ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к антителам, обладающим специфическими последовательностями, которые связываются с лигандом запрограммированной клеточной смерти 1 (PD-L1) человека и могут использоваться для обнаружения экспрессии PD-L1 в образцах ткани человека с помощью иммуногистохимического анализа (IHC). Изобретение также относится к специфическим анализам IHC, в которых используются эти антитела против PD-L1 человека.
УРОВЕНЬ ТЕХНИКИ, К КОТОРОМУ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
PD-L1 представляет собой гликопротеин клеточной поверхности, который является одним из двух известных лигандов для белка запрограммированной гибели 1 (PD-1), который распознается в качестве важного участника иммунной регуляции и поддержания периферической толерантности. Экспрессию PD-L1 наблюдали на поверхности различных иммунных клеток, включая наивные лимфоциты и активированные B- и T-клетки, моноциты и дендритные клетки (Там же). Более того, мРНК PD-L1 экспрессируется нелимфоидными тканями, включающими эндотелиальные клетки сосудов, эпителиальные клетки, мышечные клетки, и тканями миндалевидной железы и плаценты. См., например, Keir, M.E. et al, Annu Rev Immunol. 26:677-704 (2008); Sharp A.H. et al., Nature Immunol. 8:239-245 (2007); Okazaki T and Honjo T, Internat. immunol. 19:813-824 (2007).
Экспрессию PD-L1 также наблюдают в различных злокачественных опухолях человека, и взаимодействие экспрессируемого опухолевыми клетками PD-L1 с PD-1 может индуцировать ингибирование или апоптоз опухолеспецифических T-клеток. Было показано, что в больших выборках образцов, например, рака яичника, рака почки, рака толстой и прямой кишки, рака поджелудочной железы, рака печени и меланомы, экспрессия PD-L1 коррелирует с плохим прогнозом и со снижением общей выживаемости, безотносительно последующего лечения. Было показано, что моноклональные антитела против PD-1, которые блокируют связывание PD-L1 с PD-1, обладают противоопухолевой активностью против различных типов опухолей, причем ранние клинические данные у человека указывают на то, что пациенты с опухолями, которые экспрессируют PD-L1, более вероятно будут отвечать на терапию против PD-1. См., например, Iwai et al., PNAS 99: 12293-12297 (2002); Ohigashi et al, Clin Cancer Res 11: 2947-2953 (2005); Ghebeh et al, Neoplasia 8:190-198 (2006); Hamanishi, J et al, PNAS 104:3360-3365 (2007); Yang et al, Invest Ophthalmol Vis Sci. 49(6):2518-2525 (2008); Gao et al, Clin Cancer Res 15:971-979 (2009); Brahmer J.R. et al, J Clin Oncol. 28:3167-3175 (2010).
В недавнем сообщении описано сравнение 15 антител против PD-L1 человека в отношении их применимости для обнаружения экспрессии hPD-L1 в фиксированных в формалине залитых парафином (FFPE) образцах меланомы человека (Gadiot, J., et al., Cancer 117(10):2192-2201 (2011)). Критериями применимости, оцениваемыми при этом сравнении, были: (1) способность окрашивать залитые парафином ткани, (2) обеспечение низкого фонового окрашивания и (3) блокирование связывания с PD-L1 путем предварительной инкубации со слитым белком PD-L1. Авторы сделали заключение, что Ab #4059, поликлональное антитело кролика против антитела человека (полученное от ProSci, Poway, CA США), представляло собой единственное антитело против PD-L1 человека из 15 протестированных, которое удовлетворяет всем этим критериям на приемлемом уровне (Там же, на 2195, 2-ая колонка).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к моноклональным антителам против PD-L1 человека, которые обеспечивают набор характеристик окрашивания IHC в ткани миндалевидной железы, обработанной FFPE, который, как полагают авторы настоящего изобретения, является более иммунологически значимым, чем набор характеристик, обеспечиваемый ProSci Ab #4059. Как описано в примерах ниже, авторы изобретения обнаружили, что это антитело ProSci (PRS4059, Sigma-Aldrich, партия 40590604) окрашивало все гемопоэтические линии диференцировки в миндалевидной железе с равной интенсивностью, в то время как два антитела по настоящему изобретению: 22C3 и 20C3 селективно окрашивали эпителий крипт миндалевидной железы и фолликулярные CD68+ миелоидные клетки, которые морфологически соответствуют макрофагам. Более того, 22C3 и 20C3 показали закономерное различие интенсивностей между этими двумя отдельными клеточными популяциями, согласно которому интенсивность окрашивания в эпителии крипт значительно превышает интенсивность окрашивания в фолликулярных макрофагах. Все три антитела (PRS4059, 22C3 и 20C3) нейтрализуются посредством предварительной инкубации с антигеном PD-L1, что указывает на то, что их реактивность опосредуется антигенсвязывающим доменом (CDR). Таким образом, изобретение также относится к применению антител по настоящему изобретению для обнаружения экспрессии PD-L1 на поверхности клеток человека, в том числе в анализах IHC для обнаружения PD-L1 в срезах тканей, обработанных FFPE.
В одном из аспектов изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, которые специфически связываются с PD-L1 человека. Выделенное моноклональное антитело или его антигенсвязывающий фрагмент содержат три CDR легкой цепи: CDRL1, CDRL2 и CDRL3, и три CDR тяжелой цепи: CDRH1, CDRH2 и CDRH3.
CDRL1 выбрана из группы, состоящей из SEQ ID NO:1, SEQ ID NO:9, SEQ ID NO:21, варианта SEQ ID NO:9 и варианта SEQ ID NO:21. CDRL2 выбрана из группы, состоящей из SEQ ID NO:2 и варианта SEQ ID NO:2. CDRL3 выбрана из группы, состоящей из SEQ ID NO:3, SEQ ID NO:10, SEQ ID NO:22, варианта SEQ ID NO:10 и варианта SEQ ID NO:22.
CDRH1 выбрана из группы, состоящей из SEQ ID NO:5, SEQ ID NO:14, SEQ ID NO:15, SEQ ID NO:26, SEQ ID NO:27, варианта SEQ ID NO:14, варианта SEQ ID NO:15, варианта SEQ ID NO:26 и варианта SEQ ID NO:27. CDRH2 выбрана из группы, состоящей из SEQ ID NO:6, SEQ ID NO:16, SEQ ID NO:28, варианта SEQ ID NO:16 и варианта SEQ ID NO:28. CDRH3 выбрана из группы, состоящей из SEQ ID NO:7, SEQ ID NO:17, SEQ ID NO:29, варианта SEQ ID NO:17 и варианта SEQ ID NO:29.
В антителах и антигенсвязывающих фрагментах по изобретению вариант последовательности CDR (легкой цепи или тяжелой цепи) идентичен эталонной последовательности, за исключением наличия одной двух консервативных аминокислотных замен в эталонной последовательности, и предпочтительно имеет только одну консервативную аминокислотную замену в эталонной последовательности. В предпочтительных вариантах осуществления не более чем две из трех CDR легкой цепи имеют последовательность варианта, и не более чем две из трех CDR тяжелой цепи имеют последовательность варианта. В более предпочтительных вариантах осуществления только три, две или одна из шести CDR представляют собой последовательности варианта.
В одном из предпочтительных вариантов осуществления три CDR легкой цепи представляют собой SEQ ID NO:1, SEQ ID NO:2 и SEQ ID NO:3, и три CDR тяжелой цепи представляют собой SEQ ID NO:5, SEQ ID NO:6 и SEQ ID NO:7.
В другом из предпочтительных вариантов осуществления три CDR легкой цепи представляют собой SEQ ID NO:9, SEQ ID NO:2 и SEQ ID NO:10, и три CDR тяжелой цепи представляют собой SEQ ID NO:14, SEQ ID NO:16 и SEQ ID NO:17.
В другом из предпочтительных вариантов осуществления три CDR легкой цепи представляют собой SEQ ID NO:21, SEQ ID NO:2 и SEQ ID NO:22, и три CDR тяжелой цепи представляют собой SEQ ID NO:26, SEQ ID NO:28 и SEQ ID NO:29.
Некоторые антитела и антигенсвязывающие фрагменты по изобретению содержат вариабельную область легкой цепи и вариабельную область тяжелой цепи. Вариабельная область легкой цепи выбрана из группы, состоящей из SEQ ID NO:4, SEQ ID NO:13, варианта SEQ ID NO:13, SEQ ID NO:25 и варианта SEQ ID NO:25, и вариабельная область тяжелой цепи выбрана из группы, состоящей из SEQ ID NO:8, SEQ ID NO:20, варианта SEQ ID NO:20, SEQ ID NO:32 и варианта SEQ ID NO:32. В таких вариантах осуществления вариант последовательности вариабельной области легкой цепи идентичен эталонной последовательности, за исключением наличия пяти консервативных аминокислотных замен в каркасной области (т.е. вне CDR), и предпочтительно имеет менее четырех, трех или двух консервативных аминокислотных замен в каркасной области. Аналогично, вариант последовательности вариабельной области тяжелой цепи идентичен эталонной последовательности, за исключением наличия вплоть до 17 консервативных аминокислотных замен в каркасной области (т.е. вне CDR), и предпочтительно имеет менее десяти, девяти, восьми, семи, шести или пяти консервативных аминокислотных замен в каркасной области.
В одном из предпочтительных антител или антигенсвязывающих фрагментах по изобретению вариабельная область легкой цепи представляет собой SEQ ID NO:13 и вариабельная область тяжелой цепи представляет собой SEQ ID NO:20.
Другое предпочтительное антитело или антигенсвязывающий фрагмент по изобретению содержат вариабельную область легкой цепи SEQ ID NO:25 и вариабельную область тяжелой цепи SEQ ID NO:32.
В другом из варианте осуществления антитело или связывающий фрагмент по изобретению содержат вариабельную область легкой цепи SEQ ID NO:25 и вариабельную область тяжелой цепи SEQ ID NO:32, где X в SEQ ID NO:32 представляет собой pE.
В другом варианте осуществления антитело или связывающий фрагмент по изобретению содержат вариабельную область легкой цепи SEQ ID NO:25 и вариабельную область тяжелой цепи SEQ ID NO:32, где X в SEQ ID NO:32 представляет собой Q.
Во всех из описанных выше вариантов осуществления антител выделенное антитело может представлять собой полноразмерное антитело любого класса иммуноглобулинов, включая IgM, IgG, IgD, IgA и IgE. Предпочтительно, антитело представляет собой IgG-антитело, такое как IgG1, IgG2, IgG3 или IgG4. В одном варианте осуществления антитело содержит константную область IgG1 мыши.
Особенно предпочтительные антитела представляют собой моноклональные антитела 20C3 и 22C3, которые представляют собой IgG1-антитела, экспрессируемые гибридомами MEB037.20C3 и MEB037.22C3, соответственно.
Изобретение также относится к выделенному моноклональному антителу или его антигенсвязывающему фрагменту, которые специфически связываются с PD-L1 человека и блокируют связывание с PD-L1 человека антитела 20C3 или 22C3 или эталонного антитела, которое содержит SEQ ID NO:25 и SEQ ID NO:32. В одном предпочтительном варианте осуществления антитело или антигенсвязывающий фрагмент по изобретению блокирует связывание с PD-L1 человека каждого из 20C3 и 22C3, или каждого из (a) эталонного антитела, которое содержит SEQ ID NO:13 и SEQ ID NO:20, и (b) эталонного антитела, которое содержит SEQ ID NO:25 и SEQ ID NO:32.
Изобретение также относится к композиции антитела, которая содержит любое из описанных выше антител или фрагментов антител в составе. Один подходящий состав содержит 20 мМ ацетат натрия и 9% сахарозу при pH 5,0. В предпочтительном варианте осуществления композиция содержит смесь молекул антител, в которой большинство (т.е. превышающее любое из 60%, 65%, 70%, 80%, 85%, 90% или 95%) молекул антител содержит SEQ ID NO:25 и SEQ ID NO:32, где X в SEQ ID NO:32 представляет собой pE, а остальная часть молекул антител содержит SEQ ID NO:25 и SEQ ID NO:32, где X в SEQ ID NO:32 представляет собой Q.
В любом из описанных выше вариантов осуществления антигенсвязывающий фрагмент представляет собой Fab-фрагмент, Fab'-фрагмент, (Fab')2-фрагмент.
В любом из описанных выше вариантов осуществления антитело или антигенсвязывающий фрагмент могут дополнительно содержать поддающуюся обнаружению метку.
Изобретение также относится к выделенной нуклеиновой кислоте, кодирующей любую из вариабельных областей антител, описанных выше. В одном из предпочтительных вариантов осуществления нуклеиновая кислота содержит одну или обе из SEQ ID NO:33 и SEQ ID NO:34. В другом из предпочтительных вариантов осуществления нуклеиновая кислота содержит одну или обе из SEQ ID NO:35 и SEQ ID NO:36. В любом из этих вариантов осуществления выделенная нуклеиновая кислота предпочтительно представляет собой вектор экспрессии.
Изобретение также относится к клетке-хозяину, содержащей вектор экспрессии, который кодирует любую из вариабельных областей антител, описанных выше. Предпочтительно, вектор экспрессии содержит SEQ ID NO:33 и SEQ ID NO:34 или содержит SEQ ID NO:35 и SEQ ID NO:36.
Изобретение также относится к способу анализа образца ткани человека, который извлечен из человека, в отношении экспрессии PD-L1. Способ анализа включает контактирование образца ткани со связывающим PD-L1 реагентом в условиях, которые обеспечивают специфическое связывание связывающего PD-L1 реагента с PD-L1 человека, удаление не связавшегося связывающего PD-L1 реагента, и обнаружение присутствия или отсутствия связанного связывающего PD-L1 реагента. В одном из предпочтительных вариантов осуществления способ дополнительно включает количественное определение связанного связывающего реагента. Связывающий реагент представляет собой любое из моноклонального антитела или антигенсвязывающих фрагментов, описанных выше. Предпочтительно, связывающий реагент представляет собой антитело, которое содержит SEQ ID NO:13 и SEQ ID NO:20 или содержит SEQ ID NO:25 и SEQ ID NO:32. В одном из предпочтительных вариантов осуществления связывающий реагент представляет собой композицию антитела, которая содержит смесь молекул антител, содержащую SEQ ID NO:25 и SEQ ID NO:32, где большинство молекул (т.е. более чем любое из 60%, 65%, 70%, 80%, 85%, 90% или 95%) имеют pE в положении X в SEQ ID NO:32 и остальные молекулы имеют Q в положении X в SEQ ID NO:32.
В другом аспекте изобретение относится к набору для анализа образца ткани человека в отношении экспрессии PD-L1. Набор содержит связывающий PD-L1 реагент и набор реагентов для обнаружения комплекса, содержащего связующий реагент, связанный с PD-L1 человека. Связывающий PD-L1 реагент представляет собой любое моноклональные антитело или антигенсвязывающий фрагмент, описанные выше, которые специфически связываются с PD-L1 человека. Предпочтительно, антитело или связывающий фрагмент содержат SEQ ID NO:13 и SEQ ID NO:20, или содержат SEQ ID NO:25 и SEQ ID NO:32. В одном из предпочтительныхм вариантов осуществления связывающий реагент представляет собой композицию антитела, которая содержит смесь молекул антител, содержащую SEQ ID NO:25 и SEQ ID NO:32, где большинство молекул (т.е. более чем любое из 60%, 65%, 70%, 80%, 85%, 90% или 95%) имеют pE в положении X в SEQ ID NO:32, и остальные молекулы имеют Q в положении X в SEQ ID NO:32.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1 показаны нуклеотидные последовательности кДНК вариабельной области легкой и тяжелой цепей, полученной из тотальной РНК, выделенной из гибридомы MEB037.20C3 и спрогнозированные аминокислотные последовательности, кодируемые ими (полужирный шрифт), причем в скобках указаны нуклеотидные и аминокислотные последовательности для лидерного пептида и подчеркиванием указаны последовательности CDR.
На фиг. 2 представлены нуклеотидные последовательности кДНК вариабельной области легкой и тяжелой цепи, полученные из тотальной РНК, выделенной из гибридомы MEB037.22C3, и спрогнозированные аминокислотные последовательности, кодируемые ими (полужирный шрифт), причем в скобках указаны нуклеотидные и аминокислотные последовательности для лидерного пептида и подчеркиванием указаны последовательности CDR.
На фиг. 3 представлены выровненные аминокислотные последовательности для зрелых вариабельных областей легкой и тяжелой цепей антител 20C3 и 22C3, при этом полужирный шрифт указывает на положения, где последовательности отличаются, подчеркиванием указаны последовательности CDR при определении с помощью системы нумерации Kabat, и в скобках указана CDR1 тяжелой цепи при определении с помощью системы нумерации Chothia.
На фиг. 4 представлено окрашивание срезов миндалевидной железы, проведенное с помощью иммуногистохимического анализа с использованием коммерчески доступного антитела PRS4059 (фиг. 4A) или антитела 22C3 по изобретению (фиг. 4B), причем на срезах справа на фиг. 4A и 4B показаны результаты после предварительной инкубации со слитым белком PD-L1-IgG1 (R&D Systems), который конкурирует с антителами против PD-L1 человека за связывание с PD-L1 человека.
На фиг. 5 представлены фотографии соседних срезов нормальной ткани миндалевидной железы, обработанных FFPE, в которых экспрессию белка и мРНК PD-L1 человека анализировали с помощью анализа IHC с использованием антитела 22C33 (фиг. 5A) и гибридизации in-situ (ISH) (фиг. 5B), соответственно, и которые показали дифференциальное окрашивание между двумя уникальными популяциями клеток: эпителий крипт (фиг. 5A, левое увеличенное изображение, и фиг. 5B, верхнее изображение) и фолликулярные макрофаги (фиг. 5A, правое увеличенное изображение, и фиг. 5B, нижнее изображение).
На фиг. 6 проиллюстрированы результаты проточно-цитометрической оценки связывания различных антител против PD-L1 человека и изотипического контрольного антитела с клетками HT144, о которых известно, что они являются отрицательными в отношении экспрессии hPD-L1 в анализе мРНК (кПЦР) (фиг. 6A), и клетками меланомы LOX, о которых известно, что они экспрессируют высокие уровни мРНК hPD-L1 (кПЦР) (фиг. 6B).
На фиг. 7 представлено окрашивание IHC антителом 22C3 на осадках клеток, обработанных FFPE, модифицированной способами инженерии клеточной линии CHO (фиг. 7A) и клеточных линий человека (фиг. 7B, верхняя панель), и показано, что интенсивность окрашивания высоко коррелирует с уровнями экспрессии мРНК hPD-L1, измеренными в тех же линиях клеток (фиг. 7B, нижняя панель).
На фиг. 8 проиллюстрировано селективное связывание и относительная аффинность антител 22C3 и 20C3 в отношении hPD-L1, причем на графиках показаны результаты клеточного эксперимента ELISA, в котором клетки, которые не экспрессируют hPD-L1 (фиг. 8A), экспрессируют hPDL-1 (фиг. 8B и 8C), или экспрессируют PD-L2 (фиг. 8D), инкубировали с идентичным первичным антителом в указанных концентрациях, а затем определяли связывание первичного антитела со вторичным антителом козы против IgG человека, как описано в примерах.
На фиг. 9 представлены результаты конкурентных анализов связывания антител, которые показали, что антитела 22C3 и 20C3 связывают неидентичные, но перекрывающиеся эпитопы.
На фиг. 10 проиллюстрированы результаты полуколичественной гештальт-оценки интенсивности окрашивания в ICH посредством 22C3 образцов, обработанных FFPE, из указанных типов опухолей, причем степень окрашивания возрастает с увеличением числовых показателей оценки.
На фиг. 11 иллюстрируется, что экспрессия PD-L1 человека, выявленная с помощью антитела 22C3 в анализе IHC, коррелирует с ответом пациентов с меланомой на терапию антителом против PD-1 человека (MK-3475), при этом на фиг. 11A представлены репрезентативные изображения окрашивания 22C3-IHC, интерпретируемые как положительные, отрицательные или неоднозначные для экспрессии hPD-L1, а на фиг. 11B представлено количество пациентов, которые имели положительный или отрицательный ответ, которые были оценены как положительные или отрицательные (включая пациентов, оцененных как неоднозначные) в отношении экспрессии hPD-L1 в анализе IHC.
ПОДРОБНОЕ ОПИСАНИЕ
Сокращения. В подробном описании и примерах по изобретению используются следующие сокращения:
ОПРЕДЕЛЕНИЯ
Чтобы изобретение было более понятным определенные технические и научные термины конкретно определены ниже. Если нет иных конкретных указаний в настоящем документе, все другие технические и научные термины, используемые в настоящем описании, имеют значение, обычно подразумеваемое специалистом в области, к которой настоящее изобретение относится.
Как используют в рамках изобретения, включая прилагаемую формулу изобретения, форма единственного числа включает соответствующее множественное число, если контекст явно не указывает на иное.
"Активация", как используют в отношении клеток или рецепторов, относится к активации или обработке клетки или рецептора лигандом, если нет иных указаний, в зависимости от контекста или недвусмысленно. "Лиганд" охватывает природные и синтетические лиганды, например, цитокины, варианты цитокинов, аналоги, мутеины и связывающие соединения, происходящие из антител. "Лиганд" также охватывает низкомолекулярные соединения, например, пептидные миметики цитокинов и пептидные миметики антител. "Активация" может относиться к активации клеток регулируемой внутренними механизмами, а также внешними факторами и факторами окружающей среды.
"Активность" молекулы может описывать или относиться к связыванию молекулы с лигандом или с рецептором, к каталитической активности; к способности стимулировать экспрессию генов или передачу сигнала дифференцировки или созревания клеток; к антигенной активности, к модулированию активности других молекул и т.п. "Активность" молекулы также может относиться к активности модулирования или поддержания взаимодействий клетка-клетка, например адгезии, или к активности поддержания структуры клетки, например, клеточных мембран или цитоскелета. "Активность" также может означать удельную активность, например, [каталитическая активность]/[мг белка], или [иммунологическая активность]/[мг белка], концентрацию в биологическом компартменте и т.п. "Активность" может относиться к модулированию компонентов врожденной или адаптивной иммунной системой.
"Введение" и "обработка", как используют в отношении животного, человека, экспериментального субъекта, клетки, ткани, органа или биологической жидкости, относится к контакту экзогенного фармацевтического, терапевтического, диагностического средства или композиции с животным, человеком, индивидуумом, клеткой, органом или биологической жидкостью. Обработка клетки охватывает контактирование реагента с клеткой, а также контактирование реагента с жидкостью, где жидкость находится в контакте с клеткой. "Введение" и "обработка" также означают обработку in vitro и ex vivo, например клетки, реагентом, диагностическим соединением, связывающим соединением или другой клеткой. Термин "индивидуум" включает любой организм, предпочтительно животное, более предпочтительно млекопитающее (например, крыса, мышь, собака, кошка, кролик) и, наиболее предпочтительно, человека.
"Лечить" или "лечение" означает введение лекарственного средства, такого как композиция, содержащая любое из антител или антигенсвязывающих фрагментов по настоящему изобретению, внутрь или снаружи индивидууму или пациенту, имеющему один или несколько симптомов заболевания, или предположительно имеющему заболевание, против которого средство обладает терапевтической активностью. Как правило, средство вводят в количестве, эффективном для смягчения одного или нескольких симптомов заболевания у подвергаемого лечению индивидуума или популяции, либо путем индукции регрессии, либо путем ингибирования прогрессирования такого симптома(ов) на любую поддающуюся клиническому измерению степень. Количество лекарственного средства, которое является эффективным для смягчения любого конкретного симптома заболевания (также обозначаемое как "терапевтически эффективное количество"), может варьировать в зависимости от таких факторов, как болезненное состояние, возраст и масса тела пациента, и способность лекарственного средства индуцировать желаемый ответ у индивидуума. То, смягчается ли симптом заболевания, можно оценивать с помощью любого клинического измерения, как правило, используемого врачами или другими лицами, осуществляющими уход за здоровьем, для оценки тяжести или статуса прогрессирования этого симптома. В то время как вариант осуществления настоящего изобретения (например, способ лечения или изделие) может не быть эффективным в отношении смягчения заданного симптома(ов) заболевания у каждого индивидуума, он должен смягчать заданный симптом(ы) заболевания у статистически значимого числа индивидуумов при определении с помощью любого статистического критерия, известного в данной области, такого как t-критерий Стьюдента, критерий хи2, U-критерий согласно Манну и Уитни, критерий Крускала-Уоллиса (H-критерий), критерий Джонкхиера-Терпстра и критерий Вилкоксона.
"Лечение", как используют в отношении человека ветеринарного или испытуемого субъекта, относится к терапевтическому лечению, а также к исследовательским и диагностическим применениям. "Лечение", как используют в отношении человека, ветеринарного или испытуемого индивидуума, или клетки, ткани или органа, охватывает контакт антител или антигенсвязывающих фрагментов по настоящему изобретению с субъектом, являющимся человеком или животным, клеткой тканью, физиологическим компартментом или физиологической жидкостью.
Антитела против PD-L1
Антитело 20C3 представляет собой антитело продуцируемое субклоном гибридомы MEB037.20C3.116.
Антитело 22C3 представляет собой антитело, продуцируемое субклоном гибридомы MEB037.22C3.138, и оно соответствует аллотипу S414R IgG1 мыши. N-концевой остаток зрелой тяжелой цепи 22C3 представляет собой либо глутамин, либо пироглутамат (pE), который представляет собой распространенную посттрансляционную модификацию, которую часто наблюдают в моноклональных антителах, когда последовательность гена кодирует N-концевой глутамин в зрелой тяжелой или легкой цепи.
Антитела и антигенсвязывающие фрагменты по настоящему изобретению связываются со зрелой формой PD-L1 человека (лишенной пресекреторной лидерной последовательности, также обозначаемой как лидерный пептид), которая экспрессируется на поверхности определенных клеток человека. Термины "PD-L1" и "зрелый PD-L1" используют в настоящем описании взаимозаменяемо, и следует понимать, что они означают одну и ту же молекулу, если нет иных указаний или если это не очевидно из контекста. Зрелая молекула PD-L1 человека состоит из аминокислот 19-290 следующей последовательности (SEQ ID NO:37):
MRIFAVFIFMTYWHLLNAFTVTVPKDLYVVEYGSNMTIECKFPVEKQLDLAALIVYWE
MEDKNIIQFVHGEEDLKVQHSSYRQRARLLKDQLSLGNAALQITDVKLQDAGVYRCMI
SYGGADYKRITVKVNAPYNKINQRILVVDPVTSEHELTCQAEGYPKAEVIWTSSDHQVL
SGKTTTTNSKREEKLFNVTSTLRINTTTNEIFYCTFRRLDPEENHTAELVIPELPLAHPPNE
RTHLVILGAILLCLGVALTFIFRLRKGRMMDVK CGIQDTNSK QSDTHLEET.
Внеклеточный домен зрелого PD-L1 человека состоит из следующей последовательности (SEQ ID NO:38):
FTVTVPKDLYVVEYGSNMTIECKFPVEKQLDLAALIVYWEMEDKNIIQFVHGEEDLKVQ HSSYRQRARLLKDQLSLGNAALQITDVKLQDAGVYRCMISYGGADYKRITVKVNAPYN KINQRILVVDPVTSEHELTCQAEGYPKAEVIWTSSDHQVLSGKTTTTNSKREEKLFNVTS TLRINTTTNEIFYCTFRRLDPEENHTAELVIPELPLAHPPNERT.
Как используют в рамках изобретения антитело против PD-L1 человека или антитело против hPD-L1 относится к антителу, которое специфически связывается с PD-L1 человека. Антитело, которое "специфически связывается с PD-L1 человека", или антитело, которое "специфически связывается с полипептидом, содержащим аминокислотную последовательность PD-L1 человека", представляет собой антитело, которое проявляет предпочтительное связывание с PD-L1 человека по сравнению с другими антигенами, но эта специфичность не требует абсолютной специфичности связывания. Антитело против hPD-L1 считается "специфическим" в отношении PD-L1 человека, если его связывание определяет присутствие PD-L1 человека в образце, например, не вызывая нежелательных результатов, таких как ложноположительные результаты, в диагностическом анализе IHC. Степень специфичности, необходимая для антитела против hPD-L1 по изобретению, может зависеть от предполагаемого применения антитела, и при любых обстоятельствах определяется его пригодностью для применения для предполагаемой цели. Антитело или его связывающий фрагмент по изобретению связывается с PD-L1 человека с аффинностью, которая по меньшей мере в два раза превышает, предпочтительно по меньшей мере в десять раз превышает, более предпочтительно по меньшей мере в 20 раз превышает и наиболее предпочтительно по меньшей мере в 100 раз превышает аффинность в отношении любого белка, не являющегося PD-L1. Как используют в рамках изобретения антитело называют специфически связывающимся с полипептидом, содержащим данную последовательность, например зрелый PD-L1 человека (в этом случае аминокислоты 19-290 SEQ ID NO:37), если оно связывается с полипептидами, содержащими эту последовательность, но не связывается с белками лишенными этой последовательности. Например, антитело, которое специфически связывается с полипептидом, содержащим аминокислоты 19-290 SEQ ID NO:37, может связываться с меченной FLAG® формой аминокислот 19-290 SEQ ID NO:37, но не связывается с другими меченными FLAG® белками.
Как используют в рамках изобретения, термин "антитело" относится к любой форме антитела, которая проявляет желаемую биологическую активность. Таким образом, его используют в наиболее широком значении и, в частности, он охватывает, но не ограничивается ими, моноклональные антитела (включая полноразмерные моноклональные антитела), поликлональные антитела, полиспецифические антитела (например, биспецифические антитела), гуманизированные, полностью человеческие антитела, химерные антитела и камелизованные однодоменные антитела. "Родительские антитела" представляют собой антитела, получаемые при воздействии на иммунную систему антигена перед модификацией антител для предполагаемого применения, такой как гуманизация антитела для применения в качестве терапевтического антитела у человека.
Как используют в рамках изобретения, если нет иных указаний, "фрагмент антитела" или "антигенсвязывающий фрагмент" относится к антигенсвязывающим фрагментам антител, т.е. к фрагментам антитела, которые сохраняют способность специфически связываться с антигеном, связываемым полноразмерным антителом, например, к фрагментам, которые сохраняют одну или несколько областей CDR. Примеры связывающих фрагментов антител включают, но не ограничиваются ими, Fab, Fab', F(ab')2 и Fv-фрагменты; диантитела; линейные антитела; одноцепочечные молекулы антител, например, sc-Fv; наноантитела и полиспецифические антитела, образованные из фрагментов антител.
"Fab-фрагмент" содержит одну легкую цепь и CH1 и вариабельные области одной тяжелой цепи. Тяжелая цепь Fab-молекулы не может образовывать дисульфидную связь с другой молекулой тяжелой цепи. "Fab-фрагмент" может быть продуктом расщепления антитела папаином.
"Fc"-область содержит два фрагмента тяжелой цепи, содержащих CH1- и CH2-домены антитела. Два фрагмента тяжелой цепи удерживаются вместе двумя или несколькими дисульфидными связями и гидрофобными взаимодействиями CH3-доменов.
"Fab'-фрагмент" содержит одну легкую цепь и часть или фрагмент одной тяжелой цепи, которые содержат VH-домен и домен CH1, а также область между доменами CH1 и CH2, так чтобы между двумя тяжелыми цепями Fab'-фрагментов могла образовываться межцепочечная дисульфидная связь с образованием молекулы F(ab')2.
"F(ab')2-фрагмент" содержит две легких цепи и две тяжелых цепи содержащих часть константной области между доменами CH1 и CH2, так что между двумя тяжелыми цепями образуется межцепочечная дисульфидная связь. Таким образом, фрагмент F(ab')2 состоит из двух Fab'-фрагментов, которые удерживаются вместе дисульфидной связью между двумя тяжелыми цепями. "F(ab')2-фрагмент" может быть продуктом расщепления антитела пепсином.
"Fv-область" содержит вариабельные области как из тяжелой, так и из легкой цепей, но лишена константных областей.
Термин "одноцепочечное Fv" или "scFv" антитело относится к фрагментам антител, содержащим домены VH и VL антитела, причем эти домены находятся в одной полипептидной цепи. Как правило, полипептид Fv дополнительно содержит полипептидный линкер между доменами VH и VL, который обеспечивает образование из scFv желаемой структуры для связывания антигена. Для обзора scFv см. Pluckthun (1994) THE PHARMACOLOGY OF MONOCLONAL ANTIBODIES, vol. 113, Rosenburg and Moore eds. Springer-Verlag, New York, pp. 269-315. Также см. публикацию международной патентной заявки № WO 88/01649 и патенты США № 4946778 и 5260203.
"Доменное антитело" представляет собой иммунологически функциональный фрагмент иммуноглобулина, содержащий только вариабельную область тяжелой цепи или вариабельную область легкой цепи. В некоторых случаях две или более областей VH ковалентно связаны пептидным линкером с образованием двухвалентного доменного антитела. Две области VH двухвалентного доменного антитела могут быть направлены на одинаковые или на различные антигены.
"Двухвалентное антитело" содержит два антигенсвязывающих центра. В некоторых случаях два связывающих центра обладают одинаковой антигенной специфичностью. Однако двухвалентные антитела могут быть биспецифическими (см. ниже).
В определенных вариантах осуществления моноклональные антитела, описанные в настоящем описании, также включают камелизованные однодоменные антитела. См., например, Muyldermans et al. (2001) Trends Biochem. Sci. 26:230; Reichmann et al. (1999) J. Immunol. Methods 231:25; WO 94/04678; WO 94/25591; патент США № 6005079). В одном варианте осуществления настоящее изобретение относится к однодоменным антителам, содержащим два VH-домена с модификациями, так что образуются однодоменные антитела.
Как используют в рамках изобретения, термин "диантитела" относится к небольшим фрагментам антител с двумя антигенсвязывающими центрами, которые содержат вариабельный домен тяжелой цепи (VH), связанный с вариабельным доменом легкой цепи (VL), в одной и той же полипептидной цепи (VH-VL или VL-VH). С использованием линкера, который является слишком коротким, чтобы обеспечить образование пар между двумя доменами на одной цепи, домены принуждают образовывать пару с комплементарными доменами другой цепи и образовывать два антигенсвязывающих центра. Диантитела более подробно описаны, например, в EP 404097; WO 93/11161; и Holliger et al. (1993) Proc. Natl. Acad. Sci. USA 90: 6444-6448. Для обзора модифицированных способами инженерии вариантов антител, главным образом, см. Holliger and Hudson (2005) Nat. Biotechnol. 23:1126-1136.
Как правило, антитело или антигенсвязывающий фрагмент по изобретению сохраняют по меньшей мере 10% их активности связывания с PD-L1 человека (по сравнению с родительским антителом), когда эту активность выражают в молярной основе. Предпочтительно, антитело или антигенсвязывающий фрагмент по изобретению сохраняют по меньшей мере 20%, 50%, 70%, 80%, 90%, 95% или 100% или более аффинности связывания родительского антитела с PD-L1 человека. Также предполагается, что антитело или антигенсвязывающий фрагмент по изобретению могут включать консервативные или неконсервативные аминокислотные замены (обозначаемые как "консервативные варианты" или "функционально консервативные варианты" антитела), которые по существу не изменяют его биологической активности.
"Выделенное антитело" относится к состоянию очистки и в таком контексте означает, что молекула по существу свободна от других биологических молекул, таких как нуклеиновые кислоты, белки, липиды, углеводы или другой материал, такой как клеточный дебрис и среда для роста. Как правило, термин "выделенный" не предназначен для обозначения полного отсутствия такого материала или отсутствия воды, буферов или солей, если они не присутствуют в количествах, которые существенно препятствуют экспериментальному или терапевтическому применению связывающего соединение, как описано в настоящем описании.
Термин "моноклональное антитело", как используют в рамках изобретения, относится к совокупности по существу однородных антител, т.е. молекулы антитела, составляющие популяцию, являются идентичными по аминокислотной последовательности, за исключением возможных встречающихся в природе мутаций, которые могут присутствовать в небольших количествах. Напротив, препараты общепринятых (поликлональных) антител, как правило, включают множество различных антител, обладающих различными аминокислотными последовательностями в их вариабельных доменах, в частности, их CDR, которые часто являются специфичными к различным эпитопам. Определение "моноклональные" указывает на тот признак антитела, что его получают из по существу, однородной популяции антител, и его не следует истолковывать как требующий продуцирования антитела любым конкретным способом. Например, моноклональные антитела, подлежащие применению в соответствии с настоящим изобретением, можно получать способом гибридом, впервые описанным Kohler et al. (1975) Nature 256: 495, или их можно получать способами рекомбинантных ДНК (см., например, патент США № 4816567). "Моноклональные антитела" также можно выделять из библиотек фаговых антител с использованием способов, описанных, например, в Clackson et al. (1991) Nature 352: 624-628 и Marks et al. (1991) J. Mol. Biol. 222: 581-597. Также см. Presta (2005) J. Allergy Clin. Immunol. 116:731.
Как правило, основной структурный элемент антитела содержит тетрамер. Каждый тетрамер включает две идентичных пары полипептидных цепей, причем каждая пара имеет одну "легкую" (приблизительно 25 кДа) и одну "тяжелую" цепь (приблизительно 50-70 кДа). N-концевая часть каждой цепи включает вариабельную область из приблизительно от 100 до 110 или более аминокислот, в основном ответственных за распознавание антигена. C-концевая часть тяжелой цепи может определять константную область, в основном ответственную за эффекторную функцию. Как правило, легкие цепи человека классифицируют как легкие цепи каппа и лямбда. Более того, тяжелые цепи человека, как правило, классифицируют как мю, дельта, гамма, альфа или эпсилон, и они определяют изотип антитела как IgM, IgD, IgG, IgA и IgE, соответственно. В легкой и тяжелой цепях вариабельные и константные области связаны областью "j" приблизительно из 12 или более аминокислот, причем тяжелая цепь также включает "D"-область приблизительно из 10 или более аминокислот. См., главным образом, Fundamental Immunology Ch. 7 (Paul, W., ed., 2nd ed. Raven Press, N.Y. (1989).
Вариабельные области каждой паря легкая/тяжелая цепь образуют участок связывания антитела. Таким образом, как правило, интактное антитело имеет два связывающих участка. За исключением бифункциональных или биспецифических антител, два связывающих участка, как правило, являются одинаковыми.
Как правило, вариабельные домены как тяжелой, так и легкой цепей содержат три гипервариабельных области, также называемые определяющими комплементарность областями (CDR), расположенных в относительно консервативных каркасных областях (FR). CDR обычно расположены параллельно каркасным областям, что обеспечивает связывание с определенным эпитопом. Как правило, от N-конца к C-концу вариабельные домены как легкой, так и тяжелой цепей содержат FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Отнесение аминокислот к каждому домену, как правило, проводят в соответствии с определениями Sequences of Proteins of Immunological Interest, Kabat, et al.; National Institutes of Health, Bethesda, Md.; 5th ed.; NIH Publ. No. 91-3242 (1991); Kabat (1978) Adv. Prot. Chem. 32: 1-75; Kabat, et al, (1977) J. Biol. Chem. 252:6609-6616; Chothia, et al, (1987) J Mol. Biol. 196:901-917 или Chothia, et al, (1989) Nature 342:878-883.
Как используют в рамках изобретения, термин "гипервариабельная область" относится к аминокислотным остаткам антитела, которые ответственны за связывание антигена. Гипервариабельная область содержит аминокислотные остатки из "определяющей комплементарность области" или "CDR" (т.е. CDRL1, CDRL2 и CDRL3 в вариабельном домене легкой цепи и CDRH1, CDRH2 и CDRH3 в вариабельном домене тяжелой цепи). См. Kabat et al. (1991) Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (где определены области CDR антитела по последовательности); также см. Chothia and Lesk (1987) J. Mol. Biol. 196: 901-917 (где определены области CDR антитела по структуре). Как используют в рамках изобретения, термин "каркасные" остатки или остатки "FR" относится к остаткам вариабельного домена, отличным от остатков гипервариабельной области, определяемых в настоящем описании как остатки CDR.
"Гомология" относится к сходству последовательностей между двумя полинуклеотидными последовательностями или между двумя полипептидными последовательностями, когда они выровнены оптимально. Когда положение в обеих из двух сравниваемых последовательностях занято тем же основанием или мономерной субъединицей аминокислоты, например, если положение в каждой из двух молекул ДНК занято аденином, тогда молекулы являются гомологичными в этом положении. Процент гомологии представляет собой число положений, являющихся общими для двух последовательностей, деленное на общее количество сравниваемых положений × 100. Например, если 8 из 10 положений в двух последовательностях совпадают или являются гомологичными, когда последовательности выровнены оптимально, тогда две последовательности являются на 80% гомологичными. Как правило, сравнение проводят, когда две последовательности выровнены для получения максимальной процентной гомологии. Например, сравнение можно проводить с помощью алгоритма BLAST, где параметры алгоритма выбраны для наибольшего соответствия между соответствующими последовательностями на протяжении всей длины соответствующих эталонных последовательностей.
"Выделенная молекула нуклеиновой кислоты" означает ДНК или РНК геномного происхождения, происхождения из мРНК, кДНК или синтетического происхождения или некоторую их комбинацию, которая не связана со всей или частью полинуклеотида, в котором выделенный полинуклеотид встречается в природе, или связана с полинуклеотидом, с которым она не связана в природе. Для целей настоящего изобретения следует понимать, что "молекула нуклеиновой кислоты, содержащая" конкретную нуклеотидную последовательность, не охватывает интактные хромосомы. Выделенные молекулы нуклеиновой кислоты, "содержащие" указанные последовательности нуклеиновой кислоты, могут включать, в дополнение к указанным последовательностям, кодирующие последовательности для вплоть до десяти или даже вплоть до двадцати или более других белков или их участков или фрагментов, или они могут включать функционально связанные регуляторные последовательности, которые контролируют экспрессию кодирующей области указанных последовательностей нуклеиновой кислоты, и/или они могут включать векторные последовательности.
Выражение "последовательности контроля" относятся к последовательностям ДНК, необходимым для экспрессии функционально связанной кодирующей последовательности в конкретном организме-хозяине. Последовательности контроля, которые могут использоваться для прокариот, например, включают промотор, необязательно последовательность оператора и участок связывания рибосом. Известно, что эукариотические клетки используют промоторы, сигналы полиаденилирования и энхансеры.
Нуклеиновая кислота является "функционально связанной", когда она находится в функциональной взаимосвязи с другой последовательностью нуклеиновой кислоты. Например, ДНК для препоследовательности или секреторного лидера функционально связана с ДНК полипептида, если она экспрессируется в качестве пребелка, который участвует в секреции полипептида; промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию последовательности; или участок связывания рибосом функционально связан с кодирующей последовательностью, если он расположен так, чтобы способствовать трансляции. Как правило, "функционально связанный" означает, что связанные последовательности ДНК являются соседними, и, в случае секреторной лидерной последовательности, соседними и находящимися в рамке считывания. Однако энхансеры не должны быть соседними. Связывание проводят путем лигирования в удобных участках рестрикции. Если такие участки не существуют, используют синтетические олигонуклеотидные адаптеры или линкеры в соответствии с общепринятой практикой.
Как используют в рамках изобретения, выражения "клетка", "клеточная линия" и "клеточная культура" используют взаимозаменяемо и все такие обозначения включают потомков. Таким образом, слова "трансформанты" и "трансформированные клетки" включают первичные клетки индивидуума и культуры, происходящие из них, независимо от числа переносов. Также следует понимать, что не все потомство имеет точно идентичное содержание ДНК, вследствие преднамеренных или случайных мутаций. Включено мутантное потомство, которое имеет ту же функцию или биологическую активность, скрининг которой проводят в первоначально трансформированной клетке. Когда предполагаются различные обозначения, это будет очевидно из контекста.
Как используют в рамках изобретения, "полимеразная цепная реакция" или "ПЦР" относится к методике или способу, в которых определенные последовательности нуклеиновых кислот, РНК и/или ДНК, амплифицируют, как описано, например, в патенте США № 4683195. Как правило, для конструирования олигонуклеотидных праймеров используют информацию о последовательности с концов представляющей интерес области или за ее пределами. Эти праймеры являются идентичными или сходными по последовательности противоположным цепям матрицы, подлежащей амплификации. 5'-концевые нуклеотиды двух праймеров могут совпадать с концами амплифицированного материала. ПЦР можно использовать для амплификации определенных последовательностей РНК, определенных последовательностей ДНК из тотальной геномной ДНК и кДНК, транскрибированной с тотальной клеточной РНК, последовательностей бактерофага или плазмиды и т.д. См., главным образом, Mullis et al. (1987) Cold Spring Harbor Symp. Quant. Biol. 51:263; Erlich, ed., (1989) PCR TECHNOLOGY (Stockton Press, N.Y.). Как используют в рамках изобретения, ПЦР считается одним, но не единственным, примером способа полимеразной реакции нуклеиновых кислот для амплификации исследуемого образца нуклеиновой кислоты, включающим применение известной нуклеиновой кислоты в качестве праймера и полимеразы для амплификации нуклеиновой кислоты или получения конкретного фрагмента нуклеиновой кислоты.
Как используют в рамках изобретения, "последовательность эмбрионального типа" относится к нереаранжированным последовательностям ДНК иммуноглобулинов. Можно использовать любой подходящий источник нереаранжированных последовательностей иммуноглобулинов. Последовательности эмбрионального типа человека можно получать, например, из баз данных эмбриональных последовательностей JOINSOLVER® на web-сайте National Institute of Arthritis and Musculoskeletal and Skin Diseases of the United States National Institutes of Health. Последовательности эмбрионального типа мыши можно получать, например, как описано в Giudicelli et al. (2005) Nucleic Acids Res. 33:D256-D261.
Физические и функциональные свойства иллюстративных антител против PD-L1
Настоящее изобретение относится выделенным антителам против PD-L1 и к способам применения антител или их антигенсвязывающих фрагментов для обнаружения экспрессии PD-L1 на поверхности клеток. Примеры антител против PD-L1 по изобретению включают, но не ограничиваются ими: антитела 20C3 и 22C3 (см. фиг. 1 и 2). Антитела 20C3 и 22C3 связывают неидентичные, но соседние эпитопы (см. пример 2 и фиг. 9), что указывает на то, что CDR этих двух антител можно смешивать для получения дополнительных антител, которые специфически связываются с PD-L1 в одном или обоих из этих эпитопов. Таким образом, выделенное антитело или его антигенсвязывающий фрагмент, которые связывают PD-L1 человека, могут содержать три определяющих комплементарность области легкой цепи (CDR) и три CDR тяжелой цепи, представленные в таблицах 1-3 ниже.
где X=Q или pE
где X1=H или N, X2=S или T, и X3=R или S
где X1=S или T, X2=H или N, X3=S или T, X4=R или S, и X5=Q или K
где X1=D или G, X2=H или N, X3=S или N, X4=E или Q, и X5=I или M
где X1=I или V и X2=F или Y
где X=Q или pE, X1=H или Q, X2=L или V, X3=E или K, X4=I или M, X5=I или L, X6=D или G, X7=H или N, X8=N или S, X9=E или Q, X10=I или M, X11=K или R, X12=A или S, X13=S или T, X14=H или Q, X15=I или T, X16=I или V, и X17=F или Y
"Консервативно модифицированные варианты" или "консервативная замена" относятся к заменам аминокислот в белке другими аминокислотами, имеющими сходные характеристики (например заряд, размер боковой цепи, гидрофобность/гидрофильность, конформация и жесткость основной цепи, и т.д.), так что изменения часто могут быть внесены без изменения биологической активности белка. Специалистам в данной области будет понятно, что, как правило, единичные аминокислотные замены в несущественных областях полипептида по существу не изменяют биологическую активность (см., например, Watson et al. (1987) Molecular Biology of the Gene, The Benjamin/Cummings Pub. Co., p. 224 (4th Ed.)). Кроме того, замены структурно или функционально сходных аминокислот с меньшей вероятностью будут нарушать биологическую активность. Иллюстративные консервативные замены указаны в таблице 4.
Также в рамках настоящего изобретения предусматриваются варианты антител по изобретению с консервативной функцией. "Варианты с консервативной функцией", как используют в рамках изобретения, относятся к антителам или фрагментам, в которых один или несколько аминокислотных остатков заменены без изменения желаемого свойства, такого как аффинность и/или специфичность к антигену. Такие варианты включают, но не ограничиваются ими, замену аминокислоты аминокислотой, имеющей сходные свойства, такой как консервативные аминокислотные замены согласно таблице 4.
В другом варианте осуществления изобретение относится к антителу или его антигенсвязывающему фрагменту, которые специфически связывают PD-L1 и имеют VL-домены и VR-домены и обладают 100% гомологией последовательности с CDR легкой и тяжелой цепей согласно таблицам 1 или 2, и по меньшей мере 90%, 92%, 94%, 96%, 98% или 99% гомологией последовательностью со зрелыми вариабельными областями легкой и тяжелой цепей согласно таблицам 1 или 2.
Нуклеиновые кислоты
Настоящее изобретение также относится к нуклеиновым кислотам, кодирующим цепи иммуноглобулинов антител против PD-L1 и антигенсвязывающие фрагменты, описанные в настоящем описании. Например, настоящее изобретение относится к нуклеиновым кислотам, кодирующим аминокислоты, описанные в таблицах 1, 2 и 3, а также нуклеиновым кислотам, которые гибридизуются с ними.
Как правило, нуклеиновые кислоты гибридизуются в условиях низкой, умеренной или высокой жесткости и кодируют антитела, которые сохраняют способность специфически связываться с PD-L1. Первая молекула нуклеиновой кислоты является "гибридизующейся" со второй молекулой нуклеиновой кислоты, когда одноцепочечная форма первой молекулы нуклеиновой кислоты может отжигаться со второй молекулой нуклеиновой кислоты в соответствующих условиях температуры и ионной силы раствора (см. Sambrook, et al., выше). Условия температуры и ионной силы определяют "жесткость" гибридизации. Типичные условия гибридизации низкой жесткости включают 55°C, 5X SSC, 0,1% SDS и отсутствие формамида; или 30% формамид, 5X SSC, 0,5% SDS при 42°C. Типичные условия гибридизации умеренной жесткости представляют собой 40% формамид с 5X или 6X SSC и 0,1% SDS при 42°C. Условия гибридизации высокой жесткости представляют собой 50% формамид, 5X или 6X SSC при 42°C или, необязательно, при более высокой температуре (например, 57°C, 59°C, 60°C, 62°C, 63°C, 65°C или 68°C). Как правило, SSC представляет собой 0,15 M NaCl и 0,015 M Na-цитрат. Гибридизация требует, чтобы две нуклеиновых кислоты содержали комплементарные последовательности, хотя, в зависимости от жесткости гибридизации, возможны несоответствия между основаниями. Соответствующая жесткость для гибридизации нуклеиновых кислот зависит от длины нуклеиновых кислот и степени комплементарности, которые являются переменными, хорошо известными в данной области. Чем более высокой является степень сходства или гомологии между двумя нуклеотидными последовательностями, тем более высокой является жесткость, при которой нуклеиновые кислоты могут гибридизоваться. Для гибридов длиной более 100 нуклеотидов были выведены уравнения для вычисления температуры плавления (см. Sambrook, et al, выше, 9.50-9.51). Для гибридизации с более короткими нуклеиновыми кислотами, например, олигонуклеотидами, положение несоответствий оснований становится более важным, и длина олигонуклеотида определяет его специфичность (см. Sambrook, et al., выше, 11.7-11.8).
Следующие ссылки относятся к алгоритмам BLAST, часто используемым для анализа последовательностей: BLAST ALGORITHMS: Altschul, S.F., et al, (1990) J. Mol. Biol. 215:403-410; Gish, W., et al, (1993) Nature Genet. 3:266-272; Madden, T.L., et al, (1996) Meth. Enzymol. 266: 131-141; Altschul, S.F., et al, (1997) Nucleic Acids Res. 25:3389-3402; Zhang, J., et al, (1997) Genome Res. 7:649-656; Wootton, J.C., et al, (1993) Comput. Chem. 17: 149-163; Hancock, J.M. et al, (1994) Comput. Appl. Biosci. 10:67-70; ALIGNMENT SCORING SYSTEMS: Dayhoff, M.O., et al, "A model of evolutionary change in proteins", Atlas of Protein Sequence and Structure, (1978) vol. 5, suppl. 3. M.O. Dayhoff (ed.), pp. 345-352, Natl. Biomed. Res. Found., Washington, DC; Schwartz, R.M., et al, "Matrices for detecting distant relationships", Atlas of Protein Sequence and Structure, (1978) vol. 5, suppl. 3." M.O. Dayhoff (ed.), pp. 353-358, Natl. Biomed. Res. Found., Washington, DC; Altschul, S.F., (1991) J. Mol. Biol. 219:555-565; States, D.J., et al, (1991) Methods 3:66-70; Henikoff, S., et al, (1992) Proc. Natl. Acad. Sci. USA 89: 10915-10919; Altschul, S.F., et al, (1993) J. Mol. Evol. 36:290-300; ALIGNMENT STATISTICS: Karlin, S., et al, (1990) Proc. Natl. Acad. Sci. USA 87:2264-2268; Karlin, S., et al, (1993) Proc. Natl. Acad. Sci. USA 90:5873-5877; Dembo, A., et al, (1994) Ann. Prob. 22:2022-2039; и Altschul, S.F. "Evaluating the statistical significance of multiple distinct local alignments", Theoretical and Computational Methods in Genome Research (S. Suhai, ed.), (1997) pp. 1-14, Plenum, New York.
В другом варианте осуществления изобретение относится к выделенной нуклеиновой кислоте или нуклеиновым кислотам, например ДНК, кодирующей по меньшей мере одну из полипептидных цепей выделенных антител против PD-L1 или антигенсвязывающих фрагментов, описанных в настоящем описании. В некоторых вариантах осуществления выделенная нуклеиновая кислота кодирует как легкую цепь, так и тяжелую цепь на единичной молекуле нуклеиновой кислоты, и в других вариантах осуществления легкая и тяжелая цепи кодируются на разных молекулах нуклеиновой кислоты. В другом варианте осуществления нуклеиновые кислоты дополнительно кодируют сигнальную последовательность.
Настоящее изобретение также относится к векторам экспрессии, содержащим выделенные нуклеиновые кислоты по изобретению, где нуклеиновая кислота функционально связана с последовательностями контроля, которые распознаются клеткой-хозяином, когда клетка-хозяина трансфицирована вектором. Также предусматриваются клетки-хозяева, содержащие вектор экспрессии по настоящему изобретению, и способы получения антитела или его антигенсвязывающего фрагмента, описанных в настоящем описании, включающие культивирование клетки-хозяина, содержащей вектор экспрессии, кодирующий антитело или антигенсвязывающий фрагмент, в культуральной среде, и выделение антигена или его антигенсвязывающего фрагмента из клетки-хозяина или культуральной среды.
Связывание эпитопа
Кроме того, настоящее изобретение относится к антителам или их антигенсвязывающим фрагментам, которые блокируют связывание антитела 20C3 или 22C3 с PD-L1 человека путем связывания с тем же эпитопом, что и у 20C3 или 22C3, соответственно. Такие антитела и связывающие фрагменты могут быть идентифицированы с использованием любых анализов перекрестного блокирования или конкурентных анализов, известных в данной области, включая конкурентные анализы Octet, описанные в примере 2, с последующей идентификацией эпитопа на PD-L1 человека, с которым связывается перекрестно блокирующее антитело. Первое антитело считается связывающимся с перекрестным блокированием со вторым антителом, если предварительное связывание мишени с первым антителом до насыщения увеличивает концентрацию второго антитела, требуемую для достижения полумаксимального связывания мишени в 2, 3, 4, 5, 10, 20, 50, 100, 200 раз или более. Связываемый эпитоп для перекрестно блокирующего антитела можно идентифицировать с использованием способов, хорошо известных в данной области.
Одним таким способом картирования эпитопов является водородно/дейтериевый обмен, сопряженный с протеолизом и масс-спектрометрией (HDX-MS). Этот способ основан на точном измерении и сравнении степени включения дейтерия антигеном при инкубации в тяжелой воде (D2O) самостоятельно и в присутствии антитела через различные интервалы времени. Дейтерий заменяется на водород в амидном остове белков в подвергнутых воздействию областях, в то время как области антигена, связанные с антителом, будут защищены и будут демонстрировать меньший или отсутствие обмена после анализа жидкостной хроматографией-тандемной масс-спектрометрией (LC-MS/MS) протеолитических фрагментов.
Исходя из картирования эпитопа HDX-MS, описанного в примере 3, предполагаемый эпитоп на зрелом PD-L1 человека для антитела 22C3 содержит остатки в двух непрерывных аминокислотных сегментах во внеклеточном домене (SEQ ID NO:38): от 156 до 178 и от 196 до 206. Дополнительные остатки эпитопа, вероятно, присутствуют в следующих сегментах внеклеточного домена (SEQ ID NO:38): с 3 по 9; с 10 по 13; с 88 по 93 и с 135 по 147.
Таким образом, в одном варианте осуществления антитело, которое блокирует связывание антитела 22C3 с PD-L1 человека посредством связывания с тем же эпитопом что и 22C3, связывается с остатками в первом сегменте аминокислот от 156 до 178 SEQ ID NO:38 и с остатками во втором сегменте аминокислот от 196 до 206 SEQ ID NO:38, и в некоторых вариантах осуществления также связывается с остатками в одном, двух или трех, или во всех четырех из следующих сегментов SEQ ID NO:38: аминокислоты с 3 по 9; аминокислоты с 10 по 13; аминокислоты с 88 по 93 и аминокислоты с 135 по 147.
Способы получения антител и их антигенсвязывающих фрагментов
Клетки гибридомы, которые продуцируют родительские (например, грызуна) моноклональные антитела против X, можно получать способами, которые широко известны в данной области. Эти способы включают, но не ограничиваются ими, способ гибридом, первоначально разработанный Kohler, et al., (1975) (Nature 256:495-497), а также способ триом (Hering, et al., (1988) Biomed. Biochim. Acta. 47:211-216 и Hagiwara, et al., (1993) Hum. Antibod. Hybridomas 4: 15), способ B-клеточных гибридом человека (Kozbor, et al., (1983) Immunology Today 4:72 и Cote, et al., (1983) Proc. Natl. Acad. Sci. U.S.A 80:2026-2030), способ EBV-гибридомы (Cole, et al, Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., pp. 77-96, 1985) и электрослияние на основе электрического поля с использованием крупнокамерного электропоратора для слияния клеток Cyto Pulse (Cyto Pulse Sciences, Inc., Glen Burnie, MD). Предпочтительно, спленоциты мыши выделяют и подвергают слиянию с помощью PEG или путем электрослияния с клетками миеломы мыши, на основе стандартных протоколов.
Затем полученные гибридомы можно подвергать скринингу в отношении продуцирования антигенспецифических антител. Например, суспензии единичных лимфоцитов селезенки от иммунизированных мышей можно подвергать слиянию с одной шестой количества P3X63-Ag8.653 несекретирующих клеток миеломы мыши (ATCC, CRL 1580) с 50% PEG. Клетки можно высевать в количестве приблизительно 2×105 клеток/мл в микропланшете для титрования с плоским дном с последующей инкубацией в течение двух недель в селективной среде, содержащей 20% эмбриональную клональную сыворотку, 18% кондиционированную среду "653", 5% ориген (IGEN), 4 мМ L-глутамин, 1 мМ L-глутамин, 1 мМ пируват натрия, 5 мМ HEPES, 0,055 мМ 2-меркаптоэтанол, 50 единиц/мл пенициллина, 50 мг/мл стрептомицина, 50 мг/мл гентамицина и 1X HAT (Sigma; HAT добавляют через 24 часа после слияния). Через две недели клетки можно культивировать в среде, в которой HAT заменена на HT. Затем индивидуальные лунки можно подвергать скринингу способом ELISA в отношении моноклональных IgG-антител против X. После того, как произойдет экстенсивный рост гибридом, среду, как правило, можно наблюдать через 10-14 суток. Секретирующие антитело гибридомы можно повторно высевать, вновь подвергать скринингу и, если они все еще являются положительными по IgG человека, моноклональные антитела против X можно субклонировать по меньшей мере два раза способом лимитирующих разведений.
Затем стабильные субклоны можно культивировать in vitro для получения небольших количеств антитела в среде для культивирования тканей для охарактеризации. Например, можно получать приблизительно 1 грамм антитела 22C3 и очищать из клеточной линии гибридомы мыши MEB037.22C3.138 с использованием следующей методики. Замороженные клетки MEB037.22C3.138 размораживают и адаптируют во вращающейся колбе с использованием бессывороточной среды для гибридом с 2 мМ дополнительным L-глутамином с 0,18% Pluronic F-68 или без него. Присутствие Pluronic F-68 может увеличить жизнеспособность культуры во вращающихся флаконах. После полной адаптации во вращающемся флаконе проводят культивирование для продуцирования в объеме 20 литров в бессывороточной среде в биореакторе WAVE (GE Healthcare Life Sciences) с добавлением 10% CHO CD efficient Feed B (Invitrogen, каталожный номер № A10240-01). Для увеличения клеток в количестве инициируют культуру объемом 1 литр в небольшом мешке WAVE, а затем культуру WAVE в объеме 1 литр увеличивают в масштабе в культуру объемом 20 л в биореакторе WAVE. Культуру объемом 20 литров можно инициировать при плотности клеток, составляющей 0,5×106 жизнеспособных клеток/мл, подпитывать 10% CHO CD Efficient Feed B на 1 сутки, и каждые сутки корректировать pH с помощью 1 Н Na2CO3. Клетки собирают через четверо суток. Небольшие образцы можно собирать каждые сутки для анализа NOVA.
Антитела против hPD-L1 по изобретению можно очищать из культуры гибридомы с помощью следующего процесса. Культуру гибридомы очищают глубинной фильтрацией с использованием 1,2-микрометрового стеклянного волокна и 0,2-микрометрового фильтра из ацетата целлюлозы. К очищенной собранной фракции добавляют равный объем 2X буфера ProSepA (100 мМ борная кислота, 5 M NaCl, pH 8,5) и разбавленную собранную фракцию наносят на колонку с белком A с объемом слоя 170 мл. Колонку промывают 5 объемами колонки (CV) 1X буфера ProSepA (50 мМ борная кислота, 2,5 M NaCl, pH 8,5), затем промывают 2 CV 1X PBS, и антитело против hPD-L1 элюируют 5 CV буфера для элюирования (0,1 M глицин, pH 3,0). Элюированные фракции, содержащие IgG, объединяют и pH нейтрализуют добавлением 1/10 объема 1,0 M pH-буфера Tris. Затем нейтрализованную композицию антител подвергают стерилизации фильтрованием с использованием 10 кДа одноразовой кассеты TFF. Антитело можно составлять для хранения путем дифильтрации против 10 литров буфера для составления (20 мМ ацетат натрия, 9% сахароза, pH 5,0) и с использованием 20 изменений объема. С использованием этого протокола можно получать антитело 22C3 в концентрации приблизительно 5,0 мг/мл, имеющее чистоту по меньшей мере 98% при измерении с помощью SDS-PAGE, SEC ВЭЖХ и C8 ОФ-ВЭЖХ с уровнями эндотоксина менее 0,1 ЕЭ/мл и менее 0,02 ЕЭ/мг.
Антитела против PD-L1, описанные в настоящем описании, также можно получать рекомбинантными способами (например, в E. экспрессирующей системе E. coli/T7, как описано выше). В этом варианте осуществления нуклеиновые кислоты, кодирующие молекулы антитела по изобретению (например, VH или VL) можно встраивать в плазмиду на основе pET и экспрессировать в системе E. coli/T7. Существует несколько способов получения рекомбинантных антител, которые известны в данной области. Один пример способа рекомбинантной продукции антител описан в патенте США № 4816567. Трансформацию можно проводить любым известным способом введения полинуклеотидов клетку-хозяина. Способы введения гетерологичных полинуклеотидов в клетки млекопитающих хорошо известны в данной области и включают опосредуемую декстраном трансфекцию, осаждение с фосфатом кальция, опосредуемую полибреном трансфекцию, слияние протопластов, электропорацию, инкапсулирование полинуклеотида(ов) в липосомы, биолистическую инъекцию и прямую микроинъекцию ДНК в ядро. Кроме того, молекулы нуклеиновой кислоты можно вводить в клетки млекопитающих с помощью вирусных векторов. Способы трансформации клеток хорошо известны в данной области. См., например, патенты США № 4399216; 4912040; 4740461 и 4959455.
Антитела против PD-L1 также можно синтезировать любыми способами, указанными в патенте США № 6331415.
Линии клеток млекопитающих, доступные в качестве хозяев для экспрессии антител или фрагментов, описанных в настоящем описании, хорошо известны в данной области и включают множество иммортализованных клеточных линий, доступных от American Type Culture Collection (ATCC). Они включают, среди прочих, клетки яичника китайского хомячка (CHO), NSO, клетки SP2, клетки HeLa, клетки почки детеныша хомяка (BHK), клетки почки обезьяны (COS), клетки печеночно-клеточной карциномы человека (например, Hep G2), клетки A549, клетки 3T3, клетки HEK-293 и ряд других клеточных линий. Клетки-хозяева млекопитающих включают клетки человека, мыши, крысы, собаки, обезьяны, свиньи, козы, животного семейства бычьих, лошади и хомяка. Особенно предпочтительные клеточные линии выбирают путем определения того, какие клеточные линии имеют высокие уровни экспрессии. Другие клеточные линии, которые можно использовать, представляют собой клеточные линии насекомых, такие как клетки Sf9, клетки земноводных, бактериальные клетки, клетки растений и клетки грибов. Когда рекомбинантные векторы экспрессии, кодирующие тяжелую цепь или ее антигенсвязывающую часть или фрагмент, легкую цепь и/или ее антигенсвязывающий фрагмент, вводят в клетки-хозяева млекопитающих, антитела продуцируют путем культивирования клеток-хозяев в течение периода времени, достаточного для обеспечения экспрессии антитела в клетках-хозяевах или, более предпочтительно, секреции антитела в культуральную среду, в которой клетки-хозяева выращивают.
Антитела можно выделять из культуральной среды с использованием стандартных способов очистки белка. Кроме того, экспрессию антител по изобретению (или других их частей) из продуцирующих клеточных линий можно усиливать с использованием ряда известных способов. Например, система экспрессии гена глутаминсинтетазы (система GS) представляет собой распространенный подход для повышения экспрессии в определенных условиях. Система GS рассмотрена в целом или частично в европейских патентах №№ 0216846, 0256055 и 0323997 и патентной заявке Европы № 89303964.4.
Поликлональное антитело представляет собой антитело, которое продуцировано среди или в присутствии одного или нескольких других неидентичных антител. Как правило, поликлональные антитела продуцируются из коллекций различных B-лимфоцитов, например, B-лимфоцитов животного, которому вводили представляющий интерес иммуноген, которое продуцирует популяцию различных антител, все из которых направлены на иммуноген. Обычно поликлональные антитела получают прямо из иммунизированного животного, например селезенки, сыворотки или асцитной жидкости.
Кроме того, настоящее изобретение относится к фрагментам антител против PD-L1, описанным в настоящем описании. Фрагменты антител включают фрагменты F(ab)2, которые могут продуцироваться путем ферментативного расщепления IgG, например, пепсином. Fab-фрагменты можно получать, например, восстановлением F(ab)2 дитиотреитолом или меркаптоэтиламином. Fab-фрагмент представляет собой цепь VL-CL, присоединенную к цепи VH-CH1 через дисульфидную связь. F(ab)2-фрагмент представляет собой два Fab-фрагмента, которые, в свою очередь, связаны двумя дисульфидными мостиками. Fab-части молекулы F(ab)2 включают части Fc-области, между которыми расположены дисульфидные мостики. Fv-фрагмент представляет собой область VL или VR.
Иммуноглобулины можно относить к различным классам в зависимости от аминокислотных последовательностей константного домена их тяжелых цепей. Существует по меньшей мере пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и несколько из них могут быть далее подразделены на подклассы (изотипы), например IgG-1, IgG-2, IgG-3 и IgG-4; IgA-1 и IgA-2. Изобретение включает антитела и антигенсвязывающие фрагменты любого из этих классов или подклассов антител.
В одном варианте осуществления антитело или антигенсвязывающий фрагмент содержит константную область тяжелой цепи, например константную область человека, такую как константная область тяжелой цепи γ1, γ2, γ3 или γ4 человека или ее вариант. В другом варианте осуществления антитело или антигенсвязывающий фрагмент содержит константную область легкой цепи, например, константную область легкой цепи человека, такую как область легкой цепи лямбда или каппа человека. В качестве неограничивающего примера, константная область тяжелой цепи человека может представлять собой γ1 и константная область легкой цепи человека может представлять собой каппа. В альтернативном варианте осуществления Fc-область антитела представляет собой γ4 с мутацией Ser228Pro (Schuurman, J et. al., Mol. Immunol. 38: 1-8, 2001).
В некоторых вариантах осуществления различные константные домены можно присоединять к гуманизированным областям VL и VH, происходящим из CDR, описанных в настоящем описании. Например, если конкретным предполагаемым применением антитела (или фрагмента) по настоящему изобретению является обеспечение измененных эффекторных функций, можно использовать константный домен тяжелой цепи, отличный от IgG1 человека, или можно использовать гибрид IgG1/IgG4.
Инженерия антител
В конкретных вариантах осуществления является желательным изменение определенных аминокислот, содержащих экспонированные боковые цепи, на другой остаток, чтобы обеспечить более высокую химическую стабильность конечного антитела, следующим образом. Дезамидирование аспарагина может происходить в последовательностях N-G или D-G и может приводить к образованию остатка изоаспарагиновой кислоты, который вносит петлю в полипептидную цепь и снижает его стабильность (эффект изоаспарагиновой кислоты). В определенных вариантах осуществления антитела по настоящему изобретению не содержат участков изомерии аспарагина.
Например, остаток аспарагина (Asn) может быть заменен на Gln или Ala для уменьшения потенциала к образованию изоаспартата в любых последовательностях Asn-Gly, в частности в CDR. Сходная проблема может происходить в последовательности Asp-Gly. Reissner and Aswad (2003) Cell. Mol. Life Sci. 60: 1281. Образование изоаспартата может истощать или полностью устранять связывание антитела с его антигеном-мишенью. См., Presta (2005) J. Allergy Clin. Immunol. 116:731, 734. В одном варианте осуществления аспарагин заменен на глутамин (Gln). Также может быть желательным изменение аминокислоты, соседней с остатком аспарагина (Asn) или глутамина (Gln) для уменьшения вероятности дезамидирования, которое происходит с большей скоростью, когда аминокислоты меньшего размера находятся рядом с аспарагином или глутамином. См., Bischoff & Kolbe (1994) J. Chromatog. 662:261. Кроме того, любые остатки метионина (как правило, подвергаемые воздействию растворителя Met) в CDR могут быть заменены на Lys, Leu, Ala или Phe для уменьшения вероятности того, что сера метионина будет окисляться, что может уменьшить аффинность связывания антигена и также внести вклад в молекулярную гетерогенность при конечном получении антител. Там же. В одном варианте осуществления метионин заменен на аланин (Ala). Кроме того, для предупреждения или минимизации потенциального расщепления пептидных связей Asn-Pro, может быть желательным изменение любых комбинаций Asn-Pro, встречающихся в CDR на Gln-Pro, Ala-Pro или Asn-Ala. Антитела с такими заменами затем подвергают скринингу, чтобы убедиться, что замены не снижают аффинности или специфичности антитела в отношении PD-L1, или другую желаемую биологическую активность до неприемлемых уровней.
(N-G)
(Q-G), (A-G) или (N-A)
(D-G)
(E-G), (A-G) или (D-A)
(M)
(K), (L), (A) или (F)
(N)
(Q) или (A)
(N-P)
(Q-P), (A-P) или (N-A)
Конъюгаты антител
Молекулы антитела против PD-L1, описанные в настоящем описании, также можно конъюгировать с химической частью, такой как радионуклид или другая поддающаяся обнаружению метка. Радионуклиды включают 99Tc, 90Y, 111In, 32P, 14C, 125I, 3H, 131I, 11C, 15O, 13N, 18F, 35S, 51Cr, 57To, 226Ra, 60Co, 59Fe, 57Se, 152Eu, 67CU, 217Ci, 211At, 212Pb, 47Sc, 109Pd, 234Th, и 40K, 157Gd, 55Mn, 52Tr и 56Fe. Флуоресцентные или хемилюминесцентные метки включают флуорофоры, такие как хелаты редкоземельных металлов, флуоресцеин и его производные, родамин и его производные, изотиоцианат, фикоэритрин, фикоцианин, аллофикоцианин, о-фталальдегид, флуорескамин, 152Eu, дансил, умбеллиферон, люциферин, люминальную метку, изолюминальную метку, метку на основе ароматического сложного эфира акридиния, имидазольную метку, метку на основе соли акридиния, метку на основе оксалатного сложного эфира, эквориновую метку, 2,3-дигидрофталазиндионы, биотин/авидин, спиновые метки и стабильные свободные радикалы.
Для конъюгации молекул антитела с различными частями можно использовать любой способ, известный в данной области, включая способы, описанные Hunter, et al, (1962) Nature 144:945; David, et al, (1974) Biochemistry 13: 1014; Pain, et al, (1981) J. Immunol. Meth. 40:219; и Nygren, J., (1982) Histochem. и Cytochem. 30:407. Способы конъюгации антител являются общепринятыми и хорошо известны в данной области.
Экспериментальные и диагностические применения
Антитела против PD-L1 и фрагменты антител, описанные в настоящем описании, можно использовать для специфического обнаружения PD-L1 человека, экспрессируемого на поверхности клетки. Клетка может присутствовать в образце ткани или сыворотки, полученном от человека, и обнаружение экспрессии PD-L1 проводят с использованием любого из множества способов анализа in vitro, известных в данной области.
Например, конкретные варианты осуществления включают анализы ELISA (твердофазный иммуноферментный анализ), которые, как правило, включают следующие стадии:
(a) покрытие субстрата (например, поверхность лунки микропланшета для титрования, например, пластмассовый планшет) антителом против PD-L1 или его антигенсвязывающим фрагментом;
(b) нанесение образца, подлежащего исследованию в отношении присутствия PD-L1, человека, на субстрат;
(c) промывание планшета так, чтобы удалить не связавшийся материал в образце;
(d) нанесение меченных поддающихся детекции меткой антител (например, связанные с ферментом антитела), которые также являются специфичными к PD-L1 человека;
(e) промывание субстрата так, чтобы удалялись не связавшиеся меченые антитела;
(f) если меченые антитела связаны с ферментом, нанесение химического вещества, которое конвертируется ферментом во флуоресцентный сигнал; и
(g) выявление присутствия меченого антитела.
В следующем варианте осуществления меченое антитело является меченным пероксидазой, которая реагирует с ABTS (например, 2,2'-азино-бис(3-этилбензтиазолин-6-сульфоновая кислота)) или 3,3',5,5'-тетраметилбензидином для обеспечения изменения цвета, которое поддается обнаружению. Альтернативно меченое антитело метят поддающимся обнаружению радиоактивным изотопом (например, 3H), который можно обнаруживать с помощью сцинтилляционного счетчика в присутствии сцинтиллятора.
Антитела против PD-L1 и их антигенсвязывающие фрагменты по изобретению можно использовать в методике Вестерн-блоттинга или иммуноблоттинга белков. Такая методика является частью настоящего изобретения и включает, например:
(1) контактирование мембраны или другой твердой подложки, подлежащей исследованию в отношении присутствия PD-L1 человека, с антителом или антигенсвязывающим фрагментом по изобретению. Такая мембрана может иметь форму нитроцеллюлозной мембраны или мембраны на основе винила (например, поливинилиденфторид (PVDF)), на которую перенесены белки, подлежащие исследованию в отношении присутствия X в неденатурирующем геле PAGE (полиакриламидный гель-электрофорез) или геле SDS-PAGE (полиакриламидный гель-электрофорез с додецилсульфатом натрия) (например, после электрофоретического разделения в геле). Перед контактированием мембраны с антителом против PD-L1 или фрагментом, мембрану необязательно блокируют, например, обезжиренным сухим молоком и т.п., чтобы связать участки неспецифического связывания белка на мембране.
(2) промывание мембраны один или несколько раз для удаления не связавшегося антитела против PD-L1 или фрагмента и других не связавшихся веществ; и
(3) обнаружение связавшегося антитела против PD-L1 или фрагмента.
Связавшееся антитело или фрагмент можно выявлять путем инкубации связавшегося антитела или фрагмента с вторичным антителом (антитело против иммуноглобулина), которое является меченным поддающейся обнаружению меткой, а затем обнаружения присутствия вторичного антитела.
Антитела против PD-L1 и их антигенсвязывающие фрагменты, описанные в настоящем описании, также можно использовать в иммуногистохимических (IHC) анализах, которые можно проводить с использованием различных форматов IHC, известных в данной области, и они составляют варианты осуществления изобретения. В типичном анализе IHC используется срез ткани FFPE размером приблизительно 3-4 миллиметра, и предпочтительно 4 микрометра, залитый и высушенный на предметном стекле микроскопа, и он включает, например, (1) проведение депарафинизации и гидратации среза ткани, контактирование регидратированного среза ткани с антителом против PD-L1 или его антигенсвязывающим фрагментом по изобретению; и (2) обнаружение антитела против PD-L1 или его антигенсвязывающего фрагмента на поверхности одной или нескольких клеток в ткани. Если антитело или фрагмент сами по себе являются меченными поддающейся обнаружению меткой, их можно выявлять прямо. Альтернативно антитело или фрагмент могут быть связаны меченным поддающейся обнаружению меткой вторичным антителом, которое выявляют.
В предпочтительном анализе IHC используется коммерчески доступная система обнаружения Dako EnVision™ FLEX, которая предназначена для применения вместе с устройством Dako Autostainer (Dako, Agilent Technologies Company, Glostrup, Дания). При использовании этой системы с антителом 22C3 или антителом, которое содержит вариабельные области тяжелой и легкой цепей антитела 22C3, анализ IHC можно проводить следующим образом. Срезы ткани, обработанные FFPE, толщиной четыре микрометра, залитые на предметных стеклах, сушат воздухом в течение ночи, обжигают при 60°C в течение 45 минут, депарафинизируют и регидратируют. После депарафинизации предметные стекла, обработанные FFPE, подвергают индуцируемому нагреванием извлечению эпитопа с использованием раствора EnVision™ FLEX High pH Target Retrieval Solution при 97°C, а затем в течение 20 минут при комнатной температуре. Затем предметные стекла промывают, окрашивают 22C3 в концентрации 2 мкг/мл в течение 60 минут, а затем подвергают обнаружению с использованием реагентов Dako EnVision™ FLEX следующим образом: EnVision™ FLEX+ MS Linker (15 минут), EnVision™ FLEX/HRP (20 минут), EnVision™ FLEX DAB (10 минут) и DAB Enhancer (7 минут) с промежуточными стадиями промывания.
Определенные антитела против PD-L1 и их антигенсвязывающие фрагменты, описанные в настоящем описании, также можно использовать для визуализации опухоли in vivo. Такой способ может включать инъекцию радиоактивно меченного антитела против PD-L1 или его антигенсвязывающего фрагмента в организм пациента-человека, подлежащего исследованию в отношении присутствия опухоли, ассоциированной с экспрессией PD-L1, с последующей ядерной визуализацией ядер в организме пациента для обнаружения присутствия меченного антитела или фрагмента, например, в локусах, содержащих высокую концентрацию антитела или фрагмента, которые связаны с опухолью.
Способы визуализации включают визуализацию SPECT (однофотонная эмиссионная компьютерная томография) или визуализацию PET (позитронная эмиссионная томография). Метки включают, например, йод-123 (123I) и технеций-99m (99mTc), например, при визуализации SPECT или 11C, 13N, 15O или 18F, например, при визуализации PET или индий-111 (см. например, Gordon et al., (2005) International Rev. Neurobiol. 67:385-440).
Наборы для обнаружения и терапевтические наборы
Для удобства антитело или специфический связывающий агент, описанные в настоящем описании, могут быть предоставлены в наборе, т.е. упакованной комбинации реагентов в заданных количествах с инструкциями по проведению диагностического анализа или анализа для обнаружения. Когда антитело является меченным ферментом, набор включает субстраты и кофакторы, требуемые для фермента (например, предшественник субстрата, который обеспечивает поддающийся обнаружению хромофор или флуорофор). Кроме того, могут быть включены другие добавки, такие как стабилизаторы, буферы (например, блокирующий буфер или лизирующий буфер) и т.п. Относительные количества различных реагентов могут широко варьировать для обеспечения концентраций в растворе реагентов, которые существенно оптимизируют чувствительность анализа. В частности, реагенты могут быть предоставлены в качестве сухих порошков, обычно лиофилизированных, включающих эксципиенты, которые при растворении обеспечат раствор реагента, имеющий соответствующую концентрацию.
Также предусматриваются диагностические реагенты или реагенты для обнаружения и наборы, содержащие один или несколько таких реагентов, для применения в различных анализах для обнаружения, включающих, например, иммуноанализы, такие как ELISA (формата сэндвич-типа или конкурентного формата). Компоненты набора могут быть предварительно связаны с твердой подложкой или их можно наносить на поверхность твердой подложки, когда набор используют. В некоторых вариантах осуществления средства генерирования сигнала могут быть предварительно связаны с антителом по изобретению или может потребоваться комбинирование с одним или несколькими компонентами, например, буферами, конъюгатами антитело-фермент, субстратами ферментов и т.п. перед применением. Наборы также могут включать дополнительные реагенты, например, блокирующие реагенты для уменьшения неспецифического связывания с поверхностью твердой фазы, реагенты для промывания, субстраты ферментов и т.п. Поверхность твердой фазы может быть в форме пробирки, гранулы, микропланшета для титрования, микросферы или других материалов, пригодных для иммобилизации белков, пептидов или полипептидов. В конкретных аспектах фермент, который катализирует образование хемилюминесцентного или хромогенного продукта или восстановление хемилюминесцентного или хромогенного субстрата, представляет собой компонент средства для генерирования сигнала. Такие ферменты хорошо известны в данной области. Наборы могут содержать любые средства для улавливания и реагенты для обнаружения, описанные в настоящем описании. Необязательно набор также может содержать инструкции по проведению способов по изобретению.
Также можно получать наборы для обнаружения, описанные в настоящем описании, которые содержат по меньшей мере одно из антитела или антигенсвязывающего фрагмента, описанных в настоящем описании, и инструкции по применению композиции в качестве реагента для обнаружения. Контейнеры для применения в таких наборах могут включать, как правило, по меньшей мере один флакон, пробирку, колбу, бутылку, шприц или другой подходящий контейнер, в который может быть помещена одна или несколько композиций(й) для обнаружения, и предпочтительно пригодным образом распределена на аликвоты. Наборы, описанные в настоящем описании, как правило, включают средство для прочного удержания флакона(ов) для коммерческой продажи, например, формованные инжекцией или раздувом пластмассовые контейнеры, в которых находится желаемый флакон(ы). Когда в набор включена радиоактивная метка, хромогенная, флуорогенная или другая поддающаяся обнаружению метка или средство обнаружения, средство для мечения может быть предоставлено либо в том же контейнере, что и сама композиция для обнаружения, либо альтернативно оно может быть помещено в другой контейнер, в который эта вторая композиция может быть помещена и пригодным образом распределена аликвотами. Альтернативно реагент для обнаружения можно получать в одном контейнере и в большинстве случаев набор также обычно включает средство для прочного удержания флакона(ов) для коммерческой продажи и/или удобного упаковывания и доставки.
Также предусматриваются устройство или аппарат для проведения способов обнаружения или мониторинга, описанных в настоящем описании. Такое устройство может включать камеру или пробирку, в которую может быть помещен образец, систему для обработки жидкостей, необязательно включающую клапаны или насосы для прямого потока образца через устройство, необязательно фильтры для отделения плазмы или сыворотки от крови, камеры для смешения для добавления средств для улавливания или реагентов для обнаружения, и необязательно устройство для обнаружения для определения количества поддающейся обнаружению метки, связанной с иммунокомплексом средства для улавливания. Поток образца может быть пассивным (например, вследствие капиллярных, гидростатических или других сил, которые не требуют дополнительного манипулирования устройством после нанесения образца) или активным (например, с использованием силы, генерируемой с помощью механических насосов, электроосмотических насосов, центрифужной силы или увеличенного давления воздуха), или с помощью комбинации активной и пассивной сил.
Следующие варианты осуществления также относятся к процессору, считываемого компьютером устройства памяти и стандартной программе, сохраненной на считываемом компьютером устройстве памяти и адаптированной для выполнения на процессоре для выполнения любого из способов, описанных в настоящем описании. Примеры считываемых компьютером систем, оборудования и/или конфигураций включают персональные компьютеры, серверные компьютеры, ручные или портативные устройства, мультипроцессорные системы, системы на основе микропроцессора, дополнительные внешние устройства, программируемую потребительскую электронику, сетевые PC, миникомпьютеры, универсальные вычислительные машины, среды распределенных вычислений, которые включают любую из описанных выше систем или устройств, или любые другие системы, известные в данной области.
ОБЩИЕ СПОСОБЫ
Стандартные способы в молекулярной биологии описаны в Sambrook, Fritsch and Maniatis (1982 & 1989 2nd Edition, 2001 3rd Edition) Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY; Sambrook and Russell (2001) Molecular Cloning, 3rd ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY; Wu (1993) Recombinant ДНК, Vol. 217, Academic Press, San Diego, CA). Стандартные способы также представлены в Ausbel, et al. (2001) Current Protocols in Molecular Biology, Vols.1-4, John Wiley and Sons, Inc. New York, NY, где описано клонирование в бактериальных клетках и мутагенез ДНК (Vol.1), клонирование в клетках млекопитающих и дрожжах (Vol.2), гликоконъюгаты и экспрессия белка (Vol.3) и биоинформатика (Vol.4).
Способы очистки белков, включающие иммунопреципитацию, хроматографию, электрофорез, центрифугирование и кристаллизацию, описаны (Coligan, et al. (2000) Current Protocols in Protein Science, Vol. 1, John Wiley и Sons, Inc., New York). Химический анализ, химическая модификация, посттрансляционная модификация, продуцирование слитых белков, гликозилирование белков, описан (см., например, Coligan, et al. (2000) Current Protocols in Protein Science, Vol. 2, John Wiley and Sons, Inc., New York; Ausubel, et al. (2001) Current Protocols in Molecular Biology, Vol. 3, John Wiley и Sons, Inc., NY, NY, pp. 16.0.5-16.22.17; Sigma-Aldrich, Co. (2001) Products for Life Science Research, St. Louis, MO; pp. 45-89; Amersham Pharmacia Biotech (2001) BioDirectory, Piscataway, N.J., pp. 384-391). Продуцирование, очистка и фрагментация поликлональных и моноклональных антител описаны (Coligan, et al. (2001) Current Protcols in Immunology, Vol. 1, John Wiley and Sons, Inc., New York; Harlow and Lane (1999) Using Antibodies, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY; Harlow and Lane, выше). Стандартные способы охарактеризации взаимодействий лиганд/рецептор являются доступными (см., например, Coligan, et al. (2001) Current Protocols in Immunology, Vol. 4, John Wiley, Inc., New York).
Можно получать моноклональные, поликлональные и гуманизированные антитела (см., например, Sheperd and Dean (eds.) (2000) Monoclonal Antibodies, Oxford Univ. Press, New York, NY; Kontermann and Dubel (eds.) (2001) Antibody Engineering, Springer-Verlag, New York; Harlow and Lane (1988) Antibodies A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, pp. 139-243; Carpenter, et al. (2000) J. Immunol. 165:6205; He, et al. (1998) J. Immunol. 160: 1029; Tang et al. (1999) J. Biol. Chem. 274:27371-27378; Baca et al. (1997) J. Biol. Chem. 272: 10678-10684; Chothia et al. (1989) Nature 342:877-883; Foote and Winter (1992) J. Mol. Biol. 224:487-499; патент США № 6329511).
Альтернативой гуманизации является применение библиотек антител человека, экспонированных на фаге, или библиотек антител человека в трансгенных мышах (Vaughan et al. (1996) Nature Biotechnol. 14:309-314; Barbas (1995) Nature Medicine 1:837-839; Mendez et al. (1997) Nature Genetics 15: 146-156; Hoogenboom и Chames (2000) Immunol. Today 21:371-377; Barbas et al. (2001) Phage Display: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York; Kay et al. (1996) Phage Display of Peptides and Proteins: A Laboratory Manual, Academic Press, San Diego, CA; de Bruin et al. (1999) Nature Biotechnol. 17:397-399).
Очистка антигена не является необходимой для получения антител. Животных можно иммунизировать клетками, несущими представляющий интерес антиген. Затем спленоциты можно выделять из иммунизированных животных и спленоциты можно подвергать слиянию с клеточной линией миеломы с получением гибридомы (см., например, Meyaard et al. (1997) Immunity 7:283-290; Wright et al. (2000) Immunity 13:233-242; Preston et al, выше; Kaithamana et al. (1999) J. Immunol. 163:5157-5164).
Антитела можно конъюгировать, например, с низкомолекулярным лекарственным средством, ферментами, липосомами, полиэтиленгликолем (PEG). Антитела являются пригодными для терапевтических, диагностических целей и для применения в наборе, и включают антитела, связанные, например, с красителями, радиоизотопами, ферментами или металлами, например, коллоидным золотом (см., например, Le Doussal et al. (1991) J. Immunol. 146: 169-175; Gibellini et al. (1998) J. Immunol. 160:3891-3898; Hsing and Bishop (1999) J. Immunol. 162:2804-2811; Everts et al. (2002) J. Immunol. 168:883-889).
Способы проточной цитометрии, включая активированную флуоресценцией сортировку клеток (FACS), являются доступными (см., например, Owens, et al. (1994) Flow Cytometry Principles for Clinical Laboratory Practice, John Wiley and Sons, Hoboken, NJ; Givan (2001) Flow Cytometry, 2nd ed.; Wiley-Liss, Hoboken, NJ; Shapiro (2003) Practical Flow Cytometry, John Wiley and Sons, Hoboken, NJ). Флуоресцентные реагенты, пригодные для модификации нуклеиновых кислот, включая праймеры и зонды нуклеиновой кислоты, полипептиды и антитела, для применения, например, в качестве диагностических реагентов, являются доступными (Molecular Probes (2003) Catalogue, Molecular Probes, Inc., Eugene, OR; Sigma-Aldrich (2003) Catalogue, St. Louis, MO).
Стандартные способы гистологии иммунной системы описаны (см., например, Muller-Harmelink (ed.) (1986) Human Thymus: Histopathology and Pathology, Springer Verlag, New York, NY; Hiatt, et al. (2000) Color Atlas of Histology, Lippincott, Williams, and Wilkins, Phila, PA; Louis, et al. (2002) Basic Histology: Text and Atlas, McGraw-Hill, New York, NY).
Пакеты программ, базы данных для определения, например, антигенных фрагментов, лидерных последовательностей, фолдинга белков, функциональных доменов, участков гликозилирования и выравнивания последовательностей являются доступными (см., например, GenBank, Vector NTI® Suite (Informax, Inc, Bethesda, MD); GCG Wisconsin Package (Accelrys, Inc., San Diego, CA); DeCypher® (TimeLogic Corp., Crystal Bay, Nevada); Menne, et al. (2000) Bioinformatics 16: 741-742; Menne, et al. (2000) Bioinformatics Applications Note 16:741-742; Wren, et al. (2002) Comput. Methods Programs Biomed. 68: 177-181; von Heijne (1983) Eur. J. Biochem. 133: 17-21; von Heijne (1986) Nucleic Acids Res. 14:4683-4690).
ПРИМЕРЫ
Пример 1. Получение и скрининг гибридом против PD-L1
Мышей Balb/C иммунизировали слитым белком PD-L1 человека-Fc (R&D Systems®, каталожный номер № 156-B7-100) в адъюванте. Этот слитый белок содержит внеклеточный домен PD-L1 (Phel9-Thr239), слитый с фрагментом IgG1 человека (Pro100-Lys 300). После 12 иммунизаций лимфатические узлы двух мышей с высокими титрами к PD-L1 человека собирали и проводили электрослияние с получением двух партий гибридом, которым были присвоены лабораторные обозначения MEB033 и MEB037.
Супернатанты партий гибридом MEB033 и MEB037 подвергали скринингу для идентификации гибридом, которые продуцируют антитела к PD-L1 человека. В скрининге использовали белковый ELISA белка для связывания с белком hPD-L1-Fc человека; и клеточные ELISA для связывания стабильных трансформантов PD-L1 человека-CHO и родительских клеток CHO в качестве отрицательного контроля. Супернатанты из 88 исследованных клонов MEB037 и из 23 исследованных клонов гибридомы MEB033, были положительными в тестах присутствия антитела против PD-L1 (данные не представлены), и их образцы исследовали в отношении реактивности IHC на срезах ткани, обработанных FFPE, из нормальных миндалевидных желез человека (данные не представлены).
Из 88 клонов только 11 клонов из партии MEB037 обеспечивали профили окрашивания с достаточной интенсивностью и кажущейся специфичностью для обоснования дальнейшей оценки, исходя из сравнения с профилями окрашивания, полученными с помощью коммерчески доступных антител против PD-L1, приведенных в таблице ниже:
Когда авторы изобретения сравнили различные профили окрашивания, полученные с помощью экспериментальных антител, с профилями, полученными с помощью этих коммерчески доступных антител против PD-L1 человека, они выявили значимые отличия между профилями окрашивания, включающие локализацию красителя и типы окрашенных клеток. В попытках объяснить эти отличия, авторы изобретения провели ряд дополнительных экспериментов и открыли, что ни одно из этих коммерчески доступных антител не обеспечивало комбинацию признаков, требуемую для применения в IHC в отношении экспрессии PD-L1 в срезах, обработанных FFPE: (1) чувствительность - способность обнаруживать нормальную физиологическую экспрессию в тканях положительного контроля (например, миндалевидная железа человека), а также экспрессию в опухолевой ткани (например, образцы меланомы человека); (2) специфичность - набор характеристик окрашивания должен коррелировать с известным анатомическим/клеточным распределением PD-L1 и окрашивание должно быть нейтрализующимся; и (3) надежность - от небольшого варьирования до отсутствия окрашивания в профилях окрашивания при использования для анализа "дублированных" срезов тканей.
Например, авторы изобретения обнаружили, что антитело Sigma/ProSco PRS4059 продемонстрировало множество полос на лизате миндалевидной железы, ни для одной из которых не было подтверждено, что она соответствует PD-L1, не окрашивало клеточные линии меланомы LOX, для которых было показано, что они являются положительными по PD-L1 с помощью проточной цитометрии, и не нельзя было различить положительные и отрицательные клеточные линии с помощью IHC.
Некоторые из этих данных представлены на фиг. 4, на которой показано, что имуногистохимическое окрашивание срезов миндалевидной железы идентифицировало 22C3 и 20C3 в качестве двух антител с необычными и полезными иммуногистохимическими свойствами на ткани, обработанной FFPE, по сравнению с антителом PRS4059, идентифицированным Gadiot et al (выше) в качестве единственного подходящего кандидата среди 15 антител против PD-L1 человека для обнаружения экспрессии PD-L1 в срезах ткани, обработанных FFPE. В экспериментах, проведенных авторами изобретения, антитело Prosci (PRS4059, партия 40590604, использованное в концентрации 0,4 мг/мл, первичное, за которым следовала полимерная система обнаружения кролика (DAKO Envision)) окрашивало все гемопоэтические линии дифференцировки в миндалевидной железе с равной интенсивностью (фиг. 4A), в то время как антитело 22C3 селективно окрашивало эпителий крипт миндалевидной железы и фолликулярные миелоидные клетки CD68+, морфология которых соответствует макрофагам (фиг. 4B). По существу тот же набор характеристик окрашивания наблюдали в случае антитела 20C3 (данные не представлены). Более того, 22C3 и 20C3 показали единообразное различие в интенсивности окрашивания между этими двумя отдельными популяциями клеток, причем интенсивность окрашивания эпителия крипт значительно превышала интенсивность окрашивания фолликулярных макрофагов. Все три антитела можно было нейтрализовать посредством предварительной инкубации с антигеном PD-L1, что указывает на то, что реактивность опосредуется антигенсвязывающим доменом (CDR).
Пример 2. Оценка качества антител против PD-L1 20C3 и 22C3
В этом примере описаны дополнительные эксперименты, которые проводили для оценки пригодности антител 20C3 и 22C3 для применения в анализах IHC срезов тканей, обработанных FFPE.
В одном эксперименте оценивали способность этих двух антител обнаруживать диапазон экспрессии белка PD-L1 человека (hPD-L1) в анализе IHC нормальных срезов миндалевидной железы человека, обработанных FFPE, и репрезентативные изображения для 22C3 представлены на фиг. 5A. Иммуногистохимическое окрашивание посредством 22C3 в высокой степени метит эпителий крипт миндалевидной железы, а также демонстрирует окрашивание от слабого до умеренного CD68+ фолликулярной миелоидной популяции (предположительно макрофаги). Оба антитела (20C3, данные не представлены) метят клетки с точно определенной картиной мембраны/клеточной поверхности в этих двух типах клеток. Соответствие экспрессии hPD-L1 в миндалевидной железе (т.е. ограничение окрашивания IHC двумя популяциями клеток (эпителий крипт и фолликулярные макрофаги) было подтверждено с помощью независимых способов (гибридизация in-situ [ISH] для мРНК hPD-L1) на соседних срезах ткани миндалевидной железы, обработанной FFPE. Кроме того, дифференциальная экспрессия белка hPD-L1 при оценке с помощью IHC (эпителий крипт >> фолликулярные макрофаги) соответствует относительному содержанию мРНК hPD-L1, наблюдаемому с помощью ISH.
В другом эксперименте оценивали специфичность связывания 20C3 и 22C3 для клеток, экспрессирующих hPD-L1. Клетки HT144, о которых было известно, что они являются отрицательными в отношении экспрессии hPD-L1 в анализе мРНК (кПЦР), и клетки меланомы LOX, о которых било известно, что они экспрессируют высокие уровни мРНК hPD-L1 (кПЦР), окрашивали посредством 1 микрограмм/мл очищенного IgG мыши из семи гибридом, полученных в экспериментах, описанных в примере 1, выше. Также использовали несоответствующее изотипическое контрольное антитело мыши в идентичной концентрации. Флуоресцентно меченное вторичное антитело против антител мыши использовали для обнаружения первичных антител мыши. После окрашивания и повторного промывания клетки анализировали проточной цитометрией со средней интенсивностью флуоресценции, вычисленной для популяции (в совокупности >10000 событий). Результаты представлены на фиг. 6.
20C3 и 22C3, а также другие антитела против hPD-L1, использовали в качестве реагентов для проточной цитометрии для обнаружения hPD-L1 на клеточной поверхности. Значительный сдвиг вправо кривых обеих гистограмм для 20C3 и 22C3 (фиг. 6A) по сравнению с изотипическим контрольным антителом отражает селективное обнаружение hPD-L1 на положительной по hPD-L1 клеточной линии меланомы LOX. Средняя интенсивность флуоресценции, исходя из этих и других гистограмм в этом анализе, представлена на фиг. 6B. Селективность связывания 20C3 и 22C3 далее подтверждается отсутствием значительного связывания (т.е. MFI 22C3 и 20C3, сравниваемых с изотипипическим антителом) на отрицательной клеточной линии HT144. Напротив, данные на фиг. 6B показали, что как 20C3, так и 22C3, обеспечивают по меньшей мере в 10-кратное увеличение MFI по сравнению с изотипическим авнтителом на положительной по hPD-L1 клеточной линии меланомы LOX. Таким образом, как 20C3, так и 22C3 (в дополнение к клонам 5F9, 7C8, 13D2 и 31D3) показали селективное связывание с клетками, экспрессирующими hPD-L1, посредством оценки с помощью проточной цитометрии.
В другом эксперименте оценивали способность антитела 22C3 обнаруживать экспрессию hPD-L1 на полученных способами инженерии клеточных линиях и клеточных линиях человека. Клеточные линии яичника китайского хомячка (CHO), которые являются отрицательными по hPD-L1, трансфицировали экспрессирующим вектором, кодирующим PD-L1 человека, с получением модифицированной способами инженерии контрольной клеточной линии. Как показано на фиг. 7B, иммуногистохимическое окрашивание посредством 22C3 фиксированных формалином залитых парафином (FFPE) клеточных осадков исходной клеточной линии CHO (отрицательный контроль) и трансфицированной клеточной линии CHO (положительный контроль) демонстрирует соответствующее положительное и отрицательное окрашивание.
Дополнительные осадки клеточных линий человека, обработанные FFPE (A375, HS578T и меланома LOX) окрашивали 22C3, и они продемонстрировали диапазон профилей и интенсивностей окрашивания, как показано на фиг. 7B. Окрашивание 22C3 было высоким и единообразным на клетках меланомы Lox, но продемонстрировало только редкие единичные клетки в клеточных осадках A375 и HS578T. Сходное окрашивание наблюдали в случае антитела 20C3 (данные не представлены). Уровни экспрессии hPD-L1, обнаруженные в этих 3 клеточных линиях с помощью 22C3 в анализе IHC, высоко коррелировали с уровнями мРНК PD-L1 в этих клеточных линиях при оценке с использованием кПЦР с мРНК убиквитина в качестве исходного уровня.
Селективное связывание и относительную аффинность 22C3 в отношении экспрессирующих hPD-L1 клеток оценивали в клеточном эксперименте ELISA. Несколько клеточных линий (клетки hPDL1-CHOK1, родительские клетки CHOK1, клетки hPDL2-CHOKl и клетки LOX) высевали в индивидуальные лунки покрытых коллагеном 96-луночных планшетов и выращивали до смыкания монослоя. Среду удаляли и заменяли средой CHOK1 (DMEM/F12, содержащая 10% BCS), содержавшей первичное антитело в возрастающих концентрациях от 1,4 до 3000 нг/мл. Использовали следующие первичные антитела: две различных полученных партии каждого из антител 20C3 и 22C3, и в качестве контролей служили изотипическое IgG1-антитело мыши против PD-L1 (BioLegend) и против PD-L2. Первичное антитело инкубировали в течение 1 ч при 37°C, промывали 3X посредством PBS/0,01% Tween 20 и добавляли вторичное антитело, представляющее собой конъюгат антитела козы против IgG человека, специфичного к Fc, с HRP (Southern Biotech, каталожный номер #1030-05) в разведении 1:2000 в среде CHOK1. Вторичное антитело инкубировали в течение 1 ч при 37°C и промывали 5X, как описано выше. ELISA проводили с использованием TMB, останавливали 0,1 Н фосфорной кислотой, и поглощение считывали при 450 нм с вычитанием фонового уровня при 650 нм. Результаты, которые показаны на фиг. 8, показали селективное связывание 22C3 и 20C3 с клетками, экспрессирующими hPD-L1, причем аффинность связывания 22C3 превышает аффинность связывания 20C3 в отношении как модифицированных способами инженерии клеток hPD-L1 CHOK1, так и клеток LOX.
Сходный эксперимент ELISA проводили для оценки того, связывается ли любое из антител 20C3 или 22C3 с PD-L1 мыши. Не наблюдали значительного связывания любого из антител с PD-L1 мыши (данные не представлены).
Проводили конкурентные анализы связывания ("перекрестное блокирование") антител между различными парами антител против hPD-L1, идентифицированных в экспериментах, описанных в примере 1. В анализах используется платформа ForteBio® Octet®, которая основана на интерферометрии биослоя. В кратком изложении, этот способ измеряет связывание исходного антитела (mAb1) с поверхностью вершины биосенсора в качестве сдвига длины волны ( Δλ), вследствие увеличения связавшимся антителом оптической плотности (ось Y) на вершине биосенсора с течением времени (ось X). Вершина состоит из сенсора с антителом против huIgG, с которым связывается hPD-L1-Fc. Изменение оптической плотности при связывании антитела против PD-L1 отражается в восходящем наклоне кривой, начинающемся в первой вертикальной пунктирной красной линии (см. графики на фиг. 9A, 9B и 9C), который отражает добавление насыщающей концентрации mAb1 (10 микрограмм/мл) в раствор. После обеспечения уравновешивания (~1000 секунд), в раствор для анализа инжектируют второе антитело (указано второй вертикальной красной пунктирной линией на фиг. 9A, 9B и 9C). Связывание второго mAb2, показанное дополнительным смещением кривой, указывает на то, что эти два антитела связываются с неперекрывающимися эпитопами, в то время как небольшое смещение кривой или его отсутствие указывает на то, что эти два антитела связываются с перекрывающимися или идентичными эпитопами.
В целом, результаты показали, что связывание 22C3 конкурирует с дополнительным связыванием всех других клонов антител против hPDL-1, исследованных в качестве mAb2, за исключением 5H9 (фиг. 9A). Аналогично, 20C3 также не конкурирует с 5H9 за связывание, но демонстрирует промежуточную степень дополнительного связывания с 22C3, а также 4B7 (фиг. B). Взятые вместе, эти данные указывают на то, что 20C3 и 22C3 связывают перекрывающиеся эпитопы.
Способность антитела 22C3 выявлять диапазон экспрессии hPD-L1 в различных типах опухолей оценивали путем проведения анализа IHC на срезах, обработанных FFPE, полученных из следующих опухолей: опухоль мочевого пузыря, пищевода, головы и шеи, почки, HCC, молочной железы, легкого, яичника и желудка. Предварительный скрининг реактивности 22C3 с этими срезами опухолевой ткани проводили с использованием полуколичественной "гештальт"-интерпретации степени окрашивания. Как представлено на фиг. 10, 22C3 способно обнаруживать диапазон экспрессии PD-L1 от по существу, отсутствия окрашивания (показатель = 0) до выраженной высокой экспрессии (показатель = 4), демонстрируя применимость анализа IHC с 22C3 для исследования дополнительных типов опухолей, которые могут отвечать на противодействие иммунодепрессивным взаимодействиям PD-1/PD-L1.
Применимость антитела 22C3 для стратификации пациентов, которые более вероятно будут отвечать на терапию, которая блокирует взаимодействие PD-1 и PD-L1, оценивали в исследованиях с использованием иммуногистохимической оценки с помощью 22C3 архивных образцов, полученных от 18 пациентов с меланомой, включенных в испытание фазы 1 (P001) с MK-3475 - терапевтическим антителом против PD1, разработанным Merck and Co., Inc. Случаи оценивались двумя патологами и их относили к "положительным", "отрицательным" или "неоднозначным", и репрезентативные изображения, отражающие эти три категории, представлены на фиг. 11A. Согласованность между патологами в этом наборе образцов (n=18) составляла 100%.
Клинические ответы оценивали с использованием критериев обусловленного иммунной системой ответа (irRC), и они коррелировали с результатами IHC. Для этого анализа "неоднозначные случаи" считали отрицательными, что дает чувствительность анализа 72% и специфичность 86%. Результаты, которые представлены на фиг. 11B, указывают на то, что иммуногистохимическое окрашивание посредством 22C3 ткани, обработанной FFPE, обладает применимостью в качестве биомаркера отбора пациентов.
Исходя из результатов экспериментов, описанных выше, авторы изобретения, описанного в настоящем описании, определили, что антитела, продуцируемые двумя из 88 экспериментальных гибридом - MEB037.20C3.138 и MEB037.20C3.116 - имели требуемую комбинацию чувствительности, специфичности и надежности, чтобы их можно было рассматривать для разработки в качестве кандидатов для реактивных к FFPE диагностических реагентов IHC.
Пример 3. Картирование эпитопа на PD-L1 человека для антитела против PD-L1 22C3
Картирование эпитопа HDX-MS проводили с использованием антитела 22C3 и белка PD-L1-His, который содержал внеклеточный домен зрелого PD-L1 человека (SEQ ID NO:38), слитый с 11-мерной гистидиновой меткой. Сегменты с 156 по 178 и с 196 по 206 на внеклеточном домене PD-L1 человека (SEQ ID NO:38) продемонстрировали высокую защиту (отличие в среднем уровне дейтерирования >10%) при связывании с антителом 22C3. Кроме того, сегменты с 3 по 9, с 10 по 13, с 88 по 93, и с 135 по 147 продемонстрировали пограничную, но все еще значительную защиту (отличие в среднем уровне дейтерирования от 5% до 10%).
Все ссылки, цитированные в настоящем описании, включены в настоящее описание в качестве ссылок, как если бы было конкретно и индивидуально указано, что каждая индивидуальная публикация, запись в базе данных (например, последовательности Genbank или записи GeneID), патентная заявка или патент включены в качестве ссылок. Заявители подразумевают, что утверждение о включении в качестве ссылки, согласно 37 C.F.R. § 1.57(b)(1), относится к каждой индивидуальной публикации, записи в базе данных (например, последовательности Genbank или записи GeneID), патентной заявке или патенту, каждый из которых явно указан в соответствии с 37 C.F.R. § 1.57(b)(2), даже если такое цитирование не находится непосредственно рядом со специальным утверждением о включении в качестве ссылки. Включение специальных утверждений о включении в качестве ссылки, при их наличии, в описании никоим образом не уменьшает значение этого общего утверждения о включении в качестве ссылки. Цитирование ссылок в настоящем описании не предназначено для допущения того, что ссылка является соответствующим уровнем техники, а также оно не предназначено для допущения в отношении содержимого или даты этих публикаций или документов.
Объем настоящего изобретения не ограничивается конкретными вариантами осуществления, описанными в настоящем описании. Действительно, различные модификации изобретения, в дополнение к модификациям, описанным в настоящем описании, будут очевидными специалистам в данной области из предшествующего описания и прилагаемых чертежей. Подразумевают, что такие модификации входят в объем прилагаемой формулы изобретения.
Представленное выше описание считается достаточным, чтобы позволить специалисту в данной области осуществлять изобретение на практике. Различные модификация изобретения в дополнение к модификациям, показанным и описанным в настоящем описании, станут очевидными специалистам в данной области из представленного выше описания и входят в объем прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА ПРОТИВ PD-1 И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2788616C2 |
| НОВЫЕ АНТИТЕЛА ПРОТИВ PD-L1 | 2016 |
|
RU2736151C2 |
| ЛЕЧЕНИЕ РАКА КОМБИНАЦИЕЙ АНТАГОНИСТА PD-1 И ДИНАЦИКЛИБА | 2014 |
|
RU2705795C2 |
| КАНИНИЗИРОВАННЫЕ МЫШИНЫЕ АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ PD-1 | 2014 |
|
RU2676158C1 |
| НОВЫЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА К БЕЛКУ ПРОГРАММИРУЕМОЙ СМЕРТИ 1(PD-1) | 2016 |
|
RU2757316C2 |
| АНТИТЕЛА ПРОТИВ ЛИГАНДА ПРОГРАММИРУЕМОЙ СМЕРТИ (PD-L1) И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2761638C1 |
| ГУМАНИЗОВАННОЕ АНТИТЕЛО ПРОТИВ PD-1 ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ ПРИМЕНЕНИЕ | 2022 |
|
RU2829508C2 |
| АНТИТЕЛА К TIGIT И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2827445C2 |
| АНТИТЕЛА К PD-1 СОБАК | 2014 |
|
RU2761663C2 |
| АНТИ-PD-L1 АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2783685C2 |



















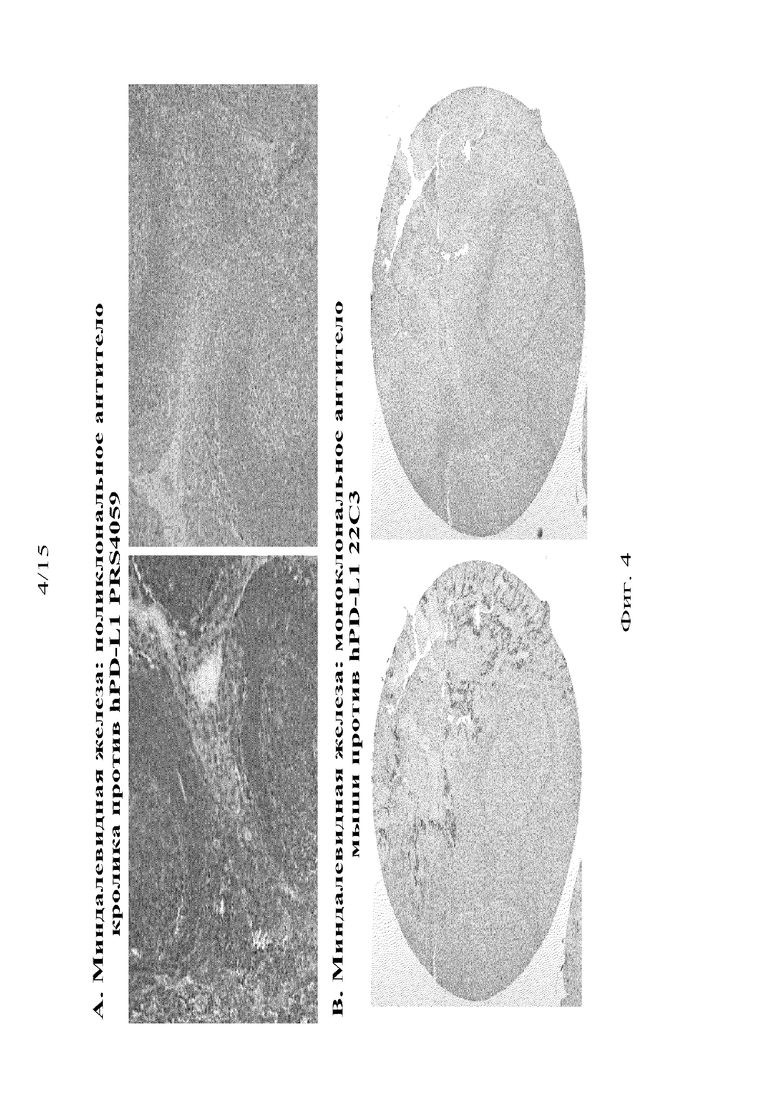
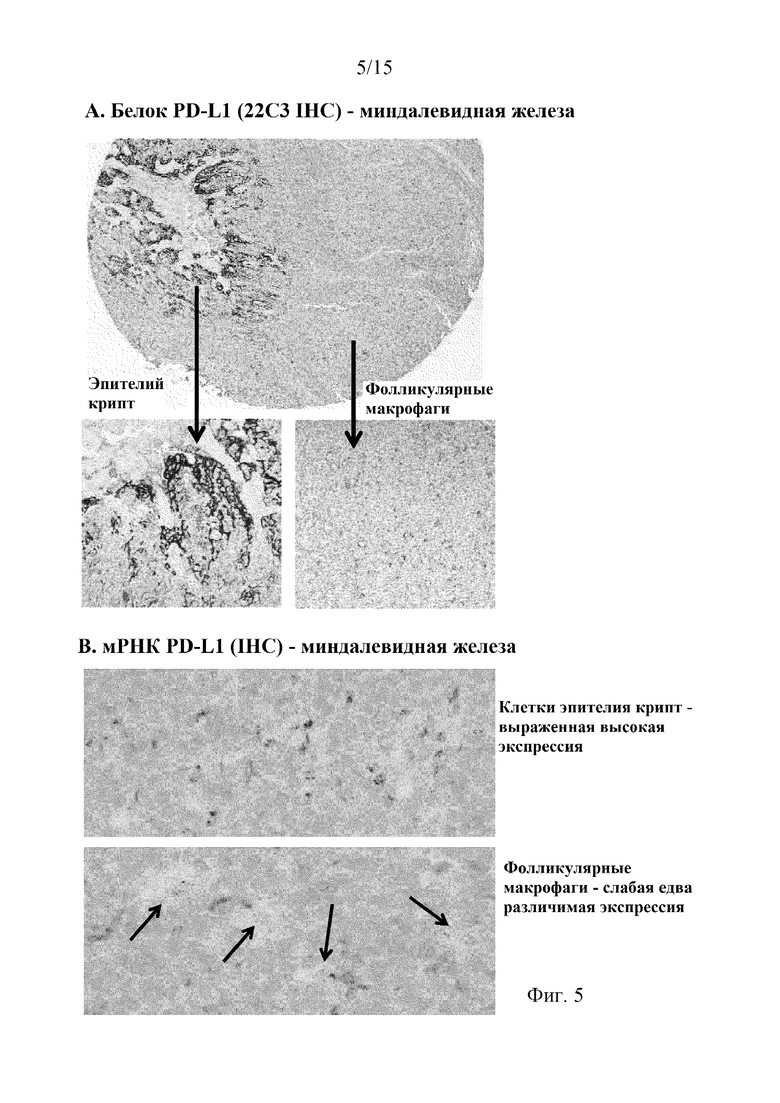
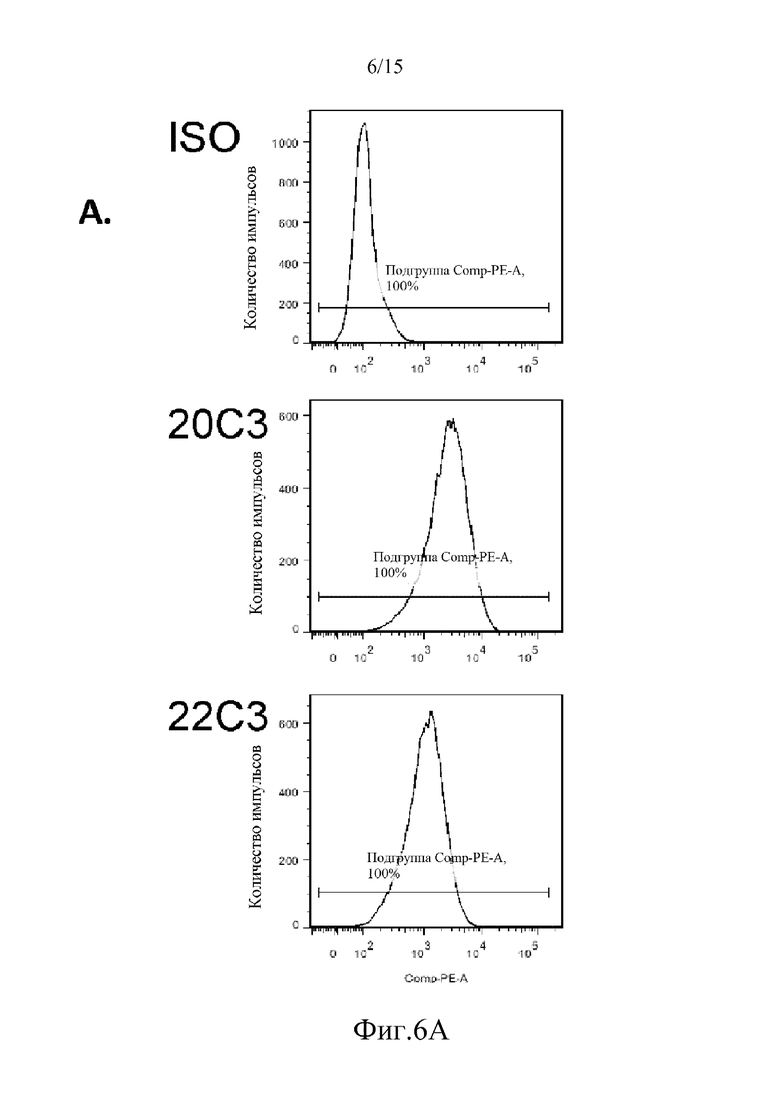

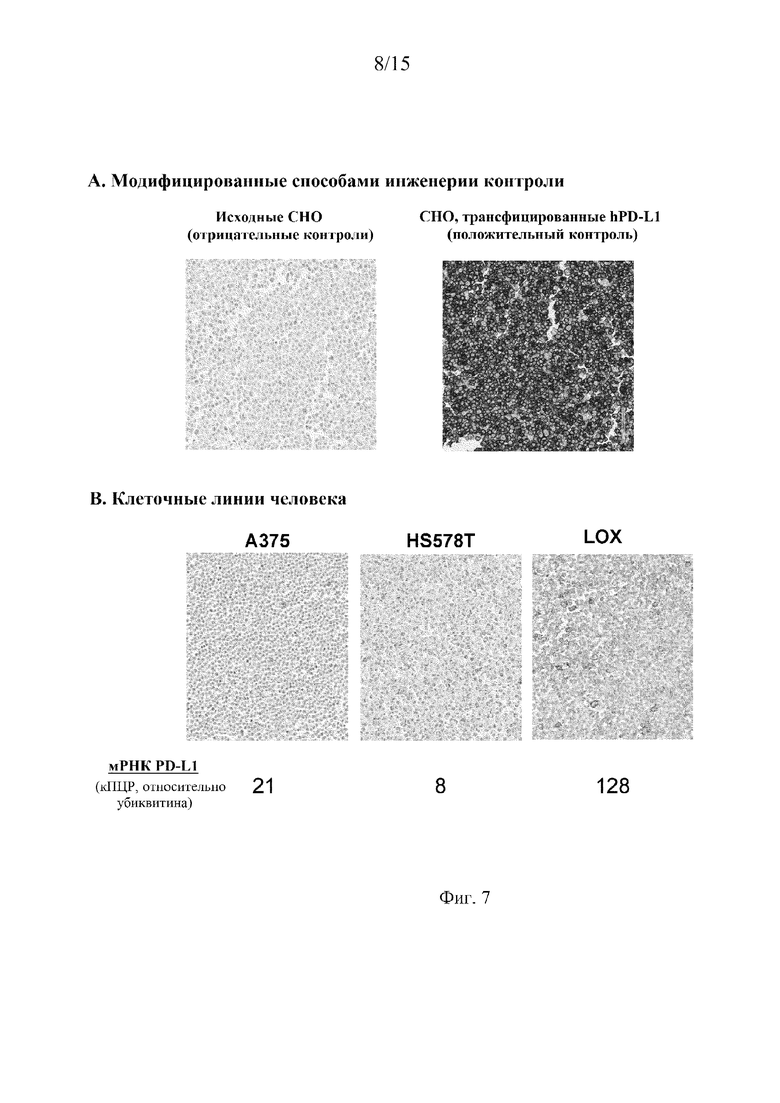
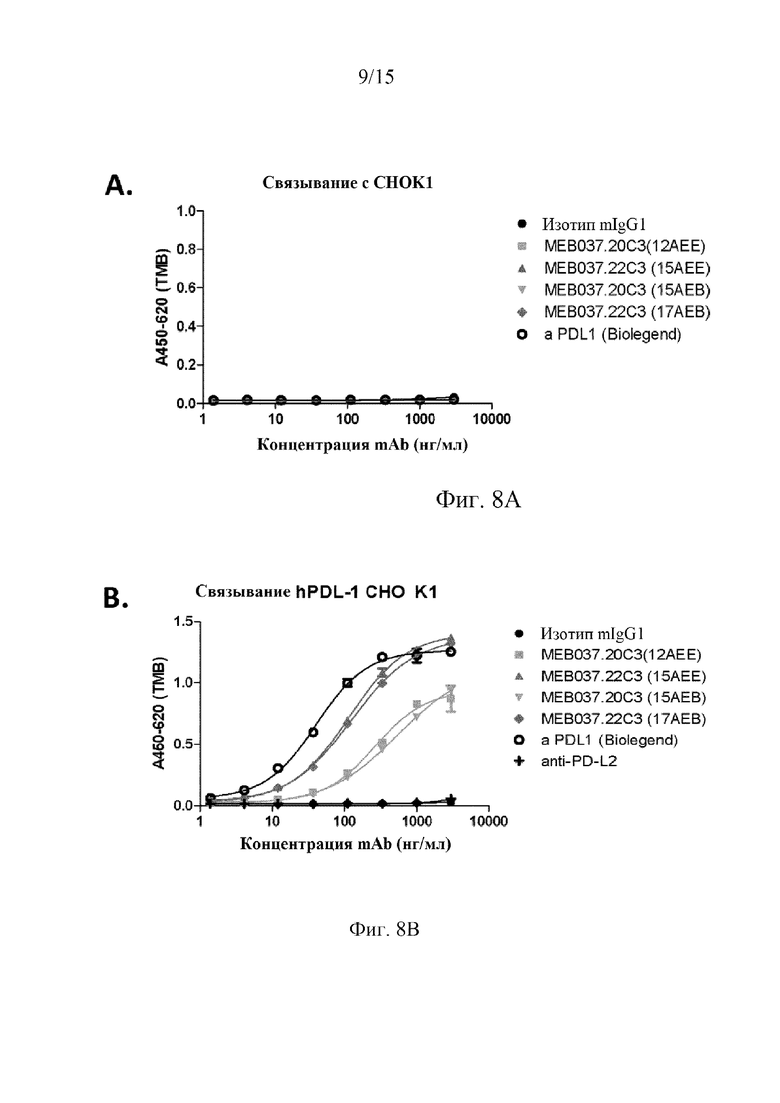
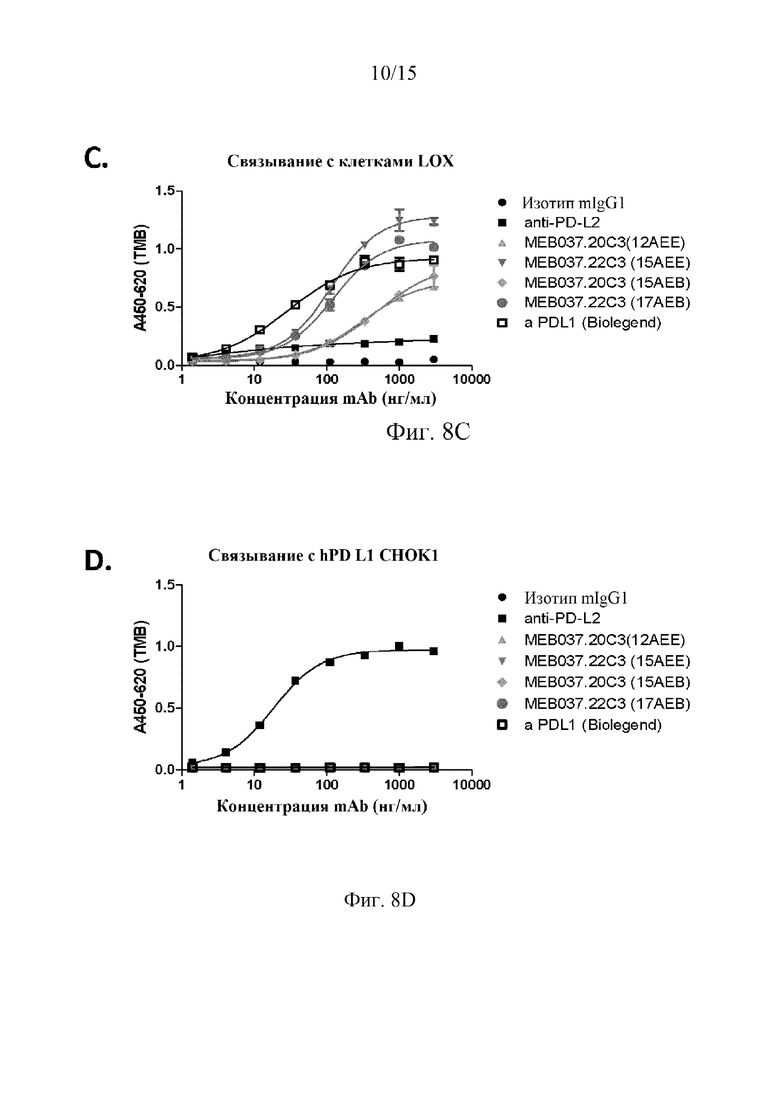
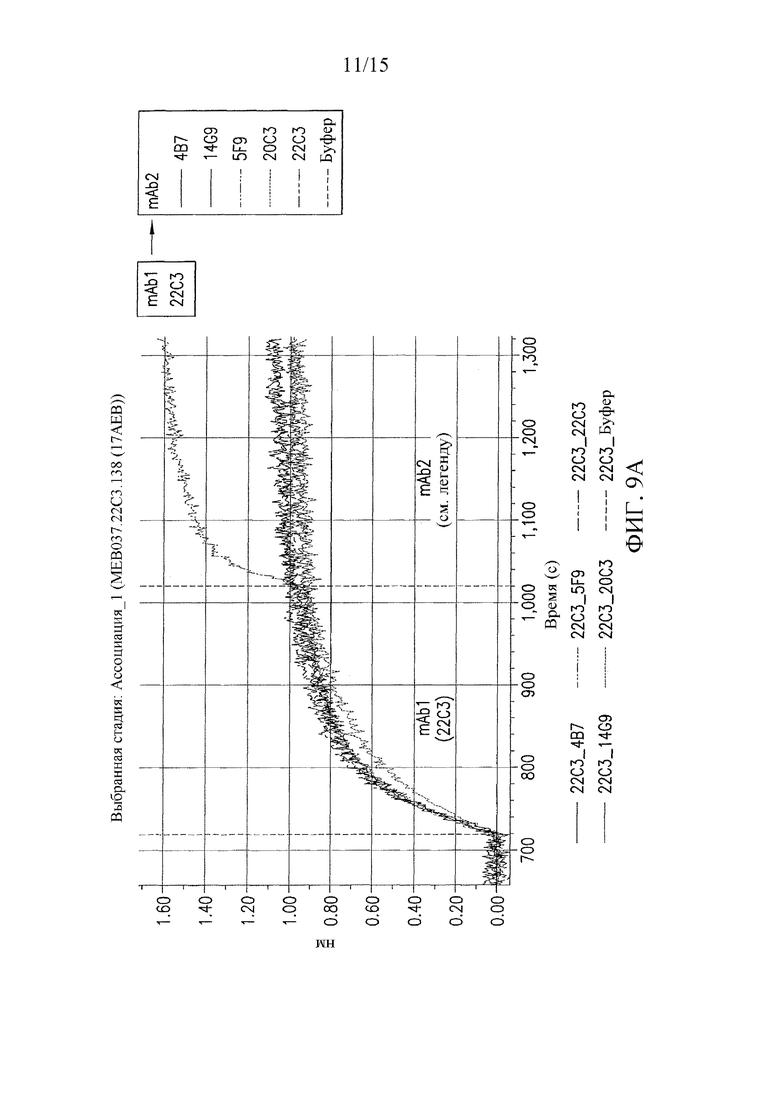
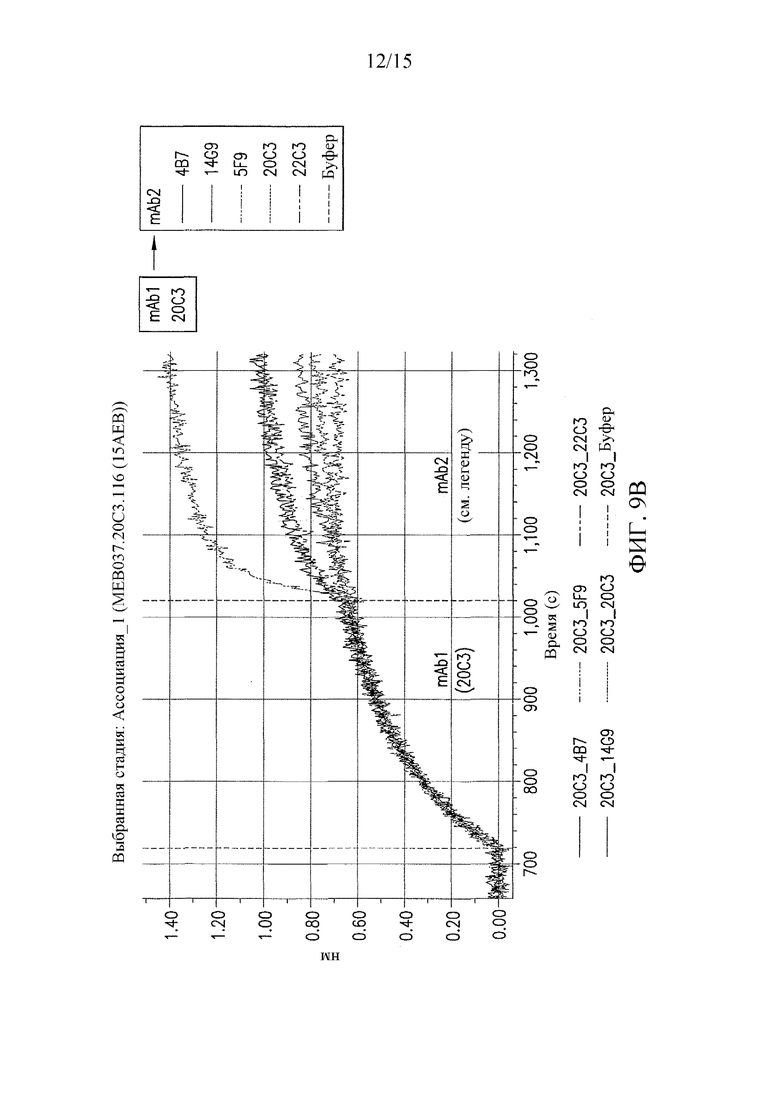
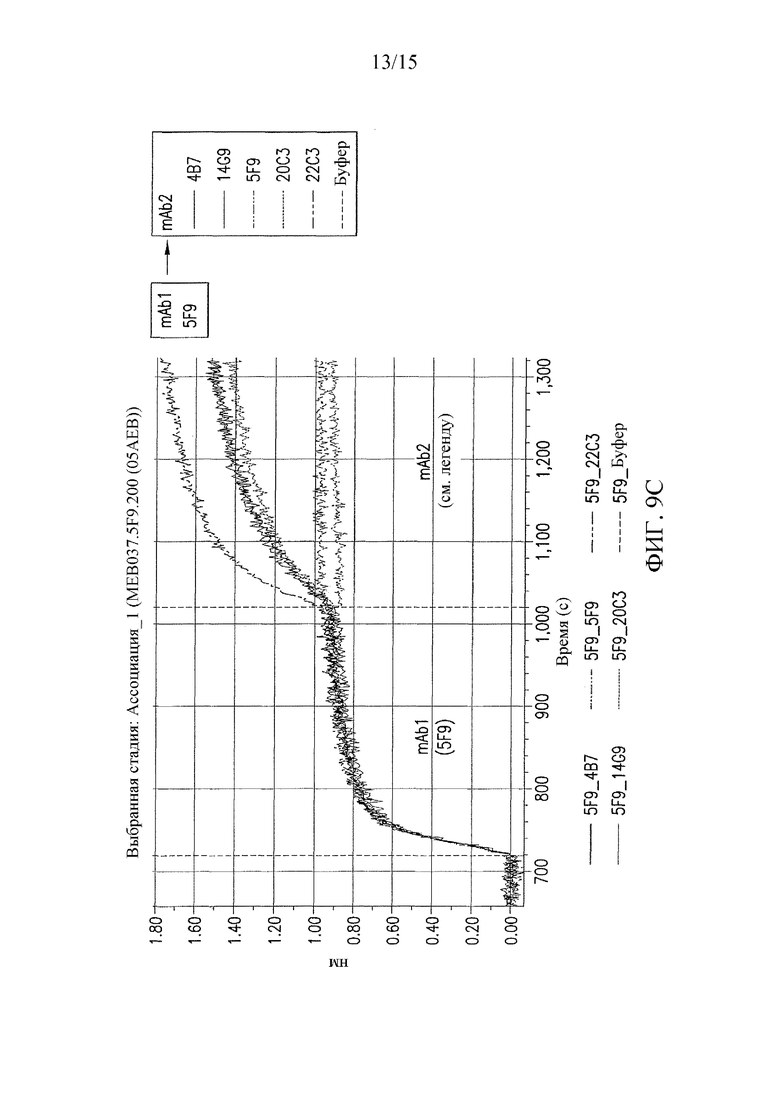

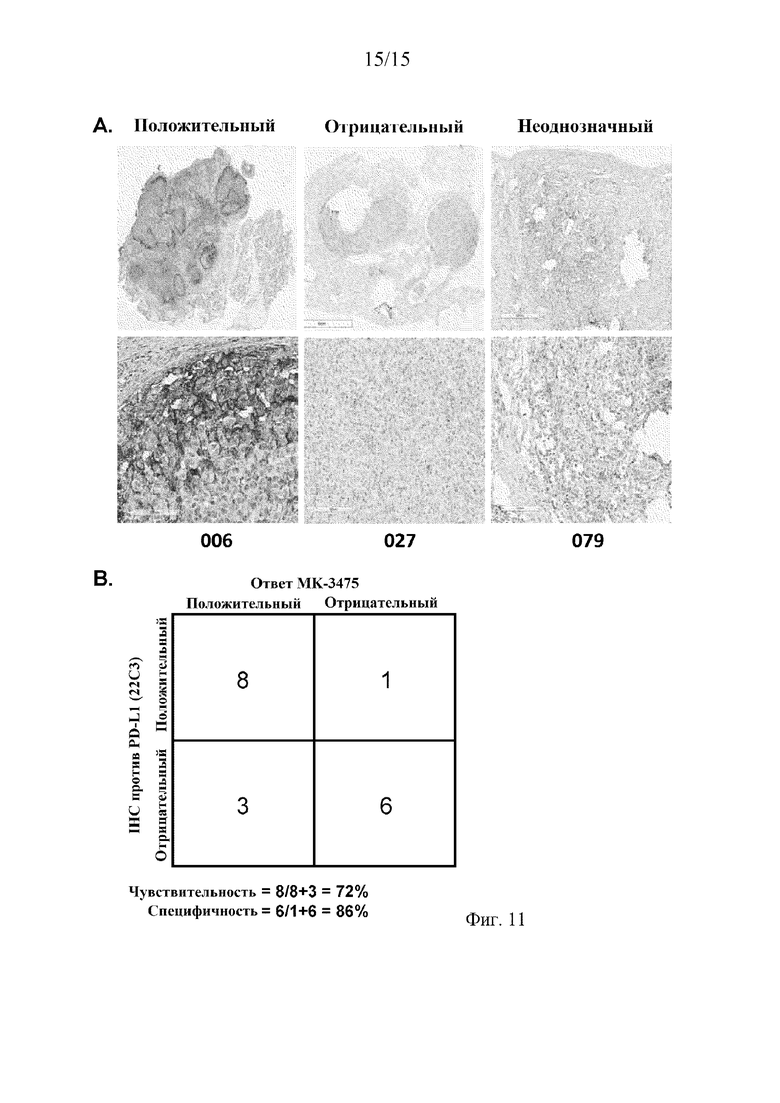
Изобретение относится к выделенным антителам, которые специфически связываются с PD-L1 человека и содержат три CDR легкой цепи: CDRL1, CDRL2 и CDRL3, и три CDR тяжелой цепи: CDRH1, CDRH2 и CDRH3, где CDRL1 представляет собой SEQ ID NO:9, CDRL2 представляет собой SEQ ID NO:2, CDRL3 представляет собой SEQ ID NO:10, CDRH1 представляет собой SEQ ID NO:14 или SEQ ID NO:15, CDRH2 представляет собой SEQ ID NO:16 и CDRН3 представляет собой SEQ ID NO:17 или CDRL1 представляет собой SEQ ID NO:21, CDRL2 представляет собой SEQ ID NO:2, CDRL3 представляет собой SEQ ID NO:22, CDRH1 представляет собой SEQ ID NO:26, CDRH2 представляет собой SEQ ID NO:28 и CDRН3 представляет собой SEQ ID NO:29, а также к антигенсвязывающим фрагментам таких антител. Представлены молекулы нуклеиновых кислот, кодирующие антитела и антигенсвязывающие фрагменты по настоящему изобретению, а также экспрессирующие векторы и клетки-хозяева для их экспрессии, способ анализа экспрессии PD-L1 человека в образце ткани, полученном у человека. Представлен набор для обнаружения комплекса антитела или антигенсвязывающего фрагмента, связавшегося с PD-L1 человека, и диагностическая композиция антител для детекции экспрессии PD-L1. Изобретение позволяет расширить ассортимент средств для детекции экспрессии PD-L1. 9 н. и 6 з.п. ф-лы, 11 ил., 5 табл., 3 пр.
1. Выделенное антитело или его антигенсвязывающий фрагмент, которые специфически связываются с PD-L1 человека и содержат три CDR легкой цепи: CDRL1, CDRL2 и CDRL3, и три CDR тяжелой цепи: CDRH1, CDRH2 и CDRH3, где:
(a) CDRL1 представляет собой SEQ ID NO:9,
CDRL2 представляет собой SEQ ID NO:2,
CDRL3 представляет собой SEQ ID NO:10,
CDRH1 представляет собой SEQ ID NO:14 или SEQ ID NO:15,
CDRH2 представляет собой SEQ ID NO:16 и
CDRН3 представляет собой SEQ ID NO:17 или
(c) CDRL1 представляет собой SEQ ID NO:21,
CDRL2 представляет собой SEQ ID NO:2,
CDRL3 представляет собой SEQ ID NO:22,
CDRH1 представляет собой SEQ ID NO:26,
CDRH2 представляет собой SEQ ID NO:28 и
CDRН3 представляет собой SEQ ID NO:29.
2. Выделенное антитело или его антигенсвязывающий фрагмент по п.1, в котором
три CDR легкой цепи представляют собой SEQ ID NO:9; SEQ ID NO:2 и SEQ ID NO:10 и
три CDR тяжелой цепи представляют собой SEQ ID NO:14, SEQ ID NO:16 и SEQ ID NO:17.
3. Выделенное антитело или его антигенсвязывающий фрагмент по п.1, в котором
три CDR легкой цепи представляют собой SEQ ID NO:21, SEQ ID NO:2 и SEQ ID NO:22 и
три CDR тяжелой цепи представляют собой SEQ ID NO:26, SEQ ID NO:28 и SEQ ID NO:29.
4. Выделенное антитело или его антигенсвязывающий фрагмент по п.1, которое содержит вариабельную область легкой цепи и вариабельную область тяжелой цепи, где:
(a) вариабельная область легкой цепи представляет собой SEQ ID NO:13 и вариабельная область тяжелой цепи представляет собой SEQ ID NO:20;
(b) вариабельная область легкой цепи представляет собой SEQ ID NO:25 и вариабельная область тяжелой цепи представляет собой SEQ ID NO:32, где X в SEQ ID NO:32 представляет собой Q; или
(c) вариабельная область легкой цепи представляет собой SEQ ID NO:25 и вариабельная область тяжелой цепи представляет собой SEQ ID NO:32, где X в SEQ ID NO:32 представляет собой pE.
5. Выделенная нуклеиновая кислота для рекомбинантной экспрессии белка, специфически связывающегося с PD-L1 человека, которая кодирует вариабельную область легкой цепи антитела и вариабельную область тяжелой цепи антитела, в которой:
(a) нуклеиновая кислота кодирует вариабельную область легкой цепи антитела, выбранную из группы, состоящей из SEQ ID NO:33 и SEQ ID NO:35; и
(b) нуклеиновая кислота кодирует вариабельную область тяжелой цепи антитела, выбранную из группы, состоящей из SEQ ID NO:34 и SEQ ID NO:36.
6. Выделенная нуклеиновая кислота для рекомбинантной экспрессии белка, специфически связывающегося с PD-L1 человека, которая кодирует вариабельную область легкой цепи антитела, в которой нуклеиновая кислота кодирует вариабельную область легкой цепи антитела, выбранную из группы, состоящей из SEQ ID NO:33 и SEQ ID NO:35.
7. Выделенная нуклеиновая кислота для рекомбинантной экспрессии белка, специфически связывающегося с PD-L1 человека, которая кодирует вариабельную область тяжелой цепи антитела, в которой нуклеиновая кислота кодирует вариабельную область тяжелой цепи антитела, выбранную из группы, состоящей из SEQ ID NO:34 и SEQ ID NO:36.
8. Вектор экспрессии, содержащий нуклеиновую кислоту по любому из пп.5-7.
9. Клетка-хозяин для продукции антитела по любому из пп.1-4, содержащая вектор экспрессии по п.8.
10. Способ анализа экспрессии PD-L1 человека в образце ткани, полученном у человека, включающий:
(a) приведение образца ткани в контакт со связывающим PD-L1 реагентом в условиях, которые обеспечивают специфическое связывание связывающего PD-L1 реагента с PD-L1 человека, где связывающий реагент содержит антитело или антигенсвязывающий фрагмент по любому из пп.1, 3 или 4,
(b) удаление не связавшегося связывающего PD-L1 реагента и
(c) обнаружение присутствия или отсутствия связавшегося связывающего PD-L1 реагента.
11. Способ по п.10, который дополнительно включает количественное определение связавшегося связывающего реагента.
12. Способ по п.10, в котором связывающий реагент содержит SEQ ID NO:13 и SEQ ID NO:20 или содержит SEQ ID NO:25 и SEQ ID NO:32.
13. Набор для обнаружения комплекса антитела или антигенсвязывающего фрагмента, связавшегося с PD-L1 человека, содержащий выделенное антитело или его антигенсвязывающий фрагмент по любому из пп.1, 3 или 4 и набор реагентов.
14. Набор по п.13, в котором антитело или антигенсвязывающий фрагмент содержат SEQ ID NO:13 и SEQ ID NO:20 или содержат SEQ ID NO:25 и SEQ ID NO:32.
15. Диагностическая композиция антител для детекции экспрессии PD-L1, которая отличается тем, что содержит смесь молекул антитела, где более 60% антител в смеси содержит SEQ ID NO:25 и SEQ ID NO:32, где X в SEQ ID NO:32 представляет собой pE, и остальная часть молекулы антитела в смеси содержит SEQ ID NO:25 и SEQ ID NO:32, где X в SEQ ID NO:32 представляет собой Q.
| OHNO et al., Antigen-binding specificities of antibodies are primarily determined by seven residues of VH, Proc | |||
| Natl | |||
| Acad | |||
| Sci | |||
| USA, 1985, vol.82 | |||
| RUDIKOFF S | |||
| et al., Single amino acid substitution altering antigen-binding specificity, Proc | |||
| Natl | |||
| Acad | |||
| Sci., 1982, 79(6) | |||
| ФИЛЛИПОВИЧ Ю.Б., Биохимические основы жизнедеятельности человека, ВЛАДОС, 2005. |

