ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Данная заявка заявляет приоритет согласно заявке на патент США №62/829,327, поданной 4 апреля 2019 г., которая включена в данный документ в полном объеме посредством ссылки для всех целей.
ССЫЛКА НА СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ ПРЕДСТАВЛЕННА В ТЕКСТОВОМ ФАЙЛЕ ПОСРЕДСТВОМ EFS WEB
[0002] Список последовательностей, записанный в файле 544999SEQLIST.txt, имеет объем 20,7 килобайт, был создан 21 марта 2020 г. и включен в данный документ посредством ссылки.
УРОВЕНЬ ТЕХНИКИ
[0003] Бесшовная конструкция ДНК имеет особое значение при создании линий трансгенных животных, поскольку швы, образованные сайтами рестрикции или путем других манипуляций, могут отрицательно повлиять на экспрессию генов, если они попадают в область, важную для регуляции. Нацеливание на геном млекопитающих часто требует конструирования больших направленных векторов с длинными плечами ДНК для прямой гомологичной рекомбинации, а также кассет устойчивости к антибиотикам для отбора клонов эмбриональных стволовых клеток. Правильно нацеленные клоны часто содержат несколько швов, необходимых для конструирования вектора, не говоря уже о самой кассете устойчивости. Для генетической абляции эти повреждения могут не иметь значения для конечного результата (нулевой аллель), но всегда есть шанс, что на экспрессию соседних генов будет оказано неблагоприятное воздействие. Для модификаций, отличных от нокаута (knock-out), таких как «нокин» (knock-in), точная экспрессия целевого локуса обычно важна для рассматриваемых исследований.
[0004] В частности, гуманизация, прямая замена мышиного гена его человеческим аналогом, требует бесшовных соединений между мышиной и человеческой последовательностью, чтобы мышиный аппарат транскрипции точно воспроизводил экспрессию нового аллеля. Следует соблюдать осторожность, чтобы скрыть швы в конструкции и кассету селекции в некодирующих областях, которые не влияют на регуляцию генов. По мере того, как животные модели становятся более сложными, к уже существующим могут быть добавлены дополнительные модификации, такие как мутации, вызывающие болезни человека, поверх гуманизированных аллелей. Дополнительные изменения могут затем добавить еще больше швов и другую кассету селекции к уже хорошо сконструированному мышиному локусу, увеличивая вероятность того, что экспрессия будет изменена, и мышиная модель не будет соответствовать заболеванию человека. С точки зрения конструирования, добавление новой кассеты к вектору, уже содержащему ее, может усложняться из-за нежелательной рекомбинации между общими элементами кассеты, такими как промоторы и поли(А)-сигналы, даже если две кассеты кодируют разные наборы. Следовательно, необходимы новые способы, чтобы упростить создание таргетинга, несущего множественные изменения (такие как гуманизированный аллель и наслоенная мутация болезни), и минимизировать швы, включенные в окончательную животную модель.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0005] Предложены способы бесшовного введения целевой генетической модификации в ранее созданный направленный вектор.
[0006] В одном аспекте некоторые такие способы включают: (а) выполнение бактериальной гомологичной рекомбинации между изначальным направленным вектором и кассетой модификации в популяции бактериальных клеток, при этом кассета модификации содержит целевую генетическую модификацию и включает вставку нуклеиновой кислоты, фланкированную 5'-плечом гомологии, соответствующим 5'-последовательности-мишени в изначальном направленном векторе и 3'-плечом гомологии, соответствующим 3'-последовательности-мишени в изначальном векторе, при этом вставка нуклеиновой кислоты содержит от 5' к 3': (i) первую повторяющуюся последовательность; (ii) первый сайт-мишень для первого нуклеазного агента; (iii) кассету селекции; (iv) второй сайт-мишень для второго нуклеазного агента; и (v) вторую повторяющуюся последовательность, идентичную первой повторяющейся последовательности; (b) отбор бактериальных клеток, содержащих модифицированный направленный вектор, содержащий кассету селекции; (с) расщепление первого сайта-мишени в модифицированном направленном векторе первым нуклеазным агентом и расщепление второго целевого сайта в модифицированном направленном векторе вторым нуклеазным агентом для удаления кассеты селекции и выявления первой повторяющейся последовательности и второй повторяющейся последовательности в модифицированном направленном векторе; и (d) соединение экспонированной первой повторяющейся последовательности с экспонированной второй повторяющейся последовательностью во внутримолекулярной реакции сборки in vitro для создания направленного вектора, содержащего бесшовную целевую генетическую модификацию, при этом ни первый сайт-мишень для первого нуклеазного агента, ни второй сайт-мишень для второго нуклеазного агента не присутствуют, и только одна копия повторяющейся последовательности присутствует в направленном векторе, содержащем бесшовную целевую генетическую модификацию.
[0007] В некоторых таких способах повторяющаяся последовательность идентична последовательности в ранее созданном направленном векторе. В некоторых таких способах целевая генетическая модификация включает вставку, а повторяющаяся последовательность идентична 5'-концу или 3'-концу вставки.
[0008] В некоторых таких способах длина повторяющейся последовательности составляет, по меньшей мере, около 20 нуклеотидов. Необязательно, длина повторяющейся последовательности составляет от около 20 нуклеотидов до около 100 нуклеотидов.
[0009] В некоторых таких способах кассета модификации представляет собой линейную двухцепочечную нуклеиновую кислоту. В некоторых таких способах кассета модификации имеет длину от около 1 т.п.н. до около 15 т.п.н. В некоторых таких способах каждое 5'-плечо гомологии и 3'-плечо гомологии имеет длину, по меньшей мере, около 35 нуклеотидов. В некоторых таких способах каждое 5'-плечо гомологии и 3'-плечо гомологии имеет длину от около 35 нуклеотидов до около 500 нуклеотидов.
[0010] В некоторых таких способах первый нуклеазный агент и/или второй нуклеазный агент представляет собой редко-расщепляющий нуклеазный агент. В некоторых таких способах первый сайт-мишень и/или второй сайт-мишень не присутствует в ранее созданном направленном векторе. В некоторых таких способах первый сайт-мишень идентичен второму сайту-мишени, и первый нуклеазный агент идентичен второму нуклеазному агенту.
[0011] В некоторых таких способах первый нуклеазный агент и/или второй нуклеазный агент включает редко-расщепляющий рестрикционный фермент. Необязательно, редко-расщепляющий рестрикционный фермент представляет собой NotI, XmaIII, SstII, Sall, NruI, NheI, Nb.BbvCI, BbvCI, AscI, AsiSI, FseI, PacI, PmeI, SbfI, SgrAI, SwaI, BspQI, SapI, SfiI, CspCI, AbsI, CciNI, FspAI, MauBI, MreI, MssI, PalAI, RgaI, RigI, SdaI, SfaAI, SgfI, SgrDI, SgsI, SmiI, SrfI, Sse2321, Sse83871, LguI, PciSI, AarI, AjuI, AloI, BarI, PpiI или PsrI.
[0012] В некоторых таких способах первый нуклеазный агент и/или второй нуклеазный агент представляет собой белок (Cas), ассоциированный с кластеризованными регулярными промежуточными короткими палиндромными повторами (CRISPR, Clustered Regularly Interspaced Short Palindromic Repeats) и направляющую РНК (нРНК, gRNA, guide RNA), нуклеазу с "цинковыми пальцами» (ZFN, zinc finger nuclease), эффекторную нуклеазу, подобную активаторам транскрипции (TALEN, Transcription Activator-Like Effector Nuclease) или сконструированную мегануклеазу. Необязательно, первый нуклеазный агент и/или второй нуклеазный агент представляет собой белок Cas и нРНК, при этом белок Cas представляет собой Cas9, и при этом нРНК содержит CRISPR-PHK (крРНК, crRNA), которая направлена, и транс-активирующую CRIPSR-PHK (тракрРНК, tracrRNA).
[0013] В некоторых таких способах целевая генетическая модификация содержит модификацию в 5'-плече гомологии или в 3'-плече гомологии. В некоторых таких способах целевая генетическая модификация содержит модификацию вставки нуклеиновой кислоты. В некоторых таких способах целевая генетическая модификация содержит точечную мутацию, делецию, вставку, замену или их комбинацию.
[0014] В некоторых таких способах кассета селекции придает устойчивость к антибиотику. Необязательно, кассета селекции придает устойчивость к ампициллину, хлорамфениколу, тетрациклину, канамицину, спектиномицину, стрептомицину, карбенициллину, блеомицину, эритромицину или полимиксину В.
[0015] В некоторых таких способах ранее созданный направленный вектор представляет собой большой направленный вектор длиной, по меньшей мере, около 10 т.п.н. Необязательно, ранее созданный направленный вектор имеет длину, по меньшей мере, около 100 т.п.н.
[0016] В некоторых таких способах ранее созданный направленный вектор содержит вторую кассету селекции. Необязательно, вторая кассета селекции придает устойчивость к антибиотику. Необязательно, кассета селекции в кассете модификации и вторая кассета селекции в ранее созданном направленном векторе придает устойчивость к другому антибиотику. Необязательно, вторая кассета селекции позволяет производить селекцию как в бактериальных клетках, так и в клетках млекопитающих.
[0017] В некоторых таких способах этап (с) выполняется in vitro.
[0018] В некоторых таких способах этап (d) включает: (i) контактирование модифицированного направленного вектора с экзонуклеазой для выявления комплементарных последовательностей между первой повторяющейся последовательностью и второй повторяющейся последовательностью; (ii) гибридизация экспонированных дополнительных последовательностей; (iii) удлинение 3'-концов гибридизированных дополнительных последовательностей; и (iv) лигирование гибридизированной комплементарной последовательности. Необязательно, этап (d) включает инкубирование модифицированного направленного вектора с экзонуклеазой, ДНК-полимеразой и ДНК-лигазой.
[0019] Некоторые такие способы дополнительно включают: (е) обработку направленного вектора первым нуклеазным агентом и вторым нуклеазным агентом после сборки in vitro на этапе (d), чтобы убедиться, что ни первый сайт-мишень для первого нуклеазного агента, ни второй сайт-мишень для второго нуклеазного агента не присутствуют.
[0020] В другом аспекте некоторые такие способы включают: (а) выполнение бактериальной гомологичной рекомбинации между ранее созданном направленным вектором и кассетой делеции в популяции бактериальных клеток, при этом кассета делеции содержит вставку нуклеиновой кислоты, фланкированную 5'-плечом гомологии, соответствующим 5'-последовательности-мишени в ранее созданном направленном векторе и 3'-плечом гомологии, соответствующим 3'-последовательности-мишени в ранее созданном векторе, при этом 5'-последовательность-мишень и 3'-последовательность-мишень фланкируют область ранее созданного направленного вектора, в которую должна быть введена целевая генетическая модификация, при этом нуклеиновая кислота вставки содержит от 5' к 3': (i) первый сайт-мишень для первого нуклеазного агента; (ii) кассету селекции; и (iii) второй сайт-мишень для второго нуклеазного агента; (b) отбор бактериальных клеток, содержащих модифицированный направленный вектор, содержащий кассету селекции; (с) расщепление первого сайта-мишени в модифицированном направленном векторе первым нуклеазным агентом и расщепление второго сайта-мишени в модифицированном направленном векторе вторым нуклеазным агентом для удаления кассеты селекции и экспонирования концевой последовательности, расположенной в направлении 5', и концевой последовательности, расположенной в направлении 3', в модифицированном направленном векторе; и (d) соединение в реакции сборки in vitro расщепленного направленного вектора с кассетой модификации, содержащей целевую генетическую модификацию, фланкированную расположенной в направлении 5' концевой последовательностью, перекрывающей расположенную в направлении 5' концевую последовательность в модифицированном направленном векторе, и расположенной в направлении 3' концевой последовательностью, перекрывающей расположенную в направлении 3' концевую последовательность в модифицированном направленном векторе для создания направленного вектора, содержащего бесшовную целевую генетическую модификацию, при этом ни первый сайт-мишень для первого нуклеазного агента, ни второй целевой сайт-мишень для второго нуклеазного агента не присутствуют в направленном векторе, содержащем бесшовную целевую генетическую модификацию.
[0021] В некоторых таких способах кассета делеции имеет длину от около 1 до около 15 т.п.н. В некоторых таких способах каждое 5'-плечо гомологии и 3'-плечо гомологии имеет длину, по меньшей мере, около 35 нуклеотидов. Необязательно, длина каждого 5'-плеча гомологии и 3'-плеча гомологии составляет от около 35 нуклеотидов до около 500 нуклеотидов. В некоторых таких способах кассета делеции представляет собой линейную двухцепочечную нуклеиновую кислоту.
[0022] В некоторых таких способах первый нуклеазный агент и/или второй нуклеазный агент представляет собой редко-расщепляющий нуклеазный агент. В некоторых таких способах первый сайт-мишень и/или второй сайт-мишень не присутствует в ранее созданном направленном векторе. В некоторых таких способах первый сайт-мишень идентичен второму сайту-мишени, и первый нуклеазный агент идентичен второму нуклеазному агенту.
[0023] В некоторых таких способах первый нуклеазный агент и/или второй нуклеазный агент включает редко-расщепляющий рестрикционный фермент. Необязательно, редко-расщепляющий рестрикционный фермент представляет собой NotI, XmaIII, SstII, Sall, NruI, NheI, Nb.BbvCI, BbvCI, AscI, AsiSI, FseI, PacI, PmeI, SbfI, SgrAI, SwaI, BspQI, SapI, SfiI, CspCI, AbsI, CciNI, FspAI, MauBI, MreI, MssI, PalAI, RgaI, RigI, SdaI, SfaAI, SgfI, SgrDI, SgsI, SmiI, SrfI, Sse2321, Sse83871, LguI, PciSI, AarI, AjuI, AloI, BarI, PpiI или PsrI.
[0024] В некоторых таких способах первый нуклеазный агент и/или второй нуклеазный агент представляет собой белок (Cas), ассоциированный с кластеризованными регулярными промежуточными короткими палиндромными повторами (CRISPR, Clustered Regularly Interspaced Short Palindromic Repeats) и направляющую РНК (нРНК, gRNA, guide RNA), нуклеазу с "цинковыми пальцами» (ZFN, zinc finger nuclease), эффекторную нуклеазу, подобную активаторам транскрипции (TALEN, Transcription Activator-Like Effector Nuclease) или сконструированную мегануклеазу. Необязательно, первый нуклеазный агент и/или второй нуклеазный агент представляет собой белок Cas и нРНК, при этом белок Cas представляет собой Cas9, и при этом нРНК содержит CRISPR-PHK (крРНК, crRNA), которая направлена, и транс-активирующую CRIPSR-PHK (тракрРНК, tracrRNA).
[0025] В некоторых таких способах кассета селекции придает устойчивость к антибиотику. Необязательно, кассета селекции придает устойчивость к ампициллину, хлорамфениколу, тетрациклину, канамицину, спектиномицину, стрептомицину, карбенициллину, блеомицину, эритромицину или полимиксину В.
[0026] В некоторых таких способах ранее созданный направленный вектор представляет собой большой направленный вектор длиной, по меньшей мере, 10 т.п.н. Необязательно, ранее созданный направленный вектор имеет длину, по меньшей мере, 100 т.п.н.
[0027] В некоторых таких способах ранее созданный направленный вектор содержит вторую кассету селекции. Необязательно, вторая кассета селекции придает устойчивость к антибиотику. Необязательно, кассета селекции в кассете делеции и вторая кассета селекции в ранее созданном направленном векторе придает устойчивость к другому антибиотику. Необязательно, вторая кассета селекции позволяет производить селекцию как в бактериальных клетках, так и в клетках млекопитающих.
[0028] В некоторых таких способах длина перекрывания между концевой последовательностью, расположенной в направлении 5' в кассете модификации и концевой последовательностью, расположенной в направлении 5' в модифицированном направленном векторе и/или длина перекрывания между концевой последовательностью, расположенной в направлении 3' в кассете модификации и концевой последовательность, расположенной в направлении 3' в модифицированном направленном векторе составляет, по меньшей мере, около 20 нуклеотидов. В некоторых таких способах длина перекрывания между концевой последовательностью, расположенной в направлении 5' в кассете модификации и концевой последовательностью, расположенной в направлении 5' в модифицированном направленном векторе и/или длина перекрывания между концевой последовательностью, расположенной в направлении 3' в кассете модификации и концевой последовательность, расположенной в направлении 3' в модифицированном направленном векторе составляет, по меньшей мере, от около 20 до около 100 нуклеотидов.
[0029] В некоторых таких способах этап (с) при этом выполняется in vitro.
[0030] В некоторых таких способах этап (d) включает: (i) контактирование расщепленного направленного вектора и кассеты модификации с экзонуклеазой для открытия комплементарных последовательностей между концевыми последовательностями в модифицированном направленном векторе и концевыми последовательностями в кассете модификации; (ii) гибридизация экспонированных дополнительных последовательностей; (iii) удлинение 3'-концов гибридизированных дополнительных последовательностей; и (iv) лигирование гибридизированной комплементарной последовательности. Необязательно, этап (d) включает инкубирование расщепленного направленного вектора и кассеты модификации с экзонуклеазой, ДНК-полимеразой и ДНК-лигазой.
[0031] В некоторых таких способах кассета модификации представляет собой линейную двухцепочечную нуклеиновую кислоту. В некоторых таких способах кассета модификации имеет длину, по меньшей мере, около 200 нуклеотидов. В некоторых таких способах кассета модификации может быть такого размера, что не может быть синтезирована напрямую или получена путем полимеразной цепной реакцией. В некоторых таких способах кассета модификации имеет длину, по меньшей мере, около 10 т.п.н.
[0032] В некоторых таких способах целевая генетическая модификация содержит точечную мутацию, делецию, вставку, замену или их комбинацию.
[0033] Некоторые такие способы дополнительно включают: (е) обработку направленного вектора первым нуклеазным агентом и вторым нуклеазным агентом после сборки in vitro на этапе (d), чтобы убедиться, что ни первый сайт-мишень для первого нуклеазного агента, ни второй сайт-мишень для второго нуклеазного агента не присутствуют.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0034] На Фиг. 1 (не в масштабе) представлена схема способа бесшовного внесения точечной мутации в большой направленный вектор посредством бактериальной гомологичной рекомбинации и внутримолекулярной сборки по Гибсону.
[0035] На Фиг. 2 (не в масштабе) представлена схема синтезированной нуклеиновой кислоты, которая будет использоваться в способе, изображенном в схеме на Фиг. 1.
[0036] На Фиг. 3 (не в масштабе) представлена схема способа бесшовного внесения точечной мутации в большой направленный вектор посредством бактериальной гомологичной рекомбинации и межмолекулярной сборки по Гибсону.
[0037] На Фиг. 4А-4В представлена традиционная целевая стратегия с использованием модифицированных мышей линии ВАС в качестве векторов и технологии самоудаления кассет, от создания вектора (Фиг. 4А) до поколения F1 мыши (Фиг. 4В). Удаление кассеты с помощью рекомбиназы Сге, управляемой мышиным протамином, оставляет шов длиной 78 п.н., содержащий единственный 1охР.
ОПРЕДЕЛЕНИЯ
[0038] Термины «белок», «полипептид» и «пептид», используемые в контексте данного документа взаимозаменяемо, включают полимерные формы аминокислот любой длины, включая кодируемые и некодируемые аминокислоты, и химически или биохимически модифицированные либо дериватизированные аминокислоты. Данные термины также включают в себя модифицированные полимеры, такие как полипептиды, имеющие модифицированные пептидные каркасы. Термин «домен» относится к любой части белка или полипептида, имеющей конкретную функцию или структуру.
[0039] Термины «нуклеиновая кислота» и «полинуклеотид», используемые в контексте данного документа взаимозаменяемо, включают полимерные формы нуклеотидов любой длины, включая рибонуклеотиды, дезоксирибонуклеотиды или их аналоги либо их модифицированные версии. Они включают одно-, двух- и многоцепочечные ДНК или РНК, геномную ДНК, кДНК, гибриды ДНК-РНК и полимеры, содержащие пуриновые основания, пиримидиновые основания или другие встречающиеся в природе, химически модифицированные, биохимически модифицированные, не встречающиеся в природе или дериватизированные нуклеотидные основания.
[0040] Термин «направленный вектор» относится к рекомбинантной нуклеиновой кислоте, которая может быть введена путем гомологичной рекомбинации, лигирования, опосредованного негомологичным соединением концов, или любым другим способом рекомбинации в целевое положение в геноме клетки.
[0041] Термин «дикий тип» включает единицы, имеющие структуру и/или активность, которые наблюдаются в нормальном (в отличие от мутантного, болезненного, измененного и т.д.) состоянии или контексте. Гены и полипептиды дикого типа часто существуют в нескольких различных формах (например, аллелях).
[0042] Термин «эндогенная последовательность» относится к последовательности нуклеиновой кислоты, которая встречается в природе внутри клетки или животного, не являющегося человеком. Например, эндогенная последовательность Rosa26 животного, не являющегося человеком, относится к нативной последовательности Rosa26, которая естественным образом встречается в локусе Rosa26 животного, не являющегося человеком.
[0043] «Экзогенные» молекулы или последовательности включают молекулы или последовательности, которые обычно не присутствуют в клетке в этой форме или локализации (например, в геномном локусе). Нормальное присутствие включает наличие в зависимости от конкретной стадии развития и условий окружающей среды клетки. Экзогенная молекула или последовательность, например, может включать мутированную версию соответствующей эндогенной последовательности внутри клетки, такую как гуманизированная версия эндогенной последовательности, или может включать последовательность, соответствующую эндогенной последовательности внутри клетки, но в другой форма (т.е. не в хромосоме). Напротив, эндогенные молекулы или последовательности включают молекулы или последовательности, которые обычно присутствуют в этой форме и локализации в конкретной клетке на конкретной стадии развития в определенных условиях окружающей среды.
[0044] Термин «гетерологичный» при использовании в контексте нуклеиновой кислоты или белка указывает на то, что нуклеиновая кислота или белок содержит, по меньшей мере, два сегмента, которые в природе не встречаются вместе в одной и той же молекуле. Например, термин «гетерологичный», когда он используется в отношении сегментов нуклеиновой кислоты или сегментов белка, указывает на то, что нуклеиновая кислота или белок содержит две или более подпоследовательностей, которые не находятся в подобной зависимости друг с другом (например, не соединены вместе) в природе. В качестве одного примера, «гетерологичная» область вектора нуклеиновой кислоты представляет собой сегмент нуклеиновой кислоты внутри молекулы или сегмент присоединенный к другой молекуле нуклеиновой кислоты, который не обнаруживается в ассоциации с другой молекулой в природе. Например, гетерологичная область вектора нуклеиновой кислоты может включать кодирующую последовательность, фланкированную последовательностями, не обнаруженными в ассоциации с кодирующей последовательностью в природе. Аналогичным образом, «гетерологичная» область белка представляет собой сегмент аминокислот внутри молекулы или присоединенный к другой молекуле пептида, который не обнаруживается в ассоциации с другой молекулой пептида в природе (например, слитый белок или белок с меткой). Точно так же нуклеиновая кислота или белок могут содержать гетерологичную метку или гетерологичную последовательность секреции или локализации.
[0045] «Оптимизация кодонов» использует преимущество вырожденности кодонов, что демонстрируется множеством комбинаций кодонов из трех пар оснований, которые определяют аминокислоту, и обычно включает процесс модификации последовательности нуклеиновой кислоты для усиления экспрессии в конкретных клетках-хозяевах путем замены, по меньшей мере, одного кодона нативной последовательности на кодон, который чаще или наиболее часто используется в генах клетки-хозяина, при сохранении нативной аминокислотной последовательности. Например, нуклеиновая кислота, кодирующая белок Cas9, может быть модифицирована для замены кодонов, имеющих более высокую частоту использования в данной прокариотической или эукариотической клетке, включая бактериальную клетку, дрожжевую клетку, человеческую клетку, клетку, которая не является человеческой, клетку млекопитающего, клетку грызуна, клетку мыши, клетку крысы, клетку хомяка или любую другую клетку-хозяин, по сравнению с встречающейся в природе последовательностью нуклеиновой кислоты. Таблицы использования кодонов легко доступны, например, в «Базе Данных Использования Кодонов» (Codon Usage Database). Эти таблицы можно адаптировать разными способами. См. Nakamura et al. (2000) Nucleic Acids Res. 28:292, включенную в данный документ в полном объеме посредством ссылки для всех целей. Также доступны компьютерные алгоритмы для оптимизации кодонов конкретной последовательности для экспрессии в конкретном хозяине (см., например, Gene Forge).
[0046] Термин «локус» относится к конкретному расположению гена (или значимой последовательности), последовательности ДНК, последовательности, кодирующей полипептид, или положению на хромосоме генома организма. Например, локус Rosa26 может относиться к определенному расположению гена Rosa26, последовательности ДНК Rosa26 или положение Rosa26 на хромосоме генома организма, которое было идентифицировано относительно того, где находится такая последовательность. «Локус Rosa26» может содержать регуляторный элемент гена Rosa26, включая, например, энхансер, промотор, 5'- и/или 3'-нетранслируемую область (UTR) или их комбинацию.
[0047] Термин «ген» относится к последовательности ДНК в хромосоме, которая кодирует продукт (например, продукт РНК и/или продукт полипептида) и содержит кодирующую область, прерванную некодирующими нитронами, и последовательность, расположенную рядом с кодирующей областью на обоих 5'- и 3'-концах, так что ген соответствует полноразмерной мРНК (включая 5' и 3' нетранс л ируемые последовательности). Термин «ген» также включает другие некодирующие последовательности, включая регуляторные последовательности (например, промоторы, энхансеры и сайты связывания факторов транскрипции), сигналы полиаденилирования, сайты внутренней посадки рибосомы, сайленсеры, изолирующие последовательности и участки прикрепления к матриксу. Эти последовательности могут быть близки к кодирующей области гена (например, в пределах 10 т.п.н.) или находиться в отдаленных сайтах, и они влияют на уровень или скорость транскрипции и трансляции гена.
[0048] «Промотор» представляет собой регуляторную область ДНК, обычно содержащую ТАТА-бокс, способный управлять РНК-полимеразой II, чтобы инициировать синтез РНК в соответствующем сайте инициации транскрипции для конкретной полинуклеотидной последовательности. Промотор может дополнительно содержать другие области, которые влияют на скорость инициации транскрипции. Описанные в данном документе промоторные последовательности модулируют транскрипцию функционально связанного полинуклеотида. Промотор может быть активным в одном или более типах клеток, описанных в данном документе (например, в прокариотической клетке или эукариотической клетке (такой как клетка млекопитающего) или их комбинации). Промотор может представлять собой, например, конститутивно активный промотор, условный промотор, индуцибельный промотор, временно ограниченный промотор (например, промотор, регулируемым развитием) или пространственно ограниченный промотор (например, клеточно-специфический или тканеспецифический промотор). Примеры промоторов можно найти, например, в WO 2013/176772, которая включена в данный документ в полном объеме посредством ссылки для всех целей.
[0049] Термин «функциональная связь» или «функционально связанный (-ая, -ые)» включает контактное расположение двух или более компонентов (например, промотора и другого элемента последовательности), таким образом, что оба компонента функционируют нормально и допускают возможность того, что, по меньшей мере, один из компонентов может опосредовать функцию, которая воздействует на, по меньшей мере, один из других компонентов. Например, промотор может быть функционально связан с кодирующей последовательностью, если промотор контролирует уровень транскрипции кодирующей последовательности в ответ на присутствие или отсутствие одного или более факторов регуляции транскрипции. Функциональная связь может включать в себя случаи, когда такие последовательности расположены смежно друг с другом или действуют в транс-положении (например, регуляторная последовательность может действовать на расстоянии, чтобы контролировать транскрипцию кодирующей последовательности).
[0050] «Комплементарность» нуклеиновых кислот означает, что нуклеотидная последовательность в одной цепи нуклеиновой кислоты из-за ориентации ее групп азотистых оснований образует водородные связи с другой последовательностью на противоположной цепи нуклеиновой кислоты. Комплементарные основания в ДНК обычно представляют собой А с Τ и С с G. В РНК они обычно представляют собой С с G и U с А. Комплементарность может быть совершенной или значительной/достаточной. Совершенная комплементарность между двумя нуклеиновыми кислотами означает, что две нуклеиновые кислоты могут образовывать дуплекс, в котором каждое основание в дуплексе связано с комплементарным основанием путем спаривания по Уотсону-Крику. «Значительная» или «достаточная» комплементарность означает, что последовательность в одной цепи не полностью и/или не совершенно комплементарна последовательности в противоположной цепи, но что между основаниями на двух цепях происходит достаточное связывание для образования стабильного гибридного комплекса в наборе условий гибридизации (например, концентрации соли и температуры). Такие условия можно предсказать с помощью последовательностей и стандартных математических расчетов для прогнозирования Тm (температуры плавления) гибридизированных цепей или путем эмпирического определения Тm с использованием обычных способов. Тm включает температуру, при которой популяция гибридизационных комплексов, образованных между двумя цепями нуклеиновой кислоты, денатурируется на 50% (т.е. популяция двухцепочечных молекул нуклеиновой кислоты становится наполовину диссоциированной на одноцепочечные). При температуре ниже Тm преимущественно происходит образование гибридизационного комплекса, тогда как при температуре выше Тm преимущественно происходит плавление или разделение цепей в гибридизационном комплексе. Тm можно оценить для нуклеиновой кислоты, имеющей известное содержание G+C в водном 1М растворе NaCl, используя, например, Tm=81,5+0,41(% G+C), хотя другие известные вычисления Тm учитывают структурные характеристики нуклеиновой кислоты.
[0051] Для гибридизации требуется, чтобы две нуклеиновые кислоты содержали комплементарные последовательности, хотя возможны несовпадения между основаниями. Условия, подходящие для гибридизации двух нуклеиновых кислот, зависят от длины нуклеиновых кислот и степени комплементарности, переменных, которые хорошо известны. Чем выше степень комплементарности между двумя нуклеотидными последовательностями, тем больше будет значение температуры плавления (Тm) для гибридов нуклеиновых кислот, имеющих эти последовательности. Для гибридизации нуклеиновых кислот с короткими участками комплементарности (например, комплементарность 35 или менее, 30 или менее, 25 или менее, 22 или менее, 20 или менее или 18 или менее нуклеотидов) положение несовпадений становится важным (см. Sambrook et al. al., цитировано выше, 11.7-11.8). Обычно длина гибридизируемой нуклеиновой кислоты составляет, по меньшей мере, около 10 нуклеотидов. Иллюстративные минимальные длины гибридизируемой нуклеиновой кислоты включают, по меньшей мере, около 15 нуклеотидов, по меньшей мере, около 20 нуклеотидов, по меньшей мере, около 22 нуклеотида, по меньшей мере, около 25 нуклеотидов и, по меньшей мере, около 30 нуклеотидов. Кроме того, температуру и концентрацию соли в промывочном растворе можно регулировать по мере необходимости в соответствии с такими факторами, как длина области комплементарности и степень комплементарности.
[0052] Последовательность полинуклеотида не обязательно должна быть на 100% комплементарной последовательности его нуклеиновой кислоты-мишени для специфической гибридизации. Более того, полинуклеотид может гибридизоваться с одним или более сегментами, так что промежуточные или соседние сегменты не участвуют в событии гибридизации (например, петлевая структура или шпилечная структура). Полинуклеотид (например, нРНК) может содержать, по меньшей мере, 70%-ную, по меньшей мере, 80%-ную, по меньшей мере, 90%-ную, по меньшей мере, 95%-ную, по меньшей мере, 99%-ную или 100%-ную комплементарность последовательности к области-мишени в последовательности нуклеиновой кислоты-мишени, на которую они нацелены. Например, нРНК, в которой 18 из 20 нуклеотидов комплементарны области-мишени и поэтому специфически гибридизуются, будет представлять 90%-ную комплементарность. В этом примере оставшиеся некомплементарные нуклеотиды могут быть сгруппированы или перемежены с комплементарными нуклеотидами и не обязательно должны быть смежными друг с другом или с комплементарными нуклеотидами.
[0053] Процент комплементарности между конкретными участками последовательностей нуклеиновых кислот внутри нуклеиновых кислот может быть определена обычным способом с использованием программ BLAST (основные инструменты поиска локального сопоставления) и программ PowerBLAST (Altschul et al. (1990) J. Mol. Biol. 215:403-410; Zhang and Madden (1997) Genome Res. 7:649-656, каждая из которых включена в данный документ в полном объеме посредством ссылки для всех целей) или с использованием программы Gap (Wisconsin Sequence Analysis Package, версия 8 для Unix, Genetics Computer Group, University Research Park, Мэдисон, Висконсин), используя настройки по умолчанию, в которых используется алгоритм Смита - Ватермана Smith and Waterman (1981) Adv. Appl. Math. 2:482-489, который включен в данный документ в полном объеме посредством ссылки для всех целей.
[0054] В способах и композициях, предложенных в данном документе, используется множество различных компонентов. Некоторые компоненты в описании могут иметь активные варианты и фрагменты. Такие компоненты включают, например, белки Cas, CRISPR-PHK, тракрРНК и направляющие РНК. Биологическая активность каждого из этих компонентов описана в данном документе в другом месте. Термин «функциональный» относится к врожденной способности белка или нуклеиновой кислоты (или их фрагмента или варианта) проявлять биологическую активность или функцию. Такие биологические активности или функции могут включать, например, способность белка Cas связываться с направляющей РНК и с последовательностью ДНК-мишени. Биологические функции функциональных фрагментов или вариантов могут быть такими же или могут фактически быть изменены (например, в отношении их специфичности, селективности или эффективности) по сравнению с исходными, но с сохранением основной биологической функции.
[0055] Термин «вариант» относится к нуклеотидной последовательности, отличающейся от последовательности, наиболее распространенной в популяции (например, на один нуклеотид), или к последовательности белка, отличной от последовательности, наиболее распространенной в популяции (например, на одну аминокислоту).
[0056] Термин «фрагмент» применительно к белку означает белок, который короче или содержит меньше аминокислот, чем полноразмерный белок. Термин «фрагмент» применительно к нуклеиновой кислоте означает нуклеиновую кислоту, которая короче или имеет меньше нуклеотидов, чем полноразмерная нуклеиновая кислота. Фрагмент может представлять собой, например, N-концевой фрагмент (т.е. образован путем удаления части С-концевого конца белка), С-концевой фрагмент (т.е. образован путем удаления части N-концевого конца белка) или внутренний фрагмент.
[0057] «Идентичность последовательностей», или «идентичность», в контексте двух полинуклеотидных или полипептидных последовательностей относится к ссылке на остатки в двух последовательностях, которые являются одинаковыми при выравнивании последовательностей для максимального соответствия в указанном окне сравнения. Когда процент идентичности последовательностей используется в отношении белков, то положения остатков, которые не идентичны, часто отличаются по консервативным аминокислотным заменам, в которых аминокислотные остатки заменены на другие аминокислотные остатки с аналогичными химическими свойствами (например, заряд или гидрофобность) и, следовательно, не меняют функциональных свойств данной молекулы. Когда последовательности отличаются по консервативным аминокислотным заменам, процент идентичности последовательностей может быть увеличен для корректировки относительно консервативного характера замены. Указывают, что последовательности, отличающиеся по таким консервативным заменам, имеют «сходство последовательностей», или «сходство». Средства для выполнения данной оценки хорошо известны. Обычно это включает в себя оценку консервативной замены как частичного, а не полного несоответствия, что тем самым увеличивает процент идентичности последовательностей. Таким образом, например, если идентичной аминокислоте присваивается оценка, равная 1, а неконсервативной замене присваивается оценка, равная нулю, то консервативной замене присваивается оценка, равная от нуля до 1. Оценка консервативных замен рассчитывается, например, так, как реализовано в программе PC/GENE (Intelligenetics, Маунтин-Вью, Калифорния, США).
[0058] «Процент идентичности последовательностей» включает значение, определенное путем сравнения двух оптимально выровненных последовательностей (наибольшее количество полностью совпадающих остатков) в окне сравнения, при этом часть полинуклеотидной последовательности в окне сравнения может содержать добавления или делеции (т.е. пробелы, гэпы) по сравнению с референтной последовательностью (которая не содержит добавлений или делеций) для оптимального выравнивания данных двух последовательностей. Данный процент рассчитывается путем определения числа положений, в которых одно и то же основание нуклеиновой кислоты или один и тот же аминокислотный остаток встречается в обеих последовательностях, чтобы получить количество совпадающих положений, с последующим делением числа совпадающих положений на общее число положений в окне сравнения и умножением полученного результата на 100, чтобы получить процент идентичности последовательностей. Если не указано иное (например, более короткая последовательность включает в себя связанную гетерологичную последовательность), окно сравнения представляет собой полную протяженность более короткой из двух сравниваемых последовательностей.
[0059] Если не указано иное, значения идентичности/сходства последовательностей включают значение, полученное с использованием программного обеспечения GAP версии 10 с использованием следующих параметров: % идентичности и % сходства для нуклеотидной последовательности с использованием штрафа в 50 за открытие гэпа и штрафа в 3 за удлинение гэпа, а также оценочной матрицы nwsgapdna.cmp; % идентичности и % сходства для нуклеотидной последовательности с использованием штрафа в 8 за открытие гэпа и штрафа в 2 за удлинение гэпа, а также оценочной матрицы BLOSUM62; или с помощью любой эквивалентной программы. «Эквивалентная программа» включает в себя любую программу для сравнения последовательностей, которая для любых двух рассматриваемых последовательностей генерирует выравнивание, имеющее идентичные совпадения нуклеотидных или аминокислотных остатков и идентичный процент идентичности последовательностей по сравнению с соответствующим выравниванием, созданным в программе GAP версии 10.
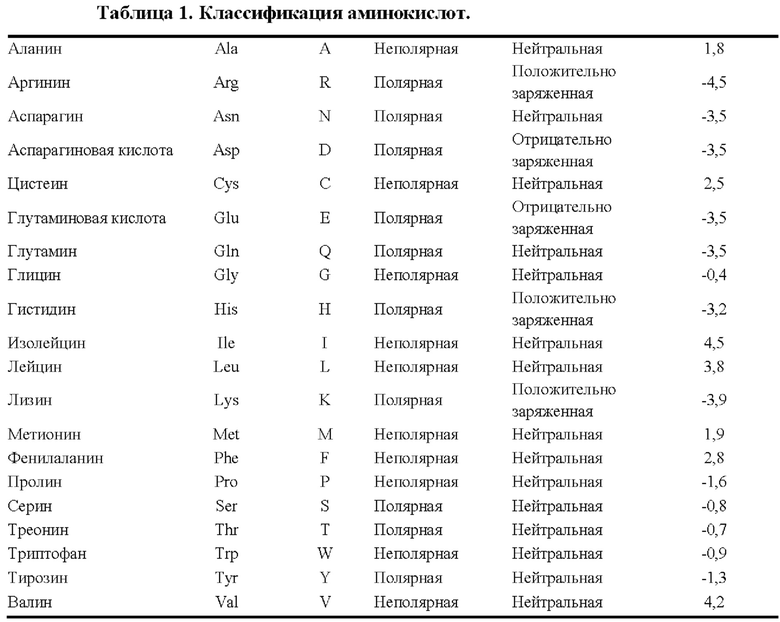
[0060] Термин «консервативная аминокислотная замена» относится к замене аминокислоты, которая обычно присутствует в данной последовательности, на другую аминокислоту аналогичного размера, заряда или полярности. Примеры консервативных замен включают в себя замену неполярного (гидрофобного) остатка, такого как изолейцин, валин или лейцин, на другой неполярный остаток. Аналогичным образом, примеры консервативных замен включают в себя замену одного полярного (гидрофильного) остатка на другой, например, замену аргинина на лизин и наоборот, замену глутамина на аспарагин и наоборот или замену глицина на серии и наоборот. Кроме того, замена основного остатка, такого как лизин, аргинин или гистидин, на другой остаток, или замена одного кислотного остатка, такого как аспарагиновая кислота или глутаминовая кислота, на другой кислотный остаток являются дополнительными примерами консервативных замен. Примеры неконсервативных замен включают в себя замену неполярного (гидрофобного) аминокислотного остатка, такого как изолейцин, валин, лейцин, аланин или метионин, на полярный (гидрофильный) остаток, такой как цистеин, глутамин, глутаминовая кислота или лизин, и (или) замену полярного остатка на неполярный остаток. Типовая классификация аминокислот обобщенно представлена в Таблице 1 ниже.
[0061]
[0062] Термин «гомологичная» последовательность (например, последовательность нуклеиновой кислоты) включает последовательность, которая либо идентична, либо по существу сходна с известной референтной последовательностью таким образом, что она, например, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентична известной референтной последовательности. Гомологичные последовательности могут включать, например, ортологическую последовательность и паралогичные последовательности. Гомологичные гены, например, обычно происходят от общей предковой последовательности ДНК либо в результате видообразования (ортологичные гены), либо в результате генетической дупликации (паралогичные гены). «Ортологические» гены включают гены разных видов, которые произошли от общего предкового гена путем видообразования. Ортологи обычно сохраняют ту же самую функцию в ходе эволюции. «Паралогичные» гены включают гены, связанные посредством дупликации в геноме. Паралоги могут развить новые функции в ходе эволюции.
[0063] Термин «in vitro» включает искусственную среду и процессы или реакции, которые происходят в искусственной среде (например, в пробирке). Термин «in vivo» включает естественную среду (например, клетку, или организм или тело) и процессы или реакции, которые происходят в естественной среде. Термин «ех vivo» включает клетки, которые были удалены из организма человека, а также процессы или реакции, происходящие в таких клетках.
[0064] Репарация в ответ на двухцепочечные разрывы (DSB) происходит главным образом через два консервативных пути репарации ДНК: гомологичную рекомбинацию (ГР, HR) и негомологичное соединение концов (NHEJ, НГР). См. Kasparek & Humphrey (2011) Seminars in Cell & Dev. Biol. 22:886-897, которая включена в полном объеме в данный документ посредством ссылки для всех целей. Точно так же репарация нуклеиновая кислоты-мишени, опосредованная экзогенной донорной нуклеиновой кислотой, может включать любой процесс обмена генетической информацией между двумя полинуклеотидами.
[0065] Термин «рекомбинация» включает любой процесс обмена генетической информацией между двумя полинуклеотидами и может происходить по любому механизму. Рекомбинация может происходить посредством направляемой гомологией репарации (НГР, HDR) или гомологичной рекомбинации (ГР). НГР или ГР включает форму репарации нуклеиновой кислоты, которая может требовать гомологии нуклеотидной последовательности, использует «донорную» молекулу в качестве матрицы для репарации молекулы «мишени» (т.е. той, которая подверглась двуцепочечному разрыву) и приводит к передаче генетической информации от донора к мишени. Не желая быть связанными какой-либо конкретной теорией, такой перенос может включать коррекцию несоответствия гетеродуплексной ДНК, которая образуется между поврежденной мишенью и донором, и/или зависимую от синтеза гибридизацию цепи, при которой донор используется для повторного синтеза генетической информации, которая станет частью мишени, и/или связанные процессы. В некоторых случаях донорный полинуклеотид, часть донорного полинуклеотида, копия донорного полинуклеотида или часть копии донорного полинуклеотида интегрируется в ДНК-мишень. См. Wang et al. (2013) Cell 153:910-918; Mandalos et al. (2012) PLOS ONE 7:e45768:1-9; и Wang et al. (2013) Nat Biotechnol. 31:530-532, каждая из которых включена в данный документ в полном объеме посредством ссылки для всех целей.
[0066] Композиции или способы, «содержащие» или «включающие» один или более число из перечисленных элементов, могут включать в себя другие элементы, не перечисленные специально. Например, композиция, которая «содержит» или «включает» белок, может содержать белок отдельно или в комбинации с другими элементами. Переходная фраза «состоящий по существу из» означает, что объем формулы изобретения следует интерпретировать как включающий в себя указанные в формуле изобретения элементы, а также те, которые существенно не влияют на основные и новые характеристики заявленного изобретения. Таким образом, термин «состоящий по существу из» при использовании в формуле данного изобретения не предназначен для интерпретации как эквивалент слову «включающий».
[0067] «По усмотрению» или «необязательно» означает, что описанное далее событие или обстоятельство может произойти, а может и не произойти, и что описание включает в себя случаи, в которых событие или обстоятельство происходит, и случаи, когда оно не происходит.
[0068] Обозначение диапазона значений включает все целые числа в пределах данного диапазона или все целые числа, определяющие данный диапазон, а также все поддиапазоны, определяемые целыми числами в пределах данного диапазона.
[0069] Если иное явным образом не следует из контекста, термин «около» охватывает значения в пределах стандартной погрешности измерения (например, SEM) указанного значения.
[0070] Термин «и/или» относится и охватывает любые и все возможные комбинации одного или более связанных перечисленных элементов, а также отсутствие комбинаций при интерпретации в альтернативе («или»).
[0071] Термин «или» относится к любому одному члену конкретного списка, а также включает любую комбинацию элементов этого списка.
[0072] Формы единственного числа включают в себя соответствующие им формы множественного числа, если из контекста явным образом не следует иное. Например, термин «белок» или «по меньшей мере, один белок» может включать множество белков, включая их смеси.
[0073] Статистически значимое означает p≤0,05.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
I. Обзор
[0074] В данном документе предложены способы дня внесения бесшовной целевой генетической модификации в ранее созданный направленный вектор. В данных способах можно использовать комбинации бактериальной гомологичной рекомбинации (БГР) и способы сборки in vitro (или внутримолекулярные или межмолекулярные) для внесения таких целевых генетических модификаций в направленный вектор без следов. Термин «бесшовный» относится к тому факту, что никакие изменения или нежелательные последовательности не вносятся в собранную ДНК в результате реакций. Комбинированная последовательность будет соответствовать точной желаемой последовательности без каких-либо изменений или артефактов, вносимых БГР или процедурами сборки in vitro.
[0075] Один из наиболее эффективных подходов к определению функции генов включает в себя преднамеренное конструирование генных мутаций в эмбриональных стволовых (ЭС, ES) клетках мыши (или других ЭС-клетках животных, кроме человека), а затем получение мышей (или других животных, кроме человека), несущих соответствующие генетические изменения. Двумя ограничивающими стадиями являются создание векторов для нацеливания на ген и последующий отбор клонов редких ЭС-клеток, в которых направленный вектор правильно изменил ген. Чтобы произвести желаемое генетическое изменение в ЭС-клетках, нужно сначала внести изменение в направленный вектор, который впоследствии используется для замены нативного гена в ЭС-клетках путем гомологичной рекомбинации.
[0076] Бесшовная конструкция ДНК имеет особое значение при создании линий трансгенных животных, поскольку швы, образованные сайтами рестрикции или другими манипуляциями, могут негативно повлиять на экспрессию генов, если они попадают в область, важную для регуляции. Нацеливание на геном млекопитающих часто требует конструирования больших направленных векторов с длинными плечами ДНК для прямой гомологичной рекомбинации, а также кассет устойчивости к антибиотикам для отбора клонов эмбриональных стволовых клеток. Правильно направленные клоны часто содержат несколько швов, необходимых для конструирования вектора и самой кассеты устойчивости. Даже при использовании технологии самоудаления кассет часто невозможно избежать «швов» (следов) экзогенных последовательностей в модифицированных локусах. См., например, Фиг. 4А-4В. Такие швы могут повлиять на правильную экспрессию локуса-мишени или даже на экспрессию соседних генов. По мере того, как животные модели становятся более сложными, к уже существующим могут быть добавлены дополнительные модификации, такие как мутации, вызывающие болезни человека, в гуманизированных аллелях. Дополнительные изменения могут затем добавить еще больше швов и другую кассету селекции к уже хорошо сконструированному мышиному локусу, увеличивая вероятность того, что экспрессия будет изменена, и мышиная модель не будет правильной. Кроме того, добавление новой кассеты к вектору, уже содержащему ее, может усложняться из-за нежелательной рекомбинации между общими элементами кассеты, такими как промоторы и поли(А)-сигналы, даже если две кассеты кодируют разные наборы. Однако такие кассеты селекции важны для того, чтобы не тратить время и ресурсы на скрининг тысяч клонов ЭС-клеток на предмет желаемой модификации.
[0077] В качестве альтернативы, использование ранее созданного направленного вектора для создания и скрининга модифицированных ЭС-клеток, содержащих модификацию ранее созданного направленного вектора, а затем повторное направленное воздействие на эти клетки вторым направленным вектором (например, ssODN) для внесения второй модификации в уже таргетированный локус, занимает много времени, и повторное нацеливание (например, с ssODN) может привести к нежелательным модификациям, таким как нежелательные вставки, нежелательные делеции, нежелательная точечная мутация или отсутствие направленного воздействия в сочетании с трансгенной вставкой в другом месте генома.
[0078] Способы, раскрытые в данном документе, обеспечивают эффективные и бесшовные способы внесения изменений в ранее созданные направленные векторы на стадии подготовки направленного вектора вместо необходимости создания и скрининга ЭС-клеток, содержащих исходный ранее созданный направленный вектор, а затем повторного направленного воздействия на эти клетки для создания вторая модификации в уже таргетирован локуса.
II. Бесшовное Внесение Целевой Модификации в Направленный Вектор с помощью Бактериальной Гомологичной Рекомбинации и Внутримолекулярной Сборки In Vitro
[0079] Некоторые описанные в данном документе способы для бесшовного внесения целевой генетической модификации в ранее созданный направленный вектор используют преимущества способов сборки in vitro для внутримолекулярной сборки. В качестве одного из примеров такие способы могут включать выполнение бактериальной гомологичной рекомбинации между ранее созданным направленным вектором и кассетой модификации в популяции бактериальных клеток. Кассета модификации может содержать вставку нуклеиновой кислоты, фланкированную 5'-плечом гомологии, соответствующим 5'-последовательности-мишени в ранее созданном направленном векторе, и 3'-плечом гомологии, соответствующим 3'-последовательности-мишени в ранее созданном векторе. Нуклеиновая кислота вставки может содержать кассету селекции, фланкированную сайтами-мишенями для одного или более нуклеазных агентов (например, редко расщепляющих нуклеазных агентов) и повторяющихся последовательностей. Например, нуклеиновая кислота вставки может содержать от 5' к 3': (1) первую повторяющуюся последовательность; (2) первый сайт-мишень для первого нуклеазного агента; (3) кассету селекции; (4) второй сайт-мишень для второго нуклеазного агента; и (5) вторую повторяющуюся последовательность.
[0080] Ранее созданный направленный вектор может представлять собой любой тип направленного вектора любого размера. В конкретном примере ранее созданный направленный вектор представляет собой большой направленный вектор (БНВЕК, LTVEC), который имеет длину, по меньшей мере, около 10 т.п.н. В другом примере длина данного вектора составляет, по меньшей мере, около 100 т.п.н. Направленные векторы и большие направленные векторы обсуждаются более подробно в другом месте в данном документе.
[0081] Кассета модификации может представлять собой линейную нуклеиновую кислоту или кольцевую нуклеиновую кислоту, это может быть одноцепочечная нуклеиновая кислота или двухцепочечная нуклеиновая кислота, и она может содержать дезоксирибонуклеиновую кислоту (ДНК) или рибонуклеиновую кислоту (РНК). В одном конкретном примере кассета модификации представляет собой линейную двухцепочечную ДНК.
[0082] Плечи гомологии в кассете модификации обозначаются в данном документе как 5'- и 3'- (т.е. расположенные в направлении 5'-конца и 3'-конца последовательности и ниже) плечи гомологии. Эта терминология относится к относительному положению плеч гомологии относительно вставки нуклеиновой кислоты в кассете модификации. 5'- и 3'-плечи гомологии соответствуют областям в пределах ранее созданного направленного вектора, подлежащего модификации, которые упоминаются в данном документе как «5'-последовательность-мишень» и «3'-последовательность-мишень», соответственно.
[0083] Плечо гомологии и последовательность-мишень «соответствуют» или являются «соответствующими» друг другу тогда, когда две области обладают достаточным уровнем идентичности последовательностей друг к другу, чтобы действовать в качестве субстратов для реакции гомологичной рекомбинации (например, бактериальной гомологичной рекомбинации). Термин «гомология» включает последовательности ДНК, которые либо идентичны, либо имеют долю идентичности последовательности с соответствующей последовательностью. Идентичность последовательности между данной последовательностью-мишенью и соответствующим плечом гомологии, обнаруженным в матрице экзогенной репарации, может иметь любую степень идентичности последовательностей, которая допускает гомологичную рекомбинацию. Например, степень идентичности последовательностей, общих для плеча гомологии экзогенной репарационной матрицы (или ее фрагмента) и последовательности-мишени (или ее фрагмента), может составлять, по меньшей мере, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательностей, так что последовательности подвергаются гомологичной рекомбинации. Кроме того, соответствующая область гомологии между плечом гомологии и соответствующей последовательностью-мишенью может иметь любую длину, достаточную для стимуляции гомологичной рекомбинации. Например, плечи гомологии могут иметь любой размер, подходящий для бактериальной гомологичной рекомбинации. Например, плечи гомологии могут состоять из, по меньшей мере, около 35 нуклеотидов, по меньшей мере, около 40 нуклеотидов, по меньшей мере, около 50 нуклеотидов, по меньшей мере, около 60 нуклеотидов, по меньшей мере, около 70 нуклеотидов, по меньшей мере, около 80 нуклеотидов, по меньшей мере, около 90 нуклеотидов, по меньшей мере, около 100 нуклеотидов. Например, плечи гомологии могут состоять из от около 35 нуклеотидов до 500 нуклеотидов, от около 75 нуклеотидов до около 500 нуклеотидов или от около 50 нуклеотидов до около 200 нуклеотидов (например, около 100 нуклеотидов). В качестве другого примера плечи гомологии могут иметь длину от около 35 нуклеотидов до около 2,5 т.п.н., иметь длину от около 35 нуклеотидов до около 1,5 т.п.н. или иметь длину от около 35 до около 500 нуклеотидов. Например, данное плечо гомологии (или каждое из плеч гомологии) и/или соответствующая последовательность-мишень могут содержать соответствующие области гомологии, которые составляют от около 35 до около 40, от около 40 до около 50, от около 50 до около 60, около 60 до около 70, от около 70 до около 80, от около 80 до около 90, от около 90 до около 100, от около 100 до около 150, от около 150 до около 200, от около 200 до около 250, от около 250 до около 300, от около 300 до около 350, от около 350 до около 400, от около 400 до около 450 или от около 450 до около 500 нуклеотидов в длину, таким образом, чтобы плечи гомологии имели достаточную гомологию для проведения гомологичной рекомбинации с соответствующими последовательностями-мишенями в нуклеиновой кислоте-мишени. Альтернативно, данное плечо гомологии (или каждое плечо гомологии) и/или соответствующая последовательность-мишень могут содержать соответствующие области гомологии, которые составляют от около 0,5 т.п.н. до около 1 т.п.н., от около 1 т.п.н. до около 1,5 т.п.н., от около 1,5 т.п.н. до около 2 т.п.н. или от около 2 т.п.н. до около 2,5 т.п.н. в длину. Например, плечи гомологии могут иметь длину около 100 нуклеотидов. Плечи гомологии могут быть симметричными (каждое плечо приблизительного одинакового размера по длине) или асимметричными (одно плечо длиннее другого).
[0084] Кассета модификации может быть любой длины. Например, кассета модификации может составлять от около 10 т.п.н. до около 400 т.п.н., от около 20 т.п.н. до около 400 т.п.н., от около 20 т.п.н. до около 30 т.п.н., от около 30 т.п.н. до около 40 т.п.н., от около 40 т.п.н. до около 50 т.п.н., от около 50 т.п.н. до около 75 т.п.н., от около 75 т.п.н. до около 100 т.п.н., от около 100 т.п.н. до 125 т.п.н., от около 125 т.п.н. до около 150 т.п.н., от около 150 т.п.н. до около 175 т.п.н., от около 175 т.п.н. до около 200 т.п.н., от около 200 т.п.н. до около 225 т.п.н., от около 225 т.п.н. до около 250 т.п.н., от около 250 т.п.н. до около 275 т.п.н. или от около 275 т.п.н. до около 300 т.п.н., от около 200 т.п.н. до около 300 т.п.н., от около 300 т.п.н. до около 350 т.п.н., или от около 350 т.п.н. до около 400 т.п.н. В одном примере кассета модификации может иметь длину, по меньшей мере, около 100 или 100 т.п.н. Кассета модификации также может составлять от около 50 т.п.н. до около 500 т.п.н., от около 100 до около 125 т.п.н., от около 300 т.п.н. до около 325 т.п.н., от около 325 т.п.н. до около 350 т.п.н., от около 350 т.п.н. до около 375 т.п.н., от около 375 т.п.н. до около 400 т.п.н., от около 400 т.п.н. до около 425 т.п.н., от около 425 т.п.н. до около 450 т.п.н., от около 450 т.п.н. до около 475 т.п.н. или от около 475 т.п.н. до около 500 т.п.н. Альтернативно, кассета модификации может составлять, по меньшей мере, 10 т.п.н., по меньшей мере, 15 т.п.н., по меньшей мере, 20 т.п.н., по меньшей мере, 30 т.п.н., по меньшей мере, 40 т.п.н., по меньшей мере, 50 т.п.н., по меньшей мере, 60 т.п.н., по меньшей мере, 70 т.п.н., по меньшей мере, 80 т.п.н., по крайней мере, 90 т.п.н., по меньшей мере, 100 т.п.н., по меньшей мере, 150 т.п.н., по меньшей мере, 200 т.п.н., по меньшей мере, 250 т.п.н., по меньшей мере, 300 т.п.н., по меньшей мере, 350 т.п.н., по меньшей мере, 400 т.п.н., по меньшей мере, 450 т.п.н., или, по меньшей мере, 500 т.п.н. или более. В одном примере кассета модификации имеет длину от около 1 т.п.н. до около 15 т.п.н. или от около 1 т.п.н. до около 10 т.п.н. (например, около 1,2 т.п.н., около 5 т.п.н., около 8 т.п.н. или около 15 т.п.н.).
[0085] Кассета модификации может содержать целевую генетическую модификацию. Например, целевая генетическая модификация может быть в 5'-плече гомологии или в 3'-плече гомологии (например, небольшая модификация, такая как точечная мутация или небольшая делеция, вставка или замена, которая не повлияет отрицательно на способность плеча гомологии рекомбинировать с последовательностью-мишенью).
Альтернативно, целевая генетическая модификация может быть в нуклеиновой кислоте вставки (например, когда целевая генетическая модификация представляет собой вставку или замену). Если единственной целевой генетической модификацией является делеция, то 5'-плечо гомологии и 3'-плечо гомологии можно сконструировать для нацеливания на 5'- и 3'-последовательности-мишени, соответственно, которые фланкируют последовательность-мишень для делеции в ранее созданном направленном векторе. В качестве одного примера целевая генетическая модификация может находиться в первой повторяющейся последовательности и/или во второй повторяющейся последовательности в нуклеиновой кислоте вставки. Типы возможных целевых генетических модификаций более подробно описаны в другом месте в данном документе. Некоторые примеры включают точечные мутации, делеции, вставки, замены или их комбинации.
[0086] Первая и вторая повторяющиеся последовательности в кассете модификации могут быть идентичны друг другу. Повторяющаяся последовательность может быть идентична последовательности в ранее созданном направленном векторе. В качестве альтернативы, в случае, если целевая генетическая модификация включает вставку (например, только вставку или вставку в сочетании с делецией (т.е. замену)), повторяющаяся последовательность может быть идентична 5'-концу или 3'-концу вставки.
[0087] Повторяющаяся последовательность может иметь любой подходящий размер для последующей сборки между первой и второй повторяющимися последовательностями в реакции сборки in vitro. В качестве одного примера, повторяющаяся последовательность может содержать, по меньшей мере, около 20 нуклеотидов, по меньшей мере, около 30 нуклеотидов, по меньшей мере, около 40 нуклеотидов или, по меньшей мере, около 50 нуклеотидов. В качестве другого примера, повторяющаяся последовательность может иметь длину от около 20 нуклеотидов до около 100 нуклеотидов, от около 20 нуклеотидов до около 90 нуклеотидов, от около 20 нуклеотидов до около 80 нуклеотидов, от около 20 нуклеотидов до около 70 нуклеотидов, от около 20 нуклеотидов и около 60 нуклеотидов, от около 20 нуклеотидов до около 50 нуклеотидов, от около 20 нуклеотидов до около 40 нуклеотидов, от около 30 нуклеотидов до около 60 нуклеотидов, или от около 40 нуклеотидов до около 50 нуклеотидов. В конкретном примере повторяющаяся последовательность может иметь длину от около 40 нуклеотидов до около 50 нуклеотидов (например, около 40 нуклеотидов или около 50 нуклеотидов).
[0088] После бактериальной гомологичной рекомбинации могут быть отобраны бактериальные клетки, содержащие модифицированный направленный вектор, содержащий кассету селекции (и содержащий целевую генетическую модификацию).
Примеры кассет селекции и способов селекции более подробно описаны в другом месте в данном документе. В конкретном примере кассета селекции придает устойчивость к антибиотику. Например, кассета может придавать устойчивость к любому из антибиотиков: ампициллину, хлорамфениколу, тетрациклину, канамицину, спектиномицину, стрептомицину, карбенициллину, блеомицину, эритромицину или полимиксину В. В некоторых способах ранее созданный направленный вектор также содержит вторую кассету селекции. Вторая кассета селекции может, например, также придавать устойчивость к антибиотику. Кассета селекции в кассете модификации и вторая кассета селекции в ранее созданном направленном векторе может придавать устойчивость к другому антибиотику. Например, кассета селекции в кассете модификации может придавать устойчивость к одному антибиотику, а вторая кассета селекции в ранее созданном направленном векторе может придавать устойчивость ко второму, другому антибиотику. В некоторых способах вторая кассета селекции может обеспечивать возможность селекции как в бактериальных клетках, так и в эукариотических клетках или клетках млекопитающих.
[0089] После селекции первый сайт-мишень в модифицированном направленном векторе может быть расщеплен первым нуклеазным агентом, а второй сайт-мишень в модифицированном направленном векторе может быть расщеплен вторым нуклеазным агентом, чтобы удалить кассету селекции и обнажить первую повторяющуюся последовательность и вторую повторяющуюся последовательность в модифицированном направленном векторе. Например, этот этап можно выполнять in vitro. В качестве примера, ДНК может быть выделена из бактериальных клеток после бактериальной гомологичной рекомбинации и селекции, после чего первый сайт-мишень в модифицированном направленном векторе может быть отщеплен первым нуклеазным агентом in vitro, а второй сайт-мишень в модифицированном направленном векторе может быть расщеплен вторым нуклеазным агентом in vitro, чтобы удалить кассету селекции и обнажить первую повторяющуюся последовательность и вторую повторяющуюся последовательность в модифицированном направленном векторе.
[0090] Первый нуклеазный агент нуклеазы и/или второй нуклеазный агент может быть редко расщепляющим нуклеазным агентом, как описано в другом месте в данном документе. Например, в некоторых способах первый сайт-мишень и/или второй сайт-мишень не присутствуют в ранее созданном направленном векторе. Первый и второй сайты-мишени могут отличаться друг от друга, или первый сайт-мишень может быть идентичен второму сайту-мишени, и первый нуклеазный агент может быть идентичен второму нуклеазному агенту. Первый нуклеазный агент и/или второй нуклеазный агент могут образовывать тупой конец, 5'-липкий конец или 3'-липкий конец. В одном примере первый нуклеазный агент и/или второй нуклеазный агент образует 3'-липкий конец.
[0091] В одном конкретном примере первый нуклеазный агент и/или второй нуклеазный агент представляют собой рестрикционный фермент или редко расщепляющий рестрикционный фермент.Примеры редко расщепляющих рестрикционных ферментов описаны в других местах данного документа, но могут включать, например, NotI, XmaIII, SstII, Sall, NruI, NheI, Nb.BbvCI, BbvCI, AscI, AsiSI, FseI, PacI, PmeI, SbfI, SgrAI, SwaI, BspQI, SapI, SfiI, CspCI, AbsI, CciNI, FspAI, MauBI, MreI, MssI, PalAI, RgaI, RigI, SdaI, SfaAI, SgfI, SgrDI, SgsI, SmiI, SrfI, Sse2321, Sse83871, LguI, PciSI, AarI, AjuI, AloI, BarI, PpiI или PsrI.
[0092] В другом конкретном примере первый нуклеазный агент и/или второй нуклеазный агент может быть сконструированным нуклеазным агентом. Например, нуклеазный агент может представлять собой белок (Cas), ассоциированный с кластеризованными регулярными промежуточными короткими палиндромными повторами (CRISPR) и направляющую РНК (нРНК), (например, Cas9 и нРНК, содержащую CRISPR-PHK (крРНК) и трансактивирующую CRISPR-PHK (тркрРНК)), нуклеазу с "цинковыми пальцами» (ZFN), эффекторную нуклеазу, подобную активаторам транскрипции (TALEN), или сконструированную мегануклеазу.
[0093] После отщепления/расщепления экспонированная первая повторяющаяся последовательность может быть собрана с экспонированной второй повторяющейся последовательностью во внутримолекулярной реакции сборки in vitro для создания направленного вектора, содержащего бесшовную целевую генетическую модификацию. Например, в некоторых таких способах ни первый сайт-мишень для первого нуклеазного агента, ни второй сайт-мишень для второго нуклеазного агента не присутствуют в направленном векторе, содержащем бесшовную целевую генетическую модификацию (т.е. после сборки in vitro). Аналогичным образом, в некоторых таких способах только одна копия повторяющейся последовательности присутствует в направленном векторе, содержащем бесшовную направленную генетическую модификацию (т.е. после сборки in vitro).
[0094] Можно использовать любой подходящий способ сборки in vitro. В одном конкретном примере этап сборки in vitro может включать инкубацию модифицированного направленного вектора с экзонуклеазой, ДНК-полимеразой и ДНК-лигазой. Например, способ сборки in vitro может включать приведение в контакт модифицированного направленного вектора с экзонуклеазой для выявления комплементарных последовательностей между первой повторяющейся последовательностью и второй повторяющейся последовательностью, гибридизация экспонированных комплементарных последовательностей, удлинение 3'-концов гибридизированных комплементарных последовательностей, и лигирование гибридизированных комплементарных последовательностей. Примеры способов сборки in vitro более подробно обсуждаются в другом месте в данном документе.
[0095] В некоторых способах для уменьшения фона вектор, полученный путем сборки in vitro, можно обработать первым нуклеазным агентом и/или вторым нуклеазным агентом для уменьшения фона (например, путем отщепления любых направленных векторов, которые не удалось собрать и, следовательно, все еще содержат сайт-мишень для первого нуклеазного агента или второго нуклеазного агента). Такой шаг может помочь проверить, что ни первый сайт-мишень для первого нуклеазного агента, ни второй сайт-мишень для второго нуклеазного агента не присутствуют.
III Бесшовное Внесение Целевой Модификации в Направленный Вектор с помощью Бактериальной Гомологичной Рекомбинации и Межмолекулярной Сборки bi Vitro
[0096] В других описанных в данном документе способах для бесшовного внесения целевой генетической модификации в ранее созданный направленный вектор используются преимущества способов сборки in vitro для межмолекулярной сборки. В качестве одного из примеров такие способы могут включать выполнение бактериальной гомологичной рекомбинации между ранее созданным направленным вектором и кассетой делеции в популяции бактериальных клеток. Кассета делеции может содержать вставку нуклеиновой кислоты, фланкированную 5'-плечом гомологии, соответствующим 5'-последовательности-мишени в ранее созданном направленном векторе, и 3'-плечом гомологии, соответствующим 3'-последовательности-мишени в ранее созданном векторе. 5'-последовательность-мишень и 3'-последовательность-мишень могут фланкировать область ранее созданного направленного вектора, в которую должна быть внесена целевая генетическая модификация. Нуклеиновая кислота вставки может содержать кассету селекции, фланкированную сайтами-мишенями для одного или более нуклеазных агентов (например, редко расщепляющих нуклеазных агентов). Например, нуклеиновая кислота вставки может содержать от 5' к 3': (1) первую повторяющуюся последовательность; (2) кассету селекции; (3) второй сайт-мишень для второго нуклеазного агента.
[0097] Ранее созданный направленный вектор может представлять собой любой тип направленного вектора любого размера. В конкретном примере ранее созданный направленный вектор представляет собой большой направленный вектор (БНВЕК, LTVEC), который имеет длину, по меньшей мере, около 10 т.п.н. В другом примере длина данного вектора составляет, по меньшей мере, около 100 т.п.н. Направленные векторы обсуждаются более подробно в другом месте в данном документе.
[0098] Кассета делеции может представлять собой линейную нуклеиновую кислоту или кольцевую нуклеиновую кислоту, это может быть одноцепочечная нуклеиновая кислота или двухцепочечная нуклеиновая кислота, и она может содержать дезоксирибонуклеиновую кислоту (ДНК) или рибонуклеиновую кислоту (РНК). В одном конкретном примере кассета модификации представляет собой линейную двухцепочечную ДНК.
[0099] Плечи гомологии в кассете делеции обозначаются в данном документе как 5'-и 3'- (т.е. расположенные в направлении 5'-конца и 3'-конца последовательности и ниже) плечи гомологии. Эта терминология относится к относительному положению плеч гомологии относительно вставки нуклеиновой кислоты в кассете делеции. 5'- и 3'-плечи гомологии соответствуют областям в пределах ранее созданного направленного вектора, подлежащего модификации, которые упоминаются в данном документе как «5'-последовательность-мишень» и «3'-последовательность-мишень», соответственно.
[00100] Плечо гомологии и последовательность-мишень «соответствуют» или являются «соответствующими» друг другу тогда, когда две области обладают достаточным уровнем идентичности последовательностей друг к другу, чтобы действовать в качестве субстратов для реакции гомологичной рекомбинации (например, бактериальной гомологичной рекомбинации). Термин «гомология» включает последовательности ДНК, которые либо идентичны, либо имеют долю идентичности последовательности с соответствующей последовательностью. Идентичность последовательности между данной последовательностью-мишенью и соответствующим плечом гомологии, обнаруженным в матрице экзогенной репарации, может иметь любую степень идентичности последовательностей, которая допускает гомологичную рекомбинацию. Например, степень идентичности последовательностей, общих для плеча гомологии экзогенной репарационной матрицы (или ее фрагмента) и последовательности-мишени (или ее фрагмента), может составлять, по меньшей мере, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательностей, так что последовательности подвергаются гомологичной рекомбинации. Кроме того, соответствующая область гомологии между плечом гомологии и соответствующей последовательностью-мишенью может иметь любую длину, достаточную для стимуляции гомологичной рекомбинации. Например, плечи гомологии могут иметь любой размер, подходящий для бактериальной гомологичной рекомбинации. Например, плечи гомологии могут состоять из, по меньшей мере, около 35 нуклеотидов, по меньшей мере, около 40 нуклеотидов, по меньшей мере, около 50 нуклеотидов, по меньшей мере, около 60 нуклеотидов, по меньшей мере, около 70 нуклеотидов, по меньшей мере, около 80 нуклеотидов, по меньшей мере, около 90 нуклеотидов, по меньшей мере, около 100 нуклеотидов. Например, плечи гомологии могут состоять из от около 35 нуклеотидов до 500 нуклеотидов, от около 75 нуклеотидов до около 500 нуклеотидов или от около 50 нуклеотидов до около 200 нуклеотидов (например, около 100 нуклеотидов). В качестве другого примера плечи гомологии могут иметь длину от около 35 нуклеотидов до около 2,5 т.п.н., иметь длину от около 35 нуклеотидов до около 1,5 т.п.н. или иметь длину от около 35 до около 500 нуклеотидов. Например, данное плечо гомологии (или каждое из плеч гомологии) и/или соответствующая последовательность-мишень могут содержать соответствующие области гомологии, которые составляют от около 35 до около 40, от около 40 до около 50, от около 50 до около 60, около 60 до около 70, от около 70 до около 80, от около 80 до около 90, от около 90 до около 100, от около 100 до около 150, от около 150 до около 200, от около 200 до около 250, от около 250 до около 300, от около 300 до около 350, от около 350 до около 400, от около 400 до около 450 или от около 450 до около 500 нуклеотидов в длину, таким образом, чтобы плечи гомологии имели достаточную гомологию для проведения гомологичной рекомбинации с соответствующими последовательностями-мишенями в нуклеиновой кислоте-мишени. Альтернативно, данное плечо гомологии (или каждое плечо гомологии) и/или соответствующая последовательность-мишень могут содержать соответствующие области гомологии, которые составляют от около 0,5 т.п.н. до около 1 т.п.н., от около 1 т.п.н. до около 1,5 т.п.н., от около 1,5 т.п.н. до около 2 т.п.н. или от около 2 т.п.н. до около 2,5 т.п.н. в длину. Например, плечи гомологии могут иметь длину около 100 нуклеотидов. Плечи гомологии могут быть симметричными (каждое плечо приблизительного одинакового размера по длине) или асимметричными (одно плечо длиннее другого).
[00101] Кассета делеции может быть любой длины. Например, кассета делеции может составлять от около 10 т.п.н. до около 400 т.п.н., от около 20 т.п.н. до около 400 т.п.н., от около 20 т.п.н. до около 30 т.п.н., от около 30 т.п.н. до около 40 т.п.н., от около 40 т.п.н. до около 50 т.п.н., от около 50 т.п.н. до около 75 т.п.н., от около 75 т.п.н. до около 100 т.п.н., от около 100 т.п.н. до 125 т.п.н., от около 125 т.п.н. до около 150 т.п.н., от около 150 т.п.н. до около 175 т.п.н., от около 175 т.п.н. до около 200 т.п.н., от около 200 т.п.н. до около 225 т.п.н., от около 225 т.п.н. до около 250 т.п.н., от около 250 т.п.н. до около 275 т.п.н. или от около 275 т.п.н. до около 300 т.п.н., от около 200 т.п.н. до около 300 т.п.н., от около 300 т.п.н. до около 350 т.п.н., или от около 350 т.п.н. до около 400 т.п.н. В одном примере кассета делеции может иметь длину, по меньшей мере, около 100 или 100 т.п.н. Кассета делеции также может составлять от около 50 т.п.н. до около 500 т.п.н., от около 100 до около 125 т.п.н., от около 300 т.п.н. до около 325 т.п.н., от около 325 т.п.н. до около 350 т.п.н., от около 350 т.п.н. до около 375 т.п.н., от около 375 т.п.н. до около 400 т.п.н., от около 400 т.п.н. до около 425 т.п.н., от около 425 т.п.н. до около 450 т.п.н., от около 450 т.п.н. до около 475 т.п.н. или от около 475 т.п.н. до около 500 т.п.н. Альтернативно, кассета делеции может составлять, по меньшей мере, 10 т.п.н., по меньшей мере, 15 т.п.н., по меньшей мере, 20 т.п.н., по меньшей мере, 30 т.п.н., по меньшей мере, 40 т.п.н., по меньшей мере, 50 т.п.н., по меньшей мере, 60 т.п.н., по меньшей мере, 70 т.п.н., по меньшей мере, 80 т.п.н., по крайней мере, 90 т.п.н., по меньшей мере, 100 т.п.н., по меньшей мере, 150 т.п.н., по меньшей мере, 200 т.п.н., по меньшей мере, 250 т.п.н., по меньшей мере, 300 т.п.н., по меньшей мере, 350 т.п.н., по меньшей мере, 400 т.п.н., по меньшей мере, 450 т.п.н., или, по меньшей мере, 500 т.п.н. или более. В одном примере кассета делеции имеет длину от около 1 т.п.н. до около 15 т.п.н. или от около 1 т.п.н. до около 10 т.п.н. (например, около 1,2 т.п.н., около 5 т.п.н., около 8 т.п.н. или около 15 т.п.н.).
[00102] После бактериальной гомологичной рекомбинации могут быть отобраны бактериальные клетки, содержащие модифицированный направленный вектор, содержащий кассету селекции. Примеры кассет селекции и способов селекции более подробно описаны в другом месте в данном документе. В конкретном примере кассета селекции придает устойчивость к антибиотику. Например, кассета может придавать устойчивость к любому из антибиотиков: ампициллину, хлорамфениколу, тетрациклину, канамицину, спектиномицину, стрептомицину, карбенициллину, блеомицину, эритромицину или полимиксину В. В некоторых способах ранее созданный направленный вектор также содержит вторую кассету селекции. Вторая кассета селекции может, например, также придавать устойчивость к антибиотику. Кассета селекции в кассете делеции и вторая кассета селекции в ранее созданном направленном векторе могут придавать устойчивость к разным антибиотикам. Например, кассета делеции в кассете модификации может придавать устойчивость к одному антибиотику, а вторая кассета селекции в ранее созданном направленном векторе может придавать устойчивость ко второму, другому антибиотику. В некоторых способах вторая кассета селекции может обеспечивать возможность селекции как в бактериальных клетках, так и в эукариотических клетках или клетках млекопитающих.
[00103] После отбора первый сайт-мишень в модифицированном направленном векторе может быть расщеплен первым нуклеазным агентом, а второй сайт-мишень в модифицированном направленном векторе может быть расщеплен вторым нуклеазным агентом, чтобы удалить кассету селекции и обнажить концевую последовательность, расположенную в направлении 5', и концевую последовательность, расположенную в направлении 3', в модифицированном направленном векторе. Например, этот этап можно выполнять in vitro. В качестве примера, ДНК может быть выделена из бактериальных клеток после бактериальной гомологичной рекомбинации и селекции, после чего первый сайт-мишень в модифицированном направленном векторе может быть отщеплен первым нуклеазным агентом in vitro, а второй сайт-мишень в модифицированном направленном векторе может быть расщеплен вторым нуклеазным агентом in vitro, чтобы удалить кассету селекции и обнажить концевую последовательность, расположенную в направлении 5', и концевую последовательность, расположенную в направлении 3', в модифицированном направленном векторе.
[00104] Первый нуклеазный агент нуклеазы и/или второй нуклеазный агент может быть редко расщепляющим нуклеазным агентом, как описано в другом месте в данном документе. Например, в некоторых способах первый сайт-мишень и/или второй сайт-мишень не присутствуют в ранее созданном направленном векторе. Первый и второй сайты-мишени могут отличаться друг от друга, или первый сайт-мишень может быть идентичен второму сайту-мишени, и первый нуклеазный агент может быть идентичен второму нуклеазному агенту. Первый нуклеазный агент и/или второй нуклеазный агент могут образовывать тупой конец, 5'-липкий конец или 3'-липкий конец. В одном примере первый нуклеазный агент и/или второй нуклеазный агент образует 3'-липкий конец.
[00105] В одном конкретном примере первый нуклеазный агент и/или второй нуклеазный агент представляют собой рестрикционный фермент или редко расщепляющий рестрикционный фермент.Примеры редко расщепляющих рестрикционных ферментов описаны в других местах данного документа, но могут включать, например, NotI, XmaIII, SstII, Sall, NruI, NheI, Nb.BbvCI, BbvCI, AscI, AsiSI, FseI, PacI, PmeI, SbfI, SgrAI, SwaI, BspQI, SapI, SfiI, CspCI, AbsI, CciNI, FspAI, MauBI, MreI, MssI, PalAI, RgaI, RigI, SdaI, SfaAI, SgfI, SgrDI, SgsI, SmiI, SrfI, Sse2321, Sse83871, LguI, PciSI, AarI, AjuI, AloI, BarI, PpiI или PsrI.
[00106] В другом конкретном примере первый нуклеазный агент и/или второй нуклеазный агент может быть сконструированным нуклеазным агентом. Например, нуклеазный агент может представлять собой белок (Cas), ассоциированный с кластеризованными регулярными промежуточными короткими палиндромными повторами (CRISPR) и направляющую РНК (нРНК), (например, Cas9 и нРНК, содержащую CRISPR-PHK (крРНК) и трансактивирующую CRISPR-PHK (тркрРНК)), нуклеазу с "цинковыми пальцами» (ZFN), эффекторную нуклеазу, подобную активаторам транскрипции (TALEN), или сконструированную мегануклеазу.
[00107] После отщепления/расщепления расщепленный направленный вектор может быть собран в реакции межмолекулярной сборки in vitro с кассетой модификации, содержащей целевую генетическую модификацию, фланкированную концевой последовательностью, расположенной в направлении 5' и перекрывающей концевую последовательность, расположенную в направлении 5', в модифицированном направленном векторе, и концевой последовательностью, расположенной в направлении 3' и перекрывающей концевую последовательность, расположенную в направлении 3', в модифицированном направленном векторе для создания направленного вектора, содержащего бесшовную целевую генетическую модификацию. Например, в некоторых таких способах ни первый сайт-мишень для первого нуклеазного агента, ни второй сайт-мишень для второго нуклеазного агента не присутствуют в направленном векторе, содержащем бесшовную целевую генетическую модификацию.
[00108] Можно использовать любой подходящий способ сборки in vitro. В одном конкретном примере этап сборки in vitro может включать инкубацию расщепленного направленного вектора и кассеты модификации с экзонуклеазой, ДНК-полимеразой и ДНК-лигазой. Например, способ сборки in vitro может включать приведение в контакт расщепленного направленного вектора и кассеты модификации с экзонуклеазой для открытия комплементарных последовательностей между концевыми последовательностями в модифицированном направленном векторе и концевыми последовательностями в кассете модификации, гибридизацию экспонированных дополнительных последовательностей, удлинение 3'-концов гибридизированных дополнительных последовательностей, и лигирование гибридизированных комплементарных последовательностей.
[00109] Кассета модификации может представлять собой линейную нуклеиновую кислоту или кольцевую нуклеиновую кислоту, это может быть одноцепочечная нуклеиновая кислота или двухцепочечная нуклеиновая кислота, и она может содержать дезоксирибонуклеиновую кислоту (ДНК) или рибонуклеиновую кислоту (РНК). В одном конкретном примере кассета модификации представляет собой линейную двухцепочечную ДНК.
[00110] Кассета модификации может быть любой длины. Например, кассета модификации может составлять от около 10 т.п.н. до около 400 т.п.н., от около 20 т.п.н. до около 400 т.п.н., от около 20 т.п.н. до около 30 т.п.н., от около 30 т.п.н. до около 40 т.п.н., от около 40 т.п.н. до около 50 т.п.н., от около 50 т.п.н. до около 75 т.п.н., от около 75 т.п.н. до около 100 т.п.н., от около 100 т.п.н. до 125 т.п.н., от около 125 т.п.н. до около 150 т.п.н., от около 150 т.п.н. до около 175 т.п.н., от около 175 т.п.н. до около 200 т.п.н., от около 200 т.п.н. до около 225 т.п.н., от около 225 т.п.н. до около 250 т.п.н., от около 250 т.п.н. до около 275 т.п.н. или от около 275 т.п.н. до около 300 т.п.н., от около 200 т.п.н. до около 300 т.п.н., от около 300 т.п.н. до около 350 т.п.н., или от около 350 т.п.н. до около 400 т.п.н. В одном примере кассета модификации может иметь длину, по меньшей мере, около 100 или 100 т.п.н. Кассета модификации также может составлять от около 50 т.п.н. до около 500 т.п.н., от около 100 до около 125 т.п.н., от около 300 т.п.н. до около 325 т.п.н., от около 325 т.п.н. до около 350 т.п.н., от около 350 т.п.н. до около 375 т.п.н., от около 375 т.п.н. до около 400 т.п.н., от около 400 т.п.н. до около 425 т.п.н., от около 425 т.п.н. до около 450 т.п.н., от около 450 т.п.н. до около 475 т.п.н. или от около 475 т.п.н. до около 500 т.п.н. Альтернативно, кассета модификации может составлять, по меньшей мере, 10 т.п.н., по меньшей мере, 15 т.п.н., по меньшей мере, 20 т.п.н., по меньшей мере, 30 т.п.н., по меньшей мере, 40 т.п.н., по меньшей мере, 50 т.п.н., по меньшей мере, 60 т.п.н., по меньшей мере, 70 т.п.н., по меньшей мере, 80 т.п.н., по крайней мере, 90 т.п.н., по меньшей мере, 100 т.п.н., по меньшей мере, 150 т.п.н., по меньшей мере, 200 т.п.н., по меньшей мере, 250 т.п.н., по меньшей мере, 300 т.п.н., по меньшей мере, 350 т.п.н., по меньшей мере, 400 т.п.н., по меньшей мере, 450 т.п.н., или, по меньшей мере, 500 т.п.н. или более. В одном конкретном примере кассета модификации имеет длину от около 400 п.н. до около 2 т.п.н. В другом примере кассета модификации имеет длину от около 1 т.п.н. до около 15 т.п.н. или от около 1 т.п.н. до около 10 т.п.н. (например, около 1,2 т.п.н., около 5 т.п.н., около 8 т.п.н. или около 15 т.п.н.). В конкретном примере кассета модификации имеет длину, по меньшей мере, около 200 нуклеотидов. В другом конкретном примере кассета модификации может быть такого размера, что не может быть синтезирована напрямую или получена путем полимеразной цепной реакцией. Например, кассета модификации может иметь длину, по меньшей мере, около 5 т.п.н., по меньшей мере, около 10 т.п.н., по меньшей мере, около 15 т.п.н., по меньшей мере, около 20 т.п.н., по меньшей мере, около 25 т.п.н. или, по меньшей мере, около 30 т.п.н.
[00111] Длина перекрывания между концевой последовательностью, расположенной в направлении 5' в кассете модификации и концевой последовательностью, расположенной в направлении 5' в модифицированном направленном векторе и/или длина перекрывания между концевой последовательностью, расположенной в направлении 3' в кассете модификации и концевой последовательность, расположенной в направлении 3' в модифицированном направленном векторе, может быть любой, подходящей для реакции сборки in vitro. В качестве одного примера, длина перекрытия может содержать, по меньшей мере, около 20 нуклеотидов, по меньшей мере, около 30 нуклеотидов, по меньшей мере, около 40 нуклеотидов или, по меньшей мере, около 50 нуклеотидов. В качестве другого примера, длина перекрытия может составлять от около 20 нуклеотидов до около 100 нуклеотидов, от около 20 нуклеотидов до около 90 нуклеотидов, от около 20 нуклеотидов до около 80 нуклеотидов, от около 20 нуклеотидов до около 70 нуклеотидов, от около 20 нуклеотидов и около 60 нуклеотидов, от около 20 нуклеотидов до около 50 нуклеотидов, от около 20 нуклеотидов до около 40 нуклеотидов, от около 30 нуклеотидов до около 60 нуклеотидов, или от около 40 нуклеотидов до около 50 нуклеотидов. В конкретном примере длина перекрытия последовательностей может составлять от около 40 нуклеотидов до около 50 нуклеотидов (например, около 40 нуклеотидов или около 50 нуклеотидов).
[00112] Кассета модификации может содержать целевую генетическую модификацию. Типы целевых генетических модификаций более подробно описаны в другом месте в данном документе. Некоторые примеры включают точечные мутации, делеции, вставки, замены или их комбинации.
[00113] В некоторых способах для уменьшения фона вектор, полученный путем сборки in vitro, можно обработать первым нуклеазным агентом и/или вторым нуклеазным агентом для уменьшения фона (например, путем отщепления любых направленных векторов, которые не удалось собрать и, следовательно, все еще содержащих сайт-мишень для первого нуклеазного агента или второго нуклеазного агента). Такой шаг может помочь проверить, что ни первый сайт-мишень для первого нуклеазного агента, ни второй сайт-мишень для второго нуклеазного агента не присутствуют.
IV. Бактериальная Гомологичная Рекомбинация
[00114] Любой подходящий способ бактериальной гомологичной рекомбинации (БГР, BHR) может быть использован в способах, описанных в данном документе. Бактериальная гомологичная рекомбинация включает временную и контролируемую экспрессию генов, которые опосредуют гомологичную рекомбинацию в бактериальных клетках, таких как Escherichia coli, тем самым позволяя бактериям опосредовать рекомбинацию между кассетой модификации и направленным вектором (например, большим направленным вектором), разделяя короткие гомологичные участки. См., например, патент США 2004/0018626 и Valenzuela et al. (2003) Biotechnol. 21(6):652-659, каждый из которых включен в данный документ в полном объеме посредством ссылки для всех целей.
[00115] Короткие гомологичные участки могут содержать область гомологии, расположенную в направлении 5', и ниже область гомологии, расположенную в направлении 3'. Области гомологии могут иметь любой размер, подходящий для бактериальной гомологичной рекомбинации. Например, области гомологии могут иметь любой размер, подходящий для бактериальной гомологичной рекомбинации. Например, области гомологии могут составлять, по меньшей мере, около 35 нуклеотидов, по меньшей мере, около 40 нуклеотидов, по меньшей мере, около 50 нуклеотидов, по меньшей мере, около 60 нуклеотидов, по меньшей мере, около 70 нуклеотидов, по меньшей мере, около 80 нуклеотидов, по меньшей мере, около 90 нуклеотидов, по меньшей мере, около 100 нуклеотидов. Например, области гомологии могут составлять от около 35 нуклеотидов до 500 нуклеотидов, от около 75 нуклеотидов до около 500 нуклеотидов или от около 50 нуклеотидов до около 200 нуклеотидов (например, около 100 нуклеотидов). В качестве другого примера области гомологии могут иметь длину от около 35 нуклеотидов до около 2,5 т.п.н., иметь длину от около 35 нуклеотидов до около 1,5 т.п.н. или иметь длину от около 35 до около 500 нуклеотидов. Например, область гомологии может иметь длину от около 35 до около 40, от около 40 до около 50, от около 50 до около 60, от около 60 до около 70, от около 70 до около 80, от около 80 до около 90, от около 90 до около 100, от около 100 до около 150, от около 150 до около 200, от около 200 до около 250, от около 250 до около 300, от около 300 до около 350, от около 350 до около 400, от около 400 до около 450 или от около 450 до около 500 нуклеотидов. Альтернативно, данная область гомологии может иметь длину от около 0,5 т.п.н. до около 1 т.п.н., от около 1 т.п.н. до около 1,5 т.п.н., от около 1,5 т.п.н. до около 2 т.п.н. или от около 2 т.п.н. до около 2,5 т.п.н. Например, область гомологии может иметь длину около 100 нуклеотидов.
[00116] Способ модификации направленного вектора с использованием бактериальной гомологичной рекомбинации может выполняться в различных системах (см., например, Yanget al. (1997) Nat.. Biotechnol. 15:859-65; Muyrers et al. (1999) Nucleic Acids Res. 27:1555-1557; Angrand et al. (1999) Nucleic Acids Res., 27:el6; Narayanan et al. (1999) Gene Ther., 6:442-447; и Yu et al. (2000) Proc. Natl. Acad. Sci. U.S.A. 97:5978-5983, каждая из которых включена в полном объеме в данный документ посредством ссылки для всех целей). Одним из примеров является ЕТ-клонирование (Zhang et al. (1998) Nat. Genet. 20:123-128 и Narayanan et al. (1999) Gene Ther., 6:442-447, каждая из которых включена в полном объеме в данный документ посредством ссылки для всех целей) и варианты этой технологии (Yu et al. (2000) Proc. Natl. Acad. Sci. U.S.A. 97:5978-5983, которая включена в полном объеме в данный документ посредством ссылки для всех целей). ЕТ относится к белкам recЕ и recТ, которые осуществляют реакцию гомологичной рекомбинации. RecE представляет собой экзонуклеазу, которая обрезает одну цепь линейной двухцепочечной ДНК от 5' к 3', оставляя таким образом линейный двухцепочечный фрагмент с 3'-одноцепочечным «липким» хвостом. Этот одноцепочечный «липкий» конец покрыт белком recТ, который обладает активностью связывания одноцепочечной ДНК (оцДНК, ssDNA). ЕТ-клонирование выполняется с использованием Е.coli, которая временно экспрессирует продукты гена Е.coli recЕ и recТ и белок лямбда (λ) бактериофага λgam. Белок λgam необходим для защиты фрагмента донорной ДНК от деградации экзонуклеазной системой recВС, и он необходим для эффективного ЕТ-клонирования в хозяевах recВС+, таких как часто используемый штамм Е. coli DH10b.
V. Сборка In Vitro
[00117] Любой способ сборки in vitro, который можно использовать для сборки, по меньшей мере, двух нуклеиновых кислот или, по меньшей мере, двух концов одной нуклеиновой кислоты в условиях, эффективных для соединения молекул ДНК с образованием по существу интактной молекулы ДНК, можно использовать в способах, описанных в данном документе. Некоторые неограничивающие примеры способов сборки in vitro включают стандартную сборку с использованием рестрикционных ферментов, сборку «in-fusion», последовательность-независимое безлигазное клонирование (SLIC, sequence and ligase independent cloning), сборку по Гибсону и сборку Golden Gate. См., например, Lee et al. (2013) Mol. Cells 35:359-370, которая включена в данный документ в полном объеме посредством ссылки для всех целей.
[00118] Одним из примеров подходящего способа сборки in vitro является изотермический способ с одной реакцией для сборки перекрывающихся молекул ДНК за счет согласованного действия экзонуклеазы (например, 5'-экзонуклеазы), ДНК-полимеразы и ДНК-лигазы. Нуклеиновые кислоты, имеющие перекрывающиеся концы (или одну линейную нуклеиновую кислоту с перекрывающимися концами), можно комбинировать с лигазой, экзонуклеазой и ДНК-полимеразой. Например, два соседних фрагмента ДНК, разделяющих перекрывающиеся концевые последовательности, могут быть объединены в ковалентно запечатанную молекулу в одностадийной изотермической реакции. В конкретном примере две или более молекулы ДНК, которые должны быть собраны, могут контактировать in vitro в одном сосуде с: (а) изолированной нетермостабильной 5'-3'-экзонуклеазой, которая лишена 3'-экзонуклеазной активности (например, непроцессивная экзонуклеаза, которая расщепляет концы двухцепочечных молекул ДНК, обнажая одноцепочечные «липкие» концы, содержащие области перекрывания); (b) краудинг-агентом (который, помимо других функций, может ускорять отжиг нуклеиновых кислот, так что одноцепочечные «липкие» концы специфически отжигаются (гибридизуются)); (с) изолированной термостабильной ДНК-полимеразой без цепь-вытесняющей активности, с 3'-экзонуклеазной активностью, или смесью указанной ДНК-полимеразы со второй ДНК-полимеразой, у которой отсутствует 3'-экзонуклеазная активность (для заполнения оставшихся одноцепочечных гэпов в гибридизированных молекулах путем удлинения 3'-концов гибридизированных областей); (d) изолированной термостабильной лигазой (которая заполняет (лигирует) образованные таким образом одноцепочечные разрывы); (е) смесью дНТФ; и (f) подходящим буфером в условиях, эффективных для соединения двух или более молекул ДНК с образованием первой собранной молекулы дцДНК в одностадийной реакции. Для одноцепочечных молекул экзонуклеаза может быть исключена, но не обязательно. В конкретном примере экзонуклеаза Т5 удаляет нуклеотиды с 5'-концов молекул двухцепочечной ДНК, «липкие» концы комплементарной одноцепочечной ДНК гибридизируются, и Phusion ДНК-полимераза заполняет гэпы, a Taq ДНК-лигаза закрывает разрывы. См., например, патент США 2010/0035768, патент США 2015/0376628, WO 2015/200334 и Gibson et al. (2009) Nat. Methods 6(5):343-345, каждый из которых включен в данный документ в полном объеме посредством ссылки для всех целей.
[00119] Первая и вторая одноцепочечные нуклеиновые кислоты имеют перекрывающиеся концы, когда их соответствующие концы комплементарны друг другу. Первая и вторая двухцепочечные нуклеиновые кислоты имеют перекрывающиеся концы, когда 5'-конец цепи первой нуклеиновой кислоты комплементарен 3'-концу цепи второй нуклеиновой кислоты, и наоборот. Например, для двухцепочечных перекрывающихся концевых последовательностей цепи одной нуклеиновой кислоты могут иметь, по меньшей мере, около 80%, по меньшей мере, около 85%, по меньшей мере, около 90%, по меньшей мере, около 95%, по меньшей мере, около 96%, по меньшей мере, около 97%, по меньшей мере, около 98%, по меньшей мере, около 99% или 100% идентичности с соответствующей цепью другой нуклеиновой кислоты. В способах, описанных в данном документе, 5'-конец цепи молекулы дцДНК, которая должна быть собрана, имеет общие перекрывающиеся концевые последовательности с 3'концом цепи другой молекулы дцДНК. Термин «перекрывающиеся концевые последовательности» включает обе цепи молекулы дцДНК. Таким образом, одна цепь из перекрывающейся области может специфически гибридизоваться со своей комплементарной цепью, когда комплементарные области перекрывающихся последовательностей представлены в виде одноцепочечных «липких» концов от 5'- и 3'-концов двух собираемых полинуклеотидов. Экзонуклеазу можно использовать для удаления нуклеотидов с 5'- или 3'-конца для создания «липких» концов последовательностей.
[00120] Длина перекрывающейся области может быть достаточной, так что область встречается только один раз в любой из собираемых нуклеиновых кислот.Таким образом, предотвращается гибридизация других полинуклеотидов с концевыми последовательностями, и сборка может быть специфичной для нуклеиновых кислот-мишеней. В качестве одного примера, длина перекрытия может содержать, по меньшей мере, около 20 нуклеотидов, по меньшей мере, около 30 нуклеотидов, по меньшей мере, около 40 нуклеотидов или, по меньшей мере, около 50 нуклеотидов. В качестве другого примера, длина перекрытия может составлять от около 20 нуклеотидов до около 100 нуклеотидов, от около 20 нуклеотидов до около 90 нуклеотидов, от около 20 нуклеотидов до около 80 нуклеотидов, от около 20 нуклеотидов до около 70 нуклеотидов, от около 20 нуклеотидов и около 60 нуклеотидов, от около 20 нуклеотидов до около 50 нуклеотидов, от около 20 нуклеотидов до около 40 нуклеотидов, от около 30 нуклеотидов до около 60 нуклеотидов, или от около 40 нуклеотидов до около 50 нуклеотидов. В конкретном примере длина перекрытия последовательностей может составлять от около 40 нуклеотидов до около 50 нуклеотидов (например, около 40 нуклеотидов или около 50 нуклеотидов).
[00121] Перекрывающиеся последовательности могут контактировать с экзонуклеазой, чтобы обнажить комплементарные последовательности (например, комплементарные одноцепочечные последовательности) между перекрывающимися последовательностями. Расщепление экзонуклеазой можно проводить в условиях, которые эффективны для удаления (отщепления) достаточного количества нуклеотидов, чтобы обеспечить возможность специфической гибридизации экспонированных одноцепочечных областей комплементарности. В общем, часть области перекрывания или вся область перекрывания отщепляется, оставляя «липкие» концы, которые составляют часть области перекрывания или всю область перекрывания. В некоторых способах расщепление экзонуклеазой может осуществляться полимеразой в отсутствие дНТФ (например, Т5 ДНК-полимеразой), тогда как в других способах расщепление экзонуклеазой может осуществляться экзонуклеазой в присутствии дНТФ, у которых отсутствует полимеразная активность (например, экзонуклеазой III).
[00122] Для отщепления концов нуклеиновых кислот в способах, описанных в данном документе, можно использовать любую из множества 5'-3'-экзодезоксирибонуклеаз, обладающих специфичностью к двухцепочечной ДНК. Термин «5'-экзонуклеаза» иногда используется в данном документе для обозначения 5'-3'-экзодезоксирибонуклеазы. Непроцессивная экзонуклеаза относится к экзонуклеазе, которая разрушает ограниченное количество (например, только несколько) нуклеотидов во время каждого события связывания ДНК. Расщепление 5'-экзонуклеазой продуцирует образование 3'-одноцепочечных «липких» хвостов в молекулах ДНК. 5'-экзонуклеазы, используемые в способах сборки in vitro, могут не обладать 3'-экзонуклеазной активностью, но могут генерировать 5'-фосфатные концы и могут инициировать деградацию как 5'-фосфорилированных, так и нефосфорилированных концов. Экзонуклеазы, используемые в описанных в данном документе способах сборки in vitro, могут инициировать расщепление с 5'-конца молекулы, будь то тупой конец или небольшой 5'- или 3'-недоступный конец. Подходящие экзонуклеазы хорошо известны и включают, например, экзонуклеазу фага Т5 (продукт гена D15 фага Т5), экзонуклеазу фага лямбда, RecE профага Rac, экзонуклеазу VIII из E.coli, экзонуклеазу фага Т7 (продукт гена 6 фага Т7) или любую из множества 5'-экзонуклеаз, которые участвуют в реакциях гомологичной рекомбинации. В качестве одного из примеров экзонуклеаза представляет собой экзонуклеазу Т5 или экзонуклеазу лямбда. В конкретном примере экзонуклеаза представляет собой экзонуклеазу Т5. В другом конкретном примере экзонуклеаза не является экзонуклеазой фага Т7.
[00123] В ситуациях, когда область перекрывания является длинной, может потребоваться отщепление только части области при условии, что образующиеся таким образом одноцепочечные «липкие» концы имеют достаточную длину и содержание оснований для гибридизации именно в условиях реакции. Термин «гибридизация (отжиг)», в частности, включает ситуации, в которых конкретная пара одноцепочечных «липких» концов будет гибридизироваться преимущественно (или исключительно) друг с другом, а не с другими одноцепочечными «липкими» концами (например, некомплементарными «липкими» концами), которые присутствуют в реакционной смеси. Под предпочтительно подразумевается, что, по меньшей мере, около 95% «липких» концов будут гибридизироваться с комплиментарными «липкими» концами. Как правило, гомологичные области перекрывания (одноцепочечные «липкие» концы или их комплементарные цепи) содержат идентичные последовательности. Однако могут использоваться частично идентичные последовательности при условии, что одноцепочечные «липкие» концы могут гибридизироваться именно в условиях реакций.
[00124] После гибридизации одноцепочечной ДНК (например, «липкие» концы, образующиеся под действием экзонуклеазы, когда молекулы ДНК, которые должны быть соединены, представляют собой дцДНК, или «липкие» концы, образующиеся путем создания разрывов в разных сайтах-мишенях на каждой цепи), одноцепочечные разрывы, оставленные экзонуклеазой могут быть заполнены подходящей ДНК-полимеразой без цепь-вытесняющей активности, и образовавшиеся таким образом разрывы могут быть закрыты лигазой. ДНК-полимераза без цепь-вытесняющей активности, как используется в данном документе, представляет собой ДНК-полимеразу, которая прекращает синтез ДНК, когда она встречает цепи ДНК, которые лежат на ее пути, когда она продолжает копировать молекулу дцДНК, или которая расщепляет встреченные нити ДНК по мере продвижения, одновременно заполняя образовавшийся разрыв, тем самым создавая движущийся одноцепочечный разрыв (ник-трансляция).
[00125] После гибридизации одной цепи первого полинуклеотида с комплементарной цепью второго полинуклеотида 3'-конец первого полинуклеотида может быть удлинен на основе матрицы второй полинуклеотидной цепи и 3'-конец второй полинуклеотидной цепи может быть удлинен на основе матрицы первой полинуклеотидной цепи. Путем удлинения комплементарного 3'-конца каждого полинуклеотида можно соединить полинуклеотиды. После сборки разрывы между удлиненным 3'-концом цепи одного фрагмента и соседним 5'-концом цепи другого фрагмента могут быть заполнены лигированием. Более конкретно, гидроксильная группа удлиненного 3'-конца первого полинуклеотида может быть лигирована с фосфатной группой 5'-конца второго полинуклеотида, и гидроксильная группа удлиненного 3'-конца второго полинуклеотида может быть лигирована с фосфатной группой 5'-конца первого полинуклеотида.
[00126] Реакцию лигирования можно проводить с помощью любой из множества подходящих термостабильных ДНК-лигаз. К подходящим лигазам относятся, например, Taq-лигаза, Термостабильная ДНК-лигаза Амплигаза или термостабильные лигазы, описанные в патенте США №6,576,453, который включен в полном объеме в данный документ посредством ссылки для всех целей.
[00127] Подходящее количество краудинг-агента, такого как ПЭГ, в реакционной смеси может способствовать, усиливать или облегчать «молекулярный краудинг». Такой краудинг-агент может позволить компонентам раствора войти в более тесный контакт друг с другом. Например, молекулы ДНК, подлежащие рекомбинации, могут оказаться ближе друг к другу; что может облегчить гибридизацию одноцепочечных «липких» концов. Подходящие краудинг-агенты известны и включают множество хорошо известных макромолекул, таких как полимеры, такие как полиэтиленгликоль (ПЭГ), фиколлы, такие как Фиколл 70, или декстраны, такие как декстран 70.
[00128] Компоненты реакции (такие как соли, буферы, подходящий источник энергии (например, АТФ или НАД), рН реакционной смеси и т.д.), которые присутствуют в реакционной смеси при сборке, могут не быть оптимальными для отдельных ферментов (экзонуклеазы, полимеразы и лигазы), но могут выступать в качестве компромиссных, которые эффективны для всего набора реакций.
VI. Направленные Векторы и Большие Направленные Векторы (БНВЕК)
[00129] Направленные векторы, используемые в описанных в данном документе способах, могут быть любыми подходящими направленными векторами. Направленные векторы могут содержать дезоксирибонуклеиновую кислоту (ДНК) или рибонуклеиновую кислоту (РНК), они могут быть одноцепочечными или двухцепочечными, и они могут иметь линейную или кольцевую форму. Направленные векторы могут представлять собой бактериальную искусственную хромосому (БИХ, ВАС), модифицированную БИХ или фрагментом БИХ. Они могут содержать ДНК человека, ДНК грызунов (например, ДНК мыши или ДНК крысы), синтетическую ДНК или любую их комбинацию.
[00130] Некоторые направленные векторы, используемые в описанных в данном документе способах, представляют собой большие направленные векторы (БНВЕК, LTVEC). БНВЕК включают направленные векторы, которые содержат плечи гомологии, которые соответствуют последовательностям нуклеиновых кислот и происходят из них, и при этом являются более крупными, чем те, которые обычно используются другими подходами, предназначенными для выполнения гомологичной рекомбинации в клетках. БНВЕК также включают направленные векторы, содержащие вставки нуклеиновых кислот, имеющие последовательности нуклеиновых кислот, большие, чем те, которые обычно используются другими подходами, предназначенными для выполнения гомологичной рекомбинации в клетках. Например, БНВЕК делают возможной модификацию больших локусов, которые не могут быть размещены традиционными направленными векторами на основе плазмид из-за их ограничений по размеру. Например, локус-мишень может представлять собой (т.е. 5'- и 3'-плечам гомологии могут соответствовать) локус клетки, на который не может быть таргетирован с помощью обычного способа или который может быть таргетирован только неправильно или только со значительно низкой эффективностью при отсутствии одноцепочечного или двухцепочечного разрыва, вызванного нуклеазным агентом (например, белком Cas). Примеры БНВЕК включают векторы, полученные из бактериальной искусственной хромосомы (БИХ), искусственной хромосомы человека или искусственной хромосомы дрожжей (ИХД, YAC). Неограничивающие примеры БНВЕК и способы их получения описаны, например, в патентах США№6,586,251; 6,596,541; и 7,105,348; и в WO 2002/036789, каждый из которых включен в данный документ в полном объеме посредством ссылки для всех целей. БНВЕК могут иметь линейную или круговую форму. БНВЕК могут иметь любую длину и обычно имеют длину, по меньшей мере, 10 т.п.н. Размер БНВЕК может быть слишком большим для возможности скрининга событий таргетирования с помощью обычных анализов, например саузерн-блоттинга и ПЦР длинных фрагментов (например, от 1 т.п.н. до 5 т.п.н.).
[00131] Направленные векторы (например, БНВЕК), используемые в описанных в данном документе способах, могут иметь любую длину. Например, направленный вектор может составлять от около 10 т.п.н. до около 400 т.п.н., от около 20 т.п.н. до около 400 т.п.н., от около 20 т.п.н. до около 30 т.п.н., от около 30 т.п.н. до около 40 т.п.н., от около 40 т.п.н. до около 50 т.п.н., от около 50 т.п.н. до около 75 т.п.н., от около 75 т.п.н. до около 100 т.п.н., от около 100 т.п.н. до 125 т.п.н., от около 125 т.п.н. до около 150 т.п.н., от около 150 т.п.н. до около 175 т.п.н., от около 175 т.п.н. до около 200 т.п.н., от около 200 т.п.н. до около 225 т.п.н., от около 225 т.п.н. до около 250 т.п.н., от около 250 т.п.н. до около 275 т.п.н. или от около 275 т.п.н. до около 300 т.п.н., от около 200 т.п.н. до около 300 т.п.н., от около 300 т.п.н. до около 350 т.п.н., или от около 350 т.п.н. до около 400 т.п.н. В одном примере направленный вектор может иметь длину, по меньшей мере, около 100 т.п.н. или 100 т.п.н. Направленный вектор также может составлять от около 50 т.п.н. до около 500 т.п.н., от около 100 до около 125 т.п.н., от около 300 т.п.н. до около 325 т.п.н., от около 325 т.п.н. до около 350 т.п.н., от около 350 т.п.н. до около 375 т.п.н., от около 375 т.п.н. до около 400 т.п.н., от около 400 т.п.н. до около 425 т.п.н., от около 425 т.п.н. до около 450 т.п.н., от около 450 т.п.н. до около 475 т.п.н. или от около 475 т.п.н. до около 500 т.п.н. Альтернативно, направленный вектор может составлять, по меньшей мере, 10 т.п.н., по меньшей мере, 15 т.п.н., по меньшей мере, 20 т.п.н., по меньшей мере, 30 т.п.н., по меньшей мере, 40 т.п.н., по меньшей мере, 50 т.п.н., по меньшей мере, 60 т.п.н., по меньшей мере, 70 т.п.н., по меньшей мере, 80 т.п.н., по крайней мере, 90 т.п.н., по меньшей мере, 100 т.п.н., по меньшей мере, 150 т.п.н., по меньшей мере, 200 т.п.н., по меньшей мере, 250 т.п.н., по меньшей мере, 300 т.п.н., по меньшей мере, 350 т.п.н., по меньшей мере, 400 т.п.н., по меньшей мере, 450 т.п.н., или, по меньшей мере, 500 т.п.н. или более.
VII. Нуклеазные Агенты
[00132] В описанных в данном документе способах можно использовать любой редко расщепляющий нуклеазный агент.Редко расщепляющий нуклеазный агент представляет собой нуклеазный агент с последовательностью-мишенью или последовательностью распознавания, которая редко встречается в геноме. Точно так же можно использовать любой нуклеазный агент с последовательностью-мишенью или последовательностью распознавания, которая не встречается за пределами желаемого сайта(-ов) расщепления в направленных векторах, описанных в данном документе. Например, может быть использован любой нуклеазный агент, который не имеет последовательности-мишени или последовательности распознавания в ранее созданных направленных векторах в способах, описанных в данном документе.
[00133] Любой нуклеазный агент, как описано выше, который вызывает одноцепочечный или двухцепочечный разрыв в желаемой последовательности-мишени, можно использовать в способах и композициях, описанных в данном документе. Можно использовать встречающийся в природе или нативный нуклеазный агент при условии, что нуклеазный агент вызывает одноцепочечный или двухцепочечный разрыв в желаемой последовательности-мишени. В качестве альтернативы можно использовать модифицированный или сконструированный нуклеазный агент. «Сконструированный нуклеазный агент» включает нуклеазу, которая сконструирована (модифицирована или получена) из ее нативной формы, чтобы специфически распознавать и индуцировать одноцепочечный или двухцепочечный разрыв в желаемой последовательности-мишени. Таким образом, сконструированный нуклеазный агент может быть получен из нативного природного нуклеазного агента или может быть создан или синтезирован искусственно. Сконструированная нуклеаза может вызывать одноцепочный или двухцепочечный разрыв в последовательности-мишени, например, при этом последовательность-мишень не является последовательностью, которая была бы распознана нативным (несконструированным или немодифицированным) нуклеазным агентом. Модификация нуклеазного агента может представлять собой изменение всего одной аминокислоты в белке расщепляющего агента или одного нуклеотида в нуклеиновой кислоте расщепляющего агента. Получение одноцепочечного или двухцепочечного разрыва в последовательности-мишени или другой ДНК может быть обозначено в данном документе как «разрезание» или «расщепление» последовательности-мишени или другой ДНК.
[00134] Также представлены активные варианты и фрагменты приведенных в качестве примеров последовательностей-мишеней. Такие активные варианты могут иметь, по меньшей мере, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичности последовательности с данной последовательностью-мишенью, при этом активные варианты сохраняют биологическую активность и, следовательно, способны распознаваться и расщепляться нуклеазным агентом специфичным для последовательности образом. Способы измерения двухцепочечного разрыва последовательности-мишени нуклеазным агентом хорошо известны. См., например, Frendewey et al. (2010) Methods in Enzymology 476:295-307, которая включена в данный документ в полном объеме посредством ссылки для всех целей.
[00135] Также предложены активные варианты и фрагменты нуклеазных агентов (т.е. сконструированные нуклеазные агенты). Такие активные варианты могут иметь, по меньшей мере, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичности последовательности с нативным нуклеазным агентом, при этом активные варианты сохраняют способность разрезать желаемую последовательность-мишень и, следовательно, сохраняют активность, индуцирующую одноцепочечный или двухцепочечный разрыв. Например, любой из описанных в данном документе нуклеазных агентов может быть модифицирован из последовательности нативной эндонуклеазы и разработан для распознавания и индуцирования одноцепочечных или двухцепочечных разрывов в последовательности-мишени, которая не распознается нативным нуклеазным агентом. Таким образом, некоторые сконструированные нуклеазы обладают специфичностью, индуцирующей одноцепочечный или двухцепочечный разрыв в последовательности-мишени, которая отличается от соответствующей последовательности-мишени нативного нуклеазного агента. Анализы на активность, индуцирующую одноцепочечный или двухцепочечный разрыв, известны и обычно измеряют общую активность и специфичность эндонуклеазы на субстратах ДНК, содержащих последовательность-мишень.
[00136] Последовательность-мишень нуклеазы содержит последовательность ДНК, в которой нуклеазный агент индуцирует одноцепочечный или двухцепочечный разрыв. Длина последовательности-мишени может варьироваться и включает, например, последовательности-мишени, которые составляют около 30-36 п.н. для пары нуклеаз с «цинковыми пальцами» (ZFN) (т.е. около 15-18 п.н. для каждой ZFN), около 36 п.н. для эффекторной нуклеазой, подобной активатору транскрипции (TALEN), или около 20 п.н. для направляющей РНК CRISPR/Cas9.
А. Рестрикционные ферменты
[00137] Нуклеазные агенты, подходящие для использования в описанных в данном документе способах, могут содержать эндонуклеазы рестрикции, которые включают эндонуклеазы Типа I, Типа II, Типа III и Типа IV. Эндонуклеазы рестрикции Типа I и Типа III распознают специфические сайты рестрикции, но обычно расщепляют в вариабельном положении от сайта связывания нуклеазы, который может находиться на расстоянии сотен пар оснований от сайта расщепления (сайта распознавания). В системах Типа II рестрикционная активность не зависит от какой-либо активности метилазы, и расщепление обычно происходит в определенных сайтах внутри или рядом с сайтом связывания. Большинство ферментов Типа II разрезают палиндромные последовательности, однако ферменты Типа IIа распознают непалиндромные сайты распознавания и расщепляют за пределами сайта распознавания, ферменты типа IIb дважды разрезают последовательности, при этом оба сайта находятся за пределами сайта распознавания, а ферменты Типа IIs распознают асимметричный сайт распознавания и расщепляют с одной стороны и на определенном расстоянии около 1-20 нуклеотидов от сайта распознавания. Рестрикционные ферменты Типа IV направлены на метилированную ДНК. Рестрикционные ферменты дополнительно описаны и классифицированы, например, в базе данных REBASE (веб-страница на rebase.neb.com; Roberts et al. (2003) Nucleic Acids Res. 31:418-20); Roberts et al. (2003) Nucleic Acids Res. 31:1805-12; и Belfort et al. (2002) in Mobile DNA II, pp.761-783, Eds. Craigie et al. (ASM Press, Вашингтон, округ Колумбия), каждая из которых включена в полном объеме в данный документ посредством ссылки для всех целей.
[00138] В некоторых способах используется редко расщепляющий рестрикционный фермент. Редко расщепляющий рестрикционный фермент относится к ферменту с сайтом-мишенью или сайтом распознавания, который редко встречается в геноме. Размер рестрикционных фрагментов, созданных путем разрезания гипотетического случайного генома рестрикционным ферментом, может быть приблизительно равен 4N, где N представляет собой количество нуклеотидов в сайте распознавания фермента. Например, фермент с сайтом распознавания, состоящим из 7 нуклеотидов, будет расщеплять геном каждые 47 п.н., продуцируя фрагменты около 16,384 п.н. Обычно редко расщепляющие ферменты имеют сайты распознавания, содержащие 6 или более нуклеотидов. Например, редко расщепляющий фермент может иметь сайт распознавания, содержащий 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 нуклеотидов или состоящий из них. Примеры редко расщепляющих рестрикционных ферментов включают NotI (GCGGCCGC), XmaIII (CGGCCG), SstII (CCGCGG), Sall (GTCGAC), NruI (TCGCGA), NheI (GCTAGC), Nb.BbvCI (CCTCAGC), BbvCI (CCTCAGC), AscI (GGCGCGCC), AsiSI (GCGATCGC), FseI (GGCCGGCC), PacI (TTAATTAA), PmeI (GTTTAAAC), SbfI (CCTGCAGG), SgrAI (CRCCGGYG), SwaI (ATTTAAAT), BspQI (GCTCTTC), SapI (GCTCTTC), SfiI (GGCCNNNNNGGCC), CspCI (CAANNNNNGTGG), AbsI (CCTCGAGG), CciNI (GCGGCCGC), FspAI (RTGCGCAY), MauBI (CGCGCGCG), MreI (CGCCGGCG), MssI (GTTTAAAC), PalAI (GGCGCGCC), RgaI (GCGATCGC), RigI (GGCCGGCC), SdaI (CCTGCAGG), SfaAI (GCGATCGC), SgfI (GCGATCGC), SgrDI (CGTCGACG), SgsI (GGCGCGCC), SmiI (ATTTAAAT), SrfI (GCCCGGGC), Sse2321 (CGCCGGCG), Sse83871 (CCTGCAGG), LguI (GCTCTTC), PciSI (GCTCTTC), AarI (CACCTGC), AjuI (GAANNNNNNNTTGG), AloI (GAACNNNNNNTCC), BarI (GAAGNNNNNNTAC), PpiI (GAACNNNNNCTC), PsrI (GAACNNNNNNTAC) и другие.
Б. Системы CRISPR/Cas
[00139] Системы (Cas) кластеризованных регулярных промежуточных коротких палиндромных повторов (CRISPR)/CRISPR-ассоциированные (Cas) системы также могут быть использованы в качестве редко расщепляющих нуклеазных агентов в описанных в данном документе способах. Системы CRISPR/Cas включают транскрипты и другие элементы, участвующие в экспрессии генов Cas или управляющие ими. Система CRISPR/Cas может быть, например, системой типа I, типа II, системой типа III или системой типа V (например, подтипом V-A или подтипом V-B). Системы CRISPR/Cas, используемые в описанных в данном документе композициях и способах, могут быть не встречающимися в природе. «Не встречающаяся в природе» система включает все, что указывает на участие руки человека, например, один или более компонентов системы, измененных или мутировавших по сравнению с их природным состоянием, по меньшей мере, практически отсутствие, по меньшей мере, одного другого компонента, с которым компоненты системы естественным образом связаны в природе, или связь, по меньшей мере, с одним другим компонентом, с которым компоненты системы не связаны в природе. Например, в некоторых системах CRISPR/Cas используются не встречающиеся в природе комплексы CRISPR, содержащие нРНК и белок Cas, которые не встречаются в природе вместе, используется белок Cas, который не встречается в природе, или используется нРНК, которая не встречается в природе.
[00140] Белки Cas и Полинуклеотиды, Кодирующие Белки Cas. Белки Cas обычно содержат, по меньшей мере, один распознающий или связывающий домен РНК, который может взаимодействовать с направляющими РНК (нРНК). Белки Cas также могут содержать нуклеазные домены (например, ДНКазные домены или РНКазные домены), ДНК-связывающие домены, геликазные домены, домены межбелкового взаимодействия, домены димеризации и другие домены. Некоторые такие домены (например, домены ДНКазы) могут происходить из природного белка Cas. Другие такие домены могут быть добавлены для получения модифицированного белка Cas. Нуклеазный домен обладает каталитической активностью в отношении расщепления нуклеиновой кислоты, которое включает разрыв ковалентных связей молекулы нуклеиновой кислоты. Расщепление может приводить к тупым или ступенчатым концам, и оно может быть одноцепочечным или двухцепочечным. Например, белок Cas9 дикого типа обычно создает тупой конец продукта расщепления. Альтернативно, белок Cpf1 дикого типа (например, FnCpf1) может приводить к продукту расщепления с 5'-«липким» концом из 5 нуклеотидов, при этом расщепление происходит после 18-й пары оснований из последовательности РАМ на нецелевой цепи и после 23-го основания на цепи-мишени. Белок Cas может обладать полной активностью расщепления для создания двухцепочечного разрыва в геномном локусе-мишени (например, двухцепочечного разрыва с тупыми концами), или это может быть никаза, которая создает одноцепочечный разрыв в геномном локусе-мишени.
[00141] Примеры белков Cas включают Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas5e (CasD), Cas6, Cas6e, Cas6f, Cas7, Cas8a1, Cas8a2, Cas8b, Cas8c, Cas9 (Csn1 или Csx12), Cas10, Cas10d, CasF, CasG, CasH, Csy1, Csy2, Csy3, Cse1 (CasA), Cse2 (CasB), Cse3 (CasE), Cse4 (CasC), Csc1, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx15, Csf1, Csf2, Csf3, Csf4 и Cu1966, и их гомологи или их модифицированные версии.
[00142] Примером белка Cas является белок Cas9 или белок, полученный из белка Cas9. Белки Cas9 происходят из системы CRISPR/Cas типа II и обычно имеют четыре ключевых мотива с консервативной архитектурой. Мотивы 1, 2 и 4 представляют собой RuvC-подобные мотивы, а мотив 3 представляет собой мотив HNH. Типовые белки Cas9 происходят из Streptococcus pyogenes, Streptococcus thermophilus, Streptococcus sp., Staphylococcus aureus, Nocardiopsis dassonvillei, Streptomyces pristinaespiralis, Streptomyces viridochromogenes, Streptomyces viridochromogenes, Streptosporangium roseum, Streptosporangium roseum, Alicyclobacillus acidocaldarius, Bacillus pseudomycoides, Bacillus selenitireducens, Exiguobacterium sibiricum, Lactobacillus delbrueckii, Lactobacillus salivarius, Microscilla marina, Burkholderiales bacterium, Polaromonas naphthalenivorans, Polaromonas sp., Crocosphaera watsonii, Cyanothece sp., Microcystis aeruginosa, Synechococcus sp.,Acetohalobium arabaticum, Ammonifex degensii, Caldicelulosiruptor becscii, Candidatus Desulforudis, Clostridium botulinum, Clostridium difficile, Finegoldia magna, Natranaerobius thermophilus, Pelotomaculum thermopropionicum, Acidithiobacillus caldus, Acidithiobacillus ferrooxidans, Allochromatium vinosum, Marinobacter sp., Nitrosococcus halophilus, Nitrosococcus watsoni, Pseudoalteromonas haloplanktis, Ktedonobacter racemifer, Methanohalobium evestigatum, Anabaena variabilis, Nodularia spumigena, Nostoc sp., Arthrospira maxima, Arthrospira platensis, Arthrospira sp., Lyngbya sp., Microcoleus chthonoplastes, Oscillatoria sp.,Petrotoga mobilis, Thermosipho africanus, Acaryochloris marina, Neisseria meningitidis или Campylobacter jejuni. Дополнительные примеры членов семейства Cas9 описаны в WO 2014/131833, включенном в данный документ в полном объеме посредством ссылки для всех целей. Cas9 из S. pyogenes (SpCas9) (которому присвоен номер доступа SwissProt Q99ZW2) представляет собой типовой белок Cas9. Cas9 из S. aureus (SaCas9) (которому присвоен номер доступа UniProt J7RUA5) представляет собой еще один пример белка Cas9. Cas9 из Campylobacter jejuni (CjCas9) (которому присвоен номер доступа UniProt Q0P897) представляет собой другой типовой белок Cas9. См., например, Kim et al. (2017) Nat. Commun. 8:14500, которая включена в данный документ в полном объеме посредством ссылки для всех целей. SaCas9 меньше, чем SpCas9, a CjCas9 меньше, чем SaCas9 и SpCas9. Cas9 из Neisseria meningitidis (Nme2Cas9) представляет собой еще один типовой белок Cas9. См., например, Edraki et al. (2019) Mol. Cell 73(4):714-726, которая включена в данный документ в полном объеме посредством ссылки для всех целей. Белки Cas9 из Streptococcus thermophilus (например, Streptococcus thermophilus LMD-9 Cas9, кодируемый локусом CRISPR1 (St1Cas9) или Streptococcus thermophilus Cas9 из локуса CRISPR3 (St3Cas9) представляют собой другие типовые белки Cas9. Cas9 из Francisella novicida (FnCas9) или вариант Cas9 RHA Francisella novicida, который распознает альтернативный РАМ (замены E1369R/E1449H/R1556A) представляют собой другие типовые белки Cas9. Эти и другие типовые белки Cas9 рассмотрены, например, в Cebrian-Serrano and Davies (2017) Mamm. Genome 28(7):247-261, которая включена в данный документ в полном объеме посредством ссылки для всех целей. Типовая последовательность белка Cas9 может содержать, состоять из или по существу из SEQ ID NO: 1. Типовая ДНК, кодирующая белок Cas9, может содержать, состоять из или по существу из SEQ ID NO: 2.
[00143] Другим примером белка Cas является белок Cpf1 (CRISPR из Prevotella и Francisella 1). Cpf1 представляет собой большой белок (около 1300 аминокислот), который содержит RuvC-подобный нуклеазный домен, гомологичный соответствующему домену Cas9, вместе с аналогом характерного богатого аргинином кластера Cas9. Однако в Cpf1 отсутствует нуклеазный домен HNH, который присутствует в белках Cas9, а RuvC-подобный домен является граничащим в последовательности Cpf1, в отличие от Cas9, где он содержит длинные вставки, включая домен HNH. См., например, Zetsche et al. (2015) Cell 163(3):759-771, которая включена в данный документ в полном объеме посредством ссылки для всех целей. Типовые белки Cpf1 происходят из Francisella tularensis 1, Francisella tularensis subsp.novicida, Prevotella albensis, Lachnospiraceae bacterium MC2017 1, Butyrivibrio proteoclasticus, Peregrinibacteria bacterium GW2011_GWA2_33_10, Parcubacteria bacterium GW2011 _GWC2_44_17, Smithella sp.SCADC, Acidaminococcus sp.BV3L6, Lachnospiraceae bacterium MA2020, Candidatus Methanoplasma termitum, Eubacterium eligens, Moraxella bovoculi 237, Leptospira inadai, Lachnospiraceae bacterium ND2006, Porphyromonas crevioricanis 3, Prevotella disiens и Ρorphyromonas macacae. Cpf1 из Francisella novicida U112 (FnCpf1; которому присвоен номер доступа UniProt A0Q7Q2) представляет собой типовой белок Cpf1.
[00144] Белки Cas могут быть белками дикого типа (т.е. белками, которые встречаются в природе), модифицированными белками Cas (т.е. вариантами белков Cas) или фрагментами белков Cas дикого типа или модифицированных белков Cas. Белки Cas также могут быть активными вариантами или фрагментами в отношении каталитической активности белков Cas дикого типа или модифицированных белков Cas. Активные варианты или фрагменты в отношении каталитической активности могут иметь, по меньшей мере, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичности последовательности с белком Cas дикого типа или модифицированным белком Cas или его частью, при этом активные варианты сохраняют способность разрезать в желаемом сайте расщепления и, следовательно, сохраняют активность, индуцирующую одноцепочечный или двухцепочечный разрыв. Анализы на активность, индуцирующую одноцепочечный или двухцепочечный разрыв, известны и обычно измеряют общую активность и специфичность белка Cas на субстратах ДНК, содержащих сайт расщепления.
[00145] Белки Cas можно модифицировать для увеличения или уменьшения одного или более из следующих параметров: аффинность связывания нуклеиновой кислоты, специфичность связывания нуклеиновой кислоты и ферментативная активность. Белки Cas также можно модифицировать для изменения любой другой активности или свойства белка, например стабильности. Например, один или более нуклеазных доменов белка Cas могут быть модифицированы, удалены или инактивированы, или белок Cas может быть усечен для удаления доменов, которые не являются существенными для функции белка, или для оптимизации (например, увеличения или уменьшения) активности или свойств белка Cas.
[00146] Одним из примеров модифицированного белка Cas является модифицированный белок SpCas9-HF1, который представляет собой высокоточный вариант Streptococcus pyogenes Cas9, несущий изменения (N497A/R661A/Q695A/Q926A), предназначенный для уменьшения неспецифических контактов ДНК. См., например, Kleinstiver et al. (2016) Nature 529(7587):490-495, которая включена в данный документ в полном объеме посредством ссылки для всех целей. Другим примером модифицированного белка Cas является модифицированный вариант eSpCas9 (K848A/K1003A/R1060A), предназначенный для уменьшения нецелевых воздействий. См., например, Slaymaker et al. (2016) Science 351(6268):84-88, которая включена в данный документ в полном объеме посредством ссылки для всех целей. Другие варианты SpCas9 включают К855А и K810A/K1003A/R1060A. Обзор этих и других модифицированных белков Cas приведен, например, в Cebrian-Serrano and Davies (2017) Mamm. Genome 28(7):247-261, которая включена в данный документ в полном объеме посредством ссылки для всех целей. Другим примером модифицированного белка Cas9 является xCas9, который представляет собой вариант SpCas9, который может распознавать расширенный диапазон последовательностей РАМ. См., например, Hu et al. (2018) Nature 556:57-63, которая включена в данный документ в полном объеме посредством ссылки для всех целей.
[00147] Белки Cas могут содержать, по меньшей мере, один нуклеазный домен, такой как ДНКазный домен. Например, белок Cpf1 дикого типа обычно содержит RuvC-подобный домен, который расщепляет обе цепи ДНК-мишени, возможно, в димерной конфигурации. Белки Cas также могут содержать, по меньшей мере, два нуклеазных домена, например, ДНКазные домены. Например, белок Cas9 дикого типа обычно содержит домен RuvC-подобный и HNH-подобный нуклеазный домен. Каждый из доменов RuvC и HNH может разрезать разные цепи двухцепочечной ДНК, чтобы образовать двухцепочечный разрыв в ДНК. См., например, Jinek et al. (2012) Science 337:816-821, которая включена в данный документ в полном объеме посредством ссылки для всех целей.
[00148] Один или более нуклеазных доменов могут быть удалены или мутированы, так что они больше не функционируют или имеют пониженную нуклеазную активность. Например, если один из нуклеазных доменов удален или мутирован в белке Cas9, полученный белок Cas9 может называться никазой и может генерировать одноцепочечный разрыв в двухцепочечной ДНК-мишени, но не двухцепочечный разрыв (т.е. он может расщеплять комплементарную цепь или некомплементарную цепь, но не обе). Примером мутации, которая превращает Cas9 в никазу, является мутация D10A (аспартат в аланин в положении 10 Cas9) в домене RuvC Cas9 из S. pyogenes. Аналогичным образом, Н939А (гистидин в аланин в положении аминокислоты 839), Н840А (гистидин в аланин в положении аминокислоты 840) или N863A (аспарагин в аланин в положении аминокислоты N863) в домене HNH Cas9 из S. pyogenes может преобразовывать Cas9 в никазу. Другие примеры мутаций, которые превращают Cas9 в никазу, включают соответствующие мутации Cas9 из S. thermophilus. См., например, Sapranauskas et al. (2011) Nucleic Acids Res. 39(21):9275-9282 и WO 2013/141680, каждая из которых включена в полном объеме в данный документ посредством ссылки для всех целей. Такие мутации могут быть получены с использованием таких способов, как сайт-направленный мутагенез, ПЦР-опосредованный мутагенез, или тотальный синтез генов. Примеры других мутаций, создающих никазы, можно найти, например, в WO 2013/176772 и WO 2013/142578, каждая из которых включена в данный документ в полном объеме посредством ссылки для всех целей.
[00149] Примеры инактивирующих мутаций в каталитических доменах xCas9 такие же, как описанные выше для SpCas9. Также известны примеры инактивирующих мутаций в каталитических доменах белков Cas9 Staphylococcus aureus. Например, фермент Cas9 Staphyloccocus aureus (SaCas9) может содержать замену в положении N580 (например, замещение N580А) для создания никазы. Альтернативно, фермент SaCas9 может содержать замену в положении D10 (например, замену D10A) для образования никазы. См., например, WO 2016/106236, которая включена в данный документ в полном объеме посредством ссылки для всех целей. Также известны примеры инактивирующих мутаций в каталитических доменах Nme2Cas9 (например, комбинация D16A и Н588А). Также известны примеры инактивирующих мутаций в каталитических доменах St1Cas9 (например, комбинация D9A, D598A, Н599А и N622A). Также известны примеры инактивирующих мутаций в каталитических доменах St3Cas9 (например, комбинация D10A и N870А). Также известны примеры инактивирующих мутаций в каталитических доменах CjCas9 (например, комбинация D8A и Н559А). Также известны примеры инактивирующих мутаций в каталитических доменах FnCas9 и RHA FnCas9 (например, N995A).
[00150] Также известны примеры инактивирующих мутаций в каталитических доменах белков Cpf1. Что касается белков Cpf1 из Francisella novicida U112 (FnCpf1), Acidaminococcus sp.BV3L6 (AsCpf1), Lachnospiraceae bacterium ND2006 (LbCpf1) и Moraxella bovoculi 237 (MbCpf1 Cpf1), такие мутации могут включать мутации в положениях 908, 993 или 1263 AsCpf1 или в соответствующих положениях в ортологах Cpf1, или в положениях 832, 925, 947 или 1180 LbCpf1 или в соответствующих положениях в ортологах Cpf1. Такие мутации могут включать, например, одну или более мутаций D908A, Е993А и D1263A AsCpf1 или соответствующие мутации в ортологах Cpf1, или D832A, Е925А, D947A и D1 180А LbCpf1 или соответствующие мутации в ортологах Cpf1. См., например, патент США 2016/0208243, который включен в данный документ в полном объеме посредством ссылки для всех целей.
[00151] Белки Cas также могут быть функционально связаны с гетерологичными полипептидами в качестве слитых белков. Например, белок Cas может быть присоединен к домену расщепления. См. WO 2014/089290, которая включена в данный документ в полном объеме посредством ссылки для всех целей. Белки Cas также могут быть слиты с гетерологичный полипептидом, обеспечивая повышенную или пониженную стабильность. Слитый домен или гетерологичный полипептид может быть расположен на N-конце, С-конце или внутри белка Cas.
[00152] В качестве одного примера, белок Cas может быть слит с одним или более гетерологичными полипептидами, которые обеспечивают субклеточную локализацию. Такие гетерологичные полипептиды могут содержать, например, один или более клеточных сигналов внутриядерной локализации (NLS), таких как однокомпонентный NLS SV40 и/или двухкомпонентный NLS альфа-импортина для нацеливания на ядро, сигнал митохондриальной локализации для нацеливания на митохондрии, сигнал ER-удержанияи т.п. См., например, Lange et al. (2007) J. Biol. Chem. 282(8):5101-5105, которая включена в данный документ в полном объеме посредством ссылки для всех целей. Такие сигналы субклеточной локализации могут быть расположены на N-конце, С-конце или где-либо в пределах белка Cas. NLS может содержать отрезок основных аминокислот и может быть однокомпонентной последовательностью или двухкомпонентной последовательностью. Необязательно, белок Cas может содержать два или более NLS, включая NLS (например, NLS альфа-импортина или однокомпонентный NLS) на N-конце и NLS (например, NLS SV40 или двухкомпонентный NLS) на С-конце. Белок Cas также может содержать два или более NLS на N-конце и/или два или более NLS на С-конце.
[00153] Белки Cas также могут быть функционально связаны с доменом проникновения в клетку, или доменом трансдукции белка. Например, домен проникновения в клетки, может происходить из белка ТАТ ВИЧ-1, TLM-мотива проникновения в клетки из вируса гепатита В человека, MPG, Рер-1, VP22, пептида проникновения в клетки из вируса простого герпеса или пептидной последовательности полиаргинина. См., например, WO 2014/089290 и WO 2013/176772, каждая из которых включена в полном объеме в данный документ посредством ссылки для всех целей. Домен проникновения в клетку может располагаться на N-конце, С-конце или где-либо в пределах белка Cas.
[00154] Белки Cas, представленные в виде мРНК, можно модифицировать для улучшения свойств стабильности и/или иммуногенности. Модификации могут быть внесены в один или более нуклеозидов внутри мРНК. Примеры химических модификаций азотистых оснований мРНК включают псевдоуридин, 1-метил-псевдоуридин и 5-метилцитидин. Например, можно использовать кэпированную и полиаденилированную мРНК Cas, содержащую N1-метил псевдоуридин. Точно так же мРНК Cas могут быть изменены за счет истощения уридина с использованием синонимичных кодонов.
[00155] Направляющие РНК. «Направляющая РНК» или «нРНК» представляет собой молекулу РНК, которая связывается с белком Cas (например, с белком Cas9) и направляет белок Cas в определенное место внутри ДНК-мишени. Направляющие РНК могут включать два сегмента: «ДНК-направляющий сегмент» и «белок-связывающий сегмент». «Сегмент» включает часть или область молекулы, такую как непрерывный участок нуклеотидов в РНК. Некоторые нРНК, например, для Cas9, могут содержать две отдельные молекулы РНК: «РНК-активатор» (например, тракрРНК) и «РНК-нацеливатель» (например, CRISPR-PHK или крРНК). Другие нРНК представляют собой одну молекулу РНК (полинуклеотид с одной РНК), которую также можно назвать «одномолекулярной нРНК», «однонаправляющей РНК» или «онРНК (sgRNA)». См., например, WO 2013/176772, WO 2014/065596, WO 2014/089290, WO 2014/093622, WO 2014/099750, WO 2013/142578 и WO 2014/131833, каждая из которых включена в полном объеме в данный документ посредством ссылки для всех целей. Для Cas9, например, однонаправляющая РНК может содержать крРНК, слитую с тракрРНК (например, через линкер). Для Cpf1, например, только крРНК необходима для достижения связывания и/или расщепления последовательности-мишени. Термины «направляющая РНК» и «нРНК» включают как двухмолекулярные (то есть модульные) нРНК, так и одномолекулярные нРНК.
[00156] Типовая двухмолекулярная нРНК включает крРНК-подобную («CRISPR-ΡΗΚ» или «РНК-нацеливатель», или «крРНК» или «повтор крРНК») молекулу и соответствующую тракрРНК-подобную («транс-активирующую CRISPR-ΡΗΚ» или «РНК-активатор» или «тракрРНК») молекулу. КрРНК содержит как ДНК-направляющий сегмент (одноцепочечный) нРНК, так и участок нуклеотидов (т.е. хвост крРНК), который формирует половину дуплекса дцРНК из белок-связывающего сегмента нРНК. Пример хвоста крРНК, расположенного ниже (направлении 3'-конца) ДНК-направляющего сегмента, включает, по существу состоит из или состоит из GUUUUAGAGCUAUGCU (SEQ ID NO: 3). Любой из описанных в данном документе ДНК-направляющих сегментов может быть присоединен к 5'-концу SEQ ID NO: 3 с образованием крРНК.
[00157] Соответствующая тракрРНК (РНК-активатор) содержит участок нуклеотидов, который формирует другую половину дуплекса дцРНК белок-связывающего сегмента нРНК. Участок нуклеотидов крРНК комплементарен участку нуклеотидов тракрРНК и гибридизуется с участком нуклеотидов тракрРНК с образованием дуплекса дцРНК белка-связывающего домена нРНК. Таким образом, можно сказать, что каждая крРНК имеет соответствующую тракрРНК. Пример последовательности тракрРНК содержит, по существу состоит из или состоит из
 Другие примеры последовательности тракрРНК содержат, по существу состоят из, или состоят из
Другие примеры последовательности тракрРНК содержат, по существу состоят из, или состоят из

[00158] В системах, в которых необходимы как крРНК, так и тракрРНК, крРНК и соответствующая тракрРНК гибридизуются с образованием нРНК. В системах, в которых необходима только крРНК, крРНК может представлять собой нРНК. КрРНК дополнительно обеспечивает одноцепочечный ДНК-направляющий сегмент, который гибридизуется с комплементарной цепью ДНК-мишени. Если используется для модификации внутри клетки, точная последовательность данной молекулы крРНК или тракрРНК может быть разработана так, чтобы быть специфичной для видов, в которых будут использоваться молекулы РНК. См., например, Mali et al. (2013) Science 339(6121):823-826; Jinek et al. (2012) Science 337(6096):816-821; Hwang et al. (2013) Nat. Biotechnol. 31(3):227-229; Jiang et al. (2013) Nat. Biotechnol. 31(3):233-239; и Cong et al. (2013) Science 339(6121): 819-823, каждая из которых включена в данный документ в полном объеме посредством ссылки для всех целей.
[00159] ДНК-направляющий сегмент (крРНК) данной нРНК содержит нуклеотидную последовательность, которая комплементарна последовательности на комплементарной цепи ДНК-мишени, как более подробно описано ниже. ДНК-направляющий сегмент нРНК взаимодействует с ДНК-мишенью специфическим для последовательности образом посредством гибридизации (то есть спаривания оснований). По существу, нуклеотидная последовательность ДНК-направляющего сегмента может варьироваться и определяет локализацию внутри ДНК-мишени, с которой будут взаимодействовать нРНК и ДНК-мишень. ДНК-направляющий сегмент заявленной нРНК может быть модифицирован для гибридизации с любой желаемой последовательностью в ДНК-мишени. Встречающиеся в природе крРНК различаются в зависимости от системы CRISPR/Cas и организма, но часто содержат направляющий сегмент длиной от 21 до 72 нуклеотидов, фланкированный двумя прямыми повторами (ПП, DR) длиной от 21 до 46 нуклеотидов (см., например, WO 2014/131833, которая включена в данный документ в полном объеме посредством ссылки для всех целей.). В случае S. pyogenes длина ПП составляет 36 нуклеотидов, а длина направляющего сегмента составляет 30 нуклеотидов. 3'-расположенный ПП комплементарен и гибридизуется с соответствующей тракрРНК, которая, в свою очередь, связывается с белком Cas.
[00160] ДНК-направляющий сегмент может иметь длину, например, по меньшей мере, около 12, 15, 17, 18, 19, 20, 25, 30, 35 или 40 нуклеотидов. Такие ДНК-направляющие сегменты могут иметь длину, например, от около 12 до около 100, от около 12 до около 80, от около 12 до около 50, от около 12 до около 40, от около 12 до около 30, от около 12 до около 25 или от около 12 до около 20 нуклеотидов. Например, ДНК-направляющий сегмент может составлять от около 15 до около 25 нуклеотидов (например, от около 17 до около 20 нуклеотидов, или около 17, 18, 19 или 20 нуклеотидов). См., например, патент США 2016/0024523, который включен в данный документ в полном объеме посредством ссылки для всех целей. Для Cas9 из S. pyogenes типичный ДНК-направляющий сегмент имеет длину от 16 до 20 нуклеотидов или от 17 до 20 нуклеотидов. Для Cas9 из S. aureus типичный ДНК-направляющий сегмент имеет длину от 21 до 23 нуклеотидов. Для Cpf1 типичный ДНК-направляющий сегмент имеет длину, по меньшей мере, 16 нуклеотидов или, по меньшей мере, 18 нуклеотидов.
[00161] тракрРНК могут быть в любой форме (например, полноразмерные тракрРНК или активные частичные тракрРНК) и различной длины. Они могут включать первичные транскрипты или формы, прошедшие процессинг.Например, тракрРНК (как часть однонаправляющей РНК или как отдельная молекула двухмолекулярной нРНК) могут содержать, состоять по существу из или состоять из всей или части последовательности тракрРНК дикого типа (например, около или более чем около 20, 26, 32, 45, 48, 54, 63, 67, 85 или более нуклеотидов последовательности тракрРНК дикого типа). Примеры последовательностей тракрРНК дикого типа из S. pyogenes включают версии, состоящие из 171 нуклеотида, 89 нуклеотидов, 75 нуклеотидов и 65 нуклеотидов. См., например, Deltcheva et al. (2011) Nature 471 (7340): 602-607; WO 2014/093661, каждая из которых полностью включена в данный документ посредством ссылки для всех целей. Примеры тракрРНК в однонаправляющих РНК (онРНК) включают сегменты тракрРНК, обнаруженные в +48, +54, +67 и +85 версиях онРНК, где «+n» означает, что до+n нуклеотида тракрРНК дикого типа включена в онРНК. См. патент США 8,697,359, который включен в данный документ посредством ссылки в полном объеме для всех целей.
[00162] Процент комплементарности между ДНК-направляющим сегментом направляющей РНК и комплементарной цепью целевой ДНК может составлять по меньшей мере 60% (например, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100%). Процент комплементарности между ДНК-направляющим сегментом и комплементарной цепью целевой ДНК может составлять по меньшей мере 60% в отношении около 20 смежных нуклеотидов. Например, процент комплементарности между ДНК-направляющим сегментом и комплементарной цепью целевой ДНК может составлять 100% в 14 смежных нуклеотидах на 5'-конце комплементарной цепи целевой ДНК и до 0% в оставшихся нуклеотидах. В таком случае можно считать, что ДНК-направляющий сегмент имеет длину 14 нуклеотидов. В качестве другого примера, процент комплементарности между ДНК-направляющим сегментом и комплементарной цепью целевой ДНК может составлять 100% в семи смежных нуклеотидах на 5'-конце комплементарной цепи целевой ДНК и составлять 0% в оставшихся нуклеотидах. В таком случае можно считать, что ДНК-направляющий сегмент имеет длину 7 нуклеотидов. В некоторых направляющих РНК по меньшей мере 17 нуклеотидов в ДНК-направляющем сегменте комплементарны комплементарной цепи целевой ДНК. Например, ДНК-направляющий сегмент может иметь длину 20 нуклеотидов и может содержать 1, 2 или 3 несовпадения с комплементарной цепью целевой ДНК. В одном примере несовпадения не примыкают к области комплементарной цепи, соответствующей последовательности мотива, примыкающего к протоспейсеру (РАМ) (т.е. обратному комплементу последовательности РАМ) (например, несовпадения находятся на 5'-конце ДНК-направляющего сегмента направляющей РНК, или несовпадения находятся по меньшей мере в 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18 или 19 пар оснований от области комплементарной цепи, соответствующей последовательности РАМ).
[00163] Белок-связывающий сегмент нРНК может содержать два участка нуклеотидов, комплементарных друг другу. Комплементарные нуклеотиды белок-связывающего сегмента гибридизуются с образованием дуплекса двухцепочечной РНК (дцРНК). Сегмент связывания с белком заявленной нРНК взаимодействует с белком Cas, и нРНК направляет связанный белок Cas к определенной нуклеотидной последовательности в целевой ДНК посредством ДНК-направляющего сегмента.
[00164] Однонаправляющие РНК могут содержать ДНК-направляющий сегмент, присоединенный к каркасной последовательности (т.е. связывающей белок или Cas-связывающей последовательности направляющей РНК). Например, такие направляющие РНК могут иметь 5'-ДНК-направляющий сегмент и 3'-каркасную последовательность. Примеры каркасных последовательностей включают, состоят по существу из или состоят из:


Другие типовые каркасные последовательности включают, состоят по существу из или состоят из:
Направляющие РНК, нацеленные на любую последовательность-мишень направляющей РНК, могут включать, например, ДНК-направляющий сегмент на 5'-конце направляющей РНК, слитый с любой из типовых каркасных последовательностей направляющей РНК на 3'-конце направляющей РНК. То есть, ДНК-направляющий сегмент может быть присоединен к 5'-концу любой из SEQ ID NO: 5-8 с образованием единой направляющей РНК (химерной направляющей РНК). Аналогичным образом, ДНК-направляющий сегмент может быть присоединен к 5'-концу любой из SEQ ID NO: 14-16 с образованием единой направляющей РНК (химерной направляющей РНК). Версии 1, 2, 3 и 4 направляющей РНК, как описано в другом месте в данном документе, относятся к ДНК-направляющим сегментам (т.е. направляющим последовательностям или направляющим), соединенным с каркасами версий 1, 2, 3 и 4, соответственно. Версии 5, 6 и 7 направляющей РНК, как описано в другом месте в данном документе, относятся к ДНК-направляющим сегментам (т.е. направляющим последовательностям или направляющим), соединенным с каркасами версий 5, 6 и 7 соответственно.
[00165] Направляющие РНК могут включать модификации или последовательности, которые обеспечивают дополнительные желательные свойства (например, модифицированную или регулируемую стабильность; субклеточное таргетирование; отслеживание с помощью флуоресцентной метки; сайт связывания для белка или белкового комплекса и т.п.). Примеры таких модификаций включают, например, 5' кэп (например, 7-метилгуанилатный кэп (m7G)); 3'-полиаденилированный хвост (т.е. 3'-поли (А) хвост); последовательность рибопереключателя (например, для обеспечения регулируемой стабильности и/или регулируемой доступности для белков и/или белковых комплексов); последовательность контроля устойчивости; последовательность, которая образует дуплекс дцРНК (т.е. шпильку); модификацию или последовательность, которая направляет РНК в субклеточную локализацию (например, ядро, митохондрии, хлоропласты и тому подобное); модификацию или последовательность, которая обеспечивает отслеживание (например, прямая конъюгация с флуоресцентной молекулой, конъюгация с фрагментом, который облегчает флуоресцентное детектирование, последовательность, которая позволяет проводить флуоресцентное детектирование и тому подобное); модификацию или последовательность, которая обеспечивает формирование сайта связывания для белков (например, белков, которые действуют на ДНК, включая ДНК-метилтрансферазы, ДНК-деметилазы, гистоновые ацетилтрансферазы, гистоновые деацетилазы и тому подобное); и их комбинации. Другие примеры модификаций включают сконструированные дуплексные структуры "петля на стержне", сконструированные области петель, сконструированные шпильки на 3' конце дуплексной структуры "петля на стержне" или любую их комбинацию. См., например, US 2015/0376586, полностью включенный в данный документ в полном объеме посредством ссылки для всех целей. Петля может представлять собой неспаренную областью нуклеотидов внутри дуплекса, состоящую из крРНК-подобной области и минимальной тракрРНК-подобной области. Петля может содержать на одной стороне дуплекса неспаренный 5'-ΧΧΧΥ-3', где X представляет собой любой пурин, a Υ может представлять собой нуклеотид, который может формировать неоднозначную пару с нуклеотидом на противоположной цепи, и регион неспаренных нуклеотидов на другой стороне дуплекса.
[00166] Немодифицированные нуклеиновые кислоты могут быть подвержены деградации. Экзогенные нуклеиновые кислоты также могут вызывать врожденный иммунный ответ. Модификации могут способствовать повышению стабильности и снижению иммуногенности. Направляющие РНК могут содержать модифицированные нуклеозиды и модифицированные нуклеотиды, включая, например, одно или более из следующего: (1) изменение или замена одного или обоих несвязанных фосфатных атомов кислорода и/или одного или более связывающих фосфатных атомов кислорода в связи фосфодиэфирного остова; (2) изменение или замена компонента сахара рибозы, такая как изменение или замена 2'-гидроксила в сахаре рибозе; (3) замена фосфатной части дефосфорилированными линкерами; (4) модификация или замена встречающегося в природе азотистого основания; (5) замена или модификация рибозо-фосфатного остова; (6) модификация 3'-конца или 5'-конца олигонуклеотида (например, удаление, модификация или замена концевой фосфатной группы или конъюгация фрагмента); и (7) модификация сахара. Другие возможные модификации направляющей РНК включают модификации или замену урацилов или полиурациловых трактов. См., например, WO 2015/048577 и патент США 2016/0237455, каждый из которых полностью включен в полном объеме в данный документ посредством ссылки для всех целей. Аналогичные модификации могут быть внесены в нуклеиновые кислоты, кодирующие Cas, такие как мРНК Cas.
[00167] В качестве одного примера, нуклеотиды на 5'- или 3'-конце направляющей РНК могут содержать фосфоротиоатные связи (например, основания могут содержать модифицированную фосфатную группу, которая представляет собой фосфоротиоатную группу). Например, направляющая РНК может содержать фосфоротиоатные связи между 2, 3 или 4 концевыми нуклеотидами на 5' или 3' конце направляющей РНК. В качестве другого примера, нуклеотиды на 5'- и/или 3'-конце направляющей РНК могут содержать 2'-O-метильные модификации. Например, направляющая РНК может содержать 2'-O-метильные модификации в 2, 3 или 4 концевых нуклеотидах на 5'- и/или 3'-конце направляющей РНК (например, на 5'-конце). См., например, WO 2017/173054 А1 и Finn et al. (2018) Cell Rep.22(9): 2227-2235, каждая из которых включена в полном объеме в данный документ посредством ссылки для всех целей. В одном конкретном примере направляющая РНК содержит 2'-O-метильные аналоги и 3'-фосфоротиоатные межнуклеотидные связи в первых трех 5'- и 3'-концевых остатках РНК. В другом конкретном примере направляющая РНК модифицируется таким образом, что все группы 2'ОН, которые не взаимодействуют с белком Cas9, заменяются 2'-О-метиловыми аналогами, а хвостовая часть направляющей РНК, которая характеризуется минимальным взаимодействием с белком Cas9, модифицирована с помощью 5'- и 3'-фосфоротиоатных межнуклеотидных связей. См., например, Yin et al. (2017) Nat. Biotech. 35(12): 1179-1187, которая включена в данный документ в полном объеме посредством ссылки для всех целей. Другие примеры модифицированных направляющих РНК представлены, например, в WO 2018/107028 А1, который включен в данный документ в полном объеме посредством ссылки для всех целей.
[00168] нРНК можно получить различными другими способами. Например, нРНК могут быть получены путем транскрипции in vitro с использованием, например, РНК-полимеразы Т7 (см., например, WO 2014/089290 и WO 2014/065596, каждый из которых включен в данный документ в полном объеме посредством ссылки для всех целей). Направляющие РНК также могут синтетически полученной молекулой, полученной путем химического синтеза. Например, направляющая РНК может быть химически синтезирована таким образом, чтобы содержать 2'-O-метильные аналоги и 3'-фосфоротиоатные межнуклеотидные связи в первых трех 5'- и 3'-концевых остатках РНК.
[00169] Последовательности-мишени направляющей РНК. ДНК-мишени для направляющих РНК включают последовательности нуклеиновых кислот, присутствующие в ДНК, с которыми будет связываться нацеленный на ДНК сегмент нРНК, при условии, что обеспечены достаточные условия для связывания. Подходящие условия связывания ДНК/РНК включают физиологические условия, обычно присутствующие в клетке. Другие подходящие условия связывания ДНК/РНК (например, условия в бесклеточной системе) известны в данной области (см., например, Molecular Cloning: A Laboratory Manual, 3rd Ed. (Sambrook et al., Harbor Laboratory Press 2001), которая включена в данный документ в полном объеме посредством ссылки для всех целей). Цепь ДНК-мишени, которая комплементарна и гибридизуется с нРНК, может быть названа «комплементарной цепью», а цепь целевой ДНК, которая комплементарна «комплементарной цепи» (и, следовательно, не комплементарна белку Cas или нРНК) может быть названа «некомплементарной цепью» или «цепью-матрицей».
[00170] ДНК-мишень содержит как последовательность на комплементарной цепи, с которой гибридизуется направляющая РНК, так и соответствующую последовательность на некомплементарной цепи (например, смежную с мотивом, примыкающим к протоспейсеру (РАМ)). Используемый в данном документе термин «последовательность-мишень направляющей РНК» относится конкретно к последовательности на некомплементарной цепи (т.е. обратному комплементу), соответствующей последовательности, с которой гибридизируется направляющая РНК на комплементарной цепи. То есть последовательность-мишень направляющей РНК относится к последовательности на некомплементарной цепи, смежной с РАМ (например, до или на 5'-конце относительно РАМ в случае Cas9). Последовательность-мишень направляющей РНК эквивалентна ДНК-направляющему сегменту направляющей РНК, но содержит тимины вместо урацилов. В качестве одного примера, последовательность-мишень направляющей РНК для фермента SpCas9 может относиться к последовательности до 5'-NGG-3' РАМ на некомплементарной цепи. Направлятарной цепи ДНК-мишени, при этом гибридизация между ДНК-направляющим сегмеющая РНК разработана таким образом, чтобы быть комплементарной к комплеменнтом направляющей РНК и комплементарной цепью ДНК-мишени способствует образованию комплекса CRISPR. Полная комплементарность не обязательно требуется при условии, что комплементарность достаточна, чтобы вызвать гибридизацию и способствовать образованию комплекса CRISPR. Если направляющая РНК упоминается в данном документе как нацеленная на последовательность-мишень направляющей РНК, это означает, что направляющая РНК гибридизируется с последовательностью комплементарной цепи ДНК-мишени, которая представляет собой обратный комплемент последовательности-мишени направляющей РНК на некомплементарной цепи.
[00171] Последовательность-мишень ДНК-мишени или направляющей РНК может содержать любой полинуклеотид и может располагаться, например, в ядре или цитоплазме клетки или в органелле клетки, такой как митохондрия или хлоропласт. Последовательность-мишень ДНК-мишени или направляющей РНК может представлять собой любую последовательность нуклеиновой кислоты, эндогенной или экзогенной для клетки. Последовательность-мишень направляющей РНК может представлять собой последовательность, кодирующую продукт гена (например, белок), или некодирующую последовательность (например, регуляторную последовательность), или может содержать и то, и другое.
[00172] Сайт-специфическое связывание и расщепление ДНК-мишени белком Cas может происходить в местах, определяемых как (i) комплементарностью спаривания оснований между направляющей РНК и комплементарной цепью ДНК-мишени, так и (ii) коротким мотивом, называемым мотивом, примыкающим к протоспейсеру (РАМ) в некомплементарной цепи ДНК-мишени. РАМ может фланкировать последовательность-мишень направляющей РНК. Необязательно, последовательность-мишень направляющей РНК может быть фланкирована на 3'-конце с помощью РАМ (например, в случае Cas9). Альтернативно, последовательность-мишень направляющей РНК может быть фланкирована на 5'-конце с помощью РАМ (например, в случае Cpf1). Например, сайт расщепления белков Cas может располагаться от около 1 до около 10 или от около 2 до около 5 пар оснований (например, 3 пары оснований) до или после последовательности РАМ (например, в пределах последовательности-мишени направляющей РНК). В случае SpCas9 последовательность РАМ (то есть на некомплементарной цепи) может представлять собой 5'-N1GG-3', где Ν1 представлять собой любой нуклеотид ДНК, а РАМ находится непосредственно на 3' конце последовательности-мишени направляющей РНК на некомплементарной цепи ДНК-мишени. Таким образом, последовательность, соответствующая РАМ на комплементарной цепи (т.е., обратный комплемент) будет представлять собой 5'-ССN2-3', где N2 представляет собой любой ДНК нуклеотид и располагается сразу на 5' конце последовательности, с которой ДНК-направляющий сегмент направляющей РНК гибридизуется на комплементарной цепи ДНК-мишени. В некоторых таких случаях Ν1 и N2 могут быть комплементарны, и пара оснований N1-N2 может представлять собой любую пару оснований (например, N1=C и N2=G; N1=G и N2=С; Ν1=Α и N2=T; или Ν1=Τ, и N2=А). В случае Cas9 из S. aureus, РАМ может представлять собой NNGRRT или NNGRR, где N может представлять собой A, G, С или Т, a R может представлять собой G или А. В случае Cas9 из С.jejuni, РАМ может представлять собой, например, NNNNACAC или NNNNRYAC, где N может представлять собой A, G, С или Т, a R может представлять собой G или А. В некоторых случаях (например, в случае FnCpf1) последовательность РАМ может располагаться до 5'-конца и иметь последовательность 5'-ΤΤΝ-3'.
[00173] Примером последовательности-мишени направляющей РНК является последовательность ДНК из 20 нуклеотидов, непосредственно предшествующая мотиву NGG, распознаваемому белком SpCas9. Например, два примера последовательностей-мишеней направляющей РНК плюс РАМ представляют собой GN19NGG (SEQ ID NO: 9) или N20NGG (SEQ ID NO: 10). См., например, WO 2014/165825, которая включена в данный документ в полном объеме посредством ссылки для всех целей Гуанин на 5'-конце может способствовать транскрипции РНК-полимеразой в клетках. Другие примеры последовательностей-мишеней направляющих РНК плюс РАМ могут содержать два гуаниновых нуклеотида на 5'-конце (например, GGN20NGG; SEQ ID NO: 11) для облегчения эффективной транскрипции полимеразой Т7 in vitro. См., например, WO 2014/065596, которая включена в данный документ в полном объеме посредством ссылки для всех целей. Другие последовательности-мишени направляющей РНК плюс РАМ могут иметь длину от 4 до 22 нуклеотидов последовательности SEQ ID NO: 9-11, включая 5' G или GG и 3' GG или NGG. Другие последовательности-мишени направляющей РНК плюс РАМ могут иметь длину от 14 до 20 нуклеотидов последовательности SEQ ID NO: 9-11.
[00174] Образование комплекса CRISPR, гибридизованного с ДНК-мишенью, может приводить к расщеплению одной или обеих цепей целевой ДНК внутри или вблизи области, соответствующей последовательности-мишени направляющей РНК (т.е. последовательности-мишени направляющей РНК на некомплементарной цепи ДНК-мишени и обратному комплементу на комплементарной цепи, с которой гибридизуется направляющая РНК). Например, сайт расщепления может располагаться в последовательности-мишени направляющей РНК (например, в определенном месте относительно последовательности РАМ). «Сайт расщепления» включает позицию целевой ДНК, в которой белок Cas вызывает одноцепочечный разрыв или двухцепочечный разрыв. Сайт расщепления может располагаться только на одной цепи (например, в случае, когда используется никаза) или на обеих цепях двухцепочечной ДНК. Сайты расщепления могут находиться в одном и том же положении на обеих цепях (образуя тупые концы; например, в случае Cas9)) или могут находиться в разных сайтах на каждой цепи (создавая липкие концы (например, выступы); например, в случае Cpf1). Липкие концы могут быть получены, например, с использованием двух белков Cas, каждый из которых вызывает одноцепочечный разрыв в отличающемся сайте расщепления на другой цепи, тем самым приводя к образованию двухцепочечного разрыва. Например, первая никаза может создавать одноцепочечный разрыв на первой цепи двухцепочечной ДНК (дцДНК), а вторая никаза может создавать одноцепочечный разрыв на второй цепи дцДНК, формируя таким образом выступающие последовательности. В некоторых случаях последовательность-мишень направляющей РНК или сайт расщепления никазы на первой цепи отделены от последовательности-мишени направляющей РНК или сайта расщепления никазы на второй цепи по меньшей мере на 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 40, 50, 75, 100, 250, 500 или 1000 пар оснований.
С. Другие Нуклеазные Агенты
[00175] В способах, описанных в данном документе, также можно использовать любой другой тип известного редко расщепляющего нуклеазного агента. Одним из примеров такого нуклеазного агента является эффекторная нуклеаза, подобная активатору транскрипции (TALEN). Эффекторные нуклеазы TAL представляют собой класс последовательность-специфичных нуклеаз, которые можно использовать для создания двухцепочечных разрывов в определенных последовательностях-мишенях в ДНК. Эффекторные нуклеазы TAL создаются путем слияния природного или сконструированного эффектора, подобного активатору транскрипции (TAL), или его функциональной части, с каталитическим доменом эндонуклеазы, такой как, например, FokI. Уникальный модульный эффекторный ДНК-связывающий домен TAL позволяет создавать белки с потенциально любой заданной специфичностью распознавания ДНК. Таким образом, ДНК-связывающие домены эффекторных нуклеаз TAL могут быть сконструированы для распознавания специфических сайтов-мишеней ДНК и, таким образом, использованы для создания двухцепочечных разрывов в желаемых последовательностях-мишенях. См. WO 2010/079430; Morbitzer et al. (2010)Proc. Natl. Acad Sci. U.S.A. 107(50):21617-21622; Scholze & Boch (2010) Virulence 1:428-432; Christian et al. Genetics (2010) 186:757-761; Li et al. (2010) Nucleic Acids Res. (2010) 39(l):359-372; и Miller et al. (2011) Nat. Biotechnol. 29:143-148, каждая из которых включена в данный документ в полном объеме посредством ссылки для всех целей.
[00176] Примеры подходящих нуклеаз TAL и способы получения подходящих нуклеаз TAL раскрыты, например, в патенте США 2011/0239315, патенте США 2011/0269234, патенте США 2011/0145940, патенте США 2003/0232410, патенте США 2005/0208489, патенте США 2005/0026157, патенте США 2005/0064474, патенте США 2006/0188987 и патенте США 2006/0063231, каждый из которых включен в данный документ в полном объеме посредством ссылки для всех целей.
[00177] В некоторых TALEN каждый мономер TALEN содержит 33-35 повторов TAL, которые распознают одну пару оснований через два гипервариабельных остатка. TALEN может представлять собой химерный белок, содержащий ДНК-связывающий домен на основе TAL-повтора, функционально связанный с независимой нуклеазой, такой как эндонуклеаза FokI. Например, нуклеазный агент может содержать ДНК-связывающий домен на основе первого TAL-повтора и второй ДНК-связывающий домен на основе TAL-повтора, при этом каждый из первого и второго ДНК-связывающих доменов на основе TAL-повторов функционально связан с нуклеазой FokI, при этом первый и второй ДНК-связывающий домен на основе TAL-повтора распознает две смежные последовательности ДНК-мишени в каждой цепи последовательности ДНК-мишени, разделенных спейсерной последовательностью различной длины (12-20 п.н.), и при этом субъединицы нуклеазы FokI димеризуются с образованием активной нуклеазы, которая делает двойной разрыв в последовательности-мишени.
[00178] Другим примером подходящего агента нуклеазы является нуклеаза с "цинковыми пальцами" (ZFN). В некоторых ZFN каждый мономер ZFN содержит 3 или более ДНК-связывающих домена с «цинковыми пальцами», при этом каждый ДНК-связывающий домен с «цинковыми пальцами» связывается с субсайтом из 3 п.н. В других ZFN, ZFN представляет собой химерный белок, содержащий ДНК-связывающий домен с «цинковыми пальцами», функционально связанный с независимой нуклеазой, такой как эндонуклеаза FokI. Например, нуклеазный агент может содержать первый ZFN и второй ZFN, при этом каждый из первого ZFN и второго ZFN функционально связан с субъединицей нуклеазы FokI, при этом первый и второй ZFN распознают две смежные последовательности ДНК-мишени в каждой цепи последовательности ДНК-мишени, разделенные спейсером около 5-7 п.н., и при этом субъединицы нуклеазы FokI димеризуются с образованием активной нуклеазы, которая делает двухцепочечный разрыв. См., например, патенте США 2006/0246567; патенте США 2008/0182332; патенте США 2002/0081614; патенте США 2003/0021776; WO 2002/057308; патенте США 2013/0123484; патенте США 2010/0291048; WO 2011/017293; и Gaj et al. (2013) Trends Biotechnol, 31 (7):397-405, каждая из которых включена в данный документ в полном объеме посредством ссылки для всех целей.
[00179] Другой тип подходящего нуклеазного агента представляет собой сконструированную мегануклеазу. Мегануклеазы были разделены на четыре семейства на основе консервативных мотивов последовательностей, семейства представляют собой семейства LAGLIDADG, GIY-YIG, HNH и His-Cys box. Эти мотивы участвуют в координации ионов металлов и гидролизе фосфодиэфирных связей. Мегануклеазы отличаются длинными последовательностями-мишенями и устойчивостью к некоторым полиморфизмам последовательностей в их ДНК-субстратах. Домены, структура и функция мегануклеаз известны, см., например, Guhan and Muniyappa (2003) Crit. Rev. Biochem.Mol. Biol. 38:199-248; Lucas et al. (2001) Nucleic Acids Res. 29:960-9; Jurica and Stoddard, (1999) Cell. Mol. Life Sci. 55:1304-26; Stoddard (2006) Q. Rev. Biophys. 38:49-95; и Moure et al. (2002) Nat. Struct. Biol. 9:764, каждая из которых включена в полном объеме в данный документ посредством ссылки для всех целей. В некоторых примерах используется встречающийся в природе вариант и/или сконструированная производная мегануклеазы. Известны способы модификации кинетики, взаимодействия кофакторов, экспрессии, оптимальных условий и/или специфичности последовательности-мишени, а также скрининга на активность. См., например, Epinat et al. (2003) Nucleic Acids Res. 31:2952-62; Chevalier et al. (2002)Mo/. Cell 10:895-905; Gimble et al. (2003)Mol. Biol. 334:993-1008; Seligman et al. (2002) Nucleic Acids Res. 30:3870-9; Sussman et al. (2004) J. Mol. Biol. 342:31-41; Rosen et al. (2006) Nucleic Acids Res. 34:4791-800; Chames et al. (2005) Nucleic Acids Res. 33:el78; Smith et al. (2006) Nucleic Acids Res. 34:el49; Gruen et al. (2002) Nucleic Acids Res. 30:e29; Chen and Zhao (2005) Nucleic Acids Res. 33:el54; WO 2005/105989; WO 2003/078619; WO 2006/097854; WO 2006/097853; WO 2006/097784 и WO 2004/031346, каждая из которых включена в полном объеме в данный документ посредством ссылки для всех целей.
[00180] Можно использовать любую мегануклеазу, включая, например, I-SceI, Ι-SceII, I-SceIII, Ι-SceIV, I-SceV, I-SceVI, I-SceVII, I-CeuI, I-CeuAIIP, I-CreI, Ι-CrepsbIP, I-CrepsbIIP, I-CrepsbIIIP, Ι-CrepsbIVP, I-TliI, I-PpoI, PI-PspI, F-SceI, F-SceII, F-SuvI, F-TevI, F-TevII, I-AmaI, I-AniI, I-ChuI, I-CmoeI, I-CpaI, Ι-CpaII, I-CsmI, I-CvuI, I-CvuAIP, I-DdiI, Ι-DdiII, I-DirI, I-DmoI, I-HmuI, I-HmuII, I-HsNIP, I-LlaI, I-MsoI, I-NaaI, I-NanI, I-NcIIP, I-NgrIP, I-NitI, I-NjaI, I-Nsp236IP, I-PakI, I-PboIP, I-PcuIP, I-PcuAI, I-PcuVI, I-PgrIP, I-PobIP, I-PorI, I-PorIIP, I-PbpIP, Ι-SpBetaIP, I-ScaI, I-SexIP, Ι-SneIP, I-SpomI, I-SpomCP, Ι-SpomIP, I-SpomIIP, I-SquIP, I-Ssp6803I, I-SthPhiJP, I-SthPhiST3P, I-SthPhiSTe3bP, Ι-TdeIP, I-TevI, I-TevII, I-TevIII, I-UarAP, I-UarHGPAIP, I-UarHGPA13P, I-VinIP, Ι-ZbiIP, PI-MtuI, PI-MtuHIP PI-MtuHIIP, PI-PfuI, ΡΙ-PfuII, PI-PkoI, ΡΙ-PkoII, PI-Rma43812IP, PI-SpBetaIP, ΡΙ-SceI, PI-TfuI, PI-TfuII, PI-ThyI, PI-TliI, PI-TliII или любые их активные варианты или фрагменты.
[00181] Мегануклеазы могут распознавать, например, двухцепочечные последовательности ДНК от 12 до 40 пар оснований. В некоторых случаях мегануклеаза распознает одну идеально подобранную последовательность-мишень в геноме. Некоторые мегануклеазы являются хоминг-нуклеазами. Один из типов хоминг-нуклеазы представляет собой семейство хоминг-нуклеаз LAGLIDADG, включая, например, I-SceI, I-CreI и I-Dmo1.
VIII. Кассеты Селекции
[00182] Любая подходящая кассета селекции может быть использована в способах, описанных в данном документе. Термин кассета селекции относится к кассете экспрессии, которая содержит одну или более последовательностей контроля экспрессии (например, промотор для экспрессии в бактериальной клетке и/или другие регуляторные последовательности, такие как энхансеры, посттранскрипционные регуляторные элементы и поли(А) последовательности), функционально связанных с нуклеиновой кислотой, кодирующей селективный маркер. Кассеты селекции могут обеспечивать селекцию в бактериальных клетках или они могут обеспечивать селекцию как в бактериальных, так и в эукариотических клетках или клетках млекопитающих. В качестве одного из примеров можно использовать такой ген, как ген неомицин-фосфотрансферазы. Неомицин-фосфотрансфераза придает устойчивость к канамицину прокариотическим клеткам и устойчивость к G418 эукариотическим клеткам. Такой ген можно использовать, например, в комбинации с системой двойного промотора, объединяющей эукариотический промотор (например, промотор эукариотической фосфоглицераткиназы (PGK)) и прокариотический промотор (например, прокариотический промотор ЕМ7).
[00183] Некоторые кассеты селекции, которые можно использовать в описанных в данном документе способах, могут придавать устойчивость к антибиотику, который в противном случае убивал бы или подавлял рост бактериальных клеток. Например, кассета селекции может придавать устойчивость к канамицину, спектиномиину, стрептомицину, ампициллину, карбенициллину, блеомицину, эритромицину, полимксину В, тетрациклину или хлорамфениколу. Такие кассеты селекции и гены, которые придают устойчивость к этим и другим антибиотикам, хорошо известны. Клетки, содержащие кассеты селекции, могут быть отобраны путем обработки клеток антибиотиком. Те клетки, которые устойчивы к антибиотику, содержат кассету селекции.
[00184] Другие кассеты селекции могут содержать репортерные гены, которые можно использовать для селекции клеток, содержащих целевую модификацию. Термин репортерный ген относится к нуклеиновой кислоте, имеющей последовательность, кодирующую продукт гена (обычно фермент), который можно легко и количественно проанализировать, когда конструкция, содержащая последовательность репортерного гена, функционально связанную с гетерологичным промоторных и/или энхансерным элементом, вносится в клетки, содержащие (или которые можно сделать так, чтобы они содержали) факторы, необходимые для активации промоторных и/или энхансерных элементов. Примеры репортерных генов включают, но не ограничиваются ими, гены, кодирующие флуоресцентные белки. Репортерный белок относится к белку, кодируемому репортерным геном.
[00185] Флуоресцентный репортерный белок представляет собой репортерный белок, который может быть обнаружен на основе флуоресценции, при этом флуоресценция может исходить либо непосредственно от репортерного белка, либо от активности репортерного белка на флуорогенном субстрате, либо от белка с аффинностью к связыванию с флуоресцентно меченным соединением. Примеры флуоресцентных белков включают зеленые флуоресцентные белки (например, GFP, GFP-2, tagGFP, turboGFP, eGFP, Emerald, Azami Green, Мономерный Azami Green, CopGFP, AceGFP и ZsGreen1), желтые флуоресцентные белки (например, YFP, eYFP, Citrine, Venus, YPet, PhiYFP и ZsYellow1), синие флуоресцентные белки (например, FP, eBFP, eBFP2, Azurite, mKalama1, GFPuv, Sapphire и T-sapphire), голубые флуоресцентные белки (например, CFP, eCFP, Cerulean, CyPet, AmCyanl и Midoriishi-Cyan), красные флуоресцентные белки (например, RFP, mKate, mKate2, mPlum, мономер DsRed, mCherry, mRFP1, DsRed-Express, DsRed2, DsRed-Monomer, HcRed-Tandem, HcRed1, AsRed2, eqFP611, mRaspberry, mStrawberry и Jred), оранжевые флуоресцентные белки (например, mOrange, mKO, Kusabira-Orange, Monomeric Kusabira-Orange, mTangerine и tdTomato) и любой другой подходящий флуоресцентный белок, присутствие которого в клетках можно определить с помощью способов проточной цитометрии. Клетки, содержащие флуоресцентный репортерный ген, могут быть отобраны, например, путем сортировки клеток, содержащих флуоресцентный репортерный белок, кодируемый этим геном.
IX. Целевые Модификации
[00186] Различные типы целевых генетических модификаций могут быть внесены с использованием описанных в данном документе способов. Такие целевые генетические модификации могут включать, например, вставку одного или более нуклеотидов, делецию одного или более нуклеотидов или замещение (замену) одного или более нуклеотидов. Такие вставки, делеции или замены могут привести, например, к точечной мутации, нокауту интересующей последовательности нуклеиновой кислоты или ее части, нокину интересующей последовательности нуклеиновой кислоты или ее части, замене последовательности эндогенной нуклеиновой кислоты на гетерологичную или экзогенную последовательность нуклеиновой кислоты, замена эндогенной последовательности нуклеиновой кислоты на гомологичную или ортологичную последовательность нуклеиновой кислоты (например, замене домена, замене экзона, замене интрона, замене регуляторной последовательности или замене гена), изменению регуляторного элемента (например, промотора или энхансера), миссенс-мутации, нонсенс-мутации, мутации со сдвигом рамки считывания генетического кода, мутации с усечением, нуль-мутации или их комбинации. Например, по меньшей мере, 1, 2, 3, 4, 5, 7, 8, 9, 10 или более нуклеотидов могут быть изменены (например, удалены, вставлены или заменены) для образования целевой генетической модификации. Делеции, вставки или замены могут быть любого размера, как описано в другом месте в данном документе. См., например, Wang et al. (2013) Cell 153:910-918; Mandalos et al. (2012) PLOS One 7:e45768; и Wang et al. (2013) Nat Biotechnol. 31:530-532, каждая из которых включена в данный документ в полном объеме посредством ссылки.
[00187] Делеции, вставки или замены могут быть любой длины. Удаленная, вставленная или замещенная нуклеиновая кислота может составлять, например, от около 1 п.н. до около 5 п.н., от около 5 п.н. до около 10 п.н., от около 10 п.н. до около 50 п.н., от около 50 п.н. до около 100 п.н., от около 100 п.н. до около 200 п.н., от около 200 до около 300 п.н., от около 300 п.н. до около 400 п.н., от около 400 п.н. до около 500 п.н., от около 500 п.н. до около 1 т.п.н., от около 1 т.п.н. до около 5 т.п.н., от около 5 т.п.н. до около 10 т.п.н., от около 10 т.п.н. до около 20 т.п.н., от около 20 т.п.н. до около 40 т.п.н., от около 40 т.п.н. до около 60 т.п.н., от около 60 т.п.н. до около 80 т.п.н., от около 80 т.п.н. до около 100 т.п.н., от около 100 т.п.н. до около 150 т.п.н. или от около 150 т.п.н. до около 200 т.п.н., от около 200 т.п.н. до около 300 т.п.н., от около 300 т.п.н. до около 400 т.п.н. или от около 400 т.п.н. до около 500 т.п.н.
[00188] Все патентные заявки, веб-сайты, другие публикации, номера доступа и т.п., указанные выше или ниже в данном документе, включены в данный документ посредством ссылки в полном объеме для всех целей в той же степени, как если бы каждый отдельный элемент был специально и отдельно указан как включенный в данный документ посредством ссылки. Если разные версии последовательности связаны с номером доступа в разное время, то подразумевается та версия, которая связана с номером доступа на действительную дату подачи данной заявки. Действительная дата подачи означает более раннюю дату из даты фактической подачи или даты подачи приоритетной заявки со ссылкой на номер доступа, если это применимо. Аналогичным образом, если разные версии публикации, веб-сайта и т.п.публикуются в разное время, то подразумевается та версия, которая была опубликована последней на действительную дату подачи данной заявки, если не указано иное. Любые признак, этап, элемент, вариант реализации или аспект данного изобретения можно применять в комбинации с любым другим, если специально не указано иное. Несмотря на то, что данного изобретение описано довольно подробно с помощью иллюстраций и примеров для ясности и понимания, будет очевидно, что могут быть осуществлены определенные изменения и модификации в пределах объема прилагаемой формулы данного изобретения.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[00189] Нуклеотидные и аминокислотные последовательности, перечисленные в сопроводительном списке последовательностей, представлены с использованием стандартных буквенных сокращений для нуклеотидных оснований и трехбуквенного кода для аминокислот. Нуклеотидные последовательности следуют стандартному соглашению: они начинаются с 5'-конца последовательности и продолжаются вперед (т.е. слева направо в каждой строке) до 3'-конца. Показана только одна цепь каждой нуклеотидной последовательности, но подразумевается, что комплементарная цепь включается любой ссылкой на отображаемую цепь. Когда предложена нуклеотидная последовательность, кодирующая аминокислотную последовательность, понятно, что также предложены ее варианты с вырожденными кодонами, которые кодируют ту же аминокислотную последовательность. Аминокислотные последовательности следуют стандартному соглашению: они начинаются с аминоконца последовательности и продолжаются вперед (т.е. слева направо в каждой строке) к карбокси-концу.
[00190]
ПРИМЕРЫ
Пример 1. Бесшовное Внесение Целевой Модификации в Большой Направленный Вектор Посредством Бактериальной Гомологичной Рекомбинации и Внутримолекулярной Сборки по Гибсону
[00191] Технология сборки по Гибсону объединяет сегменты ДНК с гомологичными концами в единую молекулу. Это отличается от традиционных лигирований между комплементарными, расположенными в ступенчатом порядке концами, созданными рестрикционными ферментами, тем, что может использоваться любая комплементарная последовательность минимального размера. Поскольку клонирование через сайты рестрикции обычно приводит к включению следов экзогенной ДНК (сайтов распознавания ферментов) в конечный продукт, сборка по Гибсону является предпочтительной, поскольку она может быть бесшовной.
[00192] Реакция сборки по Гибсону является изотермической и включает три разных фермента: экзонуклеазу Т5, ДНК-полимер азу и лигазу. См., например, патент США 2010/0035768, патент США 2015/0376628, WO 2015/200334 и Gibson et al. (2009) Nat. Methods 6(5):343-345, каждый из которых включен в данный документ в полном объеме посредством ссылки для всех целей. Реакция начинается с образования концов одноцепочечной ДНК из-за 5'-3'-экзонуклеазной активности экзонуклеазы Т5. Фрагменты ДНК с комплементарными одноцепочечными концами затем выравниваются по простым правилам спаривания оснований, и ДНК-полимераза заполняет промежутки, перемещаясь от 5' к 3'. ДНК-лигаза закрывает последний разрыв, и в результате получается бесшовная двухцепочечная молекула ДНК. Было показано, что комплементарные концы сорока пар оснований эффективны, и фактическая последовательность, как правило, не важна. Исходные фрагменты можно получить с помощью ПЦР, рестрикции или прямого синтеза.
[00193] Бесшовная конструкция ДНК имеет особое значение при создании линий трансгенных животных, поскольку швы, образованные сайтами рестрикции или путем других манипуляций, могут отрицательно повлиять на экспрессию генов, если они попадают в область, важную для регуляции. Нацеливание на геном млекопитающих часто требует конструирования больших направленных векторов с длинными плечами ДНК для прямой гомологичной рекомбинации, а также кассет устойчивости к антибиотикам для отбора клонов эмбриональных стволовых клеток. Правильно нацеленные клоны часто содержат несколько швов, необходимых для конструирования вектора, не говоря уже о самой кассете устойчивости. Для генетической абляции эти повреждения могут не иметь значения для конечного результата (нуль-аллель), но всегда есть шанс, что на экспрессию соседних генов будет оказано неблагоприятное воздействие. Для модификаций, отличных от нокаута, таких как нокин (например, репортеры или мутантные аллели), правильная экспрессия локуса-мишени обычно важна для рассматриваемых исследований. Сборка по Гибсону может устранить необходимость в некоторых из этих швов и даже облегчить конструирование самого вектора в некоторых случаях, но уникальные сайты рестрикции могут быть трудными для поиска.
[00194] Гуманизация, прямая замена мышиного гена его человеческим аналогом, в частности, требует бесшовных соединений между мышиной и человеческой последовательностью, чтобы мышиный аппарат транскрипции точно воспроизводил экспрессию нового аллеля. Необходимо проявлять осторожность, чтобы скрыть швы конструкции и кассету селекции в некодирующих областях, которые не влияют на регуляцию генов. По мере того, как животные модели становятся более сложными, к уже существующим могут быть добавлены дополнительные модификации, такие как мутации, вызывающие болезни человека, в гуманизированных аллелях. Дополнительные изменения могут затем добавить еще больше швов и другую кассету селекции к уже хорошо сконструированному мышиному локусу, увеличивая вероятность того, что экспрессия будет изменена, и мышиная модель не будет соответствовать заболеванию человека. С точки зрения конструирования, добавление новой кассеты к вектору, уже содержащему ее, может усложняться из-за нежелательной рекомбинации между общими элементами кассеты, такими как промоторы и поли(А)-сигналы, даже если две кассеты кодируют разные наборы.
[00195] Принимая во внимание эти препятствия, мы разработали способы для упрощения создания направленных векторов, несущих множественные изменения, такие как гуманизированный аллель и наслоенная мутация, вызывающая заболевание. Эти способы позволяют упростить конструирование и минимизировать количество швов, включенных в окончательную животную модель.
[00196] В первом способе синтезируется небольшой фрагмент ДНК, несущий желаемую мутацию, фланкированный короткими (<500 п.н.) плечами гомологии. Через несколько пар оснований в направлении 3' желаемой мутации выбирают область из 40-50 пар оснований и дублируют, чтобы создать прямые повторы, фланкирующие редкие сайты рестрикции или последовательности-мишени направляющей РНК Cas9, фланкирующие кассету устойчивости. Затем эту небольшую конструкцию гомологично рекомбинируют с созданным направленным мышиным вектором (таким как направленный вектор для гуманизации, с его собственной кассетой устойчивости) с помощью технологии рекомбинации. После подтверждения того, что желаемая мутация включена, новый вектор разрезают с помощью редко расщепляющего/направляющего Cas9, выпуская кассету и обнажая прямые повторы из 40-50 пар оснований. Затем сборка по Гибсону плавно закрывает разрыв во внутримолекулярной реакции. Полученный в результате направленный вектор теперь несет желаемую мутацию и не содержит дополнительных швов или кассет, кроме тех, которые изначально присутствовали при гуманизации.
[00197] В конкретном примере мы создали аллель, включающий точечную мутацию, в целевой конструкции (большом направленном векторе), содержащей гуманизированный Ген-Мишень 1. См. Фиг. 1. Цель состояла в том, чтобы эффективно и без швов создать большой направленный вектор, содержащий гуманизированный Ген-Мишень 1 с точечной мутацией, вместо того, чтобы ретаргетировать гуманизированные мышиные эмбриональные стволовые (ЭС) клетки, содержащие гуманизированный Ген-Мишень 1, чтобы внести точечную мутацию в эти гуманизированные мышиные ЭС-клетки. Первоначальная направленная конструкция содержала геномную последовательность человеческого Гена-Мишени 1 от стартового кодона до стоп-кодона, включая все интроны, для замены геномной мышиной последовательности соответствующего мышиного Гена-Мишени 1 от стартового кодона до стоп-кодона. Кроме того, нуклеиновая кислота вставки в направленной конструкции содержала само удаляющуюся кассету устойчивости к гигромицину после поли(А)-последовательности. Затем этот исходный гуманизированный вектор был модифицирован, как описано выше, с помощью точечной мутации и кассетой устойчивости к неомицину, фланкированной сайтами рестрикции AscI и прямыми повторами из 50 пар оснований последовательности Гена-Мишени 1 человека, расположенными в направлении 3' сразу после точечной мутации. Затем была синтезирована нуклеиновая кислота, содержащая кассету неомицина ЕМ7, фланкированную редкими сайтами рестрикции (AscI), и прямой повтор длиной 50 п.н. от экзона, в который должна быть внесена мутация, а также фрагменты гомологии в 5'- и 3'-направлении, включая мутацию, которая должна быть внесена во фрагмент гомологии, расположенный 5'-направлении. См. Фиг. 2. Кассета устойчивости к неомицину, была вставлена в середину экзона, который подвергался мутации, но, поскольку способ является бесшовным, экзон был повторен в конце способа. Нуклеиновую кислоту линеаризовали расщеплением с помощью HindIII, и бактериальную гомологичную рекомбинацию использовали для вставки линеаризованной синтетической нуклеиновой кислоты в большой направленный вектор, содержащий гуманизированный Ген-Мишень 1. См., например, патент США 2004/0018626 и Valenzuela et al. (2003) Biotechnol. 21(6):652-659, каждый из которых включен в данный документ в полном объеме посредством ссылки для всех целей. Кассету устойчивости к неомицину вырезали с помощью AscI, которая выпадала из неомициновой кассеты и экспонировала прямые повторы. Затем конструкция была повторно закрыта с помощью внутримолекулярной сборки по Гибсону, которая привела прямые повторы до единственной копии, оставив экзон (теперь содержащий мутацию) неповрежденным без швов. После сборки по Гибсону, реакцию снова проводили с расщеплением с помощью AscI, чтобы вырезать все, что не удалило сайты AscI во время сборки по Гибсона, тем самым уменьшая фон. Конечное секвенирование подтвердило наличие точечной мутации и отсутствие дополнительных изменений по сравнению с исходным направленным вектором. Вновь модифицированный вектор был электропорирован в эмбриональные стволовые клетки мыши, и положительные клоны были идентифицированы с помощью TAQMAN с последующим секвенированием по Сэнгеру для подтверждения внесения точечной мутации.
Пример 2. Бесшовное Внесение Целевой Модификации в Большой Направленный Вектор Посредством Бактериальной Гомологичной Рекомбинации и Межмолекулярной Сборки по Гибсону
[00198] Во втором способе желаемая мутация вводится в ДНК бактериальной искусственной хромосомы (БИХ) в два основных этапа. На первом этапе интересующая область в БИХ (область, охватывающая около 100-200 п.н. по каждую сторону мутации) удаляется бактериальной гомологичной рекомбинацией с использованием кассеты селекции, фланкированной сайтом редко расщепляющего рестрикционного фермента с каждой стороны. На втором этапе, делеция БИХ заменяется желаемой мутантной последовательностью путем сборки по Гибсону с использованием фрагмента ДНК размером около 200-500 п.н., имеющего гетерологичные 5'- и 3'-концы, гомологичные последовательности-мишени БИХ, граничащей с редко расщепляемым сайтом. Для этого БИХ-мишень на первом этапе расщепляется редко расщепляющим ферментом, обнажая два конца, гомологичные мутированному фрагменту. Рестрикционный фермент также сохраняет БИХ-мишень открытой, обеспечивая низкий фоновый эффект без необходимости добавления селективного маркера. См. Фиг. 3. Этот способ особенно полезен, например, когда в конструкцию необходимо вставить более крупные фрагменты, которые нельзя получить с помощью ПЦР (например, 15 т.п.н. или 30 т.п.н.). Например, такой большой фрагмент может быть вырезан из его источника, такого как БИХ (например, с использованием CRISPR/Cas9), а затем вставлен с помощью сборки по Гибсону в модифицированную БИХ, которая несет гомологию с 5'- и 3'-концами этого фрагмента, тем самым создавая окончательную направленную конструкцию.
[00199] В конкретном примере данный способ был использован для внесения мутации сплайсинга в большой направленный вектор, содержащий гуманизированный Ген-Мишень 2. Первоначальная направленная конструкция содержала геномную последовательность Гена-Мишени 2 дикого типа, включая интроны, предназначенную для замены соответствующей геномной последовательности Гена-Мишени 2 мыши от стартового кодона до предпоследнего экзона, включая добавление самоудаляющейся кассеты устойчивости к неомицину в интроне. Затем этот исходный гуманизированный вектор был модифицирован, как описано выше, с помощью кассеты устойчивости к гигромицину, фланкированной сайтами рестрикции AscI и гомологичной последовательностью из сорока пар оснований, расположенной в направлении 3' и 5' желаемой мутации сплайсинга. Кассету с устойчивостью к гигромицину вырезали с помощью AscI, и конструкцию повторно закрывали с помощью межмолекулярной сборки по Гибсону с фрагментом ДНК, содержащим мутацию сплайсинга, фланкированную гетерологичными 5'- и 3'-концами, гомологичными последовательности целевой направленной конструкции, граничащими с редко расщепляющимся сайтом рестрикции. Конечное секвенирование подтвердило наличие мутации сплайсинга и отсутствие дополнительных изменений по сравнению с исходным направленным вектором. Вновь модифицированный вектор был электропорирован в эмбриональные стволовые клетки мыши, и положительные клоны были идентифицированы с помощью TAQMAN с последующим секвенированием по Сэнгеру для подтверждения внесения мутации сплайсинга.
[00200] В третьем способе фрагмент ДНК человека из бактериальной искусственной хромосомы (БИХ) вырезается с помощью CRISPR/Cas9. Данный фрагмент ДНК человека сливается посредством сборки по Гибсону с БИХ мыши, на которую ранее была направлена кассета селекции. Сайт разрезания редко расщепляющим рестрикционным ферментом конструируется в области, где должен был быть интегрирован фрагмент ДНК человека. В мышиной БИХ-мишени имеется 40 п.н. гомологичных последовательностей с каждой стороны этого редко расщепляющегося сайта рестрикции. Последовательности гомологии являются гомологичными 5'- и 3'-концам фрагмента ДНК человека. Конечная конструкция выбирается из того же антибиотика, что и исходная мышиная БИХ-мишень. Несмотря на то, что в окончательную реакцию сборки по Гибсону не включается новый селекции, наблюдается низкий фон. Добавление редко расщепляющего рестрикционного фермента после сборки по Гибсону сохраняет фон на низком уровне.
[00201] В конкретном примере вышеупомянутый эксперимент заключался во внесении аллеля, содержащего область Гена-Мишени 3, кодирующего эктодомен Белка-Мишени 3, в Ген-Мишень 3 мыши. Первоначальная направленная конструкция содержала мышиную геномную последовательность Гена-Мишени 3 дикого типа, включая интроны. Самоудаляющуюся кассету устойчивости к неомицину добавляли путем бактериальной гомологичной рекомбинации, удаляя область, кодирующую эктодомен Гена-Мишени 3 мыши. В направлении 5' перед кассетой устойчивости к неомицину находился сайт рестрикции SgrDI, который разделяет 5'- и 3'-области гомологии с последовательностью человека, составляющие по 40 п.н., которые будут взаимодействовать с человеческим фрагментом. Все эти последовательности были внесены ранее описанной бактериальной гомологичной рекомбинацией. Фрагмент ДНК человека длиной 32 т.п.н. вырезали из БИХ человека с помощью CRISPR/Cas9, оставляя 5'- и 3'-концы открытыми для внутримолекулярной реакции сборки по Гибсону с мышиной направленной БИХ, которая была открыта путем расщепления SgrDI. Вновь модифицированный вектор был электропорирован в эмбриональные стволовые клетки мыши, и положительный клон был идентифицирован с помощью TAQMAN.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Regeneron Pharmaceuticals, Inc.
<120> СПОСОБЫ БЕСШОВНОГО ВНЕСЕНИЯ ЦЕЛЕВЫХ МОДИФИКАЦИЙ В НАПРАВЛЕННЫЕ ВЕКТОРЫ
<130> 057766-544999
<150> US 62/829,327
<151> 2019-04-04
<160> 16
<170> PatentIn версия 3.5
<210> 1
<211> 1370
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 1
Gly Ala Ala Ser Asp Lys Lys Tyr Ser Ile Gly Leu Asp Ile Gly Thr
1 5 10 15
Asn Ser Val Gly Trp Ala Val Ile Thr Asp Glu Tyr Lys Val Pro Ser
20 25 30
Lys Lys Phe Lys Val Leu Gly Asn Thr Asp Arg His Ser Ile Lys Lys
35 40 45
Asn Leu Ile Gly Ala Leu Leu Phe Asp Ser Gly Glu Thr Ala Glu Ala
50 55 60
Thr Arg Leu Lys Arg Thr Ala Arg Arg Arg Tyr Thr Arg Arg Lys Asn
65 70 75 80
Arg Ile Cys Tyr Leu Gln Glu Ile Phe Ser Asn Glu Met Ala Lys Val
85 90 95
Asp Asp Ser Phe Phe His Arg Leu Glu Glu Ser Phe Leu Val Glu Glu
100 105 110
Asp Lys Lys His Glu Arg His Pro Ile Phe Gly Asn Ile Val Asp Glu
115 120 125
Val Ala Tyr His Glu Lys Tyr Pro Thr Ile Tyr His Leu Arg Lys Lys
130 135 140
Leu Val Asp Ser Thr Asp Lys Ala Asp Leu Arg Leu Ile Tyr Leu Ala
145 150 155 160
Leu Ala His Met Ile Lys Phe Arg Gly His Phe Leu Ile Glu Gly Asp
165 170 175
Leu Asn Pro Asp Asn Ser Asp Val Asp Lys Leu Phe Ile Gln Leu Val
180 185 190
Gln Thr Tyr Asn Gln Leu Phe Glu Glu Asn Pro Ile Asn Ala Ser Gly
195 200 205
Val Asp Ala Lys Ala Ile Leu Ser Ala Arg Leu Ser Lys Ser Arg Arg
210 215 220
Leu Glu Asn Leu Ile Ala Gln Leu Pro Gly Glu Lys Lys Asn Gly Leu
225 230 235 240
Phe Gly Asn Leu Ile Ala Leu Ser Leu Gly Leu Thr Pro Asn Phe Lys
245 250 255
Ser Asn Phe Asp Leu Ala Glu Asp Ala Lys Leu Gln Leu Ser Lys Asp
260 265 270
Thr Tyr Asp Asp Asp Leu Asp Asn Leu Leu Ala Gln Ile Gly Asp Gln
275 280 285
Tyr Ala Asp Leu Phe Leu Ala Ala Lys Asn Leu Ser Asp Ala Ile Leu
290 295 300
Leu Ser Asp Ile Leu Arg Val Asn Thr Glu Ile Thr Lys Ala Pro Leu
305 310 315 320
Ser Ala Ser Met Ile Lys Arg Tyr Asp Glu His His Gln Asp Leu Thr
325 330 335
Leu Leu Lys Ala Leu Val Arg Gln Gln Leu Pro Glu Lys Tyr Lys Glu
340 345 350
Ile Phe Phe Asp Gln Ser Lys Asn Gly Tyr Ala Gly Tyr Ile Asp Gly
355 360 365
Gly Ala Ser Gln Glu Glu Phe Tyr Lys Phe Ile Lys Pro Ile Leu Glu
370 375 380
Lys Met Asp Gly Thr Glu Glu Leu Leu Val Lys Leu Asn Arg Glu Asp
385 390 395 400
Leu Leu Arg Lys Gln Arg Thr Phe Asp Asn Gly Ser Ile Pro His Gln
405 410 415
Ile His Leu Gly Glu Leu His Ala Ile Leu Arg Arg Gln Glu Asp Phe
420 425 430
Tyr Pro Phe Leu Lys Asp Asn Arg Glu Lys Ile Glu Lys Ile Leu Thr
435 440 445
Phe Arg Ile Pro Tyr Tyr Val Gly Pro Leu Ala Arg Gly Asn Ser Arg
450 455 460
Phe Ala Trp Met Thr Arg Lys Ser Glu Glu Thr Ile Thr Pro Trp Asn
465 470 475 480
Phe Glu Glu Val Val Asp Lys Gly Ala Ser Ala Gln Ser Phe Ile Glu
485 490 495
Arg Met Thr Asn Phe Asp Lys Asn Leu Pro Asn Glu Lys Val Leu Pro
500 505 510
Lys His Ser Leu Leu Tyr Glu Tyr Phe Thr Val Tyr Asn Glu Leu Thr
515 520 525
Lys Val Lys Tyr Val Thr Glu Gly Met Lys Pro Ala Phe Leu Ser Gly
530 535 540
Glu Gln Lys Lys Ala Ile Val Asp Leu Leu Phe Lys Thr Asn Arg Lys
545 550 555 560
Val Thr Val Lys Gln Leu Lys Glu Asp Tyr Phe Lys Lys Ile Glu Cys
565 570 575
Phe Asp Ser Val Glu Ile Ser Gly Val Glu Asp Arg Phe Asn Ala Ser
580 585 590
Leu Gly Thr Tyr His Asp Leu Leu Lys Ile Ile Lys Asp Lys Asp Phe
595 600 605
Leu Asp Asn Glu Glu Asn Glu Asp Ile Leu Glu Asp Ile Val Leu Thr
610 615 620
Leu Thr Leu Phe Glu Asp Arg Glu Met Ile Glu Glu Arg Leu Lys Thr
625 630 635 640
Tyr Ala His Leu Phe Asp Asp Lys Val Met Lys Gln Leu Lys Arg Arg
645 650 655
Arg Tyr Thr Gly Trp Gly Arg Leu Ser Arg Lys Leu Ile Asn Gly Ile
660 665 670
Arg Asp Lys Gln Ser Gly Lys Thr Ile Leu Asp Phe Leu Lys Ser Asp
675 680 685
Gly Phe Ala Asn Arg Asn Phe Met Gln Leu Ile His Asp Asp Ser Leu
690 695 700
Thr Phe Lys Glu Asp Ile Gln Lys Ala Gln Val Ser Gly Gln Gly Asp
705 710 715 720
Ser Leu His Glu His Ile Ala Asn Leu Ala Gly Ser Pro Ala Ile Lys
725 730 735
Lys Gly Ile Leu Gln Thr Val Lys Val Val Asp Glu Leu Val Lys Val
740 745 750
Met Gly Arg His Lys Pro Glu Asn Ile Val Ile Glu Met Ala Arg Glu
755 760 765
Asn Gln Thr Thr Gln Lys Gly Gln Lys Asn Ser Arg Glu Arg Met Lys
770 775 780
Arg Ile Glu Glu Gly Ile Lys Glu Leu Gly Ser Gln Ile Leu Lys Glu
785 790 795 800
His Pro Val Glu Asn Thr Gln Leu Gln Asn Glu Lys Leu Tyr Leu Tyr
805 810 815
Tyr Leu Gln Asn Gly Arg Asp Met Tyr Val Asp Gln Glu Leu Asp Ile
820 825 830
Asn Arg Leu Ser Asp Tyr Asp Val Asp His Ile Val Pro Gln Ser Phe
835 840 845
Leu Lys Asp Asp Ser Ile Asp Asn Lys Val Leu Thr Arg Ser Asp Lys
850 855 860
Asn Arg Gly Lys Ser Asp Asn Val Pro Ser Glu Glu Val Val Lys Lys
865 870 875 880
Met Lys Asn Tyr Trp Arg Gln Leu Leu Asn Ala Lys Leu Ile Thr Gln
885 890 895
Arg Lys Phe Asp Asn Leu Thr Lys Ala Glu Arg Gly Gly Leu Ser Glu
900 905 910
Leu Asp Lys Ala Gly Phe Ile Lys Arg Gln Leu Val Glu Thr Arg Gln
915 920 925
Ile Thr Lys His Val Ala Gln Ile Leu Asp Ser Arg Met Asn Thr Lys
930 935 940
Tyr Asp Glu Asn Asp Lys Leu Ile Arg Glu Val Lys Val Ile Thr Leu
945 950 955 960
Lys Ser Lys Leu Val Ser Asp Phe Arg Lys Asp Phe Gln Phe Tyr Lys
965 970 975
Val Arg Glu Ile Asn Asn Tyr His His Ala His Asp Ala Tyr Leu Asn
980 985 990
Ala Val Val Gly Thr Ala Leu Ile Lys Lys Tyr Pro Lys Leu Glu Ser
995 1000 1005
Glu Phe Val Tyr Gly Asp Tyr Lys Val Tyr Asp Val Arg Lys Met
1010 1015 1020
Ile Ala Lys Ser Glu Gln Glu Ile Gly Lys Ala Thr Ala Lys Tyr
1025 1030 1035
Phe Phe Tyr Ser Asn Ile Met Asn Phe Phe Lys Thr Glu Ile Thr
1040 1045 1050
Leu Ala Asn Gly Glu Ile Arg Lys Arg Pro Leu Ile Glu Thr Asn
1055 1060 1065
Gly Glu Thr Gly Glu Ile Val Trp Asp Lys Gly Arg Asp Phe Ala
1070 1075 1080
Thr Val Arg Lys Val Leu Ser Met Pro Gln Val Asn Ile Val Lys
1085 1090 1095
Lys Thr Glu Val Gln Thr Gly Gly Phe Ser Lys Glu Ser Ile Leu
1100 1105 1110
Pro Lys Arg Asn Ser Asp Lys Leu Ile Ala Arg Lys Lys Asp Trp
1115 1120 1125
Asp Pro Lys Lys Tyr Gly Gly Phe Asp Ser Pro Thr Val Ala Tyr
1130 1135 1140
Ser Val Leu Val Val Ala Lys Val Glu Lys Gly Lys Ser Lys Lys
1145 1150 1155
Leu Lys Ser Val Lys Glu Leu Leu Gly Ile Thr Ile Met Glu Arg
1160 1165 1170
Ser Ser Phe Glu Lys Asn Pro Ile Asp Phe Leu Glu Ala Lys Gly
1175 1180 1185
Tyr Lys Glu Val Lys Lys Asp Leu Ile Ile Lys Leu Pro Lys Tyr
1190 1195 1200
Ser Leu Phe Glu Leu Glu Asn Gly Arg Lys Arg Met Leu Ala Ser
1205 1210 1215
Ala Gly Glu Leu Gln Lys Gly Asn Glu Leu Ala Leu Pro Ser Lys
1220 1225 1230
Tyr Val Asn Phe Leu Tyr Leu Ala Ser His Tyr Glu Lys Leu Lys
1235 1240 1245
Gly Ser Pro Glu Asp Asn Glu Gln Lys Gln Leu Phe Val Glu Gln
1250 1255 1260
His Lys His Tyr Leu Asp Glu Ile Ile Glu Gln Ile Ser Glu Phe
1265 1270 1275
Ser Lys Arg Val Ile Leu Ala Asp Ala Asn Leu Asp Lys Val Leu
1280 1285 1290
Ser Ala Tyr Asn Lys His Arg Asp Lys Pro Ile Arg Glu Gln Ala
1295 1300 1305
Glu Asn Ile Ile His Leu Phe Thr Leu Thr Asn Leu Gly Ala Pro
1310 1315 1320
Ala Ala Phe Lys Tyr Phe Asp Thr Thr Ile Asp Arg Lys Arg Tyr
1325 1330 1335
Thr Ser Thr Lys Glu Val Leu Asp Ala Thr Leu Ile His Gln Ser
1340 1345 1350
Ile Thr Gly Leu Tyr Glu Thr Arg Ile Asp Leu Ser Gln Leu Gly
1355 1360 1365
Gly Asp
1370
<210> 2
<211> 4104
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 2
gataagaaat actcaatagg cttagatatc ggcacaaata gcgtcggatg ggcggtgatc 60
actgatgaat ataaggttcc gtctaaaaag ttcaaggttc tgggaaatac agaccgccac 120
agtatcaaaa aaaatcttat aggggctctt ttatttgaca gtggagagac agcggaagcg 180
actcgtctca aacggacagc tcgtagaagg tatacacgtc ggaagaatcg tatttgttat 240
ctacaggaga ttttttcaaa tgagatggcg aaagtagatg atagtttctt tcatcgactt 300
gaagagtctt ttttggtgga agaagacaag aagcatgaac gtcatcctat ttttggaaat 360
atagtagatg aagttgctta tcatgagaaa tatccaacta tctatcatct gcgaaaaaaa 420
ttggtagatt ctactgataa agcggatttg cgcttaatct atttggcctt agcgcatatg 480
attaagtttc gtggtcattt tttgattgag ggagatttaa atcctgataa tagtgatgtg 540
gacaaactat ttatccagtt ggtacaaacc tacaatcaat tatttgaaga aaaccctatt 600
aacgcaagtg gagtagatgc taaagcgatt ctttctgcac gattgagtaa atcaagacga 660
ttagaaaatc tcattgctca gctccccggt gagaagaaaa atggcttatt tgggaatctc 720
attgctttgt cattgggttt gacccctaat tttaaatcaa attttgattt ggcagaagat 780
gctaaattac agctttcaaa agatacttac gatgatgatt tagataattt attggcgcaa 840
attggagatc aatatgctga tttgtttttg gcagctaaga atttatcaga tgctatttta 900
ctttcagata tcctaagagt aaatactgaa ataactaagg ctcccctatc agcttcaatg 960
attaaacgct acgatgaaca tcatcaagac ttgactcttt taaaagcttt agttcgacaa 1020
caacttccag aaaagtataa agaaatcttt tttgatcaat caaaaaacgg atatgcaggt 1080
tatattgatg ggggagctag ccaagaagaa ttttataaat ttatcaaacc aattttagaa 1140
aaaatggatg gtactgagga attattggtg aaactaaatc gtgaagattt gctgcgcaag 1200
caacggacct ttgacaacgg ctctattccc catcaaattc acttgggtga gctgcatgct 1260
attttgagaa gacaagaaga cttttatcca tttttaaaag acaatcgtga gaagattgaa 1320
aaaatcttga cttttcgaat tccttattat gttggtccat tggcgcgtgg caatagtcgt 1380
tttgcatgga tgactcggaa gtctgaagaa acaattaccc catggaattt tgaagaagtt 1440
gtcgataaag gtgcttcagc tcaatcattt attgaacgca tgacaaactt tgataaaaat 1500
cttccaaatg aaaaagtact accaaaacat agtttgcttt atgagtattt tacggtttat 1560
aacgaattga caaaggtcaa atatgttact gaaggaatgc gaaaaccagc atttctttca 1620
ggtgaacaga agaaagccat tgttgattta ctcttcaaaa caaatcgaaa agtaaccgtt 1680
aagcaattaa aagaagatta tttcaaaaaa atagaatgtt ttgatagtgt tgaaatttca 1740
ggagttgaag atagatttaa tgcttcatta ggtacctacc atgatttgct aaaaattatt 1800
aaagataaag attttttgga taatgaagaa aatgaagata tcttagagga tattgtttta 1860
acattgacct tatttgaaga tagggagatg attgaggaaa gacttaaaac atatgctcac 1920
ctctttgatg ataaggtgat gaaacagctt aaacgtcgcc gttatactgg ttggggacgt 1980
ttgtctcgaa aattgattaa tggtattagg gataagcaat ctggcaaaac aatattagat 2040
tttttgaaat cagatggttt tgccaatcgc aattttatgc agctgatcca tgatgatagt 2100
ttgacattta aagaagacat tcaaaaagca caagtgtctg gacaaggcga tagtttacat 2160
gaacatattg caaatttagc tggtagccct gctattaaaa aaggtatttt acagactgta 2220
aaagttgttg atgaattggt caaagtaatg gggcggcata agccagaaaa tatcgttatt 2280
gaaatggcac gtgaaaatca gacaactcaa aagggccaga aaaattcgcg agagcgtatg 2340
aaacgaatcg aagaaggtat caaagaatta ggaagtcaga ttcttaaaga gcatcctgtt 2400
gaaaatactc aattgcaaaa tgaaaagctc tatctctatt atctccaaaa tggaagagac 2460
atgtatgtgg accaagaatt agatattaat cgtttaagtg attatgatgt cgatcacatt 2520
gttccacaaa gtttccttaa agacgattca atagacaata aggtcttaac gcgttctgat 2580
aaaaatcgtg gtaaatcgga taacgttcca agtgaagaag tagtcaaaaa gatgaaaaac 2640
tattggagac aacttctaaa cgccaagtta atcactcaac gtaagtttga taatttaacg 2700
aaagctgaac gtggaggttt gagtgaactt gataaagctg gttttatcaa acgccaattg 2760
gttgaaactc gccaaatcac taagcatgtg gcacaaattt tggatagtcg catgaatact 2820
aaatacgatg aaaatgataa acttattcga gaggttaaag tgattacctt aaaatctaaa 2880
ttagtttctg acttccgaaa agatttccaa ttctataaag tacgtgagat taacaattac 2940
catcatgccc atgatgcgta tctaaatgcc gtcgttggaa ctgctttgat taagaaatat 3000
ccaaaacttg aatcggagtt tgtctatggt gattataaag tttatgatgt tcgtaaaatg 3060
attgctaagt ctgagcaaga aataggcaaa gcaaccgcaa aatatttctt ttactctaat 3120
atcatgaact tcttcaaaac agaaattaca cttgcaaatg gagagattcg caaacgccct 3180
ctaatcgaaa ctaatgggga aactggagaa attgtctggg ataaagggcg agattttgcc 3240
acagtgcgca aagtattgtc catgccccaa gtcaatattg tcaagaaaac agaagtacag 3300
acaggcggat tctccaagga gtcaatttta ccaaaaagaa attcggacaa gcttattgct 3360
cgtaaaaaag actgggatcc aaaaaaatat ggtggttttg atagtccaac ggtagcttat 3420
tcagtcctag tggttgctaa ggtggaaaaa gggaaatcga agaagttaaa atccgttaaa 3480
gagttactag ggatcacaat tatggaaaga agttcctttg aaaaaaatcc gattgacttt 3540
ttagaagcta aaggatataa ggaagttaaa aaagacttaa tcattaaact acctaaatat 3600
agtctttttg agttagaaaa cggtcgtaaa cggatgctgg ctagtgccgg agaattacaa 3660
aaaggaaatg agctggctct gccaagcaaa tatgtgaatt ttttatattt agctagtcat 3720
tatgaaaagt tgaagggtag tccagaagat aacgaacaaa aacaattgtt tgtggagcag 3780
cataagcatt atttagatga gattattgag caaatcagtg aattttctaa gcgtgttatt 3840
ttagcagatg ccaatttaga taaagttctt agtgcatata acaaacatag agacaaacca 3900
atacgtgaac aagcagaaaa tattattcat ttatttacgt tgacgaatct tggagctccc 3960
gctgctttta aatattttga tacaacaatt gatcgtaaac gatatacgtc tacaaaagaa 4020
gttttagatg ccactcttat ccatcaatcc atcactggtc tttatgaaac acgcattgat 4080
ttgagtcagc taggaggtga ctga 4104
<210> 3
<211> 16
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 3
guuuuagagc uaugcu 16
<210> 4
<211> 67
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 4
agcauagcaa guuaaaauaa ggcuaguccg uuaucaacuu gaaaaagugg caccgagucg 60
gugcuuu 67
<210> 5
<211> 77
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 5
guuuuagagc uagaaauagc aaguuaaaau aaggcuaguc cguuaucaac uugaaaaagu 60
ggcaccgagu cggugcu 77
<210> 6
<211> 82
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 6
guuggaacca uucaaaacag cauagcaagu uaaaauaagg cuaguccguu aucaacuuga 60
aaaaguggca ccgagucggu gc 82
<210> 7
<211> 76
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 7
guuuuagagc uagaaauagc aaguuaaaau aaggcuaguc cguuaucaac uugaaaaagu 60
ggcaccgagu cggugc 76
<210> 8
<211> 86
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 8
guuuaagagc uaugcuggaa acagcauagc aaguuuaaau aaggcuaguc cguuaucaac 60
uugaaaaagu ggcaccgagu cggugc 86
<210> 9
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<220>
<221> misc_feature
<222> (2)..(21)
<223> n представляет собой a, c, g или t
<400> 9
gnnnnnnnnn nnnnnnnnnn ngg 23
<210> 10
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<220>
<221> misc_feature
<222> (1)..(21)
<223> n представляет собой a, c, g или t
<400> 10
nnnnnnnnnn nnnnnnnnnn ngg 23
<210> 11
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<220>
<221> misc_feature
<222> (3)..(23)
<223> n представляет собой a, c, g или t
<400> 11
ggnnnnnnnn nnnnnnnnnn nnngg 25
<210> 12
<211> 72
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 12
aaacagcaua gcaaguuaaa auaaggcuag uccguuauca acuugaaaaa guggcaccga 60
gucggugcuu uu 72
<210> 13
<211> 82
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 13
guuggaacca uucaaaacag cauagcaagu uaaaauaagg cuaguccguu aucaacuuga 60
aaaaguggca ccgagucggu gc 82
<210> 14
<211> 83
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 14
guuuuagagc uagaaauagc aaguuaaaau aaggcuaguc cguuaucaac uugaaaaagu 60
ggcaccgagu cggugcuuuu uuu 83
<210> 15
<211> 80
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 15
guuuuagagc uagaaauagc aaguuaaaau aaggcuaguc cguuaucaac uugaaaaagu 60
ggcaccgagu cggugcuuuu 80
<210> 16
<211> 92
<212> РНК
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 16
guuuaagagc uaugcuggaa acagcauagc aaguuuaaau aaggcuaguc cguuaucaac 60
uugaaaaagu ggcaccgagu cggugcuuuu uu 92
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МЫШИ, СОДЕРЖАЩИЕ МУТАЦИИ, ВСЛЕДСТВИЕ КОТОРЫХ ЭКСПРЕССИРУЕТСЯ УКОРОЧЕННЫЙ НА С-КОНЦЕ ФИБРИЛЛИН-1 | 2017 |
|
RU2721125C1 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ОЦЕНКИ ОПОСРЕДОВАННОГО CRISPR/CAS РАЗРУШЕНИЯ ИЛИ ВЫРЕЗАНИЯ И ИНДУЦИРОВАННОЙ CRISPR/CAS РЕКОМБИНАЦИИ С ЭКЗОГЕННОЙ ДОНОРСКОЙ НУКЛЕИНОВОЙ КИСЛОТОЙ IN VIVO | 2018 |
|
RU2782356C2 |
| ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА ЖИВОТНЫЕ, ВКЛЮЧАЮЩИЕ В СЕБЯ ГУМАНИЗИРОВАННЫЙ TTR ЛОКУС, И СПОСОБЫ ПРИМЕНЕНИЯ | 2018 |
|
RU2784927C1 |
| Животные, отличные от человека, содержащие гуманизированный локус альбумина | 2020 |
|
RU2815514C2 |
| ЭКСПРЕССИРУЮЩИЕ CAS МЫШИНЫЕ ЭМБРИОНАЛЬНЫЕ СТВОЛОВЫЕ КЛЕТКИ И МЫШИ И ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2782358C2 |
| СИСТЕМА РЕДАКТИРОВАНИЯ ГЕНОМА CRISPR/CAS9 II ТИПА И ЕЕ ПРИМЕНЕНИЕ | 2022 |
|
RU2794774C1 |
| СРЕДСТВО РАЗРЕЗАНИЯ ДНК НА ОСНОВЕ CAS9 БЕЛКА ИЗ БАКТЕРИИ DEMEQUINA SEDIMINICOLA | 2019 |
|
RU2722933C1 |
| ПРИМЕНЕНИЕ CAS9 БЕЛКА ИЗ БАКТЕРИИ PASTEURELLA PNEUMOTROPICA ДЛЯ МОДИФИКАЦИИ ГЕНОМНОЙ ДНК В КЛЕТКАХ | 2019 |
|
RU2724470C1 |
| СРЕДСТВО РАЗРЕЗАНИЯ ДНК НА ОСНОВЕ CAS9 БЕЛКА ИЗ БАКТЕРИИ PASTEURELLA PNEUMOTROPICA | 2019 |
|
RU2722934C1 |
| ГЕННОЕ РЕДАКТИРОВАНИЕ ГЛУБОКИХ ИНТРОННЫХ МУТАЦИЙ | 2016 |
|
RU2759335C2 |
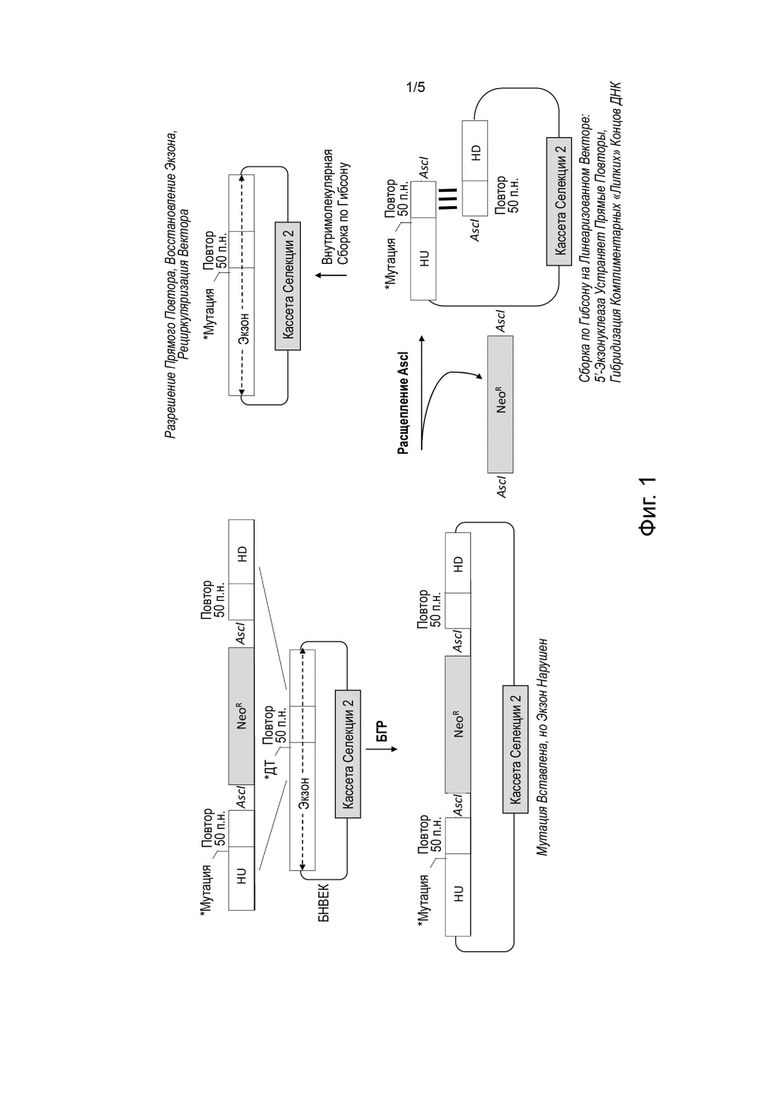
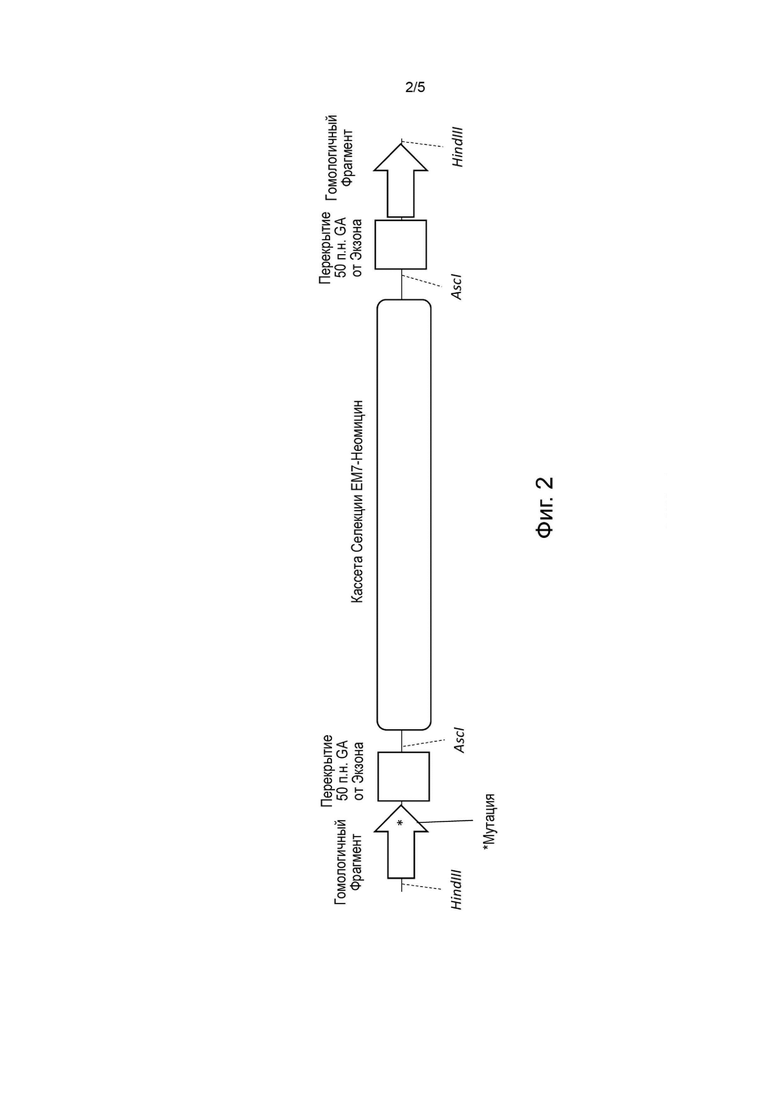

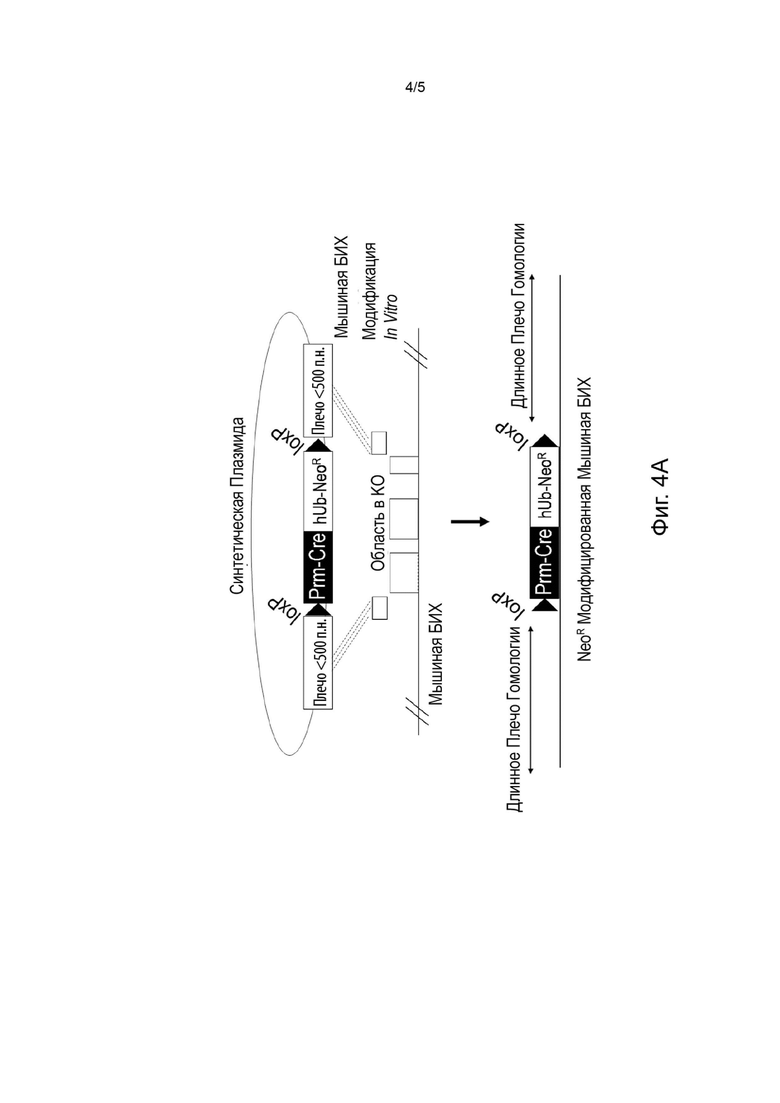

Изобретение относится к области биотехнологии. Предложены способы для внесения бесшовной целевой генетической модификации в ранее созданный направленный вектор. Изобретение может быть использовано, чтобы упростить создание таргетинга, несущего множественные изменения (такие как гуманизированный аллель и наслоенная мутация болезни), и минимизировать швы, включенные в окончательную животную модель. 2 н. и 28 з.п. ф-лы, 5 ил., 2 табл., 2 пр.
1. Способ внесения бесшовной целевой генетической модификации в ранее созданный направленный вектор, включающий:
а) выполнение бактериальной гомологичной рекомбинации между ранее созданным направленным вектором и кассетой модификации в популяции бактериальных клеток, при этом кассета модификации содержит целевую генетическую модификацию и содержит нуклеиновую кислоту вставки, фланкированную 5’-плечом гомологии, соответствующим 5’-последовательности-мишени в ранее созданном направленном векторе, и 3’-плечом гомологии, соответствующим 3’-последовательности-мишени в ранее созданном векторе, при этом нуклеиновая кислота вставки содержит от 5’- к 3’-концу:
(i) первую повторяющуюся последовательность;
(ii) первый сайт-мишень для первого нуклеазного агента;
(iii) кассету селекции;
(iv) второй сайт-мишень для второго нуклеазного агента; и
(v) вторую повторяющуюся последовательность, идентичную первой повторяющейся последовательности;
(b) отбор бактериальных клеток, содержащих модифицированный направленный вектор, содержащий кассету селекции;
(c) расщепление первого сайта-мишени в модифицированном направленном векторе первым нуклеазным агентом и расщепление второго сайта-мишени в модифицированном направленном векторе вторым нуклеазным агентом для удаления кассеты селекции и открытия первой повторяющейся последовательности и второй повторяющейся последовательности в модифицированном направленном векторе; и
(d) сборку экспонированной первой повторяющейся последовательности с экспонированной второй повторяющейся последовательностью во внутримолекулярной реакции сборки in vitro для создания направленного вектора, содержащего бесшовную целевую генетическую модификацию, при этом ни первый сайт-мишень для первого нуклеазного агента, ни второй сайт-мишень для второго нуклеазного агента не присутствуют, и только одна копия повторяющейся последовательности присутствует в направленном векторе, содержащем бесшовную направленную генетическую модификацию.
2. Способ по п. 1, отличающийся тем, что:
(I) (a) повторяющаяся последовательность идентична последовательности в ранее созданном направленном векторе или
(b) целевая генетическая модификация содержит вставку, а повторяющаяся последовательность идентична 5’-концу или 3’-концу вставки; и/или
(II) повторяющаяся последовательность имеет длину, по меньшей мере, 20 нуклеотидов, необязательно повторяющаяся последовательность имеет длину от 20 нуклеотидов до 100 нуклеотидов.
3. Способ по п. 1 или 2, отличающийся тем, что:
(I) кассета модификации представляет собой линейную двухцепочечную нуклеиновую кислоту; и/или
(II) кассета модификации имеет длину от 1 т.п.н. до 15 т.п.н.
4. Способ по любому из предшествующих пунктов, отличающийся тем, что каждое 5’-плечо гомологии и 3’-плечо гомологии имеют длину, по меньшей мере, 35 нуклеотидов или каждое 5’-плечо гомологии и 3’-плечо гомологии имеют длину от 35 нуклеотидов до 500 нуклеотидов.
5. Способ по любому из предшествующих пунктов, отличающийся тем, что первый нуклеазный агент и/или второй нуклеазный агент представляют собой редко расщепляющий нуклеазный агент.
6. Способ по любому из предшествующих пунктов, отличающийся тем, что:
(I) первый сайт-мишень и/или второй сайт-мишень отсутствуют в ранее созданном направленном векторе; и/или
(II) первый сайт-мишень идентичен второму сайту-мишени, и первый нуклеазный агент идентичен второму нуклеазному агенту.
7. Способ по любому из предшествующих пунктов, отличающийся тем, что:
(I) первый нуклеазный агент и/или второй нуклеазный агент содержат редко расщепляющий рестрикционный фермент, необязательно редко расщепляющий рестрикционный фермент представляет собой NotI, XmaIII, SstII, Sall, NruI, NheI, Nb.BbvCI, BbvCI, AscI, AsiSI, FseI, PacI, PmeI, SbfI, SgrAI, SwaI, BspQI, SapI, SfiI, CspCI, AbsI, CciNI, FspAI, MauBI, MreI, MssI, PalAI, RgaI, RigI, SdaI, SfaAI, SgfI, SgrDI, SgsI, SmiI, SrfI, Sse2321, Sse83871, LguI, PciSI, AarI, AjuI, AloI, BarI, PpiI или PsrI; или
(II) первый нуклеазный агент и/или второй нуклеазный агент представляет собой белок (Cas), ассоциированный с кластеризованными регулярными промежуточными короткими палиндромными повторами (CRISPR) и направляющую РНК (нРНК), нуклеазу с "цинковыми пальцами" (ZFN), эффекторную нуклеазу, подобную активаторам транскрипции (TALEN), или сконструированную мегануклеазу, необязательно первый нуклеазный агент и/или второй нуклеазный агент представляют собой белок Cas и нРНК, при этом белок Cas представляет собой Cas9, и при этом нРНК содержит CRISPR-РНК (крРНК) и транс-активирующую CRISPR-РНК (тракрРНК).
8. Способ по любому из предшествующих пунктов, отличающийся тем, что целевая генетическая модификация содержит:
(I) модификацию в 5’-плече гомологии или в 3’-плече гомологии или
(II) модификацию нуклеиновой кислоты вставки.
9. Способ по любому из предшествующих пунктов, отличающийся тем, что целевая генетическая модификация содержит точечную мутацию, делецию, вставку, замену или их комбинацию.
10. Способ по любому из предшествующих пунктов, отличающийся тем, что кассета селекции придает устойчивость к антибиотику, необязательно кассета селекции придает устойчивость к ампициллину, хлорамфениколу, тетрациклину, канамицину, спектиномицину, стрептомицину, карбенициллину, блеомицину, эритромицину или полимиксину B.
11. Способ по любому из предшествующих пунктов, отличающийся тем, что ранее созданный направленный вектор представляет собой большой направленный вектор, имеющий длину, по меньшей мере, 10 т.п.н. или ранее созданный направленный вектор имеет длину, по меньшей мере, 100 т.п.н.
12. Способ по любому из предшествующих пунктов, отличающийся тем, что ранее созданный направленный вектор содержит вторую кассету селекции, необязательно вторая кассета селекции придает устойчивость к антибиотику, необязательно кассета селекции в кассете модификации и вторая кассета селекции в ранее созданном направленном векторе придают устойчивость к разным антибиотикам и/или необязательно вторая кассета селекции позволяет производить селекцию как в бактериальных клетках, так и в клетках млекопитающих.
13. Способ по любому из предшествующих пунктов, отличающийся тем, что этап (с) выполняется in vitro.
14. Способ по любому из предшествующих пунктов, отличающийся тем, что этап (d) включает:
(i) контактирование модифицированного направленного вектора с экзонуклеазой для выявления комплементарных последовательностей между первой повторяющейся последовательностью и второй повторяющейся последовательностью;
(ii) гибридизацию экспонированных комплементарных последовательностей;
(iii) удлинение 3’-концов гибридизированных комплементарных последовательностей; и
(iv) лигирование гибридизированной комплементарной последовательности, и необязательно
этап (d) включает инкубирование модифицированного направленного вектора с экзонуклеазой, ДНК-полимеразой и ДНК-лигазой.
15. Способ по любому из предшествующих пунктов, дополнительно включающий:
(е) обработку направленного вектора первым нуклеазным агентом и вторым нуклеазным агентом после сборки in vitro на этапе (d), чтобы убедиться, что ни первый сайт-мишень для первого нуклеазного агента, ни второй сайт-мишень для второго нуклеазного агента не присутствуют.
16. Способ для внесения бесшовной целевой генетической модификации в ранее созданный направленный вектор, включающий:
а) выполнение бактериальной гомологичной рекомбинации между ранее созданным направленным вектором и кассетой делеции в популяции бактериальных клеток, при этом кассета делеции содержит вставку нуклеиновой кислоты, фланкированную 5’-плечом гомологии, соответствующим 5’-последовательности-мишени в ранее созданном направленном векторе, и 3’-гомологическим плечом, соответствующим 3’- последовательности-мишени в ранее созданном направленном векторе, при этом 5’-последовательность-мишень и 3’-последовательность-мишень фланкируют область ранее созданного направленного вектора, в которую должна быть внесена целевая генетическая модификация, и при этом нуклеиновая кислота вставки содержит от 5’- к 3’-концу:
(i) первый сайт-мишень для первого нуклеазного агента;
(ii) кассету селекции; и
(iii) второй сайт-мишень для второго нуклеазного агента;
(b) отбор бактериальных клеток, содержащих модифицированный направленный вектор, содержащий кассету селекции;
(c) расщепление первого сайта-мишени в модифицированном направленном векторе первым нуклеазным агентом и расщепление второго сайта-мишени в модифицированном направленном векторе вторым нуклеазным агентом для удаления кассеты селекции и экспонирования последовательности, расположенной в направлении 5’- к 3’-концу последовательности, в модифицированном направленном векторе; и
(d) соединение в реакции сборки in vitro расщепленного направленного вектора с кассетой модификации, содержащей целевую генетическую модификацию, фланкированную концевой последовательностью, расположенной в направлении 5’ и перекрывающей концевую последовательность, расположенную в направлении 5’, в модифицированном направленном векторе, и концевой последовательностью, расположенной в направлении 3’ и перекрывающей концевую последовательность, расположенную в направлении 3’, в модифицированном направленном векторе для создания направленного вектора, содержащего бесшовную целевую генетическую модификацию, при этом и первый сайт-мишень для первого нуклеазного агента, и второй сайт- мишень для второго нуклеазного агента отсутствуют в направленном векторе, содержащем бесшовную целевую генетическую модификацию.
17. Способ по п. 16, отличающийся тем, что:
(I) кассета делеции имеет длину от 1 т.п.н. до 15 т.п.н. и/или
(II) кассета делеции представляет собой линейную двухцепочечную нуклеиновую кислоту.
18. Способ по п. 16 или 17, отличающийся тем, что каждое 5’-плечо гомологии и 3’-плечо гомологии имеют длину, по меньшей мере, 35 нуклеотидов или каждое 5’-плечо гомологии и 3’-плечо гомологии имеют длину от 35 нуклеотидов до 500 нуклеотидов.
19. Способ по любому из пп. 16-18, отличающийся тем, что первый нуклеазный агент и/или второй нуклеазный агент представляют собой редко расщепляющие нуклеазные агенты.
20. Способ по любому из пп. 16-19, отличающийся тем, что:
(I) первый сайт-мишень и/или второй сайт-мишень отсутствуют в ранее созданном направленном векторе; и/или
(II) первый сайт-мишень идентичен второму сайту-мишени, и первый нуклеазный агент идентичен второму нуклеазному агенту.
21. Способ по любому из пп. 16-20, отличающийся тем, что:
(I) первый нуклеазный агент и/или второй нуклеазный агент представляют собой редко расщепляющие рестрикционные ферменты, необязательно редко расщепляющий рестрикционный фермент представляет собой NotI, XmaIII, SstII, Sall, NruI, NheI, Nb.BbvCI, BbvCI, AscI, AsiSI, FseI, PacI, PmeI, SbfI, SgrAI, SwaI, BspQI, SapI, SfiI, CspCI, AbsI, CciNI, FspAI, MauBI, MreI, MssI, PalAI, RgaI, RigI, SdaI, SfaAI, SgfI, SgrDI, SgsI, SmiI, SrfI, Sse2321, Sse83871, LguI, PciSI, AarI, AjuI, AloI, BarI, PpiI или PsrI; или
(II) первый нуклеазный агент и/или второй нуклеазный агент представляет собой белок (Cas), ассоциированный с кластеризованными регулярными промежуточными короткими палиндромными повторами (CRISPR) и направляющую РНК (нРНК), нуклеазу с «цинковыми пальцами» (ZFN), эффекторную нуклеазу, подобную активаторам транскрипции (TALEN) или сконструированную мегануклеазу, необязательно первый нуклеазный агент и/или второй нуклеазный агент представляют собой белок Cas и нРНК, при этом белок Cas представляет собой Cas9, и при этом нРНК содержит CRISPR-РНК (крРНК)и транс-активирующую CRISPR-РНК (тракрРНК).
22. Способ по любому из пп. 16-21, отличающийся тем, что кассета селекции придает устойчивость к антибиотику, необязательно кассета селекции придает устойчивость к ампициллину, хлорамфениколу, тетрациклину, канамицину, спектиномицину, стрептомицину, карбенициллину, блеомицину, эритромицину или полимиксину B.
23. Способ по любому из пп. 16-22, отличающийся тем, что ранее созданный направленный вектор представляет собой большой направленный вектор, имеющий длину, по меньшей мере, 10 т.п.н. или ранее созданный направленный вектор имеет длину, по меньшей мере, 100 т.п.н.
24. Способ по любому из пп. 16-23, отличающийся тем, что ранее созданный направленный вектор содержит вторую кассету селекции, необязательно, вторая кассета селекции придает устойчивость к антибиотику, необязательно кассета селекции в кассете делеции и вторая кассета селекции в ранее созданном направленном векторе придают устойчивость к разным антибиотикам и/или необязательно вторая кассета селекции позволяет производить селекцию как в бактериальных клетках, так и в клетках млекопитающих.
25. Способ по любому из пп. 16-24, отличающийся тем, что длина перекрывания между концевой последовательностью, расположенной в направлении 5’ в кассете модификации, и концевой последовательностью, расположенной в направлении 5’ в модифицированном направленном векторе, и/или длина перекрывания между концевой последовательностью, расположенной в направлении 3’ в кассете модификации, и концевой последовательностью, расположенной в направлении 3’ в модифицированном направленном векторе, составляет, по меньшей мере, 20 нуклеотидов, или длина перекрывания между концевой последовательностью, расположенной в направлении 5’ в кассете модификации, и концевой последовательностью, расположенной в направлении 5’ в модифицированном направленном векторе, и/или длина перекрывания между концевой последовательностью, расположенной в направлении 3’ в кассете модификации, и концевой последовательностью, расположенной в направлении 3’ в модифицированном направленном векторе, составляет от 20 до 100 нуклеотидов.
26. Способ по любому из пп. 16-25, отличающийся тем, что этап (с) выполняется in vitro.
27. Способ по любому из пп. 16-26, отличающийся тем, что этап (d) включает:
(i) контактирование расщепленного направленного вектора и кассеты модификации с экзонуклеазой для открытия комплементарных последовательностей между концевыми последовательностями в модифицированном направленном векторе и концевыми последовательностями в кассете модификации;
(ii) гибридизацию экспонированных комплементарных последовательностей;
(iii) удлинение 3’-концов гибридизированных комплементарных последовательностей; и
(iv) лигирование гибридизированной комплементарной последовательности, и необязательно этап (d) включает инкубирование расщепленного направленного вектора и кассеты модификации с экзонуклеазой, ДНК-полимеразой и ДНК-лигазой.
28. Способ по любому из пп. 16-27, отличающийся тем, что:
(I) кассета модификации представляет собой линейную двухцепочечную нуклеиновую кислоту;
(II) кассета модификации имеет длину, по меньшей мере, 200 нуклеотидов, необязательно кассета модификации имеет длину, по меньшей мере, 10 т.п.н.; и/или
(III) кассета модификации может иметь такой размер, что не может быть синтезирована напрямую или получена путем полимеразной цепной реакции.
29. Способ по любому из пп. 16-28, отличающийся тем, что целевая генетическая модификация содержит точечную мутацию, делецию, вставку, замену или их комбинацию.
30. Способ по любому из пп. 16-29, дополнительно включающий:
(е) обработку направленного вектора первым нуклеазным агентом и вторым нуклеазным агентом после сборки in vitro на этапе (d), чтобы убедиться, что ни первый сайт-мишень для первого нуклеазного агента, ни второй сайт-мишень для второго нуклеазного агента не присутствуют.
| J.YANG | |||
| High-efficiency scarless genetic modification in Escherichia coli by using lambda red recombination and I-SceI cleavage | |||
| Appl Environ Microbiol | |||
| Способ защиты переносных электрических установок от опасностей, связанных с заземлением одной из фаз | 1924 |
|
SU2014A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Способ защиты переносных электрических установок от опасностей, связанных с заземлением одной из фаз | 1924 |
|
SU2014A1 |
| Стержень для наматывания пожарного шланга | 1930 |
|
SU27289A1 |
| ПЕПТИДНЫЙ ВЕКТОР, СПОСОБ ЕГО ПОЛУЧЕНИЯ, НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК И ШТАММ ESCHERICHIA COLI B-8389 ВКПМ ДЛЯ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ГЕНЕТИЧЕСКОЙ МОДИФИКАЦИИ КЛЕТОК МЛЕКОПИТАЮЩИХ И ЧЕЛОВЕКА | 2003 |
|
RU2248983C1 |

