Область техники, к которой относится настоящее изобретение
Настоящее изобретение относится к области биотехнологии, в частности к системе редактирования генома ДНК эукариотической клетки на основе нуклеотидной последовательности, кодирующей белок SuCas9_NLS, в качестве инструмента для внесения целевых двунитевых разрывов в ДНК эукариотической клетки, в частности для редактирования генома человека. Система редактирования генома согласно настоящему изобретению имеет важное значение для генной терапии наследственных заболеваний человека.
Предшествующий уровень техники настоящего изобретения
Направленное внесение двунитевых разрывов в последовательности ДНК лежит в основе редактирования геномов эукариотических и прокариотических организмов. Таким образом, поиск новых и модификация существующих инструментов, позволяющих производить подобные манипуляции с молекулами ДНК, является актуальной задачей биотехнологии.
В настоящий момент активно используются следующие три методики: 1) искусственные нуклеазные системы, содержащей домены типа «цинковые пальцы» (ZFN); 2) эффекторные нуклеазы, похожие на активатор транскрипции (Transcription Activator-Like Effector Nuclease; TALEN-системы); 3) CRISPR-Cas системы бактерий и архей. Первые два подхода используют узнавание целевых сайтов за счет ДНК-белковых взаимодействий. Следовательно, их применение предполагает адаптацию аминокислотной последовательности для узнавания каждой новой целевой мишени (Boch J., Scholze H., Schornack S., Landgraf A., Hahn S., Kay S., Lahaye T., Nickstadt A. & Bonas U. Breaking the code of DNA binding specificity of TAL-type III effectors // Science. - 2009. - Т. 326. - №5959. - С. 1509-1512; Carroll D. Genome engineering with zinc-finger nucleases // Genetics. - 2011. - Т. 188. - №4. - С. 773-782). В случае CRISPR-Cas систем структурами, узнающими целевые последовательности, являются не белки, а короткие направляющие РНК. Таким образом, разрезание каждой новой мишени не требует синтеза нуклеазы или ее гена de novo, а обеспечивается использованием направляющих РНК, комплементарных целевой последовательности. Это делает CRISPR-Cas системы удобными и эффективными инструментами разрезания различных ДНК-последовательностей. Методика позволяет осуществлять единовременное разрезание ДНК в нескольких участках при использовании направляющих РНК разной последовательностей. Такой подход используется в том числе для одновременного изменения нескольких генов в эукариотических организмах.
CRISPR-Cas системы изначально были обнаружены как системы адаптивного иммунитета бактерий и архей. Аббревиатура CRISPR расшифровывается как «Clustered Regularly Interspaced Short Palindromic Repeats» (Jansen R., Embden J. D., Gaastra W. & Schouls L. M. Identification of genes that are associated with DNA repeats in prokaryotes // Molecular microbiology. - 2002. - Т. 43. - №. 6. - С. 1565-1575), что переводе с английского обозначает «короткие палиндромные повторы, регулярно расположенные группами». CRISPR-Cas система включает CRISPR кассету и кластер генов, кодирующих Cas белки. CRISPR кассета состоит из коротких уникальных последовательностей (спейсеров), разделенных короткими повторяющимися последовательностями (прямыми повторами; DR). В результате транскрипции CRISPR кассеты и процессинга некодирующей РНК образуются крРНК, каждая из которых содержит последовательность одного из спейсеров и фрагмент прямого повтора. Индивидуальные крРНК связываются с Cas нуклеазами, образуя эффекторные комплексы (Brouns S. J., Jore M. M., Lundgren M., Westra E. R., Slijkhuis R. J., Snijders A. P., Dickman M. J., Makarova K. S., Koonin E. V. & van der Oost J. Small CRISPR RNAs guide antiviral defense in prokaryotes // Science. - 2008. - Т. 321. - №. 5891. - С. 960-964). За счет комплементарного спаривания крРНК с целевым участком ДНК (протоспейсером) Cas нуклеаза узнает мишень и высоко специфично вносит в нее разрыв.
CRISPR-Cas системы, эффекторный комплекс которых представлен одиночным Cas белком, составляют класс 2. Класс 2 подразделяется на три типа в зависимости от Cas белков, входящих в состав систем (II, V и VI). Типы в свою очередь разделяются на подтипы (Makarova K. S., Wolf Y. I., Iranzo J., Shmakov S. A., Alkhnbashi O. S., Brouns S. J. J., Charpentier E., Cheng D., Haft D. H., Horvath P., Moineau S., Mojica F. J. M., Scott D., Shah S. A., Siksnys V., Terns M. P., Venclovas Č., White M. F., Yakunin A. F., Yan W., Zhang F., Garrett R. A., Backofen R., van der Oost J., Barrangou R. & Koonin E. V. Evolutionary classification of CRISPR-Cas systems: a burst of class 2 and derived variants // Nature Reviews Microbiology. - 2020. - Т. 18. - №2. - С. 67-83).
Наиболее изученными эффекторами класса 2 являются несколько представителей типа II. Эффекторы данного типа носят название Cas9 белков. На данный момент они уже имеют широкое практическое применение. Например, Cas эффектор CRISPR-Cas системы типа II-A из бактерии Streptococcus pyogenes (SpCas9) активно используется для генного редактирования эукариот (Cong L., Ran F. A., Cox D., Lin S., Barretto R., Habib N., Hsu P. D., Wu X., Jiang W., Marraffini L. A. & Zhang F. Multiplex genome engineering using CRISPR/Cas systems // Science. - 2013. - Т. 339. - № 6121. - С. 8197-823).
Инструменты геномного редактирования, основанные на CRISPR-Cas системах II типа, отличаются простотой и удобством работы: для их функционирования необходимо формирование эффекторного комплекса, состоящего лишь из одного Cas9 белка и двух коротких РНК: крРНК и трейсерной РНК (трРНК). Трейсерная РНК комплементарно спаривается с участком крРНК, образуя вторичную структуру, необходимую для связывания направляющих РНК с Cas эффектором (Deltcheva E., Chylinski K., Sharma C. M., Gonzales K., Chao Y., Pirzada Z. A., Eckert M. R., Vogel J. & Charpentier E. CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III // Nature. - 2011. - Т. 471. - № 7340. - С. 602-607). Также было показано, что трРНК может быть соединена с крРНК посредством короткого четырехнуклеотидного линкера, образуя единую молекулу направляющей РНК (гРНК; sgRNA) (Jinek M., Chylinski K., Fonfara I., Hauer M., Doudna J. A. & Charpentier E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity // Science. - 2012. - Т. 337. - №6096. - С. 816-821). Тем самым, использование конструкции, объединяющей в себе обе РНК, уменьшает количество компонентов инструмента редактирования на основе CRISPR-Cas системы II типа до двух.
Для распознавания и последующего гидролиза ДНК помимо комплементарного соответствия спейсера крРНК и протоспейсера эффекторному рибонуклеиновому комплексу необходимо присутствие PAM (Protospacer Adjusted Motif) на ДНК мишени (Shah, S. A., Erdmann, S., Mojica F. J. M. & Garrett R. A. Protospacer recognition motifs: mixed identities and functional diversity // RNA Biology. - 2013. - Т. 10. - №5. - С. 891-899). PAM представляет собой строго определенную последовательность из нескольких нуклеотидов, фланкирующую 3'-конец протоспейсера на нетаргетной цепи. При отсутствии PAM гидролиза связей в ДНК с образованием двунитевого разрыва не происходит. Необходимость присутствия PAM последовательности на мишени повышает специфичность узнавания, но в то же время накладывает ограничения в выборе целевых участков ДНК, в которые необходимо внести разрыв. Таким образом, наличие нужной PAM последовательности является характеристикой, ограничивающей применение Cas нуклеаз на любых участках ДНК.
На сегодняшний день известен целый ряд Cas9 нуклеаз CRISPR-Cas систем II типа, способных направлено и специфично вносить двунитевые разрывы в ДНК. Однако, лишь небольшая их часть показала эффективную работу в эукариотических клетках, в том числе в культурах клеток человека. Например, Nme1Cas9 из Neisseria meningitidis 8013 (Esvelt K. M., Mali P., Braff J. L., Moosburner M., Yaung S. J. & Church G. M. Orthogonal Cas9 Proteins for RNA-Guided Gene Regulation and Editing // Nature Methods. - 2013. - Т. 10. - №11. - С. 1116-1121), St1Cas9 из Streptococcus thermophilus LMD-9, Cje1Cas9 из Campylobacter jejuni subsp. jejuni NCTC 11168 (Fonfara I., Le Rhun A., Chylinski K., Makarova K. S., Lécrivain A.-L., Bzdrenga J., Koonin E. V. & Charpentier E. Phylogeny of Cas9 determines functional exchangeability of dual-RNA and Cas9 among orthologous type II CRISPR-Cas systems // Nucleic Acids Research. - 2014. - Т. 42. - №4. - С. 2577-2590), SaCas9 из Staphylococcus aureus subsp. aureus (Ran F. A., Cong L., Yan W. X., Scott D. A., Gootenberg J. S., Kriz A. J., Zetsche B., Shalem O., Wu X., Makarova K. S., Koonin E. V., Sharp P. A. & Zhang F. In vivo genome editing using Staphylococcus aureus Cas9 // Nature. - 2015. - Т. 520. - №7546. - С. 186-191), FnCas9 из Francisella tularensis subsp. novicida U112 (Hirano H., Gootenberg J. S., Horii T., Abudayyeh O. O., Kimura M., Hsu P. D., Nakane T., Ishitani R., Hatada I., Zhang F., Nishimasu H. & Nureki O. Structure and Engineering of Francisella novicida Cas9 // Cell. - 2016. - Т. 164. - №5. - С. 950-961), GeoCas9 из Geobacillus stearothermophilus ATCC 7953 (Harrington L. B., Paez-Espino D., Staahl B. T., Chen J. S., Ma E., Kyrpides N. C. & Doudna J. A. A thermostable Cas9 with increased lifetime in human plasma // Nature Communications. - 2017. - Т. 8. - №1. - С. 1424), ScCas9 из Streptococcus canis FSL Z3-227 (Chatterjee P., Jakimo N. & Jacobson J. M. Minimal PAM specificity of a highly similar SpCas9 ortholog // Science Advances. - 2018. - Т. 4. - №10. - С. eaau0766), SauriCas9 из Staphylococcus auricularis Ani-GT-018 (Hu Z., Wang S., Zhang C., Gao N., Li M., Wang D., Wang D., Liu D., Liu H., Ong S.-G., Wang H. & Wang Y. A compact Cas9 ortholog from Staphylococcus auricularis (SauriCas9) expands the DNA targeting scope // PLOS Biology. - 2020. - Т. 18. - №3. - С. e3000686), BlatCas9 из Brevibacillus laterosporus Rsp (Gao N., Zhang C., Hu Z., Li M., Wei J., Wang Y. & Liu H. Characterization of Brevibacillus laterosporus Cas9 (BlatCas9) for Mammalian Genome Editing // Frontiers in Cell and Developmental Biology. - 2020. - Т. 8. - С. 583164), PpCas9 из Pasteurella pneumotropica ATCC 35149 (Fedorova I., Vasileva A., Selkova P., Abramova M., Arseniev A., Pobegalov G., Kazalov M., Musharova O., Goryanin I., Artamonova D., Zyubko T., Shmakov S., Artamonova T., Khodorkovskii M. & Severinov K. PpCas9 from Pasteurella pneumotropica - a compact Type II-C Cas9 ortholog active in human cells // Nucleic Acids Research. - 2020. - Т. 48. - №21. - С. 12297-12309), FrCas9 из Faecalibaculum rodentium NYU-BL-K8 (Cui Z., Tian R., Huang Z., Jin Z., Li L., Liu J., Huang Z., Xie H., Liu D., Mo H., Zhou R., Lang B., Meng B., Weng H. & Hu Z. FrCas9 is a CRISPR/Cas9 system with high editing efficiency and fidelity // Nature Communications. - 2022. - Т. 13. - №1. - С. 1425), AtCas9 из Alicyclobacillus tengchongensis CGMCC1504 (Shi Y.-J., Duan M., Ding J.-M., Wang F.-Q., Bi L.-L., Zhang C.-X., Zhang Y.-Z., Duan J.-Y., Huang A.-H., Lei X.-L., Yin H. & Zhang Y. DNA topology regulates PAM-Cas9 interaction and DNA unwinding to enable near-PAMless cleavage by thermophilic Cas9 // Molecular Cell. - 2022. - Т. 82. - С. 1-16).
Таким образом, из уровня техники известны различные CRISPR-Cas белки, которые используют для своей работы разные оригинальные PAM последовательности, а также известно использование белка Cas в комбинации с направляющей РНК для направленного редактирования генома. Многие системы были исследованы in vitro, однако, последующее изучение данных систем редактирования для редактирования генома человека показало, что многие из предложенных систем редактирования не показали эффективного образования двунитевого разрыва в строго определенных местах последовательности ДНК клеток человека.
Таким образом, несмотря на известность ряда способов разрезания ДНК и изменения последовательности геномной ДНК на сегодняшний день сохраняется потребность в новых эффективных инструментах для модификации ДНК эукариотической клетки, в частности клетки человека, в строго определенных местах последовательности ДНК с целью генной терапии наследственных заболеваний человека.
Краткое раскрытие настоящего изобретения
Задачей настоящего изобретения является создание новых эффективных инструментов для редактирования последовательности геномной ДНК эукариотической клетки, в частности генома человека.
Для решения этой задачи авторами была создана уникальная система редактирования, содержащая:
(1) нуклеотидную последовательность SEQ ID NO: 2, кодирующую белок SuCas9_NLS с аминокислотной последовательностью SEQ ID NO: 1,
(2) нуклеотидную последовательность, кодирующую направляющую РНК, взаимодействующую с указанным белком и содержащую последовательность, образующую дуплекс с нуклеотидной последовательностью участка геномной ДНК организма, непосредственно примыкающей к нуклеотидной последовательности PAM 5’-NNAAА-3’,
где указанная система редактирования приводит к образованию двунитевого разрыва в последовательности геномной ДНК эукариотической клетки, непосредственно примыкающей к последовательности PAM 5'- NNAAА-3',
где указанная эукариотическая клетка не является клеткой зародышевой линии клеток человек.
Изменение последовательности геномной ДНК осуществляется путем введения в по меньше мере одну целевую клетку эффективного количества системы редактирования согласно настоящему изобретению, в результате чего непосредственно в редактируемых клетках синтезируется и формируется рибонуклеопротеиновый редактирующий комплекс, представляющий собой эффекторный белок SuCas9_NLS в комбинации с направляющей РНК.
Взаимодействие указанного редактирующего комплекса, направленного с помощью специфического фрагмента направляющей РНК на последовательность мишени, фланкированной последовательностью PAM 5'-NNAAА-3', приводит к образованию двунитевого разрыва в последовательности геномной ДНК эукариотической клетки, непосредственно примыкающей к последовательности PAM 5'-NNAAА-3'.
Согласно первому аспекту настоящее изобретение относится к системе редактирования последовательности геномной ДНК эукариотической клетки, содержащей
(1) нуклеотидную последовательность SEQ ID NO: 2, кодирующую белок SuCas9_NLS с аминокислотной последовательностью SEQ ID NO: 1,
(2) нуклеотидную последовательность, кодирующую направляющую РНК, взаимодействующую с указанным белком и содержащую последовательность, образующую дуплекс с нуклеотидной последовательностью участка геномной ДНК организма, непосредственно примыкающей к нуклеотидной последовательности PAM 5’-NNAAА-3’,
где указанная система приводит к образованию двунитевого разрыва в последовательности геномной ДНК эукариотической клетки, непосредственно примыкающей к последовательности PAM 5'- NNAAА-3',
где указанная эукариотическая клетка не является клеткой зародышевой линии клеток человека.
Согласно одному варианту осуществления системы редактирования согласно настоящему изобретению эукариотическая клетка представляет собой клетку человека.
Согласно второму аспекту настоящее изобретение относится к экспрессионному вектору для редактирования последовательности геномной ДНК эукариотической клетки, несущему систему редактирования согласно настоящему изобретению и регуляторный элемент, способный к функционированию в эукариотической клетке, имеющему элементы и их расположение, как представлено на фиг. 2, где указанная эукариотическая клетка не является клеткой зародышевой линии клеток человека.
Согласно третьему аспекту настоящее изобретение относится к способу редактирования последовательности геномной ДНК эукариотической клетки, предусматривающему введение в указанную клетку эффективного количества системы редактирования согласно настоящему изобретению или экспрессионного вектора согласно настоящему изобретению, где указанная эукариотическая клетка не является клеткой зародышевой линии клеток человека.
Технический результат настоящего изобретения состоит в обеспечении надежной эффективной системы редактирования для модификации ДНК эукариотической клетки, в частности клетки человека, которая позволяет проводить модификацию генома живого организма и обеспечивает внесение направленных изменений в последовательности геномной ДНК эукариотической клетки. В частности, система редактирования согласно настоящему изобретению приводит к образованию двунитевого разрыва в строго определенном месте в последовательности геномной ДНК эукариотической клетки, а именно в ее части, непосредственно примыкающей к последовательности PAM 5'- NNAAА-3'. В частности, система редактирования согласно настоящему изобретению приводит к направленному внесению двунитевых разрывов в геноме человека. Система редактирования согласно настоящему изобретению приводит к эффективной высокой экспрессии белка SuCas9_NLS в эукариотической клетке, в частности в клетке человека. Система редактирования согласно настоящему изобретению приводит к эффективному нацеливанию белка SuCas9_NLS посредством направляющей РНК для направленного редактирования и образованию двунитевого разрыва в строго определенном месте в последовательности геномной ДНК эукариотической клетки, а именно в ее части, непосредственно примыкающей к последовательности PAM 5'- NNAAА-3'.
Образование двунитевого разрыва в строго определенном месте в последовательности геномной ДНК эукариотической клетки, а именно в ее части, непосредственно примыкающей к последовательности PAM 5'- NNAAА-3', достигается посредством того, что охарактеризованная в настоящем изобретении нуклеаза SuCas9_NLS на основе SuCas9 из бактерии Streptococcus uberis обладает коротким, трехбуквенным PAM мотивом, необходимым для редактирования ДНК эукариотической клетки, в частности ДНК клетки человека. Согласно настоящему изобретению для успешного функционирования нуклеазы SuCas9_NLS in vivo достаточно присутствия короткого PAM мотива (AAА), расположенного в двух нуклеотидах от протоспейсера. Многие известные на сегодняшний день малоразмерные Cas нуклеазы, способные вносить двунитевые разрывы в геномную ДНК, имеют сложные многобуквенные PAM последовательности или PAM последовательности, которые не позволяют редактировать ДНК эукариотической клетки, в частности ДНК клетки человека
Кроме того, техническим результатом настоящего изобретения является расширение спектра доступных генетических редакторов на основе систем CRISPR-Cas II типа, что позволит использовать такие редакторы для разрезания геномной ДНК эукариотических клеток в большем количестве специфических сайтов и специфических условий.
Краткое описание чертежей
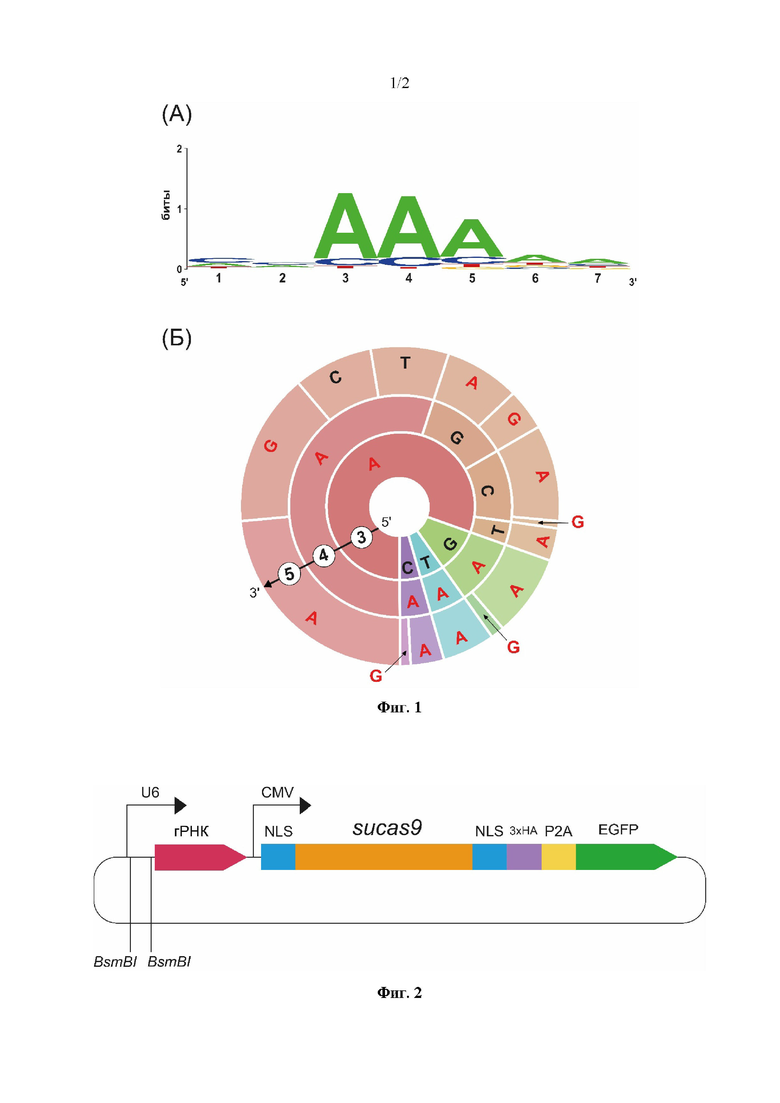
Фиг. 1. Результаты определения PAM последовательности для нуклеазы SuCas9 с использованием 7N in vitro деплеционного теста. А - визуализация результатов в виде PAM Лого; видна консервативность 3, 4, 5 позиций PAM. Б - визуализация результатов в виде PAM Колеса для 3, 4, 5 позиций; преобладающие нуклеотиды рассматриваемых позиций выделены красным цветом; в 3 и 4 позиции видно преобладание A, в 5 позиции - A или G.
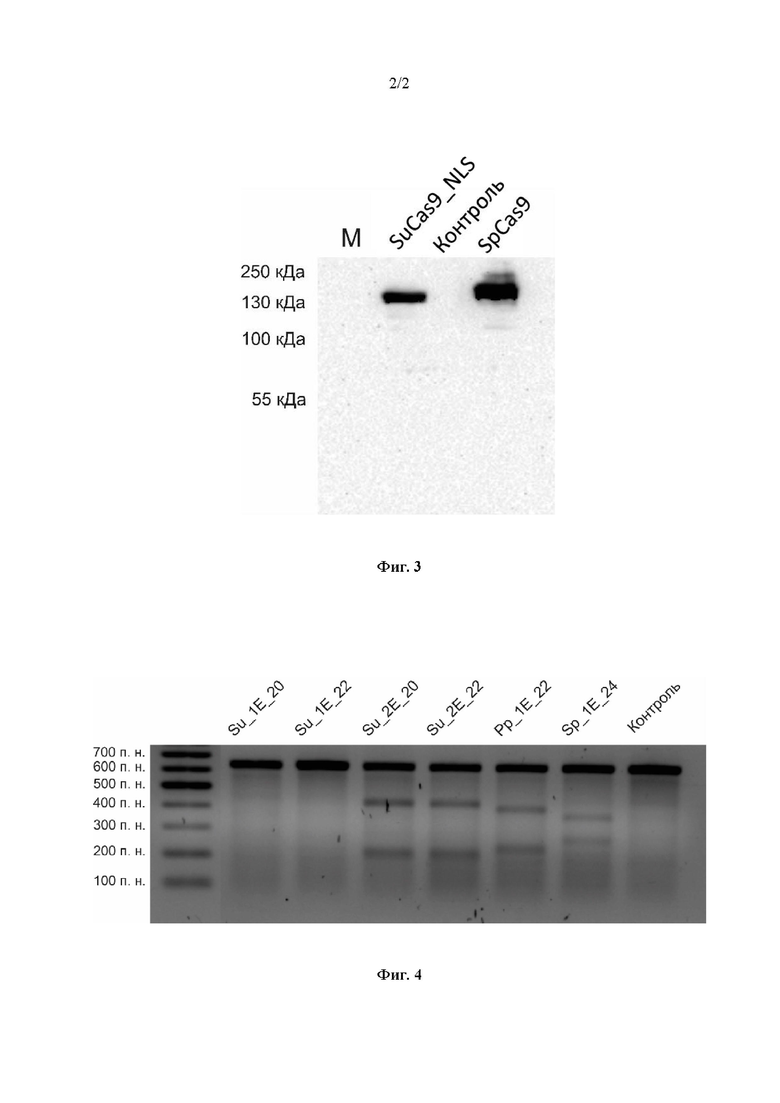
Фиг. 2. Схема экспрессионного вектора, несущего нуклеотидную последовательность SEQ ID NO: 2, кодирующую белок SuCas9_NLS с аминокислотной последовательностью SEQ ID NO: 1, и нуклеотидную последовательность, кодирующую направляющую РНК. Черными стрелками обозначено направление транскрипции направляющей РНК и нуклеазы со связанными с ним конструкциями. NLS - последовательность, кодирующая сигнал ядерной локализации, 3xHA - HA-таг, P2A - линкерный пептид, EGFP - ген зеленого флуоресцентного белка.
Фиг. 3. Проверка эффективности синтеза нуклеазы SuCas9_NLS в клетках человека HEK293T с помощью вестерн-блот анализа. Размеры белков: SuCas9_NLS - 131,78 кДа, SpCas9 (положительный контроль) - 158,46 кДа. Контроль - отрицательный контроль; клетки HEK293T, не подвергшиеся трансфекции.
Фиг. 4. Проверка наличия нуклеазной активности SuCas9_NLS в клетках человека с помощью реакции c Т7 эндонуклеазой I. «20», «22» и «24» обозначает длину участка спейсера гРНК. Ожидаемые размеры продуктов рестрикции для мишеней Su_1E_20 и Su_1E_22, фланкированных последовательностью PAM 5'-TGAAG-3': 186 п.н. + 453 п.н. - отрицательный результат, нет продуктов реакции. Ожидаемые размеры продуктов рестрикции для мишеней Su_2E_20 и Su_2E_22, фланкированных последовательностью PAM 5'-ACAAA-3': 212 п.н. + 427 п.н. Ожидаемые размеры продуктов рестрикции для мишени Pp_1E_22: 237 п.н. + 402 п.н. Ожидаемый размер продуктов рестрикции для мишени Sp_1E_24: 271 п.н. + 368 п.н.
Подробное раскрытие настоящего изобретения
Если здесь не указано иное, все используемые технические и научные термины обычно понимаются специалистами в области генной инженерии, биохимии, генетики, молекулярной биологии и имеют то же значение, что и общепринятое.
В контексте настоящего изобретения термин «процент гомологии двух последовательностей» эквивалентен термину «процент идентичности двух последовательностей». Идентичность последовательностей определяется на основании референсной последовательности. Алгоритмы для анализа последовательности известны в данной области, такие как BLAST, описанный в Altschul et al. (Basic local alignment search tool // Journal of Molecular Biology. - 1990. - Т. 215. - С. 403-410). Для целей настоящего изобретения для определения уровня идентичности и сходства между нуклеотидными последовательностями и аминокислотными последовательностями может быть использовано сравнение нуклеотидных и аминокислотных последовательностей, производимое с помощью пакета программ BLAST, предоставляемого National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/blast) с использованием содержащего разрывы выравнивания со стандартными параметрами. Процент идентичности двух последовательностей определяется числом положений идентичных аминокислот в этих двух последовательностях с учетом числа пробелов и длины каждого пробела, которые необходимо ввести для оптимального сопоставления двух последовательностей путем выравнивания. Процент идентичности равен числу идентичных аминокислот в данных положениях с учетом выравнивания последовательностей, разделенному на общее число положений и умноженному на 100.
В контексте настоящего изобретения термин «специфически гибридизуется» относится к ассоциации между двумя одноцепочечными молекулами нуклеиновых кислот или в достаточной степени комплементарными последовательностями, что разрешает такую гибридизацию в предопределенных условиях, обычно использующихся в данной области.
В контексте настоящего изобретения фраза «двунитевой разрыв, расположенный непосредственно перед нуклеотидной последовательностью РАМ» означает, что двунитевой разрыв в целевой последовательности ДНК произведен на расстоянии от 0 до 20 нуклеотидов перед нуклеотидной последовательностью РАМ.
В контексте настоящего изобретения под рибонуклеопротеиновым комплексом подразумевается совокупность эффекторного белка нуклеазы SuCas9_NLS и направляющей РНК, содержащей последовательность комплементарной ДНК мишени.
Согласно первому аспекту настоящее изобретение относится к системе редактирования последовательности геномной ДНК эукариотической клетки, содержащей
(1) нуклеотидную последовательность SEQ ID NO: 2, кодирующую белок SuCas9_NLS с аминокислотной последовательностью SEQ ID NO: 1,
(2) нуклеотидную последовательность, кодирующую направляющую РНК, взаимодействующую с указанным белком и содержащую последовательность, образующую дуплекс с нуклеотидной последовательностью участка геномной ДНК организма, непосредственно примыкающей к нуклеотидной последовательности PAM 5’-NNAAА-3’,
где указанная система приводит к образованию двунитевого разрыва в последовательности геномной ДНК эукариотической клетки, непосредственно примыкающей к последовательности PAM 5'- NNAAА-3',
где указанная эукариотическая клетка не является клеткой зародышевой линии клеток человека.
Белок SuCas9_NLS представляет собой рекомбинантный белок на основе последовательности нуклеазы Cas9 из бактерии Streptococcus uberis, имеющий аминокислотную последовательность SEQ ID NO: 1.
Нативная SuCas9 представляет собой нуклеазу Cas9, обнаруженную в бактерии Streptococcus uberis. CRISPR-Cas система из Streptococcus uberis относится к типу II-C и состоит из CRISPR кассеты, несущей прямые повторы последовательностью 5'-GTTTTTGTACTCTCAAGATTTAAGTAACTATAAAAC-3'. К CRISPR кассете прилегает ген эффекторного белка SuCas9.
Для обеспечения эффективной высокой экспрессии белка SuCas9_NLS с вводимой в эукариотическую клетку системой редактирования согласно настоящему изобретению авторы настоящего изобретения разработали уникальную нуклеотидную последовательность согласно SEQ ID NO: 2, кодирующую белок SuCas9_NLS с аминокислотной последовательностью согласно SEQ ID NO: 1, обеспечивающую эффективное редактирование ДНК эукариотической клетки, в частности редактирование генома человека. Авторы настоящего изобретения кодон-оптимизировали нуклеотидную последовательность нуклеазы SuCas9_NLS с достижением эффективной экспрессии нуклеазы SuCas9_NLS в клетках человека, предотвращения возможности появления бактериальных редких кодонов в кодирующей последовательности нуклеазы в клетках человека и эффективной работы нуклеазы SuCas9 в эукариотических клетках. В частности, нуклеотидная последовательность нуклеазы SuCas9_NLS содержит последовательность, кодирующую сигнал ядерной локализации (NLS, Nuclear Localization Signal), для обеспечения импорта этого белка внутрь ядра эукариотической клетки. Кроме того, нуклеотидная последовательность SuCas9_NLS разработана таким образом, что из нее исключены сайты узнавания распространенных рестриктаз для ускорения клонирования новых вставок, кодирующих направляющую РНК, комплементарную новым мишеням ДНК. Полученная последовательность нуклеотидов SEQ ID NO: 2 использовалась в создании системы редактирования согласно настоящему изобретению для редактирования геномной ДНК клеток человека. Разработанная нуклеотидная последовательность согласно SEQ ID NO: 2 приводит к эффективному редактированию ДНК эукариотической клетки, в частности эффективному редактированию генома человека, посредством эффективной высокой экспрессии белка SuCas9_NLS в эукариотической клетки, который в комплексе с направляющей РНК согласно настоящему изобретению, приводит к эффективному образованию двунитевого разрыва в последовательности геномной ДНК эукариотической клетки, непосредственно примыкающей к последовательности PAM 5'- NNAAА-3'.
Согласно настоящему изобретению используется направляющая РНК, взаимодействующую с указанным белком SuCas9_NLS и содержащая последовательность, образующую дуплекс с нуклеотидной последовательностью участка геномной ДНК организма, непосредственно примыкающей к нуклеотидной последовательности PAM 5’-NNAAА-3’. Согласно одному варианту осуществления настоящего изобретения в качестве направляющей РНК используется разработанная гибридная РНК, сконструированная на основе крРНК и трРНК (гРНК) из нативного организма, однако, содержащая важные отличия, необходимые для работы в эукариотическом организме. Согласно предпочтительному варианту осуществления настоящего изобретения в качестве направляющей РНК используется синтетические гибридные последовательности гРНК (гидовая РНК), примеры которых приведены в таблице 1. Такие гибридные направляющие РНК были разработаны на основе биоинформатического анализа стабильности слитых РНК структур нативных крРНК и трРНК. Кроме того, экспериментально была подобрана предпочтительная длина гибридизуемого с ДНК мишени РНК фрагмента для успешной работы рибонуклеопротеинового комплекса в эукариотичских клетках и, в частности, в клетках человека.

Таким образом, авторами настоящего изобретения была предложена длина спейсерной части направляющей РНК, меньшая по сравнению с используемыми в распространенных системах редактирования и которая составляет без ограничения по меньшей мере 20 п.н., предпочтительно от 20 до 22 п.н. и более предпочтительно 22 п.н. Указанная длина спейсерной части направляющей РНК ранее не использовалась в комплексе с белком SuCas9_NLS для редактирования ДНК эукариотической клетки.
Проводимые ранее биохимические исследования нуклеазы SuCas9 in vitro показали, что нативный белок является активной нуклеазой, способной вносить двунитевые разрывы в ДНК, ограниченные PAM 5'-NNAAR-3' (5'-NNAA A/G -3') (фиг. 1).
Однако в результате проведенных исследований по редактированию SuCas9 геномной ДНК клеток человека, авторами настоящего изобретения неожиданно было обнаружено, что достижение двухнитевого разрыва происходит в ДНК клеток человека, ограниченных PAM 5'-NNAAА-3', тогда как для ДНК клеток человека, ограниченных PAM 5'-NNAAG-3', был получен отрицательный результат, как показано на фиг.4. Таким образом, охарактеризованная в настоящем изобретении нуклеаза SuCas9_NLS на основе SuCas9 из бактерии Streptococcus uberis содержит короткий трехбуквенный PAM мотив, необходимый для редактирования ДНК эукариотической клетки, в частности ДНК клетки человека. Согласно настоящему изобретению для успешного функционирования нуклеазы SuCas9_NLS in vivo достаточно присутствия короткого PAM мотива (AAА), расположенного в двух нуклеотидах от протоспейсера. Многие известные на сегодняшний день малоразмерные Cas нуклеазы, способные вносить двунитевые разрывы в геномную ДНК, имеют сложные многобуквенные PAM последовательности или PAM последовательности, которые не позволяют редактировать ДНК эукариотической клетки, в частности ДНК клетки человека.
Кроме того, авторами настоящего изобретения было обнаружено, что разработанная нуклеотидная последовательность SEQ ID NO: 2, кодирующая белок SuCas9_NLS с аминокислотной последовательностью SEQ ID NO: 1, приводит к эффективной высокой экспрессии белка SuCas9_NLS в эукариотической клетке, в частности в клетке человека, как показано на фиг. 3, при введении в указанную клетку системы редактирования согласно настоящему изобретению или экспрессионного вектора согласно настоящему изобретению.
Кроме того, авторами настоящего изобретения было обнаружено, что надежное внесение двунитевого разрыва в ДНК, ограниченные PAM 5'-NNAAА-3', при использовании нуклеазы SuCas9_NLS в комплексе с направляющей РНК, достигается при любой длине спейсерной последовательности направляющей РНК, однако, предпочтительной является длина спейсерной последовательности от 20 п.н. до 22 п.н. При этом, использование направляющей РНК, имеющей длину спейсерной последовательности 22 п.н., неожиданно показало по меньшей мере не менее эффективные результаты направленного расщепления геномной ДНК в сравнении с использованием направляющей РНК, имеющей длину спейсерной последовательности 20 п.н., как показано на фиг. 4.
Система редактирования согласно настоящему изобретению, кодирующая рибонуклеопротеиновый комплекс, может быть введена в клетку непосредственно или опосредованно - посредством трансфекции или трансформации клеток известными специалистам способами, за счет использования рекомбинантного вируса, посредством манипуляций с клеткой, таких как микроинъекция ДНК и т. п.
Для введения системы редактирования согласно настоящему изобретению в клетки целевого организма (во все клетки или в часть клеток) авторы настоящего изобретения использовали подход с использованием известных специалистам в данной области техники способов.
Для введения системы редактирования согласно настоящему изобретению в клетки целевого организма (во все клетки или в часть клеток) могут применятся подходы доставки рибонуклепротеинового комплекса с использованием аденоассоциированных вирусов (AAV; Adeno-associated virus).
Метод доставки раскрыт в источниках (Ran F. A., Cong L., Yan W. X., Scott D. A., Gootenberg J. S., Kriz A. J., Zetsche B., Shalem O., Wu X., Makarova K. S., Koonin E. V., Sharp P. A. & Zhang F. In vivo genome editing using Staphylococcus aureus Cas9 // Nature. - 2015. - Т. 520. - №7546. - С. 186-191; Edraki A., Mir A., Ibraheim R., Gainetdinov I., Yoon Y., Song C.-Q., Cao Y., Gallant J., Xue W., Rivera-Pérez J. A. & Sontheimer E. J. A Compact, High-Accuracy Cas9 with a Dinucleotide PAM for In Vivo Genome Editing // Molecular Cell. - 2019. - Т. 73. - №4. - С. 714-726; Kim E., Koo T., Park S. W., Kim D., Kim K., Cho H.-Y., Song D. W., Lee K. J., Jung M. H., Kim S., Kim J. H., Kim J. H. & Kim J. S. In vivo genome editing with a small Cas9 orthologue derived from Campylobacter jejuni // Nature Communications. - 2017. - Т. 8. - С. 14500).
Настоящее изобретение далее проиллюстрировано посредством примера.
Пример
Пример 1. Изменение геномной ДНК клеток человека с помощью рибонуклеопротеинового комплекса SuCas9_NLS.
Для изучения активности нуклеазы SuCas9_NLS в клетках человека был создан экспрессионный вектор, несущий нуклеотидную последовательность SEQ ID NO: 2, кодирующую белок SuCas9_NLS с аминокислотной последовательностью SEQ ID NO: 1, и нуклеотидную последовательность, кодирующую направляющую РНК, под регуляцией эукариотических промоторов. Для экспрессии гРНК использовали U6 промотор с дополнительным G нуклеотидом на 3'-конце. Для возможности дальнейшей вставки различных последовательностей спейсера на 5'-конец консервативной последовательности гРНК после U6 промотора вставляли два сайта эндонуклеазы рестрикции BsmBI. Для эффективной экспрессии нуклеотидной последовательности нуклеазы использовали цитомегаловирусный CMV промотор (Cytomegalovirus). После NLS сигнала на C-конце нуклеотидной последовательности нуклеазы был в трех повторах помещен HA-таг (Human influenza hemagglutinin tag), используемый для детекции рекомбинантных белков. Для возможности проверки эффективности продукции SuCas9_NLS в клетках человека после нуклеотидной последовательности нуклеазы помещали последовательность, кодирующую зеленый флуоресцентный белок (EGFP; Enhanced Green Fluorescent Protein) через линкерный пептид P2A. Схема полученного экспрессионного вектора представлена на фиг. 2.
Созданный экспрессионный вектор был трансфицирован в клетки человека линии НЕК293Т с помощью реагента липофектамина 2000 с целью проверки внутриклеточного синтеза исследуемой нуклеазы. По истечении 72 часов после трансфекции клетки лизировали, и с полученными лизатами был проведен вестерн-блот анализ. В анализе использовались первичные антитела к HA-тагу, который был присоединен к нуклеазе SuCas9_NLS с С-конца. В качестве положительного контроля использовались клетки, трансфицированные экспрессионным вектором, обеспечивающим эффективный синтез нуклеазы SpCas9 из бактерии Streptococcus pyogenes SF370 (M1 GAS) (размер 158,46 кДа). В качестве отрицательного контроля использовались клетки, не подвергшиеся трансфекции. На фиг. 3 видно, что ген эффекторного белка SuCas9_NLS экспрессируется в клетках с созданной генетической конструкции, приводя к синтезу нуклеазы соответствующего размера (131,78 кДа).
Для проверки непосредственно нуклеазной активности рибонуклеопротеинового комплекса SuCas9_NLS в клетках человека на основе полученного экспрессионного вектора был создан ряд новых экспрессионных векторов. В них со стороны 5'-конца последовательности, кодирующей направляющую гРНК (гидовую РНК), вставляли последовательности, комплементарные выбранным мишеням в геноме человека (последовательности спейсеров). На основе данных последовательностей должны синтезироваться полноценные эффекторные рибонуклеопротеиновые комплексы SuCas9_NLS, направляемые гидовыми РНК (гРНК) на выбранные мишени. Для вставки в экспрессионные векторы спейсерных последовательностей использовался метод клонирования «Golden Gate» (Bird J. E., Marles-Wright J. & Giachino A. A User's Guide to Golden Gate Cloning Methods and Standards // ACS Synthetic Biology. - 2022. - Т. 11. - №11. - С. 3551-3563).
Для эффективной работы комплекса в клетках эукариот были разработаны гибридные гРНК с последовательностью, комплементарной ДНК-мишени длиной 20 и 22 п.н. В качестве положительных контролей использовались аналогичные экспрессионные векторы, несущие системы изменения геномной ДНК на основе нуклеаз PpCas9 и SpCas9. ДНК-мишени были фланкированы c 3'-конца PAM последовательностями SuCas9_NLS 5'-NNAAR-3', PpCas9 5'-NNNNRTT-3' или SpCas9 5'-NGG-3'.
В качестве ДНК-мишеней использовались следующие участки гена EMX1 человека (таблица 2):
Экспрессионные векторы с SuCas9_NLS, PpCas9 и SрCas9 были трансфицированы в культуру клеток человека HEK293T с помощью реагента липофектамина 2000. По истечении 72 часов после трансфекции клетки лизировали и с полученных лизатов проводили ПЦР для наработки участков, содержащих целевые сайты изменения геномной ДНК. Полученные ПЦР фрагменты подвергали in vitro реакции с T7 эндонуклеазой I для определения частоты внесения вставок и делеций в целевых сайтах геномной ДНК. Продукты реакции анализировали при помощи электрофореза на 2%-ном агарозном геле. На фиг. 4 показано, что SuCas9_NLS активно вносит изменения в ген EMX1 с эффективностью, схожей с эффективностью работы описанных в уровне техники PpCas и SpCas9.
В приведенном примере происходит двунитевый разрыв в последовательности геномной ДНК человека, непосредственно примыкающей к последовательности 5'-NNAAА-3', в то время как двунитевый разрыв в последовательности геномной ДНК человека, непосредственно примыкающей к последовательности 5'-NNAAG-3' не происходит, как показано на фиг. 4.
Также в приведенном примере эффективность генетических модификаций при использовании гРНК c последовательностью комплементарной ДНК-мишени длиной 22 п.н. сопоставима с эффективностью внесения направленного расщепления ДНК в геноме при использовании последовательности комплементарной мишени длиной 20 п.н.
Таким образом, авторами настоящего изобретения неожиданно обнаружено и подтверждено посредством описанного исследования, что:
1) достигается высокий уровень экспрессии белка SuCas9_NLS в клетках человека при применении нуклеотидной последовательности SEQ ID NO: 2, как показано на фиг. 3,
2) происходит двунитевый разрыв в последовательности геномной ДНК человека, непосредственно примыкающей к последовательности PAM 5'-NNAAА-3', в то время как двунитевый разрыв в последовательности геномной ДНК человека, непосредственно примыкающей к последовательности PAM 5'-NNAAG-3' не происходит, как показано на фиг. 4,
3) применение гРНК со спейсерной частью 22 п.н. по меньшей мере не менее эффективно, чем применение гРНК со спейсерной частью 20 п.н., как показано на фиг. 4.
Таким образом, авторами настоящего изобретения была разработана уникальная система редактирования ДНК эукариотической клетки, подходящая для редактирования генома человека, содержащая нуклеотидную последовательность SEQ ID NO: 2, кодирующую белок SuCas9_NLS с аминокислотной последовательностью SEQ ID NO: 1, и нуклеотидную последовательность, кодирующую направляющую РНК, взаимодействующую с указанным белком и содержащую последовательность, образующую дуплекс с нуклеотидной последовательностью участка геномной ДНК организма, непосредственно примыкающей к нуклеотидной последовательности PAM 5’-NNAAА-3’, где указанная система приводит к образованию двунитевого разрыва в последовательности геномной ДНК эукариотической клетки, непосредственно примыкающей к последовательности PAM 5'- NNAAА-3', где указанная эукариотическая клетка не является клеткой зародышевой линии клеток человека.
Система редактирования согласно настоящему изобретению содержит уникальную нуклеотидную последовательность SEQ ID NO: 2, кодирующую белок SuCas9_NLS с аминокислотной последовательностью SEQ ID NO: 1, обеспечивающую редактирование ДНК эукариотической клетки, в частности генома человека, с внесением двунитевого разрыва в части, непосредственно примыкающей к последовательности PAM 5'- NNAAА-3'.
Использование в системе редактирования согласно настоящему изобретению уникальной нуклеотидной последовательности SEQ ID NO: 2 приводит к надежной эффективной экспрессии белка SuCas9_NLS в эукариотической клетке, в частности в клетке человека, и обеспечивает эффективное редактирование ДНК эукариотической клетки, в частности генома человека, с внесением двунитевого разрыва в части, непосредственно примыкающей к последовательности PAM 5'- NNAAА-3'.
Использование указанной нуклеотидной последовательности SEQ ID NO: 2 вместе с нуклеотидной последовательностью, кодирующей направляющую РНК, взаимодействующей с белком SuCas9_NLS и содержащей последовательность, образующую дуплекс с нуклеотидной последовательностью участка геномной ДНК эукариотического организма, непосредственно примыкающей к нуклеотидной последовательности PAM 5’-NNAAА-3’, приводит к образованию двунитевого разрыва в последовательности геномной ДНК эукариотической клетки, непосредственно примыкающей к последовательности PAM 5'- NNAAА-3'. При этом авторы настоящего изобретения обнаружили, что эффективное нацеливание и внесение двунитевого разрыва в последовательности геномной ДНК эукариотической клетки, непосредственно примыкающей к последовательности PAM 5'- NNAAА-3', происходит при использовании любой направляющей РНК, взаимодействующей с белком SuCas9_NLS и содержащей последовательность, образующую дуплекс с нуклеотидной последовательностью участка геномной ДНК эукариотического организма, непосредственно примыкающей к нуклеотидной последовательности PAM 5’-NNAAА-3’. При этом использование синтетических гибридных последовательностей гРНК, разработанных авторами настоящего изобретения, обеспечивает дополнительные преимущества при редактировании генома эукариотической клетки. Авторы настоящего изобретения также обнаружили, что эффективное нацеливание происходит при любой длине спейсерной части направляющей РНК. При этом использование направляющей РНК со специально подобранной длиной спейсерной части по меньшей мере 20 п.н., предпочтительно от 20 п.н. до 22 п.н. и наиболее предпочтительно 22 п.н. обеспечивает дополнительные преимущества при редактировании генома эукариотической клетки.
Несмотря на то что настоящее изобретение описано со ссылкой на раскрываемые варианты воплощения, для специалистов в данной области должно быть очевидно, что конкретные подробно описанные случаи приведены лишь в целях иллюстрирования настоящего изобретения, и их не следует рассматривать как каким-либо образом ограничивающие объем изобретения. Должно быть, понятно, что возможно осуществление различных модификаций без отступления от сути настоящего изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ УЧРЕЖДЕНИЕ ИНСТИТУТ МОЛЕКУЛЯРНОЙ
ГЕНЕТИКИ НАЦИОНАЛЬНОГО ИССЛЕДОВАТЕЛЬСКОГО ЦЕНТРА «КУРЧАТОВСКИЙ ИНСТИТУТ»
(НИЦ «КУРЧАТОВСКИЙ ИНСТИТУТ» - ИМГ)
<120> СИСТЕМА РЕДАКТИРОВАНИЯ ГЕНОМной ДНК ЭУКАРИОТИЧЕСКОЙ КЛЕТКИ НА
ОСНОВЕ нуклеотидной последовательности, кодирующей БЕЛОК SUCAS9_NLS
SEQ ID NO: 1 - последовательность аминокислот нуклеазы SuCas9_NLS
MAPKKKRKVGIHGVPAAEMTNGMILGLDIGVASVGVGIIEADSGKIVHASSRIFPSANADNNVDRRKFRGSRRLLRRKKHRV
KRLQDLFDKYDIVTNFDNLNLNPYELRVKGLNEPLSNEELFASLRNITKHRGISYLDDGEDDSSGNGTEYAKAIELNQQLLK
EKTPGQIQFDRLNQYGQLRGNFDIVDENGEIHHVINVFSTSSYRKEAEQILKKQSETNTSISTDFINDFIQLLISKRKYYHG
PGNPKSRTDYGRYRTDGTDLDNIFDVLIGKCSFYPEEYRASKTSYTAQEFNFLNDLNNLTLPTETGKLSEQQKIDLVNWAKE
TKILGPKKLLQEIAKRNNCKFEDIRGYRLDNKDNPDMHVFDVYRKMNFDLETISVKDLSVDSLNQLARILTLNTEREGIEEA
IKKLMPNQFTEKQMLELIAFRKSNSSIFGKGWHSLSIKLMKELIPELYHTSDEQMTILNRFGKFKLTKLDSKRTNYIDENSV
TDEIYNPVVAKSVRQAIKIINASIKKWGDFDKIVIEMPRDKNEEEERKRIADGQKVNAKEKEQAEKHAAKLFNGKEELPSEV
FHGYKELALRIRLWYQQDQKCLYSGKEITISDLIYNRELFEIDHILPLSLSFDDSLSNKVLVYCWANQEKGQRTPFQALDSM
KSAWSYREFKNAILHNSKISRKKRDYFLTEQDISKIEVKQKFIERNLVDTRYASRTVLNVLQQSLKNLEKETKVSVVRGQFT
SQLRRKWHIDKTRDTYHHHAVDALIIAASAKLRYWKKQGDILFENYLINRHVDRVTGEIQSDDSYKEEVFTPPYDGFVQTIS
NPGFEDEILFSYQVDSKVNRKISDATIYATRSAKLEKDKKEQTYVLGKIKDIYSQTGFENFLKIYNKDKSKFLIYQKDPETW
EKIIEPILKNYREFDNKGKDIVNPFEKYRNDNGPICKYSRKGNGPEIKQFKYYDTVLGKYIEITPESSRNIVALRSLNPWRT
DVYFNEKTLKYEFLGLKYSDLNIQNKEEYGISEENYQKIKKEEGVSEESIFKFTLYRNDLIIIKDLSKGKSQIFRFWSRTGK
VNEKHKLELKPYNKSSFEKGESLEVIRNLYPSSNQFQEKLNKSDLSIYKLKTDILGNKFISKNENLFPKLIYKKIKRKFKRP
AATKKAGQAKKKK
SEQ ID NO: 2 - нуклеотидная последовательность, кодирующая белок SuCas9_NLS
с аминокислотной последовательностью SEQ ID NO: 1
ATGGCTCCCAAAAAGAAAAGGAAAGTAGGAATCCACGGCGTGCCGGCAGCCGAAATGACCAATGGCATGATTCTGGGTCTCG
ATATCGGCGTAGCGTCCGTGGGTGTTGGTATTATTGAAGCTGATTCTGGTAAGATCGTTCATGCGTCCAGCAGGATCTTTCC
GAGCGCGAACGCGGATAATAACGTTGACCGTCGCAAGTTCCGTGGTAGCCGTCGTCTGCTCCGTCGCAAGAAACACCGTGTC
AAACGCTTACAGGACTTGTTCGACAAGTACGATATCGTGACCAACTTTGATAATCTGAACCTCAACCCGTACGAGCTGCGTG
TTAAAGGCCTGAACGAACCGCTGAGCAATGAAGAGTTGTTCGCCTCGTTGCGCAACATCACGAAACACCGCGGCATTAGCTA
CCTGGACGACGGCGAGGACGACTCGTCGGGTAATGGTACAGAGTATGCGAAAGCCATTGAACTTAACCAACAATTGCTGAAA
GAAAAAACGCCGGGTCAAATCCAGTTCGACCGCCTGAATCAGTATGGTCAACTGAGAGGCAATTTCGACATCGTGGATGAGA
ACGGTGAAATACACCACGTGATTAACGTGTTTAGCACCTCTAGCTACCGGAAGGAGGCGGAACAAATTTTGAAGAAGCAGAG
CGAAACAAACACCAGCATCAGCACCGACTTTATCAACGACTTCATTCAACTGCTGATCAGCAAGCGCAAGTACTACCACGGT
CCGGGCAATCCGAAGTCCAGAACGGACTATGGTCGTTATCGGACTGATGGCACCGATTTGGACAACATTTTCGACGTGCTGA
TCGGTAAGTGCAGCTTTTATCCGGAGGAATATAGAGCAAGCAAGACCTCTTACACCGCACAGGAGTTTAATTTCCTGAACGA
TCTCAACAACTTGACCCTGCCGACCGAAACCGGCAAGCTGAGCGAGCAGCAGAAGATCGATTTGGTTAACTGGGCGAAAGAG
ACGAAAATTCTGGGACCGAAAAAACTGCTGCAAGAAATTGCCAAACGTAACAATTGTAAGTTCGAAGATATTCGCGGCTATC
GCTTGGACAACAAGGACAACCCGGACATGCATGTTTTCGACGTCTATCGTAAAATGAATTTTGACTTAGAAACGATCAGCGT
GAAAGATCTGAGCGTGGATTCCCTGAACCAGCTGGCGCGTATTCTGACCCTGAATACCGAGCGTGAGGGTATTGAAGAGGCA
ATCAAGAAGCTGATGCCGAATCAGTTCACCGAAAAGCAAATGCTGGAACTGATCGCGTTTCGTAAGTCCAACTCCAGCATTT
TCGGCAAAGGTTGGCATAGCTTATCTATTAAACTTATGAAAGAGCTTATCCCGGAACTCTACCACACCAGCGACGAGCAGAT
GACCATCTTAAACCGTTTTGGTAAGTTCAAACTGACGAAACTGGACTCTAAACGTACCAATTATATCGATGAAAATAGCGTA
ACCGACGAGATCTACAATCCCGTGGTAGCCAAATCCGTTCGTCAGGCGATTAAGATTATTAATGCGTCTATTAAAAAATGGG
GTGATTTTGATAAGATTGTGATTGAAATGCCACGTGATAAGAACGAGGAAGAGGAGCGTAAGCGCATTGCGGATGGTCAAAA
AGTCAATGCAAAAGAGAAGGAGCAAGCGGAAAAGCACGCTGCTAAGCTCTTCAACGGTAAAGAGGAGCTGCCAAGCGAGGTT
TTTCATGGCTATAAGGAACTCGCCCTGCGCATTCGTTTATGGTATCAGCAGGATCAGAAATGCCTGTATTCGGGTAAGGAGA
TTACCATTTCGGATCTAATCTATAACCGTGAACTGTTCGAAATTGATCACATTCTGCCGCTGTCTCTTAGCTTTGACGATAG
CCTGTCCAACAAGGTGCTGGTCTACTGCTGGGCAAATCAAGAAAAGGGCCAGCGTACCCCGTTTCAGGCGCTGGATAGCATG
AAATCTGCTTGGAGCTATCGCGAGTTTAAGAACGCTATATTACATAATTCCAAAATCTCCCGTAAGAAACGCGACTACTTCC
TCACTGAGCAAGATATCAGCAAAATCGAGGTTAAACAGAAGTTTATCGAGCGCAACCTGGTGGACACTCGCTATGCCTCTCG
TACGGTTCTCAACGTTTTGCAGCAGTCTTTGAAGAACCTGGAGAAAGAAACCAAAGTGAGCGTCGTGCGTGGTCAATTTACC
TCGCAATTGCGTCGTAAGTGGCACATTGATAAAACCCGTGACACATATCATCACCATGCAGTTGATGCGCTGATCATTGCAG
CGAGTGCCAAACTACGTTACTGGAAAAAACAGGGTGACATCCTGTTCGAGAACTACCTGATTAACCGTCATGTGGATCGTGT
TACCGGAGAAATCCAAAGCGACGACTCGTACAAGGAGGAAGTGTTCACCCCGCCTTATGACGGCTTTGTTCAGACCATCTCA
AACCCGGGTTTCGAGGATGAAATCCTGTTCTCCTACCAGGTTGATAGCAAGGTTAATCGTAAGATCTCCGATGCTACCATCT
ATGCTACTCGTAGCGCGAAGCTTGAAAAAGATAAGAAGGAACAAACGTACGTTCTGGGCAAGATTAAGGACATCTACAGCCA
GACCGGCTTTGAAAACTTCCTGAAGATCTATAATAAGGACAAAAGCAAGTTCTTGATTTACCAAAAGGATCCGGAAACCTGG
GAGAAAATTATCGAGCCGATTCTGAAGAACTATCGCGAATTCGACAACAAAGGCAAGGACATCGTGAACCCGTTTGAGAAAT
ACCGTAATGATAATGGCCCGATTTGTAAATATAGCCGTAAAGGCAATGGTCCGGAAATCAAACAGTTCAAATACTATGACAC
TGTTCTGGGTAAATACATTGAAATTACCCCGGAGAGCTCCCGCAACATTGTAGCACTGCGCTCTCTGAACCCGTGGAGAACG
GACGTTTATTTCAATGAGAAAACCTTGAAATACGAATTCCTGGGTCTGAAGTACTCTGATTTGAACATCCAAAATAAAGAGG
AATATGGCATCAGCGAAGAGAACTACCAGAAAATCAAGAAAGAGGAGGGCGTTAGCGAGGAAAGCATCTTTAAGTTCACTCT
GTACCGTAATGATCTAATCATCATTAAGGATCTGTCCAAAGGCAAAAGCCAGATTTTTCGTTTTTGGTCTCGTACGGGTAAA
GTTAACGAAAAGCACAAATTGGAATTGAAACCGTACAATAAGTCTTCCTTCGAAAAGGGTGAAAGCCTGGAGGTGATCCGCA
ACCTGTACCCGAGTTCCAACCAGTTTCAAGAGAAATTGAACAAGTCCGATTTGTCCATCTACAAACTGAAAACCGATATCCT
CGGCAACAAATTTATCAGTAAAAACGAGAACCTCTTCCCGAAACTGATCTATAAGAAGATCAAGCGCAAATTCAAACGTCCG
GCTGCGACCAAGAAGGCGGGTCAAGCGAAGAAAAAGAAG
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Средство разрезания ДНК на основе Cas9 белка из бактерии Streptococcus uberis NCTC3858 | 2022 |
|
RU2788197C1 |
| Средство разрезания ДНК на основе Cas9 белка из биотехнологически значимой бактерии Clostridium cellulolyticum | 2018 |
|
RU2712497C1 |
| СРЕДСТВО РАЗРЕЗАНИЯ ДНК НА ОСНОВЕ CAS9 БЕЛКА ИЗ DEFLUVIIMONAS SP. | 2018 |
|
RU2712492C1 |
| НУКЛЕАЗА CPF1 ИЗ БАКТЕРИИ Ruminococcus bromii, МОЛЕКУЛА ДНК ИЛИ РНК, КОДИРУЮЩАЯ НУКЛЕАЗУ, ВЕКТОР, СОДЕРЖАЩИЙ УКАЗАННУЮ МОЛЕКУЛУ ДНК, СИСТЕМА CRISPR/CPF1, СОДЕРЖАЩАЯ УКАЗАННУЮ НУКЛЕАЗУ И НАПРАВЛЯЮЩУЮ РНК, КЛЕТКА-ХОЗЯИН ДЛЯ ПОЛУЧЕНИЯ НУКЛЕАЗЫ CPF1, СПОСОБ ПОЛУЧЕНИЯ НУКЛЕАЗЫ CPF1 И ЕЕ ПРИМЕНЕНИЕ | 2022 |
|
RU2816876C1 |
| СРЕДСТВО РАЗРЕЗАНИЯ ДНК НА ОСНОВЕ CAS9 БЕЛКА ИЗ БАКТЕРИИ PASTEURELLA PNEUMOTROPICA | 2019 |
|
RU2722934C1 |
| ПРИМЕНЕНИЕ CAS9 БЕЛКА ИЗ БАКТЕРИИ PASTEURELLA PNEUMOTROPICA ДЛЯ МОДИФИКАЦИИ ГЕНОМНОЙ ДНК В КЛЕТКАХ | 2019 |
|
RU2724470C1 |
| СРЕДСТВО РАЗРЕЗАНИЯ ДНК НА ОСНОВЕ CAS9 БЕЛКА ИЗ БАКТЕРИИ DEMEQUINA SEDIMINICOLA | 2019 |
|
RU2722933C1 |
| Средство разрезания ДНК на основе ScCas12a белка из бактерии Sedimentisphaera cyanobacteriorum | 2022 |
|
RU2791447C1 |
| Средство разрезания ДНК на основе Cas9 белка из бактерии Capnocytophaga ochracea | 2021 |
|
RU2778156C1 |
| Средство разрезания двунитевой ДНК с помощью Cas12d белка из Katanobacteria и гибридной РНК, полученной путем слияния направляющей CRISPR РНК и scout РНК | 2020 |
|
RU2771626C1 |
Группа изобретений относится к биотехнологии. Представлены: система редактирования на основе нуклеотидной последовательности, кодирующая белок SuCas9_NLS, в качестве инструмента для внесения целевых двунитевых разрывов в ДНК эукариотической клетки для редактирования генома человека, экспрессионный вектор и способ редактирования последовательности геномной ДНК эукариотической клетки. Система редактирования генома согласно настоящему изобретению имеет важное значение для генной терапии наследственных заболеваний человека. 3 н. и 3 з.п. ф-лы, 4 ил., 2 табл., 2 пр.
1. Система редактирования последовательности геномной ДНК эукариотической клетки, содержащая
(1) нуклеотидную последовательность SEQ ID NO: 2, кодирующую белок SuCas9_NLS с аминокислотной последовательностью SEQ ID NO: 1,
(2) нуклеотидную последовательность, кодирующую направляющую РНК, взаимодействующую с указанным белком и содержащую последовательность, образующую дуплекс с нуклеотидной последовательностью участка геномной ДНК организма, непосредственно примыкающей к нуклеотидной последовательности PAM 5'-NNAAА-3',
где указанная система редактирования приводит к образованию двунитевого разрыва в последовательности геномной ДНК эукариотической клетки, непосредственно примыкающей к последовательности PAM 5'- NNAAА-3',
где указанная эукариотическая клетка не является клеткой зародышевой линии клеток человек.
2. Система редактирования по п.1, где эукариотическая клетка представляет собой клетку человека.
3. Экспрессионный вектор для редактирования последовательности геномной ДНК эукариотической клетки, несущий систему редактирования по п.1 и регуляторный элемент, способный к функционированию в эукариотической клетке, имеющий элементы и их расположение, как представлено на фиг. 2, где указанная эукариотическая клетка не является клеткой зародышевой линии клеток человека.
4. Экспрессионный вектор по п.3, где эукариотическая клетка представляет собой клетку человека.
5. Способ редактирования последовательности геномной ДНК эукариотической клетки, предусматривающий введение в указанную клетку эффективного количества системы редактирования по п.1 или экспрессионного вектора по п.3, где указанная эукариотическая клетка не является клеткой зародышевой линии клеток человека.
6. Способ по п.5, где эукариотическая клетка представляет собой клетку человека.
| RAN F | |||
| A., CONG L., YAN W | |||
| X., SCOTT D | |||
| A., GOOTENBERG J | |||
| S., KRIZ A | |||
| J., ZETSCHE B., SHALEM O., WU X., MAKAROVA K | |||
| S., KOONIN E | |||
| V., SHARP P | |||
| A | |||
| & ZHANG F | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| Устройство для закрепления лыж на раме мотоциклов и велосипедов взамен переднего колеса | 1924 |
|
SU2015A1 |
| - Т | |||
| Способ обогащения руд флотацией (отпениванием) с прибавлением масла | 1917 |
|
SU520A1 |
| АВТОМАТИЧЕСКИЙ ПРИЕМЩИК ДЛЯ ОТПЕЧАТАННЫХ ЛИСТОВ В ЛИТОГРАФСКОЙ МАШИНЕ | 1926 |
|
SU7546A1 |
| - С | |||
| Пружинная погонялка к ткацким станкам | 1923 |
|
SU186A1 |
| SHI Y.-J., DUAN M., DING J.-M., WANG F.-Q., BI L.-L., ZHANG C.-X., ZHANG Y.-Z., DUAN | |||
