Область техники
Изобретение относится к области биотехнологии, а именно к способу получения препарата рекомбинантной нуклеазы Cas путем рекомбинантной продукции в клетках Escherichia coli, очистки и доочистки нуклеазы Cas от эндотоксинов продуцента. Изобретение также относится к препаратам рекомбинантной нуклеазы Cas, полученным предложенным способом, и наборам для использования в системе CRISPR/Cas, содержащим препарат рекомбинантной нуклеазы Cas, по существу, свободный от бактериальных эндотоксинов.
Изобретение позволяет получать в значительных количествах и в ускоренные сроки рекомбинантную нуклеазу Cas, характеризующуюся минимальной токсичностью, что позволяет использовать ее в системе CRISPR/Cas в различных областях техники, связанных с редактированием генов. Начиная от создания клеточных моделей наследственных заболеваний человека и животных, функционального скрининга геномов, изучения эпигеномов и визуализации клеточных процессов до прикладного применения в пищевой промышленности для получения обогащенных продуктов питания, в сельском хозяйстве для создания новых пород животных и сортов растений, а также в медицине.
Систему CRISPR/Cas с рекомбинантной нуклеазой Cas, полученной согласно предложенному в настоящей заявке способу и отвечающей высоким требованиям к эффективности, безопасности и доступности, возможно использовать для диагностики заболеваний путем идентификации генетических последовательностей, например вирусов или онкогенов, профилактики инфекционных заболеваний за счет модификации генов не человека, а переносчиков болезней, например малярии и лечения социально значимых заболеваний, как наследственных, так и приобретенных, среди которых рак, аутоиммунные, орфанные заболевания, инфекционные заболевания, в том числе ВИЧ и многие другие.
Уровень техники
Направленное геномное редактирование с использованием программируемых нуклеаз за короткое время заняло передовые позиции среди технологий модификаций генома. На сегодняшний день существует три основных системы для направленного редактирования генома: нуклеазы с «цинковыми пальцами», TALE-нуклеазы и CRISPR/CAS нуклеазы (Nemudryi AA, Valetdinova KR, Medvedev SP and Zakian SM., TALEN and CRISPR/Cas Genome Editing Systems: Tools of Discovery, Acta Naturae, 2014, v.6, n.3, p:19-40; German DM, Mitalipov S, Mishra A and Kaul S., Therapeutic Genome Editing in Cardiovascular Diseases, JACC Basic Transl Sci., 2019, v.4, n.1, p:122-131; Kwarteng A, Ahuno ST and Kwakye-Nuako G., The therapeutic landscape of HIV-1 via genome editing, AIDS Res Ther., 2017, v.14, n.1, p:32; A programmable dual RNA-guided DNA endonuclease in adaptive bacterial immunity // Martin Jinek, Krzysztof Chylinski, Ines Fonfara, Michael Hauer, Jennifer A. Doudna and Emmanuelle Charpentier, Science, 2012, v.337, n.6096, p:816-821).
Направленное редактирование генома с использованием CRISPR/CAS нуклеаз обладает рядом преимуществ: высокая эффективность, возможность множественного редактирования, невысокая стоимость, а также скорость разработки (Loureiro A and da Silva GJ., CRISPR-Cas: Converting A Bacterial Defence Mechanism into A State-of-the-Art Genetic Manipulation Tool, Antibiotics (Basel), 2019, v.8, n.1, pii:E18; Karimian A, Azizian K, Parsian H, Rafieian S, Shafiei-Irannejad V, Kheyrollah M, Yousefi M, Majidinia M and Yousefi B., CRISPR/Cas9 technology as a potent molecular tool for gene therapy, J. Cell Physiol., 2019, doi: 10.1002/jcp.27972).
CRISPR/CAS система направленного редактирования генома состоит из СAS-нуклеазы (белка) и направляющей РНК, специфичной к последовательности-мишени. Доставку CRISPR/CAS системы в живую клетку осуществляют тремя методами: в виде комплекса плазмидных ДНК, кодирующих CAS-белок и направляющую РНК; в виде смеси мРНК, кодирующей CAS-белок, и направляющей РНК; и третий вариант доставки CRISPR/CAS системы предполагает введение в клетку готового рибонуклеопротеинового комплекса (смесь CAS-белка и направляющей РНК). Доставка CRISPR/CAS системы в виде готового рибонуклеопротеинового комплекса имеет ряд преимуществ, среди которых высокая эффективность редактирования; низкая неспецифическая активность; редактирование сразу после доставки в клетку; возможность быстрого скрининга эффективности направляющих РНК в пробирке; сниженная иммуногенность ввиду непродолжительного времени пребывания элементов системы CRISPR/CAS в клетке-мишени. Наиболее перспективным с точки зрения создания терапевтических агентов направлением в редактировании генома с использованием систем CRISPR/CAS является работа с готовыми рибонуклеопротеиновыми комплексами.
Для получения рекомбинантных белков в настоящее время широко применяют несколько гетерологичных систем экспрессии, среди которых клетки бактерий (Escherichia coli и др.); клетки дрожжей (Saccharomyces cerevisiae, Pichia pastoris и др.); клетки насекомых; клетки растений и клетки млекопитающих.
Каждая из упомянутых систем экспрессии имеет преимущества и недостатки, и окончательный выбор системы экспрессии, как правило, зависит от свойств целевого продукта (Xu J, Ge X and Dolan MC, Towards high-yield production of pharmaceutical proteins with plant cell suspension cultures, Biotechnol Adv., 2011, v.29, n.3, p:278-299).
К главным преимуществам экспрессии рекомбинантных белков в клетках Escherichia coli стоит отнести простоту и скорость получения штамма-продуцента; сравнительно невысокую стоимость процесса разработки; масштабируемость процессов экспрессии и очистки и высокий выход целевого белка.
При получении рекомбинантных белков семейства CRISPR/CAS в клетках Е. coli возникает проблема дополнительной очистки от бактериальных эндотоксинов, содержание которых регламентировано в Общей Фармакопейной Статье ОФС.1.2.4.0006.15 Бактериальные эндотоксины. Пороговой пирогенной дозой считается доза, равная 5 ЕЭ/кг в 1 час для испытуемого лекарственного препарата, если он вводится пациенту любым парентеральным путем, кроме интратекального. Белки семейства CRISPR/CAS обладают достаточно высокими значениями изоэлектрической точки (pi > 9), поэтому в растворах, имеющих физиологический pH = 6,5-8,0, имеют высокий положительный суммарный заряд. Именно высокий положительный суммарный заряд белков семейства CRISPR/CAS приводит к тому, что при выделении из клеток Е. coli, бактериальные эндотоксины разрушенных клеточных стенок бактерий, заряженные отрицательно, прочно связываются с выделяемыми белками семейства CRISPR/CAS.
Стоит отметить, что при получении рекомбинантных белков семейства CRISPR/CAS в других системах экспрессии представляется затруднительным получить препараты, свободные от бактериальных эндотоксинов (в виду их физико-химических свойств, pI > 9). Это связано с тем, что остаточные бактериальные эндотоксины могут содержаться в питательной среде и добавках, применяемых при культивировании, а также в воде, которая используется при приготовлении буферных растворов.
Рекомбинантное получение нуклеаз семейства CRISPR/CAS в клетках- продуцентах E. coli с различными подходами к очистке целевых белков описано в статьях, например Nandhakishore Rajagopalan, Sateesh Kagale, Pankaj Bhowmik and Halim Song, A Two-Step Method for Obtaining Highly Pure Cas9 Nuclease for Genome Editing, Biophysical, and Structural Studies, 2018, v.1, n.17, p.1-8; Wenqiang Li, Shungtang Li, Jie Qiao, Fei Wang, Yang Liu, Ruyi He, Yi Liu and Lixing Ma, Direct preparation of Cas9 ribonucleoprotein from E. coli for PCR-free seamless DNA assembly, doi: https://doi.org/10.1101/328468; Sojung Kim, Daesik Kim, Seung Woo Cho, Jungeun Kim and Jin-Soo Kim, Highly efficient RNA-guided genome editing in human cells via delivery of purified Cas9 ribonucleoproteins, Genome Res., 2014, v.24, n.6, p:1012-1019; Zuris JA, Thompson DB, Shu Y2, Guilinger JP, Bessen JL, Hu JH, Maeder ML, Joung JK, Chen ZY and Liu DR, Cationic lipid-mediated delivery of proteins enables efficient protein-based genome editing in vitro and in vivo, Nat. Biotechnol., 2015, v.33, n.1, p.73-80; Koziel D, Michaelis U, Kruse T., Broad application and optimization of a single wash-step for integrated endotoxin depletion during protein purification, J. Chromatogr. B Analyt. Technol. Biomed. Life Sci., 2018, v.1091, p.101-107; Lin S, Staahl BT, Alla RK and Doudna JA, Enhanced homology-directed human genome engineering by controlled timing of CRISPR/Cas9 delivery, Elife, 2014, n.3, p:e04766; Liang Z, Chen K, Li T, Zhang Y, Wang Y, Zhao Q, Liu J, Zhang H, Liu C, Ran Y and Gao C, Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes, Nat. Commun., 2017, n.8, p:14261, а также в патентных документах, например EA 201790968 A1, опубл. 31.01.2018; WO 2016205703 А1, опубл. 22.12.2016; EA 24121 B9, опубл. 30.01.2017 и WO 2016161516 A1, опубл. 13.10.2016.
Ближайшим аналогом изобретения является способ получения рекомбинантного белка семейства Cas в клетках E.coli с использованием для очистки целевого белка металл-хелатной и катионообменной хроматографии, раскрытый в статье Nandhakishore Rajagopalan, Sateesh Kagale, Pankaj Bhowmik and Halim Song, A Two-Step Method for Obtaining Highly Pure Cas9 Nuclease for Genome Editing, Biophysical, and Structural Studies, 2018, v.1, n.17, p.1-8. Способ направлен на получение Cas9 Streptococcus pyogenes (NLS-Cas9-6xHis) в клетках E.coli BL21 star (DE3)pLysS с использованием среды Лурии-Бертани (LB), содержащей ампициллин (100 μg/mL), в которую инокулировали 5 mL культуры и выращивали ночь при 37°C до достижения оптической плотности OD600≈0,6. Затем индуцировали ИПТГ (0.5 mM) и инкубировали при 18°C 16 часов. По завершении культивирования проводили центрифугирование при 5000×g 15 минут при 4°C. Ресуспендировали в ледяном буфере для лизиса LW (20 mM HEPES pH 7.5, 300 mM NaCl, 25 mM имидазол, 0.5 mM TCEP). Разрушение клеток проводили с использованием пресса Френча с последующим центрифугированием лизата при 39706×g при 4°С в течение 20 минут. Очистку целевого белка проводили двухстадийной хроматографией. На первой стадии проводилась металл-хелатная хроматография с использованием сорбента Ni-NTA агароза (Qiagen, США) c ледяным буфером LW. Элюцию осуществляли ледяным буфером EB (pH 7.5, 20 mM HEPES, 300 mM NaCl, 250 mM имидазол, 0.5 mM TCEP). На второй стадии проводилась катионообменная хроматография с использованием сорбента Resource S (GE Healthcare, Великобритания) и буфера А (pH 7.5, 20 mM HEPES, 200 mM KCl, 10 mM MgCl2, 0.5 mM TCEP). Элюцию осуществляли буфером B (pH 7.5, 20 mM HEPES, 1 M KCl, 10 mM MgCl2, 0.5 mM TCEP). Затем проводили центрифугирование, добавляли буфер А и глицерол для замораживания. Описанный способ дает возможность после двухстадийной хроматографической очистки получить порядка 1 миллиграмма высокоочищенного белка Cas9 Streptococcus pyogenes с 1 литра культуры штамма-продуцента.
Недостатками способа, описанного в ближайшем аналоге, является относительно невысокий выход целевого белка, а также отсутствие какой-либо информации о содержании в полученном препарате белка остаточных бактериальных эндотоксинов, что ставит под вопрос безопасность применения полученных таким способом нуклеаз в живых организмах.
Исходя из этого, возникает техническая проблема, заключающаяся в необходимости повышения выхода нуклеаз Cas при рекомбинантных способах получения в Е. coli и минимизации содержания токсичных примесей в препаратах рекомбинантных белков семейства CRISPR/CAS.
Краткое описание сущности изобретения
Изобретение относится к способу получения препарата рекомбинантной нуклеазы Cas путем рекомбинантной продукции в клетках Escherichia coli, очистки и доочистки нуклеазы Cas от эндотоксинов продуцента, а также к препаратам рекомбинантной нуклеазы Cas, полученным предложенным способом, и наборам для использования в системе CRISPR/Cas, содержащим препараты рекомбинантной нуклеазы Cas, по существу, свободные от бактериальных эндотоксинов.
Технической задачей предложенного изобретения является повышение степени очистки рекомбинантных белков семейства CRISPR/Cas с уменьшением содержания бактериальных эндотоксинов в растворах рекомбинантных белков семейства CRISPR/Cas для возможности их использования в составе препаратов рекомбинантной нуклеазы семейства Cas системы CRISPR/Cas, применяемых in vivo.
При осуществлении настоящего изобретения, согласно приведенной в формуле изобретения совокупности существенных признаков, достигается неожиданный технический результат - увеличение выхода целевого белка при экспрессии в E. coli, а также повышение эффективности очистки рекомбинантных нуклеаз семейства Cas с сохранением или увеличением их специфической активности и уменьшением содержания бактериальных эндотоксинов до уровня 0,3-1,5 ЕЭ/мл в препаратах полученных рекомбинантных нуклеаз семейства Cas, пригодных для применения в живых организмах (in vivo). Удалось достичь выход не менее 20 миллиграмм высокоочищенного, по существу, свободного от эндотоксинов рекомбинантного белка семейства CRISPR/Cas с 1 литра культуры штамма-продуцента.
Технический результат достигается за счет:
• Использования материнского штамма E. coli Rosetta-gami B (DE3) для продукции рекомбинантных нуклеаз семейства Cas и соблюдения протокола культивирования клеток указанного штамма с использованием бульона Лурии-Бертани (LB) с 100 мкг/мл ампициллина, дополненного MgSO4 и глюкозой для стартовой культуры с последующим культивированием до момента индукции в той же питательной среде, но с уменьшенным, по меньшей мере в 2 раза содержанием глюкозы. Отклонения от протокола приводят к отсутствию экспрессии белка;
• Индукции экспрессии по достижению оптической плотности OD600≈1,5-1,9, что значительно увеличивает выход целевого белка;
• Осуществления экспрессии белка при 22°С. Отклонение от протокола приводит к агрегации белка во время последующей очистки;
• Соблюдения разработанного авторами протокола хроматографической очистки рекомбинантных нуклеаз семейства Cas с определенным составом буферов и строгим требованием к кондуктивности раствора Cas-белка, подвергаемого катионообменной хроматографии. Отклонение от протокола приводит к потерям в количестве очищаемого белка;
• Соблюдения разработанного авторами протокола удаления бактериальных эндотоксинов из растворов рекомбинантных нуклеаз семейства CAS, экспрессированных в клетках Е. coli, с использованием детергента Triton X-114 и по меньшей мере 6-кратного повторения циклов очистки, что позволяет достичь выход не менее 20 миллиграмм с 1 литра культуры штамма-продуцента высокоочищенного с минимальным содержанием эндотоксинов рекомбинантного белка семейства CRISPR/Cas.
Подробное раскрытие сущности изобретения
Перед тем как описать способы, препараты и наборы согласно настоящему изобретению, следует отметить, что настоящее изобретение не ограничивается конкретными описанными способами, используемыми средствами и методами, продуктами или комбинациями, поскольку такие способы, средства и методы, продукты или комбинации, без сомнения, могут варьироваться. Также следует отметить, что терминология, используемая в настоящей заявке, не подразумевается как ограничивающая, поскольку объем настоящего изобретения будет ограничиваться только предлагаемой формулой изобретения.
Если не определено иное, все термины и понятия, применяемые в раскрытии настоящего изобретения, включая технические и научные термины, имеют значение, которое обычно понятно среднему специалисту в технической области, к которой принадлежит настоящее изобретение. Как правило, используемая классификация и методы клеточной биологии, молекулярной биологии, иммунологии, микробиологии, генетики, аналитической химии, химии органического синтеза, медицинской и фармацевтической химии, а также гибридизации и химии белков и нуклеиновых кислот, описанные в настоящей заявке, хорошо известны специалистам и широко применяются в данной области. Методики рекомбинантного получения и культивирования клеток-продуцентов, выделения целевого белка, очистки и проверки активности целевого белка осуществляют в соответствии с инструкциями производителя, как это обычно осуществляется в данной области, или как это подробно описано в настоящей заявке.
Далее приведены определения терминов и понятий, используемых в настоящей заявке, для лучшего понимания существа настоящего изобретения.
Используемые в настоящей заявке формы единственного числа включают объекты как в единственном, так и множественном числе, если из контекста явно не следует иное.
Используемые в настоящей заявке термины "содержащий", "содержит" и "содержащийся" являются синонимами "включающий", "включает" или "охватывающие", "охватывает", и являются включительными или неограничивающими и не исключают дополнительные, неупомянутые части, компоненты продуктов или стадии способов. Используемые в настоящей заявке термины "содержащий", "содержит" и "содержащийся" предусматривают термины "состоящий из", "состоит" и "состоит из", а также термины "состоящий, по сути, из", "состоит по сути" и "состоит, по сути, из".
Описание диапазона числовых значений с помощью крайних точек подразумевает все числа и дробные числа, относящиеся к соответствующим диапазонам, а также указанные крайние точки.
Используемые в настоящей заявке понятия "приблизительно" или "примерно" при ссылке на измеряемую величину, такую как параметр, количество, промежуток времени и т.п., охватывает изменения, составляющие +/-20% или меньше, предпочтительно +/-10% или меньше, более предпочтительно +/-5% или меньше, и еще более предпочтительно +/-1% или меньше, от указанного значения, в той степени, в которой такие изменения подходят для осуществления в раскрытом изобретении. Значение, к которому относится понятие "приблизительно" или "примерно", также раскрыто специально, и предпочтительно, само по себе.
Несмотря на то, что понятия "один или несколько" или "по меньшей мере один", как, например, один или несколько или по меньшей мере один компонент (компонентов) из группы компонентов, ясны сами по себе и следуют из приведенных в настоящей заявке примеров, данное понятие охватывает, помимо прочего, любой из указанных компонентов, или на любые два или более указанных компонентов, таких как, например, любые из 3-х, 4-х, 5-ти, 6-ти, 7-ми, 8-ми, 9-ти, 10-ти и т.п. указанных компонентов и вплоть до всех указанных компонентов.
Термин "по существу" не означает "полностью", но и не исключает такую возможность, например препарат, который «по существу очищен», может совсем не содержать примесей или может содержать примеси в незначительных количествах. В настоящей заявке предлагается препарат очищенного белка Cas, полученного рекомбинантным путем, при этом "очищенный" означает, что целевой белок присутствует в составе препарата, который, по существу, не содержит других экспрессированных белков или эндотоксинов продуцента, при этом "по существу, не содержит" означает, что в составе препарата содержатся минимальные количества примесей.
Все документы, цитируемые или приводимые в качестве ссылок в настоящей заявке, и все документы, цитируемые или приводимые в качестве ссылок в документах, цитируемых в настоящей заявке, вместе с любыми инструкциями производителя, описаниями и режимами использования способа или продуктов, упомянутых в настоящей заявке или в любом документе, включенном в настоящую заявку посредством ссылки, включены в предложенную заявку посредством ссылки и могут быть использованы при осуществлении предложенного изобретения на практике.
В следующих частях различные аспекты настоящего изобретения определены более подробно. Каждый аспект, определенный таким образом, может быть скомбинирован с любым другим аспектом или аспектами, если явно не указано обратное. В частности, любая характеристика, обозначенная как являющийся предпочтительным или преимущественным, может быть скомбинирована с любой другой характеристикой или характеристиками, обозначенными как являющиеся предпочтительными или преимущественными.
Понятие «при необходимости» следует понимать, как обозначающее дополнительное, необязательное условие, стадию, методику или компонент способа или продукта, либо как обозначающее альтернативное воплощение изобретения или варианта его выполнения, в частности его условий, стадий, методик или компонентов состава.
CRISPR (от англ. Clustered Regularly Interspaced Short Palindromic Repeats; короткие палиндромные повторы, регулярно расположенные группами) - это особые локусы бактерий и архей, состоящие из прямых повторяющихся последовательностей, которые разделены уникальными последовательностями (спейсерами).
Система направленного редактирования генома CRISPR-Cas («система CRISPR-Cas», «система CRISPR-Cas9» или «система CRISPR») состоит из СAS-нуклеазы (белка) и направляющей РНК, специфичной к целевой последовательности-мишени. В целом система CRISPR-Cas, характеризуется элементами, которые содействуют образованию комплекса CRISPR в сайте целевой последовательности (протоспейсер).
В контексте комплекса CRISPR "целевая последовательность" обозначает последовательность, относительно которой конструируется направляющая последовательность таким образом, чтобы обладать комплементарностью, при этом гибридизация между целевой последовательностью и направляющей последовательностью содействует образованию комплекса CRISPR. Целевая последовательность может содержать любой полинуклеотид, как, например, полинуклеотиды ДНК или РНК. В некоторых вариантах осуществления целевая последовательность находится в ядре или цитоплазме клетки, и она может включать нуклеиновые кислоты в митохондриях, органеллах, везикулах, липосомах или частицах, присутствующих в пределах клетки, или из них.
Для специфического нацеливания белка системы CRISPR/Cas могут быть использованы различные типы направляющих РНК, способные направлять Cas на целевой локус. Так, например, направляющая РНК может представлять собой комплекс CRISPR РНК crRNA, отвечающей за специфическое узнавание мишени, а также трансактивирующей CRISPR РНК tracrRNA, которая отвечает за связывание фермента системы CRISPR/Cas. Кроме того, направляющая РНК может представлять собой единую комбинированную РНК (sgРНК), которая сочетает в себе свойства CRISPR РНК crRNA и CRISPR РНК tracrRNA в одной молекуле.
Направляющая РНК может содержать направляющую последовательность, способную гибридизироваться с целевым локусом генома в клетке, tracr-последовательность и парную tracr-последовательность. Все они могут находиться в одиночной РНК, т.е. sgRNA (расположенной в ориентации 5' - 3'), или tracrRNA может представлять собой РНК, отличную от РНК, содержащей направляющую и tracr-последовательность. Tracr-последовательность может гибридизоваться c парной tracr-последовательностью и направлять комплекс CRISPR/Cas к целевой последовательности.
Используемый термин "crRNA", или "направляющая РНК", или "одиночная направляющая РНК", или "sgRNA" предусматривает любую полинуклеотидную последовательность, характеризующуюся достаточной комплементарностью с целевой последовательностью нуклеиновой кислоты для гибридизации с целевой последовательностью нуклеиновой кислоты и управления специфичным к последовательности связыванием комплекса нацеливания на нуклеиновую кислоту с целевой последовательностью нуклеиновой кислоты.
Способность направляющей последовательности, в рамках направляющей РНК для нацеливания на нуклеиновую кислоту, управлять специфичным к последовательности связыванием комплекса нацеливания на нуклеиновую кислоту с целевой последовательностью нуклеиновой кислоты можно оценивать с помощью любого подходящего анализа. Например, компоненты системы CRISPR для нацеливания на нуклеиновую кислоту, достаточные для образования комплекса нацеливания на нуклеиновую кислоту, в том числе направляющая последовательность, подлежащую тестированию, могут обеспечиваться в клетке-хозяине, имеющей соответствующую целевую последовательность нуклеиновой кислоты, как, например, с помощью трансфекции векторами, кодирующими компоненты комплекса нацеливания на нуклеиновую кислоту, с последующей оценкой предпочтительного нацеливания (например, расщепления) в пределах целевой последовательности нуклеиновой кислоты, как, например, с помощью анализа, описываемого в настоящей заявке.
Расщепление целевой последовательности нуклеиновой кислоты можно определять in vitro путем обеспечения целевой последовательности нуклеиновой кислоты, компонентов комплекса нацеливания на нуклеиновую кислоту, в том числе направляющей последовательности, подлежащей тестированию, и контрольной направляющей последовательности, отличной от тестируемой направляющей последовательности, и сравнения связывания или степени расщепления целевой последовательности в случае реакций с тестируемой и контрольной направляющей последовательностью. Возможны и другие анализы, и они могут быть выполнены специалистами в данной области.
Направляющая последовательность и, следовательно, направляющая РНК для нацеливания на нуклеиновую кислоту, может быть выбрана для нацеливания на любую целевую последовательность нуклеиновой кислоты. Целевая последовательность может представлять собой ДНК. Целевая последовательность может представлять собой любую последовательность РНК. В некоторых вариантах осуществления целевая последовательность может представлять собой последовательность в пределах молекулы РНК, выбранной из группы, состоящей из матричной РНК (мРНК), pre-mRNA, рибосомальной РНК (rRNA), транспортной РНК (tRNA), микро-РНК (miRNA), малой интерферирующей РНК (siRNA), малой ядерной РНК (snRNA), малой ядрышковой РНК (snoRNA), двухнитевой РНК (dsRNA), некодирующей РНК (ncRNA), длинной некодирующей РНК (lncRNA) и малой цитоплазматической РНК (scRNA).
В некоторых вариантах осуществления направляющая РНК или crRNA может содержать или состоять из последовательности прямого повтора (DR) и направляющей последовательности и/или спейсерной последовательности.
Последовательность "tracrRNA" включает любую полинуклеотидную последовательность, которая характеризуется достаточной комплементарностью с последовательностью crRNA для возможности гибридизации. В некоторых вариантах осуществления последовательность crRNA и парная tracr-последовательность содержатся в одном транскрипте, так что при гибридизация между ними двумя образуется транскрипт со вторичной структурой, такой как "шпилька", которых может быть одна или несколько.
В терминологии, используемой в данной области техники, входящий в систему CRISPR/Cas белок обозначается как «белок Cas», «белок CRISPR/Cas», «белок семейства Cas», «белок системы CRISPR/Cas», «белок семейства Cas системы CRISPR/Cas», «нуклеаза Cas», «нуклеаза CRISPR/Cas», «нуклеаза семейства CRISPR/Cas», «нуклеаза системы CRISPR/Cas», «нуклеаза семейства Cas системы CRISPR/Cas», «эндонуклеаза Cas», «эндонуклеаза CRISPR/Cas», «эндонуклеаза семейства CRISPR/Cas», «эндонуклеаза системы CRISPR/Cas», «эндонуклеаза семейства Cas системы CRISPR/Cas», в настоящей заявке, если не указано иное, перечисленные определения взаимозаменяемы.
Нуклеазы - группа ферментов, гидролизующих фосфодиэфирную связь между субъединицами нуклеиновых кислот. Различают несколько типов нуклеаз в зависимости от их специфичности и активности: экзонуклеазы и эндонуклеазы, рибонуклеазы и дезоксирибонуклеазы, никазы, рестриктазы и некоторые другие.
Раскрытый в настоящей заявке способ обеспечивает получение препаратов рекомбинантных эндонуклеаз семейства Cas системы CRISPR/Cas I типа, II типа или III типа. Подходящие белки CRISPR/Cas могут быть выбраны из Cas1, Cas1b, Cas2, Cas3, Cas4, Cas5, Cas5e (CasD), Cas6, Cas6e, Cas6f, Cas7, Cas8al, Cas8a2, Cas8b, Cas8c, Cas9, Cas9D10A, Cas12a (Cpf1), CaslO, CaslOd, CasF, CasG, CasH, Csyl, Csy2, Csy3, Csel (CasA), Cse2 (CasB), Cse3 (CasE), Cse4 (CasC), Csel, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmrl, Cmr3, Cmr4, Cmr5, Cmr6, Csbl, Csb2, Csb3, Csxl7, Csxl4, CsxlO, Csxl6, CsaX, Csx3, Cszl, Csxl5, Csfl, Csf2, Csf3, Csf4, Cul966 и других.
Нуклеаза Cas может происходить из Streptococcus pyogenes, Streptococcus thermophilus, Streptococcus sp., Nocardiopsis dassonvillei, Streptomyces pristinaespiralis, Streptomyces viridochromogenes, Streptosporangium roseum, Alicy clobacdlus acidocaldarius, Bacillus pseudomycoides, Bacillus selenitireducens, Exiguobacterium sibiricum, Lactobacillus delbrueckii, Lactobacillus salivarius, Microscilla marina, Burkholderiales bacterium, Polaromonas naphthalenivorans, Polaromonas sp., Crocosphaera watsonii, Cyanothece sp., Microcystis aeruginosa, Synechococcus sp., Acetohalobium arabaticum, Ammonifex degensii, Caldicelulosiruptor becscii, Candidatus Desulforudis, Clostridium botulinum, Clostridium difficile, Finegoldia magna, Natranaerobius thermophilus, Pelotomaculum thermopropionicum, Acidithiobacillus caldus, Acidithiobacillus ferrooxidans, Allochromatium vinosum, Marinobacter sp., Nitrosococcus halophilus, Nitrosococcus watsoni, Pseudoalteromonas haloplanktis, Ktedonobacter racemifer, Methanohalobium evestigatum, Anabaena variabilis, Nodularia spumigena, Nostoc sp., Arthrospira maxima, Arthrospira platensis, Arthrospira sp., Lyngbya sp., Microcoleus chthonoplastes, Oscillatoria sp., Petrotoga mobilis, Thermosipho africanus или Acaryochloris marina, Pseudomonas aeruginosa, Acinetobacter baumannii, Salmonella enterica, Streptococcus agalactiae, Escherichia coli, Streptococcus suis, Salinispora arenicola, Klebsiella pneumoniae, Neisseria meningitidis, Vibrio cholerae, Leptospira interrogans, Listeria monocytogenes, Fusobacterium nucleatum, Enterococcus faecalis, Salinispora pacifica, Pasteurella multocida, Moraxella catarrhalis, Leptospira santarosai, Bifidobacterium longum, Serratia marcescens, Clostridioides difficile, Bifidobacterium breve, Methanosarcina mazei, Yersinia pseudotuberculosis, Fusobacterium necrophorum, Propionibacterium freudenreichii, Campylobacter coli, Clostridium novyi, Corynebacterium ulcerans, Salinispora tropica, Lactobacillus paracasei, Aggregatibacter actinomycet., Treponema pedis, Corynebacterium striatum, Lactobacillus rhamnosus, Xenorhabdus nematophila, Campylobacter jejuni, Lachnospiraceae bacterium, Treponema denticola, Staphylococcus aureus, Bacillus halodurans.
Нуклеазы Cas согласно настоящему изобретению могут быть получены из белка Cas дикого типа или его фрагмента или из модифицированного с целью изменения одного или нескольких биологических или физико-химических свойств белка Cas путем добавления функциональных элементов для повышения активности, аффинности, растворимости, стабильности нуклеазы, для облегчения проникновения в клетку или ее компартменты и т.п. Либо домены белка Cas, не вовлеченные в РНК-направляемое расщепление, могут быть удалены из белка с целью уменьшения размера модифицированного белка Cas по сравнению с диким типом.
Предпочтительными вариантами являются РНК-направляемые эндонуклеазы системы CRISPR/Cas: SPCas9 - нуклеаза Cas9 из Streptococcus pyogenes, STCas9 - нуклеаза Cas9 из Streptococcus thermophilus, SPCas9D10A - никаза Cas9 из Streptococcus pyogenes и AsCpf1 - нуклеаза Cpf1 из Acidaminococcus, аминокислотные последовательности которых могут быть дополнены с N-конца и/или C-конца:
• 6×His - полигистидиновой последовательностью для повышения эффективности очистки;
• SV40 NLS - последовательностью сигнала ядерной локализации большого T-антигена вируса обезьян SV40 для эффективной доставки белка в ядро клетки-мишени;
• Nucleoplasmin NLS - последовательностью сигнала ядерной локализации нуклеоплазмина;
• TAT - последовательностью проникающего пептида TAT вируса иммунодефицита человека (ВИЧ-1).
Наиболее предпочтительны РНК-направляемые эндонуклеазы, характеризующиеся, имеющие или содержащие аминокислотную последовательность, выбранную из:
• SEQ ID NO: 1,
• SEQ ID NO: 2,
• SEQ ID NO: 3,
•SEQ ID NO: 4,
•SEQ ID NO: 5,
• SEQ ID NO: 6,
• SEQ ID NO: 7
• SEQ ID NO: 8
• или идентичной любой из них по меньшей мере на 95%, 96%, 97%, 98% или 99%,
а также кодируемые нуклеиновыми кислотами, характеризующимися, имеющими или содержащими нуклеотидную последовательность, выбранную из
• SEQ ID NO: 9,
• SEQ ID NO: 10,
• SEQ ID NO: 11,
• SEQ ID NO: 12,
• SEQ ID NO: 13,
• SEQ ID NO: 14,
• SEQ ID NO: 15,
• SEQ ID NO: 16
или идентичной любой из них по меньшей мере на 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%,
• или комплементарной любой из них,
• или гибридизующейся с любой из них в строгих условиях, которые могут быть
•связаны с регуляторными последовательностями,
•находится в составе кассеты экспрессии, вектора или плазмиды экспрессии, клонирования или интегрирования в геном или
• в составе клетки-продуцента.
Термин "строгие условия гибридизации" относится в данном контексте к параметрам, которые известны специалисту в данной области, включающим вариацию температуры гибридизации в зависимости от длины нуклеиновой кислоты. Параметры гибридизации нуклеиновых кислот могут быть найдены в ссылках, которые объединяют такие способы, Sambrook J. et al., Molecular Cloning, a Laboratory Manual. Cold Spring Harbor, Laboratory Press, New York, 1989 и Ausubel M. et al., Current protocols in molecular biology, edited by Volumes 1 and 2. John Wiley & Sons, Inc., Media, PA, 1988). Например, строгие условия гибридизации, в данном контексте, могут относиться к гибридизации при 65°С в буфере для гибридизации (3,5×SSC, 0,02% Фиколл, 0,02% поливинилпирролидон, 0,02% бычий сывороточный альбумин, 2,5 мМ NaH2PO4, 0,5% ДСН, 2 мМ ЭДТА, pH 7).
Согласно настоящему изобретению способ получения препарата рекомбинантной нуклеазы семейства Cas системы CRISPR/Cas, как и описанный в ближайшем аналоге (Nandhakishore Rajagopalan, Sateesh Kagale, Pankaj Bhowmik and Halim Song, A Two-Step Method for Obtaining Highly Pure Cas9 Nuclease for Genome Editing, Biophysical, and Structural Studies, 2018, v.1, n.17, p.1-8) основан на трансформации компетентных клеток E. сoli экспрессионной плазмидой, кодирующей целевой белок, с дальнейшим культивированием трансформированных клеток-продуцентов в условиях, обеспечивающих экспрессию указанной рекомбинантной нуклеазы в питательной среде Лурии-Бертани (LB) с добавлением ампициллина и использованием в качестве индуктора экспрессии целевого белка изопропил-β-D-тиогалактозида (ИПТГ), выделение целевого белка из растворимой фракции и проведение двухстадийной хроматографической очистки.
Однако достижение неожиданного технического результата обеспечивается тем, что, в отличие от способа согласно ближайшему аналогу, в настоящем изобретении:
в качестве клеток-продуцентов рекомбинантной нуклеазы семейства Cas системы CRISPR/Cas используют компетентные клетки E. сoli Rosetta-gami B (DE3);
в питательную среду добавляют MgSO4 и глюкозу и выращивают клетки-продуценты до получения стартовой культуры, после чего разводят культуральную среду до оптической плотности OD600≈0,3 и уменьшают содержание глюкозы в указанной среде, по меньшей мере, в два раза;
затем клетки-продуценты культивируют до достижения оптической плотности OD600≈1,5-1,9, добавляют ИПТГ и инкубируют при 22°С для эффективной экспрессии целевого белка;
после центрифугирования по окончании культивирования клетки ресуспендируют в буфере для лизиса на основе Na2HPO4/NaH2PO4 и разрушают методом ультразвуковой дезинтеграции с последующим центрифугированием для отделения фракции растворимой цитоплазмы, содержащей рекомбинантную нуклеазу семейства Cas системы CRISPR/Cas, от нерастворимой фракции телец включения.
Для получения стартовой культуры предпочтительно добавляют 2 mM MgSO4 и 2% глюкозы в питательную среду LB с 100 мкг/мл ампициллина, а после разводят в питательной среде LB с 100 мкг/мл ампициллина, дополненной 2 mM MgSO4 и 1% глюкозы.
Также для получения стартовой культуры предпочтительно клетки-продуценты выращивают при 37°С в течение 16-20 часов, после разведения культивируют при 37°С до достижения оптической плотности OD600≈1,5-1,9, а после добавления ИПТГ инкубируют при 22°С в течение 20-24 часов.
Предпочтительно клетки ресуспендируют в буфере для лизиса, содержащем 20 mM Na2HPO4/NaH2PO4 при pH 6,6-8,0.
Центрифугирование по окончании культивирования проводят предпочтительно при 6000 g в течение 10 минут и/или центрифугирование для отделения фракции растворимой цитоплазмы проводят при 10000 g в течение одного часа.
Достижение неожиданного технического результата обеспечивается также тем, что, в отличие от способа согласно ближайшему аналогу, в настоящем изобретении:
для очистки к полученной после центрифугирования фракции растворимой цитоплазмы, содержащей целевой белок, добавляют NaCl и имидазол и проводят первый этап хроматографической очистки путем металл-хелатной хроматографии, при этом элюцию целевого белка проводят буфером, содержащим Na2HPO4/NaH2PO4, NaCl и имидазол;
на втором этапе хроматографической очистки целевой белок в указанном буфере разбавляют не менее чем в 20 раз буфером с Tris-HCl, при этом кондуктивность раствора не превышает 5 mS/cm, и подвергают катионообменной хроматографии с последующей элюцией целевого белка буфером, содержащим Tris-HCl и NaCl, где для стабилизации целевого белка к указанному буферу добавляют DTT и EDTA.
На первом этапе хроматографической очистки к указанной фракции растворимой цитоплазмы добавляют NaCl предпочтительно до конечной концентрации 500 mM и имидазол предпочтительно до конечной концентрации 10 mM, элюцию целевого белка проводят предпочтительно буфером с pH 6,6-8,0, содержащим 20 mM Na2HPO4/NaH2PO4, 500 mM NaCl и 250 mM имидазола.
В качестве сорбента на первом этапе хроматографической очистки предпочтительно используют сорбент Chelating Sepharose FF.
На втором этапе хроматографической очистки целевой белок в указанном буфере разбавляют не менее чем в 20 раз предпочтительно буфером с pH 8,2, содержащим 20 mM Tris-HCl с кондуктивностью раствора 3-5 mS/cm, элюцию целевого белка предпочтительно проводят буфером с pH 8,2, содержащим 20 mM Tris-HCl и 600-1000 mM NaCl.
В качестве сорбента на втором этапе хроматографической очистки предпочтительно используют сорбент SP Sepharose FF.
Для стабилизации целевого белка к буферу добавляют до 2 mM DTT и 0,2 mM EDTA.
Достижение неожиданного технического результата обеспечивается также тем, что, в отличие от способа согласно ближайшему аналогу, в настоящем изобретении проводят дополнительную очистку целевого белка от эндотоксинов продуцента путем:
добавления в указанный раствор целевого белка в буфере после хроматографии детергента Triton X-114 для удаления бактериальных эндотоксинов в процессе перехода Triton X-114 из водонерастворимой фракции в водорастворимую и обратно с образованием мицелл и последующим разделением водорастворимой и водонерастворимой фракций методом центрифугирования,
затем снова добавляют Triton X-114 в водорастворимую фракцию, полученную после центрифугирования, с переводом из водонерастворимой фракции в водорастворимую и обратно с образованием мицелл и разделением путем центрифугирования,
при этом для удаления бактериальных эндотоксинов из препарата рекомбинантной нуклеазы семейства Cas системы CRISPR/Cas до уровня, позволяющего применять указанный препарат in vivo, стадии с добавлением Triton X-114 повторяют по меньшей мере 6 раз, предпочтительно повторяют 6-9 раз.
После финального центрифугирования по завершении этапа дополнительной очистки отбирают водную фазу, включающую очищенную рекомбинантную нуклеазу семейства Cas, и при необходимости проводят замораживание или лиофилизацию с добавлением вспомогательных веществ для замораживания или лиофилизации, соответственно.
Центрифугирование на этапе дополнительной очистки от эндотоксинов предпочтительно проводят при 10000 g в течение 10 мин при 23°С.
Предпочтительно дополнительную очистку рекомбинантной нуклеазы семейства Cas от эндотоксинов проводят путем выполнения следующих стадий:
добавления в раствор целевого белка в буфере, полученный после этапов хроматографической очистки, детергента Triton X-114 до 1%;
помещения полученного раствора на лед до достижения абсолютной прозрачности раствора с последующим выдерживанием при 37°С до достижения абсолютной непрозрачности раствора с целью обогащения раствора мицеллами;
последующего проведения разделения водной и мицеллярной фракций раствора с помощью центрифугирования;
добавления детергента Triton X-114 до 1% к верхней водной фазе, полученной после центрифугирования, и повторения обработки до получения препарата рекомбинантной нуклеазы семейства Cas с содержанием бактериальных эндотоксинов, сниженным до уровня, позволяющего применять указанный препарат in vivo.
При этом раствор помещают на лед и/или выдерживают при 37°С предпочтительно по меньшей мере 15 мин.
Согласно предложенному способу может быть получена рекомбинантная нуклеаза семейства Cas системы CRISPR/Cas, характеризующаяся аминокислотной последовательностью, выбранной из SEQ ID NO: 1-8.
Для получения рекомбинантной нуклеазы семейства Cas системы CRISPR/Cas может быть использована кодирующая нуклеотидная последовательность, выбранная из SEQ ID NO: 9-16.
Согласно предложенному способу получают препарат рекомбинантной нуклеазы семейства Cas системы CRISPR/Cas, по существу, свободный от бактериальных эндотоксинов, для использования в системе CRISPR/Cas.
По существу, свободный от бактериальных эндотоксинов препарат рекомбинантной нуклеазы семейства Cas системы CRISPR/Cas содержит менее 1,5 ЕЭ/мл бактериальных эндотоксинов, предпочтительно 0,3 ЕЭ/мл.
Препарат может быть в жидком виде и представляет собой водную фазу, включающую очищенную рекомбинантную нуклеазу семейства Cas, полученную после финального центрифугирования по завершении этапа дополнительной очистки.
Предпочтительно препарат рекомбинантной нуклеазы семейства Cas включает указанную нуклеазу с содержанием эндотоксинов 0,3-1,5 ЕЭ/мл, находящуюся в буферном растворе, содержащем Tris-HCl, NaCl, DTT и EDTA, наиболее предпочтительно 20 mM Tris-HCl, 600-1000 mM NaCl, 2 mM DTT, 0,2 mM EDTA при pH 8,2.
Препарат может быть в сухом виде, предпочтительно лиофильно высушенный препарат, и представляет собой водную фазу, включающую очищенную рекомбинантную нуклеазу семейства Cas, полученную после финального центрифугирования по завершении этапа дополнительной очистки и лиофильно высушенную.
Предпочтительно лиофилизированный препарат рекомбинантной нуклеазы семейства Cas получают добавлением вспомогательных веществ для лиофилизации, выбранных из глицерина или трегалозы, предпочтительно глицерина в концентрации 10-50% или трегалозы в концентрации 50-500 mM к буферному раствору, содержащему рекомбинантную нуклеазу семейства Cas с 0,3-1,5 ЕЭ/мл, Tris-HCl, NaCl, DTT и EDTA, наиболее предпочтительно 20 mM Tris-HCl, 600-1000 mM NaCl, 2 mM DTT, 0,2 mM EDTA при pH 8,2, с последующим проведением лиофилизации.
Концентрация рекомбинантной нуклеазы семейства Cas в составе препарата может варьировать в широких пределах в зависимости от целей использования от 0,3 мг/мл до 3 мг/мл.
Полученный препарат рекомбинантной нуклеазы семейства Cas системы CRISPR/Cas может быть использован в системе CRISPR/Cas в составе набора с инструкцией по применению.
Набор может дополнительно включать мишень-специфическую направляющую РНК. При этом препарат рекомбинантной нуклеазы семейства CRISPR/Cas в составе набора может находиться в комплексе с мишень-специфической направляющей РНК в одном контейнере или отдельно в разных контейнерах.
Препараты и наборы согласно настоящему изобретению возможно использовать в системе CRISPR/Cas в различных областях техники, связанных с редактированием генов: для создания клеточных моделей наследственных заболеваний, функционального скрининга геномов, изучения эпигеномов и визуализации клеточных процессов, в пищевой промышленности для получения обогащенных продуктов питания, в сельском хозяйстве для создания новых пород животных и сортов растений, а также в медицине для диагностики, предупреждения и лечения различных заболеваний.
Краткое описание чертежей
Фиг. 1. Схемы рекомбинантных белков семейства CRISPR/CAS.
6×His - полигистидиновая последовательность;
SV40 NLS - сигнал ядерной локализации большого T-антигена вируса обезьян SV40 для эффективной доставки белка в ядро клетки-мишени;
Nucleoplasmin NLS - сигнал ядерной локализации нуклеоплазмина, первого идентифицированного молекулярного шаперона;
TAT - пептид вируса иммунодефицита человека;
SPCas9 - нуклеаза Cas9 из Streptococcus pyogenes;
SPCas9D10A - никаза Cas9 из Streptococcus pyogenes;
STCas9 - нуклеаза Cas9 из Streptococcus thermophilus;
AsCpf1 - нуклеаза Cpf1 из Acidaminococcus;
TATSPCas9 - нуклеаза Cas9 из Streptococcus pyogenes, слитый белок с TAT-пептидом на N-конце;
SPCas9TAT - нуклеаза Cas9 из Streptococcus pyogenes, слитый белок с TAT-пептидом на C-конце;
TATSPCas9TAT - нуклеаза Cas9 из Streptococcus pyogenes, слитый белок с TAT-пептидом на N- и C-концах;
SPCas9D10ATAT - никаза Cas9 из Streptococcus pyogenes, слитый белок с TAT-пептидом на C-конце.
Фиг. 2. Электрофореграмма растворимых фракций, которые должны содержать рекомбинантные белки семейства CRISPR/CAS, полученных при экспрессии в клетках E. coli BL21(DE3), где цифрами 1-10 обозначены:
1 - растворимая фракция E. coli BL21(DE3), не трансформированной плазмидной ДНК pAM, несущей ген, содержащий последовательность SEQ ID NO: NO: 9-16;
2 - растворимая фракция E. coli BL21(DE3), трансформированной плазмидной ДНК pAM-SPCas9, не индуцированной добавлением ИПТГ;
3 - растворимая фракция E. coli BL21(DE3), трансформированной плазмидной ДНК pAM-SPCas9, индуцированной добавлением ИПТГ, культивированной при комнатной температуре в течение 4 часов;
4 - растворимая фракция E. coli BL21(DE3), трансформированной плазмидной ДНК pAM-SPCas9, индуцированной добавлением ИПТГ, культивированной при комнатной температуре в течение 20 часов;
5 - растворимая фракция E. coli BL21(DE3), трансформированной плазмидной ДНК pAM-SPCas9D10A, не индуцированной добавлением ИПТГ;
6 - растворимая фракция E. coli BL21(DE3), трансформированной плазмидной ДНК pAM-SPCas9D10A, индуцированной добавлением ИПТГ, культивированной при комнатной температуре в течение 4 часов;
7 - растворимая фракция E. coli BL21(DE3), трансформированной плазмидной ДНК pAM-SPCas9D10A, индуцированной добавлением ИПТГ, культивированной при комнатной температуре в течение 20 часов;
8 - растворимая фракция E. coli BL21(DE3), трансформированной плазмидной ДНК pAM-AsCpf1, не индуцированной добавлением ИПТГ;
9 - растворимая фракция E. coli BL21(DE3), трансформированной плазмидной ДНК pAM-AsCpf1, индуцированной добавлением ИПТГ, культивированной при комнатной температуре в течение 4 часов;
10 - растворимая фракция E. coli BL21(DE3), трансформированной плазмидной ДНК pAM-AsCpf1, индуцированной добавлением ИПТГ, культивированной при комнатной температуре в течение 20 часов;
буквой М обозначены стандарты молекулярных масс,
стрелкой отмечена область геля, в которой по расчетам должны были находиться рекомбинантные белки семейства CRISPR/CAS.
Фиг. 3. Электрофореграмма растворимых фракций, которые должны содержать рекомбинантные белки семейства CRISPR/CAS, полученных при экспрессии в клетках E. coli Rosetta-gami B (DE3) с использованием стартовой культуры, выращенной в бульоне Лурии-Бертани, не содержащем 2mM MgSO4 и 2% глюкозы, где цифрами 1-5 обозначены:
1 - растворимая фракция E. coli Rosetta-gami B (DE3), не трансформированной плазмидной ДНК pAM, несущей ген, содержащий последовательность SEQ ID NO: NO: 9-16;
2 - растворимая фракция E. coli Rosetta-gami B (DE3), трансформированной плазмидной ДНК pAM, несущей ген, содержащий последовательность SEQ ID NO: NO: 9, не индуцированной добавлением ИПТГ;
3 - растворимая фракция E. coli Rosetta-gami B (DE3), трансформированной плазмидной ДНК pAM, несущей ген, содержащий последовательность SEQ ID NO: NO: 9, индуцированной добавлением ИПТГ, культивированной при комнатной температуре в течение 20 часов;
4 - растворимая фракция E. coli Rosetta-gami B (DE3), трансформированной плазмидной ДНК pAM, несущей ген, содержащий последовательность SEQ ID NO: NO: 10, не индуцированной добавлением ИПТГ;
5 - растворимая фракция E. coli Rosetta-gami B (DE3), трансформированной плазмидной ДНК pAM, несущей ген, содержащий последовательность SEQ ID NO: NO: 10, индуцированной добавлением ИПТГ, культивированной при комнатной температуре в течение 20 часов;
буквой М обозначены стандарты молекулярных масс.
Стрелкой отмечена область геля, в которой по расчетам должны были находиться рекомбинантные белки семейства CRISPR/CAS.
Фиг. 4. Электрофореграмма препаратов белка, содержащих рекомбинантные белки семейства CRISPR/CAS SPCas9 и STCas9, полученных при хроматографической очистке; где цифрами 1-6 обозначены:
1 - препарат SPCas9, полученный при хроматографической очистке белка из растворимой цитоплазмы штамма-продуцента после индукции по достижению OD600=0,6-1,0;
2 - препарат SPCas9, полученный при хроматографической очистке белка из растворимой цитоплазмы штамма-продуцента после индукции по достижению OD600=1,1-1,4;
3 - препарат SPCas9, полученный при хроматографической очистке белка из растворимой цитоплазмы штамма-продуцента после индукции по достижению OD600=1,5-1,9;
4 - препарат STCas9, полученный при хроматографической очистке белка из растворимой цитоплазмы штамма-продуцента после индукции по достижению OD600=0,6-1,0;
5 - препарат STCas9, полученный при хроматографической очистке белка из растворимой цитоплазмы штамма-продуцента после индукции по достижению OD600=1,1-1,4;
6 - препарат STCas9, полученный при хроматографической очистке белка из растворимой цитоплазмы штамма-продуцента после индукции по достижению OD600=1,5-1,9;буквами M1 и M2 обозначены маркеры молекулярных масс.
Фиг. 5. Хроматографический профиль очистки рекомбинантного белка SPCas9 семейства CRISPR/CAS на Chelating sepharose при pH=6,6; где цифрами 1-5 обозначены этапы процесса:
1 - нанесение;
2 - отмывка несвязавшихся белков;
3 - элюция раствором, содержащим 50 mM имидазола;
4 - элюция раствором, содержащим 250 mM имидазола;
5 - промывка раствором, содержащим 1 M хлорида натрия.
Фиг. 6. Хроматографический профиль очистки рекомбинантного белка SPCas9 семейства CRISPR/CAS на Chelating sepharose при pH=7,3; где цифрами 1-5 обозначены этапы процесса:
1 - нанесение;
2 - отмывка несвязавшихся белков;
3 - элюция раствором, содержащим 50 mM имидазола;
4 - элюция раствором, содержащим 250 mM имидазола;
5 - промывка раствором, содержащим 1 M хлорида натрия.
Фиг. 7. Хроматографический профиль очистки рекомбинантного белка SPCas9 семейства CRISPR/CAS на Chelating sepharose при pH=8,0; где цифрами 1-5 обозначены этапы процесса:
1 - нанесение;
2 - отмывка несвязавшихся белков;
3 - элюция раствором, содержащим 50 mM имидазола;
4 - элюция раствором, содержащим 250 mM имидазола;
5 - промывка раствором, содержащим 1 M хлорида натрия.
Фиг. 8. Электрофореграмма хроматографических фракций, содержащих рекомбинантный белок SPCas9 семейства CRISPR/CAS, полученных при очистке на Chelating sepharose; где цифрами 1-4 обозначены фракции:
1 - растворимая фракция Escherichia coli Rosetta-gami B (DE3), трансформированной плазмидной ДНК pAM, несущей ген, содержащий последовательность SEQ ID NO: NO: 9;
2 - проскок (хроматографическая фракция, содержащая в себе все белки, не связавшиеся с Chelating sepharose);
3 - элюат, содержащий 50 mM имидазола;
4 - элюат, содержащий 250 mM имидазола; буквой М обозначены стандарты молекулярных масс. Стрелками отмечено положение рекомбинантного белка SPCas9 семейства CRISPR/CAS в геле.
Фиг. 9. Хроматографический профиль очистки рекомбинантного белка SPCas9 семейства CRISPR/CAS на SP Sepharose; где цифрами 1-5 обозначены этапы процесса:
1 - нанесение;
2 - отмывка несвязавшихся белков;
3 - элюция раствором, содержащим 300 mM хлорида натрия;
4 - элюция раствором, содержащим 600 mM хлорида натрия;
5 - промывка раствором, содержащим 1 M хлорида натрия.
Фиг. 10. Электрофореграмма хроматографических фракций, содержащих рекомбинантный белок SPCas9 семейства CRISPR/CAS, полученных при очистке на SP Sepharose; где цифрами 1-4 обозначены фракции:
1 - элюат с Chelating sepharose, содержащий рекомбинантный белок SPCas9 (250 mM имидазола);
2 - проскок (хроматографическая фракция, содержащая в себе все белки, не связавшиеся с SP Sepharose);
3 - элюат, содержащий 300 mM хлорида натрия;
4 - элюат, содержащий 600 mM хлорида натрия; буквой М обозначены стандарты молекулярных масс. Стрелками отмечено положение рекомбинантного белка SPCas9 семейства CRISPR/CAS в геле.
Фиг. 11. Хроматографический профиль очистки рекомбинантного белка SPCas9D10A семейства CRISPR/CAS на Chelating sepharose при pH=6,6, экспрессированного при температуре 27°С в течение 20 часов; где цифрами 1-5 обозначены этапы процесса:
1 - нанесение;
2 - отмывка несвязавшихся белков;
3 - элюция раствором, содержащим 50 mM имидазола;
4 - элюция раствором, содержащим 250 mM имидазола;
5 - промывка раствором, содержащим 1 M хлорида натрия.
Фиг. 12. Хроматографический профиль очистки рекомбинантного белка SPCas9D10A семейства CRISPR/CAS на SP Sepharose, экспрессированного при температуре 27°С в течение 20 часов; где цифрами 1-14 обозначены этапы процесса:
1 - нанесение;
2 - отмывка несвязавшихся белков;
3 - элюция раствором, содержащим 50 mM хлорида натрия;
4 - элюция раствором, содержащим 100 mM хлорида натрия;
5 - элюция раствором, содержащим 150 mM хлорида натрия;
6 - элюция раствором, содержащим 200 mM хлорида натрия;
7 - элюция раствором, содержащим 250 mM хлорида натрия;
8 - элюция раствором, содержащим 300 mM хлорида натрия;
9 - элюция раствором, содержащим 400 mM хлорида натрия;
10 - элюция раствором, содержащим 500 mM хлорида натрия;
11 - элюция раствором, содержащим 600 mM хлорида натрия;
12 - элюция раствором, содержащим 800 mM хлорида натрия;
13 - элюция раствором, содержащим 1 M хлорида натрия;
14 - промывка раствором, содержащим 2 M хлорида натрия.
Фиг. 13. Хроматографический профиль очистки рекомбинантного белка AsCpf1 семейства CRISPR/CAS на Chelating sepharose при pH=6,6; где цифрами 1-5 обозначены этапы процесса:
1 - нанесение;
2 - отмывка несвязавшихся белков;
3 - элюция раствором, содержащим 50 mM имидазола;
4 - элюция раствором, содержащим 250 mM имидазола;
5 - промывка раствором, содержащим 1 M хлорида натрия.
Фиг. 14. Электрофореграмма хроматографических фракций, содержащих рекомбинантный белок AsCpf1 семейства CRISPR/CAS, полученных при очистке на Chelating sepharose; где цифрами 1-4 обозначены фракции:
1 - растворимая фракция Escherichia coli Rosetta-gami B (DE3), трансформированной плазмидной ДНК pAM, несущей ген, содержащий последовательность SEQ ID NO: NO: 12;
2 - проскок (хроматографическая фракция, содержащая в себе все белки, не связавшиеся с Chelating sepharose);
3 - элюат, содержащий 50 mM имидазола;
4 - элюат, содержащий 250 mM имидазола;
буквой М обозначены стандарты молекулярных масс,
стрелками отмечено положение рекомбинантного белка AsCpf1 семейства CRISPR/CAS в геле.
Фиг. 15. Хроматографический профиль очистки рекомбинантного белка AsCpf1 семейства CRISPR/CAS на SP Sepharose; где цифрами 1-5 обозначены этапы процесса:
1 - нанесение;
2 - отмывка несвязавшихся белков;
3 - элюция раствором, содержащим 300 mM хлорида натрия;
4 - элюция раствором, содержащим 600 mM хлорида натрия;
5 - промывка раствором, содержащим 1 M хлорида натрия.
Фиг. 16. Электрофореграмма хроматографических фракций, содержащих рекомбинантный белок AsCpf1 семейства CRISPR/CAS, полученных при очистке на SP Sepharose; где цифрами 1-3 обозначены фракции:
1 - элюат с Chelating sepharose, содержащий рекомбинантный белок AsCpf1 (250 mM имидазола);
2 - проскок (хроматографическая фракция, содержащая в себе все белки, не связавшиеся с SP Sepharose);
3 - элюат, содержащий 600 mM хлорида натрия;
буквой М обозначены стандарты молекулярных масс,
стрелками отмечено положение рекомбинантного белка AsCpf1 семейства CRISPR/CAS в геле.
Фиг. 17. Хроматографический профиль очистки рекомбинантного белка STCas9 семейства CRISPR/CAS на Chelating sepharose при pH=6,6; где цифрами 1-5 обозначены этапы процесса:
1 - нанесение;
2 - отмывка несвязавшихся белков;
3 - элюция раствором, содержащим 50 mM имидазола;
4 - элюция раствором, содержащим 250 mM имидазола;
5 - промывка раствором, содержащим 1 M хлорида натрия.
Фиг. 18. Хроматографический профиль очистки рекомбинантного белка STCas9 семейства CRISPR/CAS на SP Sepharose; где цифрами 1-5 обозначены этапы процесса:
1 - нанесение;
2 - отмывка несвязавшихся белков;
3 - элюция раствором, содержащим 300 mM хлорида натрия;
4 - элюция раствором, содержащим 1 M хлорида натрия;
5 - промывка раствором, содержащим 2 M хлорида натрия.
Фиг. 19. Электрофореграмма хроматографических фракций, содержащих рекомбинантный белок STCas9 семейства CRISPR/CAS, полученных при очистке на Chelating sepharose и SP Sepahrose; где цифрами 1-7 обозначены фракции:
1 - растворимая фракция Escherichia coli Rosetta-gami B (DE3), трансформированной плазмидной ДНК pAM, несущей ген, содержащий последовательность SEQ ID NO: NO: 11;
2 - проскок (хроматографическая фракция, содержащая в себе все белки, не связавшиеся с Chelating sepharose);
3 - элюат c Chelating sepharose, содержащий 50 mM имидазола;
4 - элюат c Chelating sepharose, содержащий 250 mM имидазола;
5 - проскок (хроматографическая фракция, содержащая в себе все белки, не связавшиеся с SP Sepharose);
6 - элюат с SP Sepharose, содержащий 300 mM хлорида натрия;
7 - элюат c SP Sepharose, содержащий 1 M хлорида натрия;
буквой М обозначены стандарты молекулярных масс,
стрелками отмечено положение рекомбинантного белка STCas9 семейства CRISPR/CAS в геле.
Фиг. 20. Хроматографический профиль очистки рекомбинантного белка SPCas9 семейства CRISPR/CAS на Chelating sepharose (дезинтеграция и хроматография проведены в присутствии хлорида натрия в концентрации 1 М); где цифрами 1-5 обозначены этапы процесса:
1 - нанесение;
2 - отмывка несвязавшихся белков;
3 - элюция раствором, содержащим 50 mM имидазола;
4 - элюция раствором, содержащим 250 mM имидазола;
5 - промывка раствором, содержащим 2 M хлорида натрия.
Фиг. 21. Хроматографический профиль очистки рекомбинантного белка SPCas9 семейства CRISPR/CAS на SP Sepharose (дезинтеграция клеток штамма-продуцента проведена при pH=9,5); где цифрами 1-4 обозначены этапы процесса:
1 - нанесение;
2 - отмывка несвязавшихся белков;
3 - элюция раствором, содержащим 600 mM хлорида натрия;
4 - промывка раствором, содержащим 1 M хлорида натрия.
Фиг. 22. Хроматографический профиль очистки рекомбинантного белка SPCas9 семейства CRISPR/CAS на SP Sepharose (хроматография проведена при pH=4,0); где цифрами 1-5 обозначены этапы процесса:
1 - нанесение;
2 - отмывка несвязавшихся белков;
3 - элюция раствором, содержащим 600 mM хлорида натрия;
4 - элюция раствором, содержащим 1,2 M хлорида натрия;
5 - промывка раствором, содержащим 2,4 M хлорида натрия.
Фиг. 23. Электрофореграмма препаратов рекомбинантного белка SPCas9 семейства CRISPR/CAS, содержащих вспомогательные вещества для лиофилизации, такие как глицерин и трегалоза; где цифрами 1-11 обозначены препараты:
1 - содержащий 50mM трегалозы;
2 - содержащий 100mM трегалозы;
3 - содержащий 150mM трегалозы;
4 - содержащий 200mM трегалозы;
5 - содержащий 250mM трегалозы;
6 - содержащий 500mM трегалозы;
7 - содержащий 10% глицерина;
8 - содержащий 20% глицерина;
9 - содержащий 30% глицерина;
10 - содержащий 40% глицерина;
11 - содержащий 50% глицерина;
буквой М обозначены стандарты молекулярных масс,
стрелкой отмечено положение препарата рекомбинантного белка SPCas9 семейства CRISPR/Cas в геле.
Фиг. 24. Электрофореграмма фрагментов ДНК, обработанных белками семейства CRISPR/Cas в присутствии мишень-специфической направляющей РНК; где цифрами 1-10 обозначены образцы:
1 - продукты гидролиза после обработки SPCas9;
2 - продукты гидролиза после обработки TATSPCas9;
3 - продукты гидролиза после обработки SPCas9TAT;
4 - продукты гидролиза после обработки TATSPCas9TAT;
5 - продукты гидролиза после обработки EnGen Cas9 (NEB);
6 - продукты гидролиза после обработки AsCpf1;
7 - недогидролизованная ДНК-матрица, полученная методом ПЦР;
8 - продукты гидролиза после обработки SPCas9D10A;
9 - продукты гидролиза после обработки SPCas9D10ATAT;
10 - недогидролизованная ДНК-матрица (суперспирализованная плазмидная ДНК);
буквой М обозначены стандарты молекулярных масс.
Фиг. 25. Анализ фрагментов ДНК, обработанных рекомбинантными белками семейства CRISPR/Cas в присутствии мишень-специфической направляющей РНК, проведенный на анализаторе Agilent Bioanalyzer 2100; где цифрами 1-6 обозначены образцы:
1 - EnGen Cas9 (NEB),
2 - TATSPCas9,
3 - TATSPCas9TAT,
4 - SPCas9TAT,
5 - AsCpf1,
6 - SPCas9;
буквой М обозначены стандарты молекулярных масс.
Таблица 1. Характеристики полученных экспрессионных конструкций.
Таблица 2. Рекомбинантные белки семейства CRISPR/Cas.
Таблица 3. Содержание бактериальных эндотоксинов в растворах рекомбинантных белков семейства CRISPR/Cas, сниженное при помощи детергента Triton X-114, на примере белков SPCas9, STCas9, AsCpf1.
Таблица 4. Эффективность специфического гидролиза ДНК-мишени рекомбинантными белками семейства CRISPR/CAS в присутствии мишень-специфической направляющей РНК.
Примеры осуществления изобретения
ПРИМЕР 1: СОЗДАНИЕ ЭКСПРЕССИОННЫХ КОНСТРУКЦИЙ ДЛЯ ПРОДУКЦИИ РЕКОМБИНАНТНЫХ БЕЛКОВ СЕМЕЙСТВА CRISPR/CAS В Е. COLI.
Для конструирования экспрессионных плазмидных ДНК, несущих ген, содержащий одну из последовательностей SEQ ID NO: NO: 9-16, для продукции рекомбинантных белков семейства CRISPR/CAS в Е. coli в качестве векторной плазмидной ДНК использовали коммерчески доступную плазмиду pET22b (Novagen, США). Физическая карта pET22b доступна на сайте https://www.snapgene.com/resources/plasmidfiles/?set=pet_and_duet_vectors_(novagen)&plasmid=pET-22b(+). Перед использованием pET22b в качестве векторной плазмидной ДНК набором для мутагенеза QuikChange XL Site-Directed Mutagenesis Kit (Agilent, США) в сайт множественного клонирования был внесен дополнительный сайт узнавания эндонуклеазы рестрикции NsiI. В качестве источников клонируемых фрагментов использованы коммерчески доступные плазмиды из репозитория (Addgene, США), клонируемые фрагменты получали методом ПЦР с использованием разработанных праймеров (табл. 1). Схематично структура клонируемых фрагментов представлена на Фиг. 1. Праймеры, используемые для ПЦР-амплификации, содержат сайты эндонуклеаз рестрикции NsiI (на 5'-конце) и NotI (на 3'-конце), а также фрагменты ДНК, кодирующие последовательности полигистидинового тракта (на 5'-конце), сигнала ядерной локализации большого T-антигена вируса обезьян SV40 (на 5'-конце) для эффективной доставки белка в ядро клетки-мишени, и последовательность, кодирующую TAT - пептид (на 5'- и/или 3'-конце) вируса иммунодефицита человека.
Продукты амплификации, полученные в ходе ПЦР с разработанными праймерами (табл. 1), расщепляют эндонуклеазами рестрикции NsiI и NotI (кроме фрагмента, кодирующего нуклеазу STCas9 где использовали NdeI и AgeI), и затем лигируют с расщепленной по этим же сайтам (соответствующим) и очищенной элюцией из агарозного геля векторной плазмидной ДНК модифицированной pET22b(NsiI). Единичные клоны, содержащие плазмиду pET22b(NsiI) c целевой вставкой, получают методом химической трансформации клеток штамма Е. coli MACH1-T1 (Thermo Fisher Scientific, США). Клоны, содержащие вставку длиной требуемого размера, идентифицируют при помощи ПЦР с праймерами Т7 forward и Т7 reverse (ГенТерра, Россия). Выделяют плазмидную ДНК и верифицируют корректность встроенной последовательности методом капиллярного секвенирования.
В результате получены экспрессионные плазмидные ДНК, несущие ген, содержащий одну из последовательностей SEQ ID NO: NO: 9-16, для продукции рекомбинантных белков семейства CRISPR/CAS в Е. coli (табл. 1, табл. 2):
1. pAM-SPCas9 размером 9852 п.н., несущая ген, содержащий последовательность SEQ ID NO: NO: 9. Кодирует нуклеазу Cas9 из Streptococcus pyogenes с аминокислотной последовательностью SEQ ID NO: NO: 1. pAM-SPCas9 содержит 33 уникальных сайта рестрикции, среди которых BamHI (7016), EcoRI (8833), NotI (9167), NsiI (4964), SalI (5551), SmaI (6379), XbaI (4918).
2. рAM-SPCa9D10A размером 9852 п.н., несущая ген, содержащий последовательность SEQ ID NO: NO: 10. Кодирует никазу Cas9 из Streptococcus pyogenes с аминокислотной последовательностью SEQ ID NO: NO: 2. pAM-SPCas9D10A содержит 34 уникальных сайта рестрикции, среди которых BamHI (7016), EcoRI (8833), NotI (9167), NsiI (4964), SalI (5551), SmaI (6379), XbaI (4918).
3. рAM-STCas9 размером 8873 п.н., несущая ген, содержащий последовательность SEQ ID NO: NO: 11. Кодирует нуклеазу Cas9 из Streptococcus thermophilus с аминокислотной последовательностью SEQ ID NO: NO: 3. pAM-STCas9 содержит 24 уникальных сайта рестрикции, среди которых AgeI (8444), NcoI (5966), NdeI (4958), PvuI (772), XbaI (4918).
4. pAM-AsCpf1 размером 9405 п.н., несущая ген, содержащий последовательность SEQ ID NO: NO: 12. Кодирует нуклеазу Cpf1 из Acidaminococcus с аминокислотной последовательностью SEQ ID NO: NO: 4. pAM-AsCpf1 содержит 28 уникальных сайтов рестрикции, среди которых BssHII (3719), NdeI (4958), NotI (8990), NsiI (4964), XbaI (4918), XhoI (8998).
5. pAM-TATSPCas9 размером 9654 п.н., несущая ген, содержащий последовательность SEQ ID NO: NO: 13. Кодирует нуклеазу Cas9 из Streptococcus pyogenes (слитый белок с TAT-пептидом на N-конце) с аминокислотной последовательностью SEQ ID NO: NO: 5. pAM-TATSPCas9 содержит 33 уникальных сайтов рестрикции, среди которых BamHI (2094), EcoRI (3911), NotI (4275), NsiI (0), XbaI (9608).
6. pAM-SPCas9TAT размером 9612 п.н., несущая ген, содержащий последовательность SEQ ID NO: NO: 14. Кодирует нуклеазу Cas9 из Streptococcus pyogenes (слитый белок с TAT-пептидом на C-конце) с аминокислотной последовательностью SEQ ID NO: NO: 6. pAM-SPCas9TAT содержит 33 уникальных сайтов рестрикции, среди которых BamHI (7016), EcoRI (8833), NotI (9197), NsiI (4964), XbaI (4918).
7. pAM-TATSPCas9TAT размером 9654 п.н., несущая ген, содержащий последовательность SEQ ID NO: NO: 15. Кодирует нуклеазу Cas9 из Streptococcus pyogenes (слитый белок с TAT-пептидом на N- и C-концах) с аминокислотной последовательностью SEQ ID NO: NO: 7. pAM-TATSPCas9TAT содержит 33 уникальных сайтов рестрикции, среди которых BamHI (7058), EcoRI (8875), NotI (9239), NsiI (4964), XbaI (4918).
8. pAM-SPCa9D10ATAT размером 9612 п.н., несущая ген, содержащий последовательность SEQ ID NO: NO: 16. Кодирует никазу Cas9 из Streptococcus pyogenes (слитый белок с TAT-пептидом на C-конце) с аминокислотной последовательностью SEQ ID NO: NO: 8. pAM-SPCa9D10ATAT содержит 33 уникальных сайтов рестрикции, среди которых BamHI (7016), EcoRI (8833), NotI (9197), NsiI (4964), XbaI (4918).
ПРИМЕР 2: ЭКСПРЕССИЯ И ПОЛУЧЕНИЕ РАСТВОРИМОЙ ФРАКЦИИ РЕКОМБИНАНТНЫХ БЕЛКОВ СЕМЕЙСТВА CRISPR/CAS В Е. COLI.
Штаммы-продуценты рекомбинантных белков семейства CRISPR/CAS для пилотной экспрессии получают методом химической трансформации компетентных клеток E. coli BL21(DE3) с использованием плазмидной ДНК pAM (табл. 1), несущей ген, содержащий одну из последовательностей SEQ ID NO: NO: 9-16. Пилотную экспрессию проводят в бульоне Лурии-Бертани (LB), содержащем 100 мкг/мл ампициллина, 2 mM MgSO4 и 1% глюкозы. Для анализа экспрессии отбирают образцы, из которых выделяют растворимую фракцию цитоплазмы, которую затем анализируют методом электрофореза в 7,5% полиакриламидном геле. После прохождения электрофореза гель окрашивают кумасси R-250 по стандартной методике Лэммли, и сканируют с помощью сканера (Epson, Япония). В ходе пилотной экспрессии не удалось получить видимой экспрессии белков SPCas9, SPCas9D10A и AsCpf1 в штаммах-продуцентах, полученных на основе материнского штамма E. coli BL21(DE3), о чем говорит отсутствие бэндов с молекулярной массой 161836,42 Да (расчетная молекулярная масса SPCas9) в дорожках 2-4, 161792,41 Да (расчетная молекулярная масса SPCas9D10A) в дорожках 5-7 и 155028,12 Да (расчетная молекулярная масса AsCpf1) дорожках 8-10 в окрашенном акриламидном геле при соотнесении с маркером молекулярных масс (сверху вниз 180, 130, 100, 70, 55, 40, 35, 25, 15, 10 кДа, PageRuler™ Prestained Protein Ladder, 10 to 180 kDa, # 26616, США) (фиг. 2).
Ввиду отсутствия экспрессии целевых белков в штамме E. coli BL21(DE3) принято решение сменить экспрессионный штамм на E. coli Rosetta-gami B (DE3), имеющий генотип: F- ompT hsdSB (rB- mB-) gal dcm lacY1 ahpC (DE3) gor522::Tn10 trxBpRARE (CamR, KanR, TetR), обладающий способностью эффективно экспрессировать целевые белки, кодируемые последовательностями, не оптимизированными по составу кодонов для E. coli, а также обладающий способностью к образованию дисульфидных связей продуцируемого целевого белка в бактериальной цитоплазме.
Штаммы-продуценты рекомбинантных белков семейства CRISPR/CAS получают методом химической трансформации компетентных клеток E. coli Rosetta-gami B (DE3) с использованием плазмидной ДНК pAM, несущей ген, содержащий одну из последовательностей SEQ ID NO: NO: 9-16.
Получение стартовой культуры для экспрессии рекомбинантных белков семейства CRISPR/CAS проводят путем инокуляции отдельного клона штамма-продуцента в бульон Лурии-Бертани (LB), содержащий 100 мкг/мл ампициллина, 2 mM MgSO4 и 2% глюкозы. Культивирование стартовой культуры проводят при 37°С в течение ночи при постоянном перемешивании. В ходе проведенных экспериментов было показано, что отсутствие в среде для культивирования стартовой культуры MgSO4 и глюкозы приводит к отсутствию экспрессии целевого продукта, о чем говорит отсутствие бэндов с молекулярной массой 161836,42 Да (расчетная молекулярная масса SPCas9) в дорожке 3 и 161792,41 Да (расчетная молекулярная масса SPCas9D10A) в дорожке 5 в окрашенном акриламидном геле при соотнесении с маркером молекулярных масс (сверху вниз 180, 130, 100, 70, 55, 40, 35, 25, 15, 10 кДа, PageRuler™ Prestained Protein Ladder, 10 to 180 kDa, # 26616, США) (фиг. 3, стрелкой указана область полиакриламидного геля, в которой должны располагаться бэнды расчетной молекулярной массы).
На следующий день клетки разводят до оптической плотности OD600≈0,3 в питательной среде LB, содержащей 100 мкг/мл ампициллина, 2 mM MgSO4 и 1% глюкозы, и выращивают при 37°С при постоянном перемешивании до оптической плотности OD600≈1,5-1,9, так как в ходе экспериментов по экспрессии и очистке было установлено, что индукция при OD600≈1,5-1,9 позволяет достигать большего выхода белка (фиг. 4).
По достижении оптической плотности OD600≈1,5-1,9 к культуре добавляют изопропил-β-D-тиогалактозид (ИПТГ) до конечной концентрации 0,5 mM для индукции экспрессии и инкубируют при 22°С в течение 20-24 часов при постоянном перемешивании.
По окончании культивирования клетки собирают центрифугированием (6000 g, 10 мин), после чего полученные клеточные осадки ресуспендируют в буфере для лизиса, содержащем 20 mM Na2HPO4/NaH2PO4 pH 6,6-8,0.
Клетки штамма-продуцента, ресуспендированные в буфере для лизиса, содержащем 20 mM Na2HPO4/NaH2PO4 pH 6,6-8,0, разрушают методом ультразвуковой дезинтеграции (см., например, методику ультразвуковой дезинтеграции, описанную в диссертации Лебедева Л.Р., Разработка технологий получения препаратов природных и рекомбинантных белков из прокариотических клеток, Автореферат Диссертации, Бердск, 1998, 136с.).
Отделение растворимой фракции, содержащей рекомбинантный белок семейства CRISPR/CAS, от нерастворимой фракции телец включения в полученном после ультразвуковой дезинтеграции растворе проводят методом центрифугирования (10000 g, 1 час). Полученная таким образом растворимая фракция, содержащая рекомбинантный белок семейства CRISPR/CAS, подвергается хроматографической очистке.
ПРИМЕР 3. ХРОМАТОГРАФИЧЕСКАЯ ОЧИСТКА РЕКОМБИНАНТНЫХ БЕЛКОВ СЕМЕЙСТВА CRISPR/CAS ИЗ РАСТВОРИМОЙ ФРАКЦИИ Е. COLI., ПРОДУЦИРУЮЩИХ РЕКОМБИНАНТНЫЕ БЕЛКИ СЕМЕЙСТВА CRISPR/CAS
К полученной методами ультразвуковой дезинтеграции и центрифугирования растворимой фракции, содержащей рекомбинантный белок семейства CRISPR/CAS, добавляют хлорид натрия до конечной концентрации 500 mM и имидазол до конечной концентрации 10 mM.
На 1-ой стадии хроматографической очистки приготовленный таким образом раствор подвергают металл-хелатной хроматографии на сорбенте Chelating Sepharose FF (GE Healthcare, Великобритания).
Для подготовки хроматографической колонки с металл-хелатным сорбентом Chelating Sepharose FF, заряженным двухвалентными ионами никеля, 10 мл металл-хелатной смолы Chelating Sepharose FF помещают в колонку XK 16/20 Column (GE Healthcare, Великобритания) и промывают 20 объемами деионизированной воды. Заряд колонки ионами никеля проводят промывкой 5 объемами 200 mM раствора хлорида никеля с последующей промывкой 20 объемами деионизированной воды.
Перед началом процесса хроматографической очистки рекомбинантного белка семейства CRISPR/CAS из растворимой фракции E. coli, его содержащей, колонку уравновешивают 5 объемами буфера, содержащего 20 mM Na2HPO4/NaH2PO4 pH 6,6-8,0, 500 mM хлорида натрия, 10 mM имидазола. Растворимую фракцию, содержащую рекомбинантный белок семейства CRISPR/CAS, 500 mM хлорида натрия и 10 mM имидазола, наносят на металл-хелатный сорбент Chelating Sepharose FF, заряженный двухвалентными ионами никеля, со скоростью 10 мл/мин.
После нанесения все белки, не связавшиеся с металл-хелатным сорбентом Chelating Sepharose FF, удаляют путем промывки колонки 5-10 объемами буфера, содержащего 20 mM Na2HPO4/NaH2PO4 pH 6,6-8,0, 500 mM хлорида натрия, 10 mM имидазола. Удаление примесных белков, связавшихся с металл-хелатным сорбентом Chelating Sepharose FF, проводят путем промывки колонки 5-10 объемами буфера, содержащего 20 mM Na2HPO4/NaH2PO4 pH 6,6-8,0, 500 mM хлорида натрия, 50 mM имидазола. Элюцию CAS-белков проводят буфером, содержащим 20 mM Na2HPO4/NaH2PO4 pH 6,6-8,0, 500 mM хлорида натрия, 250 mM имидазола.
После элюции целевого белка хроматографическую колонку, содержащую металл-хелатный сорбент Chelating Sepharose FF, отмывают буфером, содержащим 20 mM Na2HPO4/NaH2PO4 pH 6,6-8,0, 1 M хлорида натрия. Отмытую таким образом хроматографическую колонку консервируют в растворе 20% этанола и помещают на хранение на +4°С.
Типичные хроматографические профили очистки на сорбенте Chelating Sepharose FF рекомбинантных белков семейства CRISPR/CAS приведены на примерах профилей очистки белков SPCas9, AsCpf1 и STCas9 на Фиг. 5, 6, 7, 13 и 17 соответственно.
Хроматографические фракции, содержащие рекомбинантные белки семейства CRISPR/CAS, полученные в ходе хроматографической очистки на сорбенте Chelating Sepharose FF, анализируют методом электрофореза в 7,5% полиакриламидном геле.
После прохождения электрофореза гель окрашивают кумасси R-250 по стандартной методике Лэммли, и сканируют с помощью сканера (Epson, Япония). Типичные электрофореграммы хроматографических фракций, содержащих рекомбинантные белки семейства CRISPR/CAS, приведены на примерах электрофореграмм хроматографических фракций, полученных в ходе очистки белков SPCas9, AsCpf1 и STCas9 на сорбенте Chelating Sepharose FF на Фиг. 8, 14 и 19 соответственно.
Целевые белки содержатся в хроматографических фракциях, содержащих 20 mM Na2HPO4/NaH2PO4 pH 6,6-8,0, 500 mM хлорида натрия, 250 mM имидазола, о чем говорит наличие бэндов с молекулярной массой 161836,42 Да (расчетная молекулярная масса SPCas9) в дорожке 4 на фиг. 8, 155028,12 Да (расчетная молекулярная масса AsCpf1) в дорожке 4 на фиг. 14, а также 135860,17 Да (расчетная молекулярная масса STCas9) в дорожке 4 на фиг. 19 в окрашенных акриламидных гелях при соотнесении с маркером молекулярных масс (сверху вниз 200, 150, 100, 85, 60, 50, 40, 30, 25, 20, 15, 10 кДа, Набор белковых маркеров (10-200 kDa) М35, Сибэнзим, Россия).
На 2-ой стадии хроматографической очистки рекомбинантный белок CRISPR/CAS семейства, находящийся в буфере, содержащем 20 mM Na2HPO4/NaH2PO4 pH 6,6-8,0, 500 mM хлорида натрия, 250 mM имидазола, подвергают ионообменной хроматографии на сорбенте SP Sepharose FF (GE Healthcare, Великобритания).
Для подготовки хроматографической колонки с ионообменным сорбентом SP Sepharose FF 10 мл ионообменной смолы SP Sepharose FF помещают в колонку XK 16/20 Column (GE Healthcare, Великобритания) и промывают 20 объемами деионизированной воды.
Перед началом процесса хроматографической очистки рекомбинантного белка семейства CRISPR/CAS из элюата, полученного в ходе хроматографической очистки на металл-хелатном сорбенте Chelating Sepharose FF, колонку уравновешивают 5 объемами буфера, содержащего 20 mM Tris-HCl pH 8,2. Элюат, полученный в ходе хроматографической очистки на металл-хелатном сорбенте Chelating Sepharose FF, разбавляют не менее чем в 20 раз буфером, содержащим 20 mM Tris-HCl pH 8,2 до конечной кондуктивности раствора 3-5 mS/cm и наносят на катионообменный сорбент SP Sepharose FF со скоростью 10 мл/мин.
После нанесения все белки, не связавшиеся с катионообменным сорбентом SP Sepharose FF, удаляют путем промывки колонки 5-10 объемами буфера, содержащего 20 mM Tris-HCl pH 8,2. Удаление примесных белков, связавшихся с катионообменным сорбентом SP Sepharose FF, проводят путем промывки колонки 5-10 объемами буфера, содержащего 20 mM Tris-HCl pH 8,2, 300 mM хлорида натрия.
Элюцию рекомбинантного белка CRISPR/CAS семейства проводят буфером, содержащим 20 mM Tris-HCl pH 8,2, 600 mM - 1 М хлорида натрия.
После элюции целевого белка хроматографическую колонку, содержащую катионообменный сорбент SP Sepharose FF, отмывают буфером, содержащим 20 mM Tris-HCl pH 8,2, 1-2 М хлорида натрия. Отмытую таким образом хроматографическую колонку консервируют в растворе 20% этанола, содержащего 200 mM ацетата натрия, и помещают на хранение на +4°С.
Типичные хроматографические профили очистки на сорбенте SP Sepharose FF рекомбинантных белков семейства CRISPR/CAS приведены на примерах профилей очистки белков SPCas9, AsCpf1 и STCas9 на Фиг. 9, 15 и 18 соответственно.
Хроматографические фракции, содержащие рекомбинантные белки семейства CRISPR/CAS, полученные в ходе хроматографической очистки на катионообменном сорбенте SP Sepharose FF, анализируют методом электрофореза в 7,5% полиакриламидном геле.
После прохождения электрофореза гель окрашивают кумасси R-250 по стандартной методике Лэммли, и сканируют с помощью сканера (Epson, Япония).
Типичные электрофореграммы хроматографических фракций, содержащих рекомбинантные белки семейства CRISPR/CAS, приведены на примерах электрофореграмм хроматографических фракций, полученных в ходе очистки белков SPCas9, AsCpf1 и STCas9 на сорбенте SP Sepharose FF на Фиг. 10, 16 и 19 соответственно.
Целевые белки содержатся в хроматографических фракциях, содержащих 20 mM Tris-HCl pH 8,2, 600 mM - 1 М хлорида натрия, о чем говорит наличие бэндов с молекулярной массой 161836,42 Да (расчетная молекулярная масса SPCas9) в дорожке 4 на фиг. 10, 155028,12 Да (расчетная молекулярная масса AsCpf1) в дорожке 3 на фиг. 16, а также 135860,17 Да (расчетная молекулярная масса STCas9) в дорожке 7 на фиг. 19 в окрашенных акриламидных гелях при соотнесении с маркером молекулярных масс (сверху вниз 200, 150, 100, 85, 60, 50, 40, 30, 25, 20, 15, 10 кДа, Набор белковых маркеров (10-200 kDa) М35, Сибэнзим, Россия).
При проведении экспрессии рекомбинантных белков семейства CRISPR/CAS при 27 С в течение 20-24 часов на 1-ой стадии хроматографическая очистка проходит без отклонений от протокола, наблюдаются характерные для данной стадии этапы нанесения, отмывки несвязавшихся белков, элюции раствором, содержащим 50 mM имидазола, элюции раствором, содержащим 250 mM имидазола, а также промывки раствором, содержащим 1 M хлорида натрия, что видно на фиг. 11.
На 2-ой стадии хроматографической очистки на сорбенте SP Sepharose FF происходит видимая агрегация белка (опалесценция раствора) при переводе в буфер для нанесения, содержащий 20 mM Tris-HCl pH 8,2. В результате агрегации рекомбинантный белок семейства CRISPR/CAS, экспрессированный при 27°С в течение 20-24 часов, теряет способность сорбироваться на сорбенте SP Sepharose FF, что видно на фиг. 12, когда на этапе нанесения (обозначен цифрой 1 на фиг. 12) происходит рост поглощения при длине волны 280 нм (mAu), а на этапах элюции (обозначены цифрами 3-13 на фиг. 12) такого роста не наблюдается.
ПРИМЕР 4. УДАЛЕНИЕ БАКТЕРИАЛЬНЫХ ЭНДОТОКСИНОВ ИЗ РАСТВОРОВ РЕКОМБИНАНТНЫХ БЕЛКОВ СЕМЕЙСТВА CRISPR/CAS.
Для удаления бактериальных эндотоксинов из растворов рекомбинантных белков рекомендуют различные методы. Для удаления бактериальных эндотоксинов из растворов рекомбинантных белков семейства CRISPR/CAS были применены различные подходы, такие как, изменения ионной силы раствора (повышение содержания хлорида натрия в растворе), изменения pH на различных стадиях хроматографической очискти, при помощи специализированного аффинного сорбента и с использованием детергента Triton X-114.
Белки семейства CRISPR/CAS обладают достаточно высокими значениями изоэлектрической точки (pI>9), поэтому в растворах, имеющих физиологический pH=6,5-8,0, имеют высокий положительный суммарный заряд. Именно высокий положительный суммарный заряд белков семейства CRISPR/CAS приводит к тому, что при выделении из клеток Е. coli, бактериальные эндотоксины разрушенных клеточных стенок бактерий, заряженные отрицательно, прочно связываются с выделяемыми белками семейства CRISPR/CAS.
Известно, что высокая ионная сила раствора может препятствовать прочному связыванию бактериальных эндотоксинов с поверхностью белка. Поэтому один из вариантов удаления бактериальных эндотоксинов предполагал дезинтеграцию бактериальных клеток штамма-продуцента в присутствии хлорида натрия в конечной концентрации 1 M. В протокол хроматографической очистки изменений не вносят, за исключением того, что на первой стадии очистки (на сорбенте Chelating sepharose) все используемые буферные растворы содержат хлорид натрия в конечной концентрации 1 M. Было показано, что такой модифицированный протокол не позволяет получить очищенный белок в виду потерь материала при хроматографической очистке, что видно по снижению пика (поглощение на длине волны 280 нм) при элюции белка раствором, содержащим 250 mM имидазола (фиг. 20, этап элюции раствором, содержащим 250 mM имидазола, обозначен цифрой 4).
Белки семейства CRISPR/CAS обладают достаточно высокими значениями изоэлектрической точки (pI > 9), поэтому в растворах, имеющих pH > pI (например, pH ≥ 9,5), должны иметь отрицательный суммарный заряд. Именно отрицательный суммарный заряд белка семейства CRISPR/CAS должен препятствовать связыванию с заряженными отрицательно бактериальными эндотоксинами разрушенных клеточных стенок бактерий.
Данный подход также был реализован для удаления бактериальных эндотоксинов из растворов рекомбинантных белков семейства CRISPR/CAS. Для этого дезинтеграцию бактериальных клеток штамма-продуцента проводят при значении pH = 9,5. В протокол хроматографической очистки изменений не вносят, кроме того, что на первой стадии очистки (на сорбенте Chelating sepharose) все используемые буферные растворы имеют pH = 9,5. Было показано, что такой модифицированный протокол не позволяет получить очищенный белок в виду потерь материала при хроматографической очистке, что видно по снижению пиков (поглощение на длине волны 280 нм) при элюции белка раствором, содержащим 600 mM хлорида натрия (фиг. 21, этап элюции раствором, содержащим 600 mM хлорида натрия, обозначен цифрой 3).
Известно, что хроматографическая очистка на катионообменном сорбенте SP Sepharose при низких значениях pH < 5 может способствовать удалению бактериальных эндотоксинов из растворов рекомбинантных белков. Данный подход был реализован для удаления бактериальных эндотоксинов из растворов рекомбинантных белков семейства CRISPR/CAS. В протокол дезинтеграции клеток штамма-продуцента изменений не вносят. В протокол хроматографической очистки изменений не вносят, кроме того, что на второй стадии очистки (на сорбенте SP Sepharose) все используемые буферные растворы имеют pH = 4. Было показано, что такой модифицированный протокол не позволяет получить очищенный белок в виду потерь материала при хроматографической очистке, что видно по сниженным пикам (поглощение на длине волны 280 нм) при элюции белка раствором, содержащим 600 mM хлорида натрия (фиг. 22, этап элюции раствором, содержащим 600 mM хлорида натрия, обозначен цифрой 4).
Была предпринята попытка удалить бактериальные эндотоксины из растворов рекомбинантных белков семейства CRISPR/CAS при помощи специализированного аффинного сорбента Polymyxin B-agarose (Sigma-Aldrich, США). Для этого 100 мкл Polymyxin B-agarose помещают в раствор рекомбинантного белка семейства CRISPR/CAS и инкубируют в течение 1 часа при постоянном перемешивании при комнатной температуре, после чего сорбент Polymyxin B-agarose удаляют центрифугированием. Такой способ финальной доочистки привел к полной преципитации рекомбинантного белка семейства CRISPR/CAS SPCas9 на поверхности аффинного сорбента Polymyxin B-agarose. Таким образом, было показано, что такой способ удаления бактериальных эндотоксинов не позволяет получить очищенный белок ввиду потерь материала в ходе его преципитации на поверхности аффинного сорбента Polymyxin B-agarose.
Удаление бактериальных эндотоксинов из растворов рекомбинантных белков семейства CRISPR/CAS проводят при помощи детергента Triton X-114, который обладает уникальным свойством - низкой температурой помутнения (23°C), что позволяет использовать его в ряде биохимических приложений, требующих солюбилизации и разделения белков.
Для удаления бактериальных эндотоксинов из растворов рекомбинантных белков семейства CRISPR/CAS при помощи детергента Triton X-114 к раствору белка в буфере, содержащем 20 mM Tris-HCl pH 8,2, 0,6-1 M хлорида натрия, 2 mM DTT и 0,2 mM EDTA, добавляют детергент Triton X-114 до конечной концентрации 1% и тщательно перемешивают на вортексе до видимого помутнения. После этого раствор помещают на ледяную баню на 15 мин до полного просветления (раствор должен стать абсолютно прозрачным). Затем раствор помещают в термостат на 37°С на 15 мин до полного помутнения (раствор должен стать абсолютно непрозрачным, что свидетельствует о том, что произошло обогащение раствора мицеллами, содержащими бактериальные эндотоксины). По окончании инкубации разделение водной и мицеллярной фракций осуществляют с помощью центрифугирования при 10000 g в течение 10 мин при 23°С. Верхнюю водную фазу отбирают в чистую пробирку. Процедуру очистки для удаления бактериальных эндотоксинов до уровня, позволяющего применять белковый препарат in vivo, повторяют 6-9 раз.
Содержание бактериальных эндотоксинов в растворах белков определено с помощью полуколичественного гель-тромб теста с использованием LAL-реагента с чувствительность 0,03 ЕЭ/мл (ЛАЛ-Центр, Россия). Данная методика рекомендована Министерством Здравоохранения Российской Федерации и описана в Общей Фармакопейной Статье ОФС.1.2.4.0006.15 Бактериальные эндотоксины. Пороговой пирогенной дозой считается доза, равная 5 ЕЭ/кг в 1 час для испытуемого лекарственного препарата, если он вводится пациенту любым парентеральным путем, кроме интратекального.
При применении разработанного протокола удаления бактериальных эндотоксинов растворы рекомбинантных белков семейства CRISPR/CAS содержат 0,3-1,5 ЕЭ/мл (табл. 3). Для сравнения использовали коммерчески доступный белок EnGen™ Cas9 NLS, S. pyogenes (M0646, NEB, США), информация о котором доступна на сайте “https://international.neb.com” или из опубликованных источников (см., например Philippe E. Mangeot, Valérie Risson, Floriane Fusil, Aline Marnef, Emilie Laurent, Juliana Blin, Virginie Mournetas, Emmanuelle Massouridès, Thibault J. M. Sohier, Antoine Corbin, Fabien Aubé, Marie Teixeira, Christian Pinset, Laurent Schaeffer, Gaëlle Legube, François-Loïc Cosset, Els Verhoeyen, Théophile Ohlmann and Emiliano P. Ricci, Genome editing in primary cells and in vivo using viral-derived Nanoblades loaded with Cas9-sgRNA ribonucleoproteins, Nat Commun., 2019; v.10, p:45). Содержание бактериальных эндотоксинов в препарате коммерчески доступного белка EnGen™ Cas9 NLS, S. pyogenes (M0646, NEB, США) составляет 3-30 ЕЭ/мл, что в 10-20 раз превышает содержание бактериальных эндотоксинов в растворах рекомбинантных белков семейства CRISPR/CAS, полученных по методике, описанной в данном изобретении.
В результате проведенной хроматографической очистки и удаления бактериальных эндотоксинов получены препараты рекомбинантных белков семейства CRISPR/CAS следующего состава:
1. рекомбинантная нуклеаза Cas9, Streptococcus pyogenes (SPCas9), в буферном растворе, содержащем 20 mM Tris-HCl pH 8,2, 600 mM NaCl, 2 mM DTT, 0,2 mM EDTA, 0,3-1,5 ЕЭ/мл;
2. рекомбинантная никаза Cas9, Streptococcus pyogenes (SPCas9D10A), в буферном растворе, содержащем 20 mM Tris-HCl pH 8,2, 600 mM NaCl, 2 mM DTT, 0,2 mM EDTA, 0,3-1,5 ЕЭ/мл;
3. рекомбинантная нуклеаза Cas9, Streptococcus thermophilus (STCas9), в буферном растворе, содержащем 20 mM Tris-HCl pH 8,2, 1000 mM NaCl, 2 mM DTT, 0,2 mM EDTA, 0,3-1,5 ЕЭ/мл;
4. рекомбинантная нуклеаза AsCpf1, Acidaminococcus (AsCpf1), в буферном растворе, содержащем 20 mM Tris-HCl pH 8,2, 600 mM NaCl, 2 mM DTT, 0,2 mM EDTA, 0,3-1,5 ЕЭ/мл;
5. рекомбинантная нуклеаза Cas9, Streptococcus pyogenes (TATSPCas9), слитый белок с TAT-пептидом на N-конце в буферном растворе, содержащем 20 mM Tris-HCl pH 8,2, 1000 mM NaCl, 2 mM DTT, 0,2 mM EDTA, 0,3-1,5 ЕЭ/мл;
6. рекомбинантная нуклеаза Cas9, Streptococcus pyogenes (SPCas9TAT), слитый белок с TAT-пептидом на C-конце в буферном растворе, содержащем 20 mM Tris-HCl pH 8,2, 1000 mM NaCl, 2 mM DTT, 0,2 mM EDTA, 0,3-1,5 ЕЭ/мл;
7. рекомбинантная нуклеаза Cas9, Streptococcus pyogenes (TATSPCas9TAT), слитый белок с TAT-пептидом на N- и C-концах в буферном растворе, содержащем 20 mM Tris-HCl pH 8,2, 1000 mM NaCl, 2 mM DTT, 0,2 mM EDTA, 0,3-1,5 ЕЭ/мл;
8. рекомбинантная никаза Cas9, Streptococcus pyogenes (SPCas9D10ATAT), слитый белок с TAT-пептидом на C-конце в буферном растворе, содержащем 20 mM Tris-HCl pH 8,2, 1000 mM NaCl, 2 mM DTT, 0,2 mM EDTA, 0,3-1,5 ЕЭ/мл.
Также проводили лиофилизацию полученных препаратов рекомбинантных белков семейства CRISPR/CAS (см., например методики лиофилизации, описанные в статье Блынская Е.В.,Тишков С.В. и Алексеев К.В., Технологические подходы к совершенствованию процесса лиофилизации белковых и пептидных лекарственных препаратов, Российский биотерапевтический журнал, 2017, т.16, н.1, с.6-11 или в статье Horn J, Schanda J and Friess W, Impact of fast and conservative freeze-drying on product quality of protein-mannitol-sucrose-glycerol lyophilizates, Eur J Pharm Biopharm., 2018, v.127, p:342-354). При этом к препаратам рекомбинантных белков семейства CRISPR/CAS, находящимся в буферном растворе, содержащем 20 mM Tris-HCl pH 8,2, 600-1000 mM NaCl, 2 mM DTT, 0,2 mM EDTA, 0,3-1,5 ЕЭ/мл, добавляли вспомогательные вещества для лиофилизации. Так, например, в качестве модификатора тоничности добавляли глицерин в концентрации 10-50% или в качестве крио-/лиопротектора - трегалозу в концентрации 50-500 mM и проводили лиофилизацию образцов с использованием лиофильной сушки FreeZone (Labconco, США).
Полученные таким образом препараты высокостабильны в приготовленных растворах, содержащих вспомогательные вещества для лиофилизации, что видно при проведении электрофореза в 7,5% полиакриламидном геле, об этом свидетельствует наличие бэндов с молекулярной массой 161836,42 Да (расчетная молекулярная масса SPCas9) в дорожках 1-11 на фиг. 23 в окрашенных акриламидных гелях при соотнесении с маркером молекулярных масс (сверху вниз 116, 66,2, 45, 35, 25, 18,4, 14,4 кДа, Unstained Protein Molecular Weight Marker, #26610, Thermo Fisher Scientific, США).
Лиофильно высушенные препараты рекомбинантных белков семейства CRISPR/CAS, содержащие вспомогательные вещества для лиофилизации, такие как глицерин и трегалоза, сохраняют свою специфическую активность на протяжении длительного срока (более года при хранении от 4 до 20°С).
ПРИМЕР 5. ТЕСТИРОВАНИЕ БИОЛОГИЧЕСКОЙ АКТИВНОСТИ ПОЛУЧЕННЫХ РЕКОМБИНАНТНЫХ БЕЛКОВ СЕМЕЙСТВА CRISPR/CAS
При применении разработанных протоколов хроматографической очистки и удаления бактериальных эндотоксинов растворы рекомбинантных белков семейства CRISPR/CAS не теряют своей специфической активности. Специфическая активность белков семейства CRISPR/CAS проявляется в способности вносить дву- и одноцепочечные разрывы в ДНК-матрицы, содержащие узнаваемые CAS-белками последовательности. Узнавание «мишени» происходит за счет направляющей РНК (gRNA).
Стандартный протокол тестирования специфической активности белков семейства CRISPR/CAS состоит из нескольких стадий:
1. Подготовка ДНК-матрицы для испытания специфической активности белков семейства CRISPR/CAS.
2. Подготовка направляющей РНК.
3. Создание готового рибонуклеопротеинового комплекса, содержащего белок семейства CRISPR/CAS и направляющую РНК.
4. Тестирование специфической активности белков семейства CRISPR/CAS и визуализация результатов эксперимента.
Использован стандартный протокол тестирования активности белков семейства CRISPR/CAS с некоторыми модификациями (Anders C and Jinek M., In vitro enzymology of Cas9, Methods Enzymol., 2014, v.546, p:1-20).
Первая стадия предполагает получение ДНК-матрицы, предназначенной для испытания специфической активности белков семейства CRISPR/CAS. ДНК-матрицу готовят в соответствии с выбранной для редактирования мишенью. Как правило, ДНК-матрица представляет собой продукт амплификации участка гена методом полимеразной цепной реакции, содержащего последовательность, на узнавание которой «запрограммирован» с помощью направляющей РНК CAS-белок. Гидролиз ДНК-матрицы должен приводить к образованию различимых между собой по длине фрагментов ДНК.
Для подготовки ДНК-матрицы использована плазмида pGEM-T-CCR5, содержащая фрагмент гена, кодирующий хемокиновый рецептор человека CCR5. Синтез ДНК-матрицы осуществлен методом ПЦР с использованием праймеров M13for и M13rev (ГенТерра, Россия). Длина ДНК-матрицы составляет 2000 пар нуклеотидов; длина фрагментов, образующихся в результате специфического гидролиза белками семейства CRISPR/CAS, составляет 1200 и 800 пар нуклеотидов.
Подготовка направляющей РНК осуществляется методом in vitro транскрипции с использованием коммерчески доступных наборов реагентов (HiScribe™ T7 High Yield RNA Synthesis Kit, NEB, США). Модификации протокола производителя, внесенные авторами, заключаются в способе очистки полученной in vitro транскрибированной направляющей РНК, который позволяет увеличить выход продукта реакции и получить желаемую концентрацию финального препарата направляющей РНК.
Для подготовки направляющей РНК проводят синтез фрагмента ДНК, кодирующего направляющую РНК, методом ПЦР с использованием праймеров Т7 forward и M13rev (ГенТерра, Россия), длина фрагмента составляет 250 пар нуклеотидов. Готовый ДНК фрагмент используют в качестве матрицы в реакции in vitro транскрипции. Продукты реакции in vitro транскрипции переосаждают из реакционной смеси добавлением хлорида натрия до конечной концентрации 400 mM и равного объема изопропилового спирта.
Создание готового рибонуклеопротеинового комплекса, содержащего белок семейства CRISPR/CAS и направляющую РНК проводят по стандартному протоколу с некоторыми модификациями (Anders C and Jinek M., In vitro enzymology of Cas9, Methods Enzymol., 2014, v.546, p:1-20).
Непосредственно перед объединением с CAS-белком препарат направляющей РНК (в количестве 500 нг) прогревают при 90°С в течение 5 минут и позволяют медленно остыть до комнатной температуры (инкубация при комнатной температуре не менее 10 минут). Такое прогревание необходимо для формирования корректной конформации шпильки, содержащейся в направляющей РНК. Многие производители коммерческих препаратов CAS-белков пропускают данный этап при подготовке рибонуклеопротеинового комплекса.
Для формирования готового рибонуклеопротеинового комплекса 500 нг CAS-белка и подготовленную направляющую РНК смешивают и инкубируют 15 минут при комнатной температуре. По истечении времени инкубации к смеси, содержащей готовый рибонуклеопротеиновый комплекс, добавляют ДНК-матрицу. Инкубацию рибонуклеопротеинового комплекса и ДНК-мишени проводят при температуре 37°С в течение 60 минут. По окончании реакции из смеси удаляют не связавшуюся РНК с помощью раствора РНКазы А. Продукты гидролиза визуализируют при помощи электрофореза в агарозном геле (фиг. 24).
Для тестирования биологической активности рекомбинантных белков семейства CRISPR/CAS SPCas9D10A и SPCas9D10ATAT, обладающих никазной активностью (способностью вносить однонитевые разрывы в двуцепочечные ДНК-матрицы) применяют описанный выше протокол, за исключением того, что в качестве ДНК-матрицы используют суперспирализованную плазмиду pGEM-T-CCR5, которая в результате специфического гидролиза превращается в релаксированную форму (изменяется подвижность на электрофорезе). Продукты гидролиза визуализируют при помощи электрофореза в агарозном геле (фиг. 24).
Количественную оценку степени гидролиза проводят с помощью коммерчески доступного набора High Sensitivity DNA Assay (Agilent, США) на анализаторе Bioanalyzer 2100 (Agilent, США). Результаты анализа приведены на фиг. 25; обсчет количественных данных анализа приведен в табл. 4.
Результаты проведенных экспериментов свидетельствуют о том, что рекомбинантные белки семейства CRISPR/CAS, полученные по технологии, описанной в изобретении, не теряют своей специфической активности и неожиданно даже обладают повышенной активностью по сравнению с коммерческим препаратом EnGen Cas9. Уровень активности можно расположить в следующем порядке SPCas9 > TATSPCas9TAT > EnGen Cas9 > TATSPCas9 > AsCpf1 > SPCas9TAT (табл. 4).
Таким образом, при осуществлении способа согласно настоящему изобретению удалось достичь выход не менее 20 миллиграмм рекомбинантного белка семейства CRISPR/Cas с 1 литра культуры штамма-продуцента и получить препарат рекомбинантной нуклеазы семейства Cas со сниженным до уровня 0,3-1,5 ЕЭ/мл содержанием бактериальных эндотоксинов, что позволяет применять указанный препарат in vivo.
Таблица 1.
5'-ggcagccccaagaagaagaggaa
ggtgatggacaagaagtactccattgg
gc-3'
2 этап. Cas9 (6His)
5'-tatatatgcatcatcaccaccaccat
cacggcagccccaagaagaagaggaaggtgatggacaagaagt-3'
5'-ccgtttaaactcattactgcggccgc
agggatcgaacc-3'
NotI
(класс 2, тип 2, Cas9), Streptococcus pyogenes
5'-ggcagccccaagaagaagaggaa
ggtgatggacaagaagtactccattgg
gc-3'
2 этап. Cas9 (6His) -
5'-tatatatgcatcatcaccaccaccat
cacggcagccccaagaagaagaggaaggtgatggacaagaagt-3'
5'-ccgtttaaactcattactgcggccgc
agggatcgaacc-3'
NotI
5'-ccaagaagaagaggaaggtgggcc
gcggagccaccatgagcgacctg-3’
2 этап. St1-Cas-for2(6His) -
5'-tatatatatatagcggccgcactcga
gaccggtagggatcgaacc-3'
AgeI
ccaccaccatcacggcagccccaagaagaagaggaaggtggg-3'
(класс 2, тип 5, Cas12a), Acidaminococcus
5'-ggcagccccaagaagaagaggaaggt
gatgacacagttcgagggc-3'
2 этап. AsCpf1-for 6His -
5'-tatatcatatgcatcatcaccaccacca
tcacggcagccccaagaagaagaggaagg-3'
accttatcactttttcttttttgcctggc-3'
NotI
5'-tatatatgcatggccgcaaaaaacg
ccgccagcgccgccgcccgcagggcagccatcaccaccaccatcacgg-3'
5'-gatttgggaccccagttctt-3'
NotI
5'-cagatttcagaaaggactttcagttt-3'
5'-atatatgcggccgcttatcactg
cgggcggcggcgctggcggcgttttttgcggccactgcccaccttcctcttcttcttggggtcagcc-3'
NotI
(класс 2, тип 2, Cas9), Streptococcus pyogenes, слитый белок с TAT-пептидом на N- и C-концах
cgggcggcggcgctggcggcgttttttgcggccactgcccaccttcctcttcttcttggggtcagcc-3'
5'-cagatttcagaaaggactttcagttt-3'
5'-atatatgcggccgcttatcactg
cgggcggcggcgctggcggcgttttttgcggccactgcccaccttcctcttcttcttggggtcagcc-3'
NotI
Таблица 2.
Таблица 3.
Н. т. - не тестировалось
Таблица 4.













































































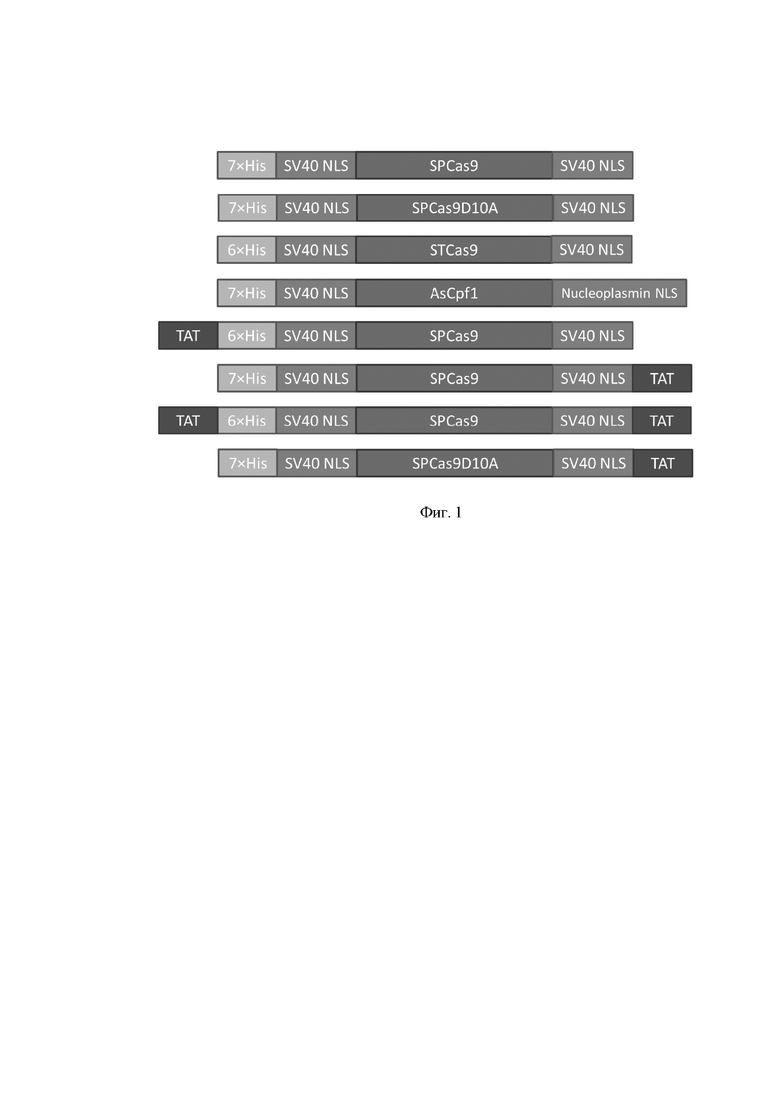
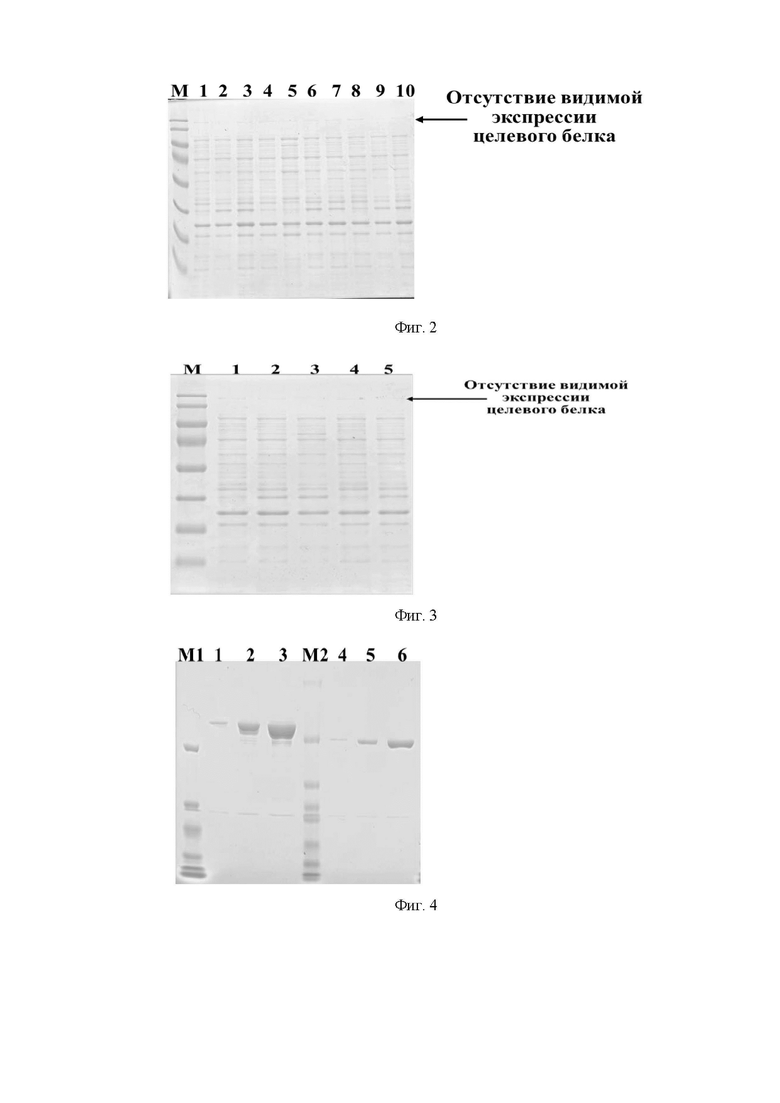
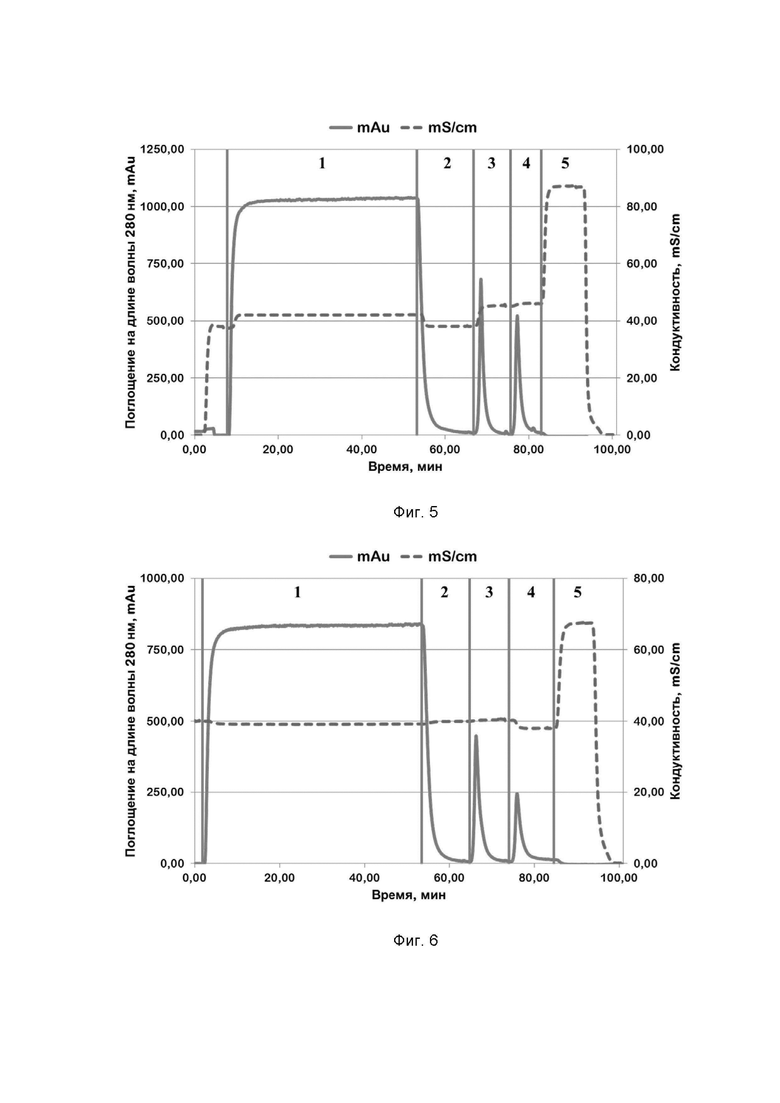
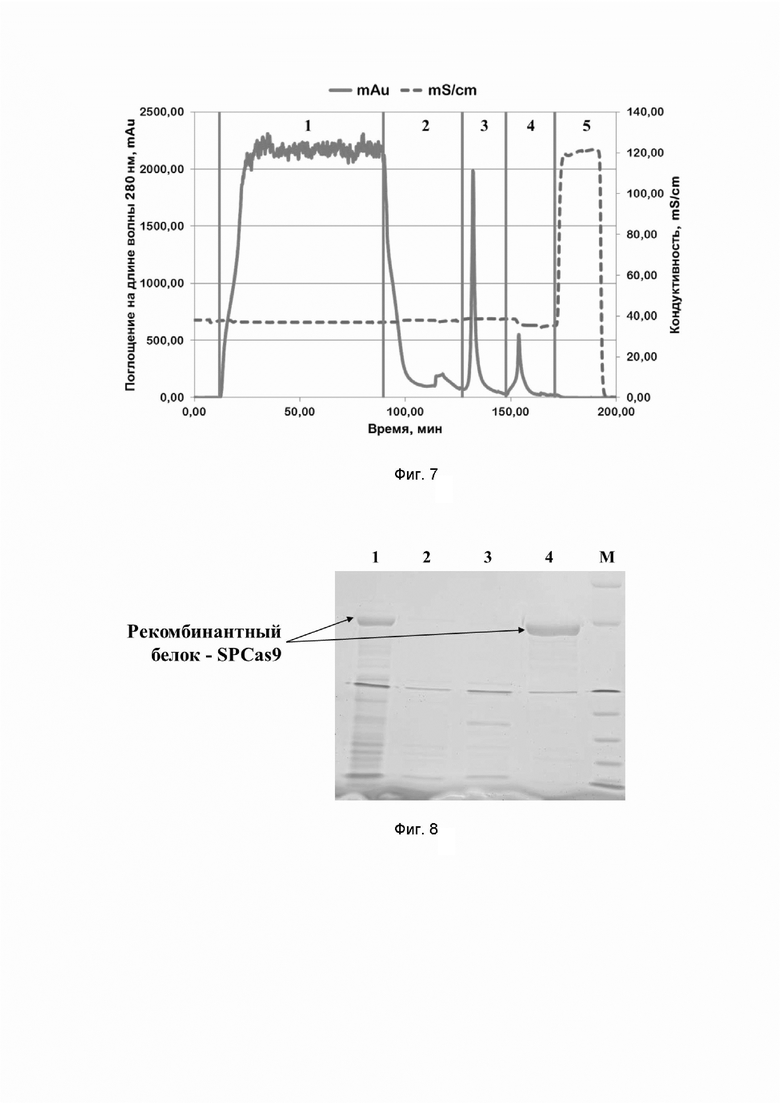
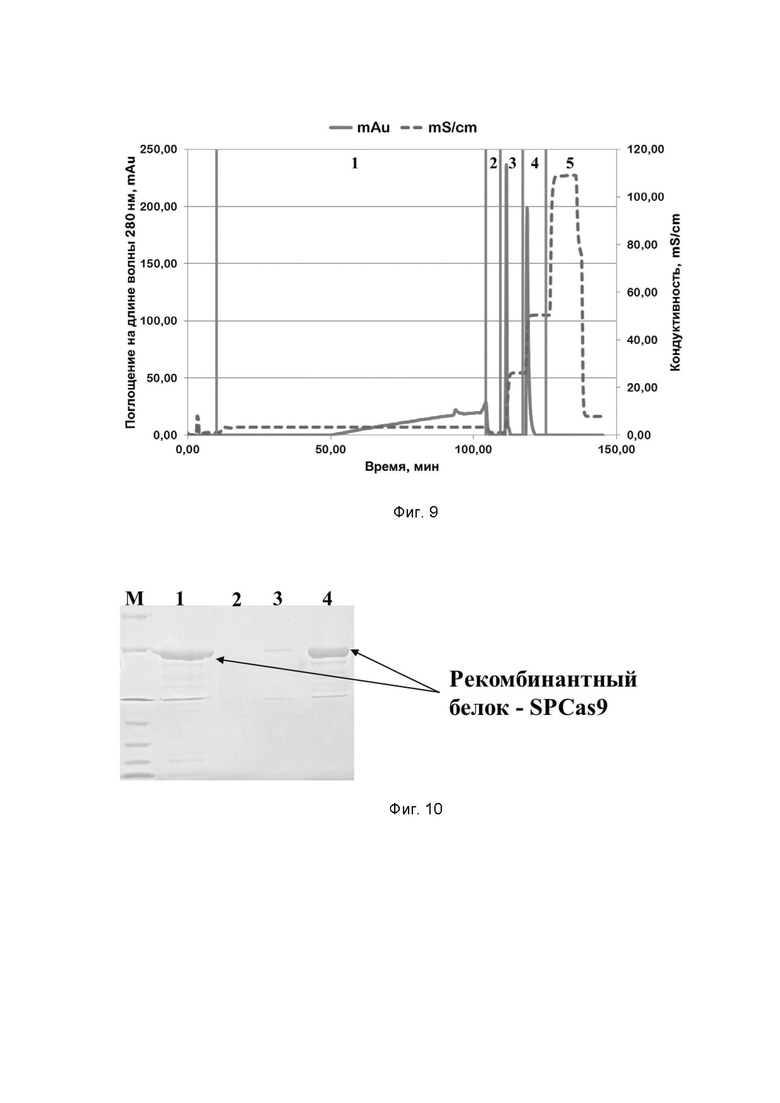
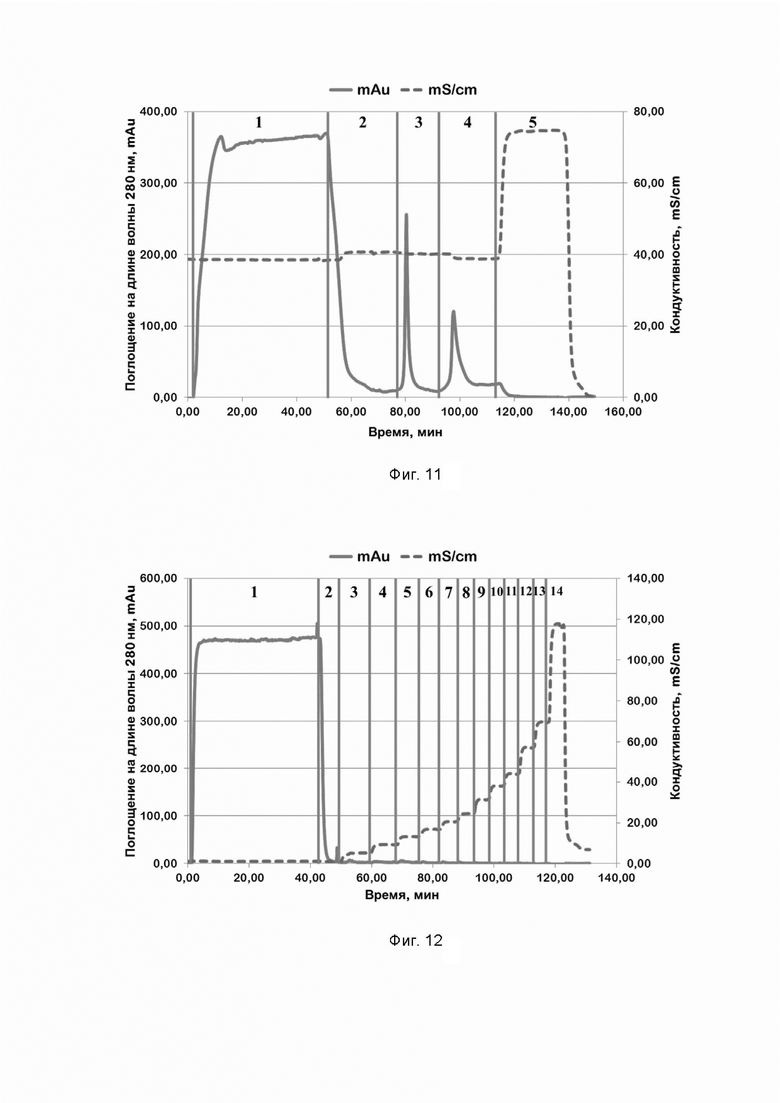
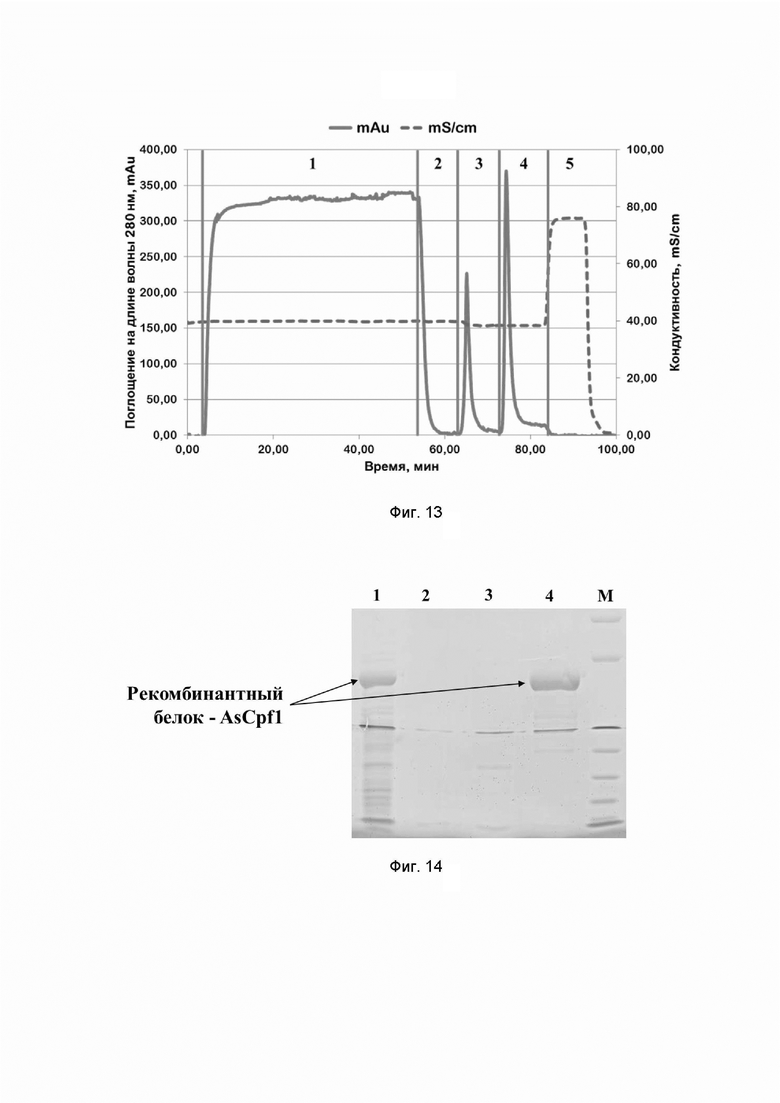
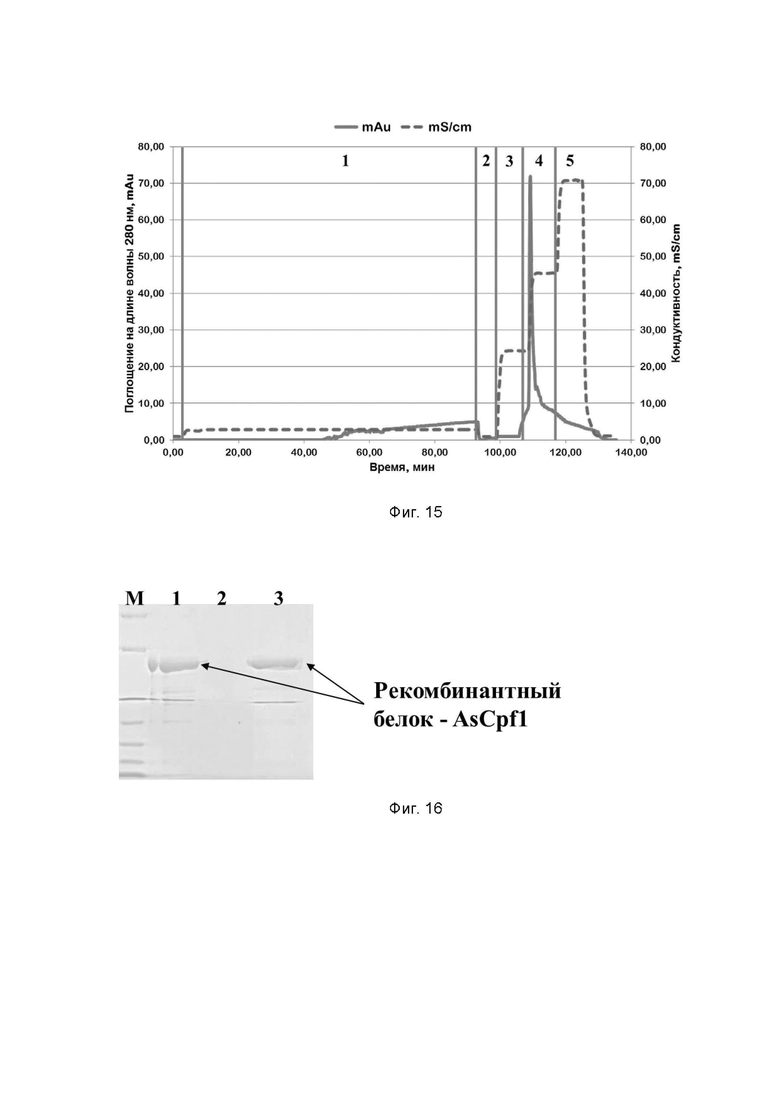
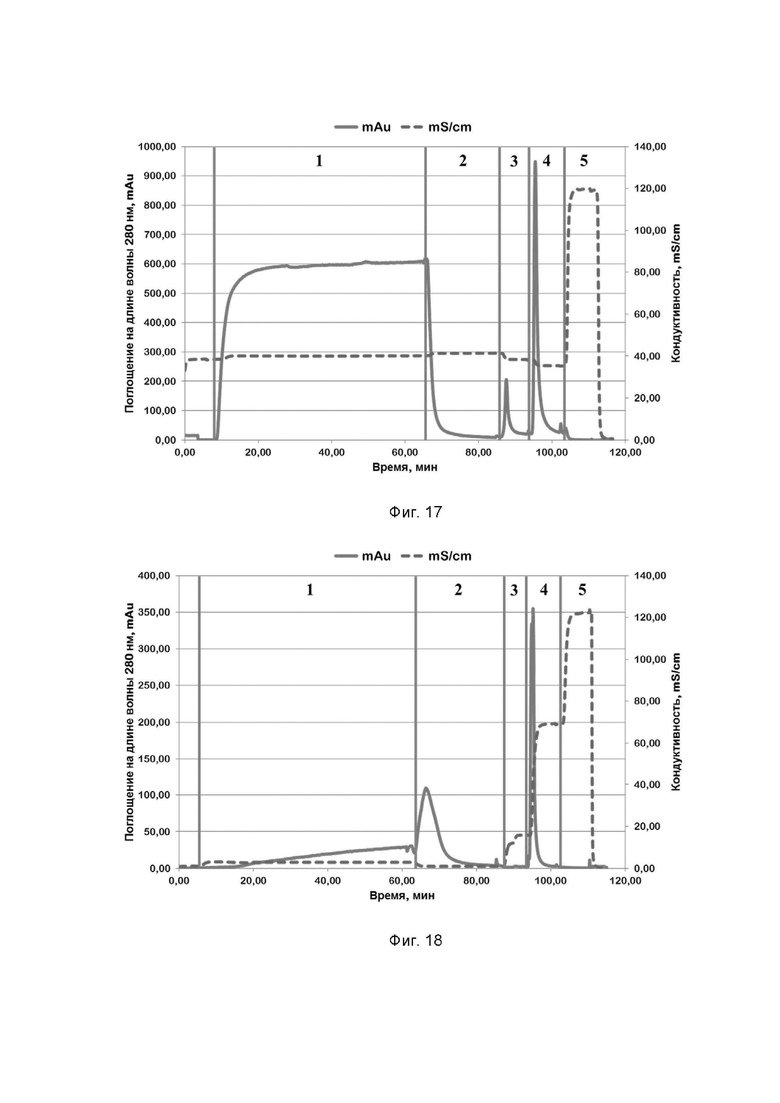
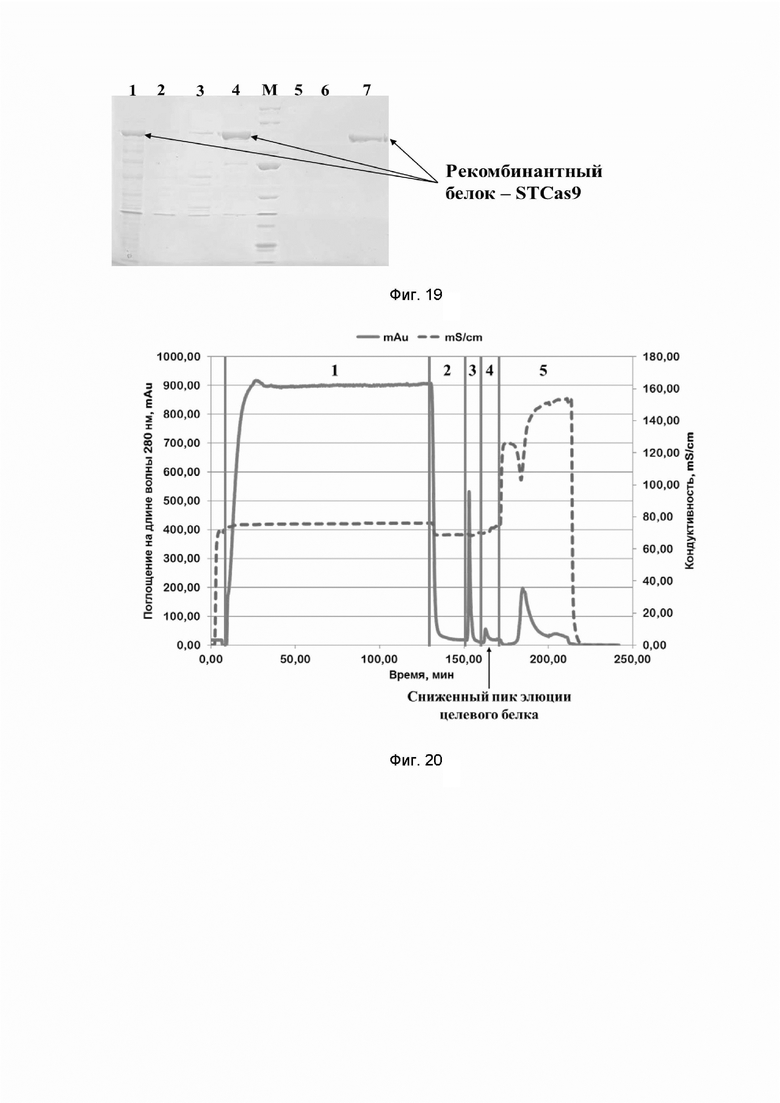
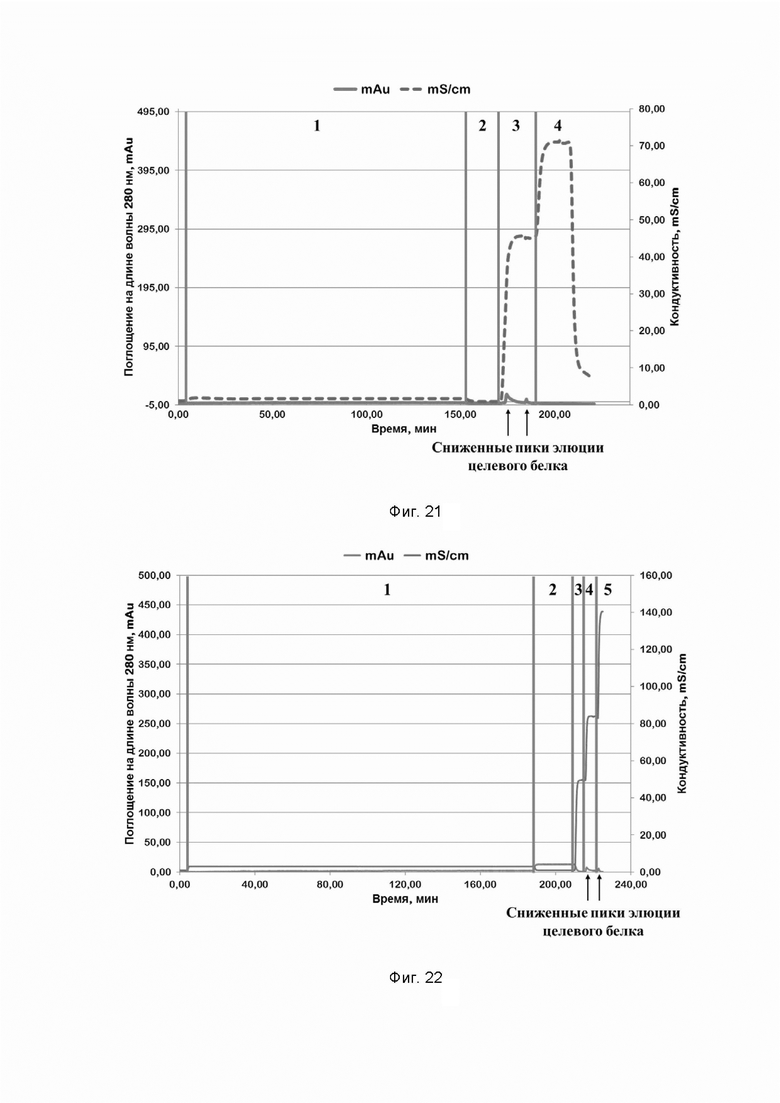
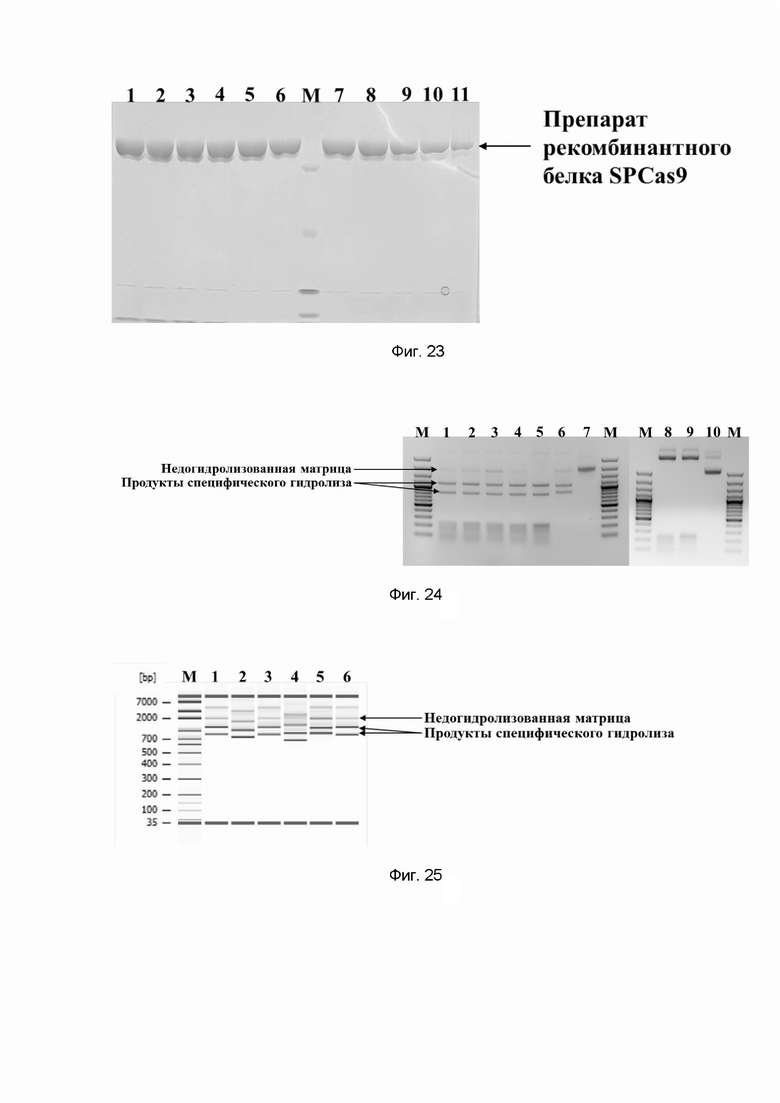
Изобретение относится к области биотехнологии, конкретно к способу получения препаратов рекомбинантной нуклеазы семейства Cas системы CRISPR/Cas в клетках штамма Escherichia coli Rosetta-gami B (DE3) и их дальнейшей очистке. Заявленный способ, включающий стадию дополнительной очистки препаратов при помощи детергента Triton X-114, позволяет уменьшить содержание бактериальных эндотоксинов до уровня 0,3-1,5 ЕЭ/мл в получаемых препаратах. 3 н. и 18 з.п. ф-лы, 25 ил., 4 табл., 5 пр.
1. Способ получения препарата рекомбинантной нуклеазы семейства Cas системы CRISPR/Cas, предусматривающий трансформацию компетентных клеток E. сoli экспрессионной плазмидой, кодирующей целевой белок, с дальнейшим культивированием трансформированных клеток-продуцентов в условиях, обеспечивающих экспрессию указанной рекомбинантной нуклеазы в питательной среде Лурии-Бертани (LB) с добавлением ампициллина и использованием в качестве индуктора экспрессии целевого белка изопропил-β-D-тиогалактозида (ИПТГ), выделение целевого белка из растворимой фракции и проведение двухстадийной хроматографической очистки, отличающийся тем, что
в качестве клеток-продуцентов используют компетентные клетки E. сoli Rosetta-gami B (DE3);
в питательную среду добавляют MgSO4 и глюкозу и выращивают клетки-продуценты до получения стартовой культуры, после чего разводят культуральную среду до оптической плотности OD600≈0,3 и уменьшают содержание глюкозы в указанной среде, по меньшей мере, в два раза;
затем клетки-продуценты культивируют до достижения оптической плотности OD600≈1,5-1,9, добавляют ИПТГ и инкубируют при 22°С для эффективной экспрессии целевого белка;
после центрифугирования по окончании культивирования клетки ресуспендируют в буфере для лизиса на основе Na2HPO4/NaH2PO4 и разрушают методом ультразвуковой дезинтеграции с последующим центрифугированием;
к полученной после центрифугирования фракции растворимой цитоплазмы, содержащей целевой белок, добавляют NaCl и имидазол и проводят первый этап хроматографической очистки путем металл-хелатной хроматографии, при этом элюцию целевого белка проводят буфером, содержащим Na2HPO4/NaH2PO4, NaCl и имидазол;
на втором этапе хроматографической очистки целевой белок в указанном буфере разбавляют не менее чем в 20 раз буфером с Tris-HCl, при этом кондуктивность раствора не превышает 5 mS/cm, и подвергают катионообменной хроматографии с последующей элюцией целевого белка буфером, содержащим Tris-HCl и NaCl, где для стабилизации целевого белка к указанному буферу добавляют DTT и EDTA;
проводят дополнительную очистку рекомбинантной нуклеазы семейства Cas от эндотоксинов клеток-продуцентов путем:
добавления в указанный раствор целевого белка в буфере после хроматографии детергента Triton X-114 для удаления бактериальных эндотоксинов в процессе перехода Triton X-114 из водонерастворимой фракции в водорастворимую и обратно с образованием мицелл и последующим разделением водорастворимой и водонерастворимой фракций методом центрифугирования,
затем снова добавляют Triton X-114 в водорастворимую фракцию, полученную после центрифугирования, с переводом из водонерастворимой фракции в водорастворимую и обратно с образованием мицелл и разделением путем центрифугирования,
при этом для удаления бактериальных эндотоксинов из препарата рекомбинантной нуклеазы семейства Cas системы CRISPR/Cas до уровня, позволяющего применять указанный препарат in vivo, стадии с добавлением Triton X-114 повторяют по меньшей мере 6 раз;
после финального центрифугирования по завершении этапа дополнительной очистки отбирают водную фазу, включающую очищенную рекомбинантную нуклеазу семейства Cas, и
при необходимости проводят лиофилизацию с добавлением вспомогательных веществ для лиофилизации.
2. Способ по п.1, отличающийся тем, что для получения стартовой культуры в питательную среду LB с 100 мкг/мл ампициллина добавляют 2 mM MgSO4 и 2% глюкозы, а после разводят в питательной среде LB с 100 мкг/мл ампициллина, дополненной 2 mM MgSO4 и 1% глюкозы.
3. Способ по п.1, отличающийся тем, что для получения стартовой культуры клетки-продуценты выращивают при 37°С в течение 16-20 часов, после разведения культивируют при 37°С до достижения оптической плотности OD600≈1,5-1,9, а после добавления ИПТГ инкубируют при 22°С в течение 20-24 часов.
4. Способ по п.1, отличающийся тем, что клетки ресуспендируют в буфере для лизиса, содержащем 20 mM Na2HPO4/NaH2PO4 при pH 6,6-8,0.
5. Способ по п.1, отличающийся тем, что центрифугирование по окончании культивирования проводят при 6000 g в течение 10 минут и/или центрифугирование для отделения фракции растворимой цитоплазмы проводят при 10000 g в течение одного часа и/или центрифугирование на этапе дополнительной очистки от эндотоксинов проводят при 10000 g в течение 10 мин при 23°С.
6. Способ по п.1, отличающийся тем, что на первом этапе хроматографической очистки к указанной фракции растворимой цитоплазмы добавляют NaCl до конечной концентрации 500 mM и имидазол до конечной концентрации 10 mM, элюцию целевого белка проводят буфером с pH 6,6-8,0, содержащим 20 mM Na2HPO4/NaH2PO4, 500 mM NaCl и 250 mM имидазола.
7. Способ по п.1, отличающийся тем, что на втором этапе хроматографической очистки целевой белок в указанном буфере разбавляют не менее чем в 20 раз буфером с pH 8,2, содержащим 20 mM Tris-HCl с кондуктивностью раствора 3-5 mS/cm, элюцию целевого белка проводят буфером с pH 8,2, содержащим 20 mM Tris-HCl и 600-1000 mM NaCl.
8. Способ по п.1, отличающийся тем, что на первом этапе хроматографической очистки используют сорбент Chelating Sepharose FF и/или на втором этапе хроматографической очистки используют сорбент SP Sepharose FF.
9. Способ по п.1, отличающийся тем, что для стабилизации целевого белка к буферу добавляют до 2 mM DTT и 0,2 mM EDTA.
10. Способ по п.1, отличающийся тем, что дополнительную очистку рекомбинантной нуклеазы семейства Cas от эндотоксинов проводят путем выполнения следующих стадий:
добавления в раствор целевого белка в буфере, полученный после этапов хроматографической очистки, детергента Triton X-114 до 1%;
помещения полученного раствора на лед до достижения абсолютной прозрачности раствора с последующим выдерживанием при 37ºС до достижения абсолютной непрозрачности раствора с целью обогащения раствора мицеллами;
последующего проведения разделения водной и мицеллярной фракций раствора с помощью центрифугирования;
добавления детергента Triton X-114 до 1% к верхней водной фазе, полученной после центрифугирования, и повторения обработки до получения препарата рекомбинантной нуклеазы семейства Cas с содержанием бактериальных эндотоксинов, сниженным до уровня, позволяющего применять указанный препарат in vivo.
11. Способ по п.10, отличающийся тем, что раствор помещают на лед и/или выдерживают при 37°С по меньшей мере 15 мин.
12. Способ по п.1, отличающийся тем, что стадии с добавлением Triton X-114 на этапе дополнительной очистки повторяют 6-9 раз.
13. Способ по п.1, отличающийся тем, что получают препарат рекомбинантной нуклеазы семейства Cas с 0,3-1,5 ЕЭ/мл, находящийся в буферном растворе, содержащем Tris-HCl, NaCl, DTT и EDTA, предпочтительно 20 mM Tris-HCl, 600-1000 mM NaCl, 2 mM DTT, 0,2 mM EDTA при pH 8,2.
14. Способ по п.1, отличающийся тем, что лиофилизированный препарат рекомбинантной нуклеазы семейства Cas получают добавлением вспомогательных веществ для лиофилизации, выбранных из глицерина или трегалозы, предпочтительно глицерина в концентрации 10-50% или трегалозы в концентрации 50-500 mM к буферному раствору, содержащему рекомбинантную нуклеазу семейства Cas с 0,3-1,5 ЕЭ/мл, Tris-HCl, NaCl, DTT и EDTA, предпочтительно 20 mM Tris-HCl, 600-1000 mM NaCl, 2 mM DTT, 0,2 mM EDTA при pH 8,2, с последующим проведением лиофилизации.
15. Способ по любому из пп.1-14, отличающийся тем, что рекомбинантная нуклеаза семейства Cas системы CRISPR/Cas характеризуется аминокислотной последовательностью, выбранной из SEQ ID NO: 1-8.
16. Способ по любому из пп.1-14, отличающийся тем, что аминокислотная последовательность рекомбинантной нуклеазы семейства Cas системы CRISPR/Cas закодирована нуклеотидной последовательностью, выбранной из SEQ ID NO: 9-16.
17. Препарат рекомбинантной нуклеазы семейства Cas системы CRISPR/Cas, по существу, свободный от бактериальных эндотоксинов, для использования в системе CRISPR/Cas, представляющий собой водную фазу, включающую очищенную рекомбинантную нуклеазу семейства Cas, полученную после финального центрифугирования по завершении этапа дополнительной очистки или лиофильно высушенную в соответствии со способом по любому из пп.1-16.
18. Препарат по п.17, где, по существу, свободный от бактериальных эндотоксинов препарат рекомбинантной нуклеазы семейства Cas системы CRISPR/Cas содержит менее 1,5 ЕЭ/мл бактериальных эндотоксинов, предпочтительно 0,3 ЕЭ/мл.
19. Препарат по любому из пп.17-18, который представляет собой лиофильно высушенный препарат.
20. Набор для использования в системе CRISPR/Cas, включающий препарат рекомбинантной нуклеазы семейства Cas системы CRISPR/Cas по любому из пп.17-19, необязательно мишень-специфическую направляющую РНК, и инструкцию по применению.
21. Набор по п.20, отличающийся тем, что включает препарат рекомбинантной нуклеазы семейства CRISPR/Cas по любому из пп.17-19 в комплексе с мишень-специфической направляющей РНК или отдельно.
| WO 2011015327, 10.02.2011 | |||
| GHOSH K | |||
| K | |||
| et al., Evaluation of a novel outer membrane surface-exposed protein, LIC13341 of Leptospira, as an adhesin and serodiagnostic candidate marker for leptospirosis, Microbiology, 2018, V | |||
| Способ получения суррогата олифы | 1922 |
|
SU164A1 |
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| RAJAGOPALAN N | |||
| et al., A Two-Step Method for Obtaining Highly Pure Cas9 Nuclease for Genome | |||

