1. ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННУЮ ЗАЯВКУ
[0001] Для настоящей заявки испрашивается приоритет в соответствии с предварительной заявкой США №62/544,718, поданной 11 августа 2017 года, и международной заявкой №PCT/US2017/046637, поданной 11 августа 2017 года, содержание которых включено в настоящий документ полностью посредством ссылки.
2. ОБЛАСТЬ ИЗОБРЕТЕНИЯ
[0002] Настоящее изобретение относится к определенным кауреноксидазам (КО), их содержащим композициям, их содержащим клеткам-хозяевам, а также способам их применения для производства ребаудиозидов, включая ребаудиозид D и ребаудиозид М.
3. ИЗВЕСТНЫЙ УРОВЕНЬ ТЕХНИКИ
[0003] Для ограничения неблагоприятных последствий потребления продуктов с высоким содержанием сахара {например, диабета и ожирения) необходимы подсластители с нулевой калорийностью, полученные из природных источников. Ребаудиозид М (RebM) - одно из множества сладких на вкус соединений, производимых растением стевия (S. rebaudiana Bertoni). Из всех ребаудиозидов RebM самый сладкий (в ~200-300 раз слаще сахарозы) и имеет самый чистый вкус. Тем не менее, RebM производится растением стевия лишь в незначительном количестве и составляет небольшую часть общего содержания стевиол-гликозидов (<1.0%). Ohta et al, 2010, J. Appl. Glycosci., 57, 199-209 (2010). В связи с этим желательно разработать биотехнологические способы производства RebM в больших количествах и с высокой степенью чистоты.
[0004] Для экономичного производства того или иного продукта при помощи биотехнологий каждый этап в биоконверсии из сырья в продукт должен иметь высокий коэффициент эффективности конверсии (в идеальных условиях >90%). В ходе нашей инженерии дрожжей для производства RebM мы выявили явное ограничение в этапе биосинтеза в начале пути производства RebM, на котором энт-каурен превращается в кауреновую кислоту (ФИГ. 1А и 1В).
[0005] Фермент КО присутствует в каждом растении, и обычно он способствует производству растительного гормона гиббереллина. Уровни гиббереллина в клетках растений на порядки ниже уровней RebM, который производится в дрожжах для промышленного производства, и в связи с этим большинство ферментов КО не обеспечивают высокую скорость потока метаболитов, необходимую для производства RebM в целях коммерческого производства. Традиционно, для превращения энт-каурена в кауреновую кислоту в инженерии дрожжей для производства RebM использовали фермент КО из Stevia rebaudiana (Sr.KO). Обычно полагали, что это растение производит высокие уровни стевиол-гликозида, поэтому фермент Sr.KO должен был обеспечивать более высокий коэффициент конверсии, или более высокий поток, по сравнению с большинством других ферментов КО.
[0006] Было обнаружено, что в штамме дрожжей с высоким потоком углерода в RebM фермент Sr.KO имеет низкий коэффициент эффективности превращения в кауреновую кислоту (25.6%), и что образовывались очень высокие уровни вышележащих промежуточных метаболитов (энт-каурена, кауренола и кауренала) (ФИГ. 1С).
[0007] Для производства RebM с высокой эффективностью и чистотой необходимы улучшенные ферменты, способные производить кауреновую кислоту с высокой эффективностью. Представленные здесь композиции и способы позволяют выполнить эту задачу и также предлагают дополнительные преимущества.
4. КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[0008] Здесь представлены композиции и способы улучшенного превращения каурена в кауреновую кислоту. Эти композиции и методы частично основаны на удивительном открытии некоторых кауреноксидаз (КО), способствующих превращать каурен в кауреновую кислоту со значительно высокой эффективностью. Даже небольшое улучшение в продуктивности штамма {например, в десять процентов) с новыми КО может потенциально сократить расходы на производство более чем на десять миллионов долларов в будущем, если предположить, что рыночный спрос на RebM составляет 5000 миллионов тон в год.
[0009] Некоторые описанные здесь КО также способны производить кауреновую кислоту с малым количеством остаточного кауренола или кауренала или вовсе без них. В связи с этим в некоторых вариантах осуществления описанные здесь композиции и способы могут сократить расходы даунстрим-процессинга для получения композиции с высоким выходом стевиол-гликозидов, например, RebM.
[0010] В одном аспекте здесь представлены генетически модифицированные клетки-хозяева и способы их применения для производства полезных в промышленном отношении соединений. В одном аспекте здесь представлена генетически модифицированная клетка-хозяин, содержащая гетерологичную нуклеиновую кислоту, кодирующую кауреноксидазу Pisum sativum. В некоторых вариантах осуществления генетически модифицированная клетка-хозяин также содержит один или более ферментных путей, способных производить стевиол и/или стевиол-гликозиды.
[0011] В некоторых вариантах осуществления здесь представлены генетически модифицированные клетки-хозяева, содержащие гетерологичную нуклеиновую кислоту, кодирующую кауреноксидазу, содержащую аминокислотную последовательность, минимум на 80%, 85%, 90% или 95% идентичную последовательности кауреноксидазы Pisum sativum (например, SEQ ID NO: 1). В некоторых вариантах осуществления генетически модифицированная клетка-хозяин способна превращать каурен в кауреновую кислоту с эффективностью более 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97% или 98%. В некоторых вариантах осуществления такими генетически модифицированными клетками-хозяевами являются дрожжевые клетки. В некоторых вариантах осуществления такими генетически модифицированными клетками-хозяевами являются клетки Saccharomyces cerevisiae.
[0012] В другом аспекте здесь представлены способы производства гетерологичного стевиол-гликозида, способ, включающий выращивание популяции представленных здесь генетически модифицированных клеток-хозяев, способных производить описанный здесь стевиол-гликозид, в среде с источником углерода в условиях, подходящих для производства указанного стевиол-гликозида; а также выделение указанного стевиол-гликозида из среды. В некоторых вариантах осуществления гетерологичный стевиол-гликозид выбирают из группы, состоящей из RebD и RebM.
[0013] В другом аспекте здесь представлены способы производства RebD, способ, включающий выращивание популяции представленных здесь генетически модифицированных клеток-хозяев, способных производить описанный здесь RebD, в среде с источником углерода в условиях, подходящих для производства указанного RebD; а также выделение указанного RebD из среды.
[0014] В другом аспекте здесь представлены способы производства RebM, способ, включающий выращивание популяции представленных здесь генетически модифицированных клеток-хозяев, способных производить описанный здесь RebM, в среде с источником углерода в условиях, подходящих для производства указанного RebM; а также выделение указанного RebM из среды.
[0015] В другом аспекте здесь представлены способы производства кауреновой кислоты, способ, включающий воздействие на каурен описанной здесь кауреноксидазой, способной превращать каурен в кауреновую кислоту в условиях, подходящих для образования кауреновой кислоты.
[0016] В некоторых вариантах осуществления такой клеткой-хозяином является дрожжевая клетка. В некоторых вариантах осуществления такими дрожжами являются Saccharomyces cerevisiae. В некоторых вариантах осуществления клетка-хозяин производит RebD или RebM с высокой эффективностью. В некоторых вариантах осуществления клетка-хозяин производит повышенное количество RebD или RebM по сравнению с дрожжевой клеткой, не содержащей фермент кауреноксидазу Pisum sativum.
5. КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
[0017] На ФИГ. 1А схематически изображено превращение фарнезилпирофосфата в стевиол.
[0018] На ФИГ. 1 В схематически изображено превращение геранилгеранилпирофосфата (GGPP) в RebM.
[0019] На ФИГ. 1С схематически изображено превращение энт-каурена в кауренол в кауренал в кауреновую кислоту.
[0020] На ФИГ. 1D представлена схема мевалонатного пути.
[0021] На ФИГ. 2 представлен пример пути получения RebM из стевиола.
[0022] На ФИГ. 3А представлена схема "посадочной площадки", используемой для того, чтобы вставить отдельные ферменты КО для скрининга в целях производства кауреновой кислоты в дрожжах.
[0023] На ФИГ. 3В представлена схема генетического конструкта КО для скрининга в целях производства кауреновой кислоты в дрожжах.
[0024] На ФИГ. 4 представлена схема, иллюстрирующая относительное повышение количества кауреновой кислоты, произведенной in vivo при использовании различных кауреноксидаз.
[0025] На ФИГ. 5 представлена столбчатая диаграмма, иллюстрирующая относительные уровни энт-каурена, кауренола и кауренала, нормированные по общему количеству кауреновой кислоты, произведенной in vivo в дрожжевом штамме с высоким потоком в RebM.
[0026] На ФИГ. 6 представлена диаграмма, иллюстрирующая относительные уровни титров RebM в штаммах с высоким потоком, содержащих Sr.KO или Ps.KO.
6. ПОДРОБНОЕ ОПИСАНИЕ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
6.1 Терминология
[0027] В контексте настоящего документа термин "гетерологичный" означает то, что обычно не встречается в природе. Термин "гетерологичная нуклеотидная последовательность" означает нуклеотидную последовательность, которая обычно не существует в указанной клетке в природе. В связи с этим гетерологичная нуклеотидная последовательность может: (а) быть чужеродной для клетки-хозяина (т.е. быть "экзогенной" для клетки); (b) естественно существовать в клетке-хозяине (т.е. быть "эндогенной"), однако присутствовать в клетке в неестественном количестве (т.е. в большем или меньшем количестве в сравнении с количеством, естественно присутствующим в клетке-хозяине); или (с) естественно существовать в клетке-хозяине, однако находиться за границами естественного локуса. Термин "гетерологичный фермент" означает фермент, который обычно не существует в указанной клетке в природе. Этот термин охватывает ферменты, которые: (а) экзогенны для указанной клетки (т.е. кодируются нуклеотидной последовательностью, которая в естественном виде не присутствует в клетке-хозяине либо обычно не присутствует в клетке-хозяине в указанных условиях); и (b) естественно присутствуют в клетке-хозяине (например, фермент кодируется нуклеотидной последовательностью, которая эндогенна для клетки), но которые производятся в неестественном количестве (например, в большем или меньшем количестве по сравнению с естественным количеством) в клетке-хозяине.
[0028] С другой стороны, термин "нативный" или "эндогенный" при его использовании в отношении молекул и, в частности, ферментов и нуклеиновых кислот обозначает молекулы, которые экспрессируются в организме, в котором они образовались или существуют в природе, независимо от уровня экспрессии, который может быть ниже, таким же или выше уровня экспрессии молекулы в нативном микроорганизме. Предполагается, что экспрессия нативных ферментов или полинуклеотидов может быть модифицирована в рекомбинантных микроорганизмах.
[0029] В контексте настоящего документа термин "родительская клетка" означает клетку, которая имеет такой же генетический фон, что и описанная здесь генетически модифицированная клетка-хозяин, за тем исключением, что она не содержит одну или более конкретных генетических модификаций модифицированной клетки-хозяина, например, одну или более модификаций, выбранных из группы, состоящей из гетерологичной экспрессии фермента пути стевиола, гетерологичной экспрессии фермента пути стевиол-гликозида, гетерологичной экспрессии геранилгеранилдифосфатсинтазы, гетерологичной экспрессии копалилдифосфатсинтазы, гетерологичной экспрессии кауренсинтазы, гетерологичной экспрессии кауреноксидазы (например, кауреноксидазы Pisum sativum), гетерологичной экспрессии стевиолсинтазы (гидроксилазы кауреновой кислоты), гетерологичной экспрессии цитохром Р450-редуктазы, гетерологичной экспрессии UGT74G1, гетерологичной экспрессии UGT76G1, гетерологичной экспрессии UGT85C2, гетерологичной экспрессии 91D и гетерологичной экспрессии UGT40087, или варианта.
[0030] В контексте настоящего документа термин "встречающийся в природе" означает то, что существует в природе. Например, кауреноксидаза, которая присутствует в организме, который можно выделить из источника в природе, и которая не была намеренно модифицирована человеком в лаборатории, является встречающейся в природе кауреноксидазой. Напротив, в контексте настоящего документа термин "не встречающийся в природе" означает то, что не существует в природе, а создано при участии человека.
[0031] Термин "среда" означает культуральную среду и/или ферментативную среду.
[0032] Термин "ферментативная композиция" означает композицию, которая содержит генетически модифицированные клетки-хозяева и продукты или метаболиты, произведенные генетически модифицированными клетками-хозяевами. Примером ферментативной композиции является цельноклеточный бульон, который может представлять собой все содержимое сосуда (например, колбы, чашки или ферментера), включая клетки, водную фазу и соединения, произведенные из генетически модифицированных клеток-хозяев.
[0033] В контексте настоящего документа термин "производство", в общем, означает количество стевиола или стевиол-гликозида, произведенного описанной здесь генетически модифицированной клеткой-хозяином. В некоторых вариантах осуществления производство выражается как выход стевиола или стевиол-гликозида для клетки-хозяина. В других вариантах осуществления производство выражается как продуктивность клетки-хозяина в плане производства стевиола или стевиол-гликозида.
[0034] В контексте настоящего документа термин "продуктивность" означает производство стевиола или стевиол-гликозида клеткой-хозяином, выраженное как количество стевиола или стевиол-гликозида, произведенное (по весу) на количество ферментативного бульона, в котором культивируется клетка-хозяин (по объему) с течением времени (в час).
[0035] В контексте настоящего документа термин "выход" означает производство стевиола или стевиол-гликозида клеткой-хозяином, выраженное как количество стевиола или стевиол-гликозида, произведенное на количество источника углерода, потребленного клеткой-хозяином, по весу.
[0036] В контексте настоящего документа термин "необнаруживаемый уровень" соединения (например, RebM2, стевиол-гликозидов или других соединений) означает уровень соединения, который слишком низкий для того, чтобы его можно было измерить и/или проанализировать при использовании стандартного метода для измерения количества соединения. Например, этот термин включает уровень соединения, который не обнаруживается аналитическими способами, описанными в Примере 6.
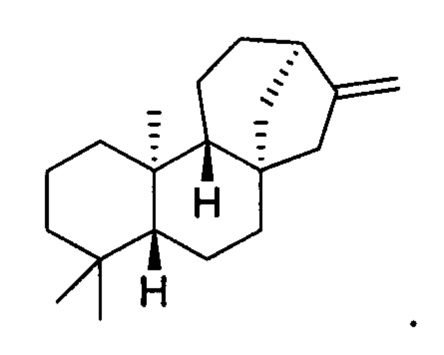
[0037] Термин "каурен" означает соединение каурен, включая любой стереоизомер каурена. В конкретных вариантах осуществления этот термин означает энантиомер, известный в данной области как энт-каурен. В конкретных вариантах осуществления этот термин означает соединение со следующей структурой:
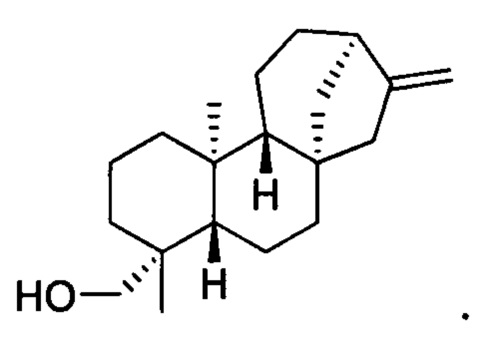
[0038] Термин "кауренол" означает соединение кауренол, включая любой стереоизомер кауренола. В конкретных вариантах осуществления этот термин означает энантиомер, известный в данной области как энт-кауренол. В конкретных вариантах осуществления этот термин означает соединение со следующей структурой:
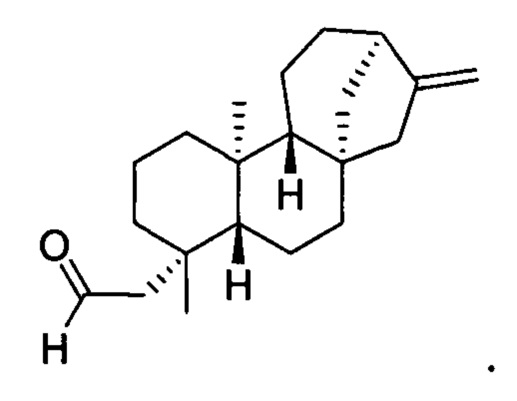
[0039] Термин "кауренал" означает соединение кауренал, включая любой стереоизомер кауренала. В конкретных вариантах осуществления этот термин означает энантиомер, известный в данной области как энт-кауренал. В конкретных вариантах осуществления этот термин означает соединение со следующей структурой:
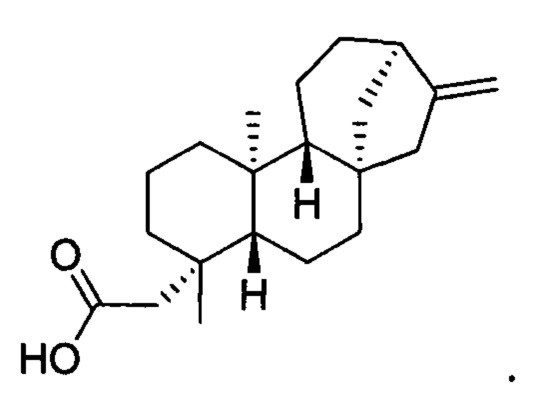
[0040] Термин "кауреновая кислота" означает соединение кауреновую кислоту, включая любой стереоизомер кауреновой кислоты. В конкретных вариантах осуществления этот термин означает энантиомер, известный в данной области как энт-кауреновая кислота. В конкретных вариантах осуществления этот термин означает соединение со следующей структурой:
[0041] В контексте настоящего документа термин "стевиол-гликозид(ы)" означает гликозид стевиола, включая, помимо прочего, встречающиеся в природе стевиол-гликозиды, например, стевиолмонозид, стевиолбиозид, рубузозид, дулкозид В, дулкозид А, ребаудиозид В, ребаудиозид G, стевиозид, ребаудиозид С, ребаудиозид F, ребаудиозид А, ребаудиозид I, ребаудиозид Е, ребаудиозид Н, ребаудиозид L, ребаудиозид К, ребаудиозид J, ребаудиозид М, ребаудиозид D, ребаудиозид N, ребаудиозид О, синтетические стевиол-гликозиды, например, глюкозилированные ферментативным путем стевиол-гликозиды и их комбинации.
[0042] В контексте настоящего документа термин "вариант" означает полипептид, который отличается от конкретного указанного "эталонного" полипептида (например, последовательности дикого типа) аминокислотными вставками, делециями, мутациями и/или заменами, но сохраняет активность, по существу аналогичную активности эталонного полипептида. В некоторых вариантах осуществления такой вариант создают при использовании технологий рекомбинантных ДНК, например, мутагенеза. В некоторых вариантах осуществления вариант полипептида отличается от эталонного полипептида заменой одного основного остатка на другой (т.е. Apr вместо Лиз), заменой одного гидрофобного остатка на другой (т.е. Лей вместо Иле) или заменой одного ароматического остатка на другой (т.е. Фен вместо Тир) и т.д. В некоторых вариантах осуществления варианты включают аналоги, когда получают консервативные замены, обуславливающие значительную структурную аналогию с эталонной последовательностью. Примеры таких консервативных замен, помимо прочего, включают замену аспарагиновой кислоты глутаминовой кислотой и наоборот; аспарагина глутамином и наоборот; треонина серином и наоборот; аргинина лизином и наоборот; или изолейцина, валина или лейцина друг другом.
[0043] В контексте настоящего документа термин "идентичность последовательностей" или "идентичность в процентах" в контексте двух или более последовательностей нуклеиновых кислот или белков означают две или более последовательности или подпоследовательности, которые одинаковы или имеют определенный процент одинаковых аминокислотных остатков или нуклеотидов. Например, последовательность может быть минимум на 60%, минимум на 65%, минимум на 70%, минимум на 75%, минимум на 80%, минимум на 85%, минимум на 90%, минимум на 91%, минимум на 92%, минимум на 93%, минимум на 94%, минимум на 95%, минимум на 96%, минимум на 97%, минимум на 98%, минимум на 99% или более идентична в заданной области эталонной последовательности при сравнении и выравнивании для максимального соответствия в окне сравнения либо в заданной области, измеренной при использовании алгоритма сравнения последовательностей или посредством выравнивания вручную и визуального контроля. Например, процент идентичности определяется вычислениями посредством деления числа идентичных нуклеотидов (или аминокислотных остатков) в последовательности на длину всех нуклеотидов (или аминокислотных остатков) за вычетом длин любых пробелов.
[0044] Для удобства степень идентичности между двумя последовательностями можно проверить при помощи компьютерной программы и математических алгоритмов, известных в данной области. Такие алгоритмы, позволяющие рассчитать идентичность последовательностей, обычно учитывают пробелы в последовательностях и несовпадения в области сравнения. Для данной цели полезны такие программы сравнения и выравнивания последовательностей, как Clustal W (Thompson et al., (1994) Nucleic Acids Res., 22: 4673-4680), ALIGN (Myers et al., (1988) CABIOS, 4: 11-17), FASTA (Pearson et al., (1988) PNAS, 85:2444-2448; Pearson (1990), Methods Enzymol., 183: 63-98) и Gapped BLAST (Altschul et al., (1997) Nucleic Acids Res., 25: 3389-3402). Программа BLAST или BLAST 2.0 (Altschul et al., J. Mol. Biol. 215:403-10, 1990) доступна из различных источников, включая Национальный центр биологической информации США (NCBI) и интернет, и может использоваться в сочетании с программами анализа последовательностей BLASTP, BLASTN, BLASTX, TBLASTN и TBLASTX. Дополнительную информацию можно узнать на веб-сайте NCBI.
[0045] В определенных вариантах осуществления выравнивание последовательностей и вычисления процента идентичности можно производить при помощи программы BLAST с использованием ее стандартных параметров (по умолчанию). Для выранивания нуклеотидных последовательностей и расчетов идентичности последовательностей используется программа BLASTN со своими параметрами по умолчанию (Параметры: Gap opening penalty=5, Gap extension penalty=2, Nucleic match=2, Nucleic mismatch=-3, Expectation value=10.0, Word size=11, Max matches in a query range=0). Для выравнивания полипептидных последовательностей и расчетов идентичности последовательностей используется программа BLASTP со своими параметрами по умолчанию (Параметры: Alignment matrix=BLOSUM62; Gap costs: Existence=11, Extension=1; Compositional adjustments=Conditional compositional score, matrix adjustment; Expectation value=10.0; Word size=6; Max matches in a query range=0. В качестве варианта используется следующая программа и параметры: Align Plus software of Clone Manager Suite, версия 5 (Sci-Ed Software); сравнение ДНК: Global comparison, Standard Linear Scoring matrix, Mismatch penalty=2, Open gap penalty=4, Extend gap penalty=1. Сравнение аминокислотных последовательностей: Global comparison, BLOSUM 62 Scoring matrix. В описанных здесь вариантах осуществления идентичность последовательностей рассчитывается при помощи программ BLASTN или BLASTP с использованием параметров по умолчанию. В описанных здесь вариантах осуществления выравнивание двух или более последовательностей выполняется при помощи Clustal W с использованием предлагаемых параметров по умолчанию (Параметры: Dealign input sequences: no; Mbed-like clustering guide-tree: yes; Mbed-like clustering iteration: yes; number of combined iterations: default(0); Max guide tree iterations: default; Max HMM iterations: default; Order: input).
6.2 Клетки-хозяева
[0046] Здесь представлены клетки-хозяева, способные производить кауреновую кислоту (КА) из каурена с высокой эффективностью. В определенных вариантах осуществления клетки-хозяева могут производить кауреновую кислоту из каурена как исходного сырья. В конкретных вариантах осуществления клетки-хозяева могут производить кауреновую кислоту из источника углерода в культуральной среде. В конкретных вариантах осуществления клетки-хозяева могут производить кауреновую кислоту из источника углерода в культуральной среде и далее могут производить RebA или RebD из кауреновой кислоты. В конкретных вариантах осуществления клетки-хозяева могут далее производить ребаудиозид М (RebM) из RebD.
[0047] В конкретных вариантах осуществления клетки-хозяева имеют ферментную активность кауреноксидазы Pisum sativum. Фермент кауреноксидаза Pisum sativum способен превращать каурен в кауреновую кислоту с высокой эффективностью. В определенных вариантах осуществления фермент кауреноксидаза Pisum sativum способен превращать каурен в кауреновую кислоту с эффективностью более 30%. В определенных вариантах осуществления фермент кауреноксидаза Pisum sativum способен превращать каурен в кауреновую кислоту с эффективностью более 35%. В определенных вариантах осуществления фермент кауреноксидаза Pisum sativum способен превращать каурен в кауреновую кислоту с эффективностью более 40%. В определенных вариантах осуществления фермент кауреноксидаза Pisum sativum способен превращать каурен в кауреновую кислоту с эффективностью более 95%. В определенных вариантах осуществления фермент кауреноксидаза Pisum sativum способен превращать каурен в кауреновую кислоту с эффективностью более 50%. В определенных вариантах осуществления фермент кауреноксидаза Pisum sativum способен превращать каурен в кауреновую кислоту с эффективностью более 55%. В определенных вариантах осуществления фермент кауреноксидаза Pisum sativum способен превращать каурен в кауреновую кислоту с эффективностью около 58%. В определенных вариантах осуществления фермент кауреноксидаза Pisum sativum способен превращать каурен в кауреновую кислоту с эффективностью более 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или 95%.
[0048] В определенных вариантах осуществления клетка-хозяин способна превращать каурен в кауреновую кислоту с эффективностью более 30%. В определенных вариантах осуществления клетка-хозяин способна превращать каурен в кауреновую кислоту с эффективностью более 35%. В определенных вариантах осуществления клетка-хозяин способна превращать каурен в кауреновую кислоту с эффективностью более 40%. В определенных вариантах осуществления клетка-хозяин способна превращать каурен в кауреновую кислоту с эффективностью более 45%. В определенных вариантах осуществления клетка-хозяин способна превращать каурен в кауреновую кислоту с эффективностью более 50%. В определенных вариантах осуществления клетка-хозяин способна превращать каурен в кауреновую кислоту с эффективностью более 55%. В определенных вариантах осуществления клетка-хозяин способна превращать каурен в кауреновую кислоту с эффективностью около 58%. В определенных вариантах осуществления клетка-хозяин способна превращать каурен в кауреновую кислоту с эффективностью более 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или 95%.
[0049] Эффективность превращения можно измерить при помощи любого метода, известного специалистам в данной области. В определенных вариантах осуществления эффективность превращения можно измерить воздействием на каурен ферментом или клеткой-хозяином в подходящих условиях для образования кауреновой кислоты. Эффективность можно измерить путем сравнения молярного количества произведенной кауреновой кислоты и общего количества каурена и кауреновой кислоты в полученной композиции. Эффективность также можно измерить посредством сравнения общего количества кауреновой кислоты и нижележащих продуктов кауреновой кислоты с общим количеством каурена, кауренола, кауренала, кауреновой кислоты и нижележащих продуктов кауреновой кислоты в полученной композиции. Например, коэффициенты эффективности конверсии штаммов, содержащих Ps.KO, показанных на ФИГ. 5, измерили посредством сравнения общего количества кауреновой кислоты и всех нижележащих соединений, показанных на ФИГ. 2, с общим количеством каурена, кауренола, кауренала, кауреновой кислоты и всех нижележащих продуктов, показанных на ФИГ.2, в полученной композиции (т.е. стевиол, 1 глюкоза + стевиол, 2 глюкоза + стевиол, 3 глюкоза + стевиол, 4 глюкоза + стевиол, 5 глюкоза + стевиол и 6 глюкоза + стевиол).
[0050] В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие кауреноксидазу, содержащую аминокислотную последовательность SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие кауреноксидазу, содержащую аминокислотную последовательность, по существу идентичную аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие кауреноксидазу, содержащую аминокислотную последовательность, которая минимум на 60% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие кауреноксидазу, содержащую аминокислотную последовательность, которая минимум на 65% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие кауреноксидазу, содержащую аминокислотную последовательность, которая минимум на 70% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие кауреноксидазу, содержащую аминокислотную последовательность, которая минимум на 75% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие кауреноксидазу, содержащую аминокислотную последовательность, которая минимум на 80% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие кауреноксидазу, содержащую аминокислотную последовательность, которая минимум на 85% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие кауреноксидазу, содержащую аминокислотную последовательность, которая минимум на 90% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие кауреноксидазу, содержащую аминокислотную последовательность, которая минимум на 95% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие кауреноксидазу, содержащую аминокислотную последовательность, которая минимум на 96% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие кауреноксидазу, содержащую аминокислотную последовательность, которая минимум на 97% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие кауреноксидазу, содержащую аминокислотную последовательность, которая минимум на 98% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие кауреноксидазу, содержащую аминокислотную последовательность, которая минимум на 99% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие кауреноксидазу, содержащую аминокислотную последовательность, которая минимум на 60%, минимум на 99% или минимум на любое число между 60% и 99% идентична аминокислотной последовательности SEQ ID NO: 1.
[0051] В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие кауреноксидазу, содержащую описанную здесь аминокислотную последовательность и способную превращать каурен в кауреновую кислоту. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие кауреноксидазу, содержащую описанную здесь аминокислотную последовательность и способную к окислению позиции 19 каурена, кауренола и кауренала. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие кауреноксидазу, способную превращать каурен в кауреновую кислоту с эффективностью более 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96% или 97%, и в которых кауреноксидаза содержит аминокислотную последовательность, которая минимум на 95% идентична аминокислотной последовательности SEQ ID №1.
[0052] В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие нуклеиновую кислоту, кодирующую кауреноксидазу Pisum sativum, содержащую аминокислотную последовательность SEQ ID №1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, по существу идентичную аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 60% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 65% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 70% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 75% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 80% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 85% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 90% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 95% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 96% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 97% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 98% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 99% идентична аминокислотной последовательности SEQ ID NO: 1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 60%, минимум на 99% или на любое число между 60% и 99% идентична аминокислотной последовательности SEQ ID NO: 1.
[0053] В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность SEQ ID №14, которая кодирует кауреноксидазу Pisum sativum, имеющую последовательность SEQ ID №1. В определенных вариантах осуществления здесь представлены клетки-хозяева, содержащие гетерологичную нуклеиновую кислоту, содержащую нуклеотидную последовательность, которая минимум на 60%, минимум на 65%, минимум на 70%, минимум на 75%, минимум на 80%, минимум на 85%, минимум на 90%, минимум на 95%, минимум на 96%, минимум на 97%, минимум на 98%, минимум на 99% идентична нуклеотидной последовательности SEQ ID №14.
[0054] В определенных вариантах осуществления клетка-хозяин содержит вариант полипептида кауреноксидазы Pisum sativum, описанного выше. В определенных вариантах осуществления этот вариант может содержать до 15, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 аминокислотной замены относительно полипептида кауреноксидазы Pisum sativum. В определенных вариантах осуществления этот вариант может содержать до 15, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 консервативной аминокислотной замены относительно полипептида кауреноксидазы Pisum sativum. В определенных вариантах осуществления любую из описанных здесь нуклеиновых кислот можно оптимизировать для клетки-хозяина, например, посредством оптимизации кодонов.
[0055] В описанных здесь вариантах осуществления можно использовать любой подходящий способ для определения соответствующих аминокислотных позиций или соответствующих локализаций петель двух полипептидов. В определенных вариантах осуществления последовательности кауреноксидазы и эталонную последовательность SEQ ID №1 можно выровнять при помощи Clustal(W) с использованием параметров по умолчанию. В другом варианте осуществления последовательности кауреноксидазы и эталонная последовательность SEQ ID №1 могут быть выравнены посредством структурных выравниваний, например, при использовании SWISS-MODEL, сервера моделирования по гомологии структуры белка, доступного через веб-сервер ExPASy или из программы Deep View (Swiss Pdb-Viewer).
[0056] В определенных вариантах осуществления каурен соответствует ФИГ. 1С. В определенных вариантах осуществления кауреноксидаза Pisum sativum или вариант кауреноксидазы Pisum sativum способны катализировать окисление каурена в С-19 для образования кауренола. В определенных вариантах осуществления кауреноксидаза Pisum sativum или вариант кауреноксидазы Pisum sativum способны катализировать окисление в С-19 для образования кауренала. В определенных вариантах осуществления кауреноксидаза Pisum sativum способна катализировать окисление кауренала в С-19 для образования кауреновой кислоты. В конкретных вариантах осуществления кауреноксидаза Pisum sativum или вариант кауреноксидазы Pisum sativum способны катализировать окисление каурена в С-19 для образования кауренола, окисление кауренола в С-19 для образования кауренала и окисление кауренала в С-19 для образования кауреновой кислоты.
[0057] В определенных вариантах осуществления RebD соответствует ФИГ.2. В определенных вариантах осуществления клетка-хозяин содержит один или более ферментов, способных превратить кауреновую кислоту в стевиол. В определенных вариантах осуществления клетка-хозяин содержит один или более ферментов, способных превратить стевиол в один или более стевиол-гликозидов. В определенных вариантах осуществления клетка-хозяин содержит один или более ферментов, способных превратить RebA в RebD. В определенных вариантах осуществления клетка-хозяин содержит один или более ферментов, способных превратить RebD в RebM.
[0058] При том, что кауреноксидаза Pisum sativum или любой вариант кауреноксидазы Pisum sativum клеток-хозяев принимает каурен как субстрат, источником каурена может быть любой источник, который специалисты в данной области считают подходящим. В определенных вариантах осуществления на кауреноксидазу Pisum sativum или любой вариант кауреноксидазы Pisum sativum можно воздействовать кауреном. В определенных вариантах осуществления на клетку-хозяина можно воздействовать кауреном. В определенных вариантах осуществления на кауреноксидазу Pisum sativum или любой вариант кауреноксидазы Pisum sativum можно воздействовать композицией, содержащей каурен, или кауренол, или кауренал, или их комбинации. В определенных вариантах осуществления эта композиция содержит каурен. В определенных вариантах осуществления эта композиция содержит кауренол. В определенных вариантах осуществления эта композиция содержит кауренал. В определенных вариантах осуществления эту композицию получают из природных продуктов, выделенных из листьев Stevia rebaudiana. В определенных вариантах осуществления эта композиция имеет микробное происхождение. В определенных вариантах осуществления на клетку-хозяина можно воздействовать композицией, содержащей один или более источников углерода.
[0059] В определенных вариантах осуществления любой вариант кауреноксидазы Pisum sativum, подходящий для катализации необходимой реакции, можно подвергнуть скринингу для любых подходящих способов, известных в данной области. Например, подходящий вариант кауреноксидазы Pisum sativum можно проанализировать in vivo посредством экспрессии гетерологичной нуклеиновой кислоты, кодирующей вариант кауреноксидазы Pisum sativum, и скрининга клеток, которые производят функциональный вариант кауреноксидазы Pisum sativum, способный катализировать окисление в необходимой локализации субстрата (например, позиции С-19 каурена, кауренола и/или кауренала). Примеры способов скрининга описаны в нижеприведенных Примерах. В другом примере подходящий вариант кауреноксидазы Pisum sativum можно подвергнуть скринингу in vitro посредством воздействия на вариант кауреноксидазы Pisum sativum таким субстратом, как каурен, кауренол и/или кауренал. В данном примере анализ присутствия кауреновой кислоты, стевиола или стевиол-гликозида, такого как RebD, можно использовать для определения того, является ли вариант кауреноксидазы Pisum sativum подходящим ферментом. Эту реакцию можно проанализировать посредством жидкостной хроматографии с масс-спектрометрией или при помощи других способов, известных в данной области. См., например, WO 2013/022989.
[0060] В определенных вариантах осуществления вариант кауреноксидазы Pisum sativum считается подходящим для превращения каурена в кауреновую кислоту, если он способен превращать каурен в кауреновую кислоту с эффективностью более 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96% или 97% in vivo.
[0061] В определенных вариантах осуществления вариант кауреноксидазы Pisum sativum считается подходящим для превращения каурена в кауренол, если он способен превращать каурен в кауренол с эффективностью более 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96% или 97% in vivo.
[0062] В определенных вариантах осуществления вариант кауреноксидазы Pisum sativum считается подходящим для превращения кауренола в кауренал, если он способен превращать кауренол в кауренал с эффективностью более 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96% или 97% in vivo.
[0063] В определенных вариантах осуществления вариант кауреноксидазы Pisum sativum считается подходящим для превращения кауренала в кауреновую кислоту, если он способен превращать кауренал в кауреновую кислоту с эффективностью более 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96% или 97% in vivo.
[0064] В определенных вариантах осуществления вариант кауреноксидазы Pisum sativum считается подходящим для превращения каурена в кауреновую кислоту, если эффективность превращения превышает 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 96% или 97% in vivo, при этом эффективность превращения рассчитывается делением общего количества кауреновой кислоты и всех нижестоящих соединений, показанных на ФИГ. 2, на общее количество каурена, кауренола, кауренала, кауреновой кислоты и всех нижестоящих соединений, показанных на ФИГ. 2, в полученной композиции (умножить на 100 процентов).
[0065] В эффективных вариантах осуществления клетка-хозяин может содержать один или более ферментных путей, способных производить каурен, при этом указанные пути берутся по-отдельности или вместе. В определенных вариантах осуществления клетки-хозяева содержат один или более ферментов, способных превращать геранилгеранилдифосфат в каурен. Специалистам в данной области известны полезные ферменты и нуклеиновые кислоты, кодирующие эти ферменты. В определенных вариантах осуществления клетки-хозяева содержат один или более ферментов, способных превращать геранилгеранилдифосфат в каурен. В других эффективных вариантах осуществления клетка-хозяин может содержать один или более ферментативных путей, способных превращать кауреновую кислоту в стевиол и/или стевиол-гликозиды, при этом указанные пути берутся по-отдельности или вместе. Специалистам в данной области известны полезные ферменты и нуклеиновые кислоты, кодирующие эти ферменты. Особенно полезные ферменты и нуклеиновые кислоты описаны в нижеприведенных разделах и также описаны, например, в US 2014/0329281 A1, US 2014/0357588 A1, US 2015/0159188, WO 2016/038095 A2 и US 2016/0198748 A1.
[0066] В дальнейших вариантах осуществления клетки-хозяева содержат один или более ферментов, способных производить геранилгеранилдифосфат из источника углерода. Они включают ферменты DXP-пути и ферменты MEV-пути. Специалистам в данной области известны полезные ферменты и нуклеиновые кислоты, кодирующие эти ферменты. Примеры ферментов каждого пути описаны далее и также описаны, например, в US 2016/0177341 A1. MEV-путь также показан на ФИГ. 1D.
[0067] В определенных вариантах осуществления эти дополнительные ферменты являются нативными. В эффективных вариантах осуществления эти дополнительные ферменты являются гетерологичными. В определенных вариантах осуществления два фермента могут быть объединены в один полипептид.
6.3 Полипептиды и нуклеиновые кислоты кауреноксидазы, не встречающейся в природе
[0068] В другом аспекте здесь представлены варианты кауреноксидазы, не встречающиеся в природе, которые включают модификацию (модификации) аминокислотных остатков в сравнении с эталонной последовательностью (например, SEQ ID №1) и при этом все равно сохраняют действие кауреноксидазы по превращению каурена в кауреновую кислоту, каурен в кауренол, кауренол в кауренал и/или кауренал в кауреновую кислоту. В определенных вариантах осуществления варианты кауреноксидазы, не встречающиеся в природе, могут включать до 20, 15, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 аминокислотной замены, делеции, добавления и/или вставки в определенных аминокислотных позициях или локализациях в сравнении с эталонной последовательностью (например, SEQ ID №1). В определенных вариантах осуществления варианты кауреноксидазы, не встречающиеся в природе, содержат любой из вариантов кауреноксидазы, которые описаны в настоящем документе.
[0069] В другом аспекте здесь представлены варианты кауреноксидазы, не встречающиеся в природе, которые включают модификацию (модификации) остатков нуклеиновых кислот в сравнении с эталонной последовательностью (например, SEQ ID №1) и при этом все равно сохраняют действие кауреноксидазы по превращению каурена в кауреновую кислоту, каурен в кауренол, кауренол в кауренал и/или кауренал в кауреновую кислоту. В определенных вариантах осуществления варианты кауреноксидазы, не встречающиеся в природе, могут кодировать любой из описанных здесь вариантов кауреноксидазы.
6.4 Клеточные штаммы
[0070] Представленные здесь полезные композиции с клетками-хозяевами и способы включают археи, прокариотические или эукариотические клетки.
[0071] Подходящие прокариотические хозяева включают, помимо прочего, любую из разновидностей грамположительных, грамотрицательных или грамвариабельных бактерий. Примеры включают, помимо прочего, клетки, принадлежащие к родам: Agrobacterium, Alicyclobacillus, Anabaena, Anacystis, Arthrobacter, Azobacter, Bacillus, Brevibacterium, Chromatium, Clostridium, Corynebacterium, Enterobacter, Erwinia, Escherichia, Lactobacillus, Lactococcus, Mesorhizobium, Methylobacterium, Microbacterium, Phormidium, Pseudomonas, Rhodobacter, Rhodopseudomonas, Rhodospirillum, Rhodococcus, Salmonella, Scenedesmun, Serratia, Shigella, Staphlococcus, Strepromyces, Synnecoccus и Zymomonas. Примеры прокариотеческих штаммов включают, помимо прочего: Bacillus subtilis, Bacillus amyloliquefacines, Brevibacterium ammoniagenes, Brevibacterium immariophilum, Clostridium beigerinckii, Enterobacter sakazakii, Escherichia coli, Lactococcus lactis, Mesorhizobium loti, Pseudomonas aeruginosa, Pseudomonas mevalonii, Pseudomonas pudica, Rhodobacter capsulatus, Rhodobacter sphaeroides, Rhodospirillum rubrum, Salmonella enterica, Salmonella typhi, Salmonella typhimurium, Shigella dysenteriae, Shigella flexneri, Shigella sonnei и Staphylococcus aureus. В конкретном варианте осуществления клеткой-хозяином является клетка Escherichia coli.
[0072] Подходящие археи-хозяева включают, помимо прочего, клетки, принадлежащие к родам: Aeropyrum, Archaeglobus, Halobacterium, Methanococcus, Methanobacterium, Pyrococcus, Sulfolobus и Thermoplasma. Примеры штаммов архей включают, помимо прочего: Archaeoglobus fulgidus, Halobacterium sp., Methanococcus jannaschii, Methanobacterium thermoautotrophicum, Thermoplasma acidophilum, Thermoplasma volcanium, Pyrococcus horikoshii, Pyrococcus abyssi и Aeropyrum pernix.
[0073] Подходящие эукариотические хозяева включают, помимо прочего, клетки грибов, клетки водорослей, клетки насекомых и клетки растений. В некоторых вариантах осуществления дрожжи, полезные в представленные способах, включают дрожжи, которые были включены в депозитарии микроорганизмов (например, IFO, АТСС и т.д.) и принадлежат к родам Aciculoconidium, Ambrosiozyma, Arthroascus, Arxiozyma, Ashbya, Babjevia, Bensingtonia, Botryoascus, Botryozyma, Brettanomyces, Bullera, Bulleromyces, Candida, Citeromyces, Clavispora, Cryptococcus, Cystofllobasidium, Debaryomyces, Dekkara, Dipodascopsis, Dipodascus, Eeniella, Endomycopsella, Eremascus, Eremothecium, Erythrobasidium, Fellomyces, Filobasidium, Galactomyces, Geotrichum, Guilliermondella, Hanseniaspora, Hansenula, Hasegawaea, Holtermannia, Hormoascus, Hyphopichia, Issatchenkia, Kloeckera, Kloeckeraspora, Kluyveromyces, Kondoa, Kuraishia, Kurtzmanomyces, Leucosporidium, Lipomyces, Lodderomyces, Malassezia, Metschnikowia, Mrakia, Myxozyma, Nadsonia, Nakazawaea, Nematospora, Ogataea, Oosporidium, Pachysolen, Phachytichospora, Phaffla, Pichia, Rhodosporidium, Rhodotorula, Saccharomyces, Saccharomycodes, Saccharomycopsis, Saitoella, Sakaguchia, Saturnospora, Schizoblastosporion, Schizosaccharomyces, Schwanniomyces, Sporidiobolus, Sporobolomyces, Sporopachydermia, Stephanoascus, Sterigmatomyces, Sterigmatosporidium, Symbiotaphrina, Sympodiomyces, Sympodiomycopsis, Torulaspora, Trichosporiella, Trichosporon, Trigonopsis, Tsuchiyaea, Udeniomyces, Waltomyces, Wickerhamia, Wickerhamiella, Williopsis, Yamadazyma, Yarrowia, Zygoascus, Zygosaccharomyces, Zygowilliopsis и Zygozyma, среди прочих.
[0074] В некоторых вариантах осуществления микробом-хозяином является Saccharomyces cerevisiae, Pichia pastoris, Schizosaccharomyces pombe, Dekkera bruxellensis, Kluyveromyces lactis (previously called Saccharomyces lactis), Kluveromyces marxianus, Arxula adeninivorans или Hansenula polymorpha (сейчас известный как Pichia angustd). В некоторых вариантах осуществления микробом-хозяином является штамм рода Candida, такого как Candida lipolytica, Candida guilliermondii, Candida krusei, Candida pseudotropicalis или Candida utilis.
[0075] В конкретном варианте осуществления микробом-хозяином является Saccharomyces cerevisiae. В некоторых вариантах осуществления хозяином является штамм Saccharomyces cerevisiae, выбранный из группы, состоящей из пекарских дрожжей, CBS 7959, CBS 7960, CBS 7961, CBS 7962, CBS 7963, CBS 7964, IZ-1904, ТА, BG-1, CR-1, SA-1, М-26, Y-904, РЕ-2, РЕ-5, VR-1, BR-1, BR-2, МЕ-2, VR-2, МА-3, МА-4, САТ-1, СВ-1, NR-1, ВТ-1 и AL-1. В некоторых вариантах осуществления микробом-хозяином является штамм Saccharomyces cerevisiae, выбранный из группы, состоящей из РЕ-2, САТ-1, VR-1, BG-1, CR-1 и SA-1. В конкретном варианте осуществления таким штаммом Saccharomyces cerevisiae является РЕ-2. В другом конкретном варианте осуществления таким штаммом Saccharomyces cerevisiae является САТ-1. В другом конкретном варианте осуществления таким штаммом Saccharomyces cerevisiae является BG-1.
[0076] В некоторых вариантах осуществления микробом-хозяином является микроб, который подходит для промышленной ферментации. В конкретных вариантах осуществления принимают меры для обеспечения выживания микроба в условиях высокой концентрации растворителя, высокой температуры, расширенного применения субстрата, лимитирования питательными веществами, осмотического стресса из-за сахара и солей, кислотности, загрязнения сульфитами и бактериями или их комбинациями, которые считаются стрессовыми условиями для среды промышленной ферментации.
6.5 Пути биосинтеза стевиола и стевиол-гликозида
[0077] В некоторых вариантах осуществления путь биосинтеза стевиола и/или путь биосинтеза стевиол-гликозида активируется в представленных здесь генетически модифицированных клетках-хозяевах посредством инженерии клеток для экспрессии полинуклеотидов и/или полипептидов, кодирующих один или более ферментов пути. На ФИГ. 1 В показан пример пути биосинтеза стевиола. На ФИГ. 2 показан пример биосинтеза стевиол-гликозида, начиная с геранилгеранилпирофосфата в различные стевиол-гликозиды.
[0078] Таким образом, в некоторых вариантах осуществления представленные здесь генетически модифицированные клетки-хозяева содержат гетерологичный полинуклеотид, кодирующий полипептид, имеющий активность геранилгеранилдифосфатсинтазы (GGPPS). В некоторых вариантах осуществления представленные здесь генетически модифицированные клетки-хозяева содержат гетерологичный полинуклеотид, кодирующий полипептид, имеющий активность копалилдифосфатсинтазы или энт-копалилпирофосфатсинтазы (CDPS; также называемой энт-копалилпирофосфатсинтазой или CPS). В некоторых вариантах осуществления представленные здесь генетически модифицированные клетки-хозяева содержат гетерологичный полинуклеотид, кодирующий полипептид, имеющий активность кауренсинтазы (KS; также называемой энт-кауренсинтазой). В конкретных вариантах осуществления представленные здесь генетически модифицированные клетки-хозяева содержат гетерологичный полинуклеотид, кодирующий полипептид, имеющий активность кауреноксидазы (КО; также называемой энт-каурен 19-оксидазой), которая описана в настоящем документе. В некоторых вариантах осуществления представленные здесь генетически модифицированные клетки-хозяева содержат гетерологичный полинуклеотид, кодирующий полипептид, имеющий активность стевиолсинтазы (также называемой 13-гидроксилазой энт-кауреновой кислоты или КАН). В некоторых вариантах осуществления представленные здесь генетически модифицированные клетки-хозяева содержат гетерологичный полинуклеотид, кодирующий полипептид, имеющий активность цитохром Р450 редуктазы (CPR).
[0079] В некоторых вариантах осуществления представленные здесь генетически модифицированные клетки-хозяева содержат гетерологичный полинуклеотид, кодирующий полипептид, имеющий активность UGT74G1. В некоторых вариантах осуществления представленные здесь генетически модифицированные клетки-хозяева содержат гетерологичный полинуклеотид, кодирующий полипептид, имеющий активность UGT76G1. В некоторых вариантах осуществления представленные здесь генетически модифицированные клетки-хозяева содержат гетерологичный полинуклеотид, кодирующий полипептид, имеющий активность UGT85C2. В некоторых вариантах осуществления представленные здесь генетически модифицированные клетки-хозяева содержат гетерологичный полинуклеотид, кодирующий полипептид, имеющий активность UGT91D. В некоторых вариантах осуществления представленные здесь генетически модифицированные клетки-хозяева содержат гетерологичный полинуклеотид, кодирующий полипептид, имеющий активность UDP-гликозилтрансферазы.
[0080] В определенных вариантах осуществления клетка-хозяин содержит вариант.В определенных вариантах осуществления такой вариант содержит 15, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 аминокислотной замены относительно релевантного полипептида. В некоторых вариантах осуществления этот вариант может содержать до 15, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 консервативной аминокислотной замены относительно эталонного полипептида. В некоторых вариантах осуществления любую из описанных здесь нуклеиновых кислот можно оптимизировать для клетки-хозяина, например, посредством оптимизации ко донов.
[0081] Далее описаны примеры нуклеиновых кислот и ферментов пути биосинтеза стевиола и/или пути биосинтеза стевиол-гликозида.
6.5.1 Герани лгеранилдифосфатсинтаза (GGPPS)
[0082] Геранилгеранилдифосфатсинтазы (ЕС 2.5.1.29) катализируют превращение фарнезилпирофосфата в геранилгеранилдифосфат.Иллюстративные примеры ферментов включают ферменты Stevia rebaudiana (учетный номер ABD92926), Gibberella fujikuroi (учетный номер САА75568), Mus musculus (учетный номер ААН69913), Thalassiosira pseudonana (учетный номер ХР_002288339), Streptomyces clavuligerus (учетный номер ZP_05004570), Sulfulobus acidocaldarius (учетный номер ВАА43200), Synechococcus sp. (учетный номер АВС98596), Arabidopsis thaliana (учетный номер NP_195399), Blakeslea trispora (учетный номер AFC92798.1) и US 2014/0329281 A1. Нуклеиновые кислоты, кодирующие эти ферменты, полезны в представленных здесь клетках и способах. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, имеющая последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одной из таких нуклеиновых кислот GGPPS. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, которая кодирует полипептид, имеющий последовательность, которая минимум на 80%, 85%, 90%, 95% идентична последовательности минимум одного из этих ферментов GGPPS.
6.5.2 Копалилдифосфатсинтаза (CDPS)
[0083] Копалилдифосфатсинтазы (ЕС 5.5.1.13) катализируют превращение фарнезилпирофосфата в геранилгеранилдифосфат. Иллюстративные примеры ферментов включают ферменты Stevia rebaudiana (учетный номер ААВ87091), Streptomyces clavuligerus (учетный номер EDY51667), Bradyrhizobium japonicum (учетный номер ААС28895.1), Zea mays (учетный номер AY562490), Arabidopsis thaliana (учетный номер NM_116512), Oryza sativa (учетный номер Q5MQ85.1) и US 2014/0329281 A1. Нуклеиновые кислоты, кодирующие эти ферменты, полезны в представленных здесь клетках и способах. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, имеющая последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одной из таких нуклеиновых кислот CDPS. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, которая кодирует полипептид, имеющий последовательность, которая минимум на 80%, 85%, 90% или 95%о идентична последовательности минимум одного из этих ферментов CDPS.
6.5.3 Кауренсинтаза (KS)
[0084] Кауренсинтазы (ЕС 4.2.3.19) катализируют превращение копалилдифосфата в каурен и дифосфат.Иллюстративные примеры ферментов включают ферменты Bradyrhizobium japonicum (учетный номер ААС28895.1), Phaeosphaeria sp.(учетный номер 013284), Arabidopsis thaliana (учетный номер Q9SAK2), Picea glauca (учетный номер ADB55711.1) и US 2014/0329281 A1. Нуклеиновые кислоты, кодирующие эти ферменты, полезны в представленных здесь клетках и способах. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, имеющая последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одной из таких нуклеиновых кислот KS. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, которая кодирует полипептид, имеющий последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одного из этих ферментов KS.
6.5.4 Бифункциональные копалилдифосфатсинтаза (CDPS) и кауренсинтаза (KS)
[0085] Также могут использоваться CDPS-KS бифункциональные ферменты (ЕС 5.5.1.13 и ЕС 4.2.3.19). Иллюстративные примеры ферментов включают ферменты Phomopsis amygdali (учетный номер BAG30962), Physcomitrella patens (учетный номер BAF61135), Gibberella fujikuroi (учетный номер Q9UVY5.1) и US 2014/0329281 A1, US 2014/0357588 A1, US 2015/0159188 и WO 2016/038095 A2. Нуклеиновые кислоты, кодирующие эти ферменты, полезны в представленных здесь клетках и способах. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, имеющая последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одной из таких нуклеиновых кислот CDPS-KS. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, которая кодирует полипептид, имеющий последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одного из этих ферментов CDPS-KS.
6.5.5 Энт-кауреноксидаза (КО)
[0086] В настоящем документе описаны энт-кауреноксидазы (ЕС 1.14.13.78; также называемые кауреноксидазы). Нуклеиновые кислоты, кодирующие эти ферменты, полезны в представленных здесь клетках и способах. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, имеющая последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одной из таких нуклеиновых кислот кауреноксидазы. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, которая кодирует полипептид, имеющий последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одного из этих ферментов кауреноксидазы.
6.5.6 Стевиолсинтаза (КАН)
[0087] Стевиолсинтазы, или гидроксилазы кауреновой кислоты (КАН), (ЕС 1.14.13) катализируют превращение кауреновой кислоты в стевиол. Иллюстративные примеры ферментов включают ферменты Stevia rebaudiana (учетный номер ACD93722), Stevia rebaudiana (SEQ ID №10) Arabidopsis thaliana (учетный номер NP_197872), Vitis vinifera (учетный номер XP_002282091), Medicago trunculata (учетный номер ABC59076) и US 2014/0329281 A1, US 2014/0357588 A1, US 2015/0159188 и WO 2016/038095 A2. Нуклеиновые кислоты, кодирующие эти ферменты, полезны в представленных здесь клетках и способах. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, имеющая последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одной из таких нуклеиновых кислот КАН. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, которая кодирует полипептид, имеющий последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одного из этих ферментов КАН.
6.5.7 Цитохром Р450 редуктаза (CPR)
[0088] Цитохром Р450 редуктазы (ЕС 1.6.2.4) способны способствовать активности вышеуказанных КО и/или КАН. Иллюстративные примеры ферментов включают ферменты Stevia rebaudiana (учетный номер АВВ88839) Arabidopsis thaliana (учетный номер NP_194183), Gibberella fujikuroi (учетный номер САЕ09055), Artemisia annua (учетный номер АВС47946.1) и US 2014/0329281 A1, US 2014/0357588 A1, US 2015/0159188 и WO 2016/038095 A2. Нуклеиновые кислоты, кодирующие эти ферменты, полезны в представленных здесь клетках и способах. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, имеющая последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одной из таких нуклеиновых кислот CPR. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, которая кодирует полипептид, имеющий последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одного из этих ферментов CPR.
6.5.8 UDP-гликозилтрансфераза 74G1 (UGT74G1)
[0089] UGT74G1 способна функционировать как уридин 5'-дифосфоглюкозил: стевиол 19-СООН трансфераза и как уридин 5'-дифосфоглюкозил: стевиол-13-О-глюкозид 19-СООН трансфераза. Как показано на ФИГ. 2, UGT74G1 способна превращать стевиол в 19-гликозид. UGT74G1 также способна превращать стевиолмонозид в рубузозид. UGT74G1 также может быть способна превращать стевиолбиозид в стевозид. Иллюстративные примеры ферментов включают ферменты Stevia rebaudiana (например, ферменты в Richman et al., 2005, Plant J. 41: 56-67 и US 2014/0329281 и WO 2016/038095 A2 и учетный номер AAR06920.1). Нуклеиновые кислоты, кодирующие эти ферменты, полезны в представленных здесь клетках и способах. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, имеющая последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одной из таких нуклеиновых кислот UGT74G1. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, которая кодирует полипептид, имеющий последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одного из этих ферментов UGT74G1.
6.5.9 UDP-гликозилтрансфераза 76G1 (UGT76G1)
[0090] UGT76G1 способна превращать остаток глюкозы в С-3' C-13-О-глюкозы акцепторной молекулы, стевиол 1,2 гликозид. Таким образом, UGT76G1 способна функционировать как уридин 5'-дифосфоглюкозил: стевиол 13-O-1,2 глюкозид С-3' глюкозилтрансферза и уридин 5'-дифосфоглюкозил: стевиол-19-О-глюкоза, 13-O-1,2 биозид С-3' глюкозилтрансфераза. Как показано на ФИГ. 2, UGT76G1 способна превращать стевиолбиозид в RebB. UGT76G1 также способна превращать стевиозид в RebA. UGT76G1 также способна превращать RebD в RebM. Иллюстративные примеры ферментов включают ферменты Stevia rebaudiana (например, ферменты в работе Richman et al., 2005, Plant J. 41: 56-67 и US 2014/0329281 A1 и WO 2016/038095 A2 и учетный номер AAR06912.1). Нуклеиновые кислоты, кодирующие эти ферменты, полезны в представленных здесь клетках и способах. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, имеющая последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одной из таких нуклеиновых кислот UGT76G1. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, которая кодирует полипептид, имеющий последовательность, которая минимум на 80%, 85%, 90%) или 95% идентична последовательности минимум одного из этих ферментов UGT76G1.
6.5.10 UDP-гликозилтрансфераза 85С2 (UGT85C2)
[0091] UGT85C2 способна функционировать как уридин 5'-дифосфоглюкозил:стевиол 13-ОН трансфераза и уридин 5'-дифосфоглюкозил:стевиол-19-O-глюкозид 13-ОН трансфераза. Таким образом, как показано на ФИГ. 2, UGT85C2 способна превращать стевиол в стевиолмонозид и также способна превращать 19-гликозид в рубузозид. Иллюстративные примеры ферментов включают ферменты Stevia rebaudiana (например, ферменты в работе Richman et al., 2005, Plant J. 41: 56-67 и US 2014/0329281 A1 и WO 2016/038095 A2 и учетный номер AAR06916.1). Нуклеиновые кислоты, кодирующие эти ферменты, полезны в представленных здесь клетках и способах. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, имеющая последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одной из таких нуклеиновых кислот UGT85C2. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, которая кодирует полипептид, имеющий последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одного из этих ферментов UGT85C2.
6.5.11 UDP-гликозилтрансфераза 91D (UGT91D)
[0092] UGT91D способна функционировать как уридин 5'-дифосфоглюкозил:стевиол-13-О-глюкозидтрансфераза, превращая остаток глюкозы в С-2' 13-О-глюкозы акцепторной молекулы, стевиол-13-О-глюкозид (стевиолмонозид) для производства стевиобиозида. UGT91D также способна функционировать как уридин 5'-дифосфоглюкозил:рубузозидтрансфераза, превращая остаток глюкозы в С-2' 13-О-глюкозы акцепторной молекулы, рубузозид, для производства стевиозида, что показано на ФИГ. 2. UGT91D также называется UGT91D2, UGT91D2e или UGT91D-like3. Иллюстративные примеры ферментов UGT91D включают ферменты Stevia rebauidana (например, ферменты UGT-последовательности с учетным номером АСЕ87855.1, US 2014/0329281 A1, WO 2016/038095 А2 и SEQ ID №7). Нуклеиновые кислоты, кодирующие эти ферменты, полезны в представленных здесь клетках и способах. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, имеющая последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одной из таких нуклеиновых кислот UGT91D. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, которая кодирует полипептид, имеющий последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одного из этих ферментов UGT91D.
6.5.12 Уридиндифосфат-зависимая гликозилтрансфераза, способная превращать RebA в RebD (UGTad)
[0093] Уридиндифосфат-зависимая гликозилтрансфераза (UGTad) способна превращать остаток глюкозы в С-2' позицию 19-О-глюкозы RebA для производства RebD, что показано на ФИГ. 2. UGTAD также способна превращать остаток глюкозы в С-2' позицию 19-О-глюкозы стевиозида для производства RebE. Полезные примеры UGT включают Os_UGT_91C1 из Oryza sativa (также называется EUGT11 в работе Houghton-Larsen et al., WO 2013/022989 A2; XP_015629141.1) и S1_UGT_101249881 из Solarium lycopersicum (также называется UGTSL2 в работе Markosyan et al., WO 2014/193888 A1; XP_004250485.1). Другие полезные UGT включают UGT40087 (XP_004982059.1), sr.UGT_9252778 (SEQ ID №16), Bd_UGT10840 (XP_003560669.1), Hv_UGT_V1 (BAJ94055.1), Bd_UGT10850 (XP_010230871.1) и Ob_UGT91Bl_like (XP_006650455.1). В описанных здесь композициях и способах может использоваться любая UGT или любой вариант UGT. Нуклеиновые кислоты, кодирующие эти ферменты, полезны в представленных здесь клетках и способах. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, имеющая последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одной из таких UGT. В определенных вариантах осуществления здесь представлены клетки и способы, в которых используется нуклеиновая кислота, которая кодирует полипептид, имеющий последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности минимум одной из этих UGT. В определенных вариантах осуществления здесь представлена нуклеиновая кислота, кодирующая описанный здесь вариант UGT.
[0094] В определенных вариантах осуществления генетически модифицированные клетки-хозяева содержат гетерологичную нуклеиновую кислоту, кодирующую UDP-гликозилтрансферазу, содержащую аминокислотную последовательность, которая минимум на 80%, 85%, 90% или 95% идентична последовательности UGT40087 (например, SEQ ID №17 или SEQ ID №18. В определенных вариантах осуществления генетически модифицированная клетка-хозяин способна превращать RebA в RebD с эффективностью более 90%, 95%, 96% или 97%. В определенных вариантах осуществления генетически модифицированная клетка-хозяин содержит UDP-гликозилтрансферазу, содержащую сахарный акцепторный домен, в котором аминокислотная последовательность сахарного акцепторного домена минимум на 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности сахарного акцепторного домена SEQ ID №17 или SEQ ID №18. В определенных вариантах осуществления генетически модифицированная клетка-хозяин содержит UDP-гликозилтрансферазу, которая содержит аминокислотную последовательность петли 1, вариант аминокислотной последовательности петли 1, аминокислотную последовательность петли 2, вариант аминокислотной последовательности петли 2, аминокислотную последовательность петли 3_1, вариант аминокислотной последовательности петли 3_1, аминокислотную последовательность петли 3_2, вариант аминокислотной последовательности петли 3_2, аминокислотную последовательность петли 4_1, вариант аминокислотной последовательности петли 4_1, аминокислотную последовательность петли 4_2 или любую их комбинацию. В определенных вариантах осуществления генетически модифицированная клетка-хозяин содержит UDP-гликозилтрансферазу, которая содержит аминокислотную последовательность, которая минимум на 61%, 65%, 70%, 75%, 80%, 85%, 90% или 95% идентична сахарному акцепторному домену SEQ ID №17 или SEQ ID №18, и также содержит аминокислотную последовательность петли 4_1 SEQ ID №17 или SEQ ID №18.
[0095] В контексте настоящего документа термин "вариант аминокислотной последовательности петли 1" означает аминокислотную последовательность, которая отличается от эталонной аминокислотной последовательности петли 1 SEQ ID №17 или 18 (или модифицированной последовательности UGT40087 петли 1, имеющей последовательность SEQ ID №28) одной, двумя, тремя, четырьмя, пятью, шестью, семью, восьмью, девятью или десятью аминокислотными вставками, делециями, мутациями и/или заменами, но допускает UDP-гликозилтрансферазу, содержащую вариант аминокислотной последовательности петли 1, вставленный в участке, который соответствует участку аминокислотной последовательности петли 1 SEQ ID №17 или 18, соответственно, для катализациии превращения RebA в RebD и/или стевиозида в RebE.
[0096] В контексте настоящего документа термин "вариант аминокислотной последовательности петли 2" означает аминокислотную последовательность, которая отличается от эталонной аминокислотной последовательности петли 2 SEQ ID №17 или 18 одной, двумя, тремя, четырьмя, пятью, шестью, семью, восьмью, девятью или десятью аминокислотными вставками, делециями, мутациями и/или заменами, но допускает UDP-гликозилтрансферазу, содержащую вариант аминокислотной последовательности петли 2, вставленный в участке, который соответствует участку аминокислотной последовательности петли 2 SEQ ID №17 или 18, соответственно, для катализациии превращения RebA в RebD и/или стевиозида в RebE.
[0097] В контексте настоящего документа термин "вариант аминокислотной последовательности петли 3_1" означает аминокислотную последовательность, которая отличается от эталонной аминокислотной последовательности петли 3_1 SEQ ID №17 или 18 одной, двумя, тремя, четырьмя, пятью, шестью, семью, восьмью, девятью или десятью аминокислотными вставками, делециями, мутациями и/или заменами, но допускает UDP-гликозилтрансферазу, содержащую вариант аминокислотной последовательности петли 3_1, вставленный в участке, который соответствует участку аминокислотной последовательности петли 3_1 SEQ ID №17 или 18, соответственно, для катализациии превращения RebA в RebD и/или стевиозида в RebE. В контексте настоящего документа термин "вариант аминокислотной последовательности петли 3_2" означает аминокислотную последовательность, которая отличается от эталонной аминокислотной последовательности петли 3_2 SEQ ID №17 или 18 одной, двумя, тремя, четырьмя, пятью, шестью, семью, восьмью, девятью или десятью аминокислотными вставками, делециями, мутациями и/или заменами, но допускает UDP-гликозилтрансферазу, содержащую вариант аминокислотной последовательности петли 3_2, вставленный в участке, который соответствует участку аминокислотной последовательности петли 3_2 SEQ ID №17 или 18, соответственно, для катализациии превращения RebA в RebD и/или стевиозида в RebE. В определенных вариантах осуществления вариант аминокислотной последовательности петли 3_2 отличается от эталонной последовательности петли 3_2 одной, двумя, тремя, четырьмя, пятью, шестью, семью, восьмью, девятью, десятью аминокислотными вставками, делециями, мутациями и/или заменами или аминокислотными вставками, делециями, мутациями и/или заменами числом до тридцати.
[0098] В контексте настоящего документа термин "вариант аминокислотной последовательности петли 4_1" означает аминокислотную последовательность, которая отличается от эталонной аминокислотной последовательности петли 4_1 SEQ ID №17 или 18 одной, двумя, тремя, четырьмя, пятью, шестью, семью, восьмью, девятью, десятью аминокислотными вставками, делециями, мутациями и/или заменами или аминокислотными вставками, делециями, мутациями и/или заменами числом до тридцати, но допускает UDP-гликозилтрансферазу, содержащую вариант аминокислотной последовательности петли 4_1, вставленный в участке, который соответствует участку аминокислотной последовательности петли 4_1 SEQ ID №17 или 18, соответственно, для катализациии превращения RebA в RebD и/или стевиозида в RebE.
[0099] В определенных вариантах осуществления клетки-хозяева содержат функциональный домен UGT40087, в котором UGT40087 содержит аминокислотную последовательность SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий N-терминальный сахарный акцепторный домен UGT40087, содержащий аминокислотную последовательность SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий С-терминальный сахарный донорный домен UGT40087, содержащий аминокислотную последовательность SEQ ID №17 или 18. В определенных вариантах осуществления сахарный акцепторный домен UGT40087 содержит аминокислотные позиции от 1 до 214 последовательности SEQ ID №18 (что соответствует аминокислотным позициям от 1 до 215 последовательности SEQ ID №17). В определенных вариантах осуществления сахарный донорный домен UGT40087 содержит аминокислотные позиции от 215 до 435 последовательности SEQ ID №18 (что соответствует аминокислотным позициям от 216 до 436 последовательности SEQ ID №17). В определенных вариантах осуществления сахарный акцепторный домен UGT40087 содержит аминокислотные позиции от 1 до 215 последовательности SEQ ID №17. В определенных вариантах осуществления сахарный донорный домен содержит аминокислотные позиции от 216 до 436 последовательности SEQ ID №17. В определенных вариантах осуществления сахарный акцепторный домен и сахарный донорный домен UGT40087 содержат более узкий диапазон аминокислотных остатков, чем от 1 до 214 или от 215 до 435, соответственно, в отношении последовательности SEQ ID №18. В определенных вариантах осуществления сахарный акцепторный домен и сахарный донорный домен UGT40087 содержат более узкий диапазон аминокислотных остатков, чем от 1 до 215 или от 216 до 436, соответственно, в отношении последовательности SEQ ID №17.
[00100] В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, в значительной степени идентичную аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 60% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 65% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 70% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 75% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 80% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 85% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 90% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 95% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 96% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 97% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 98% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 99% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления здесь представлены клетки-хозяева, которые содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 60%, минимум на 99% или на любое число от 60% до 99% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18.
[00101] В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую UGT40087: которая содержит аминокислотную последовательность N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, в значительной степени идентичную аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 60% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 65% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 70% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 75% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 80% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 85% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 90% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 95% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 96% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 97% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 98% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 99% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена последовательностей SEQ ID №17 или 18.
[00102] В определенных вариантах осуществления, когда сравнили и проанализировали смоделированные в трехмерном пространстве структуры UGT40087 и другой UDP-гликозилтрансферазы, было обнаружено четыре петли (т.е. петля 1, петля 2, петля 3 и петля 4), которые имеют значительные конформационные отличия в N-терминальном сахарном акцепторном домене. Экспериментальные результаты обмена соответствующих петлевых последовательностей между двумя UGT показали, что петлю 1, петлю 2, петлю 3_1, петлю 3_2 и петлю 4_1 UGT40087 можно заменить их соответствующими петлевыми последовательностями от других UDP-гликозилтрансфераз, которые способны превращать RebA в RebD. В этих вариантах осуществления были разработаны две версии петли 3 (т.е. петля 3_1 и петля 3_2) и петли 4 (т.е. петля 4_1 и петля 4_2), чтобы учесть две возможные длины петли.
[00103] Таким образом, в определенных вариантах осуществления клетки-хозяева содержат UDP-гликозилтрансферазу, содержащую аминокислотную последовательность, которая минимум на 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат гетерологичную нуклеиновую кислоту, кодирующую UDP-гликозилтрансферазу, содержащую аминокислотную последовательность, которая минимум на 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена SEQ ID №17 или 18. В определенных вариантах осуществления UDP-гликозилтрансфераза содержит аминокислотную последовательность петли 1 UGT40087 (т.е. SEQ ID №17 или 18), в участке UDP-гликозилтрансферазы, который соответствует участку петли 1 SEQ ID №17 или 18, соответственно. В определенных вариантах осуществления аминокислотная последовательность петли 1 SEQ ID №17 или 18 имеет аминокислотную последовательность SEQ ID №30. В определенных вариантах осуществления аминокислотная последовательность петли 1 имеет последовательность SEQ ID №28. В определенных вариантах осуществления UDP-гликозилтрансфераза содержит вариант аминокислотной последовательности петли 1 UGT40087 в участке UDP-гликозилтрансферазы, который соответствует участку петли 1 SEQ ID №17 или 18, соответственно. Вариант аминокислотной последовательности петли 1 означает аминокислотную последовательность, которая отличается от эталонной аминокислотной последовательности петли 1 SEQ ID №17 или 18 или аминокислотной последовательности петли 1 с SEQ ID №28, но допускает UDP-гликозилтрансферазу, содержащую вариант аминокислотной последовательности петли 1 для сохранения своего действия по превращению RebA в RebD и/или по превращению стевиозида в RebE.
[00104] В определенных вариантах осуществления UDP-гликозилтрансфераза содержит аминокислотную последовательность петли 2 UGT40087 (т.е. SEQ ID №17 или 18), в участке UDP-гликозилтрансферазы, который соответствует участку петли 2 SEQ ID №17 или 18, соответственно. В определенных вариантах осуществления аминокислотная последовательность петли 2 SEQ ID №17 или 18 имеет аминокислотную последовательность SEQ ID №24. В определенных вариантах осуществления UDP-гликозилтрансфераза содержит вариант аминокислотной последовательности петли 2 в участке UDP-гликозилтрансферазы, который соответствует участку петли 2 SEQ ID №17 или 18, соответственно. Вариант аминокислотной последовательности петли 2 означает аминокислотную последовательность, которая отличается от эталонной аминокислотной последовательности петли 2 SEQ ID №17 или 18, но допускает UDP-гликозилтрансферазу, содержащую вариант аминокислотной последовательности петли 2 для сохранения своего действия по превращению RebA в RebD и/или по превращению стевиозида в RebE.
[00105] В определенных вариантах осуществления UDP-гликозилтрансфераза содержит аминокислотную последовательность петли 3_1 UGT40087 (т.е. SEQ ID №17 или 18), в участке UDP-гликозилтрансферазы, который соответствует участку петли 3_1 SEQ ID №17 или 18, соответственно. В определенных вариантах осуществления аминокислотная последовательность петли 3_1 SEQ ID №17 или 18 имеет аминокислотную последовательность SEQ ID №25. В определенных вариантах осуществления UDP-гликозилтрансфераза содержит вариант аминокислотной последовательности петли 3_1 в участке UDP-гликозилтрансферазы, который соответствует участку петли 3_1 SEQ ID №17 или 18, соответственно. Вариант аминокислотной последовательности петли 31 означает аминокислотную последовательность, которая отличается от эталонной аминокислотной последовательности петли 3_1 SEQ ID №17 или 18, но допускает UDP-гликозилтрансферазу, содержащую вариант аминокислотной последовательности петли 3_1 для сохранения своего действия по превращению RebA в RebD и/или по превращению стевиозида в RebE.
[00106] В определенных вариантах осуществления UDP-гликозилтрансфераза содержит аминокислотную последовательность петли 3_2 UGT40087 (т.е. SEQ ID №17 или 18), в участке UDP-гликозилтрансферазы, который соответствует участку петли 3_2 SEQ ID №17 или 18, соответственно. В определенных вариантах осуществления аминокислотная последовательность петли 3_2 SEQ ID №17 или 18 имеет аминокислотную последовательность SEQ ID №26. В определенных вариантах осуществления UDP-гликозилтрансфераза содержит вариант аминокислотной последовательности петли 3_2 в участке UDP-гликозилтрансферазы, который соответствует участку петли 3_2 SEQ ID №17 или 18, соответственно. Вариант аминокислотной последовательности петли 3_2 означает аминокислотную последовательность, которая отличается от эталонной аминокислотной последовательности петли 3_2 SEQ ID №17 или 18, но допускает UDP-гликозилтрансферазу, содержащую вариант аминокислотной последовательности петли 3_2 для сохранения своего действия по превращению RebA в RebD и/или стевиозида по превращению в RebE.
[00107] В определенных вариантах осуществления UDP-гликозилтрансфераза содержит аминокислотную последовательность петли 4_1 UGT40087 (т.е. SEQ ID №17 или 18), в участке UDP-гликозилтрансферазы, который соответствует участку петли 4_1 SEQ ID №17 или 18, соответственно. В определенных вариантах осуществления аминокислотная последовательность петли 4_1 SEQ ID №17 или 18 имеет аминокислотную последовательность SEQ ID №27. В определенных вариантах осуществления UDP-гликозилтрансфераза содержит вариант аминокислотной последовательности петли 4_1 в участке UDP-гликозилтрансферазы, который соответствует участку петли 4_1 SEQ ID №17 или 18, соответственно. Вариант аминокислотной последовательности петли 4_1 означает аминокислотную последовательность, которая отличается от эталонной аминокислотной последовательности петли 4_1 SEQ ID №17 или 18, но допускает UDP-гликозилтрансферазу, содержащую вариант аминокислотной последовательности петли 4_1 для сохранения своего действия по превращению RebA в RebD и/или по превращению стевиозида в RebE.
[00108] В определенных вариантах осуществления UDP-гликозилтрансфераза содержит аминокислотную последовательность петли 4_2 UGT40087 (т.е. SEQ ID №17 или 18), в участке UDP-гликозилтрансферазы, который соответствует участку петли 4_2 SEQ ID №17 или 18, соответственно. Аминокислотная последовательность петли 4_2 SEQ ID №17 или 18 имеет аминокислотную последовательность SEQ ID №28.
[00109] В определенных вариантах осуществления клетки-хозяева содержат UDP-гликозилтрансферазу, содержащую аминокислотную последовательность, которая минимум на 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена SEQ ID №17 или 18, или гетерологичную нуклеиновую кислоту, кодирующую UDP-гликозилтрансферазу, и содержащую любую комбинацию из следующих компонентов:
(a) Аминокислотная последовательность петли 1 SEQ ID №17 или 18, аминокислотная последовательность SEQ ID №30 или вариант аминокислотной последовательности петли 1 в участке UDP-гликозилтрансферазы, который соответствует участку петли 1 последовательности SEQ ID №17 или 18, соответственно;
(b) аминокислотная последовательность петли 2 SEQ ID №17 или 18 или вариант аминокислотной последовательности петли 2 в участке UDP-гликозилтрансферазы, который соответствует участку петли 2 последовательности SEQ ID №17 или 18, соответственно;
(c) аминокислотная последовательность петли 3_1 SEQ ID №17 или 18 или вариант аминокислотной последовательности петли 3_1 в участке UDP-гликозилтрансферазы, который соответствует участку петли 31 последовательности SEQ ID №17 или 18, соответственно;
(d) аминокислотная последовательность петли 3_2 SEQ ID №17 или 18 или вариант аминокислотной последовательности петли 3_2 в участке UDP-гликозилтрансферазы, который соответствует участку петли 3 2 последовательности SEQ ID №17 или 18, соответственно;
(e) аминокислотная последовательность петли 4_1 SEQ ID №17 или 18 или вариант аминокислотной последовательности петли 4_1 в участке UDP-гликозилтрансферазы, который соответствует участку петли 4_1 последовательности SEQ ID №17 или 18, соответственно;
(f) аминокислотная последовательность петли 4_2 SEQ ID №17 или 18 в участке UDP-гликозилтрансферазы, который соответствует участку петли 4_2 последовательности SEQ ID №17 или 18, соответственно;
[00110] В определенных вариантах осуществления, когда сравнили и проанализировали смоделированные в трехмерном пространстве структуры UDP-гликозилтрансфераз, способных превращать RebA в RebD, было обнаружено, что петля 4_1 UGT40087, при включении в соответствующий участок петли 4_1 другой UDP-гликозилтрансферазы (и заменяющая ее нативную аминокислотную последовательность петли 4_1) обуславливала более эффективное действие варианта UDP-гликозилтрансферазы в плане ее способности превращать RebA в RebD. См. Пример 12. Эти результаты показывают, что аминокислотная последовательность петли 4_1 любой подходящей UDP-гликозилтрансферазы может быть заменена аминокислотной последовательностью петли 4_1 SEQ ID №17 или 18 для превращения RebA в RebD.
[00111] В связи с этим в определенных вариантах осуществления клетки-хозяева содержат UDP-гликозилтрансферазу, содержащую аминокислотную последовательность, которая минимум на 61%, 65%, 70%, 75%, 80%, 85%, 90% или 95% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена SEQ ID №17 или 18, и содержит аминокислотную последовательность петли 4_1 (т.е. SEQ ID №27) UGT40087 (т.е. SEQ ID №17 или 18). В определенных вариантах осуществления клетки-хозяева содержат гетерологичную нуклеиновую кислоту, кодирующую UDP-гликозилтрансферазу, содержащую аминокислотную последовательность, которая минимум на 61%, 65%, 70%, 75%, 80%, 85%, 90% или 95% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена SEQ ID №17 или 18 и содержит аминокислотную последовательность петли 4_1 (например, SEQ ID №27) последовательности SEQ ID №17 или 18. В определенных вариантах осуществления любая подходящая UDP-гликозилтрансфераза, которая содержит аминокислотную последовательность, которая минимум на 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или 95% идентична SEQ ID №17 или 18, может использоваться для включения аминокислотной последовательности петли 4_1 из SEQ ID №17 или 18 в соответствующем участке петли 4_1 (заменяющей ее нативную аминокислотную последовательность петли 4_1). Например, Ob_UGT91B_like, Hv_UGT_V1, S1_UGT_101249881, Sr.UGT_g252778, Os_UGT_91C1, Bd_UGT10840, Bd_UGT10850 или Si91Dlike могут использоваться в качестве базы для включения аминокислотной последовательности петли 4_1 из SEQ ID №17 или 18 в ее соответствующем участке петли 4_1. В определенных вариантах осуществления UDP-гликозилтрансфераза содержит аминокислотную последовательность SEQ ID №33.
[00112] В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, по существу идентичную аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 60% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 65% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 70% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 75% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 80% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 85% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 90% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 95% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 96% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 97% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 98% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат полипептид, содержащий аминокислотную последовательность, которая минимум на 99% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18.
[00113] В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую UGT40087, которая содержит аминокислотную последовательность С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, в значительной степени идентичную аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 60% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 65% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 70% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 75% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 80% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 85% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 90% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 95% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 96% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 97% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 98% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат нуклеиновую кислоту, кодирующую полипептид, содержащий аминокислотную последовательность, которая минимум на 99% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18.
[00114] В определенных вариантах осуществления клетки-хозяева содержат UDP-гликозилтрансферазу, содержащую аминокислотную последовательность, которая минимум на 60%, 65%, 70%, 75%, 80%, 85%, 90% или 95% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления клетки-хозяева содержат гетерологичную нуклеиновую кислоту, кодирующую UDP-гликозилтрансферазу, содержащую аминокислотную последовательность, которая минимум на 60%, 65%, 70%, 75%, 80%, 85%, 90% или 95% идентична аминокислотной последовательности С-терминального сахарного донорного домена последовательностей SEQ ID №17 или 18. В определенных вариантах осуществления UDP-гликозилтрансфераза содержит С-терминальный сахарный донорный домен от другой UDP-гликозилтрансферазы. Примеры других UDP-гликозилтрансфераз с подходящими С-терминальными сахарными донорными доменами включают Ob_UGT91B_like, Hv_UGT_V1, SI_UGT_101249881, Sr.UGT_g252778, Os_UGT_91C1, Bd_UGT10840, Bd_UGT10850 или Si91Dlike.
[00115] В определенных вариантах осуществления было обнаружено, что определенные аминокислотные остатки в N-терминальном сахарном акцепторном домене могут восстанавливать каталитическую активность нефункциональной предполагаемой UDP-гликозилтрансферазы в активную UDP-гликозилтрансферазу. В связи с этим клетки-хозяева содержат UDP-гликозилтрансферазу, содержащую аминокислотную последовательность, которая минимум на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентична аминокислотной последовательности N-терминального сахарного акцепторного домена SEQ ID №17 или 18 и содержит один или более следующих аминокислотных остатков:
(a) валин в аминокислотной позиции UDP-гликозилтрансферазы, которая соответствует аминокислотной позиции 11 SEQ ID №18;
(b) изолейцин в аминокислотной позиции UDP-гликозилтрансферазы, которая соответствует аминокислотной позиции 12 SEQ ID №18;
(c) пролин в аминокислотной позиции UDP-гликозилтрансферазы, которая соответствует аминокислотной позиции 55 SEQ ID №18;
(d) глутаминовая кислота в аминокислотной позиции UDP-гликозилтрансферазы, которая соответствует аминокислотной позиции 90 SEQ ID №18;
(e) серии в аминокислотной позиции UDP-гликозилтрансферазы, которая соответствует аминокислотной позиции 203 SEQ ID №18;
(f) глутаминовая кислота в аминокислотной позиции UDP-гликозилтрансферазы, которая соответствует аминокислотной позиции 223 SEQ ID №18; или
(g) валин в аминокислотной позиции UDP-гликозилтрансферазы, которая соответствует аминокислотной позиции 413 SEQ ID №18,
где аминокислотные позиции UDP-гликозилтрансферазы, соответствующие аминокислотным позициям SEQ ID №18, определяются посредством выравнивания последовательностей.
[00116] В определенных вариантах осуществления клетки-хозяева содержат UDP-гликозилтрансферазу, содержащую аминокислотную последовательность SEQ ID №32.
[00117] В некоторых вариантах осуществления клетка-хозяин содержит вариант полипептида UGT40087, описанный выше. В определенных вариантах осуществления такой вариант содержит до 15, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 аминокислотной замены относительно полипептида UGT40087. В некоторых вариантах осуществления этот вариант может содержать до 15, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 консервативной аминокислотной замены относительно полипептида UGT40087. В некоторых вариантах осуществления любую из описанных здесь нуклеиновых кислот можно оптимизировать для клетки-хозяина, например, посредством оптимизации ко донов. Полезные нуклеиновые кислоты включают SEQ ID №35 и 36.
6.6 Производство FPP и/или GGPP по MEV-пути
[00118] В некоторых вариантах осуществления представленная здесь генетически модифицированная клетка-хозяин содержит один или более гетерологичных ферментов MEV-пути, полезных для производства FPP и/или GGPP. См. ФИГ. 1D. В некоторых вариантах осуществления один или более ферментов MEV-пути содержат фермент, который конденсирует ацетил-СоА с малонил-СоА, в результате чего образуется ацетоацетил-СоА. В некоторых вариантах осуществления один или более ферментов MEV-пути содержат фермент, который конденсирует две молекулы ацетил-СоА, в результате чего образуется ацетоацетил-СоА. В некоторых вариантах осуществления один или более ферментов MEV-пути содержат фермент, который конденсирует ацетоацетил-СоА с ацетил-СоА, в результате чего образуется HMG-CoA. В некоторых вариантах осуществления один или более ферментов MEV-пути содержат фермент, который превращает HMG-CoA в мевалонат. В некоторых вариантах осуществления один или более ферментов MEV-пути содержат фермент, который фосфорилирует мевалонат в мевалонат 5-фосфат. В некоторых вариантах осуществления один или более ферментов MEV-пути содержат фермент, который превращает мевалонат 5-фосфат в мевалонат 5-пирофосфат.В некоторых вариантах осуществления один или более ферментов MEV-пути содержат фермент, который превращает мевалонат 5-пирофосфат в изопентенилпирофосфат.
[00119] В некоторых вариантах осуществления один или более ферментов MEV-пути выбираются из группы, состоящей из ацетил-СоА тиолазы, ацетоацетил-СоА синтетазы, HMG-CoA синтазы, HMG-CoA редуктазы, мевалонаткиназы, фосфомевалонаткиназы и мевалонатпирофосфатдекарбоксилазы. В некоторых вариантах осуществления, относительно фермента MEV-пути, способного катализировать образование ацетоацетил-СоА, генетически модифицированная клетка-хозяин содержит либо фермент, который конденсирует две молекулы ацетил-СоА, в результате чего образуется ацетоацетил-СоА, например, ацетил-СоА тиолаза; либо фермент, который конденсирует ацетил-СоА с малонил-СоА, в результате чего образуется ацетоацетил-СоА, например, ацетоацетил-СоА синтаза. В некоторых вариантах осуществления генетически модифицированная клетка-хозяин содержит и фермент, который конденсирует две молекулы ацетил-СоА, в результате чего образуется ацетоацетил-СоА, например, ацетил-СоА тиолаза, и фермент, который конденсирует ацетил-СоА с малонил-СоА, в результате чего образуется ацетоацетил-СоА, например, ацетоацетил-СоА синтаза.
[00120] В некоторых вариантах осуществления клетка-хозяин содержит одну или более гетерологичных нуклеотидных последовательностей, кодирующих более одного фермента MEV-пути. В некоторых вариантах осуществления клетка-хозяин содержит одну или более гетерологичных нуклеотидных последовательностей, кодирующих два фермента MEV-пути. В некоторых вариантах осуществления клетка-хозяин содержит одну или более гетерологичных нуклеотидных последовательностей, кодирующих фермент, который может превращать HMG-CoA в мевалонат, и фермент, который может превращать мевалонат в мевалонат 5-фосфат.В некоторых вариантах осуществления клетка-хозяин содержит одну или более гетерологичных нуклеотидных последовательностей, кодирующих три фермента MEV-пути. В некоторых вариантах осуществления клетка-хозяин содержит одну или более гетерологичных нуклеотидных последовательностей, кодирующих четыре фермента MEV-пути. В некоторых вариантах осуществления клетка-хозяин содержит одну или более гетерологичных нуклеотидных последовательностей, кодирующих пять ферментов MEV-пути. В некоторых вариантах осуществления клетка-хозяин содержит одну или более гетерологичных нуклеотидных последовательностей, кодирующих шесть ферментов MEV-пути. В некоторых вариантах осуществления клетка-хозяин содержит одну или более гетерологичных нуклеотидных последовательностей, кодирующих семь ферментов MEV-пути. В некоторых вариантах осуществления клетка-хозяин содержит множество гетерологичных нуклеиновых кислот, кодирующих все ферменты MEV-пути.
[00121] В некоторых вариантах осуществления генетически модифицированная клетка-хозяин содержит гетерологичную нуклеиновую кислоту, кодирующую фермент, который может превращать изопентенилпирофосфат (IPP) в диметилаллилпирофосфат (DMAPP). В некоторых вариантах осуществления генетически модифицированная клетка-хозяин содержит гетерологичную нуклеиновую кислоту, кодирующую фермент, который может конденсировать молекулы IPP и/или DMAPP, в результате чего образуется полипренильное соединение. В некоторых вариантах осуществления генетически модифицированная клетка-хозяин содержит гетерологичную нуклеиновую кислоту, кодирующую фермент, который может модифицировать IPP или полипренил, в результате чего образуется изопреноидное соединение, такое как FPP.
6.6.1 Превращение ацетил-СоА в ацетоацетил-СоА
[00122] В некоторых вариантах осуществления генетически модифицированная клетка-хозяин содержит гетерологичную нуклеотидную последовательность, кодирующую фермент, который может конденсировать две молекулы ацетилкоэнзима А, в результате чего образуется ацетоацетил-СоА, например, ацетил-СоА тиолаза. Иллюстративные последовательности нуклеотидных последовательностей, кодирующих такой фермент, включают, помимо прочего: (NC_000913 УЧАСТОК: 2324131.2325315; Escherichia coli), (D49362; Paracoccus denitriftcans) И (L20428; Saccharomyces cerevisiae).
[00123] Ацетил-СоА тиолаза катализирует обратимую конденсацию двух молекул ацетил-СоА, в результате чего получают ацетоацетил-СоА, но эта реакция является термодинамически неблагоприятной; тиолиз ацетоацетил-СоА предпочитают синтезу ацетоацетил-СоА. Ацетоацетил-СоА синтаза (AACS) (также называемая ацетил-СоА:малонил-СоА ацилтрансферазой; ЕС 2.3.1.194) конденсирует ацетил-СоА с малонил-СоА, в результате чего образуется ацетоацетил-СоА. В отличие от ацетил-СоА тиолазы, AACS-катализируемый синтез ацетоацетил-СоА, по существу, является реакцией, благоприятной в энергетическом плане, благодаря соответствующему декарбоксилированию малонил-СоА. Кроме того, AACS не демонстрирует тиолитической активности против ацетоацетил-СоА, в связи с чем эта реакция является необратимой.
[00124] В клетках-хозяевах, содержащих ацетил-СоА тиолазу и гетерологичную ADA и/или фосфотрансацетилазу (РТА), обратимая реакция, катализируемая ацетил-СоА тиолазой, которая благоприятствует тиолизу ацетоацетил-СоА, может привести к большому пулу ацетил-СоА. Ввиду обратимой активности ADA, этот пул ацетил-СоА может в свою очередь направлять ADA к обратимой реакции превращения ацетил-СоА в ацетальдегид, что уменьшает преимущества, предоставляемые ADA в плане производства ацетил-СоА. Аналогичным образом, действие РТА является необратимым, в связи с чем большой пул ацетил-СоА может привести РТА к обратимой реакции превращения ацетил-СоА в ацетилфосфат. Таким образом, в некоторых вариантах осуществления, для стимулирования того, чтобы ацетил-СоА способствовал прямой реакции ADA и РТА, в MEV-пути представленной здесь генетически модифицированной клетки-хозяина используется ацетоацетил-СоА синтаза для получения ацетоацетил-СоА из ацетил-СоА и малонил-СоА.
[00125] В некоторых вариантах осуществления AACS получают из штамма Streptomyces sp. CL190 (Okamura et al., Proc Natl Acad Sci USA 107(25):11265-70 (2010). Репрезентативные нуклеотидные последовательности AACS штамма Streptomyces sp. CL190 включают учетный номер АВ540131.1. Репрезентативные последовательности белка AACS штамма Streptomyces sp. CL190 включают учетные номера D7URV0, BAJ10048. Другие ацетоацетил-СоА синтазы, полезные для представленных здесь композиций и способов, включают, помимо прочего, Streptomyces sp.(АВ183750; КО-3988 BAD86806); штамм S. anulatus 9663 (FN178498; САХ48662); Streptomyces sp.КО-3988 (АВ212624; ВАЕ78983); Actinoplanes sp.A40644 (AB113568; BAD07381); Streptomyces sp.С (NZ_ACEW010000640; ZP_05511702); Nocardiopsis dassonvillei DSM 43111 (NZ_ABUI01000023; ZP_04335288); Mycobacterium ulcerans Agy99 (NC_008611; YP_907152); Mycobacterium marinum M (NC_010612; YP_001851502); Streptomyces sp.Mgl (NZ_DS570501; ZP_05002626); Streptomyces sp.AA4 (NZ_ACEV01000037; ZP_05478992); S. roseosporus NRRL 15998 (NZ_ABYB01000295; ZP_04696763); Streptomyces sp.ACTE (NZ_ADFD01000030; ZP_06275834); S. viridochromogenes DSM 40736 (NZ_ACEZ01000031; ZP_05529691); Frankia sp.CcI3 (NC_007777; YP_480101); Nocardia brasiliensis (NC_018681; YP_006812440.1); и Austwickia chelonae (NZ_BAGZ01000005; ZP_10950493.1). Дополнительные подходящие ацетоацетил-СоА синтазы включают те, которые описаны в Публикациях заявок на патент США под номерами 2010/0285549 и 2011/0281315, содержание которых включается в настоящий документ полностью посредством ссылки.
[00126] Ацетоацетил-СоА синтазы, также полезные в представленных здесь композициях и способах, включают молекулы, которые называют "производными" любой из описанных здесь ацетоацетил-СоА синтаз. Такое "производное" имеет следующие характеристики: (1) оно имеет значительную гомологию с любой из описанных здесь ацетоацетил-СоА синтаз; и (2) способно катализировать необратимую конденсацию ацетил-СоА с малонил-СоА, в результате чего образуется ацетоацетил-СоА. Говорят, что производное ацетоацетил-СоА синтазы имеет значительную степень гомологии с ацетоацетил-СоА синтазой, если аминокислотные последовательности производного минимум на 80%, а предпочтительнее минимум на 90%, а предпочтительнее всего минимум на 95% совпадают с аминокислотными последовательностями ацетоацетил-СоА синтазы.
6.6.2 Превращение ацетоацетил-СоА в HMG-CoA
[00127] В некоторых вариантах осуществления клетка-хозяин содержит гетерологичную нуклеотидную последовательность, кодирующую фермент, который может конденсировать ацетоацетил-СоА с другой молекулой ацетил-СоА, в результате чего образуется 3-гидрокси-3-метилглютарил-СоА (HMG-CoA), например, HMG-CoA синтаза. Иллюстративные последовательности нуклеотидных последовательностей, кодирующих такой фермент, включают, помимо прочего: (NC_001145. комплемент 19061.20536; Saccharomyces cerevisiae), (Х96617; Saccharomyces cerevisiae), (X83882; Arabidopsis thaliana), (AB037907; Kitasatospora griseola), (BT007302; Homo sapiens), и (NC_002758, номер локуса SAV2546, идент. гена 1122571; Staphylococcus aureus).
6.6.3 Превращение HMG-CoA в мевалонат
[00128] В некоторых вариантах осуществления клетка-хозяин содержит гетерологичную нуклеотидную последовательность, кодирующую фермент, который может превращать HMG-CoA в мевалонат, например, HMG-CoA редуктазу. В некоторых вариантах осуществления HMG-CoA редуктазой является использующая NADH гидроксиметилглютарил-СоА редуктаза-СоА редуктаза. HMG-CoA редуктазы (ЕС 1.1.1.34; ЕС 1.1.1.88) катализируют восстановительное деацилирование (S)-HMG-CoA в (R)-мевалонат и могут быть разделены на два класса, HMGrs класса I и класса II. Класс I включает ферменты из эукариотов и большинства архей, а класс II включает HMG-CoA редуктазы определенных прокариотов и архей. Помимо расхождения в последовательностях, ферменты этих двух классов также различаются в плане своей кофакторной специфичности. В отличие от ферментов класса I, которые используют исключительно NADPH, HMG-CoA редуктазы класса II различаются способностью к дискриминации между NADPH и NADH. См., например, Hedl et al., Journal of Bacteriology 186 (7): 1927-1932 (2004). Далее приведены кофакторные специфичности для отдельных HMG-CoA редуктаз класса II.
[0001] Кофакторные специфичности для отдельных HMG-CoA редуктаз класса II
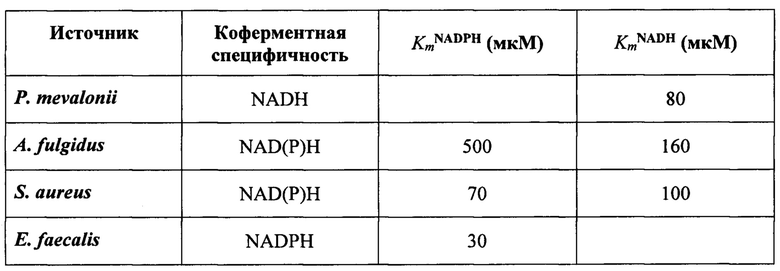
[00129] Полезные HMG-CoA редуктазы для представленных здесь композиций и способов включают HMG-CoA редуктазы, способные использовать NADH в качестве кофактора, например, HMG-CoA редуктаза из P. mevalonii, A. fulgidus или S. aureus. В конкретных вариантах осуществления HMG-CoA редуктаза способна использовать только NADH в качестве кофактора, например, HMG-CoA редуктаза из P. mevalonii, S. pomeroyi или Д acidovorans.
[00130] В некоторых вариантах осуществления использующая NADH HMG-CoA редуктаза получена из Pseudomonas mevalonii. Ранее был описан mvaA ген дикого типа Pseudomonas mevalonii, который кодирует HMG-CoA редуктазу (ЕС 1.1.1.88). См. Beach and Rodwell, J. Bacteriol. 171:2994-3001 (1989). Репрезентативные нуклеотидные последовательности mvaA Pseudomonas mevalonii включают учетный номер М24015. Репрезентативные последовательности белка HMG-CoA редуктазы Pseudomonas mevalonii включают учетные номера ААА25837, Р13702, MVAA_PSEMV.
[00131] В некоторых вариантах осуществления использующая NADH HMG-CoA редуктаза получена из Silicibacter pomeroyi. Репрезентативные нуклеотидные последовательности HMG-CoA редуктазы Silicibacter pomeroyi включают учетный номер NC_006569.1. Репрезентативные последовательности белка HMG-CoA редуктазы Silicibacter pomeroyi включают учетный номер YP_164994.
[00132] В некоторых вариантах осуществления использующая NADH HMG-CoA редуктаза получена из Delftia acidovorans. Репрезентативные нуклеотидные последовательности HMG-CoA редуктазы Delftia acidovorans включают NC_010002 УЧАСТОК: комплемент (319980..321269). Репрезентативные последовательности белка HMG-CoA редуктазы Delftia acidovorans включают учетный номер YP_001561318.
[00133] В некоторых вариантах осуществления использующая NADH HMG-CoA редуктаза получена из Solanum tuberosum (Crane et al., J. Plant Physiol. 159:1301-1307 (2002)).
[00134] Использующие NADH HMG-CoA редуктазы, также полезные в представленных здесь композициях и способах, включают молекулы, которые называются "производными" любой из описанных здесь использующих NADH HMG-CoA редуктаз, например, из P. mevalonii, S. pomeroyi и D. acidovorans. Такое "производное" имеет следующие характеристики: (1) оно имеет значительную степень гомологии с описанными здесь использующими NADH HMG-CoA редуктазами; и (2) способно катализировать восстановительное деацилирование (S)-HMG-CoA в (R)-мевалонат при предпочтительном использовании NADH в качестве кофактора. Говорят, что производное использующей NADH HMG-CoA редуктазы имеет значительную степень гомологии с использующей NADH HMG-CoA редуктазой, если аминокислотные последовательности производного минимум на 80%, а предпочтительнее минимум на 90%, а предпочтительнее всего минимум на 95% совпадают с аминокислотными последовательностями использующей NADH HMG-CoA редуктазы.
[00135] В контексте настоящего документа "использующая NADH" означает, что использующая NADH HMG-CoA редуктаза предпочитает NADH, нежели NADPH, в качестве кофактора, например, демонстрируя более высокую специфическую активность для NADH, чем для NADPH. В некоторых вариантах осуществления предпочтение к NADH в качестве кофактора выражается как соотношение kcat(NADH)/kcat(NADPH). В некоторых вариантах осуществления использующая NADH HMG-CoA редуктаза имеет соотношение kcat(NADH)/kcat(NADPH) минимум 5, 10, 15, 20, 25 или более 25. В некоторых вариантах осуществления использующая NADH HMG-CoA редуктаза использует исключительно NADH. Например, использующая NADH HMG-CoA редуктаза, которая использует исключительно NADH, демонстрирует некоторую активность с NADH, единственным кофакторомт vitro, и не демонстрирует никакой обнаруживаемой активности, когда NADPH является единственным кофактором. Для идентификации HMG-CoA редуктаз, предпочитающих NADH в качестве кофактора, можно использовать любой способ для определения кофакторной специфичности, известный в данной области, включая способы, описанные в работах Kim et al., Protein Science 9:1226-1234 (2000); и Wilding et al., J. Bacteriol. 182(18):5147-52 (2000), содержание которых включается в настоящий документ полностью посредством ссылки.
[00136] В некоторых вариантах осуществления использующую NADH HMG-CoA редуктазу модифицируют на селективность к NADH, нежели к NAPDH, например, посредством сайт-направленного мутагенеза кофактор-связывающего кармана. Способы модификации селективности к NADH описаны в работе Watanabe et al., Microbiology 153:3044-3054 (2007), а способы для определения кофакторной специфичности HMG-CoA редуктаз описаны в работе Kim et al., Protein Sci. 9:1226-1234 (2000), содержание которых включается в настоящий документ полностью посредством ссылки.
[00137] В некоторых вариантах осуществления использующую NADH HMG-CoA редуктазу получают из вида хозяев, который нативно содержит мевалонатный деструктивный путь, например, вида хозяев, который катаболизирует мевалонат как его единственный источник углерода. В таких вариантах осуществления использующая NADH HMG-CoA редуктаза, которая обычно катализирует окислительное ацилирование интернализованного (R)-мевалоната в (S)-HMG-CoA в его нативной клетке-хозяине, используется для катализации обратной реакции, то есть восстановительного деацилирования (S)-HMG-CoA в (R)-мевалонат, в генетически модифицированной клетке-хозяине, содержащей путь биосинтеза мевалоната. Прокариоты, способные к росту на мевалонате как их единственном источнике углерода, были описаны в следующих работах: Anderson et al., J. Bacteriol, 171(12):6468-6472 (1989); Beach et al., J. Bactehol 171:2994-3001 (1989); Bensch et al., J. Biol Chem. 245:3755-3762; Fimongnari et al., Biochemistry 4:2086-2090 (1965); Siddiqi et al., Biochem. Biophys. Res. Commun. 8:110-113 (1962); Siddiqi et al., J. Bacteriol 93:207-214 (1967); и Takatsuji et al., Biochem. Biophys. Res. Commun.110:187-193 (1983), содержание которых включается в настоящий документ полностью посредством ссылки.
[00138] В некоторых вариантах осуществления представленных здесь композиций и способов клетка-хозяин содержит и использующую NADH HMGr, и использующую NADPH HMG-CoA редуктазу. Иллюстративные примеры нуклеотидных последовательностей, кодирующих использующую NADPH HMG-CoA редуктазу, включают, помимо прочего: (NM_206548; Drosophila melanogaster), (NC_002758, номер локуса SAV2545, идент. гена 1122570; Staphylococcus aureus), (АВ015627; Streptomyces sp. КО 3988), (AX128213, обеспечивающая последовательность, кодирующую процессированную HMG-CoA редуктазу; Saccharomyces cerevisiae), и (NC_001145: комплемент (115734.118898; Saccharomyces cerevisiae).
6.6.4 Превращение мевалоната в мевалонат-5-фосфат
[00139] В некоторых вариантах осуществления клетка-хозяин содержит гетерологичную нуклеотидную последовательность, кодирующую фермент, который может превращать мевалонат в мевалонат 5-фосфат, например, мевалонаткиназу. Иллюстративные последовательности нуклеотидных последовательностей, кодирующих такой фермент, включают, помимо прочего: (L77688; Arabidopsis thaliana) и (Х55875; Saccharomyces cerevisiae).
6.6.5 Превращение мевалонат-5-фосфата в мевалонат-5-пирофосфат
[00140] В некоторых вариантах осуществления клетка-хозяин содержит гетерологичную нуклеотидную последовательность, кодирующую фермент, который может превращать мевалонат 5-фосфат в мевалонат 5-пирофосфат, например, фосфомевалонаткиназу. Иллюстративные последовательности нуклеотидных последовательностей, кодирующих такой фермент, включают, помимо прочего: (AF429385; Hevea brasiliensis), (NM_006556; Homo sapiens) и (NC_001145. комплемент 712315.713670; Saccharomyces cerevisiae).
6.6.6 Превращение мевалонат-5-пирофосфата в IPP
[00141] В некоторых вариантах осуществления клетка-хозяин содержит гетерологичную нуклеотидную последовательность, кодирующую фермент, который может превращать мевалонат 5-пирофосфат в изопентенилдифосфат (IPP), например, мевалонатпирофосфатдекарбоксилазу. Иллюстративные последовательности нуклеотидных последовательностей, кодирующих такой фермент, включают, помимо прочего: (Х97557; Saccharomyces cerevisiae), (AF290095; Enterococcus faecium) и (U49260; Homo sapiens).
6.6.7 Превращение IPP в DMAPP
[00142] В некоторых вариантах осуществления клетка-хозяин содержит гетерологичную нуклеотидную последовательность, кодирующую фермент, который может превращать IPP, созданный посредством MEV-пути, в диметилаллилпирофосфат (DMAPP), например, IPP изомеразу. Иллюстративные последовательности нуклеотидных последовательностей, кодирующих такой фермент, включают, помимо прочего: (NC_000913, 3031087.3031635; Escherichia coli) и (AF082326; Haematococcuspluvialis).
6.6.8 Полипренилсинтазы
[00143] В некоторых вариантах осуществления клетка-хозяин содержит гетерологичную нуклеотидную последовательность, кодирующую полипренилсинтазу, которая может конденсировать молекулы IPP и/или DMAPP, в результате чего образуются полипренильные соединения, содержащие более пяти углеродов.
[00144] В некоторых вариантах осуществления клетка-хозяин содержит гетерологичную нуклеотидную последовательность, кодирующую фермент, который может конденсировать одну молекулу IPP с одной молекулой DMAPP, в результате чего образуется одна молекула геранилпирофосфата (GPP), например, GPP синтаза. Иллюстративные последовательности нуклеотидных последовательностей, кодирующих такой фермент, включают, помимо прочего: (AF513111; Abies grandis), (AF513112; Abies grandis), (AF513113; Abies grandis), (AY534686; Antirrhinum majus), (AY534687; Antirrhinum majus), (Y17376; Arabidopsis thaliana), (AE016877, Locus API 1092; Bacillus cereus; ATCC 14579), (AJ243739; Citrus sinensis), (AY534745; Clarkia breweri), (AY953508; Ips pini), (DQ286930; Lycopersicon esculentum), (AF182828; Mentha x piperita), (AF182827; Mentha x piperita), (MPI249453; Mentha x piperita), (PZE431697, Locus CAD24425; Paracoccus zeaxanthinifaciens), (AY866498; Picrorhiza kurrooa), (AY351862; Vitis vinifera) и (AF203881, Locus AAF12843; Zymomonas mobilis).
[00145] В некоторых вариантах осуществления клетка-хозяин содержит гетерологичную нуклеотидную последовательность, кодирующую фермент, который может конденсировать две молекулы IPP с одной молекулой DMAPP или добавлять одну молекулу IPP к молекуле GPP, в результате чего образуется молекула фарнезилпирофосфата (FPP), например, FPP синтаза. Иллюстративные последовательности нуклеотидных последовательностей, кодирующих такой фермент, включают, помимо прочего: (ATU80605; Arabidopsis thaliana), (ATHFPS2R; Arabidopsis thaliana), (AAU36376; Artemisia annua), (AF461050; Bos taurus), (D00694; Escherichia coli K-12), (AE009951, Locus AAL95523; Fusobacterium nucleatum subsp.nucleatum ATCC 25586), (GFFPPSGEN; Gibberella fujikuroi), (CP000009, Locus AAW60034; Gluconobacter oxydans 621H), (AF019892; Helianthus annuus), (HUMFAPS; Homo sapiens), (KLPFPSQCR; Kluyveromyces lactis), (LAU15777; Lupinus albus), (LAU20771; Lupinus albus), (AF309508; Mus musculus), (NCFPPSGEN; Neurospora crassa), (PAFPS1; Parthenium argentatum), (PAFPS2; Parthenium argentatum), (RATFAPS; Rattus norvegicus), (YSCFPP; Saccharomyces cerevisiae), (D89104; Schizosaccharomyces pombe), (CP000003, Locus AAT87386; Streptococcus pyogenes), (CP000017, Locus AAZ51849; Streptococcus pyogenes), (NC_008022, Locus YP_598856; Streptococcus pyogenes MGAS10270), (NC_008023, Locus YP 600845; Streptococcus pyogenes MGAS2096), (NC_008024, Locus YP_602832; Streptococcus pyogenes MGAS10750), (MZEFPS; Zea mays), (AE000657, Locus AAC06913; Aquifex aeolicus VF5), (NM_202836; Arabidopsis thaliana), (D84432, Locus BAA12575; Bacillus subtilis), (U12678, Locus AAC28894; Bradyrhizobium japonicum USDA 110), (BACFDPS; Geobacillus stearothermophilus), (NC_002940, Locus NP_873754; Haemophilus ducreyi 35000HP), (L42023, Locus AAC23087; Haemophilus influenzae Rd KW20), (J05262; Homo sapiens), (YP_395294; Lactobacillus sakei subsp.sakei 23K), (NC_005823, Locus YP 000273; Leptospira interrogans serovar Copenhageni str. Fiocruz L1-130), (AB003187; Micrococcus luteus), (NC_002946, Locus YP 208768; Neisseria gonorrhoeae FA 1090), (U00090, Locus AAB91752; Rhizobium sp.NGR234), (J05091; Saccharomyces cerevisae), (CP000031, Locus AAV93568; Silicibacter pomeroyi DSS-3), (AE008481, Locus AAK99890; Streptococcus pneumoniae R6) и (NC_004556, Locus NP_779706; Xylella fastidiosa Temeculal).
[00146] В некоторых вариантах осуществления клетка-хозяин содержит гетерологичную нуклеотидную последовательность, кодирующую фермент, который может комбинировать IPP и DMAPP или IPP и FPP, в результате чего образуется геранилгеранилпирофосфат (GGPP). Иллюстративные последовательности нуклеотидных последовательностей, кодирующих такой фермент, включают, помимо прочего: (ATHGERPYRS; Arabidopsis thaliana), (ВТ005328; Arabidopsis thaliana), (NM_119845; Arabidopsis thaliana), (NZ_AAJM01000380, Locus ZP_00743052; Bacillus thuringiensis serovar israelensis, ATCC 35646 sql563), (CRGGPPS; Catharanthus roseus), (NZ_AABF02000074, Locus ZP_00144509; Fusobacterium nucleatum subsp.vincentii, ATCC 49256), (GFGGPPSGN; Gibberella fujikuroi), (AY371321; Ginkgo biloba), (AB055496; Hevea brasiliensis), (ABO 17971; Homo sapiens), (MCI276129; Mucor circinelloides f. lusitanicus), (ABO 16044; Mus musculus), (AABX01000298, Locus NCU01427; Neurospora crassa), (NCU20940; Neurospora crassa), (NZ_AAKL01000008, Locus ZP_00943566; Ralstonia solanacearum UW551), (AB118238; Rattus norvegicus), (SCU31632; Saccharomyces cerevisiae), (ABO 16095; Synechococcus elongates), (SAGGPS; Sinapis alba), (SSOGDS; Sulfolobus acidocaldarius), (NC_007759, Locus YP 461832; Syntrophus aciditrophicus SB), (NC_006840, Locus YP_204095; Vibrio fischeri ESI 14), (NM_112315; Arabidopsis thaliana), (ERWCRTE; Pantoea agglomerans), (D90087, Locus BAA14124; Pantoea ananatis), (X52291, Locus CAA36538; Rhodobacter capsulatus), (AF195122, Locus AAF24294; Rhodobacter sphaeroides) и (NC_004350, Locus NP_721015; Streptococcus mutans UA159).
[00147] При том, что ранее были описаны примеры ферментов мевалонатного пути, в определенных вариантах осуществления для альтернативного или дополнительного пути производства DMAPP и IPP в клетках-хозяевах, композициях и способах, представленных здесь, могут использоваться ферменты DXP-пути. Ферменты и нуклеиновые кислоты, кодирующие ферменты DXP-пути, хорошо известны и описаны в данной области. WO 2012/135591 А2.
6.7 Способы производства стевиол-гликозидов
[00148] В другом аспекте здесь представлен способ производства стевиол-гликозида, способ, включающий следующие этапы: (а) культивирование популяции любых из генетически модифицированных клеток-хозяев, описанных здесь, которые способны производить стевиол-гликозид в среде с источником углерода в условиях, подходящих для создания стевиол-гликозидов; и (b) выделение указанного стевиол-гликозида из среды.
[00149] В некоторых вариантах осуществления генетически модифицированная клетка-хозяин производит повышенное количество стевиол-гликозида по сравнению с родительской клеткой, не содержащей одну или более модификаций, или родительской клеткой, содержащей только одну подгруппу одной или более модификаций генетически модифицированной клетки-хозяина, а в остальном является генетически идентичной. В некоторых вариантах осуществления повышенное количество означает количество, которое минимум на 1%, 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 100% или более 100% больше, по результатам измерения, например, в выходе продукта, производстве, производительности, в граммах на литр клеточной культуры, миллиграммах на грамм сухого веса клеток, на основании удельного объема клеточной культуры, на основании удельного сухого веса клеток, на основании удельного объема клеточной культуры в единицу времени или на основании удельного сухого веса клеток в единицу времени.
[00150] В некоторых вариантах осуществления клетка-хозяин производит повышенный уровень стевиол-гликозида, который превышает значение около 10 грамм на литр ферментативной среды. В некоторых таких вариантах осуществления стевиол-гликозид производится в количестве от около 10 до около 50 грамм, более чем, около 15 грамм, более, чем около 20 грамм, более, чем около 25 грамм или более, чем около 30 грамм на литр клеточной культуры.
[00151] В некоторых вариантах осуществления клетка-хозяин производит повышенный уровень стевиол-гликозида, который превышает значение около 50 миллиграмм на грамм сухого веса клеток. В некоторых таких вариантах осуществления стевиол-гликозид производится в количестве от около 50 до около 1500 миллиграмм, более, чем около 100 миллиграмм, более, чем около 150 миллиграмм, более, чем около 200 миллиграмм, более, чем около 250 миллиграмм, более, чем около 500 миллиграмм, болееб чем около 750 миллиграмм, или более, чем около 1000 миллиграмм на грамм сухого веса клеток.
[00152] В некоторых вариантах осуществления клетка-хозяин производит повышенный уровень стевиол-гликозида, который минимум на около 10%, минимум на около 15%, минимум на около 20%, минимум на около 25%, минимум на около 30%, минимум на около 35%, минимум на около 40%, минимум на около 45%, минимум на около 50%, минимум на около 60%, минимум на около 70%, минимум на около 80%, минимум на около 90%, минимум приблизительно в 2 раза, минимум приблизительно в 2.5 раза, минимум приблизительно в 5 раз, минимум приблизительно в 10 раз, минимум приблизительно в 20 раз, минимум приблизительно в 30 раз, минимум приблизительно в 40 раз, минимум приблизительно в 50 раз, минимум приблизительно в 75 раз, минимум приблизительно в 100 раз, минимум приблизительно в 200 раз, минимум приблизительно в 300 раз, минимум приблизительно в 400 раз, минимум приблизительно в 500 раз или минимум приблизительно в 1000 раз или более превышает уровень стевиол-гликозида, производимого родительской клеткой, на основании удельного объема клеточной культуры.
[00153] В некоторых вариантах осуществления клетка-хозяин производит повышенный уровень стевиол-гликозида, который минимум на около 10%, минимум на около 15%, минимум на около 20%, минимум на около 25%, минимум на около 30%, минимум на около 35%, минимум на около 40%, минимум на около 45%, минимум на около 50%, минимум на около 60%, минимум на около 70%, минимум на около 80%, минимум на около 90%, минимум приблизительно в 2 раза, минимум приблизительно в 2.5 раза, минимум приблизительно в 5 раз, минимум приблизительно в 10 раз, минимум приблизительно в 20 раз, минимум приблизительно в 30 раз, минимум приблизительно в 40 раз, минимум приблизительно в 50 раз, минимум приблизительно в 75 раз, минимум приблизительно в 100 раз, минимум приблизительно в 200 раз, минимум приблизительно в 300 раз, минимум приблизительно в 400 раз, минимум приблизительно в 500 раз или минимум приблизительно в 1000 раз или более превышает уровень стевиол-гликозида, производимого родительской клеткой, на основании удельного сухого веса клеток.
[00154] В некоторых вариантах осуществления клетка-хозяин производит повышенный уровень стевиол-гликозида, который минимум на около 10%, минимум на около 15%, минимум на около 20%, минимум на около 25%, минимум на около 30%, минимум на около 35%, минимум на около 40%, минимум на около 45%, минимум на около 50%, минимум на около 60%, минимум на около 70%, минимум на около 80%, минимум на около 90%, минимум приблизительно в 2 раза, минимум приблизительно в 2.5 раза, минимум приблизительно в 5 раз, минимум приблизительно в 10 раз, минимум приблизительно в 20 раз, минимум приблизительно в 30 раз, минимум приблизительно в 40 раз, минимум приблизительно в 50 раз, минимум приблизительно в 75 раз, минимум приблизительно в 100 раз, минимум приблизительно в 200 раз, минимум приблизительно в 300 раз, минимум приблизительно в 400 раз, минимум приблизительно в 500 раз или минимум приблизительно в 1000 раз или более превышает уровень стевиол-гликозида, производимого родительской клеткой, на основании удельного объема клеточной культуры на единицу времени.
[00155] В некоторых вариантах осуществления клетка-хозяин производит повышенный уровень стевиол-гликозида, который минимум на около 10%, минимум на около 15%, минимум на около 20%, минимум на около 25%, минимум на около 30%, минимум на около 35%, минимум на около 40%, минимум на около 45%, минимум на около 50%, минимум на около 60%, минимум на около 70%, минимум на около 80%, минимум на около 90%, минимум приблизительно в 2 раза, минимум приблизительно в 2.5 раза, минимум приблизительно в 5 раз, минимум приблизительно в 10 раз, минимум приблизительно в 20 раз, минимум приблизительно в 30 раз, минимум приблизительно в 40 раз, минимум приблизительно в 50 раз, минимум приблизительно в 75 раз, минимум приблизительно в 100 раз, минимум приблизительно в 200 раз, минимум приблизительно в 300 раз, минимум приблизительно в 400 раз, минимум приблизительно в 500 раз или минимум приблизительно в 1000 раз или более превышает уровень стевиол-гликозида, производимого родительской клеткой, на основании удельного сухого веса клеток в единицу времени.
[00156] В большинстве вариантов осуществления производство повышенного уровня стевиол-гликозида родительской клеткой индуцируется индуцирующим соединением. Такой клеткой-хозяином можно легко манипулировать в отсутствие индуцирующего соединения. Индуцирующее соединение затем добавляют для стимулирования производства повышенного уровня стевиол-гликозида клеткой-хозяином. В других вариантах осуществления производство повышенного уровня стевиол-гликозида стимулируется меняющимися условиями культивирования, такими, как, например, температура роста, составляющие среды и т.д.
6.8 Культуральные среды и условия культивирования
[00157] Материалы и способы поддержки и роста микробных культур хорошо известны специалистам в области микробиологии или науки о ферментации (см., например, Bailey et al., Biochemical Engineering Fundamentals, second edition, McGraw Hill, New York, 1986). Необходимо учитывать подходящую культуральную среду, рН, температуру и требования к аэробным, микроаэробным или анаэробным условиям, в зависимости от конкретных требований клетки-хозяина, ферментации и процесса.
[00158] Представленные здесь способы производства стевиол-гликозидов могут применяться в подходящей культуральной среде (например, с добавлением или без добавления пантотената) в подходящем контейнере, включая, помимо прочего, планшет для культивирования клеток, колбу или ферментер. Далее эти способы можно реализовывать в любых масштабах ферментации, известных в данной области, для обеспечения промышленного производства микробных продуктов. Можно использовать любой подходящий ферментер, включая ферментер с механическим перемешиванием, ферментер с подачей воздуха, ферментер с пузырящейся колонкой или любую их комбинацию. В конкретных вариантах осуществления, в которых в качестве клетки-хозяина используется Saccharomyces cerevisiae, штаммы могут выращиваться в ферментере, подробно описанном в работе Kosaric, et al, in Ullmann's Encyclopedia of Industrial Chemistry, Sixth Edition, Volume 12, pages 398-473, Wiley-VCH Verlag GmbH & Co. KDaA, Вайнхайм, Германия.
[00159] В некоторых вариантах осуществления культуральной средой является любая культуральная среда, в которой может существовать генетически модифицированный микроорганизм, способный производить стевиол-гликозид, т.е. среда, которая поддерживает рост и жизнеспособность. В некоторых вариантах осуществления такой культуральной средой является водная среда, содержащая источники усвояемого углерода, азота и фосфата. Такая среда также может включать подходящие соли, минералы, металлы и другие питательные вещества. В некоторых вариантах осуществления источник углерода и каждое из основных питательных веществ для клеток добавляются последовательно или непрерывно к ферментативной среде, и каждое необходимое питательное вещество поддерживают на минимальном, по существу, уровне, необходимом для эффективной ассимиляции растущими клетками, например, в соответствии с заранее заданной кривой клеточного роста, основанной на метаболической или респираторной функции клеток, которые превращают источник углерода в биомассу.
[00160] Подходящие условия и подходящие среды для культивирования микроорганизмов хорошо известны в данной области. В некоторых вариантах осуществления в подходящую среду добавляют один или более дополнительных агентов, таких как, например, индуктор (например, когда одна или более нуклеотидных последовательностей, кодирующих генный продукт, контролируются индуцируемым промотором), репрессор (например, когда одна или более нуклеотидных последовательностей, кодирующих генный продукт, контролируются репрессируемым промотором) или селекционный агент (например, антибиотик для селекции микроорганизмов, содержащих генетические модификации).
[00161] В некоторых вариантах осуществления источником углерода является моносахарид (простой сахар), дисахарид, полисахарид, не поддающийся ферментации источник углерода или одна или несколько их комбинаций. Неограничивающие примеры подходящих моносахаридов включают глюкозу, галактозу, маннозу, фруктозу, ксилозу, рибозу и их комбинации. Неограничивающие примеры подходящих дисахаридов включают сукрозу, лактозу, мальтозу, трегалозу, целлобиозу и их комбинации. Неограничивающие примеры подходящих полисахаридов включают крахмал, гликоген, целлюлозу, хитин и их комбинации. Неограничивающие примеры подходящих неферментируемых источников углерода включают ацетат и глицерол.
[00162] Концентрация источника углерода, такого как глюкоза, в культуральной среде должна стимулировать клеточный рост, но не должна быть настолько высока, чтобы подавлять рост используемого микроорганизма. Обычно для культивирования используется источник углерода, такой как глюкоза, который добавляется на уровнях для достижения необходимого уровня роста и биомассы, но на необнаруживаемых уровнях (с пределами обнаружения около <0.1 г/л). В других вариантах осуществления концентрация источника углерода, такого как глюкоза, в культуральной среде выше, чем около 1 г/л, предпочтительно выше, чем около 2 г/л, и предпочтительнее выше, чем около 5 г/л. Кроме того, концентрация источника углерода, такого как глюкоза, в культуральной среде обычно менее, чем около 100 г/л, предпочтительно менее, чем около 50 г/л, и предпочтительнее менее, чем около 20 г/л. Следует отметить, что при указании концентраций компонентов культурной среды могут указываться как первоначальная, так и текущая концентрации компонентов. В некоторых случаях может быть желательно позволить культуральной среде обедняться источником углерода во время культивирования.
[00163] Источники усвояемого азота, которые могут использоваться в подходящей культуральной среде, включают, помимо прочего, источники простого азота, источники органического азота и источники сложного азота. Такие источники азота включают безводный аммиак, аммонийные соли и вещества животного, растительного и/или микробного происхождения. Подходящие источники азота включают, помимо прочего, белковые гидролизаты, гидролизаты микробной биомассы, пептон, дрожжевой экстракт, сульфат аммония, мочевину и аминокислоты. Обычно концентрация источников азота в культуральной среде выше, чем около 0.1 г/л, предпочтительно выше, чем около 0.25 г/л, и предпочтительнее выше, чем около 1.0 г/л. Тем не менее, несоблюдение определенных концентраций при добавлении источника азота к культуральной среде не является благоприятным для роста микроорганизмов. В результате, концентрация источников азота в культуральной среде менее, чем около 20 г/л, предпочтительно менее, чем около 10 г/л, и предпочтительнее менее, чем около 5 г/л. Более того, в некоторых случаях может быть желательно позволить культуральной среде обедняться источниками азота во время культивирования.
[00164] Эффективная культуральная среда может содержать другие соединения, такие как неорганические соли, витамины, микропримеси металла или стимуляторы роста. Такие другие соединения также могут присутствовать в источниках углерода, азота или минералов в эффективной среде или могут специально добавляться к среде.
[00165] Культуральная среда также может содержать подходящий источник фосфата. Такие источники фосфата включают источники как неорганического, так и органического фосфата. Предпочтительные источники фосфата включают, помимо прочего, фосфатные соли, такие как моно- или двунатрия и моно- или двукалия фосфаты, фосфат аммония и их смеси. Обычно концентрация фосфата в культуральной среде выше, чем около 1.0 г/л, предпочтительно выше, чем около 2.0 г/л, и предпочтительнее выше, чем около 5.0 г/л. Тем не менее, несоблюдение определенных концентраций при добавлении фосфата к культуральной среде не является благоприятным для роста микроорганизмов. Соответственно, концентрация фосфата в культуральной среде обычно менее, чем около 20 г/л, предпочтительно менее, чем около 15 г/л, и предпочтительнее менее, чем около 10 г/л.
[00166] Подходящая культуральная среда также может включать источник магния, предпочтительно в форме физиологически приемлемой соли, такой как гептагидрат сульфата магния, хотя также могут использоваться другие источники магния в концентрациях, которые дают аналогичные количества магния. Обычно концентрация магния в культуральной среде выше, чем около 0.5 г/л, предпочтительно выше, чем около 1.0 г/л, и предпочтительнее выше, чем около 2.0 г/л. Тем не менее, несоблюдение определенных концентраций при добавлении магния к культуральной среде не является благоприятным для роста микроорганизмов. Соответственно, концентрация магния в культурной среде менее, чем около 10 г/л, предпочтительно менее, чем около 5 г/л, и предпочтительнее менее, чем около 3 г/л. Более того, в некоторых случаях может быть желательно позволить культуральной среде обедняться источником магния во время культивирования.
[00167] В некоторых вариантах осуществления культуральная среда также может включать биологически приемлемый хелатообразующий агент, такой как тринатрий цитрат дигидрат.В таком случае концентрация хелатообразующего агента в культуральной среде выше, чем около 0.2 г/л, предпочтительно выше, чем около 0.5 г/л, и предпочтительнее выше, чем около 1 г/л. Тем не менее, несоблюдение определенных концентраций при добавлении хелатообразующего агента к культуральной среде не является благоприятным для роста микроорганизмов. Соответственно, концентрация хелатообразующего агента в культуральной среде обычно менее, чем около 10 г/л, предпочтительно менее, чем около 5 г/л, и предпочтительнее менее, чем около 2 г/л.
[00168] Культуральная среда также может первоначально включать биологически приемлемую кислоту или основание для поддержания необходимого рН культуральной среды. Биологически приемлемые кислоты включают, помимо прочего, соляную кислоту, серную кислоту, азотную кислоту, фосфорную кислоту и их смеси. Биологически приемлемые основания включают, помимо прочего, гидроксид аммония, гидроксид натрия, гидроксид калия и их смеси. В некоторых вариантах осуществления используемым основанием является гидроксид аммония.
[00169] Культуральная среда также может включать биологически приемлемый источник кальция, включая, помимо прочего, хлорид кальция. Обычно концентрация источника кальция, такого как кальция хлорид, дигидрат, в культуральной среде находится в пределах от около 5 мг/л до около 2000 мг/л, предпочтительно в пределах от около 20 мг/л до около 1000 мг/л, и предпочтительнее в пределах от около 50 мг/л до около 500 мг/л.
[00170] Культуральная среда также может включать хлорид натрия. Обычно концентрация хлорида натрия в культуральной среде находится в пределах от около 0.1 г/л до около 5 г/л, предпочтительно в пределах от около 1 г/л до около 4 г/л, и предпочтительнее в пределах от около 2 г/л до около 4 г/л.
[00171] В некоторых вариантах осуществления культуральная среда также может включать микропримеси металлов. Такие микропримеси металлов могут добавляться к культуральной среде в виде исходного раствора, который для удобства может быть приготовлен отдельно от остальной части культуральной среды. Обычно количество такого раствора микропримесей металлов, добавляемого к культуральной среде, выше, чем около 1 мл/л, предпочтительно выше, чем около 5 мл/л, и предпочтительнее выше, чем около 10 мл/л. Тем не менее, несоблюдение определенных концентраций при добавлении микропримесей металлов к культуральной среде не является благоприятным для роста микроорганизмов. Соответственно, количество такого раствора микропримесей металлов, добавляемого к культуральной среде, обычно менее, чем около 100 мл/л, предпочтительно менее, чем около 50 мл/л, и предпочтительнее менее, чем около 30 мл/л. Следует отметить, что дополнительно к добавлению микропримесей металлов к исходному раствору, отдельные компоненты могут добавляться по-отдельности, каждый в пределах, независимо соответствующих количествам компонентов, обусловленных выше указанными пределами раствора микропримесей металлов.
[00172] Культуральная среда может включать другие витамины, такие как пантотенат, биотин, кальций, пантотенат, инозитол, пиридоксин-HCl и тиамин-HCl. Такие витамины могут добавляться к культуральной среде в виде исходного раствора, который для удобства может быть приготовлен отдельно от остальной части культуральной среды. Тем не менее, несоблюдение определенных концентраций при добавлении витаминов к культуральной среде не является благоприятным для роста микроорганизмов.
[00173] Описанные здесь ферментативные способы могут использоваться в традиционных режимах культивирования, которые включают, помимо прочего, прерывный метод, метод с подпитыванием, клеточный рисайкл, непрерывный и полунепрерывный способ. В некоторых вариантах осуществления ферментирование выполняется с подпитыванием. В таком случае некоторые компоненты среды истощаются во время культивирования, включая пантотенат на производственном этапе ферментирования. В некоторых вариантах осуществления к культуре могут добавляться относительно высокие концентрации таких компонентов в начале, например, этапа производства, чтобы рост и/или производство стевиол-гликозида поддерживались в течение периода времени до появления необходимости в добавлениях. Предпочтительные диапазоны значений этих компонентов поддерживаются на протяжении культивирования посредством их добавления по мере истощения в среде. Уровни содержания компонентов в культуральной среде можно контролировать, например, посредством периодического забора проб культуральной среды и их анализа на концентрации компонентов. В качестве варианта, после разработки стандартной процедуры культивирования добавления можно производить через заданные интервалы времени в соответствии с известными уровнями концентраций в определенные моменты времени на протяжении культивирования. Как известно специалистам в данной области, скорость потребления питательных веществ повышается во время культивирования по мере роста плотности клеток в среде. Более того, во избежание попадания посторонних микроорганизмов в культуральную среду компоненты добавляются с использованием асептических способов добавления, которые известны в данной области. Кроме того, во время культивирования можно добавлять небольшое количество противовспенивающего средства.
[00174] Температурой культуральной среды может быть любая температура, подходящая для роста генетически модифицированных клеток и/или производства стевиол-гликозида. Например, перед инокуляцией культуральной среды инокулятом температуру культуральной среды можно довести до диапазона от около 20°С до около 45°С, предпочтительно до температуры в диапазоне от около 25°С до около 40°С и предпочтительнее в диапазоне от около 28°С до около 32°С и поддерживать температуру в таких диапазонах.
[00175] рН культуральной среды можно контролировать посредством добавления кислоты или основания к культуральной среде. В таких случаях, когда для контроля рН используется аммиак, он также служит источником азота в культуральной среде. Предпочтительно, рН поддерживают на уровне от около 3.0 до около 8.0, предпочтительнее от около 3.5 до около 7.0, а предпочтительнее всего от около 4.0 до около 6.5.
[00176] В некоторых вариантах осуществления концентрацию источника углерода, например, концентрацию глюкозы, культуральной среды контролируют на протяжении культивирования. Концентрацию глюкозы в культуральной среде можно контролировать при использовании известных методов, таких как, например, использование теста на фермент глюкооксидазу или жидкостную хроматографию высокого давления, которые можно использовать для контроля концентрации глюкозы в супернатанте, например, бесклеточном компоненте культуральной среды. Как указывалось ранее, концентрацию источника углерода необходимо поддерживать ниже уровня, при котором происходит ингибирование клеточного роста. Хотя такая концентрация может быть различной для разных организмов, для глюкозы как источника углерода ингибирование клеточного роста происходит при концентрациях глюкозы, превышающих значение около 60 г/л, и его можно легко определить опытным путем. Соответственно, когда глюкоза используется в качестве источника углерода, глюкозу предпочтительно подают в ферментер и поддерживают на уровне ниже пределов обнаружения. Как вариант, концентрацию глюкозы в культуральной среде поддерживают в диапазоне от около 1 г/л до около 100 г/л, предпочтительнее в диапазоне от около 2 г/л до около 50 г/л, и еще предпочтительнее в диапазоне от около 5 г/л до около 20 г/л. Хотя концентрацию источника углерода можно поддерживать в необходимом диапазоне посредством добавления, например, по существу чистого раствора глюкозы, приемлемым и, возможно, предпочтительным способом является поддержание концентрации источника углерода в культуральной среде посредством добавления аликвот исходной культуральной среды. Использование аликвот исходной культуральной среды может быть желательно, поскольку одновременно можно поддерживать концентрации других питательных веществ в среде (например, источников азота и фосфата). Аналогичным образом, концентрации микропримесей металлов можно поддерживать в культуральной среде посредством добавления аликвот раствора с микропримесями металлов.
[00177] Другая подходящая ферментативная среда и способы описаны, например, в WO 2016/196321.
6.9 Ферментативные композиции
[00178] В другом аспекте здесь представлены ферментативные композиции, содержащие описанную здесь генетически модифицированную клетку-хозяина и стевиол-гликозиды, произведенные генетически модифицированной клеткой-хозяином. Ферментативные композиции могут содержать среду. В определенных вариантах осуществления ферментативные композиции содержат генетически модифицированную клетку-хозяина и также содержат RebA, RebD и RebM. В определенных вариантах осуществления представленные здесь ферментативные композиции содержат RebM как основной компонент стевиол-гликозидов, произведенных генетически модифицированной клеткой-хозяином. В определенных вариантах осуществления ферментативные композиции содержат RebA, RebD и RebM в соотношении минимум 1:7:50. В определенных вариантах осуществления ферментативные композиции содержат RebA, RebD и RebM в соотношении минимум от 1:7:50 до 1:100:1000. В определенных вариантах осуществления ферментативные композиции содержат RebA, RebD и RebM в соотношении минимум от 1:7:50 to 1:200:2000. В определенных вариантах осуществления соотношение RebA, RebD и RebM основывается на общем содержании стевиол-гликозидов, которые ассоциируются с генетически модифицированной клеткой-хозяином и средой. В определенных вариантах осуществления соотношение RebA, RebD и RebM основывается на общем содержании стевиол-гликозидов в среде. В определенных вариантах осуществления соотношение RebA, RebD и RebM основывается на общем содержании стевиол-гликозидов, которые ассоциируются с генетически модифицированной клеткой-хозяином.
[00179] В определенных вариантах осуществления представленные здесь ферментативные композиции содержат RebM2 на необнаруживаемом уровне. В определенных вариантах осуществления представленные здесь ферментативные композиции содержат не встречающиеся в природе стевиол-гликозиды на необнаруживаемом уровне. В определенных вариантах осуществления представленные здесь ферментативные композиции, для которых проводится газовая хроматография, не дают пик "стевиол + 2 глюкоза" между пиком RebA и RebB на обнаруживаемом уровне.
6.10 Выделение стевиол-гликозидов
[00180] После того, как клетка-хозяин производит стевиол-гликозид, его можно выделить для последующего использования с помощью любых подходящих способов выделения и очистки, известных в данной области. В некоторых вариантах осуществления органическую фазу, содержащую стевиол-гликозид, отделяют от ферментативной среды посредством центрифугирования. В других вариантах осуществления органическая фаза, содержащая стевиол-гликозид, отделяется от ферментативной среды спонтанно. В некоторых вариантах осуществления органическую фазу, содержащую стевиол-гликозид, отделяют от ферментативной среды посредством добавления деэмульгатора и/или ядрообразующего агента к ферментативной реакционной смеси. Иллюстративные примеры деэмульгаторов включают флокулянты и коагулянты. Иллюстративные примеры ядрообразующих агентов включают капли самого стевиол-гликозида и орагнических растворителей, таких как додекан, изопропилмиристрат и метилолеат.
[00181] Стевиол-гликозид, произведенный в этих клетках, может присутствовать в супернатанте клеточной культуры и/или ассоциироваться с клетками-хозяевами. В вариантах осуществления, в которых стевиол-гликозид ассоциируется с клеткой-хозяином, выделение стевиол-гликозида может включать способ пермеабилизации или лизинга клеток. В альтернативном варианте или в то же самое время стевиол-гликозид в культуральной среде можно выделить с использованием процесса выделения, включающего, помимо прочего, хроматографию, экстрагирование, экстрагирование растворителем, мембранное разделение, электродиализ, обратный осмос, дистилляцию, получение химическим методом и кристаллизацию.
[00182] В некоторых вариантах осуществления стевиол-гликозид отделяют от других продуктов, которые могут присутствовать в органической фазе. В некоторых вариантах осуществления разделение обеспечивается при помощи поглощения, дистилляции, экстрагирования газ-жидкость (стриппинга), экстрагирования жидкость-жидкость (экстрагирования растворителем), вакуумного экстрагирования, испарения, ультрафильтрации и стандартных хроматографических методов. Другая подходящая ферментативная среда и способы описаны, например, в US 2016/0185813.
6.11 Способы создания генетически модифицированных клеток
[00183] Также здесь представлены способы создания генетически модифицированной клетки-хозяина, которая содержит одну или более описанных ранее модификаций, например, одну или более нуклеиновых гетерологичных нуклеиновых кислот, кодирующих кауреноксидазу Pisum sativum, и/или ферменты пути биосинтеза, например, для соединения стевиол-гликозида. Экспрессию гетерологичного фермента в клетке-хозяине можно обеспечить посредством введения в клетки-хозяева нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую фермент, под контролем регуляторных элементов, которые разрешают экспрессию в клетке-хозяине. В некоторых вариантах осуществления такой нуклеиновой кислотой является экстрахромосомная плазмида. В других вариантах осуществления такой нуклеиновой кислотой является хромосомный интеграционный вектор, который может интегрировать нуклеотидную последовательность в хромосому клетки-хозяина.
[00184] Нуклеиновые кислоты, кодирующие эти белки, можно ввести в клетку-хозяина любым способом, известным специалисту в данной области, без ограничения (см., например, Hinnen et al. (1978) Proc. Natl. Acad. Sci. USA 75:1292-3; Cregg et al. (1985) Mol. Cell. Biol. 5:3376-3385; Goeddel et al. eds, 1990, Methods in Enzymology, vol. 185, Academic Press, Inc., CA; Krieger, 1990, Gene Transfer and Expression -- A Laboratory Manual, Stockton Press, NY; Sambrook et al., 1989, Molecular Cloning -- A Laboratory Manual, Cold Spring Harbor Laboratory, NY; and Ausubel et al., eds., Current Edition, Current Protocols in Molecular Biology, Greene Publishing Associates and Wiley Interscience, NY). Примеры методов включают, помимо прочего, сферопластинг, электропорацию, трансформацию, опосредованную PEG 1000, и трансформацию, опосредованную ацетатом лития или хлоридом лития.
[00185] Число копий фермента в клетке-хозяине можно изменить посредством модификации транскрипции гена, который кодирует этот фермент. Это можно сделать, например, посредством модификации числа копий нуклеотидной последовательности, кодирующей этот фермент {например, при помощи вектора экспрессии большего или меньшего числа копий, содержащего эту нуклеотидную последовательность, или посредством введения дополнительных копий нуклеотидной последовательности в геном клетки-хозяина или посредством удаления или разрушения нуклеотидной последовательности в геноме клетки-хозяина), посредством изменения порядка кодирующих последовательностей в полицистронной мРНК оперона или разделения оперона на отдельные гены, каждый со своими собственными регуляторными элементами, или посредством повышения силы промотера или оператора, с которыми функционально связана нуклеотидная последовательность. В качестве альтернативного варианта или дополнительно число копий фермента в клетке-хозяине можно изменить посредством модификации уровня трансляции мРНК, кодирующей фермент.Это можно сделать, например, посредством модификации стабильности мРНК, модификации последовательности сайта связывания рибосомы, модификации расстояния или последовательности между сайтом связывания рибосомы и инициирующего кодона кодирующей фермент последовательности, модификации всего межцистронного участка, расположенного выше или рядом с 5' стороной инициирующего кодона кодирующего фермент участка, стабилизации 3'-конца транскрипта мРНК с использованием шпилек и специализированных последовательностей, модификации частоты использования кодона фермента, изменения экспрессии тРНК с редкими кодонами, используемых в биосинтезе фермента, и/или повышения стабильности фермента, например, посредством мутации кодирующей его последовательности.
[00186] Активность фермента в клетке-хозяине можно изменить разными способами, включая, помимо прочего, экспрессию модифицированной формы фермента, которая демонстрирует повышенную или пониженную растворимость в клетке-хозяине, экспрессию измененной формы фермента, в которой нет домена, через который ингибируется активность фермента, экспрессию модифицированной формы фермента с более высоким или низким показателем Kcat или с более высоким или низким показателем Km для субстрата, или экспрессию измененной формы фермента, на которую в большей или меньшей степени влияет регуляция по типу обратной связи или по типу прямой связи другой молекулой в пути.
[00187] В некоторых вариантах осуществления нуклеиновая кислота, используемая для генетической модификации клетки-хозяина, содержит один или более селектируемых маркеров, полезных для селекции трансформированных клеток-хозяев и для оказания селективного давления на клетку-хозяина для сохранения чужеродной ДНК.
[00188] В некоторых вариантах осуществления таким селектируемым маркером является маркер устойчивости к антибиотикам. Иллюстративные примеры маркеров устойчивости к антибиотикам включают, помимо прочего, продукты генов BLA, NAT1, PAT, AUR1-C, PDR4, SMR1, CAT, dhfr мыши, НРН, DSDA, KANR и SH BLE. Продукт гена BLA из Е. coli придает устойчивость к бета-лактамным антибиотикам {например, узкого спектра цефалоспоринам, цефамицинам и карбапенемам (эртапенемам), цефамандолу и цефоперазону) и ко всем пенициллинам против грам-отрицательных бактерий, кроме темоциллина; продукт гена NAT1 из S. noursei придает устойчивость к ноурсетрицину; продукт гена PAT из S. viridochromogenes Tu94 придает устойчивость к биалафосу; продукт гена AUR1-C из Saccharomyces cerevisiae придает устойчивость к Auerobasidin А (AbA); продукт гена PDR4 придает устойчивость к церуленину; продукт гена SMR1 придает устойчивость к сульфометуронметилу; продукт гена CAT из транспорона Tn9 придает устойчивость к хлорамфениколу; продукт гена dhfr мыши придает устойчивость к метотрексату; продукт гена НРН Klebsiella pneumonia придает устойчивость к Гигромицину В; продукт гена DSDA Е. coli позволяет клеткам расти в планшетах с D-серином в качестве единственного источника азота; ген KANR транспорона Tn903 придает устойчивость к G418; и продукт гена SH BLE из Streptoalloteichus hindustanus придает устойчивость к зеоцину (блеомицину). В некоторых вариантах осуществления маркер устойчивости к антибиотикам удаляют после того, как представленную здесь генетически модифицированную клетку выделяют.
[00189] В некоторых вариантах осуществления этот селектируемый маркер спасает от ауксотрофии (например, нутриционной ауксотрофии) в генетически модифицированном микроорганизме. В таких вариантах осуществления родительский микроорганизм содержит функциональное нарушение в одном или более генных продуктов, которые функционируют в аминокислотном или нуклеотидном пути биосинтеза и которые, будучи нефункциональными, делают родительскую клетку не способной к росту в среде без добавления одного или более питательных веществ. Такие генные продукты включают, помимо прочего, продукты генов HIS3, LEU2, LYS1, LYS2, METIS, TRP1, ADE2 и URA3 в дрожжах. Затем от ауксотрофного фенотипа можно будет спастись посредством трансформации родительской клетки с вектором экспрессии или хромосомным интеграционным конструктом, кодирующим функциональную копию нарушенного генного продукта, и созданную генетически модифицированную клетку-хозяина можно выбрать на основании утраты ауксотрофного фенотипа родительской клетки. Использование генов URA3, TRP1 и LYS2 в качестве селектируемых маркеров имеет выраженное преимущество, поскольку возможны как позитивная, так и негативная селекции. Позитивная селекция выполняется посредством ауксотрофной комплементации мутаций URA3, TRP1 и LYS2, в то время как негативная селекция основывается на специфичных ингибиторах, т.е. 5-фтороротовой кислоте (FOA), 5-фторотоаминобензойной кислоте и аминоадипиновой кислоте (аАА), соответственно, которые предотвращают рост прототрофных штаммов, но разрешают рост мутантов URA3, TRP1 и LYS2, соответственно. В других вариантах осуществления селектируемый маркер спасает от других нелетальных дефицитов или фенотипов, что можно определить при помощи известного способа селекции.
[00190] Здесь описаны конкретные гены и белки, полезные для способов, композиций и организмов данного описания изобретения; тем не менее, понятно, что абсолютная идентичность с такими генами не требуется. Например, могут выполняться и анализироваться на активность изменения в определенном гене или полинуклеотиде, содержащем последовательность, кодирующую тот или иной полипептид или фермент.Обычно такие изменения включают консервативные мутации и молчащие мутации. Такие модифицированные или мутированные полинуклеотиды и полипептиды можно подвергнуть скринингу на экспрессию функционального фермента с использованием известных в данной области методов.
[00191] Из-за присущей дегенерации генетического кода для клонирования и экспрессии полинуклеотидов, кодирующих такие ферменты, также можно использовать другие полинуклеотиды, которые кодируют, по существу, такие же или функционально эквивалентные полипептиды.
[00192] Как понятно специалистам в данной области, может быть эффективно модифицировать кодирующую последовательность для улучшения ее экспрессии в определенном хозяине. Генетический код включает 64 возможных кодона, однако большинство организмов обычно используют подгруппу таких кодонов. Кодоны, которые используются в том или ином виде чаще всего, называются оптимальными кодонами, а те, которые не используются очень часто, классифицируются как редкие или мало используемые кодоны. Кодоны могут заменяться для отражения предпочтительного использования кодонов хозяина в процессе, иногда называемом "оптимизацией кодонов" или "контролированием предпочтений кодонов". Оптимизацию кодонов для других клеток-хозяев можно легко определить при помощи таблиц частот использования кодонов либо можно выполнить при помощи имеющегося в продаже программного обеспечения, такого как CodonOp (www.idtdna.com/CodonOptfrom) от Integrated DNA Technologies.
[00193] Можно приготовить оптимизированные кодирующие последовательности, содержащие кодоны, предпочитаемые определенным прокариотическим или эукариотическим хозяином (Murray et al., 1989, Nucl Acids Res. 17: 477-508), например, чтобы повысить скорость трансляции или производить транскрипты с рекомбинантной РНК с необходимыми свойствами, такими как большее время полужизни по сравнению с транскриптами, произведенными из неоптимизированной последовательности. Для отражения предпочтений хозяина также могут быть модифицированы терминирующие трансляцию кодоны. Например, типичными стоп-кодонами для S. cerevisiae и млекопитающих являются UAA и UGA, соответственно. Типичным стоп-кодоном для однодольных растений является UGA, а насекомые и Е. coli в качестве стоп-кодона обычно используют UAA (Dalphin et al., 1996, Nucl Acids Res. 24: 216-8).
[00194] Специалистам в данной области понятно, что из-за вырожденности генетического кода для кодирования заданного фермента данного описания изобретения можно использовать различные молекулы ДНК, различающиеся своими нуклеотидными последовательностями. Здесь упоминаются последовательности нативной ДНК, кодирующие описанные выше ферменты биосинтеза, только для иллюстрации варианта осуществления изобретения, и изобретение включает молекулы ДНК любой последовательности, которая кодирует аминокислотные последовательности полипептидов и белков ферментов, используемых в способах настоящего изобретения. Аналогичным образом, полипептид обычно может допустить одну или более аминокислотных замен, делеций и вставок в своей аминокислотной последовательности без утраты или значительной утраты необходимой активности. Настоящее изобретение включает такие полипептиды с другими аминокислотными последовательностями, чем у описанных здесь конкретных белков, если модифицированные полипептиды или варианты полипептидов имеют ферментную анаболическую или катаболическую активность эталонного полипептида. Более того, указанные здесь аминокислотные последовательности, закодированные последовательностями ДНК, только иллюстрируют варианты осуществления настоящего изобретения.
[00195] Кроме того, настоящее изобретение включает гомологи ферментов, полезных для представленных здесь композиций и способов. В некоторых вариантах осуществления два белка (или участок белков) являются, по существу, гомологичными, когда аминокислотные последовательности идентичны минимум на 30%, 40%, 50%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%. Чтобы определить процент идентичности двух аминокислотных последовательностей или двух последовательностей нуклеиновых кислот, последовательности выравниваются для целей оптимального сравнения (например, для оптимального выравнивания в одну, первую или вторую, или в обе аминокислотные последовательности или последовательности нуклеиновых кислот могут быть введены пробелы и негомологичные последовательности могут не учитываться для целей сравнения). В одном варианте осуществления длина эталонной последовательности, выравненной для целей сравнения, составляет минимум 30%, обычно минимум 40%, чаще минимум 50%, еще чаще минимум 60%, и даже еще чаще 70%, 80%, 90%, 100% длины эталонной последовательности. Затем сравниваются аминокислотные остатки или нуклеотиды в соответствующих аминокислотных позициях или нуклеотидных позициях. Когда та или иная позиция в первой последовательности занята тем же аминокислотным остатком или нуклеотидом, что и соответствующая позиция во второй последовательности, тогда молекулы являются идентичными в этой позиции (в контексте настоящего документа "идентичность" аминокислот или нуклеиновых кислот эквивалентна "гомологии" аминокислот или нуклеиновых кислот). Процент идентичности между двумя последовательностями представляет собой" функцию числа идентичных позиций, общих для последовательностей, с учетом числа пробелов и длины каждого пробела, которые необходимо вводить для оптимального выравнивания этих двух последовательностей.
[00196] Когда термин "гомологичный" используется в отношении белков или пептидов, понятно, что позиции остатков, которые не идентичны, часто различаются консервативными аминокислотными заменами. "Консервативная аминокислотная замена" - это замена, в которой аминокислотный остаток заменяется другим аминокислотным остатком, имеющим боковую цепь (R группа) с аналогичными химическими свойствами (например, зарядом или гидрофобностью). В общем, консервативная аминокислотная замена существенно не изменит функциональные свойства того или иного белка. В случаях, когда две или более аминокислотных последовательности различаются друг от друга консервативными заменами, процент идентичности последовательностей или степень гомологичности можно откорректировать в направлении вверх, чтобы принять во внимание консервативную природу этой замены. Средства для такой корректировки хорошо известны специалистам в данной области (см., например, Pearson W.R., 1994, Methods in Mol Biol 25: 365-89).
[00197] Каждая из следующих шести групп содержит аминокислоты, которые являются консервативными заменами друг для друга. 1) Серии (S), треонин (Т); 2) аспарагиновая кислота (D), глутаминовая кислота (Е); 3) аспарагин (N), глутамин (Q); 4) аргинин (R), лизин (K); 5) изолейцин (I), лейцин (L), аланин (А), валин (V) и 6) фенилаланин (F), тирозин (Y), триптофан (W).
[00198] Гомология последовательностей для полипептидов, которую также называют идентичностью последовательностей в процентах, обычно измеряется при помощи программного обеспечения для анализа последовательностей. Типичным алгоритмом, используемым для сравнения последовательности молекулы с базой данных, содержащей большое количество последовательностей из различных организмов, является компьютерная программа BLAST. При выполнении поиска в базе данных, содержащей последовательности из большого числа различных организмов, обычно сравнивают аминокислотные последовательности.
[00199] Более того, любой из генов, кодирующих указанные ранее ферменты (или любые другие, упомянутые здесь (или любые из регуляторных элементов, которые контролируют или модулируют их экспрессию)) может быть оптимизирован при помощи методов генной/белковой инженерии, таких как направленная эволюция или рациональный мутагенез, которые хорошо известны специалистам в данной области. Такое действие позволяет специалистам в данной области оптимизировать ферменты для экспрессии и активности в дрожжах.
[00200] Кроме того, гены, кодирующие такие ферменты, могут быть идентифицированы из других видов грибов и бактерий и могут быть экспрессированы для модуляции этого пути. В качестве источников для таких ферментов могут служить различные организмы, включая, помимо прочего, Saccharomyces spp., включая S. cerevisiae и S. uvarum, Kluyveromyces spp., включая K. thermotolerans, K. lactis и K. marxianus, Pichia spp., Hansenula spp., включая H polymorpha, Candida spp., Trichosporon spp., Yamadazyma spp., включая Y. spp. stipitis, Torulaspora pretoriensis, Issatchenkia orientalis, Schizosaccharomyces spp., включая S. pombe, Cryptococcus spp., Aspergillus spp., Neurospora spp.или Ustilago spp. Источники генов из анаэробных грибов включают, помимо прочего, Piromyces spp, Orpinomyces spp.или Neocallimastix spp. Источники полезных прокариотических ферментов включают, помимо прочего, Escherichia, coli, Zymomonas mobilis, Staphylococcus aureus, Bacillus spp., Clostridium spp., Corynebacterium spp., Pseudomonas spp., Lactococcus spp., Enterobacter spp. и Salmonella spp.
[00201] Методы, известные специалистам в данной области, могут подходить для идентификации дополнительных гомологичных генов и гомологичных ферментов. В общем, аналогичные гены и/или аналогичные ферменты можно идентифицировать посредством функционального анализа, и они будут иметь функциональные сходства. Методы, известные специалистам в данной области, могут подходить для идентификации аналогичных генов и аналогичных ферментов. Например, методы выявления гомологичных или аналогичных UDP-гликозилтрансфераз, РТА или любых генов, белков или ферментов пути биосинтеза могут включать, помимо прочего, клонирование гена посредством ПЦР с использованием праймеров, основанных на опубликованной последовательности представляющего интерес гена/фермента, либо посредством ПЦР с использованием вырожденных праймеров, предназначенных для амплификации консервативной области в представляющем интерес гене. Более того, специалист в данной области может использовать методы для выявления гомологичных или аналогичных генов, белков или ферментов с функциональной гомологией или сходством. Методы включают контроль клетки или клеточной культуры на каталитическую активность фермента посредством in vitro ферментного анализа на указанную активность (например, как описано в настоящем документе и в работе Kiritani, K., Branched-Chain Amino Acids Methods Enzymology, 1970), последующее выделение фермента с указанной активностью посредством очистки, определение белковой последовательности фермента при помощи таких методов, как расщепление по Эдману, конструирование праймеров для ПЦР по вероятной последовательности нуклеиновой кислоты, амплификацию указанной последовательности ДНК через ПЦР и клонирование указанной последовательности нуклеиновой кислоты. Методы определения гомологичных или похожих генов и/или гомологичных или похожих ферментов, аналогичных генов и/или аналогичных ферментов или белков также включают сравнение данных, касающихся гена-кандидата или фермента-кандидата с базами данных, такими как BRENDA, KEGG или MetaCYC. Ген-кандидат или фермент-кандидат можно идентифицировать в указанных выше базах данных в соответствии с представленными здесь идеями.
7. ПРИМЕРЫ
Пример 1: Создание базового дрожжевого штамма, способного к высокому потоку в фарнезилпирофосфат (FPP) и изопреноид-фарнезен.
[00202] Штамм для производства фарнезена создали из штамма Saccharomyces cerevisiae (CEN.PK2) дикого типа посредством экспрессии генов мевалонатного пути (ФИГ. 1D) под контролем промотеров GAL1 или GAL10. Данный штамм содержал следующие хромосомно-интегрированные гены мевалонатного пути из S. cerevisiae: ацетил-СоА тиолаза, HMG-CoA синтаза, HMG-CoA редуктаза, мевалонаткиназа, фосфомевалонаткиназа, мевалонатпирофосфатдекарбоксилаза и IPP:DMAPP изомераза. Все описанные здесь гены были подвергнуты оптимизации кодонов с использованием общедоступных или других подходящих алгоритмов. Кроме того, этот штамм содержал шесть копий фарнезенсинтазы из Artemisinin annua, также под контролем одного из промотеров GAL1 или GAL10. Штамм также содержал делецию гена GAL80 и дополнительную копию GAL4 под контролем промотера GAL4oc, при этом кодирующая последовательность гена GAL4 Saccharomyces cerevisiae находится под регуляторным контролем "оперативной конститутивной" версии его нативного промотера (PGAL4oc; см., например, Griggs & Johnston (1991) PNAS 88(19):8597-8601). Наконец, ген ERG9, кодирующий скваленсинтазу, понижающе регулируется посредством замены нативного промотера промотером дрожжевого гена МЕТ3 (Westfall et al PNAS 2012).
Пример 2. Создание базового дрожжевого штамма, способного к высокому потоку в Ребаудиозид A (RebA).
[00203] На ФИГ. 1В показан пример пути биосинтеза из FPP в стевиол. На ФИГ. 2 показан пример пути биосинтеза из стевиола в гликозид RebM. Для превращения базового штамма для производства фарнезена, описанного выше, для обеспечения высокого потока в С-20 изопреноид-каурен, в геном интегрировали шесть копий геранилгеранилпирофосфатсинтазы (GGPPs), а затем по четыре копии копалилдифосфатсинтазы и кауренсинтазы. В Таблице 1 указаны все гены и промотеры, используемые для превращения FPP в RebA. На данном этапе шесть копий фарнезенсинтазы удалили из штамма. После подтверждения того, что новый штамм производит энт-каурен, в геном интегрировали оставшиеся гены для превращения энт-каурена в RebA. Каждый ген интегрировали с одной копией, за исключением фермента Sr.KAH, в случае с которым интегрировали две копии (Таблица 1.) Штамм, содержащий все гены, описанные в Таблице 1, главным образом, производил RebA. Фермент UGT91D_like3 проявлял низкую активность по превращению RebA в Ребаудиозид D (RebD). По нашим измерениям, одна копия 91D_like3 способна превращать приблизительно (3%) RebA в штамме в RebD in vivo в дрожжевом штамме, описанном выше (ФИГ. З и Таблица 2). Затем UGT76G1 может превращать RebD в конечный продукт Ребаудиозид М (RebM).
Пример 3. Скрининг ферментов кауреноксидазы (КО) для превращения каурена в кауреновую кислоту с более высокой эффективностью.
[00204] Для создания штамма с высоким потоком в RebM штамм, описанный в Примере 2, трансформировали с одной копией гена UGT40087 (описанного в Примере 8 и таблицах и Фигурах в Заявке согласно РСТ АМ-7400 РСТ, которая приложена к настоящему документу) под контролем промотера GAL1. Данный штамм производит, главным образом, RebM. Для скрининга различных аллелей КО касательно превращения каурена в кауреновую кислоту in vivo, ген КО Stevia rebuaudiana в данном штамме RebM удалили и заменили посадочной площадкой, содержащей только промотер GAL1 и терминатор, с рестрикционной последовательностью F-CphI между промотером и терминатором (ФИГ. 3). Теперь в этом скрининговом штамме нет фермента КО, и он производит только энт-каурен.
[00205] Тринадцать ферментов КО (Таблица 1) по данным литературы подвергли оптимизации кодонов для оптимальной экспрессии в S. cerevisiae и синтезировали с 60 п. о. последовательности, гомологичной PGAL1, и дрожжевым терминатором, фланкирующим последовательности F-CphI в посадочной площадке, описанной на ФИГ. 3А. Каждый синтезированный ген КО проверили по-отдельности, с одной копией, на способность превращать эн/и-каурен в кауреновую кислоту in vivo в описанном выше дрожжевом штамме. Дрожжи трансформировали донорской ДНК КО и плазмидой, содержащей эндонуклеазу F-CphI, чтобы разрезать ДНК в посадочной площадке. Правильность интеграций проверили посредством ПЦР для отбора колоний с использованием обратного праймера, внутреннего для конкретного гена КО в каждой трансформации, и универсального прямого праймера в конце промотера GAL1. На ФИГ. 3В показан итоговый генетический конструкт после правильного разрезания F-CphI и гомологичной рекомбинации ДНК КО.
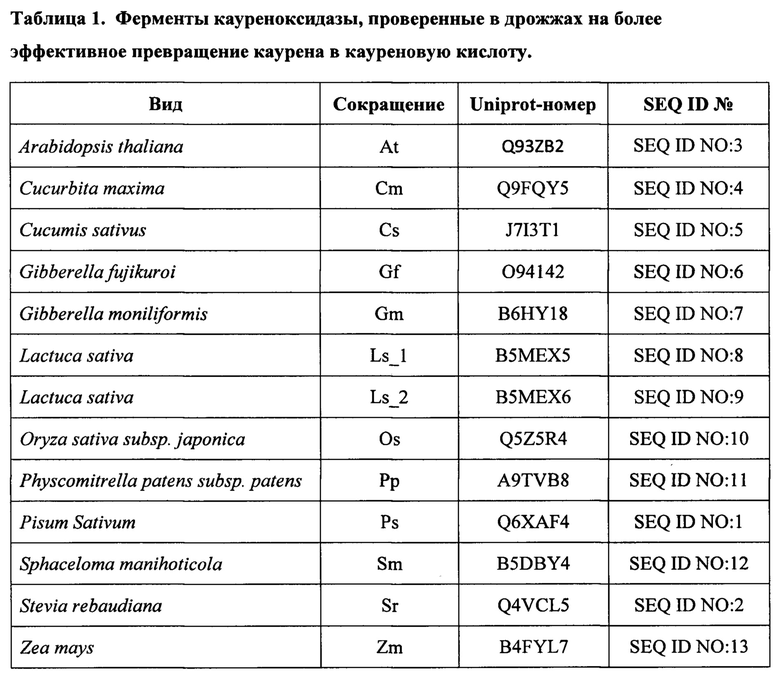
[00206] На ФИГ. 4 показаны результаты скрининга КО. Было обнаружено, что один фермент КО (Ps.KO) из растения Pisum sativum (горох огородный) имеет повышенную способность (эффективнее приблизительно в 3.5 раза) к превращению эняг-каурена в кауреновую кислоту, по сравнению с ферментом КО из Stevia rebaudiana (Sr.KO) на данном фоне штаммов. Кодон-оптимизированные последовательности нуклеиновых кислот фермента КО Pisum sativum, используемые для экспрессии в дрожжевых клетках, показаны как SEQ ID NO: 15.
Пример 4. Создание штамма с высоким потоком в RebM с более эффективным превращением энш-каурена в кауреновую кислоту.
[00207] Затем активность Ps.KO проверили в сравнении с Sr.KO в штамме с очень высоким потоком в RebM. Обычно ферменты КО в большинстве растений участвуют в производстве растительного гормона гиббереллина. Уровни гиббереллина в клетках растений на порядки ниже уровней RebM, который производится в дрожжах для промышленного производства, и в связи с этим большинство ферментов КО не обеспечивают высокий поток, необходимый для производства RebM в целях коммерческого производства. В Таблице 3 указаны все гены и промотеры, содержащиеся в штамме с более высоким потоком в RebM по сравнению со штаммом, использованным для первоначального скрининга ферментов КО (т.е. "базовым штаммом" КО). Все гены в Таблице 3 интегрировали в дрожжевой геном. Фермент КО заставляет эняг-каурен пройти через три этапа последующего окисления, в результате чего производится кауреновая кислота. Порядок реакций и промежуточные соединения следующие: в результате первого окисления энт-каурен превращается в кауренол (К-OL), в результате второго окисления кауренол превращается в кауренал (K-AL), а в результате третьего окисления кауренал превращается в кауреновую кислоту (-кислота) (ФИГ. 1С). Для достижения максимального потока из энт-каурена в RebM фермент КО должен полностью окислять эн»з-каурен в К-кислоту. Неполное превращение израсходует углерод, снизит общие титры RebM и произведет потенциально токсичные промежуточные соединения. Данные на ФИГ. 5 показывают, как в штамме с высоким потоком углерода в RebM аллель Sr.KO накапливает значительные количества вышележащих промежуточных соединения энт-каурена, кауренола (K-OL), кауренала (К-AL), в то время как фермент Ps.KO демонстрирует значительно меньшее накопление этих промежуточных соединений.
[00208] ФИГ. 6 показывает, что Ps.KO повышает количество RebM, производимого в клетке, из-а большего количества кауреновой кислоты, производимой с Ps.KO по сравнению с Sr.KO. В штамме с высоким потоком в RebM наблюдается 16% повышение титров RebM в штамме с Ps.KO по сравнению с идентичным штаммом с Sr.KO. Этот более высокий титр RebM обусловлен большим количеством кауреновой кислоты, производимой в штамме с Ps.KO.
Пример 5. Условия культивирования дрожжей
[00209] Дрожжевые колонии, проверенные на содержание предполагаемого гена кауреноксидазы, распределили в 96-луночный микротитрационный планшет, содержащий среду Bird Seed Media (BSM, первоначально описанную в работе van Hoek et al., Biotechnology and Bioengineering 68(5), 2000, pp.517-523) с 20 г/л сахарозы и 37.5 г/л сульфата аммония. Клетки культивировали при 30°С в высокопроизводительном инкубаторе микротитрационных планшетов со встряхиванием в 1000 об/мин и влажностью 80% в течение 3 дней до истощения углерода. Далее культуры пересеяли в новые планшеты, содержащие BSM с 40 г/л сахарозы и 150 г/л сульфата аммония посредством отбора 14.4 мкл из культур и разбавления в 360 мкл свежей среды. Клетки в среде культивировали при 30°С в высокопроизводительном инкубаторе-встряхивателе микротитрационных планшетов со встряхиванием 1000 об/мин и влажностью 80% еще на протяжении 3 дней с последующим экстрагированием и анализом. По завершении весь клеточный бульон разбавили 360 мкл 100% этанола, запечатали с использованием прокладки-уплотнителя с мембраной из фольги и встряхивали с 1250 об/мин в течение 30 мин для экстрагирования ребаудизиодов. 490 мкл смеси 50:50 этанол:вода добавили в новый 1.1-мл аналитический планшет и 10 мкл смеси культура/этанол добавили в аналитический планшет. Смесь центрифугировали для осаждения любых твердых частиц и 400 мкл раствора перенесли в новый 1.1-мл планшет и проанализировали посредством жидкостной хроматографии с масс-спектрометрией.
Пример 6. Аналитические методы
Обнаружение стевиола и стевиол-гликозидов при помощи масс-спектрометра:
[00210] Образцы анализируют при использовании масс-спектрометра для жидкостной хроматографии с масс-спектрометрией (АВ QTrap 4000) с применением пептида Sigma Ascentis Express Peptide ES-C18 (5 см, 2.1 мм, 2.7 мкм; артикул 53301-U) со следующим градиентом:
[00211] Масс-спектрометр использовали в режиме определения отрицательных ионов при селективной регистрации избранных реакций распада нескольких ионов (MRM). Каждый изомер ребаудиозида выявляли по времени удержания (RT), определенному на основании действующего стандарта, и по MRM переходу:
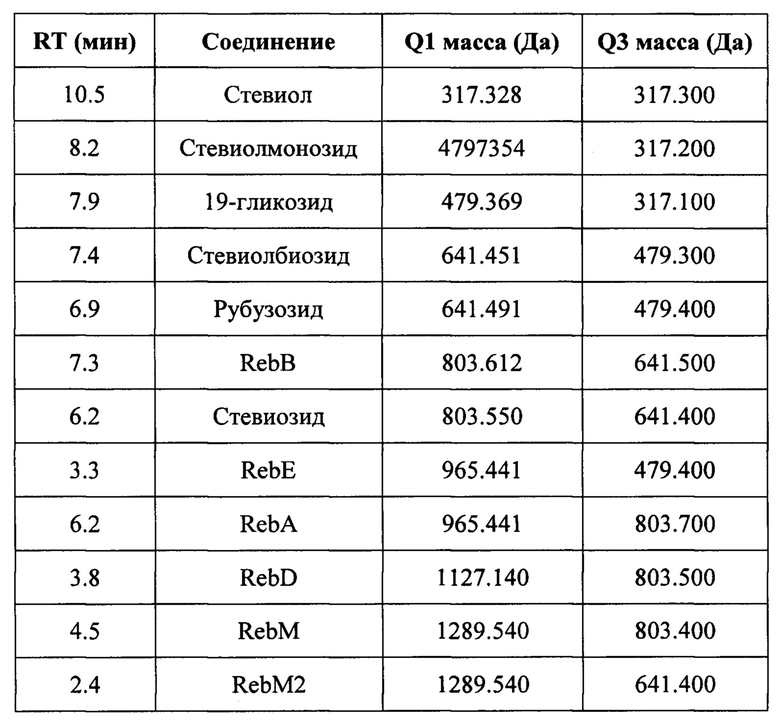
Количественное определение каурена:
[00212] Титр энт-каурена в бульонной культуре измеряют при помощи газового хроматографа, оснащенного печью с ограниченной тепловой массой и пламенно-ионизационным детектором. Бульонные образцы экстрагируют с использованием равных частей бульона и метанола и встряхивают в герметичном контейнере в течение 30 мин для выделения энт-каурена из клеток. Аликвоту 240 мкл раствора бульон:метанол затем разбавляют 1 мл этилацетата, запечатывают и встряхивают еще на протяжении 30 мин для экстрагирования энт-каурена в органическую фазу. Органическую фазу разбавляют необходимым образом, чтобы оставаться в границах линейного диапазона анализа, и разделяют на аликвоты во флаконы для образца. Образцы вводят с соответствующим индексом разведения, чтобы оставаться в пределах линейного диапазона. Разделение образца происходит на колонке Agilent DB-1MS LTM II, с использованием водорода в качестве газа-носителя в режиме постоянного давления, с использованием температурного градиента: (1) первоначальная температура 150°С в течение 0 мин, (2) повышение температуры на 25°С/мин до температуры 230°С, (3) повышение температуры на 1800°С/мин до температуры 320°С и поддержание температуры в течение 1 мин. Для определения количества энт-каурена используют внешнюю калибровку с использованием аутентичного стандарта энт-каурена.
Количественное определение кауреновой кислоты, кауренола и кауренала:
[00213] Титры кауреновой кислоты, кауренола и кауренала в бульонной культуре определяют при помощи жидкостного хроматографа высокого давления, оснащенного детектором с переменной длиной волны. Образец бульона (100 мкл) разбавляют в 300 мкл этанола и встряхивают в герметичном контейнере в течение 30 мин. 200 мкл воды добавляют к смеси бульон:этанол, смешивают и центрифугируют. Аликвоту полученного раствора (без клеточного осадка) переносят во флакон для образца и анализируют посредством высокоэффективной жидкостной хроматографии. Разделение образца происходит на Agilent Eclipse Plus C18 USP LI (4.6 мм × 50 мм × 1.8 мкм) со следующими растворителями:
• Подвижная фаза А: 0.1% муравьиная кислота в воде (об/об)
• Подвижная фаза В: 0.1% муравьиная кислота в ацетонитриле (об/об) с градиентом растворителя:

Аналиты обнаруживают с использованием УФ поглощения на 200 нм и определяют количественно при помощи внешней калибровки с коэффициентами относительной чувствительности к стандарту стевиола.
Метод количественного определения Ребаудиозида М, используемый для данных, показанных на ФИГ. 6:
[00214] Титры Ребаудидиозида М в бульоне определяют при помощи жидкостного хроматографа высокого давления, оснащенного трехквадрупольным масс-спектрометром. Образец бульона разделяют на аликвоты в пробирки Eppendorf с 200-800-кратным разбавлением в смеси 50:50 этанол:вода, смешивают в течение 20 мин, центрифугируют для удаления клеточного осадка и дебриса и аликвоту супернатанта переносят во флакон для образца для анализа. Образцы прогоняют в проточно-инжекционном режиме, и количественное определение аналитов проводят на основании интенсивности сигнала MRM переходов. Подвижная фаза 40% вода + 0.1% муравьиная кислота и 60% ацетонитрил + 0.1% муравьиная кислота с расходом в 1.1 мл/мин. Концентрацию Ребаудиозида М определяют посредством его отклика, нормализованного по отклику внутреннего стандарта (Ребаудиозид N).
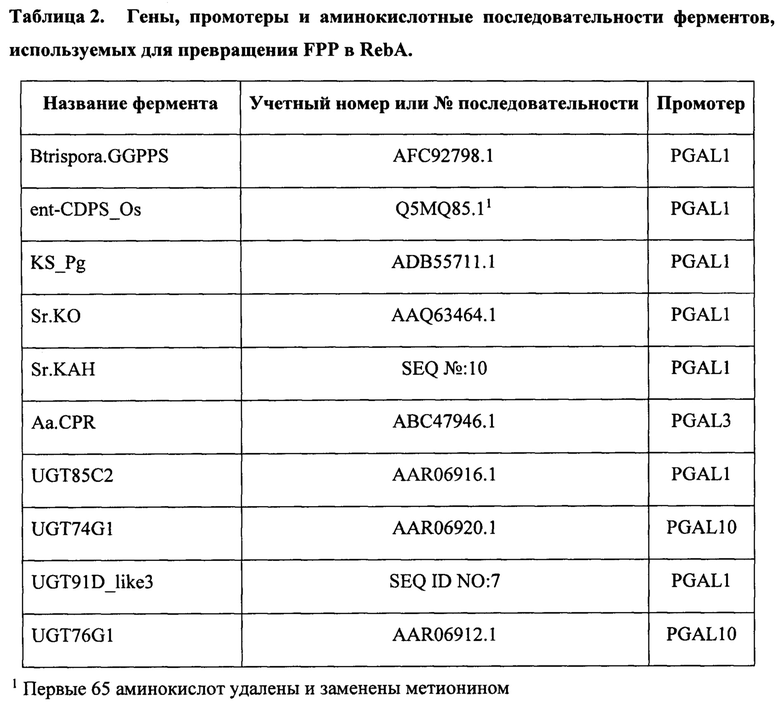
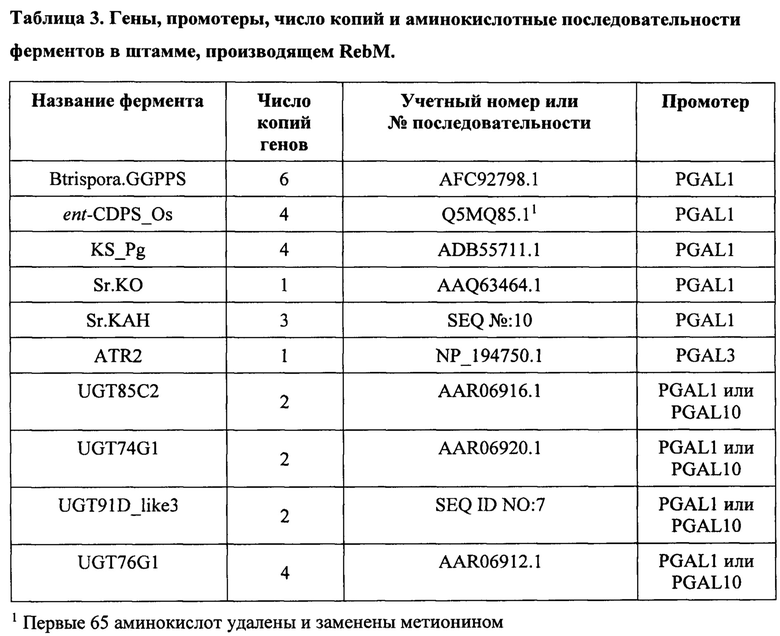
[00215] Все публикации, патенты и патентные заявки, упомянутые в настоящей спецификации, включены в настоящий документ посредством ссылки, как если бы было конкретно и по-отдельности указано, что каждая отдельная публикация, патент или патентная заявка включены в настоящий документ посредством ссылки. Несмотря на то, что описанное выше изобретение было описано довольно подробно посредством иллюстраций и примеров для упрощения понимания, специалистам в данной области должно быть ясно в свете идей данного изобретения, что в отношении данного изобретения можно выполнить некоторые изменения и модификации без отступления от духа или объема прилагаемых пунктов формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| UDP-ЗАВИСИМАЯ ГЛИКОЗИЛТРАНСФЕРАЗА ДЛЯ ВЫСОКОЭФФЕКТИВНОГО ПРОДУЦИРОВАНИЯ РЕБАУДИОЗИДОВ | 2017 |
|
RU2777901C2 |
| РЕКОМБИНАНТНОЕ ПОЛУЧЕНИЕ СТЕВИОЛ-ГЛИКОЗИДОВ | 2014 |
|
RU2706789C2 |
| БИОСИНТЕТИЧЕСКОЕ ПОЛУЧЕНИЕ СТЕВИОЛОВОГО ГЛИКОЗИДА РЕБАУДИОЗИДА D4 ИЗ РЕБАУДИОЗИДА E | 2017 |
|
RU2764803C2 |
| РЕКОМБИНАНТНОЕ ПОЛУЧЕНИЕ СТЕВИОЛ-ГЛИКОЗИДОВ | 2014 |
|
RU2741103C2 |
| АBC-ТРАНСПОРТЕРЫ ДЛЯ ВЫСОКОЭФФЕКТИВНОГО ПРОИЗВОДСТВА РЕБАУДИОЗИДОВ | 2020 |
|
RU2795855C2 |
| ГИДРОЛИЗ СТЕВИОЛОВЫХ ГЛИКОЗИДОВ С ПОМОЩЬЮ БЕТА-ГЛЮКОЗИДАЗЫ | 2018 |
|
RU2775697C2 |
| СПОСОБЫ И ВЕЩЕСТВА ДЛЯ ОСНОВАННОГО НА РЕКОМБИНАЦИИ ПОЛУЧЕНИЯ СОЕДИНЕНИЙ ШАФРАНА | 2012 |
|
RU2676730C2 |
| НОВЫЕ ФУКОЗИЛТРАНСФЕРАЗЫ И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2642307C2 |
| Способ получения ребаудиозида D и ребаудиозида M | 2022 |
|
RU2839979C2 |
| СПОСОБЫ ФЕРМЕНТАТИВНОГО СИНТЕЗА МОГРОЗИДНЫХ СОЕДИНЕНИЙ | 2012 |
|
RU2633644C2 |









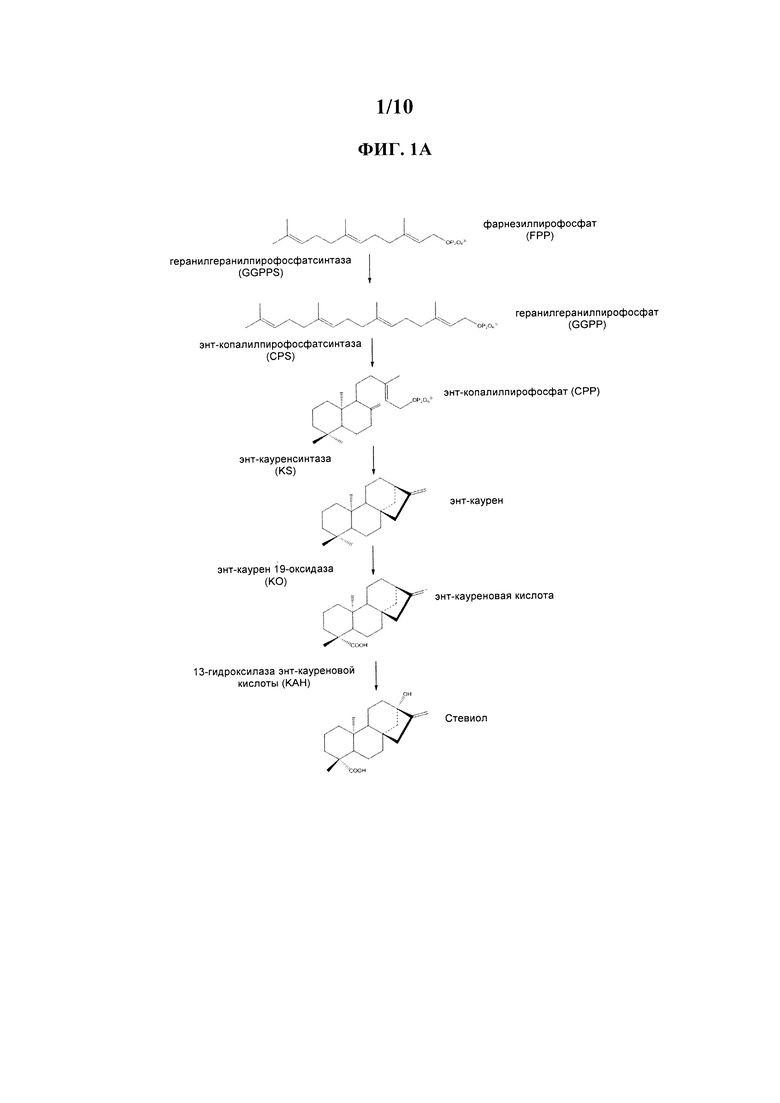
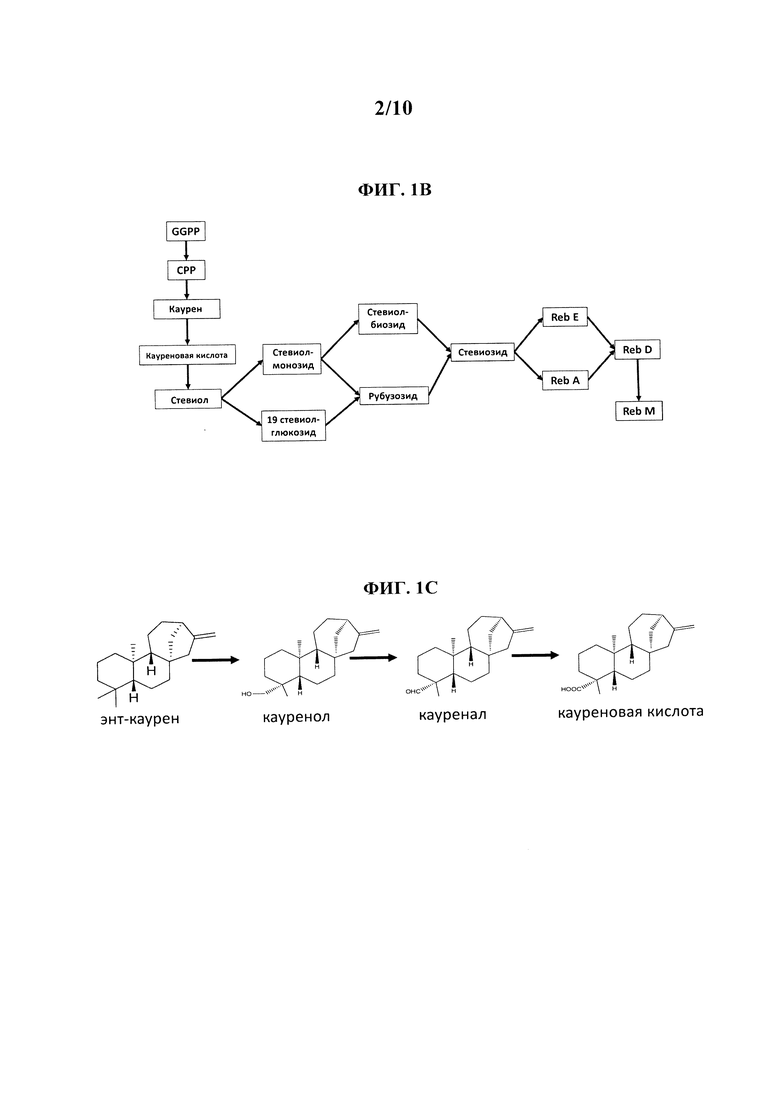
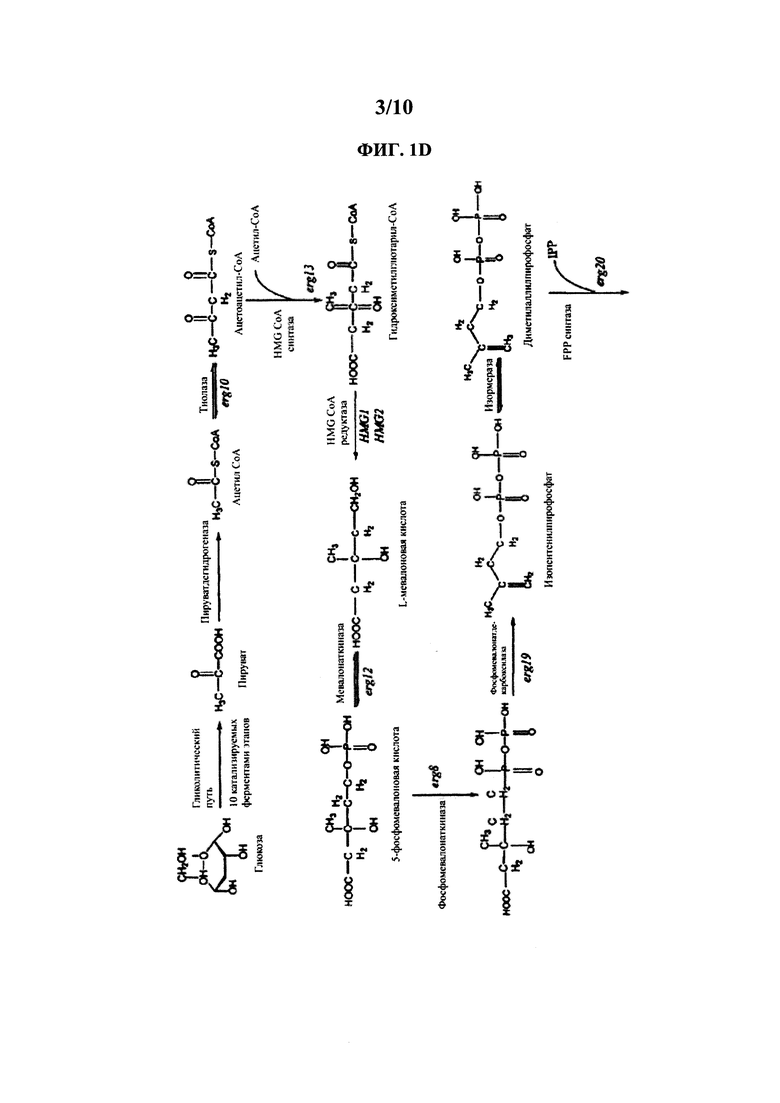
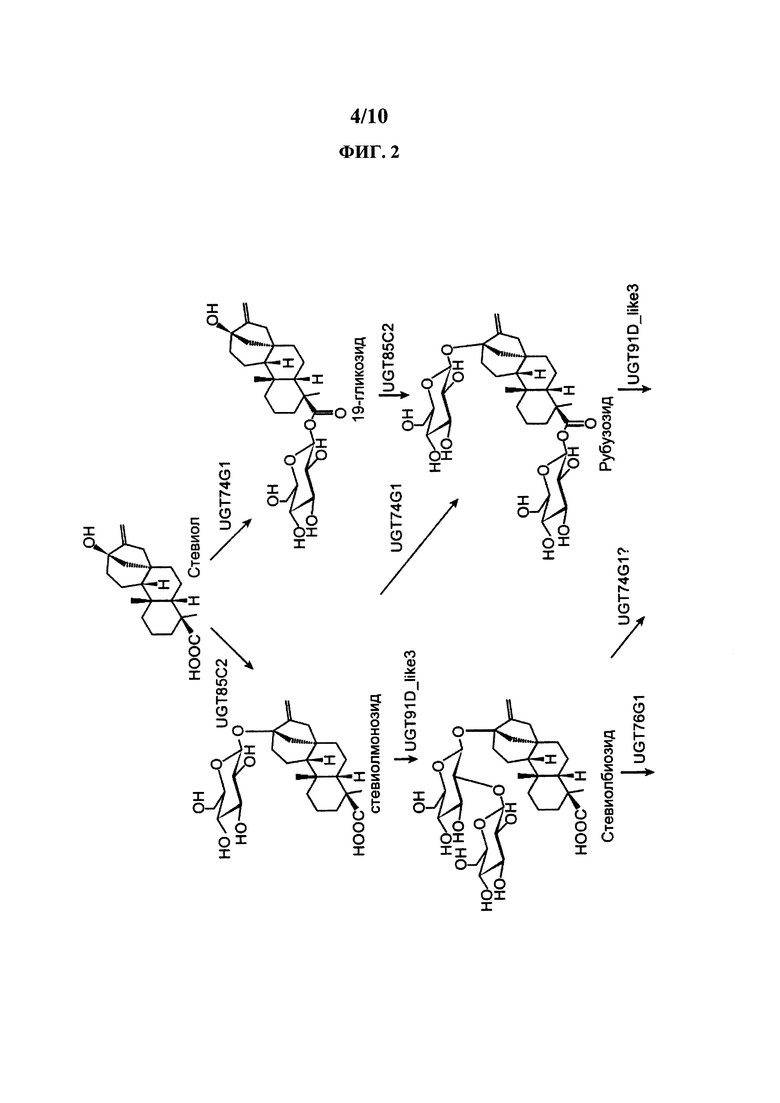
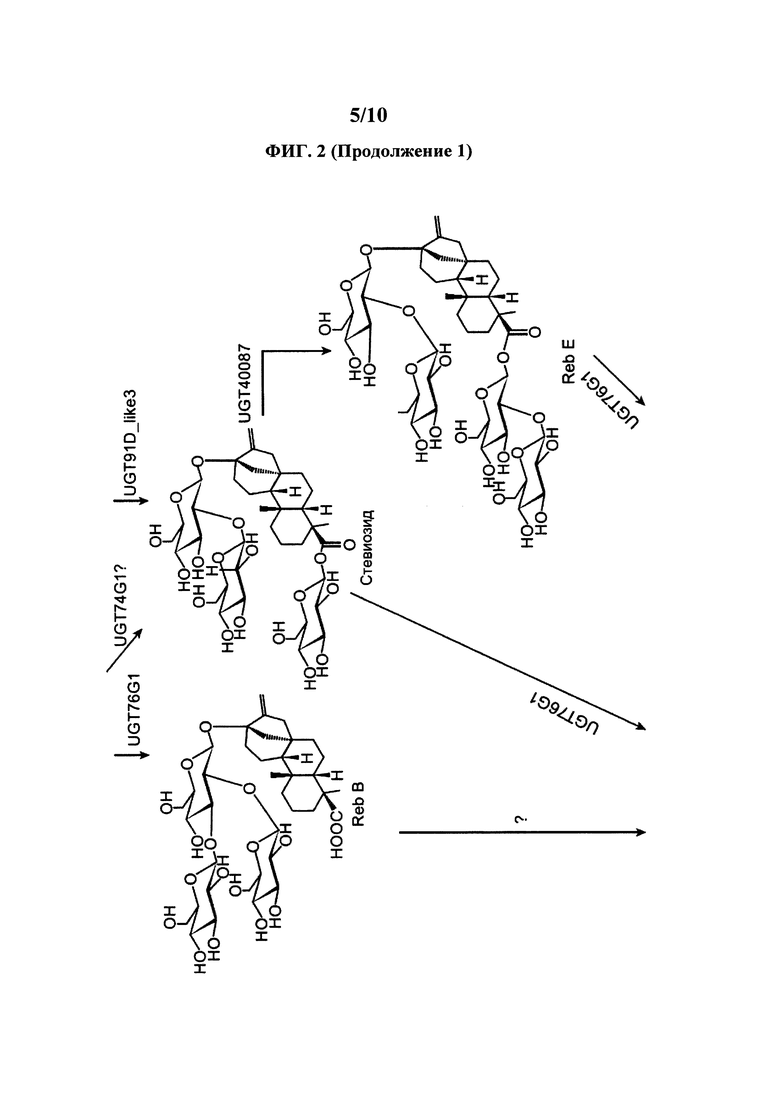
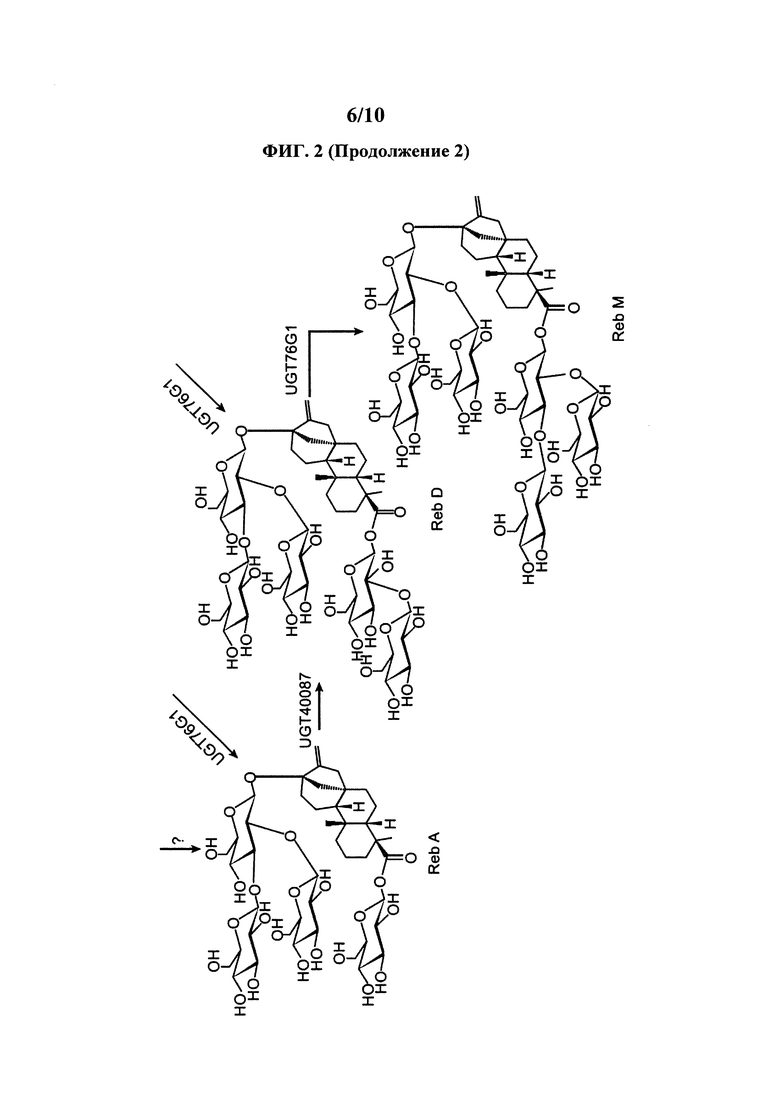
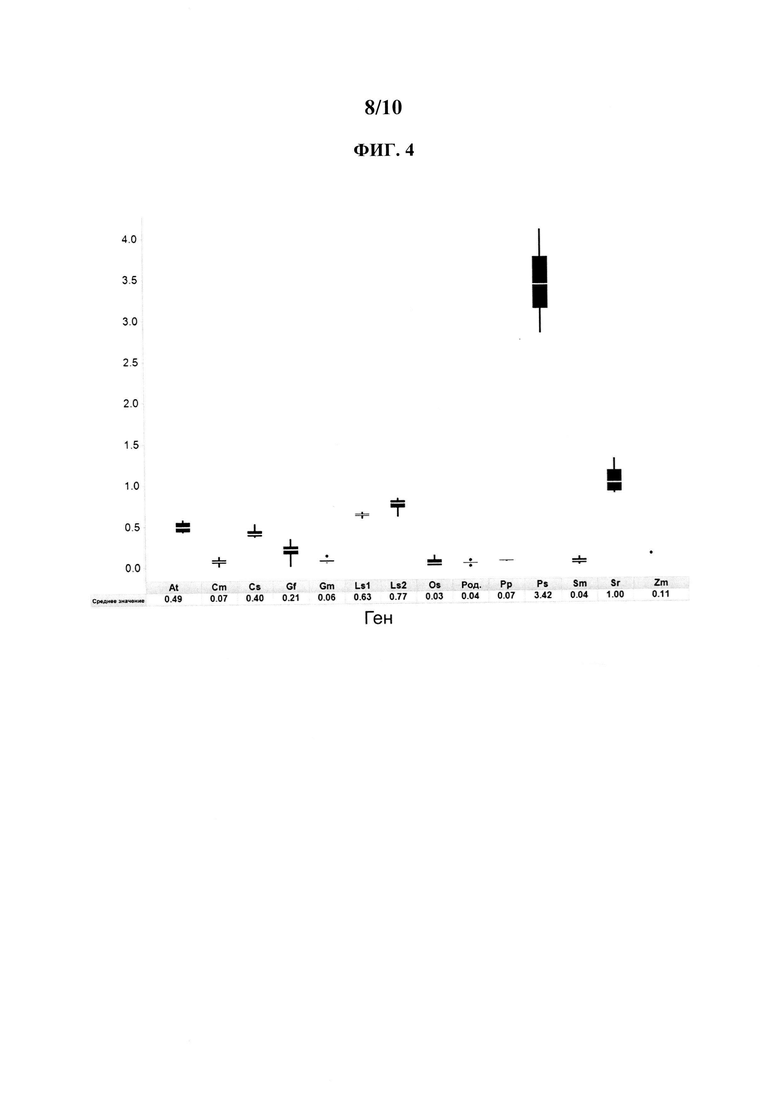
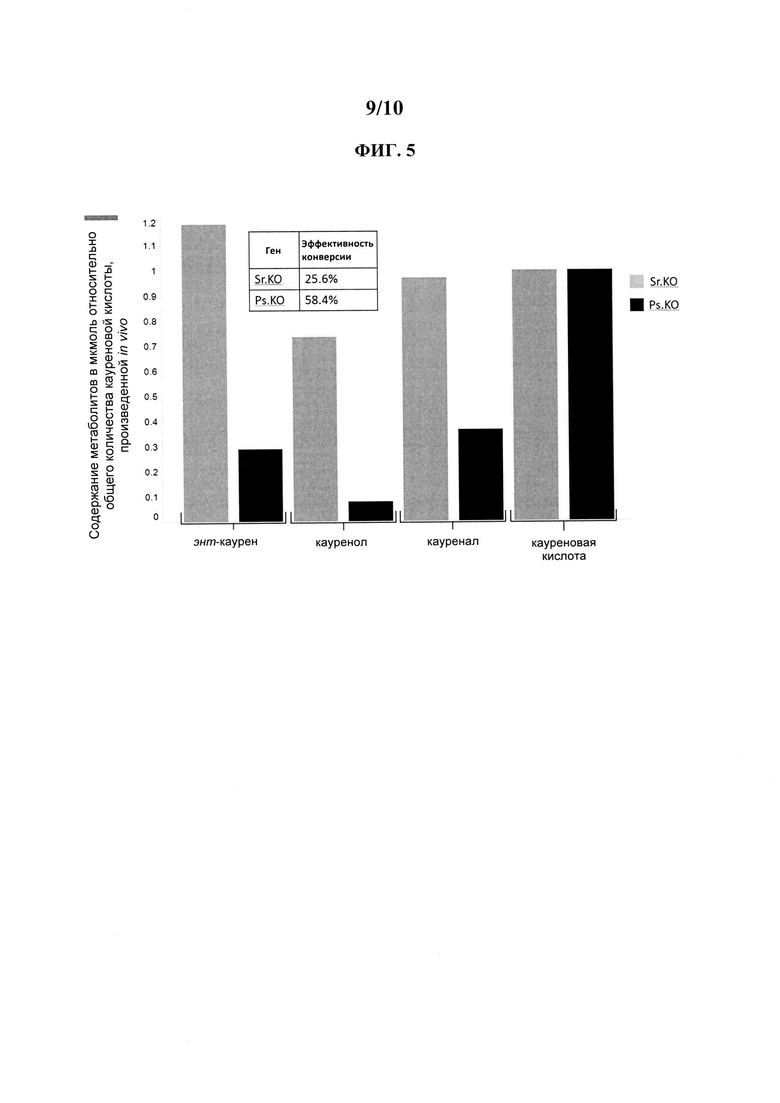

Изобретение относится к биотехнологии и представляет собой композиции и способы для более эффективного производства стевиол-гликозидов в клетке-хозяине. В некоторых вариантах осуществления данную клетку-хозяина генетически модифицируют так, чтобы она содержала гетерологичную нуклеотидную последовательность, кодирующую кауреноксидазу Pisum sativum или вариант кауреноксидазы. В некоторых вариантах осуществления данная клетка-хозяин также содержит одну или более гетерологичных нуклеотидных последовательностей, кодирующих ферменты пути, способные производить стевиол-гликозиды в клетке-хозяине. Описанные здесь композиции и способы образуют эффективный метод гетерологичного производства стевиол-гликозидов, включая, помимо прочего, ребаудиозид D и ребаудиозид М. 6 н. и 32 з.п. ф-лы, 6 ил., 3 табл., 6 пр.
1. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae, способная производить один или более стевиол-гликозидов, содержащих гетерологичную нуклеиновую кислоту, кодирующую кауреноксидазу, содержащую аминокислотную последовательность, выбранную из группы SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12 и SEQ ID NO: 13, где клетка-хозяин содержит одну или более гетерологичных нуклеиновых кислот, кодирующих один или более ферментов пути для производства стевиол-гликозида.
2. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по п. 1, способная превращать каурен в кауреновую кислоту с эффективностью более 30%, 35%, 40%, 45%, 50% или 55%.
3. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, в которой кауреноксидаза содержит аминокислотную последовательность, имеющую последовательность SEQ ID NO: 1.
4. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, в которой кауреноксидаза способна к окислению позиции C19 каурена, кауренола и/или кауренала.
5. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, в которой кауреноксидаза кодируется гетерологичной нуклеиновой кислотой, при этом гетерологичная нуклеиновая кислота содержит нуклеотидную последовательность, минимум на 80%, 85%, 90% или 95% идентичную SEQ ID NO: 15.
6. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, в которой кауреноксидаза кодируется гетерологичной нуклеиновой кислотой, имеющей последовательность SEQ ID NO: 15.
7. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, в которой кауреноксидазой является полипептид, имеющий функциональный домен SEQ ID NO: 1.
8. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, способная превращать каурен в кауреновую кислоту с эффективностью более 55%.
9. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, способная производить кауреновую кислоту.
10. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, способная производить стевиол.
11. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, способная производить RebD.
12. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, способная производить RebM.
13. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, способная производить RebM и RebM2 в соотношении минимум 10:1, 100:1 или 1000:1.
14. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, при этом генетически модифицированная клетка-хозяин производит необнаруживаемый уровень RebM2.
15. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, при этом генетически модифицированная клетка-хозяин дополнительно содержит одну или более гетерологичных нуклеиновых кислот, кодирующих один или более ферментов пути для производства стевиола.
16. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, при этом генетически модифицированная клетка-хозяин дополнительно содержит одну или более гетерологичных нуклеиновых кислот, кодирующих один или более ферментов пути для производства RebA.
17. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, при этом генетически модифицированная клетка-хозяин дополнительно содержит одну или более гетерологичных нуклеиновых кислот, кодирующих один или более ферментов пути для производства RebM.
18. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, при этом генетически модифицированная клетка-хозяин дополнительно содержит одну или более гетерологичных нуклеиновых кислот, кодирующих один или более ферментов пути для производства RebE.
19. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, в которой один или более ферментов пути включают геранилгеранилдифосфатсинтазу.
20. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, в которой один или более ферментов пути включают копалилдифосфатсинтазу.
21. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, в которой один или более ферментов пути включают энт-кауренсинтазу.
22. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, в которой один или более ферментов пути включают гидроксилазу кауреновой кислоты.
23. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, в которой один или более ферментов пути включают цитохром P450 редуктазу.
24. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, в которой один или более ферментов пути включают одну или более уридин 5'-дифосфат-зависимых гликозилтрансфераз.
25. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, в которой один или более ферментов пути включают Os_UGT_91C1, Sl_UGT_101249881, UGT40087, sr.UGT_9252778, Bd_UGT10840, Hv_UGT_V1, Bd_UGT10850 или Ob_UGT91B1_like.
26. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, в которой один или более ферментов пути включают UGT74G1, UGT76G1, UGT85C2, UGT91D или UGT40087 или вариант.
27. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, в которой один или более ферментов пути включают геранилгеранилдифосфатсинтазу, копалилдифосфатсинтазу, энт-кауренсинтазу, кауреноксидазу, гидроксилазу кауреновой кислоты, цитохром P450 редуктазу, UGTAD, UGT74G1, UGT76G1, UGT85C2 и UGT91D.
28. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, в которой один или более ферментов пути включают геранилгеранилдифосфатсинтазу, копалилдифосфатсинтазу, энт-кауренсинтазу, кауреноксидазу, гидроксилазу кауреновой кислоты, цитохром P450 редуктазу, UGT40087 или вариант, UGT74G1, UGT76G1, UGT85C2 и UGT91D.
29. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, в которой UGT40087 соответствует SEQ ID NO: 17, 18 или 33.
30. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, в которой один или более ферментов пути включают бифункциональную копалилдифосфатсинтазу и кауренсинтазу.
31. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, в которой одна или более гетерологичных нуклеиновых кислот, кодирующих один или более ферментов пути, регулируются одним транскрипционным регулятором.
32. Генетически модифицированная клетка-хозяин Saccharomyces cerevisiae по любому из предшествующих пунктов, в которой одна или более гетерологичных нуклеиновых кислот, кодирующих один или более ферментов пути, регулируются множеством гетерологичных транскрипционных регуляторов.
33. Способ производства кауреновой кислоты:
(a) культивирование популяции генетически модифицированных клеток-хозяев Saccharomyces cerevisiae по любому из предшествующих пунктов в среде с источником углерода в условиях, подходящих для производства кауреновой кислоты; и
(b) выделение указанного соединения кауреновой кислоты из среды.
34. Способ производства RebD:
(a) культивирование популяции генетически модифицированных клеток-хозяев по любому из предшествующих пунктов в среде с источником углерода в условиях, подходящих для производства RebD; и
(b) выделение указанного соединения RebD из среды.
35. Способ производства RebM:
(a) культивирование популяции генетически модифицированных клеток-хозяев по любому из предшествующих пунктов в среде с источником углерода в условиях, подходящих для производства RebM; и
(b) выделение указанного соединения RebM из среды.
36. Ферментативная композиция для производства стевиол-гликозидов, включающая:
(a) генетически модифицированную клетку-хозяина Saccharomyces cerevisiae, способную производить один или более стевиол-гликозидов, включающую гетерологичную нуклеиновую кислоту, кодирующую кауреноксидазу, включающую аминокислотную последовательность выбранную из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12 и SEQ ID NO: 13; и
(b) стевиол-глиозиды, произведенные от генетически модифицированной клетки-хозяина.
37. Ферментативная композиция по п. 36, в которой стевиол-гликозиды содержат RebA, RebD и RebM в соотношении RebA:RebD:RebM минимум 1:7:50.
38. Способ производства стевиол-гликозида, включающий
культивирование популяции генетически модифицированных клеток-хозяев по любому из предшествующих пунктов в среде с источником углерода в условиях, подходящих для производства стевиол-гликозида.
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Токарный резец | 1924 |
|
SU2016A1 |
| РЕБАУДИОЗИД D ВЫСОКОЙ СТЕПЕНИ ЧИСТОТЫ И ЕГО ПРИМЕНЕНИЕ | 2010 |
|
RU2596190C9 |

