn Phe Ala Ala Ile Pro Met Ile Phe Asp Glu Ile Ala Gln Asn Lys
485 490 495
Asp Asn Leu Ala Gln Ile Ser Ile Lys Tyr Gln Asn Gln Gly Lys Lys
500 505 510
Asp Leu Leu Gln Ala Ser Ala Glu Asp Asp Val Lys Ala Ile Lys Asp
515 520 525
Leu Leu Asp Gln Thr Asn Asn Leu Leu His Lys Leu Lys Ile Phe His
530 535 540
Ile Ser Gln Ser Glu Asp Lys Ala Asn Ile Leu Asp Lys Asp Glu His
545 550 555 560
Phe Tyr Leu Val Phe Glu Glu Cys Tyr Phe Glu Leu Ala Asn Ile Val
565 570 575
Pro Leu Tyr Asn Lys Ile Arg Asn Tyr Ile Thr Gln Lys Pro Tyr Ser
580 585 590
Asp Glu Lys Phe Lys Leu Asn Phe Glu Asn Ser Thr Leu Ala Asn Gly
595 600 605
Trp Asp Lys Asn Lys Glu Pro Asp Asn Thr Ala Ile Leu Phe Ile Lys
610 615 620
Asp Asp Lys Tyr Tyr Leu Gly Val Met Asn Lys Lys Asn Asn Lys Ile
625 630 635 640
Phe Asp Asp Lys Ala Ile Lys Glu Asn Lys Gly Glu Gly Tyr Lys Lys
645 650 655
Ile Val Tyr Lys Leu Leu Pro Gly Ala Asn Lys Met Leu Pro Lys Val
660 665 670
Phe Phe Ser Ala Lys Ser Ile Lys Phe Tyr Asn Pro Ser Glu Asp Ile
675 680 685
Leu Arg Ile Arg Asn His Ser Thr His Thr Lys Asn Gly Ser Pro Gln
690 695 700
Lys Gly Tyr Glu Lys Phe Glu Phe Asn Ile Glu Asp Cys Arg Lys Phe
705 710 715 720
Ile Asp Phe Tyr Lys Gln Ser Ile Ser Lys His Pro Glu Trp Lys Asp
725 730 735
Phe Gly Phe Arg Phe Ser Asp Thr Gln Arg Tyr Asn Ser Ile Asp Glu
740 745 750
Phe Tyr Arg Glu Val Glu Asn Gln Gly Tyr Lys Leu Thr Phe Glu Asn
755 760 765
Ile Ser Glu Ser Tyr Ile Asp Ser Val Val Asn Gln Gly Lys Leu Tyr
770 775 780
Leu Phe Gln Ile Tyr Asn Lys Asp Phe Ser Ala Tyr Ser Lys Gly Arg
785 790 795 800
Pro Asn Leu His Thr Leu Tyr Trp Lys Ala Leu Phe Asp Glu Arg Asn
805 810 815
Leu Gln Asp Val Val Tyr Lys Leu Asn Gly Glu Ala Glu Leu Phe Tyr
820 825 830
Arg Lys Gln Ser Ile Pro Lys Lys Ile Thr His Pro Ala Lys Glu Ala
835 840 845
Ile Ala Asn Lys Asn Lys Asp Asn Pro Lys Lys Glu Ser Val Phe Glu
850 855 860
Tyr Asp Leu Ile Lys Asp Lys Arg Phe Thr Glu Asp Lys Phe Phe Phe
865 870 875 880
His Cys Pro Ile Thr Ile Asn Phe Lys Ser Ser Gly Ala Asn Lys Phe
885 890 895
Asn Asp Glu Ile Asn Leu Leu Leu Lys Glu Lys Ala Asn Asp Val His
900 905 910
Ile Leu Ser Ile Asp Arg Gly Glu Arg His Leu Ala Tyr Tyr Thr Leu
915 920 925
Val Asp Gly Lys Gly Asn Ile Ile Lys Gln Asp Thr Phe Asn Ile Ile
930 935 940
Gly Asn Asp Arg Met Lys Thr Asn Tyr His Asp Lys Leu Ala Ala Ile
945 950 955 960
Glu Lys Asp Arg Asp Ser Ala Arg Lys Asp Trp Lys Lys Ile Asn Asn
965 970 975
Ile Lys Glu Met Lys Glu Gly Tyr Leu Ser Gln Val Val His Glu Ile
980 985 990
Ala Lys Leu Val Ile Glu Tyr Asn Ala Ile Val Val Phe Glu Asp Leu
995 1000 1005
Asn Phe Gly Phe Lys Arg Gly Arg Phe Lys Val Glu Lys Gln Val
1010 1015 1020
Tyr Gln Lys Leu Glu Lys Met Leu Ile Glu Lys Leu Asn Tyr Leu
1025 1030 1035
Val Phe Lys Asp Asn Glu Phe Asp Lys Thr Gly Gly Val Leu Arg
1040 1045 1050
Ala Tyr Gln Leu Thr Ala Pro Phe Glu Thr Phe Lys Lys Met Gly
1055 1060 1065
Lys Gln Thr Gly Ile Ile Tyr Tyr Val Pro Ala Gly Phe Thr Ser
1070 1075 1080
Lys Ile Cys Pro Val Thr Gly Phe Val Asn Gln Leu Tyr Pro Lys
1085 1090 1095
Tyr Glu Ser Val Ser Lys Ser Gln Glu Phe Phe Ser Lys Phe Asp
1100 1105 1110
Lys Ile Cys Tyr Asn Leu Asp Lys Gly Tyr Phe Glu Phe Ser Phe
1115 1120 1125
Asp Tyr Lys Asn Phe Gly Asp Lys Ala Ala Lys Gly Lys Trp Thr
1130 1135 1140
Ile Ala Ser Phe Gly Ser Arg Leu Ile Asn Phe Arg Asn Ser Asp
1145 1150 1155
Lys Asn His Asn Trp Asp Thr Arg Glu Val Tyr Pro Thr Lys Glu
1160 1165 1170
Leu Glu Lys Leu Leu Lys Asp Tyr Ser Ile Glu Tyr Gly His Gly
1175 1180 1185
Glu Cys Ile Lys Ala Ala Ile Cys Gly Glu Ser Asp Lys Lys Phe
1190 1195 1200
Phe Ala Lys Leu Thr Ser Val Leu Asn Thr Ile Leu Gln Met Arg
1205 1210 1215
Asn Ser Lys Thr Gly Thr Glu Leu Asp Tyr Leu Ile Ser Pro Val
1220 1225 1230
Ala Asp Val Asn Gly Asn Phe Phe Asp Ser Arg Gln Ala Pro Lys
1235 1240 1245
Asn Met Pro Gln Asp Ala Asp Ala Asn Gly Ala Tyr His Ile Gly
1250 1255 1260
Leu Lys Gly Leu Met Leu Leu Gly Arg Ile Lys Asn Asn Gln Glu
1265 1270 1275
Gly Lys Lys Leu Asn Leu Val Ile Lys Asn Glu Glu Tyr Phe Glu
1280 1285 1290
Phe Val Gln Asn Arg Asn Asn
1295 1300
<210> 60
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 60
gucuaagaac uuuaaauaau uucuacuguu guagau 36
<210> 61
<211> 71
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 61
aucuacaaaa uuauaaacua aauaaagauu cuuauaauaa cuuuauauau aaucgaaaug 60
uagagaauuu u 71
<210> 62
<211> 1300
<212> PRT
<213> Francisella ularensis
<400> 62
Met Ser Ile Tyr Gln Glu Phe Val Asn Lys Tyr Ser Leu Ser Lys Thr
1 5 10 15
Leu Arg Phe Glu Leu Ile Pro Gln Gly Lys Thr Leu Glu Asn Ile Lys
20 25 30
Ala Arg Gly Leu Ile Leu Asp Asp Glu Lys Arg Ala Lys Asp Tyr Lys
35 40 45
Lys Ala Lys Gln Ile Ile Asp Lys Tyr His Gln Phe Phe Ile Glu Glu
50 55 60
Ile Leu Ser Ser Val Cys Ile Ser Glu Asp Leu Leu Gln Asn Tyr Ser
65 70 75 80
Asp Val Tyr Phe Lys Leu Lys Lys Ser Asp Asp Asp Asn Leu Gln Lys
85 90 95
Asp Phe Lys Ser Ala Lys Asp Thr Ile Lys Lys Gln Ile Ser Glu Tyr
100 105 110
Ile Lys Asp Ser Glu Lys Phe Lys Asn Leu Phe Asn Gln Asn Leu Ile
115 120 125
Asp Ala Lys Lys Gly Gln Glu Ser Asp Leu Ile Leu Trp Leu Lys Gln
130 135 140
Ser Lys Asp Asn Gly Ile Glu Leu Phe Lys Ala Asn Ser Asp Ile Thr
145 150 155 160
Asp Ile Asp Glu Ala Leu Glu Ile Ile Lys Ser Phe Lys Gly Trp Thr
165 170 175
Thr Tyr Phe Lys Gly Phe His Glu Asn Arg Lys Asn Val Tyr Ser Ser
180 185 190
Asn Asp Ile Pro Thr Ser Ile Ile Tyr Arg Ile Val Asp Asp Asn Leu
195 200 205
Pro Lys Phe Leu Glu Asn Lys Ala Lys Tyr Glu Ser Leu Lys Asp Lys
210 215 220
Ala Pro Glu Ala Ile Asn Tyr Glu Gln Ile Lys Lys Asp Leu Ala Glu
225 230 235 240
Glu Leu Thr Phe Asp Ile Asp Tyr Lys Thr Ser Glu Val Asn Gln Arg
245 250 255
Val Phe Ser Leu Asp Glu Val Phe Glu Ile Ala Asn Phe Asn Asn Tyr
260 265 270
Leu Asn Gln Ser Gly Ile Thr Lys Phe Asn Thr Ile Ile Gly Gly Lys
275 280 285
Phe Val Asn Gly Glu Asn Thr Lys Arg Lys Gly Ile Asn Glu Tyr Ile
290 295 300
Asn Leu Tyr Ser Gln Gln Ile Asn Asp Lys Thr Leu Lys Lys Tyr Lys
305 310 315 320
Met Ser Val Leu Phe Lys Gln Ile Leu Ser Asp Thr Glu Ser Lys Ser
325 330 335
Phe Val Ile Asp Lys Leu Glu Asp Asp Ser Asp Val Val Thr Thr Met
340 345 350
Gln Ser Phe Tyr Glu Gln Ile Ala Ala Phe Lys Thr Val Glu Glu Lys
355 360 365
Ser Ile Lys Glu Thr Leu Ser Leu Leu Phe Asp Asp Leu Lys Ala Gln
370 375 380
Lys Leu Asp Leu Ser Lys Ile Tyr Phe Lys Asn Asp Lys Ser Leu Thr
385 390 395 400
Asp Leu Ser Gln Gln Val Phe Asp Asp Tyr Ser Val Ile Gly Thr Ala
405 410 415
Val Leu Glu Tyr Ile Thr Gln Gln Ile Ala Pro Lys Asn Leu Asp Asn
420 425 430
Pro Ser Lys Lys Glu Gln Glu Leu Ile Ala Lys Lys Thr Glu Lys Ala
435 440 445
Lys Tyr Leu Ser Leu Glu Thr Ile Lys Leu Ala Leu Glu Glu Phe Asn
450 455 460
Lys His Arg Asp Ile Asp Lys Gln Cys Arg Phe Glu Glu Ile Leu Ala
465 470 475 480
Asn Phe Ala Ala Ile Pro Met Ile Phe Asp Glu Ile Ala Gln Asn Lys
485 490 495
Asp Asn Leu Ala Gln Ile Ser Ile Lys Tyr Gln Asn Gln Gly Lys Lys
500 505 510
Asp Leu Leu Gln Ala Ser Ala Glu Asp Asp Val Lys Ala Ile Lys Asp
515 520 525
Leu Leu Asp Gln Thr Asn Asn Leu Leu His Lys Leu Lys Ile Phe His
530 535 540
Ile Ser Gln Ser Glu Asp Lys Ala Asn Ile Leu Asp Lys Asp Glu His
545 550 555 560
Phe Tyr Leu Val Phe Glu Glu Cys Tyr Phe Glu Leu Ala Asn Ile Val
565 570 575
Pro Leu Tyr Asn Lys Ile Arg Asn Tyr Ile Thr Gln Lys Pro Tyr Ser
580 585 590
Asp Glu Lys Phe Lys Leu Asn Phe Glu Asn Ser Thr Leu Ala Asn Gly
595 600 605
Trp Asp Lys Asn Lys Glu Pro Asp Asn Thr Ala Ile Leu Phe Ile Lys
610 615 620
Asp Asp Lys Tyr Tyr Leu Gly Val Met Asn Lys Lys Asn Asn Lys Ile
625 630 635 640
Phe Asp Asp Lys Ala Ile Lys Glu Asn Lys Gly Glu Gly Tyr Lys Lys
645 650 655
Ile Val Tyr Lys Leu Leu Pro Gly Ala Asn Lys Met Leu Pro Lys Val
660 665 670
Phe Phe Ser Ala Lys Ser Ile Lys Phe Tyr Asn Pro Ser Glu Asp Ile
675 680 685
Leu Arg Ile Arg Asn His Ser Thr His Thr Lys Asn Gly Ser Pro Gln
690 695 700
Lys Gly Tyr Glu Lys Phe Glu Phe Asn Ile Glu Asp Cys Arg Lys Phe
705 710 715 720
Ile Asp Phe Tyr Lys Gln Ser Ile Ser Lys His Pro Glu Trp Lys Asp
725 730 735
Phe Gly Phe Arg Phe Ser Asp Thr Gln Arg Tyr Asn Ser Ile Asp Glu
740 745 750
Phe Tyr Arg Glu Val Glu Asn Gln Gly Tyr Lys Leu Thr Phe Glu Asn
755 760 765
Ile Ser Glu Ser Tyr Ile Asp Ser Val Val Asn Gln Gly Lys Leu Tyr
770 775 780
Leu Phe Gln Ile Tyr Asn Lys Asp Phe Ser Ala Tyr Ser Lys Gly Arg
785 790 795 800
Pro Asn Leu His Thr Leu Tyr Trp Lys Ala Leu Phe Asp Glu Arg Asn
805 810 815
Leu Gln Asp Val Val Tyr Lys Leu Asn Gly Glu Ala Glu Leu Phe Tyr
820 825 830
Arg Lys Gln Ser Ile Pro Lys Lys Ile Thr His Pro Ala Lys Glu Ala
835 840 845
Ile Ala Asn Lys Asn Lys Asp Asn Pro Lys Lys Glu Ser Val Phe Glu
850 855 860
Tyr Asp Leu Ile Lys Asp Lys Arg Phe Thr Glu Asp Lys Phe Phe Phe
865 870 875 880
His Cys Pro Ile Thr Ile Asn Phe Lys Ser Ser Gly Ala Asn Lys Phe
885 890 895
Asn Asp Glu Ile Asn Leu Leu Leu Lys Glu Lys Ala Asn Asp Val His
900 905 910
Ile Leu Ser Ile Asp Arg Gly Glu Arg His Leu Ala Tyr Tyr Thr Leu
915 920 925
Val Asp Gly Lys Gly Asn Ile Ile Lys Gln Asp Thr Phe Asn Ile Ile
930 935 940
Gly Asn Asp Arg Met Lys Thr Asn Tyr His Asp Lys Leu Ala Ala Ile
945 950 955 960
Glu Lys Asp Arg Asp Ser Ala Arg Lys Asp Trp Lys Lys Ile Asn Asn
965 970 975
Ile Lys Glu Met Lys Glu Gly Tyr Leu Ser Gln Val Val His Glu Ile
980 985 990
Ala Lys Leu Val Ile Glu Tyr Asn Ala Ile Val Val Phe Glu Asp Leu
995 1000 1005
Asn Phe Gly Phe Lys Arg Gly Arg Phe Lys Val Glu Lys Gln Val
1010 1015 1020
Tyr Gln Lys Leu Glu Lys Met Leu Ile Glu Lys Leu Asn Tyr Leu
1025 1030 1035
Val Phe Lys Asp Asn Glu Phe Asp Lys Thr Gly Gly Val Leu Arg
1040 1045 1050
Ala Tyr Gln Leu Thr Ala Pro Phe Glu Thr Phe Lys Lys Met Gly
1055 1060 1065
Lys Gln Thr Gly Ile Ile Tyr Tyr Val Pro Ala Gly Phe Thr Ser
1070 1075 1080
Lys Ile Cys Pro Val Thr Gly Phe Val Asn Gln Leu Tyr Pro Lys
1085 1090 1095
Tyr Glu Ser Val Ser Lys Ser Gln Glu Phe Phe Ser Lys Phe Asp
1100 1105 1110
Lys Ile Cys Tyr Asn Leu Asp Lys Gly Tyr Phe Glu Phe Ser Phe
1115 1120 1125
Asp Tyr Lys Asn Phe Gly Asp Lys Ala Ala Lys Gly Lys Trp Thr
1130 1135 1140
Ile Ala Ser Phe Gly Ser Arg Leu Ile Asn Phe Arg Asn Ser Asp
1145 1150 1155
Lys Asn His Asn Trp Asp Thr Arg Glu Val Tyr Pro Thr Lys Glu
1160 1165 1170
Leu Glu Lys Leu Leu Lys Asp Tyr Ser Ile Glu Tyr Gly His Gly
1175 1180 1185
Glu Cys Ile Lys Ala Ala Ile Cys Gly Glu Ser Asp Lys Lys Phe
1190 1195 1200
Phe Ala Lys Leu Thr Ser Val Leu Asn Thr Ile Leu Gln Met Arg
1205 1210 1215
Asn Ser Lys Thr Gly Thr Glu Leu Asp Tyr Leu Ile Ser Pro Val
1220 1225 1230
Ala Asp Val Asn Gly Asn Phe Phe Asp Ser Arg Gln Ala Pro Lys
1235 1240 1245
Asn Met Pro Gln Asp Ala Asp Ala Asn Gly Ala Tyr His Ile Gly
1250 1255 1260
Leu Lys Gly Leu Met Leu Leu Gly Arg Ile Lys Asn Asn Gln Glu
1265 1270 1275
Gly Lys Lys Leu Asn Leu Val Ile Lys Asn Glu Glu Tyr Phe Glu
1280 1285 1290
Phe Val Gln Asn Arg Asn Asn
1295 1300
<210> 63
<211> 138
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 63
aaaaggccgg cggccacgaa aaaggccggc caggcaaaaa agaaaaaggg atcctaccca 60
tacgatgttc cagattacgc ttatccctac gacgtgcctg attatgcata cccatatgat 120
gtccccgact atgcctaa 138
<210> 64
<211> 1388
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 64
Met Ser Leu Asn Arg Ile Tyr Gln Gly Arg Val Ala Ala Val Glu Thr
1 5 10 15
Gly Thr Ala Leu Ala Lys Gly Asn Val Glu Trp Met Pro Ala Ala Gly
20 25 30
Gly Asp Glu Val Leu Trp Gln His His Glu Leu Phe Gln Ala Ala Ile
35 40 45
Asn Tyr Tyr Leu Val Ala Leu Leu Ala Leu Ala Asp Lys Asn Asn Pro
50 55 60
Val Leu Gly Pro Leu Ile Ser Gln Met Asp Asn Pro Gln Ser Pro Tyr
65 70 75 80
His Val Trp Gly Ser Phe Arg Arg Gln Gly Arg Gln Arg Thr Gly Leu
85 90 95
Ser Gln Ala Val Ala Pro Tyr Ile Thr Pro Gly Asn Asn Ala Pro Thr
100 105 110
Leu Asp Glu Val Phe Arg Ser Ile Leu Ala Gly Asn Pro Thr Asp Arg
115 120 125
Ala Thr Leu Asp Ala Ala Leu Met Gln Leu Leu Lys Ala Cys Asp Gly
130 135 140
Ala Gly Ala Ile Gln Gln Glu Gly Arg Ser Tyr Trp Pro Lys Phe Cys
145 150 155 160
Asp Pro Asp Ser Thr Ala Asn Phe Ala Gly Asp Pro Ala Met Leu Arg
165 170 175
Arg Glu Gln His Arg Leu Leu Leu Pro Gln Val Leu His Asp Pro Ala
180 185 190
Ile Thr His Asp Ser Pro Ala Leu Gly Ser Phe Asp Thr Tyr Ser Ile
195 200 205
Ala Thr Pro Asp Thr Arg Thr Pro Gln Leu Thr Gly Pro Lys Ala Arg
210 215 220
Ala Arg Leu Glu Gln Ala Ile Thr Leu Trp Arg Val Arg Leu Pro Glu
225 230 235 240
Ser Ala Ala Asp Phe Asp Arg Leu Ala Ser Ser Leu Lys Lys Ile Pro
245 250 255
Asp Asp Asp Ser Arg Leu Asn Leu Gln Gly Tyr Val Gly Ser Ser Ala
260 265 270
Lys Gly Glu Val Gln Ala Arg Leu Phe Ala Leu Leu Leu Phe Arg His
275 280 285
Leu Glu Arg Ser Ser Phe Thr Leu Gly Leu Leu Arg Ser Ala Thr Pro
290 295 300
Pro Pro Lys Asn Ala Glu Thr Pro Pro Pro Ala Gly Val Pro Leu Pro
305 310 315 320
Ala Ala Ser Ala Ala Asp Pro Val Arg Ile Ala Arg Gly Lys Arg Ser
325 330 335
Phe Val Phe Arg Ala Phe Thr Ser Leu Pro Cys Trp His Gly Gly Asp
340 345 350
Asn Ile His Pro Thr Trp Lys Ser Phe Asp Ile Ala Ala Phe Lys Tyr
355 360 365
Ala Leu Thr Val Ile Asn Gln Ile Glu Glu Lys Thr Lys Glu Arg Gln
370 375 380
Lys Glu Cys Ala Glu Leu Glu Thr Asp Phe Asp Tyr Met His Gly Arg
385 390 395 400
Leu Ala Lys Ile Pro Val Lys Tyr Thr Thr Gly Glu Ala Glu Pro Pro
405 410 415
Pro Ile Leu Ala Asn Asp Leu Arg Ile Pro Leu Leu Arg Glu Leu Leu
420 425 430
Gln Asn Ile Lys Val Asp Thr Ala Leu Thr Asp Gly Glu Ala Val Ser
435 440 445
Tyr Gly Leu Gln Arg Arg Thr Ile Arg Gly Phe Arg Glu Leu Arg Arg
450 455 460
Ile Trp Arg Gly His Ala Pro Ala Gly Thr Val Phe Ser Ser Glu Leu
465 470 475 480
Lys Glu Lys Leu Ala Gly Glu Leu Arg Gln Phe Gln Thr Asp Asn Ser
485 490 495
Thr Thr Ile Gly Ser Val Gln Leu Phe Asn Glu Leu Ile Gln Asn Pro
500 505 510
Lys Tyr Trp Pro Ile Trp Gln Ala Pro Asp Val Glu Thr Ala Arg Gln
515 520 525
Trp Ala Asp Ala Gly Phe Ala Asp Asp Pro Leu Ala Ala Leu Val Gln
530 535 540
Glu Ala Glu Leu Gln Glu Asp Ile Asp Ala Leu Lys Ala Pro Val Lys
545 550 555 560
Leu Thr Pro Ala Asp Pro Glu Tyr Ser Arg Arg Gln Tyr Asp Phe Asn
565 570 575
Ala Val Ser Lys Phe Gly Ala Gly Ser Arg Ser Ala Asn Arg His Glu
580 585 590
Pro Gly Gln Thr Glu Arg Gly His Asn Thr Phe Thr Thr Glu Ile Ala
595 600 605
Ala Arg Asn Ala Ala Asp Gly Asn Arg Trp Arg Ala Thr His Val Arg
610 615 620
Ile His Tyr Ser Ala Pro Arg Leu Leu Arg Asp Gly Leu Arg Arg Pro
625 630 635 640
Asp Thr Asp Gly Asn Glu Ala Leu Glu Ala Val Pro Trp Leu Gln Pro
645 650 655
Met Met Glu Ala Leu Ala Pro Leu Pro Thr Leu Pro Gln Asp Leu Thr
660 665 670
Gly Met Pro Val Phe Leu Met Pro Asp Val Thr Leu Ser Gly Glu Arg
675 680 685
Arg Ile Leu Leu Asn Leu Pro Val Thr Leu Glu Pro Ala Ala Leu Val
690 695 700
Glu Gln Leu Gly Asn Ala Gly Arg Trp Gln Asn Gln Phe Phe Gly Ser
705 710 715 720
Arg Glu Asp Pro Phe Ala Leu Arg Trp Pro Ala Asp Gly Ala Val Lys
725 730 735
Thr Ala Lys Gly Lys Thr His Ile Pro Trp His Gln Asp Arg Asp His
740 745 750
Phe Thr Val Leu Gly Val Asp Leu Gly Thr Arg Asp Ala Gly Ala Leu
755 760 765
Ala Leu Leu Asn Val Thr Ala Gln Lys Pro Ala Lys Pro Val His Arg
770 775 780
Ile Ile Gly Glu Ala Asp Gly Arg Thr Trp Tyr Ala Ser Leu Ala Asp
785 790 795 800
Ala Arg Met Ile Arg Leu Pro Gly Glu Asp Ala Arg Leu Phe Val Arg
805 810 815
Gly Lys Leu Val Gln Glu Pro Tyr Gly Glu Arg Gly Arg Asn Ala Ser
820 825 830
Leu Leu Glu Trp Glu Asp Ala Arg Asn Ile Ile Leu Arg Leu Gly Gln
835 840 845
Asn Pro Asp Glu Leu Leu Gly Ala Asp Pro Arg Arg His Ser Tyr Pro
850 855 860
Glu Ile Asn Asp Lys Leu Leu Val Ala Leu Arg Arg Ala Gln Ala Arg
865 870 875 880
Leu Ala Arg Leu Gln Asn Arg Ser Trp Arg Leu Arg Asp Leu Ala Glu
885 890 895
Ser Asp Lys Ala Leu Asp Glu Ile His Ala Glu Arg Ala Gly Glu Lys
900 905 910
Pro Ser Pro Leu Pro Pro Leu Ala Arg Asp Asp Ala Ile Lys Ser Thr
915 920 925
Asp Glu Ala Leu Leu Ser Gln Arg Asp Ile Ile Arg Arg Ser Phe Val
930 935 940
Gln Ile Ala Asn Leu Ile Leu Pro Leu Arg Gly Arg Arg Trp Glu Trp
945 950 955 960
Arg Pro His Val Glu Val Pro Asp Cys His Ile Leu Ala Gln Ser Asp
965 970 975
Pro Gly Thr Asp Asp Thr Lys Arg Leu Val Ala Gly Gln Arg Gly Ile
980 985 990
Ser His Glu Arg Ile Glu Gln Ile Glu Glu Leu Arg Arg Arg Cys Gln
995 1000 1005
Ser Leu Asn Arg Ala Leu Arg His Lys Pro Gly Glu Arg Pro Val
1010 1015 1020
Leu Gly Arg Pro Ala Lys Gly Glu Glu Ile Ala Asp Pro Cys Pro
1025 1030 1035
Ala Leu Leu Glu Lys Ile Asn Arg Leu Arg Asp Gln Arg Val Asp
1040 1045 1050
Gln Thr Ala His Ala Ile Leu Ala Ala Ala Leu Gly Val Arg Leu
1055 1060 1065
Arg Ala Pro Ser Lys Asp Arg Ala Glu Arg Arg His Arg Asp Ile
1070 1075 1080
His Gly Glu Tyr Glu Arg Phe Arg Ala Pro Ala Asp Phe Val Val
1085 1090 1095
Ile Glu Asn Leu Ser Arg Tyr Leu Ser Ser Gln Asp Arg Ala Arg
1100 1105 1110
Ser Glu Asn Thr Arg Leu Met Gln Trp Cys His Arg Gln Ile Val
1115 1120 1125
Gln Lys Leu Arg Gln Leu Cys Glu Thr Tyr Gly Ile Pro Val Leu
1130 1135 1140
Ala Val Pro Ala Ala Tyr Ser Ser Arg Phe Ser Ser Arg Asp Gly
1145 1150 1155
Ser Ala Gly Phe Arg Ala Val His Leu Thr Pro Asp His Arg His
1160 1165 1170
Arg Met Pro Trp Ser Arg Ile Leu Ala Arg Leu Lys Ala His Glu
1175 1180 1185
Glu Asp Gly Lys Arg Leu Glu Lys Thr Val Leu Asp Glu Ala Arg
1190 1195 1200
Ala Val Arg Gly Leu Phe Asp Arg Leu Asp Arg Phe Asn Ala Gly
1205 1210 1215
His Val Pro Gly Lys Pro Trp Arg Thr Leu Leu Ala Pro Leu Pro
1220 1225 1230
Gly Gly Pro Val Phe Val Pro Leu Gly Asp Ala Thr Pro Met Gln
1235 1240 1245
Ala Asp Leu Asn Ala Ala Ile Asn Ile Ala Leu Arg Gly Ile Ala
1250 1255 1260
Ala Pro Asp Arg His Asp Ile His His Arg Leu Arg Ala Glu Asn
1265 1270 1275
Lys Lys Arg Ile Leu Ser Leu Arg Leu Gly Thr Gln Arg Glu Lys
1280 1285 1290
Ala Arg Trp Pro Gly Gly Ala Pro Ala Val Thr Leu Ser Thr Pro
1295 1300 1305
Asn Asn Gly Ala Ser Pro Glu Asp Ser Asp Ala Leu Pro Glu Arg
1310 1315 1320
Val Ser Asn Leu Phe Val Asp Ile Ala Gly Val Ala Asn Phe Glu
1325 1330 1335
Arg Val Thr Ile Glu Gly Val Ser Gln Lys Phe Ala Thr Gly Arg
1340 1345 1350
Gly Leu Trp Ala Ser Val Lys Gln Arg Ala Trp Asn Arg Val Ala
1355 1360 1365
Arg Leu Asn Glu Thr Val Thr Asp Asn Asn Arg Asn Glu Glu Glu
1370 1375 1380
Asp Asp Ile Pro Met
1385
<210> 65
<211> 1108
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 65
Met Ala Thr Arg Ser Phe Ile Leu Lys Ile Glu Pro Asn Glu Glu Val
1 5 10 15
Lys Lys Gly Leu Trp Lys Thr His Glu Val Leu Asn His Gly Ile Ala
20 25 30
Tyr Tyr Met Asn Ile Leu Lys Leu Ile Arg Gln Glu Ala Ile Tyr Glu
35 40 45
His His Glu Gln Asp Pro Lys Asn Pro Lys Lys Val Ser Lys Ala Glu
50 55 60
Ile Gln Ala Glu Leu Trp Asp Phe Val Leu Lys Met Gln Lys Cys Asn
65 70 75 80
Ser Phe Thr His Glu Val Asp Lys Asp Val Val Phe Asn Ile Leu Arg
85 90 95
Glu Leu Tyr Glu Glu Leu Val Pro Ser Ser Val Glu Lys Lys Gly Glu
100 105 110
Ala Asn Gln Leu Ser Asn Lys Phe Leu Tyr Pro Leu Val Asp Pro Asn
115 120 125
Ser Gln Ser Gly Lys Gly Thr Ala Ser Ser Gly Arg Lys Pro Arg Trp
130 135 140
Tyr Asn Leu Lys Ile Ala Gly Asp Pro Ser Trp Glu Glu Glu Lys Lys
145 150 155 160
Lys Trp Glu Glu Asp Lys Lys Lys Asp Pro Leu Ala Lys Ile Leu Gly
165 170 175
Lys Leu Ala Glu Tyr Gly Leu Ile Pro Leu Phe Ile Pro Phe Thr Asp
180 185 190
Ser Asn Glu Pro Ile Val Lys Glu Ile Lys Trp Met Glu Lys Ser Arg
195 200 205
Asn Gln Ser Val Arg Arg Leu Asp Lys Asp Met Phe Ile Gln Ala Leu
210 215 220
Glu Arg Phe Leu Ser Trp Glu Ser Trp Asn Leu Lys Val Lys Glu Glu
225 230 235 240
Tyr Glu Lys Val Glu Lys Glu His Lys Thr Leu Glu Glu Arg Ile Lys
245 250 255
Glu Asp Ile Gln Ala Phe Lys Ser Leu Glu Gln Tyr Glu Lys Glu Arg
260 265 270
Gln Glu Gln Leu Leu Arg Asp Thr Leu Asn Thr Asn Glu Tyr Arg Leu
275 280 285
Ser Lys Arg Gly Leu Arg Gly Trp Arg Glu Ile Ile Gln Lys Trp Leu
290 295 300
Lys Met Asp Glu Asn Glu Pro Ser Glu Lys Tyr Leu Glu Val Phe Lys
305 310 315 320
Asp Tyr Gln Arg Lys His Pro Arg Glu Ala Gly Asp Tyr Ser Val Tyr
325 330 335
Glu Phe Leu Ser Lys Lys Glu Asn His Phe Ile Trp Arg Asn His Pro
340 345 350
Glu Tyr Pro Tyr Leu Tyr Ala Thr Phe Cys Glu Ile Asp Lys Lys Lys
355 360 365
Lys Asp Ala Lys Gln Gln Ala Thr Phe Thr Leu Ala Asp Pro Ile Asn
370 375 380
His Pro Leu Trp Val Arg Phe Glu Glu Arg Ser Gly Ser Asn Leu Asn
385 390 395 400
Lys Tyr Arg Ile Leu Thr Glu Gln Leu His Thr Glu Lys Leu Lys Lys
405 410 415
Lys Leu Thr Val Gln Leu Asp Arg Leu Ile Tyr Pro Thr Glu Ser Gly
420 425 430
Gly Trp Glu Glu Lys Gly Lys Val Asp Ile Val Leu Leu Pro Ser Arg
435 440 445
Gln Phe Tyr Asn Gln Ile Phe Leu Asp Ile Glu Glu Lys Gly Lys His
450 455 460
Ala Phe Thr Tyr Lys Asp Glu Ser Ile Lys Phe Pro Leu Lys Gly Thr
465 470 475 480
Leu Gly Gly Ala Arg Val Gln Phe Asp Arg Asp His Leu Arg Arg Tyr
485 490 495
Pro His Lys Val Glu Ser Gly Asn Val Gly Arg Ile Tyr Phe Asn Met
500 505 510
Thr Val Asn Ile Glu Pro Thr Glu Ser Pro Val Ser Lys Ser Leu Lys
515 520 525
Ile His Arg Asp Asp Phe Pro Lys Phe Val Asn Phe Lys Pro Lys Glu
530 535 540
Leu Thr Glu Trp Ile Lys Asp Ser Lys Gly Lys Lys Leu Lys Ser Gly
545 550 555 560
Ile Glu Ser Leu Glu Ile Gly Leu Arg Val Met Ser Ile Asp Leu Gly
565 570 575
Gln Arg Gln Ala Ala Ala Ala Ser Ile Phe Glu Val Val Asp Gln Lys
580 585 590
Pro Asp Ile Glu Gly Lys Leu Phe Phe Pro Ile Lys Gly Thr Glu Leu
595 600 605
Tyr Ala Val His Arg Ala Ser Phe Asn Ile Lys Leu Pro Gly Glu Thr
610 615 620
Leu Val Lys Ser Arg Glu Val Leu Arg Lys Ala Arg Glu Asp Asn Leu
625 630 635 640
Lys Leu Met Asn Gln Lys Leu Asn Phe Leu Arg Asn Val Leu His Phe
645 650 655
Gln Gln Phe Glu Asp Ile Thr Glu Arg Glu Lys Arg Val Thr Lys Trp
660 665 670
Ile Ser Arg Gln Glu Asn Ser Asp Val Pro Leu Val Tyr Gln Asp Glu
675 680 685
Leu Ile Gln Ile Arg Glu Leu Met Tyr Lys Pro Tyr Lys Asp Trp Val
690 695 700
Ala Phe Leu Lys Gln Leu His Lys Arg Leu Glu Val Glu Ile Gly Lys
705 710 715 720
Glu Val Lys His Trp Arg Lys Ser Leu Ser Asp Gly Arg Lys Gly Leu
725 730 735
Tyr Gly Ile Ser Leu Lys Asn Ile Asp Glu Ile Asp Arg Thr Arg Lys
740 745 750
Phe Leu Leu Arg Trp Ser Leu Arg Pro Thr Glu Pro Gly Glu Val Arg
755 760 765
Arg Leu Glu Pro Gly Gln Arg Phe Ala Ile Asp Gln Leu Asn His Leu
770 775 780
Asn Ala Leu Lys Glu Asp Arg Leu Lys Lys Met Ala Asn Thr Ile Ile
785 790 795 800
Met His Ala Leu Gly Tyr Cys Tyr Asp Val Arg Lys Lys Lys Trp Gln
805 810 815
Ala Lys Asn Pro Ala Cys Gln Ile Ile Leu Phe Glu Asp Leu Ser Asn
820 825 830
Tyr Asn Pro Tyr Glu Glu Arg Ser Arg Phe Glu Asn Ser Lys Leu Met
835 840 845
Lys Trp Ser Arg Arg Glu Ile Pro Arg Gln Val Ala Leu Gln Gly Glu
850 855 860
Ile Tyr Gly Leu Gln Val Gly Glu Val Gly Ala Gln Phe Ser Ser Arg
865 870 875 880
Phe His Ala Lys Thr Gly Ser Pro Gly Ile Arg Cys Ser Val Val Thr
885 890 895
Lys Glu Lys Leu Gln Asp Asn Arg Phe Phe Lys Asn Leu Gln Arg Glu
900 905 910
Gly Arg Leu Thr Leu Asp Lys Ile Ala Val Leu Lys Glu Gly Asp Leu
915 920 925
Tyr Pro Asp Lys Gly Gly Glu Lys Phe Ile Ser Leu Ser Lys Asp Arg
930 935 940
Lys Leu Val Thr Thr His Ala Asp Ile Asn Ala Ala Gln Asn Leu Gln
945 950 955 960
Lys Arg Phe Trp Thr Arg Thr His Gly Phe Tyr Lys Val Tyr Cys Lys
965 970 975
Ala Tyr Gln Val Asp Gly Gln Thr Val Tyr Ile Pro Glu Ser Lys Asp
980 985 990
Gln Lys Gln Lys Ile Ile Glu Glu Phe Gly Glu Gly Tyr Phe Ile Leu
995 1000 1005
Lys Asp Gly Val Tyr Glu Trp Gly Asn Ala Gly Lys Leu Lys Ile
1010 1015 1020
Lys Lys Gly Ser Ser Lys Gln Ser Ser Ser Glu Leu Val Asp Ser
1025 1030 1035
Asp Ile Leu Lys Asp Ser Phe Asp Leu Ala Ser Glu Leu Lys Gly
1040 1045 1050
Glu Lys Leu Met Leu Tyr Arg Asp Pro Ser Gly Asn Val Phe Pro
1055 1060 1065
Ser Asp Lys Trp Met Ala Ala Gly Val Phe Phe Gly Lys Leu Glu
1070 1075 1080
Arg Ile Leu Ile Ser Lys Leu Thr Asn Gln Tyr Ser Ile Ser Thr
1085 1090 1095
Ile Glu Asp Asp Ser Ser Lys Gln Ser Met
1100 1105
<210> 66
<211> 1108
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 66
Met Ala Ile Arg Ser Ile Lys Leu Lys Leu Lys Thr His Thr Gly Pro
1 5 10 15
Glu Ala Gln Asn Leu Arg Lys Gly Ile Trp Arg Thr His Arg Leu Leu
20 25 30
Asn Glu Gly Val Ala Tyr Tyr Met Lys Met Leu Leu Leu Phe Arg Gln
35 40 45
Glu Ser Thr Gly Glu Arg Pro Lys Glu Glu Leu Gln Glu Glu Leu Ile
50 55 60
Cys His Ile Arg Glu Gln Gln Gln Arg Asn Gln Ala Asp Lys Asn Thr
65 70 75 80
Gln Ala Leu Pro Leu Asp Lys Ala Leu Glu Ala Leu Arg Gln Leu Tyr
85 90 95
Glu Leu Leu Val Pro Ser Ser Val Gly Gln Ser Gly Asp Ala Gln Ile
100 105 110
Ile Ser Arg Lys Phe Leu Ser Pro Leu Val Asp Pro Asn Ser Glu Gly
115 120 125
Gly Lys Gly Thr Ser Lys Ala Gly Ala Lys Pro Thr Trp Gln Lys Lys
130 135 140
Lys Glu Ala Asn Asp Pro Thr Trp Glu Gln Asp Tyr Glu Lys Trp Lys
145 150 155 160
Lys Arg Arg Glu Glu Asp Pro Thr Ala Ser Val Ile Thr Thr Leu Glu
165 170 175
Glu Tyr Gly Ile Arg Pro Ile Phe Pro Leu Tyr Thr Asn Thr Val Thr
180 185 190
Asp Ile Ala Trp Leu Pro Leu Gln Ser Asn Gln Phe Val Arg Thr Trp
195 200 205
Asp Arg Asp Met Leu Gln Gln Ala Ile Glu Arg Leu Leu Ser Trp Glu
210 215 220
Ser Trp Asn Lys Arg Val Gln Glu Glu Tyr Ala Lys Leu Lys Glu Lys
225 230 235 240
Met Ala Gln Leu Asn Glu Gln Leu Glu Gly Gly Gln Glu Trp Ile Ser
245 250 255
Leu Leu Glu Gln Tyr Glu Glu Asn Arg Glu Arg Glu Leu Arg Glu Asn
260 265 270
Met Thr Ala Ala Asn Asp Lys Tyr Arg Ile Thr Lys Arg Gln Met Lys
275 280 285
Gly Trp Asn Glu Leu Tyr Glu Leu Trp Ser Thr Phe Pro Ala Ser Ala
290 295 300
Ser His Glu Gln Tyr Lys Glu Ala Leu Lys Arg Val Gln Gln Arg Leu
305 310 315 320
Arg Gly Arg Phe Gly Asp Ala His Phe Phe Gln Tyr Leu Met Glu Glu
325 330 335
Lys Asn Arg Leu Ile Trp Lys Gly Asn Pro Gln Arg Ile His Tyr Phe
340 345 350
Val Ala Arg Asn Glu Leu Thr Lys Arg Leu Glu Glu Ala Lys Gln Ser
355 360 365
Ala Thr Met Thr Leu Pro Asn Ala Arg Lys His Pro Leu Trp Val Arg
370 375 380
Phe Asp Ala Arg Gly Gly Asn Leu Gln Asp Tyr Tyr Leu Thr Ala Glu
385 390 395 400
Ala Asp Lys Pro Arg Ser Arg Arg Phe Val Thr Phe Ser Gln Leu Ile
405 410 415
Trp Pro Ser Glu Ser Gly Trp Met Glu Lys Lys Asp Val Glu Val Glu
420 425 430
Leu Ala Leu Ser Arg Gln Phe Tyr Gln Gln Val Lys Leu Leu Lys Asn
435 440 445
Asp Lys Gly Lys Gln Lys Ile Glu Phe Lys Asp Lys Gly Ser Gly Ser
450 455 460
Thr Phe Asn Gly His Leu Gly Gly Ala Lys Leu Gln Leu Glu Arg Gly
465 470 475 480
Asp Leu Glu Lys Glu Glu Lys Asn Phe Glu Asp Gly Glu Ile Gly Ser
485 490 495
Val Tyr Leu Asn Val Val Ile Asp Phe Glu Pro Leu Gln Glu Val Lys
500 505 510
Asn Gly Arg Val Gln Ala Pro Tyr Gly Gln Val Leu Gln Leu Ile Arg
515 520 525
Arg Pro Asn Glu Phe Pro Lys Val Thr Thr Tyr Lys Ser Glu Gln Leu
530 535 540
Val Glu Trp Ile Lys Ala Ser Pro Gln His Ser Ala Gly Val Glu Ser
545 550 555 560
Leu Ala Ser Gly Phe Arg Val Met Ser Ile Asp Leu Gly Leu Arg Ala
565 570 575
Ala Ala Ala Thr Ser Ile Phe Ser Val Glu Glu Ser Ser Asp Lys Asn
580 585 590
Ala Ala Asp Phe Ser Tyr Trp Ile Glu Gly Thr Pro Leu Val Ala Val
595 600 605
His Gln Arg Ser Tyr Met Leu Arg Leu Pro Gly Glu Gln Val Glu Lys
610 615 620
Gln Val Met Glu Lys Arg Asp Glu Arg Phe Gln Leu His Gln Arg Val
625 630 635 640
Lys Phe Gln Ile Arg Val Leu Ala Gln Ile Met Arg Met Ala Asn Lys
645 650 655
Gln Tyr Gly Asp Arg Trp Asp Glu Leu Asp Ser Leu Lys Gln Ala Val
660 665 670
Glu Gln Lys Lys Ser Pro Leu Asp Gln Thr Asp Arg Thr Phe Trp Glu
675 680 685
Gly Ile Val Cys Asp Leu Thr Lys Val Leu Pro Arg Asn Glu Ala Asp
690 695 700
Trp Glu Gln Ala Val Val Gln Ile His Arg Lys Ala Glu Glu Tyr Val
705 710 715 720
Gly Lys Ala Val Gln Ala Trp Arg Lys Arg Phe Ala Ala Asp Glu Arg
725 730 735
Lys Gly Ile Ala Gly Leu Ser Met Trp Asn Ile Glu Glu Leu Glu Gly
740 745 750
Leu Arg Lys Leu Leu Ile Ser Trp Ser Arg Arg Thr Arg Asn Pro Gln
755 760 765
Glu Val Asn Arg Phe Glu Arg Gly His Thr Ser His Gln Arg Leu Leu
770 775 780
Thr His Ile Gln Asn Val Lys Glu Asp Arg Leu Lys Gln Leu Ser His
785 790 795 800
Ala Ile Val Met Thr Ala Leu Gly Tyr Val Tyr Asp Glu Arg Lys Gln
805 810 815
Glu Trp Cys Ala Glu Tyr Pro Ala Cys Gln Val Ile Leu Phe Glu Asn
820 825 830
Leu Ser Gln Tyr Arg Ser Asn Leu Asp Arg Ser Thr Lys Glu Asn Ser
835 840 845
Thr Leu Met Lys Trp Ala His Arg Ser Ile Pro Lys Tyr Val His Met
850 855 860
Gln Ala Glu Pro Tyr Gly Ile Gln Ile Gly Asp Val Arg Ala Glu Tyr
865 870 875 880
Ser Ser Arg Phe Tyr Ala Lys Thr Gly Thr Pro Gly Ile Arg Cys Lys
885 890 895
Lys Val Arg Gly Gln Asp Leu Gln Gly Arg Arg Phe Glu Asn Leu Gln
900 905 910
Lys Arg Leu Val Asn Glu Gln Phe Leu Thr Glu Glu Gln Val Lys Gln
915 920 925
Leu Arg Pro Gly Asp Ile Val Pro Asp Asp Ser Gly Glu Leu Phe Met
930 935 940
Thr Leu Thr Asp Gly Ser Gly Ser Lys Glu Val Val Phe Leu Gln Ala
945 950 955 960
Asp Ile Asn Ala Ala His Asn Leu Gln Lys Arg Phe Trp Gln Arg Tyr
965 970 975
Asn Glu Leu Phe Lys Val Ser Cys Arg Val Ile Val Arg Asp Glu Glu
980 985 990
Glu Tyr Leu Val Pro Lys Thr Lys Ser Val Gln Ala Lys Leu Gly Lys
995 1000 1005
Gly Leu Phe Val Lys Lys Ser Asp Thr Ala Trp Lys Asp Val Tyr
1010 1015 1020
Val Trp Asp Ser Gln Ala Lys Leu Lys Gly Lys Thr Thr Phe Thr
1025 1030 1035
Glu Glu Ser Glu Ser Pro Glu Gln Leu Glu Asp Phe Gln Glu Ile
1040 1045 1050
Ile Glu Glu Ala Glu Glu Ala Lys Gly Thr Tyr Arg Thr Leu Phe
1055 1060 1065
Arg Asp Pro Ser Gly Val Phe Phe Pro Glu Ser Val Trp Tyr Pro
1070 1075 1080
Gln Lys Asp Phe Trp Gly Glu Val Lys Arg Lys Leu Tyr Gly Lys
1085 1090 1095
Leu Arg Glu Arg Phe Leu Thr Lys Ala Arg
1100 1105
<210> 67
<211> 1334
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 67
Met Lys Ile Ser Lys Val Asp His Thr Arg Met Ala Val Ala Lys Gly
1 5 10 15
Asn Gln His Arg Arg Asp Glu Ile Ser Gly Ile Leu Tyr Lys Asp Pro
20 25 30
Thr Lys Thr Gly Ser Ile Asp Phe Asp Glu Arg Phe Lys Lys Leu Asn
35 40 45
Cys Ser Ala Lys Ile Leu Tyr His Val Phe Asn Gly Ile Ala Glu Gly
50 55 60
Ser Asn Lys Tyr Lys Asn Ile Val Asp Lys Val Asn Asn Asn Leu Asp
65 70 75 80
Arg Val Leu Phe Thr Gly Lys Ser Tyr Asp Arg Lys Ser Ile Ile Asp
85 90 95
Ile Asp Thr Val Leu Arg Asn Val Glu Lys Ile Asn Ala Phe Asp Arg
100 105 110
Ile Ser Thr Glu Glu Arg Glu Gln Ile Ile Asp Asp Leu Leu Glu Ile
115 120 125
Gln Leu Arg Lys Gly Leu Arg Lys Gly Lys Ala Gly Leu Arg Glu Val
130 135 140
Leu Leu Ile Gly Ala Gly Val Ile Val Arg Thr Asp Lys Lys Gln Glu
145 150 155 160
Ile Ala Asp Phe Leu Glu Ile Leu Asp Glu Asp Phe Asn Lys Thr Asn
165 170 175
Gln Ala Lys Asn Ile Lys Leu Ser Ile Glu Asn Gln Gly Leu Val Val
180 185 190
Ser Pro Val Ser Arg Gly Glu Glu Arg Ile Phe Asp Val Ser Gly Ala
195 200 205
Gln Lys Gly Lys Ser Ser Lys Lys Ala Gln Glu Lys Glu Ala Leu Ser
210 215 220
Ala Phe Leu Leu Asp Tyr Ala Asp Leu Asp Lys Asn Val Arg Phe Glu
225 230 235 240
Tyr Leu Arg Lys Ile Arg Arg Leu Ile Asn Leu Tyr Phe Tyr Val Lys
245 250 255
Asn Asp Asp Val Met Ser Leu Thr Glu Ile Pro Ala Glu Val Asn Leu
260 265 270
Glu Lys Asp Phe Asp Ile Trp Arg Asp His Glu Gln Arg Lys Glu Glu
275 280 285
Asn Gly Asp Phe Val Gly Cys Pro Asp Ile Leu Leu Ala Asp Arg Asp
290 295 300
Val Lys Lys Ser Asn Ser Lys Gln Val Lys Ile Ala Glu Arg Gln Leu
305 310 315 320
Arg Glu Ser Ile Arg Glu Lys Asn Ile Lys Arg Tyr Arg Phe Ser Ile
325 330 335
Lys Thr Ile Glu Lys Asp Asp Gly Thr Tyr Phe Phe Ala Asn Lys Gln
340 345 350
Ile Ser Val Phe Trp Ile His Arg Ile Glu Asn Ala Val Glu Arg Ile
355 360 365
Leu Gly Ser Ile Asn Asp Lys Lys Leu Tyr Arg Leu Arg Leu Gly Tyr
370 375 380
Leu Gly Glu Lys Val Trp Lys Asp Ile Leu Asn Phe Leu Ser Ile Lys
385 390 395 400
Tyr Ile Ala Val Gly Lys Ala Val Phe Asn Phe Ala Met Asp Asp Leu
405 410 415
Gln Glu Lys Asp Arg Asp Ile Glu Pro Gly Lys Ile Ser Glu Asn Ala
420 425 430
Val Asn Gly Leu Thr Ser Phe Asp Tyr Glu Gln Ile Lys Ala Asp Glu
435 440 445
Met Leu Gln Arg Glu Val Ala Val Asn Val Ala Phe Ala Ala Asn Asn
450 455 460
Leu Ala Arg Val Thr Val Asp Ile Pro Gln Asn Gly Glu Lys Glu Asp
465 470 475 480
Ile Leu Leu Trp Asn Lys Ser Asp Ile Lys Lys Tyr Lys Lys Asn Ser
485 490 495
Lys Lys Gly Ile Leu Lys Ser Ile Leu Gln Phe Phe Gly Gly Ala Ser
500 505 510
Thr Trp Asn Met Lys Met Phe Glu Ile Ala Tyr His Asp Gln Pro Gly
515 520 525
Asp Tyr Glu Glu Asn Tyr Leu Tyr Asp Ile Ile Gln Ile Ile Tyr Ser
530 535 540
Leu Arg Asn Lys Ser Phe His Phe Lys Thr Tyr Asp His Gly Asp Lys
545 550 555 560
Asn Trp Asn Arg Glu Leu Ile Gly Lys Met Ile Glu His Asp Ala Glu
565 570 575
Arg Val Ile Ser Val Glu Arg Glu Lys Phe His Ser Asn Asn Leu Pro
580 585 590
Met Phe Tyr Lys Asp Ala Asp Leu Lys Lys Ile Leu Asp Leu Leu Tyr
595 600 605
Ser Asp Tyr Ala Gly Arg Ala Ser Gln Val Pro Ala Phe Asn Thr Val
610 615 620
Leu Val Arg Lys Asn Phe Pro Glu Phe Leu Arg Lys Asp Met Gly Tyr
625 630 635 640
Lys Val His Phe Asn Asn Pro Glu Val Glu Asn Gln Trp His Ser Ala
645 650 655
Val Tyr Tyr Leu Tyr Lys Glu Ile Tyr Tyr Asn Leu Phe Leu Arg Asp
660 665 670
Lys Glu Val Lys Asn Leu Phe Tyr Thr Ser Leu Lys Asn Ile Arg Ser
675 680 685
Glu Val Ser Asp Lys Lys Gln Lys Leu Ala Ser Asp Asp Phe Ala Ser
690 695 700
Arg Cys Glu Glu Ile Glu Asp Arg Ser Leu Pro Glu Ile Cys Gln Ile
705 710 715 720
Ile Met Thr Glu Tyr Asn Ala Gln Asn Phe Gly Asn Arg Lys Val Lys
725 730 735
Ser Gln Arg Val Ile Glu Lys Asn Lys Asp Ile Phe Arg His Tyr Lys
740 745 750
Met Leu Leu Ile Lys Thr Leu Ala Gly Ala Phe Ser Leu Tyr Leu Lys
755 760 765
Gln Glu Arg Phe Ala Phe Ile Gly Lys Ala Thr Pro Ile Pro Tyr Glu
770 775 780
Thr Thr Asp Val Lys Asn Phe Leu Pro Glu Trp Lys Ser Gly Met Tyr
785 790 795 800
Ala Ser Phe Val Glu Glu Ile Lys Asn Asn Leu Asp Leu Gln Glu Trp
805 810 815
Tyr Ile Val Gly Arg Phe Leu Asn Gly Arg Met Leu Asn Gln Leu Ala
820 825 830
Gly Ser Leu Arg Ser Tyr Ile Gln Tyr Ala Glu Asp Ile Glu Arg Arg
835 840 845
Ala Ala Glu Asn Arg Asn Lys Leu Phe Ser Lys Pro Asp Glu Lys Ile
850 855 860
Glu Ala Cys Lys Lys Ala Val Arg Val Leu Asp Leu Cys Ile Lys Ile
865 870 875 880
Ser Thr Arg Ile Ser Ala Glu Phe Thr Asp Tyr Phe Asp Ser Glu Asp
885 890 895
Asp Tyr Ala Asp Tyr Leu Glu Lys Tyr Leu Lys Tyr Gln Asp Asp Ala
900 905 910
Ile Lys Glu Leu Ser Gly Ser Ser Tyr Ala Ala Leu Asp His Phe Cys
915 920 925
Asn Lys Asp Asp Leu Lys Phe Asp Ile Tyr Val Asn Ala Gly Gln Lys
930 935 940
Pro Ile Leu Gln Arg Asn Ile Val Met Ala Lys Leu Phe Gly Pro Asp
945 950 955 960
Asn Ile Leu Ser Glu Val Met Glu Lys Val Thr Glu Ser Ala Ile Arg
965 970 975
Glu Tyr Tyr Asp Tyr Leu Lys Lys Val Ser Gly Tyr Arg Val Arg Gly
980 985 990
Lys Cys Ser Thr Glu Lys Glu Gln Glu Asp Leu Leu Lys Phe Gln Arg
995 1000 1005
Leu Lys Asn Ala Val Glu Phe Arg Asp Val Thr Glu Tyr Ala Glu
1010 1015 1020
Val Ile Asn Glu Leu Leu Gly Gln Leu Ile Ser Trp Ser Tyr Leu
1025 1030 1035
Arg Glu Arg Asp Leu Leu Tyr Phe Gln Leu Gly Phe His Tyr Met
1040 1045 1050
Cys Leu Lys Asn Lys Ser Phe Lys Pro Ala Glu Tyr Val Asp Ile
1055 1060 1065
Arg Arg Asn Asn Gly Thr Ile Ile His Asn Ala Ile Leu Tyr Gln
1070 1075 1080
Ile Val Ser Met Tyr Ile Asn Gly Leu Asp Phe Tyr Ser Cys Asp
1085 1090 1095
Lys Glu Gly Lys Thr Leu Lys Pro Ile Glu Thr Gly Lys Gly Val
1100 1105 1110
Gly Ser Lys Ile Gly Gln Phe Ile Lys Tyr Ser Gln Tyr Leu Tyr
1115 1120 1125
Asn Asp Pro Ser Tyr Lys Leu Glu Ile Tyr Asn Ala Gly Leu Glu
1130 1135 1140
Val Phe Glu Asn Ile Asp Glu His Asp Asn Ile Thr Asp Leu Arg
1145 1150 1155
Lys Tyr Val Asp His Phe Lys Tyr Tyr Ala Tyr Gly Asn Lys Met
1160 1165 1170
Ser Leu Leu Asp Leu Tyr Ser Glu Phe Phe Asp Arg Phe Phe Thr
1175 1180 1185
Tyr Asp Met Lys Tyr Gln Lys Asn Val Val Asn Val Leu Glu Asn
1190 1195 1200
Ile Leu Leu Arg His Phe Val Ile Phe Tyr Pro Lys Phe Gly Ser
1205 1210 1215
Gly Lys Lys Asp Val Gly Ile Arg Asp Cys Lys Lys Glu Arg Ala
1220 1225 1230
Gln Ile Glu Ile Ser Glu Gln Ser Leu Thr Ser Glu Asp Phe Met
1235 1240 1245
Phe Lys Leu Asp Asp Lys Ala Gly Glu Glu Ala Lys Lys Phe Pro
1250 1255 1260
Ala Arg Asp Glu Arg Tyr Leu Gln Thr Ile Ala Lys Leu Leu Tyr
1265 1270 1275
Tyr Pro Asn Glu Ile Glu Asp Met Asn Arg Phe Met Lys Lys Gly
1280 1285 1290
Glu Thr Ile Asn Lys Lys Val Gln Phe Asn Arg Lys Lys Lys Ile
1295 1300 1305
Thr Arg Lys Gln Lys Asn Asn Ser Ser Asn Glu Val Leu Ser Ser
1310 1315 1320
Thr Met Gly Tyr Leu Phe Lys Asn Ile Lys Leu
1325 1330
<210> 68
<211> 1120
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 68
Met Trp Ile Ser Ile Lys Thr Leu Ile His His Leu Gly Val Leu Phe
1 5 10 15
Phe Cys Asp Tyr Met Tyr Asn Arg Arg Glu Lys Lys Ile Ile Glu Val
20 25 30
Lys Thr Met Arg Ile Thr Lys Val Glu Val Asp Arg Lys Lys Val Leu
35 40 45
Ile Ser Arg Asp Lys Asn Gly Gly Lys Leu Val Tyr Glu Asn Glu Met
50 55 60
Gln Asp Asn Thr Glu Gln Ile Met His His Lys Lys Ser Ser Phe Tyr
65 70 75 80
Lys Ser Val Val Asn Lys Thr Ile Cys Arg Pro Glu Gln Lys Gln Met
85 90 95
Lys Lys Leu Val His Gly Leu Leu Gln Glu Asn Ser Gln Glu Lys Ile
100 105 110
Lys Val Ser Asp Val Thr Lys Leu Asn Ile Ser Asn Phe Leu Asn His
115 120 125
Arg Phe Lys Lys Ser Leu Tyr Tyr Phe Pro Glu Asn Ser Pro Asp Lys
130 135 140
Ser Glu Glu Tyr Arg Ile Glu Ile Asn Leu Ser Gln Leu Leu Glu Asp
145 150 155 160
Ser Leu Lys Lys Gln Gln Gly Thr Phe Ile Cys Trp Glu Ser Phe Ser
165 170 175
Lys Asp Met Glu Leu Tyr Ile Asn Trp Ala Glu Asn Tyr Ile Ser Ser
180 185 190
Lys Thr Lys Leu Ile Lys Lys Ser Ile Arg Asn Asn Arg Ile Gln Ser
195 200 205
Thr Glu Ser Arg Ser Gly Gln Leu Met Asp Arg Tyr Met Lys Asp Ile
210 215 220
Leu Asn Lys Asn Lys Pro Phe Asp Ile Gln Ser Val Ser Glu Lys Tyr
225 230 235 240
Gln Leu Glu Lys Leu Thr Ser Ala Leu Lys Ala Thr Phe Lys Glu Ala
245 250 255
Lys Lys Asn Asp Lys Glu Ile Asn Tyr Lys Leu Lys Ser Thr Leu Gln
260 265 270
Asn His Glu Arg Gln Ile Ile Glu Glu Leu Lys Glu Asn Ser Glu Leu
275 280 285
Asn Gln Phe Asn Ile Glu Ile Arg Lys His Leu Glu Thr Tyr Phe Pro
290 295 300
Ile Lys Lys Thr Asn Arg Lys Val Gly Asp Ile Arg Asn Leu Glu Ile
305 310 315 320
Gly Glu Ile Gln Lys Ile Val Asn His Arg Leu Lys Asn Lys Ile Val
325 330 335
Gln Arg Ile Leu Gln Glu Gly Lys Leu Ala Ser Tyr Glu Ile Glu Ser
340 345 350
Thr Val Asn Ser Asn Ser Leu Gln Lys Ile Lys Ile Glu Glu Ala Phe
355 360 365
Ala Leu Lys Phe Ile Asn Ala Cys Leu Phe Ala Ser Asn Asn Leu Arg
370 375 380
Asn Met Val Tyr Pro Val Cys Lys Lys Asp Ile Leu Met Ile Gly Glu
385 390 395 400
Phe Lys Asn Ser Phe Lys Glu Ile Lys His Lys Lys Phe Ile Arg Gln
405 410 415
Trp Ser Gln Phe Phe Ser Gln Glu Ile Thr Val Asp Asp Ile Glu Leu
420 425 430
Ala Ser Trp Gly Leu Arg Gly Ala Ile Ala Pro Ile Arg Asn Glu Ile
435 440 445
Ile His Leu Lys Lys His Ser Trp Lys Lys Phe Phe Asn Asn Pro Thr
450 455 460
Phe Lys Val Lys Lys Ser Lys Ile Ile Asn Gly Lys Thr Lys Asp Val
465 470 475 480
Thr Ser Glu Phe Leu Tyr Lys Glu Thr Leu Phe Lys Asp Tyr Phe Tyr
485 490 495
Ser Glu Leu Asp Ser Val Pro Glu Leu Ile Ile Asn Lys Met Glu Ser
500 505 510
Ser Lys Ile Leu Asp Tyr Tyr Ser Ser Asp Gln Leu Asn Gln Val Phe
515 520 525
Thr Ile Pro Asn Phe Glu Leu Ser Leu Leu Thr Ser Ala Val Pro Phe
530 535 540
Ala Pro Ser Phe Lys Arg Val Tyr Leu Lys Gly Phe Asp Tyr Gln Asn
545 550 555 560
Gln Asp Glu Ala Gln Pro Asp Tyr Asn Leu Lys Leu Asn Ile Tyr Asn
565 570 575
Glu Lys Ala Phe Asn Ser Glu Ala Phe Gln Ala Gln Tyr Ser Leu Phe
580 585 590
Lys Met Val Tyr Tyr Gln Val Phe Leu Pro Gln Phe Thr Thr Asn Asn
595 600 605
Asp Leu Phe Lys Ser Ser Val Asp Phe Ile Leu Thr Leu Asn Lys Glu
610 615 620
Arg Lys Gly Tyr Ala Lys Ala Phe Gln Asp Ile Arg Lys Met Asn Lys
625 630 635 640
Asp Glu Lys Pro Ser Glu Tyr Met Ser Tyr Ile Gln Ser Gln Leu Met
645 650 655
Leu Tyr Gln Lys Lys Gln Glu Glu Lys Glu Lys Ile Asn His Phe Glu
660 665 670
Lys Phe Ile Asn Gln Val Phe Ile Lys Gly Phe Asn Ser Phe Ile Glu
675 680 685
Lys Asn Arg Leu Thr Tyr Ile Cys His Pro Thr Lys Asn Thr Val Pro
690 695 700
Glu Asn Asp Asn Ile Glu Ile Pro Phe His Thr Asp Met Asp Asp Ser
705 710 715 720
Asn Ile Ala Phe Trp Leu Met Cys Lys Leu Leu Asp Ala Lys Gln Leu
725 730 735
Ser Glu Leu Arg Asn Glu Met Ile Lys Phe Ser Cys Ser Leu Gln Ser
740 745 750
Thr Glu Glu Ile Ser Thr Phe Thr Lys Ala Arg Glu Val Ile Gly Leu
755 760 765
Ala Leu Leu Asn Gly Glu Lys Gly Cys Asn Asp Trp Lys Glu Leu Phe
770 775 780
Asp Asp Lys Glu Ala Trp Lys Lys Asn Met Ser Leu Tyr Val Ser Glu
785 790 795 800
Glu Leu Leu Gln Ser Leu Pro Tyr Thr Gln Glu Asp Gly Gln Thr Pro
805 810 815
Val Ile Asn Arg Ser Ile Asp Leu Val Lys Lys Tyr Gly Thr Glu Thr
820 825 830
Ile Leu Glu Lys Leu Phe Ser Ser Ser Asp Asp Tyr Lys Val Ser Ala
835 840 845
Lys Asp Ile Ala Lys Leu His Glu Tyr Asp Val Thr Glu Lys Ile Ala
850 855 860
Gln Gln Glu Ser Leu His Lys Gln Trp Ile Glu Lys Pro Gly Leu Ala
865 870 875 880
Arg Asp Ser Ala Trp Thr Lys Lys Tyr Gln Asn Val Ile Asn Asp Ile
885 890 895
Ser Asn Tyr Gln Trp Ala Lys Thr Lys Val Glu Leu Thr Gln Val Arg
900 905 910
His Leu His Gln Leu Thr Ile Asp Leu Leu Ser Arg Leu Ala Gly Tyr
915 920 925
Met Ser Ile Ala Asp Arg Asp Phe Gln Phe Ser Ser Asn Tyr Ile Leu
930 935 940
Glu Arg Glu Asn Ser Glu Tyr Arg Val Thr Ser Trp Ile Leu Leu Ser
945 950 955 960
Glu Asn Lys Asn Lys Asn Lys Tyr Asn Asp Tyr Glu Leu Tyr Asn Leu
965 970 975
Lys Asn Ala Ser Ile Lys Val Ser Ser Lys Asn Asp Pro Gln Leu Lys
980 985 990
Val Asp Leu Lys Gln Leu Arg Leu Thr Leu Glu Tyr Leu Glu Leu Phe
995 1000 1005
Asp Asn Arg Leu Lys Glu Lys Arg Asn Asn Ile Ser His Phe Asn
1010 1015 1020
Tyr Leu Asn Gly Gln Leu Gly Asn Ser Ile Leu Glu Leu Phe Asp
1025 1030 1035
Asp Ala Arg Asp Val Leu Ser Tyr Asp Arg Lys Leu Lys Asn Ala
1040 1045 1050
Val Ser Lys Ser Leu Lys Glu Ile Leu Ser Ser His Gly Met Glu
1055 1060 1065
Val Thr Phe Lys Pro Leu Tyr Gln Thr Asn His His Leu Lys Ile
1070 1075 1080
Asp Lys Leu Gln Pro Lys Lys Ile His His Leu Gly Glu Lys Ser
1085 1090 1095
Thr Val Ser Ser Asn Gln Val Ser Asn Glu Tyr Cys Gln Leu Val
1100 1105 1110
Arg Thr Leu Leu Thr Met Lys
1115 1120
<210> 69
<211> 1152
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 69
Met Lys Val Thr Lys Val Asp Gly Ile Ser His Lys Lys Tyr Ile Glu
1 5 10 15
Glu Gly Lys Leu Val Lys Ser Thr Ser Glu Glu Asn Arg Thr Ser Glu
20 25 30
Arg Leu Ser Glu Leu Leu Ser Ile Arg Leu Asp Ile Tyr Ile Lys Asn
35 40 45
Pro Asp Asn Ala Ser Glu Glu Glu Asn Arg Ile Arg Arg Glu Asn Leu
50 55 60
Lys Lys Phe Phe Ser Asn Lys Val Leu His Leu Lys Asp Ser Val Leu
65 70 75 80
Tyr Leu Lys Asn Arg Lys Glu Lys Asn Ala Val Gln Asp Lys Asn Tyr
85 90 95
Ser Glu Glu Asp Ile Ser Glu Tyr Asp Leu Lys Asn Lys Asn Ser Phe
100 105 110
Ser Val Leu Lys Lys Ile Leu Leu Asn Glu Asp Val Asn Ser Glu Glu
115 120 125
Leu Glu Ile Phe Arg Lys Asp Val Glu Ala Lys Leu Asn Lys Ile Asn
130 135 140
Ser Leu Lys Tyr Ser Phe Glu Glu Asn Lys Ala Asn Tyr Gln Lys Ile
145 150 155 160
Asn Glu Asn Asn Val Glu Lys Val Gly Gly Lys Ser Lys Arg Asn Ile
165 170 175
Ile Tyr Asp Tyr Tyr Arg Glu Ser Ala Lys Arg Asn Asp Tyr Ile Asn
180 185 190
Asn Val Gln Glu Ala Phe Asp Lys Leu Tyr Lys Lys Glu Asp Ile Glu
195 200 205
Lys Leu Phe Phe Leu Ile Glu Asn Ser Lys Lys His Glu Lys Tyr Lys
210 215 220
Ile Arg Glu Tyr Tyr His Lys Ile Ile Gly Arg Lys Asn Asp Lys Glu
225 230 235 240
Asn Phe Ala Lys Ile Ile Tyr Glu Glu Ile Gln Asn Val Asn Asn Ile
245 250 255
Lys Glu Leu Ile Glu Lys Ile Pro Asp Met Ser Glu Leu Lys Lys Ser
260 265 270
Gln Val Phe Tyr Lys Tyr Tyr Leu Asp Lys Glu Glu Leu Asn Asp Lys
275 280 285
Asn Ile Lys Tyr Ala Phe Cys His Phe Val Glu Ile Glu Met Ser Gln
290 295 300
Leu Leu Lys Asn Tyr Val Tyr Lys Arg Leu Ser Asn Ile Ser Asn Asp
305 310 315 320
Lys Ile Lys Arg Ile Phe Glu Tyr Gln Asn Leu Lys Lys Leu Ile Glu
325 330 335
Asn Lys Leu Leu Asn Lys Leu Asp Thr Tyr Val Arg Asn Cys Gly Lys
340 345 350
Tyr Asn Tyr Tyr Leu Gln Val Gly Glu Ile Ala Thr Ser Asp Phe Ile
355 360 365
Ala Arg Asn Arg Gln Asn Glu Ala Phe Leu Arg Asn Ile Ile Gly Val
370 375 380
Ser Ser Val Ala Tyr Phe Ser Leu Arg Asn Ile Leu Glu Thr Glu Asn
385 390 395 400
Glu Asn Asp Ile Thr Gly Arg Met Arg Gly Lys Thr Val Lys Asn Asn
405 410 415
Lys Gly Glu Glu Lys Tyr Val Ser Gly Glu Val Asp Lys Ile Tyr Asn
420 425 430
Glu Asn Lys Gln Asn Glu Val Lys Glu Asn Leu Lys Met Phe Tyr Ser
435 440 445
Tyr Asp Phe Asn Met Asp Asn Lys Asn Glu Ile Glu Asp Phe Phe Ala
450 455 460
Asn Ile Asp Glu Ala Ile Ser Ser Ile Arg His Gly Ile Val His Phe
465 470 475 480
Asn Leu Glu Leu Glu Gly Lys Asp Ile Phe Ala Phe Lys Asn Ile Ala
485 490 495
Pro Ser Glu Ile Ser Lys Lys Met Phe Gln Asn Glu Ile Asn Glu Lys
500 505 510
Lys Leu Lys Leu Lys Ile Phe Lys Gln Leu Asn Ser Ala Asn Val Phe
515 520 525
Asn Tyr Tyr Glu Lys Asp Val Ile Ile Lys Tyr Leu Lys Asn Thr Lys
530 535 540
Phe Asn Phe Val Asn Lys Asn Ile Pro Phe Val Pro Ser Phe Thr Lys
545 550 555 560
Leu Tyr Asn Lys Ile Glu Asp Leu Arg Asn Thr Leu Lys Phe Phe Trp
565 570 575
Ser Val Pro Lys Asp Lys Glu Glu Lys Asp Ala Gln Ile Tyr Leu Leu
580 585 590
Lys Asn Ile Tyr Tyr Gly Glu Phe Leu Asn Lys Phe Val Lys Asn Ser
595 600 605
Lys Val Phe Phe Lys Ile Thr Asn Glu Val Ile Lys Ile Asn Lys Gln
610 615 620
Arg Asn Gln Lys Thr Gly His Tyr Lys Tyr Gln Lys Phe Glu Asn Ile
625 630 635 640
Glu Lys Thr Val Pro Val Glu Tyr Leu Ala Ile Ile Gln Ser Arg Glu
645 650 655
Met Ile Asn Asn Gln Asp Lys Glu Glu Lys Asn Thr Tyr Ile Asp Phe
660 665 670
Ile Gln Gln Ile Phe Leu Lys Gly Phe Ile Asp Tyr Leu Asn Lys Asn
675 680 685
Asn Leu Lys Tyr Ile Glu Ser Asn Asn Asn Asn Asp Asn Asn Asp Ile
690 695 700
Phe Ser Lys Ile Lys Ile Lys Lys Asp Asn Lys Glu Lys Tyr Asp Lys
705 710 715 720
Ile Leu Lys Asn Tyr Glu Lys His Asn Arg Asn Lys Glu Ile Pro His
725 730 735
Glu Ile Asn Glu Phe Val Arg Glu Ile Lys Leu Gly Lys Ile Leu Lys
740 745 750
Tyr Thr Glu Asn Leu Asn Met Phe Tyr Leu Ile Leu Lys Leu Leu Asn
755 760 765
His Lys Glu Leu Thr Asn Leu Lys Gly Ser Leu Glu Lys Tyr Gln Ser
770 775 780
Ala Asn Lys Glu Glu Thr Phe Ser Asp Glu Leu Glu Leu Ile Asn Leu
785 790 795 800
Leu Asn Leu Asp Asn Asn Arg Val Thr Glu Asp Phe Glu Leu Glu Ala
805 810 815
Asn Glu Ile Gly Lys Phe Leu Asp Phe Asn Glu Asn Lys Ile Lys Asp
820 825 830
Arg Lys Glu Leu Lys Lys Phe Asp Thr Asn Lys Ile Tyr Phe Asp Gly
835 840 845
Glu Asn Ile Ile Lys His Arg Ala Phe Tyr Asn Ile Lys Lys Tyr Gly
850 855 860
Met Leu Asn Leu Leu Glu Lys Ile Ala Asp Lys Ala Lys Tyr Lys Ile
865 870 875 880
Ser Leu Lys Glu Leu Lys Glu Tyr Ser Asn Lys Lys Asn Glu Ile Glu
885 890 895
Lys Asn Tyr Thr Met Gln Gln Asn Leu His Arg Lys Tyr Ala Arg Pro
900 905 910
Lys Lys Asp Glu Lys Phe Asn Asp Glu Asp Tyr Lys Glu Tyr Glu Lys
915 920 925
Ala Ile Gly Asn Ile Gln Lys Tyr Thr His Leu Lys Asn Lys Val Glu
930 935 940
Phe Asn Glu Leu Asn Leu Leu Gln Gly Leu Leu Leu Lys Ile Leu His
945 950 955 960
Arg Leu Val Gly Tyr Thr Ser Ile Trp Glu Arg Asp Leu Arg Phe Arg
965 970 975
Leu Lys Gly Glu Phe Pro Glu Asn His Tyr Ile Glu Glu Ile Phe Asn
980 985 990
Phe Asp Asn Ser Lys Asn Val Lys Tyr Lys Ser Gly Gln Ile Val Glu
995 1000 1005
Lys Tyr Ile Asn Phe Tyr Lys Glu Leu Tyr Lys Asp Asn Val Glu
1010 1015 1020
Lys Arg Ser Ile Tyr Ser Asp Lys Lys Val Lys Lys Leu Lys Gln
1025 1030 1035
Glu Lys Lys Asp Leu Tyr Ile Arg Asn Tyr Ile Ala His Phe Asn
1040 1045 1050
Tyr Ile Pro His Ala Glu Ile Ser Leu Leu Glu Val Leu Glu Asn
1055 1060 1065
Leu Arg Lys Leu Leu Ser Tyr Asp Arg Lys Leu Lys Asn Ala Ile
1070 1075 1080
Met Lys Ser Ile Val Asp Ile Leu Lys Glu Tyr Gly Phe Val Ala
1085 1090 1095
Thr Phe Lys Ile Gly Ala Asp Lys Lys Ile Glu Ile Gln Thr Leu
1100 1105 1110
Glu Ser Glu Lys Ile Val His Leu Lys Asn Leu Lys Lys Lys Lys
1115 1120 1125
Leu Met Thr Asp Arg Asn Ser Glu Glu Leu Cys Glu Leu Val Lys
1130 1135 1140
Val Met Phe Glu Tyr Lys Ala Leu Glu
1145 1150
<210> 70
<211> 1389
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 70
Met Gly Asn Leu Phe Gly His Lys Arg Trp Tyr Glu Val Arg Asp Lys
1 5 10 15
Lys Asp Phe Lys Ile Lys Arg Lys Val Lys Val Lys Arg Asn Tyr Asp
20 25 30
Gly Asn Lys Tyr Ile Leu Asn Ile Asn Glu Asn Asn Asn Lys Glu Lys
35 40 45
Ile Asp Asn Asn Lys Phe Ile Arg Lys Tyr Ile Asn Tyr Lys Lys Asn
50 55 60
Asp Asn Ile Leu Lys Glu Phe Thr Arg Lys Phe His Ala Gly Asn Ile
65 70 75 80
Leu Phe Lys Leu Lys Gly Lys Glu Gly Ile Ile Arg Ile Glu Asn Asn
85 90 95
Asp Asp Phe Leu Glu Thr Glu Glu Val Val Leu Tyr Ile Glu Ala Tyr
100 105 110
Gly Lys Ser Glu Lys Leu Lys Ala Leu Gly Ile Thr Lys Lys Lys Ile
115 120 125
Ile Asp Glu Ala Ile Arg Gln Gly Ile Thr Lys Asp Asp Lys Lys Ile
130 135 140
Glu Ile Lys Arg Gln Glu Asn Glu Glu Glu Ile Glu Ile Asp Ile Arg
145 150 155 160
Asp Glu Tyr Thr Asn Lys Thr Leu Asn Asp Cys Ser Ile Ile Leu Arg
165 170 175
Ile Ile Glu Asn Asp Glu Leu Glu Thr Lys Lys Ser Ile Tyr Glu Ile
180 185 190
Phe Lys Asn Ile Asn Met Ser Leu Tyr Lys Ile Ile Glu Lys Ile Ile
195 200 205
Glu Asn Glu Thr Glu Lys Val Phe Glu Asn Arg Tyr Tyr Glu Glu His
210 215 220
Leu Arg Glu Lys Leu Leu Lys Asp Asp Lys Ile Asp Val Ile Leu Thr
225 230 235 240
Asn Phe Met Glu Ile Arg Glu Lys Ile Lys Ser Asn Leu Glu Ile Leu
245 250 255
Gly Phe Val Lys Phe Tyr Leu Asn Val Gly Gly Asp Lys Lys Lys Ser
260 265 270
Lys Asn Lys Lys Met Leu Val Glu Lys Ile Leu Asn Ile Asn Val Asp
275 280 285
Leu Thr Val Glu Asp Ile Ala Asp Phe Val Ile Lys Glu Leu Glu Phe
290 295 300
Trp Asn Ile Thr Lys Arg Ile Glu Lys Val Lys Lys Val Asn Asn Glu
305 310 315 320
Phe Leu Glu Lys Arg Arg Asn Arg Thr Tyr Ile Lys Ser Tyr Val Leu
325 330 335
Leu Asp Lys His Glu Lys Phe Lys Ile Glu Arg Glu Asn Lys Lys Asp
340 345 350
Lys Ile Val Lys Phe Phe Val Glu Asn Ile Lys Asn Asn Ser Ile Lys
355 360 365
Glu Lys Ile Glu Lys Ile Leu Ala Glu Phe Lys Ile Asp Glu Leu Ile
370 375 380
Lys Lys Leu Glu Lys Glu Leu Lys Lys Gly Asn Cys Asp Thr Glu Ile
385 390 395 400
Phe Gly Ile Phe Lys Lys His Tyr Lys Val Asn Phe Asp Ser Lys Lys
405 410 415
Phe Ser Lys Lys Ser Asp Glu Glu Lys Glu Leu Tyr Lys Ile Ile Tyr
420 425 430
Arg Tyr Leu Lys Gly Arg Ile Glu Lys Ile Leu Val Asn Glu Gln Lys
435 440 445
Val Arg Leu Lys Lys Met Glu Lys Ile Glu Ile Glu Lys Ile Leu Asn
450 455 460
Glu Ser Ile Leu Ser Glu Lys Ile Leu Lys Arg Val Lys Gln Tyr Thr
465 470 475 480
Leu Glu His Ile Met Tyr Leu Gly Lys Leu Arg His Asn Asp Ile Asp
485 490 495
Met Thr Thr Val Asn Thr Asp Asp Phe Ser Arg Leu His Ala Lys Glu
500 505 510
Glu Leu Asp Leu Glu Leu Ile Thr Phe Phe Ala Ser Thr Asn Met Glu
515 520 525
Leu Asn Lys Ile Phe Ser Arg Glu Asn Ile Asn Asn Asp Glu Asn Ile
530 535 540
Asp Phe Phe Gly Gly Asp Arg Glu Lys Asn Tyr Val Leu Asp Lys Lys
545 550 555 560
Ile Leu Asn Ser Lys Ile Lys Ile Ile Arg Asp Leu Asp Phe Ile Asp
565 570 575
Asn Lys Asn Asn Ile Thr Asn Asn Phe Ile Arg Lys Phe Thr Lys Ile
580 585 590
Gly Thr Asn Glu Arg Asn Arg Ile Leu His Ala Ile Ser Lys Glu Arg
595 600 605
Asp Leu Gln Gly Thr Gln Asp Asp Tyr Asn Lys Val Ile Asn Ile Ile
610 615 620
Gln Asn Leu Lys Ile Ser Asp Glu Glu Val Ser Lys Ala Leu Asn Leu
625 630 635 640
Asp Val Val Phe Lys Asp Lys Lys Asn Ile Ile Thr Lys Ile Asn Asp
645 650 655
Ile Lys Ile Ser Glu Glu Asn Asn Asn Asp Ile Lys Tyr Leu Pro Ser
660 665 670
Phe Ser Lys Val Leu Pro Glu Ile Leu Asn Leu Tyr Arg Asn Asn Pro
675 680 685
Lys Asn Glu Pro Phe Asp Thr Ile Glu Thr Glu Lys Ile Val Leu Asn
690 695 700
Ala Leu Ile Tyr Val Asn Lys Glu Leu Tyr Lys Lys Leu Ile Leu Glu
705 710 715 720
Asp Asp Leu Glu Glu Asn Glu Ser Lys Asn Ile Phe Leu Gln Glu Leu
725 730 735
Lys Lys Thr Leu Gly Asn Ile Asp Glu Ile Asp Glu Asn Ile Ile Glu
740 745 750
Asn Tyr Tyr Lys Asn Ala Gln Ile Ser Ala Ser Lys Gly Asn Asn Lys
755 760 765
Ala Ile Lys Lys Tyr Gln Lys Lys Val Ile Glu Cys Tyr Ile Gly Tyr
770 775 780
Leu Arg Lys Asn Tyr Glu Glu Leu Phe Asp Phe Ser Asp Phe Lys Met
785 790 795 800
Asn Ile Gln Glu Ile Lys Lys Gln Ile Lys Asp Ile Asn Asp Asn Lys
805 810 815
Thr Tyr Glu Arg Ile Thr Val Lys Thr Ser Asp Lys Thr Ile Val Ile
820 825 830
Asn Asp Asp Phe Glu Tyr Ile Ile Ser Ile Phe Ala Leu Leu Asn Ser
835 840 845
Asn Ala Val Ile Asn Lys Ile Arg Asn Arg Phe Phe Ala Thr Ser Val
850 855 860
Trp Leu Asn Thr Ser Glu Tyr Gln Asn Ile Ile Asp Ile Leu Asp Glu
865 870 875 880
Ile Met Gln Leu Asn Thr Leu Arg Asn Glu Cys Ile Thr Glu Asn Trp
885 890 895
Asn Leu Asn Leu Glu Glu Phe Ile Gln Lys Met Lys Glu Ile Glu Lys
900 905 910
Asp Phe Asp Asp Phe Lys Ile Gln Thr Lys Lys Glu Ile Phe Asn Asn
915 920 925
Tyr Tyr Glu Asp Ile Lys Asn Asn Ile Leu Thr Glu Phe Lys Asp Asp
930 935 940
Ile Asn Gly Cys Asp Val Leu Glu Lys Lys Leu Glu Lys Ile Val Ile
945 950 955 960
Phe Asp Asp Glu Thr Lys Phe Glu Ile Asp Lys Lys Ser Asn Ile Leu
965 970 975
Gln Asp Glu Gln Arg Lys Leu Ser Asn Ile Asn Lys Lys Asp Leu Lys
980 985 990
Lys Lys Val Asp Gln Tyr Ile Lys Asp Lys Asp Gln Glu Ile Lys Ser
995 1000 1005
Lys Ile Leu Cys Arg Ile Ile Phe Asn Ser Asp Phe Leu Lys Lys
1010 1015 1020
Tyr Lys Lys Glu Ile Asp Asn Leu Ile Glu Asp Met Glu Ser Glu
1025 1030 1035
Asn Glu Asn Lys Phe Gln Glu Ile Tyr Tyr Pro Lys Glu Arg Lys
1040 1045 1050
Asn Glu Leu Tyr Ile Tyr Lys Lys Asn Leu Phe Leu Asn Ile Gly
1055 1060 1065
Asn Pro Asn Phe Asp Lys Ile Tyr Gly Leu Ile Ser Asn Asp Ile
1070 1075 1080
Lys Met Ala Asp Ala Lys Phe Leu Phe Asn Ile Asp Gly Lys Asn
1085 1090 1095
Ile Arg Lys Asn Lys Ile Ser Glu Ile Asp Ala Ile Leu Lys Asn
1100 1105 1110
Leu Asn Asp Lys Leu Asn Gly Tyr Ser Lys Glu Tyr Lys Glu Lys
1115 1120 1125
Tyr Ile Lys Lys Leu Lys Glu Asn Asp Asp Phe Phe Ala Lys Asn
1130 1135 1140
Ile Gln Asn Lys Asn Tyr Lys Ser Phe Glu Lys Asp Tyr Asn Arg
1145 1150 1155
Val Ser Glu Tyr Lys Lys Ile Arg Asp Leu Val Glu Phe Asn Tyr
1160 1165 1170
Leu Asn Lys Ile Glu Ser Tyr Leu Ile Asp Ile Asn Trp Lys Leu
1175 1180 1185
Ala Ile Gln Met Ala Arg Phe Glu Arg Asp Met His Tyr Ile Val
1190 1195 1200
Asn Gly Leu Arg Glu Leu Gly Ile Ile Lys Leu Ser Gly Tyr Asn
1205 1210 1215
Thr Gly Ile Ser Arg Ala Tyr Pro Lys Arg Asn Gly Ser Asp Gly
1220 1225 1230
Phe Tyr Thr Thr Thr Ala Tyr Tyr Lys Phe Phe Asp Glu Glu Ser
1235 1240 1245
Tyr Lys Lys Phe Glu Lys Ile Cys Tyr Gly Phe Gly Ile Asp Leu
1250 1255 1260
Ser Glu Asn Ser Glu Ile Asn Lys Pro Glu Asn Glu Ser Ile Arg
1265 1270 1275
Asn Tyr Ile Ser His Phe Tyr Ile Val Arg Asn Pro Phe Ala Asp
1280 1285 1290
Tyr Ser Ile Ala Glu Gln Ile Asp Arg Val Ser Asn Leu Leu Ser
1295 1300 1305
Tyr Ser Thr Arg Tyr Asn Asn Ser Thr Tyr Ala Ser Val Phe Glu
1310 1315 1320
Val Phe Lys Lys Asp Val Asn Leu Asp Tyr Asp Glu Leu Lys Lys
1325 1330 1335
Lys Phe Lys Leu Ile Gly Asn Asn Asp Ile Leu Glu Arg Leu Met
1340 1345 1350
Lys Pro Lys Lys Val Ser Val Leu Glu Leu Glu Ser Tyr Asn Ser
1355 1360 1365
Asp Tyr Ile Lys Asn Leu Ile Ile Glu Leu Leu Thr Lys Ile Glu
1370 1375 1380
Asn Thr Asn Asp Thr Leu
1385
<210> 71
<211> 1300
<212> PRT
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polypeptide"
<400> 71
Met Ser Ile Tyr Gln Glu Phe Val Asn Lys Tyr Ser Leu Ser Lys Thr
1 5 10 15
Leu Arg Phe Glu Leu Ile Pro Gln Gly Lys Thr Leu Glu Asn Ile Lys
20 25 30
Ala Arg Gly Leu Ile Leu Asp Asp Glu Lys Arg Ala Lys Asp Tyr Lys
35 40 45
Lys Ala Lys Gln Ile Ile Asp Lys Tyr His Gln Phe Phe Ile Glu Glu
50 55 60
Ile Leu Ser Ser Val Cys Ile Ser Glu Asp Leu Leu Gln Asn Tyr Ser
65 70 75 80
Asp Val Tyr Phe Lys Leu Lys Lys Ser Asp Asp Asp Asn Leu Gln Lys
85 90 95
Asp Phe Lys Ser Ala Lys Asp Thr Ile Lys Lys Gln Ile Ser Glu Tyr
100 105 110
Ile Lys Asp Ser Glu Lys Phe Lys Asn Leu Phe Asn Gln Asn Leu Ile
115 120 125
Asp Ala Lys Lys Gly Gln Glu Ser Asp Leu Ile Leu Trp Leu Lys Gln
130 135 140
Ser Lys Asp Asn Gly Ile Glu Leu Phe Lys Ala Asn Ser Asp Ile Thr
145 150 155 160
Asp Ile Asp Glu Ala Leu Glu Ile Ile Lys Ser Phe Lys Gly Trp Thr
165 170 175
Thr Tyr Phe Lys Gly Phe His Glu Asn Arg Lys Asn Val Tyr Ser Ser
180 185 190
Asn Asp Ile Pro Thr Ser Ile Ile Tyr Arg Ile Val Asp Asp Asn Leu
195 200 205
Pro Lys Phe Leu Glu Asn Lys Ala Lys Tyr Glu Ser Leu Lys Asp Lys
210 215 220
Ala Pro Glu Ala Ile Asn Tyr Glu Gln Ile Lys Lys Asp Leu Ala Glu
225 230 235 240
Glu Leu Thr Phe Asp Ile Asp Tyr Lys Thr Ser Glu Val Asn Gln Arg
245 250 255
Val Phe Ser Leu Asp Glu Val Phe Glu Ile Ala Asn Phe Asn Asn Tyr
260 265 270
Leu Asn Gln Ser Gly Ile Thr Lys Phe Asn Thr Ile Ile Gly Gly Lys
275 280 285
Phe Val Asn Gly Glu Asn Thr Lys Arg Lys Gly Ile Asn Glu Tyr Ile
290 295 300
Asn Leu Tyr Ser Gln Gln Ile Asn Asp Lys Thr Leu Lys Lys Tyr Lys
305 310 315 320
Met Ser Val Leu Phe Lys Gln Ile Leu Ser Asp Thr Glu Ser Lys Ser
325 330 335
Phe Val Ile Asp Lys Leu Glu Asp Asp Ser Asp Val Val Thr Thr Met
340 345 350
Gln Ser Phe Tyr Glu Gln Ile Ala Ala Phe Lys Thr Val Glu Glu Lys
355 360 365
Ser Ile Lys Glu Thr Leu Ser Leu Leu Phe Asp Asp Leu Lys Ala Gln
370 375 380
Lys Leu Asp Leu Ser Lys Ile Tyr Phe Lys Asn Asp Lys Ser Leu Thr
385 390 395 400
Asp Leu Ser Gln Gln Val Phe Asp Asp Tyr Ser Val Ile Gly Thr Ala
405 410 415
Val Leu Glu Tyr Ile Thr Gln Gln Ile Ala Pro Lys Asn Leu Asp Asn
420 425 430
Pro Ser Lys Lys Glu Gln Glu Leu Ile Ala Lys Lys Thr Glu Lys Ala
435 440 445
Lys Tyr Leu Ser Leu Glu Thr Ile Lys Leu Ala Leu Glu Glu Phe Asn
450 455 460
Lys His Arg Asp Ile Asp Lys Gln Cys Arg Phe Glu Glu Ile Leu Ala
465 470 475 480
Asn Phe Ala Ala Ile Pro Met Ile Phe Asp Glu Ile Ala Gln Asn Lys
485 490 495
Asp Asn Leu Ala Gln Ile Ser Ile Lys Tyr Gln Asn Gln Gly Lys Lys
500 505 510
Asp Leu Leu Gln Ala Ser Ala Glu Asp Asp Val Lys Ala Ile Lys Asp
515 520 525
Leu Leu Asp Gln Thr Asn Asn Leu Leu His Lys Leu Lys Ile Phe His
530 535 540
Ile Ser Gln Ser Glu Asp Lys Ala Asn Ile Leu Asp Lys Asp Glu His
545 550 555 560
Phe Tyr Leu Val Phe Glu Glu Cys Tyr Phe Glu Leu Ala Asn Ile Val
565 570 575
Pro Leu Tyr Asn Lys Ile Arg Asn Tyr Ile Thr Gln Lys Pro Tyr Ser
580 585 590
Asp Glu Lys Phe Lys Leu Asn Phe Glu Asn Ser Thr Leu Ala Asn Gly
595 600 605
Trp Asp Lys Asn Lys Glu Pro Asp Asn Thr Ala Ile Leu Phe Ile Lys
610 615 620
Asp Asp Lys Tyr Tyr Leu Gly Val Met Asn Lys Lys Asn Asn Lys Ile
625 630 635 640
Phe Asp Asp Lys Ala Ile Lys Glu Asn Lys Gly Glu Gly Tyr Lys Lys
645 650 655
Ile Val Tyr Lys Leu Leu Pro Gly Ala Asn Lys Met Leu Pro Lys Val
660 665 670
Phe Phe Ser Ala Lys Ser Ile Lys Phe Tyr Asn Pro Ser Glu Asp Ile
675 680 685
Leu Arg Ile Arg Asn His Ser Thr His Thr Lys Asn Gly Ser Pro Gln
690 695 700
Lys Gly Tyr Glu Lys Phe Glu Phe Asn Ile Glu Asp Cys Arg Lys Phe
705 710 715 720
Ile Asp Phe Tyr Lys Gln Ser Ile Ser Lys His Pro Glu Trp Lys Asp
725 730 735
Phe Gly Phe Arg Phe Ser Asp Thr Gln Arg Tyr Asn Ser Ile Asp Glu
740 745 750
Phe Tyr Arg Glu Val Glu Asn Gln Gly Tyr Lys Leu Thr Phe Glu Asn
755 760 765
Ile Ser Glu Ser Tyr Ile Asp Ser Val Val Asn Gln Gly Lys Leu Tyr
770 775 780
Leu Phe Gln Ile Tyr Asn Lys Asp Phe Ser Ala Tyr Ser Lys Gly Arg
785 790 795 800
Pro Asn Leu His Thr Leu Tyr Trp Lys Ala Leu Phe Asp Glu Arg Asn
805 810 815
Leu Gln Asp Val Val Tyr Lys Leu Asn Gly Glu Ala Glu Leu Phe Tyr
820 825 830
Arg Lys Gln Ser Ile Pro Lys Lys Ile Thr His Pro Ala Lys Glu Ala
835 840 845
Ile Ala Asn Lys Asn Lys Asp Asn Pro Lys Lys Glu Ser Val Phe Glu
850 855 860
Tyr Asp Leu Ile Lys Asp Lys Arg Phe Thr Glu Asp Lys Phe Phe Phe
865 870 875 880
His Cys Pro Ile Thr Ile Asn Phe Lys Ser Ser Gly Ala Asn Lys Phe
885 890 895
Asn Asp Glu Ile Asn Leu Leu Leu Lys Glu Lys Ala Asn Asp Val His
900 905 910
Ile Leu Ser Ile Asp Arg Gly Glu Arg His Leu Ala Tyr Tyr Thr Leu
915 920 925
Val Asp Gly Lys Gly Asn Ile Ile Lys Gln Asp Thr Phe Asn Ile Ile
930 935 940
Gly Asn Asp Arg Met Lys Thr Asn Tyr His Asp Lys Leu Ala Ala Ile
945 950 955 960
Glu Lys Asp Arg Asp Ser Ala Arg Lys Asp Trp Lys Lys Ile Asn Asn
965 970 975
Ile Lys Glu Met Lys Glu Gly Tyr Leu Ser Gln Val Val His Glu Ile
980 985 990
Ala Lys Leu Val Ile Glu Tyr Asn Ala Ile Val Val Phe Glu Asp Leu
995 1000 1005
Asn Phe Gly Phe Lys Arg Gly Arg Phe Lys Val Glu Lys Gln Val
1010 1015 1020
Tyr Gln Lys Leu Glu Lys Met Leu Ile Glu Lys Leu Asn Tyr Leu
1025 1030 1035
Val Phe Lys Asp Asn Glu Phe Asp Lys Thr Gly Gly Val Leu Arg
1040 1045 1050
Ala Tyr Gln Leu Thr Ala Pro Phe Glu Thr Phe Lys Lys Met Gly
1055 1060 1065
Lys Gln Thr Gly Ile Ile Tyr Tyr Val Pro Ala Gly Phe Thr Ser
1070 1075 1080
Lys Ile Cys Pro Val Thr Gly Phe Val Asn Gln Leu Tyr Pro Lys
1085 1090 1095
Tyr Glu Ser Val Ser Lys Ser Gln Glu Phe Phe Ser Lys Phe Asp
1100 1105 1110
Lys Ile Cys Tyr Asn Leu Asp Lys Gly Tyr Phe Glu Phe Ser Phe
1115 1120 1125
Asp Tyr Lys Asn Phe Gly Asp Lys Ala Ala Lys Gly Lys Trp Thr
1130 1135 1140
Ile Ala Ser Phe Gly Ser Arg Leu Ile Asn Phe Arg Asn Ser Asp
1145 1150 1155
Lys Asn His Asn Trp Asp Thr Arg Glu Val Tyr Pro Thr Lys Glu
1160 1165 1170
Leu Glu Lys Leu Leu Lys Asp Tyr Ser Ile Glu Tyr Gly His Gly
1175 1180 1185
Glu Cys Ile Lys Ala Ala Ile Cys Gly Glu Ser Asp Lys Lys Phe
1190 1195 1200
Phe Ala Lys Leu Thr Ser Val Leu Asn Thr Ile Leu Gln Met Arg
1205 1210 1215
Asn Ser Lys Thr Gly Thr Glu Leu Asp Tyr Leu Ile Ser Pro Val
1220 1225 1230
Ala Asp Val Asn Gly Asn Phe Phe Asp Ser Arg Gln Ala Pro Lys
1235 1240 1245
Asn Met Pro Gln Asp Ala Asp Ala Asn Gly Ala Tyr His Ile Gly
1250 1255 1260
Leu Lys Gly Leu Met Leu Leu Gly Arg Ile Lys Asn Asn Gln Glu
1265 1270 1275
Gly Lys Lys Leu Asn Leu Val Ile Lys Asn Glu Glu Tyr Phe Glu
1280 1285 1290
Phe Val Gln Asn Arg Asn Asn
1295 1300
<210> 72
<211> 7403
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 72
tatccggtcg aatcgagaat gacgaccgct acgtcttgga ctacgaagcc gtggcccttg 60
ccgatgctct cggtgtggat gttgccgacc tgttccgcaa gatcgattgc cccaagaacc 120
tgctgcgcag gcgggcaggg taggggagcg gtttccggcg gagattttcg gaggcgccgg 180
taacgttatg tcggggaatt tgctatacat cgacgataat tagttttgtt gattcaggat 240
cgaaatgcgc tcaaacaaag aacgttccgc gtttccctca tgcgctacta cgcccacacc 300
gccatctttc ggcacgcaaa caaagcagat gggttgcctg tcaatgggtg atcattgcct 360
gaagttacca tccatcaata atataaatca tccttactcc gaatgtccct caatcgcatc 420
tatcaaggcc gcgtggcggc cgtcgaaaca ggaacggcct tagcgaaagg taatgtcgaa 480
tggatgcctg ccgcaggagg cgacgaagtt ctctggcagc accacgaact tttccaagct 540
gccatcaact actatctcgt cgccctgctc gcactcgccg acaaaaacaa tcccgtactt 600
ggcccgctga tcagccagat ggataatccc caaagccctt accatgtctg gggaagtttc 660
cgccgccaag gacgtcagcg cacaggtctc agtcaagccg ttgcacctta tatcacgccg 720
ggcaataacg ctcccaccct tgacgaagtt ttccgctcca ttcttgcggg caacccaacc 780
gaccgcgcaa ctttggacgc tgcactcatg caattgctca aggcttgtga cggcgcgggc 840
gctatccagc aggaaggtcg ttcctactgg cccaaattct gcgatcctga ctccactgcc 900
aacttcgcgg gagatccggc catgctccgg cgtgaacaac accgcctcct ccttccgcaa 960
gttctccacg atccggcgat tactcacgac agtcctgccc ttggctcgtt cgacacttat 1020
tcgattgcta cccccgacac cagaactcct caactcaccg gccccaaggc acgcgcccgt 1080
cttgagcagg cgatcaccct ctggcgcgtc cgtcttcccg aatcggctgc tgacttcgat 1140
cgccttgcca gttccctcaa aaaaattccg gacgacgatt ctcgccttaa ccttcagggc 1200
tacgtcggca gcagtgcgaa aggcgaagtt caggcccgtc ttttcgccct tctgctattc 1260
cgtcacctgg agcgttcctc ctttacgctt ggccttctcc gttccgccac cccgccgccc 1320
aagaacgctg aaacacctcc tcccgccggc gttcctttac ctgcggcgtc cgcagccgat 1380
ccggtgcgga tagcccgtgg caaacgcagt tttgtttttc gcgcattcac cagtctcccc 1440
tgctggcatg gcggtgataa catccatccc acctggaagt cattcgacat cgcagcgttc 1500
aaatatgccc tcacggtcat caaccagatc gaggaaaaga cgaaagaacg ccaaaaagaa 1560
tgtgcggaac ttgaaactga tttcgactac atgcacggac ggctcgccaa gattccggta 1620
aaatacacga ccggcgaagc cgaaccgccc cccattctcg caaacgatct ccgcatcccc 1680
ctcctccgcg aacttctcca gaatatcaag gtcgacaccg cactcaccga tggcgaagcc 1740
gtctcctatg gtctccaacg ccgcaccatt cgcggtttcc gcgagctgcg ccgcatctgg 1800
cgcggccatg cccccgctgg cacggtcttt tccagcgagt tgaaagaaaa actagccggc 1860
gaactccgcc agttccagac cgacaactcc accaccatcg gcagcgtcca actcttcaac 1920
gaactcatcc aaaacccgaa atactggccc atctggcagg ctcctgacgt cgaaaccgcc 1980
cgccaatggg ccgatgccgg ttttgccgac gatccgctcg ccgcccttgt gcaagaagcc 2040
gaactccagg aagacatcga cgccctcaag gctccagtca aactcactcc ggccgatcct 2100
gagtattcaa gaaggcaata cgatttcaat gccgtcagca aattcggggc cggctcccgc 2160
tccgccaatc gccacgaacc cgggcagacg gagcgcggcc acaacacctt taccaccgaa 2220
atcgccgccc gtaacgcggc ggacgggaac cgctggcggg caacccacgt ccgcatccat 2280
tactccgctc cccgccttct tcgtgacgga ctccgccgac ctgacaccga cggcaacgaa 2340
gccctggaag ccgtcccttg gctccagccc atgatggaag ccctcgcccc tctcccgacg 2400
cttccgcaag acctcacagg catgccggtc ttcctcatgc ccgacgtcac cctttccggt 2460
gagcgtcgca tcctcctcaa tcttcctgtc accctcgaac cagccgctct tgtcgaacaa 2520
ctgggcaacg ccggtcgctg gcaaaaccag ttcttcggct cccgcgaaga tccattcgct 2580
ctccgatggc ccgccgacgg tgctgtaaaa accgccaagg ggaaaaccca cataccttgg 2640
caccaggacc gcgatcactt caccgtactc ggcgtggatc tcggcacgcg cgatgccggg 2700
gcgctcgctc ttctcaacgt cactgcgcaa aaaccggcca agccggtcca ccgcatcatt 2760
ggtgaggccg acggacgcac ctggtatgcc agccttgccg acgctcgcat gatccgcctg 2820
cccggggagg atgcccggct ctttgtccgg ggaaaactcg ttcaggaacc ctatggtgaa 2880
cgcgggcgaa acgcgtctct tctcgaatgg gaagacgccc gcaatatcat ccttcgcctt 2940
ggccaaaatc ccgacgaact cctcggcgcc gatccccggc gccattcgta tccggaaata 3000
aacgataaac ttctcgtcgc ccttcgccgc gctcaggccc gtcttgcccg tctccagaac 3060
cggagctggc ggttgcgcga ccttgcagaa tcggacaagg cccttgatga aatccatgcc 3120
gagcgtgccg gggagaagcc ttctccgctt ccgcccttgg ctcgcgacga tgccatcaaa 3180
agcaccgacg aagccctcct ttcccagcgt gacatcatcc ggcgatcctt cgttcagatc 3240
gccaacttga tccttcccct tcgcggacgc cgatgggaat ggcggcccca tgtcgaggtc 3300
ccggattgcc acatccttgc gcagagcgat cccggtacgg atgacaccaa gcgtcttgtc 3360
gccggacaac gcggcatctc tcacgagcgt atcgagcaaa tcgaagaact ccgtcgtcgc 3420
tgccaatccc tcaaccgtgc cctgcgtcac aaacccggag agcgtcccgt gctcggacgc 3480
cccgccaagg gcgaggaaat cgccgatccc tgtcccgcgc tcctcgaaaa gatcaaccgt 3540
ctccgggacc agcgcgttga ccaaaccgcg catgccatcc tcgccgccgc tctcggtgtt 3600
cgactccgcg ccccctcaaa agaccgcgcc gaacgccgcc atcgcgacat ccatggcgaa 3660
tacgaacgct ttcgtgcgcc cgctgatttt gtcgtcatcg aaaacctctc ccgttatctc 3720
agctcgcagg atcgtgctcg tagtgaaaac acccgtctca tgcagtggtg ccatcgccag 3780
atcgtgcaaa aactccgtca gctctgcgag acctacggca tccccgtcct cgccgtcccg 3840
gcggcctact catcgcgttt ttcttcccgg gacggctcgg ccggattccg ggccgtccat 3900
ctgacaccgg accaccgtca ccggatgcca tggagccgca tcctcgcccg cctcaaggcc 3960
cacgaggaag acggaaaaag actcgaaaag acggtgctcg acgaggctcg cgccgtccgg 4020
ggactctttg accggctcga ccggttcaac gccgggcatg tcccgggaaa accttggcgc 4080
acgctcctcg cgccgctccc cggcggccct gtgtttgtcc ccctcgggga cgccacaccc 4140
atgcaggccg atctgaacgc cgccatcaac atcgccctcc ggggcatcgc ggctcccgac 4200
cgccacgaca tccatcaccg gctccgtgcc gaaaacaaaa aacgcatcct gagcttgcgt 4260
ctcggcactc agcgcgagaa agcccgctgg cctggaggag ctccggcggt gacactctcc 4320
actccgaaca acggcgcctc tcccgaagat tccgatgcgt tgcccgaacg ggtatccaac 4380
ctgtttgtgg acatcgccgg tgtcgccaac ttcgagcgag tcacgatcga aggagtctcg 4440
caaaaattcg ccaccgggcg tggcctttgg gcctccgtca agcaacgtgc atggaaccgc 4500
gttgccagac tcaacgagac agtaacagat aacaacagga acgaagagga ggacgacatt 4560
ccgatgtaac cattgcttca ttacatctga gtctcccctc aatccctctg ccccatgcgt 4620
gatataacct ccacctcatg tcccggatcg gcgccggcaa cctgtagttc ccttccatcc 4680
tccaacactc ccgcagatcg cgatccgctg ccgccgatgc cggtgcgccg ccttcacaac 4740
tatctctact gtccgcggct tttttatctc cagtgggtcg agaatctctt tgaggaaaat 4800
gccgacacca ttgccggcag cgccgtgcat cgtcacgccg acaaacctac gcgttacgat 4860
gatgaaaaag ccgaggcact tcgcactggt ctccctgaag gcgcgcacat acgcagcctt 4920
cgcctggaaa acgcccaact cggtctcgtt ggcgtggtgg atatcgtgga gggaggcccc 4980
gacggactcg aactcgtcga ctacaaaaaa ggttccgcct tccgcctcga cgacggcacg 5040
ctcgctccca aggaaaacga caccgtgcaa cttgccgcct acgctcttct cctggctgcc 5100
gatggtgcgc gcgttgcgcc catggcgacg gtctattacg ctgccgatcg ccggcgtgtc 5160
accttcccgc tcgatgacgc cctctacgcc cgcacccgtt ccgccctcga agaggcccgc 5220
gccgttgcaa cctcggggcg catacctccg ccgctcgtct ctgacgtccg ctgcctccat 5280
tgttcctcct atgcgctttg ccttccccgc gagtccgcct ggtggtgccg ccatcgcagc 5340
acgccgcggg gagccggcca cacccccatg ttgccgggct ttgaggatga cgccgccgcc 5400
attcaccaaa tctccgaacc tgacaccgag ccaccacccg atcttgccag ccagcctccc 5460
cgtcccccgc ggctcgatgg agaattgttg gttgtccaga ctccgggagc gatgatcgga 5520
caaagcggcg gtgagtttac cgtgtccgtc aagggtgagg ttttgcgcaa gcttccggtt 5580
catcaactcc gggccattta cgtttacgga gccgtgcaac tcacggcgca tgctgtgcag 5640
accgcccttg aggaggatat cgacgtctcc tattttgcgc ccagcggccg ctttcttggc 5700
ctcctccgcg gcctgcccgc atccggcgtg gatgcgcgtc tcgggcaata caccctgttt 5760
cgcgaaccct ttggccgtct ccgtctcgcc tgcgaggcga ttcgggccaa gatccataac 5820
cagcgcgtcc tcctcatgcg taacggcgag cccggggagg gcgtcttgcg cgaactcgcc 5880
cgtctgcgcg acgccaccag tgaggcgact tcgctcgacg aactcctcgg catcgagggc 5940
atcgccgcgc atttctattt ccagtatttt cccaccatgc tgaaagaacg ggcggcctgg 6000
gcctttgatt tttccggacg caatcgccgc ccgccgcgcg acccggtcaa cgccctgctt 6060
tcgttcggtt acagcgtgtt gtccaaggaa cttgccggcg tctgccacgc tgttggccta 6120
gacccgtttt tcggcttcat gcaccagccg cgttacgggc gccccgcact cgctctcgat 6180
ctgatggagg agtttcgccc tctcatcgcc gacagtgttg ccctgaatct catcaaccgt 6240
ggcgaactcg acgaagggga ctttatccgg tcggccaatg gcaccgcgct caatgatcgg 6300
ggccgccggc gtttttggga ggcatggttc cggcgtctcg acagcgaagt cagccatcct 6360
gaatttggtt acaagatgag ctatcgacgg atgcttgaag tgcaggcgcg ccagctatgg 6420
cgctatgtgc gcggtgacgc cttccgctac cacggattca ccacccgttg attccgatgt 6480
cagatccccg ccgccgttat cttgtgtgtt acgacatcgc caatccgaag cgattgcgcc 6540
aagtggccaa gctgctggag agctatggca cgcgtctgca atactcggtt ttcgaatgtc 6600
ctttggacga tcttcgtctt gaacaggcga aggctgattt gcgcgacacg attaatgccg 6660
accaagacca ggtgttattt gtttcgcttg gccccgaagc caacgatgcc acgttgatca 6720
tcgccacgct tgggctccct tataccgtgc gctcgcgagt gacgattatc tgacccataa 6780
cccacgtgtt gaagaggctg aaaacagacg gacctctatg aagaacaatt gacgttttgg 6840
ccgaactcag cagaccttta tgcggctaag gccaatgatc atccatccta ccgccattgg 6900
gctggagacg ttttttgaaa cggcgagtgc tgcggatagc gagtttctct tggggaggcg 6960
ctcgcggcca cttttacaga ggagatgttc gggcgaactg gccgacctaa caaggcgtac 7020
ccggctcaaa atcgaggcac gctcgcacgg gatgatgtaa ttcgttgttt ttcagcatac 7080
cgtgcgagca cgggccgcag cgaatgccgt ttcacgaatc gtcaggcggc ggggagaagt 7140
catttaataa ggccactgtt aaaagccgca gcgaatgccg tttcacgaat cgtcaggcgg 7200
gcagtggatg tttttccatg aggcgaagaa tttcatcgcc gcagtgaatg ccgtttcacc 7260
attgatgaag aatgcgaggt gaaaacagag aaattgggtc aactctatca ctcttattca 7320
gccatcgttt caagaaagga tacctcgtat tggatacaac acagctcgtt cgttctctct 7380
acctccctcg acaatctcaa gga 7403
<210> 73
<211> 6789
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 73
taataaaatt gaaatatcac tatggattat tgtaatatta ccataaagat aggtgacgtt 60
tttttgaaaa ttgtaaacct aatttgaaga aaaccaatta aaaatcgctt cggctttttt 120
ttaagtgcca ggtagcattg atgctaaccc atgtgtaata aaggtttgtt ttccttcggg 180
gcacgaacac attataaggg aaacctaaag attccctttc ttgtttaata ttataaccag 240
tgaaaataag aataatgcac ctaaaactaa tatacagaaa ataagaatta aaagtactaa 300
tatatacatc atatgttatc ctccaatgct ttatttttta ataattgatg ttagtattag 360
ttttatttta atttctaaac ataagaattt gaaaaggatg tgtttattat ggcgacacgc 420
agttttattt taaaaattga accaaatgaa gaagttaaaa agggattatg gaagacgcat 480
gaggtattga atcatggaat tgcctactac atgaatattc tgaaactaat tagacaggaa 540
gctatttatg aacatcatga acaagatcct aaaaatccga aaaaagtttc aaaagcagaa 600
atacaagccg agttatggga ttttgtttta aaaatgcaaa aatgtaatag ttttacacat 660
gaagttgaca aagatgttgt ttttaacatc ctgcgtgaac tatatgaaga gttggtccct 720
agttcagtcg agaaaaaggg tgaagccaat caattatcga ataagtttct gtacccgcta 780
gttgatccga acagtcaaag tgggaaaggg acggcatcat ccggacgtaa acctcggtgg 840
tataatttaa aaatagcagg cgacccatcg tgggaggaag aaaagaaaaa atgggaagag 900
gataaaaaga aagatcccct tgctaaaatc ttaggtaagt tagcagaata tgggcttatt 960
ccgctattta ttccatttac tgacagcaac gaaccaattg taaaagaaat taaatggatg 1020
gaaaaaagtc gtaatcaaag tgtccggcga cttgataagg atatgtttat ccaagcatta 1080
gagcgttttc tttcatggga aagctggaac cttaaagtaa aggaagagta tgaaaaagtt 1140
gaaaaggaac acaaaacact agaggaaagg ataaaagagg acattcaagc atttaaatcc 1200
cttgaacaat atgaaaaaga acggcaggag caacttctta gagatacatt gaatacaaat 1260
gaataccgat taagcaaaag aggattacgt ggttggcgtg aaattatcca aaaatggcta 1320
aagatggatg aaaatgaacc atcagaaaaa tatttagaag tatttaaaga ttatcaacgg 1380
aaacatccac gagaagccgg ggactattct gtctatgaat ttttaagcaa gaaagaaaat 1440
cattttattt ggcgaaatca tcctgaatat ccttatttgt atgctacatt ttgtgaaatt 1500
gacaaaaaaa agaaagacgc taagcaacag gcaactttta ctttggctga cccgattaac 1560
catccgttat gggtacgatt tgaagaaaga agcggttcga acttaaacaa atatcgaatt 1620
ttaacagagc aattacacac tgaaaagtta aaaaagaaat taacagttca acttgatcgt 1680
ttaatttatc caactgaatc cggcggttgg gaggaaaaag gtaaagtaga tatcgttttg 1740
ttgccgtcaa gacaatttta taatcaaatc ttccttgata tagaagaaaa ggggaaacat 1800
gcttttactt ataaggatga aagtattaaa ttccccctta aaggtacact tggtggtgca 1860
agagtgcagt ttgaccgtga ccatttgcgg agatatccgc ataaagtaga atcaggaaat 1920
gttggacgga tttattttaa catgacagta aatattgaac caactgagag ccctgttagt 1980
aagtctttga aaatacatag ggacgatttc cccaagttcg ttaattttaa accgaaagag 2040
ctcaccgaat ggataaaaga tagtaaaggg aaaaaattaa aaagtggtat agaatccctt 2100
gaaattggtc tacgggtgat gagtatcgac ttaggtcaac gtcaagcggc tgctgcatcg 2160
atttttgaag tagttgatca gaaaccggat attgaaggga agttattttt tccaatcaaa 2220
ggaactgagc tttatgctgt tcaccgggca agttttaaca ttaaattacc gggtgaaaca 2280
ttagtaaaat cacgggaagt attgcggaaa gctcgggagg acaacttaaa attaatgaat 2340
caaaagttaa actttctaag aaatgttcta catttccaac agtttgaaga tatcacagaa 2400
agagagaagc gtgtaactaa atggatttct agacaagaaa atagtgatgt tcctcttgta 2460
tatcaagatg agctaattca aattcgtgaa ttaatgtata aaccctataa agattgggtt 2520
gcctttttaa aacaactcca taaacggcta gaagtcgaga ttggcaaaga ggttaagcat 2580
tggcgaaaat cattaagtga cgggagaaaa ggtctttacg gaatctccct aaaaaatatt 2640
gatgaaattg atcgaacaag gaaattcctt ttaagatgga gcttacgtcc aacagaacct 2700
ggggaagtaa gacgcttgga accaggacag cgttttgcga ttgatcaatt aaaccaccta 2760
aatgcattaa aagaagatcg attaaaaaag atggcaaata cgattatcat gcatgcctta 2820
ggttactgtt atgatgtaag aaagaaaaag tggcaggcaa aaaatccagc atgtcaaatt 2880
attttatttg aagatttatc taactacaat ccttacgagg aaaggtcccg ttttgaaaac 2940
tcaaaactga tgaagtggtc acggagagaa attccacgac aagtcgcctt acaaggtgaa 3000
atttacggat tacaagttgg ggaagtaggt gcccaattca gttcaagatt ccatgcgaaa 3060
accgggtcgc cgggaattcg ttgcagtgtt gtaacgaaag aaaaattgca ggataatcgc 3120
ttttttaaaa atttacaaag agaaggacga cttactcttg ataaaatcgc agttttaaaa 3180
gaaggagact tatatccaga taaaggtgga gaaaagttta tttctttatc aaaggatcga 3240
aagttggtaa ctacgcatgc tgatattaac gcggcccaaa atttacagaa gcgtttttgg 3300
acaagaacac atggatttta taaagtttac tgcaaagcct atcaggttga tggacaaact 3360
gtttatattc cggagagcaa ggaccaaaaa caaaaaataa ttgaagaatt tggggaaggc 3420
tattttattt taaaagatgg tgtatatgaa tggggtaatg cggggaaact aaaaattaaa 3480
aaaggttcct ctaaacaatc atcgagtgaa ttagtagatt cggacatact gaaagattca 3540
tttgatttag caagtgaact taagggagag aaactcatgt tatatcgaga tccgagtgga 3600
aacgtatttc cttccgacaa gtggatggca gcaggagtat tttttggcaa attagaaaga 3660
atattgattt ctaagttaac aaatcaatac tcaatatcaa caatagaaga tgattcttca 3720
aaacaatcaa tgtaaaagtt tgcccgtata agaacttaat taattaggat ggtaggatgt 3780
tactaaatat gtctgtaggc atcattccta ctatccgttt tgtccgaata tcagagcatt 3840
aggtgaggaa tggtaagaaa ggaaaattta tatgaaccaa ccgattccta ttcgaatgtt 3900
aaatgaaata caatattgtg agcgactttt ttactttatg catgtccaaa agctatttga 3960
tgagaatgca gatacagttg aaggaagtgc acagcatgag cgggcagaaa gaagcaaaag 4020
accaagtaaa atgggaccaa aggaattatg gggtgaggcg ccaagaagtc ttaagcttgg 4080
tgatgagctg ttaaatatta ccggtgttct tgatgccata agtcatgaag agaacagttg 4140
gatcccggtt gaatcaaaac acagttccgc accggatgga ttgaaccctt ttaaagtaga 4200
tggctttcta cttgacgggt ctgcatggcc aaacgatcaa attcaacttt gtgcacaagg 4260
cttgctcttg aatgccaatg gatacccgtg tgattatggg tatttatttt atcgtggtaa 4320
taagaaaaag gtgaaaattt attttactga agatttaatc gctgccacaa agtactatat 4380
taaaaaagca cacgagatac tagtattatc tggtgatgaa tcagctattc ctaagccttt 4440
aattgattct aataagtgtt ttcgctgttc tttaaactat atctgtcttc cggatgaaac 4500
gaactatcta ttaggggcaa gttcaacaat tcgtaaaatt gtgccttcaa ggacagatgg 4560
tggcgtttta tatgtatcag agtctggtac aaaattagga aaatcgggtg aggagttaat 4620
cattcagtat aaagatggcc aaaagcaggg tgttcctata aaagatatta ttcaagtttc 4680
gttaattgga aatgttcaat gctcaacgca attacttcat tttttaatgc aatcaaatat 4740
tcctgtaagt tatttatcat cccacggtcg tttgattggt gtcagttcat ctttagttac 4800
aaaaaatgtt ttaacaaggc agcaacagtt cattaaattt acaaatcctg agtttggact 4860
aaatctagca aaacaaattg tttatgccaa gattcgaaat caacgaactt tacttagaag 4920
aaatgggggg agtgaggtaa aggagatttt aacagattta aaatctttaa gtgacagtgc 4980
actgaacgca atatcaatag aacaattacg gggtattgaa gggatttctg caaaacatta 5040
tttcgcagga tttccgttta tgttgaaaaa tgaattacgt gaattgaatt taatgaaagg 5100
gcgtaatagg agaccgccaa aagatcctgt aaatgtactt ctttctcttg gttatacttt 5160
attgacacgt gatattcatg ctgcgtgtgg ttcagtcgga ttggatccga tgtttggttg 5220
ttaccatcgt ccagaagcag gtcgaccggc tctagtatta gatgttatgg aaacatttcg 5280
accacttatt gtagacagta ttgtcatccg agctttgaat acgggtgaaa tctcattaaa 5340
agatttttat ataggaaaag atagttgtca attattaaaa catggccgcg attccttttt 5400
tgccatttat gaaagaagaa tgcatgaaac tattaccgat ccaattttcg gctataagat 5460
tagctatcgc cgtatgctcg atttgcacat tcgaatgctt gcaaggttta ttgaagggga 5520
actgccggaa tataaaccat taatgacccg gtgagtttgt ttattaggtt aaaagaaggt 5580
gaagacatgc agcaatacgt ccttgtttct tatgatattt cggaccaaaa aagatggaga 5640
aaagtattta aactgatgaa aggatacgga gaacatgttc aatattccgt attcatatgc 5700
cagttaactg aattacagaa ggcaaaatta caagcctctt tagaagacat tatccatcat 5760
aagaatgacc aagtaatgtt tgttcacatc gggccagtga aagatggtca actatctaaa 5820
aaaatctcaa caattgggaa agaatttgtt ccattggatt taaagcggct tatattttga 5880
aaagatatag caaagaaatc ttatgaaaaa aatacaaaaa tatattgtta aaaaataggg 5940
aatattatat aatggactta cgaggttctg tcttttggtc aggacaaccg tctagctata 6000
agtgctgcag gggtgtgaga aactcctatt gctggacgat gtctctttta tttctttttt 6060
cttggatctg agtacgagca cccacattgg acatttcgca tggtgggtgc tcgtactata 6120
ggtaaaacaa acctttttaa gaagaataca aaaataacca caatattttt taaaaggaat 6180
tttgatggat ttacataacc tctcgcaaca tgcttctaaa acccaagccc accatagccc 6240
aaaaccccct gcggtccaag aaaaaagaaa tgatacgagg cattagcacc ggggagaagt 6300
catttaataa ggccactgtt aaaagtccaa gaaaaaagaa atgatacgag gcattagcac 6360
aacaatataa acgactactt taccgtgttc aagaaaaaag aaatgatatg aggcattagc 6420
acgatgggat gggagagaga ggacagttct actcttgctg tatccagctt cttttacttt 6480
atccggtatc atttcttcac ttctttctgc acataaaaaa gcacctaact atttggataa 6540
gttaagtgct tttatttccg tttgaagttg tctattgctt ttttcttcat atcttcaaat 6600
tttttctgtt tctcagagtc aactttacca actgtaatcc cttttctttt tggcattggg 6660
gtatctttcc accttagtgt gttcataagg cttatattta tcactcattg tattcctcca 6720
acacaattat aatttttccg tcatcctcaa tccaaccgtc aactgtgaca aaagacgaat 6780
ctctcttat 6789
<210> 74
<211> 6214
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 74
gtttcatttg gaaagggaga gcattggctt ttctctttgt aaataaagtg caagctttgt 60
aataagcttc tagtggagaa gtgattgttt gaatcaccca atgcacacgc actaaagtta 120
gacgaaccta taattcgtat tagtaagtat agtacatgaa gaaaaatgca acaagcattt 180
actctctttt aaataaagaa ttgatagctg ttaatattga tagtatatta taccttatag 240
atgttcgatt ttttttgaaa ttcaaaaatc atacttagta aagaaaggaa ataacgtcat 300
ggacaagcga aagcgtagaa gttacgagtt taggtgggaa gcgggaggca ccagtcatgg 360
caatccgtag cataaaacta aaactaaaaa cccacacagg cccggaagcg caaaacctcc 420
gaaaaggaat atggcggacg catcggttgt taaatgaagg cgtcgcctat tacatgaaaa 480
tgctcctgct ctttcgtcag gaaagcactg gtgaacggcc aaaagaagaa ctacaggaag 540
aactgatttg tcacatacgc gaacagcaac aacgaaatca ggcagataaa aatacgcaag 600
cgcttccgct agataaggca ctggaagctt tgcgccaact atatgaactg cttgtcccct 660
cctcggtcgg acaaagtggc gacgcccaga tcatcagccg aaagtttctc agcccgctcg 720
tcgatccgaa cagcgaaggc ggcaaaggta cttcgaaggc aggggcaaaa cccacttggc 780
agaagaaaaa agaagcgaac gacccaacct gggaacagga ttacgaaaaa tggaaaaaaa 840
gacgcgagga agacccaacc gcttctgtga ttactacttt ggaggaatac ggcattagac 900
cgatctttcc cctgtacacg aacaccgtaa cagatatcgc gtggttgcca cttcaatcca 960
atcagtttgt gcgaacctgg gacagagaca tgcttcaaca agcgattgaa agactgctca 1020
gttgggagag ctggaacaaa cgtgtccagg aagagtatgc caagctgaaa gaaaaaatgg 1080
ctcaactgaa cgagcaactc gaaggcggtc aggaatggat cagcttgcta gagcagtacg 1140
aagaaaaccg agagcgagag cttagggaaa acatgaccgc tgccaatgac aagtatcgga 1200
ttaccaagcg gcaaatgaaa ggctggaacg agctgtacga gctatggtca acctttcccg 1260
ccagtgccag tcacgagcaa tacaaagagg cgctcaagcg tgtgcagcag cgactgagag 1320
ggcggtttgg ggatgctcat ttcttccagt atctgatgga agagaagaac cgcctgatct 1380
ggaaggggaa tccgcagcgt atccattatt ttgtcgcgcg caacgaactg acgaaacggc 1440
tggaggaagc caagcaaagc gccacgatga cgttgcccaa tgccaggaag catccattgt 1500
gggtgcgctt cgatgcacgg ggaggaaatt tgcaagacta ctacttgacg gctgaagcgg 1560
acaaaccgag aagcagacgt tttgtaacgt ttagtcagtt gatatggcca agcgaatcgg 1620
gatggatgga aaagaaagac gtcgaggtcg agctagcttt gtccaggcag ttttaccagc 1680
aggtgaagtt gctgaaaaat gacaaaggca agcagaaaat cgagttcaag gataaaggtt 1740
cgggctcgac gtttaacgga cacttggggg gagcaaagct acaactggag cggggcgatt 1800
tggagaagga agaaaaaaac ttcgaggacg gggaaatcgg cagcgtttac cttaacgttg 1860
tcattgattt cgaacctttg caagaagtga aaaatggccg cgtgcaggcg ccgtatggac 1920
aagtactgca actcattcgt cgccccaacg agtttcccaa ggtcactacc tataagtcgg 1980
agcaacttgt tgaatggata aaagcttcgc cacaacactc ggctggggtg gagtcgctgg 2040
catccggttt tcgtgtaatg agcatagacc ttgggctgcg cgcggctgca gcgacttcta 2100
ttttttctgt agaagagagt agcgataaaa atgcggctga tttttcctac tggattgaag 2160
gaacgccgct ggtcgctgtc catcagcgga gctatatgct caggttgcct ggtgaacagg 2220
tagaaaaaca ggtgatggaa aaacgggacg agcggttcca gctacaccaa cgtgtgaagt 2280
ttcaaatcag agtgctcgcc caaatcatgc gtatggcaaa taagcagtat ggagatcgct 2340
gggatgaact cgacagcctg aaacaagcgg ttgagcagaa aaagtcgccg ctcgatcaaa 2400
cagaccggac attttgggag gggattgtct gcgacttaac aaaggttttg cctcgaaacg 2460
aagcggactg ggaacaagcg gtagtgcaaa tacaccgaaa agcagaggaa tacgtcggaa 2520
aagccgttca ggcatggcgc aagcgctttg ctgctgacga gcgaaaaggc atcgcaggtc 2580
tgagcatgtg gaacatagaa gaattggagg gcttgcgcaa gctgttgatt tcctggagcc 2640
gcaggacgag gaatccgcag gaggttaatc gctttgagcg aggccatacc agccaccagc 2700
gtctgttgac ccatatccaa aacgtcaaag aggatcgcct gaagcagtta agtcacgcca 2760
ttgtcatgac tgccttgggg tatgtttacg acgagcggaa acaagagtgg tgcgccgaat 2820
acccggcttg ccaggtcatt ctgtttgaaa atctgagcca gtaccgttct aacctggatc 2880
gctcgaccaa agaaaactcc accttgatga agtgggcgca tcgcagcatt ccgaaatacg 2940
tccacatgca ggcggagcca tacgggattc agattggcga tgtccgggcg gaatattcct 3000
ctcgttttta cgccaagaca ggaacgccag gcattcgttg taaaaaggtg agaggccaag 3060
acctgcaggg cagacggttt gagaacttgc agaagaggtt agtcaacgag caatttttga 3120
cggaagaaca agtgaaacag ctaaggcccg gcgacattgt cccggatgat agcggagaac 3180
tgttcatgac cttgacagac ggaagcggaa gcaaggaggt cgtgtttctc caggccgata 3240
ttaacgcggc gcacaatctg caaaaacgtt tttggcagcg atacaatgaa ctgttcaagg 3300
ttagctgccg cgtcatcgtc cgagacgagg aagagtatct cgttcccaag acaaaatcgg 3360
tgcaggcaaa gctgggcaaa gggctttttg tgaaaaaatc ggatacagcc tggaaagatg 3420
tatatgtgtg ggacagccag gcaaagctta aaggtaaaac aacctttaca gaagagtctg 3480
agtcgcccga acaactggaa gactttcagg agatcatcga ggaagcagaa gaggcgaaag 3540
gaacataccg tacactgttc cgcgatccta gcggagtctt ttttcccgaa tccgtatggt 3600
atccccaaaa agatttttgg ggcgaggtga aaaggaagct gtacggaaaa ttgcgggaac 3660
ggtttttgac aaaggctcgg taagggtgtg caaggagagt gaatggcttg tcctggatac 3720
ctgtccgcat gctaaatgaa attcagtatt gtgagcgact gtaccatatt atgcatgtgc 3780
aggggctgtt tgaggaaagc gcagacacgg tcgaaggagc agcacaacac aagcgtgcag 3840
agacacatct gcgcaaaagc aaggcagcgc cggaagagat gtggggggac gctccgttta 3900
gcttgcagct cggcgaccct gtgcttggca ttacgggaaa gctggatgcc gtctgtctgg 3960
aagaaggtaa gcagtggatt ccggtagaag gaaagcattc ggcgtcgcca gaaggcgggc 4020
agatgttcac tgtaggcgtg tattcgctgg acggttctgc ctggcccaac gaccaaatcc 4080
aattgtgtgc gcaaggcttg ctgcttcgcg cgaatggata tgaatccgat tatggctact 4140
tatactaccg tggcaataaa aagaaggttc gcattccttt ttcgcaggaa ctcatagcgg 4200
ctactcacgc ctgcattcaa aaagctcatc agcttcggga agccgaaatt ccccctccgt 4260
tgcaggagtc gaaaaagtgc tttcgatgct cgttaaatta cgtatgcatg cctgacgaga 4320
cgaattacat gttggggttg agcgcaaaca tcagaaagat tgtgcccagt cgtccagatg 4380
gcggggtact gtatgttaca gagcaggggg caaaactggg cagaagcgga gaaagcttga 4440
ccatcacctg ccggggcgaa aagatagacg aaatcccgat caaagacttg attcacgtga 4500
gcttgatggg gcatgtgcaa tgctctacgc agcttctgca caccttgatg aactgtggcg 4560
tccacgtcag ctacttgact acgcatggca cattgacagg aataatgact ccccctttat 4620
cgaaaaacat tcgaacaaga gccaagcagt ttatcaaatt tcagcacgcg gagatcgccc 4680
ttggaatcgc gagaagggtc gtgtatgcga aaatttccaa tcagcgcacg atgctgcgcc 4740
gcaatggctc accagataaa gcagttttaa aagagttaaa agagcttaga gatcgcgcgt 4800
gggaggcgcc atcactggaa atagtgagag gtatcgaggg acgtgcagca cagttgtaca 4860
tgcagttttt ccctaccatg ttaaagcacc cagtagtaga cggtatggcg atcatgaacg 4920
gtcgcaaccg tcgcccgccc aaagatccgg tcaatgcgct gctctccctc ggctatacgc 4980
ttctttcacg ggatgtttac tccgcatgtg ccaatgtcgg actcgatcca ctgttcggct 5040
ttttccatac gatggagccg ggcagaccag ctttggcact cgatctgatg gaaccgttcc 5100
gcgccttgat tgccgatagc gtagcgatac gtaccttgaa tacggaggaa ctcaccctcg 5160
gggactttta ttggggaaaa gacagttgtt atttgaaaaa ggcaggaaga caaacgtatt 5220
tcgctgccta tgaaagacgg atgaacgaga cgctgacgca tccgcaattt gggtataagc 5280
tcagctatcg ccgtatgctg gagctggaag caaggttttt ggcccggtat ctggatggag 5340
agctggtgga atatacgccg ctcatgacaa ggtaggaaat gaccatgcga caatttgttc 5400
tggtaagcta tgatattgcc gatcaaaaac gttggagaaa agtattcaag ctgatgaagg 5460
ggcaaggcga gcacgtccag tactcggtgt ttctgtgcca actcaccgag attcagcaag 5520
ccaagctaaa ggtaagcctg gcggagctgg ttcaccatgg agaagaccag gtcatgtttg 5580
taaaaatcgg cccagtgacg agagatcaac tggacaagcg gatatctact gttggcaggg 5640
agtttctgcc tcgcgatttg accaaattta tctattaagg aatgaagaaa gctagttgta 5700
acaaaagtgg aaaaagagta aaataaaggt gtcagtcgca cgctataggc cataagtcga 5760
cttacatatc cgtgcgtgtg cattatgggc ccatccacag gtctattccc acggataatc 5820
acgactttcc actaagcttt cgaattttat gatgcgagca tcctctcagg tcaaaaaagc 5880
cgggggatgc tcgaactctt tgtgggcgta ggctttccag agttttttag gggaagaggc 5940
agccgatgga taagaggaat ggcgattgaa ttttggcttg ctcgaaaaac gggtctgtaa 6000
ggcttgcggc tgtaggggtt gagtgggaag gagttcgaaa gcttagtgga aagcttcgtg 6060
gttagcaccg gggagaagtc atttaataag gccactgtta aaagttcgaa agcttagtgg 6120
aaagcttcgt ggttagcacg ctaaagtccg tctaaactac tgagatctta aatcggcgct 6180
caaataaaaa acctcgctaa tgcgaggttt cagc 6214
<210> 75
<211> 12338
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 75
gaagttatgt tgataaaatg gtttatgaaa acgtgagtct gtggtagtat tataaacaat 60
gatggaataa agtgtttttt gcgccgcacg gcatgaattc aggggttagc ttggttttgt 120
gtataaataa atgttctaca tatttatttt gttttttgcg ccgcaaaatg caactgaaag 180
ccgcatctag agcaccctgt agaagacagg gttttgagaa tagcccgaca tagagggcaa 240
tagacacggg gagaagtcat ttaataaggc cactgttaaa agttttgaga atagcccgac 300
atagagggca atagactttt gcttcgtcac ggatggactt cacaatggca acaacgtttt 360
gagaatagcc cgacatagtt atagagatgt ataaatataa ccgataaaca ttgactaatt 420
tgttgaagtc agtgtttatc ggttttttgt gtaaatatag gagttgttag aatgatactt 480
tttgcctaat tttggaactt tatgaggata taagatagac ttgataaaaa ggtaaaagaa 540
aggttaaaga gcatggcagg aatagtgacc tgtgatgaag atgatggtag aattaaaagt 600
gttcttaaag aaaaacaata ttggataagg aaaataattc aatagataaa aaatttaggg 660
ggaaaaatga aaatatcaaa agtcgatcat accagaatgg cggttgctaa aggtaatcaa 720
cacaggagag atgagattag tgggattctc tataaggatc cgacaaagac aggaagtata 780
gattttgatg aacgattcaa aaaactgaat tgttcggcga agatacttta tcatgtattc 840
aatggaattg ctgagggaag caataaatac aaaaatattg ttgataaagt aaataacaat 900
ttagataggg tcttatttac aggtaagagc tatgatcgaa aatctatcat agacatagat 960
actgttctta gaaatgttga gaaaattaat gcatttgatc gaatttcaac agaggaaaga 1020
gaacaaataa ttgacgattt gttagaaata caattgagga aggggttaag gaaaggaaaa 1080
gctggattaa gagaggtatt actaattggt gctggtgtaa tagttagaac cgataagaag 1140
caggaaatag ctgattttct ggagatttta gatgaagatt tcaataagac gaatcaggct 1200
aagaacataa aattgtctat tgagaatcag gggttggtgg tctcgcctgt atcaagggga 1260
gaggaacgga tttttgatgt cagtggcgca caaaagggaa aaagcagcaa aaaagcgcag 1320
gagaaagagg cactatctgc atttctgtta gattatgctg atcttgataa gaatgtcagg 1380
tttgagtatt tacgtaaaat tagaagactg ataaatctat atttctatgt caaaaatgat 1440
gatgttatgt ctttaactga aattccggca gaagtgaatc tggaaaaaga ttttgatatc 1500
tggagagatc acgaacaaag aaaggaagag aatggagatt ttgttggatg tccggacata 1560
cttttggcag atcgtgatgt gaagaaaagt aacagtaagc aggtaaaaat tgcagagagg 1620
caattaaggg agtcaatacg tgaaaaaaat ataaaacgat atagatttag cataaaaacg 1680
attgaaaagg atgatggaac atactttttt gcaaataagc agataagtgt attttggatt 1740
catcgcattg aaaatgctgt agaacgtata ttaggatcta ttaatgataa aaaactgtat 1800
agattacgtt taggatatct aggagaaaaa gtatggaagg acatactcaa ttttctcagc 1860
ataaaataca ttgcagtagg caaggcagta ttcaattttg caatggatga tctgcaggag 1920
aaggatagag atatagaacc cggcaagata tcagaaaatg cagtaaatgg attgacttcg 1980
tttgattatg agcaaataaa ggcagatgag atgctgcaga gagaagttgc tgttaatgta 2040
gcattcgcag caaataatct tgctagagta actgtagata ttccgcaaaa tggagaaaaa 2100
gaggatatcc ttctttggaa taaaagtgac ataaaaaaat acaaaaagaa ttcaaagaaa 2160
ggtattctga aatctatact tcagtttttt ggtggtgctt caacttggaa tatgaaaatg 2220
tttgagattg catatcatga tcagccaggt gattacgaag aaaactacct atatgacatt 2280
attcagatca tttactcgct cagaaataag agctttcatt tcaagacata tgatcatggg 2340
gataagaatt ggaatagaga actgatagga aagatgattg agcatgatgc tgaaagagtc 2400
atttctgttg agagggaaaa gtttcattcc aataacctgc cgatgtttta taaagacgct 2460
gatctaaaga aaatattgga tctcttgtat agcgattatg caggacgtgc atctcaggtt 2520
ccggcattta acactgtctt ggttcgaaag aactttccgg aatttcttag gaaagatatg 2580
ggctacaagg ttcattttaa caatcctgaa gtagagaatc agtggcacag tgcggtgtat 2640
tacctatata aagagattta ttacaatcta tttttgagag ataaagaggt aaagaatctt 2700
ttttatactt cattaaaaaa tataagaagt gaagtttcgg acaaaaaaca aaagttagct 2760
tcagatgatt ttgcatccag gtgtgaagaa atagaggata gaagtcttcc ggaaatttgt 2820
cagataataa tgacagaata caatgcgcag aactttggta atagaaaagt taaatctcag 2880
cgtgttattg aaaaaaataa ggatattttc agacattata aaatgctttt gataaagact 2940
ttagcaggtg ctttttctct ttatttgaag caggaaagat ttgcatttat tggtaaggca 3000
acacctatac catacgaaac aaccgatgtt aagaattttt tgcctgaatg gaaatccgga 3060
atgtatgcat cgtttgtaga ggagataaag aataatcttg atcttcaaga atggtatatc 3120
gtcggacgat tccttaatgg gaggatgctc aatcaattgg caggaagcct gcggtcatac 3180
atacagtatg cggaagatat agaacgtcgt gctgcagaaa ataggaataa gcttttctcc 3240
aagcctgatg aaaagattga agcatgtaaa aaagcggtca gagtgcttga tttgtgtata 3300
aaaatttcaa ctagaatatc tgcggaattt actgactatt ttgatagtga agatgattat 3360
gcagattatc ttgaaaaata tctcaagtat caggatgatg ccattaagga attgtcagga 3420
tcttcgtatg ctgcgttgga tcatttttgc aacaaggatg atctgaaatt tgatatctat 3480
gtaaatgccg gacagaagcc tatcttacag agaaatatcg tgatggcaaa gctttttgga 3540
ccagataaca ttttgtctga agttatggaa aaggtaacag aaagtgccat acgagaatac 3600
tatgactatc tgaagaaagt ttcaggatat cgggtaaggg gaaaatgtag tacagagaaa 3660
gaacaggaag atctgctaaa gttccaaaga ttgaaaaacg cagtagaatt ccgggatgtt 3720
actgaatatg ctgaggttat taatgagctt ttaggacagt tgataagttg gtcatatctt 3780
agggagaggg atctattata tttccagctg ggattccatt acatgtgtct gaaaaacaaa 3840
tctttcaaac cggcagaata tgtggatatt cgtagaaata atggtacgat tatacataat 3900
gcgatacttt accagattgt ttcgatgtat attaatggac tggatttcta tagttgtgat 3960
aaagaaggga aaacgctcaa accaattgaa acaggaaagg gcgtaggaag taagatagga 4020
caatttataa agtattccca gtatttatac aatgatccgt catataagct tgagatctat 4080
aatgcaggat tagaagtttt tgaaaacatt gatgaacatg ataatattac agatcttaga 4140
aagtatgtgg atcattttaa gtattatgca tatggtaata aaatgagcct gcttgatctg 4200
tatagtgaat tcttcgatcg tttctttaca tatgatatga agtatcagaa gaatgtagtg 4260
aatgtgttgg agaatatcct tttaaggcat tttgtaattt tctatccgaa gtttggatca 4320
ggaaaaaaag atgttggaat tagggattgt aaaaaagaaa gagctcagat tgaaataagt 4380
gagcagagcc tcacatcgga agacttcatg tttaagcttg acgacaaagc aggagaagaa 4440
gcaaagaagt ttccggcaag ggatgaacgt tatctccaga caatagccaa gttgctctat 4500
tatcctaacg aaattgagga tatgaacaga ttcatgaaga aaggagaaac gataaataaa 4560
aaagttcagt ttaatagaaa aaagaagata accaggaaac aaaagaataa ttcatcaaac 4620
gaggtattgt cttcaactat gggttattta tttaagaaca ttaaattgta aaaaagattc 4680
gttgtagata attgataggt aaaagctgac cggagccttt ggctccggac agttgtatat 4740
aagaggatat taatgactga aaatgatttt tgttggaagt cagttttttc tgtggaaagc 4800
gaaatcgaat atgatgagta tgcatatggc agaagagctg tagaaggcga gaatacatat 4860
gattacatta ctaaggaaga aagaccggaa cttaatgacg aatatgtagc gagacgttgc 4920
attttcggta aaaaagcagg aaaaatatcc aggtcggatt ttagtaggat aagatctgcg 4980
ttggatcatg cgatgataaa taatacacat acagcatttg ccagatttat cactgaaaat 5040
ctgacgagac tcaatcacaa agaacatttt ctgaatgtga cacgtgcata ttctaaacct 5100
gattctgaaa aattgataca accgagatac tggcagtcgc ctgtagttcc aaaggataaa 5160
caaatatatt atagcaagaa tgcgattaaa aaatggtgtg gttacgaaga tgatattccg 5220
cctcgttctg tgatagttca gatgtgtcta ttgtggggga ctgatcatga agaggcagat 5280
catatccttc gcagttcagg atacgcggcg cttagtcctg ttgtacttcg agatcttatc 5340
tatatgtatt atctggatca tcaggatttg caaaaaaatg agttgatatg ggaagtaaaa 5400
aagcagttgg atcacttcga tttgacaaat agaaattatg atacaaatcc ttttgatgta 5460
gggggcagcg taaatgatca tatctgtgaa ctgagcgagc atatagcgaa ggctcattat 5520
atttatgaga gggctaagga aggaccattg caaaatgtaa ttcgggatat tttgggagat 5580
acacctgccc tttattctga aatggcattt cctcagctag catctataaa caggtgtgct 5640
tgcaattcgc tttcttcata tcaaaaaaat atttttgata ctgacatagc tatatatgca 5700
gatgaaaagg acacaagagg taaatcagac cgtatccttg ttgagggcgc atcttcgaaa 5760
tggtatgaat tgaagaaacg cgatgctaat aatgtcaaaa tttctgaaaa gctgagtata 5820
ctcaatacta ttcttaaatt taatagtgtt ttttgggaag aatgttacct tgatggaaat 5880
ataaaacaat cgagcggaaa gcgatctgag gcaggaaaaa ttctttatgg tcgcgacaac 5940
ggaaaagaaa atgtcggagt ttcaaaattg gaattggtgc ggtatatgat agctgcaggt 6000
caggaacaaa atctgggaaa ttacctggtg agttcaggat tttggagaaa aaatcatatg 6060
ctgtcattta tacaaggcaa tgatatagcg cttgatgaga tggatgaatt ggatctctta 6120
gactatattc tgatatatgc atggggattt agggaaaata tcattaaaaa gaacagtaat 6180
gtgaattctt tggatgaaaa gactagaaaa gtgcagtttc cgtttataaa gttactcatg 6240
gcaattgcaa gagatatcca gatacttata tgttcagcac atgaaaaaac agtcgatgag 6300
tcatctcgaa atgcagcaaa gaagatagat atattgggaa attatattcc ttttcagatt 6360
catcttcaga gaactaaaaa agatggtgga agagtggtaa tggatacatt gtgtgctgat 6420
tggattgcgg attatgaatg gtacattgat cttgagaaag gaacacttgg atgagcagtg 6480
atgaaaggat atttaaaaaa tttttggaaa aaggatcgat ttctgagcag aaaaagatgc 6540
ttttagaaga aaagaaatgt tcggataaac taactgcact gcttgggaat tactgcatac 6600
cgatagacaa tatttcagag tcagacggaa aaatatatgc ggtctataag cttccaaaaa 6660
atgttaaacc tttgtccgaa atcattaatg atgtatcctt ttctgattgt acgatgagag 6720
tacgtttgct tctcataaag agaattctgg aactcgtgtg tgcttttcac gaaaaaaaat 6780
ggtattgtct cagtatttca ccgggaatgc tcatggttga agattttgat ataccgatgg 6840
gaaatgtcgg aaaagtattg atatatgatt tcagaaatcc tgttccgttc gagtcagtaa 6900
atgaaagaca taattttaac gtttcaaata aatacacttc accggagctg ctcatccatt 6960
caagatatga cgagtcgaaa tctgtgagtg aaaaatcaga tttgtattct gttgcaaaaa 7020
ttgcggaaac aataatagga gattttaaca gtattattgc aaatggaaat ttgatactac 7080
ttgcaatgct tagagttttt atcagtacag ggaaaagtcc ggaacctgag tatcggtttg 7140
aatcgtcgga aaatatgctt tcagtatttg aaaatttgat caaagaaaat tgtttttttg 7200
aaaaaaacga ttatacatct atgtttcatc aggcgtatga caattttttt gaatggcagg 7260
aatgtttgat atcaccggat cacttggata aaaatatgtt cgaggcagct ttatcaaatc 7320
ttgaggatca gctgcttagg gttgatattg ataagtatag agcagagtac ttctataagc 7380
ttctccgaga gttgtctaat aaatataaaa atacaattac tgatgaacaa aaggtaaggt 7440
tggcaatact tggaatcaga gcgaaaaata atctgggaaa aagttttgat gcattggaaa 7500
tatatgagtc agtacgtgat ttagaaacta tgttggagga gatggcagag cttagtcctg 7560
tcattgcttc gacatatatg gattgctacc gatatgcaga tgcgcagaaa gtggcggaag 7620
aaaacattat caggcttcat aatagtaata ttcgtatgga gaaaaaaaga atactgcttg 7680
gaaggtcata tagttcaaaa gggtgcagca tggggtttca gcatattctt ggtgcggatg 7740
agtcatttga acaggcttta tatttcttta acgaaaagga caatttttgg aaagaaatat 7800
ttgagagcag aaatttagag gacagcgata gacttataaa gtctttacga agcaatacgc 7860
atattacgct gtttcattac atgcaatatg catgtgaaac aaggagaaag gaattatatg 7920
gagcactttc agacaaatat tttataggta aagaatggac agaaagactc aaagcatata 7980
taagcaacaa ggatatatgg aaaaactatt atgagatata tattctgcta aagggtattt 8040
attgcttcta tccagaagtc atgtgttcgt ctgcgtttta tgatgaaatc caaaaaatgt 8100
acgatcttga atttgaaaag gaaaaaatgt tttacccatt gagtctgata gaactgtatc 8160
ttgctctgat agagataaaa gttaatggga gtctgacgga gaatgccgag aagttgttta 8220
aacaggcatt gacacatgac aatgaagtca aaaaaggaaa tatgaatatt cagaccgcca 8280
tttggtatcg aatatatgca ctgtataacg atgtaaaaga tgaaactgat aagaataaaa 8340
ggcttttaaa acggcttatg attctttgcc gacgatttgg ttgggcggat atgtatagtg 8400
ctttggagaa ggatgggaag ttaattgatt ttttgagatt tgaggtatgt taaatgataa 8460
cacttgcatt agatgaaaat ggcaaatttg aagatgcttt ttctaaaaaa aatgaaaaac 8520
cgataatgat tgcggggata atctatgatg acaaggggaa agagtatgat gctgagaatg 8580
aacgctacag gatatccagt tatctgcgag cagtatgtga cagtttgggt gcgaaatacc 8640
ctcaggatct acattcaaat agtaatggaa ataaggcgac tgttgggaaa gtaaaatgta 8700
aaattggtga aacactaaag gaattcttga gagaaggaac ctatgaaaaa aaggaattgc 8760
cgacaaagaa cggttattta aataagagat ctggaaaata tgtaatgttt gcagaactca 8820
ggagtagtca gggagttaaa aagcgtgtta gtggttggaa tgacaatgat ctgactcagg 8880
atgaaaaggt cagcaatctg taccttcata tggcagaaaa tgccgttgtc agaatgctct 8940
tccataatcc tatatatgaa gatgtaacag atgtaaatct ctattttccc acgcgaaaag 9000
ttgttctgaa agatagagat agagaatacg ataaacaaga tttcaaaata tatggtgata 9060
aggacaagtg cgaagcagaa agcgggagat tggtgcatta tgatatcgtg tcatcggatt 9120
tttaccgtac gataatggag aacgaatgta caagaattaa taaaaagcaa ttaaatgttc 9180
attatatgaa cacaagccca atttcgtact gggagaaaaa tgaaaaatat aatacatttt 9240
tatatttggc tgacatagtt tgttctatgc tggattatta caaaaagggt tcgagtccgg 9300
cagagtggat ggattctttt gccgaatggg gaaacaaata ttttggtgat gatcagataa 9360
tcttatttgg gtatgatgat atagatgaca aatacatgga ggctgtagat gcagtaggac 9420
agggagagta ttttcatgcg ctggatatta tatatgatgc ggaatgtagt ggaagtgaat 9480
ttgagaagca ctacaaagat tattggtttc caaagcttat aaaaaagata cgaataacag 9540
caactgtgga taatttatgc agatcgatct cagatctgga gagttttaca tatcgaagta 9600
atcttgatca gcagaaactt ttgtggattt ttgaggaaat caaagctatc gtcgataagg 9660
gagattttgg aaagaaatat catacagatc aggttatgtt tgatatgtgt aatgccggta 9720
ttgctgtgta caatcatatc ggagattttg ggactgcaaa ggaatactat gatgagtgca 9780
tgaaacacac tggggatgtg gatctggtaa agatacttcg tgcatcaaat aaaatggtgg 9840
tctttcttga cgatgctttt aggtatggtg acgcgacaga acgtgccagg aagaatgttg 9900
aataccaaaa agctttgcac gatataaaga gtgagatttg tccggaaaag aaagatgaag 9960
acttgaacta tgccatatcg ctcagtcaat ttggacaggc gcttgcgtgt gaaaaaaatt 10020
ctgatgcaga gagtgttttc ctagagtcgt tgcggcatat gaggaaaggg actgccaatt 10080
atcagattac tctttcatat ttactccatt tttatctgga tatgggaatg acagattctt 10140
atcgagaaaa aacaaaggac tattttggaa gtgaaaaacc aaaggaacag ctgaaagaat 10200
tgctgaagtt atcgggaaag gatgatagta tagttacttt caaatttgca atgtatgtct 10260
atttacgtgc actttgggta ttacaggaac cgcttactga ttttatcaga acaagattag 10320
aggacatacg tgagactctt gtaaagaaga aaatgagtga acatatggtt ggacatccgt 10380
gggagttgat ttataaatat ctggcatttc ttttttatcg tgatggaaat tgtgaagctg 10440
ctgaaaaata tattcataaa agtgaagagt gcttggaaac acaaggactg actatagatg 10500
cgattattca taatggtaag tatgaatatg cagaattgtc aggtgacgag gagatgatgg 10560
caagagagaa agcgtacttt gatgaaaaag ggatagatag aaaaaatgtt tgtactttta 10620
tgtatcattg atgtttaata agatttgacc gaggagtgac aggtaatcgc cggtatatct 10680
ggtattacct gtcatttttt gatgaaataa gctacttttt gcctaaaaaa cgaaactgtt 10740
ggtgttttat gatgattgtg tcaacaaaag agagcaaaag aagaggagaa aagtaatgtc 10800
aatgatttca tgtccgaatt gtggtggaga gatatctgaa aggtcaaaga aatgtgttca 10860
ttgtggatat gtgttagtcg aagaagctaa agtagtgtgc acagaatgtg gaactgaggt 10920
agagagtggc gctgctgtat gtccgaagtg cggctgtcct gtaaatgata gtgagacgcc 10980
tcagaaagtt gaagtgacta gggtaaatgt atcttccgta atcagcaaaa aagtcgttgt 11040
aagcatactg atcgcagtga ttacaattgc aggttttttc tatggagtga agtattcgca 11100
ggaaaagaaa gcaattgaag agtcagtaaa gcagaaggaa gactatcaaa gtacgctaga 11160
gcttgcttcg ctaatgatgc ttcaaggagc ttcggatgca gaaacttgtg ggaatttggt 11220
taggaaagtg tggagcaact gcatttataa ggagagggat gaagaaaccg acaagtatac 11280
gtgtgatagc aggggtgcag gatggtttta tgatgatttt aatgatgcat taatggctct 11340
ttacagtgac agcagttttg gcaagaagat aaatgaaatc aaaaacggtc aggaaaccgt 11400
tgcggcgatg atgaaagatc tgaaaaatcc gccggatgag atggcagatg cctatgagga 11460
tattcaaaat ttttatgtgt cctatctaac gctgacagaa atggttgtga atccaactgg 11520
aagtttgagt tctttttcat ctgatttttc cgatgcggat acggaggtgt ccaatgccta 11580
tagccggatg aagttgtatt tagattaaac tattgaggaa aaaatggagg tgctttaatg 11640
cgggggagaa actgtggagg gtcatcaggc gacggactgc tggtacttct cgtactgctt 11700
gtcctttttt ataaaatcat gccattcata ggtttatgga ttttaatttt tggtgatgct 11760
gaacgtaaag atctgggtat gggtatgatt attgtcggga tagttctata tgtattatta 11820
gaggtttttt aatgtgagtt tctgtggtaa actataaaag tacaagcttt tgcgccgcac 11880
cgcataaata gcggatttat gaccattatt tggtgaaaaa aatggtgtac acctgtgttt 11940
ttttgttttg cgccgcaaaa tgcgccacgg aaccgcatgc agagcaccct gcaagagaca 12000
gggttatgaa aacagcccga catagagggc aatagacacg gggagaagtc atttaataag 12060
gccactgtta aaagttatga aaacagcccg acatagaggg caatagacat aaagaccaaa 12120
aacaggtcat ctgcatactg tgttatgaaa acagcccgat atagagggtg tgagagatat 12180
agttctcgtc acagtgcaga aaatgaccta ttatgtgccg aaaaacaaaa tgaaaaaaga 12240
atggaaaggc gtatttaatg aaatgctgat ctgttgattt gaattaacaa aaaaaggtcg 12300
ccccacggat gacaaaaaca tccgggggcg accctttt 12338
<210> 76
<211> 6098
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 76
tactgtgtgc ataagtcttc cttagatcca taggtacagc agttttattt attagcctta 60
gaaaatggaa aatagagctt ataaatgata tgatatttat gaataaaatg attgcattct 120
cgtgcaaact ttaaatatat tgattatatc ctttacattg gttgttttaa ttactattat 180
taagtaggaa tacgatatac ctctaaatga aagaggacta aaacccgcca aaagtatcag 240
aaaatgttat tgcagtaaga gactacctct atatgaaaga ggactaaaac ttttaacagt 300
ggccttatta aatgacttct gtaagagact acctctatat gaaagaggac taaaacgtct 360
aatgtggata agtataaaaa cgcttatcca tcatttaggt gttttatttt tttgtgatta 420
tatgtacaat agaagagaga aaaaaatcat tgaggtgaaa actatgagaa ttactaaagt 480
agaggttgat agaaaaaaag tactaatttc tagggataaa aacgggggca agttagttta 540
tgaaaatgaa atgcaagata atacagaaca aatcatgcat cacaaaaaaa gttcttttta 600
caaaagtgtg gtaaacaaaa ctatttgtcg tcctgaacaa aaacaaatga aaaaattagt 660
tcatggatta ttacaagaaa atagtcaaga aaaaataaaa gtttcagatg tcactaaact 720
taatatctca aatttcttaa atcatcgttt caaaaaaagt ttatattatt ttcctgaaaa 780
tagtcctgac aaaagcgaag aatacagaat agaaataaat ctctcccaat tgttagaaga 840
tagcttaaaa aaacagcaag ggacatttat atgttgggaa tcttttagca aagacatgga 900
attatacatt aattgggcgg aaaattatat ttcatcaaaa acgaagctaa taaaaaaatc 960
cattcgaaac aatagaattc aatctactga atcaagaagt ggacaactaa tggatagata 1020
tatgaaagac attttaaata aaaacaaacc tttcgatatc caatcagtta gcgaaaagta 1080
ccaacttgaa aaattgacta gtgctttaaa agctactttt aaagaagcga agaaaaacga 1140
caaagagatt aactataagc ttaagtccac tctccaaaac catgaaagac aaataataga 1200
agaattgaag gaaaattccg aactgaacca atttaatata gaaataagaa aacatcttga 1260
aacttatttt cctattaaga aaacaaacag aaaagttgga gatataagga atttagaaat 1320
aggagaaatc caaaaaatag taaatcatcg gttgaaaaat aaaatagttc aacgcattct 1380
ccaagaaggg aaattagctt cttatgagat tgaatcaaca gttaactcta attccttaca 1440
aaaaattaaa attgaagaag catttgcctt aaagtttatc aatgcttgtt tatttgcttc 1500
taacaattta aggaatatgg tatatcctgt ttgcaaaaag gatatattaa tgataggtga 1560
atttaaaaat agttttaaag aaataaaaca caaaaaattc attcgtcaat ggtcgcaatt 1620
cttctctcaa gaaataactg ttgatgacat tgaattagct tcatgggggc tgagaggagc 1680
cattgcacca ataagaaatg aaataattca tttaaagaag catagctgga aaaaattttt 1740
taataaccct actttcaaag tgaaaaaaag taaaataata aatgggaaaa cgaaagatgt 1800
tacatctgaa ttcctttata aagaaacttt atttaaggat tatttctata gtgagttaga 1860
ttctgttcca gaattgatta ttaataaaat ggaaagtagc aaaattttag attattattc 1920
cagtgaccag cttaaccaag tttttacaat tccgaatttc gaattatctt tactgacttc 1980
ggccgttccc tttgcaccta gctttaaacg agtttatttg aaaggctttg attatcagaa 2040
tcaagatgaa gcacaaccgg attataatct taaattaaat atctataacg aaaaagcctt 2100
taattcggag gcatttcagg cgcaatattc attatttaaa atggtttatt atcaagtctt 2160
tttaccgcaa ttcactacaa ataacgattt atttaagtca agtgtggatt ttattttaac 2220
attaaacaaa gaacggaaag gttacgccaa agcatttcaa gatattcgaa agatgaataa 2280
agatgaaaag ccctcagaat atatgagtta cattcagagt caattaatgc tctatcaaaa 2340
aaagcaagaa gaaaaagaga aaattaatca ttttgaaaaa tttataaatc aagtgtttat 2400
taaaggtttc aattctttta tagaaaagaa tagattaacc tatatttgcc atccaaccaa 2460
aaacacagtg ccagaaaatg ataatataga aatacctttc cacacggata tggatgattc 2520
caatattgca ttttggctta tgtgtaaatt attagatgct aaacaactta gcgaattacg 2580
taatgaaatg ataaaattca gttgttcctt acaatcaact gaagaaataa gcacatttac 2640
caaggcgcga gaagtgattg gtttagctct tttaaatggc gaaaaaggat gtaatgattg 2700
gaaagaactt tttgatgata aagaagcttg gaaaaagaac atgtccttat atgtttccga 2760
ggaattgctt caatcattgc cgtacacaca agaagatggt caaacacctg taattaatcg 2820
aagtatcgat ttagtaaaaa aatacggtac agaaacaata ctagagaaat tattttcctc 2880
ctcagatgat tataaagttt cagctaaaga tatcgcaaaa ttacatgaat atgatgtaac 2940
ggagaaaata gcacagcaag agagtctaca taagcaatgg atagaaaagc ccggtttagc 3000
ccgtgactca gcatggacaa aaaaatacca aaatgtgatt aatgatatta gtaattacca 3060
atgggctaag acaaaggtcg aattaacaca agtaaggcat cttcatcaat taactattga 3120
tttgctttca aggttagcag gatatatgtc tatcgctgac cgtgatttcc agttttctag 3180
taattatatt ttagaaagag agaactctga gtatagagtt acaagttgga tattattaag 3240
tgaaaataaa aataaaaata aatataacga ctacgaattg tataatctaa aaaatgcctc 3300
tataaaagta tcatcaaaaa atgatcccca gttaaaagtt gatcttaagc aattacgatt 3360
aaccttagag tacttagaac tttttgataa ccgattgaaa gaaaaacgaa ataacatttc 3420
acattttaat taccttaacg gacagttagg gaactctatt ttagaattat ttgacgatgc 3480
tcgagatgta ctttcctatg atcgtaaact aaagaatgcg gtgtctaaat ctttgaaaga 3540
aattttaagc tctcatggaa tggaagtgac atttaaacca ctatatcaaa ccaatcatca 3600
tttaaaaatt gataaactcc aacctaaaaa aatacaccac ttaggtgaaa aaagtactgt 3660
ttcttcaaat caagtttcta atgaatactg tcaactagta agaacgctat taacgatgaa 3720
gtaattcttt taaagcacat taattacctc taaatgaaaa gaggactaaa actgaaagag 3780
gactaaaaca ccagatgtgg ataactatat tagtggctat taaaaattcg tcgatattag 3840
agaggaaact ttagatgaag atgaaatgga aattaaaaga aaatgacgtt cgcaaagggg 3900
tggtggtcat tgagtaaaat tgacatcgga gaagtaaccc actttttaca aggtctaaag 3960
aaaagtaacg aaaacgcccg aaaaatgata gaagacattc aatcggctgt caaagcctac 4020
gctgatgata caactttaaa aggaaaagca gtggattctt cacaaagata ctttgatgaa 4080
acgtatactg ttatttgtaa aagtatcata gaagcattag atgaaagcga agagagatta 4140
caacaatata ttcatgattt tggagatcaa gtggattctt cacctaacgc acgaattgat 4200
gcggaattac tacaagaagc aatgagtagg ttagctgaca taaagcggaa gcaagaagca 4260
cttatgcaat ccttatcttc ttctacagca acgctttacg aaggcaagca acaagcgtta 4320
cacactcaat tcacggatgc gctggagcaa gaaaaaatat tggaacgcta tattactttt 4380
gaacaaactc acgggaattt ttttgactca tttggagaac ttgtctatcg aacgggacaa 4440
gcagtgcgtg aattagctaa taacgtcaca ttcgagagcc aaacaggaag ctatcatttt 4500
gataaaatag atgcttctag attccaaact ttgcaagaaa tgttgccaaa ggcaaagaaa 4560
aaagcattta attttaatga ctaccaaata acatggaatg gcaccacgca ccttttatgg 4620
aaaaatggta aagtggatgc agaagcaacc aaagcttata acgaggcgaa actgaatgga 4680
aagctaccaa aggaaggtaa tgtagcaaca caagatgcag aactattaaa aggcattttg 4740
gcttcactga aaaacaagaa agatcctatc actggagcag atataagcag tgtgcatgta 4800
ttatctatcc ttagcgggct cgcattctcc tatacagctg ggaattataa gggaagaaaa 4860
cttactgttc caaaaagttt cttagacaaa ttaaagaaaa accgaaaatc taaagtacct 4920
aaactatcta gtttatcaga aaaacaacaa ctaaaactcg caaataaata caagaaaaaa 4980
tcacctattc caattccaga tgatgctaaa atcaaagctc agacgaaaaa ggctggttat 5040
gaacaaatat cttataaatg gaaagagaat gggataacct ttgaagttag atggcatact 5100
aggacaccag gtgcaccaaa ggaacaagga aatacgtttg ttatagaaag aaaaattcag 5160
ggtacagcag aagggaaaac aaaagttcaa caaatattgg ttggagataa taagtgggtg 5220
agtaaaagtg agtggcaaaa ggctataact gataagaaaa atggtgtaag tacctcggag 5280
caaaataaaa tgttgtctga tggacattgg aaagaataga aaggagcaaa atgatggaag 5340
attattataa aggttttgag ggatatccag agatagattt ttatacgtat atagatgata 5400
tgaaattggg tatagcaatg tgggaaggat actttgacaa cattatgaaa gaaattaatc 5460
caagtaacgg aagatggact tcattagcgt attattatca tttagatgag gggtggtatg 5520
atgaaagtcc ttgggaaata ccaagtaata cagaagcatt agaattattg gaaacaatcc 5580
atatatctaa tctagatact atcacacaag agatattact taaattaata aatttattaa 5640
agaagaatat aaatagacaa gtttatattg aatactcata aaaaagatga ttatgatata 5700
ttatagaaca aacgaacaag ccccaaatac gaggtttgtt cgtttgtttt caatataatt 5760
atttgccacc aagtgagata ttacggtttt aaatagctta tttgacgata ccaaaccctg 5820
ataagagaaa gaagaaagag aaagctggtg tagttgtttt aagtgaacta gataaaaaat 5880
taatagcaaa acttgaaaaa gatggtgtga aaatatcaaa agaagatgtt ataggaataa 5940
aataattgcc agatgatgag aaatcgtttg gctggaaaaa ggaaatccat ccgctggatt 6000
tgagcatatt cttattgaac atggtgaaca atttgctaaa tagggaattt caaaagctga 6060
gttacctgat tttttgatga ctgctttaga aaaggaaa 6098
<210> 77
<211> 6222
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 77
attctttaaa aatatctaat aatttattta ctatatactc taatacatct tttaacctat 60
ctaaaacatc atcacctaca acatcccaaa aatcatctaa aaagttaaaa aaatccatct 120
ttatcaactc ctatatctat tttttattgt gtaattcctg agttacaaaa ccattataac 180
acgtattaca cacgtagtca atacttcaaa aaaatttttt gtatattttt ttgaataagt 240
aaataaaaag agctgtgtag ctctttatta aaatcaatat ttttattttg ttaacaaact 300
tagacaacat taaatttaga aacctatata tatttcagta cttttcattt ttaggtagtc 360
taaatcagaa atggttttgt ctaaatgatg tatgtaagtt ttagtcccct tcgtttttag 420
ggtagtctaa atcagaagtc atttaataag gccactgtta aaagttttag tccccttcgt 480
ttttagggta gtctaaatcc catccaaatt atgggataat atgttacttt ttattttaat 540
atttgattat ttattgtttt tttactgatt tagattaccc ctttaattta ttttaccata 600
tttttctcat aatgcaaact aatattccaa aatttttgtt tcttttctta tgatcttttc 660
tccgatagtt atttctccag ataagatttt catttttttg aattgatctt ctgttagaat 720
taatgttctt actgatgaat tttctggaac tatcattgac aactgatttt cataggaaat 780
tattttttct tttgtgctag aacttacaat gtatactgat ttttgtacct gataatatcc 840
ttttcttata atttcttttc taaattttgc atattctttt ttttcttttc ctgtttgcat 900
tggaaaatca tacattagaa tccctacata attagtactc ataatcctct atccttaact 960
caggaatttc tacttctgac atttctcctg taaaataatt tctaatatta tctaaaaaat 1020
aatcaatcac ttgagccaat tcatattttt tatttttcca ataaactttt tgtgttaata 1080
ccaataacaa tttttgtctt aatgatttat tcaaacttac ttcttcctgt tgattaaaat 1140
atacgatata atctaccatt ggacgaaata tttcaataat atcatctgca aaattataat 1200
tattaaattg tgaactgtga tgtattccca aacttggatg aaatccttta gccacaattt 1260
ttgaagagat taagcttctc aaaaccatat acccataatt taatgccgaa tttgtcccgt 1320
cttcaccaaa tctcttaaat tttttcccaa aaagttcacc aaaatacatt cttgcagcaa 1380
ttgcttcctg atgttccgct tcttttcctt ttaatctaat attattttca tatgcttcca 1440
acttatatga tacttcctga gattttttca aaaactgcaa taaatttctt tgattttcta 1500
tttttctcat tacaattttt ctccagattt cttctttttt atcgtcaatc cagctcactt 1560
gctcattaat tcttgttgtt acttgaaaat gattatacag tcctaatgaa tgtaaaactg 1620
gctgatgttt ttcattacaa attatcagtg gaatattatg ttctgataat cttaactgta 1680
atattccgct aattttacat ctgcaatttt caactacaat tgccatgata tcatttaaag 1740
atactttatc agccttattt tcatcatctt catttatcat cacaagctgg ttatttaaaa 1800
ctgataattc attgactctt gttacatgga taatattaga catttttatt actcctttac 1860
tctaaagctt tatattcaaa cataactttc acaagttcac acaattcttc tgaatttcta 1920
tcagtcatta attttttctt ttttaaattt ttcaaatgta caattttttc cgattctaaa 1980
gtctgaattt ctattttctt atctgctcct attttaaatg ttgctacaaa accatattcc 2040
tttaatatat ccactattga tttcataatt gcatttttaa gttttctatc ataagaaagt 2100
aattttctta aattttccag cacttctaaa agtgaaattt cagcatgcgg aatatagtta 2160
aaatgtgcaa tatagtttcg tatatacaaa tcttttttct cttgttttaa tttttttact 2220
tttttatcag aatagatgct tcttttttct acattatctt tgtataattc tttataaaaa 2280
tttatatatt tttcaacaat ttgcccactt ttatatttta catttttact gttatcaaaa 2340
ttaaatattt cttcaatata atgattttca ggaaattcac ctttcaatct aaatcttaag 2400
tccctttccc agatcgaagt atatcccaca agtctgtgga gtatttttaa taacaagcct 2460
tgcaacaagt ttaattcatt aaattccact ttatttttca aatgagtata tttttgtata 2520
tttccaattg ctttttcata ttctttataa tcttcatcat taaatttttc atctttttta 2580
ggtcttgcat attttctatg taaattttgc tgcattgtat aatttttttc tatttcattt 2640
tttttattgc tgtattcttt caattctttt aaacttattt tatacttcgc tttatcagct 2700
attttttcaa gtaaatttaa catcccatat ttttttatat tataaaaagc tctatgcttt 2760
ataatatttt ctccatcaaa atatatttta tttgtgtcaa atttcttcaa ttctttccta 2820
tcttttattt tattttcatt aaaatctaaa aattttccaa tttcattcgc ttctaattca 2880
aaatcttctg ttactctatt attatctaaa tttaaaagat ttataagttc aagttcatct 2940
gaaaaagttt cttctttatt tgcactctga tatttttcaa gacttccctt caaattagtc 3000
aattctttat gattaagcaa ttttaaaatt aaataaaaca tattcaaatt ttcagtgtat 3060
tttaatatct ttcctaattt tatctctctt acaaattcat ttatttcatg tggaatttct 3120
ttattcctat tatgtttttc ataatttttt aaaattttat catatttttc tttattatct 3180
ttttttattt ttattttaga aaatatatca ttattatcat tgttattatt actttctata 3240
tattttaaat tatttttatt caaataatct ataaaacctt ttaaaaatat ttgttgtata 3300
aaatcaatgt atgtattttt ttcttcttta tcttgattat taatcatctc cctactttgt 3360
ataatagcaa gatattctac tggtacagtt ttttctatat tttcaaattt ttgatattta 3420
taatgtcctg ttttttgatt tctttgttta tttattttta ttacttcatt agttatttta 3480
aaaaaaactt tactattttt aacaaattta ttaagaaatt caccataata aatatttttc 3540
aaaagatata tttgagcatc tttttcttct ttatccttag gaacactcca aaaaaatttt 3600
aaagtatttc ttaaatcttc tattttatta tataatttcg taaaagaagg aacaaaagga 3660
atattcttat ttacaaaatt aaattttgta ttttttaaat atttaattat cacatccttt 3720
tcataataat taaatacatt tgcactattt aactgcttaa atatcttcaa tttcaatttt 3780
ttctcattta tttcattttg aaacattttt tttgaaattt cagaaggagc tatattttta 3840
aatgcaaata tatctttccc ttctaattcc aaattaaaat gcacaatccc atgtctaata 3900
ctgctaatag cttcatcaat atttgcaaaa aaatcttcta tctcattttt attatccata 3960
ttaaaatcat aactatagaa catttttaaa ttttctttta cttcattttg cttgttttca 4020
ttatatattt tatcaacttc tccagaaaca tatttttctt cgcccttatt attttttaca 4080
gtttttcctc tcattctacc tgtaatatca ttctcatttt cagtttcaag aatatttctc 4140
aatgaaaaat atgcaaccga agaaactcca attatatttc gtaaaaatgc ttcattttgt 4200
ctattcctag caataaaatc acttgttgca atctctccaa cttgtaaata ataattgtat 4260
ttcccacaat ttcttacata agtatccaat ttatttagta atttgttttc aattaatttt 4320
tttaaatttt gatattcaaa tattctctta attttatcgt tacttatgtt actcagtctt 4380
ttatacacat aatttttcaa aagctgactc atttcaattt ccacaaaatg acaaaaagca 4440
tattttatat ttttatcatt aagttcttct ttatccaaat aatatttata aaacacttgt 4500
gattttttta attcactcat atccggaatt ttttcaatta attcttttat attatttaca 4560
ttttgtattt cttcgtaaat aattttagca aaattttctt tatcattttt tcttccaatt 4620
attttgtgat agtattctct tattttatat ttttcatgtt tttttgaatt ttctattaaa 4680
aaaaataact tctcaatatc ttctttttta tacaatttat caaatgcttc ctgtacatta 4740
tttatataat cattacgctt tgctgattct ctataataat cataaataat atttcttttg 4800
ctcttccctc caactttttc aacattattt tcattaattt tctgataatt agccttattt 4860
tcttcaaatg aatattttaa agaatttatc ttattcaatt ttgcctcaac atcttttcta 4920
aatatttcta attcttcaga gttcacatct tcatttaaca atattttctt taaaactgaa 4980
aaactatttt tattttttaa atcatattct gaaatatctt cttcagaata atttttatcc 5040
tgtactgcat ttttctcttt cctattcttt aaatacagaa cactatcttt tagatgcaat 5100
actttatttg aaaaaaactt ttttaaattt tctcttctta ttctattttc ttcttcactt 5160
gcattatcag gattttttat atatatatcc agtcttatac ttaaaagctc tgacaatctc 5220
tcactagtcc tattttcttc gctcgtactt tttactaatt ttccctcttc aatatatttt 5280
ttatgcgaaa ttccatcaac ttttgtaact ttcatatata aaaacctcct aatatctata 5340
ttttttactc aatacctaat tcttttttca atgctttttg taaaatttgt gaaaaattca 5400
gatttttttc ctgtgccaat atatctaacc aaacaggaat tgttaaagtt ttctttttaa 5460
gtgcatttgt aacttttgcc acttcataca ctggatcaac agataaaata tacaaatact 5520
gattttcttt cagtttcaca tcctccactt ttgaaggctc aggaaatttt tttcttacat 5580
ccaaaaaatc agccaaatgc agacccaatg tctctctcaa attggaaaca gcctcctcca 5640
tgctatctcc aaatgtagca taataattta tctctccatc ttcaaactta tcaaaatcaa 5700
caatacaacc ataataagtc ccatcttcct tagttaccac tgctggataa aatacatcca 5760
ttttaattat ctccaatcta taccacgtgt taaatacgtg tttaaaaata tttataaaat 5820
tttttagcat ctctgctaaa ataaaacaat tatttcaaat ttttctattc cttaatcact 5880
cattgttagt gattcttttt ttacttggac aatttttcat ttaatttctt caattttttt 5940
aaaatcacat ttttttaata ttccttattt aattgcaaat tttcattact tttggggtgc 6000
tctaaatccc atccaaatta tgggataata atttttagtg aaagcaagaa gggactagaa 6060
tttaatccca acttgttttt caatacttct taatgttcct acaggtatat cttttgaata 6120
tggtactgtg accacacctt ccacacctgg gatcatccat tgataatgac tacctcttat 6180
acgcacaact tttccgccta attttctaaa tcttttttcg at 6222
<210> 78
<211> 6337
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 78
ctttctatct ttttcaaata aaattaggct ctagttagcc taatcgcata attatttatt 60
atagtataat tcttattttt tttcaaccta aaaatttaaa acatctccaa aaattttcgt 120
ttcagaacaa ccaagcaacc atattcaaaa aacaataaaa aatgagcaag aattgaaatt 180
ttattctcac tcagaagtta tttttattaa atatcacttt tcgatattgg ggtggtctat 240
atcaatttaa aagacagaat agataattct ttagagtttt agtccccttc gatattgggg 300
tggtctatat cagaagtcat ttaataaggc cactgttaaa agttttagtc cccttcgata 360
ttggggtggt ctatatccca tcctaatttc ttgctgatga gatatttatt tctaattttt 420
ctattttgtc tttattttca atactttcaa tcctattttt ctctttatta ataatataga 480
accaccctat actattatac catatttttt gatttttcaa aattccaata ttttgttttg 540
tgaaattttt tctcccattg tcacttctcc tgcaagtacc ttcatttttt gaaactgatc 600
ttctgtcagg ataatggaac ggattgatga attttctgga gcgagcattg ataactgttt 660
ttctgccagt tcgatttttt cttttgtttt cgacctcatt atatataccg atttttgaag 720
ctgataatat cccttttcta tcaatttttt cctaaaagtc ctatattcaa atctctcaac 780
atctgtctgc ataggaaaat catacataag cagaccaaaa tactcaatac tcatagtcca 840
tcacgctcaa tgtcggaatt atcacttctt catcttttac aaaataattt cgtatactat 900
ccaaataata gtctaccgct tggaaaaaat catatttctt attgttaaat aataccttct 960
gctgtgctac aagaagtatt ttttgcctta tttccttact taatttcact tcattcaaaa 1020
tatccttgta catataaaca agataatcca ccataggacg aaaaacctct attatatcat 1080
cagaaaaatt ataggcatta aactgtgact tatgatgtaa tcctaaactt ggatgaaatc 1140
cttttgctac aatctttgat gatattatag ctcttaaaat catatatcca taattaagtg 1200
cagaattcac tccatcttca tcaaatcttt taaaactatt actatacaat tcctgaaaat 1260
atatccttga agctattgct tcctgatgtt ctgcactcgc atcatctttt ttcaagtttt 1320
ccttatatgt tttcagtctt tcaatggaaa tatcactttt ttcaagatac tctaacaatg 1380
ctctttgatt ttcaatctta ttctccacta tcctgctcca caatttttcc tttttctctt 1440
tttcccactc aatctgctca tttattcgta aagtcacttg aaaatgatta aataatccca 1500
gcgaatgaat ttcaggctga tgtttctcgt tgcaaataat aatcggaatg ttattttcca 1560
ccagcctcaa ctgcaaaatc gcactaatct tacaatagca gttttcaata actatcgcag 1620
atatatcatt caaagaaatc ttatttttct catcattatt gtcttcatca accattataa 1680
gctgattatt cgatattgac aaatcatcag cccttgttat gtgaattata ttgggcattt 1740
taatcatact ccttataaat ttcattctta taacgtatca ttcgtatttt ctatttttgt 1800
taaaagttct attatcaagt ttttaatata atcagaatta taactttcta attctaaaac 1860
agaaactttt ttaggtttca ttaatctttc aagtatatca ttattaccga taagtttaaa 1920
ttttttcttt aattcatcat aatctaaatt cacatctttt ttaaatactt caaatacact 1980
tgcataagtt gaattattat aacgtgtact atatgataat aaattagaaa ctctatcaat 2040
ttgttctgca atactgtaat cagcaaacgg atttcttaca atatagaaat gtgaaatata 2100
gtttctaata ctttcatttt ccggcttatt aatttcagaa ttttcagaca aatcaattcc 2160
aaatccataa catattttct caaatttttt ataagattct tcatcaaaaa atttatagta 2220
tgctgttgtt gtataaaagc catcagatcc attacgctta ggataagctc tacttattcc 2280
agtattgtag ccacttaact taataattcc taattctctt agcccattta caatatagtg 2340
catatctctt tcaaatctag ccatttgaat agcaagtttc caatttatat ctatcaaata 2400
actttctatt ttattcaaat aattaaattc taccaaatct ctaatttttt tgtattcaga 2460
aactctatta taatcttttt caaatgattt atagttttta ttttgtatat tttttgcaaa 2520
aaagtcatca ttttctttca atttttttat atacttctct ttgtattctt tagaatatcc 2580
atttagttta tcatttagat ttttcaatat tgcatcaatt tcagatattt tattttttct 2640
aatattttta ccatcaatat taaataaaaa ttttgcatca gccattttaa tatcatttga 2700
aattaatcca taaattttat caaaatttgg atttccaata tttaaaaata aattcttttt 2760
ataaatatat aattcattct tacgttcttt aggataatat atttcttgaa atttattttc 2820
attctctgat tccatatctt ctattaaatt atctatttct tttttgtatt tttttaaaaa 2880
atcagaatta aatattattc tacacaatat tttactcttt atttcctgat ctttatcttt 2940
tatatactga tcaacctttt ttttcaaatc ctttttattt atgtttgata actttctttg 3000
ttcatcttgt aatatattcg attttttatc tatctcaaat ttagtttcat catcaaaaat 3060
tacaattttt tctaattttt tctctaaaac atcacaacca ttaatatcat ctttaaattc 3120
agttaatata ttatttttta tatcctcata ataattatta aaaatttctt ttttagtttg 3180
tattttaaaa tcatcaaagt ctttttctat ctctttcatt ttttgaataa attcttctaa 3240
attaagattc caattttcag ttatacattc atttctcaaa gtatttaatt gcattatttc 3300
atctaaaata tctataatat tttgatattc tgaagtattt aaccaaactg atgttgcaaa 3360
aaatctattt ctaattttat ttataaccgc attactattt aacagtgcaa atattgaaat 3420
tatatattca aaatcatcat ttattactat agttttatca ctagtcttta cagttattct 3480
ttcgtaagtt ttattatcat taatgtcttt tatttgtttc ttaatttctt gaatattcat 3540
tttaaaatct gaaaaatcaa aaagttcctc ataatttttt ctcaaatatc caatataaca 3600
ttctattact tttttctgat attttttaat agctttatta ttaccttttg aagcagaaat 3660
ctgagcattt ttataataat tttctataat attttcatct atttcatcaa tgtttcctaa 3720
agttttcttt aattcttgta aaaatatatt cttactttca ttttcttcta aatcatcttc 3780
taaaattaat ttcttataca attctttatt cacatatatt aaagcattta atactatttt 3840
ttctgtttct atagtatcaa atggttcatt cttaggatta ttcctatata aatttaatat 3900
ttcaggaagt actttagaaa aggatggtaa atatttaata tcattattat tttcttctga 3960
aattttaata tcatttattt tagtaattat atttttttta tctttaaata ctacatctaa 4020
atttaatgct tttgacactt cttcatctga tatttttaaa ttttgaatta tatttatgac 4080
tttattatag tcatcttgcg ttccttgtaa atctctttcc ttgctaatcg catgtaatat 4140
cctgtttctt tcatttgttc ctatctttgt aaatttccta ataaaattat ttgtaatgtt 4200
atttttatta tctataaaat ctaagtctct tattattttt atttttgaat ttaaaatttt 4260
tttatcaagt acgtaatttt tttctcgatc tcctccaaag aaatctatat tttcatcatt 4320
atttatattt tctctagaaa aaatcttatt taattccata ttggtagaag caaaaaaagt 4380
aatcaattct aaatccaatt cctctttagc gtgaagtcta gaaaaatcat cagtatttac 4440
tgttgtcata tctatatcat tatgtcttaa tttccctaaa tacataatat gctctaacgt 4500
atattgctta actcttttta aaattttttc agataatata ctttcattta aaattttttc 4560
tatttctatt ttttccattt tctttaatct gactttttgt tcatttacca atattttttc 4620
aattcttcct ttcaaatatc gatatatgat tttatatagt tctttttctt catcagattt 4680
ctttgaaaat tttttcgaat caaaattaac tttataatgt tttttaaata ttccaaaaat 4740
ttctgtatca caatttcctt tttttagttc tttttctaat ttttttatta attcatctat 4800
tttaaattct gctaaaattt tttctatttt ttcttttata ctattatttt ttatattttc 4860
tacaaaaaat tttacaattt tatctttttt attttctctt tctattttaa atttttcgtg 4920
cttatctaat agtacataag attttatata tgttctattt cttctctttt caagaaattc 4980
attattaact ttttttactt tttcaattct tttagtaata ttccaaaatt ctaactcttt 5040
tataacaaaa tcagctatat cttctactgt taaatctaca tttatattta aaattttttc 5100
aacaagcatt tttttatttt tagatttctt tttatcacca ccaacattaa gataaaattt 5160
tacaaaaccc agaatttcta aattactttt tattttttct cttatttcca taaaattagt 5220
caaaataaca tctattttat catctttcaa taatttttct cttaaatgtt cttcataata 5280
tcgattttca aatacttttt ctgtttcatt ttcaattatt ttttctataa tcttatataa 5340
actcatgtta atatttttaa aaatttcgta aattgatttt tttgtttcta attcatcatt 5400
ttctattatt cttaatatta ttgaacaatc atttagtgtt ttattagtat actcatctct 5460
gatatctatc tctatttctt cttcattctc ttgtctcttt atttctattt ttttatcatc 5520
tttagttatt ccttgcctaa ttgcttcatc tattattttc ttttttgtaa tccccaatgc 5580
tttcaatttc tcagattttc catatgcttc tatatataat acaacttctt ctgtttccaa 5640
aaaatcatca ttattttcta ttcttatgat tccttcttta cctttcaact taaatagaat 5700
atttcctgca tgaaattttc ttgtaaattc tttaagaata ttatcatttt ttttgtaatt 5760
aatatatttt ctaataaatt tattattatc aattttttct ttattattat tttcattaat 5820
atttaaaatg tatttgtttc catcatagtt ccttttaact tttactttcc gttttatttt 5880
aaaatctttt ttatcacgaa cttcatacca tctcttatgt ccaaataaat ttcccattcc 5940
aatctcctcg tttctacttt aatctaataa aatattttta aattaaatca attttacatc 6000
tttctaatca aaaatacaat tttccatttt tagtatacca catcaatatt aaatctcaaa 6060
aaaataagga gccgtcaaac atagctccct acttctattt actcataatc cccatctatc 6120
cttacttttc gtaaaatcaa tccttctttc gcctttagat ccaacttaat tttcccattt 6180
gaacctgttc taaatgttct gccttctgtt accaaatcaa taaatctttc atcctgataa 6240
tttgtttcaa attccacatt ttcccagctg ttaaacgaat tatttattac aacaataatt 6300
aaatgatcct cgattactct ttcatacaca attattt 6337
<210> 79
<211> 91
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 79
cgagguucug ucuuuugguc aggacaaccg ucuagcuaua agugcugcag gggugugaga 60
aacuccuauu gcuggacgau gucucuuuua u 91
<210> 80
<211> 16
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 80
uacgaggcau uagcac 16
<210> 81
<211> 101
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 81
ucagucgcac gcuauaggcc auaagucgac uuacauaucc gugcgugugc auuaugggcc 60
cauccacagg ucuauuccca cggauaauca cgacuuucca c 101
<210> 82
<211> 20
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 82
gaaagcuucg ugguuagcac 20
<210> 83
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 83
cccataattg ataggatcta tgaggt 26
<210> 84
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 84
aaatataaat acattaa 17
<210> 85
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 85
cccataattg ataggatcta tgaggt 26
<210> 86
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 86
atcggttata caggcta 17
<210> 87
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 87
cccataattg ataggatcta tgagga 26
<210> 88
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 88
acggcgccgg aaaacat 17
<210> 89
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 89
cccataattg ataggatcta tgaggc 26
<210> 90
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 90
agagccgcgc ctattgg 17
<210> 91
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 91
cccataattg ataggatcta tgaggt 26
<210> 92
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 92
aaaacggtcc cactaat 17
<210> 93
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 93
cccataattg ataggatcta tgaggt 26
<210> 94
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 94
aaaacggtcc cactaat 17
<210> 95
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 95
cccataattg ataggatcta tgaggt 26
<210> 96
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 96
tgagctgctg gcccgca 17
<210> 97
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 97
cccataattg ataggatcta tgagga 26
<210> 98
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 98
tggttccaat gcagtaa 17
<210> 99
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 99
cccataattg ataggatcta tgaggt 26
<210> 100
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 100
acctataacg gcaccta 17
<210> 101
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 101
cccataattg ataggatcta tgaggc 26
<210> 102
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 102
cccataattg ataggatcta tgaggt 26
<210> 103
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 103
cccataattg ataggatcta tgaggt 26
<210> 104
<211> 18
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 104
ttgccgccgt cctgcatg 18
<210> 105
<211> 27
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 105
cacgtagttt tgaagggaag ttagagg 27
<210> 106
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 106
accgaatatg agaactg 17
<210> 107
<211> 27
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 107
atcgtagttt tgaagggaag ttagagg 27
<210> 108
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 108
tacaaataaa catcaag 17
<210> 109
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 109
tcgtagtttt gaagggaagt tagagg 26
<210> 110
<211> 16
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 110
ccgctccggc cgtgat 16
<210> 111
<211> 27
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 111
atcgtagttt tgaagggaag ttagagg 27
<210> 112
<211> 27
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 112
atcgtagttt tgaagggaag ttagagg 27
<210> 113
<211> 27
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 113
atcgtagttt tgaagggaag ttagagg 27
<210> 114
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 114
gttagaattg agaggatgtt gaagga 26
<210> 115
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 115
ttacacggcc tatggtc 17
<210> 116
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 116
gttagaattg agaggatgtt gaaggt 26
<210> 117
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 117
cttatgcaca acccttt 17
<210> 118
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 118
gttagaattg agaggatgtt gaagga 26
<210> 119
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 119
aactgaacgg cttgttt 17
<210> 120
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 120
gttagaattg agaggatgtt gaagga 26
<210> 121
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 121
tcttactaat ttgccga 17
<210> 122
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 122
gttagaattg agaggatgtt gaagga 26
<210> 123
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 123
gttagaattg agaggatgtt gaagga 26
<210> 124
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 124
gttagaattg agaggatgtt gaagga 26
<210> 125
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 125
aactgaacgg cttgttt 17
<210> 126
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 126
cccataattg ataggatcta tgaggt 26
<210> 127
<211> 27
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 127
atcgtagttt tgaagggaag ttagagg 27
<210> 128
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 128
gttagaattg agaggatgtt gaagga 26
<210> 129
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 129
cccataattg ataggatcta tgaggt 26
<210> 130
<211> 27
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 130
atcgtagttt tgaagggaag ttagagg 27
<210> 131
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 131
gttagaattg agaggatgtt gaagga 26
<210> 132
<211> 27
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 132
cctctaactt cccttcaaaa ctacgat 27
<210> 133
<211> 17
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 133
cagttctcat attcggt 17
<210> 134
<211> 225
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 134
cctctaactt cccttcaaaa ctacgtgaca aaggttaaca tattgtaatt ccacttgatt 60
ttgtcatacc agggagaaaa ttggcatgga aatggatcaa tttcagcacc ccgattcggt 120
ttggacatgg aaatctaacc aacggggccg caaggcaagt ctgtggattc cctacctcga 180
tcaaataaga aaaattaagg ggactcgctg ggaatttgtt tataa 225
<210> 135
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 135
gtctcggcaa gcttggtcag tgttgggtga ttggcac 37
<210> 136
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 136
ccgcctgacg attcgtgaaa cggcattcgc tgcggc 36
<210> 137
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 137
cttcaacgaa gccacgccgc gaacggcgtg gagtg 35
<210> 138
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 138
gcttcaacga agccacgccg cgaacggcgt ggagtg 36
<210> 139
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 139
gtccaagaaa aaagaaatga tacgaggcat tagcac 36
<210> 140
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 140
gtgctaacca cgaagctttc cactaagctt tcgaac 36
<210> 141
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 141
gtgctaacca cgaagctttc cactaagctt tcgaac 36
<210> 142
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 142
gtgctaacca cgaagctttc cactaagctt tcgaac 36
<210> 143
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 143
gtgtcagtcg atcaagctgt tttcaccatc ggaacccc 38
<210> 144
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 144
gtgctaatcc cgaagctttc cactaagctt tcgaac 36
<210> 145
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 145
gtgccaacgc gctcaggatc tggcgcccac tgcgac 36
<210> 146
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 146
gtttcaatcc gcgcccccgt gagggggc 28
<210> 147
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 147
cggatcatcc ccgcatccgc gggggacac 29
<210> 148
<211> 31
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 148
gtttcaatcc gcgcccccgt gagagggcga c 31
<210> 149
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 149
gtctattgcc aactatatct ggcttttctc aatac 35
<210> 150
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 150
gtctattgcc atctttatct ggcttttctc aatac 35
<210> 151
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 151
gttattgccc tctatcttgg gctcttctca tcaac 35
<210> 152
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 152
gttattgccc tctttcttgg gctatcttct ccagc 35
<210> 153
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 153
gtttggagaa cagcccgata tagagggcaa tagac 35
<210> 154
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 154
gttttgagaa tagcccgaca tagagggcaa tagac 35
<210> 155
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 155
gttatagtcc tcttacattt agaggtagtc tttaat 36
<210> 156
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 156
gttatagtcc tcttacattt agaggtagtc tttaat 36
<210> 157
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 157
gttcttaatc taaaatagtg aaatgtaaat 30
<210> 158
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 158
gcatttacat tacattattt tagattaaga ac 32
<210> 159
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 159
actacattat agctgattct gtaaggaaac tatagc 36
<210> 160
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 160
actacattat agctgattct gtaaggaaac tatagc 36
<210> 161
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 161
tagttccctt caattttggg attatcca 28
<210> 162
<211> 26
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 162
cctttcaaac gaagggcact tacaac 26
<210> 163
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 163
gttttaacta cttattgtga aatgtaaat 29
<210> 164
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 164
gttttaatta cttattgtga aatgtaaat 29
<210> 165
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 165
gtaagagact acctctatat gaaagaggac taaaac 36
<210> 166
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 166
gatttagagt acctcaaaac agaagaggac taaaac 36
<210> 167
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 167
gatttagagt agctcaaaaa agaagaggtc taaaac 36
<210> 168
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 168
gatttagagt acctcaaaac aaaagaggac taaaac 36
<210> 169
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 169
gttttagtcc ccttcgtttt tggggtagt 29
<210> 170
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 170
tcaatcctta ttttaatgga ttcactattc ttac 34
<210> 171
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 171
gttttaatag cacaaattgt attgtaaat 29
<210> 172
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 172
gtttcaatcc ttgttttaat ggataactta ctttaac 37
<210> 173
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 173
gtttcaatcc ttgttttaat ggatactcta ctttaac 37
<210> 174
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 174
gttttagtcc ccttcgatat tggggtggtc tatatc 36
<210> 175
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 175
acctctcccg ccagtaagcg gattgagac 29
<210> 176
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 176
gcccagacac ctctcccgcc agtaagcgga ttgagac 37
<210> 177
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 177
gcccagacac ctctcccgcc agtaagcgga ttgagac 37
<210> 178
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 178
gtcgctcccc ccgcgggggc gtggatcgaa ac 32
<210> 179
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 179
gtcgctcccc ccgcgggggc gtggatcgaa ac 32
<210> 180
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 180
ggttcagtcc gccgtcgtct tggcggtgat gtga 34
<210> 181
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 181
gccctctcct cctccctggg ccg 23
<210> 182
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 182
ggttcagtcc gccgtcgtct tggcggtgat gtga 34
<210> 183
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 183
ggttcagtcc gccgtcgtct tggcggtgat gtga 34
<210> 184
<211> 3994
<212> DNA
<213> Listeria weihenstephanensis
<400> 184
tggctgctga gctgtctctt aatttatgaa aaaataatta tgttttgcta atctgtcaag 60
ctgcagattc attaataatc tggtatagtt atgttgttgc aagcgaatag gcggatatat 120
tacctcaaaa cagaagagga ctaaaaccca aacgattggt gttacattat tttcatagat 180
ttagagtacc tcaaaacaga agaggactaa aacgcactct ccgacaataa tctcgtccat 240
tttgatttag agtacctcaa aacaaaagag gactaaaaca actctgtact tgtgaagtac 300
gttaaatccg atttagagta cctcaaaaca aaagaggact aaaacctctt ttgtggataa 360
gtattcgaaa taaagccata aaaactgtga tccaaagaac tggattattg gtttttatgg 420
ctttattcaa ttcttagtat tgtagatgaa ctgtcagcga atgttgtctt gcaacgtgcc 480
ttcttgtata atgaatatat tgataaataa tagaaatttc atacacgcat gaagaaacca 540
attaaagttt cagcaataat gaagcattag gtacatgact ataaaaccaa atggagctga 600
gtagacagat gaaaatcaca aagatgagag tagatggaag aactatcgta atggagagga 660
caagtaagga aggtcaactg gtttatgaag gtatcgatgg aaataagaca acagaaatta 720
tatttgataa gaaaaaagaa tcgttttata agagtatcct caataaaact gtgagaaaac 780
ctgatgaaaa aagaaaaaaa ataggcgtaa gcaggcaatt aataaagcga ttaataaaga 840
aataacagaa ttaatgttgg cgctgttaca tcaagaagtg ccaagccaaa agttacataa 900
tttaaagagt ctaaatacgg aatctttaac taaactattt aaaccgaagt tccaaaacat 960
gatttcttat ccgcctagca aaggtgccga acatgttcaa ttttgcctta cagatatagc 1020
ggtaccagcg attcgagatt tagatgaaat taagccagat tggggcattt tttttgaaaa 1080
attgaaaccc tatacggatt gggcagaatc atacattcac tataagcaga caaccataca 1140
gaaatccatt gagcaaaaca aaatacagtc ccctgattcg ccaaggaaat tagtattgca 1200
aaaatatgtc acagcctttt tgaatggaga accgctggga ctcgatcttg tggcgaaaaa 1260
atataaactg gcagacttag cggagtcgtt taaagtagta gatttgaacg aggataaaag 1320
tgcaaactat aaaattaaag cgtgcttgca acaacatcag cgaaatattt tggatgaatt 1380
gaaagaagat ccagagttaa atcaatatgg tatagaagtg aagaagtata tacagcgata 1440
tttcccaatc aaacgtgcac cgaatagaag taaacatgcg cgagcggact ttttgaaaaa 1500
ggaattaatt gagtctacag tggagcagca atttaaaaat gctgtatatc attatgtact 1560
ggaacaagga aaaatggagg catatgagct aacagatcct aaaacaaaag acttgcagga 1620
tattagatct ggtgaggcat ttagcttcaa atttattaat gcttgcgcct tcgcatccaa 1680
taatttgaag atgattttaa accctgaatg tgaaaaggat attttaggta agggcgattt 1740
taaaaagaat ttgccaaaca gtactacgca gtctgatgtt gtgaaaaaaa tgattccttt 1800
tttctcggat gagattcaaa atgtgaattt tgatgaagct atctgggcga ttaggggctc 1860
tattcagcaa attagaaatg aggtttacca ttgcaaaaag cattcttgga aaagcatact 1920
taaaataaaa ggctttgaat ttgaacctaa caatatgaaa tatacggatt ctgatatgca 1980
aaaattgatg gataaagata tcgccaaaat tccagacttc atcgaagaaa aacttaaaag 2040
tagtgggata ataaggttct acagtcatga taaattgcag tctatctggg aaatgaagca 2100
agggttttcg ttgttgacta ctaatgcgcc gtttgtccca agctttaaac gtgtctacgc 2160
aaaagggcac gactaccaaa cttctaaaaa tagatattat gatttaggtt tgactacttt 2220
tgatattttg gaatatggag aagaagattt tcgtgcacgc tatttcctga cgaagctagt 2280
ttattatcaa caatttatgc catggtttac agctgataat aatgctttcc gagatgctgc 2340
caattttgta ttgcgattaa ataaaaatag acagcaggat gcaaaagctt ttattaacat 2400
tagagaagtt gaagaaggtg agatgcctag agactatatg ggctatgtcc aaggtcaaat 2460
agcgatacat gaggattcaa ctgaggatac accgaatcat tttgaaaaat ttattagcca 2520
ggtttttatt aagggatttg atagtcatat gagatctgct gatttaaaat ttattaaaaa 2580
tccaagaaat caggggctag aacaaagtga aattgaggaa atgagctttg atattaaagt 2640
agagccatca tttttgaaaa ataaagatga ctatattgca ttttggacat tctgcaaaat 2700
gctggatgct aggcatttaa gcgagctaag aaacgaaatg attaagtatg acggtcattt 2760
aactggagaa caagaaatca ttggtttagc attgcttgga gtggattcac gagagaatga 2820
ttggaagcaa ttttttagct cagaacggga atacgagaaa attatgaagg gctatgttgg 2880
agaggaattg tatcagcggg aaccgtaccg acaaagtgat ggcaaaacac cgattctttt 2940
tcgtggtgta gagcaagcga ggaagtatgg tactgaaaca gtgattcaac ggctttttga 3000
tgctagtcct gagtttaaag tgtcgaaatg caacataact gagtgggagc ggcaaaaaga 3060
aaccattgaa gagactattg agcgaagaaa agaattgcat aatgaatggg aaaaaaatcc 3120
caaaaaaccg caaaataatg cattttttaa agagtataaa gagtgttgtg acgctattga 3180
tgcttacaat tggcataaaa ataaaactac gcttgtatac gttaatgagc tgcaccattt 3240
gctaattgaa attctgggaa gatatgttgg ctatgtagca atagctgata gagactttca 3300
atgtatggcg aatcaatatt ttaagcattc aggaataact gagagagtgg aatattgggg 3360
cgataataga ctaaaaagta ttaaaaagct ggatacattc ttgaaaaaag aaggactgtt 3420
tgtttctgag aaaaatgcaa ggaatcatat agcgcattta aattatttat cactcaaatc 3480
tgagtgcacg ttgctgtatt tatctgagag gttgagagaa atttttaagt atgatcgtaa 3540
attaaagaat gccgtttcca agtcattaat cgatatttta gatagacatg gtatgagcgt 3600
cgtatttgct aacttgaaag aaaataaaca taggttggtg ataaaaagct tagagccaaa 3660
aaaattgaga catctaggtg agaaaaaaat cgataatggt tatatagaaa caaatcaagt 3720
ttcagaagag tattgtggta tagtaaagag actattagaa atttgatata gaatgttgca 3780
gttaatttca aaatgaaatt ggatgggaga atgcaatttc attttgctat taaaaaatac 3840
ctcaaaacag aagaggacta aaacaaatga ttgcaagccg ttacaaaaaa tatgatttag 3900
agtacctcag aacaaaagag ggctaaaact agaataatcg agtgagacgg tctacaagtg 3960
atttagagta gctcaaaaaa gaagaggtct aaaa 3994
<210> 185
<211> 4053
<212> DNA
<213> Listeria newyorkensis
<400> 185
tgtattctga gttgtctctt aatttatgaa aaaataatta tgttttggta atctgtcaag 60
atgcagatcc attaataatc tggtatagtt atgttgttgc aagtgaatat attacctcaa 120
aataaaagag gactaaaact aattggtggc gtgatacgcc gtaatgcttg atttagagta 180
cctcaaaaca aaagaggact aaaactactt gtcgatatgg tatagctttt ttcagattta 240
gagtacctca aaacaaaaga ggactaaaac taaagcttct aaatggtggc gcgttacgcc 300
gatttagaat acctcaaaac aaaagaggac taaaacctag caagccggtc gccgcgctca 360
aagtaagatt tagagtacct caaaacaaaa gaggactaaa acctcttttg tggataagta 420
ttcgaaataa agccataaaa actgtgatcc aaagagctgg attattggtt tttatggctt 480
tattcaattc ttagtattgt agatgaacca tcagtgaatg ttgtcttgca acgtgccttc 540
ttgtataatg aatatattga taaataatag aaatttcata cacgcatgaa gaaaccaatt 600
aaattttcag cattaatgaa gcattaggta catgactata aaaccaaatg gagctgagta 660
gacagatgaa aatcacaaag atgagagtag atggaagaac tatcgtaatg gagaggacaa 720
gcaaggaagg tcaactgggt tatgaaggta tcgatggaaa taagacaaca gaaattatat 780
ttgataagaa aaaagagtca ttttataaga gtatcctcaa taaaactgtg agaaaacccg 840
atgaaaaaga aaagaatagg cgtaagcagg caattaataa agcgattaat aaagaaataa 900
cagaattaat gttggcggtg ttacatcaag aagtgccaag ccaaaagtta cataatttaa 960
agagtctaaa tacggaatct ttaactaaac tatttaaacc gaagttccaa aacatgattt 1020
cttatccgcc tagcaaaggt gccgaacatg ttcagttttg ccttacagat atagcggtac 1080
cagcgattcg agatttagat gaaattaagc cagattgggg catttttttt gaaaaattga 1140
aaccctatac ggattgggca gaatcataca ttcactataa gcagacaacc atacagaaat 1200
ccattgagca aaacaaaata cagtcccctg attcgccaag gaaattagta ttgcaaaaat 1260
atgtcacagc ctttttgaat ggagaaccgc tgggactcga tcttgtggcg aaaaaatata 1320
aactggcaga cttagcggag tcgtttaaat tagtagattt gaacgaggat aaaagtgcaa 1380
actataaaat taaagcgtgc ttgcaacaac atcagcgaaa tattttggat gaattgaaag 1440
aagatccgga gttaaatcaa tatggtatag aagtgaagaa atatatacag cgatatttcc 1500
caatcaaacg tgcaccgaat agaagtaaac atgcacgagc ggactttttg aaaaaggaat 1560
taattgagtc tacagtggag caacaattta aaaatgctgt atatcattat gtactggaac 1620
aaggcaaaat ggaggcatat gagctaacag atcctaaaac aaaagacttg caggatatta 1680
gatctggtga ggcatttagc ttcaaattta ttaatgcttg cgccttcgca tcgaataatt 1740
tgaagatgat tttaaaccct gaatgtgaaa aggatatcct aggtaagggc aattttaaaa 1800
agaatttgcc aaacagtact acgcgatctg atgttgtgaa gaaaatgatt ccctttttct 1860
cggatgagct tcaaaatgtg aattttgatg aagctatctg ggcgattagg ggctctattc 1920
agcaaattag aaatgaggtt taccattgta aaaagcattc ttggaaaagc atacttaaaa 1980
taaaaggctt tgaatttgaa cctaacaata tgaaatatgc ggattctgat atgcaaaaat 2040
tgatggataa agatatcgcc aaaattccag agttcatcga agaaaaactt aaaagtagtg 2100
gagtagtaag gttctacagg catgatgagt tgcaatccat atgggaaatg aaacaaggat 2160
tttcgttgct gactactaat gcgccgtttg tcccaagctt taagcgtgtc tacgcaaaag 2220
ggcacgacta ccaaacttct aaaaatagat actataattt ggatttgact acttttgata 2280
ttttggaata tggagaagaa gattttcgtg cacgctattt cctgacgaag ctagtttatt 2340
atcagcaatt tatgccatgg tttacagctg ataataatgc tttccgagat gctgccaatt 2400
ttgtattgcg attaaataaa aatagacagc aggatgcaaa agcttttatt aacattagag 2460
aagttgaaga aggtgagatg cctagagact atatgggtta tgtccaaggt caaatagcga 2520
tacatgagga ttcaattgag gatacaccga atcattttga gaaatttatt agtcaggttt 2580
ttattaaggg ctttgatagg catatgagat ctgctaattt aaaatttatt aaaaatccaa 2640
gaaatcaggg gctagaacaa agtgagattg aggaaatgag ctttgatatt aaagtggagc 2700
cgtcattttt gaaaaataaa gatgactata ttgcattttg gatattctgc aaaatgcttg 2760
atgctaggca tttaagcgag ctaagaaacg aaatgattaa gtatgacggt catttaactg 2820
gagaacaaga aatcattggt ttagcattgc tcggagtgga ttcacgagag aatgattgga 2880
agcagttttt tagctcagaa cgggaatacg agaaaattat gaagggctat gttgtagagg 2940
aattgtatca gcgggaaccg taccgacaaa gtgatggcaa aacaccgatt ctttttcgtg 3000
gtgtagagca agcgaggaag tatggtactg aaacagtgat tcaacggctt tttgatgcta 3060
atcctgagtt caaagtgtca aaatgcaact tagcagagtg ggagcggcaa aaagaaacca 3120
ttgaagagac tattaagcga agaaaagaat tgcataatga atgggcaaaa aatccaaaaa 3180
aaccgcaaaa taatgcattt tttaaagagt ataaagagtg ttgtgacgct attgacgctt 3240
acaattggca taaaaataaa actacgcttg catacgttaa tgagctgcac catttgctaa 3300
ttgaaattct gggaagatat gttggctatg tagcaatagc tgatagagac tttcaatgta 3360
tggcgaatca atattttaag cattcaggaa taactgagag agtggaatat tggggcgata 3420
atagactaaa aagtattaaa aagctggata cattcttgaa aaaagaagga ctgtttgttt 3480
ctgagaaaaa tgcaaggaat catatagcgc atttaaatta tttatcactc aaatctgagt 3540
gcacgttgct gtatttatcc gagaggttga gagaaatttt taagtatgat cgtaaattaa 3600
agaatgccgt ttccaagtca ttaatcgata ttttagatag acatggtatg agcgtcgtat 3660
ttgctaactt gaaagaaaat aaacataggt tggtgataaa aagcttagag ccaaaaaaat 3720
tgagacatct gggtgggaaa aaaatcgatg gtggttatat agaaacaaat caagtttcag 3780
aagagtattg tggtatagtg aagagactgt tagaaatgtg atgtaggatg ttacagtcaa 3840
tctcaaaatg aaattggaag ttacaagagg acgtaatatg atgttactat taaaaatacc 3900
tcagaataga agaggactaa aacaaatgat tgcaagccgt tacaaaaaat atgatttaga 3960
gtacctcaga acaaaagagg gctaaaacta gaataatcga ttgagacggt ctacaagtga 4020
tttagagtac ctcaaaacag aagaggacta aaa 4053
<210> 186
<211> 53
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 186
auggccacuu uccagguggc aaagcccguu gacguuguga gcttctcaaa aag 53
<210> 187
<211> 32
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> modified_base
<222> (15)..(32)
<223> a, c, u, g, unknown or other
<400> 187
cugagaagug gcacnnnnnn nnnnnnnnnn nn 32
<210> 188
<211> 14
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 188
cugagaagug gcac 14
<210> 189
<211> 79
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 189
gucuagagga cagaauuuuu caacgggugu gccaauggcc acuuuccagg uggcaaagcc 60
cguugagcuu cucaaaaag 79
<210> 190
<211> 32
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> modified_base
<222> (15)..(32)
<223> a, c, u, g, unknown or other
<400> 190
cugagaagug gcacnnnnnn nnnnnnnnnn nn 32
<210> 191
<211> 39
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 191
acuuuccagg uggcaaagcc cguugagcuu cucaaaaag 39
<210> 192
<211> 53
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> source
<223> /note="Description of Combined DNA/RNA Molecule: Synthetic
oligonucleotide"
<400> 192
auggccacuu uccagguggc aaagcccguu gacguuguga gcttctcaaa aag 53
<210> 193
<211> 32
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> modified_base
<222> (15)..(32)
<223> a, c, u, g, unknown or other
<400> 193
cugagaagug gcacnnnnnn nnnnnnnnnn nn 32
<210> 194
<211> 111
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<220>
<221> modified_base
<222> (92)..(111)
<223> a, c, u, g, unknown or other
<400> 194
gucuagagga cagaauuuuu caacgggugu gccaauggcc acuuuccagg uggcaaagcc 60
cguugagcuu cucaaaucug agaaguggca cnnnnnnnnn nnnnnnnnnn n 111
<210> 195
<211> 52
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 195
aagtgctgca ggggtgtgag aaactcctat tgctggacga tgtctctttt at 52
<210> 196
<211> 33
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<220>
<221> modified_base
<222> (15)..(33)
<223> a, c, t, g, unknown or other
<400> 196
cgaggcatta gcacnnnnnn nnnnnnnnnn nnn 33
<210> 197
<211> 91
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 197
cgagguucug ucuuuugguc aggacaaccg ucuagcuaua agugcugcag gggugugaga 60
aacuccuauu gcuggacgau gucucuuuua u 91
<210> 198
<211> 14
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 198
cgaggcauua gcac 14
<210> 199
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 199
guucgaaagc uuaguggaaa gcuucguggu uagcac 36
<210> 200
<211> 101
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 200
ucagucgcac gcuauaggcc auaagucgac uuacauaucc gugcgugugc auuaugggcc 60
cauccacagg ucuauuccca cggauaauca cgacuuucca c 101
<210> 201
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 201
guucgaaagc uuaguggaaa gcuucguggu uagcac 36
<210> 202
<211> 18
<212> PRT
<213> Alicyclobacillus acidoterrestris
<400> 202
Leu Arg Val Met Ser Val Asp Leu Gly Leu Arg Thr Ser Ala Ser Ile
1 5 10 15
Ser Val
<210> 203
<211> 22
<212> PRT
<213> Alicyclobacillus acidoterrestris
<400> 203
Gln Arg Thr Leu Arg Gln Leu Arg Thr Gln Leu Ala Tyr Leu Arg Leu
1 5 10 15
Leu Val Arg Cys Gly Ser
20
<210> 204
<211> 12
<212> PRT
<213> Alicyclobacillus acidoterrestris
<400> 204
Cys Gln Leu Ile Leu Leu Glu Glu Leu Ser Glu Tyr
1 5 10
<210> 205
<211> 16
<212> PRT
<213> Alicyclobacillus acidoterrestris
<400> 205
His Gln Ile His Ala Asp Leu Asn Ala Ala Gln Asn Leu Gln Gln Arg
1 5 10 15
<210> 206
<211> 18
<212> PRT
<213> Alicyclobacillus contaminans
<400> 206
Val Arg Val Met Ser Val Asp Leu Gly Val Arg Tyr Gly Ala Ala Ile
1 5 10 15
Ser Val
<210> 207
<211> 22
<212> PRT
<213> Alicyclobacillus contaminans
<400> 207
Lys Gln Ala Leu Ala Ala Ile Arg Ala Glu Met Ser Ile Leu Arg Lys
1 5 10 15
Trp Leu Arg Val Ser Gln
20
<210> 208
<211> 12
<212> PRT
<213> Alicyclobacillus contaminans
<400> 208
Cys Asp Leu Ile Leu Phe Glu Asp Leu Ser Arg Tyr
1 5 10
<210> 209
<211> 16
<212> PRT
<213> Alicyclobacillus contaminans
<400> 209
Lys Cys Val His Ala Asp Ile Asn Ala Ala His Asn Leu Gln Arg Arg
1 5 10 15
<210> 210
<211> 18
<212> PRT
<213> Desulfovibrio inopinatus
<400> 210
Leu Arg Val Leu Ser Val Asp Leu Gly Met Arg Thr Phe Ala Ser Cys
1 5 10 15
Ser Val
<210> 211
<211> 22
<212> PRT
<213> Desulfovibrio inopinatus
<400> 211
Arg Ala Glu Ile Tyr Ala Leu Lys Arg Asp Ile Gln Arg Leu Lys Ser
1 5 10 15
Leu Leu Arg Leu Gly Glu
20
<210> 212
<211> 12
<212> PRT
<213> Desulfovibrio inopinatus
<400> 212
Cys Gln Leu Ile Leu Phe Glu Asp Leu Ala Arg Tyr
1 5 10
<210> 213
<211> 16
<212> PRT
<213> Desulfovibrio inopinatus
<400> 213
Cys Val Ile His Ala Asp Met Asn Ala Ala Gln Asn Leu Gln Arg Arg
1 5 10 15
<210> 214
<211> 18
<212> PRT
<213> Desulfonatronum thiodismutans
<400> 214
Leu Arg Val Leu Ser Val Asp Leu Gly Val Arg Ser Phe Ala Ala Cys
1 5 10 15
Ser Val
<210> 215
<211> 22
<212> PRT
<213> Desulfonatronum thiodismutans
<400> 215
Met Glu Glu Leu Arg Ser Leu Asn Gly Asp Ile Arg Arg Leu Lys Ala
1 5 10 15
Ile Leu Arg Leu Ser Val
20
<210> 216
<211> 12
<212> PRT
<213> Desulfonatronum thiodismutans
<400> 216
Cys Arg Leu Ile Leu Phe Glu Asp Leu Ala Arg Tyr
1 5 10
<210> 217
<211> 16
<212> PRT
<213> Desulfonatronum thiodismutans
<400> 217
His Val Ile His Ala Asp Ile Asn Ala Ala Gln Asn Leu Gln Arg Arg
1 5 10 15
<210> 218
<211> 18
<212> PRT
<213> Tuberibacillus calidus
<400> 218
Leu Arg Val Met Ser Val Asp Leu Gly Gln Arg Gln Ala Ala Ala Ile
1 5 10 15
Ser Ile
<210> 219
<211> 22
<212> PRT
<213> Tuberibacillus calidus
<400> 219
Asp Gln Ala Ile Arg Asp Leu Ser Arg Lys Leu Lys Phe Leu Lys Asn
1 5 10 15
Val Leu Asn Met Gln Lys
20
<210> 220
<211> 12
<212> PRT
<213> Tuberibacillus calidus
<400> 220
Cys Gln Leu Val Leu Phe Glu Asp Leu Ser Arg Tyr
1 5 10
<210> 221
<211> 16
<212> PRT
<213> Tuberibacillus calidus
<400> 221
Val Ile Thr His Ala Asp Ile Asn Ala Ala Gln Asn Leu Gln Lys Arg
1 5 10 15
<210> 222
<211> 18
<212> PRT
<213> Bacillus thermoamylovorans
<400> 222
Leu Arg Val Met Ser Ile Asp Leu Gly Gln Arg Gln Ala Ala Ala Ala
1 5 10 15
Ser Ile
<210> 223
<211> 22
<212> PRT
<213> Bacillus thermoamylovorans
<400> 223
Glu Asp Asn Leu Lys Leu Met Asn Gln Lys Leu Asn Phe Leu Arg Asn
1 5 10 15
Val Leu His Phe Gln Gln
20
<210> 224
<211> 12
<212> PRT
<213> Bacillus thermoamylovorans
<400> 224
Cys Gln Ile Ile Leu Phe Glu Asp Leu Ser Asn Tyr
1 5 10
<210> 225
<211> 16
<212> PRT
<213> Bacillus thermoamylovorans
<400> 225
Val Thr Thr His Ala Asp Ile Asn Ala Ala Gln Asn Leu Gln Lys Arg
1 5 10 15
<210> 226
<211> 18
<212> PRT
<213> Bacillus sp.
<400> 226
Phe Arg Val Met Ser Ile Asp Leu Gly Leu Arg Ala Ala Ala Ala Thr
1 5 10 15
Ser Ile
<210> 227
<211> 22
<212> PRT
<213> Bacillus sp.
<400> 227
Phe Gln Leu His Gln Arg Val Lys Phe Gln Ile Arg Val Leu Ala Gln
1 5 10 15
Ile Met Arg Met Ala Asn
20
<210> 228
<211> 12
<212> PRT
<213> Bacillus sp.
<400> 228
Cys Gln Val Ile Leu Phe Glu Asn Leu Ser Gln Tyr
1 5 10
<210> 229
<211> 16
<212> PRT
<213> Bacillus sp.
<400> 229
Val Phe Leu Gln Ala Asp Ile Asn Ala Ala His Asn Leu Gln Lys Arg
1 5 10 15
<210> 230
<211> 18
<212> PRT
<213> Methylobacterium nodulans
<400> 230
Leu Arg Val Leu Ser Ile Asp Leu Gly Val Arg Ser Phe Ala Thr Cys
1 5 10 15
Ser Val
<210> 231
<211> 22
<212> PRT
<213> Methylobacterium nodulans
<400> 231
Asp Ala Glu Leu Arg Gln Leu Arg Gly Gly Leu Asn Arg His Arg Gln
1 5 10 15
Leu Leu Arg Ala Ala Thr
20
<210> 232
<211> 12
<212> PRT
<213> Methylobacterium nodulans
<400> 232
Cys His Val Ile Leu Phe Glu Asp Leu Ser Arg Tyr
1 5 10
<210> 233
<211> 16
<212> PRT
<213> Methylobacterium nodulans
<400> 233
Ser Arg Ile His Ala Asp Ile Asn Ala Ala Gln Asn Leu Gln Arg Arg
1 5 10 15
<210> 234
<211> 18
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Candidatus Methanomethylophilus alvus sequence"
<400> 234
Leu Lys Ile Ile Gly Ile Asp Arg Gly Glu Arg Asn Leu Ile Tyr Val
1 5 10 15
Thr Met
<210> 235
<211> 22
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Candidatus Methanomethylophilus alvus sequence"
<400> 235
Arg Lys Ala Leu Asp Val Arg Glu Tyr Asp Asn Lys Glu Ala Arg Arg
1 5 10 15
Asn Trp Thr Lys Val Glu
20
<210> 236
<211> 13
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Candidatus Methanomethylophilus alvus sequence"
<400> 236
Asn Ala Ile Ile Val Met Glu Asp Leu Asn His Gly Phe
1 5 10
<210> 237
<211> 16
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Candidatus Methanomethylophilus alvus sequence"
<400> 237
Leu Pro Gln Asp Ser Asp Ala Asn Gly Ala Tyr Asn Ile Ala Leu Lys
1 5 10 15
<210> 238
<211> 18
<212> PRT
<213> Synergistes jonesii
<400> 238
Val Asn Ile Ile Gly Ile Asp Arg Gly Glu Arg Asn Leu Val Tyr Val
1 5 10 15
Ser Leu
<210> 239
<211> 22
<212> PRT
<213> Synergistes jonesii
<400> 239
His Ala Lys Leu Asn Gln Lys Glu Lys Glu Arg Asp Thr Ala Arg Lys
1 5 10 15
Ser Trp Lys Thr Ile Gly
20
<210> 240
<211> 13
<212> PRT
<213> Synergistes jonesii
<400> 240
Asn Ala Val Ile Val Met Glu Asp Leu Asn Ile Gly Phe
1 5 10
<210> 241
<211> 16
<212> PRT
<213> Synergistes jonesii
<400> 241
Leu Pro Ile Asp Ala Asp Ala Asn Gly Ala Tyr His Ile Ala Leu Lys
1 5 10 15
<210> 242
<211> 18
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 242
Pro Tyr Val Ile Gly Ile Asp Arg Gly Glu Arg Asn Leu Leu Tyr Ile
1 5 10 15
Val Val
<210> 243
<211> 22
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 243
His Ser Leu Leu Asp Lys Lys Glu Lys Glu Arg Phe Glu Ala Arg Gln
1 5 10 15
Asn Trp Thr Ser Ile Glu
20
<210> 244
<211> 13
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 244
Asp Ala Val Ile Ala Leu Glu Asp Leu Asn Ser Gly Phe
1 5 10
<210> 245
<211> 16
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 245
Leu Pro Lys Asn Ala Asp Ala Asn Gly Ala Tyr Asn Ile Ala Arg Lys
1 5 10 15
<210> 246
<211> 18
<212> PRT
<213> Francisella tularensis
<400> 246
Val His Ile Leu Ser Ile Asp Arg Gly Glu Arg His Leu Ala Tyr Tyr
1 5 10 15
Thr Leu
<210> 247
<211> 22
<212> PRT
<213> Francisella tularensis
<400> 247
His Asp Lys Leu Ala Ala Ile Glu Lys Asp Arg Asp Ser Ala Arg Lys
1 5 10 15
Asp Trp Lys Lys Ile Asn
20
<210> 248
<211> 13
<212> PRT
<213> Francisella tularensis
<400> 248
Asn Ala Ile Val Val Phe Glu Asp Leu Asn Phe Gly Phe
1 5 10
<210> 249
<211> 16
<212> PRT
<213> Francisella tularensis
<400> 249
Met Pro Gln Asp Ala Asp Ala Asn Gly Ala Tyr His Ile Gly Leu Lys
1 5 10 15
<210> 250
<211> 18
<212> PRT
<213> Moraxella caprae
<400> 250
Val Asn Val Ile Gly Ile Asp Arg Gly Glu Arg His Leu Leu Tyr Leu
1 5 10 15
Thr Val
<210> 251
<211> 22
<212> PRT
<213> Moraxella caprae
<400> 251
His Lys Ile Leu Asp Lys Arg Glu Ile Glu Arg Leu Asn Ala Arg Val
1 5 10 15
Gly Trp Gly Glu Ile Glu
20
<210> 252
<211> 13
<212> PRT
<213> Moraxella caprae
<400> 252
Asn Ala Ile Val Val Leu Glu Asp Leu Asn Phe Gly Phe
1 5 10
<210> 253
<211> 16
<212> PRT
<213> Moraxella caprae
<400> 253
Gln Pro Gln Asn Ala Asp Ala Asn Gly Ala Tyr His Ile Ala Leu Lys
1 5 10 15
<210> 254
<211> 18
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 254
Met His Ile Ile Gly Ile Asp Arg Gly Glu Arg Asn Leu Ile Tyr Leu
1 5 10 15
Cys Met
<210> 255
<211> 22
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 255
His Gln Leu Leu Lys Thr Arg Glu Asp Glu Asn Lys Ser Ala Arg Gln
1 5 10 15
Ser Trp Gln Thr Ile His
20
<210> 256
<211> 13
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 256
Asn Ala Ile Val Val Leu Glu Asp Leu Asn Phe Gly Phe
1 5 10
<210> 257
<211> 16
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 257
Met Pro Leu Asp Ala Asp Ala Asn Gly Ala Tyr Asn Ile Ala Arg Lys
1 5 10 15
<210> 258
<211> 18
<212> PRT
<213> Prevotella albensis
<400> 258
Thr His Ile Ile Gly Ile Asp Arg Gly Glu Arg His Leu Leu Tyr Leu
1 5 10 15
Ser Leu
<210> 259
<211> 22
<212> PRT
<213> Prevotella albensis
<400> 259
His Asn Leu Leu Glu Lys Arg Glu Lys Glu Arg Thr Glu Ala Arg His
1 5 10 15
Ser Trp Ser Ser Ile Glu
20
<210> 260
<211> 13
<212> PRT
<213> Prevotella albensis
<400> 260
Asn Ala Ile Val Val Leu Glu Asp Leu Asn Gly Gly Phe
1 5 10
<210> 261
<211> 16
<212> PRT
<213> Prevotella albensis
<400> 261
Phe Pro Glu Asn Ala Asp Ala Asn Gly Ala Tyr Asn Ile Ala Arg Lys
1 5 10 15
<210> 262
<211> 18
<212> PRT
<213> Smithella sp.
<400> 262
Ile Asn Ile Ile Gly Ile Asp Arg Gly Glu Arg His Leu Leu Tyr Tyr
1 5 10 15
Ala Leu
<210> 263
<211> 22
<212> PRT
<213> Smithella sp.
<400> 263
His Asn Leu Leu Asp Lys Lys Glu Gly Asp Arg Ala Thr Ala Arg Gln
1 5 10 15
Glu Trp Gly Val Ile Glu
20
<210> 264
<211> 13
<212> PRT
<213> Smithella sp.
<400> 264
Asn Ala Ile Ile Val Met Glu Asp Leu Asn Phe Gly Phe
1 5 10
<210> 265
<211> 16
<212> PRT
<213> Smithella sp.
<400> 265
Met Pro Lys Asn Ala Asp Ala Asn Gly Ala Tyr His Ile Ala Leu Lys
1 5 10 15
<210> 266
<211> 18
<212> PRT
<213> Porphyromonas crevioricanis
<400> 266
Met His Val Ile Gly Ile Asp Arg Gly Glu Arg Asn Leu Leu Tyr Ile
1 5 10 15
Cys Val
<210> 267
<211> 22
<212> PRT
<213> Porphyromonas crevioricanis
<400> 267
His Asp Leu Leu Glu Ser Arg Asp Lys Asp Arg Gln Gln Glu Arg Arg
1 5 10 15
Asn Trp Gln Thr Ile Glu
20
<210> 268
<211> 13
<212> PRT
<213> Porphyromonas crevioricanis
<400> 268
Lys Ala Val Val Ala Leu Glu Asp Leu Asn Met Gly Phe
1 5 10
<210> 269
<211> 16
<212> PRT
<213> Porphyromonas crevioricanis
<400> 269
Leu Pro Lys Asp Ala Asp Ala Asn Gly Ala Tyr Asn Ile Ala Leu Lys
1 5 10 15
<210> 270
<211> 18
<212> PRT
<213> Sulfolobus solfataricus
<400> 270
Gly Lys Val Val Ala Ile Asp Val Gly Val Glu Lys Leu Leu Ile Thr
1 5 10 15
Ser Asp
<210> 271
<211> 23
<212> PRT
<213> Sulfolobus solfataricus
<400> 271
Val Lys His Ile His Arg Glu Leu Ser Arg Lys Lys Phe Leu Ser Asn
1 5 10 15
Asn Trp Phe Lys Ala Lys Val
20
<210> 272
<211> 13
<212> PRT
<213> Sulfolobus solfataricus
<400> 272
Tyr Asp Val Val Val Met Glu Gly Ile His Ala Lys Gln
1 5 10
<210> 273
<211> 16
<212> PRT
<213> Sulfolobus solfataricus
<400> 273
Trp Ile Ala Asp Arg Asp Tyr Asn Ala Ser Leu Asn Ile Leu Arg Gly
1 5 10 15
<210> 274
<211> 18
<212> PRT
<213> Nostoc sp.
<400> 274
Leu Lys Thr Ile Gly Leu Asp Val Gly Leu Asn His Phe Leu Thr Asp
1 5 10 15
Ser Glu
<210> 275
<211> 23
<212> PRT
<213> Nostoc sp.
<400> 275
Leu Lys Arg Leu Gln Arg Arg Leu Ser Lys Thr Lys Lys Gly Ser Asn
1 5 10 15
Asn Arg Val Lys Ala Arg Asn
20
<210> 276
<211> 13
<212> PRT
<213> Nostoc sp.
<400> 276
Ser Asp Leu Val Ala Tyr Glu Asp Leu Gln Val Arg Asn
1 5 10
<210> 277
<211> 16
<212> PRT
<213> Nostoc sp.
<400> 277
His Ile Gln Asp Arg Asp Trp Asn Ala Ala Arg Asn Ile Leu Glu Leu
1 5 10 15
<210> 278
<211> 18
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Candidatus Nitrososphaera gargensis sequence"
<400> 278
Ala Lys Pro Val Gly Ile Asp Val Gly Ile Ala Lys Phe Cys His His
1 5 10 15
Ser Asp
<210> 279
<211> 23
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Candidatus Nitrososphaera gargensis sequence"
<400> 279
Leu Arg Arg Ala His Arg Arg Val Ser Arg Arg Gln Ile Gly Ser Asn
1 5 10 15
Asn Arg Lys Lys Ala Lys Arg
20
<210> 280
<211> 13
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Candidatus Nitrososphaera gargensis sequence"
<400> 280
Tyr Asp Leu Ile Phe Leu Glu Arg Leu Arg Val Met Asn
1 5 10
<210> 281
<211> 16
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Candidatus Nitrososphaera gargensis sequence"
<400> 281
Ala Ile Leu Asp Arg Asp Tyr Asn Ser Ala Ile Asn Ile Leu Lys Arg
1 5 10 15
<210> 282
<211> 18
<212> PRT
<213> Helicobacter pylori
<400> 282
Lys Lys Ala Val Gly Leu Asp Met Gly Leu Arg Thr Leu Ile Val Thr
1 5 10 15
Ser Asp
<210> 283
<211> 23
<212> PRT
<213> Helicobacter pylori
<400> 283
Leu Thr Lys Ala Gln Arg Arg Leu Ser Lys Lys Val Lys Asp Ser Asn
1 5 10 15
Asn Arg Lys Lys Gln Ala Lys
20
<210> 284
<211> 13
<212> PRT
<213> Helicobacter pylori
<400> 284
Tyr Asp Leu Ile Gly Val Glu Thr Leu Asn Val Lys Ala
1 5 10
<210> 285
<211> 16
<212> PRT
<213> Helicobacter pylori
<400> 285
Thr Thr His His Arg Asp Tyr Asn Ala Ser Val Asn Ile Arg Asn Tyr
1 5 10 15
<210> 286
<211> 18
<212> PRT
<213> Flexibacter litoralis
<400> 286
Asn Gln Ala Val Gly Ile Asp Met Gly Ile Thr Phe Phe Cys Ile Asp
1 5 10 15
Ser Asn
<210> 287
<211> 23
<212> PRT
<213> Flexibacter litoralis
<400> 287
Leu Arg Ile Ala Asn Arg Ser Leu Ser Arg Lys Lys Lys Phe Ser Asn
1 5 10 15
Gly Trp Tyr Lys Lys Lys Val
20
<210> 288
<211> 13
<212> PRT
<213> Flexibacter litoralis
<400> 288
Asn Ser Leu Val Val Val Glu Asp Leu Lys Val Lys Asn
1 5 10
<210> 289
<211> 16
<212> PRT
<213> Flexibacter litoralis
<400> 289
His Glu Thr Asn Ala Asp Glu Asn Ala Ser Lys Asn Ile Leu Ser Glu
1 5 10 15
<210> 290
<211> 18
<212> PRT
<213> Escherichia coli
<400> 290
Ala Ser Met Val Gly Leu Asp Ala Gly Val Ala Lys Leu Ala Thr Leu
1 5 10 15
Ser Asp
<210> 291
<211> 23
<212> PRT
<213> Escherichia coli
<400> 291
Leu Ala Arg Leu Gln Arg Gln Leu Ser Arg Lys Val Lys Phe Ser Asn
1 5 10 15
Asn Trp Gln Lys Gln Lys Arg
20
<210> 292
<211> 13
<212> PRT
<213> Escherichia coli
<400> 292
His Ala Met Ile Val Ile Glu Asp Leu Lys Val Ser Asn
1 5 10
<210> 293
<211> 16
<212> PRT
<213> Escherichia coli
<400> 293
Tyr Thr Ala Asn Ala Asp Val Asn Gly Ala Arg Asn Ile Leu Ala Ala
1 5 10 15
<210> 294
<211> 18
<212> PRT
<213> Clostridium botulinum
<400> 294
Asn Lys Lys Val Gly Ile Asp Val Gly Leu Lys Glu Phe Ala Thr Thr
1 5 10 15
Ser Asp
<210> 295
<211> 23
<212> PRT
<213> Clostridium botulinum
<400> 295
Leu Ala Lys Leu Gln Lys Asp Leu Ser Arg Lys Lys Lys Asn Ser Asn
1 5 10 15
Asn Arg Lys Lys Ala Arg Leu
20
<210> 296
<211> 13
<212> PRT
<213> Clostridium botulinum
<400> 296
Asn Gln Ala Ile Val Ile Glu Asn Leu Lys Val Ser Asn
1 5 10
<210> 297
<211> 16
<212> PRT
<213> Clostridium botulinum
<400> 297
Met Ile Met Asp Arg Asp Leu Asn Ala Ser Lys Asn Leu Leu Asn Leu
1 5 10 15
<210> 298
<211> 18
<212> PRT
<213> Acidaminococcus sp.
<400> 298
Met Tyr Tyr Leu Gly Leu Asp Ile Gly Thr Asn Ser Val Gly Tyr Ala
1 5 10 15
Val Thr
<210> 299
<211> 22
<212> PRT
<213> Acidaminococcus sp.
<400> 299
Ala Glu Arg Arg Ser Phe Arg Thr Ser Arg Arg Arg Leu Asp Arg Arg
1 5 10 15
Gln Gln Arg Val Lys Leu
20
<210> 300
<211> 13
<212> PRT
<213> Acidaminococcus sp.
<400> 300
Pro Lys Arg Ile Phe Ile Glu Met Ala Arg Asp Gly Glu
1 5 10
<210> 301
<211> 16
<212> PRT
<213> Acidaminococcus sp.
<400> 301
Leu His His Ala Lys Asp Ala Phe Leu Ala Ile Val Thr Gly Asn Val
1 5 10 15
<210> 302
<211> 18
<212> PRT
<213> Coprococcus catus
<400> 302
Glu Tyr Phe Leu Gly Leu Asp Met Gly Thr Gly Ser Leu Gly Trp Ala
1 5 10 15
Val Thr
<210> 303
<211> 22
<212> PRT
<213> Coprococcus catus
<400> 303
Glu Glu Arg Arg Met Phe Arg Thr Ala Arg Arg Arg Leu Asp Arg Arg
1 5 10 15
Asn Trp Arg Ile Gln Val
20
<210> 304
<211> 13
<212> PRT
<213> Coprococcus catus
<400> 304
Pro Lys Arg Val Phe Val Glu Met Ala Arg Glu Lys Gln
1 5 10
<210> 305
<211> 16
<212> PRT
<213> Coprococcus catus
<400> 305
Leu His His Ala Lys Asp Ala Tyr Leu Asn Ile Val Val Gly Asn Ala
1 5 10 15
<210> 306
<211> 18
<212> PRT
<213> Treponema denticola
<400> 306
Asp Tyr Phe Leu Gly Leu Asp Val Gly Thr Gly Ser Val Gly Trp Ala
1 5 10 15
Val Thr
<210> 307
<211> 22
<212> PRT
<213> Treponema denticola
<400> 307
Glu Val Arg Arg Leu His Arg Gly Ala Arg Arg Arg Ile Glu Arg Arg
1 5 10 15
Lys Lys Arg Ile Lys Leu
20
<210> 308
<211> 13
<212> PRT
<213> Treponema denticola
<400> 308
Pro Lys Lys Ile Phe Ile Glu Met Ala Lys Gly Ala Glu
1 5 10
<210> 309
<211> 16
<212> PRT
<213> Treponema denticola
<400> 309
Phe His His Ala His Asp Ala Tyr Leu Asn Ile Val Val Gly Asn Val
1 5 10 15
<210> 310
<211> 18
<212> PRT
<213> Mycoplasma mobile
<400> 310
Lys Val Val Leu Gly Leu Asp Leu Gly Ile Ala Ser Val Gly Trp Cys
1 5 10 15
Leu Thr
<210> 311
<211> 22
<212> PRT
<213> Mycoplasma mobile
<400> 311
Glu Thr Arg Arg Lys Lys Arg Gly Gln Arg Arg Arg Asn Arg Arg Leu
1 5 10 15
Phe Thr Arg Lys Arg Asp
20
<210> 312
<211> 13
<212> PRT
<213> Mycoplasma mobile
<400> 312
Ile Glu Lys Ile Val Val Glu Val Thr Arg Ser Ser Asn
1 5 10
<210> 313
<211> 16
<212> PRT
<213> Mycoplasma mobile
<400> 313
Gly His His Ala Glu Asp Ala Tyr Phe Ile Thr Ile Ile Ser Gln Tyr
1 5 10 15
<210> 314
<211> 18
<212> PRT
<213> Streptococcus thermophilus
<400> 314
Asp Leu Val Leu Gly Leu Asp Ile Gly Ile Gly Ser Val Gly Val Gly
1 5 10 15
Ile Leu
<210> 315
<211> 22
<212> PRT
<213> Streptococcus thermophilus
<400> 315
Leu Val Arg Arg Thr Asn Arg Gln Gly Arg Arg Leu Thr Arg Arg Lys
1 5 10 15
Lys His Arg Ile Val Arg
20
<210> 316
<211> 13
<212> PRT
<213> Streptococcus thermophilus
<400> 316
Phe Asp Asn Ile Val Ile Glu Met Ala Arg Glu Thr Asn
1 5 10
<210> 317
<211> 16
<212> PRT
<213> Streptococcus thermophilus
<400> 317
His His His Ala Val Asp Ala Leu Ile Ile Ala Ala Ser Ser Gln Leu
1 5 10 15
<210> 318
<211> 18
<212> PRT
<213> Campylobacter jejuni
<400> 318
Ala Arg Ile Leu Ala Phe Asp Ile Gly Ile Ser Ser Ile Gly Trp Ala
1 5 10 15
Phe Ser
<210> 319
<211> 22
<212> PRT
<213> Campylobacter jejuni
<400> 319
Leu Pro Arg Arg Leu Ala Arg Ser Ala Arg Lys Arg Leu Ala Arg Arg
1 5 10 15
Lys Ala Arg Leu Asn His
20
<210> 320
<211> 13
<212> PRT
<213> Campylobacter jejuni
<400> 320
Val His Lys Ile Asn Ile Glu Leu Ala Arg Glu Val Gly
1 5 10
<210> 321
<211> 16
<212> PRT
<213> Campylobacter jejuni
<400> 321
Leu His His Ala Ile Asp Ala Val Ile Ile Ala Tyr Ala Asn Asn Ser
1 5 10 15
<210> 322
<211> 18
<212> PRT
<213> Clostridium perfringens
<400> 322
Asn Tyr Ala Leu Gly Leu Asp Ile Gly Ile Thr Ser Val Gly Trp Ala
1 5 10 15
Val Ile
<210> 323
<211> 22
<212> PRT
<213> Clostridium perfringens
<400> 323
Leu Pro Arg Arg Leu Ala Arg Gly Arg Arg Arg Leu Leu Arg Arg Lys
1 5 10 15
Ala Tyr Arg Val Glu Arg
20
<210> 324
<211> 13
<212> PRT
<213> Clostridium perfringens
<400> 324
Pro Val Arg Ile Asn Ile Glu Leu Ala Arg Asp Leu Ala
1 5 10
<210> 325
<211> 16
<212> PRT
<213> Clostridium perfringens
<400> 325
Lys His His Ala Leu Asp Ala Ala Val Val Gly Val Thr Thr Gln Gly
1 5 10 15
<210> 326
<211> 18
<212> PRT
<213> Akkermansia muciniphila
<400> 326
Ser Leu Thr Phe Ser Phe Asp Ile Gly Tyr Ala Ser Ile Gly Trp Ala
1 5 10 15
Val Ile
<210> 327
<211> 22
<212> PRT
<213> Akkermansia muciniphila
<400> 327
Phe Lys Arg Arg Glu Tyr Arg Arg Leu Arg Arg Asn Ile Arg Ser Arg
1 5 10 15
Arg Val Arg Ile Glu Arg
20
<210> 328
<211> 13
<212> PRT
<213> Akkermansia muciniphila
<400> 328
Ile Ser Arg Val Cys Val Glu Val Gly Lys Glu Leu Thr
1 5 10
<210> 329
<211> 16
<212> PRT
<213> Akkermansia muciniphila
<400> 329
Leu His His Ala Leu Asp Ala Cys Val Leu Gly Leu Ile Pro Tyr Ile
1 5 10 15
<210> 330
<211> 18
<212> PRT
<213> Bifidobacterium longum
<400> 330
Arg Tyr Arg Ile Gly Ile Asp Val Gly Leu Asn Ser Val Gly Leu Ala
1 5 10 15
Ala Val
<210> 331
<211> 22
<212> PRT
<213> Bifidobacterium longum
<400> 331
Asn Met Ser Gly Val Ala Arg Arg Thr Arg Arg Met Arg Arg Arg Lys
1 5 10 15
Arg Glu Arg Leu His Lys
20
<210> 332
<211> 13
<212> PRT
<213> Bifidobacterium longum
<400> 332
Pro Val Ser Val Asn Ile Glu His Val Arg Ser Ser Phe
1 5 10
<210> 333
<211> 16
<212> PRT
<213> Bifidobacterium longum
<400> 333
Arg His His Ala Val Asp Ala Ser Val Ile Ala Met Met Asn Thr Ala
1 5 10 15
<210> 334
<211> 18
<212> PRT
<213> Wolinella succinogenes
<400> 334
Val Ser Pro Ile Ser Val Asp Leu Gly Gly Lys Asn Thr Gly Phe Phe
1 5 10 15
Ser Phe
<210> 335
<211> 22
<212> PRT
<213> Wolinella succinogenes
<400> 335
Val Gly Arg Arg Ser Lys Arg His Ser Lys Arg Asn Asn Leu Arg Asn
1 5 10 15
Lys Leu Val Lys Arg Leu
20
<210> 336
<211> 13
<212> PRT
<213> Wolinella succinogenes
<400> 336
Lys Val Pro Ile Ile Leu Glu Gln Asn Ala Phe Glu Tyr
1 5 10
<210> 337
<211> 16
<212> PRT
<213> Wolinella succinogenes
<400> 337
Ser Ser His Ala Ile Asp Ala Val Met Ala Phe Val Ala Arg Tyr Gln
1 5 10 15
<210> 338
<211> 18
<212> PRT
<213> Legionella pneumophila
<400> 338
Leu Ser Pro Ile Gly Ile Asp Leu Gly Gly Lys Phe Thr Gly Val Cys
1 5 10 15
Leu Ser
<210> 339
<211> 22
<212> PRT
<213> Legionella pneumophila
<400> 339
Ala Gln Arg Arg Ala Thr Arg His Arg Val Arg Asn Lys Lys Arg Asn
1 5 10 15
Gln Phe Val Lys Arg Val
20
<210> 340
<211> 13
<212> PRT
<213> Legionella pneumophila
<400> 340
Leu Ile Pro Ile Tyr Leu Glu Gln Asn Arg Phe Glu Phe
1 5 10
<210> 341
<211> 15
<212> PRT
<213> Legionella pneumophila
<400> 341
Pro Ser His Ala Ile Asp Ala Thr Leu Thr Met Ser Ile Gly Leu
1 5 10 15
<210> 342
<211> 18
<212> PRT
<213> Francisella tularensis
<400> 342
Ile Leu Pro Ile Ala Ile Asp Leu Gly Val Lys Asn Thr Gly Val Phe
1 5 10 15
Ser Ala
<210> 343
<211> 22
<212> PRT
<213> Francisella tularensis
<400> 343
Asn Asn Arg Thr Ala Arg Arg His Gln Arg Arg Gly Ile Asp Arg Lys
1 5 10 15
Gln Leu Val Lys Arg Leu
20
<210> 344
<211> 13
<212> PRT
<213> Francisella tularensis
<400> 344
His Ile Pro Ile Ile Thr Glu Ser Asn Ala Phe Glu Phe
1 5 10
<210> 345
<211> 16
<212> PRT
<213> Francisella tularensis
<400> 345
Tyr Ser His Leu Ile Asp Ala Met Leu Ala Phe Cys Ile Ala Ala Asp
1 5 10 15
<210> 346
<211> 18
<212> PRT
<213> Streptococcus pyogenes
<400> 346
Lys Tyr Ser Ile Gly Leu Asp Ile Gly Thr Asn Ser Val Gly Trp Ala
1 5 10 15
Val Ile
<210> 347
<211> 22
<212> PRT
<213> Streptococcus pyogenes
<400> 347
Glu Ala Thr Arg Leu Lys Arg Thr Ala Arg Arg Arg Tyr Thr Arg Arg
1 5 10 15
Lys Asn Arg Ile Cys Tyr
20
<210> 348
<211> 13
<212> PRT
<213> Streptococcus pyogenes
<400> 348
Pro Glu Asn Ile Val Ile Glu Met Ala Arg Glu Asn Gln
1 5 10
<210> 349
<211> 16
<212> PRT
<213> Streptococcus pyogenes
<400> 349
Tyr His His Ala His Asp Ala Tyr Leu Asn Ala Val Val Gly Thr Ala
1 5 10 15
<210> 350
<211> 18
<212> PRT
<213> Lactobacillus delbrueckii
<400> 350
Lys Val Ser Leu Gly Val Asp Thr Gly Gln Arg His Ile Gly Phe Ala
1 5 10 15
Ile Val
<210> 351
<211> 23
<212> PRT
<213> Lactobacillus delbrueckii
<400> 351
Tyr Thr Arg Lys Ile Tyr Arg Arg Ser Lys Arg Asn Arg Lys Thr Arg
1 5 10 15
Tyr Arg Gln Ala Arg Phe Leu
20
<210> 352
<211> 13
<212> PRT
<213> Lactobacillus delbrueckii
<400> 352
Asn Pro Asp Leu His Ile Glu Val Gly Lys Phe Asp Met
1 5 10
<210> 353
<211> 15
<212> PRT
<213> Lactobacillus delbrueckii
<400> 353
Lys Gly His Phe Met Asp Ala Ile Ala Ile Ser Gly Ile Lys Pro
1 5 10 15
<210> 354
<211> 18
<212> PRT
<213> Methanohalobium evestigatum
<400> 354
Pro Val Val Ala Gly Met Asp Ser Gly Ser Lys His Ile Gly Cys Ala
1 5 10 15
Ala Val
<210> 355
<211> 23
<212> PRT
<213> Methanohalobium evestigatum
<400> 355
Lys Asp Arg Ala Asp Tyr Arg Arg Asn Arg Arg Ser Arg Lys Thr Arg
1 5 10 15
Tyr Arg Lys Pro Arg Phe Asp
20
<210> 356
<211> 13
<212> PRT
<213> Methanohalobium evestigatum
<400> 356
Val Lys Lys Trp Ile Val Glu Thr Ala Ser Phe Asp Ile
1 5 10
<210> 357
<211> 15
<212> PRT
<213> Methanohalobium evestigatum
<400> 357
Lys Thr His Tyr Asn Asp Ala Val Ala Ile Cys Cys Asp Glu Asn
1 5 10 15
<210> 358
<211> 18
<212> PRT
<213> Clostridium botulinum
<400> 358
Pro Ile Thr Leu Gly Ile Asp Ser Gly Tyr Leu Asn Ile Gly Phe Ser
1 5 10 15
Ala Ile
<210> 359
<211> 22
<212> PRT
<213> Clostridium botulinum
<400> 359
Lys Glu Lys Ala Met Tyr Arg Arg Gln Arg Arg Ser Arg Leu Arg Tyr
1 5 10 15
Arg Lys Pro Arg Phe Asn
20
<210> 360
<211> 13
<212> PRT
<213> Clostridium botulinum
<400> 360
Ile Thr Asn Ile Ile Ile Glu Val Ala Asn Phe Asp Thr
1 5 10
<210> 361
<211> 15
<212> PRT
<213> Clostridium botulinum
<400> 361
Lys Thr His Tyr Asn Asp Ala Phe Cys Ile Ala Gly Ser Ser Asn
1 5 10 15
<210> 362
<211> 18
<212> PRT
<213> Geobacillus thermoleovorans
<400> 362
Pro Val Ser Leu Gly Val Asp Met Gly Thr Arg His Val Gly Ile Ser
1 5 10 15
Ala Thr
<210> 363
<211> 23
<212> PRT
<213> Geobacillus thermoleovorans
<400> 363
Ala Ile Arg Arg Gln Phe Arg Arg Ser Arg Arg Asn Arg Lys Thr Arg
1 5 10 15
Tyr Arg Glu Ala Arg Phe Leu
20
<210> 364
<211> 13
<212> PRT
<213> Geobacillus thermoleovorans
<400> 364
Val Thr Ser Val Thr Ile Glu Val Ala Ala Phe Asp Thr
1 5 10
<210> 365
<211> 15
<212> PRT
<213> Geobacillus thermoleovorans
<400> 365
Lys Ser His Met Val Asp Ala Arg Cys Ile Ser Gly Asn Pro Leu
1 5 10 15
<210> 366
<211> 18
<212> PRT
<213> Ammonifex degensii
<400> 366
Ser Leu Arg Ala Lys Val Asp Asp Gly Ser Arg Tyr Val Gly Ile Ala
1 5 10 15
Leu Val
<210> 367
<211> 23
<212> PRT
<213> Ammonifex degensii
<400> 367
Thr Leu Arg Arg Glu Tyr Arg Arg Gly Arg Arg Tyr Arg Ile Val Arg
1 5 10 15
His Arg Pro Cys Arg Asn Arg
20
<210> 368
<211> 13
<212> PRT
<213> Ammonifex degensii
<400> 368
Ile Ser Gly Val Asp Val Glu Leu Val Ser Ser Gly Val
1 5 10
<210> 369
<211> 15
<212> PRT
<213> Ammonifex degensii
<400> 369
Lys Ser His Thr Asn Asp Ala Leu Ser Leu Phe Leu Pro Gly Gly
1 5 10 15
<210> 370
<211> 18
<212> PRT
<213> Polaromonas sp.
<400> 370
Pro Leu Arg Ile Lys Leu Asp Pro Gly Ser Lys Thr Thr Gly Val Ala
1 5 10 15
Leu Val
<210> 371
<211> 22
<212> PRT
<213> Polaromonas sp.
<400> 371
Thr Ala Arg Arg Gln Met Arg Arg Arg Arg Arg Ser Asn Leu Arg Cys
1 5 10 15
Arg Ala Pro Arg Phe Leu
20
<210> 372
<211> 13
<212> PRT
<213> Polaromonas sp.
<400> 372
Val Arg Ala Ile Ser Ser Glu Leu Val Arg Phe Asp Met
1 5 10
<210> 373
<211> 15
<212> PRT
<213> Polaromonas sp.
<400> 373
Lys Thr His Ala Leu Asp Ala Ala Cys Val Gly Gln Val Arg Phe
1 5 10 15
<210> 374
<211> 18
<212> PRT
<213> Anabaena variabilis
<400> 374
Asp Leu Arg Ile Lys Leu Asp Pro Gly Ala Lys Ile Thr Gly Ile Ala
1 5 10 15
Leu Val
<210> 375
<211> 23
<212> PRT
<213> Anabaena variabilis
<400> 375
Ile Ser Arg Arg Gln Leu Arg Arg Thr Arg Arg Asn Arg Lys Thr Arg
1 5 10 15
Tyr Arg Lys Pro Arg Phe Leu
20
<210> 376
<211> 13
<212> PRT
<213> Anabaena variabilis
<400> 376
Ile Thr Ala Ile Ser Thr Glu Leu Val Lys Phe Asp Met
1 5 10
<210> 377
<211> 15
<212> PRT
<213> Anabaena variabilis
<400> 377
Lys Thr His Trp Leu Asp Ala Ala Cys Val Gly Gln Ser Thr Pro
1 5 10 15
<210> 378
<211> 18
<212> PRT
<213> Nostoc sp.
<400> 378
Pro Leu Arg Leu Lys Phe Asp Pro Gly Ala Lys Tyr Thr Gly Ile Ala
1 5 10 15
Leu Val
<210> 379
<211> 23
<212> PRT
<213> Nostoc sp.
<400> 379
Thr Ser Arg Arg Gln Leu Arg Arg Ser Arg Arg Ser Arg Lys Thr Arg
1 5 10 15
Tyr Arg Gln Pro Arg Phe Phe
20
<210> 380
<211> 13
<212> PRT
<213> Nostoc sp.
<400> 380
Ile Thr Ala Ile Ser Gln Glu Leu Val Lys Phe Asp Thr
1 5 10
<210> 381
<211> 15
<212> PRT
<213> Nostoc sp.
<400> 381
Lys Ser His Trp Leu Asp Ala Cys Cys Val Gly Ala Ser Thr Pro
1 5 10 15
<210> 382
<211> 18
<212> PRT
<213> Thermus thermophilus
<400> 382
Met Val Val Ala Gly Ile Asp Pro Gly Ile Thr His Leu Gly Leu Gly
1 5 10 15
Val Val
<210> 383
<211> 13
<212> PRT
<213> Thermus thermophilus
<400> 383
Pro Glu Ala Val Ala Val Glu Glu Gln Phe Phe Tyr Arg
1 5 10
<210> 384
<211> 15
<212> PRT
<213> Thermus thermophilus
<400> 384
Pro Ser His Leu Ala Asp Ala Leu Ala Ile Ala Leu Thr His Ala
1 5 10 15
<210> 385
<211> 18
<212> PRT
<213> Thermus thermophilus
<400> 385
Met Val Val Ala Gly Ile Asp Pro Gly Ile Thr His Leu Gly Leu Gly
1 5 10 15
Val Val
<210> 386
<211> 13
<212> PRT
<213> Thermus thermophilus
<400> 386
Pro Glu Ala Val Ala Val Glu Glu Gln Phe Phe Tyr Arg
1 5 10
<210> 387
<211> 15
<212> PRT
<213> Thermus thermophilus
<400> 387
Pro Ser His Leu Ala Asp Ala Leu Ala Ile Ala Leu Thr His Ala
1 5 10 15
<210> 388
<211> 18
<212> PRT
<213> Campylobacter jejuni
<400> 388
Ala Arg Ile Leu Ala Phe Asp Ile Gly Ile Ser Ser Ile Gly Trp Ala
1 5 10 15
Phe Ser
<210> 389
<211> 22
<212> PRT
<213> Campylobacter jejuni
<400> 389
Leu Pro Arg Arg Leu Ala Arg Ser Ala Arg Lys Arg Leu Ala Arg Arg
1 5 10 15
Lys Ala Arg Leu Asn His
20
<210> 390
<211> 13
<212> PRT
<213> Campylobacter jejuni
<400> 390
Val His Lys Ile Asn Ile Glu Leu Ala Arg Glu Val Gly
1 5 10
<210> 391
<211> 16
<212> PRT
<213> Campylobacter jejuni
<400> 391
Leu His His Ala Ile Asp Ala Val Ile Ile Ala Tyr Ala Asn Asn Ser
1 5 10 15
<210> 392
<211> 18
<212> PRT
<213> Clostridium perfringens
<400> 392
Asn Tyr Ala Leu Gly Leu Asp Ile Gly Ile Thr Ser Val Gly Trp Ala
1 5 10 15
Val Ile
<210> 393
<211> 22
<212> PRT
<213> Clostridium perfringens
<400> 393
Leu Pro Arg Arg Leu Ala Arg Gly Arg Arg Arg Leu Leu Arg Arg Lys
1 5 10 15
Ala Tyr Arg Val Glu Arg
20
<210> 394
<211> 13
<212> PRT
<213> Clostridium perfringens
<400> 394
Pro Val Arg Ile Asn Ile Glu Leu Ala Arg Asp Leu Ala
1 5 10
<210> 395
<211> 16
<212> PRT
<213> Clostridium perfringens
<400> 395
Lys His His Ala Leu Asp Ala Ala Val Val Gly Val Thr Thr Gln Gly
1 5 10 15
<210> 396
<211> 18
<212> PRT
<213> Akkermansia muciniphila
<400> 396
Ser Leu Thr Phe Ser Phe Asp Ile Gly Tyr Ala Ser Ile Gly Trp Ala
1 5 10 15
Val Ile
<210> 397
<211> 22
<212> PRT
<213> Akkermansia muciniphila
<400> 397
Phe Lys Arg Arg Glu Tyr Arg Arg Leu Arg Arg Asn Ile Arg Ser Arg
1 5 10 15
Arg Val Arg Ile Glu Arg
20
<210> 398
<211> 13
<212> PRT
<213> Akkermansia muciniphila
<400> 398
Ile Ser Arg Val Cys Val Glu Val Gly Lys Glu Leu Thr
1 5 10
<210> 399
<211> 16
<212> PRT
<213> Akkermansia muciniphila
<400> 399
Leu His His Ala Leu Asp Ala Cys Val Leu Gly Leu Ile Pro Tyr Ile
1 5 10 15
<210> 400
<211> 18
<212> PRT
<213> Bifidobacterium longum
<400> 400
Arg Tyr Arg Ile Gly Ile Asp Val Gly Leu Asn Ser Val Gly Leu Ala
1 5 10 15
Ala Val
<210> 401
<211> 22
<212> PRT
<213> Bifidobacterium longum
<400> 401
Asn Met Ser Gly Val Ala Arg Arg Thr Arg Arg Met Arg Arg Arg Lys
1 5 10 15
Arg Glu Arg Leu His Lys
20
<210> 402
<211> 13
<212> PRT
<213> Bifidobacterium longum
<400> 402
Pro Val Ser Val Asn Ile Glu His Val Arg Ser Ser Phe
1 5 10
<210> 403
<211> 16
<212> PRT
<213> Bifidobacterium longum
<400> 403
Arg His His Ala Val Asp Ala Ser Val Ile Ala Met Met Asn Thr Ala
1 5 10 15
<210> 404
<211> 18
<212> PRT
<213> Wolinella succinogenes
<400> 404
Val Ser Pro Ile Ser Val Asp Leu Gly Gly Lys Asn Thr Gly Phe Phe
1 5 10 15
Ser Phe
<210> 405
<211> 22
<212> PRT
<213> Wolinella succinogenes
<400> 405
Val Gly Arg Arg Ser Lys Arg His Ser Lys Arg Asn Asn Leu Arg Asn
1 5 10 15
Lys Leu Val Lys Arg Leu
20
<210> 406
<211> 13
<212> PRT
<213> Wolinella succinogenes
<400> 406
Lys Val Pro Ile Ile Leu Glu Gln Asn Ala Phe Glu Tyr
1 5 10
<210> 407
<211> 16
<212> PRT
<213> Wolinella succinogenes
<400> 407
Ser Ser His Ala Ile Asp Ala Val Met Ala Phe Val Ala Arg Tyr Gln
1 5 10 15
<210> 408
<211> 18
<212> PRT
<213> Legionella pneumophila
<400> 408
Leu Ser Pro Ile Gly Ile Asp Leu Gly Gly Lys Phe Thr Gly Val Cys
1 5 10 15
Leu Ser
<210> 409
<211> 22
<212> PRT
<213> Legionella pneumophila
<400> 409
Ala Gln Arg Arg Ala Thr Arg His Arg Val Arg Asn Lys Lys Arg Asn
1 5 10 15
Gln Phe Val Lys Arg Val
20
<210> 410
<211> 13
<212> PRT
<213> Legionella pneumophila
<400> 410
Leu Ile Pro Ile Tyr Leu Glu Gln Asn Arg Phe Glu Phe
1 5 10
<210> 411
<211> 15
<212> PRT
<213> Legionella pneumophila
<400> 411
Pro Ser His Ala Ile Asp Ala Thr Leu Thr Met Ser Ile Gly Leu
1 5 10 15
<210> 412
<211> 18
<212> PRT
<213> Francisella tularensis
<400> 412
Ile Leu Pro Ile Ala Ile Asp Leu Gly Val Lys Asn Thr Gly Val Phe
1 5 10 15
Ser Ala
<210> 413
<211> 22
<212> PRT
<213> Francisella tularensis
<400> 413
Asn Asn Arg Thr Ala Arg Arg His Gln Arg Arg Gly Ile Asp Arg Lys
1 5 10 15
Gln Leu Val Lys Arg Leu
20
<210> 414
<211> 13
<212> PRT
<213> Francisella tularensis
<400> 414
His Ile Pro Ile Ile Thr Glu Ser Asn Ala Phe Glu Phe
1 5 10
<210> 415
<211> 16
<212> PRT
<213> Francisella tularensis
<400> 415
Tyr Ser His Leu Ile Asp Ala Met Leu Ala Phe Cys Ile Ala Ala Asp
1 5 10 15
<210> 416
<211> 18
<212> PRT
<213> Streptococcus pyogenes
<400> 416
Lys Tyr Ser Ile Gly Leu Asp Ile Gly Thr Asn Ser Val Gly Trp Ala
1 5 10 15
Val Ile
<210> 417
<211> 22
<212> PRT
<213> Streptococcus pyogenes
<400> 417
Glu Ala Thr Arg Leu Lys Arg Thr Ala Arg Arg Arg Tyr Thr Arg Arg
1 5 10 15
Lys Asn Arg Ile Cys Tyr
20
<210> 418
<211> 13
<212> PRT
<213> Streptococcus pyogenes
<400> 418
Pro Glu Asn Ile Val Ile Glu Met Ala Arg Glu Asn Gln
1 5 10
<210> 419
<211> 16
<212> PRT
<213> Streptococcus pyogenes
<400> 419
Tyr His His Ala His Asp Ala Tyr Leu Asn Ala Val Val Gly Thr Ala
1 5 10 15
<210> 420
<211> 18
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Candidatus Methanomethylophilus alvus sequence"
<400> 420
Leu Lys Ile Ile Gly Ile Asp Arg Gly Glu Arg Asn Leu Ile Tyr Val
1 5 10 15
Thr Met
<210> 421
<211> 22
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Candidatus Methanomethylophilus alvus sequence"
<400> 421
Arg Lys Ala Leu Asp Val Arg Glu Tyr Asp Asn Lys Glu Ala Arg Arg
1 5 10 15
Asn Trp Thr Lys Val Glu
20
<210> 422
<211> 13
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Candidatus Methanomethylophilus alvus sequence"
<400> 422
Asn Ala Ile Ile Val Met Glu Asp Leu Asn His Gly Phe
1 5 10
<210> 423
<211> 16
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Candidatus Methanomethylophilus alvus sequence"
<400> 423
Leu Pro Gln Asp Ser Asp Ala Asn Gly Ala Tyr Asn Ile Ala Leu Lys
1 5 10 15
<210> 424
<211> 18
<212> PRT
<213> Synergistes jonesii
<400> 424
Val Asn Ile Ile Gly Ile Asp Arg Gly Glu Arg Asn Leu Val Tyr Val
1 5 10 15
Ser Leu
<210> 425
<211> 22
<212> PRT
<213> Synergistes jonesii
<400> 425
His Ala Lys Leu Asn Gln Lys Glu Lys Glu Arg Asp Thr Ala Arg Lys
1 5 10 15
Ser Trp Lys Thr Ile Gly
20
<210> 426
<211> 13
<212> PRT
<213> Synergistes jonesii
<400> 426
Asn Ala Val Ile Val Met Glu Asp Leu Asn Ile Gly Phe
1 5 10
<210> 427
<211> 16
<212> PRT
<213> Synergistes jonesii
<400> 427
Leu Pro Ile Asp Ala Asp Ala Asn Gly Ala Tyr His Ile Ala Leu Lys
1 5 10 15
<210> 428
<211> 18
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 428
Pro Tyr Val Ile Gly Ile Asp Arg Gly Glu Arg Asn Leu Leu Tyr Ile
1 5 10 15
Val Val
<210> 429
<211> 22
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 429
His Ser Leu Leu Asp Lys Lys Glu Lys Glu Arg Phe Glu Ala Arg Gln
1 5 10 15
Asn Trp Thr Ser Ile Glu
20
<210> 430
<211> 13
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 430
Asp Ala Val Ile Ala Leu Glu Asp Leu Asn Ser Gly Phe
1 5 10
<210> 431
<211> 16
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 431
Leu Pro Lys Asn Ala Asp Ala Asn Gly Ala Tyr Asn Ile Ala Arg Lys
1 5 10 15
<210> 432
<211> 18
<212> PRT
<213> Francisella tularensis
<400> 432
Val His Ile Leu Ser Ile Asp Arg Gly Glu Arg His Leu Ala Tyr Tyr
1 5 10 15
Thr Leu
<210> 433
<211> 22
<212> PRT
<213> Francisella tularensis
<400> 433
His Asp Lys Leu Ala Ala Ile Glu Lys Asp Arg Asp Ser Ala Arg Lys
1 5 10 15
Asp Trp Lys Lys Ile Asn
20
<210> 434
<211> 13
<212> PRT
<213> Francisella tularensis
<400> 434
Asn Ala Ile Val Val Phe Glu Asp Leu Asn Phe Gly Phe
1 5 10
<210> 435
<211> 16
<212> PRT
<213> Francisella tularensis
<400> 435
Met Pro Gln Asp Ala Asp Ala Asn Gly Ala Tyr His Ile Gly Leu Lys
1 5 10 15
<210> 436
<211> 18
<212> PRT
<213> Moraxella caprae
<400> 436
Val Asn Val Ile Gly Ile Asp Arg Gly Glu Arg His Leu Leu Tyr Leu
1 5 10 15
Thr Val
<210> 437
<211> 22
<212> PRT
<213> Moraxella caprae
<400> 437
His Lys Ile Leu Asp Lys Arg Glu Ile Glu Arg Leu Asn Ala Arg Val
1 5 10 15
Gly Trp Gly Glu Ile Glu
20
<210> 438
<211> 13
<212> PRT
<213> Moraxella caprae
<400> 438
Asn Ala Ile Val Val Leu Glu Asp Leu Asn Phe Gly Phe
1 5 10
<210> 439
<211> 16
<212> PRT
<213> Moraxella caprae
<400> 439
Gln Pro Gln Asn Ala Asp Ala Asn Gly Ala Tyr His Ile Ala Leu Lys
1 5 10 15
<210> 440
<211> 18
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 440
Met His Ile Ile Gly Ile Asp Arg Gly Glu Arg Asn Leu Ile Tyr Leu
1 5 10 15
Cys Met
<210> 441
<211> 22
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 441
His Gln Leu Leu Lys Thr Arg Glu Asp Glu Asn Lys Ser Ala Arg Gln
1 5 10 15
Ser Trp Gln Thr Ile His
20
<210> 442
<211> 13
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 442
Asn Ala Ile Val Val Leu Glu Asp Leu Asn Phe Gly Phe
1 5 10
<210> 443
<211> 16
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 443
Met Pro Leu Asp Ala Asp Ala Asn Gly Ala Tyr Asn Ile Ala Arg Lys
1 5 10 15
<210> 444
<211> 18
<212> PRT
<213> Prevotella albensis
<400> 444
Thr His Ile Ile Gly Ile Asp Arg Gly Glu Arg His Leu Leu Tyr Leu
1 5 10 15
Ser Leu
<210> 445
<211> 22
<212> PRT
<213> Prevotella albensis
<400> 445
His Asn Leu Leu Glu Lys Arg Glu Lys Glu Arg Thr Glu Ala Arg His
1 5 10 15
Ser Trp Ser Ser Ile Glu
20
<210> 446
<211> 13
<212> PRT
<213> Prevotella albensis
<400> 446
Asn Ala Ile Val Val Leu Glu Asp Leu Asn Gly Gly Phe
1 5 10
<210> 447
<211> 16
<212> PRT
<213> Prevotella albensis
<400> 447
Phe Pro Glu Asn Ala Asp Ala Asn Gly Ala Tyr Asn Ile Ala Arg Lys
1 5 10 15
<210> 448
<211> 18
<212> PRT
<213> Smithella sp.
<400> 448
Ile Asn Ile Ile Gly Ile Asp Arg Gly Glu Arg His Leu Leu Tyr Tyr
1 5 10 15
Ala Leu
<210> 449
<211> 22
<212> PRT
<213> Smithella sp.
<400> 449
His Asn Leu Leu Asp Lys Lys Glu Gly Asp Arg Ala Thr Ala Arg Gln
1 5 10 15
Glu Trp Gly Val Ile Glu
20
<210> 450
<211> 13
<212> PRT
<213> Smithella sp.
<400> 450
Asn Ala Ile Ile Val Met Glu Asp Leu Asn Phe Gly Phe
1 5 10
<210> 451
<211> 16
<212> PRT
<213> Smithella sp.
<400> 451
Met Pro Lys Asn Ala Asp Ala Asn Gly Ala Tyr His Ile Ala Leu Lys
1 5 10 15
<210> 452
<211> 18
<212> PRT
<213> Porphyromonas crevioricanis
<400> 452
Met His Val Ile Gly Ile Asp Arg Gly Glu Arg Asn Leu Leu Tyr Ile
1 5 10 15
Cys Val
<210> 453
<211> 22
<212> PRT
<213> Porphyromonas crevioricanis
<400> 453
His Asp Leu Leu Glu Ser Arg Asp Lys Asp Arg Gln Gln Glu Arg Arg
1 5 10 15
Asn Trp Gln Thr Ile Glu
20
<210> 454
<211> 13
<212> PRT
<213> Porphyromonas crevioricanis
<400> 454
Lys Ala Val Val Ala Leu Glu Asp Leu Asn Met Gly Phe
1 5 10
<210> 455
<211> 16
<212> PRT
<213> Porphyromonas crevioricanis
<400> 455
Leu Pro Lys Asp Ala Asp Ala Asn Gly Ala Tyr Asn Ile Ala Leu Lys
1 5 10 15
<210> 456
<211> 18
<212> PRT
<213> Alicyclobacillus acidoterrestris
<400> 456
Leu Arg Val Met Ser Val Asp Leu Gly Leu Arg Thr Ser Ala Ser Ile
1 5 10 15
Ser Val
<210> 457
<211> 22
<212> PRT
<213> Alicyclobacillus acidoterrestris
<400> 457
Gln Arg Thr Leu Arg Gln Leu Arg Thr Gln Leu Ala Tyr Leu Arg Leu
1 5 10 15
Leu Val Arg Cys Gly Ser
20
<210> 458
<211> 12
<212> PRT
<213> Alicyclobacillus acidoterrestris
<400> 458
Cys Gln Leu Ile Leu Leu Glu Glu Leu Ser Glu Tyr
1 5 10
<210> 459
<211> 16
<212> PRT
<213> Alicyclobacillus acidoterrestris
<400> 459
His Gln Ile His Ala Asp Leu Asn Ala Ala Gln Asn Leu Gln Gln Arg
1 5 10 15
<210> 460
<211> 18
<212> PRT
<213> Alicyclobacillus contaminans
<400> 460
Val Arg Val Met Ser Val Asp Leu Gly Val Arg Tyr Gly Ala Ala Ile
1 5 10 15
Ser Val
<210> 461
<211> 22
<212> PRT
<213> Alicyclobacillus contaminans
<400> 461
Lys Gln Ala Leu Ala Ala Ile Arg Ala Glu Met Ser Ile Leu Arg Lys
1 5 10 15
Trp Leu Arg Val Ser Gln
20
<210> 462
<211> 12
<212> PRT
<213> Alicyclobacillus contaminans
<400> 462
Cys Asp Leu Ile Leu Phe Glu Asp Leu Ser Arg Tyr
1 5 10
<210> 463
<211> 16
<212> PRT
<213> Alicyclobacillus contaminans
<400> 463
Lys Cys Val His Ala Asp Ile Asn Ala Ala His Asn Leu Gln Arg Arg
1 5 10 15
<210> 464
<211> 18
<212> PRT
<213> Desulfovibrio inopinatus
<400> 464
Leu Arg Val Leu Ser Val Asp Leu Gly Met Arg Thr Phe Ala Ser Cys
1 5 10 15
Ser Val
<210> 465
<211> 22
<212> PRT
<213> Desulfovibrio inopinatus
<400> 465
Arg Ala Glu Ile Tyr Ala Leu Lys Arg Asp Ile Gln Arg Leu Lys Ser
1 5 10 15
Leu Leu Arg Leu Gly Glu
20
<210> 466
<211> 12
<212> PRT
<213> Desulfovibrio inopinatus
<400> 466
Cys Gln Leu Ile Leu Phe Glu Asp Leu Ala Arg Tyr
1 5 10
<210> 467
<211> 16
<212> PRT
<213> Desulfovibrio inopinatus
<400> 467
Cys Val Ile His Ala Asp Met Asn Ala Ala Gln Asn Leu Gln Arg Arg
1 5 10 15
<210> 468
<211> 18
<212> PRT
<213> Desulfonatronum thiodismutans
<400> 468
Leu Arg Val Leu Ser Val Asp Leu Gly Val Arg Ser Phe Ala Ala Cys
1 5 10 15
Ser Val
<210> 469
<211> 22
<212> PRT
<213> Desulfonatronum thiodismutans
<400> 469
Met Glu Glu Leu Arg Ser Leu Asn Gly Asp Ile Arg Arg Leu Lys Ala
1 5 10 15
Ile Leu Arg Leu Ser Val
20
<210> 470
<211> 12
<212> PRT
<213> Desulfonatronum thiodismutans
<400> 470
Cys Arg Leu Ile Leu Phe Glu Asp Leu Ala Arg Tyr
1 5 10
<210> 471
<211> 16
<212> PRT
<213> Desulfonatronum thiodismutans
<400> 471
His Val Ile His Ala Asp Ile Asn Ala Ala Gln Asn Leu Gln Arg Arg
1 5 10 15
<210> 472
<211> 18
<212> PRT
<213> Tuberibacillus calidus
<400> 472
Leu Arg Val Met Ser Val Asp Leu Gly Gln Arg Gln Ala Ala Ala Ile
1 5 10 15
Ser Ile
<210> 473
<211> 22
<212> PRT
<213> Tuberibacillus calidus
<400> 473
Asp Gln Ala Ile Arg Asp Leu Ser Arg Lys Leu Lys Phe Leu Lys Asn
1 5 10 15
Val Leu Asn Met Gln Lys
20
<210> 474
<211> 12
<212> PRT
<213> Tuberibacillus calidus
<400> 474
Cys Gln Leu Val Leu Phe Glu Asp Leu Ser Arg Tyr
1 5 10
<210> 475
<211> 16
<212> PRT
<213> Tuberibacillus calidus
<400> 475
Val Ile Thr His Ala Asp Ile Asn Ala Ala Gln Asn Leu Gln Lys Arg
1 5 10 15
<210> 476
<211> 18
<212> PRT
<213> Bacillus thermoamylovorans
<400> 476
Leu Arg Val Met Ser Ile Asp Leu Gly Gln Arg Gln Ala Ala Ala Ala
1 5 10 15
Ser Ile
<210> 477
<211> 22
<212> PRT
<213> Bacillus thermoamylovorans
<400> 477
Glu Asp Asn Leu Lys Leu Met Asn Gln Lys Leu Asn Phe Leu Arg Asn
1 5 10 15
Val Leu His Phe Gln Gln
20
<210> 478
<211> 12
<212> PRT
<213> Bacillus thermoamylovorans
<400> 478
Cys Gln Ile Ile Leu Phe Glu Asp Leu Ser Asn Tyr
1 5 10
<210> 479
<211> 16
<212> PRT
<213> Bacillus thermoamylovorans
<400> 479
Val Thr Thr His Ala Asp Ile Asn Ala Ala Gln Asn Leu Gln Lys Arg
1 5 10 15
<210> 480
<211> 18
<212> PRT
<213> Bacillus sp.
<400> 480
Phe Arg Val Met Ser Ile Asp Leu Gly Leu Arg Ala Ala Ala Ala Thr
1 5 10 15
Ser Ile
<210> 481
<211> 22
<212> PRT
<213> Bacillus sp.
<400> 481
Phe Gln Leu His Gln Arg Val Lys Phe Gln Ile Arg Val Leu Ala Gln
1 5 10 15
Ile Met Arg Met Ala Asn
20
<210> 482
<211> 12
<212> PRT
<213> Bacillus sp.
<400> 482
Cys Gln Val Ile Leu Phe Glu Asn Leu Ser Gln Tyr
1 5 10
<210> 483
<211> 16
<212> PRT
<213> Bacillus sp.
<400> 483
Val Phe Leu Gln Ala Asp Ile Asn Ala Ala His Asn Leu Gln Lys Arg
1 5 10 15
<210> 484
<211> 18
<212> PRT
<213> Methylobacterium nodulans
<400> 484
Leu Arg Val Leu Ser Ile Asp Leu Gly Val Arg Ser Phe Ala Thr Cys
1 5 10 15
Ser Val
<210> 485
<211> 22
<212> PRT
<213> Methylobacterium nodulans
<400> 485
Asp Ala Glu Leu Arg Gln Leu Arg Gly Gly Leu Asn Arg His Arg Gln
1 5 10 15
Leu Leu Arg Ala Ala Thr
20
<210> 486
<211> 12
<212> PRT
<213> Methylobacterium nodulans
<400> 486
Cys His Val Ile Leu Phe Glu Asp Leu Ser Arg Tyr
1 5 10
<210> 487
<211> 16
<212> PRT
<213> Methylobacterium nodulans
<400> 487
Ser Arg Ile His Ala Asp Ile Asn Ala Ala Gln Asn Leu Gln Arg Arg
1 5 10 15
<210> 488
<211> 18
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 488
Lys Asn Ile Val Ser Ile Asp Gln Gly Glu Ala Gly Phe Ala Tyr Ala
1 5 10 15
Val Phe
<210> 489
<211> 22
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 489
His Ser Val Lys Lys Tyr Arg Gly Lys Lys Gln Arg Ile Gln Asn Phe
1 5 10 15
Asn Gln Lys Phe Asp Ser
20
<210> 490
<211> 13
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 490
Asn Ala Phe Pro Ile Leu Glu Lys Gln Val Gly Asn Leu
1 5 10
<210> 491
<211> 16
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 491
Lys Glu Gln His Ala Asp Val Asn Ala Ala Ile Asn Ile Gly Arg Arg
1 5 10 15
<210> 492
<211> 18
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 492
Asp His Ile Val Ala Ile Asp Leu Gly Glu Arg Ser Val Gly Phe Ala
1 5 10 15
Val Phe
<210> 493
<211> 22
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 493
Lys Ala Val Arg Ser His Arg Arg Arg Arg Gln Pro Asn Gln Lys Val
1 5 10 15
Asn Gln Thr Tyr Ser Thr
20
<210> 494
<211> 13
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 494
Asn Ala Phe Pro Val Leu Glu Phe Gln Ile Lys Asn Phe
1 5 10
<210> 495
<211> 16
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 495
Trp Thr Gly His Ala Asp Glu Asn Ala Ala Ile Asn Ile Gly Arg Arg
1 5 10 15
<210> 496
<211> 18
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Metagenome sequence"
<400> 496
Asp Arg Ile Val Ala Ile Asp Leu Gly Glu Arg Lys Ile Gly Tyr Ala
1 5 10 15
Ile Phe
<210> 497
<211> 22
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Metagenome sequence"
<400> 497
Lys Ala Val Gln Thr His Arg Asn Arg Arg Gln Pro Asn Tyr Arg Ile
1 5 10 15
Asp Gln Thr Tyr Ser Lys
20
<210> 498
<211> 13
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Metagenome sequence"
<400> 498
Gly Gly Phe Pro Val Leu Glu Ser Ser Val Arg Asn Phe
1 5 10
<210> 499
<211> 16
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Metagenome sequence"
<400> 499
His Glu Cys His Ala Asp Glu Asn Ala Ala Ile Asn Ile Gly Arg Lys
1 5 10 15
<210> 500
<211> 18
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 500
Asp His Leu Leu Ala Ile Asp Leu Gly Glu Lys Arg Val Gly Tyr Ala
1 5 10 15
Val Tyr
<210> 501
<211> 22
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 501
Lys Ala Val Arg Ser His Arg Gln Gln Arg Gln Pro Asn Gln Lys Val
1 5 10 15
Asn Gln Thr Tyr Ser Thr
20
<210> 502
<211> 13
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 502
Asn Ala Phe Pro Val Leu Glu Ser Ser Val Met Asn Phe
1 5 10
<210> 503
<211> 16
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 503
Phe Thr Gly His Ala Asp Glu Asn Ala Ala Ile Asn Ile Gly Trp Lys
1 5 10 15
<210> 504
<211> 15
<212> PRT
<213> Homo sapiens
<400> 504
Ile Pro Asp Trp Ile Val Asp Leu Arg His Glu Leu Thr His Lys
1 5 10 15
<210> 505
<211> 15
<212> PRT
<213> Homo sapiens
<400> 505
Ile Pro Asp Trp Ile Val Asp Leu Arg His Glu Leu Thr His Lys
1 5 10 15
<210> 506
<211> 15
<212> PRT
<213> Mycobacterium tuberculosis
<400> 506
Leu Gly Arg Phe Glu Ser Arg Val Arg Asn Thr Ala Ala His Glu
1 5 10 15
<210> 507
<211> 15
<212> PRT
<213> Mycobacterium tuberculosis
<400> 507
Leu Gly Arg Phe Glu Ser Arg Val Arg Asn Thr Ala Ala His Glu
1 5 10 15
<210> 508
<211> 15
<212> PRT
<213> Lactococcus lactis
<400> 508
Trp Ile Arg Ala Gly Trp Arg Ile Arg Asn Arg Ser Ala His Tyr
1 5 10 15
<210> 509
<211> 15
<212> PRT
<213> Lactococcus lactis
<400> 509
Trp Ile Arg Ala Gly Trp Phe Ile Arg Asn Arg Ser Ala His Tyr
1 5 10 15
<210> 510
<211> 15
<212> PRT
<213> Thermus thermophilus
<220>
<221> MOD_RES
<222> (5)..(5)
<223> Any amino acid
<400> 510
Leu Ala Leu Gly Xaa Val Asp Asp Arg Ser Leu Thr Val His Thr
1 5 10 15
<210> 511
<211> 15
<212> PRT
<213> Thermus thermophilus
<220>
<221> MOD_RES
<222> (5)..(5)
<223> Any amino acid
<400> 511
Leu Ala Leu Gly Xaa Val Asp Asp Arg Ser Leu Thr Val His Thr
1 5 10 15
<210> 512
<211> 22
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 512
Leu Lys Ser Met Leu Tyr Ser Met Arg Asn Ser Ser Phe His Phe Ser
1 5 10 15
Thr Glu Asn Val Asp Asn
20
<210> 513
<211> 14
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 513
Phe Arg Asn Glu Ile Asp His Phe His Tyr Phe Tyr Asp Arg
1 5 10
<210> 514
<211> 22
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 514
Leu Lys Asp Val Ile Tyr Ser Met Arg Asn Asp Ser Phe His Tyr Ala
1 5 10 15
Thr Glu Asn His Asn Asn
20
<210> 515
<211> 14
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 515
Leu Arg Asn Tyr Ile Glu His Phe Arg Tyr Tyr Ser Ser Phe
1 5 10
<210> 516
<211> 22
<212> PRT
<213> Clostridium aminophilum
<400> 516
Leu Arg Lys Ala Ile Tyr Ser Leu Arg Asn Glu Thr Phe His Phe Thr
1 5 10 15
Thr Leu Asn Lys Gly Ser
20
<210> 517
<211> 14
<212> PRT
<213> Clostridium aminophilum
<400> 517
Val Arg Lys Tyr Val Asp His Phe Lys Tyr Tyr Ala Thr Ser
1 5 10
<210> 518
<211> 22
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 518
Ile Ile Gln Ile Ile Tyr Ser Leu Arg Asn Lys Ser Phe His Phe Lys
1 5 10 15
Thr Tyr Asp His Gly Asp
20
<210> 519
<211> 14
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 519
Leu Arg Lys Tyr Val Asp His Phe Lys Tyr Tyr Ala Tyr Gly
1 5 10
<210> 520
<211> 22
<212> PRT
<213> Carnobacterium gallinarum
<400> 520
Leu Arg Gly Ser Val Gln Gln Ile Arg Asn Glu Ile Phe His Ser Phe
1 5 10 15
Asp Lys Asn Gln Lys Phe
20
<210> 521
<211> 14
<212> PRT
<213> Carnobacterium gallinarum
<400> 521
Ile Arg Asn Gln Thr Ala His Leu Ser Val Leu Gln Leu Glu
1 5 10
<210> 522
<211> 21
<212> PRT
<213> Carnobacterium gallinarum
<400> 522
Ile Arg Gly Ala Val Gln Arg Val Arg Asn Gln Ile Phe His Gln Gln
1 5 10 15
Ile Asn Lys Arg His
20
<210> 523
<211> 14
<212> PRT
<213> Carnobacterium gallinarum
<400> 523
Ile Arg Asn Asn Ile Ala His Leu His Val Leu Arg Asn Asp
1 5 10
<210> 524
<211> 20
<212> PRT
<213> Paludibacter propionicigenes
<400> 524
Ile Arg Gly Ala Val Gln Gln Ile Arg Asn Asn Val Asn His Tyr Lys
1 5 10 15
Lys Asp Ala Leu
20
<210> 525
<211> 14
<212> PRT
<213> Paludibacter propionicigenes
<400> 525
Ile Arg Asn His Ile Ala His Phe Asn Tyr Leu Thr Lys Asp
1 5 10
<210> 526
<211> 20
<212> PRT
<213> Listeria seeligeri
<400> 526
Leu Arg Gly Ala Ile Ala Pro Ile Arg Asn Glu Ile Ile His Leu Lys
1 5 10 15
Lys His Ser Trp
20
<210> 527
<211> 14
<212> PRT
<213> Listeria seeligeri
<400> 527
Lys Arg Asn Asn Ile Ser His Phe Asn Tyr Leu Asn Gly Gln
1 5 10
<210> 528
<211> 20
<212> PRT
<213> Listeria weihenstephanensis
<400> 528
Ile Arg Gly Ser Ile Gln Gln Ile Arg Asn Glu Val Tyr His Cys Lys
1 5 10 15
Lys His Ser Trp
20
<210> 529
<211> 14
<212> PRT
<213> Listeria weihenstephanensis
<400> 529
Ala Arg Asn His Ile Ala His Leu Asn Tyr Leu Ser Leu Lys
1 5 10
<210> 530
<211> 20
<212> PRT
<213> Listeria newyorkens
<400> 530
Ile Arg Gly Ser Ile Gln Gln Ile Arg Asn Glu Val Tyr His Cys Lys
1 5 10 15
Lys His Ser Trp
20
<210> 531
<211> 14
<212> PRT
<213> Listeria newyorkens
<400> 531
Ala Arg Asn His Ile Ala His Leu Asn Tyr Leu Ser Leu Lys
1 5 10
<210> 532
<211> 21
<212> PRT
<213> Leptotrichia wadei
<400> 532
Ile Ser Tyr Ser Ile Tyr Asn Val Arg Asn Gly Val Gly His Phe Asn
1 5 10 15
Lys Leu Ile Leu Gly
20
<210> 533
<211> 14
<212> PRT
<213> Leptotrichia wadei
<400> 533
Phe Arg Asn Tyr Ile Ala His Phe Leu His Leu His Thr Lys
1 5 10
<210> 534
<211> 21
<212> PRT
<213> Leptotrichia wadei
<400> 534
Met Leu Asn Ala Ile Thr Ser Ile Arg His Arg Val Val His Tyr Asn
1 5 10 15
Met Asn Thr Asn Ser
20
<210> 535
<211> 14
<212> PRT
<213> Leptotrichia wadei
<400> 535
Ile Arg Asn Tyr Ile Ala His Phe Asn Tyr Ile Pro Asp Ala
1 5 10
<210> 536
<211> 21
<212> PRT
<213> Leptotrichia wadei
<400> 536
Ile Asp Glu Ala Ile Ser Ser Ile Arg His Gly Ile Val His Phe Asn
1 5 10 15
Leu Glu Leu Glu Gly
20
<210> 537
<211> 14
<212> PRT
<213> Leptotrichia wadei
<400> 537
Ile Arg Asn Tyr Ile Ala His Phe Asn Tyr Ile Pro His Ala
1 5 10
<210> 538
<211> 22
<212> PRT
<213> Rhodobacter capsulatus
<400> 538
Leu Leu Arg Tyr Leu Arg Gly Cys Arg Asn Gln Thr Phe His Leu Gly
1 5 10 15
Ala Arg Ala Gly Phe Leu
20
<210> 539
<211> 21
<212> PRT
<213> Leptotrichia buccalis
<400> 539
Ile Asp Glu Ala Ile Ser Ser Ile Arg His Gly Ile Val His Phe Asn
1 5 10 15
Leu Glu Leu Glu Gly
20
<210> 540
<211> 14
<212> PRT
<213> Rhodobacter capsulatus
<400> 540
Thr Arg Lys Asp Leu Ala His Phe Asn Val Leu Asp Arg Ala
1 5 10
<210> 541
<211> 21
<212> PRT
<213> Leptotrichia sp.
<400> 541
Ile Asp Glu Ala Ile Ser Ser Ile Arg His Gly Ile Val His Phe Asn
1 5 10 15
Leu Glu Leu Glu Gly
20
<210> 542
<211> 14
<212> PRT
<213> Leptotrichia buccalis
<400> 542
Ile Arg Asn Tyr Ile Ala His Phe Asn Tyr Ile Pro His Ala
1 5 10
<210> 543
<211> 16
<212> PRT
<213> Leptotrichia sp.
<400> 543
Phe Gln Lys Glu Gly Tyr Leu Leu Arg Asn Lys Ile Leu His Asn Ser
1 5 10 15
<210> 544
<211> 14
<212> PRT
<213> Leptotrichia sp.
<400> 544
Ile Arg Asn Tyr Ile Ala His Phe Asn Tyr Ile Pro Asn Ala
1 5 10
<210> 545
<211> 15
<212> PRT
<213> Leptotrichia shahii
<400> 545
Phe Thr Lys Ile Gly Thr Asn Glu Arg Asn Arg Ile Leu His Ala
1 5 10 15
<210> 546
<211> 14
<212> PRT
<213> Leptotrichia sp.
<400> 546
Ile Arg Asn Tyr Ile Ser His Phe Tyr Ile Val Arg Asn Pro
1 5 10
<210> 547
<211> 14
<212> PRT
<213> Leptotrichia shahii
<400> 547
Ile Arg Asn Tyr Ile Ser His Phe Tyr Ile Val Arg Asn Pro
1 5 10
<210> 548
<211> 1129
<212> PRT
<213> Alicyclobacillus acidoterrestris
<400> 548
Met Ala Val Lys Ser Ile Lys Val Lys Leu Arg Leu Asp Asp Met Pro
1 5 10 15
Glu Ile Arg Ala Gly Leu Trp Lys Leu His Lys Glu Val Asn Ala Gly
20 25 30
Val Arg Tyr Tyr Thr Glu Trp Leu Ser Leu Leu Arg Gln Glu Asn Leu
35 40 45
Tyr Arg Arg Ser Pro Asn Gly Asp Gly Glu Gln Glu Cys Asp Lys Thr
50 55 60
Ala Glu Glu Cys Lys Ala Glu Leu Leu Glu Arg Leu Arg Ala Arg Gln
65 70 75 80
Val Glu Asn Gly His Arg Gly Pro Ala Gly Ser Asp Asp Glu Leu Leu
85 90 95
Gln Leu Ala Arg Gln Leu Tyr Glu Leu Leu Val Pro Gln Ala Ile Gly
100 105 110
Ala Lys Gly Asp Ala Gln Gln Ile Ala Arg Lys Phe Leu Ser Pro Leu
115 120 125
Ala Asp Lys Asp Ala Val Gly Gly Leu Gly Ile Ala Lys Ala Gly Asn
130 135 140
Lys Pro Arg Trp Val Arg Met Arg Glu Ala Gly Glu Pro Gly Trp Glu
145 150 155 160
Glu Glu Lys Glu Lys Ala Glu Thr Arg Lys Ser Ala Asp Arg Thr Ala
165 170 175
Asp Val Leu Arg Ala Leu Ala Asp Phe Gly Leu Lys Pro Leu Met Arg
180 185 190
Val Tyr Thr Asp Ser Glu Met Ser Ser Val Glu Trp Lys Pro Leu Arg
195 200 205
Lys Gly Gln Ala Val Arg Thr Trp Asp Arg Asp Met Phe Gln Gln Ala
210 215 220
Ile Glu Arg Met Met Ser Trp Glu Ser Trp Asn Gln Arg Val Gly Gln
225 230 235 240
Glu Tyr Ala Lys Leu Val Glu Gln Lys Asn Arg Phe Glu Gln Lys Asn
245 250 255
Phe Val Gly Gln Glu His Leu Val His Leu Val Asn Gln Leu Gln Gln
260 265 270
Asp Met Lys Glu Ala Ser Pro Gly Leu Glu Ser Lys Glu Gln Thr Ala
275 280 285
His Tyr Val Thr Gly Arg Ala Leu Arg Gly Ser Asp Lys Val Phe Glu
290 295 300
Lys Trp Gly Lys Leu Ala Pro Asp Ala Pro Phe Asp Leu Tyr Asp Ala
305 310 315 320
Glu Ile Lys Asn Val Gln Arg Arg Asn Thr Arg Arg Phe Gly Ser His
325 330 335
Asp Leu Phe Ala Lys Leu Ala Glu Pro Glu Tyr Gln Ala Leu Trp Arg
340 345 350
Glu Asp Ala Ser Phe Leu Thr Arg Tyr Ala Val Tyr Asn Ser Ile Leu
355 360 365
Arg Lys Leu Asn His Ala Lys Met Phe Ala Thr Phe Thr Leu Pro Asp
370 375 380
Ala Thr Ala His Pro Ile Trp Thr Arg Phe Asp Lys Leu Gly Gly Asn
385 390 395 400
Leu His Gln Tyr Thr Phe Leu Phe Asn Glu Phe Gly Glu Arg Arg His
405 410 415
Ala Ile Arg Phe His Lys Leu Leu Lys Val Glu Asn Gly Val Ala Arg
420 425 430
Glu Val Asp Asp Val Thr Val Pro Ile Ser Met Ser Glu Gln Leu Asp
435 440 445
Asn Leu Leu Pro Arg Asp Pro Asn Glu Pro Ile Ala Leu Tyr Phe Arg
450 455 460
Asp Tyr Gly Ala Glu Gln His Phe Thr Gly Glu Phe Gly Gly Ala Lys
465 470 475 480
Ile Gln Cys Arg Arg Asp Gln Leu Ala His Met His Arg Arg Arg Gly
485 490 495
Ala Arg Asp Val Tyr Leu Asn Val Ser Val Arg Val Gln Ser Gln Ser
500 505 510
Glu Ala Arg Gly Glu Arg Arg Pro Pro Tyr Ala Ala Val Phe Arg Leu
515 520 525
Val Gly Asp Asn His Arg Ala Phe Val His Phe Asp Lys Leu Ser Asp
530 535 540
Tyr Leu Ala Glu His Pro Asp Asp Gly Lys Leu Gly Ser Glu Gly Leu
545 550 555 560
Leu Ser Gly Leu Arg Val Met Ser Val Asp Leu Gly Leu Arg Thr Ser
565 570 575
Ala Ser Ile Ser Val Phe Arg Val Ala Arg Lys Asp Glu Leu Lys Pro
580 585 590
Asn Ser Lys Gly Arg Val Pro Phe Phe Phe Pro Ile Lys Gly Asn Asp
595 600 605
Asn Leu Val Ala Val His Glu Arg Ser Gln Leu Leu Lys Leu Pro Gly
610 615 620
Glu Thr Glu Ser Lys Asp Leu Arg Ala Ile Arg Glu Glu Arg Gln Arg
625 630 635 640
Thr Leu Arg Gln Leu Arg Thr Gln Leu Ala Tyr Leu Arg Leu Leu Val
645 650 655
Arg Cys Gly Ser Glu Asp Val Gly Arg Arg Glu Arg Ser Trp Ala Lys
660 665 670
Leu Ile Glu Gln Pro Val Asp Ala Ala Asn His Met Thr Pro Asp Trp
675 680 685
Arg Glu Ala Phe Glu Asn Glu Leu Gln Lys Leu Lys Ser Leu His Gly
690 695 700
Ile Cys Ser Asp Lys Glu Trp Met Asp Ala Val Tyr Glu Ser Val Arg
705 710 715 720
Arg Val Trp Arg His Met Gly Lys Gln Val Arg Asp Trp Arg Lys Asp
725 730 735
Val Arg Ser Gly Glu Arg Pro Lys Ile Arg Gly Tyr Ala Lys Asp Val
740 745 750
Val Gly Gly Asn Ser Ile Glu Gln Ile Glu Tyr Leu Glu Arg Gln Tyr
755 760 765
Lys Phe Leu Lys Ser Trp Ser Phe Phe Gly Lys Val Ser Gly Gln Val
770 775 780
Ile Arg Ala Glu Lys Gly Ser Arg Phe Ala Ile Thr Leu Arg Glu His
785 790 795 800
Ile Asp His Ala Lys Glu Asp Arg Leu Lys Lys Leu Ala Asp Arg Ile
805 810 815
Ile Met Glu Ala Leu Gly Tyr Val Tyr Ala Leu Asp Glu Arg Gly Lys
820 825 830
Gly Lys Trp Val Ala Lys Tyr Pro Pro Cys Gln Leu Ile Leu Leu Glu
835 840 845
Glu Leu Ser Glu Tyr Gln Phe Asn Asn Asp Arg Pro Pro Ser Glu Asn
850 855 860
Asn Gln Leu Met Gln Trp Ser His Arg Gly Val Phe Gln Glu Leu Ile
865 870 875 880
Asn Gln Ala Gln Val His Asp Leu Leu Val Gly Thr Met Tyr Ala Ala
885 890 895
Phe Ser Ser Arg Phe Asp Ala Arg Thr Gly Ala Pro Gly Ile Arg Cys
900 905 910
Arg Arg Val Pro Ala Arg Cys Thr Gln Glu His Asn Pro Glu Pro Phe
915 920 925
Pro Trp Trp Leu Asn Lys Phe Val Val Glu His Thr Leu Asp Ala Cys
930 935 940
Pro Leu Arg Ala Asp Asp Leu Ile Pro Thr Gly Glu Gly Glu Ile Phe
945 950 955 960
Val Ser Pro Phe Ser Ala Glu Glu Gly Asp Phe His Gln Ile His Ala
965 970 975
Asp Leu Asn Ala Ala Gln Asn Leu Gln Gln Arg Leu Trp Ser Asp Phe
980 985 990
Asp Ile Ser Gln Ile Arg Leu Arg Cys Asp Trp Gly Glu Val Asp Gly
995 1000 1005
Glu Leu Val Leu Ile Pro Arg Leu Thr Gly Lys Arg Thr Ala Asp
1010 1015 1020
Ser Tyr Ser Asn Lys Val Phe Tyr Thr Asn Thr Gly Val Thr Tyr
1025 1030 1035
Tyr Glu Arg Glu Arg Gly Lys Lys Arg Arg Lys Val Phe Ala Gln
1040 1045 1050
Glu Lys Leu Ser Glu Glu Glu Ala Glu Leu Leu Val Glu Ala Asp
1055 1060 1065
Glu Ala Arg Glu Lys Ser Val Val Leu Met Arg Asp Pro Ser Gly
1070 1075 1080
Ile Ile Asn Arg Gly Asn Trp Thr Arg Gln Lys Glu Phe Trp Ser
1085 1090 1095
Met Val Asn Gln Arg Ile Glu Gly Tyr Leu Val Lys Gln Ile Arg
1100 1105 1110
Ser Arg Val Pro Leu Gln Asp Ser Ala Cys Glu Asn Thr Gly Asp
1115 1120 1125
Ile
<210> 549
<211> 872
<212> PRT
<213> Alicyclobacillus contaminans
<400> 549
Met Gly Phe Asn Thr Ala Glu Leu Leu Arg Lys Val Glu Glu Glu Met
1 5 10 15
Arg Lys Thr Ser Val Gly Phe Asp Thr Asp Asn Pro Phe Ala His Arg
20 25 30
Ile Thr Arg Arg Ala Ile Arg Gly Trp Asp Arg Ile Ala Glu Ala Trp
35 40 45
Arg Arg Leu Pro Pro Asp Ala Pro Glu Ser Glu Tyr Ile Glu Ala Phe
50 55 60
Lys Asp Ile Gln Arg Lys Asn Pro Arg Lys Ile Gly Ser Glu Pro Leu
65 70 75 80
Phe Lys Asn Leu Ala Ala Pro Gly Val Arg Ser Glu Leu Leu Asn Asn
85 90 95
Pro Gln Val Leu Ile Thr Phe Ala Lys Tyr Asn Glu Leu Gln Arg Gln
100 105 110
Leu Ala Lys Ala Lys Gln Phe Ala Gln Lys Thr Leu Pro His Pro Val
115 120 125
Phe His Pro Val Trp Val Arg Tyr Asp Lys Leu Gly Gly Asn Leu His
130 135 140
His Tyr Gln Ile Glu Pro Ala Val His Ala Asn Asp Thr His Lys Val
145 150 155 160
Lys Phe Ser Ser Leu Leu Leu Pro Gln Glu Asp Gly Ser Tyr Ala Glu
165 170 175
Val Lys Asp Val Thr Val Ser Leu Ala Pro Ser Leu Gln Phe Pro Thr
180 185 190
Gly Leu Val His Pro Lys Val Thr Thr Pro Pro Arg Thr Gly Leu Val
195 200 205
Thr Val Met Asp Glu Glu Ala Gly Lys Pro Val Val Cys Tyr Arg Asp
210 215 220
Arg Gly His Asp Ala Leu Val Pro Val Ala Phe Gly Gly Ala Lys Leu
225 230 235 240
Gln Phe Asn Arg Ala His Leu Ser Ala Gly Tyr Arg Lys Gly Val Leu
245 250 255
Ser Ala Gly Gly Gly Gly Ser Ile Tyr Phe Asn Val Thr Leu Asp Val
260 265 270
Gln Val Pro Asn Glu Arg Asp Val Ser Lys Thr Phe Ser Phe Ser Arg
275 280 285
Asp Arg Asp Leu Val Ser Leu Lys Ala Glu Glu Leu Lys Arg Tyr Met
290 295 300
Glu Thr Lys Pro Leu Gly Met Pro Gly Val Arg Val Met Ser Val Asp
305 310 315 320
Leu Gly Val Arg Tyr Gly Ala Ala Ile Ser Val Phe Glu Val Lys Pro
325 330 335
Phe Ala Glu Val Arg Lys Asp Lys Leu His Tyr Pro Ile Thr Gly Cys
340 345 350
Glu Gly Phe Val Ala Glu His Glu Arg Ser Val Ile Leu Lys Leu Pro
355 360 365
Gly Glu Gly Val Arg Thr Ala Gly Lys Gln Ser Glu Arg Lys Gln Ala
370 375 380
Leu Ala Ala Ile Arg Ala Glu Met Ser Ile Leu Arg Lys Trp Leu Arg
385 390 395 400
Val Ser Gln Val Thr Glu Glu Asp Arg Ala Lys Ala Val Arg Gly Leu
405 410 415
Leu Glu Asp Glu Arg Gly Gly Gly Trp Thr Met Asp Pro Gly Glu Asp
420 425 430
Ser Asp His Gln Pro Leu Gln Gln Phe Leu His Glu Ala Arg Leu Ala
435 440 445
Val Gly Glu Leu Val Asn Leu Val His Leu Ser Pro Ala Glu Trp Glu
450 455 460
Arg Ala Val Ile Glu Arg His Arg Arg Leu Glu Arg Ile Thr Ala Ser
465 470 475 480
His Ile Arg Val Phe Gln Thr Met Arg Lys Val Trp Gly Lys Arg Arg
485 490 495
Asn Glu Asp Ala Ala His Thr Gly Gly Ile Ser Leu Ala His Ile Glu
500 505 510
His Leu Ile Gln Gln Arg Lys Leu Phe Ile Arg Trp Ser Thr His Ala
515 520 525
Arg Thr Tyr Gly Glu Val Arg Arg Leu Pro Lys His Glu Gly Phe Ala
530 535 540
Lys Arg Leu Gln Lys His Thr Asn His Val Lys Glu Asp Arg Ile Lys
545 550 555 560
Lys Leu Ala Asp Met Ile Val Met Ala Ala Arg Gly Tyr Arg Phe Leu
565 570 575
Asp Lys Arg Ala Arg Trp Val Lys Thr Arg His Ala Pro Cys Asp Leu
580 585 590
Ile Leu Phe Glu Asp Leu Ser Arg Tyr Arg Phe Thr Met Asp Arg Pro
595 600 605
Pro Thr Glu Asn Ser Gln Leu Met Asn Trp Ser His Arg Glu Leu Leu
610 615 620
Lys Thr Val Lys Met Gln Ala Ala Leu Phe Gly Ile Gly Val Gly Thr
625 630 635 640
Val Pro Ala Ala Phe Thr Ser Arg Phe Asp Ala Gln Thr Gly Ala Pro
645 650 655
Gly Leu Arg Cys Lys Arg Val Thr Lys Gln Asp Lys Glu Lys Thr Pro
660 665 670
Phe Trp Leu Ile Gln Phe Ala Glu Ile Thr Gly Val Asn Val Thr Asn
675 680 685
Val Glu Pro Gly Gln Leu Ile Pro Val Asp Gly Gly Glu Trp Phe Val
690 695 700
Ser Pro Lys Gly Pro Arg Ala Ala Asp Gly Leu Lys Cys Val His Ala
705 710 715 720
Asp Ile Asn Ala Ala His Asn Leu Gln Arg Arg Phe Trp Ile Pro Arg
725 730 735
Leu Pro Ser Val Lys Cys Arg Arg Tyr Val Glu Ala Glu Gly Phe Ala
740 745 750
Ala Val Pro Ser Ser Thr Ala Phe Met Lys Val His Gly Lys Gly Ala
755 760 765
Phe Val Ser Val Asp Gly Glu Phe Tyr Glu Tyr Gln Lys Gly Arg Arg
770 775 780
Val Ala Val Asn Arg Ala Asp Arg Thr Ser Ser Thr Leu Asp Glu Asp
785 790 795 800
Glu Gly Asp Ile Gly Glu Glu Met Leu Val Ser Ser Asn Gly Ala Gly
805 810 815
Glu Phe Val Arg Met Phe Tyr Asp Glu Ser Gly Tyr Val Gly Tyr Gly
820 825 830
Arg Trp Met Asp Ser Lys Val Phe Trp Gly Lys Val Arg Gln Ile Val
835 840 845
His Arg Ala Ile Gln Asp Gln Val Glu Lys Arg Ala Ala Ala Arg Gly
850 855 860
Glu Asn Gly Ala Thr Ser Ser Arg
865 870
<210> 550
<211> 1149
<212> PRT
<213> Desulfovibrio inopinatus
<400> 550
Met Pro Thr Arg Thr Ile Asn Leu Lys Leu Val Leu Gly Lys Asn Pro
1 5 10 15
Glu Asn Ala Thr Leu Arg Arg Ala Leu Phe Ser Thr His Arg Leu Val
20 25 30
Asn Gln Ala Thr Lys Arg Ile Glu Glu Phe Leu Leu Leu Cys Arg Gly
35 40 45
Glu Ala Tyr Arg Thr Val Asp Asn Glu Gly Lys Glu Ala Glu Ile Pro
50 55 60
Arg His Ala Val Gln Glu Glu Ala Leu Ala Phe Ala Lys Ala Ala Gln
65 70 75 80
Arg His Asn Gly Cys Ile Ser Thr Tyr Glu Asp Gln Glu Ile Leu Asp
85 90 95
Val Leu Arg Gln Leu Tyr Glu Arg Leu Val Pro Ser Val Asn Glu Asn
100 105 110
Asn Glu Ala Gly Asp Ala Gln Ala Ala Asn Ala Trp Val Ser Pro Leu
115 120 125
Met Ser Ala Glu Ser Glu Gly Gly Leu Ser Val Tyr Asp Lys Val Leu
130 135 140
Asp Pro Pro Pro Val Trp Met Lys Leu Lys Glu Glu Lys Ala Pro Gly
145 150 155 160
Trp Glu Ala Ala Ser Gln Ile Trp Ile Gln Ser Asp Glu Gly Gln Ser
165 170 175
Leu Leu Asn Lys Pro Gly Ser Pro Pro Arg Trp Ile Arg Lys Leu Arg
180 185 190
Ser Gly Gln Pro Trp Gln Asp Asp Phe Val Ser Asp Gln Lys Lys Lys
195 200 205
Gln Asp Glu Leu Thr Lys Gly Asn Ala Pro Leu Ile Lys Gln Leu Lys
210 215 220
Glu Met Gly Leu Leu Pro Leu Val Asn Pro Phe Phe Arg His Leu Leu
225 230 235 240
Asp Pro Glu Gly Lys Gly Val Ser Pro Trp Asp Arg Leu Ala Val Arg
245 250 255
Ala Ala Val Ala His Phe Ile Ser Trp Glu Ser Trp Asn His Arg Thr
260 265 270
Arg Ala Glu Tyr Asn Ser Leu Lys Leu Arg Arg Asp Glu Phe Glu Ala
275 280 285
Ala Ser Asp Glu Phe Lys Asp Asp Phe Thr Leu Leu Arg Gln Tyr Glu
290 295 300
Ala Lys Arg His Ser Thr Leu Lys Ser Ile Ala Leu Ala Asp Asp Ser
305 310 315 320
Asn Pro Tyr Arg Ile Gly Val Arg Ser Leu Arg Ala Trp Asn Arg Val
325 330 335
Arg Glu Glu Trp Ile Asp Lys Gly Ala Thr Glu Glu Gln Arg Val Thr
340 345 350
Ile Leu Ser Lys Leu Gln Thr Gln Leu Arg Gly Lys Phe Gly Asp Pro
355 360 365
Asp Leu Phe Asn Trp Leu Ala Gln Asp Arg His Val His Leu Trp Ser
370 375 380
Pro Arg Asp Ser Val Thr Pro Leu Val Arg Ile Asn Ala Val Asp Lys
385 390 395 400
Val Leu Arg Arg Arg Lys Pro Tyr Ala Leu Met Thr Phe Ala His Pro
405 410 415
Arg Phe His Pro Arg Trp Ile Leu Tyr Glu Ala Pro Gly Gly Ser Asn
420 425 430
Leu Arg Gln Tyr Ala Leu Asp Cys Thr Glu Asn Ala Leu His Ile Thr
435 440 445
Leu Pro Leu Leu Val Asp Asp Ala His Gly Thr Trp Ile Glu Lys Lys
450 455 460
Ile Arg Val Pro Leu Ala Pro Ser Gly Gln Ile Gln Asp Leu Thr Leu
465 470 475 480
Glu Lys Leu Glu Lys Lys Lys Asn Arg Leu Tyr Tyr Arg Ser Gly Phe
485 490 495
Gln Gln Phe Ala Gly Leu Ala Gly Gly Ala Glu Val Leu Phe His Arg
500 505 510
Pro Tyr Met Glu His Asp Glu Arg Ser Glu Glu Ser Leu Leu Glu Arg
515 520 525
Pro Gly Ala Val Trp Phe Lys Leu Thr Leu Asp Val Ala Thr Gln Ala
530 535 540
Pro Pro Asn Trp Leu Asp Gly Lys Gly Arg Val Arg Thr Pro Pro Glu
545 550 555 560
Val His His Phe Lys Thr Ala Leu Ser Asn Lys Ser Lys His Thr Arg
565 570 575
Thr Leu Gln Pro Gly Leu Arg Val Leu Ser Val Asp Leu Gly Met Arg
580 585 590
Thr Phe Ala Ser Cys Ser Val Phe Glu Leu Ile Glu Gly Lys Pro Glu
595 600 605
Thr Gly Arg Ala Phe Pro Val Ala Asp Glu Arg Ser Met Asp Ser Pro
610 615 620
Asn Lys Leu Trp Ala Lys His Glu Arg Ser Phe Lys Leu Thr Leu Pro
625 630 635 640
Gly Glu Thr Pro Ser Arg Lys Glu Glu Glu Glu Arg Ser Ile Ala Arg
645 650 655
Ala Glu Ile Tyr Ala Leu Lys Arg Asp Ile Gln Arg Leu Lys Ser Leu
660 665 670
Leu Arg Leu Gly Glu Glu Asp Asn Asp Asn Arg Arg Asp Ala Leu Leu
675 680 685
Glu Gln Phe Phe Lys Gly Trp Gly Glu Glu Asp Val Val Pro Gly Gln
690 695 700
Ala Phe Pro Arg Ser Leu Phe Gln Gly Leu Gly Ala Ala Pro Phe Arg
705 710 715 720
Ser Thr Pro Glu Leu Trp Arg Gln His Cys Gln Thr Tyr Tyr Asp Lys
725 730 735
Ala Glu Ala Cys Leu Ala Lys His Ile Ser Asp Trp Arg Lys Arg Thr
740 745 750
Arg Pro Arg Pro Thr Ser Arg Glu Met Trp Tyr Lys Thr Arg Ser Tyr
755 760 765
His Gly Gly Lys Ser Ile Trp Met Leu Glu Tyr Leu Asp Ala Val Arg
770 775 780
Lys Leu Leu Leu Ser Trp Ser Leu Arg Gly Arg Thr Tyr Gly Ala Ile
785 790 795 800
Asn Arg Gln Asp Thr Ala Arg Phe Gly Ser Leu Ala Ser Arg Leu Leu
805 810 815
His His Ile Asn Ser Leu Lys Glu Asp Arg Ile Lys Thr Gly Ala Asp
820 825 830
Ser Ile Val Gln Ala Ala Arg Gly Tyr Ile Pro Leu Pro His Gly Lys
835 840 845
Gly Trp Glu Gln Arg Tyr Glu Pro Cys Gln Leu Ile Leu Phe Glu Asp
850 855 860
Leu Ala Arg Tyr Arg Phe Arg Val Asp Arg Pro Arg Arg Glu Asn Ser
865 870 875 880
Gln Leu Met Gln Trp Asn His Arg Ala Ile Val Ala Glu Thr Thr Met
885 890 895
Gln Ala Glu Leu Tyr Gly Gln Ile Val Glu Asn Thr Ala Ala Gly Phe
900 905 910
Ser Ser Arg Phe His Ala Ala Thr Gly Ala Pro Gly Val Arg Cys Arg
915 920 925
Phe Leu Leu Glu Arg Asp Phe Asp Asn Asp Leu Pro Lys Pro Tyr Leu
930 935 940
Leu Arg Glu Leu Ser Trp Met Leu Gly Asn Thr Lys Val Glu Ser Glu
945 950 955 960
Glu Glu Lys Leu Arg Leu Leu Ser Glu Lys Ile Arg Pro Gly Ser Leu
965 970 975
Val Pro Trp Asp Gly Gly Glu Gln Phe Ala Thr Leu His Pro Lys Arg
980 985 990
Gln Thr Leu Cys Val Ile His Ala Asp Met Asn Ala Ala Gln Asn Leu
995 1000 1005
Gln Arg Arg Phe Phe Gly Arg Cys Gly Glu Ala Phe Arg Leu Val
1010 1015 1020
Cys Gln Pro His Gly Asp Asp Val Leu Arg Leu Ala Ser Thr Pro
1025 1030 1035
Gly Ala Arg Leu Leu Gly Ala Leu Gln Gln Leu Glu Asn Gly Gln
1040 1045 1050
Gly Ala Phe Glu Leu Val Arg Asp Met Gly Ser Thr Ser Gln Met
1055 1060 1065
Asn Arg Phe Val Met Lys Ser Leu Gly Lys Lys Lys Ile Lys Pro
1070 1075 1080
Leu Gln Asp Asn Asn Gly Asp Asp Glu Leu Glu Asp Val Leu Ser
1085 1090 1095
Val Leu Pro Glu Glu Asp Asp Thr Gly Arg Ile Thr Val Phe Arg
1100 1105 1110
Asp Ser Ser Gly Ile Phe Phe Pro Cys Asn Val Trp Ile Pro Ala
1115 1120 1125
Lys Gln Phe Trp Pro Ala Val Arg Ala Met Ile Trp Lys Val Met
1130 1135 1140
Ala Ser His Ser Leu Gly
1145
<210> 551
<211> 1194
<212> PRT
<213> Desulfonatronum thiodismutans
<400> 551
Met Val Leu Gly Arg Lys Asp Asp Thr Ala Glu Leu Arg Arg Ala Leu
1 5 10 15
Trp Thr Thr His Glu His Val Asn Leu Ala Val Ala Glu Val Glu Arg
20 25 30
Val Leu Leu Arg Cys Arg Gly Arg Ser Tyr Trp Thr Leu Asp Arg Arg
35 40 45
Gly Asp Pro Val His Val Pro Glu Ser Gln Val Ala Glu Asp Ala Leu
50 55 60
Ala Met Ala Arg Glu Ala Gln Arg Arg Asn Gly Trp Pro Val Val Gly
65 70 75 80
Glu Asp Glu Glu Ile Leu Leu Ala Leu Arg Tyr Leu Tyr Glu Gln Ile
85 90 95
Val Pro Ser Cys Leu Leu Asp Asp Leu Gly Lys Pro Leu Lys Gly Asp
100 105 110
Ala Gln Lys Ile Gly Thr Asn Tyr Ala Gly Pro Leu Phe Asp Ser Asp
115 120 125
Thr Cys Arg Arg Asp Glu Gly Lys Asp Val Ala Cys Cys Gly Pro Phe
130 135 140
His Glu Val Ala Gly Lys Tyr Leu Gly Ala Leu Pro Glu Trp Ala Thr
145 150 155 160
Pro Ile Ser Lys Gln Glu Phe Asp Gly Lys Asp Ala Ser His Leu Arg
165 170 175
Phe Lys Ala Thr Gly Gly Asp Asp Ala Phe Phe Arg Val Ser Ile Glu
180 185 190
Lys Ala Asn Ala Trp Tyr Glu Asp Pro Ala Asn Gln Asp Ala Leu Lys
195 200 205
Asn Lys Ala Tyr Asn Lys Asp Asp Trp Lys Lys Glu Lys Asp Lys Gly
210 215 220
Ile Ser Ser Trp Ala Val Lys Tyr Ile Gln Lys Gln Leu Gln Leu Gly
225 230 235 240
Gln Asp Pro Arg Thr Glu Val Arg Arg Lys Leu Trp Leu Glu Leu Gly
245 250 255
Leu Leu Pro Leu Phe Ile Pro Val Phe Asp Lys Thr Met Val Gly Asn
260 265 270
Leu Trp Asn Arg Leu Ala Val Arg Leu Ala Leu Ala His Leu Leu Ser
275 280 285
Trp Glu Ser Trp Asn His Arg Ala Val Gln Asp Gln Ala Leu Ala Arg
290 295 300
Ala Lys Arg Asp Glu Leu Ala Ala Leu Phe Leu Gly Met Glu Asp Gly
305 310 315 320
Phe Ala Gly Leu Arg Glu Tyr Glu Leu Arg Arg Asn Glu Ser Ile Lys
325 330 335
Gln His Ala Phe Glu Pro Val Asp Arg Pro Tyr Val Val Ser Gly Arg
340 345 350
Ala Leu Arg Ser Trp Thr Arg Val Arg Glu Glu Trp Leu Arg His Gly
355 360 365
Asp Thr Gln Glu Ser Arg Lys Asn Ile Cys Asn Arg Leu Gln Asp Arg
370 375 380
Leu Arg Gly Lys Phe Gly Asp Pro Asp Val Phe His Trp Leu Ala Glu
385 390 395 400
Asp Gly Gln Glu Ala Leu Trp Lys Glu Arg Asp Cys Val Thr Ser Phe
405 410 415
Ser Leu Leu Asn Asp Ala Asp Gly Leu Leu Glu Lys Arg Lys Gly Tyr
420 425 430
Ala Leu Met Thr Phe Ala Asp Ala Arg Leu His Pro Arg Trp Ala Met
435 440 445
Tyr Glu Ala Pro Gly Gly Ser Asn Leu Arg Thr Tyr Gln Ile Arg Lys
450 455 460
Thr Glu Asn Gly Leu Trp Ala Asp Val Val Leu Leu Ser Pro Arg Asn
465 470 475 480
Glu Ser Ala Ala Val Glu Glu Lys Thr Phe Asn Val Arg Leu Ala Pro
485 490 495
Ser Gly Gln Leu Ser Asn Val Ser Phe Asp Gln Ile Gln Lys Gly Ser
500 505 510
Lys Met Val Gly Arg Cys Arg Tyr Gln Ser Ala Asn Gln Gln Phe Glu
515 520 525
Gly Leu Leu Gly Gly Ala Glu Ile Leu Phe Asp Arg Lys Arg Ile Ala
530 535 540
Asn Glu Gln His Gly Ala Thr Asp Leu Ala Ser Lys Pro Gly His Val
545 550 555 560
Trp Phe Lys Leu Thr Leu Asp Val Arg Pro Gln Ala Pro Gln Gly Trp
565 570 575
Leu Asp Gly Lys Gly Arg Pro Ala Leu Pro Pro Glu Ala Lys His Phe
580 585 590
Lys Thr Ala Leu Ser Asn Lys Ser Lys Phe Ala Asp Gln Val Arg Pro
595 600 605
Gly Leu Arg Val Leu Ser Val Asp Leu Gly Val Arg Ser Phe Ala Ala
610 615 620
Cys Ser Val Phe Glu Leu Val Arg Gly Gly Pro Asp Gln Gly Thr Tyr
625 630 635 640
Phe Pro Ala Ala Asp Gly Arg Thr Val Asp Asp Pro Glu Lys Leu Trp
645 650 655
Ala Lys His Glu Arg Ser Phe Lys Ile Thr Leu Pro Gly Glu Asn Pro
660 665 670
Ser Arg Lys Glu Glu Ile Ala Arg Arg Ala Ala Met Glu Glu Leu Arg
675 680 685
Ser Leu Asn Gly Asp Ile Arg Arg Leu Lys Ala Ile Leu Arg Leu Ser
690 695 700
Val Leu Gln Glu Asp Asp Pro Arg Thr Glu His Leu Arg Leu Phe Met
705 710 715 720
Glu Ala Ile Val Asp Asp Pro Ala Lys Ser Ala Leu Asn Ala Glu Leu
725 730 735
Phe Lys Gly Phe Gly Asp Asp Arg Phe Arg Ser Thr Pro Asp Leu Trp
740 745 750
Lys Gln His Cys His Phe Phe His Asp Lys Ala Glu Lys Val Val Ala
755 760 765
Glu Arg Phe Ser Arg Trp Arg Thr Glu Thr Arg Pro Lys Ser Ser Ser
770 775 780
Trp Gln Asp Trp Arg Glu Arg Arg Gly Tyr Ala Gly Gly Lys Ser Tyr
785 790 795 800
Trp Ala Val Thr Tyr Leu Glu Ala Val Arg Gly Leu Ile Leu Arg Trp
805 810 815
Asn Met Arg Gly Arg Thr Tyr Gly Glu Val Asn Arg Gln Asp Lys Lys
820 825 830
Gln Phe Gly Thr Val Ala Ser Ala Leu Leu His His Ile Asn Gln Leu
835 840 845
Lys Glu Asp Arg Ile Lys Thr Gly Ala Asp Met Ile Ile Gln Ala Ala
850 855 860
Arg Gly Phe Val Pro Arg Lys Asn Gly Ala Gly Trp Val Gln Val His
865 870 875 880
Glu Pro Cys Arg Leu Ile Leu Phe Glu Asp Leu Ala Arg Tyr Arg Phe
885 890 895
Arg Thr Asp Arg Ser Arg Arg Glu Asn Ser Arg Leu Met Arg Trp Ser
900 905 910
His Arg Glu Ile Val Asn Glu Val Gly Met Gln Gly Glu Leu Tyr Gly
915 920 925
Leu His Val Asp Thr Thr Glu Ala Gly Phe Ser Ser Arg Tyr Leu Ala
930 935 940
Ser Ser Gly Ala Pro Gly Val Arg Cys Arg His Leu Val Glu Glu Asp
945 950 955 960
Phe His Asp Gly Leu Pro Gly Met His Leu Val Gly Glu Leu Asp Trp
965 970 975
Leu Leu Pro Lys Asp Lys Asp Arg Thr Ala Asn Glu Ala Arg Arg Leu
980 985 990
Leu Gly Gly Met Val Arg Pro Gly Met Leu Val Pro Trp Asp Gly Gly
995 1000 1005
Glu Leu Phe Ala Thr Leu Asn Ala Ala Ser Gln Leu His Val Ile
1010 1015 1020
His Ala Asp Ile Asn Ala Ala Gln Asn Leu Gln Arg Arg Phe Trp
1025 1030 1035
Gly Arg Cys Gly Glu Ala Ile Arg Ile Val Cys Asn Gln Leu Ser
1040 1045 1050
Val Asp Gly Ser Thr Arg Tyr Glu Met Ala Lys Ala Pro Lys Ala
1055 1060 1065
Arg Leu Leu Gly Ala Leu Gln Gln Leu Lys Asn Gly Asp Ala Pro
1070 1075 1080
Phe His Leu Thr Ser Ile Pro Asn Ser Gln Lys Pro Glu Asn Ser
1085 1090 1095
Tyr Val Met Thr Pro Thr Asn Ala Gly Lys Lys Tyr Arg Ala Gly
1100 1105 1110
Pro Gly Glu Lys Ser Ser Gly Glu Glu Asp Glu Leu Ala Leu Asp
1115 1120 1125
Ile Val Glu Gln Ala Glu Glu Leu Ala Gln Gly Arg Lys Thr Phe
1130 1135 1140
Phe Arg Asp Pro Ser Gly Val Phe Phe Ala Pro Asp Arg Trp Leu
1145 1150 1155
Pro Ser Glu Ile Tyr Trp Ser Arg Ile Arg Arg Arg Ile Trp Gln
1160 1165 1170
Val Thr Leu Glu Arg Asn Ser Ser Gly Arg Gln Glu Arg Ala Glu
1175 1180 1185
Met Asp Glu Met Pro Tyr
1190
<210> 552
<211> 1388
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 552
Met Ser Leu Asn Arg Ile Tyr Gln Gly Arg Val Ala Ala Val Glu Thr
1 5 10 15
Gly Thr Ala Leu Ala Lys Gly Asn Val Glu Trp Met Pro Ala Ala Gly
20 25 30
Gly Asp Glu Val Leu Trp Gln His His Glu Leu Phe Gln Ala Ala Ile
35 40 45
Asn Tyr Tyr Leu Val Ala Leu Leu Ala Leu Ala Asp Lys Asn Asn Pro
50 55 60
Val Leu Gly Pro Leu Ile Ser Gln Met Asp Asn Pro Gln Ser Pro Tyr
65 70 75 80
His Val Trp Gly Ser Phe Arg Arg Gln Gly Arg Gln Arg Thr Gly Leu
85 90 95
Ser Gln Ala Val Ala Pro Tyr Ile Thr Pro Gly Asn Asn Ala Pro Thr
100 105 110
Leu Asp Glu Val Phe Arg Ser Ile Leu Ala Gly Asn Pro Thr Asp Arg
115 120 125
Ala Thr Leu Asp Ala Ala Leu Met Gln Leu Leu Lys Ala Cys Asp Gly
130 135 140
Ala Gly Ala Ile Gln Gln Glu Gly Arg Ser Tyr Trp Pro Lys Phe Cys
145 150 155 160
Asp Pro Asp Ser Thr Ala Asn Phe Ala Gly Asp Pro Ala Met Leu Arg
165 170 175
Arg Glu Gln His Arg Leu Leu Leu Pro Gln Val Leu His Asp Pro Ala
180 185 190
Ile Thr His Asp Ser Pro Ala Leu Gly Ser Phe Asp Thr Tyr Ser Ile
195 200 205
Ala Thr Pro Asp Thr Arg Thr Pro Gln Leu Thr Gly Pro Lys Ala Arg
210 215 220
Ala Arg Leu Glu Gln Ala Ile Thr Leu Trp Arg Val Arg Leu Pro Glu
225 230 235 240
Ser Ala Ala Asp Phe Asp Arg Leu Ala Ser Ser Leu Lys Lys Ile Pro
245 250 255
Asp Asp Asp Ser Arg Leu Asn Leu Gln Gly Tyr Val Gly Ser Ser Ala
260 265 270
Lys Gly Glu Val Gln Ala Arg Leu Phe Ala Leu Leu Leu Phe Arg His
275 280 285
Leu Glu Arg Ser Ser Phe Thr Leu Gly Leu Leu Arg Ser Ala Thr Pro
290 295 300
Pro Pro Lys Asn Ala Glu Thr Pro Pro Pro Ala Gly Val Pro Leu Pro
305 310 315 320
Ala Ala Ser Ala Ala Asp Pro Val Arg Ile Ala Arg Gly Lys Arg Ser
325 330 335
Phe Val Phe Arg Ala Phe Thr Ser Leu Pro Cys Trp His Gly Gly Asp
340 345 350
Asn Ile His Pro Thr Trp Lys Ser Phe Asp Ile Ala Ala Phe Lys Tyr
355 360 365
Ala Leu Thr Val Ile Asn Gln Ile Glu Glu Lys Thr Lys Glu Arg Gln
370 375 380
Lys Glu Cys Ala Glu Leu Glu Thr Asp Phe Asp Tyr Met His Gly Arg
385 390 395 400
Leu Ala Lys Ile Pro Val Lys Tyr Thr Thr Gly Glu Ala Glu Pro Pro
405 410 415
Pro Ile Leu Ala Asn Asp Leu Arg Ile Pro Leu Leu Arg Glu Leu Leu
420 425 430
Gln Asn Ile Lys Val Asp Thr Ala Leu Thr Asp Gly Glu Ala Val Ser
435 440 445
Tyr Gly Leu Gln Arg Arg Thr Ile Arg Gly Phe Arg Glu Leu Arg Arg
450 455 460
Ile Trp Arg Gly His Ala Pro Ala Gly Thr Val Phe Ser Ser Glu Leu
465 470 475 480
Lys Glu Lys Leu Ala Gly Glu Leu Arg Gln Phe Gln Thr Asp Asn Ser
485 490 495
Thr Thr Ile Gly Ser Val Gln Leu Phe Asn Glu Leu Ile Gln Asn Pro
500 505 510
Lys Tyr Trp Pro Ile Trp Gln Ala Pro Asp Val Glu Thr Ala Arg Gln
515 520 525
Trp Ala Asp Ala Gly Phe Ala Asp Asp Pro Leu Ala Ala Leu Val Gln
530 535 540
Glu Ala Glu Leu Gln Glu Asp Ile Asp Ala Leu Lys Ala Pro Val Lys
545 550 555 560
Leu Thr Pro Ala Asp Pro Glu Tyr Ser Arg Arg Gln Tyr Asp Phe Asn
565 570 575
Ala Val Ser Lys Phe Gly Ala Gly Ser Arg Ser Ala Asn Arg His Glu
580 585 590
Pro Gly Gln Thr Glu Arg Gly His Asn Thr Phe Thr Thr Glu Ile Ala
595 600 605
Ala Arg Asn Ala Ala Asp Gly Asn Arg Trp Arg Ala Thr His Val Arg
610 615 620
Ile His Tyr Ser Ala Pro Arg Leu Leu Arg Asp Gly Leu Arg Arg Pro
625 630 635 640
Asp Thr Asp Gly Asn Glu Ala Leu Glu Ala Val Pro Trp Leu Gln Pro
645 650 655
Met Met Glu Ala Leu Ala Pro Leu Pro Thr Leu Pro Gln Asp Leu Thr
660 665 670
Gly Met Pro Val Phe Leu Met Pro Asp Val Thr Leu Ser Gly Glu Arg
675 680 685
Arg Ile Leu Leu Asn Leu Pro Val Thr Leu Glu Pro Ala Ala Leu Val
690 695 700
Glu Gln Leu Gly Asn Ala Gly Arg Trp Gln Asn Gln Phe Phe Gly Ser
705 710 715 720
Arg Glu Asp Pro Phe Ala Leu Arg Trp Pro Ala Asp Gly Ala Val Lys
725 730 735
Thr Ala Lys Gly Lys Thr His Ile Pro Trp His Gln Asp Arg Asp His
740 745 750
Phe Thr Val Leu Gly Val Asp Leu Gly Thr Arg Asp Ala Gly Ala Leu
755 760 765
Ala Leu Leu Asn Val Thr Ala Gln Lys Pro Ala Lys Pro Val His Arg
770 775 780
Ile Ile Gly Glu Ala Asp Gly Arg Thr Trp Tyr Ala Ser Leu Ala Asp
785 790 795 800
Ala Arg Met Ile Arg Leu Pro Gly Glu Asp Ala Arg Leu Phe Val Arg
805 810 815
Gly Lys Leu Val Gln Glu Pro Tyr Gly Glu Arg Gly Arg Asn Ala Ser
820 825 830
Leu Leu Glu Trp Glu Asp Ala Arg Asn Ile Ile Leu Arg Leu Gly Gln
835 840 845
Asn Pro Asp Glu Leu Leu Gly Ala Asp Pro Arg Arg His Ser Tyr Pro
850 855 860
Glu Ile Asn Asp Lys Leu Leu Val Ala Leu Arg Arg Ala Gln Ala Arg
865 870 875 880
Leu Ala Arg Leu Gln Asn Arg Ser Trp Arg Leu Arg Asp Leu Ala Glu
885 890 895
Ser Asp Lys Ala Leu Asp Glu Ile His Ala Glu Arg Ala Gly Glu Lys
900 905 910
Pro Ser Pro Leu Pro Pro Leu Ala Arg Asp Asp Ala Ile Lys Ser Thr
915 920 925
Asp Glu Ala Leu Leu Ser Gln Arg Asp Ile Ile Arg Arg Ser Phe Val
930 935 940
Gln Ile Ala Asn Leu Ile Leu Pro Leu Arg Gly Arg Arg Trp Glu Trp
945 950 955 960
Arg Pro His Val Glu Val Pro Asp Cys His Ile Leu Ala Gln Ser Asp
965 970 975
Pro Gly Thr Asp Asp Thr Lys Arg Leu Val Ala Gly Gln Arg Gly Ile
980 985 990
Ser His Glu Arg Ile Glu Gln Ile Glu Glu Leu Arg Arg Arg Cys Gln
995 1000 1005
Ser Leu Asn Arg Ala Leu Arg His Lys Pro Gly Glu Arg Pro Val
1010 1015 1020
Leu Gly Arg Pro Ala Lys Gly Glu Glu Ile Ala Asp Pro Cys Pro
1025 1030 1035
Ala Leu Leu Glu Lys Ile Asn Arg Leu Arg Asp Gln Arg Val Asp
1040 1045 1050
Gln Thr Ala His Ala Ile Leu Ala Ala Ala Leu Gly Val Arg Leu
1055 1060 1065
Arg Ala Pro Ser Lys Asp Arg Ala Glu Arg Arg His Arg Asp Ile
1070 1075 1080
His Gly Glu Tyr Glu Arg Phe Arg Ala Pro Ala Asp Phe Val Val
1085 1090 1095
Ile Glu Asn Leu Ser Arg Tyr Leu Ser Ser Gln Asp Arg Ala Arg
1100 1105 1110
Ser Glu Asn Thr Arg Leu Met Gln Trp Cys His Arg Gln Ile Val
1115 1120 1125
Gln Lys Leu Arg Gln Leu Cys Glu Thr Tyr Gly Ile Pro Val Leu
1130 1135 1140
Ala Val Pro Ala Ala Tyr Ser Ser Arg Phe Ser Ser Arg Asp Gly
1145 1150 1155
Ser Ala Gly Phe Arg Ala Val His Leu Thr Pro Asp His Arg His
1160 1165 1170
Arg Met Pro Trp Ser Arg Ile Leu Ala Arg Leu Lys Ala His Glu
1175 1180 1185
Glu Asp Gly Lys Arg Leu Glu Lys Thr Val Leu Asp Glu Ala Arg
1190 1195 1200
Ala Val Arg Gly Leu Phe Asp Arg Leu Asp Arg Phe Asn Ala Gly
1205 1210 1215
His Val Pro Gly Lys Pro Trp Arg Thr Leu Leu Ala Pro Leu Pro
1220 1225 1230
Gly Gly Pro Val Phe Val Pro Leu Gly Asp Ala Thr Pro Met Gln
1235 1240 1245
Ala Asp Leu Asn Ala Ala Ile Asn Ile Ala Leu Arg Gly Ile Ala
1250 1255 1260
Ala Pro Asp Arg His Asp Ile His His Arg Leu Arg Ala Glu Asn
1265 1270 1275
Lys Lys Arg Ile Leu Ser Leu Arg Leu Gly Thr Gln Arg Glu Lys
1280 1285 1290
Ala Arg Trp Pro Gly Gly Ala Pro Ala Val Thr Leu Ser Thr Pro
1295 1300 1305
Asn Asn Gly Ala Ser Pro Glu Asp Ser Asp Ala Leu Pro Glu Arg
1310 1315 1320
Val Ser Asn Leu Phe Val Asp Ile Ala Gly Val Ala Asn Phe Glu
1325 1330 1335
Arg Val Thr Ile Glu Gly Val Ser Gln Lys Phe Ala Thr Gly Arg
1340 1345 1350
Gly Leu Trp Ala Ser Val Lys Gln Arg Ala Trp Asn Arg Val Ala
1355 1360 1365
Arg Leu Asn Glu Thr Val Thr Asp Asn Asn Arg Asn Glu Glu Glu
1370 1375 1380
Asp Asp Ile Pro Met
1385
<210> 553
<211> 1132
<212> PRT
<213> Tuberibacillus calidus
<400> 553
Met Ala Thr Lys Ser Phe Ile Leu Lys Met Lys Thr Lys Asn Asn Pro
1 5 10 15
Gln Leu Arg Leu Ser Leu Trp Lys Thr His Glu Leu Phe Asn Phe Gly
20 25 30
Val Ala Tyr Tyr Met Asp Leu Leu Ser Leu Phe Arg Gln Lys Asp Leu
35 40 45
Tyr Met His Asn Asp Glu Asp Pro Asp His Pro Val Val Leu Lys Lys
50 55 60
Glu Glu Ile Gln Glu Arg Leu Trp Met Lys Val Arg Glu Thr Gln Gln
65 70 75 80
Lys Asn Gly Phe His Gly Glu Val Ser Lys Asp Glu Val Leu Glu Thr
85 90 95
Leu Arg Ala Leu Tyr Glu Glu Leu Val Pro Ser Ala Val Gly Lys Ser
100 105 110
Gly Glu Ala Asn Gln Ile Ser Asn Lys Tyr Leu Tyr Pro Leu Thr Asp
115 120 125
Pro Ala Ser Gln Ser Gly Lys Gly Thr Ala Asn Ser Gly Arg Lys Pro
130 135 140
Arg Trp Lys Lys Leu Lys Glu Ala Gly Asp Pro Ser Trp Lys Asp Ala
145 150 155 160
Tyr Glu Lys Trp Glu Lys Glu Arg Gln Glu Asp Pro Lys Leu Lys Ile
165 170 175
Leu Ala Ala Leu Gln Ser Phe Gly Leu Ile Pro Leu Phe Arg Pro Phe
180 185 190
Thr Glu Asn Asp His Lys Ala Val Ile Ser Val Lys Trp Met Pro Lys
195 200 205
Ser Lys Asn Gln Ser Val Arg Lys Phe Asp Lys Asp Met Phe Asn Gln
210 215 220
Ala Ile Glu Arg Phe Leu Ser Trp Glu Ser Trp Asn Glu Lys Val Ala
225 230 235 240
Glu Asp Tyr Glu Lys Thr Val Ser Ile Tyr Glu Ser Leu Gln Lys Glu
245 250 255
Leu Lys Gly Ile Ser Thr Lys Ala Phe Glu Ile Met Glu Arg Val Glu
260 265 270
Lys Ala Tyr Glu Ala His Leu Arg Glu Ile Thr Phe Ser Asn Ser Thr
275 280 285
Tyr Arg Ile Gly Asn Arg Ala Ile Arg Gly Trp Thr Glu Ile Val Lys
290 295 300
Lys Trp Met Lys Leu Asp Pro Ser Ala Pro Gln Gly Asn Tyr Leu Asp
305 310 315 320
Val Val Lys Asp Tyr Gln Arg Arg His Pro Arg Glu Ser Gly Asp Phe
325 330 335
Lys Leu Phe Glu Leu Leu Ser Arg Pro Glu Asn Gln Ala Ala Trp Arg
340 345 350
Glu Tyr Pro Glu Phe Leu Pro Leu Tyr Val Lys Tyr Arg His Ala Glu
355 360 365
Gln Arg Met Lys Thr Ala Lys Lys Gln Ala Thr Phe Thr Leu Cys Asp
370 375 380
Pro Ile Arg His Pro Leu Trp Val Arg Tyr Glu Glu Arg Ser Gly Thr
385 390 395 400
Asn Leu Asn Lys Tyr Arg Leu Ile Met Asn Glu Lys Glu Lys Val Val
405 410 415
Gln Phe Asp Arg Leu Ile Cys Leu Asn Ala Asp Gly His Tyr Glu Glu
420 425 430
Gln Glu Asp Val Thr Val Pro Leu Ala Pro Ser Gln Gln Phe Asp Asp
435 440 445
Gln Ile Lys Phe Ser Ser Glu Asp Thr Gly Lys Gly Lys His Asn Phe
450 455 460
Ser Tyr Tyr His Lys Gly Ile Asn Tyr Glu Leu Lys Gly Thr Leu Gly
465 470 475 480
Gly Ala Arg Ile Gln Phe Asp Arg Glu His Leu Leu Arg Arg Gln Gly
485 490 495
Val Lys Ala Gly Asn Val Gly Arg Ile Phe Leu Asn Val Thr Leu Asn
500 505 510
Ile Glu Pro Met Gln Pro Phe Ser Arg Ser Gly Asn Leu Gln Thr Ser
515 520 525
Val Gly Lys Ala Leu Lys Val Tyr Val Asp Gly Tyr Pro Lys Val Val
530 535 540
Asn Phe Lys Pro Lys Glu Leu Thr Glu His Ile Lys Glu Ser Glu Lys
545 550 555 560
Asn Thr Leu Thr Leu Gly Val Glu Ser Leu Pro Thr Gly Leu Arg Val
565 570 575
Met Ser Val Asp Leu Gly Gln Arg Gln Ala Ala Ala Ile Ser Ile Phe
580 585 590
Glu Val Val Ser Glu Lys Pro Asp Asp Asn Lys Leu Phe Tyr Pro Val
595 600 605
Lys Asp Thr Asp Leu Phe Ala Val His Arg Thr Ser Phe Asn Ile Lys
610 615 620
Leu Pro Gly Glu Lys Arg Thr Glu Arg Arg Met Leu Glu Gln Gln Lys
625 630 635 640
Arg Asp Gln Ala Ile Arg Asp Leu Ser Arg Lys Leu Lys Phe Leu Lys
645 650 655
Asn Val Leu Asn Met Gln Lys Leu Glu Lys Thr Asp Glu Arg Glu Lys
660 665 670
Arg Val Asn Arg Trp Ile Lys Asp Arg Glu Arg Glu Glu Glu Asn Pro
675 680 685
Val Tyr Val Gln Glu Phe Glu Met Ile Ser Lys Val Leu Tyr Ser Pro
690 695 700
His Ser Val Trp Val Asp Gln Leu Lys Ser Ile His Arg Lys Leu Glu
705 710 715 720
Glu Gln Leu Gly Lys Glu Ile Ser Lys Trp Arg Gln Ser Ile Ser Gln
725 730 735
Gly Arg Gln Gly Val Tyr Gly Ile Ser Leu Lys Asn Ile Glu Asp Ile
740 745 750
Glu Lys Thr Arg Arg Leu Leu Phe Arg Trp Ser Met Arg Pro Glu Asn
755 760 765
Pro Gly Glu Val Lys Gln Leu Gln Pro Gly Glu Arg Phe Ala Ile Asp
770 775 780
Gln Gln Asn His Leu Asn His Leu Lys Asp Asp Arg Ile Lys Lys Leu
785 790 795 800
Ala Asn Gln Ile Val Met Thr Ala Leu Gly Tyr Arg Tyr Asp Gly Lys
805 810 815
Arg Lys Lys Trp Ile Ala Lys His Pro Ala Cys Gln Leu Val Leu Phe
820 825 830
Glu Asp Leu Ser Arg Tyr Ala Phe Tyr Asp Glu Arg Ser Arg Leu Glu
835 840 845
Asn Arg Asn Leu Met Arg Trp Ser Arg Arg Glu Ile Pro Lys Gln Val
850 855 860
Ala Gln Ile Gly Gly Leu Tyr Gly Leu Leu Val Gly Glu Val Gly Ala
865 870 875 880
Gln Tyr Ser Ser Arg Phe His Ala Lys Ser Gly Ala Pro Gly Ile Arg
885 890 895
Cys Arg Val Val Lys Glu His Glu Leu Tyr Ile Thr Glu Gly Gly Gln
900 905 910
Lys Val Arg Asn Gln Lys Phe Leu Asp Ser Leu Val Glu Asn Asn Ile
915 920 925
Ile Glu Pro Asp Asp Ala Arg Arg Leu Glu Pro Gly Asp Leu Ile Arg
930 935 940
Asp Gln Gly Gly Asp Lys Phe Ala Thr Leu Asp Glu Arg Gly Glu Leu
945 950 955 960
Val Ile Thr His Ala Asp Ile Asn Ala Ala Gln Asn Leu Gln Lys Arg
965 970 975
Phe Trp Thr Arg Thr His Gly Leu Tyr Arg Ile Arg Cys Glu Ser Arg
980 985 990
Glu Ile Lys Asp Ala Val Val Leu Val Pro Ser Asp Lys Asp Gln Lys
995 1000 1005
Glu Lys Met Glu Asn Leu Phe Gly Ile Gly Tyr Leu Gln Pro Phe
1010 1015 1020
Lys Gln Glu Asn Asp Val Tyr Lys Trp Val Lys Gly Glu Lys Ile
1025 1030 1035
Lys Gly Lys Lys Thr Ser Ser Gln Ser Asp Asp Lys Glu Leu Val
1040 1045 1050
Ser Glu Ile Leu Gln Glu Ala Ser Val Met Ala Asp Glu Leu Lys
1055 1060 1065
Gly Asn Arg Lys Thr Leu Phe Arg Asp Pro Ser Gly Tyr Val Phe
1070 1075 1080
Pro Lys Asp Arg Trp Tyr Thr Gly Gly Arg Tyr Phe Gly Thr Leu
1085 1090 1095
Glu His Leu Leu Lys Arg Lys Leu Ala Glu Arg Arg Leu Phe Asp
1100 1105 1110
Gly Gly Ser Ser Arg Arg Gly Leu Phe Asn Gly Thr Asp Ser Asn
1115 1120 1125
Thr Asn Val Glu
1130
<210> 554
<211> 1108
<212> PRT
<213> Bacillus thermoamylovorans
<400> 554
Met Ala Thr Arg Ser Phe Ile Leu Lys Ile Glu Pro Asn Glu Glu Val
1 5 10 15
Lys Lys Gly Leu Trp Lys Thr His Glu Val Leu Asn His Gly Ile Ala
20 25 30
Tyr Tyr Met Asn Ile Leu Lys Leu Ile Arg Gln Glu Ala Ile Tyr Glu
35 40 45
His His Glu Gln Asp Pro Lys Asn Pro Lys Lys Val Ser Lys Ala Glu
50 55 60
Ile Gln Ala Glu Leu Trp Asp Phe Val Leu Lys Met Gln Lys Cys Asn
65 70 75 80
Ser Phe Thr His Glu Val Asp Lys Asp Val Val Phe Asn Ile Leu Arg
85 90 95
Glu Leu Tyr Glu Glu Leu Val Pro Ser Ser Val Glu Lys Lys Gly Glu
100 105 110
Ala Asn Gln Leu Ser Asn Lys Phe Leu Tyr Pro Leu Val Asp Pro Asn
115 120 125
Ser Gln Ser Gly Lys Gly Thr Ala Ser Ser Gly Arg Lys Pro Arg Trp
130 135 140
Tyr Asn Leu Lys Ile Ala Gly Asp Pro Ser Trp Glu Glu Glu Lys Lys
145 150 155 160
Lys Trp Glu Glu Asp Lys Lys Lys Asp Pro Leu Ala Lys Ile Leu Gly
165 170 175
Lys Leu Ala Glu Tyr Gly Leu Ile Pro Leu Phe Ile Pro Phe Thr Asp
180 185 190
Ser Asn Glu Pro Ile Val Lys Glu Ile Lys Trp Met Glu Lys Ser Arg
195 200 205
Asn Gln Ser Val Arg Arg Leu Asp Lys Asp Met Phe Ile Gln Ala Leu
210 215 220
Glu Arg Phe Leu Ser Trp Glu Ser Trp Asn Leu Lys Val Lys Glu Glu
225 230 235 240
Tyr Glu Lys Val Glu Lys Glu His Lys Thr Leu Glu Glu Arg Ile Lys
245 250 255
Glu Asp Ile Gln Ala Phe Lys Ser Leu Glu Gln Tyr Glu Lys Glu Arg
260 265 270
Gln Glu Gln Leu Leu Arg Asp Thr Leu Asn Thr Asn Glu Tyr Arg Leu
275 280 285
Ser Lys Arg Gly Leu Arg Gly Trp Arg Glu Ile Ile Gln Lys Trp Leu
290 295 300
Lys Met Asp Glu Asn Glu Pro Ser Glu Lys Tyr Leu Glu Val Phe Lys
305 310 315 320
Asp Tyr Gln Arg Lys His Pro Arg Glu Ala Gly Asp Tyr Ser Val Tyr
325 330 335
Glu Phe Leu Ser Lys Lys Glu Asn His Phe Ile Trp Arg Asn His Pro
340 345 350
Glu Tyr Pro Tyr Leu Tyr Ala Thr Phe Cys Glu Ile Asp Lys Lys Lys
355 360 365
Lys Asp Ala Lys Gln Gln Ala Thr Phe Thr Leu Ala Asp Pro Ile Asn
370 375 380
His Pro Leu Trp Val Arg Phe Glu Glu Arg Ser Gly Ser Asn Leu Asn
385 390 395 400
Lys Tyr Arg Ile Leu Thr Glu Gln Leu His Thr Glu Lys Leu Lys Lys
405 410 415
Lys Leu Thr Val Gln Leu Asp Arg Leu Ile Tyr Pro Thr Glu Ser Gly
420 425 430
Gly Trp Glu Glu Lys Gly Lys Val Asp Ile Val Leu Leu Pro Ser Arg
435 440 445
Gln Phe Tyr Asn Gln Ile Phe Leu Asp Ile Glu Glu Lys Gly Lys His
450 455 460
Ala Phe Thr Tyr Lys Asp Glu Ser Ile Lys Phe Pro Leu Lys Gly Thr
465 470 475 480
Leu Gly Gly Ala Arg Val Gln Phe Asp Arg Asp His Leu Arg Arg Tyr
485 490 495
Pro His Lys Val Glu Ser Gly Asn Val Gly Arg Ile Tyr Phe Asn Met
500 505 510
Thr Val Asn Ile Glu Pro Thr Glu Ser Pro Val Ser Lys Ser Leu Lys
515 520 525
Ile His Arg Asp Asp Phe Pro Lys Phe Val Asn Phe Lys Pro Lys Glu
530 535 540
Leu Thr Glu Trp Ile Lys Asp Ser Lys Gly Lys Lys Leu Lys Ser Gly
545 550 555 560
Ile Glu Ser Leu Glu Ile Gly Leu Arg Val Met Ser Ile Asp Leu Gly
565 570 575
Gln Arg Gln Ala Ala Ala Ala Ser Ile Phe Glu Val Val Asp Gln Lys
580 585 590
Pro Asp Ile Glu Gly Lys Leu Phe Phe Pro Ile Lys Gly Thr Glu Leu
595 600 605
Tyr Ala Val His Arg Ala Ser Phe Asn Ile Lys Leu Pro Gly Glu Thr
610 615 620
Leu Val Lys Ser Arg Glu Val Leu Arg Lys Ala Arg Glu Asp Asn Leu
625 630 635 640
Lys Leu Met Asn Gln Lys Leu Asn Phe Leu Arg Asn Val Leu His Phe
645 650 655
Gln Gln Phe Glu Asp Ile Thr Glu Arg Glu Lys Arg Val Thr Lys Trp
660 665 670
Ile Ser Arg Gln Glu Asn Ser Asp Val Pro Leu Val Tyr Gln Asp Glu
675 680 685
Leu Ile Gln Ile Arg Glu Leu Met Tyr Lys Pro Tyr Lys Asp Trp Val
690 695 700
Ala Phe Leu Lys Gln Leu His Lys Arg Leu Glu Val Glu Ile Gly Lys
705 710 715 720
Glu Val Lys His Trp Arg Lys Ser Leu Ser Asp Gly Arg Lys Gly Leu
725 730 735
Tyr Gly Ile Ser Leu Lys Asn Ile Asp Glu Ile Asp Arg Thr Arg Lys
740 745 750
Phe Leu Leu Arg Trp Ser Leu Arg Pro Thr Glu Pro Gly Glu Val Arg
755 760 765
Arg Leu Glu Pro Gly Gln Arg Phe Ala Ile Asp Gln Leu Asn His Leu
770 775 780
Asn Ala Leu Lys Glu Asp Arg Leu Lys Lys Met Ala Asn Thr Ile Ile
785 790 795 800
Met His Ala Leu Gly Tyr Cys Tyr Asp Val Arg Lys Lys Lys Trp Gln
805 810 815
Ala Lys Asn Pro Ala Cys Gln Ile Ile Leu Phe Glu Asp Leu Ser Asn
820 825 830
Tyr Asn Pro Tyr Glu Glu Arg Ser Arg Phe Glu Asn Ser Lys Leu Met
835 840 845
Lys Trp Ser Arg Arg Glu Ile Pro Arg Gln Val Ala Leu Gln Gly Glu
850 855 860
Ile Tyr Gly Leu Gln Val Gly Glu Val Gly Ala Gln Phe Ser Ser Arg
865 870 875 880
Phe His Ala Lys Thr Gly Ser Pro Gly Ile Arg Cys Ser Val Val Thr
885 890 895
Lys Glu Lys Leu Gln Asp Asn Arg Phe Phe Lys Asn Leu Gln Arg Glu
900 905 910
Gly Arg Leu Thr Leu Asp Lys Ile Ala Val Leu Lys Glu Gly Asp Leu
915 920 925
Tyr Pro Asp Lys Gly Gly Glu Lys Phe Ile Ser Leu Ser Lys Asp Arg
930 935 940
Lys Leu Val Thr Thr His Ala Asp Ile Asn Ala Ala Gln Asn Leu Gln
945 950 955 960
Lys Arg Phe Trp Thr Arg Thr His Gly Phe Tyr Lys Val Tyr Cys Lys
965 970 975
Ala Tyr Gln Val Asp Gly Gln Thr Val Tyr Ile Pro Glu Ser Lys Asp
980 985 990
Gln Lys Gln Lys Ile Ile Glu Glu Phe Gly Glu Gly Tyr Phe Ile Leu
995 1000 1005
Lys Asp Gly Val Tyr Glu Trp Gly Asn Ala Gly Lys Leu Lys Ile
1010 1015 1020
Lys Lys Gly Ser Ser Lys Gln Ser Ser Ser Glu Leu Val Asp Ser
1025 1030 1035
Asp Ile Leu Lys Asp Ser Phe Asp Leu Ala Ser Glu Leu Lys Gly
1040 1045 1050
Glu Lys Leu Met Leu Tyr Arg Asp Pro Ser Gly Asn Val Phe Pro
1055 1060 1065
Ser Asp Lys Trp Met Ala Ala Gly Val Phe Phe Gly Lys Leu Glu
1070 1075 1080
Arg Ile Leu Ile Ser Lys Leu Thr Asn Gln Tyr Ser Ile Ser Thr
1085 1090 1095
Ile Glu Asp Asp Ser Ser Lys Gln Ser Met
1100 1105
<210> 555
<211> 864
<212> PRT
<213> Brevibacillus sp.
<400> 555
Met Pro Lys Ile Leu Arg Gly His Lys Trp Ile Ser Leu Leu Glu Gln
1 5 10 15
Tyr Glu Glu Asn Arg Glu Arg Glu Leu Arg Glu Asn Met Thr Ala Ala
20 25 30
Asn Asp Lys Tyr Arg Ile Thr Lys Arg Gln Met Lys Gly Trp Asn Glu
35 40 45
Leu Tyr Glu Leu Trp Ser Thr Phe Pro Ala Ser Ala Ser His Glu Gln
50 55 60
Tyr Lys Glu Ala Leu Lys Arg Val Gln Gln Arg Leu Arg Gly Arg Phe
65 70 75 80
Gly Asp Ala His Phe Phe Gln Tyr Leu Met Glu Glu Lys Asn Arg Leu
85 90 95
Ile Trp Lys Gly Asn Pro Gln Arg Ile His Tyr Phe Val Ala Arg Asn
100 105 110
Glu Leu Thr Lys Arg Leu Glu Glu Ala Lys Gln Ser Ala Thr Met Thr
115 120 125
Leu Pro Asn Ala Arg Lys His Pro Leu Trp Val Arg Phe Asp Ala Arg
130 135 140
Gly Gly Asn Leu Gln Asp Tyr Tyr Leu Thr Ala Glu Ala Asp Lys Pro
145 150 155 160
Arg Ser Arg Arg Phe Val Thr Phe Ser Gln Leu Ile Trp Pro Ser Glu
165 170 175
Ser Gly Trp Met Glu Lys Lys Asp Val Glu Val Glu Leu Ala Leu Ser
180 185 190
Arg Gln Phe Tyr Gln Gln Val Lys Leu Leu Lys Asn Asp Lys Gly Lys
195 200 205
Gln Lys Ile Glu Phe Lys Asp Lys Gly Ser Gly Ser Thr Phe Asn Gly
210 215 220
His Leu Gly Gly Ala Lys Leu Gln Leu Glu Arg Gly Asp Leu Glu Lys
225 230 235 240
Glu Glu Lys Asn Phe Glu Asp Gly Glu Ile Gly Ser Val Tyr Leu Asn
245 250 255
Val Val Ile Asp Phe Glu Pro Leu Gln Glu Val Lys Asn Gly Arg Val
260 265 270
Gln Ala Pro Tyr Gly Gln Val Leu Gln Leu Ile Arg Arg Pro Asn Glu
275 280 285
Phe Pro Lys Val Thr Thr Tyr Lys Ser Glu Gln Leu Val Glu Trp Ile
290 295 300
Lys Ala Ser Pro Gln His Ser Ala Gly Val Glu Ser Leu Ala Ser Gly
305 310 315 320
Phe Arg Val Met Ser Ile Asp Leu Gly Leu Arg Ala Ala Ala Ala Thr
325 330 335
Ser Ile Phe Ser Val Glu Glu Ser Ser Asp Lys Asn Ala Ala Asp Phe
340 345 350
Ser Tyr Trp Ile Glu Gly Thr Pro Leu Val Ala Val His His Arg Ser
355 360 365
Tyr Met Leu Arg Leu Pro Gly Glu Gln Val Glu Lys Gln Val Met Glu
370 375 380
Lys Arg Asp Glu Arg Phe Gln Leu His Gln Arg Val Lys Phe Gln Ile
385 390 395 400
Arg Val Leu Ala Gln Ile Met Arg Met Ala Asn Lys Gln Tyr Gly Asp
405 410 415
Arg Trp Asp Glu Leu Asp Ser Leu Lys Gln Ala Val Glu Gln Lys Lys
420 425 430
Ser Pro Leu Asp Gln Thr Asp Arg Thr Phe Trp Glu Gly Ile Val Cys
435 440 445
Asp Leu Thr Lys Val Leu Pro Arg Asn Glu Ala Asp Trp Glu Gln Ala
450 455 460
Val Val Gln Ile His Arg Lys Ala Glu Glu Tyr Val Gly Lys Ala Val
465 470 475 480
Gln Ala Trp Arg Lys Arg Phe Ala Ala Asp Glu Arg Lys Gly Ile Ala
485 490 495
Gly Leu Ser Met Trp Asn Ile Glu Glu Leu Glu Gly Leu Arg Lys Leu
500 505 510
Leu Ile Ser Trp Ser Arg Arg Ser Arg Asn Pro Gln Glu Val Asn Arg
515 520 525
Phe Glu Arg Gly His Thr Ser His Gln Arg Leu Leu Thr His Ile Gln
530 535 540
Asn Val Lys Glu Asp Arg Leu Lys Gln Leu Ser His Ala Ile Val Met
545 550 555 560
Thr Ala Leu Gly Tyr Val Tyr Asp Glu Arg Lys Gln Glu Trp Cys Ala
565 570 575
Glu Tyr Pro Ala Cys Gln Val Ile Leu Phe Glu Asn Leu Ser Gln Tyr
580 585 590
Arg Ser Asn Leu Asp Arg Ser Thr Lys Glu Asn Ser Thr Leu Met Lys
595 600 605
Trp Ala His Arg Ser Ile Pro Lys Tyr Val His Met Gln Ala Glu Pro
610 615 620
Tyr Gly Ile Gln Ile Gly Asp Val Arg Ala Glu Tyr Ser Ser Arg Phe
625 630 635 640
Tyr Ala Lys Thr Gly Thr Pro Gly Ile Arg Cys Lys Lys Val Arg Gly
645 650 655
Gln Asp Leu Gln Gly Arg Arg Phe Glu Asn Leu Gln Lys Arg Leu Val
660 665 670
Asn Glu Gln Phe Leu Thr Glu Glu Gln Val Lys Gln Leu Arg Pro Gly
675 680 685
Asp Ile Val Pro Asp Asp Ser Gly Glu Leu Phe Met Thr Leu Thr Asp
690 695 700
Gly Ser Gly Ser Lys Glu Val Val Phe Leu Gln Ala Asp Ile Asn Ala
705 710 715 720
Ala His Asn Leu Gln Lys Arg Phe Trp Gln Arg Tyr Asn Glu Leu Phe
725 730 735
Lys Val Ser Cys Arg Val Ile Val Arg Asp Glu Glu Glu Tyr Leu Val
740 745 750
Pro Lys Thr Lys Ser Val Gln Ala Lys Leu Gly Lys Gly Leu Phe Val
755 760 765
Lys Lys Ser Asp Thr Ala Trp Lys Asp Val Tyr Val Trp Asp Ser Gln
770 775 780
Ala Lys Leu Lys Gly Lys Thr Thr Phe Thr Glu Glu Ser Glu Ser Pro
785 790 795 800
Glu Gln Leu Glu Asp Phe Gln Glu Ile Ile Glu Glu Ala Glu Glu Ala
805 810 815
Lys Gly Thr Tyr Arg Thr Leu Phe Arg Asp Pro Ser Gly Val Phe Phe
820 825 830
Pro Glu Ser Val Trp Tyr Pro Gln Lys Asp Phe Trp Gly Glu Val Lys
835 840 845
Arg Lys Leu Tyr Gly Lys Leu Arg Glu Arg Phe Leu Thr Lys Ala Arg
850 855 860
<210> 556
<211> 1108
<212> PRT
<213> Bacillus sp.
<400> 556
Met Ala Ile Arg Ser Ile Lys Leu Lys Leu Lys Thr His Thr Gly Pro
1 5 10 15
Glu Ala Gln Asn Leu Arg Lys Gly Ile Trp Arg Thr His Arg Leu Leu
20 25 30
Asn Glu Gly Val Ala Tyr Tyr Met Lys Met Leu Leu Leu Phe Arg Gln
35 40 45
Glu Ser Thr Gly Glu Arg Pro Lys Glu Glu Leu Gln Glu Glu Leu Ile
50 55 60
Cys His Ile Arg Glu Gln Gln Gln Arg Asn Gln Ala Asp Lys Asn Thr
65 70 75 80
Gln Ala Leu Pro Leu Asp Lys Ala Leu Glu Ala Leu Arg Gln Leu Tyr
85 90 95
Glu Leu Leu Val Pro Ser Ser Val Gly Gln Ser Gly Asp Ala Gln Ile
100 105 110
Ile Ser Arg Lys Phe Leu Ser Pro Leu Val Asp Pro Asn Ser Glu Gly
115 120 125
Gly Lys Gly Thr Ser Lys Ala Gly Ala Lys Pro Thr Trp Gln Lys Lys
130 135 140
Lys Glu Ala Asn Asp Pro Thr Trp Glu Gln Asp Tyr Glu Lys Trp Lys
145 150 155 160
Lys Arg Arg Glu Glu Asp Pro Thr Ala Ser Val Ile Thr Thr Leu Glu
165 170 175
Glu Tyr Gly Ile Arg Pro Ile Phe Pro Leu Tyr Thr Asn Thr Val Thr
180 185 190
Asp Ile Ala Trp Leu Pro Leu Gln Ser Asn Gln Phe Val Arg Thr Trp
195 200 205
Asp Arg Asp Met Leu Gln Gln Ala Ile Glu Arg Leu Leu Ser Trp Glu
210 215 220
Ser Trp Asn Lys Arg Val Gln Glu Glu Tyr Ala Lys Leu Lys Glu Lys
225 230 235 240
Met Ala Gln Leu Asn Glu Gln Leu Glu Gly Gly Gln Glu Trp Ile Ser
245 250 255
Leu Leu Glu Gln Tyr Glu Glu Asn Arg Glu Arg Glu Leu Arg Glu Asn
260 265 270
Met Thr Ala Ala Asn Asp Lys Tyr Arg Ile Thr Lys Arg Gln Met Lys
275 280 285
Gly Trp Asn Glu Leu Tyr Glu Leu Trp Ser Thr Phe Pro Ala Ser Ala
290 295 300
Ser His Glu Gln Tyr Lys Glu Ala Leu Lys Arg Val Gln Gln Arg Leu
305 310 315 320
Arg Gly Arg Phe Gly Asp Ala His Phe Phe Gln Tyr Leu Met Glu Glu
325 330 335
Lys Asn Arg Leu Ile Trp Lys Gly Asn Pro Gln Arg Ile His Tyr Phe
340 345 350
Val Ala Arg Asn Glu Leu Thr Lys Arg Leu Glu Glu Ala Lys Gln Ser
355 360 365
Ala Thr Met Thr Leu Pro Asn Ala Arg Lys His Pro Leu Trp Val Arg
370 375 380
Phe Asp Ala Arg Gly Gly Asn Leu Gln Asp Tyr Tyr Leu Thr Ala Glu
385 390 395 400
Ala Asp Lys Pro Arg Ser Arg Arg Phe Val Thr Phe Ser Gln Leu Ile
405 410 415
Trp Pro Ser Glu Ser Gly Trp Met Glu Lys Lys Asp Val Glu Val Glu
420 425 430
Leu Ala Leu Ser Arg Gln Phe Tyr Gln Gln Val Lys Leu Leu Lys Asn
435 440 445
Asp Lys Gly Lys Gln Lys Ile Glu Phe Lys Asp Lys Gly Ser Gly Ser
450 455 460
Thr Phe Asn Gly His Leu Gly Gly Ala Lys Leu Gln Leu Glu Arg Gly
465 470 475 480
Asp Leu Glu Lys Glu Glu Lys Asn Phe Glu Asp Gly Glu Ile Gly Ser
485 490 495
Val Tyr Leu Asn Val Val Ile Asp Phe Glu Pro Leu Gln Glu Val Lys
500 505 510
Asn Gly Arg Val Gln Ala Pro Tyr Gly Gln Val Leu Gln Leu Ile Arg
515 520 525
Arg Pro Asn Glu Phe Pro Lys Val Thr Thr Tyr Lys Ser Glu Gln Leu
530 535 540
Val Glu Trp Ile Lys Ala Ser Pro Gln His Ser Ala Gly Val Glu Ser
545 550 555 560
Leu Ala Ser Gly Phe Arg Val Met Ser Ile Asp Leu Gly Leu Arg Ala
565 570 575
Ala Ala Ala Thr Ser Ile Phe Ser Val Glu Glu Ser Ser Asp Lys Asn
580 585 590
Ala Ala Asp Phe Ser Tyr Trp Ile Glu Gly Thr Pro Leu Val Ala Val
595 600 605
His Gln Arg Ser Tyr Met Leu Arg Leu Pro Gly Glu Gln Val Glu Lys
610 615 620
Gln Val Met Glu Lys Arg Asp Glu Arg Phe Gln Leu His Gln Arg Val
625 630 635 640
Lys Phe Gln Ile Arg Val Leu Ala Gln Ile Met Arg Met Ala Asn Lys
645 650 655
Gln Tyr Gly Asp Arg Trp Asp Glu Leu Asp Ser Leu Lys Gln Ala Val
660 665 670
Glu Gln Lys Lys Ser Pro Leu Asp Gln Thr Asp Arg Thr Phe Trp Glu
675 680 685
Gly Ile Val Cys Asp Leu Thr Lys Val Leu Pro Arg Asn Glu Ala Asp
690 695 700
Trp Glu Gln Ala Val Val Gln Ile His Arg Lys Ala Glu Glu Tyr Val
705 710 715 720
Gly Lys Ala Val Gln Ala Trp Arg Lys Arg Phe Ala Ala Asp Glu Arg
725 730 735
Lys Gly Ile Ala Gly Leu Ser Met Trp Asn Ile Glu Glu Leu Glu Gly
740 745 750
Leu Arg Lys Leu Leu Ile Ser Trp Ser Arg Arg Thr Arg Asn Pro Gln
755 760 765
Glu Val Asn Arg Phe Glu Arg Gly His Thr Ser His Gln Arg Leu Leu
770 775 780
Thr His Ile Gln Asn Val Lys Glu Asp Arg Leu Lys Gln Leu Ser His
785 790 795 800
Ala Ile Val Met Thr Ala Leu Gly Tyr Val Tyr Asp Glu Arg Lys Gln
805 810 815
Glu Trp Cys Ala Glu Tyr Pro Ala Cys Gln Val Ile Leu Phe Glu Asn
820 825 830
Leu Ser Gln Tyr Arg Ser Asn Leu Asp Arg Ser Thr Lys Glu Asn Ser
835 840 845
Thr Leu Met Lys Trp Ala His Arg Ser Ile Pro Lys Tyr Val His Met
850 855 860
Gln Ala Glu Pro Tyr Gly Ile Gln Ile Gly Asp Val Arg Ala Glu Tyr
865 870 875 880
Ser Ser Arg Phe Tyr Ala Lys Thr Gly Thr Pro Gly Ile Arg Cys Lys
885 890 895
Lys Val Arg Gly Gln Asp Leu Gln Gly Arg Arg Phe Glu Asn Leu Gln
900 905 910
Lys Arg Leu Val Asn Glu Gln Phe Leu Thr Glu Glu Gln Val Lys Gln
915 920 925
Leu Arg Pro Gly Asp Ile Val Pro Asp Asp Ser Gly Glu Leu Phe Met
930 935 940
Thr Leu Thr Asp Gly Ser Gly Ser Lys Glu Val Val Phe Leu Gln Ala
945 950 955 960
Asp Ile Asn Ala Ala His Asn Leu Gln Lys Arg Phe Trp Gln Arg Tyr
965 970 975
Asn Glu Leu Phe Lys Val Ser Cys Arg Val Ile Val Arg Asp Glu Glu
980 985 990
Glu Tyr Leu Val Pro Lys Thr Lys Ser Val Gln Ala Lys Leu Gly Lys
995 1000 1005
Gly Leu Phe Val Lys Lys Ser Asp Thr Ala Trp Lys Asp Val Tyr
1010 1015 1020
Val Trp Asp Ser Gln Ala Lys Leu Lys Gly Lys Thr Thr Phe Thr
1025 1030 1035
Glu Glu Ser Glu Ser Pro Glu Gln Leu Glu Asp Phe Gln Glu Ile
1040 1045 1050
Ile Glu Glu Ala Glu Glu Ala Lys Gly Thr Tyr Arg Thr Leu Phe
1055 1060 1065
Arg Asp Pro Ser Gly Val Phe Phe Pro Glu Ser Val Trp Tyr Pro
1070 1075 1080
Gln Lys Asp Phe Trp Gly Glu Val Lys Arg Lys Leu Tyr Gly Lys
1085 1090 1095
Leu Arg Glu Arg Phe Leu Thr Lys Ala Arg
1100 1105
<210> 557
<211> 1489
<212> PRT
<213> Desulfatirhabdium butyrativorans
<400> 557
Met Pro Leu Ser Asn Asn Pro Pro Val Thr Gln Arg Ala Tyr Thr Leu
1 5 10 15
Arg Leu Arg Gly Ala Asp Pro Ser Asp Leu Ser Trp Arg Glu Ala Leu
20 25 30
Trp His Thr His Glu Ala Val Asn Lys Gly Ala Lys Val Phe Gly Asp
35 40 45
Trp Leu Leu Thr Leu Arg Gly Gly Leu Asp His Thr Leu Ala Asp Thr
50 55 60
Lys Val Lys Gly Gly Lys Gly Lys Pro Asp Arg Asp Pro Thr Pro Glu
65 70 75 80
Glu Arg Lys Ala Arg Arg Ile Leu Leu Ala Leu Ser Trp Leu Ser Val
85 90 95
Glu Ser Lys Leu Gly Ala Pro Ser Ser Tyr Ile Val Ala Ser Gly Asp
100 105 110
Glu Pro Ala Lys Asp Arg Asn Asp Asn Val Val Ser Ala Leu Glu Glu
115 120 125
Ile Leu Gln Ser Arg Lys Val Ala Lys Ser Glu Ile Asp Asp Trp Lys
130 135 140
Arg Asp Cys Ser Ala Ser Leu Ser Ala Ala Ile Arg Asp Asp Ala Val
145 150 155 160
Trp Val Asn Arg Ser Lys Val Phe Asp Glu Ala Val Lys Ser Val Gly
165 170 175
Ser Ser Leu Thr Arg Glu Glu Ala Trp Asp Met Leu Glu Arg Phe Phe
180 185 190
Gly Ser Arg Asp Ala Tyr Leu Thr Pro Met Lys Asp Pro Glu Asp Lys
195 200 205
Ser Ser Glu Thr Glu Gln Glu Asp Lys Ala Lys Asp Leu Val Gln Lys
210 215 220
Ala Gly Gln Trp Leu Ser Ser Arg Tyr Gly Thr Ser Glu Gly Ala Asp
225 230 235 240
Phe Cys Arg Met Ser Asp Ile Tyr Gly Lys Ile Ala Ala Trp Ala Asp
245 250 255
Asn Ala Ser Gln Gly Gly Ser Ser Thr Val Asp Asp Leu Val Ser Glu
260 265 270
Leu Arg Gln His Phe Asp Thr Lys Glu Ser Lys Ala Thr Asn Gly Leu
275 280 285
Asp Trp Ile Ile Gly Leu Ser Ser Tyr Thr Gly His Thr Pro Asn Pro
290 295 300
Val His Glu Leu Leu Arg Gln Asn Thr Ser Leu Asn Lys Ser His Leu
305 310 315 320
Asp Asp Leu Lys Lys Lys Ala Asn Thr Arg Ala Glu Ser Cys Lys Ser
325 330 335
Lys Ile Gly Ser Lys Gly Gln Arg Pro Tyr Ser Asp Ala Ile Leu Asn
340 345 350
Asp Val Glu Ser Val Cys Gly Phe Thr Tyr Arg Val Asp Lys Asp Gly
355 360 365
Gln Pro Val Ser Val Ala Asp Tyr Ser Lys Tyr Asp Val Asp Tyr Lys
370 375 380
Trp Gly Thr Ala Arg His Tyr Ile Phe Ala Val Met Leu Asp His Ala
385 390 395 400
Ala Arg Arg Ile Ser Leu Ala His Lys Trp Ile Lys Arg Ala Glu Ala
405 410 415
Glu Arg His Lys Phe Glu Glu Asp Ala Lys Arg Ile Ala Asn Val Pro
420 425 430
Ala Arg Ala Arg Glu Trp Leu Asp Ser Phe Cys Lys Glu Arg Ser Val
435 440 445
Thr Ser Gly Ala Val Glu Pro Tyr Arg Ile Arg Arg Arg Ala Val Asp
450 455 460
Gly Trp Lys Glu Val Val Ala Ala Trp Ser Lys Ser Asp Cys Lys Ser
465 470 475 480
Thr Glu Asp Arg Ile Ala Ala Ala Arg Ala Leu Gln Asp Asp Ser Glu
485 490 495
Ile Asp Lys Phe Gly Asp Ile Gln Leu Phe Glu Ala Leu Ala Glu Asp
500 505 510
Asp Ala Leu Cys Val Trp His Lys Asp Gly Glu Ala Thr Asn Glu Pro
515 520 525
Asp Phe Gln Pro Leu Ile Asp Tyr Ser Leu Ala Ile Glu Ala Glu Phe
530 535 540
Lys Lys Arg Gln Phe Lys Val Pro Ala Tyr Arg His Pro Asp Glu Leu
545 550 555 560
Leu His Pro Val Phe Cys Asp Phe Gly Lys Ser Arg Trp Lys Ile Asn
565 570 575
Tyr Asp Val His Lys Asn Val Gln Ala Pro Phe Tyr Arg Gly Leu Cys
580 585 590
Leu Thr Leu Trp Thr Gly Ser Glu Ile Lys Pro Val Pro Leu Cys Trp
595 600 605
Gln Ser Lys Arg Leu Thr Arg Asp Leu Ala Leu Gly Asn Asn His Arg
610 615 620
Asn Asp Ala Ala Ser Ala Val Thr Arg Ala Asp Arg Leu Gly Arg Ala
625 630 635 640
Ala Ser Asn Val Thr Lys Ser Asp Met Val Asn Ile Thr Gly Leu Phe
645 650 655
Glu Gln Ala Asp Trp Asn Gly Arg Leu Gln Ala Pro Arg Gln Gln Leu
660 665 670
Glu Ala Ile Ala Val Val Arg Asp Asn Pro Arg Leu Ser Glu Gln Glu
675 680 685
Arg Asn Leu Arg Met Cys Gly Met Ile Glu His Ile Arg Trp Leu Val
690 695 700
Thr Phe Ser Val Lys Leu Gln Pro Gln Gly Pro Trp Cys Ala Tyr Ala
705 710 715 720
Glu Gln His Gly Leu Asn Thr Asn Pro Gln Tyr Trp Pro His Ala Asp
725 730 735
Thr Asn Arg Asp Arg Lys Val His Ala Arg Leu Ile Leu Pro Arg Leu
740 745 750
Pro Gly Leu Arg Val Leu Ser Val Asp Leu Gly His Arg Tyr Ala Ala
755 760 765
Ala Cys Ala Val Trp Glu Ala Val Asn Thr Glu Thr Val Lys Glu Ala
770 775 780
Cys Gln Asn Val Gly Arg Asp Met Pro Lys Glu His Asp Leu Tyr Leu
785 790 795 800
His Ile Lys Val Lys Lys Gln Gly Ile Gly Lys Gln Thr Glu Val Asp
805 810 815
Lys Thr Thr Ile Tyr Arg Arg Ile Gly Ala Asp Thr Leu Pro Asp Gly
820 825 830
Arg Pro His Pro Ala Pro Trp Ala Arg Leu Asp Arg Gln Phe Leu Ile
835 840 845
Lys Leu Gln Gly Glu Glu Lys Asp Ala Arg Glu Ala Ser Asn Glu Glu
850 855 860
Ile Trp Ala Leu His Gln Met Glu Cys Lys Leu Asp Arg Thr Lys Pro
865 870 875 880
Leu Ile Asp Arg Leu Ile Ala Ser Gly Trp Gly Leu Leu Lys Arg Gln
885 890 895
Met Ala Arg Leu Asp Ala Leu Lys Glu Leu Gly Trp Ile Pro Ala Pro
900 905 910
Asp Ser Ser Glu Asn Leu Ser Arg Glu Asp Gly Glu Ala Lys Asp Tyr
915 920 925
Arg Glu Ser Leu Ala Val Asp Asp Leu Met Phe Ser Ala Val Arg Thr
930 935 940
Leu Arg Leu Ala Leu Gln Arg His Gly Asn Arg Ala Arg Ile Ala Tyr
945 950 955 960
Tyr Leu Ile Ser Glu Val Lys Ile Arg Pro Gly Gly Ile Gln Glu Lys
965 970 975
Leu Asp Glu Asn Gly Arg Ile Asp Leu Leu Gln Asp Ala Leu Ala Leu
980 985 990
Trp His Glu Leu Phe Ser Ser Pro Gly Trp Arg Asp Glu Ala Ala Lys
995 1000 1005
Gln Leu Trp Asp Ser Arg Ile Ala Thr Leu Ala Gly Tyr Lys Ala
1010 1015 1020
Pro Glu Glu Asn Gly Asp Asn Val Ser Asp Val Ala Tyr Arg Lys
1025 1030 1035
Lys Gln Gln Val Tyr Arg Glu Gln Leu Arg Asn Val Ala Lys Thr
1040 1045 1050
Leu Ser Gly Asp Val Ile Thr Cys Lys Glu Leu Ser Asp Ala Trp
1055 1060 1065
Lys Glu Arg Trp Glu Asp Glu Asp Gln Arg Trp Lys Lys Leu Leu
1070 1075 1080
Arg Trp Phe Lys Asp Trp Val Leu Pro Ser Gly Thr Gln Ala Asn
1085 1090 1095
Asn Ala Thr Ile Arg Asn Val Gly Gly Leu Ser Leu Ser Arg Leu
1100 1105 1110
Ala Thr Ile Thr Glu Phe Arg Arg Lys Val Gln Val Gly Phe Phe
1115 1120 1125
Thr Arg Leu Arg Pro Asp Gly Thr Arg His Glu Ile Gly Glu Gln
1130 1135 1140
Phe Gly Gln Lys Thr Leu Asp Ala Leu Glu Leu Leu Arg Glu Gln
1145 1150 1155
Arg Val Lys Gln Leu Ala Ser Arg Ile Ala Glu Ala Ala Leu Gly
1160 1165 1170
Ile Gly Ser Glu Gly Gly Lys Gly Trp Asp Gly Gly Lys Arg Pro
1175 1180 1185
Arg Gln Arg Ile Asn Asp Ser Arg Phe Ala Pro Cys His Ala Val
1190 1195 1200
Val Ile Glu Asn Leu Ala Asn Tyr Arg Pro Asp Glu Thr Arg Thr
1205 1210 1215
Arg Leu Glu Asn Arg Arg Leu Met Thr Trp Ser Ala Ser Lys Val
1220 1225 1230
His Lys Tyr Leu Ser Glu Ala Cys Gln Leu Asn Gly Leu Tyr Leu
1235 1240 1245
Cys Thr Val Ser Ala Trp Tyr Thr Ser Arg Gln Asp Ser Arg Thr
1250 1255 1260
Gly Ala Pro Gly Ile Arg Cys Gln Asp Val Ser Val Arg Glu Phe
1265 1270 1275
Met Gln Ser Pro Phe Trp Arg Lys Gln Val Lys Gln Ala Glu Ala
1280 1285 1290
Lys His Asp Glu Asn Lys Gly Asp Ala Arg Glu Arg Phe Leu Cys
1295 1300 1305
Glu Leu Asn Lys Thr Trp Lys Ala Lys Thr Pro Ala Glu Trp Lys
1310 1315 1320
Lys Ala Gly Phe Val Arg Ile Pro Leu Arg Gly Gly Glu Ile Phe
1325 1330 1335
Val Ser Ala Asp Ser Lys Ser Pro Ser Ala Lys Gly Ile His Ala
1340 1345 1350
Asp Leu Asn Ala Ala Ala Asn Ile Gly Leu Arg Ala Leu Thr Asp
1355 1360 1365
Pro Asp Trp Pro Gly Lys Trp Trp Tyr Val Pro Cys Asp Pro Val
1370 1375 1380
Ser Phe Glu Ser Lys Met Asp Tyr Val Lys Gly Cys Ala Ala Val
1385 1390 1395
Lys Val Gly Gln Pro Leu Arg Gln Pro Ala Gln Thr Asn Ala Asp
1400 1405 1410
Gly Ala Ala Ser Lys Ile Arg Lys Gly Lys Lys Asn Arg Thr Ala
1415 1420 1425
Gly Thr Ser Lys Glu Lys Val Tyr Leu Trp Arg Asp Ile Ser Ala
1430 1435 1440
Phe Pro Leu Glu Ser Asn Glu Ile Gly Glu Trp Lys Glu Thr Ser
1445 1450 1455
Ala Tyr Gln Asn Asp Val Gln Tyr Arg Val Ile Arg Met Leu Lys
1460 1465 1470
Glu His Ile Lys Ser Leu Asp Asn Arg Thr Gly Asp Asn Val Glu
1475 1480 1485
Gly
<210> 558
<211> 277
<212> PRT
<213> Alicyclobacillus herbarius
<400> 558
Met Leu Lys Gln Ala Val Leu Gly Asn Gly Pro Leu Ile Asn Trp Glu
1 5 10 15
Lys Asn Val Lys Arg Gly Lys Gly Met Ala Thr Lys Ser Ile Lys Val
20 25 30
Lys Leu Arg Leu Gly Lys His Pro Asp Ile Arg Ala Gly Ile Trp Gln
35 40 45
Leu His Lys Ala Ala Asn Ala Gly Val Arg Tyr Tyr Thr Glu Trp Leu
50 55 60
Ser Leu Met Arg Gln Lys Asn Leu Tyr Thr Arg Gly Pro Lys Gly Glu
65 70 75 80
Gln Gln Leu Tyr Arg Ser Gly Glu Gln Cys Arg Arg Glu Leu Leu Gln
85 90 95
Arg Leu Arg Glu Arg Gln Arg Leu Asn Gly Arg Thr Asp Glu Pro Gly
100 105 110
Thr Asp Glu Glu Leu Leu Lys Val Ala Arg Gln Ile Tyr Glu Val Leu
115 120 125
Val Pro Gln Ser Ile Gly Lys Ser Gly Asp Ala Gln Gln Leu Ala Ser
130 135 140
Asn Phe Leu Ser Pro Leu Val Asp Pro Asn Ser Lys Gly Gly Gln Gly
145 150 155 160
Gln Ser Asn Ala Gly Arg Lys Pro Ala Trp Gln Lys Met Arg Asp Glu
165 170 175
Gly Asn Pro Gly Trp Val Ala Ala Lys Glu Arg Tyr Glu Gln Arg Lys
180 185 190
Ala Thr Asp Pro Thr Lys Lys Met Ile Glu Met Leu Asp Gly Leu Gly
195 200 205
Leu Lys Pro Leu Phe Ser Val Phe Thr Glu Thr Tyr Thr Thr Gly Val
210 215 220
Lys Trp Lys Asp Leu Ser Lys Arg Gln Gly Val Arg Thr Trp Asp Arg
225 230 235 240
Asp Met Phe Gln Ser Leu Ser Glu Arg Ser Gly Val Ile Asp Val Gly
245 250 255
Ser His Thr Val His His Ile Asp Leu Ala Thr Ala Ser Asp Ala Gln
260 265 270
Ile Gln Tyr Glu Leu
275
<210> 559
<211> 277
<212> PRT
<213> Alicyclobacillus contaminans
<400> 559
Met Ser Val Lys Ser Ile Lys Phe Lys Leu Met Ile Gly Gly Pro Gln
1 5 10 15
Tyr Thr Arg Ile Arg Arg Gly Ile Tyr Lys Thr His Glu Val Phe Asn
20 25 30
Glu Gly Val Arg Tyr Tyr Gln Glu Trp Leu Leu Leu Met Arg Gln Gly
35 40 45
Asp Val Tyr Arg Tyr Gln Asp Asp Lys Pro Glu Ile Val Leu Ser Ala
50 55 60
Glu His Cys Lys Arg Glu Leu Leu Arg Arg Leu Arg Gln Val Gln Lys
65 70 75 80
Glu Asn Val Gly Arg Thr Ser His Thr Asp Glu Glu Leu Leu Gln Val
85 90 95
Met Arg Ala Leu Tyr Glu Leu Ile Val Pro Ser Ala Val Gly Lys Lys
100 105 110
Gly Asp Ala Ala Ser Leu Ser Arg Lys Phe Leu Ser Pro Leu Ala Trp
115 120 125
Lys Asp Ser Lys Gly Leu Thr Gly Glu Ser Lys Ala Gly Asn Lys Pro
130 135 140
Arg Trp Lys Arg Leu Gln Glu Gln Gly Leu Pro Tyr Glu Glu Glu Tyr
145 150 155 160
Asn Arg Trp Leu Arg Glu Lys Glu Ser Asp Pro Ala Lys His Ile Pro
165 170 175
Ala Gln Leu Ala Ser Met Gly Leu Lys Pro Phe Leu Lys Val Phe Thr
180 185 190
Glu Ser Thr Glu Gly Ile Ala Trp Leu Pro Leu Ala Lys Asp Gln Gly
195 200 205
Val Arg Thr Trp Asp Arg Asp Met Phe Gln Gln Ala Ile Glu Gly Leu
210 215 220
Leu Ser Trp Glu Ser Trp Asn Arg Arg Val Arg Glu Glu Tyr Asp Ala
225 230 235 240
Leu Ser Ala Arg Val Tyr Ala Tyr His Ala Lys His Phe Ala Asp Gln
245 250 255
Pro Gly Trp Ala Val Tyr Trp Pro Gln Ser Gln Pro Arg Gln Lys Gly
260 265 270
Trp Val Lys Met Lys
275
<210> 560
<211> 218
<212> PRT
<213> Citrobacter freundii
<400> 560
Met Tyr Glu Leu Trp Ser Thr Phe Pro Ala Ser Ala Ser His Glu Gln
1 5 10 15
Tyr Lys Glu Ala Leu Lys Arg Val Gln Gln Arg Leu Arg Gly Arg Phe
20 25 30
Gly Asp Ala His Phe Phe Gln Tyr Leu Met Glu Glu Lys Asn Arg Leu
35 40 45
Ile Trp Lys Gly Asn Pro Gln Arg Ile His Tyr Phe Val Ala Arg Asn
50 55 60
Glu Leu Thr Lys Arg Leu Glu Glu Ala Lys Gln Ser Ala Thr Met Thr
65 70 75 80
Leu Pro Asn Ala Arg Lys His Pro Leu Trp Val Arg Phe Asp Ala Arg
85 90 95
Gly Gly Asn Leu Gln Asp Tyr Tyr Leu Thr Ala Glu Ala Asp Lys Pro
100 105 110
Arg Ser Arg Arg Phe Val Thr Phe Ser Gln Leu Ile Trp Pro Ser Glu
115 120 125
Ser Gly Trp Met Glu Lys Lys Asp Val Glu Val Glu Leu Ala Leu Ser
130 135 140
Arg Gln Phe Tyr Gln Gln Val Lys Leu Leu Lys Asn Asp Lys Gly Lys
145 150 155 160
Gln Lys Ile Glu Phe Lys Asp Lys Gly Ser Gly Ser Thr Phe Asn Gly
165 170 175
His Leu Gly Gly Ala Lys Leu Gln Leu Glu Arg Gly Asp Leu Glu Lys
180 185 190
Glu Glu Lys Thr Ser Arg Thr Gly Lys Ser Ala Ala Phe Thr Leu Thr
195 200 205
Leu Ser Leu Ile Ser Asn Leu Cys Lys Lys
210 215
<210> 561
<211> 218
<212> PRT
<213> Citrobacter freundii
<400> 561
Leu Tyr Glu Leu Trp Ser Thr Phe Pro Ala Ser Ala Ser His Glu Gln
1 5 10 15
Tyr Lys Glu Ala Leu Lys Arg Val Gln Gln Arg Leu Arg Gly Arg Phe
20 25 30
Gly Asp Ala His Phe Phe Gln Tyr Leu Met Glu Glu Lys Asn Arg Leu
35 40 45
Ile Trp Lys Gly Asn Pro Gln Arg Ile His Tyr Phe Val Ala Arg Asn
50 55 60
Glu Leu Thr Lys Arg Leu Glu Glu Ala Lys Gln Ser Ala Thr Met Thr
65 70 75 80
Leu Pro Asn Ala Arg Lys His Pro Leu Trp Val Arg Phe Asp Ala Arg
85 90 95
Gly Gly Asn Leu Gln Asp Tyr Tyr Leu Thr Ala Glu Ala Asp Lys Pro
100 105 110
Arg Ser Arg Arg Phe Val Thr Phe Ser Gln Leu Ile Trp Pro Ser Glu
115 120 125
Ser Gly Trp Met Glu Lys Lys Asp Val Glu Val Glu Leu Ala Leu Ser
130 135 140
Arg Gln Phe Tyr Gln Gln Val Lys Leu Leu Lys Asn Asp Lys Gly Lys
145 150 155 160
Gln Lys Ile Glu Phe Lys Asp Lys Gly Ser Gly Ser Thr Phe Asn Gly
165 170 175
His Leu Gly Gly Ala Lys Leu Gln Leu Glu Arg Gly Asp Leu Glu Lys
180 185 190
Glu Glu Lys Thr Ser Arg Thr Gly Lys Ser Ala Ala Phe Thr Leu Thr
195 200 205
Leu Ser Leu Ile Ser Asn Leu Cys Lys Lys
210 215
<210> 562
<211> 482
<212> PRT
<213> Brevibacillus agri
<400> 562
Met Glu Lys Arg Asp Glu Arg Phe Gln Leu His Gln Arg Val Lys Phe
1 5 10 15
Gln Ile Arg Val Leu Ala Gln Ile Met Arg Met Ala Asn Lys Gln Tyr
20 25 30
Gly Asp Arg Trp Asp Glu Leu Asp Ser Leu Lys Gln Ala Val Glu Gln
35 40 45
Lys Lys Ser Pro Leu Asp Gln Thr Asp Arg Thr Phe Trp Glu Gly Ile
50 55 60
Val Cys Asp Leu Thr Lys Val Leu Pro Arg Asn Glu Ala Asp Trp Glu
65 70 75 80
Gln Ala Val Val Gln Ile His Arg Lys Ala Glu Glu Tyr Val Gly Lys
85 90 95
Ala Val Gln Ala Trp Arg Lys Arg Phe Ala Ala Asp Glu Arg Lys Gly
100 105 110
Ile Ala Gly Leu Ser Met Trp Asn Ile Glu Glu Leu Glu Gly Leu Arg
115 120 125
Lys Leu Leu Ile Ser Trp Ser Arg Arg Thr Arg Asn Pro Gln Glu Val
130 135 140
Asn Arg Phe Glu Arg Gly His Thr Ser His Gln Arg Leu Leu Thr His
145 150 155 160
Ile Gln Asn Val Lys Glu Asp Arg Leu Lys Gln Leu Ser His Ala Ile
165 170 175
Val Met Thr Ala Leu Gly Tyr Val Tyr Asp Glu Arg Lys Gln Glu Trp
180 185 190
Cys Ala Glu Tyr Pro Ala Cys Gln Val Ile Leu Phe Glu Asn Leu Ser
195 200 205
Gln Tyr Arg Ser Asn Leu Asp Arg Ser Thr Lys Glu Asn Ser Thr Leu
210 215 220
Met Lys Trp Ala His Arg Ser Ile Pro Lys Tyr Val His Met Gln Ala
225 230 235 240
Glu Pro Tyr Gly Ile Gln Ile Gly Asp Val Arg Ala Glu Tyr Ser Ser
245 250 255
Arg Phe Tyr Ala Lys Thr Gly Thr Pro Gly Ile Arg Cys Lys Lys Val
260 265 270
Arg Gly Gln Asp Leu Gln Gly Arg Arg Phe Glu Asn Leu Gln Lys Arg
275 280 285
Leu Val Asn Glu Gln Phe Leu Thr Glu Glu Gln Val Lys Gln Leu Arg
290 295 300
Pro Gly Asp Ile Val Pro Asp Asp Ser Gly Glu Leu Phe Met Thr Leu
305 310 315 320
Thr Asp Gly Ser Gly Ser Lys Glu Val Val Phe Leu Gln Ala Asp Ile
325 330 335
Asn Ala Ala His Asn Leu Gln Lys Arg Phe Trp Gln Arg Tyr Asn Glu
340 345 350
Leu Phe Lys Val Ser Cys Arg Val Ile Val Arg Asp Glu Glu Glu Tyr
355 360 365
Leu Val Pro Lys Thr Lys Ser Val Gln Ala Lys Leu Gly Lys Gly Leu
370 375 380
Phe Val Lys Lys Ser Asp Thr Ala Trp Lys Asp Val Tyr Val Trp Asp
385 390 395 400
Ser Gln Ala Lys Leu Lys Gly Lys Thr Thr Phe Thr Glu Glu Ser Glu
405 410 415
Ser Pro Glu Gln Leu Glu Asp Phe Gln Glu Ile Ile Glu Glu Ala Glu
420 425 430
Glu Ala Lys Gly Thr Tyr Arg Thr Leu Phe Arg Asp Pro Ser Gly Val
435 440 445
Phe Phe Pro Glu Ser Val Trp Tyr Pro Gln Lys Asp Phe Trp Gly Glu
450 455 460
Val Lys Arg Lys Leu Tyr Gly Lys Leu Arg Glu Arg Phe Leu Thr Lys
465 470 475 480
Ala Arg
<210> 563
<211> 584
<212> PRT
<213> Brevibacillus agri
<400> 563
Met Ala Ile Arg Ser Ile Lys Leu Lys Leu Lys Thr His Thr Gly Pro
1 5 10 15
Glu Ala Gln Asn Leu Arg Lys Gly Ile Trp Arg Thr His Arg Leu Leu
20 25 30
Asn Glu Gly Val Ala Tyr Tyr Met Lys Met Leu Leu Leu Phe Arg Gln
35 40 45
Glu Ser Thr Gly Glu Arg Pro Lys Glu Glu Leu Gln Glu Glu Leu Ile
50 55 60
Cys His Ile Arg Glu Gln Gln Gln Arg Asn Gln Ala Asp Lys Asn Thr
65 70 75 80
Gln Ala Leu Pro Leu Asp Lys Ala Leu Glu Ala Leu Arg Gln Leu Tyr
85 90 95
Glu Leu Leu Val Pro Ser Ser Val Gly Gln Ser Gly Asp Ala Gln Ile
100 105 110
Ile Ser Arg Lys Phe Leu Ser Pro Leu Val Asp Pro Asn Ser Glu Gly
115 120 125
Gly Lys Gly Thr Ser Lys Ala Gly Ala Lys Pro Thr Trp Gln Lys Lys
130 135 140
Lys Glu Ala Asn Asp Pro Thr Trp Glu Gln Asp Tyr Glu Lys Trp Lys
145 150 155 160
Lys Arg Arg Glu Glu Asp Pro Thr Ala Ser Val Ile Thr Thr Leu Glu
165 170 175
Glu Tyr Gly Ile Arg Pro Ile Phe Pro Leu Tyr Thr Asn Thr Val Thr
180 185 190
Asp Ile Ala Trp Leu Pro Leu Gln Ser Asn Gln Phe Val Arg Thr Trp
195 200 205
Asp Arg Asp Met Leu Gln Gln Ala Ile Glu Arg Leu Leu Ser Trp Glu
210 215 220
Ser Trp Asn Lys Arg Val Gln Glu Glu Tyr Ala Lys Leu Lys Glu Lys
225 230 235 240
Met Ala Gln Leu Asn Glu Gln Leu Glu Gly Gly Gln Glu Trp Ile Ser
245 250 255
Leu Leu Glu Gln Tyr Glu Glu Asn Arg Glu Arg Glu Leu Arg Glu Asn
260 265 270
Met Thr Ala Ala Asn Asp Lys Tyr Arg Ile Thr Lys Arg Gln Met Lys
275 280 285
Gly Trp Asn Glu Leu Tyr Glu Leu Trp Ser Thr Phe Pro Ala Ser Ala
290 295 300
Ser His Glu Gln Tyr Lys Glu Ala Leu Lys Arg Val Gln Gln Arg Leu
305 310 315 320
Arg Gly Arg Phe Gly Asp Ala His Phe Phe Gln Tyr Leu Met Glu Glu
325 330 335
Lys Asn Arg Leu Ile Trp Lys Gly Asn Pro Gln Arg Ile His Tyr Phe
340 345 350
Val Ala Arg Asn Glu Leu Thr Lys Arg Leu Glu Glu Ala Lys Gln Ser
355 360 365
Ala Thr Met Thr Leu Pro Asn Ala Arg Lys His Pro Leu Trp Val Arg
370 375 380
Phe Asp Ala Arg Gly Gly Asn Leu Gln Asp Tyr Tyr Leu Thr Ala Glu
385 390 395 400
Ala Asp Lys Pro Arg Ser Arg Arg Phe Val Thr Phe Ser Gln Leu Ile
405 410 415
Trp Pro Ser Glu Ser Gly Trp Met Glu Lys Lys Asp Val Glu Val Glu
420 425 430
Leu Ala Leu Ser Arg Gln Phe Tyr Gln Gln Val Lys Leu Leu Lys Asn
435 440 445
Asp Lys Gly Lys Gln Lys Ile Glu Phe Lys Asp Lys Gly Ser Gly Ser
450 455 460
Thr Phe Asn Gly His Leu Gly Gly Ala Lys Leu Gln Leu Glu Arg Gly
465 470 475 480
Asp Leu Glu Lys Glu Glu Lys Asn Phe Glu Asp Gly Glu Ile Gly Ser
485 490 495
Val Tyr Leu Asn Val Val Ile Asp Phe Glu Pro Leu Gln Glu Val Lys
500 505 510
Asn Gly Arg Val Gln Ala Pro Tyr Gly Gln Val Leu Gln Leu Ile Arg
515 520 525
Arg Pro Asn Glu Phe Pro Lys Val Thr Thr Tyr Lys Ser Glu Gln Leu
530 535 540
Val Glu Trp Ile Lys Ala Ser Pro Gln His Ser Ala Gly Val Glu Ser
545 550 555 560
Leu Ala Ser Gly Phe Arg Val Met Ser Ile Asp Leu Gly Leu Arg Ala
565 570 575
Ala Ala Ala Thr Ser Ile Phe Leu
580
<210> 564
<211> 259
<212> PRT
<213> Brevibacillus sp.
<400> 564
Met Ala Ile Arg Ser Ile Lys Leu Lys Leu Lys Thr His Thr Gly Pro
1 5 10 15
Glu Ala Gln Asn Leu Arg Lys Gly Ile Trp Arg Thr His Arg Leu Leu
20 25 30
Asn Glu Gly Val Ala Tyr Tyr Met Lys Met Leu Leu Leu Phe Arg Gln
35 40 45
Glu Ser Thr Gly Glu Arg Pro Lys Glu Glu Leu Gln Glu Glu Leu Ile
50 55 60
Cys His Ile Arg Glu Gln Gln Gln Arg Asn Gln Ala Asp Lys Asn Thr
65 70 75 80
Gln Ala Leu Pro Leu Asp Lys Ala Leu Glu Ala Leu Arg Gln Leu Tyr
85 90 95
Glu Leu Leu Val Pro Ser Ser Val Gly Gln Ser Gly Asp Ala Gln Ile
100 105 110
Ile Ser Arg Lys Phe Leu Ser Pro Leu Val Asp Pro Asn Ser Glu Gly
115 120 125
Gly Lys Gly Thr Ser Lys Ala Gly Ala Lys Pro Thr Trp Gln Lys Lys
130 135 140
Lys Glu Ala Asn Asp Pro Thr Trp Glu Gln Asp Tyr Glu Lys Trp Lys
145 150 155 160
Lys Arg Arg Glu Glu Asp Pro Thr Ala Ser Val Ile Thr Thr Leu Glu
165 170 175
Glu Tyr Gly Ile Arg Pro Ile Phe Pro Leu Tyr Thr Asn Thr Val Thr
180 185 190
Asp Ile Ala Trp Leu Pro Leu Gln Ser Asn Gln Phe Val Arg Thr Trp
195 200 205
Asp Arg Asp Met Leu Gln Gln Ala Ile Glu Arg Leu Leu Ser Trp Glu
210 215 220
Ser Trp Asn Lys Arg Val Gln Glu Glu Tyr Ala Lys Leu Lys Glu Lys
225 230 235 240
Met Ala Gln Leu Asn Glu Gln Leu Glu Gly Gly Gln Glu Trp Cys Thr
245 250 255
Leu Ser Arg
<210> 565
<211> 658
<212> PRT
<213> Methylobacterium nodulans
<400> 565
Met Leu Thr Lys Gln Asp Lys Gln Gln Lys Ile Thr Tyr Cys Thr Asn
1 5 10 15
Met Asn Glu Val Phe Glu Ala Lys Leu Gly Ser Ala Asp Leu Leu Leu
20 25 30
Asn Trp Asp His Leu Arg Gly Arg Ile Arg Asp Arg Val Asp Ala Gly
35 40 45
Asp Ile Gly Ser Ala Phe Leu Lys Leu Ala Leu Asp Val Ala His Val
50 55 60
Leu Pro Asp Gly Val Asp Asp Gln Leu Ala Arg Ala Ala Phe His Phe
65 70 75 80
Gln Ser Ala Lys Gly Ala Lys Ser Lys His Ala Asp Ser Val Gln Ala
85 90 95
Gly Leu Arg Val Leu Ser Ile Asp Leu Gly Val Arg Ser Phe Ala Thr
100 105 110
Cys Ser Val Phe Glu Leu Lys Asp Thr Ala Pro Thr Thr Gly Val Ala
115 120 125
Phe Pro Leu Ala Glu Phe Arg Leu Trp Ala Val His Glu Arg Ser Phe
130 135 140
Thr Leu Glu Leu Pro Gly Glu Asn Val Gly Ala Ala Gly Gln Gln Trp
145 150 155 160
Arg Ala Gln Ala Asp Ala Glu Leu Arg Gln Leu Arg Gly Gly Leu Asn
165 170 175
Arg His Arg Gln Leu Leu Arg Ala Ala Thr Val Gln Lys Gly Glu Arg
180 185 190
Asp Ala Tyr Leu Thr Asp Leu Arg Glu Ala Trp Ser Ala Lys Glu Leu
195 200 205
Trp Pro Phe Glu Ala Ser Leu Leu Ser Glu Leu Glu Arg Cys Ser Thr
210 215 220
Val Ala Asp Pro Leu Trp Gln Asp Thr Cys Lys Arg Ala Ala Arg Leu
225 230 235 240
Tyr Arg Thr Glu Phe Gly Ala Val Val Ser Glu Trp Arg Ser Arg Thr
245 250 255
Arg Ser Arg Glu Asp Arg Lys Tyr Ala Gly Lys Ser Met Trp Ser Val
260 265 270
Gln His Leu Thr Asp Val Arg Arg Phe Leu Gln Ser Trp Ser Leu Ala
275 280 285
Gly Arg Ala Ser Gly Asp Ile Arg Arg Leu Asp Arg Glu Arg Gly Gly
290 295 300
Val Phe Ala Lys Asp Leu Leu Asp His Ile Asp Ala Leu Lys Asp Asp
305 310 315 320
Arg Leu Lys Thr Gly Ala Asp Leu Ile Val Gln Ala Ala Arg Gly Phe
325 330 335
Gln Arg Asn Glu Phe Gly Tyr Trp Val Gln Lys His Ala Pro Cys His
340 345 350
Val Ile Leu Phe Glu Asp Leu Ser Arg Tyr Arg Met Arg Thr Asp Arg
355 360 365
Pro Arg Arg Glu Asn Ser Gln Leu Met Gln Trp Ala His Arg Gly Val
370 375 380
Pro Asp Met Val Gly Met Gln Gly Glu Ile Tyr Gly Ile Gln Asp Arg
385 390 395 400
Arg Asp Pro Asp Ser Ala Arg Lys His Ala Arg Gln Pro Leu Ala Ala
405 410 415
Phe Cys Leu Asp Thr Pro Ala Ala Phe Ser Ser Arg Tyr His Ala Ser
420 425 430
Thr Met Thr Pro Gly Ile Arg Cys His Pro Leu Arg Lys Arg Glu Phe
435 440 445
Glu Asp Gln Gly Phe Leu Glu Leu Leu Lys Arg Glu Asn Glu Gly Leu
450 455 460
Asp Leu Asn Gly Tyr Lys Pro Gly Asp Leu Val Pro Leu Pro Gly Gly
465 470 475 480
Glu Val Phe Val Cys Leu Asn Ala Asn Gly Leu Ser Arg Ile His Ala
485 490 495
Asp Ile Asn Ala Ala Gln Asn Leu Gln Arg Arg Phe Trp Thr Gln His
500 505 510
Gly Asp Ala Phe Arg Leu Pro Cys Gly Lys Ser Ala Val Gln Gly Gln
515 520 525
Ile Arg Trp Ala Pro Leu Ser Met Gly Lys Arg Gln Ala Gly Ala Leu
530 535 540
Gly Gly Phe Gly Tyr Leu Glu Pro Thr Gly His Asp Ser Gly Ser Cys
545 550 555 560
Gln Trp Arg Lys Thr Thr Glu Ala Glu Trp Arg Arg Leu Ser Gly Ala
565 570 575
Gln Lys Asp Arg Asp Glu Ala Ala Ala Ala Glu Asp Glu Glu Leu Gln
580 585 590
Gly Leu Glu Glu Glu Leu Leu Glu Arg Ser Gly Glu Arg Val Val Phe
595 600 605
Phe Arg Asp Pro Ser Gly Val Val Leu Pro Thr Asp Leu Trp Phe Pro
610 615 620
Ser Ala Ala Phe Trp Ser Ile Val Arg Ala Lys Thr Val Gly Arg Leu
625 630 635 640
Arg Ser His Leu Asp Ala Gln Ala Glu Ala Ser Tyr Ala Val Ala Ala
645 650 655
Gly Leu
<210> 566
<211> 464
<212> PRT
<213> Methylobacterium nodulans
<400> 566
Met Pro Val Arg Ser Leu Lys Leu Lys Ile Val Val Pro Arg His Pro
1 5 10 15
Ser Glu Leu Glu Lys Ala Gln Ala Leu Trp Ser Thr His Arg Leu Val
20 25 30
Asn Glu Ala Val Ser Phe Tyr Glu Gln Lys Leu Leu Leu Leu Arg Gly
35 40 45
Glu Thr Tyr Ser Thr Ser Asp Gly Ser Val Pro Gln Asp Glu Val Arg
50 55 60
Arg Gln Leu Leu Glu Gln Ala Arg Glu Ala Gln Ala Arg Asn Gly Gly
65 70 75 80
Ser Gly Gly Ser Asp Asp Glu Ile Val Arg Leu Cys Arg Ser Leu Tyr
85 90 95
Glu Ala Ile Val Leu Ala Asp Asp Ala Asn Ala Gln Leu Ala Asn Ala
100 105 110
Phe Leu Gly Pro Leu Thr Asp Pro Asn Ser Ala Gly Phe Leu Glu Ala
115 120 125
Phe Asn Lys Val Asp Arg Pro Ala Pro Ser Trp Leu Asp Gln Val Pro
130 135 140
Ala Ser Asp Pro Ile Asp Pro Ala Val Leu Ala Glu Ala Asn Ala Trp
145 150 155 160
Leu Asp Thr Asp Ala Gly Arg Ala Trp Leu Val Asp Thr Gly Ala Pro
165 170 175
Pro Arg Trp Arg Ser Leu Ala Ala Lys Gln Asp Pro Ile Trp Pro Arg
180 185 190
Glu Phe Ala Arg Lys Leu Gly Glu Leu Arg Lys Glu Ala Ala Ser Gly
195 200 205
Thr Ser Ala Ile Ile Lys Ala Leu Lys Arg Asp Phe Gly Val Leu Pro
210 215 220
Leu Phe Gln Pro Ser Leu Ala Pro Arg Ile Leu Gly Ser Arg Ser Ser
225 230 235 240
Leu Thr Pro Trp Asp Arg Leu Ala Phe Arg Leu Ala Val Gly His Leu
245 250 255
Leu Ser Trp Glu Ser Trp Cys Thr Arg Ala Arg Asp Glu His Thr Ala
260 265 270
Arg Val Gln Arg Leu Glu Gln Phe Ser Ser Ala His Leu Lys Gly Asp
275 280 285
Leu Ala Thr Lys Val Ser Thr Leu Arg Glu Tyr Glu Arg Ala Arg Lys
290 295 300
Glu Gln Ile Ala Gln Leu Gly Leu Pro Met Gly Glu Arg Asp Phe Leu
305 310 315 320
Ile Thr Val Arg Met Thr Arg Gly Trp Asp Asp Leu Arg Glu Lys Trp
325 330 335
Arg Arg Ser Gly Asp Lys Gly Gln Glu Ala Leu His Ala Ile Ile Ala
340 345 350
Thr Glu Gln Thr Arg Lys Arg Gly Arg Phe Gly Asp Pro Asp Leu Phe
355 360 365
Arg Trp Leu Ala Arg Pro Glu Asn His His Val Trp Ala Asp Gly His
370 375 380
Ala Asp Ala Val Gly Val Leu Ala Arg Val Asn Ala Met Glu Arg Leu
385 390 395 400
Val Glu Arg Ser Arg Asp Thr Ala Leu Met Thr Leu Pro Asp Pro Val
405 410 415
Ala His Pro Arg Ser Ala Gln Trp Glu Ala Glu Gly Gly Ser Asn Leu
420 425 430
Arg Asn Tyr Gln Leu Glu Ala Val Gly Gly Glu Leu Gln Ile Thr Leu
435 440 445
Pro Leu Leu Lys Ala Ala Asp Asp Gly Arg Cys Ile Asp Thr Pro Leu
450 455 460
<210> 567
<211> 370
<212> PRT
<213> Methylobacterium nodulans
<400> 567
Met Tyr Glu Ala Ile Val Leu Ala Asp Asp Ala Asn Ala Gln Leu Ala
1 5 10 15
Asn Ala Phe Leu Gly Pro Leu Thr Asp Pro Asn Ser Ala Gly Phe Leu
20 25 30
Glu Ala Phe Asn Lys Val Asp Arg Pro Ala Pro Ser Trp Leu Asp Gln
35 40 45
Val Pro Ala Ser Asp Pro Ile Asp Pro Ala Val Leu Ala Glu Ala Asn
50 55 60
Ala Trp Leu Asp Thr Asp Ala Gly Arg Ala Trp Leu Val Asp Thr Gly
65 70 75 80
Ala Pro Pro Arg Trp Arg Ser Leu Ala Ala Lys Gln Asp Pro Ile Trp
85 90 95
Pro Arg Glu Phe Ala Arg Lys Leu Gly Glu Leu Arg Lys Glu Ala Ala
100 105 110
Ser Gly Thr Ser Ala Ile Ile Lys Ala Leu Lys Arg Asp Phe Gly Val
115 120 125
Leu Pro Leu Phe Gln Pro Ser Leu Ala Pro Arg Ile Leu Gly Ser Arg
130 135 140
Ser Ser Leu Thr Pro Trp Asp Arg Leu Ala Phe Arg Leu Ala Val Gly
145 150 155 160
His Leu Leu Ser Trp Glu Ser Trp Cys Thr Arg Ala Arg Asp Glu His
165 170 175
Thr Ala Arg Val Gln Arg Leu Glu Gln Phe Ser Ser Ala His Leu Lys
180 185 190
Gly Asp Leu Ala Thr Lys Val Ser Thr Leu Arg Glu Tyr Glu Arg Ala
195 200 205
Arg Lys Glu Gln Ile Ala Gln Leu Gly Leu Pro Met Gly Glu Arg Asp
210 215 220
Phe Leu Ile Thr Val Arg Met Thr Arg Gly Trp Asp Asp Leu Arg Glu
225 230 235 240
Lys Trp Arg Arg Ser Gly Asp Lys Gly Gln Glu Ala Leu His Ala Ile
245 250 255
Ile Ala Thr Glu Gln Thr Arg Lys Arg Gly Arg Phe Gly Asp Pro Asp
260 265 270
Leu Phe Arg Trp Leu Ala Arg Pro Glu Asn His His Val Trp Ala Asp
275 280 285
Gly His Ala Asp Ala Val Gly Val Leu Ala Arg Val Asn Ala Met Glu
290 295 300
Arg Leu Val Glu Arg Ser Arg Asp Thr Ala Leu Met Thr Leu Pro Asp
305 310 315 320
Pro Val Ala His Pro Arg Ser Ala Gln Trp Glu Ala Glu Gly Gly Ser
325 330 335
Asn Leu Arg Asn Tyr Gln Leu Glu Ala Val Gly Gly Glu Leu Gln Ile
340 345 350
Thr Leu Pro Leu Leu Lys Ala Ala Asp Asp Gly Arg Cys Ile Asp Thr
355 360 365
Pro Leu
370
<210> 568
<211> 1050
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 568
Met Arg Ser Asn Tyr His Gly Gly Arg Asn Ala Arg Gln Trp Arg Lys
1 5 10 15
Gln Ile Ser Gly Leu Ala Arg Arg Thr Lys Glu Thr Val Phe Thr Tyr
20 25 30
Lys Phe Pro Leu Glu Thr Asp Ala Ala Glu Ile Asp Phe Asp Lys Ala
35 40 45
Val Gln Thr Tyr Gly Ile Ala Glu Gly Val Gly His Gly Ser Leu Ile
50 55 60
Gly Leu Val Cys Ala Phe His Leu Ser Gly Phe Arg Leu Phe Ser Lys
65 70 75 80
Ala Gly Glu Ala Met Ala Phe Arg Asn Arg Ser Arg Tyr Pro Thr Asp
85 90 95
Ala Phe Ala Glu Lys Leu Ser Ala Ile Met Gly Ile Gln Leu Pro Thr
100 105 110
Leu Ser Pro Glu Gly Leu Asp Leu Ile Phe Gln Ser Pro Pro Arg Ser
115 120 125
Arg Asp Gly Ile Ala Pro Val Trp Ser Glu Asn Glu Val Arg Asn Arg
130 135 140
Leu Tyr Thr Asn Trp Thr Gly Arg Gly Pro Ala Asn Lys Pro Asp Glu
145 150 155 160
His Leu Leu Glu Ile Ala Gly Glu Ile Ala Lys Gln Val Phe Pro Lys
165 170 175
Phe Gly Gly Trp Asp Asp Leu Ala Ser Asp Pro Asp Lys Ala Leu Ala
180 185 190
Ala Ala Asp Lys Tyr Phe Gln Ser Gln Gly Asp Phe Pro Ser Ile Ala
195 200 205
Ser Leu Pro Ala Ala Ile Met Leu Ser Pro Ala Asn Ser Thr Val Asp
210 215 220
Phe Glu Gly Asp Tyr Ile Ala Ile Asp Pro Ala Ala Glu Thr Leu Leu
225 230 235 240
His Gln Ala Val Ser Arg Cys Ala Ala Arg Leu Gly Arg Glu Arg Pro
245 250 255
Asp Leu Asp Gln Asn Lys Gly Pro Phe Val Ser Ser Leu Gln Asp Ala
260 265 270
Leu Val Ser Ser Gln Asn Asn Gly Leu Ser Trp Leu Phe Gly Val Gly
275 280 285
Phe Gln His Trp Lys Glu Lys Ser Pro Lys Glu Leu Ile Asp Glu Tyr
290 295 300
Lys Val Pro Ala Asp Gln His Gly Ala Val Thr Gln Val Lys Ser Phe
305 310 315 320
Val Asp Ala Ile Pro Leu Asn Pro Leu Phe Asp Thr Thr His Tyr Gly
325 330 335
Glu Phe Arg Ala Ser Val Ala Gly Lys Val Arg Ser Trp Val Ala Asn
340 345 350
Tyr Trp Lys Arg Leu Leu Asp Leu Lys Ser Leu Leu Ala Thr Thr Glu
355 360 365
Phe Thr Leu Pro Glu Ser Ile Ser Asp Pro Lys Ala Val Ser Leu Phe
370 375 380
Ser Gly Leu Leu Val Asp Pro Gln Gly Leu Lys Lys Val Ala Asp Ser
385 390 395 400
Leu Pro Ala Arg Leu Val Ser Ala Glu Glu Ala Ile Asp Arg Leu Met
405 410 415
Gly Val Gly Ile Pro Thr Ala Ala Asp Ile Ala Gln Val Glu Arg Val
420 425 430
Ala Asp Glu Ile Gly Ala Phe Ile Gly Gln Val Gln Gln Phe Asn Asn
435 440 445
Gln Val Lys Gln Lys Leu Glu Asn Leu Gln Asp Ala Asp Asp Glu Glu
450 455 460
Phe Leu Lys Gly Leu Lys Ile Glu Leu Pro Ser Gly Asp Lys Glu Pro
465 470 475 480
Pro Ala Ile Asn Arg Ile Ser Gly Gly Ala Pro Asp Ala Ala Ala Glu
485 490 495
Ile Ser Glu Leu Glu Glu Lys Leu Gln Arg Leu Leu Asp Ala Arg Ser
500 505 510
Glu His Phe Gln Thr Ile Ser Glu Trp Ala Glu Glu Asn Ala Val Thr
515 520 525
Leu Asp Pro Ile Ala Ala Met Val Glu Leu Glu Arg Leu Arg Leu Ala
530 535 540
Glu Arg Gly Ala Thr Gly Asp Pro Glu Glu Tyr Ala Leu Arg Leu Leu
545 550 555 560
Leu Gln Arg Ile Gly Arg Leu Ala Asn Arg Val Ser Pro Val Ser Ala
565 570 575
Gly Ser Ile Arg Glu Leu Leu Lys Pro Val Phe Met Glu Glu Arg Glu
580 585 590
Phe Asn Leu Phe Phe His Asn Arg Leu Gly Ser Leu Tyr Arg Ser Pro
595 600 605
Tyr Ser Thr Ser Arg His Gln Pro Phe Ser Ile Asp Val Gly Lys Ala
610 615 620
Lys Ala Ile Asp Trp Ile Ala Gly Leu Asp Gln Ile Ser Ser Asp Ile
625 630 635 640
Glu Lys Ala Leu Ser Gly Ala Gly Glu Ala Leu Gly Asp Gln Leu Arg
645 650 655
Asp Trp Ile Asn Leu Ala Gly Phe Ala Ile Ser Gln Arg Leu Arg Gly
660 665 670
Leu Pro Asp Thr Val Pro Asn Ala Leu Ala Gln Val Arg Cys Pro Asp
675 680 685
Asp Val Arg Ile Pro Pro Leu Leu Ala Met Leu Leu Glu Glu Asp Asp
690 695 700
Ile Ala Arg Asp Val Cys Leu Lys Ala Phe Asn Leu Tyr Val Ser Ala
705 710 715 720
Ile Asn Gly Cys Leu Phe Gly Ala Leu Arg Glu Gly Phe Ile Val Arg
725 730 735
Thr Arg Phe Gln Arg Ile Gly Thr Asp Gln Ile His Tyr Val Pro Lys
740 745 750
Asp Lys Ala Trp Glu Tyr Pro Asp Arg Leu Asn Thr Ala Lys Gly Pro
755 760 765
Ile Asn Ala Ala Val Ser Ser Asp Trp Ile Glu Lys Asp Gly Ala Val
770 775 780
Ile Lys Pro Val Glu Thr Val Arg Asn Leu Ser Ser Thr Gly Phe Ala
785 790 795 800
Gly Ala Gly Val Ser Glu Tyr Leu Val Gln Ala Pro His Asp Trp Tyr
805 810 815
Thr Pro Leu Asp Leu Arg Asp Val Ala His Leu Val Thr Gly Leu Pro
820 825 830
Val Glu Lys Asn Ile Thr Lys Leu Lys Arg Leu Thr Asn Arg Thr Ala
835 840 845
Phe Arg Met Val Gly Ala Ser Ser Phe Lys Thr His Leu Asp Ser Val
850 855 860
Leu Leu Ser Asp Lys Ile Lys Leu Gly Asp Phe Thr Ile Ile Ile Asp
865 870 875 880
Gln His Tyr Arg Gln Ser Val Thr Tyr Gly Gly Lys Val Lys Ile Ser
885 890 895
Tyr Glu Pro Glu Arg Leu Gln Val Glu Ala Ala Val Pro Val Val Asp
900 905 910
Thr Arg Asp Arg Thr Val Pro Glu Pro Asp Thr Leu Phe Asp His Ile
915 920 925
Val Ala Ile Asp Leu Gly Glu Arg Ser Val Gly Phe Ala Val Phe Asp
930 935 940
Ile Lys Ser Cys Leu Arg Thr Gly Glu Val Lys Pro Ile His Asp Asn
945 950 955 960
Asn Gly Asn Pro Val Val Gly Thr Val Ala Val Pro Ser Ile Arg Arg
965 970 975
Leu Met Lys Ala Val Arg Ser His Arg Arg Arg Arg Gln Pro Asn Gln
980 985 990
Lys Val Asn Gln Thr Tyr Ser Thr Ala Leu Gln Asn Tyr Arg Glu Asn
995 1000 1005
Val Ile Gly Asp Val Cys Asn Arg Ile Asp Thr Leu Met Glu Arg
1010 1015 1020
Tyr Asn Ala Phe Pro Val Leu Glu Phe Gln Ile Lys Asn Phe Gln
1025 1030 1035
Ala Gly Ala Lys Gln Leu Glu Ile Val Tyr Gly Ser
1040 1045 1050
<210> 569
<211> 1222
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 569
Met Lys Lys Phe Glu Leu Lys Gln Asn Phe Arg Asn Asn Tyr Ser Gly
1 5 10 15
Lys Thr Leu Arg Asn Phe Arg Gln Thr Leu Ala Gln Ile Ala Asn Lys
20 25 30
Lys Ser Ser Asp Ser Ile Leu Thr Ile Lys Phe Lys Leu Asp Cys Ser
35 40 45
Lys Thr Gly Lys Leu Pro Lys Tyr Glu Asn Leu Ile Ser Leu Tyr Asp
50 55 60
Thr Ile Glu Asp Ile Lys Lys Gly Thr Leu Ser Tyr Tyr Leu Phe Thr
65 70 75 80
Leu Ile Val Ser Gly Phe Lys Phe Phe Gly Ser Ala Ser Gln Ala Lys
85 90 95
Ala Phe Ser Thr Lys Asp Ile Phe Lys Asp Asn Asp Phe Tyr Asn Gln
100 105 110
Phe Lys Ile Gln Ser His Leu Asp Leu Pro Asp Phe Val Pro Ser Lys
115 120 125
Ile Tyr Gln Arg Leu Lys Lys Asn Val Arg Ser Thr Asn Gly Lys Asp
130 135 140
Asn Ala Phe Lys Ala Ser Val Ile Val Ala Glu Tyr Arg Lys Glu Ile
145 150 155 160
Gly Lys Leu Lys Asn Lys Asp Glu Ser Ser Glu His Gln Cys Glu Glu
165 170 175
Leu Phe Lys Lys Ile Gly Thr Ala Leu Glu Thr Arg Phe Ser Ser Trp
180 185 190
Gln Asp Leu Ile Asn Asn Cys Ser Thr Gly Cys Glu Ile Ile Asp Glu
195 200 205
Ile Leu Asn Asp Ser Phe Gly Thr Leu Pro Ser Ile Lys Lys Met Val
210 215 220
Leu Ala Ser Thr Thr Gln Ser Ser Asp Gly Glu Gln Asp Gly Ile Ala
225 230 235 240
Ile Ala Tyr Asp Pro Asp Ser Thr Phe Ile Lys Ser Asp Glu Leu Leu
245 250 255
Asn Pro Tyr Phe Ala Val Ala Thr Ile Leu Lys Ser Met Pro Pro Glu
260 265 270
Ile Gln Gln Asp Lys Lys Ser Ala Tyr Val Lys Ala Asn Leu Thr Thr
275 280 285
Pro Thr His Asn Ala Leu Ser Trp Ile Phe Gly Lys Gly Leu Thr Leu
290 295 300
Phe Gln Thr Glu Ser Thr Glu Lys Leu Cys Ala Met Phe Asn Val Ser
305 310 315 320
Asp Lys Arg Val Ile Glu Gln Val Gln Asp Ala Ala Lys Ala Val Lys
325 330 335
Leu Pro Ala Glu Leu Asp Leu Asn His Cys Thr Leu Lys Phe Gln Asp
340 345 350
Phe Arg Ser Ser Leu Gly Gly His Leu Asp Ser Trp Thr Thr Asn Tyr
355 360 365
Leu Lys Arg Leu Asp Glu Leu Asn Asp Leu Leu Leu Asn Leu Pro Lys
370 375 380
Asn Leu Ser Leu Pro Asp Ile Phe Met Ile Asp Gly Lys Asp Phe Ile
385 390 395 400
Glu Tyr Ser Gly Cys Asn Arg Asp Glu Ile Gln Gln Met Ile Asp Phe
405 410 415
Val Val Asn Glu Gln Asn Arg Ile Lys Leu Gln Glu Ser Leu Asn Ala
420 425 430
Leu Leu Gly Lys Gly Asn Asn Gln Ile Cys Ser Asp Asp Ile Ser Thr
435 440 445
Val Lys Asp Phe Ser Glu Ile Val Asn Ser Leu His Ser Phe Val Gln
450 455 460
Gln Ile Asp Asn Ser Leu Glu Gln Ser Ser Asn Glu Ala Asn Ser Ile
465 470 475 480
Phe Ser Glu Leu Lys Lys Lys Ile Glu Lys Asn Glu Lys Trp Asp Ile
485 490 495
Trp Lys Asn Asn Leu Lys Lys Ile Pro Lys Leu Asn Lys Leu Ser Gly
500 505 510
Gly Val Pro Asp Ala Trp Lys Glu Ile Arg Glu Ile Glu Gln Lys Phe
515 520 525
His Glu Ile Ser Glu Asn Gln Lys Lys His Phe Thr Glu Val Met Glu
530 535 540
Trp Ile Asp Ala Gly Asn Gly Thr Ile Asp Ile Phe Glu Ser Arg Phe
545 550 555 560
Lys Tyr Asp Glu Leu Leu Lys Lys Ser Lys Lys Asn Asn Leu Gln Ser
565 570 575
Ala Asp Glu Leu Ala Phe Arg Ser Val Leu Asn Lys Leu Gly Arg Phe
580 585 590
Ala Arg Gln Gly Asn Asp Leu Val Cys Glu Lys Ile Lys Asn Trp Phe
595 600 605
Lys Glu Gln Asn Ile Phe Asp Ser Ser Lys Asp Phe Asn Arg Tyr Phe
610 615 620
Ile Asn Gln Lys Gly Phe Ile Phe Lys His Pro Ser Ser Lys Lys Asp
625 630 635 640
Asn Ser Pro Tyr Asn Leu Ser Ala Asn Leu Leu Glu Lys Arg Tyr Glu
645 650 655
Val Thr Asn Thr Val Gly Ala Leu Leu Glu Gln Cys Glu Ser Asp Pro
660 665 670
Ala Ile Val Asn Asp Pro Phe Ser Met Arg Ser Leu Val Glu Phe Arg
675 680 685
Ala Leu Trp Phe Ser Ile Asn Ile Ser Gly Ile Ser Lys Glu Gln His
690 695 700
Ile Pro Thr Lys Ile Ala Gln Pro Lys Leu Asp Asp Ser Thr Tyr Gln
705 710 715 720
Glu Ser Val Ser Pro Thr Leu Lys Tyr Arg Leu Glu Lys Glu Gln Ile
725 730 735
Thr Ser Ser Glu Leu Asn Ser Ile Phe Thr Val Tyr Lys Ser Leu Leu
740 745 750
Ser Gly Leu Ser Ile Arg Leu Ser Arg Asn Ser Phe Tyr Leu Arg Thr
755 760 765
Lys Phe Ser Trp Ile Gly Asn Asn Ser Leu Ile Tyr Cys Pro Lys Glu
770 775 780
Thr Thr Trp Lys Ile Pro Ala Ala Tyr Phe Lys Ser Asp Leu Trp Asn
785 790 795 800
Glu Tyr Lys Asp Lys Gln Ile Leu Ile Val Asn Glu Glu Tyr Asp Val
805 810 815
Asp Val Val Lys Thr Phe Glu Ser Val Tyr Lys Ile Val Lys Ser Lys
820 825 830
Asp Asn Asn Glu Lys Asn Arg Ile Leu Pro Leu Leu Lys Gln Leu Pro
835 840 845
His Asp Trp Met Phe Lys Leu Pro Phe Gly Ala Ser Asn Ala Glu Lys
850 855 860
Cys Lys Val Leu Lys Leu Glu Lys Asn Asn Lys Lys Phe Lys Pro Leu
865 870 875 880
Ser Val Ser Lys Asp Ser Leu Ala Arg Leu Ser Gly Pro Ser Thr Tyr
885 890 895
Phe Asn Gln Ile Asp Glu Ile Met Met Asn Asp Glu Ser Glu Leu Ser
900 905 910
Glu Met Thr Leu Leu Ala Asp Glu Pro Val Arg Gln Gln Met Ser Asn
915 920 925
Gly Lys Ile Glu Ile Ile Pro Asp Asp Tyr Val Met Ser Leu Ala Ile
930 935 940
Pro Ile Thr Arg Ser Leu Lys Lys Gly Asn Thr Glu Ser Phe Pro Phe
945 950 955 960
Lys Asn Ile Val Ser Ile Asp Gln Gly Glu Ala Gly Phe Ala Tyr Ala
965 970 975
Val Phe Lys Leu Ser Asp Cys Gly Asn Glu Arg Ala Glu Pro Ile Ala
980 985 990
Thr Gly Leu Ile Pro Ile Pro Ser Ile Arg Arg Leu Ile His Ser Val
995 1000 1005
Lys Lys Tyr Arg Gly Lys Lys Gln Arg Ile Gln Asn Phe Asn Gln
1010 1015 1020
Lys Phe Asp Ser Thr Met Phe Thr Leu Arg Glu Asn Val Thr Gly
1025 1030 1035
Asp Ile Cys Gly Leu Ile Val Ala Leu Met Lys Lys Tyr Asn Ala
1040 1045 1050
Phe Pro Ile Leu Glu Lys Gln Val Gly Asn Leu Glu Ser Gly Ser
1055 1060 1065
Lys Gln Leu Met Leu Val Tyr Lys Ala Val Asn Ser Lys Phe Leu
1070 1075 1080
Ala Ala Lys Val Asp Met Gln Asn Asp Gln Arg Arg Ser Trp Trp
1085 1090 1095
Tyr Gln Gly Asn Ser Trp Asn Thr Pro Ile Leu Arg Ile Ser Asn
1100 1105 1110
Pro Asn Gln Ser Asn Asn Lys Asn Ile Val Lys Asn Ile Asn Gly
1115 1120 1125
Lys Lys Tyr Glu Glu Leu Lys Ile Tyr Pro Gly Tyr Ser Val Ser
1130 1135 1140
Ala Tyr Met Thr Ser Cys Ile Cys His Val Cys Gly Arg Asn Ala
1145 1150 1155
Leu Glu Leu Leu Lys Asn Asp Asp Ser Thr Gly Lys Val Lys Lys
1160 1165 1170
Tyr Gln Ile Asn Gln Asp Gly Glu Val Thr Ile Gly Gly Glu Val
1175 1180 1185
Ile Lys Leu Tyr Arg Lys Pro Asp Arg Leu Thr Pro Val Lys Asn
1190 1195 1200
Leu Ala Lys Lys Gly Asn Arg Glu Arg Thr Tyr Ala Ser Ile Asn
1205 1210 1215
Glu Arg Ala Pro
1220
<210> 570
<211> 1255
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 570
Met Arg Ser Asn Tyr His Gly Gly Arg Asn Ala Arg Gln Trp Arg Lys
1 5 10 15
Gln Ile Ser Gly Leu Ala Arg Arg Thr Lys Glu Thr Val Phe Thr Tyr
20 25 30
Lys Phe Pro Leu Glu Thr Asp Ala Ala Glu Ile Asp Phe Asp Lys Ala
35 40 45
Val Gln Thr Tyr Gly Ile Ala Glu Gly Val Gly His Gly Ser Leu Ile
50 55 60
Gly Leu Val Cys Ala Phe His Leu Ser Gly Phe Arg Leu Phe Ser Lys
65 70 75 80
Ala Gly Glu Ala Met Ala Phe Arg Asn Arg Ser Arg Tyr Pro Thr Asp
85 90 95
Ala Phe Ala Glu Lys Leu Ser Ala Ile Met Gly Ile Gln Leu Pro Thr
100 105 110
Leu Ser Pro Glu Gly Leu Asp Leu Ile Phe Gln Ser Pro Pro Arg Ser
115 120 125
Arg Asp Gly Ile Ala Pro Val Trp Ser Glu Asn Glu Val Arg Asn Arg
130 135 140
Leu Tyr Thr Asn Trp Thr Gly Arg Gly Pro Ala Asn Lys Pro Asp Glu
145 150 155 160
His Leu Leu Glu Ile Ala Gly Glu Ile Ala Lys Gln Val Phe Pro Lys
165 170 175
Phe Gly Gly Trp Asp Asp Leu Ala Ser Asp Pro Asp Lys Ala Leu Ala
180 185 190
Ala Ala Asp Lys Tyr Phe Gln Ser Gln Gly Asp Phe Pro Ser Ile Ala
195 200 205
Ser Leu Pro Ala Ala Ile Met Leu Ser Pro Ala Asn Ser Thr Val Asp
210 215 220
Phe Glu Gly Asp Tyr Ile Ala Ile Asp Pro Ala Ala Glu Thr Leu Leu
225 230 235 240
His Gln Ala Val Ser Arg Cys Ala Ala Arg Leu Gly Arg Glu Arg Pro
245 250 255
Asp Leu Asp Gln Asn Lys Gly Pro Phe Val Ser Ser Leu Gln Asp Ala
260 265 270
Leu Val Ser Ser Gln Asn Asn Gly Leu Ser Trp Leu Phe Gly Val Gly
275 280 285
Phe Gln His Trp Lys Glu Lys Ser Pro Lys Glu Leu Ile Asp Glu Tyr
290 295 300
Lys Val Pro Ala Asp Gln His Gly Ala Val Thr Gln Val Lys Ser Phe
305 310 315 320
Val Asp Ala Ile Pro Leu Asn Pro Leu Phe Asp Thr Thr His Tyr Gly
325 330 335
Glu Phe Arg Ala Ser Val Ala Gly Lys Val Arg Ser Trp Val Ala Asn
340 345 350
Tyr Trp Lys Arg Leu Leu Asp Leu Lys Ser Leu Leu Ala Thr Thr Glu
355 360 365
Phe Thr Leu Pro Glu Ser Ile Ser Asp Pro Lys Ala Val Ser Leu Phe
370 375 380
Ser Gly Leu Leu Val Asp Pro Gln Gly Leu Lys Lys Val Ala Asp Ser
385 390 395 400
Leu Pro Ala Arg Leu Val Ser Ala Glu Glu Ala Ile Asp Arg Leu Met
405 410 415
Gly Val Gly Ile Pro Thr Ala Ala Asp Ile Ala Gln Val Glu Arg Val
420 425 430
Ala Asp Glu Ile Gly Ala Phe Ile Gly Gln Val Gln Gln Phe Asn Asn
435 440 445
Gln Val Lys Gln Lys Leu Glu Asn Leu Gln Asp Ala Asp Asp Glu Glu
450 455 460
Phe Leu Lys Gly Leu Lys Ile Glu Leu Pro Ser Gly Asp Lys Glu Pro
465 470 475 480
Pro Ala Ile Asn Arg Ile Ser Gly Gly Ala Pro Asp Ala Ala Ala Glu
485 490 495
Ile Ser Glu Leu Glu Glu Lys Leu Gln Arg Leu Leu Asp Ala Arg Ser
500 505 510
Glu His Phe Gln Thr Ile Ser Glu Trp Ala Glu Glu Asn Ala Val Thr
515 520 525
Leu Asp Pro Ile Ala Ala Met Val Glu Leu Glu Arg Leu Arg Leu Ala
530 535 540
Glu Arg Gly Ala Thr Gly Asp Pro Glu Glu Tyr Ala Leu Arg Leu Leu
545 550 555 560
Leu Gln Arg Ile Gly Arg Leu Ala Asn Arg Val Ser Pro Val Ser Ala
565 570 575
Gly Ser Ile Arg Glu Leu Leu Lys Pro Val Phe Met Glu Glu Arg Glu
580 585 590
Phe Asn Leu Phe Phe His Asn Arg Leu Gly Ser Leu Tyr Arg Ser Pro
595 600 605
Tyr Ser Thr Ser Arg His Gln Pro Phe Ser Ile Asp Val Gly Lys Ala
610 615 620
Lys Ala Ile Asp Trp Ile Ala Gly Leu Asp Gln Ile Ser Ser Asp Ile
625 630 635 640
Glu Lys Ala Leu Ser Gly Ala Gly Glu Ala Leu Gly Asp Gln Leu Arg
645 650 655
Asp Trp Ile Asn Leu Ala Gly Phe Ala Ile Ser Gln Arg Leu Arg Gly
660 665 670
Leu Pro Asp Thr Val Pro Asn Ala Leu Ala Gln Val Arg Cys Pro Asp
675 680 685
Asp Val Arg Ile Pro Pro Leu Leu Ala Met Leu Leu Glu Glu Asp Asp
690 695 700
Ile Ala Arg Asp Val Cys Leu Lys Ala Phe Asn Leu Tyr Val Ser Ala
705 710 715 720
Ile Asn Gly Cys Leu Phe Gly Ala Leu Arg Glu Gly Phe Ile Val Arg
725 730 735
Thr Arg Phe Gln Arg Ile Gly Thr Asp Gln Ile His Tyr Val Pro Lys
740 745 750
Asp Lys Ala Trp Glu Tyr Pro Asp Arg Leu Asn Thr Ala Lys Gly Pro
755 760 765
Ile Asn Ala Ala Val Ser Ser Asp Trp Ile Glu Lys Asp Gly Ala Val
770 775 780
Ile Lys Pro Val Glu Thr Val Arg Asn Leu Ser Ser Thr Gly Phe Ala
785 790 795 800
Gly Ala Gly Val Ser Glu Tyr Leu Val Gln Ala Pro His Asp Trp Tyr
805 810 815
Thr Pro Leu Asp Leu Arg Asp Val Ala His Leu Val Thr Gly Leu Pro
820 825 830
Val Glu Lys Asn Ile Thr Lys Leu Lys Arg Leu Thr Asn Arg Thr Ala
835 840 845
Phe Arg Met Val Gly Ala Ser Ser Phe Lys Thr His Leu Asp Ser Val
850 855 860
Leu Leu Ser Asp Lys Ile Lys Leu Gly Asp Phe Thr Ile Ile Ile Asp
865 870 875 880
Gln His Tyr Arg Gln Ser Val Thr Tyr Gly Gly Lys Val Lys Ile Ser
885 890 895
Tyr Glu Pro Glu Arg Leu Gln Val Glu Ala Ala Val Pro Val Val Asp
900 905 910
Thr Arg Asp Arg Thr Val Pro Glu Pro Asp Thr Leu Phe Asp His Ile
915 920 925
Val Ala Ile Asp Leu Gly Glu Arg Ser Val Gly Phe Ala Val Phe Asp
930 935 940
Ile Lys Ser Cys Leu Arg Thr Gly Glu Val Lys Pro Ile His Asp Asn
945 950 955 960
Asn Gly Asn Pro Val Val Gly Thr Val Ala Val Pro Ser Ile Arg Arg
965 970 975
Leu Met Lys Ala Val Arg Ser His Arg Arg Arg Arg Gln Pro Asn Gln
980 985 990
Lys Val Asn Gln Thr Tyr Ser Thr Ala Leu Gln Asn Tyr Arg Glu Asn
995 1000 1005
Val Ile Gly Asp Val Cys Asn Arg Ile Asp Thr Leu Met Glu Arg
1010 1015 1020
Tyr Asn Ala Phe Pro Val Leu Glu Phe Gln Ile Lys Asn Phe Gln
1025 1030 1035
Ala Gly Ala Lys Gln Leu Glu Ile Val Tyr Gly Ser Val Leu His
1040 1045 1050
Arg Tyr Thr Phe Ser Gly Val Asp Ala His Lys Ala Lys Arg Arg
1055 1060 1065
Glu Tyr Trp Tyr Asn Gly Glu Leu Trp Glu His Pro Tyr Leu Met
1070 1075 1080
Ala Lys Lys Trp Asn Glu Glu Thr Asn Ser Met Ser Gly Ala Pro
1085 1090 1095
Lys Pro Val Ser Leu Phe Pro Gly Val Thr Val Asn Ala Ala Arg
1100 1105 1110
Thr Ser Gln Ile Cys His Gln Cys Gln Arg Asn Pro Met Ser His
1115 1120 1125
Leu Arg Gly Leu Thr Gly Thr Ile Glu Ile Ser Ser Asp Gly Leu
1130 1135 1140
Leu Glu Leu Asp Asp Gly Thr Ile Arg Leu Phe Glu Thr Ser Asp
1145 1150 1155
Tyr Asp Glu Asp Lys Phe Lys Gln Ser Arg Arg Glu Lys Arg Arg
1160 1165 1170
Leu Asp Ala Asn Val Leu Leu Ser Gly Arg His Arg Ala Glu Tyr
1175 1180 1185
Ile Tyr Thr Val Ala Lys Arg Asn Leu Arg Arg Pro Pro Lys Asn
1190 1195 1200
Val Met Thr Lys Asp Thr Thr Gln Ser Arg Tyr Thr Cys Leu Tyr
1205 1210 1215
Lys Asn Cys Ser Trp Thr Gly His Ala Asp Glu Asn Ala Ala Ile
1220 1225 1230
Asn Ile Gly Arg Arg Tyr Leu Ala Glu Arg Ile Asp Met Pro Ala
1235 1240 1245
Ser Lys Thr Lys Ala Ala Val
1250 1255
<210> 571
<211> 1258
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Metagenome sequence"
<400> 571
Met Asn Ala Arg Asp Trp Arg Lys His Val Gly Val Leu Ala Gln Gln
1 5 10 15
His Lys Glu Thr Thr Arg Thr Tyr Thr Phe Pro Leu Asp Thr Thr Gly
20 25 30
Ser Ala Ile Asp Phe Asp Ala Ala Leu Gln Ala Tyr Asn Ala Val Glu
35 40 45
Gly Val Gly Tyr Gly Ser Leu Leu Gly Leu Ala Cys Ala Val His Leu
50 55 60
Ser Gly Phe Arg Leu Phe Ser Thr Gly Lys Glu Ala Ala Thr Phe Arg
65 70 75 80
Asn Arg Ala Arg Tyr Pro Asn Ala Ala Phe Gln Ala Ala Leu Arg Lys
85 90 95
Glu Leu Gly Thr Thr Ile Thr Thr Leu Thr Pro Glu Thr Leu Asp Arg
100 105 110
Leu Phe Ser Ser Arg Pro Lys Arg Arg Asn Gly Val Pro Leu Pro Trp
115 120 125
Asn Gln Asp Ser Ile Arg Asp Arg Leu Tyr Thr Asn Trp Val Lys Pro
130 135 140
Arg Pro Gly Asp Thr Pro Asp Ala Val Leu Phe Gln Ile Ala Thr Gly
145 150 155 160
Ile Ala Gln Glu Ile Thr Glu Asp Val Ser Ser Trp Thr Asp Leu Ala
165 170 175
Lys Asn Ser Asp Arg Gly Leu Lys Ala Ala His Arg Tyr Phe Ala Arg
180 185 190
Val Gly Gly Phe Pro Ala Phe Asp Asn Leu Thr Pro Pro Ala Thr Val
195 200 205
Gln Pro Thr Asp Thr Thr Ile Asp Tyr Asp Pro Asn Ala Pro Phe His
210 215 220
Leu Val Ser His Ala Asp Gln Thr Leu Ile His Gln Ser Ile Ser Leu
225 230 235 240
Cys Ala His Arg Ile Arg Gln Glu Asp Pro Ala Leu Asp Pro Asn Lys
245 250 255
Ser Gly Phe Ile Lys Gln Leu Gln Asn Asn Phe Leu Ser Gln Thr Phe
260 265 270
Tyr Gly Leu Ser Trp Leu Phe Gly Ala Gly Tyr Val His Phe Arg Glu
275 280 285
Cys Thr Ala Asn Asp Leu Ala Ile Gln Tyr Gly Ile Pro Asn Asn Cys
290 295 300
Arg Asp Gly Ile His Gln Ile Lys Ser Phe Ala Asp Ala Ile Leu Pro
305 310 315 320
Asn Thr Phe Phe Glu Lys Lys His Tyr Arg Lys Asp Ser Arg Ser Val
325 330 335
Gly Lys Lys Ala Lys Ser Trp Ile Ser Asn Tyr Trp Gln Arg Leu Leu
340 345 350
Gln Leu Gln Thr Trp Val Asp Asp His Thr Trp Val Thr Leu Pro Gln
355 360 365
Glu Leu Thr Glu Ala Gln Phe Lys Pro Leu Phe Arg Gly Leu Leu Val
370 375 380
Asp Ala Val Glu Leu Met Ala Ile Ala Glu Arg Leu Pro Gln Arg Leu
385 390 395 400
Ala Asp Cys Arg Asp Ser Leu Asp Cys Leu Met Gly Lys Gly Pro Gln
405 410 415
Ala Ala Thr Lys Asn Asp Val Glu Ile Val Glu Lys Val Arg Glu Glu
420 425 430
Ile Glu Ser Phe Val Gly Gln Ile Glu Gln Leu Gly Asn Gln Leu Arg
435 440 445
His Gln Leu Glu Asn Glu Asn Asn Asp Gln Val His Arg Asp Asn Leu
450 455 460
His Gln Leu Lys Asn Arg Leu Pro Leu Asp Leu Arg Arg Pro Gln Ala
465 470 475 480
Leu Asn Lys Ile Ser Gly Gly Val Pro Asp Val Ala Lys Ser Ile Arg
485 490 495
Gly Leu Glu Thr Gln Leu Asp Gln Val Leu Lys Glu Arg Arg Ser His
500 505 510
Phe Gly Arg Leu Thr Lys Trp Ala Lys Glu Cys Gly Ile Thr Leu Asp
515 520 525
Pro Leu Gln Pro Leu Ile Glu Ser Glu Lys Gln Arg Val Ala Glu Arg
530 535 540
Gly Ser Ala His Asp Ala Lys Glu Leu Ala Ile Arg Leu Leu Leu Gln
545 550 555 560
Arg Ile Gly Arg Leu Gly His Arg Leu Ser Pro Thr Asn Ala Thr Ala
565 570 575
Ile Gln Glu Leu Leu Arg Pro Val Phe Ala Val Lys Arg Glu Phe Asn
580 585 590
Leu Phe Phe His Asn His Met Gly Ala Leu Tyr Arg Ser Pro Tyr Ser
595 600 605
Thr Ser Arg His Gln Pro Phe Gln Ile Asn Val Asp Val Ala His Gly
610 615 620
Thr Asp Trp Ile Gly Thr Ile Glu Thr Leu Ile Gln Asn Leu Phe Thr
625 630 635 640
Gln Ile Gln Asp Asp Ala Leu Leu Arg Asp Leu Val Gln Leu Glu Gly
645 650 655
Phe Val Phe Ser His Lys Leu Arg Ala Leu Pro Gly Val Ile Pro Ser
660 665 670
Glu Leu Ala Arg Pro Asn Asn Leu Gln Gln Met Gly Leu Pro Ala Leu
675 680 685
Leu Leu Val Leu Leu Gln Ala Asp Gln Val His Arg Glu Thr Val Leu
690 695 700
Arg Val Phe Asn Leu Tyr Gly Ser Ala Ile Asn Gly Tyr Leu Phe Gln
705 710 715 720
Ala Leu Arg Pro Gly Phe Ile Val Arg Ala Gly Phe Gln Arg Leu Glu
725 730 735
Thr Lys Lys Leu Arg Tyr Val Pro Lys Ala Gln Ser Trp Gln Tyr Pro
740 745 750
Asp Arg Leu His His Ala Lys Ser Ala Ile Lys Asn Ser Leu Ser Ala
755 760 765
Gly Trp Ile Lys Lys Asn His Gln Gly Ala Ile Leu Pro Gln Lys Thr
770 775 780
Leu Thr Ala Leu Val Lys Gln Lys Ser Leu Lys Asp Thr Gly Val Pro
785 790 795 800
Glu Tyr Leu Val Gln Ala Pro His Asp Trp Tyr Val Pro Ile Asp Leu
805 810 815
Arg Gly Pro Ala Ile Pro Ile Glu Gly Leu Thr Val Gly Thr Glu Gly
820 825 830
Pro Glu Leu Thr Gln Leu Gly Pro Met Lys Asp Asp Cys Ala Phe Arg
835 840 845
Ala Ile Gly Pro Ser Ser Phe Lys Ser Lys Ile Asp Ala Gly Leu Leu
850 855 860
Pro Gln Asp Val Lys Tyr Gly Asp Met Thr Leu Ile Phe Asp Gln His
865 870 875 880
Tyr Gln Gln Ser Ile Ser Phe Ala Asn Gly Thr Phe Ser Ile Gln Tyr
885 890 895
Gln Pro Thr Ser Leu Gln Val Lys Ala Ala Ile Pro Val Val Asp Lys
900 905 910
Arg Pro Arg Asp Thr Arg Asn Asn Ser His Leu Tyr Asp Arg Ile Val
915 920 925
Ala Ile Asp Leu Gly Glu Arg Lys Ile Gly Tyr Ala Ile Phe Asp Leu
930 935 940
Lys Gln Val Leu Lys Ser Glu Gln Leu Glu Pro Met Arg Glu Asp Gly
945 950 955 960
Lys Pro Leu Ile Gly Ser Ile Ser Ile Arg Ser Ile Arg Gly Leu Met
965 970 975
Lys Ala Val Gln Thr His Arg Asn Arg Arg Gln Pro Asn Tyr Arg Ile
980 985 990
Asp Gln Thr Tyr Ser Lys Ala Leu Met His Tyr Arg Glu Ser Val Ile
995 1000 1005
Gly Asp Val Cys Asn Ala Ile Asp Thr Leu Cys Ala Arg Tyr Gly
1010 1015 1020
Gly Phe Pro Val Leu Glu Ser Ser Val Arg Asn Phe Glu Val Gly
1025 1030 1035
Ser Ala Gln Leu Lys Thr Val Tyr Gly Ser Val Ser Arg Arg Tyr
1040 1045 1050
Thr Trp Ser Ala Val Asp Ala His Lys Asn Gln Arg Gln Gln Tyr
1055 1060 1065
Trp Leu Gly Gly Thr Lys Asp Lys Ile Pro Ile Trp Thr His Pro
1070 1075 1080
Tyr Leu Met Thr Arg Glu Trp Asp Glu Lys Asn Ser Lys Trp Ser
1085 1090 1095
Asn Arg Ser Lys Pro Leu Lys Met His Pro Gly Val Glu Val His
1100 1105 1110
Pro Ala Gly Thr Ser Gln Ile Cys His Gln Cys Lys Arg Asn Pro
1115 1120 1125
Ile Gly Ala Leu Trp Asn Val Ala Asp Thr Val Val Leu Asp Asp
1130 1135 1140
Gln Gly Gln Leu Asp Leu Asp Asp Gly Thr Ile Arg Leu Asn Ser
1145 1150 1155
Gly Tyr Ile Asp Thr Thr Glu Ile Lys Arg Ala Arg Arg Lys Lys
1160 1165 1170
Ile Arg Leu Pro Glu Asn Lys Pro Leu Thr Gly Ser His Lys Thr
1175 1180 1185
Ser His Val Arg Ala Val Ala Arg Arg Asn Leu Arg Gln Pro Pro
1190 1195 1200
Lys Ser Thr Arg Ala Lys Asp Thr Thr Gln Ser Arg Tyr Thr Cys
1205 1210 1215
Leu Tyr Val Asp Cys Gly His Glu Cys His Ala Asp Glu Asn Ala
1220 1225 1230
Ala Ile Asn Ile Gly Arg Lys Tyr Leu Gln Glu Arg Ile His Ile
1235 1240 1245
Glu Ala Ser Arg Gln Ala Leu Ser Thr Arg
1250 1255
<210> 572
<211> 1269
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Marine metagenome sequence"
<400> 572
Met Val Ala Gly Leu Lys Lys Ile Lys Arg Asp Gly Val Thr Met Lys
1 5 10 15
Ser Asn Tyr His Gly Gly Val Lys Ala Arg Ala Trp Arg Lys Arg Ile
20 25 30
Gly Gly Leu Ala Arg Arg Gln Lys Glu Thr Val Phe Thr Tyr Lys Phe
35 40 45
Pro Leu Glu Thr Glu Glu Ala Gly Ile Asp Phe Asp Lys Ala Val Gln
50 55 60
Thr Tyr Gly Ile Ala Glu Gly Ile Ser Gln Gly Ser Leu Ile Gly Leu
65 70 75 80
Val Cys Ala Phe His Leu Ser Gly Phe Arg Leu Phe Ser Lys Ala Asp
85 90 95
Glu Thr Lys Ala Phe Cys Asn Gln Gly Arg Tyr Pro Asn Gln Ala Phe
100 105 110
Ala Glu Lys Leu Arg Asn Glu Leu Ser Val Thr Leu Pro Lys Leu Ser
115 120 125
Pro Gln Ser Leu Asp Val Leu Phe Gln Ser Ser Pro Lys Ser Lys Asn
130 135 140
Gly Val Ala Pro Glu Trp Ser Lys Asn Ala Ile Arg Asn Arg Leu Tyr
145 150 155 160
Thr Asn Trp Thr Gly Lys Gly Ala Gly Thr Asn Pro Asp Glu His Leu
165 170 175
Leu Glu Ile Ala Glu Asp Ile Ala Ala Glu Ile Asp Ser Asp Leu Asp
180 185 190
Gly Trp Lys Asp Leu Glu Glu His Pro Glu Lys Gly Leu Ser Ala Ala
195 200 205
Asp Arg Tyr Phe Gln Ala Gln Gly Asp Phe Pro Ser Leu Thr Gly Leu
210 215 220
Pro Pro Ser Val Pro Leu Thr Pro Gln Asn Ser Thr Val Ala Phe Glu
225 230 235 240
Gly Asp Pro Val Cys Leu Asn Pro Ser Asp Asn Thr Leu Leu His Gln
245 250 255
Ala Val Ala Arg Cys Ala Gly Arg Ile Leu Gln Glu Gln Pro Asn Leu
260 265 270
Ser Pro Asp Lys Asn Arg Phe Ile Asn Gln Leu Gln Asp Glu Leu Val
275 280 285
Ser Ser Gln Asn Asn Gly Leu Ser Trp Leu Phe Gly Val Gly Phe Lys
290 295 300
Tyr Trp Lys Glu Met Ser Val Asp Gln Leu Ala Asp Asp Tyr Lys Val
305 310 315 320
Lys Ser Thr Asp Leu Asp Ala Leu Lys Gln Val Lys Ser Phe Ile Asp
325 330 335
Ala Ile Pro Leu Asn Pro Leu Phe Asp Thr Pro His Tyr Gly Glu Phe
340 345 350
Arg Ala Ser Val Ala Gly Lys Met Arg Ser Trp Val Lys Asn Tyr Trp
355 360 365
Lys Arg Leu Leu Asp Leu Lys Ser Gln Leu Gly Thr Ala Asn Ile Asn
370 375 380
Leu Pro Glu Gly Leu Asp Glu Gln Arg Ala Glu Asn Leu Phe Ser Gly
385 390 395 400
Leu Leu Ile Asp Ser Lys Gly Leu Arg Gln Val Thr Asp Lys Leu Pro
405 410 415
Ser Arg Leu Lys Lys Ala Glu Asp Thr Ile Asp Arg Leu Met Gly Asp
420 425 430
Gly Asn Pro Thr Ser Asp Asp Ile Glu Gln Val Glu Thr Val Ala Ala
435 440 445
Glu Ile Ser Ala Phe Ile Gly Gln Val Glu Gln Phe Asn Asn Gln Leu
450 455 460
Glu Gln Arg Leu Glu Asn Pro Leu Glu Gly Asp Asp Glu Thr Phe Leu
465 470 475 480
Lys Gln Leu Lys Ile Asp Leu Pro Ala Glu Phe Lys Lys Pro Pro Ala
485 490 495
Ile Asn Arg Ile Ser Gly Gly Ser Pro Asp Pro Thr Ala Glu Ile Ala
500 505 510
Glu Leu Glu Glu Lys Leu Asp Arg Leu Met Ser Ala Arg Lys Glu His
515 520 525
Tyr Glu Thr Ile Ala Glu Trp Ala Ser Ala Asn Lys Val Thr Leu Asp
530 535 540
Pro Met Glu Ala Met Thr Thr Leu Glu Ala Gln Arg Leu Thr Glu Arg
545 550 555 560
Gly Ala Glu Gly Asp Gln Glu Glu Phe Ala Leu Arg Leu Leu Leu Gln
565 570 575
Arg Ile Gly Arg Leu Ala Asn Arg Leu Ser Pro Gln Gly Ala Thr Ala
580 585 590
Ile Arg Asp Leu Leu Arg Pro Val Phe Thr Glu Lys Arg Glu Phe Asn
595 600 605
Leu Phe Phe His Asn Arg Met Gly Ser Leu Tyr Arg Ser Pro Tyr Ser
610 615 620
Thr Ser Arg His Gln Pro Phe Thr Ile Asp Val Ala Val Ala Lys Asn
625 630 635 640
Thr Asp Trp Met Asp Ala Leu Asp Gly Ile Ala Glu Thr Ile Met Lys
645 650 655
Gly Leu Ser Gln Ala Gly Asp Glu Leu Ser Leu Arg Leu Arg Asp Trp
660 665 670
Ile Asn Ile Ser Gly Phe Ser Leu Ser Gln Arg Leu Arg Gly Leu Pro
675 680 685
Asp Thr Val Pro Gly Glu Leu Ala Leu Val Arg Ser Ala Asp Asp Val
690 695 700
Arg Ile Pro Pro Met Leu Ala Leu Gln Leu Glu Glu Asp Glu Val Ser
705 710 715 720
Arg Glu Val Cys Leu Lys Ala Phe Asn Leu Tyr Val Ser Ala Ile Asn
725 730 735
Gly Cys Leu Phe Arg Ala Leu Arg Glu Gly Phe Ile Val Arg Thr Lys
740 745 750
Phe Gln Arg Leu Glu Arg Asp Val Leu Ser Tyr Val Pro Lys Thr Lys
755 760 765
Leu Trp Asn Tyr Pro Gln Arg Leu Asp Thr Ala Arg Gly Pro Ile His
770 775 780
Ser Ala Leu Ala Ala Ala Trp Ile Asn Lys Glu Gly Ser Val Ile Asp
785 790 795 800
Pro Val Glu Thr Val Thr Ala Leu Ser Asp Thr Gly Phe Ser Asp Asp
805 810 815
Gly Ile Pro Glu Tyr Leu Val Gln Ala Pro His Asp Trp Tyr Thr Pro
820 825 830
Ile Asp Leu Arg Asp Ile Ser Lys Pro Val Ser Gly Leu Pro Val Lys
835 840 845
Lys Asn Ile Thr Gly Leu Lys Arg Gln Lys Lys Gln Thr Ala Phe Arg
850 855 860
Met Val Gly Pro Ser Ser Phe Lys Ser His Leu Asp Ser Thr Leu Leu
865 870 875 880
Ser Glu Glu Val Lys Leu Gly Asp Phe Thr Leu Ile Phe Asp Gln Tyr
885 890 895
Tyr Lys Gln Arg Val Ser Tyr Asn Gly Arg Val Lys Ile Thr Phe Glu
900 905 910
Pro Asp Arg Leu His Val Glu Ala Ala Val Pro Val Ile Asp Lys Arg
915 920 925
Val Arg Pro Ser Thr Glu Glu Asp Ala Leu Phe Asp His Leu Leu Ala
930 935 940
Ile Asp Leu Gly Glu Lys Arg Val Gly Tyr Ala Val Tyr Asp Ile Lys
945 950 955 960
Ala Cys Leu Arg Thr Gly Asp Ile Lys Pro Leu Glu Asp Gly Asp Gly
965 970 975
Lys Pro Ile Val Gly Ser Val Ala Val Pro Ser Ile Arg Arg Leu Met
980 985 990
Lys Ala Val Arg Ser His Arg Gln Gln Arg Gln Pro Asn Gln Lys Val
995 1000 1005
Asn Gln Thr Tyr Ser Thr Ala Leu Met Asn Tyr Arg Glu Asn Val
1010 1015 1020
Ile Gly Asp Val Cys Asn Arg Ile Asp Thr Leu Met Glu Lys Tyr
1025 1030 1035
Asn Ala Phe Pro Val Leu Glu Ser Ser Val Met Asn Phe Glu Ala
1040 1045 1050
Gly Ser Arg Gln Leu Glu Met Val Tyr Gly Ser Val Leu His Arg
1055 1060 1065
Tyr Thr Tyr Ser Lys Ile Asp Ala His Thr Ala Lys Arg Lys Glu
1070 1075 1080
Tyr Trp Tyr Thr Gly Glu Tyr Trp Asp His Pro Tyr Leu Met Ala
1085 1090 1095
His Lys Trp Asn Glu Arg Thr Arg Ser Tyr Ser Gly Ser Leu Ser
1100 1105 1110
Ala Leu Thr Leu Tyr Pro Gly Val Met Val His Pro Ala Gly Thr
1115 1120 1125
Ser Gln Arg Cys His Gln Cys Lys Arg Asn Pro Met Val Glu Ile
1130 1135 1140
Lys Gln Leu Thr Gly Gln Val Glu Ile Asn Ala Asp Gly Ser Leu
1145 1150 1155
Glu Leu Asp Asp Gly Thr Ile Cys Leu Tyr Glu Gly Tyr Asp Tyr
1160 1165 1170
Ser Pro Glu Glu Tyr Lys Lys Ala Lys Arg Glu Lys Arg Arg Leu
1175 1180 1185
Asp Pro Asn Val Pro Leu Ser Gly Arg His Gln Ala Lys His Val
1190 1195 1200
Ser Ala Val Ala Lys Arg Asn Leu Arg Arg Pro Thr Val Ser Met
1205 1210 1215
Met Ser Gly Asp Thr Thr Gln Ala Arg Tyr Val Cys Leu Tyr Thr
1220 1225 1230
Asp Cys Asp Phe Thr Gly His Ala Asp Glu Asn Ala Ala Ile Asn
1235 1240 1245
Ile Gly Trp Lys Tyr Leu Thr Glu Arg Ile Ala Leu Ser Glu Ser
1250 1255 1260
Lys Asp Lys Ala Gly Val
1265
<210> 573
<211> 1285
<212> PRT
<213> Rhodobacter capsulatus
<400> 573
Met Gln Ile Gly Lys Val Gln Gly Arg Thr Ile Ser Glu Phe Gly Asp
1 5 10 15
Pro Ala Gly Gly Leu Lys Arg Lys Ile Ser Thr Asp Gly Lys Asn Arg
20 25 30
Lys Glu Leu Pro Ala His Leu Ser Ser Asp Pro Lys Ala Leu Ile Gly
35 40 45
Gln Trp Ile Ser Gly Ile Asp Lys Ile Tyr Arg Lys Pro Asp Ser Arg
50 55 60
Lys Ser Asp Gly Lys Ala Ile His Ser Pro Thr Pro Ser Lys Met Gln
65 70 75 80
Phe Asp Ala Arg Asp Asp Leu Gly Glu Ala Phe Trp Lys Leu Val Ser
85 90 95
Glu Ala Gly Leu Ala Gln Asp Ser Asp Tyr Asp Gln Phe Lys Arg Arg
100 105 110
Leu His Pro Tyr Gly Asp Lys Phe Gln Pro Ala Asp Ser Gly Ala Lys
115 120 125
Leu Lys Phe Glu Ala Asp Pro Pro Glu Pro Gln Ala Phe His Gly Arg
130 135 140
Trp Tyr Gly Ala Met Ser Lys Arg Gly Asn Asp Ala Lys Glu Leu Ala
145 150 155 160
Ala Ala Leu Tyr Glu His Leu His Val Asp Glu Lys Arg Ile Asp Gly
165 170 175
Gln Pro Lys Arg Asn Pro Lys Thr Asp Lys Phe Ala Pro Gly Leu Val
180 185 190
Val Ala Arg Ala Leu Gly Ile Glu Ser Ser Val Leu Pro Arg Gly Met
195 200 205
Ala Arg Leu Ala Arg Asn Trp Gly Glu Glu Glu Ile Gln Thr Tyr Phe
210 215 220
Val Val Asp Val Ala Ala Ser Val Lys Glu Val Ala Lys Ala Ala Val
225 230 235 240
Ser Ala Ala Gln Ala Phe Asp Pro Pro Arg Gln Val Ser Gly Arg Ser
245 250 255
Leu Ser Pro Lys Val Gly Phe Ala Leu Ala Glu His Leu Glu Arg Val
260 265 270
Thr Gly Ser Lys Arg Cys Ser Phe Asp Pro Ala Ala Gly Pro Ser Val
275 280 285
Leu Ala Leu His Asp Glu Val Lys Lys Thr Tyr Lys Arg Leu Cys Ala
290 295 300
Arg Gly Lys Asn Ala Ala Arg Ala Phe Pro Ala Asp Lys Thr Glu Leu
305 310 315 320
Leu Ala Leu Met Arg His Thr His Glu Asn Arg Val Arg Asn Gln Met
325 330 335
Val Arg Met Gly Arg Val Ser Glu Tyr Arg Gly Gln Gln Ala Gly Asp
340 345 350
Leu Ala Gln Ser His Tyr Trp Thr Ser Ala Gly Gln Thr Glu Ile Lys
355 360 365
Glu Ser Glu Ile Phe Val Arg Leu Trp Val Gly Ala Phe Ala Leu Ala
370 375 380
Gly Arg Ser Met Lys Ala Trp Ile Asp Pro Met Gly Lys Ile Val Asn
385 390 395 400
Thr Glu Lys Asn Asp Arg Asp Leu Thr Ala Ala Val Asn Ile Arg Gln
405 410 415
Val Ile Ser Asn Lys Glu Met Val Ala Glu Ala Met Ala Arg Arg Gly
420 425 430
Ile Tyr Phe Gly Glu Thr Pro Glu Leu Asp Arg Leu Gly Ala Glu Gly
435 440 445
Asn Glu Gly Phe Val Phe Ala Leu Leu Arg Tyr Leu Arg Gly Cys Arg
450 455 460
Asn Gln Thr Phe His Leu Gly Ala Arg Ala Gly Phe Leu Lys Glu Ile
465 470 475 480
Arg Lys Glu Leu Glu Lys Thr Arg Trp Gly Lys Ala Lys Glu Ala Glu
485 490 495
His Val Val Leu Thr Asp Lys Thr Val Ala Ala Ile Arg Ala Ile Ile
500 505 510
Asp Asn Asp Ala Lys Ala Leu Gly Ala Arg Leu Leu Ala Asp Leu Ser
515 520 525
Gly Ala Phe Val Ala His Tyr Ala Ser Lys Glu His Phe Ser Thr Leu
530 535 540
Tyr Ser Glu Ile Val Lys Ala Val Lys Asp Ala Pro Glu Val Ser Ser
545 550 555 560
Gly Leu Pro Arg Leu Lys Leu Leu Leu Lys Arg Ala Asp Gly Val Arg
565 570 575
Gly Tyr Val His Gly Leu Arg Asp Thr Arg Lys His Ala Phe Ala Thr
580 585 590
Lys Leu Pro Pro Pro Pro Ala Pro Arg Glu Leu Asp Asp Pro Ala Thr
595 600 605
Lys Ala Arg Tyr Ile Ala Leu Leu Arg Leu Tyr Asp Gly Pro Phe Arg
610 615 620
Ala Tyr Ala Ser Gly Ile Thr Gly Thr Ala Leu Ala Gly Pro Ala Ala
625 630 635 640
Arg Ala Lys Glu Ala Ala Thr Ala Leu Ala Gln Ser Val Asn Val Thr
645 650 655
Lys Ala Tyr Ser Asp Val Met Glu Gly Arg Ser Ser Arg Leu Arg Pro
660 665 670
Pro Asn Asp Gly Glu Thr Leu Arg Glu Tyr Leu Ser Ala Leu Thr Gly
675 680 685
Glu Thr Ala Thr Glu Phe Arg Val Gln Ile Gly Tyr Glu Ser Asp Ser
690 695 700
Glu Asn Ala Arg Lys Gln Ala Glu Phe Ile Glu Asn Tyr Arg Arg Asp
705 710 715 720
Met Leu Ala Phe Met Phe Glu Asp Tyr Ile Arg Ala Lys Gly Phe Asp
725 730 735
Trp Ile Leu Lys Ile Glu Pro Gly Ala Thr Ala Met Thr Arg Ala Pro
740 745 750
Val Leu Pro Glu Pro Ile Asp Thr Arg Gly Gln Tyr Glu His Trp Gln
755 760 765
Ala Ala Leu Tyr Leu Val Met His Phe Val Pro Ala Ser Asp Val Ser
770 775 780
Asn Leu Leu His Gln Leu Arg Lys Trp Glu Ala Leu Gln Gly Lys Tyr
785 790 795 800
Glu Leu Val Gln Asp Gly Asp Ala Thr Asp Gln Ala Asp Ala Arg Arg
805 810 815
Glu Ala Leu Asp Leu Val Lys Arg Phe Arg Asp Val Leu Val Leu Phe
820 825 830
Leu Lys Thr Gly Glu Ala Arg Phe Glu Gly Arg Ala Ala Pro Phe Asp
835 840 845
Leu Lys Pro Phe Arg Ala Leu Phe Ala Asn Pro Ala Thr Phe Asp Arg
850 855 860
Leu Phe Met Ala Thr Pro Thr Thr Ala Arg Pro Ala Glu Asp Asp Pro
865 870 875 880
Glu Gly Asp Gly Ala Ser Glu Pro Glu Leu Arg Val Ala Arg Thr Leu
885 890 895
Arg Gly Leu Arg Gln Ile Ala Arg Tyr Asn His Met Ala Val Leu Ser
900 905 910
Asp Leu Phe Ala Lys His Lys Val Arg Asp Glu Glu Val Ala Arg Leu
915 920 925
Ala Glu Ile Glu Asp Glu Thr Gln Glu Lys Ser Gln Ile Val Ala Ala
930 935 940
Gln Glu Leu Arg Thr Asp Leu His Asp Lys Val Met Lys Cys His Pro
945 950 955 960
Lys Thr Ile Ser Pro Glu Glu Arg Gln Ser Tyr Ala Ala Ala Ile Lys
965 970 975
Thr Ile Glu Glu His Arg Phe Leu Val Gly Arg Val Tyr Leu Gly Asp
980 985 990
His Leu Arg Leu His Arg Leu Met Met Asp Val Ile Gly Arg Leu Ile
995 1000 1005
Asp Tyr Ala Gly Ala Tyr Glu Arg Asp Thr Gly Thr Phe Leu Ile
1010 1015 1020
Asn Ala Ser Lys Gln Leu Gly Ala Gly Ala Asp Trp Ala Val Thr
1025 1030 1035
Ile Ala Gly Ala Ala Asn Thr Asp Ala Arg Thr Gln Thr Arg Lys
1040 1045 1050
Asp Leu Ala His Phe Asn Val Leu Asp Arg Ala Asp Gly Thr Pro
1055 1060 1065
Asp Leu Thr Ala Leu Val Asn Arg Ala Arg Glu Met Met Ala Tyr
1070 1075 1080
Asp Arg Lys Arg Lys Asn Ala Val Pro Arg Ser Ile Leu Asp Met
1085 1090 1095
Leu Ala Arg Leu Gly Leu Thr Leu Lys Trp Gln Met Lys Asp His
1100 1105 1110
Leu Leu Gln Asp Ala Thr Ile Thr Gln Ala Ala Ile Lys His Leu
1115 1120 1125
Asp Lys Val Arg Leu Thr Val Gly Gly Pro Ala Ala Val Thr Glu
1130 1135 1140
Ala Arg Phe Ser Gln Asp Tyr Leu Gln Met Val Ala Ala Val Phe
1145 1150 1155
Asn Gly Ser Val Gln Asn Pro Lys Pro Arg Arg Arg Asp Asp Gly
1160 1165 1170
Asp Ala Trp His Lys Pro Pro Lys Pro Ala Thr Ala Gln Ser Gln
1175 1180 1185
Pro Asp Gln Lys Pro Pro Asn Lys Ala Pro Ser Ala Gly Ser Arg
1190 1195 1200
Leu Pro Pro Pro Gln Val Gly Glu Val Tyr Glu Gly Val Val Val
1205 1210 1215
Lys Val Ile Asp Thr Gly Ser Leu Gly Phe Leu Ala Val Glu Gly
1220 1225 1230
Val Ala Gly Asn Ile Gly Leu His Ile Ser Arg Leu Arg Arg Ile
1235 1240 1245
Arg Glu Asp Ala Ile Ile Val Gly Arg Arg Tyr Arg Phe Arg Val
1250 1255 1260
Glu Ile Tyr Val Pro Pro Lys Ser Asn Thr Ser Lys Leu Asn Ala
1265 1270 1275
Ala Asp Leu Val Arg Ile Asp
1280 1285
<210> 574
<211> 1340
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 574
Met Gln Ile Ser Lys Val Asn His Lys His Val Ala Val Gly Gln Lys
1 5 10 15
Asp Arg Glu Arg Ile Thr Gly Phe Ile Tyr Asn Asp Pro Val Gly Asp
20 25 30
Glu Lys Ser Leu Glu Asp Val Val Ala Lys Arg Ala Asn Asp Thr Lys
35 40 45
Val Leu Phe Asn Val Phe Asn Thr Lys Asp Leu Tyr Asp Ser Gln Glu
50 55 60
Ser Asp Lys Ser Glu Lys Asp Lys Glu Ile Ile Ser Lys Gly Ala Lys
65 70 75 80
Phe Val Ala Lys Ser Phe Asn Ser Ala Ile Thr Ile Leu Lys Lys Gln
85 90 95
Asn Lys Ile Tyr Ser Thr Leu Thr Ser Gln Gln Val Ile Lys Glu Leu
100 105 110
Lys Asp Lys Phe Gly Gly Ala Arg Ile Tyr Asp Asp Asp Ile Glu Glu
115 120 125
Ala Leu Thr Glu Thr Leu Lys Lys Ser Phe Arg Lys Glu Asn Val Arg
130 135 140
Asn Ser Ile Lys Val Leu Ile Glu Asn Ala Ala Gly Ile Arg Ser Ser
145 150 155 160
Leu Ser Lys Asp Glu Glu Glu Leu Ile Gln Glu Tyr Phe Val Lys Gln
165 170 175
Leu Val Glu Glu Tyr Thr Lys Thr Lys Leu Gln Lys Asn Val Val Lys
180 185 190
Ser Ile Lys Asn Gln Asn Met Val Ile Gln Pro Asp Ser Asp Ser Gln
195 200 205
Val Leu Ser Leu Ser Glu Ser Arg Arg Glu Lys Gln Ser Ser Ala Val
210 215 220
Ser Ser Asp Thr Leu Val Asn Cys Lys Glu Lys Asp Val Leu Lys Ala
225 230 235 240
Phe Leu Thr Asp Tyr Ala Val Leu Asp Glu Asp Glu Arg Asn Ser Leu
245 250 255
Leu Trp Lys Leu Arg Asn Leu Val Asn Leu Tyr Phe Tyr Gly Ser Glu
260 265 270
Ser Ile Arg Asp Tyr Ser Tyr Thr Lys Glu Lys Ser Val Trp Lys Glu
275 280 285
His Asp Glu Gln Lys Ala Asn Lys Thr Leu Phe Ile Asp Glu Ile Cys
290 295 300
His Ile Thr Lys Ile Gly Lys Asn Gly Lys Glu Gln Lys Val Leu Asp
305 310 315 320
Tyr Glu Glu Asn Arg Ser Arg Cys Arg Lys Gln Asn Ile Asn Tyr Tyr
325 330 335
Arg Ser Ala Leu Asn Tyr Ala Lys Asn Asn Thr Ser Gly Ile Phe Glu
340 345 350
Asn Glu Asp Ser Asn His Phe Trp Ile His Leu Ile Glu Asn Glu Val
355 360 365
Glu Arg Leu Tyr Asn Gly Ile Glu Asn Gly Glu Glu Phe Lys Phe Glu
370 375 380
Thr Gly Tyr Ile Ser Glu Lys Val Trp Lys Ala Val Ile Asn His Leu
385 390 395 400
Ser Ile Lys Tyr Ile Ala Leu Gly Lys Ala Val Tyr Asn Tyr Ala Met
405 410 415
Lys Glu Leu Ser Ser Pro Gly Asp Ile Glu Pro Gly Lys Ile Asp Asp
420 425 430
Ser Tyr Ile Asn Gly Ile Thr Ser Phe Asp Tyr Glu Ile Ile Lys Ala
435 440 445
Glu Glu Ser Leu Gln Arg Asp Ile Ser Met Asn Val Val Phe Ala Thr
450 455 460
Asn Tyr Leu Ala Cys Ala Thr Val Asp Thr Asp Lys Asp Phe Leu Leu
465 470 475 480
Phe Ser Lys Glu Asp Ile Arg Ser Cys Thr Lys Lys Asp Gly Asn Leu
485 490 495
Cys Lys Asn Ile Met Gln Phe Trp Gly Gly Tyr Ser Thr Trp Lys Asn
500 505 510
Phe Cys Glu Glu Tyr Leu Lys Asp Asp Lys Asp Ala Leu Glu Leu Leu
515 520 525
Tyr Ser Leu Lys Ser Met Leu Tyr Ser Met Arg Asn Ser Ser Phe His
530 535 540
Phe Ser Thr Glu Asn Val Asp Asn Gly Ser Trp Asp Thr Glu Leu Ile
545 550 555 560
Gly Lys Leu Phe Glu Glu Asp Cys Asn Arg Ala Ala Arg Ile Glu Lys
565 570 575
Glu Lys Phe Tyr Asn Asn Asn Leu His Met Phe Tyr Ser Ser Ser Leu
580 585 590
Leu Glu Lys Val Leu Glu Arg Leu Tyr Ser Ser His His Glu Arg Ala
595 600 605
Ser Gln Val Pro Ser Phe Asn Arg Val Phe Val Arg Lys Asn Phe Pro
610 615 620
Ser Ser Leu Ser Glu Gln Arg Ile Thr Pro Lys Phe Thr Asp Ser Lys
625 630 635 640
Asp Glu Gln Ile Trp Gln Ser Ala Val Tyr Tyr Leu Cys Lys Glu Ile
645 650 655
Tyr Tyr Asn Asp Phe Leu Gln Ser Lys Glu Ala Tyr Lys Leu Phe Arg
660 665 670
Glu Gly Val Lys Asn Leu Asp Lys Asn Asp Ile Asn Asn Gln Lys Ala
675 680 685
Ala Asp Ser Phe Lys Gln Ala Val Val Tyr Tyr Gly Lys Ala Ile Gly
690 695 700
Asn Ala Thr Leu Ser Gln Val Cys Gln Ala Ile Met Thr Glu Tyr Asn
705 710 715 720
Arg Gln Asn Asn Asp Gly Leu Lys Lys Lys Ser Ala Tyr Ala Glu Lys
725 730 735
Gln Asn Ser Asn Lys Tyr Lys His Tyr Pro Leu Phe Leu Lys Gln Val
740 745 750
Leu Gln Ser Ala Phe Trp Glu Tyr Leu Asp Glu Asn Lys Glu Ile Tyr
755 760 765
Gly Phe Ile Ser Ala Gln Ile His Lys Ser Asn Val Glu Ile Lys Ala
770 775 780
Glu Asp Phe Ile Ala Asn Tyr Ser Ser Gln Gln Tyr Lys Lys Leu Val
785 790 795 800
Asp Lys Val Lys Lys Thr Pro Glu Leu Gln Lys Trp Tyr Thr Leu Gly
805 810 815
Arg Leu Ile Asn Pro Arg Gln Ala Asn Gln Phe Leu Gly Ser Ile Arg
820 825 830
Asn Tyr Val Gln Phe Val Lys Asp Ile Gln Arg Arg Ala Lys Glu Asn
835 840 845
Gly Asn Pro Ile Arg Asn Tyr Tyr Glu Val Leu Glu Ser Asp Ser Ile
850 855 860
Ile Lys Ile Leu Glu Met Cys Thr Lys Leu Asn Gly Thr Thr Ser Asn
865 870 875 880
Asp Ile His Asp Tyr Phe Arg Asp Glu Asp Glu Tyr Ala Glu Tyr Ile
885 890 895
Ser Gln Phe Val Asn Phe Gly Asp Val His Ser Gly Ala Ala Leu Asn
900 905 910
Ala Phe Cys Asn Ser Glu Ser Glu Gly Lys Lys Asn Gly Ile Tyr Tyr
915 920 925
Asp Gly Ile Asn Pro Ile Val Asn Arg Asn Trp Val Leu Cys Lys Leu
930 935 940
Tyr Gly Ser Pro Asp Leu Ile Ser Lys Ile Ile Ser Arg Val Asn Glu
945 950 955 960
Asn Met Ile His Asp Phe His Lys Gln Glu Asp Leu Ile Arg Glu Tyr
965 970 975
Gln Ile Lys Gly Ile Cys Ser Asn Lys Lys Glu Gln Gln Asp Leu Arg
980 985 990
Thr Phe Gln Val Leu Lys Asn Arg Val Glu Leu Arg Asp Ile Val Glu
995 1000 1005
Tyr Ser Glu Ile Ile Asn Glu Leu Tyr Gly Gln Leu Ile Lys Trp
1010 1015 1020
Cys Tyr Leu Arg Glu Arg Asp Leu Met Tyr Phe Gln Leu Gly Phe
1025 1030 1035
His Tyr Leu Cys Leu Asn Asn Ala Ser Ser Lys Glu Ala Asp Tyr
1040 1045 1050
Ile Lys Ile Asn Val Asp Asp Arg Asn Ile Ser Gly Ala Ile Leu
1055 1060 1065
Tyr Gln Ile Ala Ala Met Tyr Ile Asn Gly Leu Pro Val Tyr Tyr
1070 1075 1080
Lys Lys Asp Asp Met Tyr Val Ala Leu Lys Ser Gly Lys Lys Ala
1085 1090 1095
Ser Asp Glu Leu Asn Ser Asn Glu Gln Thr Ser Lys Lys Ile Asn
1100 1105 1110
Tyr Phe Leu Lys Tyr Gly Asn Asn Ile Leu Gly Asp Lys Lys Asp
1115 1120 1125
Gln Leu Tyr Leu Ala Gly Leu Glu Leu Phe Glu Asn Val Ala Glu
1130 1135 1140
His Glu Asn Ile Ile Ile Phe Arg Asn Glu Ile Asp His Phe His
1145 1150 1155
Tyr Phe Tyr Asp Arg Asp Arg Ser Met Leu Asp Leu Tyr Ser Glu
1160 1165 1170
Val Phe Asp Arg Phe Phe Thr Tyr Asp Met Lys Leu Arg Lys Asn
1175 1180 1185
Val Val Asn Met Leu Tyr Asn Ile Leu Leu Asp His Asn Ile Val
1190 1195 1200
Ser Ser Phe Val Phe Glu Thr Gly Glu Lys Lys Val Gly Arg Gly
1205 1210 1215
Asp Ser Glu Val Ile Lys Pro Ser Ala Lys Ile Arg Leu Arg Ala
1220 1225 1230
Asn Asn Gly Val Ser Ser Asp Val Phe Thr Tyr Lys Val Gly Ser
1235 1240 1245
Lys Asp Glu Leu Lys Ile Ala Thr Leu Pro Ala Lys Asn Glu Glu
1250 1255 1260
Phe Leu Leu Asn Val Ala Arg Leu Ile Tyr Tyr Pro Asp Met Glu
1265 1270 1275
Ala Val Ser Glu Asn Met Val Arg Glu Gly Val Val Lys Val Glu
1280 1285 1290
Lys Ser Asn Asp Lys Lys Gly Lys Ile Ser Arg Gly Ser Asn Thr
1295 1300 1305
Arg Ser Ser Asn Gln Ser Lys Tyr Asn Asn Lys Ser Lys Asn Arg
1310 1315 1320
Met Asn Tyr Ser Met Gly Ser Ile Phe Glu Lys Met Asp Leu Lys
1325 1330 1335
Phe Asp
1340
<210> 575
<211> 1437
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 575
Met Lys Ile Ser Lys Val Arg Glu Glu Asn Arg Gly Ala Lys Leu Thr
1 5 10 15
Val Asn Ala Lys Thr Ala Val Val Ser Glu Asn Arg Ser Gln Glu Gly
20 25 30
Ile Leu Tyr Asn Asp Pro Ser Arg Tyr Gly Lys Ser Arg Lys Asn Asp
35 40 45
Glu Asp Arg Asp Arg Tyr Ile Glu Ser Arg Leu Lys Ser Ser Gly Lys
50 55 60
Leu Tyr Arg Ile Phe Asn Glu Asp Lys Asn Lys Arg Glu Thr Asp Glu
65 70 75 80
Leu Gln Trp Phe Leu Ser Glu Ile Val Lys Lys Ile Asn Arg Arg Asn
85 90 95
Gly Leu Val Leu Ser Asp Met Leu Ser Val Asp Asp Arg Ala Phe Glu
100 105 110
Lys Ala Phe Glu Lys Tyr Ala Glu Leu Ser Tyr Thr Asn Arg Arg Asn
115 120 125
Lys Val Ser Gly Ser Pro Ala Phe Glu Thr Cys Gly Val Asp Ala Ala
130 135 140
Thr Ala Glu Arg Leu Lys Gly Ile Ile Ser Glu Thr Asn Phe Ile Asn
145 150 155 160
Arg Ile Lys Asn Asn Ile Asp Asn Lys Val Ser Glu Asp Ile Ile Asp
165 170 175
Arg Ile Ile Ala Lys Tyr Leu Lys Lys Ser Leu Cys Arg Glu Arg Val
180 185 190
Lys Arg Gly Leu Lys Lys Leu Leu Met Asn Ala Phe Asp Leu Pro Tyr
195 200 205
Ser Asp Pro Asp Ile Asp Val Gln Arg Asp Phe Ile Asp Tyr Val Leu
210 215 220
Glu Asp Phe Tyr His Val Arg Ala Lys Ser Gln Val Ser Arg Ser Ile
225 230 235 240
Lys Asn Met Asn Met Pro Val Gln Pro Glu Gly Asp Gly Lys Phe Ala
245 250 255
Ile Thr Val Ser Lys Gly Gly Thr Glu Ser Gly Asn Lys Arg Ser Ala
260 265 270
Glu Lys Glu Ala Phe Lys Lys Phe Leu Ser Asp Tyr Ala Ser Leu Asp
275 280 285
Glu Arg Val Arg Asp Asp Met Leu Arg Arg Met Arg Arg Leu Val Val
290 295 300
Leu Tyr Phe Tyr Gly Ser Asp Asp Ser Lys Leu Ser Asp Val Asn Glu
305 310 315 320
Lys Phe Asp Val Trp Glu Asp His Ala Ala Arg Arg Val Asp Asn Arg
325 330 335
Glu Phe Ile Lys Leu Pro Leu Glu Asn Lys Leu Ala Asn Gly Lys Thr
340 345 350
Asp Lys Asp Ala Glu Arg Ile Arg Lys Asn Thr Val Lys Glu Leu Tyr
355 360 365
Arg Asn Gln Asn Ile Gly Cys Tyr Arg Gln Ala Val Lys Ala Val Glu
370 375 380
Glu Asp Asn Asn Gly Arg Tyr Phe Asp Asp Lys Met Leu Asn Met Phe
385 390 395 400
Phe Ile His Arg Ile Glu Tyr Gly Val Glu Lys Ile Tyr Ala Asn Leu
405 410 415
Lys Gln Val Thr Glu Phe Lys Ala Arg Thr Gly Tyr Leu Ser Glu Lys
420 425 430
Ile Trp Lys Asp Leu Ile Asn Tyr Ile Ser Ile Lys Tyr Ile Ala Met
435 440 445
Gly Lys Ala Val Tyr Asn Tyr Ala Met Asp Glu Leu Asn Ala Ser Asp
450 455 460
Lys Lys Glu Ile Glu Leu Gly Lys Ile Ser Glu Glu Tyr Leu Ser Gly
465 470 475 480
Ile Ser Ser Phe Asp Tyr Glu Leu Ile Lys Ala Glu Glu Met Leu Gln
485 490 495
Arg Glu Thr Ala Val Tyr Val Ala Phe Ala Ala Arg His Leu Ser Ser
500 505 510
Gln Thr Val Glu Leu Asp Ser Glu Asn Ser Asp Phe Leu Leu Leu Lys
515 520 525
Pro Lys Gly Thr Met Asp Lys Asn Asp Lys Asn Lys Leu Ala Ser Asn
530 535 540
Asn Ile Leu Asn Phe Leu Lys Asp Lys Glu Thr Leu Arg Asp Thr Ile
545 550 555 560
Leu Gln Tyr Phe Gly Gly His Ser Leu Trp Thr Asp Phe Pro Phe Asp
565 570 575
Lys Tyr Leu Ala Gly Gly Lys Asp Asp Val Asp Phe Leu Thr Asp Leu
580 585 590
Lys Asp Val Ile Tyr Ser Met Arg Asn Asp Ser Phe His Tyr Ala Thr
595 600 605
Glu Asn His Asn Asn Gly Lys Trp Asn Lys Glu Leu Ile Ser Ala Met
610 615 620
Phe Glu His Glu Thr Glu Arg Met Thr Val Val Met Lys Asp Lys Phe
625 630 635 640
Tyr Ser Asn Asn Leu Pro Met Phe Tyr Lys Asn Asp Asp Leu Lys Lys
645 650 655
Leu Leu Ile Asp Leu Tyr Lys Asp Asn Val Glu Arg Ala Ser Gln Val
660 665 670
Pro Ser Phe Asn Lys Val Phe Val Arg Lys Asn Phe Pro Ala Leu Val
675 680 685
Arg Asp Lys Asp Asn Leu Gly Ile Glu Leu Asp Leu Lys Ala Asp Ala
690 695 700
Asp Lys Gly Glu Asn Glu Leu Lys Phe Tyr Asn Ala Leu Tyr Tyr Met
705 710 715 720
Phe Lys Glu Ile Tyr Tyr Asn Ala Phe Leu Asn Asp Lys Asn Val Arg
725 730 735
Glu Arg Phe Ile Thr Lys Ala Thr Lys Val Ala Asp Asn Tyr Asp Arg
740 745 750
Asn Lys Glu Arg Asn Leu Lys Asp Arg Ile Lys Ser Ala Gly Ser Asp
755 760 765
Glu Lys Lys Lys Leu Arg Glu Gln Leu Gln Asn Tyr Ile Ala Glu Asn
770 775 780
Asp Phe Gly Gln Arg Ile Lys Asn Ile Val Gln Val Asn Pro Asp Tyr
785 790 795 800
Thr Leu Ala Gln Ile Cys Gln Leu Ile Met Thr Glu Tyr Asn Gln Gln
805 810 815
Asn Asn Gly Cys Met Gln Lys Lys Ser Ala Ala Arg Lys Asp Ile Asn
820 825 830
Lys Asp Ser Tyr Gln His Tyr Lys Met Leu Leu Leu Val Asn Leu Arg
835 840 845
Lys Ala Phe Leu Glu Phe Ile Lys Glu Asn Tyr Ala Phe Val Leu Lys
850 855 860
Pro Tyr Lys His Asp Leu Cys Asp Lys Ala Asp Phe Val Pro Asp Phe
865 870 875 880
Ala Lys Tyr Val Lys Pro Tyr Ala Gly Leu Ile Ser Arg Val Ala Gly
885 890 895
Ser Ser Glu Leu Gln Lys Trp Tyr Ile Val Ser Arg Phe Leu Ser Pro
900 905 910
Ala Gln Ala Asn His Met Leu Gly Phe Leu His Ser Tyr Lys Gln Tyr
915 920 925
Val Trp Asp Ile Tyr Arg Arg Ala Ser Glu Thr Gly Thr Glu Ile Asn
930 935 940
His Ser Ile Ala Glu Asp Lys Ile Ala Gly Val Asp Ile Thr Asp Val
945 950 955 960
Asp Ala Val Ile Asp Leu Ser Val Lys Leu Cys Gly Thr Ile Ser Ser
965 970 975
Glu Ile Ser Asp Tyr Phe Lys Asp Asp Glu Val Tyr Ala Glu Tyr Ile
980 985 990
Ser Ser Tyr Leu Asp Phe Glu Tyr Asp Gly Gly Asn Tyr Lys Asp Ser
995 1000 1005
Leu Asn Arg Phe Cys Asn Ser Asp Ala Val Asn Asp Gln Lys Val
1010 1015 1020
Ala Leu Tyr Tyr Asp Gly Glu His Pro Lys Leu Asn Arg Asn Ile
1025 1030 1035
Ile Leu Ser Lys Leu Tyr Gly Glu Arg Arg Phe Leu Glu Lys Ile
1040 1045 1050
Thr Asp Arg Val Ser Arg Ser Asp Ile Val Glu Tyr Tyr Lys Leu
1055 1060 1065
Lys Lys Glu Thr Ser Gln Tyr Gln Thr Lys Gly Ile Phe Asp Ser
1070 1075 1080
Glu Asp Glu Gln Lys Asn Ile Lys Lys Phe Gln Glu Met Lys Asn
1085 1090 1095
Ile Val Glu Phe Arg Asp Leu Met Asp Tyr Ser Glu Ile Ala Asp
1100 1105 1110
Glu Leu Gln Gly Gln Leu Ile Asn Trp Ile Tyr Leu Arg Glu Arg
1115 1120 1125
Asp Leu Met Asn Phe Gln Leu Gly Tyr His Tyr Ala Cys Leu Asn
1130 1135 1140
Asn Asp Ser Asn Lys Gln Ala Thr Tyr Val Thr Leu Asp Tyr Gln
1145 1150 1155
Gly Lys Lys Asn Arg Lys Ile Asn Gly Ala Ile Leu Tyr Gln Ile
1160 1165 1170
Cys Ala Met Tyr Ile Asn Gly Leu Pro Leu Tyr Tyr Val Asp Lys
1175 1180 1185
Asp Ser Ser Glu Trp Thr Val Ser Asp Gly Lys Glu Ser Thr Gly
1190 1195 1200
Ala Lys Ile Gly Glu Phe Tyr Arg Tyr Ala Lys Ser Phe Glu Asn
1205 1210 1215
Thr Ser Asp Cys Tyr Ala Ser Gly Leu Glu Ile Phe Glu Asn Ile
1220 1225 1230
Ser Glu His Asp Asn Ile Thr Glu Leu Arg Asn Tyr Ile Glu His
1235 1240 1245
Phe Arg Tyr Tyr Ser Ser Phe Asp Arg Ser Phe Leu Gly Ile Tyr
1250 1255 1260
Ser Glu Val Phe Asp Arg Phe Phe Thr Tyr Asp Leu Lys Tyr Arg
1265 1270 1275
Lys Asn Val Pro Thr Ile Leu Tyr Asn Ile Leu Leu Gln His Phe
1280 1285 1290
Val Asn Val Arg Phe Glu Phe Val Ser Gly Lys Lys Met Ile Gly
1295 1300 1305
Ile Asp Lys Lys Asp Arg Lys Ile Ala Lys Glu Lys Glu Cys Ala
1310 1315 1320
Arg Ile Thr Ile Arg Glu Lys Asn Gly Val Tyr Ser Glu Gln Phe
1325 1330 1335
Thr Tyr Lys Leu Lys Asn Gly Thr Val Tyr Val Asp Ala Arg Asp
1340 1345 1350
Lys Arg Tyr Leu Gln Ser Ile Ile Arg Leu Leu Phe Tyr Pro Glu
1355 1360 1365
Lys Val Asn Met Asp Glu Met Ile Glu Val Lys Glu Lys Lys Lys
1370 1375 1380
Pro Ser Asp Asn Asn Thr Gly Lys Gly Tyr Ser Lys Arg Asp Arg
1385 1390 1395
Gln Gln Asp Arg Lys Glu Tyr Asp Lys Tyr Lys Glu Lys Lys Lys
1400 1405 1410
Lys Glu Gly Asn Phe Leu Ser Gly Met Gly Gly Asn Ile Asn Trp
1415 1420 1425
Asp Glu Ile Asn Ala Gln Leu Lys Asn
1430 1435
<210> 576
<211> 1385
<212> PRT
<213> Clostridium aminophilum
<400> 576
Met Lys Phe Ser Lys Val Asp His Thr Arg Ser Ala Val Gly Ile Gln
1 5 10 15
Lys Ala Thr Asp Ser Val His Gly Met Leu Tyr Thr Asp Pro Lys Lys
20 25 30
Gln Glu Val Asn Asp Leu Asp Lys Arg Phe Asp Gln Leu Asn Val Lys
35 40 45
Ala Lys Arg Leu Tyr Asn Val Phe Asn Gln Ser Lys Ala Glu Glu Asp
50 55 60
Asp Asp Glu Lys Arg Phe Gly Lys Val Val Lys Lys Leu Asn Arg Glu
65 70 75 80
Leu Lys Asp Leu Leu Phe His Arg Glu Val Ser Arg Tyr Asn Ser Ile
85 90 95
Gly Asn Ala Lys Tyr Asn Tyr Tyr Gly Ile Lys Ser Asn Pro Glu Glu
100 105 110
Ile Val Ser Asn Leu Gly Met Val Glu Ser Leu Lys Gly Glu Arg Asp
115 120 125
Pro Gln Lys Val Ile Ser Lys Leu Leu Leu Tyr Tyr Leu Arg Lys Gly
130 135 140
Leu Lys Pro Gly Thr Asp Gly Leu Arg Met Ile Leu Glu Ala Ser Cys
145 150 155 160
Gly Leu Arg Lys Leu Ser Gly Asp Glu Lys Glu Leu Lys Val Phe Leu
165 170 175
Gln Thr Leu Asp Glu Asp Phe Glu Lys Lys Thr Phe Lys Lys Asn Leu
180 185 190
Ile Arg Ser Ile Glu Asn Gln Asn Met Ala Val Gln Pro Ser Asn Glu
195 200 205
Gly Asp Pro Ile Ile Gly Ile Thr Gln Gly Arg Phe Asn Ser Gln Lys
210 215 220
Asn Glu Glu Lys Ser Ala Ile Glu Arg Met Met Ser Met Tyr Ala Asp
225 230 235 240
Leu Asn Glu Asp His Arg Glu Asp Val Leu Arg Lys Leu Arg Arg Leu
245 250 255
Asn Val Leu Tyr Phe Asn Val Asp Thr Glu Lys Thr Glu Glu Pro Thr
260 265 270
Leu Pro Gly Glu Val Asp Thr Asn Pro Val Phe Glu Val Trp His Asp
275 280 285
His Glu Lys Gly Lys Glu Asn Asp Arg Gln Phe Ala Thr Phe Ala Lys
290 295 300
Ile Leu Thr Glu Asp Arg Glu Thr Arg Lys Lys Glu Lys Leu Ala Val
305 310 315 320
Lys Glu Ala Leu Asn Asp Leu Lys Ser Ala Ile Arg Asp His Asn Ile
325 330 335
Met Ala Tyr Arg Cys Ser Ile Lys Val Thr Glu Gln Asp Lys Asp Gly
340 345 350
Leu Phe Phe Glu Asp Gln Arg Ile Asn Arg Phe Trp Ile His His Ile
355 360 365
Glu Ser Ala Val Glu Arg Ile Leu Ala Ser Ile Asn Pro Glu Lys Leu
370 375 380
Tyr Lys Leu Arg Ile Gly Tyr Leu Gly Glu Lys Val Trp Lys Asp Leu
385 390 395 400
Leu Asn Tyr Leu Ser Ile Lys Tyr Ile Ala Val Gly Lys Ala Val Phe
405 410 415
His Phe Ala Met Glu Asp Leu Gly Lys Thr Gly Gln Asp Ile Glu Leu
420 425 430
Gly Lys Leu Ser Asn Ser Val Ser Gly Gly Leu Thr Ser Phe Asp Tyr
435 440 445
Glu Gln Ile Arg Ala Asp Glu Thr Leu Gln Arg Gln Leu Ser Val Glu
450 455 460
Val Ala Phe Ala Ala Asn Asn Leu Phe Arg Ala Val Val Gly Gln Thr
465 470 475 480
Gly Lys Lys Ile Glu Gln Ser Lys Ser Glu Glu Asn Glu Glu Asp Phe
485 490 495
Leu Leu Trp Lys Ala Glu Lys Ile Ala Glu Ser Ile Lys Lys Glu Gly
500 505 510
Glu Gly Asn Thr Leu Lys Ser Ile Leu Gln Phe Phe Gly Gly Ala Ser
515 520 525
Ser Trp Asp Leu Asn His Phe Cys Ala Ala Tyr Gly Asn Glu Ser Ser
530 535 540
Ala Leu Gly Tyr Glu Thr Lys Phe Ala Asp Asp Leu Arg Lys Ala Ile
545 550 555 560
Tyr Ser Leu Arg Asn Glu Thr Phe His Phe Thr Thr Leu Asn Lys Gly
565 570 575
Ser Phe Asp Trp Asn Ala Lys Leu Ile Gly Asp Met Phe Ser His Glu
580 585 590
Ala Ala Thr Gly Ile Ala Val Glu Arg Thr Arg Phe Tyr Ser Asn Asn
595 600 605
Leu Pro Met Phe Tyr Arg Glu Ser Asp Leu Lys Arg Ile Met Asp His
610 615 620
Leu Tyr Asn Thr Tyr His Pro Arg Ala Ser Gln Val Pro Ser Phe Asn
625 630 635 640
Ser Val Phe Val Arg Lys Asn Phe Arg Leu Phe Leu Ser Asn Thr Leu
645 650 655
Asn Thr Asn Thr Ser Phe Asp Thr Glu Val Tyr Gln Lys Trp Glu Ser
660 665 670
Gly Val Tyr Tyr Leu Phe Lys Glu Ile Tyr Tyr Asn Ser Phe Leu Pro
675 680 685
Ser Gly Asp Ala His His Leu Phe Phe Glu Gly Leu Arg Arg Ile Arg
690 695 700
Lys Glu Ala Asp Asn Leu Pro Ile Val Gly Lys Glu Ala Lys Lys Arg
705 710 715 720
Asn Ala Val Gln Asp Phe Gly Arg Arg Cys Asp Glu Leu Lys Asn Leu
725 730 735
Ser Leu Ser Ala Ile Cys Gln Met Ile Met Thr Glu Tyr Asn Glu Gln
740 745 750
Asn Asn Gly Asn Arg Lys Val Lys Ser Thr Arg Glu Asp Lys Arg Lys
755 760 765
Pro Asp Ile Phe Gln His Tyr Lys Met Leu Leu Leu Arg Thr Leu Gln
770 775 780
Glu Ala Phe Ala Ile Tyr Ile Arg Arg Glu Glu Phe Lys Phe Ile Phe
785 790 795 800
Asp Leu Pro Lys Thr Leu Tyr Val Met Lys Pro Val Glu Glu Phe Leu
805 810 815
Pro Asn Trp Lys Ser Gly Met Phe Asp Ser Leu Val Glu Arg Val Lys
820 825 830
Gln Ser Pro Asp Leu Gln Arg Trp Tyr Val Leu Cys Lys Phe Leu Asn
835 840 845
Gly Arg Leu Leu Asn Gln Leu Ser Gly Val Ile Arg Ser Tyr Ile Gln
850 855 860
Phe Ala Gly Asp Ile Gln Arg Arg Ala Lys Ala Asn His Asn Arg Leu
865 870 875 880
Tyr Met Asp Asn Thr Gln Arg Val Glu Tyr Tyr Ser Asn Val Leu Glu
885 890 895
Val Val Asp Phe Cys Ile Lys Gly Thr Ser Arg Phe Ser Asn Val Phe
900 905 910
Ser Asp Tyr Phe Arg Asp Glu Asp Ala Tyr Ala Asp Tyr Leu Asp Asn
915 920 925
Tyr Leu Gln Phe Lys Asp Glu Lys Ile Ala Glu Val Ser Ser Phe Ala
930 935 940
Ala Leu Lys Thr Phe Cys Asn Glu Glu Glu Val Lys Ala Gly Ile Tyr
945 950 955 960
Met Asp Gly Glu Asn Pro Val Met Gln Arg Asn Ile Val Met Ala Lys
965 970 975
Leu Phe Gly Pro Asp Glu Val Leu Lys Asn Val Val Pro Lys Val Thr
980 985 990
Arg Glu Glu Ile Glu Glu Tyr Tyr Gln Leu Glu Lys Gln Ile Ala Pro
995 1000 1005
Tyr Arg Gln Asn Gly Tyr Cys Lys Ser Glu Glu Asp Gln Lys Lys
1010 1015 1020
Leu Leu Arg Phe Gln Arg Ile Lys Asn Arg Val Glu Phe Gln Thr
1025 1030 1035
Ile Thr Glu Phe Ser Glu Ile Ile Asn Glu Leu Leu Gly Gln Leu
1040 1045 1050
Ile Ser Trp Ser Phe Leu Arg Glu Arg Asp Leu Leu Tyr Phe Gln
1055 1060 1065
Leu Gly Phe His Tyr Leu Cys Leu His Asn Asp Thr Glu Lys Pro
1070 1075 1080
Ala Glu Tyr Lys Glu Ile Ser Arg Glu Asp Gly Thr Val Ile Arg
1085 1090 1095
Asn Ala Ile Leu His Gln Val Ala Ala Met Tyr Val Gly Gly Leu
1100 1105 1110
Pro Val Tyr Thr Leu Ala Asp Lys Lys Leu Ala Ala Phe Glu Lys
1115 1120 1125
Gly Glu Ala Asp Cys Lys Leu Ser Ile Ser Lys Asp Thr Ala Gly
1130 1135 1140
Ala Gly Lys Lys Ile Lys Asp Phe Phe Arg Tyr Ser Lys Tyr Val
1145 1150 1155
Leu Ile Lys Asp Arg Met Leu Thr Asp Gln Asn Gln Lys Tyr Thr
1160 1165 1170
Ile Tyr Leu Ala Gly Leu Glu Leu Phe Glu Asn Thr Asp Glu His
1175 1180 1185
Asp Asn Ile Thr Asp Val Arg Lys Tyr Val Asp His Phe Lys Tyr
1190 1195 1200
Tyr Ala Thr Ser Asp Glu Asn Ala Met Ser Ile Leu Asp Leu Tyr
1205 1210 1215
Ser Glu Ile His Asp Arg Phe Phe Thr Tyr Asp Met Lys Tyr Gln
1220 1225 1230
Lys Asn Val Ala Asn Met Leu Glu Asn Ile Leu Leu Arg His Phe
1235 1240 1245
Val Leu Ile Arg Pro Glu Phe Phe Thr Gly Ser Lys Lys Val Gly
1250 1255 1260
Glu Gly Lys Lys Ile Thr Cys Lys Ala Arg Ala Gln Ile Glu Ile
1265 1270 1275
Ala Glu Asn Gly Met Arg Ser Glu Asp Phe Thr Tyr Lys Leu Ser
1280 1285 1290
Asp Gly Lys Lys Asn Ile Ser Thr Cys Met Ile Ala Ala Arg Asp
1295 1300 1305
Gln Lys Tyr Leu Asn Thr Val Ala Arg Leu Leu Tyr Tyr Pro His
1310 1315 1320
Glu Ala Lys Lys Ser Ile Val Asp Thr Arg Glu Lys Lys Asn Asn
1325 1330 1335
Lys Lys Thr Asn Arg Gly Asp Gly Thr Phe Asn Lys Gln Lys Gly
1340 1345 1350
Thr Ala Arg Lys Glu Lys Asp Asn Gly Pro Arg Glu Phe Asn Asp
1355 1360 1365
Thr Gly Phe Ser Asn Thr Pro Phe Ala Gly Phe Asp Pro Phe Arg
1370 1375 1380
Asn Ser
1385
<210> 577
<211> 1334
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 577
Met Lys Ile Ser Lys Val Asp His Thr Arg Met Ala Val Ala Lys Gly
1 5 10 15
Asn Gln His Arg Arg Asp Glu Ile Ser Gly Ile Leu Tyr Lys Asp Pro
20 25 30
Thr Lys Thr Gly Ser Ile Asp Phe Asp Glu Arg Phe Lys Lys Leu Asn
35 40 45
Cys Ser Ala Lys Ile Leu Tyr His Val Phe Asn Gly Ile Ala Glu Gly
50 55 60
Ser Asn Lys Tyr Lys Asn Ile Val Asp Lys Val Asn Asn Asn Leu Asp
65 70 75 80
Arg Val Leu Phe Thr Gly Lys Ser Tyr Asp Arg Lys Ser Ile Ile Asp
85 90 95
Ile Asp Thr Val Leu Arg Asn Val Glu Lys Ile Asn Ala Phe Asp Arg
100 105 110
Ile Ser Thr Glu Glu Arg Glu Gln Ile Ile Asp Asp Leu Leu Glu Ile
115 120 125
Gln Leu Arg Lys Gly Leu Arg Lys Gly Lys Ala Gly Leu Arg Glu Val
130 135 140
Leu Leu Ile Gly Ala Gly Val Ile Val Arg Thr Asp Lys Lys Gln Glu
145 150 155 160
Ile Ala Asp Phe Leu Glu Ile Leu Asp Glu Asp Phe Asn Lys Thr Asn
165 170 175
Gln Ala Lys Asn Ile Lys Leu Ser Ile Glu Asn Gln Gly Leu Val Val
180 185 190
Ser Pro Val Ser Arg Gly Glu Glu Arg Ile Phe Asp Val Ser Gly Ala
195 200 205
Gln Lys Gly Lys Ser Ser Lys Lys Ala Gln Glu Lys Glu Ala Leu Ser
210 215 220
Ala Phe Leu Leu Asp Tyr Ala Asp Leu Asp Lys Asn Val Arg Phe Glu
225 230 235 240
Tyr Leu Arg Lys Ile Arg Arg Leu Ile Asn Leu Tyr Phe Tyr Val Lys
245 250 255
Asn Asp Asp Val Met Ser Leu Thr Glu Ile Pro Ala Glu Val Asn Leu
260 265 270
Glu Lys Asp Phe Asp Ile Trp Arg Asp His Glu Gln Arg Lys Glu Glu
275 280 285
Asn Gly Asp Phe Val Gly Cys Pro Asp Ile Leu Leu Ala Asp Arg Asp
290 295 300
Val Lys Lys Ser Asn Ser Lys Gln Val Lys Ile Ala Glu Arg Gln Leu
305 310 315 320
Arg Glu Ser Ile Arg Glu Lys Asn Ile Lys Arg Tyr Arg Phe Ser Ile
325 330 335
Lys Thr Ile Glu Lys Asp Asp Gly Thr Tyr Phe Phe Ala Asn Lys Gln
340 345 350
Ile Ser Val Phe Trp Ile His Arg Ile Glu Asn Ala Val Glu Arg Ile
355 360 365
Leu Gly Ser Ile Asn Asp Lys Lys Leu Tyr Arg Leu Arg Leu Gly Tyr
370 375 380
Leu Gly Glu Lys Val Trp Lys Asp Ile Leu Asn Phe Leu Ser Ile Lys
385 390 395 400
Tyr Ile Ala Val Gly Lys Ala Val Phe Asn Phe Ala Met Asp Asp Leu
405 410 415
Gln Glu Lys Asp Arg Asp Ile Glu Pro Gly Lys Ile Ser Glu Asn Ala
420 425 430
Val Asn Gly Leu Thr Ser Phe Asp Tyr Glu Gln Ile Lys Ala Asp Glu
435 440 445
Met Leu Gln Arg Glu Val Ala Val Asn Val Ala Phe Ala Ala Asn Asn
450 455 460
Leu Ala Arg Val Thr Val Asp Ile Pro Gln Asn Gly Glu Lys Glu Asp
465 470 475 480
Ile Leu Leu Trp Asn Lys Ser Asp Ile Lys Lys Tyr Lys Lys Asn Ser
485 490 495
Lys Lys Gly Ile Leu Lys Ser Ile Leu Gln Phe Phe Gly Gly Ala Ser
500 505 510
Thr Trp Asn Met Lys Met Phe Glu Ile Ala Tyr His Asp Gln Pro Gly
515 520 525
Asp Tyr Glu Glu Asn Tyr Leu Tyr Asp Ile Ile Gln Ile Ile Tyr Ser
530 535 540
Leu Arg Asn Lys Ser Phe His Phe Lys Thr Tyr Asp His Gly Asp Lys
545 550 555 560
Asn Trp Asn Arg Glu Leu Ile Gly Lys Met Ile Glu His Asp Ala Glu
565 570 575
Arg Val Ile Ser Val Glu Arg Glu Lys Phe His Ser Asn Asn Leu Pro
580 585 590
Met Phe Tyr Lys Asp Ala Asp Leu Lys Lys Ile Leu Asp Leu Leu Tyr
595 600 605
Ser Asp Tyr Ala Gly Arg Ala Ser Gln Val Pro Ala Phe Asn Thr Val
610 615 620
Leu Val Arg Lys Asn Phe Pro Glu Phe Leu Arg Lys Asp Met Gly Tyr
625 630 635 640
Lys Val His Phe Asn Asn Pro Glu Val Glu Asn Gln Trp His Ser Ala
645 650 655
Val Tyr Tyr Leu Tyr Lys Glu Ile Tyr Tyr Asn Leu Phe Leu Arg Asp
660 665 670
Lys Glu Val Lys Asn Leu Phe Tyr Thr Ser Leu Lys Asn Ile Arg Ser
675 680 685
Glu Val Ser Asp Lys Lys Gln Lys Leu Ala Ser Asp Asp Phe Ala Ser
690 695 700
Arg Cys Glu Glu Ile Glu Asp Arg Ser Leu Pro Glu Ile Cys Gln Ile
705 710 715 720
Ile Met Thr Glu Tyr Asn Ala Gln Asn Phe Gly Asn Arg Lys Val Lys
725 730 735
Ser Gln Arg Val Ile Glu Lys Asn Lys Asp Ile Phe Arg His Tyr Lys
740 745 750
Met Leu Leu Ile Lys Thr Leu Ala Gly Ala Phe Ser Leu Tyr Leu Lys
755 760 765
Gln Glu Arg Phe Ala Phe Ile Gly Lys Ala Thr Pro Ile Pro Tyr Glu
770 775 780
Thr Thr Asp Val Lys Asn Phe Leu Pro Glu Trp Lys Ser Gly Met Tyr
785 790 795 800
Ala Ser Phe Val Glu Glu Ile Lys Asn Asn Leu Asp Leu Gln Glu Trp
805 810 815
Tyr Ile Val Gly Arg Phe Leu Asn Gly Arg Met Leu Asn Gln Leu Ala
820 825 830
Gly Ser Leu Arg Ser Tyr Ile Gln Tyr Ala Glu Asp Ile Glu Arg Arg
835 840 845
Ala Ala Glu Asn Arg Asn Lys Leu Phe Ser Lys Pro Asp Glu Lys Ile
850 855 860
Glu Ala Cys Lys Lys Ala Val Arg Val Leu Asp Leu Cys Ile Lys Ile
865 870 875 880
Ser Thr Arg Ile Ser Ala Glu Phe Thr Asp Tyr Phe Asp Ser Glu Asp
885 890 895
Asp Tyr Ala Asp Tyr Leu Glu Lys Tyr Leu Lys Tyr Gln Asp Asp Ala
900 905 910
Ile Lys Glu Leu Ser Gly Ser Ser Tyr Ala Ala Leu Asp His Phe Cys
915 920 925
Asn Lys Asp Asp Leu Lys Phe Asp Ile Tyr Val Asn Ala Gly Gln Lys
930 935 940
Pro Ile Leu Gln Arg Asn Ile Val Met Ala Lys Leu Phe Gly Pro Asp
945 950 955 960
Asn Ile Leu Ser Glu Val Met Glu Lys Val Thr Glu Ser Ala Ile Arg
965 970 975
Glu Tyr Tyr Asp Tyr Leu Lys Lys Val Ser Gly Tyr Arg Val Arg Gly
980 985 990
Lys Cys Ser Thr Glu Lys Glu Gln Glu Asp Leu Leu Lys Phe Gln Arg
995 1000 1005
Leu Lys Asn Ala Val Glu Phe Arg Asp Val Thr Glu Tyr Ala Glu
1010 1015 1020
Val Ile Asn Glu Leu Leu Gly Gln Leu Ile Ser Trp Ser Tyr Leu
1025 1030 1035
Arg Glu Arg Asp Leu Leu Tyr Phe Gln Leu Gly Phe His Tyr Met
1040 1045 1050
Cys Leu Lys Asn Lys Ser Phe Lys Pro Ala Glu Tyr Val Asp Ile
1055 1060 1065
Arg Arg Asn Asn Gly Thr Ile Ile His Asn Ala Ile Leu Tyr Gln
1070 1075 1080
Ile Val Ser Met Tyr Ile Asn Gly Leu Asp Phe Tyr Ser Cys Asp
1085 1090 1095
Lys Glu Gly Lys Thr Leu Lys Pro Ile Glu Thr Gly Lys Gly Val
1100 1105 1110
Gly Ser Lys Ile Gly Gln Phe Ile Lys Tyr Ser Gln Tyr Leu Tyr
1115 1120 1125
Asn Asp Pro Ser Tyr Lys Leu Glu Ile Tyr Asn Ala Gly Leu Glu
1130 1135 1140
Val Phe Glu Asn Ile Asp Glu His Asp Asn Ile Thr Asp Leu Arg
1145 1150 1155
Lys Tyr Val Asp His Phe Lys Tyr Tyr Ala Tyr Gly Asn Lys Met
1160 1165 1170
Ser Leu Leu Asp Leu Tyr Ser Glu Phe Phe Asp Arg Phe Phe Thr
1175 1180 1185
Tyr Asp Met Lys Tyr Gln Lys Asn Val Val Asn Val Leu Glu Asn
1190 1195 1200
Ile Leu Leu Arg His Phe Val Ile Phe Tyr Pro Lys Phe Gly Ser
1205 1210 1215
Gly Lys Lys Asp Val Gly Ile Arg Asp Cys Lys Lys Glu Arg Ala
1220 1225 1230
Gln Ile Glu Ile Ser Glu Gln Ser Leu Thr Ser Glu Asp Phe Met
1235 1240 1245
Phe Lys Leu Asp Asp Lys Ala Gly Glu Glu Ala Lys Lys Phe Pro
1250 1255 1260
Ala Arg Asp Glu Arg Tyr Leu Gln Thr Ile Ala Lys Leu Leu Tyr
1265 1270 1275
Tyr Pro Asn Glu Ile Glu Asp Met Asn Arg Phe Met Lys Lys Gly
1280 1285 1290
Glu Thr Ile Asn Lys Lys Val Gln Phe Asn Arg Lys Lys Lys Ile
1295 1300 1305
Thr Arg Lys Gln Lys Asn Asn Ser Ser Asn Glu Val Leu Ser Ser
1310 1315 1320
Thr Met Gly Tyr Leu Phe Lys Asn Ile Lys Leu
1325 1330
<210> 578
<211> 1175
<212> PRT
<213> Carnobacterium gallinarum
<400> 578
Met Arg Ile Thr Lys Val Lys Ile Lys Leu Asp Asn Lys Leu Tyr Gln
1 5 10 15
Val Thr Met Gln Lys Glu Glu Lys Tyr Gly Thr Leu Lys Leu Asn Glu
20 25 30
Glu Ser Arg Lys Ser Thr Ala Glu Ile Leu Arg Leu Lys Lys Ala Ser
35 40 45
Phe Asn Lys Ser Phe His Ser Lys Thr Ile Asn Ser Gln Lys Glu Asn
50 55 60
Lys Asn Ala Thr Ile Lys Lys Asn Gly Asp Tyr Ile Ser Gln Ile Phe
65 70 75 80
Glu Lys Leu Val Gly Val Asp Thr Asn Lys Asn Ile Arg Lys Pro Lys
85 90 95
Met Ser Leu Thr Asp Leu Lys Asp Leu Pro Lys Lys Asp Leu Ala Leu
100 105 110
Phe Ile Lys Arg Lys Phe Lys Asn Asp Asp Ile Val Glu Ile Lys Asn
115 120 125
Leu Asp Leu Ile Ser Leu Phe Tyr Asn Ala Leu Gln Lys Val Pro Gly
130 135 140
Glu His Phe Thr Asp Glu Ser Trp Ala Asp Phe Cys Gln Glu Met Met
145 150 155 160
Pro Tyr Arg Glu Tyr Lys Asn Lys Phe Ile Glu Arg Lys Ile Ile Leu
165 170 175
Leu Ala Asn Ser Ile Glu Gln Asn Lys Gly Phe Ser Ile Asn Pro Glu
180 185 190
Thr Phe Ser Lys Arg Lys Arg Val Leu His Gln Trp Ala Ile Glu Val
195 200 205
Gln Glu Arg Gly Asp Phe Ser Ile Leu Asp Glu Lys Leu Ser Lys Leu
210 215 220
Ala Glu Ile Tyr Asn Phe Lys Lys Met Cys Lys Arg Val Gln Asp Glu
225 230 235 240
Leu Asn Asp Leu Glu Lys Ser Met Lys Lys Gly Lys Asn Pro Glu Lys
245 250 255
Glu Lys Glu Ala Tyr Lys Lys Gln Lys Asn Phe Lys Ile Lys Thr Ile
260 265 270
Trp Lys Asp Tyr Pro Tyr Lys Thr His Ile Gly Leu Ile Glu Lys Ile
275 280 285
Lys Glu Asn Glu Glu Leu Asn Gln Phe Asn Ile Glu Ile Gly Lys Tyr
290 295 300
Phe Glu His Tyr Phe Pro Ile Lys Lys Glu Arg Cys Thr Glu Asp Glu
305 310 315 320
Pro Tyr Tyr Leu Asn Ser Glu Thr Ile Ala Thr Thr Val Asn Tyr Gln
325 330 335
Leu Lys Asn Ala Leu Ile Ser Tyr Leu Met Gln Ile Gly Lys Tyr Lys
340 345 350
Gln Phe Gly Leu Glu Asn Gln Val Leu Asp Ser Lys Lys Leu Gln Glu
355 360 365
Ile Gly Ile Tyr Glu Gly Phe Gln Thr Lys Phe Met Asp Ala Cys Val
370 375 380
Phe Ala Thr Ser Ser Leu Lys Asn Ile Ile Glu Pro Met Arg Ser Gly
385 390 395 400
Asp Ile Leu Gly Lys Arg Glu Phe Lys Glu Ala Ile Ala Thr Ser Ser
405 410 415
Phe Val Asn Tyr His His Phe Phe Pro Tyr Phe Pro Phe Glu Leu Lys
420 425 430
Gly Met Lys Asp Arg Glu Ser Glu Leu Ile Pro Phe Gly Glu Gln Thr
435 440 445
Glu Ala Lys Gln Met Gln Asn Ile Trp Ala Leu Arg Gly Ser Val Gln
450 455 460
Gln Ile Arg Asn Glu Ile Phe His Ser Phe Asp Lys Asn Gln Lys Phe
465 470 475 480
Asn Leu Pro Gln Leu Asp Lys Ser Asn Phe Glu Phe Asp Ala Ser Glu
485 490 495
Asn Ser Thr Gly Lys Ser Gln Ser Tyr Ile Glu Thr Asp Tyr Lys Phe
500 505 510
Leu Phe Glu Ala Glu Lys Asn Gln Leu Glu Gln Phe Phe Ile Glu Arg
515 520 525
Ile Lys Ser Ser Gly Ala Leu Glu Tyr Tyr Pro Leu Lys Ser Leu Glu
530 535 540
Lys Leu Phe Ala Lys Lys Glu Met Lys Phe Ser Leu Gly Ser Gln Val
545 550 555 560
Val Ala Phe Ala Pro Ser Tyr Lys Lys Leu Val Lys Lys Gly His Ser
565 570 575
Tyr Gln Thr Ala Thr Glu Gly Thr Ala Asn Tyr Leu Gly Leu Ser Tyr
580 585 590
Tyr Asn Arg Tyr Glu Leu Lys Glu Glu Ser Phe Gln Ala Gln Tyr Tyr
595 600 605
Leu Leu Lys Leu Ile Tyr Gln Tyr Val Phe Leu Pro Asn Phe Ser Gln
610 615 620
Gly Asn Ser Pro Ala Phe Arg Glu Thr Val Lys Ala Ile Leu Arg Ile
625 630 635 640
Asn Lys Asp Glu Ala Arg Lys Lys Met Lys Lys Asn Lys Lys Phe Leu
645 650 655
Arg Lys Tyr Ala Phe Glu Gln Val Arg Glu Met Glu Phe Lys Glu Thr
660 665 670
Pro Asp Gln Tyr Met Ser Tyr Leu Gln Ser Glu Met Arg Glu Glu Lys
675 680 685
Val Arg Lys Ala Glu Lys Asn Asp Lys Gly Phe Glu Lys Asn Ile Thr
690 695 700
Met Asn Phe Glu Lys Leu Leu Met Gln Ile Phe Val Lys Gly Phe Asp
705 710 715 720
Val Phe Leu Thr Thr Phe Ala Gly Lys Glu Leu Leu Leu Ser Ser Glu
725 730 735
Glu Lys Val Ile Lys Glu Thr Glu Ile Ser Leu Ser Lys Lys Ile Asn
740 745 750
Glu Arg Glu Lys Thr Leu Lys Ala Ser Ile Gln Val Glu His Gln Leu
755 760 765
Val Ala Thr Asn Ser Ala Ile Ser Tyr Trp Leu Phe Cys Lys Leu Leu
770 775 780
Asp Ser Arg His Leu Asn Glu Leu Arg Asn Glu Met Ile Lys Phe Lys
785 790 795 800
Gln Ser Arg Ile Lys Phe Asn His Thr Gln His Ala Glu Leu Ile Gln
805 810 815
Asn Leu Leu Pro Ile Val Glu Leu Thr Ile Leu Ser Asn Asp Tyr Asp
820 825 830
Glu Lys Asn Asp Ser Gln Asn Val Asp Val Ser Ala Tyr Phe Glu Asp
835 840 845
Lys Ser Leu Tyr Glu Thr Ala Pro Tyr Val Gln Thr Asp Asp Arg Thr
850 855 860
Arg Val Ser Phe Arg Pro Ile Leu Lys Leu Glu Lys Tyr His Thr Lys
865 870 875 880
Ser Leu Ile Glu Ala Leu Leu Lys Asp Asn Pro Gln Phe Arg Val Ala
885 890 895
Ala Thr Asp Ile Gln Glu Trp Met His Lys Arg Glu Glu Ile Gly Glu
900 905 910
Leu Val Glu Lys Arg Lys Asn Leu His Thr Glu Trp Ala Glu Gly Gln
915 920 925
Gln Thr Leu Gly Ala Glu Lys Arg Glu Glu Tyr Arg Asp Tyr Cys Lys
930 935 940
Lys Ile Asp Arg Phe Asn Trp Lys Ala Asn Lys Val Thr Leu Thr Tyr
945 950 955 960
Leu Ser Gln Leu His Tyr Leu Ile Thr Asp Leu Leu Gly Arg Met Val
965 970 975
Gly Phe Ser Ala Leu Phe Glu Arg Asp Leu Val Tyr Phe Ser Arg Ser
980 985 990
Phe Ser Glu Leu Gly Gly Glu Thr Tyr His Ile Ser Asp Tyr Lys Asn
995 1000 1005
Leu Ser Gly Val Leu Arg Leu Asn Ala Glu Val Lys Pro Ile Lys
1010 1015 1020
Ile Lys Asn Ile Lys Val Ile Asp Asn Glu Glu Asn Pro Tyr Lys
1025 1030 1035
Gly Asn Glu Pro Glu Val Lys Pro Phe Leu Asp Arg Leu His Ala
1040 1045 1050
Tyr Leu Glu Asn Val Ile Gly Ile Lys Ala Val His Gly Lys Ile
1055 1060 1065
Arg Asn Gln Thr Ala His Leu Ser Val Leu Gln Leu Glu Leu Ser
1070 1075 1080
Met Ile Glu Ser Met Asn Asn Leu Arg Asp Leu Met Ala Tyr Asp
1085 1090 1095
Arg Lys Leu Lys Asn Ala Val Thr Lys Ser Met Ile Lys Ile Leu
1100 1105 1110
Asp Lys His Gly Met Ile Leu Lys Leu Lys Ile Asp Glu Asn His
1115 1120 1125
Lys Asn Phe Glu Ile Glu Ser Leu Ile Pro Lys Glu Ile Ile His
1130 1135 1140
Leu Lys Asp Lys Ala Ile Lys Thr Asn Gln Val Ser Glu Glu Tyr
1145 1150 1155
Cys Gln Leu Val Leu Ala Leu Leu Thr Thr Asn Pro Gly Asn Gln
1160 1165 1170
Leu Asn
1175
<210> 579
<211> 1164
<212> PRT
<213> Carnobacterium gallinarum
<400> 579
Met Arg Met Thr Lys Val Lys Ile Asn Gly Ser Pro Val Ser Met Asn
1 5 10 15
Arg Ser Lys Leu Asn Gly His Leu Val Trp Asn Gly Thr Thr Asn Thr
20 25 30
Val Asn Ile Leu Thr Lys Lys Glu Gln Ser Phe Ala Ala Ser Phe Leu
35 40 45
Asn Lys Thr Leu Val Lys Ala Asp Gln Val Lys Gly Tyr Lys Val Leu
50 55 60
Ala Glu Asn Ile Phe Ile Ile Phe Glu Gln Leu Glu Lys Ser Asn Ser
65 70 75 80
Glu Lys Pro Ser Val Tyr Leu Asn Asn Ile Arg Arg Leu Lys Glu Ala
85 90 95
Gly Leu Lys Arg Phe Phe Lys Ser Lys Tyr His Glu Glu Ile Lys Tyr
100 105 110
Thr Ser Glu Lys Asn Gln Ser Val Pro Thr Lys Leu Asn Leu Ile Pro
115 120 125
Leu Phe Phe Asn Ala Val Asp Arg Ile Gln Glu Asp Lys Phe Asp Glu
130 135 140
Lys Asn Trp Ser Tyr Phe Cys Lys Glu Met Ser Pro Tyr Leu Asp Tyr
145 150 155 160
Lys Lys Ser Tyr Leu Asn Arg Lys Lys Glu Ile Leu Ala Asn Ser Ile
165 170 175
Gln Gln Asn Arg Gly Phe Ser Met Pro Thr Ala Glu Glu Pro Asn Leu
180 185 190
Leu Ser Lys Arg Lys Gln Leu Phe Gln Gln Trp Ala Met Lys Phe Gln
195 200 205
Glu Ser Pro Leu Ile Gln Gln Asn Asn Phe Ala Val Glu Gln Phe Asn
210 215 220
Lys Glu Phe Ala Asn Lys Ile Asn Glu Leu Ala Ala Val Tyr Asn Val
225 230 235 240
Asp Glu Leu Cys Thr Ala Ile Thr Glu Lys Leu Met Asn Phe Asp Lys
245 250 255
Asp Lys Ser Asn Lys Thr Arg Asn Phe Glu Ile Lys Lys Leu Trp Lys
260 265 270
Gln His Pro His Asn Lys Asp Lys Ala Leu Ile Lys Leu Phe Asn Gln
275 280 285
Glu Gly Asn Glu Ala Leu Asn Gln Phe Asn Ile Glu Leu Gly Lys Tyr
290 295 300
Phe Glu His Tyr Phe Pro Lys Thr Gly Lys Lys Glu Ser Ala Glu Ser
305 310 315 320
Tyr Tyr Leu Asn Pro Gln Thr Ile Ile Lys Thr Val Gly Tyr Gln Leu
325 330 335
Arg Asn Ala Phe Val Gln Tyr Leu Leu Gln Val Gly Lys Leu His Gln
340 345 350
Tyr Asn Lys Gly Val Leu Asp Ser Gln Thr Leu Gln Glu Ile Gly Met
355 360 365
Tyr Glu Gly Phe Gln Thr Lys Phe Met Asp Ala Cys Val Phe Ala Ser
370 375 380
Ser Ser Leu Arg Asn Ile Ile Gln Ala Thr Thr Asn Glu Asp Ile Leu
385 390 395 400
Thr Arg Glu Lys Phe Lys Lys Glu Leu Glu Lys Asn Val Glu Leu Lys
405 410 415
His Asp Leu Phe Phe Lys Thr Glu Ile Val Glu Glu Arg Asp Glu Asn
420 425 430
Pro Ala Lys Lys Ile Ala Met Thr Pro Asn Glu Leu Asp Leu Trp Ala
435 440 445
Ile Arg Gly Ala Val Gln Arg Val Arg Asn Gln Ile Phe His Gln Gln
450 455 460
Ile Asn Lys Arg His Glu Pro Asn Gln Leu Lys Val Gly Ser Phe Glu
465 470 475 480
Asn Gly Asp Leu Gly Asn Val Ser Tyr Gln Lys Thr Ile Tyr Gln Lys
485 490 495
Leu Phe Asp Ala Glu Ile Lys Asp Ile Glu Ile Tyr Phe Ala Glu Lys
500 505 510
Ile Lys Ser Ser Gly Ala Leu Glu Gln Tyr Ser Met Lys Asp Leu Glu
515 520 525
Lys Leu Phe Ser Asn Lys Glu Leu Thr Leu Ser Leu Gly Gly Gln Val
530 535 540
Val Ala Phe Ala Pro Ser Tyr Lys Lys Leu Tyr Lys Gln Gly Tyr Phe
545 550 555 560
Tyr Gln Asn Glu Lys Thr Ile Glu Leu Glu Gln Phe Thr Asp Tyr Asp
565 570 575
Phe Ser Asn Asp Val Phe Lys Ala Asn Tyr Tyr Leu Ile Lys Leu Ile
580 585 590
Tyr His Tyr Val Phe Leu Pro Gln Phe Ser Gln Ala Asn Asn Lys Leu
595 600 605
Phe Lys Asp Thr Val His Tyr Val Ile Gln Gln Asn Lys Glu Leu Asn
610 615 620
Thr Thr Glu Lys Asp Lys Lys Asn Asn Lys Lys Ile Arg Lys Tyr Ala
625 630 635 640
Phe Glu Gln Val Lys Leu Met Lys Asn Glu Ser Pro Glu Lys Tyr Met
645 650 655
Gln Tyr Leu Gln Arg Glu Met Gln Glu Glu Arg Thr Ile Lys Glu Ala
660 665 670
Lys Lys Thr Asn Glu Glu Lys Pro Asn Tyr Asn Phe Glu Lys Leu Leu
675 680 685
Ile Gln Ile Phe Ile Lys Gly Phe Asp Thr Phe Leu Arg Asn Phe Asp
690 695 700
Leu Asn Leu Asn Pro Ala Glu Glu Leu Val Gly Thr Val Lys Glu Lys
705 710 715 720
Ala Glu Gly Leu Arg Lys Arg Lys Glu Arg Ile Ala Lys Ile Leu Asn
725 730 735
Val Asp Glu Gln Ile Lys Thr Gly Asp Glu Glu Ile Ala Phe Trp Ile
740 745 750
Phe Ala Lys Leu Leu Asp Ala Arg His Leu Ser Glu Leu Arg Asn Glu
755 760 765
Met Ile Lys Phe Lys Gln Ser Ser Val Lys Lys Gly Leu Ile Lys Asn
770 775 780
Gly Asp Leu Ile Glu Gln Met Gln Pro Ile Leu Glu Leu Cys Ile Leu
785 790 795 800
Ser Asn Asp Ser Glu Ser Met Glu Lys Glu Ser Phe Asp Lys Ile Glu
805 810 815
Val Phe Leu Glu Lys Val Glu Leu Ala Lys Asn Glu Pro Tyr Met Gln
820 825 830
Glu Asp Lys Leu Thr Pro Val Lys Phe Arg Phe Met Lys Gln Leu Glu
835 840 845
Lys Tyr Gln Thr Arg Asn Phe Ile Glu Asn Leu Val Ile Glu Asn Pro
850 855 860
Glu Phe Lys Val Ser Glu Lys Ile Val Leu Asn Trp His Glu Glu Lys
865 870 875 880
Glu Lys Ile Ala Asp Leu Val Asp Lys Arg Thr Lys Leu His Glu Glu
885 890 895
Trp Ala Ser Lys Ala Arg Glu Ile Glu Glu Tyr Asn Glu Lys Ile Lys
900 905 910
Lys Asn Lys Ser Lys Lys Leu Asp Lys Pro Ala Glu Phe Ala Lys Phe
915 920 925
Ala Glu Tyr Lys Ile Ile Cys Glu Ala Ile Glu Asn Phe Asn Arg Leu
930 935 940
Asp His Lys Val Arg Leu Thr Tyr Leu Lys Asn Leu His Tyr Leu Met
945 950 955 960
Ile Asp Leu Met Gly Arg Met Val Gly Phe Ser Val Leu Phe Glu Arg
965 970 975
Asp Phe Val Tyr Met Gly Arg Ser Tyr Ser Ala Leu Lys Lys Gln Ser
980 985 990
Ile Tyr Leu Asn Asp Tyr Asp Thr Phe Ala Asn Ile Arg Asp Trp Glu
995 1000 1005
Val Asn Glu Asn Lys His Leu Phe Gly Thr Ser Ser Ser Asp Leu
1010 1015 1020
Thr Phe Gln Glu Thr Ala Glu Phe Lys Asn Leu Lys Lys Pro Met
1025 1030 1035
Glu Asn Gln Leu Lys Ala Leu Leu Gly Val Thr Asn His Ser Phe
1040 1045 1050
Glu Ile Arg Asn Asn Ile Ala His Leu His Val Leu Arg Asn Asp
1055 1060 1065
Gly Lys Gly Glu Gly Val Ser Leu Leu Ser Cys Met Asn Asp Leu
1070 1075 1080
Arg Lys Leu Met Ser Tyr Asp Arg Lys Leu Lys Asn Ala Val Thr
1085 1090 1095
Lys Ala Ile Ile Lys Ile Leu Asp Lys His Gly Met Ile Leu Lys
1100 1105 1110
Leu Thr Asn Asn Asp His Thr Lys Pro Phe Glu Ile Glu Ser Leu
1115 1120 1125
Lys Pro Lys Lys Ile Ile His Leu Glu Lys Ser Asn His Ser Phe
1130 1135 1140
Pro Met Asp Gln Val Ser Gln Glu Tyr Cys Asp Leu Val Lys Lys
1145 1150 1155
Met Leu Val Phe Thr Asn
1160
<210> 580
<211> 1154
<212> PRT
<213> Paludibacter propionicigenes
<400> 580
Met Arg Val Ser Lys Val Lys Val Lys Asp Gly Gly Lys Asp Lys Met
1 5 10 15
Val Leu Val His Arg Lys Thr Thr Gly Ala Gln Leu Val Tyr Ser Gly
20 25 30
Gln Pro Val Ser Asn Glu Thr Ser Asn Ile Leu Pro Glu Lys Lys Arg
35 40 45
Gln Ser Phe Asp Leu Ser Thr Leu Asn Lys Thr Ile Ile Lys Phe Asp
50 55 60
Thr Ala Lys Lys Gln Lys Leu Asn Val Asp Gln Tyr Lys Ile Val Glu
65 70 75 80
Lys Ile Phe Lys Tyr Pro Lys Gln Glu Leu Pro Lys Gln Ile Lys Ala
85 90 95
Glu Glu Ile Leu Pro Phe Leu Asn His Lys Phe Gln Glu Pro Val Lys
100 105 110
Tyr Trp Lys Asn Gly Lys Glu Glu Ser Phe Asn Leu Thr Leu Leu Ile
115 120 125
Val Glu Ala Val Gln Ala Gln Asp Lys Arg Lys Leu Gln Pro Tyr Tyr
130 135 140
Asp Trp Lys Thr Trp Tyr Ile Gln Thr Lys Ser Asp Leu Leu Lys Lys
145 150 155 160
Ser Ile Glu Asn Asn Arg Ile Asp Leu Thr Glu Asn Leu Ser Lys Arg
165 170 175
Lys Lys Ala Leu Leu Ala Trp Glu Thr Glu Phe Thr Ala Ser Gly Ser
180 185 190
Ile Asp Leu Thr His Tyr His Lys Val Tyr Met Thr Asp Val Leu Cys
195 200 205
Lys Met Leu Gln Asp Val Lys Pro Leu Thr Asp Asp Lys Gly Lys Ile
210 215 220
Asn Thr Asn Ala Tyr His Arg Gly Leu Lys Lys Ala Leu Gln Asn His
225 230 235 240
Gln Pro Ala Ile Phe Gly Thr Arg Glu Val Pro Asn Glu Ala Asn Arg
245 250 255
Ala Asp Asn Gln Leu Ser Ile Tyr His Leu Glu Val Val Lys Tyr Leu
260 265 270
Glu His Tyr Phe Pro Ile Lys Thr Ser Lys Arg Arg Asn Thr Ala Asp
275 280 285
Asp Ile Ala His Tyr Leu Lys Ala Gln Thr Leu Lys Thr Thr Ile Glu
290 295 300
Lys Gln Leu Val Asn Ala Ile Arg Ala Asn Ile Ile Gln Gln Gly Lys
305 310 315 320
Thr Asn His His Glu Leu Lys Ala Asp Thr Thr Ser Asn Asp Leu Ile
325 330 335
Arg Ile Lys Thr Asn Glu Ala Phe Val Leu Asn Leu Thr Gly Thr Cys
340 345 350
Ala Phe Ala Ala Asn Asn Ile Arg Asn Met Val Asp Asn Glu Gln Thr
355 360 365
Asn Asp Ile Leu Gly Lys Gly Asp Phe Ile Lys Ser Leu Leu Lys Asp
370 375 380
Asn Thr Asn Ser Gln Leu Tyr Ser Phe Phe Phe Gly Glu Gly Leu Ser
385 390 395 400
Thr Asn Lys Ala Glu Lys Glu Thr Gln Leu Trp Gly Ile Arg Gly Ala
405 410 415
Val Gln Gln Ile Arg Asn Asn Val Asn His Tyr Lys Lys Asp Ala Leu
420 425 430
Lys Thr Val Phe Asn Ile Ser Asn Phe Glu Asn Pro Thr Ile Thr Asp
435 440 445
Pro Lys Gln Gln Thr Asn Tyr Ala Asp Thr Ile Tyr Lys Ala Arg Phe
450 455 460
Ile Asn Glu Leu Glu Lys Ile Pro Glu Ala Phe Ala Gln Gln Leu Lys
465 470 475 480
Thr Gly Gly Ala Val Ser Tyr Tyr Thr Ile Glu Asn Leu Lys Ser Leu
485 490 495
Leu Thr Thr Phe Gln Phe Ser Leu Cys Arg Ser Thr Ile Pro Phe Ala
500 505 510
Pro Gly Phe Lys Lys Val Phe Asn Gly Gly Ile Asn Tyr Gln Asn Ala
515 520 525
Lys Gln Asp Glu Ser Phe Tyr Glu Leu Met Leu Glu Gln Tyr Leu Arg
530 535 540
Lys Glu Asn Phe Ala Glu Glu Ser Tyr Asn Ala Arg Tyr Phe Met Leu
545 550 555 560
Lys Leu Ile Tyr Asn Asn Leu Phe Leu Pro Gly Phe Thr Thr Asp Arg
565 570 575
Lys Ala Phe Ala Asp Ser Val Gly Phe Val Gln Met Gln Asn Lys Lys
580 585 590
Gln Ala Glu Lys Val Asn Pro Arg Lys Lys Glu Ala Tyr Ala Phe Glu
595 600 605
Ala Val Arg Pro Met Thr Ala Ala Asp Ser Ile Ala Asp Tyr Met Ala
610 615 620
Tyr Val Gln Ser Glu Leu Met Gln Glu Gln Asn Lys Lys Glu Glu Lys
625 630 635 640
Val Ala Glu Glu Thr Arg Ile Asn Phe Glu Lys Phe Val Leu Gln Val
645 650 655
Phe Ile Lys Gly Phe Asp Ser Phe Leu Arg Ala Lys Glu Phe Asp Phe
660 665 670
Val Gln Met Pro Gln Pro Gln Leu Thr Ala Thr Ala Ser Asn Gln Gln
675 680 685
Lys Ala Asp Lys Leu Asn Gln Leu Glu Ala Ser Ile Thr Ala Asp Cys
690 695 700
Lys Leu Thr Pro Gln Tyr Ala Lys Ala Asp Asp Ala Thr His Ile Ala
705 710 715 720
Phe Tyr Val Phe Cys Lys Leu Leu Asp Ala Ala His Leu Ser Asn Leu
725 730 735
Arg Asn Glu Leu Ile Lys Phe Arg Glu Ser Val Asn Glu Phe Lys Phe
740 745 750
His His Leu Leu Glu Ile Ile Glu Ile Cys Leu Leu Ser Ala Asp Val
755 760 765
Val Pro Thr Asp Tyr Arg Asp Leu Tyr Ser Ser Glu Ala Asp Cys Leu
770 775 780
Ala Arg Leu Arg Pro Phe Ile Glu Gln Gly Ala Asp Ile Thr Asn Trp
785 790 795 800
Ser Asp Leu Phe Val Gln Ser Asp Lys His Ser Pro Val Ile His Ala
805 810 815
Asn Ile Glu Leu Ser Val Lys Tyr Gly Thr Thr Lys Leu Leu Glu Gln
820 825 830
Ile Ile Asn Lys Asp Thr Gln Phe Lys Thr Thr Glu Ala Asn Phe Thr
835 840 845
Ala Trp Asn Thr Ala Gln Lys Ser Ile Glu Gln Leu Ile Lys Gln Arg
850 855 860
Glu Asp His His Glu Gln Trp Val Lys Ala Lys Asn Ala Asp Asp Lys
865 870 875 880
Glu Lys Gln Glu Arg Lys Arg Glu Lys Ser Asn Phe Ala Gln Lys Phe
885 890 895
Ile Glu Lys His Gly Asp Asp Tyr Leu Asp Ile Cys Asp Tyr Ile Asn
900 905 910
Thr Tyr Asn Trp Leu Asp Asn Lys Met His Phe Val His Leu Asn Arg
915 920 925
Leu His Gly Leu Thr Ile Glu Leu Leu Gly Arg Met Ala Gly Phe Val
930 935 940
Ala Leu Phe Asp Arg Asp Phe Gln Phe Phe Asp Glu Gln Gln Ile Ala
945 950 955 960
Asp Glu Phe Lys Leu His Gly Phe Val Asn Leu His Ser Ile Asp Lys
965 970 975
Lys Leu Asn Glu Val Pro Thr Lys Lys Ile Lys Glu Ile Tyr Asp Ile
980 985 990
Arg Asn Lys Ile Ile Gln Ile Asn Gly Asn Lys Ile Asn Glu Ser Val
995 1000 1005
Arg Ala Asn Leu Ile Gln Phe Ile Ser Ser Lys Arg Asn Tyr Tyr
1010 1015 1020
Asn Asn Ala Phe Leu His Val Ser Asn Asp Glu Ile Lys Glu Lys
1025 1030 1035
Gln Met Tyr Asp Ile Arg Asn His Ile Ala His Phe Asn Tyr Leu
1040 1045 1050
Thr Lys Asp Ala Ala Asp Phe Ser Leu Ile Asp Leu Ile Asn Glu
1055 1060 1065
Leu Arg Glu Leu Leu His Tyr Asp Arg Lys Leu Lys Asn Ala Val
1070 1075 1080
Ser Lys Ala Phe Ile Asp Leu Phe Asp Lys His Gly Met Ile Leu
1085 1090 1095
Lys Leu Lys Leu Asn Ala Asp His Lys Leu Lys Val Glu Ser Leu
1100 1105 1110
Glu Pro Lys Lys Ile Tyr His Leu Gly Ser Ser Ala Lys Asp Lys
1115 1120 1125
Pro Glu Tyr Gln Tyr Cys Thr Asn Gln Val Met Met Ala Tyr Cys
1130 1135 1140
Asn Met Cys Arg Ser Leu Leu Glu Met Lys Lys
1145 1150
<210> 581
<211> 1120
<212> PRT
<213> Listeria seeligeri
<400> 581
Met Trp Ile Ser Ile Lys Thr Leu Ile His His Leu Gly Val Leu Phe
1 5 10 15
Phe Cys Asp Tyr Met Tyr Asn Arg Arg Glu Lys Lys Ile Ile Glu Val
20 25 30
Lys Thr Met Arg Ile Thr Lys Val Glu Val Asp Arg Lys Lys Val Leu
35 40 45
Ile Ser Arg Asp Lys Asn Gly Gly Lys Leu Val Tyr Glu Asn Glu Met
50 55 60
Gln Asp Asn Thr Glu Gln Ile Met His His Lys Lys Ser Ser Phe Tyr
65 70 75 80
Lys Ser Val Val Asn Lys Thr Ile Cys Arg Pro Glu Gln Lys Gln Met
85 90 95
Lys Lys Leu Val His Gly Leu Leu Gln Glu Asn Ser Gln Glu Lys Ile
100 105 110
Lys Val Ser Asp Val Thr Lys Leu Asn Ile Ser Asn Phe Leu Asn His
115 120 125
Arg Phe Lys Lys Ser Leu Tyr Tyr Phe Pro Glu Asn Ser Pro Asp Lys
130 135 140
Ser Glu Glu Tyr Arg Ile Glu Ile Asn Leu Ser Gln Leu Leu Glu Asp
145 150 155 160
Ser Leu Lys Lys Gln Gln Gly Thr Phe Ile Cys Trp Glu Ser Phe Ser
165 170 175
Lys Asp Met Glu Leu Tyr Ile Asn Trp Ala Glu Asn Tyr Ile Ser Ser
180 185 190
Lys Thr Lys Leu Ile Lys Lys Ser Ile Arg Asn Asn Arg Ile Gln Ser
195 200 205
Thr Glu Ser Arg Ser Gly Gln Leu Met Asp Arg Tyr Met Lys Asp Ile
210 215 220
Leu Asn Lys Asn Lys Pro Phe Asp Ile Gln Ser Val Ser Glu Lys Tyr
225 230 235 240
Gln Leu Glu Lys Leu Thr Ser Ala Leu Lys Ala Thr Phe Lys Glu Ala
245 250 255
Lys Lys Asn Asp Lys Glu Ile Asn Tyr Lys Leu Lys Ser Thr Leu Gln
260 265 270
Asn His Glu Arg Gln Ile Ile Glu Glu Leu Lys Glu Asn Ser Glu Leu
275 280 285
Asn Gln Phe Asn Ile Glu Ile Arg Lys His Leu Glu Thr Tyr Phe Pro
290 295 300
Ile Lys Lys Thr Asn Arg Lys Val Gly Asp Ile Arg Asn Leu Glu Ile
305 310 315 320
Gly Glu Ile Gln Lys Ile Val Asn His Arg Leu Lys Asn Lys Ile Val
325 330 335
Gln Arg Ile Leu Gln Glu Gly Lys Leu Ala Ser Tyr Glu Ile Glu Ser
340 345 350
Thr Val Asn Ser Asn Ser Leu Gln Lys Ile Lys Ile Glu Glu Ala Phe
355 360 365
Ala Leu Lys Phe Ile Asn Ala Cys Leu Phe Ala Ser Asn Asn Leu Arg
370 375 380
Asn Met Val Tyr Pro Val Cys Lys Lys Asp Ile Leu Met Ile Gly Glu
385 390 395 400
Phe Lys Asn Ser Phe Lys Glu Ile Lys His Lys Lys Phe Ile Arg Gln
405 410 415
Trp Ser Gln Phe Phe Ser Gln Glu Ile Thr Val Asp Asp Ile Glu Leu
420 425 430
Ala Ser Trp Gly Leu Arg Gly Ala Ile Ala Pro Ile Arg Asn Glu Ile
435 440 445
Ile His Leu Lys Lys His Ser Trp Lys Lys Phe Phe Asn Asn Pro Thr
450 455 460
Phe Lys Val Lys Lys Ser Lys Ile Ile Asn Gly Lys Thr Lys Asp Val
465 470 475 480
Thr Ser Glu Phe Leu Tyr Lys Glu Thr Leu Phe Lys Asp Tyr Phe Tyr
485 490 495
Ser Glu Leu Asp Ser Val Pro Glu Leu Ile Ile Asn Lys Met Glu Ser
500 505 510
Ser Lys Ile Leu Asp Tyr Tyr Ser Ser Asp Gln Leu Asn Gln Val Phe
515 520 525
Thr Ile Pro Asn Phe Glu Leu Ser Leu Leu Thr Ser Ala Val Pro Phe
530 535 540
Ala Pro Ser Phe Lys Arg Val Tyr Leu Lys Gly Phe Asp Tyr Gln Asn
545 550 555 560
Gln Asp Glu Ala Gln Pro Asp Tyr Asn Leu Lys Leu Asn Ile Tyr Asn
565 570 575
Glu Lys Ala Phe Asn Ser Glu Ala Phe Gln Ala Gln Tyr Ser Leu Phe
580 585 590
Lys Met Val Tyr Tyr Gln Val Phe Leu Pro Gln Phe Thr Thr Asn Asn
595 600 605
Asp Leu Phe Lys Ser Ser Val Asp Phe Ile Leu Thr Leu Asn Lys Glu
610 615 620
Arg Lys Gly Tyr Ala Lys Ala Phe Gln Asp Ile Arg Lys Met Asn Lys
625 630 635 640
Asp Glu Lys Pro Ser Glu Tyr Met Ser Tyr Ile Gln Ser Gln Leu Met
645 650 655
Leu Tyr Gln Lys Lys Gln Glu Glu Lys Glu Lys Ile Asn His Phe Glu
660 665 670
Lys Phe Ile Asn Gln Val Phe Ile Lys Gly Phe Asn Ser Phe Ile Glu
675 680 685
Lys Asn Arg Leu Thr Tyr Ile Cys His Pro Thr Lys Asn Thr Val Pro
690 695 700
Glu Asn Asp Asn Ile Glu Ile Pro Phe His Thr Asp Met Asp Asp Ser
705 710 715 720
Asn Ile Ala Phe Trp Leu Met Cys Lys Leu Leu Asp Ala Lys Gln Leu
725 730 735
Ser Glu Leu Arg Asn Glu Met Ile Lys Phe Ser Cys Ser Leu Gln Ser
740 745 750
Thr Glu Glu Ile Ser Thr Phe Thr Lys Ala Arg Glu Val Ile Gly Leu
755 760 765
Ala Leu Leu Asn Gly Glu Lys Gly Cys Asn Asp Trp Lys Glu Leu Phe
770 775 780
Asp Asp Lys Glu Ala Trp Lys Lys Asn Met Ser Leu Tyr Val Ser Glu
785 790 795 800
Glu Leu Leu Gln Ser Leu Pro Tyr Thr Gln Glu Asp Gly Gln Thr Pro
805 810 815
Val Ile Asn Arg Ser Ile Asp Leu Val Lys Lys Tyr Gly Thr Glu Thr
820 825 830
Ile Leu Glu Lys Leu Phe Ser Ser Ser Asp Asp Tyr Lys Val Ser Ala
835 840 845
Lys Asp Ile Ala Lys Leu His Glu Tyr Asp Val Thr Glu Lys Ile Ala
850 855 860
Gln Gln Glu Ser Leu His Lys Gln Trp Ile Glu Lys Pro Gly Leu Ala
865 870 875 880
Arg Asp Ser Ala Trp Thr Lys Lys Tyr Gln Asn Val Ile Asn Asp Ile
885 890 895
Ser Asn Tyr Gln Trp Ala Lys Thr Lys Val Glu Leu Thr Gln Val Arg
900 905 910
His Leu His Gln Leu Thr Ile Asp Leu Leu Ser Arg Leu Ala Gly Tyr
915 920 925
Met Ser Ile Ala Asp Arg Asp Phe Gln Phe Ser Ser Asn Tyr Ile Leu
930 935 940
Glu Arg Glu Asn Ser Glu Tyr Arg Val Thr Ser Trp Ile Leu Leu Ser
945 950 955 960
Glu Asn Lys Asn Lys Asn Lys Tyr Asn Asp Tyr Glu Leu Tyr Asn Leu
965 970 975
Lys Asn Ala Ser Ile Lys Val Ser Ser Lys Asn Asp Pro Gln Leu Lys
980 985 990
Val Asp Leu Lys Gln Leu Arg Leu Thr Leu Glu Tyr Leu Glu Leu Phe
995 1000 1005
Asp Asn Arg Leu Lys Glu Lys Arg Asn Asn Ile Ser His Phe Asn
1010 1015 1020
Tyr Leu Asn Gly Gln Leu Gly Asn Ser Ile Leu Glu Leu Phe Asp
1025 1030 1035
Asp Ala Arg Asp Val Leu Ser Tyr Asp Arg Lys Leu Lys Asn Ala
1040 1045 1050
Val Ser Lys Ser Leu Lys Glu Ile Leu Ser Ser His Gly Met Glu
1055 1060 1065
Val Thr Phe Lys Pro Leu Tyr Gln Thr Asn His His Leu Lys Ile
1070 1075 1080
Asp Lys Leu Gln Pro Lys Lys Ile His His Leu Gly Glu Lys Ser
1085 1090 1095
Thr Val Ser Ser Asn Gln Val Ser Asn Glu Tyr Cys Gln Leu Val
1100 1105 1110
Arg Thr Leu Leu Thr Met Lys
1115 1120
<210> 582
<211> 970
<212> PRT
<213> Listeria weihenstephanensis
<400> 582
Met Leu Ala Leu Leu His Gln Glu Val Pro Ser Gln Lys Leu His Asn
1 5 10 15
Leu Lys Ser Leu Asn Thr Glu Ser Leu Thr Lys Leu Phe Lys Pro Lys
20 25 30
Phe Gln Asn Met Ile Ser Tyr Pro Pro Ser Lys Gly Ala Glu His Val
35 40 45
Gln Phe Cys Leu Thr Asp Ile Ala Val Pro Ala Ile Arg Asp Leu Asp
50 55 60
Glu Ile Lys Pro Asp Trp Gly Ile Phe Phe Glu Lys Leu Lys Pro Tyr
65 70 75 80
Thr Asp Trp Ala Glu Ser Tyr Ile His Tyr Lys Gln Thr Thr Ile Gln
85 90 95
Lys Ser Ile Glu Gln Asn Lys Ile Gln Ser Pro Asp Ser Pro Arg Lys
100 105 110
Leu Val Leu Gln Lys Tyr Val Thr Ala Phe Leu Asn Gly Glu Pro Leu
115 120 125
Gly Leu Asp Leu Val Ala Lys Lys Tyr Lys Leu Ala Asp Leu Ala Glu
130 135 140
Ser Phe Lys Val Val Asp Leu Asn Glu Asp Lys Ser Ala Asn Tyr Lys
145 150 155 160
Ile Lys Ala Cys Leu Gln Gln His Gln Arg Asn Ile Leu Asp Glu Leu
165 170 175
Lys Glu Asp Pro Glu Leu Asn Gln Tyr Gly Ile Glu Val Lys Lys Tyr
180 185 190
Ile Gln Arg Tyr Phe Pro Ile Lys Arg Ala Pro Asn Arg Ser Lys His
195 200 205
Ala Arg Ala Asp Phe Leu Lys Lys Glu Leu Ile Glu Ser Thr Val Glu
210 215 220
Gln Gln Phe Lys Asn Ala Val Tyr His Tyr Val Leu Glu Gln Gly Lys
225 230 235 240
Met Glu Ala Tyr Glu Leu Thr Asp Pro Lys Thr Lys Asp Leu Gln Asp
245 250 255
Ile Arg Ser Gly Glu Ala Phe Ser Phe Lys Phe Ile Asn Ala Cys Ala
260 265 270
Phe Ala Ser Asn Asn Leu Lys Met Ile Leu Asn Pro Glu Cys Glu Lys
275 280 285
Asp Ile Leu Gly Lys Gly Asp Phe Lys Lys Asn Leu Pro Asn Ser Thr
290 295 300
Thr Gln Ser Asp Val Val Lys Lys Met Ile Pro Phe Phe Ser Asp Glu
305 310 315 320
Ile Gln Asn Val Asn Phe Asp Glu Ala Ile Trp Ala Ile Arg Gly Ser
325 330 335
Ile Gln Gln Ile Arg Asn Glu Val Tyr His Cys Lys Lys His Ser Trp
340 345 350
Lys Ser Ile Leu Lys Ile Lys Gly Phe Glu Phe Glu Pro Asn Asn Met
355 360 365
Lys Tyr Thr Asp Ser Asp Met Gln Lys Leu Met Asp Lys Asp Ile Ala
370 375 380
Lys Ile Pro Asp Phe Ile Glu Glu Lys Leu Lys Ser Ser Gly Ile Ile
385 390 395 400
Arg Phe Tyr Ser His Asp Lys Leu Gln Ser Ile Trp Glu Met Lys Gln
405 410 415
Gly Phe Ser Leu Leu Thr Thr Asn Ala Pro Phe Val Pro Ser Phe Lys
420 425 430
Arg Val Tyr Ala Lys Gly His Asp Tyr Gln Thr Ser Lys Asn Arg Tyr
435 440 445
Tyr Asp Leu Gly Leu Thr Thr Phe Asp Ile Leu Glu Tyr Gly Glu Glu
450 455 460
Asp Phe Arg Ala Arg Tyr Phe Leu Thr Lys Leu Val Tyr Tyr Gln Gln
465 470 475 480
Phe Met Pro Trp Phe Thr Ala Asp Asn Asn Ala Phe Arg Asp Ala Ala
485 490 495
Asn Phe Val Leu Arg Leu Asn Lys Asn Arg Gln Gln Asp Ala Lys Ala
500 505 510
Phe Ile Asn Ile Arg Glu Val Glu Glu Gly Glu Met Pro Arg Asp Tyr
515 520 525
Met Gly Tyr Val Gln Gly Gln Ile Ala Ile His Glu Asp Ser Thr Glu
530 535 540
Asp Thr Pro Asn His Phe Glu Lys Phe Ile Ser Gln Val Phe Ile Lys
545 550 555 560
Gly Phe Asp Ser His Met Arg Ser Ala Asp Leu Lys Phe Ile Lys Asn
565 570 575
Pro Arg Asn Gln Gly Leu Glu Gln Ser Glu Ile Glu Glu Met Ser Phe
580 585 590
Asp Ile Lys Val Glu Pro Ser Phe Leu Lys Asn Lys Asp Asp Tyr Ile
595 600 605
Ala Phe Trp Thr Phe Cys Lys Met Leu Asp Ala Arg His Leu Ser Glu
610 615 620
Leu Arg Asn Glu Met Ile Lys Tyr Asp Gly His Leu Thr Gly Glu Gln
625 630 635 640
Glu Ile Ile Gly Leu Ala Leu Leu Gly Val Asp Ser Arg Glu Asn Asp
645 650 655
Trp Lys Gln Phe Phe Ser Ser Glu Arg Glu Tyr Glu Lys Ile Met Lys
660 665 670
Gly Tyr Val Gly Glu Glu Leu Tyr Gln Arg Glu Pro Tyr Arg Gln Ser
675 680 685
Asp Gly Lys Thr Pro Ile Leu Phe Arg Gly Val Glu Gln Ala Arg Lys
690 695 700
Tyr Gly Thr Glu Thr Val Ile Gln Arg Leu Phe Asp Ala Ser Pro Glu
705 710 715 720
Phe Lys Val Ser Lys Cys Asn Ile Thr Glu Trp Glu Arg Gln Lys Glu
725 730 735
Thr Ile Glu Glu Thr Ile Glu Arg Arg Lys Glu Leu His Asn Glu Trp
740 745 750
Glu Lys Asn Pro Lys Lys Pro Gln Asn Asn Ala Phe Phe Lys Glu Tyr
755 760 765
Lys Glu Cys Cys Asp Ala Ile Asp Ala Tyr Asn Trp His Lys Asn Lys
770 775 780
Thr Thr Leu Val Tyr Val Asn Glu Leu His His Leu Leu Ile Glu Ile
785 790 795 800
Leu Gly Arg Tyr Val Gly Tyr Val Ala Ile Ala Asp Arg Asp Phe Gln
805 810 815
Cys Met Ala Asn Gln Tyr Phe Lys His Ser Gly Ile Thr Glu Arg Val
820 825 830
Glu Tyr Trp Gly Asp Asn Arg Leu Lys Ser Ile Lys Lys Leu Asp Thr
835 840 845
Phe Leu Lys Lys Glu Gly Leu Phe Val Ser Glu Lys Asn Ala Arg Asn
850 855 860
His Ile Ala His Leu Asn Tyr Leu Ser Leu Lys Ser Glu Cys Thr Leu
865 870 875 880
Leu Tyr Leu Ser Glu Arg Leu Arg Glu Ile Phe Lys Tyr Asp Arg Lys
885 890 895
Leu Lys Asn Ala Val Ser Lys Ser Leu Ile Asp Ile Leu Asp Arg His
900 905 910
Gly Met Ser Val Val Phe Ala Asn Leu Lys Glu Asn Lys His Arg Leu
915 920 925
Val Ile Lys Ser Leu Glu Pro Lys Lys Leu Arg His Leu Gly Glu Lys
930 935 940
Lys Ile Asp Asn Gly Tyr Ile Glu Thr Asn Gln Val Ser Glu Glu Tyr
945 950 955 960
Cys Gly Ile Val Lys Arg Leu Leu Glu Ile
965 970
<210> 583
<211> 1051
<212> PRT
<213> Listeria newyorkensis
<400> 583
Met Lys Ile Thr Lys Met Arg Val Asp Gly Arg Thr Ile Val Met Glu
1 5 10 15
Arg Thr Ser Lys Glu Gly Gln Leu Gly Tyr Glu Gly Ile Asp Gly Asn
20 25 30
Lys Thr Thr Glu Ile Ile Phe Asp Lys Lys Lys Glu Ser Phe Tyr Lys
35 40 45
Ser Ile Leu Asn Lys Thr Val Arg Lys Pro Asp Glu Lys Glu Lys Asn
50 55 60
Arg Arg Lys Gln Ala Ile Asn Lys Ala Ile Asn Lys Glu Ile Thr Glu
65 70 75 80
Leu Met Leu Ala Val Leu His Gln Glu Val Pro Ser Gln Lys Leu His
85 90 95
Asn Leu Lys Ser Leu Asn Thr Glu Ser Leu Thr Lys Leu Phe Lys Pro
100 105 110
Lys Phe Gln Asn Met Ile Ser Tyr Pro Pro Ser Lys Gly Ala Glu His
115 120 125
Val Gln Phe Cys Leu Thr Asp Ile Ala Val Pro Ala Ile Arg Asp Leu
130 135 140
Asp Glu Ile Lys Pro Asp Trp Gly Ile Phe Phe Glu Lys Leu Lys Pro
145 150 155 160
Tyr Thr Asp Trp Ala Glu Ser Tyr Ile His Tyr Lys Gln Thr Thr Ile
165 170 175
Gln Lys Ser Ile Glu Gln Asn Lys Ile Gln Ser Pro Asp Ser Pro Arg
180 185 190
Lys Leu Val Leu Gln Lys Tyr Val Thr Ala Phe Leu Asn Gly Glu Pro
195 200 205
Leu Gly Leu Asp Leu Val Ala Lys Lys Tyr Lys Leu Ala Asp Leu Ala
210 215 220
Glu Ser Phe Lys Leu Val Asp Leu Asn Glu Asp Lys Ser Ala Asn Tyr
225 230 235 240
Lys Ile Lys Ala Cys Leu Gln Gln His Gln Arg Asn Ile Leu Asp Glu
245 250 255
Leu Lys Glu Asp Pro Glu Leu Asn Gln Tyr Gly Ile Glu Val Lys Lys
260 265 270
Tyr Ile Gln Arg Tyr Phe Pro Ile Lys Arg Ala Pro Asn Arg Ser Lys
275 280 285
His Ala Arg Ala Asp Phe Leu Lys Lys Glu Leu Ile Glu Ser Thr Val
290 295 300
Glu Gln Gln Phe Lys Asn Ala Val Tyr His Tyr Val Leu Glu Gln Gly
305 310 315 320
Lys Met Glu Ala Tyr Glu Leu Thr Asp Pro Lys Thr Lys Asp Leu Gln
325 330 335
Asp Ile Arg Ser Gly Glu Ala Phe Ser Phe Lys Phe Ile Asn Ala Cys
340 345 350
Ala Phe Ala Ser Asn Asn Leu Lys Met Ile Leu Asn Pro Glu Cys Glu
355 360 365
Lys Asp Ile Leu Gly Lys Gly Asn Phe Lys Lys Asn Leu Pro Asn Ser
370 375 380
Thr Thr Arg Ser Asp Val Val Lys Lys Met Ile Pro Phe Phe Ser Asp
385 390 395 400
Glu Leu Gln Asn Val Asn Phe Asp Glu Ala Ile Trp Ala Ile Arg Gly
405 410 415
Ser Ile Gln Gln Ile Arg Asn Glu Val Tyr His Cys Lys Lys His Ser
420 425 430
Trp Lys Ser Ile Leu Lys Ile Lys Gly Phe Glu Phe Glu Pro Asn Asn
435 440 445
Met Lys Tyr Ala Asp Ser Asp Met Gln Lys Leu Met Asp Lys Asp Ile
450 455 460
Ala Lys Ile Pro Glu Phe Ile Glu Glu Lys Leu Lys Ser Ser Gly Val
465 470 475 480
Val Arg Phe Tyr Arg His Asp Glu Leu Gln Ser Ile Trp Glu Met Lys
485 490 495
Gln Gly Phe Ser Leu Leu Thr Thr Asn Ala Pro Phe Val Pro Ser Phe
500 505 510
Lys Arg Val Tyr Ala Lys Gly His Asp Tyr Gln Thr Ser Lys Asn Arg
515 520 525
Tyr Tyr Asn Leu Asp Leu Thr Thr Phe Asp Ile Leu Glu Tyr Gly Glu
530 535 540
Glu Asp Phe Arg Ala Arg Tyr Phe Leu Thr Lys Leu Val Tyr Tyr Gln
545 550 555 560
Gln Phe Met Pro Trp Phe Thr Ala Asp Asn Asn Ala Phe Arg Asp Ala
565 570 575
Ala Asn Phe Val Leu Arg Leu Asn Lys Asn Arg Gln Gln Asp Ala Lys
580 585 590
Ala Phe Ile Asn Ile Arg Glu Val Glu Glu Gly Glu Met Pro Arg Asp
595 600 605
Tyr Met Gly Tyr Val Gln Gly Gln Ile Ala Ile His Glu Asp Ser Ile
610 615 620
Glu Asp Thr Pro Asn His Phe Glu Lys Phe Ile Ser Gln Val Phe Ile
625 630 635 640
Lys Gly Phe Asp Arg His Met Arg Ser Ala Asn Leu Lys Phe Ile Lys
645 650 655
Asn Pro Arg Asn Gln Gly Leu Glu Gln Ser Glu Ile Glu Glu Met Ser
660 665 670
Phe Asp Ile Lys Val Glu Pro Ser Phe Leu Lys Asn Lys Asp Asp Tyr
675 680 685
Ile Ala Phe Trp Ile Phe Cys Lys Met Leu Asp Ala Arg His Leu Ser
690 695 700
Glu Leu Arg Asn Glu Met Ile Lys Tyr Asp Gly His Leu Thr Gly Glu
705 710 715 720
Gln Glu Ile Ile Gly Leu Ala Leu Leu Gly Val Asp Ser Arg Glu Asn
725 730 735
Asp Trp Lys Gln Phe Phe Ser Ser Glu Arg Glu Tyr Glu Lys Ile Met
740 745 750
Lys Gly Tyr Val Val Glu Glu Leu Tyr Gln Arg Glu Pro Tyr Arg Gln
755 760 765
Ser Asp Gly Lys Thr Pro Ile Leu Phe Arg Gly Val Glu Gln Ala Arg
770 775 780
Lys Tyr Gly Thr Glu Thr Val Ile Gln Arg Leu Phe Asp Ala Asn Pro
785 790 795 800
Glu Phe Lys Val Ser Lys Cys Asn Leu Ala Glu Trp Glu Arg Gln Lys
805 810 815
Glu Thr Ile Glu Glu Thr Ile Lys Arg Arg Lys Glu Leu His Asn Glu
820 825 830
Trp Ala Lys Asn Pro Lys Lys Pro Gln Asn Asn Ala Phe Phe Lys Glu
835 840 845
Tyr Lys Glu Cys Cys Asp Ala Ile Asp Ala Tyr Asn Trp His Lys Asn
850 855 860
Lys Thr Thr Leu Ala Tyr Val Asn Glu Leu His His Leu Leu Ile Glu
865 870 875 880
Ile Leu Gly Arg Tyr Val Gly Tyr Val Ala Ile Ala Asp Arg Asp Phe
885 890 895
Gln Cys Met Ala Asn Gln Tyr Phe Lys His Ser Gly Ile Thr Glu Arg
900 905 910
Val Glu Tyr Trp Gly Asp Asn Arg Leu Lys Ser Ile Lys Lys Leu Asp
915 920 925
Thr Phe Leu Lys Lys Glu Gly Leu Phe Val Ser Glu Lys Asn Ala Arg
930 935 940
Asn His Ile Ala His Leu Asn Tyr Leu Ser Leu Lys Ser Glu Cys Thr
945 950 955 960
Leu Leu Tyr Leu Ser Glu Arg Leu Arg Glu Ile Phe Lys Tyr Asp Arg
965 970 975
Lys Leu Lys Asn Ala Val Ser Lys Ser Leu Ile Asp Ile Leu Asp Arg
980 985 990
His Gly Met Ser Val Val Phe Ala Asn Leu Lys Glu Asn Lys His Arg
995 1000 1005
Leu Val Ile Lys Ser Leu Glu Pro Lys Lys Leu Arg His Leu Gly
1010 1015 1020
Gly Lys Lys Ile Asp Gly Gly Tyr Ile Glu Thr Asn Gln Val Ser
1025 1030 1035
Glu Glu Tyr Cys Gly Ile Val Lys Arg Leu Leu Glu Met
1040 1045 1050
<210> 584
<211> 1182
<212> PRT
<213> Leptotrichia wadei
<400> 584
Met Tyr Met Lys Ile Thr Lys Ile Asp Gly Val Ser His Tyr Lys Lys
1 5 10 15
Gln Asp Lys Gly Ile Leu Lys Lys Lys Trp Lys Asp Leu Asp Glu Arg
20 25 30
Lys Gln Arg Glu Lys Ile Glu Ala Arg Tyr Asn Lys Gln Ile Glu Ser
35 40 45
Lys Ile Tyr Lys Glu Phe Phe Arg Leu Lys Asn Lys Lys Arg Ile Glu
50 55 60
Lys Glu Glu Asp Gln Asn Ile Lys Ser Leu Tyr Phe Phe Ile Lys Glu
65 70 75 80
Leu Tyr Leu Asn Glu Lys Asn Glu Glu Trp Glu Leu Lys Asn Ile Asn
85 90 95
Leu Glu Ile Leu Asp Asp Lys Glu Arg Val Ile Lys Gly Tyr Lys Phe
100 105 110
Lys Glu Asp Val Tyr Phe Phe Lys Glu Gly Tyr Lys Glu Tyr Tyr Leu
115 120 125
Arg Ile Leu Phe Asn Asn Leu Ile Glu Lys Val Gln Asn Glu Asn Arg
130 135 140
Glu Lys Val Arg Lys Asn Lys Glu Phe Leu Asp Leu Lys Glu Ile Phe
145 150 155 160
Lys Lys Tyr Lys Asn Arg Lys Ile Asp Leu Leu Leu Lys Ser Ile Asn
165 170 175
Asn Asn Lys Ile Asn Leu Glu Tyr Lys Lys Glu Asn Val Asn Glu Glu
180 185 190
Ile Tyr Gly Ile Asn Pro Thr Asn Asp Arg Glu Met Thr Phe Tyr Glu
195 200 205
Leu Leu Lys Glu Ile Ile Glu Lys Lys Asp Glu Gln Lys Ser Ile Leu
210 215 220
Glu Glu Lys Leu Asp Asn Phe Asp Ile Thr Asn Phe Leu Glu Asn Ile
225 230 235 240
Glu Lys Ile Phe Asn Glu Glu Thr Glu Ile Asn Ile Ile Lys Gly Lys
245 250 255
Val Leu Asn Glu Leu Arg Glu Tyr Ile Lys Glu Lys Glu Glu Asn Asn
260 265 270
Ser Asp Asn Lys Leu Lys Gln Ile Tyr Asn Leu Glu Leu Lys Lys Tyr
275 280 285
Ile Glu Asn Asn Phe Ser Tyr Lys Lys Gln Lys Ser Lys Ser Lys Asn
290 295 300
Gly Lys Asn Asp Tyr Leu Tyr Leu Asn Phe Leu Lys Lys Ile Met Phe
305 310 315 320
Ile Glu Glu Val Asp Glu Lys Lys Glu Ile Asn Lys Glu Lys Phe Lys
325 330 335
Asn Lys Ile Asn Ser Asn Phe Lys Asn Leu Phe Val Gln His Ile Leu
340 345 350
Asp Tyr Gly Lys Leu Leu Tyr Tyr Lys Glu Asn Asp Glu Tyr Ile Lys
355 360 365
Asn Thr Gly Gln Leu Glu Thr Lys Asp Leu Glu Tyr Ile Lys Thr Lys
370 375 380
Glu Thr Leu Ile Arg Lys Met Ala Val Leu Val Ser Phe Ala Ala Asn
385 390 395 400
Ser Tyr Tyr Asn Leu Phe Gly Arg Val Ser Gly Asp Ile Leu Gly Thr
405 410 415
Glu Val Val Lys Ser Ser Lys Thr Asn Val Ile Lys Val Gly Ser His
420 425 430
Ile Phe Lys Glu Lys Met Leu Asn Tyr Phe Phe Asp Phe Glu Ile Phe
435 440 445
Asp Ala Asn Lys Ile Val Glu Ile Leu Glu Ser Ile Ser Tyr Ser Ile
450 455 460
Tyr Asn Val Arg Asn Gly Val Gly His Phe Asn Lys Leu Ile Leu Gly
465 470 475 480
Lys Tyr Lys Lys Lys Asp Ile Asn Thr Asn Lys Arg Ile Glu Glu Asp
485 490 495
Leu Asn Asn Asn Glu Glu Ile Lys Gly Tyr Phe Ile Lys Lys Arg Gly
500 505 510
Glu Ile Glu Arg Lys Val Lys Glu Lys Phe Leu Ser Asn Asn Leu Gln
515 520 525
Tyr Tyr Tyr Ser Lys Glu Lys Ile Glu Asn Tyr Phe Glu Val Tyr Glu
530 535 540
Phe Glu Ile Leu Lys Arg Lys Ile Pro Phe Ala Pro Asn Phe Lys Arg
545 550 555 560
Ile Ile Lys Lys Gly Glu Asp Leu Phe Asn Asn Lys Asn Asn Lys Lys
565 570 575
Tyr Glu Tyr Phe Lys Asn Phe Asp Lys Asn Ser Ala Glu Glu Lys Lys
580 585 590
Glu Phe Leu Lys Thr Arg Asn Phe Leu Leu Lys Glu Leu Tyr Tyr Asn
595 600 605
Asn Phe Tyr Lys Glu Phe Leu Ser Lys Lys Glu Glu Phe Glu Lys Ile
610 615 620
Val Leu Glu Val Lys Glu Glu Lys Lys Ser Arg Gly Asn Ile Asn Asn
625 630 635 640
Lys Lys Ser Gly Val Ser Phe Gln Ser Ile Asp Asp Tyr Asp Thr Lys
645 650 655
Ile Asn Ile Ser Asp Tyr Ile Ala Ser Ile His Lys Lys Glu Met Glu
660 665 670
Arg Val Glu Lys Tyr Asn Glu Glu Lys Gln Lys Asp Thr Ala Lys Tyr
675 680 685
Ile Arg Asp Phe Val Glu Glu Ile Phe Leu Thr Gly Phe Ile Asn Tyr
690 695 700
Leu Glu Lys Asp Lys Arg Leu His Phe Leu Lys Glu Glu Phe Ser Ile
705 710 715 720
Leu Cys Asn Asn Asn Asn Asn Val Val Asp Phe Asn Ile Asn Ile Asn
725 730 735
Glu Glu Lys Ile Lys Glu Phe Leu Lys Glu Asn Asp Ser Lys Thr Leu
740 745 750
Asn Leu Tyr Leu Phe Phe Asn Met Ile Asp Ser Lys Arg Ile Ser Glu
755 760 765
Phe Arg Asn Glu Leu Val Lys Tyr Lys Gln Phe Thr Lys Lys Arg Leu
770 775 780
Asp Glu Glu Lys Glu Phe Leu Gly Ile Lys Ile Glu Leu Tyr Glu Thr
785 790 795 800
Leu Ile Glu Phe Val Ile Leu Thr Arg Glu Lys Leu Asp Thr Lys Lys
805 810 815
Ser Glu Glu Ile Asp Ala Trp Leu Val Asp Lys Leu Tyr Val Lys Asp
820 825 830
Ser Asn Glu Tyr Lys Glu Tyr Glu Glu Ile Leu Lys Leu Phe Val Asp
835 840 845
Glu Lys Ile Leu Ser Ser Lys Glu Ala Pro Tyr Tyr Ala Thr Asp Asn
850 855 860
Lys Thr Pro Ile Leu Leu Ser Asn Phe Glu Lys Thr Arg Lys Tyr Gly
865 870 875 880
Thr Gln Ser Phe Leu Ser Glu Ile Gln Ser Asn Tyr Lys Tyr Ser Lys
885 890 895
Val Glu Lys Glu Asn Ile Glu Asp Tyr Asn Lys Lys Glu Glu Ile Glu
900 905 910
Gln Lys Lys Lys Ser Asn Ile Glu Lys Leu Gln Asp Leu Lys Val Glu
915 920 925
Leu His Lys Lys Trp Glu Gln Asn Lys Ile Thr Glu Lys Glu Ile Glu
930 935 940
Lys Tyr Asn Asn Thr Thr Arg Lys Ile Asn Glu Tyr Asn Tyr Leu Lys
945 950 955 960
Asn Lys Glu Glu Leu Gln Asn Val Tyr Leu Leu His Glu Met Leu Ser
965 970 975
Asp Leu Leu Ala Arg Asn Val Ala Phe Phe Asn Lys Trp Glu Arg Asp
980 985 990
Phe Lys Phe Ile Val Ile Ala Ile Lys Gln Phe Leu Arg Glu Asn Asp
995 1000 1005
Lys Glu Lys Val Asn Glu Phe Leu Asn Pro Pro Asp Asn Ser Lys
1010 1015 1020
Gly Lys Lys Val Tyr Phe Ser Val Ser Lys Tyr Lys Asn Thr Val
1025 1030 1035
Glu Asn Ile Asp Gly Ile His Lys Asn Phe Met Asn Leu Ile Phe
1040 1045 1050
Leu Asn Asn Lys Phe Met Asn Arg Lys Ile Asp Lys Met Asn Cys
1055 1060 1065
Ala Ile Trp Val Tyr Phe Arg Asn Tyr Ile Ala His Phe Leu His
1070 1075 1080
Leu His Thr Lys Asn Glu Lys Ile Ser Leu Ile Ser Gln Met Asn
1085 1090 1095
Leu Leu Ile Lys Leu Phe Ser Tyr Asp Lys Lys Val Gln Asn His
1100 1105 1110
Ile Leu Lys Ser Thr Lys Thr Leu Leu Glu Lys Tyr Asn Ile Gln
1115 1120 1125
Ile Asn Phe Glu Ile Ser Asn Asp Lys Asn Glu Val Phe Lys Tyr
1130 1135 1140
Lys Ile Lys Asn Arg Leu Tyr Ser Lys Lys Gly Lys Met Leu Gly
1145 1150 1155
Lys Asn Asn Lys Phe Glu Ile Leu Glu Asn Glu Phe Leu Glu Asn
1160 1165 1170
Val Lys Ala Met Leu Glu Tyr Ser Glu
1175 1180
<210> 585
<211> 1180
<212> PRT
<213> Leptotrichia wadei
<400> 585
Met Lys Ile Thr Lys Ile Asp Gly Val Ser His Tyr Lys Lys Gln Asp
1 5 10 15
Lys Gly Ile Leu Lys Lys Lys Trp Lys Asp Leu Asp Glu Arg Lys Gln
20 25 30
Arg Glu Lys Ile Glu Ala Arg Tyr Asn Lys Gln Ile Glu Ser Lys Ile
35 40 45
Tyr Lys Glu Phe Phe Arg Leu Lys Asn Lys Lys Arg Ile Glu Lys Glu
50 55 60
Glu Asp Gln Asn Ile Lys Ser Leu Tyr Phe Phe Ile Lys Glu Leu Tyr
65 70 75 80
Leu Asn Glu Lys Asn Glu Glu Trp Glu Leu Lys Asn Ile Asn Leu Glu
85 90 95
Ile Leu Asp Asp Lys Glu Arg Val Ile Lys Gly Tyr Lys Phe Lys Glu
100 105 110
Asp Val Tyr Phe Phe Lys Glu Gly Tyr Lys Glu Tyr Tyr Leu Arg Ile
115 120 125
Leu Phe Asn Asn Leu Ile Glu Lys Val Gln Asn Glu Asn Arg Glu Lys
130 135 140
Val Arg Lys Asn Lys Glu Phe Leu Asp Leu Lys Glu Ile Phe Lys Lys
145 150 155 160
Tyr Lys Asn Arg Lys Ile Asp Leu Leu Leu Lys Ser Ile Asn Asn Asn
165 170 175
Lys Ile Asn Leu Glu Tyr Lys Lys Glu Asn Val Asn Glu Glu Ile Tyr
180 185 190
Gly Ile Asn Pro Thr Asn Asp Arg Glu Met Thr Phe Tyr Glu Leu Leu
195 200 205
Lys Glu Ile Ile Glu Lys Lys Asp Glu Gln Lys Ser Ile Leu Glu Glu
210 215 220
Lys Leu Asp Asn Phe Asp Ile Thr Asn Phe Leu Glu Asn Ile Glu Lys
225 230 235 240
Ile Phe Asn Glu Glu Thr Glu Ile Asn Ile Ile Lys Gly Lys Val Leu
245 250 255
Asn Glu Leu Arg Glu Tyr Ile Lys Glu Lys Glu Glu Asn Asn Ser Asp
260 265 270
Asn Lys Leu Lys Gln Ile Tyr Asn Leu Glu Leu Lys Lys Tyr Ile Glu
275 280 285
Asn Asn Phe Ser Tyr Lys Lys Gln Lys Ser Lys Ser Lys Asn Gly Lys
290 295 300
Asn Asp Tyr Leu Tyr Leu Asn Phe Leu Lys Lys Ile Met Phe Ile Glu
305 310 315 320
Glu Val Asp Glu Lys Lys Glu Ile Asn Lys Glu Lys Phe Lys Asn Lys
325 330 335
Ile Asn Ser Asn Phe Lys Asn Leu Phe Val Gln His Ile Leu Asp Tyr
340 345 350
Gly Lys Leu Leu Tyr Tyr Lys Glu Asn Asp Glu Tyr Ile Lys Asn Thr
355 360 365
Gly Gln Leu Glu Thr Lys Asp Leu Glu Tyr Ile Lys Thr Lys Glu Thr
370 375 380
Leu Ile Arg Lys Met Ala Val Leu Val Ser Phe Ala Ala Asn Ser Tyr
385 390 395 400
Tyr Asn Leu Phe Gly Arg Val Ser Gly Asp Ile Leu Gly Thr Glu Val
405 410 415
Val Lys Ser Ser Lys Thr Asn Val Ile Lys Val Gly Ser His Ile Phe
420 425 430
Lys Glu Lys Met Leu Asn Tyr Phe Phe Asp Phe Glu Ile Phe Asp Ala
435 440 445
Asn Lys Ile Val Glu Ile Leu Glu Ser Ile Ser Tyr Ser Ile Tyr Asn
450 455 460
Val Arg Asn Gly Val Gly His Phe Asn Lys Leu Ile Leu Gly Lys Tyr
465 470 475 480
Lys Lys Lys Asp Ile Asn Thr Asn Lys Arg Ile Glu Glu Asp Leu Asn
485 490 495
Asn Asn Glu Glu Ile Lys Gly Tyr Phe Ile Lys Lys Arg Gly Glu Ile
500 505 510
Glu Arg Lys Val Lys Glu Lys Phe Leu Ser Asn Asn Leu Gln Tyr Tyr
515 520 525
Tyr Ser Lys Glu Lys Ile Glu Asn Tyr Phe Glu Val Tyr Glu Phe Glu
530 535 540
Ile Leu Lys Arg Lys Ile Pro Phe Ala Pro Asn Phe Lys Arg Ile Ile
545 550 555 560
Lys Lys Gly Glu Asp Leu Phe Asn Asn Lys Asn Asn Lys Lys Tyr Glu
565 570 575
Tyr Phe Lys Asn Phe Asp Lys Asn Ser Ala Glu Glu Lys Lys Glu Phe
580 585 590
Leu Lys Thr Arg Asn Phe Leu Leu Lys Glu Leu Tyr Tyr Asn Asn Phe
595 600 605
Tyr Lys Glu Phe Leu Ser Lys Lys Glu Glu Phe Glu Lys Ile Val Leu
610 615 620
Glu Val Lys Glu Glu Lys Lys Ser Arg Gly Asn Ile Asn Asn Lys Lys
625 630 635 640
Ser Gly Val Ser Phe Gln Ser Ile Asp Asp Tyr Asp Thr Lys Ile Asn
645 650 655
Ile Ser Asp Tyr Ile Ala Ser Ile His Lys Lys Glu Met Glu Arg Val
660 665 670
Glu Lys Tyr Asn Glu Glu Lys Gln Lys Asp Thr Ala Lys Tyr Ile Arg
675 680 685
Asp Phe Val Glu Glu Ile Phe Leu Thr Gly Phe Ile Asn Tyr Leu Glu
690 695 700
Lys Asp Lys Arg Leu His Phe Leu Lys Glu Glu Phe Ser Ile Leu Cys
705 710 715 720
Asn Asn Asn Asn Asn Val Val Asp Phe Asn Ile Asn Ile Asn Glu Glu
725 730 735
Lys Ile Lys Glu Phe Leu Lys Glu Asn Asp Ser Lys Thr Leu Asn Leu
740 745 750
Tyr Leu Phe Phe Asn Met Ile Asp Ser Lys Arg Ile Ser Glu Phe Arg
755 760 765
Asn Glu Leu Val Lys Tyr Lys Gln Phe Thr Lys Lys Arg Leu Asp Glu
770 775 780
Glu Lys Glu Phe Leu Gly Ile Lys Ile Glu Leu Tyr Glu Thr Leu Ile
785 790 795 800
Glu Phe Val Ile Leu Thr Arg Glu Lys Leu Asp Thr Lys Lys Ser Glu
805 810 815
Glu Ile Asp Ala Trp Leu Val Asp Lys Leu Tyr Val Lys Asp Ser Asn
820 825 830
Glu Tyr Lys Glu Tyr Glu Glu Ile Leu Lys Leu Phe Val Asp Glu Lys
835 840 845
Ile Leu Ser Ser Lys Glu Ala Pro Tyr Tyr Ala Thr Asp Asn Lys Thr
850 855 860
Pro Ile Leu Leu Ser Asn Phe Glu Lys Thr Arg Lys Tyr Gly Thr Gln
865 870 875 880
Ser Phe Leu Ser Glu Ile Gln Ser Asn Tyr Lys Tyr Ser Lys Val Glu
885 890 895
Lys Glu Asn Ile Glu Asp Tyr Asn Lys Lys Glu Glu Ile Glu Gln Lys
900 905 910
Lys Lys Ser Asn Ile Glu Lys Leu Gln Asp Leu Lys Val Glu Leu His
915 920 925
Lys Lys Trp Glu Gln Asn Lys Ile Thr Glu Lys Glu Ile Glu Lys Tyr
930 935 940
Asn Asn Thr Thr Arg Lys Ile Asn Glu Tyr Asn Tyr Leu Lys Asn Lys
945 950 955 960
Glu Glu Leu Gln Asn Val Tyr Leu Leu His Glu Met Leu Ser Asp Leu
965 970 975
Leu Ala Arg Asn Val Ala Phe Phe Asn Lys Trp Glu Arg Asp Phe Lys
980 985 990
Phe Ile Val Ile Ala Ile Lys Gln Phe Leu Arg Glu Asn Asp Lys Glu
995 1000 1005
Lys Val Asn Glu Phe Leu Asn Pro Pro Asp Asn Ser Lys Gly Lys
1010 1015 1020
Lys Val Tyr Phe Ser Val Ser Lys Tyr Lys Asn Thr Val Glu Asn
1025 1030 1035
Ile Asp Gly Ile His Lys Asn Phe Met Asn Leu Ile Phe Leu Asn
1040 1045 1050
Asn Lys Phe Met Asn Arg Lys Ile Asp Lys Met Asn Cys Ala Ile
1055 1060 1065
Trp Val Tyr Phe Arg Asn Tyr Ile Ala His Phe Leu His Leu His
1070 1075 1080
Thr Lys Asn Glu Lys Ile Ser Leu Ile Ser Gln Met Asn Leu Leu
1085 1090 1095
Ile Lys Leu Phe Ser Tyr Asp Lys Lys Val Gln Asn His Ile Leu
1100 1105 1110
Lys Ser Thr Lys Thr Leu Leu Glu Lys Tyr Asn Ile Gln Ile Asn
1115 1120 1125
Phe Glu Ile Ser Asn Asp Lys Asn Glu Val Phe Lys Tyr Lys Ile
1130 1135 1140
Lys Asn Arg Leu Tyr Ser Lys Lys Gly Lys Met Leu Gly Lys Asn
1145 1150 1155
Asn Lys Phe Glu Ile Leu Glu Asn Glu Phe Leu Glu Asn Val Lys
1160 1165 1170
Ala Met Leu Glu Tyr Ser Glu
1175 1180
<210> 586
<211> 1197
<212> PRT
<213> Leptotrichia wadei
<400> 586
Met Lys Val Thr Lys Ile Asp Gly Leu Ser His Lys Lys Phe Glu Asp
1 5 10 15
Glu Gly Lys Leu Val Lys Phe Arg Asn Asn Lys Asn Ile Asn Glu Ile
20 25 30
Lys Glu Arg Leu Lys Lys Leu Lys Glu Leu Lys Leu Asp Asn Tyr Ile
35 40 45
Lys Asn Pro Glu Asn Val Lys Asn Lys Asp Lys Asp Ala Glu Lys Glu
50 55 60
Thr Lys Ile Arg Arg Thr Asn Leu Lys Lys Tyr Phe Ser Glu Ile Ile
65 70 75 80
Leu Arg Lys Glu Asp Glu Lys Tyr Ile Leu Lys Lys Thr Lys Lys Phe
85 90 95
Lys Asp Ile Asn Gln Glu Ile Asp Tyr Tyr Asp Val Lys Ser Lys Lys
100 105 110
Asn Gln Gln Glu Ile Phe Asp Val Leu Lys Glu Ile Leu Glu Leu Lys
115 120 125
Ile Lys Glu Thr Glu Lys Glu Glu Ile Ile Thr Phe Asp Ser Glu Lys
130 135 140
Leu Lys Lys Val Phe Gly Glu Asp Phe Val Lys Lys Glu Ala Lys Ile
145 150 155 160
Lys Ala Ile Glu Lys Ser Leu Lys Ile Asn Lys Ala Asn Tyr Lys Lys
165 170 175
Asp Ser Ile Lys Ile Gly Asp Asp Lys Tyr Ser Asn Val Lys Gly Glu
180 185 190
Asn Lys Arg Ser Arg Ile Tyr Glu Tyr Tyr Lys Lys Ser Glu Asn Leu
195 200 205
Lys Lys Phe Glu Glu Asn Ile Arg Glu Ala Phe Glu Lys Leu Tyr Thr
210 215 220
Glu Glu Asn Ile Lys Glu Leu Tyr Ser Lys Ile Glu Glu Ile Leu Lys
225 230 235 240
Lys Thr His Leu Lys Ser Ile Val Arg Glu Phe Tyr Gln Asn Glu Ile
245 250 255
Ile Gly Glu Ser Glu Phe Ser Lys Lys Asn Gly Asp Gly Ile Ser Ile
260 265 270
Leu Tyr Asn Gln Ile Lys Asp Ser Ile Lys Lys Glu Glu Asn Phe Ile
275 280 285
Glu Phe Ile Glu Asn Thr Gly Asn Leu Glu Leu Lys Glu Leu Thr Lys
290 295 300
Ser Gln Ile Phe Tyr Lys Tyr Phe Leu Glu Asn Glu Glu Leu Asn Asp
305 310 315 320
Glu Asn Ile Lys Phe Ala Phe Cys Tyr Phe Val Glu Ile Glu Val Asn
325 330 335
Asn Leu Leu Lys Glu Asn Val Tyr Lys Ile Lys Arg Phe Asn Glu Ser
340 345 350
Asn Lys Lys Arg Ile Glu Asn Ile Phe Glu Tyr Gly Lys Leu Lys Lys
355 360 365
Leu Ile Val Tyr Lys Leu Glu Asn Lys Leu Asn Asn Tyr Val Arg Asn
370 375 380
Cys Gly Lys Tyr Asn Tyr His Met Glu Asn Gly Asp Ile Ala Thr Ser
385 390 395 400
Asp Ile Asn Met Arg Asn Arg Gln Thr Glu Ala Phe Leu Arg Ser Ile
405 410 415
Ile Gly Val Ser Ser Phe Gly Tyr Phe Ser Leu Arg Asn Ile Leu Gly
420 425 430
Val Asn Asp Asp Asp Phe Tyr Glu Thr Glu Glu Asp Leu Thr Lys Lys
435 440 445
Glu Arg Arg Asn Leu Glu Lys Ala Lys Glu Asp Ile Thr Ile Lys Asn
450 455 460
Thr Phe Asp Glu Val Val Val Lys Ser Phe Gln Lys Lys Gly Ile Tyr
465 470 475 480
Asn Ile Lys Glu Asn Leu Lys Met Phe Tyr Gly Asp Ser Phe Asp Asn
485 490 495
Ala Asp Lys Asp Glu Leu Lys Gln Phe Phe Val Asn Met Leu Asn Ala
500 505 510
Ile Thr Ser Ile Arg His Arg Val Val His Tyr Asn Met Asn Thr Asn
515 520 525
Ser Glu Asn Ile Phe Asn Phe Ser Gly Ile Glu Val Ser Lys Leu Leu
530 535 540
Lys Ser Ile Phe Glu Lys Glu Thr Asp Lys Arg Glu Leu Lys Leu Lys
545 550 555 560
Ile Phe Arg Gln Leu Asn Ser Ala Gly Val Phe Asp Tyr Trp Glu Asn
565 570 575
Arg Lys Ile Asp Lys Tyr Leu Glu Asn Ile Glu Phe Lys Phe Val Asn
580 585 590
Lys Asn Ile Pro Phe Val Pro Ser Phe Thr Lys Leu Tyr Asn Arg Ile
595 600 605
Asp Asn Leu Lys Gly Asn Asn Ala Leu Asn Leu Gly Tyr Ile Asn Ile
610 615 620
Pro Lys Arg Lys Glu Ala Arg Asp Ser Gln Ile Tyr Leu Leu Lys Asn
625 630 635 640
Ile Tyr Tyr Gly Glu Phe Val Glu Lys Phe Val Asn Asn Asn Asp Asn
645 650 655
Phe Glu Lys Ile Phe Arg Glu Ile Ile Glu Ile Asn Lys Lys Asp Gly
660 665 670
Thr Asn Thr Lys Thr Lys Phe Tyr Lys Leu Glu Lys Phe Glu Thr Leu
675 680 685
Lys Ala Asn Ala Pro Ile Glu Tyr Leu Glu Lys Leu Gln Ser Leu His
690 695 700
Gln Ile Asn Tyr Asn Arg Glu Lys Val Glu Glu Asp Lys Asp Ile Tyr
705 710 715 720
Val Asp Phe Val Gln Lys Ile Phe Leu Lys Gly Phe Ile Asn Tyr Leu
725 730 735
Gln Gly Ser Asp Leu Leu Lys Ser Leu Asn Leu Leu Asn Leu Lys Lys
740 745 750
Asp Glu Ala Ile Ala Asn Lys Lys Ser Phe Tyr Asp Glu Lys Leu Lys
755 760 765
Leu Trp Gln Asn Asn Gly Ser Asn Leu Ser Lys Met Pro Glu Glu Ile
770 775 780
Tyr Asp Tyr Ile Lys Lys Ile Lys Ile Asn Lys Ile Asn Tyr Ser Asp
785 790 795 800
Arg Met Ser Ile Phe Tyr Leu Leu Leu Lys Leu Ile Asp His Lys Glu
805 810 815
Leu Thr Asn Leu Arg Gly Asn Leu Glu Lys Tyr Val Ser Met Asn Lys
820 825 830
Asn Lys Ile Tyr Ser Glu Glu Leu Asn Ile Val Asn Leu Val Ser Leu
835 840 845
Asp Asn Asn Lys Val Arg Ala Asn Phe Asn Leu Lys Pro Glu Asp Ile
850 855 860
Gly Lys Phe Leu Lys Thr Glu Thr Ser Ile Arg Asn Ile Asn Gln Leu
865 870 875 880
Asn Asn Phe Ser Glu Ile Phe Ala Asp Gly Glu Asn Val Ile Lys His
885 890 895
Arg Ser Phe Tyr Asn Ile Lys Lys Tyr Gly Ile Leu Asp Leu Leu Glu
900 905 910
Lys Ile Val Asp Lys Ala Asp Leu Lys Ile Thr Lys Glu Glu Ile Lys
915 920 925
Lys Tyr Glu Asn Leu Gln Asn Glu Leu Lys Arg Asn Asp Phe Tyr Lys
930 935 940
Ile Gln Glu Arg Ile His Arg Asn Tyr Asn Gln Lys Pro Phe Leu Ile
945 950 955 960
Lys Asn Asn Glu Lys Asp Phe Asn Asp Tyr Lys Lys Ala Ile Glu Asn
965 970 975
Ile Gln Asn Tyr Thr Gln Leu Lys Asn Lys Ile Glu Phe Asn Asp Leu
980 985 990
Asn Leu Leu Gln Ser Leu Leu Phe Arg Ile Leu His Arg Leu Ala Gly
995 1000 1005
Tyr Thr Ser Leu Trp Glu Arg Asp Leu Gln Phe Lys Leu Lys Gly
1010 1015 1020
Glu Tyr Pro Glu Asn Lys Tyr Ile Asp Glu Ile Phe Asn Phe Asp
1025 1030 1035
Asn Ser Lys Asn Lys Ile Tyr Asn Glu Lys Asn Glu Arg Gly Gly
1040 1045 1050
Ser Val Val Ser Lys Tyr Gly Tyr Phe Leu Val Glu Lys Asp Gly
1055 1060 1065
Glu Ile Gln Arg Lys Asn Ala Arg Asp Lys Lys Lys Asn Lys Ile
1070 1075 1080
Ile Lys Lys Glu Gly Leu Glu Ile Arg Asn Tyr Ile Ala His Phe
1085 1090 1095
Asn Tyr Ile Pro Asp Ala Thr Lys Ser Ile Leu Glu Ile Leu Glu
1100 1105 1110
Glu Leu Arg Asn Leu Leu Lys Tyr Asp Arg Lys Leu Lys Asn Ala
1115 1120 1125
Val Met Lys Ser Ile Lys Asp Ile Phe Lys Glu Tyr Gly Leu Ile
1130 1135 1140
Ile Glu Phe Lys Ile Ser His Val Asn Asn Ser Glu Lys Ile Glu
1145 1150 1155
Val Leu Asn Val Asp Ser Glu Lys Ile Lys His Leu Lys Asn Asn
1160 1165 1170
Gly Leu Val Thr Thr Arg Asn Ser Glu Asp Leu Cys Glu Leu Ile
1175 1180 1185
Lys Met Met Leu Glu Tyr Lys Lys Ser
1190 1195
<210> 587
<211> 1152
<212> PRT
<213> Leptotrichia wadei
<400> 587
Met Lys Val Thr Lys Val Asp Gly Ile Ser His Lys Lys Tyr Ile Glu
1 5 10 15
Glu Gly Lys Leu Val Lys Ser Thr Ser Glu Glu Asn Arg Thr Ser Glu
20 25 30
Arg Leu Ser Glu Leu Leu Ser Ile Arg Leu Asp Ile Tyr Ile Lys Asn
35 40 45
Pro Asp Asn Ala Ser Glu Glu Glu Asn Arg Ile Arg Arg Glu Asn Leu
50 55 60
Lys Lys Phe Phe Ser Asn Lys Val Leu His Leu Lys Asp Ser Val Leu
65 70 75 80
Tyr Leu Lys Asn Arg Lys Glu Lys Asn Ala Val Gln Asp Lys Asn Tyr
85 90 95
Ser Glu Glu Asp Ile Ser Glu Tyr Asp Leu Lys Asn Lys Asn Ser Phe
100 105 110
Ser Val Leu Lys Lys Ile Leu Leu Asn Glu Asp Val Asn Ser Glu Glu
115 120 125
Leu Glu Ile Phe Arg Lys Asp Val Glu Ala Lys Leu Asn Lys Ile Asn
130 135 140
Ser Leu Lys Tyr Ser Phe Glu Glu Asn Lys Ala Asn Tyr Gln Lys Ile
145 150 155 160
Asn Glu Asn Asn Val Glu Lys Val Gly Gly Lys Ser Lys Arg Asn Ile
165 170 175
Ile Tyr Asp Tyr Tyr Arg Glu Ser Ala Lys Arg Asn Asp Tyr Ile Asn
180 185 190
Asn Val Gln Glu Ala Phe Asp Lys Leu Tyr Lys Lys Glu Asp Ile Glu
195 200 205
Lys Leu Phe Phe Leu Ile Glu Asn Ser Lys Lys His Glu Lys Tyr Lys
210 215 220
Ile Arg Glu Tyr Tyr His Lys Ile Ile Gly Arg Lys Asn Asp Lys Glu
225 230 235 240
Asn Phe Ala Lys Ile Ile Tyr Glu Glu Ile Gln Asn Val Asn Asn Ile
245 250 255
Lys Glu Leu Ile Glu Lys Ile Pro Asp Met Ser Glu Leu Lys Lys Ser
260 265 270
Gln Val Phe Tyr Lys Tyr Tyr Leu Asp Lys Glu Glu Leu Asn Asp Lys
275 280 285
Asn Ile Lys Tyr Ala Phe Cys His Phe Val Glu Ile Glu Met Ser Gln
290 295 300
Leu Leu Lys Asn Tyr Val Tyr Lys Arg Leu Ser Asn Ile Ser Asn Asp
305 310 315 320
Lys Ile Lys Arg Ile Phe Glu Tyr Gln Asn Leu Lys Lys Leu Ile Glu
325 330 335
Asn Lys Leu Leu Asn Lys Leu Asp Thr Tyr Val Arg Asn Cys Gly Lys
340 345 350
Tyr Asn Tyr Tyr Leu Gln Val Gly Glu Ile Ala Thr Ser Asp Phe Ile
355 360 365
Ala Arg Asn Arg Gln Asn Glu Ala Phe Leu Arg Asn Ile Ile Gly Val
370 375 380
Ser Ser Val Ala Tyr Phe Ser Leu Arg Asn Ile Leu Glu Thr Glu Asn
385 390 395 400
Glu Asn Asp Ile Thr Gly Arg Met Arg Gly Lys Thr Val Lys Asn Asn
405 410 415
Lys Gly Glu Glu Lys Tyr Val Ser Gly Glu Val Asp Lys Ile Tyr Asn
420 425 430
Glu Asn Lys Gln Asn Glu Val Lys Glu Asn Leu Lys Met Phe Tyr Ser
435 440 445
Tyr Asp Phe Asn Met Asp Asn Lys Asn Glu Ile Glu Asp Phe Phe Ala
450 455 460
Asn Ile Asp Glu Ala Ile Ser Ser Ile Arg His Gly Ile Val His Phe
465 470 475 480
Asn Leu Glu Leu Glu Gly Lys Asp Ile Phe Ala Phe Lys Asn Ile Ala
485 490 495
Pro Ser Glu Ile Ser Lys Lys Met Phe Gln Asn Glu Ile Asn Glu Lys
500 505 510
Lys Leu Lys Leu Lys Ile Phe Lys Gln Leu Asn Ser Ala Asn Val Phe
515 520 525
Asn Tyr Tyr Glu Lys Asp Val Ile Ile Lys Tyr Leu Lys Asn Thr Lys
530 535 540
Phe Asn Phe Val Asn Lys Asn Ile Pro Phe Val Pro Ser Phe Thr Lys
545 550 555 560
Leu Tyr Asn Lys Ile Glu Asp Leu Arg Asn Thr Leu Lys Phe Phe Trp
565 570 575
Ser Val Pro Lys Asp Lys Glu Glu Lys Asp Ala Gln Ile Tyr Leu Leu
580 585 590
Lys Asn Ile Tyr Tyr Gly Glu Phe Leu Asn Lys Phe Val Lys Asn Ser
595 600 605
Lys Val Phe Phe Lys Ile Thr Asn Glu Val Ile Lys Ile Asn Lys Gln
610 615 620
Arg Asn Gln Lys Thr Gly His Tyr Lys Tyr Gln Lys Phe Glu Asn Ile
625 630 635 640
Glu Lys Thr Val Pro Val Glu Tyr Leu Ala Ile Ile Gln Ser Arg Glu
645 650 655
Met Ile Asn Asn Gln Asp Lys Glu Glu Lys Asn Thr Tyr Ile Asp Phe
660 665 670
Ile Gln Gln Ile Phe Leu Lys Gly Phe Ile Asp Tyr Leu Asn Lys Asn
675 680 685
Asn Leu Lys Tyr Ile Glu Ser Asn Asn Asn Asn Asp Asn Asn Asp Ile
690 695 700
Phe Ser Lys Ile Lys Ile Lys Lys Asp Asn Lys Glu Lys Tyr Asp Lys
705 710 715 720
Ile Leu Lys Asn Tyr Glu Lys His Asn Arg Asn Lys Glu Ile Pro His
725 730 735
Glu Ile Asn Glu Phe Val Arg Glu Ile Lys Leu Gly Lys Ile Leu Lys
740 745 750
Tyr Thr Glu Asn Leu Asn Met Phe Tyr Leu Ile Leu Lys Leu Leu Asn
755 760 765
His Lys Glu Leu Thr Asn Leu Lys Gly Ser Leu Glu Lys Tyr Gln Ser
770 775 780
Ala Asn Lys Glu Glu Thr Phe Ser Asp Glu Leu Glu Leu Ile Asn Leu
785 790 795 800
Leu Asn Leu Asp Asn Asn Arg Val Thr Glu Asp Phe Glu Leu Glu Ala
805 810 815
Asn Glu Ile Gly Lys Phe Leu Asp Phe Asn Glu Asn Lys Ile Lys Asp
820 825 830
Arg Lys Glu Leu Lys Lys Phe Asp Thr Asn Lys Ile Tyr Phe Asp Gly
835 840 845
Glu Asn Ile Ile Lys His Arg Ala Phe Tyr Asn Ile Lys Lys Tyr Gly
850 855 860
Met Leu Asn Leu Leu Glu Lys Ile Ala Asp Lys Ala Lys Tyr Lys Ile
865 870 875 880
Ser Leu Lys Glu Leu Lys Glu Tyr Ser Asn Lys Lys Asn Glu Ile Glu
885 890 895
Lys Asn Tyr Thr Met Gln Gln Asn Leu His Arg Lys Tyr Ala Arg Pro
900 905 910
Lys Lys Asp Glu Lys Phe Asn Asp Glu Asp Tyr Lys Glu Tyr Glu Lys
915 920 925
Ala Ile Gly Asn Ile Gln Lys Tyr Thr His Leu Lys Asn Lys Val Glu
930 935 940
Phe Asn Glu Leu Asn Leu Leu Gln Gly Leu Leu Leu Lys Ile Leu His
945 950 955 960
Arg Leu Val Gly Tyr Thr Ser Ile Trp Glu Arg Asp Leu Arg Phe Arg
965 970 975
Leu Lys Gly Glu Phe Pro Glu Asn His Tyr Ile Glu Glu Ile Phe Asn
980 985 990
Phe Asp Asn Ser Lys Asn Val Lys Tyr Lys Ser Gly Gln Ile Val Glu
995 1000 1005
Lys Tyr Ile Asn Phe Tyr Lys Glu Leu Tyr Lys Asp Asn Val Glu
1010 1015 1020
Lys Arg Ser Ile Tyr Ser Asp Lys Lys Val Lys Lys Leu Lys Gln
1025 1030 1035
Glu Lys Lys Asp Leu Tyr Ile Arg Asn Tyr Ile Ala His Phe Asn
1040 1045 1050
Tyr Ile Pro His Ala Glu Ile Ser Leu Leu Glu Val Leu Glu Asn
1055 1060 1065
Leu Arg Lys Leu Leu Ser Tyr Asp Arg Lys Leu Lys Asn Ala Ile
1070 1075 1080
Met Lys Ser Ile Val Asp Ile Leu Lys Glu Tyr Gly Phe Val Ala
1085 1090 1095
Thr Phe Lys Ile Gly Ala Asp Lys Lys Ile Glu Ile Gln Thr Leu
1100 1105 1110
Glu Ser Glu Lys Ile Val His Leu Lys Asn Leu Lys Lys Lys Lys
1115 1120 1125
Leu Met Thr Asp Arg Asn Ser Glu Glu Leu Cys Glu Leu Val Lys
1130 1135 1140
Val Met Phe Glu Tyr Lys Ala Leu Glu
1145 1150
<210> 588
<211> 1159
<212> PRT
<213> Leptotrichia buccalis
<400> 588
Met Lys Val Thr Lys Val Gly Gly Ile Ser His Lys Lys Tyr Thr Ser
1 5 10 15
Glu Gly Arg Leu Val Lys Ser Glu Ser Glu Glu Asn Arg Thr Asp Glu
20 25 30
Arg Leu Ser Ala Leu Leu Asn Met Arg Leu Asp Met Tyr Ile Lys Asn
35 40 45
Pro Ser Ser Thr Glu Thr Lys Glu Asn Gln Lys Arg Ile Gly Lys Leu
50 55 60
Lys Lys Phe Phe Ser Asn Lys Met Val Tyr Leu Lys Asp Asn Thr Leu
65 70 75 80
Ser Leu Lys Asn Gly Lys Lys Glu Asn Ile Asp Arg Glu Tyr Ser Glu
85 90 95
Thr Asp Ile Leu Glu Ser Asp Val Arg Asp Lys Lys Asn Phe Ala Val
100 105 110
Leu Lys Lys Ile Tyr Leu Asn Glu Asn Val Asn Ser Glu Glu Leu Glu
115 120 125
Val Phe Arg Asn Asp Ile Lys Lys Lys Leu Asn Lys Ile Asn Ser Leu
130 135 140
Lys Tyr Ser Phe Glu Lys Asn Lys Ala Asn Tyr Gln Lys Ile Asn Glu
145 150 155 160
Asn Asn Ile Glu Lys Val Glu Gly Lys Ser Lys Arg Asn Ile Ile Tyr
165 170 175
Asp Tyr Tyr Arg Glu Ser Ala Lys Arg Asp Ala Tyr Val Ser Asn Val
180 185 190
Lys Glu Ala Phe Asp Lys Leu Tyr Lys Glu Glu Asp Ile Ala Lys Leu
195 200 205
Val Leu Glu Ile Glu Asn Leu Thr Lys Leu Glu Lys Tyr Lys Ile Arg
210 215 220
Glu Phe Tyr His Glu Ile Ile Gly Arg Lys Asn Asp Lys Glu Asn Phe
225 230 235 240
Ala Lys Ile Ile Tyr Glu Glu Ile Gln Asn Val Asn Asn Met Lys Glu
245 250 255
Leu Ile Glu Lys Val Pro Asp Met Ser Glu Leu Lys Lys Ser Gln Val
260 265 270
Phe Tyr Lys Tyr Tyr Leu Asp Lys Glu Glu Leu Asn Asp Lys Asn Ile
275 280 285
Lys Tyr Ala Phe Cys His Phe Val Glu Ile Glu Met Ser Gln Leu Leu
290 295 300
Lys Asn Tyr Val Tyr Lys Arg Leu Ser Asn Ile Ser Asn Asp Lys Ile
305 310 315 320
Lys Arg Ile Phe Glu Tyr Gln Asn Leu Lys Lys Leu Ile Glu Asn Lys
325 330 335
Leu Leu Asn Lys Leu Asp Thr Tyr Val Arg Asn Cys Gly Lys Tyr Asn
340 345 350
Tyr Tyr Leu Gln Asp Gly Glu Ile Ala Thr Ser Asp Phe Ile Ala Arg
355 360 365
Asn Arg Gln Asn Glu Ala Phe Leu Arg Asn Ile Ile Gly Val Ser Ser
370 375 380
Val Ala Tyr Phe Ser Leu Arg Asn Ile Leu Glu Thr Glu Asn Glu Asn
385 390 395 400
Asp Ile Thr Gly Arg Met Arg Gly Lys Thr Val Lys Asn Asn Lys Gly
405 410 415
Glu Glu Lys Tyr Val Ser Gly Glu Val Asp Lys Ile Tyr Asn Glu Asn
420 425 430
Lys Lys Asn Glu Val Lys Glu Asn Leu Lys Met Phe Tyr Ser Tyr Asp
435 440 445
Phe Asn Met Asp Asn Lys Asn Glu Ile Glu Asp Phe Phe Ala Asn Ile
450 455 460
Asp Glu Ala Ile Ser Ser Ile Arg His Gly Ile Val His Phe Asn Leu
465 470 475 480
Glu Leu Glu Gly Lys Asp Ile Phe Ala Phe Lys Asn Ile Ala Pro Ser
485 490 495
Glu Ile Ser Lys Lys Met Phe Gln Asn Glu Ile Asn Glu Lys Lys Leu
500 505 510
Lys Leu Lys Ile Phe Arg Gln Leu Asn Ser Ala Asn Val Phe Arg Tyr
515 520 525
Leu Glu Lys Tyr Lys Ile Leu Asn Tyr Leu Lys Arg Thr Arg Phe Glu
530 535 540
Phe Val Asn Lys Asn Ile Pro Phe Val Pro Ser Phe Thr Lys Leu Tyr
545 550 555 560
Ser Arg Ile Asp Asp Leu Lys Asn Ser Leu Gly Ile Tyr Trp Lys Thr
565 570 575
Pro Lys Thr Asn Asp Asp Asn Lys Thr Lys Glu Ile Ile Asp Ala Gln
580 585 590
Ile Tyr Leu Leu Lys Asn Ile Tyr Tyr Gly Glu Phe Leu Asn Tyr Phe
595 600 605
Met Ser Asn Asn Gly Asn Phe Phe Glu Ile Ser Lys Glu Ile Ile Glu
610 615 620
Leu Asn Lys Asn Asp Lys Arg Asn Leu Lys Thr Gly Phe Tyr Lys Leu
625 630 635 640
Gln Lys Phe Glu Asp Ile Gln Glu Lys Ile Pro Lys Glu Tyr Leu Ala
645 650 655
Asn Ile Gln Ser Leu Tyr Met Ile Asn Ala Gly Asn Gln Asp Glu Glu
660 665 670
Glu Lys Asp Thr Tyr Ile Asp Phe Ile Gln Lys Ile Phe Leu Lys Gly
675 680 685
Phe Met Thr Tyr Leu Ala Asn Asn Gly Arg Leu Ser Leu Ile Tyr Ile
690 695 700
Gly Ser Asp Glu Glu Thr Asn Thr Ser Leu Ala Glu Lys Lys Gln Glu
705 710 715 720
Phe Asp Lys Phe Leu Lys Lys Tyr Glu Gln Asn Asn Asn Ile Lys Ile
725 730 735
Pro Tyr Glu Ile Asn Glu Phe Leu Arg Glu Ile Lys Leu Gly Asn Ile
740 745 750
Leu Lys Tyr Thr Glu Arg Leu Asn Met Phe Tyr Leu Ile Leu Lys Leu
755 760 765
Leu Asn His Lys Glu Leu Thr Asn Leu Lys Gly Ser Leu Glu Lys Tyr
770 775 780
Gln Ser Ala Asn Lys Glu Glu Ala Phe Ser Asp Gln Leu Glu Leu Ile
785 790 795 800
Asn Leu Leu Asn Leu Asp Asn Asn Arg Val Thr Glu Asp Phe Glu Leu
805 810 815
Glu Ala Asp Glu Ile Gly Lys Phe Leu Asp Phe Asn Gly Asn Lys Val
820 825 830
Lys Asp Asn Lys Glu Leu Lys Lys Phe Asp Thr Asn Lys Ile Tyr Phe
835 840 845
Asp Gly Glu Asn Ile Ile Lys His Arg Ala Phe Tyr Asn Ile Lys Lys
850 855 860
Tyr Gly Met Leu Asn Leu Leu Glu Lys Ile Ala Asp Lys Ala Gly Tyr
865 870 875 880
Lys Ile Ser Ile Glu Glu Leu Lys Lys Tyr Ser Asn Lys Lys Asn Glu
885 890 895
Ile Glu Lys Asn His Lys Met Gln Glu Asn Leu His Arg Lys Tyr Ala
900 905 910
Arg Pro Arg Lys Asp Glu Lys Phe Thr Asp Glu Asp Tyr Glu Ser Tyr
915 920 925
Lys Gln Ala Ile Glu Asn Ile Glu Glu Tyr Thr His Leu Lys Asn Lys
930 935 940
Val Glu Phe Asn Glu Leu Asn Leu Leu Gln Gly Leu Leu Leu Arg Ile
945 950 955 960
Leu His Arg Leu Val Gly Tyr Thr Ser Ile Trp Glu Arg Asp Leu Arg
965 970 975
Phe Arg Leu Lys Gly Glu Phe Pro Glu Asn Gln Tyr Ile Glu Glu Ile
980 985 990
Phe Asn Phe Glu Asn Lys Lys Asn Val Lys Tyr Lys Gly Gly Gln Ile
995 1000 1005
Val Glu Lys Tyr Ile Lys Phe Tyr Lys Glu Leu His Gln Asn Asp
1010 1015 1020
Glu Val Lys Ile Asn Lys Tyr Ser Ser Ala Asn Ile Lys Val Leu
1025 1030 1035
Lys Gln Glu Lys Lys Asp Leu Tyr Ile Arg Asn Tyr Ile Ala His
1040 1045 1050
Phe Asn Tyr Ile Pro His Ala Glu Ile Ser Leu Leu Glu Val Leu
1055 1060 1065
Glu Asn Leu Arg Lys Leu Leu Ser Tyr Asp Arg Lys Leu Lys Asn
1070 1075 1080
Ala Val Met Lys Ser Val Val Asp Ile Leu Lys Glu Tyr Gly Phe
1085 1090 1095
Val Ala Thr Phe Lys Ile Gly Ala Asp Lys Lys Ile Gly Ile Gln
1100 1105 1110
Thr Leu Glu Ser Glu Lys Ile Val His Leu Lys Asn Leu Lys Lys
1115 1120 1125
Lys Lys Leu Met Thr Asp Arg Asn Ser Glu Glu Leu Cys Lys Leu
1130 1135 1140
Val Lys Ile Met Phe Glu Tyr Lys Met Glu Glu Lys Lys Ser Glu
1145 1150 1155
Asn
<210> 589
<211> 913
<212> PRT
<213> Leptotrichia sp.
<400> 589
Met Lys Glu Leu Ile Glu Lys Val Pro Asn Val Ser Glu Leu Lys Lys
1 5 10 15
Ser Gln Val Phe Tyr Lys Tyr Tyr Leu Asn Lys Glu Lys Leu Asn Asp
20 25 30
Glu Asn Ile Lys Tyr Val Phe Cys His Phe Val Glu Ile Glu Met Ser
35 40 45
Lys Leu Leu Lys Asn Tyr Val Tyr Lys Lys Pro Ser Asn Ile Ser Asn
50 55 60
Asp Lys Val Lys Arg Ile Phe Glu Tyr Gln Ser Leu Lys Lys Leu Ile
65 70 75 80
Glu Asn Lys Leu Leu Asn Lys Leu Asp Thr Tyr Ile Arg Asn Cys Gly
85 90 95
Lys Tyr Ser Phe Tyr Leu Gln Asp Gly Glu Ile Ala Thr Ser Asp Phe
100 105 110
Ile Val Gly Asn Arg Gln Asn Glu Ala Phe Leu Arg Asn Ile Ile Gly
115 120 125
Val Ser Ser Ala Ala Tyr Phe Ser Leu Arg Asn Ile Leu Glu Thr Glu
130 135 140
Asn Glu Asn Asp Ile Thr Gly Lys Met Arg Gly Lys Thr Val Lys Asn
145 150 155 160
Lys Lys Gly Glu Glu Lys Tyr Ile Ser Gly Glu Ile Asp Lys Leu Tyr
165 170 175
Asp Asn Asn Lys Gln Asn Glu Val Lys Lys Asn Leu Lys Met Phe Tyr
180 185 190
Ser Tyr Asp Phe Asn Met Asn Ser Lys Lys Glu Ile Glu Asp Phe Phe
195 200 205
Ser Asn Ile Asp Glu Ala Ile Ser Ser Ile Arg His Gly Ile Val His
210 215 220
Phe Asn Leu Glu Leu Glu Gly Lys Asp Ile Phe Thr Phe Lys Asn Ile
225 230 235 240
Val Pro Ser Gln Ile Ser Lys Lys Met Phe His Asp Glu Ile Asn Glu
245 250 255
Lys Lys Leu Lys Leu Lys Ile Phe Lys Gln Leu Asn Ser Ala Asn Val
260 265 270
Phe Arg Tyr Leu Glu Lys Tyr Lys Ile Leu Asn Tyr Leu Asn Arg Thr
275 280 285
Arg Phe Glu Phe Val Asn Lys Asn Ile Pro Phe Val Pro Ser Phe Thr
290 295 300
Lys Leu Tyr Ser Arg Ile Asp Asp Leu Lys Asn Ser Leu Cys Ile Tyr
305 310 315 320
Trp Lys Ile Pro Lys Ala Asn Asp Asn Asn Lys Thr Lys Glu Ile Thr
325 330 335
Asp Ala Gln Ile Tyr Leu Leu Lys Asn Ile Tyr Tyr Ser Glu Phe Leu
340 345 350
Asn Tyr Phe Met Ser Asn Asn Gly Asn Phe Phe Glu Ile Ile Lys Glu
355 360 365
Ile Ile Glu Leu Asn Lys Asn Asp Lys Arg Asn Leu Lys Thr Gly Phe
370 375 380
Tyr Lys Leu Gln Lys Phe Glu Asn Leu Gln Glu Lys Thr Pro Lys Glu
385 390 395 400
Tyr Leu Ala Asn Ile Gln Ser Phe Tyr Met Ile Asp Ala Gly Asn Lys
405 410 415
Asp Glu Glu Glu Lys Asp Ala Tyr Ile Asp Phe Ile Gln Lys Ile Phe
420 425 430
Leu Lys Gly Phe Met Thr Tyr Leu Ala Asn Asn Gly Arg Leu Ser Leu
435 440 445
Met Tyr Ile Gly Asn Asp Glu Gln Ile Asn Thr Ser Leu Ala Glu Lys
450 455 460
Lys Gln Glu Phe Asp Lys Phe Leu Lys Lys Tyr Glu Gln Asn Asn Asn
465 470 475 480
Ile Lys Ile Pro Tyr Glu Ile Asn Glu Phe Leu Arg Glu Ile Lys Leu
485 490 495
Gly Asn Ile Leu Lys Tyr Thr Glu Arg Leu Asn Met Phe Tyr Leu Ile
500 505 510
Leu Lys Leu Leu Asn His Lys Glu Leu Thr Asn Leu Lys Gly Ser Leu
515 520 525
Glu Lys Tyr Gln Ser Ala Asn Lys Glu Glu Ala Phe Ser Asp Gln Leu
530 535 540
Glu Leu Ile Asn Leu Leu Asn Leu Asp Asn Asn Arg Val Thr Glu Asp
545 550 555 560
Phe Glu Leu Glu Ala Asp Glu Ile Gly Lys Phe Leu Asp Phe Asn Gly
565 570 575
Asn Lys Val Lys Asp Asn Lys Glu Leu Lys Lys Phe Asp Thr Asn Lys
580 585 590
Ile Tyr Phe Asp Gly Glu Asn Ile Ile Lys His Arg Ala Phe Tyr Asn
595 600 605
Ile Lys Lys Tyr Gly Met Leu Asn Leu Leu Glu Lys Ile Ser Asp Glu
610 615 620
Ala Lys Tyr Lys Ile Ser Ile Glu Glu Leu Lys Asn Tyr Ser Asn Lys
625 630 635 640
Lys Asn Glu Ile Glu Lys Asn His Thr Asn Gln Glu Asn Leu His Arg
645 650 655
Lys Tyr Ala Arg Pro Arg Lys Asp Glu Lys Phe Asn Asp Glu Asp Tyr
660 665 670
Lys Lys Tyr Glu Lys Ala Ile Arg Asn Ile Gln Gln Tyr Thr His Leu
675 680 685
Lys Asn Lys Val Glu Phe Asn Glu Leu Asn Leu Leu Gln Ser Leu Leu
690 695 700
Leu Arg Ile Leu His Arg Leu Val Gly Tyr Thr Ser Ile Trp Glu Arg
705 710 715 720
Asp Leu Arg Phe Arg Leu Lys Gly Glu Phe Pro Glu Asn Gln Tyr Ile
725 730 735
Glu Glu Ile Phe Asn Phe Asn Asn Ser Lys Asn Val Lys Tyr Lys Asn
740 745 750
Gly Gln Ile Val Glu Lys Tyr Ile Ser Phe Tyr Lys Glu Leu Tyr Lys
755 760 765
Asp Asp Thr Glu Lys Ile Ser Ile Tyr Ser Asp Lys Lys Val Lys Glu
770 775 780
Leu Lys Lys Glu Lys Lys Asp Leu Tyr Ile Arg Asn Tyr Ile Ala His
785 790 795 800
Phe Asn Tyr Ile Pro Asn Ala Glu Ile Ser Leu Leu Glu Val Leu Glu
805 810 815
Asn Leu Arg Lys Leu Leu Ser Tyr Asp Arg Lys Leu Lys Asn Ala Ile
820 825 830
Met Lys Ser Ile Val Asp Ile Leu Lys Glu Tyr Gly Phe Val Val Thr
835 840 845
Phe Lys Ile Glu Lys Asp Lys Lys Ile Arg Ile Glu Ser Leu Lys Ser
850 855 860
Glu Glu Val Val His Leu Lys Lys Leu Lys Leu Lys Asp Asn Asp Lys
865 870 875 880
Lys Lys Glu Pro Ile Lys Thr Tyr Arg Asn Ser Lys Glu Leu Cys Glu
885 890 895
Leu Val Lys Val Met Phe Glu Tyr Lys Met Lys Glu Lys Lys Ser Glu
900 905 910
Asn
<210> 590
<211> 1385
<212> PRT
<213> Leptotrichia sp.
<400> 590
Met Gly Asn Leu Phe Gly His Lys Arg Trp Tyr Glu Val Arg Asp Lys
1 5 10 15
Lys Asp Phe Lys Ile Lys Arg Lys Val Lys Val Lys Arg Asn Tyr Asp
20 25 30
Gly Asn Lys Tyr Ile Leu Asn Ile Asn Glu Asn Asn Asn Lys Glu Lys
35 40 45
Ile Asp Asn Asn Lys Phe Ile Gly Glu Phe Val Asn Tyr Lys Lys Asn
50 55 60
Asn Asn Val Leu Lys Glu Phe Lys Arg Lys Phe His Ala Gly Asn Ile
65 70 75 80
Leu Phe Lys Leu Lys Gly Lys Glu Glu Ile Ile Arg Ile Glu Asn Asn
85 90 95
Asp Asp Phe Leu Glu Thr Glu Glu Val Val Leu Tyr Ile Glu Val Tyr
100 105 110
Gly Lys Ser Glu Lys Leu Lys Ala Leu Glu Ile Thr Lys Lys Lys Ile
115 120 125
Ile Asp Glu Ala Ile Arg Gln Gly Ile Thr Lys Asp Asp Lys Lys Ile
130 135 140
Glu Ile Lys Arg Gln Glu Asn Glu Glu Glu Ile Glu Ile Asp Ile Arg
145 150 155 160
Asp Glu Tyr Thr Asn Lys Thr Leu Asn Asp Cys Ser Ile Ile Leu Arg
165 170 175
Ile Ile Glu Asn Asp Glu Leu Glu Thr Lys Lys Ser Ile Tyr Glu Ile
180 185 190
Phe Lys Asn Ile Asn Met Ser Leu Tyr Lys Ile Ile Glu Lys Ile Ile
195 200 205
Glu Asn Glu Thr Glu Lys Val Phe Glu Asn Arg Tyr Tyr Glu Glu His
210 215 220
Leu Arg Glu Lys Leu Leu Lys Asp Asn Lys Ile Asp Val Ile Leu Thr
225 230 235 240
Asn Phe Met Glu Ile Arg Glu Lys Ile Lys Ser Asn Leu Glu Ile Met
245 250 255
Gly Phe Val Lys Phe Tyr Leu Asn Val Ser Gly Asp Lys Lys Lys Ser
260 265 270
Glu Asn Lys Lys Met Phe Val Glu Lys Ile Leu Asn Thr Asn Val Asp
275 280 285
Leu Thr Val Glu Asp Ile Val Asp Phe Ile Val Lys Glu Leu Lys Phe
290 295 300
Trp Asn Ile Thr Lys Arg Ile Glu Lys Val Lys Lys Phe Asn Asn Glu
305 310 315 320
Phe Leu Glu Asn Arg Arg Asn Arg Thr Tyr Ile Lys Ser Tyr Val Leu
325 330 335
Leu Asp Lys His Glu Lys Phe Lys Ile Glu Arg Glu Asn Lys Lys Asp
340 345 350
Lys Ile Val Lys Phe Phe Val Glu Asn Ile Lys Asn Asn Ser Ile Lys
355 360 365
Glu Lys Ile Glu Lys Ile Leu Ala Glu Phe Lys Ile Asn Glu Leu Ile
370 375 380
Lys Lys Leu Glu Lys Glu Leu Lys Lys Gly Asn Cys Asp Thr Glu Ile
385 390 395 400
Phe Gly Ile Phe Lys Lys His Tyr Lys Val Asn Phe Asp Ser Lys Lys
405 410 415
Phe Ser Asn Lys Ser Asp Glu Glu Lys Glu Leu Tyr Lys Ile Ile Tyr
420 425 430
Arg Tyr Leu Lys Gly Arg Ile Glu Lys Ile Leu Val Asn Glu Gln Lys
435 440 445
Val Arg Leu Lys Lys Met Glu Lys Ile Glu Ile Glu Lys Ile Leu Asn
450 455 460
Glu Ser Ile Leu Ser Glu Lys Ile Leu Lys Arg Val Lys Gln Tyr Thr
465 470 475 480
Leu Glu His Ile Met Tyr Leu Gly Lys Leu Arg His Asn Asp Ile Val
485 490 495
Lys Met Thr Val Asn Thr Asp Asp Phe Ser Arg Leu His Ala Lys Glu
500 505 510
Glu Leu Asp Leu Glu Leu Ile Thr Phe Phe Ala Ser Thr Asn Met Glu
515 520 525
Leu Asn Lys Ile Phe Asn Gly Lys Glu Lys Val Thr Asp Phe Phe Gly
530 535 540
Phe Asn Leu Asn Gly Gln Lys Ile Thr Leu Lys Glu Lys Val Pro Ser
545 550 555 560
Phe Lys Leu Asn Ile Leu Lys Lys Leu Asn Phe Ile Asn Asn Glu Asn
565 570 575
Asn Ile Asp Glu Lys Leu Ser His Phe Tyr Ser Phe Gln Lys Glu Gly
580 585 590
Tyr Leu Leu Arg Asn Lys Ile Leu His Asn Ser Tyr Gly Asn Ile Gln
595 600 605
Glu Thr Lys Asn Leu Lys Gly Glu Tyr Glu Asn Val Glu Lys Leu Ile
610 615 620
Lys Glu Leu Lys Val Ser Asp Glu Glu Ile Ser Lys Ser Leu Ser Leu
625 630 635 640
Asp Val Ile Phe Glu Gly Lys Val Asp Ile Ile Asn Lys Ile Asn Ser
645 650 655
Leu Lys Ile Gly Glu Tyr Lys Asp Lys Lys Tyr Leu Pro Ser Phe Ser
660 665 670
Lys Ile Val Leu Glu Ile Thr Arg Lys Phe Arg Glu Ile Asn Lys Asp
675 680 685
Lys Leu Phe Asp Ile Glu Ser Glu Lys Ile Ile Leu Asn Ala Val Lys
690 695 700
Tyr Val Asn Lys Ile Leu Tyr Glu Lys Ile Thr Ser Asn Glu Glu Asn
705 710 715 720
Glu Phe Leu Lys Thr Leu Pro Asp Lys Leu Val Lys Lys Ser Asn Asn
725 730 735
Lys Lys Glu Asn Lys Asn Leu Leu Ser Ile Glu Glu Tyr Tyr Lys Asn
740 745 750
Ala Gln Val Ser Ser Ser Lys Gly Asp Lys Lys Ala Ile Lys Lys Tyr
755 760 765
Gln Asn Lys Val Thr Asn Ala Tyr Leu Glu Tyr Leu Glu Asn Thr Phe
770 775 780
Thr Glu Ile Ile Asp Phe Ser Lys Phe Asn Leu Asn Tyr Asp Glu Ile
785 790 795 800
Lys Thr Lys Ile Glu Glu Arg Lys Asp Asn Lys Ser Lys Ile Ile Ile
805 810 815
Asp Ser Ile Ser Thr Asn Ile Asn Ile Thr Asn Asp Ile Glu Tyr Ile
820 825 830
Ile Ser Ile Phe Ala Leu Leu Asn Ser Asn Thr Tyr Ile Asn Lys Ile
835 840 845
Arg Asn Arg Phe Phe Ala Thr Ser Val Trp Leu Glu Lys Gln Asn Gly
850 855 860
Thr Lys Glu Tyr Asp Tyr Glu Asn Ile Ile Ser Ile Leu Asp Glu Val
865 870 875 880
Leu Leu Ile Asn Leu Leu Arg Glu Asn Asn Ile Thr Asp Ile Leu Asp
885 890 895
Leu Lys Asn Ala Ile Ile Asp Ala Lys Ile Val Glu Asn Asp Glu Thr
900 905 910
Tyr Ile Lys Asn Tyr Ile Phe Glu Ser Asn Glu Glu Lys Leu Lys Lys
915 920 925
Arg Leu Phe Cys Glu Glu Leu Val Asp Lys Glu Asp Ile Arg Lys Ile
930 935 940
Phe Glu Asp Glu Asn Phe Lys Phe Lys Ser Phe Ile Lys Lys Asn Glu
945 950 955 960
Ile Gly Asn Phe Lys Ile Asn Phe Gly Ile Leu Ser Asn Leu Glu Cys
965 970 975
Asn Ser Glu Val Glu Ala Lys Lys Ile Ile Gly Lys Asn Ser Lys Lys
980 985 990
Leu Glu Ser Phe Ile Gln Asn Ile Ile Asp Glu Tyr Lys Ser Asn Ile
995 1000 1005
Arg Thr Leu Phe Ser Ser Glu Phe Leu Glu Lys Tyr Lys Glu Glu
1010 1015 1020
Ile Asp Asn Leu Val Glu Asp Thr Glu Ser Glu Asn Lys Asn Lys
1025 1030 1035
Phe Glu Lys Ile Tyr Tyr Pro Lys Glu His Lys Asn Glu Leu Tyr
1040 1045 1050
Ile Tyr Lys Lys Asn Leu Phe Leu Asn Ile Gly Asn Pro Asn Phe
1055 1060 1065
Asp Lys Ile Tyr Gly Leu Ile Ser Lys Asp Ile Lys Asn Val Asp
1070 1075 1080
Thr Lys Ile Leu Phe Asp Asp Asp Ile Lys Lys Asn Lys Ile Ser
1085 1090 1095
Glu Ile Asp Ala Ile Leu Lys Asn Leu Asn Asp Lys Leu Asn Gly
1100 1105 1110
Tyr Ser Asn Asp Tyr Lys Ala Lys Tyr Val Asn Lys Leu Lys Glu
1115 1120 1125
Asn Asp Asp Phe Phe Ala Lys Asn Ile Gln Asn Glu Asn Tyr Ser
1130 1135 1140
Ser Phe Gly Glu Phe Glu Lys Asp Tyr Asn Lys Val Ser Glu Tyr
1145 1150 1155
Lys Lys Ile Arg Asp Leu Val Glu Phe Asn Tyr Leu Asn Lys Ile
1160 1165 1170
Glu Ser Tyr Leu Ile Asp Ile Asn Trp Lys Leu Ala Ile Gln Met
1175 1180 1185
Ala Arg Phe Glu Arg Asp Met His Tyr Ile Val Asn Gly Leu Arg
1190 1195 1200
Glu Leu Gly Ile Ile Lys Leu Ser Gly Tyr Asn Thr Gly Ile Ser
1205 1210 1215
Arg Ala Tyr Pro Lys Arg Asn Gly Ser Asp Gly Phe Tyr Thr Thr
1220 1225 1230
Thr Ala Tyr Tyr Lys Phe Phe Asp Glu Glu Ser Tyr Lys Lys Phe
1235 1240 1245
Glu Lys Ile Cys Tyr Gly Phe Gly Ile Asp Leu Ser Glu Asn Ser
1250 1255 1260
Glu Ile Asn Lys Pro Glu Asn Glu Ser Ile Arg Asn Tyr Ile Ser
1265 1270 1275
His Phe Tyr Ile Val Arg Asn Pro Phe Ala Asp Tyr Ser Ile Ala
1280 1285 1290
Glu Gln Ile Asp Arg Val Ser Asn Leu Leu Ser Tyr Ser Thr Arg
1295 1300 1305
Tyr Asn Asn Ser Thr Tyr Ala Ser Val Phe Glu Val Phe Lys Lys
1310 1315 1320
Asp Val Asn Leu Asp Tyr Asp Glu Leu Lys Lys Lys Phe Arg Leu
1325 1330 1335
Ile Gly Asn Asn Asp Ile Leu Glu Arg Leu Met Lys Pro Lys Lys
1340 1345 1350
Val Ser Val Leu Glu Leu Glu Ser Tyr Asn Ser Asp Tyr Ile Lys
1355 1360 1365
Asn Leu Ile Ile Glu Leu Leu Thr Lys Ile Glu Asn Thr Asn Asp
1370 1375 1380
Thr Leu
1385
<210> 591
<211> 1389
<212> PRT
<213> Leptotrichia shahii
<400> 591
Met Gly Asn Leu Phe Gly His Lys Arg Trp Tyr Glu Val Arg Asp Lys
1 5 10 15
Lys Asp Phe Lys Ile Lys Arg Lys Val Lys Val Lys Arg Asn Tyr Asp
20 25 30
Gly Asn Lys Tyr Ile Leu Asn Ile Asn Glu Asn Asn Asn Lys Glu Lys
35 40 45
Ile Asp Asn Asn Lys Phe Ile Arg Lys Tyr Ile Asn Tyr Lys Lys Asn
50 55 60
Asp Asn Ile Leu Lys Glu Phe Thr Arg Lys Phe His Ala Gly Asn Ile
65 70 75 80
Leu Phe Lys Leu Lys Gly Lys Glu Gly Ile Ile Arg Ile Glu Asn Asn
85 90 95
Asp Asp Phe Leu Glu Thr Glu Glu Val Val Leu Tyr Ile Glu Ala Tyr
100 105 110
Gly Lys Ser Glu Lys Leu Lys Ala Leu Gly Ile Thr Lys Lys Lys Ile
115 120 125
Ile Asp Glu Ala Ile Arg Gln Gly Ile Thr Lys Asp Asp Lys Lys Ile
130 135 140
Glu Ile Lys Arg Gln Glu Asn Glu Glu Glu Ile Glu Ile Asp Ile Arg
145 150 155 160
Asp Glu Tyr Thr Asn Lys Thr Leu Asn Asp Cys Ser Ile Ile Leu Arg
165 170 175
Ile Ile Glu Asn Asp Glu Leu Glu Thr Lys Lys Ser Ile Tyr Glu Ile
180 185 190
Phe Lys Asn Ile Asn Met Ser Leu Tyr Lys Ile Ile Glu Lys Ile Ile
195 200 205
Glu Asn Glu Thr Glu Lys Val Phe Glu Asn Arg Tyr Tyr Glu Glu His
210 215 220
Leu Arg Glu Lys Leu Leu Lys Asp Asp Lys Ile Asp Val Ile Leu Thr
225 230 235 240
Asn Phe Met Glu Ile Arg Glu Lys Ile Lys Ser Asn Leu Glu Ile Leu
245 250 255
Gly Phe Val Lys Phe Tyr Leu Asn Val Gly Gly Asp Lys Lys Lys Ser
260 265 270
Lys Asn Lys Lys Met Leu Val Glu Lys Ile Leu Asn Ile Asn Val Asp
275 280 285
Leu Thr Val Glu Asp Ile Ala Asp Phe Val Ile Lys Glu Leu Glu Phe
290 295 300
Trp Asn Ile Thr Lys Arg Ile Glu Lys Val Lys Lys Val Asn Asn Glu
305 310 315 320
Phe Leu Glu Lys Arg Arg Asn Arg Thr Tyr Ile Lys Ser Tyr Val Leu
325 330 335
Leu Asp Lys His Glu Lys Phe Lys Ile Glu Arg Glu Asn Lys Lys Asp
340 345 350
Lys Ile Val Lys Phe Phe Val Glu Asn Ile Lys Asn Asn Ser Ile Lys
355 360 365
Glu Lys Ile Glu Lys Ile Leu Ala Glu Phe Lys Ile Asp Glu Leu Ile
370 375 380
Lys Lys Leu Glu Lys Glu Leu Lys Lys Gly Asn Cys Asp Thr Glu Ile
385 390 395 400
Phe Gly Ile Phe Lys Lys His Tyr Lys Val Asn Phe Asp Ser Lys Lys
405 410 415
Phe Ser Lys Lys Ser Asp Glu Glu Lys Glu Leu Tyr Lys Ile Ile Tyr
420 425 430
Arg Tyr Leu Lys Gly Arg Ile Glu Lys Ile Leu Val Asn Glu Gln Lys
435 440 445
Val Arg Leu Lys Lys Met Glu Lys Ile Glu Ile Glu Lys Ile Leu Asn
450 455 460
Glu Ser Ile Leu Ser Glu Lys Ile Leu Lys Arg Val Lys Gln Tyr Thr
465 470 475 480
Leu Glu His Ile Met Tyr Leu Gly Lys Leu Arg His Asn Asp Ile Asp
485 490 495
Met Thr Thr Val Asn Thr Asp Asp Phe Ser Arg Leu His Ala Lys Glu
500 505 510
Glu Leu Asp Leu Glu Leu Ile Thr Phe Phe Ala Ser Thr Asn Met Glu
515 520 525
Leu Asn Lys Ile Phe Ser Arg Glu Asn Ile Asn Asn Asp Glu Asn Ile
530 535 540
Asp Phe Phe Gly Gly Asp Arg Glu Lys Asn Tyr Val Leu Asp Lys Lys
545 550 555 560
Ile Leu Asn Ser Lys Ile Lys Ile Ile Arg Asp Leu Asp Phe Ile Asp
565 570 575
Asn Lys Asn Asn Ile Thr Asn Asn Phe Ile Arg Lys Phe Thr Lys Ile
580 585 590
Gly Thr Asn Glu Arg Asn Arg Ile Leu His Ala Ile Ser Lys Glu Arg
595 600 605
Asp Leu Gln Gly Thr Gln Asp Asp Tyr Asn Lys Val Ile Asn Ile Ile
610 615 620
Gln Asn Leu Lys Ile Ser Asp Glu Glu Val Ser Lys Ala Leu Asn Leu
625 630 635 640
Asp Val Val Phe Lys Asp Lys Lys Asn Ile Ile Thr Lys Ile Asn Asp
645 650 655
Ile Lys Ile Ser Glu Glu Asn Asn Asn Asp Ile Lys Tyr Leu Pro Ser
660 665 670
Phe Ser Lys Val Leu Pro Glu Ile Leu Asn Leu Tyr Arg Asn Asn Pro
675 680 685
Lys Asn Glu Pro Phe Asp Thr Ile Glu Thr Glu Lys Ile Val Leu Asn
690 695 700
Ala Leu Ile Tyr Val Asn Lys Glu Leu Tyr Lys Lys Leu Ile Leu Glu
705 710 715 720
Asp Asp Leu Glu Glu Asn Glu Ser Lys Asn Ile Phe Leu Gln Glu Leu
725 730 735
Lys Lys Thr Leu Gly Asn Ile Asp Glu Ile Asp Glu Asn Ile Ile Glu
740 745 750
Asn Tyr Tyr Lys Asn Ala Gln Ile Ser Ala Ser Lys Gly Asn Asn Lys
755 760 765
Ala Ile Lys Lys Tyr Gln Lys Lys Val Ile Glu Cys Tyr Ile Gly Tyr
770 775 780
Leu Arg Lys Asn Tyr Glu Glu Leu Phe Asp Phe Ser Asp Phe Lys Met
785 790 795 800
Asn Ile Gln Glu Ile Lys Lys Gln Ile Lys Asp Ile Asn Asp Asn Lys
805 810 815
Thr Tyr Glu Arg Ile Thr Val Lys Thr Ser Asp Lys Thr Ile Val Ile
820 825 830
Asn Asp Asp Phe Glu Tyr Ile Ile Ser Ile Phe Ala Leu Leu Asn Ser
835 840 845
Asn Ala Val Ile Asn Lys Ile Arg Asn Arg Phe Phe Ala Thr Ser Val
850 855 860
Trp Leu Asn Thr Ser Glu Tyr Gln Asn Ile Ile Asp Ile Leu Asp Glu
865 870 875 880
Ile Met Gln Leu Asn Thr Leu Arg Asn Glu Cys Ile Thr Glu Asn Trp
885 890 895
Asn Leu Asn Leu Glu Glu Phe Ile Gln Lys Met Lys Glu Ile Glu Lys
900 905 910
Asp Phe Asp Asp Phe Lys Ile Gln Thr Lys Lys Glu Ile Phe Asn Asn
915 920 925
Tyr Tyr Glu Asp Ile Lys Asn Asn Ile Leu Thr Glu Phe Lys Asp Asp
930 935 940
Ile Asn Gly Cys Asp Val Leu Glu Lys Lys Leu Glu Lys Ile Val Ile
945 950 955 960
Phe Asp Asp Glu Thr Lys Phe Glu Ile Asp Lys Lys Ser Asn Ile Leu
965 970 975
Gln Asp Glu Gln Arg Lys Leu Ser Asn Ile Asn Lys Lys Asp Leu Lys
980 985 990
Lys Lys Val Asp Gln Tyr Ile Lys Asp Lys Asp Gln Glu Ile Lys Ser
995 1000 1005
Lys Ile Leu Cys Arg Ile Ile Phe Asn Ser Asp Phe Leu Lys Lys
1010 1015 1020
Tyr Lys Lys Glu Ile Asp Asn Leu Ile Glu Asp Met Glu Ser Glu
1025 1030 1035
Asn Glu Asn Lys Phe Gln Glu Ile Tyr Tyr Pro Lys Glu Arg Lys
1040 1045 1050
Asn Glu Leu Tyr Ile Tyr Lys Lys Asn Leu Phe Leu Asn Ile Gly
1055 1060 1065
Asn Pro Asn Phe Asp Lys Ile Tyr Gly Leu Ile Ser Asn Asp Ile
1070 1075 1080
Lys Met Ala Asp Ala Lys Phe Leu Phe Asn Ile Asp Gly Lys Asn
1085 1090 1095
Ile Arg Lys Asn Lys Ile Ser Glu Ile Asp Ala Ile Leu Lys Asn
1100 1105 1110
Leu Asn Asp Lys Leu Asn Gly Tyr Ser Lys Glu Tyr Lys Glu Lys
1115 1120 1125
Tyr Ile Lys Lys Leu Lys Glu Asn Asp Asp Phe Phe Ala Lys Asn
1130 1135 1140
Ile Gln Asn Lys Asn Tyr Lys Ser Phe Glu Lys Asp Tyr Asn Arg
1145 1150 1155
Val Ser Glu Tyr Lys Lys Ile Arg Asp Leu Val Glu Phe Asn Tyr
1160 1165 1170
Leu Asn Lys Ile Glu Ser Tyr Leu Ile Asp Ile Asn Trp Lys Leu
1175 1180 1185
Ala Ile Gln Met Ala Arg Phe Glu Arg Asp Met His Tyr Ile Val
1190 1195 1200
Asn Gly Leu Arg Glu Leu Gly Ile Ile Lys Leu Ser Gly Tyr Asn
1205 1210 1215
Thr Gly Ile Ser Arg Ala Tyr Pro Lys Arg Asn Gly Ser Asp Gly
1220 1225 1230
Phe Tyr Thr Thr Thr Ala Tyr Tyr Lys Phe Phe Asp Glu Glu Ser
1235 1240 1245
Tyr Lys Lys Phe Glu Lys Ile Cys Tyr Gly Phe Gly Ile Asp Leu
1250 1255 1260
Ser Glu Asn Ser Glu Ile Asn Lys Pro Glu Asn Glu Ser Ile Arg
1265 1270 1275
Asn Tyr Ile Ser His Phe Tyr Ile Val Arg Asn Pro Phe Ala Asp
1280 1285 1290
Tyr Ser Ile Ala Glu Gln Ile Asp Arg Val Ser Asn Leu Leu Ser
1295 1300 1305
Tyr Ser Thr Arg Tyr Asn Asn Ser Thr Tyr Ala Ser Val Phe Glu
1310 1315 1320
Val Phe Lys Lys Asp Val Asn Leu Asp Tyr Asp Glu Leu Lys Lys
1325 1330 1335
Lys Phe Lys Leu Ile Gly Asn Asn Asp Ile Leu Glu Arg Leu Met
1340 1345 1350
Lys Pro Lys Lys Val Ser Val Leu Glu Leu Glu Ser Tyr Asn Ser
1355 1360 1365
Asp Tyr Ile Lys Asn Leu Ile Ile Glu Leu Leu Thr Lys Ile Glu
1370 1375 1380
Asn Thr Asn Asp Thr Leu
1385
<210> 592
<211> 10
<212> PRT
<213> Lactococcus lactis
<400> 592
Phe Leu Val Asn His Asn Tyr Tyr Ser Phe
1 5 10
<210> 593
<211> 16
<212> PRT
<213> Lactococcus lactis
<400> 593
Leu Gln Lys Phe Thr Gly Asp Ile Glu Asn Leu Val Lys Ala Ser Leu
1 5 10 15
<210> 594
<211> 9
<212> PRT
<213> Lactococcus lactis
<400> 594
Val Ile Val Pro Glu Leu Thr Phe Gly
1 5
<210> 595
<211> 15
<212> PRT
<213> Lactococcus lactis
<400> 595
Trp Ile Arg Ala Gly Trp Phe Ile Arg Asn Arg Ser Ala His Tyr
1 5 10 15
<210> 596
<211> 13
<212> PRT
<213> Lactococcus lactis
<400> 596
Asn Lys Asp Leu Phe Ala Phe Met Leu Ser Ile Lys Gln
1 5 10
<210> 597
<211> 10
<212> PRT
<213> Lactococcus lactis
<400> 597
Phe Leu His Lys Asn Ser Tyr Phe Arg Phe
1 5 10
<210> 598
<211> 16
<212> PRT
<213> Lactococcus lactis
<400> 598
Leu Phe Ile Phe Ser Thr Arg Leu Glu Ile Phe Trp Lys Lys Lys Ile
1 5 10 15
<210> 599
<211> 9
<212> PRT
<213> Lactococcus lactis
<400> 599
Ala Leu Val Glu Glu Leu Thr Phe Gly
1 5
<210> 600
<211> 15
<212> PRT
<213> Lactococcus lactis
<400> 600
Trp Met Asn Val Val Arg Leu Tyr Arg Asn Lys Ser Ala His Gly
1 5 10 15
<210> 601
<211> 13
<212> PRT
<213> Lactococcus lactis
<400> 601
Lys Ser Tyr Leu Tyr Gly Ala Leu Tyr Val Phe Lys His
1 5 10
<210> 602
<211> 10
<212> PRT
<213> Corynebacterium diphtheriae
<400> 602
Leu Leu Ala Gln Leu Asn Tyr Tyr Arg Leu
1 5 10
<210> 603
<211> 16
<212> PRT
<213> Corynebacterium diphtheriae
<400> 603
Val Phe Ile Glu Leu Asp Arg Val Glu Leu Ala Ile Gln Thr Arg Leu
1 5 10 15
<210> 604
<211> 9
<212> PRT
<213> Corynebacterium diphtheriae
<400> 604
Ala Ala Val Glu Val Met Asp Trp Gly
1 5
<210> 605
<211> 15
<212> PRT
<213> Corynebacterium diphtheriae
<400> 605
Trp Leu Lys Ser Leu Asn Ile Leu Arg Asn Tyr Ala Ala His His
1 5 10 15
<210> 606
<211> 13
<212> PRT
<213> Corynebacterium diphtheriae
<400> 606
Gly Gln Leu Ser Met Ile Gln Tyr Leu His His Gln Leu
1 5 10
<210> 607
<211> 12
<212> PRT
<213> Shewanella baltica
<400> 607
Met Leu Ile Glu Asn Asp Leu Asp Gly Ile Glu Asn
1 5 10
<210> 608
<211> 16
<212> PRT
<213> Shewanella baltica
<400> 608
Asn Tyr Gln Leu Phe Tyr Phe Leu Glu Lys Thr Ile Arg Asn Gln Ile
1 5 10 15
<210> 609
<211> 15
<212> PRT
<213> Shewanella baltica
<400> 609
Val Met Phe Asn Leu Asn Thr Leu Arg Asn Pro Ile Ala His Cys
1 5 10 15
<210> 610
<211> 13
<212> PRT
<213> Shewanella baltica
<400> 610
Asp Glu Lys Leu Arg Leu Glu Ile Ser Leu Arg Asp Trp
1 5 10
<210> 611
<211> 12
<212> PRT
<213> Lactococcus lactis
<400> 611
Leu Arg Glu Ile Asn Ile Lys Ala Ser Lys Ser Arg
1 5 10
<210> 612
<211> 16
<212> PRT
<213> Lactococcus lactis
<400> 612
Leu Leu Pro Leu Leu His Lys Tyr Glu Trp Ser Leu Arg Lys Leu Ile
1 5 10 15
<210> 613
<211> 9
<212> PRT
<213> Lactococcus lactis
<400> 613
Tyr Asp Phe Glu Glu Tyr Leu Phe Gly
1 5
<210> 614
<211> 15
<212> PRT
<213> Lactococcus lactis
<400> 614
Asp Met Arg Leu Ile Arg Asp Gly Arg Asn Ile Val Gly His Asn
1 5 10 15
<210> 615
<211> 13
<212> PRT
<213> Lactococcus lactis
<400> 615
Leu Ser Lys Gly Leu Lys Lys Tyr Ile Lys Lys Leu Asp
1 5 10
<210> 616
<211> 12
<212> PRT
<213> Geobacter bemidjiensis
<400> 616
Arg Leu Pro Leu Thr Ser His Ile Gln Lys Gln Asp
1 5 10
<210> 617
<211> 16
<212> PRT
<213> Geobacter bemidjiensis
<400> 617
Ile Tyr Pro Lys Leu Asn Arg Ile Glu Asn Arg Leu Arg His Tyr Leu
1 5 10 15
<210> 618
<211> 9
<212> PRT
<213> Geobacter bemidjiensis
<400> 618
Phe Glu Leu Gly Lys Ile Val Tyr Ala
1 5
<210> 619
<211> 15
<212> PRT
<213> Geobacter bemidjiensis
<400> 619
Lys Trp Ile Arg Leu Glu Glu Ile Arg His Lys Val Ala His Asn
1 5 10 15
<210> 620
<211> 13
<212> PRT
<213> Geobacter bemidjiensis
<400> 620
Ala Asn Glu Tyr Ile Asp Ser Leu Gln Ser Ile Ile Asp
1 5 10
<210> 621
<211> 12
<212> PRT
<213> Salmonella enterica
<400> 621
Phe Val Thr Ser Leu Glu His Leu Arg Gln Gln Gln
1 5 10
<210> 622
<211> 16
<212> PRT
<213> Salmonella enterica
<400> 622
Ala Gln Arg Gln Leu Arg Ala Ile Glu Leu Thr Leu Lys Ala Leu Ile
1 5 10 15
<210> 623
<211> 9
<212> PRT
<213> Salmonella enterica
<400> 623
Asn His Tyr Leu Lys Gln His Phe Gly
1 5
<210> 624
<211> 15
<212> PRT
<213> Salmonella enterica
<400> 624
Phe Leu Asp Asp Cys Arg Leu Ala Arg Asn Glu Val Ile Ala Arg
1 5 10 15
<210> 625
<211> 13
<212> PRT
<213> Salmonella enterica
<400> 625
Leu Met Leu Leu Asn Val Gln Tyr Gln Gln Ile Val Arg
1 5 10
<210> 626
<211> 12
<212> PRT
<213> Shigella flexneri
<400> 626
Phe Leu Trp Gln Leu Glu Tyr Leu Arg Glu Lys Gln
1 5 10
<210> 627
<211> 16
<212> PRT
<213> Shigella flexneri
<400> 627
Ser Leu Gln Gln Val Arg Ala Leu Glu Leu Thr Ile Arg Ser Leu Ile
1 5 10 15
<210> 628
<211> 9
<212> PRT
<213> Shigella flexneri
<400> 628
Leu Glu His Leu Asn Lys Leu Phe Gly
1 5
<210> 629
<211> 15
<212> PRT
<213> Shigella flexneri
<400> 629
Phe Leu Asp Asp Ile Arg Val Ile Arg Asn Arg Leu Ala His His
1 5 10 15
<210> 630
<211> 13
<212> PRT
<213> Shigella flexneri
<400> 630
Thr Thr Leu Val Asn Tyr Tyr Tyr Arg Glu Ile Thr Glu
1 5 10
<210> 631
<211> 16
<212> PRT
<213> Streptomyces avermitilis
<400> 631
Ala Tyr Ile Trp Leu Asn Leu Val Glu Gln Arg Leu Arg Ala Val Val
1 5 10 15
<210> 632
<211> 9
<212> PRT
<213> Streptomyces avermitilis
<400> 632
Asn Val Leu Ser Phe Leu Thr Leu Pro
1 5
<210> 633
<211> 11
<212> PRT
<213> Streptomyces avermitilis
<400> 633
Leu Glu Val Thr Arg Asn Val Val Ser Arg Asn
1 5 10
<210> 634
<211> 13
<212> PRT
<213> Streptomyces avermitilis
<400> 634
Arg Tyr Gly Asp Val Val Gly Val His Pro Asp Arg Val
1 5 10
<210> 635
<211> 10
<212> PRT
<213> Helicobacter pylori
<400> 635
Ser Ile Ser Val Leu His Tyr Asp Tyr Leu
1 5 10
<210> 636
<211> 16
<212> PRT
<213> Helicobacter pylori
<400> 636
Leu Phe Leu Trp Ile His Phe Phe Glu Thr Ala Leu Arg Ser Lys Met
1 5 10 15
<210> 637
<211> 9
<212> PRT
<213> Helicobacter pylori
<400> 637
Gln Ile Leu Asn Leu Phe Thr Leu Gly
1 5
<210> 638
<211> 15
<212> PRT
<213> Helicobacter pylori
<400> 638
Thr Phe Ser Leu Ile Arg Lys Ala Arg Asn Asp Leu Phe His Asn
1 5 10 15
<210> 639
<211> 13
<212> PRT
<213> Helicobacter pylori
<400> 639
Thr Leu Lys Leu Glu Arg Ala Ile Phe Phe Lys Thr Ile
1 5 10
<210> 640
<211> 11
<212> PRT
<213> Methylomirabilis oxyfera
<400> 640
Gly Pro Pro Glu Tyr Tyr Tyr Arg Leu Cys Arg
1 5 10
<210> 641
<211> 16
<212> PRT
<213> Methylomirabilis oxyfera
<400> 641
Ala Asp Ser Lys Leu Lys Asp Thr Val Ser Glu Met Arg Lys Phe Ile
1 5 10 15
<210> 642
<211> 15
<212> PRT
<213> Methylomirabilis oxyfera
<400> 642
Trp Met Asn Arg Ile Asn Glu Leu Arg Arg Ile Pro Ala His Pro
1 5 10 15
<210> 643
<211> 13
<212> PRT
<213> Methylomirabilis oxyfera
<400> 643
Asp Phe Glu Tyr Ile Asp Phe Ile Tyr Asp Glu Leu Met
1 5 10
<210> 644
<211> 12
<212> PRT
<213> Novosphingobium aromaticivorans
<400> 644
Thr Ala Val Lys Gln Gln Ser Phe Gly Met Glu Ala
1 5 10
<210> 645
<211> 16
<212> PRT
<213> Novosphingobium aromaticivorans
<400> 645
Ala Ala Ala Lys Val Thr Gln Ile His Lys Lys Leu Phe Asn Tyr Val
1 5 10 15
<210> 646
<211> 15
<212> PRT
<213> Novosphingobium aromaticivorans
<400> 646
Trp Ile Lys Val Leu Asn Asp Ile Arg Gln Tyr Thr Ala His Pro
1 5 10 15
<210> 647
<211> 13
<212> PRT
<213> Novosphingobium aromaticivorans
<400> 647
Gln Val Ser Phe Val Asn Glu Val Tyr Glu Lys Val Glu
1 5 10
<210> 648
<211> 11
<212> PRT
<213> Elizabethkingia anophelis
<400> 648
Gly Glu Ile Lys Tyr Trp Arg Thr Phe Gln Lys
1 5 10
<210> 649
<211> 16
<212> PRT
<213> Elizabethkingia anophelis
<400> 649
Ala Ile Ala Tyr Ile Arg Asp Ile Glu Thr Glu Phe Lys Ser Asp Phe
1 5 10 15
<210> 650
<211> 15
<212> PRT
<213> Elizabethkingia anophelis
<400> 650
Trp Met Val Lys Leu Glu Arg Ile Arg Asn Gln Asn Phe His Ser
1 5 10 15
<210> 651
<211> 13
<212> PRT
<213> Elizabethkingia anophelis
<400> 651
Glu Leu Ser Phe Leu Glu Glu Leu His Asp Trp Ile Tyr
1 5 10
<210> 652
<211> 12
<212> PRT
<213> Escherichia coli
<400> 652
Phe Ser Ala Leu Pro Arg Ile Ile Glu Tyr Ala Tyr
1 5 10
<210> 653
<211> 16
<212> PRT
<213> Escherichia coli
<400> 653
Pro Phe Leu Leu Leu Ser Glu Ile Glu Asn His Ile Arg Lys Leu Ile
1 5 10 15
<210> 654
<211> 9
<212> PRT
<213> Escherichia coli
<400> 654
Glu Ser Val Ala Asp Leu Thr Phe Gly
1 5
<210> 655
<211> 15
<212> PRT
<213> Escherichia coli
<400> 655
Glu Leu Asp Lys Val Arg Ile Ile Arg Asn Asp Val Met His Phe
1 5 10 15
<210> 656
<211> 13
<212> PRT
<213> Escherichia coli
<400> 656
Asn His Glu Leu Leu His Asn Phe Val Arg Phe Ile His
1 5 10
<210> 657
<211> 12
<212> PRT
<213> Haloarcula marismortui
<400> 657
Phe Glu Leu Phe Asp Thr Leu Ala Glu Asp Asp Tyr
1 5 10
<210> 658
<211> 16
<212> PRT
<213> Haloarcula marismortui
<400> 658
Pro Phe Leu Gln Ile Gly Glu Ile Glu Glu Ser Leu Arg His Leu Phe
1 5 10 15
<210> 659
<211> 9
<212> PRT
<213> Haloarcula marismortui
<400> 659
Asp Arg Pro Glu Asp Phe Ser Phe Asp
1 5
<210> 660
<211> 15
<212> PRT
<213> Haloarcula marismortui
<400> 660
Leu Leu Glu Asp Ile Arg Glu Thr Arg Asn Ala Leu Leu His Phe
1 5 10 15
<210> 661
<211> 13
<212> PRT
<213> Haloarcula marismortui
<400> 661
Asp Arg Asp Gln Leu Asp Met Ala His Gly Tyr Phe Thr
1 5 10
<210> 662
<211> 12
<212> PRT
<213> Nostoc sp.
<400> 662
Met Lys Leu Leu Pro Ile Leu Gln Gln Asn Pro Arg
1 5 10
<210> 663
<211> 15
<212> PRT
<213> Nostoc sp.
<400> 663
Phe Gly Leu Val Thr Leu Leu Glu Met Asn Leu Leu Arg Leu Val
1 5 10 15
<210> 664
<211> 9
<212> PRT
<213> Nostoc sp.
<400> 664
Asp Leu Leu Asp Tyr Leu Gln Phe Cys
1 5
<210> 665
<211> 15
<212> PRT
<213> Nostoc sp.
<400> 665
Phe Leu Lys Ser Ala Glu Gln Leu Arg Asn Arg Leu Ala His Ala
1 5 10 15
<210> 666
<211> 13
<212> PRT
<213> Nostoc sp.
<400> 666
Ser Trp Asn Asp Leu Ile Ser Leu Ala Glu Ala Met Glu
1 5 10
<210> 667
<211> 10
<212> PRT
<213> Xanthobacter autotrophicus
<400> 667
Val Phe Glu Gly Met Glu Leu Leu Pro Ala
1 5 10
<210> 668
<211> 14
<212> PRT
<213> Xanthobacter autotrophicus
<400> 668
Ala Leu Ile Pro Phe Val Glu Lys Arg Leu Glu Thr Ser Leu
1 5 10
<210> 669
<211> 8
<212> PRT
<213> Xanthobacter autotrophicus
<400> 669
Glu Ala Phe Lys Ala Val Leu Gly
1 5
<210> 670
<211> 15
<212> PRT
<213> Xanthobacter autotrophicus
<400> 670
Leu Val Asn Glu Leu Gly Asp Val Arg Asn Lys Leu Ser His Asn
1 5 10 15
<210> 671
<211> 13
<212> PRT
<213> Xanthobacter autotrophicus
<400> 671
Tyr Asp Asp Ala Glu Arg Ala Leu Asp Thr Met Arg Arg
1 5 10
<210> 672
<211> 10
<212> PRT
<213> Methanospirillum hungatei
<400> 672
Val Gly Arg Ala Met Asp Gln Leu Lys Thr
1 5 10
<210> 673
<211> 14
<212> PRT
<213> Methanospirillum hungatei
<400> 673
Gly Leu Met Arg Phe Val Glu Arg Glu Met Lys Ser Ala Tyr
1 5 10
<210> 674
<211> 8
<212> PRT
<213> Methanospirillum hungatei
<400> 674
Lys Val Phe Ser Gln Ile Leu Gly
1 5
<210> 675
<211> 15
<212> PRT
<213> Methanospirillum hungatei
<400> 675
Leu Val Ser Glu Leu Arg Glu Thr Arg Asn Gln Trp Ala His Gln
1 5 10 15
<210> 676
<211> 13
<212> PRT
<213> Methanospirillum hungatei
<400> 676
Thr Asn Asp Thr Leu Arg Ala Leu Asp Ser Thr Ala Arg
1 5 10
<210> 677
<211> 10
<212> PRT
<213> Roseiflexus sp.
<400> 677
Ile Gly Lys Ala Leu Asp Leu Leu Arg Gln
1 5 10
<210> 678
<211> 14
<212> PRT
<213> Roseiflexus sp.
<400> 678
Gly Leu Gln Pro Phe Ile Glu Arg Glu Leu Gln Asn His Tyr
1 5 10
<210> 679
<211> 8
<212> PRT
<213> Roseiflexus sp.
<400> 679
Asp Val Phe Arg Lys Thr Leu Gly
1 5
<210> 680
<211> 15
<212> PRT
<213> Roseiflexus sp.
<400> 680
Leu Val Ser Glu Leu Arg Glu Trp Arg Asn Lys Trp Ala His Gln
1 5 10 15
<210> 681
<211> 13
<212> PRT
<213> Roseiflexus sp.
<400> 681
Thr Asp Asp Thr Tyr Arg Val Leu Asp Ser Ala Ala Arg
1 5 10
<210> 682
<211> 10
<212> PRT
<213> Plasmodium yoelii
<400> 682
Ile Leu Asn Ile Phe His Ile Leu Ser Ala
1 5 10
<210> 683
<211> 14
<212> PRT
<213> Plasmodium yoelii
<400> 683
His Leu Ser Pro Ile Ile Glu Gln Ile Met Glu Met Glu Tyr
1 5 10
<210> 684
<211> 7
<212> PRT
<213> Plasmodium yoelii
<400> 684
Asp Ile Phe Glu Asn Arg Ile
1 5
<210> 685
<211> 15
<212> PRT
<213> Plasmodium yoelii
<400> 685
Ile Leu Glu Asn Leu Gln Lys Ala Ser Ile Phe Trp Ala Asn Gln
1 5 10 15
<210> 686
<211> 13
<212> PRT
<213> Plasmodium yoelii
<400> 686
Glu Phe Phe Leu Ser Asn Leu Val Ser Ser Tyr Phe Phe
1 5 10
<210> 687
<211> 10
<212> PRT
<213> Theileria parva
<400> 687
Val Val Met Ile Phe Gln Cys Val Cys Asp
1 5 10
<210> 688
<211> 14
<212> PRT
<213> Theileria parva
<400> 688
Ala Phe Gln Pro Phe Ile Ser Lys Cys Met Leu Lys Lys Phe
1 5 10
<210> 689
<211> 7
<212> PRT
<213> Theileria parva
<400> 689
Asp Ile Phe Glu Gln Val Leu
1 5
<210> 690
<211> 15
<212> PRT
<213> Theileria parva
<400> 690
His Leu Asn Thr Ile Gln Thr Ala Ser Ile Tyr Trp Ala Asn Gln
1 5 10 15
<210> 691
<211> 8
<212> PRT
<213> Theileria parva
<400> 691
Asn Tyr Gly Lys Cys Arg Lys Ile
1 5
<210> 692
<211> 10
<212> PRT
<213> Daphnia pulex
<400> 692
Ser Ser Lys Glu Ser Ala Ala Ile Ala Ile
1 5 10
<210> 693
<211> 14
<212> PRT
<213> Daphnia pulex
<400> 693
Gly His Ile Val Phe Asp Thr Phe Leu Glu Asp Val Ala Pro
1 5 10
<210> 694
<211> 8
<212> PRT
<213> Daphnia pulex
<400> 694
Asp Cys Phe Ile Ile Pro Pro Gly
1 5
<210> 695
<211> 15
<212> PRT
<213> Daphnia pulex
<400> 695
Ile Leu Glu Arg Ala Met Asp Gly Arg His Ala Val Ser His His
1 5 10 15
<210> 696
<211> 13
<212> PRT
<213> Daphnia pulex
<400> 696
Trp Glu Gln His Leu Lys Asp Tyr Val Tyr Ile Leu Thr
1 5 10
<210> 697
<211> 10
<212> PRT
<213> Homo sapiens
<400> 697
Ala Gly His Cys Leu Leu Leu Leu Arg Ser
1 5 10
<210> 698
<211> 16
<212> PRT
<213> Homo sapiens
<400> 698
Cys Leu Gln Gly Phe Val Gly Arg Glu Val Leu Ser Phe His Arg Gly
1 5 10 15
<210> 699
<211> 15
<212> PRT
<213> Homo sapiens
<400> 699
Lys Val Thr Glu Val Ile Lys Cys Arg Asn Glu Ile Met His Ser
1 5 10 15
<210> 700
<211> 13
<212> PRT
<213> Homo sapiens
<400> 700
Ser Ser Thr Trp Leu Arg Asp Phe Gln Met Lys Ile Gln
1 5 10
<210> 701
<211> 10
<212> PRT
<213> Branchiostoma floridae
<400> 701
Val Gly Ile Ala Leu Leu Thr Thr Arg Asp
1 5 10
<210> 702
<211> 16
<212> PRT
<213> Branchiostoma floridae
<400> 702
Gly Leu Thr Asn Val Thr Glu Gln Ala Ala Lys Glu Leu Gln Ala Glu
1 5 10 15
<210> 703
<211> 15
<212> PRT
<213> Branchiostoma floridae
<400> 703
Pro Leu Lys Asn Val Ile Glu Val Arg Asn Lys Thr Met His Ser
1 5 10 15
<210> 704
<211> 13
<212> PRT
<213> Branchiostoma floridae
<400> 704
Asp Arg Gln Thr Phe Asn Glu Tyr Met Asp Lys Met Glu
1 5 10
<210> 705
<211> 10
<212> PRT
<213> Homo sapiens
<400> 705
Val Ser Asp Leu Glu Lys Ser Leu Gly Thr
1 5 10
<210> 706
<211> 14
<212> PRT
<213> Homo sapiens
<400> 706
Gly Leu Ser Ser Ile Leu Glu Thr Glu Met Lys Ile Ala Phe
1 5 10
<210> 707
<211> 8
<212> PRT
<213> Homo sapiens
<400> 707
Lys His Trp Leu Ala Val Phe Gly
1 5
<210> 708
<211> 16
<212> PRT
<213> Homo sapiens
<400> 708
Thr Ile Glu Ser Leu Tyr Lys Asn Leu Arg Lys Ala Asn Lys Ala Val
1 5 10 15
<210> 709
<211> 13
<212> PRT
<213> Homo sapiens
<400> 709
Ser Arg Ser Leu Leu His Ala Phe Ser Thr Arg Ser Asn
1 5 10
<210> 710
<211> 10
<212> PRT
<213> Ostreococcus lucimarinus
<400> 710
Met Glu Arg Leu Met Met Val Leu Asp His
1 5 10
<210> 711
<211> 14
<212> PRT
<213> Ostreococcus lucimarinus
<400> 711
Val Leu Ala Ile Val Leu Glu Gly Gly Leu Arg Ala Glu Phe
1 5 10
<210> 712
<211> 8
<212> PRT
<213> Ostreococcus lucimarinus
<400> 712
Ala Asn Trp Gly Ser Leu Phe Ser
1 5
<210> 713
<211> 14
<212> PRT
<213> Ostreococcus lucimarinus
<400> 713
Glu Ile Glu Val Leu Leu Asp Ala Ala Ile Arg Gln Arg Lys
1 5 10
<210> 714
<211> 13
<212> PRT
<213> Ostreococcus lucimarinus
<400> 714
Ala Arg Asp Val Ser Ser Ala Ala Val Ala Leu Leu Asn
1 5 10
<210> 715
<211> 10
<212> PRT
<213> Branchiostoma floridae
<400> 715
Leu Cys Gly Met Lys Thr Leu Leu Lys Ala
1 5 10
<210> 716
<211> 14
<212> PRT
<213> Branchiostoma floridae
<400> 716
Val Leu Ala Val Val Leu Glu Thr Glu Met Lys Ala Val Phe
1 5 10
<210> 717
<211> 8
<212> PRT
<213> Branchiostoma floridae
<400> 717
Lys His Trp Ile Ala Val Phe Gly
1 5
<210> 718
<211> 17
<212> PRT
<213> Branchiostoma floridae
<400> 718
His Leu Asp Ser Leu Val Lys His Phe Thr Arg Gly Arg Ser Tyr Gly
1 5 10 15
Val
<210> 719
<211> 13
<212> PRT
<213> Branchiostoma floridae
<400> 719
Ala Leu Gln Leu Val Arg Gln Leu His Asn His Ser Thr
1 5 10
<210> 720
<211> 10
<212> PRT
<213> Microcystis aeruginosa
<400> 720
Leu Asn Trp Leu Asp Gln Leu His Asp Asp
1 5 10
<210> 721
<211> 16
<212> PRT
<213> Microcystis aeruginosa
<400> 721
Leu Ile Glu Leu Cys Gly Trp Ile Glu Glu Thr Met Asp Asp Ile Val
1 5 10 15
<210> 722
<211> 9
<212> PRT
<213> Microcystis aeruginosa
<400> 722
Phe Arg Lys Met Leu Met Met Val Ile
1 5
<210> 723
<211> 15
<212> PRT
<213> Microcystis aeruginosa
<400> 723
Tyr Leu Gly Asn Leu Lys Asp Ser Arg Asn Arg Ala Ala His Thr
1 5 10 15
<210> 724
<211> 13
<212> PRT
<213> Microcystis aeruginosa
<400> 724
Phe Asp Lys Ile Tyr Gly Leu Leu Lys Glu Leu Asp Ala
1 5 10
<210> 725
<211> 12
<212> PRT
<213> Lactococcus lactis
<400> 725
Leu Ser Glu Leu His Glu Phe Ile Lys Lys Leu Asn
1 5 10
<210> 726
<211> 16
<212> PRT
<213> Lactococcus lactis
<400> 726
Val Ile Arg Ser Cys Gly Ile Ile Glu Gln Leu Thr Lys Thr Leu Ile
1 5 10 15
<210> 727
<211> 9
<212> PRT
<213> Lactococcus lactis
<400> 727
Ile Asn Gly Leu Ile Asp Thr Phe Asp
1 5
<210> 728
<211> 15
<212> PRT
<213> Lactococcus lactis
<400> 728
His Ile Asp Ser Leu Arg Gln Leu Arg Asn Ser Ile Ala His Gly
1 5 10 15
<210> 729
<211> 13
<212> PRT
<213> Lactococcus lactis
<400> 729
Met Gly Tyr Phe Asp Ser Cys Ile Ile Leu Met Phe Arg
1 5 10
<210> 730
<211> 12
<212> PRT
<213> Frankia sp.
<400> 730
Leu Ser Glu Leu Ala Ala Leu Val Gln Asp Gln Ala
1 5 10
<210> 731
<211> 16
<212> PRT
<213> Frankia sp.
<400> 731
Val Ile Arg Ser Cys Gly Tyr Leu Glu Gln Thr Val Ala Gly Thr Phe
1 5 10 15
<210> 732
<211> 9
<212> PRT
<213> Frankia sp.
<400> 732
Leu Glu Thr Leu Ala Gly Arg Phe Asp
1 5
<210> 733
<211> 15
<212> PRT
<213> Frankia sp.
<400> 733
Glu Leu Ala Thr Leu Val Asp Arg Arg Asn Arg Ile Ala His Gly
1 5 10 15
<210> 734
<211> 13
<212> PRT
<213> Frankia sp.
<400> 734
Leu Glu Leu His Arg Val Ala Cys Glu Ala Ala Asp Trp
1 5 10
<210> 735
<211> 10
<212> PRT
<213> Neisseria meningitidis
<400> 735
Cys Cys Ser Ile Phe Ser Asp Phe Arg Met
1 5 10
<210> 736
<211> 16
<212> PRT
<213> Neisseria meningitidis
<400> 736
Leu Phe His Val Val Ser Ile Phe Glu Ile Val Leu Arg Asn Lys Ile
1 5 10 15
<210> 737
<211> 9
<212> PRT
<213> Neisseria meningitidis
<400> 737
Gln Leu Val Ala Gly Leu Gly Phe Gly
1 5
<210> 738
<211> 15
<212> PRT
<213> Neisseria meningitidis
<400> 738
Glu Leu Ser Asn Ile Asn Lys Phe Arg Asn Arg Leu Ala His His
1 5 10 15
<210> 739
<211> 13
<212> PRT
<213> Neisseria meningitidis
<400> 739
Asp Val Asp Thr Ala Ser Val Phe Ser His Phe Ser Asp
1 5 10
<210> 740
<211> 10
<212> PRT
<213> Pseudomonas syringae
<400> 740
Leu Glu Lys His Phe Ser Ser Ala Arg Leu
1 5 10
<210> 741
<211> 16
<212> PRT
<213> Pseudomonas syringae
<400> 741
Met Met Pro Met Leu Ser Val Leu Glu Ile Ala Leu Lys Asn Gly Ile
1 5 10 15
<210> 742
<211> 9
<212> PRT
<213> Pseudomonas syringae
<400> 742
Lys Ile Val Ala Glu Leu Ala Phe Gly
1 5
<210> 743
<211> 15
<212> PRT
<213> Pseudomonas syringae
<400> 743
Ala Leu Asn Leu Ile Arg Asn Leu Arg Asn Arg Val Phe His His
1 5 10 15
<210> 744
<211> 13
<212> PRT
<213> Pseudomonas syringae
<400> 744
Asp Pro Gln Leu Val Pro Trp Leu Ala Gln Tyr Asp Arg
1 5 10
<210> 745
<211> 10
<212> PRT
<213> Geobacter uraniireducens
<400> 745
Leu Arg Arg Ala Ile Ser His Glu Arg Leu
1 5 10
<210> 746
<211> 16
<212> PRT
<213> Geobacter uraniireducens
<400> 746
Leu Tyr Thr Pro Leu Gln Cys Leu Glu Val Cys Leu Arg Asn Ser Ile
1 5 10 15
<210> 747
<211> 9
<212> PRT
<213> Geobacter uraniireducens
<400> 747
Arg Ile Ile Pro Glu Leu Thr Phe Gly
1 5
<210> 748
<211> 15
<212> PRT
<213> Geobacter uraniireducens
<400> 748
Arg Phe Asn His Ile Arg Thr Leu Arg Asn Arg Ile Phe His His
1 5 10 15
<210> 749
<211> 13
<212> PRT
<213> Geobacter uraniireducens
<400> 749
Asn Pro Ala Met Met Thr Phe Val Glu Pro Phe Asp Ser
1 5 10
<210> 750
<211> 16
<212> PRT
<213> Sulfuricurvum kujiense
<400> 750
Glu Glu Lys Ser Glu Phe Ile Arg Glu Phe Phe Lys Arg Thr Leu His
1 5 10 15
<210> 751
<211> 9
<212> PRT
<213> Sulfuricurvum kujiense
<400> 751
Thr Gln Thr Ile Asn Ser Phe Leu Gly
1 5
<210> 752
<211> 14
<212> PRT
<213> Sulfuricurvum kujiense
<400> 752
Phe Arg Asn Tyr Leu Lys Arg Leu Arg Asn Ala Val Ser His
1 5 10
<210> 753
<211> 13
<212> PRT
<213> Sulfuricurvum kujiense
<400> 753
Val Asn Leu Leu Ile Thr Leu Leu Ser Arg Asn Ile Leu
1 5 10
<210> 754
<211> 13
<212> PRT
<213> Dethiobacter alkaliphilus
<400> 754
Gln Val Val Glu Lys Asp Phe Val Ala Arg Thr Met His
1 5 10
<210> 755
<211> 9
<212> PRT
<213> Dethiobacter alkaliphilus
<400> 755
Thr Leu Leu Ile Asn Cys Leu Leu Gly
1 5
<210> 756
<211> 14
<212> PRT
<213> Dethiobacter alkaliphilus
<400> 756
Ala Ser Arg Phe Leu Gln Cys Met Arg Asn Ser Val Ala His
1 5 10
<210> 757
<211> 10
<212> PRT
<213> Dethiobacter alkaliphilus
<400> 757
Leu Ala Thr Lys Leu Ala Gln Tyr Val Gln
1 5 10
<210> 758
<211> 13
<212> PRT
<213> Klebsiella pneumoniae
<400> 758
Ser Asp Phe Glu Thr Asp Phe Val Gln Arg Thr Leu Ala
1 5 10
<210> 759
<211> 9
<212> PRT
<213> Klebsiella pneumoniae
<400> 759
Thr Leu Thr Leu Asn Cys Leu Leu Gly
1 5
<210> 760
<211> 14
<212> PRT
<213> Klebsiella pneumoniae
<400> 760
Leu Arg Gln Leu Ile His Lys Met Arg Asn Ser Val Ala His
1 5 10
<210> 761
<211> 13
<212> PRT
<213> Klebsiella pneumoniae
<400> 761
Leu Leu Pro Phe Leu Lys Tyr Tyr Ala Thr Leu Leu Leu
1 5 10
<210> 762
<211> 10
<212> PRT
<213> Lactobacillus casei
<400> 762
Lys Ile Asp Arg Glu Met Phe Trp Arg Arg
1 5 10
<210> 763
<211> 16
<212> PRT
<213> Lactobacillus casei
<400> 763
Tyr Leu Leu Leu Tyr Ser Ser Trp Glu Gly Phe Ile Arg Ser Ile Ala
1 5 10 15
<210> 764
<211> 9
<212> PRT
<213> Lactobacillus casei
<400> 764
Leu Ala Arg Ile Val Ser Val Leu Asp
1 5
<210> 765
<211> 14
<212> PRT
<213> Lactobacillus casei
<400> 765
Asp Arg Asp Leu Leu Lys Val Arg Asn Glu Ile Ala His Gly
1 5 10
<210> 766
<211> 13
<212> PRT
<213> Lactobacillus casei
<400> 766
Thr Val Ser His Val Leu Glu Met Met Asp Leu Phe Ser
1 5 10
<210> 767
<211> 10
<212> PRT
<213> Caulobacter sp.
<400> 767
Asp Leu Asp Ala Ala Arg Leu Arg Arg Ala
1 5 10
<210> 768
<211> 16
<212> PRT
<213> Caulobacter sp.
<400> 768
Ile Val Leu Ala Tyr Ser His Trp Glu Gly Phe Tyr Asn Glu Cys Ile
1 5 10 15
<210> 769
<211> 9
<212> PRT
<213> Caulobacter sp.
<400> 769
Leu Lys Glu Asn Phe Arg Ile Leu Gly
1 5
<210> 770
<211> 14
<212> PRT
<213> Caulobacter sp.
<400> 770
Asn Lys Glu Leu Val Gly Trp Arg His Ser Ile Ala His Gly
1 5 10
<210> 771
<211> 13
<212> PRT
<213> Caulobacter sp.
<400> 771
His Ile Ile Leu Thr Asn Ser Leu Leu Leu Thr Leu Ser
1 5 10
<210> 772
<211> 10
<212> PRT
<213> Microcystis aeruginosa
<400> 772
Asn Leu Asp Glu Asp Met Ala Trp Arg Ile
1 5 10
<210> 773
<211> 16
<212> PRT
<213> Microcystis aeruginosa
<400> 773
Ile Thr Thr Leu Tyr Ala His Trp Glu Gly Phe Ile Lys Tyr Ala Ala
1 5 10 15
<210> 774
<211> 9
<212> PRT
<213> Microcystis aeruginosa
<400> 774
Phe Thr Asp Ile Cys Thr Ile Leu Gly
1 5
<210> 775
<211> 14
<212> PRT
<213> Microcystis aeruginosa
<400> 775
Asp Glu Gln Leu Leu Thr Gln Arg Asn Lys Ile Ala His Gly
1 5 10
<210> 776
<211> 13
<212> PRT
<213> Microcystis aeruginosa
<400> 776
Thr Tyr Asn Leu Val Ile Lys Leu Ile Arg Asp Phe Lys
1 5 10
<210> 777
<211> 11
<212> PRT
<213> Arabidopsis thaliana
<400> 777
Pro Trp Leu Ser Trp Glu Glu Trp Asp Ser Val
1 5 10
<210> 778
<211> 15
<212> PRT
<213> Arabidopsis thaliana
<400> 778
Gly Ser Leu Pro Ala Pro Val Asp Val Thr Cys Ser Leu Ile Glu
1 5 10 15
<210> 779
<211> 9
<212> PRT
<213> Arabidopsis thaliana
<400> 779
Ile Ala Asp Ala Ala Arg Ala Ile Gly
1 5
<210> 780
<211> 15
<212> PRT
<213> Arabidopsis thaliana
<400> 780
Ile Pro Arg Lys Leu Ile Asp Leu Arg His Glu Gly Ser His Arg
1 5 10 15
<210> 781
<211> 13
<212> PRT
<213> Arabidopsis thaliana
<400> 781
Ala Ala Asp Glu Ala Leu Glu Trp Leu Lys Ser Tyr Tyr
1 5 10
<210> 782
<211> 11
<212> PRT
<213> Homo sapiens
<400> 782
Ala Trp Leu Ser Arg Ala Glu Trp Asp Gln Val
1 5 10
<210> 783
<211> 16
<212> PRT
<213> Homo sapiens
<400> 783
Gly Asn Glu Leu Pro Leu Ala Val Ala Ser Thr Ala Asp Leu Ile Arg
1 5 10 15
<210> 784
<211> 9
<212> PRT
<213> Homo sapiens
<400> 784
Leu Lys Cys Leu Ala Gln Glu Val Asn
1 5
<210> 785
<211> 15
<212> PRT
<213> Homo sapiens
<400> 785
Ile Pro Asp Trp Ile Val Asp Leu Arg His Glu Leu Thr His Lys
1 5 10 15
<210> 786
<211> 13
<212> PRT
<213> Homo sapiens
<400> 786
Gly Cys Tyr Phe Val Leu Asp Trp Leu Gln Lys Thr Tyr
1 5 10
<210> 787
<211> 11
<212> PRT
<213> Saccharomyces cerevisiae
<400> 787
Pro Trp Arg Asp Phe Ala Glu Leu Glu Glu Leu
1 5 10
<210> 788
<211> 16
<212> PRT
<213> Saccharomyces cerevisiae
<400> 788
Ser Gln Tyr Leu Pro His Val Val Asp Ser Thr Ala Gln Ile Thr Cys
1 5 10 15
<210> 789
<211> 9
<212> PRT
<213> Saccharomyces cerevisiae
<400> 789
Leu His Thr Leu Ala Ala Lys Ile Gly
1 5
<210> 790
<211> 15
<212> PRT
<213> Saccharomyces cerevisiae
<400> 790
Leu Pro Ser Trp Phe Val Asp Leu Arg His Trp Gly Thr His Glu
1 5 10 15
<210> 791
<211> 13
<212> PRT
<213> Saccharomyces cerevisiae
<400> 791
Ala Ala Asn Glu Ala Leu Ser Trp Leu Tyr Asp His Tyr
1 5 10
<210> 792
<211> 11
<212> PRT
<213> Streptococcus pneumoniae
<400> 792
Ser Lys Pro Cys Ile Glu Ala Glu Asn Met Ile
1 5 10
<210> 793
<211> 16
<212> PRT
<213> Streptococcus pneumoniae
<400> 793
Ala Phe Met Ala Arg Arg Ala Leu Glu Gln Ala Val His Trp Ile Tyr
1 5 10 15
<210> 794
<211> 8
<212> PRT
<213> Streptococcus pneumoniae
<400> 794
Ser Ser Leu Val Trp Asp Asp Asp
1 5
<210> 795
<211> 15
<212> PRT
<213> Streptococcus pneumoniae
<400> 795
Gln Ile Val Leu Leu Ile Arg Trp Gly Asn His Ala Ala His Gly
1 5 10 15
<210> 796
<211> 13
<212> PRT
<213> Streptococcus pneumoniae
<400> 796
Ala Leu His His Leu Tyr Gln Phe Val Asn Phe Ile Asp
1 5 10
<210> 797
<211> 11
<212> PRT
<213> Microcystis aeruginosa
<400> 797
Tyr Asp His Ala Ser Gln Ala Glu Gly Leu Val
1 5 10
<210> 798
<211> 16
<212> PRT
<213> Microcystis aeruginosa
<400> 798
Cys Phe Tyr Thr Arg Phe Val Leu Glu Gln Met Val Cys Trp Leu Tyr
1 5 10 15
<210> 799
<211> 8
<212> PRT
<213> Microcystis aeruginosa
<400> 799
Gly Ala Leu Ile His Glu Gln Thr
1 5
<210> 800
<211> 15
<212> PRT
<213> Microcystis aeruginosa
<400> 800
Lys Ile Arg Thr Ile His Lys Val Gly Asn Asn Ala Ala His Asp
1 5 10 15
<210> 801
<211> 13
<212> PRT
<213> Microcystis aeruginosa
<400> 801
Leu Ile Glu Glu Leu Phe His Leu Thr Tyr Trp Leu Val
1 5 10
<210> 802
<211> 11
<212> PRT
<213> Escherichia coli
<400> 802
Tyr Ala Ile Ala Cys Ala Ala Glu Asn Asn Tyr
1 5 10
<210> 803
<211> 16
<212> PRT
<213> Escherichia coli
<400> 803
Leu Ile Lys Met Arg Met Phe Gly Glu Ala Thr Ala Lys His Leu Gly
1 5 10 15
<210> 804
<211> 8
<212> PRT
<213> Escherichia coli
<400> 804
His Asp Leu Leu Arg Glu Leu Gly
1 5
<210> 805
<211> 15
<212> PRT
<213> Escherichia coli
<400> 805
Val Phe His Lys Leu Arg Arg Ile Gly Asn Gln Ala Val His Glu
1 5 10 15
<210> 806
<211> 13
<212> PRT
<213> Escherichia coli
<400> 806
Cys Leu Arg Leu Gly Phe Arg Leu Ala Val Trp Tyr Tyr
1 5 10
<210> 807
<211> 11
<212> PRT
<213> Bradyrhizobium japonicum
<400> 807
Val Gln Lys Leu Ile Lys Ala Ser Gln Leu Ala
1 5 10
<210> 808
<211> 16
<212> PRT
<213> Bradyrhizobium japonicum
<400> 808
Leu Thr Glu Val Arg Arg Ala Met Lys Ala Ala Ala Asp Leu Phe Trp
1 5 10 15
<210> 809
<211> 10
<212> PRT
<213> Bradyrhizobium japonicum
<400> 809
Leu Asn Arg Leu Gln Glu Phe Ala Arg Val
1 5 10
<210> 810
<211> 13
<212> PRT
<213> Bradyrhizobium japonicum
<400> 810
Arg Arg Leu Asn Asp Leu Ala Ser Lys Gly Val His Ala
1 5 10
<210> 811
<211> 13
<212> PRT
<213> Bradyrhizobium japonicum
<400> 811
Ala Glu Ala Arg Gln Gly Leu Val Gly Leu Tyr Phe Phe
1 5 10
<210> 812
<211> 11
<212> PRT
<213> Leptospira meyeri
<400> 812
Leu Pro Lys Phe Ser Ala Ile Tyr Ser Asn Leu
1 5 10
<210> 813
<211> 16
<212> PRT
<213> Leptospira meyeri
<400> 813
Val His Ser Cys Arg Arg Leu Leu Gln Ser Val Ala Asp Lys Leu Met
1 5 10 15
<210> 814
<211> 10
<212> PRT
<213> Leptospira meyeri
<400> 814
Ile Asn Arg Leu Ile Tyr Tyr Ile Glu Thr
1 5 10
<210> 815
<211> 13
<212> PRT
<213> Leptospira meyeri
<400> 815
Asp Ser Val Phe Gln Ala Ser Gln Lys Gly Ser His Ser
1 5 10
<210> 816
<211> 13
<212> PRT
<213> Leptospira meyeri
<400> 816
Gln Glu Ala Asp Arg Tyr Val Ile His Thr Phe Leu Leu
1 5 10
<210> 817
<211> 11
<212> PRT
<213> Bacteroides coprosuis
<400> 817
Val Val Asp Asp Arg Asp Phe Ser Leu Leu Ala
1 5 10
<210> 818
<211> 16
<212> PRT
<213> Bacteroides coprosuis
<400> 818
Leu Asp Arg Leu His Thr Tyr Val Ile Lys Phe Ile Arg Gln Leu Cys
1 5 10 15
<210> 819
<211> 9
<212> PRT
<213> Bacteroides coprosuis
<400> 819
Phe Gly Lys Tyr Val Lys Phe Ile Val
1 5
<210> 820
<211> 16
<212> PRT
<213> Bacteroides coprosuis
<400> 820
Ile Glu Ala Phe Asn Asp Ile Arg Asn Asn Lys Ser Phe Ala His Asp
1 5 10 15
<210> 821
<211> 13
<212> PRT
<213> Bacteroides coprosuis
<400> 821
Tyr Ala Glu Ser Val Leu Ile Phe Asn Asn Val Thr Asn
1 5 10
<210> 822
<211> 10
<212> PRT
<213> Escherichia coli
<400> 822
Asn Val Asn Glu Asn Ile Tyr Gln Ala Leu
1 5 10
<210> 823
<211> 16
<212> PRT
<213> Escherichia coli
<400> 823
Tyr Asp Arg Val His Thr Ala Leu His Ala Ser Leu Arg Gln Met Cys
1 5 10 15
<210> 824
<211> 9
<212> PRT
<213> Escherichia coli
<400> 824
Leu Ser Leu Ile Thr Ala His Leu Lys
1 5
<210> 825
<211> 16
<212> PRT
<213> Escherichia coli
<400> 825
Leu His Gly Ile Asn Asn Leu Arg Asn Asn Tyr Ser Met Ala His Pro
1 5 10 15
<210> 826
<211> 13
<212> PRT
<213> Escherichia coli
<400> 826
Glu Ala Asp Ala Arg Phe Ala Ile Asn Leu Val Arg Ser
1 5 10
<210> 827
<211> 11
<212> PRT
<213> Lactococcus lactis
<400> 827
Ile Met Asn Ile Gly Tyr Val Glu Lys Ile Leu
1 5 10
<210> 828
<211> 16
<212> PRT
<213> Lactococcus lactis
<400> 828
Val Thr Lys Ser Arg Thr Ile Ile Glu Thr Val Phe Ile Ala Ile Leu
1 5 10 15
<210> 829
<211> 9
<212> PRT
<213> Lactococcus lactis
<400> 829
Arg Ser Leu Val Asn Lys Thr Leu Gly
1 5
<210> 830
<211> 16
<212> PRT
<213> Lactococcus lactis
<400> 830
Val Asp Ser Ile Thr Thr Met Arg Asn Ile Asn Ser Asp Ser His Gly
1 5 10 15
<210> 831
<211> 13
<212> PRT
<213> Lactococcus lactis
<400> 831
Glu Ala Glu Ala Glu Leu Ile Leu Asn Ser Ala Val Asn
1 5 10
<210> 832
<211> 11
<212> PRT
<213> Peptoniphilus indolicus
<400> 832
Phe Leu Tyr Leu Lys Thr Leu Lys Asn Lys Glu
1 5 10
<210> 833
<211> 16
<212> PRT
<213> Peptoniphilus indolicus
<400> 833
Arg Gly Ile Thr Pro Leu Val Thr Glu Leu Phe Ile Leu Ile Ile Asp
1 5 10 15
<210> 834
<211> 9
<212> PRT
<213> Peptoniphilus indolicus
<400> 834
Leu Ile Glu Ile Ile Lys Asn Glu Arg
1 5
<210> 835
<211> 15
<212> PRT
<213> Peptoniphilus indolicus
<400> 835
Ile Arg Asp Val Glu Gly Lys Leu Arg Asn Arg Ala Ala His Glu
1 5 10 15
<210> 836
<211> 13
<212> PRT
<213> Peptoniphilus indolicus
<400> 836
Gly Asn Asn His Tyr Asp Ser Tyr Asp Leu Met Asn Lys
1 5 10
<210> 837
<211> 11
<212> PRT
<213> Mycobacterium tuberculosis
<400> 837
Ile Ser Ala Leu Ala Leu Leu Ala Lys Arg Glu
1 5 10
<210> 838
<211> 16
<212> PRT
<213> Mycobacterium tuberculosis
<400> 838
Arg Ser Ala Thr Pro Ala Ile Thr Ile Val Leu Arg Ala Ala Val Ala
1 5 10 15
<210> 839
<211> 9
<212> PRT
<213> Mycobacterium tuberculosis
<400> 839
Trp Leu Ala Leu Leu Arg Gln Phe Ala
1 5
<210> 840
<211> 15
<212> PRT
<213> Mycobacterium tuberculosis
<400> 840
Leu Gly Arg Phe Glu Ser Arg Val Arg Asn Thr Ala Ala His Glu
1 5 10 15
<210> 841
<211> 11
<212> PRT
<213> Mycobacterium tuberculosis
<400> 841
Ala Asp Leu Thr Leu Tyr Asp Arg Leu Asn Asp
1 5 10
<210> 842
<211> 12
<212> PRT
<213> Streptococcus thermophilus
<400> 842
Tyr Leu Met Ile Asp Val Leu Lys Glu Arg Glu His
1 5 10
<210> 843
<211> 16
<212> PRT
<213> Streptococcus thermophilus
<400> 843
Ile Glu Glu Ile Ile Lys Lys Asp His Glu Gly Leu Ile Val Phe Asp
1 5 10 15
<210> 844
<211> 9
<212> PRT
<213> Streptococcus thermophilus
<400> 844
Tyr Leu Asn Ile Leu Glu Phe Tyr Glu
1 5
<210> 845
<211> 14
<212> PRT
<213> Streptococcus thermophilus
<400> 845
Ile Leu Ser Leu Asn Gly Glu Arg Asn Lys Val Ala His Gly
1 5 10
<210> 846
<211> 13
<212> PRT
<213> Streptococcus thermophilus
<400> 846
Asp Ser Ser Tyr Phe Asn Tyr Tyr Asp Lys Gln Asn Lys
1 5 10
<210> 847
<211> 10
<212> PRT
<213> Synechocystis sp.
<400> 847
Leu Ile Ser Val Val Ala Phe Arg Leu Gly
1 5 10
<210> 848
<211> 16
<212> PRT
<213> Synechocystis sp.
<400> 848
Ile Leu Asp His Arg Lys Gln Ile Asn Phe Ala Leu Asn Asn Gly Gly
1 5 10 15
<210> 849
<211> 10
<212> PRT
<213> Synechocystis sp.
<400> 849
Thr Glu Ile Arg Asn Asp Leu Ala His Cys
1 5 10
<210> 850
<211> 12
<212> PRT
<213> Synechocystis sp.
<400> 850
Asn Lys Ile Phe Pro Gln Leu Glu Glu Ile Ala Asn
1 5 10
<210> 851
<211> 5
<212> PRT
<213> Methanocaldococcus jannaschii
<400> 851
Lys Asn Thr Leu Phe
1 5
<210> 852
<211> 14
<212> PRT
<213> Methanocaldococcus jannaschii
<400> 852
Lys Glu Asn Pro Asn Ser Gln Tyr Ile Lys Asn Glu Ile Ser
1 5 10
<210> 853
<211> 15
<212> PRT
<213> Methanocaldococcus jannaschii
<400> 853
Glu Asn Ile Asp Lys Phe Lys Ile Arg Asn Phe Leu Ala His Ala
1 5 10 15
<210> 854
<211> 13
<212> PRT
<213> Methanocaldococcus jannaschii
<400> 854
Ser Glu Lys Thr Ser Leu Arg Tyr Asn Lys Asn Tyr Ile
1 5 10
<210> 855
<211> 11
<212> PRT
<213> Pyrococcus furiosus
<400> 855
Ser Lys Ile Phe Glu Ser Leu Pro Arg Ile Gly
1 5 10
<210> 856
<211> 15
<212> PRT
<213> Pyrococcus furiosus
<400> 856
Arg Gln Val Glu Trp Leu Arg Asn Leu Val Tyr Gly Arg Leu Trp
1 5 10 15
<210> 857
<211> 15
<212> PRT
<213> Pyrococcus furiosus
<400> 857
Thr Ile Glu Ser Pro Asn Val Val Arg Asn Phe Ile Ala His Ser
1 5 10 15
<210> 858
<211> 13
<212> PRT
<213> Pyrococcus furiosus
<400> 858
Asp Lys Glu Lys Ala Ala Asn Leu Ala Tyr Glu Ala Leu
1 5 10
<210> 859
<211> 5
<212> PRT
<213> Sulfolobus solfataricus
<400> 859
Ala Glu Thr Tyr Ala
1 5
<210> 860
<211> 13
<212> PRT
<213> Sulfolobus solfataricus
<400> 860
Asp Lys Val Thr Arg Ala Ile Ile Glu Asn Glu Val Asp
1 5 10
<210> 861
<211> 13
<212> PRT
<213> Sulfolobus solfataricus
<400> 861
Gly Lys Gly Phe Asp Lys Arg Ile Leu Tyr Ala His Gly
1 5 10
<210> 862
<211> 10
<212> PRT
<213> Sulfolobus solfataricus
<400> 862
Asp Lys Ile Asp Glu Ile Glu Arg Gln Ile
1 5 10
<210> 863
<211> 12
<212> PRT
<213> Desulfococcus oleovorans
<400> 863
Phe Ala Asn Ala Glu Arg Arg Phe Asp Glu Gly Lys
1 5 10
<210> 864
<211> 16
<212> PRT
<213> Desulfococcus oleovorans
<400> 864
Val Leu Arg Leu Tyr Arg Ile Val Glu Met Ala Gly Gln Gln Arg Leu
1 5 10 15
<210> 865
<211> 9
<212> PRT
<213> Desulfococcus oleovorans
<400> 865
Gly Tyr Ser Leu Leu Lys Glu Met Gly
1 5
<210> 866
<211> 17
<212> PRT
<213> Desulfococcus oleovorans
<400> 866
Ser Phe Leu Lys Ile Gln Asp Ser Arg Asn His Ser Phe Leu Ala His
1 5 10 15
Gly
<210> 867
<211> 13
<212> PRT
<213> Desulfococcus oleovorans
<400> 867
Tyr Met Ser Leu Arg Asp Phe Ile Val Ser Leu Asn Ile
1 5 10
<210> 868
<211> 12
<212> PRT
<213> Oscillochloris trichoides
<400> 868
Leu Arg Asn Ala Glu Arg Arg Ala Ala Arg Ala Arg
1 5 10
<210> 869
<211> 16
<212> PRT
<213> Oscillochloris trichoides
<400> 869
Val Ala Arg Leu Tyr Arg Ala Thr Glu Leu Phe Ala Gln Ile Arg Leu
1 5 10 15
<210> 870
<211> 9
<212> PRT
<213> Oscillochloris trichoides
<400> 870
Ser Tyr Ala Leu Leu Gly Lys Leu Asp
1 5
<210> 871
<211> 17
<212> PRT
<213> Oscillochloris trichoides
<400> 871
Pro Leu Asn Asn Ala Leu Thr Arg Arg Asn Gln Ser Ile Leu Ala His
1 5 10 15
Gly
<210> 872
<211> 13
<212> PRT
<213> Oscillochloris trichoides
<400> 872
Tyr His Asp Leu Ala Ser His Leu Tyr Thr Leu Ile Asn
1 5 10
<210> 873
<211> 12
<212> PRT
<213> Homo sapiens
<400> 873
Phe Pro Glu Ile Phe Asp Ala Leu Glu Ser Leu Gln
1 5 10
<210> 874
<211> 14
<212> PRT
<213> Homo sapiens
<400> 874
Lys Leu Thr Ser Cys Leu Glu Arg Ala Leu Gly Asp Val Phe
1 5 10
<210> 875
<211> 9
<212> PRT
<213> Homo sapiens
<400> 875
Ser Glu Glu Leu Ala Gln Val Phe Ser
1 5
<210> 876
<211> 15
<212> PRT
<213> Homo sapiens
<400> 876
Gly Ser Pro Cys Gly Leu Asn Leu Arg Asn Val Leu Trp His Gly
1 5 10 15
<210> 877
<211> 13
<212> PRT
<213> Homo sapiens
<400> 877
Tyr Cys Ser Met Met Ile Leu Leu Thr Ala Gly Leu Gly
1 5 10
<210> 878
<211> 12
<212> PRT
<213> Entamoeba histolytica
<400> 878
Trp Phe Glu Ser Phe Gln Glu Ile Ile Gln Thr Pro
1 5 10
<210> 879
<211> 14
<212> PRT
<213> Entamoeba histolytica
<400> 879
Leu Leu Ser Val Gln Phe Asn Val His Leu Lys Asp Asn Ile
1 5 10
<210> 880
<211> 9
<212> PRT
<213> Entamoeba histolytica
<400> 880
Lys Met Tyr Glu Glu His Thr Val Pro
1 5
<210> 881
<211> 15
<212> PRT
<213> Entamoeba histolytica
<400> 881
Gly Pro Pro Thr Gly Leu Asn Leu Arg Asn Leu Leu Trp His Gly
1 5 10 15
<210> 882
<211> 13
<212> PRT
<213> Entamoeba histolytica
<400> 882
His Ile Cys Leu Leu Ile Ile Leu Tyr Gln Thr Ile Gln
1 5 10
<210> 883
<211> 12
<212> PRT
<213> Staphylococcus aureus
<400> 883
Ile Glu His Gly Ile Ser Arg Phe Leu Glu Lys Asp
1 5 10
<210> 884
<211> 14
<212> PRT
<213> Staphylococcus aureus
<400> 884
Ile Leu Val Pro Gln Phe Glu Ser Thr Val Arg Arg Met Phe
1 5 10
<210> 885
<211> 9
<212> PRT
<213> Staphylococcus aureus
<400> 885
Arg Asp Asp Val Lys Ser Thr Leu Gly
1 5
<210> 886
<211> 15
<212> PRT
<213> Staphylococcus aureus
<400> 886
Val Glu Gln Ser Gly Leu Asn Leu Arg Asn Glu Ile Ala His Gly
1 5 10 15
<210> 887
<211> 12
<212> PRT
<213> Staphylococcus aureus
<400> 887
Lys Cys Ile Leu Val Ile Tyr Leu Phe Leu Ile Leu
1 5 10
<210> 888
<211> 12
<212> PRT
<213> Cyanothece sp.
<400> 888
Leu Leu Lys Gly Ile Gln Ala Tyr Leu Glu Glu Asp
1 5 10
<210> 889
<211> 14
<212> PRT
<213> Cyanothece sp.
<400> 889
Leu Leu Ile Pro Gln Ile Glu Ala Ala Ile Arg Asn Leu Val
1 5 10
<210> 890
<211> 9
<212> PRT
<213> Cyanothece sp.
<400> 890
Ser Glu Gln Val Lys Gln Ser Leu Gly
1 5
<210> 891
<211> 15
<212> PRT
<213> Cyanothece sp.
<400> 891
Thr Asp Gln Arg Gly Trp Asn Val Arg Asn Asn Val Cys His Gly
1 5 10 15
<210> 892
<211> 12
<212> PRT
<213> Cyanothece sp.
<400> 892
Leu Thr Glu Arg Leu Ile His Ile Leu Leu Ile Leu
1 5 10
<210> 893
<211> 9
<212> PRT
<213> Sulfolobus solfataricus
<400> 893
Ile Ser Thr Ser Ala Glu Val Tyr Tyr
1 5
<210> 894
<211> 16
<212> PRT
<213> Sulfolobus solfataricus
<400> 894
Cys Glu Lys Tyr Tyr Lys Ala Ala Glu Glu Ala Ile Lys Leu Leu Val
1 5 10 15
<210> 895
<211> 8
<212> PRT
<213> Sulfolobus solfataricus
<400> 895
Lys Leu Leu Arg Ser Asn Asn Thr
1 5
<210> 896
<211> 15
<212> PRT
<213> Sulfolobus solfataricus
<400> 896
Leu Trp Lys Ser Ala Trp Thr Leu His Val Glu Gly Phe His Glu
1 5 10 15
<210> 897
<211> 13
<212> PRT
<213> Sulfolobus solfataricus
<400> 897
Leu Lys Glu Asp Val Arg Lys Leu Val Ile Phe Ala Val
1 5 10
<210> 898
<211> 9
<212> PRT
<213> Pyrobaculum aerophilum
<400> 898
Tyr Ala Glu Ala Ala Arg Glu Leu Leu
1 5
<210> 899
<211> 16
<212> PRT
<213> Pyrobaculum aerophilum
<400> 899
Ser Glu Lys Ala Trp Gly Ala Ala Ala Leu Ala Val Lys Ala Tyr Ala
1 5 10 15
<210> 900
<211> 7
<212> PRT
<213> Pyrobaculum aerophilum
<400> 900
Lys Ile Ala Gly Glu Leu Gly
1 5
<210> 901
<211> 14
<212> PRT
<213> Pyrobaculum aerophilum
<400> 901
Ala Trp Ala Gln Ala Asn Ala Met His Ile Asn Phe Tyr Glu
1 5 10
<210> 902
<211> 13
<212> PRT
<213> Pyrobaculum aerophilum
<400> 902
Ala Leu Lys Lys Val Ser Arg Leu Val Glu Glu Leu Thr
1 5 10
<210> 903
<211> 11
<212> PRT
<213> Homo sapiens
<400> 903
Arg Arg Trp Leu Arg Gln Ala Arg Ala Asn Phe
1 5 10
<210> 904
<211> 16
<212> PRT
<213> Homo sapiens
<400> 904
Asn Glu Trp Val Cys Phe Lys Cys Tyr Leu Ser Thr Lys Leu Ala Leu
1 5 10 15
<210> 905
<211> 8
<212> PRT
<213> Homo sapiens
<400> 905
Ala Gln Lys Ile Glu Glu Tyr Ser
1 5
<210> 906
<211> 15
<212> PRT
<213> Homo sapiens
<400> 906
Val His Thr Leu Glu Ala Tyr Gly Val Asp Ser Leu Lys Thr Arg
1 5 10 15
<210> 907
<211> 13
<212> PRT
<213> Homo sapiens
<220>
<221> MOD_RES
<222> (2)..(2)
<223> Any amino acid
<400> 907
Val Xaa Glu Cys Thr Ala Cys Ile Ile Ile Lys Leu Glu
1 5 10
<210> 908
<211> 11
<212> PRT
<213> Heamophilus influenzae
<400> 908
Lys Leu Asn Leu Asn Val Leu Asp Ala Ala Phe
1 5 10
<210> 909
<211> 16
<212> PRT
<213> Heamophilus influenzae
<220>
<221> MOD_RES
<222> (14)..(15)
<223> Any amino acid
<400> 909
Ile Gln Lys Phe Glu Phe Val Tyr Glu Leu Ser Leu Lys Xaa Xaa Lys
1 5 10 15
<210> 910
<211> 8
<212> PRT
<213> Heamophilus influenzae
<400> 910
Leu Arg Glu Ala Leu Arg Phe Gly
1 5
<210> 911
<211> 15
<212> PRT
<213> Heamophilus influenzae
<220>
<221> MOD_RES
<222> (8)..(8)
<223> Any amino acid
<400> 911
Lys Trp Val Ala Tyr Arg Asp Xaa Arg Asn Ile Thr Ser His Thr
1 5 10 15
<210> 912
<211> 13
<212> PRT
<213> Heamophilus influenzae
<400> 912
Asp Phe Leu Ile Glu Ser Ser Phe Leu Leu Glu Gln Leu
1 5 10
<210> 913
<211> 6
<212> PRT
<213> Thermus thermophilus
<220>
<221> MOD_RES
<222> (1)..(1)
<223> Any amino acid
<400> 913
Xaa Ala Glu Lys Ala Leu
1 5
<210> 914
<211> 16
<212> PRT
<213> Thermus thermophilus
<400> 914
Ile Gln Arg Phe Glu Tyr Thr Phe Glu Ala Phe Trp Lys Ala Leu Gln
1 5 10 15
<210> 915
<211> 8
<212> PRT
<213> Thermus thermophilus
<400> 915
Ile Arg Leu Ala Arg Glu Val Gly
1 5
<210> 916
<211> 15
<212> PRT
<213> Thermus thermophilus
<220>
<221> MOD_RES
<222> (5)..(5)
<223> Any amino acid
<400> 916
Leu Ala Leu Gly Xaa Val Asp Asp Arg Ser Leu Thr Val His Thr
1 5 10 15
<210> 917
<211> 13
<212> PRT
<213> Thermus thermophilus
<220>
<221> MOD_RES
<222> (12)..(12)
<223> Any amino acid
<400> 917
Ile Phe Arg Arg Leu Pro Asp Tyr Ala Arg Leu Xaa Glu
1 5 10
<210> 918
<211> 11
<212> PRT
<213> Rhodococcus equi
<400> 918
Val Asn Leu Leu Arg Arg Ala Asp Gly Leu Leu
1 5 10
<210> 919
<211> 16
<212> PRT
<213> Rhodococcus equi
<400> 919
Phe Cys Ala Ala Tyr Val Gly Ala Leu Arg Gly Ala Ala Ala Val Leu
1 5 10 15
<210> 920
<211> 8
<212> PRT
<213> Rhodococcus equi
<400> 920
Trp Val Leu Met Ala Arg Ala Glu
1 5
<210> 921
<211> 15
<212> PRT
<213> Rhodococcus equi
<400> 921
Tyr Phe Ala Gly Tyr Ser Gly Leu Arg Ala Asp Leu Glu Ala Gly
1 5 10 15
<210> 922
<211> 13
<212> PRT
<213> Rhodococcus equi
<400> 922
Asp Ala Glu Glu Val Asp Gly Phe Tyr Ala Glu Val Gly
1 5 10
<210> 923
<211> 11
<212> PRT
<213> Streptomyces avermitilis
<400> 923
Leu Asp Leu Leu Ala Gln Ala Arg Ala Gly Leu
1 5 10
<210> 924
<211> 16
<212> PRT
<213> Streptomyces avermitilis
<400> 924
Tyr Ala Thr Ala His Leu Ala Ala Leu Arg Thr Ala Ala Ala Val Leu
1 5 10 15
<210> 925
<211> 8
<212> PRT
<213> Streptomyces avermitilis
<400> 925
Trp Glu Val Leu Pro Glu Ile Ala
1 5
<210> 926
<211> 15
<212> PRT
<213> Streptomyces avermitilis
<400> 926
Leu Phe Ala Ser Gly Ala Gly Arg Arg Ala Arg Ala Glu Ala Gly
1 5 10 15
<210> 927
<211> 13
<212> PRT
<213> Streptomyces avermitilis
<400> 927
Ser Asn Arg Asp Ala Asp Asp Leu Ile Arg Asp Val Ala
1 5 10
<210> 928
<211> 11
<212> PRT
<213> Staphylococcus aureus
<400> 928
Ala Leu Ile Val Glu Glu Leu Phe Glu Tyr Ala
1 5 10
<210> 929
<211> 16
<212> PRT
<213> Staphylococcus aureus
<400> 929
Pro Ser Leu Thr Val Gln Val Ala Met Ala Gly Ala Met Leu Ile Gly
1 5 10 15
<210> 930
<211> 8
<212> PRT
<213> Staphylococcus aureus
<400> 930
Thr Glu Ala Val Lys Gln Ser Asp
1 5
<210> 931
<211> 10
<212> PRT
<213> Staphylococcus aureus
<400> 931
His Leu Cys Gln Phe Val Met Ser Gly Gln
1 5 10
<210> 932
<211> 13
<212> PRT
<213> Staphylococcus aureus
<400> 932
Ser Glu Lys Leu Leu Glu Ser Leu Glu Asn Phe Trp Asn
1 5 10
<210> 933
<211> 11
<212> PRT
<213> Enterococcus faecium
<400> 933
Asn Phe Leu Leu Cys Asn Phe Ser Asn Leu Trp
1 5 10
<210> 934
<211> 16
<212> PRT
<213> Enterococcus faecium
<400> 934
Leu Glu Leu Leu Ser Gln Leu Gln Lys Asn Thr Leu Gln Leu Ile Arg
1 5 10 15
<210> 935
<211> 10
<212> PRT
<213> Enterococcus faecium
<400> 935
Lys Lys Phe Ala Lys Thr Thr Ala Arg Leu
1 5 10
<210> 936
<211> 13
<212> PRT
<213> Enterococcus faecium
<400> 936
Lys Val Glu Leu Phe Glu Ala Tyr Lys Asn Ser Leu Leu
1 5 10
<210> 937
<211> 9
<212> PRT
<213> Escherichia coli
<400> 937
Gly Val Tyr Ala Asn Glu Leu Arg Ala
1 5
<210> 938
<211> 16
<212> PRT
<213> Escherichia coli
<400> 938
Gly Gly Ile Arg Glu Ile Glu Phe Ile Val Gln Val Phe Gln Leu Ile
1 5 10 15
<210> 939
<211> 8
<212> PRT
<213> Escherichia coli
<400> 939
Thr Leu Ser Ala Ile Ala Glu Leu
1 5
<210> 940
<211> 15
<212> PRT
<213> Escherichia coli
<400> 940
Glu Gln Leu Arg Val Ala Tyr Leu Phe Leu Arg Arg Leu Glu Asn
1 5 10 15
<210> 941
<211> 13
<212> PRT
<213> Escherichia coli
<400> 941
Leu Thr Gly His Met Thr Asn Val Arg Arg Val Phe Asn
1 5 10
<210> 942
<211> 11
<212> PRT
<213> Sebaldella termitidis
<400> 942
Ser Arg Cys Met Lys Ile Ala Gln Ser Gly Gln
1 5 10
<210> 943
<211> 16
<212> PRT
<213> Sebaldella termitidis
<400> 943
Ile Ala Glu Ala Glu Phe Ile Asn Glu Ser Ile Tyr Met Ile Tyr Leu
1 5 10 15
<210> 944
<211> 8
<212> PRT
<213> Sebaldella termitidis
<400> 944
Lys Asp Met Gln Phe Leu Pro Ile
1 5
<210> 945
<211> 14
<212> PRT
<213> Sebaldella termitidis
<400> 945
Asn Leu Leu Asn Asn Leu Ile Ser Ile Gln Asn Ser Glu Lys
1 5 10
<210> 946
<211> 13
<212> PRT
<213> Sebaldella termitidis
<400> 946
Ala Glu Lys Ile Cys Gly Leu Ile Ile Asn Glu Leu Lys
1 5 10
<210> 947
<211> 10
<212> PRT
<213> Streptomyces coelicolor
<400> 947
Ala Arg Leu Asp Ala Tyr Ala Asn Ser His
1 5 10
<210> 948
<211> 16
<212> PRT
<213> Streptomyces coelicolor
<400> 948
Leu Asp Ala Ala Asp Ser Ile Gly Phe Leu Leu Glu Leu Leu Phe Ala
1 5 10 15
<210> 949
<211> 8
<212> PRT
<213> Streptomyces coelicolor
<400> 949
Trp Glu Leu Asp Arg Phe Pro Leu
1 5
<210> 950
<211> 14
<212> PRT
<213> Streptomyces coelicolor
<400> 950
Glu Leu Leu Ala Thr Leu Gly Arg Ile Thr Gly Ala Gly Gly
1 5 10
<210> 951
<211> 13
<212> PRT
<213> Streptomyces coelicolor
<400> 951
Gln Arg Glu Leu Phe Gly Arg Val Glu Ala Ala Ala Arg
1 5 10
<210> 952
<211> 11
<212> PRT
<213> Flavobacterium psychrophilum
<400> 952
Tyr Ser Ile Tyr Lys Asn Ala Arg Gln Leu Arg
1 5 10
<210> 953
<211> 16
<212> PRT
<213> Flavobacterium psychrophilum
<400> 953
Thr Ser Leu Leu Ile Leu Ser Ser Glu Glu Val Ile Lys Ser Ile Leu
1 5 10 15
<210> 954
<211> 8
<212> PRT
<213> Flavobacterium psychrophilum
<400> 954
Gln Leu Ile Glu Leu Ser Ile Gly
1 5
<210> 955
<211> 15
<212> PRT
<213> Flavobacterium psychrophilum
<400> 955
Lys Leu Thr Glu Phe Asp Asp Lys Lys Asn Gln Gly Phe Tyr Val
1 5 10 15
<210> 956
<211> 13
<212> PRT
<213> Flavobacterium psychrophilum
<400> 956
Lys Thr Glu Phe Thr Glu Thr Lys Val Val Val Asp Arg
1 5 10
<210> 957
<211> 10
<212> PRT
<213> Lactococcus lactis
<400> 957
Lys Cys Ile Asp His Ile Ser Val Leu Ile
1 5 10
<210> 958
<211> 16
<212> PRT
<213> Lactococcus lactis
<400> 958
Thr Phe Ile Ser Ile Thr Ile Ile Glu Glu Val Gly Lys Thr His Ile
1 5 10 15
<210> 959
<211> 8
<212> PRT
<213> Lactococcus lactis
<400> 959
Ser Leu Pro Thr Ile Lys Met Gly
1 5
<210> 960
<211> 14
<212> PRT
<213> Lactococcus lactis
<400> 960
Thr Gly Glu Leu Ile Ser Ile Arg Glu Ser Ser Leu Tyr Ala
1 5 10
<210> 961
<211> 13
<212> PRT
<213> Lactococcus lactis
<400> 961
Lys Glu Gln Ser Arg Ala Leu Leu Leu Tyr Ala Ile Glu
1 5 10
<210> 962
<211> 11
<212> PRT
<213> Pseudomonas putida
<400> 962
Asp Ala Leu Leu Thr Asn Ala Ala Ser Leu Ile
1 5 10
<210> 963
<211> 16
<212> PRT
<213> Pseudomonas putida
<400> 963
Phe Ala Leu Ala His Leu Ala Arg Glu Glu Ile Ala Lys Thr Leu Met
1 5 10 15
<210> 964
<211> 8
<212> PRT
<213> Pseudomonas putida
<400> 964
Thr Ile Asn Ser Ile Val Phe Cys
1 5
<210> 965
<211> 12
<212> PRT
<213> Pseudomonas putida
<400> 965
Phe Arg Asn Asp Leu Lys Asn Asn Ser Leu Tyr Val
1 5 10
<210> 966
<211> 13
<212> PRT
<213> Pseudomonas putida
<400> 966
Ala Glu Arg Ala Leu Arg Thr Ile Thr Leu Ala Trp Asp
1 5 10
<210> 967
<211> 11
<212> PRT
<213> Selenomonas sputigena
<400> 967
Gln Ile Ala Tyr Tyr Leu Tyr Phe Met Tyr Leu
1 5 10
<210> 968
<211> 16
<212> PRT
<213> Selenomonas sputigena
<400> 968
Met Thr Ser Phe Ala Tyr Tyr Lys Ser Tyr Phe Asp Arg Val Thr Ala
1 5 10 15
<210> 969
<211> 10
<212> PRT
<213> Selenomonas sputigena
<400> 969
Arg Leu Cys Glu Phe Tyr Glu Glu Phe Asp
1 5 10
<210> 970
<211> 17
<212> PRT
<213> Selenomonas sputigena
<400> 970
Ile Ile Asp Lys Ala Gln Ala Leu Arg Tyr Ala Asn Pro Leu Thr His
1 5 10 15
Ser
<210> 971
<211> 13
<212> PRT
<213> Selenomonas sputigena
<400> 971
Ile Arg Glu Leu Ser Thr Leu Leu Asp Arg Tyr Ile Ala
1 5 10
<210> 972
<211> 11
<212> PRT
<213> Lactobacillus helveticus
<400> 972
Trp Ile Ser Tyr Tyr Leu Tyr Phe Glu Ser Ile
1 5 10
<210> 973
<211> 16
<212> PRT
<213> Lactobacillus helveticus
<400> 973
Leu Thr Ser Tyr Ala Phe Phe Lys Asn Tyr Phe Asp Arg Thr Thr Ala
1 5 10 15
<210> 974
<211> 10
<212> PRT
<213> Lactobacillus helveticus
<400> 974
Gln Leu Gln Lys Val Tyr Arg Ile Leu Asn
1 5 10
<210> 975
<211> 17
<212> PRT
<213> Lactobacillus helveticus
<400> 975
Ile Ile Ser Lys Ala Asn Asp Leu Arg Asn Asn Asn Pro Leu Ser His
1 5 10 15
Ala
<210> 976
<211> 13
<212> PRT
<213> Lactobacillus helveticus
<400> 976
Ile Ala Thr Met Arg Ser Leu Phe Lys Leu Leu Val Glu
1 5 10
<210> 977
<211> 11
<212> PRT
<213> Lactococcus lactis
<400> 977
Lys Ile Leu Asn Phe Ile Tyr Phe Arg Ala Lys
1 5 10
<210> 978
<211> 16
<212> PRT
<213> Lactococcus lactis
<400> 978
Leu Glu Ser Phe Ala Tyr Tyr Lys Asn Tyr Phe Asp Arg Phe Val Ala
1 5 10 15
<210> 979
<211> 10
<212> PRT
<213> Lactococcus lactis
<400> 979
Lys Leu Ile Asp Gly Leu Lys Gln Leu Asn
1 5 10
<210> 980
<211> 17
<212> PRT
<213> Lactococcus lactis
<400> 980
Ile Ile Asn Glu Ala His Lys Ile Arg Asn Ser Asn Pro Val Ser His
1 5 10 15
Ser
<210> 981
<211> 13
<212> PRT
<213> Lactococcus lactis
<400> 981
Leu Asn Asp Leu Lys Ile Ile Ile Glu Gln Leu Ser Thr
1 5 10
<210> 982
<211> 11
<212> PRT
<213> Pseudomonas sp.
<400> 982
Lys Trp Leu Phe Ile Asp Gln Met Val Asp Leu
1 5 10
<210> 983
<211> 12
<212> PRT
<213> Pseudomonas sp.
<400> 983
Phe Lys Phe Arg Glu Ile Arg Ile Glu Tyr Ser Gln
1 5 10
<210> 984
<211> 9
<212> PRT
<213> Pseudomonas sp.
<400> 984
Tyr Glu Tyr Ala Gln Glu Ile Arg Ser
1 5
<210> 985
<211> 14
<212> PRT
<213> Pseudomonas sp.
<400> 985
Arg Lys Ile Pro Asp Phe Arg Gly Lys Tyr Ala Ala His Ile
1 5 10
<210> 986
<211> 13
<212> PRT
<213> Pseudomonas sp.
<400> 986
Lys Ala Leu Glu Phe Tyr Asn Trp Ile His Ser Asn Glu
1 5 10
<210> 987
<211> 11
<212> PRT
<213> Vibrio paracholerae
<400> 987
Glu Glu Ile Leu Ser Gly Leu Ile Gly Asp Leu
1 5 10
<210> 988
<211> 12
<212> PRT
<213> Vibrio paracholerae
<400> 988
Arg Lys Tyr Val Glu Leu Asn Gln Lys Tyr Gly Lys
1 5 10
<210> 989
<211> 10
<212> PRT
<213> Vibrio paracholerae
<400> 989
Gly Val Tyr Asn Asn Glu Ile Asn Lys Asn
1 5 10
<210> 990
<211> 14
<212> PRT
<213> Vibrio paracholerae
<400> 990
Thr Ala Ile Lys Lys Leu Arg Asn His Cys Val Ala His Val
1 5 10
<210> 991
<211> 13
<212> PRT
<213> Vibrio paracholerae
<400> 991
Phe Ala Asp Glu Phe Leu Asp Trp Ile Cys Pro Asp Asn
1 5 10
<210> 992
<211> 11
<212> PRT
<213> Escherichia coli
<400> 992
Thr Met Ala Asp His Met Val Asn Glu Ala Trp
1 5 10
<210> 993
<211> 16
<212> PRT
<213> Escherichia coli
<400> 993
Phe Asn Leu Ile Leu Gln Ser Ile Glu Phe Arg Leu Lys Gly Leu Ile
1 5 10 15
<210> 994
<211> 10
<212> PRT
<213> Escherichia coli
<400> 994
Lys Val Tyr Asn Thr Phe Ala Ser Lys Ser
1 5 10
<210> 995
<211> 14
<212> PRT
<213> Escherichia coli
<400> 995
Trp Phe Asn Ser Met Arg Ile Leu Arg Asn Arg Phe Met His
1 5 10
<210> 996
<211> 13
<212> PRT
<213> Escherichia coli
<400> 996
Asp Ile Met Pro Glu Leu Ile Phe Thr Ser Val Val Arg
1 5 10
<210> 997
<211> 11
<212> PRT
<213> Geobacter sulfurreducens
<400> 997
Leu Asn Tyr Glu Ala Leu Tyr Val Lys Ser Lys
1 5 10
<210> 998
<211> 16
<212> PRT
<213> Geobacter sulfurreducens
<400> 998
Gln Leu Trp Ala Ser Met Ala Leu Glu Leu Leu Ala Lys Ser Ser Leu
1 5 10 15
<210> 999
<211> 9
<212> PRT
<213> Geobacter sulfurreducens
<400> 999
Gln Arg Leu Gly His Ile Ser Lys Leu
1 5
<210> 1000
<211> 15
<212> PRT
<213> Geobacter sulfurreducens
<400> 1000
Phe Cys Glu Gln Leu Ser Leu Arg Arg Asn Ser Glu Ile His Ser
1 5 10 15
<210> 1001
<211> 13
<212> PRT
<213> Geobacter sulfurreducens
<400> 1001
Asp Ala Trp Glu Val Lys Tyr Trp Tyr Ala Ile Glu Val
1 5 10
<210> 1002
<211> 11
<212> PRT
<213> Streptomyces coelicolor
<400> 1002
Asp Val Ser Tyr Thr Pro Val Ser Asn Gly Met
1 5 10
<210> 1003
<211> 16
<212> PRT
<213> Streptomyces coelicolor
<400> 1003
Val Leu His Leu Gln Ala Ala Thr Glu Val Leu Leu Lys Ala Arg Leu
1 5 10 15
<210> 1004
<211> 10
<212> PRT
<213> Streptomyces coelicolor
<400> 1004
Asp Arg Leu Arg Asp Ile Ala Arg Leu Asp
1 5 10
<210> 1005
<211> 15
<212> PRT
<213> Streptomyces coelicolor
<400> 1005
Arg Ile Lys Glu Pro Gly Glu Ser Arg Asn Ala Leu Gln His Tyr
1 5 10 15
<210> 1006
<211> 13
<212> PRT
<213> Streptomyces coelicolor
<400> 1006
Tyr Ala Ile Glu Ser Arg Ala Ala Arg Val Leu Asp Phe
1 5 10
<210> 1007
<211> 10
<212> PRT
<213> Leptospira interrogans
<400> 1007
Cys Thr Arg Leu Tyr Asn Gln Ile Leu Glu
1 5 10
<210> 1008
<211> 16
<212> PRT
<213> Leptospira interrogans
<400> 1008
Tyr Thr Lys Leu Phe Asn Ile Leu Asp Lys Val Ala Ala Ile Val Tyr
1 5 10 15
<210> 1009
<211> 6
<212> PRT
<213> Leptospira interrogans
<400> 1009
Phe Pro Ser Thr Phe Gly
1 5
<210> 1010
<211> 13
<212> PRT
<213> Leptospira interrogans
<400> 1010
His His Leu Arg Val Arg Arg Asn Asn Ile Val His Trp
1 5 10
<210> 1011
<211> 13
<212> PRT
<213> Leptospira interrogans
<400> 1011
Glu Glu Asp Val Gln Arg Leu Phe Leu Ile Ser Lys Ala
1 5 10
<210> 1012
<211> 10
<212> PRT
<213> Shigella boydii
<400> 1012
Met Glu Met Val Leu Asn Arg Leu Lys Ser
1 5 10
<210> 1013
<211> 16
<212> PRT
<213> Shigella boydii
<400> 1013
Phe Arg Leu Cys Phe Gly Ile Leu Asp Lys Ile Ala Val Ala Ile Cys
1 5 10 15
<210> 1014
<211> 8
<212> PRT
<213> Shigella boydii
<400> 1014
Pro Gln Lys Asn Ile Tyr Phe Gln
1 5
<210> 1015
<211> 15
<212> PRT
<213> Shigella boydii
<400> 1015
Glu Leu Ala Phe Tyr Lys Glu Trp Arg Asn Gly Leu Glu His Lys
1 5 10 15
<210> 1016
<211> 13
<212> PRT
<213> Shigella boydii
<400> 1016
Ile His His Phe Glu His Leu Leu Gln Ile Thr Arg Ser
1 5 10
<210> 1017
<211> 10
<212> PRT
<213> Enterococcus faecalis
<400> 1017
Phe Tyr Ser Leu Phe Asn Gln Ile Lys Gln
1 5 10
<210> 1018
<211> 16
<212> PRT
<213> Enterococcus faecalis
<400> 1018
Tyr Arg Ser Val Tyr Ser Ile Phe Asp Lys Ile Ala Tyr Phe Leu Asn
1 5 10 15
<210> 1019
<211> 8
<212> PRT
<213> Enterococcus faecalis
<400> 1019
Pro Lys Asn Leu Ile Thr Phe His
1 5
<210> 1020
<211> 15
<212> PRT
<213> Enterococcus faecalis
<400> 1020
Asn Leu Glu Lys Ile Ala Glu Ile Arg Asn Ala Met Glu His Lys
1 5 10 15
<210> 1021
<211> 13
<212> PRT
<213> Enterococcus faecalis
<400> 1021
Glu Lys Ile Thr Leu Glu Leu Phe Lys Leu Thr Arg Glu
1 5 10
<210> 1022
<211> 10
<212> PRT
<213> Clostridium thermocellum
<400> 1022
Phe Asn Asn Arg Ala Phe Asp Leu Ile Val
1 5 10
<210> 1023
<211> 16
<212> PRT
<213> Clostridium thermocellum
<400> 1023
Tyr Thr Arg Phe Glu Gly Leu Ile Asp Thr Ile Tyr His Ile Ile Asn
1 5 10 15
<210> 1024
<211> 7
<212> PRT
<213> Clostridium thermocellum
<400> 1024
Lys Pro Ser Ser Glu Phe Arg
1 5
<210> 1025
<211> 15
<212> PRT
<213> Clostridium thermocellum
<400> 1025
Val Tyr Lys Lys Ile Asn Lys Phe Arg Asn Asn Ile Val His Asn
1 5 10 15
<210> 1026
<211> 13
<212> PRT
<213> Clostridium thermocellum
<400> 1026
Tyr Thr Thr Ser Thr Glu Phe Leu Asn Asn Ile Lys Asp
1 5 10
<210> 1027
<211> 10
<212> PRT
<213> Bacillus cereus
<400> 1027
Leu Asn Asn Arg Ile Phe Gln Leu Asp Leu
1 5 10
<210> 1028
<211> 16
<212> PRT
<213> Bacillus cereus
<400> 1028
Phe Pro Lys Ala Phe Thr Ala Leu Asp Leu Leu Ala His Leu Leu Phe
1 5 10 15
<210> 1029
<211> 7
<212> PRT
<213> Bacillus cereus
<400> 1029
Lys Thr Glu Lys Lys Ile Lys
1 5
<210> 1030
<211> 15
<212> PRT
<213> Bacillus cereus
<400> 1030
Glu Phe Gln Lys Ala Ser Lys Val Arg Asn Asp Ile Ile His Asn
1 5 10 15
<210> 1031
<211> 13
<212> PRT
<213> Bacillus cereus
<400> 1031
Tyr Thr Pro Ser Lys Glu Ile Leu Asn Ile Ala Arg Gly
1 5 10
<210> 1032
<211> 10
<212> PRT
<213> Pseudomonas syringae
<400> 1032
Glu Tyr Leu Arg Cys Lys Asp Ala Phe Glu
1 5 10
<210> 1033
<211> 16
<212> PRT
<213> Pseudomonas syringae
<400> 1033
Ser Ser Phe Ile His His Leu Tyr Glu Leu Tyr Met Ala Leu Phe Ala
1 5 10 15
<210> 1034
<211> 8
<212> PRT
<213> Pseudomonas syringae
<400> 1034
Ser Ile Asp Arg Gly Ala Val Ser
1 5
<210> 1035
<211> 16
<212> PRT
<213> Pseudomonas syringae
<400> 1035
Phe Gly Pro Ala Phe Arg Ser Met Arg Asn Lys Ile Ala Gly His Val
1 5 10 15
<210> 1036
<211> 13
<212> PRT
<213> Pseudomonas syringae
<400> 1036
Val Lys Leu Thr Glu Phe Phe Gln Lys Tyr His Pro Tyr
1 5 10
<210> 1037
<211> 10
<212> PRT
<213> Burkholderia xenovorans
<400> 1037
Glu Tyr Leu Arg Cys Asp Asp Ala Leu His
1 5 10
<210> 1038
<211> 16
<212> PRT
<213> Burkholderia xenovorans
<400> 1038
Ala Arg Phe Ile His His Leu Tyr Glu Phe Asn Ile Ala Cys Ala Gln
1 5 10 15
<210> 1039
<211> 8
<212> PRT
<213> Burkholderia xenovorans
<400> 1039
Arg Val Arg Arg Gln Ala Tyr Asn
1 5
<210> 1040
<211> 16
<212> PRT
<213> Burkholderia xenovorans
<400> 1040
Phe Ala Lys Ala Phe Arg Thr Ala Arg Asn Thr Thr Asn Gly His Ala
1 5 10 15
<210> 1041
<211> 13
<212> PRT
<213> Burkholderia xenovorans
<400> 1041
Leu Asn Leu Ser Asp Phe Phe Thr Arg Tyr His Arg Phe
1 5 10
<210> 1042
<211> 10
<212> PRT
<213> Microcystis aeruginosa
<400> 1042
Glu His Leu Asp Cys Glu Leu Trp Glu Arg
1 5 10
<210> 1043
<211> 16
<212> PRT
<213> Microcystis aeruginosa
<400> 1043
Ile Arg Asn Ala Thr Val Ile Leu Glu Asp Arg Met Arg Lys Leu Gly
1 5 10 15
<210> 1044
<211> 8
<212> PRT
<213> Microcystis aeruginosa
<400> 1044
Gly Ile Val Asn Leu Ile Phe Gly
1 5
<210> 1045
<211> 15
<212> PRT
<213> Microcystis aeruginosa
<400> 1045
Tyr Ser Gly Thr Met Lys Ile Phe Arg Asn Arg Tyr Ala His Arg
1 5 10 15
<210> 1046
<211> 13
<212> PRT
<213> Microcystis aeruginosa
<400> 1046
Ile Ile Val Phe Ile Asp Leu Leu Leu Lys Met Leu Asp
1 5 10
<210> 1047
<211> 11
<212> PRT
<213> Vibrio parahaemolyticus
<400> 1047
Ser Arg Asn Val His Pro Asp Val Leu Lys Tyr
1 5 10
<210> 1048
<211> 16
<212> PRT
<213> Vibrio parahaemolyticus
<400> 1048
Val Phe Glu Ala Thr Lys Ser Val Ala Asp Lys Ile Arg Asn Lys Thr
1 5 10 15
<210> 1049
<211> 8
<212> PRT
<213> Vibrio parahaemolyticus
<400> 1049
Val Leu Val Asp Glu Ala Phe Ser
1 5
<210> 1050
<211> 15
<212> PRT
<213> Vibrio parahaemolyticus
<400> 1050
Leu Lys Gly Leu Phe Gly Thr Phe Arg Asn Thr Thr Ala His Ala
1 5 10 15
<210> 1051
<211> 13
<212> PRT
<213> Vibrio parahaemolyticus
<400> 1051
Ile Leu Ser Met Val Ser Leu Val His Arg Arg Leu Asp
1 5 10
<210> 1052
<211> 11
<212> PRT
<213> Lactococcus lactis
<400> 1052
Ala Leu Glu Leu His Ser Glu Val Thr Lys Tyr
1 5 10
<210> 1053
<211> 16
<212> PRT
<213> Lactococcus lactis
<400> 1053
Val Phe Glu Ser Cys Lys Gly Leu Phe Asp Arg Ile Arg Leu Ile Ser
1 5 10 15
<210> 1054
<211> 8
<212> PRT
<213> Lactococcus lactis
<400> 1054
Thr Leu Ile Asn Gln Ala Phe Asn
1 5
<210> 1055
<211> 15
<212> PRT
<213> Lactococcus lactis
<400> 1055
Ile Lys Thr Cys Leu Tyr Leu Tyr Arg Asn His Gln Ala His Val
1 5 10 15
<210> 1056
<211> 13
<212> PRT
<213> Lactococcus lactis
<400> 1056
Gly Leu Met Ser Ile Ser Leu Ala His Glu Leu Leu Asp
1 5 10
<210> 1057
<211> 11
<212> PRT
<213> Nematostella vectensis
<400> 1057
Ser Thr Thr Leu Thr Thr Phe Leu Asn Leu His
1 5 10
<210> 1058
<211> 16
<212> PRT
<213> Nematostella vectensis
<400> 1058
Glu Asp Tyr Asp Ile Thr Leu Leu Thr Cys Leu Leu Arg Asn Ile Cys
1 5 10 15
<210> 1059
<211> 8
<212> PRT
<213> Nematostella vectensis
<400> 1059
Asp Lys Leu Pro Pro Ala Tyr Asp
1 5
<210> 1060
<211> 15
<212> PRT
<213> Nematostella vectensis
<400> 1060
Val Val Arg Leu Arg His Tyr Arg Asn Asp Leu Tyr Ala His Ile
1 5 10 15
<210> 1061
<211> 13
<212> PRT
<213> Nematostella vectensis
<400> 1061
Trp Ala Asp Ile Ser Ala Ala Leu Leu Ser Leu Gly Gly
1 5 10
<210> 1062
<211> 11
<212> PRT
<213> Branchiostoma floridae
<400> 1062
Pro Pro Ser Leu Pro Ala Gln Leu Lys Lys His
1 5 10
<210> 1063
<211> 16
<212> PRT
<213> Branchiostoma floridae
<400> 1063
Glu Glu Phe Asp Ile Ser Leu Leu Leu Leu Leu Leu Lys Glu Leu Val
1 5 10 15
<210> 1064
<211> 8
<212> PRT
<213> Branchiostoma floridae
<400> 1064
Gly Arg Asp Ala Pro Tyr Ser Asp
1 5
<210> 1065
<211> 13
<212> PRT
<213> Branchiostoma floridae
<400> 1065
Lys Leu Gly Gln Phe Arg Asn Lys Asn Tyr Gly His Ile
1 5 10
<210> 1066
<211> 13
<212> PRT
<213> Branchiostoma floridae
<400> 1066
Trp Asp Glu Leu Thr Glu Ile Leu Val Asp Leu Gly Gly
1 5 10
<210> 1067
<211> 11
<212> PRT
<213> Homo sapiens
<400> 1067
Pro Pro Leu Leu Lys Lys Glu Leu Leu Ile His
1 5 10
<210> 1068
<211> 16
<212> PRT
<213> Homo sapiens
<400> 1068
Lys Gln Phe Asp Leu Cys Leu Leu Leu Ala Leu Ile Lys His Leu Asn
1 5 10 15
<210> 1069
<211> 8
<212> PRT
<213> Homo sapiens
<400> 1069
Asn Met Glu Pro Pro Ser Ser Asp
1 5
<210> 1070
<211> 15
<212> PRT
<213> Homo sapiens
<400> 1070
Ile Leu Arg Leu Cys Lys Tyr Arg Asp Ile Leu Leu Ser Glu Ile
1 5 10 15
<210> 1071
<211> 13
<212> PRT
<213> Homo sapiens
<400> 1071
Trp Lys Lys Val Ser Asp Ile Leu Leu Arg Leu Gly Met
1 5 10
<210> 1072
<211> 11
<212> PRT
<213> Escherichia coli
<400> 1072
Val Thr Ala Glu Lys Leu Leu Val Ser Gly Leu
1 5 10
<210> 1073
<211> 16
<212> PRT
<213> Escherichia coli
<400> 1073
Leu Tyr Pro Glu Leu Arg Thr Ile Glu Gly Val Leu Lys Ser Lys Met
1 5 10 15
<210> 1074
<211> 8
<212> PRT
<213> Escherichia coli
<400> 1074
Tyr Ile Leu Lys Pro Gln Phe Ala
1 5
<210> 1075
<211> 14
<212> PRT
<213> Escherichia coli
<400> 1075
Ala Tyr Thr Phe Phe Asn Val Glu Arg His Ser Leu Phe His
1 5 10
<210> 1076
<211> 13
<212> PRT
<213> Escherichia coli
<400> 1076
Met Ile Ser Asp Met Ala Arg Leu Met Gly Lys Ala Thr
1 5 10
<210> 1077
<211> 11
<212> PRT
<213> Photobacterium profundum
<400> 1077
Asp Thr Tyr Arg Ser Leu Leu Ser Ser Ser Tyr
1 5 10
<210> 1078
<211> 16
<212> PRT
<213> Photobacterium profundum
<400> 1078
Ile Tyr Pro Asp Leu Arg Val Leu Glu Gly Val Ile Lys Glu Ala Met
1 5 10 15
<210> 1079
<211> 8
<212> PRT
<213> Photobacterium profundum
<400> 1079
Thr Glu Leu Lys Thr Glu Tyr Asn
1 5
<210> 1080
<211> 14
<212> PRT
<213> Photobacterium profundum
<400> 1080
Cys Tyr Ala Tyr Phe Lys Ala His Arg His Ser Leu Phe His
1 5 10
<210> 1081
<211> 13
<212> PRT
<213> Photobacterium profundum
<400> 1081
Thr Thr Asp Thr Ile Gly Glu Val Met Gln Met Ser Glu
1 5 10
<210> 1082
<211> 11
<212> PRT
<213> Geobacillus thermoglucosidasius
<400> 1082
Leu Tyr Asp Arg Asp Arg Ile Glu Ala Ser Glu
1 5 10
<210> 1083
<211> 16
<212> PRT
<213> Geobacillus thermoglucosidasius
<400> 1083
Val Ser Gly Thr Leu Arg Ala Phe Glu Gly Phe Phe Lys Lys Leu Leu
1 5 10 15
<210> 1084
<211> 8
<212> PRT
<213> Geobacillus thermoglucosidasius
<400> 1084
Asp Ile Ser Glu Lys Val Phe Asn
1 5
<210> 1085
<211> 14
<212> PRT
<213> Geobacillus thermoglucosidasius
<400> 1085
Met Leu Asn His Met Ser Gln Asp Arg Asn Pro Tyr Ser His
1 5 10
<210> 1086
<211> 13
<212> PRT
<213> Geobacillus thermoglucosidasius
<400> 1086
Pro Leu Arg Thr Leu Asn Gln Ala Ile Ser Leu His Asn
1 5 10
<210> 1087
<211> 12
<212> PRT
<213> Teredinibacter turnerae
<400> 1087
Cys Arg Ser Ile Arg Lys Leu Leu Asn Met Asn Ala
1 5 10
<210> 1088
<211> 16
<212> PRT
<213> Teredinibacter turnerae
<400> 1088
Ser Tyr Pro Leu Ile Tyr Glu Ile Glu Asn Leu Val Arg Lys Leu Ile
1 5 10 15
<210> 1089
<211> 9
<212> PRT
<213> Teredinibacter turnerae
<400> 1089
Ile Gln Leu Ser Asn Phe Leu Phe Asp
1 5
<210> 1090
<211> 15
<212> PRT
<213> Teredinibacter turnerae
<400> 1090
Arg Trp Gly Lys Leu Tyr Lys Leu Arg Cys Lys Ile Ala His Asn
1 5 10 15
<210> 1091
<211> 13
<212> PRT
<213> Teredinibacter turnerae
<400> 1091
Thr Thr Lys Leu Val Glu Glu Val Lys Leu Lys Ile Leu
1 5 10
<210> 1092
<211> 5
<212> PRT
<213> Methanococcus maripaludis
<400> 1092
Phe Arg Leu Met Tyr
1 5
<210> 1093
<211> 16
<212> PRT
<213> Methanococcus maripaludis
<400> 1093
Phe Leu Asp Ser Val Leu Ala Leu Glu Ile Tyr His Thr Leu Lys Phe
1 5 10 15
<210> 1094
<211> 10
<212> PRT
<213> Methanococcus maripaludis
<400> 1094
Phe Ile Asn Lys Met Lys Asp Val Phe Asn
1 5 10
<210> 1095
<211> 15
<212> PRT
<213> Methanococcus maripaludis
<400> 1095
Ile Cys Arg Ile Ile Arg Asp Thr Arg Asn Lys Leu Val His Asp
1 5 10 15
<210> 1096
<211> 13
<212> PRT
<213> Methanococcus maripaludis
<400> 1096
Pro Tyr Phe Leu Ile Glu Leu Leu Lys Asn Ile Phe Lys
1 5 10
<210> 1097
<211> 11
<212> PRT
<213> Novosphingobium pentaromativorans
<400> 1097
Val His Arg Ala Leu Ser Trp Leu Arg Arg Ala
1 5 10
<210> 1098
<211> 14
<212> PRT
<213> Novosphingobium pentaromativorans
<400> 1098
Phe Ile Leu Leu Trp Ile Gly Phe Asn Ala Ala Tyr Ala Gly
1 5 10
<210> 1099
<211> 10
<212> PRT
<213> Novosphingobium pentaromativorans
<400> 1099
Glu Arg Ser Arg Thr Ala Ile Asn Tyr Ala
1 5 10
<210> 1100
<211> 15
<212> PRT
<213> Novosphingobium pentaromativorans
<400> 1100
Leu Phe Asp Arg Leu Tyr Val Leu Arg Asn Gln Leu Val His Gly
1 5 10 15
<210> 1101
<211> 13
<212> PRT
<213> Novosphingobium pentaromativorans
<400> 1101
Arg Asp Gln Val Arg Asp Gly Ala Ser Leu Leu Gly Cys
1 5 10
<210> 1102
<211> 12
<212> PRT
<213> Chlorobium chlorochromatii
<400> 1102
Ile Met Glu Gln Arg Lys Ala Ile Leu Glu Pro Leu
1 5 10
<210> 1103
<211> 16
<212> PRT
<213> Chlorobium chlorochromatii
<400> 1103
Ala Val Ala Tyr Asn His Phe Val Pro Leu Leu Ala Gln Asp Leu Ile
1 5 10 15
<210> 1104
<211> 6
<212> PRT
<213> Chlorobium chlorochromatii
<400> 1104
Lys Ile Ser Asn Lys Lys
1 5
<210> 1105
<211> 15
<212> PRT
<213> Chlorobium chlorochromatii
<400> 1105
Ser Glu Lys Leu Lys Thr Phe Arg Asp Lys Tyr Tyr Ala His Leu
1 5 10 15
<210> 1106
<211> 13
<212> PRT
<213> Chlorobium chlorochromatii
<400> 1106
Phe Leu Gly Ile His Arg Lys Ser Ala Asn Glu Met Trp
1 5 10
<210> 1107
<211> 12
<212> PRT
<213> Lactococcus lactis
<400> 1107
Asp Ala Tyr Asn Lys Leu Ile Leu Leu Lys Gln Tyr
1 5 10
<210> 1108
<211> 15
<212> PRT
<213> Lactococcus lactis
<400> 1108
Phe Phe Tyr Asn Asn Leu Leu Asp Ser Leu Val Ile Ala Ile Phe
1 5 10 15
<210> 1109
<211> 6
<212> PRT
<213> Lactococcus lactis
<400> 1109
Asn Tyr Thr Asn Phe Pro
1 5
<210> 1110
<211> 15
<212> PRT
<213> Lactococcus lactis
<400> 1110
Leu Glu Tyr Leu Tyr Ala Gln Arg Asn Lys Ile Tyr Val His Asn
1 5 10 15
<210> 1111
<211> 13
<212> PRT
<213> Lactococcus lactis
<400> 1111
Asn Tyr Ala Trp Glu Pro Thr Asn Ile Asn Asp Trp Glu
1 5 10
<210> 1112
<211> 16
<212> PRT
<213> Escherichia coli
<400> 1112
Glu Ser Val Ile Ala His Met Asn Glu Leu Leu Ile Ala Leu Ser Asp
1 5 10 15
<210> 1113
<211> 15
<212> PRT
<213> Escherichia coli
<400> 1113
Arg Tyr Thr Gln Gln Gln Arg Leu Arg Thr Ala Ile Ala His His
1 5 10 15
<210> 1114
<211> 12
<212> PRT
<213> Escherichia coli
<400> 1114
Glu Ala Arg His Glu Gln Leu Thr Lys Gly Gly Thr
1 5 10
<210> 1115
<211> 16
<212> PRT
<213> Cronobacter sakazakii
<400> 1115
Gln His Val Ile Ala Pro Met Asn Glu Leu Leu Ile Ala Leu Ser Asp
1 5 10 15
<210> 1116
<211> 15
<212> PRT
<213> Cronobacter sakazakii
<400> 1116
Arg Tyr Asp Leu Gln Gln Gln Leu Arg Thr Ala Ile Ala His His
1 5 10 15
<210> 1117
<211> 13
<212> PRT
<213> Cronobacter sakazakii
<400> 1117
Ala Ala Glu Arg Leu Ala Glu Leu Thr Arg Gly Gly Thr
1 5 10
<210> 1118
<211> 16
<212> PRT
<213> Homo sapiens
<400> 1118
Glu Ser Arg Tyr Arg Thr Leu Arg Asn Val Gly Asn Glu Ser Asp Ile
1 5 10 15
<210> 1119
<211> 10
<212> PRT
<213> Homo sapiens
<400> 1119
Leu Gln Pro Gly Pro Ser Glu His Ser Lys
1 5 10
<210> 1120
<211> 14
<212> PRT
<213> Homo sapiens
<400> 1120
Val Gly Asp Leu Leu Lys Phe Ile Arg Asn Leu Gly Glu His
1 5 10
<210> 1121
<211> 13
<212> PRT
<213> Homo sapiens
<400> 1121
Ile Gly Asp Pro Ser Leu Tyr Phe Gln Lys Thr Phe Pro
1 5 10
<210> 1122
<211> 16
<212> PRT
<213> Arabidopsis thaliana
<400> 1122
Glu Met Arg Leu Ser Phe Leu Arg Asp Ala Ser Asp Arg Val Glu Leu
1 5 10 15
<210> 1123
<211> 10
<212> PRT
<213> Arabidopsis thaliana
<400> 1123
Met Glu Ser Thr Ala Pro Val Ala Ile Gly
1 5 10
<210> 1124
<211> 15
<212> PRT
<213> Arabidopsis thaliana
<400> 1124
Ile Arg Asp Leu Leu Arg Val Ile Arg Asn Lys Leu Asn His His
1 5 10 15
<210> 1125
<211> 13
<212> PRT
<213> Arabidopsis thaliana
<400> 1125
Pro Glu Gly Phe Asp Glu Tyr Phe Ala Val Arg Phe Pro
1 5 10
<210> 1126
<211> 12
<212> PRT
<213> Helicobacter pylori
<400> 1126
Tyr Glu Leu Leu Trp Gln Glu Val Ile Arg Ala Lys
1 5 10
<210> 1127
<211> 15
<212> PRT
<213> Helicobacter pylori
<400> 1127
Trp Val Ser Leu Gln Asn Val Met Arg Arg Ile Ile Glu Tyr Tyr
1 5 10 15
<210> 1128
<211> 5
<212> PRT
<213> Helicobacter pylori
<400> 1128
Phe Arg Ile Leu Gly
1 5
<210> 1129
<211> 17
<212> PRT
<213> Helicobacter pylori
<400> 1129
Lys Gln Val Phe Ser Ser Phe Ile Ser Trp Phe Asn Asp Gly Ser His
1 5 10 15
Gly
<210> 1130
<211> 13
<212> PRT
<213> Helicobacter pylori
<400> 1130
Ile Glu Thr Tyr Leu Lys Val Phe Glu Asn Ile Phe Lys
1 5 10
<210> 1131
<211> 12
<212> PRT
<213> Streptococcus mutans
<400> 1131
His Leu Met Leu Val Asp Glu Leu Lys Lys Ala Ile
1 5 10
<210> 1132
<211> 15
<212> PRT
<213> Streptococcus mutans
<400> 1132
Glu Lys Tyr His Phe Asn Leu Leu Arg Asn Leu Leu Glu Lys Thr
1 5 10 15
<210> 1133
<211> 5
<212> PRT
<213> Streptococcus mutans
<400> 1133
Ala Thr Phe Leu Gly
1 5
<210> 1134
<211> 14
<212> PRT
<213> Streptococcus mutans
<400> 1134
Pro Ala Pro Tyr Ile Arg Arg Ile Asn Leu His Ser His Ser
1 5 10
<210> 1135
<211> 13
<212> PRT
<213> Streptococcus mutans
<400> 1135
Lys Lys Val Leu Glu Arg Val Phe Asn Gln Phe Leu Gln
1 5 10
<210> 1136
<211> 12
<212> PRT
<213> Escherichia coli
<400> 1136
His Leu His Leu Lys Gln Thr Ile Glu Gln Ala Ile
1 5 10
<210> 1137
<211> 15
<212> PRT
<213> Escherichia coli
<400> 1137
Glu Arg Tyr His Phe Thr Leu Leu Arg Asn Leu Tyr Glu Lys Thr
1 5 10 15
<210> 1138
<211> 5
<212> PRT
<213> Escherichia coli
<400> 1138
Ala Ser Phe Leu Gly
1 5
<210> 1139
<211> 13
<212> PRT
<213> Escherichia coli
<400> 1139
Leu Tyr Leu Ser Arg Ile Ile Asn Phe Thr Ser His Ser
1 5 10
<210> 1140
<211> 13
<212> PRT
<213> Escherichia coli
<400> 1140
Lys Ala Thr Val Lys Leu Leu Leu Asp His Leu Lys Asn
1 5 10
<210> 1141
<211> 12
<212> PRT
<213> Bacteroides fragilis
<400> 1141
Lys Glu Ile Glu Glu Glu Arg Thr Val Gln Asn Ile
1 5 10
<210> 1142
<211> 16
<212> PRT
<213> Bacteroides fragilis
<400> 1142
Thr Ser Phe Gly Glu Val Thr Glu Glu Tyr His Asp Glu Leu Tyr Ser
1 5 10 15
<210> 1143
<211> 8
<212> PRT
<213> Bacteroides fragilis
<400> 1143
Tyr Ile Lys Glu Leu Ser Asn Gly
1 5
<210> 1144
<211> 15
<212> PRT
<213> Bacteroides fragilis
<400> 1144
Gln Lys Thr Leu Thr Glu Lys Ile Arg His Gln Ile His His Pro
1 5 10 15
<210> 1145
<211> 13
<212> PRT
<213> Bacteroides fragilis
<400> 1145
Glu Thr Glu Ile Arg Gln Ser Ile Glu Asp Met Arg Ala
1 5 10
<210> 1146
<211> 12
<212> PRT
<213> Methylobacillus flagellates
<400> 1146
Ser Asn Gln Ile Pro Thr Arg Val Ser Pro Val Leu
1 5 10
<210> 1147
<211> 16
<212> PRT
<213> Methylobacillus flagellates
<400> 1147
Ser Ala Phe Gly Glu Ala Ser Tyr Glu Tyr His Asn Glu Leu Tyr Gly
1 5 10 15
<210> 1148
<211> 8
<212> PRT
<213> Methylobacillus flagellates
<400> 1148
Tyr Asn Arg Leu Arg Arg Asp Gly
1 5
<210> 1149
<211> 15
<212> PRT
<213> Methylobacillus flagellates
<400> 1149
Gln Val Ile Leu Thr Glu Tyr Ile Arg His Gln Ile His His Pro
1 5 10 15
<210> 1150
<211> 13
<212> PRT
<213> Methylobacillus flagellates
<400> 1150
Thr Ala Glu Leu Thr Glu Ser Ile Glu Thr Met Arg Leu
1 5 10
<210> 1151
<211> 12
<212> PRT
<213> Campylobacter hominis
<400> 1151
Lys Asp Gly Glu Gln Lys Lys Glu Val Lys Asn Val
1 5 10
<210> 1152
<211> 16
<212> PRT
<213> Campylobacter hominis
<400> 1152
Met Ala Phe Gly Glu Ile Thr Glu Glu Tyr His Asn Glu Leu Tyr Gly
1 5 10 15
<210> 1153
<211> 8
<212> PRT
<213> Campylobacter hominis
<400> 1153
Tyr Lys Lys Leu Lys Lys Asp Gly
1 5
<210> 1154
<211> 15
<212> PRT
<213> Campylobacter hominis
<400> 1154
Lys Leu Thr Leu Thr Glu Tyr Ile Arg His Gln Ile His His Pro
1 5 10 15
<210> 1155
<211> 13
<212> PRT
<213> Campylobacter hominis
<400> 1155
Leu Ser Glu Leu Lys Asp Ser Ile Glu Met Met Arg Asn
1 5 10
<210> 1156
<211> 15
<212> PRT
<213> Homo sapiens
<400> 1156
Ile Pro Asp Trp Ile Val Asp Leu Arg His Glu Leu Thr His Lys
1 5 10 15
<210> 1157
<211> 15
<212> PRT
<213> Mycobacterium tuberculosis
<400> 1157
Leu Gly Arg Phe Glu Ser Arg Val Arg Asn Thr Ala Ala His Glu
1 5 10 15
<210> 1158
<211> 15
<212> PRT
<213> Lactococcus lactis
<400> 1158
Trp Ile Arg Ala Gly Trp Phe Ile Arg Asn Arg Ser Ala His Tyr
1 5 10 15
<210> 1159
<211> 15
<212> PRT
<213> Thermus thermophilus
<220>
<221> MOD_RES
<222> (5)..(5)
<223> Any amino acid
<400> 1159
Leu Ala Leu Gly Xaa Val Asp Asp Arg Ser Leu Thr Val His Thr
1 5 10 15
<210> 1160
<211> 22
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1160
Leu Lys Ser Met Leu Tyr Ser Met Arg Asn Ser Ser Phe His Phe Ser
1 5 10 15
Thr Glu Asn Val Asp Asn
20
<210> 1161
<211> 22
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1161
Leu Lys Asp Val Ile Tyr Ser Met Arg Asn Asp Ser Phe His Tyr Ala
1 5 10 15
Thr Glu Asn His Asn Asn
20
<210> 1162
<211> 22
<212> PRT
<213> Clostridium aminophilum
<400> 1162
Leu Arg Lys Ala Ile Tyr Ser Leu Arg Asn Glu Thr Phe His Phe Thr
1 5 10 15
Thr Leu Asn Lys Gly Ser
20
<210> 1163
<211> 22
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1163
Ile Ile Gln Ile Ile Tyr Ser Leu Arg Asn Lys Ser Phe His Phe Lys
1 5 10 15
Thr Tyr Asp His Gly Asp
20
<210> 1164
<211> 22
<212> PRT
<213> Carnobacterium gallinarum
<400> 1164
Leu Arg Gly Ser Val Gln Gln Ile Arg Asn Glu Ile Phe His Ser Phe
1 5 10 15
Asp Lys Asn Gln Lys Phe
20
<210> 1165
<211> 21
<212> PRT
<213> Carnobacterium gallinarum
<400> 1165
Ile Arg Gly Ala Val Gln Arg Val Arg Asn Gln Ile Phe His Gln Gln
1 5 10 15
Ile Asn Lys Arg His
20
<210> 1166
<211> 20
<212> PRT
<213> Paludibacter propionicigenes
<400> 1166
Ile Arg Gly Ala Val Gln Gln Ile Arg Asn Asn Val Asn His Tyr Lys
1 5 10 15
Lys Asp Ala Leu
20
<210> 1167
<211> 20
<212> PRT
<213> Listeria seeligeri
<400> 1167
Leu Arg Gly Ala Ile Ala Pro Ile Arg Asn Glu Ile Ile His Leu Lys
1 5 10 15
Lys His Ser Trp
20
<210> 1168
<211> 20
<212> PRT
<213> Listeria weihenstephanensis
<400> 1168
Ile Arg Gly Ser Ile Gln Gln Ile Arg Asn Glu Val Tyr His Cys Lys
1 5 10 15
Lys His Ser Trp
20
<210> 1169
<211> 20
<212> PRT
<213> Listeria newyorkensis
<400> 1169
Ile Arg Gly Ser Ile Gln Gln Ile Arg Asn Glu Val Tyr His Cys Lys
1 5 10 15
Lys His Ser Trp
20
<210> 1170
<211> 21
<212> PRT
<213> Leptotrichia wadei
<400> 1170
Ile Ser Tyr Ser Ile Tyr Asn Val Arg Asn Gly Val Gly His Phe Asn
1 5 10 15
Lys Leu Ile Leu Gly
20
<210> 1171
<211> 21
<212> PRT
<213> Leptotrichia wadei
<400> 1171
Met Leu Asn Ala Ile Thr Ser Ile Arg His Arg Val Val His Tyr Asn
1 5 10 15
Met Asn Thr Asn Ser
20
<210> 1172
<211> 21
<212> PRT
<213> Leptotrichia wadei
<400> 1172
Ile Asp Glu Ala Ile Ser Ser Ile Arg His Gly Ile Val His Phe Asn
1 5 10 15
Leu Glu Leu Glu Gly
20
<210> 1173
<211> 22
<212> PRT
<213> Rhodobacter capsulatus
<400> 1173
Leu Leu Arg Tyr Leu Arg Gly Cys Arg Asn Gln Thr Phe His Leu Gly
1 5 10 15
Ala Arg Ala Gly Phe Leu
20
<210> 1174
<211> 21
<212> PRT
<213> Leptotrichia buccalis
<400> 1174
Ile Asp Glu Ala Ile Ser Ser Ile Arg His Gly Ile Val His Phe Asn
1 5 10 15
Leu Glu Leu Glu Gly
20
<210> 1175
<211> 21
<212> PRT
<213> Leptotrichia sp.
<400> 1175
Ile Asp Glu Ala Ile Ser Ser Ile Arg His Gly Ile Val His Phe Asn
1 5 10 15
Leu Glu Leu Glu Gly
20
<210> 1176
<211> 16
<212> PRT
<213> Leptotrichia sp.
<400> 1176
Phe Gln Lys Glu Gly Tyr Leu Leu Arg Asn Lys Ile Leu His Asn Ser
1 5 10 15
<210> 1177
<211> 15
<212> PRT
<213> Leptotrichia shahii
<400> 1177
Phe Thr Lys Ile Gly Thr Asn Glu Arg Asn Arg Ile Leu His Ala
1 5 10 15
<210> 1178
<211> 14
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1178
Phe Arg Asn Glu Ile Asp His Phe His Tyr Phe Tyr Asp Arg
1 5 10
<210> 1179
<211> 14
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1179
Leu Arg Asn Tyr Ile Glu His Phe Arg Tyr Tyr Ser Ser Phe
1 5 10
<210> 1180
<211> 14
<212> PRT
<213> Clostridium aminophilum
<400> 1180
Val Arg Lys Tyr Val Asp His Phe Lys Tyr Tyr Ala Thr Ser
1 5 10
<210> 1181
<211> 14
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1181
Leu Arg Lys Tyr Val Asp His Phe Lys Tyr Tyr Ala Tyr Gly
1 5 10
<210> 1182
<211> 14
<212> PRT
<213> Carnobacterium gallinarum
<400> 1182
Ile Arg Asn Gln Thr Ala His Leu Ser Val Leu Gln Leu Glu
1 5 10
<210> 1183
<211> 14
<212> PRT
<213> Carnobacterium gallinarum
<400> 1183
Ile Arg Asn Asn Ile Ala His Leu His Val Leu Arg Asn Asp
1 5 10
<210> 1184
<211> 14
<212> PRT
<213> Paludibacter propionicigenes
<400> 1184
Ile Arg Asn His Ile Ala His Phe Asn Tyr Leu Thr Lys Asp
1 5 10
<210> 1185
<211> 14
<212> PRT
<213> Listeria seeligeri
<400> 1185
Lys Arg Asn Asn Ile Ser His Phe Asn Tyr Leu Asn Gly Gln
1 5 10
<210> 1186
<211> 14
<212> PRT
<213> Listeria weihenstephanensis
<400> 1186
Ala Arg Asn His Ile Ala His Leu Asn Tyr Leu Ser Leu Lys
1 5 10
<210> 1187
<211> 14
<212> PRT
<213> Listeria newyorkensis
<400> 1187
Ala Arg Asn His Ile Ala His Leu Asn Tyr Leu Ser Leu Lys
1 5 10
<210> 1188
<211> 14
<212> PRT
<213> Leptotrichia wadei
<400> 1188
Phe Arg Asn Tyr Ile Ala His Phe Leu His Leu His Thr Lys
1 5 10
<210> 1189
<211> 14
<212> PRT
<213> Leptotrichia wadei
<400> 1189
Ile Arg Asn Tyr Ile Ala His Phe Asn Tyr Ile Pro Asp Ala
1 5 10
<210> 1190
<211> 14
<212> PRT
<213> Leptotrichia wadei
<400> 1190
Ile Arg Asn Tyr Ile Ala His Phe Asn Tyr Ile Pro His Ala
1 5 10
<210> 1191
<211> 14
<212> PRT
<213> Rhodobacter capsulatus
<400> 1191
Thr Arg Lys Asp Leu Ala His Phe Asn Val Leu Asp Arg Ala
1 5 10
<210> 1192
<211> 14
<212> PRT
<213> Leptotrichia buccalis
<400> 1192
Ile Arg Asn Tyr Ile Ala His Phe Asn Tyr Ile Pro His Ala
1 5 10
<210> 1193
<211> 14
<212> PRT
<213> Leptotrichia sp.
<400> 1193
Ile Arg Asn Tyr Ile Ala His Phe Asn Tyr Ile Pro Asn Ala
1 5 10
<210> 1194
<211> 14
<212> PRT
<213> Leptotrichia sp.
<400> 1194
Ile Arg Asn Tyr Ile Ser His Phe Tyr Ile Val Arg Asn Pro
1 5 10
<210> 1195
<211> 14
<212> PRT
<213> Leptotrichia shahii
<400> 1195
Ile Arg Asn Tyr Ile Ser His Phe Tyr Ile Val Arg Asn Pro
1 5 10
<210> 1196
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1196
gugccacuuc ucagaucgcu cgcucaguga uccgac 36
<210> 1197
<211> 105
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1197
gucagaacac ugagcgagcg uucuuuuuga gaagcucaac gggcuuugcc accuggaaag 60
uggccauugg cacacccguu gaaaaaauuc uguccucuag acaga 105
<210> 1198
<211> 105
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1198
gucagaacac ugagcgagcg uucuuuuuga gaagcucaac gggcuuugcc accuggaaag 60
uggccauugg cacacccguu gaaaaaauuc uguccucuag acaga 105
<210> 1199
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1199
gugccacuuc ucagaucgcu cgcucaguga uccgac 36
<210> 1200
<211> 37
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1200
gugccaauca cccaacacug accaagcuug ccgagac 37
<210> 1201
<211> 64
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1201
cuuggggaaa gcuaggcaag uuuuggauga uaagaaauaa ucaugucaca aggagggagu 60
uuuu 64
<210> 1202
<211> 64
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1202
cuuggggaaa gcuaggcaag uuuuggauga uaagaaauaa ucaugucaca aggagggagu 60
uuuu 64
<210> 1203
<211> 37
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1203
gugccaauca cccaacacug accaagcuug ccgagac 37
<210> 1204
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1204
gccgcagcga augccguuuc acgaaucguc aggcgg 36
<210> 1205
<211> 75
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1205
gcuggagacg uuuuuugaaa cggcgagugc ugcggauagc gaguuucucu uggggaggcg 60
cucgcggcca cuuuu 75
<210> 1206
<211> 75
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1206
gcuggagacg uuuuuugaaa cggcgagugc ugcggauagc gaguuucucu uggggaggcg 60
cucgcggcca cuuuu 75
<210> 1207
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1207
gccgcagcga augccguuuc acgaaucguc aggcgg 36
<210> 1208
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1208
guccaagaaa aaagaaauga uacgaggcau uagcac 36
<210> 1209
<211> 107
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1209
cuggacgaug ucucuuuuau uucuuuuuuc uuggaucuga guacgagcac ccacauugga 60
cauuucgcau ggugggugcu cguacuauag guaaaacaaa ccuuuuu 107
<210> 1210
<211> 107
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1210
cuggacgaug ucucuuuuau uucuuuuuuc uuggaucuga guacgagcac ccacauugga 60
cauuucgcau ggugggugcu cguacuauag guaaaacaaa ccuuuuu 107
<210> 1211
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1211
guccaagaaa aaagaaauga uacgaggcau uagcac 36
<210> 1212
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1212
guucgaaagc uuaguggaaa gcuucguccu uagcac 36
<210> 1213
<211> 69
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1213
cacggauaau cacgacuuuc cacuaagcuu ucgaauuuua ugaugcgagc auccucucag 60
gucaaaaaa 69
<210> 1214
<211> 69
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1214
cacggauaau cacgacuuuc cacuaagcuu ucgaauuuua ugaugcgagc auccucucag 60
gucaaaaaa 69
<210> 1215
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1215
guucgaaagc uuaguggaaa gcuucguggu uagcac 36
<210> 1216
<211> 35
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1216
guauugagaa aagccagaua uaguuggcaa uagac 35
<210> 1217
<211> 62
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1217
auauuuugau ucccauuuau gguuauuuac cauaaauggg aaucaacuaa aaaauauuuu 60
uu 62
<210> 1218
<211> 35
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1218
guauugagaa aagccagaua uaguuggcaa uagac 35
<210> 1219
<211> 62
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1219
auauuuugau ucccauuuau gguuauuuac cauaaauggg aaucaacuaa aaaauauuuu 60
uu 62
<210> 1220
<211> 35
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1220
guugaugaga agagcccaag auagagggca auaac 35
<210> 1221
<211> 35
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1221
gcuggagaag auagcccaag aaagagggca auaac 35
<210> 1222
<211> 78
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1222
auuauuacca uuuugguugg aaugcuauua uaaaggauca uucgauuauu accucuaccu 60
cccuucccac gauuucuu 78
<210> 1223
<211> 78
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1223
attattacca ttttggttgg aatgctatta taaaggatca ttcgattatt acctctacct 60
cccttcccac gatttctt 78
<210> 1224
<211> 35
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1224
guugaugaga agagcccaag auagagggca auaac 35
<210> 1225
<211> 78
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1225
attattacca ttttggttgg aatgctatta taaaggatca ttcgattatt acctctacct 60
cccttcccac gatttctt 78
<210> 1226
<211> 35
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1226
gcuggagaag auagcccaag aaagagggca auaac 35
<210> 1227
<211> 35
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1227
guuuggagaa cagcccgaua uagagggcaa uagac 35
<210> 1228
<211> 81
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1228
gucuuacgac cucaguauua ggaagauuuc aaccaagaaa acuuaguuuc aggcuuaaug 60
aucgagucau gcagccaaag u 81
<210> 1229
<211> 81
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1229
gucuuacgac cucaguauua ggaagauuuc aaccaagaaa acuuaguuuc aggcuuaaug 60
aucgagucau gcagccaaag u 81
<210> 1230
<211> 35
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1230
guuuggagaa cagcccgaua uagagggcaa uagac 35
<210> 1231
<211> 35
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1231
guuuugagaa uagcccgaca uagagggcaa uagac 35
<210> 1232
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1232
guuaugaaaa cagcccgaca uagagggcaa uagaca 36
<210> 1233
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1233
guuauagucc ucuuacauuu agagguaguc uuuaau 36
<210> 1234
<211> 98
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1234
ucuuaagaac uucucuaccu gaaguuggau uauaaaugac ucuugcucuc auagauaucc 60
uccuuugaaa auauacacug ccgauuaauu accguuuu 98
<210> 1235
<211> 98
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1235
ucuuaagaac uucucuaccu gaaguuggau uauaaaugac ucuugcucuc auagauaucc 60
uccuuugaaa auauacacug ccgauuaauu accguuuu 98
<210> 1236
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1236
guuauagucc ucuuacauuu agagguaguc uuuaau 36
<210> 1237
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1237
guuauagucc ucuuacauuu agagguaguu uauauu 36
<210> 1238
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1238
guuauagucc ccuuacauuu agggguaguc uuuaau 36
<210> 1239
<211> 102
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1239
aauauaaauu cucccuaaau auaagagaau aauaacucaa ucucuucauu cguauuuugu 60
cuaguuaaga uaaguaccac caaauacaau caauccaaaa aa 102
<210> 1240
<211> 102
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1240
aauauaaauu cucccuaaau auaagagaau aauaacucaa ucucuucauu cguauuuugu 60
cuaguuaaga uaaguaccac caaauacaau caauccaaaa aa 102
<210> 1241
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1241
guuauagucc ucuuacauuu agagguaguu uauauu 36
<210> 1242
<211> 102
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1242
aauauaaauu cucccuaaau auaagagaau aauaacucaa ucucuucauu cguauuuugu 60
cuaguuaaga uaaguaccac caaauacaau caauccaaaa aa 102
<210> 1243
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1243
guuauagucc ccuuacauuu agggguaguc uuuaau 36
<210> 1244
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1244
guuguaguuc ccuucaauuu ugggauaauc cacaag 36
<210> 1245
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1245
guuuuagucc ucuuucauau agagguaguc ucuuac 36
<210> 1246
<211> 99
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1246
augaaaagag gacuaaaacu gaaagaggac uaaaacacca gauguggaua acuauauuag 60
uggcuauuaa aaauucgucg auauuagaga ggaaacuuu 99
<210> 1247
<211> 99
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1247
augaaaagag gacuaaaacu gaaagaggac uaaaacacca gauguggaua acuauauuag 60
uggcuauuaa aaauucgucg auauuagaga ggaaacuuu 99
<210> 1248
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1248
guuuuagucc ucuuucauau agagguaguc ucuuac 36
<210> 1249
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1249
guuuuagacc ucuucuauuu ugagguacuc uaaauc 36
<210> 1250
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1250
guuuuagucc ucuuuuguuu ugagguacuc uaaauc 36
<210> 1251
<211> 147
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1251
aagucagcgc acaacaaaga agaugacgaa caaaaucucu cgccaucuuc uuaaaauuau 60
uugccacaca gccaacauua uaagcguuaa aaccagcacc augaguacau uucacccaac 120
aaucagaauc cccguuucuc cguuuuu 147
<210> 1252
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1252
guuuuagucc ucuuuuguuu ugagguacuc uaaauc 36
<210> 1253
<211> 147
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1253
aagucagcgc acaacaaaga agaugacgaa caaaaucucu cgccaucuuc uuaaaauuau 60
uugccacaca gccaacauua uaagcguuaa aaccagcacc augaguacau uucacccaac 120
aaucagaauc cccguuucuc cguuuuu 147
<210> 1254
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1254
guuuuagauc ccuucguuuu ugggguuauc uauauc 36
<210> 1255
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1255
guuuuagucc ccuucguuuu ugggguaguc uaaauc 36
<210> 1256
<211> 113
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1256
gauuuagagc accccaaaag uaaugaaaau uugcaauuaa auaaggaaua uuaaaaaaau 60
gugauuuuaa aaaaauugaa gaaauuaaau gaaaaauugu ccaaguaaaa aaa 113
<210> 1257
<211> 70
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1257
auuuagauua ccccuuuaau uuauuuuacc auauuuuucu cauaaugcaa acuaauauuc 60
caaaauuuuu 70
<210> 1258
<211> 113
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1258
gauuuagagc accccaaaag uaaugaaaau uugcaauuaa auaaggaaua uuaaaaaaau 60
gugauuuuaa aaaaauugaa gaaauuaaau gaaaaauugu ccaaguaaaa aaa 113
<210> 1259
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1259
guuuuagucc ccuucguuuu ugggguaguc uaaauc 36
<210> 1260
<211> 70
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1260
auuuagauua ccccuuuaau uuauuuuacc auauuuuucu cauaaugcaa acuaauauuc 60
caaaauuuuu 70
<210> 1261
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1261
guuuuagucc ccuucguuuu ugggguaguc uaaauc 36
<210> 1262
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1262
guuuuagucc ccuucgauau uggggugguc uauauc 36
<210> 1263
<211> 95
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1263
auugaugugg uauacuaaaa auggaaaauu guauuuuuga uuagaaagau guaaaauuga 60
uuuaauuuaa aaauauuuua uuagauuaaa guaga 95
<210> 1264
<211> 95
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1264
auugaugugg uauacuaaaa auggaaaauu guauuuuuga uuagaaagau guaaaauuga 60
uuuaauuuaa aaauauuuua uuagauuaaa guaga 95
<210> 1265
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1265
guuuuagucc ccuucgauau uggggugguc uauauc 36
<210> 1266
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1266
guucaguccg ccgucgucuu ggcggugaug ugaggc 36
<210> 1267
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1267
guucaguccg ccgucauuuu ggcggugaug ugcucc 36
<210> 1268
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1268
guucaguccg ccgucgucuu ggcggugaug ugaggc 36
<210> 1269
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1269
guucaguccg ccgucauuuu ggcggugaug ugcucc 36
<210> 1270
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1270
guucaguccg ccgucgucuu ggcggugaug ugaggc 36
<210> 1271
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1271
guucaguccg ccgucauuuu ggcggugaug ugcucc 36
<210> 1272
<211> 81
<212> DNA
<213> Alicyclobacillus acidoterrestris
<400> 1272
agcgcccgct gtcggatcac tgagcgagcg atctgagaag tggcactgtt tggtaaaggt 60
aaaaagacga atgatgcatc c 81
<210> 1273
<211> 83
<212> DNA
<213> Alicyclobacillus acidoterrestris
<400> 1273
tgatgcatcc gtcggatcac tgagcgagcg atctgagaag tggcaccctt tataaaaagg 60
ggcgtccttt agtaccgtgt act 83
<210> 1274
<211> 84
<212> DNA
<213> Alicyclobacillus acidoterrestris
<400> 1274
accgtgtact gtcggatcac tgagcgagcg atctgagaag tggcacaagc cttgagtaat 60
tcgccgtggg attccccgcc gtat 84
<210> 1275
<211> 80
<212> DNA
<213> Alicyclobacillus acidoterrestris
<400> 1275
cccgccgtat gtcggatcac tgagcgagcg atctgagaag tggcactaat gaagttaaag 60
gagatgagac aatgaaagaa 80
<210> 1276
<211> 83
<212> DNA
<213> Alicyclobacillus acidoterrestris
<400> 1276
aatgaaagaa gtcggatcac tgagcgagcg atctgagaag tggcactgca atgcgttgga 60
ttatgacgat gcaggccaag gaa 83
<210> 1277
<211> 56
<212> DNA
<213> Alicyclobacillus acidoterrestris
<400> 1277
ggccaaggaa gtcggatcac tgagcgagcg atctgagaag tggcactgat gtcagc 56
<210> 1278
<211> 36
<212> DNA
<213> Alicyclobacillus acidoterrestris
<400> 1278
gtcggatcac tgagcgagcg atctgagaag tggcac 36
<210> 1279
<211> 36
<212> DNA
<213> Alicyclobacillus acidoterrestris
<400> 1279
gtcggatcac tgagcgagcg atctgagaag tggcac 36
<210> 1280
<211> 84
<212> DNA
<213> Desulfonatronum thiodismutans
<400> 1280
ccggctcgag gtctcggcaa gcttggtcag tgttgggtga ttggcacatc caggtggttg 60
gatgcgggac ataccttccg cctt 84
<210> 1281
<211> 84
<212> DNA
<213> Desulfonatronum thiodismutans
<400> 1281
cttccgcctt gtctcggcaa gcttggtcag tgttgggtga ttggcactgc ttcccggcga 60
acggcgagct gacctcctag atgt 84
<210> 1282
<211> 83
<212> DNA
<213> Desulfonatronum thiodismutans
<400> 1282
tcctagatgt gtctcggcaa gcttggtcag tgttgggtga ttggcaccgt ctgctcggtc 60
tcggacttca ccaccacgtc cac 83
<210> 1283
<211> 83
<212> DNA
<213> Desulfonatronum thiodismutans
<400> 1283
ccacgtccac gtctcggcaa gcttggtcag tgttgggtga ttggcacggg tgcggcattt 60
gcgggtgttg ggggagtggc agg 83
<210> 1284
<211> 37
<212> DNA
<213> Desulfonatronum thiodismutans
<400> 1284
gtctcggcaa gcttggtcag tgttgggtga ttggcac 37
<210> 1285
<211> 15
<212> DNA
<213> Desulfonatronum thiodismutans
<400> 1285
gtctcggcaa gcttg 15
<210> 1286
<211> 22
<212> DNA
<213> Desulfonatronum thiodismutans
<400> 1286
gtcagtgttg ggtgattggc ac 22
<210> 1287
<211> 80
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1287
acatccactg ccgcctgacg attcgtgaaa cggcattcgc tgcggcaata gtctctggaa 60
atgttatagt agctcctaca 80
<210> 1288
<211> 81
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1288
agctcctaca ccgcctgacg attcgtgaaa cggcattcgc tgcggcaaga aaaagagtcg 60
tggtgttggc gcgggtcaga c 81
<210> 1289
<211> 80
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1289
cgggtcagac ccgcctgacg attcgtgaaa cggcattcgc tgcggcaatg taacgcctgg 60
agcatggctt gacccgaacc 80
<210> 1290
<211> 82
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1290
gacccgaacc ccgcctgacg attcgtgaaa cggcattcgc tgcggcaata tacgtctgat 60
taaaggtatg ggattccctg tt 82
<210> 1291
<211> 80
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1291
attccctgtt ccgcctgacg attcgtgaaa cggcattcgc tgcggctctc agtcaattcg 60
aatatgatgc ggggtactgg 80
<210> 1292
<211> 80
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1292
ggggtactgg ccgcctgacg attcgtgaaa cggcattcgc tgcggcgctc cacaaaagcg 60
attatcattt cccggttata 80
<210> 1293
<211> 82
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1293
cccggttata ccgcctgacg attcgtgaaa cggcattcgc tgcggcgtgc ccggccatgc 60
ggttatcggt ctcgatggcc tt 82
<210> 1294
<211> 79
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1294
cgatggcctt ccgcctgacg attcgtgaaa cggcattcgc tgcggctcgc ggggaaacga 60
gtgcgtagtc gatcgtcac 79
<210> 1295
<211> 79
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1295
cgatcgtcac ccgcctgacg attcgtgaaa cggcattcgc tgcggcgtag ctgtcgccgt 60
ctttcttgta ttctttttt 79
<210> 1296
<211> 82
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1296
attctttttt ccgcctgacg attcgtgaaa cggcattcgc tgcggcgatc ggacaatcac 60
gccagacatt gccggtcatg at 82
<210> 1297
<211> 83
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1297
cggtcatgat ccgcctgacg attcgtgaaa cggcattcgc tgcggcttgg cgaccttcag 60
gcgagcgtta ttggcggcat aga 83
<210> 1298
<211> 81
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1298
gcggcataga ccgcctgacg attcgtgaaa cggcattcgc tgcggcaaaa atggcgaaac 60
cgaagccgcc gacgcgatac a 81
<210> 1299
<211> 80
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1299
acgcgataca ccgcctgacg attcgtgaaa cggcattcgc tgcggcatgt caattttggt 60
aacacttcgc cttggcacca 80
<210> 1300
<211> 80
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1300
cttggcacca ccgcctgacg attcgtgaaa cggcattcgc tgcggcatgt caattttggt 60
aacacttcgc cttggcacca 80
<210> 1301
<211> 80
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1301
cttggcacca ccgcctgacg attcgtgaaa cggcattcgc tgcggcagca cgtggggttt 60
ttgctctcac aaagtaaatt 80
<210> 1302
<211> 79
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1302
aaagtaaatt ccgcctgacg attcgtgaaa cggcattcgc tgcggcatag catcggcgag 60
tgtgtctgac agtcctact 79
<210> 1303
<211> 83
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1303
cagtcctact ccgcctgacg attcgtgaaa cggcattcgc tgcggcggcg aaccctcgtc 60
aaccgcgacc tcaagatggc aca 83
<210> 1304
<211> 81
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1304
agatggcaca ccgcctgacg attcgtgaaa cggcattcgc tgcggcaacg catcacgcgc 60
ctccggctcc atttccttgc c 81
<210> 1305
<211> 80
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1305
tttccttgcc ccgcctgacg attcgtgaaa cggcattcgc tgcggctccg ctcacagggg 60
caatctacgc tcaggagatg 80
<210> 1306
<211> 81
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1306
tcaggagatg ccgcctgacg attcgtgaaa cggcattcgc tgcggcgaaa acgccccgaa 60
acacagttcc cgaattcaaa t 81
<210> 1307
<211> 79
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1307
gaattcaaat ccgcctgacg attcgtgaaa cggcattcgc tgcggcagca caattccgag 60
aaacggtttc aggatcata 79
<210> 1308
<211> 81
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1308
caggatcata ccgcctgacg attcgtgaaa cggcattcgc tgcggcgcca gcgttccctg 60
cgatgctgtt gcgaaatccc c 81
<210> 1309
<211> 80
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1309
cgaaatcccc ccgcctgacg attcgtgaaa cggcattcgc tgcggcttgc actgcgtggt 60
gaaagagttc aaagcgtcca 80
<210> 1310
<211> 56
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1310
aaagcgtcca ccgcctgacg attcgtgaaa cggcattcgc tgcggcccgt gctcgc 56
<210> 1311
<211> 36
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1311
ccgcctgacg attcgtgaaa cggcattcgc tgcggc 36
<210> 1312
<211> 36
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1312
ccgcctgacg attcgtgaaa cggcattcgc tgcggc 36
<210> 1313
<211> 10
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1313
ccgcctgacg 10
<210> 1314
<211> 12
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1314
attcgtgaaa cg 12
<210> 1315
<211> 14
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1315
gcattcgctg cggc 14
<210> 1316
<211> 10
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1316
ccgcctgacg 10
<210> 1317
<211> 12
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1317
attcgtgaaa cg 12
<210> 1318
<211> 14
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1318
gcattcgctg cggc 14
<210> 1319
<211> 36
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1319
ccgcctgacg attcgtgaaa cggcattcgc tgcggc 36
<210> 1320
<211> 10
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1320
ccgcctgacg 10
<210> 1321
<211> 12
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1321
attcgtgaaa cg 12
<210> 1322
<211> 14
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1322
gcattcgctg cggc 14
<210> 1323
<211> 36
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1323
ccgcctgacg attcgtgaaa cggcattcgc tgcggc 36
<210> 1324
<211> 10
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1324
ccgcctgacg 10
<210> 1325
<211> 12
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1325
attcgtgaaa cg 12
<210> 1326
<211> 14
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1326
gcattcgctg cggc 14
<210> 1327
<211> 81
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1327
accccctgcg gtccaagaaa aaagaaatga tacgaggcat tagcaccatg caaacggatt 60
gttatataaa tcttcttgaa c 81
<210> 1328
<211> 82
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1328
cttcttgaac gtccaagaaa aaagaaatga tacgaggcat tagcacattg ttccggcggc 60
taatttgtct gcggtaatcg aa 82
<210> 1329
<211> 87
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1329
ggtaatcgaa gtccaagaaa aaagaaatga tacgaggcat tagcaccttc gccatcctca 60
tcccttatca gttgattgcc tagcgtt 87
<210> 1330
<211> 83
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1330
gcctagcgtt gtccaagaaa aaagaaatga tacgaggcat tagcacacac agaaaccaaa 60
tgggaacacg ttttcgttaa taa 83
<210> 1331
<211> 84
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1331
tcgttaataa gtccaagaaa aaagaaatga tacgaggcat tagcacgaca ttaaaaaatt 60
ccaaccaagc gagttattga gtgg 84
<210> 1332
<211> 80
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1332
tattgagtgg gtccaagaaa aaagaaatga tacgaggcat tagcacttat gagcttaaaa 60
gcttgttagc gacattaaac 80
<210> 1333
<211> 83
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1333
gacattaaac gtccaagaaa aaagaaatga tacgaggcat tagcacgttt tgttacagct 60
ttattcctta cttgatcgac tct 83
<210> 1334
<211> 83
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1334
gatcgactct gtccaagaaa aaagaaatga tacgaggcat tagcaccaga acctatctca 60
agaggatgca ttctggaaag gaa 83
<210> 1335
<211> 86
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1335
tggaaaggaa gtccaagaaa aaagaaatga tacgaggcat tagcacctta taacaataat 60
ttaaaagcaa tttatgactg tataga 86
<210> 1336
<211> 82
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1336
actgtataga gtccaagaaa aaagaaatga tacgaggcat tagcactttt aagggacatc 60
agaaacacta taagctcact tg 82
<210> 1337
<211> 81
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1337
agctcacttg gtccaagaaa aaagaaatga tacgaggcat tagcacataa tcgactttgc 60
atttctatag tgtcgttcat c 81
<210> 1338
<211> 85
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1338
gtcgttcatc gtccaagaaa aaagaaatga tacgaggcat tagcacaaaa tggaacaagg 60
aacaatagac gtttataagt atgga 85
<210> 1339
<211> 83
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1339
taagtatgga gtccaagaaa aaagaaatga tacgaggcat tagcactttc aataaagcat 60
caaactcttt ttgcattttt tca 83
<210> 1340
<211> 83
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1340
cattttttca gtccaagaaa aaagaaatga tacgaggcat tagcacaccg agtaaggaat 60
cgtttaatca acaaacttga aac 83
<210> 1341
<211> 84
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1341
aacttgaaac gtccaagaaa aaagaaatga tacgaggcat tagcacggta gtcggcggct 60
aagtgtcgca gggttaacac cgat 84
<210> 1342
<211> 85
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1342
taacaccgat gtccaagaaa aaagaaatga tacgaggcat tagcacacgc tgaacaaaac 60
tcacgaaacc aaaagtttat aaaat 85
<210> 1343
<211> 81
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1343
tttataaaat gtccaagaaa aaagaaatga tacgaggcat tagcacggaa aggatttact 60
agatctcgca agaaaggtaa c 81
<210> 1344
<211> 82
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1344
gaaaggtaac gtccaagaaa aaagaaatga tacgaggcat tagcacattt agtatattgt 60
tgttttcatt tgcttttttc gc 82
<210> 1345
<211> 83
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1345
cttttttcgc gtccaagaaa aaagaaatga tacgaggcat tagcacttat accgtaaaaa 60
attttggatt tgatgtcacc gtc 83
<210> 1346
<211> 82
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1346
tgtcaccgtc gtccaagaaa aaagaaatga tacgaggcat tagcacagaa cacaaaaagc 60
ggaaaaattg cacttatttt cg 82
<210> 1347
<211> 82
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1347
cttattttcg gtccaagaaa aaagaaatga tacgaggcat tagcacaata ctcgtctaca 60
aactttttct gcttttctgt ta 82
<210> 1348
<211> 84
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1348
ttttctgtta gtccaagaaa aaagaaatga tacgaggcat tagcaccata caggacactt 60
aaactctact ttacgatttt caaa 84
<210> 1349
<211> 82
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1349
gattttcaaa gtccaagaaa aaagaaatga tatgaggcat tagcacgatt taaaacttct 60
tctggcatcc agtacataga tt 82
<210> 1350
<211> 82
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1350
tacatagatt gtccaagaaa aaagaaatga tatgaggcat tagcacgaaa aaactgtttc 60
cttatcacac ctatagcaat aa 82
<210> 1351
<211> 84
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1351
atagcaataa gtccaagaaa aaagaaatga tatgaggcat tagcactcta gatatgttca 60
agaaaatatg cagtctatcg gtca 84
<210> 1352
<211> 85
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1352
ctatcggtca gtccaagaaa aaagaaatga tatgaggcat tagcacgaat caattaatcc 60
aatttgttgc aacttaggta gagag 85
<210> 1353
<211> 84
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1353
aggtagagag gtccaagaaa aaagaaatga tatgaggcat tagcacccga attagcatcg 60
taccaattac atccatatga tgct 84
<210> 1354
<211> 82
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1354
atatgatgct gtccaagaaa aaagaaatga tatgaggcat tagcactttt ctatttttat 60
attactgttt gtttggtgat aa 82
<210> 1355
<211> 84
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1355
ttggtgataa gtccaagaaa aaagaaatga tatgaggcat tagcactgaa tctaaccaag 60
gaagaaataa aagatattta taag 84
<210> 1356
<211> 81
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1356
tatttataag gtccaagaaa aaagaaatga tatgaggcat tagcacaaaa aggagtgttt 60
caaaatggca aatgaaaaaa g 81
<210> 1357
<211> 82
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1357
atgaaaaaag gtccaagaaa aaagaaatga tatgaggcat tagcacagct tggattgaat 60
caaacaatgg tggtcgaggt tt 82
<210> 1358
<211> 82
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1358
gtcgaggttt gtccaagaaa aaagaaatga tatgaggcat tagcacttga gttgtaatta 60
atcaccgttc tacctgaaaa ct 82
<210> 1359
<211> 80
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1359
cctgaaaact gtccaagaaa aaagaaatga tatgaggcat tagcactaga tgttgaaaac 60
ggcggttttg tctattatcc 80
<210> 1360
<211> 84
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1360
tctattatcc gtccaagaaa aaagaaatga tatgaggcat tagcacaaac ccgcaaaaaa 60
taagctgttg acgagtctag ttag 84
<210> 1361
<211> 85
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1361
gtctagttag gtccaagaaa aaagaaatga tacgaggcat tagcacttgt caatcctcga 60
aaatgccggg cggacgggct tgcca 85
<210> 1362
<211> 81
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1362
gggcttgcca gtccaagaaa aaagaaatga aacgaggcat tagcacccac caccgacggc 60
gtccacgcca attgcctgct t 81
<210> 1363
<211> 72
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1363
ttgcctgctt gtccaagaaa aaagaaatga tacgaggcat tagcacaaca atataaacga 60
ctactttacc gt 72
<210> 1364
<211> 56
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1364
actttaccgt gttcaagaaa aaagaaatga tatgaggcat tagcacgatg ggatgg 56
<210> 1365
<211> 36
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1365
gtccaagaaa aaagaaatga tacgaggcat tagcac 36
<210> 1366
<211> 36
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1366
gtccaagaaa aaagaaatga tacgaggcat tagcac 36
<210> 1367
<211> 36
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1367
gtccaagaaa aaagaaatga tacgaggcat tagcac 36
<210> 1368
<211> 83
<212> DNA
<213> Bacillus sp.
<400> 1368
ggactttagc gtgctaacca cgaagctttc cactaagctt tcgaacaccg gattggtgga 60
aagacgccgc tgccgaccga aaa 83
<210> 1369
<211> 83
<212> DNA
<213> Bacillus sp.
<400> 1369
cgaccgaaaa gtgctaacca cgaagctttc cactaagctt tcgaacaccg gattggtgga 60
aagacgccgc tgccgaccga aaa 83
<210> 1370
<211> 87
<212> DNA
<213> Bacillus sp.
<400> 1370
cgaccgaaaa gtgctaacca cgaagctttc cactaagctt tcgaacggga aaacgtgggc 60
gcaactttgc tattctggca tgtttgc 87
<210> 1371
<211> 83
<212> DNA
<213> Bacillus sp.
<400> 1371
gcatgtttgc gtgctaacca cgaagctttc cactaagctt tcgaacaagt ttggaagggg 60
gcgatagcga tgggttttaa gat 83
<210> 1372
<211> 81
<212> DNA
<213> Bacillus sp.
<400> 1372
gttttaagat gtgctaacca cgaagctttc cactaagctt tcgaacccga tcaccgtcaa 60
aaccgtagta gtaagactcg c 81
<210> 1373
<211> 81
<212> DNA
<213> Bacillus sp.
<400> 1373
taagactcgc gtgctaacca cgaagctttc cactaagctt tcgaacaagg cggcctaaat 60
cacttgggcc gcccttaaga t 81
<210> 1374
<211> 86
<212> DNA
<213> Bacillus sp.
<400> 1374
cccttaagat gtgctaacca cgaagctttc cactaagctt tcgaactgct tttatttcca 60
tgctaccatt ggctgaatcg ggactg 86
<210> 1375
<211> 82
<212> DNA
<213> Bacillus sp.
<400> 1375
atcgggactg gtgctaacca cgaagctttc cactaagctt tcgaacaaac aggtatctgc 60
ttatcaacgt gcagcacagg cg 82
<210> 1376
<211> 82
<212> DNA
<213> Bacillus sp.
<400> 1376
agcacaggcg gtgctaacca cgaagctttc cactaagctt tcgaacctat atgagtagaa 60
cgtctctcaa taagcgtaga at 82
<210> 1377
<211> 81
<212> DNA
<213> Bacillus sp.
<400> 1377
agcgtagaat gtgctaacca cgaagctttc cactaagctt tcgaacgcgc cttgttgcta 60
tcattagggt cgcgatcaac a 81
<210> 1378
<211> 88
<212> DNA
<213> Bacillus sp.
<400> 1378
gcgatcaaca gtgctaacca cgaagctttc cactaagctt tcgaacgtat aacttttaga 60
taacaaatgt tatacaaaat gcttgacg 88
<210> 1379
<211> 81
<212> DNA
<213> Bacillus sp.
<400> 1379
atgcttgacg gtgctaacca cgaagctttc cactaagctt tcgaacagcc caagctttac 60
atacacctat gcgtatgctt t 81
<210> 1380
<211> 56
<212> DNA
<213> Bacillus sp.
<400> 1380
cgtatgcttt gtgctaacca cgaagctttc cactaagctt tcgaactcct tcccac 56
<210> 1381
<211> 36
<212> DNA
<213> Bacillus sp.
<400> 1381
gtgctaacca cgaagctttc cactaagctt tcgaac 36
<210> 1382
<211> 84
<212> DNA
<213> Bacillus sp.
<400> 1382
tggacaagct gtgctaacca cgaagctttc cactaagctt tcgaacctct ggtgtaaggc 60
ggtttcttgc ttgtaagtgc gcgg 84
<210> 1383
<211> 84
<212> DNA
<213> Bacillus sp.
<400> 1383
aagtgcgcgg gtgctaatcc cgaagctttc cactaagctt tcgaacaaaa gagtataagt 60
accactctta gccgagtagt tcaa 84
<210> 1384
<211> 80
<212> DNA
<213> Bacillus sp.
<400> 1384
agtagttcaa gtgctaatcc cgaagctttc cactaagctt tcgaacactg gagaaccttc 60
caagtgtgcg atagcgtatc 80
<210> 1385
<211> 80
<212> DNA
<213> Bacillus sp.
<400> 1385
atagcgtatc gtgctaatcc cgaagctttc cactaagctt tcgaactttt ttgttccgcc 60
aaggtaaaac gtaccgagtt 80
<210> 1386
<211> 83
<212> DNA
<213> Bacillus sp.
<400> 1386
gtaccgagtt gtgctaatcc cgaagctttc cactaagctt tcgaactttc cgtaaaactt 60
tttgagaaaa caggagggcg act 83
<210> 1387
<211> 86
<212> DNA
<213> Bacillus sp.
<400> 1387
gagggcgact gtgctaatcc cgaagctttc cactaagctt tcgaaccagg ctttttgtgc 60
ataggtcatg cagtaaccta atccgc 86
<210> 1388
<211> 85
<212> DNA
<213> Bacillus sp.
<400> 1388
cctaatccgc gtgctaatcc cgaagctttc cactaagctt tcgaacgacc atgatctcga 60
aaagaaaatt acgcgtgcat cgcaa 85
<210> 1389
<211> 84
<212> DNA
<213> Bacillus sp.
<400> 1389
tgcatcgcaa gtgctaatcc cgaagctttc cactaagctt tcgaactggg ccgctccgat 60
tcgataaaga cgggtatctg gaga 84
<210> 1390
<211> 56
<212> DNA
<213> Bacillus sp.
<400> 1390
tatctggaga gtgctaatcc cgaagctttc cactaagctt tcgaaccagg gtaccc 56
<210> 1391
<211> 36
<212> DNA
<213> Bacillus sp.
<400> 1391
gtgctaatcc cgaagctttc cactaagctt tcgaac 36
<210> 1392
<211> 82
<212> DNA
<213> Bacillus sp.
<400> 1392
gctacctctg gtacatcccc ttcaatttcc actaagcttt cgaacgttcg gcgtagtagt 60
gtacacgctc ttgaatatga tg 82
<210> 1393
<211> 81
<212> DNA
<213> Bacillus sp.
<400> 1393
gaatatgatg gtacatcccc ttcaatttcc actaagcttt cgaacgccgc aacgtgctcc 60
gggagaggcg cacgttcaga g 81
<210> 1394
<211> 80
<212> DNA
<213> Bacillus sp.
<400> 1394
acgttcagag gtacatcccc ttcaatttcc actaagcttt cgaacgcgac cagtccgagc 60
agggcaatcc cgacaatgcg 80
<210> 1395
<211> 81
<212> DNA
<213> Bacillus sp.
<400> 1395
cgacaatgcg gtacatcccc ttcaatttcc actaagcttt cgaacaaccc gaaggactac 60
accatttccc gtgataaagc g 81
<210> 1396
<211> 81
<212> DNA
<213> Bacillus sp.
<400> 1396
tgataaagcg gtacatcccc ttcaatttcc actaagcttt cgaacaaaag cttggggagg 60
gcttcgagtc gggtatacag g 81
<210> 1397
<211> 79
<212> DNA
<213> Bacillus sp.
<400> 1397
ggtatacagg gtacatcccc ttcaatttcc actaagcttt cgaacccttg tccgcgataa 60
atctgtacgt cgaatccgt 79
<210> 1398
<211> 81
<212> DNA
<213> Bacillus sp.
<400> 1398
tcgaatccgt gtacatcccc ttcaatttcc actaagcttt cgaactgttt ggttgctgcg 60
cggccgaagt tgtctgacag t 81
<210> 1399
<211> 84
<212> DNA
<213> Bacillus sp.
<400> 1399
gtctgacagt gtacatcccc ttcaatttcc actaagcttt cgaacccgtt attaagttac 60
ctctttctag cataacctga gtgt 84
<210> 1400
<211> 88
<212> DNA
<213> Bacillus sp.
<400> 1400
acctgagtgt gtacatcccc ttcaatttcc actaagcttt cgaacatgta catcctgcat 60
cagttcgaag gaagcgggat cagctagt 88
<210> 1401
<211> 86
<212> DNA
<213> Bacillus sp.
<400> 1401
atcagctagt gtacatcccc ttcaatttcc actaagcttt cgaaccaaga gacgtgttga 60
cgatcatctt ttttgagatc tgccgt 86
<210> 1402
<211> 87
<212> DNA
<213> Bacillus sp.
<400> 1402
gatctgccgt gtacatcccc ttcaatttcc actaagcttt cgaacgcgca actcttgcaa 60
agttttttgc agcaacattt tttccgt 87
<210> 1403
<211> 82
<212> DNA
<213> Bacillus sp.
<400> 1403
ttttttccgt gtacatcccc ttcaatttcc actaagcttt cgaactcaac catacttatt 60
tattttagca ctgccccaaa gt 82
<210> 1404
<211> 56
<212> DNA
<213> Bacillus sp.
<400> 1404
gccccaaagt gtacatcccc ttcaatttcc actaagcttt cgaacaacct ctgctg 56
<210> 1405
<211> 35
<212> DNA
<213> Bacillus sp.
<400> 1405
gtacatcccc ttcaatttcc actaagcttt cgaac 35
<210> 1406
<211> 36
<212> DNA
<213> Bacillus sp.
<400> 1406
gtgctaacca cgaagctttc cactaagctt tcgaac 36
<210> 1407
<211> 83
<212> DNA
<213> Desulfatirhabdium butyrativorans
<400> 1407
cccatccatt tgtccgtcga tcaagctgct ttcaccatcg gaaccccggc caccgaggca 60
gccaacacca cccaggccga gct 83
<210> 1408
<211> 83
<212> DNA
<213> Desulfatirhabdium butyrativorans
<400> 1408
aggccgagct gtgtcagtcg atcaagctgt tttcaccatc ggaacccctt gaattgagac 60
ggttccggag cgcaatatgg tga 83
<210> 1409
<211> 59
<212> DNA
<213> Desulfatirhabdium butyrativorans
<400> 1409
aatatggtga gtgtcagtcg atcaagctgt tttcaccatc ggaacccccc ccggccatc 59
<210> 1410
<211> 38
<212> DNA
<213> Desulfatirhabdium butyrativorans
<400> 1410
gtgtcagtcg atcaagctgt tttcaccatc ggaacccc 38
<210> 1411
<211> 38
<212> DNA
<213> Desulfatirhabdium butyrativorans
<400> 1411
gtgtcagtcg atcaagctgt tttcaccatc ggaacccc 38
<210> 1412
<211> 19
<212> DNA
<213> Desulfatirhabdium butyrativorans
<400> 1412
gtgtcagtcg atcaagctg 19
<210> 1413
<211> 19
<212> DNA
<213> Desulfatirhabdium butyrativorans
<400> 1413
ttttcaccat cggaacccc 19
<210> 1414
<211> 82
<212> DNA
<213> Brevibacillus agri
<400> 1414
cgaggatttt gtgctaacca cgaagctttc cactaagctt tcgaaccata ccgccgttta 60
ctttacctgg tacggcgact at 82
<210> 1415
<211> 81
<212> DNA
<213> Brevibacillus agri
<400> 1415
cggcgactat gtgctaacca cgaagctttc cactaagctt tcgaactctg caagtgcgcg 60
tctatgaaaa gaactgggaa a 81
<210> 1416
<211> 86
<212> DNA
<213> Brevibacillus agri
<400> 1416
aactgggaaa gtgctaacca cgaagctttc cactaagctt tcgaacgttc aaccttcttc 60
gtgagaaaat tgatgatatt ttgaag 86
<210> 1417
<211> 85
<212> DNA
<213> Brevibacillus agri
<400> 1417
tattttgaag gtgctaacca cgaagctttc cactaagctt tcgaaccgga tcactctcaa 60
cagctccatt acaacgatac ttatt 85
<210> 1418
<211> 83
<212> DNA
<213> Brevibacillus agri
<400> 1418
gatacttatt gtgctaacca cgaagctttc cactaagctt tcgaacgaag ttatgaagaa 60
agtctagcat gtattcccga tgt 83
<210> 1419
<211> 84
<212> DNA
<213> Brevibacillus agri
<400> 1419
ttcccgatgt gtgctaacca cgaagctttc cactaagctt tcgaaccatc aacagcaaca 60
tcaacacgag atcgaggttg aggc 84
<210> 1420
<211> 81
<212> DNA
<213> Brevibacillus agri
<400> 1420
aggttgaggc gtgctaacca cgaagctttc cactaagctt tcgaacctct gtttgcagca 60
gcatacttca aaaagtcttc a 81
<210> 1421
<211> 85
<212> DNA
<213> Brevibacillus agri
<400> 1421
aaagtcttca gtgctaacca cgaagctttc cactaagctt tcgaacataa tccgtttgct 60
gtacggatat aaaaatcttg tatag 85
<210> 1422
<211> 80
<212> DNA
<213> Brevibacillus agri
<400> 1422
tcttgtatag gtgctaacca cgaagctttc cactaagctt tcgaaccgta tgcgaagtct 60
gttatacacc gcattttatg 80
<210> 1423
<211> 57
<212> DNA
<213> Brevibacillus agri
<400> 1423
gcattttatg gtgctaacca cgaagctttc cactaagctt tcgaactcct tcccact 57
<210> 1424
<211> 36
<212> DNA
<213> Brevibacillus agri
<400> 1424
gtgctaacca cgaagctttc cactaagctt tcgaac 36
<210> 1425
<211> 87
<212> DNA
<213> Brevibacillus agri
<400> 1425
tggacaagct gtgctaacca cgaagctttc cactaagctt tcgaacttct taacacgcga 60
cgactctttc cccacacgat ttcgatg 87
<210> 1426
<211> 84
<212> DNA
<213> Brevibacillus agri
<400> 1426
gatttcgatg gtgctaatcc cgaagctttc cactaagctt tcgaacaact ataacataac 60
ccaagcaacc gcttatagtt tctg 84
<210> 1427
<211> 86
<212> DNA
<213> Brevibacillus agri
<400> 1427
atagtttctg gtgctaatcc cgaagctttc cactaagctt tcgaacacca gcgccttgtc 60
tgccatgtat tgctgtgcca aagcca 86
<210> 1428
<211> 81
<212> DNA
<213> Brevibacillus agri
<400> 1428
gccaaagcca gtgctaatcc cgaagctttc cactaagctt tcgaactaac cgtaagcctg 60
taacgaagcg ttcgctccaa c 81
<210> 1429
<211> 81
<212> DNA
<213> Brevibacillus agri
<400> 1429
tcgctccaac gtgctaatcc cgaagctttc cactaagctt tcgaactggg acggtgcaca 60
gcagaccgta aaagatgcgt t 81
<210> 1430
<211> 82
<212> DNA
<213> Brevibacillus agri
<400> 1430
aagatgcgtt gtgctaatcc cgaagctttc cactaagctt tcgaacatct agtcacccca 60
aggataattg tcgtcgtagt cc 82
<210> 1431
<211> 84
<212> DNA
<213> Brevibacillus agri
<400> 1431
gtcgtagtcc gtgctaatcc cgaagctttc cactaagctt tcgaacctct agcttgctgt 60
cgaagttagc cctactgttg atga 84
<210> 1432
<211> 84
<212> DNA
<213> Brevibacillus agri
<400> 1432
ctgttgatga gtgctaatcc cgaagctttc cactaagctt tcgaacatct gcatcacggg 60
tagcaacgcc cgcagtcagg tctt 84
<210> 1433
<211> 82
<212> DNA
<213> Brevibacillus agri
<400> 1433
gtcaggtctt gtgctaatcc cgaagctttc cactaagctt tcgaacgagg atcgtgtctg 60
tcaacgggta ttgcacgctt cc 82
<210> 1434
<211> 84
<212> DNA
<213> Brevibacillus agri
<400> 1434
gcacgcttcc gtgctaatcc cgaagctttc cactaagctt tcgaacacgg agtggatttt 60
agagaggggt ggtttttgtt tcta 84
<210> 1435
<211> 81
<212> DNA
<213> Brevibacillus agri
<400> 1435
tttgtttcta gtgctaatcc cgaagctttc cactaagctt tcgaacattt tcgttgccca 60
gacgggagag aggaagagag t 81
<210> 1436
<211> 82
<212> DNA
<213> Brevibacillus agri
<400> 1436
ggaagagagt gtgctaatcc cgaagctttc cactaagctt tcgaactctc ctgatacgcc 60
ccatcagact gcttttctag ca 82
<210> 1437
<211> 83
<212> DNA
<213> Brevibacillus agri
<400> 1437
ttttctagca gtgctaatcc cgaagctttc cactaagctt tcgaactcat atttatcctg 60
taagcttccc tcactcattt tta 83
<210> 1438
<211> 82
<212> DNA
<213> Brevibacillus agri
<400> 1438
ctcattttta gtgctaatcc cgaagctttc cactaagctt tcgaacttga ttgttcggct 60
aaggcgaagg acagagctaa gg 82
<210> 1439
<211> 82
<212> DNA
<213> Brevibacillus agri
<400> 1439
agagctaagg gtgctaatcc cgaagctttc cactaagctt tcgaacgtcc tgacataccg 60
atgttcggcg ctcggtcgaa gt 82
<210> 1440
<211> 86
<212> DNA
<213> Brevibacillus agri
<400> 1440
cggtcgaagt gtgctaatcc cgaagctttc cactaagctt tcgaacgcca ttcgccgagc 60
ctgcggacgt cttcttttcc ggatac 86
<210> 1441
<211> 81
<212> DNA
<213> Brevibacillus agri
<400> 1441
ttccggatac gtgctaatcc cgaagctttc cactaagctt tcgaacccca ggaaacttat 60
atcggcaata gtagctgtct g 81
<210> 1442
<211> 82
<212> DNA
<213> Brevibacillus agri
<400> 1442
tagctgtctg gtgctaatcc cgaagctttc cactaagctt tcgaacgtgc tgcgagtacg 60
agatcaagca ccgcaaggct aa 82
<210> 1443
<211> 85
<212> DNA
<213> Brevibacillus agri
<400> 1443
gcaaggctaa gtgctaatcc cgaagctttc cactaagctt tcgaacttgc cacatttctc 60
tacacggggc tgcgggtcag cgagc 85
<210> 1444
<211> 86
<212> DNA
<213> Brevibacillus agri
<400> 1444
gtcagcgagc gtgctaatcc cgaagctttc cactaagctt tcgaacgtaa cttgcttact 60
caaagtaagc gtgtagtcac ccgccg 86
<210> 1445
<211> 85
<212> DNA
<213> Brevibacillus agri
<400> 1445
tcacccgccg gtgctaatcc cgaagctttc cactaagctt tcgaacggca cgaattcgac 60
ttcgtccgta tgggcgacta tatga 85
<210> 1446
<211> 84
<212> DNA
<213> Brevibacillus agri
<400> 1446
gactatatga gtgctaatcc cgaagctttc cactaagctt tcgaacacta tggcaggatt 60
ggcagaactt tcgccgtttt ttca 84
<210> 1447
<211> 83
<212> DNA
<213> Brevibacillus agri
<400> 1447
cgttttttca gtgctaatcc cgaagctttc cactaagctt tcgaaccccc tttttgacca 60
tgccaagagg aaaagatagc gga 83
<210> 1448
<211> 85
<212> DNA
<213> Brevibacillus agri
<400> 1448
agatagcgga gtgctaatcc cgaagctttc cactaagctt tcgaacttat tttgggtctc 60
gtgagtcgat gcagttgcgt atcta 85
<210> 1449
<211> 82
<212> DNA
<213> Brevibacillus agri
<400> 1449
tgcgtatcta gtgctaatcc cgaagctttc cactaagctt tcgaacccac gtggcgaaag 60
caagttttcc atatcgtcat aa 82
<210> 1450
<211> 81
<212> DNA
<213> Brevibacillus agri
<400> 1450
atcgtcataa gtgctaatcc cgaagctttc cactaagctt tcgaaccttc tattttggga 60
gttggtgctg tgatggtggc g 81
<210> 1451
<211> 82
<212> DNA
<213> Brevibacillus agri
<400> 1451
gatggtggcg gtgctaatcc cgaagctttc cactaagctt tcgaacttct ttggtattca 60
ctccccatcc cgtctcatgg ct 82
<210> 1452
<211> 83
<212> DNA
<213> Brevibacillus agri
<400> 1452
tctcatggct gtgctaatcc cgaagctttc cactaagctt tcgaacgcta gccgtggaca 60
aggaagaaat aaaggaactc att 83
<210> 1453
<211> 82
<212> DNA
<213> Brevibacillus agri
<400> 1453
ggaactcatt gtgctaatcc cgaagctttc cactaagctt tcgaactacc actacctcaa 60
cgctcccgtc gaaaccatca tg 82
<210> 1454
<211> 80
<212> DNA
<213> Brevibacillus agri
<400> 1454
aaccatcatg gtgctaatcc cgaagctttc cactaagctt tcgaactata caggcccgct 60
tttttgatgt cgatgctgga 80
<210> 1455
<211> 82
<212> DNA
<213> Brevibacillus agri
<400> 1455
cgatgctgga gtgctaatcc cgaagctttc cactaagctt tcgaacggtt tcttgatgaa 60
acgcagcgcg tccgtaaagg at 82
<210> 1456
<211> 56
<212> DNA
<213> Brevibacillus agri
<400> 1456
cgtaaaggat gtgctaatcc cgaagctttc cactaagctt tcgaacccct gtcttc 56
<210> 1457
<211> 36
<212> DNA
<213> Brevibacillus agri
<400> 1457
gtgctaatcc cgaagctttc cactaagctt tcgaac 36
<210> 1458
<211> 36
<212> DNA
<213> Brevibacillus agri
<400> 1458
gtgctaacca cgaagctttc cactaagctt tcgaac 36
<210> 1459
<211> 397
<212> DNA
<213> Brevibacillus sp.
<400> 1459
gtgctaacca cgaagctttc cactaagctt tcgaacgctt acagcttccg gtcaaccccg 60
catccatccg catagtgcta atcccgaagc tttccactaa gctttcgaac tcaagatcga 120
ggcactgtac ccgggcacga aagggtgcta atcccgaagc tttccactaa gctttcgaac 180
ttaatatctt ggttgtagtt gcgcaaacct tttatgtgct aatcccgaag ctttccacta 240
agctttcgaa ccttactcag tttgcgtcct ccgaagaagt cgtccataag tgctaatccc 300
gaagctttcc actaagcttt cgaacgcgac acgaccgtat tcgtggacga gccggtcttt 360
gtgtgctaat cccgaagctt tccactaagc tttcgaa 397
<210> 1460
<211> 36
<212> DNA
<213> Brevibacillus sp.
<400> 1460
gtgctaacca cgaagctttc cactaagctt tcgaac 36
<210> 1461
<211> 80
<212> DNA
<213> Methylobacterium nodulans
<400> 1461
aacagatcgt gtgccaacgc gatcaggatc tggcgcccac tgcgaccccc cgtgtccaca 60
tcgacggccg cgatgcacca 80
<210> 1462
<211> 81
<212> DNA
<213> Methylobacterium nodulans
<400> 1462
cgatgcacca gtgccaacgc gctcaggatc tggcgcccac tgcgacttcc ggcgaccctg 60
gatctgcacg gctacacaac t 81
<210> 1463
<211> 81
<212> DNA
<213> Methylobacterium nodulans
<400> 1463
ctacacaact gtgccaacgc gctcaggatc tggcgcccac tgcgacttgg ccccgccggc 60
accgggtagg agacccgcca t 81
<210> 1464
<211> 79
<212> DNA
<213> Methylobacterium nodulans
<400> 1464
gacccgccat gtgccaacgc gctcaggatc tggcgcccac tgcgacatag ttctggctgt 60
tggaggccag attgttctg 79
<210> 1465
<211> 81
<212> DNA
<213> Methylobacterium nodulans
<400> 1465
gattgttctg gtgccaacgc gctcaggatc tggcgcccac tgcgacctga ccttcacgcg 60
tgtctactcg ggcgtggcca a 81
<210> 1466
<211> 81
<212> DNA
<213> Methylobacterium nodulans
<400> 1466
gcgtggccaa gtgccaacgc gctcaggatc tggcgcccac tgcgacgtcc aggccgagtt 60
gggcaaggtc cgccggatcc t 81
<210> 1467
<211> 56
<212> DNA
<213> Methylobacterium nodulans
<400> 1467
gccggatcct gtgccaacgc gctcaggatc tggcgcccac tgcgacggcg ggcacg 56
<210> 1468
<211> 36
<212> DNA
<213> Methylobacterium nodulans
<400> 1468
gtgccaacgc gctcaggatc tggcgcccac tgcgac 36
<210> 1469
<211> 36
<212> DNA
<213> Methylobacterium nodulans
<400> 1469
gtgccaacgc gctcaggatc tggcgcccac tgcgac 36
<210> 1470
<211> 12
<212> DNA
<213> Methylobacterium nodulans
<400> 1470
gtgccaacgc gc 12
<210> 1471
<211> 24
<212> DNA
<213> Methylobacterium nodulans
<400> 1471
tcaggatctg gcgcccactg cgac 24
<210> 1472
<211> 36
<212> DNA
<213> Alicyclobacillus acidoterrestris
<400> 1472
gtcggatcac tgagcgagcg atctgagaag tggcac 36
<210> 1473
<211> 37
<212> DNA
<213> Desulfonatronum thiodismutans
<400> 1473
gtctcggcaa gcttggtcag tgttgggtga ttggcac 37
<210> 1474
<211> 36
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1474
ccgcctgacg attcgtgaaa cggcattcgc tgcggc 36
<210> 1475
<211> 36
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1475
gtccaagaaa aaagaaatga tacgaggcat tagcac 36
<210> 1476
<211> 36
<212> DNA
<213> Bacillus sp.
<400> 1476
gtgctaacca cgaagctttc cactaagctt tcgaac 36
<210> 1477
<211> 38
<212> DNA
<213> Desulfatirhabdium butyrativorans
<400> 1477
gtgtcagtcg atcaagctgt tttcaccatc ggaacccc 38
<210> 1478
<211> 36
<212> DNA
<213> Brevibacillus agri
<400> 1478
gtgctaacca cgaagctttc cactaagctt tcgaac 36
<210> 1479
<211> 36
<212> DNA
<213> Brevibacillus sp.
<400> 1479
gtgctaacca cgaagctttc cactaagctt tcgaac 36
<210> 1480
<211> 36
<212> DNA
<213> Methylobacterium nodulans
<400> 1480
gtgccaacgc gctcaggatc tggcgcccac tgcgac 36
<210> 1481
<211> 36
<212> DNA
<213> Alicyclobacillus acidoterrestris
<400> 1481
gtcggatcac tgagcgagcg atctgagaag tggcac 36
<210> 1482
<211> 37
<212> DNA
<213> Desulfonatronum thiodismutans
<400> 1482
gtctcggcaa gcttggtcag tgttgggtga ttggcac 37
<210> 1483
<211> 36
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Opitutaceae bacterium sequence"
<400> 1483
ccgcctgacg attcgtgaaa cggcattcgc tgcggc 36
<210> 1484
<211> 36
<212> DNA
<213> Bacillus thermoamylovorans
<400> 1484
gtccaagaaa aaagaaatga tacgaggcat tagcac 36
<210> 1485
<211> 36
<212> DNA
<213> Bacillus sp.
<400> 1485
gtgctaacca cgaagctttc cactaagctt tcgaac 36
<210> 1486
<211> 38
<212> DNA
<213> Desulfatirhabdium butyrativorans
<400> 1486
gtgtcagtcg atcaagctgt tttcaccatc ggaacccc 38
<210> 1487
<211> 36
<212> DNA
<213> Brevibacillus agri
<400> 1487
gtgctaacca cgaagctttc cactaagctt tcgaac 36
<210> 1488
<211> 36
<212> DNA
<213> Brevibacillus sp.
<400> 1488
gtgctaacca cgaagctttc cactaagctt tcgaac 36
<210> 1489
<211> 36
<212> DNA
<213> Methylobacterium nodulans
<400> 1489
gtcgcagtgg gcgccagatc ctgagcgcgt tggcac 36
<210> 1490
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1490
gtcggatcac tgagcgagcg atctgagaag tggcac 36
<210> 1491
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1491
gtctcggcaa gcttggtcag tgttgggtga ttggcac 37
<210> 1492
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1492
gccgcagcga atgccgtttc acgaatcgtc aggcgg 36
<210> 1493
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1493
gtccaagaaa aaagaaatga tacgaggcat tagcac 36
<210> 1494
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1494
gttcgaaagc ttagtggaaa gcttcgtggt tagcac 36
<210> 1495
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1495
gttcgaaagc ttagtggaaa ttgaagggga tgtacca 37
<210> 1496
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1496
ggggttccga tggtgaaaac agcttgatcg actgac 36
<210> 1497
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1497
gttcgaaagc ttagtggaaa gcttcgtggt tagcac 36
<210> 1498
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1498
gttcgaaagc ttagtggaaa gcttcgtggt tagcac 36
<210> 1499
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1499
gtctcaacgg gcgccagttc ctgagctcgt tggcac 36
<210> 1500
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1500
tgatgtcagc gaaacgacca ccatcggggt tataac 36
<210> 1501
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1501
gggtgcggca tttgcgggtg ttgggggagt ggcagg 36
<210> 1502
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1502
cagtggatgt ttttccatga ggcgaagaat ttcatc 36
<210> 1503
<211> 26
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1503
aacaatataa acgactactt taccgt 26
<210> 1504
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1504
agcttgtcca acttgatgct ccttttcatc 30
<210> 1505
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1505
atcatattca agagcgtgta cactactacg ccgaac 36
<210> 1506
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1506
acagctcggc ctgggtggtg ttggctgcct cggtggcc 38
<210> 1507
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1507
agcttgtcca acttgatgct ccttttcatc 30
<210> 1508
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1508
agcttgtcca acttgatgct ccttttcatc 30
<210> 1509
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1509
tgcatgcgct cctcgaggcg gaggatccga tcgc 34
<210> 1510
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1510
gtcggatcac tgagtgatct aacctatcaa atgcccaaac cacctcctgc ggggttcgaa 60
tcccttcgag tgcgccacaa acataagtaa aacgccggat tcagtttatc gagtccagcg 120
tttttctttt ctactgcagt gttctacgca tcttactatg ccgttgtaat acgaagattt 180
gcgaacatct tggaatacag 200
<210> 1511
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1511
gtctcggcaa gcttggtcag tgatgggtga ttgacatcag tgatgggtga ttgacactag 60
aagcgcacgg cctaggttac gttcttgggg aaagctaggc aagttttgga tgataagaaa 120
taatcatgtc acaaggaggg agtttttgtg gcgagaatca ccatgtatcg aattcaaatc 180
aatacagaag ctataagaaa 200
<210> 1512
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1512
gccgcagtga atgccgtttc accattgatg aagaatgcga ggtgaaaaca gagaaattgg 60
gtcaactcta tcactcttat tcagccatcg tttcaagaaa ggatacctcg tattggatac 120
aacacagctc gttcgttctc tctacctccc tcgacaatct caaggactat ggcgatgcca 180
gcgatggaac agccgaggca 200
<210> 1513
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1513
gttcaagaaa aaagaaatga tatgaggcat tagcacgatg ggatgggaga gagaggacag 60
ttctactctt gctgtatcca gcttctttta ctttatccgg tatcatttct tcacttcttt 120
ctgcacataa aaaagcacct aactatttgg ataagttaag tgcttttatt tccgtttgaa 180
gttgtctatt gcttttttct 200
<210> 1514
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1514
gttcgaaaac ttggtgaaat accttgaaaa ttaacacatc aaagatgccc ctgctttacg 60
ttaggggcat ctttctatta aataacctat agtgaacttc aaaatcattg atctcttgat 120
aataatatat accaaattca atagtcatag tgagaggttt acaggtgctc gatacgtttt 180
cttgcaaaaa agaagcagtc 200
<210> 1515
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1515
gttcgaaagc ttagtggaaa ttgaagggga tgtaccagag gtagcactta gcaagttttt 60
tgagaaacaa gtatctggcc ccaatagcta ggcttatgtg aaatgtagca tgagtggtgt 120
gcaatgaaaa aatagggagg atattagcac tatgaaaaag cagcttaaaa aaacagttaa 180
atgcgtgatg gcgtctatat 200
<210> 1516
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1516
ggggttccga tggtgaaagc agcttgatcg acggacaaat ggatgggcat gcgtaggggc 60
ggttcgcgaa ccgcccctac aactacaccg aaagataccg cgactgaatc tcggtgttgg 120
ccagaagctc ctgcctggtg ccttcgtagc ggatgaggcc tttgtcgatg acatagcccc 180
gatcggagat ggagagggcg 200
<210> 1517
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1517
gttcgaaaac ttggtgaaat accttgaaaa ttaacacatc aaagatgccc ctgctttacg 60
ttaggggcat ctttctatta aataacctat agtgaacttc aaaatcattg atctcttgat 120
aataatatat accaaattca atagtcatag tgagaggttt acaggtgctc gatacgtttt 180
cttgcaaaaa agaagcagtc 200
<210> 1518
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1518
gttcgaaaac ttggtgaaat accttgaaaa ttaacacatc aaagatgccc ctgctttacg 60
ttaggggcat ctttctatta aataacctat agtgaacttc aaaatcattg atctcttgat 120
aataatatat accaaattca atagtcatag tgagaggttt acaggtgctc gatacgtttt 180
cttgcaaaaa agaagcagtc 200
<210> 1519
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1519
gtcgcaacgg atcccagatc ctgatctcgc tggcaggtgc cctggtagat gacgactcgc 60
gctcgttgca gggaggctcg cgcagacgag cgggaaactg cttgtcaggc aatcgcctgt 120
gctgggtccg ctggattctg tgttcgttgg cacgggggag ccgtgggaat gcctactgcc 180
tggatggggt cgcaccaagt 200
<210> 1520
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1520
gttcgaaagc ttagtggaaa ttgaagggga tgtaccc 37
<210> 1521
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1521
gucggaucac ugagcgagcg aucugagaag uggcac 36
<210> 1522
<211> 158
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1522
ucuagaggac agaauuuuuc aacgggugug ccaauggcca cuuuccaggu ggcaaagccc 60
guugagcuuc ucaaaaagaa cgcucgcuca guguucugac cuuucgagcg ccuguucagg 120
gcgaaaaccc ugggaggcgc ucgaaucaua ggugggac 158
<210> 1523
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1523
gccgcagcga augccguuuc acgaaucguc aggcgg 36
<210> 1524
<211> 75
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1524
gcuggagacg uuuuuugaaa cggcgagugc ugcggauagc gaguuucucu uggggaggcg 60
cucgcggcca cuuuu 75
<210> 1525
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1525
guccaagaaa aaagaaauga uacgaggcau uagcac 36
<210> 1526
<211> 107
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1526
cuggacgaug ucucuuuuau uucuuuuuuc uuggaucuga guacgagcac ccacauugga 60
cauuucgcau ggugggugcu cguacuauag guaaaacaaa ccuuuuu 107
<210> 1527
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1527
guucgaaagc uuaguggaaa gcuucguccu uagcac 36
<210> 1528
<211> 69
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1528
cacggauaau cacgacuuuc cacuaagcuu ucgaauuuua ugaugcgagc auccucucag 60
gucaaaaaa 69
<210> 1529
<211> 79
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1529
gaagaaatga ctattgccaa ctatatctgg cttttctcaa tacccttagc gcgaaaatac 60
cccctcgcca taaccaacc 79
<210> 1530
<211> 84
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1530
ataaccaacc gtctattgcc atctttatct ggcttttctc aatactatca aggtacagca 60
aatatgctca tcagtgctat gaag 84
<210> 1531
<211> 82
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1531
tgctatgaag gtctattgcc atctttatct ggcttttctc aatacagtag aaatgattat 60
acctgacaac gttaaagaga gt 82
<210> 1532
<211> 79
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1532
taaagagagt gtctattgcc atctttatct ggcttttctc aatacaaaag aaaccaaaaa 60
attcattgcg taacaccat 79
<210> 1533
<211> 80
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1533
gtaacaccat gtctattgcc aactatatct ggcttttctc aatacaaaat acgcacagcc 60
tcattaccac tcagtttatt 80
<210> 1534
<211> 80
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1534
tcagtttatt gtctattgcc aactatatct ggcttttctc aatactaaaa tggatgattg 60
gtggctcgat gacactgtcc 80
<210> 1535
<211> 82
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1535
gacactgtcc gtctattgcc aactatatct ggcttttctc aatacagctt ctagtggtat 60
gccttcggga ccatcaaagg aa 82
<210> 1536
<211> 81
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1536
atcaaaggaa gtctattgcc aactatatct ggcttttctc aatacgcata tcttgaggat 60
tgggattctg aactcttaga g 81
<210> 1537
<211> 82
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1537
actcttagag gtctattgcc aactatatct ggcttttctc aatacctcac ttattcacct 60
gggttaactg catgaactca ac 82
<210> 1538
<211> 55
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1538
tgaactcaac gtctattgcc aactttatct ggcttttctc aataccctac cttct 55
<210> 1539
<211> 35
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1539
gtctattgcc aactatatct ggcttttctc aatac 35
<210> 1540
<211> 35
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1540
gtctattgcc aactatatct ggcttttctc aatac 35
<210> 1541
<211> 78
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1541
accttccaga gttattgccc tctatcttgg actcttctca tcaactcata ataccaagaa 60
gaaatcaagc atattatc 78
<210> 1542
<211> 80
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1542
gcatattatc gttattgccc tctatcttgg gctcttctca tcaacacgag ataggcaaac 60
cgaagaacat gatatatata 80
<210> 1543
<211> 78
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1543
gatatatata gttattgccc tctatcttgg gctcttctca tcaacgaata cgatacaact 60
ggcgactggt ttgattaa 78
<210> 1544
<211> 55
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1544
gtttgattaa gttattgccc tctatcttgg gctcttctca tcaacaccgt cctct 55
<210> 1545
<211> 35
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1545
gttattgccc tctatcttgg gctcttctca tcaac 35
<210> 1546
<211> 35
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1546
gttattgccc tctatcttgg gctcttctca tcaac 35
<210> 1547
<211> 80
<212> DNA
<213> Clostridium aminophilum
<400> 1547
aaagagcagg gtttggagaa cagcccgata tagagggcaa tagacgcctt ttgtcgcaag 60
tgttgccgta ccggataacg 80
<210> 1548
<211> 80
<212> DNA
<213> Clostridium aminophilum
<400> 1548
ccggataacg gtttggagaa cagcccgata tagagggcaa tagacgcaac atatgcttca 60
tcggggaaga gttctttccc 80
<210> 1549
<211> 80
<212> DNA
<213> Clostridium aminophilum
<400> 1549
gttctttccc gtttggagaa cagcccgata tagagggcaa tagacccgga agcaattccc 60
ccaatggcct gtccaagcca 80
<210> 1550
<211> 79
<212> DNA
<213> Clostridium aminophilum
<400> 1550
gtccaagcca gtttggagaa cagcccgata tagagggcaa tagacttaat tgccggatac 60
ttgagatgga acttgtcac 79
<210> 1551
<211> 78
<212> DNA
<213> Clostridium aminophilum
<400> 1551
aacttgtcac gtttggagaa cagcccgata tagagggcaa tagacatatc atcatcaaga 60
gtctgccttg catcacaa 78
<210> 1552
<211> 79
<212> DNA
<213> Clostridium aminophilum
<400> 1552
tgcatcacaa gtttggagaa cagcccgata tagagggcaa tagacgttta tgtaaactct 60
taacgagtaa ccatcttaa 79
<210> 1553
<211> 81
<212> DNA
<213> Clostridium aminophilum
<400> 1553
accatcttaa gtttggagaa cagcccgata aagagggcaa tagaccagac catacctttt 60
cagacgggat ctttaccatc t 81
<210> 1554
<211> 79
<212> DNA
<213> Clostridium aminophilum
<400> 1554
tttaccatct gtttggagaa cagcccgata aagagggcaa tagacctgat gctgcgaatc 60
tgccagaact cttgcgacc 79
<210> 1555
<211> 56
<212> DNA
<213> Clostridium aminophilum
<400> 1555
tcttgcgacc gtttggagaa cagcccgata tagagggcaa tagacggtgt acttgt 56
<210> 1556
<211> 35
<212> DNA
<213> Clostridium aminophilum
<400> 1556
gtttggagaa cagcccgata tagagggcaa tagac 35
<210> 1557
<211> 35
<212> DNA
<213> Clostridium aminophilum
<400> 1557
gtttggagaa cagcccgata tagagggcaa tagac 35
<210> 1558
<211> 81
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1558
agaagacagg gttttgagaa tagcccgaca tagagggcaa tagacaacgg ttttaccatt 60
gtcaaactcg cctgctgtcg t 81
<210> 1559
<211> 84
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1559
ctgctgtcgt gttttgagaa tagcccgaca tagagggcaa tagacttttg cttcgtcacg 60
gatggacttc acaatggcaa caac 84
<210> 1560
<211> 79
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1560
tggcaacaac gttttgagaa tagcccgaca tagagggcaa tagaccttca taaatccaag 60
atacggatgc atgattacg 79
<210> 1561
<211> 77
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1561
catgattacg gttttgagaa tagcccgaca tagagggcaa tagactttta aagggatgat 60
aaggaagccg tatactc 77
<210> 1562
<211> 82
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1562
ccgtatactc gttttgagaa tagcccgaca tagagggcaa tagacctaag ctggtttttc 60
catgttgaca ccttgcctag tt 82
<210> 1563
<211> 81
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1563
ttgcctagtt gttttgagaa tagcccgaca tagagggcaa tagacttctt ctcttgtcat 60
tcttctacct ctaaaatctc a 81
<210> 1564
<211> 35
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1564
gttttgagaa tagcccgaca tagagggcaa tagac 35
<210> 1565
<211> 35
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1565
gttttgagaa tagcccgaca tagagggcaa tagac 35
<210> 1566
<211> 35
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1566
gttttgagaa tagcccgaca tagagggcaa tagac 35
<210> 1567
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1567
aaaatggaaa gttatagtcc tcttacattt agaggtagtc tttaattaaa agattacaat 60
gaaggttatc cacttc 76
<210> 1568
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1568
tatccacttc gttatagtcc tcttacattt agaggtagtc tttaatttaa ctcatgggat 60
atgaattatc atttag 76
<210> 1569
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1569
tatcatttag gttatagtcc tcttacattt agaggtagtc tttaatgtat taggaacaat 60
aaacgactct ttttta 76
<210> 1570
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1570
ctctttttta gttatagtcc tcttacattt agaggtagtc tttaattcaa attaatcgtt 60
ctctttatat ctggga 76
<210> 1571
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1571
atatctggga gttatagtcc tcttacattt agaggtagtc tttaattgat tatatcgaaa 60
atcaaataaa tgcgct 76
<210> 1572
<211> 37
<212> DNA
<213> Carnobacterium gallinarum
<400> 1572
gttatagtcc tcttacattt agaggtagtc tttaatt 37
<210> 1573
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1573
ccaataaacg gttatagtcc tcttacattt agaggtagtc tttaattctc ctttttcatg 60
aatggccgtt aaccct 76
<210> 1574
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1574
cgttaaccct gttatagtcc tcttacattt agaggtagtc tttaattgcg acaatcagta 60
tgattacgat gctgac 76
<210> 1575
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1575
cgatgctgac gttatagtcc tcttacattt agaggtagtc tttaattttt acataaaaaa 60
aggagggtat aatcat 76
<210> 1576
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1576
gtataatcat gttatagtcc tcttacattt agaggtagtc tttaatttac gccaagaatt 60
attaatgttg atgcaa 76
<210> 1577
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1577
gttgatgcaa gttatagtcc tcttacattt agaggtagtc tttaatcttt atccatgaat 60
taactcatgc gattgc 76
<210> 1578
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1578
atgcgattgc gttatagtcc tcttacattt agaggtagtc tttaatagaa atatattaat 60
agcgacttat attaca 76
<210> 1579
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1579
ttatattaca gttatagtcc tcttacattt agaggtagtc tttaatatat tggagaaaaa 60
agaaaaagat tagtca 76
<210> 1580
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1580
agattagtca gttatagtcc tcttacattt agaggtagtc tttaattcct ctattgtact 60
taacatcatc tactga 76
<210> 1581
<211> 56
<212> DNA
<213> Carnobacterium gallinarum
<400> 1581
catctactga gttatagtcc tcttacattt agaggtagtc tttaataccc ttctat 56
<210> 1582
<211> 36
<212> DNA
<213> Carnobacterium gallinarum
<400> 1582
gttatagtcc tcttacattt agaggtagtc tttaat 36
<210> 1583
<211> 37
<212> DNA
<213> Carnobacterium gallinarum
<400> 1583
gttatagtcc tcttacattt agaggtagtc tttaatt 37
<210> 1584
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1584
tctagttctg attaaagact acccctaaat gtaaggggac tataactcct caatgctttt 60
attttgagcg ttcgct 76
<210> 1585
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1585
agcgttcgct attaaagact acccctaaat gtaaggggac tataactacc gactagagaa 60
ctctttagtc acttga 76
<210> 1586
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1586
agtcacttga attaaagact acccctaaat gtaaggggac tataactgct tttgttaaat 60
tgattattgg gtcttc 76
<210> 1587
<211> 37
<212> DNA
<213> Carnobacterium gallinarum
<400> 1587
attaaagact acccctaaat gtaaggggac tataact 37
<210> 1588
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1588
gttgaggggt aatataaact acctctaaat gtaagaggac tataacttta ttagttgttc 60
atctgttttt ttttaa 76
<210> 1589
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1589
ttttttttaa aatataaact acctctaaat gtaagaggac tataacctac aaaaacgact 60
aaaaaagcgt ttaaag 76
<210> 1590
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1590
gcgtttaaag aatataaact acctctaaat gtaagaggac tataactttg gttgtacccc 60
tttttgaaca tatagg 76
<210> 1591
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1591
aacatatagg aatataaact acctctaaat gtaagaggac tataactgcg taatgatgag 60
gtggcagaat caatgc 76
<210> 1592
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1592
gaatcaatgc aatataaact acctctaaat gtaagaggac tataaccata aaaccttgat 60
tttcataaga tatatc 76
<210> 1593
<211> 76
<212> DNA
<213> Carnobacterium gallinarum
<400> 1593
aagatatatc aatataaact acctctaaat gtaagaggac tataacccaa tagtcaattc 60
tgttagccag taggga 76
<210> 1594
<211> 57
<212> DNA
<213> Carnobacterium gallinarum
<400> 1594
ccagtaggga aatataaact acctctaaat gtaagaggac tataactgct atataaa 57
<210> 1595
<211> 36
<212> DNA
<213> Carnobacterium gallinarum
<400> 1595
aatataaact acctctaaat gtaagaggac tataac 36
<210> 1596
<211> 37
<212> DNA
<213> Carnobacterium gallinarum
<400> 1596
attaaagact acccctaaat gtaaggggac tataact 37
<210> 1597
<211> 75
<212> DNA
<213> Paludibacter propionicigenes
<400> 1597
gcatgtcctt gttttagttc ccttcaattt tgggataatc cacaagtatt actgctgcaa 60
tgcagattgg cgtat 75
<210> 1598
<211> 75
<212> DNA
<213> Paludibacter propionicigenes
<400> 1598
attggcgtat gttgtagttc ccttcaattt tgggataatc cacaagtatt actgctgcaa 60
tgcagattgg cgtat 75
<210> 1599
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1599
attggcgtat gttgtagttc ccttcaattt tgggataatc cacaaggaaa acgaaaatta 60
taacttcaat aaattc 76
<210> 1600
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1600
caataaattc gttgtagttc cctttaattt tgggataatc cacaagagta aaaccgtaca 60
atggagaaga taacgt 76
<210> 1601
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1601
aagataacgt gatgtagttc ccttcaattt tgggatagtc cacaagagcg gtgcggattt 60
gagcgatgcg aatttg 76
<210> 1602
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1602
tgcgaatttg gttgtagttc ccttcaatat tgggataatc cacaaggaat taggattatg 60
agtttatata gtttga 76
<210> 1603
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1603
tatagtttga gttgtagttc ccttcaatat tgggataatt cacaagtaaa cattaaaaca 60
ataaacggaa gttcga 76
<210> 1604
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1604
ggaagttcga gttgtagttc ccttcaattt tgggataatc cataagaaag caaaaatgaa 60
cacaaaacaa tttgaa 76
<210> 1605
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1605
acaatttgaa gttttagttc ccttcgattt tgggataatc cacaagtaca gcagtaagcg 60
gaaacactca ggcaat 76
<210> 1606
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1606
ctcaggcaat gttgtagttc ccttcaattt tgggataatc cacaaggccg taaaaaatcc 60
acacaacgat aagatg 76
<210> 1607
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1607
cgataagatg gttttagttc ccttcaattt tgggataatc cataagctct tactggctgg 60
tgccgggctt gctgca 76
<210> 1608
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1608
gcttgctgca gttgtagttc ccttcaattt tgggattatc cacaagagaa agagtgatga 60
acgacgagca ggatat 76
<210> 1609
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1609
agcaggatat gttgtagttc ccttcaattt tgggattatc cacaagttac acctgatcaa 60
tggaaattgc ttagtg 76
<210> 1610
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1610
ttgcttagtg gttgtagttc ccttcaattt tgggataatc cacaagtgtg attaatgata 60
ttaccgtagc taacgg 76
<210> 1611
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1611
tagctaacgg gttgtagttc ccttcattat tgggatagtc cacaagaaaa caacctgcat 60
ggataaatta tatgac 76
<210> 1612
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1612
attatatgac gttctagctc ccttcaattt tgggataatc cacaagtaaa aacatatttg 60
ctgaagacta tggaag 76
<210> 1613
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1613
actatggaag gttgtagttc ccttcaatat tgggataatc cacaagtgct attcggaata 60
gatttttacc ccacca 76
<210> 1614
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1614
taccccacca gttgtagttc ccttcaattt tgggataatt cacaagtggg attatgaaat 60
aaccgattat aaacta 76
<210> 1615
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1615
ttataaacta gttgtagttc ccttcaattt tgggataatt cacaagagaa atcaaaaatt 60
gattcttcag tagctt 76
<210> 1616
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1616
tcagtagctt gttgtagttc ccttcaattt tgggataatt cacaagcgaa tcctaatggt 60
tatattactg cttcat 76
<210> 1617
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1617
actgcttcat gttgtagttc ccttcaattt tgggataatt cacaagtaaa ttgaaaattt 60
ctggacaagc tccagt 76
<210> 1618
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1618
aagctccagt gttgtagttc ccttcaatat tgggataatc cacaagaaac gaatcacaga 60
acatgtgaac ctggta 76
<210> 1619
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1619
gaacctggta gttgtagttc ccttcaattt tgggataatc cacaagagat gataacgtag 60
gcaacatccc tgataa 76
<210> 1620
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1620
tccctgataa gttttagttc ccttcaattt tgggataatc cacaagcgtt attcaggaga 60
gagtttcaaa gagtat 76
<210> 1621
<211> 76
<212> DNA
<213> Paludibacter propionicigenes
<400> 1621
caaagagtat gttttagttc ccttcaatat tgggataatc cacaagtaaa acattggcta 60
ataacagaac tagggc 76
<210> 1622
<211> 56
<212> DNA
<213> Paludibacter propionicigenes
<400> 1622
gaactagggc gttttagttc ccttcaatat tgggataatc cacaagatag tatctg 56
<210> 1623
<211> 36
<212> DNA
<213> Paludibacter propionicigenes
<400> 1623
gttgtagttc ccttcaattt tgggataatc cacaag 36
<210> 1624
<211> 36
<212> DNA
<213> Paludibacter propionicigenes
<400> 1624
gttgtagttc ccttcaattt tgggataatc cacaag 36
<210> 1625
<211> 36
<212> DNA
<213> Paludibacter propionicigenes
<400> 1625
gttgtagttc ccttcaattt tgggataatc cacaag 36
<210> 1626
<211> 36
<212> DNA
<213> Paludibacter propionicigenes
<400> 1626
gttgtagttc ccttcaattt tgggataatc cacaag 36
<210> 1627
<211> 76
<212> DNA
<213> Listeria seeligeri
<400> 1627
tgttattgca gtaagagact acctctatat gaaagaggac taaaaccata tttccaaact 60
ccactttgac tacacc 76
<210> 1628
<211> 76
<212> DNA
<213> Listeria seeligeri
<400> 1628
tgactacacc gtaagagact acctctatat gaaagaggac taaaacggtc ccactacttg 60
aggtacgaac atatca 76
<210> 1629
<211> 76
<212> DNA
<213> Listeria seeligeri
<400> 1629
gaacatatca gtaagagact acctctatat gaaagaggac taaaacttag tcaacccctc 60
gctgcatttt cacatt 76
<210> 1630
<211> 76
<212> DNA
<213> Listeria seeligeri
<400> 1630
ttttcacatt gtaagagact acctctatat gaaagaggac taaaacgatg gataataggg 60
atagatcatt agtccg 76
<210> 1631
<211> 56
<212> DNA
<213> Listeria seeligeri
<400> 1631
cattagtccg gtaagagact acctctatat gaaagaggac taaaacgtct aatgtg 56
<210> 1632
<211> 36
<212> DNA
<213> Listeria seeligeri
<400> 1632
gtaagagact acctctatat gaaagaggac taaaac 36
<210> 1633
<211> 36
<212> DNA
<213> Listeria seeligeri
<400> 1633
gtaagagact acctctatat gaaagaggac taaaac 36
<210> 1634
<211> 76
<212> DNA
<213> Listeria weihenstephanensis
<400> 1634
tattttcata gatttagagt acctcaaaac agaagaggac taaaacgcac tctccgacaa 60
taatctcgtc catttt 76
<210> 1635
<211> 76
<212> DNA
<213> Listeria weihenstephanensis
<400> 1635
cgtccatttt gatttagagt acctcaaaac aaaagaggac taaaacaact ctgtacttgt 60
gaagtacgtt aaatcc 76
<210> 1636
<211> 56
<212> DNA
<213> Listeria weihenstephanensis
<400> 1636
cgttaaatcc gatttagagt acctcaaaac aaaagaggac taaaacctct tttgtg 56
<210> 1637
<211> 36
<212> DNA
<213> Listeria weihenstephanensis
<400> 1637
gatttagagt acctcaaaac aaaagaggac taaaac 36
<210> 1638
<211> 76
<212> DNA
<213> Listeria weihenstephanensis
<400> 1638
gtctacaagt gatttagagt agctcaaaaa agaagaggtc taaaacagag atactgatac 60
tattgagccc agtcat 76
<210> 1639
<211> 76
<212> DNA
<213> Listeria weihenstephanensis
<400> 1639
gcccagtcat gatttagagt acctcaaaat agaagaggtc taaaactgtt ccaaggcttg 60
ctaactcacc tttttc 76
<210> 1640
<211> 76
<212> DNA
<213> Listeria weihenstephanensis
<400> 1640
cacctttttc gatttagagt acctcaaaat agaagaggtc taaaacatct tgtttatatc 60
cgtatcggca cccaaa 76
<210> 1641
<211> 76
<212> DNA
<213> Listeria weihenstephanensis
<400> 1641
ggcacccaaa gatttagagt gcctcaaaat agaagaggtc taaaactcac tctccatcca 60
ctcaatcaga tcattt 76
<210> 1642
<211> 76
<212> DNA
<213> Listeria weihenstephanensis
<400> 1642
cagatcattt gatttagagt acctcaaaat agaagaggtc taaaactatc aatttccttt 60
tgatttctta aatcag 76
<210> 1643
<211> 76
<212> DNA
<213> Listeria weihenstephanensis
<400> 1643
cttaaatcag gatttagagt acctcaaaat agaagaggtc taaaacgttc caatatggtt 60
ttttccacat cttccg 76
<210> 1644
<211> 57
<212> DNA
<213> Listeria weihenstephanensis
<400> 1644
acatcttccg gatttagagt acctcaaaat agaagaggtc taaaacctca aattgaa 57
<210> 1645
<211> 36
<212> DNA
<213> Listeria weihenstephanensis
<400> 1645
gatttagagt acctcaaaat agaagaggtc taaaac 36
<210> 1646
<211> 36
<212> DNA
<213> Listeria weihenstephanensis
<400> 1646
gatttagagt acctcaaaac aaaagaggac taaaac 36
<210> 1647
<211> 36
<212> DNA
<213> Listeria weihenstephanensis
<400> 1647
gatttagagt acctcaaaac aaaagaggac taaaac 36
<210> 1648
<211> 75
<212> DNA
<213> Listeria newyorkensis
<400> 1648
cgtaatgctt gatttagagt acctcaaaac aaaagaggac taaaactact tgtcgatatg 60
gtatagcttt tttca 75
<210> 1649
<211> 76
<212> DNA
<213> Listeria newyorkensis
<400> 1649
gcttttttca gatttagagt acctcaaaac aaaagaggac taaaactaaa gcttctaaat 60
ggtggcgcgt tacgcc 76
<210> 1650
<211> 76
<212> DNA
<213> Listeria newyorkensis
<400> 1650
gcgttacgcc gatttagaat acctcaaaac aaaagaggac taaaacctag caagccggtc 60
gccgcgctca aagtaa 76
<210> 1651
<211> 57
<212> DNA
<213> Listeria newyorkensis
<400> 1651
ctcaaagtaa gatttagagt acctcaaaac aaaagaggac taaaacctct tttgtgg 57
<210> 1652
<211> 36
<212> DNA
<213> Listeria newyorkensis
<400> 1652
gatttagagt acctcaaaac aaaagaggac taaaac 36
<210> 1653
<211> 36
<212> DNA
<213> Listeria newyorkensis
<400> 1653
gatttagagt acctcaaaac aaaagaggac taaaac 36
<210> 1654
<211> 36
<212> DNA
<213> Listeria newyorkensis
<400> 1654
gatttagagt acctcaaaac aaaagaggac taaaac 36
<210> 1655
<211> 78
<212> DNA
<213> Leptotrichia wadei
<400> 1655
atgatgtatg taagttttag tccccttcgt ttttggggta gtctaaatcc tggtaaacca 60
atccacatcg aagaaaag 78
<210> 1656
<211> 76
<212> DNA
<213> Leptotrichia wadei
<400> 1656
cgaagaaaag aaagttttag tccccttcgt ttttggggta gtctaaatca tgtagaagaa 60
gttattgtat ctattt 76
<210> 1657
<211> 59
<212> DNA
<213> Leptotrichia wadei
<400> 1657
gtatctattt tacgttttag tccccttcgt ttttggggta gtctaaatct ttatatcaa 59
<210> 1658
<211> 39
<212> DNA
<213> Leptotrichia wadei
<400> 1658
taagttttag tccccttcgt ttttggggta gtctaaatc 39
<210> 1659
<211> 39
<212> DNA
<213> Leptotrichia wadei
<400> 1659
taagttttag tccccttcgt ttttggggta gtctaaatc 39
<210> 1660
<211> 39
<212> DNA
<213> Leptotrichia wadei
<400> 1660
taagttttag tccccttcgt ttttggggta gtctaaatc 39
<210> 1661
<211> 78
<212> DNA
<213> Leptotrichia shahii
<400> 1661
attctttaga gttttagtcc ccttcgatat tggggtggtc tatatcgaaa aagaagagtt 60
tattcagata gatttgtc 78
<210> 1662
<211> 77
<212> DNA
<213> Leptotrichia shahii
<400> 1662
tagatttgtc gttttagtcc ccttcgatat tggggtggtc tatatcaata tggattactt 60
ggtagaacag caatcta 77
<210> 1663
<211> 57
<212> DNA
<213> Leptotrichia shahii
<400> 1663
cagcaatcta gttttagtcc ccttcgatat tggggtggtc tatatcccat cctaatt 57
<210> 1664
<211> 36
<212> DNA
<213> Leptotrichia shahii
<400> 1664
gttttagtcc ccttcgatat tggggtggtc tatatc 36
<210> 1665
<211> 36
<212> DNA
<213> Leptotrichia shahii
<400> 1665
gttttagtcc ccttcgatat tggggtggtc tatatc 36
<210> 1666
<211> 80
<212> DNA
<213> Rhodobacter capsulatus
<400> 1666
gaacatcatg ggttcagtcc gccgtcgtct tggcggtgat gtgaggctct cccagcatac 60
caaaccgctg gcgaccatca 80
<210> 1667
<211> 78
<212> DNA
<213> Rhodobacter capsulatus
<400> 1667
gcgaccatca ggttcagtcc gccgtcgtct tggcggtgat gtgatgcaca ggagagaccg 60
atgaaaatca cggccttc 78
<210> 1668
<211> 79
<212> DNA
<213> Rhodobacter capsulatus
<400> 1668
cacggccttc ggttcagtcc gccgtcgtct tggcggtgac gtgaggcacc taaacaagag 60
gttctacgat gccgaaagg 79
<210> 1669
<211> 58
<212> DNA
<213> Rhodobacter capsulatus
<400> 1669
tgccgaaagg ggttcagtcc gccgtcgtct tggcggtgat gtgaggctca ggtcccgc 58
<210> 1670
<211> 37
<212> DNA
<213> Rhodobacter capsulatus
<400> 1670
ggttcagtcc gccgtcgtct tggcggtgat gtgaggc 37
<210> 1671
<211> 37
<212> DNA
<213> Rhodobacter capsulatus
<400> 1671
ggttcagtcc gccgtcgtct tggcggtgat gtgaggc 37
<210> 1672
<211> 37
<212> DNA
<213> Rhodobacter capsulatus
<400> 1672
ggttcagtcc gccgtcgtct tggcggtgat gtgaggc 37
<210> 1673
<211> 37
<212> DNA
<213> Rhodobacter capsulatus
<400> 1673
ggttcagtcc gccgtcgtct tggcggtgat gtgaggc 37
<210> 1674
<211> 80
<212> DNA
<213> Rhodobacter capsulatus
<400> 1674
gaacatcatg ggttcagtcc gccgtcgtct tggcggtgat gtgaggctct cccagcatac 60
caaaccgctg gcgaccatca 80
<210> 1675
<211> 78
<212> DNA
<213> Rhodobacter capsulatus
<400> 1675
gcgaccatca ggttcagtcc gccgtcgtct tggcggtgat gtgatgcaca ggagagaccg 60
atgaaaatca acggcttc 78
<210> 1676
<211> 79
<212> DNA
<213> Rhodobacter capsulatus
<400> 1676
caacggcttc ggttcagtcc gccgtcgtct tggcggtgac gtgaggcacc taaacaagag 60
gttctacgat gccgaaagg 79
<210> 1677
<211> 58
<212> DNA
<213> Rhodobacter capsulatus
<400> 1677
tgccgaaagg ggttcagtcc gccgtcgtct tggcggtgat gtgaggctca ggtcccgc 58
<210> 1678
<211> 37
<212> DNA
<213> Rhodobacter capsulatus
<400> 1678
ggttcagtcc gccgtcgtct tggcggtgat gtgaggc 37
<210> 1679
<211> 37
<212> DNA
<213> Rhodobacter capsulatus
<400> 1679
ggttcagtcc gccgtcgtct tggcggtgat gtgaggc 37
<210> 1680
<211> 37
<212> DNA
<213> Rhodobacter capsulatus
<400> 1680
ggttcagtcc gccgtcgtct tggcggtgat gtgaggc 37
<210> 1681
<211> 37
<212> DNA
<213> Rhodobacter capsulatus
<400> 1681
ggttcagtcc gccgtcgtct tggcggtgat gtgaggc 37
<210> 1682
<211> 80
<212> DNA
<213> Rhodobacter capsulatus
<400> 1682
gaacatcatg ggttcagtcc gccgtcgtct tggcggtgat gtgaggctct cccagcatac 60
caaaccgctg gcgaccatca 80
<210> 1683
<211> 78
<212> DNA
<213> Rhodobacter capsulatus
<400> 1683
gcgaccatca ggttcagtcc gccgtcgtct tggcggtgat gtgatgcaca ggagagaccg 60
atgaaaatca acggcttc 78
<210> 1684
<211> 79
<212> DNA
<213> Rhodobacter capsulatus
<400> 1684
caacggcttc ggttcagtcc gccgtcgtct tggcggtgac gtgaggcacc taaacaagag 60
gttctacgat gccgaaagg 79
<210> 1685
<211> 58
<212> DNA
<213> Rhodobacter capsulatus
<400> 1685
tgccgaaagg ggttcagtcc gccgtcgtct tggcggtgat gtgaggctca ggtcccgc 58
<210> 1686
<211> 37
<212> DNA
<213> Rhodobacter capsulatus
<400> 1686
ggttcagtcc gccgtcgtct tggcggtgat gtgaggc 37
<210> 1687
<211> 37
<212> DNA
<213> Rhodobacter capsulatus
<400> 1687
ggttcagtcc gccgtcgtct tggcggtgat gtgaggc 37
<210> 1688
<211> 37
<212> DNA
<213> Rhodobacter capsulatus
<400> 1688
ggttcagtcc gccgtcgtct tggcggtgat gtgaggc 37
<210> 1689
<211> 37
<212> DNA
<213> Rhodobacter capsulatus
<400> 1689
ggttcagtcc gccgtcgtct tggcggtgat gtgaggc 37
<210> 1690
<211> 35
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1690
gtctattgcc aactatatct ggcttttctc aatac 35
<210> 1691
<211> 35
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1691
gttattgccc tctatcttgg gctcttctca tcaac 35
<210> 1692
<211> 35
<212> DNA
<213> Clostridium aminophilum
<400> 1692
gtttggagaa cagcccgata tagagggcaa tagac 35
<210> 1693
<211> 35
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1693
gttttgagaa tagcccgaca tagagggcaa tagac 35
<210> 1694
<211> 37
<212> DNA
<213> Carnobacterium gallinarum
<400> 1694
gttatagtcc tcttacattt agaggtagtc tttaatt 37
<210> 1695
<211> 36
<212> DNA
<213> Carnobacterium gallinarum
<400> 1695
aatataaact acctctaaat gtaagaggac tataac 36
<210> 1696
<211> 36
<212> DNA
<213> Paludibacter propionicigenes
<400> 1696
gttgtagttc ccttcaattt tgggataatc cacaag 36
<210> 1697
<211> 36
<212> DNA
<213> Listeria seeligeri
<400> 1697
gtaagagact acctctatat gaaagaggac taaaac 36
<210> 1698
<211> 36
<212> DNA
<213> Listeria weihenstephanensis
<400> 1698
gatttagagt acctcaaaac aaaagaggac taaaac 36
<210> 1699
<211> 36
<212> DNA
<213> Listeria newyorkensis
<400> 1699
gatttagagt acctcaaaac aaaagaggac taaaac 36
<210> 1700
<211> 39
<212> DNA
<213> Leptotrichia wadei
<400> 1700
taagttttag tccccttcgt ttttggggta gtctaaatc 39
<210> 1701
<211> 36
<212> DNA
<213> Leptotrichia shahii
<400> 1701
gttttagtcc ccttcgatat tggggtggtc tatatc 36
<210> 1702
<211> 37
<212> DNA
<213> Rhodobacter capsulatus
<400> 1702
ggttcagtcc gccgtcgtct tggcggtgat gtgaggc 37
<210> 1703
<211> 37
<212> DNA
<213> Rhodobacter capsulatus
<400> 1703
ggttcagtcc gccgtcgtct tggcggtgat gtgaggc 37
<210> 1704
<211> 37
<212> DNA
<213> Rhodobacter capsulatus
<400> 1704
ggttcagtcc gccgtcgtct tggcggtgat gtgaggc 37
<210> 1705
<211> 35
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1705
gtctattgcc aactatatct ggcttttctc aatac 35
<210> 1706
<211> 35
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1706
gttattgccc tctatcttgg gctcttctca tcaac 35
<210> 1707
<211> 35
<212> DNA
<213> Clostridium aminophilum
<400> 1707
gtttggagaa cagcccgata tagagggcaa tagac 35
<210> 1708
<211> 35
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Lachnospiraceae bacterium sequence"
<400> 1708
gttttgagaa tagcccgaca tagagggcaa tagac 35
<210> 1709
<211> 37
<212> DNA
<213> Carnobacterium gallinarum
<400> 1709
gttatagtcc tcttacattt agaggtagtc tttaatt 37
<210> 1710
<211> 36
<212> DNA
<213> Carnobacterium gallinarum
<400> 1710
aatataaact acctctaaat gtaagaggac tataac 36
<210> 1711
<211> 36
<212> DNA
<213> Paludibacter propionicigenes
<400> 1711
gttgtagttc ccttcaattt tgggataatc cacaag 36
<210> 1712
<211> 36
<212> DNA
<213> Listeria seeligeri
<400> 1712
gtaagagact acctctatat gaaagaggac taaaac 36
<210> 1713
<211> 36
<212> DNA
<213> Listeria weihenstephanensis
<400> 1713
gatttagagt acctcaaaac aaaagaggac taaaac 36
<210> 1714
<211> 36
<212> DNA
<213> Listeria newyorkensis
<400> 1714
gatttagagt acctcaaaac aaaagaggac taaaac 36
<210> 1715
<211> 39
<212> DNA
<213> Leptotrichia wadei
<400> 1715
taagttttag tccccttcgt ttttggggta gtctaaatc 39
<210> 1716
<211> 36
<212> DNA
<213> Leptotrichia shahii
<400> 1716
gttttagtcc ccttcgatat tggggtggtc tatatc 36
<210> 1717
<211> 37
<212> DNA
<213> Rhodobacter capsulatus
<400> 1717
ggttcagtcc gccgtcgtct tggcggtgat gtgaggc 37
<210> 1718
<211> 37
<212> DNA
<213> Rhodobacter capsulatus
<400> 1718
ggttcagtcc gccgtcgtct tggcggtgat gtgaggc 37
<210> 1719
<211> 37
<212> DNA
<213> Rhodobacter capsulatus
<400> 1719
ggttcagtcc gccgtcgtct tggcggtgat gtgaggc 37
<210> 1720
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1720
gtattgagaa aagccagata tagttggcaa tagtc 35
<210> 1721
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1721
attaaagact acctctaaat gtaagaggac tataac 36
<210> 1722
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1722
gttatagtcc tcttacattt agaggtagtc tttaat 36
<210> 1723
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1723
attaaagact acctctaaat gtaagaggac tataac 36
<210> 1724
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1724
gttttagtcc tcttctgttt tgaggtactc taaatc 36
<210> 1725
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1725
gttttagccc tcttttgttc tgaggtactc taaatc 36
<210> 1726
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1726
gttttgagaa tagcccgaca tagagggcaa tagac 35
<210> 1727
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1727
gtttggagaa cagcccgata tagagggcaa tagac 35
<210> 1728
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1728
tcacatcacc gccaagacga cggcggactg aacc 34
<210> 1729
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1729
tcacatcacc gccaagacga cggcggactg aacc 34
<210> 1730
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1730
tcacatcacc gccaagacga cggcggactg aacc 34
<210> 1731
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1731
gctggagaag atagcccaag aaagagggca ataac 35
<210> 1732
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1732
gttttagtcc tcttttgttt tgaggtactc taaatc 36
<210> 1733
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1733
gatatagacc accccaatat cgaaggggac taaaac 36
<210> 1734
<211> 28
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1734
tggattatcc caaaattgaa gggaacta 28
<210> 1735
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1735
gttttagtcc tctttcatat agaggtagtc tcttac 36
<210> 1736
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1736
atttcttctc ttgtcattct tctacctttc gcacaa 36
<210> 1737
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1737
tttccatttt cccactactt tctaaaaaga 30
<210> 1738
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1738
tgattatatc gaaaatcaaa taaatgcgct 30
<210> 1739
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1739
cgtttattgg tcagagtaaa ctcaactccg 30
<210> 1740
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1740
tatgaaaata atgtaacacc aatcgtttgg 30
<210> 1741
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1741
atattttttg taacggcttg caatcattt 29
<210> 1742
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1742
ttcttctctt gtcattcttc tacctctaaa atctca 36
<210> 1743
<211> 35
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1743
ggtgtacttg ttccactcaa tccacttatc atctt 35
<210> 1744
<211> 33
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1744
catgatgttc ctttcttggg tatggggtaa gcc 33
<210> 1745
<211> 33
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1745
catgatgttc ctttcttggg tatggggtaa gcc 33
<210> 1746
<211> 33
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1746
catgatgttc ctttcttggg tatggggtaa gcc 33
<210> 1747
<211> 34
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1747
gctaacatct ccggtgttat taccactcca ttct 34
<210> 1748
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1748
aagcattacg gcgtatcacg ccaccaatta 30
<210> 1749
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1749
tctaaagaat tatctattct gtcttttaaa tt 32
<210> 1750
<211> 38
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1750
aaacaaggac atgcacatac ccacatgttt ttctcttg 38
<210> 1751
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1751
tgcaataaca ttttctgata cttttggcgg 30
<210> 1752
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1752
gtattgagaa aagccagata tagcatgaac acttcggtgt ttgtgctttt ttagtatgac 60
gggcatgccg tcagtctgtg gtgaaagtcc acaaggggcg tagttgccaa cgaaccccaa 120
agcaactccc aaggtttaca ccgtgaggtg taggcgagaa gaagggatag caaaatcgta 180
gcctgacgaa cagaaacctg 200
<210> 1753
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1753
attaaagact acctctaaat gtaagaaaaa taaaaaaata aaagagttac atatagttaa 60
gaaaaagaag taggaatatt tattcctact tctttttcgt tgtatttaat ttatttatat 120
gaaaatatga taagatagat atatagtatt agaatggagg gtattgttaa gatgcgtata 180
acaaaagtga aaataaaatt 200
<210> 1754
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1754
gttatagtcc tcttacattt agaggttgga atggcaacag tttttttgac aaattttata 60
aggtgcagaa cttctttccg tatgctattc cgattgtcct tgacaatgag cctcctcgga 120
tgtgacgatc ttcggcagga gctaacagtt aaagttagac tgcctcgcta gcgttagcac 180
atccgagctc attatcaagg 200
<210> 1755
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1755
attaaagact acctggatag gctacaatta actagacaga aaaattaagg tgtgtagact 60
agaagaaaac ataccaggag gaatttttat gtctaagaga acacgaagaa ctttttcaca 120
agaattcaag caacaaatcg tcaatcttta cttagctgga aagccacgtg tagaaatcat 180
tcgagaatat gaactaacgg 200
<210> 1756
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1756
gttttagtcc tcttctgttt tgaggtaata tatccgccta ttcgcttgca acaacataac 60
tataccagat tattaatgaa tctgcagctt gacagattag caaaacataa ttattttttc 120
ataaattaag agacagctca gcagccatct cttaaattca accaattatc ttctgctaac 180
aaaatacctt ccttcttgaa 200
<210> 1757
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1757
gttttagtcc tcttctgttt tgaggtattt tttaatagca aaatgaaatt gcattctccc 60
atccaatttc attttgaaat taactgcaac attctatatc aaatttctaa tagtctcttt 120
actataccac aatactcttc tgaaacttga tttgtttcta tataaccatt atcgattttt 180
ttctcaccta gatgtctcaa 200
<210> 1758
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1758
gttttgagaa tagcccgaca tagttataga gatgtataaa tataaccgat aaacattgac 60
taatttgttg aagtcagtgt ttatcggttt tttgtgtaaa tataggagtt gttagaatga 120
tactttttgc ctaattttgg aactttatga ggatataaga tagacttgat aaaaaggtaa 180
aagaaaggtt aaagagcatg 200
<210> 1759
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1759
gtttgtagaa cagcctgata tagagggcga taggactttg gctgcatgac tcgatcatta 60
agcctgaaac taagttttct tgtgttgaaa tcttcctaat actgaggtcg taagaccatc 120
ttgattattc acgaatctgt gactctgttc tgagaacaat ctcatactat aaggacaatg 180
ttttttgaaa tggaggattt 200
<210> 1760
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1760
tcacatcacc gccaagatga cggcgggaac ccaatgcaaa cggaggtgcg gcaacagcaa 60
ggttgcaccg gctggacttc ggcggcagtc tggcgcaacg gggcgcagga cacggaagat 120
gtggcggggg caagatggac ctgtttttca aggccacgga atatgagacc ctgcaggcct 180
catggctcaa ggtccagcaa 200
<210> 1761
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1761
tcacatcacc gccaagatga cggcgggaac ccaatgcaaa cggaggtgcg gcaacagcaa 60
ggttgcaccg gctggacttc ggcggcagtc tggcgcaacg gggcgcagga cacggaagat 120
gtggcggggg caagatggac ctgtttttca aggccacgga atatgagacc ctgcaggcct 180
catggctcaa ggtccagcaa 200
<210> 1762
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1762
tcacatcacc gccaagatga cggcgggaac ccaatgcaaa cggaggtgcg gcaacagcaa 60
ggttgcaccg gctggacttc ggcggcagtc tggcgcaacg gggcgcagga cacggaagat 120
gtggcggggg caagatggac ctgtttttca aggccacgga atatgagacc ctgcaggcct 180
catggctcaa ggtccagcaa 200
<210> 1763
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1763
gttggagaag agagcccaag atagaggaga ttgacattta ttacaagcgg agattaaaac 60
gataactgag aaaaaatgaa taacgctgat gaaaacggcc ggattcttgg ccgttttttt 120
gtctatttgc taagtgcaca aagattgtga aataacatct gctactatgt atttatcgag 180
gtacgtaaat ctaggtggtg 200
<210> 1764
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1764
gttttagtcc tcttttattt tgaggtaata tattcacttg caacaacata actataccag 60
attattaatg gatctgcatc ttgacagatt accaaaacat aattattttt tcataaatta 120
agagacaact cagaatacag aattgcctct acatgtctaa ttttcccact gtcaattcct 180
ctgcgcataa tatctccacc 200
<210> 1765
<211> 200
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1765
gatatagacc accccaatat cgaaaagtga tatttaataa aaataacttc tgagtgagaa 60
taaaatttca attcttgctc attttttatt gttttttgaa tatggttgct tggttgttct 120
gaaacgaaaa tttttggaga tgttttaaat ttttaggttg aaaaaaaata agaattatac 180
tataataaat aattatgcga 200
<210> 1766
<211> 250
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1766
tggattatcc caaaattgaa gggtaacact acagctgaca tcaaagcaca aacaaccccg 60
aatgaaattc atcattcggg gttgttttta taaaggttag cttagctaat tgcagtccta 120
cagcaaatca ctacttcttc aaacgcaata tctccggatt ttctgcaata aatttattgg 180
cagcttcgta gccctgatga tagatagtga agtaagagtc gtagctaaac tcatgaaagg 240
tttcgatggc 250
<210> 1767
<211> 250
<212> DNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1767
gttttagtcc tctttcattt agaggtatat cgtattccta cttaataata gtaattaaaa 60
caaccaatgt aaaggatata atcaatatat ttaaagtttg cacgagaatg caatcatttt 120
attcataaat atcatatcat ttataagctc tattttccat tttctaaggc taataaataa 180
aactgctgta cctatggatc taaggaagac ttatgcacac agtacagcaa cttttcagca 240
tgatttgtgt 250
<210> 1768
<211> 78
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1768
auuauuacca uuuugguugg aaugcuauua uaaaggauca uucgauuauu accucuaccu 60
cccuucccac gauuucuu 78
<210> 1769
<211> 35
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1769
gcuggagaag auagcccaag aaagagggca auaac 35
<210> 1770
<211> 81
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1770
gucuuacgac cucaguauua ggaagauuuc aaccaagaaa acuuaguuuc aggcuuaaug 60
aucgagucau gcagccaaag u 81
<210> 1771
<211> 35
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1771
guuuggagaa cagcccgaua uagagggcaa uagac 35
<210> 1772
<211> 99
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1772
augaaaagag gacuaaaacu gaaagaggac uaaaacacca gauguggaua acuauauuag 60
uggcuauuaa aaauucgucg auauuagaga ggaaacuuu 99
<210> 1773
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1773
guuuuagucc ucuuucauau agagguaguc ucuuac 36
<210> 1774
<211> 106
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
polynucleotide"
<400> 1774
uuaguauacc acaucaauau uaaaucucaa aaaaauaagg agccgucaaa cauagcuccc 60
uacuucuauu uacucauaau ccccaucuau ccuuacuuuu cguaaa 106
<210> 1775
<211> 36
<212> RNA
<213> Artificial Sequence
<220>
<221> source
<223> /note="Description of Artificial Sequence: Synthetic
oligonucleotide"
<400> 1775
guuuuagucc ccuucgauau uggggugguc uauauc 36
<210> 1776
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 1776
cccataattg ataggatcta tgaggt 26
<210> 1777
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 1777
ctcccgaaaa gccttgt 17
<210> 1778
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 1778
cccatgattg ataggatcta tgaggt 26
<210> 1779
<211> 18
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 1779
tttcccccga caggcgta 18
<210> 1780
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 1780
cccataattg ataggatcta tgaggt 26
<210> 1781
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 1781
tccatatgaa tggcgcg 17
<210> 1782
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 1782
cccataattg ataggatcta tgaggt 26
<210> 1783
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 1783
tgccgccgtc ctgcatg 17
<210> 1784
<211> 26
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 1784
cccataattg ataggatcta tgaggt 26
<210> 1785
<211> 17
<212> DNA
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 1785
gcccggacca catgcac 17
<210> 1786
<211> 51
<212> PRT
<213> Unknown
<220>
<221> source
<223> /note="Description of Unknown:
Gut metagenome sequence"
<400> 1786
Met Ser Lys Asp Thr Thr Gln Ser Arg Tyr Phe Cys Val Phe Lys Asn
1 5 10 15
Cys Pro Cys His Asn Lys Glu Gln His Ala Asp Val Asn Ala Ala Ile
20 25 30
Asn Ile Gly Arg Arg Phe Leu Lys Asp Cys Ile Leu Asp Asp Asn Lys
35 40 45
Glu Lys Asp
50
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЕ ФЕРМЕНТЫ CRISPR И СИСТЕМЫ | 2016 |
|
RU2771826C2 |
| НОВЫЕ ФЕРМЕНТЫ И СИСТЕМЫ CRISPR | 2016 |
|
RU2737537C2 |
| НОВЫЕ ФЕРМЕНТЫ И СИСТЕМЫ CRISPR | 2023 |
|
RU2832308C2 |
| ВАКЦИНА И СПОСОБЫ ОБНАРУЖЕНИЯ И ПРОФИЛАКТИКИ ФИЛЯРИОЗА | 2021 |
|
RU2832185C1 |
| КОНСТРУИРОВАНИЕ СИСТЕМ, СПОСОБЫ И ОПТИМИЗИРОВАННЫЕ НАПРАВЛЯЮЩИЕ КОМПОЗИЦИИ ДЛЯ МАНИПУЛЯЦИИ С ПОСЛЕДОВАТЕЛЬНОСТЯМИ | 2013 |
|
RU2796017C2 |
| БИОЛОГИЧЕСКИЕ МАТЕРИАЛЫ ДЛЯ МОДУЛЯЦИИ ИММУННЫХ ОТВЕТОВ | 2017 |
|
RU2770060C2 |
| ЛЕЧЕНИЕ ЗЛОКАЧЕСТВЕННОГО НОВООБРАЗОВАНИЯ С ИСПОЛЬЗОВАНИЕМ ГУМАНИЗИРОВАННОГО ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА ПРОТИВ ВСМА | 2015 |
|
RU2751660C2 |
| ДОСТАВКА, КОНСТРУИРОВАНИЕ И ОПТИМИЗАЦИЯ СИСТЕМ, СПОСОБОВ И КОМПОЗИЦИЙ ДЛЯ МАНИПУЛЯЦИИ С ПОСЛЕДОВАТЕЛЬНОСТЯМИ И ПРИМЕНЕНИЯ В ТЕРАПИИ | 2013 |
|
RU2721275C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЕГРАДАЦИИ НЕПРАВИЛЬНО УПАКОВАННЫХ БЕЛКОВ | 2016 |
|
RU2761564C2 |
| КОМПОНЕНТЫ СИСТЕМЫ CRISPR-CAS, СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ МАНИПУЛЯЦИИ С ПОСЛЕДОВАТЕЛЬНОСТЯМИ | 2013 |
|
RU2796549C2 |
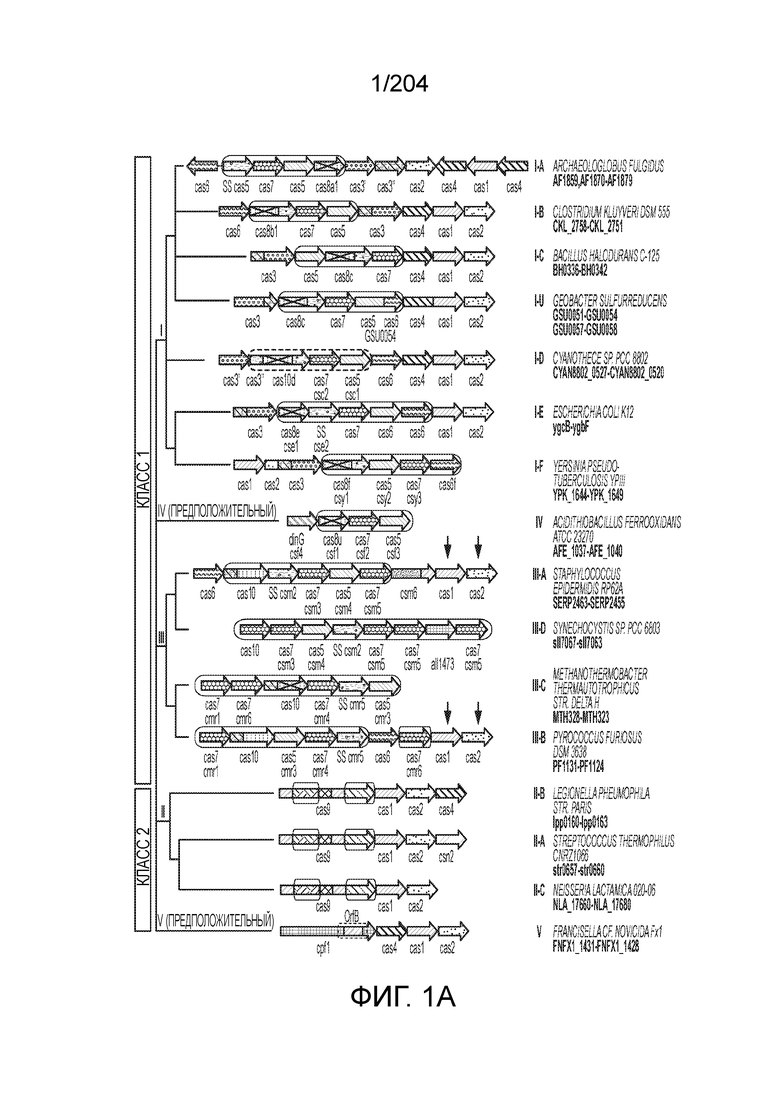
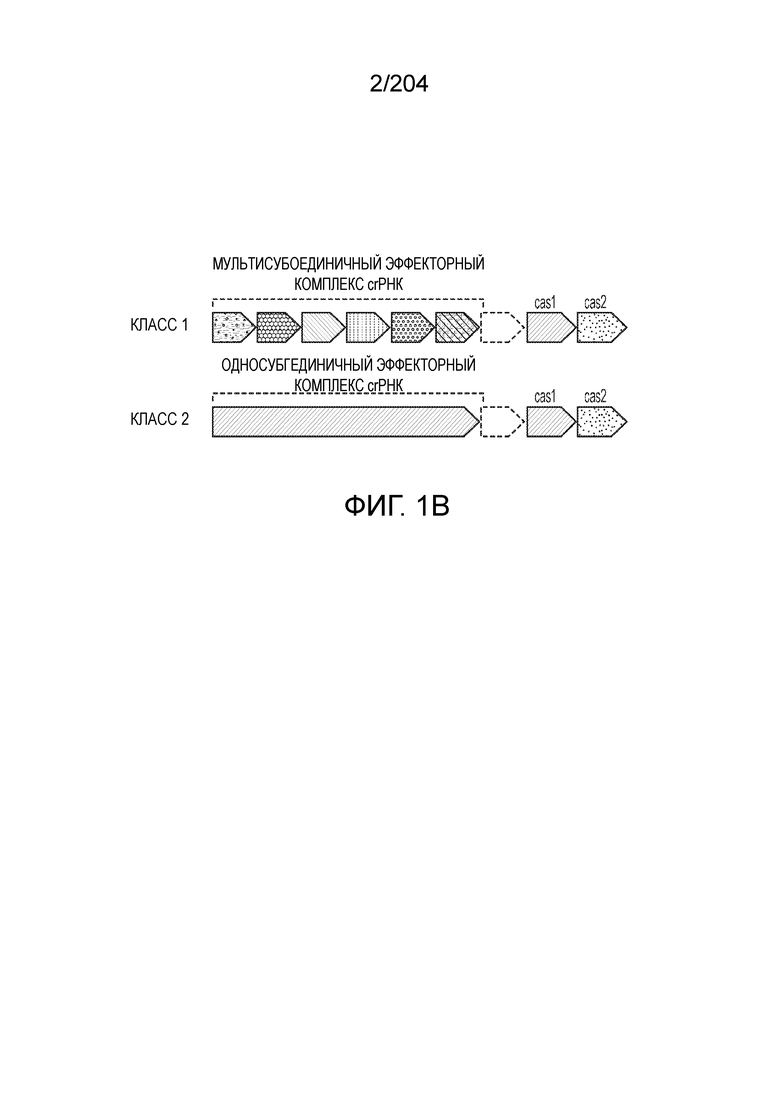
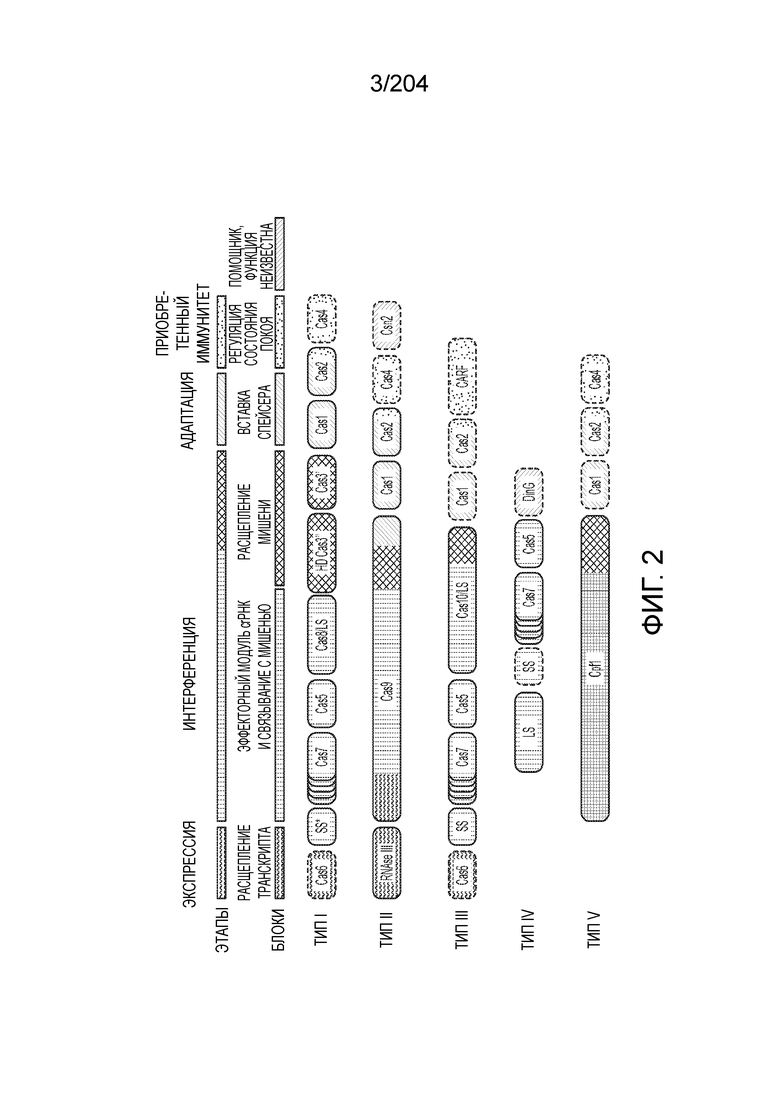
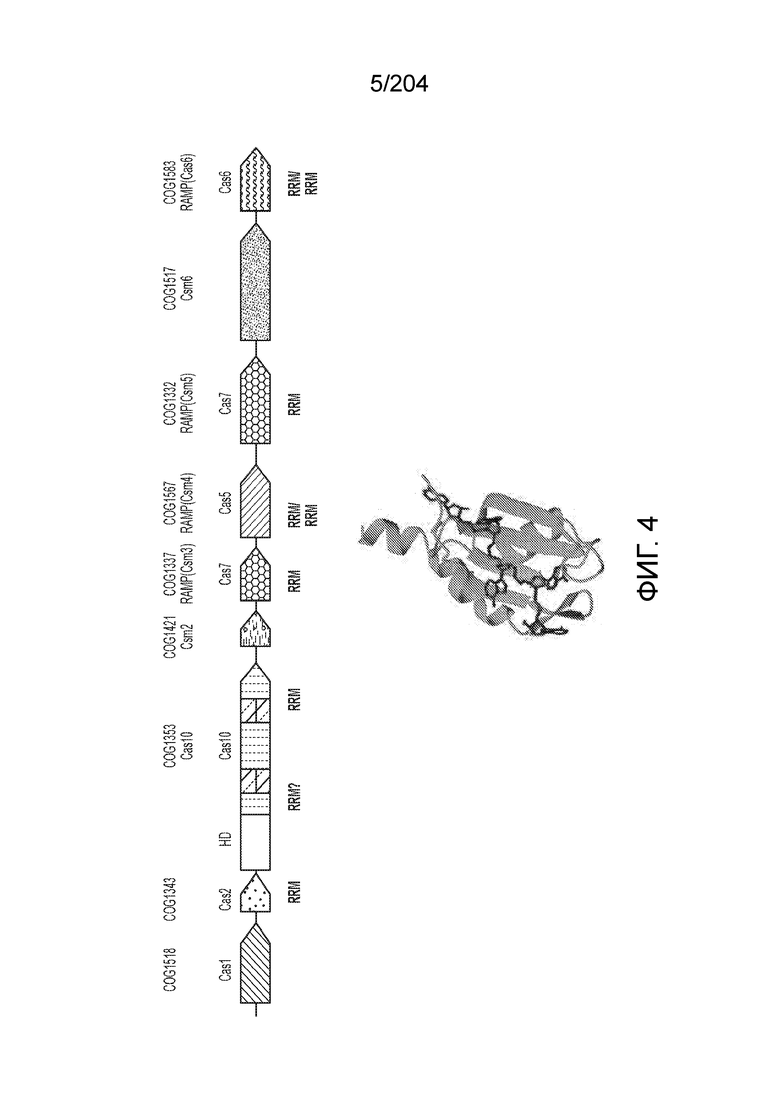
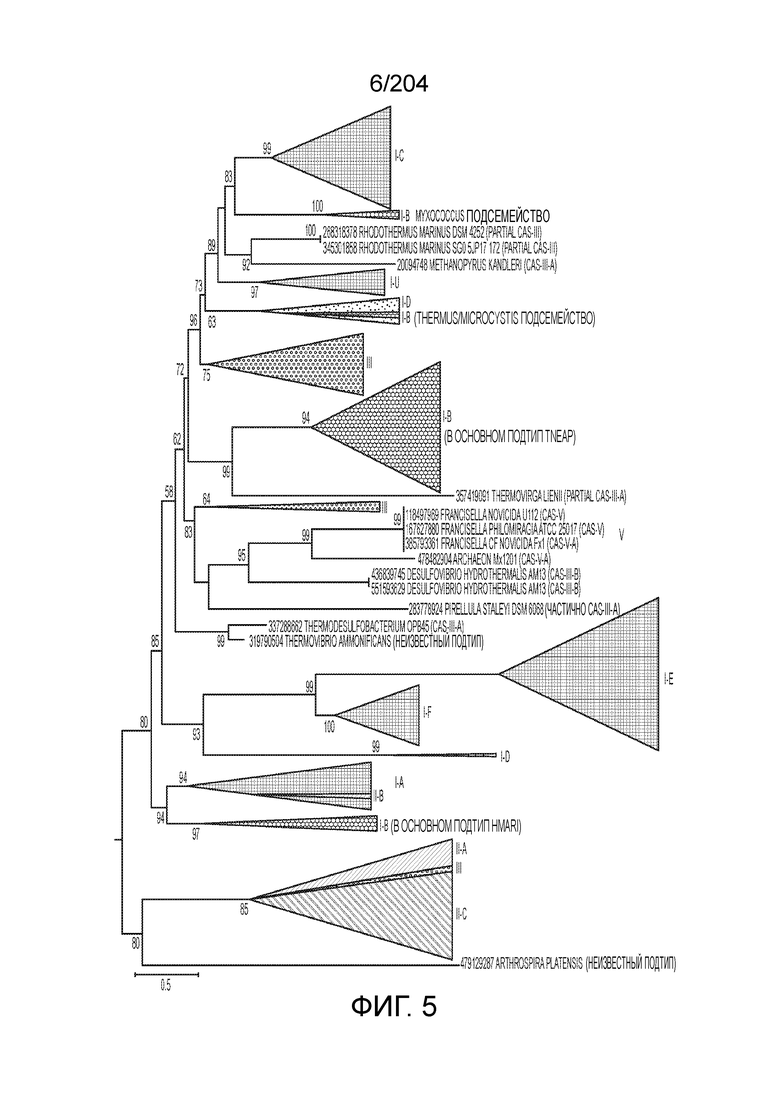

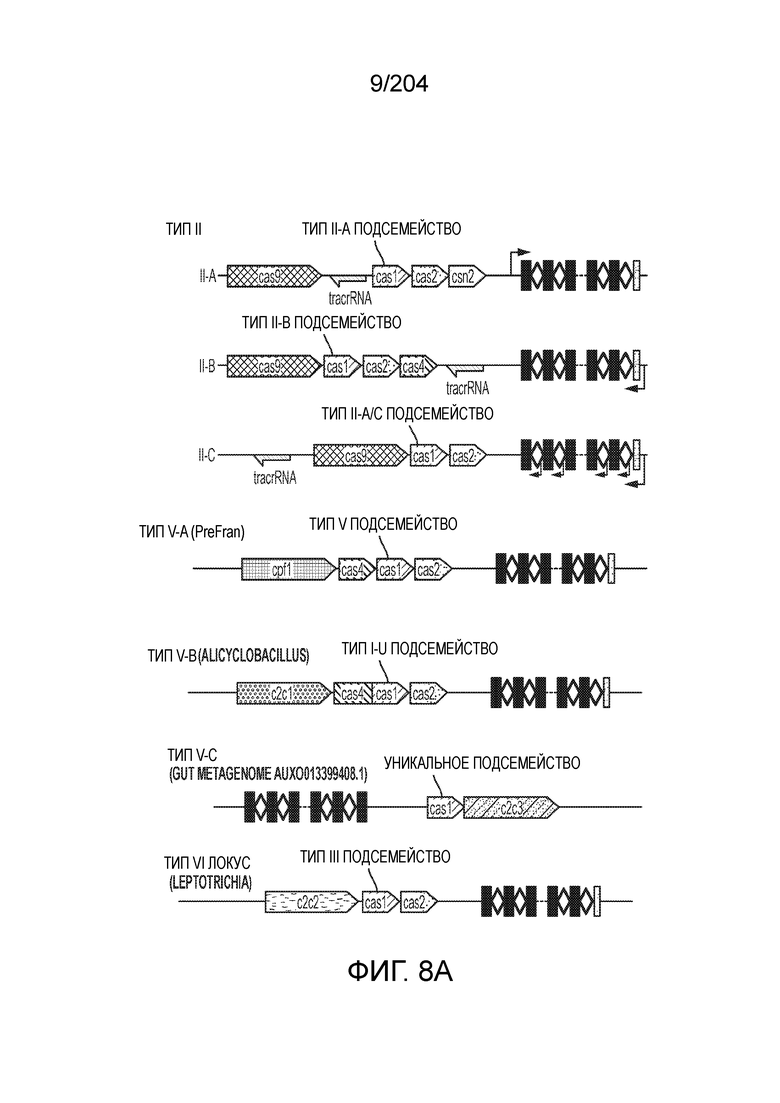
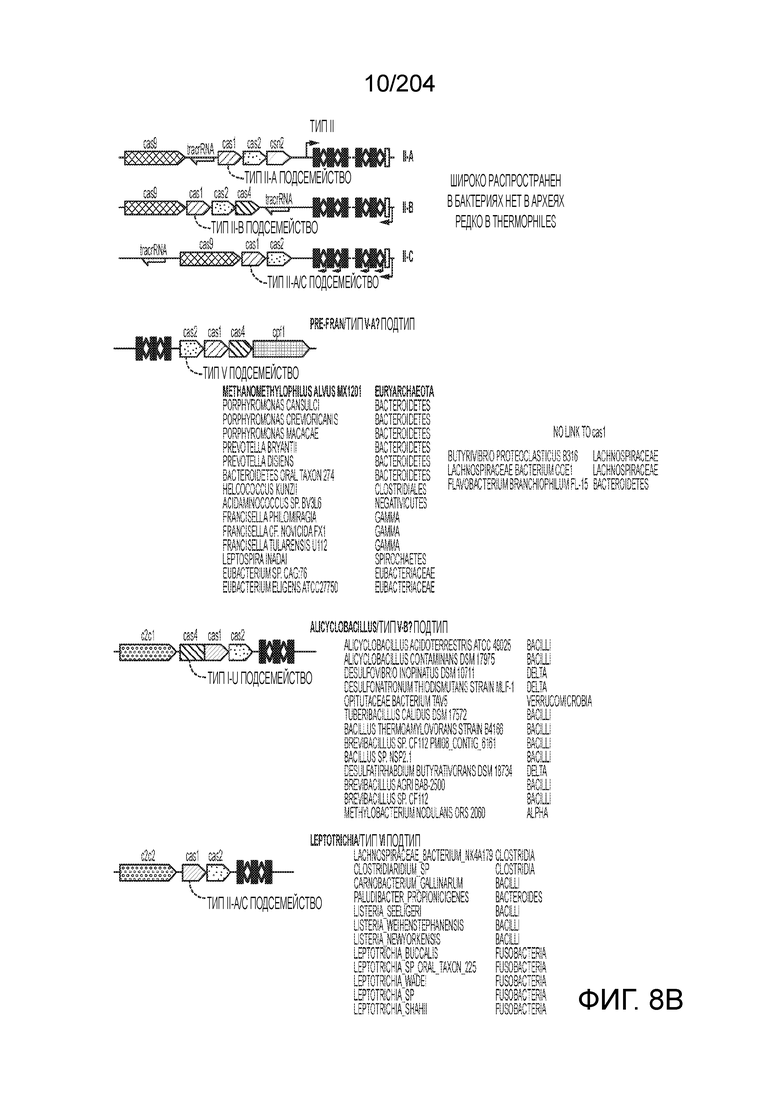
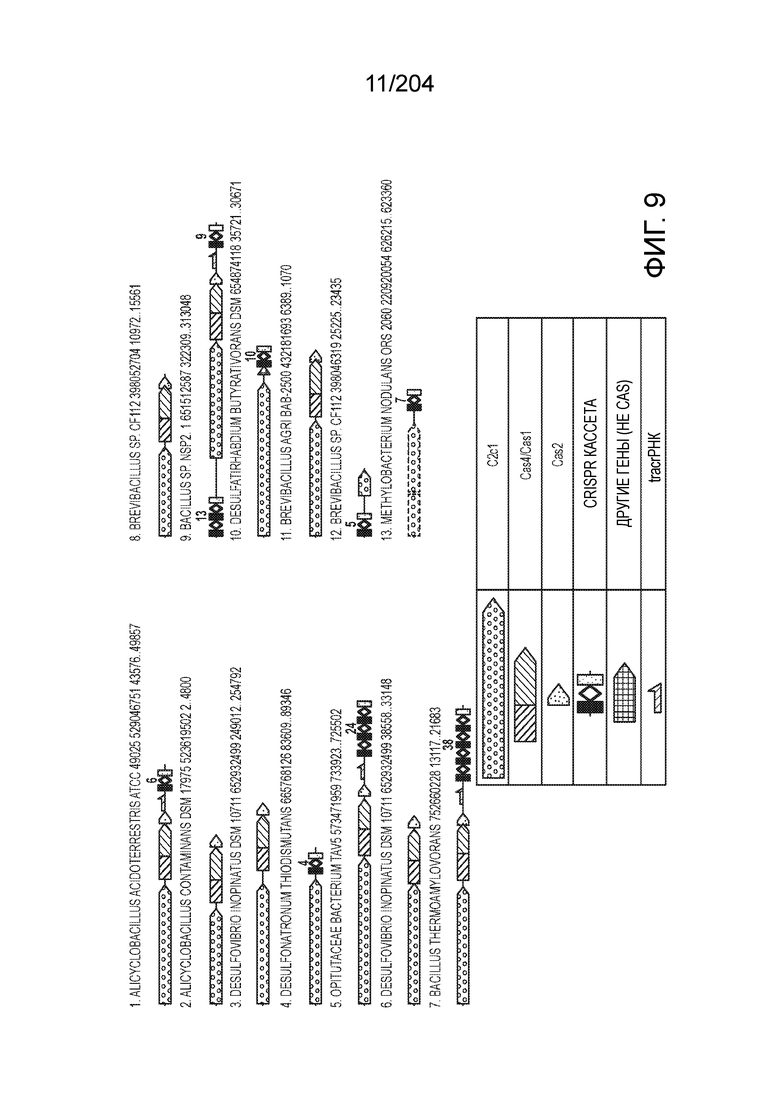
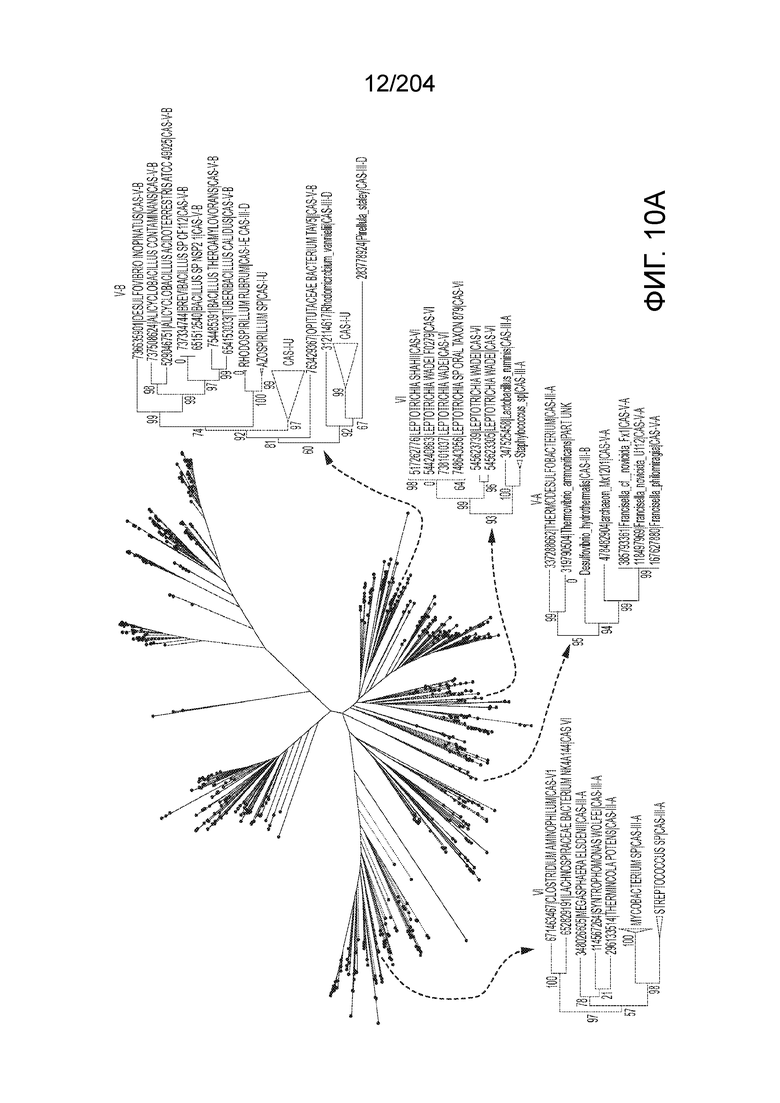
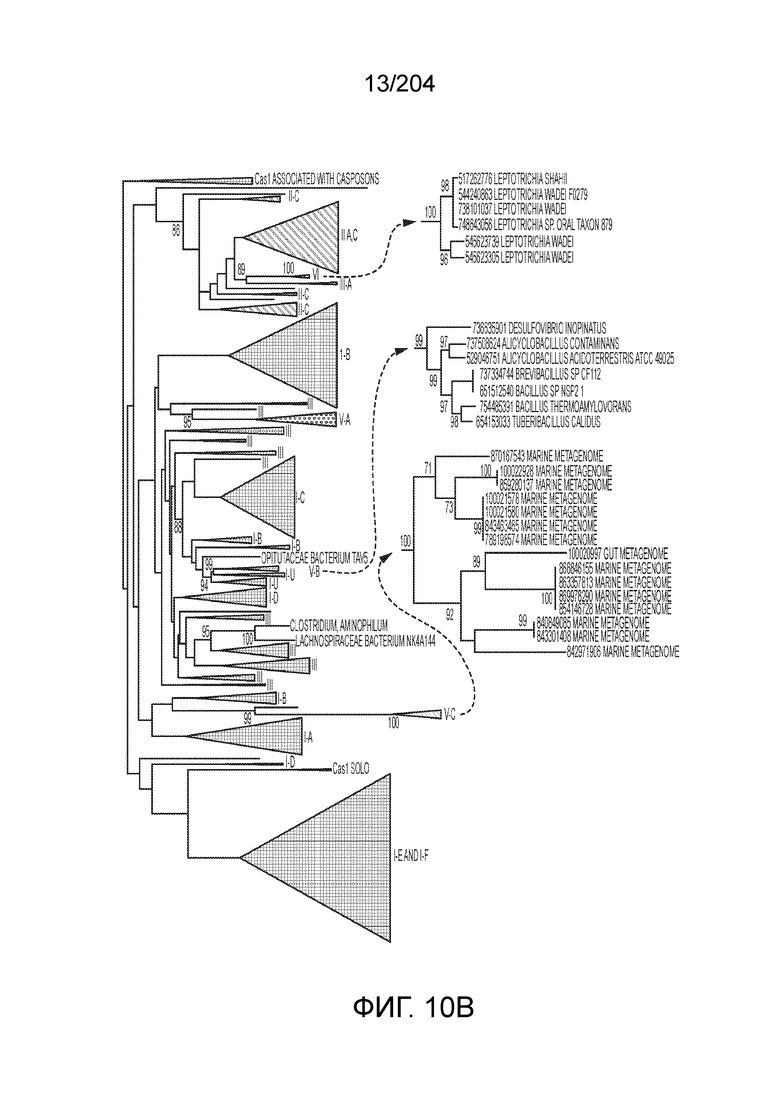









































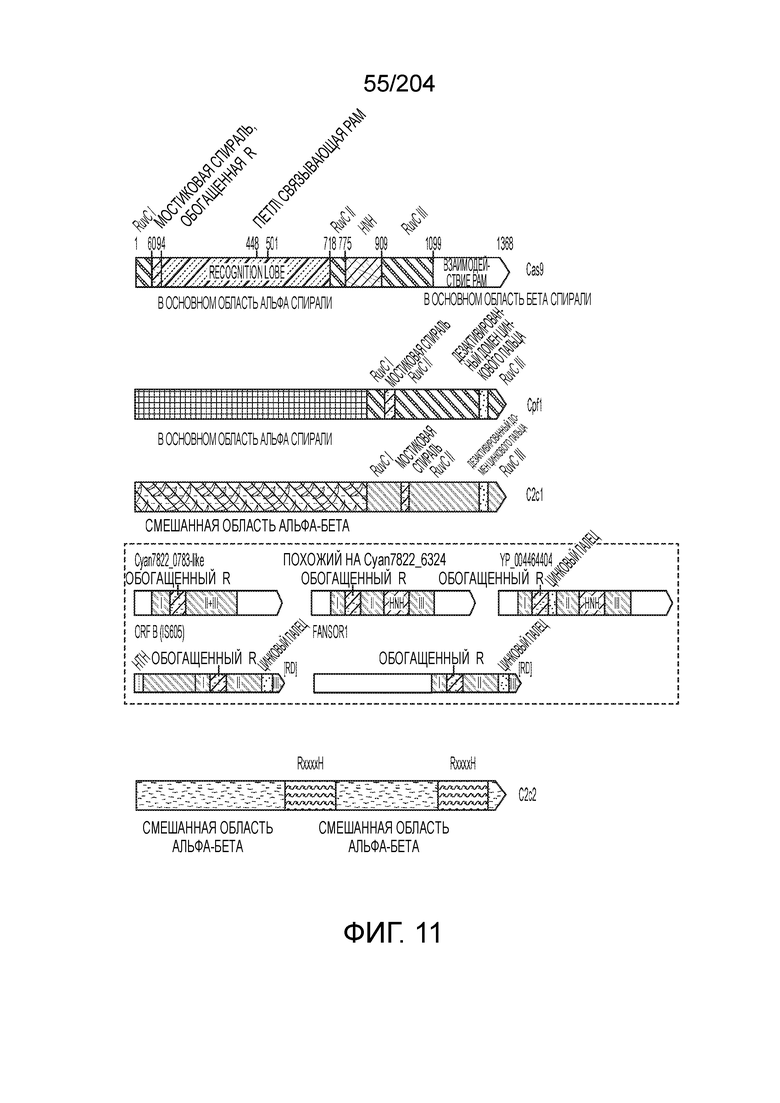


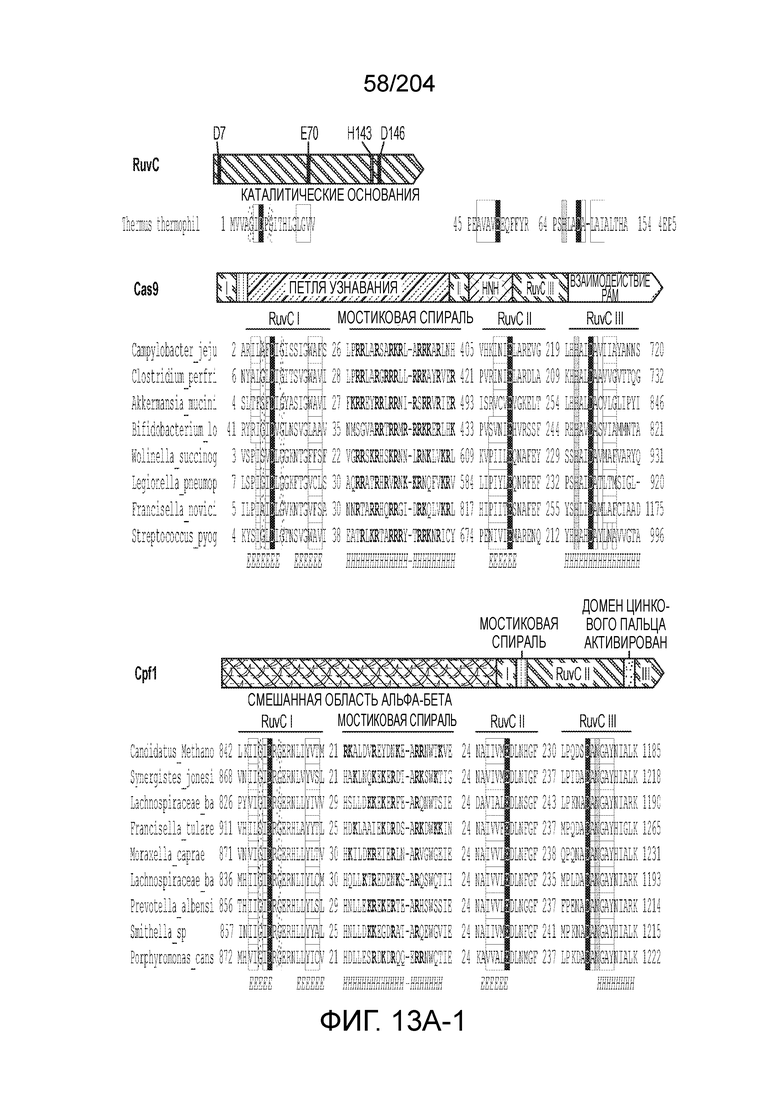
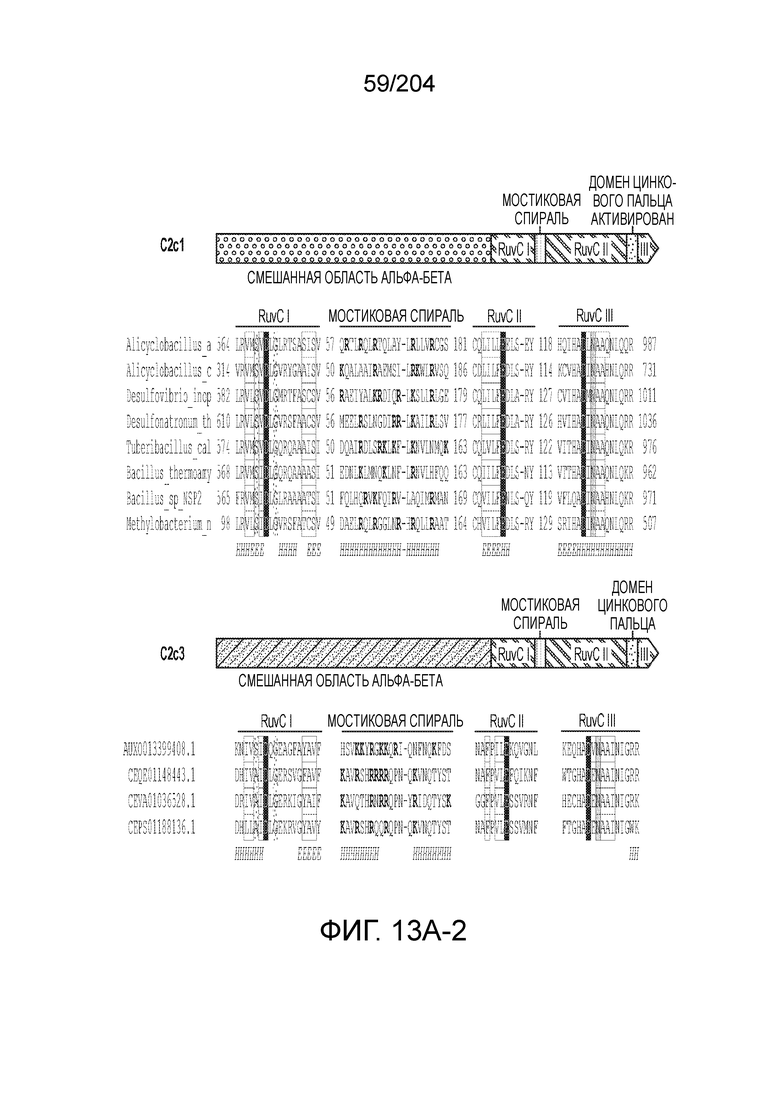
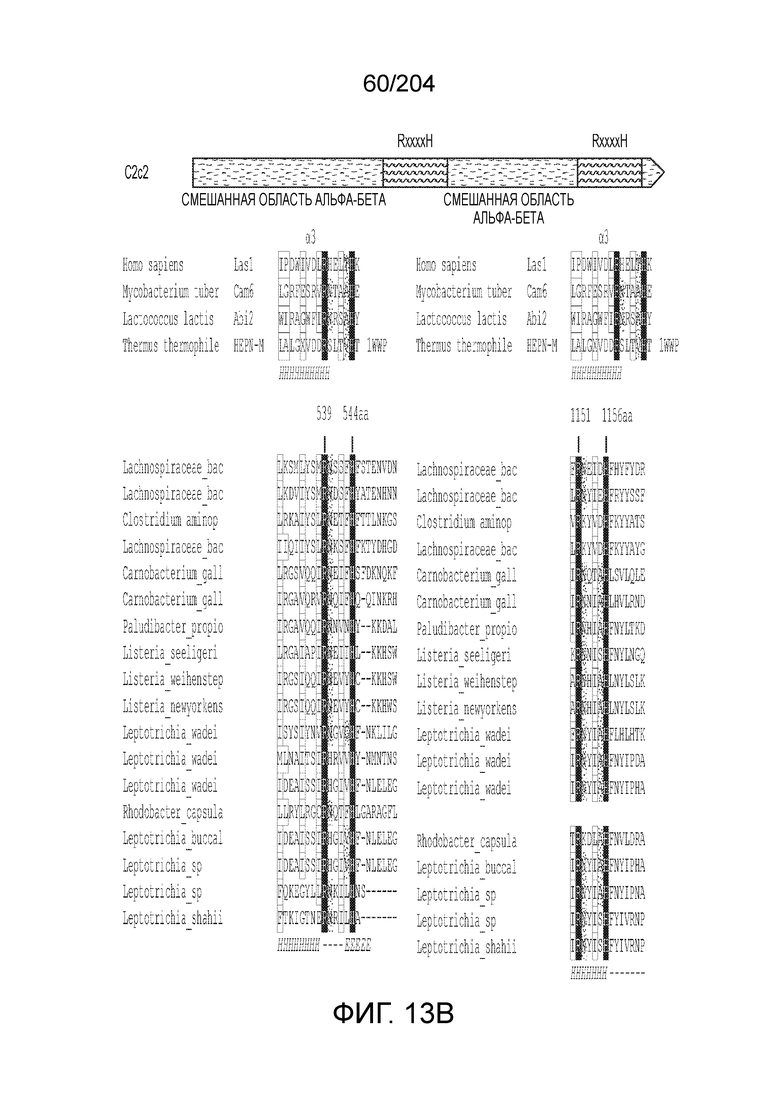
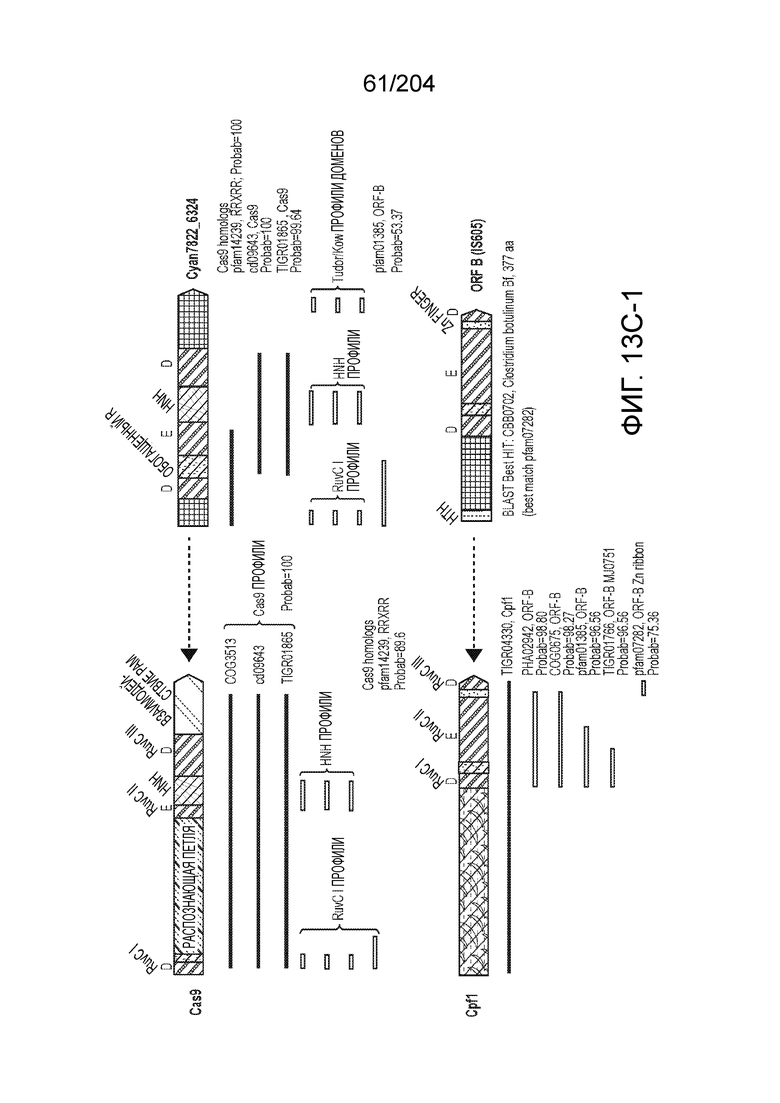
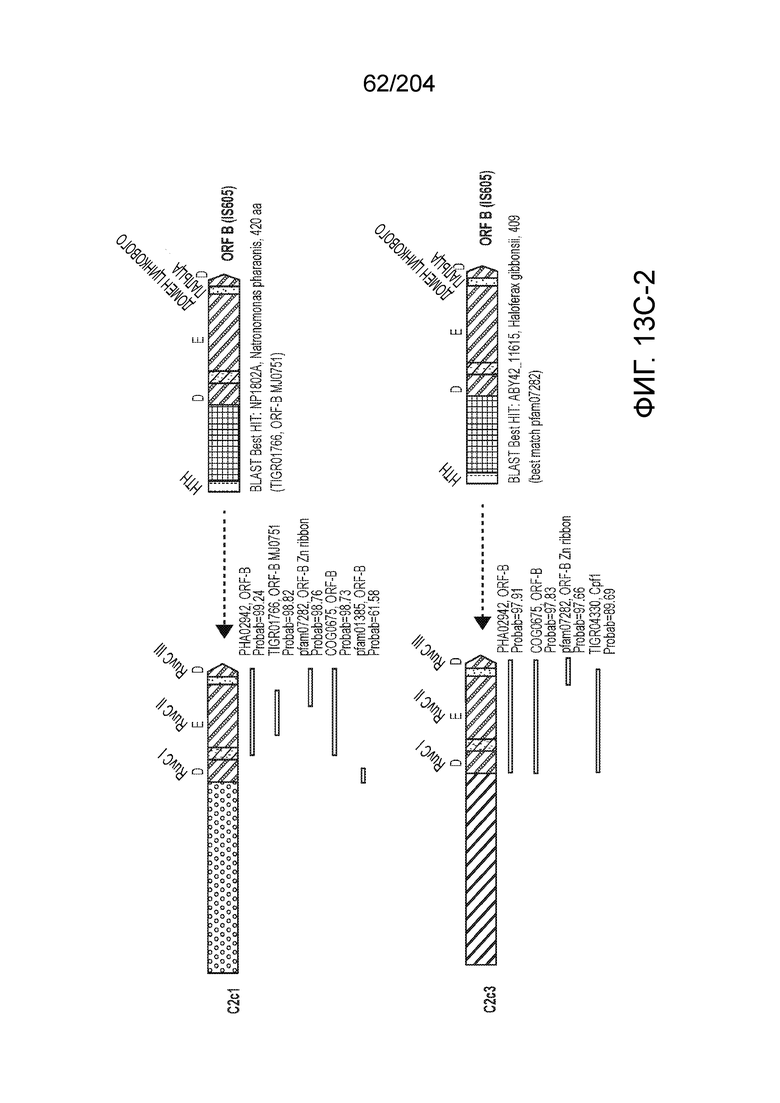

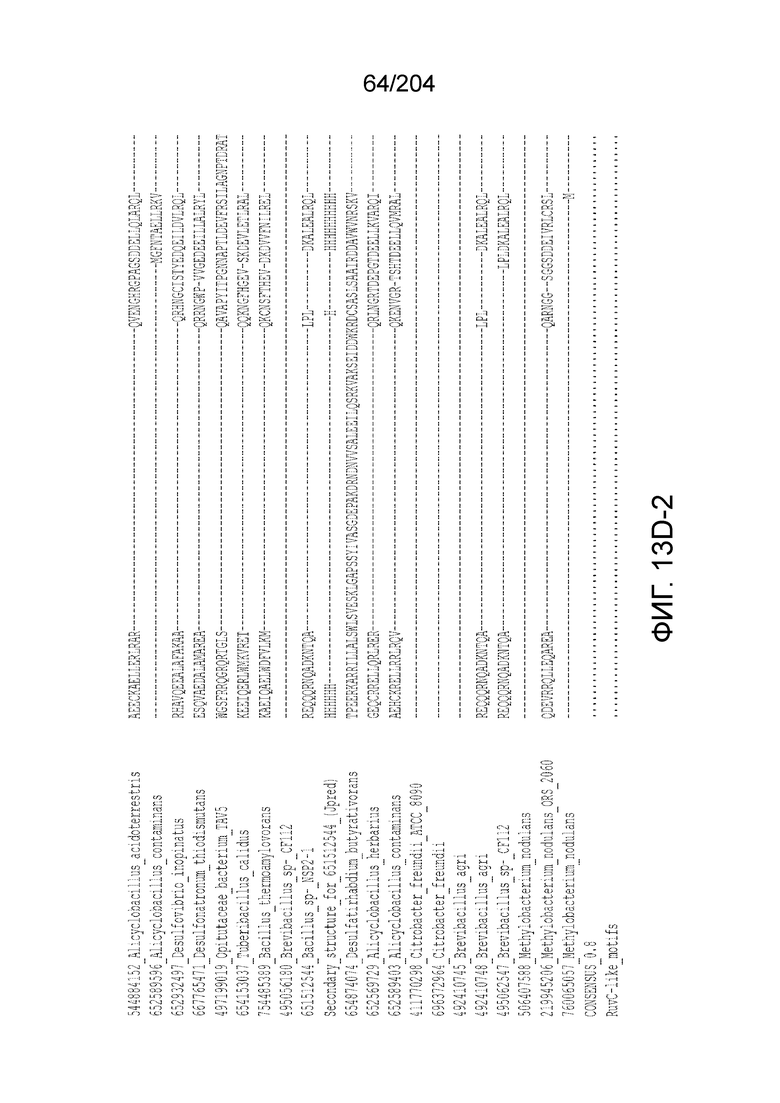

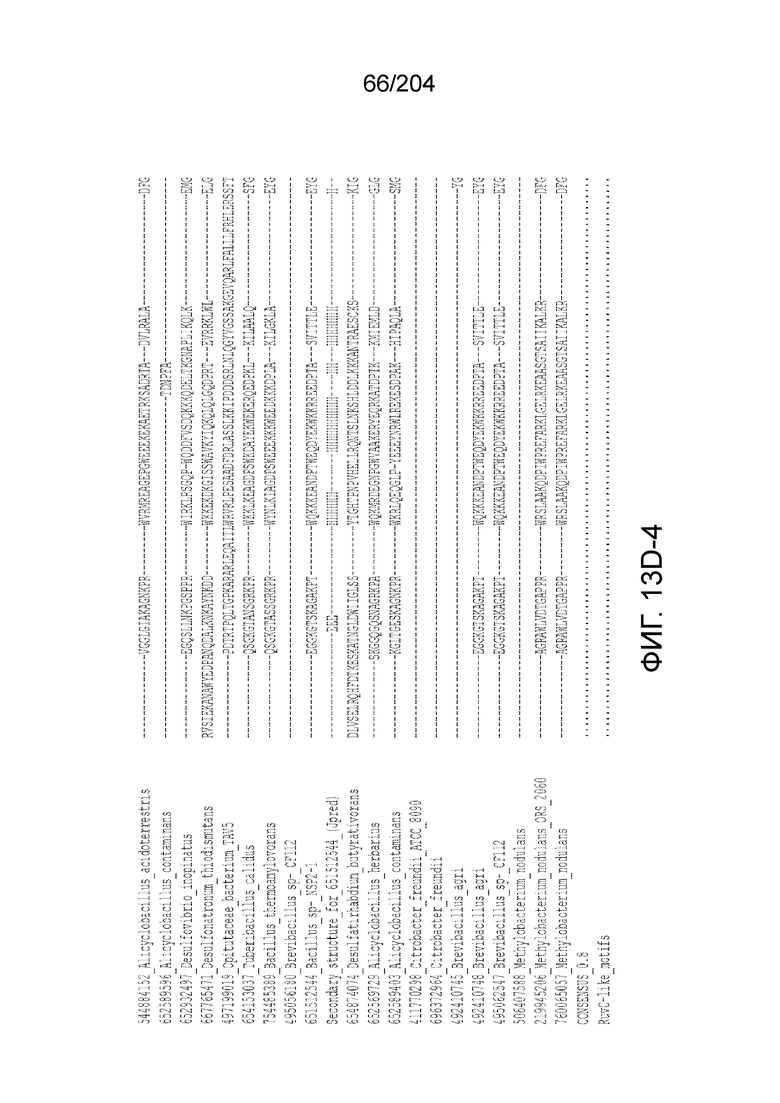










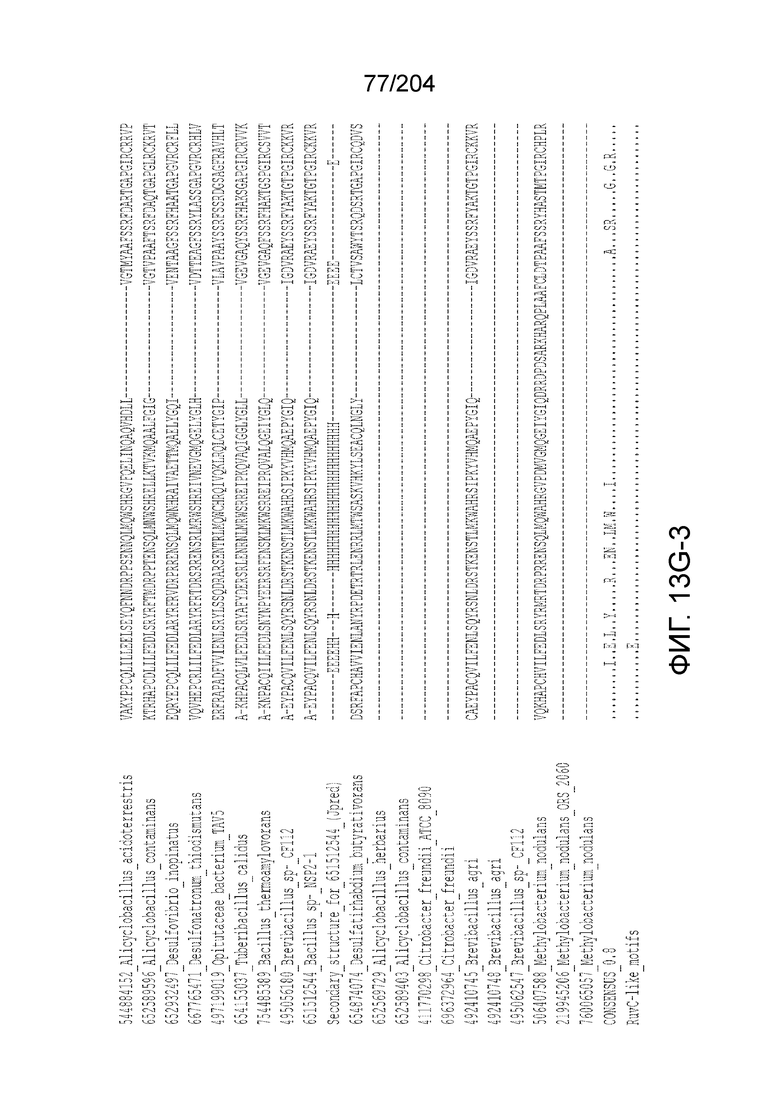
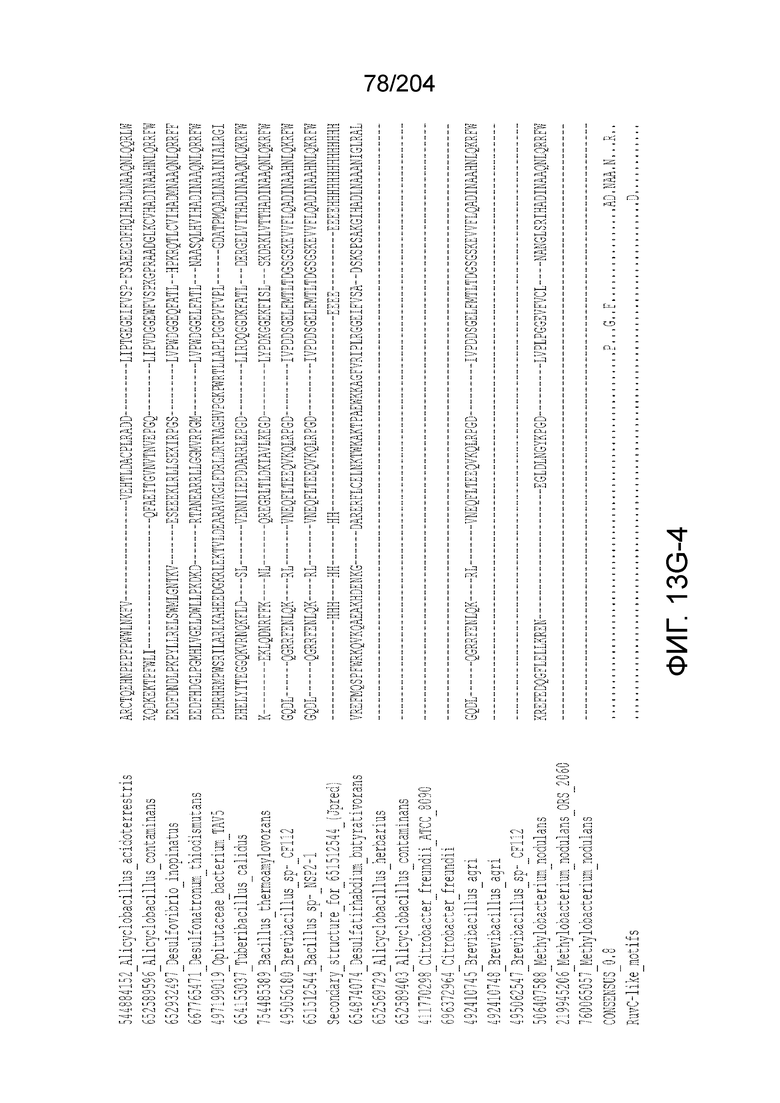


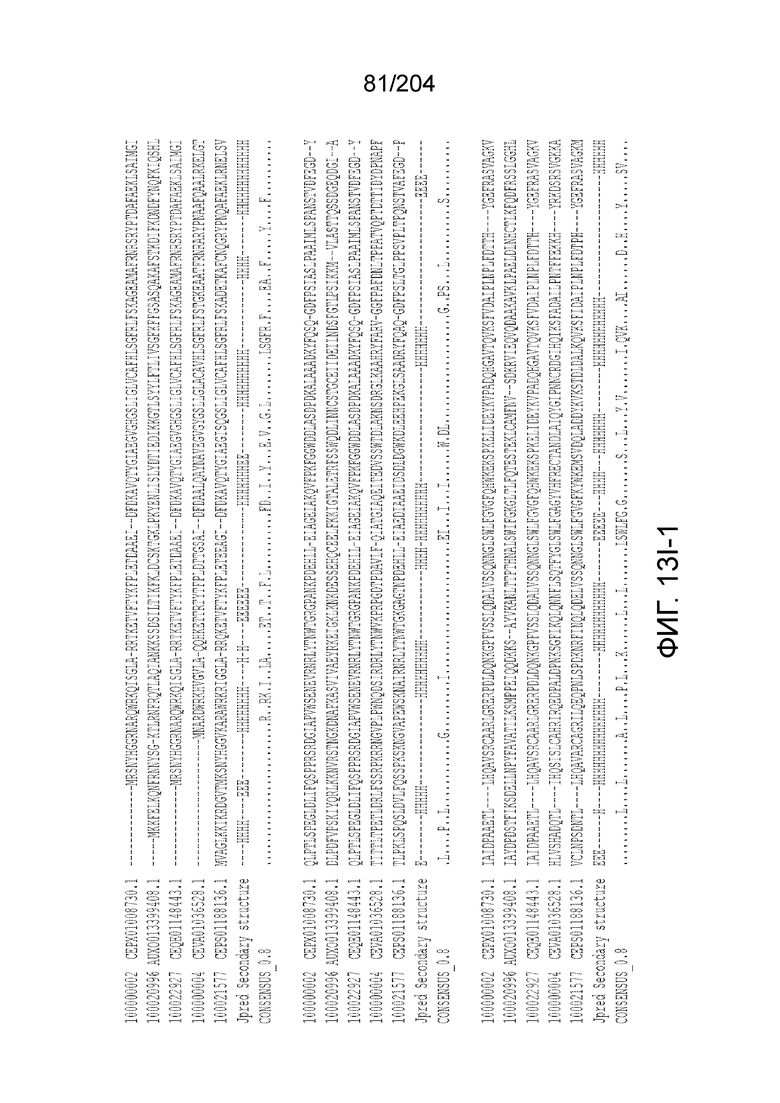























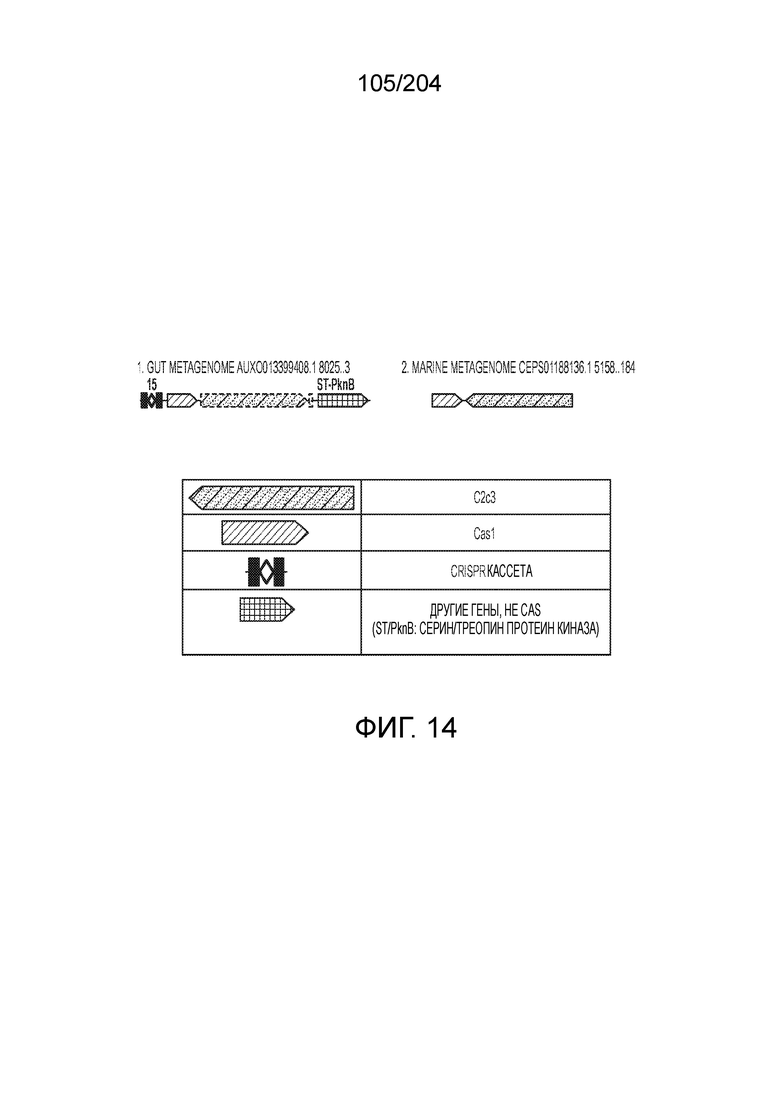
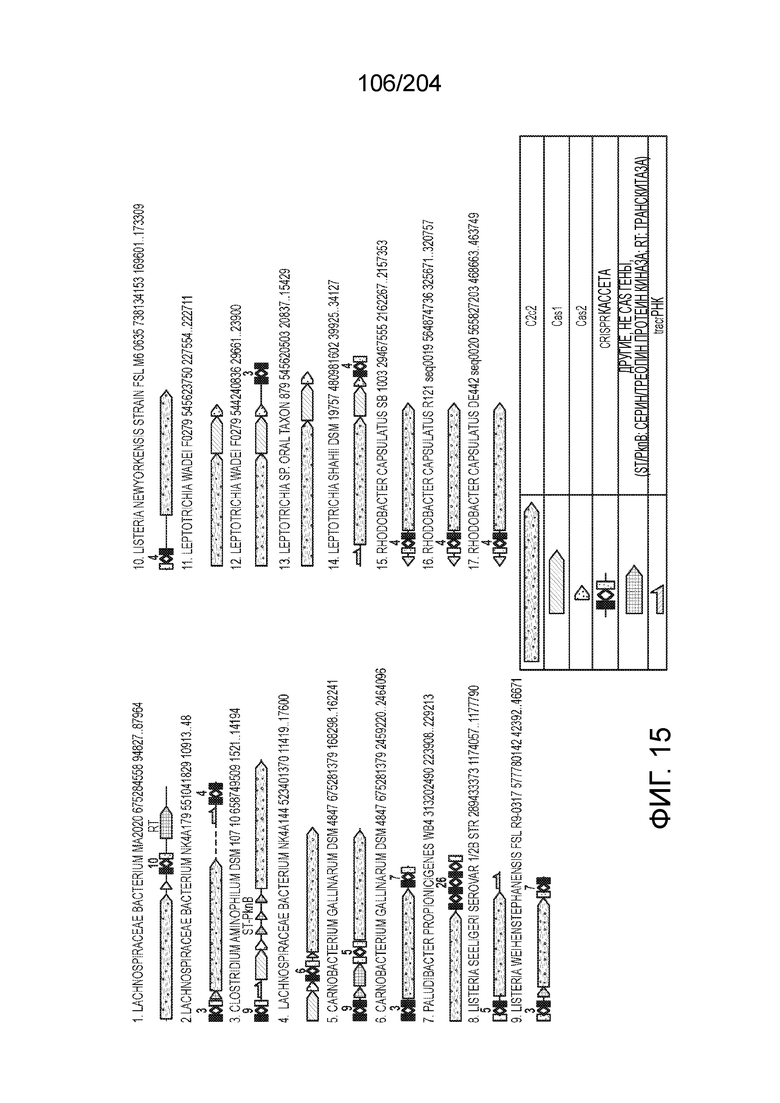






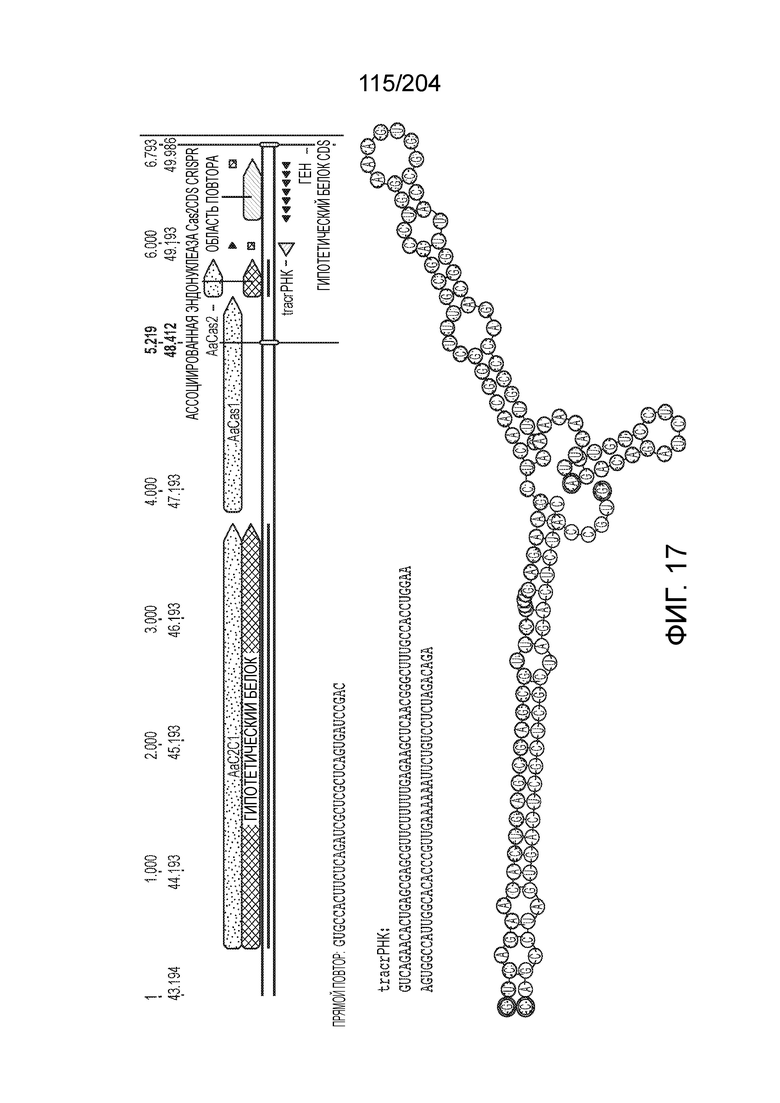
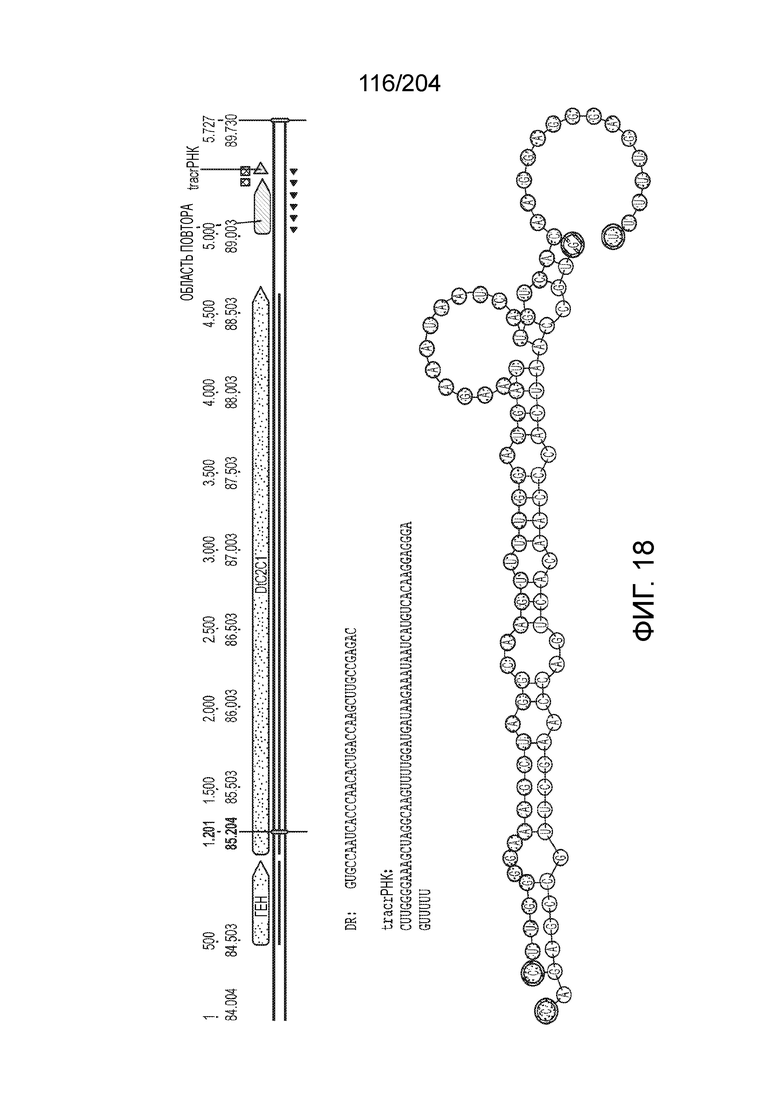
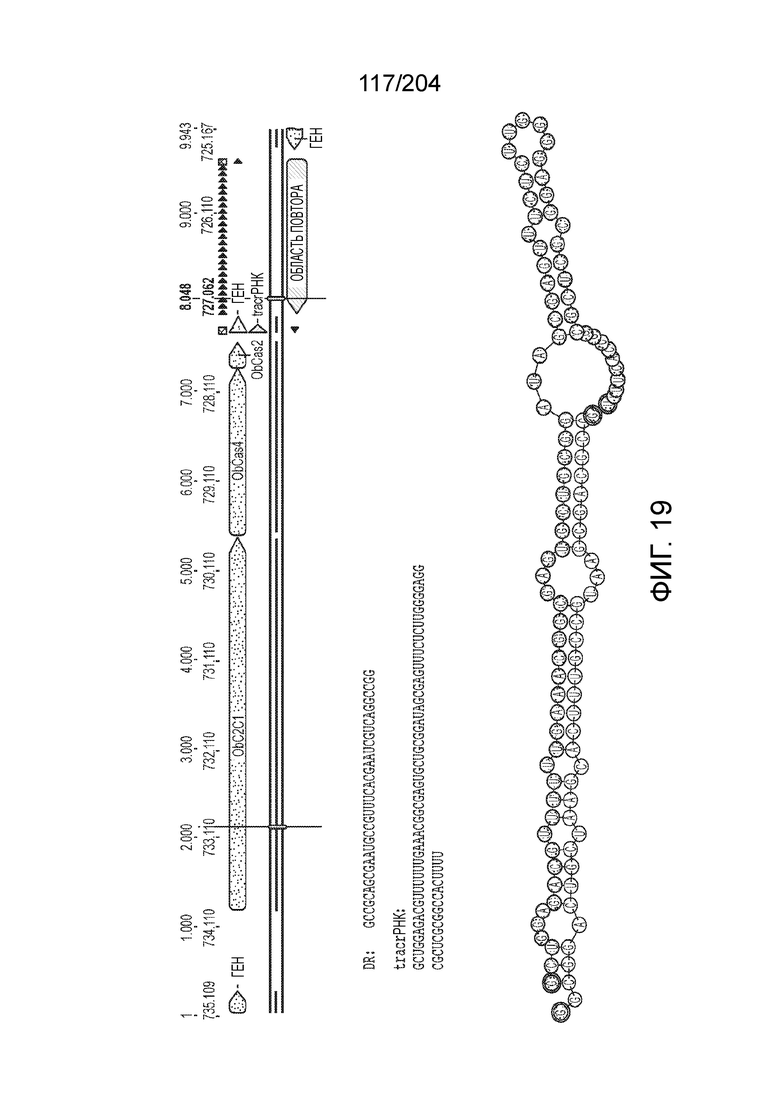
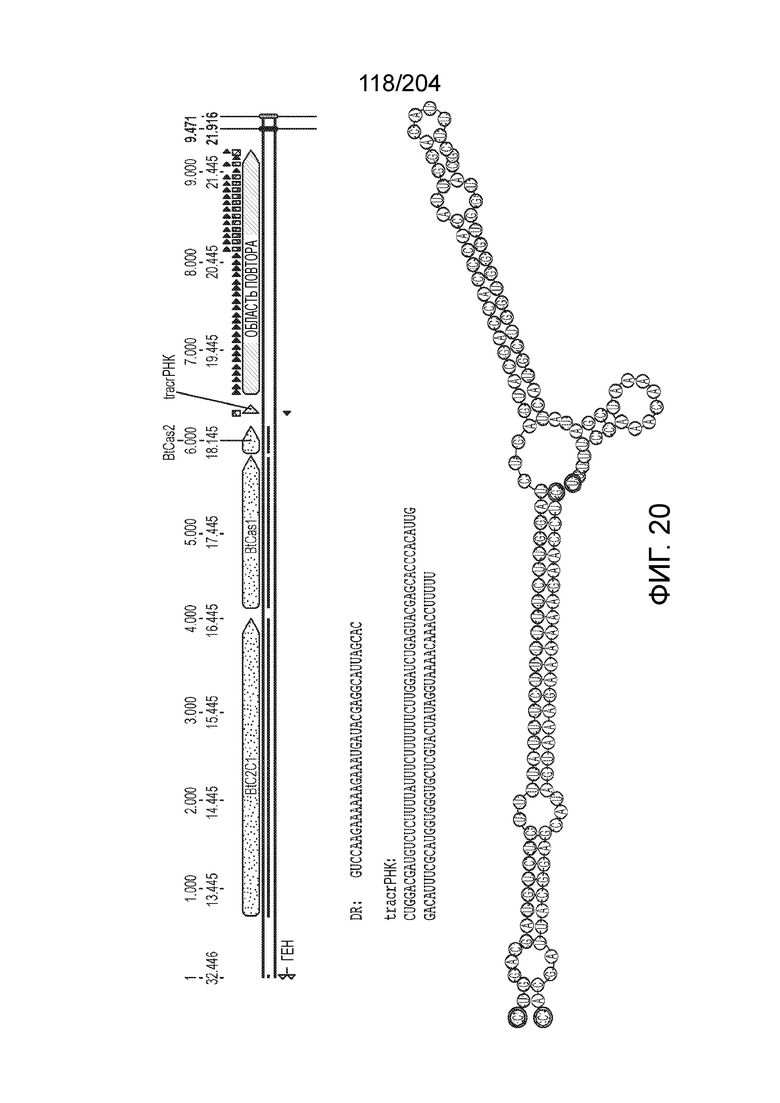
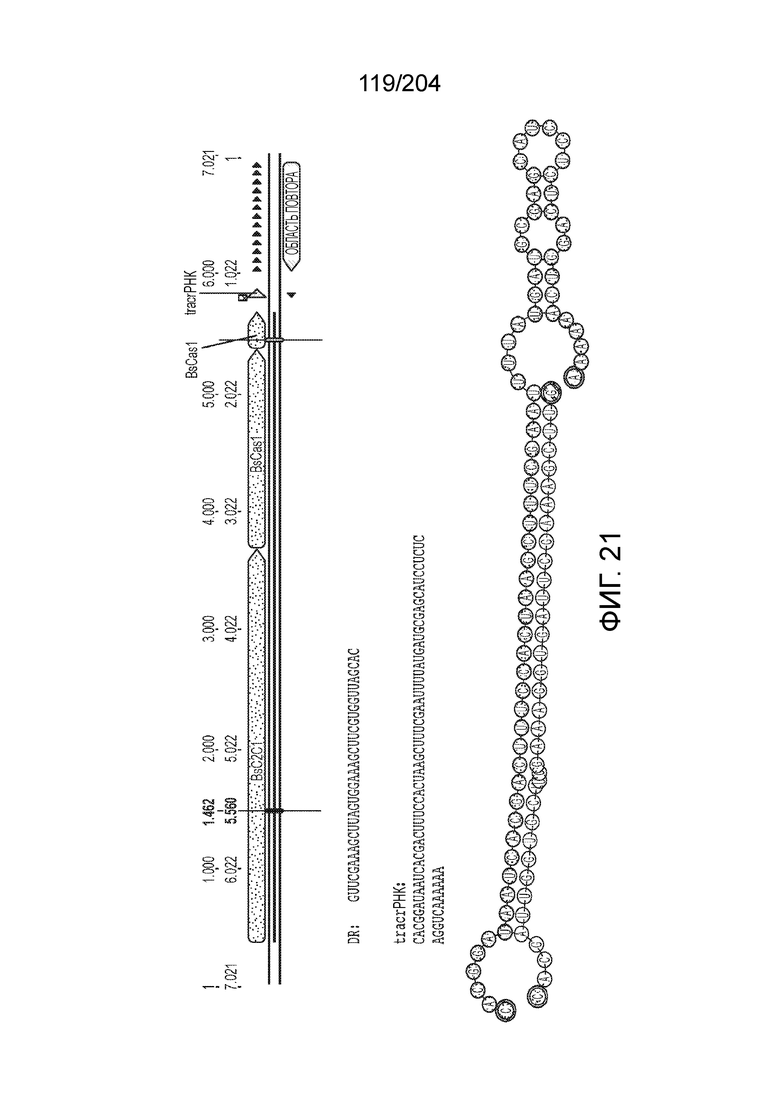
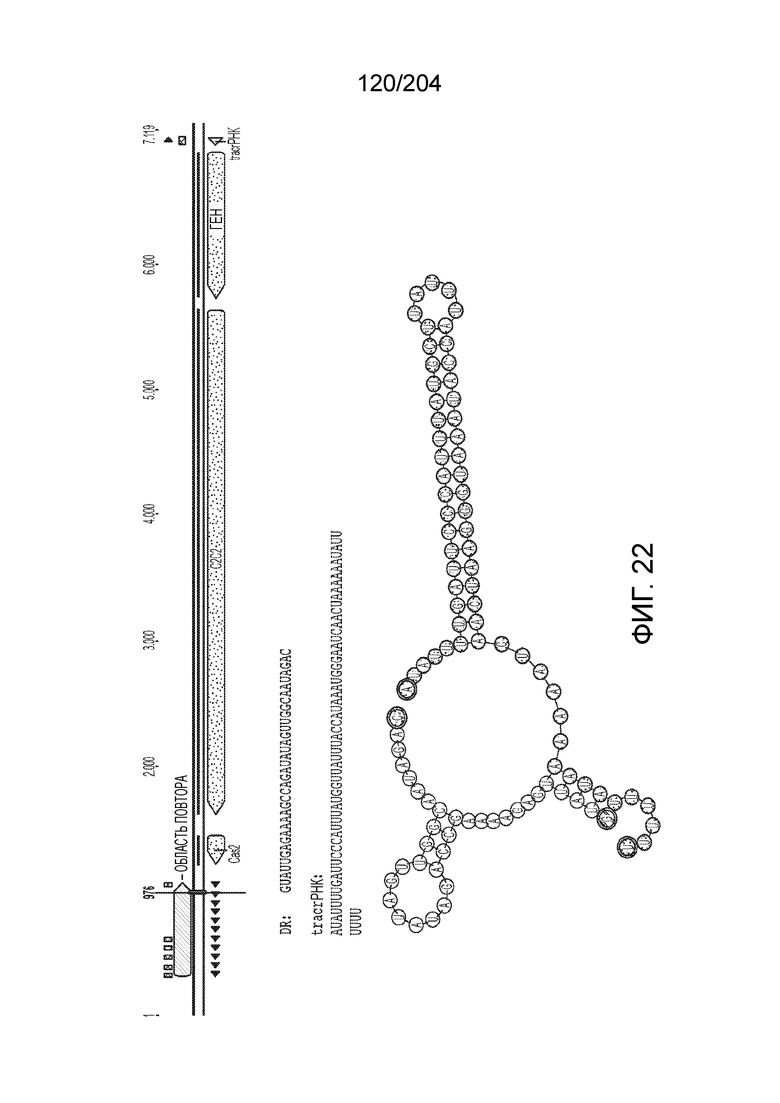
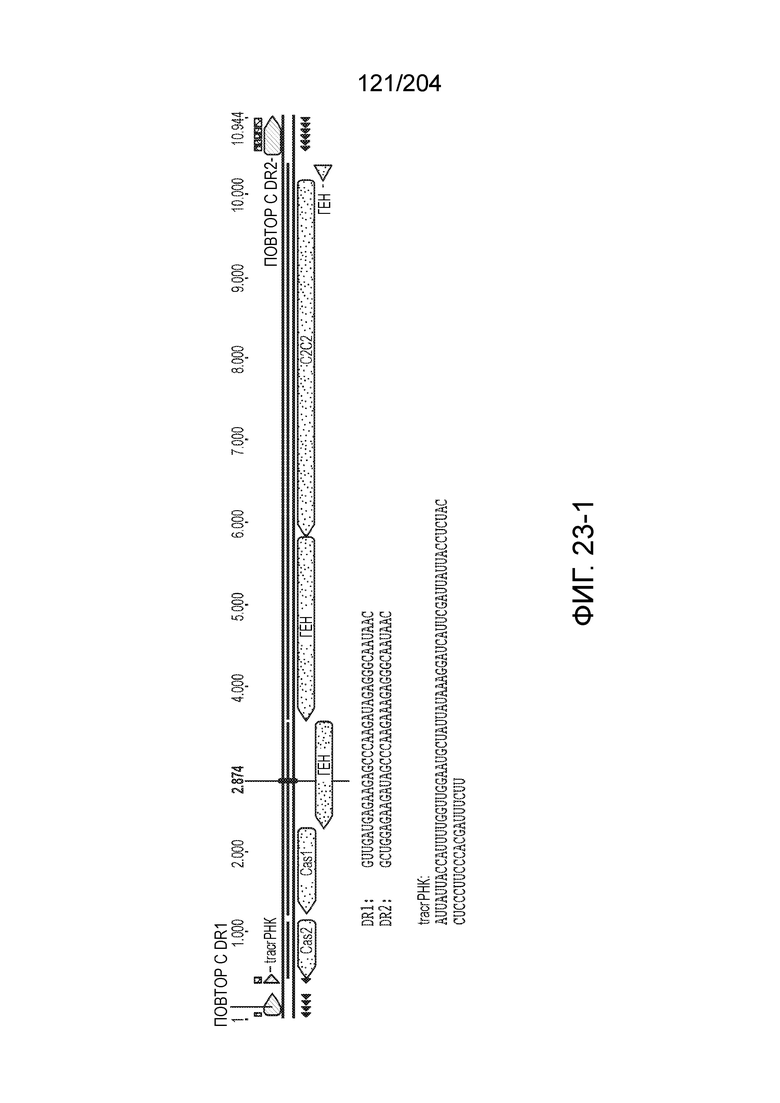
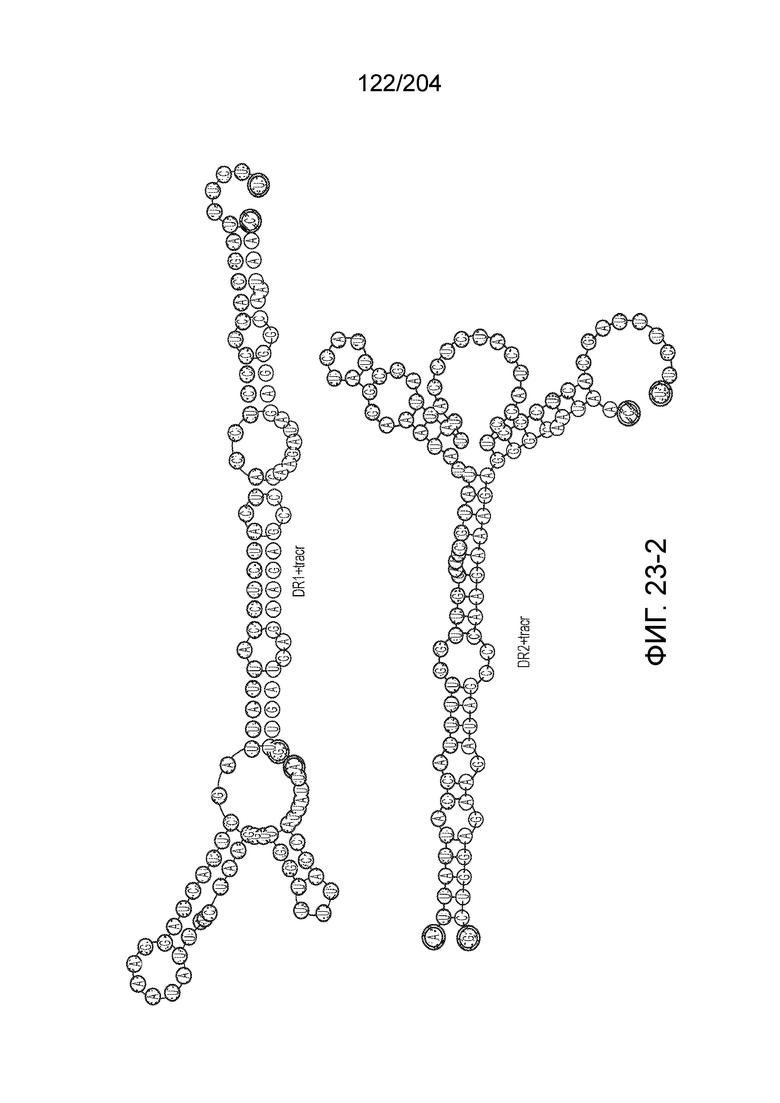
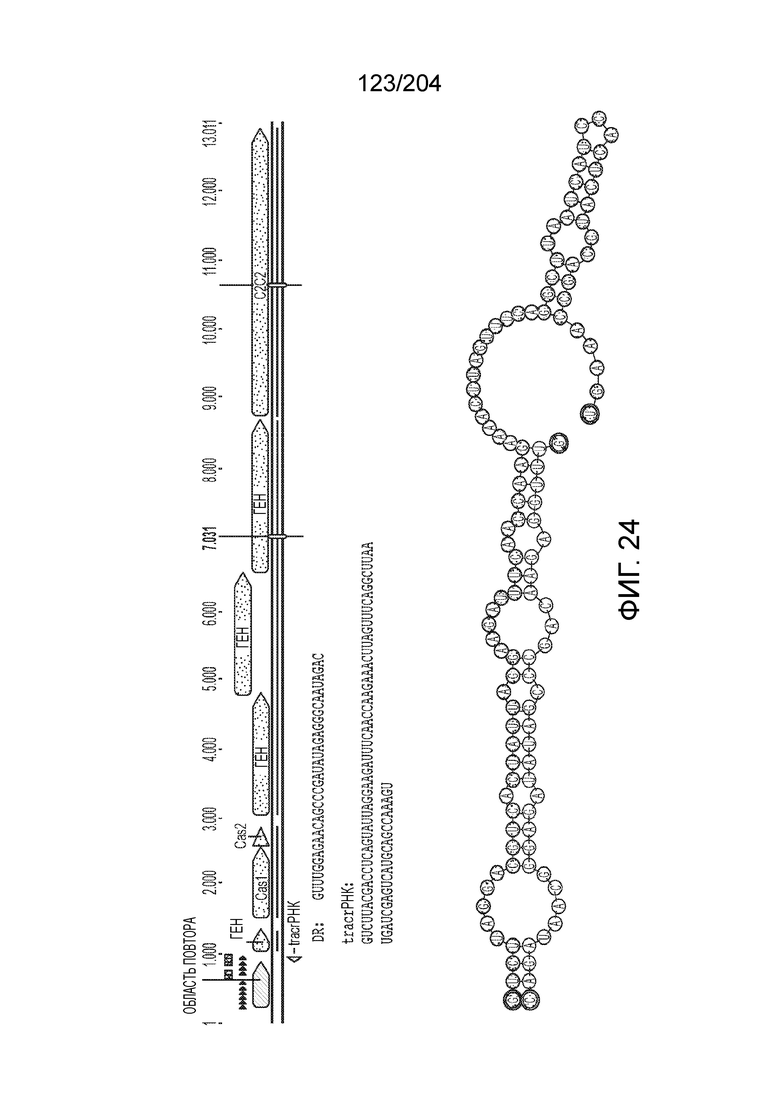
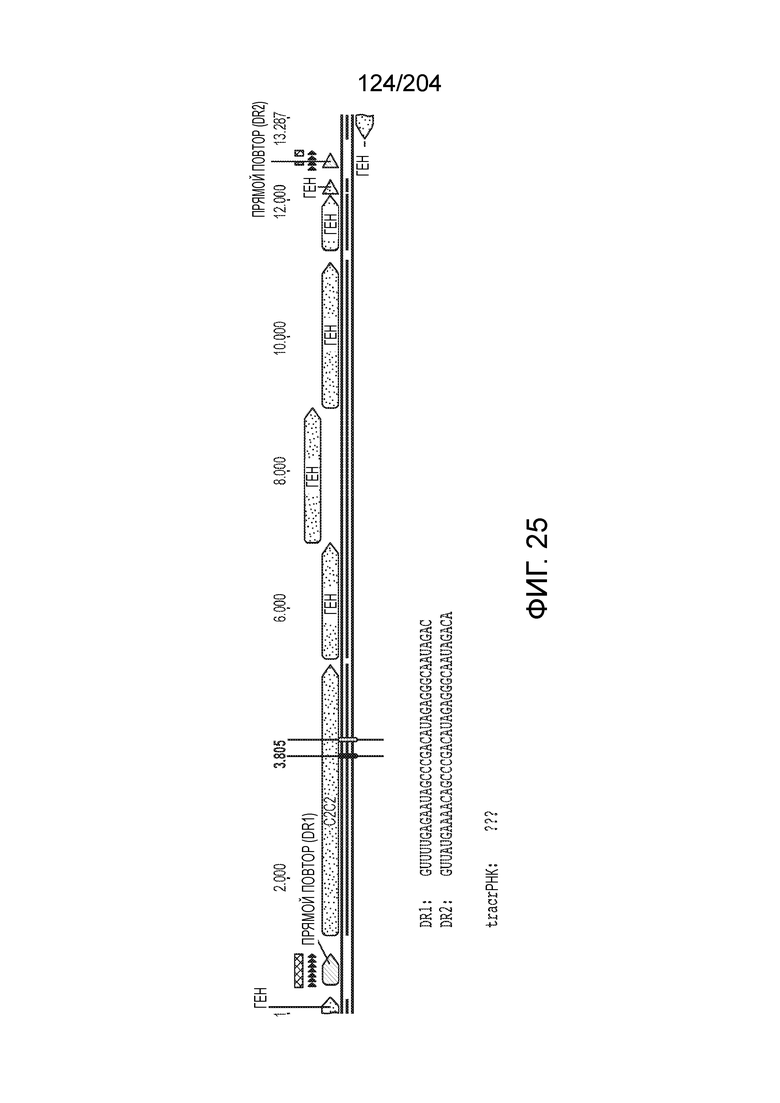
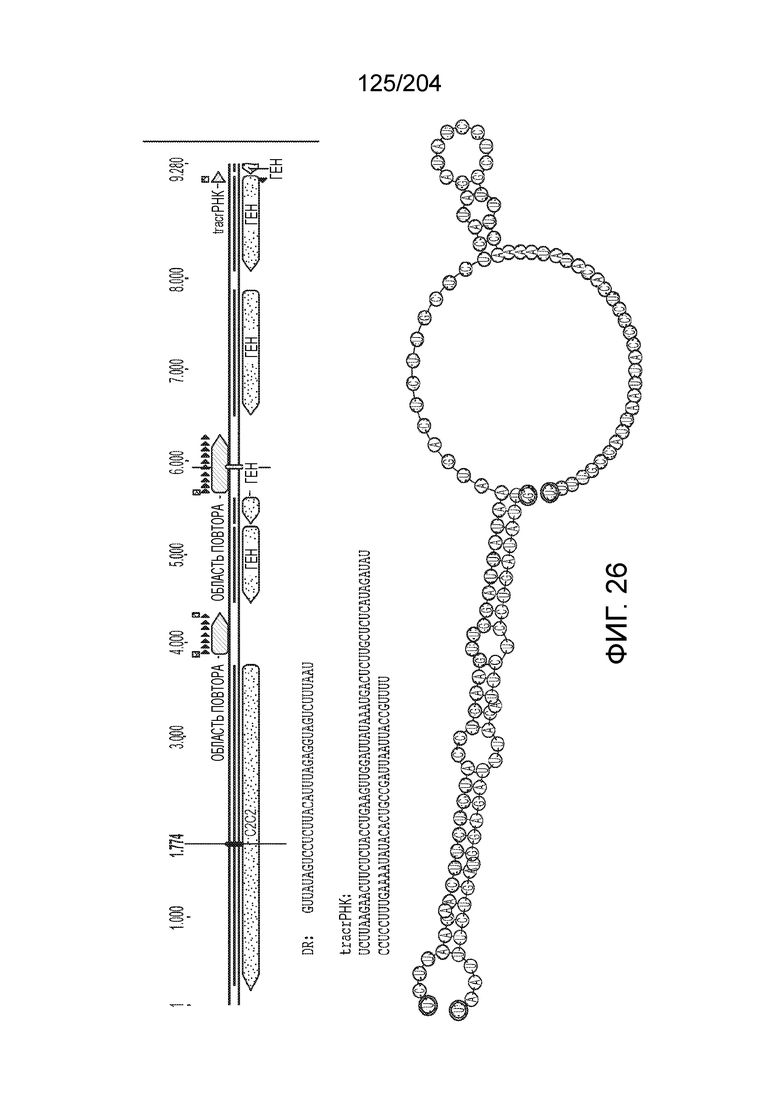
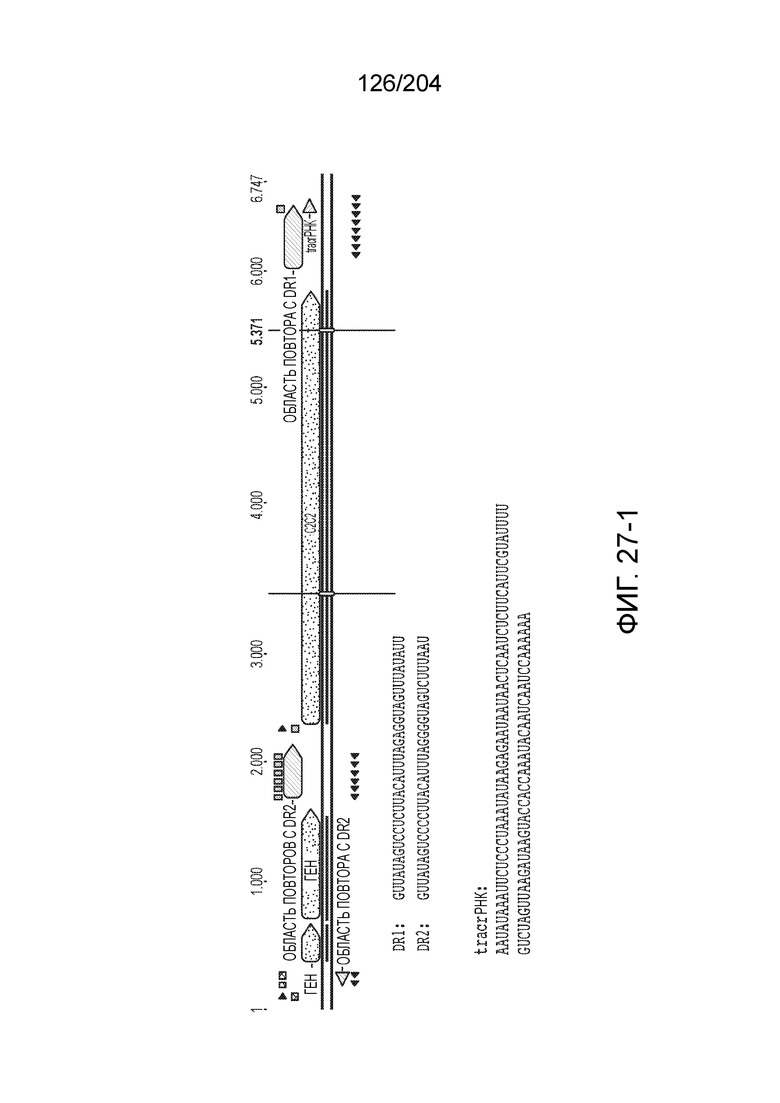
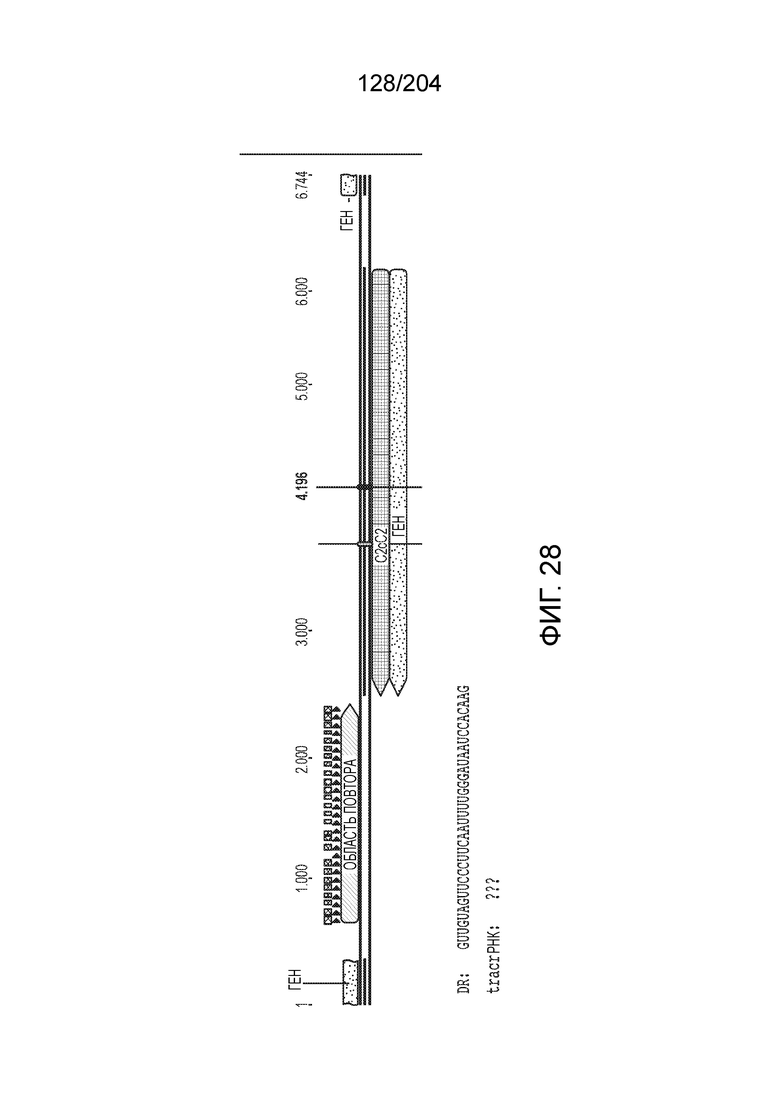
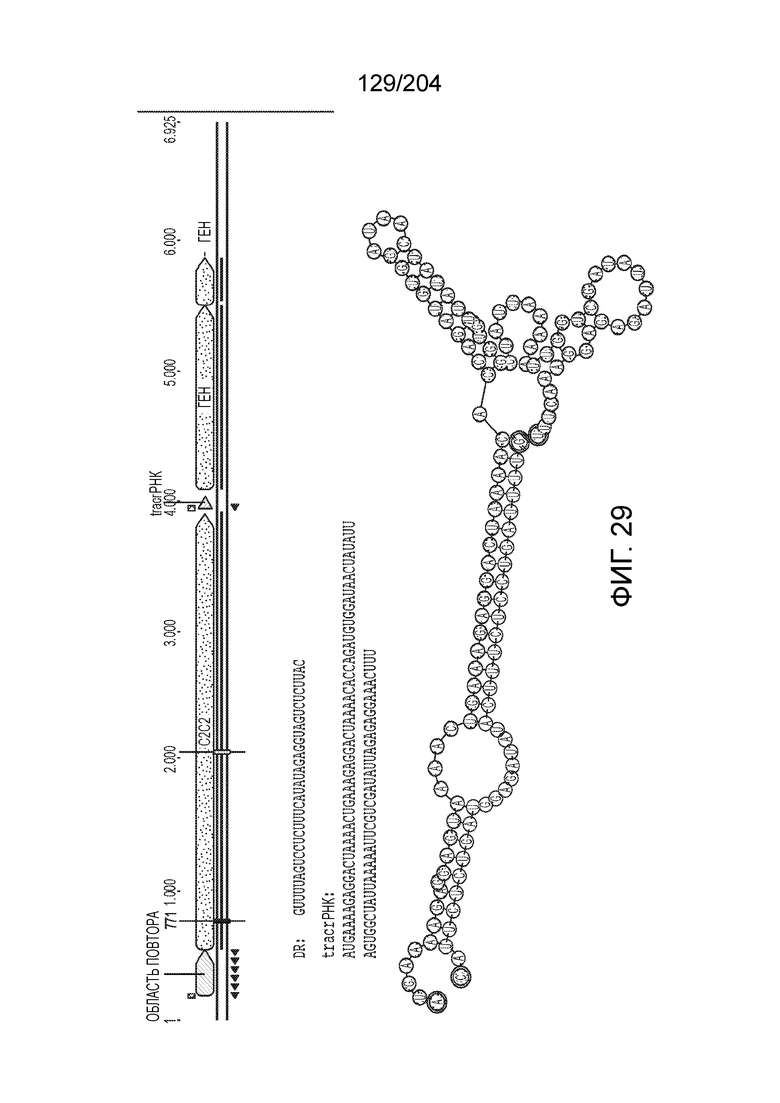
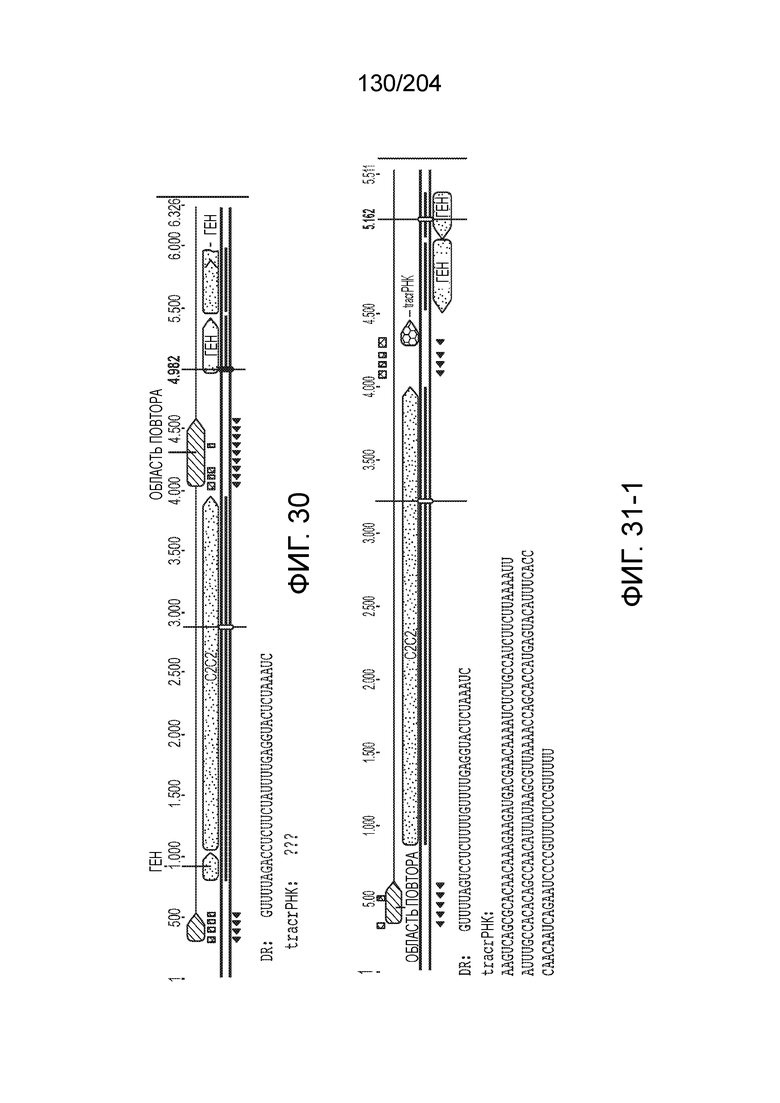
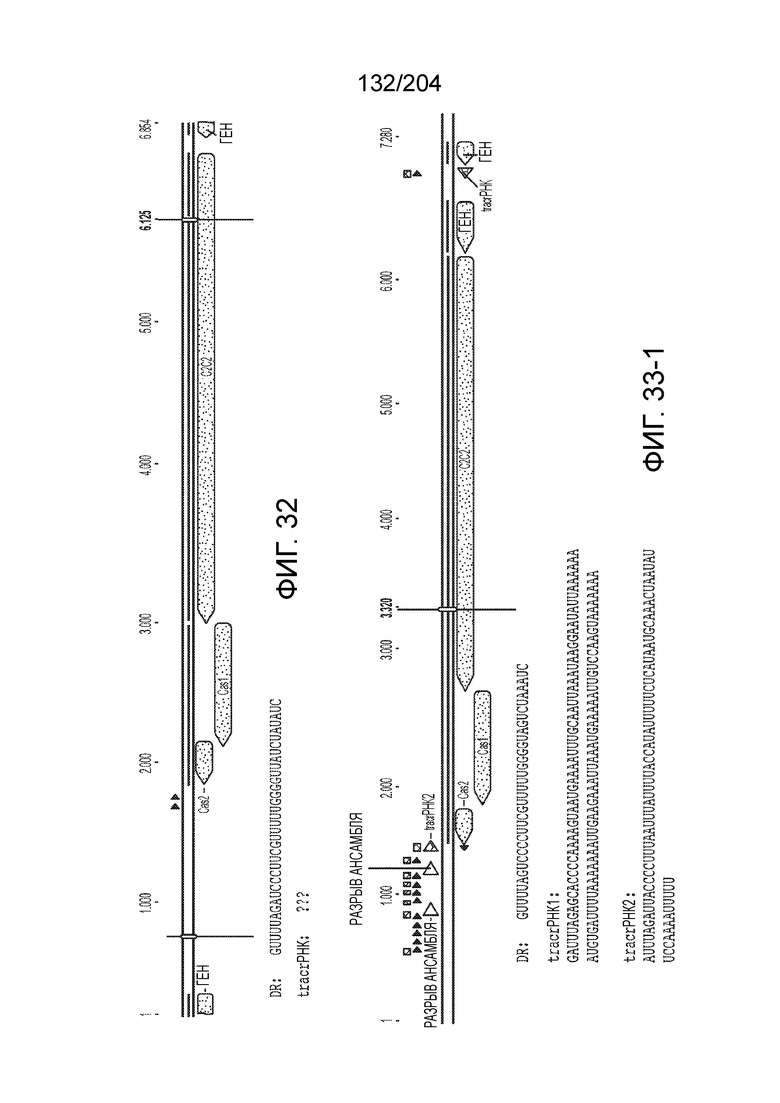
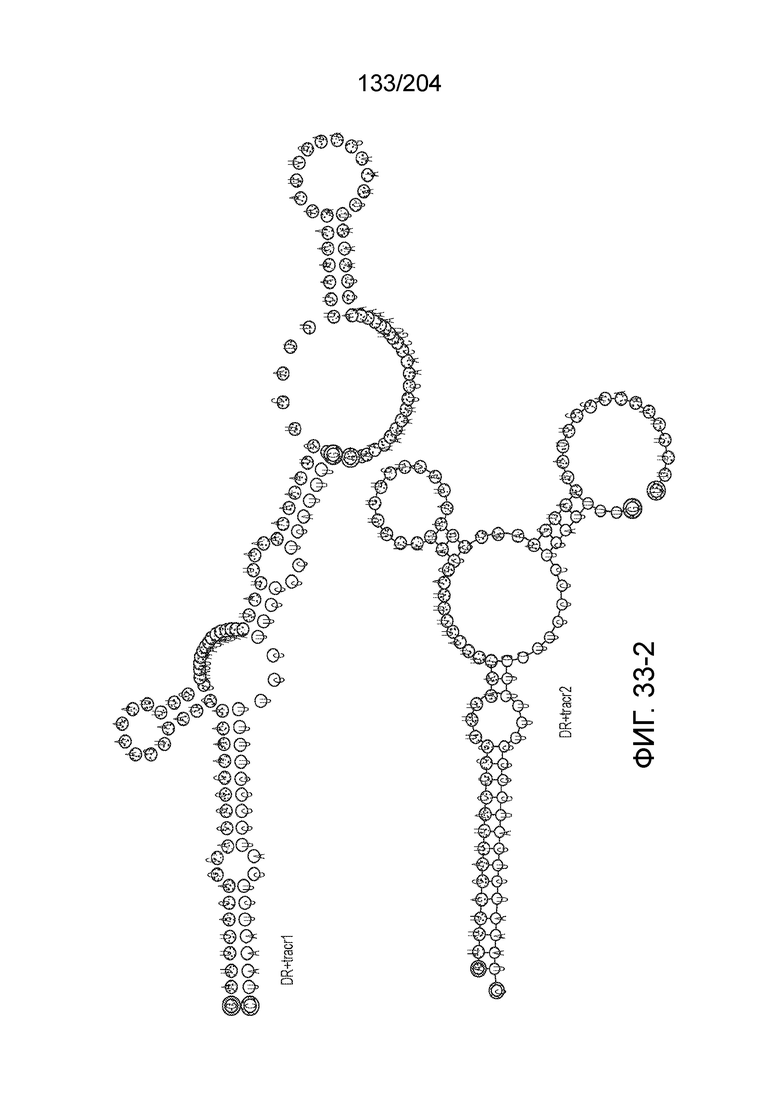
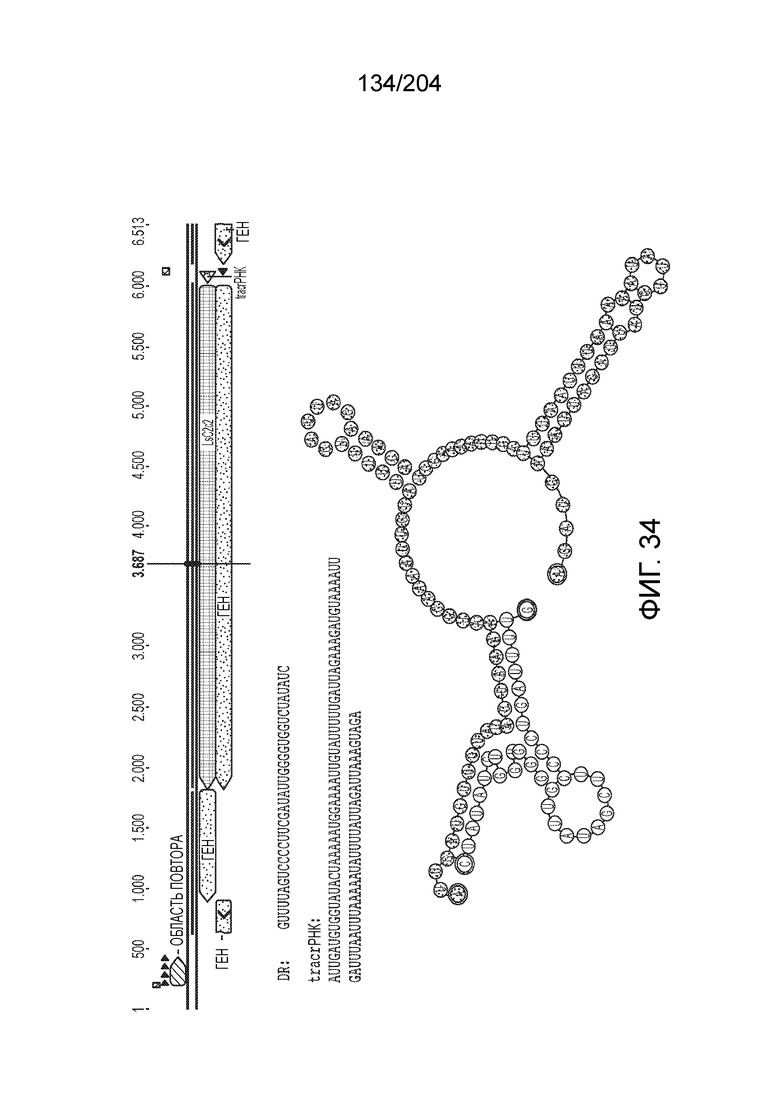
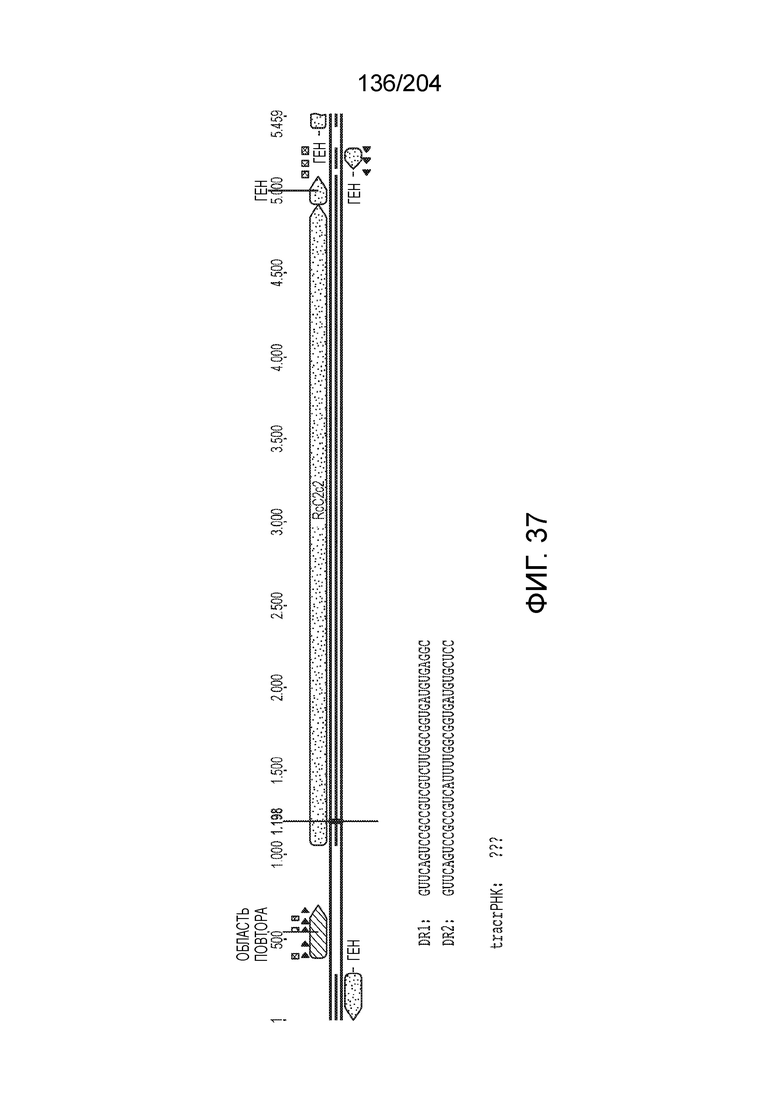

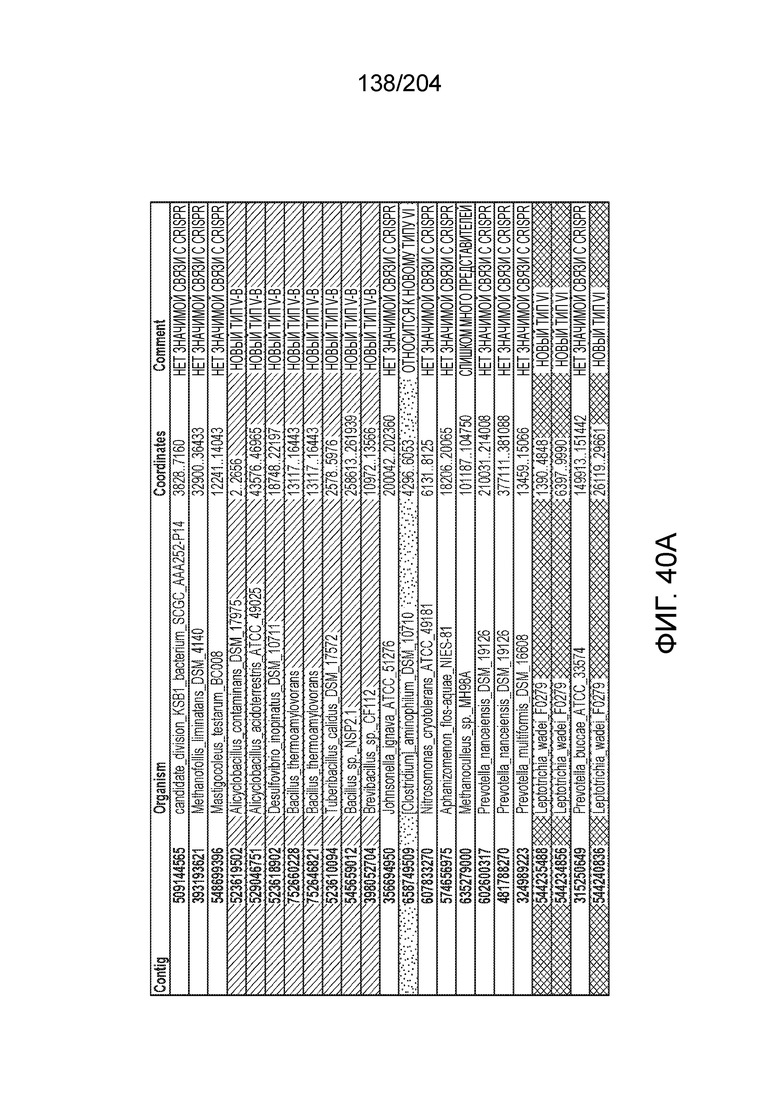











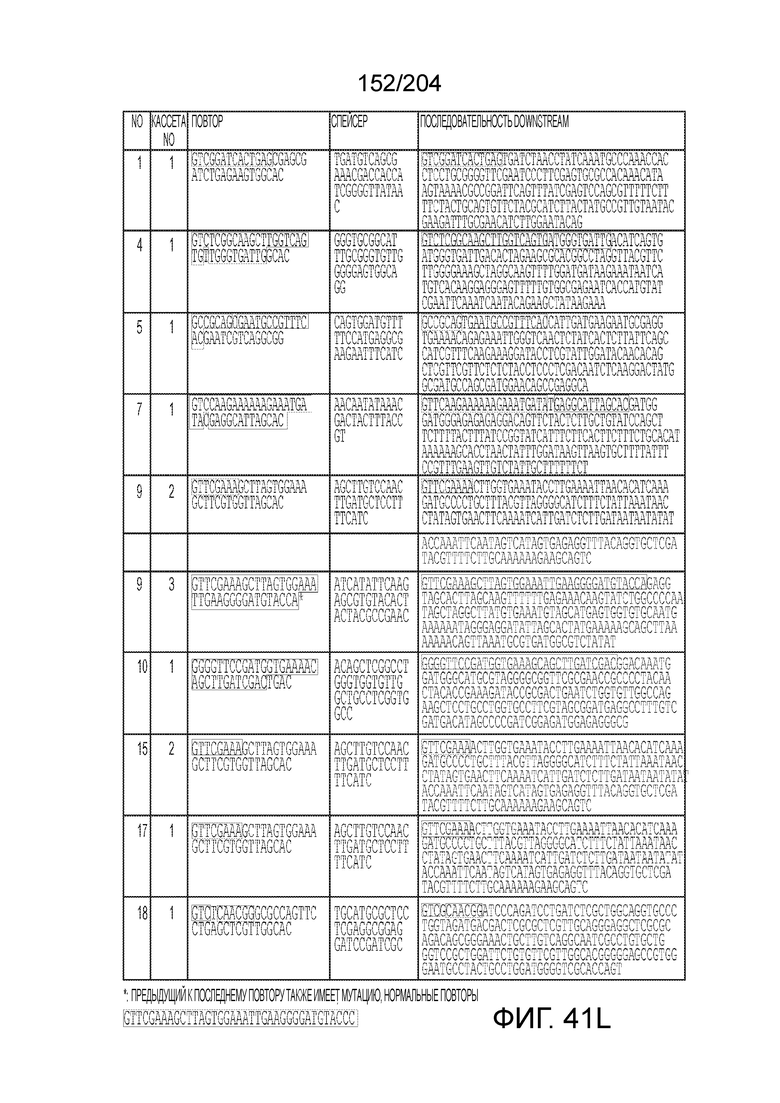
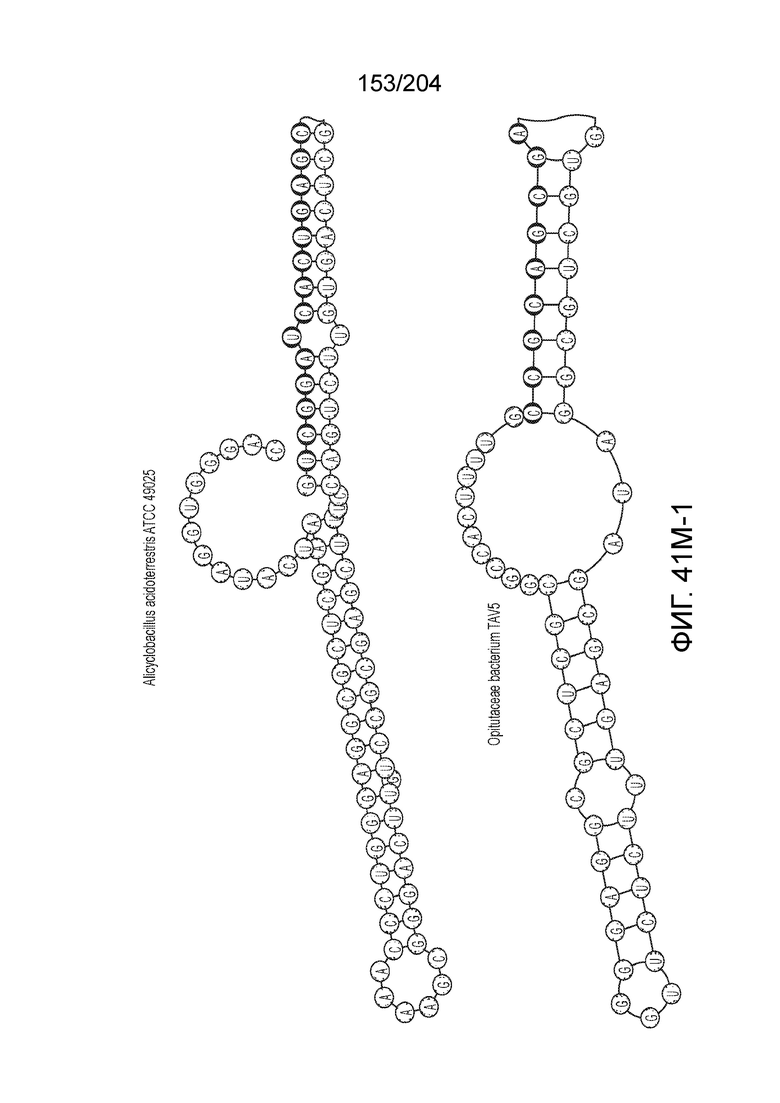
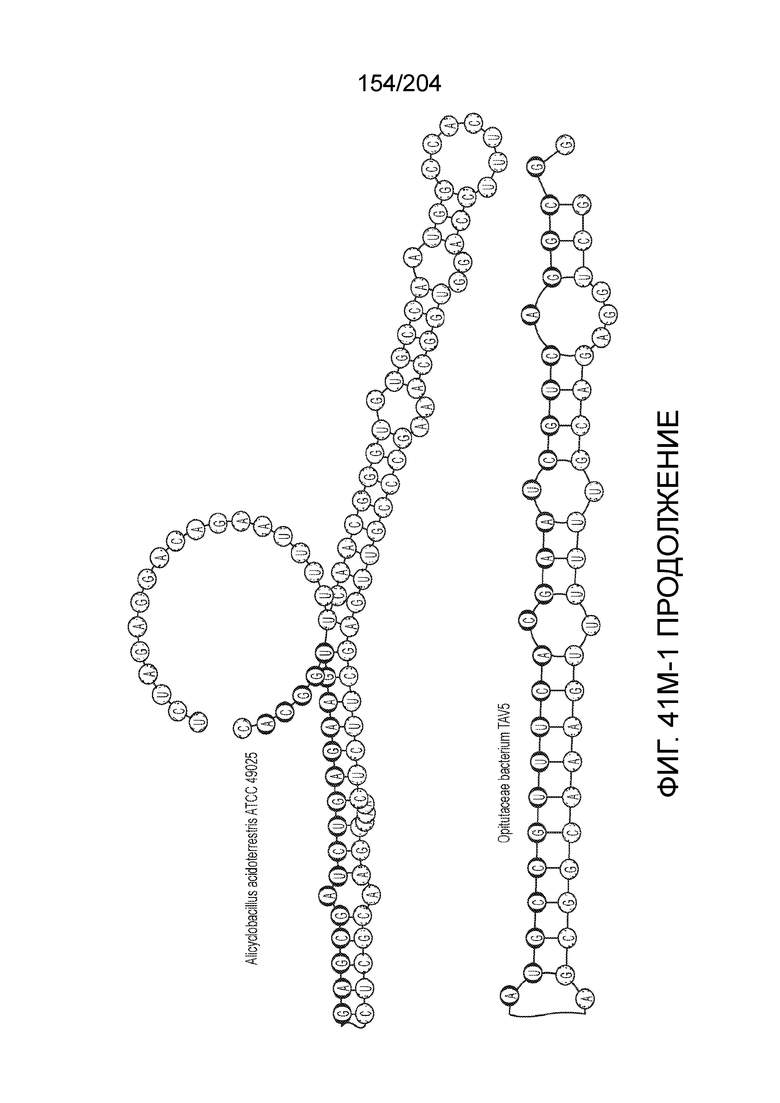
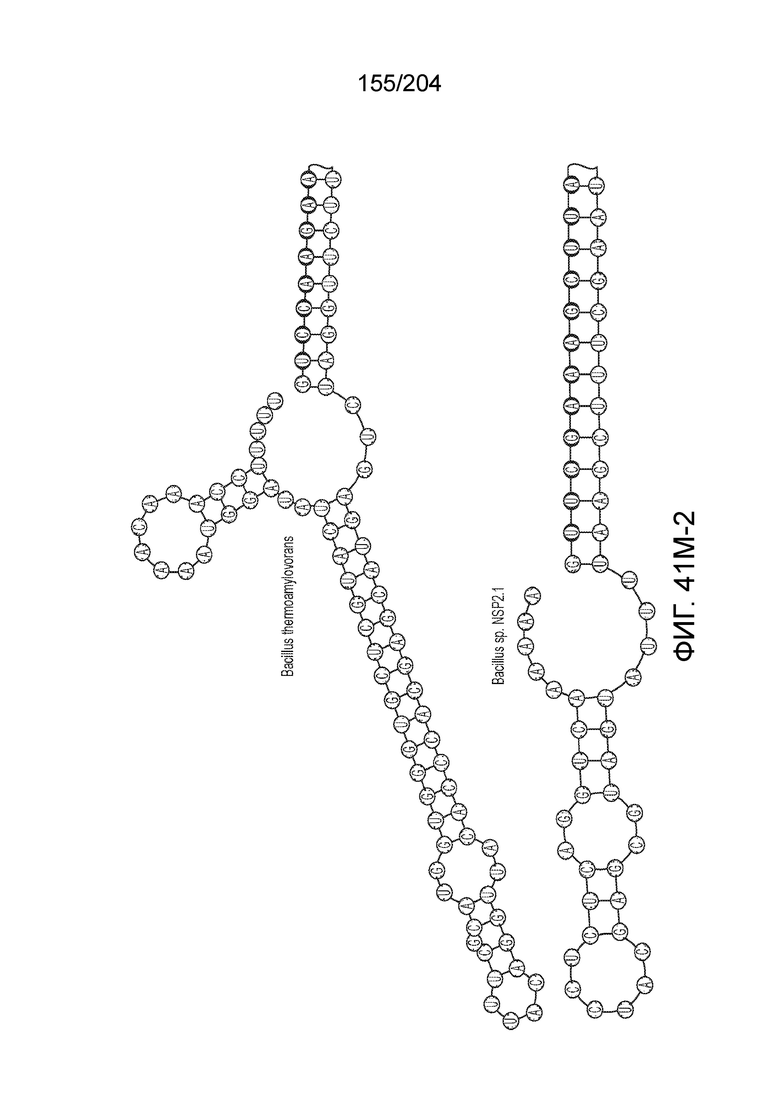
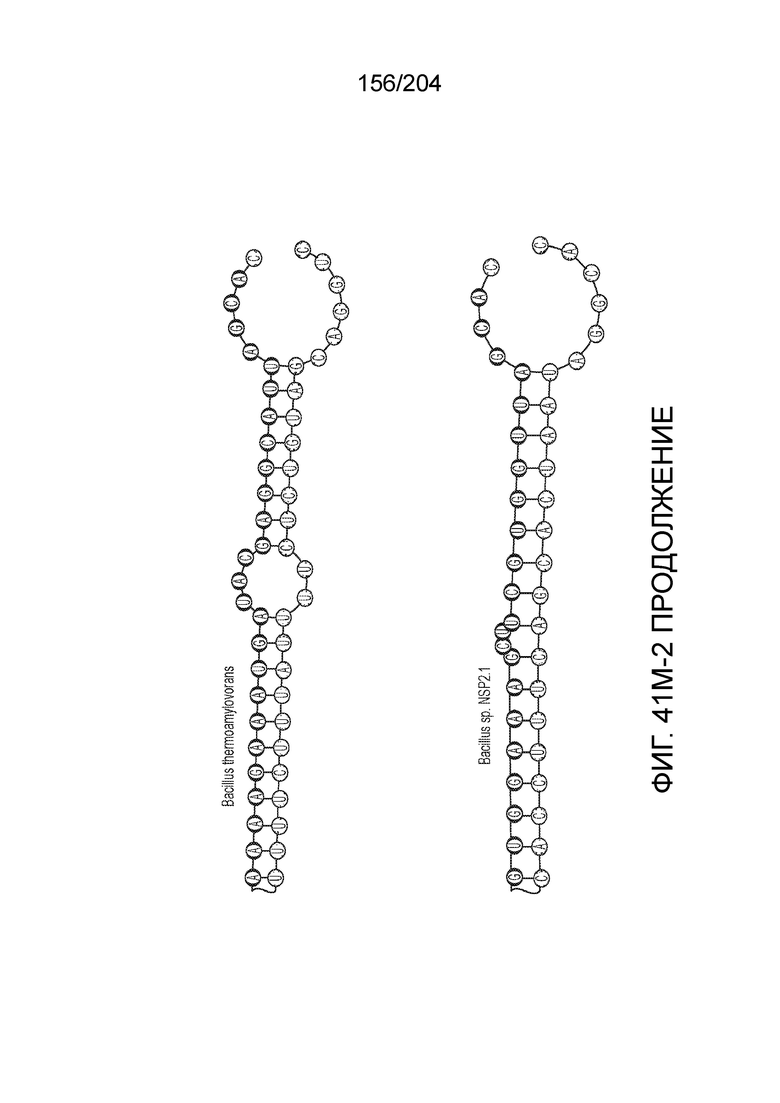














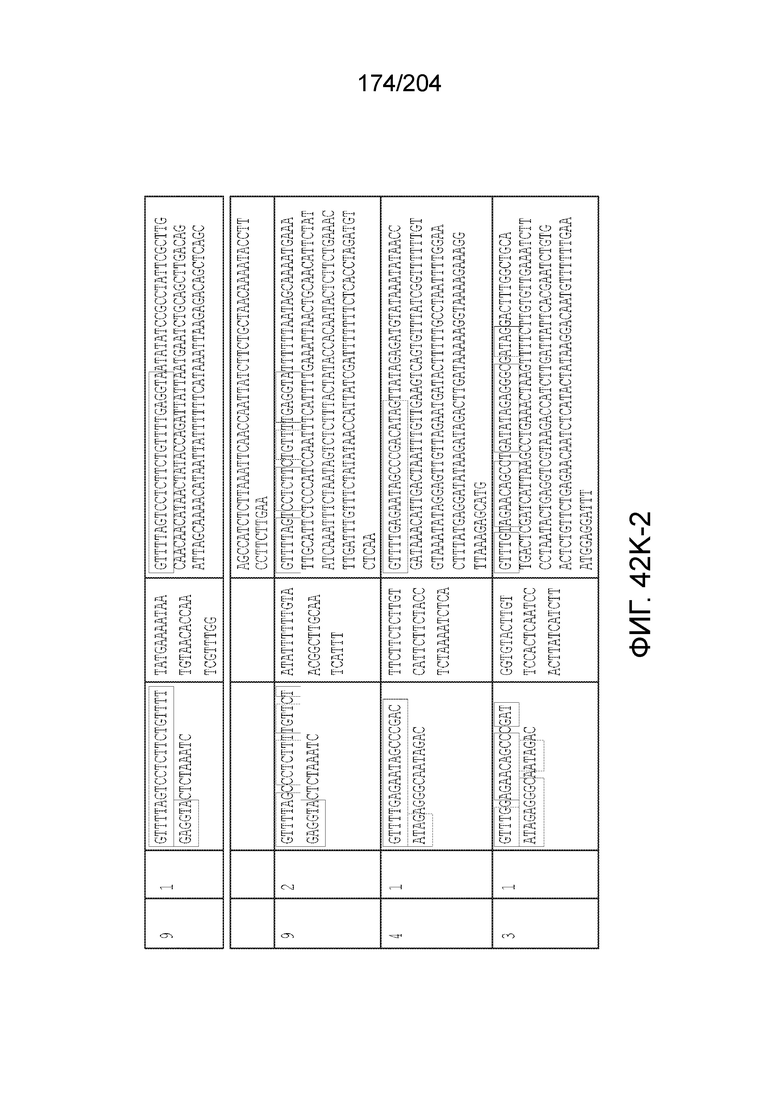
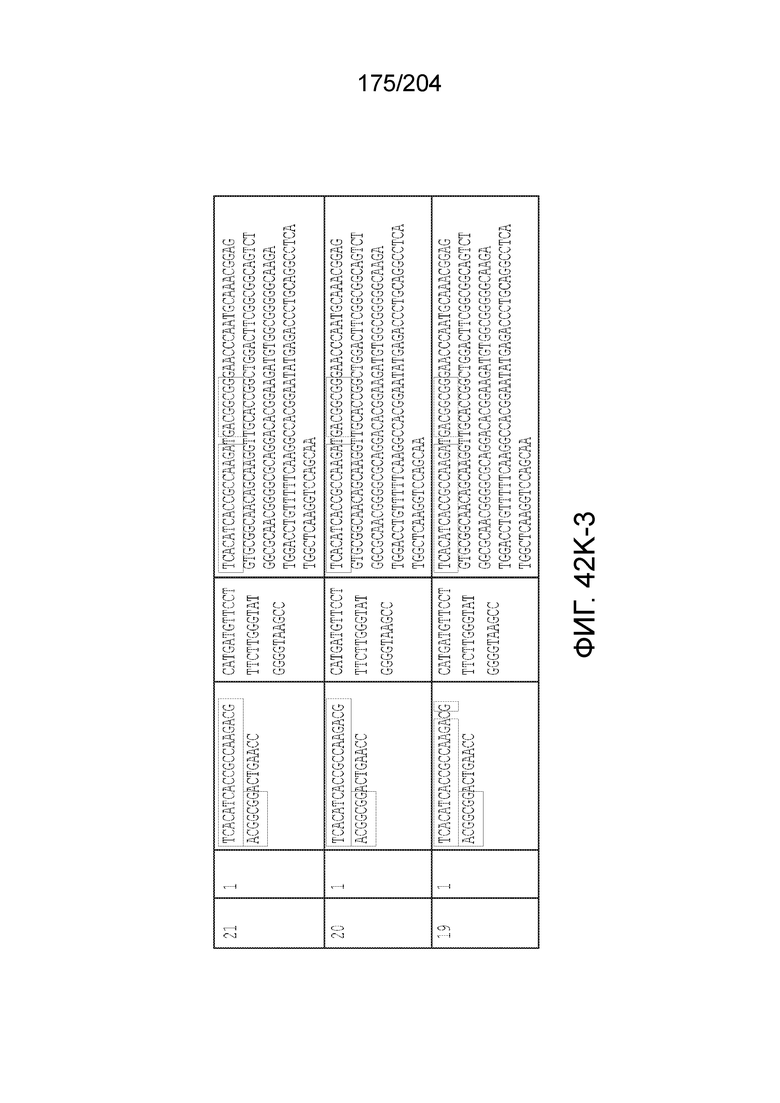
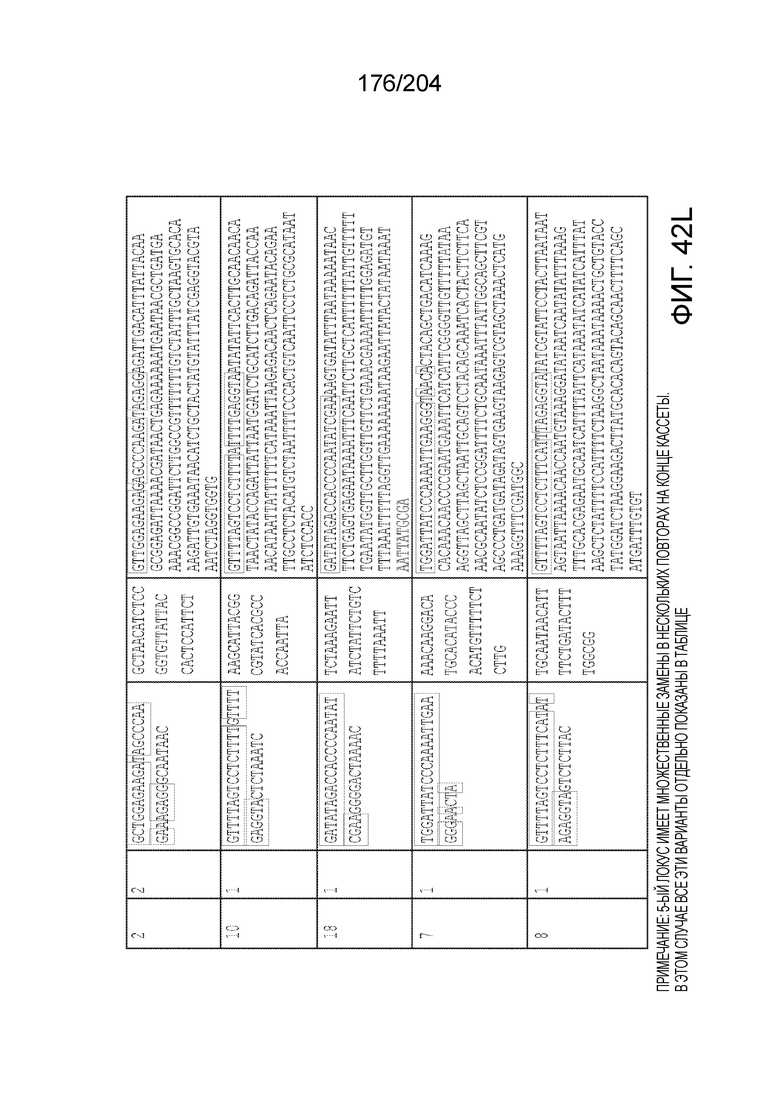
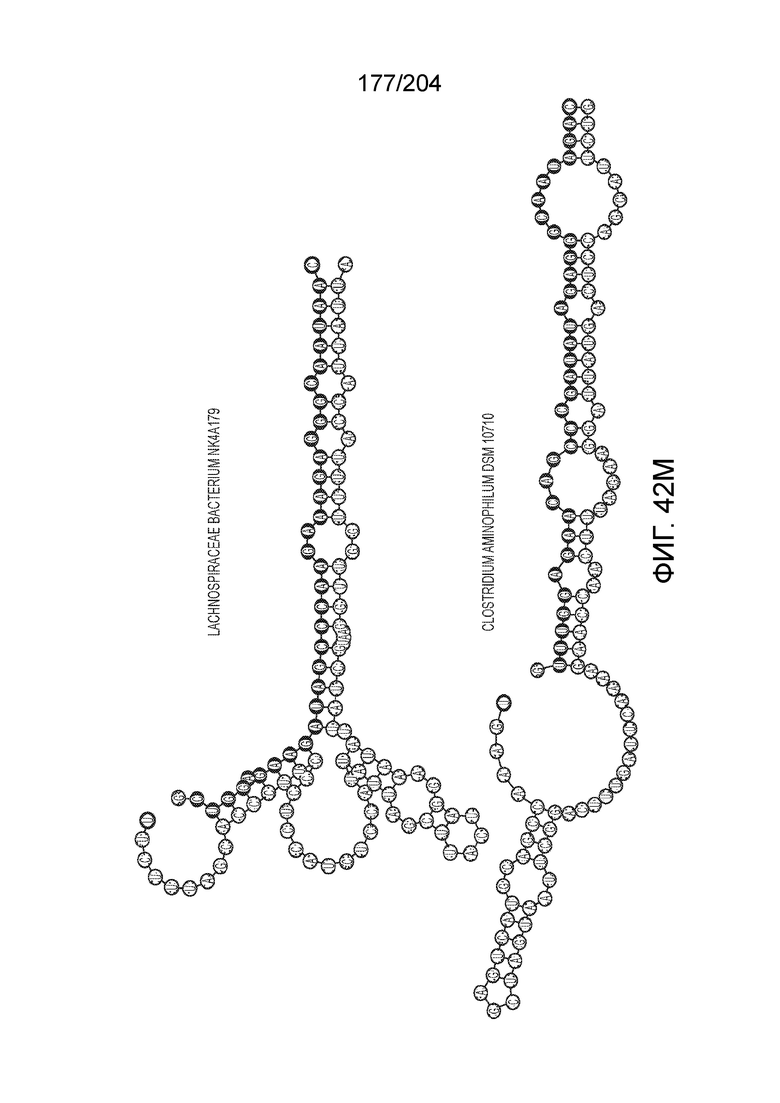
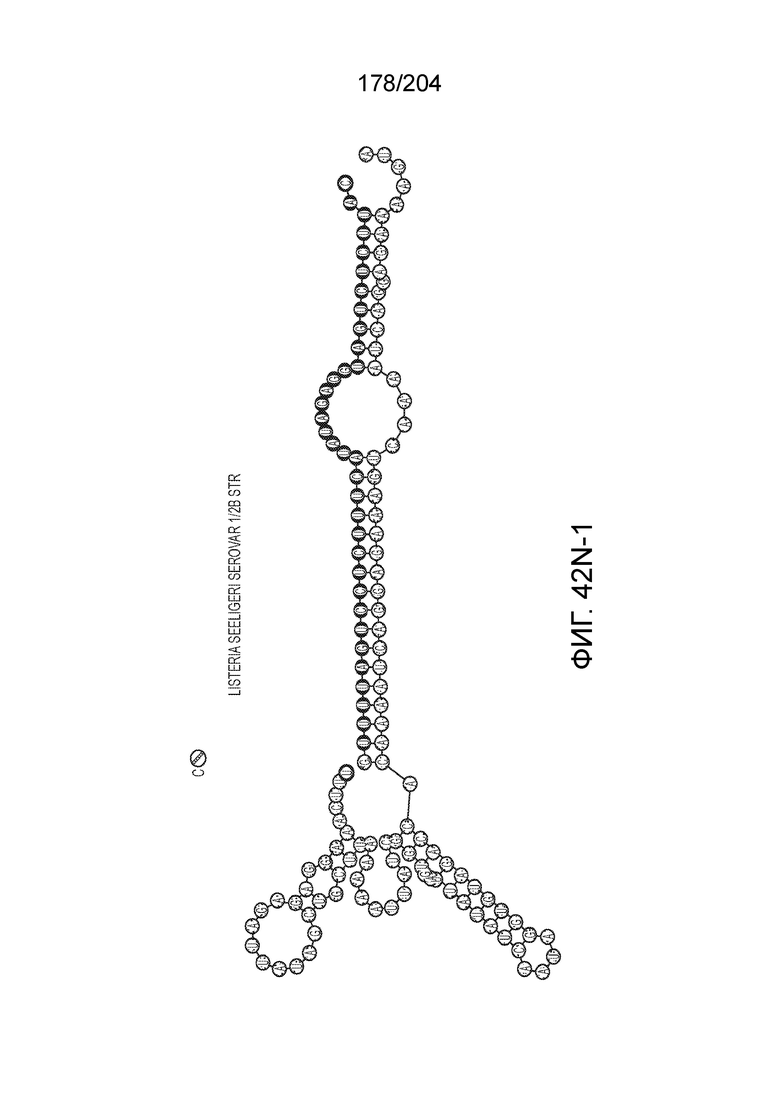
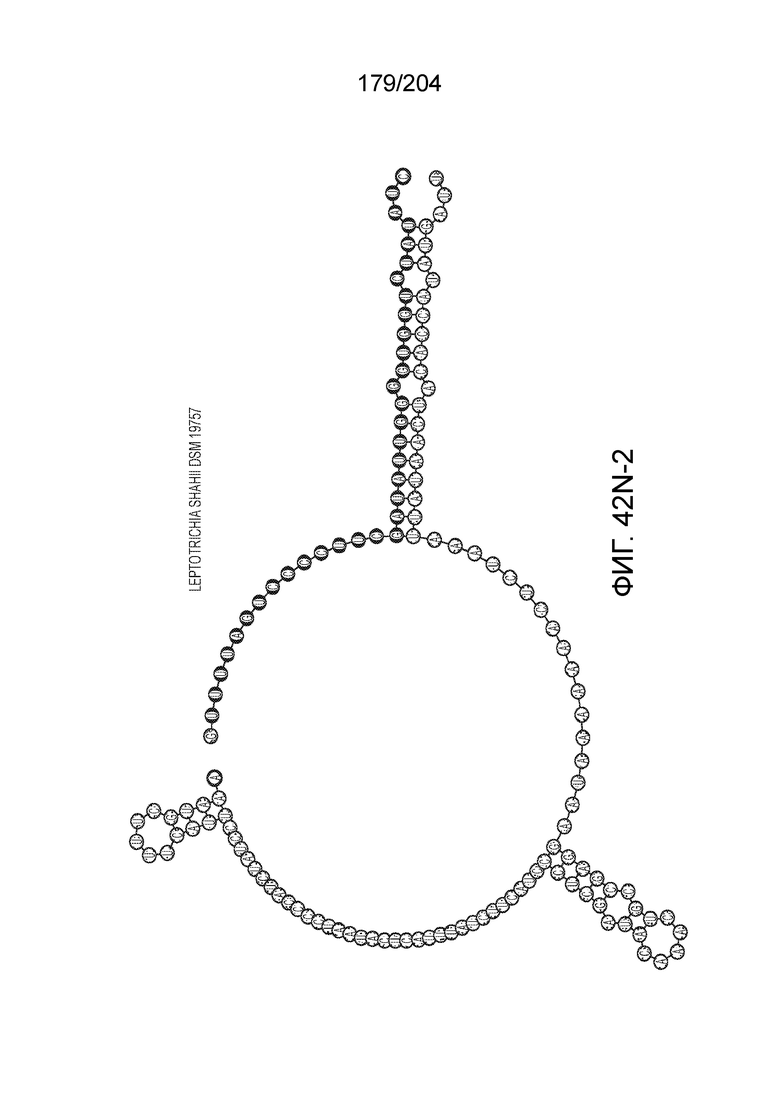
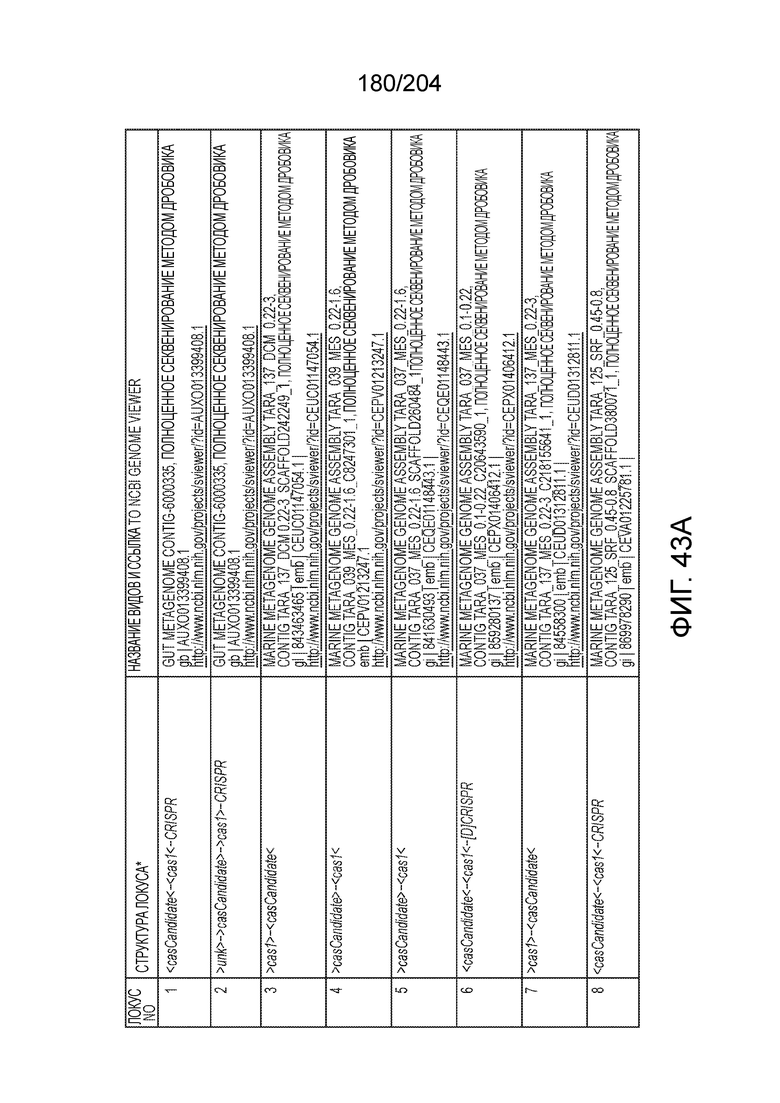





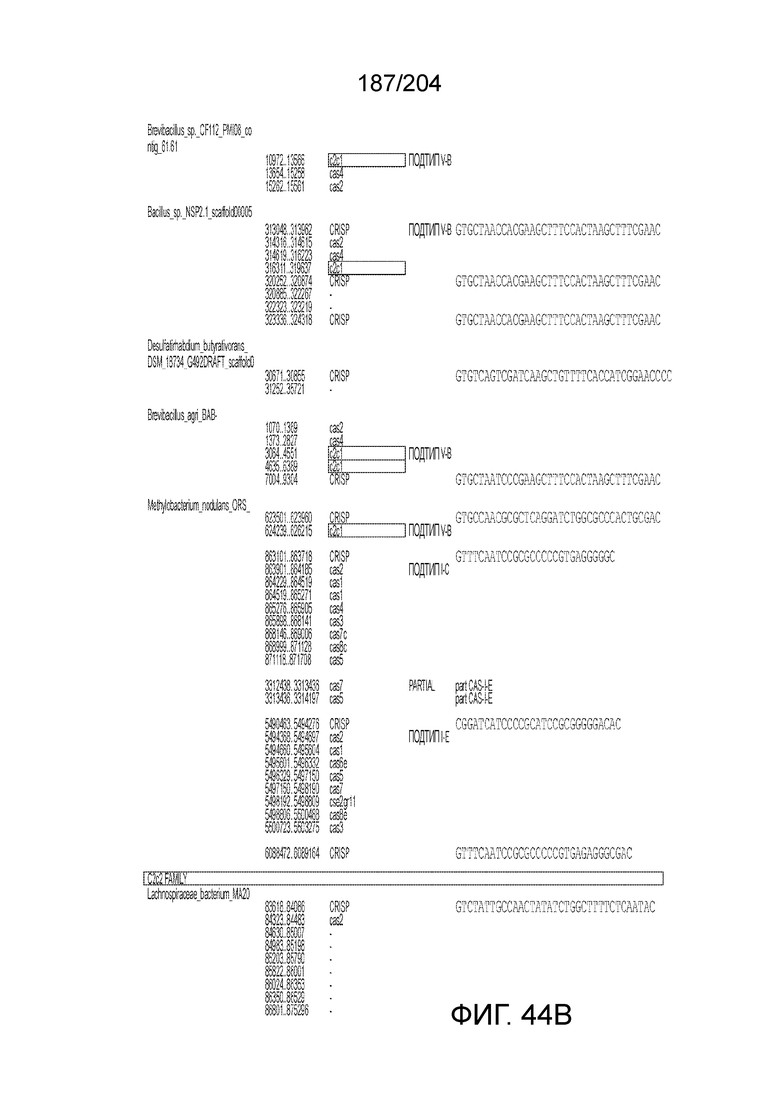

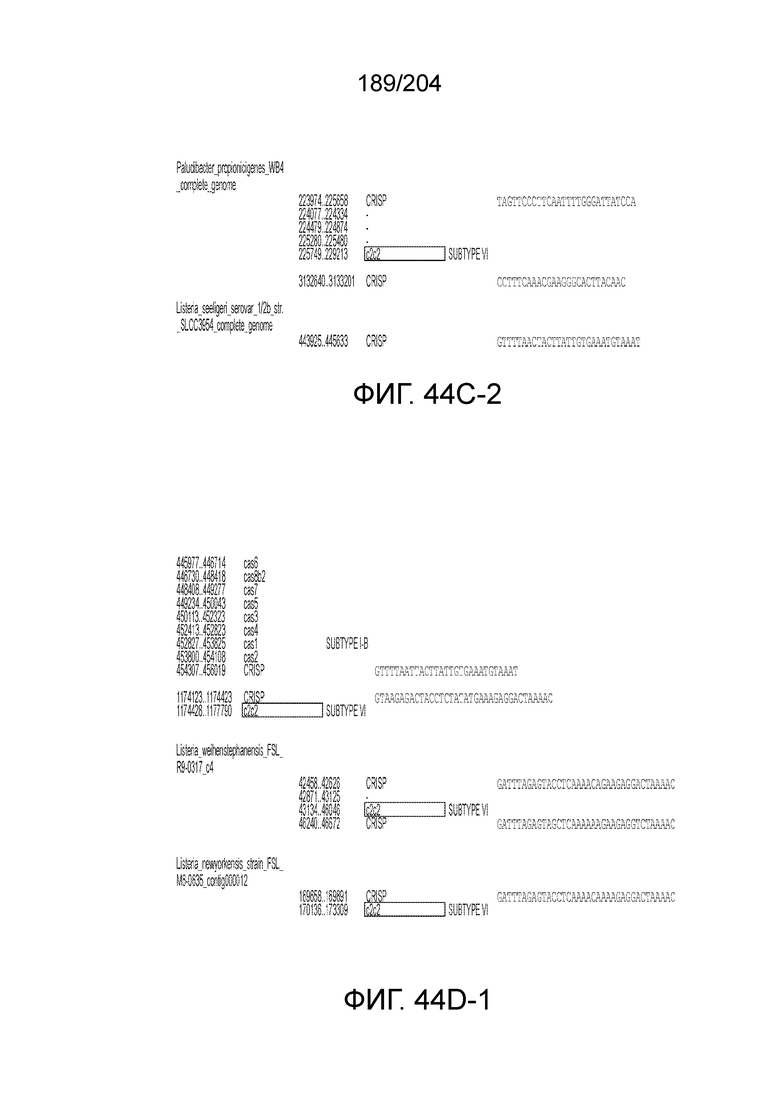
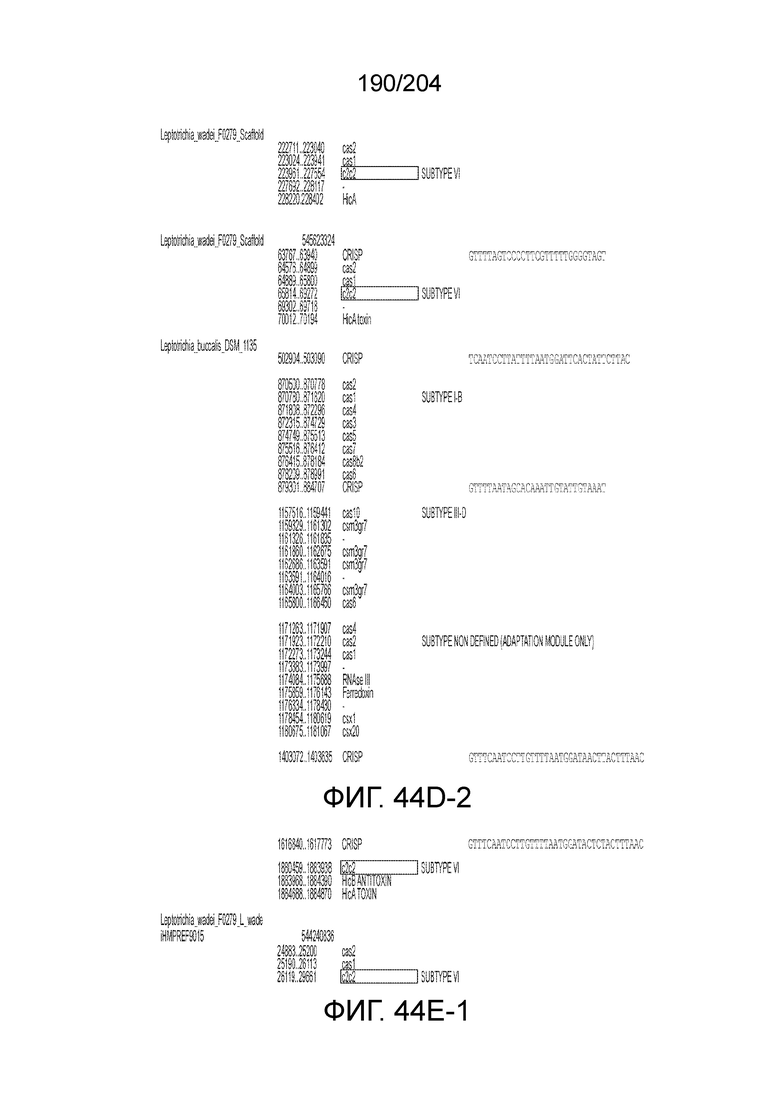
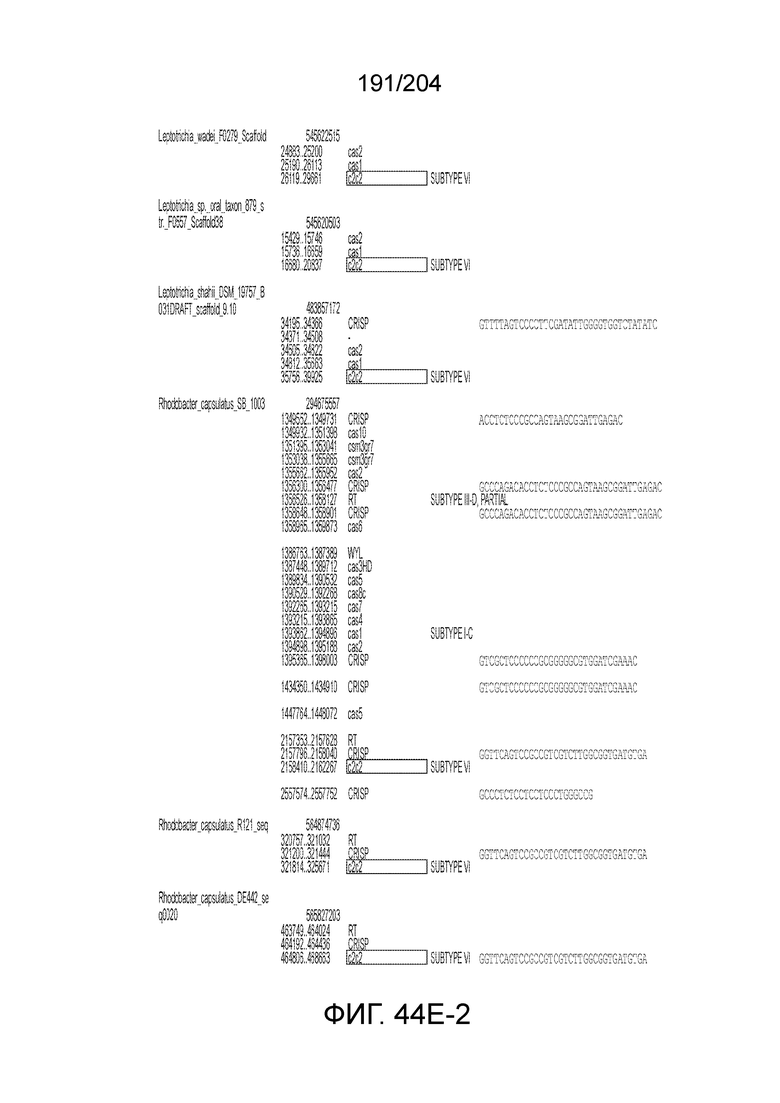
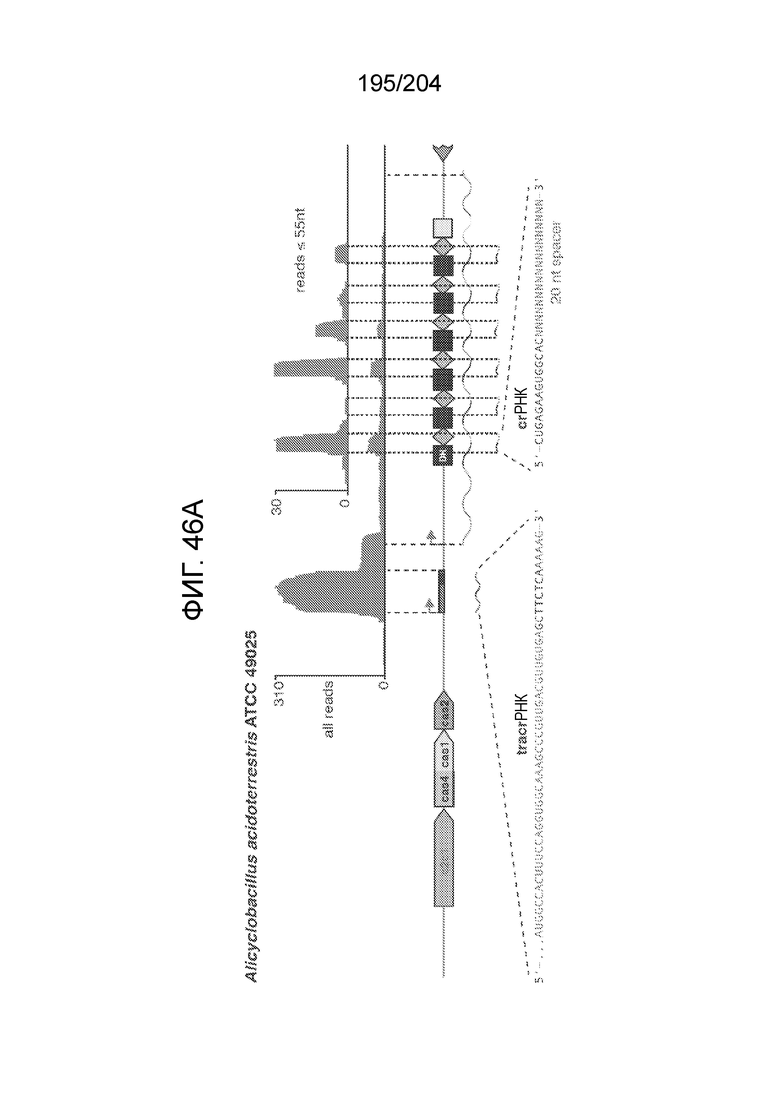
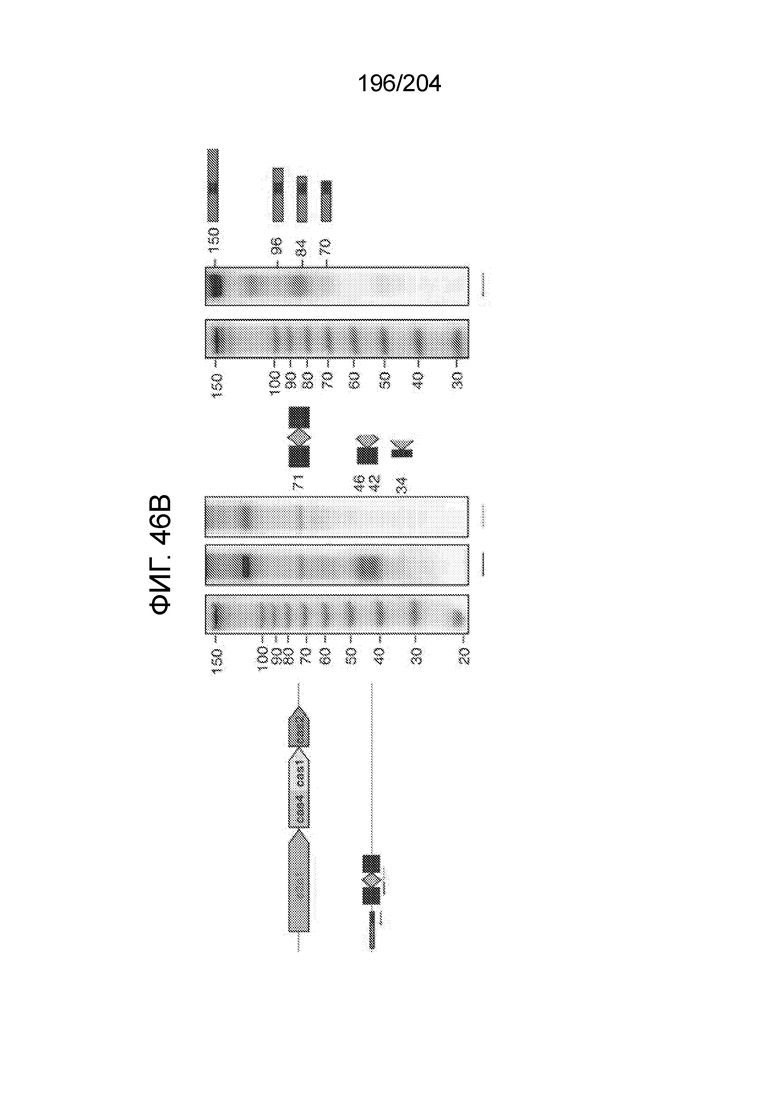
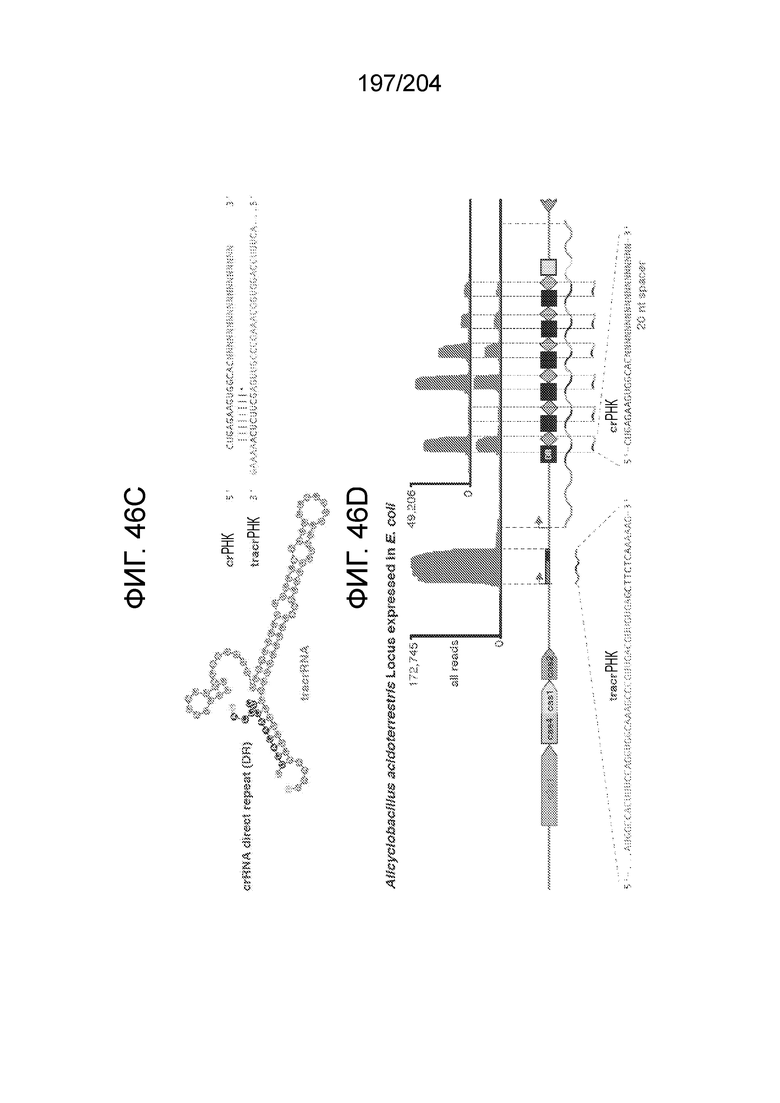
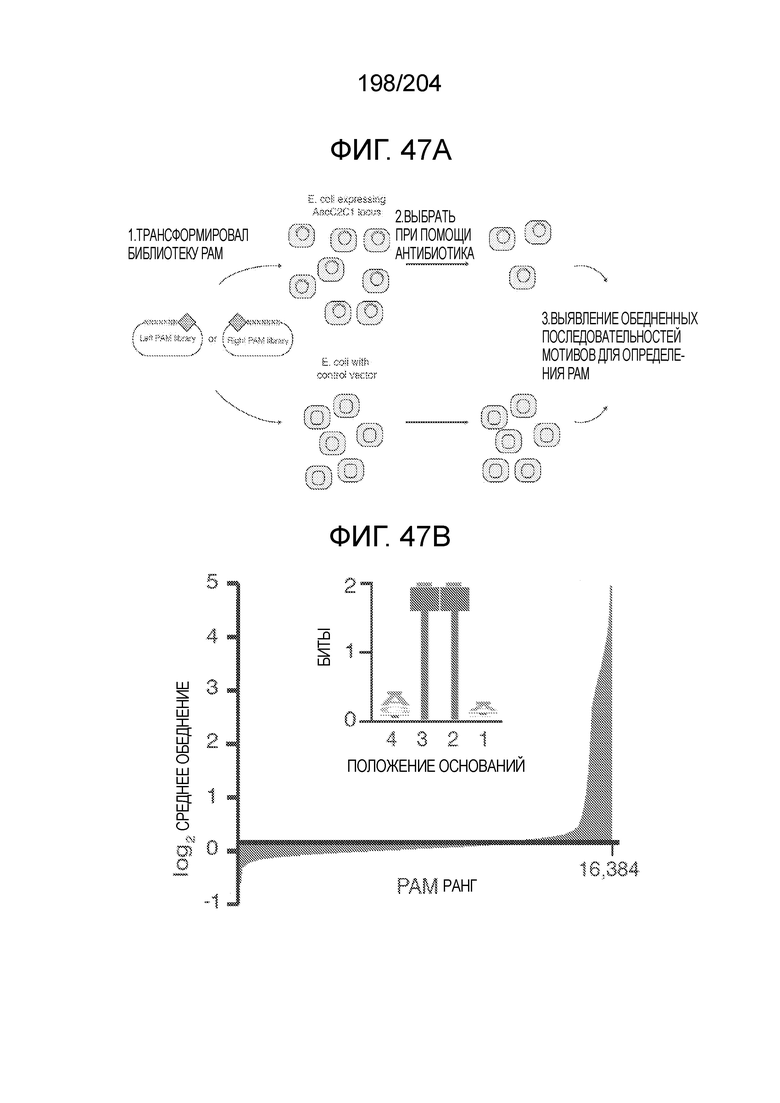
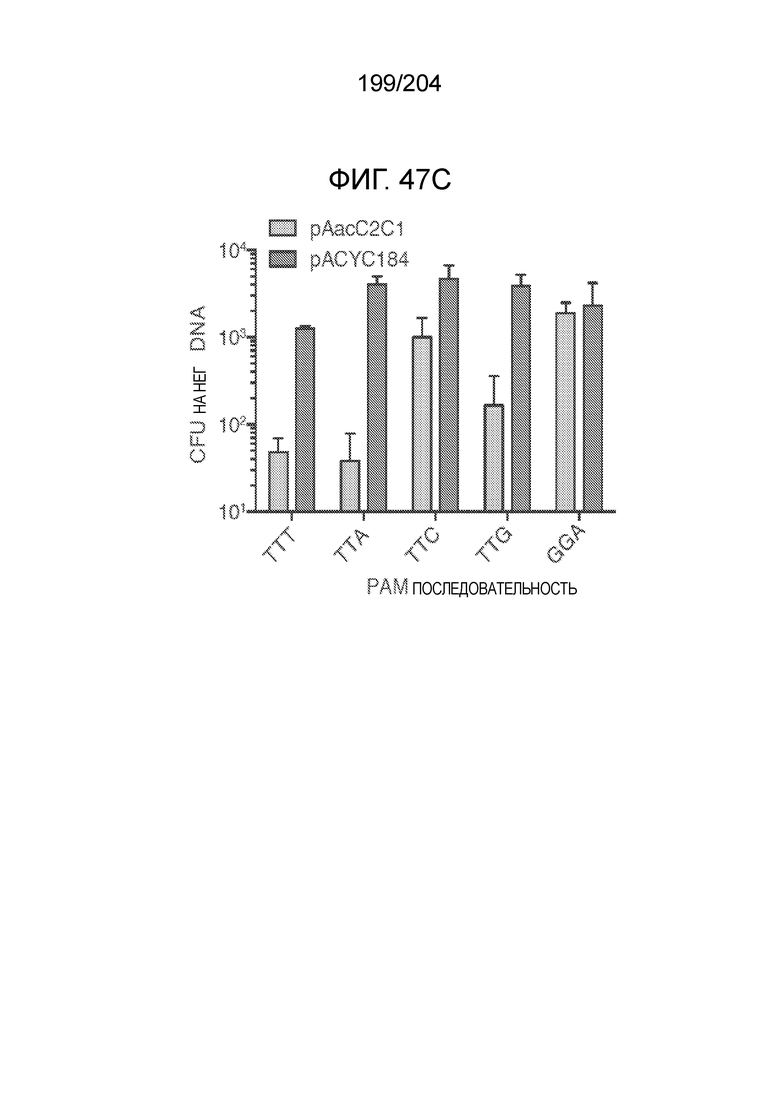
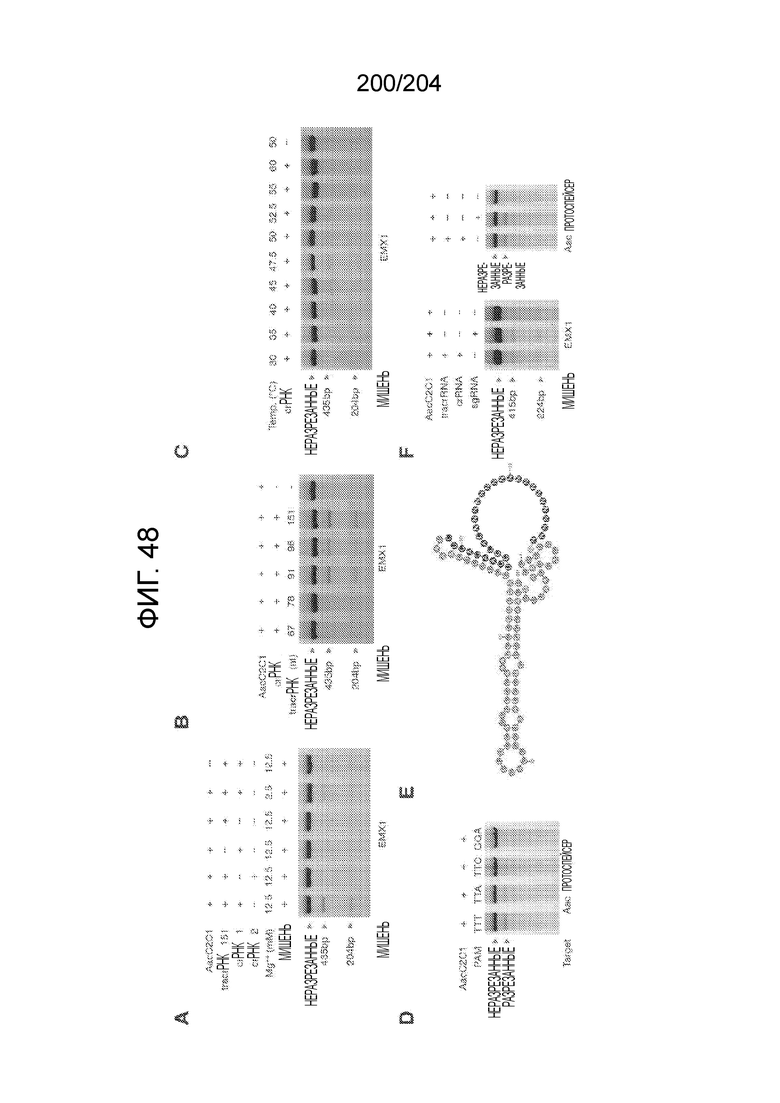
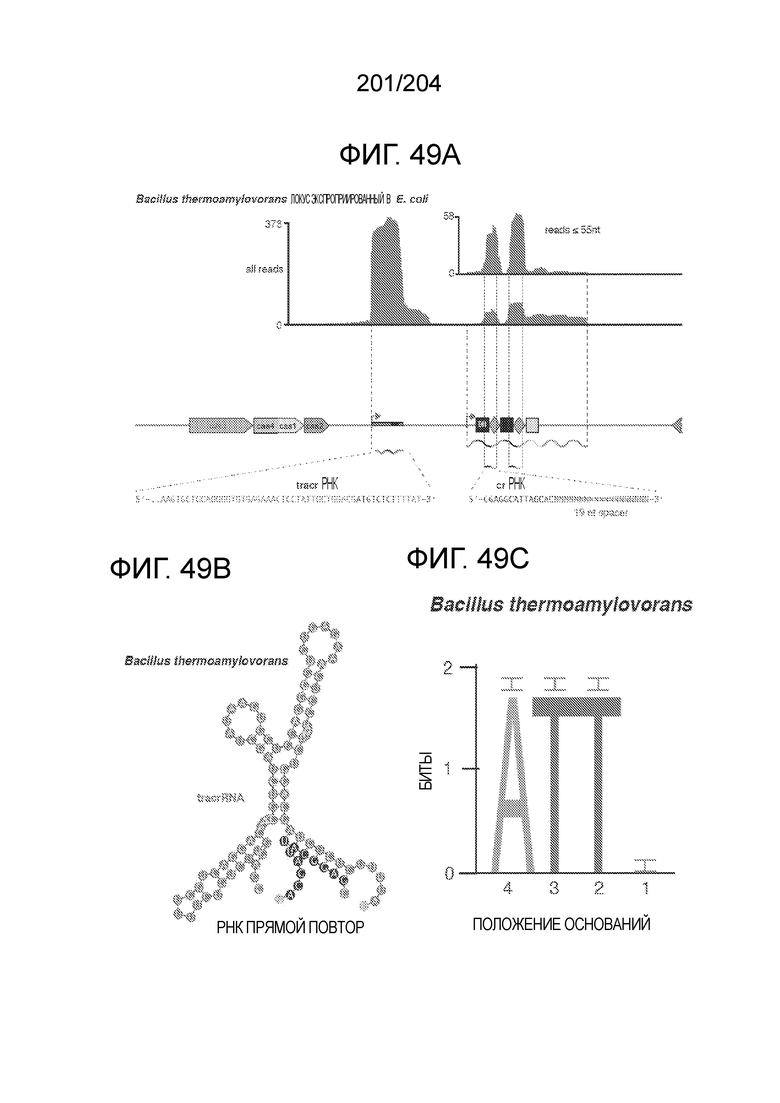
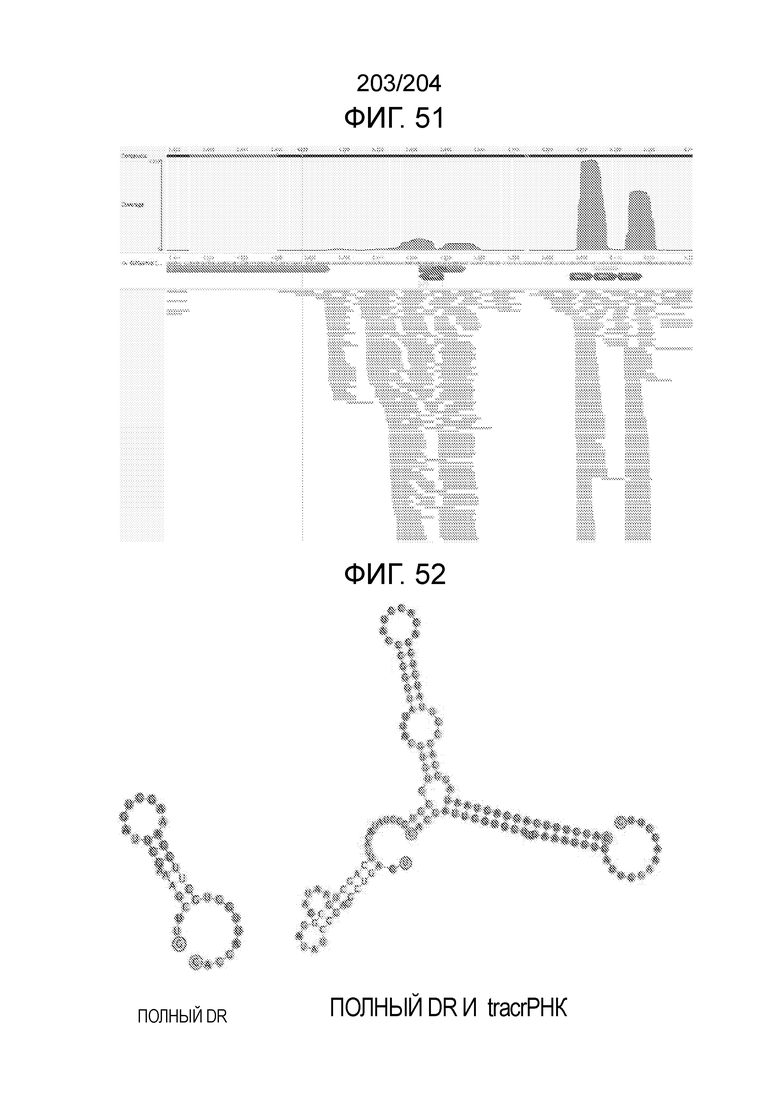
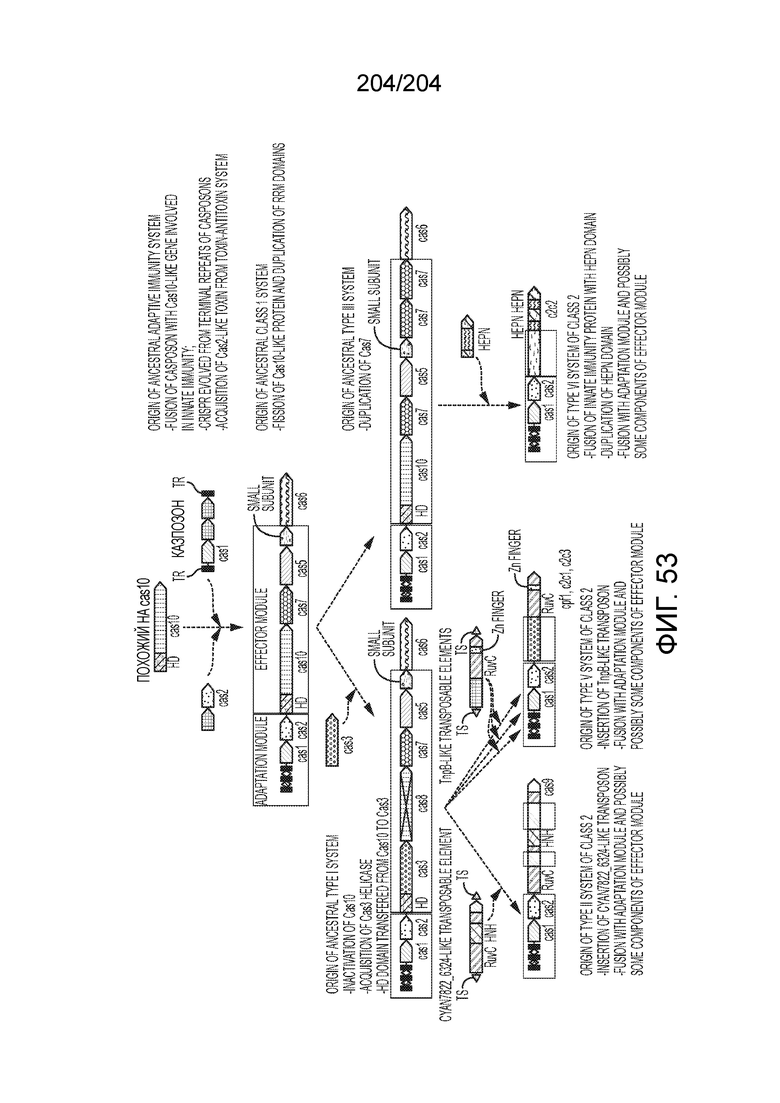
Изобретение относится к области биотехнологии, в частности к способу модификации представляющего интерес локуса-мишени, включающему доставку в указанный локус не встречающейся в природе или сконструированной композиции, включающей одномолекулярный эффекторный белок CRISPR-Cas, содержащий три каталитических домена нуклеазы RuvC, но не содержащий домен HNH, и один или более компонентов, являющихся нуклеиновыми кислотами, включающих гетерологичную направляющую последовательность и последовательность tracr-PHK. Также раскрыты не встречающаяся в природе или сконструированная композиция и система для модификации представляющей интерес последовательности-мишени, а также содержащая ее система доставки и векторная система. Изобретение также относится к частице, клетке, содержащей ее клеточной линии, а также к способу получения растения и способу ингибирования роста клеток. Изобретение эффективно для модификации представляющего интерес локуса-мишени. 12 н. и 49 з.п. ф-лы, 53 ил., 12 табл., 5 пр.
1. Способ модификации представляющего интерес локуса-мишени, включающий доставку в указанный локус не встречающейся в природе или сконструированной композиции, включающей одномолекулярный эффекторный белок CRISPR-Cas, содержащий три каталитических домена нуклеазы RuvC, но не содержащий домен HNH, и один или более компонентов, являющихся нуклеиновыми кислотами, включающих гетерологичную направляющую последовательность и последовательность tracr-PHK, где эффекторный белок образует комплекс с одним или более компонентами, являющимися нуклеиновыми кислотами, и при связывании указанного комплекса с представляющим интерес локусом-мишенью эффекторный белок индуцирует модификацию представляющего интерес локуса-мишени,
и где способ модификации не включает модификацию генетической целостности клетки зародышевой линии человека.
2. Способ по п. 1, где представляющий интерес локус-мишень представляет собой ДНК.
3. Способ по п. 1 или 2, где модификация представляющего интерес локуса-мишени представляет собой разрыв цепи.
4. Способ по любому из предшествующих пунктов, где представляющий интерес локус-мишень находится в молекуле ДНК in vitro.
5. Способ по любому из предшествующих пунктов, где представляющий интерес локус-мишень находится в ДНК в клетке.
6. Способ по п. 5, где клетка является прокариотической клеткой.
7. Способ по п. 5, где клетка является эукариотической клеткой.
8. Способ по любому из предшествующих пунктов, где представляющий интерес локус-мишень является представляющим интерес геномным локусом.
9. Способ по любому из предшествующих пунктов, где в комплексе с эффекторным белком компонент(ы), являющийся нуклеиновой кислотой, способен осуществлять или осуществляет специфическое для последовательности связывание комплекса с последовательностью представляющего интерес локуса-мишени.
10. Способ по любому из предшествующих пунктов, где компонент(ы), являющийся нуклеиновой кислотой, представляет собой последовательность РНК CRISPR (cr-РНК) и/или последовательность трансактивирующей РНК CRISPR (tracr-PHK).
11. Способ по любому из пп. 3-10, где разрыв цепи представляет собой одноцепочечный разрыв цепи.
12. Способ по любому из пп. 3-10, где разрыв цепи представляет собой двухцепочечный разрыв цепи.
13. Способ по любому из предшествующих пунктов, где эффекторный белок и компонент(ы), являющийся нуклеиновой кислотой, предоставляются посредством одной или более полинуклеотидных молекул, кодирующих полипептид и/или компонент(ы), являющийся нуклеиновой кислотой, и где одна или более полинуклеотидных молекул функционально организованы таким образом, чтобы происходила экспрессия полипептидов и/или компонента(ов), являющегося нуклеиновой кислотой.
14. Способ по п. 13, где одна или более полинуклеотидных молекул содержат один или более регуляторных элементов, функционально организованных таким образом, чтобы происходила экспрессия полипептидов и/или компонента(ов), являющегося нуклеиновой кислотой, в котором необязательно один или более регуляторных элементов включают индуцибельные промоторы.
15. Способ по п. 13 или 14, где одна или более полинуклеотидных молекул находятся в одном или более векторах.
16. Способ по любому из пп. 13-15, где одна или более полинуклеотидных молекул находятся в системе доставки.
17. Способ по п. 15, где один или более векторов находятся в системе доставки.
18. Способ по любому из предшествующих пунктов, где не встречающуюся в природе или сконструированную композицию доставляют посредством носителя для доставки, включающего липосому(ы), частицу(ы), экзосому(ы), микровезикулу(ы), генную пушку или один или более вирусных векторов.
19. Не встречающаяся в природе или сконструированная композиция для модификации представляющего интерес локуса-мишени, включающая одномолекулярный эффекторный белок CRISPR-Cas, содержащий три каталитических домена нуклеазы RuvC, но не содержащий домен HNH, и один или более компонентов, являющихся нуклеиновыми кислотами, включающих гетерологичную направляющую последовательность и последовательность tracr-PHK,
где эффекторный белок образует комплекс с одним или более компонентами, являющимися нуклеиновыми кислотами, и при связывании указанного комплекса с представляющим интерес локусом-мишенью эффекторный белок индуцирует модификацию представляющего интерес локуса-мишени,
и при этом модификация не включает модификацию генетической целостности клетки зародышевой линии человека.
20. Композиция по п. 19, где представляющий интерес локус-мишень представляет собой ДНК.
21. Композиция по п. 19 или 20, где модификация представляющего интерес локуса-мишени представляет собой разрыв цепи.
22. Композиция по любому из пп. 19-21, где представляющий интерес локус-мишень находится в молекуле ДНК in vitro.
23. Композиция по любому из пп. 19-21, где представляющий интерес локус-мишень находится в ДНК в клетке.
24. Композиция по п. 23, где клетка является прокариотической клеткой.
25. Композиция по п. 23, где клетка является эукариотической клеткой.
26. Композиция по любому из пп. 19-25, где представляющий интерес локус-мишень является представляющим интерес геномным локусом.
27. Композиция по любому из пп. 19-26, где в комплексе с эффекторным белком компонент(ы), являющийся нуклеиновой кислотой, способен осуществлять или осуществляет специфическое для последовательности связывание комплекса с последовательностью представляющего интерес локуса-мишени.
28. Композиция по любому из пп. 19-27, где компонент(ы), являющийся нуклеиновой кислотой, представляет собой последовательность РНК CRISPR (сг-РНК) и/или последовательность трансактивирующей РНК CRISPR (tracr-PHK).
29. Композиция по любому из пп. 21-28, где разрыв цепи представляет собой одноцепочечный разрыв цепи.
30. Композиция по любому из пп. 21-28, где разрыв цепи представляет собой двухцепочечный разрыв цепи.
31. Композиция по любому из пп. 19-30, где эффекторный белок и компонент(ы), являющийся нуклеиновой кислотой, предоставляются посредством одной или более полинуклеотидных молекул, кодирующих полипептид и/или компонент(ы), являющийся нуклеиновой кислотой, и где одна или более полинуклеотидных молекул функционально организованы таким образом, чтобы происходила экспрессия полипептидов и/или компонента(ов), являющегося нуклеиновой кислотой.
32. Композиция по п. 31, где одна или более полинуклеотидных молекул содержат один или более регуляторных элементов, функционально организованных таким образом, чтобы происходила экспрессия полипептидов и/или компонента(ов), являющегося нуклеиновой кислотой, в котором необязательно один или более регуляторных элементов включают индуцибельные промоторы.
33. Композиция по п. 31 или 32, где одна или более полинуклеотидных молекул находятся в одном или более векторах.
34. Композиция по любому из пп. 31, 32, где одна или более полинуклеотидных молекул находятся в системе доставки.
35. Композиция по п. 33, где один или более векторов находятся в системе доставки.
36. Композиция по любому из пп. 19-35, где не встречающуюся в природе или сконструированную композицию доставляют посредством носителя для доставки, включающего липосому(ы), частицу(ы), экзосому(ы), микровезикулу(ы), генную пушку или один или более вирусных векторов.
37. Векторная система для модификации представляющего интерес локуса-мишени, включающая один или более векторов, причем один или более векторов включают одну или более полинуклеотидных молекул, кодирующих компоненты не встречающейся в природе или сконструированной композиции по любому из пп. 19-36, где один или более векторов содержат: а) первый регуляторный элемент, функционально связанный с одной или более нуклеотидными последовательностями, кодирующими одну или более полинуклеотидных последовательностей CRISPR-Cas, включающих направляющую РНК, которая содержит направляющую последовательность, соединенную с последовательностью прямого повтора, где направляющая последовательность способна гибридизоваться с локусом-мишенью; б) второй регуляторный элемент, функционально связанный с нуклеотидной последовательностью, кодирующей одномолекулярный эффекторный белок CRISPR-Cas, содержащий три каталитических домена нуклеазы RuvC, но не содержащий домен HNH; где компоненты (а) и (б) находятся на одном и том же или различных векторах системы, где при транскрипции одна или более направляющих последовательностей гибридизуются с указанным локусом-мишенью, локус-мишень находится с 3'-стороны от прилегающего к протоспэйсеру мотива (РАМ) и указанная направляющая РНК образует комплекс с указанным эффекторным белком.
38. Система доставки для модификации представляющей интерес последовательности-мишени, включающая один или более векторов или одну или более полинуклеотидных молекул, причем один или более векторов или полинуклеотидных молекул включают одну или более полинуклеотидных молекул, кодирующих компоненты не встречающейся в природе или сконструированной композиции, которая представляет собой композицию по любому из пп. 19-36, где один или более векторов содержат: (а) первый регуляторный элемент, функционально связанный с одной или более нуклеотидными последовательностями, кодирующими одну или более полинуклеотидных последовательностей CRISPR-Cas, включающих направляющую РНК, которая содержит направляющую последовательность, соединенную с последовательностью прямого повтора, где направляющая последовательность способна гибридизоваться с локусом-мишенью; (б) второй регуляторный элемент, функционально связанный с нуклеотидной последовательностью, кодирующей одномолекулярный эффекторный белок CRISPR-Cas, содержащий три каталитических домена нуклеазы RuvC, но не содержащий домен HNH; где компоненты (а) и (б) находятся на одном и том же или различных векторах системы, где при транскрипции одна или более направляющих последовательностей гибридизуются с указанным локусом-мишенью, локус-мишень находится с 3'-стороны от прилегающего к протоспэйсеру мотива (РАМ) и указанная направляющая РНК образует комплекс с указанным эффекторным белком.
39. Не встречающаяся в природе или сконструированная композиция по любому из пп. 19-36, векторная система по п. 37 или система доставки по п. 38 для применения в терапевтическом способе лечения.
40. Не встречающаяся в природе или сконструированная композиция по любому из пп. 19-36 для применения в терапевтическом способе лечения, где указанный терапевтический способ лечения включает редактирование гена или генома или генную терапию.
41. Не встречающаяся в природе или сконструированная композиция по любому из пп. 19-36, векторная система по п. 37 или система доставки по п. 38 для применения в:
- сайт-специфическом нокауте гена;
- сайт-специфическом редактировании генома;
- специфической для последовательности ДНК-интерференции; или
- мультиплексной модификации генома способами инженерии.
42. Клетка, не являющаяся эмбриональной клеткой человека, модифицированная способами инженерии таким образом, чтобы она содержала или экспрессировала не встречающуюся в природе или сконструированную композицию по любому из пп. 19-36 для модификации представляющего интерес локуса-мишени, включающую одномолекулярный эффекторный белок CRISPR-Cas, содержащий три каталитических домена нуклеазы RuvC, но не содержащий домен HNH, и один или более компонентов, являющихся нуклеиновыми кислотами, включающих гетерологичную направляющую последовательность и последовательность tracr-PHK, где эффекторный белок образует комплекс с одним или более компонентами, являющимися нуклеиновыми кислотами, и при связывании указанного комплекса с представляющим интерес локусом-мишенью эффекторный белок индуцирует модификацию представляющего интерес локуса-мишени.
43. Клеточная линия, состоящая из клеток по п. 42, модифицированных способами инженерии таким образом, чтобы содержать или экспрессировать не встречающуюся в природе или сконструированную композицию по любому из пп. 19-36, для модификации представляющего интерес локуса-мишени.
44. Сконструированная, не встречающаяся в природе система CRISPR-Cas для модификации представляющего интерес локуса-мишени, включающая:
а. одну или более полинуклеотидных последовательностей CRISPR-Cas, включающих направляющую РНК (гРНК), которая содержит направляющую последовательность, соединенную с последовательностью прямого повтора, где направляющая последовательность способна гибридизоваться с последовательностью-мишенью, или одну или более нуклеотидных последовательностей, кодирующих одну или более полинуклеотидных последовательностей CRISPR-Cas; и
b. одномолекулярный эффекторный белок CRISPR-Cas, содержащий три каталитических домена нуклеазы RuvC, но не содержащий домен HNH, или одну или более нуклеотидных последовательностей, кодирующих указанный одномолекулярный эффекторный белок; где одна или более направляющих последовательностей гибридизуются с указанным локусом-мишенью, указанный локус-мишень находится с 3'-стороны от прилегающего к протоспэйсеру мотива (РАМ) и указанная направляющая РНК образует комплекс с указанным эффекторным белком.
45. Сконструированная, не встречающаяся в природе векторная система CRISPR-Cas для модификации представляющего интерес локуса-мишени, включающая один или более векторов, содержащих:
a. первый регуляторный элемент, функционально связанный с одной или более нуклеотидными последовательностями, кодирующими одну или более полинуклеотидных последовательностей CRISPR-Cas, включающих направляющую РНК, которая содержит направляющую последовательность, соединенную с последовательностью прямого повтора, где направляющая последовательность способна гибридизоваться с локусом-мишенью;
b. второй регуляторный элемент, функционально связанный с нуклеотидной последовательностью, кодирующей одномолекулярный эффекторный белок CRISPR-Cas, содержащий три каталитических домена нуклеазы RuvC, но не содержащий домен HNH; где компоненты (а) и (b) находятся на одном и том же или различных векторах системы, где при транскрипции одна или более направляющих последовательностей гибридизуются с указанным локусом-мишенью, локус-мишень находится с 3'-стороны от прилегающего к протоспэйсеру мотива (РАМ) и указанная направляющая РНК образует комплекс с указанным эффекторным белком.
46. Система по п. 44 или 45, где локус-мишень находится в клетке.
47. Система по п. 45, где клетка представляет собой эукариотическую клетку.
48. Система по п. 44 или 45, где при транскрипции одна или более направляющих последовательностей гибридизуются с локусом-мишенью и направляющая РНК образует комплекс с одномолекулярный эффекторным белком CRISPR-Cas, содержащим три каталитических домена нуклеазы RuvC, но не содержащим домен HNH, что вызывает расщепление дистальнее локуса-мишени.
49. Система по п. 48, где указанное расщепление образует ступенчатый двухцепочечный разрыв с 5'-выступающим концом из 4 или 5 нуклеотидов.
50. Система по п. 44 или 45, где РАМ содержит 5'-тимин-богатый мотив, выбранный из TTN, TTTV или ATTN, где N представляет собой А, С, G или Т, а V представляет собой А, С или G.
51. Система по п. 44 или 45, где эффекторный белок представляет собой эффекторный белок, полученный из видов бактерий, выбранных из группы, состоящей из Alicyclobacillus acidoterrestris, Alicyclobacillus contaminans, Desulfovibrio inopinaius, Desulfonatronum ihiodismutans, бактерий Opitutaceae TAV5, Tuberibacillus calidus, Bacillus thermoamyiovorans, Brevibacillus sp.CF112, Bacillus sp.NSP2.1, Desulfatirhabdium butyrativorans, Alicyclobacillus herbarius, Citrobacier freundii, Brevibacillus agri, Methylobacterium nodularis.
52. Система по п. 51, в которой РАМ-последовательность представляет собой TTN, где N представляет собой А, С, G или Т или где РАМ-последовательность представляет собой TTTV, где V представляет собой А, С или G, а эффекторный белок получен из бактерий Alicyclobacillus acidoterrestris.
53. Система по п. 44 или 45, где эффекторный белок содержит один или более сигналов ядерной локализации.
54. Система по п. 44 или 45, где последовательности нуклеиновых кислот, кодирующие эффекторный белок, являются кодон-оптимизированными для экспрессии в эукариотической клетке.
55. Система по п. 44 или 45, где компоненты (а) и (б) или нуклеотидные последовательности расположены на одном векторе.
56. Способ получения растения, имеющего модифицированный, представляющий интерес признак, кодируемый представляющим интерес геном, причем указанный способ включает приведение клетки растения в контакт с системой по п. 44 или 45 или воздействие на клетку растения способом по п. 1, тем самым либо модифицируя, либо внося указанный, представляющий интерес ген, и регенерацию растения из указанной клетки растения.
57. Частица для доставки системы в клетку, содержащая систему по п. 44 или 45.
58. Частица по п. 57, где частица содержит эффекторный белок в комплексе с направляющей РНК.
59. Система по п. 44 или 45 или способ по п. 1, где комплекс, направляющая РНК или белок конъюгированы по меньшей мере с одним сахарным фрагментом, необязательно с N-ацетилгалактозамином (GalNAc), в частности с трехантенным GalNAc.
60. Сконструированная, не встречающаяся в природе композиция для модификации представляющего интерес локуса-мишени, содержащая систему CRISPR-Cas, причем указанная система содержит функциональный одномолекулярный эффекторный белок системы CRISPR-Cas и направляющую РНК (гРНК),
где гРНК содержит нефункциональную направляющую последовательность; где гРНК способна гибридизоваться с локусом-мишенью;
где система CRISPR-Cas направлена с уменьшенной инсерционно-делеционной активностью на локус-мишень, и
где функциональный одномолекулярный эффекторный белок системы CRISPR-Cas представляет собой белок, содержащий три каталитических домена нуклеазы RuvC, но не содержащий домен HNH.
61. Способ ингибирования роста клеток, причем способ включает доставку в клетку не встречающейся в природе или сконструированной композиции, содержащей функциональный одномолекулярный эффекторный белок системы CRISPR-Cas и направляющую РНК (гРНК),
где гРНК способна гибридизоваться с последовательностью ДНК-мишени клетки;
где система CRISPR-Cas направлена с уменьшенной инсерционно-делеционной активностью на последовательность ДНК-мишени; и
где фунциональный одномолекулярный эффекторный белок системы CRISPR-Cas представляет собой белок, содержащий три каталитических домена нуклеазы RuvC, но не содержащий домен HNH.
| WO2015071474 A2, 21.05.2015 | |||
| US2015079680 A1, 19.03.2015 | |||
| MAKAROVA KIRA S | |||
| et al | |||
| Evolution and classification of the CRISPR-Cas systems, Nature Reviews Microbiology, 2011, 9(6), pp | |||
| Детекторный радиоприемник гетеродин | 1923 |
|
SU467A1 |
| WO2014204724 A1, 24.12.2014 | |||
| US2010076057 A1, 25.03.2010 | |||
| ПУГАЧ К.С | |||
| и др | |||
| CRISPR-системы адаптивного иммунитета прокариот, Молекулярная биология, 2012, | |||

