Настоящее изобретение относится к нуклеиновой кислоте, которая кодирует аутоактивирующийся белок устойчивости для достижения устойчивости к патогенам у растений, к применению нуклеиновой кислоты для получения трансгенного растения, а также к трансгенным растениям.
Грибы, вирусы, нематоды и бактерии, вызывающие известные заболевания растений, являются причиной большой потери урожая по всему миру, причиняют ущерб качеству продуктов урожая и делают необходимым дорогостоящее применение химических средств защиты для растений, поскольку естественные защитные средства растений, которые защищают их от большого количества потенциальных возбудителей заболеваний или могут замедлять их распространение, часто недостаточны. К таким защитным средствам относятся реакция гиперчувтвительности, которая контролирует гибель клеток тканей хозяина в очаге инфекции, усиление растительной клеточной стенки с помощью лигнификации и образования каллуса, образование фитоалексинов и продукцию СП-(связанных с патогенезом)-белков. Ключевыми молекулами для активации индуцируемых средств защиты являются гены устойчивости (резистентности) растений (R-гены). В соответствии с гипотезой "ген-на-ген" Флора (Flor), белок R-гена взаимодействует с соответствующим белком микробного гена авирулентности (Avr-ген) и таким образом происходит индуцируемая защитная реакция.
Большее число R-генов может в зависимости от структуры кодируемого ими Р-белка быть разделено на 5 классов (Martin et al., 2003). Класс 1 содержит только Pto-ген томата, который кодирует серин/треониновую киназу. Основное число R-генов растений принадлежит к суперсемейству NBS-LRR-генов, которые кодируют "нуклеотид-связывающий сайт" (NBS - nucleotide-binding site) и "лейцин-богатый повтор" (LRR - leucine rich repeat). NBS-LRR-гены, содержащие на своем N-конце структуру "биспирали" (coiled-coil) (CC), как, например, "лейциновая молния", будут принадлежать к CC-NBS-LRR-генам класса 2. R-гены CC-NBS-LRR-типа были найдены у всех покрытосеменных. К классу 3 относят R-гены TIR-NBS-LRR-типа, которые несут на N-конце последовательность, имеющую гомологию с TIR-участком ("toll-interleukin-1-receptor - toll-интерлейкин-1-рецептор") животных вместо СС-домена. TIR-NBS-LRR-гены составляют до 75% от всех R-генов у Arabidopsis thaliana, но не обнаружены у злаковых и сахарной свеклы (Tian et al., 2004).
4 класс R-генов составляют Cf-гены томатов. У CF-белков нет NBS-доменов, но они несут трансмембранный домен (ТМ) и внеклеточный лейцин-богатый повтор (LRR). К 5 классу относятся Ха21-белок риса, состоящий из внеклеточного LRR-домена, трансмембранного домена и внутриклеточного киназного домена.
Хотя R-гены слабо экспрессируются под промоторами R-генов, сильная конститутивная экспрессия R-генов 1, 2 и 3 классов приводит к активации защиты от патогенов самих растений в отсутствии соответствующих продуктов генов авирулентности и, таким образом, к аутоактивации Р-белков (Tang et al., 1999; Oldroyd and Staskawicz, 1998; Bendahmane et al., 2002).
Обычно конститутивная сверхэкспрессия R-генов в трансгенных растениях связана с нежелательными с точки зрения агрономии свойствами, такими как микронекрозы (Tang et al., 1999) или низкорослость растений (Frost et al., 2004).
Еще одной возможностью для обеспечения аутоактивации Р-белков классов 2 и 3 является мутагенез в специальных консервативных аминокислотных мотивах в полноразмерных CC-NBS-LRR или TIR-NBS-LRR белках. Мутагенез последовательности в NBS- или LRR-домене Rx-гена картофеля (Bendahmane et al., 2002) и NBS-домена LRR-гена льна (Howles et al., 2005) приводит к образованию мутантов, у которых в отсутствии соответствующих генов авирулентности после транзитной экспрессии запускается гибель клеток.
Эксперименты с делениями у Rx-гена показали, что продукты делеции, состоящие из СС-домена и частей NBS-домена, после их транзитной сверхэкспрессии также могут вызвать клеточную гибель, которая наступает быстрее, чем при применении полноразмерных R-генов. Эти продукты делеции помимо СС-домена нуждаются еще в Р-петле, киназе-2 и полноразмерной киназе-3 NBS-домена. В противном случае дальнейшее укорочение NBS-домена приведет к более медленному по сравнению с полноразмерным R-геном включению реакции гиперчувствительности (Bendahname et al., 2002).
Аутоактивация гена L10, одного из R-генов, принадлежащих к классу 3, может быть достигнута с помощью получения укороченного TIR-NBS-LRR-белка, состоящего из TIR-домена и 34 аминокислот смежного NBS-домена, включающего Р-петлю (Frost et al., 2004).
Хотя известно множество способов аутоактивации R-генов, до настоящего времени не было описано никаких трансгенных растений, у которых аутоактивация R-белков приводила бы к повышенной противогрибковой устойчивости без одновременного нанесения ущерба агрономическим свойствам. Попытка стабильно трансформировать два аутоактивируемых полноразмерных варианта L6-генов под контролем нативных L6-промоторов устойчивости или промоторов, индуцируемых грибами у льна, каждый раз приводила к появлению растений с нормальным ростом, восприимчивых к грибковой инфекции, либо к появлению низкорослых растений, устойчивых к грибковой инфекции (Howles et al., 2005).
Задача настоящего изобретения заключается в том, чтобы изменить защитную функцию растения против патогенов таким образом, чтобы защитная реакция растения при атаке патогенов надежно активировалась бы без негативного влияния на агрономические свойства растения.
В соответствии с изобретением поставленная задача решается с помощью нуклеиновой кислоты, включающей ограниченный участок NBS-LRR-гена устойчивости, который простирается от 5'-конца кодирующей области NBS-LRR-гена устойчивости в направлении 5'-3' до начала NBS-домена, причем NBS-LRR-ген устойчивости не является TIR-NBS-LRR-геном устойчивости. Такая нуклеиновая кислота может быть выделена из растения или получена синтетически.
Ограниченная область NBS-LRR-гена устойчивости, начиная со старт-кодона для трансляции (ATG-кодон) и до NBS-домена, известна как базовая благодаря Р-петле (мотив киназы-1a). Для того чтобы область NBS-LRR-гена устойчивости по изобретению могла функционировать, она не должна включать Р-петлю. Также не должны присутствовать и другие отрезки NBS- и LRR-доменов NBS-LRR-гена устойчивости. Однако отдельные нуклеотиды NBS-домена, включающего нуклеотиды Р-петли, могут быть оставлены, если они не будут отрицательно влиять на проявление реакции гиперчувствительности.
Под аутоактивирующимся белком устойчивости будет пониматься такой белок, который в отсутствии соответствующего продукта гена авирулентности будет приводить к активированию защиты от патогенов у растения. В этом отношении изобретение обладает тем преимуществом, что при возникновении устойчивости против патогенов не требуется никакого взаимодействия между белком устойчивости и белком авирулентности, благодаря чему защитная реакция растения является более направленной и в конечном итоге может протекать безопаснее.
Аутоактивация может быть вызвана, например, с помощью транзитной сверхэкспрессии гена устойчивости. Сверхэкспрессия означает, что сила экспрессии нативного промотора гена устойчивости будет настолько повышена, что каскад передачи сигнала, регулируемый R-белком, будет активирован при отсутствии соответствующего микробного продукта гена авирулентности. Это приведет к активации защиты от патогенов, которая приведет в результате к частичной или полной устойчивости к болезни.
Аутоактивация белков устойчивости может быть достигнута также с помощью укорочения полноразмерных R-генов BvKWS3_165, BvKWS3_135, Bv13033 и Bv12069 сахарной свеклы, а также StR3a гена картофеля в 5'-области, при котором в N-конце белка, лишенном NBS- и LRR- доменов, кодируется в конечном итоге случайный СС-домен. N-конец, лишенный NBS-домена, означает в данном контексте, что 5'-конец кодирующей области NBS-LRR-гена устойчивости распространяется настолько далеко к 3'-концу, что функциональная структура Р-петли NBS-LRR-гена устойчивости не включается в состав гена. В простых случаях Р-петля отсутствует полностью. Однако в укороченном гене устойчивости могут также присутствовать отдельные нуклеотиды Р-петли, если из-за них проявление реакции гиперчувствительности не будет замедлено. При укорочении NBS-LRR-гена устойчивости с N-конца будут также удалены последовательности киназы-2, киназы-3, GLPC и MHD, включая фланкирующие аминокислоты в соответствии с информацией банков данных "Просайт" (Prosite) (Bairoch et al., 1996) и "Пфам" (Pfam) (Sonnhammer et al., 1997), а также в соответствии с определением мотивов согласно Бендамане и др. (Bendahmane et al., 2002).
Применение укороченных R-генов 165_#176, 135_#147, 13033_#159 и Bv12069 и StR3a-#1-155 приводит, в сравнении с полноразмерными R-генами, к более быстрому запуску клеточной гибели в тканях растения. В комбинации с патоген-индуцируемым промотором это приводит к улучшенной индуцированной защите от патогенов. Это действует также для таких R-белков, которые не могут быть аутоактивированы из-за известных мутаций в MHD- или VHD-домене, проявляющихся в экспрессии гена 135_#147 BvKWS3 и 135-D480V.
Поскольку укороченные R-гены в состоянии вызвать клеточную гибель существенно раньше по сравнению с полноразмерными R-генами, для укороченных R-генов достаточно наличия слабой экспрессии для достижения концентрации белка, критической для защиты от патогенов, как было показано для R-гена 135_#147.
Р-петля или мотив киназы-1a совместно с мотивами киназы-2 и киназы-3 являются характерными для АТФ- или ГТФ-гидролизующих белков (Traut, 1994) и обнаружены в NBS-доменах NBS-LRR-генов. Р-петля характеризует N-концевую область NBS-домена (Bendahmane et al., 2002). Консенсусной последовательностью Р-петли для R-генов Prf, Rx, Rpm1, BvKWS3_135, BvKWS3_133 и BvKWS3_165 является (I/V)VG(M/I)GG(L/I/S)GKTT(L/V).
Неожиданно было обнаружено, что особенно хорошей активностью обладают нуклеиновые кислоты, кодирующие аминокислотную последовательность, включающую мотив DAE, и в особенности нуклеиновые кислоты, кодирующие мотив последовательности AVLXDAE. Мотивы последовательности DAE и AVLXDAE представлены, например. в SEQ ID NOS: 13 и 15.
Предпочтительными последовательностями нуклеиновых кислот являются выбранные из следующей группы:
а) нуклеотидная последовательность в соответствии с SEQ ID NO: 1, или нуклеотидная последовательность, комплементарная нуклеотидной последовательности в соответствии с SEQ ID NO: 1, или нуклеотидная последовательность, которая может быть гибридизована с нуклеотидной последовательностью в соответствии с SEQ ID NO: 1 или с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности в соответствии с SEQ ID NO: 1;
б) нуклеотидная последовательность в соответствии с SEQ ID NO: 2, или нуклеотидная последовательность, комплементарная нуклеотидной последовательности в соответствии с SEQ ID NO: 2, или нуклеотидная последовательность, которая может быть гибридизована с нуклеотидной последовательностью в соответствии с SEQ ID NO: 2 или с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности в соответствии с SEQ ID NO: 2;
в) нуклеотидная последовательность в соответствии с SEQ ID NO: 3, или нуклеотидная последовательность, комплементарная нуклеотидной последовательности в соответствии с SEQ ID NO: 3, или нуклеотидная последовательность, которая может быть гибридизована с нуклеотидной последовательностью в соответствии с SEQ ID NO: 3 или с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности в соответствии с SEQ ID NO: 3;
г) нуклеотидная последовательность в соответствии с SEQ ID NO: 4, или нуклеотидная последовательность, комплементарная нуклеотидной последовательности в соответствии с SEQ ID NO: 4, или нуклеотидная последовательность, которая может быть гибридизована с нуклеотидной последовательностью в соответствии с SEQ ID NO: 4 или с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности в соответствии с SEQ ID NO: 4;
д) нуклеотидная последовательность в соответствии с SEQ ID NO: 16, или нуклеотидная последовательность, комплементарная нуклеотидной последовательности в соответствии с SEQ ID NO: 16, или нуклеотидная последовательность, которая может быть гибридизована с нуклеотидной последовательностью в соответствии с SEQ ID NO: 16 или с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности в соответствии с SEQ ID NO: 16.
Ограниченный участок NBS-LRR-гена устойчивости находится в предпочтительных нуклеотидных последовательностях, указанных ниже:
SEQ ID NO: 1 c 124 по 654
SEQ ID NO: 2 с 155 по 598
SEQ ID NO: 3 с 94 по 573
SEQ ID NO: 4 c 194 по 694
Применяемый здесь термин "гибридизировать" обозначает гибридизацию в общепринятых условиях, в таких как описанные у Сембрук и др. (Sambrook et al., 1989), предпочтительно в строгих условиях. Строгими условиями гибридизации является, например, гибридизация в 4-кратном растворе хлорида и цитрата натрия (SSC) при 65°C с последующим многократным отмыванием в 0,1-кратном SSC при 65°C в течение примерно 1 часа. Менее строгими условиями гибридизации является, например, гибридизация в 4-кратном SSC при 37°C с последующим многократным отмыванием в 0,1-кратном SSC при комнатной температуре. "Строгие условия гибридизации" могут также означать гибридизацию при 68°C в 0,25 М фосфате натрия, рН 7,2, 7% додецилсульфате натрия (SDS), 1 мМ этилендиаминуксусной кислоте (ЭДТА) и 1% бычьем сывороточном альбумине (БСА) в течение 16 часов с последующим двукратным отмыванием в 2-кратном SSC и 0,2% SDS при 68°C.
Предпочтительно ген устойчивости, кодирующий аутоактивирующийся белок устойчивости, происходит из сахарной свеклы или картофеля.
Предпочтительно, чтобы нуклеиновая кислота по изобретению кодировала аминокислотную последовательность в соответствии с SEQ ID NO: 13 по 15. Внутри консенсусных последовательностей функциональные равнозначные аминокислоты могут быть взаимозаменяемы, например Asp вместо Glu, Leu вместо Ile, Ala или Val, Arg вместо Lys, Phe вместо Trp.
Обе консенсусные последовательности в соответствии с SEQ ID NOS: 13 и 14 представляют собой два функциональных блока, которые могут располагаться относительно друг друга на не очень жестко установленном расстоянии. Предпочтительно промежуток между обоими блоками представляет собой консенсусную последовательность в соответствии с SEQ ID NO: 15, а также консенсусную последовательность в соответствии с фиг.10.
Нуклеиновую кислоту по изобретению предпочтительно комбинировать с промотором, индуцируемым патогенами. Индуцируемый патогенами промотор активируется в ответ на инфекцию тканей хозяина возбудителем болезни, например повреждающим грибом, бактерией, вирусом или нематодами. Индуцируемый патогенами промотор более активен в растительных тканях во время проникновения в них инфекции или в успешно инфицированных тканях, чем в неинфицированных растительных тканях.
Промоторы, индуцируемые патогенами, в основном известны специалистам. Примеры промоторов, индуцируемых патогенами, включают хитиназный промотор (Samac and Shah, 1991), глюканазный промотор (Hennig et al., 1993) и prp-1 промотор (Martini et al., 1993).
С помощью индуцированной патогенами сверхэкспрессии R-гена можно предотвратить негативные последствия конститутивной экспрессии, такие как, например, низкорослость или дефицит растений.
Особенно подходящими показали себя синтетические промоторы. В данном изобретении описаны промоторы, полученные с помощью молекулярно-биологических методов, которые в таком виде не были найдены в природе. Синтетический промотор представляет собой минимальный промотор, который помимо минимального промотора содержит только один или более определенным образом подобранный цис-элемент. Эти цис-элементы представляют собой сайты связывания ДНК-связывающих белков, таких как транскрипционные факторы, и они выделены из природных промоторов, отобраны из готовых выделенных цис-элементов или получены искусственно при помощи способа случайной рекомбинации и отобраны при помощи подходящего способа. По сравнению с природным промотором синтетический промотор ввиду своего менее сложного строения активируется только с помощью небольшого количества экзогенных и эндогенных факторов, поэтому регулируется более специфически.
Минимальный промотор или "коровый" промотор представляет собой последовательность нуклеотидов, содержащую сайты связывания основных комплексов транскрипционных факторов и обеспечивающую точную инициацию транскрипции с помощью РНК-полимеразы II. Характерный мотив последовательности минимального промотора представляет собой ТАТА-бокс, инициаторный элемент (Inr), "элемент, распознающий TFBII" (BRE - ТАТА binding factor II recognition element) и "нижележащий элемент корового промотора" (DPE - downstream core promoter element). Эти элементы могут встречаться в минимальном промоторе вместе или по отдельности. В продаже имеются минимальные промоторы или мотивы их последовательности, выделенные из любого растительного или вирусного гена.
В объеме настоящего изобретения разработаны новые синтетические промоторы, которые в комбинации с известными генами устойчивости, не обязательно кодирующими аутоактивирующиеся белки устойчивости, можно применять для получения растений, устойчивых к патогенам. В данном изобретении описаны промоторы типа n×S-m×D-минимальных промоторов, n×W2-m×D-минимальных промоторов и n×Gst1-m×D-минимальных промоторов, таким образом, синтетический промотор содержит в себе одну или более чем одну следующую комбинацию цис-элементов:
а) n×S-m×D-бокс;
б) n×W2-m×D-бокс;
в) n×Gst1-m×D-бокс,
где n и m представляют собой натуральное число от 1 до 10
S-бокс (CAGCCACCAAAGAGGACCCAGAAT), имеющий последовательность нуклеотидов SEQ ID NO: 6, W2-бокс (TTATTCAGCCATCAAAAGTTGACCAATAAT), имеющий последовательность нуклеотидов SEQ ID NO: 7,
D-бокс (TACAATTCAAACATTGTTCAAACAAGGAACC),
имеющий последовательность нуклеотидов SEQ ID NO: 8 и Gst-бокс (TTCTAGCCACCAGATTTGACCAAAC), имеющий последовательность нуклеотидов SEQ ID NO: 9, описаны Раштоном в 2002 (Rushton et al., 2002), включая функции важных ядерных последовательностей.
Промоторы различаются в зависимости от набора элементов (n×S-m×D, n×W2-m×D или n×Gst1-m×D) по их базовой активности, способности быть индуцированными патогенами, кинетике активации и силе промотора, как, например, было показано для промоторов с комбинациями цис-элементов 2×S-2×D с нуклеотидной последовательностью SEQ ID NO: 10, 2×W2-2×D с нуклеотидной последовательностью SEQ ID NO: 11 и 2×Gst1-2×D с нуклеотидной последовательностью SEQ ID NO: 12. Свойства синтетического промотора дают возможность модифицировать генную экспрессию путем изменения числа цис-элементов (n, m=1…10) в соответствии с конкретными требованиями. Сравнение промоторов 2×S-2×D с вариантами 2×S-4×D, 4×S-2×D и 4×S-4×D показало, что в среднем сила промотора при применении тетрамеров повышается по сравнению с промоторами, состоящими из димеров. Кроме того, способность к индуцированию патогенами возрастает от димер-димерного промотора (2×S-2×D) к тетрамер-димерному и димер-тетрамерному промоторам (4×S-2×D, 2×S-4×D) и тетрамер-тетрамерному промотору (4×S-4×D) во всех временных точках измерения. Одновременно с возрастанием силы промотора и способности к индуцированию патогенами в случае описанных примеров тетрамерсодержащих промоторов обнаруживается усиление базовой активности. Эти примеры показывают, что важные свойства промоторов регулируются количеством цис-элементов и оптимальные варианты промоторов могут быть получены и идентифицированы для соответствующих искусственных реаранжировок.
Соответствующие результаты могут быть также получены с комбинациями цис-элементов, представляющими собой производные нуклеотидных последовательностей SEQ ID NO: 10, SEQ ID NO: 11 или SEQ ID NO: 12 и обладающими свойствами, сравнимыми со свойствами комбинаций цис-элементов SEQ ID NO: 10, SEQ ID NO: 11 или SEQ ID NO: 12.
Промоторы 2×S-2×D-минимальный промотор и 2×W2-2×D-минимальный промотор являются, например, комбинированными соответственно с четырьмя полноразмерными R-генами BvKWS3_133, BvKWS3_123, BvKWS3_135 и BvKWS3_165, и ими может быть трансформирована сахарная свекла. Испытание на устойчивость к грибам трансгенных растений с участием важнейшего гриба-паразита сахарной свеклы Cercospora beticola, возбудителя пятнистости листьев, в случае каждой из конструкций приводило к улучшению устойчивости к грибам, и при этом трансгенные растения не отличались от нетрансгенных растений по росту или другим агрономическим свойствам. Этот результат показывает, что при применении промотора, способного индуцироваться патогенами, в целом можно достичь сверхэкспрессии R-генов, запускающих клеточную гибель, и, следовательно, улучшенной устойчивости к болезням без негативного влияния на успешное развитие растения. При применении оптимальных промоторов в результате выбора наиболее подходящего количества повторов цис-элементов устойчивость к болезням дополнительно улучшается.
Настоящее изобретение также относится к трансгенным растениям, трансформированным новыми конструкциями нуклеиновых кислот, в особенности к растениям сахарной свеклы, частям, таким как семена или посевной материал этих растений, а также применению новых конструкций нуклеиновых кислот для получения трансгенных растений.
Изобретение далее будет пояснено со ссылкой на чертежи и примеры.
Изобретение, описанное в качестве примера с сахарной свеклой, также может без проблем быть применено и к другим пищевым растениям, у которых выделены гены устойчивости.
Графические материалы
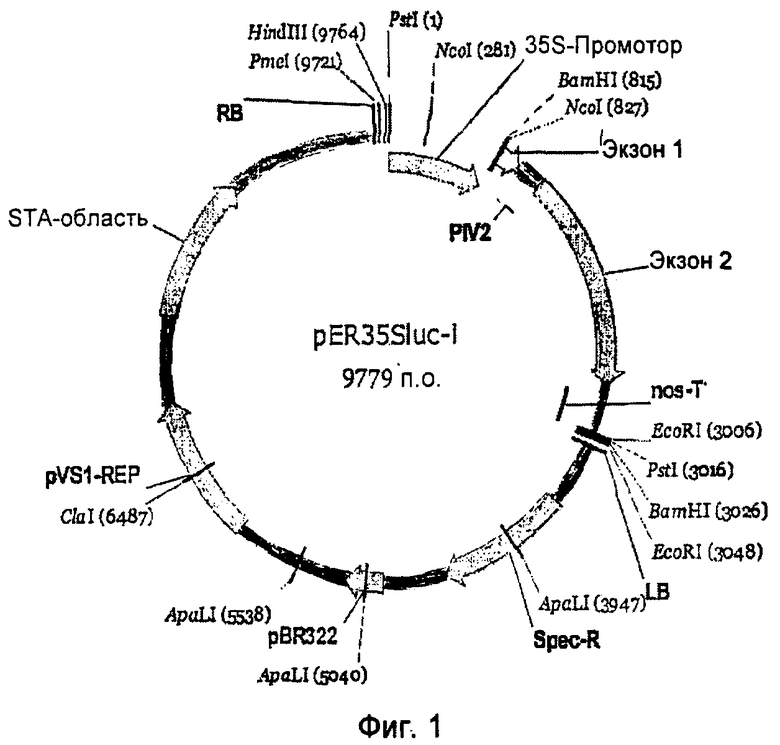
На Фиг.1 показана карта бинарного вектора pER-35Sluci, применяемого для опосредованной Agrobacterium tumefaciens транзитной экспрессии R-генов в листьях сахарной свеклы. Вектор несет один ген люциферазы с одним интроном Photinus pyralis, который не может экспрессироваться в A. tumefaciens.
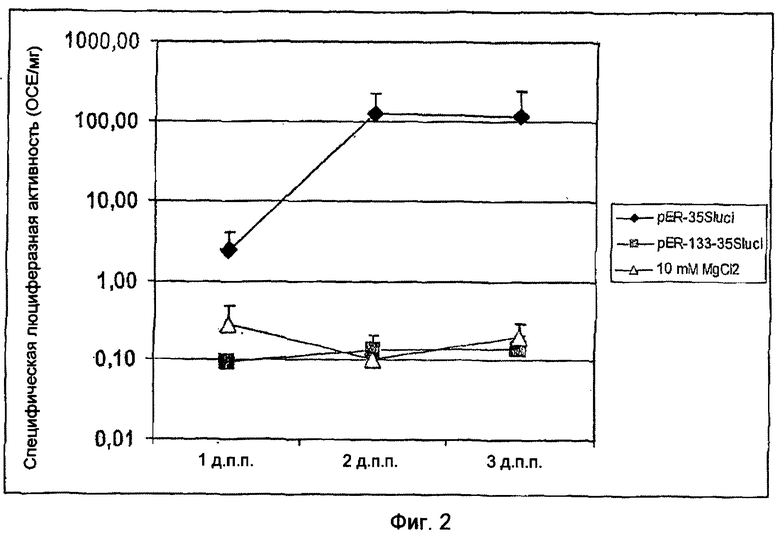
На Фиг.2 показан запуск клеточной гибели в листьях сахарной свеклы после транзитной экспрессии R-гена BvKWS3_133 с помощью Agrobacterium tumefaciens. В то время как транзитная экспрессия конструкций pER-35Sluci приводит к усиленной активности генов-репортеров в листьях свеклы, экспрессия конструкций pER133-35Sluci приводит к клеточной гибели таким образом, что невозможно выявить какую-либо активность генов-репортеров.
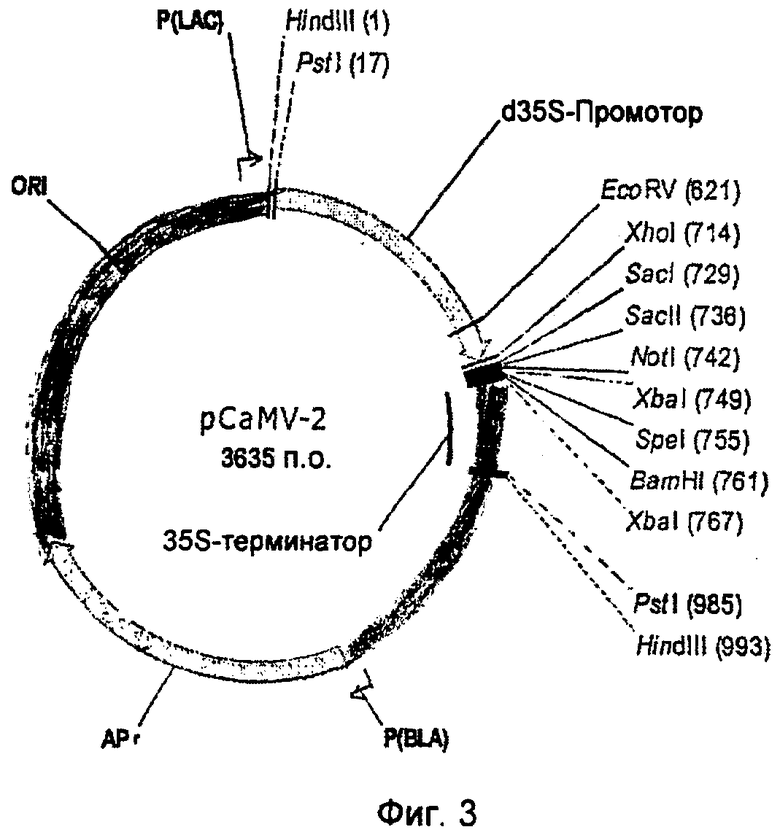
На Фиг.3 показан вектор pCaMV-2, который применялся для транзитной биолистической трансформации листьев сахарной свеклы. Полноразмерные и укороченные R-гены доставляли под контролем двойного 35S промотора, как было описано выше.
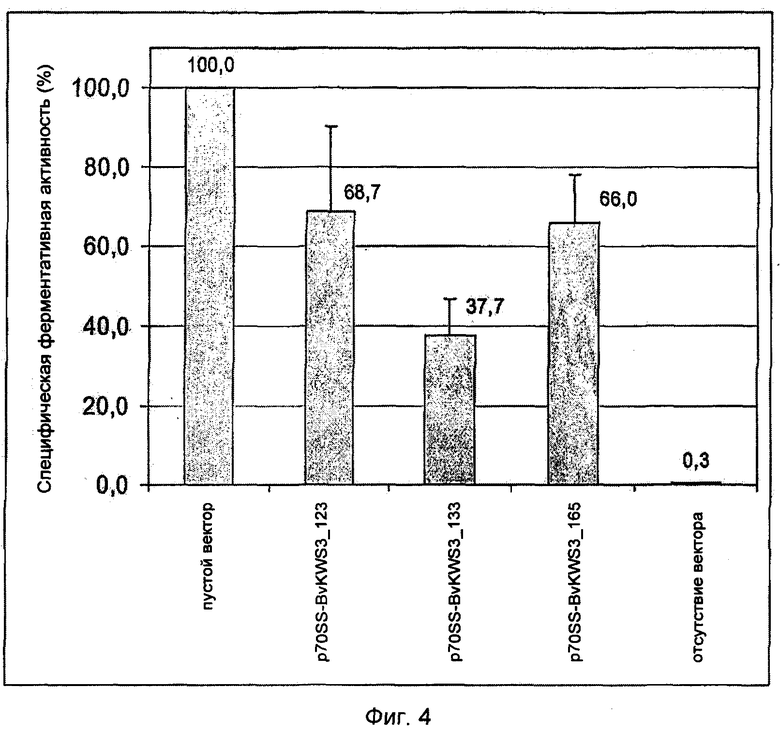
На Фиг.4 показан запуск клеточной гибели в листьях сахарной свеклы после транзитной экспрессии R-генов BvKWS3_123, BvKWS3_133 и BvKWS3_165 посредством биолистической трансформации. Проводили совместную трансформацию генами BvKWS3_123, BvKWS3_133 и BvKWS3_165, находящимися под контролем двойного 35S-промотора (d35S), и конструкцией гена-репортера p70S-luc. Активность гена-репортера измеряли через 20 ч после трансформации. Как видно по запуску реакции гиперчувствительности, активность гена-репортера уменьшена по сравнению с контролем (пустой вектор pCaMV-2 и p70S-luc). Представлено среднее значение 3-х независимых повторяющихся параллелей по меньшей мере с 9 единичными экспериментами на конструкцию. Погрешность обозначена стандартной ошибкой.
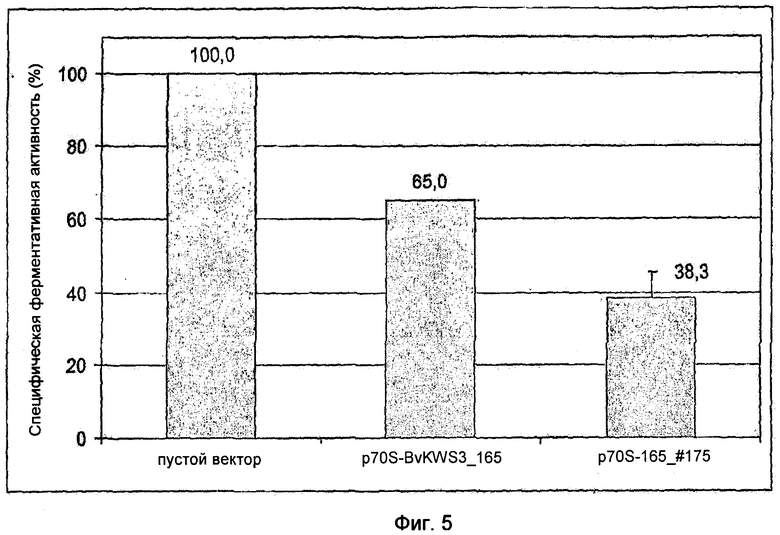
На Фиг.5 показано усиление запуска клеточной гибели в результате экспрессии 5'-концевых областей R-гена BvKWS3_165 по сравнению с экспрессией полноразмерного R-гена BvKWS3_165. Проводили биолистическую трансформацию листьев сахарной свеклы N-концевой областью и полноразмерным R-геном под контролем d35S промотора (p70S-165_#175 и p70S-BvKWS3_165) с конструкцией p70S-luc. Представлено среднее значение 3-х независимых повторяющихся параллелей по меньшей мере с 9-12 единичными экспериментами на конструкцию.
На Фиг.6 показано усиление запуска клеточной гибели в результате экспрессии полноразмерного R-гена BvKWS3_135 по сравнению с усилением запуска клеточной гибели с помощью 5'-концевой области 135_#147 R-гена BvKWS3_135. Проводили биолистическую трансформацию листьев сахарной свеклы полноразмерным R-геном и n-концевой областью 135_#147 под контролем d35S промотора (p70S-BvKWS3_135 и p70S-135_#147) с конструкцией p70S-luc. Представлено среднее значение 2-х независимых повторяющихся параллелей по меньшей мере с 9-12 единичными экспериментами на конструкцию.
На Фиг.7 показано усиление запуска клеточной гибели в результате экспрессии 5'-концевых областей 13033_#159 R-гена Bv13033 по сравнению с экспрессией полноразмерного R-гена Bvl3033. Проводили биолистическую трансформацию листьев сахарной свеклы полноразмерным R-геном и N-концевой областью 13033_#159 под контролем d35S промотора (p70S-13033 и p70S-13033_#159) с конструкцией p70S-luc. Представлено среднее значение 2-х независимых повторяющихся параллелей по меньшей мере с 9-12 экспериментами на конструкцию.
На Фиг.8 показано усиление запуска клеточной гибели в результате экспрессии R-гена Bv12069.
На Фиг.9 показана аутоактивация белков BvKWS3_135 в результате укорочения 5'-области клона кДНК 135_#147 по сравнению с мутацией VHD мотива NBS-домена.
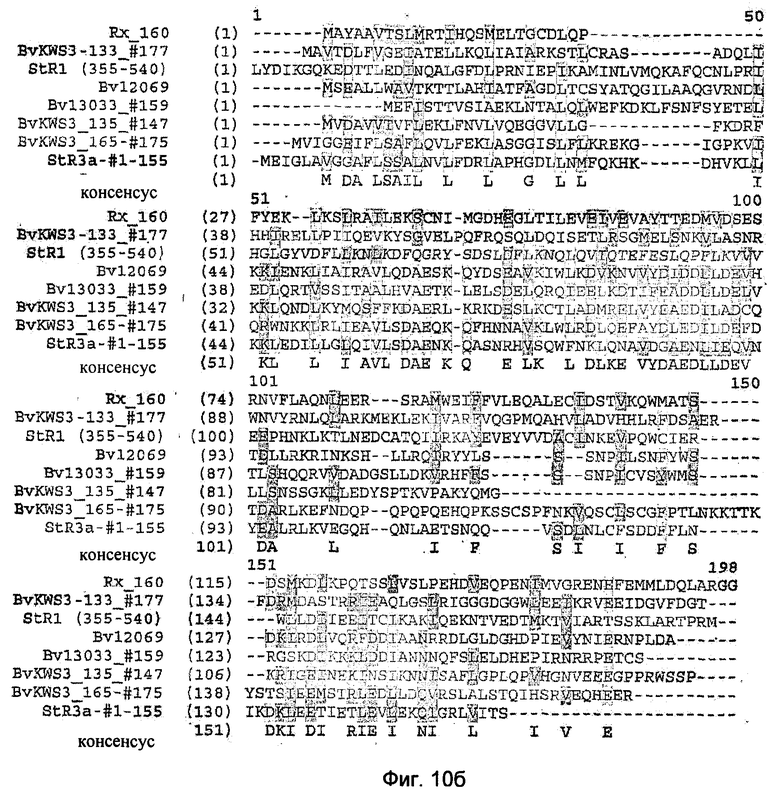
На Фиг.10 а)-в) показано сравнение аминокислотных последовательностей укороченных аутоактивных белков Bv12069, Bv13033_#159, BvKWS135_#147, BvKWS3_165_#175 и StR3a-#l-155 между собой со сравнительными последовательностями не обладающих аутоактивностью укороченных белков устойчивости картофеля (RX-160) и StR1(355-540), а также с полноразмерными R-белками NBS-LRR-типа Arabidopsis thaliana (AtAB028617), фасоли (Р.vulgarisJ71), риса (O.sativaАР003073), сои (G.maxKR4) томатов (Tomato-I2). Консенсусные последовательности подчеркнуты.
На Фиг.11 показано, что делеция аминокислот 147-175 достоверно снижает способность белков к аутоактивации.
На Фиг.12 показана активация синтетических промоторов 2×S-2×D в трансгенной сахарной свекле после атаки Cercospora beticola.
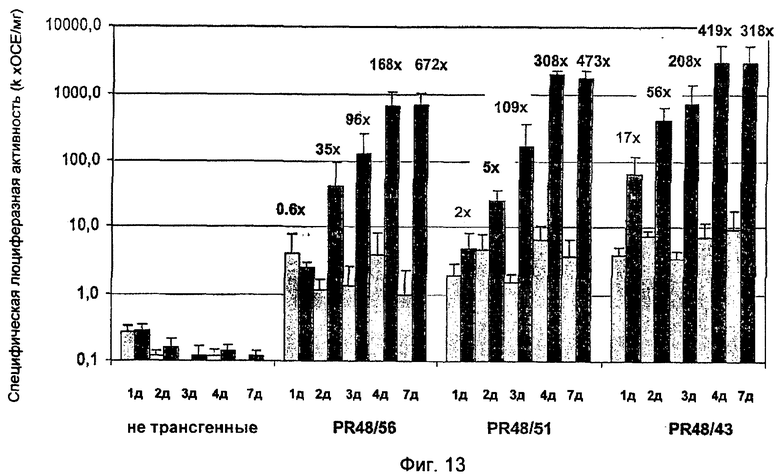
На Фиг.13 показана активация синтетических промоторов 2×W2-2×D в трансгенной сахарной свекле после атаки Cercospora beticola.
На Фиг.14 показано сравнение активностей промоторов генов-репортеров 2×S-2×D, 4×S-2×D, 2×S-4×D и 4×S-4×D в трансгенной сахарной свекле после атаки Cercospora beticola.
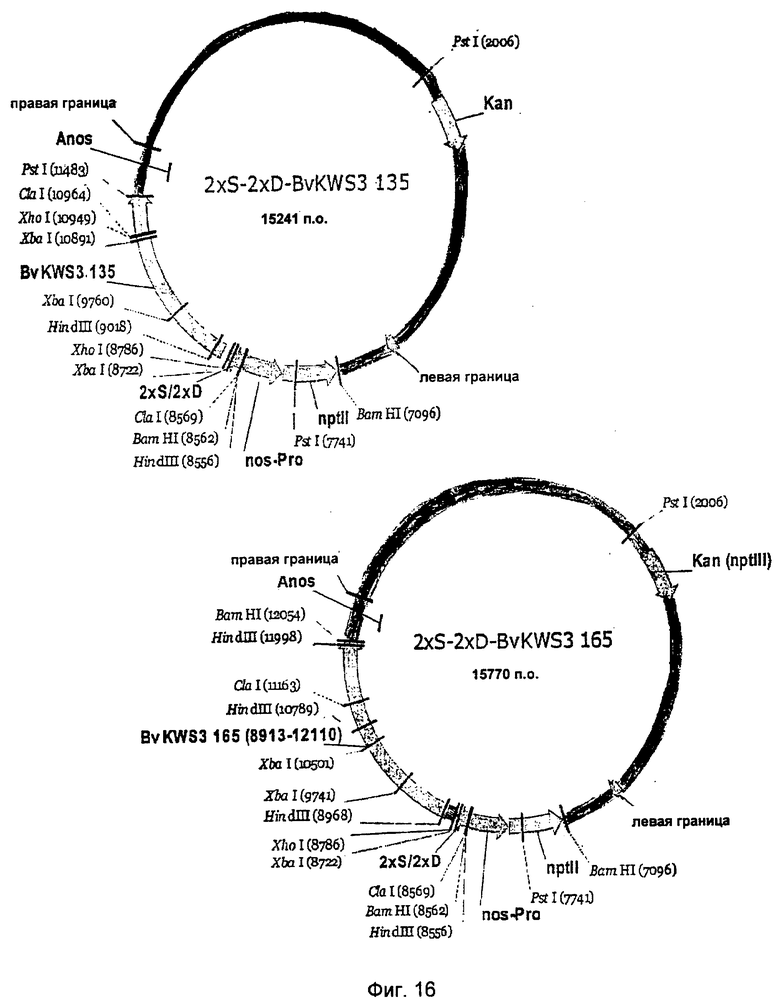
На Фиг.15 и 16 показаны комбинации полноразмерных R-генов 123, 133, 135, 165 с синтетическим промотором 2×S-2×D.
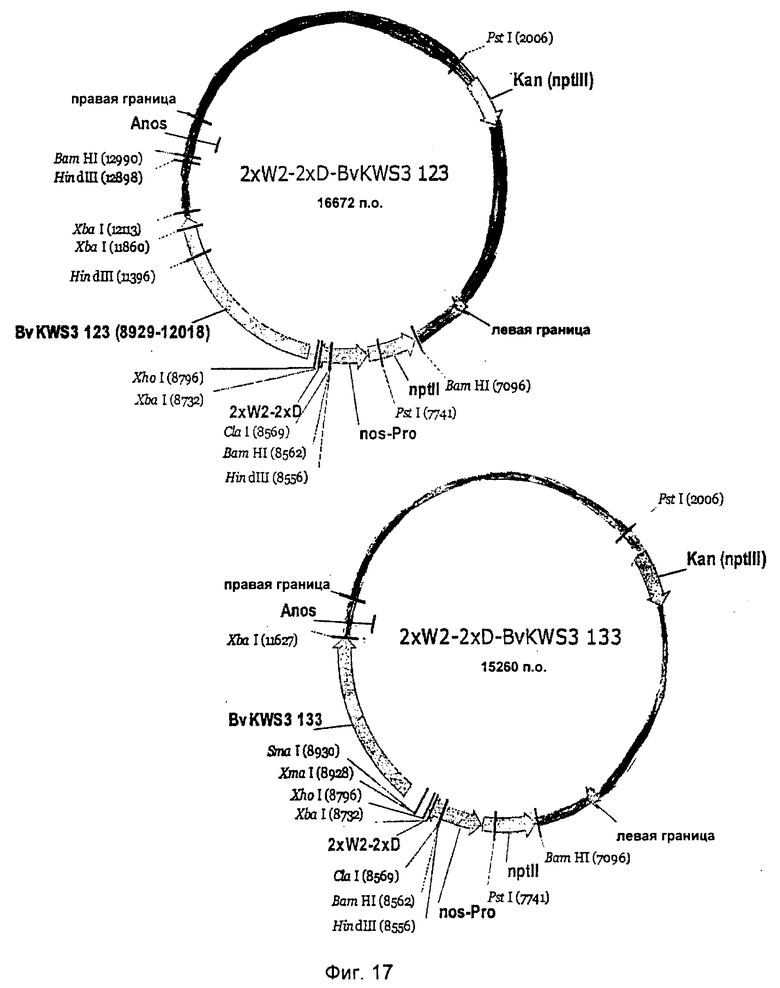
На Фиг.17 и 18 показаны комбинации полноразмерных R-генов 123, 133, 135, 165 с синтетическим промотором 2×W2-2×D.
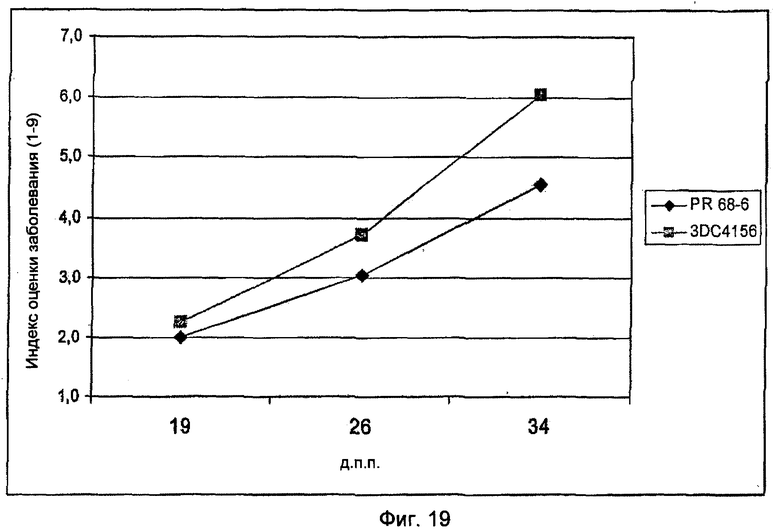
На Фиг.19 показано повышение устойчивости трансгенной сахарной свеклы линии PR68-6 против гриба-паразита Cercospora beticola по сравнению с нетрансгенным контролем 3DC4156.
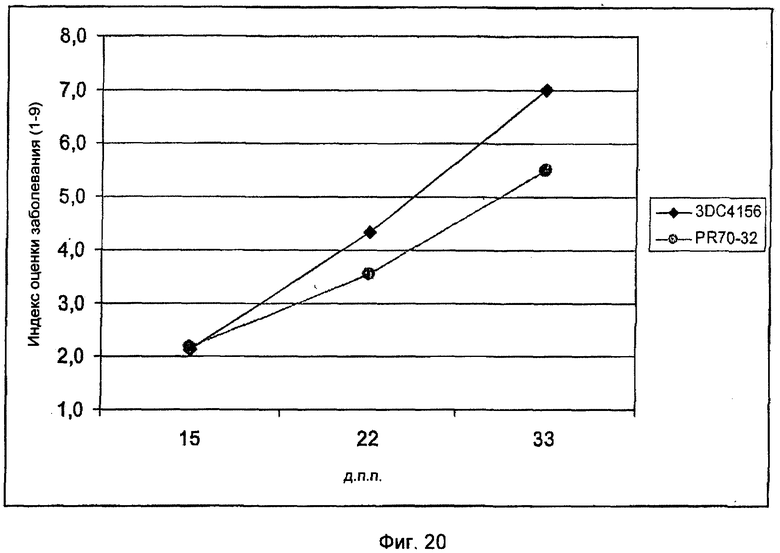
На Фиг.20 показано повышение устойчивости трансгенной сахарной свеклы линии PR70-32 против гриба-паразита Cercospora beticola по сравнению с нетрансгенным контролем 3DC4156.
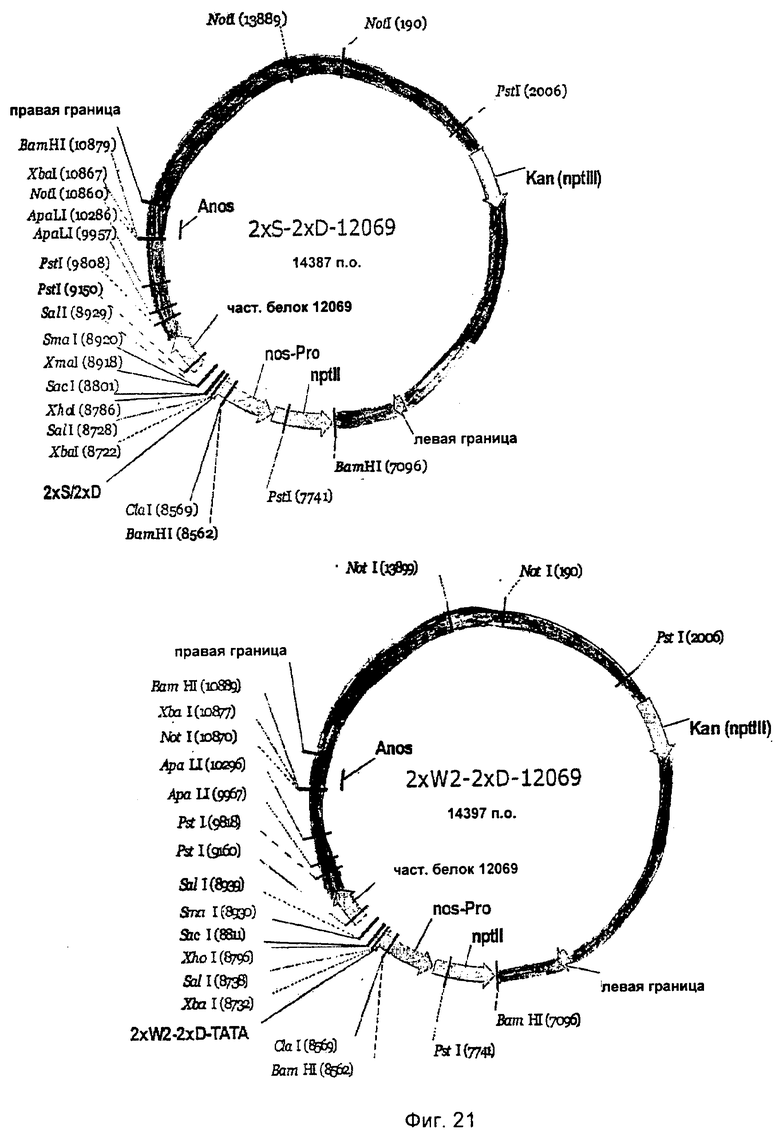
На Фиг.21 и 22 показана комбинация N-концевых областей R-генов 165_#176 и 12069 с синтетическими промоторами 2×S-2×D и 2×W2-2×D.
Примеры
Доказательство проявления быстрой реакции устойчивости в листьях сахарной свеклы в результате сверхэкспрессии гена BvKWS3_133
Транзитная сверхэкспрессия полноразмерного клона кДНК гена BvKWS3_133 в листьях сахарной свеклы с помощью Agrobacterium tumefaciens вызывает быструю клеточную гибель без видимого формирования некроза. кДНК-клон BvKWS3_133 комбинировали с d35S-промотором и встраивали в бинарный вектор pER-35Sluci (Фиг.1). Векторами pER-35Sluci и pER133-35Sluci трансформировали штамм С58С1 Agrobacterium (An, 1987). Положительные агробактерии помещали для транзитной экспрессии в 50 мл LB-среды с 100 мг/мл стрептомицина и 20 мкМ ацетосирингона на 4-5 ч. После этого бактерии центрифугировали, осадок помещали в раствор 10 мМ MgCl2, 10 мМ MES, 100 мкМ ацетосириногена и определяли плотность бактерий при OD600=0,1. Бактериальную суспензию оставляли на 2-3 часа и затем с помощью 2,5 мл шприца инъецировали через нижнюю сторону листа 10-недельных растений сахарной свеклы внутрь листа. После инкубации при 25°C в культуральном шкафу измеряли активность гена-репортера люциферазы Photinus pyralis в трансформированных листьях через 1, 2 и 3 дня после прививки. Для этого определили активность люциферазы с помощью системы для люциферазного анализа Luciferase Assay System (Promega, Mannheim, Deutschland) в люминометре Sirius (Berthold Detection System GmbH, Pforzheim, Deutschland) в соответствии с инструкциями изготовителя. Для получения ферментативного экстракта, подходящего для измерения, для каждого измерения вырезали из листьев по меньшей мере 2 круглых фрагмента. На каждую конструкцию проводили по 8 измерений в течение дня. Образцы листьев гомогенизировали в ступке с добавлением морского песка в 10 кратном объеме (об./мас.) пассивного литического буфера (ПЛБ). Жидкий супернатант переносили в 1,5 мл пробирки типа "Эппендорф" и центрифугировали в течение 5 мин при 4°C и 20000 g. Прозрачный супернатант отбирали и для измерения активности люциферазы Photinus использовали по 10 мкл центрифугированного экстракта. Листья сахарной свеклы, которые трансформировали контрольной конструкцией pER-35Sluci, в первый день показали слабую активность люциферазы, а на 2 и 3 день активность люциферазы составляла 124000 или 116000 относительных световых единиц (ОСЕ)/мг ткани листа. Листья свеклы, трансформированные конструкцией pER133-35Sluci, на всех 3-х временных точках измерения не продемонстрировали активности, которая была бы выше, чем в листьях, привитых MgCl2 (Фиг.2). При этом транзитная экспрессия клона кДНК BvKWS3_133 проявилась в очень быстрой клеточной гибели в привитых листьях свеклы.
Конститутивная экспрессия R-генов BvKWS3_123, BvKWS3_133 и BvKWS3_165 вызывает клеточную гибель в листьях сахарной свеклы
R-ген BvKWS3_133, также как R-ген BvKWS3_165 с нуклеотидной последовательностью в соответствии с SEQ ID NO: 5 и R-ген BvKWS3_123 комбинировали с двойным 35S промотором вектора pCaMV-2 (Фиг.3). Созданные векторы несли метки p70S-BvKWS3_133, p70S-BvKWS3_165 и p70S-BvKWS3_123. Чтобы проверить функциональность R-генов в листьях сахарной свеклы, осуществили транзитную биолистическую трансформацию по Шмидту и др. (Schmidt et al., 2004), в результате чего осуществили экспрессию конструкции p70S-BvKWS3_133, p70S-BvKWS3_165 и p70S-BvKWS3_123 с вектором гена-репортера p70S-luc. В качестве положительного контроля использовали пустой вектор pCaMV-2 в комбинации с вектором гена-репортера p70S-luc. От использования нормализированного вектора отказались, в отличие от Шмидта и др. (Schmidt et al., 2004). Люциферазную активность определяли через 20 ч после трансформации с помощью системы для люциферазного анализа Luciferase Assay System (Promega, Mannheim, Deutschland). Эксперименты по трансформации повторяли в 3-х параллелях, и каждый эксперимент включал 9 повторных испытаний на конструкцию. Получив среднее значение для 3-х экспериментов, выяснили, что по сравнению с практически 100% активностью положительного контроля люциферазы (пустого вектора), активность гена-репортера составляет при p70S-BvKWS3_133 только 37,7%, при p70S-BvKWS3_165 только 66% и при p70S-BvKWS3_123 только 68,7% (Фиг.4). Сильная экспрессия R-генов BvKWS3_133, BvKWS3_165 и BvKWS3_123, опосредованная d35S промотором, приводит либо к клеточной гибели, либо к реакции гиперчувствительности в части трансформированных клеток, которой препятствует коэкспрессия векторов генов-репортеров, которые совместно были внесены в клетки. При этом показано, что сильная экспрессия трех R-генов приводит к клеточной гибели или реакции гиперчувствительности в отсутствии соответствующих продуктов гена авирулентности.
5'-Область гена BvKWS3_165 вызывает более быструю клеточную гибель по сравнению с полноразмерным клоном кДНК BvKWS3_165
5'-Область полноразмерного кДНК клона BvKWS3_165 с последовательностью нуклеотидов в соответствии с SEQ ID NO: 5 амплифицировали в конструкции p70S-BvKWS3_165 с помощью Pfu-полимеразы (Stratagene) и использованием праймеров S316 (CTCGAGAATTCGAGCTCCACCGCGG) и S318 (CTGGATCCTCACCTCCGTTCTTCATGTTGCTCTACC) и одновременно ввели стоп-кодон в кодируемую область. Амплифицированная область соответствовала последовательности нуклеотидов в соответствии с SEQ ID NO: 1 и кодировала аминокислотную последовательность 1-174 BvKWS3_165 (Фиг.10). Аминокислотная последовательность содержала только N-концевую область BvKWS3_165 и не включала в себя никаких NBS- и LRR-доменов (Фиг.10). Продукт полимеразной цепной реакции (ПЦР-продукт) резали с помощью ферментов рестрикции SacII и BamHI и клонировали в вектор pCaMV-2. Получившийся вектор содержал метку p70S-165_#175. Способность конструкций p70S-BvKWS3_165 и p70S-165_#175 вызывать клеточную гибель в листьях сахарной свеклы количественно тестировали с помощью транзитной биолистической трансформации. Для этого осуществляли котрансформацию каждым из векторов совместно с вектором гена-репортера p70S-luc. В качестве положительного контроля использовали пустой вектор pCaMV-2 в комбинации с вектором гена-репортера p70S-luc. По сравнению с трансформацией пустым вектором (pCaMV-2) трансформация p70S-BvKWS3_165 дает активность, составляющую 65% активности гена-репортера, а трансформация p70S-165_#175 к активности, составляющей только 38% активности гена-репортера (Фиг.5). Этот результат показывает, что экспрессия только N-конца 165_#175 размером 175 аминокислот приводит к более интенсивному запуску клеточной гибели в трансформированных листьях сахарной свеклы, чем использование полноразмерного белка BvKWS3_165, состоящего из 1066 аминокислот. При экспрессии 165_#175 погибают больше клеток трансформированных листьев, чем в случае экспрессии BvKWS3_165. Причиной этого различия является новая, более интенсивная форма аутоактивирования R-белков, полученная в результате укорочения N-конца.
5'-Область гена BvKWS3_135 вызывает более быструю клеточную гибель по сравнению с полноразмерным клоном кДНК BvKWS3_135
5'-Область полноразмерного кДНК клона BvKWS3_135 в конструкции p70S-BvKWS3_135 амплифицировали с помощью Pfu-полимеразы (Stratagene) с использованием праймеров S316 (CTCGAGAATTCGAGCTCCACCGCGG) и S330 (CTGGATCCTCAGGGAGAACTCCATCTGGGTGGTCC) и одновременно ввели стоп-кодон в кодируемую область. Амплифицированная область соответствовала последовательности нуклеотидов в соответствии с SEQ ID NO: 2 и кодировала аминокислотную последовательность 1-174 BvKWS3_135 (Фиг.10). Аминокислотная последовательность содержала только N-концевую область BvKWS3_135 и не включала в себя никаких NBS- и LRR-доменов или мотивов этих доменов. Продукт полимеразной цепной реакции (ПЦР-продукт) разрезали с помощью ферментов рестрикции SacII и BamHI и клонировали в векторе pCaMV-2. Получившийся вектор содержал метку p70S-135_#147. Способность конструкций p70S-BvKWS3_135 и p70S-135_#147 вызывать клеточную гибель в листьях сахарной свеклы количественно тестировали с помощью транзитной биолистической трансформации. Для этого проводили совместную трансформацию каждым вектором с вектором гена репортера p70S-luc. В качестве положительного контроля использовали пустой вектор pCaMV-2 в комбинации с вектором гена-репортера p70S-luc. По сравнению с трансформацией пустым вектором (pCaMV-2) трансформация p70S-BvKWS3_135 приводит к активности, составляющей 74,5% активности гена-репортера, а трансформация p70S-135_#147 - к активности, составляющей только 58,5% активности гена-репортера (Фиг.6). Результаты показывают, что экспрессия полноразмерного клона BvKWS3_135 вызывает запуск клеточной гибели в трансформированных тканях. Однако экспрессия только N-конца 135_#147 размером 147 аминокислот приводит к более интенсивному запуску клеточной гибели в трансформированных листьях сахарной свеклы по сравнению с использованием полноразмерного белка BvKWS3_135, состоящего из 844 аминокислот. При экспрессии 135_#147 погибают больше клеток трансформированных листьев, чем в случае экспрессии BvKWS3_135. Причиной этого различия является новая более интенсивная форма аутоактивации R-белков, полученная в результате укорочения N-конца.
5'-Область гена Bv13033 вызывает более быструю клеточную гибель по сравнению с полноразмерным клоном кДНК Bv13033
5'-Область полноразмерного кДНК клона BvKWS3_135 в конструкции p70S-BvKWS3_135 амплифицировали с помощью Pfu-полимеразы (Stratagene) с использованием праймеров S316 (CTCGAGAATTCGAGCTCCACCGCGG) и S333 (CTGGATCCTCAAGAACAAGTCTCAGGCCTTCTGTT) и одновременно ввели стоп-кодон в кодируемую область. Амплифицированная область соответствовала последовательности нуклеотидов в соответствии с SEQ ID NO: 3 и кодировала аминокислотную последовательность 1-159 Bv13033 (Фиг.10). Аминокислотная последовательность содержала только N-концевую область Bv13033 и не включала в себя никаких NBS- и LRR-доменов или мотивов этих доменов. Продукт полимеразной цепной реакции (ПЦР-продукт) разрезали с помощью ферментов рестрикции SacII и BamHI и клонировали в векторе pCaMV-2. Получившийся вектор содержал метку p70S-13033_#159. Способность конструкций p70S-13033 и p70S-13033_#159 вызывать клеточную гибель в листьях сахарной свеклы количественно тестировали с помощью транзитной биолистической трансформации. Для этого проводили совместную трансформацию каждым из векторов с вектором гена репортера p70S-luc. В качестве положительного контроля использовали пустой вектор pCaMV-2. По сравнению с трансформацией пустым вектором (pCaMV-2) трансформация p70S-13033 приводит к активности, составляющей 95% активности гена-репортера, а трансформация p70S-165_#175 - к активности, составляющей только 68% активности гена-репортера (Фиг.7). Эти результаты показывают, что экспрессия полноразмерного клона Bvl3033 приводит только к слабому запуску клеточной гибели в трансформированных тканях. Экспрессия только N-конца 13033_#159 размером 159 аминокислот приводит, напротив, к более интенсивному запуску клеточной гибели в трансформированных листьях сахарной свеклы. Причиной этого различия является новая более интенсивная форма аутоактивации R-белков, полученная в результате укорочения N-конца.
Запуск клеточной гибели в листьях сахарной свеклы с помощью 5'-области гена Bv12069
R-Ген Bv12069 с последовательностью нуклеотидов в соответствии с SEQ ID NO: 4 кодирует N-конец R-белка размером 166 аминокислот. Белок Bv12069, не содержащий никаких NBS- и LRR-доменов, обладает отчетливой гомологией с N-концами аутоактивируемых R-белков 165_#175, 135_#147, 13033_#159 размером 175, 147 и 159 аминокислот соответственно (Фиг.10). Клон кДНК с помощью двойного промотора 35S вектора pCaMV-2 (Фиг.3) комбинировали с вектором p70S-12069. Для проверки функциональности гена Bv12069 осуществляли экспрессию конструкции p70S-12069 в комбинации с вектором гена-репортера p70S-luc в листьях сахарной свеклы с помощью транзитной биолистической трансформации. Активность гена-репортера в листьях, трансформированных p70S-12069 и p70S-luc, составила в трех независимых параллелях всего 51% от активности, измеренной для положительного контроля (пустых векторов pCaMV-2 и p70S-luc) (Фиг.8). Таким образом, экспрессия белка Bv12069 размером 166 аминокислот вызывает клеточную гибель в листьях сахарной свеклы.
Укорочение гена BvKWS3_135 приводит к образованию аутоактивирующегося белка, а не к мутагенезу в МНD-домене
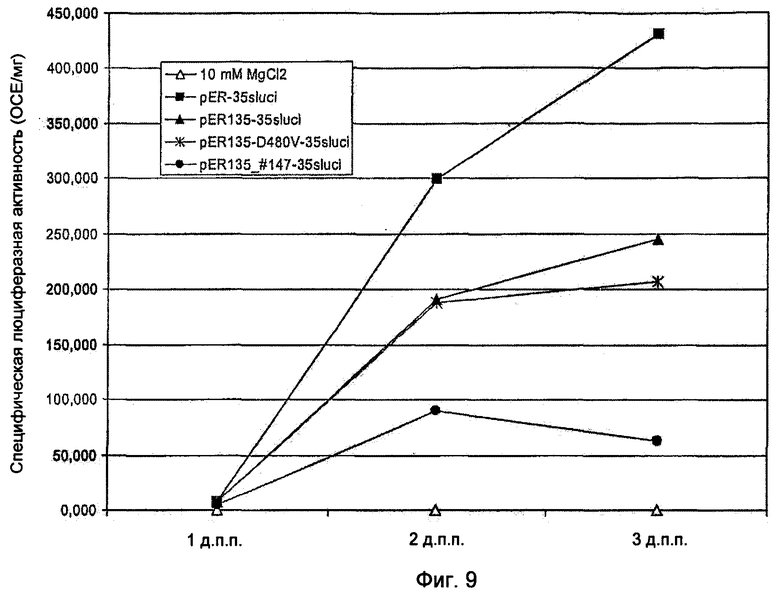
Механизм аутоактивации по изобретению с помощью укорочения R-белка NBS-LRR типа на конце, не имеющем NBS и LRR, сравнивали со способом аутоактивации с помощью мутагенеза MHD-мотива. Мутагенез MHD-мотива Rx-гена картофеля и L6-гена льна приводит к аутоактивации данных генов (Bendahmane et al., 2002; Howles et al., 2005). Клон кДНК BvKWS3_135 кодирует VHD-мотив, соответствующий MHD-мотиву, который также часто обнаруживают рядом с MHD-мотивом в R-генах (Howles et al., 2005). Получили соответствующую мутацию полноразмерного BvKWS3_135, как описано у Bendahmane et al. (2002). Для этого аминокислоту аспартат в мотиве VHD гена BvKWS3_135 заменили на аминокислоту валин. Полученный ген нес метку BvKWS3J35_D480V. Эффективность генов 135_#147, BvKWS3_135_D480V и немодифицированного гена BvKWS3_135 проверяли с помощью транзитной опосредованной Agrobacterium tumefaciens сверхэкспрессии в листьях сахарной свеклы. Для этого клон кДНК BvKWS3_135 комбинировали с d35S промотором и встраивали в бинарный вектор pER-35Sluci. Полученный вектор нес метку pER135-35Sluci. Соответствующим образом поступали и с укороченным клоном кДНК 135_#147 с последовательностью нуклеотидов в соответствии с SEQ ID NO: 2 и мутированным клоном кДНК BvKWS3_135_D480V. Полученные векторы несли метки pER135_#147-35Sluci и pER135_D480V-35Sluci. Векторы трансформировали, как было описано для штамма Agrobacterium C58C1, и вместе с положительным контролем pER-35Sluci инъецировали в листья сахарной свеклы. Активность гена-репортера люциферазы Photinus pyralis в трансформированных листьях измеряли через 1, 2 и 3 дня после прививки. Листья сахарной свеклы, трансформированные контрольной конструкцией pER-35Sluci, в первый день показали слабую активность люциферазы, а на 2-й и 3-й день показали активность люциферазы, составляющую 190000 и 245000 ОСЕ/мг ткани листа, и, таким образом, измеримую клеточную гибель по сравнению с положительным контролем. Активность гена-репортера в конструкции pER135_D480V-35Sluci составила на 2-й и 3-й день 188000 и 206000 ОСЕ/мг (Фиг.9). При этом введение MHD-мутации в ген BvKWS3_135 не приводило совсем или приводило только к едва детектируемой аутоактивации. В соответствии с этим способ с использованием укороченного R-гена 135_#147, напротив, на 2-й и 3-й день показал активность гена-репортера, составляющую 90000 и 63000 ОСЕ/мг (Фиг.9), и, следовательно, отчетливо более сильное проявление клеточной гибели и аутоактивацию по сравнению с конструкцией pER-35Sluci и pER135_D480V-35Sluci.
Идентификация общих аминокислотных мотивов в N-концах R-белков BvKWS3_165, BvKWS3_135, Bv13033 и Bv12069 и StR3a
Установление гомологии между N-концами R-белков BvKWS3_165, BvKWS3_135, Bv13033 и Bv12069 длиной 175, 147, 159 и 166 аминокислот, а также N-концом R3a гена картофеля длиной 155 аминокислот (Huang et al., 2005) проводили для идентификации общих мотивов последовательности. Сравнение привело к идентификации большего числа консенсусных последовательностей в N-концах аутоактивируемых R-белков. Общие мотивы в последовательностях представлены в виде консенсусных последовательностей на Фиг.10а.
Консенсусная последовательность соответствует аминокислотной последовательности в соответствии с SEQ ID NO: 13: AVLXDAEXKQXX XXXLXXWLXDLKDXVYDXDDILDE. Другая консенсусная последовательность соответствует аминокислотной последовательности в соответствии с SEQ ID NO: 14: IXEIXXKLDDL.
Буква X обозначает здесь произвольную аминокислоту.
Обе консенсусные последовательности в описанных формах содержатся только в таких N-концах CC-NBS-LRR R-белков, экспрессия которых приводит к аутоактивации. Таким образом, СС-домен RX-гена длиной 160 аминокислот не в состоянии вызвать клеточную гибель или реакцию гиперчувствительности (Bendahmane et al., 2002). Транзитная экспрессия N-конца R-гена BvKWS3_133_e08 сахарной свеклы длиной 177 аминокислот, а также N-конца R1 гена картофеля длиной 540 аминокислот (Ballvora et al., 2002) не приводит к усилению клеточной гибели по сравнению с полноразмерным R-геном BvKWS3_133_e08, а в случае полноразмерного R1 гена не приводит вообще к клеточной гибели (данные не представлены). Сравнение аминокислот N-концов аутоактивируемых белков BvKWS3_165_#176, BvKWS3_135_#147, Bv13033_#159, Bv12069 и StR3a-, #1-155 с аминокислотными последовательностями СС-доменов Rx-, StR1- и BvKWS3_133_#177-белков показало отсутствие описанных выше консенсусных последовательностей в неаутоакитивируемых N-концах (Фиг.10б). В особенности важным вспомогательным средством для идентификации R-белков, чьи N-концы аутоактивны, является последовательный мотив DAE. С помощью мотива DAE в консенсусной последовательности обнаружили подходящие для аутоактивации R-гены у многих видов растений, например те, которые показаны для примера на Фиг.10 для Arabidopsis thaliana (AtAB028617), фасоли (PvulgarisJ71), риса (osativaAp003073), сои (GmaxKR4) и томатов (Tomato-I2).
Аминокислотная последовательность 147-175 важна для аутоактивации R-белка 165 #175
Для идентификации аминокислотного отрезка в белке 165_#175, важного для аутоактивации N-конца NBS-LRR-белков, укоротили кодирующую область клона кДНК 165_#175. Клоны кДНК 165_#93 и 165_#146 кодируют аминокислоты 1-93 или 1-146 белка 165_#175. Транзитное биолитическое тестирование конструкций p70S_165_#93, p70S_165_#146 и p70S_165_#175 показало, что только белок 165_#175, но не 165_#93 и не 165_#146 приводит к заметной клеточной гибели (Фиг.11). Поэтому область последовательности 146-175 существенна для аутоактивации NBS-LRR-белков. В этой области находится мотив последовательности, консервативной для всех тестированных белков (Фиг.10а).
Быстрая активация синтетических индуцируемых патогенами промоторов 2×S-2×D и 2×W2-2×D в результате грибковой инфекции
Для индуцированной патогенами сверхэкспрессии полноразмерных или неполных генов устойчивости особенно подходят промоторы типа n×S-m×D, n×W2-m×D и n×Gst1-m×D, где n=1, 2, 3, 4, 5, 6, 7, 8, 9, 10 и m=1, 2, 3, 4, 5, 6, 7, 8, 9, 10. Например, промоторы типа 2×S-2×D в соответствии с SEQ ID No. 10, 2×W2-2×D в соответствии с SEQ ID No. 11, а также 2×Gst1-2×D в соответствии с SEQ ID No. 12 комбинировали с геном люциферазы Photinus pyralis, трансформировали ими сахарную свеклу и анализировали реакцию на грибковую инфекцию.
Для трансформации растений применяли бинарные векторы 2×S-2×D-luc-kan, 2×W2-2×D-luc-kan и 2×Gst1-2×D-luc-kan. Бинарными векторами трансформировали штамм Agrobacterium tumefaciens C58C1 с резидентной плазмидой pGV2260 способом прямой трансформации ДНК (An, 1987). Отбор рекомбинантных клонов A. tumefaciens проводили с применением антибиотика канамицина (50 мг/л).
Трансформацию сахарной свеклы проводили по Линдсею и др. (Lindsey et al., 1991) с применением антибиотика канамицина. Трансгенность растений проверяли с помощью ПЦР. Использование праймеров GTGGAGAGGCTATTCGGTA и CCACCATGATATTCGGCAAG привело к амплификации фрагмента ДНК размером 553 пар оснований (п.о.) гена nptII. ПЦР проводили с использованием 10 нг геномной ДНК, концентрации праймеров 0,2 мкМ при температуре отжига 55°C в мультициклере РТС-200 (MJ Research, Watertown, USA).
Для анализа способности промотора быть индуцированным патогенами сахарную свеклу инфецировали Cercospora beticola, возбудителем пятнистости листьев сахарной свеклы в условиях in vitro. В каждом случае 4 растения каждой трансгенной линии погружали в суспензию фрагментов мицелия С. beticola (400.000 фрагментов/мл) и контрольных 4 растения погружали в разбавленный овощной сок. Инфицированные растения и контрольные растения инкубировали затем в культуральном шкафу при 25°C на свету в течение 16 ч. Инфицированный и неинфицированный листовой материал брали через 1, 2, 3, 4 и 6-7 дней после прививки и определяли активность гена-репортера люциферазы с помощью Luciferase Assay System (Promega, Mannheim, Deutschland), как было описано выше.
Промотор 2×S-2×D, так же как и промотор 2×W2-2×D, проявлял быструю и сильную способность быть индуцированными патогенами на ранней фазе инфекции, различаясь между собой только в базовой активности и силе промотора (Фиг.12-13). Промотор 2×S-2×D в случае трансгенных линий PR39/11, PR39/48 и PR39/49 индуцировался очень быстро - индукция была в 11-59 раз сильнее уже на 1-й день после прививки и в 21-380 раз сильнее на 2-й день по сравнению с неинфицированными растениями (Фиг.12). В то время как в 1-й день отмечали прорастание гифов грибов на эпидермисе, на 2-й день происходило проникновение в лист через строму и затем проникновение в ткань листа. На поздней стадии инфекции на 7-й день обнаруживали индукцию промотора в 113-792 раз более сильную при видимом развитии некроза. Базовая активность промотора 2×S-2×D, измеренная как активность гена-репортера у неинфицированных растений, очень слаба, и ее значение только в 1-10 раз превышает значение активности люциферазы в нетрансгенных растениях.
Активация промотора 2×W2-2×D протекает немного медленнее по сравнению с активацией промотора 2×S-2×D. В первый день инфекции промотор 2×W2-2×D проявляет в 2-17 раз более сильную индукцию в ответ на патогены, на второй день инфекции - в 5-56 раз. При проявлении некроза на 7 день индукция в ответ на патогены достигает максимального значения в 318-672 ед. (Фиг.13). Базовая активность 2×W2-2×D промотора, измеренная как активность гена-репортера у нетрансгенных растений, в 10-50 раз выше, чем в случае 2×S-2×D промотора. 2×W2-2×D промотор также отчетливо отличается от 2×S-2×D промотора, будучи примерно в 10 раз более сильным.
Оптимизация свойств промотора с помощью изменения количества цис-элементов
Свойства синтетического промотора типа n×S-m×D, n×W2-m×D и n×Gst1-m×D с n=1, 2, 3, 4, 5, 6, 7, 8, 9, 10 и m=1, 2, 3, 4, 5, 6, 7, 8, 9, 10 модулировали и оптимизировали, варьируя количество цис-элементов в соответствии с потребностями генной экспрессии. Это показано, например, для промоторов типа n×S-m×D. Рядом с бинарным вектором 2×S-2×D-luc-kan встраивали бинарные векторы 4×S-2×D-luc-kan, 2×S-4×D-luc-kan и 4×S-4×D-luc-kan и трансформировали ими сахарную свеклу. Трансгенные растения инфицировали С. beticola, как было описано, и ежедневно измеряли активность гена-репортера после внедрения гриба. Результаты измерений для 13 независимых 2×S-2×D-luc линий, 14 независимых 4×S-2×D-luc линий, 15 независимых 2×S-4×D-luc линий, а также 15 независимых 4×S-4×D-luc линий усреднили и сравнили средние значения силы промотора, индукции патогенами и базовой активности.
Сравнение свойств промотора 2×S-2×D с вариантами 2×S-4×D, 4×S-2×D и 4×S-4×D показало, что в среднем сила промотора при использовании тетрамеров по сравнению с промотором, сконструированным из тримеров, повышается (Фиг.14). К тому же, в ряду димер-димерного промотора (2×S-2×D), тетрамер-димерного, димер-тетрамерного промоторов (4×S-2×D, 4×S-2×D) и тетрамер-тетрамерного промотора (4×S-4×D) повышается способность быть индуцированным патогенами во всех временных точках измерения (табл.1).
Здесь представлено среднее значение индукции патогеном для 13-15 независимых трансформантов (линий) на промоторную конструкцию через 1-4 дня после прививки.
Параллельно с усилением силы промотора и способности индуцироваться патогенами повышается базовая активность промоторов, содержащих тетрамеры (табл. 2).
Здесь представлено среднее значение базовой активности 13-15 независимых трансформантов (линий) на промоторную конструкцию, которые измеряли в качестве неинфицированных контролей в течение четырехдневного эксперимента по инфицированию. Базовая активность показывала относительную величину активности гена-репортера трансгенных растений в сравнении с неспецифической фоновой активностью нетрансгенных растений.
Этот пример показал, что, используя концепцию важных свойств промотора, таких как сила промотора, способность индуцироваться патогенами и базовая активность, которые регулируются с помощью количества цис-элементов, для соответствующей технической реализации можно изготовить оптимальные варианты промоторов. Оптимальное количество цис-элементов индуцируемых патогенами промоторов в изученных примерах с учетом способности индуцироваться патогенами больше, чем у раствора димеров, описанного Раштоном и др., 2002 (Rushton et al., 2002).
Получение сахарной свеклы, устойчивой к грибам, с помощью трансформации индуцируемых патогенами генов устойчивости
Для усиления устойчивости к грибам сахарной свеклы промоторы 2×S-2×D или 2×W2-2×D комбинировали соответственно с каждым из четырех R-генов BvKWS3_123, BvKWS3_133, BvKWS3_135 и BvKWS3_165 и трансформировали ими сахарную свеклу. Для этого бинарные векторы 2×S-2×D-luc-kan и 2×W2-2×D-luc-kan размером 13959 и 13969 п.о. разрезали с помощью Sacl и места разреза заполняли с помощью обработки Т4-ДНК полимеразой. Затем векторы разрезали Xhol, разделяли электрофорезом и векторы размером 12284 и 12294 отделяли от гена люциферазы размером 1675 п.о. и выделяли их.
Выделение генов устойчивости сахарной свеклы осуществляли из векторов p70S-BvKWS3_123, p70S-BvKWS3_133, p70S-BvKWS3_135 и p70S-BvKWS3_165. Для этого векторы сначала делали линейными с помощью Notl и места разреза заполняли с помощью обработки фрагментом Кленова. Затем векторы разрезали Xhol и изолировали R-гены. Полученные векторы несли метку 2×S-2×D-BvKWS3_123, 2×S-2×D-BvKWS3_133, 2×S-2×D-BvKWS3_135 и 2×S-2×D-BvKWS3_165, а также 2×W2-2×D-BvKWS3_123, 2×W2-2×D-BvKWS3J33, 2×W2-2×D-BvKWS3_135 и 2×W2-2×D-BvKWS3_J65 (Фиг.15-18). Бинарные векторы использовали для получения трансгенной сахарной свеклы, как было описано.
Идентификация сахарной свеклы, устойчивой к грибам, с помощью испытания устойчивости к грибу-паразиту Cercospora beticola
Повышенную устойчивость к грибам растений наблюдали при испытании устойчивости к грибам, как описано в качестве примера, для проверки устойчивости сахарной свеклы к Cercospora beticola.
Чтобы инфицировать сахарную свеклу возбудителем листовых пятен С. beticola, в теплице рядом с трансгенными растениями посадили сахарную свеклу с генотипом 3DC4156, используемую для трансформации. За 2 недели до запланированной инокуляции планшеты с овощным соком (40% овощной сок Albani) инокулировали агрессивным изолятом Ahlburg С. beticola и инкубировали при 25°C. Непосредственно перед инокуляцией агар, заросший грибами, соскабливали с помощью предметного стекла и небольшого количества воды. Концентрацию фрагментов мицелия и грибных спор определяли с помощью счетной камеры. Плотность возбудителя доводили до концентрации 20000 фрагментов/мл, разбавляя водой. Для инфицирования 10-12 недельные растения верхушками окунали в 5 л стаканы, заполненные возбудителем. В каждой исследуемой линии инокулировали 30 растений и растения расставляли в теплице в произвольном порядке.
После инокуляции растения инкубировали в теплице в течение 4-х дней при 28°C и 95% влажности воздуха. Спустя 4 дня влажность воздуха снижали до 60%-70%. Через две, три и четыре недели после прививки поражение листьев оценивали оптически с помощью девятибальной схемы оценки компании Кляйнванцлебенер Заатцзухт (Kleinwanzlebener Saatzucht (KWS)) (1970) (1 = здоровые листья, 9 = 100% разрушенные листья). Трансгенные линии, трансформированные одной из конструкций 2×S-2×D-BvKWS3_123, 2×S-2×D-BvKWS3_133, 2×S-2×D-BvKWS3_165, 2×W2-2×D-BvKWS3_123, 2×W2-2×D-BvKWS3_133, 2×W2-2×D-BvKWS3_135 или 2×W2-2×D-BvKWS3_165, продемонстрировали повышенную устойчивость к грибам (табл. 3) по сравнению с контролем.
Анализ развития во времени поражения у трансформантов PR68-6 и PR70-32 в 3-х точках показал, что в процессе испытания различие в развитии поражения между контролем и трансгенной линией усиливается (Фиг.19 и 20). Эти результаты демонстрируют, что индуцированная экспрессия различных R-генов сахарной свеклы с помощью патогенспецифических промоторов приводит к повышению устойчивости к грибам.
Получение растений, устойчивых к грибам, с помощью трансформации N-концевых областей R-генов под контролем патогенчувствительных промоторов
Для получения растений, устойчивых к грибам, с применением N-концевых отрезков R-генов, укороченные R-гены 13033_#159, 135_#147, 165_#175 и Bv12069 комбинировали с промоторами 2×S-2×D и 2×W2-2×D и трансформировали ими сахарную свеклу.
Для этого бинарные векторы 2×S-2×D-luc-kan и 2×W2-2×D-luc-kan размером 13959 и 13969 п.о. разрезали Sacl и заполняли разрезанные части с помощью Т4-обработки ДНК-полимеразой. Затем векторы разрезали Xhol, разделяли с помощью гель-электрофореза и отделяли и изолировали векторы размером 12284 и 12294 п.о. от гена люциферазы размером 1675 п.о.
Выделение укороченных R-генов осуществляли из векторов p70S-12069, p70S-13033_#159, p70S-135_#147 и p70S-165_#175. Векторы затем делали линейными с помощью Xbal, места разреза заполняли с помощью обработки фрагментом Кленова и затем разрезали векторы с помощью Xhol. Выделенные фрагменты R-генов после этого клонировали в подготовленных бинарных векторах. Созданные векторы содержали метку 2×S-2×D-12069, 2×S-2×D-13033_#159, 2×S-2×D-135_#147, 2×S-2×D-165_#175 и 2×W2-2×D-12069, 2×W2-2×D-13033_#159, 2×W2-2×D-135_#147, 2×W2-2×D-165_#175 (Фиг.21-22). Бинарными векторами трансформировали сахарную свеклу так, как было описано, и устойчивые к грибам растения идентифицировали с помощью испытания на устойчивость с Cercospora beticola.
Источники информации
Altschul, S.F. et al. (1990). Basic Local Alignment search tool, J.Mol. Biol. 215: 403-410.
An, G. (1987). Binary Ti vectors for plant transformation and promoter analysis. Methods Enzymol. 153, 292-305.
Bairoch et al., (1996). The PROSITE database, its status in 1995. Nucleic Acids Res 24:189-96.
Ballavora A., Ercolana M.R., Weiss J., Meksem K., Bormann C.A.I., Oberhagemann P., Salamini F., Gebhardt C. (2002). The Rl gene for potato resistance to late blight (Phytophthora infestans) belongs to the leucine zipper/NBS/LRR class of plant resistance genes. Plant J.30(3):361-71.
Bendahmane A., Famham G., Moffett P., and Baulcombe D.C. (2002). Constitutive gain-of-runction mutants in a nucleotide binding site-leucine rich repeat protein encoded at the Rx locus of potato. Plant J. Oct; 32(2):195-204.
Frost D., Way H., Howles P., Luck J., Manners J., Hardham A., Finnegan J., and Ellis J. (2004). Tobacco transgenic for the flax rust resistance gene L expresses allele-specific activation of defense responses. Mol Plant Microbe Interact. 17(2):224-32.
Hennig, J., Dewey, R.E., Cutt, J.R, and Klessig, D.F. (1993). Pathogen, salicylic acid and developmental dependent expression of a beta-1,3-glucanase/GUS gene fusion in transgenic tobacco plants. Plant J. 4(3):481-93.
Howles P., Lawrence G., Finnegan J., McFadden H., Ayliffe M., Dodds D., and Ellis J. (2005). Autoactive Alleles of the Flax L6 Rust Resistance Gene Induce Non-Race-Specific Rust Resistance Associated with the Hypersensitive Response. Mol Plant Microbe Interact 18(6):570-582.
Huang S., an der Vossen E.A., Kuang H., Vlesshouwers V.G., Zhang N., Borm, T.J., van Eck HJ., Baker В., Jacobsen E., and Visser R.G. (2005). Comparative genomics enabled the isolation of the R3a late blight resistance gene in potato. Plant J. 42(2):251-61.
Lindsey, K., Gallois, P., and Eady, C. (1991). Regeneration and transformation of sugar beet by Agrobacterium tumefaciens. Plant Tissue Culture Manual B7:1-13; Kluwer Academic Publishers.
Lupas, A., Van Dyke M., and Stock J. (1991) Predicting coiled coils from protein sequences. Science 252:1162-4.
Martin, G. В., Bogdanove, A. J., and Sessa, G. (2003). Understanding the functions of plant disease resistance proteins. Annu. Rev. Plant Biol. 54:23-61.
Martini, N., Egen, M., Rüntz, I., and Strittmatter, G. (1993). Promoter sequences of a potato pathogenesis-related gene mediate transcriptional activation selectively upon fungal infection. Mol Gen Genet 236:179-186.
Oldroyd, G. E. D., and. Staskawicz, B. J. (1998). Genetically engineered broad-spectrum disease resistance in tomato. Proc Natl Acad Sei USA. 95(17):10300-10305.
Rushton P. J., Reinstadler A., Lipka V., Lippok В., Somssich I.E. (2002). Synthetic plant promoters containing defined regulatory elements provide novel insights into pathogen-and wound-induced signaling. Plant Cell 14(4):749-62.
Samac, D. A. and Shah, D.M. (1991). Developmental and Pathogen-Induced Activation of the Arabidopsis Acidic Chitinase Promoter. Plant Cell. 3(10):1063-1072.
Schmidt К., Heberle В., Kurrasch J., Nehls R., and Stahl D. J. (2004). Suppression of phenylalanine ammonia lyase expression in sugar beet by the fungal pathogen Cercospora beticola is mediated at the core promoter of the gene. Plant Mol. Biol., 55: 835-852.
Sonnhammer, E. L, Eddy, S. R., and Durbin, R. D. (1997) Pfam: A comprehensive database of protein domain families based on seed alignments. Proteins 28:405-20.
Tang X., Xie M., Kirn Y.J., Zhou J., Klessig D.F., Martin G.B. (1999). Overexpression of Pto activates defense responses and confers broad resistance. Plant Cell 11(1):15-29.
Tian Y., Fan L, Thurau Т., Jung C, and Cai D. (2004). The absence of TIR-type resistance gene analogues in the sugar beet (Beta vulgaris L.) genome. J. Mol. Evol. 58(1):40-53.
Traut, T. W. (1994). The functions and consensus motifs of nine types of peptide segments that form different types of nucleotide binding Sites. Eur. J. Biochem. 222:9-19.





| название | год | авторы | номер документа |
|---|---|---|---|
| ПРОМОТОР, ИНДУЦИРУЕМЫЙ ПРИ ХРАНЕНИИ | 2005 |
|
RU2433183C2 |
| КЛОНИРОВАНИЕ И ИСПОЛЬЗОВАНИЕ ФУНКЦИОНАЛЬНОГО R-ГЕНА ИЗ SOLANUM CHACOENSE | 2010 |
|
RU2586509C2 |
| СПОСОБ ПОВЫШЕНИЯ РЕЗИСТЕНТНОСТИ ТРАНСГЕННЫХ РАСТЕНИЙ К ПАТОГЕННЫМ ВОЗДЕЙСТВИЯМ | 2007 |
|
RU2428480C9 |
| РАСТИТЕЛЬНЫЕ КЛЕТКИ И ОРГАНИЗМЫ РАСТЕНИЙ С МОДИФИЦИРОВАННЫМ КЛЕТОЧНЫМ РОСТОМ, РАЗВИТИЕМ И ДИФФЕРЕНЦИРОВКОЙ | 2004 |
|
RU2349642C2 |
| РЕГУЛЯТОРНЫЕ МОЛЕКУЛЫ НУКЛЕИНОВЫХ КИСЛОТ ДЛЯ УСИЛЕНИЯ СЕМЯ-СПЕЦИФИЧНОЙ И/ИЛИ СЕМЯ-ПРЕДПОЧТИТЕЛЬНОЙ ГЕННОЙ ЭКСПРЕССИИ В РАСТЕНИЯХ | 2010 |
|
RU2559534C2 |
| СКОНСТРУИРОВАННЫЕ ПЕСТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2017 |
|
RU2772947C2 |
| НОВЫЕ РАСТИТЕЛЬНЫЕ ЦИС-РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ ДЛЯ РАЗРАБОТКИ ПАТОГЕН-ЧУВСТВИТЕЛЬНЫХ ХИМЕРНЫХ ПРОМОТОРОВ | 2012 |
|
RU2660569C2 |
| ПАТОГЕН-ИНДУЦИРУЕМЫЙ СИНТЕТИЧЕСКИЙ ПРОМОТОР | 2007 |
|
RU2406761C2 |
| БОЛЕЗНЕУСТОЙЧИВЫЕ РАСТЕНИЯ | 2006 |
|
RU2393227C2 |
| ЭНХАНСЕР ПАЛОЧКОВИДНОГО ВИРУСА САХАРНОГО ТРОСТНИКА (SCBV) И ЕГО ПРИМЕНЕНИЕ В ФУНКЦИОНАЛЬНОЙ ГЕНОМИКЕ РАСТЕНИЙ | 2013 |
|
RU2639517C2 |










Нуклеиновая кислота, кодирующая аутоактивирующийся белок устойчивости, включает ограниченный участок NBS-LRR-гена, который простирается от 5'-конца кодирующей области NBS-LRR-гена в направлении 5'-3' до начала NBS-домена, причем NBS-LRR-ген не является TIR-NBS-LRR-геном. Растения, включающие аутоактивирующийся белок устойчивости, приобретают устойчивость к патогенам. 5 н. и 14 з.п. ф-лы, 24 ил., 3 табл.
1. Аутоактивирующийся белок устойчивости для выработки устойчивости к патогенам у растений, отличающийся тем, что его кодирует нуклеиновая кислота, включающая ограниченный участок NBS-LRR-гена устойчивости, который простирается от 5'-конца кодирующей области NBS-LRR-гена устойчивости в направлении 5'-3' до начала NBS-домена NBS-LRR-гена устойчивости, но не включая Р-петлю, причем NBS-LRR-ген устойчивости не является TIR-NBS-LRR-геном устойчивости.
2. Аутоактивирующийся белок устойчивости по п.1, включающий аминокислотную последовательность с мотивом последовательности DAE.
3. Аутоактивирующийся белок устойчивости по п.1, включающий аминокислотную последовательность с мотивом последовательности AVLXDAE.
4. Аутоактивирующийся белок устойчивости по п.1, отличающийся тем, что нуклеиновая кислота включает нуклеотидную последовательность, выбранную из следующей группы:
а) нуклеотидная последовательность в соответствии с SEQ ID NO:1 или нуклеотидная последовательность, комплементарная нуклеотидной последовательности в соответствии с SEQ ID NO:1, или нуклеотидная последовательность, которая может быть гибридизована с нуклеотидной последовательностью в соответствии с SEQ ID NO:1 или с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности в соответствии с SEQ ID NO:1;
б) нуклеотидная последовательность в соответствии с SEQ ID NO:2 или нуклеотидная последовательность, комплементарная нуклеотидной последовательности в соответствии с SEQ ID NO:2, или нуклеотидная последовательность, которая может быть гибридизована с нуклеотидной последовательностью в соответствии с SEQ ID NO:2 или с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности в соответствии с SEQ ID NO:2;
в) нуклеотидная последовательность в соответствии с SEQ ID NO:3 или нуклеотидная последовательность, комплементарная нуклеотидной последовательности в соответствии с SEQ ID NO:3, или нуклеотидная последовательность, которая может быть гибридизована с нуклеотидной последовательностью в соответствии с SEQ ID NO:3 или с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности в соответствии с SEQ ID NO:3;
г) нуклеотидная последовательность в соответствии с SEQ ID NO:4 или нуклеотидная последовательность, комплементарная нуклеотидной последовательности в соответствии с SEQ ID NO:4, или нуклеотидная последовательность, которая может быть гибридизована с нуклеотидной последовательностью в соответствии с SEQ ID NO:4 иди с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности в соответствии с SEQ ID NO:4;
д) нуклеотидная последовательность в соответствии с SEQ ID NO:16 или нуклеотидная последовательность, комплементарная нуклеотидной последовательности в соответствии с SEQ ID NO:16, или нуклеотидная последовательность, которая может быть гибридизована с нуклеотидной последовательностью в соответствии с SEQ ID NO:16 или с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности в соответствии с SEQ ID NO:16.
5. Аутоактивирующийся белок устойчивости по п.1, отличающийся тем, что ген устойчивости NBS-LRR представляет собой ген устойчивости сахарной свеклы или картофеля.
6. Аутоактивирующийся белок устойчивости по п.1, включающий аминокислотную последовательность, выбранную из следующей группы:
a) SEQ ID NO:13
б) SEQ ID NO:14
B) SEQ ID NO:15
7. Трансгенное растение, включающее аутоактивирующийся белок устойчивости по любому из пп.1-6.
8. Часть трансгенного растения, включающая аутоактивирующийся белок устойчивости по любому из пп.1-6.
9. Семя или посевной материал трансгенного растения, включающие аутоактивирующийся белок устойчивости по любому из пп.1-6.
10. Способ получения трансгенного растения с увеличенной устойчивостью к патогенам, а также трансгенного семени или трансгенного посевного материала такого растения, при котором применяют нуклеиновую кислоту, где нуклеиновая кислота включает ограниченный участок NBS-LRR-гена устойчивости, который простирается от 5'-конца кодирующей области NBS-LRR-гена устойчивости в направлении 5'-3' до начала NBS-домена NBS-LRR-гена устойчивости, но не включая Р-петлю, причем NBS-LRR-ген устойчивости не является TIR-NBS-LRR-геном устойчивости.
11. Способ по п.10, отличающийся тем, что нуклеиновая кислота кодирует аминокислотную последовательность с мотивом последовательности DAE.
12. Способ по п.10, отличающийся тем, что нуклеиновая кислота кодирует аминокислотную последовательность с мотивом последовательности AVLXDAE.
13. Способ по п.10, отличающийся тем, что нуклеиновая кислота включает нуклеотидную последовательность, выбранную из следующей группы:
а) нуклеотидная последовательность в соответствии с SEQ ID NO:1 или нуклеотидная последовательность, комплементарная нуклеотидной последовательности в соответствии с SEQ ID NO:1, или нуклеотидная последовательность, которая может быть гибридизована с нуклеотидной последовательностью в соответствии с SEQ ID NO:1 или с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности в соответствии с SEQ ID NO:1;
б) нуклеотидная последовательность в соответствии с SEQ ID NO:2 или нуклеотидная последовательность, комплементарная нуклеотидной последовательности в соответствии с SEQ ID NO:2, или нуклеотидная последовательность, которая может быть гибридизована с нуклеотидной последовательностью в соответствии с SEQ ID NO:2 или с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности в соответствии с SEQ ID NO:2;
в) нуклеотидная последовательность в соответствии с SEQ ID NO:3 или нуклеотидная последовательность, комплементарная нуклеотидной последовательности в соответствии с SEQ ID NO:3, или нуклеотидная последовательность, которая может быть гибридизована с нуклеотидной последовательностью в соответствии с SEQ ID NO:3 или с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности в соответствии с SEQ ID NO:3,
г) нуклеотидная последовательность в соответствии с SEQ ID NO:4 или нуклеотидная последовательность, комплементарная нуклеотидной последовательности в соответствии с SEQ ID NO:4, или нуклеотидная последовательность, которая может быть гибридизована с нуклеотидной последовательностью в соответствии с SEQ ID NO:4 или с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности в соответствии с SEQ ID NO:4;
д) нуклеотидная последовательность в соответствии с SEQ ID NO:16 или нуклеотидная последовательность, комплементарная нуклеотидной последовательности в соответствии с SEQ ID NO:16, или нуклеотидная последовательность, которая может быть гибридизована с нуклеотидной последовательностью в соответствии с SEQ ID NO:16 или с нуклеотидной последовательностью, комплементарной нуклеотидной последовательности в соответствии с SEQ ID NO:16.
14. Способ по п.10, отличающийся тем, что ген устойчивости NBS-LRR представляет собой ген устойчивости сахарной свеклы или картофеля.
15. Способ по п.10, отличающийся тем, что нуклеиновая кислота кодирует аминокислотную последовательность, выбранную из следующей группы:
а) SEQ ID NO:13
б) SEQ ID NO: 14
в) SEQ ID NO: 15
16. Способ по п.10, отличающийся тем, что применяют конструкцию
нуклеиновой кислоты, включающую
а) промотор, индуцируемый патогенами, а также
б) нуклеиновую кислоту по любому из пп.10-15, контролируемую промотором.
17. Способ по п.16, отличающийся тем, что патоген-индуцируемый промотор представляет собой синтетический промотор.
18. Способ по п.17, отличающийся тем, что синтетический промотор включает одну или более чем одну следующую комбинацию цис-элементов, где n и m представляют собой натуральное число от 1 до 10:
а) n×S-m×D-бокс
б) n×W2-m×D-бокс
в) n×Gstl-m×D-бокс
19. Способ по. 18, отличающийся тем, что комбинация цис-элементов включает:
а) последовательность нуклеотидов SEQ ID NO:10, или
б) последовательность нуклеотидов SEQ ID NO:11, или
в) последовательность нуклеотидов SEQ ID NO:12.
| DATADASE EMBL, Accession # AV827853, 21.03.2002 | |||
| UniProtKB/Swiss-Prot, Accession # Q38834, 11.04.2003 | |||
| PLANT JOURNAL, 2002 Oct, 32(2): 195-204 | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

