Область техники, к которой относится изобретение
Настоящее изобретение относится к химерным антигенным рецепторам (CAR). CAR способны перенаправлять специфичность и реактивность иммунной клетки в направлении выбранной мишени с помощью свойств лигандсвязывающего домена. В частности, настоящее изобретение относится к многоцепочечному химерному антигенному рецептору, в котором внеклеточные лигандсвязывающие и сигнальные домены разделены по различным трансмембранным полипептидам для улучшения их функций. Различные трансмембранные полипептиды, составляющие многоцепочечный CAR по настоящему изобретению, когда собраны вместе, могут специфически связываться с одним или несколькими лигандами в мишени и индуцировать активацию иммунных клеток, в которых они экспрессируются, и иммунный ответ. Настоящее изобретение также относится к полинуклеотидам, векторам, кодирующим такие трансмембранные полипептиды, и выделенным клеткам, экспрессирующим указанный многоцепочечный CAR на своей поверхности, для иммунотерапии. Настоящее изобретение также относится к способам получения иммунных клеток способами генной инженерии, экспрессирующих многоцепочечный CAR на своей поверхности. Изобретение открывает путь эффективным стратегиям адоптивной иммунотерапии для лечения злокачественной опухоли и вирусных инфекций.
Уровень техники изобретения
Адоптивная иммунотерапия, которая включает перенос аутологичных антиген-специфических Т-клеток, полученных ex vivo, является многообещающей стратегией для лечения вирусных инфекций и злокачественных опухолей. T-клетки, используемые для адоптивной иммунотерапии, могут быть получены или путем размножения антиген-специфических T-клеток, или путем перенаправления T-клеток посредством генной инженерии (Park, Rosenberg et al., 2011). Перенос специфичных к вирусному антигену T-клеток является хорошо отлаженной процедурой, используемой для лечения связанных с трансплантатами вирусных инфекций и редких связанных с вирусами злокачественных опухолей. Кроме того, было показано, что выделение и перенос специфичных к опухоли T-клеток являются успешными при лечении меланомы.
Новые специфичности T-клеток были успешно созданы посредством генетического переноса трансгенных Т-клеточных рецепторов или химерных антигенных рецепторов (CAR) (Jena, Dotti et al., 2010). CAR представляют собой синтетические рецепторы, состоящие из направляющего участка, который связан с одним или несколькими сигнальными доменами в одной молекуле слияния. Обычно связывающий участок CAR состоит из антигенсвязывающего домена одноцепочечного антитела (scFv), содержащего легкие и вариабельные фрагменты моноклонального антитела, присоединенные посредством гибкого линкера. Также успешно применялись связывающие участки, основанные на доменах рецептора или лиганда. Сигнальные домены для первого поколения CAR получают из цитоплазматического участка цепей CD3-дзета или Fc-рецептора-гамма. Было показано, что CAR первого поколения успешно перенаправляют цитотоксичность T-клеток, однако они не могли обеспечивать продолжительное размножение и противоопухолевую активность in vivo. Для повышения выживаемости и улучшения пролиферации модифицированных CAR T-клеток присоединяли сигнальные домены из костимулирующих молекул, включая CD28, OX-40 (CD134) и 4-1BB (CD137), по отдельности (второе поколение) или в комбинации (третье поколение). CAR успешно позволяли перенаправление T-клеток против антигенов, экспрессированных на поверхности опухолевых клеток из различных злокачественных опухолей, включая лимфомы и солидные опухоли (Jena, Dotti et al., 2010).
Современные архитектуры CAR построены на конструкции, в которой все нужные домены содержатся в одном полипептиде (US 7741465). Такая конструкция требует последовательного присоединения сигнальных доменов, что требует переместить некоторые домены из их природных околомембранных положений, удаленных от плазматической мембраны. Тем не менее, архитектуры, в которых лиганды и сигнальные домены отделены в своих нормальных околомембранных положениях (т.е. поблизости от клеточной мембраны на ее внутренней стороне), были бы более желательными, и считается, что они позволят улучшенное функционирование костимулирующих доменов. Высокоаффинный рецептор для IgE (FcεRI) мог бы обеспечивать такую архитектуру. В самом деле, FcεRI, который присутствует на тучных клетках и базофилах, представляет собой тетрамерный комплекс, состоящий из лигандсвязывающей альфа-субъединицы, бета-субъединицы и гомодимера двух передающих сигнал гамма-субъединиц (Metzger, Alcaraz et al., 1986). FcεRI-альфа домен состоит из внеклеточного домена, содержащего два Ig-подобных домена, которые связывают IgE, трансмембранный домен и короткий цитоплазматический хвост. Бета-субъединица содержит четыре трансмембранных сегмента, разделяющих амино- и карбоксиконцевые цитоплазматические хвосты. Гамма-цепь состоит по существу из трансмембранного участка и цитоплазматического хвоста, содержащего один иммунорецепторный тирозиновый активирующий мотив (ITAM) (Cambier, 1995).
Современный протокол для лечения пациентов с помощью адоптивной иммунотерапии основан на переносе аутологичных клеток. При таком подходе T-лимфоциты получают от пациентов, генетически модифицируют или отбирают ex vivo, культивируют in vitro, для того чтобы увеличить количество клеток, если необходимо, и, наконец, вводят в пациента. В дополнение к введению лимфоцитов на хозяина можно воздействовать другими способами, которые поддерживают приживление Т-клеток или их участие в иммунном ответе, например, посредством предварительного кондиционирования (с помощью излучения или химиотерапии) и введения факторов роста лимфоцитов (таких как IL-2). Каждый пациент получает индивидуально подобранное лечение с использованием собственных лимфоцитов пациента (т.е. аутологическую терапию).
Аутологические терапии сталкиваются со значительными техническими и логистическими препятствиями для практического применения, их создание требует дорогостоящих специализированных технических средств и высококвалифицированного персонала, они должны быть созданы в течение короткого времени после диагностики пациента, и, во многих случаях, предварительная обработка пациента приводит к ухудшению иммунной функции, так что лимфоциты пациента могут становиться малофункциональными и присутствовать в очень малых количествах. Из-за этих препятствий каждый препарат аутологичных клеток пациента является по существу эффективным новым продуктом, что ведет к существенной вариативности в эффективности и безопасности. В идеале, хотелось бы использовать стандартизированную терапию, в которой аллогенные терапевтические клетки можно было бы создавать заранее, подробно характеризовать и делать доступными для немедленного введения пациентам. Под аллогенными подразумевается, что клетки получают от индивидуумов, принадлежащих к одному и тому же виду, но генетически разнородных. Однако, использование аллогенных клеток имеет в настоящее время много недостатков. В иммунокомпетентных хозяевах аллогенные клетки быстро отторгаются, процесс называется отторжением хозяин против трансплантата (HvG), и это существенно ограничивает эффективность перенесенных клеток. В иммунонекомпетентных хозяевах аллогенные клетки могут приживаться, но их эндогенные специфичности TCR распознают ткань хозяина как чужие, что приводит к заболеванию трансплантат против хозяина (GvHD), которое может приводить к серьезному повреждению тканей и смерти. Для того, чтобы эффективно использовать аллогенные клетки, нужно преодолеть обе эти проблемы.
В иммунокомпетентных хозяевах аллогенные клетки быстро отторгаются иммунной системой хозяина. Было продемонстрировано, что аллогенные лейкоциты, присутствующие в необлученных продуктах крови, сохраняются в течение не больше чем от 5 до 6 дней. (Boni, Muranski et al., 2008). Таким образом, чтобы предотвратить отторжение аллогенных клеток, иммунная система хозяина должна быть эффективно подавлена. Глюкокортикостероиды широко используются терапевтически для иммуносупрессии (Coutinho and Chapman, 2011). Этот класс стероидных гормонов связывается с глюкокортикоидным рецептором (GR), присутствующим в цитозоле T-клеток, что ведет к транслокации в ядро и связыванию специфических мотивов ДНК, которые регулируют экспрессию ряда генов, участвующих в иммунологическом процессе. Обработка T-клеток с помощью глюкокортикоидных стероидов приводит к пониженному уровню производства цитокинов, что ведет к Т-клеточной анергии и препятствию к активации Т-клеток. Алемтузумаб, также известный как CAMPATH1-H, представляет собой гуманизированное моноклональное антитело, направленное на CD52, 12-аминокислотный гликопротеин, соединенный с гликозилфосфатидилинозитолом (GPI) (Waldmann and Hale, 2005). CD52 экспрессируется в больших количествах на T и B-лимфоцитах и в малых количествах на моноцитах, при этом отсутствуя на гранулоцитах и предшественниках костного мозга. Было показано, что лечение с помощью алемтузумаба, гуманизированного моноклонального антитела, направленного против CD52, индуцирует быстрое истощение циркулирующих лимфоцитов и моноцитов. Его часто используют при лечении Т-клеточных лимфом и в некоторых случаях как часть режима кондиционирования для трансплантации. Однако, в случае адоптивной иммунотерапии использование иммуносупрессивных препаратов также будет иметь отрицательное воздействие на введенные терапевтические T-клетки. Следовательно, для эффективного использования подхода адоптивной иммунотерапии в данных условиях введенные клетки должны быть резистенстными к иммуносупрессивной обработке.
С другой стороны, T-клеточные рецепторы (TCR) представляют собой рецепторы клеточной поверхности, которые участвуют в активации Т-клеток в ответ на презентацию антигена. TCR, как правило, состоит из двух цепей, альфа и бета, которые собираются с образованием гетеродимера и связываются с передающими субъединицами CD3 с образованием комплекса T-клеточного рецептора, присутствующего на клеточной поверхности. Каждая альфа и бета-цепь TCR состоит из иммуноглобулин-подобного N-концевого вариабельного (V) и константного (C) участка, гидрофобного трансмембранного домена и короткого цитоплазматического участка. Что касается молекул иммуноглобулина, вариабельный участок альфа и бета-цепей создается посредством V(D)J-рекомбинации, создающей большое разнообразие антиген-специфичностей в популяции T-клеток. Однако, в отличие от иммуноглобулинов, которые распознают интактный антиген, T-клетки активируются процессированными пептидными фрагментами, связанными с молекулой MHC, что вводит дополнительное измерение в распознавание антигена T-клетками, известное как MHC-рестрикция. Распознавание несовместимостей MHC между донором и реципиентом посредством T-клеточного рецептора приводит к пролиферации T-клеток и потенциальному развитию GVHD. Было показано, что нормальная поверхностная экспрессия TCR зависит от согласованного синтеза и сборки всех семи компонентов комплекса (Ashwell and Klusner, 1990). Инактивация TCR-альфа или TCR-бета может приводить к элиминации TCR с поверхности T-клеток, что предотвращает распознавание аллоантигена и, следовательно, GVHD. Однако, разрушение TCR приводит к элиминации сигнального компонента CD3 и изменяет способы дальнейшего размножения T-клеток.
Опосредованный T-клетками иммунитет включает несколько последовательных стадий, регулируемых равновесием между костимулирующими и ингибирующими сигналами, которые точно настраивают иммунный ответ. Ингибирующие сигналы, называемые иммунными контрольными точками, являются существенными для поддержания аутотолерантности, а также для ограничения иммунно-опосредованного сопутствующего повреждения ткани. Экспрессия белка иммунных контрольных точек может быть дерегулирована опухолями. Способность опухолей присваивать данные ингибирующие пути представляет собой важный механизм в иммуннорезистентности и ограничивает успех иммунотерапии. Одним из перспективных подходов к активации терапевтического T-клеточного иммунного ответа является блокада этих иммунных контрольных точек (Pardoll, 2012). Иммунные контрольные точки представляют значительные препятствия для активации функционального клеточного иммунитета при раке, и антагонистические антитела, специфичные для ингибирующих лигандов на T-клетках, включая CTLA4 и белок программируемой смерти-1 (PD-1), являются примерами направленных средств, исследуемых в клиниках.
Антиген 4 цитотоксических T-лимфоцитов (CTLA-4; также известный как CD152) снижает амплитуду активации T-клеток, и было показано, что лечение с помощью антагониста антител к CTLA4 (ипилимумаба) дает эффект выживаемости пациентов с меланомой (Robert and Mateus, 2011). Белок программируемой клеточной смерти 1 (PD1 или PDCD1, также известный как CD279) представляет собой другую очень перспективную мишень для иммунотерапии (Pardoll and Drake, 2012; Pardoll, 2012). В отличие от CTLA-4, PD1 ограничивает T-клеточные эффекторные функции в периферической ткани в момент воспалительного ответа на инфекцию и для ограничения аутоиммунитета. Первое клиническое испытание антитела PD1 продемонстрировало несколько случаев регрессии опухоли (Brahmer, Drake et al., 2010). Многие дополнительные белки иммунных контрольных точек, судя по недавним исследованиям, представляют собой перспективные мишени для терапевтической блокады.
В нормальных T-клетках T-клеточные рецепторы получаются из рецепторов пре-T-клеток (pTCR), которые экспрессируются незрелыми тимоцитами и имеют решающее значение для развития Т-клеток из двойной отрицательной (CD4- CD8-) в двойную положительную (CD4+ CD8+) стадии. Пре-T-клетки, которые успешно прошли продуктивные реаранжировки локуса TCR-бета, экспрессируют функциональную цепь TCR-бета, которая образует пару с инвариантной цепью preT-альфа и сигнальными компонентами CD3 с образованием комплекса пре-TCR. Экспрессия пре-TCR на клеточной поверхности является необходимой для запуска бета-селекции, процесса, который индуцирует размножение развивающихся T-клеток, обеспечивает аллельное исключение локуса TCR-бета и приводит к индукции реаранжировок в локусе TCR-альфа (von Boehmer, 2005). После продуктивных реаранжировок TCR-альфа и замещения pT-альфа на TCR-альфа с образованием зрелых TCR, тимоциты подвергаются второй стадии селекции, называемой положительной или TCR-альфа/бета селекцией при связывании комплексов аутопептида и MHC, экспрессированных на эпителиальных клетках тимуса. Таким образом, зрелые T-клетки распознают и отвечают на комплекс антиген/MHC посредством своих TCR. Наиболее непосредственным следствием активации TCR является инициирование сигнальных путей посредством связанных CD3-субъединиц, которые приводят к нескольким событиям, включая клональное размножение T-клеток, повышающую регуляцию активации маркеров на клеточной поверхности и индукцию цитотоксичности или секрецию цитокинов.
Благодаря характеру селекции цепей TCR-бета посредством образования пары с preT-альфа во время развития в тимусе в T-клетках, в которых TCR-альфа была инактивирована, гетерологичное введение трансгена pT-альфа может приводить к образованию пре-TCR. Этот pTCR может служить в качестве средства активации или стимуляции Т-клеток таким образом, который не является зависимым от MHC, тем самым позволяя, например, непрерывное размножение альфа/бета T-клеток после инактивации TCR-альфа. Важно отметить, что комплекс pTCR демонстрирует такой же биохимический состав, что и TCR, в отношении связанных CD3-субъединиц (Carrasco, Ramiro et al., 2001). Кроме того, в отличие от TCR, сигнализация пре-TCR может происходить частично в связи с независимыми от лигандов событиями. Кристаллическая структура внеклеточного домена pTCR обеспечивает структурную основу для возможной независимости сигнализации pTCR от лигандов. Было показано, что pTCR образует димер голова к хвосту, в котором связаны два гетеродимера pT-альфа-TCR-бета (Pang, Berry et al., 2010).
В контексте разработки сконструированных иммунных клеток терапевтического качества, которые могут быть направлены на злокачественные или инфицированные клетки, изобретатели стремились к улучшенным архитектурам CAR, который были бы близки к природным и, вероятно, вели бы себя соответственно, используя любые внеклеточные моно- или мультиспецифические лигандсвязывающие домены.
В результате, авторы разработали новое поколение CAR, использующих отдельные полипептидные субъединицы в соответствии с настоящим изобретением, именуемых "многоцепочечные CAR".
Сущность изобретения
Химерные антигенные рецепторы (CAR) в предшествующем уровне техники присутствуют в виде одиночных молекул слияния, которые требуют последовательного присоединения сигнальных доменов. Однако, удаление сигнальных доменов из их природного околомембранного положения препятствует их функции. Таким образом, чтобы преодолеть этот недостаток, изобретатели смогли сконструировать так называемые многоцепочечные CAR, которые делают возможным околомембранное положение всех соответствующих сигнальных доменов. В многоцепочечных CAR сигнальные домены помещены на различных полипептидных цепях, так что они размещены в околомембранных положениях. Например, многоцепочечный CAR может быть получен из FcεRI посредством замены высокоаффинного IgE-связывающего домена альфа-цепи FcεRI на внеклеточный лигандсвязывающий домен, такой как scFv, в то время как N- и/или C-концевые хвосты бета- и/или гамма-цепи FcεRI используются для помещения передающего сигнал домена в нормальные околомембранные положения. Внеклеточный лигандсвязывающий домен выполняет задачу перенаправления специфичности T-клеток в направлении клеток-мишеней, тогда как передающие сигнал домены, оставленные в околомембранных положениях, активируют иммунный клеточный ответ.
Тот факт, что сигнальные домены в околомембранном положении присутствуют на полипептиде(ах), отличающемся от несущего внеклеточный лигандсвязывающий домен, обеспечивает более гибкую архитектуру для CAR. Соответственно, дополнительные сигнальные домены или костимулирующие домены могут быть присоединены или отсоединены от CAR в зависимости от интенсивности взаимодействия, наблюдаемого между лигандсвязывающим доменом CAR и его рецептором. Такая гибкость в архитектуре CAR также позволяет вводить дополнительный внеклеточный лигандсвязывающий домен(ы). Это может быть сделано путем слияния второго лигандсвязывающего домена с полипептидом, несущим первый лигандсвязывающий домен, или путем введения дополнительных внеклеточных лигандсвязывающих доменов. Если данный второй лигандсвязывающий домен имеет другую специфичность, это образует биспецифический многоцепочечный CAR, а если имеют место дополнительные специфичности, то мультиспецифический многоцепочечный CAR. Это особенно выгодно с точки зрения нацеливания на заданные клетку или вирус, несущие два или более различных рецептора, присутствующих на их поверхности. Это увеличивает специфичность такого многоцепочечного CAR в направлении указанных клетки или вируса. С другой стороны, количество сигнальных доменов может позволять модуляцию сигнала в соответствии с различными связывающими доменами. Кроме того, каждый рекомбинантный полипептид, образующий CAR, может быть экспрессирован независимо с различными уровнями экспрессии для дополнительной модуляции активности CAR.
Настоящее изобретение относится к данной новой архитектуре CAR, к рекомбинантным полинуклеотидам, кодирующим их, а также к способам конструирования иммунных клеток для специфичного распознавания клеток, какими бы ни были цель и природа иммунных клеток.
Настоящее изобретение дополнительно предлагает способы получения таких иммунных клеток, более подходящих для целей иммунотерапии.
Например, иммунные клетки могут быть дополнительно сконструированы так, чтобы быть неаллореактивными и/или резистентными к иммуносупрессивным средствам, в частности, посредством инактивации генов TCR-альфа или бета в первичных клетках и/или посредством соединения инактивации генов, кодирующих мишени для различных иммуносупрессивных средств, в частности, CD52 и GR (глюкокортикоидных рецепторов), и последующей селекции клеток, устойчивых к упомянутым иммуносупрессивным средствам.
В дополнение к вышеуказанной концепции генетически модифицированных иммунных клеток настоящее изобретение также относится к вариантам осуществления, в которых иммунные клетки конструируют так, чтобы позволять пролиферацию, когда TCR-альфа инактивирована, например, путем временной экспрессии preT-альфа в клетках, посредством чего восстанавливается функциональный комплекс CD3 в отсутствие функциональной альфа/бета-TCR.
Для того, чтобы сконструировать генетически высоко активные модифицированные иммунные клетки, изобретение также предлагает способы, в которых иммунные контрольные точки блокированы недостаточной экспрессией генов, таких как PD1 и CTLA-4.
Настоящая заявка дополнительно раскрывает сконструированные иммунные клетки, в частности, T-клетки, для применения в качестве лекарственного средства, более конкретно, для лечения или профилактики злокачественных опухолей посредством введения таких иммунных клеток в живой организм.
Краткое описание фигур и таблиц
В дополнение к вышеуказанным признакам изобретение дополнительно содержит другие признаки, которые будут следовать из следующего описания, а также из прилагаемых чертежей. Более полное понимание настоящего изобретения и многих его сопутствующих преимуществ будет легко получено, поскольку оно станет более понятным с помощью ссылки на следующие фигуры в сочетании с подробным описанием ниже.
Фигура 1: Схематическое представление нормального отношения между T-клетками и антиген-презентирующей клеткой.
Фигура 2: Схематическое представление генетически модифицированных терапевтических T-клеток в соответствии с настоящим изобретением и опухолевых клеток пациента.
Фигура 3: Схематическое представление многоцепочечного CAR.
Фигура 4: Схема различных вариантов многоцепочечных CAR. A. Схема рецептора FcεRI. B-C. Различные варианты многоцепочечных CAR (от csm1 до csm10), содержащие scFv и участок в виде ножки CD8, слитый с трансмембранным доменом альфа-цепи FcεRI. По меньшей мере один из доменов 41BB, CD28 и/или CD3-дзета может быть слит с FcεRI-альфа, бета и/или гамма-цепью.
Фигура 5: Схематическое представление одного примера способа получения аллогенных клеток человека способами генной инженерии для иммунной терапии.
Фигура 6: Концентрация в клетках на миллилитр живых CD52-положительных или CD52-отрицательных клеток после лечения с помощью анти-CD52 антитела (CAMPATH1-H) с комплементом или контролем.
Фигура 7: Сравнение распределения прямого светорассеяния (FSC), индикатора размера клеток, между TCR-положительными и TCR-отрицательными клетками или между CD52-положительными и CD52-отрицательными клетками и не активированными клетками в качестве контроля.
Фигура 8: Анализ с помощью проточной цитометрии экспрессии CD107a (маркер дегрануляции) на целевых инактивированных по CD52 и TCR-альфа T-клетках. Экспрессию CD107 анализировали на CD52+TCRαβ+ клетках (первая колонка), CD52-TCRαβ- клетках (вторая колонка), CD52-TCRαβ+ клетках (третья колонка) и CD52+TCRαβ- клетках (четвертая колонка) перед (A) и после инкубации с клетками Дауди (B); C) представляет анализ с помощью проточной цитометрии T-клеток, дополнительно трансфицированных CAR и инкубированных с клетками Дауди; D) представляет анализ с помощью проточной цитометрии T-клеток, трансфицированных CAR, но не инкубированных с клетками Дауди, и E) представляет анализ с помощью проточной цитометрии T-клеток, трансфицированных CAR и обработанных PMA/иономицином (положительный контроль).
Фигура 9: Анализ с помощью глубокого секвенирования потенциальных мишеней вне сайта TALE-нуклеаз CD52 и TRAC. Фигура 9 раскрывает последовательности "левой полумишени" как SEQ ID NO:149-165 и последовательности "правой полумишени" как SEQ ID NO:166-182, все, соответственно, в порядке изображения.
Фигура 10: Анализ геномного локуса PDCD1 и CTLA-4 посредством T7-эндонуклеазного анализа. Стрелки показывают на расщепленные продукты ПЦР.
Фигура 11: Схематическое представление некоторых примеров конструктов preT-альфа.
Фигура 12: Анализ с помощью проточной цитометрии эффективности трансдукции (% BFP+ клеток) и активность конструктов FL, Δ18, Δ48 pT-альфа (% поверхностной экспрессии CD3) в TCR-альфа-инактивированных клетках Jurkat.
Фигура 13: Схематическое представление лентивирусного конструкта, кодирующего белок pT-альфа (пре-TCRα).
Фигура 14: A. Изображение экспериментального протокола. B. Анализ с помощью проточной цитометрии экспрессии TCR-альфа/бета, CD3 и экспрессии BFP на TCR-альфа-инактивированных T-клетках (KO), трансдуцированных или BFP-2A-pT-альфа-Δ48 (KO/Δ48), или контрольным лентивирусным вектором BFP (KO/BFP), до и после очистки. C. Анализ с помощью проточной цитометрии экспрессии TCR-альфа/бета и CD3 на очищенных TCR-альфа-инактивированных клетках, трансдуцированных (BFPpos) или не трансдуцированных (BFPneg) лентивирусным вектором BFP-2A-pT-альфа-Δ48. NEP представляет не электропорированные клетки с TRAC-TALE-нуклеазами.
Фигура 15: A-B. Анализ с помощью проточной цитометрии экспрессии маркера ранней активации CD69 (A), маркера поздней активации CD25 (B) через 24 и 48 часов после реактивации с помощью гранул с анти-CD3/CD28, соответственно, на не электропорированных клетках (NEP) и TCR-альфа-инактивированных клетках (KO), трансдуцированных лентивирусным вектором BFP-2A-pΤα-Δ48 (pΤα-Δ48), лентивирусным вектором BFP-2A-pΤα-Δ48.41BB (pΤα-Δ48.ΒΒ) или контрольным вектором BFP (BFP). Гистограммы pΤα-Δ48 соответствуют сигналу, детектируемому в TCR-инактивированных клетках, экспрессирующих pΤα-Δ48 (BFP+ клетки), тогда как гистограммы KO соответствуют TCR-альфа-инактивированным клеткам, которые не экспрессируют pΤα-Δ48 (BFP- клетки). Гистограммы pΤα-Δ48.ΒΒ соответствуют сигналу, детектируемому в TCR-инактивированных клетках, экспрессирующих pΤα-Δ48.41ΒΒ (BFP+ клетки), тогда как гистограммы KO соответствуют TCR-альфа-инактивированным клеткам, которые не экспрессируют pΤα-Δ48.41ΒΒ (BFP-клетки). Гистограммы NEP (не электропорированные) соответствуют сигналу, детектируемому в неинженерных клетках. C. Анализ с помощью проточной цитометрии размера клеток через 72 часа после реактивации с помощью гранул с анти-CD3/CD28 на не электропорированных клетках (NEP) и TCR-альфа-инактивированных клетках (KO), трансдуцированных лентивирусным вектором BFP-2A-pΤα-Δ48 (pΤα-Δ48), лентивирусным вектором BFP-2A-pΤα-Δ48.41BB (pΤα-Δ48.ΒΒ) или контрольным вектором BFP (BFP). Значения, указанные в верхней части каждого графика, соответствуют среднему геометрическому флуоресценции каждой популяции.
Фигура 16: Анализ клеточного роста TCR-альфа-инактивированных клеток (KO), трансдуцированных pT-альфа-Δ48 (pTaΔ48) или контрольным вектором BFP (BFP), выдерживаемых в IL2 или в IL2 с гранулами анти-CD3/CD28, в различных временных точках (ось x). Количество BFP+ клеток определяли в различных временных точках для каждого варианта и определяли, во сколько раз изменится индукция данных клеток (ось y) по отношению к значению, полученному на день 2 после реактивации. Результаты получали с двумя независимыми донорами. Для второго донора клеточный рост также определяли для клеток, трансдуцированных pT-альфа-Δ48.41BB (pΤα-Δ48.BB) и полноразмерным pT-альфа (pΤα-FL).
Фигура 17: Анализ с помощью проточной цитометрии GFP-положительных клеток на PBMC, электропорированных с помощью пяти различных программ CytoPulse. A. NEP, EP#1 (GFP) и EP#2 (GFP); B. EP#3 (pUC), EP#4 (GFP) и EP#5 (GFP). Верхняя линия соответствует трансфекции 6×106 клеток на кювету, в то время как нижняя линия соответствует трансфекции 3×106 клеток на кювету.
Фигура 18: Анализ с помощью проточной цитометрии смертности очищенных T-клеток с помощью красителя для определения жизнеспособности (eFluor-450) и GFP-положительных клеток среди жизнеспособной популяции после электропорации мРНК GFP, ДНК GFP и контрольной ДНК pUC. A. NT и NEP; B. EP (нет ДНК) и ДНК GFP; C. ДНК pUC и мРНК GFP. NEP соответствует клеткам, которые выдерживали в буфере для электропорации, но не электропорировали, и NT соответствует не электропорированным клеткам, выдерживаемым в культуральной среде.
Фигура 19: Анализ с помощью проточной цитометрии экспрессии TCR-альфа/бета и CD3 на первичных T-клетках человека после электропорации мРНК TRAC-TALE-нуклеазы (сверху). Анализ с помощью глубокого секвенирования геномной ДНК, экстрагированной из первичных T-клеток человека после электропорации мРНК TRAC-TALE-нуклеазы (снизу) (SEQ ID NO:183-192, соответственно, в порядке изображения).
Фигура 20: A. Анализ с помощью проточной цитометрии экспрессии CAR (анти-F(ab')2) после электропорации T-клеток с или без мРНК, кодирующей одноцепочечный CAR. B. Анализ с помощью проточной цитометрии экспрессии CD107a (маркер дегрануляции) на электропорированных T-клетках, совместно культивированных с клетками Дауди.
Фигура 21: A. Изображение мРНК, кодирующей многоцепочечный CAR. B. Анализ с помощью проточной цитометрии экспрессии CAR (анти-F(ab')2) на жизнеспособных T-клетках, электропорированных с или без полицистронной мРНК, кодирующей многоцепочечный CAR. C. Анализ с помощью проточной цитометрии экспрессии CD107a (маркер дегрануляции) на электропорированных T-клетках, совместно культивированных с клетками Дауди.
Фигура 22: Экспрессия многоцепочечных CAR в T-клетках человека после электропорации полицистронных мРНК.
Фигура 23: Экспрессия мультисубъединичных CAR обусловливается экспрессией трех цепей: α, β и γ.
Фигура 24: T-клетки человека, временно экспрессирующие многоцепочечные CAR, подвергаются дегрануляции после совместного культивирования с клетками-мишенями.
Фигура 25: T-клетки человека, временно экспрессирующие многоцепочечные CAR, секретируют цитокины после совместного культивирования с клетками-мишенями.
Фигура 26: T-клетки человека, временно экспрессирующие многоцепочечные CAR, лизируют клетки-мишени.
Таблица 1: Описание GR-TALE-нуклеаз и последовательности сайтов-мишеней TALE-нуклеаз в гене GR человека.
Таблица 2: Расщепляющая активность GR-TALE-нуклеаз в дрожжах. Значения лежат между 0 и 1. Максимальное значение составляет 1.
Таблица 3: Процентная доля направленного мутагенеза в сайтах-мишенях эндогенных TALE-нуклеаз в клетках 293.
Таблица 4: Процентная доля направленного мутагенеза в сайтах-мишенях эндогенных TALE-нуклеаз в первичных T-лимфоцитах.
Таблица 5: Описание TALE-нуклеаз CD52, TRAC и TRBC и последовательности сайтов-мишеней TALE-нуклеаз в соответствующих генах человека.
Таблица 6: Дополнительные последовательности-мишени для TALE-нуклеаз TRAC и CD52.
Таблица 7: Процентная доля вставок-делеций для TALE-нуклеазы, нацеленной на мишени CD52_T02, TRAC_T01, TRBC_T01 и TRBC_T02.
Таблица 8: Процентные доли CD52-отрицательных, TCR-отрицательных и CD52/TCR-двойных отрицательных T-лимфоцитов после трансфекции соответствующих экспрессирующих TALE-нуклеазу полинуклеотидов.
Таблица 9: Процентные доли TCR-отрицательных T-лимфоцитов после трансфекции экспрессирующих TRBC-TALE-нуклеазу полинуклеотидов.
Таблица 10: Описание TALE-нуклеаз CTLA4 и PDCD1 и последовательности сайтов-мишеней TALE-нуклеаз в соответствующих генах человека.
Таблица 11: Описание выборки конструктов pT-альфа.
Таблица 12: Активность различных конструктов pT-альфа в TCR-альфа-инактивированных клетках Jurkat. Активность измеряли посредством анализа с помощью проточной цитометрии экспрессии CD3 на TCR-альфа-инактивированных клетках Jurkat, трансфицированных различными конструктами preT-альфа.
Таблица 13: Различные программы CytoPulse, использованные для определения минимального напряжения, требуемого для электропорации в происходящих от PBMC T-клетках.
Таблица 14: Программа CytoPulse, использованная для электропорации очищенных T-клеток.
Таблица 15: Описание составов цепей FcR вариантов многоцепочечных CAR.
Подробное описание изобретения
Если специально не определено в настоящем документе, все используемые технические и научные термины имеют то значение, которое обычно понимается специалистом в областях генной терапии, биохимии, генетики и молекулярной биологии.
Все способы и материалы, аналогичные или эквивалентные тем, которые описаны в настоящем документе, могут быть использованы в осуществлении на практике или тестировании настоящего изобретения, причем подходящие способы и материалы описаны в настоящем документе. Все публикации, патентные заявки, патенты и другие ссылки, указанные в настоящем документе, включены в качестве ссылки во всей их полноте. В случае конфликта, настоящее описание изобретения, включая определения, будет иметь преимущество. Кроме того, материалы, способы и примеры являются только иллюстративными и не предназначены для ограничения, если не указано иное.
Осуществление настоящего изобретения на практике будет использовать, если не указано иное, обычные методы клеточной биологии, клеточной культуры, молекулярной биологии, трансгенной биологии, микробиологии, рекомбинантной ДНК и иммунологии, которые известны специалистам в данной области. Такие методы подробно описаны в литературе. См., например, Current Protocols in Molecular Biology (Frederick M. AUSUBEL, 2000, Wiley and son Inc, Library of Congress, США); Molecular Cloning: A Laboratory Manual, Third Edition, (Sambrook et al, 2001, Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press); Oligonucleotide Synthesis (M. J. Gait ed., 1984); Mullis et al., патент США № 4683195; Nucleic Acid Hybridization (B. D. Harries & S. J. Higgins eds. 1984); Transcription And Translation (B. D. Hames & S. J. Higgins eds. 1984); Culture Of Animal Cells (R. I. Freshney, Alan R. Liss, Inc., 1987); Immobilized Cells And Enzymes (IRL Press, 1986); B. Perbal, A Practical Guide To Molecular Cloning (1984); серия Methods In ENZYMOLOGY (J. Abelson and M. Simon, eds.-in-chief, Academic Press, Inc., New York), особенно тома 154 и 155 (Wu et al. eds.) и том 185 "Gene Expression Technology" (D. Goeddel, ed.); Gene Transfer Vectors For Mammalian Cells (J. H. Miller and M. P. Calos eds., 1987, Cold Spring Harbor Laboratory); Immunochemical Methods In Cell And Molecular Biology (Mayer and Walker, eds., Academic Press, London, 1987); Handbook Of Experimental Immunology, Volumes I-IV (D. M. Weir and C. C. Blackwell, eds., 1986); и Manipulating the Mouse Embryo, (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1986).
Многоцепочечный химерный антигенный рецептор (CAR)
Настоящее изобретение относится к многоцепочечному химерному антигенному рецептору (CAR), в частности, к приспособленному к иммунным клеткам, применяемым в иммунотерапии.
Многоцепочечный CAR в соответствии с настоящим изобретением обычно содержит по меньшей мере:
- один трансмембранный полипептид, содержащий по меньшей мере один внеклеточный лигандсвязывающий домен и;
- один трансмембранный полипептид, содержащий по меньшей мере один передающий сигнал домен;
так что указанные полипептиды собирают вместе с образованием многоцепочечного химерного антигенного рецептора.
Термин "внеклеточный лигандсвязывающий домен", как используется в настоящем документе, определяется как олиго- или полипептид, который способен к связыванию с лигандом. Предпочтительно, данный домен будет способен к взаимодействию с молекулой клеточной поверхности. Например, внеклеточный лигандсвязывающий домен может быть выбран для распознавания лиганда, который действует в качестве маркера клеточной поверхности на клетках-мишенях, связанных с конкретным состоянием заболевания. Таким образом, примеры маркеров клеточной поверхности, которые могут действовать как лиганды, включают связанные с вирусными, бактериальными и паразитарными инфекциями, аутоиммунным заболеванием и раковыми клетками. В частности, внеклеточный лигандсвязывающий домен может содержать антигенсвязывающий домен, происходящий от антитела против антигена мишени. В качестве неограничивающих примеров, антиген мишени может представлять собой опухолеассоциированный поверхностный антиген, такой как ErbB2 (HER2/neu), карциноэмбриональный антиген (CEA), молекула адгезии эпителиальных клеток (EpCAM), рецептор эпидермального фактора роста (EGFR), EGFR вариант III (EGFRvIII), CD19, CD20, CD30, CD40, дисиалоганглиозид GD2, дуктально-эпителиальный муцин, gp36, TAG-72, гликосфинголипиды, глиома-ассоциированный антиген, β-хорионический гонадотропин человека, альфафетопротеин (AFP), лектин-реактивный AFP, тиреоглобулин, RAGE-1, MN-CA IX, обратная транскриптаза теломеразы человека, RU1, RU2 (AS), кишечная карбоксилэстераза, mut hsp70-2, M-CSF, простаза, простатический специфический антиген (PSA), PAP, NY-ESO-1, LAGA-1a, p53, простеин, PSMA, выживание и теломераза, опухолевый антиген-1 карциномы простаты (PCTA-1), MAGE, ELF2M, нейтрофил-эластаза, эфрин B2, CD22, инсулиноподобный фактор роста (IGF1)-I, IGF-II, рецептор IGFI, мезотелин, молекула главного комплекса гистосовместимости (MHC), презентирующая эпитоп опухолеспецифического пептида, 5T4, ROR1, Nkp30, NKG2D, опухолевые стромальные антигены, экстра домен A (EDA) и экстра домен B (EDB) фибронектина, и домен A1 тенасцина-C (TnC A1), и связанный с фибробластами белок (fap); линиеспецифический или тканеспецифический антиген, такой как CD3, CD4, CD8, CD24, CD25, CD33, CD34, CD133, CD138, CTLA-4, B7-1 (CD80), B7-2 (CD86), эндоглин, молекула главного комплекса гистосовместимости (MHC), BCMA (CD269, TNFRSF 17), или вирусоспецифический поверхностный антиген, такой как ВИЧ-специфический антиген (такой как HIV gp120); EBV-специфический антиген, CMV-специфический антиген, HPV-специфический антиген, специфический к вирусу Ласса антиген, специфический к вирусу гриппа антиген, а также любое производное или вариант этих поверхностных маркеров.
Внеклеточный лигандсвязывающий домен может также содержать пептид, связывающий антиген мишени, пептид или белок, связывающий антитело, которое связывает антиген мишени, пептидный или белковый лиганд, такой как фактор роста, цитокин или гормон в качестве неограничивающих примеров, связывающий рецептор на мишени, или домен, происходящий от рецептора, такого как рецептор фактора роста, цитокиновый рецептор или рецептор гормона в качестве неограничивающих примеров, связывающий пептидный или белковый лиганд на мишени. Предпочтительно, мишень представляет собой клетку или вирус.
В предпочтительном варианте осуществления указанный внеклеточный лигандсвязывающий домен представляет собой одноцепочечный фрагмент антитела (scFv), содержащий легкий (VL) и тяжелый (VH) вариабельный фрагмент специфического к антигену мишени моноклонального антитела, соединенные гибким линкером. В предпочтительном варианте осуществления указанный scFv представляет собой анти-CD19 scFV, предпочтительно, scFV-4G7 (Peipp et al., J Immunol Methods., 2004, Feb 15;285(2):265-80) (VH: SEQ ID NO:193 и VL: SEQ ID NO:194, scFV: SEQ ID NO:195).
Можно также использовать связывающий домен, отличный от scFv, для заранее определенного нацеливания на лимфоциты, такой как фрагменты однодоменных антител верблюда или лиганды рецепторов, такие как полипептид фактор роста сосудистого эндотелия, интегрин-связывающий пептид, херегулин или мутеин IL-13, антителосвязывающие домены, гипервариабельные петли или CDR антител в качестве неограничивающих примеров.
В предпочтительном варианте осуществления указанный трансмембранный полипептид дополнительно содержит участок в виде ножки между упомянутым внеклеточным лигандсвязывающим доменом и упомянутым трансмембранным доменом. Термин "участок в виде ножки", используемый в настоящем документе, в общем означает любой олиго- или полипептид, который функционирует так, чтобы связывать трансмембранный домен с внеклеточным лигандсвязывающим доменом. В частности, участок в виде ножки используют для обеспечения большей гибкости и доступности для внеклеточного лигандсвязывающего домена. Участок в виде ножки может содержать вплоть до 300 аминокислот, предпочтительно, от 10 до 100 аминокислот и, наиболее предпочтительно, от 25 до 50 аминокислот. Участок в виде ножки может быть получен из всей или части природной молекулы, как, например, из всего или части внеклеточного участка CD8, CD4 или CD28 или из всего или части константного участка антитела. Альтернативно, участок в виде ножки может представлять собой синтетическую последовательность, которая соответствует природной последовательности ножки, или может представлять собой полностью синтетическую последовательность ножки. В предпочтительном варианте осуществления указанный участок в виде ножки является частью альфа-цепи CD8 человека (например, NP_001139345.1) (SEQ ID NO:196).
Таким образом, экспрессия многоцепочечного CAR в иммунных клетках приводит к модифицированным клеткам, которые селективно элиминируют определенные мишени, включая, но без ограничения, злокачественные клетки, несущие соответствующий опухолеассоциированный поверхностный антиген, или инфицированные вирусом клетки, несущий вирусоспецифический поверхностный антиген, или клетки-мишени, несущие линиеспецифический или тканеспецифический поверхностный антиген.
Обычно в злокачественных клетках наблюдается понижающая регуляция или мутация антигенов мишени, создающая варианты ускользания благодаря потере антигенов. Таким образом, для компенсации ускользания опухоли и придания иммунной клетке большей специфичности к мишени многоцепочечный CAR может содержать несколько внеклеточных лигандсвязывающих доменов для одновременного связывания различных элементов в мишени, посредством этого увеличивая активацию и функционирование иммунных клеток. В одном из вариантов осуществления внеклеточные лигандсвязывающие домены могут быть размещены в тандеме на одном трансмембранном полипептиде и, необязательно, могут быть разделены с помощью линкера. В другом варианте осуществления указанные различные внеклеточные лигандсвязывающие домены могут быть размещены на различных трансмембранных полипептидах, составляя многоцепочечный CAR. В другом варианте осуществления настоящее изобретение относится к популяции многоцепочечных CAR, причем все они содержат различные внеклеточные лигандсвязывающие домены. В частности, настоящее изобретение относится к способу получения иммунных клеток способом генной инженерии, включающему наличие иммунной клетки и экспрессию на поверхности указанной клетки популяции многоцепочечных CAR, причем все они содержат различные внеклеточные лигандсвязывающие домены. В другом конкретном варианте осуществления настоящее изобретение относится к способу получения иммунной клетки способами генной инженерии, включающему наличие иммунной клетки и введение в указанную клетку полинуклеотидов, кодирующих полипептиды, составляющие популяцию многоцепочечных CAR, причем все они содержат различные внеклеточные лигандсвязывающие домены. В конкретном варианте осуществления способ получения иммунной клетки способами генной инженерии включает экспрессию на поверхности клетки по меньшей мере части бета и/или гамма-цепи FcεRI, слитой с передающим сигнал доменом, и нескольких частей альфа-цепей FcsRI, слитых с различными внеклеточными лигандсвязывающими доменами. В более конкретном варианте осуществления указанный способ включает введение в указанную клетку по меньшей мере одного полинуклеотида, который кодирует часть бета и/или гамма-цепи FcsRI, слитой с передающим сигнал доменом, и несколько альфа-цепей FcεRI, слитых с различными внеклеточными лигандсвязывающими доменами. Под популяцией многоцепочечных CAR имеется в виду по меньшей мере два, три, четыре, пять, шесть или больше многоцепочечных CAR, причем все они содержат различные внеклеточные лигандсвязывающие домены. Различные внеклеточные лигандсвязывающие домены в соответствии с настоящим изобретением могут, предпочтительно, одновременно связывать различные элементы в мишени, посредством этого увеличивая активацию и функционирование иммунных клеток.
Настоящее изобретение также относится к выделенной иммунной клетке, которая содержит популяцию многоцепочечных CAR, причем все они содержат различные внеклеточные лигандсвязывающие домены.
Передающий сигнал домен или внутриклеточный сигнальный домен многоцепочечного CAR по настоящему изобретению отвечает за внутриклеточную сигнализацию после связывания внеклеточного лигандсвязывающего домена с мишенью, что приводит к активации иммунной клетки и иммунному ответу. Другими словами, передающий сигнал домен отвечает за активацию по меньшей мере одной из нормальных эффекторных функций иммунной клетки, в которой экспрессируется многоцепочечный CAR. Например, эффекторная функция T-клетки может представлять собой цитолитическую активность или хелперную активность, включая секрецию цитокинов. Таким образом, термин "передающий сигнал домен" относится к части белка, которая передает сигнал эффекторной сигнальной функции и направляет клетку к выполнению специализированной функции.
Предпочтительные примеры передающего сигнал домена для использования в многоцепочечном CAR могут представлять собой цитоплазматические последовательности Fc-рецептора или T-клеточного рецептора и корецепторов, которые действуют согласованно, чтобы инициировать передачу сигнала после контакта антигенного рецептора, а также любое производное или вариант данных последовательностей и любую синтетическую последовательность, которая обладает той же функциональной способностью. Передающий сигнал домен содержит два различных класса цитоплазматической сигнальной последовательности, те, которые инициируют антиген-зависимую первичную активацию, и те, которые действуют антиген-независимым образом для обеспечения вторичного или костимулирующего сигнала. Первичная цитоплазматическая сигнальная последовательность может содержать сигнальные мотивы, который известны как иммунорецепторные тирозиновые активирующие мотивы ITAM. ITAM представляют собой хорошо определенные сигнальные мотивы, обнаруженные во внутрицитоплазматическом хвосте множества рецепторов, которые служат в качестве сайтов связывания для тирозинкиназ класса syk/zap70. Примеры ITAM, используемых в настоящем изобретении, могут включать в качестве неограничивающих примеров происходящие от TCR-дзета, FcR-гамма, FcR-бета, FcR-эпсилон, CD3-гамма, CD3-дельта, CD3-эпсилон, CD5, CD22, CD79a, CD79b и CD66d. В предпочтительном варианте осуществления передающий сигнал домен многоцепочечного CAR может содержать сигнальный домен CD3-дзета или внутрицитоплазматический домен бета или гамма-цепей FcεRI.
В конкретном варианте осуществления передающий сигнал домен многоцепочечного CAR по настоящему изобретению содержит костимулирующую сигнальную молекулу. Костимулирующая молекула представляет собой молекулу клеточной поверхности, отличную от антигенного рецептора или его лигандов, которая требуется для эффективного иммунного ответа. "Костимулирующий лиганд" обозначает молекулу на антиген-презентирующей клетке, которая специфически связывает распознанную костимулирующую молекулу на T-клетке, посредством этого обеспечивая сигнал, который, в дополнение к первичному сигналу, обеспечиваемому, например, связыванием комплекса TCR/CD3 с молекулой MHC, нагруженной пептидом, опосредует T-клеточный ответ, включая, но без ограничения, активацию пролиферации, дифференцировку и тому подобное. Костимулирующий лиганд может включать, но без ограничения, CD7, B7-1 (CD80), B7-2 (CD86), PD-L1, PD-L2, 4-1BBL, OX40L, индуцируемый костимулирующий лиганд (ICOS-L), молекулу межклеточной адгезии (ICAM), CD30L, CD40, CD70, CD83, HLA-G, MICA, M1CB, HVEM, рецептор лимфотоксина-бета, 3/TR6, ILT3, ILT4, агонист или антитело, которое связывает рецептор толл-лиганда, и лиганд, который специфически связывается с B7-H3. Костимулирующий лиганд также охватывает, в числе прочего, антитело, которое специфически связывается с костимулирующей молекулой, присутствующей на T-клетке, такой как, но без ограничения, CD27, CD28, 4-1BB, OX40, CD30, CD40, PD-1, ICOS, функционально-связанный антиген-1 лимфоцитов (LFA-1), CD2, CD7, LTGHT, NKG2C, B7-H3, лиганд, который специфически связывается с CD83.
"Костимулирующая молекула" обозначает распознанного партнера по связыванию на T-клетке, который специфически связывается с костимулирующим лигандом, тем самым опосредуя костимулирующий ответ клетки, такой как, но без ограничения, пролиферация. Костимулирующие молекулы включают, но без ограничения, молекулу MHC класса I, BTLA и рецептор толл-лиганда. Примеры костимулирующих молекул включают CD27, CD28, CD8, 4-1BB (CD137), OX40, CD30, CD40, PD-1, ICOS, функционально-связанный антиген-1 лимфоцитов (LFA-1), CD2, CD7, LIGHT, NKG2C, B7-H3 и лиганд, который специфически связывается с CD83, и тому подобное.
В другом конкретном варианте осуществления указанный передающий сигнал домен представляет собой связывающие мотивы TNFR-ассоциированного фактора 2 (TRAF2), внутрицитоплазматический хвост костимулирующего члена семейства TNFR. Цитоплазматический хвост костимулирующего члена семейства TNFR содержит связывающие мотивы TRAF2, состоящие из основного консервативного мотива (P/S/A)X(Q/E)E) или минорного мотива (PXQXXD), где X представляет собой любую аминокислоту. Белки TRAF рекрутируются во внутриклеточные хвосты многих TNFR в ответ на тримеризацию рецептора.
В предпочтительном варианте осуществления домен передачи сигнала многоцепочечного CAR по настоящему изобретению содержит часть костимулирующей сигнальной молекулы, выбранной из группы, состоящей из 4-1BB (GenBank: AAA53133) и CD28 (NP_006130.1). В частности, домен передачи сигнала многоцепочечного CAR по настоящему изобретению содержит аминокислотную последовательность, выбранную из SEQ ID NO:200 и SEQ ID NO:201.
Отличительные признаки соответствующих трансмембранных полипептидов включают способность к экспрессии на поверхности иммунной клетки, в частности, клетках лимфоцитов или клетках природных киллеров (NK), и к совместному действию для направления клеточного ответа иммунной клетки против заранее определенной клетки-мишени. Различные трансмембранные полипептиды многоцепочечного CAR по настоящему изобретению, содержащие внеклеточный лигандсвязывающий домен и/или передающий сигнал домен, совместно действуют для участия в передаче сигнала после связывания с целевым лигандом и индуцируют иммунный ответ. Трансмембранный домен может происходить или из природного, или из синтетического источника. Трансмембранный домен может происходить от любого мембраносвязанного или трансмембранного белка. В качестве неограничивающих примеров, трансмембранный полипептид может представлять собой субъединицу T-клеточного рецептора, такую как α, β, γ или δ, полипептид, образующий комплекс CD3, p55 (α-цепь), p75 (β-цепь) или γ-цепь IL2-рецептора, субъединичную цепь Fc-рецепторов, в частности, Fcγ рецептор III или CD-белки. Альтернативно, трансмембранный домен может быть синтетическим и может содержать преимущественно гидрофобные остатки, такие как лейцин и валин. В предпочтительном варианте осуществления трансмембранный полипептид, происходящий от цепей рецептора Fcε или их варианта, содержит, в частности, часть цепей α (SEQ ID NO:202), β (SEQ ID NO:203) и/или γ (SEQ ID NO:204) FcεRI или их вариант.
Термин "происходящий от" обозначает полипептид, имеющий аминокислотную последовательность, которая эквивалентна последовательности рецептора Fcε, которая включают одну или несколько аминокислотных модификаций последовательности рецептора Fcε. Такая аминокислотная модификация(и) может включать аминокислотную замену(ы), удаление(я), добавление(я) или комбинацию любых таких модификаций, и может изменять биологическую активность связывающего участка Fc по отношению к Fc-рецептору. С другой стороны, связывающие участки Fc, происходящие от конкретного Fc-рецептора, могут включать одну или несколько аминокислотных модификаций, которые по существу не изменяют биологическую активность связывающего участка Fc по отношению к Fc-рецептору. Аминокислотная модификация(и) такого типа, как правило, содержит замену(ы) консервативной аминокислоты.
В конкретном варианте осуществления многоцепочечный CAR содержит трансмембранный полипептид, происходящий от цепи FcεRI. В более специальном варианте осуществления FcεRI цепь представляет собой α-цепь FcεRI, в которой внеклеточный домен заменен внеклеточным лигандсвязывающим доменом, предпочтительно scFV, более предпочтительно scFv-4G7 (SEQ ID NO:195).
В более специальном варианте осуществления указанный многоцепочечный CAR может содержать часть альфа-цепи FcεRI и часть бета-цепи FcεRI или их вариант, так что указанные цепи FcεRI спонтанно димеризуются вместе с образованием димерного химерного антигенного рецептора. В другом варианте осуществления многоцепочечный химерный антиген может содержать часть альфа-цепи FcεRI и часть гамма-цепи FcεRI или их вариант, так что указанные цепи FcεRI спонтанно тримеризуются вместе с образованием тримерного химерного антигенного рецептора, и в другом варианте осуществления многоцепочечный химерный антигенный рецептор может содержать часть альфа-цепи FcεRI, часть бета-цепи FcεRI и часть гамма-цепи FcεRI или их варианты, так что указанные цепи FcεRI спонтанно тетрамеризуются вместе с образованием тетрамерного химерного антигенного рецептора.
Другими словами, многоцепочечный CAR, содержащий по меньшей мере два из следующих компонентов:
a) один полипептид, содержащий часть альфа-цепи FcεRI и внеклеточный лигандсвязывающий домен,
b) один полипептид, содержащий часть бета-цепи FcεRI, и/или
c) один полипептид, содержащий часть гамма-цепь FcεRI, в результате чего различные полипептиды мультимеризуются вместе спонтанно с образованием димерного, тримерного или тетрамерного CAR.
В предпочтительном варианте осуществления многоцепочечный CAR по настоящему изобретению содержит часть полипептида с аминокислотной последовательностью, выбранной из группы, состоящей из от SEQ ID NO:202 до SEQ ID NO:204.
Термин "часть", используемый в настоящем документе, относится к любому сокращенному варианту молекулы, который представляет собой более короткий пептид. Альтернативно, функциональные варианты аминокислотной последовательности полипептида могут быть получены посредством мутаций в ДНК, которая кодирует полипептид. Такие варианты или функциональные варианты включают, например, удаления, или вставки, или замены остатков в аминокислотной последовательности. Любая комбинация удаления, вставки и замены может также приводить к конечному конструкту, при условии, что конечный конструкт обладает желаемой активностью, в частности, проявляет специфическую клеточную иммунную активность против мишени. Функциональность многоцепочечного CAR по настоящему изобретению в клетке-хозяине может быть детектирована в анализе, подходящем для демонстрации сигнального потенциала указанного многоцепочечного CAR после связывания конкретной мишени. Такие анализы доступны специалисту в данной области. Например, данный анализ позволяет детектировать сигнальный путь, запускаемый при связывании мишени, как, например, анализ, включающий измерение повышения высвобождения ионов кальция, внутриклеточного фосфорилирования тирозина, оборота инозитолфосфата или производства интерлейкина (IL) 2, интерферона-γ, GM-CSF, IL-3, IL-4, вызываемых при этом.
В качестве неограничивающего примера, различные варианты многоцепочечного CAR проиллюстрированы на фигуре 4. В более предпочтительном варианте осуществления многоцепочечный CAR по настоящему изобретению содержит полипептид с аминокислотной последовательностью, выбранной из группы, состоящей из от SEQ ID NO:206 до SEQ ID NO:213. В предпочтительном варианте осуществления многоцепочечный CAR содержит полипептид с аминокислотной последовательностью, которая имеет идентичность последовательности по меньшей мере 70%, предпочтительно, по меньшей мере 80%, более предпочтительно, по меньшей мере 90%, 95%, 97% или 99% с аминокислотной последовательностью, выбранной из группы, состоящей из от SEQ ID NO:206 до SEQ ID NO:213.
Полинуклеотиды, векторы:
Настоящее изобретение также относится к полинуклеотидам, векторам, кодирующим вышеописанный многоцепочечный CAR в соответствии с настоящим изобретением. Настоящее изобретение предлагает полинуклеотиды, включая молекулы ДНК и РНК, которые кодируют трансмембранные полипептиды, раскрытые в настоящем документе, которые могут быть включены в многоцепочечный CAR. В частности, настоящее изобретение относится к полинуклеотиду, содержащему последовательность нуклеиновой кислоты, кодирующую по меньшей мере один трансмембранный полипептид, составляющий многоцепочечный CAR, как описано выше. Более конкретно, настоящее изобретение относится к полинуклеотиду, содержащему две или более последовательности нуклеиновой кислоты, кодирующие трансмембранные полипептиды, составляющие многоцепочечный CAR, как описано выше. В предпочтительном варианте осуществления настоящее изобретение относится к полинуклеотиду, выбранному из группы, состоящей из от SEQ ID NO:214 до SEQ ID NO:223. В предпочтительном варианте осуществления полинуклеотид имеет идентичность последовательности по меньшей мере 70%, предпочтительно, по меньшей мере 80%, более предпочтительно, по меньшей мере 90%, 95%, 97% или 99% с последовательностью нуклеиновой кислоты, выбранной из группы, состоящей из от SEQ ID NO:214 до SEQ ID NO:223.
Полинуклеотид может содержаться в кассете экспрессии или векторе экспрессии (например, плазмиде для введения в бактериальную клетку-хозяина, или вирусном векторе, таком как бакуловирусный вектор, для трансфекции клетки-хозяина из насекомых, или плазмиде или вирусном векторе, таком как лентивирусный, для трансфекции клетки-хозяина млекопитающих).
В конкретном варианте осуществления различные последовательности нуклеиновой кислоты могут быть включены в один полинуклеотид или вектор, который содержит последовательность нуклеиновой кислоты, кодирующую последовательность рибосомного прыжка, такую как последовательность, кодирующую пептид 2A. Пептиды 2A, которые были идентифицированы в подгруппе афтовирусы пикорнавирусов, вызывают рибосомный "прыжок" от одного кодона к следующему без образования пептидной связи между двумя аминокислотами, кодируемыми данными кодонами (см. Donnelly et al., J. of General Virology 82: 1013-1025 (2001); Donnelly et al., J. of Gen. Virology 78: 13-21 (1997); Doronina et al., Mol. And. Cell. Biology 28(13): 4227-4239 (2008); Atkins et al., RNA 13: 803-810 (2007)). Под "кодоном" понимаются три нуклеотида на мРНК (или на смысловой цепи молекулы ДНК), которые транслируются рибосомой в один аминокислотный остаток. Таким образом, два полипептида могут быть синтезированы из одной сплошной открытой рамки считывания в мРНК, когда полипептиды разделены последовательностью олигопептида 2A, которая находится внутри рамки. Такие механизмы рибосомного прыжка хорошо известны в данной области, и известно, что они используются некоторыми векторами для экспрессии нескольких белков, кодируемых одной матричной РНК. В качестве неограничивающего примера, в настоящем изобретении пептиды 2A применялись для экспрессии в клетке различных полипептидов многоцепочечного CAR. В более предпочтительном варианте осуществления настоящее изобретение относится к полинуклеотидам, выбранным из группы, состоящей из от SEQ ID NO:224 до SEQ ID NO:232.
Для направления трансмембранного полипептида, такого как FcεR, на секреторный путь клетки-хозяина, в полинуклеотидной последовательности или векторной последовательности предусмотрена последовательность секреторного сигнала (также известная как лидерная последовательность, препро-последовательность или пре-последовательность). Последовательность секреторного сигнала может представлять собой последовательность из FcεR или может происходить от другого секретируемого (например, t-PA) или впервые синтезированного белка. Последовательность секреторного сигнала функционально связана с последовательностью трансмембранной нуклеиновой кислоты, т.е. данные две последовательности соединены в правильной рамке считывания и расположены так, чтобы направлять вновь синтезированный полипептид на секреторный путь клетки-хозяина. Последовательности секреторного сигнала обычно расположены со стороны 5' от последовательности нуклеиновой кислоты, кодирующей представляющий интерес полипептид, хотя некоторые последовательности секреторного сигнала могут быть расположены в другом месте представляющей интерес последовательности нуклеиновой кислоты (см., например, Welch et al., патент США № 5037743; Holland et al., патент США № 5143830). В предпочтительном варианте осуществления сигнальный пептид содержит остатки с 1 по 25 альфа-цепи FcεRI (NP_001992,1) и имеет аминокислотную последовательность SEQ ID NO:205.
Специалистам в данной области будет понятно, что, с учетом вырожденности генетического кода, возможна значительная изменчивость последовательности среди данных полинуклеотидных молекул. Предпочтительно, последовательности нуклеиновой кислоты настоящего изобретения являются кодон-оптимизированными для экспрессии в клетках млекопитающих, предпочтительно, для экспрессии в клетках человека. Кодонная оптимизация обозначает замену в представляющей интерес последовательности кодонов, которые, как правило, редки в высоко экспрессированных генах данного вида, на кодоны, которые, как правило, часты в высоко экспрессированных генах таких видов, причем такие кодоны кодируют те же аминокислоты, что и кодоны, которые подвергают замене.
В предпочтительном варианте осуществления полинуклеотид в соответствии с настоящим изобретением содержит последовательность нуклеиновой кислоты, выбранную из группы, состоящей из от SEQ ID NO:214 до SEQ ID NO:223. Настоящее изобретение относится к полинуклеотидам, содержащим последовательность нуклеиновой кислоты, которая имеет идентичность последовательности по меньшей мере 70%, предпочтительно, по меньшей мере 80%, более предпочтительно, по меньшей мере 90%, 95%, 97% или 99% с последовательностью нуклеиновой кислоты, выбранной из группы, состоящей из от SEQ ID NO:214 до SEQ ID NO:222.
Способы получения иммунной клетки способами генной инженерии:
В охватываемом конкретном варианте осуществления настоящее изобретение относится к способу получения иммунных клеток для иммунотерапии, включающему введение в указанные иммунные клетки полипептидов, составляющих указанный многоцепочечный CAR, и размножение указанных клеток. В конкретном варианте осуществления настоящее изобретение относится к способу получения иммунной клетки способами генной инженерии, включающему наличие клетки и экспрессию на поверхности указанной клетки по меньшей мере одного многоцепочечного CAR, как описано выше. В конкретном варианте осуществления данный способ включает трансформацию клетки с помощью по меньшей мере одного полинуклеотида, кодирующего полипептиды, составляющие по меньшей мере один многоцепочечный CAR, как описано выше, и экспрессию указанных полинуклеотидов в указанной клетке.
В другом варианте осуществления настоящее изобретение относится к способу получения клеток для иммунотерапии, включающему введение в указанные клетки различных полипептидов, составляющих указанный многоцепочечный CAR, и размножение указанных клеток. В предпочтительном варианте осуществления указанные полинуклеотиды включают в лентивирусные векторы, поскольку они стабильно экспрессируются в клетках.
В другом варианте осуществления указанный способ дополнительно включает стадию генетической модификации указанных клеток посредством инактивации по меньшей мере одного гена, экспрессирующего один компонент TCR, мишень для иммуносупрессивного средства, ген HLA и/или ген иммунной контрольной точки, такой как PDCD1 или CTLA-4. В предпочтительном варианте осуществления указанный ген выбирают из группы, состоящей из TCR-альфа, TCR-бета, CD52, GR, PD1 и CTLA-4. В предпочтительном варианте осуществления указанный способ дополнительно включает введение в указанные T-клетки редкощепящей эндонуклеазы, способной к селективной инактивации посредством расщепления ДНК указанных генов. В более предпочтительном варианте осуществления указанная редкощепящая эндонуклеаза представляет собой TALE-нуклеазу. Предпочтительными TALE-нуклеазами в соответствии с настоящим изобретением являются распознающие и расщепляющие последовательность-мишень, выбранную из группы, состоящей из: SEQ ID NO:1-6 (GR), SEQ ID NO:37, 57-60 (TCR-альфа), SEQ ID NO:38 или 39 (TCR-бета) и SEQ ID NO:40, от SEQ ID NO:61 до SEQ ID NO:65 (CD52), от SEQ ID NO:74 до SEQ ID NO:78 (PD1 и CTLA-4).
Предполагается, что благодаря инактивации гена представляющий интерес ген не экспрессируется в форме функционального белка. В конкретном варианте осуществления генетическая модификация данного способа основана на экспрессии в клетках, предлагаемых для получения способами генной инженерии, одной редкощепящей эндонуклеазы, так что указанная редкощепящая эндонуклеаза специфически катализирует расщепление в одном целевом гене, посредством этого инактивируя указанный целевой ген. Разрывы нити нуклеиновой кислоты, вызываемые редкощепящей эндонуклеазой, обычно исправляются посредством отличающихся механизмов гомологичной рекомбинации или негомологичного соединения концов (NHEJ). Однако, NHEJ является несовершенным процессом репарации, который часто приводит к изменениям в последовательности ДНК в сайте расщепления. Механизмы включают повторное соединение того, что остается от двух концов ДНК, посредством прямого повторного лигирования (Critchlow and Jackson, 1998) или посредством так называемого опосредованного микрогомологией соединения концов (Ma, Kim et al., 2003). Репарация посредством негомологичного соединения концов (NHEJ) часто приводит к небольшим вставкам или делециям и может быть использована для создания специфических генных нокаутов. Указанная модификация может представлять собой замену, делецию или присоединение по меньшей мере одного нуклеотида. Клетки, в которых происходит событие индуцированного расщеплением мутагенеза, т.е. событие мутагенеза, следующее за событием NHEJ, могут быть идентифицированы и/или отобраны посредством хорошо известного в данной области способа.
В другом варианте осуществления дополнительный каталитический домен может быть дополнительно введен в клетку с указанной редкощепящей эндонуклеазой для усиления мутагенеза, для того чтобы усилить способность инактивировать целевые гены, описанные в настоящем описании. В частности, указанный дополнительный каталитический домен представляет собой обрабатывающий концы ДНК фермент. Неограничивающие примеры обрабатывающих концы ДНК ферментов включают 5-3' экзонуклеазы, 3-5' экзонуклеазы, 5-3' щелочные экзонуклеазы, 5' флэп-эндонуклеазы, геликазы, фосфатазы, гидролазы и независимые от шаблона ДНК-полимеразы. Неограничивающие примеры такого каталитического домена содержат белковый домен или каталитически активное производное белкового домена, выбранного из группы, состоящей из hExol (EXO1_HUMAN), Exol дрожжей (EXO1_YEAST), Exol E.coli, TREX2 человека, TREX1 мыши, TREX1 человека, бычьего TREX1, TREX1 крысы, TdT (концевой дезоксинуклеотидтрансферазы) DNA2 человека, DNA2 дрожжей (DNA2_YEAST). В предпочтительном варианте осуществления указанный дополнительный каталитический домен обладает 3'-5'-экзонуклеазной активностью, и в более предпочтительном варианте осуществления указанный дополнительный каталитический домен представляет собой каталитический домен TREX, более предпочтительно TREX2 (WO2012/058458). В другом предпочтительном варианте осуществления указанный каталитический домен кодируется одноцепочечным TREX-полипептидом. Указанный дополнительный каталитический домен может быть слит с нуклеазным слитым белком или химерным белком в соответствии с настоящим изобретением, необязательно с помощью пептидного линкера.
Известно, что эндонуклеолитические разрывы увеличивают скорость гомологичной рекомбинации. Таким образом, в другом варианте осуществления стадия генетической модификации данного способа дополнительно включает стадию введения в клетки экзогенной нуклеиновой кислоты, содержащей по меньшей мере последовательность, гомологичную части целевой последовательности нуклеиновой кислоты, так что происходит гомологичная рекомбинация между целевой последовательностью нуклеиновой кислоты и экзогенной нуклеиновой кислотой. В конкретных вариантах осуществления указанная экзогенная нуклеиновая кислота содержит первую и вторую части, которые, соответственно, гомологичны участку 5' и 3' целевой последовательности нуклеиновой кислоты. Указанная экзогенная нуклеиновая кислота в данных вариантах осуществления также содержит третью часть, расположенную между первой и второй частями, которая не содержит гомологии с участками 5' и 3' целевой последовательности нуклеиновой кислоты. После расщепления целевой последовательности нуклеиновой кислоты вызывается событие гомологичной рекомбинации между целевой последовательностью нуклеиновой кислоты и экзогенной нуклеиновой кислотой. Предпочтительно, в указанной донорной матрице используют гомологичные последовательности из по меньшей мере 50 п.о., предпочтительно, больше, чем 100 п.о., и, более предпочтительно, больше, чем 200 п.о. Следовательно, экзогенная нуклеиновая кислота составляет, предпочтительно, от 200 п.о. до 6000 п.о., более предпочтительно от 1000 п.о. до 2000 п.о. Действительно, общие гомологии нуклеиновой кислоты расположены в участках, фланкирующих в прямом и в обратном направлении сайт разрыва, и подлежащая введению последовательность нуклеиновой кислоты должна быть расположена между двумя плечами.
В частности, указанная экзогенная нуклеиновая кислота последовательно содержит первый участок гомологии с последовательностями, расположенными в обратном направлении от указанного расщепления, последовательность для инактивации одного целевого гена, выбранного из группы, состоящей из TCR-альфа, TCR-бета, CD52, GR, генов иммунных контрольных точек, и второй участок гомологии с последовательностями, расположенными в прямом направлении от расщепления. Указанная стадия введения полинуклеотида может выполняться одновременно, до или после введения или экспрессии указанной редкощепящей эндонуклеазы. В зависимости от расположения целевой последовательности нуклеиновой кислоты, в которой произошло событие разрыва, такая экзогенная нуклеиновая кислота может использоваться для нокаута гена, например, когда экзогенная нуклеиновая кислота расположена в пределах открытой рамки считывания указанного гена, или для введения новых последовательностей или генов, представляющих интерес. Вставки последовательностей с использованием такой экзогенной нуклеиновой кислоты можно использовать для модификации целевого существующего гена посредством коррекции или замены указанного гена (обмен аллелями в качестве неограничивающего примера) или регулировать экспрессию целевого гена повышающим или понижающим образом (обмен промоторами в качестве неограничивающего примера) посредством коррекции или замены указанного целевого гена. В предпочтительном варианте осуществления инактивация генов из группы, состоящей из TCR-альфа, TCR-бета, CD52, GR, генов иммунных контрольных точек, может быть осуществлена в точной геномной локализации, на которую воздействует специфическая TALE-нуклеаза, причем указанная специфическая TALE-нуклеаза катализирует расщепление, и причем указанная экзогенная нуклеиновая кислота последовательно содержит по меньшей мере участок гомологии и последовательность для инактивации одного целевого гена, выбранного из группы, состоящей из TCR-альфа, TCR-бета, CD52, GR, генов иммунных контрольных точек, которые интегрированы посредством гомологичной рекомбинации. В другом варианте осуществления несколько генов могут быть, последовательно или в одно и то же время, инактивированы с использованием нескольких TALE-нуклеаз, соответственно и специфически направленных на один определенный ген, и нескольких специфических полинуклеотидов для специфической инактивации генов.
Посредством стадии дополнительной геномной модификации может быть предусмотрена также инактивация другого гена, выбранного из группы, состоящей из TCR-альфа, TCR-бета, CD52, GR, генов иммунных контрольных точек. Как упомянуто выше, указанная стадия дополнительной геномной модификации может представлять собой стадию инактивации, включающую:
(a) введение в указанные клетки по меньшей мере одной редкощепящей эндонуклеазы, так что указанная редкощепящая эндонуклеаза специфически катализирует расщепление в одной целевой последовательности генома указанной клетки.
(b) необязательно, введение в указанные клетки экзогенной нуклеиновой кислоты, последовательно содержащей первый участок гомологии с последовательностями в обратном направлении от указанного расщепления, последовательность, подлежащую вставке в геном указанной клетки, и второй участок гомологии с последовательностями в прямом направлении от указанного расщепления,
причем указанная вводимая экзогенная нуклеиновая кислота инактивирует ген и интегрирует по меньшей мере одну экзогенную полинуклеотидную последовательность, кодирующую по меньшей мере один представляющий интерес рекомбинантный белок. В другом варианте осуществления указанная экзогенная полинуклеотидная последовательность интегрирована в ген, выбранный из группы, состоящей из TCR-альфа, TCR-бета, CD52, GR, генов иммунных контрольных точек.
- Резистентные к иммуносупрессивным средствам T-клетки:
В конкретном аспекте одна из стадий генетической модификации клеток может представлять собой способ, включающий:
(a) модификацию T-клеток посредством инактивации по меньшей мере одного гена, экспрессирующего мишень для иммуносупрессивного средства, и
(b) размножение указанных клеток, необязательно в присутствии указанного иммуносупрессивного средства.
Иммуносупрессивное средство представляет собой средство, которое подавляет иммунную функцию посредством одного из нескольких механизмов действия. Другими словами, иммуносупрессивное средство представляет собой роль, исполняемую соединением, которая проявляется в способности уменьшать степень и/или ненасытность иммунного ответа. В качестве неограничивающего примера иммуносупрессивное средство может представлять собой ингибитор кальциневрина, мишень рапамицина, блокатор α-цепи интерлейкина-2, ингибитор инозинмонофосфатдегидрогеназы, ингибитор редуктазы дигидрофолиевой кислоты, кортикостероид или иммуносупрессивный антиметаболит. Классические цитотоксические иммуносупрессанты действуют посредством ингибирования синтеза ДНК. Другие могут действовать посредством активации T-клеток или посредством ингибирования активации хелперных клеток. Способ в соответствии с настоящим изобретением позволяет придавать T-клеткам резистентность к иммуносупрессивным средствам для иммунотерапии посредством инактивации мишени иммуносупрессивного средства в T-клетках. В качестве неограничивающих примеров, мишени для иммуносупрессивного средства могут представлять собой рецептор для иммуносупрессивного средства, такой как: CD52, глюкокортикоидный рецептор (GR), ген члена семейства FKBP и ген члена семейства циклофилинов.
Предполагается, что благодаря инактивации гена представляющий интерес ген не экспрессируется в форме функционального белка. В конкретном варианте осуществления генетическая модификация данного способа основана на экспрессии в клетках, предлагаемых для получения способами генной инженерии, одной редкощепящей эндонуклеазы, так что указанная редкощепящая эндонуклеаза специфически катализирует расщепление в одном целевом гене, посредством этого инактивируя указанный целевой ген. В конкретном варианте осуществления указанный способ получения клеток способами генной инженерии включает по меньшей мере одну из следующих стадий:
(a) Наличие T-клетки, предпочтительно, из клеточной культуры или из образца крови;
(b) Выбор гена в указанной T-клетке, экспрессирующего мишень для иммуносупрессивного средства;
(c) Введение в указанную T-клетку редкощепящей эндонуклеазы, способной селективно инактивировать посредством расщепления ДНК, предпочтительно, посредством двухнитевого разрыва, указанный ген, кодирующий мишень для указанного иммуносупрессивного средства, и
(d) Размножение указанных клеток, необязательно в присутствии указанного иммуносупрессивное средства.
В более предпочтительном варианте осуществления указанный способ включает:
(a) Наличие T-клетки, предпочтительно, из клеточной культуры или из образца крови;
(b) Выбор гена в указанной T-клетке, экспрессирующего мишень для иммуносупрессивного средства;
(c) Трансформацию указанной T-клетки с помощью нуклеиновой кислоты, кодирующей редкощепящую эндонуклеазу, способную селективно инактивировать посредством расщепления ДНК, предпочтительно, посредством двухнитевого разрыва, указанный ген, кодирующий мишень для указанного иммуносупрессивного средства, и
(d) Экспрессию указанных редкощепящих эндонуклеаз в указанных T-клетках;
(e) Размножение указанных клеток, необязательно в присутствии указанного иммуносупрессивного средства.
В конкретном варианте осуществления указанная редкощепящая эндонуклеаза специфически воздействует на один ген, выбранный из группы, состоящей из CD52, GR. В другом варианте осуществления указанный ген стадии (b), определенный для иммуносупрессивной обработки, представляет собой CD52, и иммуносупрессивная обработка стадии (d) или (e) содержит гуманизированное антитело, направленное на антиген CD52.
В другом варианте осуществления указанный ген стадии (b), определенный для иммуносупрессивной обработки, представляет собой глюкокортикоидный рецептор (GR), и иммуносупрессивная обработка стадии d) или (e) содержит кортикостероид, такой как дексаметазон.
В другом варианте осуществления указанный целевой ген стадии (b), определенный для иммуносупрессивной обработки, представляет собой ген члена семейства FKBP или его варианта, и иммуносупрессивная обработка стадии (d) или (e) содержит FK506, также известный как такролимус или фуджимицин. В другом варианте осуществления указанный ген члена семейства FKBP представляет собой FKBP12 или его вариант.
В другом варианте осуществления указанный ген стадии (b), определенный для иммуносупрессивной обработки, представляет собой ген члена семейства циклофилинов или его варианта, и иммуносупрессивная обработка стадии (d) или (e) содержит циклоспорин.
В другом варианте осуществления указанная редкощепящая эндонуклеаза может представлять собой мегануклеазу, цинковопальцевую нуклеазу или TALE-нуклеазу. В предпочтительном варианте осуществления указанная редкощепящая эндонуклеаза представляет собой TALE-нуклеазу. Предпочтительными TALE-нуклеазами в соответствии с настоящим изобретением являются распознающие и расщепляющие последовательность-мишень, выбранную из группы, состоящей из:
- SEQ ID NO:1-6 (GR), и
- SEQ ID NO:40, 61-65 (CD52).
Упомянутые TALE-нуклеазы, предпочтительно, содержат полипептидную последовательность, выбранную из от SEQ ID NO:7 до SEQ ID NO:18 и от SEQ ID NO:47 до SEQ ID NO:48, для того чтобы расщеплять соответствующие последовательности-мишени SEQ ID NO:1-6 и SEQ ID NO:40.
- Высокоактивные T-клетки для иммунотерапии
В конкретном аспекте одна конкретная стадия генетической модификации клетки может представлять собой способ, включающий:
(a) модификацию T-клеток посредством инактивации по меньшей мере одного гена иммунных контрольных точек; и
(b) размножение указанных клеток.
Опосредованный T-клетками иммунитет включает несколько последовательных стадий, включающих клональную селекцию антиген-специфических клеток, их активацию и пролиферацию во вторичную лимфоидную ткань, их направленную миграцию к участкам антигена и воспаления, выполнение прямой эффекторной функции и предоставление помощи (посредством цитокинов и мембранных лигандов) для множества эффекторных иммунных клеток. Каждая из данных стадий регулируется посредством уравновешивания стимулирующего и ингибирующего сигнала, что точно регулирует ответ. Средним специалистам в данной области будет понятно, что термин "иммунные контрольные точки" обозначает группу молекул, экспрессируемых T-клетками. Данные молекулы эффективно служат в качестве "тормозов" для понижающей модуляции или ингибирования иммунного ответа. Молекулы иммунных контрольных точек включают, но без ограничения, белок программируемой смерти 1 (PD-1, также известный как PDCD1 или CD279, номер доступа: M_005018), антиген 4 цитотоксических T-лимфоцитов (CTLA-4, также известный как CD152, номер доступа GenBank AF414120.1), LAG3 (также известный как CD223, номер доступа: NM_002286,5), Tim3 (также известный как HAVCR2, номер доступа GenBank: JX049979.1), BTLA (также известный как CD272, номер доступа: NM_181780.3), BY55 (также известный как CD160, номер доступа GenBank: CR541888.1), TIGIT (также известный как VSTM3, номер доступа: NM_173799), B7H5 (также известный как C10orf54, гомолог мышиного гена vista, номер доступа: NM_022153.1), LAIR1 (также известный как CD305, номер доступа GenBank: CR542051.1), SIGLEC10 (номер доступа GeneBank: AY358337.1), 2B4 (также известный как CD244, номер доступа: NM_001166664.1), которые непосредственно ингибируют иммунные клетки. Например, CTLA-4 представляет собой белок клеточной поверхности, экспрессируемый на некоторых CD4 и CD8 T-клетках; при контакте с его лигандами (B7-1 и B7-2) на антиген-презентирующих клетках ингибируется активация и эффекторная функция T-клеток. Таким образом, настоящее изобретение относится к способу получения T-клеток, особенно для иммунотерапии, включающему генетическую модификацию T-клеток посредством инактивации по меньшей мере одного белка, задействованного в иммунной контрольной точке, в частности, PD1 и/или CTLA-4.
В конкретном варианте осуществления указанный способ получения клеток способами генной инженерии включает по меньшей мере одну из следующих стадий:
(a) наличие T-клетки, предпочтительно, из клеточной культуры или из образца крови;
(b) ведение в указанную T-клетку редкощепящей эндонуклеазы, способной селективно инактивировать посредством расщепления ДНК, предпочтительно, посредством двухнитевого разрыва, один ген, кодирующий белок иммунных контрольных точек,
(c) размножение указанных клеток.
В более предпочтительном варианте осуществления указанный способ включает:
(a) наличие T-клетки, предпочтительно из клеточной культуры или из образца крови;
(b) трансформацию указанной T-клетки с помощью нуклеиновой кислоты, кодирующей редкощепящую эндонуклеазу, способную селективно инактивировать посредством расщепления ДНК, предпочтительно посредством двухнитевого разрыва, ген, кодирующий белок иммунных контрольных точек;
(c) экспрессию указанных редкощепящих эндонуклеаз в указанных T-клетках;
(d) размножение указанных клеток.
В конкретном варианте осуществления указанная редкощепящая эндонуклеаза специфически воздействует на один ген, выбранный из группы, состоящей из: PD1, CTLA-4, LAG3, Tim3, BTLA, BY55, TIGIT, B7H5, LAIR1, SIGLEC10, 2B4, TCR-альфа и TCR-бета. В другом варианте осуществления указанная редкощепящая эндонуклеаза может представлять собой мегануклеазу, цинковопальцевую нуклеазу или TALE-нуклеазу. В предпочтительном варианте осуществления указанная редкощепящая эндонуклеаза представлять собой TALE-нуклеазу. Под TALE-нуклеазой понимается слитый белок, состоящий из ДНК-связывающего домена, происходящего от подобного активатору транскрипции эффектора (TALE), и одного нуклеазного каталитического домена для расщепления целевой последовательности нуклеиновой кислоты. (Boch, Scholze et al., 2009; Moscou and Bogdanove, 2009; Christian, Cermak et al., 2010; Cermak, Doyle et al., 2011; Geissler, Scholze et al., 2011; Huang, Xiao et al., 2011; Li, Huang et al., 2011; Mahfouz, Li et al., 2011; Miller, Tan et al., 2011; Morbitzer, Romer et al., 2011; Mussolino, Morbitzer et al., 2011; Sander, Cade et al., 2011; Tesson, Usal et al., 2011; Weber, Gruetzner et al., 2011; Zhang, Cong et al., 2011; Deng, Yan et al., 2012; Li, Piatek et al., 2012; Mahfouz, Li et al., 2012; Mak, Bradley et al., 2012).
В настоящем изобретении были разработаны новые TALE-нуклеазы для точного нацеливания на соответствующие гены для стратегий адоптивной иммунотерапии. Предпочтительными TALE-нуклеазами в соответствии с настоящим изобретением являются распознающие и расщепляющие последовательность-мишень, выбранную из группы, состоящей из: SEQ ID NO:77 и SEQ ID NO:78 (PD1), от SEQ ID NO:74 до SEQ ID NO:76 (CTLA-4). Настоящее изобретение также относится к полипептидам TALE-нуклеаз, которые содержат аминокислотную последовательность, выбранную из группы, состоящей из от SEQ ID NO:79 до SEQ ID NO:88.
Настоящее изобретение также относится к полипептидам, содержащим аминокислотную последовательность, которая имеет идентичность последовательности по меньшей мере 70%, предпочтительно, по меньшей мере 80%, более предпочтительно, по меньшей мере 90%, 95%, 97% или 99% с аминокислотной последовательностью, выбранной из группы, состоящей из от SEQ ID NO:79 до SEQ ID NO:88. Также в объем настоящего изобретения включены полинуклеотиды, векторы, кодирующие вышеописанные редкощепящие эндонуклеазы в соответствии с настоящим изобретением. Данный способ может быть связан с любым из различных способов, описанных в настоящем раскрытии.
- Неаллореактивные T-клетки:
В другом варианте осуществления настоящее изобретение может особенно подходить для аллогенной иммунотерапии. В таком случае одна из стадий генетической модификации клетки может представлять собой способ, включающий:
(a) модификацию T-клеток посредством инактивации по меньшей мере одного гена, кодирующего компонент T-клеточного рецептора (TCR)
(b) размножение указанных клеток.
В конкретном варианте осуществления генетическая модификация способа основана на экспрессии в клетках, предлагаемых для получения способами генной инженерии, одной редкощепящей эндонуклеазы, так что указанная редкощепящая эндонуклеаза специфически катализирует расщепление в одном целевом гене, посредством этого инактивируя указанный целевой ген. В конкретном варианте осуществления указанный способ получения клеток способами генной инженерии включает по меньшей мере одну из следующих стадий:
(a) наличие T-клетки, предпочтительно, из клеточной культуры или из образца крови;
Введение в указанную T-клетку редкощепящей эндонуклеазы, способной селективно инактивировать посредством расщепления ДНК, предпочтительно посредством двухнитевого разрыва, по меньшей мере один ген, кодирующий компонент T-клеточного рецептора (TCR).
(b) Размножение указанных клеток.
В более предпочтительном варианте осуществления указанный способ включает:
(a) наличие T-клетки, предпочтительно, из клеточной культуры или из образца крови;
(b) Трансформацию указанной T-клетки с помощью нуклеиновой кислоты, кодирующей редкощепящую эндонуклеазу, способную селективно инактивировать посредством расщепления ДНК, предпочтительно, посредством двухнитевого разрыва, по меньшей мере один ген, кодирующий компонент T-клеточного рецептора (TCR);
(c) Экспрессию указанных редкощепящих эндонуклеаз в указанных T-клетках;
(d) Сортировку трансформированных T-клеток, которые не экспрессируют TCR на своей клеточной поверхности;
(e) Размножение указанных клеток.
В другом варианте осуществления указанная редкощепящая эндонуклеаза может представлять собой мегануклеазу, цинковопальцевую нуклеазу или TALE-нуклеазу. В предпочтительном варианте осуществления указанная редкощепящая эндонуклеаза представляет собой TALE-нуклеазу. Предпочтительными TALE-нуклеазами в соответствии с настоящим изобретением являются распознающие и расщепляющие последовательность-мишень, выбранную из группы, состоящей из:
- SEQ ID NO:37, 57-60 (TCR-альфа),
- SEQ ID NO:38 или 39 (TCR-бета).
Упомянутые TALE-нуклеазы, предпочтительно, содержат полипептидную последовательность, выбранную из от SEQ ID NO:41 до SEQ ID NO:48, для того чтобы расщеплять соответствующие последовательности-мишени SEQ ID NO:37-39.
- PreT-альфа
В другом аспекте еще одна стадия генетической модификации клеток может представлять собой способ размножения TCR-альфа-дефицитных T-клеток, включающий введение в указанную T-клетку pT-альфа (также называемой пре-TCRα) или ее функционального варианта и размножение указанных клеток, необязательно посредством стимуляции комплекса CD3. В предпочтительном варианте осуществления данный способ включает:
a) Трансформацию указанных клеток с помощью нуклеиновой кислоты, кодирующей по меньшей мере фрагмент pT-альфа, для поддержания поверхностной экспрессии CD3
b) Экспрессию указанной pT-альфа в указанных клетках
c) Размножение указанных клеток, необязательно посредством стимуляции комплекса CD3.
Настоящее изобретение также относится к способу получения T-клеток для иммунотерапии, содержащему стадии способа размножения T-клеток.
В конкретном варианте осуществления полинуклеотидная последовательность pT-альфа может быть введена случайным образом или же посредством гомологичной рекомбинации, в частности, вставка может быть связана с инактивацией гена TCR-альфа.
В соответствии с настоящим изобретением используют различные функциональные варианты pT-альфа. "Функциональный вариант" пептида обозначает молекулу, по существу аналогичную или всему пептиду, или его фрагменту. "Фрагмент" pT-альфа или ее функционального варианта настоящего изобретения относится к любому сокращенному варианту молекулы, то есть более короткому пептиду. Предпочтительные pT-альфа или функциональный варианты могут представлять собой полноразмерную pT-альфа или C-концевой усеченный вариант pT-альфа. В C-концевой усеченной pT-альфа с C-концевого конца отсутствуют один или несколько остатков. В качестве неограничивающих примеров, в C-концевом усеченном варианте pT-альфа отсутствуют 18, 48, 62, 78, 92, 110 или 114 остатков с C-конца белка (от SEQ ID NO:107 до SEQ ID NO:114). Кроме того, варианты аминокислотной последовательности пептида могут быть получены посредством мутаций в ДНК, которая кодирует пептид. Такие функциональные варианты включают, например, удаления, или вставки, или замены остатков в аминокислотной последовательности. Любая комбинация удаления, вставки и замены может также приводить к конечному конструкту, при условии, что конечный конструкт обладает желаемой активностью, в частности, восстанавливает функциональный комплекс CD3. В предпочтительном варианте осуществления по меньшей мере одну мутацию вводят в различные варианты pT-альфа, как описано выше, для воздействия на димеризацию. В качестве неограничивающего примера, мутировавший остаток может представлять собой по меньшей мере W46R, D22A, K24A, R102A или R117A белка pT-альфа человека или выровненные с помощью метода CLUSTALW положения на члене семейства pT-альфа или гомологе. Предпочтительно, pT-альфа или ее вариант, как описано выше, содержат мутировавший остаток W46R (SEQ ID NO:123) или мутировавшие остатки D22A, K24A, R102A и R117A (SEQ ID NO:124). В конкретном варианте осуществления указанные pT-альфа или варианты также слиты с передающим сигнал доменом, таким как CD28, OX40, ICOS, CD27, CD137 (4-1BB) и CD8, в качестве неограничивающих примеров (от SEQ ID NO:115 до SEQ ID NO:120). Внеклеточный домен pT-альфа или вариантов, как описано выше, может быть слит с фрагментом белка TCR-альфа, в частности, с трансмембранным и внутриклеточным доменом TCR-альфа (SEQ ID NO:122). Варианты pT-альфа также могут быть слиты с внутриклеточным доменом TCR-альфа (SEQ ID NO:121).
В другом варианте осуществления указанные варианты pT-альфа слиты с внеклеточным лигандсвязывающим доменом, и, более предпочтительно, pT-альфа или ее функциональный вариант слиты с одноцепочечным фрагментом антитела (scFV), содержащим легкий (VL) и тяжелый (VH) вариабельный фрагмент специфического к антигену мишени моноклонального антитела, соединенные гибким линкером. В качестве неограничивающего примера, аминокислотную последовательность pT-альфа или ее функционального варианта выбирают из группы, состоящей из от SEQ ID NO:107 до SEQ ID NO:124.
Поскольку некоторая изменчивость может возникать из геномных данных, из которых эти полипептиды происходят, а также принимая во внимание возможность замены некоторых из аминокислот, присутствующих в этих полипептидах, без значительной потери активности (функциональные варианты), настоящее изобретение охватывает полипептидные варианты вышеуказанных полипептидов, которые имеют идентичность по меньшей мере 70%, предпочтительно, по меньшей мере 80%, более предпочтительно, по меньшей мере 90% и, еще более предпочтительно, по меньшей мере 95% с последовательностями, предлагаемыми в данной патентной заявке.
Настоящее изобретение, таким образом, обращается к полипептидам, содержащим полипептидную последовательность, которая имеет идентичность по меньшей мере 70%, предпочтительно, по меньшей мере 80%, более предпочтительно, по меньшей мере 90%, 95% 97% или 99% с последовательностями с аминокислотной последовательностью, выбранной из группы, состоящей из от SEQ ID NO:107 до SEQ ID NO:124.
Под TCR-альфа-дефицитной T-клеткой понимается выделенная T-клетка, в которой отсутствует экспрессия функциональной цепи TCR-альфа. Это может быть достигнуто с помощью различных средств, в качестве неограничивающих примеров, посредством получения способом генной инженерии T-клетки таким образом, что она не экспрессирует никакой функциональной TCR-альфа на своей клеточной поверхности, или посредством получения T-клетки таким образом, что она производит очень мало функциональной цепи TCR-альфа на своей поверхности, или посредством получения T-клетки для экспрессии мутировавшей или усеченной формы цепи TCR-альфа.
TCR-альфа-дефицитные клетки больше не могут распространяться посредством комплекса CD3. Таким образом, для преодоления данной проблемы и для предоставления возможности пролиферации TCR-альфа-дефицитным клеткам в указанные клетки вводят pT-альфа или ее функциональный вариант, тем самым восстанавливая функциональный комплекс CD3. В предпочтительном варианте осуществления данный способ дополнительно включает введение в указанные T-клетки редкощепящих эндонуклеаз, способных селективно инактивировать посредством расщепления ДНК один ген, кодирующий один компонент T-клеточного рецептора (TCR). В конкретном варианте осуществления указанная редкощепящая эндонуклеаза представляет собой TALE-нуклеазу. В качестве неограничивающих примеров, TALE-нуклеаза направлена против одной из последовательностей-мишеней гена TCR-альфа, выбранных из группы, состоящей из SEQ ID NO:37 и SEQ ID NO:57-60. Предпочтительно, TALE-нуклеазы выбирают из группы, состоящей из SEQ ID NO:41 и SEQ ID NO:42.
Также в настоящее изобретение включены полипептиды, кодирующие pT-альфа, в частности, функциональные варианты, описанные выше. В предпочтительном варианте осуществления настоящее изобретение относится к pT-альфа или ее функциональному варианту, слитому с передающим сигнал доменом, таким как CD28, OX40, ICOS, CD137 и CD8. Более конкретно, настоящее изобретение относится к функциональному варианту pT-альфа, содержащему аминокислотную последовательность, выбранную из группы, состоящей из от SEQ ID NO:107 до SEQ ID NO:124. Также в настоящее изобретение включены полинуклеотиды, векторы, кодирующие pT-альфа или ее функциональные варианты, описанные выше.
Объем настоящего изобретения также включает выделенные клетки или клеточные линии, допускающие получение посредством указанного способа. В частности, указанные выделенные клетки или клеточные линии получают посредством введения в указанные клетки pT-альфа или ее функционального варианта для поддержания поверхностной экспрессии CD3. В предпочтительном варианте осуществления указанная выделенная клетка или клеточная линия дополнительно генетически модифицированы посредством инактивации гена TCR-альфа. Данный ген, предпочтительно, инактивируется посредством по меньшей мере одной редкощепящей эндонуклеазы. В предпочтительном варианте осуществления указанная редкощепящая эндонуклеаза представляет собой TALE-нуклеазу.
Объем настоящего изобретения также включает выделенные клетки или клеточные линии, допускающие получение посредством указанного способа получения клеток способами генной инженерии, в частности, T-клеток, в которых инактивирован по меньшей мере один ген, выбранный из группы, состоящей из TCR-альфа, TCR-бета, CD52, GR, генов иммунных контрольных точек.
- Биспецифические антитела
В соответствии со следующим вариантом осуществления сконструированные T-клетки, полученные различными способами, как описано выше, могут быть дополнительно подвергнуты воздействию биспецифических антител. Упомянутые T-клетки могут быть подвергнуты воздействию биспецифических антител ex vivo до введения пациенту или in vivo после введения пациенту. Упомянутые биспецифические антитела содержат два вариабельных участка с различными антигенными свойствами, которые позволяют приводить сконструированные клетки в непосредственную близость к антигену мишени. В качестве неограничивающего примера, упомянутое биспецифическое антитело направлено против опухолевого маркера и лимфоцитарного антигена, такого как CD3, и имеет возможность перенаправлять и активировать любые циркулирующие T-клетки против опухолей.
Способы доставки
Различные способы, описанные выше, включают введение многоцепочечного CAR, pT-альфа или ее функциональных вариантов, редкощепящей эндонуклеазы, TALE-нуклеазы, CAR, необязательно с обрабатывающим концы ДНК ферментом или экзогенной нуклеиновой кислотой, в клетку.
В качестве неограничивающего примера, указанные многоцепочечный CAR, pT-альфа или ее функциональный вариант, редкощепящие эндонуклеазы, TALE-нуклеазы или CAR, необязательно с обрабатывающим концы ДНК ферментом или экзогенной нуклеиновой кислотой, могут быть введены в виде трансгенов, кодируемых одним или различными плазмидными векторами. Различные трансгены могут быть включены в один вектор, который содержит последовательность нуклеиновой кислоты, кодирующую последовательность рибосомного прыжка, такую как последовательность, кодирующую пептид 2A. Пептиды 2A, которые были идентифицированы в подгруппе афтовирусы пикорнавирусов, вызывают рибосомный "прыжок" от одного кодона к следующему без образования пептидной связи между двумя аминокислотами, кодируемыми данными кодонами (см. Donnelly et al., J. of General Virology 82: 1013-1025 (2001); Donnelly et al., J. of Gen. Virology 78: 13-21 (1997); Doronina et al., Mol. And. Cell. Biology 28(13): 4227-4239 (2008); Atkins et al., RNA 13: 803-810 (2007)). Под "кодоном" понимаются три нуклеотида на мРНК (или на смысловой цепи молекулы ДНК), которые транслируются рибосомой в один аминокислотный остаток. Таким образом, два полипептида могут быть синтезированы из одной сплошной открытой рамки считывания в мРНК, когда полипептиды разделены последовательностью олигопептида 2A, которая находится внутри рамки. Такие механизмы рибосомного прыжка хорошо известны в данной области, и известно, что они используются некоторыми векторами для экспрессии нескольких белков, кодируемых одной матричной РНК. В качестве неограничивающего примера, в настоящем изобретении пептиды 2A применялись для экспрессии в клетке редкощепящей эндонуклеазы и обрабатывающего концы ДНК фермента или различных полипептидов многоцепочечного CAR.
Указанный плазмидный вектор может также содержать маркер селекции, который предназначен для идентификации и/или селекции клеток, которые получили указанный вектор.
Полипептиды могут быть синтезированы in situ в клетке в результате введения полинуклеотидов, кодирующих указанные полипептиды, в клетку. Альтернативно, указанные полипептиды могут быть получены вне клетки и затем введены в нее. Способы введения полинуклеотидного конструкта в клетки животных известны в данной области и включают, в качестве неограничивающих примеров, способы стабильной трансформации, когда полинуклеотидный конструкт интегрируют в геном клетки, способы временной трансформации, когда полинуклеотидный конструкт не интегрируют в геном клетки, и способы, опосредуемые вирусами. Упомянутые полинуклеотиды могут быть введены в клетку с помощью, например, рекомбинантных вирусных векторов (например, ретровирусов, аденовирусов), липосом и тому подобного. Например, способы временной трансформации включают, например, микроинъекцию, электропорацию или бомбардировку частицами. Упомянутые полинуклеотиды могут быть включены в векторы, более конкретно в плазмиды или вирус, для экспрессии в клетках.
- Электропорация
В конкретном варианте осуществления настоящего изобретения полинуклеотиды, кодирующие полипептиды в соответствии с настоящим изобретением, могут представлять собой мРНК, которую вводят непосредственно в клетки, например, посредством электропорации. Изобретатели определили оптимальные условия для электропорации мРНК в T-клетку.
Изобретатель использовал метод CytoPulse, который позволяет посредством использования импульсных электрических полей временно делать проницаемыми живые клетки для доставки материала в данные клетки. Метод, основанный на использовании форм сигналов электропорации PulseAgile (собственность Cellectis) обеспечивает точный контроль продолжительности, интенсивности импульсов, а также интервала между импульсами (патент США 6010613 и международная заявка PCT WO2004083379). Все эти параметры могут быть изменены, для того чтобы достигать наилучших условий для высокой эффективности трансфекции с минимальной смертностью. В основном, первые сильные импульсы электрического поля делают возможным образование пор, тогда как последующие слабые импульсы электрического поля делают возможным движение полинуклеотида в клетку. В одном из аспектов настоящего изобретения изобретатель описывает стадии, которые ведут к достижению эффективности трансфекции мРНК в T-клетки >95%, и использование протокола электропорации для временной экспрессии различных типов белков в T-клетках. В частности, настоящее изобретение относится к способу трансформации T-клетки, включающему приведение указанной T-клетки в контакт с РНК и прикладывание к T-клетке адаптивной импульсной последовательности, состоящей из:
(a) одного электрического импульса с напряжением в диапазоне от 2250 до 3000 В на сантиметр, шириной импульса, равной 0,1 мс, и интервалом между импульсами, равным от 0,2 до 10 мс, между электрическими импульсами стадий (a) и (b);
(b) одного электрического импульса с напряжением в диапазоне от 2250 до 3000 В, с шириной импульса, равной 100 мс, и интервалом между импульсами, равным 100 мс, между электрическим импульсом стадии (b) и первым электрическим импульсом стадии (c); и
(c) 4 электрических импульсов с напряжением, равным 325 В, с шириной импульса, равной 0,2 мс, и интервалом между импульсами, равным 2 мс, между всеми 4 электрическими импульсами.
В конкретном варианте осуществления способ трансформации T-клетки, включающий приведение указанной T-клетки в контакт с РНК и прикладывание к T-клетке адаптивной импульсной последовательности, состоящей из:
(a) одного электрического импульса с напряжением, равным 2250, 2300, 2350, 2400, 2450, 2500, 2550, 2400, 2450, 2500, 2600, 2700, 2800, 2900 или 3000 В на сантиметр, шириной импульса, равной 0,1 мс, и интервалом между импульсами, равным 0,2, 0,5, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 мс, между электрическими импульсами стадий (a) и (b);
(b) одного электрического импульса с напряжением в диапазоне от 2250, равным 2250, 2300, 2350, 2400, 2450, 2500, 2550, 2400, 2450, 2500, 2600, 2700, 2800, 2900 или 3000 В, с шириной импульса, равной 100 мс, и интервалом между импульсами, равным 100 мс, между электрическим импульсом стадии (b) и первым электрическим импульсом стадии (c); и
(c) 4 электрических импульсов с напряжением, равным 325 В, с шириной импульса, равной 0,2 мс, и интервалом между импульсами, равным 2 мс, между всеми 4 электрическими импульсами.
Любые значения, включенные в диапазон значений, описанный выше, раскрыты в настоящей заявке. Среда для электропорации может представлять собой любую подходящую среду, известную в данной области. Предпочтительно, среда для электропорации имеет проводимость в диапазоне, охватывающем от 0,01 до 1,0 миллисименс.
В конкретных вариантах осуществления, в качестве неограничивающих примеров, указанная РНК кодирует редкощепящую эндонуклеазу, один мономер редкощепящей эндонуклеазы, такой как половина TALE-нуклеазы, химерный антигенный рецептор, по меньшей мере один компонент многоцепочечного химерного антигенного рецептора, pT-альфа или ее функциональный вариант, экзогенную нуклеиновую кислоту, один дополнительный каталитический домен.
Модифицированные T-клетки
Настоящее изобретение также относится к выделенным клеткам или клеточным линиям, допускающим получение посредством указанного способа получения клеток способами генной инженерии. В частности, указанная выделенная клетка содержит по меньшей мере один многоцепочечный CAR, как описано выше. В другом варианте осуществления указанная выделенная клетка содержит популяцию многоцепочечных CAR, причем все они содержат различные внеклеточные лигандсвязывающие домены. В частности, указанная выделенная клетка содержит экзогенные полинуклеотидные последовательности, кодирующие полипептиды, составляющие по меньшей мере один многоцепочечный CAR. Упомянутые клетки могут также дополнительно содержать по меньшей мере один инактивированный ген, выбранный из группы, состоящей из CD52, GR, TCR-альфа, TCR-бета, гена HLA, генов иммунных контрольных точек, таких как PD1 и CTLA-4, или может экспрессировать трансген pT-альфа.
Объем настоящего изобретения также включает выделенную иммунную клетку, предпочтительно, T-клетку, полученную в соответствии с любым из способов, описанных выше. Указанная иммунная клетка обозначает клетку гемопоэтического происхождения, функционально участвующую в инициировании и/или осуществлении врожденного и/или адаптивного иммунного ответа. Указанная иммунная клетка в соответствии с настоящим изобретением может происходить от стволовой клетки. Стволовые клетки могут представлять собой взрослые стволовые клетки, эмбриональные стволовые клетки, более конкретно нечеловеческие стволовые клетки, стволовые клетки пуповинной крови, клетки-предшественники, стволовые клетки костного мозга, индуцированные плюрипотентные стволовые клетки, тотипотентные стволовые клетки или гемопоэтические стволовые клетки. Типичными клетками человека являются CD34+ клетки. Указанная выделенная клетка может также представлять собой дендритную клетку, дендритную клетку-киллер, тучную клетку, NK-клетку, B-клетку или T-клетку, выбранную из группы, состоящей из воспалительных T-лимфоцитов, цитотоксических T-лимфоцитов, регуляторных T-лимфоцитов или хелперных T-лимфоцитов. В другом варианте осуществления указанная клетка может происходить из группы, состоящей из CD4+ T-лимфоцитов и CD8+ T-лимфоцитов. Перед размножением и генетической модификацией клеток настоящего изобретения источник клеток может быть получен от индивида посредством множества неограничивающих способов. Клетки могут быть получены из ряда неограничивающих источников, включая мононуклеарные клетки периферической крови, костный мозг, ткань лимфатических узлов, пуповинную кровь, ткань вилочковой железы, ткань из участка инфекции, асциты, плевральный выпот, ткань селезенки и опухоли. В определенных вариантах осуществления настоящего изобретения можно использовать любой ряд T-клеточных линий, доступных и известных специалистам в данной области. В другом варианте осуществления указанная клетка может быть получена от здорового донора, от пациента с диагнозом рака или от пациента с диагнозом инфекции. В другом варианте осуществления указанная клетка является частью смешанной популяции клеток, которые представляют различные фенотипические характеристики. Объем настоящего изобретения также охватывает клеточную линию, получаемую из трансформированной T-клетки в соответствии со способом, описанным выше. Модифицированные клетки, резистентные к иммуносупрессивной обработке и допускающие получение указанным выше способом, включены в объем настоящего изобретения.
В другом варианте осуществления указанная выделенная клетка в соответствии с настоящим изобретением содержит один инактивированный ген, выбранный из группы, состоящей из CD52, GR, PD1, CTLA-4, LAG3, Tim3, BTLA, BY55, TIGIT, B7H5, LAIR1, SIGLEC10, 2B4, HLA, TCR-альфа и TCR-бета, и/или экспрессирует CAR, многоцепочечный CAR и/или трансген pT-альфа. В другом конкретном варианте осуществления указанная выделенная клетка содержит полинуклеотиды, кодирующие указанные полипептиды, составляющие многоцепочечный CAR.
В другом варианте осуществления указанная выделенная клетка в соответствии с настоящим изобретением содержит два инактивированных гена, выбранных из группы, состоящей из CD52 и GR, CD52 и TCR-альфа, CDR52 и TCR-бета, GR и TCR-альфа, GR и TCR-бета, TCR-альфа и TCR-бета, PD1 и TCR-альфа, PD1 и TCR-бета, CTLA-4 и TCR-альфа, CTLA-4 и TCR-бета, LAG3 и TCR-альфа, LAG3 и TCR-бета, Tim3 и TCR-альфа, Tim3 и TCR-бета, BTLA и TCR-альфа, BTLA и TCR-бета, BY55 и TCR-альфа, BY55 и TCR-бета, TIGIT и TCR-альфа, TIGIT и TCR-бета, B7H5 и TCR-альфа, B7H5 и TCR-бета, LAIR1 и TCR-альфа, LAIR1 и TCR-бета, SIGLEC10 и TCR-альфа, SIGLEC10 и TCR-бета, 2B4 и TCR-альфа, 2B4 и TCR-бета, и/или экспрессирует CAR, многоцепочечный CAR и/или трансген pT-альфа.
В другом варианте осуществления TCR делают не функциональным в клетках в соответствии с настоящим изобретением посредством инактивации гена TCR-альфа и/или гена(ов) TCR-бета. Вышеуказанные стратегии используют, в частности, для избежания GvHD. В конкретном аспекте настоящего изобретения предлагается способ получения модифицированных клеток, происходящих от индивидуума, причем указанные клетки могут пролиферировать независимо от сигнального пути главного комплекса гистосовместимости. Указанный способ включает следующие стадии:
(a) Получение клеток от указанного индивидуума;
(b) Генетическую модификацию указанных клеток ex vivo посредством инактивации генов TCR-альфа или TCR-бета;
(c) Культивирование генетически модифицированных T-клеток in vitro в соответствующих условиях для амплификации указанных клеток.
Модифицированные клетки, которые могут пролиферировать независимо от сигнального пути главного комплекса гистосовместимости, допускающие получение посредством данного способа, включены в объем настоящего изобретения. Упомянутые модифицированные клетки можно использовать в конкретном аспекте настоящего изобретения для лечения пациентов от отторжения хозяин против трансплантата (HvG) и заболевания трансплантат против хозяина (GvHD); следовательно, в объем настоящего изобретения попадает способ лечения пациентов от отторжения хозяин против трансплантата (HvG) и заболевания трансплантат против хозяина (GvHD), включающий лечение указанного пациента посредством введения указанному пациенту эффективного количества модифицированных клеток, содержащих инактивированные гены TCR-альфа и/или TCR-бета.
Активация и размножение T-клеток
Или до, или после генетической модификации T-клеток T-клетки могут быть активированы и размножены, как правило, с помощью способов, описанных, например, в патентах США 6352694; 6534055; 6905680; 6692964; 5858358; 6887466; 6905681; 7144575; 7067318; 7172869; 7232566; 7175843; 5883223; 6905874; 6797514; 6867041; и публикации патентной заявки США № 20060121005. T-клетки могут быть размножены in vitro или in vivo.
Обычно T-клетки настоящего изобретения размножают посредством контакта со средством, которое имитирует комплекс CD3 TCR, и костимулирующей молекулой на поверхности T-клеток для создания сигнала активации для T-клетки.
Например, для создания сигнала активации для T-клетки можно использовать химические вещества, такие как ионофор кальция A23187, форбол-12-миристат-13-ацетат (PMA) или митогенные лектины вроде фитогемагглютинина (PHA).
В качестве неограничивающих примеров, популяции Т-клеток могут быть стимулированы in vitro, как, например, посредством контакта с анти-CD3 антителом или его антигенсвязывающим фрагментом или анти-CD2 антителом, иммобилизованным на поверхности, или посредством контакта с активатором протеинкиназы C (например, бриостатином) в сочетании с ионофором кальция. Для костимуляции вспомогательной молекулы на поверхности T-клеток используют лиганд, который связывает вспомогательную молекулу. Например, популяция T-клеток может быть приведена в контакт с анти-CD3 антителом и анти-CD28 антителом в условиях, подходящих для стимуляции пролиферации T-клеток. Для стимуляции пролиферации или CD4+ T-клеток, или CD8+ T-клеток анти-CD3 антитело и анти-CD28 антитело. Например, средства, обеспечивающие каждый сигнал, могут находиться в растворе или быть связаны с поверхностью. Как легко могут понять средние специалисты в данной области, отношение частиц к клеткам может зависеть от размера частиц по отношению к клетке-мишени. В других вариантах осуществления настоящего изобретения клетки, такие как T-клетки, объединяют с покрытыми средством гранулами, после этого гранулы и клетки разделяют, и затем клетки культивируют. В альтернативном варианте осуществления перед культивированием покрытые средством гранулы и клетки не разделяют, но культивируют вместе. Условия, подходящие для культивирования T-клеток, включают подходящие среды (например, минимальные питательные среды, или среды RPMI-1640, или X-vivo 5 (Lonza)), который могут содержать факторы, необходимые для пролиферации и жизнеспособности, включая сыворотку (например, фетальную бычью или человеческую сыворотку), интерлейкин-2 (IL-2), инсулин, IFN-g, 1L-4, 1L-7, GM-CSF, -10, -2, 1L-15, TGFp и TNF- или любые другие добавки для роста клеток, известные специалисту. Другие добавки для роста клеток включают, но без ограничения, поверхностно-активное вещество, плазманат и восстанавливающие средства, такие как N-ацетилцистеин и 2-меркаптоэтанол. Среды могут включать RPMI 1640, A1M-V, DMEM, MEM, a-MEM, F-12, X-Vivo 1 и X-Vivo 20, Optimizer, с добавленными аминокислотами, пируватом натрия и витаминами, или свободные от сыворотки, или дополненные соответствующим количеством сыворотки (или плазмы), или определенным набором гормонов, и/или количеством цитокина(ов), достаточным для роста и размножения T-клеток. Антибиотики, например, пенициллин и стрептомицин, включают только в экспериментальные культуры, не в культуры клеток, которые предназначены для введения индивиду. Клетки-мишени выдерживают в условиях, необходимых для поддержания роста, например, подходящих температуре (например, 37°C) и атмосфере (например, воздух плюс 5% CO2). T-клетки, которые подвергались стимуляции в течение различного времени, могут проявлять различные характеристики.
В другом конкретном варианте осуществления указанные клетки могут быть размножены посредством совместного культивирования с тканью или клетками. Упомянутые клетки могут также быть размножены in vivo, например, в крови индивида после введения указанной клетки индивиду.
Терапевтические применения
В другом варианте осуществления выделенная клетка, полученная посредством различных способов, или клеточная линия, происходящая от указанной выделенной клетки, как описано выше, может быть использована в качестве лекарственного средства. В другом варианте осуществления упомянутое лекарственное средство может быть использовано для лечения злокачественных опухолей или инфекции у пациента. В другом варианте осуществления указанная выделенная клетка в соответствии с настоящим изобретением или клеточная линия, происходящая от указанной выделенной клетки, могут быть использованы в изготовлении лекарственного средства для лечения рака, вирусной инфекции или аутоиммунного заболевания у пациента.
В другом аспекте настоящее изобретение основано на способах лечения пациентов, причем указанный способ включает по меньшей мере одну из следующих стадий:
(a) наличие иммунной-клетки, получаемой посредством любого из способов, описанных выше;
(b) введение указанных трансформированных иммунных клеток указанному пациенту.
В одном из вариантов осуществления указанные T-клетки по настоящему изобретению могут подвергаться устойчивому размножению T-клеток in vivo и могут существовать в течение длительного периода времени.
Упомянутое лечение может быть улучшающим, радикальным или профилактическим. Оно может быть или частью аутологичной иммунотерапии, или частью аллогенного иммунотерапевтического лечения. Под аутологичным понимается, что клетки, клеточная линия или популяция клеток, используемые для лечения пациентов, происходят от указанного пациента или от совместимого по лейкоцитарному антигену человека (HLA) донора. Под аллогенным понимается, что клетки или популяция клеток, используемые для лечения пациентов, происходят не от указанного пациента, но от донора.
Настоящее изобретение особенно подходит для аллогенной иммунотерапии, поскольку она дает возможность трансформации T-клеток, как правило полученных от доноров, в неаллореактивные клетки. Это может быть сделано в рамках стандартных протоколов и воспроизведено столько раз, сколько необходимо. Итоговые модифицированные T-клетки могут быть объединены и введены одному или нескольким пациентам, будучи доступны в виде "готового" терапевтического продукта.
Клетки, которые могут быть использованы с раскрытыми способами, описаны в предыдущем разделе. Упомянутое лечение может быть использовано для лечения пациентов с диагнозом рака, вирусной инфекции, аутоиммунных нарушений или заболевания трансплантат против хозяина (GvHD). Злокачественные опухоли, которые можно лечить, включают опухоли, которые не васкуляризированы или еще по существу не васкуляризированы, а также васкуляризированные опухоли. Злокачественные опухоли могут включать несолидные опухоли (такие как гематологические опухоли, например, лейкемии и лимфомы) или могут включать солидные опухоли. Виды рака, подлежащие лечению с помощью многоцепочечных CAR по настоящему изобретению, включают, но без ограничения, карциному, бластому и саркому и определенную лейкемию или лимфоидные злокачественные опухоли, доброкачественные и злокачественные опухоли и злокачественные опухоли, например, саркомы, карциномы и меланомы. Также включены опухоли/раки взрослых и опухоли/раки детей.
Может иметь место лечение в комбинации с одной или несколькими терапиями против рака, выбранными из группы из терапии антителами, химиотерапии, терапии цитокинами, терапии дендритными клетками, генной терапии, гормональной терапии, терапии лазерным светом и лучевой терапии.
В соответствии с предпочтительным вариантом осуществления настоящего изобретения упомянутое лечение может вводиться пациентам, проходящим иммуносупрессивное лечение. Действительно, настоящее изобретение, предпочтительно, основано на клетках или популяции клеток, которые были сделаны резистентными к по меньшей мере одному иммуносупрессивному средству вследствие инактивации гена, кодирующего рецептор для такого иммуносупрессивного средства. В данном аспекте иммуносупрессивное лечение должно способствовать селекции и размножению T-клеток в соответствии с настоящим изобретением в пациенте.
Введение клеток или популяции клеток в соответствии с настоящим изобретением может быть осуществлено любым удобным способом, в том числе посредством аэрозольной ингаляции, инъекции, приема внутрь, переливания, имплантации или трансплантации. Композиции, описанные в настоящем документе, могут быть введены пациенту подкожно, внутрикожно, внутрь опухоли, интранодально, интрамедуллярно, внутримышечно, посредством внутривенной или внутрилимфатической инъекции или внутрибрюшинно. В одном из вариантов осуществления клеточные композиции настоящего изобретения, предпочтительно, вводят посредством внутривенной инъекции.
Введение клеток или популяции клеток может состоять из введения 104-109 клеток на кг веса тела, предпочтительно, от 105 до 106 клеток/кг веса тела, включая все целые значения числа клеток в пределах данных диапазонов. Клетки или популяция клеток могут быть введены в одной или нескольких дозах. В другом варианте осуществления упомянутое эффективное количество клеток вводят в виде однократной дозы. В другом варианте осуществления упомянутое эффективное количество клеток вводят в виде больше чем одной дозы в течение периода времени. График введения остается на усмотрение управляющего врача и зависит от клинического состояния пациента. Клетки или популяция клеток могут быть получены из любого источника, такого как банк крови или донор. При том, что индивидуальные потребности различаются, определение оптимальных интервалов эффективных количеств данного типа клеток для конкретного заболевания или условий лежит в пределах компетентности специалиста в данной области. Эффективное количество означает количество, которое обеспечивает терапевтический или профилактический эффект. Применяемая дозировка будет зависеть от возраста, состояния здоровья и веса реципиента, вида сопутствующего лечения, если таковое имеется, частоты лечения и природы желаемого эффекта.
В другом варианте осуществления упомянутое эффективное количество клеток или композицию, содержащую данные клетки, вводят парентерально. Упомянутое введение может представлять собой внутривенное введение. Упомянутое введение может быть осуществлено непосредственно путем инъекции в опухоль.
В определенных вариантах осуществления настоящего изобретения клетки вводят пациенту в сочетании (например, до, одновременно или после) с любым количеством соответствующих модальностей лечения, включая, но без ограничения, лечение с помощью средств, таких как противовирусная терапия, цидофовир и интерлейкин-2, цитарабин (также известный как ARA-C), или лечение с помощью натализумаба для пациентов с MS, или лечение с помощью эфализумаба для пациентов с псориазом, или другие методы лечения для пациентов с PML. В других вариантах осуществления T-клетки настоящего изобретения могут использоваться в комбинации с химиотерапией, излучением, иммуносупрессивными средствами, такими как циклоспорин, азатиоприн, метотрексат, микофенолат и FK506, антителами или другими иммунодеструктивными средствами, такими как CAM PATH, анти-CD3 антитела или другие терапии антителами, цитоксином, флударибином, циклоспорином, FK506, рапамицином, микофеноловой кислотой, стероидами, FR901228, цитокинами и облучением. Данные препараты или ингибируют кальций-зависимую фосфатазу кальциневрин (циклоспорин и FK506), или ингибируют киназу p70S6, что важно для индуцируемой факторами роста сигнализации (рапамицин) (Liu et al., Cell 66:807-815, 1 1; Henderson et al., Immun. 73:316-321, 1991; Bierer et al., Citrr. Opin. mm n. 5:763-773, 93). В другом варианте осуществления клеточные композиции настоящего изобретения вводят пациенту в сочетании (например, до, одновременно или после) с трансплантацией костного мозга, T-клеточной аблативной терапией, используя или химиотерапевтические средства, такие как флударабин, наружная дистанционная лучевая терапия (XRT), циклофосфамид, или антитела, такие как OKT3 или CAMPATH. В другом варианте осуществления клеточные композиции настоящего изобретения вводят после B-клеточной аблативной терапии, такой как средства, которые реагируют с CD20, например, ритуксан. Например, в одном из вариантов осуществления индивиды могут проходить стандартное лечение с помощью высоких доз химиотерапии с последующей трансплантации стволовых клеток периферической крови. В определенных вариантах осуществления после трансплантата индивиды получали введение размноженных иммунных клеток настоящего изобретения. В дополнительном варианте осуществления размноженные клетки вводят до или после хирургии. Упомянутые модифицированные клетки, полученные посредством любого из способов, описанных в настоящем документе, могут использоваться в конкретном аспекте настоящего изобретения для лечения пациентов от отторжения хозяин против трансплантата (HvG) и заболевания трансплантат против хозяина (GvHD); следовательно, в объем настоящего изобретения попадает способ лечения пациентов от отторжения хозяин против трансплантата (HvG) и заболевания трансплантат против хозяина (GvHD), включающий лечение указанного пациента посредством введения указанному пациенту эффективного количества модифицированных клеток, содержащих инактивированные гены TCR-альфа и/или TCR-бета.
Пример способа получения аллогенных клеток человека способами генной инженерии для иммунотерапии
Для лучшего понимания настоящего изобретения один пример способа получения аллогенных клеток человека для иммунотерапии проиллюстрирован на фигуре 5. Данный способ включает комбинацию одной или нескольких из следующих стадий:
1. Наличие T-клеток из клеточной культуры или из образца крови от одного отдельного пациента или из банка крови и активация указанных T-клеток с помощью гранул активатора анти-CD3/C28. Гранулы обеспечивают как первичный, так и костимулирующий сигналы, которые требуются для активации и размножения T-клеток.
2. a) Трансдукция указанных клеток трансгеном pT-альфа или ее функционального варианта для поддержания поверхностной экспрессии CD3 и предоставления клетке возможности размножения посредством стимуляции комплекса CD3. Ожидается, что разрушение TCR элиминирует комплекс TCR и удалит аллореактивность (GvHD), но оно может изменить размножение аллогенных клеток из-за потери сигнального компонента CD3. Ожидается, что трансдуцированные клетки будут экспрессировать цепь pT-альфа или ее функциональный вариант. Данная цепь pT-альфа образует пару с цепью TCR-бета и сигнальными компонентами CD3 для образования комплекса пре-TCR и, посредством этого, восстановления функционального комплекса CD3 и поддержания активации или стимуляции инактивированных по TCR-альфа клеток. Трансдукция T-клеток с помощью лентивирусного вектора pT-альфа может быть осуществлена до или после инактивации TCR-альфа.
b) Трансдукция указанных клеток многоцепочечными CAR позволяет перенаправлять T-клетки против антигенов, экспрессированных на поверхности клеток-мишеней от различных злокачественных опухолей, включая лимфомы и солидные опухоли. Для улучшения функции костимулирующего домена изобретатели разработали многоцепочечный CAR, происходящий от FcεRI, как описано выше. Трансдукция может быть осуществлена до или после инактивации TCR-альфа и других генов, таких как гены CD52.
3. Инженерия не аллореактивных и иммуносупрессивно резистентных T-клеток:
a) Существует возможность инактивировать TCR-альфа в указанных клетках, для того чтобы элиминировать TCR с поверхности клетки и предотвратить распознавание посредством TCR аллогенных клеток ткани хозяина как чужой и, таким образом, избежать GvHD.
b) Также существует возможность инактивировать один ген, кодирующий мишень для иммуносупрессивного средства, для того чтобы сделать указанные клетки резистентными к иммуносупрессивному лечению для предотвращения отторжения трансплантата без воздействия на трансплантированные T-клетки. В данном примере мишенью иммуносупрессивных средств является CD52, и иммуносупрессивное средство представляет собой гуманизированное моноклональное анти-CD52 антитело.
Изобретателями было показано, что использование TALE-нуклеазы, делающее возможным более высокий уровень событий DSB в T-клетках, оказалось особенно выгодным для достижения вышеуказанной двойной инактивации в T-клетках. Предпочтительно, гены TCR-альфа и CD52 инактивируют посредством электропорирования T-клеток мРНК, кодирующей TALE-нуклеазу, нацеленную на указанные гены. Изобретателями было обнаружено, что использование мРНК, приводившее к высоким степеням трансформации, оказалось менее вредным для T-клеток и, таким образом, оказалось существенным в процессе инженерии T-клеток. Затем инактивированные T-клетки сортируют с помощью магнитных гранул. Например, T-клетки, экспрессирующие CD52, удаляли посредством фиксации на твердой поверхности, и инактивированные клетки не подвергали стрессу прохождения через колонку. Такой мягкий способ повышает концентрацию правильно сконструированных T-клеток.
4. Размножение in vitro сконструированных T-клеток перед введением пациенту или in vivo после введения пациенту посредством стимуляции комплекса CD3. Перед стадией введения пациенты могут подвергаться иммуносупрессивному лечению, как например, CAMPATH1-H, гуманизированным моноклональным анти-CD52 антителом.
5. Необязательно, указанные клетки подвергали воздействию биспецифических антител ex vivo до введения пациенту или in vivo после введения пациенту для приведения сконструированных клеток в непосредственную близость к антигену мишени.
Другие определения
- Если не указано иное, "по меньшей мере один" используется взаимозаменяемо и обозначает один или больше, чем один. - Аминокислотные остатки в полипептидной последовательности обозначены в настоящем документе в соответствии с однобуквенным кодом, в котором, например, Q обозначает Gln или остаток глутамина, R обозначает Arg или остаток аргинина, и D обозначает Asp или остаток аспарагиновой кислоты.
- Аминокислотная замена означает замещение одного аминокислотного остатка другим, например, замещение остатка аргинина остатком глутамина в пептидной последовательности представляет собой аминокислотную замену.
- Нуклеотиды обозначаются следующим образом: однобуквенный код используется для обозначения основания нуклеозида: a представляет собой аденин, t представляет собой тимин, c представляет собой цитозин, и g представляет собой гуанин. Для вырожденных нуклеотидов r обозначает g или a (пуриновые нуклеотиды), k обозначает g или t, s обозначает g или c, w обозначает a или t, m обозначает a или c, y обозначает t или c (пиримидиновые нуклеотиды), d обозначает g, a или t, v обозначает g, a или c, b обозначает g, t или c, h обозначает a, t или c, и n обозначает g, a, t или c.
- Как используется в настоящем документе, "нуклеиновая кислота" или "полинуклеотиды" обозначает нуклеотиды и/или полинуклеотиды, такие как дезоксирибонуклеиновую кислоту (ДНК) или рибонуклеиновую кислоту (РНК), олигонуклеотиды, фрагменты, создаваемые полимеразной цепной реакцией (ПЦР), и фрагменты, создаваемые любым из лигирования, расщепления, действия эндонуклеазы и действия экзонуклеазы. Молекулы нуклеиновой кислоты могут быть составлены из мономеров, которые представляют собой природные нуклеотиды (как, например, ДНК и РНК), или аналоги природных нуклеотидов (например, энантиомерные формы природных нуклеотидов), или комбинацию тех и других. Модифицированные нуклеотиды могут иметь изменения во фрагментах сахаров и/или во фрагментах пиримидиновых или пуриновых оснований. Модификации сахаров включают, например, замену одной или нескольких гидроксильных групп галогенами, алкильными группами, аминами и азидогруппами, или сахара могут быть функционализированы как простые эфиры или сложные эфиры. Кроме того, весь фрагмент сахара может быть заменен стерически и электронно аналогичными структурами, такими как азасахара и карбоциклические аналоги сахаров. Примеры модификаций во фрагменте основания включают алкилированные пурины и пиримидины, ацилированные пурины или пиримидины или другие хорошо известные гетероциклические заместители. Мономеры нуклеиновой кислоты могут быть связаны фосфодиэфирными связями или аналогами таких связей. Нуклеиновые кислоты могут быть или одноцепочечными или двухцепочечными.
- Под "полинуклеотидом, последовательно содержащим первый участок гомологии с последовательностями в обратном направлении от указанного двухнитевого разрыва, последовательность, подлежащую вставке в геном указанной клетки, и второй участок гомологии с последовательностями в прямом направлении от от указанного двухнитевого разрыва" понимается конструкт или матрица ДНК, содержащие первую и вторую часть, которые гомологичны участкам 5' и 3' ДНК-мишени in situ. Конструкт ДНК также содержит третью часть, расположенную между первой и второй частью, которая имеет некоторую гомологию с соответствующей последовательностью ДНК in situ или, альтернативно, не имеет гомологии с участками 5' и 3' ДНК-мишени in situ. После расщепления ДНК-мишени возникает событие гомологичной рекомбинации между геномом, содержащим целевой ген, содержащийся в представляющем интерес локусе, и матрицей, причем геномная последовательность, содержащая ДНК-мишень, замещается третьей частью матрицы и вариабельной частью первой и второй частей указанной матрицы.
- Под "ДНК-мишенью", "последовательностью ДНК-мишени", "целевой последовательностью ДНК", "целевой последовательностью нуклеиновой кислоты", "последовательностью-мишенью" или "сайтом процессинга" понимается полинуклеотидная последовательность, на которую может быть направлена редкощепящая эндонуклеаза в соответствии с настоящим изобретением, и которая может быть ей обработана. Эти термины относятся к определенному местоположению ДНК, предпочтительно, местоположению генома в клетке, но также к части генетического материала, которая может существовать независимо от основной части генетического материала, как например, плазмиды, эписомы, вирус, транспозоны, или в органеллах, таких как митохондрии в качестве неограничивающего примера. В качестве неограничивающих примеров мишеней TALE-нуклеаз, целевые геномные последовательности обычно состоят из двух последовательностей длиной 17 п.о. (называемых полумишенями), разделенных спейсером 15 п.о. Каждая полумишень распознается повторами TALE-нуклеаз, перечисленных в таблицах 1, 5, 6 и 10 в качестве неограничивающих примеров, кодируемых в плазмидах, под контролем промотора EF1-альфа или промотора T7. Целевая последовательность нуклеиновой кислоты определяется последовательностью от 5' до 3' одной нити указанной мишени, как показано в таблицах 1, 5, 6 и 10.
- Под химерным антигенным рецептором (CAR) понимаются молекулы, которые объединяют связывающий домен против компонента, присутствующего на клетке-мишени, например, основанную на антителах специфичность к желаемому антигену (например, опухолевому антигену), с активирующим T-клеточный рецептор внутриклеточным доменом для создания химерного белка, который проявляет специфическую клеточную иммунную активность против мишени. Как правило, CAR состоит из внеклеточного одноцепочечного антитела (scFvFc), слитого с внутриклеточным сигнальным доменом дзета-цепи комплекса T-клеточного антигенного рецептора (scFvFc:ζ), и обладает способностью при экспрессии в T-клетках перенаправлять распознавание антигена на основании специфичности моноклонального антитела. Одним примером CAR, используемого в настоящем изобретении, является CAR, направленный против антигена CD19, и он может содержать в качестве неограничивающего примера аминокислотную последовательность: SEQ ID NO:73.
- Под "вектором доставки" или "векторами доставки" понимается любой вектор доставки, который можно использовать в настоящем изобретении для того, чтобы ввести в контакт с клеткой (т.е. "контактирование") или доставить внутрь клеток или внутриклеточных компартментов (т.е. "введение") средства/химические вещества и молекулы (белки или нуклеиновые кислоты), нужные в настоящем изобретении. Он включает, но без ограничения, липосомные векторы доставки, вирусные векторы доставки, лекарственные векторы доставки, химические носители, полимерные носители, липоплексы, полиплексы, дендримеры, микропузырьки (ультразвуковые контрастные средства), наночастицы, эмульсии или другие соответствующие векторы переноса. Данные векторы доставки делают возможной доставку молекул, химических веществ, макромолекул (генов, белков) или других векторов, таких как плазмиды, пептиды, разработанные Diatos. В этих случаях векторы доставки представляют собой молекулярные носители. Под "вектором доставки" или "векторами доставки" также понимаются способы доставки для осуществления трансфекции.
- Термины "вектор" или "векторы" относятся к молекуле нуклеиновой кислоты, способной транспортировать другую нуклеиновую кислоту, с которой она связана. "Вектор" в настоящем изобретении включает, но без ограничения, вирусный вектор, плазмиду, РНК-вектор или линейную или кольцевую молекулу ДНК или РНК, которая может состоять из хромосомных, не хромосомных, полусинтетических или синтетических нуклеиновых кислот. Предпочтительными векторами являются способные к автономной репликации (эписомальный вектор) и/или экспрессии нуклеиновых кислот, с которыми они связаны (векторы экспрессии). Большое количество подходящих векторов известно специалистам в данной области и коммерчески доступно.
Вирусные векторы включают ретровирус, аденовирус, парвовирус (например, аденоассоциированные вирусы), коронавирус, РНК-вирусы с отрицательной нитью, такие как ортомиксовирус (например, вирус гриппа), рабдовирус (например, вирус бешенства и везикулярного стоматита), парамиксовирус (например, кори и Сендай), РНК-вирусы с положительной нитью, такие как пикорнавирус и альфавирус, и вирусы с двухнитевой ДНК, включая аденовирус, вирус герпеса (например, вирус простого герпеса типов 1 и 2, вирус Эпштейна-Барр, цитомегаловирус) и поксвирус (например, осповакцины, оспы кур и канарипокс). Другие вирусы включают, например, вирус Норфолк, тогавирус, флавивирус, реовирусы, паповавирус, гепаднавирус и вирус гепатита. Примеры ретровирусов включают: лейкоза-саркомы птиц, типа C млекопитающих, вирусы типа B, вирусы типа D, группы HTLV-BLV, лентивирус, спумавирус (Coffin, J. M., Retroviridae: The viruses and their replication, Fundamental Virology, Third Edition, B. N. Fields, et al., Eds., Lippincott-Raven Publishers, Philadelphia, 1996).
- Под "лентивирусным вектором" понимаются основанные на ВИЧ лентивирусные векторы, которые являются очень перспективными для доставки генов из-за их относительно большой емкости упаковки, уменьшенной иммуногенности и их способности к стабильной трансдукции с высокой эффективностью большого диапазона различных типов клеток. Лентивирусные векторы обычно создают после временной трансфекции трех (упаковывающей, оболочки и переноса) или более плазмид в клетки-продуценты. Как и ВИЧ, лентивирусные векторы входят в клетку-мишень посредством взаимодействия вирусных поверхностных гликопротеинов с рецепторами на клеточной поверхности. При входе вирусная РНК подвергается обратной транскрипции, которая опосредуется комплексом вирусной обратной транскриптазы. Продуктом обратной транскрипции является двухцепочечная линейная вирусная ДНК, которая является субстратом для вирусной интеграции в ДНК инфицированных клеток. Под "интегративными лентивирусными векторами (или LV)" понимаются такие векторы, в качестве неограничивающего примера, которые способны интегрироваться в геном клетки-мишени. Напротив, под "неинтегративными лентивирусными векторами (или NILV)" понимаются эффективные векторы доставки генов, которые не интегрируются в геном клетки-мишени посредством действия интегразы вируса.
- Векторы доставки и векторы могут быть связаны или объединены с любыми методами клеточной пермеабилизации, такими как сонопорация, или электропорация, или производные данных методов.
- Под клеткой или клетками понимаются любые эукариотические живые клетки, первичные клетки и клеточные линии, происходящие от этих организмов, для культур in vitro.
- Под "первичной клеткой" или "первичными клетками" понимаются клетки, взятые непосредственно из живой ткани (т.е. биопсийного материала) и адаптированные для роста in vitro, которые подвергались очень небольшому количеству удвоений популяции и, следовательно, лучше представляют главные функциональный компоненты и характеристики тканей, из которых они происходят, по сравнению с непрерывными опухолеродными или искусственно иммортализованными клеточными линиями.
В качестве неограничивающих примеров, клеточные линии могут быть выбраны из группы, состоящей из клеток CHO-K1; клеток HEK293; клеток Caco2; клеток U2-OS; клеток NIH 3T3; клеток NSO; клеток SP2; клеток CHO-S; клеток DG44; клеток K-562, клеток U-937; клеток MRC5; клеток IMR90; клеток Jurkat; клеток HepG2; клеток HeLa; клеток HT-1080; клеток HCT-116; клеток Hu-h7; клеток Huvec; клеток Molt 4.
Все эти клеточные линии могут быть модифицированы с помощью способа настоящего изобретения для получения моделей клеточных линий для производства, экспрессии, количественного анализа, детектирования, исследования представляющего интерес гена или белка; данные модели могут быть также использованы для скрининга представляющих интерес биологически активных молекул при исследовании и производстве и в различных областях, таких как химические продукты, биотопливо, терапевтические средства и агрономия в качестве неограничивающих примеров.
- под "мутацией" понимается замена, удаление, вставка вплоть до одного, двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти, одиннадцати, двенадцати, тринадцати, четырнадцати, пятнадцати, двадцати, двадцати пяти, тридцати, сорока, пятидесяти или более нуклеотидов/аминокислот в полинуклеотидной (кДНК, ген) или полипептидной последовательности. Мутация может воздействовать на кодирующую последовательность гена или его регуляторную последовательность. Она может также воздействовать на структуру геномной последовательности или структуру/стабильность кодируемой мРНК.
- под "вариантом(ами)" понимается вариант повтора, вариант, вариант связывания ДНК, вариант TALE-нуклеазы, вариант полипептида, полученный посредством мутации или замены по меньшей мере одного остатка в аминокислотной последовательности родительской молекулы.
- под "функциональным вариантом" понимается каталитически активный мутант белка или белкового домена; такой мутант может иметь такую же активность как и его родительский белок или белковый домен или дополнительные свойства или более высокую или более низкую активность.
- Под "геном" понимается основная единица наследственности, состоящая из сегмента ДНК, расположенного линейным образом вдоль хромосомы, который кодирует специфический белок или сегмент белка. Ген обычно включает промотор, 5'-нетранслируемую область, одну или несколько кодирующих последовательностей (экзоны), необязательно, интроны, 3'-нетранслируемую область. Ген может дополнительно содержать терминатор, энхансеры и/или сайленсеры.
- Как используется в настоящем документе, термин "локус" обозначает определенное физическое местоположение последовательности ДНК (например, гена) на хромосоме. Термин "локус" может относиться к определенному физическому местоположению последовательности-мишени редкощепящей эндонуклеазы на хромосоме. Такой локус может содержать последовательность-мишень, которая распознается и/или расщепляется редкощепящей эндонуклеазой в соответствии с настоящим изобретением. Следует понимать, что представляющий интерес локус настоящего изобретения может не только определять последовательность нуклеиновой кислоты, которая существует в основной части генетического материала (т.е. в хромосоме) клетки, но также часть генетического материала, которая может существовать независимо от указанной основной части генетического материала, такую как плазмиды, эписомы, вирус, транспозоны, или в органеллах, таких как митохондрии в качестве неограничивающих примеров.
- Термин "эндонуклеаза" относится к любому ферменту дикого типа или его варианту, способному катализировать гидролиз (расщепление) связей между нуклеиновыми кислотами в молекуле ДНК или РНК, предпочтительно молекуле ДНК. Эндонуклеазы не расщепляют молекулу ДНК или РНК независимо от ее последовательности, но распознают и расщепляют молекулу ДНК или РНК у определенных полинуклеотидных последовательностей, далее называемых "последовательности-мишени" или "сайты-мишени". Эндонуклеазы могут быть классифицированы как редкощепящие эндонуклеазы, когда они имеют, как правило, полинуклеотидный сайт распознавания, больший, чем 12 пар оснований (п.о.) в длину, более предпочтительно, равный 14-55 п.о. Редкощепящие эндонуклеазы значительно увеличивают HR посредством индуцирования двухнитевых разрывов ДНК (DSB) в определенном локусе (Rouet, Smih et al., 1994; Choulika, Perrin et al., 1995; Pingoud and Silva, 2007). Редкощепящие эндонуклеазы могут, например, представлять собой хоминг-эндонуклеазу (Paques and Duchateau, 2007), химерную цинковопальцевую нуклеазу (ZFN), получающуюся от слияния сконструированных цинковопальцевых доменов с каталитическим доменом рестрикционного фермента, такого как FokI (Porteus and Carroll, 2005), или химическую эндонуклеазу (Eisenschmidt, Lanio et al., 2005; Arimondo, Thomas et al., 2006). В химических эндонуклеазах химический или пептидный расщепитель конъюгирован или с полимером нуклеиновых кислот, или с другой ДНК, распознающей определенную последовательность-мишень, посредством этого направляя расщепляющую активность на определенную последовательность. Химические эндонуклеазы также включают синтетические нуклеазы, такие как конъюгаты ортофенантролина, ДНК-расщепляющую молекулу, и триплексформирующие олигонуклеотиды (TFO), которые, как известно, связывают определенные последовательности ДНК (Kalish and Glazer, 2005). Такие химические эндонуклеазы заключены в термине "эндонуклеаза" в соответствии с настоящим изобретением.
Редкощепящие эндонуклеазы могут также представлять собой, например, TALE-нуклеазы, новый класс химерных нуклеаз, использующий каталитический домен FokI и ДНК-связывающий домен, происходящий от подобного активатору транскрипции эффектора (TALE), семейства белков, используемых в инфекционном процессе патогенами растений рода Xanthomonas (Boch, Scholze et al., 2009; Moscou and Bogdanove, 2009; Christian, Cermak et al., 2010; Li, Huang et al.). Функциональная компоновка основанной на FokI TALE-нуклеазы (TALE-нуклеазы) по существу такая же, как у ZFN, но цинковопальцевый ДНК-связывающий домен заменен на домен TALE. Как таковое, расщепление ДНК TALE-нуклеазой требует двух распознающих ДНК участков, фланкирующих неспецифический центральный участок. Редкощепящие эндонуклеазы, охватываемые настоящим изобретением, могут также происходить от TALE- нуклеаз.
Редкощепящая эндонуклеаза может представлять собой хоминг-эндонуклеазу, также известную под названием мегануклеазы. Такие хоминг-эндонуклеазы хорошо известны в данной области (Stoddard, 2005). Хоминг-эндонуклеазы распознают целевую последовательность ДНК и создают одно- или двухнитевой разрыв. Хоминг-эндонуклеазы высоко специфичны, распознавая сайты-мишени ДНК в диапазоне от 12 до 45 пар оснований (п.о.) в длину, обычно в диапазоне от 14 до 40 п.о. в длину. Хоминг-эндонуклеаза в соответствии с настоящим изобретением может, например, соответствовать эндонуклеазе LAGLIDADG ("LAGLIDADF" раскрыто как SEQ ID NO:127), эндонуклеазе HNH или эндонуклеазе GIY-YIG. Предпочтительная хоминг-эндонуклеаза в соответствии с настоящим изобретением может представлять собой вариант I-Crel.
- Под "TALE-нуклеазой" (TALEN) понимается слитый белок, состоящий из связывающего нуклеиновую кислоту домена, обычно происходящего от подобного активатору транскрипции эффектора (TALE), и одного нуклеазного каталитического домена для расщепления целевой последовательности нуклеиновой кислоты. Каталитический домен, предпочтительно, представляет собой нуклеазный домен и, более предпочтительно, домен, обладающий эндонуклеазной активностью, такой как, например, I-Tevl, ColE7, NucA и Fok-I. В конкретном варианте осуществления домен TALE может быть слит с мегануклеазой, такой как, например, I-Crel и I-Onul, или ее функциональным вариантом. В более предпочтительном варианте осуществления указанная нуклеаза представляет собой мономерную TALE-нуклеазу. Мономерная TALE-нуклеаза представляет собой TALE-нуклеазу, которая не требует димеризации для специфического распознавания и расщепления, такого как слияния сконструированных TAL-повторов с каталитическим доменом I-Tevl, описанные в WO2012138927. Подобный активатору транскрипции эффектор (TALE) представляет собой белки из вида бактерий Xanthomonas, которые содержат множество повторяющихся последовательностей, причем каждый повтор содержит два остатка в положении 12 и 13 (RVD), которые специфичны для каждого нуклеотидного основания целевой последовательности нуклеиновой кислоты. Связывающие домены со схожими свойствами модульного связывания нуклеиновых кислот основание за основанием (MBBBD) могут также происходить от новых модульных белков, недавно обнаруженных заявителем в другом виде бактерий. Новые модульные белки имеют преимущество в том, что демонстрируют большую изменчивость последовательности, чем TAL-повторы. Предпочтительно, RVD, связанные с распознаванием различных нуклеотидов, представляют собой HD для распознавания C, NG для распознавания T, NI для распознавания A, NN для распознавания G или A, NS для распознавания A, C, G или T, HG для распознавания T, IG для распознавания T, NK для распознавания G, HA для распознавания C, ND для распознавания C, HI для распознавания C, HN для распознавания G, NA для распознавания G, SN для распознавания G или A и YG для распознавания T, TL для распознавания A, VT для распознавания A или G и SW для распознавания A. В другом варианте осуществления существенные аминокислоты 12 и 13 могут мутировать в направлении других аминокислотных остатков, для того чтобы модулировать их специфичность в направлении нуклеотидов A, T, C и G и, в частности, усиливать эту специфичность. TALE-нуклеаза уже была описана и использована для стимулирования нацеливания на гены и модификаций генов (Boch, Scholze et al., 2009; Moscou and Bogdanove, 2009; Christian, Cermak et al., 2010; Li, Huang et al.). Сконструированные TAL-нуклеазы коммерчески доступны под торговым наименованием TALEN™ (Cellectis, 8 rue de la Croix Jarry, 75013 Paris, Франция).
- Термин "расщепление" относится к разрыву ковалентного остова полинуклеотида. Расщепление может быть инициировано множеством способов, включая, но без ограничения, ферментативный или химический гидролиз фосфодиэфирной связи. Возможны как однонитевое расщепление, так и двухнитевое расщепление, и двухнитевое расщепление может происходить в результате двух различных событий однонитевого расщепления. Двухнитевое расщепление ДНК, РНК или гибрида ДНК/РНК может приводить к получению или тупых концов, или ступенчатых концов.
- Под "слитым белком" понимается результат хорошо известного в данной области процесса, состоящего в соединении двух или более генов, которые исходно кодируют отдельные белки или их часть, причем трансляция указанного "слитого гена" приводит к одному полипептиду с функциональными свойствами, происходящими от каждого из исходных белков.
- "Идентичность" относится к идентичности последовательностей между двумя молекулами нуклеиновой кислоты или полипептидами. Идентичность можно определять посредством сравнения положения в каждой последовательности, которое с целью сравнения может быть выровнено. Когда положение в сравниваемой последовательности занято тем же основанием, то молекулы идентичны в этом положении. Степень сходства или идентичности между нуклеиновокислотными или аминокислотными последовательностями является функцией количества идентичных или совпадающих нуклеотидов в положениях, общих для данных последовательностей нуклеиновой кислоты. Можно использовать различные алгоритмы и/или программы выравнивания для вычисления идентичности между двумя последовательностями, включая FASTA или BLAST, который доступен как часть пакета для анализа последовательностей GCG (University of Wisconsin, Madison, Wis.) и может быть использован, например, с установками по умолчанию. Например, рассматриваются полипептиды, имеющие идентичность по меньшей мере 70%, 85%, 90%, 95%, 98% или 99% с определенными полипептидами, описанными в настоящем документе, и, предпочтительно, проявляющие по существу те же функции, а также полинуклеотид, кодирующий такие полипептиды.
- "сходство" описывает отношение между аминокислотными последовательностями двух или более полипептидов. Можно также использовать BLASTP для идентификации аминокислотной последовательности, имеющей сходство последовательности по меньшей мере 70%, 75%, 80%, 85%, 87,5%, 90%, 92,5%, 95%, 97,5%, 98%, 99% с эталонной аминокислотной последовательностью, с помощью матрицы сходства, такой как BLOSUM45, BLOSUM62 или BLOSUM80. Если не указано иное, оценка сходства будет основываться на использовании BLOSUM62. Когда используется BLASTP, процентное сходство основывается на оценке положительных результатов BLASTP, и процентная идентичность последовательности основывается на оценке идентичности BLASTP. "Идентичность" в BLASTP показывает количество и долю всех остатков в парах последовательностей с высокой оценкой, которые идентичны; и "число положительных результатов" BLASTP показывает количество и долю остатков, для которых оценки выравнивания имеют положительные значения, и которые схожи друг с другом. Аминокислотные последовательности, имеющие такие степени идентичности или сходства или любую промежуточную степень идентичности или сходства с аминокислотными последовательностями, раскрытыми в настоящем документе, рассматриваются и охвачены настоящим раскрытием. Полинуклеотидные последовательности схожих полипептидов определяют с использованием генетического кода, и они могут быть получены обычными средствами. Например, функциональный вариант pT-альфа может иметь сходство последовательности 70%, 75%, 80%, 85%, 87,5%, 90%, 92,5%, 95%, 97,5%, 98%, 99% с аминокислотной последовательностью SEQ ID NO:107. Полинуклеотид, кодирующий такой функциональный вариант, может быть получен посредством обратной трансляции его аминокислотной последовательности с помощью генетического кода.
- "передающий сигнал домен" или "костимулирующий лиганд" обозначает молекулу на антиген-презентирующей клетке, которая специфически связывает распознанную костимулирующую молекулу на T-клетке, посредством этого обеспечивая сигнал, который, в дополнение к первичному сигналу, обеспечиваемому, например, связыванием комплекса TCR/CD3 с молекулой MHC, нагруженной пептидом, опосредует T-клеточный ответ, включая, но без ограничения, активацию пролиферации, дифференцировку и тому подобное. Костимулирующий лиганд может включать, но без ограничения, CD7, B7-1 (CD80), B7-2 (CD86), PD-L1, PD-L2, 4-1BBL, OX40L, индуцируемый костимулирующий лиганд (ICOS-L), молекулу межклеточной адгезии (ICAM), CD30L, CD40, CD70, CD83, HLA-G, MICA, M1CB, HVEM, рецептор лимфотоксина-бета, 3/TR6, ILT3, ILT4, агонист или антитело, которое связывает рецептор толл-лиганда, и лиганд, который специфически связывается с B7-H3. Костимулирующий лиганд также охватывает, в числе прочего, антитело, которое специфически связывается с костимулирующей молекулой, присутствующей на T-клетке, такой как, но без ограничения, CD27, CD28, 4-1BB, OX40, CD30, CD40, PD-1, ICOS, функционально-связанный антиген-1 лимфоцитов (LFA-1), CD2, CD7, LTGHT, NKG2C, B7-H3, лиганд, который специфически связывается с CD83.
"Костимулирующая молекула" обозначает распознанного партнера по связыванию на T-клетке, который специфически связывается с костимулирующим лигандом, тем самым опосредуя костимулирующий ответ клетки, такой как, но без ограничения, пролиферация. Костимулирующие молекулы включают, но без ограничения, молекулу MHC класса I, BTLA и рецептор толл-лиганда.
"Костимулирующий сигнал", как используется в настоящем документе, обозначает сигнал, который в комбинации с первичным сигналом, таким как лигирование TCR/CD3, приводит к пролиферации и/или повышающей регуляции T-клеток или понижающей регуляции основных молекул.
- "биспецифическое антитело" обозначает антитело, которое имеет связывающие сайты для двух различных антигенов в одной молекуле антитела. Специалистам в данной области понятно, что в дополнение к канонической структуре антитела могут быть сконструированы другие молекулы с двумя специфичностями связывания. Кроме того, следует понимать, что связывание антигена биспецифическими антителами может быть одновременным или последовательным. Биспецифические антитела могут быть получены с помощью химических методов (см., например, Kranz et al. (1981) Proc. Natl. Acad. Sci. USA 78, 5807), методами "polydoma" (см. патент США № 4474893) или методами рекомбинантной ДНК, все из которых известны per se. В качестве неограничивающего примера, каждый связывающий домен содержит по меньшей мере один вариабельный участок от тяжелой цепи антитела ("участок VH или H"), причем участок VH первого связывающего домена специфически связывается с маркером лимфоцитов, таким как CD3, и участок VH второго связывающего домена специфически связывается с опухолевым антигеном.
- Термин "внеклеточный лигандсвязывающий домен", как используется в настоящем документе, определяют как олиго- или полипептид, который способен к связыванию лиганда. Предпочтительно, данный домен будет способен к взаимодействию с молекулой клеточной поверхности. Например, внеклеточный лигандсвязывающий домен может быть выбран для распознавания лиганда, который действует в качестве маркера клеточной поверхности на клетках-мишенях, связанных с конкретным состоянием заболевания. Таким образом, примеры маркеров клеточной поверхности, которые могут действовать как лиганды, включают связанные с вирусными, бактериальными и паразитарными инфекциями, аутоиммунным заболеванием и раковыми клетками.
Термин "индивид" или "пациент", как используется в настоящем документе, включает всех членов царства животных, включая отличных от человека приматов и людей.
Приведенное выше описание настоящего изобретения предлагает способ и процесс его осуществления и применения, так что любой специалист в данной области может его осуществлять и применять, причем данная возможность обеспечена, в частности, для содержания прилагаемой формулы изобретения, которое составляет часть исходного описания.
Там, где в настоящем документе установлен числовой предел или диапазон, включены концевые точки. Кроме того, все значения и поддиапазоны в границах числового предела или диапазона прямо включены, как если бы они были явно указаны.
Приведенное выше описание представлено, чтобы дать возможность специалисту в данной области осуществлять и применять настоящее изобретение, и приведено в контексте конкретного применения и его требований. Различные модификации предпочтительных вариантов осуществления будут очевидны специалистам в данной области, и общие принципы, определенные в настоящем документе, могут быть применены к другим вариантам осуществления и применениям без отступления от сущности и объема настоящего изобретения. Таким образом, не предполагается, что настоящее изобретение ограничено показанными вариантами осуществления, но должно соответствовать самому широкому объему, совместимому с принципами и признаками, раскрытыми в настоящем документе.
После общего описания настоящего изобретения дополнительное понимание может быть получено посредством ссылки на определенные конкретные примеры, которые приведены в настоящем документе только с целью иллюстрации, и не предназначены для ограничения, если не указано иное.
Примеры
Пример 1: TALE-нуклеазы, расщепляющие ген GR человека
Было разработано и получено 6 гетеродимерных TALE-нуклеаз, нацеленных на экзоны гена GR человека. Таблица 1 ниже указывает последовательности-мишени, расщепляемые каждой TALE-нуклеазой. TALE-нуклеаза GR состоит из двух независимых единиц (называемых половинами TALE-нуклеазы), причем каждая содержит повторную последовательность, сконструированную для связи и расщепления последовательности-мишени GR, состоящей из двух последовательностей длиной 17 п.о. (называемых полумишенями), разделенных спейсером 15 п.о.


Аминокислотные последовательности N-концевого, C-концевого доменов и повтора основаны на TALE AvrBs3 (ref: GenBank: X16130.1). C-концевой и N-концевой домены разделены двумя сайтами рестрикции BsmBI. Наборы повторов (SEQ ID NO:7-18), нацеленных на желаемые последовательности (SEQ ID NO:1-6), синтезировали с помощью способа твердой подложки, состоящего из последовательных стадий рестрикции/лигирования/промывания (международная заявка PCT WO2013/017950). Кратко говоря, первый блок (кодирующий двойной повтор) иммобилизовывали на твердой подложке посредством взаимодействия биотин/стрептавидин, затем второй блок (тройной повтор) лигировали к первому и после расщепления SfaNI присоединяли третий блок (тройной повтор). Данный процесс повторяли, используя блоки тройных или двойных повторов до получения желаемого набора повторов. Затем продукт клонировали в классической плазмиде для клонирования pAPG10 для амплификации в E. coli и секвенировали. Последовательности наборов повторов, полученные таким образом, субклонировали в дрожжевом TALE-векторе экспрессии, используя ферменты рестрикции BsmBI типа IIS для получения плазмиды и BbvI и SfaNI для вставляемой последовательности повтора. ДНК, кодирующую половину TALE-нуклеазы, содержащую происходящий от TALE ДНК-связывающий домен, слитый с каталитическим доменом фермента рестрикции FokI, амплифицировали в E. Coli, выделяли с помощью стандартных минипрепаративных методов и секвенировали для определения целостности вставки.
Активность TALE-нуклеазы GR в дрожжах:
Нуклеазную активность шести GR-TALE-нуклеаз проверяли при 37°C и 30°C в нашем SSA-анализе в дрожжах, описанном ранее (международные заявки PCT WO 2004/067736 и в Epinat, Arnould et al., 2003; Chames, Epinat et al., 2005; Arnould, Chames et al., 2006; Smith, Grizot et al., 2006), на мишенях, содержащих две последовательности-мишени TALE, обращенные друг к другу на нити ДНК, разделенные спейсером в 15 п.о., что давало SEQ ID NO:1-6. Все дрожжевые целевые репортерные плазмиды, содержащие целевые последовательности ДНК TALE-нуклеазы, конструировали, как описано выше (международные заявки PCT WO 2004/067736 и в Epinat, Arnould et al., 2003; Chames, Epinat et al., 2005; Arnould, Chames et al., 2006; Smith, Grizot et al., 2006). Уровень TALE-нуклеазной расщепляющей активности в дрожжах отдельных клонов на мишенях представлен в таблице 2.

Значения содержатся между 0 и 1. Максимальное значение составляет 1.
Активность TALE-нуклеаз GR в клетках HEK293:
Каждый TALE-нуклеазный конструкт субклонировали, используя расщепление ферментом рестрикции в векторе экспрессии млекопитающих под контролем длинного промотора pEF1-альфа.
Один миллион клеток HEK293 высевали за один день до трансфекции. Клетки котрансфицировали 2,5 мкг каждой из двух плазмид, кодирующих левую и правую половину TALE-нуклеаз GRex2, GRex3T2, GRex3T4, GRex5T1, GRex5T2 или GRex5T3, распознающих представляющие интерес геномные последовательности двух полумишеней в гене GR под контролем промотора EF1-альфа, используя 25 мкл липофектамина (Invitrogen) в соответствии с инструкциями изготовителя. В качестве контроля клетки котрансфицировали 2,5 мкг каждой из двух плазмид, кодирующих левую и правую половины TALE-нуклеаз, нацеленных на сайт-мишень ((TALE-нуклеаза TRAC_T01-L и -R (SEQ ID NO:41 и SEQ ID NO:42), сайт-мишень TRAC_T01 (SEQ ID NO:37)) участка константной альфа-цепи (TRAC_T01) T-клеточного рецептора под контролем промотора EF1-альфа. Двухнитевой разрыв, создаваемый TALE-нуклеазами в последовательности, кодирующей GR, индуцировал негомологичное соединение концов (NHEJ), которое представляет собой механизм, подверженный ошибкам. Активность TALE-нуклеаз измеряли по частоте вставок или делеций в целевом геномном локусе.
Через 2 или 7 дней после трансфекции клетки собирали, и осуществляли локус-специфические ПЦР на геномной ДНК, экстрагированной с использованием следующих составных праймеров: 5'-CCATCTCATCCCTGCGTGTCTCCGACTCAG-3' (прямая адапторная последовательность, SEQ ID NO:126) -10N (TAG)- локус-специфическая прямая последовательность для экзона 2 GR: 5'-GGTTCATTTAACAAGCTGCC-3' (SEQ ID NO:127; составной праймер, раскрытый как SEQ ID NO:31), для экзона 3 GR: 5'-GCATTCTGACTATGAAGTGA-3' (SEQ ID NO:128; составной праймер, раскрытый как SEQ ID NO:32) и для экзона 5 GR: 5'-TCAGCAGGCCACTACAGGAGTCTCACAAG-3' (SEQ ID NO:129; составной праймер, раскрытый как SEQ ID NO:33) и обратный составной праймер 5'-CCTATCCCCTGTGTGCCTTGGCAGTCTCAG-3' (обратная адапторная последовательность, SEQ ID NO:130))- локус-специфическая обратная последовательность для экзона 2 GR: 5'-AGCCAGTGAGGGTGAAGACG-3' (SEQ ID NO:131; составной праймер, раскрытый как SEQ ID NO:34), для экзона 3 GR: 5'-GGGCTTTGCATATAATGGAA-3' (SEQ ID NO:132; составной праймер, раскрытый как SEQ ID NO:35) и для экзона 5 GR: 5'-CTGACTCTCCCCTTCATAGTCCCCAGAAC-3' (SEQ ID NO:133; составной праймер, раскрытый как SEQ ID NO:36).
Продукты ПЦР секвенировали с помощью системы секвенирования 454 (454 Life Sciences). Получали приблизительно 10000 последовательностей на продукт ПЦР, и затем их анализировали на присутствие событий сайт-специфических вставок или делеций. Таблица 3 указывает процентную долю последовательностей, демонстрирующих вставки или делеции в сайте-мишени TALE-нуклеазы, от общего числа последовательностей в образце. В таблице 3 приведены для GRex2, GRex3T2 и GRex3T4 результаты типичного эксперимента.
Во всех исследованных случаях % мутагенеза был на день 7 близок при сравнении с образцом на день 2 после трансфекции. Природу событий мутагенеза также проанализировали, обнаружив во всех случаях преимущество делеций по сравнению со вставками.
Процентная доля направленного мутагенеза в сайтах-мишенях эндогенных TALE-нуклеаз клеток HEK293
Активность TALE-нуклеаз GR в первичных T-лимфоцитах:
Каждый TALE-нуклеазный конструкт субклонировали, используя расщепление ферментом рестрикции в векторе экспрессии под контролем промотора T7.
мРНК, кодирующую TALE-нуклеазы, расщепляющие геномные последовательности GR, синтезировали из каждой плазмиды, несущей кодирующие последовательности в прямом направлении от промотора T7. T-лимфоциты, выделенные из периферической крови, активировали в течение 5 дней с использованием гранул активатора анти-CD3/CD28 (Life technologies), и 5 миллионов клеток трансфицировали посредством электропорации 10 мкг каждой из 2 мРНК, кодирующих обе половины TALE-нуклеазы, используя прибор CytoLVT-P (BTX-Harvard apparatus). T-клетки, трансфицированные 10 мкг каждой из 2 мРНК, кодирующих обе половины TALE-нуклеазы, нацеленные на ген CD52 (CD52_T02-L и -R TALEN (SEQ ID NO:55 и 56), последовательность-мишень CD52_T02 SEQ ID NO:40) использовали в качестве контроля.
Через 3 и 7 дней после трансфекции геномную ДНК выделели из трансфицированных клеток и осуществляли локус-специфические ПЦР, используя праймеры, описанные ранее. Продукты ПЦР секвенировали с помощью системы секвенирования 454 (454 Life Sciences). Получали приблизительно 10000 последовательностей на продукт ПЦР, и затем их анализировали на присутствие событий сайт-специфических вставок или делеций; результаты приведены в таблице 4.
Процентная доля направленного мутагенеза в сайтах-мишенях эндогенных TALE-нуклеаз в первичных T-лимфоцитах
Пример 2: TALE-нуклеазы, расщепляющие ген CD52 человека, константной цепи альфа T-клеточного рецептора человека (TRAC) и константных цепей бета 1 и 2 T-клеточного рецептора человека (TRBC)
Как описано в примере 1, были разработаны и получены гетеродимерные TALE-нуклеазы, нацеленные на гены CD52, TRAC и TRBC, соответственно. Целевые геномные последовательности состоят из двух последовательностей длиной 17 п.о. (называемых полумишенями), разделенных спейсером в 11 или 15 п.о. Каждая полумишень распознается повторами половин TALE-нуклеаз, приведенных в таблице 5. Геном человека содержит две функциональные бета-цепи T-клеточного рецептора (TRBC1 и TRBC2). Во время развития альфа/бета T-лимфоцитов одна из этих двух константных цепей выбирается в каждой клетке для сплайсига в вариабельный участок TCR-бета и образует функциональную полноразмерную бета-цепь. 2 мишени TRBC были выбраны в последовательностях, консервативных между TRBC1 и TRBC2, так что соответствующая TALE-нуклеаза будет расщеплять как TRBC1, так в то же время и TRBC2.
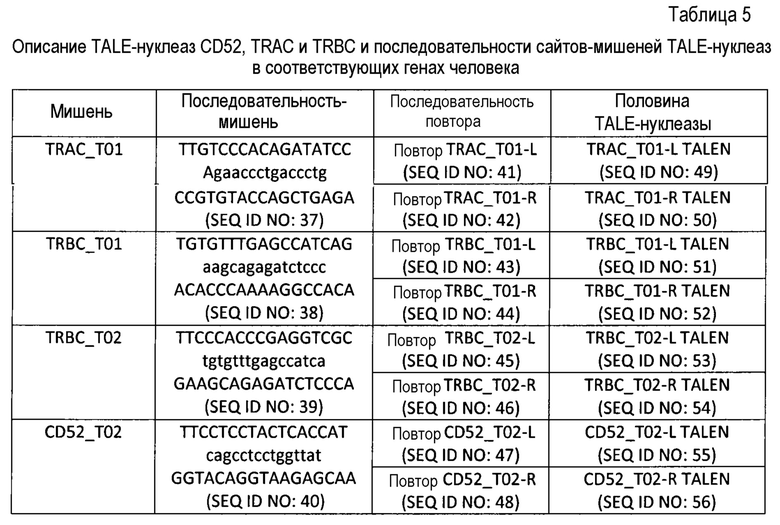
Были разработаны и другие последовательности-мишени в генах TRAC и CD52, которые приведены в таблице 6.

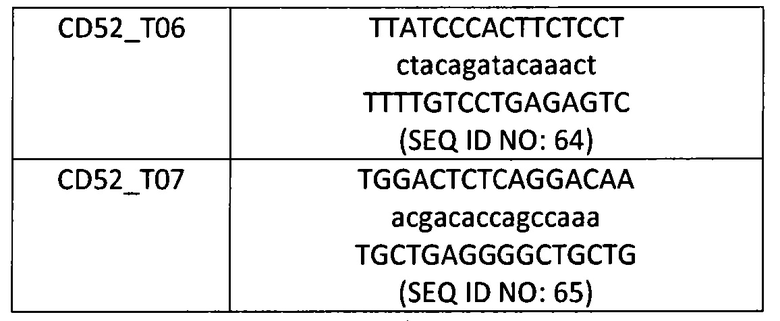
Активность CD52-TALE-нуклеазы, TRAC-TALE-нуклеазы и TRBC-TALE-нуклеазы в клетках HEK293
Каждый TALE-нуклеазный конструкт субклонировали, используя расщепление ферментом рестрикции в векторе экспрессии млекопитающих под контролем длинного промотора pEF1-альфа. Один миллион клеток HEK293 высевали за один день до трансфекции. Клетки котрансфицировали 2,5 мкг каждой из двух плазмид, кодирующих TALE-нуклеазы, распознающие две полумишени в представляющей интерес геномной последовательности в гене CD52, участка константной цепи альфа T-клеточного рецептора (TRAC) или участка константной цепи бета T-клеточного рецептора (TRBC) под контролем промотора EF1-альфа, или 5 мкг контрольного вектора pUC (pCLS0003), используя 25 мкл липофектамина (Invitrogen) в соответствии с инструкциями изготовителя. Двухнитевое расщепление, создаваемое TALE-нуклеазами в последовательности, кодирующей CD52 или TRAC, восстанавливалось в живых клетках посредством негомологичного соединения концов (NHEJ), которое представляет собой механизм, подверженный ошибкам. Активность TALE-нуклеаз в живых клетках измеряли по частоте вставок или делеций в целевом геномном локусе. Через 48 часов после трансфекции геномную ДНК выделели из трансфицированных клеток, и осуществляли локус-специфические ПЦР с использованием следующих составных праймеров: 5'-CCATCTCATCCCTGCGTGTCTCCGACTCAG (прямая адапторная последовательность, SEQ ID NO:126)- 10N (TAG)- локус-специфическая прямая последовательность для CD52: 5'-CAGATCTGCAGAAAGGAAGC-3' (SEQ ID NO:134; составной праймер, раскрытый как SEQ ID NO:66), для TRAC: 5'-ATCACTGGCATCTGGACTCCA-3' (SEQ ID NO:135; составной праймер, раскрытый как SEQ ID NO:67), для TRBC1: 5'-AGAGCCCCTACCAGAACCAGAC-3' (SEQ ID NO:136; составной праймер, раскрытый как SEQ ID NO:68) или для TRBC2: 5'-GGACCTAGTAACATAATTGTGC-3' (SEQ ID NO:137; составной праймер, раскрытый как SEQ ID NO:69) и обратный составной праймер 5'-CCTATCCCCTGTGTGCCTTGGCAGTCTCAG (обратная адапторная последовательность, SEQ ID NO:130)-эндогенная локус-специфическая обратная последовательность для CD52: 5'-CCTGTTGGAGTCCATCTGCTG-3' (SEQ ID NO:138; составной праймер, раскрытый как SEQ ID NO:70), для TRAC: 5'-CCTCATGTCTAGCACAGTTT-3' (SEQ ID NO:139; составной праймер, раскрытый как SEQ ID NO:71), для TRBC1 и TRBC2: 5'-ACCAGCTCAGCTCCACGTGGT-3' (SEQ ID NO:140; составной праймер, раскрытый как SEQ ID NO:72). Продукты ПЦР секвенировали с помощью системы секвенирования ПЦР 454 (454 Life Sciences). Получали приблизительно 10000 последовательностей на продукт ПЦР, и затем их анализировали на присутствие событий сайт-специфических вставок или делеций; результаты приведены в таблице 7.
Процентные доли вставок-делеций для TALE-нуклеазы, нацеленной на мишени CD52_T02, TRAC_T01, TRBC_T01 и TRBC_T02
Активность CD52-TALE-нуклеазы, TRBC-TALE-нуклеазы и TRAC-TALE-нуклеазы в первичных T-лимфоцитах
Каждый TALE-нуклеазный конструкт субклонировали, используя расщепление ферментом рестрикции в векторе экспрессии млекопитающих под контролем промотора T7.
мРНК, кодирующую TALE-нуклеазу, расщепляющую геномные последовательности CD52 TRAC и TRBC, синтезировали из плазмиды, несущей кодирующие последовательности в прямом направлении от промотора T7. T-лимфоциты, выделенные из периферической крови, активировали в течение 5 дней с использованием гранул активатора анти-CD3/CD28 (Life technologies), и затем 5 миллионов клеток трансфицировали посредством электропорации 10 мкг каждой из 2 мРНК, кодирующих обе половины TALE-нуклеазы (или некодирующей РНК в качестве контроля), используя прибор CytoLVT-P. В результате вставок и делеций, индуцированных NHEJ, кодирующая последовательность для CD52 и/или TRAC оказывается в части клеток вне пределов рамки, что ведет к нефункциональным генам. Через 5 дней после электропорации клетки помечали конъюгированным с флуорохромом анти-CD52 или анти-TCR антителом посредством проточной цитометрии на присутствие CD52 или TCR на их клеточной поверхности. Поскольку все T-лимфоциты, размноженные из периферической крови, обычно экспрессируют CD52 и TCR, соотношение CD52-отрицательных или TCR-отрицательных клеток является прямой мерой TALE-нуклеазной активности. В таблице 8 приведены результаты типичного эксперимента. Таблица 9 показывает результаты типичного эксперимента для определения эффективности TALE-нуклеазы TRBC.
Процентные доли CD52-отрицательных, TCR-отрицательных и CD52/TCR-двойных отрицательных T-лимфоцитов после трансфекции соответствующих экспрессирующих TALE-нуклеазу полинуклеотидов.
Процентные доли TCR-отрицательных T-лимфоцитов после трансфекции экспрессирующих TALE-нуклеазу TRBC полинуклеотидов
Функциональный анализ T-клеток с целевым геном CD52
Целью инактивации гена CD52 является получение T-лимфоцитов, резистентных к опосредованной анти-CD52 антителом иммуносупрессии. Как описано в предыдущем абзаце, T-лимфоциты трансфицировали мРНК, кодирующей TALE-нуклеазу, расщепляющую CD52. Через 7 дней после трансфекции клетки обрабатывали 50 мкг/мл моноклонального анти-CD52 антитела (или крысиным IgG в качестве контроля) с или без 30% кроличьего комплемента (Cedarlane). После 2 часов инкубации при 37°C клетки помечали конъюгированным с флуорохромом анти-CD52 антителом вместе с флуоресцентным красителем для определения жизнеспособности (eBioscience) и анализировали посредством проточной цитометрии для измерения частоты CD52-положительных и CD52-отрицательных клеток среди живых клеток. Фигура 6 показывает результат типичного эксперимента, демонстрирующего, что CD52- отрицательные клетки полностью резистентны к комплемент-опосредованной токсичности анти-CD52 антител.
Функциональный анализ T-клеток с целевым геном TRAC
Целью инактивации гена TRAC является получение T-лимфоцитов, нечувствительных к стимуляции T-клеточного рецептора. Как описано в предыдущем абзаце, T-лимфоциты трансфицировали мРНК, кодирующей TALE-нуклеазу, расщепляющую TRAC или CD52. Через 16 дней после трансфекции клетки обрабатывали вплоть до 5 г/мл фитогемагглютинина (PHA, Sigma-Aldrich), Т-клеточного митогена, действующего через T-клеточный рецептор. Клетки с функциональным T-клеточным рецептором должны увеличиваться в размерах после обработки PHA. После трех дней инкубации клетки помечали конъюгированным с флуорохромом анти-CD52 или анти-TCR антителом и анализировали посредством проточной цитометрии для сравнения распределения размера клеток между TCR-положительными и TCR-отрицательными клетками или между CD52-положительными и CD52-отрицательными клетками. На фигуре 7 показано, что TCR-положительные клетки значительно увеличиваются в размерах после обработки PHA, тогда как TCR-отрицательные клетки имеют тот же размер, что и необработанные клетки, что указывает на то, что инактивация TRAC делает их нечувствительными к TCR-сигнализации. Напротив, CD52-положительные и CD52-отрицательные увеличиваются в размерах в одинаковой степени.
Функциональный анализ T-клеток с целевыми генами CD52 и TRAC
Для того, чтобы убедиться, что генная инженерия не влияет на способность Т-клеток демонстрировать противоопухолевую активность при снабжении химерным антигенным рецептором (CAR), авторы трансфицировали T-клетки, которые были целевыми для CD52-TALE-нуклеазы и TRAC-TALE-нуклеазы, 10 мкг РНК, кодирующей анти-CD19 CAR (SEQ ID NO:73). Через 24 часа T-клетки инкубировали в течение 4 часов с клетками Дауди, экспрессирующими CD19. Повышающую регуляцию CD107a, маркера цитотоксической гранулосекреции T-лимфоцитов (называемой дегрануляцией), на клеточной поверхности измеряли посредством анализа с помощью проточной цитометрии (Betts, Brenchley et al., 2003). Результаты представлены на фигуре 8 и показывают, что CD52-отрицательные/TCRαβ-отрицательные клетки и CD52-положительные/TCRαβ-положительные обладают одинаковой способностью к дегрануляции в ответ на PMA/иономицин (положительный контроль) или CD19+ клетки Дауди. Повышающая регуляция CD107 зависит от присутствия CD19+. Эти данные свидетельствуют о том, что генная инженерия не оказывает отрицательного воздействия на способность T-клеток к осуществлению контролируемого противоопухолевого ответа.
Безопасность CD52-TALE-нуклеазы и TRAC-TALE-нуклеазы для генома в первичных T-лимфоцитах
Поскольку наши конструкты включают нуклеазные субъединицы, важным вопросом является, может ли множественная TALE-нуклеазная трансфекция приводить к генотоксичности и расщеплению вне мишени при "близком соответствии" последовательностей-мишеней или посредством ошибочного спаривания половин TALE-нуклеазы. Для определения воздействия TRAC-TALE-нуклеазы и CD52-TALE-нуклеазы на целостность клеточных геномов авторы составили список последовательностей в геноме человека, которые дают возможность для расщепления вне сайта. Для создания данного списка авторы идентифицировали все последовательности в геноме с вплоть до 4 замен по сравнению с оригинальными полумишенями и затем идентифицировали пары потенциальных полумишеней в ориентации голова к голове со спейсером от 9 до 30 п.о. друг между другом. Данный анализ включал сайты, потенциально целевые для гомодимеров одной молекулы половины TALE-нуклеазы или гетеродимеров, образованных одной половиной TALE-нуклеазы CD52 и одной половиной TALE-нуклеазы TRAC. Авторы оценивали потенциальные мишени вне сайта на основании данных о специфичности, принимая во внимание стоимость отдельных замен и положение замен (причем несовпадения лучше переносятся основаниями с 3'-конца полумишени). Авторы получили 173 уникальные последовательности с оценкой, отражающей определение вероятности расщепления. Авторы выбрали 15 лучших оценок и проанализировали посредством глубокого секвенирования частоты мутаций, обнаруженных в данных локусах в T-клетках, одновременно трансфицированных TALE-нуклеазой CD52 и TRAC и очищенных посредством магнитного разделения на CD52-отрицательные, TCRαβ-отрицательные. Результаты приведены на фигуре 9. Самая высокая частота вставок/делеций составила 7×10-4. Данные результаты делают мутацию в гипотетической мишени вне сайта по меньшей мере в 600 раз менее вероятной, чем в предполагаемой мишени. Следовательно, TALE-нуклеазные реагенты, используемые в данном исследовании, оказываются чрезвычайно специфическими.
Пример 3: TALE-нуклеазы, расщепляющие ген CTLA4 человека и ген PDCD1 человека.
Как описано в примере 1, были разработаны и получены гетеродимерные TALE-нуклеазы, нацеленные соответственно на гены PDCD1 и CTLA4. Целевые геномные последовательности состоят из двух последовательностей длиной 17 п.о. (называемых полумишенями), разделенных спейсером в 11 или 15 п.о. Каждая полумишень распознается повторами половин TALE-нуклеаз, перечисленных в таблице 10.

Активность CTLA4-TALE-нуклеазы и PDCD1-TALE-нуклеазы в клетках HEK293
Каждый TALE-нуклеазный конструкт субклонировали, используя расщепление ферментом рестрикции в векторе экспрессии млекопитающих под контролем длинного промотора pEF1-альфа. Один миллион клеток HEK293 высевали за один день до трансфекции. Клетки котрансфицировали 2,5 мкг каждой из двух плазмид, кодирующих TALE-нуклеазы, распознающие две полумишени в представляющей интерес геномной последовательности в гене PDCD1 и CTLA-4 под контролем промотора EF1-альфа, или 5 мкг контрольного вектора pUC (pCLS0003), используя 25 мкл липофектамина (Invitrogen) в соответствии с инструкциями изготовителя. Двухнитевое расщепление, создаваемое TALE-нуклеазами в последовательностях, кодирующих PDCD1 или CTLA-4, восстанавливалось в живых клетках посредством негомологичного соединения концов (NHEJ), которое представляет собой механизм, подверженный ошибкам. Активность TALE-нуклеаз в живых клетках измеряли по частоте вставок или делеций в целевом геномном локусе. Через 48 часов после трансфекции геномную ДНК выделели из трансфицированных клеток, и осуществляли локус-специфические ПЦР с использованием следующих составных праймеров: 5'-CCATCTCATCCCTGCGTGTCTCCGACTCAG (прямая адапторная последовательность, SEQ ID NO:126)- 10N (TAG)- локус-специфическая прямая последовательность для CTLA4_T01: 5'-CTCTACTTCCTGAAGACCTG-3' (SEQ ID NO:141; составной праймер, раскрытый как SEQ ID NO:99), для CTLA4_T03/T04: 5'-ACAGTTGAGAGATGGAGGGG-3' (SEQ ID NO:142; составной праймер, раскрытый как SEQ ID NO:100), для PDCD1_T01: 5'-CCACAGAGGTAGGTGCCGC-3' (SEQ ID NO:143; составной праймер, раскрытый как SEQ ID NO:101) или для PDCD1_T03: 5'-GACAGAGATGCCGGTCACCA-3' (SEQ ID NO:144; составной праймер, раскрытый как SEQ ID NO:102) и обратный составной праймер 5'-CCTATCCCCTGTGTGCCTTGGCAGTCTCAG (обратная адапторная последовательность, SEQ ID NO:130)- эндогенная локус-специфическая обратная последовательность для CTLA4_T01: 5'-TGGAATACAGAGCCAGCCAA-3' (SEQ ID NO:145; составной праймер, раскрытый как SEQ ID NO:103), для CTLA4_T03/T04: 5'-GGTGCCCGTGCAGATGGAAT-3' (SEQ ID NO:146; составной праймер, раскрытый как SEQ ID NO:104), для PDCD1_T01: 5'-GGCTCTGCAGTGGAGGCCAG-3' (SEQ ID NO:147; составной праймер, раскрытый как SEQ ID NO:105) или для PDCD1_T03: 5'-GGACAACGCCACCTTCACCT-3' (SEQ ID NO:148; составной праймер, раскрытый как SEQ ID NO:106).
Продукты ПЦР анализировали посредством T7-эндонуклеазного анализа: кратко говоря, после денатурации и повторного отжига продукта ПЦР эндонуклеаза T7 специфически расщепляет ошибочную ДНК, состоящую из дикого типа и мутировавшей нитей. Продукт расщепления затем получают посредством электрофореза в полиакриламидном геле. Присутствие расщепленного продукта указывает на мутировавшие последовательности, индуцированные TALE-нуклеазной активностью. Результаты изображены на фигуре 10, где стрелки указывают на расщепленные продукты ПЦР. Они демонстрируют, что все TALE-нуклеазы PDCD1_T1, PDCD1_T3, CTLA4_T1, CTLA4_T3 и CTLA4_T4 проявляют мутагенную нуклеазную активность на своих сайтах-мишенях.
Пример 4: pT-альфа делает возможной поверхностную экспрессию CD3 в TCR-альфа-инактивированных T-лимфоцитах:
Описание различных вариантов preT-альфа:
Ген pT-альфа человека кодирует трансмембранный гликопротеин, содержащий внеклеточный Ig-подобный домен, гидрофобный трансмембранный домен и большой C-концевой внутрицитоплазматический хвост. Различные варианты, происходящие от pT-альфа гликопротеина человека, были разработаны и описаны в таблице 11 и представлены на фигуре 11.
Описание выборки конструктов pT-альфа
Различные исследованные конструкты preT-альфа включают:
1) Делеционные мутанты pT-альфа: Различные делеции были созданы во внутриклеточном цитоплазматическом хвосте белка pT-альфа человека (который содержит 114 аминокислот) (SEQ ID NO:107). Исследованные конструкты включают полноразмерный вариант белка (FL) и мутанты, в которых было удалено 18, 48, 62, 78, 92, 110 и 114 аминокислот с C-конца белка (от SEQ ID NO:108 до SEQ ID NO:114).
2) Мутанты pT-альфа, содержащие внутриклеточные активационные домены: Варианты FL и Δ48, слитые с внутриклеточными доменами активации CD8, CD28 или 41BB со своих C-концов (от SEQ ID NO:115 до SEQ ID NO:120).
3) Химерные мутанты pT-альфа/TCRα: В одном из конструктов внутриклеточный домен TCRα (IC) был слит с лишенным хвоста вариантом (Δ114) pT-альфа (SEQ ID NO:121). Также был создан второй конструкт, в котором внеклеточный домен pT-альфа был слит с трансмембранным (TM) и IC доменами от TCRα (SEQ ID NO:122).
4) Мутанты димеризации pT-альфа: Некоторые мутации были описаны в литературе как способные изменять способность к олигомеризации/димеризации комплекса пре-TCR. Предполагается, что данные мутанты делают возможной экспрессию пре-TCR на клеточной поверхности, не индуцируя конститутивную сигнализацию (как предполагается, индуцируемую при олигомеризации пре-TCR). Данные мутации были введены в вариант pT-альфа-Δ48 и представляют собой:
- 1×MUT: W46R (SEQ ID NO:123)
- 4×MUT: D22A, K24A, R102A, R117A (SEQ ID NO:124).
Активность различных конструктов preT-альфа в TRAC-инактивированных клетках Jurkat:
Для скрининга различных вариантов pT-альфа на их способность восстанавливать поверхностную экспрессию CD3 в TCR-альфа-инактивированных клетках была создана клеточная линия, в которой ген TCR-альфа был разрушен с помощью TALEN, нацеленной на TRAC. Клетки Jurkat (T-клеточная клеточная линия лейкемии) трансфицировали плазмидами, кодирующими для TALEN, расщепляющую TRAC, с помощью электропорации CytoPulse, и затем были очищены клетки KO (TCRα/βNEG; CD3NEG) посредством отрицательной селекции с помощью магнитных гранул CD3. Популяцию KO (JKT_KOx3 клетки) амплифицировали и использовали для скрининга различных вариантов pT-альфа. Скрининг проводили посредством трансфекции одного миллиона клеток JKT_KOx3 15 мкг плазмиды, кодирующей различные варианты pT-альфа, под контролем промотора EF1α, с последующим анализом посредством проточной цитометрии клеточной поверхностной экспрессии CD3 через 48 часов после трансфекции. Фигура 12 представляет собой типичный пример эффективности трансфекции (% BFP+ клеток) и активности конструктов pT-альфа FL, Δ18 и Δ48 в клетках JKT_KOx3 на основании % CD3+ клеток, определенного посредством проточной цитометрии. Результаты от различных конструктов сгруппированы в таблице 12.
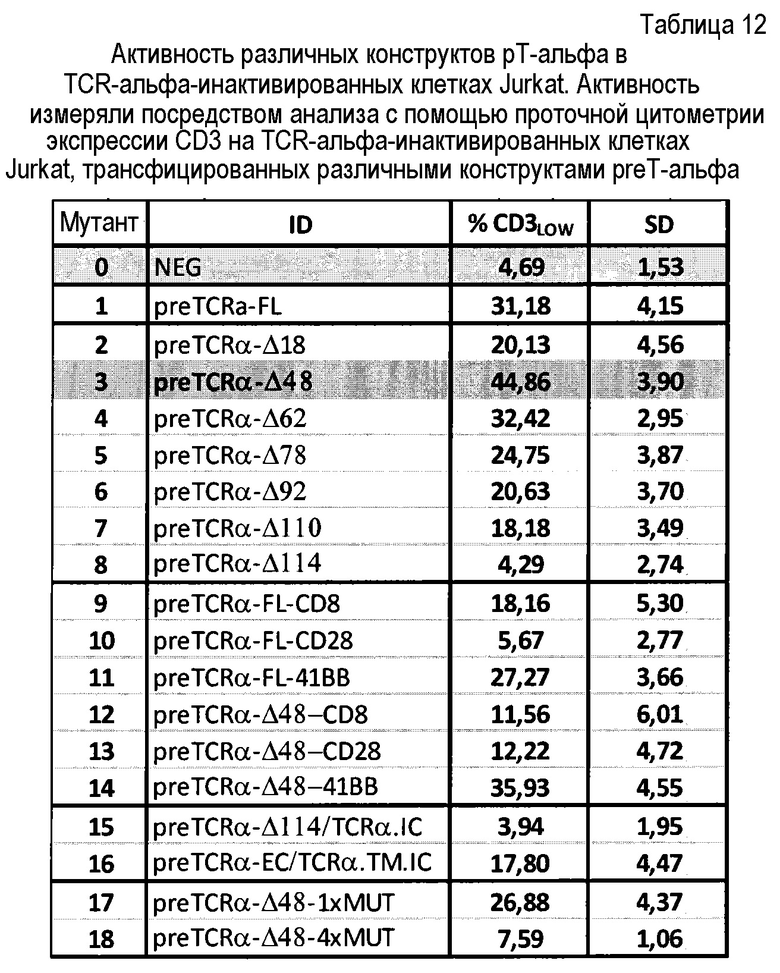
Активность pT-альфа-FL и pT-альфа-Δ48 в TCR-альфа-инактивированных первичных T-лимфоцитах:
Для того, чтобы проверить способность вариантов pT-альфа-FL и pT-альфа-Δ48 индуцировать поверхностную экспрессию CD3 в TCR-альфа-инактивированных T-лимфоцитах, последовательности, кодирующие pT-альфа-FL и pT-альфа-Δ48 клонировали в самоинактивирующийся лентивирусный вектор pLV-SFFV-BFP-2A-PCTRA, который кодирует синий флуоресцентный белок (BFP) под контролем промотора SFFV, за которым следует саморасщепляющийся пептид T2A (фигура 13).
T-лимфоциты, выделенные из периферической крови, активировали в течение 72 часов с использованием гранул активатора анти-CD3/CD28 (Life technologies), и 4,5 миллиона клеток трансфицировали посредством электропорации 10 мкг мРНК, кодирующей TALE-нуклеазу, нацеленную на участок константной цепи TCR-альфа (TRAC), используя прибор CytoLVT-S (BTX-Harvard Harbour). Через два дня после электропорации T-клетки трансдуцировали лентивирусными векторами или LV-SFFV-BFP-2A-pT-альфа-Δ48, или LV-SFFV-BFP-2A-control. CD3-отрицательные и с низким содержанием CD3 T-клетки затем очищали с помощью магнитных гранул анти-CD3 (Miltenyi Biotech). Этот экспериментальный протокол представлен на фигуре 14A.
Фигура 14B изображает анализ с помощью проточной цитометрии клеточной поверхностной экспрессии TCR-альфа/бета, CD3 и экспрессии BFP на TCR-альфа-инактивированных T-клетках (KO), трансдуцированных лентивирусным вектором или BFP-2A-pT-альфа-Δ48 (KO/Δ48), или контрольным BFP (KO/BFP), до и после очистки с помощью гранул CD3. TCR-альфа-инактивированные клетки, трансдуцированные вектором BFP-T2A-pT-альфа-Δ48 (BFP+ клетки), демонстрируют более высокий уровень CD3 по сравнению с нетрансдуцированными клетками (BFP- клетки). Никаких различий не наблюдается среди клеток, трансдуцированных контрольным вектором BFP. Эти результаты показывают, что pT-альфа опосредует восстановление экспрессии CD3 на клеточной поверхности TCR-альфа-инактивированных клеток. Напротив, окрашивание TCR-альфа/бета остается, как и ожидалось, без изменений в клетках, трансдуцированных или нет вектором экспрессии pT-альфа-Δ48.
pT-альфа-опосредованная экспрессия CD3 поддерживает активацию TCR-дефицитных T-клеток:
Для определения способности pT-альфа передавать сигналы активации клеток, анализировали экспрессию маркеров ранней и поздней активации на TCR-альфа-инактивированных T-клетках, трансдуцированных pT-альфа-Δ48 и pT-альфа-Δ48.41BB. TCR-альфа-инактивированные T-клетки, трансдуцированные pT-альфа-Δ48 и pT-альфа-Δ48.41BB, создавали из первичных T-клеток человека, как описано в предыдущем разделе и на фигуре 14A.
Для детектирования сигнализации посредством CD3 клетки реактивировали с помощью покрытых анти-CD3/CD28 гранул через 3 дня после очистки TCR-альфа-инактивированных T-клеток с помощью гранул CD3 (фигура 14A). Клетки окрашивали конъюгированными с флуорохромом анти-CD69 (маркер ранней активации) и анти-CD25 (маркер поздней активации), соответственно, через 24 и 48 часов после реактивации и анализировали посредством проточной цитометрии (фигура 15A-B). Как показано на фигуре 15A-B, TCR-альфа-инактивированные клетки, экспрессирующие pT-альфа-Δ48 (KO/pΤα-Δ48) или pT-альфа-Δ48.41BB (KO/pΤα-Δ48.ΒΒ), демонстрируют повышающую регуляцию маркеров активации до уровней, аналогичных наблюдаемым в экспрессирующих TCR-альфа/бета клетках (NEP: не электропорированные клетки).
Другим индикатором активации T-клеток является увеличение размера клеток, которые иногда называют "взрывом". Способность комплексов пре-TCR индуцировать "взрыв" измеряли посредством анализа с помощью проточной цитометрии размера клеток через 72 часа после реактивации с помощью гранул анти-CD3/CD28 (фигура 15C). Стимуляция с помощью гранул анти-CD3/CD28 индуцировала сопоставимое увеличение размера клеток в клетках, экспрессирующих комплексы TCR-альфа/бета, по сравнению с клетками, экспрессирующими pT-альфа-Δ48 или pT-альфа-Δ48.41BB. Взятые вместе, эти результаты показывают, что комплексы пре-TCR могут передавать сигналы, которые эффективно связываются с механизмами, опосредующими повышающую регуляцию маркеров активации.
Экспрессия CD3, опосредованная pT-альфа, поддерживает размножение TCR-дефицитных первичных T-клеток с помощью стимулирующих анти-CD3/CD28 антител
Для определения способности комплексов пре-TCR поддерживать долгосрочную клеточную пролиферацию измеряли пролиферацию клеток, созданных, как описано выше. Десять дней после начальной активации клетки выдерживали в IL2 (нет реактивации) или в IL2 с гранулами анти-CD3/CD28 (реактивация). Для всех условий клетки подсчитывали и анализировали с помощью проточной цитометрии в различных временных точках для определения количества BFP+ клеток. Сравнивали рост TCR-альфа-инактивированных клеток (KO), трансдуцированных векторами BFP или BFP-T2A-пре-TCRα-Δ48, и определяли, во сколько раз изменится индукция данных клеток по отношению к значению, полученному на день 2 после реактивации. Фигура 16 показывает результаты, полученные с двумя независимыми донорами. В обоих случаях TCR-альфа-инактивированные клетки, экспрессирующие pT-альфа-Δ48, демонстрировали большее размножение, чем TCR-альфа-инактивированные клетки, экспрессирующие только контрольный вектор BFP. Для второго донора также рассмотрели TCR-альфа-инактивированные клетки, экспрессирующие pT-альфа-Δ48.41BB или полноразмерный pT-альфа, продемонстрировавшие также большее размножение, чем TCR-альфа-инактивированные клетки, экспрессирующие только контрольный вектор BFP.
Пример 5: оптимизация трансфекции мРНК в T-клетках с помощью метода CytoPulse.
Определение оптимизированной программы CytoPulse
Первую серию экспериментов проводили на не активированных PBMC, для того чтобы определить диапазон напряжения, в котором клетки могут быть трансфицированы. Было протестировано пять различных программ, как описано в таблице 13.
Различные программы CytoPulse, использованные для определения минимального напряжения, требуемого для электропорации в происходящих от PBMC T-клетках
3 или 6 миллионов клеток электропорировали в кювете с зазором 0,4 см (30 или 15×106 клеток/мл) с 20 мкг плазмид, кодирующих GFP, и контрольных плазмид pUC, используя различные программы CytoPulse. Чрез 24 часа после электропорации анализировали экспрессию GFP в электропорированных клетках с помощью проточной цитометрии для определения эффективности трансфекции. Данные, показанные на фигуре 17, указывают минимальное напряжение, требуемое для электропорации плазмид в происходящих от PBMC T-клетках. Эти результаты демонстрируют, что программы CytoPulse 3 и 4 делают возможной эффективную трансформацию T-клеток (EP#3 и #4).
Электропорация мРНК в очищенных активированных T-клетках
После определения лучшей программы CytoPulse, которая делает возможной эффективную электропорацию ДНК в T-клетах, авторы проверили, применим ли данный способ к электропорации мРНК.
5×106 очищенных T-клеток, предварительно активированных в течение 6 дней с помощью PHA/IL2, ресуспендировали в буфере cytoporation T (BTX-Harvard apparatus) и электропорировали в кюветах 0,4 см 10 мкг мРНК, кодирующей GFP, или 20 мкг плазмид, кодирующих GFP или pUC, используя предпочтительную программу CytoPulse, как определено в предыдущем разделе (таблица 14).
Программа CytoPulse, использованная для электропорации очищенных T-клеток
Через 48 часов после трансфекции клетки окрашивали красителем для определения жизнеспособности (eFluor-450) и определяли клеточную жизнеспособность и % жизнеспособных GFP+ клеток посредством анализа с помощью проточной цитометрии (фигура 18).
Данные, представленные на фигуре 18, указывают что электропорация РНК при определенных здесь оптимальных условиях является не токсичной и делает возможной трансфекцию больше чем 95% жизнеспособных клеток.
В итоге, весь набор данных показывает, что Т-клетки могут быть эффективно трансфицированы или ДНК, или РНК. В частности, трансфекция РНК не оказывает влияния на клеточную жизнеспособность и делает возможным однородный уровень экспрессии представляющего интерес трансфицированного гена в клеточной популяции.
Эффективная трансфекция может быть достигнута вскоре после клеточной активации, независимо от используемого способа активации (PHA/IL-2 или покрытые CD3/CD28 гранулы). Изобретатели добились успеха в трансфекции клеток от 72 часов после активации с эффективностью >95%. Кроме того, также может быть получена эффективная трансфекция T-клеток после оттаивания и активации с помощью того же протокола электропорации.
Электропорация мРНК в первичных T-клетках человека для функциональной экспрессии TALE-нуклеазы
После демонстрации того, что электропорация мРНК делает возможной эффективную экспрессию GFP в первичных T-клетках человека, авторы проверили, применим ли данный способ к экспрессии других представляющих интерес белков. Нуклеазы на основе подобных активатору транскрипции эффекторов (TALE-нуклеазы) представляют собой сайт-специфические нуклеазы, создаваемые посредством слияния ДНК-связывающего домена TAL с ДНК-расщепляющим доменом. Они являются мощными инструментами редактирования генома, поскольку они индуцируют двухнитевые разрывы практически в любой желаемой последовательности ДНК. Эти двухнитевые разрывы активируют негомологичное соединение концов (NHEJ), подверженный ошибкам механизм репарации ДНК, потенциально ведущий к инактивации любого желаемого представляющего интерес гена. Альтернативно, если в то же время в клетки ввести адекватный шаблон репарации, индуцируемые TALE-нуклеазой разрывы ДНК могут быть восстановлены посредством гомологичной рекомбинации, что предлагает возможность модификации последовательности генов по желанию.
Авторы использовали электропорацию мРНК для экспрессии TALE-нуклеазы, разработанной так, чтобы специфически расщеплять последовательность в гене человека, кодирующем альфа-цепь T-клеточного антигенного рецептора (TRAC). Ожидается, что мутации, индуцируемые в данной последовательности, приведут к инактивации гена и исчезновению комплекса TCRαβ с клеточной поверхности. РНК TALE-нуклеазы TRAC или не кодирующую РНК в качестве контроля трансфицировали в активированные первичные T-лимфоциты человека с помощью метода CytoPulse. Электропорационная последовательность состояла из 2 импульсов по 1200 В с последующими четырьмя импульсами по 130 В, как описано в таблице 14.
Посредством анализа с помощью проточной цитометрии поверхностной экспрессии TCR через 7 дней после электропорации (фигура 19, верхняя панель) авторы обнаружили, что 44% T-клеток лишились экспрессии TCRαβ. Авторы проанализировали геномную ДНК трансфицированных клеток с помощью ПЦР-амплификации локуса TRAC с последующим высокопроизводительным секвенированием 454. 33% секвенированных аллелей (727 из 2153) содержали вставку или делецию в сайте расщепления TALE-нуклеазы. Фигура 19 (нижняя панель) показывает примеры мутировавших аллелей.
Эти данные показывают, что электропорация мРНК с помощью метода CytoPulse приводит к функциональной экспрессии TALE-нуклеазы TRAC.
Электропорация T-клеток моноцистронной мРНК, кодирующей одноцепочечный химерный антигенный анти-CD19 рецептор (CAR):
5×106 T-клеток, предварительно активированных в течение нескольких дней (3-5) с помощью покрытых анти-CD3/CD28 гранул и IL2, ресуспендировали в буфере cytoporation T и электропорировали в кюветах 0,4 см без мРНК или с 10 мкг мРНК, кодирующей одноцепочечный CAR (SEQ ID NO:73), используя программу, описанную в таблице 14.
Через 24 часа после электропорации клетки окрашивали с помощью фиксируемого красителя для определения жизнеспособности eFluor-780 и PE-конъюгированного фрагмента F(ab')2 козьего антимышиного IgG, специфического для определения клеточной поверхностной экспрессии данного CAR на живых клетках. Данные показаны на фигуре 20. Они демонстрируют, что подавляющее большинство живых Т-клеток, электропорированных моноцистронной мРНК, описанной выше, экспрессируют CAR на своей поверхности.
Через 24 часа после электропорации T-клетки совместно культивировали с клетками Дауди (CD19+) в течение 6 часов и анализировали с помощью проточной цитометрии для детектирования экспрессии маркера дегрануляции CD107a на своей поверхности (Betts, Brenchley et al., 2003).
Данные, показанные на фигуре 20, демонстрируют, что большинство клеток, электропорированных моноцистронной мРНК, описанной выше, подвергаются дегрануляции в присутствии клеток-мишеней, экспрессирующих CD19. Эти результаты ясно демонстрируют, что CAR, экспрессированный на поверхности электропорированных T-клеток, активен.
Электропорация T-клеток полицистронной мРНК, кодирующей мультисубъединичный химерный антигенный анти-CD19 рецептор (CAR):
5×106 T-клеток, предварительно активированных в течение нескольких дней (3-5) с помощью покрытых анти-CD3/CD28 гранул и IL2, электропорировали в буфере cytoporation T и электропорировали в кюветах 0,4 см без мРНК или с 45 мкг мРНК, кодирующей многоцепочечный CAR (SEQ ID NO:226) (фигура 21A и фигура 4B (csm4)), используя программу, описанную в таблице 14.
Через 24 часа после электропорации клетки окрашивали с помощью фиксируемого красителя для определения жизнеспособности eFluor-780 и PE-конъюгированного фрагмента F(ab')2 козьего антимышиного IgG, специфического для определения клеточной поверхностной экспрессии данного CAR на живых клетках. Данные, показанные на фигуре 21, демонстрируют, что подавляющее большинство живых Т-клеток, электропорированных полицистронной мРНК, описанной выше, экспрессируют CAR на своей поверхности.
Через 24 часа после электропорации T-клетки совместно культивировали с клетками Дауди (CD19+) в течение 6 часов и анализировали с помощью проточной цитометрии для детектирования экспрессии маркера дегрануляции CD107a на своей поверхности. Данные, показанные на фигуре 21 демонстрируют, что большинство клеток, электропорированных полицистронной мРНК, описанной выше, подвергаются дегрануляции в присутствии клеток-мишеней, экспрессирующих CD19. Эти результаты ясно демонстрируют, что CAR, экспрессированный на поверхности электропорированных T-клеток, активен.
Пример 6: Многоцепочечные CAR
A. Конструирование многоцепочечных CAR
Девять многоцепочечных CAR, нацеленных на антиген CD19, были разработаны на основании высокоаффинного рецептора для IgE (FcεRI). FcεRI, экспрессированный на тучных клетках и базофилах, вызывает аллергические реакции. Он представляет собой тетрамерный комплекс, состоящий из одной α-субъединицы (SEQ ID NO:202), одной β-субъединицы (SEQ ID NO:203) и двух дисульфидно-сшитых γ-субъединиц (SEQ ID NO:204). α-субъединица содержит IgE-связывающий домен. β и γ-субъединицы содержат ITAM (SEQ ID NO:198 и SEQ ID NO:199), которые опосредуют передачу сигнала (фигура 4A). Многоцепочечные CAR были разработаны, как описано на фигурах 4B, C и в таблице 15. В каждом многоцепочечном CAR внеклеточный домен цепи FcRα удаляли и заменяли на 4G7scFv и шарнир CD8α (SEQ ID NO:206). В многоцепочечных CAR csm2, csm4, csm5, csm8 и csm10 удаляли ITAM цепи FcRβ и/или цепи FcRγ. В многоцепочечных CAR csm2, csm4, csm5, csm6, csm8, csm9 и csm10 добавляли 3 ITAM CD3ζ (SEQ ID NO:197) к цепи FcRβ или цепи FcRγ. В многоцепочечных CAR csm2, csm4, csm5, csm6, csm7, csm8, csm9 и csm10 добавляли внутриклеточный домен 4-1BB (SEQ ID NO:200) к цепи FcRα, цепи FcRβ или цепи FcRγ. В многоцепочечном CAR csm10 добавляли внутриклеточный домен CD28 к цепи FcRα (SEQ ID NO:201).
Описание составов цепей вариантов многоцепочечных CAR
(SEQ ID NO:206)
(SEQ ID NO:203)
(SEQ ID NO:204)
(SEQ ID NO:206)
(SEQ ID NO:203)
(SEQ ID NO:212)
(SEQ ID NO:206)
(SEQ ID NO:208)
(SEQ ID NO:213)
(SEQ ID NO:206)
(SEQ ID NO:209)
(SEQ ID NO:204)
(SEQ ID NO:206)
(SEQ ID NO:210)
(SEQ ID NO:214)
(SEQ ID NO:207)
(SEQ ID NO:203)
(SEQ ID NO:204)
(SEQ ID NO:207)
(SEQ ID NO:211)
(SEQ ID NO:204)
(SEQ ID NO:207)
(SEQ ID NO:203)
(SEQ ID NO:214)
(SEQ ID NO:206)
(SEQ ID NO:208)
(SEQ ID NO:213)
B. Временно экспрессированные многоцепочечные CAR могут вызывать активацию T-клеток
Многоцепочечные CAR могут быть экспрессированы в T-клетках человека после электропорации полицистронной мРНК.
T-клетки электропорировали кэпированной и полиаденилированой полицистронной мРНК (фигура 21A), которую получали с помощью набора mMESSAGE mMACHINE и линеаризованных плазмид в качестве шаблона. Плазмиды, используемые в качестве шаблона, содержали РНК-полимеразный промотор T7, за которым следовала полицистронная последовательность ДНК, кодирующая csm1, csm2, csm4, csm5, csm6, csm7, csm8, csm9 или csm10 (SEQ ID NO:224-232).
Электропорацию полицистронных мРНК в T-клетки человека осуществляли с помощью устройства CytoLVT-S (Cellectis) в соответствии со следующим протоколом: 5×106 T-клеток, предварительно активированных в течение нескольких дней (3-5) с помощью покрытых анти-CD3/CD28 гранул и IL2, ресуспендировали в буфере cytoporation T и электропорировали в кюветах 0,4 см 45 мкг мРНК, используя программу PBMC3 (таблица 14).
Через 24 часа после электропорации T-клетки человека, сконструированные с помощью полицистронных мРНК, кодирующих многоцепочечные CAR, помечали с помощью фиксируемого красителя для определения жизнеспособности eFluor-780 и специфического PE-конъюгированного фрагмента F(ab')2 козьего антимышиного IgG и анализировали с помощью проточной цитометрии. Данные, показанные на фигуре 22, демонстрируют, что живые T-клетки, сконструированные с помощью полицистронных мРНК, экспрессируют многоцепочечные CAR csm1, csm2, csm4, csm5, csm6, csm7, csm8, csm9 и csm10.
Как описано в литературе для FcεRI, экспрессия мультисубъединичных CAR обуславливается экспрессией 3 цепей. Одна только цепь α экспрессируется меньше, чем комплекс цепей α+γ, и комплекс цепей α+γ экспрессируется меньше, чем комплекс цепей α+β+γ (фигура 23).
T-клетки человека, временно экспрессирующие многоцепочечные CAR, подвергаются дегрануляции после совместного культивирования с клетками-мишенями
Через 24 часа после электропорации T-клетки человека, сконструированные с помощью полицистронных мРНК, кодирующих многоцепочечные CAR, культивировали совместно с целевыми (Дауди) или контрольными (K562) клетками в течение 6 часов. Затем CD8+ T-клетки анализировали с помощью проточной цитометрии для детектирования экспрессии маркера дегрануляции CD107a на своей поверхности. Данные, показанные на фигуре 24, демонстрируют, что CD8+ T-клетки человека, экспрессирующие многоцепочечные CAR csm1, csm2, csm4, csm5, csm6, csm7, csm8, csm9 и csm10, подвергаются дегрануляции при совместном культивировании с экспрессирующими CD19 клетками-мишенями, но не при совместном культивировании с контрольными клетками.
T-клетки человека, временно экспрессирующие многоцепочечные CAR, секретируют цитокины после совместного культивирования с клетками-мишенями
Через 24 часа после электропорации T-клетки человека, сконструированные с помощью полицистронных мРНК, кодирующих многоцепочечные CAR, культивировали совместно с целевыми (Дауди) или контрольными (K562) клетками в течение 24 часов. Затем супернатанты собирали и анализировали с помощью цитометрического набора с множеством гранул для цитокинов TH1/TH2, для того чтобы определить количество цитокинов, производимых T-клетками. Данные, показанные на фигуре 25A-C, демонстрируют, что T-клетки человека, экспрессирующие многоцепочечные CAR csm1, csm2, csm4, csm5, csm6, csm7, csm8, csm9 и csm10, производят IFNγ, IL8 и IL5 при совместном культивировании с экспрессирующими CD19 клетками-мишенями, но не при совместном культивировании с контрольными клетками.
T-клетки человека, временно экспрессирующие многоцепочечные CAR, лизируют клетки-мишени
Через 24 часа после электропорации T-клетки человека, сконструированные с помощью полицистронных мРНК, кодирующих многоцепочечные CAR, культивировали совместно с целевыми (Дауди) или контрольными (K562) клетками в течение 4 часов. Затем клетки-мишени анализировали с помощью проточной цитометрии для анализа их жизнеспособности. Данные, показанные на фигуре 26, демонстрируют, что клетки, экспрессирующие многоцепочечные CAR csm1, csm2, csm4, csm5, csm6, csm7, csm8, csm9 и csm10, лизируют экспрессирующие CD19 клетки-мишени, но не контрольные клетки.
Список ссылок, цитированных в настоящем описании
Arimondo, P. B., C. J. Thomas, et al. (2006). "Exploring the cellular activity of camptothecin-triple-helix-forming oligonucleotide conjugates." Mol Cell Biol 26(1): 324-33.
Arnould, S., P. Chames, et al. (2006). "Engineering of large numbers of highly specific homing endonucleases that induce recombination on novel DNA targets." J Mol Biol 355(3): 443-58.
Ashwell, J. D. and R. D. Klusner (1990). "Genetic and mutational analysis of the T-cell antigen receptor." Annu Rev Immunol 8: 139-67.
Betts, M. R., J. M. Brenchley, et al. (2003). "Sensitive and viable identification of antigen-specific CD8+ T cells by a flow cytometric assay for degranulation." J Immunol Methods 281(1-2): 65-78.
Boch, J., H. Scholze, et al. (2009). "Breaking the code of DNA binding specificity of TAL-type III effectors." Science 326(5959): 1509-12.
Boni, A., P. Muranski, et al. (2008). "Adoptive transfer of allogeneic tumor-specific T cells mediates effective regression of large tumors across major histocompatibility barriers." Blood 112(12): 4746-54.
Brahmer, J. R., C. G. Drake, et al. (2010). "Phase I study of single-agent anti-programmed death-1 (MDX-1106) in refractory solid tumors: safety, clinical activity, pharmacodynamics, and immunologic correlates." J Clin Oncol 28(19): 3167-75.
Cambier, J. C. (1995). "Antigen and Fc receptor signaling. The awesome power of the immunoreceptor tyrosine-based activation motif (ITAM)." J Immunol 155(7): 3281-5.
Carrasco, Y. R., A. R. Ramiro, et al. (2001). "An endoplasmic reticulum retention function for the cytoplasmic tail of the human pre-T cell receptor (TCR) alpha chain: potential role in the regulation of cell surface pre-TCR expression levels." J Exp Med 193(9): 1045- 58.
Cermak, T., E. L. Doyle, et al. (2011). "Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting." Nucleic Acids Res 39(12): e82.
Chames, P., J. C. Epinat, et al. (2005). "In vivo selection of engineered homing endonucleases using double-strand break induced homologous recombination." Nucleic Acids Res 33(20): e178.
Choulika, A., A. Perrin, et al. (1995). "Induction of homologous recombination in mammalian chromosomes by using the l-Scel system of Saccharomyces cerevisiae." Mol Cell Biol 15(4): 1968-73.
Christian, M., T. Cermak, et al. (2010). "Targeting DNA double-strand breaks with TAL effector nucleases." Genetics 186(2): 757-61.
Coutinho, A. E. and K. E. Chapman (2011). "The anti-inflammatory and immunosuppressive effects of glucocorticoids, recent developments and mechanistic insights." Mol Cell Endocrinol 335(1): 2-13.
Critchlow, S. E. and S. P. Jackson (1998). "DNA end-joining: from yeast to man." Trends Biochem Sci 23(10): 394-8.
Deng, D., C. Yan, et al. (2012). "Structural basis for sequence-specific recognition of DNA by TAL effectors." Science 335(6069): 720-3.
Eisenschmidt, K., T. Lanio, et al. (2005). "Developing a programmed restriction endonuclease for highly specific DNA cleavage." Nucleic Acids Res 33(22): 7039-47.
Epinat, J. C, S. Arnould, et al. (2003). "A novel engineered meganuclease induces homologous recombination in yeast and mammalian cells." Nucleic Acids Res 31(11):2952-62.
Geissler, R., H. Scholze, et al. (2011). "Transcriptional activators of human genes with programmable DNA-specificity." PLoS One 6(5): e19509.
Howard, F. D., H. R. Rodewald, et al. (1990). "CD3 zeta subunit can substitute for the gamma subunit of Fc epsilon receptor type I in assembly and functional expression of the high-affinity IgE receptor: evidence for interreceptor complementation." Proc Natl Acad Sci USA 87(18): 7015-9.
Huang, P., A. Xiao, et al. (2011). "Heritable gene targeting in zebrafish using customized TALENs." Nat Biotechnol 29(8): 699-700.
Jena, B., G. Dotti, et al. (2010). "Redirecting T-cell specificity by introducing a tumor-specific chimeric antigen receptor." Blood 116(7): 1035-44.
Kalish, J. M. and P. M. Glazer (2005). "Targeted genome modification via triple helix formation." Ann N Y Acad Sci 1058: 151-61.
Li, L., M. J. Piatek, et al. (2012). "Rapid and highly efficient construction of TALE-based transcriptional regulators and nucleases for genome modification." Plant Mol Biol 78(4-5): 407-16.
Li, T., S. Huang, et al. (2011). "TAL nucleases (TALNs): hybrid proteins composed of TAL effectors and Fokl DNA-cleavage domain." Nucleic Acids Res 39(1): 359-72.
Li, T., S. Huang, et al. (2011). "Modularly assembled designer TAL effector nucleases for targeted gene knockout and gene replacement in eukaryotes." Nucleic Acids Res 39(14): 6315-25.
Ma, J. L., E. M. Kim, et al. (2003). "Yeast Mre11 and Rad1 proteins define a Ku-independent mechanism to repair double-strand breaks lacking overlapping end sequences." Mol Cell Biol 23(23): 8820-8.
Mahfouz, M. M., L. Li, et al. (2012). "Targeted transcriptional repression using a chimeric TALE-SRDX repressor protein." Plant Mol Biol 78(3): 311-21.
Mahfouz, M. M., L. Li, et al. (2011). "De novo-engineered transcription activator-like effector (TALE) hybrid nuclease with novel DNA binding specificity creates double-strand breaks." Proc Natl Acad Sci USA 108(6): 2623-8.
Mak, A. N., P. Bradley, et al. (2012). "The crystal structure of TAL effector PthXo1 bound to its DNA target." Science 335(6069): 716-9.
Metzger, H., G. Alcaraz, et al. (1986). "The receptor with high affinity for immunoglobulin E." Annu Rev Immunol 4: 419-70.
Miller, J. C., S. Tan, et al. (2011). "A TALE nuclease architecture for efficient genome editing." Nat Biotechnol 29(2): 143-8.
Morbitzer, R., P. Romer, et al. (2011). "Regulation of selected genome loci using de novo- engineered transcription activator-like effector (TALE)-type transcription factors." Proc Natl Acad Sci USA 107(50): 21617-22.
Moscou, M. J. and A. J. Bogdanove (2009). "A simple cipher governs DNA recognition by TAL effectors." Science 326(5959): 1501.
Mussolino, C, R. Morbitzer, et al. (2011). "A novel TALE nuclease scaffold enables high genome editing activity in combination with low toxicity." Nucleic Acids Res 39(21):9283-93.
Pang, S. S., R. Berry, et al. (2010). "The structural basis for autonomous dimerization of the pre-T-cell antigen receptor." Nature 467(7317): 844-8.
Paques, F. and P. Duchateau (2007). "Meganucleases and DNA double-strand break-induced recombination: perspectives for gene therapy." Curr Gene Ther 7(1): 49-66.
Pardoll, D. and C. Drake (2012). "Immunotherapy earns its spot in the ranks of cancer therapy." J Exp Med 209(2): 201-9.
Pardoll, D. M. (2012). "The blockade of immune checkpoints in cancer immunotherapy." Nat Rev Cancer 12(4): 252-64.
Park, T. S., S. A. Rosenberg, et al. (2011). "Treating cancer with genetically engineered T cells." Trends Biotechnol 29(11): 550-7.
Pingoud, A. and G. H. Silva (2007). "Precision genome surgery." Nat Biotechnol 25(7): 743-4.
Porteus, M. H. and D. Carroll (2005). "Gene targeting using zinc finger nucleases." Nat Biotechnol 23(8): 967-73.
Robert, C. and C. Mateus (2011). "[Anti-CTLA-4 monoclonal antibody: a major step in the treatment of metastatic melanoma]." Med Sci (Paris) 27(10): 850-8.
Rouet, P., F. Smih, et al. (1994). "Introduction of double-strand breaks into the genome of mouse cells by expression of a rare-cutting endonuclease." Mol Cell Biol 14(12):8096-106.
Saint-Ruf, C, O. Lechner, et al. (1998). "Genomic structure of the human pre-T cell receptor alpha chain and expression of two mRNA isoforms." Eur J Immunol 28(11): 3824-31.
Sander, J. D., L. Cade, et al. (2011). "Targeted gene disruption in somatic zebrafish cells using engineered TALENs." Nat Biotechnol 29(8): 697-8.
Smith, J., S. Grizot, et al. (2006). "A combinatorial approach to create artificial homing endonucleases cleaving chosen sequences." Nucleic Acids Res 34(22): e149.
Stoddard, B. L. (2005). "Homing endonuclease structure and function." Q Rev Biophys 38(1):49-95.
Tesson, L, C. Usal, et al. (2011). "Knockout rats generated by embryo microinjection of TALENs." Nat Biotechnol 29(8): 695-6.
von Boehmer, H. (2005). "Unique features of the pre-T-cell receptor alpha-chain: not just a surrogate." Nat Rev Immunol 5(7): 571-7.
Waldmann, H. and G. Hale (2005). "CAMPATH: from concept to clinic." Philos Trans R Soc Lond B Biol Sci 360(1461): 1707-11.
Weber, E., R. Gruetzner, et al. (2011). "Assembly of designer TAL effectors by Golden Gate cloning." PLoS One 6(5): e19722.
Yamasaki, S., E. Ishikawa, et al. (2006). "Mechanistic basis of pre-T cell receptor-mediated autonomous signaling critical for thymocyte development." Nat Immunol 7(1): 67-75.
Zhang, F., L. Cong, et al. (2011). "Efficient construction of sequence-specific TAL effectors for modulating mammalian transcription." Nat Biotechnol 29(2): 149-53.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ КОНСТРУИРОВАНИЯ АЛЛОГЕННЫХ И ВЫСОКОАКТИВНЫХ Т-КЛЕТОК ДЛЯ ИММУНОТЕРАПИИ | 2014 |
|
RU2736616C2 |
| СПОСОБ СОЗДАНИЯ Т-КЛЕТОК, ПРИГОДНЫХ ДЛЯ АЛЛОГЕННОЙ ТРАНСПЛАНТАЦИИ | 2015 |
|
RU2752933C2 |
| СПОСОБЫ КОНСТРУИРОВАНИЯ ВЫСОКОАКТИВНЫХ Т-КЛЕТОК ДЛЯ ИММУНОТЕРАПИИ | 2014 |
|
RU2725542C2 |
| Химерные антигенные рецепторы, нацеленные на антиген созревания B-клеток | 2016 |
|
RU2706582C2 |
| СПОСОБ КОНСТРУИРОВАНИЯ АЛЛОГЕННЫХ И УСТОЙЧИВЫХ К ЛЕКАРСТВЕННЫМ ПРЕПАРАТАМ Т-КЛЕТОК ДЛЯ ИММУНОТЕРАПИИ | 2014 |
|
RU2689558C1 |
| ИНСЕРЦИЯ ТАРГЕТНОГО ГЕНА ДЛЯ УЛУЧШЕННОЙ КЛЕТОЧНОЙ ИММУНОТЕРАПИИ | 2017 |
|
RU2824204C2 |
| CD33-СПЕЦИФИЧЕСКИЕ ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ ДЛЯ ИММУНОТЕРАПИИ РАКА | 2015 |
|
RU2701341C2 |
| НАЦЕЛИВАНИЕ НА КЛЕТКИ, ОПОСРЕДУЕМОЕ ХИМЕРНЫМ АНТИГЕННЫМ РЕЦЕПТОРОМ | 2018 |
|
RU2783316C2 |
| CD19-СПЕЦИФИЧЕСКИЙ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР И ЕГО ПРИМЕНЕНИЯ | 2014 |
|
RU2727447C2 |
| СПОСОБЫ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ ИСТОЩЕНИЯ TCRαβ КЛЕТОК | 2020 |
|
RU2829624C2 |


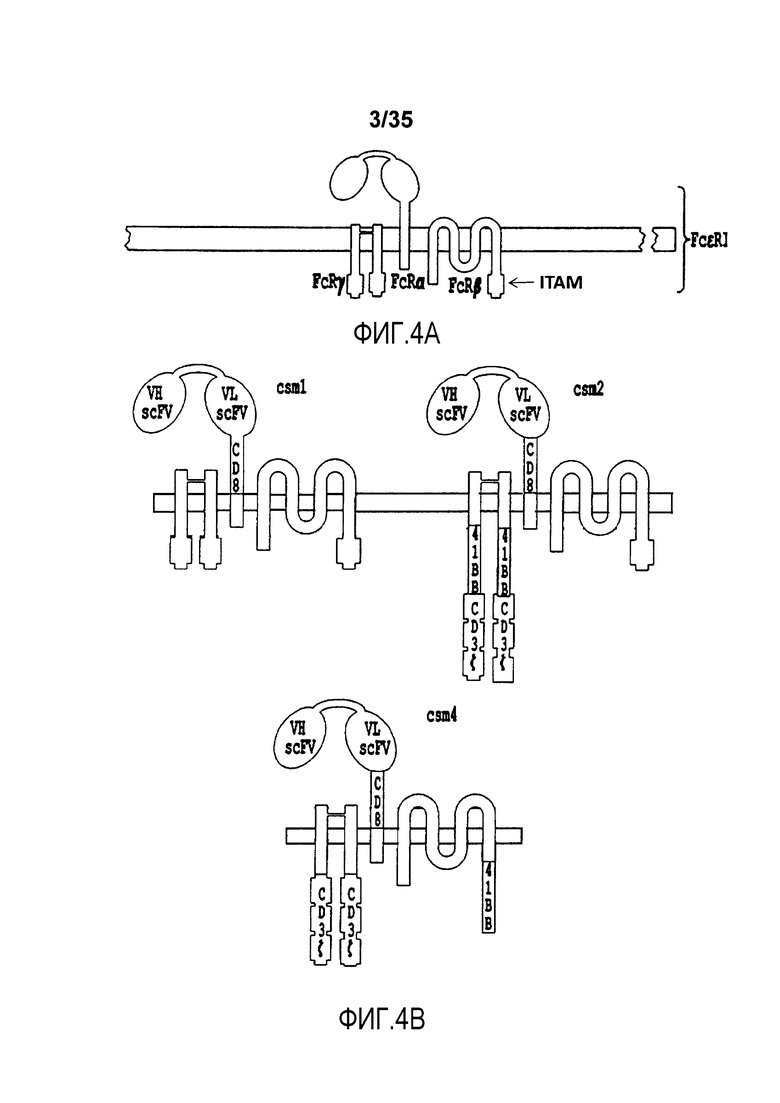

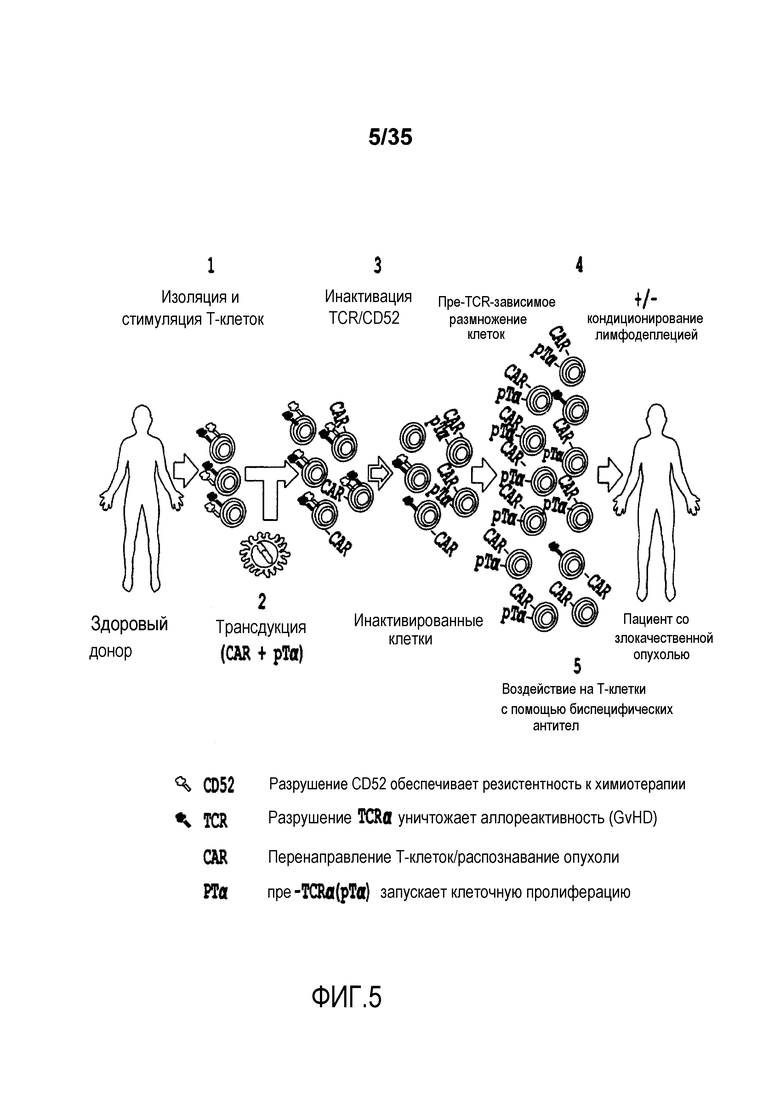
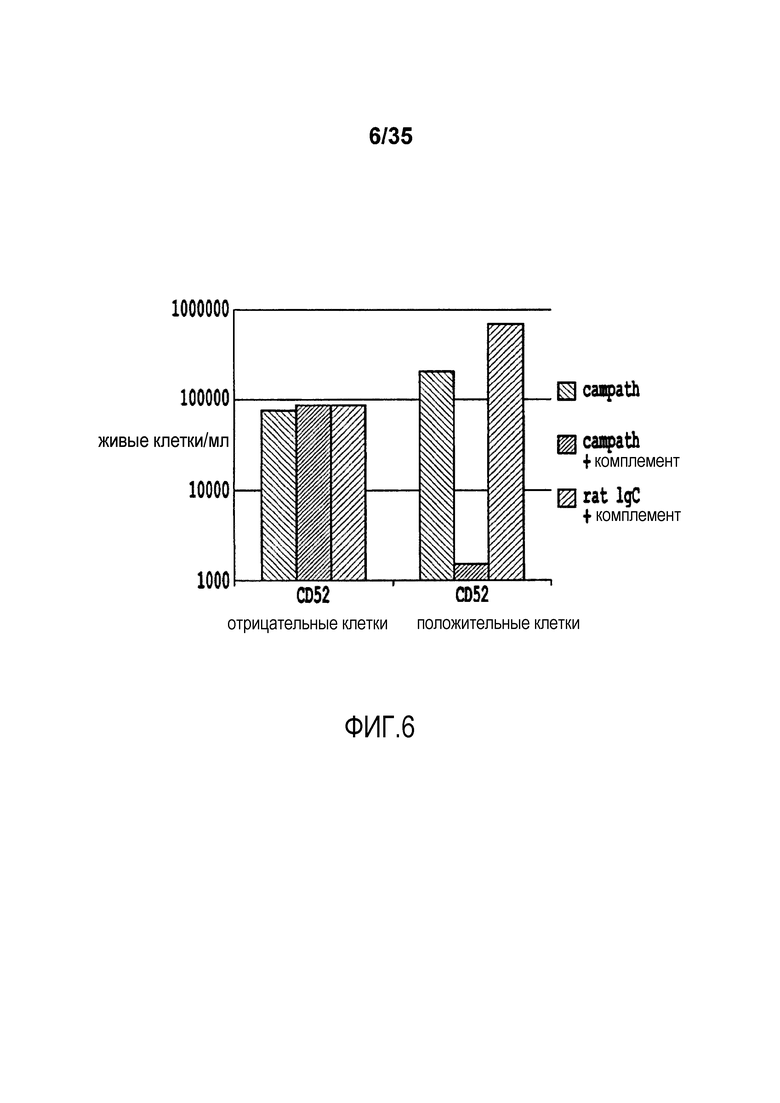

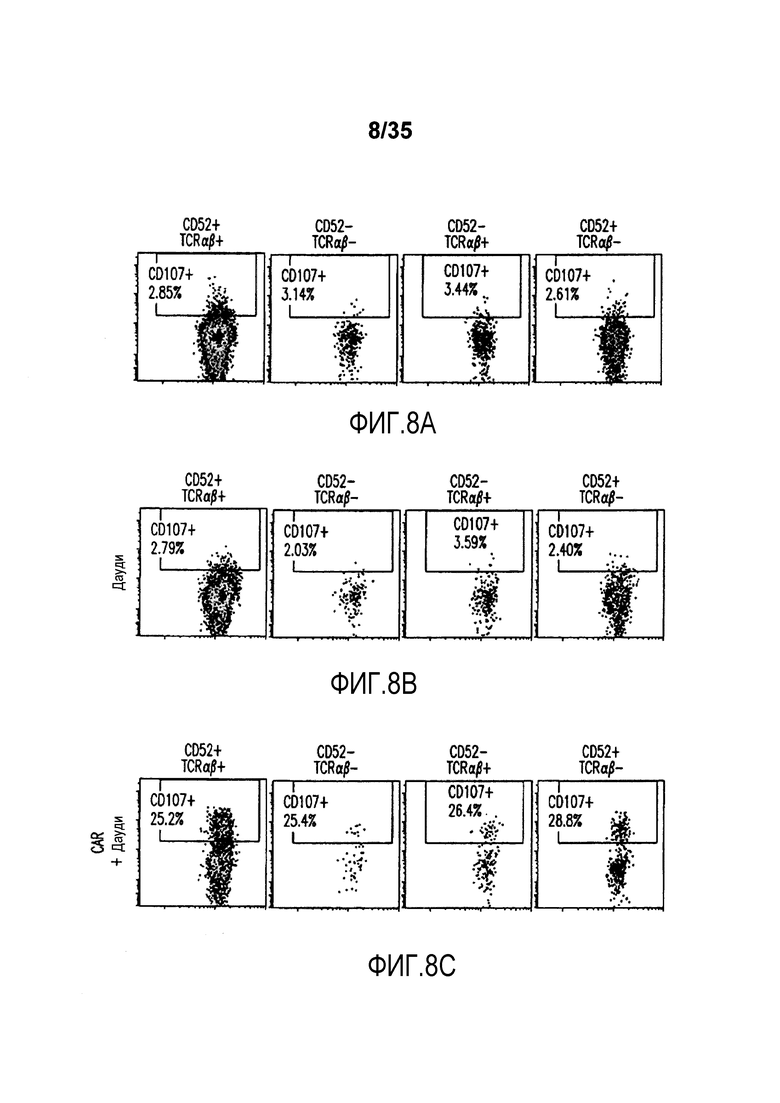
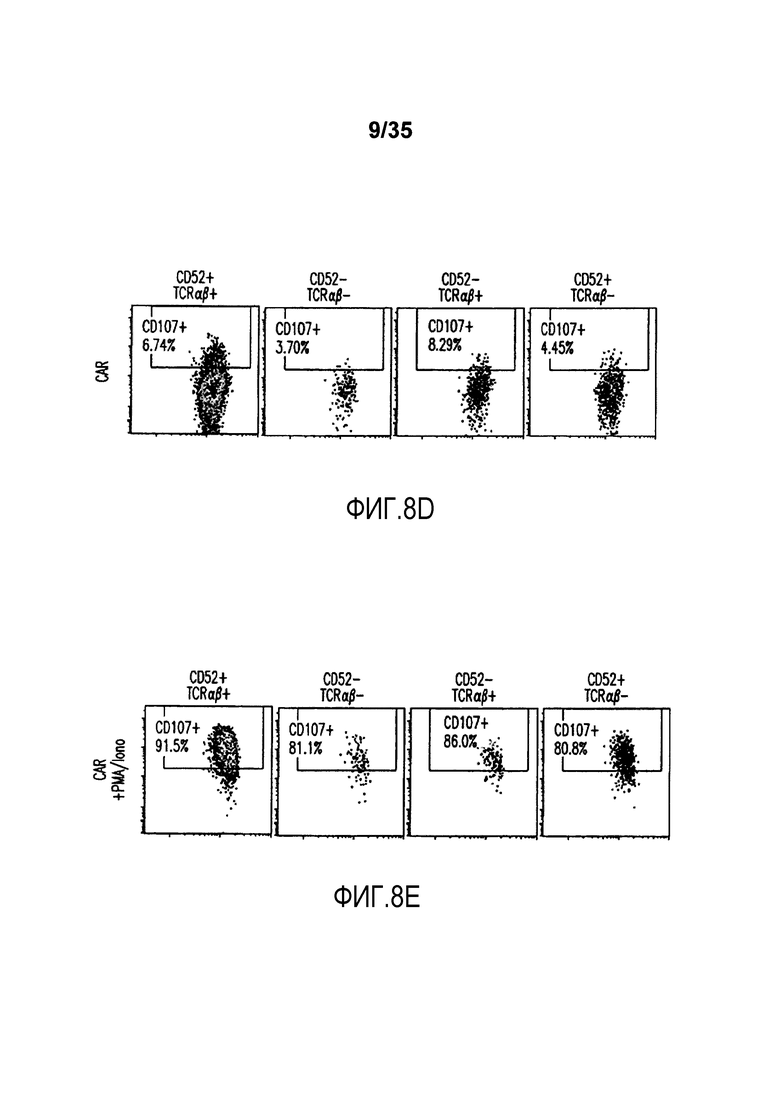






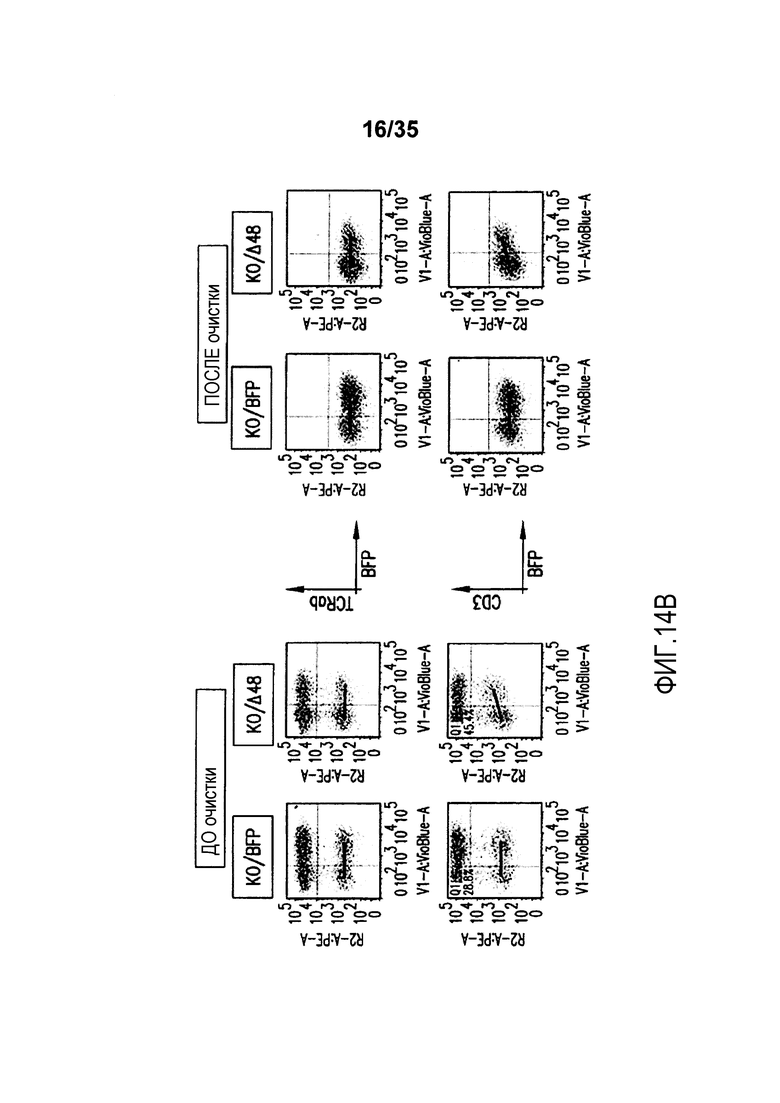

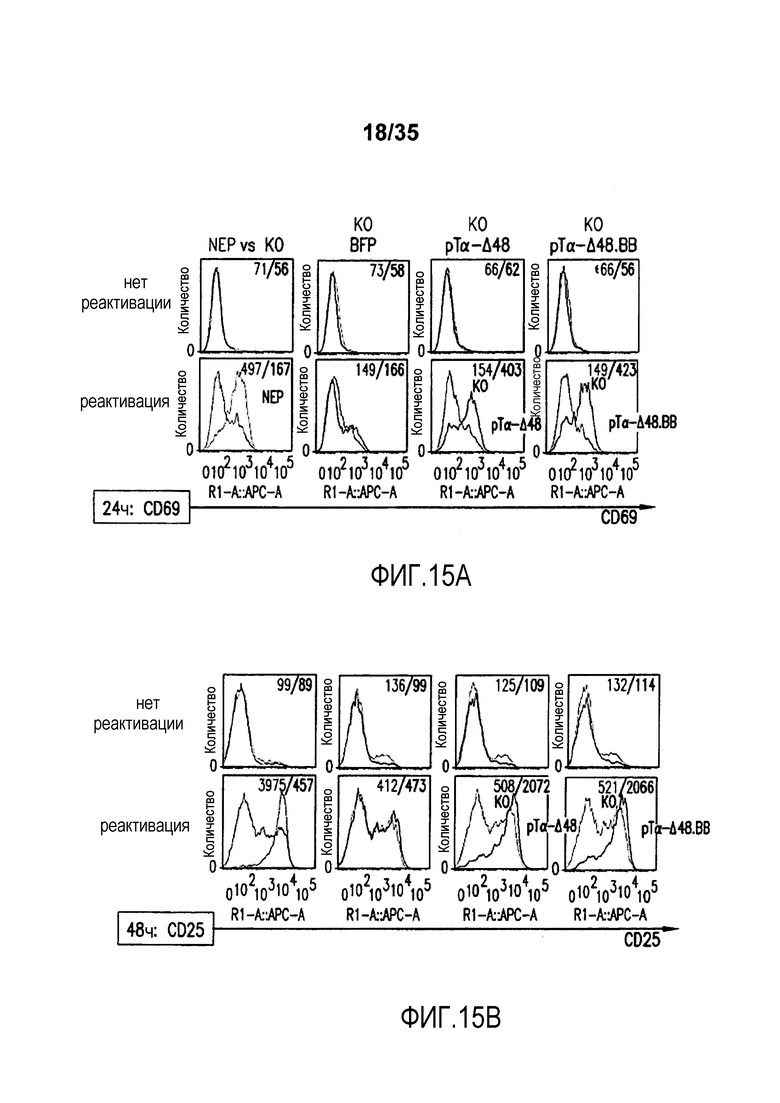
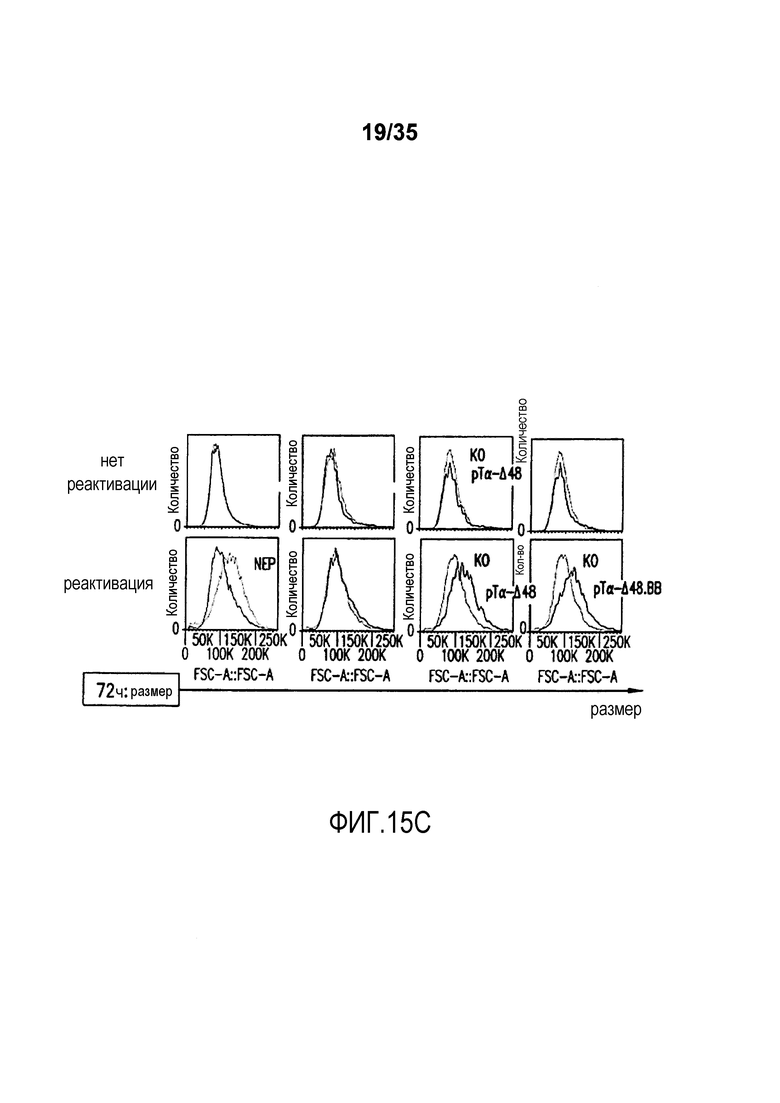


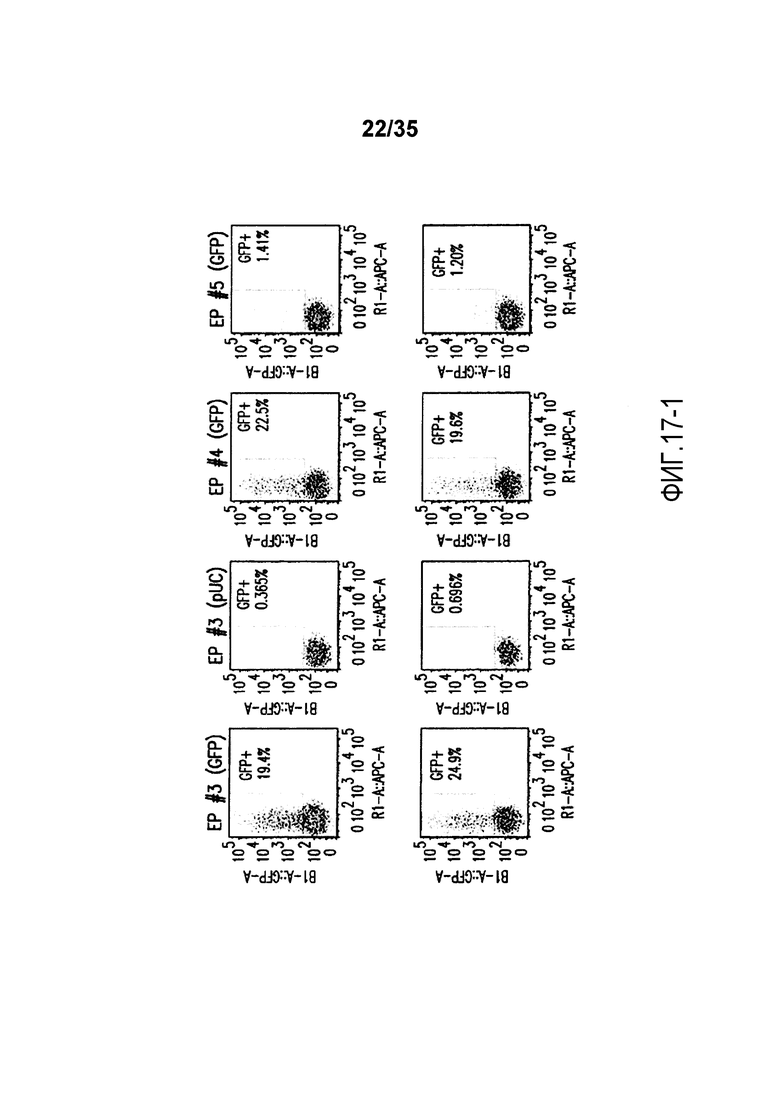

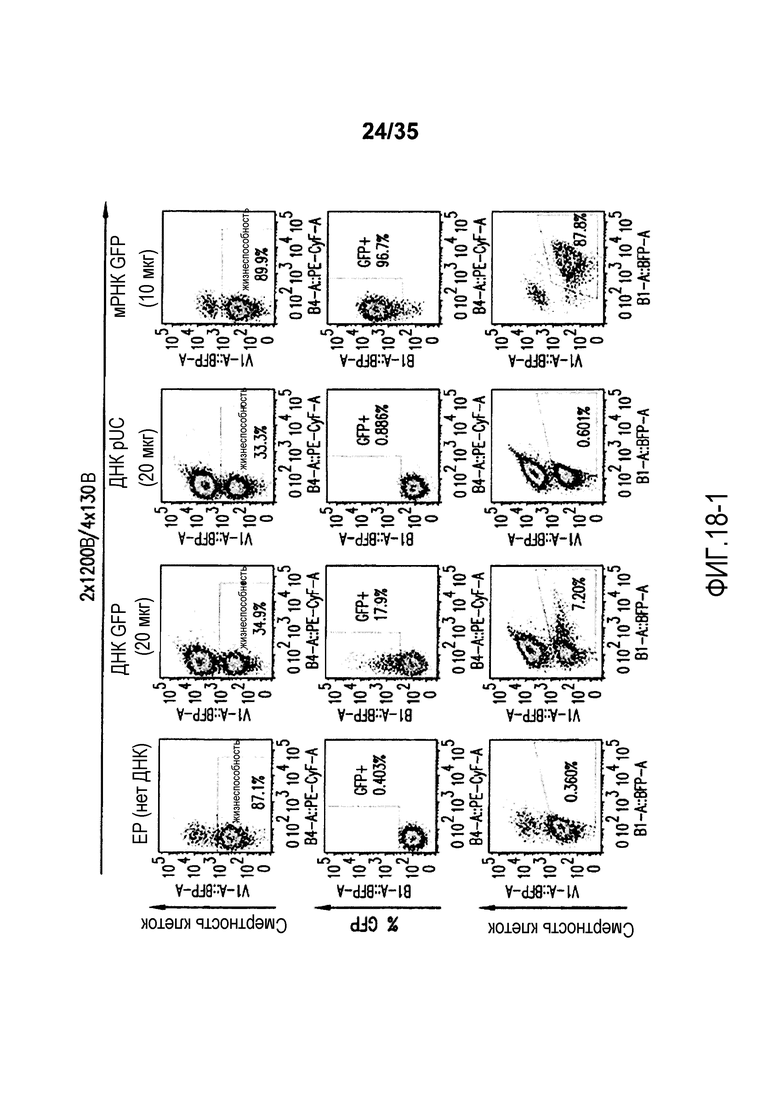
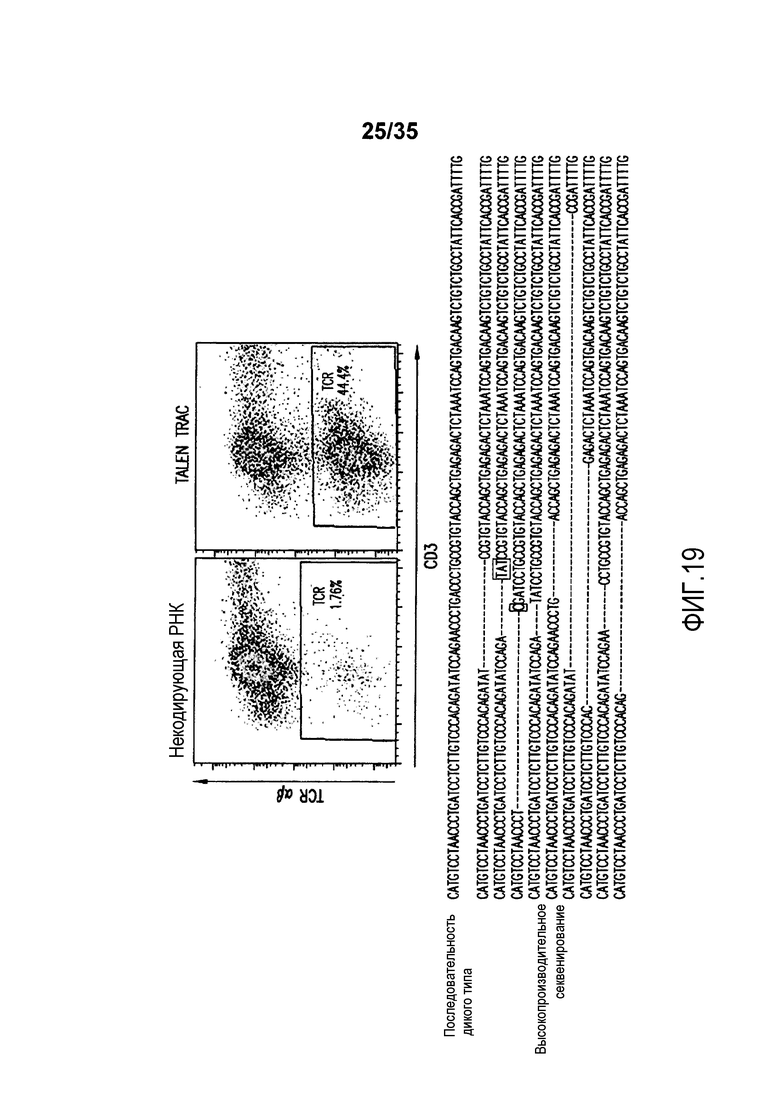

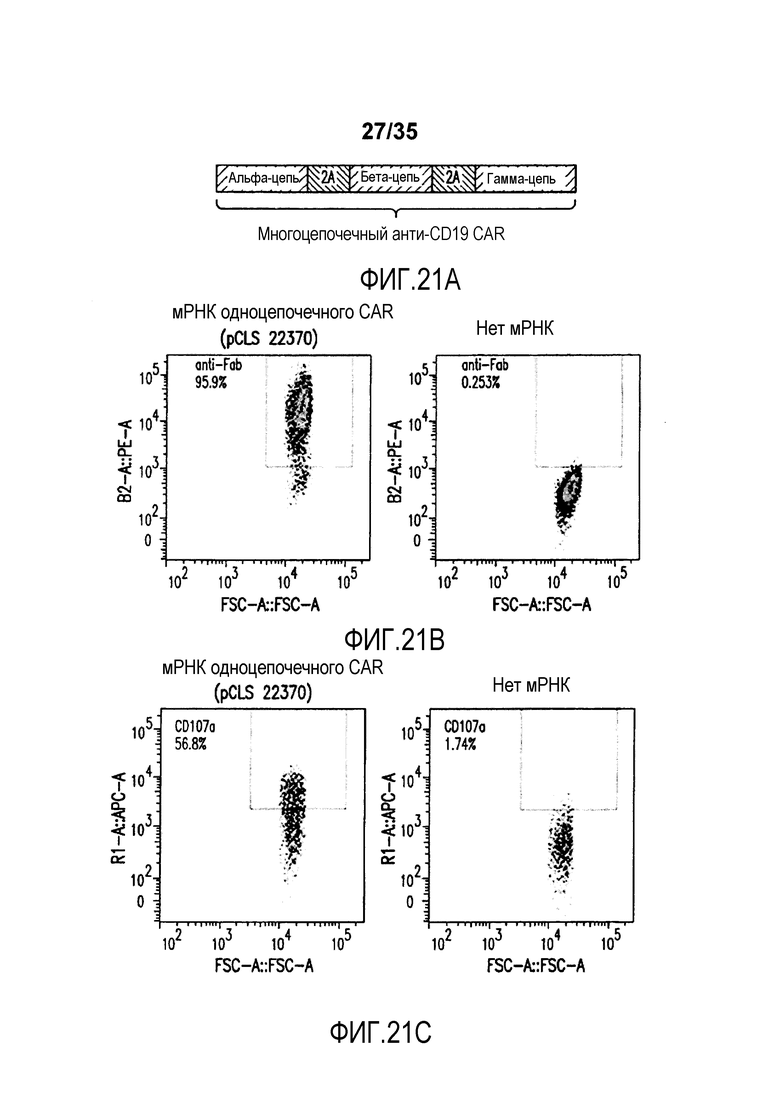

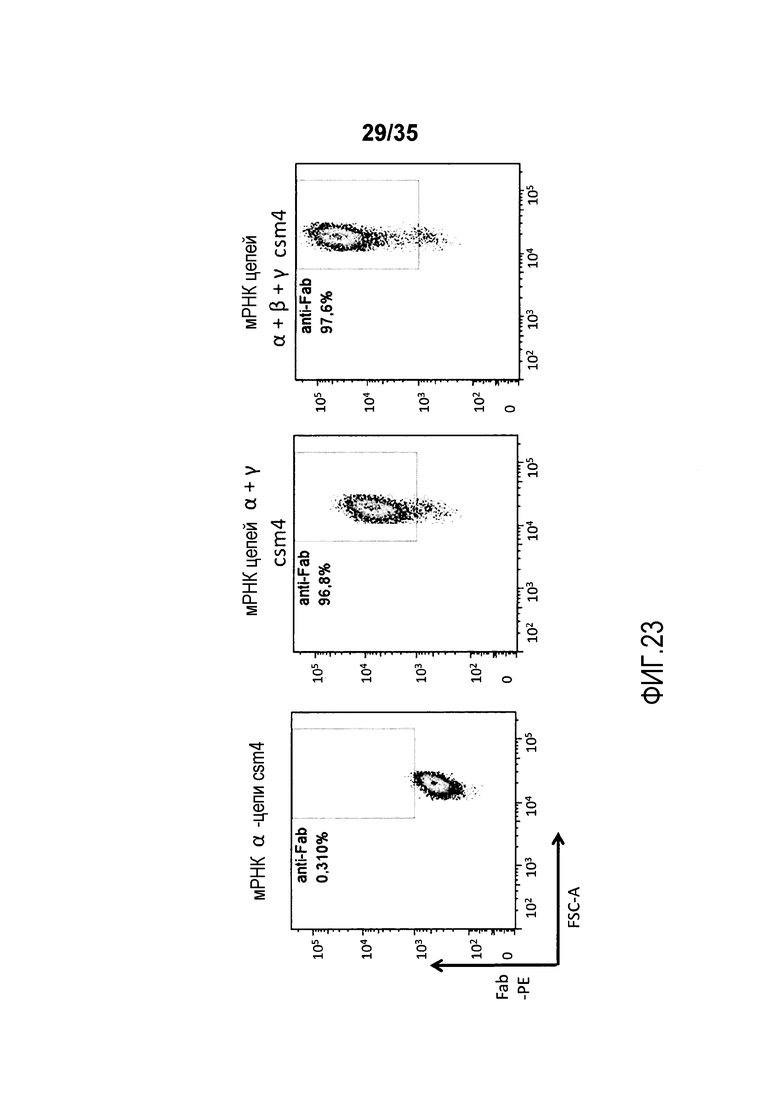

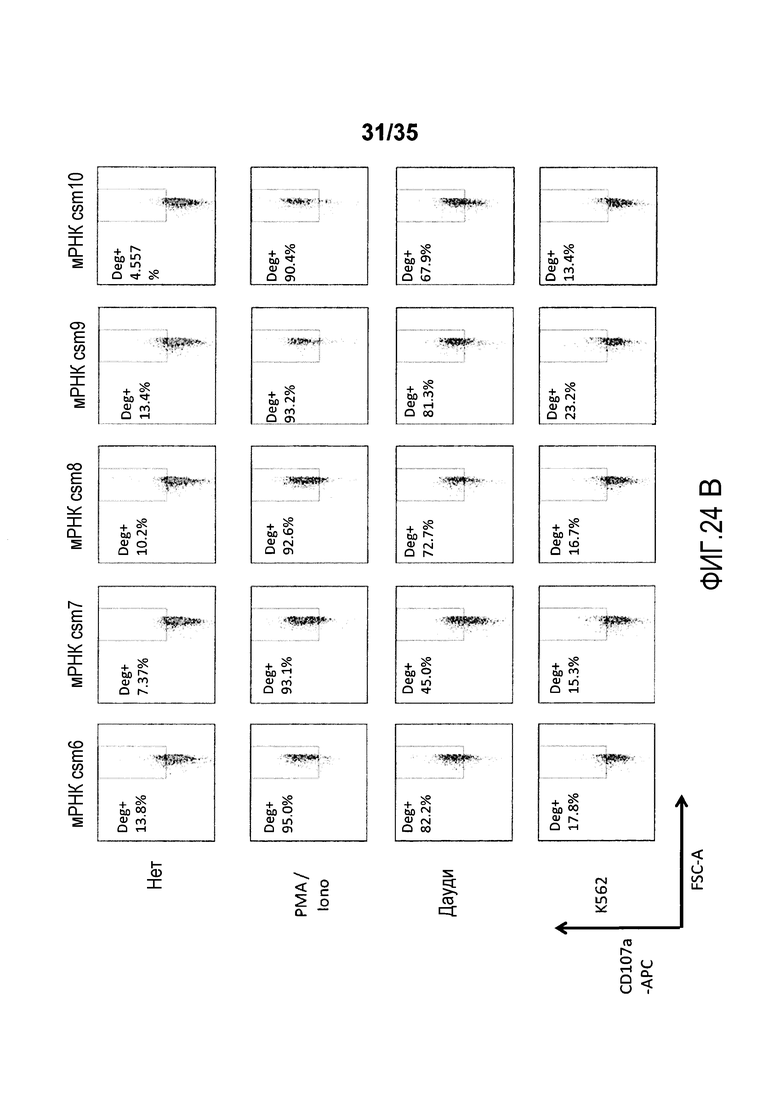

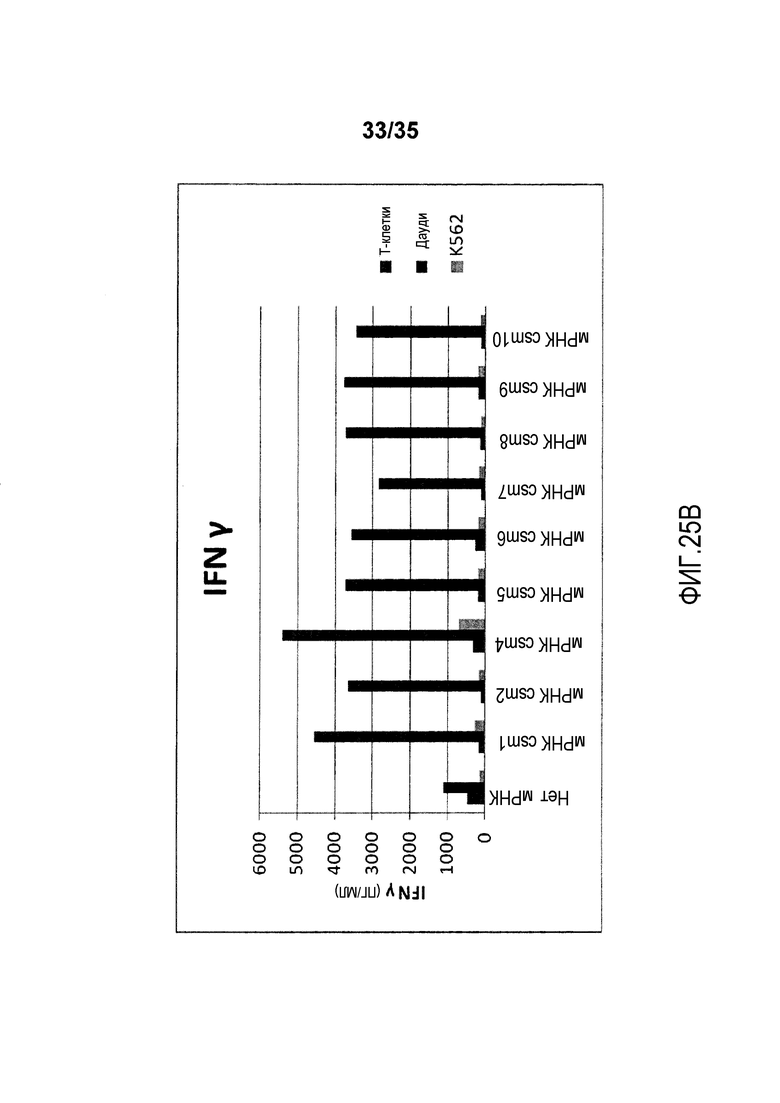
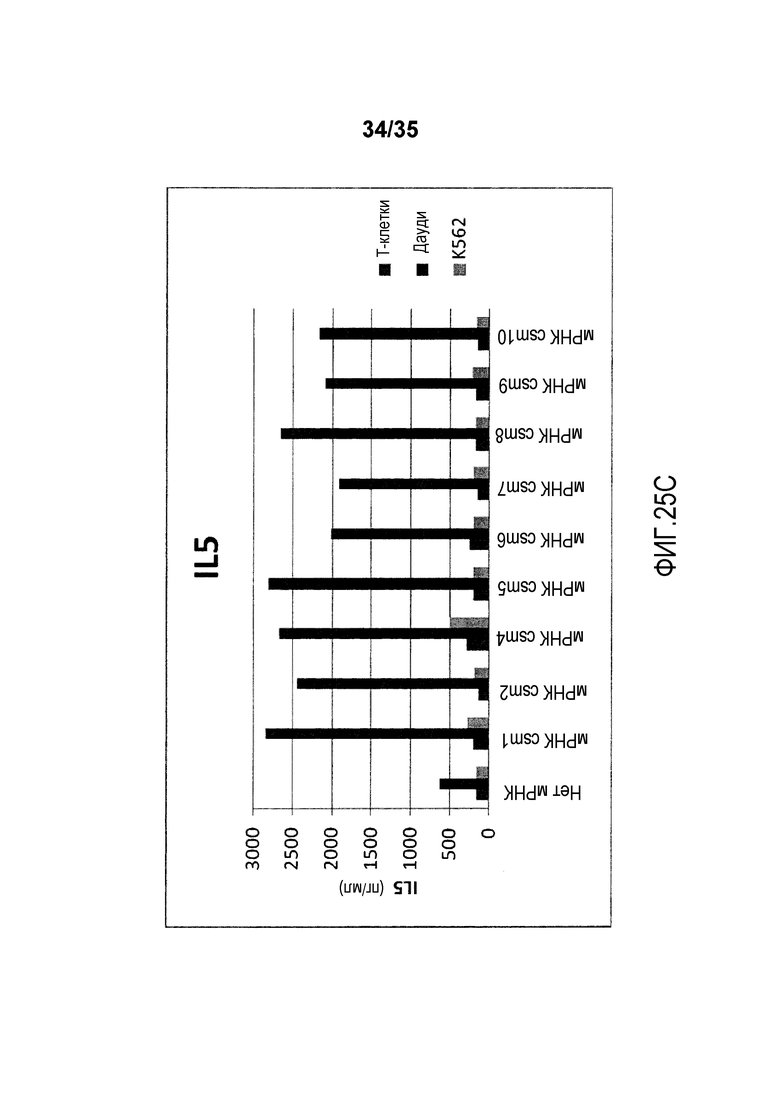

Изобретение относится к области биотехнологии, конкретно к получению химерных антигенных рецепторов (CAR), называемых многоцепочечными CAR, и может быть применено в медицине для противоопухолевой терапии. Многоцепочечный CAR направлен против маркера клеточной поверхности, представленного на целевой клетке и содержит два нековалентно связанных трансмембранных полипептида. Предложенная структура CAR обеспечивает оптимальную передачу сигнала. Первый трансмембранный полипептид содержит трансмембранный домен и одноцепочечный фрагмент антитела (scFv), который способен связываться с маркером клеточной поверхности, представленным на целевой клетке. Второй трансмембранный полипептид содержит трансмембранный домен и передающий сигнал домен. Изобретение позволяет получить CAR, перенаправляющий специфичность и реактивность иммунных клеток в направлении выбранной мишени с использованием свойств лигандсвязывающего домена. 6 н. и 16 з.п. ф-лы, 26 ил., 15 табл., 6 пр.
1. Многоцепочечный химерный антигенный рецептор (CAR), направленный против маркера клеточной поверхности, представленного на целевой клетке, где многоцепочечный CAR содержит:
первый трансмембранный полипептид, содержащий трансмембранный домен и по меньшей мере один одноцепочечный фрагмент антитела (scFv), причем данный по меньшей мере один одноцепочечный фрагмент антитела (scFv) способен связываться с маркером клеточной поверхности, представленным на целевой клетке, и
по меньшей мере один другой трансмембранный полипептид, содержащий трансмембранный домен и по меньшей мере один передающий сигнал домен,
причем указанный первый трансмембранный полипептид и указанный по меньшей мере один другой трансмембранный полипептид являются нековалентно связанными.
2. Многоцепочечный химерный антигенный рецептор по п. 1, в котором по меньшей мере один трансмембранный полипептид содержит часть Fc-рецептора, субъединицы Т-клеточного рецептора, рецептора IL-2, p55 (α-цепи) или CD-белка.
3. Многоцепочечный химерный антигенный рецептор по п. 1, где по меньшей мере один трансмембранный полипептид содержит часть рецептора Fc.
4. Многоцепочечный химерный антигенный рецептор по п. 2, в котором указанную часть Fc-рецептора выбирают из группы, состоящей из: (a) альфа-цепи FcεRI, (b) бета-цепи FcεRI и (c) гамма-цепи FcεRI.
5. Многоцепочечный химерный антигенный рецептор по п. 4, содержащий часть альфа-цепи FcεRI и часть гамма-цепи FcεRI, так что указанные цепи FcεRI димеризуются вместе с образованием димерного химерного антигенного рецептора.
6. Многоцепочечный химерный антигенный рецептор по п. 4, содержащий часть альфа-цепи FcεRI, часть бета-цепи FcεRI и часть гамма-цепи FcεRI, так что указанные цепи FcεRI тетрамеризуются вместе с образованием тетрамерного химерного антигенного рецептора.
7. Многоцепочечный химерный антигенный рецептор по п. 1, где целевая клетка представляет собой клетку злокачественной опухоли.
8. Многоцепочечный химерный антигенный рецептор по п. 1, в котором указанный первый трансмембранный полипептид дополнительно содержит шарнир между указанным одноцепочечным фрагментом антитела (scFv) и указанным трансмембранным доменом.
9. Многоцепочечный химерный антигенный рецептор по п. 1, в котором указанный передающий сигнал домен выбирают из группы, состоящей из дзета-цепи TCR, цепи FcεRIβ, цепи FcεRIγ, иммунорецепторного тирозинового активирующего мотива (ITAM).
10. Многоцепочечный химерный антигенный рецептор по п. 1, в котором указанный CAR содержит костимулирующий домен.
11. Многоцепочечный химерный антигенный рецептор по п. 1, дополнительно содержащий костимулирующую молекулу, выбранную из группы, состоящей из CD28, OX40, ICOS, CD137 и CD8.
12. Многоцепочечный химерный антигенный рецептор по п. 1, содержащий полипептиды, выбранные из группы, состоящей из от SEQ ID NO: 202 до SEQ ID NO: 204 и от SEQ ID NO: 206 до SEQ ID NO: 213.
13. Полинуклеотид, кодирующий многоцепочечный CAR по любому из пп. 1-12, причем указанный полинуклеотид содержит две или более последовательности нуклеиновой кислоты, кодирующие трансмембранные полипептиды, составляющие многоцепочечный CAR.
14. Вектор для экспрессии многоцепочечного CAR по любому из пп. 1-12 в клетке, причем указанный вектор содержит полинуклеотид по п. 13.
15. Способ получения генетически измененной иммунной клетки, направленной на маркер клеточной поверхности, представленный на целевой клетке, причем указанный способ включает:
(a) наличие иммунной клетки;
(b) введение в указанную иммунную клетку по меньшей мере одного полинуклеотида по п. 13 и
(c) экспрессию на поверхности иммунной клетки по меньшей мере одного многоцепочечного CAR по любому из пп. 1-12,
где указанная иммунная клетка представляет собой Т-клетку, дендритную клетку, дендритную клетку-киллер, тучную клетку, клетку естественного киллера (NK-клетку) или B-клетку.
16. Способ получения генетически измененной иммунной клетки по п. 15, дополнительно включающий стадию (d) инактивации в указанной иммунной клетке по меньшей мере одного гена, выбранного из группы, состоящей из TCR-альфа, TCR-бета, CD52, GR и генов иммунных контрольных точек.
17. Способ по п. 15 или 16, где указанная иммунная клетка представляет собой Т-клетку, дендритную клетку-киллер или клетку естественного киллера (NK-клетку).
18. Выделенная генетически измененная иммунная клетка, направленная против маркера клеточной поверхности, представленного на целевой клетке, полученная согласно способу по п. 15, причем указанная иммунная клетка экспрессирует на своей поверхности по меньшей мере один многоцепочечный CAR по любому из пп. 1-12, и где указанная иммунная клетка представляет собой Т-клетку, дендритную клетку, дендритную клетку-киллер, тучную клетку, клетку естественного киллера (NK-клетку) или B-клетку.
19. Выделенная генетически измененная иммунная клетка по п. 18, где указанная иммунная клетка представляет собой Т-клетку, дендритную клетку-киллер или клетку естественного киллера (NK-клетку).
20. Выделенная генетически измененная иммунная клетка по п. 18, где по меньшей мере один ген, выбранный из группы, состоящей из TCR альфа, TCR бета, CD52, GR и генов иммунных контрольных точек, инактивирован.
21. Выделенная генетически измененная иммунная клетка по п. 18 или 19, где Т-клетка выбрана из воспалительных T-лимфоцитов, цитотоксических T-лимфоцитов, регуляторных T-лимфоцитов или хелперных T-лимфоцитов.
22. Применение выделенной генетически измененной иммунной клетки по любому из пп. 18-21 в качестве лекарственного средства для лечения злокачественной опухоли, при этом указанный по меньшей мере один CAR, экспрессируемый на поверхности указанной иммунной клетки, направлен против маркера клеточной поверхности, представленного на клетке указанной злокачественной опухоли.
| RA C | |||
| ET AL., Complete Structure of the Mouse Mast Cell Receptor for IgE (FcRI) and Surface Expression of Chimeric Receptors (Rat-Mouse-Human) on Transfected Cells, THE JOURNAL OF BIOLOCICAL CHEMISTRY, 1989, v.264, n.26, p.15323-15327 | |||
| US 5770396, 23.06.1998 | |||
| BIPULENDU JENA ET AL., Redirecting T-cell specificity by introducing a tumor-specific chimeric antigen receptor, BLOOD, 2010, v.116, p.1035-1044 | |||
| Способ изготовления электродов для гальванических элементов | 1927 |
|
SU9956A1 |

