ОБЛАСТЬ ТЕХНИКИ
[0001] Настоящее изобретение относится к новой лизиндекарбоксилазе, микроорганизму, трансформированному геном, кодирующим белок, имеющий соответствующую активность, и способу получения кадаверина с их использованием.
УРОВЕНЬ ТЕХНИКИ
[0002] Общий способ получения полиамидов (nylon) с использованием диамина представляет собой химический процесс с использованием 1,4-диаминобутана и гексаметилендиамина в качестве исходных материалов. Эти исходные материалы получают из органических соединений нефтяного происхождения. Поэтому по мере ужесточения природоохранительного законодательства растет коммерческая потребность в альтернативных материалах, полученных биологическим путем.
[0003] Между тем кадаверин представляет собой диамин - органическое соединение, состоящее из 5 атомов углерода, с молекулярной формулой NH2(CH2)5NH2, и он может быть исходным материалом для nylon 5,6. Ожидается, что при осуществлении получения кадаверина биологическим путем, можно будет получать разнообразные полиамиды, при этом удовлетворяя коммерческий спрос на полученные из биологического сырья материалы.
[0004] Что касается производства кадаверина биологическим путем, исследования переработки лизина с помощью микроорганизмов (биоконверсии) были широко известны еще до 1940-х гг. (Gale E.F., Epps Н.М. 1944. Studies on bacterial amino-acid decarboxylases. Biochem J. 38, 232-242). Ключевой стадией биоконверсии является образование кадаверина из лизина с помощью фермента лизиндекарбоксилазы (ФИГ. 1). Сообщалось об активности лизиндекарбоксилазы во многих различных микроорганизмах, и лизиндекарбоксилазу с известной удельной активностью (ммоль/мин/мг) получают из четырех типов микроорганизмов (Escherichia coli, Bacterium cadaveris, Glycine max и Selenomonas ruminantium). Было показано, что среди этих лизиндекарбоксилаз лизиндекарбоксилаза, полученная из Escherichia coli, обладает самой высокой активностью, и фермент, используемый в практическом производстве, также сводится к единственному CadA, который получают из Escherichia coli (патент Японии №2005-147171, Европейский патент №2004-010711 и патент Японии №2002-257374). Однако при получении кадаверина реакцией лизина с лизиндекарбоксилазой образуется диоксид углерода вследствие декарбоксилирования лизина и двухвалентный катион - кадаверин из одновалентного катиона - лизина, тем самым повышая рН в ходе реакции. Таким образом, при ферментативной реакции лизиндекарбоксилазы меняется рН, что создает проблему, связанную со снижением производительности. Кроме того, может происходить денатурация фермента кислотой, образующейся в реакционном растворе, или основанием, что приводит к потере его активности.
[0005] Соответственно, изобретателями данного изобретения были открыты новая лизиндекарбоксилаза, обладающая устойчивостью к высоким температурам и рН, и то, что лизиндекарбоксилазу можно экспрессировать в микроорганизме, относящемуся к виду Escherichia sp., и тем самым выполнить настоящее изобретение.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ ТЕХНИЧЕСКАЯ ЗАДАЧА
[0006] Задачей настоящего изобретения является предоставить новую лизиндекарбоксилазу и полинуклеотид, кодирующий белок, обладающий соответствующей активностью.
[0007] Другой задачей настоящего изобретения является создание микроорганизма, который трансформирован для экспрессии лизиндекарбоксилазы.
[0008] Еще одной задачей настоящего изобретения является предложить способ получения кадаверина путем использования фермента или микроорганизма, содержащего его.
ТЕХНИЧЕСКОЕ РЕШЕНИЕ
[0009] В конкретном аспекте данного изобретения предложен белок, обладающий новой лизиндекарбоксилазной активностью, включающий в себя аминокислотную последовательность SEQ ID NO: 1 или аминокислотную последовательность, гомологичную не менее чем 75% этой последовательности.
[0010] В данном контексте термин «белок, обладающий лизиндекарбоксилазной активностью» относится к белку, обладающему каталитической активностью в реакции декарбоксилирования лизина при использовании пиридоксаль-5'-фосфата в качестве кофермента при декарбоксилировании лизина, тем самым производящему кадаверин и диоксид углерода.
[0011] Белок, обладающий лизиндекарбоксилазной активностью, включающий в себя аминокислотную последовательность SEQ ID NO: 1 или аминокислотную последовательность, гомологичную не менее чем 75% этой последовательности, может представлять собой белок, обладающий лизиндекарбоксилазной активностью, который впервые обнаружен в микроорганизме из Pseudomonas sp., и этот белок может включать любые белки при условии, что они обладают соответствующей активностью и обнаруживаются в микроорганизме из Pseudomonas sp. Например, микроорганизм из Pseudomonas sp. может представлять собой Pseudomonas thermotolerans, Pseudomonas alcaligenes, Pseudomonas resinovorans, Pseudomonas putida и Pseudomonas synxantha.
[0012] В частности, белок, обладающий новой лизиндекарбоксилазной активностью, и полученный из микроорганизма Pseudomonas thermotolerans, может иметь аминокислотную последовательность SEQ ID NO: 1 или аминокислотную последовательность, гомологичную не менее чем примерно 75%, не менее чем примерно 80%, не менее чем примерно 85%, не менее чем примерно 90% или не менее чем примерно 95% последовательности SEQ ID NO: 1. Белок, обладающий лизиндекарбоксилазной активностью, и полученный из микроорганизма Pseudomonas alcaligenes, может иметь аминокислотную последовательность SEQ ID NO: 3 или аминокислотную последовательность, гомологичную не менее чем примерно 75%, не менее чем примерно 80%, не менее чем примерно 90% или не менее чем примерно 95% последовательности SEQ ID NO: 3. Белок, обладающий лизиндекарбоксилазной активностью, и полученный из микроорганизма Pseudomonas resinovorans, может иметь аминокислотную последовательность SEQ ID NO: 5 или аминокислотную последовательность, гомологичную не менее чем примерно 75%, не менее чем примерно 80%, не менее чем примерно 90% или не менее чем примерно 95% последовательности с SEQ ID NO: 5. Белок, обладающий лизиндекарбоксилазной активностью, и полученный из микроорганизма Pseudomonas putida, может иметь аминокислотную последовательность SEQ ID NO: 7 или аминокислотную последовательность, гомологичную не менее чем примерно 75%, не менее чем примерно 80%, не менее чем примерно 90% или не менее чем примерно 95% последовательности SEQ ID NO: 7. Белок, обладающий лизиндекарбоксилазной активностью, и полученный из микроорганизма Pseudomonas synxantha, может иметь аминокислотную последовательность SEQ ID NO: 9 или аминокислотную последовательность, гомологичную не менее чем примерно 75%, не менее чем примерно 80%, не менее чем примерно 90% или не менее чем примерно 95% последовательности SEQ ID NO: 9. Однако эти белки не ограничиваются приведенными выше аминокислотными последовательностями, и белки могут иметь любую аминокислотную последовательность при условии, что аминокислотная последовательность способна стабильно проявлять лизиндекарбоксилазную активность.
[0013] Кроме того, в другом конкретном аспекте настоящего изобретения предложен полинуклеотид, кодирующий новый белок, обладающий лизиндекарбоксилазной активностью, в частности, полинуклеотид, последовательность которого гомологична не менее чем 75% нуклеотидной последовательности SEQ ID NO: 2.
[0014] Нуклеотидную последовательность, кодирующую белок, обладающий лизиндекарбоксилазной активностью, можно получать из известной геномной последовательности, полученной из микроорганизма Pseudomonas sp. В частности, нуклеотидную последовательность можно получать из геномных последовательностей, полученных из одного или нескольких микроорганизмов, выбранных из группы, состоящей из Pseudomonas thermotolerans, Pseudomonas alcaligenes, Pseudomonas resinovorans, Pseudomonas putida и Pseudomonas synxantha. Нуклеотидная последовательность, кодирующая лизиндекарбоксилазу, и получаемая из микроорганизма Pseudomonas thermotolerans, может иметь нуклеотидную последовательность SEQ ID NO: 2, а также может иметь нуклеотидную последовательность, гомологичную не менее чем примерно 75%, не менее чем примерно 80%, не менее чем примерно 90% или не менее чем примерно 95% нуклеотидной последовательности SEQ ID NO: 2. Нуклеотидная последовательность, кодирующая лизиндекарбоксилазу, и получаемая из микроорганизма Pseudomonas alcaligenes, может иметь нуклеотидную последовательность SEQ ID NO: 4, а также может иметь нуклеотидную последовательность, гомологичную не менее чем примерно 75%, не менее чем примерно 80%, не менее чем примерно 90% или не менее чем примерно 95% нуклеотидной последовательности SEQ ID NO: 4. Нуклеотидную последовательность, кодирующую лизиндекарбоксилазу, и получаемую из микроорганизма Pseudomonas resinovorans можно получать из известной геномной последовательности Pseudomonas resinovorans, и, в частности, она может иметь нуклеотидную последовательность SEQ ID NO: 6, а также может иметь нуклеотидную последовательность, гомологичную не менее чем примерно 75%, не менее чем примерно 80%, не менее чем примерно 90% или не менее чем примерно 95% нуклеотидной последовательности SEQ ID NO: 6. Нуклеотидная последовательность, кодирующая лизиндекарбоксилазу, и получаемая из микроорганизма Pseudomonas putida, может иметь нуклеотидную последовательность SEQ ID NO: 8, а также может иметь нуклеотидную последовательность, гомологичную не менее чем примерно 75%, не менее чем примерно 80%, не менее чем примерно 90% или не менее чем примерно 95% нуклеотидной последовательности SEQ ID NO: 8. Нуклеотидная последовательность, кодирующая L-лизиндекарбоксилазу, и получаемая из микроорганизма Pseudomonas synxantha, может иметь нуклеотидную последовательность SEQ ID NO: 10, а также может иметь нуклеотидную последовательность, гомологичную не менее чем примерно 75%, не менее чем примерно 80%, не менее чем примерно 90% или не менее чем примерно 95% нуклеотидной последовательности SEQ ID NO: 10. Однако полинуклеотиды, кодирующие белки, обладающие лизиндекарбоксилазной активностью, не ограничиваются этим, и полинуклеотиды могут включать любые полинуклеотиды без ограничения при условии, что эти полинуклеотиды способны кодировать новый белок, обладающий лизиндекарбоксилазной активностью согласно настоящему изобретению.
[0015] В данном контексте термин «гомология», «гомологичный» относится к степени совпадения с выбранной аминокислотной последовательностью или нуклеотидной последовательностью, и гомология может выражаться в процентах. В настоящем раскрытии гомологичная последовательность, обладающая активностью, которые идентична или схожа с выбранной аминокислотной последовательностью или нуклеотидной последовательностью, выражаются как «% гомологии». Например, гомологию можно определять, используя стандартную программу, рассчитывающую такие параметры, как балл, идентичность, схожесть и т.п., в частности, BLAST 2.0, или путем сравнения последовательностей, как определено в эксперименте по Southern гибридизации в жестких условиях, а подбор подходящих условий гибридизации может быть выполнен широко известным специалистам в данной области способом, (например, см. Sambrook et al., 1989, ниже).
[0016] Более конкретно, лизиндекарбоксилаза может содержать одну или несколько аминокислотных последовательностей, выбранных из группы, включающей SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5, SEQ ID NO: 7 и SEQ ID NO: 9. Кроме того, полинуклеотид, кодирующий белок, обладающий L-лизиндекарбоксилазной активностью, может содержать одну или несколько нуклеотидных последовательностей, выбранных из группы, состоящей из SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6, SEQ ID NO: 8 и SEQ ID NO: 10.
[0017] В варианте осуществления настоящего изобретения подтверждается, что лизиндекарбоксилазы, полученные из вышеуказанных микроорганизмов Pseudomonas sp., не проявляют существенных изменений своей активности при высоких рН, и вследствие этого они устойчивы к рН.
[0018] В еще одном конкретном аспекте настоящего изобретения предложен микроорганизм, трансформированный для экспрессии нового белка, обладающего лизиндекарбоксилазной активностью. Трансформированный микроорганизм может представлять собой любой из прокариотических и эукариотических микроорганизмов при условии, что он трансформируется для экспрессии белка, обладающего соответствующей декарбоксилазной активностью. Например, трансформированный микроорганизм может включать в себя микроорганизмы Escherichia sp., Erwinia sp., Serratia sp., Providencia sp. и Corynebacterium sp. Микроорганизм может быть, в частности, микроорганизмом, принадлежащим, без ограничения, к Escherichia sp.или Corynebacterium sp., и более конкретно - к Е. coli или Corynebacterium glutamicum.
[0019] Кроме того, родительский штамм трансформированного микроорганизма может представлять собой микроорганизм, но не ограничиваясь им, обладающий улучшенной способностью продуцировать лизин по сравнению с диким типом. В данном контексте выражение «микроорганизм, обладающий улучшенной способностью продуцировать лизин по сравнению с диким типом» относится к микроорганизму с улучшенной способностью продуцировать лизин по сравнению с природным микроорганизмом или родительским штаммом, и микроорганизм, обладающий улучшенной способностью продуцировать лизин конкретно не ограничивается при условии, что он представляет собой микроорганизм, обладающий улучшенной способностью продуцировать лизин по сравнению с родительским штаммом.
[0020] Для придания улучшенной способности продуцировать лизин по сравнению с диким типом можно применять общий способ выращивания микроорганизмов, такой как способ создания ауксотрофных мутантных штаммов, аналог-устойчивых штаммов или мутантных штаммов с регулируемым обменом, способных продуцировать лизин, и способ получения рекомбинантных штаммов с повышенной ферментативной активностью биосинтеза лизина. При выращивании продуцирующих лизин микроорганизмов, такие свойства, как ауксотрофия, аналог-устойчивость и мутации регулирования обмена, могут быть приданы отдельно или в комбинации. Повышенную ферментативную активность биосинтеза лизина можно придать отдельно или в комбинации. Кроме того, ферментативную активность биосинтеза лизина можно повышать одновременно с приданием, например, следующих свойств: ауксотрофии, аналог-устойчивости и мутации регулирования обмена. В частности, ген, кодирующий фермент биосинтеза лизина, может включать, но не ограничиваться ими, ген дигидродипиколинатсинтазы (dapA), ген аспартокиназы (lysC), ген дигидродипиколинатредуктазы (dapB), ген диаминопимелатдекарбоксилазы (lysA), ген диаминопимелатдегидрогеназы (ddh), ген фосфоэнолпируваткарбоксилазы (ррс), ген аспартатаминотрансферазы (aspC) и ген аспартат полуальдегиддегидрогеназы (asd). Способ придания или повышения способности продуцировать лизин путем усиления ферментативной активности биосинтеза лизина можно осуществлять путем введения мутаций в гены, кодирующие соответствующие ферменты, или усиления генов для повышения внутриклеточной активности ферментов. Эти способы можно выполнять с помощью генетической рекомбинации, но не ограничиваясь ею.
[0021] Микроорганизм может представлять собой любой из прокариотических и эукариотических микроорганизмов при условии, что он обладает улучшенной способностью продуцировать лизин по сравнению с диким типом. В частности, микроорганизм может представлять собой микроорганизм Escherichia sp. или микроорганизм Coryneform. Микроорганизм Escherichia может представлять собой, но не ограничиваться ими, Escherichia coli, Escherichia albertii, Escherichia blattae, Escherichia fergusonii, Escherichia hermannii или Escherichia vulneris. Более конкретно, микроорганизм Escherichia sp. может представлять собой Escherichia coli. Микроорганизм Coryneform может включать в себя микроорганизм Corynebacterium или Brevibacterium sp. Кроме того, микроорганизм Coryneform может представлять собой, но не ограничиваться ими, в частности, Corynebacterium glutamicum, Corynebacterium thermoaminogenes, Brevibacterium flavum или Brevibacterium lactofermentum.
[0022] Для преобразования микроорганизма таким образом, чтобы микроорганизм экспрессировал белок, обладающий активностью лизиндекарбоксилазы, ген лизиндекарбоксилазы по настоящему изобретению можно включать в виде белка лизиндекарбоксилазы или в виде блока экспрессии гена в микроорганизм, подлежащий преобразованию. Блок экспрессии гена лизиндекарбоксилазы может быть функционально связан с вектором, а затем трансформирован в микроорганизм или вставлен в хромосому микроорганизма. В частности, ген лизиндекарбоксилазы может быть функционально связан так, чтобы ген сверхэкспрессировался промотором выше от инициирующего кодона.
[0023] В данном контексте термин «блок экспрессии» относится к фрагменту, включающему промотор, который функционально связан с полинуклеотидом, кодирующим белок, и может дополнительно включать 3'-UTL, 5'-UTL, поли(А)-хвост и т.п. В данном контексте выражение «блок экспрессии» может быть взаимозаменяемым с выражением «экспрессионная кассета».
[0024] В данном контексте термин «функционально связанный» относится к функциональной связи между нуклеотидной последовательностью гена и нуклеотидной последовательностью, имеющей промоторную активность, посредством чего транскрипция гена, кодирующего лизиндекарбоксилазу, инициируется и опосредуется, указывая на то, что нуклеотидная последовательность с активностью промотора функционально связана с геном лизиндекарбоксилазы для регулирования транскрипционной активности гена лизиндекарбоксилазы.
[0025] В данном контексте термин «трансформация» означает общее действие по введению гена лизиндекарбоксилазы, полученного из микроорганизма Pseudomonas sp., в клетку-хозяина, в частности, микроорганизм Escherichia sp. или микроорганизм Coryneform, для экспрессии гена в клетке-хозяине. В этом отношении ген лизиндекарбоксилазы представляет собой полинуклеотид, включая ДНК и РНК, способный кодировать лизиндекарбоксилазу. Ген может быть введен в любом виде при условии, что его можно ввести в клетку-хозяина и экспрессировать в ней. Например, ген можно вводить в клетку-хозяина в экспрессионной кассете, которая представляет собой полинуклеотидную структуру, включающую в себя полные элементы для экспрессии гена. Экспрессионная кассета включает промотор, функционально связанный с геном, сигнал окончания транскрипции, участок связывания рибосомы и сигнал окончания трансляции. Экспрессионная кассета может быть самовоспроизводимым вектором экспрессии. Ген также может вводиться в клетку-хозяина самостоятельно или в полинуклеотидной структуре для функционального связывания с последовательностью, необходимой для экспрессии в клетке-хозяине. Рекомбинантный вектор представляет собой средство, с помощью которого ДНК вводится в клетку-хозяина для экспрессии белка, и можно использовать такой известный вектор экспрессии, как плазмидный вектор, космидный вектор, вектор бактериофага и т.п. Вектор может быть легко приготовлен специалистами в данной области согласно любому известному способу использования метода рекомбинации ДНК, но не ограничивается ими.
[0026] Способ трансформации может быть любым способом введения полинуклеотида в клетки и может быть выполнен путем выбора подходящего стандартного метода, известного в данной области техники. Например, способ может включать, но не ограничивается ими, электропорацию, совместное осаждение с фосфатом кальция, ретровирусную инфекцию, микроинъекцию, метод DEAE-декстрана, катионный липосомный метод и т.д.
[0027] В конкретном варианте осуществления микроорганизм, имеющий улучшенную способность продуцировать лизин, трансформируется таким образом, что экспрессируется белок, обладающий лизиндекарбоксилазной активностью по настоящему изобретению, тем самым обладающий превосходной способностью продуцировать кадаверин.
[0028] В еще одном конкретном варианте осуществления настоящего изобретения предложено применение новой лизиндекарбоксилазы или микроорганизма, трансформированного для экспрессии нового белка, обладающего лизиндекарбоксилазной активностью, в производстве кадаверина.
[0029] Новая лизиндекарбоксилаза и микроорганизм, трансформированный для экспрессии нового белка, обладающего лизиндекарбоксилазной активностью, такие же, как и те, что описаны выше. В конкретном варианте осуществления продемонстрировано, что лизиндекарбоксилаза настоящего изобретения обладает более высокой устойчивостью к изменению температуры или рН, чем полученная из Е. Coli лизиндекарбоксилаза, которая широко используется при производстве кадаверина. В частности, новая лизиндекарбоксилаза настоящего изобретения обладает более высокой устойчивостью к рН, чем полученная из Е. coli лизиндекарбоксилаза, и, следовательно, это является преимуществом в реакции, превращающей лизин в кадаверин. Соответственно, новая лизиндекарбоксилаза настоящего изобретения и микроорганизм, трансформированный для экспрессии нового белка, обладающего лизиндекарбоксилазной активностью, можно применять для получения кадаверина.
[0030] В еще одном аспекте настоящего изобретения предложен способ получения кадаверина.
[0031] В конкретном варианте осуществления способа получения кадаверина по настоящему изобретению, способ представляет собой способ получения кадаверина, включающий в себя стадии конверсии лизина в кадаверин путем использования нового белка, обладающего лизиндекарбоксилазной активностью, или микроорганизма, трансформированного для экспрессии белка, обладающего лизиндекарбоксилазной активностью; и извлечения полученного кадаверина.
[0032] Новый белок, обладающий лизиндекарбоксилазной активностью, и трансформированный микроорганизм такие же, как и те, что описаны выше. Трансформированный микроорганизм может, в частности, представлять собой микроорганизм Escherichia sp.
[0033] На стадии превращения лизина в кадаверин новый белок, обладающий лизиндекарбоксилазной активностью, экстрагируют из микроорганизма, экспрессирующего белок, фермент очищают и используют для декарбоксилирования лизина, тем самым получая кадаверин. Альтернативно, лизин добавляют к культуре, полученной культивированием трансформированного микроорганизма, и микроорганизм используется без дальнейшей переработки для декарбоксилирования лизина, тем самым превращая лизин в кадаверин.
[0034] В другом конкретном варианте осуществления способа получения кадаверина по настоящему изобретению предложен способ получения кадаверина, включающий стадии культивирования в среде микроорганизма, способного продуцировать кадаверин, причем микроорганизм с улучшенной способностью продуцировать лизин по сравнению с диким типом, трансформирован для экспрессии нового белка, обладающего лизиндекарбоксилазной активностью; и извлечения кадаверина из микроорганизма или культуры.
[0035] Новый белок, обладающий L-лизиндекарбоксилазной активностью, и микроорганизм с улучшенной способностью продуцировать лизин по сравнению с диким типом такие же, как и те, что описаны выше.
[0036] Культивирование можно проводить в подходящей среде в условиях культивирования, которые хорошо известны в данной области. Условия среды и культуры могут быть легко изменены любым специалистом в данной области в зависимости от выбранного микроорганизма. Способ культивирования может включать в себя, без ограничения, периодическую культуру, непрерывную культуру, культуру с подпиткой или их комбинацию.
[0037] Среда может включать в себя множество источников углерода, источников азота и микроэлементов.
[0038] Например, в частности, источники углерода могут включать углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, мальтоза, крахмал и целлюлоза; масла, такие как соевое масло, подсолнечное масло, касторовое масло и кокосовое масло; жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота; спирты, такие как глицерин и этанол; органические кислоты, такие как уксусная кислота, или их комбинации. В частности, в качестве источника углерода, без ограничения, может использоваться глюкоза. Источники азота, без ограничения, могут включать источники органического азота, такие как пептон, дрожжевой экстракт, мясной бульон, солодовый экстракт, кукурузный крутой ликер (CSL) и соевый шрот, неорганические источники азота, такие как мочевина, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония или их комбинации. Среда может включать в качестве источников фосфора, но без ограничения, например, вторичный кислый фосфат калия, первичный кислый фосфат калия и соответствующие натрий-содержащие соли, и такие соли металлов, как сульфат магния и сульфат железа. Кроме того, в среду могут входить аминокислоты, витамины и подходящие предшественники. Среду или индивидуальные составляющие можно добавлять в среду в периодическом или непрерывном режиме. Эти примеры приведены только целях иллюстрации, и настоящее изобретение этими примерами не ограничивается.
[0039] Кроме того, рН среды можно регулировать в ходе культивирования путем добавления такого соединения, как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота или серная кислота подходящим образом. Выделение пузырьков воздуха можно подавлять в ходе культивирования, используя пеногасящую добавку, такую как полигликолевый эфир жирной кислоты. Для поддержания аэробных условий в среде в культуру можно впрыскивать кислород или кислородсодержащий газ (например, воздух). Температура среды в общем может быть от 20°С до 45°С, например, от 25°С до 40°С. Культивирование может продолжаться пока продуцирование лизиндекарбоксилазы не достигнет требуемого уровня, например, от 10 часов до 160 часов.
[0040] Способ извлечения кадаверина можно осуществлять, например, путем сбора или извлечения произведенного кадаверина из среды с помощью подходящего способа, известного в уровне техники, согласно периодическому режиму, непрерывному режиму или периодическому режиму с подпиткой средой. В способе извлечения можно использовать центрифугирование, фильтрацию, ионообменную хроматографию, кристаллизацию и т.п. Например, биомассу можно удалять из культуры путем низкоскоростного центрифугирования, а полученную надосадочную жидкость очищать ионообменной хроматографией.
[0041] Кроме того, способ получения кадаверина может дополнительно включать стадию извлечения лизиндекарбоксилазы из микроорганизма или среды.
[0042] Способ извлечения лизиндекарбоксилазы из микроорганизма или среды можно осуществлять, например, путем сбора или извлечения произведенной лизиндекарбоксилазы из микроорганизма или среды с помощью подходящего способа, известного в уровне техники, согласно периодическому режиму, непрерывному режиму или периодическому режиму с подпиткой средой. В данном способе извлечения можно использовать центрифугирование, фильтрацию, ионообменную хроматографию, кристаллизацию и т.п. Например, биомассу можно удалять из культуры путем низкоскоростного центрифугирования, а полученную надосадочную жидкость очищать ионообменной хроматографией. Кроме того, лизиндекарбоксилазу можно извлекать из клеточного лизата, который получают разрушением микроорганизма в среде. Клеточный лизат можно получать, используя подходящий способ, известный в области техники. Например, можно использовать механический гомогенизатор или поступающий в продажу буфер для лизиса клеток. Лизиндекарбоксилазу можно получать из клеточного лизата соответствующим способом, известным в области техники, таким как центрифугирование и т.п.
ПОЛЕЗНЫЕ ТЕХНИЧЕСКИЕ ЭФФЕКТЫ ИЗОБРЕТЕНИЯ
[0043] Новый белок, обладающий лизиндекарбоксилазной активностью, и полученный из микроорганизма Pseudomonas sp., предложенный в настоящем изобретении, может обладать стабильной активностью даже при изменениях рН, и, следовательно, белок может быть эффективно применен в реакции конверсии лизина в кадаверин, и поэтому может широко применяться в производстве кадаверина.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0044] На ФИГ. 1 показан механизм реакции лизиндекарбоксилазы, в которой из лизина образуется кадаверин;
[0045] ФИГ. 2 представляет собой изображение SDS-PAGE геля, показывающего результаты экспрессии PtLDC и PtLDC с N-концевыми his-tag;
[0046] На ФИГ. 3 показана реакционная способность PtLDC при конверсии лизина в кадаверин;
[0047] На ФИГ. 4 показана относительная ферментативная активность PtLDC при разных температурах;
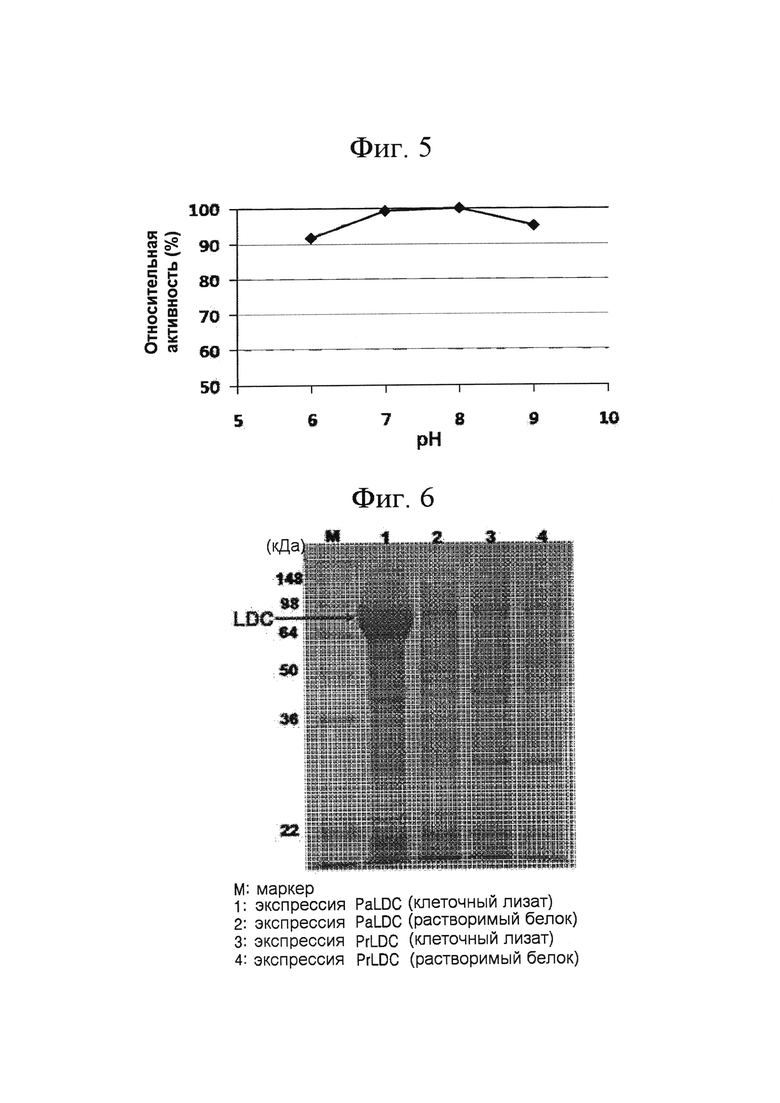
[0048] На ФИГ. 5 показана относительная ферментативная активность PtLDC при разных рН;
[0049] ФИГ. 6 представляет собой изображение SDS-PAGE геля, показывающего экспрессию PaLDC и PrLDC;
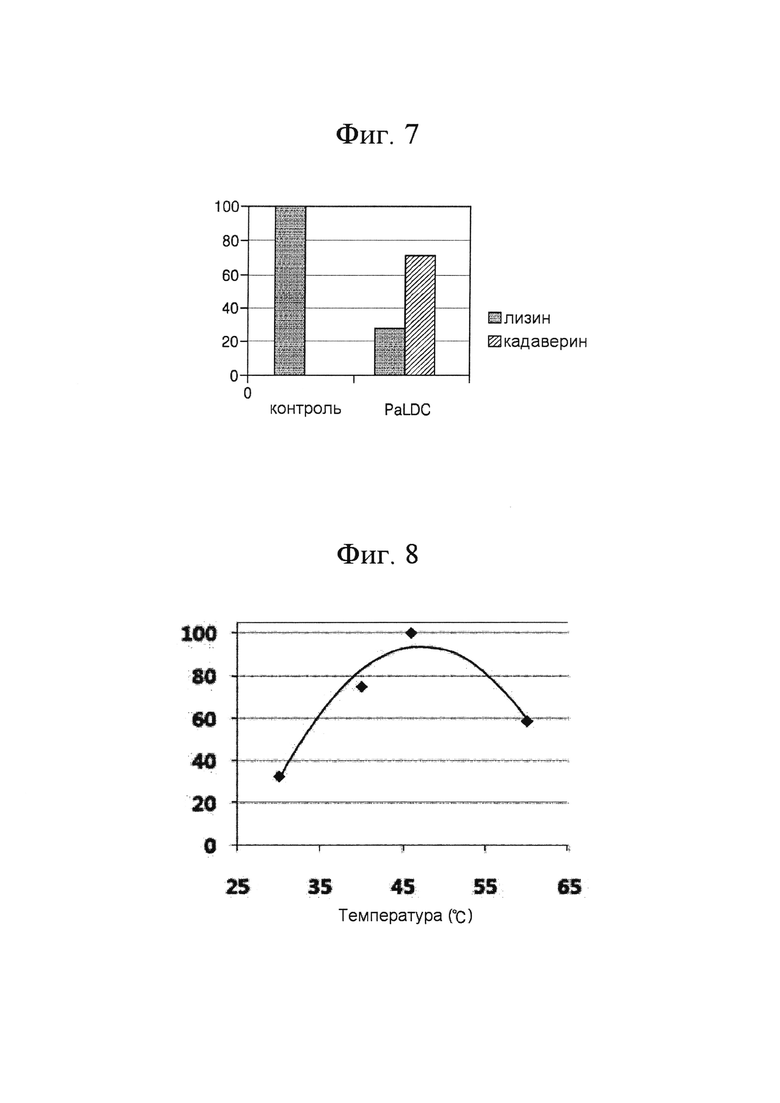
[0050] На ФИГ. 7 показана реакционная способность PaLDC при конверсии лизина в кадаверин;
[0051] На ФИГ. 8 показана относительная ферментативная активность PaLDC при разных температурах;
[0052] На ФИГ. 9 показана относительная ферментативная активность PaLDC при разных рН;
[0053] На ФИГ. 10 показана реакционная способность PrLDC при конверсии лизина в кадаверин;
[0054] На ФИГ. 11 показана относительная ферментативная активность PrLDC при разных температурах;
[0055] На ФИГ. 12 показана относительная ферментативная активность PrLDC при разных рН;
[0056] ФИГ. 13 представляет собой изображение SDS-PAGE геля, показывающего результаты экспрессии EcLDC, PpLDC, PtLDC и PxLDC;
[0057] На ФИГ. 14 показана реакционная способность PpLDC при конверсии лизина в кадаверин;
[0058] На ФИГ. 15 показана относительная ферментативная активность PpLDC при разных температурах;
[0059] На ФИГ. 16 показана относительная ферментативная активность PpLDC при разных рН;
[0060] На ФИГ. 17 показана реакционная способность PxLDC при конверсии лизина в кадаверин;
[0061] На ФИГ. 18 показана относительная ферментативная активность PxLDC при разных рН;
[0062] На ФИГ. 19 показаны соответствующие относительные ферментативные активности EcLDC и PtLDC при разных температурах; и
[0063] На ФИГ. 20 показаны соответствующие относительные ферментативные активности EcLDC и PtLDC при разных рН.
ТЕХНИЧЕСКОЕ РЕШЕНИЕ ИЗОБРЕТЕНИЯ
[0064] Далее в данном документе настоящее изобретение будет описано более подробно со ссылками на Примеры. Однако эти примеры приведены только в целях иллюстрации, и объем настоящего изобретения этими примерами не ограничивается.
[0065] Пример 1. Отбор новых лизиндекарбоксилаз для получения кадаверина
[0066] 1-1. Отбор лизиндекарбоксилазы, полученной из Pseudomonas thermotolerans
[0067] Чтобы отобрать новую лизиндекарбоксилазу для применения в получении кадаверина, использовали программное обеспечение BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastn&PAGE_TYPE=BlastSearc h&LINK_LOC=blasthome), предоставленное Национальным центром биотехнологической информации (NCBI) США, для поиска лизиндекарбоксилазы, полученной из термофильной бактерии, которая имеет высокое сходство с пептидной последовательностью активного сайта лизиндекарбоксилазы, полученной из Е. coli. А именно, проводили поиск с помощью BLAST, по в общей сложности 31 пептидной последовательности (GRVEGKVIYETQSTHKLLAAFSQASMIHVKG: SEQ ID NO: 12), каждая из которых включала 15 аминокислот на N-конце и С-конце, сосредоточенных на 367-м лизине, который является основной аминокислотой, полученной из Е. Coli лизиндекарбоксилазы. В результате подтвердилось, что микроорганизмы Escherichia, Shigella, Enterobacteria, Edwardsiella, Klebsiella, Serratia, Yersinia, Yokenella, Raoultella, Ceratitis, Salmonella, Sutterella, Shimwellia, Vibrio и Pseudomonas sp. имеют высокую степень гомологии. Исследование было направлено на поиск лизиндекарбоксилазы, имеющей высокую термическую стабильность, и при этом обладающей активностью, сходной с активностью лизиндекарбоксилазы из Е. coli. Как правило, обнаруживаемые в термофильных бактериях белки известны своей высокой термической стабильностью, и поэтому среди микроорганизмов, обнаруженных при поиске, был выбран Pseudomonas thermotolerans, который характеризуется как термофильный (46-60°С) микроорганизм.
[0068] 1-2. Отбор лизиндекарбоксилазы, полученной из различных микроорганизмов Pseudomonas sp.
[0069] Для отбора лизиндекарбоксилаз, полученных из микроорганизмов Pseudomonas sp., отличающихся от Pseudomonas thermotolerans, были отобраны четыре микроорганизма (Pseudomonas alcaligenes, Pseudomonas resinovorans, Pseudomonas putida и Pseudomonas synxantha), продемонстрировавших низкую гомологию с Pseudomonas sp. Для идентификации нуклеотидов и аминокислотных последовательностей лизиндекарбоксилаз, полученных из четырех микроорганизмов Pseudomonas sp., отобранных как указано выше, использовали программы для нуклеотидов и геномов, предоставленные Национальным центром биотехнологической информации (NCBI) США (http://www.ncbi.nlm.nih.gov/).
[0070] В следующей Таблице 1 показана гомология аминокислотных последовательностей лизиндекарбоксилаз, полученных из Pseudomonas sp.

[0071] PtLDC: лизиндекарбоксилаза, полученная из Pseudomonas thermotolerans (P. thermotolerans)
[0072] PaLDC: лизиндекарбоксилаза, полученная из Pseudomonas alcaligenes (P. alcaligenes)
[0073] PrLDC: лизиндекарбоксилаза, полученная из Pseudomonas resinovorans (P. resinovorans)
[0074] PpLDC: лизиндекарбоксилаза, полученная из Pseudomonas putida (P. putida)
[0075] PxLDC: лизиндекарбоксилаза, полученная из Pseudomonas synxantha (P. synxantha)
[0076] Пример 2. Получение Е. coli со вставленным геном лизиндекарбоксилазы, полученным из Pseudomonas Thermotolerans, и анализ активности лизиндекарбоксилазы, экспрессированной в нем
[0077] 2-1. Трансформация Е. coli геном лизиндекарбоксилазы, полученным из Pseudomonas thermotolerans
[0078] Для введения гена лизиндекарбоксилазы, полученного из Pseudomonas thermotolerans, в Е. coli и экспрессии в нем гена выполняли клонирование рекомбинантного гена. Генетическую информацию о Pseudomonas thermotolerans получали из геномных данных в NCBI (http://www.ncbi.nlm.nih.gov/genome/).
[0079] Геномную ДНК Pseudomonas thermotolerans получали и затем использовали в качестве матрицы для амплификации гена лизиндекарбоксилазы, полученного из Pseudomonas thermotolerans- (ptldc), используя полимеразную цепную реакцию (ПЦР). Для выполнения ПЦР использовали праймеры 5_LDC_NdeI (AATATACATATGTACAAAGACCTCCAATTCCCC) (SEQ ID NO: 13) и 3_LDC_XhoI (AATATACTCGAGTCAGATCTTGATGCAGTCCACCG) (SEQ ID NO: 14) и PfuUltraTM ДНК полимеразу (Stratagene, США), ПЦР выполняли за 30 циклов в следующих условиях: 94°С: 30 сек, 55°С: 30 сек и 72°С: 2 мин. В результате получали амплифицированный ptldc (SEQ ID NO: 2). Далее, для экспрессии полученной из Pseudomonas thermotolerans лизиндекарбоксилазы с N-концевым His-tag, использовали праймеры 5_LDC_BamHI (AATATAGGATCCGTACAAAGACCTCCAATTCCCC) (SEQ ID NO: 15) и 3_LDC_SacI (AATATAGAGCTCTCAGATCTTGATGCAGTCCACCG) (SEQ ID NO: 16) для выполнения ПЦР таким же образом, как и в описанном выше способе ПЦР. Затем каждый ген ptldc, полученный в ПЦР, вводили в экспрессионный вектор Е. coli - pET-Deut1. После этого, каждую плазмиду, клонированную с геном ptldc, вводили в E. coli Rosetta методом трансформации тепловым шоком. Каждый из трансформированых E. coli культивировали в 50 мл жидкой среды LB (содержащей 50 мг/мл ампициллина) при 37°С. Когда значение OD600 достигало 0,8, туда добавляли 0,4 мМ изопропил-β-D-1-тиогалактопиранозид (ИПТГ) и инкубировали при 18°С в течение 48 часов для стимулирования экспрессии. Каждую полученную из Pseudomonas thermotolerans лизиндекарбоксилазу (PtLDC), полностью экспрессированную таким образом, идентифицировали, используя SDS-PAGE (ФИГ. 2). Результаты SDS-PAGE показали, что PtLDC и PtLDC с His-tag, экспрессированные при низких температурах, были свехэкспрессированы в виде растворимых белков (линии 2 и 4 на ФИГ. 2).
[0080] E. coli Rosetta, трансформированный плазмидой, включающей ptldc (SEQ ID NO: 2), обозначали как 'Escherichia coli CC04-0055', и депонировали в Корейском центре культур микроорганизмов (Korean Culture Center of Microorganisms, KCCM) 24 июля 2014 г. под Инвентарным № KCCM11559P.
[0081] 2-2. Исследование активности полученной из Pseudomonas thermotolerans лизиндекарбоксилазы, экспрессированной в E. coli
[0082] (1) Исследование реакционной способности лизиндекарбоксилазы
[0083] Для исследования реакционной способности PtLDC и PtLDC с His-tag, проводили реакцию 50 мл растворимого белка, 100 мМ пиридоксаль-фосфата (пиридоксаль-фосфат, PLP) и 250 мМ лизина в объеме 200 мл при 46°С в течение 2 часов. Реакционный буферный раствор представлял собой 50 мМ фосфата натрия с рН 6,2. В качестве контроля использовали микроорганизм с введенным пустым вектором и анализировали количества лизина и кадаверина (ФИГ. 3). Для точного анализа количеств лизина и кадаверина использовали высокоэффективную жидкостную хроматографию (Waters, Milford, MA) с использованием детектора показателя преломления 2414 Refractive Indes Detector (Waters, Milford, MA). Реагент лизин-HCl и реагент 1,5-диаминпентан (кадаверин) приобретали у Sigma (St. Louis, МО) и подвижную фазу, состоящую из 1 мМ лимонной кислоты, 10 мМ винной кислоты, 24 мМ этилендиамина и 5% ацетонитрила, использовали для разделения и количественного анализа двух материалов в колонке lonoSpher С3-100 мм, 5 мм. Продуцирования кадаверина в контрольном образце обнаружено не было. PtLDC с N-концевым His-tag показал 72% конверсию лизина, a PtLDC показал 100% конверсию лизина, что указывает на образование кадаверина.
[0084] (2) Исследование активности лизиндекарбоксилазы в зависимости от температуры и рН
[0085] Для анализа ферментативных свойств PtLDC в различных температурных условиях (30°С, 42°С, 50°С, 60°С, 70°С и 80°С) сравнивали относительную активность. При разведении PtLDC и реакции с 250 мМ лизинового субстрата при 60°С в течение 30 минут было найдено, что образовалось 42 мМ кадаверина. С учетом этого анализировали концентрации кадаверина, используя 50 мМ натрийфосфатного буфера (рН 6,2) в качестве буферного раствора, и эквивалентное количество фермента в тех же реакционных условиях, за исключением температуры реакции, которая составляла 30°С, 42°С, 50°С, 70°С и 80°С, и сравнивали в количеством кадаверина, продуцированного при температуре реакции 60°С (ФИГ. 4). Как видно из ФИГ. 4, PtLDC продемонстрировал самую высокую активность при 60°С. Помимо этого, активность PtLDC поддерживалась на уровне не менее 80% от активности при температуре 55°С~65°С.
[0086] Кроме того, активность лизиндекарбоксилазы была оценена при различных значениях рН (6,2, 7,0, 8,0 и 9,0). Температуру реакции фиксировали при 60°С, и использовали равные объемы фермента в тех же реакционных условиях, за исключением того, что использовали 50 мМ натрийфосфатного буфера (рН 6,2), 50 мМ трис-буфера (рН 7,0), 100 мМ калийфосфатного буфера (рН 8,0) и 50 мМ трис-буфера (рН 9,0). Сравнивали реакционную способность лизиндекарбоксилазы при различных рН (ФИГ. 5). PtLDC продемонстрировал наивысшую активность при рН 8,0, и его активность поддерживалась на уровне не менее 90% при значениях рН 6 - рН 9. Количество кадаверина, полученного при каждом значении рН, сравнивали с количеством кадаверина, полученного при рН 8 (ФИГ. 5). Экспериментальный результат показал, что PtLDC обладает высокой стабильностью при изменении рН или при высоких значениях рН.
[0087] Пример 3. Получение Е. coli со вставленным геном лизиндекарбоксилазы, полученным из Pseudomonas Alcaligenes, и анализ активности лизиндекарбоксилазы. экспрессированной в нем
[0088] 3-1. Трансформация Е. coli геном лизиндекарбоксилазы, полученным из Pseudomonas alcaligenes
[0089] Для клонирования гена лизиндекарбоксилазы (paldc), полученного из Pseudomonas alcaligenes, использовали праймеры 5_PaLDC_NdeI (AATATACATATGTACAAAGACCTGAA GTTCCCCATCC) (SEQ ID NO: 17) и 3_PaLDC_XhoI (AATATACTCGAGTCACTCCCTTATGCAATCAACGGTATAGC) (SEQ ID NO: 18) и очищенную геномную ДНК Pseudomonas alcaligenes в качестве матрицы для выполнения ПЦР. Pfu ДНК полимерузу использовали в качестве полимеразы, и ПЦР выполняли за 30 циклов в следующих условиях: 94°С: 30 сек, 55°С: 30 сек и 72°С: 2 мин. В результате получали амплифицированный ген paldc (SEQ ID NO: 4).
[0090] Полученный ген paldc экспрессировали при низкой температуре в E.coli таким же образом, как и в Примерах 2-1, и идентифицировали, используя SDS-PAGE (ФИГ. 6). Как видно из ФИГ. 6, полученная из Pseudomonas alcaligenes лизиндекарбоксилаза (PaLDC) была в основном экспрессирована в виде нерастворимого белка, а растворимый белок на SDS-PAGE геле обнаружен не был (ФИГ. 6, см. линии 1 и 2).
[0091] 3-2. Исследование активности полученной из Pseudomonas alcaligenes лизиндекарбоксилазы, экспрессированной в E. coli
[0092] (1) Исследование реакционной способности лизиндекарбоксилазы
[0093] Для исследования реакционной способности PaLDC, клеточный лизат PaLDC, полученный в Примерах 3-1, центрифугировали при 13000 об/мин в течение 15 минут с получением надосадочной жидкости (растворимый белок), которую использовали для конверсии лизина. Проводили реакцию 50 мкл растворимого белка, 100 мМ PLP и 250 мМ лизина в 50 мМ натрийфосфатного буфера (рН 6,2) в реакционном объеме 200 мкл при 46°С в течение 2 часов. В результате была обнаружена 70% конверсия лизина в кадаверин под действием PaLDC (ФИГ. 7).
[0094] (2) Исследование активности лизиндекарбоксилазы в зависимости от температуры и рН
[0095] Для того, чтобы найти оптимальные температурные условия для активности полученной из Pseudomonas alcaligenes лизиндекарбоксилазы, определяли ферментативную активность при следующих температурах: 30°С, 40°С, 46°С и 60°С таким же образом, как и в Примере 2-2. В результате было обнаружено, что наивысшая активность PaLDC наблюдается при 50°С (ФИГ. 8).
[0096] Активность полученной из Pseudomonas alcaligenes лизиндекарбоксилазы при различных значениях рН определяли таким же образом, как и в Примере 2-2. В результате было показано, что наивысшая стабильность PaLDC наблюдается при рН 8 и рН 9, и она поддерживается на уровне не менее 95% при рН 6 (ФИГ. 9).
[0097] Пример 4. Получение Е. coli со вставленным геном лизиндекарбоксилазы, полученным из Pseudomonas Resinovorans, и анализ активности лизиндекарбоксилазы, экспрессированной в нем
[0098] 4-1. Трансформация Е. coli геном лизиндекарбоксилазы, полученным из Pseudomonas resinovorans
[0099] Для клонирования гена лизиндекарбоксилазы (pride), полученного из Pseudomonas resinovorans, использовали праймеры 5_PrLDC_NdeI (AATATACATATGTACAAAGAGCTC AAGTTCCCCGTCCTC) (SEQ ID NO: 19) и 3_PrLDC_XhoI (AATATACTCGAG TTATTCCCTGATGCAGTCCACTGTA TAGC) (SEQ ID NO: 20) и очищенную геномную ДНК Pseudomonas resinovorans в качестве матрицы для выполнения ПЦР. ПЦР выполняли, используя такую же полимеразу в тех же условиях ПЦР, как и в Примере 3-1. В результате получали амплифицированный pride (SEQ ID NO: 6).
[00100] Полученный ген pride экспрессировали при низкой температуре в E.coli таким же образом, как и в Примере 2-1, и идентифицировали, используя SDS-PAGE (ФИГ. 6; линии 3 и 4). В результате было обнаружено, что PrLDC практически не экспрессируется при низкой температуре.
[00101] 4-2. Исследование активности полученной из Pseudomonas resinovorans лизиндекарбоксилазы, экспрессированной в E. coli
[00102] (1) Исследование реакционной способности лизиндекарбоксилазы
[00103] Для исследования реакционной способности лизиндекарбоксилазы (PrLDC), полученной из Pseudomonas resinovorans, клеточный лизат PrLDC, полученный в Примере 4-1, центрифугировали при 13000 об/мин в течение 15 минут с получением надосадочной жидкости, которую использовали для конверсии лизина. Проводили реакцию 50 мкл растворимого белка, 100 мМ PLP и 250 мМ лизина в 50 мМ натрийфосфатного буфера (рН 6,2) в реакционном объеме 200 мкл при 46°С в течение 2 часов. В результате получили 66% кадаверина (ФИГ. 10).
[00104] (2) Исследование активности лизиндекарбоксилазы в зависимости от температуры и рН
[00105] Для того, чтобы найти оптимальные температурные условия для активности PrLDC, определяли ферментативную активность при следующих температурах: 30°С, 40°С, 46°С и 60°С таким же образом, как и в Примере 2-2. В результате было обнаружено, что наивысшая активность PrLDC наблюдается при 60°С (ФИГ. 11).
[00106] Активность PrLDC при различных значениях рН определяли таким же образом, как и в Примере 2-2. В результате было показано, что наивысшая стабильность PrLDC наблюдается при рН 6, и она поддерживается на уровне не менее 90% при рН 9 (ФИГ. 12).
[00107] Пример 5. Получение Е. coli со вставленным геном лизиндекарбоксилазы, полученным из Pseudomonas putida, и анализ активности лизиндекарбоксилазы, экспрессированной в нем
[00108] 5-1. Трансформация Е. coli геном лизиндекарбоксилазы, полученным из Pseudomonas putida
[00109] Для клонирования гена лизиндекарбоксилазы (ppldc), полученного из Pseudomonas putida, использовали праймеры 5_PpLDC_NdeI (AATATACATATGTACAAAGACCTCCAA ТТСССС) (SEQ ID NO: 21) и 3_PpLDC_XhoI (AATATACTCGAGTCACTCCCTTATGCAATCAACGGTATAGC) (SEQ ID NO: 22) и очищенную геномную ДНК Pseudomonas putida в качестве матрицы для выполнения ПЦР. Pfu ДНК полимерузу использовали в качестве полимеразы, и ПЦР выполняли за 30 циклов в следующих условиях: 94°С: 30 сек, 55°С: 30 сек и 72°С: 2 мин. В результате получали амплифицированный ген ppldc (SEQ ID NO: 8).
[00110] Полученный ген ppldc экспрессировали при низкой температуре в E. coli таким же образом, как и в Примере 2-1, и идентифицировали, используя SDS-PAGE (ФИГ. 13). Как видно по Линиям 3 и 4 на ФИГ. 13, полученная из Pseudomonas putida лизиндекарбоксилаза (PpLDC) практически не экспрессируется при низких температурах.
[00111] Клеточный лизат центрифугировали при 13000 об/мин в течение 15 минут и использовали надосадочную жидкость для реакции конверсии лизина.
[00112] 5-2. Исследование активности полученной из Pseudomonas putida лизиндекарбоксилазы, экспрессированной в E. coli
[00113] (1) Исследование реакционной способности лизиндекарбоксилазы
[00114] Для исследования реакционной способности PpLDC, клеточный лизат PpLDC, полученный в Примере 5-1, центрифугировали при 13000 об/мин в течение 15 минут с получением надосадочной жидкости, которую использовали для конверсии лизина. Проводили реакцию 50 мкл растворимого белка, 100 мМ PLP и 250 мМ лизина в 50 мМ натрийфосфатного буфера (рН 6,2) в реакционном объеме 200 мкл при 46°С в течение 2 часов. В результате получили 16% кадаверина (ФИГ. 14).
[00115] (2) Исследование активности лизиндекарбоксилазы в зависимости от температуры и рН
[00116] Для того, чтобы найти оптимальные температурные условия для активности PpLDC, определяли ферментативную активность при следующих температурах: 50°С, 60°С и 70°С таким же образом, как и в Примере 2-2. В результате было обнаружено, что наивысшая активность PpLDC наблюдается при 50°С (ФИГ. 15).
[00117] Активность PpLDC при различных значениях рН определяли таким же образом, как и в Примере 2-2. В результате наивысшая активность PpLDC наблюдалась при рН 6, и его реакционная способность падала с повышением значения рН (ФИГ. 16).
[00118] Пример 6. Получение Е. coli со вставленным геном лизиндекарбоксилазы, полученным из Pseudomonas synxantha, и анализ активности лизиндекарбоксилазы, экспрессированной в нем
[00119] 6-1. Трансформация Е. coli геном лизиндекарбоксилазы, полученным из Pseudomonas synxantha
[00120] Для клонирования гена лизиндекарбоксилазы (pxldc), полученного из Pseudomonas synxantha, использовали праймеры 5_PxLDC_NdeI (AATATACATATGTACAAAGACCTCCAA ТТСССС) (SEQ ID NO: 23) и 3_PxLDC_XhoI (AATATACTCGAGTCACTCCCTTATGCAATCAACGGTATAGC) (SEQ ID NO: 24) и очищенную геномную ДНК Pseudomonas synxantha в качестве матрицы для выполнения ПЦР. Pfu ДНК полимеразу использовали для амплификации гена, и ПЦР выполняли за 30 циклов в следующих условиях: 94°С: 30 сек, 45°С: 30 сек и 72°С: 2 мин с получением амплифицированного pxldc (SEQ ID NO: 10).
[00121] Полученный ген pxldc экспрессировали при низкой температуре в E. coli таким же образом, как и в Примере 2-1, и идентифицировали, используя SDS-PAGE (ФИГ. 13). Как видно по Линиям 7 и 8 на ФИГ. 13, полученная из Pseudomonas synxantha лизиндекарбоксилаза (PxLDC) сверхэкспрессируется в виде растворимого белка при низких температурах.
[00122] 6-2. Исследование активности полученной из Pseudomonas synxantha лизиндекарбоксилазы, экспрессированной в E. coli
[00123] (1) Исследование реакционной способности PxLDC
[00124] Для исследования реакционной способности PxLDC, клеточный лизат PxLDC, полученный в Примере 6-1, центрифугировали при 13000 об/мин в течение 15 минут с получением надосадочной жидкости, которую использовали для конверсии лизина. Проводили реакцию 50 мкл растворимого белка, 100 мМ PLP и 250 мМ лизина в 50 мМ натрийфосфатного буфера (рН 6,2) в реакционном объеме 200 мкл при 46°С в течение 2 часов. В результате получили 25% кадаверина (ФИГ. 17).
[00125] (2) Исследование активности лизиндекарбоксилазы в зависимости от рН
[00126] Для того, чтобы найти оптимальные условия рН для PxLDC, определяли ферментативную активность при различных значениях рН таким же образом, как и в Примере 2-2 (ФИГ. 18). В результате наивысшая активность PxLDC наблюдалась при рН 6, и его реакционная способность падала с повышением значения рН.
[00127] Пример 7. Сравнение активности полученной из E. coli лизиндекарбоксилазы и полученной из Pseudomonas thermotolerans лизиндекарбоксилазы
[00128] 7-1. Клонирование и экспрессия полученной из E. coli лизиндекарбоксилазы
[00129] Ген лизиндекарбоксилазы E. coli - cadA - клонировали для экспрессии EcLDC (SEQ ID NO: 11). Гомология между аминокислотными последовательностями PtLDC и EcLDC составляла 36%. Клонированную с геном cadA плазмиду вводили в E. coli K-12 BL21 и инкубировали при 37°С в течение 4 часов для стимулирования экспрессии. Полностью экспрессированную таким образом EcLDC идентифицировали, используя SDS-PAGE (ФИГ. 13; Линии 1 и 2). В результате EcLDC сверхэкспрессировался в виде растворимого белка.
[00130] 7-2. Сравнение относительной ферментативной активности EcLDC и PtLDC
[00131] (1) Сравнение активности в зависимости от температуры
[00132] Относительную ферментативную активность (относительную активность) сравнивали для EcLDC и PtLDC при различных температурах (37°С, 42°С, 50°С, 60°С, 70°С и 80°С) таким же образом, как и в Примере 2-2 (ФИГ. 19).
[00133] В результате было найдено, что наивысшая активность у обоих EcLDC и PtLDC наблюдается при 60°С. Относительная активность EcLDC при 50°С составляла 54% (при этом активность EcLDC при 60°С была принята за 100%), а относительная активность EcLDC при 80°С составляла 12%. Относительная активность PtLDC при 50°С составляла 76% (при этом активность PtLDC при 60°С была принята за 100%), а относительная активность PtLDC при 80°С составляла 19%. Было найдено, что активность PtLDC хорошо сохранялась при высоких температурах. В заключение, оба фермента продемонстрировали большое различие в уровнях их активности в зависимости от температуры, и относительная активность PtLDC сохранялась лучше, чем у EcLDC.
[00134] (2) Сравнение активности в зависимости от рН
[00135] Кроме того, активность оценивали при разных значениях рН (6,2, 7,4, 8,0 и 9,0) таким же образом, как и в Примере 2-2 (ФИГ. 20). В результате наивысшая активность EcLDC наблюдалась при рН 6, и ферментативная активность EcLDC сильно падала с повышением значения рН. При рН 9 сохранялось 50% активности EcLDC. В отличие от этого, PtLDC не проявлял существенных изменений в своей активности в зависимости от рН, и активность PtLDC поддерживалась на уровне не менее 90% при значениях рН 6,2~9. Соответственно, оценка показала, что PtLDC обладает более высокой устойчивостью к температуре или изменению рН, чем EcLDC.
[00136] (3) Сравнение активности у PtLDC и EcLDC
[00137] После количественного анализа белков PtLDC и EcLDC для оценки удельной активности (ед./мг), PtLDC продемонстрировал значение, равное 10060 (ед./мг), a EcLDC продемонстрировал значение, равное 36335 (ед./мг). При сравнении их реакционных способностей оказалось, что активность EcLDC примерно в 3,6 раза выше активности PtLDC. Кроме того, при сравнении оптимальных температур для этих ферментов, оба фермента продемонстрировали оптимальное действие при 60°С, а их активность сильно снижалась при перемене температуры. Однако при сравнении оптимальных значений рН оказалось, что у EcLDC наблюдается более высокая специфическая инактивация при повышении рН, а у PtLDC существенного изменения ферментативной активности при изменении рН не наблюдалось.
[00138] Активность EcLDC выше, чем у PtLDC. Однако активность EcLDC может быть более подвержена влиянию изменения рН, поскольку в ходе реакции с участием лизиндекарбоксилазы рН повышается. PtLDC более устойчив к рН, чем EcLDC, что является преимуществом в реакции конверсии лизина. Коммерческое производство кадаверина биоконверсией лизина требует регулирования рН путем обработки кислотой, но при применении PtLDC необходимость в титровании рН может ослабнуть. Ожидается, что затраты на производство кадаверина биоконверсией могут быть снижены.
[00139] Депозитарий: Корейский центр культур микроорганизмов (KCCM)
[00140] Инвентарный номер: KCCM11559P
[00141] Дата депонирования: 20140724.

































Группа изобретений относится к области биотехнологии. Предложен микроорганизм, трансформированный полинуклеотидом, кодирующим белок, обладающий лизиндекарбоксилазной активностью, содержащий аминокислотную последовательность SEQ ID NO: 1 или аминокислотную последовательность, гомологичную не менее чем на 75% этой последовательности, для экспрессии белка, обладающего лизиндекарбоксилазной активностью, где белок, обладающий лизиндекарбоксилазной активностью, происходит из Pseudomonas sp. и микроорганизм представляет собой микроорганизм Escherichia sp. Предложен микроорганизм, способный продуцировать кадаверин, где микроорганизм обладает улучшенной способностью продуцировать лизин по сравнению с диким типом и трансформирован полинуклеотидом, кодирующим белок, обладающий лизиндекарбоксилазной активностью, содержащий аминокислотную последовательность SEQ ID NO: 1 или аминокислотную последовательность, гомологичную не менее чем на 75% этой последовательности, для экспрессии белка, обладающего лизиндекарбоксилазной активностью, где белок, обладающий лизиндекарбоксилазной активностью, происходит из Pseudomonas sp. и микроорганизм представляет собой микроорганизм Escherichia sp. Предложены варианты способа получения кадаверина с использованием указанных микроорганизмов. Группа изобретений позволяет снизить затраты на производство кадаверина биоконверсией лизина. 4 н. и 6 з.п. ф-лы, 20 ил., 1 табл., 7 пр.
1. Микроорганизм, трансформированный полинуклеотидом, кодирующим белок, обладающий лизиндекарбоксилазной активностью, содержащий аминокислотную последовательность SEQ ID NO: 1 или аминокислотную последовательность, гомологичную не менее чем на 75% этой последовательности, для экспрессии белка, обладающего лизиндекарбоксилазной активностью, где белок, обладающий лизиндекарбоксилазной активностью, происходит из Pseudomonas sp. и микроорганизм представляет собой микроорганизм Escherichia sp.
2. Микроорганизм, способный продуцировать кадаверин, где микроорганизм обладает улучшенной способностью продуцировать лизин по сравнению с диким типом и трансформирован полинуклеотидом, кодирующим белок, обладающий лизиндекарбоксилазной активностью, содержащий аминокислотную последовательность SEQ ID NO: 1 или аминокислотную последовательность, гомологичную не менее чем на 75% этой последовательности, для экспрессии белка, обладающего лизиндекарбоксилазной активностью, где белок, обладающий лизиндекарбоксилазной активностью, происходит из Pseudomonas sp. и микроорганизм представляет собой микроорганизм Escherichia sp.
3. Микроорганизм по п. 1 или 2, где аминокислотная последовательность, гомологичная не менее чем на 75%, представляет собой аминокислотную последовательность SEQ ID NO: 3, 5, 7 или 9.
4. Микроорганизм по п. 1 или 2, где полинуклеотид имеет нуклеотидную последовательность SEQ ID NO: 2 или нуклеотидную последовательность, гомологичную не менее чем на 75% этой последовательности.
5. Микроорганизм по п. 4, где нуклеотидная последовательность, гомологичная не менее чем на 75%, представляет собой последовательность SEQ ID NO: 4, 6, 8 или 10.
6. Способ получения кадаверина, включающий стадии:
конверсии лизина в кадаверин путем использования микроорганизма по п. 1; и
извлечения полученного кадаверина.
7. Способ получения кадаверина, включающий стадии:
культивирования микроорганизма по п. 2 в среде; и
извлечения кадаверина из микроорганизма или среды.
8. Способ по п. 6 или 7, где аминокислотная последовательность, гомологичная не менее чем на 75%, представляет собой аминокислотную последовательность SEQ ID NO: 3, 5, 7 или 9.
9. Способ по п. 6 или 7, где полинуклеотид имеет нуклеотидную последовательность SEQ ID NO: 2 или нуклеотидную последовательность, гомологичную не менее чем 75% этой последовательности.
10. Способ по п. 9, где нуклеотидная последовательность, гомологичная не менее чем на 75%, представляет собой последовательность SEQ ID NO: 4, 6, 8 или 10.
| Data Basa Online GenBank WP_017938426 от 28.06.2013 [Найдено онлайн 24.05.2018] | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| US 20110039313 A1, 17.02.2011 | |||
| WO 2014113999 A1, 31.07.2014 | |||
| СПОСОБ ПОЛУЧЕНИЯ ХИМИЧЕСКОГО ПРОДУКТА И АППАРАТ ДЛЯ НЕПРЕРЫВНОЙ ФЕРМЕНТАЦИИ | 2009 |
|
RU2513694C2 |
| СПОСОБ ПОЛУЧЕНИЯ САХАРНОГО РАСТВОРА | 2009 |
|
RU2516792C2 |

