Изобретение относится к области биотехнологии и может быть использовано для элиминации раковых стволовых клеток и предотвращения рецидивов опухолей при нацеленной иммунотерапии злокачественных заболеваний.
На сегодняшний день, одним из самых безопасных и перспективных методов лечения онкологических заболеваний считается иммунотерапия на основе адоптивного Т-клеточного переноса. Данный подход базируется на стандартной методике клеточного переноса, суть которой заключается в получении популяции иммунных клеток пациента (в данном случае CD8+ Т-лимфоцитов), увеличении их количества in vitro и введении обратно пациенту. Для повышения цитотоксической активности Т-клеток и преодоления явления толерантности был разработан подход, заключающийся в создании популяции генетически модифицированных цитотоксических Т-клеток, способных экспрессировать последовательность химерного Т-клеточного рецептора. Благодаря наличию в составе химерного рецептора последовательности антитела, специфичного к определенному опухолевому рецептору, Т-клетка приобретает способность распознавать необходимый антиген на поверхности опухолевой клетки без необходимости активации через систему главного комплекса гистосовместимости.
В уровне техники раскрываются различные генетически модифицированные цитотоксические Т-клетки.
Известен химерный антигенспецифический рецептор на основе гуманизированного антитела против EGFRvIII. Описана выделенная молекула нуклеиновой кислоты, кодирующая химерный антигенспецифический рецептор (CAR), причем указанный CAR включает связывающий домен антитела против EGFRvIII, трансмембранный домен и внутриклеточный домен сигнализации, включающий стимулирующий домен. Кодируемый CAR включает трансмембранный домен, который включает домен белка, выбираемого из группы, состоящей из альфа-, бета- или дзета-цепи Т-клеточного рецептора, CD28, CD3 эпсилон, CD45, CD4, CD5, CD8, CD9, CD16, CD22, CD33, CD37, CD64, CD80, CD86, CD134, CD137 и CD154, а кодируемый внутриклеточный домен сигнализации включает функциональный домен сигнализации CD3 дзета (заявка RU 2015139874).
Известна изолированная молекула нуклеиновой кислоты, кодирующая химерный антигенный рецептор a (CAR) и/или RIAD или эзрин-полипептид, где CAR содержит антигенсвязывающий домен (например, антитело или фрагмент антитела, TCR или фрагмент TCR), который связывается с опухолевым антигеном, трансмембранный домен и внутриклеточный сигнальный домен, например, внутриклеточный сигнальный домен, содержащий костимулирующий домен. В некоторых вариантах осуществления опухолевой антиген выбирают из одного или более: CD19, CD123, CD22, CD30, CD171, CS-1 (также называемый CD2 подмножеством 1, CRACC, SLAMF7, CD319 и 19А24), лектин-подобная молекула-1 С-типа (CLL-1 или CLECL1), CD33, вариант III рецептора эпидермального фактора роста (EGFRvIII); ганглиозид G2 (GD2), ганглиозид GD3 (международная заявка WO 2016172583 (А1), опубл. 2016-10-27).
Известен химерный мономолекулярный Т-клеточный рецептор, специфичный к раково-эмбриональному антигену, включающий одноцепочечное антитело, кодируемое нуклеотидной последовательностью SEQ ID NO: 1 или SEQ ID NO: 2, в котором эффекторный фрагмент Т-клеточного рецептора объединен с антиген-распознающей частью, представляющей собой вариабельные фрагменты двух различных антител к раково-эмбриональному антигену (РЭА) (патент RU 2522004 (С2), опубл. 2014-07-10).
Известен генно-инженерный, CD19-специфический химерный рецептор Т-клеток к иммунным клеткам, экспрессирующим химерный рецептор. Настоящее изобретение также относится к применению таких клеток для клеточной иммунотерапии злокачественных новообразований. Химерный рецептор представляет собой одноцепочечный scFvFc: zeta-рецептор, где scFvFc обозначает внеклеточный домен, scFv обозначает цепочки VH и VL одноцепочечного моноклонального антитела к CD19, Fc представляет собой по меньшей мере часть константной области IgG1, а zeta представляет собой внутриклеточный сигнальный домен дзета-цепи человеческого CD3. Внеклеточный домен scFvFc и дзета внутриклеточного домена связаны трансмембранным доменом, таким как трансмембранный домен CD4 (патент US 7446179 (В2), опубл. 2008-11-04).
Известен химерный антигенный рецептор (CAR), специфичный для CD33, вектора, кодирующие их, и рекомбинантные Т-клетки, содержащие CD33 CAR. Изобретение может быть взято в качестве наиболее близкого аналога (прототипа). Раскрытие также включает в себя способы введения генно-модифицированной Т-клетки, экспрессирующей CAR, которая содержит CD33-связывающий домен. Примеры фрагментов антител включают, но не ограничиваются ими, Fab, Fab', F(ab')2, фрагменты scFv-антитела, линейные антитела, однодоменные антитела, такие как sdAb (VL или VH), VHH верблюда (заявка WO 2016014576, опубл. 28.01.2016).
Указанные аналоги имеют ряд принципиальных недостатков, связанных, главным образом, с антигенраспознающей частью химерного рецептора. Антигенраспознающая часть большинства современных химерных рецепторов представлена одноцепочечными scFv-моноклональными антителами. По своей структуре они представляют собой комплекс из тяжелой и легкой вариабельных цепей стандартного антитела, объединенных линкерной последовательностью, от выбора которой, зависит формирование правильной пространственной ориентации вариабельных антиген-распознающих доменов и, как следствие, эффективность распознавания целевых антигенов. Практическое применение существующих аналогов предполагает необходимость проведения расчетов пространственной ориентации вариабельных цепей в зависимости от того или иного линкера, что значительно усложняет рабочий процесс. Другим важным недостатком является высокая молекулярная масса scFv-моноклональных антител, препятствующая эффективной экспрессии последовательности трансгена и презентации рецептора на поверхности Т-клеток. Высокая молекулярная масса антигенраспознающей части химерного рецептора может также являться причиной иммуногенности всей структуры рецептора и приводить к быстрому удалению трансплантированных лимфоцитов из организма пациента.
Решением подобных проблем является замена антигенраспознающего домена на основе scFv-антител верблюжьими одноцепочечными моноклональными VHH-антителами. VHH-антитела характеризуются крайне низкой молекулярной массой и высоким уровнем гомологии с вариабельной тяжелой цепью иммуноглобулина человека, что позволяет избежать иммуногенности химерных рецепторов и значительно увеличить время циркуляции модифицированных лимфоцитов в организме пациента. За счет наличия в составе тяжелой цепи иммуноглобулина единственного вариабельного домена, обладающего большей протяженностью, чем у обычных антител, VHH-антитела отличаются высокой аффинностью к своим лигандам. Этот тип антител также гораздо более удобен для создания иммунных библиотек вариабельных фрагментов и последующего отбора высоко специфичных вариантов антител при помощи фагового или рибосомального дисплея.
Хотя в ближайшем аналоге упомянута возможность включения фрагмента верблюжьих антител, конкретные конструкции не описаны.
Технической задачей изобретения является создание генетической конструкции - лентивирусного вектора, включающего полинуклеотид антигенраспознающего домена VHH, кодирующий Т-клеточный химерный рецептор, специфичный к опухолевому рецептору CD47. Задача решается тем, что разработана генетическая конструкция на основе экспрессионного плазмидного лентивирусного вектора, имеющая в составе последовательность одноцепочечных VHH антител к опухолевому рецептору CD47, кодирующих структуру Т-клеточного химерного рецептора, В состав вектора также входят полинуклеотиды, кодирующие N-концевую сигнальную последовательность рецептора CD8, антиген-распознающую часть, представляющую собой последовательность моноклонального одноцепочечного VHH-антитела, специфичного к CD47, спейсерную область, представленную 12 аминокислотной последовательностью шарнирного домена иммуноглобулина G4; трансмембранный домен рецептора CD28, сигнальный домен, включающий последовательности цитоплазматических доменов: CD28, CD137 и CD3ζ, а также ряд регуляторных элементов: 5' и 3' LTR (long terminal repeat), Rev (RRE или Rev-response element), Env - регуляторную последовательность, обеспечивающую упаковку вирусного генома в структуру вирусной частицы с образованием вириона, сРРТ - центральный полипуриновый тракт, WPRE (посттранскрипционный регуляторный элемент вируса гепатита сурков), MCS (сайт множественного клонирования) и промотор EF1alpha.
Технический результат, обеспечиваемый предложенной конструкцией: эффективная экспрессия и последующая презентация на поверхности клеток Т-клеточного химерного рецептора на основе одноцепочечных VHH-антител, характеризующегося высокой аффинностью к опухолевым антигенам (рецептору CD47), простотой создания иммунных библиотек и более высокой результативностью при использовании в нацеленной иммунотерапии злокачественных новообразований. Изобретение иллюстрируют следующие примеры:
Пример 1. Получение генетической конструкции, кодирующей структуру Т-клеточного химерного рецептора на основе одноцепочечных VHH-антител, специфичных к опухолевому рецептору CD47.
В качестве основы для создания генетической конструкции, содержащей последовательность химерного Т-клеточного рецептора, используют классический лентивирусный вектор следующей структуры (фиг. 1):
В представленный вектор методом клонирования встраивают следующие элементы:
 Последовательность сигнального домена рецептора CD8
Последовательность сигнального домена рецептора CD8
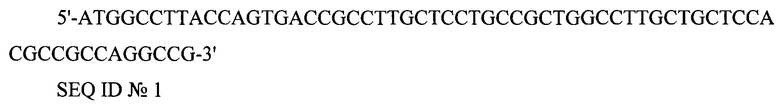
 Последовательность шарнирного домена IgG4 (12 аминокислот)
Последовательность шарнирного домена IgG4 (12 аминокислот)
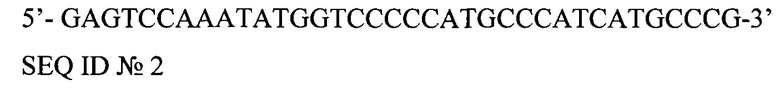
 Последовательность трансмембранного домена рецептора CD8
Последовательность трансмембранного домена рецептора CD8
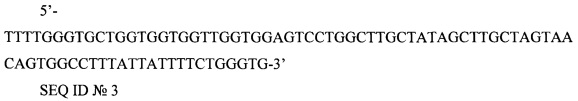
 Сигнальный домен рецептора CD28
Сигнальный домен рецептора CD28
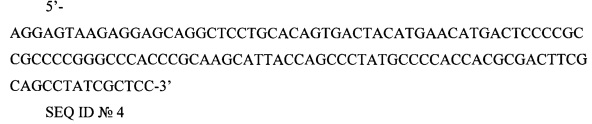
 Сигнальный домен рецептора CD137
Сигнальный домен рецептора CD137
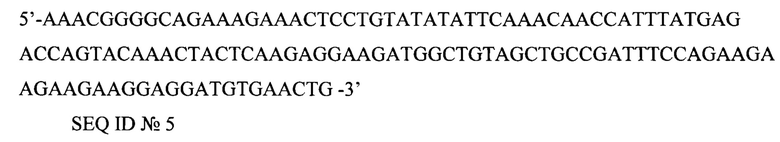
Сигнальный домен рецептора CD3ζ

 Антигенраспознающий домен (VHH-19)
Антигенраспознающий домен (VHH-19)
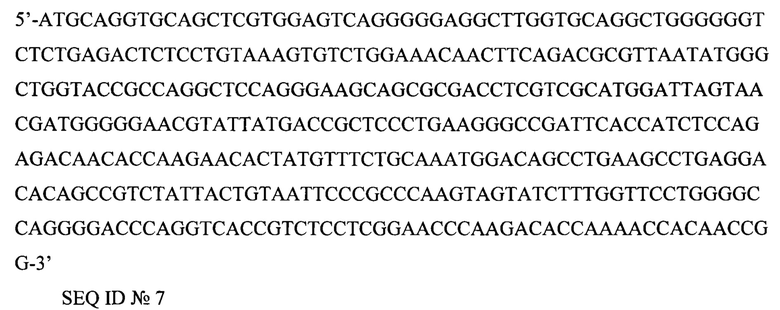
 5' LTR (long terminal repeat)
5' LTR (long terminal repeat)
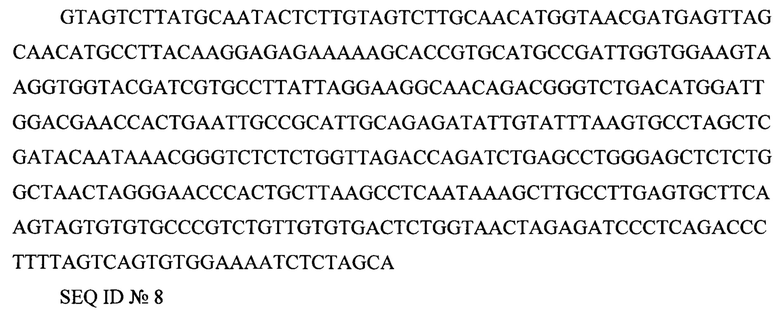
 3' LTR (long terminal repeat)
3' LTR (long terminal repeat)
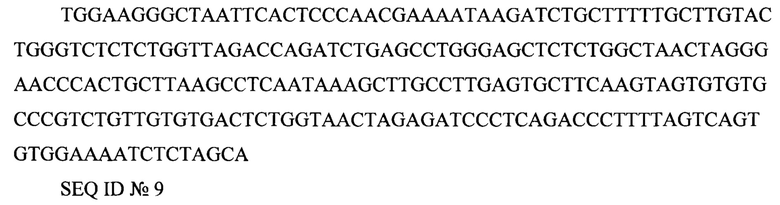
 Rev (RRE или Rev-response element)
Rev (RRE или Rev-response element)
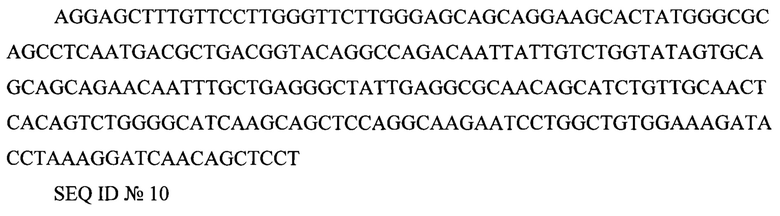
 Env - регуляторная последовательность, обеспечивающая упаковку вирусного генома в структуру вирусной частицы с образованием вириона
Env - регуляторная последовательность, обеспечивающая упаковку вирусного генома в структуру вирусной частицы с образованием вириона

 сРРТ - центральный полипуриновый тракт
сРРТ - центральный полипуриновый тракт
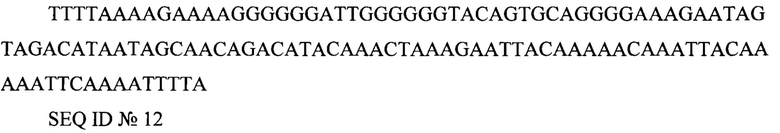
 WPRE (посттранскрипционный регуляторный элемент вируса гепатита сурков)
WPRE (посттранскрипционный регуляторный элемент вируса гепатита сурков)

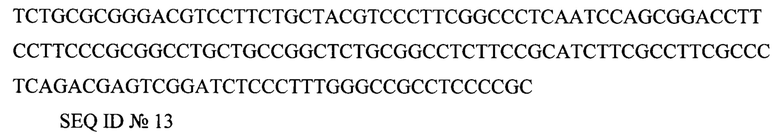
 MCS (сайт множественного клонирования)
MCS (сайт множественного клонирования)
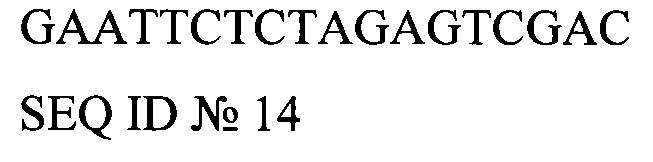
 промотор EF1alpha
промотор EF1alpha

В силу того, что промоторная область цитомегаловируса обеспечивает слабый уровень экспрессии в популяции цитотоксических Т-лимфоцитов человека, ее заменяют на последовательность промотора EF1alpha.
Рецептор CD47 экспрессируется на многих типах клеток и участвует во множестве клеточных процессов, включая такие механизмы, как пролиферация, дифференцировка, клеточная миграция, адгезия клеток и проведение сигналов, обеспечивающих апоптоз. Исследования показали, что CD47 гиперэкспрессирован в различных типах опухолевых клеток таких, например, как, клетки рака мочевого пузыря или клетки острого миелобластного лейкоза. Таким образом, CD47 представляется одной из перспективных мишеней для направленной терапии. Блокирование этого рецептора нарушает супрессирующее действие опухолевых клеток на компоненты врожденного иммунитета, что способствует развитию противоопухолевого иммунного ответа.
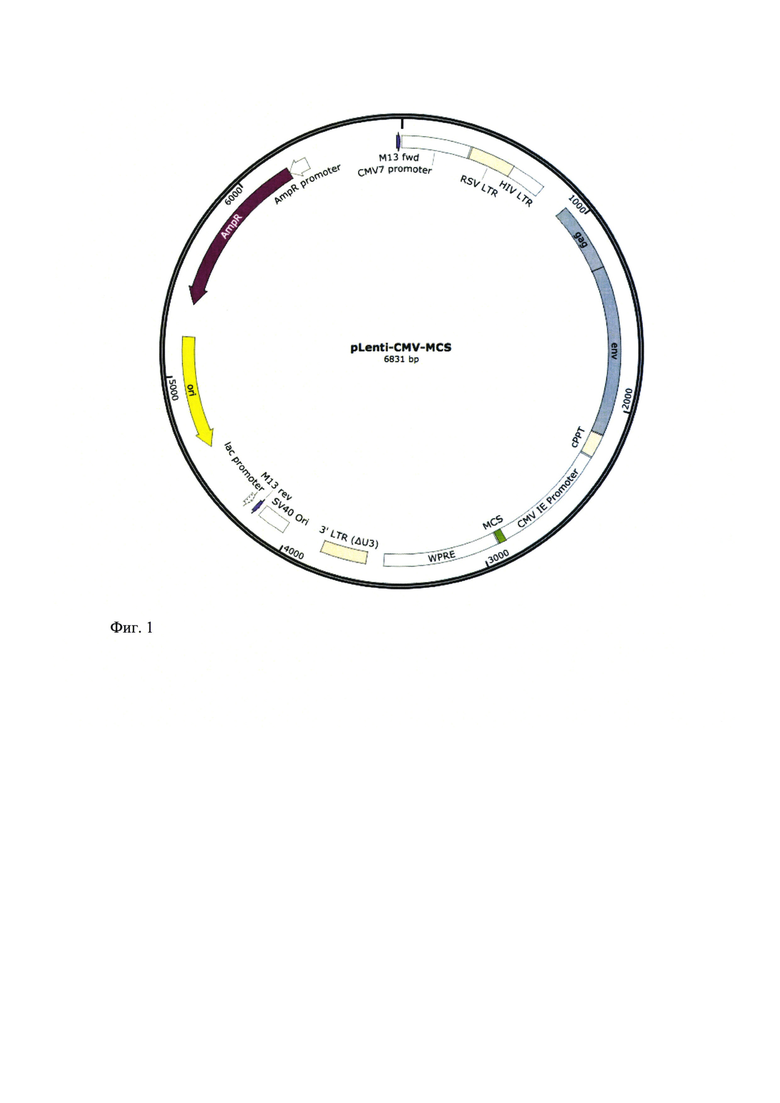
Предварительно приведенные последовательности функциональных элементов химерного рецептора нарабатывают в ходе ПЦР. Разделение амплифицированных фрагментов проводят в ходе горизонтального электрофореза в агарозном геле. Для очистки ДНК-фрагментов используют набор CleanUp Mini (Евроген, Россия). Очищенные фрагменты ДНК и лентивирусный вектор pLenti-CMV-MCS рестрицируют соответствующими эндонуклеазами рестрикции, после чего проводят еще один этап очистки рестрикционных смесей в агарозном геле. На завершающем этапе проводят лигирование полученных генетических последовательностей и экспрессионного вектора. Анализ нуклеотидной последовательности полученной генетической конструкции анализируют методом секвенирования, доказавшим успешность проведенного клонирования. Результатом работы стало получение генетической конструкции, кодирующей структуру Т-клеточного химерного рецептора на основе одноцепочечных VHH-антител, специфичных к опухолевому рецептору CD47; структура генетической конструкции представлена на фиг. 2.
Пример 2. Структура лентивирусного вектора, кодирующего структуру Т-клеточного химерного рецептора на основе одноцепочечных VHH-антител, специфичных к опухолевому рецептору CD47
В состав заявленного вектора входят ряд базовых элементов, кодирующих функциональные компоненты структуры Т-клеточного химерного рецептора, а также ряд регуляторных мотивов, обеспечивающих эффективную работу генетической конструкции.
Регуляторные элементы
 5' и 3' LTR (long terminal repeat) - длинные концевые повторы, последовательности ДНК, обеспечивающие внедрение вирусного генома в геном клетки-хозяина при участии фермента интегразы.
5' и 3' LTR (long terminal repeat) - длинные концевые повторы, последовательности ДНК, обеспечивающие внедрение вирусного генома в геном клетки-хозяина при участии фермента интегразы.
 Rev (RRE или Rev-response element) - регуляторная последовательность, обеспечивающая транспортировку молекулы вирусной мРНК из ядра в цитоплазму, где происходит ее экспрессия.
Rev (RRE или Rev-response element) - регуляторная последовательность, обеспечивающая транспортировку молекулы вирусной мРНК из ядра в цитоплазму, где происходит ее экспрессия.
 Env - регуляторная последовательность, обеспечивающая упаковку вирусного генома в структуру вирусной частицы с образованием вириона.
Env - регуляторная последовательность, обеспечивающая упаковку вирусного генома в структуру вирусной частицы с образованием вириона.
 сРРТ - центральный полипуриновый тракт - последовательность, которая способствует проникновению обратно-транскрибируемой вирусной ДНК в ядро клетки, обеспечивая тем самым заражение митотически неактивных клеток.
сРРТ - центральный полипуриновый тракт - последовательность, которая способствует проникновению обратно-транскрибируемой вирусной ДНК в ядро клетки, обеспечивая тем самым заражение митотически неактивных клеток.
 WPRE (woodchuck hepatitis virus posttranscriptional regulatory element-посттранскрипционный регуляторный элемент вируса гепатита сурков) - энхансер - последовательность, обеспечивающая формирование особой пространственной структуры трансгена и увеличивающая вероятность распознавания промоторного региона факторами траскрипции. Кроме того, увеличивает стабильность мРНК трансгена.
WPRE (woodchuck hepatitis virus posttranscriptional regulatory element-посттранскрипционный регуляторный элемент вируса гепатита сурков) - энхансер - последовательность, обеспечивающая формирование особой пространственной структуры трансгена и увеличивающая вероятность распознавания промоторного региона факторами траскрипции. Кроме того, увеличивает стабильность мРНК трансгена.
 Промотор EF1alpha. Эффективный промотор, обеспечивающий высокий уровень экспрессии во многих типах клеток.
Промотор EF1alpha. Эффективный промотор, обеспечивающий высокий уровень экспрессии во многих типах клеток.
Базовые функциональные элементы
 Внеклеточная часть или эктодомен, включающий следующие структуры:
Внеклеточная часть или эктодомен, включающий следующие структуры:
 N-концевая сигнальная последовательность рецептора CD8, обеспечивающая секрецию химерного белка через мембрану клетки.
N-концевая сигнальная последовательность рецептора CD8, обеспечивающая секрецию химерного белка через мембрану клетки.
 Антиген-распознающая часть, представляющая собой последовательность моноклонального одноцепочечного VHH-антитела, специфичного к CD47.
Антиген-распознающая часть, представляющая собой последовательность моноклонального одноцепочечного VHH-антитела, специфичного к CD47.
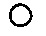 Спейсерная область, представленная минимальной 12-ти аминокислотной последовательностью шарнирного домена иммуноглобулина G4; обеспечивает присоединение антиген-распознающего участка с трансмембранным доменом.
Спейсерная область, представленная минимальной 12-ти аминокислотной последовательностью шарнирного домена иммуноглобулина G4; обеспечивает присоединение антиген-распознающего участка с трансмембранным доменом.
 Внутриклеточная часть (эндодомен) включает следующие элементы:
Внутриклеточная часть (эндодомен) включает следующие элементы:
 Трансмембранный домен рецептора CD28. По структуре представляет собой гидрофобную альфа-спираль и обеспечивает заякоривание химерного рецептора в мембране клетки.
Трансмембранный домен рецептора CD28. По структуре представляет собой гидрофобную альфа-спираль и обеспечивает заякоривание химерного рецептора в мембране клетки.
 Цитоплазматическая часть, представленная сигнальным доменом третьего поколения, включающим последовательности цитоплазматических доменов: CD28, CD137 и CD3ζ, комбинация которых обеспечивает генерацию активирующего сигнала в Т-лимфоците после взаимодействия антиген-распознающей части с антигеном на поверхности раковой клетки.
Цитоплазматическая часть, представленная сигнальным доменом третьего поколения, включающим последовательности цитоплазматических доменов: CD28, CD137 и CD3ζ, комбинация которых обеспечивает генерацию активирующего сигнала в Т-лимфоците после взаимодействия антиген-распознающей части с антигеном на поверхности раковой клетки.
Пример 3. Анализ эффективности элиминирования раковых клеток Т-лимфоцитами, трансфецированными лентивирусным вектором, кодирующим Т-клеточный химерный рецептора на основе одноцепочечных VHH-антител, специфичных к опухолевому рецептору CD47.
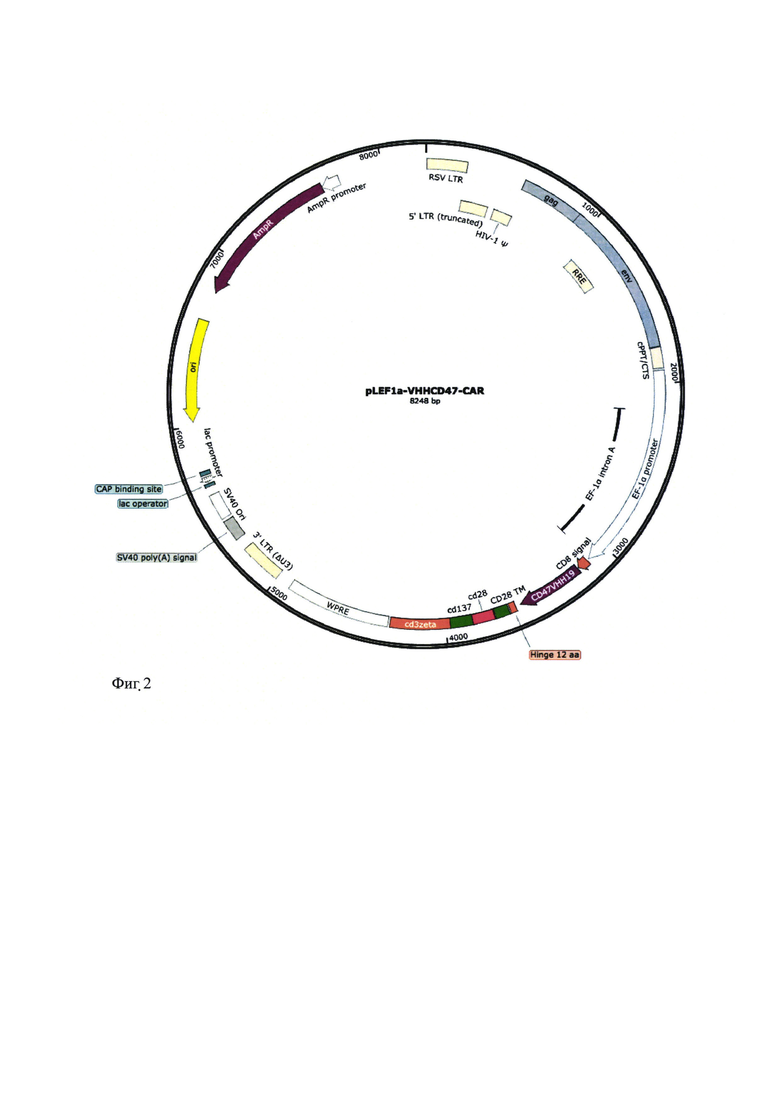
В ходе in vitro анализа проводят культивирование клеток С047-позититивных (1 - U937, 2 - MDA-MB-468) и CD47-негативных (3 - MDA-MB-231, 4 - SW527) линий в присутствии Т-лимфоцитов, трансфецированных генетической конструкцией, кодирующей Т-клеточный химерный рецептора на основе одноцепочечных VHH-антител. В качестве контроля используют Т-лимфоциты, трансфецированные исходным лентивирусным вектором, не кодирующим структуру химерного рецептора. По завершении инкубации (24 часа) анализируют количество живых раковых клеток в ходе количественной проточной цитометрии; полученные результаты представлены на фиг. 3.
Полученные результаты свидетельствуют об эффективности элиминирования раковых клеток Т-лимфоцитами, трансфецированными генетической конструкцией, кодирующей Т-клеточный химерный рецептора на основе одноцепочечных VHH-антител, специфичных к опухолевому рецептору CD47 и позволяют рекомендовать разработанную генетическую конструкцию для использования в иммунотерапии злокачественных заболеваний.
Изобретение иллюстрируют следующие графические материалы:
Фиг. 1 - Схематическое изображение лентивирусного вектора, взятого за основу при создании генетической конструкции, содержащей последовательность химерного Т-клеточного рецептора.
Фиг 2 - Схематичное изображение лентивирусного вектора, кодирующего структуру Т-клеточного химерного рецептора на основе одноцепочечных VHH-антител, специфичных к опухолевому рецептору CD47.
Фиг. 3 - Результаты анализа эффективности элиминирования раковых клеток Т-лимфоцитами, трансфецированными лентивирусным вектором, кодирующим Т-клеточный химерный рецептора на основе одноцепочечных VHH-антител, специфичных к опухолевому рецептору CD47. Представлены данные для CD47-позититивных (1 - U937, 2 - MDA-MB-468) и CD47-негативных (3 - MDA-MB-231, 4 - SW527) линий.
Последовательность сигнального домена рецептора CD8
5'-ATGGCCTTACCAGTGACCGCCTTGCTCCTGCCGCTGGCCTTGCTGCTCCACGCCGCCAGGCCG-3'
SEQ ID №1
Последовательность шарнирного домена IgG4 (12 аминокислот)
5'-GAGTCCAAATATGGTCCCCCATGCCCATCATGCCCG-3'
SEQ ID №2
Последовательность трансмембранного домена рецептора CD8
5'-TTTTGGGTGCTGGTGGTGGTTGGTGGAGTCCTGGCTTGCTATAGCTTGCTAGTAACAGTGGCCTTTATTATTTTCTGGGTG-3'
SEQ ID №3
Сигнальный домен рецептора CD28
5'-AGGAGTAAGAGGAGCAGGCTCCTGCACAGTGACTACATGAACATGACTCCCCGCCGCCCCGGGCCCACCCGCAAGCATTACCAGCCCTATGCCCCACCACGCGACTTCGCAGCCTATCGCTCC-3'
SEQ ID №4
Сигнальный домен рецептора CD137
5'-AAACGGGGCAGAAAGAAACTCCTGTATATATTCAAACAACCATTTATGAGACCAGTACAAACTACTCAAGAGGAAGATGGCTGTAGCTGCCGATTTCCAGAAGAAGAAGAAGGAGGATGTGAACTG-3'
SEQ ID №5
Сигнальный домен рецептора CD3ζ
5'-AGAGTGAAGTTCAGCAGGAGCGCAGACGCCCCCGCGTACCAGCAGGGCCAGAACCAGCTCTATAACGAGCTCAATCTAGGACGAAGAGAGGAGTACGATGTTTTGGACAAGAGACGTGGCCGGGACCCTGAGATGGGGGGAAAGCCGCAGAGAAGGAAGAACCCTCAGGAAGGCCTGTACAATGAACTGCAGAAAGATAAGATGGCGGAGGCCTACAGTGAGATTGGGATGAAAGGCGAGCGCCGGAGGGGCAAGGGGCACGATGGCCTTTACCAGGGTCTCAGTACAGCCACCAAGGACACCTACGACGCCCTTCACATGCAGGCCCTGCCCCCTCGC-3'
SEQ ID №6
Антигенраспознающий домен (VHH-19)
5'-ATGCAGGTGCAGCTCGTGGAGTCAGGGGGAGGCTTGGTGCAGGCTGGGGGGTCTCTGAGACTCTCCTGTAAAGTGTCTGGAAACAACTTCAGACGCGTTAATATGGGCTGGTACCGCCAGGCTCCAGGGAAGCAGCGCGACCTCGTCGCATGGATTAGTAACGATGGGGGAACGTATTATGACCGCTCCCTGAAGGGCCGATTCACCATCTCCAGAGACAACACCAAGAACACTATGTTTCTGCAAATGGACAGCCTGAAGCCTGAGGACACAGCCGTCTATTACTGTAATTCCCGCCCAAGTAGTATCTTTGGTTCCTGGGGCCAGGGGACCCAGGTCACCGTCTCCTCGGAACCCAAGACACCAAAACCACAACCGG-3'
SEQ ID №7
5' LTR (long terminal repeat) GTAGTCTTATGCAATACTCTTGTAGTCTTGCAACATGGTAACGATGAGTTAGCAACATGCCTTACAAGGAGAGAAAAAGCACCGTGCATGCCGATTGGTGGAAGTAAGGTGGTACGATCGTGCCTTATTAGGAAGGCAACAGACGGGTCTGACATGGATTGGACGAACCACTGAATTGCCGCATTGCAGAGATATTGTATTTAAGTGCCTAGCTCGATACAATAAACGGGTCTCTCTGGTTAGACCAGATCTGAGCCTGGGAGCTCTCTGGCTAACTAGGGAACCCACTGCTTAAGCCTCAATAAAGCTTGCCTTGAGTGCTTCAAGTAGTGTGTGCCCGTCTGTTGTGTGACTCTGGTAACTAGAGATCCCTCAGACCCTTTTAGTCAGTGTGGAAAATCTCTAGCA
SEQ ID №8
3' LTR (long terminal repeat) TGGAAGGGCTAATTCACTCCCAACGAAAATAAGATCTGCTTTTTGCTTGTACTGGGTCTCTCTGGTTAGACCAGATCTGAGCCTGGGAGCTCTCTGGCTAACTAGGGAACCCACTGCTTAAGCCTCAATAAAGCTTGCCTTGAGTGCTTCAAGTAGTGTGTGCCCGTCTGTTGTGTGACTCTGGTAACTAGAGATCCCTCAGACCCTTTTAGTCAGTGTGGAAAATCTCTAGCA
SEQ ID №9
Rev (RRE или Rev-response element) AGGAGCTTTGTTCCTTGGGTTCTTGGGAGCAGCAGGAAGCACTATGGGCGCAGCCTCAATGACGCTGACGGTACAGGCCAGACAATTATTGTCTGGTATAGTGCAGCAGCAGAACAATTTGCTGAGGGCTATTGAGGCGCAACAGCATCTGTTGCAACTCACAGTCTGGGGCATCAAGCAGCTCCAGGCAAGAATCCTGGCTGTGGAAAGATACCTAAAGGATCAACAGCTCCT
SEQ ID №10
Env - регуляторная последовательность, обеспечивающая упаковку вирусного генома в структуру вирусной частицы с образованием вириона GCGGCCGCTGATCTTCAGACCTGGAGGAGGAGATATGAGGGACAATTGGAGAAGTGAATTATATAAATATAAAGTAGTAAAAATTGAACCATTAGGAGTAGCACCCACCAAGGCAAAGAGAAGAGTGGTGCAGAGAGAAAAAAGAGCAGTGGGAATAGGAGCTTTGTTCCTTGGGTTCTTGGGAGCAGCAGGAAGCACTATGGGCGCAGCCTCAATGACGCTGACGGTACAGGCCAGACAATTATTGTCTGGTATAGTGCAGCAGCAGAACAATTTGCTGAGGGCTATTGAGGCGCAACAGCATCTGTTGCAACTCACAGTCTGGGGCATCAAGCAGCTCCAGGCAAGAATCCTGGCTGTGGAAAGATACCTAAAGGATCAACAGCTCCTGGGGATTTGGGGTTGCTCTGGAAAACTCATTTGCACCACTGCTGTGCCTTGGAATGCTAGTTGGAGTAATAAATCTCTGGAACAGATTGGAATCACACGACCTGGATGGAGTGGGACAGAGAAATTAACAATTACACAAGCTTAATACACTCCTTAATTGAAGAATCGCAAAACCAGCAAGAAAAGAATGAACAAGAATTATTGGAATTAGATAAATGGGCAAGTTTGTGGAATTGGTTTAACATAACAAATTGGCTGTGGTATATAAAATTATTCATAATGATAGTAGGAGGCTTGGTAGGTTTAAGAATAGTTTTTGCTGTACTTTCTATAGTGAATAGAGTTAGGCAGGGATATTCACCATTATCGTTTCAGACCCACCTCCCAACCCCGAGGGGACCCGACAGGCCCGAAGGAATAGAAGAAGAAGGTGGAGAGAGAGACAGAGACAGATCCATTCGATTAGTGAACGGATCTCGACG
SEQ ID №11
Env - регуляторная последовательность, обеспечивающая упаковку вирусного генома в структуру вирусной частицы с образованием вириона GCGGCCGCTGATCTTCAGACCTGGAGGAGGAGATATGAGGGACAATTGGAGAAGTGAATTATATAAATATAAAGTAGTAAAAATTGAACCATTAGGAGTAGCACCCACCAAGGCAAAGAGAAGAGTGGTGCAGAGAGAAAAAAGAGCAGTGGGAATAGGAGCTTTGTTCCTTGGGTTCTTGGGAGCAGCAGGAAGCACTATGGGCGCAGCCTCAATGACGCTGACGGTACAGGCCAGACAATTATTGTCTGGTATAGTGCAGCAGCAGAACAATTTGCTGAGGGCTATTGAGGCGCAACAGCATCTGTTGCAACTCACAGTCTGGGGCATCAAGCAGCTCCAGGCAAGAATCCTGGCTGTGGAAAGATACCTAAAGGATCAACAGCTCCTGGGGATTTGGGGTTGCTCTGGAAAACTCATTTGCACCACTGCTGTGCCTTGGAATGCTAGTTGGAGTAATAAATCTCTGGAACAGATTGGAATCACACGACCTGGATGGAGTGGGACAGAGAAATTAACAATTACACAAGCTTAATACACTCCTTAATTGAAGAATCGCAAAACCAGCAAGAAAAGAATGAACAAGAATTATTGGAATTAGATAAATGGGCAAGTTTGTGGAATTGGTTTAACATAACAAATTGGCTGTGGTATATAAAATTATTCATAATGATAGTAGGAGGCTTGGTAGGTTTAAGAATAGTTTTTGCTGTACTTTCTATAGTGAATAGAGTTAGGCAGGGATATTCACCATTATCGTTTCAGACCCACCTCCCAACCCCGAGGGGACCCGACAGGCCCGAAGGAATAGAAGAAGAAGGTGGAGAGAGAGACAGAGACAGATCCATTCGATTAGTGAACGGATCTCGACG
SEQ ID №11
cPPT - центральный полипуриновый тракт TTTTAAAAGAAAAGGGGGGATTGGGGGGTACAGTGCAGGGGAAAGAATAGTAGACATAATAGCAACAGACATACAAACTAAAGAATTACAAAAACAAATTACAAAAATTCAAAATTTTA
SEQ ID №12
WPRE (посттранскрипционный регуляторный элемент вируса гепатита сурков) AATCAACCTCTGGATTACAAAATTTGTGAAAGATTGACTGGTATTCTTAACTATGTTGCTCCTTTTACGCTATGTGGATACGCTGCTTTAATGCCTTTGTATCATGCTATTGCTTCCCGTATGGCTTTCATTTTCTCCTCCTTGTATAAATCCTGGTTGCTGTCTCTTTATGAGGAGTTGTGGCCCGTTGTCAGGCAACGTGGCGTGGTGTGCACTGTGTTTGCTGACGCAACCCCCACTGGTTGGGGCATTGCCACCACCTGTCAGCTCCTTTCCGGGACTTTCGCTTTCCCCCTCCCTATTGCCACGGCGGAACTCATCGCCGCCTGCCTTGCCCGCTGCTGGACAGGGGCTCGGCTGTTGGGCACTGACAATTCCGTGGTGTTGTCGGGGAAGCTGACGTCCTTTCCATGGCTGCTCGCCTGTGTTGCCACCTGGATTCTGCGCGGGACGTCCTTCTGCTACGTCCCTTCGGCCCTCAATCCAGCGGACCTTCCTTCCCGCGGCCTGCTGCCGGCTCTGCGGCCTCTTCCGCATCTTCGCCTTCGCCCTCAGACGAGTCGGATCTCCCTTTGGGCCGCCTCCCCGC
SEQ ID №13
MCS (сайт множественного клонирования)
GAATTCTCTAGAGTCGAC
SEQ ID №14
Промотор EF1alpha GGCTCCGGTGCCCGTCAGTGGGCAGAGCGCACATCGCCCACAGTCCCCGAGAAGTTGGGGGGAGGGGTCGGCAATTGAACCGGTGCCTAGAGAAGGTGGCGCGGGGTAAACTGGGAAAGTGATGTCGTGTACTGGCTCCGCCTTTTTCCCGAGGGTGGGGGAGAACCGTATATAAGTGCAGTAGTCGCCGTGAACGTTCTTTTTCGCAACGGGTTTGCCGCCAGAACACAGGTAAGTGCCGTGTGTGGTTCCCGCGGGCCTGGCCTCTTTACGGGTTATGGCCCTTGCGTGCCTTGAATTACTTCCACCTGGCTGCAGTACGTGATTCTTGATCCCGAGCTTCGGGTTGGAAGTGGGTGGGAGAGTTCGAGGCCTTGCGCTTAAGGAGCCCCTTCGCCTCGTGCTTGAGTTGAGGCCTGGCCTGGGCGCTGGGGCCGCCGCGTGCGAATCTGGTGGCACCTTCGCGCCTGTCTCGCTGCTTTCGATAAGTCTCTAGCCATTTAAAATTTTTGATGACCTGCTGCGACGCTTTTTTTCTGGCAAGATAGTCTTGTAAATGCGGGCCAAGATCTGCACACTGGTATTTCGGTTTTTGGGGCCGCGGGCGGCGACGGGGCCCGTGCGTCCCAGCGCACATGTTCGGCGAGGCGGGGCCTGCGAGCGCGGCCACCGAGAATCGGACGGGGGTAGTCTCAAGCTGGCCGGCCTGCTCTGGTGCCTGGCCTCGCGCCGCCGTGTATCGCCCCGCCCTGGGCGGCAAGGCTGGCCCGGTCGGCACCAGTTGCGTGAGCGGAAAGATGGCCGCTTCCCGGCCCTGCTGCAGGGAGCTCAAAATGGAGGACGCGGCGCTCGGGAGAGCGGGCGGGTGAGTCACCCACACAAAGGAAAAGGGCCTTTCCGTCCTCAGCCGTCGCTTCATGTGACTCCACGGAGTACCGGGCGCCGTCCAGGCACCTCGATTAGTTCTCGAGCTTTTGGAGTACGTCGTCTTTAGGTTGGGGGGAGGGGTTTTATGCGATGGAGTTTCCCCACACTGAGTGGGTGGAGACTGAAGTTAGGCCAGCTTGGCACTTGATGTAATTCTCCTTGGAATTTGCCCTTTTTGAGTTTGGATCTTGGTTCATTCTCAAGCCTCAGACAGTGGTTCAAAGTTTTTTTCTTCCATTTCAGGTGTCGTGA
SEQ ID №15
Изобретение относится к области биотехнологии, конкретно к получению лентивирусного вектора для экспрессии химерного антигенного рецептора (CAR) на основе VHH-антитела к CD47 в Т-клетках человека, и может быть использовано в медицине. Полученные нацеленные Т-клетки, презентирующие CAR к CD47, характеризуются высоким уровнем экспрессии CAR, низкой иммуногенностью и увеличенным временем циркуляции в организме и могут быть использованы для эффективной иммунотерапии злокачественных новообразований. 3 ил., 3 пр.
Лентивирусный вектор для экспрессии химерного антигенного рецептора (CAR) к CD47 в Т-клетках человека, включающий полинуклеотид с SEQ ID NO: 7, кодирующий антиген-распознающую часть, представляющую собой последовательность VHH к CD47; полинуклеотид с SEQ ID NO: 2, кодирующий шарнирный домен IgG4; полинуклеотид с SEQ ID NO: 4, кодирующий трансмембранный домен рецептора CD28; полинуклеотид с SEQ ID NO: 5, кодирующий CD137, полинуклеотид с SEQ ID NO: 6, кодирующий CD3ζ, SEQ ID NO: 8 и NO: 9 - 5'LTR и 3' LTR соответственно, полинуклеотид SEQ ID NO: 10, кодирующий Rev, полинуклеотид SEQ ID NO: 11, кодирующий регуляторную последовательность Env, обеспечивающую упаковку вирусного генома, SEQ ID NO: 12 - центральный полинуклеотид, кодирующий сРРТ полипуриновый тракт, SEQ ID NO: 13 - последовательность, кодирующая посттранскрипционный регуляторный элемент вируса гепатита сурков WPRE, SEQ ID NO: 14 - сайт множественного клонирования MCS и SEQ ID NO: 15 - промотор EF1alpha, где сама конструкция лентиновирусного вектора представлена на Фигуре 2.
| RU 2015139874, 24.03.2017 | |||
| ЭКСПРЕССИОННЫЙ ПЛАЗМИДНЫЙ ЛЕНТИВИРУСНЫЙ ВЕКТОР ДЛЯ ГЕТЕРОЛОГИЧНОЙ ЭКСПРЕССИИ РЕКОМБИНАНТНОГО ЧЕЛОВЕЧЕСКОГО БЕЛКА CD47 | 2015 |
|
RU2627181C2 |
| RU 2015156509, 05.07.2017 | |||
| WEISKOPF K | |||
| et al | |||
| Способ очищения сернокислого глинозема от железа | 1920 |
|
SU47A1 |
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| КУЛУМЗИН С | |||
| В | |||
| и др.,Основы дизайна химерных антигенных рецепторов, ACTA NATURAE, | |||

