Изобретение относится к медицине, биотехнологии, фармакологии, а именно, к способам оценки сродства лигандов к биомишени и может быть использовано для оценки эффективности связывания новых синтезированных олигонуклеотидов с мишенями.
В современной медицине одним из приоритетных направлений является поиск таргетных лекарственных средств, которые действуют избирательно на определенные типы клеток, не затрагивая клетки, не вовлеченные в патологический процесс. Особенно важна селективность при создании противоопухолевых препаратов, когда необходимо действовать на малигнизированные клетки. При этом желательна невысокая токсичность для всего организма. Одним из классов веществ, способных распознавать молекулы-мишени на нужных клетках являются олигонуклеотидные аптамеры. Технология их получения основана на отборе по связыванию с мишенью. И поэтому способы, позволяющие оценить связывание олигонуклеотидного аптамера, весьма востребованы [Jing, М.; Bowser, М.Т. Anal Chim Acta 2011, 686, 9-18].
Известен способ оценки специфичности связывания молекул с мишенью с применением радиоактивной метки [Международная заявка WO 00/52473 от 08.09.2000 «Набор для радиоактивного мечения и анализ связывания»]. Предметом изобретения является набор реактивов, позволяющий помимо диагностических целей оценивать специфичность связывания антител с клетками мишенями, положительными по антигену.
Однако, набор и способ предназначен для анализа антител и не предполагает использование для оценки сродства олигонуклеотидов. Кроме того, применение радиоактивной метки сопряжено с осуществлением мер безопасности и затратами на утилизацию радиоактивных отходов, что делает способ трудновыполнимым в лаборатории без изотопного блока.
Известно устройство, применяемое для детектирования связывания [Патент RU 2609184 от 17.01.2013, «Устройство, применяемое для детектирования сродства»], в котором осуществлен способ оценки сродства, основанный на рассеивании затухающего поля когерентного света, распространяющегося через планарный волновод по поверхности, модифицированной молекулами-мишенями, связывающимися с испытуемым веществом.
К недостаткам способа следует отнести то, что мишень, привязанная к подложке может изменить свои конформационные свойства и результаты оценки специфичности связывания будут искажены. Кроме того, модель не вполне адекватна реальному событию связывания олигонуклеотида с рецептором-мишенью на поверхности клетки.
Известен способ оценки сродства, на основе селектора и системы количественной оценки [Заявка RU 2014135576 А от 30.01.2013, «Распознавание на основе селектора и системы количественной оценки, а также способ единого анализа множества анализируемых веществ»]. Способ использует принцип разделения иммунных комплексов с помощью технологий селективной адсорбции на колонке или картридже.
К недостаткам способа относится использование предварительно подготовленных колонок и картриджей, необходимых для разделения связавшегося и несвязанного селектора, которые зачастую недоступны или требуют дополнительных исследований для их создания.
Наиболее адекватной моделью оценки афинности на наш взгляд является непосредственный контакт олигонуклеотида с клеткой-мишенью, имеющей большое число рецепторов целевого белка на внешней мембране. Этот способ оценки афинности аптамеров описан в статье «Dynamics and Visualization of MCF7 Adenocarcinoma Cell Death by Aptamer-C1q-Mediated Membrane Attack» [J.R. Stecker, A.A. Savage, J.G. Bruno, D.M.  and J.R. Koke. Dynamics and Visualization of MCF7 Adenocarcinoma Cell Death by Aptamer-C1q-Mediated Membrane Attack. 2012 Nucleic Acid Ther; 22(4): 275-282] Этот способ выбран в качестве наиболее близкого к представленному изобретению. Способ осуществляют следующим образом.
and J.R. Koke. Dynamics and Visualization of MCF7 Adenocarcinoma Cell Death by Aptamer-C1q-Mediated Membrane Attack. 2012 Nucleic Acid Ther; 22(4): 275-282] Этот способ выбран в качестве наиболее близкого к представленному изобретению. Способ осуществляют следующим образом.
Известный дезоксирибонуклеотидный аптамер MUC1-5TR-1, разработанный ранее [Ferreira CS, Papamichael K, Guilbault G, Schwarzacher T, Gariepy J, Missailidis S. DNA aptamers against the MUC1 tumour marker: design of aptamer-antibody sandwich ELISA for the early diagnosis of epithelial tumours. Anal. Bioanal. Chem. 2008;390:1039-1050] был выбран. Его константа диссоциации с мишенью составляла 47.3 nM. Для исследования использовали указанный олигонуклеотидный аптамер с 5'-биотин- или 5'-Alexa Fluor 546 (AF546) меткой, нуклеотидная последовательность: 5'-GGGAGACAAGAATAAACGCTCAAGAAGTGAAAATGACAGAACACAACATTCGACAGGAGGCTCACAACAGGC-3'.
Исследуемый аптамер метили по флангу 5'-биотин- или 5'-Alexa Fluor 546 (AF546) меткой.
Мишенью для исследуемого аптамера служил маркер MUC1, в большом количестве представленный на поверхности клеток линии MCF7. Клетки линии MCF7 рака груди человека были выращены на стеклянных чашках до 80-90% покрытия монослоя чашки. Клетки промывали дважды 1х буфером для связывания (1× ВВ; 0.5 М NaCl, 10 мМ Tris-HCl, и 1 мМ MgCl2, в воде, свободной от нуклеаз, рН 7.5-7.6), затем к клеткам добавляли 2,4 мл свежей стерилизованной фильтрованием среды и 200 мкл исследуемого олигонуклеотида (1.5 мг/мл), инкубировали 5 мин при 37°С с 95% воздушной смеси с добавлением 5% СО2.
Клетки дважды отмывали 1× ВВ в течение 5 мин, фиксировали в метаноле при -20°С в течение 1 мин. После высушивания, клетки трижды промывали 1× ВВ, каждая отмывка в течение 10 мин, инкубировали с 1:20 моноклональным мышиным анти-человеческим антителом С9 (United States Biological, Inc.) в течение ночи при 4°С. Далее клетки трижды промывали 1× ВВ, каждая отмывка в течение 10 мин, блокировали 20% сухим молоком в течение 2 часов при комнатной температуре, после чего еще трижды промывали 1× ВВ, каждая отмывка в течение 10 мин. Затем клетки инкубировали с анти-мышиным вторичным антителом, содержащим Су5-метку (Invitrogen) в течение 2 часов при комнатной температуре. Далее клетки снова трижды промывали 1× ВВ, каждая отмывка в течение 10 мин, в течение 20 мин инкубировали с 1:2,000 красителем ядер Хехста, снова отмывали, регистрировали изображение с помощью флуоресцентной микроскопии.
Оценку аффинности аптамера олигонуклеотидной природы с мишенью осуществляли визуально, разглядывая фотографии светящихся клеток.
Тем не менее к недостаткам способа следует отнести отсутствие количественных характеристик визуальной оценки результата. Это сильно затрудняет сравнение исследуемых аптамеров по сродству. Именно на решение этой задачи направлено данное изобретение.
Сущность изобретения состоит в том, что испытуемый олигонуклеотид используют с ковалентно пришитым флуоресцентным красителем.
Выбирают культуру клеток, которая содержит на поверхности клеток богато представленную мишень для исследуемого олигонуклеотида. В качестве контрольной культуры выбирают культуру клеток с низкой представленностью целевого рецептора на внешней поверхности клеточной мембраны.
Испытуемый олигонуклеотид инкубируют с положительными клетками, содержащими целевой белковый рецептор в большем количестве. Одновременно в тех же условиях проводят инкубацию испытуемого образца олигонуклеотида с культурой контрольных клеток (не обогащенных целевым рецептором).
Культуры клеток отмывают от не связявшегося олигонуклеотида. Осуществляют флуоресцентную микроскопию и фотографирование образцов клеток (положительной и контрольной линии).
Фотографию контрольного образца вместе с изображением опытного образца объединяют в один графический растровый файл. Инвертируют изображение и измеряют оптическую плотность окрашенных клеток при помощи программы анализа графических файлов. Сродство олигонуклеотида к рецептору рассчитывают по формуле:
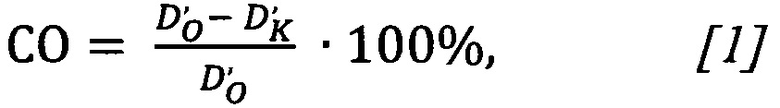
где СО - сродство олигонуклеотида;
D'о - оптическая плотность опытного образца за вычетом оптической плотности фона;
D'к - оптическая плотность контрольного образца за вычетом оптической плотности фона.
Следует отметить, что в аналогах сродство олигонуклеотида оценивают с помощью кажущейся константы диссоциации (KD), а в заявляемом способе - по формуле [1]. Расчет KD связан с необходимостью получения точных данных о текущей концентрации каждого компонента в смеси. Это усложняет задачу и не помогает достигать практических целей, а именно оценки сродства олигонуклеотида с многокомпонентной живой структурой - клеткой. В отличие от прототипа в заявленном способе используют контрольную культуру клеток и предложенный количественный критерий. Экспериментальным путем подобраны условия экспозиции (инкубации и отмывки) олигонуклеотида с клетками, при которых наблюдается различие в связывании контрольных и опытных образцов. Подбор условий инкубации включал использование клеток иммобилизованных на поверхности луночных планшетов и в суспензии.
Приведенные ниже примеры раскрывают конкретную реализацию выполнения способа, описанного в изобретении, но никоем образом не ограничивают сам способ.
Пример 1
Испытан олигонуклеотид TuTu22 описанный в [D.-L.Wang, Y.-L. Songc, Z. Zhuc, X.-L. Lic, Y. Zouc, H.-T. Yanga, J.-J. Wanga, P.-S. Yaob, R.-J. Pana, C.J. Yangc, D.-Z. Kang, Selection of DNA aptamers against epidermal growth factor receptor with high affinity and specificity, BBRC, 453, 4, 681-685] к рецептору эпидермального фактора роста EGFR. Нуклеотидная последовательность: 5'-FAM-TACCAGTGCGATGCTCAGTCCGTATGCTCAGTCTTACTATAACTTCTCTGCTCGTTGCTGACGCATTCGGTTGAC-3'
Этот олигонуклеотид был помечен по флангу ковалентно конъюгированным флуоресцентным красителем флуоресцеином (FAM).
В качестве опытной культуры клеток использовали стандартную линию клеток А431 (культура клеток карциномы почек человека). В качестве контроля использовали линию С6 (линия клеток крысы). Ранее с помощью ПЦР в реальном времени нами был показан высокий уровень экспрессии белка EGFR (60-ти кратно более высокий уровень экспрессии EGFR, чем в клетках фибробластов человека). В контрольных клетках С6 наблюдалась нулевая экспрессия человеческого EGFR.
Клетки культивировали в стандартных условиях для выращивания, количество клеток составляло 3×106 клеток на лунку стандартного четырехлуночного планшета. Опытные и контрольные культуры клеток выращивали в лунках одного и того же планшета.
Готовили раствор аптамера с концентрацией 100 нМ в смеси 50% фосфатного буферного раствора и 50% бессывороточной среды для культивирования клеток. Среду для культивирования анализируемых клеточных линий заменяли на 200 мкл раствора аптамера. Инкубировали испытуемый аптамер с опытной и контрольной культурами клеток в течение 60 мин при бережном перемешивании на термостатированном шейкере при температуре 37°С. Все манипуляции проводили в условиях затемнения для предотвращения «выцветания» флуоресцентной метки аптамера.
Первичную отмывку проводили дважды раствором фосфатного буфера: предварительно удалили раствор аптамера и добавили 1000 мкл буфера к клеткам. После двукратной отмывки буфером, раствор сливали, добавляли 1000 мкл бессывороточной среды инкубации, клетки выдерживали 10 мин при бережном перемешивании на термостатированном шейкере при температуре 37°С.
После окончания отмывки среду удаляли, к клеткам приливали 200 мкл свежей среды для инкубации и помещали четырехлуночный планшет на предметный столик флуоресцентного микроскопа.
Чтобы убедиться, что зоны окрашивания совпадают с очертаниями клеток, проводили визуализацию зоны наблюдения при видимом свете. Фоторегистрацию проводили с помощью флуоресцентного микроскопа Olympus IX.
Визуализацию опытных и контрольных клеток проводили при одних и тех же условиях фоторегистрации.
Фотографии опытного и контрольного образцов объединяли в один графический файл для синхронизации обработки и анализа изображений.
На Фигуре 1 представлено цветное (А) и черно-белое (В) изображение на левой части которого - культура С6, на правой - А431.
С помощью программного обеспечения PhotoM 1.31 полученное изображение клеток опытного и контрольного образца инвертировали и определяли оптические плотности участков изображений, соответствующих флуоресцентно-окрашенных исследуемым олигонуклеотидом клеток-мишеней в опытных и контрольных образцах и вычитали оптическую плотность участков вне клеток для опытного и контрольного изображения. Для повышения точности оценки измерение характеристик проводили по пяти различным зонам с последующим усреднением результатов.
Указанное программное обеспечение позволяет вычитать фоновую оптическую плотность. На Фигуре 2 представлен скриншот (изображение) программы PhotoM 1.31 демонстрирующий вышеназванное свойство для культур С6 и А431 изображенных на Фигуре 1.
Были получены следующие значения:
Для С6 оптическая плотность по выделенным областям:
1. 1942 пкс (0.07%). Плотность 0.0137.
2. 2127 пкс (0.08%). Плотность 0.00913.
3. 2081 пкс (0.07%). Плотность 0.00508.
4. 2092 пкс (0.08%). Плотность 0.00426.
5. 2331 пкс (0.08%). Плотность 0.00794.
Средняя по объекту 0.00796
Для А431 оптическая плотность по выделенным областям:
1. 984 пкс (0.04%). Плотность 0.00678.
2. 743 пкс (0.03%). Плотность 0.00615.
3. 627 пкс (0.02%). Плотность 0.0104.
4. 775 пкс (0.03%). Плотность 0.0104.
5. 914 пкс (0.03%). Плотность 0.00548.
Средняя по объекту 0.00799.
D'о=0.00799;
D'к=0.00796.
При подстановке полученных значений оптической плотности в формулу [1]
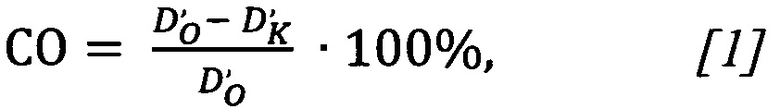
получили значение сродства олигонуклеотида СО=(0.00799-0.00796)/(0.00799)⋅100%=0,4%
На основании полученного результата делаем вывод, что испытуемый аптамер не обладает выраженным сродством к выбранной мишени.
Пример 2.
С помощью заявленного способа оценивали аффинность олигонуклеотида GREF18, меченного Су3 к клеткам ФЭЧ-6. Контрольной культурой клеток служила линия клеток А431. Способ осуществляли аналогично примеру 1.
На Фигуре 3 представлена фоторегистрация окрашенных клеток в цветном (А) и черно-белом варианте (В), где в левой части изображения - культура А431, в правой - ФЭЧ6. На Фигуре 4 представлен скриншот (изображение) программы PhotoM 1.31 с перенесенными в нее данными по культурам А4631 и ФЭЧ6 изображенных на Фигуре 2.
Были получены следующие значения:
Для А431 оптическая плотность по выделенным областям:
1. 22426 пкс (0.80%). Плотность 0.0222.
2. 10714 пкс (0.38%). Плотность 0.0228.
3. 14440 пкс (0.52%). Плотность 0.0288.
4. 16055 пкс (0.58%). Плотность 0.0104.
5. 9979 пкс (0.36%). Плотность 0.0125.
Средняя по объекту 0.0197.
Для ФЭЧ6 оптическая плотность по выделенным областям:
1. 11617 пкс (0.42%). Плотность 0.186.
2. 9528 пкс (0.34%). Плотность 0.0701.
3. 9729 пкс (0.35%). Плотность 0.203.
4. 6376 пкс (0.23%). Плотность 0.128.
5. 7629 пкс (0.27%). Плотность 0.154.
Средняя по объекту 0.151.
D'o=0.151;
D'к=0.020.
При подстановке полученных значений оптической плотности в формулу [1]
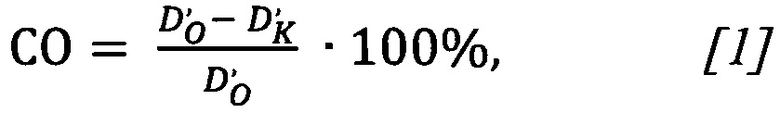
получили значение сродства олигонуклеотида СО=(0.151-0.020)/(0.151)⋅100%=87%
Делаем вывод о сильном сродстве испытуемого олигонуклеотида к культуре клеток ФЭЧ6.
Изобретение позволяет сравнивать олигонуклеотиды по сродству к клеткам, поверхность которых обогащена целевыми молекулами-мишенями.
Изобретение вводит количественные критерии для отбора наиболее высокоаффинных олигонуклеотидов к молекулам-мишеням на поверхности клеток. С другой стороны, изобретение может быть применено для маркирования клеток и тканей определенного типа, например, при росте раковой опухоли.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ВЫДЕЛЕНИЯ И АНАЛИЗА ЭКЗОСОМ | 2022 |
|
RU2788198C1 |
| АПТАМЕР, СПЕЦИФИЧНЫЙ К ОПУХОЛЕВЫМ ТКАНЯМ ЛЕГКОГО ЧЕЛОВЕКА | 2012 |
|
RU2528870C2 |
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК-АПТАМЕРОВ, СВЯЗЫВАЮЩАЯ ШИГА-ТОКСИН ТИПА 2 | 2014 |
|
RU2566552C1 |
| СПОСОБ СОЗДАНИЯ И СЕЛЕКЦИИ БИБЛИОТЕКИ МОДИФИЦИРОВАННЫХ АПТАМЕРОВ | 2016 |
|
RU2652952C1 |
| ФОТОАКТИВИРУЕМОЕ ХИМИЧЕСКОЕ ОБЕСЦВЕЧИВАНИЕ КРАСИТЕЛЕЙ | 2012 |
|
RU2623880C2 |
| Способ термической диссоциации для проведения селекции ДНК-аптамеров | 2019 |
|
RU2723373C1 |
| Способ терапии глиомы человека за счет снижения пролиферации и индукции дифференцировки опухолевых клеток глиомы человека | 2022 |
|
RU2820200C2 |
| СПОСОБ СЕЛЕКЦИИ АПТАМЕРОВ К ЗАДАННЫМ БЕЛКОВЫМ МИШЕНЯМ НА ПОВЕРХНОСТИ КЛЕТОК | 2012 |
|
RU2518368C1 |
| АПТАМЕРЫ, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ TLR-4, И ИХ ПРИМЕНЕНИЕ | 2015 |
|
RU2709718C2 |
| ДНК-аптамеры к ДНК-полимеразе эта человека | 2020 |
|
RU2737285C1 |
Изобретение относится к медицине, а именно к фармакологии, и может быть использовано для оценки сродства олигонуклеотида к мишени. Для этого проводят мечение олигонуклеотида флуорохромом. Затем осуществляют контакт меченного олигонуклеотида с мишенью-рецептором на поверхности клеток. После чего проводят фоторегистрацию полученного образца с использованием флуоресцентного микроскопа. При этом измеряют оптическую плотность области окрашивания инвертированных изображений и сравнивают с отрицательным контролем. Затем рассчитывают сродство олигонуклеотида к мишени. Изобретение обеспечивает отбор наиболее высокоаффинных олигонуклеотидов к молекулам-мишеням на поверхности клеток для маркирования клеток и тканей. 4 ил., 2 пр.
Способ оценки сродства олигонуклеотида к мишени, включающий мечение олигонуклеотида флуорохромом, контакт меченного олигонуклеотида с мишенью-рецептором на поверхности клеток, фоторегистрацию результата с использованием флуоресцентного микроскопа, отличающийся тем, что измеряют оптическую плотность области окрашивания инвертированных изображений, сравнивают с отрицательным контролем, а расчет сродства осуществляют по формуле:
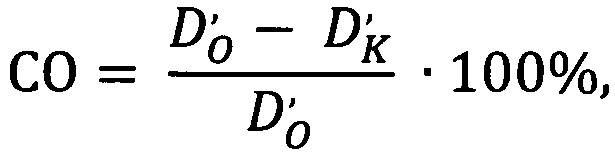
где СО - сродство олигонуклеотида;
D'о - оптическая плотность опытного образца за вычетом оптической плотности фона;
D'к - оптическая плотность контрольного образца за вычетом оптической плотности фона.
| ДВИГАТЕЛЬ ДИЗЕЛЯ ДЛЯ ТЕПЛОВОЗА | 1926 |
|
SU6512A1 |
| RU 2014133427 A, 10.03.2016 | |||
| УСТРОЙСТВО ДЛЯ ИССЛЕДОВАНИЯ ПРОЦЕССА ВОЛОЧЕНИЯ | 1936 |
|
SU52473A1 |
| STECKER J.R | |||
| et al., Dynamics and visualization of MCF7 adenocarcinoma cell death by aptamer-C1q-mediated membrane attack, Nucleic Acid Ther., 2012 Aug; 22 (4):275-82 | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |

