ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Данная заявка содержит объект изобретения, который частично относится к предварительной заявке США № 61/546028, поданной 11 октября 2011 года, предварительной заявке США № 61/323708, поданной 13 апреля 2010 года; международной заявке № PCT/US2011/032184, поданной 12 апреля 2011 года; международной заявке № PCT/US2011/032188, поданной 12 апреля 2011 года; и PCT заявке № PCT/US2008/012398, поданной 10 октября 2008 года и опубликованной в виде международной публикации № WO 2009/058379 A2. Каждая из вышеуказанных заявок включена в данный документ посредством ссылки во всей ее полноте.
CCЫЛКА НА ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Данная заявка включает посредством ссылки перечень последовательностей, поданный с заявкой с помощью EFS-Web в виде текста, представленного под названием ”CD40L-101WO1_SL.txt”, созданного 3 октября 2012 года и имеющего размер 232 килобайта.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Область изобретения
[0003] Настоящее изобретение в целом относится к области миметиков антител, а именно к каркасным структурам, происходящим из третьего домена фибронектина типа III тенасцина С человека, пригодным, например, для создания продуктов, обладающих новыми характеристиками связывания. В частности, настоящее изобретение относится к CD40L-специфичным Tn3-каркасным структурам, способам получения подобных каркасных структур и способам применения для диагностики и лечения системной красной волчанки и других аутоиммунных и/или воспалительных нарушений.
Уровень техники
[0004] Настоящее изобретение относится к CD40L-специфичным белковым каркасным структурам, которые связываются с CD40L, пригодным, например, для лечения аутоиммунных и/или воспалительных нарушений.
[0005] Биомолекулы, способные к специфическому связыванию с желаемым целевым эпитопом, имеют большое значение в качестве терапевтических средств, научно-исследовательских и медицинских диагностических средств. Хорошо известным примером данного класса молекул является антитело. Антитела можно выбрать таким образом, чтобы они специфически связывались и являлись аффинными к практически любому структурному эпитопу. Однако, природные антитела представляют собой гетеротетрамерные, сложные по своей структуре молекулы, которые с трудом экспрессируются в простых эукариотических системах. В результате большинство антител получают с использованием сложных и дорогостоящих экспрессирующих систем на основе клеток млекопитающих.
[0006] Белки, имеющие относительно определенные трехмерные структуры, обычно называемые белковыми каркасными структурами, можно использовать в качестве реагентов для создания сконструированных продуктов. Одной специфической сферой, в которой используют подобные каркасные структуры, является область создания миметиков антител. Миметики антител, т.е., малые, не являющиеся антителами белковые терапевтические средства, обладающие преимуществами антител и фрагментов антител, такими как высокая аффинность связывания с мишенями и низкие иммуногенность и токсичность, избегают при этом некоторых недостатков, таких как склонность фрагментов антител к агрегации и меньшая стабильность, чем у полноразмерных IgG.
[0007] Данные недостатки можно устранить путем использования фрагментов антител, созданных путем удаления частей в нативной структуре антитела. Однако, это часто вызывает агрегацию, если аминокислотные остатки, которые в норме погружены в гидрофобное окружение, такое как граница раздела между вариабельным и константным доменом, подвергаются воздействию растворителя. Один пример миметика антитела на основе каркасной структуры основан на структуре домена фибронектина типа III (FnIII), домена, который широко распространен среди типов и классов белков, таких как белки крови млекопитающих и структурные белки. Конструирование и применение каркасных структур FnIII, полученных из третьего домена FnIII человеческого тенасцина C, описано в PCT заявках PCT/US2011/032184 и PCT/US2011/032188, обе из которых включены в данный документ посредством ссылки во всей их полноте.
[0008] CD40L является представителем семейства молекул TNF, которые первично экспрессируются на активированных T-клетках (в том числе подтипах Th0, Th1 и Th2), и образуют гомотримеры, сходные с другими представителями данного семейства. Кроме того, обнаружили, что CD40L экспрессируется на Mast-клетках и активированных базофилах и эозинофилах. CD40L связывается с рецептором CD40 (CD40R) на антиген-презентирующих клетках (APC), что приводит к различным эффектам в зависимости от типа клетки-мишени. В целом, CD40L играет роль в качестве костимулирующей молекулы и индуцирует активацию APC в ассоциации со стимуляцией T-клеточного рецептора молекулами MHC на APC.
[0009] Передача сигнала посредством рецептора CD40 с помощью CD40L запускает каскад реакций, что приводит в результате к активации клеток, несущих рецептор CD40, и оптимальному примированию CD4+ T-клеток. Более конкретно, когнатное взаимодействие между CD40L и рецептором CD40 стимулирует дифференциацию B-клеток в антитело-секретирующие клетки и В-клетки памяти (Burkly, In Adv. Exp. Med. Bio., Vol. 489., D. M. Monroe, U. Hedner, M. R. Hoffman, C. Negrier, G. F. Savidge, and G. C. I. White, eds. Klower Academic/Plenum Publishers, 2001, p. 135). Дополнительно, взаимодействие между CD40L и рецептором CD40 стимулирует клеточно-опосредованный иммунитет посредством активации макрофагов и дендритных клеток и образования естественных клеток-киллеров, а также цитотоксических Т-лимфоцитов (смотри Burkly выше).
[0010] Было показано, что взаимодействие между CD40L и рецептором CD40 играет важную роль в нескольких экспериментально индуцированных аутоиммунных заболеваниях, таких как коллаген-индуцированный артрит, экспериментальный аллергический энцефаломиелит, оофорит, колит, медикаментозный волчаночный нефрит. В частности, было показано, что индукцию заболевания во всех из этих моделей можно блокировать CD40L-антагонистами во время введения антигена. Прекращение развития заболевания с использованием антитела, являющегося антагонистом к CD40L, также было показано в моделях спонтанного аутоиммунного заболевания на животных, включая формы инсулинозависимого диабета и волчаночный нефрит, а также заболевание "трансплантат против хозяина", трансплантацию, фиброз легких и модели заболевания атеросклерозом.
[0011] Было показано, что нарушение каскада реакций CD40L/CD40R посредством блокирования CD40L является благоприятным при многих аутоиммунно-опосредованных заболеваниях (например, но без ограничения, системная красная волчанка (SLE), ревматоидный артрит (RA), рассеянный склероз (MS), воспалительные заболевания кишечника (IBD) и отторжение аллотрансплантата. Например, лечение антителами к CD40L обеспечивает предупреждение или улучшение течения нефрита в мышиной модели коллаген-индуцированного артрита (Mohan et al. J. Immuno. 154:1470). Кроме того, антитела к CD40L обеспечивают сохранение почечной функции у мышей SNF1 с тяжелым нефритом. (Kalled et al. J. Immuno. 160:2158). Уровни CD40L тесно коррелируют с клинической тяжестью заболевания (т.е., уменьшение воспаления), и повреждением ткани-мишени как у субъектов, не относящихся к людям, так и у людей.
[0012] SLE представляет собой прогрессирующее и в ряде случаев приводящее к смерти заболевание. Различные проявления волчанки варьируют от сыпи и артрита вплоть до анемии и тромбоцитопении и даже психоза. Существует явное свидетельство, демонстрирующее, что многие механизмы иммунной системы вовлечены в воспалительный процесс, приводящий к заболеваниям почек, кожи, головного мозга и тромбозу. Одним характерным признаком SLE является утрата В-клеточной толерантности и выраженность наличия аутоантител у пациентов с этим заболеванием. При волчаночном заболевании почек аутоантитела к двухцепочечной ДНК могут образовывать комплексы антитело-нуклеосома и осаждаться на почечной клубочковой базальной мембране. Данные иммунные комплексы, в свою очередь, активируют комплемент, что может привести к гломерулонефриту.
[0013] Как было выявлено, у пациентов с SLE повышена экспрессия CD40R, а также CD40L. Усиленный костимулирующий сигнал, вероятно, способствует патологическому воспалительному ответу, выявляемому при SLE. T-клетки при SLE характеризуются спонтанным повышением активации, связанным с пониженным порогом активации на аутоантигены. Кроме того, эти клетки имеют пониженную чувствительность к дополнительной антигенной стимуляции, устойчивы к апоптозу, характеризуются увеличенной продолжительностью существования после активации и имеют множество измененных внутриклеточных сигнальных путей. После активации CD40R на APC T-клеточными CD40L, как APC, так и T-клетки становятся активными, продуцируют цитокины и при SLE способствуют продуцированию патогенных аутоантител и повреждению ткани (волчаночный нефрит). Блокирование сигнального пути CD40R/CD40L отдельно или в комбинации является эффективным при прекращении развития заболевания у предрасположенных к волчанке мышей. У пациентов с SLE гуманизированное антитело к CD40L приводило к уменьшению количества антител к двухцепочечной ДНК и В-клеток, протеинурии и облегчало тяжесть заболевания при SLE.
[0014] Однако, нацеленное воздействие на CD40L естественными антителами существенно повышало факторы опасности. Например, исследование с использованием антитела к CD40L 5c8 (BIOGEN®) у пациентов, страдающих от хронической резистентной идиопатической тромбоцитопенической пурпуры (ITP) были приостановлены по причине зарегистрированных тромбоэмболических осложнений (Davidson et al. Arth Rheu, 43:S271). Кроме того, дополнительные испытания с использованием альтернативных антител, направленных против CD40L, привели к другим связанным с тромбозом осложнениям (Davis et al. Arth Rheu, 43:S281; Schuler, Transplantation, 77:717). С учетом трудностей, связанных с антагонизмом по отношению к CD40L, направляемым антителом, существует неудовлетворенная потребность в нацеленном воздействии и антагонизме по отношению к CD40L при помощи альтернативного средства, не являющегося антителом. Таким образом, направленное воздействие на CD40L при помощи каркасной структуры на основе Tn3 представляет собой эффективную альтернативу с предотвращением Fab2 и/или Fc-опосредованной агрегации тромбоцитов и последующих побочных эффектов.
[0015] Цитирование или обсуждение ссылки в данном документе не следует истолковывать как допущение того, что таковая относится к предшествующему уровню техники для настоящего изобретения.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0016] Настоящее изобретение предлагает Tn3-каркасную структуру, содержащую CD40L-специфичные мономерные субъединицы, где мономерная субъединица содержит семь бета-цепей, обозначенных A, B, C, D, E, F и G, и шесть петлевых участков, обозначенных AB, BC, CD, DE, EF и FG, и где Tn3-каркасная структура специфически связывается с CD40L. В некоторых вариантах осуществления Tn3-каркасная структура содержит одну CD40L-специфичную мономерную субъединицу. В других вариантах осуществления Tn3-каркасная структура содержит две CD40L-специфичные мономерные субъединицы, соединенные в тандем. В некоторых конкретных вариантах осуществления Tn3-каркасная структура содержит две CD40L-специфичные мономерные субъединицы, которые соединены непосредственно.
[0017] В некоторых вариантах осуществления две CD40L-специфичные мономерные субъединицы соединены линкером. В других вариантах осуществления линкер содержит пептидный линкер, который может представлять собой гибкий пептидный линкер. В некоторых вариантах осуществления пептидный линкер содержит последовательность (GmX)n, где X представляет собой cерин (S), аланин (A), глицин (G), лейцин (L), изолейцин (I) или валин (V); m и n представляют собой целые значения; m равно 1, 2, 3 или 4; и n равно 1, 2, 3, 4, 5, 6 или 7. В некоторых вариантах осуществления пептидный линкер содержит SEQ ID NO: 131, SEQ ID NO: 132, SEQ ID NO: 142 или SEQ ID NO: 143.
[0018] В некоторых вариантах осуществления связывание Tn3-каркасной структуры, содержащей две CD40L-специфичные мономерные субъединицы, с CD40L улучшено по сравнению с Tn3-каркасной структурой, содержащей одну CD40L-специфичную мономерную субъединицу. В других вариантах осуществления связывание Tn3-каркасной структуры, содержащей две CD40L-специфичные мономерные субъединицы, с CD40L обеспечивает улучшение воздействия на мишень по сравнению с Tn3-каркасной структурой, содержащей одну CD40L-специфичную мономерную субъединицу. В других вариантах осуществления улучшение связывания Tn3-каркасной структуры с CD40L представляет собой улучшение аффинности связывания, улучшение авидности связывания, или и то и другое. В конкретных вариантах осуществления аффиность связывания Tn3-каркасной структуры, содержащей две CD40L-специфичные мономерные субъединицы, с CD40L и стабильность белка Tn3-каркасной структуры улучшены по сравнению с Tn3-каркасной структурой, содержащей одну CD40L-специфичную мономерную субъединицу. В некоторых вариантах осуществления авидность связывания Tn3-каркасной структуры, содержащей две CD40L-специфичные мономерные субъединицы, по отношению к CD40L и стабильность белка Tn3-каркасной структуры улучшены по сравнению с Tn3-каркасной структурой, содержащей одну CD40L-специфичную мономерную субъединицу.
[0019] В некоторых вариантах осуществления по меньшей мере одна CD40L-специфичная мономерная субъединица в Tn3-каркасной структуре связана с линкером или гетерологичным фрагментом. В других вариантах осуществления линкер или гетерологичный фрагмент в Tn3-каркасной структурe конъюгирован с N-концом или C-концом CD40L-специфичной мономерной субъединицы. В конкретных вариантах осуществления линкер, связанный с CD40L-специфичной мономерной субъединицей в Tn3-каркасной структуре, содержит пептидный линкер, который в некоторых вариантах осуществления может представлять собой гибкий пептидный линкер. Данный пептидный линкер в определенных вариантах осуществления может содержать последовательность (GmX)n, где X представляет собой серин (S), аланин (A), глицин (G), лейцин (L), изолейцин (I) или валин (V); m и n представляют собой целые числа; m равно 1, 2, 3 или 4; и n равно 1, 2, 3, 4, 5, 6 или 7. В некоторых вариантах осуществления пептидный линкер, связанный с CD40L-специфичной мономерной субъединицей в Tn3-каркасной структуре, содержит SEQ ID NO: 131, SEQ ID NO: 132, SEQ ID NO: 142 или SEQ ID NO: 143.
[0020] В некоторых вариантах осуществления Tn3-каркасная структура содержит линкер, который содержит фунциональную группу. В некоторых вариантах осуществления данная функциональная группа представляет собой иммуноглобулин или его фрагмент. В конкретных вариантах осуществления данный иммуноглобулин или его фрагмент содержит Fc-домен. В некоторых вариантах осуществления данный Fc-домен не индуцирует по меньшей мере одну FcγR-опосредованную эффекторную функцию. В некоторых вариантах осуществления данная по меньшей мере одна FcγR-опосредованная эффекторная функция представляет собой ADCC.
[0021] В некоторых вариантах осуществления Tn3-каркасная структура содержит по меньшей мере одну CD40L-специфичную мономерную субъединицу, связанную с гетерологичным фрагментом. В некоторых вариантах осуществления данный гетерологичный фрагмент содержит структуру, выбранную из группы, состоящей из следующего: белок, пептид, белковый домен, линкер, лекарственное средство, токсин, цитотоксическое средство, радиофармацевтическое средство, радионуклид, радиоактивное соединение, органический полимер, неорганический полимер, полиэтиленгликоль (PEG), биотин, альбумин, человеческий сывороточный альбумин (HSA), участок, отвечающий за связывание HSA c FcRn, антитело, домен антитела, фрагмент антитела, одноцепочечное антитело, антитело на основе домена, альбумин-связывающий домен, фермент, лиганд, рецептор, связывающий пептид, каркасная структура, отличающаяся от FnIII, эпитопная метка, рекомбинантный полипептидный полимер, цитокин и комбинация двух или более из указанных фрагментов.
[0022] В некоторых вариантах осуществления Tn3-каркасная структура содержит по меньшей мере одну CD40L-специфичную мономерную субъединицу, конъюгированную с PEG. В некоторых вариантах осуществления Tn3-каркасная структура содержит по меньшей мере одну CD40L-специфичную мономерную субъединицу, конъюгированную с альбумином. В конкретных вариантах осуществления данный альбумин представляет собой человеческий сывороточный альбумин (HSA). В других вариантах осуществления данный HSA представляет собой вариант HSA. В некоторых конкретных вариантах осуществления аминокислотная последовательность варианта HSA представляет собой SEQ ID NO: 133. В других вариантах осуществления вариант HSA имеет по меньшей мере одно улучшенное свойство по сравнению с нативным HSA или фрагментом нативного HSA. В конкретных вариантах осуществления улучшенное свойство представляет собой измененное время полужизни в плазме по сравнению с временем полужизни в плазме нативного HSA или фрагмента нативного HSA. В некоторых вариантах осуществления измененное время полужизни в плазме представляет собой более длительное время полужизни в плазме по сравнению с временем полужизни в плазме нативного HSA или фрагмента нативного HSA. В других вариантах осуществления измененное время полужизни в плазме представляет собой более короткое время полужизни в плазме по сравнению с временем полужизни в плазме нативного HSA или фрагмента нативного HSA.
[0023] В некоторых вариантах осуществления Tn3-каркасная структура является слитой с вариантом HSA, содержащим по меньшей мере одну аминокислотную замену в домене III HSA. В других вариантах осуществления Tn3-каркасная структура является слитой с вариантом HSA, содержащим последовательность полноразмерного зрелого HSA (SEQ ID NO: 133 или 138) или ее фрагмент, за исключением по меньшей мере одной аминокислотной замены, пронумерованной относительно положения в полноразмерном зрелом HSA, в положении, выбранном из группы, состоящей из 407, 415, 463, 500, 506, 508, 509, 511, 512, 515, 516, 521, 523, 524, 526, 535, 550, 557, 573, 574 и 580; где по меньшей мере одна аминокислотная замена не включает замену лизина (K) на глутаминовую кислоту (E) в положении 573, и где Tn3-каркасная структура имеет время полужизни в плазме больше, чем время полужизни в плазме Tn3-каркасной структуры, которая не является конъюгированной с таким вариантом HSA.
[0024] В некоторых вариантах осуществления Tn3-каркасная структура является слитой с вариантом HSA, где по меньшей мере одна аминокислотная замена, пронумерованная относительно положения в полноразмерном зрелом HSA, находится в положении, выбранном из группы, состоящей из 463, 508, 523 и 524, где указанная Tn3-каркасная структура имеет более длительное время полужизни в плазме, чем время полужизни в плазме Tn3-каркасной структуры, которая не является конъюгированной с указанным вариантом HSA. В некоторых вариантах осуществления вариант HSA, содержит последовательность полноразмерного зрелого HSA (SEQ ID NO: 133 или 138) или ее фрагмент, за исключением по меньшей мере одной аминокислотной замены, пронумерованной относительно положения в полноразмерном зрелом HSA, выбранной из группы, состоящей из: (a) замены лейцина (L) в положении 407 на аспарагин (N) или тирозин (Y); (b) замены валина (V) в положении 415 на треонин (T); (c) замены лейцина (L) в положении 463 на аспарагин (N); (d) замены лизина (K) в положении 500 на аргинин (R); (e) замены треонина (T) в положении 506 на тирозин (Y); (f) замены треонина (T) в положении 508 на аргинин (R); (g) замены фенилаланина (F) в положении 509 на метионин (M) или триптофан (W); (h) замены аланина (A) в положении 511 на фенилаланин (F); (i) замены аспарагиновой кислоты (D) в положении 512 на тирозин (Y); (j) замены треонина (T) в положении 515 на глутамин (Q); (k) замены лейцина (L) в положении 516 на треонин (T) или триптофан (W); (l) замены аргинина (R) в положении 521 на триптофан (W); (m) замены изолейцина (I) в положении 523 на аспарагиновую кислоту (D), глутаминовую кислоту (E), глицин (G), лизин (K) или аргинин (R); (n) замены лизина (K) в положении 524 на лейцин (L); (o) замены глутамина (Q) в положении 526 на метионин (M); (p) замены гистидина (H) в положении 535 на пролин (P); (q) замены аспарагиновой кислоты (D) в положении 550 на глутаминовую кислоту (E); (r) замены лизина (K) в положении 557 на глицин (G); (s) замены лизина (K) в положении 573 на фенилаланин (F), гистидин (H), пролин (P), триптофан (W) или тирозин (Y); (t) замены лизина (K) в положении 574 на аспарагин (N); (u) замены глутамина (Q) в положении 580 на лизин (K) и (v) комбинации из двух или более указанных замен, где указанная Tn3-каркасная структура имеет более длительное время полужизни в плазме, чем время полужизни Tn3-каркасной структуры, которая не является конъюгированной с указанным вариантом HSA.
[0025] В некоторых вариантах осуществления Tn3-каркасная структура является слитой с вариантом HSA, содержащим последовательность полноразмерного зрелого HSA (SEQ ID NO: 133 или 138) или ее фрагмент, за исключением по меньшей мере одной аминокислотной замены, пронумерованной относительно положения в полноразмерном зрелом HSA, выбранной из группы, состоящей из: (a) замены лейцина (L) в положении 463 на аспарагин (N); (b) замены треонина (T) в положении 508 на аргинин (R); (c) замены изолейцина (I) в положении 523 на аспарагиновую кислоту (D), глутаминовую кислоту (E), глицин (G), лизин (K) или аргинин (R); (d) замены лизина (K) в положении 524 на лейцин (L) и (e) комбинации из двух или более указанных замен, где указанная Tn3-каркасная структура имеет более длительное время полужизни в плазме, чем время полужизни в плазме Tn3-каркасной структуры, которая не является конъюгированной с указанным вариантом HSA.
[0026] В некоторых вариантах осуществления Tn3-каркасная структура содержит по меньшей мере две идентичные CD40L-специфичные мономерные субъединицы. В других вариантах осуществления Tn3-каркасная структура содержит по меньшей мере две различные CD40L-специфичные мономерные субъединицы. В некоторых вариантах осуществления Tn3-каркасная структура представляет собой агонист рецептора CD40L. В некоторых вариантах осуществления Tn3-каркасная структура представляет собой антагонист рецептора CD40.
[0027] В некоторых вариантах осуществления Tn3-каркасная структура содержит по меньшей мере две CD40L-специфичные мономерные субъединицы, которые специфически связываются с одним и тем же эпитопом CD40L. В некоторых вариантах осуществления Tn3-каркасная структура содержит по меньшей мере две CD40L-специфичные мономерные субъединицы, которые специфически связываются с различными эпитопами CD40L. В некоторых вариантах осуществления данные различные эпитопы CD40L представляют собой неперекрывающиеся эпитопы. В других вариантах осуществления данные различные эпитопы CD40L представляют собой перекрывающиеся эпитопы.
[0028] В некоторых вариантах осуществления Tn3-каркасная структура связывается по меньшей мере с двумя молекулами CD40L. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, которая связывается по меньшей мере c двумя молекулами CD40L.
[0029] В некоторых вариантах осуществления бета-цепи по меньшей мере одной CD40L-специфичной мономерной субъединицы из Tn3-каркасной структуры имеют по меньшей мере 90% идентичность последовательности с бета-цепями с SEQ ID NO: 3. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, содержащую бета-цепь A, которая содержит SEQ ID NO: 11, или содержащую бета-цепь A, которая содержит SEQ ID NO: 11, за исключением по меньшей мере одной мутации. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, содержащую бета-цепь B, которая содержит SEQ ID NO: 12, или содержащую бета-цепь B, которая содержит SEQ ID NO: 12, за исключением по меньшей мере одной мутации. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, содержащую бета-цепь C, которая содержит SEQ ID NO: 13 или 14, или содержащую бета-цепь С, которая содержит SEQ ID NO: 13 или 14, за исключением по меньшей мере одной мутации, и где цистеин в SEQ ID NO: 13 или 14 не заменен. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, содержащую бета-цепь D, которая содержит SEQ ID NO: 15, или содержащую бета-цепь D, которая содержит SEQ ID NO: 15, за исключением по меньшей мере одной мутации.
[0030] В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, содержащую бета-цепь E, которая содержит SEQ ID NO: 16, или содержащую бета-цепь E, которая содержит SEQ ID NO: 16, за исключением по меньшей мере одной мутации. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, содержащую бета-цепь F, которая содержит SEQ ID NO: 17, или содержащую бета-цепь F, которая содержит SEQ ID NO: 17, за исключением по меньшей мере одной мутации, и где цистеин в SEQ ID NO: 17 не заменен. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, содержащую бета-цепь G, которая содержит SEQ ID NO: 18, или содержащую бета-цепь G, которая содержит SEQ ID NO: 18, за исключением по меньшей мере одной мутации.
[0031] В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, которая содержит аминокислотную последовательность:
IEV(XAB)nALITW(XBC)nCELX1YGI(XCD)nTTIDL(XDE)nYSI(XEF)nYEVSLIC(XFG)nKETFTT,
где XAB, XBC, XCD, XDE, XEF и XFG представляют собой аминокислотные остатки, присутствующие в последовательностях петель AB, BC, CD, DE, EF и FG, соответственно; X1 представляет собой аминокислотный остаток A или T; и длина петли n представляет собой целое число от 2 до 26.
[0032] В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли AB содержит SEQ ID NO: 4 или SEQ ID NO: 136, последовательность петли CD содержит SEQ ID NO: 6, и последовательность петли EF содержит SEQ ID NO: 8 или SEQ ID NO: 137. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли BC содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93 и 168. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли DE содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 94, 95, 96, 97, 98 и 169. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли FG содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 9, 99, 139 и 170.
[0033] В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли BC содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116, 117 и 174. В определенных вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли DE содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 118, 119, 120, 121, 122, 123, 124, 125, 126, 127, 128 и 175. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли FG содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 129, 130 и 177. В определенных вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли АВ содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 4 и 136.
[0034] В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 83, последовательность петли DЕ содержит SEQ ID NO: 94 и последовательность петли FG содержит SEQ ID NO: 9 или 139. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 83, последовательность петли DЕ содержит SEQ ID NO: 94 и последовательность петли FG содержит SEQ ID NO: 99. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 84, последовательность петли DЕ содержит SEQ ID NO: 95 и последовательность петли FG содержит SEQ ID NO: 9 или 139.
[0035] В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 85, последовательность петли DЕ содержит SEQ ID NO: 94 и последовательность петли FG содержит SEQ ID NO: 9 или 139. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 86, последовательность петли DЕ содержит SEQ ID NO: 96 и последовательность петли FG содержит SEQ ID NO: 9 или 139. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 87, последовательность петли DЕ содержит SEQ ID NO: 97 и последовательность петли FG содержит SEQ ID NO: 9 или 139.
[0036] В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 88, последовательность петли DЕ содержит SEQ ID NO: 95 и последовательность петли FG содержит SEQ ID NO: 9 или 139. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 89, последовательность петли DЕ содержит SEQ ID NO: 94 и последовательность петли FG содержит SEQ ID NO: 9 или 139. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 90, последовательность петли DЕ содержит SEQ ID NO: 94 и последовательность петли FG содержит SEQ ID NO: 9 или 139.
[0037] В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 91, последовательность петли DЕ содержит SEQ ID NO: 95 и последовательность петли FG содержит SEQ ID NO: 9 или 139. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 92, последовательность петли DЕ содержит SEQ ID NO: 98, последовательность петли FG содержит SEQ ID NO: 9 или 139. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 93, последовательность петли DЕ содержит SEQ ID NO: 94 и последовательность петли FG содержит SEQ ID NO: 9 или 139.
[0038] В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 100, последовательность петли DЕ содержит SEQ ID NO: 118 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 101, последовательность петли DЕ содержит SEQ ID NO: 119 и последовательность петли FG содержит SEQ ID NO: 129. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 102, последовательность петли DЕ содержит SEQ ID NO: 120 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 103, последовательность петли DЕ содержит SEQ ID NO: 121 и последовательность петли FG содержит SEQ ID NO: 129. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли АВ содержит SEQ ID NO: 136.
[0039] В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 104, последовательность петли DЕ содержит SEQ ID NO: 122 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 105, последовательность петли DЕ содержит SEQ ID NO: 121 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 106, последовательность петли DЕ содержит SEQ ID NO: 123 и последовательность петли FG содержит SEQ ID NO: 129. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли АВ содержит SEQ ID NO: 136.
[0040] В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 107, последовательность петли DЕ содержит SEQ ID NO: 123 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 108, последовательность петли DЕ содержит SEQ ID NO: 118 и последовательность петли FG содержит SEQ ID NO: 129. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 109, последовательность петли DЕ содержит SEQ ID NO: 123 и последовательность петли FG содержит SEQ ID NO: 129. В некоторых вариантах осуществления последовательность петли BC содержит SEQ ID NO: 110, последовательность петли DЕ содержит SEQ ID NO: 121 и последовательность петли FG содержит SEQ ID NO: 129. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли АВ содержит SEQ ID NO: 136.
[0041] В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 111, последовательность петли DЕ содержит SEQ ID NO: 123 и последовательность петли FG содержит SEQ ID NO: 130. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 108, последовательность петли DЕ содержит SEQ ID NO: 121 и последовательность петли FG содержит SEQ ID NO: 129. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 112, последовательность петли DЕ содержит SEQ ID NO: 124 и последовательность петли FG содержит SEQ ID NO: 129. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 113, последовательность петли DЕ содержит SEQ ID NO: 125 и последовательность петли FG содержит SEQ ID NO: 129. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли АВ содержит SEQ ID NO: 136.
[0042] В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 114, последовательность петли DЕ содержит SEQ ID NO: 118 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 115, последовательность петли DЕ содержит SEQ ID NO: 126 и последовательность петли FG содержит SEQ ID NO: 129. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 116, последовательность петли DЕ содержит SEQ ID NO: 127 и последовательность петли FG содержит SEQ ID NO: 129. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли ВС содержит SEQ ID NO: 117, последовательность петли DЕ содержит SEQ ID NO: 128 и последовательность петли FG содержит SEQ ID NO: 129. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где последовательность петли АВ содержит SEQ ID NO: 136.
[0043] В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42 и 146. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, которая содержит аминокислотную последовательность:
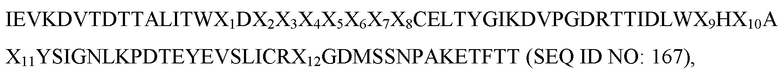
где (a) X1 представляет собой аминокислотный остаток серин (S) или лейцин (L); (b) X2 представляет собой аминокислотный остаток аспарагиновую кислоту (D) или глутаминовую кислоту (E); (c) X3 представляет собой аминокислотный остаток гистидин (H), изолейцин (I), валин (V), фенилаланин (F) или триптофан (W); (d) X4 представляет собой аминокислотный остаток аланин (A), глицин (G), глутаминовую кислоту (E) или аспарагиновую кислоту (D); (e) X5 представляет собой аминокислотный остаток глутаминовую кислоту (E), лейцин (L), глутамин (Q), серин (S), аспарагиновую кислоту (D) или аспарагин (N); (f) X6 представляет собой аминокислотный остаток фенилаланин (F) или тирозин (Y); (g) X7 представляет собой аминокислотный остаток изолейцин (I), валин (V), гистидин (H), глутаминовую кислоту (E) или аспарагиновую кислоту (D); (h) X8 представляет собой аминокислотный остаток глицин (G), триптофан (W) или валин (V); (i) X9 представляет собой аминокислотный остаток триптофан (W), фенилаланин (F) или тирозин (Y); (j) X10 представляет собой аминокислотный остаток серин (S), глутамин (Q), метионин (M) или гистидин (H); (k) X11 представляет собой аминокислотный остаток триптофан (W) или гистидин (H) и (l) X12 представляет собой аминокислотный остаток аргинин (R) или серин (S).
[0044] В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, содержащую последовательность, выбранную из группы, состоящей из SEQ ID NO: 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80 и 82.
[0045] В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, которая содержит аминокислотную последовательность:
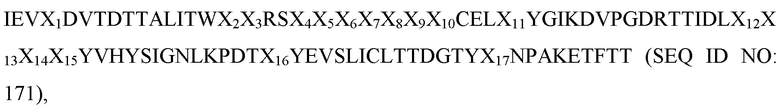
где
(a) X1 представляет собой аминокислотный остаток лизин (K) или глутаминовую кислоту (E);
(b) X2 представляет собой аминокислотный остаток треонин (T) или изолейцин (I);
(c) X3 представляет собой аминокислотный остаток аспарагин (N) или аланин (A);
(d) X4 представляет собой аминокислотный остаток серин (S), лейцин (L), аланин (A), фенилаланин (F) или тирозин (Y);
(e) X5 представляет собой аминокислотный остаток тирозин (Y), аланин (A), глицин (G), валин (V), изолейцин (I) или серин (S);
(f) X6 представляет собой аминокислотный остаток тирозин (Y), серин (S), аланин (A) или гистидин (H);
(g) X7 представляет собой аминокислотный остаток аспарагин (N), аспарагиновую кислоту (D), гистидин (H) или тирозин (Y);
(h) X8 представляет собой аминокислотный остаток лейцин (L), фенилаланин (F), гистидин (H) или тирозин (Y);
(i) X9 представляет собой аминокислотный остаток гистидин (H), пролин (P), серин (S), лейцин (L) или аспарагиновую кислоту (D);
(j) X10 представляет собой аминокислотный остаток глицин (G), фенилаланин (F), гистидин (H) или тирозин (Y);
(k) X11 представляет собой аминокислотный остаток аланин (A) или треонин (T);
(l) X12 представляет собой аминокислотный остаток серин (S), аспарагин (N), глутаминовую кислоту (E), аспарагин (R) или аспарагиновую кислоту (D);
(m) X13 представляет собой аминокислотный остаток серин (S), глутамин (Q), треонин (T), аспарагин (N) или аланин (A);
(n) X14 представляет собой аминокислотный остаток пролин (P), валин (V), изолейцин (I) или аланин (A) или не является аминокислотой;
(o) X15 представляет собой аминокислотный остаток изолейцин (I) или не является аминокислотой;
(p) X16 представляет собой аминокислотный остаток глутаминовую кислоту (E) или лизин (K); и
(q) X17 представляет собой аминокислотный остаток серин (S) или аспарагин (N).
[0046] В некоторых вариантах осуществления Tn3-каркасная структура содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 134, 135, 205, 206, 207 и 208. В некоторых вариантах осуществления Tn3-каркасная структура состоит из последовательности, выбранной из группы, состоящей из SEQ ID NO: 134, 135, 205, 206, 207 и 208.
[0047] В некоторых вариантах осуществления Tn3-каркасная структура содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 201, 202, 203 и 204. В некоторых вариантах осуществления Tn3-каркасная структура состоит из последовательности, выбранной из группы, состоящей из SEQ ID NO: 201, 202, 203 и 204. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где указанная специфичность по отношению к CD40L относится к CD40L человека. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, где указанная специфичность по отношению к CD40L относится к мембраносвязанному CD40L (SEQ ID NO: 1), растворимому CD40L (SEQ ID NO: 2) или его фрагменту. В некоторых конкретных вариантах осуществления Tn3-каркасная структура связывается с CD40L и предупреждает связывание CD40L с CD40.
[0048] Настоящее изобретение также предлагает способ изменения активности в клетке, экспрессирующей CD40L, включающий приведение в контакт клетки с Tn3-каркасной структурой, где Tn3-каркасная структура связывается с CD40L и предупреждает связывание CD40L с CD40. В некоторых вариантах осуществления Tn3-каркасная структура связывается с CD40L с аффинностью (Kd) приблизительно 1 мкМ или менее, или приблизительно 500 нМ или менее, или приблизительно 100 нМ или менее, или приблизительно 50 нМ или менее, или приблизительно 25 нМ или менее, или приблизительно 10 нМ или менее, или приблизительно 5 нМ или менее, или приблизительно 2 нМ или менее.
[0049] В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, которая специфично связывается с эпитопом CD40L, содержащим аминокислоты, расположенные в положениях 142-155, 200-230 или 247-251 в SEQ ID NO: 2. В некоторых вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, которая взаимодействует с аминокислотами CD40L E142, Y146, M148, N151, L155, R200, R203 и E230. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, которая взаимодействует с аминокислотами CD40L R203, I204, V247, H249 и T251. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичная мономерную субъединицу, которая взаимодействует с аминокислотами CD40L E142, Y146, M148, N151, L155, которые расположены в первой молекуле CD40L, и с аминокислотами CD40L R200, R203 и E230, которые расположены во второй молекуле CD40L. В других вариантах осуществления Tn3-каркасная структура содержит CD40L-специфичную мономерную субъединицу, которая взаимодействует с аминокислотами CD40L R203 и I204, которые расположены в первой молекуле CD40L, и с аминокислотами CD40L V247, H249 и T251, которые расположены во второй молекуле CD40L.
[0050] Настоящее изобретение также предлагает полипептиды, содержащие один или несколько CD40L-специфичных Tn3-мономеров, включая, но без ограничения, слияния сывороточного альбумина, описанные в данном документе.
[0051] Настоящее изобретение также предлагает выделенную молекулу нуклеиновой кислоты, кодирующую CD40L-специфичную Tn3-каркасную структуру, вектор экспрессии, содержащий молекулу нуклеиновой кислоты, кодирующую CD40L-специфичную Tn3-каркасную структуру, и клетку-хозяина, содержащую такой вектор. Настоящее изобретение также предлагает способ получения Tn3-каркасной структуры, включающий культивирование клетки-хозяина в условиях, при которых экспрессируется CD40L-специфичная Tn3-каркасная структура, кодируемая молекулой нуклеиновой кислоты.
[0052] Настоящее изобретение также предлагает фармацевтическую композицию, содержащую CD40L-специфичную Tn3-каркасную структуру и фармацевтически приемлемый наполнитель. Настоящее изобретение также предлагает способ предупреждения, лечения, уменьшения интенсивности, управления течением аутоиммунного заболевания у пациента, нуждающегося в этом, включающий введение эффективного количества фармацевтической композиции, содержащей CD40L-специфичную Tn3-каркасную структуру.
[0053] Настоящее изобретение также предлагает способ снижения частоты или количества кортикостероидов, вводимых пациенту с аутоиммунным заболеванием, включающий введение пациенту терапевтически эффективного количествa фармацевтической композиции, содержащей CD40L-специфичную Tn3-каркасную структуру.
[0054] Аутоиммунным заболеванием, которое подлежит лечению путем введения CD40L-специфичной Tn3-каркасной структуры, может быть очаговая алопеция, анкилозирующий спондилит, антифосфолипидный синдром, аутоиммунная болезнь Аддисона, аутоиммунные заболевания надпочечников, аутоиммунная гемолитическая анемия, аутоиммунный гепатит, аутоиммунный оофорит и орхит, синдром Шегрена, псориаз, атеросклероз, диабетическая и другие ретинопатии, ретролентальная фиброплазия, возрастная макулярная дегенерация, неоваскулярная глаукома, гемангиомы, тиреоидные гиперплазии (в том числе болезнь Грейвса), трансплантация ткани роговицы и другой ткани, а также хроническое воспаление, сепсис, ревматоидный артрит, перитонит, болезнь Крона, реперфузионное повреждение, септицемия, эндотоксический шок, муковисцидоз, эндокардит, псориаз, артрит (например, псориатический артрит), анафилактический шок, ишемия органа, реперфузионное повреждение, повреждение спинного мозга и отторжение аллотрансплантата, аутоиммунная тромбоцитопения, болезнь Бехчета, булезный пемфигоид, кардиомиопатия, дерматит, связанный с целиакией спру, синдром хронический усталости и иммунной дисфункции (CFIDS), хроническая воспалительная демиелинизирующая полинейропатия, синдром Черджа-Стросса, рубцовый пемфигоид, CREST-синдром, болезнь холодовых агглютининов, болезнь Крона, дискоидная волчанка, идиопатическая криоглобулинемия смешанного типа, фибромиалгия-фибромиозит, гломерулонефрит, болезнь Грейвса, Гийена-Барре, тиреоидит Хашимото, идиопатический фиброз легких, идиопатическая тромбоцитопеническая пурпура (ITP), IgA-нейропатия, болезнь Стилла, красный плоский лишай, красная волчанка, болезнь Меньера, смешанное заболевание соединительной ткани, рассеянный склероз, сахарный диабет 1 типа или иммуноопосредованный, миастения, вульгарный пемфигус, пернициозная анемия, нодозный полиартериит, полихондрит, полигландулярные синдромы, ревматическая полимиалгия, полимиозит и дерматомиозит, первичная агаммаглобулинемия, первичный билиарный цирроз, псориаз, псориатический артрит, болезнь Рейно, синдром Рейтера, ревматоидный артрит, саркоидоз, склеродермия, синдром Шегрена, синдром мышечной скованности, системная красная волчанка, красная волчанка, артериит Такаясу, височный артериит/гигантоклеточный артериит, язвенный колит, увеит, васкулит, такой как герпетиформный дерматит, витилиго и гранулематоз Вегенера.
[0055] В некоторых конкретных вариантах осуществления аутоиммунное заболевание, которое подлежит лечению путем введения CD40L-специфичной Tn3-каркасной структуры, представляет собой системную красную волчанку (SLE).
[0056] Кроме того, способы лечения с помощью CD40L-специфичной Tn3-каркасной структуры могут предусматривать дополнительную терапию, такую как иммунотерапия, биологическая терапия, химиотерапия, лучевая терапия или терапия с помощью низкомолекулярного лекарственного средства.
[0057] Настоящее изобретение также предлагает белковый кристалл, содержащий Tn3-каркасную структуру, состоящую из SEQ ID NO: 20, в комплексе с растворимым CD40L (SEQ ID NO: 2), где данный кристалл имеет кристаллическую решетку с P212121-орторомбической пространственной группой и постоянными решетки, +/- 0,1%, a=85,69 Å, b=90,64 Å, c=95,56 Å. В некоторых вариантах осуществления асимметрическая единица кристалла включает тример CD40L и три молекулы Tn3-каркасной структуры. В других вариантах осуществления кристалл характеризуется рассеиванием рентгеновского излучения при определении структурных координат до разрешения со значением, равным 3,2 Å или менее.
[0058] Настоящее изобретение также предлагает белковый кристалл, содержащий Tn3-каркасную структуру, состоящую из SEQ ID NO: 68, в комплексе с растворимым CD40L (SEQ ID NO: 2), где данный кристалл имеет кристаллическую решетку с P213-кубической пространственной группой и постоянными решетки, +/- 0,1%, a=b=c=97,62 Å. В некоторых вариантах осуществления асимметрическая единица кристалла содержит одну молекулу CD40L и одну молекулу Tn3-каркасной структуры. В других вариантах осуществления кристалл характеризуется рассеиванием рентгеновского излучения при определении структурных координат до разрешения со значением, равным 2,7 Å или менее.
[0059] Настоящее изобретение также предлагает белковый кристалл, содержащий Tn3-каркасную структуру, состоящую из SEQ ID NO: 28 или 146, в комплексе с растворимым CD40L (SEQ ID NO: 2), где данный кристалл имеет кристаллическую решетку с P321-пространственной группой и постоянными решетки, +/- 0,1%, a=95,53 Å, b=93,53 Å, c=66,69 Å. В некоторых вариантах осуществления асимметрическая единица кристалла содержит одну молекулу CD40L и одну молекулу Tn3-каркасной структуры. В других вариантах осуществления кристалл характеризуется рассеиванием рентгеновского излучения при определении структурных координат до разрешения со значением, равным 2,8 Å или менее.
[0060] Настоящее изобретение также предлагает белковый кристалл, содержащий две различные Tn3-каркасные структуры, состоящие из SEQ ID NO: 68 и SEQ ID NO: 28 или 146, в комплексе с растворимым CD40L (SEQ ID NO: 2), где данный кристалл имеет кристаллическую решетку с P21-кубической пространственной группой и постоянными решетки, +/- 0,1%, a=80,32 Å, b=143,48 Å, c=111,27 Å, β=98,22 Å. В некоторых вариантах осуществления асимметрическая единица кристалла содержит два тримера CD40L и шесть молекул каждой Tn3-каркасной структуры. В других вариантах осуществления кристалл характеризуется рассеиванием рентгеновского излучения при определении структурных координат до разрешения со значением, равным 1,9 Å или менее.
[0061] В некоторых вариантах осуществления белковый кристалл получают с использованием диффузии паров в сидячей капле. Настоящее изобретение также предлагает способ получения белкового кристалла, предусматривающий: (a) смешивание объема раствора, содержащего Tn3-каркасную структуру, которая содержит CD40L-специфичную мономерную субъединицу, в комплексе с CD40L, с объемом резервуарного раствора, содержащего осаждающее средство; и (b) инкубирование смеси, полученной на этапе (a) в закрытом контейнере в пригодных для кристаллизации условиях до образования белкового кристалла. В некоторых вариантах осуществления способ получения белкового кристалла предусматривает применение диффузии паров в сидячей капле.
[0062] В некоторых вариантах осуществления способ создания белкового кристалла применяют для получения кристаллов, содержащих CD40L-специфичные Tn3-мономерные субъединицы с SEQ ID NO: 20, SEQ ID NO: 28, SEQ ID NO: 68 или SEQ ID NO: 146.
[0063] Настоящее изобретение также предлагает машиночитаемый накопитель данных, содержащий материал для хранения данных, закодированный машиночитаемыми командами для: (a) преобразования данных в графическое трехмерное изображение структуры части белкового кристалла Tn3-каркасной структуры, содержащей CD40L-специфичную мономерную субъединицу, образующую комплекс с CD40L; и (b) обеспечения отображения указанного графического трехмерного изображения. В некоторых вариантах осуществления такая Tn3-каркасная структура содержит SEQ ID NO: 20, SEQ ID NO: 28, SEQ ID NO: 68 или SEQ ID NO: 146. В других вариантах осуществления такой белковый кристалл представляет собой
[0064] (a) белковый кристалл, содержащий Tn3-каркасную структуру, состоящую из SEQ ID NO: 20, в комплексе с растворимым CD40L (SEQ ID NO: 2), где данный кристалл имеет кристаллическую решетку с P212121-орторомбической пространственной группой и постоянными решетки, +/- 0,1%, a=85,69 Å, b=90,64 Å, c=95,56 Å;
[0065] (b) белковый кристалл, содержащий Tn3-каркасную структуру, состоящую из SEQ ID NO: 68, в комплексе с растворимым CD40L (SEQ ID NO: 2), где данный кристалл имеет кристаллическую решетку с P213-кубической пространственной группой и постоянными решетки, +/- 0,1%, a=b=c=97,62 Å; или
[0066] (c) белковый кристалл, содержащий Tn3-каркасную структуру, состоящую из SEQ ID NO: 20, и Tn3-каркасную структуру, состоящую из SEQ ID NO: 68, где обе Tn3-каркасные структуры находятся в комплексе с растворимым CD40L (SEQ ID NO: 2);
[0067] (d) белковый кристалл, содержащий Tn3-каркасную структуру, состоящую из SEQ ID NO: 28 или 146, в комплексе с растворимым CD40L (SEQ ID NO: 2), где данный кристалл имеет кристаллическую решетку с P321-пространственной группой и постоянными решетки, +/- 0,1%, a=95,53 Å, b=93,53 Å, c=66,69 Å;
[0068] (е) белковый кристалл, содержащий две различные Tn3-каркасные структуры, состоящие из SEQ ID NO: 68 и SEQ ID NO: 28 или 146, в комплексе с растворимым CD40L (SEQ ID NO: 2), где данный кристалл имеет кристаллическую решетку с P21-кубической пространственной группой и постоянными решетки, +/- 0,1%, a=80,32 Å, b=143,48 Å, c=111,27 Å, β=98,22 Å.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0069] С целью иллюстрации настоящего изобретения в графических материалах изображены определенные варианты осуществления настоящего изобретения. Тем не менее, настоящее изобретение не ограничено точными структурами и средствами из вариантов осуществления, изображенных на графических материалах.
[0070] На Фиг. 1A показано ингибирование экспрессии CD86, которая индуцирована мышиным CD40L (MuCD40L), измеренное с помощью анализа D10G4.1/PBMC (мононуклеарные клетки периферической крови). Анализировали мышиную CD40L-специфичную Tn3-каркасную структуру M13, ее вариант M31 с оптимизированной аффинностью (приблизительно 20x улучшение аффинности), моноклональное антитело MR1 к CD40L, а также негативный контроль. Также показаны значения IC50.
[0071] На Фиг. 1B показано ингибирование CD40L в анализе с мышиным NfkB. В данном анализе используют NIHT3T-клетки, экспрессирующие мышиный CD40R и содержащие репортерный конструкт NfkB-люцифераза. Добавление CD40L приводит к возникновению сигнала (измеряется по люциферазной активности), который ингибируется как антителом MR1 к CD40L, так и CD40L-специфичной Tn3-каркасной структурой M31.
[0072] На Фиг. 2A показано конструирование конструктов слияния CD40L-специфичных тандемных бивалентных Tn3-каркасных структур и сывороточного альбумина (SA).
[0073] На Фиг. 2B показано анализ в SDS-PAGE очищенного моновалентного конструкта M13 (CD40L-специфичный Tn3-конструкт) или тандемных бивалентных каркасных структур с линкерами, содержащими 1, 3, 5 или 7 звеньев Gly4Ser (обозначены как GS), соединяющими две мономерные субъединицы Tn3 M13. Моновалентный конструкт M13 разгоняли на дорожке 2, димерный конструкт с 1 звеном GS (C1) разгоняли на дорожках 3 и 7, димерный конструкт с 3 звеньями GS (C2) разгоняли на дорожках 4 и 8, димерный конструкт с 5 звеньями GS (C3) разгоняли на дорожках 5 и 9, а димерный конструкт с 7 звеньями GS (C4) разгоняли на дорожках 6 и 10. Образцы разгоняли либо при невосстанавливающих условиях (дорожки 2-6), либо при восстанавливающих условиях (дорожки 7-10).
[0074] На Фиг. 2C показано конкурентное ингибирование связывания мышиного CD40L с мышиным рецептором CD40, иммобилизированном на биосенсорном чипе, под воздействием мышиных CD40L-специфичных моновалентных (M13) или бивалентных тандемных каркасных структур (M13-xGS-M13, где x составляет 1, 3, 5 или 7, соответствующих бивалентным каркасным структурам с линкерами, содержащими 1, 3, 5 или 7 звеньев Gly4Ser). Также указана полумаксимальная ингибирующая концентрация (IC50) для различных конструктов.
[0075] На Фиг. 2D показан ингибирующий эффект мышиных CD40L-специфичных Tn3 моновалентных (M13) и бивалентных тандемных каркасных структур на экспрессию CD86, которая индуцирована мышиными CD40L, в В-клетках. Приведены значения IC50 для всех Tn3-конструктов и для антитела MR1 к мышиному CD40L.
[0076] На Фиг. 3A показаны высокие уровни экспрессии мышиной CD40L-специфичной тандемной бивалентной Tn3-каркасной структуры, слитой с мышиным сывороточным альбумином (MSA), в клетках HEK 293. Данные конструкты имеют 1 (G4S) повтор в линкере между звеньями Tn3-каркасной структуры и 3 (G4S) повторами в линкере между Tn3-каркасной структурой и MSA. Кроме того, данный конструкт содержит мутацию N49Q в каждой из каркасных структур M13 и M31 для удаления потенциального сайта N-связанного гликозилирования. 10 мкл культурального супернатанта, отобранного 3 или 6 дней спустя после трансфекции, разгоняли в геле в SDS-PAGE параллельно с известными количествами очищенного белка. Уровень экспрессии оценивали до 200 мг/л на 6 день после после трансфекции. Очистку осуществляли с помощью IMAC с использованием C-концевой His-метки.
[0077] На Фиг. 3B показано ингибирование экспрессии CD86, которая индуцирована мышиным CD40L (MuCD40L), измеренное с помощью клеточного анализа D10G4.1/PBMC. Анализировали CD40L-специфичную тандемную бивалентную Tn3-каркасную структуру (M13-1GS-M13), такой же конструкт, слитый с мышиным сывороточным альбумином (MSA) (M13-1GS-M13-MSA), а также моноклональное антитело MR1 к мышиному CD40L. Для всех конструктов приведены значения IC50.
[0078] На Фиг. 3C показано ингибирование экспрессии CD86, которая индуцирована мышиным CD40L (MuCD40L), измеренное с помощью анализа D10G4.1/PBMC. Aнализировали CD40L-специфичную тандемную бивалентную Tn3-каркасную структуру (M13-1GS-M13), слитую с мышиным сывороточным альбумином (MSA)(M13-1GS-M13-MSA), вариант каркасной структуры с созревшей аффинностью M13, конъюгированный с MSA (M31Mono-MSA), тандемная бивалентная каркасная структура, содержащая вариант M31 с оптимизированной аффинностью, конъюгированный с MSA (M31-1GS-M31-MSA), тандемную бивалентную каркасную структуру в качестве негативного контроля, которая не связывается с мышиным CD40L (D1-1GS-D1-MSA), а также моноклональное антитело MR1. Также приведены значения IC50.
[0079] На Фиг. 4A показаны фармакокинетические данные по отношению к нескольким мышиным CD40L-специфичных конструктам у мышей, определенные с помощью ELISA. Для каждого конструкта указаны значения полужизни в плазме (t1/2).
[0080] На Фиг. 4B показаны фармакокинетические данные для 342-HSA, специфичного по отношению к CD40L человека, и варианта 342-HSA, содержащего замену Leu в положении 463 на Asn (L463N) и замену Lys в положении 524 на Leu (K524L), у яванского макака, определенные с помощью ELISA.
[0081] На Фиг. 5A показано созревание B-клеток в зародышевых центрах (GC) в анализе после иммунизации бараньими эритроцитами (SRBC). Анализировали моноклональное антитело MR1 к CD40L.
[0082] На Фиг. 5B показано созревание B-клеток в зародышевых центрах (GC) в анализе после иммунизации бараньими эритроцитами (SRBC). Анализировали моновалентный и бивалентный конструкты, полученные из M31, слитые с MSA. В качестве негативного контроля использовали бивалентный конструкт D1-D1, конъюгированный с MSA.
[0083] На Фиг. 5C показано созревание B-клеток на периферии (не в GC) в анализе после иммунизации бараньими эритроцитами (SRBC). Анализировали моновалентный и бивалентный конструкты, полученные из M31, слитые с MSA. В качестве негативного контроля использовали бивалентный конструкт D1-D1, конъюгированный с MSA.
[0084] На Фиг. 5D показан процент (% CD4) и число (#CD4) CD4-положительных клеток в анализе после иммунизации бараньими эритроцитами (SRBC). Анализировали моновалентный и бивалентный конструкты, полученные из M31, слитые с MSA, а также моноклональные антитела MR1 к CD40L. В качестве негативного контроля использовали бивалентный конструкт D1-D1, конъюгированный с MSA.
[0085] На Фиг. 5E показан процент (% CD44hi) и число (#CD44hi) CD44hi-положительных клеток в анализе после иммунизации бараньими эритроцитами (SRBC). Анализировали моновалентный и бивалентный конструкты, полученные из M31, слитые с MSA, а также моноклональные антитела MR1 к CD40L. В качестве негативного контроля использовали бивалентный конструкт D1-D1, конъюгированный с MSA.
[0086] На Фиг. 5F показано количество IgG к SRBC в анализе после иммунизации бараньими эритроцитами (SRBC). Анализировали моновалентный и бивалентный конструкты, полученные из M31, слитые с MSA. В качестве негативного контроля использовали бивалентный конструкт D1-D1, конъюгированный с MSA.
[0087] На Фиг. 5G показаны титры IgM к KLH в модели KLH-специфичного T-клеточно-зависимого гуморального иммунного ответа (TDAR). Анализировали моновалентный и бивалентный конструкты, полученные из 342, слитые с HSA.
[0088] На Фиг. 5H показаны титры IgG к KLH в модели KLH-специфичного T-клеточно-зависимого гуморального иммунного ответа (TDAR). Анализировали моновалентный и бивалентный конструкты, полученные из 342, слитые с HSA.
[0089] На Фиг. 6A показан ингибирующий эффект моновалентных мономерных Tn3-каркасных структур 309 и 311, специфичных по отношению к CD40L человека, на экспрессию CD86 человека, которая индуцирована CD40L, в CD19-положительных PBMC человека, стимулированных клетками Jurkat D1.1.
[0090] На Фиг. 6B показан ингибирующий эффект моновалентных мономерных Tn3-каркасных структур 309 и 311, специфичных по отношению к CD40L человека, на пролиферацию В-клеток, стимулированную CD40L человека.
[0091] На Фиг. 6C показан ингибирующий эффект моновалентных мономерных Tn3-каркасных структур 309 и 311, специфичных по отношению к CD40L человека, на количество плазматических клеток при совместном культивировании T/B-клеток. Также с помощью FACS было показано, что Tn3-каркасная структура 309 связывается с активированными первичными T-клетками (данные не показаны). Каркасную структуру D1 ("Neg Tn3") использовали в качестве контроля. Также в качестве контролей использовали два моноклональных антитела к CD40L, обозначенных aCD40L(RE) и aCD40L(Bio) (моноклональное антитело Biogen 5c8 к CD40L человека).
[0092] На Фиг. 7A показано, что моновалентные Tn3-каркасные структуры 309 и 311, специфичные по отношению к CD40L человека, имеют сходные биофизические характеристики. Обе каркасные структуры являются монодисперсными, как определено с помощью SEC.
[0093] На Фиг. 7B показано, что моновалентные Tn3-каркасные структуры 309 и 311, специфичные по отношению к CD40L человека, имеют сходные биофизические характеристики. Обе каркасные структуры характеризуются сходной термостабильностью, так как исходная Tn3-каркасная структура (обозначена Tn3 (дикий тип) на графическом изображении), как измерено при помощи дифференциальной сканирующей калориметрии (DSC).
[0094] На Фиг. 8A показано ингибирование экспрессии CD86, которая индуцирована CD40L человека, в CD19-положительных PBMC человека, стимулированных клетками Jurkat D1.1. Анализировали моновалентные (311) и бивалентные (311_3GS и 311_7GS) Tn3-каркасные структуры, специфичные по отношению к CD40L человека. Показаны значения IC50 для каждого конструкта.
[0095] На Фиг. 8B показано ингибирование экспрессии CD86, которая индуцирована CD40L человека, в CD19-положительных PBMC человека, стимулированных клетками Jurkat D1.1. Анализировали моновалентные (309) и бивалентные (309_3GS и 309_7GS) Tn3-каркасные структуры, специфичные по отношению к CD40L человека, также как моноклональное антитело Biogen 5c8 к CD40L человека. Показаны значения IC50 для каждого конструкта и антитела.
[0096] На Фиг. 9A показано конструирование типичной тандемной бивалентной Tn3-каркасной структуры, специфичной по отношению к CD40L человека, слитой с человеческим сывороточным альбумином (HSA). “GGGGG” (SEQ ID NO: 148) и “GGGGA” (SEQ ID NO: 149) представляют собой линкеры, альтернативные по отношению к линкерам "GGGGS" (SEQ ID NO: 147).
[0097] На Фиг. 9B показана тестовая очистка из 293F-клеток на колонке IEX. Плечевая фракция (<10% основного пика) содержит O-гликозилированный белок, соединенный с остатками серина, присутствующими в линкерах.
[0098] На Фиг. 9С показано ингибирование экспрессии CD86, которая индуцирована CD40L человека, в CD19-положительных PBMC человека, стимулированных клетками Jurkat D1.1. Анализировали бивалентную Tn3-каркасную структуру (309), специфичную по отношению к CD40L человека, такую же каркасную структуру, слитую с HSA, а также моноклональное антитело Biogen 5c8 к CD40L человека.
[0099] На Фиг. 9D показано ингибирование экспрессии CD86, которая индуцирована CD40L человека, в CD19-положительных PBMC человека, стимулированных клетками Jurkat D1.1. Тестировали три бивалентные (309) Tn3-каркасные структуры, специфичные по отношению к CD40L человека. Три (G4S) повтора присутствовали в линкере между субъединицами, специфичными по отношению к CD40L человека (309 в данном примере), тогда как линкер между субъединицами 309 и HSA варьировал от 1 до 3 (G4S) повторов. Также анализировали моноклональное антитело Biogen 5c8 к CD40L человека.
[0100] На Фиг. 10A показано влияние мутации в петлевых последовательностях в 309 (левая панель) и 311 (правая панель) на связывание CD40L. На связывание указывает усиление сигнала в анализе связывания. WT представляет собой вариант с первоначальной лидерной последовательностью (исходной Tn3-последовательностью), где BC, DE и FG означает варианты, в которых петлевую последовательность BC, DE или FG изменяли к исходной Tn3-последовательности, как представлено в тенасцине C человека.
[0101] На Фиг. 10B показаны профили ингибирования панели каркасных структур с оптимизированной аффинностью, определенные по экспрессии CD86, которая индуцирована CD40L человека, в CD19-положительных PBMC человека, стимулированных клетками Jurkat D1.1. Мономеры из клона 309 Tn3, специфичного по отношению к CD40L человека, были оптимизированы по аффинности. Оптимизированные по аффинности мономеры обозначены как клон 340 - клон 349. В конструкте клон 309wtFG целая петля FG была заменена на петлю FG исходной Tn3-каркасной структуры. Также анализировали моноклональное антитело 5c8 к CD40L.
[0102] На Фиг. 10С показаны профили ингибирования, определенные по экспрессии CD86, которая индуцирована CD40L человека, в CD19-положительных PBMC человека, стимулированных клетками Jurkat D1.1. Показаны профиль мономера 311 Tn3, специфичного по отношению к CD40L человека, его вариант K4E, а также негативный контроль.
[0103] На Фиг. 10D показаны профили ингибирования, определенные по экспрессии CD86, которая индуцирована CD40L человека, в CD19-положительных PBMC человека, стимулированных клетками Jurkat D1.1. Показаны профили мономера 311K4E Tn3, специфичного по отношению к CD40L человека, мономера с оптимизированной аффиностью 311K4E_12, а также моноклонального антитела 5c8 к CD40L. Также представлены IC50 для двух конструктов и антитела.
[0104] На Фиг. 11A и Фиг. 11B показаны множественные выравнивания последовательностей исходной CD40L-специфичной Tn3-каркасной структуры 309, варианта 309FGwt, а также вариантов 340-349 с оптимизированной аффинностью. Аминокислотные остатки 1-42 показаны на Фиг. 11A, а аминокислотные остатки 43-83 показаны на Фиг. 11B. Варианты петель затемнены. Консенсусная аминокислотная последовательность представлена ниже множественного выравнивания последовательностей. Выровненные последовательности соответствуют аминокислотным последовательностям клонов 309 Tn3-каркасной структуры (SEQ ID NO: 20), 309FGwt (SEQ ID NO: 22), 340 (SEQ ID NO: 24), 341 (SEQ ID NO: 26), 342 (SEQ ID NO: 28), 343 (SEQ ID NO: 30), 344 (SEQ ID NO: 32), 345 (SEQ ID NO: 34), 346 (SEQ ID NO: 36), 347 (SEQ ID NO: 38), 348 (SEQ ID NO: 40) и 349 (SEQ ID NO: 42).
[0105] На Фиг. 12A и Фиг. 12B показаны множественные выравнивания последовательностей исходной CD40L-специфичной Tn3-каркасной структуры 311, варианта 311K4E, а также вариантов 311K4E_1 - 311K4E_21 с оптимизированной аффинностью. Аминокислотные остатки 1-44 показаны на Фиг. 12A, а аминокислотные остатки 45-87 показаны на Фиг. 12B. Варианты петель затемнены. Аминокислотные вариации вне затемненных петель помещены в рамку. Консенсусная последовательность представлена ниже множественного выравнивания последовательностей. Выровненные последовательности соответствуют аминокислотным последовательностям клонов 311 Tn3-каркасной структуры (SEQ ID NO: 44), 311K4E (SEQ ID NO: 46), 311K4E_1 (SEQ ID NO: 48), 311K4E_2 (SEQ ID NO: 50), 311K4E_2 (SEQ ID NO: 52), 311K4E_3 (SEQ ID NO: 54), 311K4E_4 (SEQ ID NO: 56), 311K4E_5 (SEQ ID NO: 58), 311K4E_7 (SEQ ID NO: 60), 311K4E_8 (SEQ ID NO: 62), 311K4E_9 (SEQ ID NO: 64), 311K4E_10 (SEQ ID NO: 66), 311K4E_11 (SEQ ID NO: 68), 311K4E_12 (SEQ ID NO: 70), 311K4E_13 (SEQ ID NO: 72), 311K4E_14 (SEQ ID NO: 74), 311K4E_15 (SEQ ID NO: 76), 311K4E_16 (SEQ ID NO: 78), 311K4E_19 (SEQ ID NO: 80), 311K4E_20 (SEQ ID NO: 82) и 311K4E_21 (SEQ ID NO: 84).
[0106] На Фиг. 13 показан анализ ингибирования NfkB человека, в котором используют клетки HEK293, экспрессирующие рецептор CD40 человека и содержащие репортерный конструкт NfkB-люцифераза. Добавление CD40L человека приводит к возникновению сигнала (измеряется по люциферазной активности), который ингибируется молекулой, связывающей CD40L. Анализировали CD40L-специфичные каркасные структуры 340 и 342 из Tn3, а также моноклональное антитело 5c8 к CD40L.
[0107] На Фиг. 14 показано связывание Tn3-каркасных структур, специфичных по отношению CD40L человека, с CD4+ T-клетками человека, активированными антителом к CD3/28 в течение 24 ч. Анализировали моновалентную каркасную структуру 342, слитую с HSA (обозначена 342-HSA) и бивалентную каркасную структуру 342, слитую с HSA (342-342-HSA).
[0108] На Фиг. 15 показано ингибирование первичной T/B-клеточной пролиферации у человека в день 3. Анализировали моновалентную каркасную структуру 340, слитую с HSA (340-HSA), моновалентную каркасную структуру 342, слитую с HSA (342-HSA), а также бивалентную каркасную структуру 342, слитую с HSA (342-342-HSA). Показаны значения IC50 для каждого конструкта.
[0109] На Фиг. 16A показан агрегационный анализ с использованием отмытых тромбоцитов. На графике показан иллюстративный позитивный контроль АDP-индуцированной агрегации для донора (три верхних записанных кривых, соответственно) АDP: 0,5 мкМ, 1 мкМ и 2 мкМ вместе с иммунным комплексом (IC) из моноклонального антитела 5c8 (600 нМ) и растворимого CD40L человека (200 нМ).
[0110] На Фиг. 16B показан агрегационный анализ с использованием отмытых тромбоцитов. На графике показан недостаток агрегации, если использовали преформированные иммунные комплексы из бивалентных каркасных структур 309-309 (не являются слитыми с HSA) и растворимого CD40L человека. Концентрацию CD40L человека (растворимая форма) поддерживали постоянной при 600 нМ, а концентрацию конструктов каркасных структур изменяли от 200 нМ до 800 нМ.
[0111] На Фиг. 16С показан агрегационный анализ с использованием отмытых тромбоцитов. На графике показан недостаток агрегации при использовании преформированных иммунных комплексов из моновалентных каркасных структур 342, слитых с HSA, и растворимого CD40L человека. Концентрацию CD40L человека (растворимая форма) поддерживали постоянной при 600 нМ, а концентрацию конструктов каркасных структур изменяли от 100 нМ до 400 нМ. На графике также показана быстрая агрегация, индуцированная иммунным комплексом из моноклонального антитела Biogen 5c8 и растворимого CD40L человека.
[0112] На Фиг. 17A показано ленточное представление кристаллической структуры растворимого CD40L в комплексе с CD40L-специфичной мономерной Tn3-каркасной структурой 309. CD40L образует тример (полипептиды A, B и C). Каждая каркасная структура 309 (полипептиды D, E и F обведены кругом) осуществляет контакт с двумя полипептидами CD40L. Перечислены специфичные контакты между мономерной каркасной структурой 309 и первым и вторым полипептидами CD40L. Это вид "сверху вниз" данной структуры.
[0113] На Фиг. 17B показано ленточное представление кристаллической структуры растворимого CD40L в комплексе с CD40L-специфичной мономерной Tn3-каркасной структурой 311K4E_12. CD40L образует тример (полипептиды A, B и C). Каждая мономерная каркасная структура 311K4E_12 (полипептиды D, E и F обведены кругом) осуществляет контакт с двумя полипептидами CD40L. Перечислены специфичные контакты между каждой мономерной каркасной структурой 311K4E_12 и первым и вторым полипептидами CD40L. Это вид "сверху вниз" данной структуры.
[0114] На Фиг. 17С показано ленточное представление, которое демонстрирует, что каркасные структуры 311K4E_12 и 309 (обведены кругом) связываются с различными эпитопами, расположенными в различных частях тримерного комплекса CD40L. Обе каркасные структуры связываются в одном и том же кармане для того, чтобы взаимодействовать с рецептором CD40. Это "вид сбоку" данной структуры.
[0115] На Фиг. 17D показано ленточное представление структуры кристалла растворимого CD40L в комплексе с CD40L-специфичной мономерной Tn3-каркасной структурой 342. Показаны лишь один CD40L и одна мономерная каркасная структура 342. Перечислены специфичные контакты между мономерной каркасной структурой 342 и первыми полипептидами CD40L.
[0116] На Фиг. 17E показано ленточное представление, которое демонстрирует, что каркасные структуры 342 и 311K4E_12 могут одновременно связываться с различными эпитопами, расположенными в различных частях тримерного комплекса CD40L. Обе каркасные структуры связываются в одной и той же бороздке, которая будет взаимодействовать с рецептором CD40. Это "вид сбоку" данной структуры.
[0117] На Фиг. 18A показано расположение контактов между аминокислотами в CD40L-специфичной мономерной Tn3-каркасной структуре 311K4E_12 (SEQ ID NO: 68) и тримером, образованным растворимой молекулой CD40L (SEQ ID NO: 2), как показано на Фиг. 17A. Каждая каркасная структура осуществляет контакт с 2 молекулами CD40L. Показана последовательность CD40L (SEQ ID NO: 2). Пунктирная подчеркивающая линия = цитоплазматический домен; сплошная подчеркивающая линия = сигнальный якорный мембранный белок типа II; двойная подчеркивающая линия = область, coвместно кристаллизированная с Tn3-каркасной структурой; затемнение = остатки на 1-м CD40L, которые контактируют с Tn3; ненасыщенное затемнение = остатки на 2-м CD40L, которые контактируют с Tn3.
[0118] На Фиг. 18B показано расположение контактов между аминокислотами CD40L-специфичной мономерной Tn3-каркасной структуры 309 и тримера CD40L, как показано на Фиг. 17B. Каждая каркасная структура осуществляет контакт с 2 молекулами CD40L. Показана последовательность CD40L (SEQ ID NO: 2). Пунктирная подчеркивающая линия = цитоплазматический домен; сплошная подчеркивающая линия = сигнальный якорный мембранный белок типа II; двойная подчеркивающая линия = область, coвместно кристаллизированная с Tn3-каркасной структурой; затемнение = остатки на 1-м CD40L, которые контактируют с Tn3; ненасыщенное затемнение = остатки на 2-м CD40L, которые контактируют с Tn3; остатки в двойной рамке контактируют с петлей FG из каркасной структуры 309, которая, по всей видимости, не является консервативной в клонах, имеющих петлю FG дикого типа.
[0119] Фиг. 19. На панели A показана иллюстративная хроматограмма элюирования Tn3-каркасной структуры (309 или 311K4E_12), CD40L и комплекса между ними в колонке для гель-хроматографии Superdex 200 10/300 GL. На панели B показаны кристаллы комплекса 309-CD40L. Кристаллы демонстрируют увеличение размеров до 0,15 × 0,15 × 0,1 мм. На панели C показаны кристаллы комплекса 311K4E_12-CD40L.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Определения
[0120] Прежде чем продолжить описание настоящего изобретения более подробно, следует понимать, что настоящее изобретение не ограничивается конкретными композициями или стадиями процесса, поскольку таковые могут изменяться. Следует отметить, что употребляемые в данном описании и прилагаемой формуле изобретения формы единственного числа включают определяемые объекты во множественном числе, если из контекста явно не следует иное. Выражения “один или несколько” и “по меньшей мере один” могут употребляться в данном документе взаимозаменяемо.
[0121] Кроме того, выражение “и/или” там, где оно употребляется в данном документе, следует рассматривать как конкретное раскрытие каждого из двух указанных признаков или компонентов с другим или без него. Таким образом, выражение "и/или" при употреблении в данном документе в такой фразе, как "А и/или В", должно включать "А и В", "А или В", "А" (отдельно) и "В" (отдельно). Аналогично, выражение "и/или", при употреблении в такой фразе, как "А, В и/или С" должно охватывать каждый из следующих вариантов осуществления: А, В и С; А, В или С; А или С; А или В; В или С; А и С; А и В; В и С; А (отдельно); В (отдельно) и С (отдельно).
[0122] Если не определено иначе, все технические и научные выражения, используемые в данном документе, имеют то же значение, которое обычно понимает специалист с обычной квалификацией в области техники, к которой относится это раскрытие. Например, the Concise Dictionary of Biomedicine and Molecular Biology, Juo, Pei-Show, 2nd ed., 2002, CRC Press; The Dictionary of Cell and Molecular Biology, 3rd ed., 1999, Academic Press и the Oxford Dictionary Of Biochemistry And Molecular Biology, Revised, 2000, Oxford University Press, обеспечивают специалиста общим словарем многих выражений, используемых в настоящем изобретении.
[0123] Единицы измерения, префиксы и символы обозначены в их форме, принятой в Système International de Unites (SI). Числовые диапазоны включают числа, определяющие диапазон. Если не указано иное, то аминокислотные последовательности записаны слева направо в направлении от амино к карбокси. Приведенные в данном документе заголовки не ограничивают различные аспекты или варианты осуществления настоящего раскрытия, которые могут обеспечиваться ссылкой на описание в целом. Соответственно, выражения, которые приведены непосредственно ниже, более полно определены посредством ссылки на описание во всей его полноте.
[0124] Ясно, что какие бы варианты осуществления ни описывались в данном документе формулировкой "содержащий", другие аналогичные варианты осуществления, описываемые выражениями "состоящий из" и/или "практически состоящий из", также представлены.
[0125] В данном документе аминокислоты указаны либо с помощью их общеизвестных трехбуквенных символов, либо с помощью однобуквенных символов, рекомендованных Комиссией по биохимической номенклатуре IUPAC-IUB. Аналогично, нуклеотиды указаны с помощью их общепринятых однобуквенных кодов.
[0126] Выражение "эпитоп", как используется в данном документе, относится к белковой детерминанте, способной связываться с каркасной структурой по настоящему изобретению. Эпитопы обычно состоят из химически активных поверхностных групп молекул, таких как аминокислоты или боковые цепи сахаров, и обычно обладают специфическими трехмерными структурными характеристиками, а также специфическими характеристиками заряда. Конформационные и неконформационные эпитопы отличаются тем, что связывание с первым, но не с последним теряется в присутствии денатурирующих растворителей.
[0127] Выражения "домен фибронектина типа III (FnIII)," "FnIII-домен" и "FnIII-каркасная структура" относятся к полипептидам, гомологичным домену фибронектина типа III, имеющим по меньшей мере 7 бета-цепей, которые расположены между двумя бета-складчатыми слоями, которые сами по себе упакованы между собой с образованием кора белка, и дополнительно содержащим доступные для растворителя петли, соединяющие бета-цепи друг с другом. Имеется по меньшей мере три таких петли на каждом краю бета-складчатого сэндвича, где край представляет собой границу белка, перпендикулярную направлению бета-цепей. В конкретных вариантах осуществления FnIII-домен содержит 7 бета-цепей, обозначенных A, B, C, D, E, F и G, соединенных с шестью петлевыми участками, обозначенными AB, BC, CD, DE, EF и FG, где петлевой участок соединен с каждой бета-цепью.
[0128] Выражение "Tn3-каркасная структура", как используется в данном документе, относится к молекулам, содержащим по меньшей мере одну FnIII-каркасную структуру, где бета-цепь A содержит SEQ ID NO: 11, бета-цепь B содержит SEQ ID NO: 12, бета-цепь С содержит SEQ ID NO: 13 или 14, бета-цепь D содержит SEQ ID NO: 15, бета-цепь Е содержит SEQ ID NO: 16, бета-цепь F содержит SEQ ID NO: 17, и бета-цепь G содержит SEQ ID NO: 18, где по меньшей мере одна петля представляет собой не встречающийся в природе вариант петель в “исходной Tn3-каркасной структуре.” В конкретных вариантах осуществления одна или несколько бета-цепей Tn3-модуля включают по меньшей мере одну аминокислотную замену, за исключение того, что остатки цистеина в бета-цепи C (например, цистеин в SEQ ID NO: 13 или 14) и бета-цепях F (SEQ ID NO: 17) не заменены.
[0129] Выражение “исходный Tn3”, как используется в данном документе, относится к FnIII-каркасной структуре, содержащей SEQ ID NO: 3, т.е термально стабилизированная и созданная по методике цистеиновой инженерии FnIII-каркасная структура, полученная из 3-го FnIII-домена тенасцина C человека.
[0130] Выражения "мультимер" или "мультимерная каркасная структура" относятся к молекуле, которая содержит по меньшей мере две FnIII-каркасные структуры в комплексе. Каркасные структуры, образующие мультимерную каркасную структуру могут быть соединены посредством линкера, что позволяет каждой каркасной структуре функционировать независимо.
[0131] Выражения "мономер", "мономерная субъединица" или "мономерная каркасная структура" относятся к молекуле, которая содержит только одну FnIII-каркасную структуру.
[0132] Выражение "CD40L-специфичная мономерная субъединица", как используется в данном документе, относится к Tn3-мономеру, происходящему из "исходного Tn3", где Tn3-мономер специфически связывается с CD40L или его фрагментом, например, растворимой формой CD40L.
[0133] Выражение "ДНК" относится к последовательности из двух или более ковалентно связанных, встречающихся в природе или модифицированных дезоксирибонуклеотидов.
[0134] Выражение "белок слияния" относится к белку, который содержит в своем составе (i) одну или несколько каркасных структур по настоящему изобретению, соединенных со (ii) вторым отличным белком (т.е "гетерологичный" белок).
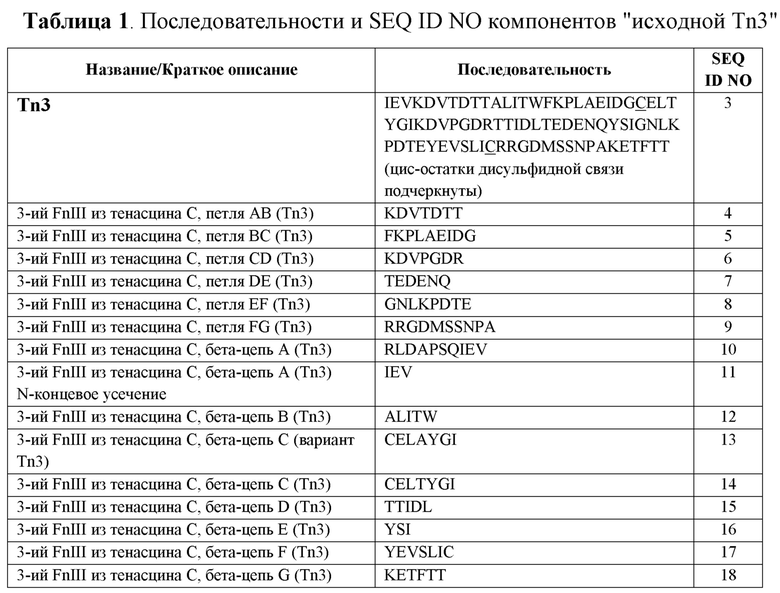
[0135] Выражение "гетерологичный фрагмент" используют в данном документе для указания добавления структуры к Tn3-каркасной структуре по настоящему изобретению, где структура, как правило, не является частью FnIII-домена. Иллюстративные гетерологичные фрагменты включают белки, пептиды, белковые домены, линкеры, лекарственные средства, токсины, радиофармацевтические средства, радиоактивные соединения, органические и неорганические полимеры, а также любые другие структуры, которые могут обеспечивать активность, не свойственную FnIII-домену самому по себе, в том числе, но без ограничения, полиэтиленгликоль (PEG), цитотоксическое средство, радионуклид, радиофармацевтическое средство, биотин, домен димеризации (например, домен "лейциновая застежка"), человеческий сывороточный альбумин (HSA) или его часть, отвечающая за связывание с FcRn, домен или фрагмент антитела (например, вариабельный домен антитела, CH1-домен, Cкаппа-домен, Cлямбда-домен, CH2- или CH3-домен), одноцепочечное антитело, антитело на основе домена, альбумин-связывающий домен, молекула IgG, фермент, лиганд, рецептор, связывающий пептид, каркасная структура, отличающаяся от FnIII, эпитопная метка, полимер на основе рекомбинантного полипептида, цитокин и тому подобные.
[0136] Выражение "линкер", как используется в данном документе, относится к любому молекулярному ансамблю, который объединяет или соединяет две или более каркасные структуры. Линкер может быть молекулой, которая функционирует в качестве "спейсера" между модулями в каркасной структуре, или также может быть молекулой с дополнительной функцией (т.е "функциональный фрагмент"). Молекула, включенная в определение "гетерологичного фрагмента", также может функционировать в качестве линкера.
[0137] Выражения "соединенный" и "слитый" используют взаимозаменямо. Данные выражения относятся к объединению двух или более каркасных структур, гетерологичных фрагментов или линкеров с помощью любых средств, включающих химическое конъюгирование или рекомбинантные способы.
[0138] Выражения "домен" или "белковый домен" относятся к участку белка, который могут укладываться в стабильную трехмерную структуру, часто независимо от остальной части белка, и которые могут быть наделены конкретной функцией. Данная структура обеспечивает определенную функцию, ассоциированную с функцией домена в пределах первоначального белка, например, ферментативная активность, образование узнающего мотива для другой молекулы или обеспечение необходимыми структурными компонентами белка для его существования в конкретной среде белков. В пределах как семейства белков, так и суперсемейств родственных белков белковые домены могут представлять собой эволюционно консервативные области. При описании компонентов мультимерной каркасной структуры выражения "домен", "мономерная каркасная структура", "мономерная субъединица" и "модуль" можно использовать взаимозаменяемо. Под “нативным FnIII-доменом” подразумевают любой нерекомбинантный FnIII-домен, который закодирован в живом организме.
[0139] "Последовательность белка" или "аминокислотная последовательность" означает линейное представление аминокислотных составляющих элементов в полипептиде в направлении от амино-конца до карбокси-конца, в котором остатки, которые граничат друг с другом в представлении являются смежными в первичной структуре полипептида.
[0140] Выражение "нуклеиновая кислота" относится к любым двум или более ковалентно связанным нуклеотидам, или аналогам нуклеотидов, или производным. Как используется в данном документе, данное выражение без ограничения включает ДНК, РНК и ПНК. “Нуклеиновая кислота” и “полинуклеотид” используют в данном документе взаимозаменяемо.
[0141] Выражение "полинуклеотид" предназначено для охвата единичной нуклеиновой кислоты, а также множественных нуклеиновых кислот, и относится к выделенной молекуле нуклеиновой кислоты или конструкту, например, информационной РНК (иРНК) или плазмидной ДНК (пДНК). Выражение "выделенная" нуклеиновая кислота или полинуклеотид относится к молекуле нуклеиновой кислоты, ДНК или РНК, которую удалили из ее нативного окружения. Например, рекомбинатный полинуклеотид, кодирующий, к примеру, каркасную структуру по настоящему изобретению, входящий в состав вектора, считают выделенным для целей настоящего изобретения. Дополнительные примеры выделенного полинуклеотида включают рекомбинантные полинуклеотиды, содержащиеся в гетерологичных клетках-хозяевах или очищенные (частично или в значительной степени) полинуклеотиды в растворе. Выделенные молекулы РНК включают in vivo или in vitro РНК-транскрипты полинуклеотидов по настоящему изобретению. Выделенные полинуклеотиды или нуклеиновые кислоты в соответствии с настоящим изобретением дополнительно включают такие молекулы, полученные синтетически. Кроме того, полинуклеотид или нуклеиновая кислота могут представлять собой или иметь в своем составе регуляторный элемент, такой как промотор, сайт связывания рибосом или терминатор транскрипции.
[0142] Выражение "фармацевтически приемлемый" относится к соединению или белку, которое можно вводить животному (например, млекопитающему) без существенных нежелательных медицинских последствий.
[0143] Выражение "физиологически приемлемый носитель" относится к носителю, который не характеризуется существенным вредным воздействием на хозяина, подлежащего обработке, и который сохраняет терапевтические свойства соединения, с которым его вводят. Одним иллюстративным физиологически приемлемым носителем является физиологический солевой раствор. Другие физиологически приемлемые носители и их составы, известные специалисту в данной области техники, и описанные, например, в Remington's Pharmaceutical Sciences, (18th edition), ed. A. Gennaro, 1990, Mack Publishing Company, Easton, Pa., включены в данный документ посредством ссылки.
[0144] Под "полипептидом" подразумевают любую последовательность из двух или более аминокислот, линейно связанных амидными связями (пептидными связями), вне зависимости от длины, посттрансляционной модификации или функции. "Полипептид", "пептид" и "белок" используют в данном документе взаимозаменяемо. Таким образом, пептиды, дипептиды, трипептиды или олигопептиды включены в определение "полипептида", и выражение "полипептид" можно использовать вместо или взаимозаменяемо с любым из данных выражений. Выражение "полипептид" также предназначено для обозначения продуктов пост-экспрессионных модификаций полипептида, включая, но без ограничения, гликозилирование, ацетилирование, фосфорилирование, амидирование, получение производных с помощью известных защитных/блокирующих групп, протеолитическое расщепление или модификацию с помощью не встречающихся в природе аминокислот. Полипептид может быть получен из природного биологического источника или создан с помощью рекомбинантной технологии, но не обязательно осуществляется трансляция из определенной последовательности нуклеиновой кислоты. Полипептид может быть получен любым способом, в том числе химическим синтезом.
[0145] Также в качестве полипептидов по настоящему изобретению предусмотрены фрагменты, производные, аналоги или варианты вышеприведенных полипептидов и любые их комбинации. Варианты могут быть встречающимися в природе или не встречающимися в природе. Не встречающиеся в природе варианты можно получить при помощи методик мутагенеза, известных в данной области техники. Варианты полипептидов могут включать консервативные или неконсервативные аминокислотные замены, делеции или добавления. Также в качестве "производных" включены те пептиды, которые содержат одну или несколько встречающихся в природе аминокислотных производных двадцати стандартных аминокислот.
[0146] Под "рандомизированной" или "мутированной" подразумевают включение одной или нескольких аминокислотных изменений, в том числе делеции, замены или добавления, относительно матричной последовательности. Под "рандомизированием" или "внедрением мутации" подразумевают процесс введения в последовательность такого аминокислотного изменения. Рандомизация или мутация может быть достигнута посредством преднамеренного, слепого или спонтанного изменения последовательности, обычно всей кодирующей последовательности нуклеиновой кислоты, и ее можно осуществить с помощью любой методики, например, ПЦР, ПЦР с использованием ошибающейся полимеразы или химического синтеза ДНК. Выражения “рандомизирование”, “рандомизированный”, “внедрение мутации”, “мутированный” и тому подобные используют взаимозаменяемо в данном документе.
[0147] Под “когнатным” или "когнатным немутированным белком" подразумевают белок, который является идентичным по последовательности варианту белка, за исключением аминокислотных мутаций, введенных в вариант белка, где вариант белка является рандомизированным или мутированным.
[0148] Под выражением "РНК" подразумевают последовательность из двух или более ковалентно связанных, встречающихся в природе или модифицированных рибонуклеотидов. Один пример модифицированной РНК, включенный в рамки данного выражения, представляет собой фосфоротиоатную РНК.
[0149] Выражения "каркасная структура по настоящему изобретению" или "каркасные структуры по настоящему изобретению", как используется в данном документе, относятся к мультимерным Tn3-каркасным структурам, а также мономерным Tn3-каркасным структурам. Выражение “мишень” относится к соединению, узнаваемому специфической каркасной структурой по настоящему изобретению. Выражения “мишень” и “антиген” используются взаимозаменяемо в данном документе. Выражение "специфичность", как используется в данном документе, например, в выражениях "специфически связывается" или "специфическое связывание", относится к относительной аффинности, при помощи которой Tn3-каркасная структура по настоящему изобретению связывается с одним или несколькими антигенами посредством одного или нескольких антиген-связывающих доменов, и тому, что связывание предусматривает некоторую комплементарность между одним или несколькими антиген-связывающими доменами и одним или несколькими антигенами. Согласно данному определению говорят, что Tn3-каркасная структура по настоящему изобретению "специфически связывается" с эпитопом, когда она связывается с данным эпитопом более легко, чем если бы она связывалась со случайным, неродственным эпитопом.
[0150] Каркасная структура со “зрелой аффинностью” представляет собой каркасную структуру с одним или несколькими изменениями, обычно в петле, результатом чего является улучшение аффинности Tn3-каркасной структуры по отношению к эпитопу в сравнении с исходной Tn3-каркасной структурой, которая не имеет такого изменения(ий).
[0151] Выражение "аффинность", как используется в данном документе, относится к измерению силы связывания конкретной Tn3-каркасной структуры по настоящему изобретению с отдельным эпитопом.
[0152] Выражение "авидность", как используется в данном документе, относится к общей стабильности комплекса между популяцией Tn3-каркасных структур по настоящему изобретению и определенным эпитопом, т.е функционально объединенная сила связывания множества Tn3-каркасных структур с антигеном. Авидность относится как к аффинности отдельных антиген-связывающих доменов в отношении конкретных эпитопов, так и также к валентности каркасной структуры по настоящему изобретению.
[0153] Выражение "воздействие на мишень" относится к связыванию Tn3-каркасной структуры по настоящему изобретению с одной или несколькими мишенями и к биологическим эффектам, являющимся результатом такого связывания. В этом отношении, множественные единицы, связывающие антиген, в Tn3-каркасной структуре могут взаимодействовать с рядом мишеней и/или эпитопов и, например, физически сближать две мишени, инициировать каскады метаболических реакций посредством взаимодействия с отдельными мишенями и т.д. В отношении CD40L, "воздействие на мишень" относится к эффекту, достигнутому, например, путем усиления, стимуляции или активации одной или нескольких биологических активностей CD40L.
[0154] Выражение "валентность", как используется в данном документе, относится к числу потенциальных антиген-связывающих модулей, например, числу FnIII-модулей в каркасной структуре по настоящему изобретению. Если Tn3-каркасная структура по настоящему изобретению содержит более одного антиген-связывающего модуля, каждый связывающий модуль может специфически связываться с тем же эпитопом или другим эпитопом в той же мишени или других мишенях.
[0155] Выражение "дисульфидная связь", как используется в данном документе, включает ковалентную связь, образованную между двумя атомами серы. Аминокислота цистеин содержит тиоловую группу, которая может образовывать дисульфидную связь или мостик со второй тиоловой группой.
[0156] Выражение “иммуноглобулин” и "антитело" включает различные широкие классы полипептидов, которые могут биохимически отличаться. Специалисты в данной области примут во внимание, что тяжелые цепи классифицируют как гамма, мю, альфа, дельта или эпсилон. Именно природа этой цепи определяет "класс" антитела как IgG, IgM, IgA, IgG или IgE, соответственно. Модифицированные версии каждого из данных классов без труда различит специалист в данной области техники. Как используется в данном документе, выражение "антитело" включает, но без ограничения, интактное антитело, модифицированное aнтитело, VL- или VL-домен, CHl-домен, Cкаппа-домен, Cлямбда-домен, Fc-домен (см. ниже), CH2- или CH3-домен.
[0157] Как используется в данном документе, выражение "Fc-домен" относится к части константной области антитела. Традиционно, выражение Fc-домен относится к продукту расщепления протеазой (например, папаином), охватывающему парные CH2, CH3 и шарнирную области антитела. В контексте данного раскрытия выражение Fc-домен или Fc относится к любому полипептиду (или нуклеиновой кислоте, кодирующей такой полипептид), независимо от способа получения, который охватывает все или часть CH2, CH3 и шарнирной областей полипептида в составе иммуноглобулина.
[0158] Как используется в данном документе, выражение "модифицированное антитело" включает синтетические формы антитела, измененные таким образом, что они не являются встречающимися в природе, например, антитела, которые содержат по меньшей мере части двух тяжелых цепей, но не две полные тяжелые цепи (как, например, антитела с удаленным доменом или минитела); мультиспецифические формы антител (например, биспецифические, триспецифические и т.д.), измененные так, чтобы связываться с двумя или более антигенами или с различными эпитопами отдельного антигена. Кроме того, выражение "модифицированное антитело" включает поливалентные формы антител (например, трехвалентные, четырехвалентные и т.д антитела к трем или более копиям одного и того же антигена). (См., например, Antibody Engineering, Kontermann & Dubel, eds., 2010, Springer Protocols, Springer).
[0159] Выражение “время полужизни in vivo” используют в его нормальном значении, т.е время, при котором 50% биологической активности полипептида все еще обнаруживают в теле/органе-мишени, или время, при котором активность полипептида представляет собой 50% от его исходного значения. В качестве альтернативы, к определению функционального времени полужизни in vivo, можно определить "время полужизни в сыворотке", т.е время, при котором 50% полипептидных молекул циркулируют в плазме или кровотоке до очищения. Определение времени полужизни в сыворотке зачастую является более простым, чем определение функционального времени полужизни in vivo, и показатель времени полужизни в сыворотке обычно является хорошим признаком показателя функционального времени полужизни. Альтернативные выражения для времени полужизни в сыворотке включают "время полужизни в плазме," время полувыведения из кровотока, время полувыведения из крови, сывороточный клиренс, плазматический клиренс и время полувыведения. Функциональность, подлежащую сохранению, обычно выбирают из прокоагулянтной, протеолитической, кофактор-связывающей, рецептор-связывающей активности или другого типа биологической активности, связанной с конкретным белком.
[0160] Выражение "повышено" в отношении функционального времени полужизни in vivo или времени полужизни в плазме используют для указания того, что относительное время полужизни полипептида представляет собой статистически значимое повышение относительно такового для контрольной молекулы (например, немодифицированный полипептид), как это определено в сопоставимых условиях.
[0161] Выражение "понижено" в отношении функционального времени полужизни in vivo или времени полужизни в плазме используют для указания того, что относительное время полужизни полипептида представляет собой статистически значимое понижение относительно такового для контрольной молекулы (например, немодифицированный полипептид), как это определено в сопоставимых условиях.
[0162] Выражение "экспрессия", как используется в данном документе, относится к способу, с помощью которого исходя из гена продуцируется биохимическое вещество, например, каркасная структура по настоящему изобретению или ее фрагмент. Способ предусматривает любое проявление функционального присутствия гена в клетке, включая, без ограничения, нокдаун гена, а также как транзиентную экспрессию, так и стабильную экспрессию. Он включает, без ограничений, транскрипцию гена в одну или несколько иРНК и трансляцию таких иРНК в один или несколько полипептидов. Если конечный желаемый продукт представляет собой биохимическое вещество, экспрессия включает создание таких биохимических веществ и любых предшественников.
[0163] “Продукт экспрессии” может представлять собой либо нуклеиновую кислоту, например, информационную РНК, полученную посредством транскрипции гена, либо полипептид. Продукты экспрессии, описываемые в данном документе, дополнительно включают нуклеиновые кислоты с посттранскрипционными модификациями, например, полиаденилированием, или полипептиды с посттрансляционными модификациями, например, метилированием, гликозилированием, добавлением липидов, ассоциацией с другими белковыми субъединицами, протеолитическим расщеплением и т.п.
[0164] Выражение "вектор" или "вектор экспрессии" применяют в данном документе для обозначения векторов, применяемых в соответствии с настоящим изобретением в качестве носителей для введения в клетку и экспрессии желаемого гена в клетке-хозяине. Как известно специалистам в данной области техники, такие векторы можно без труда выбрать из группы, состоящей из плазмид, фагов, вирусов и ретровирусов. Как правило, векторы, совместимые с настоящим изобретением, как правило, будут содержать селектируемый маркер, соответствующие сайты рестрикции для облегчения клонирования желаемого гена и обладать способностью проникать в эукариотические или прокариотические клетки и/или реплицироваться в них.
[0165] Выражение "клетки-хозяева" относится к клеткам, которые содержат векторы, сконструированные с помощью методик рекомбинантных ДНК и кодирующие по меньшей мере один гетерологичный ген. В описаниях способов выделения продукта экспрессии из рекомбинантных хозяев выражения "клетка" и "культура клеток" используют взаимозаменяемо для указания источника продукта экспрессии, если четко не указано иное, т.е извлечение продукта экспрессии из "клеток" означает либо извлечение из отцентрифугированных цельных клеток, либо извлечение из культуры клеток, содержащих как среду, так и суспендированные клетки.
[0166] Выражения "лечить" или "лечение", как используются в данном документе, относятся как к терапевтическому лечению, так и к профилактическим или превентивным мерам, где целью является предупреждение или замедление (уменьшение) нежелательного физиологического изменения или нарушения, такого как прогрессирование воспалительного заболевания или состояния. Благоприятные или желаемые клинические результаты включают, но без ограничения, облегчение симптомов, уменьшение степени заболевания, стабилизированное (т.е. не ухудшающееся) состояние заболевания, задержку или замедление прогрессирования заболевания, улучшение или временное облегчение болезненного состояния и ремиссию (либо частичную, либо полную), либо поддающуюся, либо не поддающуюся обнаружению.
[0167] Выражение "лечение" может также означать увеличение выживаемости по сравнению с ожидаемой выживаемостью в случае без получения лечения. К тем, кто нуждается в лечении, относятся те, у кого уже есть состояние или нарушение, а также те, кто склонен иметь состояние или нарушение, или те, у кого нужно предотвратить состояние или нарушение.
[0168] Выражения "субъект", "индивидуум" "животное", "пациент" или "млекопитающее" относятся к любому индивидууму, пациенту или животному, в частности, субъекту-млекопитающему, для которого необходимы диагностирование, прогнозирование или терапия. Субъекты-млекопитающие включают людей, домашних животных, сельскохозяйственных животных и зоопарковых, используемых в спорте или комнатных животных, таких как собаки, кошки, морские свинки, кролики, крысы, мыши, лошади, крупный рогатый скот, коровы и т.д.
[0169] Выражение “CD40L”, как используется в данном документе, относится без ограничений к CD40L, экспрессированному на поверхности T-клеток, рекомбинантно экспрессированному CD40L, CD40L, экспрессированному в E.coli и очищенному из них или других пригодных систем экспрессии рекомбинантных белков, негликозилированному CD40L и растворимым фрагментам CD40L. Как используется в данном документе, "CD40L" также относится к MegaCD40L. MegaCD40L™ представляет собой высокоактивный конструкт, в котором два тримерных CD40-лиганда искусственно соединены через коллагеновый домен домен из ACRP30/адипонектина. Данный конструкт очень эффективно стимулирует естественную мембранносвязанную агрегацию CD40L in vivo. Он обеспечивает простую и в равной степени эффективную альтернативу комбинациям [CD40L+энхансер] (Alexis biochemicals). Выражение "CD40L" относится к мономерным формам CD40L, а также к олигомерным формам, например, тримерный CD40L.
[0170] Выражение “CD40L” относится как к полноразмерному CD40L, так и к растворимым фрагментам, например, формам внеклеточного домена CD40L, возникающим в результате протеолиза. Аминокислотные последовательности мембранносвязанных и растворимых форм CD40L человека (Swissprot: P29965) показаны в SEQ ID NO: 1 и SEQ ID NO: 2, соответственно.
[0171] Выражения “CD40L-антагонист” или “антагонист” используют в самом широком смысле, и они охватывают любую молекулу, которая частично или полностью ингибирует, снижает или инактивирует одну или несколько биологических активностей CD40L и его биологически активных вариантов in vitro, in situ или in vivo. К примеру, CD40L-антагонист может функционировать для того, чтобы частично или полностью ингибировать, снижать или инактивировать одну или несколько биологических активностей одной или нескольких молекул CD40L или одной или нескольких молекул CD40L, связанных с CD40 или другими мишенями, in vivo, in vitro или in situ, как результат его связывания с CD40L.
[0172] Выражение “CD40L-агонист” или “агонист” используют в самом широком смысле, и оно охватывает любую молекулу, которая частично или полностью усиливает, стимулирует или активирует одну или несколько биологических активностей CD40L и его биологически активных вариантов in vitro, in situ или in vivo. К примеру, CD40L-агонист может функционировать для того, чтобы частично или полностью усиливать, стимулировать или активировать одну или несколько биологических активностей одной или нескольких молекул CD40L или одной или нескольких молекул CD40L, связанных с CD40R или другими мишенями, in vivo, in vitro или in situ, как результат его связывания с CD40L.
[0173] Выражение “кристалл”, как используется в данном документе, относится к одной форме твердого состояния вещества, в которой атомы упорядочены в структуру, которая периодически повторяется в трех координатах, как правило, с образованием кристаллической решетки.
[0174] Выражение “симметрия пространственной структуры”, как используется в данном документе, относится к полной симметрии кристалла, которая объединяет симметрию переноса кристаллической решетки с симметрией точечной группы. “Пространственная группа” обозначается заглавной буквой, идентифицирующей группу кристаллической решетки (P, A, F и т.д), за которой следует символ точечной группы, в котором элементы вращения и отражения расширены для включения осей винтовой симметрии и плоскостей скользящего отражения. Заметьте, что точечная группа симметрии для данной пространственной группы может быть определена удалением символа центрировки ячейки пространственной группы и замещением всех осей винтовой симметрии осями сходной симметрии и замещением всех плоскостей скользящего отражения плоскостями зеркального отражения. Точечная группа симметрии для пространственной группы описывает истинную симметрию ее обратной кристаллической решетки.
[0175] Выражение “базисная ячейка”, как используется в данном документе, означает атомы в кристалле, которые упорядочены в кристалле в регулярно повторяющуюся структуру, в которой наименьшая повторяющаяся единица называется базисной ячейкой. Целостную структуру можно воссоздать из данных о базисной ячейке, которая характеризуется тремя размерностями (a, b и c) и тремя углами (α, β, и γ). Величины a и b представляют собой размерности сторон основания ячейки, а γ представляет собой угол между двумя этими сторонами. Величина с представляет собой высоту базисной ячейки. Углы α и β описывают углы между основанием и вертикальными сторонами базисной ячейки.
[0176] Выражение “машиночитаемый накопитель данных”, как используется в данном документе, означает материал для хранения информации, закодированный машиночитаемыми командами, где устройство программируют с использованием команд для использования таких данных и оно способно отображать данные в необходимом формате, например, графическом трехмерном представлении молекул или молекулярных комплексов.
[0177] Выражение “дифракционная рентгенограмма” означает рентгенограмму, полученную в результате рассеяния рентгеновского излучения периодическим ансамблем молекул или атомов в кристалле. Рентгеноструктурная кристаллография представляет собой методику, в которой используется тот факт, что рентгеновское излучение рассеивается кристаллами. Рентгеновское излучение характеризуется определенной длиной волны (в диапазоне в ангстремах, приблизительно 10-8 см), подлежащей рассеиванию облаком электронов атома сопоставимого размера. Исходя из дифракционной рентгенограммы, полученной в результате рассеяния рентгеновского излучения периодическим ансамблем молекул или атомов в кристалле можно воссоздать распределение электронной плотности. Дополнительную фазовую информацию можно получить либо из данных дифракции, либо из дополняющих экспериментов по дифракции для завершения воссоздания (фазовая проблема в кристаллографии). Модель прогрессивно встроена в экспериментальную плотность электронов, уточненную относительно данных для получения точной молекулярной структуры. Рентгеновская структура координат определяет уникальную конфигурацию точек в пространстве. Специалисты в данной области поймут, что совокупность структурных координат для белка или комплекса белок-лиганд или его части определяет относительную совокупность точек, что в свою очередь определяет конфигурацию в трех измерениях. Сходную или идентичную конфигурацию можно определить с помощью совершенно различного набора координат, при условии, что расстояния и углы между координатами остаются практически одинаковыми. Кроме того, конфигурацию точек можно определить путем увеличения или уменьшения расстояний между координатами посредством скалярного множителя с сохранением при этом практически таких же углов.
[0178] Выражение “кристаллическая структура”, как используется в данном документе, относится к пространственной или кристаллической сборке повторяющихся атомных или молекулярных единиц в кристаллическом материале. Кристаллическую структуру кристаллического материала можно определить с помощью способов рентгеноструктурной кристаллографии, см., например, “Principles of Protein X-Ray Crystallography” by Jan Drenth, Springer Advanced Texts in Chemistry, Springer Verlag, 2nd ed., February 199, ISBN: 0387985875, и “Introduction to Macromolecular Crystallography” by Alexander McPherson, Wiley-Liss, Oct. 18, 2002, ISBN: 0471251224.
[0179] Выражение “эффекторная функция” относится к таковым биологическим активностям антитела или фрагмента антитела, присущим Fc-участку (нативный Fc-участок или вариант аминокислотной последовательности Fc-участка) антитела, и которые изменяются в зависимости от изотипа антитела. Примеры эффекторных функций антител включают связывание C1q и комплементзависимая цитотоксичность; связывание Fc-рецептора; антителозависимая клеточноопосредованная цитотоксичность (ADCC); фагоцитоз; понижающая регуляция клеточных поверхностных рецепторов (например, рецепторов B-клеток) и активация B-клеток.
[0180] Выражение “антителозависимая клеточноопосредованная цитотоксичность” или “ADCC” относится к форме цитотоксичности, при которой секретируемый Ig связывается с Fc-рецепторами (FcR), представленными на определенных цитотоксических клетках (например, естественные клетки-киллеры (NK), нейтрофилы и макрофаги), что дает возможность данным цитотоксическим эффекторным клеткам специфически связываться с антиген-несущей клеткой-мишенью и в дальнейшем уничтожать клетки-мишени с помощью цитотоксинов.
[0181] Выражение “Fc-рецептор” или “FcR” описывает рецептор, который связывается с Fc-участком антитела. FcR может быть нативной последовательностью FcR человека. FcR может связывать IgG-антитело (гамма-рецептор) и включает рецепторы подклассов FcγRI, FcγRII и FcγRIII, в том числе аллельные варианты и подвергнутые альтернативному сплайсингу формы данных рецепторов. Выражение также включает неонатальный рецептор FcRn.
[0182] Выражение “консенсусная последовательность” относится к белковой последовательности, которая демонстрирует наиболее часто встречающиеся аминокислоты в конкретном положении после множественного выравнивания последовательностей. Консенсусная последовательность представляет собой способ представления результатов множественного выравнивания последовательностей при сравнении друг с другом родственных последовательностей. Консенсусная последовательность показывает, какие остатки являются наиболее распространенными при выравнивании в каждом положении, и степень вариабельности в каждом положении.
Введение
[0183] CD40L (также известный как CD154, CD40-лиганд, gp39 или TBAM) представляет собой 33 кДа мембранный гликопротеин типа II (Swiss-ProtAcc-No P29965). Дополнительно, существуют более короткие 18 кДа растворимые формы CD40L (также известны как sCD40L или растворимый CD40L). Эти растворимые формы CD40L получают с помощью протеолитического процессинга мембраносвязанного белка, но клеточная активность растворимой разновидности является слабой при отсутствии олигомеризации более высокого порядка (например, тримеризации).
[0184] Настоящее изобретение предлагает семейство рекомбинантных, не встречающихся в природе белковых каркасных структур (Tn3-каркасные структуры), способных связываться с CD40L. В частности, белки, описанные в данном документе, можно использовать для выявления определенных петель, которые являются аналогичными по отношению к участкам, определяющим комплементарность ("CDR") вариабельного участка антител. Эти петли могут подвергаться рандомизации или перегруппровке с ограничениями для получения разнообразия, обеспечивающего способность к связыванию с множеством соединений-мишеней. Tn3-каркасные структуры можно использовать в виде мономеров или составить в мультимерные каркасные структуры, способные к связыванию с CD40L.
[0185] В конкретных вариантах осуществления настоящее изобретение предлагает средства для CD40L-специфичного связывания, которые пригодны для предупреждения, уменьшения интенсивности, выявления, диагностики или контроля заболеваний, как, например, но без ограничения, аутоиммунное заболевание. В других конкретных вариантах осуществления CD40L-специфичные Tn3-каркасные структуры по настоящему изобретению пригодны для лечения аутоиммунных заболеваний и состояний. В некоторых вариантах осуществления, аутоиммунные заболевания могут включать, но без ограничения, системную красную волчанку (SLE), ревматоидный артрит (RA), рассеянный склероз (MS), воспалительное заболевание кишечника (IBD) и отторжение аллотрансплантата.
[0186] Tn3-каркасные структуры по настоящему изобретению содержат CD40L-специфичные мономерные субъединицы, происходящие из третьего FnIII-домена тенасцина C человека, в которых сконструировали по меньшей мере одну не встречающуюся в природе внутримолекулярную дисульфидную связь. Мономерные субъединицы, которые составляют Tn3-каркасные структуры по настоящему изобретению, точно уложены независимо друг от друга с сохранением их специфичности связывания и аффинности, и каждая из мономерных каркасных структур сохраняет свои функциональные свойства. Если мономерные субъединицы собраны в мультимерные Tn3-каркасные структуры с высокой валентностью, мономерные субъединицы точно уложены независимо друг от друга с сохранением их специфичности связывания и аффинности, и каждый из мономеров сохраняет свои функциональные свойства.
[0187] Tn3-каркасные структуры по настоящему изобретению, содержащие более одной мономерной субъединицы, могут связываться со множеством эпитопов, например, (i) связываться со множеством эпитопов в одной мишени, (ii) связываться с одним эпитопом во множестве мишеней, (iii) связываться со множеством эпитопов, расположенных на различных субъединицах одной мишени, или (iv) связываться со множеством эпитопов на множестве мишеней с повышением, таким образом, авидности.
[0188] Кроме того, в связи с возможностью варьировать расстояние между множеством мономеров посредством линкеров мультимерные Tn3-каркасные структуры являются способными к связыванию со множеством молекул-мишеней на поверхности (или на одной и той же клетке/поверхности, или на различных-клетках/поверхностях). Вследствие их способности связываться одновременно с более, чем одной мишенью, мультимерную Tn3-каркасную структуру по настоящему изобретению можно использовать для модуляции множества каскадов реакций, перекрестного связывания с рецепторами на поверхности клетки, связывания поверхностных клеточных рецепторов на отдельных клетках и/или связывания молекул-мишеней или клеток с субстратом.
[0189] Дополнительно, настоящее изобретение предлагает каркасные структуры со зрелой аффинностью, где аффинность каркасной структуры для конкретной мишени модулируют посредством мутации. Также, настоящее изобретение предлагает способы получения каркасных структур по настоящему изобретению, а также способы конструирования каркасных структур с желаемыми физико-химическими, фармакологическими или иммунологическими свойствами. Более того, настоящее изобретение предлагает применения таких каркасных структур и способы терапевтического, профилактического и диагностического применения.
Структурный мотив FnIII
[0190] Tn3-каркасные структуры по настоящему изобретению основаны на структуре модуля фибронектина (FnIII) типа III, домена, широко распространенного среди всех трех надцарств живых организмов и вирусов и во множестве классов белков. В конкретных вариантах осуществления каркасные структуры по настоящему изобретению происходят из третьего FnIII-домена тенасцина C человека (см. международную заявку №, международную заявку № PCT/US2008/012398, опубликованную в качестве WO 2009/058379; PCT/US2011/032184, опубликованную в качестве WO 2011/130324; и международную заявку № PCT/US2011/032188, опубликованную в качестве WO2011130328).
[0191] В одном конкретном варианте осуществления Tn3-каркасные структуры по настоящему изобретению содержат CD40L-специфичную мономерную субъединицу, происходящую из исходной Tn3-каркасной структуры. Общая трехмерная укладка мономера тесно связана с таковой у наименьшего функционального фрагмента антитела, вариабельного участка тяжелой цепи (VH), который в однодоменных антителах, полученных от верблюдов и верблюдовых (например, лам), содержит целую антиген-распознающую единицу.
[0192] Tn3-мономерные субъединицы по настоящему изобретению и нативный FnIII-домен из тенасцина C характеризуются одинаковой третичной структурой, называемой структура "бета-сэндвич" с тремя бета-цепями (A, B, и E) с одной стороны и четырьмя бета-цепями (C, D, F и G) с другой стороны, соединенными шестью петлевыми участками. Эти петлевые участки обозначены в соответствии с бета-цепями, соединенными с N- и C-концами каждой петли. Соответственно, петля AB расположена между бета-цепями A и B, петля BC расположена между бета-цепями B и C, петля CD расположена между бета-цепями C и D, петля DE расположена между бета-цепями D и E, петля EF расположена между бета-цепями E и F, и петля FG расположена между бета-цепями F и G. FnIII-домены содержат открытую для растворителя петлю, толерантную к рандомизации, что облегчает образование различных пулов белковых каркасных структур, способных связываться со специфическими мишенями с высокой аффинностью.
[0193] В одном аспекте настоящего изобретения Tn3-мономерные субъединицы, подвергнутые прямой перегруппировке, конструируют для рандомизации одной или нескольких петель, которые аналогичны участкам, определяющим комплементарность (CDR), в вариабельном участке антитела. Такой принцип прямой перегруппировки в результате приводит к получению антителоподобных молекул с высокими аффинностями к мишеням, представляющим интерес, например, CD40L.
[0194] Кроме того, в некоторых вариантах осуществления Tn3-каркасные структуры, описанные в данном документе, можно использовать для выявления открытых петель (например, предварительно рандомизированные петли и выбранные на основе мишени для связывания) для прямой перегруппировки молекул, которые связываются с такими введенными петлями. Этот тип отбора можно осуществлять для идентификации молекул для узнавания применительно к любой отдельной CDR-подобной петле или, в качестве альтернативы, для узнавания двух или трех CDR-подобных петель, объединенных в нелинейном эпитоп-связывающем фрагменте. Группа из трех петель (обозначенных BC, DE и FG), которые могут обеспечивать мишень-специфическое связывание, уложена между цепями B и C; цепями D и E и бета-цепями F и G, соответственно. Петли BC, DE и FG третьего FnIII-домена тенасцина C человека имеют длину 9, 6 и 10 аминокислотных остатков, соответственно. Длина этих петель находится в узком пределе когнатных антиген-распознающих петель, обнаруженных в тяжелых цепях антител, и составляет 7-10, 4-8 и 4-28 аминокислот в длину, соответственно. Подобным образом, вторая группа петель, петли AB, CD и EF (7, 7 и 8 аминокислот в длину, соответственно) уложены между бета-цепями A и B; бета-цепями C и D и бета-цепями E и F, соответственно.
[0195] После того, как рандомизированы и отобраны по высокой аффинности связывания с мишенью, петли в Tn3-мономерной каркасной структуре могут образовывать контакты с мишенями, эквивалентные контактам когнатных CDR-петель у антител. Соответственно, в некоторых вариантах осуществления петли AB, CD и EF рандомизированы и отобраны по высокой аффинности связывания с одной или несколькими мишенями, например, CD40L. В некоторых вариантах осуществления данный способ рандомизации и отбора можно осуществлять параллельно с рандомизацией петель BC, DE и FG, при этом в других вариантах осуществления данный способ рандомизации и отбора проводят последовательно.
CD40L-специфичные мономерные субъединицы
[0196] Настоящее изобретение предлагает CD40L-специфичные рекомбинантные, не встречающиеся в природе Tn3-каркасные структуры, содержащие множество доменов с бета-цепями, связанными со множеством петлевых участков, где в один или несколько указанных петлевых участков вносят изменения путем делеции, замены или добавления по меньшей мере одной аминокислоты из когнатных петель в Tn3 дикого типа (SEQ ID NO: 3) (см. Таблицу 1).
[0197] Для получения улучшенных CD40L-специфичных Tn3-мономерных субъединиц с новыми характеристиками связывания исходный Tn3 подвергают добавлениям, делециям или заменам аминокислот. Будет понятно, что при сравнении последовательности CD40L-специфичной Tn3-мономерной субъединицы с последовательностью исходной Tn3 используют одинаковое определение бета-цепей и петель. В некоторых вариантах осуществления CD40L-специфичные Tn3-мономерные субъединицы по настоящему изобретению содержат аминокислотную последовательность:
IEV(XAB)nALITW(XBC)nCELX1YGI(XCD)nTTIDL(XDE)nYSI(XEF)nYEVSLIC(XFG)nKETFTT,
где
(a) XAB, XBC, XCD, XDE, XEF, и XFG представляют собой аминокислотные остатки, присутствующие в последовательностях петель AB, BC, CD, DE, EF и FG, соответственно;
(b) X1 представляет собой аминокислотный остаток аланин (A) или треонин (T); и
(c) длина петли n представляет собой целое число от 2 до 26.
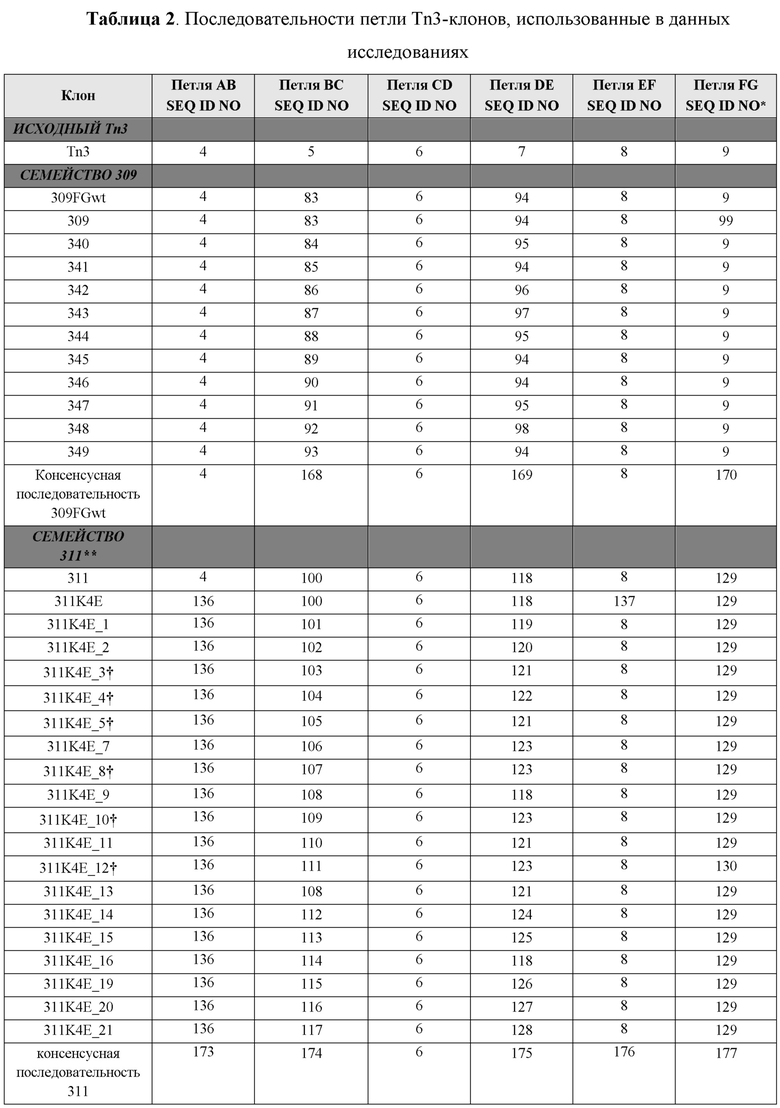
† Клоны, содержащие бета-цепь C, имеющую последовательность CELAYGI (SEQ ID NO: 14), при этом все другие клоны содержат бета-цепь C, имеющую последовательность CELTYGI (SEQ ID NO: 13).
* В некоторых вариантах в семействе 309, например, 342, петлю FG можно заменить на SEQ ID NO: 139.
** В некоторых вариантах в семействе 311 петлю BC можно сконструировать с заменой тирозина в положении 21. Специально предусмотрено, что замещающие аминокислотные остатки могут иметь малую боковую цепь.
[0198] В некоторых вариантах осуществления CD40L-специфичные Tn3-мономерные субъединицы по настоящему изобретению состоят из аминокислотной последовательности:
IEV(XAB)nALITW(XBC)nCELX1YGI(XCD)nTTIDL(XDE)nYSI(XEF)nYEVSLIC(XFG)nKETFTT,
где
(a) XAB, XBC, XCD, XDE, XEF и XFG представляют собой аминокислотные остатки, присутствующие в последовательностях петель AB, BC, CD, DE, EF и FG, соответственно;
(b) X1 представляет собой аминокислотный остаток аланин (A) или треонин (T); и
(c) длина петли n представляет собой целое число от 2 до 26.
[0199] В одном варианте осуществления бета-цепи CD40L-специфичной Tn3-мономерной каркасной структуры имеют по меньшей мере 90% идентичность последовательности с бета-цепями исходной Tn3-каркасной структуры (SEQ ID NO: 3). Для вычисления такого процента идентичности последовательности аминокислотные последовательности выравнивают с помощью способов, известных в данной области техники. Процент идентичности аминокислотной последовательности определяют как соотношение между (a) числом аминокислот, расположенных в бета-цепях, которые идентичны в выравнивании последовательности, и (b) общим числом аминокислот, расположенных в бета-цепи.
[0200] В одном варианте осуществления последовательность петли АB содержит SEQ ID NO: 4 или SEQ ID NO: 136. В другом варианте осуществления последовательность петли CD содержит SEQ ID NO: 6. В другом варианте осуществления последовательность петли EF содержит SEQ ID NO: 8 или SEQ ID NO: 137. В одном варианте осуществления последовательность петли АB состоит из SEQ ID NO: 4 или SEQ ID NO: 136. В другом варианте осуществления последовательность петли СD состоит из SEQ ID NO: 6. В другом варианте осуществления последовательность петли EF состоит из SEQ ID NO: 8 или SEQ ID NO: 137.
[0201] В одном варианте осуществления последовательность петли BC содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 83, 84, 85, 86, 87, 88, 89, 90, 91, 92 и 93. В другом варианте осуществления последовательность петли BC состоит из последовательности, выбранной из группы, состоящей из SEQ ID NO: 83, 84, 85, 86, 87, 88, 89, 90, 91, 92 и 93.
[0202] В одном варианте осуществления последовательность петли DE содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 94, 95, 96, 97 и 98. В другом варианте осуществления последовательность петли DE состоит из последовательности, выбранной из группы, состоящей из SEQ ID NO: 94, 95, 96, 97 и 98.
[0203] В одном варианте осуществления последовательность петли FG содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 9, 99 и 139. В другом варианте осуществления последовательность петли FG состоит из последовательности, выбранной из группы, состоящей из SEQ ID NO: 9, 99 и 139.
[0204] В одном варианте осуществления последовательность петли BC содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116 и 117. В другом варианте осуществления последовательность петли BC состоит из последовательности, выбранной из группы, состоящей из SEQ ID NO: 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116 и 117.
[0205] В некоторых вариантах осуществления последовательность петли DE содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 118, 119, 120, 121, 122, 123, 124, 125, 126, 127 и 128. В других вариантах осуществления последовательность петли DE состоит из последовательности, выбранной из группы, состоящей из SEQ ID NO: 118, 119, 120, 121, 122, 123, 124, 125, 126, 127 и 128.
[0206] В некоторых вариантах осуществления последовательность петли FG содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 129 и 130. В других вариантах осуществления последовательность петли FG состоит из последовательности, выбранной из группы, состоящей из SEQ ID NO: 129 и 130.
[0207] В некоторых вариантах осуществления последовательность петли BC содержит SEQ ID NO: 83, последовательность петли DЕ содержит SEQ ID NO: 94 и последовательность петли FG содержит SEQ ID NO: 9 или 139. В некоторых вариантах осуществления последовательность петли BC состоит из SEQ ID NO: 83, последовательность петли DЕ состоит из SEQ ID NO: 94 и последовательность петли FG состоит из SEQ ID NO: 9 или 139.
[0208] В некоторых вариантах осуществления последовательность петли BC содержит SEQ ID NO: 83, последовательность петли DЕ содержит SEQ ID NO: 94 и последовательность петли FG содержит SEQ ID NO: 99. В других вариантах осуществления последовательность петли BC состоит из SEQ ID NO: 83, последовательность петли DЕ состоит из SEQ ID NO: 94 и последовательность петли FG состоит из SEQ ID NO: 99.
[0209] В некоторых вариантах осуществления последовательность петли BC содержит SEQ ID NO: 84, последовательность петли DЕ содержит SEQ ID NO: 95 и последовательность петли FG содержит SEQ ID NO: 9 или 139. В других вариантах осуществления последовательность петли BC состоит из SEQ ID NO: 84, последовательность петли DЕ состоит из SEQ ID NO: 95 и последовательность петли FG состоит из SEQ ID NO: 9 или 139.
[0210] В некоторых вариантах осуществления последовательность петли BC содержит SEQ ID NO: 85, последовательность петли DЕ содержит SEQ ID NO: 94 и последовательность петли FG содержит SEQ ID NO: 9 или 139. В других вариантах осуществления последовательность петли BC состоит из SEQ ID NO: 85, последовательность петли DЕ состоит из SEQ ID NO: 94 и последовательность петли FG состоит из SEQ ID NO: 9 или 139.
[0211] В некоторых вариантах осуществления последовательность петли BC содержит SEQ ID NO: 86, последовательность петли DЕ содержит SEQ ID NO: 96 и последовательность петли FG содержит SEQ ID NO: 9 или 139. В других вариантах осуществления последовательность петли BC состоит из SEQ ID NO: 86, последовательность петли DЕ состоит из SEQ ID NO: 96 и последовательность петли FG состоит из SEQ ID NO: 9 или 139.
[0212] В некоторых вариантах осуществления последовательность петли BC содержит SEQ ID NO: 87, последовательность петли DЕ содержит SEQ ID NO: 97 и последовательность петли FG содержит SEQ ID NO: 9 или 139. В других вариантах осуществления последовательность петли BC состоит из SEQ ID NO: 87, последовательность петли DЕ состоит из SEQ ID NO: 97 и последовательность петли FG состоит из SEQ ID NO: 9 или 139.
[0213] В некоторых вариантах осуществления последовательность петли BC содержит SEQ ID NO: 88, последовательность петли DЕ содержит SEQ ID NO: 95 и последовательность петли FG содержит SEQ ID NO: 9 или 139. В других вариантах осуществления последовательность петли BC состоит из SEQ ID NO: 88, последовательность петли DЕ состоит из SEQ ID NO: 95 и последовательность петли FG состоит из SEQ ID NO: 9 или 139.
[0214] В некоторых вариантах осуществления последовательность петли BC содержит SEQ ID NO: 89, последовательность петли DЕ содержит SEQ ID NO: 94 и последовательность петли FG содержит SEQ ID NO: 9 или 139. В других вариантах осуществления последовательность петли BC состоит из SEQ ID NO: 89, последовательность петли DЕ состоит из SEQ ID NO: 94 и последовательность петли FG состоит из SEQ ID NO: 9 или 139.
[0215] В некоторых вариантах осуществления последовательность петли BC содержит SEQ ID NO: 90, последовательность петли DЕ содержит SEQ ID NO: 94 и последовательность петли FG содержит SEQ ID NO: 9 или 139. В других вариантах осуществления последовательность петли BC состоит из SEQ ID NO: 90, последовательность петли DЕ состоит из SEQ ID NO: 94 и последовательность петли FG состоит из SEQ ID NO: 9 или 139.
[0216] В некоторых вариантах осуществления последовательность петли BC содержит SEQ ID NO: 91, последовательность петли DЕ содержит SEQ ID NO: 95 и последовательность петли FG содержит SEQ ID NO: 9 или 139. В других вариантах осуществления последовательность петли BC состоит из SEQ ID NO: 91, последовательность петли DЕ состоит из SEQ ID NO: 95 и последовательность петли FG состоит из SEQ ID NO: 9 или 139.
[0217] В некоторых вариантах осуществления последовательность петли BC содержит SEQ ID NO: 92, последовательность петли DЕ содержит SEQ ID NO: 98 и последовательность петли FG содержит SEQ ID NO: 9 или 139. В других вариантах осуществления последовательность петли BC состоит из SEQ ID NO: 92, последовательность петли DЕ состоит из SEQ ID NO: 98 и последовательность петли FG состоит из SEQ ID NO: 9 или 139.
[0218] В некоторых вариантах осуществления последовательность петли BC содержит SEQ ID NO: 93, последовательность петли DЕ содержит SEQ ID NO: 94 и последовательность петли FG содержит SEQ ID NO: 9 или 139. В других вариантах осуществления последовательность петли BC состоит из SEQ ID NO: 93, последовательность петли DЕ состоит из SEQ ID NO: 94 и последовательность петли FG состоит из SEQ ID NO: 9 или 139.
[0219] В некоторых вариантах осуществления последовательность петли BC содержит SEQ ID NO: 168, последовательность петли DЕ содержит SEQ ID NO: 169 и последовательность петли FG содержит SEQ ID NO: 170. В других вариантах осуществления последовательность петли BC состоит из SEQ ID NO: 168, последовательность петли DЕ состоит из SEQ ID NO: 169 и последовательность петли FG состоит из SEQ ID NO: 170.
[0220] В некоторых вариантах осуществления последовательность петли BC содержит SEQ ID NO: 100, последовательность петли DЕ содержит SEQ ID NO: 118 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления последовательность петли BC состоит из SEQ ID NO: 100, последовательность петли DЕ состоит из SEQ ID NO: 118 и последовательность петли FG состоит из SEQ ID NO: 129.
[0221] В некоторых вариантах осуществления последовательность петли АB содержит SEQ ID NO: 136 последовательность петли BC содержит SEQ ID NO: 101, последовательность петли DЕ содержит SEQ ID NO: 119 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления последовательность петли АB состоит из SEQ ID NO: 136, последовательность петли ВС состоит из SEQ ID NO: 101, последовательность петли DЕ состоит из SEQ ID NO: 119 и последовательность петли FG состоит из SEQ ID NO: 129.
[0222] В некоторых вариантах осуществления последовательность петли АB содержит SEQ ID NO: 136, последовательность петли BC содержит SEQ ID NO: 102, последовательность петли DЕ содержит SEQ ID NO: 120 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления последовательность петли АB состоит из SEQ ID NO: 136, последовательность петли ВС состоит из SEQ ID NO: 102, последовательность петли DЕ состоит из SEQ ID NO: 120 и последовательность петли FG состоит из SEQ ID NO: 129.
[0223] В некоторых вариантах осуществления последовательность петли АB содержит SEQ ID NO: 136, последовательность петли BC содержит SEQ ID NO: 103, последовательность петли DЕ содержит SEQ ID NO: 121 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления последовательность петли АB состоит из SEQ ID NO: 136, последовательность петли ВС состоит из SEQ ID NO: 103, последовательность петли DЕ состоит из SEQ ID NO: 121 и последовательность петли FG состоит из SEQ ID NO: 129.
[0224] В некоторых вариантах осуществления последовательность петли АB содержит SEQ ID NO: 136, последовательность петли BC содержит SEQ ID NO: 104, последовательность петли DЕ содержит SEQ ID NO: 122 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления последовательность петли АB состоит из SEQ ID NO: 136, последовательность петли ВС состоит из SEQ ID NO: 104, последовательность петли DЕ состоит из SEQ ID NO: 122 и последовательность петли FG состоит из SEQ ID NO: 129.
[0225] В некоторых вариантах осуществления последовательность петли АB содержит SEQ ID NO: 136, последовательность петли BC содержит SEQ ID NO: 105, последовательность петли DЕ содержит SEQ ID NO: 121 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления последовательность петли АB состоит из SEQ ID NO: 136, последовательность петли ВС состоит из SEQ ID NO: 105, последовательность петли DЕ состоит из SEQ ID NO: 121 и последовательность петли FG состоит из SEQ ID NO: 129.
[0226] В некоторых вариантах осуществления последовательность петли АB содержит SEQ ID NO: 136, последовательность петли BC содержит SEQ ID NO: 106, последовательность петли DЕ содержит SEQ ID NO: 123 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления последовательность петли АB состоит из SEQ ID NO: 136, последовательность петли ВС состоит из SEQ ID NO: 106, последовательность петли DЕ состоит из SEQ ID NO: 123 и последовательность петли FG состоит из SEQ ID NO: 129.
[0227] В некоторых вариантах осуществления последовательность петли АB содержит SEQ ID NO: 136, последовательность петли BC содержит SEQ ID NO: 107, последовательность петли DЕ содержит SEQ ID NO: 123 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления последовательность петли АB состоит из SEQ ID NO: 136, последовательность петли ВС состоит из SEQ ID NO: 107, последовательность петли DЕ состоит из SEQ ID NO: 123 и последовательность петли FG состоит из SEQ ID NO: 129.
[0228] В некоторых вариантах осуществления последовательность петли АB содержит SEQ ID NO: 136, последовательность петли BC содержит SEQ ID NO: 108, последовательность петли DЕ содержит SEQ ID NO: 118 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления последовательность петли АB состоит из SEQ ID NO: 136, последовательность петли ВС состоит из SEQ ID NO: 108, последовательность петли DЕ состоит из SEQ ID NO: 118 и последовательность петли FG состоит из SEQ ID NO: 129.
[0229] В некоторых вариантах осуществления последовательность петли АB содержит SEQ ID NO: 136, последовательность петли BC содержит SEQ ID NO: 109, последовательность петли DЕ содержит SEQ ID NO: 123 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления последовательность петли АB состоит из SEQ ID NO: 136, последовательность петли ВС состоит из SEQ ID NO: 109, последовательность петли DЕ состоит из SEQ ID NO: 123 и последовательность петли FG состоит из SEQ ID NO: 129.
[0230] В некоторых вариантах осуществления последовательность петли АB содержит SEQ ID NO: 136, последовательность петли BC содержит SEQ ID NO: 110, последовательность петли DЕ содержит SEQ ID NO: 121 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления последовательность петли АB состоит из SEQ ID NO: 136, последовательность петли ВС состоит из SEQ ID NO: 110, последовательность петли DЕ состоит из SEQ ID NO: 121 и последовательность петли FG состоит из SEQ ID NO: 129.
[0231] В некоторых вариантах осуществления последовательность петли АB содержит SEQ ID NO: 136, последовательность петли BC содержит SEQ ID NO: 111, последовательность петли DЕ содержит SEQ ID NO: 123 и последовательность петли FG содержит SEQ ID NO: 130. В других вариантах осуществления последовательность петли АB состоит из SEQ ID NO: 136, последовательность петли ВС состоит из SEQ ID NO: 111, последовательность петли DЕ состоит из SEQ ID NO: 123 и последовательность петли FG состоит из SEQ ID NO: 130.
[0232] В некоторых вариантах осуществления последовательность петли АB содержит SEQ ID NO: 136, последовательность петли BC содержит SEQ ID NO: 108, последовательность петли DЕ содержит SEQ ID NO: 121 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления последовательность петли АB состоит из SEQ ID NO: 136, последовательность петли ВС состоит из SEQ ID NO: 108, последовательность петли DЕ состоит из SEQ ID NO: 121 и последовательность петли FG состоит из SEQ ID NO: 129.
[0233] В некоторых вариантах осуществления последовательность петли АB содержит SEQ ID NO: 136, последовательность петли BC содержит SEQ ID NO: 112, последовательность петли DЕ содержит SEQ ID NO: 124 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления последовательность петли АB состоит из SEQ ID NO: 136, последовательность петли ВС состоит из SEQ ID NO: 112, последовательность петли DЕ состоит из SEQ ID NO: 124 и последовательность петли FG состоит из SEQ ID NO: 129.
[0234] В некоторых вариантах осуществления последовательность петли АB содержит SEQ ID NO: 136, последовательность петли BC содержит SEQ ID NO: 113, последовательность петли DЕ содержит SEQ ID NO: 125 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления последовательность петли АB состоит из SEQ ID NO: 136, последовательность петли BC состоит из SEQ ID NO: 113, последовательность петли DЕ состоит из SEQ ID NO: 125 и последовательность петли FG состоит из SEQ ID NO: 129.
[0235] В некоторых вариантах осуществления последовательность петли АB содержит SEQ ID NO: 136, последовательность петли BC содержит SEQ ID NO: 114, последовательность петли DЕ содержит SEQ ID NO: 118 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления последовательность петли АB состоит из SEQ ID NO: 136, последовательность петли ВС состоит из SEQ ID NO: 114, последовательность петли DЕ состоит из SEQ ID NO: 118 и последовательность петли FG состоит из SEQ ID NO: 129.
[0236] В некоторых вариантах осуществления последовательность петли АB содержит SEQ ID NO: 136, последовательность петли BC содержит SEQ ID NO: 115, последовательность петли DЕ содержит SEQ ID NO: 126 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления последовательность петли АB состоит из SEQ ID NO: 136, последовательность петли ВС состоит из SEQ ID NO: 115, последовательность петли DЕ состоит из SEQ ID NO: 126 и последовательность петли FG состоит из SEQ ID NO: 129.
[0237] В некоторых вариантах осуществления последовательность петли АB содержит SEQ ID NO: 136, последовательность петли BC содержит SEQ ID NO: 116, последовательность петли DЕ содержит SEQ ID NO: 127 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления последовательность петли АB состоит из SEQ ID NO: 136, последовательность петли ВС состоит из SEQ ID NO: 116, последовательность петли DЕ состоит из SEQ ID NO: 127 и последовательность петли FG состоит из SEQ ID NO: 129.
[0238] В некоторых вариантах осуществления последовательность петли АB содержит SEQ ID NO: 136, последовательность петли BC содержит SEQ ID NO: 117, последовательность петли DЕ содержит SEQ ID NO: 128 и последовательность петли FG содержит SEQ ID NO: 129. В других вариантах осуществления последовательность петли АB состоит из SEQ ID NO: 136, последовательность петли ВС состоит из SEQ ID NO: 117, последовательность петли DЕ состоит из SEQ ID NO: 128 и последовательность петли FG состоит из SEQ ID NO: 129.
[0239] В некоторых вариантах осуществления последовательность петли BC содержит SEQ ID NO: 174, последовательность петли DЕ содержит SEQ ID NO: 175 и последовательность петли FG содержит SEQ ID NO: 177. В других вариантах осуществления последовательность петли BC состоит из SEQ ID NO: 174, последовательность петли DЕ состоит из SEQ ID NO: 175 и последовательность петли FG состоит из SEQ ID NO: 177.
[0240] В некоторых вариантах осуществления CD40L-специфичная мономерная субъединица содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42 и 146. В других вариантах осуществления CD40L-специфичная мономерная субъединица состоит из последовательности, выбранной из группы, состоящей из SEQ ID NO: 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42 и 146.
[0241] В некоторых вариантах осуществления CD40L-специфичная мономерная субъединица содержит SEQ ID NO: 28 или 146. В других вариантах осуществления CD40L-специфичная мономерная субъединица состоит из SEQ ID NO: 28 или 146.
[0242] В некоторых вариантах осуществления CD40L-специфичные Tn3-мономерные субъединицы по настоящему изобретению содержат аминокислотную последовательность:
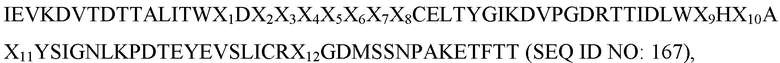
где
(a) X1 представляет собой аминокислотный остаток серин (S) или лейцин (L);
(b) X2 представляет собой аминокислотный остаток аспарагиновую кислоту (D) или глутаминовую кислоту (E);
(c) X3 представляет собой аминокислотный остаток гистидин (H), изолейцин (I), валин (V), фенилаланин (F) или триптофан (W);
(d) X4 представляет собой аминокислотный остаток аланин (A), глицин (G), глутаминовую кислоту (E) или аспарагиновую кислоту (D);
(e) X5 представляет собой аминокислотный остаток глутаминовую кислоту (E), лейцин (L), глутамин (Q), серин (S), аспарагиновую кислоту (D) или аспарагин (N);
(f) X6 представляет собой аминокислотный остаток фенилаланин (F) или тирозин (Y);
(g) X7 представляет собой аминокислотный остаток изолейцин (I), валин (V), гистидин (H), глутаминовую кислоту (E) или аспарагиновую кислоту (D);
(h) X8 представляет собой аминокислотный остаток глицин (G), триптофан (W) или валин (V);
(i) X9 представляет собой аминокислотный остаток триптофан (W), фенилаланин (F) или тирозин (Y);
(j) X10 представляет собой аминокислотный остаток серин (S), глутамин (Q), метионин (M) или гистидин (H);
(k) X11 представляет собой аминокислотный остаток триптофан (W) или гистидин (H); и
(l) X12 представляет собой аминокислотный остаток аргинин (R) или серин (S).
[0243] В некоторых вариантах осуществления CD40L-специфичные Tn3-мономерные субъединицы по настоящему изобретению состоят из аминокислотной последовательности:
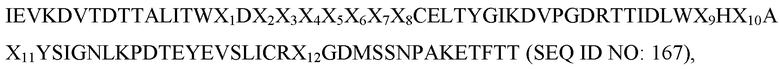
где
(a) X1 представляет собой аминокислотный остаток серин (S) или лейцин (L);
(b) X2 представляет собой аминокислотный остаток аспарагиновую кислоту (D) или глутаминовую кислоту (E);
(c) X3 представляет собой аминокислотный остаток гистидин (H), изолейцин (I), валин (V), фенилаланин (F) или триптофан (W);
(d) X4 представляет собой аминокислотный остаток аланин (A), глицин (G), глутаминовую кислоту (E) или аспарагиновую кислоту (D);
(e) X5 представляет собой аминокислотный остаток глутаминовую кислоту (E), лейцин (L), глутамин (Q), серин (S), аспарагиновую кислоту (D) или аспарагин (N);
(f) X6 представляет собой аминокислотный остаток фенилаланин (F) или тирозин (Y);
(g) X7 представляет собой аминокислотный остаток изолейцин (I), валин (V), гистидин (H), глутаминовую кислоту (E) или аспарагиновую кислоту (D);
(h) X8 представляет собой аминокислотный остаток глицин (G), триптофан (W) или валин (V);
(i) X9 представляет собой аминокислотный остаток триптофан (W), фенилаланин (F) или тирозин (Y);
(j) X10 представляет собой аминокислотный остаток серин (S), глутамин (Q), метионин (M) или гистидин (H);
(k) X11 представляет собой аминокислотный остаток триптофан (W) или гистидин (H); и
(l) X12 представляет собой аминокислотный остаток аргинин (R) или серин (S).
[0244] В некоторых вариантах осуществления CD40L-специфичная мономерная субъединица содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80 и 82. В некоторых вариантах осуществления CD40L-специфичная мономерная субъединица состоит из последовательности, выбранной из группы, состоящей из SEQ ID NO: 44, 46, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76, 78, 80 и 82.
[0245] В некоторых вариантах осуществления CD40L-специфичные Tn3-мономерные субъединицы по настоящему изобретению содержат аминокислотную последовательность:

где
(a) X1 представляет собой аминокислотный остаток лизин (K) или глутаминовую кислоту (E);
(b) X2 представляет собой аминокислотный остаток треонин (T) или изолейцин (I);
(c) X3 представляет собой аминокислотный остаток аспарагин (N) или аланин (A);
(d) X4 представляет собой аминокислотный остаток серин (S), лейцин (L), аланин (A), фенилаланин (F) или тирозин (Y);
(e) X5 представляет собой аминокислотный остаток тирозин (Y), аланин (A), глицин (G), валин (V), изолейцин (I) или серин (S);
(f) X6 представляет собой аминокислотный остаток тирозин (Y), серин (S), аланин (A) или гистидин (H);
(g) X7 представляет собой аминокислотный остаток аспарагин (N), аспарагиновую кислоту (D), гистидин (H) или тирозин (Y);
(h) X8 представляет собой аминокислотный остаток лейцин (L), фенилаланин (F), гистидин (H) или тирозин (Y);
(i) X9 представляет собой аминокислотный остаток гистидин (H), пролин (P), серин (S), лейцин (L) или аспарагиновую кислоту (D);
(j) X10 представляет собой аминокислотный остаток глицин (G), фенилаланин (F), гистидин (H) или тирозин (Y);
(k) X11 представляет собой аминокислотный остаток аланин (A) или треонин (T);
(l) X12 представляет собой аминокислотный остаток серин (S), аспарагин (N), глутаминовую кислоту (E), аспарагин (R) или аспарагиновую кислоту (D);
(m) X13 представляет собой аминокислотный остаток серин (S), глутамин (Q), треонин (T), аспарагин (N) или аланин (A);
(n) X14 представляет собой аминокислотный остаток пролин (P), валин (V), изолейцин (I) или аланин (A) или не является аминокислотой;
(o) X15 представляет собой аминокислотный остаток изолейцин (I) или не является аминокислотой;
(p) X16 представляет собой аминокислотный остаток глутаминовую кислоту (E) или лизин (K); и
(q) X17 представляет собой аминокислотный остаток серин (S) или аспарагин (N).
[0246] В некоторых вариантах осуществления CD40L-специфичные Tn3-мономерные субъединицы по настоящему изобретению состоят из аминокислотной последовательности:
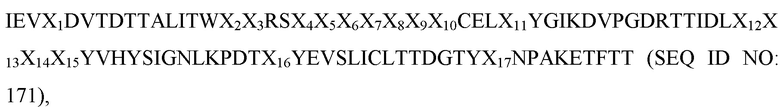
где
(a) X1 представляет собой аминокислотный остаток лизин (K) или глутаминовую кислоту (E);
(b) X2 представляет собой аминокислотный остаток треонин (T) или изолейцин (I);
(c) X3 представляет собой аминокислотный остаток аспарагин (N) или аланин (A);
(d) X4 представляет собой аминокислотный остаток серин (S), лейцин (L), аланин (A), фенилаланин (F) или тирозин (Y);
(e) X5 представляет собой аминокислотный остаток тирозин (Y), аланин (A), глицин (G), валин (V), изолейцин (I) или серин (S);
(f) X6 представляет собой аминокислотный остаток тирозин (Y), серин (S), аланин (A) или гистидин (H);
(g) X7 представляет собой аминокислотный остаток аспарагин (N), аспарагиновую кислоту (D), гистидин (H) или тирозин (Y);
(h) X8 представляет собой аминокислотный остаток лейцин (L), фенилаланин (F), гистидин (H) или тирозин (Y);
(i) X9 представляет собой аминокислотный остаток гистидин (H), пролин (P), серин (S), лейцин (L) или аспарагиновую кислоту (D);
(j) X10 представляет собой аминокислотный остаток глицин (G), фенилаланин (F), гистидин (H) или тирозин (Y);
(k) X11 представляет собой аминокислотный остаток аланин (A) или треонин (T);
(l) X12 представляет собой аминокислотный остаток серин (S), аспарагин (N), глутаминовую кислоту (E), аспарагин (R) или аспарагиновую кислоту (D);
(m) X13 представляет собой аминокислотный остаток серин (S), глутамин (Q), треонин (T), аспарагин (N) или аланин (A);
(n) X14 представляет собой аминокислотный остаток пролин (P), валин (V), изолейцин (I) или аланин (A) или не является аминокислотой;
(o) X15 представляет собой аминокислотный остаток изолейцин (I) или не является аминокислотой;
(p) X16 представляет собой аминокислотный остаток глутаминовую кислоту (E) или лизин (K); и
(q) X17 представляет собой аминокислотный остаток серин (S) или аспарагин (N).
[0247] В некоторых вариантах осуществления CD40L-специфичная мономерная каркасная структура содержит Tn3-модуль, где одна или несколько бета-цепей содержат по меньшей мере одну аминокислотную замену за исключением того, что остатки цистеина в бета-цепях C и F (SEQ ID NO: 13 или 14 и SEQ ID NO: 17, соответственно) могут не быть замененными.
[0248] Петли, соединяющие различные бета-цепи CD40L-специфичной мономерной субъединицы, можно рандомизировать по длине и/или вариабельности последовательности. В одном варианте осуществления CD40L-специфичная мономерная субъединица имеет по меньшей мере одну петлю, которая рандомизирована по длине и/или вариабельности последовательности. В одном варианте осуществления по меньшей мере одна, по меньшей мере две, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять или по меньшей мере шесть петель CD40L-специфичной мономерной субъединицы рандомизированы по длине и/или вариабельности последовательности. В одном варианте осуществления по меньшей мере одна петля CD40L-специфичной мономерной субъединицы сохранена неизменной, тогда как по меньшей мере одна дополнительная петля рандомизирована по длине и/или вариабельности последовательности. В другом варианте осуществления по меньшей мере одна, по меньшей мере две или все три петли AB, CD и EF сохранены неизменными, тогда как по меньшей мере одна, по меньшей мере две или все три петли BC, DE и FG рандомизированы по длине или вариабельности последовательности. В другом варианте осуществления по меньшей мере одна, по меньшей мере две или все три петли AB, CD и EF рандомизированы, тогда как по меньшей мере одна, по меньшей мере две или все три петли BC, DE и FG рандомизированы по длине или вариабельности последовательности. В еще одном варианте осуществления по меньшей мере одна, по меньшей мере две, по меньшей мере три, по меньшей мере 4, по меньшей мере пять или все шесть петель AB, CD, EF, BC, DE и FG рандомизированы по длине и/или вариабельности последовательности.
[0249] В некоторых вариантах осуществления один или несколько остатков в пределах петли оставлены неизменными, тогда как другие остатки рандомизированы по длине и/или вариабельности последовательности. В некоторых вариантах осуществления один или несколько остатков в пределах петли ограничены заранее определенным и ограниченным числом различных аминокислот, тогда как другие остатки рандомизированы по длине и/или вариабельности последовательности. Соответственно, CD40L-специфичная мономерная субъединица по настоящему изобретению может содержать одну или несколько петель, имеющих вырожденную консенсусную последовательность и/или один или несколько инвариантных аминокислотных остатков.
[0250] В одном варианте осуществления CD40L-специфичная мономерная субъединица по настоящему изобретению содержит петлю AB, которая является рандомизированной. В другом варианте осуществления CD40L-специфичная мономерная субъединица по настоящему изобретению содержит петлю BC, которая является рандомизированной. В одном варианте осуществления CD40L-специфичная мономерная субъединица по настоящему изобретению содержит петлю CD, которая является рандомизированной. В одном варианте осуществления CD40L-специфичная мономерная субъединица по настоящему изобретению содержит петлю DE, которая является рандомизированной. В одном варианте осуществления CD40L-специфичная мономерная субъединица по настоящему изобретению содержит петлю EF, которая является рандомизированной.
[0251] В конкретных вариантах осуществления CD40L-специфичная мономерная субъединица по настоящему изобретению содержит петлю FG, которая является по меньшей мере на один аминокислотный остаток более короткой, чем когнатная петля FG третьего FnIII-домена тенасцина C человека, и дополнительно рандомизирована по одному или нескольким положениям.
[0252] В конкретных вариантах осуществления по меньшей мере одна из петель BC, DE и FG рандомизирована, причем бета-цепь A содержит SEQ ID NO:10 или 11, бета-цепь B содержит SEQ ID NO:12, бета-цепь C содержит SEQ ID NO:13 или 14, бета-цепь D содержит SEQ ID NO:15, бета-цепь E содержит SEQ ID NO:16, бета-цепь F содержит SEQ ID NO:17 и бета-цепь G содержит SEQ ID NO:18, петля AB содержит SEQ ID NO:4 или 136, петля CD содержит SEQ ID NO:6 и петля EF содержит SEQ ID NO:8 или 137.
[0253] В других конкретных вариантах осуществления по меньшей мере одна из петель AB, CD и EF рандомизирована, причем бета-цепь A содержит SEQ ID NO:10 или 11, бета-цепь B содержит SEQ ID NO:12, бета-цепь C содержит SEQ ID NO:13 или 14, бета-цепь D содержит SEQ ID NO:15, бета-цепь E содержит SEQ ID NO:16, бета-цепь F содержит SEQ ID NO:17 и бета-цепь G содержит SEQ ID NO:18, петля BC содержит SEQ ID NO:5, петля DE содержит SEQ ID NO:7 и петля FG содержит SEQ ID NO:9 или 139.
Повышенная стабильность каркасной структуры
[0254] Стабильность Tn3-каркасной структуры по настоящему изобретению можно увеличить с помощью различных подходов. В некоторых вариантах осуществления Tn3-каркасные структуры по настоящему изобретению можно стабилизировать путем удлинения N- и/или C-концевых участков. N- и/или C-концевые участки можно удлинить на 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более 10 аминокислот. В других вариантах осуществления Tn3-каркасные структуры по настоящему изобретению можно стабилизировать путем введения изменения, которое увеличивает время полужизни в сыворотке, как описано в данном документе. В еще одном варианте осуществления Tn3-каркасные структуры по настоящему изобретению содержат добавление, делецию или замену по меньшей мере одного аминокислотного остатка для стабилизации гидрофобного кора каркасной структуры.
[0255] Tn3-каркасные структуры по настоящему изобретению могут быть эффективно стабилизированы путем конструирования дисульфидных связей, не относящихся к природным, как раскрыто в международной заявке на патент № PCT/US2011/032184. В некоторых вариантах осуществления каркасные структуры по настоящему изобретению содержат не встречающиеся в природе дисульфидные связи, как описано в PCT публикации № WO 2009/058379. Биоинформационные подходы можно использовать для выявления кандидатных положений, пригодных для конструирования дисульфидных связей.
[0256] В одном варианте осуществления Tn3-мономерная субъединица по настоящему изобретению содержит по меньшей мере одну, по меньшей мере две, по меньшей мере три, по меньшей мере четыре или по меньшей мере пять не встречающихся в природе внутримолекулярных дисульфидных связей. В одном варианте осуществления Tn3-мономерная субъединица по настоящему изобретению содержит по меньшей мере одну не встречающуюся в природе внутримолекулярную дисульфидную связь, где указанная по меньшей мере одна не встречающаяся в природе дисульфидная связь стибилизирует мономер. В еще одном варианте осуществления Tn3-каркасные структуры по настоящему изобретению содержат по меньшей мере одну не встречающуюся в природе дисульфидную связь, где данная связь расположена между двумя отдельными мономерными или мультимерными Tn3-каркасными структурами, т.е данная дисульфидная связь является внутримолекулярной дисульфидной связью. Например, дисульфидная связь может соединять отдельные каркасные структуры (например, две CD40L-специфичные мономерные каркасные структуры), Tn3-каркасную структуру и линкер, или Tn3-каркасную структуру и Fc-домен, или Tn3-каркасную структуру и антитело или его фрагмент.
[0257] В некоторых вариантах осуществления Tn3-каркасные структуры по настоящему изобретению содержат по меньшей мере одну не встречающуюся в природе внутримолекулярную дисульфидную связь, которая соединяет Tn3-мономерную субъединицу и выделенный гетерологичный фрагмент, Tn3-мономерную субъединицу и гетерологичный фрагмент, слитый или конъюгированный с такой же Tn3-каркасной структурой, или Tn3-мономерную субъединицу и гетерологичный фрагмент, слитый или конъюгированный с другой Tn3-каркасной структурой.
[0258] В некоторых вариантах осуществления Tn3-каркасные структуры по настоящему изобретению содержат дисульфидную связь, которая образует Tn3-мультимерную каркасную структуру из по меньшей мере 2, по меньшей мере 3, по меньшей мере 4 или более мономерных субъединиц.
[0259] В другом варианте осуществления Tn3-каркасные структуры по настоящему изобретению могут предусматривать удлинение N- и/или C-концевых участков. В одном варианте осуществления Tn3-каркасная структура по настоящему изобретению содержит изменение для увеличения времени полужизни в сыворотке, как описано в данном документе. В еще одном варианте осуществления Tn3-каркасные структуры по настоящему изобретению содержат добавление, делецию или замену по меньшей мере одного аминокислотного остатка для стабилизации гидрофобного кора каркасной структуры.
Измерения стабильности
[0260] Стабильность Tn3-мономерных субъединиц по настоящему изобретению, выделенных или в виде части мультимерной Tn3-каркасной структуры, можно легко измерять с помощью методик, хорошо известных в данной области техники, таких как термическая денатурация (Tпл) и хаотропная денатурация (например, обработка мочевиной или солями гуанидина), обработка протеазами (например, обработка термолизином) или другим способом, принятым в данной области техники, для определения стабильности белка. Исчерпывающий обзор методик, используемых для измерения стабильности белка, можно найти, например, в "Current Protocols in Molecular Biology" и "Current Protocols in Protein Science" by John Wiley and Sons. 2007.
Мультимерные Tn3-каркасные структуры
[0261] Один аспект настоящего изобретения предлагает мультимерные Tn3-каркасные структуры, содержащие по меньшей мере две Tn3-мономерные субъединицы по настоящему изобретению, соединенные в тандем, и при этом по меньшей мере один из мономеров представляет собой CD40L-специфичную мономерную субъединицу. Такие мультимерные Tn3-каркасные структуры могут быть собраны во множество конфигураций. В конкретном аспекте настоящее изобретение предлагает мультимерные Tn3-каркасные структуры, где по меньшей мере две CD40L-специфичные мономерные субъединицы соединены в тандем посредством пептидного линкера. В некоторых вариантах осуществления мультимерная Tn3-каркасная структура характеризуется повышением валентности и/или авидности в отношении связывания с мишенью или другим воздействием на мишень(и). В некоторых вариантах осуществления повышение валентности и/или авидности в отношении связывания мишени достигается, если множественные мономерные субъединицы связываются с одной и той же мишенью. В некоторых вариантах осуществления повышение валентности улучшает специфическое воздействие на мишень, такое как повышение димеризации белка-мишени.
[0262] В конкретном варианте осуществления, мультимерная Tn3-каркасная структура по настоящему изобретению содержит по меньшей мере две CD40L-специфичные мономерные субъединицы, соединенные в тандем, причем каждая CD40L-специфичная мономерная субъединица связывает по меньшей мере одну мишень, и при этом каждая CD40L-специфичная мономерная субъединица содержит множество бета-цепей, соединенных с множеством петлевых участков, где по меньшей мере одна петля представляет собой не встречающийся в природе вариант когнатной петли в исходной Tn3-каркасной структуре (SEQ ID NO: 3).
[0263] В одном варианте осуществления мультимерные Tn3-каркасные структуры получают посредством ковалентного связывания между CD40L-специфичными мономерными субъединицами, например, путем непосредственного связывания CD40L-специфичных мономерных субъединиц или путем введения линкера, например, пептидного линкера. В конкретных примерах ковалентно связанные Tn3-каркасные структуры получают путем создания генов слияния, которые кодируют CD40L-специфичные мономерные субъединицы или, в качестве альтернативы, путем конструирования кодонов для остатков цистеина в CD40L-специфичных мономерных субъединицах, что обеспечивает образование дисульфидной связи между продуктами экспрессии.
[0264] В одном варианте осуществления мультимерные Tn3-каркасные структуры по настоящему изобретению содержат по меньшей мере две CD40L-специфичные мономерные субъединицы, которые непосредственно соединены друг с другом без какого-либо дополнительного вовлечения аминокислот. В другом варианте осуществления мультимерные Tn3-каркасные структуры по настоящему изобретению содержат по меньшей мере две CD40L-специфичные мономерные субъединицы, которые соединены в тандем посредством линкера, например, пептидного линкера.
[0265] В конкретном варианте осуществления мультимерные Tn3-каркасные структуры по настоящему изобретению содержат по меньшей мере две CD40L-специфичные мономерные субъединицы, которые соединены в тандем посредством пептидного линкера, причем данный пептидный линкер содержит от 1 до приблизительно 1000, или от 1 до приблизительно 500, или от 1 до приблизительно 250, или от 1 до приблизительно 100, или от 1 до приблизительно 50, или от 1 до приблизительно 25 аминокислот. В конкретном варианте осуществления мультимерная Tn3-каркасная структура содержит по меньшей мере две CD40L-специфичные мономерные субъединицы, которые соединены в тандем посредством пептидного линкера, причем данный пептидный линкер содержит от 1 до приблизительно 20, или от 1 до приблизительно 15, или от 1 до приблизительно 10, или от 1 до приблизительно 5 аминокислот.
[0266] В конкретном варианте осуществления мультимерная Tn3-каркасная структура содержит по меньшей мере две CD40L-специфичные мономерные субъединицы, которые соединены в тандем посредством линкера, например, пептидного линкера, причем данный линкер представляет собой функциональный фрагмент. Данный функциональный фрагмент будут выбирать исходя из требуемых функций и/или характеристик мультимерной Tn3-каркасной структуры. Например, функциональный фрагмент, пригодный для очистки (например, гистидиновая метка) можно использовать в качестве линкера. Функциональные фрагменты, пригодные в качестве линкеров, включают, но без ограничения, полиэтиленгликоль (PEG), цитотоксическое средство, радионуклид, радиофармацевтическое средство, биотин, домен димеризации, человеческий сывороточный альбумин (HSA) или его участок, отвечающий за связывание с FcRn, домен или фрагмент антитела, одноцепочечное антитело, антитело на основе домена, альбумин-связывающий домен, молекулу IgG, фермент, лиганд, рецептор, связывающий пептид, каркасную структуру, отличающуюся от Tn3, эпитопную метку, рекомбинантный полипептидный полимер, цитокин и тому подобные. Конкретные пептидные линкеры и функциональные фрагменты, которые можно использовать в качестве линкеров, раскрыты ниже.
[0267] В конкретных вариантах осуществления данная функциональная группа представляет собой иммуноглобулин или его фрагмент. В некоторых вариантах осуществления данный иммуноглобулин или его фрагмент содержит Fc-домен. В некоторых вариантах осуществления данный Fc-домен не индуцирует по меньшей мере одну FcγR-опосредованную эффекторную функцию, такую как ADCC (антителозависимую клеточноопосредованную цитотоксичность). В данной области техники известно, что Fc-домен может быть подвергнут изменению для снижения или подавления по меньшей мере одной FcγR-опосредованной эффекторной функции, см., например, патенты США №№ 5624821 и 6737056.
[0268] В некоторых вариантах осуществления мультимерная Tn3-каркасная структура содержит по меньшей мере две CD40L-специфичные мономерные субъединицы, которые связаны посредством одного или нескольких линкеров, где эти линкеры, помещенные между каждой CD40L-специфичной мономерной субъединицей, могут быть одинаковыми линкерами или различными линкерами. В некоторых вариантах осуществления линкер может содержать множествo линкеров, которые могут быть одинаковыми линкерами или различными линкерами. В некоторых вариантах осуществления, если множество линкеров взаимозависимо соединены, некоторые или все линкеры могут быть функциональными фрагментами.
Стехиометрия связывания каркасной структуры
[0269] В некоторых вариантах осуществления мономерная или мультимерная Tn3-каркасная структура может содержать CD40L-специфичную мономерную субъединицу, специфическую по отношению к различным эпитопам, которые могут представлять собой различные эпитопы на одной молекуле CD40L или на различных молекулах-мишенях CD40L. В некоторых вариантах осуществления мультимерная Tn3-каркасная структура может содержать CD40L-специфичные мономерные субъединицы, где каждая субъединица нацеливается на один или несколько различных эпитопов на одной или нескольких молекулах CD40L.
[0270] В других вариантах осуществления мономерная или мультимерная Tn3-каркасная структура может связывать два или более различных эпитопов на одной и той же молекуле CD40L. В некоторых вариантах осуществления данные различные эпитопы представляют собой неперекрывающиеся эпитопы. В других вариантах осуществления данные различные эпитопы представляют собой перекрывающиеся эпитопы.
[0271] В еще одном конкретном варианте осуществления мономерная или мультимерная Tn3-каркасная структура может связывать один или несколько эпитопов на молекуле CD40L и дополнительно связывать один или несколько эпитопов на второй молекуле CD40L. В некоторых вариантах осуществления молекулы-мишени являются частью олигомерного комплекса, например, тримерного комплекса CD40L.
[0272] В еще одном конкретном варианте осуществления мономерная или мультимерная Tn3-каркасная структура может связываться с одним эпитопом на CD40L-тримере. В еще одном варианте осуществления мономерная или мультимерная Tn3-каркасная структура может связываться с одним и тем же эпитопом по меньшей мере на двух CD40L-тримерах.
[0273] В конкретных вариантах осуществления мономерная или мультимерная Tn3-каркасная структура может связываться с одним и тем же эпитопом на двух или более копиях молекулы CD40L на поверхности смежных клеток. В конкретных вариантах осуществления мономерная или мультимерная Tn3-каркасная структура может связываться с одним и тем же эпитопом на двух или более копиях молекулы CD40L в растворе. В некоторых вариантах осуществления мономерная или мультимерная Tn3-каркасная структура может связываться с одним и тем же эпитопом или различными эпитопами на CD40L с одной и той же или различными аффинностями связывания и/или авидностями.
[0274] В другом варианте осуществления мономерные или мультимерные Tn3-каркасные структуры могут связываться с эпитопом на одной или нескольких копиях CD40L и достигать или усиливать (например, синергетически) необходимое воздействие на мишень, например, препятствовать связыванию с рецептором или препятствовать олигомеризации.
[0275] Кроме того, если мономерная или мультимерная Tn3-каркасная структура по настоящему изобретению содержит множество CD40L-специфичных мономерных субъединиц, например, различные мономеры, причем каждый мономер нацелен на различные эпитопы на CD40L, такие мономерные субъединицы могут быть упорядочены в соответствии с определенной структурой или специальной ориентацией для достижения или усиления определенного биологического эффекта. Такие комбинации мономерных субъединиц можно собрать и в дальнейшем оценить с использованием способов, известных в данной области техники.
Слияния
[0276] Настоящее изобретение предлагает Tn3-каркасные структуры, где по меньшей мере одна CD40L-специфичная мономерная субъединица может быть слитой с гетерологичным фрагментом. В данном контексте гетерологичный фрагмент не используют для соединения каркасных структур в качестве спейсера, но он может обеспечивать Tn3-каркасную структуру дополнительной функциональностью. В некоторых вариантах осуществления гетерологичный фрагмент также может выполнять функцию линкера. Настоящее изобретение охватывает применение Tn3-каркасных структур, конъюгированных или слитых с одним или несколькими гетерологичными фрагментами, включая, но без ограничения, пептиды, полипептиды, белки, белки слияния, молекулы нуклеиновой кислоты, малые молекулы, средства-миметики, синтетические лекарственные средства, неорганические молекулы и органические молекулы. Соответственно, настоящее изобретение предлагает полипептиды, содержащие один или несколько CD40L-специфичных Tn3-мономеров, включая, но без ограничения, белки слияния, описанные в данном документе.
[0277] Настоящее изобретение охватывает применение Tn3-каркасных структур, рекомбинантно слитых или химически конъюгированных с гетерологичным белком, или полипептидом, или их фрагментом. Конъюгация включает как ковалентную, так и нековалентную конъюгацию. В некоторых вариантах осуществления Tn3-каркасная структура может быть слитой или химически конъюгированной с полипептидом из по меньшей мере 10, по меньшей мере 20, по меньшей мере 30, по меньшей мере 40, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 200, по меньшей мере 300, по меньшей мере 500 или по меньшей мере 1000 аминокислот для получения белков слияния.
[0278] Слияние или конъюгирование Tn3-каркасной структуры с одним или несколькими гетерологичными фрагментами может быть непосредственным, т.е без линкера, уложенного между Tn3-каркасной структурой и гетерологичным фрагментом, или посредством одной или нескольких линкерных последовательностей, описанных в данном документе. В некоторых вариантах осуществления каркасные структуры можно применять для нацеливания гетерологичных полипептидов на конкретные типы клеток, либо in vitro, либо in vivo, путем слияния или конъюгирования Tn3-каркасных структур с антителами, специфическими по отношению к конкретным рецепторам на поверхности клетки у целевых клеток.
[0279] Tn3-каркасные структуры, которые являются слитыми или конъюгированными с гетерологичными полипептидами, также можно использовать в иммунологических анализах in vitro и способах очистки с использованием способов, известных в данной области техники. См., например, международную публикацию № WO 93/21232; европейский патент № EP 439095; Naramura et al., Immunol. Lett. 39:91-99, 1994; патент США № 5474981; Gillies et al., PNAS 89:1428-1432, 1992 и Fell et al., J. Immunol. 146:2446-2452, 1991, которые включены в данный документ посредством ссылки во всей их полноте.
[0280] В некоторых вариантах осуществления Tn3-каркасные структуры могут быть сопряжены с иммунным ответом человека путем слияния или конъюгирования каркасной структуры с иммуноглобулином или его доменом, включая, но без ограничения, константный участок IgG (Fc), например, через N- или C-конец. Подобным образом, слияние между Tn3-каркасной структурой и белком комплемента, таким как CIq, можно использовать для нацеливания на клетки.
[0281] Различные публикации, в которых описываются способы получения физиологически активных молекул, время полужизни которых модифицировано путем введения в молекулы FcRn-связывающего полипептида (см., например, WO 97/43316; патент США № 5869046; патент США № 5747035; WO 96/32478 и WO 91/14438) путем связывания молекул с антителами, FcRn-связывающие аффинности которых сохранены, но аффинности для других Fc-рецепторов были в значительной степени снижены (см., например, WO 99/43713), или путем слияния молекул с FcRn-связывающими доменами антител (смотри, например, WO 00/09560; патент США № 4703039). Специальные методики и способы увеличения времени полужизни физиологически активных молекул также можно найти в патенте США № 7083784. Более конкретно, полагают, что Tn3-каркасные структуры могут быть слиты с Fc-участком из IgG, где данный Fc-участок содержит мутации аминокислотного остатка M252Y/S254T/T256E или H433K/N434F/Y436H, где положения аминокислот обозначены в соответствии со схемой нумерации по Kabat. В особенности, предполагают, что слияние Tn3-каркасной структуры с вариантом Fc-домена не способно индуцировать ADCC.
[0282] В некоторых вариантах осуществления время полужизни Tn3-каркасной структуры может быть увеличено путем слияния при помощи методик генной инженерии Tn3-каркасной структуры, по сути, с неструктурированным рекомбинантным полипептидом (например, полипептид XTEN™) или путем конъюгации с полиэтиленгликолем (PEG).
[0283] В некоторых вариантах осуществления Tn3-каркасная структура может быть слитой с молекулами, которые увеличивают или удлиняют in vivo время полужизни в сыворотке. В некоторых вариантах осуществления данная каркасная структура может быть слитой или конъюгированной с альбумином, таким как человеческий сывороточный альбумин (HSA), неонатальным Fc-рецептором (FcRn) или его связывающем фрагментом, PEG, полисахаридами, антителами, комплементом, гемоглобином, связывающим пептидом, липопротеинами и другими факторами для увеличения ее времени полужизни в кровотоке и/или ее тканевой проницаемости. Любое из данных слияний можно получить с помощью стандартных методик, например, путем экспрессии белка слияния из рекомбинантного гена слияния, сконструированного с использованием общедоступных последовательностей генов.
[0284] В некоторых вариантах осуществления, свойство Tn3-каркасной структуры может быть улучшено путем конъюгации или слияния с вариантом HSA, т.е. молекула, полученная из полноразмерного HSA (SEQ ID NO: 139), содержащая по меньшей мере аминокислотную замену, делецию или усечение последовательности.
[0285] В некоторых вариантах осуществления свойство, улучшенное при помощи конъюгации с вариантом HSA, представляет собой время полужизни в плазме. Улучшение времени полужизни в плазме Tn3-каркасной структуры может представлять собой изменение в таком смысле, как увеличение или уменьшение времени полужизни в плазме, или изменения в других фармакокинетических параметрах. В некоторых вариантах осуществления вариант HSA представляет собой мутант, полученный из полноразмерного HSA (SEQ ID NO: 138). В конкретном варианте осуществления вариант HSA включает замену цистеина в положении 34 на серин (SEQ ID NO: 133). Варианты HSA, которые можно использовать для модифицирования времени полужизни в плазме Tn3-каркасной структуры, описаны, например, в международных публикациях WO 2011/103076 и WO 2011/051489, обе из которых включены в данный документ посредством ссылки во всей их полноте. В некоторых вариантах осуществления время полужизни в плазме Tn3-каркасной структуры по настоящему изобретению увеличивают путем слияния с вариантом HSA, содержащим по меньшей мере одну аминокислотную замену в домене III HSA.
[0286] В некоторых вариантах осуществления Tn3-каркасная структура по настоящему изобретению содержит вариант HSA, содержащий последовательность полноразмерного зрелого HSA (SEQ ID NO: 138) или ее фрагмент, за исключением по меньшей мере одной аминокислотной замены, пронумерованной относительно положения в полноразмерном зрелом HSA, в положении, выбранном из группы, состоящей из 407, 415, 463, 500, 506, 508, 509, 511, 512, 515, 516, 521, 523, 524, 526, 535, 550, 557, 573, 574 и 580; при этом по меньшей мере одна аминокислотная замена не предусматривает замену лизина (K) на глутаминовую кислоту (E) в положении 573, и при этом Tn3-каркасная структура характеризуется большим временем полужизни в плазме, чем время полужизни в плазме Tn3-каркасной структуры, не являющейся конъюгированной с вариантом HSA.
[0287] В некоторых вариантах осуществления по меньшей мере одна аминокислотная замена, пронумерованная относительно положения в полноразмерном зрелом HSA, находится в положении, выбранном из группы, состоящей из 463, 508, 523 и 524, причем указанная Tn3-каркасная структура имеет более длительное время полужизни в плазме, чем время полужизни в плазме Tn3-каркасной структуры, не являющейся конъюгированной с вариантом HSA.
[0288] В некоторых вариантах осуществления Tn3-каркасная структура по настоящему изобретению содержит вариант HSA, содержащий последовательность полноразмерного зрелого HSA (SEQ ID NO: 133 или 138) или ее фрагмент, за исключением по меньшей мере одной аминокислотной замены, пронумерованной относительно положения в полноразмерном зрелом HSA, выбранной из группы, состоящей из:
(a) замены лейцина (L) в положении 407 на аспарагин (N) или тирозин (Y);
(b) замены валина (V) в положении 415 на треонин (T);
(c) замены лейцина (L) в положении 463 на аспарагин (N);
(d) замены лизина (K) в положении 500 на аргинин (R);
(e) замены треонина (T) в положении 506 на тирозин (Y);
(f) замены треонина (T) в положении 508 на аргинин (R);
(g) замены фенилаланина (F) в положении 509 на метионин (M) или триптофан (W);
(h) замены аланина (A) в положении 511 на фенилаланин (F);
(i) замены аспарагиновой кислоты (D) в положении 512 на тирозин (Y);
(j) замены треонина (T) в положении 515 на глутамин (Q);
(k) замены лейцина (L) в положении 516 на треонин (T) или триптофан (W);
(l) замены аргинина (R) в положении 521 на триптофан (W);
(m) замены изолейцина (I) в положении 523 на аспарагиновую кислоту (D), глутаминовую кислоту (E), глицин (G), лизин (K) или аргинин (R);
(n) замены лизина (K) в положении 524 на лейцин (L);
(o) замены глутамина (Q) в положении 526 на метионин (M);
(p) замены гистидина (H) в положении 535 на пролин (P);
(q) замены аспарагиновой кислоты (D) в положении 550 на глутаминовую кислоту (E);
(r) замены лизина (K) в положении 557 на глицин (G);
(s) замены лизина (K) в положении 573 на фенилаланин (F), гистидин (H), пролин (P), триптофан (W) или тирозин (Y);
(t) замены лизина (K) в положении 574 на аспарагин (N);
(u) замены глутамина (Q) в положении 580 на лизин (K); и
(v) комбинации двух или более указанных замен,
где указанная Tn3-каркасная структура имеет более длительное время полужизни в плазме, чем время полужизни в плазме Tn3-каркасной структуры, не являющейся конъюгированной с указанным вариантом HSA.
[0289] В некоторых вариантах осуществления Tn3-каркасная структура по настоящему изобретению содержит вариант HSA, содержащий последовательность полноразмерного зрелого HSA (SEQ ID NO: 133 или 138) или ее фрагмент, за исключением по меньшей мере одной аминокислотной замены, пронумерованной относительно положения в полноразмерном зрелом HSA, выбранной из группы, состоящей из:
(a) замены лейцина (L) в положении 463 на аспарагин (N);
(b) замены треонина (Т) в положении 508 на аргинин (R);
(c) замены изолейцина (I) в положении 523 на аспарагиновую кислоту (D), глутаминовую кислоту (E), глицин (G), лизин (K) или аргинин (R);
(d) замены лизина (K) в положении 524 на лейцин (L); и
(e) комбинации двух или более указанных замен,
где указанная Tn3-каркасная структура имеет более длительное время полужизни в плазме, чем время полужизни в плазме Tn3-каркасной структуры, не являющейся конъюгированной с указанным вариантом HSA.
[0290] Более того, Tn3-каркасные структуры по настоящему изобретению могут быть слиты с маркерными последовательностями, такими как пептид, для облегчения очистки. В некоторых вариантах осуществления маркерная аминокислотная последовательность представляет собой полигистидиновый пептид (His-метка), например, oктагистидин-метка (His-8-метка) или гексагистидин-метка (His-6-метка), как, например, метка, помещенная в вектор экспрессии pQE (QIAGEN, Inc., 9259 Eton Avenue, Чэтсуорт, Калифорния, 91311), наряду с прочими векторами, многие их которых являются коммерчески доступными. Как описано у Gentz et al., Proc. Natl. Acad. Sci. USA 86:821-824, 1989, к примеру, полигистидин вносят для подходящей очистки белка слияния. Другие пептидные метки, пригодные для очистки включают, но без ограничения, гемагглютининовую ("HA") метку, которая соответствует эпитопу, полученному из белка гемагглютинина вируса гриппа (смотри, например, Wilson et al., Cell 37:767, 1984), FLAG-метку, Strep-метку, myc-метку, V5-метку, GFP-метку, AU1-метку, AU5-метку, ECS-метку, GST-метку или OLLAS-метку.
[0291] Дополнительные белки слияния, содержащие Tn3-каркасные структуры по настоящему изобретению, могут быть получены известными методиками перестановки генов, перестановки мотивов, перестановки экзонов и/или перестановки кодонов (вместе названных “перестановкой ДНК”).
[0292] Перестановку ДНК можно использовать для внесения изменения для воздействия Tn3-каркасных структур на мишень (например, получение каркасных структур с более высокими аффинностями и более низкими скоростями диссоциации). Внесение изменений в Tn3-каркасные структуры можно осуществить с помощью случайного мутагенеза посредством ПЦР с использованием ошибающейся полимеразы, произвольной вставки нуклеотида или других способов перед рекомбинацией. Одна или несколько частей полинуклеотида, кодирующего каркасную структуру, которая связывается со специфической мишенью, можно рекомбинировать с одним или несколькими компонентами, мотивами, сегментами, частями, доменами, фрагментами и т.д. одной или нескольких гетерологичных молекул.
Слияния антитела и Fc-домена
[0293] В некоторых вариантах осуществления Tn3-каркасная структура по настоящему изобретению содержит CD40L-специфичную мономерную субъединицу, слитую с доменом или фрагментом антитела (например, IgG), в том числе, но без ограничения, Fc-доменом.
[0294] В некоторых вариантах осуществления только одна CD40L-специфичная мономерная субъединица конъюгирована или слита с доменом или фрагментом антитела. К примеру, одна CD40L-специфичная мономерная субъединица может быть слитой с N-концом полипептида домена или фрагмента антитела (например, тяжелая цепь или легкая цепь антитела). В других вариантах осуществления Tn3-каркасные структуры создают путем слияния или конъюигирования одной или нескольких CD40L-специфичных мономерных субъединиц с N-концом и/или C-концом полипептида домена или фрагмента антитела (например, тяжелая цепь, и/или легкая цепь антитела, или Fc-домен).
[0295] В некоторых вариантах осуществления несколько или все CD40L-специфичные мономерные субъединицы, слитые с доменом или фрагментом антитела, являются идентичными. В некоторых других вариантах осуществления несколько или все CD40L-специфичные мономерные субъединицы, слитые с доменом или фрагментом антитела, являются различными.
[0296] В конкретном варианте осуществления Tn3-каркасная структура по настоящему изобретению содержит одну CD40L-специфичную мономерную субъединицу, слитую с Fc-доменом. В других вариантах осуществления Tn3-каркасная структура по настоящему изобретению содержит по меньшей мере две CD40L-специфичные мономерные субъединицы, слитые с Fc-доменом. В одном конкретном варианте осуществления две из CD40L-специфичных мономерных субъединиц, слитых с Fc-доменом, являются идентичными. В одном конкретном варианте осуществления две из CD40L-специфичных мономерных субъединиц, слитых с Fc-доменом, являются различными. В одном конкретном варианте осуществления две CD40L-специфичные мономерные субъединицы, слитые с Fc-доменом, соединены друг с другом в тандем, и одна из CD40L-специфичных мономерных субъединиц является слитой с Fc-доменом.
[0297] В некоторых вариантах осуществления можно получить димеры различных Tn3-каркасных структур по настоящему изобретению путем использования Fc-домена с мутациями, которые способствуют образованию гетеродимеров. В данной области техники известно, что варианты Fc-участка (например, аминокислотные замены, и/или добавления, и/или делеции) усиливают или ослабляют эффекторную функцию антитела и могут изменять фармакокинетические свойства (например, время полужизни) антитела. Таким образом, в определенных вариантах осуществления Tn3-каркасные структуры по настоящему изобретению содержат Fc-домен (домены), которые содержат измененный Fc-участок, в котором одно или несколько изменений были осуществлены в Fc-участке для изменения функциональных и/или фармакокинетических свойств Tn3-каркасной cтуктуры. В определенных вариантах осуществления Tn3-каркасные структуры по настоящему изобретению содержат Fc-домен (домены), которые содержат измененный Fc-участок, в котором одно или несколько изменений были осуществлены в Fc-участке для уменьшения или устранения по меньшей мере одной FcγR-опосредованной эффекторной функции.
[0298] Также известно, что гликозилирование Fc-участка можно модифицировать для повышения или понижения эффекторной функции и/или противовоспалительной активности. Соответственно, в одном варианте осуществления Tn3-каркасная структура по настоящему изобретению содержит Fc-участок с измененным гликозилированием аминокислотных остатков для изменения цитотоксических и/или противовоспалительных свойств Tn3-каркасных структур.
Топологии Tn3-каркасной структуры
[0299] Tn3-каркасные структуры по настоящему изобретению могут быть слиты с C-концом Fc-доменов, легкими цепями антитела и тяжелыми цепями антитела в любом подходящем пространственном расположении. См., например, международную публикацию PCT/US2011/032184 для подробного описания топологий предусмотренных каркасных структур.
Получение каркасных структур по настоящему изобретению
[0300] Tn3-каркасные структуры, описанные в данном документе, могут быть использованы в любой методике для создания новых или улучшенных мишень-связывающих белков. В одном конкретном варианте осуществления мишень является иммобилизированной на твердой подложке, такой как смола для колоночной хроматографии или лунка титровального микроплашета, и мишень контактировала с библиотекой кандидатных связывающих белков на основе каркасной структуры. Такая библиотека может состоять из клонов, сконструированных из Tn3-каркасной структуры, посредством рандомизации последовательности и/или длины CDR-подобных петель.
[0301] В связи с этим, бактериофаговый (фаговый) дисплей является одной хорошо известной методикой, которая позволяет осуществлять скрининг больших олигопептидных библиотек для идентификации представителя(ей) из данных библиотек, которые способны специфически связываться с мишенью. Фаговый дисплей представляет собой методику, с помощью которой вариантные полипептиды представляют в качестве белков слияния с белком оболочки на поверхности частиц бактериофага (Scott, J. K. and Smith, G. P. (1990) Science 249: 386). Биоинформационные подходы можно использовать для определения длины петли и предпочтительных характеристик вариабельности встречающихся в природе FnIII-доменов. При помощи этого анализа предпочтительные характеристики для длины петли и вариабельности последовательности можно использовать для разработки подхода "рандомизация с ограничением". При данной рандомизации с ограничением предпочтительные характеристики относительной длины петли и последовательности предусмотрены для разработки стратегии библиотеки. Включение анализа длины петли и вариабельности последовательности в разработку библиотеки дает в результате рандомизацию с ограничением (т.е. определенные положения в рандомизированной петле ограничены в том, что определенная аминокислота может находиться в таком положении).
[0302] Настоящее изобретение также предлагает рекомбинантные библиотеки, включающие разнообразные популяции не встречающихся в природе Tn3-каркасных структур. В одном варианте осуществления библиотеки включают не встречающиеся в природе Tn3-каркасные структуры, содержащие множество доменов с бета-цепями, связанными со множеством петлевых участков, где в один или несколько указанных петлевых участков вносят изменения путем делеции, замены или добавления по меньшей мере одной аминокислоты. В конкретном варианте осуществления библиотеки включают Tn3-каркасные структуры, полученные из Tn3-каркасной структуры дикого типа.
[0303] Как подробно описано выше, петли, соединяющие различные бета-цепи каркасных структур, можно рандомизировать по длине и/или вариабельности последовательности. В одном варианте осуществления библиотеки по настоящему изобретению содержат Tn3-каркасные структуры, имеющие по меньшей мере одну петлю, которая рандомизирована по длине и/или вариабельности последовательности. В одном варианте осуществления по меньшей мере одна, по меньшей мере две, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять или по меньшей мере шесть петель Tn3-каркасных структур рандомизированы по длине и/или вариабельности последовательности. В одном варианте осуществления по меньшей мере одна петля сохранена неизменной, тогда как по меньшей мере одна дополнительная петля рандомизирована по длине и/или вариабельности последовательности. В другом варианте осуществления по меньшей мере одна, по меньшей мере две или все три петли AB, CD и EF сохранены неизменными, тогда как по меньшей мере одна, по меньшей мере две или все три петли BC, DE и FG рандомизированы по длине или вариабельности последовательности. В другом варианте осуществления по меньшей мере одна, по меньшей мере две или по меньшей мере все три петли AB, CD и EF рандомизированы, тогда как по меньшей мере одна, по меньшей мере две или все три петли BC, DE и FG рандомизированы по длине и/или вариабельности последовательности.
[0304] В конкретном варианте осуществления библиотеки по настоящему изобретению содержат FnIII-каркасные структуры, где бета-цепь A содержит SEQ ID NO: 10 или 11, бета-цепь В содержит SEQ ID NO: 12, бета-цепь С содержит SEQ ID NO: 13 или 14, бета-цепь D содержит SEQ ID NO: 15, бета-цепь Е содержит SEQ ID NO: 16, бета-цепь F содержит SEQ ID NO: 17, и бета-цепь G содержит SEQ ID NO: 18.
[0305] В конкретном варианте осуществления библиотеки по настоящему изобретению содержат FnIII-каркасные структуры, где бета-цепь A состоит из SEQ ID NO: 10 или 11, бета-цепь В состоит из SEQ ID NO: 12, бета-цепь С состоит из SEQ ID NO: 13 или 14, бета-цепь D состоит из SEQ ID NO: 15, бета-цепь E состоит из SEQ ID NO: 16, бета-цепь F состоит из SEQ ID NO: 17, и бета-цепь G состоит из SEQ ID NO: 18.
[0306] В конкретном варианте осуществления библиотеки по настоящему изобретению содержат FnIII-каркасные структуры, где бета-цепь A практически состоит из SEQ ID NO: 10 или 11, бета-цепь В практически состоит из SEQ ID NO: 12, бета-цепь С практически состоит из SEQ ID NO: 13 или 14, бета-цепь D практически состоит из SEQ ID NO: 15, бета-цепь E практически состоит из SEQ ID NO: 16, бета-цепь F практически состоит из SEQ ID NO: 17, и бета-цепь G практически состоит из SEQ ID NO: 18.
[0307] Как подробно описано выше, один или несколько остатков в пределах петли оставлены неизменными, тогда как другие остатки рандомизированы по длине и/или вариабельности последовательности. Необязательно или в качестве альтернативы, один или несколько остатков в пределах петли ограничены заранее определенным и ограниченным числом различных аминокислот, тогда как другие остатки рандомизированы по длине и/или вариабельности последовательности. Соответственно, библиотеки по настоящему изобретению содержат Tn3-каркасные структуры, которые могут содержать одну или несколько петель, имеющих вырожденную консенсусную последовательность и/или один или несколько инвариантных аминокислотных остатков. В другом варианте осуществления библиотеки по настоящему изобретению содержат Tn3-каркасные структуры, имеющие петли BC, которые являются рандомизированными. В другом варианте осуществления библиотеки по настоящему изобретению содержат Tn3-каркасные структуры, имеющие петли BC, которые являются рандомизированными. В еще одном варианте осуществления библиотеки по настоящему изобретению содержат Tn3-каркасные структуры, имеющие петли BC, которые являются рандомизированными.
[0308] В одном варианте осуществления библиотеки по настоящему изобретению содержат Tn3-каркасные структуры, имеющие петли DE, которые являются рандомизированными. В одном варианте осуществления библиотеки по настоящему изобретению содержат Tn3-каркасные структуры, имеющие петли FG, которые являются рандомизированными. В другом варианте осуществления библиотеки по настоящему изобретению содержат FnIII-каркасные структуры, имеющие петли FG, которые являются рандомизированными.
[0309] В конкретном варианте осуществления библиотеки по настоящему изобретению содержат каркасные структуры, причем каркасные структуры содержат аминокислотную последовательность:
IEV(XAB)nALITW(XBC)nCELX1YGI(XCD)nTTIDL(XDE)nYSI(XEF)nYEVSLIC(XFG)nKETFTT,
где
(a) XAB, XBC, XCD, XDE, XEF и XFG представляют собой аминокислотные остатки, присутствующие в последовательностях петель AB, BC, CD, DE, EF и FG, соответственно;
(b) X1 представляет собой аминокислотный остаток A или T; и
(c) длина петли n представляет собой целое число от 2 до 26.
[0310] В некоторых вариантах осуществления библиотеки по настоящему изобретению содержат CD40L-специфичные Tn3-мономерные субъединицы по настоящему изобретению, содержащие аминокислотную последовательность:
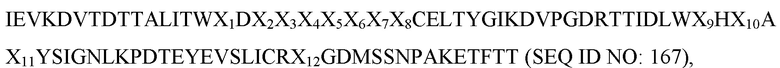
где
(a) X1 представляет собой аминокислотный остаток серин (S) или лейцин (L);
(b) X2 представляет собой аминокислотный остаток аспарагиновую кислоту (D) или глутаминовую кислоту (E);
(c) X3 представляет собой аминокислотный остаток гистидин (H), изолейцин (I), валин (V), фенилаланин (F) или триптофан (W);
(d) X4 представляет собой аминокислотный остаток аланин (A), глицин (G), глутаминовую кислоту (E) или аспарагиновую кислоту (D);
(e) X5 представляет собой аминокислотный остаток глутаминовую кислоту (E), лейцин (L), глутамин (Q), серин (S), аспарагиновую кислоту (D) или аспарагин (N);
(f) X6 представляет собой аминокислотный остаток фенилаланин (F) или тирозин (Y);
(g) X7 представляет собой аминокислотный остаток изолейцин (I), валин (V), гистидин (H), глутаминовую кислоту (E) или аспарагиновую кислоту (D);
(h) X8 представляет собой аминокислотный остаток глицин (G), триптофан (W) или валин (V);
(i) X9 представляет собой аминокислотный остаток триптофан (W), фенилаланин (F) или тирозин (Y);
(j) X10 представляет собой аминокислотный остаток серин (S), глутамин (Q), метионин (M) или гистидин (H);
(k) X11 представляет собой аминокислотный остаток триптофан (W) или гистидин (H); и
(l) X12 представляет собой аминокислотный остаток аргинин (R) или серин (S).
[0311] В некоторых вариантах осуществления библиотеки по настоящему изобретению содержат CD40L-специфичные Tn3-мономерные субъединицы по настоящему изобретению, содержащие аминокислотную последовательность:
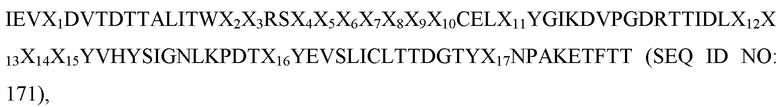
где
(a) X1 представляет собой аминокислотный остаток лизин (K) или глутаминовую кислоту (E);
(b) X2 представляет собой аминокислотный остаток треонин (T) или изолейцин (I);
(c) X3 представляет собой аминокислотный остаток аспарагин (N) или аланин (A);
(d) X4 представляет собой аминокислотный остаток серин (S), лейцин (L), аланин (A), фенилаланин (F) или тирозин (Y);
(e) X5 представляет собой аминокислотный остаток тирозин (Y), аланин (A), глицин (G), валин (V), изолейцин (I) или серин (S);
(f) X6 представляет собой аминокислотный остаток тирозин (Y), серин (S), аланин (A) или гистидин (H);
(g) X7 представляет собой аминокислотный остаток аспарагин (N), аспарагиновую кислоту (D), гистидин (H) или тирозин (Y);
(h) X8 представляет собой аминокислотный остаток лейцин (L), фенилаланин (F), гистидин (H) или тирозин (Y);
(i) X9 представляет собой аминокислотный остаток гистидин (H), пролин (P), серин (S), лейцин (L) или аспарагиновую кислоту (D);
(j) X10 представляет собой аминокислотный остаток глицин (G), фенилаланин (F), гистидин (H) или тирозин (Y);
(k) X11 представляет собой аминокислотный остаток аланин (A) или треонин (T);
(l) X12 представляет собой аминокислотный остаток серин (S), аспарагин (N), глутаминовую кислоту (E), аспарагин (R) или аспарагиновую кислоту (D);
(m) X13 представляет собой аминокислотный остаток серин (S), глутамин (Q), треонин (T), аспарагин (N) или аланин (A);
(n) X14 представляет собой аминокислотный остаток пролин (P), валин (V), изолейцин (I) или аланин (A) или не является аминокислотой;
(o) X15 представляет собой аминокислотный остаток изолейцин (I) или не является аминокислотой;
(p) X16 представляет собой аминокислотный остаток глутаминовую кислоту (E) или лизин (K); и
(q) X17 представляет собой аминокислотный остаток серин (S) или аспарагин (N).
[0312] Настоящее изобретение дополнительно предлагает способы для идентифицирования рекомбинантной Tn3-каркасной структуры, которая связывает мишень, например, CD40L, и характеризуется повышенной стабильностью или улучшенным воздействием на мишень, например, CD40L при сравнении с исходной Tn3-каркасной структурой, путем скрининга библиотек по настоящему изобретению.
[0313] В конкретных вариантах осуществления способ для идентифицирования рекомбинантной Tn3-каркасной структуры, имеющей повышенную стабильность белка по сравнению с исходной Tn3-каркасной структурой, и специфично связывающей мишень, предусматривает
[0314] приведение в контакт лиганда-мишени с библиотекой по настоящему изобретению при условиях, подходящих для образования комплекса каркасная структура:лиганд-мишень;
[0315] получение из комплекса каркасной структуры, которая связывает лиганд-мишень;
[0316] определение, является ли стабильность каркасной структуры, полученной на этапе (b), большей, чем у Tn3-каркасной структуры дикого типа.
[0317] Тот же способ можно использовать для идентификации рекомбинантной Tn3-каркасной структуры с улучшенной аффинностью связывания, авидностью и т.д в отношении мишени. В одном варианте осуществления на этапе (a) библиотеку каркасных структур по настоящему изобретению инкубируют с иммобилизированой мишенью. В одном варианте осуществления на этапе (b) комплекс каркасная структура:лиганд-мишень отмывают для удаления неспецифических связывающих веществ, и наиболее устойчивые связывающие вещества элюируют при очень жестких условиях, и осуществляют ПЦР для получения информации о последовательности. Специально предусмотрено, что связывающие вещества и/или информация о последовательности, полученные на этапе (b), могут быть использованы для создания новой библиотеки с использованием способов, раскрытых в данном документе, или известных специалисту в данной области техники, которые могут быть использованы для повтора способа отбора с или без дополнительного введения мутации в последовательность. В некоторых вариантах осуществления количество циклов отбора можно проводить до получения связывающих веществ с достаточной аффинностью к антигену.
[0318] Дополнительным вариантом осуществления настоящего изобретения является накопление молекул выделенной нуклеиновой кислоты, кодирующей библиотеку, содержащую каркасные структуры по настоящему изобретению, как описано выше.
[0319] Каркасные структуры по настоящему изобретению можно подвергнуть созреванию аффинности. При данном способе, принятом в данной области техники, специфический связывающий белок подвергают процедуре, при которой осуществляется отбор по повышенной аффинности к специфичной мишени (см. Wu et al., Proc. Natl. Acad. Sci. USA. 95(11):6037-42). Полученные в результате каркасные структуры настоящего изобретения могут проявлять по меньшей мере настолько же высокие характеристики связывания по сравнению с каркасными структурами перед созреванием аффинности.
[0320] Настоящее изобретение также предлагает способы идентифицирования аминокислотной последовательности, кодирующей белок каркасной структуры, способной связываться с мишенью с тем, чтобы образовывать комплекс каркасная структура:мишень. В одном варианте осуществления данный способ предусматривает (a) приведение в контакт библиотеки настоящего изобретения с иммобилизированной или отделяемой мишенью; (b) отделение комплексов каркасная структура:мишень от свободных каркасных структур; (c) инициирование репликации отделенных каркасных структур из (b) с тем, чтобы привести к образованию новой полипептидной библиотеки дисплея, отличной от таковой в (a), при наличии пониженной вариабельности и будучи обогащенной в представленных каркасных структурах, способных к связыванию мишени; d) необязательного повтора этапов (a) и (b) с новой библиотекой из (c); и e) определение последовательности нуклеиновых кислот участка, кодирующего представленную каркасную структуру вида из (d), и, следовательно, установление последовательности пептидов, способной связываться с мишенью.
[0321] В другом варианте осуществления Tn3-каркасные структуры настоящего изобретения можно дополнительно рандомизировать после идентификации в скринированной библиотеке. В одном варианте осуществления способы настоящего изобретения включают дополнительное рандомизирование по меньшей мере одной, по меньшей мере двух, по меньшей мере трех, по меньшей мере четырех, по меньшей мере пяти или по меньшей мере шести петель из каркасной структуры, идентифицированной в библиотеке, с использованием способа, описанного в данном документе. В другом варианте осуществления дополнительно рандомизированную каркасную структуру подвергают следующему способу идентифицирования каркасной структуры, способной к связыванию мишени. Данный способ включает (a) приведение в контакт дополнительно рандомизированной каркасной структуры с иммобилизированной или отделяемой мишенью, (b) отделение комплексов дополнительно рандомизированная каркасная структура:мишень от свободных каркасных структур, (c) инициирование репликации отделенных каркасных структур из (b), необязательный повтор этапов (a)-(c), и (d) определение последовательности нуклеиновых кислот участка, кодирующего указанную дополнительно рандомизированную каркасная структуру, и, следовательно, установление последовательности пептидов, способной связываться с мишенью.
[0322] В дополнительном варианте осуществления дополнительно рандомизированные каркасные структуры содержат по меньшей мере одну, по меньшей мере две, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять или по меньшей мере шесть рандомизированных петель, которые предварительно рандомизировали в первой библиотеке. В альтернативном дополнительном варианте осуществления дополнительно рандомизированные каркасные структуры содержат по меньшей мере одну, по меньшей мере две, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять или по меньшей мере шесть рандомизированных петель, которые не были предварительно рандомизированы в первой библиотеке.
[0323] Настоящее изобретение также предлагает способ получения по меньшей мере двух Tn3-каркасных структур, которые связываются с по меньшей мере одной или несколькими мишенями. Данный способ позволяет проводить скрининг средств, которые совместно воздействуют для активации определенного ответа. Может быть преимущественным использование такого скрининга, когда необходима активность агонистов, требующая кооперативного взаимодействия более чем одной каркасной структуры. Данный способ позволяет проводить скрининг кооперативно взаимодействующих средств с суммарной активностью без изменения формата библиотеки с образованием мультимерных комплексов. В одном варианте осуществления способ настоящего изобретения включает приведение в контакт лиганда-мишени с библиотекой настоящего изобретения при условиях, которые позволяют формироваться комплексу каркасная структура:лиганд-мишень, приводя в контакт указанные каркасные структуры со средством для перекрестного связывания (определено как средство, которое приводит в непосредственную близость по меньшей мере идентичные или отличающиеся каркасные структуры), где перекрестное связывание каркасных структур активирует поддающийся детекции ответ и получение из комплекса указанных каркасных структур, которые связывают мишень. В дополнительном варианте осуществления средство для перекрестного связывания представляет собой специфичное к каркасной структуре антитело или его фрагмент, специфичное к эпитопной метке антитело или его фрагмент, домен димеризации, такой как Fc-участок, суперспиральный мотив (например, но не ограничивая, "лейциновая застежка"), химическое перекрестно-сшивающее средство или другой домен димеризации, известный в данной области.
[0324] Настоящее изобретение также предлагает способы детектирования соединения, использующего Tn3-каркасные структуры по настоящему изобретению. Исходя из специфичностей связывания Tn3-каркасных структур, полученных с помощью скрининга библиотеки, возможным является использование таких Tn3-каркасных структур в анализах для детекции специфичной мишени в образце, таких как диагностические способы. В одном варианте осуществления данный способ детектирования соединения включает приведение в контакт указанного соединения в образце с Tn3-каркасной структурой настоящего изобретения при условиях, которые позволяют формирование комплекса соединение: каркасная структура и детектирование каркасной структуры, тем самым детектируя указанное соединение в образце. В дополнительных вариантах осуществления каркасную структуру метят (т.е радиоизотопная метка, флуоресцентная, связанная с ферментом или колориметрическая метка) для облегчения обнаружения соединения.
[0325] Настоящее изобретение также предлагает способы фиксации соединения, использующего Tn3-каркасные структуры по настоящему изобретению. Исходя из специфичностей связывания Tn3-каркасных структур, полученных с помощью скрининга библиотеки, возможным является использование таких Tn3-каркасных структур в анализах для связывания специфичной мишени в образце, таких как для способов очистки. В одном варианте осуществления данный способ связывания соединения включает приведение в контакт указанного соединения в образце с каркасной структурой по настоящему изобретению при условиях, которые позволяют образование комплекса соединение: каркасная структура и удаление данного комплекса из образца, тем самым обеспечивая связывание указанного соединение в указанном образце. В дополнительных вариантах осуществления Tn3-каркасная структура является иммобилизированной для облегчения удаления комплекса соединение:каркасная структура.
[0326] В некоторых вариантах осуществления Tn3-каркасные структуры, выделенные из библиотек настоящего изобретения, содержат по меньшей мере одну, по меньшей мере две, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть или более рандомизированных петель. В некоторых вариантах осуществления выделенные петлевые последовательности Tn3-каркасной структуры можно переместить из донорной каркасной структуры в любую петлю принимающей FnIII-каркасной структуры, включая, но не ограничивая, принимающую Tn3-каркасную структуру (например, петлевую последовательность FG из донорной каркасной структуры можно переместить из донорной каркасной структуры в любую петлю в принимающей FnIII-каркасной структуре). В конкретных вариантах осуществления выделенные петлевые последовательности можно переместить в когнатную петлю в принимающей каркасной структуре (например, петлевую последовательность FG из донорной каркасной структуры можно переместить в принимающую FnIII-каркасную структуру в положение петли FG). В некоторых вариантах осуществления выделенные петлевые последовательности могут быть "перемешаны и совмещены" случайным образом с различным принимающими каркасными структурами.
[0327] В других вариантах осуществления выделенные последовательности, кодирующие Tn3-каркасные структуры, можно идентифицировать с помощью петлевой последовательности. Например, библиотеку используют для реакции связывания с определенной мишенью и выделяют коллекцию специфических связывающих веществ. Рандомизированные петлевые последовательности могут характеризоваться как специфичные последовательности, независимо от окружения Tn3-каркасной структуры (т.е каркасная структура, которая связывает мишень X, где указанная каркасная структура содержит петлевую последовательность FG из SEQ ID NO:X). В альтернативных вариантах осуществления, где каркасная структура имеет петлевые последовательности, которые связывают мишень X, петлевые последовательности могут характеризоваться как связывающие мишень X в отсутствие последовательности каркасной структуры. Иными словами, предполагается, что каркасные структуры, выделенные из библиотеки, которые связывают конкретную мишень, могут экспрессироваться в виде вариабельных последовательностей петли, которые связывают ту мишень, независимую от остова каркасной структуры. Данный процесс будет аналогичен концепции CDR в вариабельных участках антител.
Созревание аффинности
[0328] Разработка Tn3-каркасных структур настоящего изобретения может включать один или несколько этапов созревания аффинности in vitro или in vivo. В некоторых вариантах осуществления Tn3-мономерные субъединицы могут подвергнуть одиночному этапу созревания аффинности. В других вариантах осуществления Tn3-мономерные субъединицы можно подвергнуть двум или более этапам созревания аффинности. Может использоваться любой подход созревания аффинности, что в результате приводит, как правило, к аминокислотным изменениям в исходной Tn3-каркасной структуре, или, более конкретно, к аминокислотным изменениям в петлях исходной Tn3-каркасной структуры, что улучшает связывание Tn3-каркасной структуры со зрелой аффинностью с необходимым антигеном.
[0329] Данных аминокислотных изменений можно, например, достигнуть посредством случайного мутагенеза, "прогонного" мутагенеза и "сканирующего" мутагенеза. Подобный мутагенез может быть достигнут использованием, например, ПЦР с использованием ошибающейся полимеразы, штаммов-“мутаторов” дрожжей или бактерий, внедрение случайных или определенных замен в нуклеиновых кислотах во время ab initio синтеза все или части связывающей молекулы на основе FnIII. Способы осуществления созревания аффинности и/или мутагенеза описаны, например, в патентах США №№ 7195880; 6951725; 7078197; 7022479; 5922545; 5830721; 5605793, 5830650; 6194550; 6699658; 7063943; 5866344 и PCT публикации WO06023144.
[0330] Такие способы созревания аффинности могут дополнительно требовать того, чтобы жесткость антиген-связывающего скринингового исследования повышали для отбора Tn3-каркасных структур с улучшенной аффинностью к антигену. Принятые в данной области техники способы повышения жесткости анализа белок-белкового взаимодействия, можно использовать в данном случае. В одном варианте осуществления одно или несколько условия для анализа варьируют (например, концентрация солей в буфере для анализа) для уменьшения аффинности Tn3-каркасной структуры к требуемому антигену. В другом варианте осуществления период времени, который дает возможность Tn3-каркасной структуре связываться с требуемым антигеном, сокращен.
[0331] В другом варианте осуществления к анализу белок-белкового взаимодействия может быть добавлен этап конкурентного связывания. Например, сперва дают возможность Tn3-каркасной структуре связаться с требуемым иммобилизированным антигеном. Затем добавляют неиммобилизированный антиген в определенной концентрации, который служит в качестве конкурента за связывание с иммобилизированным антигеном, чтобы таким образом Tn3-каркасные структуры с наиболее низкой аффинностью к антигену извлекать из иммобилизированного антигена, что приводит к отбору Tn3-каркасных структур с улучшенной антиген-связывающей аффинностью. Жесткость условий анализа можно дополнительно повысить путем увеличения концентрации неиммобилизированного антиген, который добавляют в анализ.
[0332] Способы скрининга также могут требовать множества циклов отбора для обогащения одной или нескольких Tn3-каркасных структур улучшенным антиген-связыванием. В одном варианте осуществления в каждом цикле отбора дополнительные аминокислотные мутации вводят в Tn3-каркасную структуру. В другом варианте осуществления в каждом цикле отбора жесткость связывания связывание с требуемым антигеном повышают для отбора Tn3-каркасных структур с повышенной аффинностью к антигену.
[0333] В некоторых вариантах осуществления созревание аффинности осуществляют путем насыщающего мутегенеза частей петель BC, DE и FG из Tn3. В некоторых вариантах осуществления насыщающий мутагенез проводят с использованием мутагенеза по Кункелю. В других вариантах осуществления насыщающий мутагенез проводят с использованием ПЦР.
[0334] В некоторых вариантах осуществления используют по меньшей мере один, по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять или более пяти циклов созревания аффинности. В некоторых вариантах осуществления насыщающий мутагенез применяют относительно только одной петли, тогда как в некоторых других вариантах осуществления только одну петлю или часть петли подвергают мутации на протяжении одного цикла созревания аффинности. В некоторых вариантах осуществления более чем одну петлю или части одной или нескольких петель подвергают мутации на протяжении одного цикла созревания аффинности.
[0335] В других вариантах осуществления петли BC, DE и FG одновременно подвергают мутации на протяжении одного и того же цикла созревания аффинности.
[0336] В случае мономеров для сборки в мультимерной Tn3-каркасной структуре, связывающейся с различными эпитопами одной и той же мишени, каждую специфичность связывания можно независимо подвергнуть скринингу.
[0337] В некоторых вариантах осуществления петли рандомизируют с использованием библиотеки фагового дисплея. В некоторых вариантах осуществления связывание Tn3-каркасной структуры с требуемой мишенью можно определить, используя способы, принятые в данной области техники. Также аминокислотные последовательности Tn3-каркасных структур, идентифицированные в скринингах, можно определять, используя способы, принятые в данной области техники.
[0338] В некоторых вариантах осуществления мономерные каркасные структуры со зрелой аффинностью по настоящему изобретению проявляют повышенную аффинности к CD40L в по меньшей мере 5 раз, по меньшей мере 10 раз, по меньшей мере 20 раз, по меньшей мере 40раз, по меньшей мере 6o раз, по меньшей мере 80 раз или по меньшей мере 100 раз или более по сравнению с той же самой Tn3-каркасной структурой до созреванием аффинности, как определено с помощью поверхностного плaзмонного резонанса или с помощью других анализов, известных в данной области техники. В некоторых вариантах осуществления мономерные каркасные структуры со зрелой аффинностью по настоящему изобретению имеют константу диссоциации (Kd) менее 5 мкМ, менее 1 мкМ, менее 500 мкМ, менее 250 мкМ, менее 100 мкМ или менее 50 мкМ, как определено с помощью поверхностного плaзмонного резонанса или с помощью других анализов, известных в данной области техники.
[0339] Данные способы созревания аффинности можно использовать для разработки Tn3-каркасных структур с требуемыми улучшенными свойствами связывания, такими как повышенная аффинность или другими требуемыми характеристиками, такими как благоприятные фармакокинетические свойства, высокая активность, низкая иммуногенность, повышенная или пониженная перекрестная реактивность и т.д.
Получение тандемных повторов
[0340] Связывание тандемных конструктов, димера, образованного путем связывания двух CD40L-специфичных мономерных субъединиц, может быть получено путем лигирования олигонуклеотидов в сайтах рестрикции с использованием рестриктаз, известных в данной области техники, включая, но без ограничения, рестриктазы типа II и типа IIS.
[0341] Мультимерные Tn3-каркасные структуры по настоящему изобретению могут содержать линкер на C-конце и/или N-конце и/или между доменами, как описано в данном документе. Дополнительно, каркасные структуры настоящего изобретения содержат по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8 или более полипептидов, при этом каркасные структуры могут быть слитыми или конъюгированными с доменом димеризации, включая, но без ограничения, фрагмент антитела, выбранный из следующего:
(i) Fab фрагмент, имеющий VL-, CL-, VH- и CH1-домены;
(ii) Fab' фрагмент, который представляет собой Fab фрагмент, имеющий один или несколько остатков цистеина на C-конце CH1-домена;
(iii) Fd фрагмент, имеющий VH- и CH1-домены;
(iv) Fd' фрагмент, имеющий VH- и CH1-домены и один или несколько остатков цистеина на C-конце CH1-домена;
(v) Fv фрагмент, имеющий VL- и VH-домены одного плеча антитела;
(vi) dAb фрагмент, который состоит из VH-домена;
(vii) выделенные CDR-участки;
(viii) F(ab')2 фрагменты, бивалентный фрагмент, включающий два Fab' фрагмента, соединенных посредством дисульфидного мостика в шарнирной области;
(ix) молекулы одноцепочечных антител (например, одноцепочечный Fv; scFv);
(x) "диатело" с двумя антиген-связывающими сайтами, содержащее вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) в той же самой полипептидной цепи;
(xi) "линейное антитело", содержащее пару тандемных Fd-сегментов (VH-CH1-VH-CH1), которые вместе с комплементарными полипептидами легкой цепи образуют пару антиген-связывающих участков;
(xii) полноразмерное антитело; и
(xiii) Fc участок, содержащий CH2-CH3, который может дополнительно содержать все или часть шарнирного участка и/или CH1-участок.
Получение Tn3-каркасной структуры
[0342] Рекомбинантная экспрессия Tn3-каркасной структуры по настоящему изобретению требует конструирования вектора экспрессии, содержащего полинуклеотид, который кодирует Tn3-каркасную структуру. Как только был получен полинуклеотид, кодирующий Tn3-каркасную структуру, вектор для получения Tn3-каркасной структуры может быть получен с помощью технологии рекомбинантной ДНК, используя методики, хорошо известные в данной области техники. Таким образом, способы получения белка посредством экспрессии полинуклеотида, содержащего Tn3-каркасную структуру, кодирующую нуклеотидную последовательность, описаны в данном документе. Способы, которые хорошо известны специалистам в данной области техники, можно применять для конструирования векторов экспрессии, содержащих последовательность, кодирующую полипептиды каркасных структур, и соответствующие сигналы регуляции транскрипции и трансляции. Эти способы включают, например, in vitro методики рекомбинантных ДНК, методики синтеза и in vivo генетическую рекомбинацию. Настоящее изобретение, таким образом, предлагает реплицируемые векторы, содержащие нуклеотидную последовательность, кодирующую Tn3-каркасную структуру настоящего изобретения, функционально связанную с промотором.
[0343] Вектор экспрессии переносят в клетку-хозяина с помощью стандартных методик, и трансфицированные клетки затем культивируют с помощью стандартных методик с получением Tn3-каркасной структуры настоящего изобретения. Таким образом, настоящее изобретение включает клетки-хозяева, содержащие полинуклеотид, кодирующий каркасную структуру настоящего изобретения, функционально связанную с гетерологичным промотором. Пригодные клетки-хозяева включают, но без ограничения, микроорганизмы, такие бактерии (например, E. coli и B. subtilis).
[0344] Множество систем экспрессии хозяин-вектор могут быть использованы для экспрессии Tn3-каркасных структур настоящего изобретения. Такие системы экспрессии хозяин-вектор представляют собой носители, с помощью которых кодирующие последовательности, представляющие интерес, можно получать и впоследствии очищать, но также представляют собой клетки, которые могут, будучи трансформированными или трансфицированными с помощью подходящих нуклеотидных кодирующих последовательностей, экспрессируют каркасную структуру по настоящему изобретению in situ. Такие включают, но без ограничения, микроорганизмы, такие как бактерии (например, E. coli и B. subtilis), трансформированные векторами экспрессии на основе ДНК рекомбинантного бактериофага, плазмидной ДНК или космидной ДНК, содержащими последовательности, кодирующие каркасную структуру, или системы на основе клеток млекопитающих (например, клетки COS, CHO, BHK, 293, NSO и 3T3).
[0345] Раскрыты способы, пригодные для получения Tn3-каркасной структуры настоящего изобретения, например, в международной патентной публикации № WO 2009/058379. Как только была получена каркасная структура настоящего изобретения путем рекомбинантной экспрессии, она может быть очищена с помощью любого способа, известного в данной области техники, для очистки белка.
[0346] В некоторых вариантах осуществления каркасные структуры настоящего изобретения можно получить в негликозилированной форме путем замещения аминокислотных остатков, которые могут быть гликозилированы во время рекомбинантной экспрессии. В одном конкретном варианте осуществления аминокислоты серин в линкере глицин-серин (например, SEQ ID NO: 131 или SEQ ID NO: 132) можно заменить на другие аминокислотные остатки, такие как аланин, глицин, лейцин, изолейцин или валин (смотри, например, SEQ ID NO: 140, 141, 142 и 143) для предупреждения гликозилирования во время рекомбинантной экспрессии. В некоторых конкретных вариантах осуществления сайт N-гликозилирования удаляют из Tn3-каркасных структур настоящего изобретения. В других вариантах осуществления каркасную структуру настоящего изобретения можно дегликозилировать после рекомбинантной экспрессии. Способы дегликозилирования in vitro после рекомбинантной экспрессии с использованием, например, ферментных смесей известны в данной области техники (например, наборы для дегликозилирования PFGase F, Enodo F Multi, Orela O-linked Glycan Release, Enzymatic CarboRelease и Enzymatic DeGlycoMx, поставляемые QA-bio, Палм-Дезерт, Калифорния).
[0347] Получение Tn3-каркасных структур настоящего изобретения в научно-исследовательской лаборатории можно увеличить в масштабе для получения каркасных структур в реакторах аналитического масштаба или реакторах промышленного масштаба, как описано в публикации патента США № 2010-0298541 A1.
Масштабируемое производство секретированных Tn3-каркасных структур
[0348] Tn3-каркасные структуры настоящего изобретения могут быть получены внутриклеточно или в виде секретированной формы. В некоторых вариантах осуществления секретированные каркасные структуры надлежащим образом уложены и полностью функциональны. Tn3 каркасные структуры настоящего изобретения можно получить с помощью масштабируемого способа. В некоторых вариантах осуществления каркасные структуры могут быть получены с помощью масштабируемого способа настоящего изобретения в научно-исследовательской лаборатории, при котором может быть увеличен масштаб для получения Tn3-каркасных структур настоящего изобретения в биореакторах аналитического масштаба (например, но не ограничивая 5 л, 10 л, 15 л, 30 л или 50 л биореакторах). В других вариантах осуществления Tn3-каркасные структуры могут быть получены с помощью масштабируемого способа настоящего изобретения в научно-исследовательской лаборатории, при котором может быть увеличен масштаб для получения Tn3-каркасных структур настоящего изобретения в биореакторах промышленного масштаба (например, но не ограничивая, 75 л, 100 л, 150 л, 300 л или 500 л). В некоторых вариантах осуществления масштабируемый способ настоящего изобретения в результате приводит к небольшому уменьшению или не приводит к уменьшению эффективности производства в сравнении со способом получения, выполненном в научно-исследовательской лаборатории.
Линкеры
[0349] Мономерные субъединицы в мультимерной Tn3-каркасной структуре могут быть соединены белковыми и/или небелковыми линкерами, где каждый линкер является слитым по меньшей мере с двумя мономерными субъединицами. Подходящий линкер может состоять из белкового линкера, небелкового линкера и их комбинаций. Комбинации линкеров могут быть гомомерными или гетеромерными. В некоторых вариантах осуществления мультимерная Tn3-каркасная структура настоящего изобретения содержит множество мономерных субъединиц, где все линкеры идентичны. В других вариантах осуществления мультимерная Tn3-каркасная структура содержит множество мономерных субъединиц, где по меньшей мере один из линкеров функционально или структурно отличается от остальных линкеров. В некоторых вариантах осуществления линкеры могут сами по себе содействовать активности мультимерной Tn3-каркасной структуры путем непосредственного или опосредственного участия в связывании с мишенью.
[0350] В некоторых вариантах осуществления белковый линкер представляет собой полипептид. Линкер-полипептид должен иметь длину, которая достаточна для связывания двух или более мономерных субъединиц таким способом, что они предполагают правильную конформацию относительно друг друга, сохраняя таким образом требуемую активность.
[0351] В одном варианте осуществления полипептидный линкер содержит от 1 до приблизительно 1000 аминокислотных остатков, от 1 до приблизительно 50 аминокислотных остатков, 1-25 аминокислотных остатков, 1-20 аминокислотных остатков, 1-15 аминокислотных остатков, 1-10 аминокислотных остатков, 1-5 аминокислотных остатков, 1-3 аминокислотных остатков. Настоящее изобретение дополнительно предлагает нуклеиновые кислоты, такие как ДНК, РНК или комбинации и той, и другой, кодирующие последовательность полипептидного линкера. Аминокислотные остатки, выбранные для включения в полипептидный линкер, должны проявлять свойства, которые существенно не затрагивают активность или функцию мультимерной Tn3-каркасной структуры настоящего изобретения. Таким образом, полипептидный линкер должен в целом не проявлять заряд, который был бы несовместим с активностью или функцией Tn3-мультимерной каркасной структуры настоящего изобретения, или затрагивать внутренние укладки, или образовывать связи или другие взаимодействия с аминокислотными остатками в одной или нескольких мономерных субъединицах, которые бы серьезно препятствовали связыванию мультимерной Tn3-каркасной структуры настоящего изобретения с CD40L.
[0352] Использование встречающихся в природе, а также искусственных пептидных линкеров для соединения полипептидов в новые связанные полипептиды слияния хорошо известно в литературе. Соответственно, линкеры, обеспечивающие слияние двух или более мономерных субъединиц, представляют собой природные линкеры, искусственные линкеры или их комбинации. В некоторых вариантах осуществления аминокислотные последовательности всех пептидных линкеров, представленных в Tn3-мультимерной каркасной структуре настоящего изобретения, являются идентичными. В некоторых вариантах осуществления аминокислотные последовательности по меньшей мере двух из пептидных линкеров, представленных в мультимерной Tn3-каркасной структуре настоящего изобретения, являются различными.
[0353] В некоторых вариантах осуществления полипептидный линкер обладает конформационной гибкостью. В некоторых вариантах осуществления последовательность полипептидного линкера содержит аминокислотную последовательность (G-G-G-G-X)m, где X представляет собой аланин (A), cерин (S), глицин (G), изолейцин (I), лейцин (L) или валин (V) и m представляет собой положительное целое число (смотри, например, SEQ ID NO: 209). В конкретном варианте осуществления последовательность полипептидного линкера содержит аминокислотную последовательность (G-G-G-G-S)m, где m представляет собой положительное целое число (смотри, например, SEQ ID NO: 147). В другом конкретном варианте осуществления последовательность полипептидного линкера содержит аминокислотную последовательность (G-G-G-G-G)m, где m представляет собой положительное целое число (смотри, например, SEQ ID NO: 148). В еще одном конкретном варианте осуществления последовательность полипептидного линкера содержит аминокислотную последовательность (G-G-G-G-A)m, где m представляет собой положительное целое число (cмотри, например, SEQ ID NO: 149). В некоторых вариантах осуществления полипептидный линкер представляет собой по своей природе неструктурированный природный или искусственный полипептид (смотри, например, Schellenberger et al., Nature Biotechnol. 27:1186-1190, 2009; смотри также Sickmeier et al., Nucleic Acids Res. 35:D786-93, 2007).
[0354] Пептидный линкер может быть модифицирован таким способом, что вводится аминокислотный остаток, содержащий присоединенную группу для неполипептидного фрагмента. Примером таких аминокислотных остатков может быть остаток цистеина (к которому в дальнейшем присоединяется неполипептидный фрагмент), или аминокислотная последовательность может включать in vivo сайт N-гликозилирования (тем самым присоединяя остаток сахара (in vivo) к пептидному линкеру).
[0355] В некоторых вариантах осуществления, аминокислотные последовательности всех пептидных линкеров, представленных в полипептидном мультимере, являются идентичными. В качестве альтернативы, аминокислотные последовательности всех пептидных линкеров, представленных в полипептидном мультимере, являются различными.
Мечение или конъюгирование Tn3-каркасных структур
[0356] Tn3-каркасные структуры настоящего изобретения можно использовать в неконъюгированной форме или конъюгированной по меньшей мере с одним из ряда гетерологичных фрагментов для облегчения обнаружения мишени, для диагностической визуализации или терапии. При проведении очистки Tn3-каркасные структуры могут быть мечеными или конъюгированными либо перед, либо после очистки.
[0357] Многие гетерологичные фрагменты не имеют в достаточном количестве подходящих функциональных групп, с которыми Tn3-каркасные структуры настоящего изобретения могут быть соединены. Таким образом, в некоторых вариантах осуществления эффекторная молекула присоединена к каркасной структуре посредством линкера, где данный линкер содержит реакционноспособные группы для конъюгирования. В некоторых вариантах осуществления гетерологичный фрагмент, конъюгированный с Tn3-каркасной структурой настоящего изобретения, может выполнять функцию линкера. В других вариантах осуществления фрагмент является конъюгированным с Tn3-каркасной структурой через линкер, который может быть расщепляемым или нерасщепляемым. В одном варианте осуществления расщепляемая связывающая молекула представляет собой расщепляемую при изменении окислительно-восстановительного потенциала связывающую молекулу, вследствие чего связывающая молекула расщепляется в средах с более низким окислительно-восстановительным потенциалом, таких как цитоплазма и другие участки с более высокими концентрациями молекул со свободными сульфгидрильными группами. Примеры соединяющих молекул, которые могут расщепляться в результате изменения окислительно-восстановительного потенциала, включают такие, которые содержат дисульфиды.
[0358] В некоторых вариантах осуществления, Tn3-каркасные структуры настоящего изобретения сконструированы для обеспечения реакционноспособных групп для конъюгирования. В таких каркасных структурах N-конец и/или C-конец служит для обеспечения реакционноспособных групп для конъюгирования. В других вариантах осуществления N-конец может быть конъюгированным с одним фрагментом (таким как, но без ограничения, PEG), тогда как C-конец является конъюгированным с другим фрагментом (таким как, но без ограничения, биотин) или наоборот.
[0359] Выражение "полиэтиленгликоль" или "PEG" означает соединение полиэтиленгликоля или его производное с или без связывающих средств, связывающих или активирующих фрагментов (например, с тиоловым, трифлатовым, трезилатовым, азиридиновым, оксирановым, N-гидроксисукцинимидным или малеинимидным фрагментом). Выражение "PEG" предназначено для обозначения полиэтиленгликоля с молекулярной массой от 500 до 150000 Да, в том числе его аналогов, где, к примеру, концевую OH-группу заменяли метоксигруппой (обозначен как mPEG).
[0360] Tn3-каркасные структуры настоящего изобретения могут быть дериватизированы полиэтиленгликолем (PEG). PEG представляет собой линейный, водорастворимый полимер из повторяющихся единиц этиленоксида с двумя концевыми гидроксильными группами. PEG классифицирую по их молекулярным массам, которые, как правило, варьируют от приблизительно 500 дальтон до приблизительно 40000 дальтон. В конкретном варианте осуществления используемые PEG имеют молекулярные массы в пределах от 5000 дальтон до приблизительно 20000 дальтон. PEG, связанные с каркасными структурами настоящего изобретения, могут быть либо разветвленными, либо неразветвленными. Смотри, например, Monfardini, C. et al. 1995 Bioconjugate Chem 6:62-69. PEG являются коммерчески доступными от Nektar Inc., Sigma Chemical Co. и других компаний. Такие PEG включают, но без ограничения, монометоксиполиэтиленгликоль (MePEG-OH), монометоксиполиэтиленгликоль-сукцинат(MePEG-S), монометоксиполиэтиленгликоль-сукцинимидилсукцинат (MePEG-S- NHS), монометоксиполиэтиленгликоль-амин (MePEG-NH2), монометоксиполиэтиленгликоль-трезилат (MePEG-TRES) и монометоксиолиэтиленгликоль-имидазолилкарбонил (MePEG-IM).
[0361] Вкратце, гидрофильный полимер, который используют, например, PEG, кэппируют на одном конце нереакционноспособной группой, такой как метокси или этоксигруппа. Затем полимер активируют на другом конце реакцией с подходящим активирующим средством, таким как циануровые галогениды (например, циануровый хлорид, бромид или фторид), карбонилимидазол, ангидридный реактив (например, дигалогенсукциновый ангидрид, такой как дибромсукциновый ангидрид), ацилазид, п-диазонийбензиловый эфир, 3-(п-диазонийфенокси)-2-гидроксипропиловый эфир) и тому подобные. Активированный полимер затем вступает в реакцию с полипептидом, как описано в данном документе, для получения полипептида, дериватизированного с полимером. В качестве альтернативы, функциональная группа в Tn3-каркасных структурах настоящего изобретения может быть активирована для реакции с полимером, или две группы могут быть соединены в согласованной реакции сочетания, используя известные способы сочетания. Будет легко понять, что полипептиды настоящего изобретения могут быть дериватизированы с PEG, используя большое число других реакционных схем известных и используемых специалистами в данной области техники. PEG может быть соединен с каркасной структурой настоящего изобретения в одной или нескольких функциональных группах либо на конце Tn3-каркасной структуры, либо внутри Tn3-каркасной структуры. В конкретных вариантах осуществления PEG присоединен либо на N-конце, либо на C-конце.
[0362] В других вариантах осуществления Tn3-каркасные структуры настоящего изобретения, их аналоги или производные могут быть конъюгированными с диагностическим или рентгеноконтрастным средством. Такие Tn3-каркасные структуры могут быть пригодны для мониторинга или прогнозирования развития или прогрессирования заболевания как части процедуры клинического испытания, такой как определение эффективности конкретной терапии.
[0363] Настоящее изобретение дополнительно охватывает применения Tn3-каркасных структур, конъюгированных с терапевтическим фрагментом. Tn3-каркасная структура может быть конъюгирована с терапевтическим фрагментом, таким как цитотоксин, например, цитостатик или средство, разрушающее клетки, терапевтическое средство или ион радиоактивного металла, например, вещество, излучающее альфа-частицы. Цитотоксин или цитотоксическое средство включает любое средство, которое является вредным для клеток.
Анализ Tn3-каркасных структур
[0364] Аффинность связывания и другие свойства связывания каркасной структуры Tn3 с антигеном можно определить с помощью разнообразных способов анализа in vitro, известных в данной области техники, в том числе, например, равновесных способов (например, твердофазного иммуноферментного анализа (ELISA) или кинетического анализа (например, анализа BIACORE®)) и других способов, таких как анализы непрямого связывания, анализы конкурентного связывания, гель-электрофорез и хроматография (например, гель-фильтрация). В этих и других способах может быть задействована метка на одном или нескольких исследуемых компонентах и/или могут использоваться разнообразные способы выявления, в том числе, без ограничений, с применением хромогенных, флуоресцентных, люминесцентных или изотопных меток. Подробное описание аффинности и кинетики связывания можно найти в Paul, W.E., ed., Fundamental Immunology, 4th Ed., Lippincott-Raven, Philadelphia (1999).
[0365] В некоторых вариантах осуществления Tn3-каркасные структуры по настоящему изобретению специфически связываются с мишенью при специфических кинетических показателях. В некоторых вариантах осуществления Tn3-каркасные структуры по настоящему изобретению могут характеризоваться константой диссоциации или Kd (koff/kon), составляющей менее 1×10−2 M, 1×10−3 M, 1×10−4 M, 1×10−5 M, 1×10−6 M, 1×10−7 M, 1×10−8 M, 1×10−9 M, 1×10−10 M, 1×10−11 M, 1×10−12 M, 1×10−13 M, 1×10−14 M или менее 1×10−15 M. В конкретных вариантах осуществления Tn3-каркасные структуры по настоящему изобретению характеризуются Kd, составляющей 500 мкМ, 100 мкМ, 500 нМ, 100 нМ, 1 нМ, 500 пM, 100 пM или менее, определенной в анализе BIAcore® или в других анализах, известных в данной области техники.
[0366] В альтернативном варианте осуществления аффинность каркасных структур Tn3 по настоящему изобретению описывается относительно константы ассоциации (Ka), вычисляемой как отношение kon/koff, которая составляет по меньшей мере 1×102 M-1, 1×103 M-1, 1×104 M-1, 1×105 M-1, 1×106 M-1, 1×107 M-1, 1×108 M-1, 1×109 M-1, 1×1010 M-1, 1×1011 M-1, 1×1012 M-1, 1×1013 M-1, 1×1014 M-1, 1×1015 M-1 или по меньшей мере 5×1015 M-1.
[0367] В определенных вариантах осуществления скорость, с которой происходит диссоциация комплекса каркасных структур Tn3 по настоящему изобретению и целевого эпитопа, может быть более значимой, чем величина Kd или Ka. В некоторых вариантах осуществления каркасные структуры Tn3 по настоящему изобретению характеризуются koff, составляющей менее 10-3 с-1, менее 5x10-3 с-1, менее 10-4 с-1, менее 5x10-4 с-1, менее 10-5 с-1, менее 5x10-5 с-1, менее 10-6 с-1, менее 5x10-6 с-1, менее 10-7 с-1, менее 5x10-7 с-1, менее 10-8 с-1, менее 5x10-8 с-1, менее 10-9 с-1, менее 5x10-9 с-1 или менее 10-10 с-1.
[0368] В определенных других вариантах осуществления скорость, с которой происходит ассоциация комплекса каркасных структур Tn3 по настоящему изобретению и целевого эпитопа, может быть более значимой, чем величина Kd или Ka. В этом случае каркасные структуры Tn3 по настоящему изобретению связываются с мишенью при скорости kon, составляющей по меньшей мере 105 M-1с-1, по меньшей мере 5x105 M-1с-1, по меньшей мере 106 M-1с-1, по меньшей мере 5 x 106 M-1с-1, по меньшей мере 107 M-1с-1, по меньшей мере 5 x 107 M-1с-1, или по меньшей мере 108 M-1с-1, или по меньшей мере 109 M-1с-1.
[0369] Каркасные структуры Tn3 по настоящему изобретению могут также быть прикреплены к твердым подложкам, которые в особенности пригодны для иммунологических анализов или очищения целевого антигена. Такие твердые подложки включают, без ограничений, стекло, целлюлозу, полиакриламид, нейлон, полистирол, поливинилхлорид или полипропилен.
CD40L-специфичные Tn3-каркасные структуры
[0370] Настоящее изобретение предлагает Tn3-каркасные структуры, специфически связывающиеся с CD40L. В конкретных вариантах осуществления каркасные структуры по настоящему изобретению специфически связываются с CD40L человека. В других конкретных вариантах осуществления Tn3-каркасные структуры по настоящему изобретению связываются с гомологами CD40L от мыши, курицы, макака-резуса, макака-крабоеда, крысы или кролика. В некоторых вариантах осуществления Tn3-каркасные структуры по настоящему изобретению связываются с доступным эпитопом CD40L. Такие варианты осуществления предусматривают CD40L, эндогенная экспрессия которого происходит в клетках, и/или клетки, подвергнутые трансфекции в целях эктопической экспрессии рецептора.
[0371] В некоторых вариантах осуществления Tn3-каркасные структуры по настоящему изобретению распознают эпитопы, экспонируемые на мономерном CD40L. В других вариантах осуществления Tn3-каркасные структуры по настоящему изобретению распознают эпитопы, экспонируемые на тримерной форме CD40L. В других вариантах осуществления Tn3-каркасные структуры по настоящему изобретению распознают эпитопы, экспонируемые на CD40L, связанном с мембраной. В других вариантах осуществления Tn3-каркасные структуры по настоящему изобретению распознают эпитопы, экспонируемые на растворимом CD40L.
[0372] В еще одних вариантах осуществления Tn3-каркасные структуры по настоящему изобретению связываются с мономерным CD40L и предупреждают олигомеризацию молекул CD40L или препятствуют ей. В еще одних вариантах осуществления каркасные структуры по настоящему изобретению ослабляют или ингибируют взаимодействие CD40L с CD40. В других вариантах осуществления Tn3-каркасные структуры по настоящему изобретению оказывают агонистическое влияние на передачу сигналов в клетке, опосредованную CD40L. В еще одних вариантах осуществления Tn3-каркасные структуры по настоящему изобретению оказывают антагонистическое влияние на передачу сигналов в клетке, опосредованную CD40L.
[0373] Настоящее изобретение также предлагает способы модулирования активности CD40L при помощи Tn3-каркасных структур, описываемых в данном документе. В некоторых вариантах осуществления способы по настоящему изобретению включают приведение CD40L в контакт с CD40L-специфичными каркасными структурами и блокирование взаимодействия между CD40 и CD40L. В других вариантах осуществления способы по настоящему изобретению включают приведение клетки, в которой происходит экспрессия CD40L, в контакт с CD40L-специфичной каркасной структурой Tn3 и предупреждение протеолитического отщепления CD40L от поверхности клетки. В других вариантах осуществления способы по настоящему изобретению включают приведение мономерного CD40L в контакт с CD40L-специфичной Tn3-каркасной структурой и предупреждение олигомеризации CD40L. В других вариантах осуществления димеризации или олигомеризации CD40L можно достичь посредством применения мультимерных Tn3-каркасных структур.
[0374] В некоторых вариантах осуществления способы по настоящему изобретению предусматривают введение CD40L-специфичной каркасной структуры, которая ослабляет CD40-опосредованный иммунный ответ (см., например, Elqueta et al. 229: 152-172, 2009) или ингибирует путь передачи сигналов в нисходящем направлении, инициируемый связыванием CD40 с CD40L, что измеряется с помощью обычных анализов, известных в данной области техники.
[0375] Не желая ограничиваться какой-либо конкретной теорией, полагают, что CD40L-специфичные каркасные структуры по настоящему изобретению могут функционировать путем предупреждения связывания CD40L с CD40, путем связывания и блокирования растворимого CD40L, путем изменения характера взаимодействия CD40L с CD40, но не предотвращения связывания, путем предотвращения или усиления опосредованного металлопротеиназами ферментативного отщепления CD40L от поверхности клетки с получением растворимого CD40L, путем предотвращения или усиления эндоцитоза, опосредованного CD40L на поверхности клетки, и т.д.
Последовательности, специфически связывающиеся с CD40L
[0376] В некоторых вариантах осуществления Tn3-каркасная структура по настоящему изобретению содержит CD40L-специфичные мономерные субъединицы, содержащие по меньшей мере одну, по меньшей мере две, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять или по меньшей мере шесть петлевых последовательностей, которые связываются с CD40L.
[0377] В некоторых вариантах осуществления CD40L-специфичные мономерные субъединицы содержат по меньшей мере одну, по меньшей мере две, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять или по меньшей мере шесть петлевых последовательностей мономерных клонов, связывающихся с CD40L, выбранных из: 309 (исходный клон из семейства 309, выделенный из наивной библиотеки Tn3; SEQ ID NO: 20), 309FGwt (исходный клон 309 с гуманизированной FG-петлей; SEQ ID NO: 22), 340 (клон 309 с созревшей аффинностью; SEQ ID NO: 24), 341 (клон 309 с созревшей аффинностью; SEQ ID NO: 26), 342 (клон 309 с созревшей аффинностью; SEQ ID NO: 28 или SEQ ID NO: 146), 343 (клон 309 с созревшей аффинностью; SEQ ID NO: 30), 344 (клон 309 с созревшей аффинностью; SEQ ID NO: 32), 345 (клон 309 с созревшей аффинностью; SEQ ID NO: 34), 346 (клон 309 с созревшей аффинностью; SEQ ID NO: 36), 347 (клон 309 с созревшей аффинностью; SEQ ID NO: 38), 348 (клон 309 с созревшей аффинностью; SEQ ID NO: 40), 349 (клон 309 с созревшей аффинностью; SEQ ID NO: 42), 311 (исходный клон из семейства 311, выделенный из наивной библиотеки Tn3; SEQ ID NO: 44), 311K4E (вариант клона из семейства 311 из первого цикла созревания аффинности; SEQ ID NO: 46); 311K4E_1 (вариант клона из семейства 311 из второго цикла созревания аффинности; SEQ ID NO: 48), 311K4E_2 (вариант клона из семейства 311 из второго цикла созревания аффинности; SEQ ID NO: 50), 311K4E_3 (вариант клона из семейства 311 из второго цикла созревания аффинности; SEQ ID NO: 52), 311K4E_4 (вариант клона из семейства 311 из второго цикла созревания аффинности; SEQ ID NO: 54), 311K4E_5 (вариант клона из семейства 311 из второго цикла созревания аффинности; SEQ ID NO: 56), 311K4E_7 (вариант клона из семейства 311 из второго цикла созревания аффинности; SEQ ID NO: 58), 311K4E_8 (вариант клона из семейства 311 из второго цикла созревания аффинности; SEQ ID NO: 60), 311K4E_9 (вариант клона из семейства 311 из второго цикла созревания аффинности; SEQ ID NO: 62), 311K4E_10 (вариант клона из семейства 311 из второго цикла созревания аффинности; SEQ ID NO: 64), 311K4E_11 (вариант клона из семейства 311 из второго цикла созревания аффинности; SEQ ID NO: 66), 311K4E_12 (вариант клона из семейства 311 из второго цикла созревания аффинности; SEQ ID NO: 68), 311K4E_13 (вариант клона из семейства 311 из второго цикла созревания аффинности; SEQ ID NO: 70), 311K4E_14 (вариант клона из семейства 311 из второго цикла созревания аффинности; SEQ ID NO: 72), 311K4E_15 (вариант клона из семейства 311 из второго цикла созревания аффинности; SEQ ID NO: 74), 311K4E_16 (вариант клона из семейства 311 из второго цикла созревания аффинности; SEQ ID NO: 76), 311K4E_19 (вариант клона из семейства 311 из второго цикла созревания аффинности; SEQ ID NO: 78), 311K4E_20 (вариант клона из семейства 311 из второго цикла созревания аффинности; SEQ ID NO: 80) и 311K4E_21 (вариант клона из семейства 311 из второго цикла созревания аффинности; SEQ ID NO: 82).
[0378] В некоторых вариантах осуществления CD40L-специфичные мономерные субъединицы содержат по меньшей мере одну петлевую последовательность, выбранную из петлевых последовательностей, перечисленных в Таблице 2. В других вариантах осуществления CD40L-специфичные мономерные субъединицы содержат по меньшей мере одну последовательность петли BC, выбранную из последовательностей петли BC, перечисленных в Таблице 2. В других вариантах осуществления CD40L-специфичные мономерные субъединицы содержат по меньшей мере одну последовательность петли DE, выбранную из последовательностей петли DE, перечисленных в Таблице 2. В других вариантах осуществления CD40L-специфичные мономерные субъединицы содержат по меньшей мере одну последовательность петли FG, выбранную из последовательностей петли FG, перечисленных в Таблице 2.
[0379] В некоторых вариантах осуществления CD40L-специфичные мономерные субъединицы содержат последовательность петли BC, выбранную из последовательностей петли BC, перечисленных в Таблице 2; и последовательность петли DE, выбранную из последовательностей петли DE, перечисленных в Таблице 2. В других вариантах осуществления CD40L-специфичные мономерные субъединицы содержат последовательность петли BC, выбранную из последовательностей петли BC, перечисленных в Таблице 2; и последовательность петли FG, выбранную из последовательностей петли FG, перечисленных в Таблице 2. В других вариантах осуществления CD40L-специфичные мономерные субъединицы содержат последовательность петли DE, выбранную из последовательностей петли DE, перечисленных в Таблице 2; и последовательность петли FG, выбранную из последовательностей петли FG, перечисленных в Таблице 2. В некоторых вариантах осуществления CD40L-специфичные мономерные субъединицы содержат петлевые последовательности, соответствующие петлевым последовательностям одного, двух или трех различных клонов Tn3.
[0380] В конкретных вариантах осуществления, в случае, если последовательность CD40L-специфичной мономерной каркасной структуры содержит линкер и/или гистидиновую метку (например, His-8-метку) на C-конце последовательности или дополнительные N-концевые аминокислоты, этот C-концевой линкер и/или гистидиновую метку и дополнительные N-концевые аминокислоты можно удалить, при этом соответствующая аминокислотная последовательность содержит, таким образом, делецию последовательностей C-концевого линкера и His-метки и N-концевой дополнительной аминокислоты или аминокислот.
[0381] В некоторых вариантах осуществления CD40L-специфичная Tn3-каркасная структура содержит одну мономерную субъединицу, например, последовательность клона 342 (клон 309 с созревшей аффинностью; SEQ ID NO: 28 и/или SEQ ID NO: 146). В других вариантах осуществления CD40L-специфичная каркасная структура содержит более одной мономерной субъединицы, например, две мономерные субъединицы клона 342 (SEQ ID NO: 28 и/или SEQ ID NO: 146) в тандеме (см., например, SEQ ID NO: 135). В конкретных вариантах осуществления Tn3-каркасные структуры по настоящему изобретению конъюгированы с вариантом HSA (см., например, SEQ ID NO: 134 и SEQ ID NO: 135). В дополнительных вариантах осуществления HSA может быть конъюгированным с N-концом или с C-концом мультимерной Tn3-каркасной структуры.
[0382] В конкретном варианте осуществления CD40L-специфичная Tn3-каркасная структура содержит одну мономерную субъединицу 311K4E_12, линкер GS и вариант C34S HSA (см., например, SEQ ID NO: 201). В другом конкретном варианте осуществления CD40L-специфичная Tn3-каркасная структура содержит одну мономерную субъединицу 311K4E_12 с вариантом CELTYG бета-цепи C, линкером, полностью состоящим из остатков глицина, и вариантом C34S HSA (см., например, SEQ ID NO: 202). В другом конкретном варианте осуществления CD40L-специфичная Tn3-каркасная структура содержит две субъединицы 311K4E_12 в тандеме и два линкера GS, где один линкер GS соединяет субъединицы друг с другом, а второй линкер GS соединяет одну субъединицу с вариантом C34S HSA (см., например, SEQ ID NO: 203). В еще одном конкретном варианте осуществления CD40L-специфичная Tn3-каркасная структура содержит две субъединицы 311K4E_12 в тандеме и два линкера, полностью состоящих из остатков глицина, где один линкер, полностью состоящий из остатков глицина, соединяет субъединицы друг с другом, а второй линкер, полностью состоящий из остатков глицина, соединяет одну субъединицу с вариантом C34S HSA (см., например, SEQ ID NO: 204).
[0383] В одном конкретном варианте осуществления CD40L-специфичная Tn3-каркасная структура содержит две субъединицы 309, соединенные в тандем посредством линкера GS (см., например, SEQ ID NO: 205). В другом конкретном варианте осуществления CD40L-специфичная Tn3-каркасная структура содержит одну субъединицу 309, соединенную с вариантом C34S HSA (см., например, SEQ ID NO: 206). В другом конкретном варианте осуществления CD40L-специфичная Tn3-каркасная структура содержит две субъединицы 309 в тандеме и два линкера GS, где один линкер GS соединяет субъединицы друг с другом, а второй линкер GS соединяет одну субъединицу с вариантом C34S HSA (см., например, SEQ ID NO: 207).
[0384] В конкретном варианте осуществления CD40L-специфичная Tn3-каркасная структура содержит одну мономерную субъединицу 342, линкер GS и вариант C34S HSA (см., например, SEQ ID NO: 134). В другом конкретном варианте осуществления CD40L-специфичная Tn3-каркасная структура содержит одну мономерную субъединицу 342, линкер, полностью состоящий из остатков глицина, и вариант C34S HSA (см., например, SEQ ID NO: 144). В другом конкретном варианте осуществления CD40L-специфичная Tn3-каркасная структура содержит две субъединицы 342 в тандеме и два линкера GS, где один линкер GS соединяет субъединицы друг с другом, а второй линкер GS соединяет одну субъединицу с вариантом C34S HSA (см., например, SEQ ID NO: 135). В еще одном конкретном варианте осуществления CD40L-специфичная Tn3-каркасная структура содержит две субъединицы 342 в тандеме и два линкера, полностью состоящих из остатков глицина, где один линкер, полностью состоящий из остатков глицина, соединяет субъединицы друг с другом, а второй линкер, полностью состоящий из остатков глицина, соединяет одну субъединицу с вариантом C34S HSA (см., например, SEQ ID NO: 145). В еще одном конкретном варианте осуществления CD40L-специфичная Tn3-каркасная структура содержит две субъединицы 342, соединенные в тандем посредством линкера GS (см., например, SEQ ID NO: 208).
[0385] В другом конкретном варианте осуществления CD40L-специфичная Tn3-каркасная структура содержит субъединицу 311 или субъединицу, происходящую из 311 (например, 311K4E_12), и субъединицу 309 или субъединицу, происходящую из 309 (например, 342), в тандеме и два линкера GS, где один линкер GS соединяет субъединицы друг с другом, а второй линкер GS соединяет одну субъединицу с вариантом C34S HSA (см., например, SEQ ID NO: 135). В еще одном конкретном варианте осуществления CD40L-специфичная Tn3-каркасная структура содержит субъединицу 311 или субъединицу, происходящую из 311 (например, 311K4E_12), и субъединицу 309 или субъединицу, происходящую из 309 (например, 342), в тандеме и два линкера, полностью состоящих из остатков глицина, где один линкер, полностью состоящий из остатков глицина, соединяет субъединицы друг с другом, а второй линкер, полностью состоящий из остатков глицина, соединяет одну субъединицу с вариантом C34S HSA (см., например, SEQ ID NO: 145).
[0386] Примеры CD40L-специфичных тандемных бивалентных Tn3-каркасных структур и конструктов слияния на основе сывороточного альбумина (SA) показаны на Фиг. 2A (также см. фиг. 9A). Хотя конкретные линкеры представлены на Фиг. 2A, рассматриваются и другие линкеры, представленные в данном документе. Хотя можно применять зрелый SA дикого типа, например, мышиный сывороточный альбумин (MSA) или человеческий сывороточный альбумин (HSA), предполагается, что один или несколько аминокислотных остатков цистеина (C) в зрелом SA могут быть заменены, например, на серин (S), аланин (A), глицин (G) и т.д.
[0387] Типичные конструкты показаны ниже. Последовательность SA подчеркнута. Линкеры заключены в рамку. Будет понятно, что многочисленные изменения находятся в пределах объема настоящего изобретения. Например, линкеры могут быть изменены (некоторые неограничивающие примеры представлены в данном документе), первые один или два N-концевых аминокислотных остатка (SQ) могут отсутствовать и/или быть заменены на альтернативные аминокислотные остатки, может быть включена метка (например, 6xHis-метка), альтернативные CD40L-специфичные каркасные структуры (например, таковые на основе 10-го домена Fn3 фибронектина) можно использовать в подобном конструкте, и т.д.
Моновалентный 342—HSA, конструкт 1 (SEQ ID NO: 134),
[мономерный 342]-(G4S)2 линкер-HSAC34S

Моновалентный 342—HSA, конструкт 2 (SEQ ID NO: 144),
[мономерный 342]-G10 линкер-HSAC34S:

Бивалентный 342—HSA, конструкт 1 (SEQ ID NO: 135),
[мономерный 342]-линкер (G4S)3-[мономерный 342]-линкер (G4S)2-HSAC34S:

Бивалентный 342—HSA, конструкт 2 (SEQ ID NO: 145),
[мономерный 342]-линкер G15-[мономерный 342]-линкер G10-HSAC34S:
 Моновалентный 311K4E_12—HSA, конструкт 1 (SEQ ID NO: 201),
Моновалентный 311K4E_12—HSA, конструкт 1 (SEQ ID NO: 201),
[мономерный 311K4E_12]-линкер (G4S)2-HSAC34S:

Моновалентный 311K4E_12—HSA, конструкт 2 (SEQ ID NO: 202),
[мономерный 311K4E_12]-линкер G10-HSAC34S:

Бивалентный 311K4E_12—HSA, конструкт 1 (SEQ ID NO: 203),
[мономерный 311K4E_12]-линкер G4S3-[мономерный 311K4E_12]-линкер (G4S)2-HSAC34S:

Бивалентный 311K4E_12—HSA, конструкт 2 (SEQ ID NO: 204),
[мономерный 311K4E_12]-линкер G15-[мономерный 311K4E_12]-линкер G10-HSAC34S:

Фармацевтические композиции
[0388] В другом аспекте настоящее изобретение предлагает композицию, например, без ограничений, фармацевтическую композицию, содержащую одну Tn3-каркасную структуру по настоящему изобретению или комбинацию таковых, составленные вместе с фармацевтически приемлемым носителем. Такие композиции могут включать одну Tn3-каркасную структуру по настоящему изобретению или комбинацию из, например, без ограничения, двух или более различных таковых. Например, фармацевтическая композиция по настоящему изобретению может содержать комбинацию Tn3-каркасных структур, которые связываются с различными эпитопами целевого антигена или которые характеризуются взаимодополняющими активностями. В конкретном варианте осуществления фармацевтическая композиция содержит одну мономерную Tn3-каркасную структуру по настоящему изобретению. В конкретном варианте осуществления фармацевтическая композиция содержит мультимерную Tn3-каркасную структуру по настоящему изобретению. В еще одном конкретном варианте осуществления фармацевтическая композиция содержит димерную Tn3-каркасную структуру по настоящему изобретению.
[0389] Фармацевтические композиции по настоящему изобретению также можно вводить при комбинированной терапии, как, например, в комбинации с другими средствами. Например, комбинированная терапия может включать применение Tn3-каркасной структуры по настоящему изобретению в комбинации по меньшей мере с одним другим видом терапии, где терапия может представлять собой иммунотерапию, химиотерапию, лучевую терапию или фармакотерапию. Фармацевтические композиции по настоящему изобретению могут включать одну или несколько фармацевтически приемлемых солей.
Способы применения каркасных структур
[0390] Tn3-каркасные структуры по настоящему изобретению обладают диагностической и терапевтической значимостью в условиях in vitro и in vivo. Например, эти молекулы можно вводить в клетки в культуре, например, in vitro или ex vivo, или субъекту, например, in vivo, для лечения, предупреждения или диагностики разнообразных нарушений.
[0391] Настоящее изобретение также предлагает способы применения Tn3-каркасных структур по настоящему изобретению. Настоящее изобретение также охватывает применение Tn3-каркасных структур по настоящему изобретению для предупреждения, диагностики, управления проявлением, лечения или уменьшения интенсивности одного или нескольких симптомов, ассоциированных с заболеваниями, нарушениями, обусловленными заболеваниями, или нарушениями, включая, без ограничений, раковые, воспалительные и аутоиммунные заболевания, инфекционные заболевания, в отдельности или в комбинации с другими видами терапии. Настоящее изобретение также охватывает применение Tn3-каркасных структур по настоящему изобретению, конъюгированных или слитых с фрагментом (например, терапевтическим средством или лекарственным средством), для предупреждения, управления проявлением, лечения или уменьшения интенсивности одного или нескольких симптомов, ассоциированных с заболеваниями, нарушениями или инфекциями, включая, без ограничений, раковые, воспалительные и аутоиммунные заболевания, инфекционные заболевания, в отдельности или в комбинации с другими видами терапии.
[0392] Настоящее изобретение также предлагает способы нацеливания на эпитоп, трудные для осуществления с традиционными антителами. Например, в одном варианте осуществления Tn3-каркасные структуры по настоящему изобретению можно применять для первоначального нацеливания на прилегающий антиген, и во время связывания другой связывающий домен может вступать в контакт с криптантигеном.
[0393] Настоящее изобретение также предлагает способы применения Tn3-каркасных структур для сближения клеток различных типов. В одном варианте осуществления белки по настоящему изобретению могут связываться с целевой клеткой посредством одного связывающего домена и рекрутировать другую клетку посредством другого связывающего домена. В другом варианте осуществления первая клетка может представлять собой раковую клетку, а вторая клетка представляет собой эффекторную клетку иммунной системы, такую как NK-клетка. В другом варианте осуществления можно применять Tn3-каркасные структуры по настоящему изобретению для укрепления взаимодействия между двумя различными клетками, такими как антиген-презентирующая клетка и T-клетка, с возможным усилением иммунного ответа.
[0394] Настоящее изобретение также предлагает способы применения каркасных структур Tn3 для элиминации клеток из популяции. В одном варианте осуществления способы по настоящему изобретению пригодны для элиминации клеток следующих типов: эозинофилы, базофилы, нейтрофилы, T-клетки, B-клетки, мастоциты, моноциты и опухолевые клетки.
[0395] Настоящее изобретение также предлагает способы применения Tn3-каркасных структур в качестве диагностических реагентов. Такие диагностические реагенты можно применять для тестирования в отношении наличия или отсутствия CD40L, наличия рецептора CD40, эффективности связывания CD40L с рецептором CD40, свободного CD40L у пациента, свободного CD40L в образце или CD40L, связанного с рецептором CD40 в образце.
[0396] Tn3-каркасные структуры по настоящему изобретению и композиции, содержащие их, пригодны для многих целей, например, в качестве терапевтических средств против широкого диапазона хронических и острых заболеваний и нарушений, включая, без ограничений, аутоиммунные и/или воспалительные заболевания. Композиции и способы настоящего изобретения, описанные в данном документе, пригодны для предупреждения или лечение аутоиммунных нарушений и/или воспалительных нарушений.
[0397] Примеры аутоиммунных и/или воспалительных нарушений включают, без ограничений, очаговую алопецию, анкилозирующий спондилит, антифосфолипидный синдром, аутоиммунную болезнь Аддисона, аутоиммунные заболевания надпочечников, аутоиммунную гемолитическую анемию, аутоиммунный гепатит, аутоиммунные оофорит и орхит, синдром Шегрена, псориаз, атеросклероз, диабетическую и другие формы ретинопатии, ретролентальную фиброплазию, возрастную макулярную дегенерацию, неоваскулярную глаукому, формы гемангиомы, формы тиреоидной гиперплазии (в том числе болезнь Грейвса), трансплантацию ткани роговицы и другой ткани, а также хроническое воспаление, сепсис, ревматоидный артрит, перитонит, болезнь Крона, реперфузионное повреждение, септицемию, эндотоксический шок, муковисцидоз, эндокардит, псориаз, артрит (например, псориатический артрит), анафилактический шок, ишемию органа, реперфузионное повреждение, повреждение спинного мозга и отторжение аллотрансплантата, аутоиммунную тромбоцитопению, болезнь Бехчета, буллезный пемфигоид, кардиомиопатию, дерматит, связанный с целиакией-спру, синдром хронической усталости и иммунной дисфункции (CFIDS), хроническую воспалительную демиелинизирующую полинейропатию, синдром Черджа-Стросса, рубцовый пемфигоид, CREST-синдром, болезнь холодовых агглютининов, болезнь Крона, дискоидную волчанку, идиопатическую криоглобулинемию смешанного типа, фибромиалгию/фибромиозит, гломерулонефрит, болезнь Грейвса, синдром Гийена-Барре, тиреоидит Хашимото, идиопатический фиброз легких, идиопатическую тромбоцитопеническую пурпуру (ITP), IgA-нейропатию, болезнь Стилла, красный плоский лишай, красную волчанку, болезнь Меньера, смешанное заболевание соединительной ткани, рассеянный склероз, сахарный диабет 1 типа или иммуноопосредованный сахарный диабет, миастению, вульгарный пемфигус, пернициозную анемию, нодозный полиартериит, полихондрит, полигландулярные синдромы, ревматическую полимиалгию, полимиозит и дерматомиозит, первичную агаммаглобулинемию, первичный билиарный цирроз, псориаз, псориатический артрит, болезнь Рейно, синдром Рейтера, ревматоидный артрит, саркоидоз, склеродермию, синдром Шегрена, синдром мышечной скованности, системную красную волчанку, красную волчанку, артериит Такаясу, височный артериит/гигантоклеточный артериит, язвенный колит, увеит, васкулит, такой как герпетиформный дерматит, витилиго и гранулематоз Вегенера.
[0398] Примеры воспалительных заболеваний включают, без ограничений, астму, энцефалит, воспалительные заболевания кишечника, хроническое обструктивное заболевание легких (COPD), аллергические нарушения, септический шок, фиброз легких, недифференцированную спондилоартропатию, недифференцированную артропатию, артрит, воспалительный остеолиз и хроническое воспаление, возникающее в результате хронических вирусных или бактериальных инфекций. Композиции и способы по настоящему изобретению можно применять с одним или несколькими видами традиционной терапии, применяемыми для предупреждения, управления течением или лечения вышеупомянутых заболеваний.
[0399] Настоящее изобретение предлагает способы предупреждения, управления течением, лечения или уменьшения интенсивности раковых, аутоиммунных, воспалительных или инфекционных заболеваний или одного или нескольких их симптомов, при этом указанные способы включают введение субъекту, нуждающемуся в этом, одной или нескольких каркасных структур Tn3 по настоящему изобретению в комбинации с одним или несколькими терапевтическими средствами, которые не являются терапевтическими средствами против рака (также называемые средствами терапии заболеваний, не относящихся к раковым).
[0400] Примеры таких средств включают, без ограничений, противорвотные средства, противогрибковые средства, антибактериальные средства, такие как антибиотики, противовоспалительные средства и противовирусные средства. Неограничивающие примеры противорвотных средств включают метопимазин и метоклопрамид. Неограничивающие примеры противогрибковых средств включают азольные лекарственные средства, имидазол, триазолы, полиен, амфотерицин и пиримидин. Неограничивающие примеры антибактериальных средств включают дактиномицин, блеомицин, эритромицин, пенициллин, митрамицин, цефалоспорин, имипенем, азтреонам, ванкомицин, циклосерин, бацитрацин, хлорамфеникол, клиндамицин, тетрациклин, стрептомицин, тобрамицин, гентамицин, амикацин, канамицин, неомицин, спектиномицин, триметоприм, норфлоксацин, рефампин, полимиксин, амфотерицин B, нистатин, кетоконазол, изониазид, метронидазол и пентамидин. Неограничивающие примеры противовирусных средств включают аналоги нуклеозидов (например, зидовудин, ацикловир, ганцикловир, видарабин, идоксуридин, трифлуридин и рибавирин), фоскарнет, амантадин, римантадин, саквинавир, индинавир, ритонавир, интерферон ("IFN")-α,β или γ и AZT. Неограничивающие примеры противовоспалительных средств включают нестероидные противовоспалительные лекарственные средства ("NSAID"), стероидные противовоспалительные лекарственные средства, бета-агонисты, антихолинергические средства и метилксантины.
[0401] В одном варианте осуществления настоящее изобретение включает композиции, при помощи которых возможно лечить хроническое воспаление. В одном варианте осуществления композиции пригодны для нацеливания на клетки иммунной системы для разрушения или инактивации. В одном варианте осуществления композиции пригодны для нацеливания на активированные T-клетки, "дремлющие" T-клетки, В-клетки, нейтрофилы, эозинофилы, базофилы, мастоциты или дендритные клетки. В другом варианте осуществления настоящее изобретение включает композиции, способные к ослаблению функции клеток иммунной системы. В другом варианте осуществления композиции способны к нивелированию функции клеток иммунной системы.
[0402] В другом варианте осуществления настоящее изобретение включает композиции, пригодные для лечения заболеваний желудочно-кишечного тракта. Каркасные структуры согласно настоящему изобретению проявляют высокий уровень стабильности в условиях низкого pH. Стабильность при низких pH позволяет предположить, что композиция будет подходящей для перорального введения при разнообразных расстройствах желудочно-кишечного тракта, таких как синдром раздраженной толстой кишки, желудочно-пищеводный рефлюкс, формы псевдонепроходимости кишечника, демпинг-синдром, устойчивая тошнота, пептическая язва, аппендицит, ишемический колит, неспецифический язвенный колит, гастрит, заболевание, вызываемое Helicobacter pylori, болезнь Крона, болезнь Уиппла, целиакия-спру, дивертикулит, дивертикулез, дисфагия, грыжа пищеводного отверстия диафрагмы, инфекционные расстройства пищевода, икота, руминация и другие.
[0403] Настоящее изобретение дополнительно предлагает комбинированные композиции и способы применения таких композиций в предупреждении, лечении, ослаблении или уменьшении интенсивности заболевания или его симптомов. Применение Tn3-каркасных структур по настоящему изобретению можно комбинировать с видами традиционной терапии, подходящими для предупреждения, лечения, ослабления или уменьшения интенсивности заболевания или его симптомов. Приводимые в качестве примера виды традиционной терапии можно найти в Physician’s Desk Reference (56th ed., 2002 и 57th ed., 2003). В некоторых вариантах осуществления применение каркасных структур Tn3 по настоящему изобретению можно комбинировать с химиотерапией, лучевой терапией, хирургическим вмешательством, иммунотерапией с помощью биологического препарата (антитела или пептида), терапией с помощью низкомолекулярных средств или другим видом терапии, известным в данной области техники. В некоторых вариантах осуществления компоненты комбинированной терапии применяют совместно. В других вариантах осуществления компоненты комбинированной терапии применяют по отдельности.
[0404] Настоящее изобретение также предлагает способы диагностирования заболеваний. Tn3-каркасные структуры по настоящему изобретению, которые связываются со специфичной мишенью, ассоциированной с заболеванием, можно внедрить в способ, применяемый для диагностики указанного заболевания. В одном варианте осуществления Tn3-каркасные структуры по настоящему изобретению применяют в способе диагностики заболевания у субъекта, при этом указанный способ включает получение образца от субъекта, приведение мишени в контакт с Tn3-каркасной структурой в указанном образце в условиях, обеспечивающих возможность образования взаимодействия мишень:каркасная структура, идентификацию комплекса мишень:каркасная структура и выявление, таким образом, мишени в образце. В других вариантах осуществления заболевание, которое следует диагностировать, описано в данном документе.
[0405] Настоящее изобретение также предлагает способы визуализации специфических мишеней. В одном варианте осуществления Tn3-каркасные структуры по настоящему изобретению, конъюгированные с визуализирующими средствами, такими как зеленые флуоресцентные белки, другие флуоресцентные метки (Cy3, Cy5, родамин и другие), биотин или радионуклиды, можно применять в способах визуализации наличия, месторасположения или продвижения специфической мишени. В некоторых вариантах осуществления способ визуализации мишени, включающий применение Tn3-каркасной структуры по настоящему изобретению, осуществляют in vitro. В других вариантах осуществления способ визуализации мишени, включающий применение Tn3-каркасной структуры по настоящему изобретению, осуществляют in vivo. В других вариантах осуществления способ визуализации мишени, включающий применение Tn3-каркасной структуры по настоящему изобретению, осуществляют с помощью MRI, PET-сканирования, рентгенографии, флуоресцентной детекции или с помощью других способов детекции, известных в данной области техники.
[0406] Настоящее изобретение также предлагает способы контроля прогрессирования, рецидива, лечения или уменьшения интенсивности заболевания при помощи каркасных структур по настоящему изобретению. В одном варианте осуществления способы контроля прогрессирования, рецидива, лечения или уменьшения интенсивности заболевания выполняют с помощью способов визуализации, диагностики или приведения в контакт соединения/мишени с Tn3-каркасной структурой по настоящему изобретению, как представлено в данном документе.
Дозировка и введение фармацевтических композиций
[0407] Для получения фармацевтических или стерильных композиций, содержащих Tn3-каркасную структуру по настоящему изобретению, каркасную структуру смешивают с фармацевтически приемлемым носителем или наполнителем. Композиции, предназначенные для введения, предпочтительно являются апирогенными, что означает, что они практически не содержат эндотоксинов и/или связанных пирогенных веществ. Выбор схемы введения терапевтического средства зависит от нескольких факторов, в том числе от скорости циркуляции объекта в сыворотке крови или ткани, уровня проявления симптомов, иммуногенности объекта и доступности клеток-мишеней в биологической среде. В конкретных вариантах осуществления схема введения максимально увеличивает количество терапевтического средства, доставляемого пациенту, что согласуется с приемлемым уровнем побочных эффектов.
[0408] Фактические уровни дозировки активных ингредиентов в фармацевтических композициях по настоящему изобретению можно изменять таким образом, чтобы получить количество активного ингредиента, эффективное для достижения необходимого терапевтического эффекта для конкретного пациента, композиции и способа введения, которое не является токсичным для пациента. Выбранный уровень дозировки будет зависеть от разнообразных фармакокинетических факторов, включая активность конкретных используемых композиций по настоящему изобретению или их сложных эфиров, солей или амидов, способ введения, время введения, скорость экскреции конкретного используемого соединения, длительность лечения, другие лекарственные средства, соединения и/или материалы, применяемые в комбинации с конкретными используемыми композициями, возраст, пол, вес, состояние, общее состояние здоровья и предшествующую историю болезни пациента, подлежащего лечению, и подобных факторов, хорошо известных в области медицины.
[0409] Композицию по настоящему изобретению можно также вводить посредством одного или нескольких путей введения при помощи одного или нескольких разнообразных способов, известных в данной области техники. В конкретных вариантах осуществления Tn3-каркасные структуры по настоящему изобретению можно составлять с обеспечением надлежащего распределения in vivo.
Эквиваленты
[0410] Специалисты в данной области признают или будут способны установить, используя не более, чем обычные эксперименты, многие эквиваленты конкретных вариантов осуществления настоящего изобретения, описанных в данном документе. Такие эквиваленты предназначены для охвата нижеследующей формулой изобретения.
[0411] Все публикации, патенты и заявки на патенты, упоминаемые в настоящем описании, включены в данное описание посредством ссылки в такой же степени, как если бы каждые отдельные публикация, патент или заявка на патент были конкретно и отдельно указаны как включенные в данный документ посредством ссылки.
ПРИМЕРЫ
[0412] Настоящее изобретение теперь описано со ссылкой на следующие примеры. Эти примеры являются только иллюстративными, и настоящее изобретение никоим образом не следует толковать как ограниченное этими примерами, а скорее следует толковать как охватывающее любые возможные варианты, которые становятся очевидными в результате реализации идей, изложенных в данном документе.
Пример 1
Конструирование библиотеки из 3 петель на основе исходной Tn3-каркасной структуры
[0413] Библиотеку конструировали на основе исходной Tn3-каркасной структуры, описанной в публикации международной заявки на патент № WO 2009/058379, где она обозначена как "Tn3 SS4." Библиотека содержит рандомизированные участки петель BC, DE и FG. В данной конструкции в библиотеку Tn3 включено разнообразие охарактеризованных последовательностей и длин петель, что согласуется с характером разнообразия, описанным для природных доменов FnIII, три различных длины петель BC и FG, и применяется схема, описывающая совокупность кодонов “NHT”, для внедрения разнообразия в библиотеку (H = A, T, C). По данной схеме образуют 12 кодонов, кодирующих 12/20 аминокислот (см. таблицу 3), т.е. каждый кодон кодирует уникальную аминокислоту. Более того, стоп-кодоны или кодоны, кодирующие цистеин (Cys), отсутствуют.
[0414] Разнообразие библиотеки создавали при помощи вырожденных олигонуклеотидов, показанных в Таблице 4.

Коды нуклеотидов: N = G/A/T/C; H = A/T/C; R = A/G; S = G/C; B = T/C/G; V = A/C/G; M = A/C; K = G/T.
[0415] Сборку библиотеки проводили при помощи олигонуклеотидов, показанных в Таблице 5.

[0416] Производили сборку смеси вырожденных олигонуклеотидов (в эквимолярных соотношениях олигонуклеотидов, соответствующих петлям BC и FG, соответственно), BCX-DE bridge v2, DE-FGX bridge v2 и KpnI amp rev v2, в 20-цикловой ПЦР-реакции без избытка внешних праймеров. Данный продукт разбавляли и амплифицировали в ходе обычной ПЦР-реакции при помощи праймеров BC library amp v2 и KpnI reverse v2. Из полученного в результате продукта ПЦР создавали полный ген Tn3, который затем расщепляли с помощью NcoI и KpnI и лигировали в вектор для фагового дисплея (описанный в WO 2009/058379). E. coli трансформировали путем введения ДНК с помощью электропорации. Конечное разнообразие библиотеки согласно оценкам составляло приблизительно 7,9 x 1010 представителей.
[0417] После электропорации библиотеку инкубировали в течение 1 часа при 37oC со встряхиванием. Добавляли хелперный фаг M13K07, и через час клетки разбавляли до большего объема и выращивали при 37oC со встряхиванием в течение ночи. На следующий день фаги удаляли и концентрировали из надосадочной жидкости путем осаждения с PEG 8000.
Пример 2
Пэннинг библиотек для идентификации Tn3-каркасных структур, специфичных по отношению к CD40L человека
[0418] Фаг-дисплейные библиотеки Tn3, содержащие >1010 уникальных последовательностей, подвергали пэннингу с CD40L. Разнообразие данных библиотек происходило из изменчивости последовательностей и длины петель BC, DE и FG, аналогичных трем петлям CDR в вариабельном домене антитела. Селекцию лидерных белков Tn3 проводили путем пэннинга библиотек на рекомбинантных биотинилированных CD40L человека и клеточной линии CHO, в которой происходит сверхэкспрессия CD40L. Чередующиеся циклы пэннинга с этими двумя реагентами применяли для обеспечения того, чтобы лидерные клоны могли распознать рекомбинантный внеклеточный домен, а также нативный CD40L, заякоренный на мембране.
[0419] Рекомбинантный CD40L человека (MegaCD40L человека; Axxora) биотинилировали с помощью сульфо-NHS-биотина EZ-Link (Pierce, Рокфорд, Иллинойс), применяя реагент для биотинилирования в 5–кратном молярном избытке. После инкубирования в течение 1 часа при комнатной температуре образец подвергали диализу в PBS в течение ночи с удалением неконъюгированного биотина. 10 мкг биотинилированного CD40L иммобилизировали на гранулах со стрептавидином M280 (Dynal, Карлсбад, Калифорния) с последующим блокированием в PBS, содержащем 10 мг/мл BSA, в течение 2 часов. Поступающий материал состоял из библиотек, разработанных согласно описанному в примере 1 или, дополнительно, библиотек, разработанных при помощи стандартных методик конструирования, таких как описанные в WO 2009/058379.
[0420] Фаги блокировали в PBS, содержащем 10 мг/мл BSA, в течение 2 часов. Блокированный поступающий материал добавляли к контрольным гранулам со стрептавидином M280 (без мишени) и инкубировали во встряхивателе-качалке в течение 2 часов при комнатной температуре для элиминации из библиотеки молекул, связывающихся с гранулами. Библиотеку, подвергнутую элиминации, затем добавляли к гранулам, покрытым CD40L, и инкубировали в течение 2 часов при комнатной температуре на качающейся платформе. После трех промываний с помощью PBST (PBS + 0,1% Tween) для удаления несвязанных фагов гранулы добавляли к клеткам XL-1 Blue E. coli в экспоненциальной фазе роста, которые затем совместно инфицировали хелперным фагом M13KO7 в 60 мл среды 2xYT, содержащей 50 мкг/мл карбенициллина. После выращивания в течение ночи при 37oC со встряхиванием фаги собирали путем осаждения с PEG из суточной культуральной среды.
[0421] Второй цикл пэннинга (цикл 2) проводили на клеточной линии CHO, в которой происходит сверхэкспрессия CD40L. Фаговую библиотеку блокировали в 3% BSA/PBS с покачиванием при комнатной температуре в течение 1 часа. Клетки разделяли с помощью Accutase (Invitrogen), промывали 2x с помощью 5 мл PBS, и 107 клеток блокировали в 1 мл 3% BSA/PBS с покачиванием при комнатной температуре в течение 30 минут. Блокированные клетки осаждали на центрифуге при 500 x g в течение 5 минут, аккуратно ресуспендировали в растворе блокированной фаговой библиотеки и инкубировали в течение 1 часа при комнатной температуре. Несвязанные фаги удаляли путем аккуратного промывания клеток 3 раза в 1 мл 3% BSA/PBS и один раз в PBS, осаждая клетки путем центрифугирования при 500 x g в течение 5 минут в микроцентрифуге и с использованием новых пробирок Eppendorf для каждого промывания. Клеточный осадок непосредственно добавляли к XL-1 Blue E. coli в экспоненциальной фазе роста, которые затем обрабатывали согласно описанному для цикла 1.
[0422] Цикл 3 пэннинга проводили согласно описанному для цикла 1, за исключением того, что связанные фаги элюировали путем добавления 100 мМ HCl с последующей нейтрализацией с помощью 1M Tris-HCl, pH 8. Элюированный нейтрализованный фаг применяли для инфицирования клеток XL1 Blue E. coli согласно описанному для цикла 1.
[0423] Цикл 4 пэннинга также выполняли на клетках согласно описанному для цикла 2, за исключением того, что производили 5 промываний в 3% BSA/PBS. Цикл 5 осуществляли при помощи 5 мкг биотинилированного MegaCD40L, но в остальном согласно описанному для цикла 3.
[0424] После 5 циклов пэннинга проводили скрининг полученных в результате вариантов Tn3 в виде растворимого белка. Исходные растворы фагов, амплифицированных и осажденных с помощью PEG, применяли в ПЦР для амплификации совокупности фрагментов, охватывающей кодируемые последовательности Tn3. Эту совокупность фрагментов расщепляли с помощью NcoI + KpnI и клонировали в соответствующие сайты NcoI - KpnI плазмиды pSec-oppA(L25M)-Tn3 (см., например, WO 2009/058379). В самоиндуцирующую среду MagicMedia (Invitrogen), содержащую карбенициллин (100 мкг/мл), в 96-луночных планшетах с глубокими лунками инокулировали клетки BL21 DE3 E. coli, трансформированные с помощью конструктов, происходящих из pSec-oppA(L25M)-Tn3. Культуры выращивали в течение 18 часов, встряхивая при 37ºC, и клетки отделяли от среды путем центрифугирования. Среду, содержащую секретируемые растворимые варианты Tn3, применяли непосредственно в скрининговом анализе связывания с CD40L.
[0425] Десять наборов из 96 клонов подвергали скринингу для идентификации белков Tn3, специфически связывающихся с рекомбинантными CD40L. Вкратце, в скрининговом анализе использовали захват растворимых вариантов Tn3 с His-меткой, секретируемых в среду, посредством связывания с антителом к His, иммобилизованным в лунках микропланшетов. После захвата среду и избыточный белок вымывали, и взаимодействие между захваченными вариантами Tn3 и CD40L отслеживали путем использования биотинилированного MegaCD40L человека и измерения уровня оставшейся мишени (после промывания планшета) с помощью SA-HRP и реагентов для традиционного ELISA.
[0426] На этапе захвата иммобилизованное антитело к His насыщали с помощью Tn3, и молярное количество захваченного Tn3 в каждой лунке становилось практически идентичным независимо от уровней экспрессии отдельных клонов. Данная нормализация уровней Tn3 давала в результате уровни в анализе, пропорциональные эффективности взаимодействия с мишенью и не подвергаемые влиянию возможных различий в уровнях экспрессии белка.
[0427] Объекты данного анализа, дающие положительный результат, секвенировали для идентификации 34 уникальных последовательностей Tn3, которые связываются с рекомбинантными CD40L. Из панели уникальных последовательностей Tn3, связывающихся с CD40L, подмножество из 24 клонов, характеризовавшихся устойчивым сигналом в анализе и удовлетворительными уровнями экспрессии, что оценивается посредством SDS-PAGE надосадочных жидкостей культуры, подвергали реэкспрессии и незначительной очистке.
[0428] Вкратце, в среду на основе супербульона, содержащую карбенициллин (100 мкг/мл) с 1% глюкозой, инокулировали клетки BL21 DE3 E. coli, трансформированные с помощью конструктов, происходящих из pSec-oppA(L25M)-Tn3. Культуры выращивали при 37ºC до достижения оптической плотности (O.D.) 0,5-0,8, затем индуцировали с помощью 0,2 мМ IPTG. После встряхивания при 37ºC в течение 5 часов клетки отделяли от среды путем центрифугирования. Очистку Tn3-каркасных структур от среды производили путем очистки на установке периодического действия с помощью Ni-NTA Superflow (Qiagen), промывания в 2x PBS с 20 мМ имидазола и элюирования с помощью 2x PBS с 250 мМ имидазола. Образцы подвергали диализу в PBS, и концентрации определяли по поглощению УФ-излучения при 280 нм согласно Gill and von Hippel (Anal. Biochem. 182: 319, 1989).
[0429] На основании ранжирования в анализе и поведения при SEC уровень экспрессии 8 лидерных клонов повышали, и их очищали до низкого уровня эндотоксинов (< 1 ЕЭ/мг) для тестирования в функциональном клеточном анализе.
[0430] Два клона Tn3 (обозначенные как 309 и 311) демонстрировали сходную активность в биохимическом и клеточном анализах (Фиг. 6A, 6B, 6C) и были в 3-5 раз более действенными, чем ближайший конкурирующий клон.
[0431] Моновалентные Tn3-каркасные структуры 309 и 311, специфичные по отношению к CD40L человека, уменьшали общее количество B-клеток, количество плазматических клеток и переключение классов Ig (Фиг. 6A, 6B и 6C). Фиг. 6A показан ингибирующий эффект 309 и 311 в отношении экспрессии CD86, индуцированной HuCD40L, на CD19-положительных PBMC человека, стимулированных клетками Jurkat D1.1; на Фиг. 6B показано ингибирование пролиферации B-клеток, стимулированной HuCD40L, с помощью 309 и 311; и на Фиг. 6C показано уменьшение количества плазматических клеток в совместных культурах T/B-клеток. Также было показано, что 309 связывается с активированными первичными T-клетками с помощью FACS (данные не показаны). PBMC стимулировали с помощью рекомбинантных MegaCD40L человека (Axxora) или клеток Jurkat, в которых происходит экспрессия CD40L человека (D1.1, ATCC), и процентную долю клеток CD19+/CD86+ измеряли с помощью FACS через 24 часа.
[0432] Лидерные клоны 309 и 311 находились в монодисперсном состоянии и не проявляли какой-либо тенденции к агрегации или образованию олигомеров высшего порядка в растворе, что определяли с помощью анализа очищенных образцов посредством эксклюзионной хроматографии (SEC) (фиг. 7A).
[0433] Значения температурной стабильности лидерных клонов 309 и 311 определяли с помощью дифференциальной сканирующей калориметрии (DSC) с применением образцов белка при 1 мг/мл в PBS с pH 7,2 и сравнивали с температурной стабильностью исходного белка Tn3 (фиг. 7B). Лидерные клоны 309 и 3011 характеризовались Tm, составляющей 70 ± 1°C, которая была лишь немного более низкой, чем Tm исходного Tn3 (72°C).
[0434] Поскольку не были идентифицированы клоны, проявляющие перекрестную реактивность в отношении лигандов мыши, осуществляли способ пэннинга, подобный описанному выше, для идентификации Tn3, специфичного по отношению к лигандам мыши, обозначенного M13. M13 также демонстрировал активность в клеточном анализе PBMC (см. Фиг. 1A).
Пример 3
Оптимизация лидерных CD40L-специфичных Tn3-каркасных структур
[0435] Для повышения действенности выбранных лидерных Tn3 применяли оптимизацию аффинности. В целом, проводили один или несколько циклов мутагенеза в пределах петель Tn3, контактирующих с мишенью, с селекцией улучшенных вариантов из комбинаторных фаг-дисплейных библиотек.
3.1 Перестановка в петлях
[0436] С целью определения того, какие из 3 петель в двух лидерных молекулах участвуют во взаимодействии с CD40L, создавали конструкты, в которых каждая отдельная последовательность петли была заменена на последовательность петли исходного Tn3, обнаруживаемую в тенасцине C человека. Показатели активности мутантных вариантов сравнивали с таковыми первоначальных вариантов в анализе связывания, проводимом в соответствии с описанным для скринингового анализа, описанного выше (Фиг. 10A). Для обеих лидерных молекул мутации в петлях BC и DE приводили в результате к полной утрате связывания с CD40L, тогда как замена петли FG на последовательность исходного Tn3 не оказывала эффект (для 309), либо оказывала ограниченный эффект (311) на связывание. Таким образом, петли BC и DE, по-видимому, содержали последовательности, несущие главную ответственность за контактирование с CD40L, и поэтому были в первую очередь выбраны для оптимизации аффинности.
3.2 Оптимизация лидерной CD40L-специфичной Tn3-каркасной структуры 309
[0437] Поскольку в эксперименте по перестановке в петлях выявили, что последовательность петли FG 309 можно заменить на последовательность петли FG исходного Tn3 без значительной утраты действенности связывания, было решено применять данный конструкт (называемый 309FGwt) в качестве остова для созревания аффинности. Это будет устранять несущественные мутации, проявляющиеся в отклонении от последовательности исходного тенасцина C, в целях снижения возможного риска иммуногенности. Следует отметить, что последовательность петли FG исходного Tn3 содержала RGD-мотив, который затем устраняли путем мутации в конечных лидерных молекулах. Создавали три библиотеки петель BC и одну библиотеку петель DE.
[0438] Для трех библиотек петель BC выполняли три цикла ПЦР с применением вырожденных олигонуклеотидов BC9 PCR, BC 9-loop NNK и 309 BC-loop NNKdope (таблица 6) вместе с обратным праймером Kpn1 amp rev v2 (таблица 5) при помощи матрицы, происходящей из 309FGwt, в которой кодоны петли BC были заменены на стоп-кодоны. Последующая ПЦР-амплификация этих фрагментов с помощью праймеров BC library amp v2 (таблица 5) и KpnI reverse v2 давала фрагмент библиотеки Tn3 полной длины.
[0439] Для библиотеки петель DE ПЦР-амплификация с помощью DE PCR и KpnI amp reverse v2 на матрице, происходящей из 309FGwt (в которой кодоны петли DE были заменены на стоп-кодоны), давала фрагмент, содержащий рандомизированную петлю DE и петлю FG Tn3 дикого типа, который объединяли с фрагментом, кодирующим участок Tn3 выше петли DE, образованным путем ПЦР с помощью BC library amp v2 и BCX-DE bridge v2 на матрице из 309. Два фрагмента соединяли в ходе ПЦР с перекрывающимися праймерами с помощью внешних праймеров BC library amp v2 и KpnI reverse v2.

1= кодоны для всех 19aa(-cys) 2= кодоны для Ala/Pro 3= кодоны для Ala/Gly;
4= 70%G10%A10%C10%T 5= 10%G,70%A,10%C,10%T
6=10%G,10%A,70%C, 10%T 7= 10%G,10%A,10%C,70%T
8= 70%A15%C15%T K = 50%G/50%T
[0440] Фрагменты NcoI-KpnI клонировали в вектор для фагового дисплея, и фаговую библиотеку создавали согласно описанному в примере 1.
[0441] Четыре библиотеки по отдельности подвергали пэннингу на биотинилированном MegaCD40L человека согласно описанному для первого цикла в примере 2 с применением 4 мкг CD40L в цикле 1 и 1 мкг в цикле 2. После амплификации фагов в выходном материале после цикла 2 одноцепочечную ДНК выделяли при помощи набора Qiagen Spin M13 (Qiagen, Валенсия, Калифорния), и пулы фрагментов, содержащих петли BC, из библиотек петель BC амплифицировали при помощи BC lib amp v2 и BC shuffle rev, а пул фрагментов, содержащих петли DE, амплифицировали из библиотеки петель DE при помощи праймеров DE-shuffle FWD и KpnI reverse v2. Фрагменты для ПЦР очищали из геля, и проводили их сборку за счет использования их перекрывающейся последовательности при помощи внешних праймеров BC lib amp v2 и KpnI reverse v2. Полученный в результате ПЦР фрагмент применяли для создания библиотеки в фаговом векторе согласно описанному ранее. Эту библиотеку подвергали пэннингу, состоящему в общей сложности из 5 циклов, на биотинилированном MegaCD40L человека согласно описанному в примере 2, за исключением того, что вначале обеспечивали контакт библиотек с мишенью в концентрации 50 нМ, 20 нМ, 20 нМ и 10 нМ (в общем объеме 50 мкл) в ходе циклов 1-4 в течение 2 часов перед инкубированием с блокированными магнитными гранулами со стрептавидином M280 в течение 10 минут с последующим промыванием.
[0442] Выходные образцы клонировали в пуле в сайты NcoI - KpnI плазмиды pSec-oppA(L25M)-Tn3 pSec, и шестнадцать 96-луночных планшетов подвергали скринингу в отношении связывания с CD40L при помощи растворимого белка в скрининговом анализе, описанном выше. 270 клонов с наивысшими количественными показателями тщательно отбирали, повторно анализировали и секвенировали. Десять клонов отбирали для дополнительной характеристики на основе анализа связывания и анализа последовательностей. Она включала оценивание действенности в анализе PBMC, определение Kd в отношении связывания с CD40L в анализе с использованием биосенсоров, определение термодинамической стабильности с помощью дифференциальной сканирующей калориметрии и выявление тенденции к агрегации или образованию олигомеров высшего порядка в растворе с помощью анализа посредством эксклюзионной хроматографии. Результаты обобщены в таблице 7. Последовательности клонов 309 и 309FGwt, выравненные с последовательностями десяти оптимизированных клонов (обозначенных 340, 341, 342, 343, 344, 345, 346, 347, 348 и 349), показаны на фиг. 11A и 11B.
[0443] Варианты с созревшей аффинностью демонстрировали на 1-3 порядка более высокую действенность, чем клон 309, сохраняли высокую стабильность, измеряемую с помощью DSC, и большинство из них находилось в монодисперсном состоянии, что измеряли с помощью SEC.
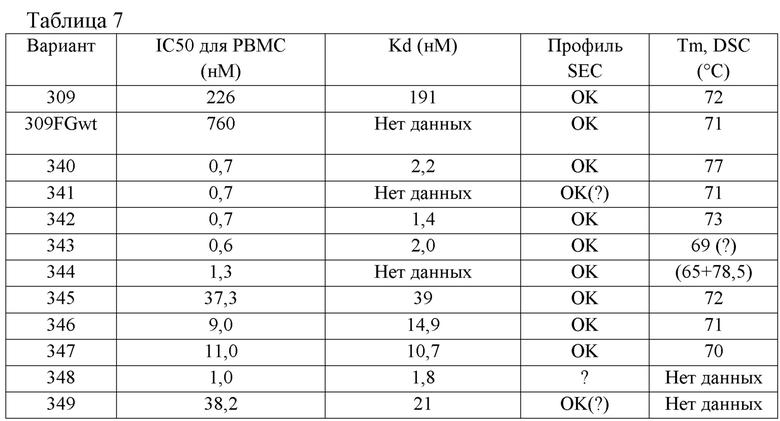
[0444] Анализы PBMC осуществляли путем стимуляции PBMC с помощью клеток Jurkat, в которых происходит экспрессия CD40L человека (D1.1, ATCC), и процентную долю клеток CD19+/CD86+ измеряли с помощью FACS через 24 часа. Данный анализ применяли для тестирования и ранжирования панели лидерных Tn3-каркасных структур, переходя к этому от установления приоритетов на основе биохимических критериев. Результаты анализов PBMC показаны на Фиг. 10B и обобщены в таблице 7.
[0445] Измерения аффинности осуществляли на матричной системе для исследования взаимодействия белков ProteOn XPR36 (Bio-Rad, Геркулес, Калифорния) с сенсорным чипом GLC при 25ºC. Фосфатно-солевой буферный раствор ProteOn с 0,005% Tween 20, pH 7,4 (PBS/Tween) применяли в качестве подвижного буфера. MegaCD40L человека иммобилизировали на чипе при поверхностной плотности приблизительно 2300 резонансных единиц. Варианты Tn3 (340, 342, 343, 345, 346, 347, 348 и 349) в двукратных разведениях получали в PBS/Tween/0,5 мг/мл BSA, pH 7,4 (от 150 до 4,7 нМ). Образцы в каждой концентрации впрыскивали в шесть каналов для анализируемых веществ при скорости потока 30 мкл/мин. в течение 300 секунд. Kd определяли путем применения анализа равновесия, настройка параметров которого осуществляется в программном обеспечении ProteOn. Результаты показаны в Таблице 7.
[0446] Десять TCD40L-специфичных вариантов Tn3 анализировали в отношении стабильности с помощью DSC. Вкратце, измерения при DSC производили на VP-Capillary DSC (MicroCal). Производили замену буфера для белков на PBS (pH 7,2) посредством обширного диализа, и их концентрацию доводили до 0,25-0,5 мг/мл для анализа по методу DSC. Образцы сканировали при 20-95°C при скорости сканирования 90°C/час без повторного сканирования. Результаты показаны в Таблице 7.
[0447] Для наилучших клонов получали улучшение IC50 по сравнению с 309 вплоть до 300-кратного и более чем 1000-кратное улучшение по сравнению с остовом 309FGwt, применяемым для создания библиотеки оптимизированных лидерных молекул. Семь клонов характеризовались Kd в диапазоне одноразрядных чисел нМ.
3.3 Оптимизация лидерных CD40L-специфичных Tn3-каркасных структур 311
Перед проведением эксперимента с использованием петель, упомянутого ранее (см. Фиг. 10A).
[0448] была предпринята первоначальная попытка создания библиотеки оптимизированных лидерных молекул, ориентированная на внедрение разнообразия в петлю FG 311, создание и скрининг полученной в результате фаг-дисплейной библиотеки. Скрининг проводили после 4 циклов пэннинга на биотинилированном MegaCD40L человека согласно описанному ранее, и было обнаружено, что большинство положительных попаданий содержало случайную мутацию замены остатка K4 на E в N-концевом константном участке Tn3 выше петли BC. Не был выявлен явный консенсус между последовательностями положительных попаданий в петле FG. Внедрение одной мутации K4E в 311 приводило в результате к приблизительно 100-кратному повышению действенности (с приблизительно 4 мкМ до 36 нМ) в анализе PBMC (см. Фиг. 10C).
[0449] Поскольку в эксперименте по перестановке в петлях выявили, что последовательности петель BC и DE были необходимы для связывания с CD40L, эти петли затем предназначались для использования в остове 311K4E для дополнительного созревания аффинности. Создавали две отдельные библиотеки. Одна библиотека предназначалась для включения петли BC согласно стратегии, где каждый остаток с 50% вероятностью был в последовательности 311 дикого типа и с приблизительно 50% вероятностью был одним из остальных 11 остатков, кодируемых NHT. Другая библиотека включала полностью рандомизированную петлю DE из 6 остатков.
[0450] Что касается библиотеки петель BC, в ПЦР-реакциях применяли олигонуклеотиды BC11-311Gly и BC11-311NHT (таблица 8) с обратным праймером KpnI amp rev v2 (таблица 5) на матрице, происходящей из 311, в которой кодоны петли BC были заменены на стоп-кодоны, с образованием фрагментов, содержащих петли BC, DE и FG. Амплификация смеси 1:1 этих фрагментов с помощью праймеров BC library amp K4E и KpnI amp rev v2 в конечном счете давала фрагмент библиотеки Tn3 полной длины.
[0451] Библиотеку петель DE создавали как библиотеку петель DE 309FGwt, описанную выше, за исключением того, что в ПЦР-реакциях применяли матрицу, происходящую из 311, и в конечной ПЦР-амплификации применяли праймер BC library amp K4E.

1=70%G, 10%A, 10%C, 10%T 2=10%G, 70%A, 10%C, 10%T
3=10%G, 10%A, 70%C, 10%T 4=10%G, 10%A, 10%C, 70%T
5=70%A, 15%C, 15%T 6=15%A, 70%C, 15%T
7=15%A, 15%C, 70%T V=33%A, 33%C, 33%G
H=33%A, 33%C, 33%T
[0452] Фрагменты библиотеки полной длины расщепляли с помощью NcoI и KpnI, клонировали в вектор для фагового дисплея, и фаговые библиотеки создавали согласно описанному в примере 1.
[0453] Две библиотеки по отдельности подвергали пэннингу на биотинилированном MegaCD40L человека согласно описанному в примере 1 с применением 10 мкг белка в цикле 1, 5 мкг в цикле 2 и 5 мкг в цикле 3. После амплификации фагов в выходном материале после цикла 3 одноцепочечную ДНК выделяли при помощи набора Qiagen (Qiagen, Валенсия, Калифорния), и две библиотеки подвергали шаффлингу согласно описанному для библиотек 309FGwt. После шаффлинга библиотеку подвергали пэннингу, состоящему в общей сложности из 5 циклов, на биотинилированном MegaCD40L человека согласно описанному выше для подвергнутой шаффлингу библиотеки 309FGwt, применяя 100 нМ, 20 нМ, 4 нМ, 1 нМ и 1 нМ мишени в ходе циклов 1-5.
[0454] Выходные образцы клонировали в пуле в растворимый вектор секреции согласно описанному ранее, и пять 96-луночных планшетов подвергали скринингу в отношении связывания с CD40L при помощи скринингового анализа растворимых белков, описанного ранее. Положительные попадания идентифицировали относительно сигнала, получаемого при использовании варианта-остова 311K4E. 18 уникальных клонов с наивысшими количественными показателями, обозначенные 311K4E_1, 311K4E_2, 311K4E_3, 311K4E_4, 311K4E_5, 311K4E_7, 311K4E_8, 311K4E_9, 311K4E_10, 311K4E_11, 311K4E_12, 311K4E_13, 311K4E_14, 311K4E_15, 311K4E_16, 311K4E_19, 311K4E_20 и 311K4E_21 (последовательности, показанные на Фиг. 12A и Фиг. 12B), анализировали в качестве сырых неочищенных белков для ранжирования по скорости диссоциации. Определение оценки скорости диссоциации неочищенных Tn3-каркасных структур осуществляли на матричной системе для исследования взаимодействия белков ProteOn XPR36 (Bio-Rad, Геркулес, Калифорния) в анализе с использованием биосенсоров с CD40L, иммобилизованным на чипе. MegaCD40L человека иммобилизировали на чипе GLC (BioRad), и все варианты разбавляли до расчетной концентрации 80 нМ, впрыскивали при скорости потока 30 мкл/мин. в течение 300 секунд при времени диссоциации, установленном на 1200 секунд. В качестве буфера подвижной фазы применяли PBS, 0,005% Tween 20, 3 мМ EDTA, pH 7,4. Скорости диссоциации ранжировали с помощью визуального изучения сенсограмм. Подмножество из четырех вариантов, демонстрирующих наиболее низкие скорости диссоциации, 311K4E_3, 311K4E_11, 311K4E_12 и 311K4E_15, очищали, и значения Kd определяли как составляющие от 1,1 до 6,4 нМ (Таблица 9).
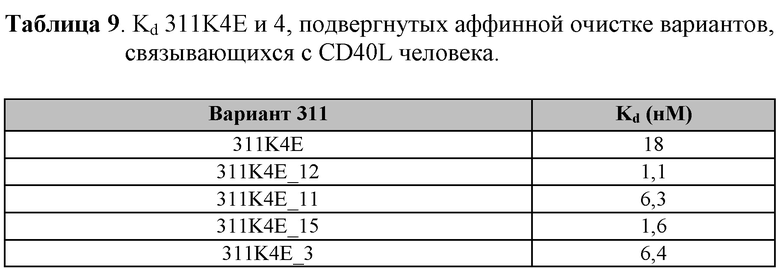
[0455] Как указано на Фиг. 10D, понижение Kd (с 18 нМ до 1 нМ) 311K4E_12 соответствовало 12-кратному повышению действенности в анализе PBMC относительно остова 311K4E.
[0456] В заключение, операция оптимизации лидерных молекул по исходным попаданиям 309 и 311 из наивных библиотек дает одноразрядное число нМ для молекул, связывающихся с CD40L.
[0457] Подобную операцию оптимизации производили в отношении молекулы M13, специфичной по отношению к лигандам мыши (данные не показаны). Полученная в результате оптимизированная молекула, специфичная по отношению к CD40L мыши (обозначенная M31), демонстрировала приблизительно 20-кратное повышение действенности в анализе PBMC по сравнению с исходной молекулой (Фиг. 1A).
Пример 4
Экспрессия и очистка конструктов слияния CD40L-специфичный Tn3—HSA без меток
[0458] Tn3-конструкты, слитые с HSA, описанные в общих чертах на Фиг. 2A и 9A, экспрессировали в клеточной линии млекопитающих 293F посредством транзиентной трансфекции. Экспрессионные конструкты слияния Tn3-HSA создавали на основе вектора экспрессии для млекопитающих собственного производства. Для повышения однородности продукта использовали мутантную форму HSA (обозначенную HSA C34S), в которой неспаренный частично доступный цистеин 34 в результате мутации был заменен на серин (Zhao et al., 2009, Eur. J. Pharm. Biopharm. 72: 405-11).
[0459] Белок слияния можно очистить за один этап очистки с помощью ионообменной хроматографии (IEX). Пример элюирования 309-309-HSA из колонки Q-HP (GE HealthCare) с помощью солевого градиента показан на Фиг. 9B. В дополнение к основному пику наблюдались минорные более поздние пики элюирования (составляющие менее 10% общей площади пика). В анализе по методу масс-спектрометрии выявили, что материал, соответствующий этим минорным побочным пикам, был обогащен O-гликозилированными видами 309-309-HSA. Фракции, содержащие материал, соответствующий основному пику, объединяли и применяли в последующих анализах активности.
[0460] В случае очистки в более крупном масштабе этапу IEX, упомянутому выше, предшествовал захват конструктов слияния Tn3-HSA из надосадочной жидкости культуры с помощью аффинной хроматографии с применением аффинных матриц для HSA, например, HiTrap Blue HP (GE HealthCare). После промывания белок слияния на основе HSA можно было элюировать буфером, содержащим октиленовую кислоту. Элюат загружали в колонку Q-HP после 3-кратного разведения в фосфатном буфере.
[0461] Анализ минорного(минорных) пика(пиков) выявил наличие углеводных фрагментов с O-связями. Предположили, что O-гликан представляет собой однородную смесь углеводов, происходящую из структуры, представляющей собой O-ксилозилированное ядро, о которой сообщалось ранее (Wakabayashi et al., 1999, J. Biol. Chem. 274:5436-5442) и которая показана ниже:
[0462] Было определено, что сайт преобладающего присоединения находится в линкере GGGGS между доменами Tn3. Было также обнаружено, что гликан присутствует в меньшей степени в линкере GGGGS, который находится между Tn3 и HSA. Уровни O-гликана, таким образом, были более высокими в бивалентных конструктах по сравнению с моновалентными конструктами и были более высокими в материале, вырабатываемом клетками HEK, по сравнению с таковым в клетках CHO. Уровни также варьировали среди различных Tn3-конструктов. Таким образом, уровень O-гликана можно понизить посредством тщательного отбора клеток-хозяев, например, применения клеток CHO или других клеток, в отношении которых было обнаружено, что они вырабатывают материал с более низкими уровнями O-гликана. В дополнение, материал, содержащий O-гликан, можно удалить посредством способов очистки с получением более однородного продукта, в котором отсутствует O-гликан. Альтернативно, линкер можно модифицировать с удалением основного(основных) сайта(сайтов) присоединения O-гликана, например, путем замены остатка Ser на Gly в результате мутации. Линкеры в некоторых конструктах можно повторно сконструировать с получением одного или нескольких линкеров GGGGG. В материале с линкером(линкерами) GGGGG не были выявлены O-гликаны какого-либо типа, и различий по активности не наблюдали (данные не показаны).
Пример 5
Увеличение периода полужизни в сыворотке CD40L-специфичных Tn3-каркасных структур
[0463] Изучали слияние с сывороточным альбумином в качестве стратегии увеличения периода полужизни в сыворотке CD40L-специфичных Tn3-каркасных структур. В целях определения фармакокинетических (PK) свойств конструктов слияния Tn3, специфичный по отношению к CD40L мыши—MSA проводили PK-анализ на мышах. Конструкты слияния на основе MSA выбирали для изучения суррогатных молекул, отдавая им преимущество перед соответствующими конструктами слияния на основе HSA, поскольку FcRn мыши связывается с HSA значительно слабее, чем он связывается с MSA, что приводит в результате к понижению рециркуляции из эндосом и, следовательно, повышению скорости циркуляции (Andersen et al. J. Biol. Chem. 285, 4826-4836, 2010).
[0464] Клетки HEK 293 применяли для экспрессии тандемной бивалентной Tn3-каркасной структуры, специфичной по отношению к CD40L мыши, слитой с MSA. Наблюдали высокие уровни экспрессии (Фиг. 3A). Данные конструкты имели линкер (G4S) между единицами Tn3-каркасной структуры и 3 повтора (G4S) в линкере между каркасной структурой и MSA. В дополнение, в каждую из каркасных структур M13 и M31 внедряли мутацию N49Q с удалением возможного сайта N-связанного гликозилирования. Данная мутация не влияла на действенность данных каркасных структур (данные не показаны). Через 6 дней после трансфекции уровень экспрессии оценивали как 200 мг/л. Очистку осуществляли с помощью IMAC с использованием C-концевой His-метки. Выход очищенного белка оценивали как 125 мг/л надосадочной жидкости культуры.
[0465] После слияния MSA с бивалентными каркасными структурами M13 наблюдали 8-кратное повышение действенности по сравнению с димерной каркасной структурой M13 без MSA (фиг. 3B). Бивалентная каркасная структура, содержащая M31 с созревшей аффинностью, слитая с MSA, была в 140 раз более действенной, чем соответствующая бивалентная каркасная структура M13, слитая с MSA, приблизительно в 900 раз более действенной, чем конструкт слияния моновалентный M31—MSA, и обладала действенностью, сравнимой с таковой для моноклонального антитела MR1 к CD40L мыши (Фиг. 3C).
[0466] Для определения PK-свойств конструктов слияния CD40L-специфичный Tn3—MSA осуществляли PK-анализ на мышах. Белковые конструкты вводили внутривенно при 10 мг/кг самкам мышей CD-1 в возрасте 5-7 недель. У каждой мыши производили забор 150 мкл крови в различные моменты времени, и с помощью анализа при помощи ELISA определяли концентрацию конструкта слияния Tn3-HSA в сыворотке крови. Вкратце, планшеты Nunc MaxiSorp покрывали антителом M2 к FLAG (Agilent), блокировали в 4% молоке в PBS+0,1% Tween (PBST) и инкубировали с MegaCD40L мыши (Axxora). MegaCD40L иммобилизировали посредством его FLAG-метки. Образцы сыворотки крови и белковые стандарты разбавляли в 4% молоке в PBST и добавляли после промывания планшета в PBST. После инкубирования планшет промывали в PBST, и для выявления конструктов слияния Tn3-HSA применяли поликлональное антитело кролика к TN3 (Covance) при помощи антитела козы к иммуноглобулинам кролика, конъюгированного с HRP (Jackson ImmunoResearch), при стандартном протоколе ELISA. Концентрации в образцах сыворотки крови определяли на основе градуировочных кривых в модели линейной регрессии, образованных в ходе анализов разведения того же самого конструкта слияния Tn3-MSA. Концентрации определяли как среднее значение для 3 различных мышей.
[0467] Как видно из Фиг. 4A, M31-MSA и M13-M13-MSA характеризовались временем полужизни, составляющими 38 часов и 31 час, соответственно, тогда как конструкт M31-M31-HSA характеризовался временем полужизни 12 часов. Для сравнения, тандемный конструкт M13-M13 сам по себе (не слитый с MSA) проявлял время полужизни, составляющее 30 минут (не показано).
[0468] В отличие от наблюдений в отношении каркасных структур мышей, при слиянии HSA с каркасными структурами, специфичными по отношению к CD40L человека, значительного понижения действенности не было. На Фиг. 9C показано, что при слиянии бивалентной каркасной структуры Tn3, специфичной по отношению к CD40L человека, содержащей два мономера 309, с HSA не было значительного понижения действенности согласно измерениям в анализе PMBC.
[0469] PK-свойства конструкта мономерный 342, специфичный по отношению к CD40L человека—HSA сравнивали с таковыми варианта 342-HSA, содержащего две замены (L463N и K524L) для увеличения времени полужизни в сыворотке у макака-крабоеда после однократной внутривенной инъекции. Белковые конструкты вводили посредством медленной болюсной инъекции при 10 мг/кг самцам макаков-крабоедов весом 2-5 кг. У каждого животного из периферического сосуда отбирали 1 мл крови в каждый из моментов времени перед дозированием, через 5 минут и через 30 минут после дозирования; через 2, 12, 24 и 48 часов после дозирования и в дни 4, 8, 11, 15, 22, 29, 36, 43 и 57. Концентрацию конструкта слияния Tn3-HSA в сыворотке определяли с помощью анализа при помощи ELISA. Вкратце, планшеты Nunc MaxiSorp покрывали антителом M2 к FLAG (Agilent), блокировали в 4% молоке в PBS+0,1% Tween (PBST) и инкубировали с MegaCD40L человека (Axxora). MegaCD40L иммобилизировали посредством его FLAG-метки. Образцы сыворотки крови и белковые стандарты разбавляли в 4% молоке в PBST и добавляли после промывания планшета в PBST. После инкубирования планшет промывали в PBST, и для выявления конструктов слияния Tn3-HSA применяли поликлональное антитело кролика к TN3 (Covance) при помощи антитела козы к иммуноглобулинам кролика, конъюгированного с HRP (Jackson ImmunoResearch), при стандартном протоколе ELISA. Концентрации в образцах сыворотки крови определяли на основе градуировочных кривых в модели линейной регрессии, образованных в ходе анализов разведения того же самого конструкта слияния Tn3-HSA. Концентрации отложены на графике на Фиг. 4B. Время полужизни конструкта 342-HSA составляло приблизительно 7 дней, тогда как конструкт, представляющий собой вариант L463N/K524L 342-HSA, демонстрировал увеличенное время полужизни 13-17 дней в ходе начальной линейной фазы (Фиг. 4B). Через 30 дней концентрации конструкта, представляющего собой вариант L463N/K524L 342-HSA, падали быстрее по сравнению с конструктом на основе HSA дикого типа. Данные наблюдения могут указывать на некоторую иммуногенность данного конструкта для обезьян.
Пример 6
Характеристика CD40L-специфичных Tn3-каркасных структур
6.1 Экспериментальные способы
[0470] 6.1.1 Анализ стимуляции PBMC. Кровь получали от здоровых доноров согласно руководствам по безопасности от MedImmune. PBMC выделяли посредством пробирок CPT (центрифугирование при 1500 g в течение 20 минут), и 1 x 106 PBMC (на условие) стимулировали с помощью рекомбинантного MegaCD40L человека (Axxora) или клеток Jurkat, в которых происходит экспрессия CD40L человека (D1.1, ATCC). Процентную долю клеток CD19+/CD86+ измеряли с помощью FACS через 24 часа после стимуляции. Данный анализ применяли для тестирования и ранжирования панели лидерных Tn3-каркасных структур, переходя к этому от установления приоритетов на основе биохимических критериев. Анализ также можно осуществлять с использованием клеточной линии, в которой происходит экспрессия CD40L мыши (D10.G4), или с MegaCD40L мыши (Axxora ALX522120) вместо стимуляции клетками человека или рекомбинантным белком, поскольку лиганд мыши дает перекрестную реакцию с рецептором человека.
[0471] 6.1.2 Анализ CD40R/NFΚB мыши. Клетки NIH3T3 с репортерами, контролируемыми NFкB (репортерная система под контролем NFΚB от Panomics и система трансфекции с помощью mCD40R собственного производства) стимулировали с помощью рекомбинантного белка MegaCD40L мыши (Axxora, № по каталогу ALX522120) или клеток D10.G4 (ATCC), в которых происходит сверхэкспрессия CD40L, в течение 24 часов в присутствии или в отсутствие Tn3-каркасных структур. Bright-Glow (Promega E2610) добавляли согласно инструкциям производителя. Регистрируемым показателем была люминесценция (700), возникающая посредством активации репортеров, контролируемых NFΚB, осуществляемой в системе EnVision (Perkin Elmer).
[0472] 6.1.3 Анализ CD40R/NFΚB человека. Клетки HEK293 с репортерами (от Panomics и собственного производства) стимулировали с помощью рекомбинантного белка MegaCD40L (Axxora ALX522110) или клеток Jurkat субклона D1.1 (ATCC), в которых происходит сверхэкспрессия CD40L, в течение 24 часов в присутствии или в отсутствие Tn3-каркасных структур. Bright-Glow (Promega E2610) добавляли согласно инструкциям производителя. Регистрируемым показателем была люминесценция (700), возникающая посредством активации репортеров, контролируемых NFΚB, осуществляемой в системе EnVision (Perkin Elmer).
[0473] 6.1.4 Анализ двух типов клеток. Первичные T/В-клетки выделяли из различных доноров. T-клетки человека CD4+ (1x105), стимулированные антителом к CD3 и обработанные митомицином C, культивировали с очищенными В-клетками (5x104). Регистрируемые показатели были следующими. День 2: маркеры активации (FACS), день 5: пролиферация B-клеток (метаболит ATP, Cell-Titer Glo, Invitrogen), день 7: дифференцировка плазматических клеток (FACS), день 7: выработка Ig (ELISA, R&D Systems).
[0474] 6.1.5 Анализы агрегации тромбоцитов. Аденозиндифосфат (ADP) был получен от Chrono-Log (Хэвертаун, Пенсильвания, США). Все остальные продукты были по меньшей мере чистыми для анализа. Образцы крови собирали от здоровых добровольцев, помещали в 12,9 мМ цитрат натрия и центрифугировали при 150 x g в течение 15 минут с получением PRP (плазмы, обогащенной тромбоцитами). После разделения PRP пробирки вновь центрифугировали при 1200 x g в течение 15 минут с получением PPP (плазмы, обедненной тромбоцитами). Тромбоциты промывали с помощью способа, описанного в Mustard et al. (Br. J. Haematol. 22:193-204, 1972), и ресуспендировали в растворе Тироде, содержащем 2 мM CaCl2, 1 мM MgCl2, 0,1% декстрозу, 0,35% бычий сывороточный альбумин, 0,05 ед./мл апиразу, pH 7,35. Агрегацию тромбоцитов изучали с помощью светового трансмиссионного агрегометра (Chrono-Log 700-4DR, Chrono-Log Corporation, Хэвертаун, Пенсильвания, США) и регистрировали в течение 10 мин. после стимуляции тромбоцитов указанными агонистами тромбоцитов согласно описанному. Tn3-каркасные структуры предварительно инкубировали с растворимым CD40L (sCD40L) с образованием иммунных комплексов перед добавлением.
[0475] 6.1.6 Анализы иммунизации. Эритроциты овцы (SRBC) приобретали у Colorado Serum (Денвер, Колорадо) и разводили в 10 раз в среде HBSS непосредственно перед применением. Мышей иммунизировали с помощью 0,2 мл разведенных SRBC в день 0. Первичный ответ зародышевого центра (GC) у мышей, подвергнутых антигенной стимуляции, оценивали через 14 дней после иммунизации посредством FACS (В-клетки из GC, B-клетки не из GC и все субпопуляции T-клеток). Tn3-каркасные структуры и контрольные образцы вводили в дни 9-13 с повышением дозы каждые 24 часа согласно указанному.
[0476] 6.1.7 Анализы зависимого от T-клеток образования KLH-специфичных антител (TDAR). Макакам-крабоедам (Macaca fascicularis) китайского происхождения весом 3,1 - 4,6 кг (Covance Research Products, Элис, Техас) вводили внутривенно (в подкожную или головную вену) один раз в неделю указанную дозу (0,5, 5, 40 мг/кг) ингибитора (342-мономер-Tn3-HSA и 342-342-бивалент-Tn3-HSA) или контрольного раствора/среды. KLH (№ партии MD158678A, Supplier Pierce Biotechnologies, Рокфорд, Иллинойс) разбавляли с помощью соответствующего количества стерильной воды для инъекций (Supplier Midwest Veterinary Supply, Норристаун, Пенсильвания) в стерильных условиях. Флаконы взбалтывали с перемешиванием, и их содержимое объединяли в стерильном флаконе. 1 мл раствора KLH (10 мг/мл) вводили подкожно в спину каждого животного слева от срединной линии дважды (в день 1 и в день 29) в течение 1 часа после окончания введения тестируемого или контрольного препарата. Образцы крови для дополнительного анализа получали от всех животных в следующие моменты времени: до тестирования, в дни 4, 6, 8, 11, 15, 22, 25, 32, 34, 36, 39, 43, 46, 50 и 57. Забор образцов, которые собирали в дни 8, 15 и 22, производили перед введением дозы. Оценивание титров KLH-специфичных антител IgM и IgG производили в дни 8, 11 и 15. Титры KLH-специфичных антител IgM и IgG в день 15 показаны на фигурах 5G и 5H, соответственно. Способ титрования с точкой отсечения использовали в формате ELISA для выявления антител к KLH в сыворотке крови обезьян. Образцы инкубировали с KLH, иммобилизованным на планшете для ELISA. После инкубирования планшеты промывали и связанные антитела выявляли с помощью антител IgG-HRP или IgM-HRP козы к иммуноглобулинам обезьяны, а затем визуализировали с помощью тетраметилбензидина (TMB).
[0477] Во всех экспериментах с использованием животных соблюдали принятые в настоящее время практические принципы надлежащего содержания животных, например, изложенные в Guide for the Care and Use of Laboratory Animals; National Academy Press, 2011. Huntingdon Life Sciences, Ист-Миллстон, Нью-Джерси, полностью аккредитованные Международной ассоциацией по аттестации и аккредитации содержания лабораторных животных (AAALAC). За животными наблюдал технический персонал в отношении любых состояний, требующих возможного ветеринарного ухода, и при необходимости их лечили.
6.2 CD40L-специфичные Tn3-каркасные структуры осуществляют функциональную нейтрализацию CD40L. Взаимодействия с CD40.
[0478] T-клетки, в которых происходит экспрессия CD40L, вступают в контакт с В-клетками, в которых происходит экспрессия CD40, что в результате приводит к активации сигнального пути NFΚB (Zangani, 2009). Таким образом, клеточную линию с репортерным конструктом NFΚB-люцифераза применяли для определения того, могли ли молекулы Tn3, связывающиеся с CD40L, ингибировать передачу сигналов в нисходящем направлении после вступления в контакт с CD40. Клетки HEK293, в которых происходит экспрессия CD40L человека и репортера, стимулировали MegaCD40L человека или мыши при EC90 (эффективная концентрация, приводящая в результате к 90% ингибированию, т.е. 1,5 мкг/мл для MegaCD40L человека и 3 мкг/мл для MegaCD40L мыши).
[0479] Молекула 342, специфичная по отношению к лигандам человека, ингибировала активность NFΚB, индуцируемую CD40L человека, при IC50 = 1,5 нМ (Фиг. 13). Молекула M31, специфичная по отношению к лигандам мыши, нейтрализовала активность NFΚB, индуцируемую CD40L мыши, при IC50 = 1,6 нМ (Фиг. 1B). Моноклональные антитела как 5c8 (антитела к CD40L человека), так и MR1 (антитела к CD40L мыши), представляющие собой антитела к CD40L из группы положительного контроля, функционировали приблизительно в 10 раз лучше, чем мономерные каркасные структуры Tn3 с IC50 0,200 нМ +/- SD (самый низкий порог в анализе). Это может быть отчасти связано с бивалентной природой моноклональных антител, которая вносит вклад в авидность взаимодействия с их соответствующими CD40L.
6.3 Димерные CD40L-специфичные Tn3-каркасные структуры проявляют улучшенное связывание.
[0480] Экспериментальные данные указывают на то, что связывание CD40L-специфичной бивалентной Tn3-каркасной структуры было улучшено по сравнению с таковым CD40L-специфичной мономерной Tn3-каркасной структуры. Связывание CD40L-специфичной бивалентной Tn3-каркасной структуры с CD40L улучшало воздействие на мишень в некоторых случаях приблизительно на 3 порядка по сравнению с таковым для CD40L-специфичной мономерной Tn3-каркасной структуры in vitro, как показано на Фиг. 2C и Фиг. 2D (для лигандов мыши) и на Фиг. 8A и Фиг. 8B (для лигандов человека).
[0481] На Фиг. 2C показано конкурентное ингибирование связывания CD40L мыши с рецептором CD40 мыши, иммобилизованным на биосенсорном чипе с помощью моновалентных (M13) или бивалентных тандемных каркасных структур, специфичных по отношению к CD40L мыши. Мономеры M13 связывались с пептидными линкерами различной длины, содержащими один (1GS), три (3GS), пять (5GS) или семь (7GS) повторов "GGGGS". IC50 каркасной структуры M13-1GS-M13 составляла 29 нМ, тогда как IC50 мономерной каркасной структуры M13 составляла 71 нМ. IC50 бивалентных каркасных структур M13 с более длинными линкерами была значительно более низкой (5-6 нМ).
[0482] На Фиг. 2D показан ингибирующий эффект моновалентных (M13) или бивалентных тандемных каркасных структур, специфичных по отношению к CD40L мыши, в отношении экспрессии CD86, индуцируемой CD40L мыши, в В-клетках. Бивалентные каркасные структуры были приблизительно на 3 порядка более действенными, чем моновалентные каркасные структуры.
[0483] На Фиг. 8A и 8B показано, что Tn3-каркасные структуры 309 и 311, специфичные по отношению к CD40L человека, проявляли увеличенную действенность в формате бивалентного тандема. Бивалентные каркасные структуры 311 (Фиг. 8A) и бивалентные каркасные структуры 309 (Фиг. 8B) демонстрировали приблизительно 7-кратное и 500-1000-кратное улучшение, соответственно, при ингибировании экспрессии, индуцируемой CD40L человека, на CD19-положительных PBMC человека, стимулированных с помощью клеток Jurkat D1.1. Бивалентные каркасные структуры 309 были сравнимы по действенности с моноклональным антителом 5c8 к CD40L человека от Biogen.
[0484] Растворимость, стабильность и легкость очистки не нарушались при добавлении пептидных линкеров различной длины, содержащих один (1GS), три (3GS), пять (5GS) или семь (7GS) повторов "GGGGS" (см. Фиг. 2B).
6.4 Связывание и функционирование CD40L-специфичных Tn3-каркасных структур.
[0485] В дополнение к биохимическому связыванию, описанному выше, было важно удостовериться в том, что эти новые Tn3-каркасные структуры были способны связываться с эндогенными CD40L, экспрессируемыми на первичных T-клетках после активации. T-клетки выделяли из нескольких доноров и активировали согласно описанному. Через 24 часа экспрессия CD40L повышалась, что определяли путем окрашивания моноклональным антителом 5c8 (специфичным по отношению к лигандам человека) и моноклональным антителом MR1 (специфичным по отношению к лигандам мыши) (данные не показаны). С помощью молекул CD40L-специфичных Tn3-каркасных структур, так же как и моноклональных антител, можно было выявлять сопоставимые уровни экспрессии CD40L, что подтверждает, что эти молекулы способны связываться с нативным белком.
[0486] Одним из функциональных последствий взаимодействия CD40L:CD40 является повышение экспрессии костимулирующих молекул на В-клетках (Yellin et al., J. Exp. Med. 6:1857-1864, 1995). Молекулы Tn3, направленные на CD40L, тестировали в отношении их способности предупреждать это явление. Клеточные линии, в которых происходит эндогенная экспрессия CD40L человека или мыши (субклон Jurkat D1.1 или D10.G4, соответственно), применяли для стимуляции мононуклеарных клеток периферической крови (PBMC). После стимуляции активацию В-клеток оценивали путем измерения процентной доли В-клеток CD19+, в которых происходит повышенная экспрессия CD86, посредством проточной цитометрии. В этом анализе моноклональные антитела из группы положительного контроля были способны уменьшать процентную долю клеток CD19+ с экспрессией CD86 при IC50, составляющей 0,170 нМ (5c8) и 0,230 нМ (MR1). Оптимизированный Tn3 342, специфичный по отношению к лигандам человека, был способен оказывать антагонистическое влияние на повышение экспрессии CD86 при значениях IC50 = 0,700 нМ (n = 5 доноров) (см. Фиг. 10B и Таблицу 7).
[0487] Оптимизированный Tn3 M31, специфичный по отношению к лигандам мыши, характеризовался IC50 1,5 нМ. Эти подобные результаты наблюдали в случае, когда для стимуляции PBMC применяли рекомбинантный белок Mega-CD40L. Экспериментальные данные демонстрировали, что обе молекулы, специфичные либо по отношению к лигандам мыши, либо по отношению к лигандам человека, могут ингибировать не только главный сигнальный путь в клетке (NFΚB), но также одну из его наиболее важных функциональных ролей: взаимодействия T- и B-клеток. Это ингибирующее действие может препятствовать внесению вклада CD40L во многие аутоиммунные заболевания и состояния.
6.5 Tn3, связывающиеся с CD40L, ингибируют пролиферацию B-клеток и дифференцировку плазматических клеток после совместного культивирования T/B-клеток.
[0488] Взаимодействия CD40L на T-клетках с В-клетками, в которых происходит экспрессия CD40, представляют собой фундаментальный аспект хелперного действия T-клеток, которое способствует развитию форм адаптивного иммунного ответа (Banchereau, 1994; Oxenius, 1996, van Kooten & Banchereau, 1997). Для моделирования этого явления связывающиеся с hCD40L конструкты слияния Tn3-HSA, содержащие 340, 342 и димер 342-342, оценивали в совместных культурах первичных клеток, содержащих T-клетки и В-клетки, где стимулированные с помощью антитела к CD3, обработанные митомицином C T-клетки CD4+ человека культивировали с очищенными В-клетками человека. Измеряли способность В-клеток к пролиферации на четвертый день, к дифференцировке в плазматические клетки (PC) к седьмому дню и к переключению классов вырабатываемых ими антител (данные относительно PC и антител не показаны) (Фиг. 15) (Ettinger, 2007). CD40L-специфичная Tn3-каркасная структура 342-342-HSA была способна снижать темпы пролиферации, индуцируемой T-клетками, по меньшей мере на 50% по сравнению с клеточной пролиферацией B-клеток в отсутствие каркасных структур или в присутствии неспецифичной контрольной каркасной структуры. Пролиферация является предшественником и первым сигналом дифференцировки плазматических клеток после лигандирования CD40L:CD40. Также наблюдали ингибирование дифференцировки плазматических клеток и переключения классов антител (данные не показаны).
6.6 In vivo разрушение оси CD40:CD40L.
[0489] Центральная роль взаимодействий CD40L:CD40R в формах T-зависимого иммунного ответа была хорошо охарактеризована (Noelle, 1992; Renshaw, 1994, Wykes, 2003). Tn3-каркасную структуру M31, специфичную по отношению к CD40L мыши (M31-MSA и M31-M31-MSA), применяли для оценивания эффектов этих новых молекул в модели T-зависимой иммунизации путем иммунизации мышей (внутривенно) в нулевой день эритроцитами овцы (SRBC).
[0490] В дни 9-13 мышам внутрибрюшинно ежедневно инъецировали указанную дозу ингибитора, и в день 14 подсчитывали количество В-клеток из GC в селезенке и в лимфатических узлах. Ежедневное дозирование было необходимо с учетом короткого T1/2 данной молекулы in vivo, составляющего 31 час (Фиг. 4). На основании предыдущих данных результатов исследований точно установлено, что CD40L регулирует такие формы гуморального ответа, как образование зародышевых центров в анатомических участках, таких как селезенка и лимфатические узлы (Jacob, 1991). В данном случае наблюдали дозозависимое разрушение оси CD40L:CD40, которая вносит вклад в такое образование, при помощи M31-MSA, в отличие от "наивной" молекулы или неспецифичного контроля, D1-MSA, как видно из процентной доли В-клеток из GC.
[0491] Даже при 10 мг/кг M31-MSA был способен свести к нулю процент В-клеток из GC (Фиг. 5B), также как и моноклональное антитело MR1 (Фиг. 5A). Другие субпопуляции клеток выглядели нормальными, в том числе специфические популяции T-клеток, что позволяет убедиться в том, что наблюдаемые результаты не были связаны с элиминацией T-клеток (Фиг. 5C, Фиг. 5D, Фиг. 5E). В дополнение, результаты по данным ELISA с Ig, представляющим собой антитела к SRBC, отражали таковые по данным относительно B-клеток из зародышевых центров (Фиг. 5F). Взятые вместе, эти данные указывают на то, что Tn3-каркасная структура M31-MSA, специфичная по отношению к лигандам мыши, может прекращать реакции, управляемые посредством сигнального пути CD40.
[0492] Подобным образом, Tn3-каркасную структуру 342, специфичную по отношению к CD40L лигандам человека (342-HSA и 342-342-HSA), применяли для оценивания эффектов этих новых молекул в модели зависимого от T-клеток образования KLH-специфичных антител (TDAR) на макаках-крабоедах. В данном случае разрушение оси CD40L:CD40 приводит в результате к дозозависимому подавлению образования антител к антигену KLH. Как показано на фигурах 5G и 5H, бивалентный конструкт 342 подавлял уровни антител IgM и IgG при 0,5 мг/кг (миллиграммов на килограмм), а при 5 мг/кг наблюдалось почти полное подавление. Мономерный конструкт 342 также подавлял уровни IgM и IgG, но при более высоких концентрациях, при этом при 40 мг/кг наблюдалось почти полное подавление. Эти данные указывают на то, что оба специфичных для лигандов человека конструкта, представляющие собой каркасные структуры Tn3 342-HSA и 342-342-HSA, могут прекращать реакции, управляемые посредством сигнального пути CD40.
6.7 Tn3-каркасные структуры, специфичные по отношению к CD40L человека, не индуцируют агрегацию тромбоцитов.
[0493] Клинические испытания с участием людей с использованием моноклональных антител к CD40L приостанавливали, когда у некоторых пациентов возникали случаи тромбоэмболии (Davidson et al. Arth Rheu, 43:S271). Последующие доклинические анализы позволили предположить, что это явление представляет собой свойственный классу эффект моноклональных антител к CD40L в отношении мишеней. Таким образом, было важно провести тестирование Tn3-каркасных структур, специфичных по отношению к CD40L человека, в анализах агрегации тромбоцитов.
[0494] В случае применения соотношения трех молекул физиологического CD40L к одной молекуле моноклонального антитела к CD40L в цитратной плазме, обогащенной тромбоцитами (PRP), промытых тромбоцитах и цельной крови наблюдали проагрегантные эффекты (Фиг. 16A). Эти эффекты были опосредованы взаимодействиями, зависимыми от Fc-домена моноклонального антитела, после связывания с CD40L (данные не показаны). В отсутствие слияния с Fc-доменом агрегацию не наблюдали. У нескольких доноров при любых специфичных по отношению к CD40L человека Tn3-каркасных структурах в качестве димеров или в качестве белков слияния на основе HSA не наблюдали агрегацию (Фиг. 16B и Фиг. 16C).
[0495] Пагубные побочные эффекты, наблюдаемые в клинических испытаниях, наблюдали при образовании растворимого иммунного комплекса CD40L/моноклональное антитело к CD40L в присутствии тромбоцитов (Фиг. 16A и кривая 5C8 на Фиг. 16C). Другой пример этого явления можно наблюдать в гистологическом исследовании трансгенного FcγRIIa человека на мышах (Francis et al., 2010). После введения растворимых иммунных комплексов CD40L/моноклональное антитело в легочной ткани в течение нескольких минут после введения наблюдали большое количество тромбов. Однако, при повторении с использованием Tn3-каркасных структур, связывающихся с CD40L, легкое характеризовалось нормальным гистологическим строением, что находилось в соответствии с контрольными образцами (данные не показаны).
Пример 7
Домены фибронектина типа III, сконструированные для связывания с CD40L: клонирование, экспрессия, очистка, кристаллизация и предварительный рентгеновский дифракционный анализ двух комплексов.
[0496] Рекомбинантный растворимый CD40L человека подвергали совместной кристаллизации с CD40L-специфичными мономерными Tn3-каркасными структурами , 309 и 311K4E-12, обе из которых были выделены в качестве молекул, связывающихся с CD40L, из фаг-дисплейных библиотек. Дифракция на кристаллах происходила при разрешении до 3,1 и 2,9 Å, соответственно. В дополнение, рекомбинантный растворимый CD40L человека подвергали совместной кристаллизации с оптимизированным мономерным Tn3 342 в отдельности и вместе, как с мономерным 342, так и с мономерным 311K4E_12. Дифракция на кристаллах, соответствующих этим структурам, происходила при разрешении до 2,8 и 1,9 Å, соответственно. Соответствующие кристаллические структуры помогают понять взаимодействие между каркасными структурами Tn3 и CD40L и могут применяться для разработки молекул, связывающихся с CD40L с более высокой аффинностью, и тандемных конструктов, связывающихся с несколькими эпитопами.
7.1 Экспрессия и очистка молекул Tn3 и растворимого CD40L человека
[0497] Для получения молекул Tn3 без меток для кристаллизации белки экспрессировали в E. coli при помощи IPTG-индуцируемого вектора собственного производства, предназначенного для секреции белков, экспрессируемых рекомбинантным путем, в периплазматическое пространство. Этот вектор имел промотор Ptac, мутантную форму L25/M сигнального пептида OppA (MTNITKRSLVAAGVLAALMAGNVAMA) (SEQ ID NO: 210), C-концевую 8xHis-метку в дополнение к сайту расщепления тромбина. Последовательности Tn3 субклонировали между сигнальным пептидом и сайтом расщепления тромбина.
[0498] Экспрессируемые секретируемые белки с His-метками очищали при помощи смолы Ni-NTA согласно инструкциям производителя (Qiagen, Валенсия, Калифорния, США) и затем расщепляли с помощью тромбина с последующей повторной аффинной очисткой при помощи Ni-NTA для удаления неразрезанного интактного белка и отрезанного фрагмента с His-меткой. За этим этапом очистки следовал этап ионного обмена при помощи колонок HiTrap Q (GE Healthcare, Пискатавэй, Нью-Джерси, США), который выполняли на Äkta Purifier (GE Healthcare, Пискатавэй, Нью-Джерси, США). Очищенные белки Tn3 без меток демонстрируют более чем 95% чистоту и однородность на основе результатов SDS-PAGE и SEC.
[0499] Ген растворимого CD40L человека (113-261, UNIPROT: P29965) синтезировали с помощью GeneArt с N-концевой 6xHis-меткой и клонировали в вектор экспрессии у млекопитающих собственного производства под контролем основного немедленно-раннего промотора цитомегаловируса (hCMVie) (Boshart et al., Cell 41: 521-530,1985). Ген CD40L клонировали внутри рамки считывания с сигнальным пептидом CD33. Гены EBNA и Ori P в векторе применяли для повышения уровня экспрессии белка. Ген CD40L также включал последовательность поли-A SV40, обеспечивающую возможность должного процессинга 3’-конца его мРНК. Конструкт подвергали транзиентной трансфекции в клетки из суспензионной культуры 293F (эмбриональные клетки почек человека [HEK], выращиваемые в среде 293 Freestyle с применением 293 Fectin, Invitrogen, Карлсбад, Калифорния, США). Клетки выращивали с применением стандартного протокола, и со среды осуществляли сбор через 4 и 8 дней. Растворимый белок CD40L затем очищали при помощи смолы Ni-NTA с последующим этапом ионного обмена с помощью колонки Hi-Trap SP FF (GE Healthcare, Пискатавэй, Нью-Джерси, США) и диализом против 50 мМ Tris с pH 7,5, 50 мМ NaCl.
[0500] Для получения комплексов молекулу Tn3, 309, или 311K4E_12, или 342, смешивали с CD40L в соотношении 1,1:1, концентрировали при помощи концентраторов Vivaspin (предельное значение 30000 Да; GE Healthcare, Пискатавэй, Нью-Джерси, США) до приблизительно 10 мг/мл и подвергали эксклюзионной хроматографии (SEC) при помощи колонки Superdex 200 10/300 GL (GE Healthcare, Пискатавэй, Нью-Джерси, США), предварительно уравновешенной с помощью 50 мМ Tris-HCl, pH 7,5, 100 мМ NaCl, 0,02% NaN3 (Фиг. 19, панель A). После этапа разделения комплекс концентрировали до 18 мг/мл и подвергали кристаллизации. Комплекс 342-311K4E_12-CD40L получали в точности так, как описано выше, с применением трех компонентов в соотношении 1,1:1,1:1.
7.2 Скрининг и оптимизация условий кристаллизации
[0501] Организацию экспериментов по кристаллизации в "сидячей капле" осуществляли в 96-луночных планшетах Intelli (Art Robbins Instruments, Саннивэйл, Калифорния, США) при помощи робота для создания кристаллизационных растворов Phoenix (Art Robbins Instruments, Саннивэйл, Калифорния, США) путем смешивания луночного раствора и раствора белкового комплекса, имеющих объемы 300 нл, в ячейке для капель и предоставления им возможности уравновешивания против 50 мкл луночного раствора. Применяли коммерческие наборы для скрининга условий кристаллизации от Hampton Research (Алисо-Вьехо, Калифорния, США), Emerald BioSystems (Бэйнбридж-Айленд, Вашингтон, США) и Molecular Dimensions (Апопка, Флорида, США).
[0502] Для кристаллизации каждого из комплексов 309-CD40L, 342-CD40L и 342-311K4E_12-CD40L требовался этап оптимизации, который включал дополнительный этап скрининга с помощью Additive Screen HT (Hampton Research, Алисо-Вьехо, Калифорния, США). На этапе оптимизации луночный раствор, которым заполняли 96-луночный планшет, состоял на 80% из раствора, дающего нужный эффект на начальном этапе скрининга, и на 20% из соответствующей добавки. Каплю создавали из 300 нл белкового раствора и 300 нл нового луночного раствора после тщательного перемешивания последнего. Кристаллы, обладающие должным качеством для дифракции, собирали непосредственно из 96-луночного планшета. В целях криоконсервации кристалл последовательно переносили в три раствора маточной жидкости с повышающейся концентрацией глицерина.
[0503] Кристаллы 311-CD40L, обладающие должным качеством для дифракции, выращивали в растворе из начального этапа скрининга, который не требует добавления криопротектора.
7.3 Рентгеновская дифракция и сбор данных
[0504] Картины рентгеновской дифракции для комплекса 309-CD40L получали от монокристалла на Beamline 5.0.3 в Усовершенствованном источнике света, Национальная лаборатория имени Лоуренса в Беркли (Калифорнийский университет, Беркли), оборудованном рентгеновским детектором ADSC Q315R CCD (Area Detector Systems Corporation, Повей, Калифорния, США). Собирали 360 последовательных изображений с диапазоном колебаний 0,5º при расстоянии между кристаллом и детектором 300 мм и времени облучения 0,8 секунд.
[0505] Картины рентгеновской дифракции для комплексов 311K4E_12-CD40L, 342-CD40L и 342-311K4E_12-CD40L получали от монокристаллов на Beamline 31-ID-D в Усовершенствованном источнике фотонов, Аргоннская национальная лаборатория (Чикагский университет, Чикаго, Иллинойс), оборудованном детектором Rayonix 225 HE (Rayonix LLC, Эванстон, Иллинойс, США). Собирали 180 последовательных изображений с диапазоном колебаний 1º при расстоянии между кристаллом и детектором 300 мм и времени облучения 0,8 секунд.
[0506] Уменьшение и увеличение размера всех наборов данных осуществляли при помощи пакета программ HKL2000 (Otwinowski & Minor, Methods in Enzymology, 276:307-326. 1997).
7.4 Результаты и обсуждение
[0507] Наиболее воспроизводимым условием кристаллизации комплекса 309-CD40L, по-видимому, являлось B5 (0,2 M NaNO3, 20% PEG 3350) в случае набора для скрининга Peg/Ion (Hampton Research). При дополнительной оптимизации с помощью Additive Screen получали кристалл, обладающий должным качеством для дифракции, получаемый при условии A1 (0,1 M BaCl2·2H2O). Кристалл, показанный на Фиг. 19, панель B, собирали с 96-луночного планшета и охлаждали в жидком азоте после переноса в раствор маточной жидкости, дополненный 20% глицерином.
[0508] Симметрия пространственной группы. Кристалл принадлежал к орторомбической пространственной группе P212121 с параметрами ячейки a=85,69 Å, b=90,64 Å, c=95,56 Å, и дифракция на нем происходила при разрешении до 3,1 Å. Ожидается, что асимметричная единица содержит тример CD40L и три молекулы 309 со значением VM приблизительно 2,3 Å3/Да.
[0509] Что касается кристаллизации 311K4E_12-CD40L, в случае набора для скрининга Cryo I & II (Emerald BioStructures) получали ряд условий, которые не требовали ни оптимизации, ни криоконсервации. Монокристалл (Фиг. 19, панель C), получаемый при условии F7 (40% PEG 600, 0,1 M CH3COONa, 0,2 M MgCl2) применяли для сбора данных.
[0510] Симметрия пространственной группы. Кристалл принадлежал к кубической пространственной группе P213 с параметром ячейки 97,62 Å, и дифракция на нем происходила при разрешении до 2,6 Å. Асимметричная единица содержит одну молекулу CD40L и одну молекулу 311K4E_12 со значением VM приблизительно 2,9 Å3/Да.
[0511] Симметрия пространственной группы 342-CD40L. Кристалл принадлежал к пространственной группе P321 с параметрами ячейки a=93,53 Å, b=93,53 Å, c=66,69 Å, разрешением 2,8 Å. Асимметричная единица содержит один мономер CD40L и один мономер 342.
[0512] Симметрия пространственной группы 342-311K4E_12-CD40L. Кристалл принадлежал к пространственной группе P21 с параметрами ячейки a=80,32 Å, b=143,48 Å, c=111,27 Å, β=98,22º, разрешением 1,9 Å. Асимметричная единица содержит два тримера CD40, шесть мономеров 342 и шесть мономеров 311K4E-12.
[0513] Статистические данные для всех структур показаны в таблице 10.
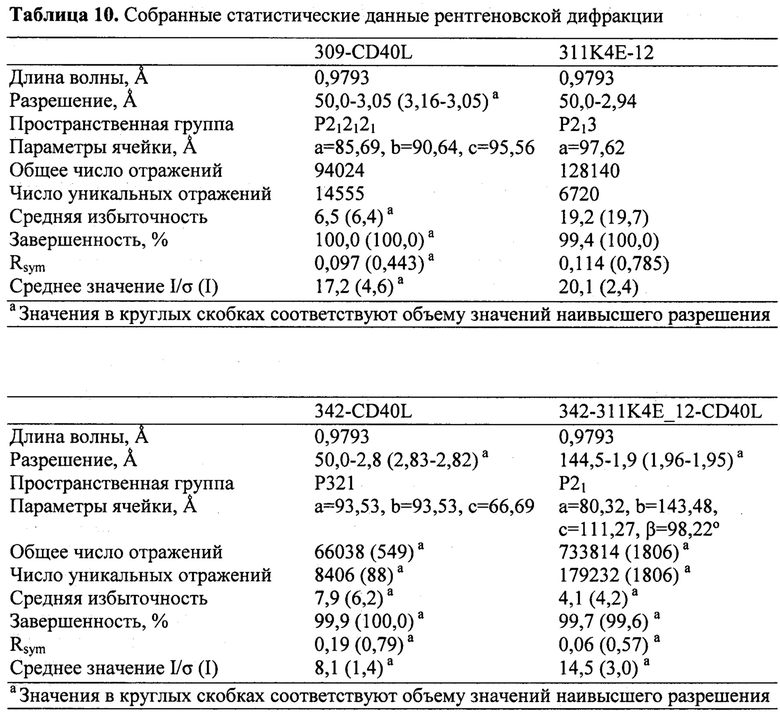
[0514] CD40L образовывал тример (полипептиды A, B и C на Фиг. 17A). Каждая Tn3-каркасная структура 309 (полипептиды D, E и F на Фиг. 17A) образовывала контакт с двумя полипептидами CD40L. В кристаллической структуре выявили, что между каждой каркасной структурой 309 и первым и вторым полипептидами CD40L существовали шесть специфических контактов. Аспарагиновая кислота 17 в BC образует контакт с треонином 251 в первом CD40L. Глутаминовая кислота 18 в петле BC образует контакт с аргинином 203 в первом CD40L и с изолейцином 204 во втором CD40L. Серин 47 в петле DE образует контакт с гистидином 249 в первом CD40L. Триптофан 49 в петле DE образует контакт с валином 247 в первом CD40L. Аспарагиновая кислота 70 в петле FG образует контакт с серином 185 во втором CD40L (см. Фиг. 17A). Аминокислотные остатки CD40L, контактирующие с каркасной структурой 311, также показаны на Фиг. 18A.
[0515] Как и в случае 309, каждая мономерная каркасная структура 311K4E_12 (полипептиды A, B и C на Фиг. 17B) образует контакт с двумя полипептидами CD40L. В кристаллической структуре выявили, что между каждой каркасной структурой 311K4E_12 и первым и вторым полипептидами CD40L существовали 19 специфических контактов. Аспарагин 17 в петле BC образует контакты с тирозином 146 и глутаминовой кислотой 142 в первом CD40L. Аргинин 18 в петле BC образует контакт с глутаминовой кислотой 142, тирозином 146 и метионином 148 в первом CD40L. Серин 19 в петле BC образует контакт с глутаминовой кислотой 142 и лейцином 155 в первом CD40L. Серин 22 в петле BC образует контакт с аспарагином 151 в первом CD40L. Гистидин 15 в петле BC образует контакт с тирозином 146 в первом CD40L. Гистидин 51 в петле DE образует контакт с тирозином 146 в первом CD40L и с глутаминовой кислотой 230 во втором CD40L. Валин 50 в петле DE образует контакт с глутаминовой кислотой 230 во втором CD40L. N-концевой участок мономерной каркасной структуры 311K4E_12 соединен со вторым CD40L. Аргинин 200 во втором CD40L образует контакт с треонином 7, аспарагиновой кислотой 8 и треонином 10 в N-концевом участке 311K4E_12. Аргинин 203 во втором CD40L образует контакт с глутаминовой кислотой 4 и аспарагиновой кислотой 5. Аминокислотные остатки CD40L, контактирующие с каркасной структурой 309, также показаны на Фиг. 18B.
[0516] Из кристаллических структур CD40L в комплексах с 309 и 311K4E_12 было видно, что мономерные каркасные структуры 311K4E_12 и 309 связываются с различными эпитопами, расположенными в различных частях тримерного комплекса CD40L (Фиг. 17C). Из структур было видно, что обе каркасные структуры связываются в одной и той же бороздке, которая будет взаимодействовать с рецептором CD40.
[0517] Кристаллическая структура 342 с CD40L представлена на фигуре 20, и из нее видно, что хотя 342 связывается с той же частью CD40L, наблюдаются специфические изменения в остатках, образующих контакты, по сравнению с исходным клоном 309. Конкретно, в 342 аспартат 18 в петле BC формирует контакт с треонином 251 CD40L, а гистидин 47 петли DE образует контакт с гистидином 249 CD40L, гистидин 48 петли DE образует контакт с гистидином 249, серином 245 и серином 248 CD40L, а гистидин 50 петли DE образует контакт с валином 247 CD40L.
[0518] Кристаллическая структура 342 и 311K4E_12 с CD40L демонстрирует, что обе каркасные структуры могут одновременно связываться с их соответствующими эпитопами, расположенными в различных частях тримерного комплекса CD40L (Фиг. 21). Контакты с каждой отдельной каркасной структурой (как описано выше) сохраняются.
[0519] Примеры, показанные выше, иллюстрируют различные аспекты настоящего изобретения и практическое осуществление способов по настоящему изобретению. Эти примеры не предназначены для обеспечения полного описания многих различных вариантов осуществления настоящего изобретения. Таким образом, хотя настоящее изобретение было довольно подробно описано посредством иллюстрации и примеров в целях ясности понимания, средние специалисты в данной области без труда осознают, что могут быть внесены многие изменения и модификации без отступления от сущности или объема прилагаемой формулы изобретения.
[0520] Все публикации, патенты и заявки на патенты, упоминаемые в настоящем описании, включены в данное описание посредством ссылки в такой же степени, как если бы каждые отдельные публикация, патент или заявка на патент были конкретно и отдельно указаны как включенные в данный документ посредством ссылки.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO: 1
CD40L sp|P29965|CD40L_ЧЕЛОВЕКА – мембранная форма
Цитоплазматический домен = 1-20
участок сигнального якорного мембранного белка типа II = 21-46
растворимая форма = 113-261
MIETYNQTSPRSAATGLPISMKIFMYLLTVFLITQMIGSALFAVYLHRRLDKIEDERNLH
EDFVFMKTIQRCNTGERSLSLLNCEEIKSQFEGFVKDIMLNKEETKKENSFEMQKGDQNP
QIAAHVISEASSKTTSVLQWAEKGYYTMSNNLVTLENGKQLTVKRQGLYYIYAQVTFCSN
REASSQAPFIASLCLKSPGRFERILLRAANTHSSAKPCGQQSIHLGGVFELQPGASVFVN
VTDPSQVSHGTGFTSFGLLKL
SEQ ID NO: 2
CD40L– растворимая форма, также соответствующая конструкту, полученному в результате совместной кристаллизации
MQKGDQNPQIAAHVISEASSKTTSVLQWAEKGYYTMSNNLVTLENGKQLTVKRQGLYYIY
AQVTFCSNREASSQAPFIASLCLKSPGRFERILLRAANTHSSAKPCGQQSIHLGGVFELQ
PGASVFVNVTDPSQVSHGTGFTSFGLLKL
SEQ ID NO: 3
Tn3 (с немодифицированными петлями)
IEVKDVTDTTALITWFKPLAEIDGCELTYGIKDVPGDRTTIDLTEDENQYSIGNLKPDTE
YEVSLICRRGDMSSNPAKETFTT
SEQ ID NO: 4
3-ий FnIII из тенасцина C, петля AB (Tn3)
KDVTDTT
SEQ ID NO: 5
3-ий FnIII из тенасцина C, петля BC (Tn3)
FKPLAEIDG
SEQ ID NO: 6
3-ий FnIII из тенасцина C, петля CD (Tn3)
KDVPGDR
SEQ ID NO: 7
3-ий FnIII из тенасцина C, петля DE (Tn3)
TEDENQ
SEQ ID NO: 8
3-ий FnIII из тенасцина C, петля EF (Tn3)
GNLKPDTE
SEQ ID NO: 9
3-й FnIII тенасцина C, FG-петля (Tn3); также в клонах 309FGwt, 340,341, 342, 343, 344, 345, 346, 347, 348 и 349
RRGDMSSNPA
SEQ ID NO: 10
3-ий FnIII из тенасцина C, бета-цепь A (Tn3)
RLDAPSQIEV
SEQ ID NO: 11
3-й FnIII тенасцина C, N-концевое усечение бета-цепи A (Tn3)
IEV
SEQ ID NO: 12
3-ий FnIII из тенасцина C, бета-цепь B (Tn3)
ALITW
SEQ ID NO: 13
3-ий FnIII из тенасцина C, бета-цепь C (вариант Tn3)
CELAYGI
SEQ ID NO: 14
3-ий FnIII из тенасцина C, бета-цепь C (Tn3)
CELTYGI
SEQ ID NO: 15
3-ий FnIII из тенасцина C, бета-цепь D (Tn3)
TTIDL
SEQ ID NO: 16
3-ий FnIII из тенасцина C, бета-цепь E (Tn3)
YSI
SEQ ID NO: 17
3-ий FnIII из тенасцина C, бета-цепь F (Tn3)
YEVSLIC
SEQ ID NO: 18
3-ий FnIII из тенасцина C, бета-цепь G (Tn3)
KETFTT
SEQ ID NO: 19
Клон 309 - исходный клон, выделенный из наивной библиотеки Tn3
AIEVKDVTDTTALITWSDEFGHYDGCELTYGIKDVPGDRTTIDLWWHSAWYSIGNLKPDT
EYEVSLICYTDQEAGNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 20
Клон 309 - исходный клон, выделенный из наивной библиотеки Tn3 (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVKDVTDTTALITWSDEFGHYDGCELTYGIKDVPGDRTTIDLWWHSAWYSIGNLKPDTE
YEVSLICYTDQEAGNPAKETFTT
SEQ ID NO: 21
Клон 309FGwt - исходный клон с "гуманизированной" петлей FG
AIEVKDVTDTTALITWSDEFGHYDGCELTYGIKDVPGDRTTIDLWWHSAWYSIGNLKPDT
EYEVSLICRRGDMSSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 22
Клон 309FGwt - исходный клон с "гуманизированной" петлей FG (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVKDVTDTTALITWSDEFGHYDGCELTYGIKDVPGDRTTIDLWWHSAWYSIGNLKPDTE
YEVSLICRRGDMSSNPAKETFTT
SEQ ID NO: 23
Клон 340 - вариант с созревшей аффинностью
AIEVKDVTDTTALITWSDDFDNYEWCELTYGIKDVPGDRTTIDLWYHMAWYSIGNLKPDT
EYEVSLICRRGDMSSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 24
Клон 340 - вариант с созревшей аффинностью (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVKDVTDTTALITWSDDFDNYEWCELTYGIKDVPGDRTTIDLWYHMAWYSIGNLKPDTE
YEVSLICRRGDMSSNPAKETFTT
SEQ ID NO: 25
Клон 341 - вариант с созревшей аффинностью
AIEVKDVTDTTALITWSDDFADYVWCELTYGIKDVPGDRTTIDLWWHSAWYSIGNLKPDT
EYEVSLICRRGDMSSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 26
Клон 341 - вариант с созревшей аффинностью (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVKDVTDTTALITWSDDFADYVWCELTYGIKDVPGDRTTIDLWWHSAWYSIGNLKPDTE
YEVSLICRRGDMSSNPAKETFTT
SEQ ID NO: 27
Клон 342 - вариант с созревшей аффинностью (с петлей FG дикого типа) AIEVKDVTDTTALITWSDDFGEYVWCELTYGIKDVPGDRTTIDLWYHHAHYSIGNLKPDT
EYEVSLICRRGDMSSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 28
Клон 342 - вариант с созревшей аффинностью (с петлей FG дикого типа; без N-концевого A, и C-концевого линкера, и His8-метки)
IEVKDVTDTTALITWSDDFGEYVWCELTYGIKDVPGDRTTIDLWYHHAHYSIGNLKPDTE
YEVSLICRRGDMSSNPAKETFTT
SEQ ID NO: 29
Клон 343 - вариант с созревшей аффинностью AIEVKDVTDTTALITWLDDWGSYHVCELTYGIKDVPGDRTTIDLWYHQAWYSIGNLKPDT
EYEVSLICRRGDMSSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 30
Клон 343 - вариант с созревшей аффинностью (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVKDVTDTTALITWLDDWGSYHVCELTYGIKDVPGDRTTIDLWYHQAWYSIGNLKPDTE
YEVSLICRRGDMSSNPAKETFTT
SEQ ID NO: 31
Клон 344 - вариант с созревшей аффинностью AIEVKDVTDTTALITWSDEVGDYVVCELTYGIKDVPGDRTTIDLWYHMAWYSIGNLKPDT
EYEVSLICRRGDMSSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 32
Клон 344 - вариант с созревшей аффинностью (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVKDVTDTTALITWSDEVGDYVVCELTYGIKDVPGDRTTIDLWYHMAWYSIGNLKPDTE
YEVSLICRRGDMSSNPAKETFTT
SEQ ID NO: 33
Клон 345 - вариант с созревшей аффинностью AIEVKDVTDTTALITWSDDFAEYVGCELTYGIKDVPGDRTTIDLWWHSAWYSIGNLKPDT
EYEVSLICRRGDMSSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 34
Клон 345 - вариант с созревшей аффинностью (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVKDVTDTTALITWSDDFAEYVGCELTYGIKDVPGDRTTIDLWWHSAWYSIGNLKPDTE
YEVSLICRRGDMSSNPAKETFTT
SEQ ID NO: 35
Клон 346 - вариант с созревшей аффинностью AIEVKDVTDTTALITWSDDFEEYVVCELTYGIKDVPGDRTTIDLWWHSAWYSIGNLKPDT
EYEVSLICRRGDMSSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 36
Клон 346 - вариант с созревшей аффинностью (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVKDVTDTTALITWSDDFEEYVVCELTYGIKDVPGDRTTIDLWWHSAWYSIGNLKPDTE
YEVSLICRRGDMSSNPAKETFTT
SEQ ID NO: 37
Клон 347 - вариант с созревшей аффинностью AIEVKDVTDTTALITWSDEVGQYVGCELTYGIKDVPGDRTTIDLWYHMAWYSIGNLKPDT
EYEVSLICRRGDMSSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 38
Клон 347 - вариант с созревшей аффинностью (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVKDVTDTTALITWSDEVGQYVGCELTYGIKDVPGDRTTIDLWYHMAWYSIGNLKPDTE
YEVSLICRRGDMSSNPAKETFTT
SEQ ID NO: 39
Клон 348 - вариант с созревшей аффинностью AIEVKDVTDTTALITWSDDIGLYVWCELTYGIKDVPGDRTTIDLWFHQAWYSIGNLKPDT
EYEVSLICRRGDMSSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 40
Клон 348 - вариант с созревшей аффинностью (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVKDVTDTTALITWSDDIGLYVWCELTYGIKDVPGDRTTIDLWFHQAWYSIGNLKPDTE
YEVSLICRRGDMSSNPAKETFTT
SEQ ID NO: 41
Клон 349 - вариант с созревшей аффинностью AIEVKDVTDTTALITWSDEHAEFIGCELTYGIKDVPGDRTTIDLWWHSAWYSIGNLKPDT
EYEVSLICRRGDMSSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 42
Клон 349 - вариант с созревшей аффинностью (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVKDVTDTTALITWSDEHAEFIGCELTYGIKDVPGDRTTIDLWWHSAWYSIGNLKPDTE
YEVSLICRRGDMSSNPAKETFTT
SEQ ID NO: 43
Клон 311 - исходный клон, выделенный из наивной библиотеки Tn3
AIEVKDVTDTTALITWTNRSSYYNLHGCELTYGIKDVPGDRTTIDLSSPYVHYSIGNLKP
DTEYEVSLICLTTDGTYSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 44
Клон 311 - исходный клон, выделенный из наивной библиотеки Tn3 (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVKDVTDTTALITWTNRSSYYNLHGCELTYGIKDVPGDRTTIDLSSPYVHYSIGNLKPD
TEYEVSLICLTTDGTYSNPAKETFTT
SEQ ID NO: 45
Клон 311K4E - вариант из первого цикла созревания аффинности
AIEVEDVTDTTALITWTNRSSYYNLHGCELTYGIKDVPGDRTTIDLSSPYVHYSIGNLKP
DTEYEVSLICLTTDGTYSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 46
Клон 311K4E - вариант из первого цикла созревания аффинности (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVEDVTDTTALITWTNRSSYYNLHGCELTYGIKDVPGDRTTIDLSSPYVHYSIGNLKPD
TEYEVSLICLTTDGTYSNPAKETFTT
SEQ ID NO: 47
Клон 311K4E_1 - вариант клона из второго цикла созревания аффинности
AIEVEDVTDTTALITWINRSYYADLHGCELTYGIKDVPGDRTTIDLDQIYVHYSIGNLKP
DTKYEVSLICLTTDGTYSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 48
Клон 311K4E_1 - вариант клона из второго цикла созревания аффинности (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVEDVTDTTALITWINRSYYADLHGCELTYGIKDVPGDRTTIDLDQIYVHYSIGNLKPD
TKYEVSLICLTTDGTYSNPAKETFTT
SEQ ID NO: 49
Клон 311K4E_2 – вариант клона из второго цикла созревания аффинности AIEVEDVTDTTALITWTNRSSYSHLDGCELTYGIKDVPGDRTTIDLSAAIYVHYSIGNLK
PDTEYEVSLICLTTDGTYSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 50
Клон 311K4E_2 – вариант клона из второго цикла созревания аффинности (без N-концевого A, и C-концевого линкера, и His8-метки) IEVEDVTDTTALITWTNRSSYSHLDGCELTYGIKDVPGDRTTIDLSAAIYVHYSIGNLKP
DTEYEVSLICLTTDGTYSNPAKETFTT
SEQ ID NO: 51
Клон 311K4E_3 – вариант клона из второго цикла созревания аффинности
AIEVEDVTDTTALITWINRSSYHNFPHCELAYGIKDVPGDRTTIDLNSPYVHYSIGNLKP
DTEYEVSLICLTTDGTYSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 52
Клон 311K4E_3 – вариант клона из второго цикла созревания аффинности (без N-концевого A, и C-концевого линкера, и His8-метки) IEVEDVTDTTALITWINRSSYHNFPHCELAYGIKDVPGDRTTIDLNSPYVHYSIGNLKPD
TEYEVSLICLTTDGTYSNPAKETFTT
SEQ ID NO: 53
Клон 311K4E_4 – вариант клона из второго цикла созревания аффинности
AIEVEDVTDTTALITWTNRSSYSNHLGCELAYGIKDVPGDRTTIDLNNIYVHYSIGNLKP
DTEYEVSLICLTTDGTYSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 54
Клон 311K4E_4 - вариант клона из второго цикла созревания аффинности (без N-концевого A, и C-концевого линкера, и His8-метки) IEVEDVTDTTALITWTNRSSYSNHLGCELAYGIKDVPGDRTTIDLNNIYVHYSIGNLKPD
TEYEVSLICLTTDGTYSNPAKETFTT
SEQ ID NO: 55
Клон 311K4E_5 - вариант клона из второго цикла созревания аффинности
AIEVEDVTDTTALITWTNRSSYSNFHGCELAYGIKDVPGDRTTIDLNSPYVHYSIGNLKP
DTEYEVSLICLTTDGTYSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 56
Клон 311K4E_5 - вариант клона из второго цикла созревания аффинности (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVEDVTDTTALITWTNRSSYSNFHGCELAYGIKDVPGDRTTIDLNSPYVHYSIGNLKPD
TEYEVSLICLTTDGTYSNPAKETFTT
SEQ ID NO: 57
Клон 311K4E_7 - вариант клона из второго цикла созревания аффинности
AIEVEDVTDTTALITWTNRSFYSNLHGCELTYGIKDVPGDRTTIDLNQPYVHYSIGNLKP
DTEYEVSLICLTTDGTYSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 58
Клон 311K4E_7 - вариант клона из второго цикла созревания аффинности (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVEDVTDTTALITWTNRSFYSNLHGCELTYGIKDVPGDRTTIDLNQPYVHYSIGNLKPD
TEYEVSLICLTTDGTYSNPAKETFTT
SEQ ID NO: 59
Клон 311K4E_8 - вариант клона из второго цикла созревания аффинности
AIEVEDVTDTTALITWTNRSSYAYLHGCELAYGIKDVPGDRTTIDLNQPYVHYSIGNLKP
DTEYEVSLICLTTDGTYSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 60
Клон 311K4E_8 - вариант клона из второго цикла созревания аффинности (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVEDVTDTTALITWTNRSSYAYLHGCELAYGIKDVPGDRTTIDLNQPYVHYSIGNLKPD
TEYEVSLICLTTDGTYSNPAKETFTT
SEQ ID NO: 61
Клон 311K4E_9 - вариант клона из второго цикла созревания аффинности
AIEVEDVTDTTALITWINRSSYANLHGCELTYGIKDVPGDRTTIDLSSPYVHYSIGNLKP
DTEYEVSLICLTTDGTYSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 62
Клон 311K4E_9 - вариант клона из второго цикла созревания аффинности (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVEDVTDTTALITWINRSSYANLHGCELTYGIKDVPGDRTTIDLSSPYVHYSIGNLKPD
TEYEVSLICLTTDGTYSNPAKETFTT
SEQ ID NO: 63
Клон 311K4E_10 - вариант клона из второго цикла созревания аффинности
AIEVEDVTDTTALITWTNRSSYANYHGCELAYGIKDVPGDRTTIDLNQPYVHYSIGNLKP
DTEYEVSLICLTTDGTYSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 64
Клон 311K4E_10 - вариант клона из второго цикла созревания аффинности (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVEDVTDTTALITWTNRSSYANYHGCELAYGIKDVPGDRTTIDLNQPYVHYSIGNLKPD
TEYEVSLICLTTDGTYSNPAKETFTT
SEQ ID NO: 65
Клон 311K4E_11 - вариант клона из второго цикла созревания аффинности
AIEVEDVTDTTALITWTNRSSYANLPGCELTYGIKDVPGDRTTIDLNSPYVHYSIGNLKP
DTEYEVSLICLTTDGTYSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 66
Клон 311K4E_11 - вариант клона из второго цикла созревания аффинности (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVEDVTDTTALITWTNRSSYANLPGCELTYGIKDVPGDRTTIDLNSPYVHYSIGNLKPD
TEYEVSLICLTTDGTYSNPAKETFTT
SEQ ID NO: 67
Клон 311K4E_12 - вариант клона из второго цикла созревания аффинности
AIEVEDVTDTTALITWTNRSSYSNLHGCELAYGIKDVPGDRTTIDLNQPYVHYSIGNLKP
DTEYEVSLICLTTDGTYNNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 68
Клон 311K4E_12 - вариант клона из второго цикла созревания аффинности (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVEDVTDTTALITWTNRSSYSNLHGCELAYGIKDVPGDRTTIDLNQPYVHYSIGNLKPD
TEYEVSLICLTTDGTYNNPAKETFTT
SEQ ID NO: 69
Клон 311K4E_13 - вариант клона из второго цикла созревания аффинности
AIEVEDVTDTTALITWINRSSYANLHGCELTYGIKDVPGDRTTIDLNSPYVHYSIGNLKP
DTEYEVSLICLTTDGTYSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 70
Клон 311K4E_13 - вариант клона из второго цикла созревания аффинности (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVEDVTDTTALITWINRSSYANLHGCELTYGIKDVPGDRTTIDLNSPYVHYSIGNLKPD
TEYEVSLICLTTDGTYSNPAKETFTT
SEQ ID NO: 71
Клон 311K4E_14 - вариант клона из второго цикла созревания аффинности
AIEVEDVTDTTALITWTARSAYSHHHYCELTYGIKDVPGDRTTIDLRQPYVHYSIGNLKP
DTEYEVSLICLTTDGTYSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 72
Клон 311K4E_14 - вариант клона из второго цикла созревания аффинности (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVEDVTDTTALITWTARSAYSHHHYCELTYGIKDVPGDRTTIDLRQPYVHYSIGNLKPD
TEYEVSLICLTTDGTYSNPAKETFTT
SEQ ID NO: 73
Клон 311K4E_15 - вариант клона из второго цикла созревания аффинности
AIEVEDVTDTTALITWTNRSSYANYHHCELTYGIKDVPGDRTTIDLELYVHYSIGNLKPD
TEYEVSLICLTTDGTYSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 74
Клон 311K4E_15 - вариант клона из второго цикла созревания аффинности (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVEDVTDTTALITWTNRSSYANYHHCELTYGIKDVPGDRTTIDLELYVHYSIGNLKPDT
EYEVSLICLTTDGTYSNPAKETFTT
SEQ ID NO: 75
Клон 311K4E_16 - вариант клона из второго цикла созревания аффинности
AIEVEDVTDTTALITWTNRSSYSDLPGCELTYGIKDVPGDRTTIDLSSPYVHYSIGNLKP
DTEYEVSLICLTTDGTYSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 76
Клон 311K4E_16 - вариант клона из второго цикла созревания аффинности (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVEDVTDTTALITWTNRSSYSDLPGCELTYGIKDVPGDRTTIDLSSPYVHYSIGNLKPD
TEYEVSLICLTTDGTYSNPAKETFTT
SEQ ID NO: 77
Клон 311K4E_19 - вариант клона из второго цикла созревания аффинности
AIEVEDVTDTTALITWTHRSAYSNHSFCELTYGIKDVPGDRTTIDLNTPYVHYSIGNLKP
DTEYEVSLICLTTDGTYSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 78
Клон 311K4E_19 - вариант клона из второго цикла созревания аффинности (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVEDVTDTTALITWTHRSAYSNHSFCELTYGIKDVPGDRTTIDLNTPYVHYSIGNLKPD
TEYEVSLICLTTDGTYSNPAKETFTT
SEQ ID NO: 79
Клон 311K4E_20 - вариант клона из второго цикла созревания аффинности
AIEVEDVTDTTALITWTNRSLYANFHGCELTYGIKDVPGDRTTIDLEQVYVHYSIGNLKP
DTEYEVSLICLTTDGTYSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 80
Клон 311K4E_20 - вариант клона из второго цикла созревания аффинности (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVEDVTDTTALITWTNRSLYANFHGCELTYGIKDVPGDRTTIDLEQVYVHYSIGNLKPD
TEYEVSLICLTTDGTYSNPAKETFTT
SEQ ID NO: 81
Клон 311K4E_21 - вариант клона из второго цикла созревания аффинности
AIEVEDVTDTTALITWTNRSSYSNLPGCELTYGIKDVPGDRTTIDLNQVYVHYSIGNLKP
DTEYEVSLICLTTDGTYSNPAKETFTTGGGTLGHHHHHHHH
SEQ ID NO: 82
Клон 311K4E_21 - вариант клона из второго цикла созревания аффинности (без N-концевого A, и C-концевого линкера, и His8-метки)
IEVEDVTDTTALITWTNRSSYSNLPGCELTYGIKDVPGDRTTIDLNQVYVHYSIGNLKPD
TEYEVSLICLTTDGTYSNPAKETFTT
SEQ ID NO: 83
Клоны 309 и 309FGwt - петля BC
SDEFGHYDG
SEQ ID NO: 84
Клон 340 - петля BC
SDDFDNYEW
SEQ ID NO: 85
Клон 341 - петля BC
SDDFADYVW
SEQ ID NO: 86
Клон 342 - петля BC
SDDFGEYVW
SEQ ID NO: 87
Клон 343 - петля BC
LDDWGSYHV
SEQ ID NO: 88
Клон 344 - петля BC
SDEVGDYVV
SEQ ID NO: 89
Клон 345 - петля BC
SDDFAEYVG
SEQ ID NO: 90
Клон 346 - петля BC
SDDFEEYVV
SEQ ID NO: 91
Клон 347 – петля BC
SDEVGQYVG
SEQ ID NO: 92
Клон 348 – петля BC
SDDIGLYVW
SEQ ID NO: 93
Клон 349 - петля BC
SDEHAEFIG
SEQ ID NO: 94
Клоны 309, 309FGwt, 341, 345, 346, 349 – петля DE
WWHSAW
SEQ ID NO: 95
Клоны 340, 344, 347 – петля DE
WYHMAW
SEQ ID NO: 96
Клон 342 - петля DE
WYHHAH
SEQ ID NO: 97
Клон 343 – петля DE
WYHQAW
SEQ ID NO: 98
Клон 348 – петля DE
WFHQAW
SEQ ID NO: 99
Клон 309 – петля FG
YTDQEAGNPA
SEQ ID NO: 100
Клоны 311, 311K4E - петля BC
TNRSSYYNLHG
SEQ ID NO: 101
Клон 311K4E_1 - петля BC
INRSYYADLHG
SEQ ID NO: 102
Клон 311K4E_2 - петля BC
TNRSSYSHLDG
SEQ ID NO: 103
Клон 311K4E_3 - петля BC
INRSSYHNFPH
SEQ ID NO: 104
Клон 311K4E_4 - петля BC
TNRSSYSNHLG
SEQ ID NO: 105
Клон 311K4E_5 - петля BC
TNRSSYSNFHG
SEQ ID NO: 106
Клон 311K4E_7 - петля BC
TNRSFYSNLHG
SEQ ID NO: 107
Клон 311K4E_8 - петля BC
TNRSSYAYLHG
SEQ ID NO: 108
Клоны 311K4E_9, 311K4E_13 - петля BC
INRSSYANLHG
SEQ ID NO: 109
Клон 311K4E_10 - петля BC
TNRSSYANYHG
SEQ ID NO: 110
Клон 311K4E_11 - петля BC
TNRSSYANLPG
SEQ ID NO: 111
Клон 311K4E_12 - петля BC
TNRSSYSNLHG
SEQ ID NO: 112
Клон 311K4E_14 - петля BC
TARSAYSHHHY
SEQ ID NO: 113
Клон 311K4E_15 - петля BC
TNRSSYANYHH
SEQ ID NO: 114
Клон 311K4E_16 - петля BC
TNRSSYSDLPG
SEQ ID NO: 115
Клон 311K4E_19 - петля BC
THRSAYSNHSF
SEQ ID NO: 116
Клон 311K4E_20 - петля BC
TNRSLYANFHG
SEQ ID NO: 117
Клон 311K4E_21 - петля BC
TNRSSYSNLPG
SEQ ID NO: 118
Клоны 311, 311K4E, 311K4E_9, 311K4E_16 - петля DE
SSPYVH
SEQ ID NO: 119
Клон 311K4E_1 - петля DE
DQIYVH
SEQ ID NO: 120
Клон 311K4E_2 - петля DE
SAAIYVH
SEQ ID NO: 121
Клон 311K4E_3, 311K4E_5, 311K4E_11, 311K4E_13 - петля DE
NSPYVH
SEQ ID NO: 122
Клон 311K4E_4 - петля DE
NNIYVH
SEQ ID NO: 123
Клон 311K4E_7, 311K4E_8, 311K4E_10, 311K4E_12 - петля DE
NQPYVH
SEQ ID NO: 124
Клон 311K4E_14 - петля DE
RQPYVH
SEQ ID NO: 125
Клон 311K4E_15 - петля DE
ELYVH
SEQ ID NO: 126
Клон 311K4E_19 - петля DE
NTPYVH
SEQ ID NO: 127
Клон 311K4E_20 - петля DE
EQVYVH
SEQ ID NO: 128
Клон 311K4E_21 - петля DE
NQVYVH
SEQ ID NO: 129
Клоны 311, 311K4E, 311K4E_1, 311K4E_2, 311K4E_3, 311K4E_4, 311K4E_5, 311K4E_7, 311K4E_8, 311K4E_9, 311K4E_10, 311K4E_11, 311K4E_13, 311K4E_14, 311K4E_15, 311K4E_16, 311K4E_19, 311K4E_20, 311K4E_21 - петля FG
LTTDGTYSNPA
SEQ ID NO: 130
Клон 311K4E_12 - петля FG
LTTDGTYNNPA
SEQ ID NO: 131
Линкер 2GS – (Gly4Ser)2
GGGGSGGGGS
SEQ ID NO: 132
Линкер 3GS – (Gly4Ser)3
GGGGSGGGGSGGGGS
SEQ ID NO: 133
Мутант HSA C34S
Место локализации мутации Cys->Ser подчеркнуто
DAHKSEVAHRFKDLGEENFKALVLIAFAQYLQQSPFEDHVKLVNEVTEFAKTCVADESAE
NCDKSLHTLFGDKLCTVATLRETYGEMADCCAKQEPERNECFLQHKDDNPNLPRLVRPEV
DVMCTAFHDNEETFLKKYLYEIARRHPYFYAPELLFFAKRYKAAFTECCQAADKAACLLP
KLDELRDEGKASSAKQRLKCASLQKFGERAFKAWAVARLSQRFPKAEFAEVSKLVTDLTK
VHTECCHGDLLECADDRADLAKYICENQDSISSKLKECCEKPLLEKSHCIAEVENDEMPA
DLPSLAADFVESKDVCKNYAEAKDVFLGMFLYEYARRHPDYSVVLLLRLAKTYETTLEKC
CAAADPHECYAKVFDEFKPLVEEPQNLIKQNCELFEQLGEYKFQNALLVRYTKKVPQVST
PTLVEVSRNLGKVGSKCCKHPEAKRMPCAEDYLSVVLNQLCVLHEKTPVSDRVTKCCTES
LVNRRPCFSALEVDETYVPKEFNAETFTFHADICTLSEKERQIKKQTALVELVKHKPKAT
KEQLKAVMDDFAAFVEKCCKADDKETCFAEEGKKLVAASQAALGL
SEQ ID NO: 134
Моновалентный конструкт 342-2GS-HSAC34S
HSAC34S подчеркнут
SQIEVKDVTDTTALITWSDDFGEYVWCELTYGIKDVPGDRTTIDLWYHHAHYSIGNLKPD
TEYEVSLICRSGDMSSNPAKETFTTGGGGSGGGGSDAHKSEVAHRFKDLGEENFKALVLI
AFAQYLQQSPFEDHVKLVNEVTEFAKTCVADESAENCDKSLHTLFGDKLCTVATLRETYG
EMADCCAKQEPERNECFLQHKDDNPNLPRLVRPEVDVMCTAFHDNEETFLKKYLYEIARR
HPYFYAPELLFFAKRYKAAFTECCQAADKAACLLPKLDELRDEGKASSAKQRLKCASLQK
FGERAFKAWAVARLSQRFPKAEFAEVSKLVTDLTKVHTECCHGDLLECADDRADLAKYIC
ENQDSISSKLKECCEKPLLEKSHCIAEVENDEMPADLPSLAADFVESKDVCKNYAEAKDV
FLGMFLYEYARRHPDYSVVLLLRLAKTYETTLEKCCAAADPHECYAKVFDEFKPLVEEPQ
NLIKQNCELFEQLGEYKFQNALLVRYTKKVPQVSTPTLVEVSRNLGKVGSKCCKHPEAKR
MPCAEDYLSVVLNQLCVLHEKTPVSDRVTKCCTESLVNRRPCFSALEVDETYVPKEFNAE
TFTFHADICTLSEKERQIKKQTALVELVKHKPKATKEQLKAVMDDFAAFVEKCCKADDKE
TCFAEEGKKLVAASQAALGL
SEQ ID NO: 135
Бивалентный конструкт 342-3GS-342-2GS-HSAC34S
HSA подчеркнут
SQIEVKDVTDTTALITWSDDFGEYVWCELTYGIKDVPGDRTTIDLWYHHAHYSIGNLKPD
TEYEVSLICRSGDMSSNPAKETFTTGGGGSGGGGSGGGGSRLDAPSQIEVKDVTDTTALI
TWSDDFGEYVWCELTYGIKDVPGDRTTIDLWYHHAHYSIGNLKPDTEYEVSLICRSGDMS
SNPAKETFTTGGGGSGGGGSDAHKSEVAHRFKDLGEENFKALVLIAFAQYLQQSPFEDHV
KLVNEVTEFAKTCVADESAENCDKSLHTLFGDKLCTVATLRETYGEMADCCAKQEPERNE
CFLQHKDDNPNLPRLVRPEVDVMCTAFHDNEETFLKKYLYEIARRHPYFYAPELLFFAKR
YKAAFTECCQAADKAACLLPKLDELRDEGKASSAKQRLKCASLQKFGERAFKAWAVARLS
QRFPKAEFAEVSKLVTDLTKVHTECCHGDLLECADDRADLAKYICENQDSISSKLKECCE
KPLLEKSHCIAEVENDEMPADLPSLAADFVESKDVCKNYAEAKDVFLGMFLYEYARRHPD
YSVVLLLRLAKTYETTLEKCCAAADPHECYAKVFDEFKPLVEEPQNLIKQNCELFEQLGE
YKFQNALLVRYTKKVPQVSTPTLVEVSRNLGKVGSKCCKHPEAKRMPCAEDYLSVVLNQL
CVLHEKTPVSDRVTKCCTESLVNRRPCFSALEVDETYVPKEFNAETFTFHADICTLSEKE
RQIKKQTALVELVKHKPKATKEQLKAVMDDFAAFVEKCCKADDKETCFAEEGKKLVAASQ
AALGL
SEQ ID NO: 136
Вариант петли AB с N-концевым E, характерный для клонов из семейства 311
EDVTDTT
SEQ ID NO: 137
Вариант петли EF с C-концевым K, характерный для клонов из семейства 311
GNLKPDTK
SEQ ID NO: 138
HSA человека полной длины
DAHKSEVAHRFKDLGEENFKALVLIAFAQYLQQCPFEDHVKLVNEVTEFAKTCVADESAE
NCDKSLHTLFGDKLCTVATLRETYGEMADCCAKQEPERNECFLQHKDDNPNLPRLVRPEV
DVMCTAFHDNEETFLKKYLYEIARRHPYFYAPELLFFAKRYKAAFTECCQAADKAACLLP
KLDELRDEGKASSAKQRLKCASLQKFGERAFKAWAVARLSQRFPKAEFAEVSKLVTDLTK
VHTECCHGDLLECADDRADLAKYICENQDSISSKLKECCEKPLLEKSHCIAEVENDEMPA
DLPSLAADFVESKDVCKNYAEAKDVFLGMFLYEYARRHPDYSVVLLLRLAKTYETTLEKC
CAAADPHECYAKVFDEFKPLVEEPQNLIKQNCELFEQLGEYKFQNALLVRYTKKVPQVST
PTLVEVSRNLGKVGSKCCKHPEAKRMPCAEDYLSVVLNQLCVLHEKTPVSDRVTKCCTES
LVNRRPCFSALEVDETYVPKEFNAETFTFHADICTLSEKERQIKKQTALVELVKHKPKAT
KEQLKAVMDDFAAFVEKCCKADDKETCFAEEGKKLVAASQAALGL
SEQ ID NO: 139
Вариант петли FG 309 (с мутацией RR->RS); может присутствовать в конструктах 342
RSGDMSSNPA
SEQ ID NO: 140
Линкер 2GX – (Gly4X)2; X = Ala, Gly, Leu, Ile, Val
GGGGXGGGGX
SEQ ID NO: 141
Линкер 3GX – (Gly4X)3; X = Ala, Gly, Leu, Ile, Val
GGGGXGGGGXGGGGX
SEQ ID NO: 142
Линкер G10 – (Gly4Gly)2
GGGGGGGGGG
SEQ ID NO: 143
Линкер G15 – (Gly4Gly)3
GGGGGGGGGGGGGGG
SEQ ID NO: 144
342-G10-HSAC34S, моновалентный конструкт 2, линкеры полностью состоят из Gly
HSA подчеркнут
SQIEVKDVTDTTALITWSDDFGEYVWCELTYGIKDVPGDRTTIDLWYHHAHYSIGNLKPD
TEYEVSLICRSGDMSSNPAKETFTTGGGGGGGGGGDAHKSEVAHRFKDLGEENFKALVLI
AFAQYLQQSPFEDHVKLVNEVTEFAKTCVADESAENCDKSLHTLFGDKLCTVATLRETYG
EMADCCAKQEPERNECFLQHKDDNPNLPRLVRPEVDVMCTAFHDNEETFLKKYLYEIARR
HPYFYAPELLFFAKRYKAAFTECCQAADKAACLLPKLDELRDEGKASSAKQRLKCASLQK
FGERAFKAWAVARLSQRFPKAEFAEVSKLVTDLTKVHTECCHGDLLECADDRADLAKYIC
ENQDSISSKLKECCEKPLLEKSHCIAEVENDEMPADLPSLAADFVESKDVCKNYAEAKDV
FLGMFLYEYARRHPDYSVVLLLRLAKTYETTLEKCCAAADPHECYAKVFDEFKPLVEEPQ
NLIKQNCELFEQLGEYKFQNALLVRYTKKVPQVSTPTLVEVSRNLGKVGSKCCKHPEAKR
MPCAEDYLSVVLNQLCVLHEKTPVSDRVTKCCTESLVNRRPCFSALEVDETYVPKEFNAE
TFTFHADICTLSEKERQIKKQTALVELVKHKPKATKEQLKAVMDDFAAFVEKCCKADDKE
TCFAEEGKKLVAASQAALGL
SEQ ID NO: 145
342-G15-342-G10-HSAC34S, бивалентный конструкт 2, линкеры полностью состоят из Gly
HSA подчеркнут
SQIEVKDVTDTTALITWSDDFGEYVWCELTYGIKDVPGDRTTIDLWYHHAHYSIGNLKPD
TEYEVSLICRSGDMSSNPAKETFTTGGGGGGGGGGGGGGGRLDAPSQIEVKDVTDTTALI
TWSDDFGEYVWCELTYGIKDVPGDRTTIDLWYHHAHYSIGNLKPDTEYEVSLICRSGDMS
SNPAKETFTTGGGGGGGGGGDAHKSEVAHRFKDLGEENFKALVLIAFAQYLQQSPFEDHV
KLVNEVTEFAKTCVADESAENCDKSLHTLFGDKLCTVATLRETYGEMADCCAKQEPERNE
CFLQHKDDNPNLPRLVRPEVDVMCTAFHDNEETFLKKYLYEIARRHPYFYAPELLFFAKR
YKAAFTECCQAADKAACLLPKLDELRDEGKASSAKQRLKCASLQKFGERAFKAWAVARLS
QRFPKAEFAEVSKLVTDLTKVHTECCHGDLLECADDRADLAKYICENQDSISSKLKECCE
KPLLEKSHCIAEVENDEMPADLPSLAADFVESKDVCKNYAEAKDVFLGMFLYEYARRHPD
YSVVLLLRLAKTYETTLEKCCAAADPHECYAKVFDEFKPLVEEPQNLIKQNCELFEQLGE
YKFQNALLVRYTKKVPQVSTPTLVEVSRNLGKVGSKCCKHPEAKRMPCAEDYLSVVLNQL
CVLHEKTPVSDRVTKCCTESLVNRRPCFSALEVDETYVPKEFNAETFTFHADICTLSEKE
RQIKKQTALVELVKHKPKATKEQLKAVMDDFAAFVEKCCKADDKETCFAEEGKKLVAASQ
AALGL
SEQ ID NO: 146
Клон 342 - вариант с созревшей аффинностью (с вариантом петли FG, RR->RS подчеркнуто)
IEVKDVTDTTALITWSDDFGEYVWCELTYGIKDVPGDRTTIDLWYHHAHYSIGNLKPDTE
YEVSLICRSGDMSSNPAKETFTT
SEQ ID NO: 147
Линкерный модуль Gly-Ser, (G4S)n, где n = 1-7; показан модуль (G4S)n, где n=1
GGGGS
SEQ ID NO: 148
Линкерный модуль Gly, (G5)n, где n = 1-7; показан модуль (G5)n, где n=1
GGGGG
SEQ ID NO: 149
Линкерный модуль Gly-Ala, (G4A)n, где n = 1-7; показан модуль (G4A)n, где n=1
GGGGA
SEQ ID NO: 150
Полигистидиновая метка (H8) - необязательный компонент Tn3-каркасных структур, пригодный для очистки, можно объединять с дополнительными остатками линкера.
HHHHHHHH
SEQ ID NO: 151
Линкер, соединенный с полигистидиновой меткой - необязательный компонент Tn3-каркасных структур, пригодный для очистки
GGGGSHHHHHHHH
SEQ ID NO: 152
Зрелый MSA дикого типа
EAHKSEIAHRYNDLGEQHFKGLVLIAFSQYLQKCSYDEHAKLVQEVTDFAKTCVADESAA
NCDKSLHTLFGDKLCAIPNLRENYGELADCCTKQEPERNECFLQHKDDNPSLPPFERPEA
EAMCTSFKENPTTFMGHYLHEVARRHPYFYAPELLYYAEQYNEILTQCCAEADKESCLTP
KLDGVKEKALVSSVRQRMKCSSMQKFGERAFKAWAVARLSQTFPNADFAEITKLATDLTK
VNKECCHGDLLECADDRAELAKYMCENQATISSKLQTCCDKPLLKKAHCLSEVEHDTMPA
DLPAIAADFVEDQEVCKNYAEAKDVFLGTFLYEYSRRHPDYSVSLLLRLAKKYEATLEKC
CAEANPPACYGTVLAEFQPLVEEPKNLVKTNCDLYEKLGEYGFQNAILVRYTQKAPQVST
PTLVEAARNLGRVGTKCCTLPEDQRLPCVEDYLSAILNRVCLLHEKTPVSEHVTKCCSGS
LVERRPCFSALTVDETYVPKEFKAETFTFHSDICTLPEKEKQIKKQTALAELVKHKPKAT
AEQLKTVMDDFAQFLDTCCKAADKDTCFSTEGPNLVTRCKDALA
SEQ ID NO: 153
Зрелый MSA-C34S/C579S с мутацией по Cys; остатки, появившиеся в результате мутации, подчеркнуты
EAHKSEIAHRYNDLGEQHFKGLVLIAFSQYLQKSSYDEHAKLVQEVTDFAKTCVADESAA
NCDKSLHTLFGDKLCAIPNLRENYGELADCCTKQEPERNECFLQHKDDNPSLPPFERPEA
EAMCTSFKENPTTFMGHYLHEVARRHPYFYAPELLYYAEQYNEILTQCCAEADKESCLTP
KLDGVKEKALVSSVRQRMKCSSMQKFGERAFKAWAVARLSQTFPNADFAEITKLATDLTK
VNKECCHGDLLECADDRAELAKYMCENQATISSKLQTCCDKPLLKKAHCLSEVEHDTMPA
DLPAIAADFVEDQEVCKNYAEAKDVFLGTFLYEYSRRHPDYSVSLLLRLAKKYEATLEKC
CAEANPPACYGTVLAEFQPLVEEPKNLVKTNCDLYEKLGEYGFQNAILVRYTQKAPQVST
PTLVEAARNLGRVGTKCCTLPEDQRLPCVEDYLSAILNRVCLLHEKTPVSEHVTKCCSGS
LVERRPCFSALTVDETYVPKEFKAETFTFHSDICTLPEKEKQIKKQTALAELVKHKPKAT
AEQLKTVMDDFAQFLDTCCKAADKDTCFSTEGPNLVTRSKDALA
SEQ ID NO: 154
Клон M13; петли BC, DE, FG подчеркнуты
IEVKDVTDTTALITWHDAFGYDFGCELTYGIKDVPGDRTTIDLPDHFHNYSIGNLKPDT
EYEVSLICANDHGFDSNPAKETFTT
SEQ ID NO: 155
Клон M13N49Q; мутация N49Q подчеркнута
IEVKDVTDTTALITWHDAFGYDFGCELTYGIKDVPGDRTTIDLPDHFHQYSIGNLKPDT
EYEVSLICANDHGFDSNPAKETFTT
SEQ ID NO: 156
Бивалентный конструкт M13N49Q-1GS-M13N49Q; мутация N49Q подчеркнута
SQIEVKDVTDTTALITWHDAFGYDFGCELTYGIKDVPGDRTTIDLPDHFHQYSIGNLKPD
TEYEVSLICANDHGFDSNPAKETFTTTGGGGSRLDAPSQIEVKDVTDTTALITWHDAFGY
DFGCELTYGIKDVPGDRTTIDLPDHFHQYSIGNLKPDTEYEVSLICANDHGFDSNPAKET
FTT
SEQ ID NO: 157
Моновалентный конструкт M13N49Q-3GS-MSA-C34S/C579S; мутации подчеркнуты
SQIEVKDVTDTTALITWHDAFGYDFGCELTYGIKDVPGDRTTIDLPDHFHQYSIGNLKPD
TEYEVSLICANDHGFDSNPAKETFTTGGGGSGGGGSGGGGSEAHKSEIAHRYNDLGEQHF
KGLVLIAFSQYLQKSSYDEHAKLVQEVTDFAKTCVADESAANCDKSLHTLFGDKLCAIPN
LRENYGELADCCTKQEPERNECFLQHKDDNPSLPPFERPEAEAMCTSFKENPTTFMGHYL
HEVARRHPYFYAPELLYYAEQYNEILTQCCAEADKESCLTPKLDGVKEKALVSSVRQRMK
CSSMQKFGERAFKAWAVARLSQTFPNADFAEITKLATDLTKVNKECCHGDLLECADDRAE
LAKYMCENQATISSKLQTCCDKPLLKKAHCLSEVEHDTMPADLPAIAADFVEDQEVCKNY
AEAKDVFLGTFLYEYSRRHPDYSVSLLLRLAKKYEATLEKCCAEANPPACYGTVLAEFQP
LVEEPKNLVKTNCDLYEKLGEYGFQNAILVRYTQKAPQVSTPTLVEAARNLGRVGTKCCT
LPEDQRLPCVEDYLSAILNRVCLLHEKTPVSEHVTKCCSGSLVERRPCFSALTVDETYVP
KEFKAETFTFHSDICTLPEKEKQIKKQTALAELVKHKPKATAEQLKTVMDDFAQFLDTCC
KAADKDTCFSTEGPNLVTRSKDALA
SEQ ID NO: 158
Бивалентный конструкт M13N49Q-1GS-M13N49Q-3GS-MSA-C34S/C579S; мутации подчеркнуты
SQIEVKDVTDTTALITWHDAFGYDFGCELTYGIKDVPGDRTTIDLPDHFHQYSIGNLKPD
TEYEVSLICANDHGFDSNPAKETFTTTGGGGSRLDAPSQIEVKDVTDTTALITWHDAFGY
DFGCELTYGIKDVPGDRTTIDLPDHFHQYSIGNLKPDTEYEVSLICANDHGFDSNPAKET
FTTGGGGSGGGGSGGGGSEAHKSEIAHRYNDLGEQHFKGLVLIAFSQYLQKSSYDEHAKL
VQEVTDFAKTCVADESAANCDKSLHTLFGDKLCAIPNLRENYGELADCCTKQEPERNECF
LQHKDDNPSLPPFERPEAEAMCTSFKENPTTFMGHYLHEVARRHPYFYAPELLYYAEQYN
EILTQCCAEADKESCLTPKLDGVKEKALVSSVRQRMKCSSMQKFGERAFKAWAVARLSQT
FPNADFAEITKLATDLTKVNKECCHGDLLECADDRAELAKYMCENQATISSKLQTCCDKP
LLKKAHCLSEVEHDTMPADLPAIAADFVEDQEVCKNYAEAKDVFLGTFLYEYSRRHPDYS
VSLLLRLAKKYEATLEKCCAEANPPACYGTVLAEFQPLVEEPKNLVKTNCDLYEKLGEYG
FQNAILVRYTQKAPQVSTPTLVEAARNLGRVGTKCCTLPEDQRLPCVEDYLSAILNRVCL
LHEKTPVSEHVTKCCSGSLVERRPCFSALTVDETYVPKEFKAETFTFHSDICTLPEKEKQ
IKKQTALAELVKHKPKATAEQLKTVMDDFAQFLDTCCKAADKDTCFSTEGPNLVTRSKDA
LA
SEQ ID NO: 159
Клон M31; петли BC, DE, FG подчеркнуты
IEVKDVTDTTALITWHDPSGYDFWCELTYGIKDVPGDRTTIDLPDHFHNYSIGNLKPDTE
YEVSLICANDHGFDSYPAKETFTT
SEQ ID NO: 160
Клон M31N49Q; мутация N49Q подчеркнута
IEVKDVTDTTALITWHDPSGYDFWCELTYGIKDVPGDRTTIDLPDHFHQYSIGNLKPDTE
YEVSLICANDHGFDSYPAKETFTT
SEQ ID NO: 161
Бивалентный конструкт M31N49Q-1GS-M31N49Q; мутация N49Q подчеркнута
SQIEVKDVTDTTALITWHDPSGYDFWCELTYGIKDVPGDRTTIDLPDHFHQYSIGNLKPD
TEYEVSLICANDHGFDSYPAKETFTTTGGGGSRLDAPSQIEVKDVTDTTALITWHDPSGY
DFWCELTYGIKDVPGDRTTIDLPDHFHQYSIGNLKPDTEYEVSLICANDHGFDSYPAKET
FTT
SEQ ID NO: 162
Моновалентный конструкт M31N49Q-3GS-MSA-C34S/C579S; мутации подчеркнуты
SQIEVKDVTDTTALITWHDPSGYDFWCELTYGIKDVPGDRTTIDLPDHFHQYSIGNLKPD
TEYEVSLICANDHGFDSYPAKETFTTGGGGSGGGGSGGGGSEAHKSEIAHRYNDLGEQHF
KGLVLIAFSQYLQKSSYDEHAKLVQEVTDFAKTCVADESAANCDKSLHTLFGDKLCAIPN
LRENYGELADCCTKQEPERNECFLQHKDDNPSLPPFERPEAEAMCTSFKENPTTFMGHYL
HEVARRHPYFYAPELLYYAEQYNEILTQCCAEADKESCLTPKLDGVKEKALVSSVRQRMK
CSSMQKFGERAFKAWAVARLSQTFPNADFAEITKLATDLTKVNKECCHGDLLECADDRAE
LAKYMCENQATISSKLQTCCDKPLLKKAHCLSEVEHDTMPADLPAIAADFVEDQEVCKNY
AEAKDVFLGTFLYEYSRRHPDYSVSLLLRLAKKYEATLEKCCAEANPPACYGTVLAEFQP
LVEEPKNLVKTNCDLYEKLGEYGFQNAILVRYTQKAPQVSTPTLVEAARNLGRVGTKCCT
LPEDQRLPCVEDYLSAILNRVCLLHEKTPVSEHVTKCCSGSLVERRPCFSALTVDETYVP
KEFKAETFTFHSDICTLPEKEKQIKKQTALAELVKHKPKATAEQLKTVMDDFAQFLDTCC
KAADKDTCFSTEGPNLVTRSKDALA
SEQ ID NO: 163
Бивалентный конструкт M31N49Q-1GS-M31N49Q-3GS-MSA-C34S/C579S; мутации подчеркнуты
SQIEVKDVTDTTALITWHDPSGYDFWCELTYGIKDVPGDRTTIDLPDHFHQYSIGNLKPD
TEYEVSLICANDHGFDSYPAKETFTTTGGGGSRLDAPSQIEVKDVTDTTALITWHDPSGY
DFWCELTYGIKDVPGDRTTIDLPDHFHQYSIGNLKPDTEYEVSLICANDHGFDSYPAKET
FTTGGGGSGGGGSGGGGSEAHKSEIAHRYNDLGEQHFKGLVLIAFSQYLQKSSYDEHAKL
VQEVTDFAKTCVADESAANCDKSLHTLFGDKLCAIPNLRENYGELADCCTKQEPERNECF
LQHKDDNPSLPPFERPEAEAMCTSFKENPTTFMGHYLHEVARRHPYFYAPELLYYAEQYN
EILTQCCAEADKESCLTPKLDGVKEKALVSSVRQRMKCSSMQKFGERAFKAWAVARLSQT
FPNADFAEITKLATDLTKVNKECCHGDLLECADDRAELAKYMCENQATISSKLQTCCDKP
LLKKAHCLSEVEHDTMPADLPAIAADFVEDQEVCKNYAEAKDVFLGTFLYEYSRRHPDYS
VSLLLRLAKKYEATLEKCCAEANPPACYGTVLAEFQPLVEEPKNLVKTNCDLYEKLGEYG
FQNAILVRYTQKAPQVSTPTLVEAARNLGRVGTKCCTLPEDQRLPCVEDYLSAILNRVCL
LHEKTPVSEHVTKCCSGSLVERRPCFSALTVDETYVPKEFKAETFTFHSDICTLPEKEKQ
IKKQTALAELVKHKPKATAEQLKTVMDDFAQFLDTCCKAADKDTCFSTEGPNLVTRSKDA
LA
SEQ ID NO: 164
Клон D1 - отрицательный контроль Tn3
IEVKDVTDTTALITWSPGERIWMFTGCELTYGIKDVPGDRTTIDLTEDENQYSIGNLKPD
TEYEVSLICPNYERISNPAKETFTTT
SEQ ID NO: 165
Бивалентный конструкт D1-1GS-D1-3G-MSA-C34S/C579S; мутации подчеркнуты
SQIEVKDVTDTTALITWSPGERIWMFTGCELTYGIKDVPGDRTTIDLTEDENQYSIGNLK
PDTEYEVSLICPNYERISNPAKETFTTTGGGGSRLDAPSQIEVKDVTDTTALITWSPGER
IWMFTGCELTYGIKDVPGDRTTIDLTEDENQYSIGNLKPDTEYEVSLICPNYERISNPAK
ETFTTGGGGSGGGGSGGGGSEAHKSEIAHRYNDLGEQHFKGLVLIAFSQYLQKSSYDEHA
KLVQEVTDFAKTCVADESAANCDKSLHTLFGDKLCAIPNLRENYGELADCCTKQEPERNE
CFLQHKDDNPSLPPFERPEAEAMCTSFKENPTTFMGHYLHEVARRHPYFYAPELLYYAEQ
YNEILTQCCAEADKESCLTPKLDGVKEKALVSSVRQRMKCSSMQKFGERAFKAWAVARLS
QTFPNADFAEITKLATDLTKVNKECCHGDLLECADDRAELAKYMCENQATISSKLQTCCD
KPLLKKAHCLSEVEHDTMPADLPAIAADFVEDQEVCKNYAEAKDVFLGTFLYEYSRRHPD
YSVSLLLRLAKKYEATLEKCCAEANPPACYGTVLAEFQPLVEEPKNLVKTNCDLYEKLGE
YGFQNAILVRYTQKAPQVSTPTLVEAARNLGRVGTKCCTLPEDQRLPCVEDYLSAILNRV
CLLHEKTPVSEHVTKCCSGSLVERRPCFSALTVDETYVPKEFKAETFTFHSDICTLPEKE
KQIKKQTALAELVKHKPKATAEQLKTVMDDFAQFLDTCCKAADKDTCFSTEGPNLVTRSK
DALA
SEQ ID NO: 166
Клон 342 с мутацией замены RDG на SDG; мутация подчеркнута
IEVKDVTDTTALITWSDDFGEYVWCELTYGIKDVPGDRTTIDLWYHHAHYSIGNLKPDTE
YEVSLICRSGDMSSNPAKETFTT
SEQ ID NO: 167
Консенсусная последовательность 309FGwt
Все цепи представляют собой цепи исходного Tn3; бета-цепь C представляет собой вариант CELTYGI (SEQ ID NO: 14); петли AB, CD, EF представляют собой петли исходного Tn3
X1= Ser или Leu
X2= Asp или Glu
X3= His, Ile, Val, Phe или Trp
X4= Ala, Gly, Glu или Asp
X5= Glu, Leu, Gln, Ser, Asp или Asn
X6= Phe или Tyr
X7= Ile, Val, His, Glu или Asp
X8= Gly, Trp или Val
X9= Trp, Phe или Tyr
X10= Ser, Gln, Met или His
X11= Trp или His
X12= Arg или Ser
IEVKDVTDTTALITWX1DX2X3X4X5X6X7X8CELTYGIKDVPGDRTTIDLWX9HX10AX11YSIGNLKPDTEYEVSLICRX12GDMSSNPAKETFTT
SEQ ID NO: 168
Консенсусная последовательность 309FGwt, петля BC
X1= Ser или Leu
X2= Asp или Glu
X3= His, Ile, Val, Phe или Trp
X4= Ala, Gly, Glu или Asp
X5= Glu, Leu, Gln, Ser, Asp или Asn
X6= Phe или Tyr
X7= Ile, Val, His, Glu или Asp
X8= Gly, Trp или Val
X1DX2X3X4X5X6X7X8
SEQ ID NO: 169
Консенсусная последовательность 309FGwt, петля DE
X9= Trp, Phe или Tyr
X10= Ser, Gln, Met или His
X11= Trp или His
WX9HX10AX11
SEQ ID NO: 170
Консенсусная последовательность 309FGwt, петля FG
X12= Arg или Ser
RX12GDMSSNPA
SEQ ID NO: 171
Консенсусная последовательность 311; все цепи представляют собой цепи исходного Tn3; два варианта бета-цепи C (SEQ ID NO: 13 и 14); петля CD представляет собой петлю исходного Tn3
X1= Lys или Glu
X2= Thr или Ile
X3= Asn или Ala
X4= Ser, Leu, Ala, Phe или Tyr
X5= Tyr, Ala, Gly, Val, Ile или Ser (контакт петли BC и N-конца)
X6= Tyr, Ser, Ala или His
X7= Asn, Asp, His или Tyr
X8= Leu, Phe, His или Tyr
X9= His, Pro, Ser, Leu или Asp
X10= Gly, Phe, His или Tyr
X11= Ala или Thr
X12= Ser, Asn, Glu, Arg или Asp
X13= Ser, Gln, Thr, Asn или Ala
X14= Pro, Val, -, Ile или Ala (- отсутствие аминокислоты)
X15= - или Ile (- отсутствие аминокислоты)
X16= Glu или Lys
X17= Ser или Asn
IEVX1DVTDTTALITWX2X3RSX4X5X6X7X8X9X10CELX11YGIKDVPGDRTTIDLX12X13X14X15YVHYSIGNLKPDTX16YEVSLICLTTDGTYX17NPAKETFTT
SEQ ID NO: 172
Консенсусная последовательность 311; бета-цепь C клонов из семейства 311
X11= Ala или Thr
CELX11YGI
SEQ ID NO: 173
Консенсусная последовательность 311; петля AB
X1= Lys или Glu
X1DVTDTT
SEQ ID NO: 174
Консенсусная последовательность 311; петля BC
X2= Thr или Ile
X3= Asn или Ala
X4= Ser, Leu, Ala, Phe или Tyr, X5= Tyr, Ala, Gly, Val, Ile или Ser (контакт петли BC и N-конца)
X6= Tyr, Ser, Ala или His
X7= Asn, Asp, His или Tyr
X8= Leu, Phe, His или Tyr
X9= His, Pro, Ser, Leu или Asp
X10= Gly, Phe, His или Tyr
X2X3RSX4X5X6X7X8X9X10
SEQ ID NO: 175
Консенсусная последовательность 311; петля DE
X12= Ser, Asn, Glu, Arg или Asp, X13= Ser, Gln, Thr, Asn или Ala
X14= Pro, Val, -, Ile или Ala (- отсутствие аминокислоты)
X15= - или Ile (- отсутствие аминокислоты)
X12X13X14X15YVH
SEQ ID NO: 176
Консенсусная последовательность 311; петля EF
X16= Glu или Lys
GNLKPDTX16
SEQ ID NO: 177
Консенсусная последовательность 311; петля FG
X17= Ser или Asn
LTTDGTYX17NPA
SEQ ID NO: 178
BC9 NHT oligo; петля BC
Коды нуклеотидов: N = G/A/T/C; H = A/T/C; R = A/G; S = G/C; B = T/C/G; V = A/C/G; M = A/C; K = G/T.
ACCGCGCTGATTACCTGGNHTNHTSCGNHTGSTNHTNHTNHTGGCTGTGAACTGACCTAT
GGCATTAAA
SEQ ID NO: 179
BC11 NHT oligo; петля BC
Коды нуклеотидов: N = G/A/T/C; H = A/T/C; R = A/G; S = G/C; B = T/C/G; V = A/C/G; M = A/C; K = G/T.
ACCGCGCTGATTACCTGGNHTNHTBSTNHTNHTNHTNHTNHTNHTNHTGGCTGTGAACTG
ACCTATGGCATTAAA
SEQ ID NO: 180
BC12 NHT oligo; петля BC
Коды нуклеотидов: N = G/A/T/C; H = A/T/C; R = A/G; S = G/C; B = T/C/G; V = A/C/G; M = A/C; K = G/T.
ACCGCGCTGATTACCTGGNHTVMACCGNHTNHTNHTRRCRGCNHTVTTNHTGGCTGTGAA
CTGACCTATGGCATTAAA
SEQ ID NO: 181
DE NHT oligo; петля DE
Коды нуклеотидов: N = G/A/T/C; H = A/T/C; R = A/G; S = G/C; B = T/C/G; V = A/C/G; M = A/C; K = G/T.
CGATCGCACCACCATAGATCTGNHTNHTNHTNHTNHTNHTTATAGCATTGGTAACCTGAA
ACCG
SEQ ID NO: 182
FG9 NHT oligo; петля FG
Коды нуклеотидов: N = G/A/T/C; H = A/T/C; R = A/G; S = G/C; B = T/C/G; V = A/C/G; M = A/C; K = G/T.
GAATATGAAGTGAGCCTGATTTGCNHTAMSNHTNHTGGTNHTNHTNHTKCGAAAGAAACC
TTTACCACCGGTG
SEQ ID NO: 183
FG10 NHT oligo; петля FG
Коды нуклеотидов: N = G/A/T/C; H = A/T/C; R = A/G; S = G/C; B = T/C/G; V = A/C/G; M = A/C; K = G/T.
GAATATGAAGTGAGCCTGATTTGCNHTAMSNHTNHTNHTNHTRGCNHTCCGGCGAAAGAA
ACCTTTACCACCGGTG
SEQ ID NO: 184
Олигонуклеотид FG11 NHT; петля FG
Коды нуклеотидов: N = G/A/T/C; H = A/T/C; R = A/G; S = G/C; B = T/C/G; V = A/C/G; M = A/C; K = G/T.
GAATATGAAGTGAGCCTGATTTGCNHTAMSNHTNHTGGTNHTNHTAGCAACCCGGCGAAA
GAAACCTTTACCACCGGTG
SEQ ID NO: 185
Олигонуклеотид BCX-DE bridge v2
CAGATCTATGGTGGTGCGATCGCCCGGCACATCTTTAATGCCATAGGTCAGTTCACA
SEQ ID NO: 186
Олигонуклеотид DE-FGX bridge v2
GCAAATCAGGCTCACTTCATATTCGGTATCCGGTTTCAGGTTACCAATGCTAT
SEQ ID NO: 187
Олигонуклеотид KpnI amp rev v2
CGGGTCGGTTGGGGTACCGCCACCGGTGGTAAAGGTTTCTTT
SEQ ID NO: 188
Олигонуклеотид KpnI reverse v2
CGGGTCGGTTGGGGTA
SEQ ID NO: 189
Олигонуклеотид BC library amp v2
GGCCCAGCCGGCCATGGCCGCCATTGAAGTGAAAGATGTGACCGATACCACCGCGCTGAT
TACCTGG
SEQ ID NO: 190
Олигонуклеотид BC9 PCR
Коды нуклеотидов: 1= кодоны для всех 19aa(-cys); 2=кодоны для Ala/Pro 50/50; 3= кодоны для Ala/Gly
ACCGCGCTGATTACCTGGTCT1213111GGCTGTGAACTGACCTATGGCATTAAAGATG
SEQ ID NO: 191
Олигонуклеотид BC 9-loop NNK
Коды нуклеотидов: K= 50%G/50%T
ACCGCGCTGATTACCTGGNNKNNKSMGNNKGSTNNKNNKNNKGGCTGTGAACTGACCTA
TGGCATTAAA
SEQ ID NO: 192
Олигонуклеотид 309 BC-loop NNKdope
Коды нуклеотидов: 4= 70%G10%A10%C10%T; 5= 10%G,70%A,10%C,10%T; 6=10%G,10%A,70%C, 10%T; 7= 10%G,10%A,10%C,70%T; 8= 70%A15%C15%T и K= 50%G/50%T
ACCGCGCTGATTACCTGG76K45K45K77K44K65K78T45K44KTGTGAACTGACCTA
TGGCATTAAA
SEQ ID NO: 193
Олигонуклеотид DE PCR
Коды нуклеотидов: 1= кодоны для всех 19aa(-cys)
GATGTGCCGGGCGATCGCACCACCATAGATCTG111111TATAGCATTGGTAACCTGAAA
CCGG
SEQ ID NO: 194
Олигонуклеотид Upstr BCloop Rev
CCAGGTAATCAGCGCGGTGGTAT
SEQ ID NO: 195
Олигонуклеотид BC shuffle rev
CAGATCTATGGTGGTGCGATCGC
SEQ ID NO: 196
Олигонуклеотид DE shuffle FWD
TGTGAACTGACCTATGGCATTAAAGATGT
SEQ ID NO: 197
Олигонуклеотид BC11-311Gly
Коды нуклеотидов: 1=70%G, 10%A, 10%C, 10%T; 2=10%G, 70%A, 10%C, 10%T; 3=10%G, 10%A, 70%C, 10%T; 4=10%G, 10%A, 10%C, 70%T; 5=70%A, 15%C, 15%T; 6=15%A, 70%C, 15%T; 7=15%A, 15%C, 70%T; V=33%A, 33%C, 33%G.
ACCGCGCTGATTACCTGG26T25TV1T46T46T45T45T25T37T35TGGCTGTGAACTG
ACCTATGGCATTAAA
SEQ ID NO: 198
Олигонуклеотид BC11-311NHT
Коды нуклеотидов: 1=70%G, 10%A, 10%C, 10%T; 2=10%G, 70%A, 10%C, 10%T; 3=10%G, 10%A, 70%C, 10%T; 4=10%G, 10%A, 10%C, 70%T; 5=70%A, 15%C, 15%T; 6=15%A, 70%C, 15%T; 7=15%A, 15%C, 70%T; V=33%A, 33%C, 33%G; и H=33%A, 33%C, 33%T
ACCGCGCTGATTACCTGG26T25TV1T46T46T45T45T25T37T35TNHTTGTGAACTG
ACCTATGGCATTAAA
SEQ ID NO: 199
Олигонуклеотид BC library amp K4E
GGCCCAGCCGGCCATGGCCGCCATTGAAGTGGAAGATGTGACCGATACCACCGCGCTGAT
TACCTGG
SEQ ID NO: 200
Вариант HSA с увеличенным временем полужизни (C34S, L463N, K524L); мутации подчеркнуты
DAHKSEVAHRFKDLGEENFKALVLIAFAQYLQQSPFEDHVKLVNEVTEFAKTCVADESAE
NCDKSLHTLFGDKLCTVATLRETYGEMADCCAKQEPERNECFLQHKDDNPNLPRLVRPEV
DVMCTAFHDNEETFLKKYLYEIARRHPYFYAPELLFFAKRYKAAFTECCQAADKAACLLP
KLDELRDEGKASSAKQRLKCASLQKFGERAFKAWAVARLSQRFPKAEFAEVSKLVTDLTK
VHTECCHGDLLECADDRADLAKYICENQDSISSKLKECCEKPLLEKSHCIAEVENDEMPA
DLPSLAADFVESKDVCKNYAEAKDVFLGMFLYEYARRHPDYSVVLLLRLAKTYETTLEKC
CAAADPHECYAKVFDEFKPLVEEPQNLIKQNCELFEQLGEYKFQNALLVRYTKKVPQVST
PTLVEVSRNLGKVGSKCCKHPEAKRMPCAEDYLSVVLNQLCVNHEKTPVSDRVTKCCTES
LVNRRPCFSALEVDETYVPKEFNAETFTFHADICTLSEKERQILKQTALVELVKHKPKAT
KEQLKAVMDDFAAFVEKCCKADDKETCFAEEGKKLVAASQAALGL
SEQ ID NO: 201
Моновалентный конструкт, представляющий собой вариант 311K4E_12-I (содержит линкер GS и HSA C34S); линкер и серин, появившийся в результате мутации, подчеркнуты
SQIEVEDVTDTTALITWTNRSSYSNLHGCELAYGIKDVPGDRTTIDLNQPYVHYSIGNLK
PDTEYEVSLICLTTDGTYNNPAKETFTTGGGGSGGGGSDAHKSEVAHRFKDLGEENFKAL
VLIAFAQYLQQSPFEDHVKLVNEVTEFAKTCVADESAENCDKSLHTLFGDKLCTVATLRE
TYGEMADCCAKQEPERNECFLQHKDDNPNLPRLVRPEVDVMCTAFHDNEETFLKKYLYEI
ARRHPYFYAPELLFFAKRYKAAFTECCQAADKAACLLPKLDELRDEGKASSAKQRLKCAS
LQKFGERAFKAWAVARLSQRFPKAEFAEVSKLVTDLTKVHTECCHGDLLECADDRADLAK
YICENQDSISSKLKECCEKPLLEKSHCIAEVENDEMPADLPSLAADFVESKDVCKNYAEA
KDVFLGMFLYEYARRHPDYSVVLLLRLAKTYETTLEKCCAAADPHECYAKVFDEFKPLVE
EPQNLIKQNCELFEQLGEYKFQNALLVRYTKKVPQVSTPTLVEVSRNLGKVGSKCCKHPE
AKRMPCAEDYLSVVLNQLCVLHEKTPVSDRVTKCCTESLVNRRPCFSALEVDETYVPKEF
NAETFTFHADICTLSEKERQIKKQTALVELVKHKPKATKEQLKAVMDDFAAFVEKCCKAD
DKETCFAEEGKKLVAASQAALGL
SEQ ID NO: 202
Моновалентный конструкт, представляющий собой вариант 311K4E_12-I (содержит вариант CELTYG бета-цепи C, линкер, полностью состоящий из G, и HSA C34S); линкер и серин, появившийся в результате мутации, подчеркнуты
SQIEVEDVTDTTALITWTNRSSYSNLHGCELTYGIKDVPGDRTTIDLNQPYVHYSIGNLK
PDTEYEVSLICLTTDGTYNNPAKETFTTGGGGGGGGGGDAHKSEVAHRFKDLGEENFKAL
VLIAFAQYLQQSPFEDHVKLVNEVTEFAKTCVADESAENCDKSLHTLFGDKLCTVATLRE
TYGEMADCCAKQEPERNECFLQHKDDNPNLPRLVRPEVDVMCTAFHDNEETFLKKYLYEI
ARRHPYFYAPELLFFAKRYKAAFTECCQAADKAACLLPKLDELRDEGKASSAKQRLKCAS
LQKFGERAFKAWAVARLSQRFPKAEFAEVSKLVTDLTKVHTECCHGDLLECADDRADLAK
YICENQDSISSKLKECCEKPLLEKSHCIAEVENDEMPADLPSLAADFVESKDVCKNYAEA
KDVFLGMFLYEYARRHPDYSVVLLLRLAKTYETTLEKCCAAADPHECYAKVFDEFKPLVE
EPQNLIKQNCELFEQLGEYKFQNALLVRYTKKVPQVSTPTLVEVSRNLGKVGSKCCKHPE
AKRMPCAEDYLSVVLNQLCVLHEKTPVSDRVTKCCTESLVNRRPCFSALEVDETYVPKEF
NAETFTFHADICTLSEKERQIKKQTALVELVKHKPKATKEQLKAVMDDFAAFVEKCCKAD
DKETCFAEEGKKLVAASQAALGL
SEQ ID NO: 203
Бивалентный конструкт, представляющий собой вариант 311K4E_12-I (содержит линкеры GS и HSA C34S); линкеры и серин, появившийся в результате мутации, подчеркнуты
SQIEVEDVTDTTALITWTNRSSYSNLHGCELAYGIKDVPGDRTTIDLNQPYVHYSIGNLK
PDTEYEVSLICLTTDGTYNNPAKETFTTGGGGSGGGGSGGGGSRLDAPSQIEVEDVTDTT
ALITWTNRSSYSNLHGCELAYGIKDVPGDRTTIDLNQPYVHYSIGNLKPDTEYEVSLICL
TTDGTYNNPAKETFTTGGGGSGGGGSDAHKSEVAHRFKDLGEENFKALVLIAFAQYLQQS
PFEDHVKLVNEVTEFAKTCVADESAENCDKSLHTLFGDKLCTVATLRETYGEMADCCAKQ
EPERNECFLQHKDDNPNLPRLVRPEVDVMCTAFHDNEETFLKKYLYEIARRHPYFYAPEL
LFFAKRYKAAFTECCQAADKAACLLPKLDELRDEGKASSAKQRLKCASLQKFGERAFKAW
AVARLSQRFPKAEFAEVSKLVTDLTKVHTECCHGDLLECADDRADLAKYICENQDSISSK
LKECCEKPLLEKSHCIAEVENDEMPADLPSLAADFVESKDVCKNYAEAKDVFLGMFLYEY
ARRHPDYSVVLLLRLAKTYETTLEKCCAAADPHECYAKVFDEFKPLVEEPQNLIKQNCEL
FEQLGEYKFQNALLVRYTKKVPQVSTPTLVEVSRNLGKVGSKCCKHPEAKRMPCAEDYLS
VVLNQLCVLHEKTPVSDRVTKCCTESLVNRRPCFSALEVDETYVPKEFNAETFTFHADIC
TLSEKERQIKKQTALVELVKHKPKATKEQLKAVMDDFAAFVEKCCKADDKETCFAEEGKK
LVAASQAALGL
SEQ ID NO: 204
Бивалентный конструкт, представляющий собой вариант 311K4E_12-I (содержит линкеры, полностью состоящие из G, и HSA S34); линкеры и серин, появившийся в результате мутации, подчеркнуты
SQIEVEDVTDTTALITWTNRSSYSNLHGCELTYGIKDVPGDRTTIDLNQPYVHYSIGNLK
PDTEYEVSLICLTTDGTYNNPAKETFTTGGGGGGGGGGGGGGGRLDAPSQIEVEDVTDTT
ALITWTNRSSYSNLHGCELAYGIKDVPGDRTTIDLNQPYVHYSIGNLKPDTEYEVSLICL
TTDGTYNNPAKETFTTGGGGGGGGGGDAHKSEVAHRFKDLGEENFKALVLIAFAQYLQQS
PFEDHVKLVNEVTEFAKTCVADESAENCDKSLHTLFGDKLCTVATLRETYGEMADCCAKQ
EPERNECFLQHKDDNPNLPRLVRPEVDVMCTAFHDNEETFLKKYLYEIARRHPYFYAPEL
LFFAKRYKAAFTECCQAADKAACLLPKLDELRDEGKASSAKQRLKCASLQKFGERAFKAW
AVARLSQRFPKAEFAEVSKLVTDLTKVHTECCHGDLLECADDRADLAKYICENQDSISSK
LKECCEKPLLEKSHCIAEVENDEMPADLPSLAADFVESKDVCKNYAEAKDVFLGMFLYEY
ARRHPDYSVVLLLRLAKTYETTLEKCCAAADPHECYAKVFDEFKPLVEEPQNLIKQNCEL
FEQLGEYKFQNALLVRYTKKVPQVSTPTLVEVSRNLGKVGSKCCKHPEAKRMPCAEDYLS
VVLNQLCVLHEKTPVSDRVTKCCTESLVNRRPCFSALEVDETYVPKEFNAETFTFHADIC
TLSEKERQIKKQTALVELVKHKPKATKEQLKAVMDDFAAFVEKCCKADDKETCFAEEGKK
LVAASQAALGL
SEQ ID NO: 205
Бивалентный конструкт 309-3GS-309
SQIEVKDVTDTTALITWSDEFGHYDGCELTYGIKDVPGDRTTIDLWWHSAWYSIGNLKPD
TEYEVSLICYTDQEAGNPAKETFTTGGGGSGGGGSGGGGSRLDAPSQIEVKDVTDTTALI
TWSDEFGHYDGCELTYGIKDVPGDRTTIDLWWHSAWYSIGNLKPDTEYEVSLICYTDQEA
GNPAKETFTT
SEQ ID NO: 206
Моновалентный конструкт 309-2GS-HSAC34S
SQIEVKDVTDTTALITWSDEFGHYDGCELTYGIKDVPGDRTTIDLWWHSAWYSIGNLKPD
TEYEVSLICYTDQEAGNPAKETFTTGGGGSGGGGSDAHKSEVAHRFKDLGEENFKALVLI
AFAQYLQQSPFEDHVKLVNEVTEFAKTCVADESAENCDKSLHTLFGDKLCTVATLRETYG
EMADCCAKQEPERNECFLQHKDDNPNLPRLVRPEVDVMCTAFHDNEETFLKKYLYEIARR
HPYFYAPELLFFAKRYKAAFTECCQAADKAACLLPKLDELRDEGKASSAKQRLKCASLQK
FGERAFKAWAVARLSQRFPKAEFAEVSKLVTDLTKVHTECCHGDLLECADDRADLAKYIC
ENQDSISSKLKECCEKPLLEKSHCIAEVENDEMPADLPSLAADFVESKDVCKNYAEAKDV
FLGMFLYEYARRHPDYSVVLLLRLAKTYETTLEKCCAAADPHECYAKVFDEFKPLVEEPQ
NLIKQNCELFEQLGEYKFQNALLVRYTKKVPQVSTPTLVEVSRNLGKVGSKCCKHPEAKR
MPCAEDYLSVVLNQLCVLHEKTPVSDRVTKCCTESLVNRRPCFSALEVDETYVPKEFNAE
TFTFHADICTLSEKERQIKKQTALVELVKHKPKATKEQLKAVMDDFAAFVEKCCKADDKE
TCFAEEGKKLVAASQAALGL
SEQ ID NO: 207
Бивалентный конструкт 309-3GS-309-2GS-HSAC34S
SQIEVKDVTDTTALITWSDEFGHYDGCELTYGIKDVPGDRTTIDLWWHSAWYSIGNLKPD
TEYEVSLICYTDQEAGNPAKETFTTGGGGSGGGGSGGGGSRLDAPSQIEVKDVTDTTALI
TWSDEFGHYDGCELTYGIKDVPGDRTTIDLWWHSAWYSIGNLKPDTEYEVSLICYTDQEA
GNPAKETFTTGGGGSGGGGSDAHKSEVAHRFKDLGEENFKALVLIAFAQYLQQSPFEDHV
KLVNEVTEFAKTCVADESAENCDKSLHTLFGDKLCTVATLRETYGEMADCCAKQEPERNE
CFLQHKDDNPNLPRLVRPEVDVMCTAFHDNEETFLKKYLYEIARRHPYFYAPELLFFAKR
YKAAFTECCQAADKAACLLPKLDELRDEGKASSAKQRLKCASLQKFGERAFKAWAVARLS
QRFPKAEFAEVSKLVTDLTKVHTECCHGDLLECADDRADLAKYICENQDSISSKLKECCE
KPLLEKSHCIAEVENDEMPADLPSLAADFVESKDVCKNYAEAKDVFLGMFLYEYARRHPD
YSVVLLLRLAKTYETTLEKCCAAADPHECYAKVFDEFKPLVEEPQNLIKQNCELFEQLGE
YKFQNALLVRYTKKVPQVSTPTLVEVSRNLGKVGSKCCKHPEAKRMPCAEDYLSVVLNQL
CVLHEKTPVSDRVTKCCTESLVNRRPCFSALEVDETYVPKEFNAETFTFHADICTLSEKE
RQIKKQTALVELVKHKPKATKEQLKAVMDDFAAFVEKCCKADDKETCFAEEGKKLVAASQ
AALGL
SEQ ID NO: 208
Бивалентный конструкт 342-3GS-342
SQIEVKDVTDTTALITWSDDFGEYVWCELTYGIKDVPGDRTTIDLWYHHAHYSIGNLKPD
TEYEVSLICRSGDMSSNPAKETFTTGGGGSGGGGSGGGGSRLDAPSQIEVKDVTDTTALI
TWSDDFGEYVWCELTYGIKDVPGDRTTIDLWYHHAHYSIGNLKPDTEYEVSLICRSGDMS
SNPAKETFTT
SEQ ID NO: 209
Линкерный модуль Gly-Ser, (G4X)n, где X=G, S, A, L, I или V и n = 1-7; показан модуль (G4X)n, где n=1
GGGGX
SEQ ID NO: 210
Мутантная форма L25/M сигнального пептида OppA
MTNITKRSLVAAGVLAALMAGNVAMA
| название | год | авторы | номер документа |
|---|---|---|---|
| CD40L-СПЕЦИФИЧНЫЕ КАРКАСНЫЕ СТРУКТУРЫ, ПРОИСХОДЯЩИЕ ИЗ TN3, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2826189C2 |
| TRAIL R2-СПЕЦИФИЧЕСКИЕ МУЛЬТИМЕРНЫЕ СКАФФОЛДЫ | 2011 |
|
RU2628699C2 |
| КОМПОЗИЦИИ НА ОСНОВЕ СТАБИЛИЗИРОВАННЫХ ФИБРОНЕКТИНОВЫХ ДОМЕНОВ, СПОСОБЫ И ОБЛАСТИ ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2603272C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИЕ К PD1 И TIM3 | 2016 |
|
RU2729371C1 |
| КОМПОЗИЦИИ IL-12, НАЦЕЛЕННЫЕ НА EDB | 2019 |
|
RU2758143C1 |
| НОВЫЕ БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, ОБЛАДАЮЩИЕ СПОСОБНОСТЬЮ СПЕЦИФИЧЕСКИ СВЯЗЫВАТЬСЯ С CD40 И FAP | 2018 |
|
RU2766234C2 |
| АНТИТЕЛА ПРОТИВ CD19 ЧЕЛОВЕКА, ОБЛАДАЮЩИЕ ВЫСОКОЙ АФФИННОСТЬЮ | 2016 |
|
RU2761077C1 |
| НОВЫЕ ПОЛИПЕПТИДЫ СО СПЕЦИФИЧЕСКИМ СВЯЗЫВАНИЕМ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2723034C2 |
| ПОЛИПЕПТИДЫ, СОДЕРЖАЩИЕ ОДИНОЧНЫЕ ВАРИАБЕЛЬНЫЕ ДОМЕНЫ ИММУНОГЛОБУЛИНОВ, НАЦЕЛИВАЮЩИЕСЯ НА IL-13 И TSLP | 2020 |
|
RU2836310C1 |
| АНТАГОНИСТЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2005 |
|
RU2401842C2 |























































































































































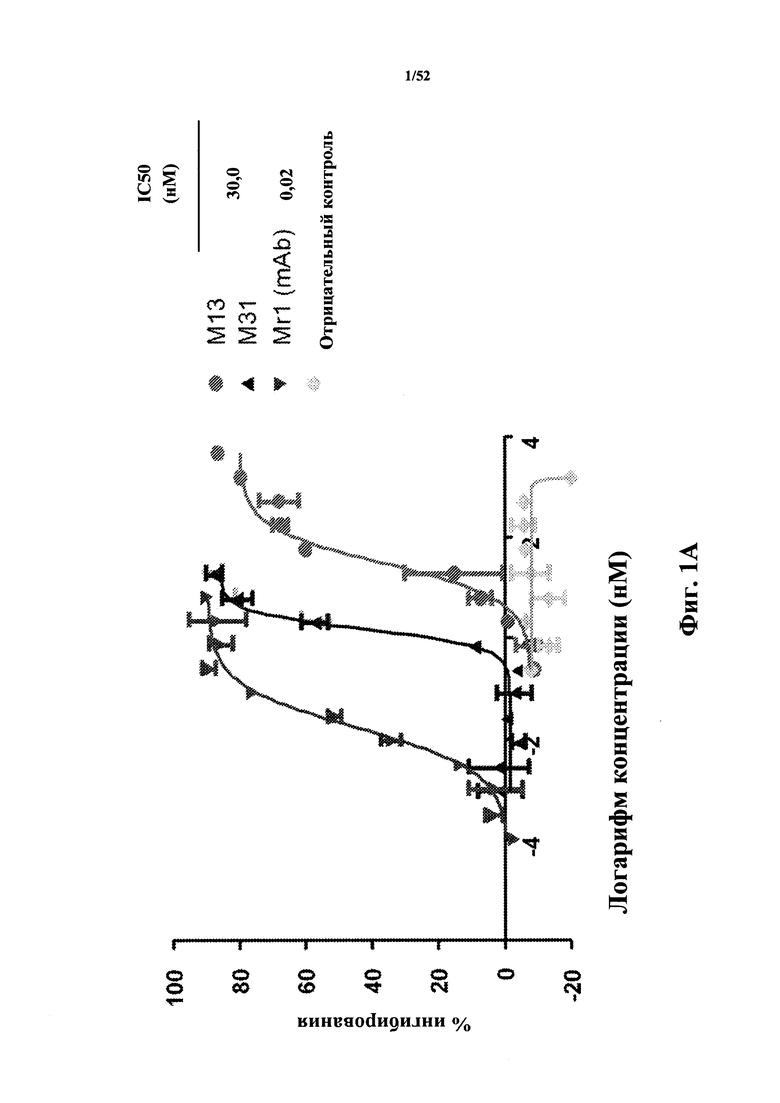
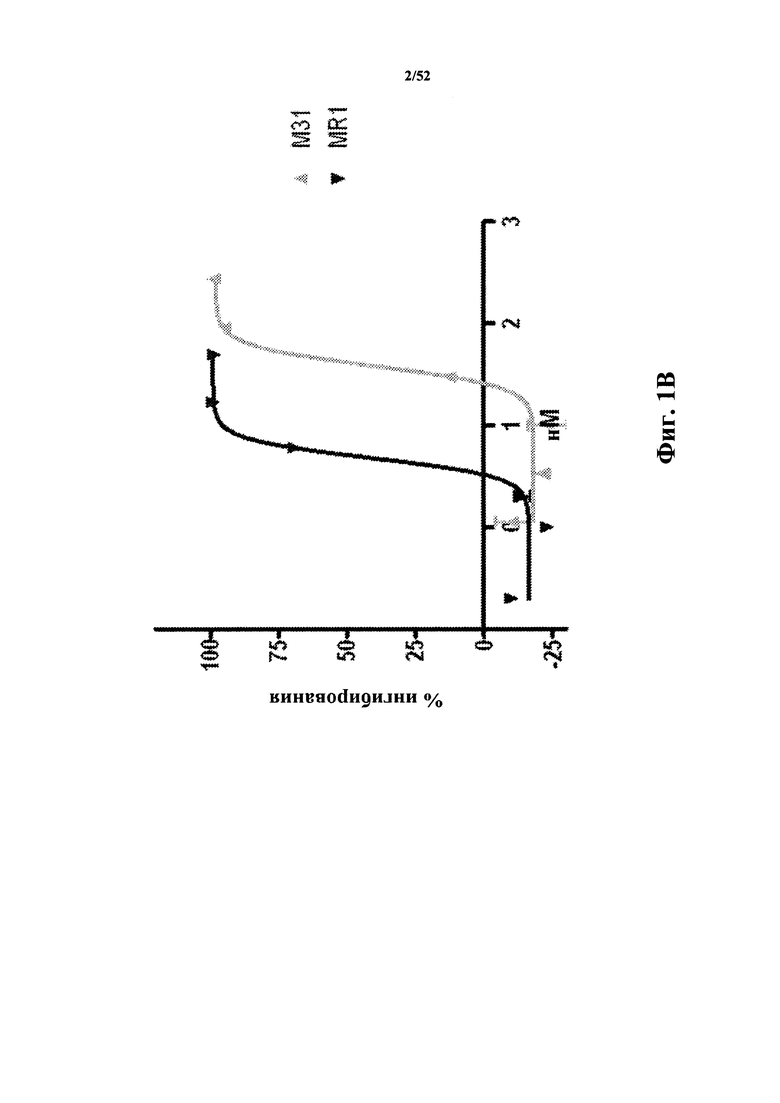
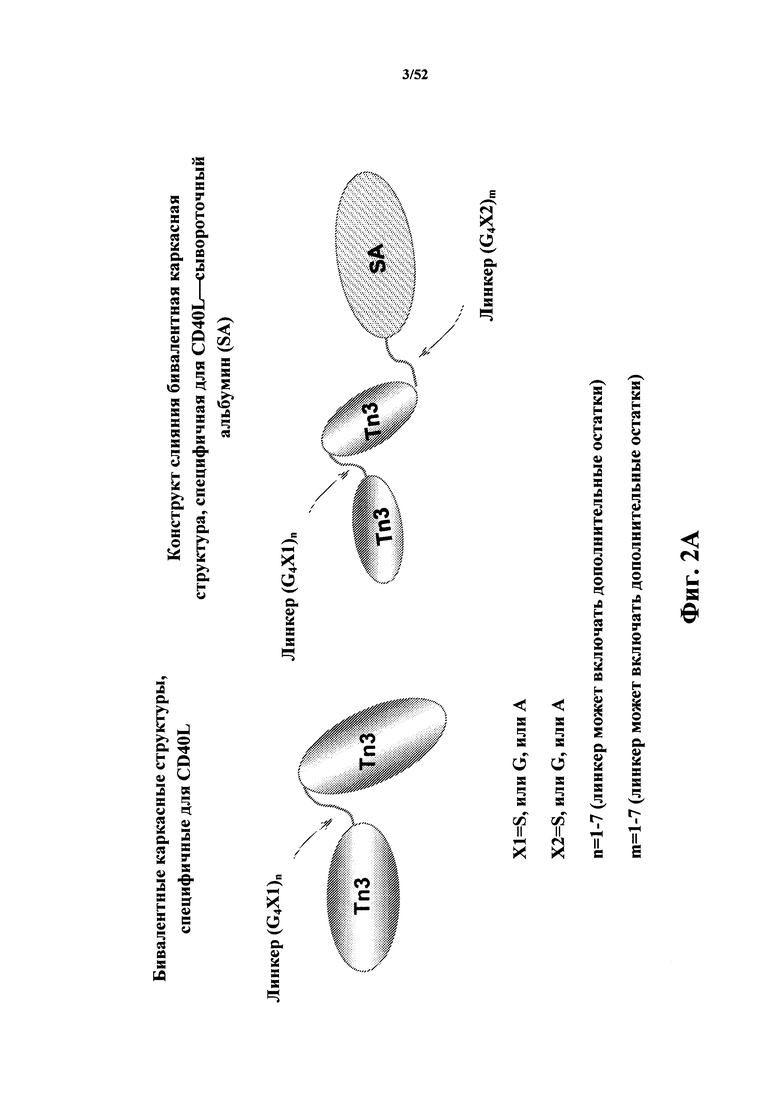

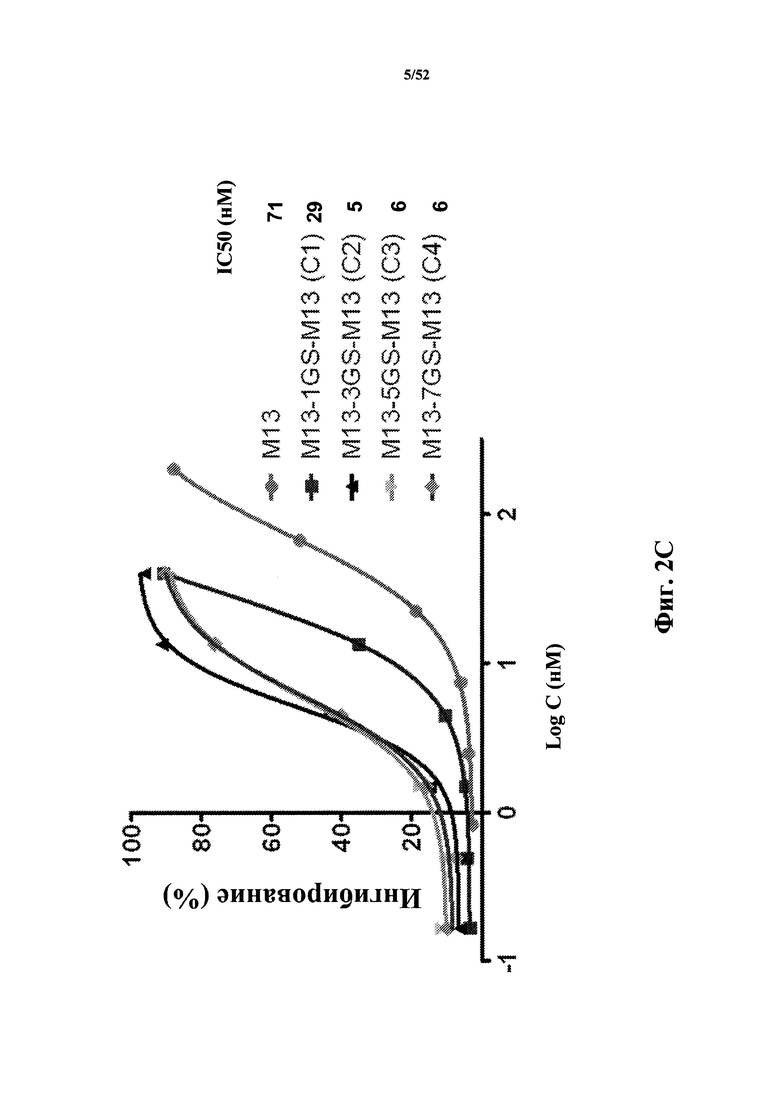
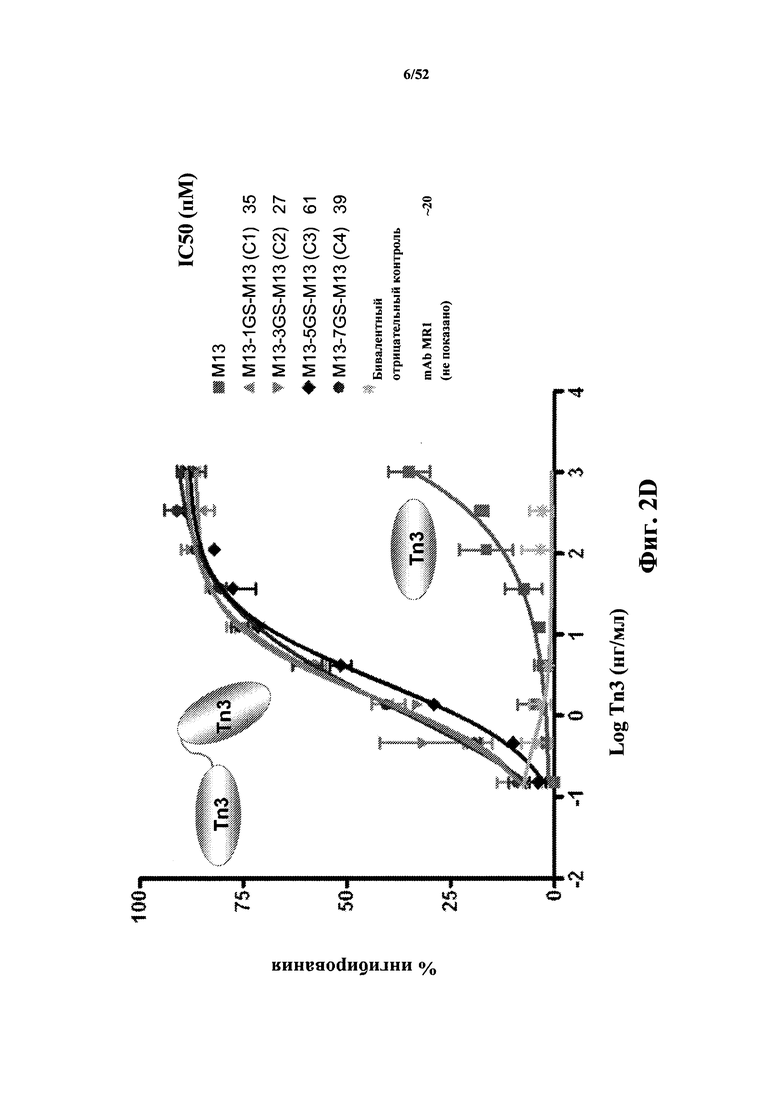
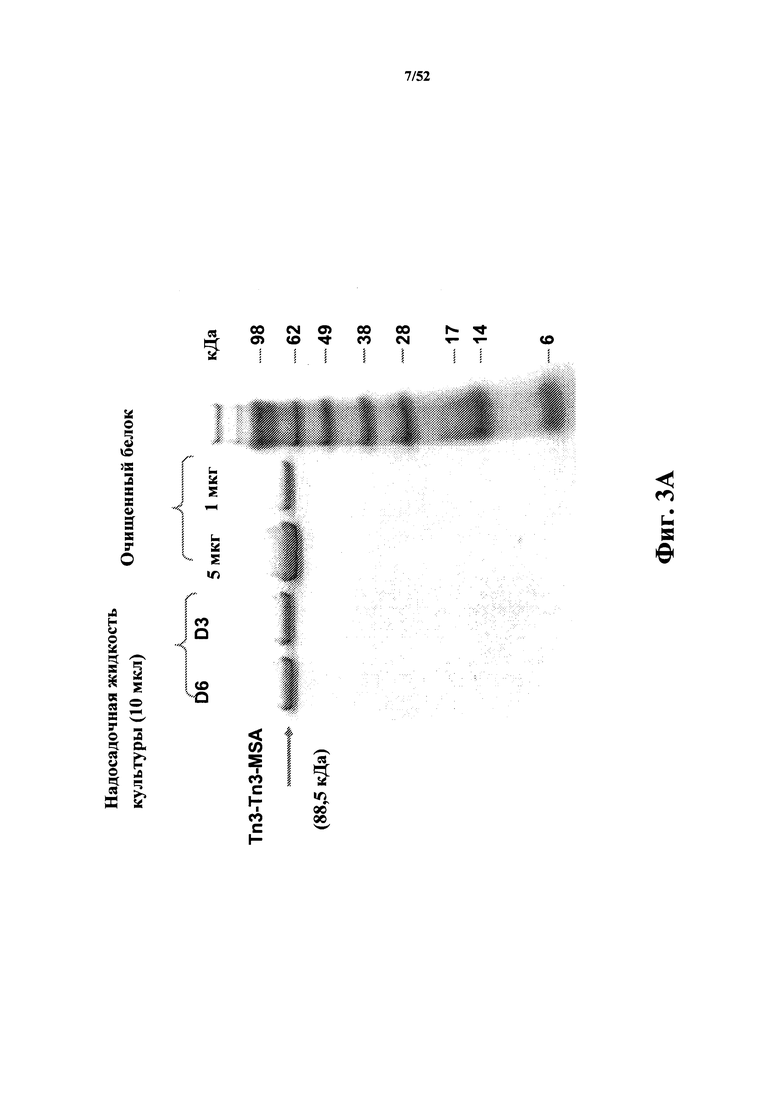
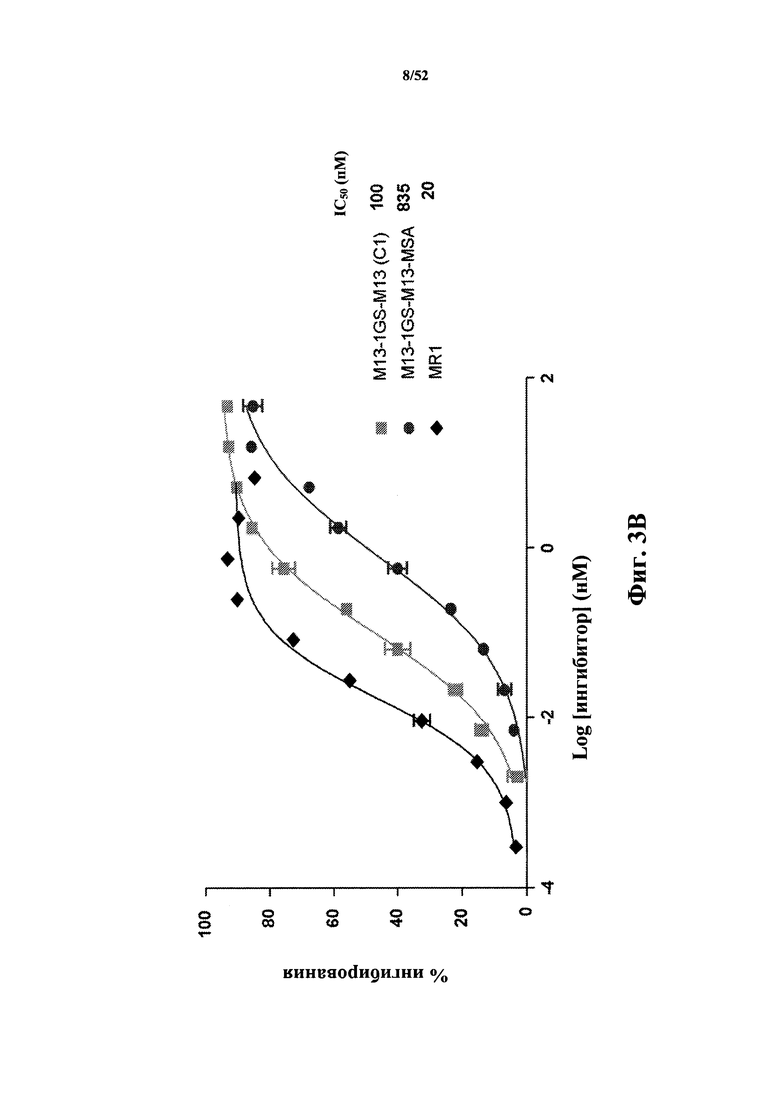
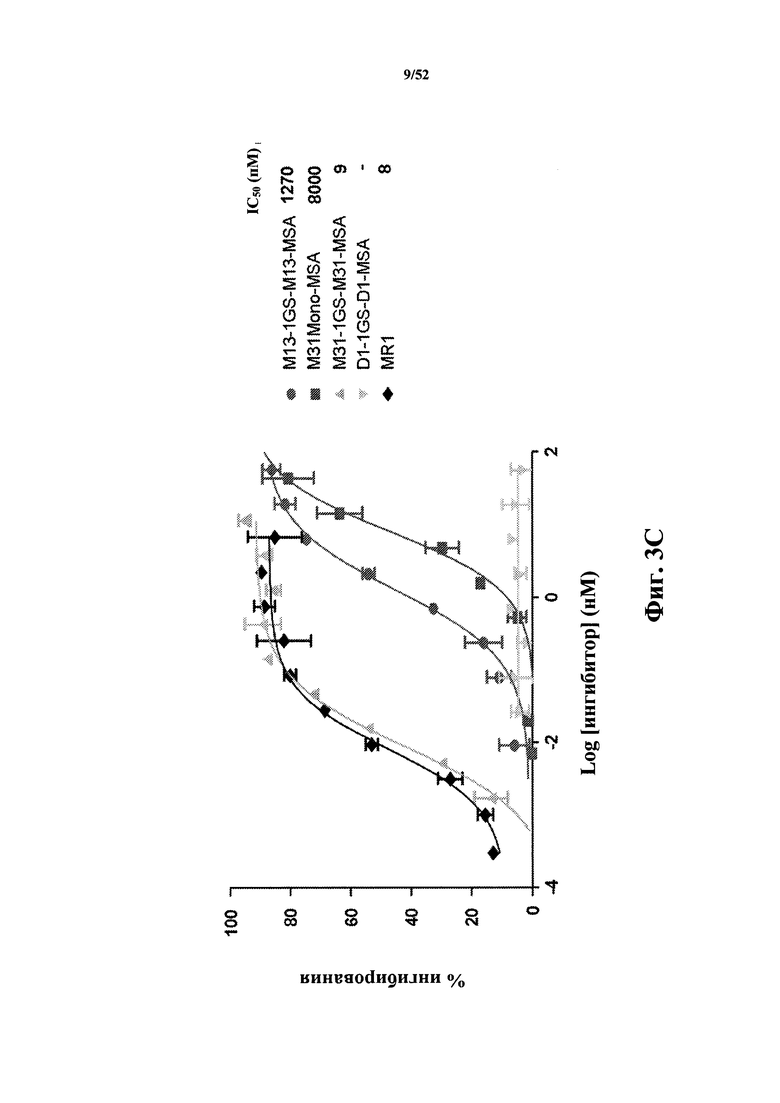

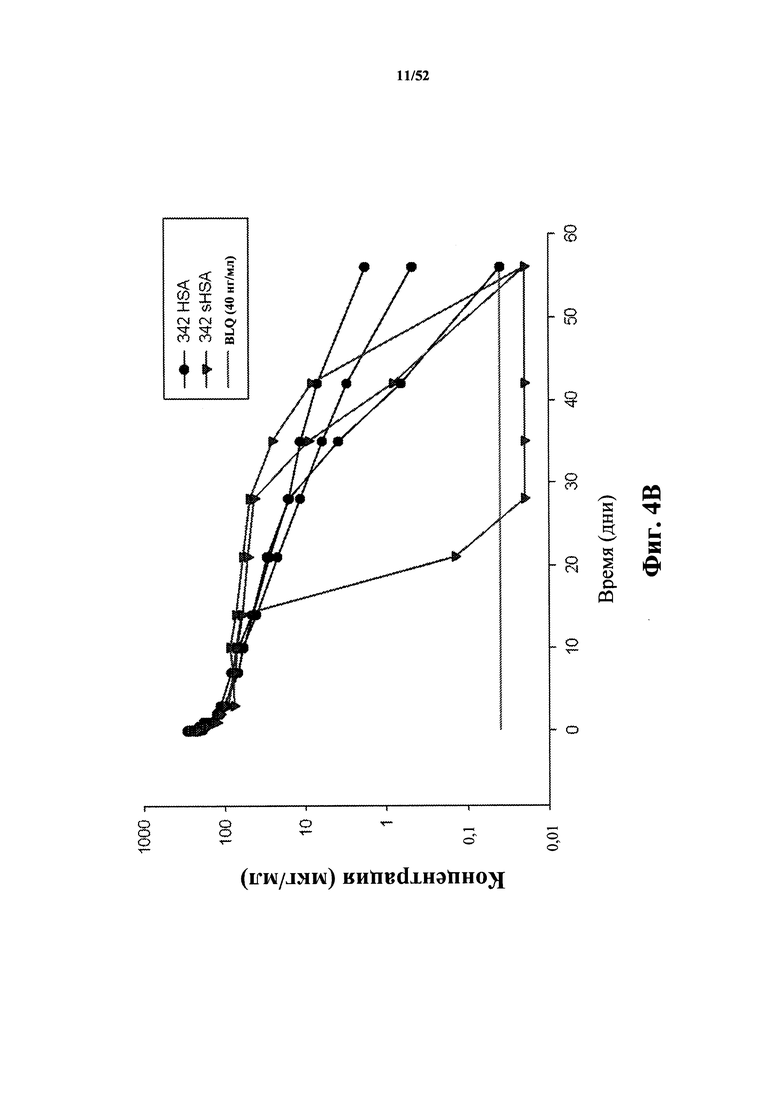
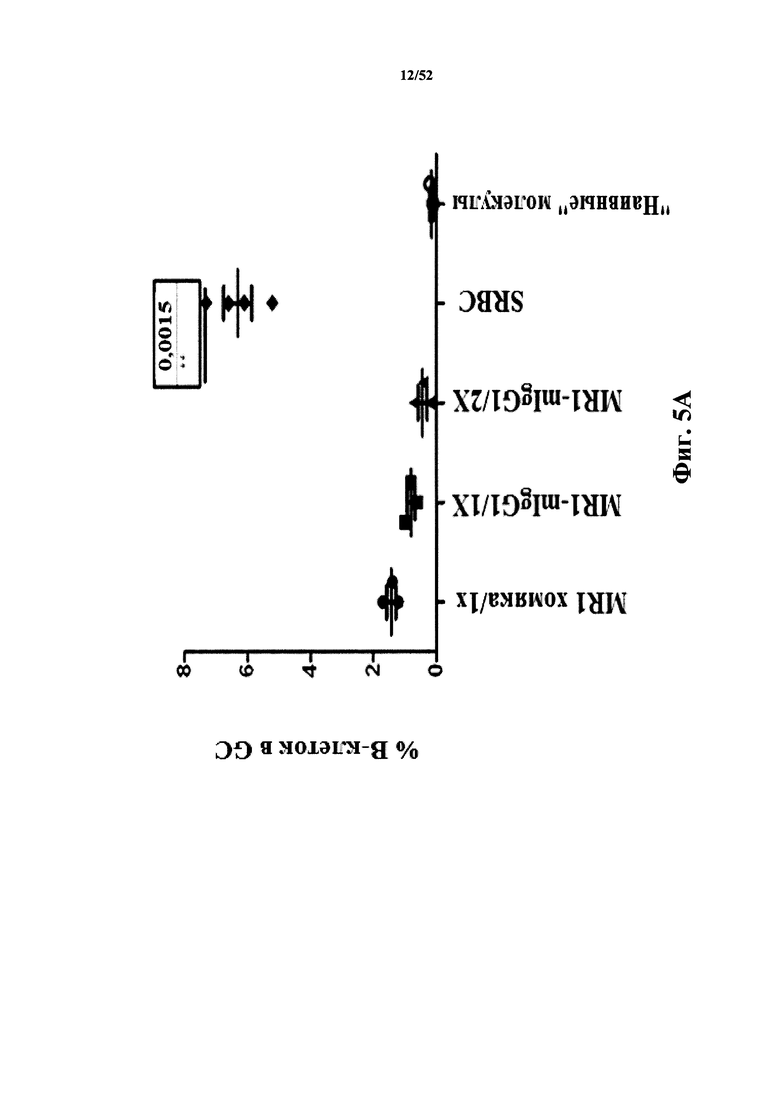
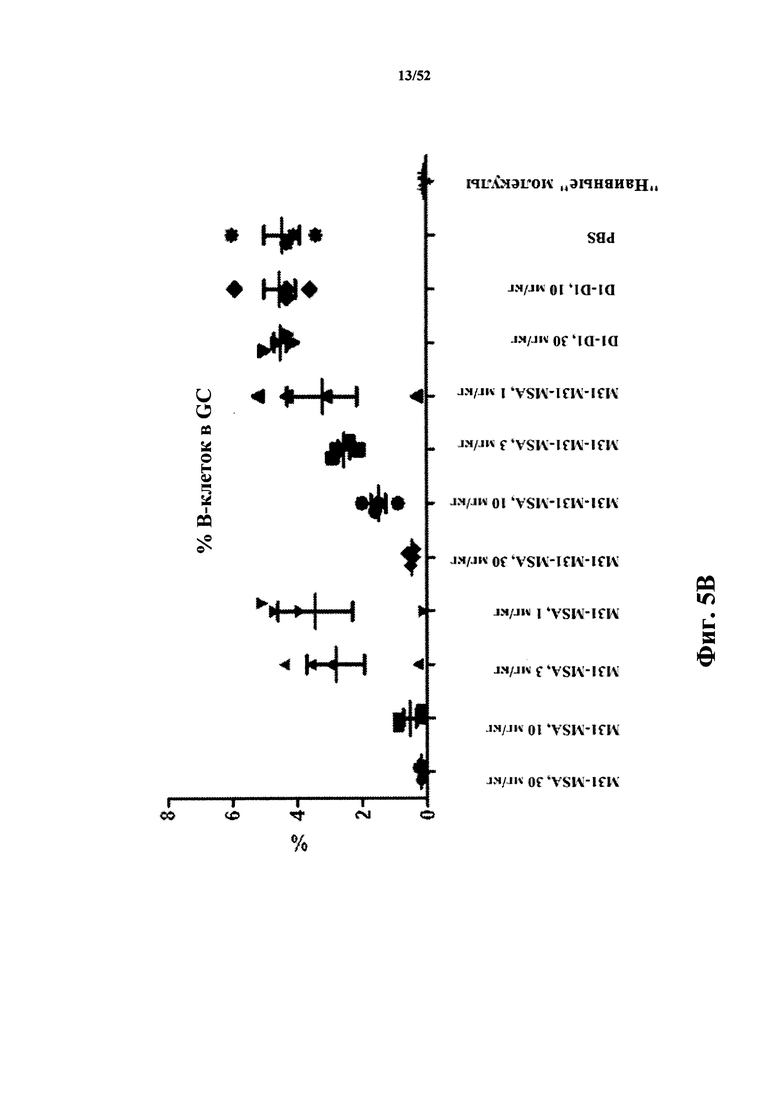
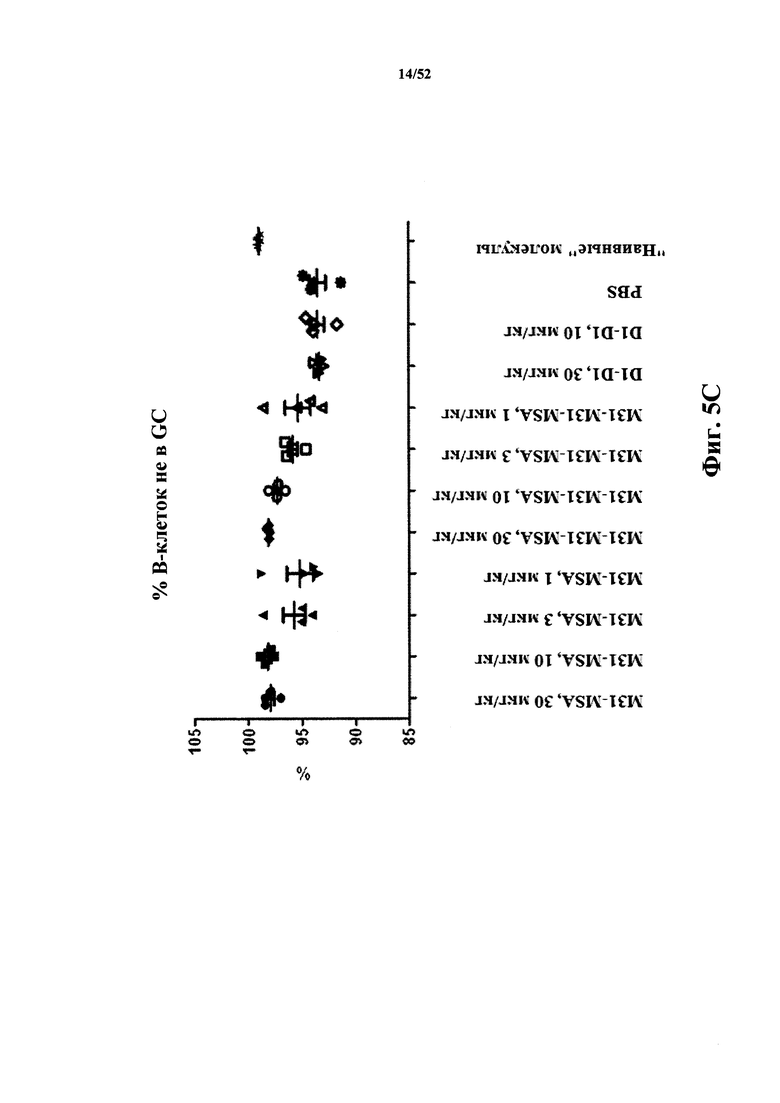
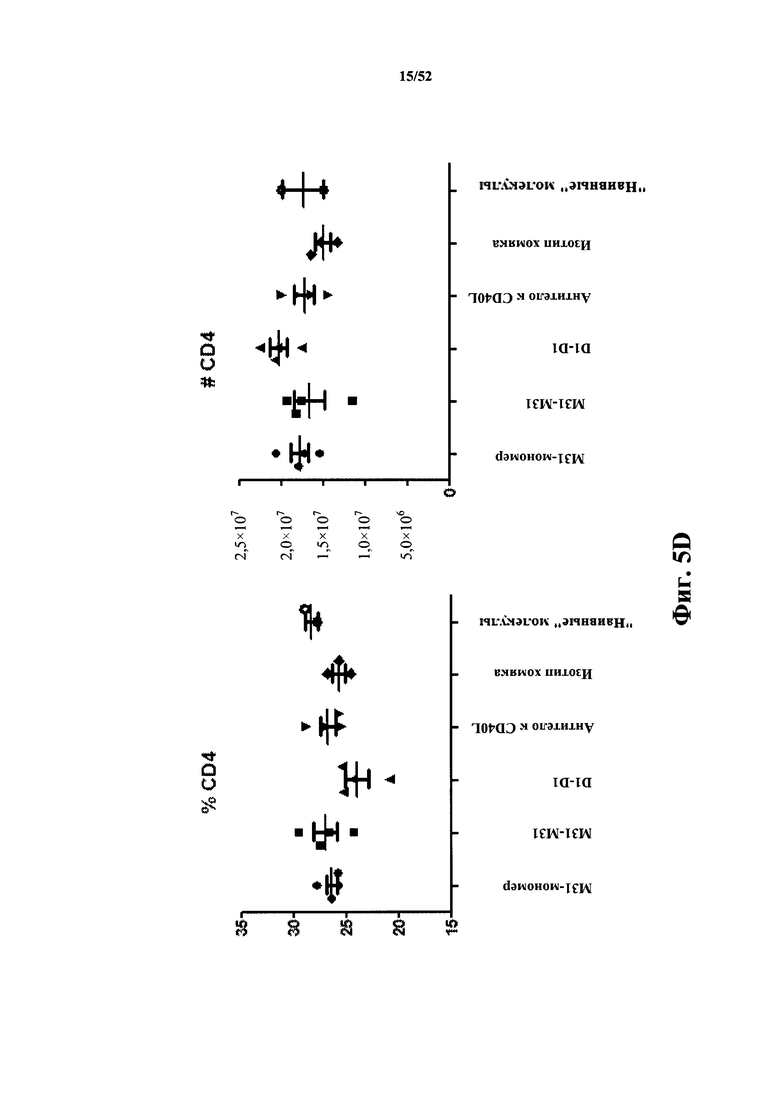
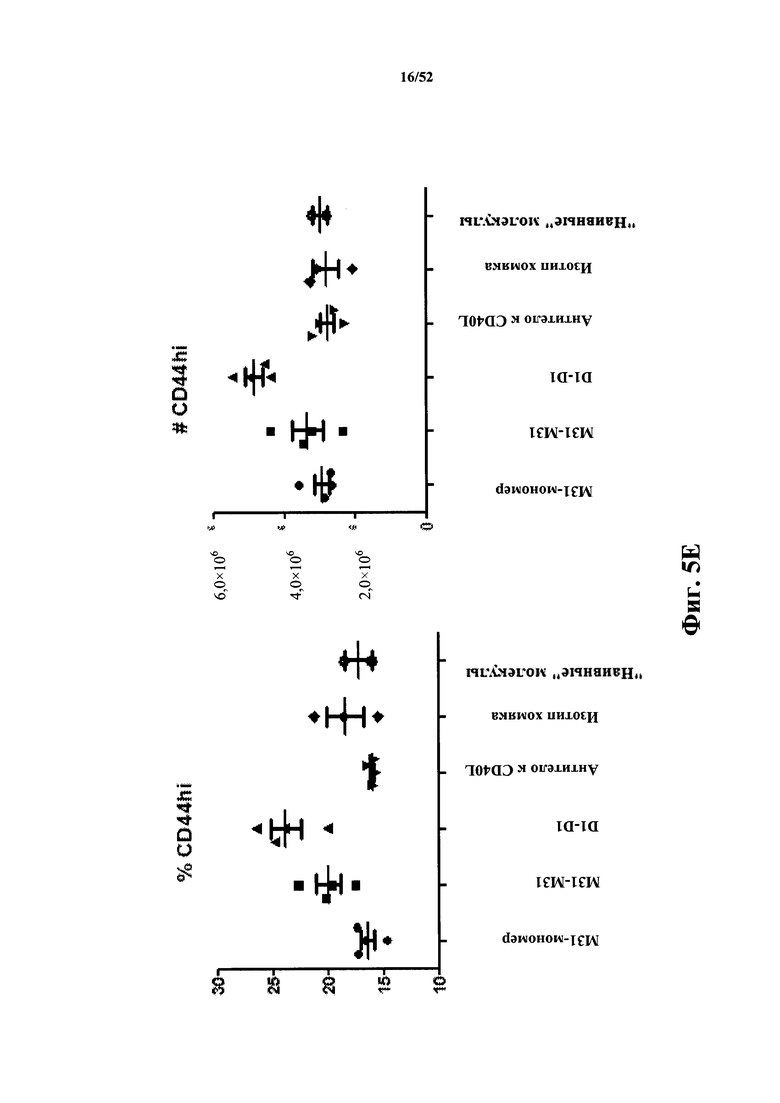
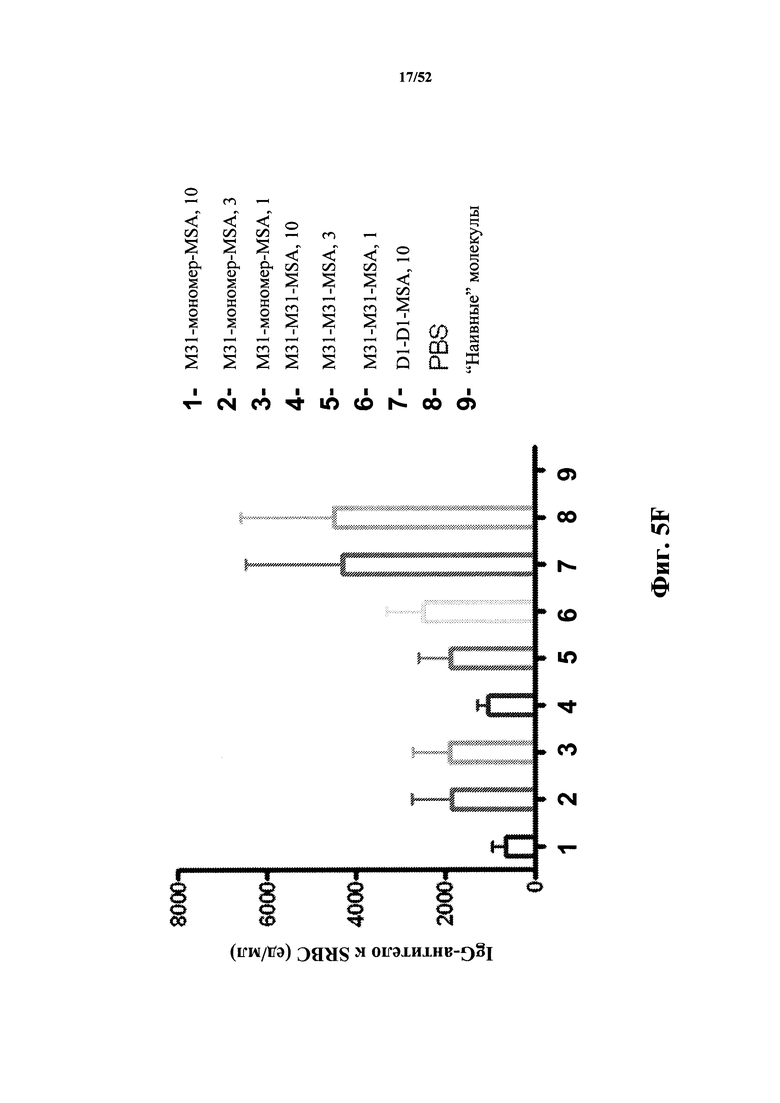
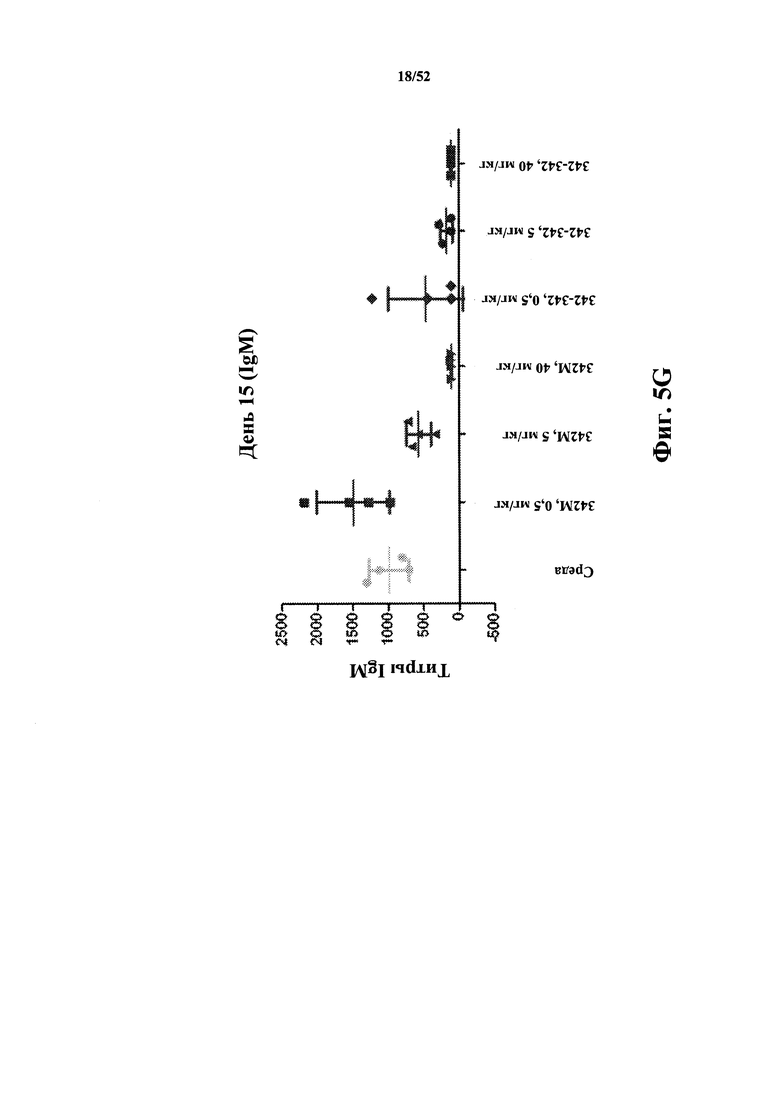
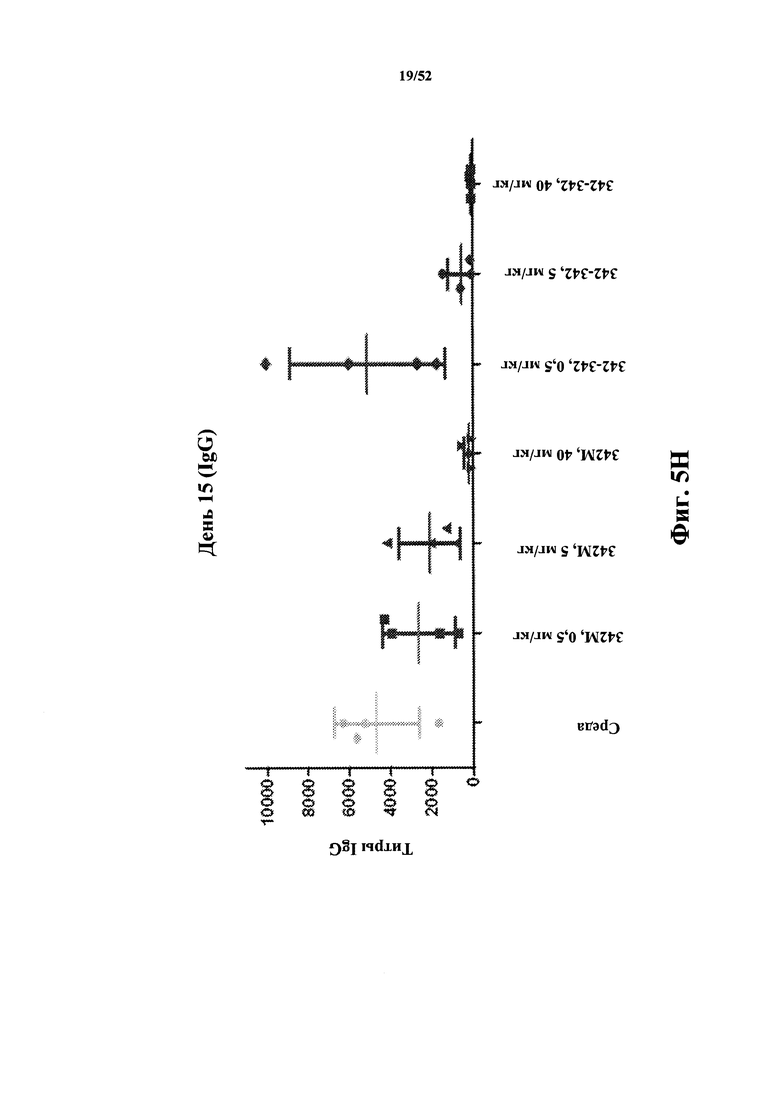
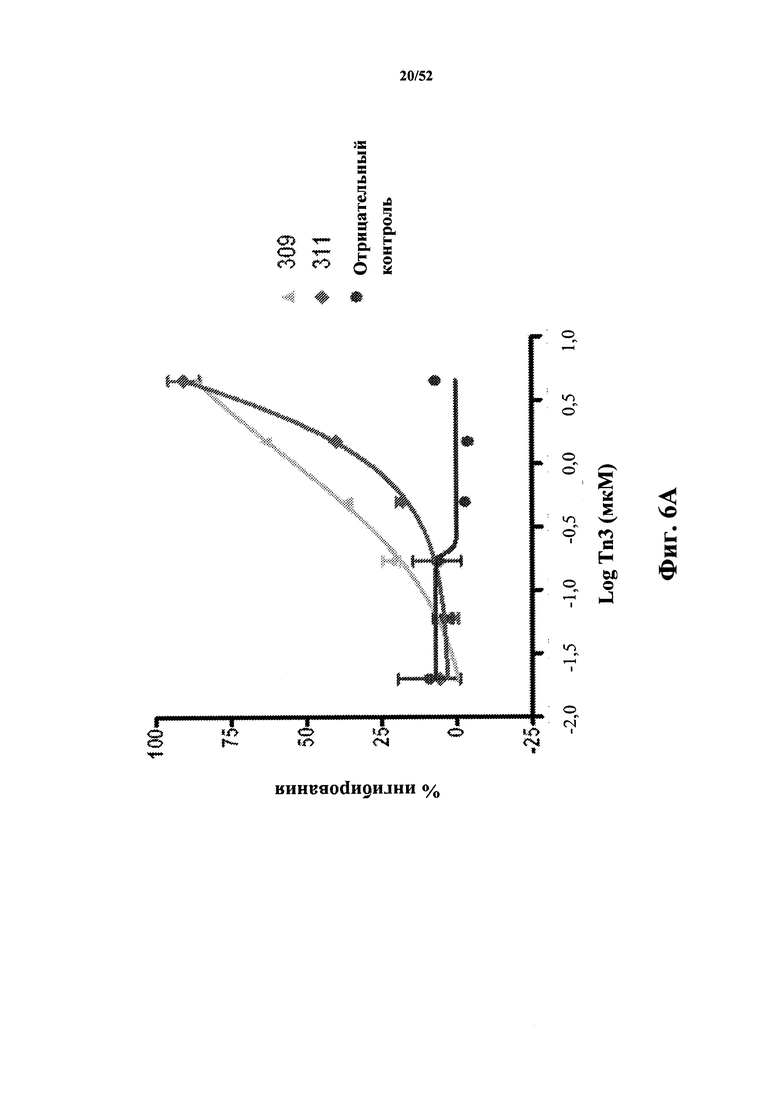
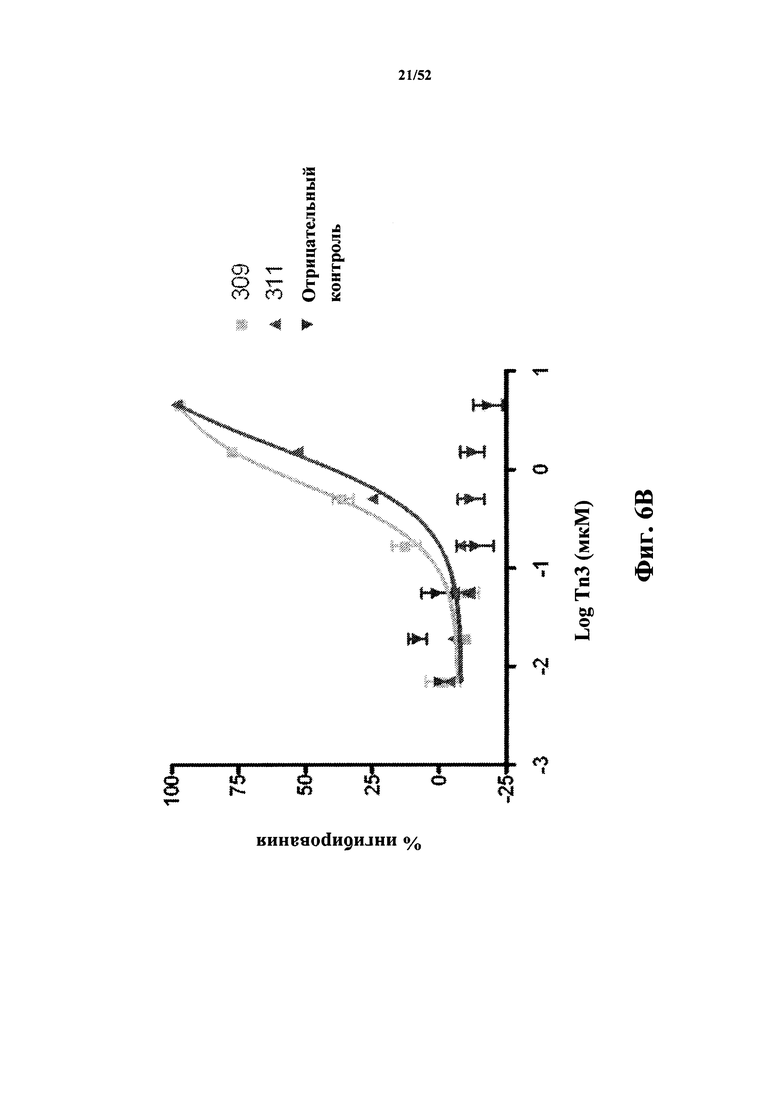
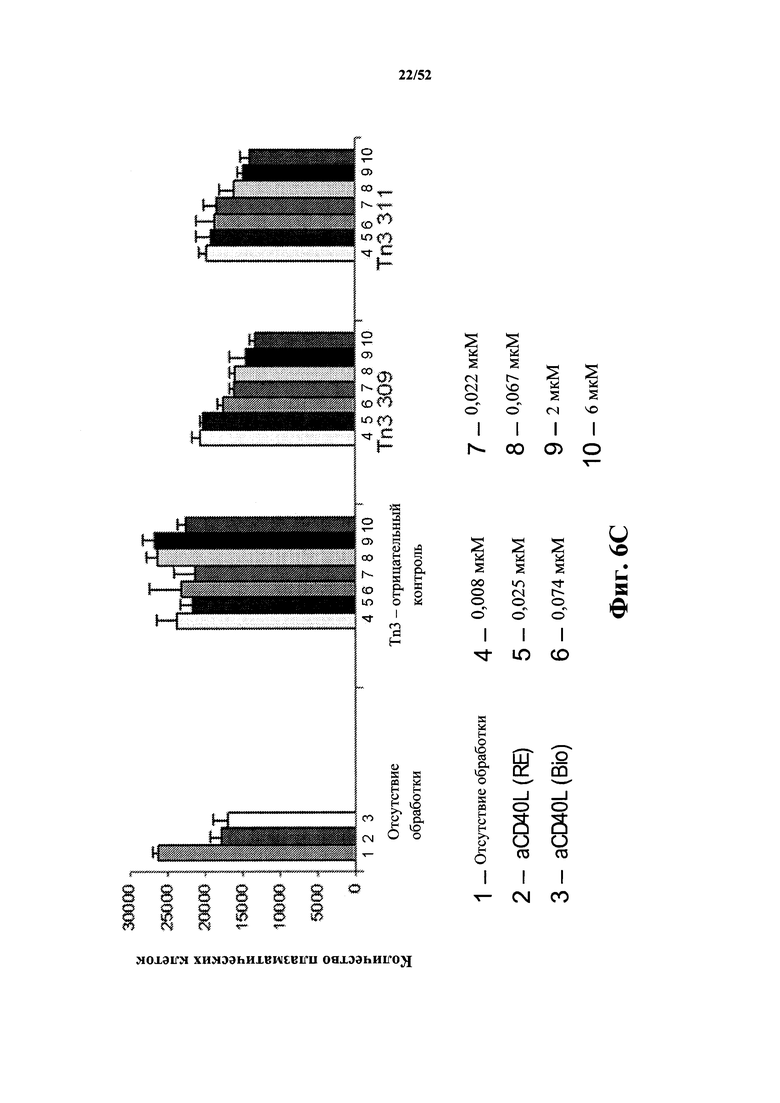
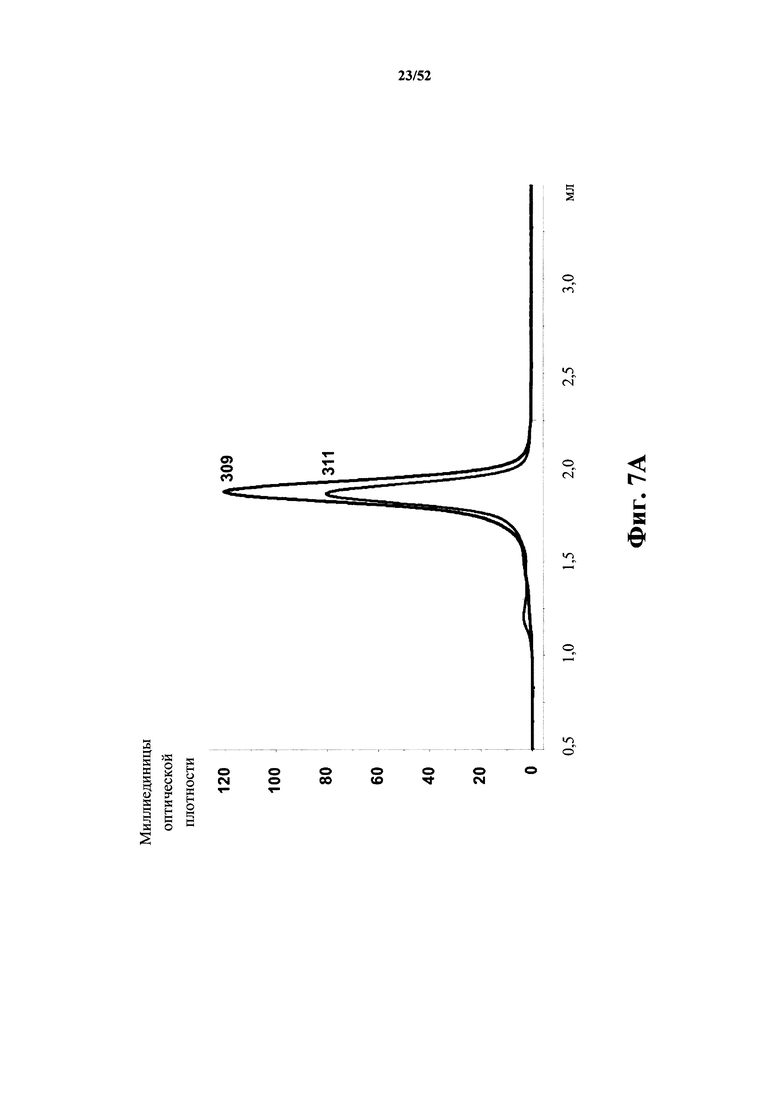
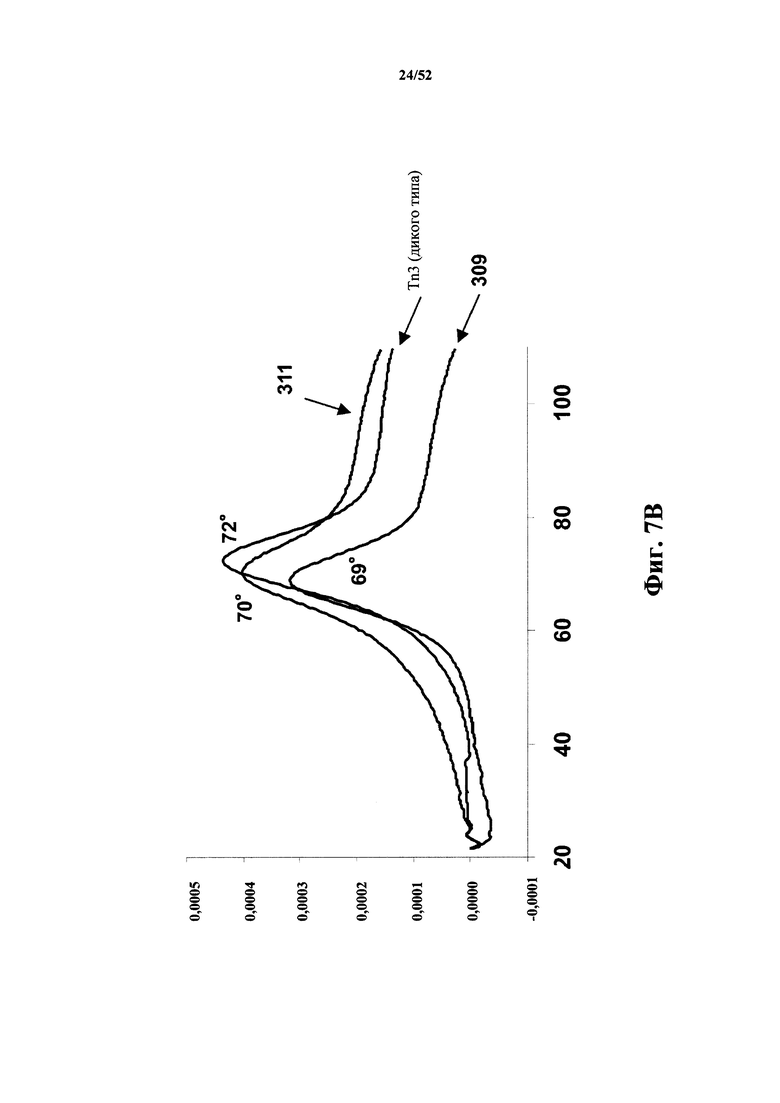
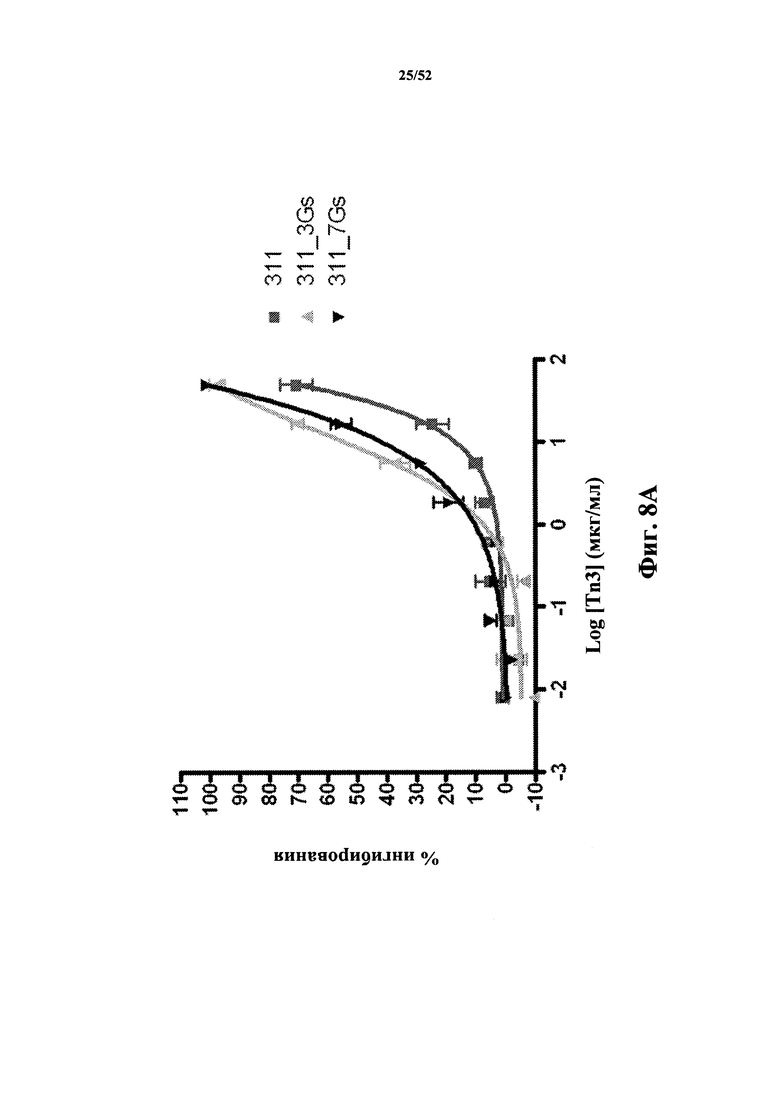
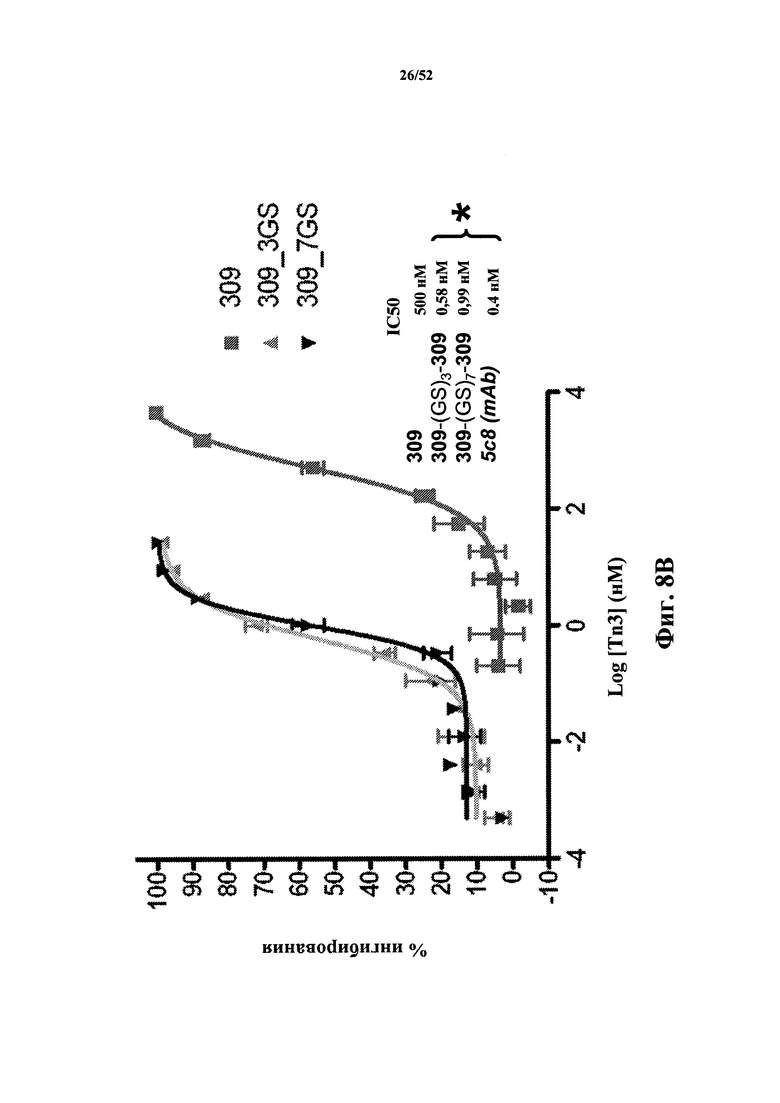

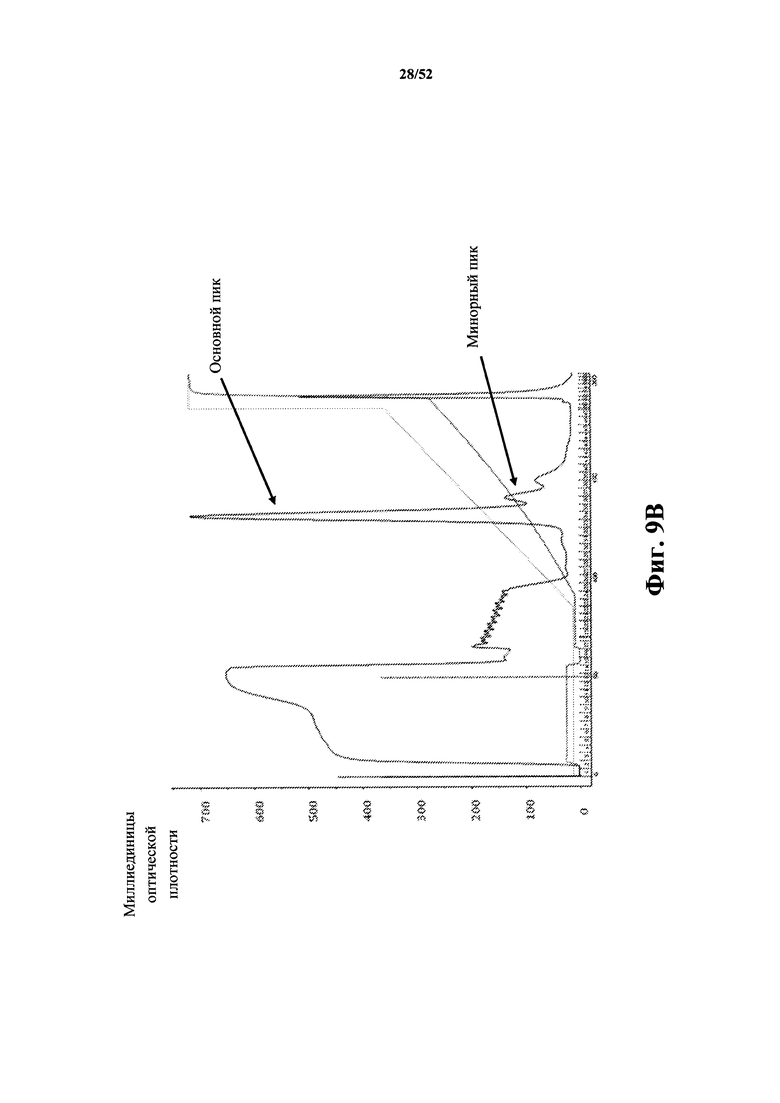

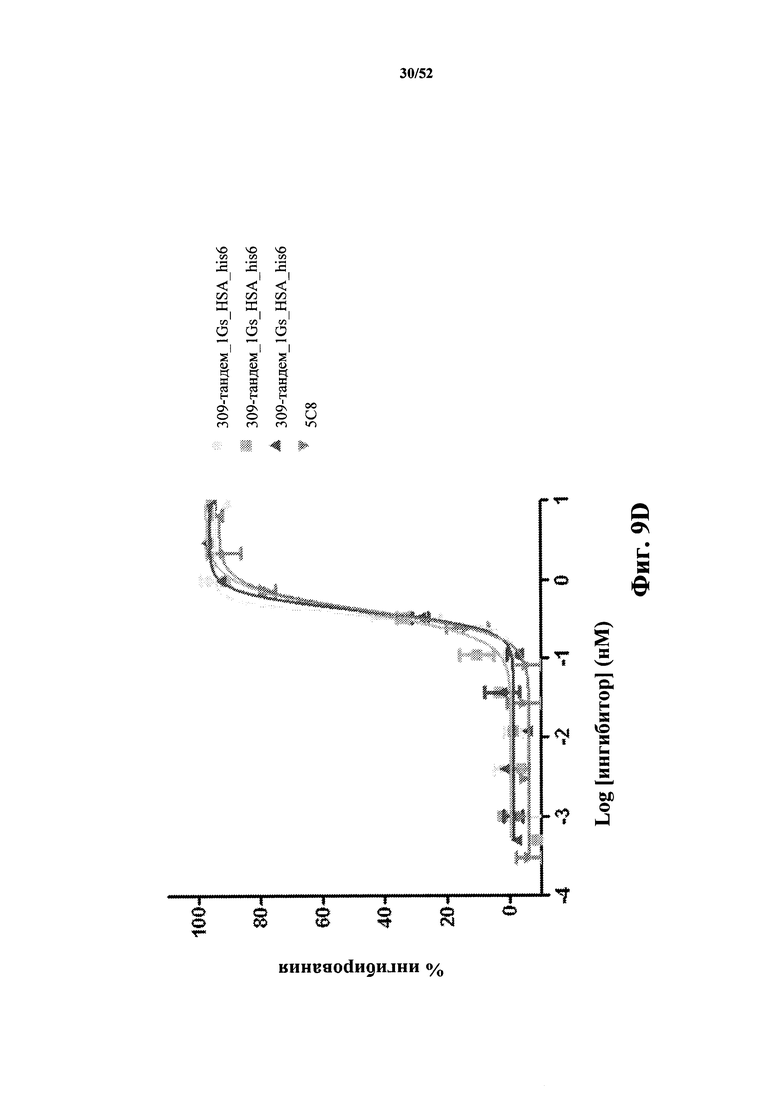
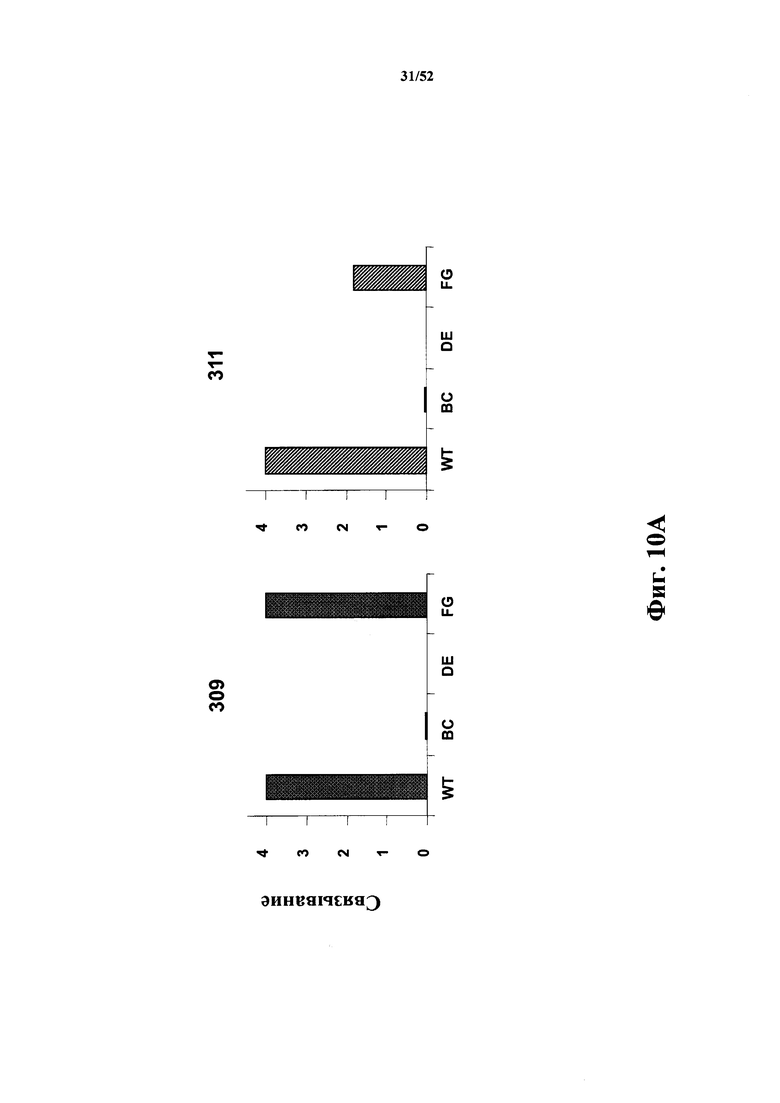
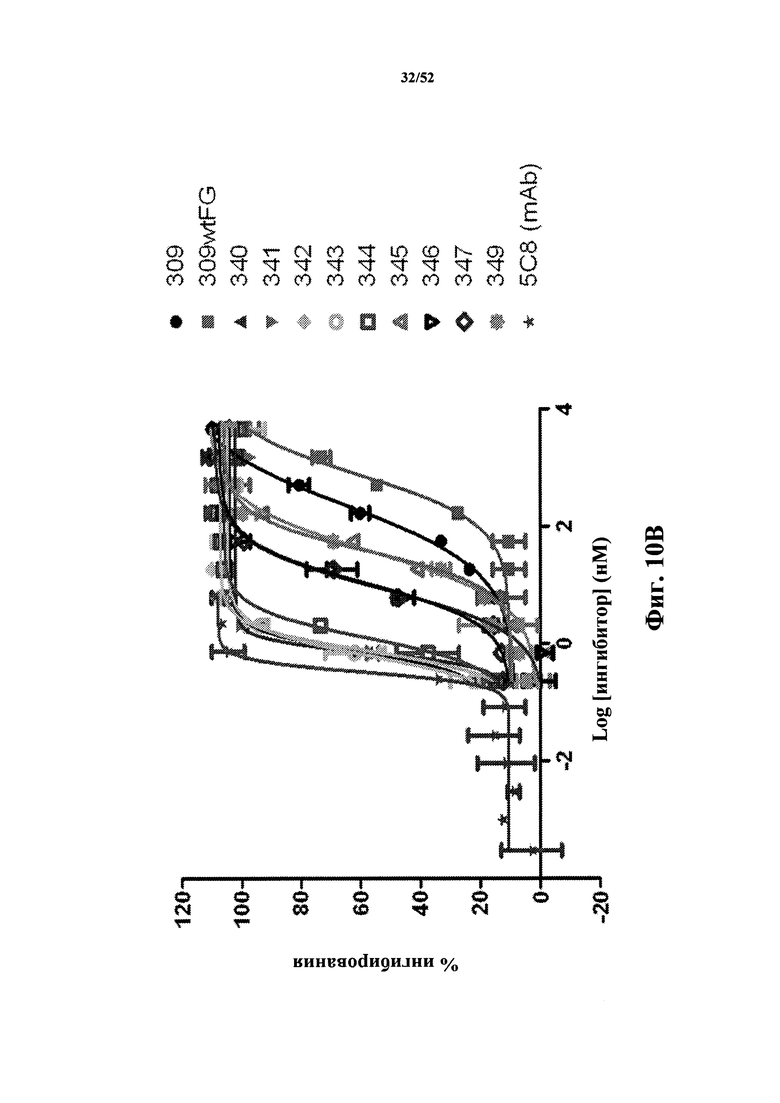
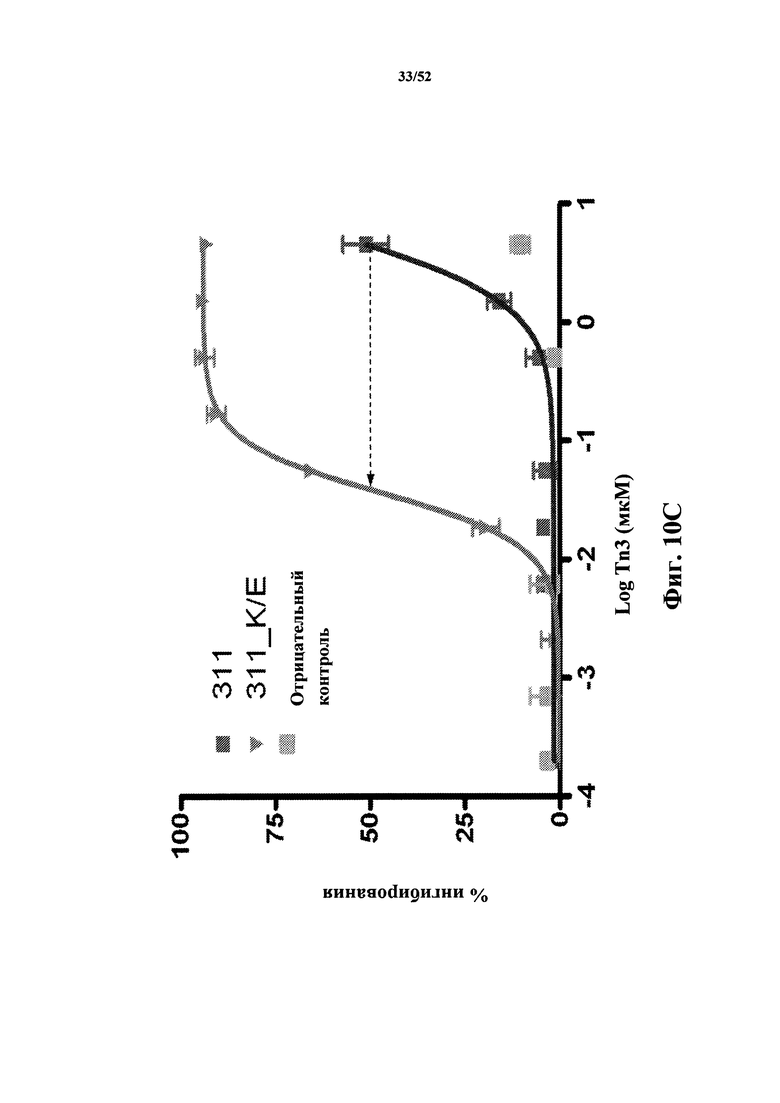
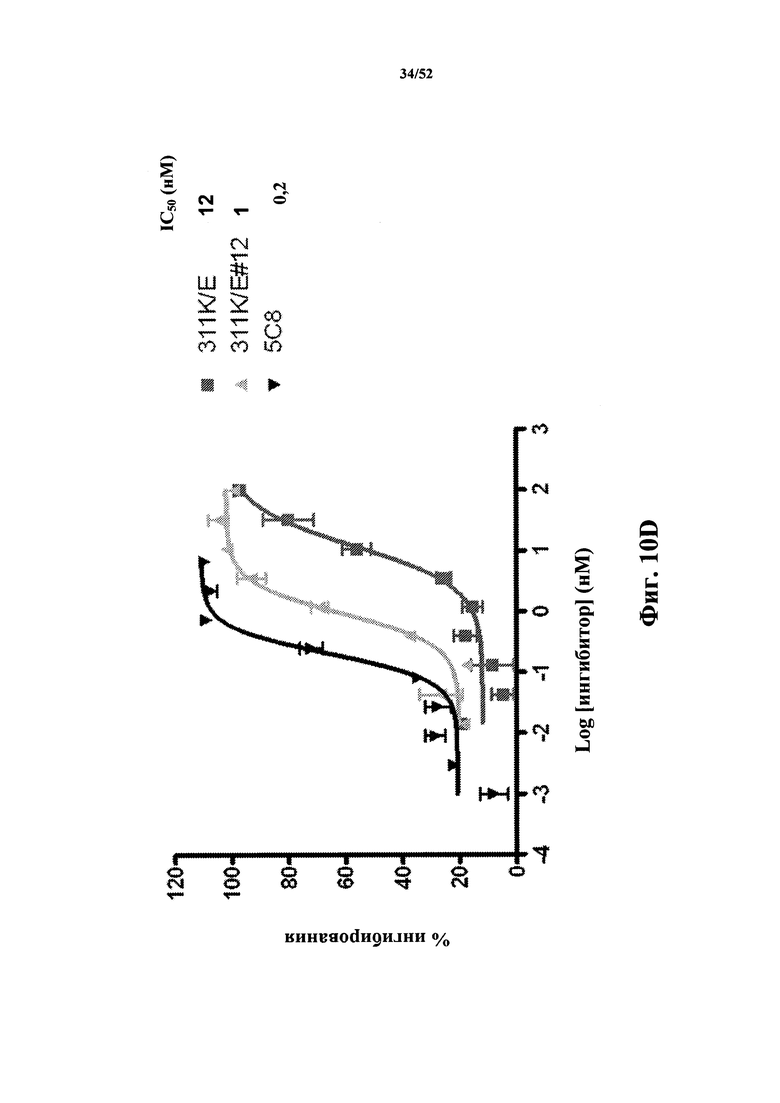
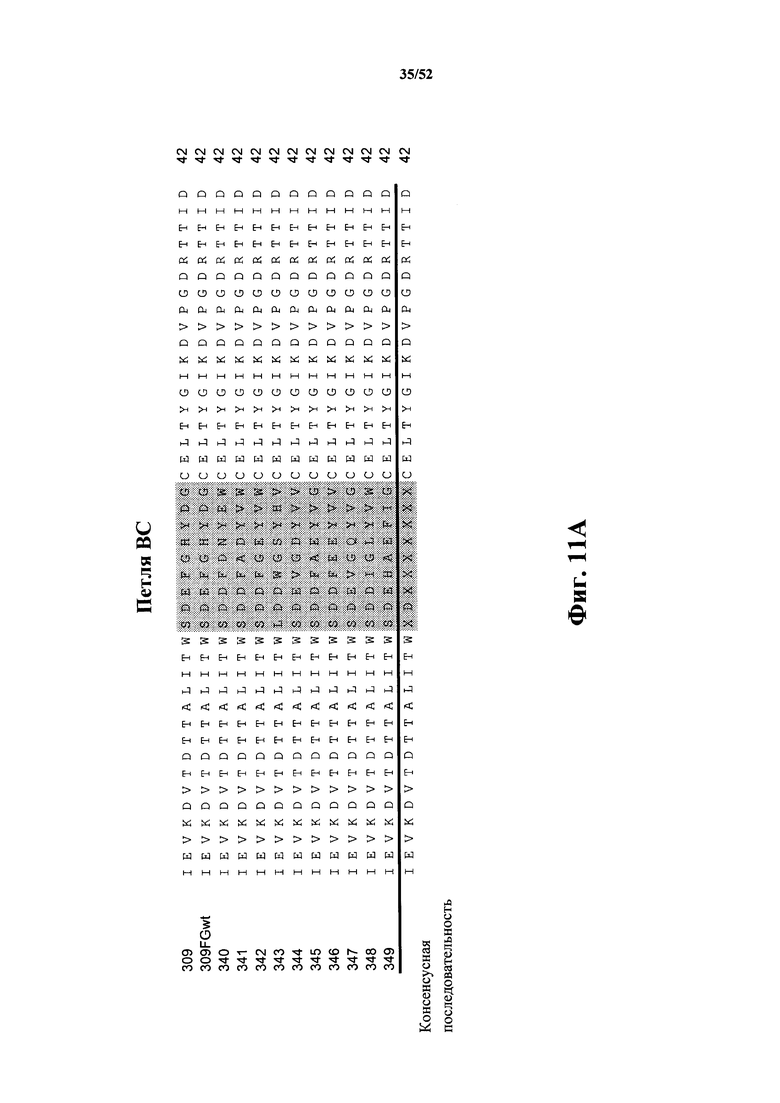
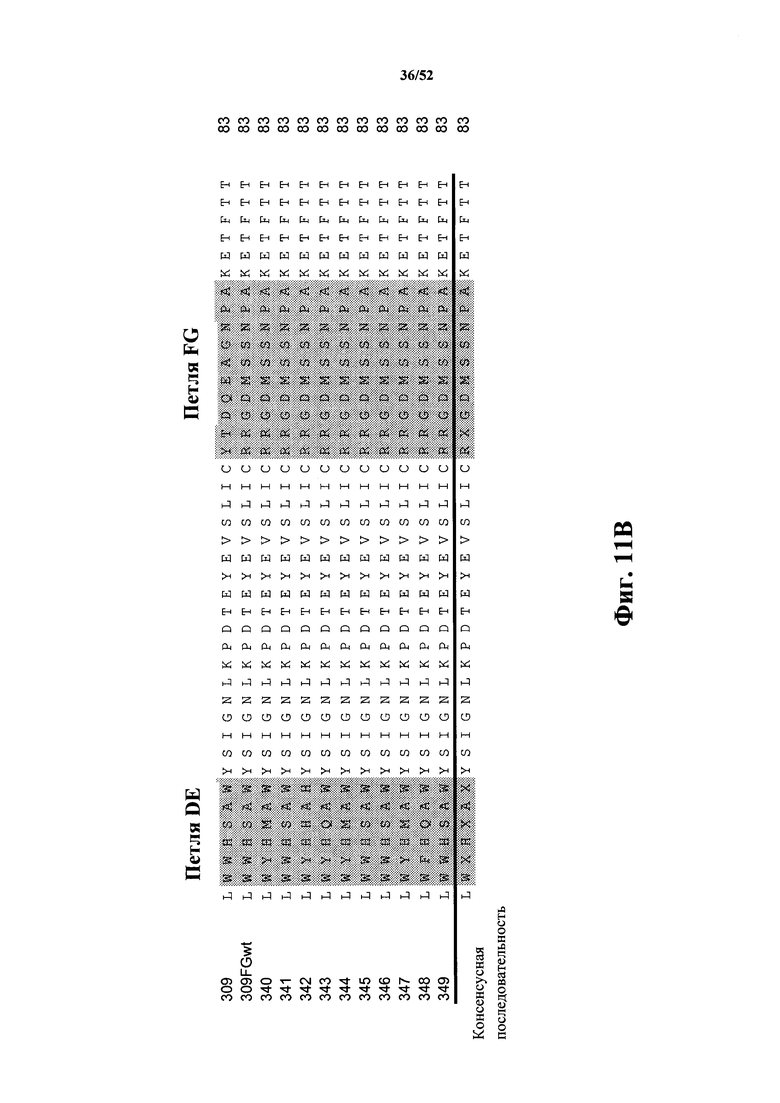
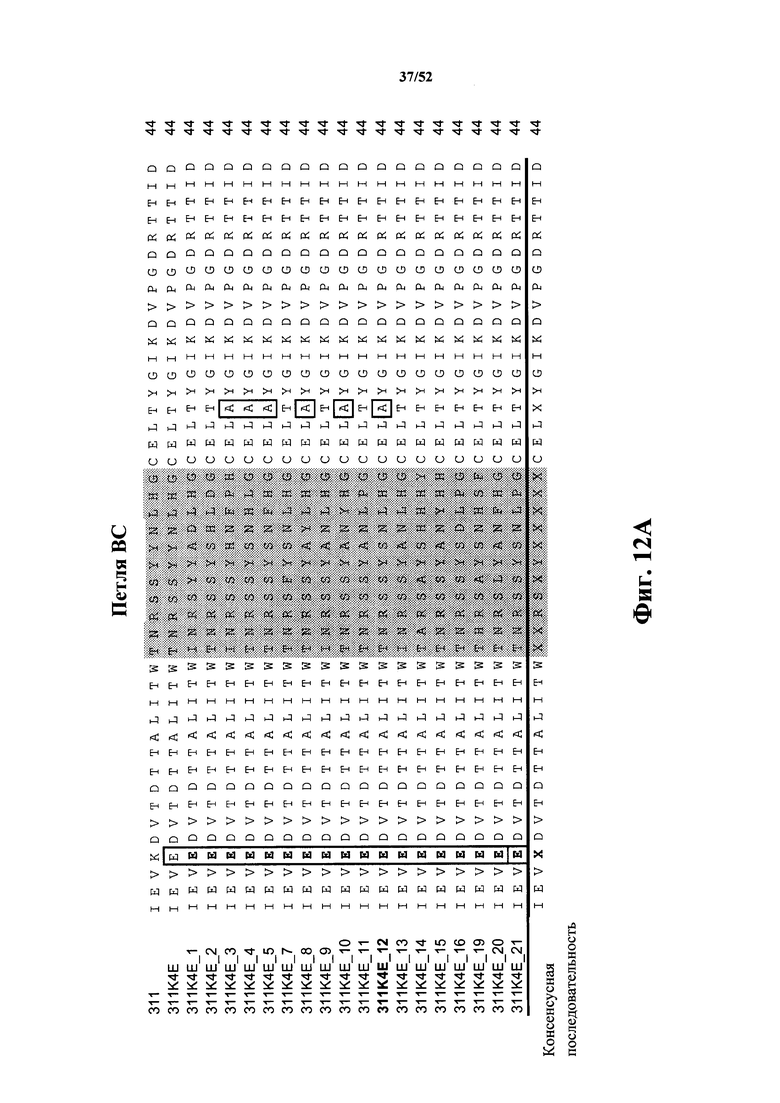
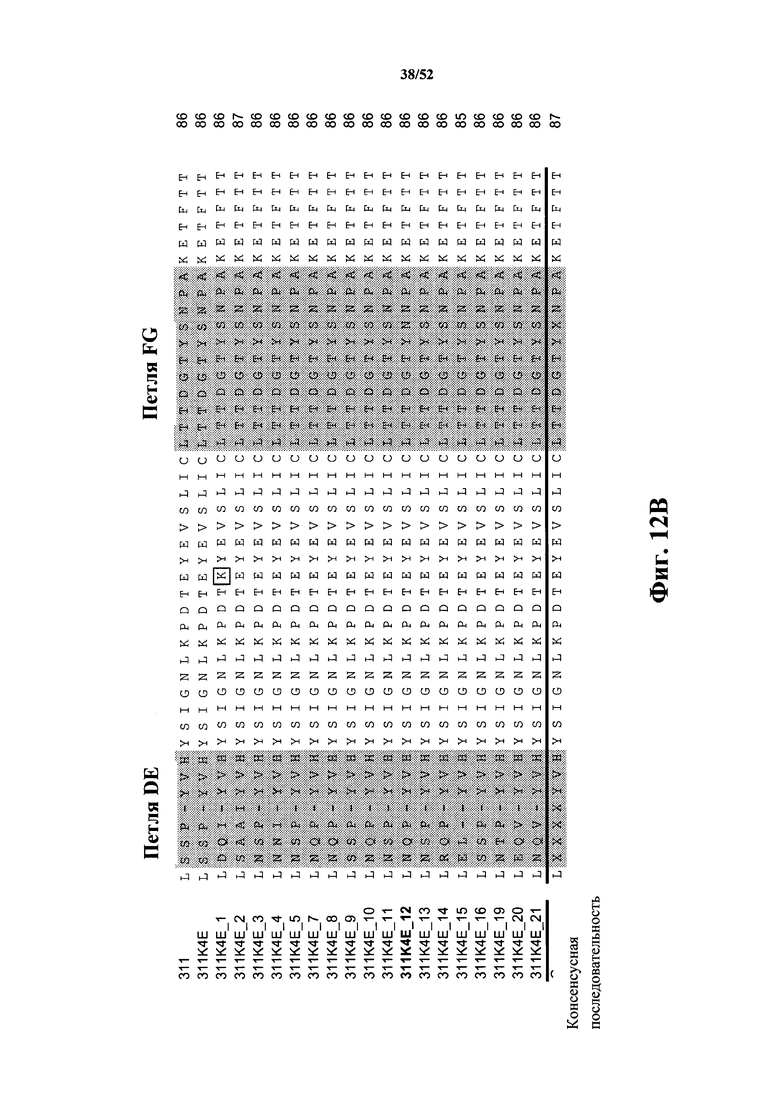
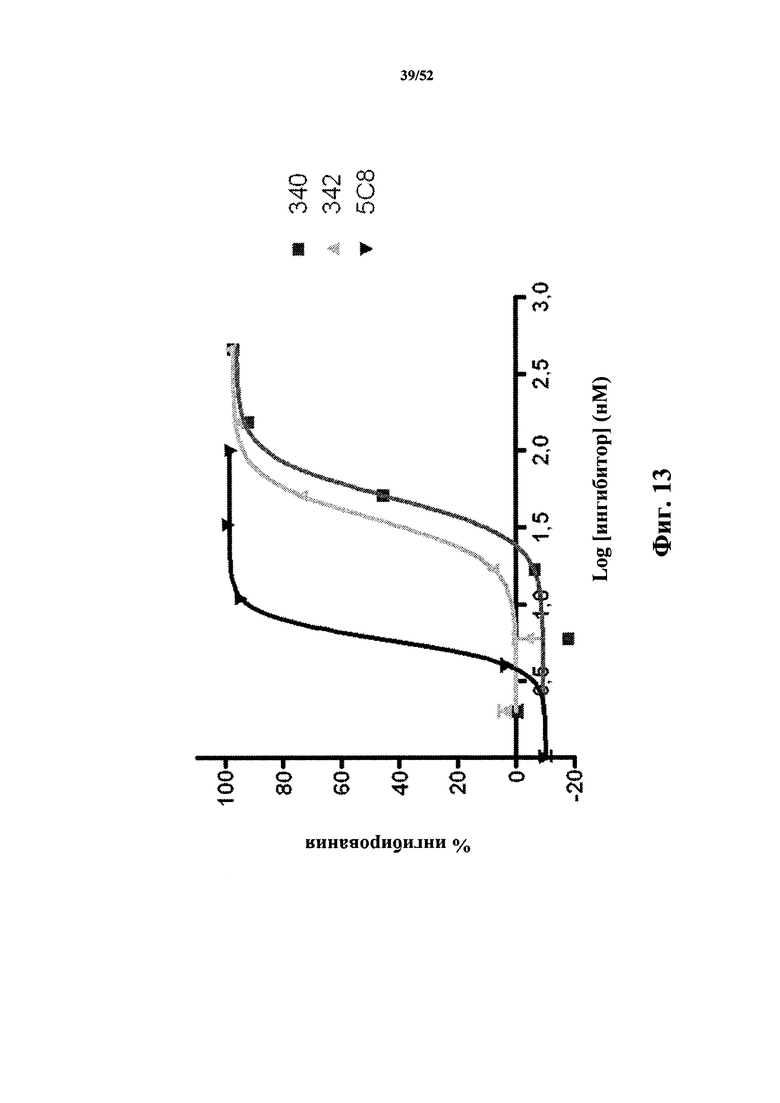
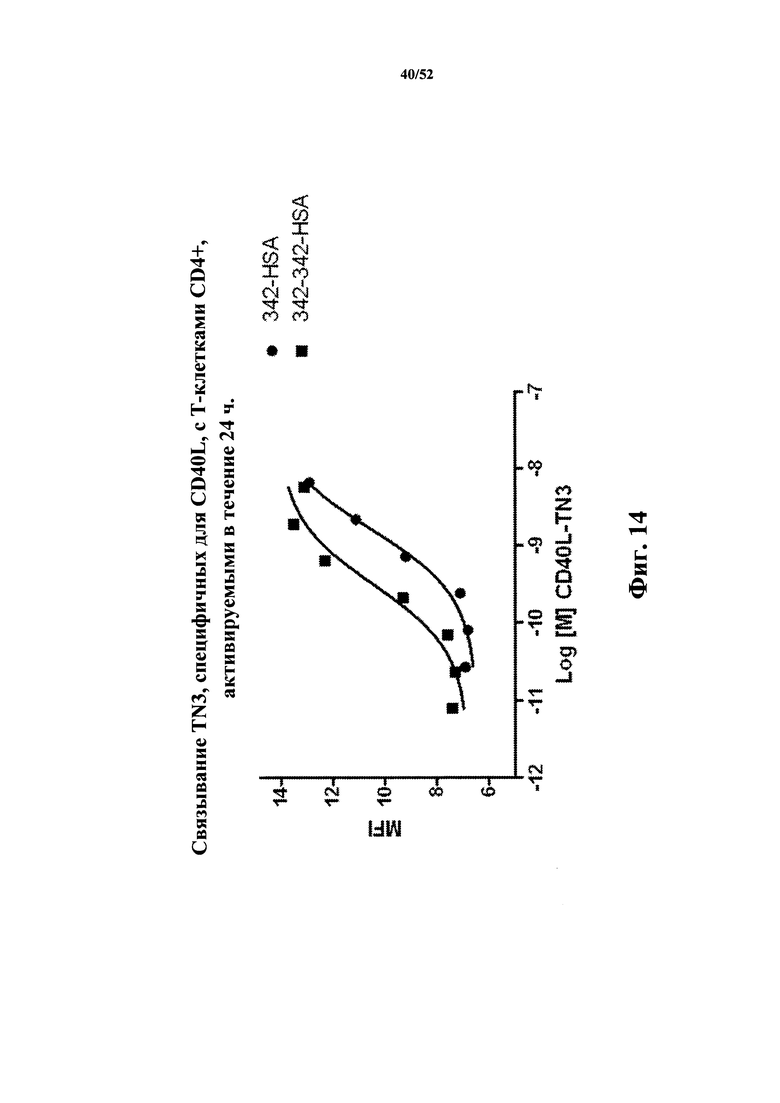
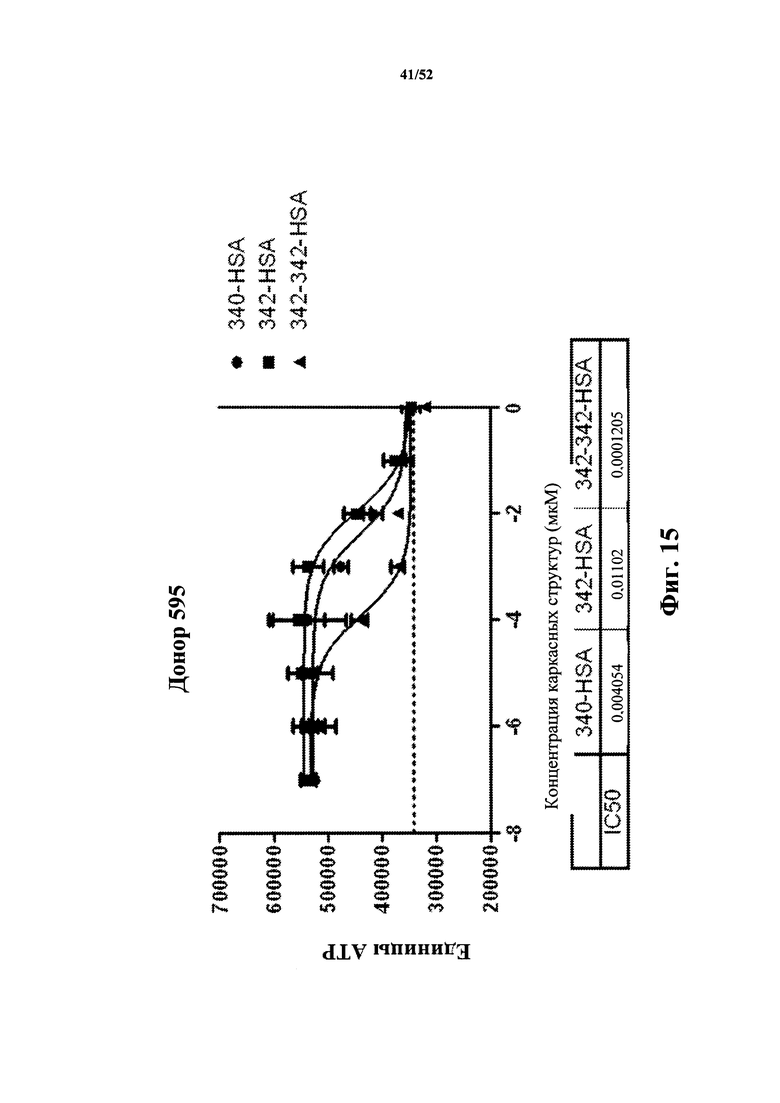
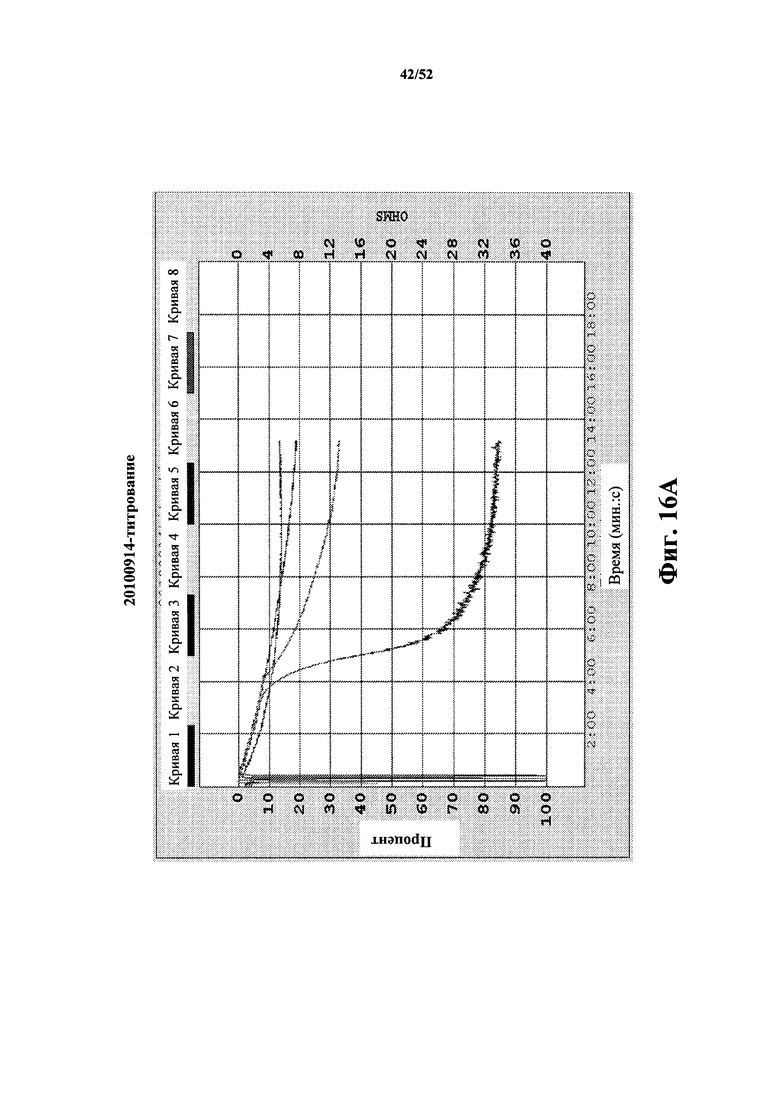

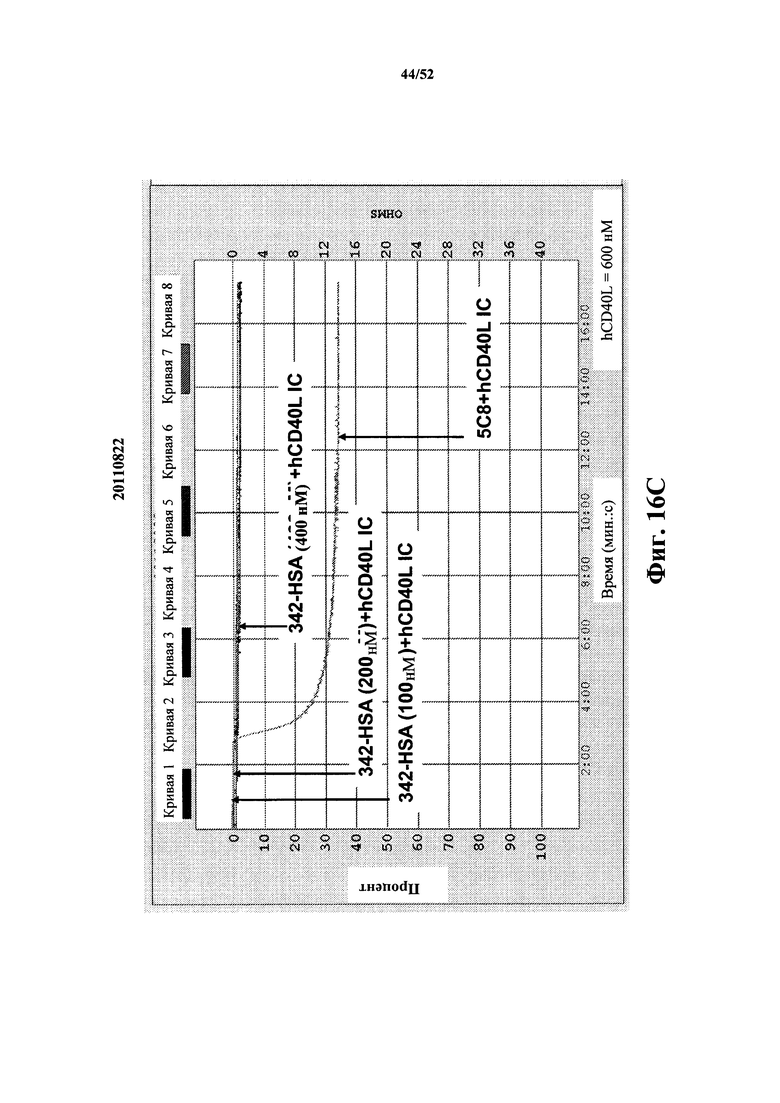
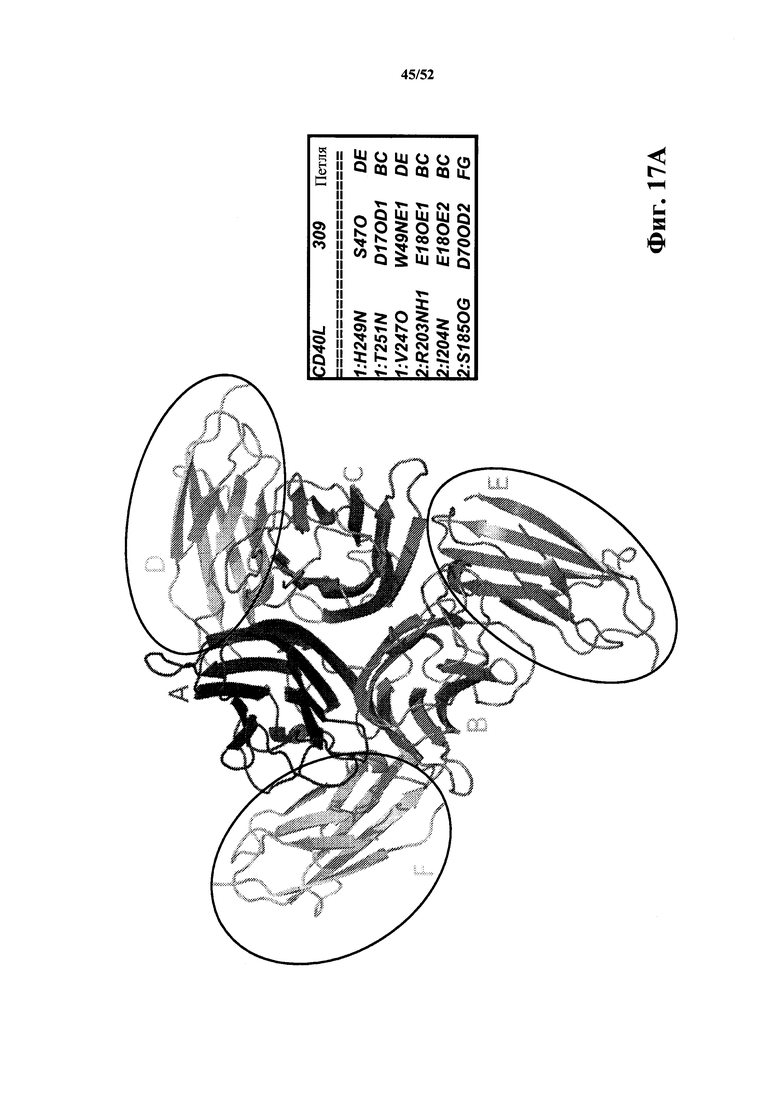
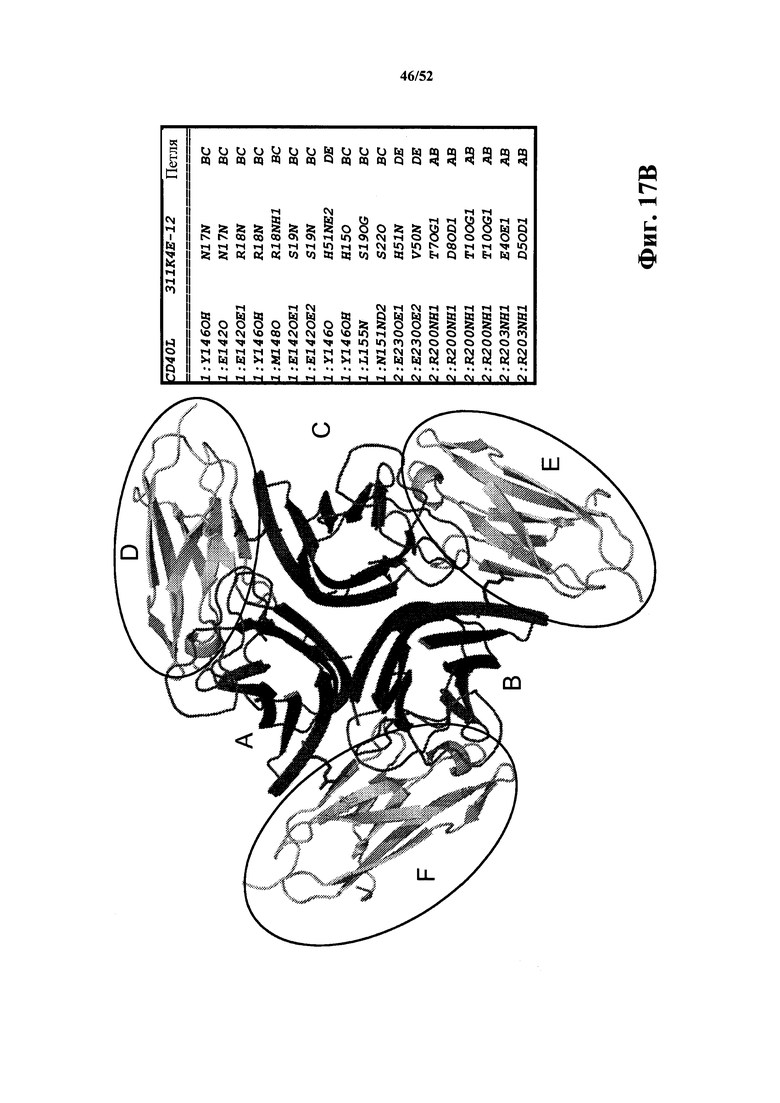
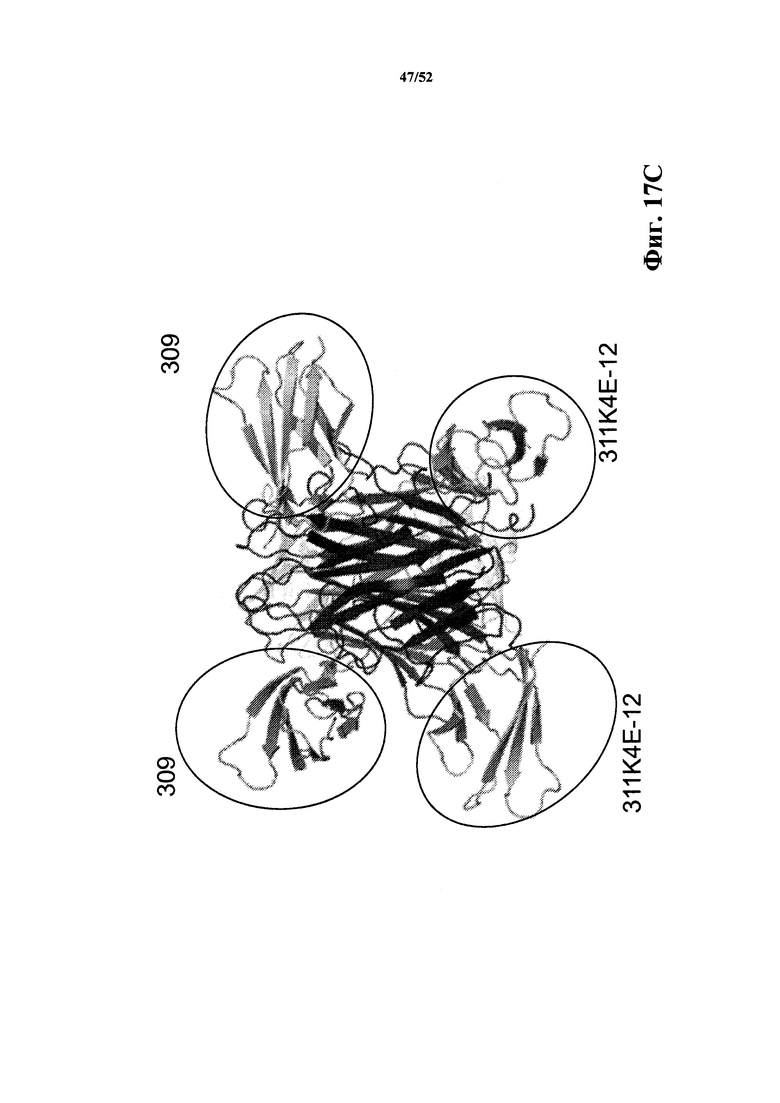

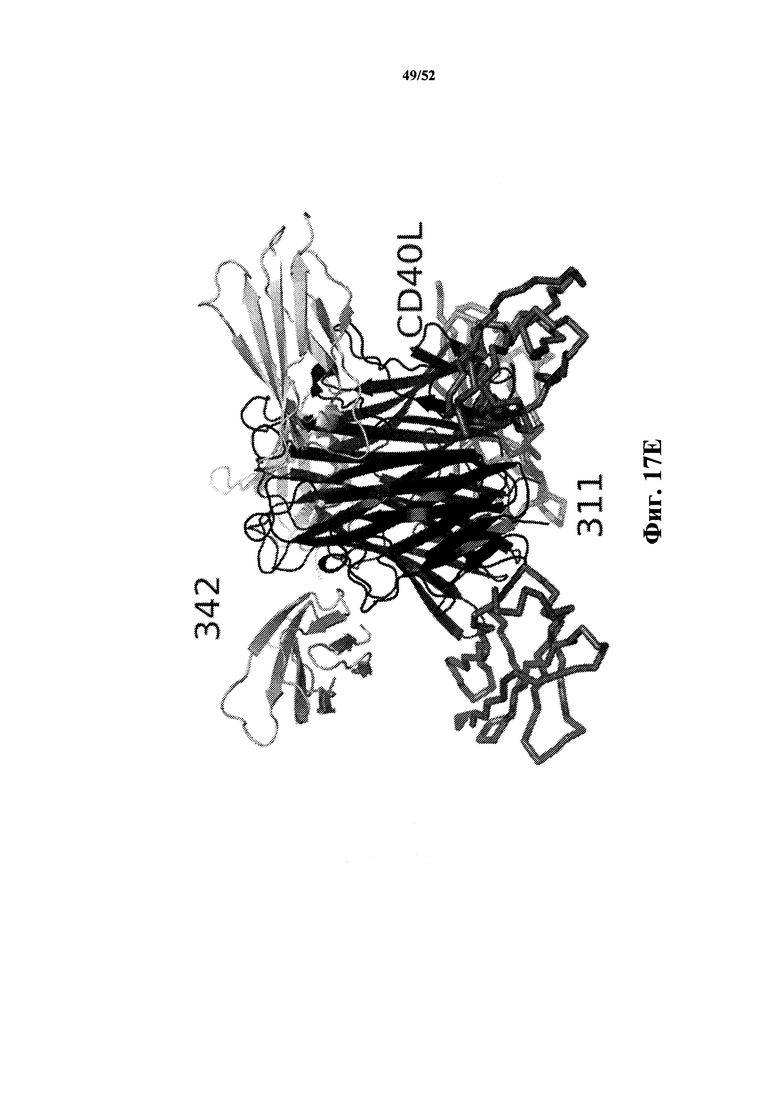


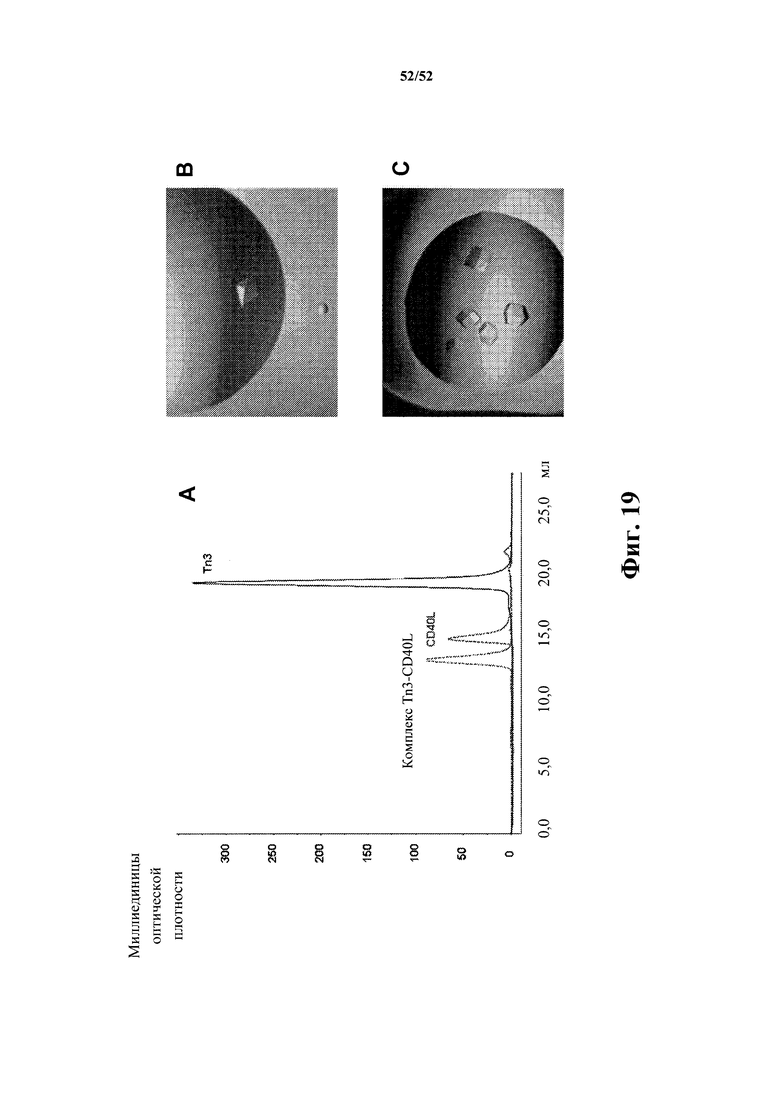
Настоящее изобретение относится к области биотехнологии, конкретно к каркасным структурам на основе FnIII-домена тенасцина-3 (Tn3). Изобретение позволяет получить каркасную структуру, способную специфически связываться с CD40L и обладающую повышенной аффинностью в отношении мишени в сравнении с известными аналогами. Сконструированные каркасные структуры по настоящему изобретению могут применяться в качестве профилактических, диагностических или терапевтических средств, в частности, для терапевтических применений против SLE и других аутоиммунных заболеваний и состояний. 16 н. и 25 з.п. ф-лы, 19 ил., 10 табл., 7 пр.
1. Tn3-каркасная структура, специфически связывающаяся с CD40L и содержащая две CD40L-специфичные мономерные субъединицы,
где каждая мономерная субъединица содержит семь бета-цепей, обозначенных A, B, C, D, E, F и G, и шесть петлевых участков, обозначенных AB, BC, CD, DE, EF и FG,
где бета-цепи и петлевые участки каждой мономерной субъединицы находятся в порядке от N-конца до C-конца: A-бета-цепь, петля AB, B-бета-цепь, петля BC, C-бета-цепь, петля CD, D-бета-цепь, петля DE, E-бета-цепь, петля EF, F-бета-цепь, петля FG, G-бета-цепь,
где петля AB представляет собой SEQ ID NO: 4, петля BC представляет собой SEQ ID NO: 86, петля CD представляет собой SEQ ID NO: 6, петля DE представляет собой SEQ ID NO: 96, петля EF представляет собой SEQ ID NO: 8, а петля FG представляет собой SEQ ID NO: 9 или 139,
где две мономерные субъединицы соединены в тандем посредством пептидного линкера, причем указанный пептидный линкер содержит аминокислотную последовательность (GmX)n, где X представляет собой серин (S), аланин (A), глицин (G), лейцин (L), изолейцин (I) или валин (V); m и n представляют собой целые числа, где m равно 1, 2, 3 или 4, n равно 1, 2, 3, 4, 5, 6 или 7.
2. Tn3-каркасная структура по п.1, где пептидный линкер содержит SEQ ID NO: 131, SEQ ID NO: 132, SEQ ID NO: 142 или SEQ ID NO: 143.
3. Tn3-каркасная структура по п.1, где
(a) A-бета-цепь содержит SEQ ID NO: 11;
(b) B-бета-цепь содержит SEQ ID NO: 12;
(c) C-бета-цепь содержит SEQ ID NO: 13 или 14;
(d) D-бета-цепь содержит SEQ ID NO: 15;
(e) E-бета-цепь содержит SEQ ID NO: 16;
(f) F-бета-цепь содержит SEQ ID NO: 17 и
(g) G-бета-цепь содержит SEQ ID NO: 18.
4. Tn3-каркасная структура по п.1, где каждая мономерная субъединица содержит SEQ ID NO: 146.
5. Tn3-каркасная структура по п.1, где каждая мономерная субъединица содержит SEQ ID NO: 208.
6. Tn3-каркасная структура по п.1 или 2, где каждая мономерная субъединица содержит аминокислотную последовательность:
IEV(XAB)nALITW(XBC)nCELX1YGI(XCD)nTTIDL(XDE)nYSI(XEF)n
YEVSLIC(XFG)nKETFTT,
где
(a) XAB, XBC, XCD, XDE, XEF и XFG представляют собой аминокислотные остатки, присутствующие в последовательностях петель AB, BC, CD, DE, EF и FG соответственно;
(b) X1 представляет собой аминокислотный остаток A или T и
(c) длина петли n представляет собой целое число от 2 до 26.
7. Конъюгат Tn3-каркасной структуры, который специфически связывается с CD40L и содержит Tn3-каркасный белок, соединенный с гетерологичным фрагментом,
где Tn3-каркасный белок содержит:
две CD40L-специфичные мономерные субъединицы, где каждая мономерная субъединица содержит семь бета-цепей, обозначенных A, B, C, D, E, F и G, и шесть петлевых участков, обозначенных AB, BC, CD, DE, EF и FG,
где бета-цепи и петлевые участки каждой мономерной субъединицы находятся в порядке от N-конца до C-конца: A-бета-цепь, петля AB, B-бета-цепь, петля BC, C-бета-цепь, петля CD, D-бета-цепь, петля DE, E-бета-цепь, петля EF, F-бета-цепь, петля FG, G-бета-цепь,
где петля AB представляет собой SEQ ID NO: 4, петля BC представляет собой SEQ ID NO: 86, петля CD представляет собой SEQ ID NO: 6, петля DE представляет собой SEQ ID NO: 96, петля EF представляет собой SEQ ID NO: 8, а петля FG представляет собой SEQ ID NO: 9 или 139,
где две мономерные субъединицы соединены в тандем посредством пептидного линкера, причем указанный пептидный линкер содержит аминокислотную последовательность (GmX)n, где X представляет собой серин (S), аланин (A), глицин (G), лейцин (L), изолейцин (I) или валин (V); m и n представляют собой целые числа, где m равно 1, 2, 3 или 4, n равно 1, 2, 3, 4, 5, 6 или 7; и
где гетерологичный фрагмент представляет собой белок, пептид, белковый домен, линкер, лекарственное средство, токсин, цитотоксическое средство, визуализирующее средство, радионуклид, радиоактивное соединение, органический полимер, неорганический полимер, полиэтиленгликоль (PEG), биотин, альбумин, человеческий сывороточный альбумин (HSA), участок, отвечающий за связывание HSA c FcRn, антитело, домен антитела, фрагмент антитела, одноцепочечное антитело, антитело на основе домена, альбумин-связывающий домен, фермент, лиганд, рецептор, связывающий пептид, каркасную структуру, отличающуюся от FnIII, эпитопную метку, рекомбинантный полипептидный полимер или цитокин.
8. Конъюгат Tn3-каркасной структуры по п.7, где гетерологичный фрагмент соединен с N-концом или C-концом каркасной структуры.
9. Конъюгат Tn3-каркасной структуры по п.8, где гетерологичный фрагмент соединен с каркасом вторым пептидным линкером.
10. Конъюгат Tn3-каркасной структуры по п.9, где второй пептидный линкер представляет собой гибкий пептидный линкер.
11. Конъюгат Tn3-каркасной структуры по п.9 или 10, где второй пептидный линкер содержит последовательность (GmX)n, где
(a) X представляет собой cерин (S), аланин (A), глицин (G), лейцин (L), изолейцин (I) или валин (V);
(b) m и n представляют собой целые числа;
(c) m равно 1, 2, 3 или 4 и
(d) n равно 1, 2, 3, 4, 5, 6 или 7.
12. Конъюгат Tn3-каркасной структуры по п.11, где второй пептидный линкер содержит SEQ ID NO: 131, SEQ ID NO: 132, SEQ ID NO: 142 или SEQ ID NO: 143.
13. Конъюгат Tn3-каркасной структуры по п.7, где гетерологичный фрагмент представляет собой PEG.
14. Конъюгат Tn3-каркасной структуры по п.9, где гетерологичный фрагмент представляет собой альбумин.
15. Конъюгат Tn3-каркасной структуры по п.14, где альбумин представляет собой сывороточный альбумин человека (HSA).
16. Конъюгат Tn3-каркасной структуры по п.15, где указанный HSA представляет собой вариант HSA.
17. Конъюгат Tn3-каркасной структуры по п.16, где аминокислотная последовательность указанного варианта HSA представляет собой SEQ ID NO: 133.
18. Конъюгат Tn3-каркасной структуры по п.16, где вариант HSA содержит последовательность полноразмерного зрелого HSA (SEQ ID NO: 133 или 138) или ее фрагмент за исключением по меньшей мере одной аминокислотной замены, пронумерованной относительно положения в полноразмерном зрелом HSA, выбранной из группы, состоящей из:
(a) замены лейцина (L) в положении 407 на аспарагин (N) или тирозин (Y);
(b) замены валина (V) в положении 415 на треонин (T);
(c) замены лейцина (L) в положении 463 на аспарагин (N);
(d) замены лизина (K) в положении 500 на аргинин (R);
(e) замены треонина (T) в положении 506 на тирозин (Y);
(f) замены треонина (T) в положении 508 на аргинин (R);
(g) замены фенилаланина (F) в положении 509 на метионин (M) или триптофан (W);
(h) замены аланина (A) в положении 511 на фенилаланин (F);
(i) замены аспарагиновой кислоты (D) в положении 512 на тирозин (Y);
(j) замены треонина (T) в положении 515 на глутамин (Q);
(k) замены лейцина (L) в положении 516 на треонин (T) или триптофан (W);
(l) замены аргинина (R) в положении 521 на триптофан (W);
(m) замены изолейцина (I) в положении 523 на аспарагиновую кислоту (D), глутаминовую кислоту (E), глицин (G), лизин (K) или аргинин (R);
(n) замены лизина (K) в положении 524 на лейцин (L);
(o) замены глутамина (Q) в положении 526 на метионин (M);
(p) замены гистидина (H) в положении 535 на пролин (P);
(q) замены аспарагиновой кислоты (D) в положении 550 на глутаминовую кислоту (E);
(r) замены лизина (K) в положении 557 на глицин (G);
(s) замены лизина (K) в положении 573 на фенилаланин (F), гистидин (H), пролин (P), триптофан (W) или тирозин (Y);
(t) замены лизина (K) в положении 574 на аспарагин (N);
(u) замены глутамина (Q) в положении 580 на лизин (K) и
(v) комбинации двух или более указанных замен,
где указанная Tn3-каркасная структура имеет более длительное время полужизни в плазме, чем время полужизни в плазме Tn3-каркасной структуры, не являющейся конъюгированной с указанным вариантом HSA.
19. Конъюгат Tn3-каркасной структуры по п.18, где вариант HSA содержит последовательность полноразмерного зрелого HSA (SEQ ID NO: 133 или 138) или ее фрагмент за исключением по меньшей мере одной аминокислотной замены, пронумерованной относительно положения в полноразмерном зрелом HSA, выбранной из группы, состоящей из:
(a) замены лейцина (L) в положении 463 на аспарагин (N);
(a) замены треонина (Т) в положении 508 на аргинин (R);
(c) замены изолейцина (I) в положении 523 на аспарагиновую кислоту (D), глутаминовую кислоту (E), глицин (G), лизин (K) или аргинин (R);
(d) замены лизина (K) в положении 524 на лейцин (L) и
(e) комбинации двух или более указанных замен,
где указанная Tn3-каркасная структура имеет более длительное время полужизни в плазме, чем время полужизни в плазме Tn3-каркасной структуры, не являющейся конъюгированной с указанным вариантом HSA.
20. Конъюгат Tn3-каркасной структуры по любому из пп.7-19, где
(a) бета-цепь A содержит SEQ ID NO: 11;
(b) бета-цепь B содержит SEQ ID NO: 12;
(c) бета-цепь C содержит SEQ ID NO: 13 или 14;
(d) бета-цепь D содержит SEQ ID NO: 15;
(e) бета-цепь E содержит SEQ ID NO: 16;
(f) бета-цепь F содержит SEQ ID NO: 17 и
(g) бета-цепь G содержит SEQ ID NO: 18.
21. Конъюгат Tn3-каркасной структуры по любому из пп.7-19, где каждая мономерная субъединица содержит аминокислотную последовательность:
IEV(XAB)nALITW(XBC)nCELX1YGI(XCD)nTTIDL(XDE)nYSI(XEF)n
YEVSLIC(XFG)nKETFTT,
где
(a) XAB, XBC, XCD, XDE, XEF и XFG представляют собой аминокислотные остатки, присутствующие в последовательностях петель AB, BC, CD, DE, EF и FG соответственно;
(b) X1 представляет собой аминокислотный остаток A или T и
(c) длина петли n представляет собой целое число от 2 до 26.
22. Конъюгат Tn3-каркасной структуры по любому из пп.7-19, где каждая мономерная субъединица содержит SEQ ID NO: 146.
23. Конъюгат Tn3-каркасной структуры по п.7, где белок Tn3-каркасной структуры содержит последовательность, выбранную из группы, состоящей из SEQ ID NO: 134, 135, 144, 145, 146 и 208.
24. Конъюгат Tn3-каркасной структуры по п.7, где белок Tn3-каркасной структуры состоит из последовательности, выбранной из группы, состоящей из SEQ ID NO: 135, 145 и 208.
25. Конъюгат Tn3-каркасной структуры по п.7, где указанный CD40L представляет собой CD40L человека.
26. Конъюгат Tn3-каркасной структуры по п.7, где указанный CD40L представляет собой мембраносвязанный CD40L (SEQ ID NO: 1), растворимый CD40L (SEQ ID NO: 2) или его фрагмент.
27. Способ ингибирования взаимодействия между CD40L и CD40 в клетке, экспрессирующей CD40L, предусматривающий приведение в контакт клетки с Tn3-каркасной структурой по любому из пп.1-6 или конъюгатом Tn3-каркасной структуры по любому из пп.7-26.
28. Способ снижения CD40-опосредованного сигнального ответа, предусматривающий приведение в контакт клетки, экспрессирующей CD40L, с Tn3-каркасной структурой по любому из пп.1-6 или конъюгатом Tn3-каркасной структуры по любому из пп.7-26,
где Tn3-каркасная структура связывается с CD40L и нарушает CD40-опосредованный сигнальный путь.
29. Способ по п.28, отличающийся тем, что CD40-опосредованный сигнальный ответ представляет собой T-зависимый иммунный ответ.
30. Выделенная молекула нуклеиновой кислоты, кодирующая Tn3-каркасную структуру по любому из пп.1-6.
31. Вектор экспрессии, содержащий нуклеиновую кислоту по п.30.
32. Клетка-хозяин для экспрессии Tn3-каркасной структуры по любому из пп.1-6,
где клетка-хозяин является одной из: бактериальной клеткой, клеткой COS, клеткой CHO, клеткой 3T3, клеткой NSO или клеткой BHK и
где указанная клетка-хозяин содержит вектор по п.31.
33. Способ получения Tn3-каркасной структуры, предусматривающий культивирование клетки-хозяина по п.32 в условиях, при которых экспрессируется Tn3-каркасная структура, кодируемая молекулой нуклеиновой кислоты.
34. Композиция для уменьшения CD40-опосредованного иммунного ответа, содержащая Tn3-каркасную структуру по любому из пп.1-6 в количестве, достаточном для достижения терапевтического ответа, и фармацевтически приемлемый наполнитель.
35. Способ лечения, уменьшения интенсивности, управления течением аутоиммунного заболевания у нуждающегося пациента, предусматривающий введение эффективного количества композиции по п.34.
36. Выделенная молекула нуклеиновой кислоты, кодирующая конъюгат Tn3-каркасной структуры по любому из пп.14-19.
37. Вектор экспрессии, содержащий нуклеиновую кислоту по п.36.
38. Клетка-хозяин для экспрессии конъюгата Tn3-каркасной структуры по любому из пп.14-19,
где клетка-хозяин является одной из: бактериальной клеткой, клеткой COS, клеткой CHO, клеткой 3T3, клеткой NSO или клеткой BHK и
где клетка-хозяин содержит вектор по п.37.
39. Способ получения конъюгата Tn3-каркасной структуры, предусматривающий культивирование клетки-хозяина по п.38 в условиях, при которых экспрессируется конъюгат Tn3-каркасной структуры, кодируемой молекулой нуклеиновой кислоты.
40. Композиция для уменьшения CD40-опосредованного иммунного ответа, содержащая конъюгат Tn3-каркасной структуры по любому из пп.7-24 в количестве, достаточном для достижения терапевтического ответа, и фармацевтически приемлемый наполнитель.
41. Способ лечения, уменьшения интенсивности, управления течением аутоиммунного заболевания у нуждающегося пациента, предусматривающий введение эффективного количества композиции по п.40.
| US 20100298541 A1, 25.11.2010 | |||
| WO 2011051466 A1, 05.05.2011 | |||
| US 20100098730 A1, 22.04.2010 | |||
| US 2008004206 A1, 03.01.2008 | |||
| US 20090176654, 09.07.2009 | |||
| ГУМАНИЗИРОВАННЫЕ АНТИ-CD40-АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2006 |
|
RU2407544C2 |

