ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к очищенному экстракту грибов (SsE1), который выявляет иммунный ответ растений, и к идентификации рецептора растительного происхождения AtRLP30, который опосредует распознавание SsE1. В настоящем изобретении предусматриваются также способы получения экстракта SsE1 и применение SsE1 и AtRLP30 в растениях для модуляции иммунного ответа растений, в частности, против грибковых инфекций Sclerotinia spp. или Botrytis spp.
УРОВЕНЬ ТЕХНИКИ
Современная модель иммунной системы растений подтверждает существование двух направлений. Одно направление основано на распознавании патоген-ассоциированных высококонсервативных молекулярных паттернов (PAMPs, ПАМП) локализованными на мембране паттерн-распознающими рецепторами (PRRs), результатом чего является активация внутриклеточных сигнальных путей, приводящая к укреплению клеточной стенки и продуцированию антимикробных соединений. PAMPs по определению являются незаменимыми молекулами, которые типичны для целого класса, и, следовательно, их трудно видоизменить или удалить, не вызывая серьезного снижения уровня приспособленности микроба. PAMPs также известны как микроб-ассоциированные молекулярные паттерны (MAMPs), так как они не ограничиваются одними патогенными (болезнетворными) микроорганизмами. Хорошо известным примером является Pep-13, экспонированный на поверхности антигенный эпитоп на кальций-зависимой тканевой трансглутаминазе (клеточной стенки), который широко распространен в патогенных оомицетах, относящихся к порядку Perenosporales (Пероноспоровые), но присутствует также в светящихся морских бактериях Vibrio. Pep-13 активирует иммунитет (защиту) петрушки и картофеля, из чего можно сделать вывод, что он функционирует как родоспецифическая детерминанта для активации иммунитета растения как у растений-хозяев, так и у растений, не являющихся хозяевами.
Позднее бактериальный пептидогликан был идентифицирован как новый РАМР/МАМР, инициирующий иммунные ответы в Arabidopsis thaliana (резуховидке Таля). Обработка пептидогликаном индуцирует поток ионов, повышение внутриклеточной концентрации ионов Ca2+, продуцирование реакционноспособных форм кислорода и фитоалксина камалексина. Эксперименты по профилированию транскриптов показали, что пептидогликан влияет на экспрессию многих генов и что перепрограммирование транскриптома Arabidopsis хорошо совпадает с изменениями, вызываемыми пептидом flg22, (N-концевым) участком флагеллина, натуральным РАМР/МАМР. Обратный генетический метод позволил идентифицировать содержащие лизиновый мотив рецептор-подобные белки (RLP) LYM1 и LYM3 в качестве рецепторов пептидогликанов, для которых дополнительно требуется действие третьего LysM белка, LysM-RLK CERK1.
Паттерн-распознающие рецепторы (PRRs) на поверхности растительной клетки распознают MAMPs и передают сигнал в клетку. До настоящего времени было идентифицировано несколько сигналов, относящихся к богатым лейциновыми повторами рецептор-подобным киназам (LRR-RLK) и к семейству белков (LRR-RLP) и к LysM-RLKs/RLPs. Лучше всего изученным рецептором РАМР/МАМР в растениях является FLS2, рецептор-подобный белок-киназа растения Arabidopsis с внеклеточным доменом, богатым лейциновыми повторами (LRR-RLK), который распознает и связывает flg22, состоящий из 22 аминокислотных остатков фрагмент бактериального флагеллина. Ортологи FLS2 присутствуют в томатах, табаке, ячмене и рисе, из чего можно сделать вывод, что распознавание флагеллина представляет собой древний эволюционный механизм обнаружения патогенов. Важно отметить, что растения Arabidopsis Т-ДНК линий с нефункциональным FLS2 более восприимчивы к инфекциям, вызванным патогенными бактериями, такими как Pto DC3000. Недавние исследования показали, что FLS2, по механизму, опосредуемому flg22, образует комплекс с BAK1, другим членом семейства LRR-RLK. Интересно отметить, что ВАК1 первоначально был идентифицирован как партнер BRI1, LRR-RLK, который распознает растительный гормон брассинолид, который принимает участие в регуляции процессов роста и развития. Действие BAK1 необходимо для реакции на некоторые PAMPs/MAMPs, включая flg22 и бактериальный фактор элонгации EF-Tu в Arabidopsis, или бактериальный белок холодового шока и раздражитель белковой природы элицитин 1NF1 из оомицетов в растении табака. Лежащая в основе молекулярного механизма активация рецептора включает индуцируемые лигандом конформационные изменения в рецепторном комплексе с последующими событиями ауто- и транс-фосфорилирования в киназном домене взаимодействующих партнеров, инициирующими нисходящие сигналы. Хотя разные PAMPs/MAMPs распознаются разными рецепторами, они индуцируют общие ранние события передачи сигнала, включающие повышенные уровни цитоплазматического кальция, активацию MAP киназ, продуцирование реакционноспособных форм кислорода и индукцию биосинтеза салициловой кислоты, жасмоната и этилена в растениях Arabidopsis.
Важно отметить, что в биологических исследованиях стимулирование с применением PAMPs/MAMPs предоставляет способ инициирования защитной системы растений без образования трансгенных продуктов и уже предлагается в сельском хозяйстве для повышения жизнеспособности растений.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
С учетом вышеизложенного целью настоящего изобретения является предоставление новых PAMPs/MAMPs в растениях и их соответствующих рецепторов, которые способствуют повышению устойчивости растений, конкретно, устойчивости к инфицированию грибковыми патогенами семейства Sclerotiniaceae.
В первом аспекте настоящего изобретения вышеуказанная цель достигается с помощью иммуногенного в отношении растений экстракта грибов, который получают способом, включающим стадии:
(а) предоставление культурального фильтрата клеток Sclerotinia sclerotiorum (склеротинии склероциевидной),
(б) введение указанного фильтрата из п. (а) в первую катионообменную колонку, уравновешенную низкосолевым буфером А,
(в) элюирование экстракта высокосолевым буфером В,
(г) разведение элюированной фракции из п. (в) в буфере А с целью сделать возможным связывание со второй катионообменной колонкой,
(д) введение разведенной фракции из п. (г) во вторую катионообменную колонку и элюирование иммуногенного в отношении растений экстракта грибов буферным солевым раствором, проводимость которого составляет 5-20 мС/см, предпочтительно, 8-16 мС/см.
В другом аспекте проблема, поставленная в настоящем изобретении, разрешается способом получения иммуногенного экстракта грибов, включающим стадии (а) предоставление культурального фильтрата клеток Sclerotinia sclerotiorum, (б) введение указанного фильтрата из п. (а) в первую катионообменную колонку, уравновешенную низкосолевым буфером А, (в) элюирование экстракта высокосолевым буфером В, (г) разведение элюированной фракции из п. (в) в буфере А с целью сделать возможным связывание со второй катионообменной колонкой, (д) введение разведенной фракции из п. (г) во вторую катионообменную колонку и элюирование иммуногенного в отношении растений экстракта грибов буферным солевым раствором, проводимость которого составляет 5-20 мС/см, предпочтительно, 8-16 мС/см.
Согласно одному предпочтительному варианту изобретения культуральный фильтрат, предоставляемый на стадии (а) вышеописанного способа, готовят фильтрованием культуральной среды 2-3-недельной культуры Sclerotinia sclerotiorum через нейлоновые сита и последующей лиофилизацией среды в течение 3-4 дней. Затем лиофилизированный материал ресуспендируют в буфере А (около 6 мл/г сухой массы) и центрифугируют, чтобы отделить нерастворимые частицы. Супернатант затем используют в качестве культурального фильтрата в способе по настоящему изобретению. Настоящее изобретение охватывает также другие способы предоставления культурального фильтрата грибов, известные специалисту в данной области техники.
Для иммуногенного в отношении растений экстракта грибов и способа его получения по настоящему изобретению предпочтительно, чтобы буфер А представлял собой 100 мМ Mes буфер с pH 5.4, и/или чтобы буфер В представлял собой 100 мМ Mes буфер с pH 5.4, 0.5 М KCl. Mes - это общепринятое (стандартное) название 2-(N-морфолино)этансульфоновой кислоты. Mes буфер готовят по стандартным методикам, известным специалисту в данной области техники.
Также предпочтительным является вышеуказанный иммуногенный в отношении растений экстракт грибов и способ его получения, где матрицей в указанной первой катионообменной колонке является GE Healthcare Sepharose SP™ FastFlow. Более предпочтительно, чтобы объем слоя указанной матрицы в колонке GE Healthcare XK16 составлял около 15 мл.
Согласно одному варианту изобретения культуральный фильтрат вводится в указанную колонку при скорости потока от 3 до 5 мл/мин. Затем, необязательно, колонку промывают буфером А.
Предпочтительно, элюирование на стадии (в) осуществляют буфером В при скорости потока 3-5 мл/мин. В данном случае предпочтительно, чтобы отбиралась одна фракция, соответствующая пику элюирования, с OD280nm и OD215nm. Необязательно, элюированную фракцию можно проверять на растениях - таких как Arabidopsis thaliana - на этилен-индуцирующую активность.
Или же активность экстракта по изобретению можно проверять по выявлению типичных индуцированных РАМР защитных реакций растений, таких как посттрансляционная активация MAP киназы и транскрипционная активация маркерных иммунных генов, например, например, белка, связанного с патогенезом, 1 (PR1), или флагеллин-отвечающей киназы 1 (FRK1) (FLG-индуцируемой (индуцибельной) рецептороподобной киназы). Однако не следует считать, что изобретение ограничивается тестированием с помощью вышеуказанных защитных реакций. Для проверки экстракта по изобретению на активность можно также проводить мониторинг экспрессии других иммунных генов или биохимических реакций, типичных для инфекции, вызываемой патогенами.
Согласно другому предпочтительному варианту изобретения указанная вторая катионообменная колонка представляет собой колонку GE Healthcare Source 15S 4.6/100 РЕ, которая, предпочтительно, заранее уравновешена буфером А.
Предпочтительным является иммуногенный в отношении растений экстракт грибов по изобретению и способ его получения, где фракция, элюированная на стадии (г), разводится в буфере А, примерно, в 10 раз.
Согласно другому предпочтительному варианту разведенную фракцию, полученную на стадии (г), вводят во вторую катионообменную колонку при скорости потока от 0.5 до 1.5 мл/мин. После ввода разведенной фракции во вторую катионообменную колонку ее, предпочтительно, промывают буфером А.
Предпочтительно вводить указанный элюирующий буфер на последней стадии элюирования (д) при скорости потока от 0.5 до 1 мл/мин.
Согласно одному предпочтительному варианту элюирование осуществляют в линейном градиенте концентрации соли в буфере В (например, от 0% до 60% в 40 объемах колонки) и в этом градиенте отбирают ряд фракций по 500 мкл. Соответственно, все фракции, элюированные буферным солевым раствором, проводимость которого составляет 5-20 мС/см, предпочтительно, 8-16 мС/см, содержат экстракт по изобретению. Или же все фракции, элюированные буферным солевым раствором, проводимость которого составляет 5-20 мС/см, предпочтительно, 8-16 мС/см, объединяют, получая экстракт по изобретению.
После стадии (д) элюированную фракцию, которая представляет собой экстракт по изобретению, можно, необязательно, проверять на этилен-индуцирующую активность, например, методами, описанными в разделе Примеры настоящего изобретения.
В контексте настоящего изобретения предпочтительно, чтобы хроматографию осуществляли на системе GE Healthcare ÄKTA Explorer FPLC при охлаждении до 4°С и при использовании программы GE Healthcare Unicorn.
Помимо этого предусматривается иммуногенная композиция, полученная способом по настоящему изобретению.
Согласно другому варианту иммуногенный экстракт грибов по изобретению содержит по меньшей мере один из белков, выбранный из Цитохрома С (A7E6R4), N-ацетилтрансферазы (A7F941), ингибитора диссоциации Rho-GDP (А7ЕТ57), полиубиквитина (А7Е4Е9), белка дисульфид-изомеразы (A7EDH2) или пектин-эстеразы (A7EXV0). Еще более предпочтительно, когда указанный экстракт грибов содержит по меньшей мере два указанных выше белка, предпочтительно по меньшей мере три указанных белка и, наиболее предпочтительно, все указанные выше белки. В скобках указан номер доступа для входа в банк данных UniProtKB/TrEMBL.
Цель настоящего изобретения достигается также с помощью иммуногенного экстракта грибов, который содержит по меньшей мере один из белков, выбранный из Цитохрома С (A7E6R4), N-ацетилтрансферазы (A7F941), ингибитора диссоциации Rho-GDP (А7ЕТ57), полиубиквитина (А7Е4Е9), белка дисульфид-изомеразы (A7EDH2) или пектин-эстеразы (A7EXV0). Еще более предпочтительно, когда указанный экстракт грибов содержит по меньшей мере два указанных выше белка, предпочтительно по меньшей мере три указанных белка и, наиболее предпочтительно, все указанные выше белки.
Один аспект изобретения включает иммуногенные экстракты грибов по изобретению, применяемые для активации, усиления или инициирования иммунного ответа растения. В другом аспекте цель настоящего изобретения достигается за счет применения выделенного белка, выбранного из Цитохрома С (A7E6R4), N-ацетилтрансферазы (A7F941), ингибитора диссоциации Rho-GDP (А7ЕТ57), полиубиквитина (А7Е4Е9), белка дисульфид-изомеразы (A7EDH2) или пектин-эстеразы (A7EXV0) или их активного фрагмента или гомолога, для активации, усиления или инициирования иммунного ответа растения.
В контексте настоящего изобретения гомологичный белок означает белок, имеющий аминокислотную последовательность, на 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или, предпочтительно, на 99% идентичную последовательности одного из белков, выбранных из Цитохрома С (A7E6R4), N-ацетилтрансферазы (A7F941), ингибитора диссоциации Rho-GDP (А7ЕТ57), полиубиквитина (А7Е4Е9), белка дисульфид-изомеразы (A7EDH2) или пектин-эстеразы (A7EXV0). Согласно настоящему изобретению активные фрагменты этих белков или их гомологов представляют собой такие участки белков, которые обладают активностью, выражающейся в способности индуцировать иммунный ответ растения на грибковую инфекцию, предпочтительно, на инфицирование грибами семейства Склеротиниевых (Sclerotiniaceae), например, на инфицирование растения грибами Sclerotinia sclerotiorum или Botrytis cinerea.
Соответственно, другой аспект изобретения также относится к применению нуклеиновой кислоты, кодирующей белок, выбранный из Цитохрома С (A7E6R4), N-ацетилтрансферазы (A7F941), ингибитора диссоциации Rho-GDP (А7ЕТ57), полиубиквитина (А7Е4Е9), белка дисульфид-изомеразы (A7EDH2) или пектин-эстеразы (A7EXV0) или их активного фрагмента или гомолога, для активации, усиления или инициирования иммунного ответа растения.
Гомологичная нуклеиновая кислота в контексте настоящего изобретения означает нуклеиновую кислоту, имеющую нуклеотидную последовательность, на 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или, предпочтительно, на 99% идентичную последовательности одной из нуклеиновых кислот, кодирующей белок, выбранный из Цитохрома С (A7E6R4), N-ацетилтрансферазы (A7F941), ингибитора диссоциации Rho-GDP (А7ЕТ57), полиубиквитина (А7Е4Е9), белка дисульфид-изомеразы (A7EDH2) или пектин-эстеразы (A7EXV0). Согласно настоящему изобретению активные фрагменты этих белков или их гомологов представляют собой такие участки белков, которые обладают активностью, выражающейся в способности индуцировать иммунный ответ растения на грибковую инфекцию, предпочтительно, на инфицирование грибами семейства Склеротиниевых (Sclerotiniaceae), например, на инфицирование растения грибами Sclerotinia sclerotiorum или Botrytis cinerea.
Еще один аспект изобретения относится к способу модуляции устойчивости растения к инфицированию патогенами (к болезнетворной инфекции), включающий модуляцию в указанном растении экспрессии белка, содержащего аминокислотную последовательность, по меньшей мере на 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или, предпочтительно, на 100% идентичной AtRLP30. Предпочтительно, AtRLP30 представляет собой белок, содержащий аминокислотную последовательность, показанную в SEQ ID No. 1.
Согласно некоторым вариантам изобретения выражение "модуляция устойчивости растения к инфицированию патогенами (к инфекции, вызываемой патогенами)" означает либо повышение, либо снижение устойчивости растения. С другой стороны выражение "модуляцию в указанном растении экспрессии белка" означает либо увеличение количества экспрессируемого белкового продукта в указанном растении по сравнению с необработанным контрольным растением, либо уменьшение количества экспрессируемого белкового продукта в указанном растении по сравнению с необработанным контрольным растением. Или же модулируется не экспрессия указанного белка, а активность белка. При этом модуляция активности белка означает либо усиление, либо ослабление биохимической/биологической функции указанного белка. Такую модуляцию активности можно стимулировать, например, вводя в растение мутировавший белок, который выявляет измененные биохимические характеристики по сравнению с белком дикого типа.
В данном аспекте повышение экспрессии/активности указанного белка приводит к повышению устойчивости указанного растения к инфекции, вызываемой патогенами. Снижение экспрессии/активности приводит к снижению устойчивости указанного растения к инфекции, вызываемой патогенами.
Помимо этого неожиданно было обнаружено, что в передаче сигнала через AtRLP30 дополнительно участвует корецепторный белок киназа, ассоциированная с BRI1- 1 (BAK1). По этой причине один предпочтительный способ по изобретению помимо модуляции AtRLP30 или его гомологов включает также модуляцию в указанном растении экспрессии белка, содержащего аминокислотную последовательность, по меньшей мере на 50%, 60%, 70%, 80%, 90%, 95%, 98%, 99% или 100% идентичную аминокислотной последовательности BAK1, причем повышение экспрессии указанного белка приводит к повышению устойчивости указанного растения к болезнетворной инфекции, а снижение экспрессии приводит к снижению устойчивости указанного растения к болезнетворной инфекции. Таким образом, в соответствии со способами по настоящему изобретению модуляция экспрессии или активности как BAK1, так и AtRLP30, или их гомологов, позволяет осуществлять сходное (в равной степени) усиление модуляции иммунного ответа растения, конкретно, иммунного ответа на грибковую инфекцию.
В контексте настоящего изобретения предпочтительный белок ВАК1 представляет собой белок BAK1 растения Arabidopsis thaliana.
Согласно предпочтительным вариантам изобретения экспрессия описанного выше указанного белка в указанном растении повышается при использовании эктопической экспрессии указанного белка. С другой стороны экспрессия указанного белка в указанном растении снижается, например, посредством направленного мутагенеза, РНК интерференции или РНК-опосредуемого метилирования ДНК.
Опосредуемые РНК механизмы сайленсинга могут препятствовать экспрессии генов на разных уровнях: некоторые РНК-управляемые механизмы действуют на посттрансляционном уровне с использованием расщепления целевых матричных РНК. Однако производные дцРНК также могут управлять изменениями в хроматиновой структуре областей ДНК, с которыми их последовательности идентичны. Например, растения используют такие РНК для того, чтобы "наложить" отпечатки (импринты) метилирования цитозина на идентичные последовательности ДНК, создавая основную модель для образования транскрипционно-молчащего гетерохроматина. Этот процесс обычно называют направляемым РНК метилированием ДНК (RdDM).
RdDM инициируется молекулами двухцепочечной РНК (дцРНК, dsRNA), находящейся в клеточном ядре. Возможно (теоретически) они инициируют de novo (первичное) метилирование всех цитозиновых оснований, которые локализованы в областях ДНК, комплементарных последовательности двойной цепи РНК. В результате этого, в организме млекопитающих и в растениях, метилированные положения в ДНК служат в качестве меток (маркеров, указателей) для перестройки (модификации) соседнего (прилегающего) хроматина таким образом, чтобы в этих локусах мог образоваться плотный гетерохроматин. Благодаря плотной среде хроматина другие белки не могут контактировать с ДНК. Так, в частности, транскрипционные факторы и компоненты механизма транскрипции нельзя собрать на метилированных последовательностях промоторов, и, следовательно, на этих участках никакая транскрипция не может происходить. Фактически гены, имеющие метилированные регуляторные последовательности, в меньшей степени транскрибируются и, следовательно, в меньшей степени экспрессируются.
Предпочтительно, модуляция экспрессии AtRLP30 и/или BAK1 или их гомологов по изобретению осуществляется с использованием опосредуемого РНК метилирования ДНК, нацеленного на последовательность, выбранную, но без ограничения, из эндогенной регуляторной последовательности, которая контролирует транскрипцию ДНК растения. Термин "регуляторная последовательность" в данном контексте означает нуклеотидную последовательность, которая, будучи функционально связана с кодирующей областью гена, влияет на транскрипцию кодирующей области таким образом, что молекула рибонуклеиновой кислоты (РНК) транскрибируется с помощью кодирующей области. Как правило, регуляторный элемент может повышать или понижать уровень транскрипции нуклеотидной последовательности, например, кодирующей последовательности, функционально связанной с элементом, по сравнению с уровнем транскрипции нуклеотидной последовательности в отсутствие регуляторного элемента. Регуляторные элементы хорошо известны в уровне техники и, предпочтительно, включают промоторы, энхансеры, сайленсеры-ослабители транскрипции, инактивированные последовательности интронных сайленсеров, 3'-нетранслируемые и 5'-нетранслируемые последовательности транскрибируемой последовательности, предпочтительно, поли-А сигнальную последовательность или другие элементы, стабилизирующие белок или РНК, инсуляторы, которые подавляют регуляторный эффект этих последовательностей в отношении заданных областей, или другие элементы контроля экспрессии генов, заведомо регулирующие экспрессию генов или количество экспрессии генного продукта. Регуляторный элемент может быть выделен из последовательности природной геномной ДНК или может быть синтетическим, например, синтетическим промотором.
В контексте настоящего изобретения термины "полинуклеотид", "олигонуклеотид" и "нуклеотидная последовательность" употребляются как синонимы по отношению к полимерной (состоящей из двух или более мономеров) форме нуклеотидов любой длины (протяженности), либо рибонуклеотидов, либо дезоксирибонуклеотидов. Хотя нуклеотиды обычно связаны фосфодиэфирными связями, термин также включает полимеры, содержащие нейтральные амидные связи в основной цепи, состоящие из аминоэтилглициновых звеньев (мономеров). Эти термины применяются только по отношению к первичной структуре молекулы. Соответственно, термин включает двухцепочечные и одноцепочечные молекулы ДНК, как отмечалось выше. Следует учитывать, что такие полинуклеотиды можно модифицировать, например, включением метки, такой как радиоактивная, флуоресцентная или другая метка, метилированием, включением кэпа, заменой одного или более натуральных нуклеотидов на нуклеотидный аналог, введением межнуклеотидной модификации, такой как нейтральные (не имеющие заряда) связи (например, метилфосфонатные, фосфотриэфирные, фосфорамидатные, карбаматные и т.п. связи), введением дополнительной молекулы, такой как молекула белка (например, молекулы нуклеазы, токсина, антитела, сигнального пептида, поли-L-лизина и т.п.), введением интеркалятора, такого как акридин или псорален, введением хелатирующего агента, который может представлять собой металл, такой как бор, окислитель-ион металла или радиоактивный металл, введением алкилирующего агента или наличием модифицированной связи (например, альфа-аномерная нуклеиновая кислота).
Предпочтительные полинуклеотиды по настоящему изобретению выбраны из оцДНК (онДНК), дцДНК (днДНК), мДНК (tDNA), оцРНК (онРНК), дцРНК (днРНК), кшРНК (shRNA), киРНК (siRNA) и мРНК. Предпочтительно, полинуклеотид представляет собой ДНК, которая кодирует молекулу дцРНК, предпочтительно, дцРНК-шпильку (шпилечную дцРНК). ДцРНК-шпильки, предпочтительно, образуются посредством экспрессии конструкции ДНК, которая кодирует смежные смысловую и антисмысловую последовательности, разделенные спейсером. После транскрипции такой конструкции полученная молекула оцРНК образует двойную цепь посредством спаривания оснований смысловой и антисмысловой последовательностей.
В контексте данного изобретения инфекция, вызываемая патогенами, предпочтительно, означает грибковую инфекцию, предпочтительно, инфекцию Sclerotinia или Botryotiona, например, инфекцию в результате заражения (инфицирования) растения с помощью Sclerotinia sclerotiorum или Botrytis cinerea.
Растения, которые, предпочтительно, используются в контексте настоящего изобретения или которые являются предпочтительными мишенями для иммуногенного экстракта, включают кукурузу (Zea mays), Brassica sp. (род Капуста, например, B. napus (брюква), B. rapa (репа) B. juncea (горчица сарепская)), люцерну (Medicago sativa), рожь (Secale cereal, рожь посевная), сорго (Sorghum bicolor (сорго двухцветное), Sorghum vulgare (сорго техническое)), подсолнечник (Helianthus annum, подсолнечник масличный), сафлор (Carthamus tinctorius, сафлор красильный)), пшеницу (Triticum aestivum, пшеница мягкая), сою (Glycine max, соя обыкновенная), табак (Nicotiana tabacum, табак настоящий), картофель (Solanum tuberosum, картофель европейский), арахис (Arachis hypogaea, земляной орех), хлопок, хлопчатник (Gossypium barbadense, хлопок американский, Gossypium hirsutum, хлопок травянистый), батат, сладкий картофель (Ipomoea batatus), кассаву (Manihot esculenta, маниок съедобный), кофе (Cofea spp.), кокосовую пальму (Cocos nucifera), ананас (Ananas comosus, ананас крупнохохолковый), цитрусовые деревья (Citrus spp.), шоколадное дерево (Theobroma cacao), чайный куст (Camellia sinensis, камелия китайская), банан (Musa spp.), авокадо (Perseaultilane), инжир, фиговое дерево (Ficus carica), гуаву (Psidium guava), манго (Mangifera indica, манго индийское), оливковое дерево (Olea europaea), папайю (Carica papaya, дынное дерево), кешью (Anacardium occidentale, анакардий западный), макадамию (Macadamia integrifolia), миндаль (Prunus amygdalus), сахарную свеклу (Beta vulgaris), сахарный тростник (Saccharum spp.), овес, ряску (Lemna), ячмень, томаты (Lycopersicon esculentum), латук (например, Lactuca sativa, салат, латук посевной), зеленую фасоль (Phaseolus vulgaris), фасоль лимскую (Phaseoluslimensis), чину (Lathyrus spp.) и членов рода Cucumis, таких как огурец (C sativus, огурец посевной), канталупа (C. cantalupensis), и мускусная дыня (C. melo). Также используются декоративные растения, такие как азалия (Rhododendron spp.), гортензия (Macrophylla hydrangea), гибискус (Hibiscus rosasinensis, китайская роза), розы (Rosa spp.), тюльпаны (Tulipa spp.), нарциссы (Narcissus spp.), петунии (Petunia hybrida), гвоздика (Dianthus caryophyllus, гвоздика голландская), пуансеттия (Euphorbia pulcherrima) и хризантемы. Другие декоративные растения, которые входят в объем изобретения, включают растения рода недотрог (бальзамин), бегонию, пеларгонию, фиалки, цикламен, вербену, барвинок, бархатцы, первоцвет (примулу), сенполию (узанбарскую фиалку), агератум, амарант, львиный зев, аквилегию, пепельник (цинерарию), клевер, растения рода Cosmos, вигну китайскую, георгины, дурман, шпорник (дельфиниум), герберы, глоксинии, гиппеаструм, растения рода мезембриантемум, сальпиглоссис и циннии. Хвойные деревья для практического применения по настоящему изобретению включают, например, сосну, такую как ладанная сосна (Pinus taeda), сосна Эллиота (Pinus elliotii), сосна желтая (Pinus ponderosa), сосна скрученная широкохвойная (Pinus contorta) и сосна лучистая (Pinus radiata), ель Дугласа (Pseudotsuga menziesii); тсугу западную (Tsugaultilane); серебристую ель (Picea glauca); секвойю вечнозеленую (Sequoia sempervirens); настоящие пихты, такие как пихта благородная (Abies amabilis) и пихта бальзамическая (Abies balsamea); и кедры, такие как туя (Thuja plicata) и аляскинский желтый кедр (Chamaecyparis nootkatensis), предпочтительно, в тех случаях, когда растение восприимчиво к инфекции Sclerotinia spp или Botrytis spp.
Цель настоящего изобретения позволяет также достичь способ получения трансгенного растения с повышенной устойчивостью к грибковой инфекции, включающий стадии (i) трансформацию растения или растительной клетки с помощью нуклеотидной последовательности, кодирующей AtRLP30 или AtRLP30-подобный белок, содержащий аминокислотную последовательность, по меньшей мере на 50%, 60%, 70%, 80%, 90%, 95%, 98%, 99% или 100% идентичную SEQ ID No 1. Согласно предпочтительным вариантам изобретения нуклеотидная последовательность, кодирующая AtRLP30 или AtRLP30-подобный белок, содержащий аминокислотную последовательность, по меньшей мере на 50%, 60%, 70%, 80%, 90%, 95%, 98%, 99% или 100% идентичную SEQ ID No 1, включена в экспрессирующий вектор, который способствует экспрессии полинуклеотида в растении.
Согласно предпочтительным вариантам изобретения любые способы получения растений по настоящему изобретению по существу не являются биологическими процессами. Более предпочтительно, способы получения растений по настоящему изобретению не содержат какие-либо стадии или не состоят из каких-либо стадий полового скрещивания полных геномов растений и последующего отбора растений.
В другом аспекте настоящего изобретения предусматривается растение, полученное способом получения трансгенного растения, обладающее повышенной устойчивостью к грибковой инфекции, см. выше в данном описании.
Еще в одном аспекте настоящего изобретения цель достигается посредством гена, содержащего нуклеотидную последовательность, кодирующую AtRLP30 или AtRLP30-подобный белок (гомолог), содержащий аминокислотную последовательность, по меньшей мере на 50%, 60%, 70%, 80%, 90%, 95%, 98%, 99% или 100% идентичную SEQ ID No 1.
Термин "ген" в контексте настоящего изобретения означает любой элемент последовательности ДНК, который может транскрибироваться в РНК и который мог бы кодировать наследственные признаки в организме. Большинство генов является генами, кодирующими белки, при этом нуклеотидная последовательность гена кодирует аминокислотную последовательность белкового продукта. Однако другие гены могут кодировать РНК, которые не транслируются в белки - так называемые некодирующие РНК (ncRNAs, нкРНК). Например, гены нкРНК кодируют транспортные РНК (тРНК) или структурные РНК, такие как обнаруженные в больших белковых комплексах, подобных рибосомам (рРНК, rRNA). Термин "ген" включает также кодирующие области для малых некодирующих РНК. "Гены" малых некодирующих РНК включают малые ядрышковые (малые нуклеолярные) РНК (мякРНК, мнРНК, snoRNAs), микроРНК (microRNAs), киРНК (siRNAs) и piPHK (пиРНК, piRNAs), и длинные нкРНК, которые включают такие примеры, как Xist и HOTAIR.
Еще в одном аспекте изобретение относится к экспрессирующей кассете, характеризующейся тем, что эта экспрессирующая кассета позволяет экспрессировать ген по изобретению.
В другом предпочтительном аспекте настоящее изобретение относится к вектору, содержащему нуклеиновую кислоту по настоящему изобретению, например, ген или экспрессирующую кассету по настоящему изобретению. Вектор по определению в настоящем изобретении означает белок или нуклеиновую кислоту или их смесь, которые могут быть введены или способны вводить содержащиеся в них полинуклеотиды в клетку. Предпочтительно, чтобы белки, кодируемые вводимой нуклеиновой кислотой, экспрессировались в клетке после введения вектора.
Согласно предпочтительному варианту вектор по настоящему изобретению включает рекомбинантный вектор, плазмиды, фагмиды, фаги, космиды, вирусы, в частности, но без ограничения, вирусные векторы на основе ампликона, векторы на основе Х-вируса картофеля, векторы на основе вируса погремковости табака, векторы на основе геминивирусов, например, векторы на основе вируса курчавости (скручивания) листьев капусты и вируса штриховатой мозаики ячменя и векторы на основе вирусов-сателлитов (см. обзор Curtin, S.J., Wang, М.-В., Watson, J.M., Roffey, P., Blanchard, C.L. and Waterhouse, P.M.. (2007), chapter 12, p 291-332 в "Rice Functional Genomics; Challenges, Progress and Prospects". Upadhyaya, Narayana M. (Ed.), ISBN: 978-0-387-48903-2), виросомы и покрытые нуклеиновыми кислотами частицы, в частности, шарики из золота.
Термин "молекула рекомбинантной нуклеиновой кислоты" ("рекомбинантная нуклеотидная молекула") относится к полинуклеотиду, получаемому при участии человека. Молекула рекомбинантной нуклеиновой кислоты может содержать две или более нуклеотидных последовательности, которые связаны таким образом, что получается продукт, в природе не встречающийся в клетке. В частности, две или более нуклеотидные последовательности могут быть функционально связаны и, например, могут кодировать слитый (гибридный) полипептид, или могут содержать нуклеотидную последовательность и регуляторный элемент. Рекомбинантная нуклеотидная молекула может быть на основе природного полинуклеотида, но может отличаться от него, например, полинуклеотид может иметь одну или более нуклеотидных замен, например, первый кодон, который обычно содержится в полинуклеотиде, заменяется на вырожденный кодон, который кодирует ту же самую или консервативную аминокислоту, или, например, в полинуклеотид вводится соответствующая последовательность, например, сайт распознавания рестриктаз или сайт сплайсинга, промотор, сайт инициации репликации ДНК и т.п.
Предпочтительным является рекомбинантный вектор по настоящему изобретению, который представляет собой экспрессирующий вектор, необязательно содержащий один или более генов, которые должны экспрессироваться. Предпочтительно, указанная экспрессия запускается регуляторной последовательностью (или последовательностями). Регуляторная последовательность может быть выделенной из последовательности натуральной геномной ДНК или может быть синтетической, например, синтетическим промотором.
Такие экспрессирующие векторы по настоящему изобретению, предпочтительно, используются согласно тем вариантам изобретения, в которых экспрессия белка по изобретению повышается с целью модуляции устойчивости указанного трансформированного растения. Предпочтительно, устойчивость означает устойчивость к инфекции, вызываемой патогенами, наиболее предпочтительно, инфекцию вызывает грибковый патоген, описанный выше.
Регуляторные последовательности могут представлять собой конститутивно экспрессируемые регуляторные последовательности, которые поддерживают экспрессию генов при относительном уровень активности (основном уровне), или могут являться регулируемыми регуляторными последовательностями. Конститутивно экспрессируемая регуляторная последовательность может экспрессироваться в клетках любых типов или может быть тканеспецифической, т.е. экспрессируется только в клетках определенных типов, фазоспецифической, т.е. экспрессируется только на определенных стадиях развития или роста растительной клетки, и т.п. Регуляторная последовательность, такая как тканеспецифическая или фазоспецифическая регуляторные последовательности, или индуцибельная (индуцируемая) регуляторная последовательность, используемая для создания рекомбинантного полинуклеотида или для практического применения способа по изобретению, может представлять собой регуляторную последовательность, которая обычно, в природе, имеется в геноме растения. Однако регуляторная последовательность также может представлять собой последовательность другого организма, отличного от растения, включая, например, вирус растений, вирус животных или клетку животного или другого многоклеточного организма.
Предпочтительной регуляторной последовательностью для экспрессии полинуклеотидов по изобретению является промоторный элемент. Применимые промоторы включают, но без ограничения, конститутивные, индуцибельные (индуцируемые), регулируемые во времени, регулируемые в процессе развития, пространственно регулируемые, химически регулируемые, отвечающие на стресс, тканеспецифические, вирусные и синтетические промоторы. Известно, что промоторные последовательности бывают сильными или слабыми. Сильный промотор обеспечивает высокий уровень экспрессии гена, тогда как слабый промотор обусловливает очень низкий уровень экспрессии гена. Индуцибельный промотор означает промотор, который обеспечивает включение и выключение экспрессии генов в ответ на агент, поступающий извне, или на экологический или дифференцировочный стимул. Бактериальный промотор можно индуцировать, варьируя уровни экспрессии генов в зависимости от уровня изотиопропил-галактозида, добавляемого к трансформируемым бактериальным клеткам. Выделенная промоторная последовательность, которая представляет собой сильный промотор для гетерологичной нуклеиновой кислоты, является предпочтительной, так как она обеспечивает достаточный уровень экспрессии генов, что позволяет легко обнаруживать и отбирать трансформированные клетки, и при необходимости обеспечивает высокий уровень экспрессии генов.
Выбор промотора в значительной степени зависит от временных и пространственных требований к экспрессии, а также от целевого объекта ("вида"). В отдельных случаях желательной является экспрессия в различных (нескольких) тканях. В то время как в других случаях целесообразной является тканеспецифическая, например, лист-специфическая, семя-специфическая, лепесток-специфическая, пыльник-специфическая или сердцевина-специфическая экспрессия. Хотя показано, что многие промоторы двудольных являются функциональными ("работают") в однодольных и наоборот, по возможности промоторы двудольных выбирают для экспрессии в двудольных, а промоторы однодольных для экспрессии в однодольных. Однако никаких ограничений относительно происхождения или источника выбранного промотора не существует. Достаточно того, чтобы промоторы были функциональными и запускали экспрессию заданной нуклеотидной последовательности в конкретной клетке.
Были найдены другие последовательности, которые повышают (усиливают) экспрессию генов в трансгенных растениях; они включают интронные последовательности (например, интрон Adh 1, bronze 1, актина 1, актина 2 (Международная заявка WO 00/760067) или сахарозосинтазы), сигналы полиаденилирования в первичной 3'-нетранслируемой области и вирусные лидерные последовательности (например, из TMV, MCMV и AMV). Например, известно, что целый ряд нетранслируемых лидерных последовательностей вирусов повышает экспрессию. Конкретно, было найдено, что лидерные последовательности вируса табачной мозаики (TMV), вируса хлорозной пятнистости кукурузы (MCMV) и вируса мозаики люцерны (AMV) эффективно повышают экспрессию (например, Gallie et al., 1987; Skuzeski et al., 1990). Другие лидерные последовательности, известные в уровне техники, включают, но без ограничения, лидерные последовательности топи-коронавирусов, например, лидерную последовательность EMCV (5'-некодирующей области вируса энцефаломиокардита; Elroy-Stein et al., 1989); лидерные последовательности потивирусов, например, лидерную последовательность TEV (вируса гравировки табака); лидерную последовательность MDMV (вируса карликовой мозаики кукурузы); лидерную последовательность человеческого белка, связывающего тяжелую цепь иммуноглобулинов (BiP), (Macejak et al., 1991); нетранслируемую лидерную последовательность мРНК обол очечного белка AMV (AMV RNA 4; Jobling et al., 1987), TMV (Gallie et al., 1989) и MCMV (Lommel et al., 1991; см. также della Cioppa etal., 1987).
Для экспрессии любых конструкций по настоящему изобретению в растении или в растительной клетке изобретение, предпочтительно, воплощается таким образом, чтобы описанные полинуклеотиды были функционально связаны с промотором и с сайтом полиаденилирования, при этом указанный промотор характеризуется тем, что он является функциональным в указанной клетке указанного растения. В данном контексте в качестве промотора подходит любой элемент последовательности, который индуцирует транскрипцию последовательности в прямом (5'-3') направлении. Минимальные требования к промоторам общеизвестны из уровня техники, и многие из таких промоторов обычно применяются для экспрессии генов в растениях. Согласно предпочтительному варианту изобретения трансформацию растения или растительной клетки при использовании любого полинуклеотида по настоящему изобретению осуществляют способом, выбранным из стандартных методов, известных из уровня техники. Трансформацию растительной ткани, предпочтительно, можно осуществлять методом бомбардировки (микро)частицами (Klein et al., "High-Velocity Microprojectiles for Delivering Nucleic Acids Into Living Cells," Nature 327:70-73 (1987)), называемым также методом баллистической трансформации клетки-хозяина, который раскрывается в патентах США №4,945,050, 5,036,006 и 5,100,792, все на имя Sanford et al., и в статье Emerschad et al., "Somatic Embryogenesis and Plant Development from Immature Zygotic Embryos of Seedless Grapes (Vitis vinifera)" Plant Cell Reports 14:6-12 (1995). При бомбардировке (микро)частицами микрочастицы вольфрама или золота (диаметром от 1 до 2 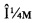 ) покрывают соответствующей ДНК, а затем бомбардируют ткань с применением газа под высоким давлением. Таким способом можно доставлять чужеродные нуклеотиды в ядро. Биологически активные частицы (например, высушенные бактериальные клетки, содержащие вектор и гетерологичную ДНК) можно доставлять в растительные клетки таким же образом. Можно также применять другие варианты бомбардировки частицами, неизвестные или которые будут разработаны в дальнейшем. Другим предпочтительным методом стабильного введения конструкции нуклеиновой кислоты в растительные клетки является заражение растительной клетки с использованием Agrobacterium tumefaciens (агробактерии опухолеобразующей) или Agrobacterium rhizogenes (агробактерии "корнеродной"), предварительно трансформированных с помощью полинуклеотидной конструкции. Как описано выше, Ti (или RT) плазмида агробактерии (Agrobacterium) делает возможным в высшей степени успешный перенос молекулы чужеродной нуклеиновой кислоты в растительные клетки. В предпочтительном варианте трансформации Agrobacterium применяют вакуумную инфильтрацию, в которой используют цельные растения (Senior, "Uses of Plant Gene Silencing," Biotechnology and Genetic Engineering Reviews 15:79-119 (1998)). Еще одним таким методом введения является слияние протопластов с другими частицами, либо с мини-клетками, клетками, лизосомами, либо с другими способными к слиянию тельцами с липидной оболочкой (Fraley et al., Proc. Natl. Acad. Sci. USA 79:1859-63 (1982)). Также предпочтительным является метод, в котором нуклеотидная молекула вводится в растительные клетки электропорацией (Fromm et al., Proc. Natl. Acad. Sci. USA 82:5824 (1985)). Согласно этому методу протопласты растений подвергают электропорации в присутствии плазмид, содержащих экспрессирующую кассету. Электрические импульсы тока большой силы обратимо пермеабилизуют биомембраны, делая возможной введение плазмид. Электропорированные протопласты растений реформируют (преобразуют) клеточную стенку, делятся и регенерируют. Другие предпочтительные методы трансформации включают обусловленную химическими соединениями трансформацию растения, микроинъекцию, физические абразивы, вирусную трансдукцию и лазерные пучки (Senior, "Uses of Plant Gene Silencing," Biotechnology and Genetic Engineering Reviews 15:79-119 (1998)). Конкретный метод трансформации не особенно важен для практического применения настоящего изобретения. Любой метод, который приводит к эффективной трансформации выбранной клетки-хозяина, является подходящим для осуществления настоящего изобретения на практике.
) покрывают соответствующей ДНК, а затем бомбардируют ткань с применением газа под высоким давлением. Таким способом можно доставлять чужеродные нуклеотиды в ядро. Биологически активные частицы (например, высушенные бактериальные клетки, содержащие вектор и гетерологичную ДНК) можно доставлять в растительные клетки таким же образом. Можно также применять другие варианты бомбардировки частицами, неизвестные или которые будут разработаны в дальнейшем. Другим предпочтительным методом стабильного введения конструкции нуклеиновой кислоты в растительные клетки является заражение растительной клетки с использованием Agrobacterium tumefaciens (агробактерии опухолеобразующей) или Agrobacterium rhizogenes (агробактерии "корнеродной"), предварительно трансформированных с помощью полинуклеотидной конструкции. Как описано выше, Ti (или RT) плазмида агробактерии (Agrobacterium) делает возможным в высшей степени успешный перенос молекулы чужеродной нуклеиновой кислоты в растительные клетки. В предпочтительном варианте трансформации Agrobacterium применяют вакуумную инфильтрацию, в которой используют цельные растения (Senior, "Uses of Plant Gene Silencing," Biotechnology and Genetic Engineering Reviews 15:79-119 (1998)). Еще одним таким методом введения является слияние протопластов с другими частицами, либо с мини-клетками, клетками, лизосомами, либо с другими способными к слиянию тельцами с липидной оболочкой (Fraley et al., Proc. Natl. Acad. Sci. USA 79:1859-63 (1982)). Также предпочтительным является метод, в котором нуклеотидная молекула вводится в растительные клетки электропорацией (Fromm et al., Proc. Natl. Acad. Sci. USA 82:5824 (1985)). Согласно этому методу протопласты растений подвергают электропорации в присутствии плазмид, содержащих экспрессирующую кассету. Электрические импульсы тока большой силы обратимо пермеабилизуют биомембраны, делая возможной введение плазмид. Электропорированные протопласты растений реформируют (преобразуют) клеточную стенку, делятся и регенерируют. Другие предпочтительные методы трансформации включают обусловленную химическими соединениями трансформацию растения, микроинъекцию, физические абразивы, вирусную трансдукцию и лазерные пучки (Senior, "Uses of Plant Gene Silencing," Biotechnology and Genetic Engineering Reviews 15:79-119 (1998)). Конкретный метод трансформации не особенно важен для практического применения настоящего изобретения. Любой метод, который приводит к эффективной трансформации выбранной клетки-хозяина, является подходящим для осуществления настоящего изобретения на практике.
Другой аспект настоящего изобретения относится к способу скрининга на микроб-ассоциированные молекулярные паттерны (PAMPs/MAMPs), включающему стадии: (i) экспрессирование в растении или в растительной клетке белка, содержащего аминокислотную последовательность, по меньшей мере на 50%, 60%, 70%, 80%, 90%, 95%, 98%, 99% или 100% идентичную SEQ ID No. 1 (AtRLP30), (ii) контактирование указанного растения или растительной клетки с соединением-кандидатом, (iii) количественное определение иммунного ответа указанного растения или указанной растительной клетки по сравнению с контрольным растением или с контрольной растительной клеткой, при этом повышенный иммунный ответ указанного растения или указанной иммунной клетки показывает, что указанное соединение-кандидат представляет собой МАМР. Предпочтительным является вышеприведенный способ, в котором указанный МАМР означает ассоциированный с грибами молекулярный паттерн, предпочтительно, имеющийся в Sclerotinia spp. или Botrytis ssp.
Предпочтительно, на стадии (iii) вышеприведенного способа по изобретению иммунный ответ указанного растения или указанной растительной клетки измеряют, определяя продуцирование этилена и/или экспрессию генов, отвечающих за иммунные реакции, или репортерных генов. Репортерные гены, применимые в контексте настоящего изобретения, состоят из промотора, отвечающего за иммунные реакции, функционально связанного с репортерным геном, который позволяет легко считывать данные экспрессии репортерного гена, например, люциферазные ферменты или флуоресцентные белки. Специалисту в данной области техники доступен широкий выбор ферментов, которые можно использовать в качестве репортерных генов. Еще один аспект изобретения относится к способу очистки МАМР, включающий применение AtRLP30 белка, или его внеклеточных участков, содержащего последовательность, по меньшей мере на 50%, 60%, 70%, 80%, 90%, 95%, 98%, 99% или 100% идентичную аминокислотной последовательности, показанной в SEQ ID No. 1.
Например, для очистки МАМР вышеуказанный белок можно связывать с твердым носителем, предпочтительно, с мембраной или гранулами, которые затем делают возможным связывание МАМР с указанным белком. Затем осуществляют контакт неочищенного или очищенного экстракта культуры клеток патогенов с носителем, связанным с белком по изобретению. С помощью последующей отмывки и элюирования на завершающей стадии можно выделить МАМР, который селективно связывается с белком по изобретению.
В другом аспекте изобретения предусматривается способ сенсибилизации растения к грибковым инфекциям, причем этот способ включает стадии обработки растения, растительной клетки или растительной ткани экстрактом грибов согласно настоящему изобретению, обладающим иммуногенными свойствами в отношении растения, или осуществление на указанном растении способа по изобретению, описанного выше. Далее для описания настоящего изобретения приводятся, но без ограничения, прилагающиеся фигуры и последовательности. Для целей настоящего изобретения все ссылочные материалы, цитируемые в настоящей заявке, включены в настоящее изобретение посредством отсылки во всей полноте.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1: Продуцирование (образование) этилена - иммунный ответ Arabidopsis на экстракты патогенов из основной фракции грибов. Фрагменты листьев Arabidopsis обрабатывали культуральным фильтратом или растворимым экстрактом мицелия восьми видов грибов. Через 3 часа методом газовой хроматографии определяли количество продуцированного этилена. Сила иммунного ответа на экстракты показана цветом, меняющимся от белого (количество образовавшегося этилена (продукция) аналогично количеству этилена, образующегося в случае необработанного контроля) через желтый и оранжевый (количество от низкого до среднего) до красного (высокое количество).
Фигура 2: Продуцирование этилена - иммунный ответ Arabidopsis на SsE1-содержащую фракцию, элюируемую с катионообменной хроматографической колонки с SP сефарозой. Фрагменты листьев Arabidopsis обрабатывали посредством 10 мкл фракции, связанной с SP-сефарозой, которая элюировалась 0.5 М раствором KCl (Буфер В). Через 3 часа методом газовой хроматографии определяли количество продуцированного этилена. В качестве негативного контроля использовали 10 мкл Буфера А (50/100 мМ Mes pH 5.4) и 10 мкл Буфера В (50/100 мМ Mes pH 5.4, 0,5 М KCl). В качестве позитивного контроля использовали 10 мкл неразведенного и 10х разведенного культурального фильтрата (неочищенный фильтрат). Фракция, не связанная с SP- сефарозой, не проявляет этилен-индуцирующей активности (элюат).
Фигура 3: Продуцирование этилена - иммунный ответ Arabidopsis на SsE1-содержащие фракции, элюируемые с катионообменной хроматографической колонки с сорбентом (смолой) Source 15S. Фрагменты листьев Arabidopsis обрабатывали с помощью 15 мкл фракции, связанной с Source 15S, которую элюировали в градиенте 0-0.5 М KCl. Отбирали образцы по 500 мкл. Через 3 часа методом газовой хроматографии определяли количество продуцированного этилена. В качестве негативного контроля использовали 15 мкл Буфера А (50/100 мМ Mes pH 5.4) и 15 мкл Буфера В (50/100 мМ Mes pH 5.4, 0,5 М KCl). В качестве позитивного контроля для определения количества образовавшегося этилена использовали 15 мкл неразведенного культурального фильтрата (неочищенный фильтрат), а также 15 мкл активной фракции, элюируемой с SP-сефарозы (SP элюирование), обе фракции без разведения и в разведении 10х. SsE1-содержащие фракции (В7-D7) элюируют при величине проводимости солевого раствора от 8 до 16 мС/см.
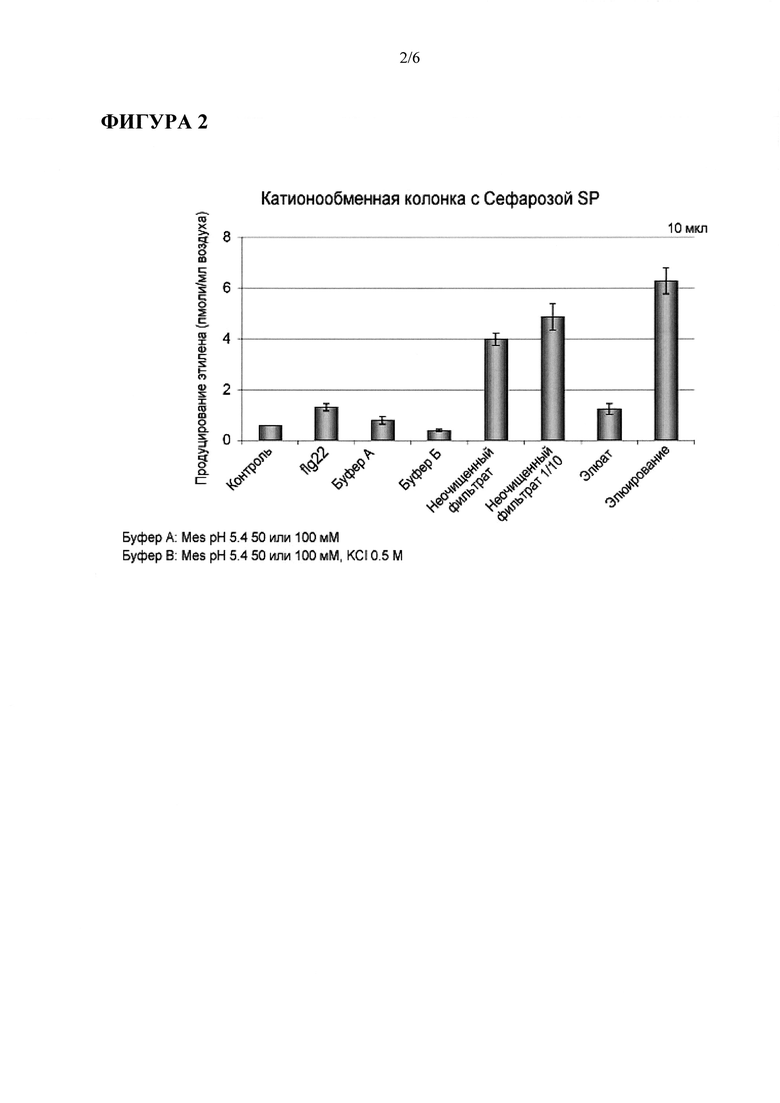
Фигура 4: Элиситор Sclerotinia sclerotiorum, фракция SsE1, вызывает (генерирует) иммунный ответ в Arabidopsis. (А) Иммуноблоттинг активированной MAPK (митоген-активируемой протеинкиназы) с помощью антитела к фосфо-р44/р42 MAPK в экстрактах листьев Arabidopsis. Образцы листьев собирали через 10 минут после инфильтрации с использованием flg22 или SsE1. В качестве контроля нагрузки служило окрашивание красителем Ponceau S Red (пунцовым С). (В) Анализ активности GUS в pPR1::GUS трансгенных растениях Arabidopsis. Листья пропитывали (инфильтрация) неочищенным фильтратом (CF) Sclerotinia, SsE1 фракцией или буфером и собирали через 24 часа для гистохимического окрашивания на активность GUS. (С) Транскрипционное профилирование FRK1 с использованием количественной ПЦР в реальном времени. Листья Arabidopsis пропитывали (инфильтрация) flg22, CF, SsE1, буфером или водой и собирали через 6 часов после обработки. Данные экспрессии нормализованы по уровням Actin транскрипта и представлены в виде х-кратной индукции по сравнению с обработанным водой контролем. Буфер А = 50 мМ Mes pH 5.4; Буфер В = 50 мМ Mes pH 5.4, 500 мМ KCI.
Фигура 5: Продуцирование этилена - иммунный ответ на SsE1 в bakl мутантах и различных экотипах Arabidopsis. Диски листьев 5-недельных растений Arabidopsis обрабатывали, применяя 500 нМ flg22 или частично очищенные экстракты Penicillium (PEN) и Sclerotinia sclerotiorum (SsE1). После инкубации в течение 3 часов методом газовой хроматографии определяли количество образовавшегося этилена. (А) Продуцирование этилена в Arabidopsis Col-0 и трех растениях линии bakl с различными Т-ДНК-инсерциями. (В) Продуцирование этилена в Arabidopsis Col-0 и пяти типах с другими кодами доступа, нечувствительных к SsE1.
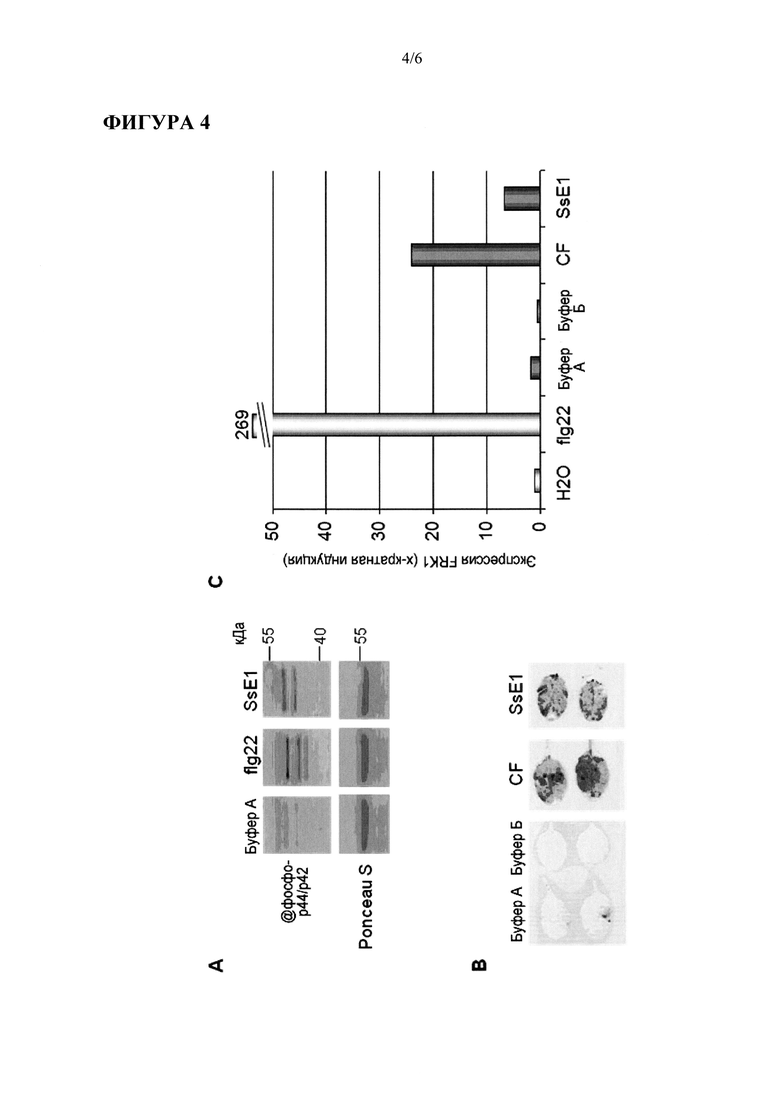
Фигура 6: Продуцирование этилена - в ответ на SsE1 в atrlp30 мутантах. Три независимых линии с Т-ДНК-инсерциями для AtRLP30 обрабатывали flg22, неочищенным препаратом элиситора из Penicillium (PEN) и SsE1.
SEQ ID NO. 1 показывает аминокислотную последовательность AtRLP30:

SEQ ID NO. 2 показывает последовательность геномной нуклеиновой кислоты AtRLP30:


ПРИМЕРЫ
Пример 1: Скрининг грибковых патогенов
В контексте настоящего изобретения основные патогенные грибы подвергались скринингу на наличие триггеров типичных РАМР-индуцируемых защитных реакций в растениях Arabidopsis. В тестировании участвовали патогены Ustilago maydis (пузырчатая головня кукурузы); Magnaporthe oryzae (пирикуляриозный гриб, возбудитель пирикуляриоза риса); Mycosphaerella graminicola (возбудитель септориозной пятнистости (STB) листьев пшеницы); Fusarium graminearum (возбудитель фузариоза (парши) колосьев пшеницы); Cercospora beticola (возбудитель церкоспороза свеклы); Sclerotinia sclerotiorum (склеротиния склероциевидная, возбудитель гнили стеблей сои и капусты); Rhizopus oryzae (возбудитель послеуборочного гниения); Rhizoctonia solani (возбудитель "черной парши" (ризоктиниоза) у многих растений). Эти патогены были выбраны потому, что их геномы были секвенированы или их секвенирование осуществляется в настоящее время.
Авторы изобретения методично проводили испытания неочищенных и не полностью очищенных препаратов как культурального фильтрата, так и мицелия грибов, выращенных без сопутствующих микроорганизмов, на их способность стимулировать продуцирование этилена во фрагментах листьев растений Arabidopsis. Наиболее сильную элиситорную активность в биологических анализах обнаруживали в экстрактах мицелия и культуральных фильтратах грибов Sclerotinia sclerotiorum, Rhizoctonia solani и Rhizopus oryzae (Фигура 1).
Пример 2: Очистка иммуногенного экстракта грибов
Сначала в качестве источника элиситора использовали культуральные фильтраты. Обладающий активностью культуральный экстракт Sclerotinia sclerotiorum очищали катионообменной хроматографией в две стадии на соединенных между собой хроматографических колонках, получали одну фракцию, содержащую элиситор, названную SsE1 (Sclerotinia sclerotiorum Elicitor 1).
Sclerotinia sclerotiorum штамм 1946 получали из Коллекции микроорганизмов и клеточных культур ФРГ (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (DSMZ)). Гриб выращивали на солодово-пептонном агар-агаре (10 г солода, 2.5 г пептона, 15 г агар-агара/л) в течение 3 дней при комнатной температуре в темноте. Затем колбы Эрленмейера на 20-25 1л, содержащие 400 мл солодово-пептонной среды (10 г солода, 2.5 г пептона) каждая, инокулируют с использованием 2-3 агар-агаровых пробок. Грибы выращивали в течение 3 недель при комнатной температуре в темноте без встряхивания. Культуральную среду фильтровали через нейлоновые сита и лиофилизировали в течение 3-4 дней. Лиофилизированный материал ресуспендировали в 100 мМ Mes буфере при pH 5.4 (около 6 мл/г сухой массы) и центрифугировали дважды по 20 мин при 10.000 g и 4°С для удаления нерастворимых частиц. Полученный супернатант использовали для выделения SsE1.
SsE1 частично очищали от концентрированного культурального фильтрата катионообменной хроматографией в две стадии. Хроматографию проводили на системе GE Healthcare ÄKTA Explorer FPLC, охлажденной до 4°С, с использованием программы GE Healthcare Unicorn. На первой стадии колонку GE Healthcare XK16 заполняли GE Healthcare Sepharose SP FastFlow сорбентом до объема насадки около 15 мл. Колонку уравновешивали буфером А (100 мМ Mes буфер, pH 5.4). Затем наносили концентрированный культуральный фильтрат при скорости потока 3-5 мл/мин. Колонку промывали буфером А, а элюирование осуществляли буфером В (100 мМ Mes буфера с pH 5.4, 500 мМ KCl) при скорости потока 3-5 мл/мин. Одну фракцию, соответствующую пику элюирования, с OD280nm и OD215nm, отбирали вручную и проверяли на этилен-индуцирующую активность (Фигура 2). На второй стадии общий объем элюата разводили 10х в буфере А и при скорости потока от 0.5 до 1.5 мл/мин наносили на колонку GE Healthcare Source 15S 4.6/100 РЕ, уравновешенную буфером А. Колонку промывали буфером А. Затем элюировали SsE1 в линейном градиенте буфера В (от 0% до 60% в 40 объемах колонки) при скорости потока от 0.5 до 1 мл/мин. Автоматически отбирали фракции по 500 мкл, соответствующие элюированию всего пика, контролируемому по OD280nm и OD215nm. Активные фракции, содержащие SsE1, идентифицировали, определяя их способность выявлять продуцирование этилена в листьях Arabidopsis thaliana Col-0 (Фигура 3).
После пропитки листьев Arabidopsis SsE1 мог индуцировать типичные РАМР-индуцируемые защитные реакции, такие как посттрансляционную активацию MAP киназы и транскрипционную активацию маркерных генов иммунного ответа, кодирующих белок, связанный с патогенезом 1 (PR1), и флагеллин-отвечающую киназу 1 (FRK1) (FRK1) (Фигура 4А-С).
Пример 3: Получение характеристик иммуногенного экстракта грибов
Для определения характеристик собирали SsE1 активные фракции, элюируемые со смолы Source 15S, и проводили масс-спектрометрический анализ методом nano-LC MS/MS (И. Macek, Proteome Centre Tubingen). Этим методом в образце было обнаружено несколько белков, которые могут отвечать за иммуногенную активность SsE1. Эти белки приводятся в Таблице 1.

Пример 4: Идентификация рецептора SsE1
Для идентификации паттерн-распознающего рецептора, опосредующего иммунный ответ на обработку иммуногенным экстрактом грибов по изобретению, на растениях Arabidopsis проводили обратный генетический анализ на рецепторные белки, отвечающие за SsE1. Увеличивающийся объем информации о структуре и функциях PRRs растений показывает, что в основном существует два типа рецепторов, опосредующих распознавание микробных паттернов различных классов. Полагают, что LRR белки и LRR-рецепторные киназы участвуют в активации иммунной системы хозяина, предпочтительно, за счет распознавания паттерна белков, тогда как белки с LysM доменами и LysM-рецепторные киназы, по-видимому, опосредуют восприятие (распознавание) микробов и иммунитет (защитные реакции) хозяина, предпочтительно, за счет распознавания паттерна гликанов. Поэтому класс подвергшихся скринингу белков бы ограничен членами семейства LRR-RLK, которые с большой степенью вероятности содержат SsE1 рецептор.
С этой целью был проведен скрининг на стойкость к SsE1 группы гомозиготных по Т-ДНК инсерции линий для 40 РАМР-индуцируемых LRR-RLKs (В. Kemmerling, ZMBP Tübingen). Кроме того, авторы изобретения получили и тестировали мутанты с единичной мутацией, fls2 efr cerkl мутанты с тройной мутацией и мутанты с четверной мутацией fls2 efr cerkl и каждый член LRR-RLK подсемейства XII, который содержит flg22 рецептор FLS2 и EF-Tu рецептор EFR (С. Zipfel, the Sainsbury laboratory Norwich, UK). Ни одна из тестированных LRR-RLKs нокаутированных линий не показала заметного уменьшения количества продуцированного этилена после обработки с помощью SsE1, за исключением bak1 мутанта. Имеется несколько мутантных аллелей BAK1: у мутантов bak1-3 и bak1-4 частично нарушены передача сигнала брассиностероидов и контроль клеточной смерти, тогда как у описанного недавно аллеля bakl-5 нарушена только РАМР передача сигнала, но не затронуты эффекты брассиностероидов и клеточная смерть. В то время как реакция в растениях bak1-3 м bak1-4 была отменена не полностью, растения bak1-5 были совершенно устойчивы к обработке с помощью SsE1 (Фигура 5А).
В параллельно проводимом исследовании изучалось продуцирование этилена в ответ на обработку с помощью SsE1 на 60 различных образцах растений Arabidopsis (Nordborg collection, Nottingham Arabidopsis Stock Centre). Естественные генетические различия между различными экотипами растений Arabidopsis могут помочь идентифицировать экотипы, которые частично или полностью нечувствительны (т.е. устойчивы) к SsE1. Такие экотипы были бы полезны для идентификации локуса, отвечающего за распознавание SsE1, с помощью позиционного клонирования. Лучшим примером успешного применения данного метода является идентификация устойчивого к флагеллину экотипа Ws-0, в котором отсутствует функциональный FLS2 рецептор. Пять экотипов (Mt-0, Lov-1, Br-0, Lov-5 и Sd-1) из 60 проверенных образцов оказались устойчивыми к SsE1. У этих экотипов не была нарушена способность продуцировать этилен, так как у них сохранялась "полная" реакция на flg22 или PEN элиситор (Фигура 3В). Скрещивание устойчивых экотипов Mt-0, Lov-1 и Sd-1 и анализ полученных F1 популяций показал, что один и тот же рецессивный ген отвечает (одни и те же рецессивные гены отвечают) на SsE1 у всех трех экотипов (данные не показаны).
Продуцирование этилена использовали в качестве признака для генетического анализа восприимчивости к SsE1 у продуктов скрещивания Lov-1 X Col-0. При этом было обнаружено, что единственный локус рецессивного аллеля, который картирован в верхнем плече хромосомы 3, отвечает за восприимчивость (чувствительность) к SsE1 (64 из 270 тестированных F2 растений были устойчивы к элиситору). Последующее более тонкое картирование с применением маркеров SSLP (Simple Sequence Length Polymorphism, "полиморфизм длин простых последовательностей") и различных маркеров CAPS (Cleaved Amplified Polymorphic Sequence, полиморфизм амплифицированных и отрестриктированных последовательностей) дало область, содержащую четыре LRR-рецептороподобных белка (RLPs), но ни одного из членов других семейств LRR- содержащих белков. Независимые Т-ДНК нокаутированные линии (растений) для каждого из LRR-RLP кандидатов обрабатывали с помощью SsE1. Результаты показали, что AtRLP30 (At3g05360) участвует в распознавании SsE1, тогда как реакции на flg22 и PEN не изменялись (Фигура 6). В нокаутированных линиях, соответствующих двум другим кандидатам, AtRLP32 (At3g05650) и AtRLP33 (Af3g05660), также наблюдалась обычная реакция на SsE1.








Изобретение относится к области биохимии, в частности к способу увеличения устойчивости растения к грибковой инфекции, вызываемой грибами семейства Sclerotiniaceae, включающему повышение в указанном растении экспрессии белка AtRLP30, а также к способу получения трансгенного растения с повышенной устойчивостью к грибковой инфекции, вызываемой грибами семейства Sclerotiniaceae, включающему трансформацию растения или растительной клетки с помощью нуклеотидной последовательности, кодирующей AtRLP30 или AtRLP30-подобный белок. Также раскрыты способ скрининга на микроб-ассоциированные молекулярные паттерны (PAMPs/MAMPs), имеющиеся в Sclerotinia spp или Botrytis ssp., предусматривающий экспрессирование в растении или в растительной клетке белка AtRLP30, а также способ очистки МАМР, имеющегося в Sclerotinia spp или Botrytis ssp, включающий применение AtRLP30 белка, или его внеклеточных участков. Изобретение также относится к способу сенсибилизации растения к грибковой инфекции, вызываемой грибами семейства Sclerotiniaceae, включающему осуществление на указанном растении любого из вышеуказанного способа. Изобретение позволяет эффективно получать растение с повышенной устойчивостью к грибковой инфекции, вызываемой грибами семейства Sclerotiniaceae. 5 н. и 5 з.п. ф-лы, 6 ил., 1 табл., 4 пр.
1. Способ увеличения устойчивости растения к грибковой инфекции, вызываемой грибами семейства Sclerotiniaceae, включающий повышение в указанном растении экспрессии белка, содержащего аминокислотную последовательность, по меньшей мере на 60% идентичную последовательности SEQ ID No. 1.
2. Способ по п. 1, включающий также модуляцию в указанном растении экспрессии белка, содержащего аминокислотную последовательность, по меньшей мере на 60% идентичную BAK1, причем повышение экспрессии указанного белка приводит к повышению устойчивости указанного растения к грибковой инфекции, вызываемой грибами семейства Sclerotiniaceae,
и причем снижение экспрессии приводит к снижению устойчивости указанного растения к грибковой инфекции, вызываемой грибами семейства Sclerotiniaceae.
3. Способ по п. 2, в котором экспрессия указанного белка в указанном растении повышается при использовании эктопической экспрессии указанного белка.
4. Способ по п. 2, в котором экспрессия указанного белка в указанном растении снижается посредством мутагенеза, РНК интерференции или РНК-опосредуемого метилирования ДНК.
5. Способ по любому из пп. 1-4, в котором указанная грибковая инфекция, вызываемая грибами семейства Sclerotiniaceae, представляет собой инфекцию растения, вызываемую Sclerotinia sclerotiorum или Botrytis cinerea.
6. Способ получения трансгенного растения с повышенной устойчивостью к грибковой инфекции, вызываемой грибами семейства Sclerotiniaceae, по сравнению с растением, нетрансформированным AtRLP30, включающий стадии (i) трансформации растения или растительной клетки с помощью нуклеотидной последовательности, кодирующей AtRLP30 или AtRLP30-подобный белок, содержащий аминокислотную последовательность, по меньшей мере на 60% идентичную SEQ ID No 1.
7. Способ скрининга на микроб-ассоциированные молекулярные паттерны (PAMPs/MAMPs), имеющиеся в Sclerotinia spp или Botrytis ssp, включающий стадии: (i) экспрессирование в растении или в растительной клетке белка, содержащего аминокислотную последовательность, по меньшей мере на 60% идентичную SEQ ID No. 1 (AtRLP30), (ii) контактирование указанного растения или указанной растительной клетки с соединением-кандидатом, (iii) количественное определение иммунного ответа указанного растения или указанной растительной клетки по сравнению с растением, нетрансформированным AtRLP30, или с растительной клеткой, нетрансформированной AtRLP30, при этом повышенный иммунный ответ указанного растения или указанной клетки показывает, что указанное соединение-кандидат представляет собой МАМР, имеющийся в Sclerotinia spp или Botrytis ssp.
8. Способ скрининга по п. 7, в котором на стадии (iii) иммунный ответ указанного растения или указанной растительной клетки измеряют, определяя продуцирование этилена и/или экспрессию генов, отвечающих за иммунные реакции, или репортерных генов.
9. Способ очистки МАМР, имеющегося в Sclerotinia spp или Botrytis ssp, включающий применение AtRLP30 белка, или его внеклеточных участков, содержащего последовательность, по меньшей мере на 60% идентичную последовательности SEQ ID No. 1, с выделением МАМР, который селективно связывается с указанным белком.
10. Способ сенсибилизации растения к грибковой инфекции, вызываемой грибами семейства Sclerotiniaceae, который включает осуществление на указанном растении способа по любому из пп. 1-6.
| GUODONG WANG, A Genome-Wide Functional Investigation into the Roles of Receptor-Like Proteins in Arabidopsis, Plant Physiology, 2008, Vol | |||
| Раздвижной паровозный золотник со скользящими по его скалке поршнями и упорными для них шайбами | 1922 |
|
SU147A1 |
| Электрический фонарь - испытательный прибор | 1912 |
|
SU503A1 |
| LOURDES GOMEZ-GOMEZ et al., FLS2: An LRR Receptor-like Kinase Involved in the Perception of the Bacterial Elicitor Flagellin in Arabidopsis, Molecular Cell, 2000, Vol | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ, УСТОЙЧИВЫХ К ВИРУСНОЙ ИНФЕКЦИИ | 2005 |
|
RU2300567C1 |

