Настоящее изобретение касается способа повышения резистентности к патогенным воздействиям в трансгенных растениях и/или растительных клетках, причем в растение или растительную клетку вводят и обеспечивают там экспрессию последовательности ДНК, кодирующей белок с лейцинобогащенным повторным (LRR) доменом и/или киназной активностью. Также настоящее изобретение касается применения нуклеиновых кислот, кодирующих такой белок, для создания трансгенных растений или растительных клеток с повышенной резистентностью к патогенным воздействиям. Кроме того, настоящее изобретение касается последовательностей нуклеиновых кислот, кодирующих белок, обеспечивающий повышенную резистентность к патогенным воздействиям у растений.
Болезни растений, вызываемые различными патогенными агентами, например, вирусами, бактериями и грибками, могут вызывать существенные потери урожая при разведении культурных растений, что, с одной стороны, имеет экономические следствия, но также и угрожает питанию людей. С прошлого века для воздействия на грибковые заболевания применяют химические фунгициды. Хотя применение этих веществ позволило снизить масштабы болезней растений, но до сих пор нельзя исключить, что эти соединения оказывают вредное воздействие на человека, животных и окружающую среду. Поэтому, чтобы снизить потребление обычных средств защиты растений до минимума, важно исследовать естественные механизмы защиты различных растений от патогенного воздействия различных возбудителей и целенаправленно использовать их, применяя методы генной инженерии, например, вводя внешние гены резистентности или изменяя внутреннюю экспрессию генов в растениях, для создания растений, обладающих резистентностью против патогенных воздействий.
Способов придать растениям резистентность к патогенным воздействиям, прежде всего патогенным грибкам, немного. Недостаток их частично обусловлен сложностью рассматриваемых биологических систем. Достижению резистентности против патогенных воздействий препятствует также то, что о взаимодействиях между патогенным агентом и растением известно немного. Большое количество различных патогенных агентов, развитые ими механизмы инфицирования, и развившиеся у отрядов, семейств и видов растений механизмы защиты взаимодействуют между собой сложным образом.
Патогенные грибки выработали в основном две стратегии инфицирования. Некоторые грибки проникают в ткань хозяина через устьица (например, ржавчинные грибки, виды Septoria, Fusarium) и проникают в ткань мезофилла, в то время как другие проникают через кутикулу в лежащие под ней клетки эпидермиса (например, виды Blumeria).
Инфекции, вызванные патогенными грибками, вызывают в пораженных растениях активацию защитных механизмов. Так, удалось показать, что защитные реакции против грибков, проникающих через эпидермис, часто начинаются с формирования резистентности против пенетрации (формирование сосочков, утолщение клеточной стенки, главный компонент - каллусная ткань) под проникающей грибковой гифой (Elliott et al. (2002) Mol. Plant Microbe Interact. 15: 1069-77).
Защитные механизмы растений, однако, во многих случаях не обеспечивают достаточной защиты от поражения патогенными агентами.
Формирование резистентности против пенетрации патогенных агентов, механизм развития инфекции которых включает в себя проникновение в клетки эпидермиса или мезофилла, имеет большое значение как для однодольных, так и для двудольных растений. В отличие от других способов придания резистентности, этот вариант может позволить сформировать резистентность широкого спектра против облигатно-биотрофных, гемибиотрофных и некротрофных грибков.
До сих пор для создания растений, обладающих резистентностью к грибкам, часто проводили скрещивание с введением количественных признаков резистентности (QTL). Этот способ, однако, обладает тем недостатком, что нередко параллельно происходит введение нежелательных признаков. Кроме того, методы разведения, необходимые для этого, очень сложны и требуют много времени. Поэтому задача настоящего изобретения состояла в том, чтобы предложить способ повышения резистентности растений против пенетрирующих патогенных агентов.
Задачу решают посредством форм исполнения, охарактеризованных в пунктах формулы изобретения.
Следовательно, настоящее изобретение касается способа повышения резистентности к патогенным воздействиям в трансгенных растениях и/или растительных клетках, причем в растение или растительную клетку вводят и обеспечивают там экспрессию последовательности ДНК, кодирующей белок, который сообщает повышенную резистентность к патогенам, предпочтительно - повышенную резистентность к патогенным грибкам.
В рамках анализа TIGS (Transient Induced Gene Silencing, преходящего вызванного "молчания" генов), проведенного на ячмене по методу Schweizer et al. (2001), было обнаружено, что "молчание" гена RNR8, опосредованное dsRNAi (интерференцией двухцепочечной РНК), повышает восприимчивость растения к патогенному грибку Blumeria graminis, и, следовательно, ген RNR8 играет роль в придании растениям ячменя резистентности к патогенам.
RNR8 относится к семейству белков, содержащих лейцинобогащенные повторы (LRR) с доменом киназы или активностью киназы. Белки, принадлежащие к этому семейству, играют важную роль в различных клеточных процессах, как то: регуляция развития эндосперма и пыльцы (Li and Wurtzel (1998) Plant Mol. Biol. 37: 749-761; Muschietti et al. (1998) Plant Cell 10: 319-330), регуляции развития меристемы и цветка (Torii et al. (1996) Plant Cell 8: 735-746; Dark et al. (1997) Cell 89: 575-585; Kim et al. (2000) Plant Sci. 152: 17-26) и в росте побегов, вызванном гиббереллином (van der Knapp et al. (1999) Plant Physiol. 120: 559-569). Домены LRR включают в себя от 2 до 45 повторений аминокислотной последовательности, в состав которой входят от 20 до 30 аминокислот, и обычно сворачивается, принимая форму подковы.
В отношении другого члена этого семейства, ОsХа21 из риса, удалось показать, что он сообщает резистентность в отношении патогенных бактерий Xanthomonas oryzae (Song et al. (1995) Science 270(5243): 1804-1806). Белок согласно изобретению гомологичен белку OsXa21 по последовательностям лишь в очень незначительной степени, примерно на 23%.
Белки согласно изобретению обеспечивают повышение резистентности растений к патогенным воздействиям, предпочтительно - резистентности к патогенным грибкам. Предпочтительно они отличаются тем, что либо содержат лейцинобогащенные повторы (LRR), либо обладают активностью киназы, особо предпочтительно, чтобы белки согласно изобретению обладали обоими признаками. В настоящее время предполагают (не фиксируясь, однако, этой гипотезой), что за сообщение резистентности против патогенных воздействий отвечает либо активность киназы, либо домен LRR, либо оба эти признака вместе. В целях простоты изложения ниже используют понятие "белок согласно изобретению".
Последовательность нуклеиновых кислот, которую вводят в растение или в растительную клетку при реализации способа согласно изобретению, и которая кодирует белок согласно изобретению, выбирают из группы, которую образуют:
i) Последовательности нуклеиновых кислот, в состав которых входят последовательности нуклеотидов, соответствующие кодирующим последовательностям SEQ ID NO 1 или их фрагментам,
ii) Последовательности нуклеиновых кислот, в состав которых входят последовательности нуклеотидов, кодирующие белок с последовательностью аминокислот, приведенной под SEQ ID NO 2, или ее фрагментами,
iii) Последовательности нуклеиновых кислот, в состав которых входят последовательности нуклеотидов, идентичные кодирующим последовательностям SEQ ID NO 1 по меньшей мере на 60% и/или
iv) Последовательности нуклеиновых кислот, в состав которых входят последовательности нуклеотидов, гибридизирующиеся с комплементарной нитью нуклеотидной последовательности от i) до iii) в обязательных условиях.
"Резистентность" означает воспрепятствование, исключение, снижение или ослабление симптомов заболевания растения, развивающихся вследствие поражения патогенным агентом. Симптомы могут быть различного рода, предпочтительно к ним, однако, относятся те, что прямым или косвенным образом ведут к снижению качества растения, количества урожая, пригодности к использованию в пищу или корм, либо же осложняют посев, выращивание, сбор урожая или обработку собранного урожая.
В предпочтительной форме исполнения происходит ослабление, снижение проявления или предотвращение следующих симптомов: образования пустул и скопления спор на поверхности пораженных тканей, мацерации тканей, распространяющегося некроза ткани, накопления микотоксинов, например, из Fusarium graminearum или F.culmorum.
"Повышенная резистентность к патогенным воздействиям" означает, что защитные механизмы определенного растения или в некоторой части растения, например, в органе, ткани, клетке или органелле, благодаря применению способа согласно изобретению при прочих равных условиях (например, климатических условиях, условиях выращивания, типе патогенного воздействия и т.д.) обладают повышенной по сравнению с подходящим контролем, например, с диким типом растения ("исходным растением", "контрольным растением"), резистентностью к одному или более патогенным воздействиям. Предпочтительно, чтобы в растении по меньшей мере эпидермис и/или ткань мезофилла или органы, имеющие эпидермис и/или ткань мезофилла, обладали повышенной резистентностью к патогенным воздействиям. В качестве примера можно указать повышение резистентности в листьях.
В одной из форм исполнения повышают резистентность в нижней цветковой чешуе (lemma), верхней цветковой чешуе (palea) или колосковой чешуе (плева, glume).
Повышенная резистентность предпочтительно выражается в снижении проявлений болезненных симптомов, причем к симптомам - помимо вышеупомянутых поражений - также относятся эффективность проникновения патогенного агента в растение или растительную клетку или эффективность пролиферации патогенного агента в таковых или на таковых. При этом предпочтительно ослабление болезненных симптомов по сравнению с контрольным растением по меньшей мере на 10% или по меньшей мере на 20%, особо предпочтительно - по меньшей мере на 40% или 60%, чрезвычайно предпочтительно - по меньшей мере на 70% или 80%, а наиболее предпочтительно - по меньшей мере на 90% или 95%.
В рамках изобретения "патоген" в рамках изобретения означает организмы, взаимодействие которых с растением ведут к вышеописанным болезненным симптомам, в частности, под "патогеном" подразумевают организмы из царства грибов. При этом предпочтительно, чтобы патоген представлял собой патогенный агент, проникающий в клетки эпидермиса или мезофилла, особо предпочтительно - патогены, входящие в растения через устьица, а затем проникающие в клетки мезофилла. При этом предпочтительно упомянуть организмы отделов Ascomycota и Basidomycota. Особо предпочтительны при этом семейства Blumeriaceae, Pucciniaceae, Mycosphaerellaceae и Hypocreaceae.
Особо предпочтительны организмы тех семейств, которые принадлежат к родам Blumeria, Puccinia, Fusarium или Mycosphaerella.
Крайне предпочтительны виды Blumeria graminis, Puccinia triticina, Puccinia striiformis, Mycosphaerella graminicola, Stagonospora nodorum, Fusarium graminearum, Fusarium culmorum, Fusarium avenaceum, Fusarium poae и Microdochium nivale.
В особо предпочтительных формах исполнения способ согласно изобретению ведет к следующим видам резистентности:
- ячменя против патогена Puccinia graminis f.sp. hordei (стеблевая ржавчина ячменя, barley stem rust),
- пшеницы против патогенов Fusarium graminearum, Fusarium avenaceum, Fusarium culmorum, Puccinia graminis f.sp. tritici (стеблевая ржавчина пшеницы, wheat stem rust), Puccinia recondita f.sp. tritici, Puccinia striiformis, Septoria nodorum, Septoria tritici и/или Septoria avenae,
- кукурузы против патогенов Fusarium moniliforme var. subglutinans, Puccinia sorghi и/или Puccinia polysora,
- сорго против патогенов Puccinia purpurea, Fusarium moniliforme, Fusarium graminearum и/или Fusarium oxysporum,
- сои против патогена Phakopsora pachyrhizi и/или Phakopsora meibromae.
Равным образом предметом изобретения является изолированная молекула нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты, выбранную из группы, которую образуют:
i) Последовательности нуклеиновых кислот, в состав которых входят последовательности нуклеотидов, соответствующие кодирующим последовательностям SEQ ID NO 1 или их фрагментам,
ii) Последовательности нуклеиновых кислот, в состав которых входят последовательности нуклеотидов, кодирующие белок с последовательностью аминокислот, приведенной под SEQ ID NO 2, или ее фрагментами,
iii) Последовательности нуклеиновых кислот, в состав которых входят последовательности нуклеотидов, идентичные кодирующим последовательностям SEQ ID NO 1 по меньшей мере на 60% и/или
iv) Последовательности нуклеиновых кислот, в состав которых входят последовательности нуклеотидов, гибридизирующиеся с комплементарной нитью нуклеотидной последовательности от i) до iii) в обязательных условиях,
кодирующие белок, который сообщает растениям повышенную резистентность к патогенам, предпочтительно - повышенную резистентность к патогенным грибкам.
Понятие "нуклеиновая кислота (молекула нуклеиновой кислоты)", в том смысле, в котором его здесь применяют, в предпочтительной форме исполнения включает в себя расположенную на 3'- и на 5'-конце кодирующего участка гена и не прошедшую трансляцию последовательность: по меньшей мере 500, предпочтительно 200, особо предпочтительно 100 нуклеотидов вверх от 5'-конца кодирующего участка и по меньшей мере 100, предпочтительно 50, особо предпочтительно 20 нуклеотидов вниз от 3'-конца кодирующего участка гена.
"Изолированную" молекулу нуклеиновой кислоты отделяют от других молекул нуклеиновой кислоты, имеющихся в естественном источнике нуклеиновых кислот. Предпочтительно, чтобы "изолированная" нуклеиновая кислота не содержала последовательностей, естественным образом фланкирующих нуклеиновые кислоты в геномной ДНК организма, из которого происходит нуклеиновая кислота (например, последовательности, находящиеся на 5'- и 3'-концах нуклеиновой кислоты). В различных формах исполнения изолированная молекула может содержать, например, менее чем 5 тысяч пар оснований (тыс. п.о., kb), 4 тыс. п.о., 3 тыс. п.о., 2 тыс. п.о., 1 тыс. п.о., 0,5 тыс. п.о. или 0,1 тыс. п.о. нуклеотидных последовательностей, естественным образом фланкирующих нуклеиновые кислоты в геномной ДНК организма, из которого происходит нуклеиновая кислота. Все упомянутые здесь молекулы нуклеиновых кислот могут представлять собой, например, РНК, ДНК или кДНК.
Применяемые при реализации способа молекулы нуклеиновых кислот, например, молекула нуклеиновой кислоты с последовательностью нуклеотидов SEQ ID NO 1 или ее частью, можно изолировать, применяя стандартные технологии молекулярной биологии и предоставленную здесь информацию о последовательностях. Кроме того, с помощью алгоритмов сравнения, которые представлены, например, на странице Национального центра биотехнологической информации (NCBI) http://www.ncbi.nlm.nih.gov, определить, например, гомологичную последовательность или гомологичные консервированные участки последовательностей на уровне ДНК или аминокислот. Основные части этой последовательности или всю гомологичную последовательность можно, применяя стандартные методики гибридизации (как описано, например, в Sambrook et al., см. выше), использовать для изоляции прочих полезных для реализации способа последовательностей нуклеиновых кислот из других организмов путем скрининга банков кДНК и/или генома. Кроме того, молекулу нуклеиновой кислоты, содержащую всю последовательность, соответствующую SEQ ID NO 1, или ее часть, можно выделить с помощью полимеразной цепной реакции, причем для этого применяют олигонуклеотидный праймер на основе приведенных здесь последовательностей или их частей (например, молекулу нуклеиновой кислоты, содержащую всю последовательность или ее часть, можно выделить с применением олигонуклеотидного праймера, созданного на основе такой же последовательности). Например, из клеток можно выделить иРНК (например, методом экстракции с гуанидинтиоцианатом по Chirgwin et al. (1979) Biochemistry 18: 5294-5299), а на ее основе с помощью обратной транскриптазы (например, обратной транскриптазы вируса лейкемии мышей (MLV) Молони, распространяется Gibco/BRL, Bethesda, MD или обратной транскриптазы вируса миелобластоза птиц (AMV), распространяется Seikagaku Amerika, Inc., St. Petersburg, FL) синтезировать кДНК. Синтетические олигонуклеотидные праймеры для амплификации с помощью полимеразной цепной реакции можно создать на основе последовательности нуклеиновой кислоты, представленной в SEQ ID NO 1, или с помощью последовательности аминокислот, представленной в SEQ ID NO 2. Нуклеиновую кислоту согласно изобретению можно подвергнуть амплификации с применением кДНК или, в качестве альтернативы - геномной ДНК в качестве матрицы и надлежащего олигонуклеотидного праймера посредством стандартных технологий амплификации ПЦР. Амплифицированную таким образом нуклеиновую кислоту можно клонировать с созданием подходящего вектора и охарактеризовать с помощью анализа последовательностей ДНК (секвенирования). Олигонуклеотиды, соответствующие последовательности нуклеотидов, кодирующей белок согласно изобретению, можно синтезировать с помощью стандартных способов синтеза, например, с помощью автоматического устройства синтеза ДНК.
Под понятием "идентичности последовательностей" двух нуклеиновых кислот подразумевают идентичность последовательностей нуклеиновых кислот в каждом случае по всей длине последовательности, в предпочтительной форме исполнения - по всей длине экспрессируемой последовательности, еще более предпочтительно - по кодирующей последовательности (предпочтительно CDS), которую рассчитывают с помощью программного алгоритма GAP (Wisconsin Package Version 10.0, University of Wisconsin, Genetics Computer Group (GCG), Madison, USA; Altschul et al. (1997) Nucleic Acids Res. 25: 3389ff), задавая следующие параметры:
значимость пробела (Gap Weight): 50
значимость длины (Length Weight): 3
среднее соответствие: 10
среднее несоответствий: 0.
Например, последовательность, гомологичная последовательности нуклеиновой кислоты с SEQ ID NO 1 по меньшей мере на 80%, при сравнении с последовательностью SEQ ID NO 1 с использованием вышеуказанного программного алгоритма с приведенными выше параметрами демонстрирует гомологичность, составляющую по меньшей мере 80%.
В одной из форм исполнения настоящее изобретение касается последовательностей нуклеиновых кислот, демонстрирующих идентичность последовательности, приведенной под SEQ ID NO 1, составляющую по меньшей мере 60%, предпочтительно - по меньшей мере 65, 70, 75 или 80%, особо предпочтительно - по меньшей мере 82, 84, 86, 88 или 90%, а наиболее предпочтительно - по меньшей мере 92, 94, 96, 98 или 99%.
Под "идентичностью двух белков" подразумевают идентичность аминокислот на определенном участке белка, предпочтительно - по всей длине белка, в особенности - идентичность, которую рассчитывают сравнением с применением программного обеспечения, например, программного пакета Lasergene производства фирмы DNA Star Inc., Madison, Wisconsin (США), используя методику CLUSTAL (Higgins et al. (1989) Comput. Appl. Biosci. 5(2): 151). Гомологию также можно рассчитывать с применением программного пакета Lasergene производства фирмы DNA Star Inc., Madison, Wisconsin (США), используя методику CLUSTAL (Higgins et al. (1989) Comput. Appl. Biosci. 5(2): 151).
Предпочтительно под "идентичностью двух белков" подразумевают идентичность аминокислотных последовательностей, предпочтительно - в каждом случае по всей длине последовательности, которую рассчитывают путем сравнения с помощью программного алгоритма GAP (Wisconsin Package Version 10.0, University of Wisconsin, Genetics Computer Group (GCG), Madison, USA), задавая следующие параметры:
значимость пробела (Gap Weight): 8
значимость длины (Length Weight): 2
среднее соответствие: 2,912
среднее несоответствий: -2,003.
"Стандартные условия гибридизации" следует понимать в широком смысле, в зависимости от применения это означает обязательные или менее обязательные условия гибридизации. В числе прочего такие условия гибридизации описаны у Sambrook and Russell, Molecular Cloning - A Laboratory Manual, 3. Auflage, Cold Spring Harbor Laboratory Press, 2001) или в Current Protocols in Molecular Biology, John Wiley & Sons, N.Y. (1989), 6.3.1-6.3.6..
Специалист выбирает условия гибридизации, позволяющие ему различать специфическую и неспецифическую гибридизацию.
Например, условия на этапе отмывки можно выбирать из условий меньшей строгости (примерно с 2Х SSC (Na-цитратный буфер) при 50°С) и таковыми высокой строгости (примерно 0.2Х SSC при 50°С, предпочтительно при 65°С) (20Х SSC: 0,3 М цитрата натрия, 3 М NaCl, pH 7,0). Кроме того, во время этапа отмывки можно повышать температуру: от менее строгих условий - примерно при 22°С - до более строгих условий, примерно при 65°С. Оба параметра - концентрацию соли и температуру - можно варьировать одновременно или по отдельности, причем в каждом случае другой параметр поддерживают на постоянном уровне. Во время гибридизации можно также применять денатурирующие агенты, как, например, формамид или ДСН. В присутствии 50% формамида гибридизацию предпочтительно проводить при 42°С. Ниже приведены примеры условий для гибридизации и этапа отмывки:
(1) Условия гибридизации можно, например, выбирать из нижеследующих вариантов условий:
а) 4Х SSC при 65°С,
b) 6X SSC при 45°С,
c) 6X SSC, 100 мкг/мл денатурированной фрагментированной ДНК из рыбьих молок при 68°С,
d) 6X SSC, 0,5% ДСН 100 мкг/мл денатурированной фрагментированной ДНК из рыбьих молок при 68°С,
e) 6X SSC, 0,5% ДСН 100 мкг/мл денатурированной фрагментированной ДНК из рыбьих молок, 50% формамида при 42°С,
f) 50% формамида, 4Х SSC при 42°С,
g) 50% (об./об.) формамида, 0,1% бычьего сывороточного альбумина, 0,1% фиколла, 0,1% поливинилпирролидона, 50 мМ натриево-фосфатного буфера, рН 6,5, 750 мМ NaCl, 75 мМ цитрата натрия при 42°С,
h) 2X или 4Х SSC при 50°С (нестрогие условия),
i) от 30 до 40% формамида, 2X или 4Х SSC при 42°С (нестрогие условия).
j) 500 мМ натриево-фосфатного буфера, рН 7,2, 7% ДСН (додецилсульфат натрия) (г/об.), 1 мМ ЭДТА, 10 мкг/мл одночепочечной ДНК, 0,5% БСА (г/об.) (Church and Gilbert (1984) Proc. Natl. Acad. Sci. U.S.A. 81(7): 1991-1995).
(2) Условия отмывки можно, например, выбирать из нижеследующих вариантов условий:
a) 0,015 М NaCl/0,0015 М цитрата натрия/0,1% ДСН при 50°С,
b) 0,1 Х SCC при 65°С,
c) 0,1Х SSC, 0,5% ДСН при 68°С,
d) 0,1Х SSC, 0,5% ДСН, 50% формамида при 42°С,
e) 0,2Х SSC, 0,1% ДСН при 42°С,
f) 2X SSC при 65°С (нестрогие условия).
В одном из вариантов исполнения условия гибридизации выбирают следующим образом.
Выбирают буфер гибридизации, содержащий формамид, NaCl и ПЭГ 6000. Присутствие формамида в буфере гибридизации дестабилизирует двухцепочечные молекулы нуклеиновых кислот, благодаря чему температуру гибридизации можно понизить до 42°С, не снижая этим строгость условий. Применение соли в буфере гибридизации повышает степень ренатурации дуплекса или эффективность гибридизации. Хотя ПЭГ повышает вязкость раствора, что оказывает отрицательное влияние на степень ренатурации, но благодаря присутствию в растворе полимера возрастает концентрация зонда в оставшейся среде, что повышает степень гибридизации. Состав буфера следующий:
Гибридизацию проводят в течение ночи при 42°С. Фильтры отмывают на следующее утро трижды 3х, используя 2×SSC и 0,1% ДСН - по 10 минут.
Последовательности нуклеиновых кислот, отличающиеся от последовательности, приведенной под SEQ ID NO 1, можно получить, осуществляя в последовательности нуклеотидов SEQ ID NO 1 замещение, добавление или делеции нуклеотидов так, чтобы возникали белки, в которых по сравнению с последовательностью, приведенной под номером SEQ ID NO 2, имеют место замещение, добавление или делеция одной или нескольких аминокислот. Мутации можно вводить в последовательность SEQ идент. №1 посредством стандартных методик, например, мутагенеза, специфичного по положению, и мутагенеза, опосредованного ПЦР. Предпочтительно осуществлять консервативные замещения аминокислот в одном или нескольких предсказанных остатках несущественных аминокислот, т.е. остатков аминокислот, не влияющих на активность киназы и/или на домен LRR. При "консервативном замещении аминокислот" остаток аминокислоты заменяют на остаток аминокислоты со схожей боковой цепью. В этой области работы определены семейства аминокислотных остатков со схожими боковыми цепями. К этим семействам относятся аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислыми боковыми цепями (например, аспарагиновая кислота и глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан). Таким образом, в белке, применяемом согласно изобретению, предпочтительно заменять предсказанный несущественный аминокислотный остаток на другой аминокислотный остаток из того же семейства по признаку боковых цепей. В качестве альтернативы при другой форме исполнения мутаций можно случайным образом вводить мутации по всей длине или части длины последовательности, кодирующей белок согласно изобретению, например, проверяя их с точки зрения способности сообщать резистентность к патогенным воздействиям.
Под понятием "фрагменты ДНК" в том смысле, в котором его применяют здесь, подразумевают части ДНК, кодирующие белок согласно изобретению, биологическая активность которых состоит в том, чтобы обеспечивать повышение резистентности к патогенным воздействиям. Предпочтительно, чтобы фрагменты ДНК кодировали белок с активностью киназы или с доменом LRR, особо предпочтительно - с обоими признаками, причем чтобы белки, кодируемые фрагментами ДНК, в основном обладали той же активностью киназы и/или имели тот же домен LRR, что и белки, кодируемые полноценной последовательностью ДНК, и чтобы с помощью этих фрагментов можно было добиться повышения резистентности трансгенных растений к патогенным воздействиям согласно изобретению.
Под понятием "фрагменты белка" в том смысле, в котором его применяют здесь, подразумевают части белка, биологическая активность которых состоит в том, чтобы обеспечивать повышение резистентности к патогенным воздействиям (предпочтительно - резистентности к патогенным грибкам) у растений. Предпочтительно, чтобы фрагменты белка обладали активностью киназы или содержали домен LRR, особо предпочтительно - обладали бы обоими признаками, причем чтобы фрагменты белка в основном обладали той же активностью киназы и/или имели тот же домен LRR, что и белки, имеющие полную длину, и чтобы с помощью этих фрагментов можно было добиться повышения резистентности трансгенных растений к патогенным воздействиям согласно изобретению.
Под "в основном одинаковой ферментативной активностью" применяемого в способе согласно изобретению белка с киназной активностью подразумевают, что ферментативная активность составляет по сравнению с ферментами, кодируемыми последовательностью с номером SEQ ID NO 1 или ее производными, по меньшей мере, 50%, предпочтительно - по меньшей мере, 60%, особо предпочтительно - по меньшей мере, 70%, в частности предпочтительно - по меньшей мере, 80%, а наиболее предпочтительно - по меньшей мере 90%. Таким образом, белки с активностью киназы, имеющие в основном одинаковую ферментативную активность, также пригодны для формирования повышенной резистентности трансгенных растений к патогенным воздействиям.
Киназную активность белков можно определить простыми методами, которые известны специалисту и которые обозначают как количественный анализ киназы (Kinase-assay). Для этого, например, подлежащий исследованию белок, который очищают, например, иммунопреципитацией, инкубируют в подходящем буферном растворе с АТФ, снабженным радиоактивной меткой, в качестве донора фосфата и надлежащим субстратом, а затем отделяют субстрат с помощью полиакриламидного геля с ДСН. Если белок обладает активностью киназы, то субстрат получает метку фосфата (в свою очередь, имеющего радиоактивную метку), а радиоактивность можно обнаружить и количественно определить с помощью соответствующих методик.
Повышения резистентности к патогенным воздействиям согласно изобретению можно также добиться, изменяя экспрессию эндогенного, собственного белка растения, который соответствует белку согласно изобретению. Таким образом, это собственный белок растения, сообщающий повышение резистентности к патогенным воздействиям и предпочтительно обладающий активностью киназы или доменом LRR, а особо предпочтительно - обоими признаками. Это изменение экспрессии белка можно обеспечить, например, изменяя последовательность промоторной ДНК гена, кодирующего белок. Такое изменение, имеющее следствием измененный, предпочтительно - повышенный уровень экспрессии эндогенного гена согласно изобретению, можно осуществить путем делеции или инсерции последовательностей ДНК. Изменение промоторной последовательности эндогенных генов согласно изобретению, как правило, ведет к количественному изменению экспрессии гена и, таким образом, например, к изменению регистрируемой в клетке или в растениях активности киназы (если белок обладает киназной активностью). Изменение промоторной последовательности эндогенного гена согласно изобретению может также вести к изменению в клетке количества белка с доменом LRR.
Еще одна возможность для повышения активности и содержания эндогенных белков согласно изобретению состоит в том, чтобы регуляторными средствами повысить активность факторов транскрипции, участвующих в транскрипции эндогенных генов, например, повышенной экспрессией. Способы повышения экспрессии факторов транскрипции известны специалисту, а также изложены в рамках настоящего изобретения для белков согласно изобретению.
Кроме того, повышенной экспрессии эндогенного гена согласно изобретению можно также добиться, вызывая взаимодействие с промотором таких генов регуляторного белка, не встречающегося в исходном (неизмененном) организме. Такой регулятор может представлять собой химерный белок, состоящий из домена связывания ДНК и домена активатора транскрипции, как это описано, например, в международной заявке WO 96/06166.
"Трансгенный" в отношении, например, последовательности нуклеиновых кислот, кассеты экспрессии или вектора, содержащих указанную последовательность нуклеиновых кислот, или организма, преобразованного указанной последовательностью нуклеиновых кислот, кассетой экспрессии или вектора, означает все возникшие с помощью генно-инженерных методик конструкции или организмы, в которых либо
a) последовательность нуклеиновых кислот RNR8, или
b) генетическая контрольная последовательность, например промотор, функционально связанная с последовательностью нуклеиновых кислот RNR8, или
с) (а) и (b)
находятся не в своем естественном генетическом окружении или были изменены генно-инженерными методами, причем к примерам изменения относятся замещение, добавление, делеция или инсерция одного или нескольких нуклеотидных остатков. "Естественное генетическое окружение" означает естественный хромосомный локус в организме происхождения или наличие в геномной библиотеке. В случае геномной библиотеки естественное генетическое окружение последовательности нуклеиновых кислот предпочтительно, по меньшей мере, частично сохранено. Окружение фланкирует последовательность нуклеиновых кислот, по меньшей мере, с одной стороны, а длина его последовательности составляет, по меньшей мере, 50 пар оснований (bp), предпочтительно, по меньшей мере, 500 bp, особо предпочтительно, по меньшей мере, 1000 bp, крайне предпочтительно - по меньшей мере, 5000 пар оснований. Природную кассету экспрессии - например, встречающееся в природе сочетание промотора RNR8 с соответствующим геном RNR8 - превращается в трансгенную кассету экспрессии, если ее изменяют иными, нежели природные, синтетическими ("искусственными") способами, например, мутагенезом. Соответствующие способы описаны (патент США US 5565350; международная заявка WO 00/15815).
В рамках изобретения "введение" означает все способы, пригодные для того, чтобы ввести последовательность нуклеиновых кислот RNR8 прямым или непрямым образом в растение или в клетку, компартмент, ткань, орган или зародыш такового, либо же сформировать ее там. Введение может вести к временному (преходящему) или постоянному (стабильному) присутствию последовательности нуклеиновых кислот RNR8.
Под "введением" подразумевают, например, такие способы как трансфекция, трансдукция или трансформация.
Введение кассеты экспрессии согласно изобретению в организм или его клетки, ткани, органы, части или зародыш (предпочтительно в растения или растительные клетки, ткани, органы, части или зародыш) целесообразно осуществлять с применением векторов, содержащих кассеты экспрессии. Кассету экспрессии можно вводить в вектор (например, в плазмиду) через надлежащий участок рестрикции. Полученную плазмиду сначала вводят в клетки E.coli. Клетки E.coli, корректно прошедшие трансформацию, отбирают, выращивают и получают рекомбинантную плазмиду с помощью знакомых специалисту методов. Для проверки этапа клонирования можно использовать рестрикционный анализ и секвенирование.
Векторы могут представлять собой, например, плазмиды, космиды, фаги, вирусы или же агробактерии. В предпочтительной форме исполнения введение кассеты экспрессии осуществляют с помощью плазмидных векторов. Предпочтительны такие векторы, которые дают возможность стабильно интегрировать кассету экспрессии в геном хозяина.
Создание измененного организма (или измененной клетки) требует введения соответствующих молекул ДНК в соответствующую клетку хозяина, а затем образования соответствующих РНК и белков посредством экспрессии гена.
Имеется множество методов, позволяющих осуществить этот процесс, называемый трансформацией или трансдукцией либо же трансфекцией (Keown et al. (1990) Methods in Enzymology 185: 527-537). Так, в качестве примера, ДНК или РНК можно ввести непосредственно - посредством микроинъекции или бомбардировки микрочастицами, покрытыми ДНК. Также возможно повысить проницаемость [стенок] клетки (пермебилизация), например, с помощью полиэтиленгликоля, так что ДНК может проникнуть в клетку путем диффузии. Также можно ввести ДНК в клетку путем слияния протопластов с другими объектами, содержащими ДНК, например, миниклетками, клетками, лизосомами или липосомами. Еще один подходящий метод введения ДНК - это электропорация, при которой с помощью электрического импульса вызывают обратимую пермеабилизацию клетки. Описания соответствующих методик имеются (например, в Bilang et al. (1991) Gene 100: 247-250; Scheid et al. (1991) Mol. Gen. Genet. 228: 104-112; Guerche et al. (1987) Plant Science 52: 111-116; Neuhause et al. (1987) Theor. Appl. Genet. 75: 30-36; Klein et al. (1987) Nature 327: 70-73; Howell et al. (1980) Science 208: 1265; Horsch et al. (1985) Science 227: 1229-1231; DeBlock et al. (1989) Plant Physiology 91: 694-701; Methods for Plant Molecular Biology (Weissbach and Weissbach, eds.) Academic Press Inc. (1988); and Methods in Plant Molecular Biology (Schuler and Zielinski, eds.) Academic Press Inc. (1989)).
При этом в растениях описанные методы трансформации и регенерации растений из растительных тканей или растительных клеток используют для преходящей или стабильной трансформации. Подходящие методы - это прежде всего трансформация протопластов путем индуцированного полиэтиленгликолем поглощения ДНК, биолистический способ с генетической пушкой, так называемая "бомбардировка частицами", электропорация, инкубация сухих эмбрионов в содержащем ДНК растворе и микроинъекция.
Помимо использования этих "прямых" методик трансформации, можно также провести трансформацию с помощью бактериальной инфекции посредством Agrobacterium tumefaciens или Agrobacterium rhizogenes. Методики описаны, например, Horsch et al. (1985) Science 225: 1229f.
Если применяют агробактерии, то кассету экспрессии необходимо интегрировать в специальные плазмиды, которые представляют собой либо промежуточный вектор (английское название: shuttle or intermediate vector), либо бинарный вектор. Если для трансформации используют Ti- или Ri-плазмиду, то по крайней мере правая граница, а в большинстве случаев, однако, как правая, так и левая границы Т-ДНК Ti- или Ri-плазмиды соединены с подлежащей вводу кассетой экспрессии в качестве фланкирующих регионов.
Предпочтительно применять бинарные векторы. Репликация бинарных (челночных) векторов возможна как в E.coli, так и в агробактерии. Как правило, они содержат ген-маркер отбора и линкер или полилинкер, фланкированный левой или правой ограничивающей последовательностью Т-ДНК. Их трансформация возможна непосредственно в агробактерии (Holsters et al. (1978) Mot. Gen. Genet. 163: 181-187). Ген-маркер отбора, например, ген nptII, сообщающий резистентность к канамицину, дает возможность отбора агробактерий, прошедших трансформацию. Агробактерия, играющая в данном случае роль организма-хозяина, должна уже содержать Ti-плазмиду-хелпер с регионом vir, необходимую для переноса Т-ДНК в растительную клетку. Агробактерию, преобразованную таким образом, можно применять для трансформации растительных клеток. Применение Т-ДНК для трансформации растительных клеток подробно исследовано и описано (европейский патент ЕР 120 516; Hoekema, в книге: The Binary Plant Vector System, Offsetdrukkerij Kanters B.V., Alblasserdam, Chapter V; An et al. (1985) EMBO J 4: 277-287). Разнообразные бинарные векторы известны и отчасти представлены в торговле, как, например, рВI101.2 или pBIN19 (Clontech Laboratories, Inc. США).
В случае инъекции или электропорации ДНК или РНК в растительные клетки каких-либо особых требований к применяемой плазмиде не предъявляют. Можно применять простые плазмиды, например, таковые из ряда pUC. Если из преобразованных клеток необходимо получать целые растения, то необходимо, чтобы в плазмиде находился дополнительный ген-маркер, позволяющий осуществлять отбор.
Прошедшие трансформацию и стабильные клетки, т.е. те, что содержат введенную ДНК в интегрированном в ДНК хозяина виде, можно отличить от не прошедших трансформацию, если компонентом введенной ДНК является пригодный к селекции маркер (McCormick et al. (1986) Plant Cell Reports 5: 81-84). В качестве примера маркера можно использовать любой ген, который способен сообщать резистентность к антибиотикам или гербицидам, (как, например, канамицину, G418, блеомицину, гигромицину или фосфинотрицину). Прошедшие трансформацию клетки, в которых имеет место экспрессия такого гена-маркера, в состоянии выжить в присутствии соответствующего антибиотика или гербицида в такой концентрации, которая губительна для дикого типа, не прошедшего трансформацию. Примеры включают в себя ген bar, придающий резистентность к гербициду фосфинотрицину (Rathore et al. (1993) Plant Mol Biol 21(5): 871-884), ген nptll, придающий резистентность к канамицину, ген hpt, придающий резистентность к гигромицину, или ген EPSP, придающий резистентность к гербициду глифосату. Полученные растения можно культивировать и скрещивать обычным образом. Чтобы убедиться, что интеграция в геном стабильна и наследуется, необходима культивация в двух или более поколениях.
Вышеуказанные способы описаны, например, в Jenes et al. (1993) Techniques for Gene Transfer, в книге: Transgenic Plants, Vol.1, Engineering and Utilization, изданной SD Kung и R Wu, Academic Press, S. 128-143, а также в Potrykus (1991) Annu. Rev. Plant Physiol. Plant Molec. Biol. 42: 205-225).
Подлежащую экспрессии конструкцию целесообразно клонировать в вектор, способный трансформировать Agrobacterium tumefaciens, например, в pBin 19 (Bevan et al. (1984) Nucl. Acids Res. 12: 8711f).
Когда создана преобразованная растительная клетка, целое растение можно получить с применением известных специалисту методов. Можно работать, например, с культурами каллуса. В этой массе недифференцированных клеток можно известным способом вызвать формирование побега и корня. Полученные проростки можно высаживать и культивировать.
Специалисту также известны способы для получения из растительных клеток частей растения и целого растения (регенерации). Для этого применяют, например, способы, описанные Fennell et al. (1992) Plant Cell Rep. 11: 567-570; Stoeger et al. (1995) Plant Cell Rep. 14: 273-278; Jahne et al. (1994) Theor. Appl. Genet. 89: 525-533.
Еще одним предметом настоящего изобретения является рекомбинантная молекула нуклеиновой кислоты, включающая, в последовательности 5'-3', следующие элементы:
- регуляторные последовательности промотора, активного в растительных клетках,
- последовательность ДНК согласно изобретению, оперативно с ними связанную,
- при необходимости - оперативно связанные с ними регуляторные последовательности, способные служить в растительной клетке сигналами транскрипции, терминации и/или полиаденилирования.
"Оперативно связанные" означает, что промотор и подлежащая экспрессии последовательность нуклеиновых кислот, а при необходимости - и прочие регуляторные элементы расположены таким образом, что каждый из регуляторных элементов способен выполнять свои функции при экспрессии последовательности нуклеиновых кислот. Прямая связь в химическом смысле для этого необязательна. Генетические контрольные последовательности, как, например, последовательности-энхансеры способны осуществлять свое воздействие на последовательность-цель, и, находясь в более удаленном положении, либо вообще из другой молекулы ДНК. Предпочтительны варианты размещения, при которых подлежащая экспрессии в трансгенном виде последовательность нуклеиновых кислот располагается следом за последовательностью, играющей роль промотора, так что обе последовательности ковалентно связаны друг с другом. При этом предпочтительно, чтобы расстояние между последовательностью-промотором и подлежащей экспрессии в трансгенном виде последовательностью нуклеиновых кислот составляло менее 200 пар оснований, особо предпочтительно - менее 100 пар оснований, а наиболее предпочтительно - менее 50 пар оснований.
Создание такой оперативной связи и синтез рекомбинантной молекулы нуклеиновой кислоты можно осуществлять с помощью общепринятых методик рекомбинации и клонирования, которые описаны, например, в Sambrook and Russell (2001) Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor (NY), в Silhavy TJ, Berman ML und Enquist LW (1984) Experiments with Gene Fusions, Cold Spring Harbor Laboratory, Cold Spring Harbor (NY), в Ausubel FM et al. (1987) Current Protocols in Molecular Biology, Greene Publishing Assoc. and Wiley Interscience и в Gelvin et al. (1990) в книге: Plant Molecular Biology Manual. Между промотором и подлежащей экспрессии последовательности нуклеиновых кислот можно, однако, размещать и другие последовательности, которые играют роль, например, линкера с определенными участками разреза для рестрикционных ферментов или сигнального пептида. Инсерция последовательностей может также вести к экспрессии химерного белка. Предпочтительно, чтобы рекомбинантная молекула нуклеиновой кислоты, в состав которой входят по меньшей мере промотор и подлежащая экспрессии последовательность нуклеиновой кислоты в оперативной связи, находилась в интегрированном в вектор состоянии, и чтобы ее можно было ввести инсерцией в растительный геном путем, например, трансформации.
Под специфичными для растений промоторами в принципе подразумевают любой промотор, который способен управлять экспрессией генов, в особенности, чужеродных генов в растениях или частях, клетках, тканях, культурах растений. Экспрессия при этом может иметь конституциональный характер, быть индуцируемой или зависеть от развития.
Предпочтительны:
а) Конституциональные промоторы
Под "конституциональными" промоторами подразумевают такие промоторы, которые обеспечивают экспрессию в многочисленных, предпочтительно во всех тканях, на протяжении длительного периода развития растения, предпочтительно во все моменты развития растения. В особенности целесообразно применять растительный промотор или промотор, происходящий от растительного вируса. Особенно предпочтителен промотор транскрипта 35S из CaMV (вируса мозаики цветной капусты) (Franck et al. (1980) Cell 21: 285-294; Odell et al. (1985) Nature 313: 810-812; Shewmaker et al. (1985) Virology 140: 281-288; Gardner et al. (1986) Plant Mol. Biol. 6: 221-228) или промотор 19S CaMV (патент США US 5352605; международная заявка WO 84/02913; Benfey et al. (1989) EMBO J. 8: 2195-2202). Другой пригодный к применению конституциональный промотор - это промотор "Rubisco small subunit (малой субъединицы рибулозо-1,5-бифосфаткарбоксилазы/оксигеназы, SSU)" (патент США US 4962028), промотор нопалин-синтазы из Agrobacterium, двойной промотор TR, промотор OCS (октопин-синтазы) из Agrobacterium, промотор убиквитина (Holtorf S et al. (1995) Plant Mol Biol 29: 637-649), промотор убиквитина 1 (Chris-tensen et al. (1992) Plant Mol Biol 18: 675-689; Bruce et al. (1989) Proc. Natl. Acad. Sci. USA 86: 9692-9696), промотор Smas, промотор циннамилалкогольдегидрогеназы (патент США US 5683439), промоторы субъединиц АТФазы или промотор пролин-обогащенного белка из пшеницы (международная заявка WO 91/13991), а также прочие промоторы генов, конституциональная экспрессия которых в растениях известна специалисту. Особо предпочтителен в качестве конституционального промотора промотор гена нитрилазы-1 (nit1) из A. thaliana (№ поступления в GenBank: Y07648.2, нуклеотиды 2456-4340, Hillebrand et al. (1996) Gene 170: 197-200).
b) Тканеспецифичные промоторы
В одной из форм исполнения применяют промоторы, обладающие специфичностью к антеридиям, архегониям, цветам, листьям, стеблю, корням и семенам.
Специфичные к семенам промоторы - это, например, промотор фазеолина (патент США 5504200; Bustos et al. (1989) Plant Cell 1(9): 839-53), гена 2S-альбумина (Joseffson et al. (1987) J. Biol. Chem. 262: 12196-12201), легумина (Shirsat et al. (1989) Mol. Gen. Genet. 215(2): 326-331), USP (неизвестного семенного белка, "unknown seed protein"; Bäumlein et al. (1991) Mol. Gen. Genet. 225(3): 459-67), гена напина (US 5608152; Stalberg et al. (1996) L. Planta 199: 515-519), гена, кодирующего белок, связывающий сахарозу (международная заявка WO 00/26388), или промотор легумина В4 (LeB4; Bäumlein et al. (1991) Mol. Gen. Genet. 225: 121-128; Bäumlein et al. (1992) Plant Journal 2(2): 233-9; Fiedler et al. (1995) Biotechnology (NY) 13(10): 1090f), промотор олеозина из арабидопсиса (резуховидки)(международная заявка WO 98/45461), промотор Все4 из Brassica (капусты) (международная заявка WO 91/13980). Прочие пригодные к использованию промоторы со специфичностью к семенам - это гены, кодирующие "High Molecular Weight Glutenin" (HMWG, глютенин высокого молекулярного веса), глиадин, фермент разветвления, АДФ-глюкозопирофосфатазу (АГФазу) или синтазу крахмала. Кроме того, предпочтительны промоторы, позволяющие специфичную по семенам экспрессию в однодольных растениях, таких как кукуруза, ячмень, пшеница, рожь, рис и т.д. Можно с выгодой применять промотор гена lpt2 или lpt1 (международные заявки WO 95/15389, WO 95/23230) или промоторы, описанные в международной заявке WO 99/16890 (промоторы гена хордеина, гена глютеина, гена оризина, гена проламина, гена глиадина, гена зеина, гена казирина, гена секалина).
Промоторы, специфичные для клубней, корнеплодов или корней, - это, например, промотор пататина класса I (B33) и промотор ингибитора катепсина D из картофеля.
Промоторы, специфичные для листьев, - это, например, промотор цитолитической ФБФазы из картофеля (международная заявка WO 97/05900), промотор малой субъединицы рибулозо-1,5-бифосфаткарбоксилазы/оксигеназы (SSU) или промотор ST-LSI из картофеля (Stockhaus et al. (1989) EMBO J. 8: 2445-2451). Промоторы, специфичные для эпидермиса, - это, например, промотор гена ОХL-Р (белка, подобного оксалат-оксидазе, "Oxalat-Oxidase like protein"); Wei et al. (1998) Plant Mol. Biol. 36: 101-112), промотор, состоящий из промотора GSTA1 и интрона WIR1a (международная заявка WO 2005/035766) и промотор GLP4-(европейская заявка РСТ/ЕР 2006/062747).
Другие тканеспецифичные промоторы - это, например, специфичные для цветка промоторы, как то: промотор фитоен-синтазы (международная заявка WO 92/16635) или промотор гена Prr (международная заявка WO 98/22593) и специфичные для пыльцевых мешков промоторы, как, например, промотор 5126 (патенты США 5689049, US 5689051), промотор globl и промотор γ-зеина.
с) Промоторы с химической индукцией
Кассеты экспрессии могут также содержать промотор с химической индукцией (обзорная статья: Gatz et al. (1997) Annu. Rev. Plant Physiol. Plant Mol. Biol. 48: 89-108), посредством которого можно в определенный момент управлять экспрессией экзогенного гена в растении. Такие промоторы также можно применять, как, например, промотор PRP1 (Ward et al. (1993) Plant Mol. Biol. 22: 361-366), промотор, индуцируемый салициловой кислотой (международная заявка WO 95/19443), промотор, индуцируемый бензолсульфонамидом (европейский патент ЕР 0388186), промотор, индуцируемый тетрациклином (Gatz et al. (1992) Plant J. 2: 397-404), промотор, индуцируемый абсцизиновой кислотой (европейский патент ЕР 0335528) или же промотор, индуцируемый этанолом или циклогексаноном (международная заявка WO 93/21334).
d) Промоторы, индуцируемые стрессом или патогенным агентом
Особо выгодно применение промоторов с индукцией патогеном, поскольку они позволяют осуществлять экспрессию только в случае потребности (т.е. поражения патогеном).
Поэтому при реализации способа согласно изобретению в одной из форм исполнения в растениях применяют активные промоторы, представляющие собой промотор, индуцируемый патогенным воздействием.
К промоторам, индуцируемым патогенным воздействием, относят промоторы генов, индуцируемых вследствие поражения патогеном, как, например, гены PR-белков, SAR-белков, β-1,3-глюканазы, хитиназы и т.д. (например, Redolfi et al. (1983) Neth. J. Plant Pathol. 89: 245-254; Uknes et al. (1992) Plant Cell 4: 645-656; Van Loon (1985) Plant Mol. Virol. 4: 111-116; Marineau et al. (1987) Plant Mol. Biol. 9: 335-342; Matton et al. (1987) Molecular Plant-Microbe Interactions 2: 325-342; Somssich et al. (1986) Proc. Natl. Acad. Sci. USA 83: 2427-2430; Somssich et al. (1988) Mol. Gen. Genetics 2: 93-98; Chen et al. (1996) Plant J. 10: 955-966; Zhang and Sing (1994) Proc. Natl. Acad. Sci. USA 91: 2507-2511; Warner et al. (1993) Plant J. 3: 191-201; Siebertz et al. (1989) Plant Cell 1: 961-968).
Это охватывает также промоторы, индуцируемые механическим повреждением (ранением), как, например, промотор гена pinll (Ryan (1990) Ann. Rev. Phytopath. 28: 425-449; Duan et al. (1996) Nat. Biotech. 14: 494-498), гена wun1 и wun2 (патент США 5428148), гена win1 и win2 (Stanford et al. (1989) Mol. Gen. Genet. 215: 200-208), гена системина (McGurl et al. (1992) Science 225: 1570-1573), гена WIP1 (Rohmeier et al. (1993) Plant Mol. Biol. 22: 783-792; Eckelkamp et al. (1993) FEBS Letters 323: 73-76), гена MPI (Corderok et al. (1994) Plant J. 6(2): 141-150) и им подобных.
Один из источников других промоторов, индуцируемых патогенным воздействием, представляет собой семейство генов PR. Ряд элементов в этих промоторах оказались полезны. Так участок от нуклеотида -364 до нуклеотида -288 в промоторе PR-2d обеспечивает специфичность по салицилату (Buchel et al. (1996) Plant Mol. Biol. 30: 493-504). Последовательность 5'-ТСАТСТТСТТ-3' многократно встречается в промоторе β-1,3-глюканазы ячменя и более чем в 30 других индуцируемых стрессом генах. В табаке этот регион связывает ядерный протеин, количество которого повышается салицилатом. В качестве промоторов с индукцией патогенным воздействием можно также применять промоторы PR-1 из табака и арабидопсиса (европейская заявка ЕР-А 0332104, международная заявка WO 98/03536). Особо предпочтительны промоторы "acidic PR-5" (aPR5) из ячменя (Schweizer et al. (1997) Plant Physiol. 114: 79-88) и пшеницы (Rebmann et al. (1991) Plant Mol. Biol. 16: 329-331), поскольку их индукция патогенным воздействием особо специфична. За 4-6 часов после поражения патогеном происходит накопление белков aPR5, а их фоновая экспрессия очень низка (международная заявка WO 99/66057). Подход для достижения повышенной специфичности индукции патогеном основывается на получении синтетических промоторов из сочетаний известных элементов, отвечающих на патогенное воздействие (Rushton et al. (2002) Plant Cell 14: 749-762; международные заявки WO 00/01830; WO 99/66057). Специалисту известны другие индуцируемые патогенным воздействием промоторы, полученные от различных видов (европейские заявки ЕР-А 1165794; ЕР-А 1062356; ЕР-А 1041148; ЕР-А 1032684).
К прочим промоторам, индуцируемым патогенным воздействием, относятся промотор Flachs Fis1 (международная заявка WO 96/34949), промотор Vst1 (Schubert et al. (1997) Plant Mol. Biol. 34: 417-426), а также промотор сесквитерпен-циклазы EAS4 из табака (патент США 6100451).
Кроме того, предпочтительны промоторы, индукцию которых обеспечивает биотический или абиотический стресс, например, индуцируемый патогенным воздействием промотор гена PRP1 (или промотор gst1), например, из картофеля (международная заявка WO 96/28561; Ward et al. (1993) Plant Mol. Biol. 22: 361-366), индуцируемый нагревом промотор hsp70 или hsp80 из томата (патент США 5187267), индуцируемый холодом промотор альфа-амилазы из картофеля (международная заявка WO 96/12814), индуцируемый светом промотор PPDK или индуцируемый механическим повреждением промотор pinll (европейская заявка ЕР-А 0375091).
е) Промоторы, специфичные для ткани мезофилла
"Ткань мезофилла" означает ткань листа, расположенную между слоями эпидермиса и состоящую из столбчатой ткани, губчатой ткани и жилок листа.
При реализации способа согласно изобретению в одной из форм исполнения применяют промоторы, специфичные для ткани мезофилла, как, например, промотор гена пшеницы Germin 9f-3.8 (№ поступления в GenBank: М63224) или промотор ячменя GerA (международная заявка WO 02/057412). Эти промоторы особо выгодны, поскольку они специфичны для ткани мезофилла, и одновременно их индуцирует патогенное воздействие. Кроме того, возможно применение специфичного для ткани мезофилла промотора из арабидопсиса САВ-2 (№ поступления в GenBank: Х15222), а также промотора Zea mays PPCZml (№ поступления в GenBank: X63869) или их гомологов. "Специфичный для ткани мезофилла" означает обусловленное специфическим взаимодействием между содержащимися в последовательности промотора цис-элементами и связывающимися с ними факторами транскрипции ограничение транскрипции гена только растительной тканью, по возможности малым ее количеством, содержащим ткань мезофилла, предпочтительно подразумевают транскрипцию, ограниченную тканью мезофилла.
Прочие специфичные для мезофилла промоторы - это PPCZm1 (=РЕРС; Kausch (2001) Plant Mol. Biol. 45: 1-15); OsrbcS (Kyozuka et al. (1993) Plant Phys. 102: 991-1000); OsPPDK, поступл. АС 099041; TaGF-2.8, поступл. M63223 (Schweizer (1999) Plant J. 20: 541-552); TaFBPase, поступл. X53957; TaWIS1, поступл. AF467542 (US 2002/115849); HvBIS1, поступл. AF467539 (US 2002/115849); ZmMIS1, поступл. AF467514 (US 2002/115849); HvPR1a, поступл. Х74939 (Bryngelsson et al. (1994) Molecular Plant-Microbe Interactions 7(2): 267-75; HvPR1b, поступл. Х74940 (Bryngelsson et al. (1994) Molecular Plant-Microbe Interactions 7(2): 267-75); HvB1, 3gluc; поступл. AF479647; HvPrx8, поступл. AJ276227 (Kristensen et al. (2001) Molecular Plant Pathology 2(6): 311-317; und HvPAL, поступл. X97313 (Wei (1998) Plant Molecular Biology 36: 101-112).
f) Промоторы, специфичные для эпидермиса
Под "тканью эпидермиса" или эпидермисом подразумевают внешние тканевые слои растений. Эпидермис может быть однослойным или многослойным; существует экспрессия генов, "обогащенная" в эпидермисе, как, например, гена Cer3, которая может служить маркером (Hannoufa. (1996) Plant J. 10(3): 459-467).
Специалист предпочтительно подразумевает под "эпидермисом" преобладающую покровную ткань первичных надземных частей растения, например, побега, листьев, цветов, плодов и семян.
Специфичные для эпидермиса промоторы - это, например, WIR5 (=GstA1), поступл. Х56012 (Dudler & Schweizer, неопубл.); GLP4, поступл. AJ310534 (Wei (1998) Plant Molecular Biology 36: 101-112); GLP2a, поступл. AJ237942 (Schweizer (1999). Plant J 20: 541-552); Prx7, поступл. AJ003141 (Kristensen (2001) Molecular Plant Pathology 2(6): 311-317); GerA, поступл. AF250933 (Wu (2000) Plant Phys. Biochem. 38: 685-698); OsRCl, поступл. АР004656; RTBV, поступл. AAV62708, AAV62707 (Klöti (1999) PMB 40: 249-266) и Cer3 (Hannoufa (1996) Plant J. 10 (3): 459-467).
g) Промоторы, зависящие от развития
Прочие пригодные к применению промоторы - это, например, промоторы, специфичные для созревания плодов, как, например, специфичный для созревания плода промотор из томата (международная заявка WO 94/21794, европейский патент ЕР 409625). Зависящие от развития промоторы частично включают в себя тканеспецифичные промоторы, поскольку образование отдельных тканей естественным образом зависит от развития.
Особо предпочтительны конституциональные, а также специфичные для листа и/или стебля, индуцируемые патогенным воздействием, специфичные для корня, специфичные для ткани мезофилла промоторы, причем конституциональные, индуцируемые патогенным воздействием, специфичные для ткани мезофилла и специфичные для корня промоторы наиболее предпочтительны.
Кроме того, в оперативной связи с подлежащей экспрессии последовательностью нуклеиновой кислоты могут находиться другие промоторы, дающие возможность экспрессии в других тканях растений или в других организмах, как, например, бактериях E.coli. В принципе, в качестве промоторов растений можно использовать все вышеописанные промоторы.
Описания других промоторов, пригодных к экспрессии в растениях, имеются (Rogers et al. (1987) Meth. in Enzymol. 153: 253-277; Schardl et al. (1987) Gene 61: 1-11; Bergeret al. (1989) Proc. Natl. Acad. Sci. USA 86: 8402-8406).
Кроме того, обычный специалист в состоянии рутинными методами выделить прочие подходящие промоторы. Так, с помощью общепринятых методик молекулярной биологии, например, экспериментов по гибридизации или исследования связывания ДНК с белком, специалист может определить, например, дальнейшие регуляторные элементы нуклеиновой кислоты, специфичные для эпидермиса. При этом, например, на первом этапе нужную ткань изолируют из желательного организма, из которого необходимо выделить регуляторные последовательности, выделяют из нее всю поли(А)+-РНК и создают банк кДНК. На втором этапе с помощью клонов кДНК, основанных на молекулах поли(A)+-PHK из другой ткани, определяют посредством гибридизации те клоны из первого банка, соответствующие молекулы поли(A)+-PHK которых накапливаются только в нужной ткани. Затем с помощью идентифицированных таким образом кДНК выделяют промоторы, обладающие тканеспецифичными регуляторными элементами. Кроме того, в распоряжении специалиста имеются другие основывающиеся на ПЦР методы изоляции надлежащих тканеспецифичных промоторов.
Последовательности нуклеиновых кислот, содержащиеся в кассетах экспрессии или векторах согласно изобретению, могут быть оперативно связаны, кроме промоторов, с другими контрольными генетическими последовательностями. Термин "контрольные генетические последовательности" следует понимать в широком смысле, под ним подразумевают все те последовательности, которые оказывают влияние на формирование или функцию рекомбинантной молекулы нуклеиновой кислоты согласно изобретению. Например, контрольные генетические последовательности модифицируют транскрипцию и трансляцию в организмах прокариот и эукариот. Предпочтительно, чтобы в состав кассет экспрессии согласно изобретению входил расположенный вверх от 5'-конца конкретной последовательности нуклеиновой кислоты, подлежащей экспрессии в генетически модифицированном виде, промотор с описанной выше специфичностью, а вниз от 3'-конца - терминаторная последовательность в качестве дополнительной контрольной генетической последовательности, а также - при необходимости - прочие обычные регуляторные элементы, причем чтобы они в каждом случае были функционально связаны с последовательностью нуклеиновой кислоты, подлежащей экспрессии в трансгенном виде.
К контрольным генетическим последовательностям также относятся другие промоторы, элементы промоторов или минимальные промоторы, способные модифицировать свойства, управляющие экспрессией. Так, благодаря контрольным генетическим последовательностям, например, тканеспецифичную экспрессию можно осуществлять с дополнительной зависимостью от определенных стресс-факторов. Соответствующие элементы описаны, например, для водного стресса, абсцизиновой кислоты (Lam E and Chua NH (1991) J. Biol. Chem. 266(26): 17131-17135) и теплового стресса (Schoffl F et al. (1989) Mol. Gen. Genet. 217(2-3): 246-53).
В принципе, при реализации способа согласно изобретению можно использовать все естественные промоторы со своими регуляционными последовательностями, как, например, вышеописанные. Кроме того, можно с выгодой использовать и синтетические промоторы.
Также к контрольным генетическим последовательностям относятся нетранслируемые 5'-регионы, интроны или не кодирующие 3'-регионы генов, как, например, интрон Actin-1, или интроны Adh1-S 1, 2 и 6 (общее описание: The Maize Handbook, Chapter 116, Freeling and Walbot, Eds., Springer, New York (1994)). Было показано, что они могут играть значительную роль при регуляции экспрессии генов. Так, было показано, что нетранслируемые 5'-последовательности могут усиливать преходящую экспрессию гетерологичных генов. В качестве примеров усилителей трансляции следует упомянуть лидерную 5'-последовательность из вируса табачной мозаики (Gallie et al. (1987) Nucl. Acids Res. 15: 8693-8711) и ей подобные. Кроме того, они могут усиливать тканевую специфичность (Rouster J et al. (1998) Plant J. 15: 435-440).
Целесообразно, чтобы в состав рекомбинантной молекулы нуклеиновой кислоты входила одна или несколько так называемых "энхансерных" последовательностей, функционально связанных с промотором, которые дают возможность повышения трансгенной экспрессии последовательности нуклеиновой кислоты. На 3'-конце последовательности нуклеиновой кислоты, подлежащей экспрессии в трансгенном виде, также можно инсерцией ввести полезные дополнительные последовательности, как то: прочие регуляторные элементы или терминаторы. Конструкция гена может содержать последовательности нуклеиновой кислоты, подлежащей экспрессии в трансгенном виде, в одной или нескольких копиях.
Сигналы полиаденилирования, пригодные для использования в качестве контрольных последовательностей, - это растительные сигналы полиаденилирования, предпочтительно те, которые соответствуют сигналам полиаденилирования Т-ДНК из Agrobacterium tumefaciens, в особенности гена 3 Т-ДНК (октопин-синтазы) Ti-плазмиды pTiACHS (Gielen et al. (1984) EMBO J. 3: 835 ff), или их функциональные эквиваленты. Примеры особо удобных терминаторных последовательностей - это терминатор OCS (октопин-синтазы) и терминатор NOS (нопалин-синтазы).
Под контрольными последовательностями также подразумевают те, что дают возможность гомологичной рекомбинации или инсерции в геном организма-хозяина или удаление из генома. При гомологичной рекомбинации возможна, например, замена естественного промотора определенного гена на промотор со специфичностью к эмбриональному эпидермису и/или цветку.
Рекомбинантная молекула нуклеиновой кислоты и полученный из нее вектор могут содержать другие функциональные элементы. Понятие "функциональный элемент" следует понимать в широком смысле, под ним подразумевают все элементы, оказывающие влияние на синтез, размножение или функционирование молекул нуклеиновых кислот, векторов или генетически модифицированных организмов согласно изобретению. В качестве примеров, не налагающих, однако, каких-либо ограничений, следует указать:
а) Маркеры отбора, сообщающие резистентность к ингибиторам метаболизма, например, 2-дезоксиглюкозо-6-фосфату (международная заявка WO 98/45456), антибиотикам или биоцидам, предпочтительно - гербицидам, как, например, канамицину, G418, блеомицину, гигромицину или фосфинотрицину. Особо предпочтительные маркеры отбора - это те, которые придают резистентность к гербицидам. В качестве примеров следует назвать; последовательности ДНК, кодирующие фосфинотрицинацетилтрансферазы (PAT) и инактивирующие ингибиторы глутамин-синтазы (ген bar и pat), гены 5-енолпирувилшикимат-3-фосфатсинтазы (гены EPSP-синтазы), придающие резистентность к Glyphosat® (N-(фосфонометил)глицину), кодирующий разлагающие Glyphosat® ферменты ген дох (глифосатоксидоредуктаза), ген deh (кодирующий дегалогеназу, которая инактивирует далапон), ацетолактатсинтазы, инактивирующие сульфонилмочевину и имидазолинон, а также гены bxn, кодирующие нитралазные ферменты, разлагающие бромоксинил, ген aasa, сообщающий резистентность к антибиотику апектиномицину, ген стрептомицинофосфотрансферазы (SPT), который придает резистентность к стрептомицину, ген неомицинофосфотрансферазы (NPTII), сообщающий резистентность к канамицину и генетицидину, ген гигромицинфосфотрансферазы (НРТ), сообщающий резистентность к гигромицину, ген ацетолактат-синтазы (ALS), придающий резистентность к гербицидам на основе сульфонилмочевины (например, мутированные варианты ALS с мутациями, например, S4 и/или Hra).
b) Репортерные гены, кодирующие легко квантифицируемые белки и обеспечивающие посредством собственного окрашивания или активности фермента оценку эффективности трансформации или места или времени экспрессии. При этом крайне предпочтительны белки-репортеры (Schenborn and Groskreutz (1999) Mol. Biotechnol. 13(1): 29-44), как то: "зеленый флуоресцентный белок" (green fluorescence protein, GFP) (Sheen et al. (1995) Plant Journal 8(5): 777-784; Haseloff et al. (1997) Proc. Natl. Acad. Sci. USA 94(6): 2122-2127; Reichel et al. (1996) Proc. Natl. Acad. Sci. USA 93(12): 5888-5893; Tian et al. (1997) Plant Cell Rep. 16: 267-271; WO 97/41228; Chui et al. (1996) Curr Biol 6: 325-330; Leffel et al. (1997) Biotechniques 23(5): 912-8), хлорамфениколтрансфераза, люцифераза (Ow et al. (1986) Science 234: 856-859; Millar et al. (1992) Plant Mol. Biol. Rep. 10: 324-414), ген экворина (Prasher et al. (1985) Biochem. Biophys. Res. Commun. 126(3): 1259-1268), β-галактозидаза, ген R-локуса (кодирует белок, регулирующий продукцию антоцианиновых пигментов (красное окрашивание) в растительных тканях и позволяющий таким образом прямо анализировать активность промотора без добавления дополнительных вспомогательных веществ или хромогенных субстратов; Dellaporta et al., в публикации: Chromosome Structure and Function: Impact of New Concepts, 18th Stadler Genetics Symposium, 11:263-282, 1988), крайне предпочтительна β-глюкуронидаза (Jefferson et al. (1987) EMBO J. 6: 3901-3907).
c) Исходные материалы репликации, обеспечивающие размножение кассет экспрессии или векторов согласно изобретению, например, в E.coli. В качестве примера следует назвать ORI (origin of DNA replication), pBR322 ori или Р15А ori (Sambrook and Russell, см. выше).
d) Элементы, которые необходимы для трансформации растений, опосредованной агробактериями, как, например, правое или левое отграничение Т-ДНК или региона vir.
Для отбора клеток, успешно прошедших трансформацию, как правило, необходимо дополнительно ввести маркер отбора, который сообщает успешно прошедшим трансформацию клеткам резистентность к биоцидному веществу (например, гербициду), ингибитору метаболизма, как то: 2-дезоксиглюкозу-6-фосфату (международная заявка WO 98/45456) или антибиотику и позволяет таким образом провести отбор, отделяя трансформированные клетки от нетрансформированных (McCormick et al. (1986) Plant Cell Reports 5: 81-84).
Кроме того, настоящее изобретение касается генетически модифицированных растительных клеток и генетически модифицированных растений, которые включают в себя последовательность нуклеиновых кислот согласно изобретению или рекомбинантную молекулу нуклеиновой кислоты согласно изобретению, а также частей растений, трансгенных продуктов урожая и трансгенного генетического материала этих растений, как то: протопластов, растительных клеток, каллусов, семян, клубней, черенков, а также трансгенного потомства этих растений.
Предпочтительно - эти растения, принадлежащие к семейству Роасеае, особо предпочтительно - растения, выбранные из родов Hordeum, Avena, Secale, Triticum, Sorghum, Zea, Saccharum и Oryza, крайне предпочтительно - растения видов Hordeum vulgare (ячмень), Triticum aestivum (пшеница), Triticum aestivum subsp.spelta (спельта), тритикале, Avena sativa (овес), Secale cereale (рожь), Sorghum bicolor (просо), Zea mays (кукуруза), Saccharum officinarum (сахарный тростник) и Oryza sativa (рис).
Способ согласно настоящему изобретению, однако, можно применять и для обработки двудольных сельскохозяйственных растений, например, хлопчатника, бобовых, стручковых и особенно люцерны, соевых бобов, рапса, томатов, сахарной свеклы, картофеля, подсолнечника, декоративных растений, а также деревьев. К другим сельскохозяйственным растениям могут относиться фрукты (в частности, яблоня, груша, вишня, виноград, цитрусовые, ананасы и бананы), масличная пальма, чай, какао и кофейное дерево, табак, сизаль, а также лекарственные растения раувольфия и дигиталис. Особо предпочтительны двудольные растения сахарная свекла, рапс, соя, томат, картофель и табак. Список прочих сельскохозяйственных растений содержится в патенте США №6137030.
Специфическую экспрессию белка согласно изобретению в растениях согласно изобретению или в растительных клетках согласно изобретению можно обнаружить и наблюдать с помощью обычных методов молекулярной биологии и биохимии. Специалисту известны эти методики, он может без ограничений выбирать подходящий метод обнаружения, например, анализ методом нозерн-блоттинга для обнаружения специфичной для белка РНК или для определения степени накопления специфичной для белка РНК или методом саузерн-блоттинга или ПЦР для обнаружения последовательностей ДНК, кодирующих белок согласно изобретению. Зондовые или праймерные последовательности, применяемые для этого, могут быть идентичны последовательности, приведенной под номером SEQ ID NO 1, или иметь незначительные отличия от этой последовательности.
Само собой разумеется, что способ согласно изобретению можно сочетать также с другими способами повышения резистентности трансгенных растений к патогенным воздействиям. Так, например, можно подходящими методами повышать количество полипептида, активность или функциональные возможности одного или нескольких факторов резистентности, выбранных из группы, которую составляют белок Вах Inhibitor 1 из Hordeum vulgare (№ поступления в GenBank: AJ290421), из Nicotiana tabacum (№ поступления в GenBank: AF390556), из риса (№ поступления в GenBank: AB025926), из арабидопсиса (резуховидки) (№ поступления в GenBank: AB025927) или табака и рапса (№ поступления в GenBank: AF390555, Bolduc et al. (2003) Planta 216: 377-386), ROR2 (например, из ячменя (№ поступления в GenBank: AY246906)), SnAP34 (например, из ячменя (№ поступления в GenBank: AY247208)) и/или связывающего белка Lumenal Binding Protein BiP, например, из риса (№ поступления в GenBank AF006825). Также можно надлежащими методами снижать количество полипептида, активность или функциональные возможности одного или нескольких факторов резистентности, выбранных из группы, которую образуют белок RacB (например, из ячменя (№ поступления в GenBank: AJ344223)), CSL1 (например, из арабидопсиса (резуховидки) (№ поступления в GenBank: NM116593)), HvNaOX (например, из ячменя (№ поступления в GenBank: AJ251717); европейский патент ЕР 1525315), MLO (например, из ячменя (№ поступления в GenBank Z83834); международные заявки WO 98/04586, WO 00/01722, WO 99/47552), ARM1 (повторяющийся белок броненосца, "armadillo repeat protein"; № европейской заявки на патент 05110468.5).
Еще один предмет изобретения касается применения трансгенных организмов согласно изобретению и происходящих от них клеток, клеточных культур, частей - как, например, в случае трансгенных растительных организмов, корней, листьев и т.д. - и трансгенного наследственного материала, как то: семян или плодов, для производства кормов или пищи, фармацевтических средств или химических реактивов высокой чистоты.
В одной из форм исполнения изобретение, кроме того, касается рекомбинантного способа производства фармацевтических агентов или химических реактивов высокой степени очистки в организме хозяина, причем организм хозяина или его часть изменены с помощью одной из вышеописанных рекомбинантных молекул нуклеиновых кислот, а эта молекула нуклеиновой кислоты содержит один или несколько структурных генов, которые кодируют желательный химический реактив или катализируют биосинтез желательного химического реактива, прошедший трансформацию организм хозяина культивируют, а желательный химикат изолируют из среды культивации. Этот способ можно широко применять для получения таких химических реактивов высокой степени очистки, как ферменты, витамины, аминокислоты, сахара, жирные кислоты, натуральные и синтетические модификаторы вкуса, ароматизаторы и красители. Особо предпочтительно производство токоферолов и токотриенолов, а также каротиноидов. Культивацию прошедших трансформацию организмов-хозяев, а также выделение из организмов хозяев или из среды культивации осуществляют известными специалисту методами. Производство фармацевтических агентов, как, например, антител или вакцин, описано у Hood and Jilka (1999). Curr. Opin. Biotechnol. 10(4): 382-6; Ma and Vine (1999) Curr. Top. Microbiol. Immunol. 236: 275-92.
Ниже представлена идентификация белка, содержащего лейцинобогащенный повтор с доменом киназы, RNR8, как гена из ячменя, сообщающего ячменю резистентность изолятам Blumeria graminis, и его применение для формирования в трансгенных растениях или растительных клетках резистентности к патогенам. Не следует считать, что нижеследующие примеры налагают какие-либо ограничения. Содержание всех литературных источников, заявок на патент, патентов и опубликованных заявок на патент, процитированных в настоящей заявке на патент, включено в нее посредством ссылок.
Примеры
Пример 1: Общий процесс клонирования
Процедуры клонирования, как, например, рестрикционное расщепление, гель-электрофорез в агарозе, очистку фрагментов ДНК, перенос нуклеиновых кислот на нитроцеллюлозные и нейлоновые мембраны, соединение фрагментов ДНК, трансформацию клеток Е.coli, культивирование бактерий и анализ последовательностей рекомбинантных ДНК проводили так, как описано у Sambrook et al. (2001), см. выше.
Пример 2: Анализ последовательности рекомбинантной ДНК
Секвенирование рекомбинантных молекул ДНК осуществляли с помощью лазерно-флуоресцентного секвенатора производства фирмы ABI по методике Sanger (Sanger et al. (1977) Proc. Natl. Acad. Sci. USA 74: 5463-5467).
Пример 3: Скрининг ВАС для идентификации клона, содержащего последовательность согласно изобретению
(ВАС - Bacterial Artificial Chromosome, искусственная бактериальная хромосома). Для идентификации гена, кодирующего в ячмене последовательность согласно изобретению, применяли пулы ДНК из библиотеки ВАС ячменя (Yu et. al. (2000) TAG 101: 1093-99). Клоны ВАС, которые несут последовательность согласно изобретению, определяли посредством ПЦР с применением праймеров 5' СТТ ТСG TGC ТТА TGT GGG TGT GАС (SEQ ID NO 4) и 5' CAT GAG GAG TCT GCA ATA AGG A (SEQ ID NO 5).
ПЦР выбрали для обнаружения искомой последовательности ДНК ввиду ее высокой чувствительности. Анализ проводили в реакционном объеме 20 мкл. Реакционная смесь состояла из 10 мМ трис-HCl, рН 9,0; 50 мМ KCl; 0,1% Тритона Х-100, 0,2 мМ дНТФ; 2 мМ MgCl2, по 0,6 мкМ олигонуклеотида и Taq-полимеразы (концентрация в реакционной смеси: ~1 ЕД мкл-1). На порцию реакционной смеси применяли либо 10 нг ДНК из пула ВАС, либо 2 мкл культуры бактерий (для ПЦР колонии). Имеющиеся последовательности сДНК служили шаблоном для получения олигонуклеотида 5'GGA TTT GTC ACG TCC AAC CT (SEQ ID NO 6) и 5'АТТ GGC ААТ TGT GAT AGC CC (SEQ ID NO 7).
Подлежащую амплификации ВАС-ДНК и праймер поместили в реакционный сосуд, а затем смешали с реакционным раствором для ПЦР. Для уничтожения и раскрытия бактерий при ПЦР колонии предварительную смесь перед добавлением реакционной смеси ПЦР в течение 5 минут нагревали до 95°С. Для денатурации двухцепочечной ДНК использовали начальный этап 5 минут при 95°С. В первые 10 циклов проводили Touchdown ПЦР с периодами 30 с 95°С; 30 с 60 к 55°С и 60 с 72°С.При этом в каждом цикле температуру снижали на 0,5°С (с 60 до 55°С). Дальнейшие 30 циклов проходили с периодами 30 с 95°С; 30 с 55°С и 60 с 72°С. Для завершающего удлинения цепи в течение 5 минут проводили инкубацию при 72°С, после чего реакционную смесь охлаждали до 20°С и поддерживали температуру на этом уровне. Анализ экспериментов с ПЦР проводили на основе короткого, как и ожидалось, продукта реакции длиной 189 п.о. с использованием 2,5%-ных агарозных гелей в 0,5х буфера ТВЕ (Трис-борат-ЭДТА).
Для идентификации гена и промотора проводили субклонирование идентифицированных отдельных клонов в два этапа. Сначала выделяли ВАС-ДНК отдельного клона с помощью колонки Qiagen (Maxi-Kit; Qiagen; изоляция по протоколу изготовителя). С помощью бокового сдвига (Hydroshear: Genomic Solutions) из этой ВАС-ДНК сформировали фрагменты длиной 5-10 тыс. п.о., а полученные концы дополнили до ровного положения с помощью фрагментов Кленов (реакция по протоколу изготовителя). Отбор фрагментов по длине проводили с помощью 0,8%-ного агарозного геля в 0,5% ТВЕ. Из геля вырезали участок с фрагментами соответствующей длины и провели элюцию ДНК из агарозного геля с помощью набора Qiagen Gel Extraction Kit (элюция по протоколу изготовителя). Прошедшие элюцию фрагменты длиной 5-10 тыс. п.о. подвергли лигации в вектор pBluescript II SK(-), линеаризованный EcoRV (рестрикция и дефосфорилирование согласно данным изготовителя), и химико-термическим способом трансформировали в высококомпетентные клетки Е.coli. Затем преобразованные клетки разместили произвольным образом с помощью робота для пикинга (Qpick, Genetix) и перенесли на микротитровальные пластины со средой LB (лизогенный бульон).
С помощью ПЦР выбрали субфрагмент, который несет представляющий интерес ген, и максимизировали длину потенциального участка вверх от 5' (5'-upstream). Отобранный субфрагмент снова разделили сдвигом на фрагменты длиной 1-2 тыс. п.о. провели лигацию, трансформацию и выложили клоны на микротитровальные пластины (см. выше). Из прошедших пикинг клонов произвольным образом выбрали 96 колоний и провели их секвенирование с помощью протокола TempliPhi (по протоколу изготовителя). Провели сбор последовательностей. Полученную информацию о последовательностях использовали для аннотации кодирующих экзонов в сравнении с уже известными последовательностями других организмов, чтобы определить последовательность согласно изобретению и ее потенциальные промоторы.
Пример 4: Субклонирование гена RNR8 в р1РКТА9
В целях более тщательной проверки того, отвечает ли идентифицированный ген RNR8 за придание резистентности к патогенным воздействиям, провели субклонирование гена в вектор pIPKTA9 по следующему протоколу.
1. Расщепление ("переваривание") NBS-LRR ВАС (№027N11) с помощью Psp 1406 I и XmaJI. Это позволяет вырезать почти всю кодирующую часть гена RNR8.
2. Формирование тупых концов с помощью фрагментов Кленов.
3. Элюция из геля полос 11 тыс. п.о. с этапа 2 с помощью набора Qiagen для гель-экстракции.
4. Лигация полос 11 тыс. п.о. NBS_LRR (этап 3) в ТА38 в присутствии Swa I (Douchkov et al. (2005) Molecular Plant-Microbe Interactions 18: 755-761).
5. ПЦР фрагмента NBS-LRR 1 тыс. п.о. в целях дополнения кодирующего участка гена на 5'-конце (upstream). Полимераза Thermal Асе (формирует фрагменты с тупыми концами).
Праймер NBS-LRR F1: GCT GAA ССА АСС CGG GGA GAA АТА (SEQ ID NO 8)
Праймер NBS-LRR F1: AGA TGA ТСG GAA GAA CAG TGC AAC (SEQ ID NO 9)
6. ПЦР для очистки (этап 5) на пластинах MinElute.
7. Клонирование фрагмента ПЦР 5' NBS-LRR (этап 6) в pIPKTA9, разрезанный Sma I.
8. Расщепление 11 тыс. п.о. LRR в ТА38 (этап 4) с помощью Aar и Not.
9. Гель-элюция 11 тыс. п.о. LRR с этапа 8.
10. Расщепление pIPKTA9_NBS_LRR_5' (этап 7) посредством Aar I и Not I.
11. Гель-элюция полос вектора ~3,5 тыс. п.о. с этапа 10.
12. Лигация фрагментов с этапов 7 и 11 ДНК-лигазой Т4 (Fermentas).
13. Подтверждение итоговой конструкции pIPKTA9_NBS-LRR секвенированием.
Поскольку при использовании конструкции pIPKTA9_NBS-LRR добились тех же эффектов, что и с клоном ВАС, из этого можно заключить, что RNR8 на ВАС №027N11 представляет собой ген, ответственный за наблюдаемую резистентность к патогенным воздействиям (см. также иллюстрацию 1).
Пример 5: Преходящая экспрессия в пшенице посредством бомбардировки частицами
В листья пшеницы с помощью "генетической пушки" (Bio-Rad, модель PDS-1000/Не, адаптер Hepta) посредством биолистической трансформации по методу Douchkov et al. (2005) Mol. Plant-Microbe Interact. 18: 755-761 ввели следующую смесь конструкций:
Для нанесения покрытия с ДНК на выстрел 2,18 мг частиц золота (диаметр 1,0 мкм, плотность частиц 25 мг мл-1 в глицерине 50% (об./об.)) смешали с 14-21 мкг "суперспирализованной" ДНК и добавили 1 М Са(NO3)2, рН 10, так что конечная концентрация Са(NO3)2 составляла 0,5 М. После центрифугирования и отмывки этанолом 70% (об./об.) частицы ресуспендировали в этаноле 96% (об./об.) и распределили по 7 макроскопическим носителям. В вакууме (3,6×103 Па) под воздействием гелиевой волны давления 7,6×106 Па частицы в каждом случае ввели в 7 сегментов листа растений пшеницы в возрасте 7 суток (сорт Kanzler). Для обстрела сегменты листьев поместили в чашку Петри на фитоагар 0,5% (в./об.), в который добавили 20 мкг мл-1 бензимидазола. Затем листья инкубировали при +20°С и непрямом освещении дневным светом в течение 4 часов.
Пример 6: Инокуляция сегментов листьев
Листья, прошедшие бомбардировку, перенесли на фитоагар 1% (в./об.) с 20 мкг мл-1 бензимидазола в поликарбонатных кюветах размером 20×20 см. В стойке для инокуляции провели инфицирование спорами мучнистой росы пшеницы, для чего в стойку насыпали споры с зараженных (в сильной степени) листьев пшеницы. Плотность инокулята составляла около 200 спор/мм2. По прошествии 5 минут кюветы изъяли, закрыли и инкубировали при +20°С и непрямом освещении дневным светом в течение 40-48 часов.
Пример 7: Гистохимическое обнаружение глюкуронидазы
Листья инфильтрировали в вакууме раствором для обнаружения глюкуронидазы (10 мМ ЭДТА, 1.4 мМ K3[Fe(CN)6], 1.4 мМ K4[Fe(CN)6], 0.1% (об./об.) Тритон Х-100, 20% (об./об.) метанола, 1 мг/мл 3-бромо-4-хлоро-3-индолил-β-D-глюкуроновая кислота, 100 мМ натриево-фосфатный буфер, рН 7.0) и инкубировали при +37°С на протяжении ночи. После удаления раствора для обнаружения листья в течение 15 минут обесцветили раствором 7.5% (мас./об.) трихлоруксусной кислоты и 50% (об./об.) метанола при +20°С.
Для оптической микроскопии применяли микроскоп Zeiss Axiolab при 200-кратном увеличении. Содержимое клеток с экспрессией глюкуронидазы имеет синее окрашивание. С помощью количественной микроскопии подсчитали количество клеток с окрашиванием глюкуронидазы и количество клеток с окрашиванием глюкуронидазы, несущих по меньшей мере 1 гаусторию грибка мучнистой росы пшеницы. Индекс восприимчивости рассчитывали путем деления количества положительных по глюкуронидазе клеток, несущих гаустории, на количество всех положительных по глюкуронидазе клеток.
Результаты эксперимента по инокуляции представлены на иллюстрации 1. Было показано, что экспрессия белка согласно изобретению из клона ВАС или вектора экспрессии pIPKTA9_NBS-LRR снижает восприимчивость растений пшеницы к мучнистой росе пшеницы примерно на 50% по сравнению с растениями, которые прошли трансформацию пустым вектором.
Иллюстрации
Иллюстрация 1: Эксперимент по преходящей комплементации в пшеницу с клоном ячменя ВАС 027N11 (банк Morex ВАС), содержащим лейцинобогащенный повтор протеинкиназы Rnr8. Листья пшеницы также бомбардировали pUbiGUS и либо pIPKTA9 (пустой вектор избыточной экспрессии), клоном ВАС 027N11, pIPKTA9_NBS LRR (см. пример 4), либо pIPKTA9:TaPERO (конструкция избыточной экспрессии пероксидазы, играющая роль положительного контроля). Представлены средние значения и стандартные отклонения двух независимых опытов, каждый с 2 параллельными бомбардировками.
| название | год | авторы | номер документа |
|---|---|---|---|
| БОЛЕЗНЕУСТОЙЧИВЫЕ РАСТЕНИЯ | 2006 |
|
RU2393227C2 |
| ПРИМЕНЕНИЕ ДЕНДРИМЕРНОЙ НАНОТЕХНОЛОГИИ ДЛЯ ДОСТАВКИ БИОМОЛЕКУЛ В РАСТИТЕЛЬНЫЕ КЛЕТКИ | 2010 |
|
RU2571928C2 |
| АУТОАКТИВИРУЮЩИЙСЯ БЕЛОК УСТОЙЧИВОСТИ | 2006 |
|
RU2375453C2 |
| ПРОМОТОР ДЛЯ СПЕЦИФИЧЕСКОЙ ЭКСПРЕССИИ ТРАНСГЕНОВ В ЭПИДЕРМИСЕ РАСТЕНИЙ | 2004 |
|
RU2382079C2 |
| СПОСОБЫ И ВЕЩЕСТВА ДЛЯ ПРИДАНИЯ РАСТЕНИЯМ УСТОЙЧИВОСТИ К ЗАБОЛЕВАНИЯМ | 1996 |
|
RU2203320C2 |
| УСТОЙЧИВЫЕ К БАКТЕРИЯМ ТРАНСГЕННЫЕ РАСТЕНИЯ | 2012 |
|
RU2640246C2 |
| КЛОНИРОВАНИЕ И ИСПОЛЬЗОВАНИЕ ФУНКЦИОНАЛЬНОГО R-ГЕНА ИЗ SOLANUM CHACOENSE | 2010 |
|
RU2586509C2 |
| ПОЛУЧЕНИЕ ФУНКЦИОНАЛИЗИРОВАННОЙ ЛИНЕЙНОЙ ДНК-КАССЕТЫ И ОПОСРЕДОВАННАЯ КВАНТОВЫМИ ТОЧКАМИ/НАНОЧАСТИЦАМИ ДОСТАВКА В РАСТЕНИЯ | 2011 |
|
RU2574785C2 |
| КОНСТРУКЦИЯ И СПОСОБ КОНСТРУИРОВАНИЯ СИНТЕТИЧЕСКОГО ДВУНАПРАВЛЕННОГО РАСТИТЕЛЬНОГО ПРОМОТОРА UBI1 | 2012 |
|
RU2639538C2 |
| РАСТИТЕЛЬНЫЙ ПЕПТИД ГАММА-ЗЕИН ДЛЯ ДОСТАВКИ БИОМОЛЕКУЛ В РАСТИТЕЛЬНЫЕ КЛЕТКИ | 2011 |
|
RU2563805C2 |








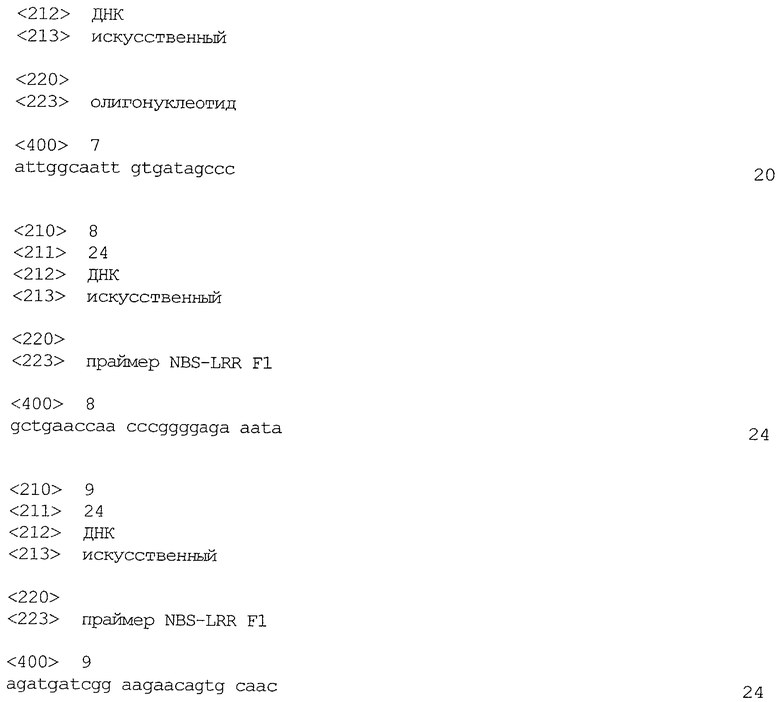

В растение или растительную клетку вводят и обеспечивают там экспрессию последовательности ДНК, кодирующей белок с лейцинобогащенным повторяющимся доменом (LRR) и/или киназной активностью. Экспрессия такой последовательности ДНК обеспечивает повышение резистентности трансгенных растений или растительных клеток к воздействиям патогенов. 2 н. и 16 з.п. ф-лы, 1 ил.
1. Способ повышения резистентности к патогенным воздействиям в трансгенных растениях, отличающийся тем, что изолированную молекулу нуклеиновой кислоты, содержащую последовательность нуклеиновой кислоты, выбранную из группы, включающей:
i) последовательности нуклеиновых кислот, в состав которых входят последовательности нуклеотидов, соответствующие кодирующей последовательности SEQ ID NO: 1,
ii) последовательности нуклеиновых кислот, в состав которых входят последовательности нуклеотидов, кодирующие белок с последовательностью аминокислот, приведенной под SEQ ID NO: 2,
iii) последовательности нуклеиновых кислот, в состав которых входят последовательности нуклеотидов, идентичные кодирующей последовательности SEQ ID NO: 1, по меньшей мере, на 60%, и/или
iv) последовательности нуклеиновых кислот, в состав которых входят последовательности нуклеотидов, гибридизирующиеся с комплементарной нитью нуклеотидной последовательности от i) до iii) в строгих условиях,
или
рекомбинантный белок, который кодируется указанными последовательностями нуклеиновых кислот, вводят в растение или в клетки растения, где его экспрессируют.
2. Способ по п.1, отличающийся тем, что последовательность нуклеиновой кислоты происходит от Hordeum vulgare.
3. Способ по п.1, отличающийся тем, что он включает в себя следующие этапы:
a) синтез рекомбинантной молекулы нуклеиновой кислоты, включающий в 5'-3'-ориентации, следующие элементы:
- регуляторные последовательности промотора, активного в растительных клетках,
- указанную в п.1 последовательность ДНК, оперативно с ними связанную,
- при необходимости - оперативно связанные с ними регуляторные последовательности, способные служить в растительной клетке сигналами транскрипции, терминации и/или полиаденилирования,
b) перенос рекомбинантной молекулы нуклеиновой кислоты из а) в растительные клетки и при необходимости интеграцию в растительный геном, и
c) регенерацию растений из трансформированных растительных клеток.
4. Способ по п.3, отличающийся тем, что экспрессию последовательности ДНК проводят под контролем конституционального промотора, предпочтительно - промотора 35S CaMV или промотора убиквитина.
5. Способ по п.3, отличающийся тем, что экспрессию последовательности ДНК проводят под контролем тканеспецифичного промотора.
6. Способ по п.3, отличающийся тем, что тканеспецифичный промотор представляет собой промотор, специфичный для эпидермиса, мезофилла или листа.
7. Способ по п.3, отличающийся тем, что экспрессию последовательности ДНК проводят под контролем индуцируемого промотора.
8. Способ по п.7, отличающийся тем, что индуцируемый промотор представляет собой промотор, индуцируемый патогеном или раной.
9. Способ по одному из пп.1-8, отличающийся тем, что патоген представляет собой патогенный грибок.
10. Способ по п.9, отличающийся тем, что патогенный грибок представляет собой грибок, выбранный из группы, состоящей из грибков мучнистой росы, ржавчинных грибков, грибков Fusarium и/или Septoria.
11. Трансгенное растение, включающее в себя растительную клетку, включающую в себя указанную в п.1 последовательность нуклеиновой кислоты или указанную в п.3 рекомбинантную молекулу нуклеиновой кислоты, или полученную в способе по одному из пп.1-10, либо созданное способом по одному из пп.1-10, а также части этих растений, трансгенные продукты урожая и трансгенный материал размножения этих растений, такой как: протопласты, растительные клетки, каллусы, семена, клубни и черенки.
12. Трансгенное растение по п.11, отличающееся тем, что трансгенная растительная клетка по сравнению с клетками дикого типа имеет повышенное количество указанного в п.1 белка.
13. Трансгенное растение по п.11, отличающееся тем, что трансгенная растительная клетка по сравнению с клетками дикого типа имеет повышенную резистентность к воздействию патогенов.
14. Трансгенное растение по п.11, отличающееся тем, что трансгенная растительная клетка по сравнению с клетками дикого типа имеет повышенную резистентность к грибкам.
15. Трансгенное растение по п.14, отличающееся тем, что трансгенная растительная клетка имеет повышенную резистентность к грибкам мучнистой росы, ржавчинным грибкам и/или грибкам Septoria.
16. Трансгенное растение по п.15, отличающееся тем, что трансгенная растительная клетка имеет повышенную резистентность к Formae speciales грибков мучнистой росы.
17. Трансгенное растение по одному из пп.11-16, отличающееся тем, что трансгенное растение представляет собой однодольное растение, предпочтительно - растение, принадлежащее к родам Avena, Triticum, Secale, Hordeum, Oryza, Panicum, Pennisetum, Setaria, Sorghum, Zea и им подобным.
18. Трансгенное растение по одному из пп.11-16, отличающееся тем, что трансгенное растение представляет собой двудольное растение, предпочтительно растение хлопчатника, бобовых, таких как стручковых и особенно люцерны, соевых бобов, рапса, канолы, томатов, сахарной свеклы, картофеля, подсолнечника, декоративных растений, табака, а также деревья.
| DATABASE EMBL, EMBL Accession no | |||
| КАМЕРА ДЛЯ СМЕСКИ ШЕРСТИ | 1926 |
|
SU5700A1 |
| PLANT CELL, 2004, vol.16, 3480-3495 | |||
| PLANT JOURNAL, 2004, vol.37, 517-527. | |||

