Данное изобретение относится к антигенсвязывающим белкам, содержащим две Fc-части, к способам их получения, фармацевтическим композициям, содержащим указанные антигенсвязывающие белки, и к их применению.
Уровень техники
В последние два десятилетия были разработаны и оценены различные инженерные производные антител, моно- или полиспецифические, либо моно- или поливалентные (см., например, Holliger, P., et al., Nature Biotech. 23 (2005) 1126-1136; Fischer, N., and Leger O., Pathobiology 74 (2007) 3-14).
US 2004/0033561 относится к ДНК и производству одновалентных моноантител путем совместной экспрессии тяжелой цепи и модифицированной тяжелой цепи. Тем не менее, при экспрессии образуется значительное число нежелательных гомодимеров в качестве побочного продукта, который трудно отделить от нужных гетеродимерных моноантител, так как гомодимер и гетеродимер имеют такие же или аналогичные молекулярные массы. WO 2007/048037 относится к одновалентным IgG, что соответствует гетеродимерным моноантителам из US 2004/0033561, но может иметь меченую группировку, присоединенную к тяжелой цепи для облегчения очистки гетеродимера от трудного для отделения гомодимерного побочного продукта.
Сущность изобретения
Изобретение включает антигенсвязывающий белок, содержащий
а) две модифицированные тяжелые цепи антитела, которое специфически связывается с антигеном, где VH каждой тяжелой цепи заменен на VL указанного антитела, причем указанные модифицированные тяжелые цепи связаны друг с другом через свои СН-домены Fc-части;
б) две модифицированные тяжелые цепи указанного антитела, где СН1 каждой тяжелой цепи заменен на CL указанного антитела, причем указанные модифицированные тяжелые цепи связаны друг с другом через свои СН-домены Fc-части;
и где VL-домены тяжелых цепей а) связаны с VH-доменами тяжелых цепей б), и CH1-домены тяжелых цепей а) связаны с CL-доменами тяжелых цепей б).
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что
СН3-домены Fc-части модифицированных тяжелых цепей а) и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют один и тот же изотип.
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что
СН2- и СН3-домены Fc-части модифицированных тяжелых цепей а) и СН2-и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют один и тот же изотип.
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что
СН2- и СН3-домены Fc-части модифицированных тяжелых цепей а) и СН2-и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют изотип IgG.
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что
СН2- и СН3-домены Fc-части модифицированных тяжелых цепей а) и СН2-и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют изотип IgG1.
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что содержит
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №1; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №2;
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №3; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №4; или
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №5; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №6.
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что
либо две модифицированные тяжелые цепи а),
либо две модифицированные тяжелые цепи б)
также модифицированы путем аминокислотных замен S364G, L368F, D399K и K409D (где аминокислотные позиции нумеруются в соответствии с индексом ЕС Кабата).
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №1, и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №2;
где либо две модифицированные тяжелые цепи а),
либо две модифицированные тяжелые цепи б)
также модифицированы путем аминокислотных замен S364G, L368F, D399K и K409D (где аминокислотные позиции нумеруются в соответствии с индексом ЕС Кабата);
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №3; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №4;
где либо две модифицированные тяжелые цепи а),
либо две модифицированные тяжелые цепи б)
также модифицированы путем аминокислотных замен S364G, L368F, D399K и K409D (где аминокислотные позиции нумеруются в соответствии с индексом ЕС Кабата);
или
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №5; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №6,
где либо две модифицированные тяжелые цепи а),
либо две модифицированные тяжелые цепи б)
также модифицированы путем аминокислотных замен S364G, L368F, D399K и K409D (где аминокислотные позиции нумеруются в соответствии с индексом ЕС Кабата).
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что
СН3-домены Fc-части модифицированных тяжелых цепей а) и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют различные изотипы.
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что
СН3-домены Fc-части модифицированных тяжелых цепей а) имеют изотип IgG1;
и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют изотип IgA.
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что содержит
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №7; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №4.
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что содержит
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №3; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №8.
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что
СН2- и СН3-домены Fc-части модифицированных тяжелых цепей а) имеют изотип IgG1;
СН2- и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют изотип IgA.
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что СН2-домены Fc-частей а) и б) имеют изотип IgG1, а антигенсвязывающий белок является афукозилированным, где количество фукозы составляет 80% или менее (предпочтительно от 65% до 5%) от общего количества олигосахаридов (сахаров) на Asn297, и имеет человеческий изотип IgG1.
Кроме того, изобретение включает способ получения антигенсвязывающего белка в соответствии с изобретением,
включающий этапы
а) трансформации клетки-хозяина векторами, содержащими нуклеиновокислотные молекулы, кодирующие антигенсвязывающий белок в соответствии с изобретением;
б) культивирования клетки-хозяина в условиях, обеспечивающих синтез указанной антигенсвязывающей белковой молекулы; и
в) выделения указанной антигенсвязывающей белковой молекулы из указанной культуры.
Изобретение также включает нуклеиновую кислоту, кодирующую антигенсвязывающий белок в соответствии с изобретением.
Кроме того, изобретение включает векторы, содержащие нуклеиновую кислоту, кодирующую антигенсвязывающий белок в соответствии с изобретением.
Кроме того, изобретение включает клетку-хозяина, содержащую указанные векторы.
Кроме того, изобретение включает композицию, предпочтительно фармацевтическую или диагностическую композицию антигенсвязывающего белка в соответствии с изобретением.
Кроме того, изобретение включает фармацевтическую композицию, содержащую антигенсвязывающий белок в соответствии с изобретением.
Кроме того, изобретение включает способ лечения пациента, нуждающегося в терапии, который характеризуется введением пациенту терапевтически эффективного количества антигенсвязывающего белка в соответствии с изобретением.
Было обнаружено, что антигенсвязывающие белки в соответствии с изобретением обладают ценными характеристиками, такими как биологическая или фармакологическая активность (например, усиленная ADCC по сравнению с родительскими антителами). Они могут быть использованы, например, для лечения таких заболеваний как рак. Кроме того, антигенсвязывающие белки в соответствии с изобретением обладают высокоценными фармакокинетическими свойствами (такими как, например, AUC0-inf, Cmax или С0).
Описание графических материалов
Фиг.1А и В: А) Схематическая структура антигенсвязывающего белка в соответствии с изобретением (сокращенно MoAb-димер) с CH1-CL-переключением. В) Схема основного побочного продукта - одновалентного мономера антитела (MoAb) с CH1-CL-переключением (сокращенно MoAb).
Фиг.1C: С) Ассоциация двух модифицированных тяжелых цепей а и b: гетеродимеризация двух различных цепей (а с b) непосредственно приводит к одновалентному антителу В (путь 2). Гомодимеризация двух идентичных цепей (а с а и b с b) приводит к предполагаемым промежуточным продуктам аа и bb (по пути 1), которые могут ассоциировать с формированием «MoAb-димера» А. Модификация контактов СН3-СН3 может влиять на распределение продуктов А (MoAb-димер) и В (MoAb). Модификации, которые способствуют гетеродимеризации (например, «ключи в замках»), будут увеличивать относительное количество соединения В по пути 2, в то время как модификации, которые поддерживают притягивающие взаимодействия между идентичными цепочками, но приводят к отталкиванию различных цепочек (например, СН3-доменов из а и b, взятых от разных изотипов), будут способствовать пути 1 и тем самым увеличивать количество А. Белые: домены легкой цепи. Пунктирные: домены тяжелой цепи.
Фиг.2: Биохимическая характеризация MoAb-димера c-Met (5D5 MoAb-димер («СН3-wt»)) (СН3-wt относится к неизмененному СН3-домену дикого типа). (А) Очищенное на белке А антитело разделяли на колонке Superdex 200 26/60. (В) Пиковые фракции (1, 2, 3) объединяли и подвергали SDS-PAGE в невосстанавливающих и восстанавливающих условиях. Полиакриламидные гели окрашивали красителем Кумасси синим. Индивидуальные пики соответствуют MoAb (3), MoAb-димеру (2) и более высокомолекулярному агрегату (1).
Фиг.3: Биохимическая характеризация MoAb-димера IGF-1R (IGF-1R AK18 MoAb-димер («СН3-wt»)) (СН3-wt относится к неизмененному СН3-домену дикого типа). (А) Очищенное на белке А антитело разделяли на колонке Superdex 200 26/60. (В) Пиковые фракции (1, 2) объединяли и подвергали SDS-PAGE в невосстанавливающих и восстанавливающих условиях. Полиакриламидные гели окрашивали красителем Кумасси синим. Индивидуальные пики соответствуют MoAb (2) и MoAb-димеру (1). (С) Молекулярную массу фракций пиков 1 и 2 исследовали путем SEC-MALLS.
Фиг.4: Биохимическая характеризация Her3 205 MoAb-димера («СН3-wt») (СН3-wt относится к неизмененному СН3-домену дикого типа). (А) Очищенное на белке А антитело разделяли на колонке Superdex 200 26/60. (В) Пиковые фракции (1, 2) объединяли и подвергали SDS-PAGE в невосстанавливающих и восстанавливающих условиях. Полиакриламидные гели окрашивали красителем Кумасси синим. Индивидуальные пики соответствуют MoAb (3), MoAb-димеру (2) и более высокомолекулярному агрегату (1).
Фиг.5: Схематическое изображение анализа поверхностного плазменного резонанса, применяемого для анализа аффинности связывания IGF-1R. Противочеловеческое IgG-антитело (JIR 109-005-098) иммобилизовали на поверхности биосенсорного чипа СМ5 и затем захватывали антитела MoAb или MoAb-димеры. Дальнейшее введение эктодомена рекомбинантного IGF-1R подтверждало функциональность антигенсвязывающих сайтов в молекулах MoAb и MoAb-димера.
Фиг.6. Клеточное связывание MoAb-димера (IGF-1R AK18 MoAb-димера («СН3-wt») (В) и родительского антитела Mab IGF-1R (А) с клетками А549 в анализе путем проточной цитометрии. Клетки А549 инкубировали с серией разведении указанных антител. Связанные антитела визуализировали с помощью Fc-связывающего вторичного антитела, связанного с флуорофором.
Фиг.7: ADCC-анализ негликоинженерного (не-GE) и гликоинженерного (GE) IGF1R-MKA и негликоинженерного IGF-1R-MoAb-димера (IGF1R AK18 MoAb-димер («СН3-wt»)). Полученные от доноров мононуклеарные клетки периферической крови (РВМС) инкубировали с клетками рака предстательной железы (DU145) в присутствии не-GE-IGF1R МКА (1), GE-IGF1R-MKA (2) и не-GE-IGF1R AK18 MoAb-димера («СН3-wt») (3).
Фиг.8: Интернализацию IGF-1R оценивали в клетках НТ29 после инкубации с антителом IGF-1R IgG1 (МКА IGF-1R) и IGF-1R MoAb-димером (IGF1R AK18 MoAb-димер («СН3-wt»)). На графике изображены общие уровни IGF-1R, на которых определяли воздействие антитела в анализе на основе ELISA.
Фиг.9: Аутофосфорилирование IGF-1R оценивали после инкубации клеток 3T3-IGF-1R с антителом IGF-1R IgG1 и IGF-1R MoAb-димером (IGF1R AK18 MoAb-димер («СН3-wt»)) в присутствии 10 нМ IGF-1. На графике изображены уровни фосфо-IGF-1R, на которых воздействие антитела определяли в анализе с установками на основе ELISA.
Фиг.10: Анализ соотношения полученного MoAb-димера (= антигенсвязывающего белка в соответствии с изобретением) и MoAb-мономера (= одновалентного побочного продукта), определенного с помощью ВЭЖХ. Различные антитела с СН3-доменами дикого типа (СН3-wt) и модифицированными СН3-доменами временно экспрессировали и определяли соотношение димера и мономера.
Фиг.11: ESI-MS-спектр IGF-1R-MoAb-димера (SEC-фракция 1) в невосстанавливающих условиях и после дегликозилирования.
Фиг.12: ESI-MS-спектр IGF-1R-MoAb-димера (SEC-фракция 1) после дегликозилирования и восстановления.
Подробное описание изобретения
Изобретение включает антигенсвязывающий белок, содержащий:
а) две модифицированные тяжелые цепи антитела, которое специфически связывается с антигеном, где VH каждой тяжелой цепи заменен на VL указанного антитела, причем указанные модифицированные тяжелые цепи связаны друг с другом через свои СН-домены Fc-части;
б) две модифицированные тяжелые цепи указанного антитела, где СН1 каждой тяжелой цепи заменен на CL указанного антитела, причем указанные модифицированные тяжелые цепи связаны друг с другом через свои СН-домены Fc-части;
и где VL-домены тяжелых цепей а) связаны с VH-доменами тяжелых цепей б), и СН1-домены тяжелых цепей а) связаны с CL-доменами тяжелых цепей б).
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что
СН3-домены Fc-части модифицированных тяжелых цепей а) и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют один и тот же изотип.
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что
СН2- и СН3-домены Fc-части модифицированных тяжелых цепей а) и СН2-и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют один и тот же изотип.
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что
СН2- и СН3-домены Fc-части модифицированных тяжелых цепей а) и СН2-и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют изотип IgG.
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что
СН2- и СН3-домены Fc-части модифицированных тяжелых цепей а) и СН2-и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют изотип IgG1.
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что содержит
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №1; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №2;
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №3; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №4; или
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №5; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №6.
Для улучшения выхода антигенсвязывающего белка в соответствии с изобретением (т.е. для повышения соотношения MoAb-димера и MoAb-мономера (см. пример 9)), СН3-домены IgG1 могут быть также модифицированы путем мутаций, вследствие чего СН3-домены IgG1 а) и природные (wt) СН3-домены IgG1 б) отличаются. Модификация/мутация должны осуществляться таким образом, чтобы поддерживать притягивающие взаимодействия между идентичными цепями, но приводить к отталкиванию различных цепей (см. также фиг.1C).
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что
либо две модифицированные тяжелые цепи а),
либо две модифицированные тяжелые цепи б)
также модифицированы путем аминокислотных замен S364G, L368F, D399K и K409D (где аминокислотные позиции нумеруются в соответствии с индексом ЕС Кабата).
Система нумерации Кабата описана в Kabat, et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD(1991).
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №1; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №2;
где либо две модифицированные тяжелые цепи а),
либо две модифицированные тяжелые цепи б)
также модифицированы путем аминокислотных замен S364G, L368F, D399K и K409D (где аминокислотные позиции нумеруются в соответствии с индексом ЕС Кабата);
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №3; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №4;
где либо две модифицированные тяжелые цепи а),
либо две модифицированные тяжелые цепи б)
также модифицированы путем аминокислотных замен S364G, L368F, D399K и K409D (где аминокислотные позиции нумеруются в соответствии с индексом ЕС Кабата);
или
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №5; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №6,
где либо две модифицированные тяжелые цепи а),
либо две модифицированные тяжелые цепи б)
также модифицированы путем аминокислотных замен S364G, L368F, D399K и K409D (где аминокислотные позиции нумеруются в соответствии с индексом ЕС Кабата).
Другая возможность улучшить выход антигенсвязывающего белка в соответствии с изобретением (т.е. повысить соотношение MoAb-димера и MoAb-мономера (см. пример 9)), СН3-домены а) и б) берут от разных изотипов. Таким образом, притягивающие взаимодействия между идентичными цепями поддерживаются, а различные цепи отталкиваются (см. также фиг.1C).
Таким образом, в одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что
СН3-домены Fc-части модифицированных тяжелых цепей а) и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют различные изотипы.
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что
СН3-домены Fc-части модифицированных тяжелых цепей а) имеют изотип IgG;
и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют изотип IgA.
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что содержит
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №7; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №4.
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что содержит
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №3; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №8.
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что
СН2- и СН3-домены Fc-части модифицированных тяжелых цепей а) имеют изотип IgG1;
и СН2- и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют изотип IgA1.
В одном воплощении антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что СН2-домены Fc-частей а) и б) имеют изотип IgG1, а антигенсвязывающий белок афукозилирован с количеством фукозы 80% или менее от общего количества олигосахаридов (сахаров) на Asn297 и имеет человеческий изотип IgG1.
Термин «антитело», используемый в данном документе, означает антитело полной длины, состоящее из двух тяжелых цепей антитела и двух легких цепей антитела (см. фиг.1). Тяжелая цепь антитела полной длины представляет собой полипептид, содержащий в направлении от N-конца к C-концу вариабельный домен тяжелой цепи антитела (VH), константный домен 1 тяжелой цепи антитела (СН1), шарнирную область антитела (HR), константный домен 2 тяжелой цепи антитела (СН2) и константный домен 3 тяжелой цепи антитела (СН3), сокращенно VH-CH1-HR-CH2-CH3; и, возможно, константный домен 4 тяжелой цепи антитела (СН4) в случае антитела класса IgE. Предпочтительно тяжелая цепь антитела полной длины представляет собой полипептид, содержащий от N-конца по направлению к С-концу VH, СН1, HR, СН2 и СН3. Легкая цепь антитела полной длины представляет собой полипептид, содержащий в направлении от N-конца к С-концу вариабельный домен легкой цепи антитела (VL) и константный домен легкой цепи антитела (CL), сокращенно VL-CL. Константный домен легкой цепи антитела (CL) может быть κ (каппа) или λ (лямбда). Цепи антитела полной длины связаны друг с другом посредством межполипептидных дисульфидных связей между доменом CL и доменом СН1 (т.е. между легкой и тяжелой цепями) и между шарнирными областями тяжелых цепей антитела полной длины. Примерами типичных антител полной длины являются природные антитела, такие как IgG (например, IgG1 и IgG2), IgM, IgA, IgD и IgE. Антитела в соответствии с изобретением могут быть от одного вида, например, человека, либо они могут быть химерными или гуманизированными антителами. Антитела полной длины в соответствии с изобретением содержат два антигенсвязывающих сайта, каждый из которых образован парой VH и VL, которые специфически связываются с одним и тем же (первым) антигеном.
Из этих антител полной длины антигенсвязывающий белок в соответствии с изобретением получают путем:
а) модификации двух тяжелых цепей антитела, которое специфически связывается с антигеном, заменяя VH-домен каждой тяжелой цепи на VL-домен указанного антитела;
б) модификации двух тяжелых цепей указанного антитела, заменяя СН1-домен каждой тяжелой цепи на CL-домен указанного антитела.
«Fc-часть» антитела или антигенсвязывающего белка непосредственно не участвует в связывании антитела с антигеном, но отвечает а) за ассоциацию цепей (модифицированного) антитела друг с другом (например, через их СН3-домены) и б) за различные эффекторные функции. «Fc-часть антитела» является термином, хорошо известным специалистам, и определяется на основании расщепления антител папаином. В зависимости от аминокислотной последовательности константной области их тяжелых цепей антитела или иммуноглобулины делятся на классы: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть разделены на подклассы (изотипы), например, IgG1, IgG2, IgG3 и IgG4, IgA1 и IgA2. В соответствии с константными областями тяжелой цепи различные классы иммуноглобулинов называются α, δ, ε, γ и μ, соответственно.
Существуют пять видов тяжелых цепей антител млекопитающих, которые обозначаются греческими буквами: α, δ, ε, γ и μ (Janeway, С.A., Jr. et al., Immunobiology, 5th ed., Garland Publishing (2001)). Тип присутствующей тяжелой цепи определяет класс антитела; эти цепи находятся в антителах IgA, IgD, IgE, IgG и IgM, соответственно (Rhoades, R.A., and Pflanzer, R.G., Human Physiology, 4th ed., Thomson Learning (2002)). Отдельные тяжелые цепи отличаются по размеру и составу; α и γ содержат приблизительно 450 аминокислот, а μ и ε имеют примерно 550 аминокислот.
Каждая тяжелая цепь имеет две области: константную область и вариабельную область. Константная область одинакова во всех антителах одного и того же изотипа, но отличается в антителах различных изотипов. Тяжелые цепи γ, α и δ имеют константную область, состоящую из трех константных доменов СН1, СН2 и СН3 (в линии) и шарнирной области для дополнительной гибкости (Woof, J., and Burton, D., Nat. Rev. Immunol. 4 (2004) 89-99); тяжелые цепи μ и ε имеют константную область, состоящую из четырех константных доменов СН1, СН2, СН3 и СН4 (Janeway, C.A., Jr. et al., Immunobiology., 5th ed., Garland Publishing (2001)). Вариабельная область тяжелой цепи отличается в антителах, продуцируемых различными В-клетками, но является одинаковой для всех антител, продуцируемых одной В-клеткой или B-клеточным клоном. Вариабельная область каждой тяжелой цепи имеет примерно 110 аминокислот в длину и составляет один домен антитела.
«СН-домены Fc-части» представляют собой константный домен 2 тяжелой цепи антитела (СН2) и константный домен 3 тяжелой цепи антитела (СН3) и, возможно, константный домен 4 тяжелой цепи антитела (СН4) в случае антитела класса IgE.
Термин «указанные модифицированные тяжелые цепи, связанные друг с другом через их СН-домены Fc-частей» относится к межцепочечному доменному спариванию константных доменов тяжелой цепи антитела (СН) из двух модифицированных тяжелых цепей друг с другом, например двух СН3-доменов обеих цепей друг с другом, например, через межцепочечное ионное взаимодействие, взаимодействие Ван-Дер-Ваальса или водородные связи (см. фиг.1А). В одном воплощении указанные модифицированные тяжелые цепи связаны друг с другом посредством, по меньшей мере, их СН3-доменов Fc-части (и, возможно, посредством их СН2-доменов или, возможно, посредством их СН2-доменов и СН4-доменов (если они присутствуют)).
Термин «где VL-домены тяжелых цепей а) связаны с VH-доменами тяжелых цепей б), и СН1-домены тяжелых цепей а) связаны с CL-доменами тяжелых цепей б)» относится к доменному спариванию указанных доменов антител (всегда одного из а) и одного из б)), как они находятся, например, в природных антителах (VL/VH и CH1/CL), например, через межцепочечное ионное взаимодействие, взаимодействие Ван-Дер-Ваальса, водородные связи или дисульфидное взаимодействие (см. фиг.1А).
«Антигенсвязывающий белок» в соответствии с изобретением содержит два антигенсвязывающих сайта и является двухвалентным. Термины «сайт связывания» или «антигенсвязывающий сайт», используемые в данном документе, обозначают область(и) антигенсвязывающего белка в соответствии с изобретением, с которой обычно связывается лиганд (например, антиген или фрагмент антигена), и которая получена из молекулы антитела или ее фрагмента (например, Fab-фрагмента). Антигенсвязывающий сайт в соответствии с изобретением состоит из вариабельных доменов тяжелой цепи антитела (VH) и вариабельных доменов легкой цепи антитела (VL).
Антигенсвязывающие сайты (т.е. пары VH/VL), которые специфически связываются с нужным антигеном, могут быть получены а) из известных антител к антигену или б) из новых антител или фрагментов антител, полученных с помощью способов иммунизации de novo с использованием, помимо прочего, антигенного белка, или нуклеиновой кислоты, или их фрагментов, или с помощью фагового дисплея.
Антигенсвязывающий сайт антигенсвязывающего белка изобретения содержит шесть определяющих комплементарность областей (CDR), которые в разной степени способствуют аффинности сайта связывания к антигену. Существуют три CDR вариабельного домена тяжелой цепи (CDRH1, CDRH2 и CDRH3) и три CDR вариабельного домена легкой цепи (CDRL1, CDRL2 и CDRL3). Протяженность CDR и каркасных областей (FR) определяют путем сравнения с базой данных аминокислотных последовательностей, в которых эти области были определены в соответствии с вариабельностью среди последовательностей.
Специфичность антитела относится к селективному распознаванию антителом определенного эпитопа антигена. Природные антитела, например, являются моноспецифическими. Биспецифические антитела представляют собой антитела, которые имеют две различные антигенсвязывающие специфичности. Антигенсвязывающие белки в соответствии с изобретением являются по меньшей мере моноспецифическими и специфически связываются с эпитопом соответствующего антигена.
Термин «валентность», используемый в данной заявке, означает наличие определенного числа сайтов связывания в молекуле антитела. Природное антитело, например, имеет два сайта связывания и является двухвалентным. Также антигенсвязывающий белок в соответствии с изобретением является по меньшей мере двухвалентным.
Используемые в данном документе термины «моноклональное антитело» или «композиция с моноклональным антителом» относятся к препарату молекул антитела единого аминокислотного состава.
Термин «химерное антитело» относится к антителу, содержащему вариабельную область, т.е. связывающую область, из одного источника или вида, и по меньшей мере часть константной области, полученной из другого источника или вида, как правило полученному с помощью методик рекомбинантной ДНК. Предпочтительными являются химерные антитела, содержащие мышиную вариабельную область и человеческую константную область. Другими предпочтительными формами «химерных антител», охватываемыми данным изобретением, являются те, в которых константную область модифицировали или изменяли по сравнению с исходным антителом, создавая свойства в соответствии с изобретением, особенно в связи с C1q-связыванием и/или Fc-рецепторным (FcR) связыванием. Такие химерные антитела также называют «антителами с переключением класса». Химерные антитела являются продуктом экспрессированных иммуноглобулиновых генов, содержащих сегменты ДНК, кодирующие вариабельные области иммуноглобулинов, и сегменты ДНК, кодирующие константные области иммуноглобулинов. Способы получения химерных антител включают обычные методики рекомбинантной ДНК и генной трансфекции, в настоящее время хорошо известные в данной области. См., например, Morrison, S.L., et al., Proc. Natl. Acad Sci. USA 81 (1984) 6851-6855; патенты US 5202238 и 5204244.
Термин «гуманизированное антитело» относится к антителам, в которых каркасные области или «области, определяющие комплементарность» (CDR), были изменены так, чтобы они содержали CDR иммуноглобулина другой специфичности, отличной от родительского иммуноглобулина. В предпочтительном воплощении мышиный CDR прививают каркасной области человеческого антитела для получения «гуманизированного антитела». См., например, Riechmann, L., et al., Nature 332 (1988) 323-327; и Neuberger, M.S., et al., Nature 314 (1985) 268-270. Особенно предпочтительные CDR соответствуют тем, которые представляют последовательности, распознающие антигены, отмеченные выше для химерных антител. Другими формами «гуманизированных антител», предусмотренными данным изобретением, являются такие антитела, которые были дополнительно модифицированы или изменены в константной области по сравнению с исходным антителом для создания свойств в соответствии с изобретением, особенно в отношении C1q-связывания и/или Fc-рецепторного (FcR) связывания.
Термин «человеческое антитело», используемый в данном документе, включает антитела с вариабельными и константными областями, полученными из человеческих зародышевых последовательностей иммуноглобулина. Человеческие антитела хорошо известны в данной области (van Dijk, M.A., and van de Winkel, J.G., Curr. Opin. Pharmacol. 5 (2001) 368-374). Человеческие антитела могут быть также получены на трансгенных животных (например, мышах), которые способны после иммунизации продуцировать полный репертуар или выборку человеческих антител в отсутствие эндогенной продукции иммуноглобулина. Перенос совокупности человеческих зародышевых генов иммуноглобулинов в такую мышь, мутантную по зародышевой линии, приведет к продукции человеческих антител в ответ на введение антигена (см., например, Jakobovits, A., et al., Proc. Natl. Acad. Sci. USA 90 (1993) 2551-2555; Jakobovits, A., et al., Nature 362 (1993) 255-258; Brueggemann, M.D., et al., Year Immunol. 7 (1993) 33-40). Человеческие антитела могут быть также получены в библиотеках фагового дисплея (Hoogenboom, H.R., and Winter, G., J. Mol. Biol. 227 (1992) 381-388; Marks, J.D., et al., J. Mol. Biol. 222 (1991) 581-597). Методики Cole, et al. и Boerner, et al. также доступны для изготовления человеческих моноклональных антител (Cole, et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p.77 (1985); и Boerner, P., et al., J. Immunol. 147 (1991) 86-95). Как уже было упомянуто в отношении химерных и гуманизированных антител в соответствии с изобретением, термин «человеческое антитело», используемый в данном документе, включает также такие антитела, которые были модифицированы в константной области для создания свойств в соответствии с изобретением, особенно в отношении C1q-связывания и/или FcR-связывания, например, путем «переключения класса», т.е. путем изменения или мутации Fc-частей (например, из IgG1 в IgG4 и/или мутации IgG1/IgG4).
Термин «рекомбинантное человеческое антитело», используемый в данном документе, охватывает все человеческие антитела, которые получают, экспрессируют, создают или выделяют с помощью рекомбинантных средств, например антитела, выделенные из клетки-хозяина, такой как клетка NS0 или СНО, или животного (например, мыши), которое является трансгенным для человеческих генов иммуноглобулинов или антител, экспрессируемых с помощью рекомбинантного экспрессионного вектора, трансфицированного в клетку-хозяина. Такие рекомбинантные человеческие антитела имеют вариабельные и константные области в перегруппированной форме. Рекомбинантные человеческие антитела в соответствии с изобретением подвергали соматическому гипермутированию in vivo. Таким образом, аминокислотные последовательности областей VH и VL рекомбинантных антител являются последовательностями, которые, будучи полученными из и связанными с человеческими зародышевыми последовательностями VH и VL, не могут естественным образом существовать в репертуаре человеческих зародышевых антител in vivo.
Термин «вариабельный домен» (вариабельный домен легкой цепи (VL), вариабельный домен тяжелой цепи (VH)), используемый в данном документе, означает каждую пару легкой и тяжелой цепей, которые непосредственно участвуют в связывании антитела с антигеном. Вариабельные домены легкой и тяжелой цепей человека имеют одинаковую общую структуру и каждый домен содержит четыре каркасные области (framework region, FR), последовательности которых в значительной степени консервативны, соединенные тремя «гипервариабельными областями» (или областями, определяющими комплементарность, CDR). Каркасные области принимают конформацию β-складчатости, а CDR могут образовывать петли, связывающие β-складчатую структуру. CDR в каждой цепи удерживаются в их трехмерной структуре с помощью каркасных областей и образуют вместе с CDR из другой цепи антигенсвязывающий сайт. CDR3-области тяжелой и легкой цепей антитела играют особенно важную роль в специфичности/аффинности связывания антител в соответствии с изобретением и, следовательно, обеспечивают дальнейший предмет изобретения.
Термины «гипервариабельная область» или «антигенсвязывающая часть антитела», используемые в данном документе, относятся к аминокислотным остаткам антитела, которые отвечают за связывание антигена. Гипервариабельная область содержит аминокислотные остатки из «областей, определяющих комплементарность», или «CDR». «Каркасные участки», или «FR-области», являются такими областями вариабельных доменов, которые отличаются от остатков гипервариабельных областей, определенных выше. Таким образом, легкая и тяжелая цепи антитела содержат от N-конца к С-концу домены FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. CDR в каждой цепи разделены такими аминокислотами каркасного участка. В частности, CDR3 тяжелой цепи является областью, которая вносит наибольший вклад в связывание антигена. CDR и FR-области определяют в соответствии со стандартным определением Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991).
Используемые в данном документе термины «связывающийся» или «специфически связывающийся» относятся к связыванию антигенсвязывающего белка с эпитопом антигена в анализе in vitro, предпочтительно в анализе плазмонного резонанса (BIAcore, GE-Healthcare, Упсала, Швеция) с очищенным антигеном дикого типа. Аффинность связывания определяется значениями ka (константа скорости ассоциации антитела из комплекса антитело/антиген), kD (константа диссоциации) и KD (kD/ka). Связывание или специфическое связывание означает аффинность связывания (KD) в 10-8 моль/л или менее (например от 10-8 М до 10-13 моль/л), предпочтительно от 10-9 М до 10-13 моль/л. Таким образом, антигенсвязывающий белок в соответствии с изобретением специфически связывается с каждым антигеном, для которого он является специфическим, с аффинностью связывания (KD) 10-8 моль/л или менее (например от 10-8 М до 10-13 моль/л), предпочтительно от 10-9 М до 10-13 моль/л.
Связывание антигенсвязывающего белка с FcγRIII может быть исследовано в анализе BIAcore (GE Healthcare, Упсала, Швеция). Аффинность связывания определяется значениями ka (константа скорости ассоциации антитела из комплекса антитело/антиген), kD (константа диссоциации) и KD (kD/ka).
Термин «эпитоп» включает любую полипептидную детерминанту, способную специфически связываться с антигенсвязывающими белками. В некоторых воплощениях эпитопная детерминанта включает химически активные поверхностные группировки молекул, такие как аминокислоты, сахарные боковые цепочки, фосфорил или сульфонил, а в некоторых воплощениях может иметь специфические трехмерные структурные характеристики и/или специфические характеристики заряда. Эпитоп является областью антигена, которая связывается антигенсвязывающим белком.
В некоторых воплощениях говорят, что антитело специфически связывается с антигеном, когда оно преимущественно распознает свой антиген-мишень в сложной смеси белков и/или макромолекул.
Термин «константная область», используемый в данной заявке, обозначает сумму доменов антитела помимо вариабельной области. Константная область непосредственно не участвует в связывании с антигеном, но проявляет различные эффекторные функции. В зависимости от аминокислотной последовательности константной области их тяжелых цепей антитела делятся на классы (также называемые изотипами): IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть разделены на подклассы (также называемые изотипами), такие как IgG1, IgG2, IgG3 и IgG4, IgA1 и IgA2. Константные области тяжелой цепи, которые соответствуют различным классам антител, называют α, δ, ε, γ и μ, соответственно. Константные области легкой цепи (CL), которые можно найти во всех пяти классах антител, называют κ (каппа) и λ (лямбда).
Термин «константная область человеческого происхождения», используемый в данной заявке, обозначает константную область тяжелой цепи человеческого антитела изотипов IgG1, IgG2, IgG3 или IgG4 и/или константную область легкой цепи каппа или лямбда. Такие константные области хорошо известны в данной области и описаны, например, в Kabat, E.A., (см., например, Johnson, G. and Wu, T.T., Nucleic Acids Res. 28 (2000) 214-218; Kabat, E.A., et al., Proc. Natl. Acad. Sci. USA 72 (1975) 2785-2788).
Термин «комплементзависимая цитотоксичность (CDC, complement-dependent cytotoxicity)» означает процесс, начинающийся при связывании фактора комплемента C1q с Fc-частью большинства подклассов IgG-антител. Связывание C1q с антителом обусловлено определенными белок-белковыми взаимодействиями на так называемом сайте связывания. Такие сайты связывания Fc-части известны в данной области (см. выше). Такими сайтами связывания Fc-части являются сайты, например, характеризующиеся аминокислотами L234, L235, D270, N297, Е318, K320, K322, Р331 и Р329 (нумерация в соответствии с EU индексом Кабата). Антитела подклассов IgG1, IgG2 и IgG3 обычно демонстрируют активацию комплемента, включающую связывание C1q и С3, в то время как IgG4 не активирует систему комплемента и не связывает C1q и/или С3.
В то время как антитела подкласса IgG4 демонстрируют уменьшение связывания Fc-рецептора (FcγRIIIa), антитела других подклассов IgG демонстрируют сильное связывание. Тем не менее, Pro238, Asp265, Asp270, Asn297 (с потерей Fc-углевода), Pro329, Leu234, Leu235, Gly236, Gly237, Ile253, Ser254, Lys288, Thr307, Gln311, Asn434 и His435 являются остатками, которые, если они изменены, также снижают связывание Fc-рецептора (Shields, R.L., et al., J. Biol. Chem. 276 (2001) 6591-6604; Lund, J., et al., FASEB J. 9(1995) 115-119; Morgan, A., et al., Immunology 86 (1995) 319-324; EP 0307434).
В одном воплощении антитело в соответствии с изобретением обладает пониженным связыванием FcR по сравнению с антителом IgG1 и родительским антителом полной длины в связи с FcR-связыванием подкласса IgG4 или подкласса IgG1 или IgG2 с мутацией в S228, L234, L235 и/или D265 и/или содержит мутацию PVA236. В одном воплощении мутациями в родительском антителе полной длины являются S228P, L234A, L235A, L235E и/или PVA236. В другом воплощении мутациями в родительском антителе полной длины являются S228P и L235E в IgG4 и L234A и L235A в IgG1.
Константная область антитела непосредственно участвует в ADCC (антителозависимой клеточной цитотоксичности) и CDC (комплементзависимой цитотоксичности). Активация комплемента (CDC) начинается при связывании фактора комплемента C1q с константной областью большинства подклассов IgG-антител. Связывание C1q с антителом вызвано определенными белок-белковыми взаимодействиями на так называемом сайте связывания. Такие сайты связывания константных областей известны в данной области и описаны, например, Lukas, T.J., et al., J. Immunol. 127 (1981) 2555-2560; Bunkhouse, R. and Cobra, J.J., Mol. Immunol. 16 (1979) 907-917; Burton, D.R., et al., Nature 288 (1980) 338-344; Thomason, J.E., et al., Mol. Immunol. 37 (2000) 995-1004; Idiocies, E.E., et al., J. Immunol. 164 (2000) 4178-4184; Hearer, M., et al., J. Virol. 75 (2001) 12161-12168; Morgan, A., et al., Immunology 86 (1995) 319-324; и ЕР 0307434. Такие сайты связывания константных областей характеризуются, например, аминокислотами L234, L235, D270, N297, Е318, K320, K322, Р331 и Р329 (нумерация в соответствии с EU индексом Кабата, которая описана в Kabat, Е.А., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD(1991)).
Термин «антителозависимая клеточная цитотоксичность (ADCC, antibody-dependent cellular cytotoxicity)» относится к лизису человеческих клеток-мишеней антителом в соответствии с изобретением в присутствии эффекторных клеток. ADCC предпочтительно измеряют путем обработки препарата клеток, экспрессирующих антиген, антителом в соответствии с изобретением в присутствии эффекторных клеток, таких как свежевыделенные РВМС или очищенные эффекторные клетки из лейкоцитной пленки, такие как моноциты, или натуральные киллеры (NK, natural killers), или постоянно растущая клеточная линия NK.
Неожиданно было обнаружено, что антигенсвязывающий белок в соответствии с изобретением обладает повышенными ADCC-свойствами по сравнению с родительским антителом полной длины, особенно в области высоких концентраций антитела. Эти повышенные ADCC-эффекты достигаются без дальнейшей модификации Fc-части, такой как гликоинженерия.
Таким образом, в одном воплощении антигенсвязывающие белки в соответствии с изобретением обладают повышенной ADCC (измеренной, как описано в примере 4) в сравнении с его родительским антителом полной длины.
У млекопитающих есть только два типа легких цепей, которые называются лямбда (λ) и каппа (κ). Легкая цепь имеет два последовательных домена: один константный домен CL и один вариабельный домен VL. Приблизительная длина легкой цепи составляет от 211 до 217 аминокислот. Предпочтительно легкая цепь является легкой каппа (κ)-цепью, а константный CL-домен предпочтительно получен из легкой каппа (κ)-цепи (константный домен Сκ).
Клеточно-опосредованные эффекторные функции моноклональных антител могут быть усилены путем инженерии их олигосахаридного компонента, как описано в Umana, P., et al., Nature Biotechnol. 17 (1999) 176-180 и патенте US 6602684. Антитела типа IgG1, наиболее часто используемые терапевтические антитела, являются гликопротеинами, которые имеют консервативный N-связанный сайт гликозилирования на Asn297 в каждом СН2-домене. Два сложных двухантенных олигосахарида, присоединенные к Asn297, скрыты между СН2-доменами, образуя обширные контакты с полипептидным остовом, и их присутствие имеет важное значение для антитела, чтобы опосредовать эффекторные функции, такие как антителозависимую клеточную цитотоксичность (ADCC) (Lifely, M.R., et al., Glycobiology 5 (1995) 813-822; Jefferis, R., et al., Immunol. Rev. 163 (1998) 59-76; Wright, A., and Morrison, S.L., Trends Biotechnol. 15 (1997) 26-32). В Umana, P., et al. Nature Biotechnol. 17 (1999) 176-180 и WO 99/54342 показано, что сверхэкспрессия в клетках яичника китайского хомячка (СНО) β(1,4)-N-ацетилглюкозаминилтрансферазы III («GnTIII»), гликозилтрансферазы, катализирующей формирование олигосахаридов с симметричным разветвлением, значительно увеличивает in vitro ADCC-активность антител. Изменения в составе Asn297-углевода или его ликвидация также влияет на связывание с FcγR и C1q (Umana, P., et al., Nature Biotechnol. 17 (1999) 176-180; Davies, J., et al., Biotechnol. Bioeng. 74 (2001) 288-294; Mimura, Y., et al., J. Biol. Chem. 276 (2001) 45539-45547; Radaev, S., et al., J. Biol. Chem. 276 (2001) 16478-16483; Shields, R.L., et al., J. Biol. Chem. 276 (2001) 6591-6604; Shields, R.L., et al., J. Biol. Chem. 277 (2002) 26733-26740; Simmons, L.C., et al., J. Immunol. Methods 263 (2002) 133-147).
В одном аспекте изобретения антигенсвязывающий белок в соответствии с изобретением характеризуется тем, что СН2-домены Fc-частей а) и б) имеют изотип IgG1, и антигенсвязывающий белок афукозилирован с количеством фукозы 80% или ниже от общего количества олигосахаридов (сахаров) на Asn297.
В одном воплощении антигенсвязывающий белок афукозилирован с количеством фукозы от 65% до 5% от общего количества олигосахаридов (сахаров)на Asn297.
Термин «афукозилированный антигенсвязывающий белок» относится к антигенсвязывающим белкам изотипа IgG1 или IgG3 (предпочтительно изотипа IgG1) с измененным характером гликозилирования в Fc-области на Asn297 со сниженным уровнем остатков фукозы. Гликозилирование человеческого IgG1 или IgG3 происходит на Asn297 как гликозилирование сердцевинного фукозилированного двухантенного сложного олигосахарида, заканчивающегося двумя остатками Gal. Эти структуры обозначаются как G0-, G1- (α-1,6- или α-1,3-) или G2-гликанановые остатки в зависимости от количества концевых Gal-остатков (Raju, T.S., Bioprocess Int. 1 (2003) 44-53). Тип гликозилирования Fc-частей антитела в СНО описан, например, Routier, F.H., Glycoconjugate J. 14 (1997) 201-207. Антитела, которые рекомбинантно экспрессируются в негликомодифицированных клетках-хозяевах СНО, обычно фукозилированы на Asn297 в количестве по меньшей мере 85%. Следует понимать, что термин «антигенсвязывающий белок», используемый в данном документе, включает антигенсвязывающий белок, не имеющий фукозы в паттерне гликозилирования. Общеизвестно, что типичной позицией гликозилированных остатков в антителе является аспарагин в позиции 297 в соответствии с системой нумерации ЕС («Asn297»).
Таким образом, афукозилированный антигенсвязывающий белок в соответствии с изобретением означает антигенсвязывающий белок изотипа IgG1 или IgG3 (предпочтительно изотипа IgG1), в котором количество фукозы составляет 80% или менее (например, от 80% до 1%) от общего количества олигосахаридов (сахаров) на Asn297 (что означает, что по меньшей мере 20% или более олигосахаридов в Fc-области являются Asn297-афукозилированными). В одном воплощении количество фукозы составляет 65% или менее (например, от 65% до 1%), в одном воплощении от 65% до 5%, в одном воплощении от 40% до 20% олигосахаридов в Fc-области на Asn297. В соответствии с изобретением «количество фукозы» означает количество указанного олигосахарида (фукозы) в олигосахаридной (сахарной) цепочке на Asn297, связанное с суммой всех олигосахаридов (сахаров), присоединенных к Asn297 (например, сложных, гибридных и высокоманнозных структур), которое измеряется масс-спектрометрией MALDI-TOF и рассчитывается как среднее значение (подробное описание процедуры определения количества фукозы см., например, в WO 2008/077546). Кроме того, в одном воплощении олигосахариды Fc-области являются симметрично разветвленными.
Афукозилированный антигенсвязывающий белок в соответствии с изобретением может быть экспрессирован в гликомодифицированной клетке-хозяине, созданной для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид с активностью GnTIII, в количестве, достаточном для частично фукозилированных олигосахаридов в Fc-области. В одном воплощении полипептид с активностью GnTIII является гибридным полипептидом. Альтернативно α1,6-фукозилтрансферазная активность клетки-хозяина может быть уменьшена или устранена в соответствии с патентом US 6946292 для создания гликомодифицированных клеток-хозяев. Степень фукозилирования антигенсвязывающего белка может быть предопределена, например, в ферментативных условиях (например, по времени ферментирования) или при сочетании по меньшей мере двух антигенсвязывающих белков с различной степенью фукозилирования. Такие способы образования афукозилированных антигенсвязывающих белков описаны в WO 2005/044859, WO 2004/065540, W02007/031875, Umana, P., et al., Nature Biotechnol. 17 (1999) 176-180, WO 99/154342, WO 2005/018572, WO 2006/116260, WO 2006/114700, WO 2005/011735, WO 2005/027966, WO 97/028267, US 2006/0134709, US 2005/0054048, US 2005/0152894, WO 2003/035835, WO 2000/061739. Эти гликоинженерные антигенсвязывающие белки в соответствии с изобретением обладают повышенной ADCC (по сравнению с родительскими антигенсвязывающими белками). Другие гликоинженерные способы, дающие афукозилированные антигенсвязывающие белки в соответствии с изобретением, описаны, например, в Niwa, R., et al., J. Immunol. Methods 306 (2005) 151-160; Shinkawa, Т., et al., J Biol Chem, 278 (2003) 3466-3473; WO 03/055993 или US 2005/0249722.
Таким образом, один аспект изобретения представляет собой афукозилированный антигенсвязывающий белокв соответствии с изобретением, который имеет изотип IgG1 или IgG3 (предпочтительно изотип IgG1) с количеством фукозы 60% или менее (например, от 60% до 1%) от общего количества олигосахаридов (сахаров)на Asn297.
Таким образом, один аспект изобретения представляет собой афукозилированный антигенсвязывающий белок в соответствии с изобретением, который имеет изотип IgG1 или IgG3 (предпочтительно изотип IgG1) с количеством фукозы 60% или менее (например, от 60% до 1%) от общего количества олигосахаридов (сахаров) на Asn297 для лечения рака.
Другой аспект изобретения представляет собой применение афукозилированного антигенсвязывающего белка в соответствии с изобретением, который имеет изотип IgG1 или IgG3 (предпочтительно изотип IgG1) с количеством фукозы 60% или менее от общего количества олигосахаридов (сахаров) на Asn297 для изготовления лекарственного препарата для лечения рака.
В одном воплощении количество фукозы составляет от 60% до 20% от общего количества олигосахаридов (сахаров) на Asn297. В одном воплощении количество фукозы составляет от 60% до 40% от общего количества олигосахаридов (сахаров) на Asn297. В одном воплощении количество фукозы составляет 0% от общего количества олигосахаридов (сахаров) на Asn297.
Понятия «система нумерации ЕС» или «индекс ЕС» обычно используются, когда речь идет об остатке в константной области тяжелой цепи иммуноглобулина (например, индекс ЕС по Kabat et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991), включенной в данный документ посредством ссылки).
Термин «сахарные цепочки демонстрируют характеристики N-связанных гликанов, прикрепленных к Asn297 антитела, рекомбинантно экспрессированного в СНО-клетке» означает, что сахарная цепочка на Asn297 родительского антитела полной длины в соответствии с изобретением имеет ту же структуру и последовательность сахарных остатков, за исключением остатка фукозы, как и в таком же антителе, экспрессированном в немодифицированных СНО-клетках, например, как сообщалось в WO 2006/103100.
Термин «NGNA», используемый в данной заявке, обозначает остаток сахара N-гликолилнейраминовой кислоты.
Гликозилирование человеческого IgG1 или IgG3 осуществляется на Asn297 как гликозилирование сердцевинного фукозилированного двухантенного сложного олигосахарида, заканчивающегося двумя остатками Gal. Человеческие константные области тяжелой цепи подкласса IgG1 или IgG3 подробно описаны в Kabat, E., A., et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991), и в Brueggemann, M., et al., J. Exp. Med. 166 (1987) 1351-1361; Love, Т., W., et al., Methods Enzymol. 178 (1989) 515-527. Эти структуры обозначаются как G0-, G1- (α-1,6- или α-1,3-) или G2-гликанановые остатки в зависимости от количества концевых Gal-остатков (Raju, T.S., Bioprocess Int. 1 (2003) 44-53). Тип гликозилирования Fc-частей антитела в СНО описан, например, Routier, F.H., Glycoconjugate J. 14 (1997) 201-207. Антитела, которые рекомбинантно экспрессируются в негликомодифицированных клетках-хозяевах СНО, обычно фукозилированы на Asn297 в количестве по меньшей мере 85%. Модифицированные олигосахариды родительского антитела полной длины могут быть гибридными или сложными. Предпочтительно восстановленные/нефукозилированные олигосахариды с симметричным разветвлением являются гибридными. В другом воплощении восстановленные/нефукозилированные олигосахариды с симметричным разветвлением являются сложными.
В соответствии с изобретением «количество фукозы» означает количество указанного сахара в сахарной цепочке на Asn297, связанное с суммой всех гликоструктур, присоединенных к Asn297 (например, сложных, гибридных и высокоманнозных структур), которое измеряется масс-спектрометрией MALDI-TOF и рассчитывается как среднее значение. Относительное количество фукозы представляет собой процент фукозосодержащих структур, связанных со всеми гликоструктурами, выявленными в образце, обработанном N-гликозидазой F (например, сложными, гибридными и олиго- и высокоманнозными структурами, соответственно) при MALDI-TOF.
Антитело в соответствии с данным изобретением получают с помощью рекомбинантных средств. Таким образом, один аспект данного изобретения представляет собой нуклеиновую кислоту, кодирующую антитело в соответствии с изобретением, а другой аспект представляет собой клетку, содержащую указанную нуклеиновую кислоту, кодирующую антитело в соответствии с изобретением. Способы рекомбинантной продукции широко известны в данной области и включают экспрессию белка в прокариотических или эукариотических клетках с последующим выделением антитела и, как правило, очисткой до фармацевтически приемлемой чистоты. Для экспрессии вышеупомянутых антител в клетке-хозяине нуклеиновые кислоты, кодирующие соответственно модифицированные легкие и тяжелые цепи, вставляют в соответствующие векторы экспрессии с помощью стандартных способов. Экспрессию осуществляют в соответствующих прокариотических или эукариотических клетках-хозяевах, таких как клетки СНО, клетки NS0, клетки SP2/0, клетки HEK293, клетки COS, клетки PER.C6, дрожжевые клетки, клетки E.coli, и выделяют антитело из клеток (из супернатанта или из клеток после лизиса). Основные способы рекомбинантной продукции антител широко известны в данной области и описаны, например, в статьях Makrides, S.C., Protein Expr. Purif. 17 (1999) 183-202; Geisse, S., et al., Protein Expr. Purif. 8 (1996) 271-282; Kaufman, R.J., Mol. Biotechnol. 16 (2000) 151-161; Werner, R.G., Drug Res. 48 (1998) 870-880.
Антигенсвязывающие белки в соответствии с изобретением соответственно выделяют из культуральной среды с помощью обычных процедур очистки иммуноглобулинов, таких как, например, с помощью белка А-сефарозы, гидроксиапатитной хроматографии, гель-электрофореза, диализа или аффинной хроматографии. ДНК и РНК, кодирующие моноклональные антитела, можно легко выделить и секвенировать с помощью обычных процедур. Гибридомные клетки могут служить источником таких ДНК и РНК. После выделения ДНК может быть вставлена в экспресионные векторы, которыми затем трансфицируют клетки-хозяева, например, клетки HEK293, клетки СНО или клетки миеломы, которые в противном случае не продуцируют иммуноглобулиновый белок, чтобы получить синтез рекомбинантных моноклональных антител в клетках-хозяевах.
Варианты (или мутанты) аминокислотной последовательности антигенсвязывающего белка получают путем внесения соответствующих нуклеотидных замен в ДНК антитела или путем нуклеотидного синтеза. Тем не менее, такие модификации могут быть выполнены лишь в очень ограниченном диапазоне, например как описано выше. Например, модификации не меняют приведенные выше характеристики антител, такие как изотип IgG и связывание антигена, но могут улучшать выход рекомбинантной продукции, стабильность белка или облегчать очистку.
Термин «клетка-хозяин», используемый в данной заявке, обозначает любой вид клеточной системы, который может быть разработан для создания антител в соответствии с данным изобретением. В одном воплощении в качестве клеток-хозяев используют клетки HEK293 и клетки СНО. Используемые в данном документе выражения «клетка», «клеточная линия» и «клеточная культура» используются взаимозаменяемо, и все такие обозначения включают потомство. Таким образом, слова «трансформанты» и «трансформированные клетки» включают первичную клетку-объект и культуры, полученные из нее, независимо от количества переносов. Также понятно, что все потомство может не быть абсолютно идентичным по содержанию ДНК из-за преднамеренных или случайных мутаций. Включается вариантное потомство, которое имеет такие же функции или биологическую активность, как и у исходной трансформированной клетки.
Экспрессия в клетках NSO описана, например, в Barnes, L.M., et al., Cytotechnology 32 (2000) 109-123; Barnes, L.M., et al., Biotech. Bioeng. 73 (2001) 261-270. Временная экспрессия описана, например, в Durocher, Y., et al., Nucl. Acids. Res. 30 (2002) E9. Клонирование вариабельных доменов описано в Orlandi, R., et al., Proc. Natl. Acad. Sci. USA 86 (1989) 3833-3837; Carter, P., et al., Proc. Natl. Acad. Sci. USA 89 (1992) 4285-4289; и Norderhaug, L, et al., J. Immunol. Methods 204 (1997) 77-87. Предпочтительная система для временной экспрессии (HEK293) описана в Schlaeger, E.-J., and Christensen, K., Cytotechnology 30 (1999) 71-83, и в Schlaeger, E.-J., in J. Immunol. Methods 194 (1996) 191-199.
Контрольные последовательности, которые подходят для прокариот, например, включают промотор, возможно операторную последовательность и сайт связывания рибосомы. Эукариотические клетки, как известно, используют промоторы, энхансеры и сигналы полиаденилирования.
Нуклеиновая кислота является «функционально связанной», когда она находится в функциональной взаимосвязи с другой нуклеиновокислотной последовательностью. Например, ДНК предпоследовательности или секреторного лидера функционально связана с ДНК полипептида, если он экспрессируется как пребелок, который участвует в секреции полипептида; промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию последовательности; или сайт связывания рибосомы функционально связан с кодирующей последовательностью, если он расположен так, чтобы облегчить трансляцию. Как правило, «функционально связан» означает, что последовательности ДНК являются смежными и, в случае секреторного лидера, смежными и одной рамке считывания. Тем не менее, энхансеры не обязательно должны быть смежными. Связывание осуществляется путем лигирования на подходящих сайтах рестрикции. Если таких сайтов не существует, то в соответствии с обычной практикой используются синтетические олигонуклеотидные адаптеры или линкеры.
Очистку антигенсвязывающих белков проводят с целью устранения других клеточных компонентов или других загрязняющих веществ, например, других клеточных нуклеиновых кислот или белков (например, пробочных продуктов на фиг.1В), с помощью стандартных методик, в том числе щелочной/SDS-обработки, CsCl, колоночной хроматографии, электрофореза в агарозном геле и других хорошо известных в данной области методик (см. Ausubel, F., et al., ed. Current Protocols in Molecular Biology, Greene Publishing and Wiley Interscience, New York (1987)). Для очистки белков приняты и широко применяются различные способы, такие как, например, аффинная хроматография с микробными белками (например, аффинная хроматография с белком А или белком G), ионообменная хроматография (например, катионообменная (с карбоксиметиловыми смолами), анионообменная (с аминоэтиловыми смолами) и со смешанным режимом обмена), тиофильная адсорбционная хроматография (например, с бета-меркаптоэтанолом и другими SH-лигандами), хроматография гидрофобных взаимодействий или адсорбционная хроматография с ароматическими группами (например, с фенил-сефарозой, аза-аренофильными смолами или м-аминофенилбороновой кислотой), металл-хелатная аффинная хроматография (например, с носителем со сродством к Ni(II)- и Cu(II)), эксклюзионная хроматография и электрофоретические способы (такие как гель-электрофорез, капиллярный электрофорез) (Vijayalakshmi, M.A., Appl. Biochem. Biotech. 75 (1998) 93-102). Один пример очистки описан в примере 1 и на соответствующих фигурах.
Одним из аспектов изобретения является фармацевтическая композиция, содержащая антигенсвязывающий белок в соответствии с изобретением. Другим аспектом изобретения является применение антигенсвязывающего белка в соответствии с изобретением для изготовления фармацевтической композиции. Еще один аспект изобретения представляет собой способ изготовления фармацевтической композиции, содержащей антигенсвязывающий белок в соответствии с изобретением. В другом аспекте данное изобретение предусматривает композицию, например фармацевтическую композицию, содержащую антигенсвязывающий белок в соответствии с данным изобретением, собранную в состав вместе с фармацевтическим носителем.
Одно воплощение изобретения представляет собой антигенсвязывающий белок в соответствии с изобретением для лечения рака.
Другой аспект изобретения представляет собой указанную фармацевтическую композицию для лечения рака.
Одно воплощение изобретения представляет собой антигенсвязывающий белок в соответствии с изобретением для применения в лечении рака.
Другим аспектом изобретения является применение антигенсвязывающего белка в соответствии с изобретением для изготовления лекарственного препарата для лечения рака.
Другим аспектом изобретения является способ лечения пациента, страдающего от рака, путем введения антигенсвязывающего белка в соответствии с изобретением пациенту, нуждающемуся в таком лечении.
Упоминаемый в данном документе «фармацевтический носитель» включает любые и все растворители, дисперсные среды, покрытия, антибактериальные и противогрибковые агенты, изотонические и задерживающие абсорбцию агенты и т.п., которые являются физиологически совместимыми. Предпочтительно, носитель подходит для внутривенного, внутримышечного, подкожного, парентерального, спинномозгового или эпидермального введения (например, путем инъекции или инфузии).
Фармацевтическая композиция данного изобретения может быть введена различными способами, известными в данной области. Как будет понятно специалисту, путь и/или режим введения будет варьировать в зависимости от желаемого результата. Для введения соединения изобретения определенным путем может потребоваться покрытие соединения или одновременное введение соединения с материалом, предотвращающим его инактивацию. Например, соединение может быть введено субъекту в соответствующем носителе, например, в липосомах или разбавителе. Фармацевтически приемлемые разбавители включают физиологический раствор и водные буферные растворы. Фармацевтические носители включают стерильные водные растворы или дисперсии и стерильные порошки для приготовления стерильных инъекционных растворов или дисперсий непосредственно перед применением. Применение таких сред и агентов для фармацевтически активных веществ известно в данной области.
Фразы «парентеральное введение» и «вводят парентерально», используемые в данном документе, означают способы введения, отличные от энтерального и местного введения, как правило, путем инъекций, и включают, но не ограничиваясь ими, внутривенные, внутримышечные, внутриартериальные, интратекальные, внутрикапсулярные, интраорбитальные, внутрисердечные, внутрикожные, внутрибрюшинные, транстрахеальные, подкожные, субкутикулярные, внутрисуставные, субкапсулярные, субарахноидальные, спинномозговые, эпидуральные и интрастенальные инъекции и инфузии.
Термин «рак», используемый в данном документе, относится к пролиферативным заболеваниям, таким как лимфомы, лимфоцитарные лейкозы, рак легкого, немелкоклеточный рак легкого (NSCL), бронхиолоальвеолярный рак легкого, рак кости, рак поджелудочной железы, рак кожи, рак головы или шеи, кожная или внутриглазная меланома, рак матки, рак яичников, рак прямой кишки, рак анальной области, рак желудка, рак толстой кишки, рак молочной железы, рак матки, карцинома фаллопиевых труб, карцинома эндометрия, карцинома шейки матки, карцинома влагалища, карцинома вульвы, болезнь Ходжкина, рак пищевода, рак тонкой кишки, рак эндокринной системы, рак щитовидной железы, рак паращитовидной железы, рак надпочечника, саркома мягких тканей, рак уретры, рак полового члена, рак предстательной железы, рак мочевого пузыря, рак почек или мочеточников, почечно-клеточная карцинома, карцинома почечной лоханки, мезотелиома, гепатоцеллюлярный рак, рак желчного пузыря, опухоли центральной нервной системы (ЦНС), опухоли спинного мозга, глиома головного мозга, мультиформная глиобластома, астроцитомы, шванномы, эпендимомы, медуллобластомы, менингиомы, плоскоклеточные карциномы, аденома гипофиза и саркома Юинга, включая рефракторные версии любого из вышеперечисленных видов рака, или комбинацию одного или более чем одного из вышеперечисленных видов рака.
Эти композиции также могут содержать вспомогательные вещества, такие как консерванты, смачивающие агенты, эмульгаторы и диспергирующие агенты. Профилактика наличия микроорганизмов может быть обеспечена как за счет стерилизации, как сказано выше, так и за счет включения различных антибактериальных и противогрибковых агентов, например парабена, хлорбутанола, фенола, сорбиновой кислоты и т.п. Также может быть целесообразным включение в композиции изотонических агентов, таких как сахара, хлорид натрия и т.п. Кроме того, удлиненную абсорбцию инъекционной фармацевтической формы можно получить путем включения агентов, которые задерживают абсорбцию, таких как моностеарат алюминия и желатин.
Независимо от выбранного пути введения соединения данного изобретения, которые могут быть использованы в подходящей гидратированной форме, и/или фармацевтические композиции данного изобретения составляют в фармацевтически приемлемые лекарственные формы с помощью традиционных способов, известных специалистам в данной области.
Фактические уровни доз активных ингредиентов в фармацевтических композициях данного изобретения можно изменить таким образом, чтобы получить количество активного ингредиента, эффективное для достижения желаемого терапевтического эффекта для конкретного пациента, композиции и способа введения, но не токсичное для пациента. Выбранный уровень доз будет зависеть от ряда фармакокинетических факторов, включая активность конкретных используемых композиций данного изобретения, путь введения, время введения, скорость выведения конкретного используемого соединения, продолжительность лечения, другие лекарственные препараты, соединения и/или материалы, используемые в сочетании с конкретными используемыми композициями, возраст, пол, вес, состояние, общее состояние здоровья и историю болезни пациента, подлежащего лечению, и т.п. факторов, хорошо известных в медицине.
Композиция должна быть стерильной и жидкой при условии, что она доставляется с помощью шприца. Кроме воды, предпочтительным носителем является изотонический буферный солевой раствор.
Соответствующая текучесть может быть обеспечена, например, путем использования покрытий, таких как лецитин, путем поддержания требуемого размера частиц в случае дисперсии и путем применения поверхностно-активных веществ. Во многих случаях в композицию предпочтительно включать изотонические агенты, например сахара, многоатомные спирты, такие как маннит или сорбит, и хлорид натрия.
Используемые в данном документе выражения «клетка», «клеточная линия» и «клеточная культура» используются взаимозаменяемо, и все такие обозначения включают потомство. Таким образом, слова «трансформанты» и «трансформированные клетки» включают первичную клетку-объект и культуры, полученные из нее, независимо от числа переносов. Также понятно, что все потомство может не быть абсолютно идентичным по содержанию ДНК из-за преднамеренных или случайных мутаций. Включено вариантное потомство, которое имеет такие же функции или биологическую активность, как и у исходной трансформированной клетки. Если имеются в виду другие значения, то это будет ясно из контекста.
Термин «трансформация», используемый в данном документе, относится к процессу переноса векторов/нуклеиновых кислот в клетку-хозяина. Если в качестве клеток-хозяев используются клетки без труднопреодолимой клеточной стенки, то трансфекцию осуществляют, например, путем осаждения фосфатом кальция, как описано в Graham, F.L., and van der Eb, A.J., Virology 52 (1978) 546. Тем не менее, также могут быть использованы другие способы введения ДНК в клетки, например введение в ядро или слияние протопластов. Если используются прокариотические клетки или клетки, которые содержат значительные конструкции клеточной стенки, то одним из способов трансфекции является, например, обработка кальцием с использованием хлорида кальция, как описано в Cohen, F.N, et al, PNAS 69 (1972) 7110.
Используемое в данном документе выражение «экспрессия» относится к процессу, посредством которого нуклеиновая кислота транскрибируется в мРНК, и/или к процессу, в котором транскрибированная мРНК (также известная как транскрипт) впоследствии транслируется в пептиды, полипептиды или белки. Транскрипты и кодируемые полипептиды в совокупности называются продуктом гена. Если полинуклеотид является производным геномной ДНК, то экспрессия в эукариотической клетке может включать сплайсинг мРНК.
«Вектор» представляет собой нуклеиновокислотную молекулу, в частности самореплицирующуюся, которая переносит вставленную нуклеиновокислотную молекулу в и/или между клетками-хозяевами. Этот термин включает векторы, которые функционируют в основном для введения ДНК или РНК в клетку (например, для хромосомной интеграции), репликации векторов, которые функционируют в основном для репликации ДНК или РНК, и экспрессионные векторы, которые отвечают за транскрипцию и/или трансляцию ДНК или РНК. Кроме того, включены векторы, которые выполняют более чем одну из описанных функций.
«Экспрессионный вектор» представляет собой полинуклеотид, который, будучи введенным в соответствующую клетку-хозяина, может быть транскрибирован и транслирован в полипептид. Понятие «экспрессионной системы» обычно относится к подходящей клетке-хозяину, содержащей экспрессионный вектор, который может функционировать для получения нужного продукта экспрессии.
Следующие примеры, список последовательностей и графические материалы приведены для облегчения понимания данного изобретения, полный объем которого изложен в прилагаемой формуле изобретения. Понятно, что в изложенных процедурах могут быть сделаны модификации без отхода от сущности данного изобретения.
Процедура эксперимента
А. Материалы и методы
Методики рекомбинантной ДНК
Были использованы стандартные способы манипуляций с ДНК, описанные в Sambrook, J. et al., Molecular cloning: A laboratory manual; Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York (1989). Молекулярно-биологические реагенты использовали в соответствии с инструкциями изготовителей.
Анализ последовательности ДНК и белка и управление данными о последовательностях
Общая информация о нуклеотидных последовательностях легких и тяжелых цепей человеческих иммуноглобулинов приведена в: Kabat, E.A., et al., (1991) Sequences of Proteins of Immunological Interest, Fifth Ed., NIH Publication No 91-3242. Аминокислоты цепей антител пронумерованы в соответствии с нумерацией ЕС (Edelman, G.M., et al., PNAS 63 (1969) 78-85; Kabat, E.A., et al., (1991) Sequences of Proteins of Immunological Interest, Fifth Ed., NIH Publication No 91-3242). Пакет программного обеспечения GCG (Genetics Computer Group, Мэдисон, Висконсин) версии 10.2 и программу от Infomax Vector NTI Advance Suite версии 8.0 использовали для создания, картирования, анализа, аннотации и иллюстрации последовательности.
Секвенирование ДНК
Последовательности ДНК определяли путем секвенирования двойной цепи, осуществляемого на SequiServe (Vaterstetten, Германия) и Geneart AG (Регенсбург, Германия).
Синтез генов
Требуемые сегменты генов были изготовлены с помощью Geneart AG (Регенсбург, Германия) из синтетических олигонуклеотидов и ПЦР-продуктов путем автоматизированного синтеза генов. Сегменты генов, фланкированные сайтами рестрикции уникальных эндонуклеаз клонировали в плазмиды pGA18 (ampR). Плазмидные ДНК очищали из трансформированных бактерий и определяли концентрацию путем УФ-спектроскопии. Последовательность ДНК субклонированных фрагментов генов подтверждали путем секвенирования ДНК. Последовательности ДНК, кодирующие две цепи антитела (VH-CL-CH2-CH3 и VL-СН1-СН2-СН3) получали в виде целых фрагментов путем синтеза генов с фланкирующими сайтами рестрикции 5'Hpal и 3'Nael. Сегменты генов, кодирующие точечные мутации в СН3-домене, которые предпочтительно давали продукт MoAb-димер, а также замену сегментов генов, кодирующих часть IgG1 с IgA-СН3-доменом (например, SEQ ID №7), получали путем синтеза генов. Эти сегменты фланкировали уникальными сайтами рестрикции, которые позволяли заменять соответствующие последовательности IgG1 дикого типа. Все конструкции были разработаны с 5'-концевой последовательностью ДНК, кодирующей лидерный пептид, который ориентирует белки на секрецию в эукариотических клетках.
Конструирование экспрессионных плазмид
Экспрессионный вектор Roche использовали для конструирования всех цепей антител. Вектор состоит из следующих элементов:
- сайта начала репликации, oriP, вируса Эпштейна-Барр (EBV),
- сайта начала репликации из вектора pUC18, который обеспечивает репликацию этой плазмиды в E.coli
- гена бета-лактамазы, который придает устойчивость к ампициллину в E.coli,
- немедленного раннего энхансера и промотора цитомегаловируса человека (HCMV),
- человеческой 1-иммуноглобулиновой сигнальной последовательности полиаденилирования («полиА»), и
- уникальных сайтов рестрикции Hpal, Bell и Nael.
Гены иммуноглобулинов в порядке VH-CL-CH2-CH3 и VL-CH1-CH2-CH3, а также конструкции, модифицированные в 3'-области, кодирующие С-конец цепи антитела (СН3), получали путем генного синтеза и клонировали в плазмиды pGA18 (ampR), как было описано. Плазмиды pG18 (ampR), несущие синтезированные ДНК-сегменты, и экспрессионный вектор Roche расщепляли с помощью рестрикционных ферментов Hpal и Nael или Bell и Nael (Roche Molecular Biochemicals) и подвергали электрофорезу в агарозном геле. Затем сегменты ДНК лигировали с выделенным фрагментом Hpal/Nael или Bcll/Nael экспрессионного вектора Roche, получая окончательные экспрессионные векторы. Окончательными экспрессионными векторами трансформировали клетки E.coli, выделяли ДНК экспрессионных плазмид (Miniprep) и подвергали рестрикционному анализу и секвенированию ДНК. Правильные клоны выращивали в 150 мл среды LB-Amp, снова выделяли плазмидную ДНК (Maxiprep) и подтверждали целостность последовательности путем секвенирования ДНК.
Временная экспрессия иммуноглобулиновых вариантов в клетках HEK293
Рекомбинантные иммуноглобулиновые варианты экспрессировали путем временной трансфекции человеческих эмбриональных почечных клеток 293-F с помощью системы для экспрессии FreeStyle™ 293 Expression System в соответствии с инструкцией производителя (Invitrogen, США). Вкратце, суспензию клеток FreeStyle™ 293-F культивировали в среде для экспрессии FreeStyle™ 293 при 37°С/8% CO2. В день трансфекции клетки высевали в свежую среду с плотностью 1-2×106 жизнеспособных клеток/мл. Комплексы ДНК-293fectin™ получали в среде Opti-MEM® I (Invitrogen, США) с использованием 325 мкл 293fectin™ (Invitrogen, Германия) и 250 мкг каждой плазмидной ДНК в молярном соотношении 1:1 в конечном объеме для трансфекции 250 мл. Клеточные культуральные супернатанты, содержащие антитела, собирали через 7 дней после трансфекции путем центрифугирования при 14000 g в течение 30 минут и фильтровали через стерильный фильтр (с размером пор 0,22 мкм). Супернатанты хранили при температуре -20°С до очистки.
Альтернативно, антитела получали путем временной трансфекции в клетках HEK293-EBNA. Антитела экспрессировали путем временной котрансфекции соответствующих экспрессионных плазмид в прикрепленных клетках HEK293-EBNA (человеческая эмбриональная почечная клеточная линия 293, экспрессирующая ядерный антиген вируса Эпштейна-Барр; депозитный номер в Американской коллекции типовых культур АТСС № CRL-10852, 959218), культивируемых в DMEM (среде Игла, модифицированной Дульбекко, Gibco) с добавлением 10% Ultra Low IgG FCS (фетальной телячьей сыворотки, Gibco), 2 мМ L-глутамина (Gibco) и 250 мкг/мл генетицина (Gibco). Для трансфекции использовали FuGENE™ 6 Transfection Reagent (Roche Molecular Biochemicals) в соотношении реагента FuGENE™ (мкл) к ДНК (мкг) 4:1 (в диапазоне от 3:1 до 6:1). Белки экспрессировали из соответствующих плазмид с использованием эквимолярного соотношения плазмид. Клетки подпитывали на 3-й день L-глутамином до 4 мМ, глюкозой [Sigma] и NAA [Gibco]. Клеточные культуральные супернатанты, содержащие иммуноглобулиновые варианты, собирали с 5-го по 11-й день после трансфекции путем центрифугирования и сохраняли при -80°С. Общая информация о рекомбинантной экспрессии человеческих иммуноглобулинов, например в клетках HEK293, дана в Meissner, P., et al., Biotechnol. Bioeng. 75 (2001) 197-203.
Очистка антител
Антитела очищали из клеточных культуральных супернатантов с помощью аффинной хроматографии с использованием белка A-Sepharose™ (GE Healthcare, Швеция) и эксклюзионной хроматографии на Superdex200. Вкратце, стерильные профильтрованные клеточные культуральные супернатанты очищали на колонке HiTrap ProteinA HP (5 мл), уравновешенной буфером PBS (10 мМ Na2HPO4, 1 мМ KH2PO4, 137 мМ NaCl и 2,7 мМ KCl, рН 7,4). Несвязанные белки отмывали с помощью уравновешивающего буфера. Антитело и варианты антитела элюировали 0,1 М цитратным буфером, рН 2,8, и фракции, содержащие белок, нейтрализовали 0,1 мл 1 М Tris, рН 8,5. Затем элюированные белковые фракции объединяли, концентрировали с помощью устройства с центробежным фильтром Amicon Ultra (MWCO: 30 K, Millipore) до объема 3 мл и наносили на гель-фильтрационную колонку Superdex200 HiLoad 120 мл 16/60 (GE Healthcare, Швеция), уравновешенную 20 мМ гистидином, 140 мМ NaCl, рН 6,0. Фракции, содержащие очищенные антитела с менее чем 5% высокомолекулярных агрегатов, собирали и хранили в виде аликвот 1,0 мг/мл при -80°С.
Анализ очищенных белков
Концентрацию белка в очищенных образцах белка определяли путем измерения оптической плотности (OD) при 280 нм, используя коэффициент молярной экстинкции, рассчитанный на основе аминокислотной последовательности. Чистоту и молекулярную массу антител анализировали с помощью SDS-PAGE в присутствии и в отсутствие восстанавливающего агента (5 мМ 1,4-дитиотреитола) и окрашивания бриллиантовым синим Кумасси. Систему гелей NuPAGE® Pre-Cast (Invitrogen, США) использовали в соответствии с инструкцией изготовителя (4-12% гели Tris-глицин). Содержание агрегатов антител в образцах анализировали с помощью высокопроизводительной SEC с использованием аналитической эксклюзиоонной колонки Superdex 200 (GE Healthcare, Швеция) в рабочем буфере 200 мМ KH2PO4, 250 мМ KCl, рН 7,0, при 25°С. 25 мкг белка наносили на колонку со скоростью потока 0,5 мл/мин и изократически элюировали более 50 минут. Для анализа стабильности очищенные белки с концентрацией 1 мг/мл инкубировали при 4°С и 40°С в течение 7 дней, а затем оценивали с помощью высокопроизводительной SEC (например, HP SEC Analysis (Purified Protein)). Целостность аминокислотного остова восстановленных иммуноглобулиновых вариантных цепей проверяли с помощью масс-спектрометрии NanoElectrospray Q-TOF после удаления N-гликанов путем ферментативной обработки пептид-N-глюкозидазой F (Roche Molecular Biochemicals).
Масс-спектрометрия
Общую дегликозилированную массу антител определяли и подтверждали путем масс-спектрометрии с ионизацией электрораспылением (ESI-MS). Вкратце, 100 мкг очищенных антител дегликозилировали с помощью 50 мЕд N-гликозидазы F (PNGaseF, Prozyme) в 100 мМ KH2PO4/K2HPO4, рН 7, при 37°С в течение 12-24 ч при концентрации белка до 2 мг/мл и затем обессоливали путем ВЭЖХ на колонке Sephadex G25 (GE Healthcare). Массу соответствующих цепей антител определяли путем ESI-MS после дегликозилирования и восстановления. Вкратце, 50 мкг антитела в 115 мкл инкубировали с 60 мкл 1М ТСЕР и 50 мкл 8М гуанидина гидрохлорида, а затем обессоливали. Общую массу и массу восстановленных цепей антител определяли путем ESI-MS на системе Q-Star Elite MS, оснащенной источником ионизации NanoMate. Записанный диапазон масс зависит от молекулярной массы образцов. В целом для восстановленных антител диапазон масс был установлен как 600-2000 m/z, а для невосстановленных антител как 1000-3600 m/z.
SEC-MALLS
SEC-MALLS (эксклюзионную хроматографию с многоугловым рассеянием лазерного света) использовали для определения приблизительной молекулярной массы белков в растворе. Согласно теории рассеяния света MALLS позволяет оценивать молекулярную массу макромолекул независимо от их молекулярной формы или других предположений. SEC-MALLS основана на разделении белков по их размеру (гидродинамический радиус) с помощью SEC-хроматографии с последующим использованием детекторов, чувствительных к концентрации и рассеянию света. SEC-MALLS, как правило, дает оценку молекулярной массы с точностью, которая позволяет ясно разделять мономеры, димеры, тримеры и т.д., если SEC-разделение является достаточным.
В этой работе были использованы следующие приборы: Dionex Ultimate 3000 HPLC; колонка: Superose 6 10/300 (GE Healthcare); элюент: 1 × PBS; скорость потока: 0,25 мл/мин; детекторы: OptiLab REX (Wyatt Inc., Dernbach), MiniDawn Treos (Wyatt Inc., Dernbach). Молекулярные массы рассчитывали с помощью программного обеспечения Astra, версия 5.3.2.13. Белки в количестве от 50 до 150 мкг наносили на колонку, a BSA (Sigma Aldrich) использовали в качестве референсного белка.
Динамическое рассеяние света (Dynamic Light Scattering, DLS) с течением времени
Образцы (30 мкл) в концентрации примерно 1 мг/мл в 20 мМ His/HisCl, 140 мМ NaCl, pH 6,0, фильтровали через 384-луночный фильтр-планшет (с размером пор 0,45 мкм) в 384-луночный оптический планшет (Corning) и покрывали 20 мкл парафинового масла (Sigma). Данные о динамическом рассеянии света повторно собирали в течение 5 дней с помощью планшетного ридера DynaPro DLS (Wyatt) при постоянной температуре 40°С. Данные обрабатывали с помощью Dynamics V6.10 (Wyatt).
Поверхностный плазменный резонанс
Связывающие свойства анти-IGF-IR-антигенсвязывающих белков и антител определяли путем методики поверхностного плазменного резонанса (SPR) с помощью инструмента BIAcore (BIAcore®, GE Healthcare, Упсала, Швеция). Эта система хорошо известна для изучения взаимодействий молекул. Она позволяет осуществлять в реальном времени непрерывный мониторинг связываний лиганда/анализируемого вещества и, таким образом, определять константы скорости ассоциации (ka), константы скорости диссоциации (kd) и константы равновесия (KD) в различных условиях анализа. SPR-технология основана на измерении показателя преломления вблизи поверхности покрытого золотом биосенсорного чипа. Изменения показателя преломления отражают изменения массы на поверхности, вызванные взаимодействием иммобилизованного лиганда с анализируемым веществом, введенным в растворе. Если молекулы связывают иммобилизованный лиганд на поверхности, то масса увеличивается, в случае диссоциации масса уменьшается. Для захвата противочеловеческое IgG-антитело иммобилизировали на поверхности биосенсорного чипа СМ5 путем аминного связывания. Проточные кюветы активировали смесью 0,1 М N-гидроксисукцинимида и 0,1 М 3-(N,N-диметиламино)пропил-N-этилкарбодиимида в соотношении 1:1 со скоростью потока 5 мкл/мин. Противочеловеческое IgG-антитело вводили в ацетате натрия, pH 5,0, в концентрации 10 мкг/мл. Референсную контрольную проточную кювету обрабатывали таким же образом, но буферами только с носителями без антитела захвата. Поверхности блокировали путем введения 1М этаноламина/HCl, pH 8,5. IGF-1R-антитела разводили в HBS-P и вводили. Все взаимодействия проводили при 25°С (стандартная температура). Регенерационный раствор 3 М хлорида магния вводили в течение 60 со скоростью потока 5 мкл/мин, чтобы удалить любые нековалентно связанные белки после каждого цикла связывания. Сигналы регистрировали в режиме одного сигнала в секунду. Образцы вводили в возрастающих концентрациях. Фиг.17 изображает примененный формат анализа. Низкая плотность загрузки антитела захвата и IGF-1R-антител была выбрана для обеспечения связывания.
Для измерения аффинности человеческое FcgIIIa иммобилизировали на поверхность сенсорного чипа СМ-5 путем захвата помеченным His-меткой рецептором анти-His-антитела (Penta-His, Qiagen), которое связывалось с поверхностью за счет стандартного аминного связывания и химической реакции блокировки на инструменте SPR (Biacore T100). После захвата FcgRIIIa вводили IGF-1R-антитело в количестве 50 нМ со скоростью потока 5 мкл/мин при 25°С, затем чип регенерировали с помощью 60-секундного импульса 10 мМ раствора глицин-HCl, рН 2,0.
Анализ антителозависимой клеточной цитотоксичности (ADCC)
Определение опосредованных антителом эффекторных функций с помощью aHTH-IGF-IR-антител. Для того чтобы определить способность полученных антител вызывать иммунные эффекторные механизмы, были проведены исследования антителозависимой клеточной цитотоксичности (ADCC). Для изучения влияний антител в ADCC, клетки DU145, экспрессирующие IGF-IR (1×106 клеток/мл), метили с помощью 1 мкл раствора BATDA (Perkin Elmer) на 1 мл в течение 25 минут при 37°С в клеточном инкубаторе. После этого клетки промывали четыре раза с помощью 10 мл RPMI-FM/PenStrep и центрифугировали в течение 10 минут при 200 g. Перед последним этапом центрифугирования определяли число клеток и после этого из осадка разводили клетки до плотности 1×105 клеток/мл в среде RPMI-FM/PenStrep. Клетки высевали по 5000 на лунку в круглодонный планшет в объеме 50 мкл. Антитела HuMAb добавляли до конечной концентрации 25-0,1 мкг/мл в клеточной культуральной среде объемом 50 мкл к 50 мкл суспензии клеток. Затем 50 мкл эффекторных клеток, свежевыделенных РВМС, добавляли в соотношении Е:Т 25:1. Планшеты центрифугировали в течение 1 минуты при 200 g с последующим этапом инкубации в течение 2 часов при 37°С. После инкубации клетки центрифугировали в течение 10 минут при 200 g, 20 мкл супернатанта собирали и переносили в планшет Optiplate 96-F. Добавляли 200 мкл раствора европия (Perkin Elmer, при комнатной температуре) и инкубировали планшеты в течение 15 минут на шейкере. Флуоресценцию количественно оценивали с помощью флуориметра временного разрешения (Victor 3, Perkin Elmer) с использованием протокола Eu-TDA от Perkin Elmer. Величина лизиса клеток в ADCC выражается как % от максимального высвобождения усилителя флуоресценции TDA из клеток-мишеней, лизированных детергентом, скорректированный с учетом спонтанного высвобождения TDA из соответствующих клеток-мишеней.
Анализ интернализации IGF-1R
Связывание антител и антигенсвязывающего белка в соответствии с изобретением с IGF-1R приводит к интернализации и деградации рецептора. Этот процесс можно контролировать путем инкубации клеток НТ29 CRC, экспрессирующих IGF-1R, с антителами, нацеленными на IGF-1R, с последующей количественной оценкой оставшихся уровней белка IGF-1R в клеточных лизатах путем ELISA.
С этой целью клетки НТ29 с плотностью 1,5×104 клеток/лунка инкубировали в 96-луночных МТР в RPMI с 10% FCS в течение ночи при 37°С и 5% CO2 для прикрепления клеток. На следующее утро среду аспирировали и добавляли 100 мкл анти-IGF-1R-антитела, разведенного в RPMI + 10% FCS до концентраций от 10 нМ до 2 пМ с шагом разведения 1:3. Клетки инкубировали с антителом в течение 18 часов при 37°С. После этого среду снова удаляли и вносили 120 мкл MES-буфера для лизиса (25 мМ MES, рН 6,5 + Complete).
Для ELISA в 96-луночные покрытые стрептавидином планшеты из полистирола (Nunc) загружали 100 мкл MAK<hu IGF-1Rα>hu-1a-IgG-Bi (Ch.10), разведенного 1:200 в 3% BSA/PBST (конечная концентрация 2,4 мкг/мл), и инкубировали при постоянном перемешивании в течение 1 ч при комнатной температуре. Затем содержимое лунок удаляли и каждую лунку промывали три раза с помощью 200 мкл PBST. В каждую лунку вносили 100 мкл раствора лизата клеток, снова инкубировали в течение 1 ч при комнатной температуре на планшетном шейкере и промывали три раза с помощью 200 мкл PBST. После удаления супернатанта добавляли 100 мкл/лунка PAK<human IGF-1Rα>Ra-C20-IgG (Santa Cruz № sc-713), разведенного 1:750 в 3% BSA/PBST с последующими такими же интервалами инкубации и промываниями, как было описано выше. Для обнаружения специфического связывания антитела с IGF-1R добавляли 100 мкл/лунка поликлонального кроличьего антитела, конъюгированного с пероксидазой хрена (Cell Signaling №7074), разведенного 1:4000 в 3% BSA/PBST. Еще через 1 ч несвязанное антитело снова удаляли путем тщательного промывания 6 раз, как описано выше. Для количественного определения связанного антитела вносили 100 мкл/лунка 3,3'-5,5'-тетраметилбензидина (Roche, BM-Blue №11484281) и инкубировали в течение 30 минут при комнатной температуре. Наконец, цветную реакцию останавливали путем добавления 25 мкл/лунка 1 М H2SO4 и измеряли поглощение света при длине волны 450 нм. Клетки, не обработанные антителом, использовали в качестве контроля для 0% понижающей регуляции, а лизирующий буфер для контроля фона.
Анализ аутофосфорилирования IGF-1R (IGF-1-стимуляция)
Нацеливание IGF-1R-антител на IGF-1R приводит к ингибированию IGF-1-индуцированного аутофосфорилирования. Мы исследовали ингибирование аутофосфорилирования MoAb-димера IGF-1R дикого типа по сравнению с родительским IGF-1R-антителом IgG1. Для этой цели клетки 3T3-IGF-1R, клеточную линию мышиных фибробластов со сверхэкспрессией человеческого IGF-1R, обрабатывали в течение 10 минут рекомбинантным человеческим IGF-1 с концентрацией 10 нМ в присутствии различных концентраций IGF-1R-антитела или IGF-1R-антигенсвязывающего белка. После лизиса клеток уровни фосфорилированного белка IGF-1R определяли с помощью фосфо-IGF-1R-специфического ELISA, сочетающего человеческое IGF-1R-специфическое антитело захвата и фосфотирозин-специфическое антитело детекции.
Определение фармакокинетических свойств: кинетика одной дозы у мышей
Методы
Животные
Мыши NMRI, самки, кормленые, вес тела 23-32 г в момент введения соединения.
Протокол исследования:
Для внутривенного введения одной дозы 10 мг/кг мышей делили на три группы по 2-3 животных в каждой. Образцы крови забирали в группе 1 через 0,5, 168 и 672 часа, в группе 2 через 24 и 336 часов, а в группе 3 через 48 и 504 часа после введения дозы.
Образцы крови объемом примерно 100 мкл получали путем ретробульбарной пункции. Образцы сыворотки объемом по меньшей мере 40 мкл получали из крови через 1 час инкубации при комнатной температуре путем центрифугирования (9300 g) в течение 2,5 мин при комнатной температуре. Образцы сыворотки замораживали непосредственно после центрифугирования и хранили в замороженном виде при -20°С до анализа.
Анализ
Концентрации человеческих антител в сыворотке мышей определяли в ферментном иммунособентном анализе (ELISA) с использованием 1% мышиной сыворотки. На первом этапе биотинилированное моноклональное антитело против человеческого Fcγ (mAb<hFcγPAN>IgG-Bi) связывалось с покрытыми стрептавидином микротитровальными планшетами. На следующем этапе вносились образцы сыворотки (в различных разведениях) и референсные стандарты, соответственно, и связывались с иммобилизованным mAb<hFcγPAN>IgG-Bi. Затем добавляли дигоксигенилированное моноклональное антитело против человеческого Fcγ (mAb<hFcγPAN>IgG-Dig). Человеческие антитела обнаруживали с помощью анти-Dig-антитела, конъюгированного с пероксидазой хрена. ABTS-раствор использовали в качестве субстрата для пероксидазы хрена. Специфичность используемых антител захвата и детекции, которые не реагируют перекрестно с IgG мыши, дает возможность количественно оценить человеческие антитела в образцах мышиной сыворотки.
Расчеты
Фармакокинетические параметры рассчитывали путем анализа без учета компартментов с использованием программы для фармакокинетической оценки WinNonlinTM, версия 5.2.1.
Следующие фармакокинетические параметры использовали для оценки человеческих антител:
- Начальная концентрация, предполагаемая только для моделей с внутривенным болюсным введением (С0).
- Максимальная наблюдаемая концентрация (Cmax), имеющая место в момент времени (Tmax).
- Время максимальной наблюдаемой концентрации (Tmax).
- площадь под кривой концентрация/время AUC (0-inf) рассчитывается по линейному правилу трапеций (с линейной интерполяцией) с момента времени 0 до бесконечности.
- Кажущееся время полужизни (T1/2) было получено из уравнения: T1/2=ln2/λz.
- общий клиренс (CL) рассчитывается как доза/AUC(0-inf).
- Объем распределения в равновесном состоянии (Vss), рассчитанный как MRT(0-inf) × CL (MRT(0-inf), определяемый как AUMC(0-inf)/AUC(0-inf).
В. Примеры
Пример 1
Создание антигенсвязывающих белков MoAb-димер
Мы разработали антигенсвязывающие белки в соответствии с изобретением против c-Met (SEQ ID №1 и SEQ ID №2), IGF-1R (SEQ ID №3 и SEQ ID №4) и HERS (SEQ ID №5 и SEQ ID №6) на основе принципа дизайна, показанного на фиг.1А. Соответствующие конструкции временно экспрессировали в клетках HEK293, как описано выше, а затем очищали путем аффинной хроматографии на белке А с последующей эксклюзионной хроматографией. Фиг.2-4 изображают хроматограммы эксклюзионной хроматографии с тремя различными полученными антигенсвязывающими белками, а также соответствующие SDS-PAGE в невосстанавливающих и восстанавливающих условиях. В дополнение к пику 3 на фиг.2, пику 2 на фиг.3, пику 3 на фиг.4, наблюдались белки, элюирующиеся из колонки раньше, пик 2 на фиг.2, пик 1 на фиг.3, пик 4 на фиг.7. На основе их времени удерживания было рассчитано, что они демонстрируют двойную молекулярную массу одновалентного антитела (фиг.1В, MoAb), соответствующего антигенсвязывающему белку в соответствии с фиг.1А (MoAb-димер).
Размер двух пиков 1 и 2 для антител IGF-1R был подтвержден посредством SEC-MALLS (фиг.3С), и было показано, что действительно белок, соответствующий пику 1, демонстрирует примерно двойную молекулярную массу одновалентного антитела (пик 2). Существование MoAb-димера и идентичность изолированных белков впоследствии были подтверждены путем масс-спектрометрии. На основании этих выводов мы получили модель структуры MoAb-димера, которая изображена на фиг.1А.
Мы провели эксперименты, такие как масс-спектрометрия, восстановление и протеазное расщепление, чтобы подтвердить предполагаемую структуру, показанную на фиг.1А.
Стабильность IGF1R AK18 MoAb-димера («СН3-wt») была изучена путем динамического рассеяния света, как описано выше. Вкратце, тенденцию IGF1R AK18 MoAb-димера («СН3-wt») к агрегации оценивали с помощью эксперимента DLS с течением времени при 40°С.
Пример 2
Аффинность связывания IGF-1R
Связывание внеклеточного домена IGF1R с AK18 MoAb-димером («СН3-wt») сравнивали со связыванием родительского <IGF-1R> IgG1-антитела с помощью поверхностного плазмонного резонанса (SPR). Фиг.5 изображает схему анализа SPR для определения аффинности. Анализ (двойное определение) показал, что аффинность связывания IGF-1R сохраняется у IGF1R AK18 MoAb-димера («СН3-wt»).
Пример 3
Клеточное связывание с клеточными линиями, экспрессирующими IGF-1R
Клеточное связывание IGF-1R MoAb-димера было продемонстрировано на клетках А549. Клетки А549 в логарифмической фазе роста отделяли с помощью аккутазы (Sigma), и 2×105 клеток использовали для каждой инкубации антитела. IGF-1R-антитело и MoAb-димер добавляли в серии трехкратных разведении (100-0,0003 мкг/мл). Связанные антитела визуализировали с помощью вторичного Alexa488-связанного антитела (5 мкг/мл), связывающегося с константной области человеческого иммуноглобулина. Мертвые клетки окрашивали с помощью 7-AAD (BD) и исключали из анализа. Интенсивность флуоресценции отдельных клеток измеряли на проточном цитометре FACS Canto (BD Biosciences). Данные показали, что MoAb-димер демонстрирует очень похожую величину полумаксимального связывания с клетками, сравнимую с таковой для родительского IGF-1R IgG1-антитела. Это означает, что MoAb-димер может связываться двумя ветвями с IGF-1R на клетках и демонстрирует эффект авидности. Тем не менее, общая MFI (средняя интенсивность флуоресценции) выше для MoAb-димера, чем для МКА IGF-1R. Это, вероятно, связано с большим числом Fc-частей на одну молекулу IGF-1R на поверхности клетки. Результаты показаны на фиг.6 и ниже.
Пример 4
Индукция ADCC
Полученные от доноров мононуклеарные клетки периферической крови (РВМС) могут быть использованы для измерения привлечения эффекторных клеток негликоинженерными и гликоинженерными антителами к раковым клеткам. Лизис раковых клеток коррелирует с опосредованной МК-клетками цитотоксичностью и пропорционален способности антитела привлекать МК-клетки. В этой конкретной ситуации клетки рака предстательной железы DU145 инкубировали в соотношении 1:25 (DU145:PBMC) в отсутствие или в присутствии соответствующих антител. Через 2 часа клеточный лизис определяли с помощью системы BATDA/европий, как описано выше. Значение клеточного лизиса из-за ADCC выражается как % от максимального высвобождения усилителя флуоресценции TDA из клеток-мишеней, лизированных детергентом, скорректированный с учетом спонтанного высвобождения TDA из соответствующих клеток-мишеней. Данные показывают, что негликоинженерный двухвалентный IGF-1R MoAb-димер превосходит негликоинженерное IGF-1R-антитело в отношении индукции ADCC. Удивительно, что негликоинженерный IGF-1R MoAb-димер даже превосходит гликоинженерное IGF-1R-антитело в отношении индукции ADCC при высокой концентрации, что показывается снижением ADCC в анализе при высоких концентрациях. Повышенную индукцию ADCC MoAb-димером в отсутствие гликоинженерии можно объяснить более высокой аффинностью конструкции к FcgRIIIa на NK-клетках из-за двухвалентности для FcgRIIIa (эффект авидности).
Результаты для негликоинженерного МКА IGF1R (1), гликоинженерного афукозилированного МКА IGF1R (2) и негликоинженерного IGF1R AK18 MoAb-димера («СН3-wt») (3) представлены на фиг.7, показывая, что IGF1R AK18 MoAb-димер («СН3-wt») имеет повышенную ADCC по сравнению с МКА IGF1R, а в более высоких концентрациях даже по сравнению с гликоинженерным афукозилированным МКА IGF1R (2).
Пример 5
Анализ интернализации IGF-1R
Нацеливание двухвалентных IGF-1R-антител на IGF-1R на опухолевых клетках приводит к интернализации и лизосомальной деградации IGF-1R. Мы исследовали свойства интернализации IGF-1R MoAb-димера по сравнению с родительским IGF-1R-антителом. С этой целью клетки рака толстой кишки НТ29 обрабатывали в течение 18 часов различными концентрациями МоА-димера и родительского IGF-1R-антитела. После лизиса клеток оставшиеся уровни IGF-1R-белка определяли путем IGF-1R-специфического ELISA.
Данные на фиг.8 показывают, что интернализация IGF-1R MoAb-димером практически идентична таковой двухвалентным родительским IGF-1R-антителом. Максимальная интернализация составляла 82,99% (IgG1) по сравнению с 83,7% (MoAb-димер), концентрация, необходимая для полумаксимального ингибирования, составляла 0,027 нМ (IgG1) по сравнению с 0,027 нМ (MoAb-димер).
Пример 6
IGF-1R-аутофосфорилирование (IGF-1-стимуляция)
Нацеливание IGF-1R-антител на IGF-1R приводит к ингибированию IGF-1-индуцированного аутофосфорилирования. Мы исследовали ингибирование аутофосфорилирования MoAb-димерного IGF-1R-антитела по сравнению с родительским IGF-1R IgG1-антителом. Для этой цели клетки 3T3-IGF-1R, клеточную линию мышиных фибробластов со сверхэкспрессией человеческого IGF-1R, обрабатывали в течение 10 минут 10 нМ рекомбинантного человеческого IGF-1 в присутствии различных концентраций IGF-1R-антитела и антигенсвязывающего белка. После лизиса клеток уровни фосфорилированного IGF-1R-белка определяли с помощью фосфо-IGF-1R-специфического ELISA, сочетая человеческое IGF-1R-специфическое антитело захвата и фосфотирозин-специфическое антитело детекции.
Данные на фиг.9 показывают, что IGF-1R MoAb-димер может ингибировать IGF-1-индуцированное аутофосфорилирование аналогично или даже немного лучше, чем родительская молекула IGF-1R IgG1. Концентрация, необходимая для полумаксимального ингибирования, составила 1,44 нМ (родительский IGF-1R IgG1) по сравнению с 3,52 нМ (MoAb-димер), максимальное наблюдаемое ингибирование составило 80,3% (родительский IGF-1R IgG1) по сравнению с 89,1% (MoAb-димер).
Пример 7
Определение фармакокинетических свойств
Фармакокинетические свойства антигенсвязывающих белков в соответствии с изобретением определяли у мышей NMRI, самок, кормленных, 23-32 г масса тела в момент введения мыши дозы соединения, в исследовании PK с однократным введением дозы, как описано выше (в разделах Методы).
PK-свойства приведены в последующей таблице и показывают, что антигенсвязывающий белок IGF1R AK18 MoAb-димер («СН3-wt») имеет весьма ценные PK-свойства по сравнению с родительским <IGF-1R> IgG1-антителом (такие как, например, AUC0-inf, Cmax или С0).
Пример 8
ESI-MS-эксперимент с IGF-1R MoAb-димером
IGF-1R-MoAb-димер (IGF1R AK18 MoAb («СН3-wt»)) (SEQ ID №3 и SEQ ID №4) временно экспрессировали и очищали путем аффинной хроматографии на белке А и эксклюзионной хроматографии. После препаративной SEC (см. фиг.3А) антитело элюировали из двух отдельных пиков, которые собирали. Аналитическая SEC первого пика (фракция 1) показывает кажущуюся молекулярную массу примерно 200 кДа, где второй пик (фракция 2) соответствует молекулярной массе 100 кДа. Различные молекулярные массы могут быть отнесены к определенному мономеру как побочному продукт (фракция 2) и как к нужному димеру (фракция 1), соответственно. SEC-MALS подтвердила первоначальный результат SEC и показала для фракции 2 (мономера) молекулярную массу 99,5 кДа, а для фракции 1 (димера) молекулярную массу 193,8 кДа.
SDS-PAGE-анализ (см. фиг.3В) двух фракций в денатурирующих и восстанавливающих условиях показал одну основную полосу с кажущейся молекулярной массой 50-60 кДа. В невосстанавливающих условиях фракция 2 показывает основную полосу с молекулярной массой примерно 100 кДа, а фракция 1 показывает очень широкий диапазон примерно 100-200 кДа.
Фракция 1=165 мл
Фракция 2=190 мл
Эти исходные данные показывают образование димера.
ESI-MS-спектры (образцы инкубировали с 60 мкл 1М ТСЕР и 50 мкл 8 М гуанидина гидрохлорида) дегликозилированного MoAb-димера из фракции 1 и MoAb-мономера из фракции 2 показали отличия. Фракция 2 показала только одну серию пиков, соответствующую мономеру с массой 98151 Да, в то время как фракция 1 показала две различные кривые, содержащие две основные серии пиков, соответствующие массе 98155 Да (мономер) и массе 196319 Да (димер).
Присутствие мономера во фракции 1 может быть объяснено условиями инкубации (инкубировали с 60 мкл 1М ТСЕР и 50 мкл 8 М гуанидина-гидрохлорида) и потенциально открытым S-S-мостиком между CH1-Ck. В этом случае перенос из водного в кислый органический растворитель в процессе подготовки образца для MS может привести к диссоциации димера на два мономера.
Это соответствует результатам анализа CE-SDS (BioAnalyzer). Фракция 1 содержит 71% димера и 16% мономера в невосстанавливающих условиях. Применение SDS отделяет нековалентно связанные цепи. Фракция 2 показала только один мономерный сигнал (98%) в невосстанавливающих условиях.
MS-измерения в восстанавливающих условиях фракции 1 и фракции 2 показали правильную последовательность и экспрессию конструкций. MS-данные от фракции 1 показали две различные тяжелые цепи с молекулярной массой 47960 Да и 50208 Да в приблизительно равных количествах. Эти две тяжелые цепи также были обнаружены во фракции 2 с похожей интенсивностью.
Пример 9
Анализ соотношений MoAb-димера и одновалентного мономера в СН3 wt и СН3-модифицированных IGF1R AK18 MoAb-димер-антигенсвязывающих белках
Модификации в СН3-доменах цепей антитела могут изменять соотношения MoAb-димера и MoAb-мономера. В качестве примера, различные модификации тяжелой цепи временно экспрессировали в клетках HEK293F вместе или в сочетании с немодифицированной тяжелой цепью. Для сравнения антитело, содержащее немодифицированные тяжелые цепи, временно трансфицировали в параллелях. Супернатант, содержащий антитела, собирали через семь дней после трансфекции. Антитело осаждали с помощью белок А-сефарозы и элюировали из гранул по стандартному протоколу с шоковым рН. Полученный элюат подвергали анализу ВЭЖХ с использованием колонки G3000SW (TSKgel). Соотношения антител количественно оценивали, рассчитывая площади под соответствующими пиками. Термин «СН3-wt» относится к СН3-домену IgG1 с природной последовательностью (дикий тип = wt.)
Пример 10
Производство гликоинженерных антител
Для производства гликоинженерного MoAb-антигенсвязывающего белка клетки HEK-EBNA трансфицировали четырьмя плазмидами с использованием кальций-фосфатного способа. Две плазмиды, кодирующие цепи антитела, одна для экспрессии гибридного полипептида GnTIII (экспрессионный вектор GnT-III) и одна для экспрессии маннозидазы II (экспрессионный вектор маннозидазы II комплекса Гольджи) в соотношении 4:4:1:1, соответственно. Клетки выращивали в виде прикрепленных однослойных культур в Т-образных сосудах с использованием культуральной среды DMEM с добавлением 10% FCS и подвергали трансфекции, когда они имели конфлюэнтность от 50 до 80%. Для трансфекции в колбу Т150 за 24 часа до трансфекции высевали 15 млн клеток в 25 мл культуральной среды DMEM, дополненной FCS (конечная концентрация 10% объем/объем), и клетки помещали в инкубатор при 37°C с атмосферой 5% СО2 на ночь. Для каждой колбы Т150, в которой проводили трансфекцию, готовили раствор ДНК, CaCl2 и воды путем смешивания 94 мкг общей плазмидной векторной ДНК, разделенной поровну между экспрессионными векторами с легкой и тяжелой цепью, воды до конечного объема 469 мкл и 469 мкл 1 М раствора CaCl2. К этому раствору добавляли 938 мкл 50 мМ HEPES, 280 мМ NaCl, 1,5 мМ Na2HPO4 при рН 7,05, немедленно перемешивали в течение 10 секунд и инкубировали при комнатной температуре в течение 20 секунд. Суспензию разводили 10 мл DMEM, дополненной 2% FCS, и вносили в Т150 вместо находящейся там среды. Затем добавляли еще 13 мл среды для трансфекции. Клетки инкубировали при 37°С, 5% CO2 в течение примерно 17-20 часов, а затем среду заменяли 25 мл DMEM, 10% FCS. Кондиционированную культуральную среду собирали примерно через 7 дней после замены среды путем центрифугирования в течение 15 минут при 210 g, раствор стерилизовали путем фильтрации (фильтр с размером пор 0,22 мкм), добавляли азид натрия до конечной концентрации 0,01% вес/объем и сохраняли при 4°С.
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2011 |
|
RU2573588C2 |
| ТРИ- ИЛИ ТЕТРАСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2010 |
|
RU2570633C2 |
| ДВУХВАЛЕНТНЫЕ БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2008 |
|
RU2547615C2 |
| ДВУХВАЛЕНТНЫЕ БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2008 |
|
RU2587616C2 |
| ЛЕГКОВЫДЕЛЯЕМЫЕ БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА С ПРИРОДНЫМ ИММУНОГЛОБУЛИНОВЫМ ФОРМАТОМ | 2015 |
|
RU2647758C2 |
| ЛЕГКОВЫДЕЛЯЕМЫЕ БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА С ПРИРОДНЫМ ИММУНОГЛОБУЛИНОВЫМ ФОРМАТОМ | 2014 |
|
RU2569157C2 |
| ДВУХВАЛЕНТНЫЕ БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2008 |
|
RU2575066C2 |
| ПОЛИСПЕЦИФИЧНЫЕ АНТИТЕЛА, ВКЛЮЧАЮЩИЕ АНТИТЕЛА ПОЛНОЙ ДЛИНЫ И ОДНОЦЕПОЧЕЧНЫЕ ФРАГМЕНТЫ FAB | 2010 |
|
RU2598248C2 |
| БИСПЕЦИФИЧЕСКИЕ ЧЕТЫРЕХВАЛЕНТНЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ | 2010 |
|
RU2604189C2 |
| МОДИФИЦИРОВАННЫЕ АСИММЕТРИЧНЫЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕ Fc-РЕЦЕПТОР, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2687043C2 |

















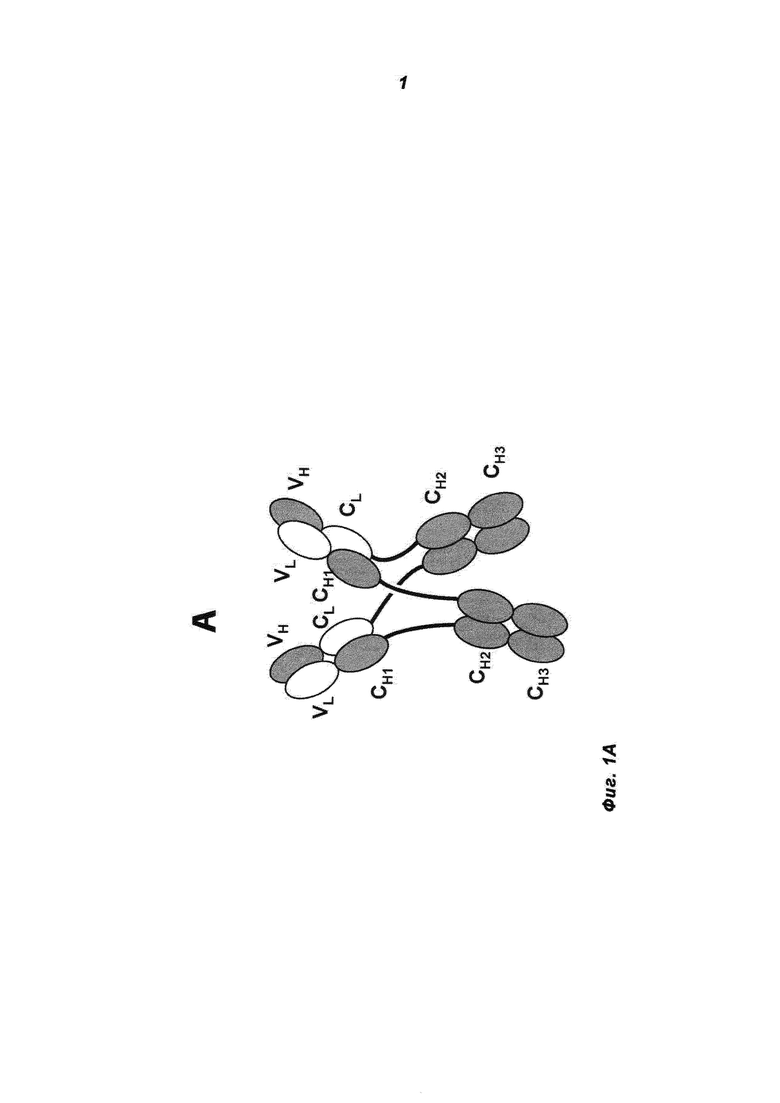
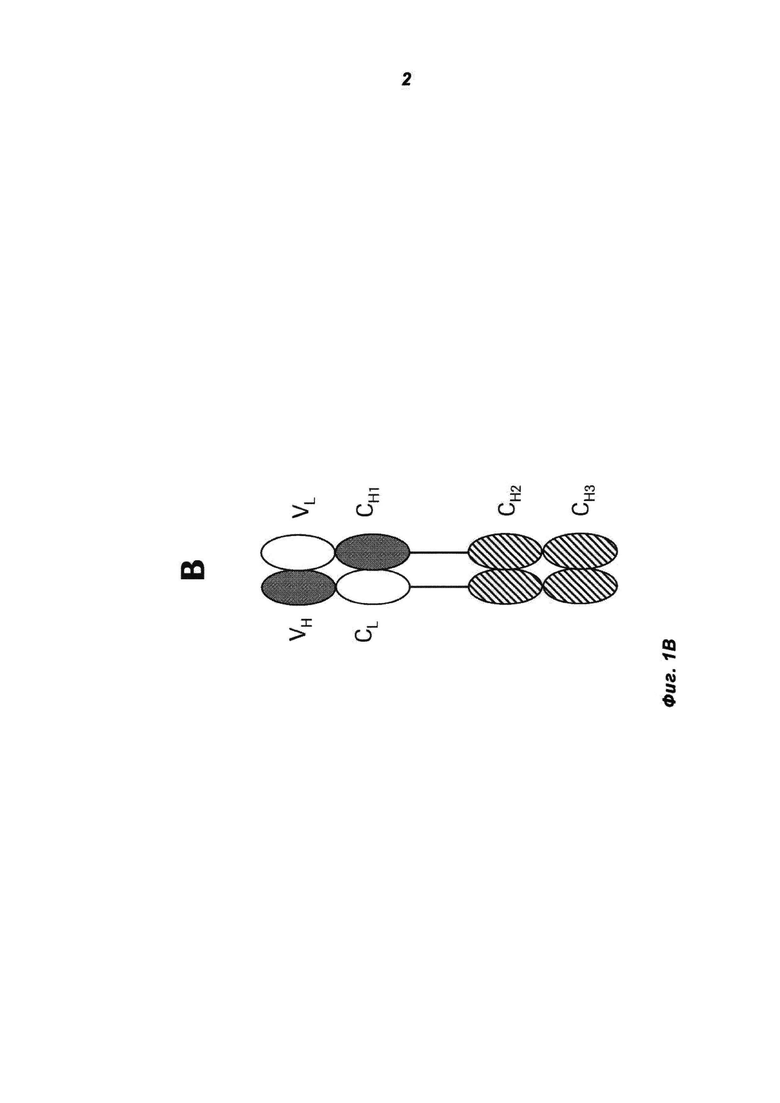
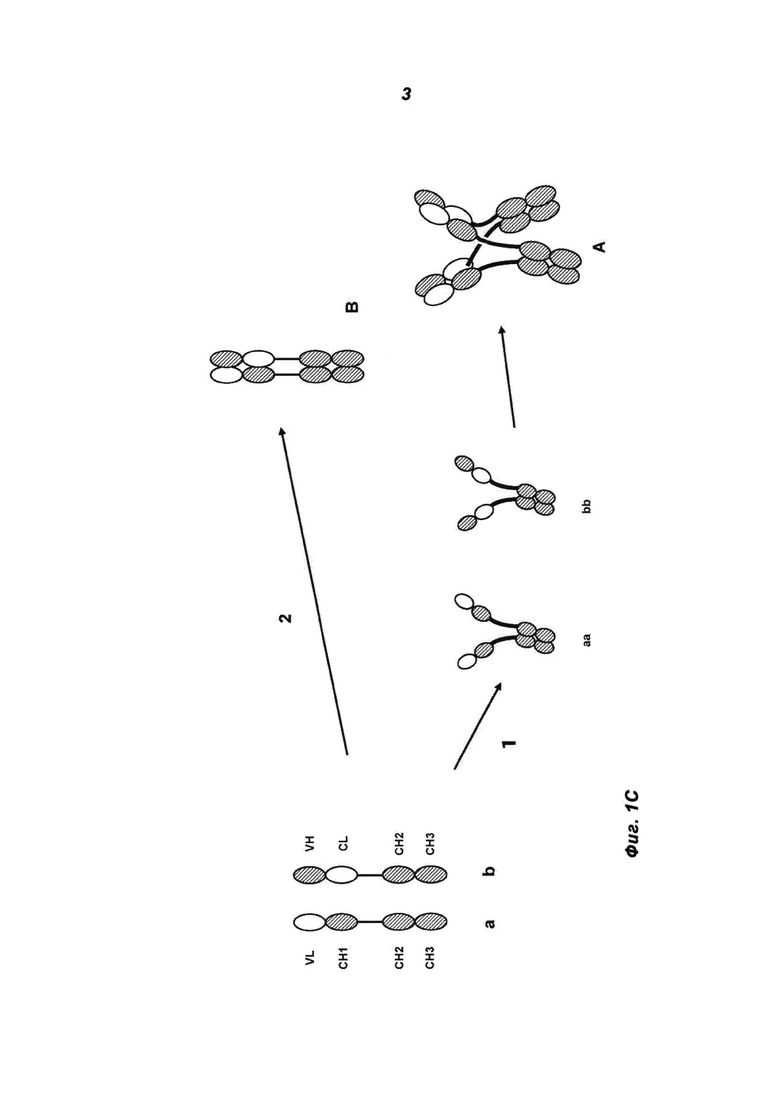
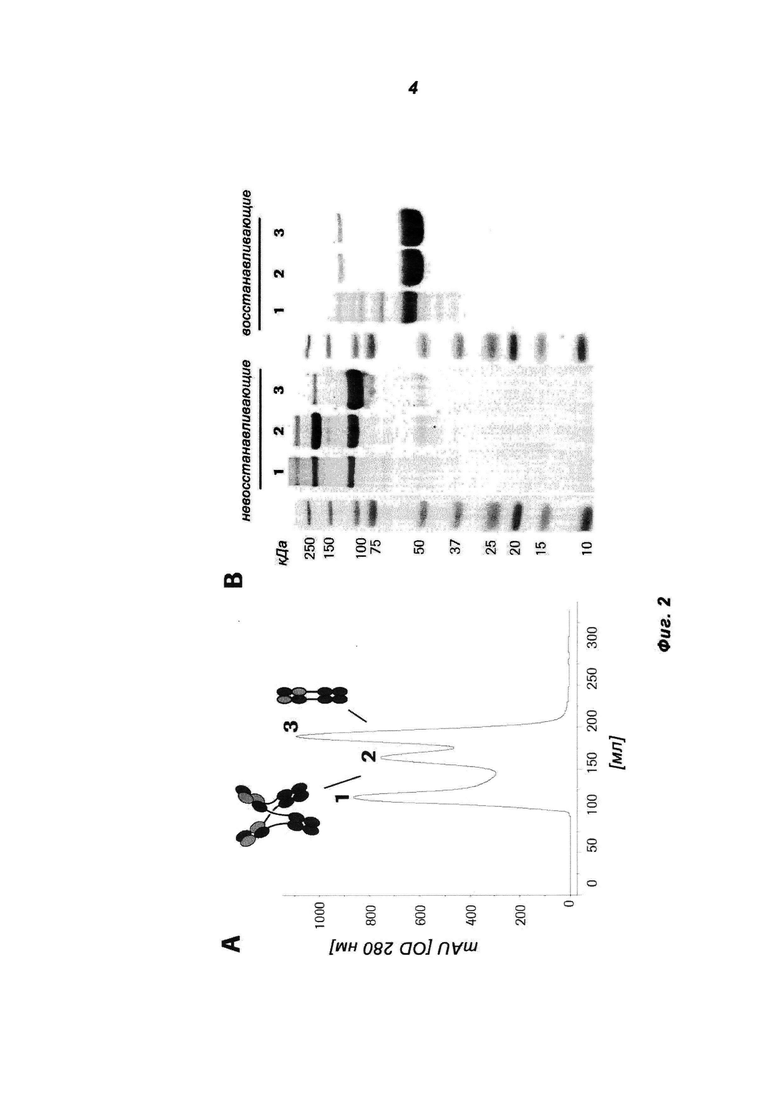
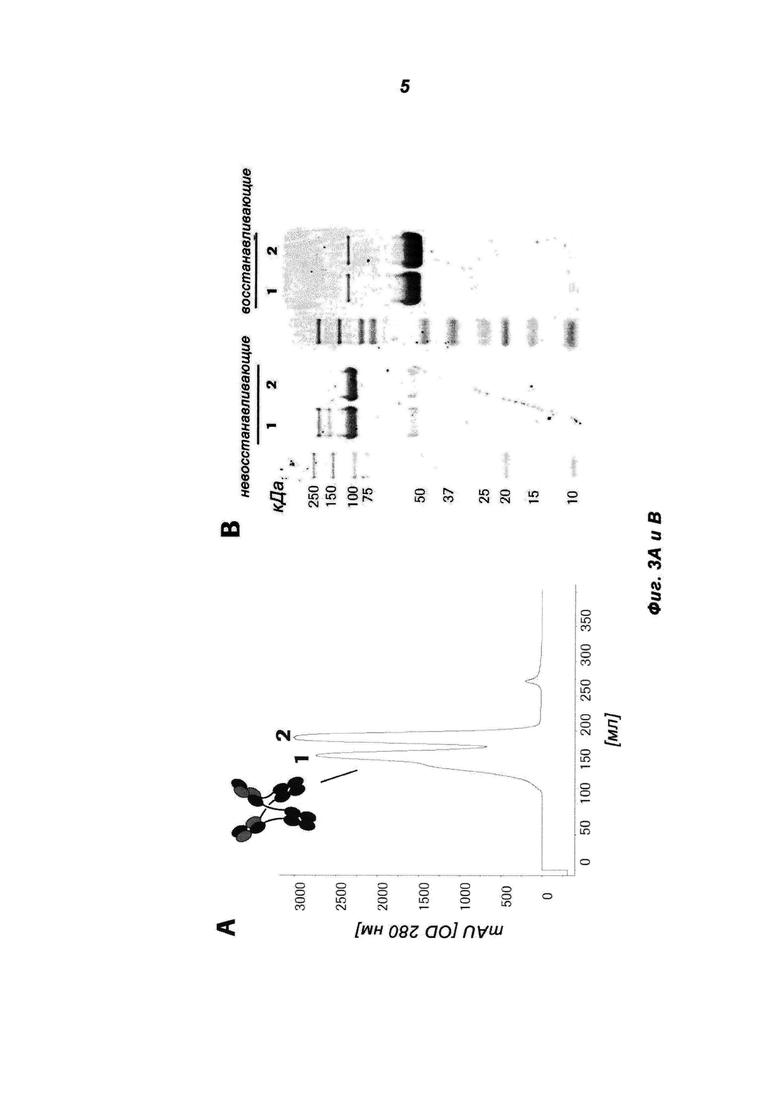
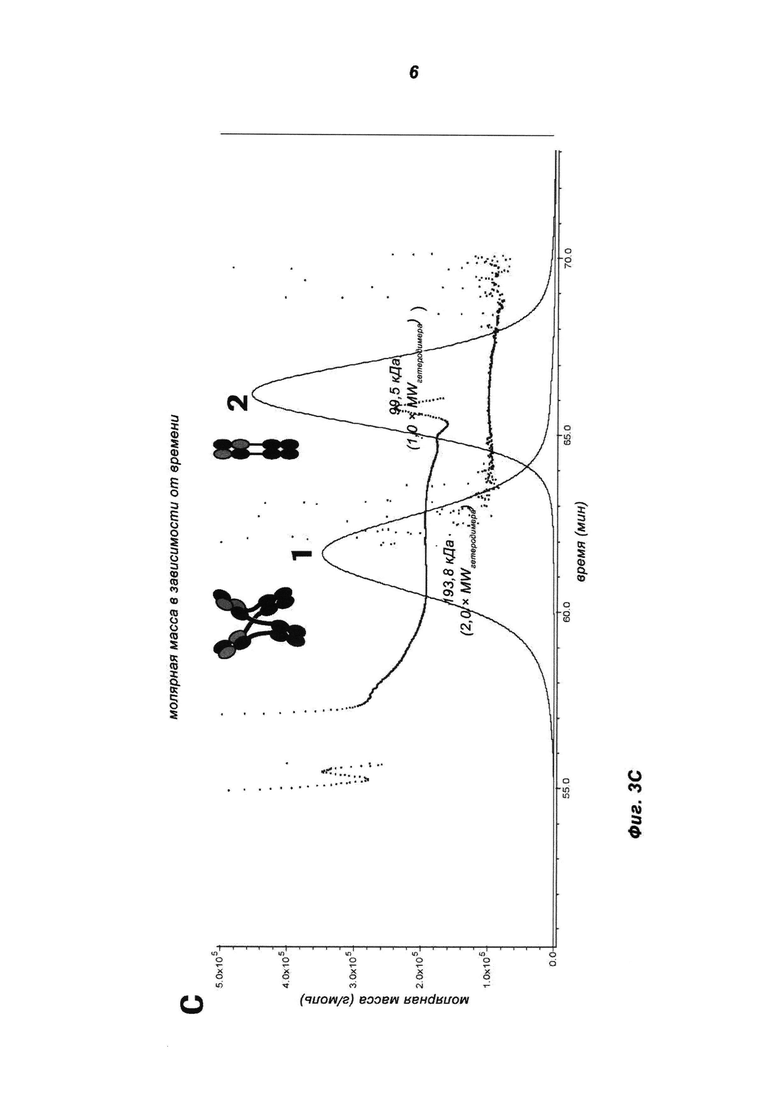
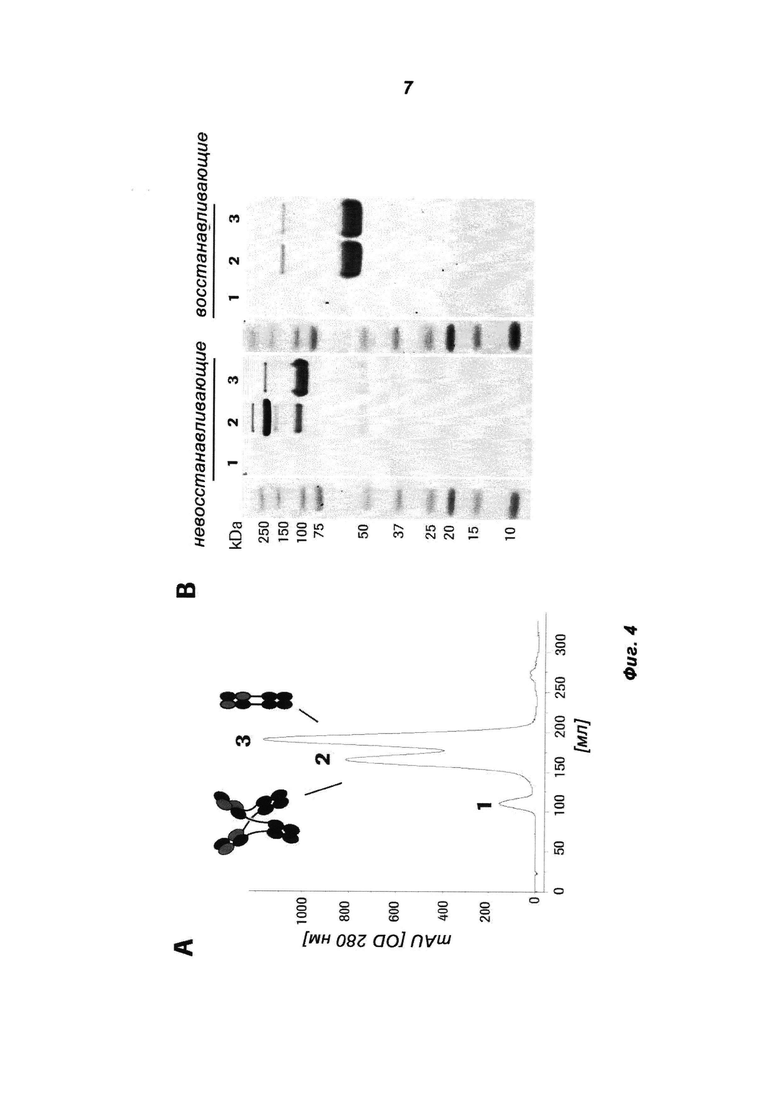
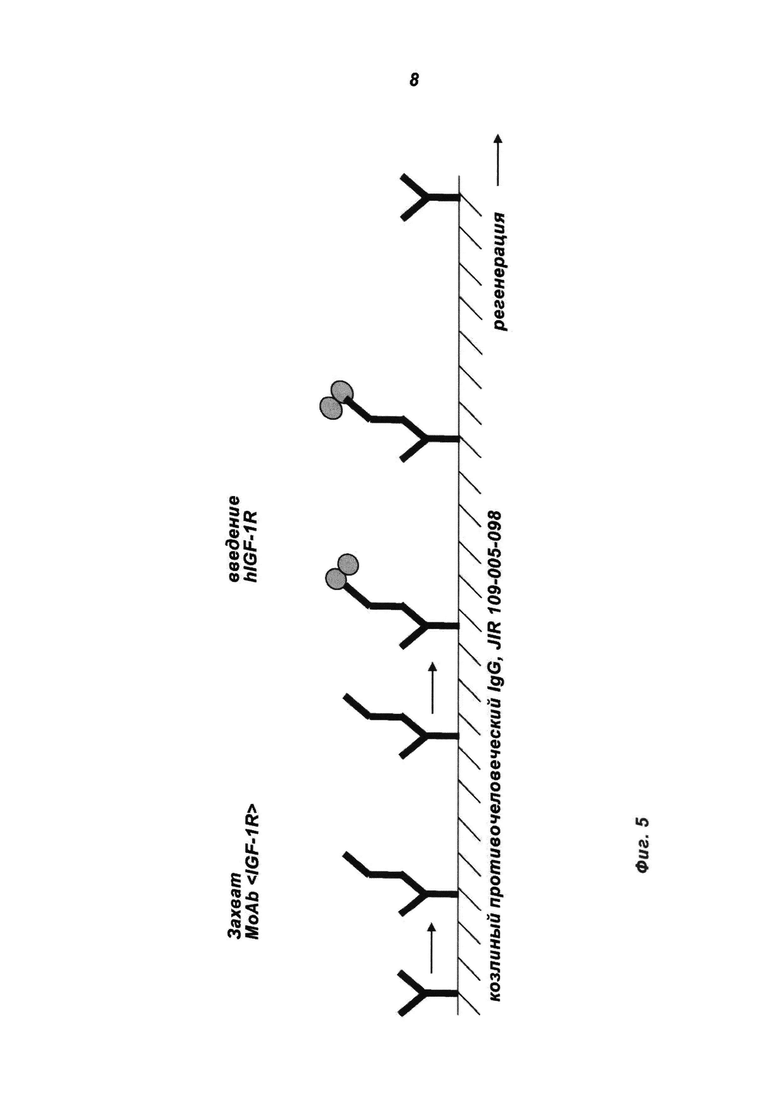
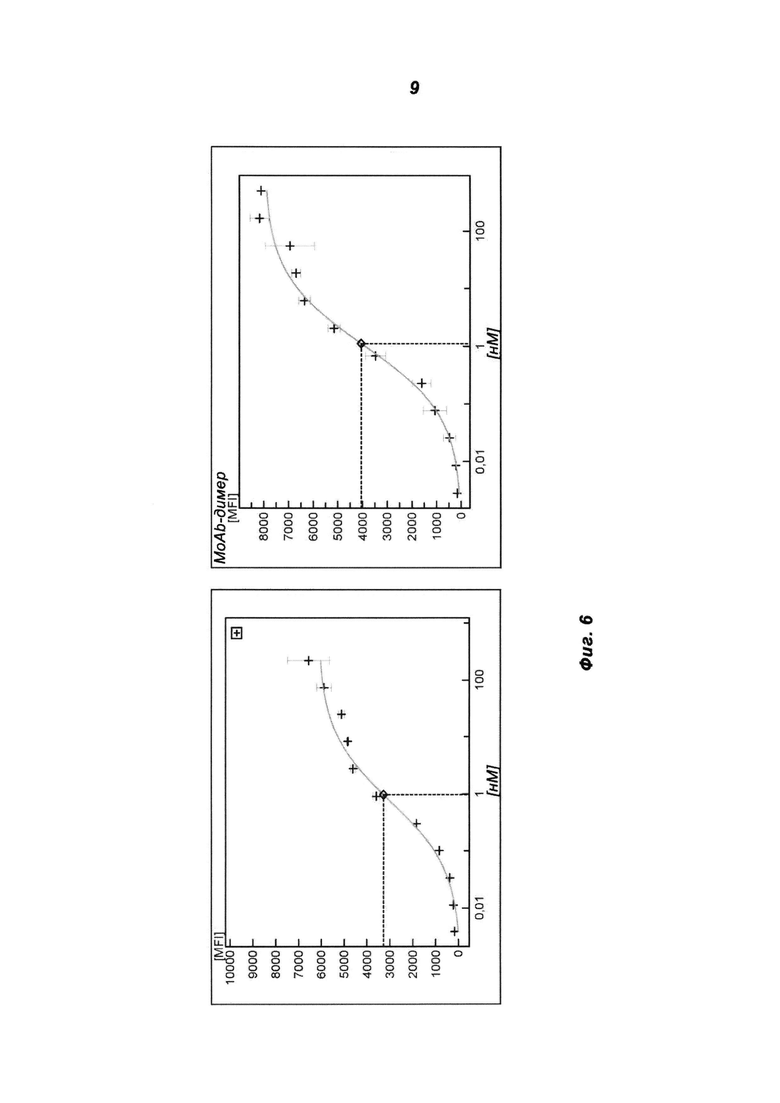
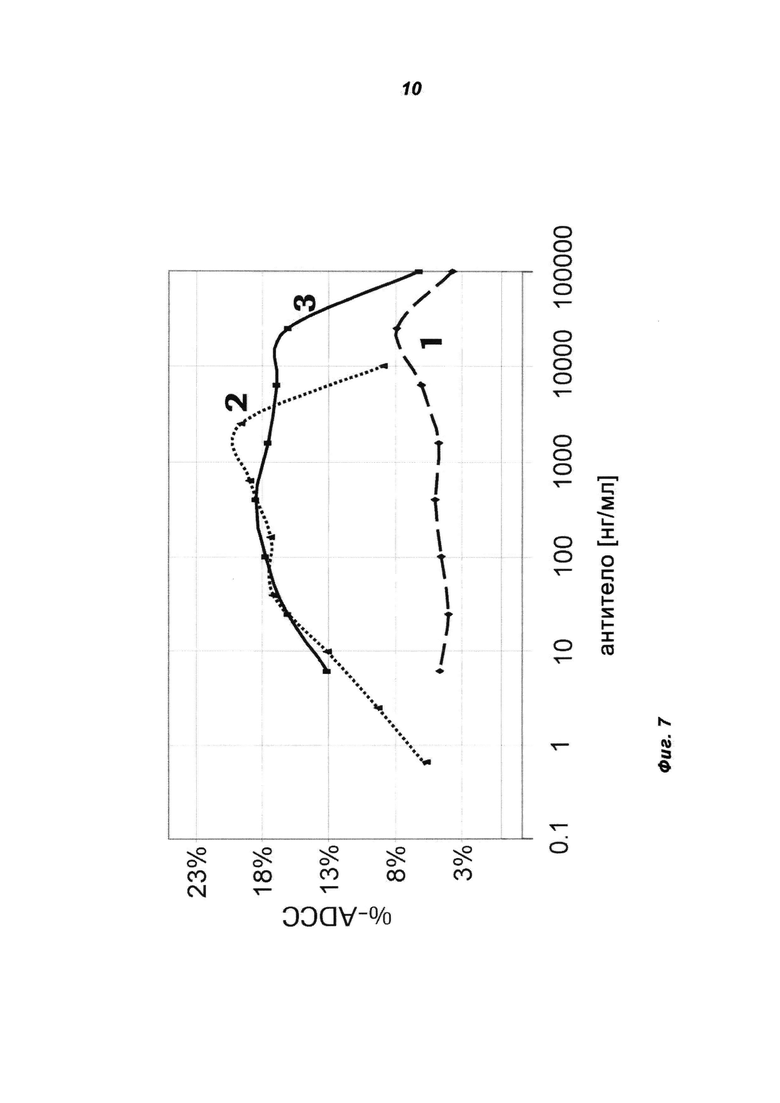
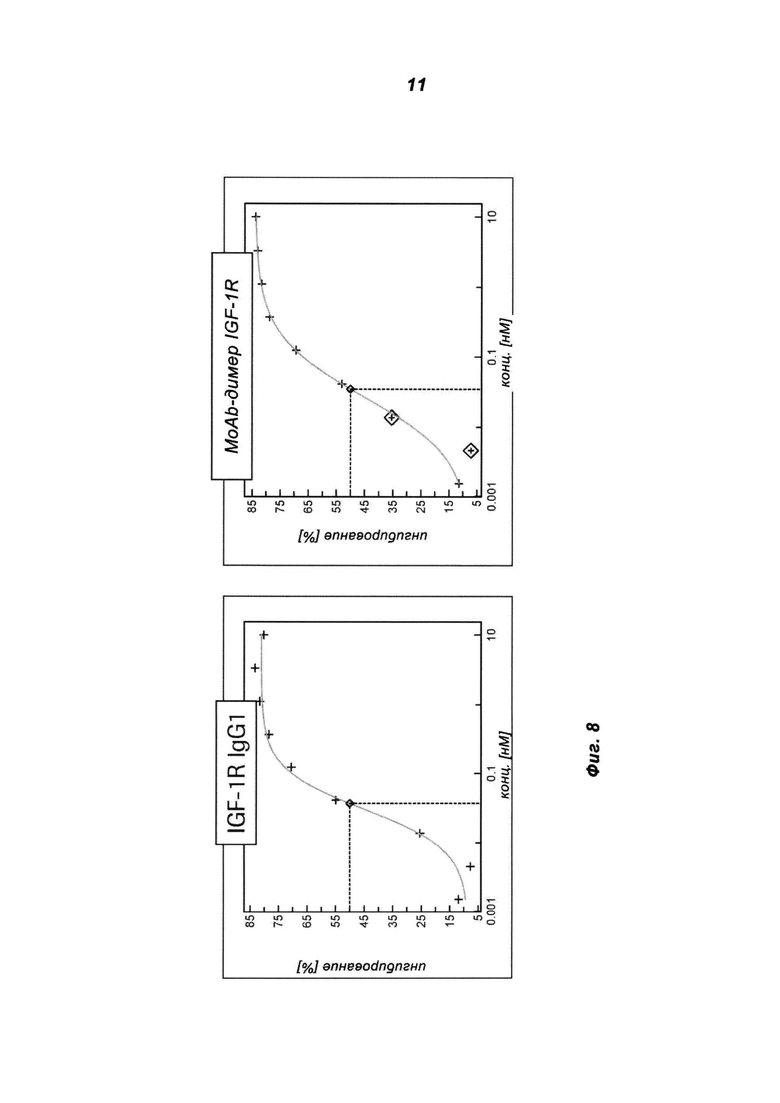
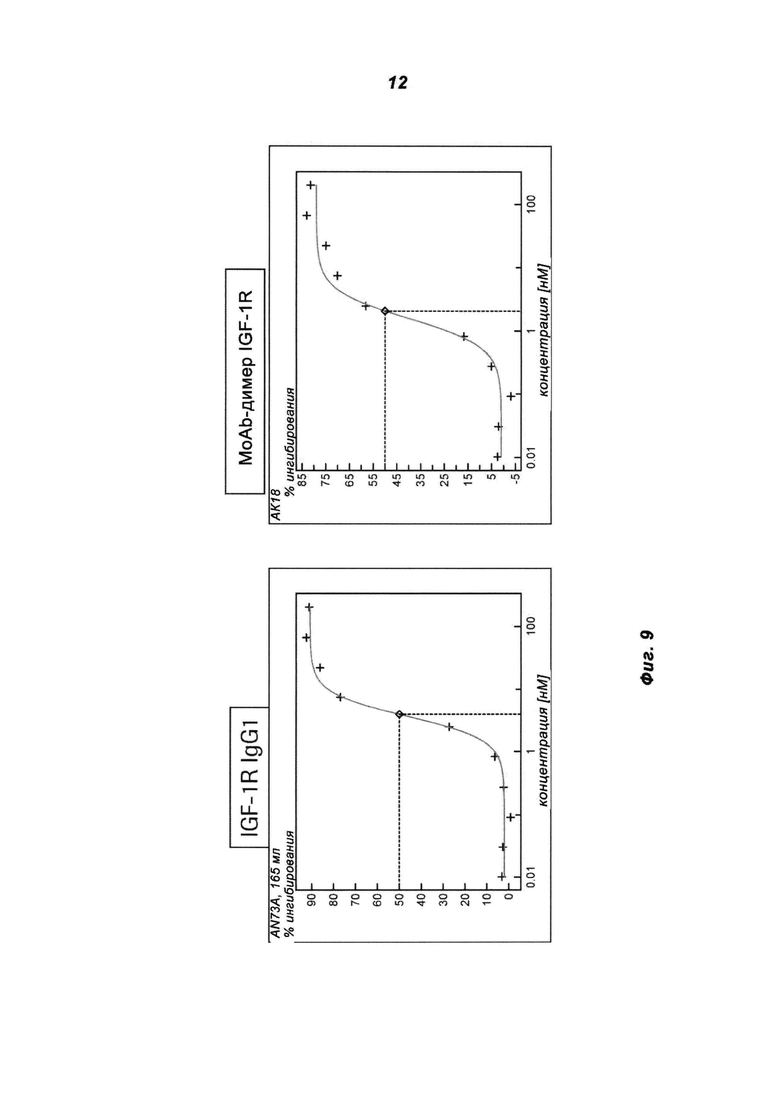
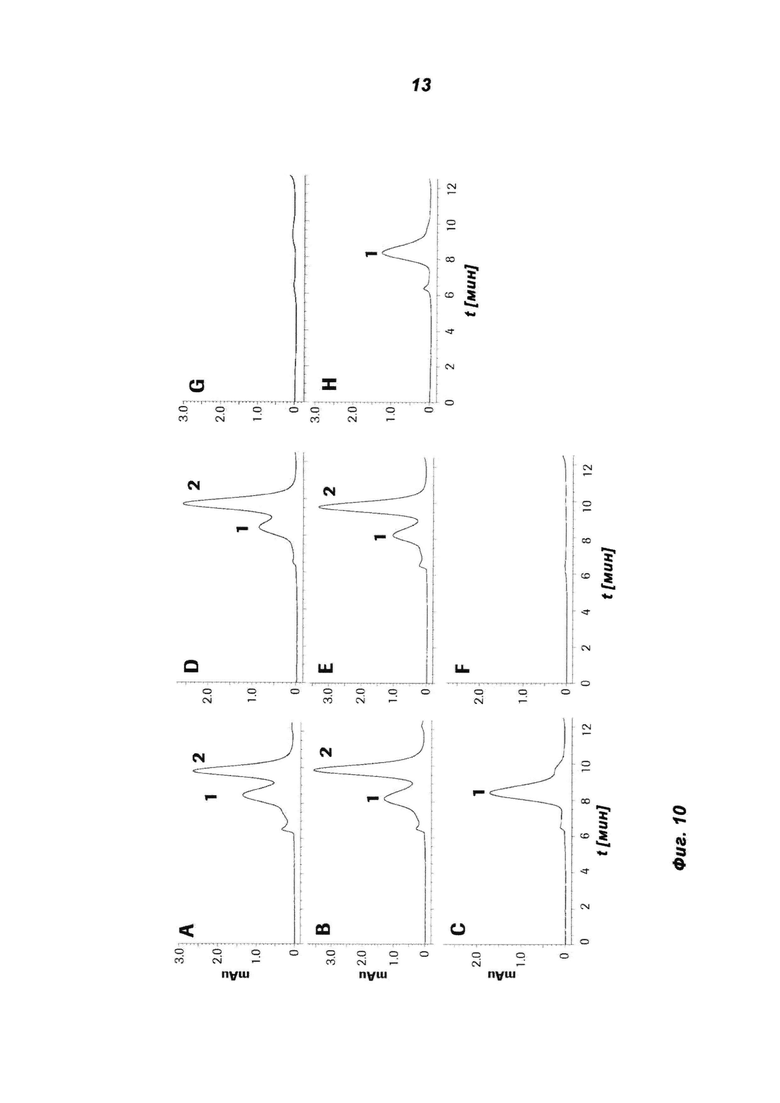
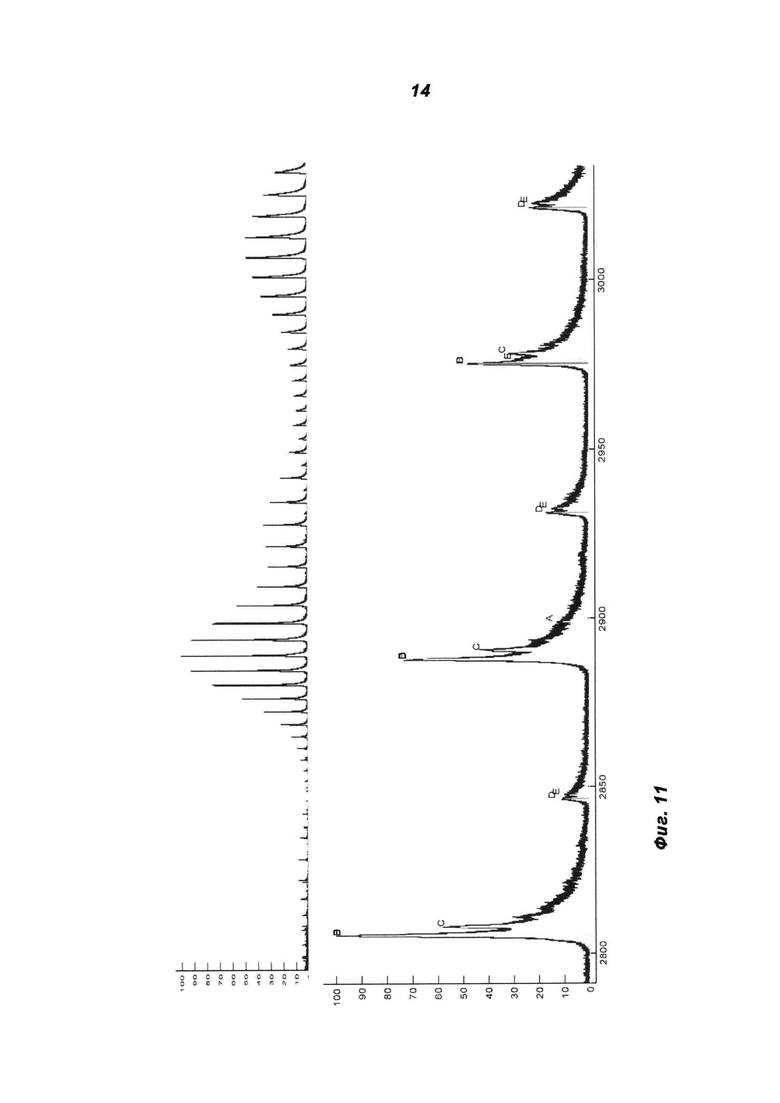
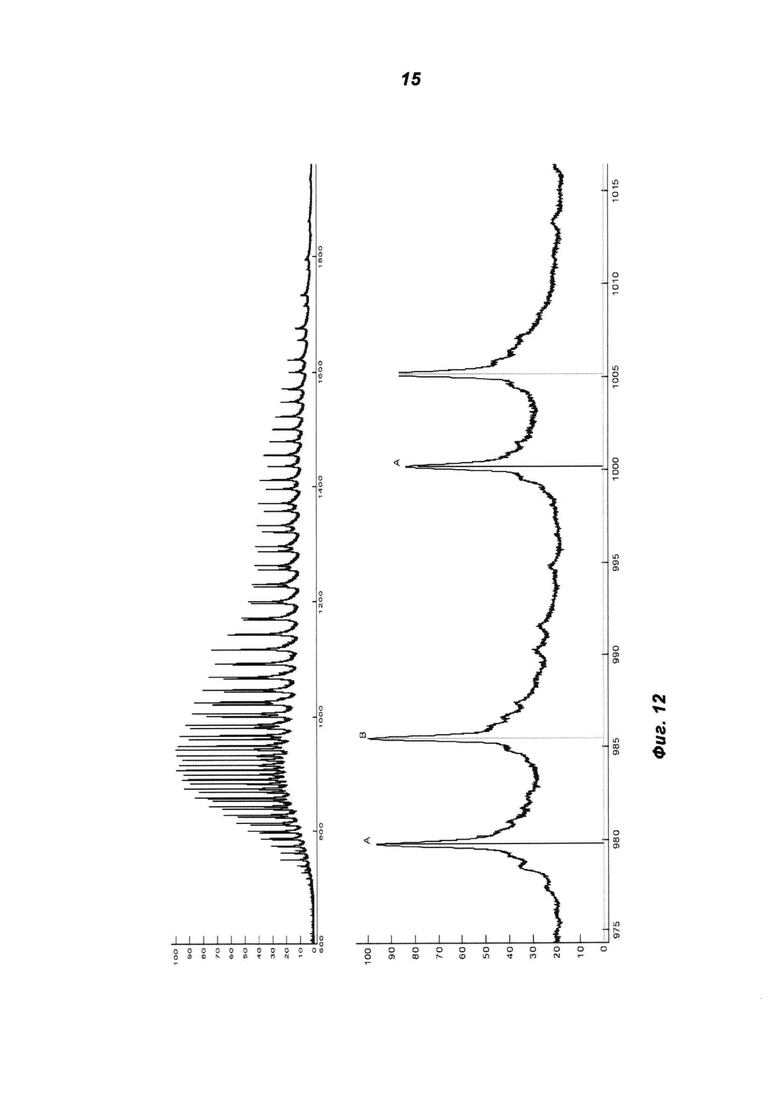
Изобретение относится к области иммунологии. Предложены способ получения антигенсвязывающего белка и клетка-хозяин для его получения. Антигенсвязывающий белок, получаемый способом по настоящему изобретению, содержит: а) две модифицированные тяжелые цепи антитела, в которых VH каждой тяжелой цепи заменен на VL указанного антитела и которые связаны друг с другом через свои СН3-домены Fc-части; и б) две модифицированные тяжелые цепи антитела, в которых СН1 каждой тяжелой цепи заменен на CL указанного антитела и которые связаны друг с другом через свои СН3-домены Fc-части; причем VL-домены тяжелых цепей а) связаны с VH-доменами тяжелых цепей б), а СН1-домены тяжелых цепей а) связаны с CL-доменами тяжелых цепей б). Получаемый антигенсвязывающий белок проявляет повышенную антителозависимую клеточноопосредованную цитотоксичность (ADCC). 2 н. и 12 з.п. ф-лы, 12 ил., 5 табл., 10 пр.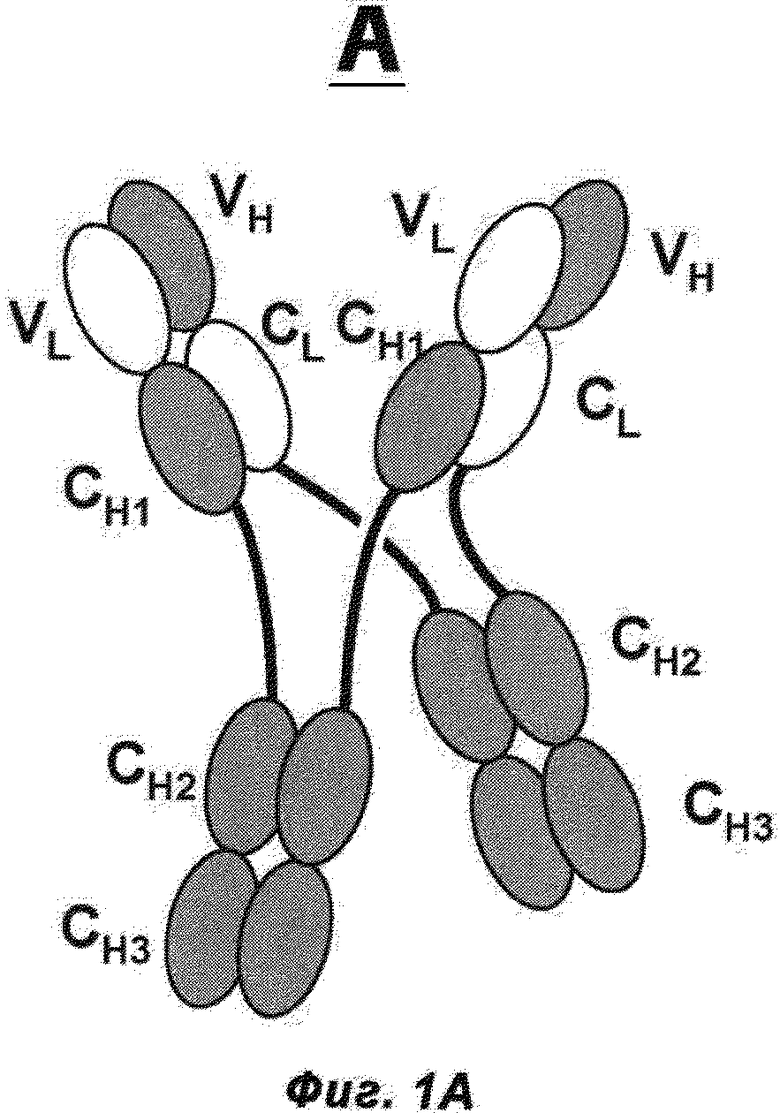
1. Способ получения антигенсвязывающего белка, где антигенсвязывающий белок содержит:
а) две модифицированные тяжелые цепи антитела, которое специфически связывается с антигеном, где VH каждой тяжелой цепи заменен на VL указанного антитела, причем указанные модифицированные тяжелые цепи связаны друг с другом через свои СН3-домены Fc-части;
б) две модифицированные тяжелые цепи указанного антитела, где СН1 каждой тяжелой цепи заменен на CL указанного антитела, причем указанные модифицированные тяжелые цепи связаны друг с другом через свои СН3-домены Fc-части;
и где VL-домены тяжелых цепей а) связаны с VH-доменами тяжелых цепей б), и СН1-домены тяжелых цепей а) связаны с CL-доменами тяжелых цепей б);
и где домены СН2-СН3 а) и б) могут иметь разный изотип;
и способ включает стадии:
i) трансформации клетки-хозяина экспрессионными векторами, содержащими молекулы нуклеиновых кислот, кодирующие
- модифицированную тяжелую цепь антитела, которое специфически связывается с антигеном, где VH-домен заменен на VL-домен указанного антитела; и
- модифицированную тяжелую цепь указанного антитела, где СН1-домен заменен на CL-домен указанного антитела,
ii) культивирования клетки-хозяина в условиях, обеспечивающих синтез указанной антигенсвязывающей белковой молекулы; и
iii) выделения указанной антигенсвязывающей белковой молекулы из указанной культуры.
2. Способ по п.1, который характеризуется тем, что в составе антигенсвязывающего белка СН3-домены Fc-части модифицированных тяжелых цепей а) и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют один и тот же изотип.
3. Способ по п.2, который характеризуется тем, что в составе антигенсвязывающего белка СН2- и СН3-домены Fc-части модифицированных тяжелых цепей а) и СН2- и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют один и тот же изотип.
4. Способ по п.3, который характеризуется тем, что в составе антигенсвязывающего белка СН2- и СН3-домены Fc-части модифицированных тяжелых цепей а) и СН2- и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют изотип IgG.
5. Способ по п.4, который характеризуется тем, что в составе антигенсвязывающего белка СН2- и СН3-домены Fc-части модифицированных тяжелых цепей а) и СН2- и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют изотип IgG1.
6. Способ по п.1, который характеризуется тем, что антигенсвязывающий белок содержит:
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №1; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №2;
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №3; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №4; или
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №5; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №6.
7. Способ по п.5, который характеризуется тем, что в составе антигенсвязывающего белка
либо две модифицированные тяжелые цепи а),
либо две модифицированные тяжелые цепи б)
также модифицированы путем аминокислотных замен S364G, L368F, D399K и K409D (где аминокислотные позиции нумеруются в соответствии с индексом ЕС Кабата).
8. Способ по п.1, который характеризуется тем, что антигенсвязывающий белок содержит:
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №1; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №2;
где либо две модифицированные тяжелые цепи а),
либо две модифицированные тяжелые цепи б)
также модифицированы путем аминокислотных замен S364G, L368F, D399K и K409D (где аминокислотные позиции нумеруются в соответствии с индексом ЕС Кабата);
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №3; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №4;
где либо две модифицированные тяжелые цепи а),
либо две модифицированные тяжелые цепи б)
также модифицированы путем аминокислотных замен S364G, L368F, D399K и K409D (где аминокислотные позиции нумеруются в соответствии с индексом ЕС Кабата);
или
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №5; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №6,
где либо две модифицированные тяжелые цепи а),
либо две модифицированные тяжелые цепи б)
также модифицированы путем аминокислотных замен S364G, L368F, D399K и K409D (где аминокислотные позиции нумеруются в соответствии с индексом ЕС Кабата).
9. Способ по п.1, который характеризуется тем, что в составе антигенсвязывающего белка СН3-домены Fc-части модифицированных тяжелых цепей а) и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют разные изотипы.
10. Способ по п.9, который характеризуется тем, что в составе антигенсвязывающего белка
СН3-домены Fc-части модифицированных тяжелых цепей а) имеют изотип IgG1;
и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют изотип IgA.
11. Способ по п.10, который характеризуется тем, что антигенсвязывающий белок содержит:
а) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №7; и
б) две модифицированные тяжелые цепи, содержащие аминокислотную последовательность SEQ ID №4.
12. Способ по п.10, который характеризуется тем, что в составе антигенсвязывающего белка
СН2- и СН3-домены Fc-части модифицированных тяжелых цепей а) имеют изотип IgG1;
и СН2- и СН3-домены Fc-части модифицированных тяжелых цепей б) имеют изотип IgA.
13. Способ по любому из пп.1-11, который характеризуется тем, что в составе антигенсвязывающего белка СН2-домены Fc-частей а) и б) имеют изотип IgG1 и антигенсвязывающий белок афукозилирован с количеством фукозы 80% или менее от общего количества олигосахаридов (сахаров) на Asn297 и имеет изотип IgG1 человека.
14. Клетка-хозяин для получения антигенсвязывающего белка, где антигенсвязывающий белок содержит:
а) две модифицированные тяжелые цепи антитела, которое специфически связывается с антигеном, где VH каждой тяжелой цепи заменен на VL указанного антитела, причем указанные модифицированные тяжелые цепи связаны друг с другом через свои СН3-домены Fc-части;
б) две модифицированные тяжелые цепи указанного антитела, где СН1 каждой тяжелой цепи заменен на CL указанного антитела, причем указанные модифицированные тяжелые цепи связаны друг с другом через свои СН3-домены Fc-части;
и где VL-домены тяжелых цепей а) связаны с VH-доменами тяжелых цепей б), и СН1-домены тяжелых цепей а) связаны с CL-доменами тяжелых цепей б);
где клетка-хозяин содержит экспрессионные векторы, содержащие молекулы нуклеиновых кислот, кодирующие:
- модифицированную тяжелую цепь антитела, которое специфически связывается с антигеном, где VH-домен заменен на VL-домен указанного антитела; и
- модифицированную тяжелую цепь указанного антитела, где СН1-домен заменен на CL-домен указанного антитела.
| GUNASEKARAN KANNAN et al | |||
| "Enhancing Antibody Fc Heterodimer Formation through Electrostatic Steering Effects | |||
| APPLICATIONS TO BISPECIFIC MOLECULES AND MONOVALENT IgG." Journal of Biological Chemistry, 2010, 285(25): 19637-19646 | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| MORRISON SHERIE L | |||
| et al | |||
| "Variable region domain exchange influences the functional properties of IgG." The Journal of Immunology, 1998, 160(6): 2802-2808 | |||
| МОДИФИЦИРОВАННОЕ АГОНИСТИЧЕСКОЕ АНТИТЕЛО | 2001 |
|
RU2295537C2 |

