ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[1] По настоящей заявке испрашивается приоритет временной заявки на патент США 61/898,571, поданной 1 ноября 2013 года, которая настоящим полностью включена посредством отсылки.
ВКЛЮЧЕНИЕ СПИСКА ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[2] Бумажная копия Списка последовательностей и машиночитаемая форма последовательности, содержащая файл под названием 32559-17_ST25.txt, который имеет размер 46751 байт (согласно измерению в Microsoft WINDOWS® Explorer), представлена в настоящей заявке и включена посредством отсылки. Указанный Список последовательностей состоит из SEQ ID NO: 1-12.
УРОВЕНЬ ТЕХНИКИ
[3] Настоящее описание в целом относится к биосинтезу стевиол-гликозидов. В частности, настоящее описание относится к рекомбинантному полипептиду, который катализирует продукцию стевиол-гликозидов, таких как ребаудиозид D, ребаудиозид E и новый ребаудиозид (ребаудиозид Z).
[4] Стевиол-гликозиды представляют собой природные продукты, выделенные из листьев Stevia rebaudiana, и широко применяются в качестве высокоинтенсивных, низкокалорийных подсластителей. Природные стевиол-гликозиды имеют одинаковую основную структуру (стевиол) и отличаются содержанием углеводных остатков (например, остатки глюкозы, рамнозы и ксилозы) в положениях C13 и C19. Стевиол-гликозиды с известной структурой включают стевиозид, ребаудиозид A, ребаудиозид B, ребаудиозид C, ребаудиозид D, ребаудиозид E, ребаудиозид F и дулькозид A.
[5] В пересчете на сухой вес стевиозид, ребаудиозид A, ребаудиозид C и дулькозид A составляют 9,1, 3,8, 0,6 и 0,3% от общего веса стевиол-гликозидов в листьях, соответственно, в то время как другие стевиол-глюкозиды присутствуют в намного более низком количестве. Экстракты из растения Stevia rebaudiana доступны коммерчески и обычно содержат стевиозид и ребаудиозид в качестве основных соединений. Другие стевиол-гликозиды обычно присутствуют в экстракте стевии в качестве минорных компонентов. Например, количество ребаудиозида A в коммерческих препаратах может изменяться от приблизительно 20% до более чем 90% от общего количества содержащихся стевиол-гликозидов, тогда как количество ребаудиозида B может составлять приблизительно 1-2%, количество ребаудиозида C может составлять приблизительно 7-15%, и количество ребаудиозида D может составлять приблизительно 2% от общего количества стевиол-гликозидов.
[6] Как натуральные подсластители, различные стевиол-глюкозиды имеют различные уровни сладости и послевкусие. Сладость стевиол-гликозидов значительно превышает сладость сахарозы. Например, стевиозид в 100-150 раз слаще сахарозы и имеет горькое послевкусие, тогда как ребаудиозид A и E в 250-450 раз слаще сахарозы и их послевкусие намного лучше, чем у стевиозида. Соответственно, на вкусовой профиль любого экстракта стевии существенно влияет относительное содержание стевиол-гликозидов в экстракте, на которое в свою очередь может влиять источник растения, факторы окружающей среды (такие как состав почвы и климат) и процесс экстракции. В частности, изменения условий экстракции могут привести к непостоянному содержанию стевиол-гликозидов в экстрактах стевии, в результате чего вкусовой профиль различается у разных партий продуктов экстракции. На вкусовой профиль экстрактов стевии также могут влиять содержащиеся в растении примеси (такие как пигменты, липиды, белки, фенольные соединения и сахариды), которые остаются в продукте после процесса экстракции. Такие примеси обычно имеют посторонние привкусы, нежелательные в случае применения экстракта стевии в качестве подсластителя.
[7] Большинство стевиол-гликозидов образуются в результате нескольких реакций гликозилирования стевиола, которые обычно катализируют УДФ-гликозилтрансферазы (УГТ) при использовании уридин-5'-дифосфоглюкозы (УДФ-глюкозы) в качестве донора сахарной группы. В растениях УГТ представляют крайне неоднородную группу ферментов, которые переносят остаток глюкозы от УДФ-глюкозы к стевиолу. Стевиозид является промежуточным соединением в биосинтезе ребаудиозидных соединений. Например, гликозилирование C-3' C-13-O-глюкозы стевиозида дает ребаудиозид A; а гликозилирование C-2' 19-O-глюкозы стевиозида дает ребаудиозид E. Дальнейшее гликозилирование ребаудиозида A (по 19-O-глюкозе) или ребаудиозида E (по C-13-O-глюкозе) дает ребаудиозид D (Фигура 1).
[8] Практический подход к улучшению вкусовых качеств экстракта стевии состоит в повышении выхода ребаудиозидных соединений в результате дальнейшего гликозилирования стевиозида. УГТ с 1,2-19-O-глюкоза гликозилирующей активностью являются важными ферментами для получения ребаудиозида D и E.
[9] Сахарозосинтазы (СС) катализируют превращение УДФ в УДФ-глюкозу в присутствии сахарозы. Таким образом, для реакции гликозилирования, использующей УДФ-глюкозу (такой как реакции, катализируемые УГТ), СС может использоваться для регенерации УДФ-глюкозы из УДФ, повышая эффективность такой реакции (Фигура 2).
[10] Таким образом, существует потребность в стевиол-гликозидах с постоянным вкусовым профилем и менее интенсивным посторонним привкусом, чем у существующих коммерческих продуктов. Как описано в настоящей заявке, в настоящем описании предложен рекомбинантный полипептид, который может применяться для получения стевиол-гликозидов (таких как ребаудиозид D и ребаудиозид E). В настоящем описании также предложен способ получения композиции стевиол-гликозида (ребаудиозида Z) с применением такого рекомбинантного полипептида.
КРАТКОЕ ОПИСАНИЕ
[11] Рассматриваемая технология в общем относится к рекомбинантным полипептидам, которые обладают активностями УДФ-гликозилтрансферазы. В частности, предложены полипептиды, обладающие 1,2-19-O-глюкоза-гликозилирующей активностью в отношении стевиол-гликозидных соединений. В одном аспекте рассматриваемая технология относится к рекомбинантному полипептиду, включающему аминокислотную последовательность, обладающую по меньшей мере 80% идентичностью с SEQ ID NO:6. В примере осуществления аминокислотная последовательность рекомбинантного полипептида, описанного в настоящей заявке, обладает по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или даже 100% идентичностью с SEQ ID NO:6.
[12] В другом аспекте рассматриваемая технология относится к выделенной нуклеиновой кислоте, включающей нуклеотидную последовательность, кодирующую рекомбинантный полипептид, описанный в настоящей заявке. В другом аспекте рассматриваемая технология относится к вектору, включающему нуклеиновую кислоту, описанную в настоящей заявке, и клетке-хозяину, включающей вектор, описанный в настоящей заявке. В примере осуществления клетка-хозяин рассматриваемой технологии выбрана из группы, состоящей из клетки бактерий, дрожжей, нитчатых грибов, цианобактерий и растений.
[13] В другом аспекте рассматриваемая технология также относится к способу получения композиции стевиол-гликозида, включающему инкубирование субстрата (такого как стевиозид, ребаудиозид A, ребаудиозид E или их комбинация) с рекомбинантным полипептидом, включающим аминокислотную последовательность, обладающую по меньшей мере 80% идентичностью с SEQ ID NO:6. В примере осуществления аминокислотная последовательность рекомбинантного полипептида, применяемого в способе, описанном в настоящей заявке, обладает по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или даже 100% идентичностью с SEQ ID NO:6.
[14] В другом аспекте рассматриваемая технология также относится к способу получения композиции стевиол-гликозида, включающему инкубирование субстрата (такого как стевиозид и ребаудиозид E) с рекомбинантным полипептидом, включающим аминокислотную последовательность, обладающую по меньшей мере 80% идентичностью с SEQ ID NO:11. В примере осуществления аминокислотная последовательность рекомбинантного полипептида, применяемого в способе, описанном в настоящей заявке, обладает по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или даже 100% идентичностью с SEQ ID NO:11.
[15] В одном варианте осуществления способ дополнительно включает инкубирование рекомбинантной сахарозосинтазы (такой как сахарозосинтаза с аминокислотной последовательностью, обладающей по меньшей мере 80% идентичностью с аминокислотной последовательностью AtSUS1, представленной в SEQ ID NO:9) с субстратом и рекомбинантным полипептидом, описанным в настоящей заявке. В другом варианте осуществления способ дополнительно включает инкубирование рекомбинантной УДФ-гликозилтрансферазы (такой как УДФ-гликозилтрансфераза с аминокислотной последовательностью, обладающей по меньшей мере 80% идентичностью с аминокислотной последовательностью UGT76G1, представленной в SEQ ID NO:11) с рекомбинантной сахарозосинтазой, субстратом и рекомбинантным полипептидом, описанным в настоящей заявке. В другом варианте осуществления способ, описанный в настоящей заявке, включает инкубирование субстрата с клеткой-хозяином, экспрессирующей рекомбинантный полипептид.
[16] Рассматриваемая технология также относится к новому стевиол-гликозиду, называемому ребаудиозид Z, который отличается временем удержания приблизительно 6,68 минут на ВЭЖХ при условиях, описанных в настоящей заявке. Рассматриваемая технология также относится к способу получения ребаудиозида Z, описанного в настоящей заявке, включающему инкубирование субстрата с рекомбинантным полипептидом, включающим аминокислотную последовательность, обладающую по меньшей мере 80% идентичностью с SEQ ID NO:6. При использовании в настоящем описании, термины "ребаудиозид Z" или "Reb Z" относятся к смеси соединений, в частности смеси ребаудиозида Z1 ("Reb Z1") и ребаудиозида Z2 ("Reb Z2").
[17] В одном варианте осуществления настоящее описание также направлено на способ синтеза ребаудиозида Z из ребаудиозида E. Способ включает: получение реакционной смеси, включающей ребаудиозид E, субстрат, выбранный из группы, состоящей из сахарозы, уридиндифосфата (УДФ) и уридиндифосфат-глюкозы (УДФ-глюкозы), и HV1, и сахарозосинтазу, инкубирование реакционной смеси в течение времени, достаточного для получения ребаудиозида Z, где глюкоза ковалентно присоединяется к ребаудиозиду E с получением ребаудиозида Z, глюкоза ковалентно присоединяется к C2'-13-O-глюкозе ребаудиозида E с получением ребаудиозида Z1, и глюкоза ковалентно присоединяется к C2'-19-O-глюкозе ребаудиозида E с получением ребаудиозида Z2.
[18] В одном варианте осуществления соединение ребаудиозид Z является ребаудиозидом Z1 (Reb Z1), имеющим следующую структуру:
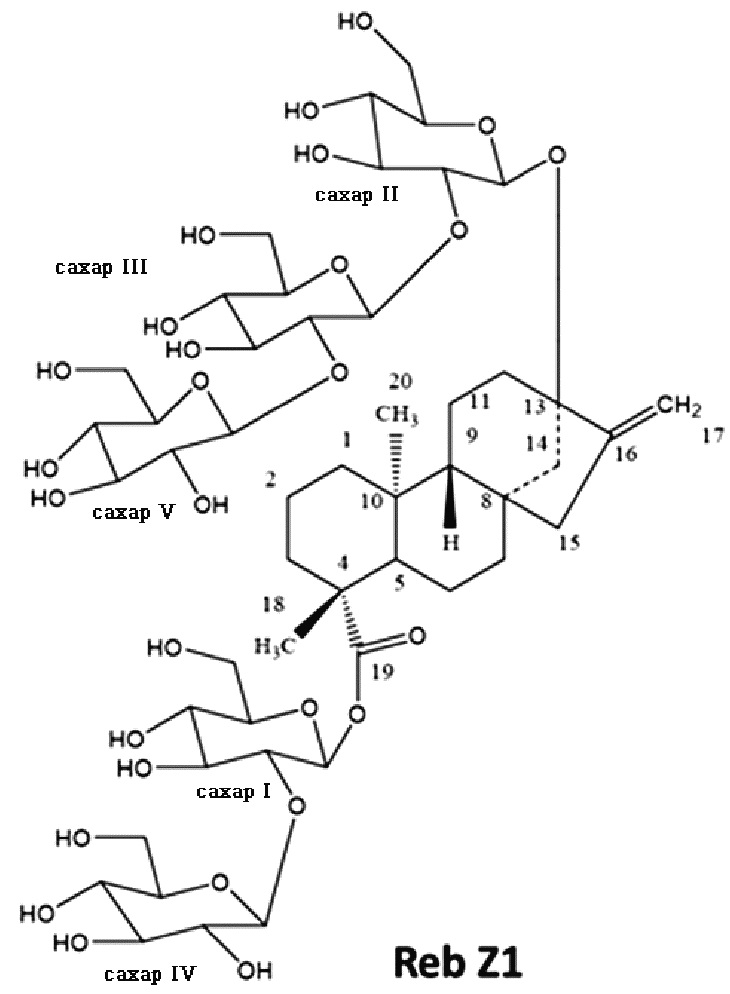 .
.
[19] В одном варианте осуществления соединение ребаудиозид Z является ребаудиозидом Z2 (Reb Z2), имеющим следующую структуру:
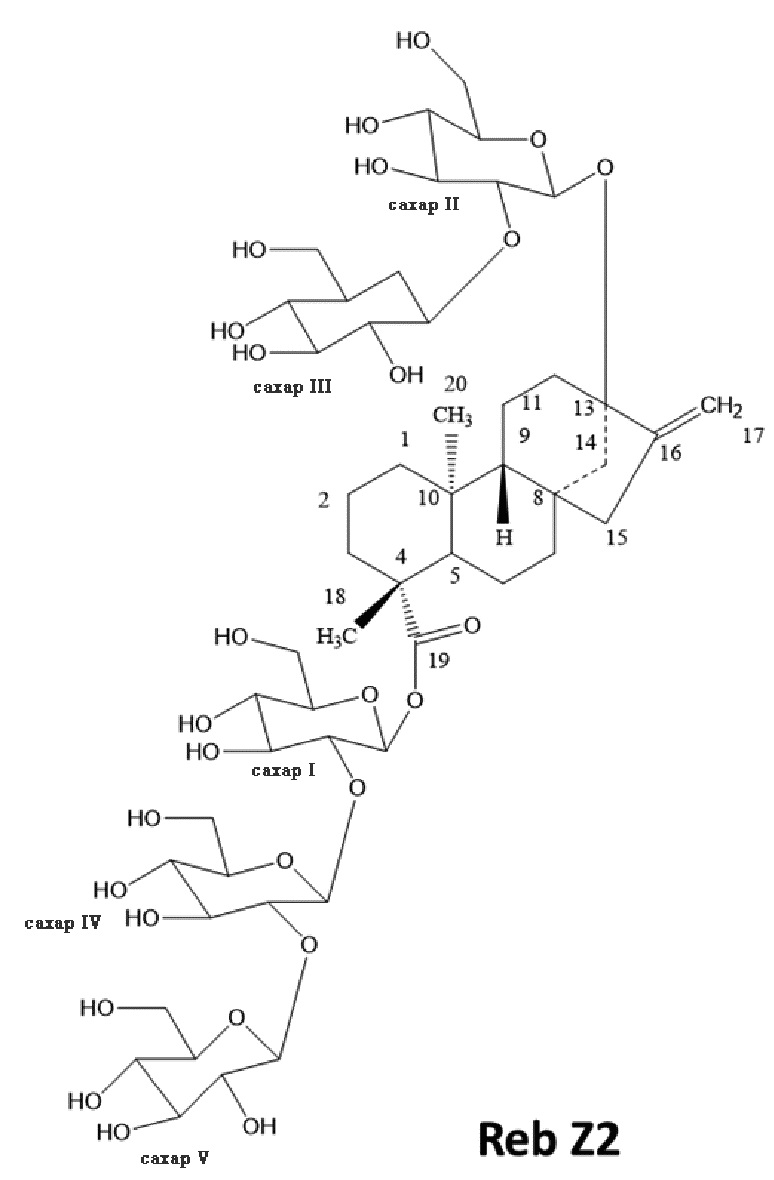 .
.
[20] Как описано в настоящей заявке, рекомбинантные полипептиды согласно настоящей технологии могут использоваться для разработки биосинтетического способа получения стевиол-гликозидов, которые обычно мало распространены в природных источниках, например, ребаудиозида D и ребаудиозида E. Таким образом, настоящая технология также обеспечивает композицию стевиол-гликозида, полученную биосинтетическим способом, описанным в настоящей заявке. Такая композиция может включать стевиол-гликозидное соединение, выбранное из группы, состоящей из ребаудиозида D, ребаудиозида E, нового ребаудиозида (указанного в настоящей заявке как "ребаудиозид Z" и "Reb Z") и их комбинаций. Кроме того, также предложен подсластитель, включающий композицию стевиол-гликозидов, описанную в настоящей заявке.
[21] В одном варианте осуществления настоящее описание направлено на подсластитель, включающий соединение, имеющее следующую химическую структуру:
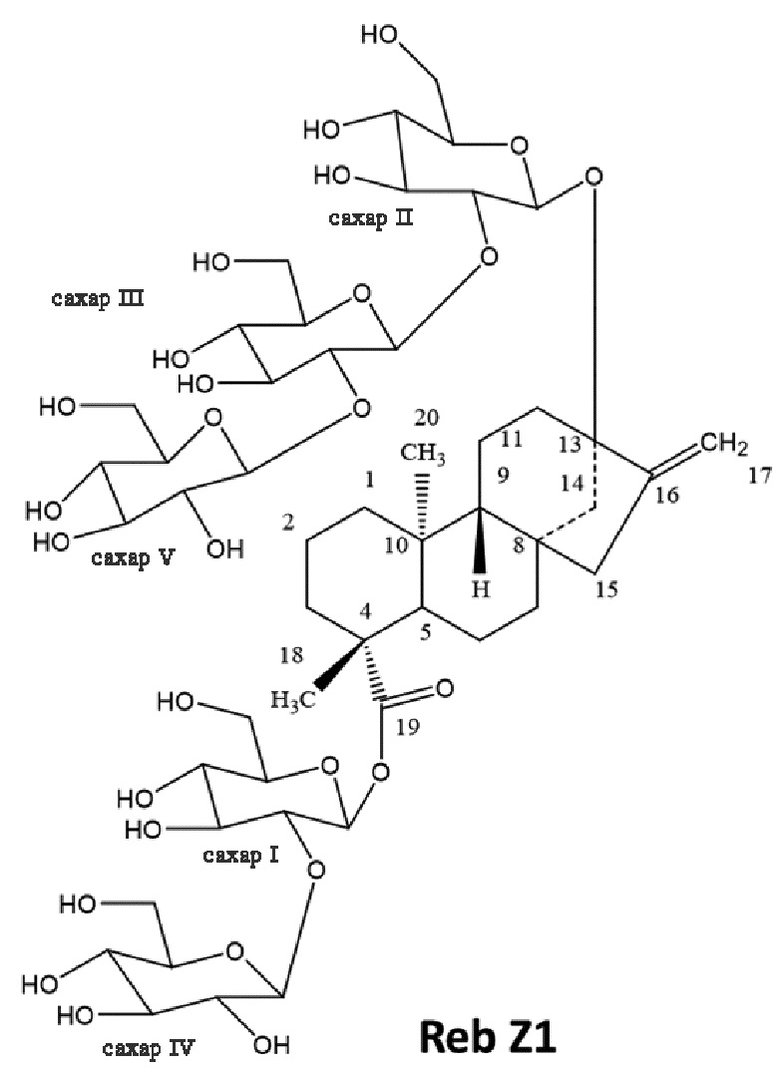 .
.
[22] В другом варианте осуществления подсластитель включает соединение, имеющее следующую химическую структуру:
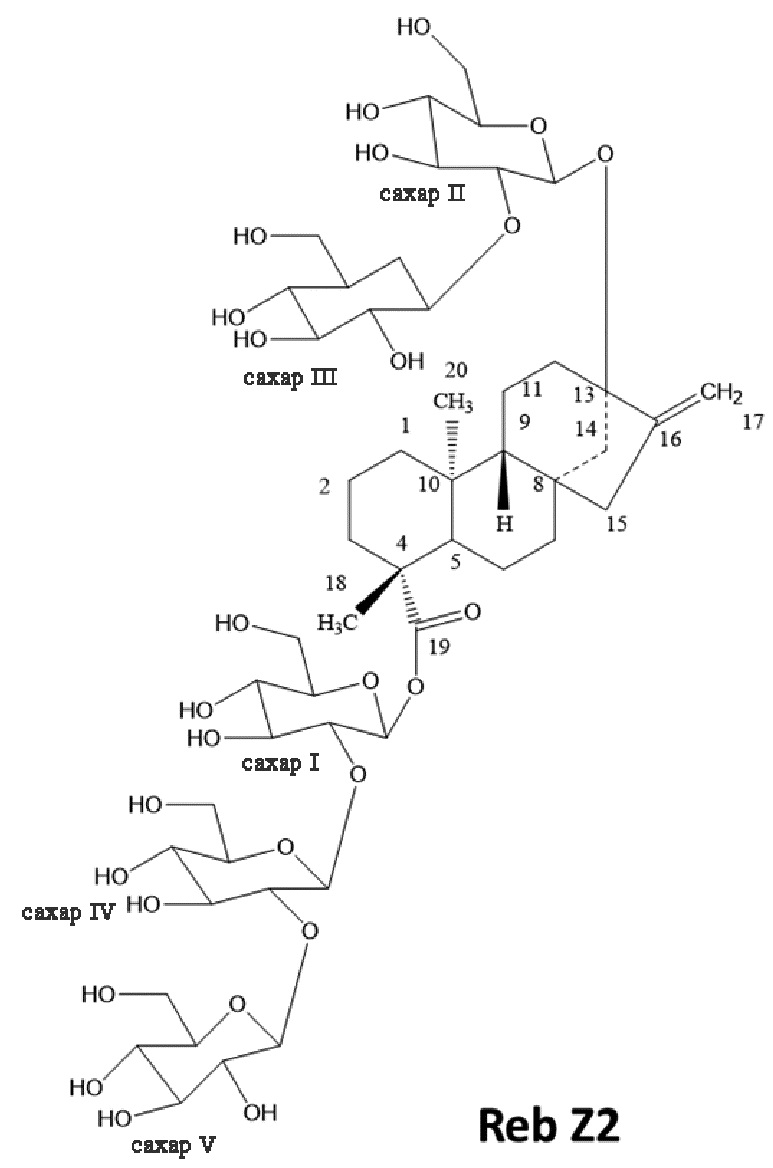 .
.
[23] Настоящее описание также направлено на применение подсластителей в потребительских продуктах, таких как напитки, кондитерские изделия, хлебобулочные изделия, печенье и жевательные резинки.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[24] Описание будет более понятным, и особенности, аспекты и преимущества, помимо представленных выше, станут ясны при рассмотрении их подробного описания, следующего далее. Такое подробное описание ссылается на следующие чертежи, где:
[25] Фигура 1 является схемой, на которой показаны пути биосинтеза стевиол-гликозида из стевиозида. Как описано в настоящей заявке, рекомбинантный полипептид HV1 ("HV1") содержит 1,2-19-O-глюкоза-гликозилирующую активность, которая переносит вторую сахарную группу на C-2' 19-O-глюкозы стевиозида с получением ребаудиозида E ("Reb E") или, аналогично, с получением ребаудиозида D ("Reb D") из ребаудиозида A ("Reb A"). На Фигуре 1 также показано, что рекомбинантный фермент UGT76G1 ("UGT76G1", отличающийся от рекомбинантного полипептида HV1) катализирует реакцию, которая переносит сахарную группу на C-3' C-13-O-глюкозы стевиозида с получением ребаудиозида A или, аналогично, с получением ребаудиозида D из ребаудиозида E.
[26] На Фигуре 2 показана примерная схема комбинированной реакционной системы УДФ-гликозилтрансферазы ("УГТ") и сахарозосинтазы ("СС"). В Реакции 1 показана катализируемая УГТ реакция, превращающая ребаудиозид A ("Reb A") в ребаудиозид D ("Reb D"), в которой УДФ-глюкоза используется в качестве донора глюкозы и приводит к получению УДФ. В Реакции 2 показана катализируемая СС реакция, превращающая УДФ в УДФ-глюкозу, в которой сахароза используется в качестве донора глюкозы. В Реакции 2 также показано, что катализируемая СС реакция может быть объединена с реакцией, катализируемой УГТ.
[27] На Фигуре 3 показано in vitro получение ребаудиозида D ("Reb D") из ребаудиозида ("Reb A"), катализируемое рекомбинантным полипептидом HV1 (SEQ ID NO:6) и рекомбинантной AtSUS1 (SEQ ID NO:9) в комбинированной реакционной системе HV1-AtSUS1, как описано в настоящей заявке. На Фигуре 3A показаны стандарты стевиозида ("Ste"), Ребаудиозида A ("Reb A") и Ребаудиозида D ("Reb D"). Результаты на 6, 9, 12 и 24 часа показаны на Фигурах 3B-E, соответственно. Результаты реакции без рекомбинантной AtSUS1 (то есть некомбинированной реакции) на 12 и 24 часа показаны на Фигурах 3F и 3G, соответственно.
[28] На Фигуре 4 показано in vitro получение ребаудиозида E ("Reb E") из стевиозида, катализируемое рекомбинантным полипептидом HV1 (SEQ ID NO:6) и рекомбинантной AtSUS1 (SEQ ID NO:9) в комбинированной реакционной системе HV1-AtSUS1, как описано в настоящей заявке. На Фигуре 4A показаны стандарты стевиозида ("Ste"), Ребаудиозида A ("Reb A") и Ребаудиозида D ("Reb D"). Результат на 20 часов показан на Фигурах 4B, который включает соединение ребаудиозид Z ("Reb Z").
[29] На Фигуре 5 показано in vitro получение ребаудиозида D ("Reb D") из стевиозида, катализируемое комбинацией рекомбинантного полипептида HV1 (SEQ ID NO:6), рекомбинантной UGT76G1 (SEQ ID NO:11) и рекомбинантной AtSUS1 (SEQ ID NO:9). На Фигуре 5A показаны стандарты стевиозида ("Ste"), Ребаудиозида A ("Reb A") и Ребаудиозида D ("Reb D"). Результаты на 6, 9, 12 и 24 часов показаны на Фигурах 5B-E, соответственно.
[30] На Фигуре 6 показан электрофоретический анализ в ДСН-ПААГ очищенного рекомбинантного полипептида HV1.
[31] На Фигуре 7 показан электрофоретический анализ в ДСН-ПААГ очищенного рекомбинантного полипептида AtSUS1.
[32] На Фигуре 8 показано in vitro получение ребаудиозида Z ("Reb Z") из ребаудиозида E ("Reb E"), катализируемое рекомбинантным полипептидом HV1 (SEQ ID NO:6) и рекомбинантной AtSUS1 (SEQ ID NO:9) в комбинированной реакционной системе HV1-AtSUS1, как описано в настоящей заявке. На Фигуре 8A показаны стандарты ребаудиозида E ("Reb E"). Результат на 24 часа показан на Фигурах 8B, который включает соединение ребаудиозид Z ("Reb Z").
[33] На Фигуре 9 показан электрофоретический анализ в ДСН-ПААГ очищенного рекомбинантного полипептида UGT76G1.
[34] Фигура 10: Структуры Ребаудиозида Z (включая Reb Z1 и Reb Z2) и Ребаудиозида E.
[35] Фигура 11: Ключевые TOCSY и HMBC корреляции Reb Z1 и Reb Z2.
[36] Фигура 12: показано in vitro получение ребаудиозида D ("Reb D") из ребаудиозида E ("Reb E"), катализируемое рекомбинантной UGT76G1 (SEQ ID NO:11). На Фигуре 12A-12B показаны стандарты Ребаудиозида E ("Reb E"), Ребаудиозида D ("Reb D"). Показаны результаты реакции без рекомбинантной AtSUS1 (Фигура 12C) (то есть некомбинированной реакции) и с рекомбинантной AtSUS1 (Фигура 12D) (то есть комбинированной УГТ-СС реакции) на 6 часов, соответственно.
[37] Хотя в отношении настоящего описания допускаются различные модификации и альтернативные формы, определенные варианты его осуществления показаны в качестве примера на чертежах и подробно описаны ниже. Впрочем, следует понимать, что описание конкретных вариантов осуществления не должно ограничивать описание и охватывает все модификации, эквиваленты и альтернативы, включенные в сущность и объем описания, как определено прилагаемой формулой изобретения.
ПОДРОБНОЕ ОПИСАНИЕ
[38] Рассматриваемая технология обеспечивает рекомбинантный полипептид, который обладает УДФ-гликозилтрансферазной активностью, такой как 1,2-19-O-глюкоза-гликозилирующая активность и 1,2-13-O-глюкоза-гликозилирующая активность, для синтеза стевиол-гликозидов. Рекомбинантный полипептид рассматриваемой технологии (который в дальнейшем может также именоваться как "рекомбинантный полипептид HV1") может применяться для биосинтеза стевиол-гликозидных соединений. В настоящем описании УДФ-гликозилтрансфераза (УГТ) относится к ферменту, который переносит остаток сахара с активированной донорной молекулы (обычно УДФ-глюкозы) на акцепторную молекулу. 1,2-19-O-глюкоза-гликозилирующая активность относится к ферментной активности, которая переносит сахарную группу на C-2' 19-O-глюкозной группы стевиозида, ребаудиозида A или ребаудиозида E (Фигура 1 и Фигура 10). 1,2-13-O-глюкоза-гликозилирующая активность относится к ферментной активности, которая переносит сахарную группу на C-2' 13-O-глюкозной группы ребаудиозида E (Фигура 10).
[39] Названия УГТ ферментов, используемые в настоящем описании, соответствуют системе номенклатуры, принятой Комитетом по номенклатуре УГТ (Mackenzie et al., "The UDP glycosyltransferase gene super family: recommended nomenclature updated based on evolutionary divergence", Pharmacogenetics, 1997, vol. 7, pp. 255-269), которая классифицирует гены УГТ комбинацией номера семейства, буквы, обозначающей подсемейство, и номера индивидуального гена. Например, название "UGT76G1" соответствует ферменту УГТ, кодируемому геном, который относится к семейству УГТ номер 76 (растительного происхождения), подсемейству G и гену номер 1.
[40] В растениях существует большое семейство генов УГТ. Впрочем, биологические функции большинства этих УГТ остаются неизвестными.
Определения
[41] При использовании в настоящем описании, формы единственного числа включают формы множественного числа, если из текста прямо не следует иное.
[42] В тех случаях, когда термин "включает", "содержит" или подобный используется в описании или формуле изобретения, предполагается, что такой термин является включительным так же, как термин "включать", при интерпретации "включать" при использовании в качестве переходного слова в формуле изобретения.
[43] Слово "примерный" используется в настоящем описании как "служащий в качестве примера, образца или иллюстрации". Любой вариант осуществления, описанный в настоящей заявке как "примерный", не должен обязательно рассматриваться как предпочтительный или преимущественный в отношении других вариантов осуществления.
[44] Термин "комплементарный" должен иметь свое обычное и общепринятое значение, известное среднему специалисту в данной области техники, и использоваться без ограничения для описания отношения между нуклеотидными основаниями, которые способны к гибридизации друг с другом. Например, в отношении ДНК, аденозин комплементарен тимину, а цитозин комплементарен гуанину. Таким образом, рассматриваемая технология также включает выделенные фрагменты нуклеиновых кислот, которые комплементарны полным последовательностям, представленным в сопровождающем Списке последовательностей, а также те, которые по существу подобны последовательностям нуклеиновых кислот.
[45] Термины "нуклеиновая кислота" и "нуклеотид" должны иметь свои соответствующие обычные и общепринятые значения, известные среднему специалисту в данной области техники, и использоваться без ограничения для обозначения дезоксирибонуклеотидов или рибонуклеотидов и их полимеров в одно- или в двухцепочечной форме. Если нет специальных ограничений, термин охватывает нуклеиновые кислоты, содержащие известные аналоги природных нуклеотидов, которые обладают подобными характеристиками связывания, как и референсная нуклеиновая кислота, и метаболизируются аналогично природным нуклеотидам. Если не указано иное, конкретная последовательность нуклеиновой кислоты также в подразумеваемой форме охватывает соответствующие консервативно модифицированные или вырожденные варианты (например, замены вырожденных кодонов) и комплементарные последовательности, а также последовательность, указанную в явной форме.
[46] Термин "выделенный" должен иметь свое обычное и общепринятое значение, известное среднему специалисту в данной области техники, и при использовании в отношении выделенной нуклеиновой кислоты или выделенного полипептида должен использоваться без ограничения для обозначения нуклеиновой кислоты или полипептида, которые, благодаря человеку, существуют отдельно от своего нативного окружения и поэтому не являются продуктом природы. Выделенная нуклеиновая кислота или полипептид могут существовать в очищенной форме или могут существовать в ненативном окружении, в таком как, например, трансгенная клетка-хозяин.
[47] Термин "инкубирование", при использовании в настоящем описании, означает процесс смешивания двух или более химических или биологических единиц (таких как химическое соединение и фермент) и обеспечения их взаимодействия при условиях, благоприятных для получения композиции стевиол-гликозидов.
[48] Термин "вырожденный вариант" относится к последовательности нуклеиновой кислоты, имеющей последовательность остатков, которая отличается от референсной последовательности нуклеиновой кислоты на одну или более вырожденных замен кодонов. Вырожденные замены кодонов могут быть получены при создании последовательностей, в которых третье положение одного или более выбранных (или всех) кодонов заменено смешанным основанием и/или остатками дезоксиинозина. Последовательность нуклеиновой кислоты и все ее вырожденные варианты будут экспрессировать ту же аминокислоту или полипептид.
[49] Термины "полипептид", "белок" и "пептид" должны иметь свое обычное и общепринятое значение, известное среднему специалисту в данной области техники; эти три термина иногда используются попеременно и используются без ограничения для обозначения полимера из аминокислот или аналогов аминокислот, независимо от его размера или функции. Хотя "белок" часто используется в отношении относительно крупных полипептидов, а "пептид" часто используется в отношении малых полипептидов, применение данных терминов в уровне техники совпадает и изменяется. Термин "полипептид", при использовании в настоящем описании, относится к пептидам, полипептидам и белкам, если не отмечено иное. Термины "белок", "полипептид" и "пептид" используются в настоящем описании попеременно при обозначении продукта полинуклеотида. Таким образом, примерные полипептиды включают продукты полинуклеотидов, природные белки, гомологи, ортологи, паралоги, фрагменты и другие эквиваленты, варианты и аналоги предыдущего.
[50] Термины "полипептидный фрагмент" и "фрагмент", при использовании в отношении референсного полипептида, должны иметь свои обычные и общепринятые значения, известные среднему специалисту в данной области техники, и использоваться без ограничения для обозначения полипептида, в котором аминокислотные остатки удалены по сравнению с референсным полипептидом непосредственно, но где остальная аминокислотная последовательность обычно идентична соответствующим положениям в референсном полипептиде. Такие делеции могут происходить на N-конце или C-конце референсного полипептида или, в альтернативном варианте, на обоих концах.
[51] Термин "функциональный фрагмент" полипептида или белка относится к пептидному фрагменту, который является частью полноразмерного полипептида или белка и обладает по существу такой же биологической активностью или имеет по существу такую же функцию, что и полноразмерный полипептид или белок (например, выполняет такую же ферментативную реакцию).
[52] Термины "вариант полипептида", "модифицированная аминокислотная последовательность" или "модифицированный полипептид", которые используются попеременно, относятся к аминокислотной последовательности, которая отличается от референсного полипептида одной или более аминокислотами, например, одной или более аминокислотными заменами, делециями и/или добавлениями. В одном из аспектов вариант является "функциональным вариантом", который сохраняет некоторые или все свойства референсного полипептида.
[53] Термин "функциональный вариант" также включает консервативно замененные варианты. Термин "консервативно замененный вариант" относится к пептиду, имеющему аминокислотную последовательность, которая отличается от референсного пептида одной или более консервативными аминокислотными заменами и сохраняет некоторую часть или всю активность референсного пептида. "Консервативная аминокислотная замена" является заменой остатка аминокислоты функционально подобным остатком. Примеры консервативных замен включают замену одного неполярного (гидрофобного) остатка, такого как изолейцин, валин, лейцин или метионин, другим; замену одного заряженного или полярного (гидрофильного) остатка другим, например, между аргинином и лизином, между глутамином и аспарагином, между треонином и серином; замену одного основного остатка, такого как лизин или аргинин, другим; или замену одного кислотного остатка, такого как аспарагиновая кислота или глутаминовая кислота, другим; или замену одного ароматического остатка, такого как фенилаланин, тирозин или триптофан, другим. Такие замены, как ожидают, будут оказывать малое или не оказывать никакого воздействия на кажущуюся молекулярную массу или изоэлектрическую точку белка или полипептида. Фраза "консервативно замененный вариант" также включает пептиды, в которых остаток заменен химически дериватизированным остатком, при условии, что полученный в результате пептид сохраняет некоторую часть или всю активность референсного пептида, как описано в настоящей заявке.
[54] Термин "вариант", применительно к полипептидам рассматриваемой технологии, также включает функционально активный полипептид, имеющий аминокислотную последовательность, которая по меньшей мере на 75%, по меньшей мере на 76%, по меньшей мере на 77%, по меньшей мере на 78%, по меньшей мере на 79%, по меньшей мере на 80%, по меньшей мере на 81%, по меньшей мере на 82%, по меньшей мере на 83%, по меньшей мере на 84%, по меньшей мере на 85%, по меньшей мере на 86%, по меньшей мере на 87%, по меньшей мере на 88%, по меньшей мере на 89%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% и даже на 100% идентична аминокислотной последовательности референсного полипептида.
[55] Термин "гомологичный", во всех его грамматических формах и вариантах написания, относится к зависимости между полинуклеотидами или полипептидами, которые обладают "общим эволюционным происхождением", включая полинуклеотиды или полипептиды из суперсемейств, а также гомологичные полинуклеотиды или белки из различных биологических видов (Reeck и другие, Клетка 50:667, 1987). Такие полинуклеотиды или полипептиды обладают гомологией последовательности, что отражается подобием их последовательностей, либо в процентах идентичности, либо в присутствии определенных аминокислот или мотивов в консервативных положениях. Например, два гомологичных полипептида могут иметь аминокислотные последовательности, которые являются по меньшей мере на 75%, по меньшей мере на 76%, по меньшей мере на 77%, по меньшей мере на 78%, по меньшей мере на 79%, по меньшей мере на 80%, по меньшей мере на 81%, по меньшей мере на 82%, по меньшей мере на 83%, по меньшей мере на 84%, по меньшей мере на 85%, по меньшей мере на 86%, по меньшей мере на 87%, по меньшей мере на 88%, по меньшей мере на 89%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% и даже на 100% идентичными.
[56] "Процент (%) идентичности аминокислотных последовательностей" в отношении последовательностей различных полипептидов рассматриваемой технологии определяется как процент аминокислотных остатков в кандидатной последовательности, которые идентичны аминокислотным остаткам референсного полипептида (такого как, например, SEQ ID NO:6), после выравнивания последовательностей и введения пропусков, при необходимости, для достижения максимального процента идентичности последовательностей, без учета каких-либо консервативных замен в качестве части идентичности последовательности.
[57] Выравнивание в целях определения процента идентичности аминокислотных последовательностей может быть выполнено различными способами, которые известны в уровне техники, например, при использовании общедоступных программ, таких как программы BLAST, BLAST-2, ALIGN, ALIGN-2 или Megalign (DNASTAR). Специалисты в данной области техники сумеют определить подходящие параметры для измерения выравнивания, включая любые алгоритмы, требуемые для получения максимального выравнивания по всей длине сравниваемых последовательностей. Например, % идентичности аминокислотных последовательностей можно определить при использовании программы сравнения последовательностей NCBI-BLAST2. Программа сравнения последовательностей NCBI-BLAST2 может быть загружена с сайта ncbi.nlm.nih.gov. В NCBI BLAST2 используется несколько параметров поиска, где все параметры поиска установлены со значениями по умолчанию, включая, например, unmask yes, strand=all, expected occurrences 10, minimum low complexity length=15/5, multi-pass e-value=0.01, constant for multi-pass=25, dropoff for final gapped alignment=25 и scoring matrix=BLOSUM62. В случаях использования NCBI-BLAST2 для сравнения аминокислотных последовательностей, % идентичности аминокислотных последовательностей для данной аминокислотной последовательности A и данной аминокислотной последовательности B (что можно альтернативно сформулировать как данная аминокислотная последовательность A, которая имеет или включает некоторый % идентичности аминокислотной последовательности по отношению к данной аминокислотной последовательности B) вычисляют следующим образом: 100 умножить на частное X/Y, где X является количеством аминокислотных остатков, оцененных как идентичные совпадения программой выравнивания последовательностей NCBI-BLAST2 при выравнивании с помощью этой программы последовательностей A и B, и где Y является общим количеством аминокислотных остатков в B. Следует понимать, что если длина аминокислотной последовательности A не равна длине аминокислотной последовательности B, % идентичности аминокислотных последовательностей A и B не будет равен % идентичности аминокислотных последовательностей B и A.
[58] В этом смысле методики определения "подобия" аминокислотных последовательностей известны в уровне техники. В общем, "подобие" означает точное сравнение аминокислоты с аминокислотой в двух или более полипептидах в подходящем положении, где аминокислоты идентичны или обладают подобными химическими и/или физическими свойствами, такими как заряд или гидрофобность. При этом между сравниваемыми полипептидными последовательностями может быть определен так называемый "процент подобия". Методики определения идентичности последовательностей нуклеиновых кислот и аминокислот также известны в уровне техники и включают определение нуклеотидной последовательности мРНК для данного гена (обычно через промежуточную кДНК) и определение аминокислотной последовательности, кодируемой в ней, и ее сравнение со второй аминокислотной последовательностью. В общем, "идентичность" относится к точному соответствию нуклеотида с нуклеотидом или аминокислоты с аминокислотой в двух полинуклеотидных или полипептидных последовательностях, соответственно. Две или более полинуклеотидных последовательности можно сравнить при определении их "процента идентичности", как и в случае двух или более аминокислотных последовательностей. Программы, доступные в пакете Wisconsin Sequence Analysis Package, Version 8 (доступном от Genetics Computer Group, Madison, Wis.), например программа GAP, позволяют вычислять идентичность между двумя полинуклеотидами и идентичность и подобие между двумя полипептидными последовательностями, соответственно. Другие программы для вычисления идентичности или подобия между последовательностями известны специалистам в данной области.
[59] Положение аминокислоты, "соответствующее" референсному положению, является положением, которое выравнивается с референсной последовательностью, как идентифицируют при выравнивании аминокислотных последовательностей. Такие выравнивания могут быть выполнены вручную или при использовании известных программ выравнивания последовательностей, таких как ClustalW2, Blast 2 и т.д.
[60] Если не определено иное, процент идентичности двух полипептидных или полинуклеотидных последовательностей именуется как процент идентичных аминокислотных или нуклеотидных остатков по всей длине более короткой из двух данных последовательностей.
[61] "Кодирующая последовательность" должна иметь свое обычное и общепринятое значение, известное среднему специалисту в данной области техники, и использоваться без ограничения для обозначения последовательности ДНК, которая кодирует определенную аминокислотную последовательность.
[62] "Подходящие регуляторные последовательности" должны иметь свое обычное и общепринятое значение, известное среднему специалисту в данной области техники, и используются без ограничения для обозначения нуклеотидных последовательностей, которые расположены перед (5' некодирующие последовательности), в или после (3' некодирующие последовательности) кодирующей последовательности, и которые влияют на транскрипцию, процессинг РНК или стабильность, или трансляцию связанной кодирующей последовательности. Регуляторные последовательности могут включать промоторы, лидерные последовательности трансляции, интроны и последовательности распознавания полиаденилирования.
[63] "Промотор" должен иметь свое обычное и общепринятое значение, известное среднему специалисту в данной области техники, и используется без ограничения для обозначения последовательности ДНК, способной регулировать экспрессию кодирующей последовательности или функциональной РНК. В общем, кодирующая последовательность расположена 3' относительно последовательности промотора. Промоторы могут быть получены полностью из нативного гена или собраны из различных элементов, полученных из различных промоторов, существующих в природе, или даже могут включать синтетические сегменты ДНК. Специалистам в данной области техники известно, что различные промоторы могут направлять экспрессию гена в различных тканях или типах клеток, или на различных стадиях развития, или в ответ на различные условия окружающей среды. Промоторы, которые вызывают экспрессию гена в большинстве типов клеток большую часть времени, обычно называются "конститутивными промоторами". Также известно, что поскольку в большинстве случаев точные границы регуляторных последовательностей не были полностью определены, фрагменты ДНК различной длины могут обладать идентичной промоторной активностью.
[64] Термин "функционально связанный" относится к ассоциации последовательностей нуклеиновых кислот на одном фрагменте нуклеиновой кислоты, в результате чего на функцию одной из последовательностей влияет другая. Например, промотор функционально связан с кодирующей последовательностью, когда он способен воздействовать на экспрессию данной кодирующей последовательности (то есть кодирующая последовательность находится под транскрипционным контролем промотора). Кодирующие последовательности могут быть функционально связаны с регуляторными последовательностями в смысловой или антисмысловой ориентации.
[65] Термин "экспрессия", при использовании в настоящем описании, должен иметь свое обычное и общепринятое значение, известное среднему специалисту в данной области техники, и используется без ограничения для обозначения транскрипции и устойчивого накопления смысловой (мРНК) или антисмысловой РНК, полученной из фрагмента нуклеиновой кислоты рассматриваемой технологии. "Оверэкспрессия" относится к синтезу продукта гена в трансгенных или рекомбинантных организмах, уровни продукции которого превышают уровни продукции в нормальных или нетрансформированных организмах.
[66] "Трансформация" должна иметь свое обычное и общепринятое значение, известное среднему специалисту в данной области техники, и используется без ограничения для обозначения переноса полинуклеотида в клетку-мишень. Перенесенный полинуклеотид может быть включен в геном или хромосомную ДНК клетки-мишени, что приводит к генетически стабильному наследованию, или он может реплицироваться независимо от хромосом хозяина. Организмы-хозяева, содержащие трансформированные фрагменты нуклеиновых кислот, называются "трансгенными" или "рекомбинантными", или "трансформированными" организмами.
[67] Термины "трансформированный", "трансгенный" и "рекомбинантный", при использовании в настоящем описании в отношении клеток-хозяев, должны иметь свои соответствующие обычные и общепринятые значения, известные среднему специалисту в данной области техники, и используются без ограничения для обозначения клетки организма-хозяина, такой как растительная или микробная клетка, в которую была введена гетерологичная молекула нуклеиновой кислоты. Молекула нуклеиновой кислоты может быть стабильно интегрирована в геном клетки-хозяина, или молекула нуклеиновой кислоты может присутствовать как внехромосомная молекула. Такая внехромосомная молекула может быть самореплицирующейся. Трансформированные клетки, ткани или субъекты, как понимают, охватывают не только конечный продукт процесса трансформации, но и его трансгенное потомство.
[68] Термины "рекомбинантный", "гетерологичный" и "экзогенный", при использовании в настоящем описании в отношении полинуклеотидов, должны иметь свои обычные и общепринятые значения, известные среднему специалисту в данной области техники, и используются без ограничения для обозначения полинуклеотида (например, последовательности ДНК или гена), который происходит из источника, чужеродного для конкретной клетки-хозяина или, если он происходит из того же источника, изменен относительно своей исходной формы. Таким образом, гетерологичный ген в клетке-хозяине включает ген, который является эндогенным по отношению к конкретной клетке-хозяину, но был изменен, например, при использовании сайт-направленного мутагенеза или других рекомбинантных технологий. Указанные термины также включают не встречающиеся в природе множественные копии природной последовательности ДНК. Таким образом, термины относятся к сегменту ДНК, который является чужеродным или гетерологичным по отношению к клетке или гомологичным по отношению к клетке, но находящимся в таком положении или форме в клетке-хозяине, в котором данный элемент обычно не находится.
[69] Аналогичным образом термины "рекомбинантный", "гетерологичный" и "экзогенный", при использовании в настоящем описании в отношении полипептидной или аминокислотной последовательности, означают полипептидную или аминокислотную последовательность, которая происходит из источника, чужеродного по отношению к конкретной клетке-хозяину или, если она происходит из того же источника, изменена относительно своей исходной формы. Таким образом, рекомбинантные сегменты ДНК могут экспрессироваться в клетке-хозяине с получением рекомбинантного полипептида.
[70] Термины "плазмида", "вектор" и "кассета" должны иметь свои соответствующие обычные и общепринятые значения, известные среднему специалисту в данной области техники, и используются без ограничения для обозначения внехромосомного элемента, часто несущего гены, которые не являются частью центрального метаболизма клетки, и который обычно находится в форме кольцевых молекул двухцепочечной ДНК. Такие элементы могут быть автономно реплицирующимися последовательностями, интегрирующимися в геном последовательностями, фаговыми или нуклеотидными последовательностями, линейными или кольцевыми, одно- или двухцепочечными ДНК или РНК, полученными из любого источника, в котором несколько нуклеотидных последовательностей были соединены или рекомбинированы с получением уникальной конструкции, которая способна вводить промоторный фрагмент и последовательность ДНК для выбранного продукта гена вместе с подходящими 3' нетранслируемыми последовательностями в клетку. "Трансформационная кассета" относится к определенному вектору, содержащему чужеродный ген и имеющему другие элементы в дополнение к чужеродному гену, которые способствуют трансформации конкретной клетки-хозяина. "Кассета экспрессии" относится к определенному вектору, содержащему чужеродный ген и имеющему другие элементы в дополнение к чужеродному гену, которые обеспечивают повышенную экспрессию данного гена в чужеродном хозяине.
[71] Стандартные методики генной инженерии и молекулярного клонирования, используемые в настоящей заявке, известны в уровне техники и описаны, например, в Sambrook, J., Fritsch, E. F. and Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory: Cold Spring Harbor, N.Y., 1989 (в дальнейшем "Maniatis"); и в Silhavy, T. J., Bennan, M. L. and Enquist, L. W. Experiments with Gene Fusions; Cold Spring Harbor Laboratory: Cold Spring Harbor, N.Y., 1984; а также в Ausubel, F. M. et al., в Current Protocols in Molecular Biology, издательство Greene Publishing and Wiley-Interscience, 1987;, которые полностью включены в настоящее описание посредством отсылки.
[72] Если не определено иное, все технические и научные термины, используемые в настоящем описании, имеют такое же значение, под которым их обычно понимает средний специалист в области техники, к которой относится настоящее описание. Хотя любые методы и материалы, подобные или эквивалентные описанным в настоящей заявке, могут использоваться при осуществлении или тестировании настоящего описания, ниже описаны предпочтительные материалы и методы.
[73] Настоящее описание будет понято более полно при рассмотрении следующих неограничивающих Примеров. Следует понимать, что данные Примеры, притом, что в них указываются предпочтительные варианты осуществления рассматриваемой технологии, даны лишь в качестве иллюстрации. Из приведенного выше обсуждения и этих Примеров, специалист в данной области техники сумеет установить существенные характеристики рассматриваемой технологии и без отступления от ее сущности и объема сможет внести различные изменения и модификации рассматриваемой технологии, чтобы адаптировать ее к различным применениям и условиям.
Рекомбинантные полипептиды
[74] В одном аспекте настоящее описание относится к рекомбинантному полипептиду, имеющему аминокислотную последовательность, обладающую по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% и даже 100% идентичностью с аминокислотной последовательностью, представленной в SEQ ID NO:6. Предпочтительно аминокислотная последовательность рекомбинантного полипептида обладает по меньшей мере 80% идентичностью с SEQ ID NO:6. Более предпочтительно аминокислотная последовательность рекомбинантного полипептида обладает по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% и даже 100% идентичностью с SEQ ID NO:6. В примере осуществления аминокислотная последовательность рекомбинантного полипептида состоит из SEQ ID NO:6. Соответственно, рекомбинантный полипептид, описанный в настоящей заявке, включает функциональные фрагменты SEQ ID NO:6, функциональные варианты SEQ ID NO:6 и другие гомологичные полипептиды, которые обладают, например, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% и даже 100% идентичностью последовательности с SEQ ID NO:6.
[75] В другом аспекте настоящее описание относится к выделенной нуклеиновой кислоте, имеющей нуклеотидную последовательность, кодирующую рекомбинантный полипептид, описанный в настоящей заявке. Например, выделенная нуклеиновая кислота может включать нуклеотидную последовательность, кодирующую полипептид, имеющий аминокислотную последовательность, которая обладает по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% и даже 100% идентичностью с аминокислотной последовательностью, представленной в SEQ ID NO:6. Предпочтительно выделенная нуклеиновая кислота включает нуклеотидную последовательность, кодирующую полипептид, имеющий аминокислотную последовательность, которая по меньшей мере на 80% идентична аминокислотной последовательности, представленной в SEQ ID NO:6. Более предпочтительно выделенная нуклеиновая кислота включает нуклеотидную последовательность, кодирующую полипептид, имеющий аминокислотную последовательность, которая обладает по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% и даже 100% идентичностью последовательности с аминокислотной последовательностью, представленной в SEQ ID NO:6. Выделенная нуклеиновая кислота, таким образом, включает нуклеиновые кислоты, которые кодируют функциональные фрагменты SEQ ID NO:6, функциональные варианты SEQ ID NO:6 или другие гомологичные полипептиды, которые обладают, например, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% и даже 100% идентичностью последовательности с SEQ ID NO:6.
[76] В варианте осуществления настоящее описание относится к выделенной нуклеиновой кислоте, имеющей нуклеотидную последовательность, которая обладает по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% и даже 100% идентичностью с нуклеотидной последовательностью, представленной в SEQ ID NO:7. Предпочтительно выделенная нуклеиновая кислота включает нуклеотидную последовательность, которая обладает по меньшей мере 80% идентичностью с нуклеотидной последовательностью, представленной в SEQ ID NO:7. Более предпочтительно выделенная нуклеиновая кислота включает нуклеотидную последовательность, которая обладает по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% и даже 100% идентичностью с последовательностью нуклеиновой кислоты, представленной в SEQ ID NO:7.
[77] В другом аспекте рассматриваемая технология относится к вектору, содержащему нуклеиновые кислоты, описанные в настоящей заявке, и клетке-хозяину, содержащей вектор, описанный в настоящей заявке. В некоторых вариантах осуществления настоящее описание относится к вектору экспрессии, включающему по меньшей мере один полинуклеотид рассматриваемой технологии, и где вектор экспрессии при трансфекции в клетку-хозяина способен к экспрессии по меньшей мере одного рекомбинантного полипептида HV1, описанного в настоящей заявке. В варианте осуществления вектор экспрессии включает нуклеотидную последовательность, представленную в SEQ ID NO:7, или ее вариант.
[78] Конструкция вектора экспрессии зависит от таких факторов как выбор трансформируемой клетки-хозяина, требуемый уровень экспрессии белка и т.п. Векторы экспрессии могут быть введены в клетку-хозяина для получения в результате рекомбинантного полипептида рассматриваемой технологии, такого как рекомбинантный полипептид HV1, имеющий аминокислотную последовательность SEQ ID NO:6 или ее вариант.
[79] Экспрессию белков в прокариотах чаще всего проводят в бактериальной клетке-хозяине с использованием векторов, содержащих конститутивные или индуцируемые промоторы, направляющие экспрессию слитых или неслитых белков. Слитые векторы добавляют некоторое количество аминокислот к белку, кодируемому в них, обычно на N-конец рекомбинантного белка. Такие слитые векторы обычно служат трем целям: 1) повысить экспрессию рекомбинантного белка; 2) повысить растворимость рекомбинантного белка; и 3) способствовать очистке рекомбинантного белка, действуя как лиганд при аффинной очистке. Часто в место соединения слитого фрагмента и рекомбинантного белка вводят сайт протеолитического расщепления, позволяющий отделить рекомбинантный белок от слитого фрагмента после очистки слитого белка. Такие векторы включены в объем настоящего описания.
[80] В варианте осуществления вектор экспрессии включает такие генетические элементы для экспрессии рекомбинантного полипептида в бактериальных клетках. Элементы для транскрипции и трансляции в бактериальной клетке могут включать промотор, кодирующую область белкового комплекса и терминатор транскрипции.
[81] В варианте осуществления векторы экспрессии рассматриваемой технологии включают бактериальные векторы экспрессии, например, рекомбинантную ДНК бактериофага, плазмидную ДНК или космидную ДНК, дрожжевые векторы экспрессии например, рекомбинантные дрожжевые векторы экспрессии, векторы для экспрессии в клетках насекомых, например, рекомбинантные вирусные векторы экспрессии, например бакуловирус, или векторы для экспрессии в клетках растений, например, рекомбинантные вирусные векторы экспрессии, такие как вирус мозаики цветной капусты (CaMV), вирус табачной мозаики (ВТМ), или рекомбинантные плазмидные векторы экспрессии, такие как Ti-плазмиды.
[82] В варианте осуществления вектор включает бактериальный вектор экспрессии. В другом варианте осуществления вектор экспрессии включает высококопийный вектор экспрессии; в альтернативе вектор экспрессии включает низкокопийный вектор экспрессии, например, плазмиду Mini-F.
[83] Среднему специалисту в данной области техники будут известны методики молекулярной биологии, доступные для получения векторов экспрессии. Полинуклеотид, используемый для включения в вектор экспрессии рассматриваемой технологии, как описано выше, может быть получен с помощью стандартных методик, таких как полимеразная цепная реакция (ПЦР).
[84] Было разработано много методик молекулярной биологии для функционального связывания ДНК с векторами через комплементарные липкие концы. В одном варианте осуществления дополнительные гомополимерные участки могут быть добавлены к молекуле нуклеиновой кислоты, которую предполагается вставить в векторную ДНК. Вектор и молекулу нуклеиновой кислоты затем соединяют с образованием водородной связи между дополнительными гомополимерными концевыми участками, получая рекомбинантные молекулы ДНК.
[85] В альтернативном варианте осуществления синтетические линкеры, содержащие один или более введенных сайтов рестрикции, используют для функционального связывания полинуклеотида рассматриваемой технологии с вектором экспрессии. В варианте осуществления полинуклеотид получают при обработке эндонуклеазой рестрикции. В варианте осуществления молекулу нуклеиновой кислоты обрабатывают ДНК-полимеразой бактериофага T4 или ДНК полимеразой I E. coli, ферментами, которые удаляют выступающие 3'-одноцепочечные концы с помощью своей 3'-5'-экзонуклеазной активности и заполняют углубленные 3'-концы с помощью своей полимеразной активности, с получением в результате сегментов ДНК с тупыми концами. Затем сегменты с тупыми концами инкубируют с большим молярным избытком молекул линкера в присутствии фермента, который способен катализировать лигирование молекул ДНК с тупыми концами, такого как ДНК-лигаза бактериофага T4. Таким образом, продуктом реакции является полинуклеотид, несущий полимерные линкерные последовательности на своих концах. Затем эти полинуклеотиды расщепляют подходящей рестриктазой и лигируют в вектор экспрессии, который был разрезан ферментом, который дает концы, совместимые с концами полинуклеотида.
[86] В альтернативе может использоваться вектор, имеющий сайты безлигазного клонирования (LIC). Затем требуемый ПЦР амплифицированный полинуклеотид можно клонировать в вектор LIC без рестрикции или лигирования (Aslanidis and de Jong, Nucl. Acid. Res. 18, 6069-6074, (1990), Haun, et al., Biotechniques 13, 515-518 (1992), которая включена в настоящее описание в той мере, в которой это соответствует настоящему описанию).
[87] В варианте осуществления для выделения и/или модификации целевого полинуклеотида, предназначенного для вставки в выбранную плазмиду, предпочтительно использовать ПЦР. Подходящие праймеры для использования при получении последовательности с помощью ПЦР могут быть подобраны для выделения требуемой кодирующей области молекулы нуклеиновой кислоты, добавления сайтов эндонуклеаз рестрикции или LIC, помещения кодирующей области в нужную рамку считывания.
[88] В варианте осуществления полинуклеотид для включения в вектор экспрессии рассматриваемой технологии получают при помощи ПЦР с использованием подходящих олигонуклеотидных праймеров. Кодирующую область амплифицируют, при этом сами праймеры включаются в последовательность амплифицируемого продукта. В варианте осуществления амплификационные праймеры содержат сайты узнавания эндонуклеаз рестрикции, которые позволяют клонировать последовательность амплифицированного продукта в подходящий вектор.
[89] В варианте осуществления полинуклеотид SEQ ID NO:7 или его вариант получают с помощью ПЦР и включают в вектор экспрессии при использовании расщепления эндонуклеазами рестрикции и лигирования согласно методикам, которые известны в уровне техники.
[90] Настоящее описание также относится к клетке-хозяину, включающей вектор экспрессии, описанный в настоящей заявке. Подходящие хозяева рассматриваемой технологии обычно включают микробные хозяева или растения-хозяева. Например, клетка-хозяин рассматриваемой технологии выбрана из группы, состоящей из клеток бактерий, дрожжей, нитчатых грибов, цианобактерий и растений.
[91] Микробные хозяева могут включать любой организм, способный к экспрессии полинуклеотида (такого как SEQ ID NO:7) с получением рекомбинантного полипептида HV1, описанного в настоящей заявке. Микроорганизмы, применимые в рассматриваемой технологии, включают бактерии, такие как кишечные бактерии (например, Escherichia и Salmonella), а также Bacillus, Acinetobacter, актиномицеты, такие как Streptomyces, Corynebacterium, метанотрофы, такие как Methylosinus, Methylomonas, Rhodococcus и Pseudomona; цианобактерии, такие как Rhodobacter и Synechocystis; дрожжи, такие как Saccharomyces, Zygosaccharomyces, Kluyveromyces, Candida, Hansenula, Debaryomyces, Mucor, Pichia и Torulopsis; и нитчатые грибы, такие как Aspergillus и Arthrobotrys, а также водоросли, и Escherichia, Klebsiella, Pantoea, Salmonella, Corynebacterium, Clostridium и Clostridium acetobutylicum, например. Предпочтительно, микробный хозяин является бактериями (такой как Escherichia) или дрожжами (такими как Saccharomyces). Векторы экспрессии могут быть включены в эти и другие микробные хозяева для получения большого, коммерчески применимого количества стевиол-гликозидов.
[92] В варианте осуществления рекомбинантный полипептид может быть экспрессирован в клетке-хозяине, которая является растительной клеткой. При использовании в настоящем описании, термин "растительная клетка", как подразумевают, означает любую клетку, полученную из однодольного или двудольного растения и способную формировать недифференцированные ткани, такие как каллусы, дифференцированные ткани, такие как эмбрионы, части однодольных растений, однодольные растения или семя. Термин "растение", как подразумевают, означает любой дифференцированный многоклеточный организм, способный к фотосинтезу, включая однодольные и двудольные. В некоторых вариантах осуществления растительная клетка может быть растительной клеткой Arabidopsis, растительной клеткой табака, растительной клеткой сои, растительной клеткой петунии или клеткой из другой масличной культуры, включающей, без ограничения, растительную клетку канолы, растительную клетку рапса, растительную клетку пальмы, растительную клетку подсолнечника, растительную клетку хлопка, растительную клетку кукурузы, растительную клетку арахиса, растительную клетку льна и растительную клетку кунжута.
[93] Применимые растения-хозяева могут включать любое растение, которое обеспечивает продукцию рекомбинантных полипептидов рассматриваемой технологии. Подходящие зеленые растения для использования в качестве хозяев включают, без ограничения перечисленными, сою, рапс (Brassica napus, B. campestris), подсолнечник (Helianthus annus), хлопок (Gossypium hirsutum), кукурузу, табак (Nicotiana tabacum), люцерну (Medicago sativa), пшеницу (Triticum sp.), ячмень (Hordeum vulgare), овес (Avena sativa), сорго (Sorghum bicolor), рис (Oryza sativa), Arabidopsis, крестоцветные овощи (брокколи, цветную капусту, капусту, пастернак и т.д.), дыню, морковь, сельдерей, петрушку, томаты, картофель, землянику, арахис, виноград, кормовые посевные культуры, сахарную свеклу, сахарный тростник, бобы, горох, рожь, лен, деревья лиственных пород, деревья хвойных пород и кормовые травы. Виды водорослей включают, без ограничения перечисленными, коммерчески значимых хозяев, таких как Spirulina, Haemotacoccus и Dunaliella. Подходящие растения для способа рассматриваемой технологии также включают культуры для получения биотоплива, биомассы и биоэнергии. Примеры растений включают Arabidopsis thaliana, рис (Oryza sativa), Hordeum vulgare, просо прутьевидное (Panicum vigratum), Brachypodium spp., Brassica spp. и Crambe abyssinica.
[94] В некоторых вариантах осуществления настоящее описание включает трансгенные клетки-хозяева или хозяева, которые были трансформированы одним или более векторами, раскрытыми в настоящей заявке.
[95] В альтернативе клетки-хозяева могут быть клетками-хозяевами, подходящими для продукции биосинтеза включая одноклеточные организмы, микроорганизмы, многоклеточные организмы, растения, грибы, бактерии, водоросли, сельскохозяйственные культуры, несельскохозяйственные культуры и/или подобные.
[96] Векторы экспрессии могут быть введены в растение или микробные клетки-хозяева с помощью стандартных методик трансформации или трансфекции. Трансформацию подходящих клеток вектором экспрессии рассматриваемой технологии осуществляют способами, известными в уровне техники, при этом обычно она зависит от типа вектора и клетки. Подходящие методики включают соосаждение с фосфатом кальция или хлоридом кальция, трансфекцию, опосредованную DEAE-декстраном, липофекцию, хемопорацию или электропорацию.
[97] Успешно трансформированные клетки, то есть клетки, содержащие вектор экспрессии, могут быть идентифицированы с помощью методик, известных в уровне техники. Например, клетки, трансфицированные вектором экспрессии рассматриваемой технологии, можно культивировать с получением полипептидов, описанных в настоящей заявке. Клетки можно проверить на наличие ДНК вектора экспрессии с помощью методик, известных в уровне техники.
[98] Клетки-хозяева могут содержать одну копию вектора экспрессии, описанного ранее, или, альтернативно, множество копий вектора экспрессии.
[99] В некоторых вариантах осуществления трансформированная клетка является клеткой животного, клеткой насекомого, клеткой растения, клеткой водоросли, клеткой гриба или клеткой дрожжей. В некоторых вариантах осуществления клетка является растительной клеткой, выбранной из группы, состоящей из: растительной клетки канолы, растительной клетки рапса, растительной клетки пальмы, растительной клетки подсолнечника, растительной клетки хлопка, растительной клетки кукурузы, растительной клетки арахиса, растительной клетки льна, растительной клетки кунжута, растительной клетки сои и растительной клетки петунии.
[100] Системы экспрессии на основе микробных клеток-хозяев и векторы экспрессии, содержащие регуляторные последовательности, которые направляют экспрессию с высоким уровнем чужеродных белков, известны специалистам в данной области техники. Любые из них могут применяться для создания векторов для экспрессии рекомбинантного полипептида рассматриваемой технологии в микробной клетке-хозяине. Такие векторы могут быть затем введены в подходящие микроорганизмы с помощью трансформации для обеспечения экспрессии с высоким уровнем рекомбинантного полипептида рассматриваемой технологии.
[101] Векторы или кассеты, применимые для трансформации подходящих микробных клеток-хозяев, известны в уровне техники. Как правило, вектор или кассета содержат последовательности, направляющие транскрипцию и трансляцию соответствующего полинуклеотида, селективного маркера и последовательностей, обеспечивающих автономную репликацию или хромосомную интеграцию. Подходящие векторы включают 5' область полинуклеотида, которая содержит области регуляции инициации транскрипции и 3' область фрагмента ДНК, которая регулирует терминацию транскрипции. Предпочтительно, чтобы обе области регуляции были получены из генов, гомологичных трансформированной клетке-хозяину, хотя следует понимать, что такие области регуляции не должны быть обязательно получены из генов, нативных по отношению к определенным видам, выбранным в качестве хозяина.
[102] Области регуляции инициации или промоторы, которые могут применяться для направления экспрессии рекомбинантного полипептида в требуемой микробной клетке-хозяине, являются многочисленными и известны специалистам в данной области техники. Фактически любой промотор, способный к направлению этих генов, подходит для рассматриваемой технологии, в том числе, без ограничения перечисленными, CYC1, HIS3, GAL1, GAL10, ADH1, PGK, PHO5, GAPDH, ADC1, TRP1, URA3, LEU2, ENO, TPI (применимый для экспрессии в Saccharomyces); AOX1 (применимый для экспрессии в Pichia); а также lac, trp, IPL, IPR, T7, tac и trc (применимые для экспрессии в Escherichia coli).
[103] Области регуляции терминации могут быть также получены из различных генов, нативных по отношению к микробным хозяевам. Необязательно может быть включен участок терминации для микробных хозяев, описанных в настоящей заявке.
[104] В растительных клетках векторы экспрессии рассматриваемой технологии могут включать кодирующую область, функционально связанную с промоторами, способными направлять экспрессию рекомбинантного полипептида рассматриваемой технологии в требуемых тканях на нужной стадии развития. По причинам удобства экспрессируемые полинуклеотиды могут включать промоторные последовательности и лидерные последовательности трансляции, полученные из одного и того же полинуклеотида. Также должны присутствовать 3' некодирующие последовательности, кодирующие сигналы терминации транскрипции. Векторы экспрессии могут также включать один или более интронов, способствующих экспрессии полинуклеотида.
[105] В отношении растительных клеток-хозяев, любая комбинация любого промотора и любого терминатора, способная вызывать экспрессию кодирующей области, может использоваться в векторных последовательностях рассматриваемой технологии. Некоторые подходящие примеры промоторов и терминаторов включают промоторы и терминаторы генов нопалинсинтазы (nos), октопинсинтазы (ocs) и вируса мозаики цветной капусты (CaMV). Одним из типов эффективного растительного промотора, который может использоваться, является высокоэффективный растительный промотор. Такие промоторы, функционально связанные с вектором экспрессии рассматриваемой технологии, должны быть способны активировать экспрессию вектора. Высокоэффективные растительные промоторы, которые могут использоваться в рассматриваемой технологии, включают промотор малой субъединицы (ss) рибулозо-1,5-бисфосфаткарбоксилазы, например, из сои (Berry-Lowe et al., J. Molecular and App. Gen., 1:483 498 (1982), которая полностью включена в настоящее описание в той мере, в которой это соответствует настоящей заявке), и промотор хлорофилл a/b-связывающего белка. Эти два промотора, как известно, индуцирует свет в растительных клетках (см., например, Genetic Engineering of Plants, an Agricultural Perspective, A. Cashmore, Plenum, N.Y. (1983), страницы 29 38; Coruzzi, G. et al., The Journal of Biological Chemistry, 258:1399 (1983), и Dunsmuir, P. et al., Journal of Molecular and Applied Genetics, 2:285 (1983), которые полностью включены в настоящее описание в той мере, в которой это соответствует настоящей заявке).
[106] Выбор плазмидного вектора зависит от способа, который будет использоваться для трансформации растений-хозяев. Специалисту хорошо известны генетические элементы, которые должны присутствовать в плазмидном векторе для успешной трансформации, отбора и размножения клеток-хозяев, содержащих химерный полинуклеотид. Специалисту также известно, что различные независимые события трансформации приводят к различным уровням и профилям экспрессии (Jones et al., EMBO J. 4:2411 2418 (1985); De Almeida et al., Mol. Gen. Genetics 218:78 86 (1989), которые полностью включены в настоящее описание в той мере, в которой это соответствует настоящей заявке) и, следовательно, что множество событий требуется подвергнуть скринингу для получения линий, демонстрирующих требуемый уровень и профиль экспрессии. Такой скрининг может быть выполнен с помощью Саузерн-анализа ДНК-блотов, Нозерн-анализая экспрессии мРНК, Вестерн-анализа экспрессии белка или фенотипического анализа.
[107] Введение вектора экспрессии рассматриваемой технологии в растительную клетку может быть выполнено множеством способов, известных средним специалистам в данной области техники, включающих вставку целевой последовательности нуклеиновой кислоты в Ri-плазмиду Agrobacterium rhizogenes или Ti-плазмиду Agrobacterium tumefaciens, микроинъекцию, электропорацию или прямое осаждение. В качестве примера, в некоторых вариантах осуществления транзиентная экспрессия целевого полинуклеотида может быть выполнена методами агро-инфильтрации. В этом отношении, суспензия Agrobacterium tumefaciens, содержащая целевой полинуклеотид, может быть выращена в культуре и затем введена в растение с помощью шприца, конец которого прижимают к нижней стороне листа, мягко надавливая при этом на другую сторону листа. При этом раствор агробактерии поступает в воздушные полости внутри листа через устьица. Попав внутрь листа, агробактерия трансформирует целевой ген в части растительных клеток, где ген затем транзиентно экспрессируется.
[108] В качестве другого примера, трансформация целевой плазмиды в растительную клетку может быть выполнена с помощью методик бомбардировки ускоренными частицами (то есть биолистики). В этом отношении, суспензия зародышей растения может быть выращена в жидкой культуре и затем подвергнута бомбардировке плазмидами или полинуклеотидами, которые прикреплены к частицам золота, где частицы золота, связанные с целевой плазмидой или нуклеиновой кислотой, могут проникать через мембраны растительных тканей, таких как зародышевая ткань. После бомбардировки трансформированные эмбрионы могут быть отобраны при использовании подходящего антибиотика с получением новых, клонально размноженных, трансформированных эмбриогенных суспензионных культур.
[109] Клетки-хозяева могут быть немодифицированными клетками или клеточными линиями, или клеточными линиями, которые были генетически модифицированы. В некоторых вариантах осуществления клетка-хозяин является клеточной линией, которая была модифицирована с целью выращивания в требуемых условиях, например, при более низкой температуре.
[110] Стандартные методики генной инженерии могут использоваться для получения нуклеиновой кислоты, которая кодирует рекомбинантный полипептид, описанный в настоящей заявке, включения нуклеиновой кислоты в вектор экспрессии и введения вектора в клетку-хозяина, как описано в Sambrook, et al. (eds), Molecular Cloning; A Laboratory Manual, Third Edition, Cold Spring Harbor, (2001); и Ausubel, F.M. et al. (eds.) Current Protocols in Molecular Biology, John Wiley & Sons (1995). Нуклеиновая кислота, кодирующая полипептид, может быть вставлена в вектор или векторы экспрессии таким образом, чтобы нуклеиновые кислоты были функционально связаны с последовательностями регуляции транскрипции и трансляции (такими как последовательность промотора, последовательность терминации транскрипции и т.д.). Вектор экспрессии и последовательности регуляции экспрессии обычно выбирают так, чтобы они были совместимы с используемой для экспрессии клеткой-хозяином.
[111] Экспрессия полипептида в хозяине, описанном в настоящей заявке, может быть дополнительно улучшена путем оптимизации кодонов. Например, замена менее распространенного кодона более распространенным кодоном может влиять на полупериод существования мРНК или изменять ее структуру посредством введения вторичной структуры, которая препятствует трансляции мРНК. Вся или часть кодирующей области может быть оптимизирована. В некоторых случаях требуемую модуляцию экспрессии достигают путем оптимизации по существу всего гена. В других случаях требуемую модуляцию достигают при оптимизации части, а не всей последовательности гена.
[112] Использование кодонов в любой кодирующей последовательности можно регулировать для достижения требуемого свойства, например, высоких уровней экспрессии в определенном типе клеток. Исходной точкой для такой оптимизации может быть кодирующая последовательность со 100% распространенных кодонов или кодирующая последовательностью, которая содержит смесь распространенных и малораспространенных кодонов.
[113] Две или более кандидатных последовательностей, которые отличаются по своей частоте использования кодонов, могут быть получены и проверены с целью определения, обладают ли они нужным свойством. Кандидатные последовательности могут быть оценены при использовании компьютера для поиска присутствия регуляторных элементов, таких как сайленсеры или энхансеры, и поиска присутствия областей кодирующей последовательности, которая могла быть преобразована в такие регуляторные элементы при изменении использования кодонов. Дополнительные критерии могут включать обогащение специфическими нуклеотидами, например, A, C, G или U, смещение кодонов по определенной аминокислоте или присутствие или отсутствие специфической вторичной или третичной структуры мРНК. Регулирование кандидатной последовательности может быть выполнено на основе ряда таких критериев.
[114] В некоторых вариантах осуществления оптимизированная по кодонам последовательность нуклеиновой кислоты может экспрессировать свой белок на уровне, который составляет приблизительно 110%, приблизительно 150%, приблизительно 200%, приблизительно 250%, приблизительно 300%, приблизительно 350%, приблизительно 400%, приблизительно 450% или приблизительно 500% от уровня белка, экспрессируемого последовательностью нуклеиновой кислоты, которая не была оптимизирована по кодонам.
[115] В дополнение к нуклеиновой кислоте, которая кодирует рекомбинантный полипептид рассматриваемой технологии, вектор экспрессии рассматриваемой технологии может дополнительно нести регуляторные последовательности, которые регулируют экспрессию белка в клетке-хозяине, такие как промоторы, энхансеры или другие элементы регуляции экспрессии, которые регулируют транскрипцию или трансляцию нуклеиновой кислоты (кислот). Такие регуляторные последовательности известны в уровне техники. Специалистам в данной области техники будет очевидно, что конструкция вектора экспрессии, в том числе выбор регуляторных последовательностей, может зависеть от таких факторов как выбор клетки-хозяина, которую предполагается трансформировать, требуемый уровень экспрессии белка и т.д. Кроме того, рекомбинантные векторы экспрессии рассматриваемой технологии могут нести дополнительные последовательности, такие как последовательности, которые регулируют репликацию вектора в клетках-хозяевах (например, точки начала репликации) и селективные маркерные гены.
Биосинтез стевиол-гликозидов
[116] Как описано в настоящей заявке, рекомбинантные полипептиды настоящей технологии обладают УДФ-гликозилтрансферазной активностью, более конкретно включающей 1,2-19-O-глюкоза-гликозилирующую активность, и могут применяться для разработки биосинтетических способов получения стевиол-гликозидов, которые обычно мало распространены в природных источниках, таких как ребаудиозид D и ребаудиозид E. Рекомбинантные полипептиды настоящей технологии обладают УДФ-гликозилтрансферазной активностью, могут применяться для разработки биосинтетических способов получения новых стевиол-гликозидов, таких как ребаудиозид Z1 и ребаудиозид Z2.
[117] Таким образом, в одном аспекте рассматриваемая технология также относится к способу получения композиции стевиол-гликозидов, включающему инкубирование субстрата с рекомбинантным полипептидом, включающим аминокислотную последовательность, обладающую по меньшей мере 80% идентичностью с SEQ ID NO:6.
[118] Субстрат может быть любым природным или синтетическим соединением, способным к превращению в стевиол-гликозидное соединение в реакции, катализируемой одной или более УДФ-гликозилтрансферазами. Например, субстрат может быть натуральным экстрактом стевии, стевиолом, стевиол-13-O-глюкозидом, стевиол-19-O-глюкозидом, стевиол-1,2-биозидом, рубузозидом, стевиозидом, ребаудиозидом A, ребаудиозидом G или ребаудиозидом E. Субстрат может быть чистым соединением или смесью различных соединений. Предпочтительно субстрат включает соединение, выбранное из группы, состоящей из рубузозида, стевиозида, ребаудиозида A, ребаудиозида E и их комбинаций.
[119] Способ, описанный в настоящей заявке, также обеспечивает комбинированную реакционную систему, в которой рекомбинантные пептиды, описанные в настоящей заявке, могут функционировать в комбинации с одним или более дополнительными ферментами для повышения эффективности или изменения результата общего биосинтеза стевиол-гликозидных соединений. Например, дополнительный фермент может регенерировать УДФ-глюкозу, необходимую для реакции гликозилирования, превращая УДФ, образующийся в реакции гликозилирования, обратно в УДФ-глюкозу (при использовании, например, сахарозы в качестве донора остатка глюкозы), что повышает эффективность реакции гликозилирования. В другом примере рекомбинантный полипептид рассматриваемой технологии может давать промежуточный стевиол-гликозидный продукт (например, ребаудиозид E), который затем превращается в другой стевиол-гликозид (например, ребаудиозид D) в реакции, катализируемой другой УДФ-гликозилтрансферазой, такой как UGT76G1. В другом примере рекомбинантный полипептид рассматриваемой технологии может давать промежуточный стевиол-гликозидный продукт (например, ребаудиозид E), который затем превращается в другой стевиол-гликозид (например, ребаудиозид Z1 и ребаудиозид Z2) в реакции, катализируемой УДФ-гликозилтрансферазой, такой как HV1.
[120] Таким образом, в одном варианте осуществления способ рассматриваемой технологии дополнительно включает инкубирование рекомбинантной сахарозосинтазы (СС) с субстратом и рекомбинантным полипептидом, описанным в настоящей заявке. Рекомбинантная сахарозосинтаза превращает УДФ в УДФ-глюкозу, используя сахарозу в качестве источника глюкозы. Подходящая сахарозосинтаза включает сахарозосинтазы, полученные из генов СС Arabidopsis thaliana и Vigna radiate, или из любого гена, который кодирует функциональный гомолог сахарозосинтазы, кодируемой последовательностью SUS1 Arabidopsis thaliana и Vigna radiate, или их функциональные гомологи. Подходящими сахарозосинтазами может быть, например, сахарозосинтаза 1 Arabidopsis; сахарозосинтаза 3 Arabidopsis; и сахарозосинтаза Vigna radiate. Наиболее подходящей сахарозосинтазой может быть, например, сахарозосинтаза 1 Arabidopsis. Например, рекомбинантная СС включает аминокислотную последовательность, которая обладает по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или даже 100% идентичностью с аминокислотной последовательности AtSUS1, представленной в SEQ ID NO:9. Предпочтительно рекомбинантная СС рассматриваемой технологии включает аминокислотную последовательность, которая обладает по меньшей мере 80% идентичностью с аминокислотной последовательностью AtSUS1, представленной в SEQ ID NO:9.
[121] Рекомбинантная сахарозосинтаза рассматриваемой технологии может быть получена при экспрессии нуклеиновой кислоты, имеющей нуклеотидную последовательность, кодирующую целевую аминокислотную последовательность (например, аминокислотную последовательность, которая обладает по меньшей мере 80% идентичностью с аминокислотной последовательности, представленной в SEQ ID NO:9), в клетке-хозяине, как описано выше. Например, вектор, включающий нуклеотидную последовательность, представленную в SEQ ID NO:10, может быть введен в микробного хозяина (такого как E. coli) с использованием стандартных методик трансформации, с получением рекомбинантной сахарозосинтазы.
[122] В другом варианте осуществления способ рассматриваемой технологии дополнительно включает инкубирование рекомбинантной УДФ-гликозилтрансферазы с рекомбинантной сахарозосинтазой, субстратом и рекомбинантным полипептидом, описанным в настоящей заявке. Рекомбинантная УДФ-гликозилтрансфераза может катализировать другую реакцию гликозилирования, отличную от реакции, катализируемой рекомбинантным полипептидом рассматриваемой технологии. Например, рекомбинантная УДФ-гликозилтрансфераза может катализировать реакцию, которая переносит сахарную группу на C-3' C-13-O-глюкозы стевиозида с получением ребаудиозида A (или аналогично с получением ребаудиозида D из ребаудиозида E), тогда как рекомбинантный полипептид рассматриваемой технологии переносит вторую сахарную группу на C-2' 19-O-глюкозы стевиозида с получением ребаудиозида E (или аналогично с получением ребаудиозида D из ребаудиозида A).
[123] Подходящая УДФ-гликозилтрансфераза включает любую УГТ, известную в уровне техники как способную катализировать одну или более реакций в биосинтезе стевиол-гликозидных соединений, таких как UGT85C2, UGT74G1, UGT76G1 или их функциональные гомологи. Например, УДФ-гликозилтрансфераза, как описано в настоящей заявке, может включать аминокислотную последовательность, которая обладает по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или даже 100% идентичностью с аминокислотной последовательностью UGT76G1, представленной в SEQ ID NO:11. Предпочтительно УДФ-гликозилтрансфераза включает аминокислотную последовательность, которая обладает по меньшей мере 80% идентичностью с аминокислотной последовательностью UGT76G1, представленной в SEQ ID NO:11.
[124] Рекомбинантная УДФ-гликозилтрансфераза может быть получена при экспрессии нуклеиновой кислоты, имеющей нуклеотидную последовательность, кодирующую целевую аминокислотную последовательность (например, аминокислотную последовательность, которая обладает по меньшей мере 80% идентичностью с аминокислотной последовательностью, представленной в SEQ ID NO:11), в клетке-хозяине, как описано выше. Например, вектор, включающий нуклеотидную последовательность, представленную в SEQ ID NO:12, может быть введен в микробного хозяина (такого как E. coli) с помощью стандартных методик трансформации с получением рекомбинантной УДФ-гликозилтрансферазы рассматриваемой технологии.
[125] Рассматриваемая технология охватывает как in vitro, так и in vivo получение стевиол-гликозидных соединений.
[126] Например, инкубирование может быть in vitro процессом, в котором субстрат взаимодействует с рекомбинантным полипептидом рассматриваемой технологии. Предпочтительно в in vitro процессе рекомбинантный полипептид очищают перед инкубированием с субстратом. Стандартные методики очистки полипептидов, такие как центрифугирование, лизис клеток и хроматография, включены в способы рассматриваемой технологии. Например, нуклеиновая кислота, кодирующая рекомбинантный полипептид рассматриваемой технологии, может быть клонирована в вектор экспрессии с гистидиновой меткой, в результате чего экспрессированный рекомбинантный полипептид может быть очищен с помощью аффинной колоночной хроматографии.
[127] In vitro способ рассматриваемой технологии включает любую буферную систему, которая является подходящей для получения стевиол-гликозида с применением одного или более рекомбинантных полипептидов рассматриваемой технологии. Как правило, буферная система является водным раствором, таким как буфер Трис, буфер HEPES, буфер MOPS, фосфатный буфер, с pH от приблизительно 6,0 до приблизительно 8,0. Более предпочтительно pH составляет от приблизительно 6,5 до приблизительно 7,5. Еще более предпочтительно pH составляет от приблизительно 7,0 до приблизительно 7,5.
[128] Как правило, в in vitro способе рассматриваемой технологии субстрат присутствует в буфере в концентрации от приблизительно 0,2 мг/мл до приблизительно 5 мг/мл, предпочтительно от приблизительно 0,5 мг/мл до приблизительно 2 мг/мл, более предпочтительно от приблизительно 0,7 мг/мл до приблизительно 1,5 мг/мл.
[129] Как правило, в in vitro способе рассматриваемой технологии УДФ-глюкоза включена в буфер в концентрации от приблизительно 0,2 мМ до приблизительно 5 мМ, предпочтительно от приблизительно 0,5 мМ до приблизительно 2 мМ, более предпочтительно от приблизительно 0,7 мМ до приблизительно 1,5 мМ. В варианте осуществления, в случае включения рекомбинантной сахарозосинтазы в реакцию, сахарозу также включают в буфер в концентрации от приблизительно 100 мМ до приблизительно 500 мМ, предпочтительно от приблизительно 200 мМ до приблизительно 400 мМ, более предпочтительно от приблизительно 250 мМ до приблизительно 350 мМ.
[130] Как правило, в in vitro способе рассматриваемой технологии весовое соотношение рекомбинантного полипептида к субстрату, в расчете на сухой вес, составляет от приблизительно 1:100 до приблизительно 1:5, предпочтительно от приблизительно 1:50 до приблизительно 1:10, более предпочтительно от приблизительно 1:25 до приблизительно 1:15.
[131] Как правило, температура реакции in vitro способа составляет от приблизительно 20°C до приблизительно 40°C, предпочтительно от 25°C до приблизительно 37°C, более предпочтительно от 28°C до приблизительно 32°C.
[132] В настоящем описании также предусмотрена композиция стевиол-гликозидов, полученная с применением биосинтетического способа, описанного в настоящей заявке. Точная природа композиции стевиол-гликозидов, полученной с применением способа, описанного в настоящей заявке, например, типы соединений и их процентное содержание в конечном продукте, зависит от используемого субстрата, условий инкубирования и ферментативной активности, включенных в реакционную систему. Например, в случае использования стевиозида в качестве субстрата, полученная композиция стевиол-гликозидов может включать ребаудиозид A, ребаудиозид D, ребаудиозид E, ребаудиозид Z1 и ребаудиозид Z2 и их комбинации.
[133] Рассматриваемая технология обеспечивает способ превращения преобладающих стевиол-гликозидных соединений (то есть стевиозида и ребаудиозида A) в натуральном экстракте стевии в ребаудиозид D и ребаудиозид E, которые в иных случаях мало распространены в натуральном экстракте. Таким образом, рассматриваемая технология также обеспечивает способ обогащения содержания одного или более определенных стевиол-гликозидов (таких как ребаудиозид D и ребаудиозид E), включающий инкубирование субстрата (такого как натуральный экстракт стевии) с рекомбинантным полипептидом, включающим аминокислотную последовательность, обладающую по меньшей мере 80% идентичностью с SEQ ID NO:6. Например, при использовании натурального экстракта стевии в качестве субстрата, получаемая композиция стевиол-гликозидов может быть обогащена ребаудиозидом D и/или ребаудиозидом E, которые мало распространены в натуральном экстракте стевии.
[134] Специалисту в данной области будет известно, что композиция стевиол-гликозидов, полученная способом, описанным в настоящей заявке, может быть затем очищена и смешана с другими стевиол-гликозидами, вкусоароматическими веществами или подсластителями, с получением требуемого вкуса и аромата или композиции подсластителя. Например, композиция, обогащенная ребаудиозидом D, получанная, как описано в настоящей заявке, может быть смешана с натуральным экстрактом стевии, содержащим ребаудиозид A в качестве преобладающего стевиол-гликозида, или с другими синтетическими или натуральными стевиол-гликозидными продуктами, с получением требуемой композиции подсластителя. В альтернативе по существу очищенный стевиол-гликозид (например, ребаудиозид D), полученный из композиции стевиол-гликозидов, описанной в настоящей заявке, может быть объединен с другими подсластителями, такими как сахароза, мальтодекстрин, аспартам, сукралоза, неотам, ацесульфам калия и сахарин. Количество стевиол-гликозида относительно других подсластителей можно регулировать для получения требуемого вкуса, как известно в уровне техники. Композиция стевиол-гликозидов, описанная в настоящей заявке (включающая ребаудиозид D, ребаудиозид E, ребаудиозид Z1, ребаудиозид Z2 или их комбинацию), может быть включена в пищевые продукты (такие как напитки, безалкогольные напитки, мороженое, молочные продукты, кондитерские изделия, изделия из дробленого зерна, жевательную резинку, хлебобулочные изделия и т.д.), биологически активные пищевые добавки, диетическое питание, а также фармацевтические продукты.
ПРИМЕРЫ
ПРИМЕР 1: Выбор кандидатных генов УГТ
[135] Филогенетический и белковый BLAST анализ использовали для идентификации 7 кандидатных генов, которые относятся к подсемейству UGT91, с 1,2-19-O-глюкоза-гликозилирующей активностью (Таблица 1).
Список кандидатных генов УГТ
[Brachypodium distachyon]
[Brachypodium distachyon]
[Brachypodium distachyon]
[Brachypodium distachyon]
[Brachypodium distachyon]
Пример 2: Скрининг ферментной активности кандидатных генов УГТ
[136] Фрагменты полноразмерной ДНК всех кандидатных генов УГТ синтезировали коммерчески. Почти все кодоны кДНК были заменены кодонами, предпочтительными для E. coli (Genscript, NJ). Синтезированную ДНК клонировали в бактериальный вектор экспрессии pETite N-His SUMO Kan Vector (Lucigen).
[137] Каждую экспрессионную конструкцию трансформировали в E. coli BL21 (DE3), которую затем выращивали в среде LB, содержащей 50 мкг/мл канамицина, при 37°C до достижения OD600 0,8-1,0. Экспрессию белка индуцировали добавлением 1 мМ изопропил-β-D-1-тиогалактопиранозида (IPTG) и выращивали культуру далее при 16°C в течение 22 ч. Клетки собирали центрифугированием (3000×g; 10 мин; 4°C). Осадки клеток собирали и использовали немедленно или хранили при -80°C.
[138] Осадки клеток обычно ресуспендировали в лизисном буфере (50 мМ калий фосфатный буфер, pH 7,2, 25 мкг/мл лизоцима, 5 мкг/мл ДНКазы I, 20 мМ имидазола, 500 мМ NaCl, 10% глицерина и 0,4% Triton X-100). Клетки разрушали ультразвуком при 4°C и удаляли клеточный дебрис центрифугированием (18000×g; 30 мин). Супернатант наносили на уравновешенную (уравновешивающий буфер: 50 мМ калий фосфатный буфер, pH 7,2, 20 мМ имидазола, 500 мМ NaCl, 10% глицерина) аффинную колонку Ni-NTA (Qiagen). После нанесения образца белка колонку промывали уравновешивающим буфером для удаления не связавшихся белковых примесей. His-меченные рекомбинантные полипептиды УГТ элюировали уравновешивающим буфером, содержащим 250 мМ имидазола.
[139] Очищенные кандидатные УГТ рекомбинантные полипептиды исследовали на наличие 1,2-19-O-глюкоза-гликозилирующей активности при использовании стевиозида или Reb A в качестве субстрата (Фигура 1). Как правило, рекомбинантный полипептид (10 мкг) тестировали в 200 мкл in vitro реакционной системе. Реакционная система содержит 50 мМ калий фосфатный буфер, pH 7,2, 3 мМ MgCl2, 1 мг/мл стевиозида или ребаудиозида A, 1 мМ УДФ-глюкозы. Реакцию проводили при 30°C и останавливали добавлением 200 мкл 1-бутанола. Образцы экстрагировали три раза 200 мкл 1-бутанола. Смешанную фракцию сушили и растворяли в 70 мкл 80% метанола для анализа с помощью высокоэффективной жидкостной хроматографии (ВЭЖХ). Экстракт стевии (Blue California, CA), содержащий 95% стевиозида, использовали в качестве стевиозид-содержащего субстрата. Ребаудиозид (чистота 99%) был также поставлен Blue California.
[140] Катализируемую УГТ реакцию гликозилирования объединяли с реакцией образования УДФ-глюкозы, катализируемой сахарозосинтазой (такой как AtSUS1 из SEQ ID NO:9). В этом способе УДФ-глюкоза образовывалась из сахарозы и УДФ (Фигура 2), при этом можно исключить добавление дополнительных количеств УДФ-глюкозы. Последовательность AtSUS1 (Bieniawska et al., Plant J. 2007, 49: 810-828) синтезировали и встраивали в бактериальный вектор экспрессии. Рекомбинантный белок AtSUS1 экспрессировали и очищали с помощью аффинной хроматографии. Очищенный рекомбинантный полипептид AtSUS1 анализировали с помощью электрофореза в ДСН-ПААГ (молекулярная масса: 106,3 кДа, Фигура 7).
[141] Таким образом, активности рекомбинантных полипептидов УГТ тестировали без AtSUS1 (50 мМ калий фосфатный буфер, pH 7,2, 3 мМ MgCl2, 1 мг/мл стевиозида или ребаудиозида A, 1 мМ УДФ) или при объединении с AtSUS (50 мМ калий фосфатный буфер, pH 7,2, 3 мМ MgCl2, 1 мг/мл стевиозида или ребаудиозида A, 1 мМ УДФ и 285 мМ сахарозы). Как правило, 10 мкг AtSUS1 использовали для 200 мкл in vitro реакции. Реакцию in vitro инкубировали при 30°C и останавливали экстракцией при использовании 1-бутанола.
[142] Затем проводили анализ ВЭЖХ при использовании системы Dionex UPLC ultimate 3000 (Sunnyvale, CA), включающей насос для четырехкомпонентных смесей, термостатируемое колоночное отделение, автодозатор и УФ-детектор. Для исследования стевиол-гликозидов использовали Phenomenex Luna NH2 с предколонкой. Ацетонитрил в воде использовали для изократического элюирования в анализе ВЭЖХ. Продукты ребаудиозид D, ребаудиозид E, ребаудиозид Z идентифицировали с помощью ЯМР-анализа.
[143] Рекомбинантный полипептид (SEQ ID NO:6), кодируемый SEQ ID NO:7, показал 1,2-19-O-глюкоза-гликозилирующую активность и был подвергнут дополнительному анализу. Ген был получен из Hordeum vulgare subsp. vulgare (в настоящей заявке сокращенно обозначен "HV1"). Очищенный рекомбинантный полипептид HV1 анализировали с помощью электрофореза в ДСН-ПААГ (Фигура 6). Как показано на Фигуре 6, рекомбинантный белок HV1 (молекулярная масса: 61,4 кДа) очищали с помощью аффинной хроматографии. Полипептиды, кодируемые другими кандидатными генами (Таблица 1), не показали обнаруживаемой активности в анализах, описанных в настоящей заявке, несмотря на то, что они обладали приблизительно 62-74% идентичностью последовательности с рекомбинантным полипептидом HV1.
[144] Как описано в настоящей заявке, рекомбинантный полипептид HV1 переносил сахарную группу на ребаудиозид A с получением ребаудиозида D при любых условиях реакции, с или без AtSUS1. Ребаудиозид A полностью превращался в ребаудиозид D под действием рекомбинантного полипептида HV1 в УГТ-СС комбинированной реакционной системе (Фигура 3B-E). Однако ребаудиозид A лишь частично превращался в ребаудиозид D через 24 часа под действием только одного рекомбинантного полипептида HV1 без объединения с AtSUS1 (Фигура 3F-G). Таким образом, рекомбинантный полипептид HV1 показал 1,2-19-O-глюкоза-гликозилирующую активность с получением ребаудиозида D из ребаудиозида A, при этом AtSUS1 повышала эффективность превращения в УГТ-СС комбинированной системе.
[145] Кроме того, рекомбинантный полипептид HV1 в комбинации с AtSUS1 (SEQ ID NO:9) превращал стевиозид в ребаудиозид E in vitro (Фигура 4). Было получено неожиданное соединение ("Reb Z"), имеющее время удержания ВЭЖХ 6,68 минуты (см. Фигуру 4), которое отличалось от ребаудиозидов D и E. Это соединение представляло собой новый стевиол-гликозид и было названо "ребаудиозид Z" ("Reb Z"). Для подтверждения превращения Reb E в Reb Z, субстрат Reb E (0,5 мг/мл) инкубировали с рекомбинантным полипептидом HV1 (20 мкг) и AtSUS1 (20 мкг) в УГТ-СС комбинированной реакционной системе (200 мкл) при условиях, аналогичных используемым в примерах выше. Как показано на Фигуре 8, Reb Z был получен с применением комбинации рекомбинантного полипептида HV1 и AtSUS1. Эти результаты показали, что HV1 может переносить глюкозную группу на Reb E с получением Reb Z.
ПРИМЕР 3: Биосинтез стевиол-гликозида с применением рекомбинантного полипептида HV1
[146] Как показано на Фигуре 1, ребаудиозид D также может образовываться при гликозилировании C-3' C-13-O-глюкозы ребаудиозида E. Таким образом, ребаудиозид D может быть получен различными биосинтетическими методами (например, через ребаудиозид, а не ребаудиозид E), в зависимости от порядка, в котором проходят реакции гликозилирования. Например, гликозилирование по C-3' C-13-O-глюкозы стевиозида может проходить сначала, с образованием промежуточного ребаудиозида A, с последующим гликозилированием по C-2' 19-O-глюкозы ребаудиозида A с получением ребаудиозида D. На настоящий момент, UGT76G1 (SEQ ID NO:11) из стевии был идентифицирован как фермент, который переносит сахарный остаток на C-3' C-13-O-глюкозы стевиозида с образованием ребаудиозида A.
[147] Кодон-оптимизированную кДНК UGT76G1 встраивали в бактериальный вектор экспрессии и рекомбинантный белок UGT76G1 экспрессировали и очищали с помощью аффинной хроматографии. Очищенный рекомбинантный полипептид UGT76G1 анализировали с помощью электрофореза в ДСН-ПААГ (молекулярная масса: 65,4 кДа, Фигура 9). Субстрат ребаудиозид E инкубировали с рекомбинантной UGT76G1, с или без AtSUS1, при условиях, аналогичных используемым в Примерах выше. Продукты анализировали с помощью ВЭЖХ. Как показано на Фигуре 12, ребаудиозид D образовывался под действием рекомбинантной UGT76G1. Добавление рекомбинантной AtSUS в реакцию повышало эффективность превращения в УГТ-СС комбинированной системе. Таким образом, рекомбинантный полипептид UGT76G1 показал 1,3-13-O-глюкоза-гликозилирующую активность с получением Reb D из Reb E.
[148] Таким образом, каталитическую активность рекомбинантного полипептида HV1 для биосинтеза стевиол-гликозидов (например, получения ребаудиозида D) также определяли в комбинации с UGT76G1. Субстрат стевиозид инкубировали с рекомбинантным полипептидом HV1 (10 мкг), UGT76G1 (10 мкг) и AtSUS1 (10 мкг) в УГТ-СС комбинированной реакционной системе (200 мкл) при условиях, аналогичных используемым в примерах выше. Продукты анализировали с помощью ВЭЖХ. Как показано на Фигуре 5, ребаудиозид D образовывался под действием комбинации рекомбинантного полипептида HV1, UGT76G1 и AtSUS1. Таким образом, рекомбинантный полипептид HV1, который показал, по меньшей мере, 1,2-19-O-глюкоза-гликозилирующую активность, может применяться в комбинации с другими УГТ ферментами (такими как UGT76G1) для комплексного, многостадийного биосинтеза стевиол-гликозидов.
Пример 4: ЯМР-анализ структуры Reb Z
[149] Материал, используемый для исследования ребаудиозида Z (Reb Z), получали при использовании ферментативного превращения ребаудиозида E и очищали с помощью ВЭЖХ.
[150] Данные МСВР были получены с использованием масс-спектрометра высокого разрешения LTQ Orbitrap Discovery, с установкой разрешения на 30000; сканирования m/z от 150 до 1500 в режиме детектирования положительно заряженных ионов с ионизацией электрораспылением. Напряжение на игле устанавливали на 4 кВ; другие исходные условия были следующими: защитный газ = 25, вспомогательный газ = 0, продувочный газ = 5 (все потоки газа указаны в условных единицах), напряжение на капилляре = 30 В, температура капилляра = 300°C и напряжение на цилиндрической линзе = 75. Образец разбавляли смесью ацетонитрила:метанола:воды 2:2:1 (такой же, как элюент для введения) и вводили 50 микролитров.
[151] Спектры ЯМР получали на спектрометрах Bruker Avance DRX 500 МГц или Varian INOVA 600 МГц при использовании стандартных последовательностей импульсов. Одномерные (1H и 13C) и двумерные (COSY, TOCSY, HMQC и HMBC) спектры ЯМР получали в C5D5N.
[152] Соединение Reb Z, показанное как смесь Reb Z1 и Reb Z2, показано на Фигуре 10. Молекулярная формула соединения Reb Z была выведена как C50H80O28 на основе его положительного масс-спектра высокого разрешения (ВР), который показал присутствие ионов-аддуктов, соответствующих [M+Na]+ при m/z 1151,4713; такой состав подтверждался данными 13C-ЯМР спектра. Данные 1H-ЯМР спектра Reb Z показали присутствие смеси двух соединений (Reb Z1 и Reb Z2) в соотношении от 60:40 до 70:30. Следовательно, данные 1H и 13C-ЯМР спектров Reb Z показали наборы пиков для каждого протона и углерода, присутствующих в его структуре. Кислотный гидролиз Reb Z 5% H2SO4 дал D-глюкозу, которую идентифицировали при непосредственном сравнении с аутентичным образцом с помощью ТСХ. Ферментативный гидролиз Reb Z дал агликон, который идентифицировали как стевиол при сравнении 1H-ЯМР и ТСХ со стандартным соединением. Значения 1H и 13C-ЯМР для соединения Reb Z были установлены на основе данных TOCSY, HMQC и HMBC. Высокие константы взаимодействия, наблюдаемые для пяти аномерных протонов глюкозных групп, указывали на их β-ориентацию, как сообщали для стевиол-гликозидов.
Данные 1H и 13C-ЯМР спектров (химические сдвиги и константы взаимодействия) ребаудиозида Z ("Reb Z") и ребаудиозида E a-c.
2,922 д (13,2)/2,79 д (12,8)
6,35 д (7,8)
5,14 д (7,4)/
5,46 д (7,8)/
5,40 д (7,6)
5,34 д (7,5)/
[153] На основе результатов из данные ЯМР-спектров и экспериментов гидролиза Reb Z, а также тщательного сравнения значений 1H и 13C ЯМР Reb Z с ребаудиозидом E предположили, что смесь двух соединений, получаемая при ферментативном превращении, была установлена как 13-[(2-O-β-D-глюкопиранозил-2-O-β-D-глюкопиранозил-β-D-глюкопиранозил)окси]энт-каур-16-ен-19-овой кислоты 2-O-β-D-глюкопиранозил-β-D-глюкопиранозиловый эфир (Reb Z1) или 13-[(2-O-β-D-глюкопиранозил-β-D-глюкопиранозил)-окси]энт-каур-16-ен-19-овой кислоты [(2-O-β-D-глюкопиранозил-2-O-β-D-глюкопиранозил-β-D-глюкопиранозиловый) эфир (Reb Z2).
[154] Кислотный гидролиз соединения Reb Z. К раствору соединения Reb Z (5 мг) в MeOH (10 мл) добавляли 3 мл 5% H2SO4 и нагревали смесь с обратным холодильником в течение 24 часов. Затем реакционную смесь нейтрализовали насыщенным карбонатом натрия и экстрагировали этилацетатом (EtOAc) (2×25 мл), получив водную фракцию, содержащую сахар, и фракцию EtOAc, содержащую часть агликона. Водную фазу выпаривали и сравнивали со стандартным сахаром при использовании системы ТСХ EtOAc/н-бутанол/вода (2:7:1) и CH2Cl2/MeOH/вода (10:6:1); сахар идентифицировали как D-глюкоза.
[155] Ферментативный гидролиз соединения Reb Z. Соединение Reb Z (1 мг) растворяли в 10 мл 0,1М натрий ацетатного буфера, pH 4,5, и добавляли неочищенную пектиназу из Aspergillus niger (50 мкл, Sigma-Aldrich, P2736). Смесь перемешивали при 50°C в течение 96 ч. В ходе реакции осаждался продукт, который фильтровали и затем кристаллизовали. Полученный продукт, полученный в результате гидролиза 1, идентифицировали как стевиол при сравнении его ТСХ со стандартным соединением и данными 1H ЯМР спектра (Фигура 11).
[156] Смесь двух соединений, названную Reb Z, получили путем биоконверсии Ребаудиозида E с использованием ферментативной методики, и структуры охарактеризовали как 13-[(2-O-β-D-глюкопиранозил-2-O-β-D-глюкопиранозил-β-D-глюкопирано-зил)окси]энт-каур-16-ен-19-овой кислоты 2-O-β-D-глюкопиранозил-β-D-глюкопиранозиловый эфир (Reb Z1) или 13-[(2-O-β-D-глюкопиранозил-β-D-глюкопиранозил)окси]энт-каур-16-ен-19-овой кислоты [(2-O-β-D-глюкопиранозил-2-O-β-D-глюкопиранозил-β-D-глюкопиранозиловый) эфир (Reb Z2), на основе обширных исследований с помощью одно и двухмерного ЯМР, а также масс-спектральных данных высокого разрешения и гидролитических исследований.
[157] Таким образом, 1,2-19-O-глюкоза-гликозилирующая активность рекомбинантного полипептида HV1 была подтверждена его способностью переносить вторую сахарную группу на C-2' 19-O-глюкозы стевиозида с получением Ребаудиозида E. Рекомбинантный полипептид HV1 также обладает способностью переносить третью группу глюкозы на C-2' 13-O-глюкозы или C-2' 19-O-глюкозы ребаудиозида E с получением Reb Z1 или Reb Z2.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНОЕ ПОЛУЧЕНИЕ СТЕВИОЛ-ГЛИКОЗИДОВ | 2014 |
|
RU2741103C2 |
| БИОСИНТЕТИЧЕСКОЕ ПОЛУЧЕНИЕ СТЕВИОЛОВОГО ГЛИКОЗИДА РЕБАУДИОЗИДА D4 ИЗ РЕБАУДИОЗИДА E | 2017 |
|
RU2764803C2 |
| ГИДРОЛИЗ СТЕВИОЛОВЫХ ГЛИКОЗИДОВ С ПОМОЩЬЮ БЕТА-ГЛЮКОЗИДАЗЫ | 2018 |
|
RU2775697C2 |
| UDP-ЗАВИСИМАЯ ГЛИКОЗИЛТРАНСФЕРАЗА ДЛЯ ВЫСОКОЭФФЕКТИВНОГО ПРОДУЦИРОВАНИЯ РЕБАУДИОЗИДОВ | 2017 |
|
RU2777901C2 |
| ПРИМЕНЕНИЕ КАУРЕНОКСИДАЗЫ PISUM SATIVUM ДЛЯ ВЫСОКОЭФФЕКТИВНОГО ПРОИЗВОДСТВА РЕБАУДИОЗИДОВ | 2018 |
|
RU2795550C2 |
| Способ получения ребаудиозида D и ребаудиозида M | 2022 |
|
RU2839979C2 |
| АBC-ТРАНСПОРТЕРЫ ДЛЯ ВЫСОКОЭФФЕКТИВНОГО ПРОИЗВОДСТВА РЕБАУДИОЗИДОВ | 2020 |
|
RU2795855C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕБАУДИОЗИДА M С ИСПОЛЬЗОВАНИЕМ ФЕРМЕНТАТИВНОГО СПОСОБА | 2013 |
|
RU2658436C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕБАУДИОЗИДА C С ПРИМЕНЕНИЕМ ФЕРМЕНТАТИВНОГО СПОСОБА | 2016 |
|
RU2736155C1 |
| СПОСОБЫ И ВЕЩЕСТВА ДЛЯ ОСНОВАННОГО НА РЕКОМБИНАЦИИ ПОЛУЧЕНИЯ СОЕДИНЕНИЙ ШАФРАНА | 2012 |
|
RU2676730C2 |























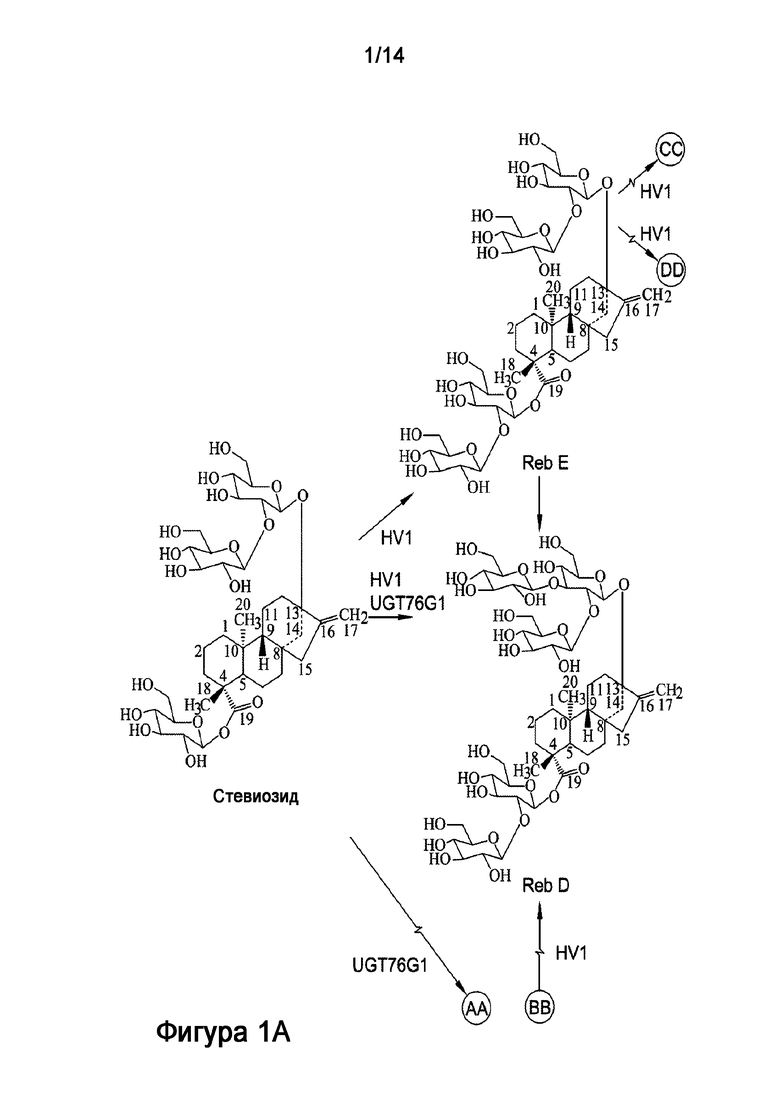
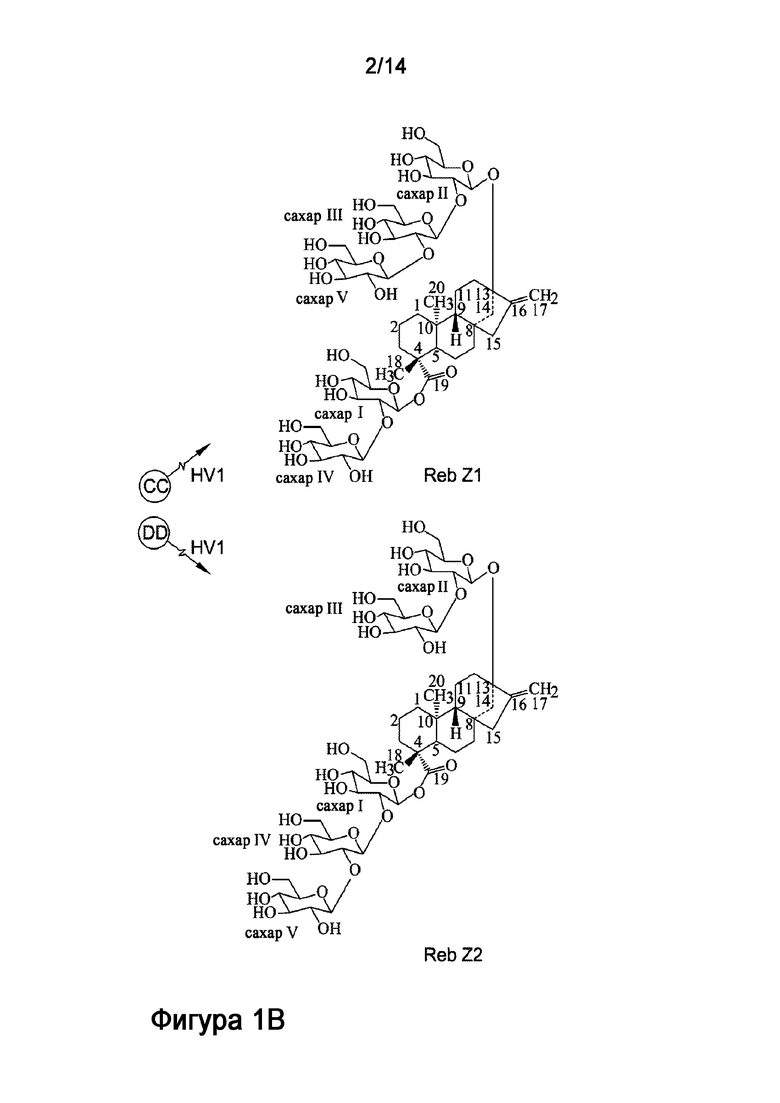
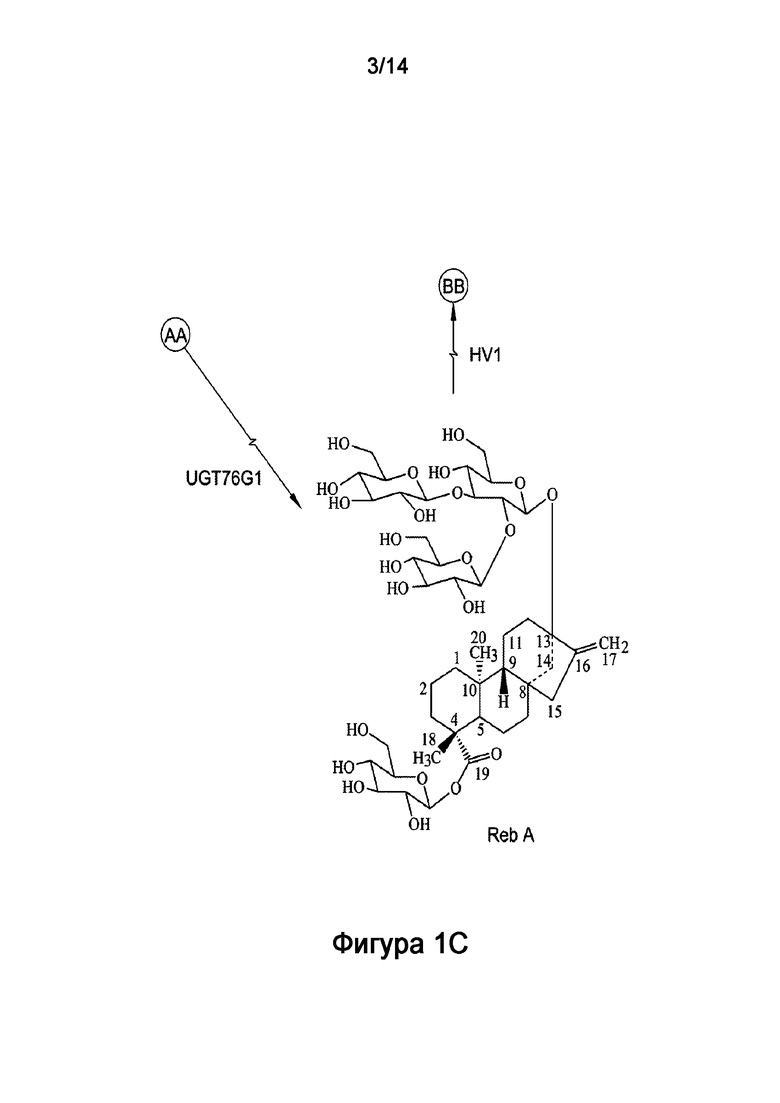
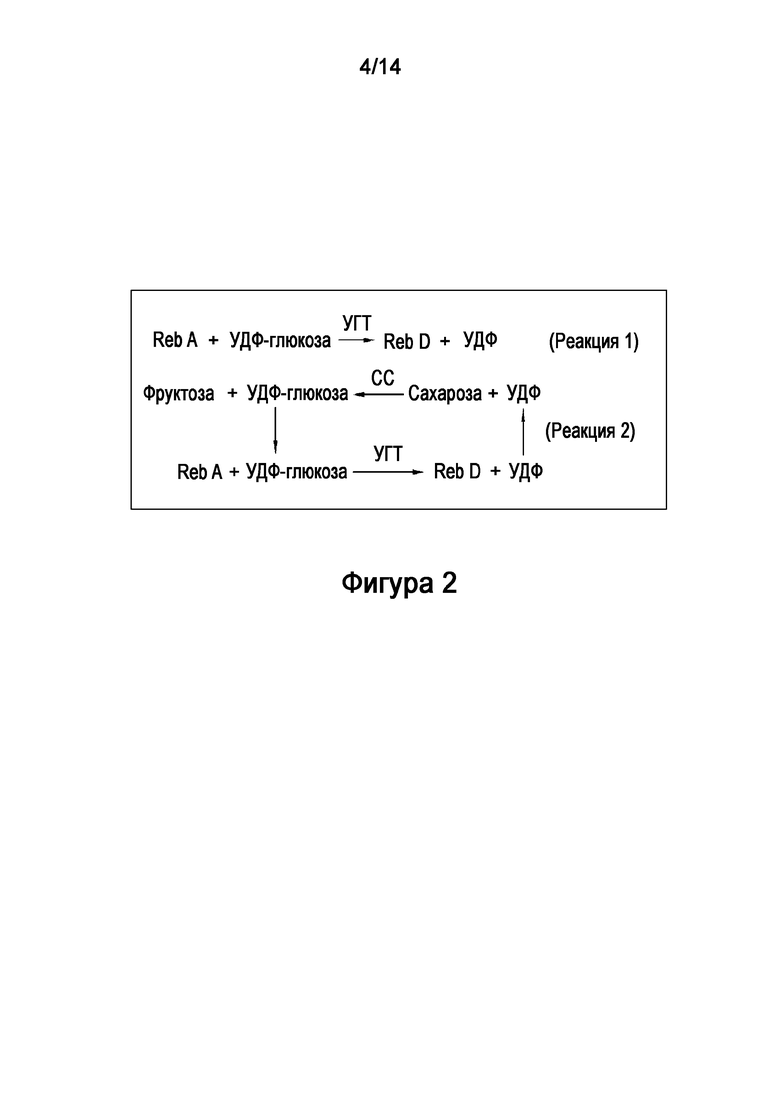
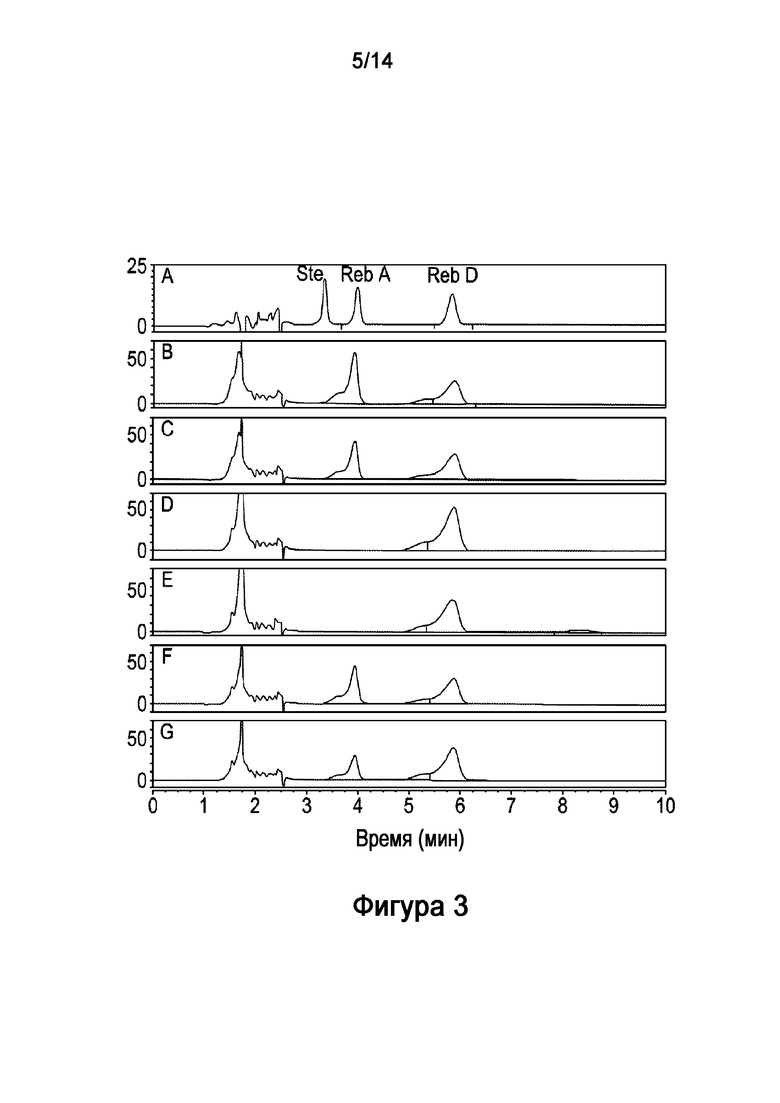
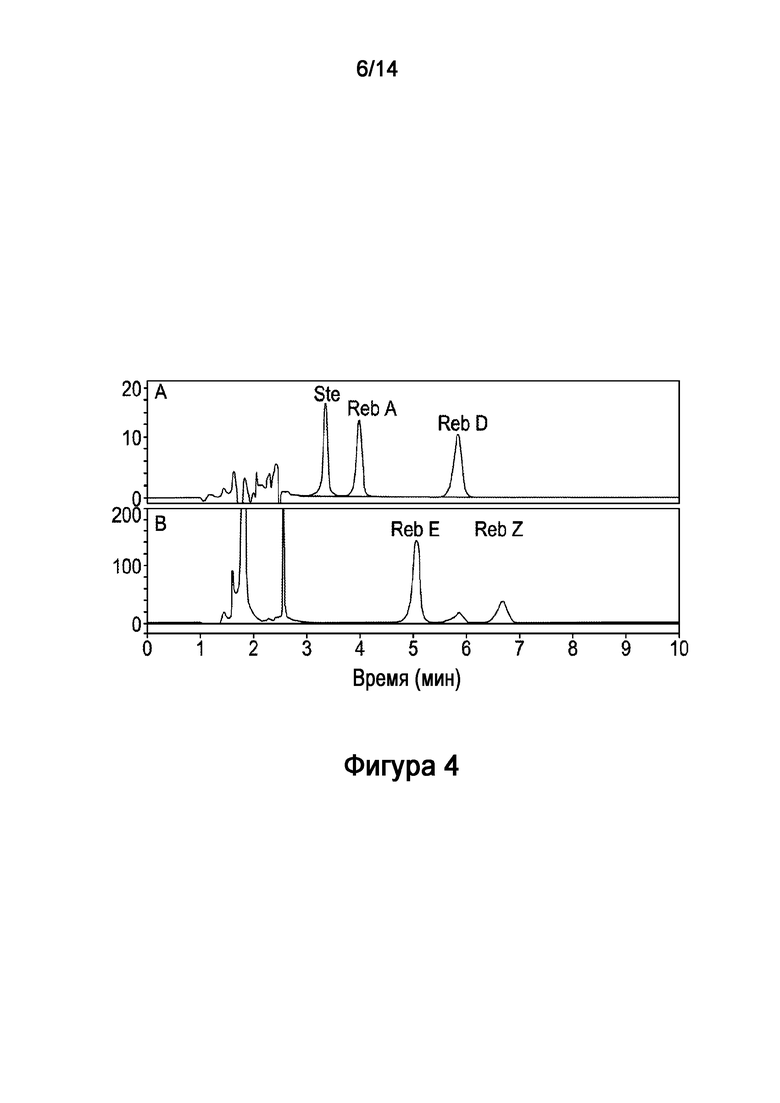
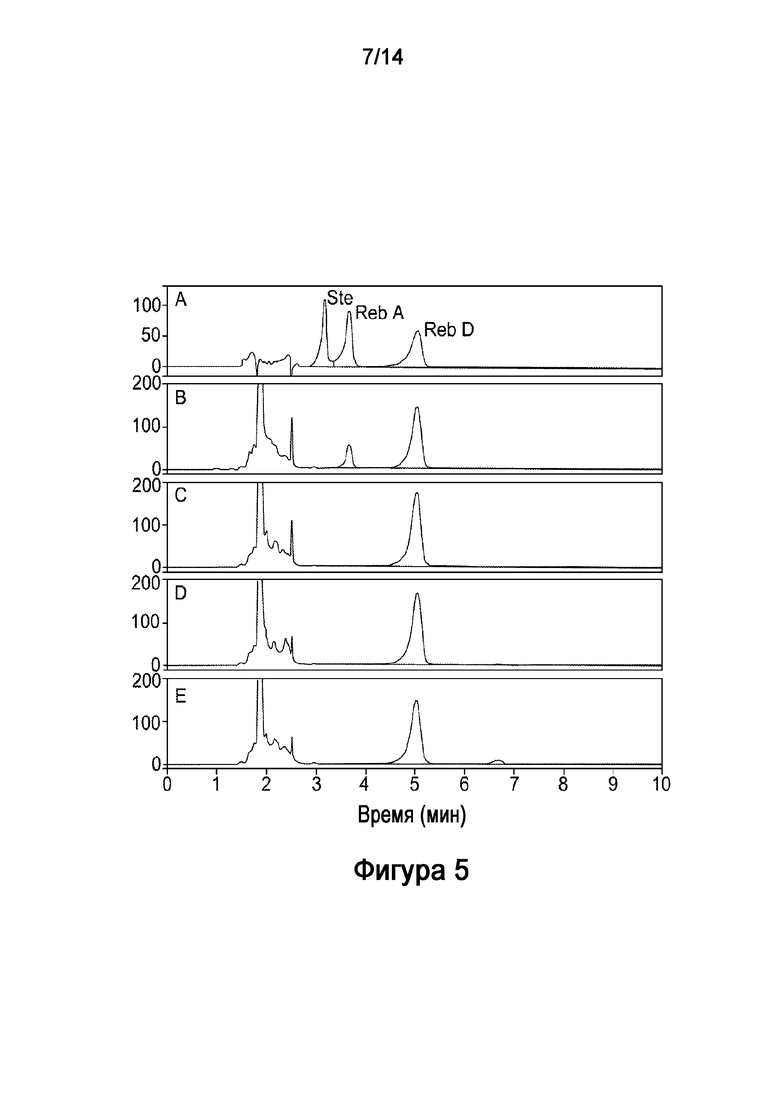
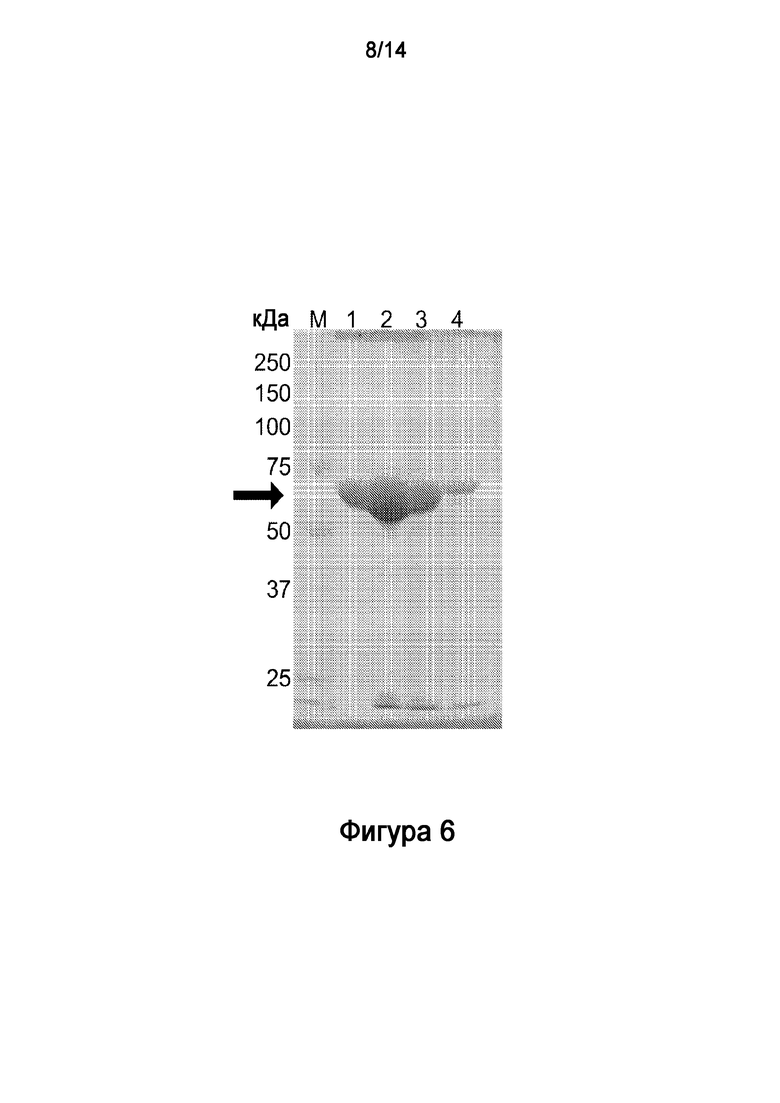
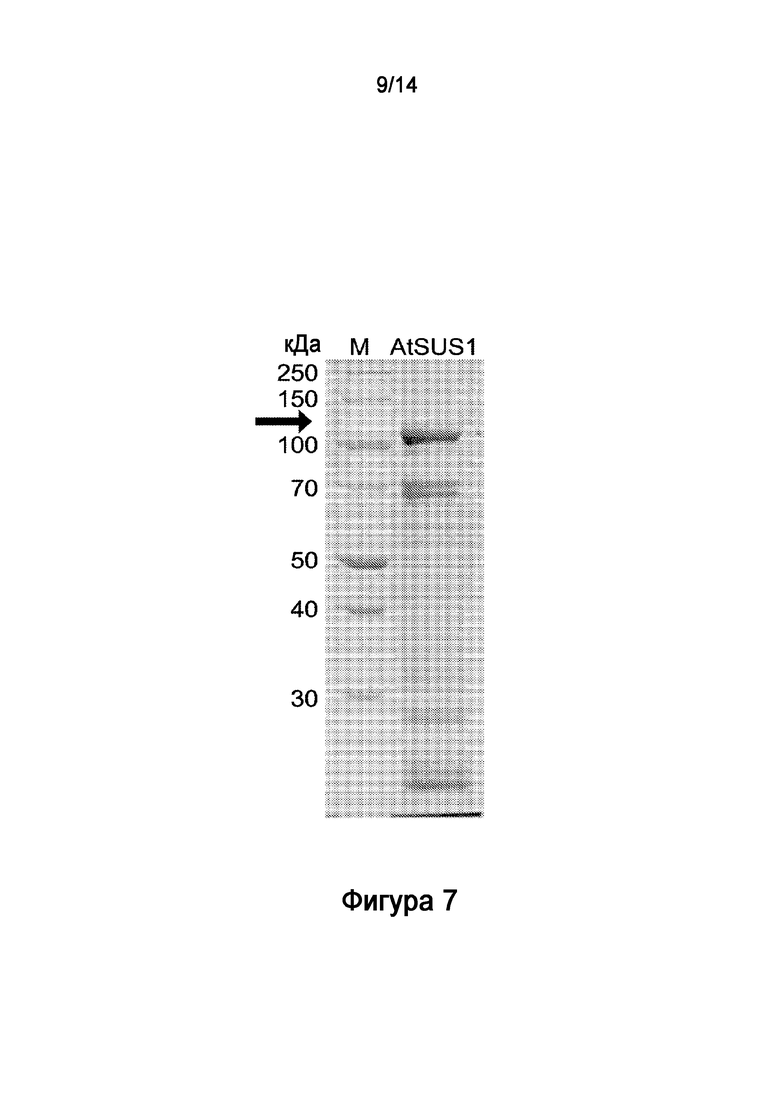
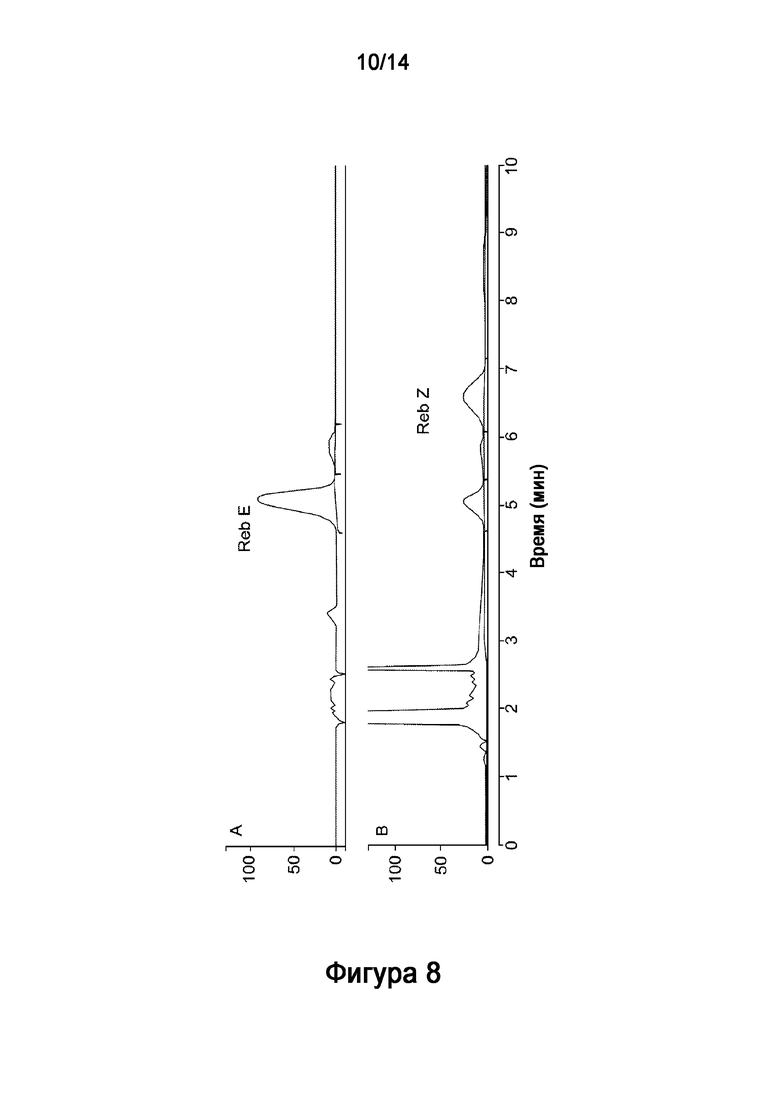
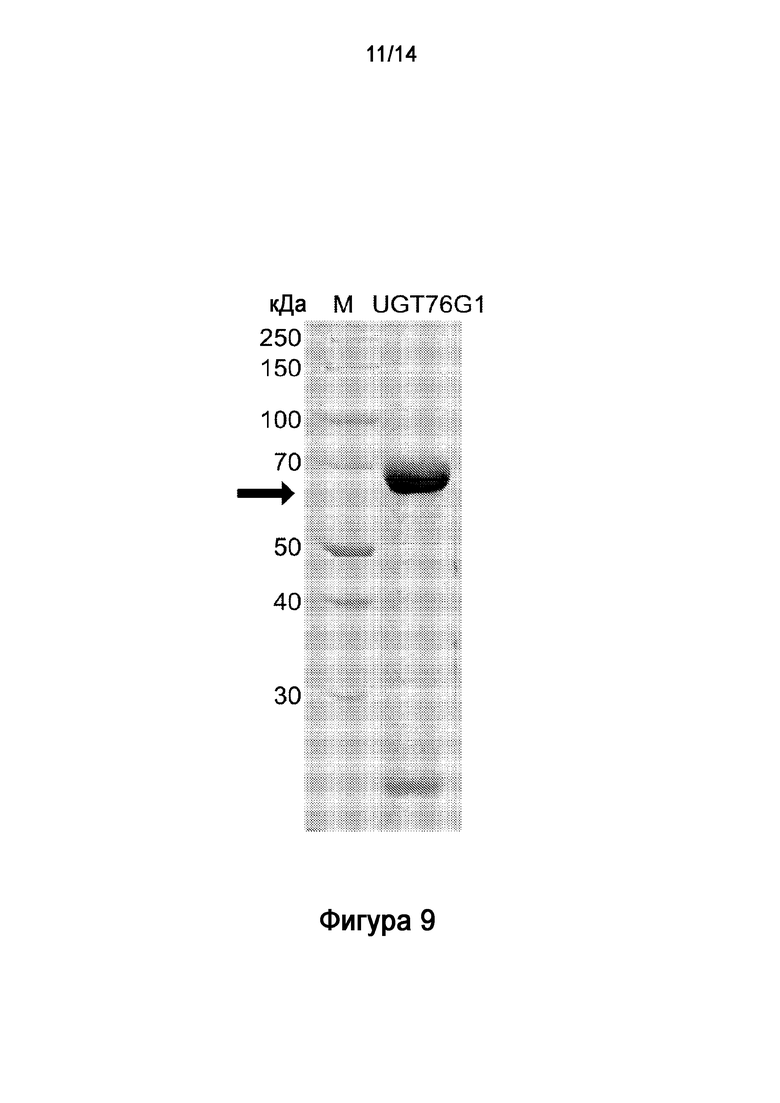
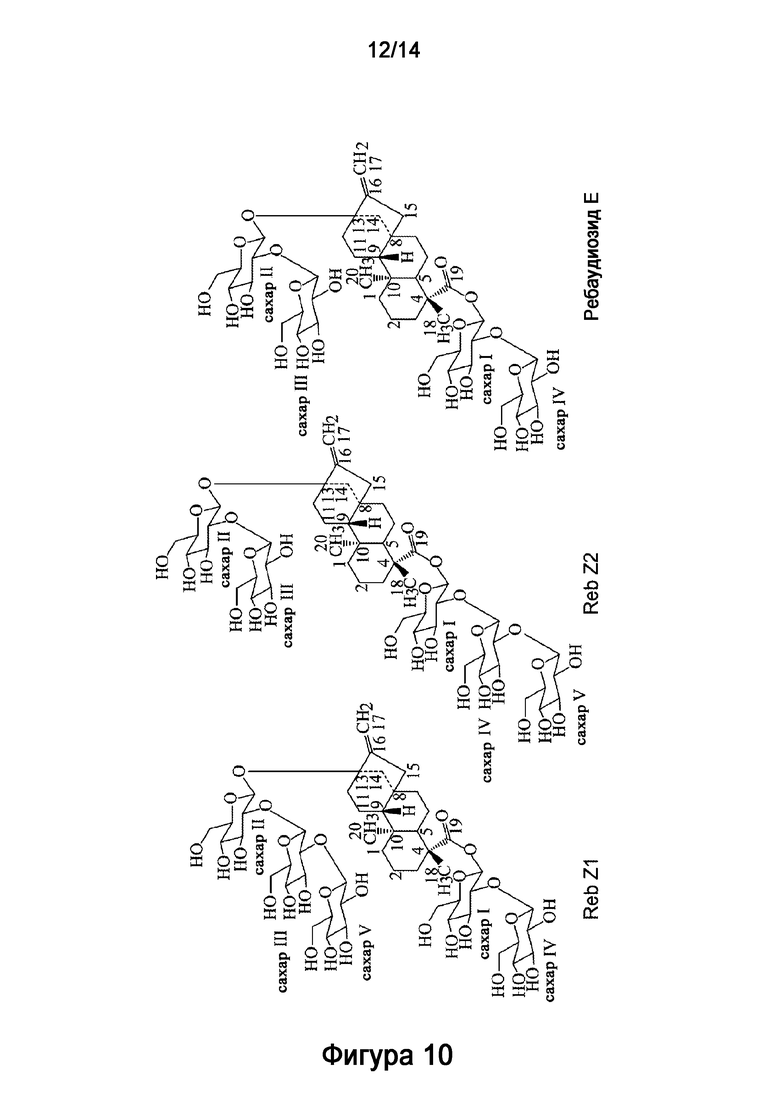
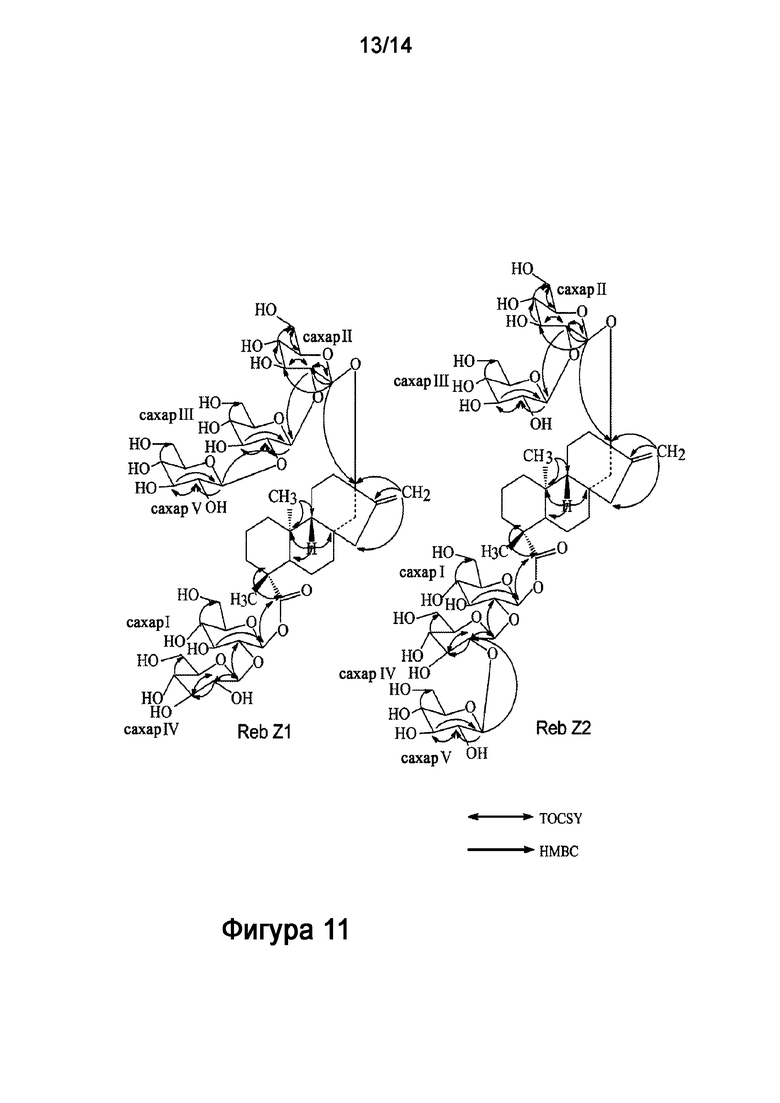
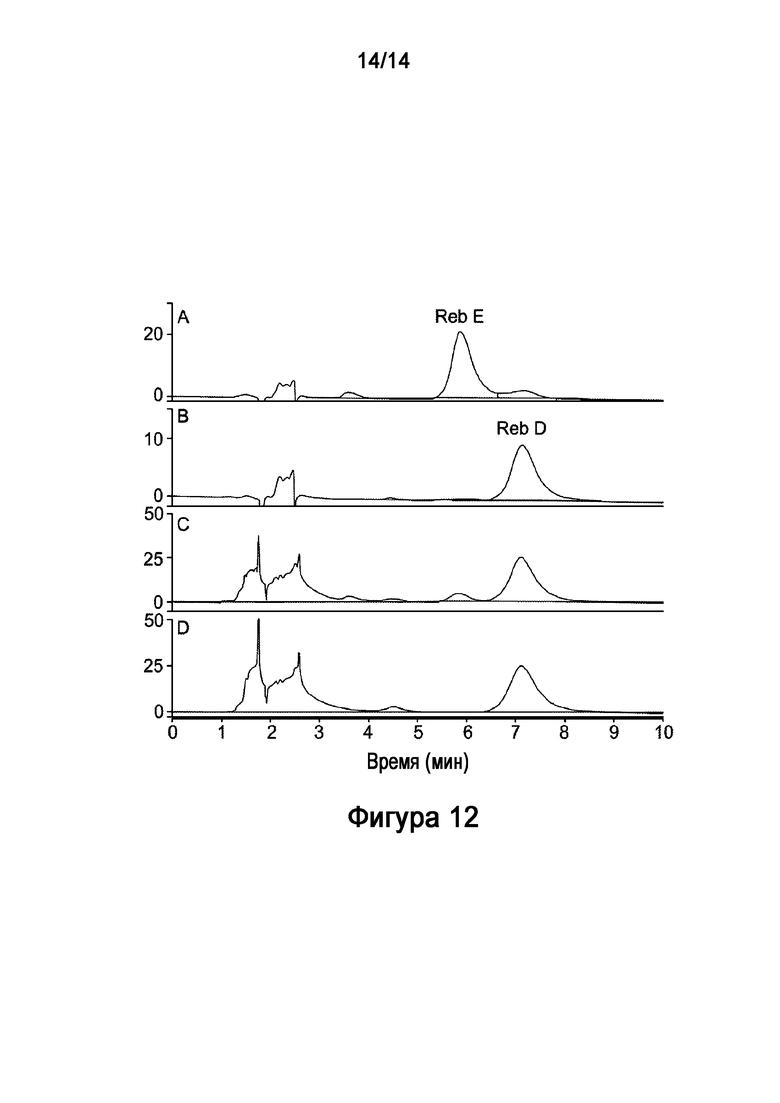
Изобретение относится к области биотехнологии, конкретно к стевиол-гликозиду ребаудиозида Z1, и может быть использовано в пищевой промышленности. Предложен способ получения композиции стевиол-гликозидов с применением рекомбинантных полипептидов, обладающих УДФ-гликозилтрансферазной активностью, включая 1,2-19-O-глюкоза-гликозилирующую активность и 1,2-13-O-глюкоза-гликозилирующую активность. Раскрытый стевиол-гликозид может выступать в качестве подсластителя. 4 н. и 7 з.п. ф-лы, 12 ил., 2 табл., 4 пр.
1. Соединение ребаудиозид Z1, имеющее структуру:
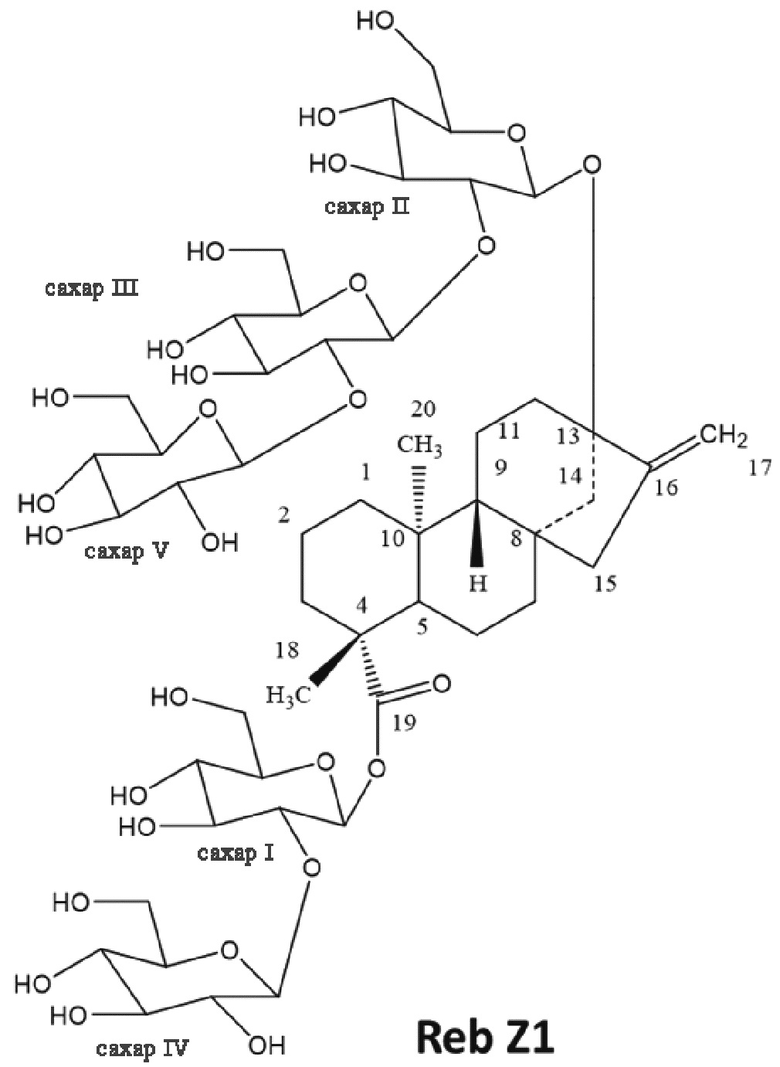
2. Подсластитель, включающий соединение, имеющее химическую структуру:
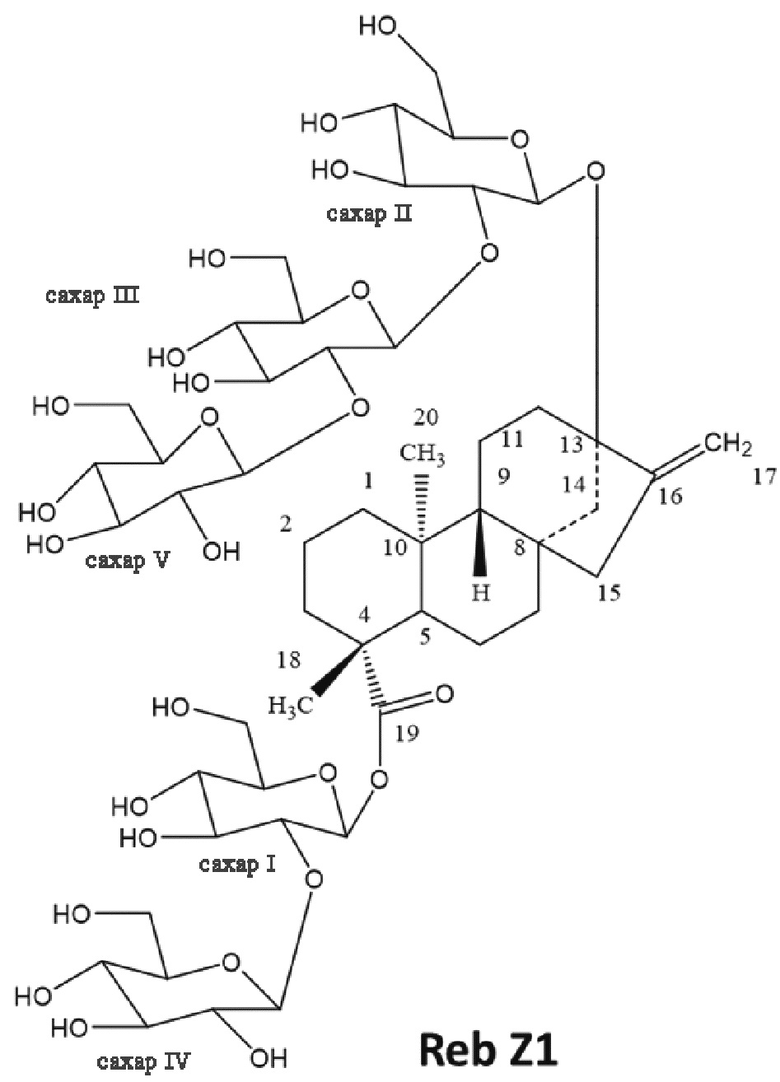
3. Подсластитель по п.2, дополнительно включающий по меньшей мере один компонент из числа наполнителя, объемообразующего вещества и ингибитора слеживания.
4. Пищевой продукт, включающий подслащивающее количество подсластителя по п.2.
5. Пищевой продукт по п.4, выбранный из группы, состоящей из напитков, кондитерских изделий, хлебобулочных изделий, печенья и жевательной резинки.
6. Способ получения соединения по п.1 из ребаудиозида Е, включающий: получение реакционной смеси, содержащей ребаудиозид Е и либо (А) рекомбинантную УДФ-гликозилтрансферазу, содержащую аминокислотную последовательность, имеющую по меньшей мере 80% идентичности с SEQ ID No:6, и УДФ-глюкозу в качестве субстрата, либо (В) сахарозу, УДФ и УДФ-глюкозу в качестве субстрата, и УДФ-гликозилтрансферазу, содержащую аминокислотную последовательность, имеющую по меньшей мере 80% идентичности с SEQ ID No:6, вместе с сахарозосинтазой; и инкубацию реакционной смеси, посредством чего глюкоза ковалентно присоединяется к C2’ 13-O-глюкозы ребаудиозида E с получением соединения по п.1.
7. Способ по п.6, в котором используют сахарозосинтазу, выбранную из группы, состоящей из сахарозосинтазы 1 Arabidopsis, сахарозосинтазы 3 Arabidopsis и сахарозосинтазы Vigna radiate и их функциональных гомологов.
8. Способ по п.7, в котором указанная сахарозосинтаза имеет по меньшей мере 80% идентичности с аминокислотной последовательностью сахарозосинтазы 1 Arabidopsis thaliana, представленной в SEQ ID No:9.
9. Способ по п.6, в котором рекомбинантная УДФ-гликозилтрансфераза содержит аминокислотную последовательность, имеющую по меньшей мере 90% идентичности с SEQ ID No:6.
10. Способ по п.6, в котором рекомбинантная УДФ-гликозилтрансфераза содержит аминокислотную последовательность, имеющую по меньшей мере 95% идентичности с SEQ ID No:6.
11. Способ по п.6, в котором рекомбинантная УДФ-гликозилтрансфераза состоит из аминокислотной последовательности гликозилтрансферазы HV1 согласно SEQ ID No:6.
| База данных "UNIPROT", последовательность F2DT21, размещена 31.05.2011 | |||
| WO 2013022989 A2, 14.02.2013 | |||
| US 2013171328 A1, 04.07.2013 | |||
| MEECH R | |||
| et al., The glycosidation of xenobiotics and endogenous compounds: Versatility and redundancy in the UDP glucpsialtransferase superfamily, PHARM & THER., 2012, v | |||
| Халат для профессиональных целей | 1918 |
|
SU134A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Мяльно-трепальный станок для обработки тресты лубовых растений | 1922 |
|
SU200A1 |
| КОМПОЗИЦИЯ ПОДСЛАСТИТЕЛЯ | 2012 |
|
RU2499427C1 |

