Данная заявка претендует на приоритет заявок на английский патент №№1422511.4, 1512467.0, 1512595.8 и 1521987.6 (поданных 17го декабря 2014 года, 16го июля 2015 года, 17го декабря 2015 года и 14го декабря 2015, соответственно), полное содержание каждой из которых включено в данное описание посредством отсылки для всех целей.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Данное изобретение относится к области редактирования РНК, при этом нуклеотидная последовательность целевой РНК модифицируется, например, с целью корректирования мутации.
УРОВЕНЬ ТЕХНИКИ
Редактирование РНК представляет собой естественный процесс, посредством которого эукариотические клетки изменяют последовательность молекул РНК, причем изменение часто является сайт-специфическим и точным, тем самым увеличивая репертуар кодируемых геномом РНК на несколько порядков. РНК, кодирующая ферменты, была описана для эукариотических организмов как в царстве животных, так и в царстве растений, и эти процессы играют важную роль в контроле клеточного гомеостаза у многоклеточных организмов, начиная от простейших форм жизни, например, таких как Caenorhabditis elegans, до человека. Примерами редактирования РНК являются превращение аденозина в инозин и цитидина в уридин с помощью ферментов, называемых аденозиндезаминаза и цитидиндезаминаза, соответственно. Наиболее активно изучаемой системой редактирования РНК является фермент аденозиндезаминаза. Аденозиндезаминаза представляет собой многодоменный белок, содержащий узнающий домен и каталитический домен. Узнающий домен распознает специфическую последовательность или конформацию дцРНК, тогда как каталитический домен превращает аденозин в инозин в ближайшем более или менее предопределенном положении целевой РНК посредством дезаминирования нуклеинового основания. Механизмы трансляции клетки обусловливают считывание инозина как гуанина, это означает, что если редактируемый аденозин находится в кодирующей области мРНК или пре-мРНК, его можно перекодировать в белковую последовательность. Превращение А в I может происходить также в 5' не-кодирующей последовательности целевой мРНК, создавая новые сайты инициации трансляции 5' (выше) относительно исходного сайта инициации трансляции, что приводит к белкам с удлиненным N-концом. Кроме того превращения А в I могут происходить в элементах сплайсинга в интронах или экзонах в пре-мРНК, тем самым изменяя паттерн сплайсинга. В результате такого редактирования РНК экзоны могут включаться или пропускаться. Аденозиндезаминазы являются частью обширного семейства ферментов, называемых РНК-зависимыми (РНК-овыми) аденозиндезаминазами (ADAR), которые включат человеческие дезаминазы hADAR1, hADAR2 и hADAR3.
Применение олигонуклеотидов для редактирования целевой РНК известно в уровне техники, см., например, Montiel-Gonzalez et al. (Proceedings of the National Academy of Sciences 2013 November 5, 2013, vol. 110, no. 45, pp. 18285-18290). Авторы описали адресное редактирование целевой РНК с использованием рекомбинантного гибридного белка, содержащего аденозиндезаминазный домен белка hADAR1, слитый с так называемым В-бокс связывающим доменом белка N бактериофага лямбда. Природный узнающий домен hADAR1 удаляли с целью исключить свойства природного ADAR узнавать субстрат и заменить его на В-бокс узнающий домен лямбда N-белка. В-бокс представляет собой короткий фрагмент РНК из 17 нуклеотидов, который распознается В-бокс-связывающим доменом N-белка. Авторы создали антисмысловой олигонуклеотид, содержащий участок гидовой РНК (гРНК), комплементарный целевой последовательности, для редактирования слитого с В- боксом участка с целью специфического узнавания последовательности гибридным белком N-домен дезаминазой. Авторы наглядно показали, что олигонуклеотид 'руководящей' РНК точно направляет гибридный белок аденозиндезаминазы на сайт-мишень, что приводит к гРНК-направленному сайт-специфическому редактированию А в I целевой РНК.
Недостатком предлагаемого способа является необходимость в гибридном белке, состоящем из В-бокс-связывающего домена N-белка бактериофага лямбда, слитого генной инженерии с аденозиндезаминазным доменом процессированного природного белка ADAR. Для этого требуется, чтобы клетки-мишени либо трансдуцировались с использованием гибридного белка, что представляет собой основную проблему, либо чтобы клетки-мишени трансфицировались с использованием нуклеотидной конструкции, кодирующей созданный методами рекомбинантной ДНК гибридный белок аденозиндезаминазы, для экспрессии в клетках-мишенях. Последнее является немалым препятствием, когда редактирование надлежит осуществлять в многоклеточном организме, например, при лечении заболевания человека.
Vogel et al. (Angewandte Chemie. Int. Ed. 2014, 53, 6267-71) раскрывают редактирование РНК, кодирующих eCFP и Фактор V Лейдена, с использованием бензилгуанин-замещенной РНК-гида и рекомбинантного гибридного белка, содержащего аденозиндезаминазные домены ADAR1 или 2, связанные методами рекомбинантной ДНК с доменом SNAP-хвоста (полученная рекомбинантными методами О6-алкилгуанин-ДНК-алкилтрансфераза). Хотя полученный методами рекомбинантной ДНК искусственный гибридный белок дезаминазы можно нацеливать на заданный сайт редактирования в целевых РНК в клетках Hela в культуре, используя ковалентно связанную (через бензилгуанин) РНК-гид, эта система имеет недостатки, сходные с недостатками рекомбинантных (генно-инженерных) ADARs, описанных выше, тем, что неясно, как применять систему, чтобы не надо было сначала генетически модифицировать ADAR, а затем трансфицировать или трансдуцировать клетки, содержащие целевую РНК, чтобы получать клетки с этим генно-инженерным белком. Ясно, что эту систему не просто адаптировать для применения на людях, например, в плане лечения.
Другой способ редактирования, в котором используются олигонуклеотиды, известен как CRISPR/Cas9 система, но этот комплекс редактирования РНК воздействует на ДНК. Последний способ страдает тем же недостатком, что и системы на основе рекомбинантных ADAR, описанные выше, поскольку в нем требуется совместная доставка к клетке-мишени CRISPR/Cas9 фермента или экспрессионной конструкции, кодирующей его, совместно с 'руководящим' олигонуклеотидом (гидом).
Следовательно, остается необходимость в новых методах, в которых могут использоваться эндогенные клеточные пути для редактирования эндогенных нуклеиновых кислот в клетках млекопитающих, даже в организме в целом, которые не сопровождаются проблемами, ассоциированными с методами из уровня техники.
РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение устраняет недостатки способов согласно известному уровню техники, предоставляя адресный подход к редактированию РНК с применением олигонуклеотидных конструкций, содержащих нацеливающий участок, специфический к целевой нуклеотидной последовательности, подлежащей редактированию, и рекрутирующий участок, способный связывать и рекрутировать редактирующую нуклеотидную структуру, естественно присутствующую в клетке. Функция рекрутирующего участка олигонуклеотидной конструкции состоит в том, чтобы селективно, с достаточной аффинностью, связываться с редактирующей РНК структурой, эндогенной к и свойственной клетке, перенацеливая такую структуру на заданный сайт-мишень с помощью нацеливающего участка олигонуклеотидной конструкции по изобретению, тем самым инициируя редактирование заданных нуклеотидных остатков в области целевой РНК, соответствующей нацеливающему участку олигонуклеотидной конструкции.
Нацеливающий участок олигонуклеотидной конструкции обычно содержит антисмысловую олигонуклеотидную последовательность, комплементарную сайту-мишени в подлежащей редактированию последовательности РНК. Один предпочтительный вариант такого адресного подхода к редактированию целевой РНК представляет собой олигонуклеотидную конструкцию, содержащую два участка, нацеливающий участок, содержащий антисмысловую последовательность, комплементарную целевой последовательности РНК, и рекрутирующий участок, содержащий последовательность, узнающую фермент для редактирования РНК.
Рекрутирующий участок может содержать дцРНК в виде шпилечной структуры со стеблем и петлей. Шпилечная структура может находиться выше (5') или ниже (3') нацеливающего участка (предпочтительно, выше). Или же рекрутирующий участок олигонуклеотидной конструкции может разрывать нацеливающий участок таким образом, что часть нацеливающего участка лежит выше (5') рекрутирующего участка, а часть нацеливающего участка лежит ниже (3') рекрутирующего участка, что заставляет рекрутирующий участок олигонуклеотидной конструкции образовывать петлю после отжига этой олигонуклеотидной конструкции с целевой РНК.
Согласно другому варианту рекрутирующий участок содержит сегмент дцРНК, имитирующий РНК-последовательность, т.е. является идентичным или сходным по структуре РНК-последовательности, которая заведомо редактируется природными структурами, редактирующими РНК. Эта РНК-последовательность, заведомо являющаяся природным субстратом для редактирования РНК, содержат сегмент дцРНК, предпочтительно, включающий один РНК-сегмент, который складывается (назад) сам с собой за счет спаривания с комплементарным нуклеиновым основанием, при этом образуется шпилечная структура или структура типа "стебель-петля". Два примера известных редактируемых РНК-последовательностей, которые охарактеризованы очень подробно, находятся в В-субъединице глутаматных рецепторов (GluR-B) AMP А (3-амино-3-гидрокси-5-метил-4-изоксазол пропионовой кислоты) подтипа. Эта модельная система содержит два часто редактируемых сайта, причем ДНК-кодируемый AGA редактируется в IGA, что приводит к замене аргинина на глицин (R/G сайт) и отдельной замене глутамина на аргинин (Q/R сайт). Известно, что GluR-B (R/G) сайт содержит структуру типа стебель-петля, состоящую из 71 нуклеотида, содержащую 3 пары некомплементарных оснований (мисмэтчи), 2 А⋅С и одну G⋅U пары оснований с нестрогим соответствием. Интересно отметить, что петля состоит из высококонсервативной структуры "пентапетли" GCUAA, которая соответствует филогенетически консервативной последовательности GCUMA, где М обозначает А или С (Aruscavage P.J. & Bass B.L. RNA. 2000; 6: 257-269). По-видимому, имеется некоторая область редактирования двух остатков аденозина с нестрогим соответствием, при этом эффективность повышается, когда основание, противостоящее редактируемому аденозину (напротив него) выбрано из цитидина или уридина, причем цитидин является предпочтительным.
Эту структуру можно просто применять без изменений или ее можно адаптировать для использования в олигонуклеотидной конструкции по изобретению в качестве рекрутирующего участка, посредством уменьшения или увеличения числа пар нуклеиновых оснований неполного соответствия в структуре стебля, чтобы модифицировать специфичность редактирования и/или перенацелить редактирование на предпочтительный(-е) сайт(-ы) в целевой РНК-последовательности. Помимо этого, или в качестве альтернативы, сайт узнавания GluR-B для hADAR1 можно модифицировать, укорачивая 'стебель', но не отменяя полностью узнавание. Такое укорочение может быть удобно с точки зрения технологичности или стоимости продукта в перспективе и т.п.
Пример рекрутирующего участка GluR-B домена содержит последовательность: 5'-(AUANa)nUAUAACAAUAUgcuaaAUGUUGUUAUA(NbUAU)n-3', где Na и Nb, каждый, обозначает единичные нуклеотиды, которые могут представлять собой A, G, С или U, при условии, что Na и Nb образуют пару некомплементарных (неспаренных) оснований (мисмэтч) при образовании структуры 'стебель-петля', и n равно 1 или 0 (т.е. SEQ ID NOs: 6 и 7). Подходящий пример такого рекрутирующего участка включает эту последовательность, где n=1, с дополнительным удлинением в направлениях 5' и 3', например, где каждое удлинение содержит от 1 до 10 нуклеотидов (или более, например, от 1 до 20 нуклеотидов или более). Например, удлинение на 3 дополнительных нуклеотида в каждом направлении дает (SEQ ID NO: 23) 5'-GGAAUANaUAUAACAAUAUgcuaaAUGUUGUUAUANbUAUCCC-3', как видно в SEQ ID NOs: 20-22 (в которых Na=Nb=G). Это дополнительное удлинение может повысить эффективность корректировки.
Другим примером рекрутирующего участка на основе субстрата полноразмерного природного GluR-B рецептора является 5'-GUGGAAUANaUAUAACAAUAUgcuaaAUGUUGUUAUANbUAUCCCAC-3' (SEQ ID NO: 24; остатки в удлинении по сравнению с последовательностью в предыдущем абзаце подчеркнуты), применяемый ниже, в разделе Примеры. Рекрутирующий участок полноразмерного GluR-B рецептора в комбинации с нацеливающими участками для A1AT-транскрипта с мутацией G в А в положении 9989, ассоциированный с недостаточностью А1АТ (альфа-1-антитрипсина), описан в Примере 4. Рекрутирующий участок полноразмерного GluR-B рецептора в комбинации с нацеливающими участками для LRRK2 транскрипта с мутацией G2019S, ассоциированный с болезнью Паркинсона, описан в Примере 5.
Рекрутирующий участок может быть связан по 5' или по 3' концу нацеливающего участка, необязательно с помощью линкера "L", который содержит один или более нуклеотидов, олигопептид или другой химический линкер, такой как полиэтиленгликоль (PEG, ПЭГ).
Нацеливающий участок может содержать последовательность, комплементарную целевой РНК-последовательности, представленной общей формулой:
N1N2N3N4N5N6N7N8N9N10N11N12N13N14N15N16N17CN18N19N20 (SEQ ID NO: 8),
где N, от N1 до N20, в зависимости от комплементарной последовательности в целевой РНК-последовательности, каждый независимо, обозначают A, G, С или U, причем N4 предпочтительно образует пару некомплементарных (неспаренных) оснований (мисмэтч) с противостоящим нуклеотидом в целевой РНК-последовательности, когда нацеливающий участок связывается с целевой РНК-последовательностью, и N10 и N16 образуют пару оснований нестрогого соответствия с противостоящими нуклеотидами в целевой РНК-последовательности, когда нацеливающий участок связывается с целевой РНК-последовательностью, и С обозначает цитидин, противостоящий аденозину в целевой РНК-последовательности, который является мишенью для дезаминирования.
Пары мисмэтч (некомплементарных) оснований представляют собой пары оснований G-A, С-A, U-C, A-A, G-G, С-С, U-U. Парами оснований нестрогого соответствия являются пары оснований: G-U, I-U, I-A и I-C. Нацеливающий участок может иметь в длину более 20 нуклеотидов, не менее, чем 200 нуклеотидов или более, хотя полагают, что более 50 нуклеотидов не является обязательным, и предпочтительно, менее 40 нуклеотидов. Еще более предпочтительно, когда рекрутирующие участки имеют длину менее 30 нуклеотидов, предпочтительно, менее 25 нуклеотидов.
Предпочтительно, нацеливающий участок содержит 2'-O метальные группы в каждом положении, которое противостоит аденозину, когда нацеливающий участок связывается с целевой РНК-последовательностью, если этот аденозин в целевой РНК-последовательности не является мишенью редактирования. Вообще для защиты нацеливающего участка от деградации нуклеазами предпочтительно, чтобы все нуклеотиды содержали 2'-O-метильные группы, за исключением нуклеотида, противостоящего целевому аденозину и нуклеотидам, соседним с указанным противостоящим нуклеотидом (один 5' и один 3'), которым следует содержать 2'-ОН группы.
Согласно предпочтительному варианту рекрутирующий участок содержит последовательность ДНК: (CG)3N1-Nn(CG)3, где с N1 до Nn могут быть одинаковыми или различными и могут быть выбраны из гуанозина, аденозина, тимидина, цитидина и инозина, 'n' имеет значение от 2 до 20, предпочтительно, от 2 до 10, более предпочтительно, от 2 до 5, еще более предпочтительно, от 3 до 5, наиболее предпочтительно, 4 или 5. Таким образом, имеется три CG повтора, фланкирующие до 20 промежуточных нуклеотидов. Эта ДНК-последовательность способна образовывать структуру типа 'стебель-петля'. Согласно другому предпочтительному варианту рекрутирующий участок олигонуклеотидной конструкции представляет собой ДНК структуру, содержащую последовательность: (CG)3Tn(CG)3, где n обозначает целое число от 3 до 5, предпочтительно, 4, (CGCGCGTTTTCGCGCG; SEQ ID NO: 5). Эта ДНК-последовательность способна образовывать структуру типа 'стебель-петля'. Также в уровне техники было описано, что последовательность (CG)3T4(CG)3 в физиологических условиях принимает конформацию Z-ДНК и что эта Z-ДНК структура узнается и связывается с белком hADAR1 (FEBS letters 458:1 1999 Sep 10 pg 27-31). Как и вышеописанный рекрутирующий участок дцРНК, рекрутирующий участок этой Z-ДНК может лежать выше или ниже нацеливающего участка или разрывать нацеливающий участок, разделяя нацеливающий участок на 5' сегмент (выше) и на 3' сегмент (ниже), при этом рекрутирующий участок ДНК образует петлю, когда нацеливающие отжигаются связываются с целевой РНК. Согласно этому варианту цитидиновые основания представляют собой, предпочтительно, 5-метилцитидин, с целью снижения потенциальной иммуногенности, обусловленной CpG последовательностями.
Олигонуклеотиды по настоящему изобретению содержат нацеливающий участок (т.е. участок, который нацеливает олигонуклеотид на корректное положение в целевой РНК-последовательности) и рекрутирующий участок (т.е. участок, основная функция которого заключается в рекрутировании редактирующей структуры, например, ADAR, и который не обязательно является комплементарным, предпочтительно, не является комплементарным целевой РНК в области остатка(-ов) аденозина, который(-ые) является (являются) мишенью редактирования). Эта двухчастная структура четко отличает олигонуклеотиды по изобретению от известных олигонуклеотидов, например, таких как олигонуклеотиды, известные из уровня техники (см. например, Международные заявки WO 2014/011053, WO 2005/094370, и Woolf et al, 1995. PNAS USA 92, 8298-8302), которые по существу комплементарны мишени по всей своей длине и не содержат рекрутирующего участка (естественно, ни один из них не является комплементарным целевой РНК, но вместо этого обладает аффинностью к редактирующей структуре, как в настоящем изобретении). Следовательно, в предпочтительном варианте по настоящему изобретению предусматривается предоставление олигонуклеотидной конструкции для сайт-направленного редактирования нуклеотида в целевой РНК-последовательности в эукариотической клетке, причем указанная олигонуклеотидная конструкция содержит:
(a) нацеливающий участок, содержащий антисмысловую последовательность, комплементарную участку целевой РНК; и
(b) рекрутирующий участок, который не является комплементарным целевой RNA-последователъности и способен связывать и рекрутировать редактирующую РНК структуру, естественно присутствующую в указанной клетке и способную осуществлять редактирование указанного нуклеотида.
Согласно другому варианту в изобретении предусматривается олигонуклеотидная конструкция, содержащая:
(a) нацеливающий участок, содержащий антисмысловую последовательность, комплементарную участку целевой РНК; и
(b) рекрутирующий участок, способный образовывать внутримолекулярную структуру типа 'стебель-петля', связывать и рекрутировать редактирующую РНК структуру, естественно присутствующую в указанной клетке и способную осуществлять редактирование указанного нуклеотида.
Согласно еще одному варианту рекрутирующий участок может являться аптамером, выбранным для связывания с редактирующей структурой, присущей клетке. Методы селекции аптамеров хорошо известны в уровне техники. Аптамеры, которые связываются с редактирующей структурой, не вызывая отмены дезаминазной активности, можно выбрать в качестве рекрутирующего участка и без всяких сложностей связать с нацеливающим участком олигонуклеотидной конструкции согласно изобретению с помощью линкера любого типа обычной (фосфодиэфирной) или модифицированной (например, фосфоротиоатной или фосфородитиоатной) межнуклеозидной связью, пептидильной связью или другой химической связью, например, с использованием полиэтиленгликоля.
Согласно еще одному варианту изобретения антитело, фрагмент антитела, его связывающий домен или антитело семейства верблюдовых, которое связывается с редактирующей структурой, постоянно присутствующей в клетке, не вызывая отмены редактирующей активности, можно выбрать в качестве рекрутирующего участка и соединить с нацеливающим участком олигонуклеотидной конструкции согласно изобретению.
Термин "олигонуклеотидная конструкция" может относиться к одному олигонуклеотиду, комплексу из двух или более олигонуклеотидов (включая аптамер) с аффинностью друг к другу (антисмысловая комплементарность или иное) или комплексу олигонуклеотида и белкового связывающего участка (например, антитела, фрагмента антитела или связывающего домена), которые могут связываться непосредственно или с помощью ПЭГ (PEG) или другого линкера.
Таким образом, в настоящем изобретении предусматриваются олигонуклеотидные конструкции и способы сайт-специфического редактирования целевых РНК-последовательностей в клетке без необходимости трансдуцировать или трансфицировать клетку с использованием рекомбинантных (генно-инженерных) редактирующих ферментов. Благодаря дизайну олигонуклеотидных конструкций редактирующие структуры, такие как ADARs, рекрутируются и нацеливаются на сайты редактирования, выбранные экспериментатором. Эти олигонуклеотидные конструкции и способы по изобретению можно просто использовать для осуществления изменений в целевых РНК-последовательностях, например, для реверсии мутаций, которые способствуют заболеванию или вызывают заболевание, тем самым ослабляя симптомы заболевания. Известно, что структуры, редактирующие РНК, очень эффективно редактируют их субстраты, с частотой, намного превосходящей опосредуемую олигонуклеотидами репарацию ДНК или РНК. Это дает большие преимущества при лечении заболевания.
Нацеливающий участок и рекрутирующий участок в олигонуклеотидной конструкции согласно изобретению могут непосредственно прилегать друг к другу. Или же нацеливающий участок и рекрутирующий участок могут быть связаны ковалентно с помощью линкера. Линкер может включать неспецифические нуклеотидные остатки (неспецифические в том смысле, что они необязательно комплементарны целевой РНК-последовательности, а также не обладают аффинностью к редактирующей структуре, постоянно находящейся в клетке), (олиго)пептидные линкеры или другие химические линкеры. Согласно еще одному варианту нацеливающий участок и рекрутирующий участок могут предусматриваться в виде двух разделенных - т.е. не связанных ковалентной связью - олигонуклеотидных последовательностей, содержащих 'антисмысловую комплементарность', способную образовывать дцРНК или гибридную структуру ДНК : РНК. Образование такой дцРНК или гибридной олигонуклеотидной конструкции может происходить до введения в клетку или субъекту, подлежащему лечению, или после введения двух отдельных олигонуклеотидов, in vitro или in vivo, например, субъекту, подлежащему лечению.
В изобретении предусматривается способ введения изменения в целевую РНК-последовательность в эукариотической клетке, предпочтительно, в клетке млекопитающего, включающий стадии: (i) введение в указанную клетку олигонуклеотидной конструкции, содержащей нацеливающий участок, который включает последовательность в достаточной степени комплементарную к целевой РНК-последовательности, чтобы связываться посредством спаривания нуклеиновых оснований с указанной целевой РНК, и рекрутирующий участок, содержащий последовательность, узнаваемую редактирующей структурой РНК, которая естественно присутствует в указанной эукариотической клетке, предпочтительно, клетке млекопитающего; (ii) выдерживание в течение времени, достаточного для того, чтобы структура, редактирующая РНК, осуществила реакцию редактирования целевой РНК-последовательности; и (iii) идентификация наличия изменения в РНК-последовательности. Согласно предпочтительному варианту реакция редактирования проводится указанной редактирующей структурой на одном или более нуклеиновых оснований в области перекрывания между целевой РНК-последовательностью и нацеливающим участком олигонуклеотидной конструкции. Согласно предпочтительному варианту нацеливающий участок олигонуклеотидной конструкции содержит мисмэтч, противостоящий нуклеиновому(-ым) основанию(-ям), подлежащему(-им) редактированию. Согласно другому предпочтительному варианту реакция редактирования включает превращение А в I посредством дезаминирования нуклеинового основания аденозина в целевой РНК-последовательности. Согласно этому последнему способу предпочтительно, чтобы олигонуклеотидная конструкция содержала С напротив подлежащего редактированию остатка аденозина. Согласно другому предпочтительному способу реакция редактирования представляет собой превращение С в U посредством дезаминирования цитидинового нуклеинового основания; согласно последнему способу предпочтительно, чтобы нацеливающий участок олигонуклеотидной конструкции содержал А напротив С в подлежащей редактированию целевой РНК-последовательности.
В изобретении также предусматривается способ редактирования мутантной CFTR целевой РНК-последовательности в человеческой клетке, включающий стадии: (i) предоставление указанной клетке олигонуклеотидной конструкции, содержащей нацеливающий участок, комплементарный CFTR целевой РНК-последовательности, и рекрутирующий участок, способный рекрутировать hADAR редактирующую структуру; и (ii) выдерживание в течение времени, достаточного для того, чтобы редактирующая структура hADAR осуществила редактирование нуклеиновых оснований в или близ области перекрывания между целевой РНК-последовательностью и нацеливающим участком олигонуклеотидной конструкции. Согласно предпочтительному варианту по изобретению мутантная CFTR целевая РНК (пре-мРНК или мРНК) содержит мутацию G551D, и реакция редактирования вызывает превращение аденозина в инозин, тем самым вызывая обращение мутации G551D (обратная мутация) в указанной целевой РНК-последовательности. Для аспарагиновой кислоты (D) имеется два кодона: GAU и GAC. Следовательно, больной муковисцидозом с мутацией G551D может иметь либо мутацию GAU или GAC в положении, соответствующем ко дону 551 белка CFTR. Дезаминирование А во втором положении кодона приведет к образованию I, который будет считываться в процессе трансляции, как G. Следовательно, дезаминирование А во втором положении мутантного кодона создает GIU или GIC, соответственно, которые фактически считываются как GGU и GGC. Как GGU, так и GGC кодируют глицин, в результате чего РНК редактирование обоих мутантных ко донов G551D с помощью аденозиндезаминазы даст нужный триплет, кодирующий глицин, эффективно обращая мутантный CFTR белок в нормальный.
Специалисту в данной области техники будет понятно, что мутация G551D используется лишь в качестве примера и никоим образом не ограничивает объем изобретения. Существуют буквально тысячи генетических заболеваний, вызванных заменами одной пары оснований, которые поддаются реверсии с применением олигонуклеотидных конструкций и способов по изобретению, посредством рекрутирования либо дезаминазы, например, такой как аденозиндезаминаза, подробно описанная в данной заявке, либо цитидиндезаминазы.
Рекрутирование цитидиндезаминазы в сайт-мишень работает таким же образом, как и в случае аденозиндезаминаз hADAR1 и hADAR2. Однако у цитидиндезаминаз другие требования к связыванию, и они узнают другие структуры в целевых РНК-последовательностях, которые определяют редактирование цитидина. Особенно хорошо изученной цитидиндезаминазой является человеческая Apobec1. Основной принцип редактирования РНК с использованием олигонуклеотидной конструкции- нацеливание на сайт редактирования и рекрутирование резидентной естественно присутствующей редактирующей структуры остается таким же для цитидиндезаминаз и является частью раскрываемого и заявленного в данном описании изобретения.
ОСУЩЕСТВЛЕНИЕ ИЗОБРЕТЕНИЯ
Олигонуклеотидные конструкции по изобретению уникальны тем, что они совмещают две важные функции: они связываются с естественно присутствующими структурами, редактирующими РНК, с определенной аффинностью и они связываются посредством комплементарного спаривания (Уотсон-Крик) антисмысловых оснований с сайтом в целевой РНК-последовательности, подлежащим редактированию, тем самым рекрутируя структуры, редактирующие РНК, к сайту редактирования. "Естественно присутствующая" структура присутствует в клетке без необходимости предшествующего вмешательства человека. Следовательно, процессированный или рекомбинантный фермент (например, такой как ферменты, описанные в публикациях Montiel-Gonzalez et al. и Vogel et al.) не является естественно присутствующим в клетке; эти ферменты могут присутствовать в клетке, но только после человеческого вмешательства (трансдукции или трансфекции). Таким образом, данное изобретение можно осуществлять с использованием РНК-редактирующих структур дикого типа, эндогенных для клетки.
Понятно, что такое рекрутирование не обязательно должно быть количественным, при котором все РНК-редактирующие структуры, резидентные в клетке, будут рекрутироваться для наиболее эффективного редактирования целевой РНК-последовательности. Предполагается, и даже считается желательным, чтобы содержание резидентных редактирующих структур оставалось подходящим для того, чтобы задействовать свои натуральные субстраты. Кроме того, согласно некоторым вариантам, например, тем, в которых рекрутирующий участок олигонуклеотидной конструкции представляет собой дцРНК-последовательность, содержащую аденозины, каталитический домен редактирующей РНК структуры после его рекрутирования олигонуклеотидной конструкцией может действовать на рекрутирующий участок олигонуклеотидной конструкции, а также на редактирующий субстрат на участке дцРНК, созданном путем связывания нацеливающего участка с комплементарной последовательностью в целевой РНК-последовательности. В любом варианте это не является проблемой, поскольку могут быть случаи применения, когда желательно дополнительное редактирование. В тех случаях, когда следует избегать дополнительного редактирования, нацеливающий участок можно полностью модифицировать химическими методами, например, вводя во все нуклеотиды 2'-O-метилированный сахар, за исключением нуклеотида(-ов), противостоящего(-их) целевому(-ым) остатку(-ам) аденозина, и двух нуклеотидов (один 5' и один 3'), фланкирующих каждый нуклеотид, противостоящий целевому аденозину. Вообще, аденозин в целевой РНК можно защищать от редактирования, вводя в противостоящий нуклеотид 2'-ОМе группу или предусматривая в качестве противостоящего основания гуанин или аденин, поскольку эти два нуклеиновых основания способны предотвращать редактирование противостоящего аденозина.
Олигонуклеотидные конструкции
Рекрутирующий участок олигонуклеотидных конструкций по изобретению характеризуется дцРНК или дцДНК структурой. Одним из способов создания двухцепочечной олигонуклеотидной структуры в единичной молекуле является создание палиндромной последовательности, способной раскручиваться по меньшей мере на протяжении части своей длины. Появление таких структур типа 'стебель-петля' может быть обусловлено (1) искусственными РНК-последовательностями, способными образовывать структуру типа 'стебель-петля', (2) искусственными ДНК-последовательностями, способными образовывать структуру типа 'стебель-петля', (3) РНК-последовательностями из известных РНК-субстратов для редактирующих структур, резидентных в клетке. Например, олигонуклеотидная конструкция по изобретению может содержать рекрутирующий участок с последовательностью, аналогичной последовательности природного сайта узнавания для редактирующей активности, или он может имитировать сайт узнавания подобно аптамеру. Каждый из этих вариантов более подробно будет описан ниже. Помимо этого, специалисты в данной области могут создать нацеливающий (проектный) и рекрутирующий участки на основе каждого из подробно описанных вариантов. Способы создания и тестирования структур нуклеиновых кислот, обладающих аффинностью к белкам, собственно, хорошо известны в уровне техники.
Нацеливающий участок олигонуклеотидной конструкции по изобретению должен иметь достаточную область перекрывания с сайтом-мишенью и быть комплементарным этому сайту, чтобы создать возможность последовательность-специфической гибридизации олигонуклеотидной конструкции с целевой РНК-последовательностью. Протяженность и степень перекрывания может меняться от мишени к мишени, но средний специалист в данной области может их определить обычным способом. В целом более длинные последовательности предусматривают более высокую степень специфичности -и, следовательно, меньше побочных эффектов, например, за счет неспецифического связывания - и более прочную связь с сайтом-мишенью. Нацеливающий участок олигонуклеотидной конструкции по изобретению обычно должен иметь в длину более 10 нуклеотидов, предпочтительно, более 11, 12, 13, 14, 15, 16, еще более предпочтительно, более 17 нуклеотидов. Нацеливающий участок, предпочтительно, короче 200 нуклеотидов, более предпочтительно, короче 100 нуклеотидов, еще более предпочтительно, короче 50 нуклеотидов, еще более предпочтительно, 25 или менее нуклеотидов.
Согласно одному варианту в изобретении предусматривается олигонуклеотидная конструкция для осуществления заданного изменения в одном или более специфических положений целевой РНК-последовательности в клетке посредством рекрутирования редактирующей РНК структуры, естественно присутствующей в указанной клетке, имеющей последовательность 5'-X-(Y-X')n-L-Z-3', где X комплементарен целевой РНК-последовательности ниже (3') специфического положения, X' комплементарен целевой РНК-последовательности выше (5') специфического положения, Y содержит один или более нуклеотидов (например, вплоть до 10, предпочтительно, от 1 до 5, более предпочтительно, от 1 до 3, например, 1 или 2), который/которые не является/не являются комплементарным/комплементарными целевой РНК-последовательности, n обозначает целое число от 1 до 10 (предпочтительно, от 1 до 5, более предпочтительно, от 1 до 3, или 1), L обозначает линкерную последовательность, которая является необязательной и может содержать любое число нуклеотидов, включая ноль, и Z обозначает последовательность, которая распознается и связывается с указанной редактирующей РНК структурой. L может также включать различные химические связи, например (олиго)пептидную связь или ПЭГ (PEG) связь.
Когда n больше или равен 1, нацеливающий участок олигонуклеотидной конструкции не является абсолютно комплементарным целевой РНК-последовательности, но на самом деле содержит один или более мисмэтчей (некомплементарных (неспаренных) оснований) или оснований неполного соответствия, которые служат для повышения специфичности, посредством повышения частоты редактирования противостоящего нуклеотида в целевой РНК-последовательности. Когда n равен двум или более, X' встречается более одного раза и два или более X' могут быть идентичными в зависимости от последовательности комплементарных оснований в целевой РНК-последовательности, но скорее всего, два или более X' не являются идентичными. Когда n обозначает 0, нацеливающий участок абсолютно комплементарен целевой РНК-последовательности на всем протяжении нацеливающего участка, т.е. без каких-либо мисмэтчей (оснований, некомплементарных основаниям целевой РНК-последовательности). Этот вариант, по-видимому, вызывает неспецифическое редактирование РНК с помощью аденозиндезаминазы, это означает, что все остатки аденозина в области перекрывания между нацеливающим участком олигонуклеотидной конструкции и целевой РНК-последовательностью имеют равную вероятность превратиться в инозин. Любое неспецифическое редактирование аденозина можно ограничить, если принять меры, чтобы остатки аденозина, которые не должны становиться мишенью, или по меньшей мере с пониженной частотой, сталкивались с противостоящим нуклеотидом с 2'-O-модифицированным рибозным фрагментом, например, с 2'-ОМе, поскольку известно, что последний снижает эффективность редактирования противостоящего аденозина. Или же, или помимо этого, можно в качестве противостоящего основания использовать гуанин или аденин, поскольку эти нуклеиновые основания обычно препятствуют дезаминированию противостоящего основания.
Согласно другому варианту в изобретении предусматривается олигонуклеотидная конструкция для осуществления заданного изменения в специфическом положении в целевой РНК-последовательности в клетке посредством рекрутирования редактирующей РНК структуры, естественно присутствующей в указанной клетке, имеющая последовательность 5'-Z-L-(X'-Y)n-X-3', где X комплементарен целевой РНК-последовательности выше (5') специфического положения, X' комплементарен целевой РНК-последовательности ниже (3') специфического положения, Y содержит один или более нуклеотидов, который/которые не является/не являются комплементарным/комплементарными целевой РНК-последовательности (например, вплоть до 10, предпочтительно, от 1 до 5, предпочтительно, от 1 до 3, например, 1 или 2), n обозначает целое число от 1 до 10 (предпочтительно, от 1 до 5, более предпочтительно, от 1 до 3, или 1), L обозначает линкерную последовательность, которая является необязательной и может содержать любое число нуклеотидов, включая ноль, и Z обозначает последовательность, которая распознается и связывается с указанной редактирующей РНК структурой. L может также включать различные химические связи, например (олиго)пептидную связь.
Когда n больше или равен 1, нацеливающий участок олигонуклеотидной конструкции не является абсолютно комплементарным целевой РНК-последовательности, но на самом деле содержит один или более мисмэтчей или оснований неполного соответствия, которые служат для повышения специфичности посредством повышения частоты редактирования противостоящего нуклеотида в целевой РНК-последовательности. Когда n равен двум или более, X' встречается более одного раза и два или более X' могут быть идентичными в зависимости от последовательности комплементарных оснований в целевой РНК-последовательности, но скорее всего, два или более X' не являются идентичными. Когда n обозначает 0, нацеливающий участок абсолютно комплементарен целевой РНК-последовательности на всем протяжении нацеливающего участка, т.е. без каких-либо мисмэтчей относительно целевой РНК-последовательности. Этот вариант, по-видимому, вызывает неспецифическое редактирование РНК при использовании аденозиндезаминазы, это означает, что все остатки аденозина в области перекрывания между нацеливающим участком олигонуклеотидной конструкции и целевой РНК-последовательностью имеют равную вероятность превратиться в инозин. Любое неспецифическое редактирование остатков аденозина можно ограничить, если принять меры, чтобы остатки аденозина, которые не должны становиться мишенью, или по меньшей мере с пониженной частотой, сталкивались с противостоящим нуклеотидом с 2'-O-модифицированным рибозным фрагментом, например, с 2'-ОМе, поскольку известно, что последний снижает эффективность редактирования противостоящего аденозина.
Олигонуклеотид по изобретению может представлять собой гибридную ДНК/РНК молекулу, т.е. включает как дезоксирибо-, так и рибо-нуклеотиды в одном и том же олигонуклеотиде.
Рекрутирующий участок: предпочтительные варианты
Рекрутирующий участок должен быть достаточно длинным, чтобы обеспечить создание такой структуры, как РНК- или ДНК-структура типа 'стебель-петля', предпочтительно, в Z-PHK или Z-ДНК конформации, которая узнается редактирующей структурой по изобретению. Если редактирующая структура представляет собой hADAR1, то в этом случае известны нуклеотидные последовательности, которые обеспечивают узнавание и связывание посредством Z-альфа домена варианта hADAR1 с молекулярной массой 150 кДа.
Рекрутирующие участки из искусственных структур типа 'стебель-петля' РНК или ДНК
Пример искусственной РНК- или ДНК-структуры типа 'стебель-петля', которая является предпочтительным рекрутирующим участком по изобретению, содержит последовательность (RY или YR)nNm(RY или YR)n, где R, Y и N представляют собой либо рибонуклеотиды, либо дезоксирибонуклеотиды, и где R обозначает А или G, Y обозначает Т, U или С, N обозначает A, G, С, Т или U, n обозначает 3 или более, m обозначает 1 или более (предпочтительно, m обозначает 2 или более, более предпочтительно, m обозначает 3 или более, еще более предпочтительно, m обозначает 4 или более) и где N образует петлю, а две (RY)n или (YR)n последовательности либо образуют дцРНК структуру типа 'стебель' (когда все нуклеотиды представляют собой рибонуклеотиды), либо дцДНК структуру (когда все нуклеотиды представляют собой дезоксирибонуклеотиды) комплементарным спариванием нуклеиновых оснований по Уотсону-Крику. Рекрутирующий участок, предпочтительно, содержит либо одни рибонуклеотиды, либо одни дезоксирибонуклеотиды, хотя не исключается рекрутирующий участок, содержащий как рибо-, так и дезоксирибонуклеотиды.
Особенно предпочтительные примеры рекрутирующих участков, заведомо связывающихся в hADAR1, представляют собой (i) ДНК структуры, представленные формулой (CG)nTm(CG)n, где n обозначает 3 и m обозначает 4 или более, предпочтительно, 4 или 5, которые имеют тенденцию образовывать так называемые Z-ДНК структуры, (ii) РНК структуры, представленные формулой (RY)nNm(RY)n, где R обозначает А или G, Y обозначает С или U, N обозначает любой из A, G, С или U, и где все N могут быть одинаковыми или различными, n обозначает 3 или более, m обозначает 4 или более, предпочтительно, 4 или 5, которые имеют тенденцию образовывать Z-PHK структуры. Z-ДНК и Z-RNA структуры отличаются от их более распространенных аналогов (для дцДНК наиболее распространенной формой является В конформация, тогда как для дцРНК наиболее распространенной является А-форма) тем, что двойная спираль является левосторонней в противоположность А и В формам, которые, обе, являются правосторонними, и нуклеиновые основания в каркасе Z-ДНК и Z-PHK расположены в пространстве в виде зигзага (отсюда приставка "Z").
Рекрутирующие участки из природных РНК-субстратов
Отличные от Z-PHK-образующих дцРНК-последовательности, образующие структуры типа 'стебель-петля' и петли, также выступают в роли рекрутирующих участков. В уровне техники описаны различные дцРНК структуры, содержащие мисмэтчи или пары оснований с нестрогим соответствием, отличные от GluR-B, довольно подробно описанной выше, которые взаимодействуют со связывающим доменом редактирующих структур, такие как GluR-C и GluR-D, 5-НТ2с серотониновые рецепторы и некоторые первичные (pri-) и пре-микроРНК и микроРНК. Предпочтительные петли в рекрутирующих участках олигонуклеотидных конструкций по изобретению соответствуют консервативным последовательностям тетра- или пентапетли UNCG, где N может обозначать любой из A, G, С или U, или GCUMA, где М обозначает А или С, соответственно. Если рекрутирующий участок содержит сегмент 'стебель-петля' из известного в уровне техники редактирующего РНК сайта, стебель можно использовать без изменений или можно изменять последовательность или длину, делать короче или модифицировать другим способом, чтобы изменять его характеристики, например, аффинность к редактирующей РНК структуре, или на основании соображений технологичности или обработки, стоимости, или по любой другой причине, до тех пор, пока функция рекрутирования еще не совсем ослабла. Эти структуры можно легко получать in vitro и тестировать их способность связывать, рекрутировать и перенацеливать редактирующие структуры. В уровне техники известны некоторые редактирующие структуры, которые можно получать в промышленных масштабах, включая hADARs, и тестировать в анализах на связывание с дцРНК и дцДНК структурами в олигонуклеотидных конструкциях по изобретению. Такие анализы легко доступны для средних специалистов в области белок-нуклеотидных взаимодействий и включают анализ сдвига электрофоретической подвижности (EMSA).
Два примера известных редактируемых РНК-последовательностей, которые охарактеризованы очень подробно, находятся в В-субъединице глутаматных рецепторов (GluR-B) АМРА (3-амино-3-гидрокси-5-метил-4-изоксазол пропионовой кислоты) подтипа. Эта модельная система содержит два часто редактируемых сайта, причем ДНК-кодируемый AGA редактируется в IGA, что приводит к замене аргинина на глицин (R/G сайт) и отдельной замене глутамина на аргинин (Q/R сайт). Известно, что GluR-B (R/G) сайт содержит структуру типа 'стебель-петля', состоящую из 71 нуклеотида, содержащую 3 пары некомплементарных (неспаренных) оснований (мисмэтчей), 2 А⋅С и одну G⋅U пары оснований с нестрогим соответствием. Интересно отметить, что петля состоит из высококонсервативной структуры "пентапетли" GCUAA, которая соответствует филогенетически консервативной последовательности GCUMA, где М обозначает А или С (Aruscavage P.J. & Bass B.L. RNA. 2000; 6: 257-269). По-видимому, отдается некоторое предпочтение редактированию двух остатков аденозина с нестрогим соответствием, при этом эффективность повышается, когда основание, противостоящее редактируемому остатку аденозина, выбрано из цитидина или уридина, причем цитидин является предпочтительным.
Эту структуру можно просто применять без изменений или ее можно адаптировать для использования в олигонуклеотидной конструкции по изобретению в качестве рекрутирующего участка, посредством уменьшения или увеличения числа пар нестрогого соответствия в структуре стебля, чтобы модифицировать специфичность редактирования и/или перенацелить редактирование на предпочтительный(-е) сайт(-ы) в целевой РНК-последовательности. Помимо этого, или в качестве альтернативы, сайт узнавания GluR-B для hADAR1 можно модифицировать, укорачивая 'стебель', но не отменяя полностью узнавание. Такое укорочение может быть удобно с точки зрения технологичности или стоимости продукта в перспективе и т.п.
Пример рекрутирующего участка из GluR-B домена, являющегося предпочтительным вариантом по изобретению, содержит последовательность: 5'-(AUANa)nUAUAACAAUAUgcuaaAUGUUGUUAUA(NbUAU)n-3', где Na и Nb, каждый, обозначает единичные нуклеотиды, которые могут представлять собой A, G, С или U, при условии, что Na и Nb образуют мисмэтч пару оснований (некомплементарных) при образовании структуры типа 'стебель-петля', и n равно 1 или 0 (т.е. SEQ ID NOs: 6 и 7).
Другой предпочтительный рекрутирующий участок содержит последовательность или состоит из последовательности 5'-GUGGNcAUANaUAUAACAAUAUgcuaaAUGUUGUUAUANbUAUNdCCAC-3' (SEQ ID NO: 25), где: каждый из Na, Nb, Nc HNd может обозначать нуклеотид A, G, С или U, при условии, что Na и Nb образуют мисмэтч пару оснований и Nc и Nd образуют мисмэтч пару оснований при образовании структуры типа 'стебель-петля'; и при этом пентануклеотид gcuaa образует петлю и последовательности, прилегающие к пентануклеотиду gcuaa выше (5') и ниже (3'), образуют стебель посредством спаривания оснований.
Рекрутирующий участок может быть связан на 5' или 3' конце с нацеливающим участком, необязательно с помощью линкера "L", который содержит один или более нуклеотидов, олигопептид или другой химический линкер, например, полиэтиленгликоль (PEG, ПЭГ).
Химическая модификация олигонуклеотидных конструкций
Известны различные химические реакции и модификация олигонуклеотидов, которые без труда можно применять согласно изобретению. Регулярные межнуклеозидные связи между нуклеотидами можно изменять путем введения моно- или дитиоэфирных групп в фосфодиэфирные связи с образованием тиофосфорных сложных эфиров (фосфоротиоатов) и дитиофосфорных сложных эфиров (фосфородитиоатов), соответственно. Возможны другие модификации межнуклеозидных связей, включая амидирование и пептидные линкеры. Сахар рибозу можно модифицировать заменой 2'-O фрагмента на низший алкил (С1-4, такой как 2'-O-Ме), алкенил (С2-4), алкинил (С2-4), метоксиэтил (2'-MOE) или другой заместитель. Предпочтительными заместителями 2' ОН группы являются метальная, метоксиэтильная или 3,3'-диметилаллильная группа. Последняя известна своей способностью ингибировать чувствительность к нуклеазам вследствие ее большого объема, в то же время повышая эффективность гибридизации (Angus & Sproat FEBS 1993 Vol. 325, no. 1, 2, 123-7). Или же можно использовать последовательности закрытых нуклеиновых кислот (LNA), содержащие 2'-4' внутримолекулярную мостиковую (обычно метиленовый мостик между 2' кислородным и 4' углеродным атомами) связь внутри рибозного кольца. Пуриновые нуклеиновые основания и/или пиримидиновые нуклеиновые основания можно модифицировать с целью изменения их свойств, например, посредством аминирования или дезаминирования гетероциклических колец. Конкретный химизм и формат могут меняться от олигонуклеотидной конструкции к олигонуклеотидной конструкции и от применения к применению и могут детально разрабатываться в соответствии с желаниями и предпочтениями специалистов в данной области.
Длина
Олигонуклеотидные конструкции по изобретению могут содержать от 20 до нескольких сотен нуклеотидов. По практическим соображениям, таким как технологичность и стоимость, длина олигонуклеотидных конструкций, предпочтительно, должна быть короче, чем 200 нуклеотидов. Предпочтительно, длина олигонуклеотидных конструкций составляет от 20 до 100 нуклеотидов, более предпочтительно, от 24 до 60 нуклеотидов, еще более предпочтительно, от 30 и 50 нуклеотидов. Нацеливающий участок олигонуклеотидной конструкции, предпочтительно, содержит более 10 нуклеотидов, предпочтительно, более 11, 12, 13, 14, 15, 16, еще более предпочтительно, более 17 нуклеотидов. Более длинные нацеливающие участки обеспечивают более высокую специфичность к сайту-мишени в подлежащей редактированию РНК-последовательности, вызывают меньше побочных эффектов вследствие случайных (побочных) связей, а также предоставляют больше возможностей для создания вторичных структур, например, таких как структуры 'стебель-петля', в самом нацеливающем участке, мисмэтчи или основания нестрогого соответствия (с учетом мисмэтчей с одним или более комплементарным(-х) основанием(-й) в целевой РНК-последовательности в подлежащем редактированию сайте или вблизи этого сайта) и т.д. Предпочтительные нацеливающие участки являются комплементарными по всей длине нацеливающего участка за исключением мисмэтча, противостоящего подлежащему редактированию нуклеотиду, и необязательно одного или двух оснований нестрогого соответствия.
Конформация
Из уровня техники известно, что редактирующие РНК структуры, например, такие как hADARs, редактируют структуры дцРНК с различной специфичностью в зависимости от ряда факторов. Одним важным фактором является степень комплементарности двух цепей, образующих последовательность дцРНК. Абсолютная комплементарность двух цепей обычно побуждает каталитический домен hADAR неизбирательно дезаминировать остатки аденозина, реагируя в той или иной степени с любым остатком аденозина, с которым он сталкивается. Специфичность hADAR1 в отношении превращения только конкретных аденозинов может быть повышена обеспечением мисмэтча в дцРНК за счет предоставления нацеливающего участка, который содержит мисмэтч напротив подлежащего редактированию аденозина. Мисмэтч, предпочтительно, создается посредством предоставления нацеливающего участка, имеющего цитидин или уридин, предпочтительно, цитидин, напротив подлежащего редактированию аденозина. В результате дезаминирования аденозина в целевой цепи целевая цепь получит инозин, который в большинстве химических процессов "считывается", в соответствии с биологическими механизмами клетки, как G. Следовательно, после превращения А в I мисмэтч разрешается, потому что I абсолютно способен к спариванию с противостоящим основанием С в нацеливающем участке олигонуклеотидной конструкции согласно изобретению. После исправления мисмэтча в результате редактирования субстрат высвобождается и комплекс олигонуклеотидная конструкция-редактирующая структура избавляется от целевой РНК-последовательности, которая затем становится доступной для последующих биохимических процессов, таких как сплайсинг и трансляция.
Желаемая степень специфичности редактирования целевой РНК-последовательности может зависеть от применения. Следуя инструкциям в настоящей заявке на патент, специалисты в данной области смогут создать нацеливающий участок олигонуклеотидной конструкции в соответствии со своими требованиями и экспериментально, методом проб и ошибок, получить нужный результат.
Нацеливающий участок олигонуклеотидных конструкций по изобретению, обычно будет содержать стандартные нуклеотиды A, G, U и С, но может также включать инозин (I), например, вместо одного или более G нуклеотидов. В рекрутирующем участке олигонуклеотидной конструкции согласно изобретению G может также заменяться на I, однако, следует обратить внимание на то, что I в стебле или петле не мешает образованию конформаций Z-ДНК или Z-PHK в тех вариантах, когда это желательно.
Специфичность редактирования
Для предупреждения нежелательного редактирования аденозинов в целевой РНК-последовательности в области перекрывания с олигонуклеотидной конструкцией нацеливающий участок олигонуклеотидной конструкции можно модифицировать химическими методами. В уровне техники показано, что 2'-O-метилирование рибозильного фрагмента нуклеозида, противостоящего аденозину в целевой РНК-последовательности, резко уменьшает дезаминирование этого аденозина с использованием ADAR (Vogel et al. 2014 Angewandte Chemie Int. Ed. 53, 6267-71). Следовательно, при включении 2-метокси (2'-OMe) нуклеотидов в нужном положении олигонуклеотидной конструкции специфичность редактирования может резко повыситься. Предполагается, что другие 2'-О замены рибозильного фрагмента, например, такие как 2'-метоксиэтильная (2'-МОЕ) и 2'-O-диметилаллильная группы, также могут уменьшить нежелательное редактирование соответствующего (противостоящего) аденозина в целевой РНК-последовательности. Другие химические модификации легкодоступны для рядового специалиста в области синтеза и дизайна олигонуклеотидов. Синтез модифицированных такими химическими методами олигонуклеотидных конструкций и их тестирование методами по изобретению не является непосильной задачей, а другие модификации охватываются настоящим изобретением.
Редактирующие структуры
Редактирующие структуры обычно являются белковыми по своей природе, например, такие как ADAR ферменты, обнаруженные в многоклеточных организмах, включая млекопитающих. Редактирующие структуры могут также содержать комплексы нуклеиновой(-ых) кислоты (кислот) и белков или пептидов, например, такие, как рибонуклеопротеины. Редактирующие ферменты могут содержать только нуклеиновую(-ые) кислоту(-ы) или состоять только из нуклеиновой(-ых) кислоты (кислот), например, рибозимы. Все такие редактирующие структуры охватываются настоящим изобретением при условии, что они рекрутируются олигонуклеотидными конструкциями по изобретению. Предпочтительно, редактирующая структура представляет собой фермент, более предпочтительно, аденозиндезаминазу или цитидиндезаминазу, еще более предпочтительно, аденозиндезаминазу. Если редактирующая структура представляет собой аденозиндезаминазу, Y, предпочтительно, обозначает цитидин или уридин, наиболее предпочтительно, цитидин. Наибольший интерес представляют человеческие ADARs, hADAR1 и hADAR2, включая любые их изоформы, например, такие как hADAR1 p110 и р150.
Известные в уровне техники редактирующие РНК ферменты, для которых можно легко создать олигонуклеотидные конструкции, включают аденозиндезаминазы, воздействующие на РНК (ADARs), такие как hADAR1 и hADAR2, у человека или в человеческих клетках, и цитидиндезаминазы. Человеческие ADAR3 (hADAR3) были описаны в уровне техники, но, согласно имеющимся сведениям, не обладают дезаминазной активностью.
Известно, что hADAR1 существует в двух изоформах: длинный, 150 кДа, интерферон-индуцибельный вариант и более короткий, 100 кДа, вариант, которые получают альтернативным сплайсингом из общей пре-мРНК. Интересно отметить, что только длинная изоформа способна связываться с Z-ДНК структурой, которая может содержаться в рекрутирующем участке олигонуклеотидной конструкции по изобретению. Соответственно, на уровень изоформы 150 кДа, присутствующей в клетке, может влиять интерферон, в частности, интерферон-гамма (IFN-гамма). hADAR1 также индуцируется с помощью TNF-альфа. Это дает возможность разрабатывать комбинированную терапию, согласно которой пациенту вводят интерферон-гамма или TNF-альфа и олигонуклеотидные конструкции, содержащие Z-ДНК в качестве рекрутирующего участка по изобретению, либо в виде комбинированного лекарственного препарата, либо в виде отдельных лекарственных препаратов, или одновременно, или последовательно, в любом порядке. Некоторые болезненные состояния могут уже сопровождаться повышенными уровнями IFN-гамма или TNF-альфа в некоторых тканях пациента, что создает дополнительные возможности для того, чтобы сделать редактирование пораженных тканей более специфическим.
Как нацеливающий участок, так и рекрутирующий участок могут содержать нуклеотиды или состоять из нуклеотидов, имеющие (имеющих) химические модификации, которые изменяют устойчивость к нуклеазам, изменяют аффинность связывания (выражающуюся температурой плавления) или другие свойства. Примерами химических модификаций являются модификации остатка сахара, в том числе посредством сшивания заместителей в остатке сахара (рибозы) (например, как в LNA, или в закрытых нуклеиновых кислотах), введением в качестве заместителя при 2'-O атоме алкильной (например, 2'-O-метил), алкинильной (2'-O-алкинил), алкенильной (2'-O-алкенил), алкоксиалкильной (например, метоксиэтильной, 2'-МОЕ) групп, имеющих длину по описанию выше, и т.п. Также фосфодиэфирную группу в скелете молекулы можно модифицировать путем введения моно- или дитиоэфирных групп, амидированием и т.п. с образованием фосфоротиоатных, фосфородитиоатных, фосфороамидатных и т.д. межнуклеозидных связей. Межнуклеотидные связи можно заменять, полностью или частично, на пептидные связи, получая последовательности пептидо-нуклеиновых кислот, и т.п. В качестве альтернативы, или помимо этого, нуклеиновые основания можно модифицировать посредством (дез)аминирования, получая инозин или 2'6'-диаминопурины и т.п.
Другая модификация может представлять собой метилирование С5 в цитидиновом остатке нуклеотида с целью ослабить потенциальные иммуногенные свойства, заведомо ассоциируемые с CpG последовательностями.
Структура олигонуклеотидных конструкций по изобретению может изменяться, переходя от "одноногих" шпилек (Фиг. 1 и 2) к "двуногим" шпилькам (Фиг. 3), причем ножки содержат нацеливающий участок или нацеливающие участки, а корпус или центральная часть шпильки создают рекрутирующий участок (Фиг. 1-3). Ножка или ножки олигонуклеотидной конструкции обеспечивают мисмэтчи (некомплементарные нуклеиновые основания) или нуклеиновые основания нестрогого соответствия, представляющие собой сайт, противостоящий сайту редактирования, например, подлежащим редактированию остаткам аденозина или цитидина, в целевой РНК. Например, в случае, когда олигонуклеотидная конструкция рекрутирует ADAR активность для редактирования превращения А в I в целевой РНК, мисмэтч или нуклеиновое основание неполного соответствия может включать остаток аденозина, гуанина, уридина или цитидина, предпочтительно, остаток цитидина. За исключением мисмэтча и нуклеинового основания неполного соответствия напротив сайта редактирования нацеливающий участок обычно абсолютно комплементарен целевой РНК, хотя ограниченное число неидеальных пар, таких как основания нестрогого соответствия или мисмэтчи (некомплементарные основания), могут быть допустимы при отсутствии неприемлемого ослабления специфичности и/или прочности связи между олигонуклеотидной конструкцией и целевой РНК-последовательностью. Стебель шпильки может состоять из абсолютно комплементарных отрезков нуклеотидов, образующих двухцепочечную РНК-структуру по всей своей длине. Или же стебель шпильки может содержать один или более противостоящих нуклеотидов нестрогого соответствия или некомплементарных нуклеотидов, при условии, что узнавание олигонуклеотидной конструкции редактирующей активностью не станет недопустимо слабым. Следует понимать, что функция редактирующей активности в клетке в ее природных сайтах редактирования может быть ослаблена вследствие рекрутирования структур, отвечающих за редактирующую активность, при использовании олигонуклеотидных конструкций по изобретению Рядовому специалисту в данной области понятно, что степень перенацеливания редактирующих структур внутри клетки на другие сайты-мишени можно регулировать, изменяя аффинность рекрутирующего участка олигонуклеотидных конструкций по изобретению к распознающему домену редактирующей структуры. Это можно осуществлять, снижая аффинность рекрутирующего участка олигонуклеотидной конструкции к редактирующей структуре любым способом или совокупностью способов, включая изменение последовательности стебля, размера структуры (последовательности, химической структуры скелета, рибозила или нуклеинового основания) петли или их комбинацию. Точную модификацию можно определить эмпирически и/или вычислительными методами с учетом структурных взаимодействий между рекрутирующим участком олигонуклеотидной конструкции и распознающим доменом редактирующей структуры.
Также, или в качестве альтернативы, степень рекрутирования и перенацеливания редактирующей структуры, резидентной в клетке, можно регулировать путем подбора и схемы введения доз олигонуклеотидной конструкции. Это важную вещь должен определить экспериментатор (in vitro) или врач-клиницист, обычно в фазе I и/или II клинических испытаний.
Предпочтительно, в изобретении предусматривается применение олигонуклеотидной конструкции, которая состоит из одного олигонуклеотида (Фиг. 1, Фиг. 2А, Фиг. 3), содержащего как нацеливающий участок, так и рекрутирующий участок, для редактирования нуклеотидных последовательностей. Несомненно, однако, что в объем настоящего изобретения входит применение олигонуклеотидной конструкции, содержащей два нуклеотида (Фиг. 2В), например, один, содержащий нацеливающий участок, и один, содержащий рекрутирующий участок. Следовательно, согласно другому варианту, в изобретении предусматривается два отдельных олигонуклеотида, причем один из них содержит нацеливающий участок, а второй содержит рекрутирующий участок, при этом два олигонуклеотида рассчитаны на обладание аффинностью друг к другу, например, за счет спаривания оснований по Уотсону-Крику между двумя функциональными участками последовательности, не имеющей специфической нацеливающей или рекрутирующей функции, например, линкерной последовательности. Преимущества такой двухкомпонентной системы могут заключаться в гибкости, применимости (например, в области персонализованной медицины), технологичности, стоимости готовых лекарственных препаратов и т.д. Два (или более) олигонуклеотида в соответствии с многокомпонентной системой не обязательно должны явно разделять различные функции (нацеливание и рекрутирование). Например, олигонуклеотиды могут способствовать возникновению нацеливающей и рекрутирующей функции только после объединения в комплекс. Если происходит отжиг двух олигонуклеотидов, образующих участок дцРНК, они действительно в результате отжига могут создать рекрутирующую функцию. Отжиг (ренатурирование) является одним из способов образования комплекса, но понятно, что существуют и другие способы, посредством которых могут объединиться два (или более) олигонуклеотидных компонента, объединив тем самым нацеливающую и рекрутирующую функции. Олигонуклеотидные конструкции, содержащие более двух олигонуклеотидов, не исключаются из объема настоящего изобретения, хотя чем больше число олигонуклеотидов в конструкции, тем более сложной является система с точки зрения производства, методов анализа, приготовления, введения или других аспектов обращения с ней, включая логистику и затраты.
Клетка млекопитающего
Данное изобретение относится к модификации целевых РНК-последовательностей в эукариотических клетках, предпочтительно, в клетках многоклеточных организмов, более предпочтительно, в клетках млекопитающих. В принципе данное изобретение можно применять в отношении клеток из любых видов млекопитающих, но предпочтительно использовать его по отношению к человеческим клеткам.
Данное изобретение можно применять в отношении клеток из любого органа, например, кожи, легкого, сердца, почки, печени, поджелудочной железы, кишечника, мышцы, железы, глаза, головного мозга, крови и т.п. Данное изобретение особенно применимо для модификации последовательностей в клетках, тканях или органах, имеющих отношение к болезненному состоянию субъекта (человека). Такие клетки включают, но без ограничения, эпителиальные клетки легкого или желудочно-кишечного тракта, клетки репродуктивных органов, мышечные клетки, клетки глаза, клетки кожи, клетки тканей и органов, например, таких как печень, почки, поджелудочная железа, иммунные клетки, раковые клетки, клетки желез, клетки головного мозга и т.п.
Данное изобретение можно также применять в отношении клеток млекопитающих, которые естественным образом не присутствуют в организме, например, в отношении линий клеток или в отношении эмбриональных стволовых (ES) клеток.
Данное изобретение можно применять в отношении различных типов стволовых клеток, включая плюрипотентные стволовые клетки, тотипотентные стволовые клетки, эмбриональные стволовые клетки, индуцируемые плюрипотентные стволовые клетки и т.д.
Клетка может находиться in vitro или in vivo. Одним из преимуществ данного изобретения является то, что его можно применять в отношении клеток in situ в живом организме, но его можно применять также в отношении клеток в культуре. Согласно некоторым вариантам клетки обрабатываются ex vivo, а затем вводятся в живой организм (например, их снова вводят в организм, из которого они были первоначально извлечены).
Изобретение можно также применять для редактирования целевых РНК-последовательностей в клетках так называемого органоида. Органоиды можно рассматривать как трехмерные полученные in vitro ткани, тем не менее стимулируемые в определенных условиях вырабатывать индивидуальные выделенные ткани (например, см. Lancaster & Knoblich, Science 2014, vol. 345 no. 6194 1247125). В терапевтическом плане они применяются, потому что их можно получать in vitro из клеток пациентов, а затем органоиды можно снова вводить пациенту в качестве аутологичного материала, который отторгается с меньшей долей вероятности, чем обычный трансплантат. Таким образом, согласно другому предпочтительному варианту изобретение может применяться на практике на органоидах, выращенных с использованием образцов ткани, взятой у пациента (например, из его желудочно-кишечного тракта; см. Sala et al. J Surg Res. 2009; 156(2):205-12, а также Sato et al. Gastroenterology 2011; 141:1762-72); после редактирования РНК в соответствии с изобретением органоиды или стволовые клетки, находящиеся в органоидах, могут использоваться для трансплантации обратно пациенту с целью усиления функции органа.
Клетка, подвергающаяся воздействию, обычно содержит генетическую мутацию. Мутация может быть гетерозиготной или гомозиготной. Обычно данное изобретение применяется для модификации точковых (точечных) мутаций, например, таких как мутации N в А, где N может обозначать G, С, U (Т на уровне ДНК), предпочтительно, мутации G в А, или мутации N в С, где N может обозначать A, G, U (Т на уровне ДНК), предпочтительно, мутации U в С. Гены, содержащие мутации, представляющие особый интерес, обсуждаются ниже. Однако согласно некоторым вариантам возможно применение изобретения в противоположном направлении путем введения мутации, ассоциированной с заболеванием, в линию клеток или животному, чтобы создать полезное средство исследования изучаемого заболевания. В качестве примера создания модели заболевания мы получили олигонуклеотидную последовательность, которая предусматривает рекрутирование редактирующей активности в человеческой клетке, чтобы создать мутацию в гене СЕР290, создающую скрытый сайт сплайсинга, который лежит в основе формы врожденного амавроза Лебера, самой распространенной формы врожденной детской слепоты.
Мутация, подлежащая реверсии посредством редактирования РНК, может возникать на уровне хромосомы или какой-либо другой формы ДНК, например, такой как митохондриальная ДНК, или РНК, включая пре-мРНК, рибосомную РНК или митохондриальную РНК. Изменение, которое надлежит сделать, может быть в целевой РНК патогена, включая грибы, дрожжи, паразиты, кинетопластиды, бактерии, бактериофаги, вирусы и т.д., которыми были инфицированы клетка или субъект. Соответственно, редактирование может осуществляться на уровне РНК на целевой последовательности внутри такой клетки, такого субъекта или патогена. Некоторые патогены, например, такие как вирусы, высвобождают свою нуклеиновую кислоту, ДНК или РНК, в клетку инфицированного хозяина (клетку). Другие патогены локализованы или циркулируют в организме инфицированного хозяина. Олигонуклеотидные конструкции по изобретению можно применять для редактирования целевых РНК-последовательностей внутри клетки патогена, локализованного или циркулирующего в эукариотическом хозяине, при условии, что клетки, в которых должно происходить редактирование, содержат структуру, совместимую с вводимой в них олигонуклеотидной конструкцией.
Без связи с какой-либо теорией полагают, что редактирование РНК с использованием hADAR1 и hADAR2 происходит на пре-мРНК в ядре в процессе транскрипции или сплайсинга. Полагают, что редактирование РНК с применением цитидиндезаминаз происходит на уровне мРНК. Не исключено редактирование кодонов митохондриальной РНК или некодирующих последовательностей в зрелых мРНК.
Целевая последовательность и изменение
Данное изобретение применяется для осуществления изменений в целевой РНК-последовательности в эукариотической клетке посредством применения олигонуклеотидной конструкции, способной нацеливать на сайт, подлежащий редактированию, и рекрутировать редактирующие РНК структуры, резидентные в клетке, осуществлять реакцию(-и) редактирования. Предпочтительными реакциями редактирования являются реакции дезаминирования аденозина и реакции дезаминирования цитидина, превращение аденозина в инозин, а цитидина в уридин, соответственно. Изменения могут осуществляться в 5' или 3' нетранслируемых областях целевой РНК, в (скрытых) сайтах сплайсинга, в экзонах (изменение аминокислот в белках, транслируемых с использованием целевой РНК, частоты использования кодонов или протекания сплайсинга посредством изменения экзонных сайленсеров или энхансеров сплайсинга, посредством введения или удаления стартовых или стоп-кодонов), в интронах (изменение сплайсинга посредством изменения интронных сайленсеров сплайсинга или интронных энхансеров сплайсинга, точек разветвления) и вообще в любой области, влияющей на устойчивость, структуру или функции РНК. Целевая РНК-последовательность может содержать мутацию, которую, например, можно пожелать скорректировать или изменить (транзиция или трансверсия). Или же мутацию в целевой РНК-последовательности осуществляют с намерением иметь измененный фенотип (или генотип, в случае РНК-организмов, таких как РНК-содержащие вирусы) в случаях, когда мутации раньше не было. Например, можно создать клеточные линии или животных, которые несут изменения (мутации) в целевой РНК-последовательности и которые можно использовать в анализах или в качестве (животных, органоидов и т.д.) модельных систем для изучения заболевания, тестирования испытуемых соединений против заболеваний и т.п. Олигонуклеотидные конструкции и способы по изобретению можно применять в системах высокопроизводительного скрининга (в упорядоченном виде) для создания банка клеток с достаточным разнообразием целевых РНК, например, кодирующих большое разнообразие изоформ белка, для дополнительного экспериментального исследования, включая скрининг соединений, белковой инженерии и т.п.
Целевая РНК может представлять собой любую клеточную или вирусную РНК-последовательность, в большинстве случаев она является пре-мРНК или мРНК с функцией кодирования белков.
Исключительно для простоты поиска, но без намерения ограничить изобретение, ниже приводится таблица для иллюстрации возможных изменений в кодонах, которые можно осуществлять редактированием с использованием аденозиндезаминазы, управляемым олигонуклеотидами по изобретению. Тем более не следует полагать, что таблица ограничивает применимость изобретения к кодирующим последовательностям в любой РНК; как уже указывалось, изобретение можно применять на практике на любой РНК-мишени, содержащей аденозин, как в кодирующей области, интроне, некодирующем экзоне (например, в 5'- или 3' нетранслируемой области), в миРНК, тРНК, рРНК и т.д. Во избежание какого-либо недопонимания в том, что касается широты применения, изменения, которые являются несущественными ('молчащими') с точки зрения кодирования, могут все же менять экспрессию генов определенного белка, учитывая, что некоторые кодоны для одной. и той же аминокислоты могут быть более предпочтительными, чем другие, и могут привести, например, к различной устойчивости транскрипции или эффективности трансляции, в связи с чем кодированный белок становится более или менее распространенным, чем белок без изменения.
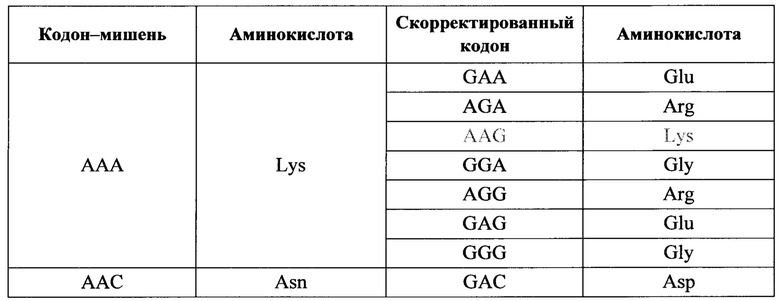

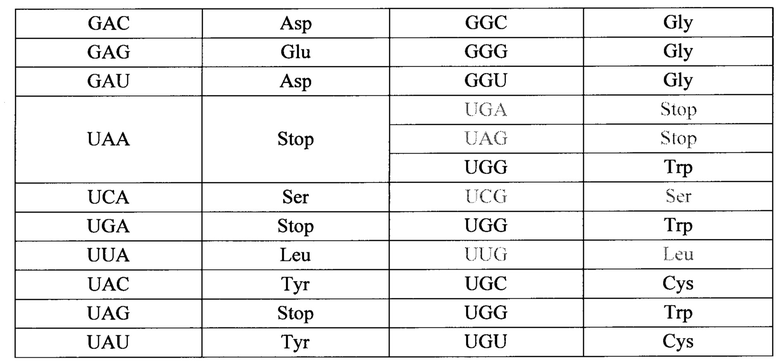
Особый интерес при редактирования с применением олигонуклеотидов по изобретению вызывают остатки аденозина-мишени, являющиеся частью кодонов для аминокислотных остатков, которые определяют важнейшие функции или характеристики, такие как каталитические сайты, сайты связывания с другими белками, связывание с субстратами, домены локализации, для ко- и пост-трансляционной модификации, например, такой как гликозилирование, гидроксилирование, миристоилирование, расщепление белков протеазами (для созревания белка и/или как часть определения внутриклеточной маршрутизации), и т.п.
Организм-носитель генетических заболеваний обусловлен мутациями G в А, и эти генетические заболевания являются предпочтительными целевыми заболеваниями, потому что дезаминирование аденозина в мутантной целевом аденозине реверсирует мутацию к состоянию дикого типа. Однако, реверсия к состоянию дикого типа не всегда необходима для достижения благоприятного эффекта. Модификация А в G в мишени может также быть благотворной, если нуклеотид дикого типа отличен от G. В некоторых случаях можно предсказать, что это случится, в других обстоятельствах это может потребовать некоторых экспериментов. В некоторых случаях модификация из А в целевой РНК в G, где нуклеотидом дикого типа является не G, может быть молчащей (не транслируемой в иную аминокислоту), или несущественной по иным причинам (например, аминокислота заменяется, но она является консервативной заменой, которая не нарушает структуру и функцию белка), или аминокислота является частью функционального домена, который обладает определенной устойчивостью к изменению. Если переход от А к G посредством редактирования согласно изобретению осуществляли в некодирующей РНК, или в некодирующем участке РНК, результат также может быть несущественным или менее серьезным, чем первоначальная мутация. Специалисты в данной области понимают, что применение настоящего изобретения является очень широким и не ограничивается даже предупреждением или лечением заболевания. Изобретение можно также применять для модификации транскриптов с целью изучения их эффектов, даже если, или в особенности если, такие модификации вызывают болезненное состояние, например, в клетке или на животной модели, причем животное отлично от человека.
Предпочтительные примеры генетических заболеваний, которые можно предупреждать и/или лечить с использованием олигонуклеотидов согласно изобретению, включают любые заболевания, при котором модификация одного или более остатков аденозина в целевой РНК (теоретически, вероятно) приведет к благотворному изменению.
Транскрибируемые РНК-последовательности, которые являются потенциальными целевыми РНК-последовательностями по изобретению, содержащие мутации, заслуживающие особого внимания, включают, но без ограничения, РНК-последовательности, транскрибируемые при использовании гена CFTR (регулятора трансмембранной проводимости кистозного фиброза), дистрофина, хантингтина, нейрофибромина 1, нейрофибромина 2, β-цепи гемоглобина, СЕР290 (центросомального белка 290 кДа), гена НЕХА β-гексозаминидазы А, и любого из генов (синдрома) Ашера (Ушера) (например, USH2B, кодирующего белок ушерин), отвечающего за форму генетической слепоты, называемую синдромом Ашера (Ушера). Более обширный список представлен ниже. Таким образом, выбирают целевую последовательность, а олигонуклеотидная конструкция будет включать нужную модификацию, чтобы скорректировать мутацию.
Специалисты в области CF мутаций знают, что в гене CFTR известно от 1000 до 2000 мутаций CFTR, включая R117H, G542X, G551D, R553X, W1282X и N1303K.
Вообще, мутациями в любой целевой РНК, которые можно реверсировать, используя олигонуклеотидные конструкции по изобретению, являются мутации G в А, в случае рекрутирования аденозиндезаминазы, и мутации U в С, в случае рекрутирования цитидиндезаминазы, и соответственным образом создавать дизайн олигонуклеотидной конструкции. Мутации, на которые можно нацеливаться с использованием олигонуклеотидных конструкций по изобретению, включают мутации С в А, U в А (Т в А на уровне ДНК) в случае рекрутирования аденозиндезаминаз, и мутации A в C и G в C в случае рекрутирования цитидиндезаминаз. Хотя редактирование РНК в последних случаях не обязательно может реверсировать мутацию к состоянию дикого типа, редактированный нуклеотид может обусловить улучшение по сравнению первоначальной мутацией. Например, мутацию, которая обусловливает появление терминирующего (стоп-) кодона в рамке считывания - что при трансляции приводит к процессированному белку - можно заменить на кодон, кодирующий аминокислоту, которая может не являться оригинальной аминокислотой в этом положении, но которая приводит к (полноразмерному) белку по меньшей мере с некоторыми функциональными свойствами, по меньшей мере повышенными по сравнению с процессированным белком.
Целевая последовательность является эндогенной для эукариотической клетки, предпочтительно, клетки млекопитающего, более предпочтительно, для человеческой клетки. Таким образом, целевая последовательность не является, например, трансгеном или маркерным геном, который искусственно введен в какой-то момент истории клетки, но скорее геном, естественно присутствующим в клетке (либо в мутантной, либо не в мутантной форме).
Данное изобретение не ограничено корректировкой мутаций, поскольку, напротив, его можно применять для изменения последовательности дикого типа на мутантную последовательность с применением олигонуклеотидов по изобретению. Одним из примеров, в котором может быть целесообразно модифицировать аденозин дикого типа, является пропуск/перепрыгивание экзона, например, посредством модификации аденозина, который оказался точкой разветвления, необходимой для сплайсинга указанного экзона. В другом примере аденозин определяет последовательность узнавания или является частью последовательности узнавания для связывания белка или входит в состав вторичной структуры, определяющей устойчивость РНК. Таким образом, как отмечается выше, данное изобретение можно применять для предоставления средства исследования заболеваний, введения новых мутаций, которые являются менее опасными, чем имеющаяся мутация, и т.д.
Области применения олигонуклеотидных конструкций
Количество вводимых олигонуклеотидных конструкций, дозировка и схема применения могут меняться в зависимости от типа клеток, подлежащего лечению заболевания, целевой популяции, способа введения (например, системного или местного), тяжести заболевания и приемлемого уровня побочного действия, но все это можно и нужно оценивать эмпирическим методом в процессе исследования in vitro, в преклинических и клинических испытаниях. Особенно эффективными являются испытания, в которых в результате модифицирования последовательности наблюдается легко детектируемое фенотипическое изменение. Возможно, что в более высоких дозах олигонуклеотид мог бы конкурировать за связывание с нуклеотидной редактирующей структурой (например, ADAR) внутри клетки, тем самым сокращая количество структуры, которая свободно участвует в редактировании РНК, но испытания с обычными дозами выявят любые такие эффекты для данного олигонуклеотида и данной мишени.
Один пригодный метод испытания включает доставку олигонуклеотидной конструкции к линиям клеток или в тест-организм, а затем отбор биоптатов в различные контрольные моменты времени. Можно проанализировать последовательность целевой РНК в биоптате, а затем легко определить долю клеток, имеющих модификацию. Такое испытание можно провести один раз, затем результаты можно сохранить и последующую доставку можно осуществлять без необходимости отбирать биоптаты.
Таким образом, способ по изобретению может включать стадию определения присутствия заданного изменения в целевой РНК-последовательности в клетке, тем самым подтверждается, что целевая РНК-последовательность модифицирована. Эта стадия обычно включает секвенирование релевантного участка целевой РНК, или ее кДНК копии (или кДНК копии ее продукта сплайсинга в случае, если целевая РНК представляет собой пре-мРНК), обсуждаемой выше, и таким образом можно легко проверить изменение последовательности. Или же изменение можно оценить на уровне белка (длину, гликозилирование, функцию и т.п.), или по результатам определения некоторых функций, например в виде (индуцируемого) тока, если белок, кодируемый целевой РНК-последовательностью, представляет собой ионный канал. В случае CFTR функции специалисту в данной области хорошо известен анализ в камере Уссинга или NPD-тест на млекопитающих, включая людей, для оценки восстановления или (мутации) приобретения функции.
После осуществления редактирования РНК в клетке концентрация модифицированной РНК может со временем уменьшаться, например, вследствие клеточного деления, ограниченного периода полужизни редактированной РНК и т.п. Поэтому при практическом применении в терапии способ по изобретению включать повторную доставку олигонуклеотидной конструкции до тех пор, пока не будет модифицировано количество РНК, достаточное для того, чтобы обеспечить пациенту ощутимый лечебный эффект и/или сохранить этот эффект во времени.
Доставка олигонуклеотидной конструкции
Олигонуклеотидные конструкции по изобретению особенно пригодны для терапевтического применения, и поэтому в изобретении предусматривается фармацевтическая композиция, содержащая олигонуклеотидную конструкцию по изобретению и фармацевтически приемлемый носитель. Согласно некоторым вариантам согласно изобретению фармацевтически приемлемый носитель может представлять собой просто физиологический солевой раствор. Эффективным может быть изотонический или гипотонический физиологический раствор, в особенности для доставки в легкие.
В изобретении также предусматривается устройство доставки (например, шприц, ингалятор, небулайзер), которое содержит фармацевтическую композицию по изобретению.
Также в изобретении предусматривается олигонуклеотидная конструкция по изобретению для применения в способе осуществления изменения в целевой РНК-последовательности в клетке млекопитающего, предпочтительно, в человеческой клетке, как описано в данной заявке. Сходным образом в данном изобретении предусматривается применение олигонуклеотидной конструкции по изобретению для изготовления лекарственного средства для введения изменений в целевую РНК-последовательность в клетке млекопитающего, предпочтительно, в человеческой клетке, как описано в данной заявке.
Композиция, подбор доз и способ введения для применения в терапии
Олигонуклеотидные конструкции по изобретению удобно вводить в виде водного раствора, например, в физиологическом растворе, или в виде суспензии, которые необязательно содержат добавки, эксципиенты и другие ингредиенты, совместимые с применением в фармации, в концентрациях в диапазоне от 1 нг/мл до 1 г/мл, предпочтительно, от 10 нг/мл до 500 мг/мл, более предпочтительно, от 100 нг/мл до 100 мг/мл. Доза колеблется от примерно 1 мкг/кг до примерно 100 мг/кг, предпочтительно, от примерно 10 мкг/кг до примерно 10 мг/кг, более предпочтительно, от примерно 100 мкг/кг до примерно 1 мг/кг. Введение можно осуществлять в виде ингаляции (например, с применением небулайзера), интраназально, перорально, с помощью инъекции или инфузии, внутривенно, подкожно, интрадермально, интракраниально, внутримышечно, интратрахеально, интраперитонеально, интраректально, непосредственной инъекцией в опухоль и т.п. Возможно введение в твердом виде, в виде порошка, в виде пилюль или в любой другой форме, совместимой с применением фармацевтической композиции у человека.
Данное изобретение особенно подходит для лечения генетических заболеваний, например, таких как муковисцидоз, альбинизм, дефицит альфа-1-антитрипсина, болезнь Альцгеймера, боковой амиотрофический склероз, астма, β-талассемия, CADASIL-синдром, болезнь Шарко-Мари-Тута, хроническая обструктивная болезнь легких (COPD, ХОБЛ), дистальная спинальная мышечная атрофия (DSMA), мышечная дистрофия Дюшенна/Беккера, дистрофический буллезный эпидермолиз, буллезный эпидермолиз, болезнь Фабри, расстройства, ассоциированные с Фактором V Лейдена, семейный аденоматозный полипоз, галактоземия, болезнь Гаухера, болезнь Гоше, дефицит активности глюкозо-6-фосфатдегидрогеназы, гемофилия, врожденный гемохроматоз, синдром Хантера, болезнь Хантингтона, синдром Гурлер, воспалительное заболевание кишечника (IBD), врожденный синдром полиагглютинации, врожденный амавроз Лебера, синдром Леша-Нихана, синдром Линча, синдром Марфана, мукополисахаридоз, мышечная дистрофия, дистрофическая миотония типа I и типа II, нейрофиброматоз, болезнь Ниманна-Пика типа А, В и С, рак, ассоциированный с антигеном NY-ESO1, болезнь Паркинсона, синдром Пейтца-Егерса, фенилкетонурия, болезнь Помпе, первичная цилиарная дискинезия, расстройства, связанные с мутацией в гене протромбина, такой как мутация в гене протромбина G20210A, легочная гипертензия, пигментная дистрофия сетчатки, болезнь Сандхоффа, синдром тяжелого комбинированного иммунодефицита (SCID), серповидноклеточная анемия, спинальная мышечная атрофия, болезнь Штаргардта, болезнь Тая-Сакса, Х-сцепленный тяжелый комбинированный иммунодефицит, различные формы рака (например, BRCA1 и 2-связанный синдром наследственного рака яичников и молочной железы) и т.п.
Согласно некоторым вариантам олигонуклеотидную конструкцию можно доставлять системно, но чаще доставляют олигонуклеотидную конструкцию к клеткам, обладающим фенотипом целевой последовательности. Например, мутации в CFTR вызывают муковисцидоз (кистозный фиброз), наблюдаемый главным образом в эпителиальной ткани легких, поэтому в случае CFTR целевой последовательности предпочтительно следует доставлять олигонуклеотидную конструкцию конкретно и непосредственно в легкие. Это удобно осуществлять посредством ингаляции, например, порошка или аэрозоля, обычно с применением небулайзера. Особенно предпочтительны небулайзеры (МЕШ-небулайзеры), в которых применяются так называемые вибрирующие сетки-мембраны, включая PARI eFlow (Rapid) или i-neb от компании Respironics. Авторы изобретения нашли, что вдыхание олигонуклеотидных конструкций может привести к системному распределению олигонуклеотидной конструкции и всасыванию, в том числе, клетками тканей кишечника, печени, поджелудочной железы, почки и слюнных желез в том числе. Поэтому следует ожидать, что доставка олигонуклеотидных конструкций по изобретению посредством вдыхания также может эффективно нацеливать на эти клетки, что в случае нацеливания на ген CFTR могло бы привести к ослаблению симптомов в желудочно-кишечном тракте, также ассоциируемых с муковисцидозом. В случае других целевых последовательностей, в зависимости от заболевания и/или органа-мишени, введение может быть топическим (например, на коже), интрадермальным, подкожным, внутримышечным, внутривенным, может являться глазной инъекцией и т.д.
При некоторых заболеваниях наблюдается повышенная толщина слизистого слоя, что приводит к уменьшению всасывания лекарственных средств в легких. Одним из таких заболеваний является хронический бронхит, а другим примером является муковисцидоз. Имеются различные лекарственные формы для нормализации слизистой оболочки, например, такие как ДНКазы, гипертонический солевой раствор или маннит, который продается под названием Бронхитол (Bronchitol). Если средства нормализации слизистой оболочки применяются в комбинации с редактирующими РНК олигонуклеотидными конструкциями, например, такими, как олигонуклеотидные конструкции по изобретению, они могут повысить эффективность этих лекарственных средств. Поэтому олигонуклеотидные конструкции по изобретению вводят субъекту, предпочтительно, человеку, предпочтительно, совместно со средствами нормализации слизистой, предпочтительно, со средствами нормализации слизистой, описанными в данной заявке. Помимо этого введение олигонуклеотидных конструкций по изобретению можно проводить совместно с введением синтетического препарата для лечения CF, такого как потенцирующие соединения, например калидеко (ивакафтор; VX-770), или корректирующие соединения (корректоры), например, VX-809 (лумакафтор) и/или VX-661. Другие комбинированные лекарственные средства при CF могут включать олигонуклеотидную конструкцию по изобретению в комбинации со стимулятором аденозиндезаминазы, в качестве которого применяется IFN-гамма или TNF-альфа.
В качестве альтернативы, или в комбинации со средствами нормализации слизистой, для эффективной доставки редактирующих РНК молекул к эпителиальным клеткам, например, легкого или кишечника, можно применять частицы или наночастицы, проникающие через слизистую. Соответственно, при введении олигонуклеотидной конструкции по изобретению субъекту, предпочтительно, человеку, предпочтительно, применяют доставку в виде частиц или наночастиц, проникающих через слизистую.
Хронические или острые легочные инфекции часто наблюдаются у пациентов с такими болезнями, как муковисцидоз. Лечение антибиотиками подавляет бактериальные инфекции и такие их симптомы, как утолщение слизистой и/или образование биопленки. Применение антибиотиков в комбинации с олигонуклеотидными конструкциями по изобретению повысило бы эффективность редактирования РНК благодаря более легкому доступу олигонуклеотидной конструкции к клеткам-мишеням. Соответственно, введение олигонуклеотидной конструкции по изобретению субъекту, предпочтительно, человеку, предпочтительно совмещать с лечением антибиотиками для подавления бактериальных инфекций и их симптомов, таких как утолщение слизистой и/или образование биопленки. Антибиотики можно вводить системно и/или местно.
Для применения по отношению к больным муковисцидозом олигонуклеотидные конструкции по изобретению, либо расфасованные, либо связанные олигонуклеотидные конструкции по изобретению, можно объединять с любыми средствами нормализации слизистой, например, такими как ДНКаза, маннит, гипертонический солевой раствор и/или с синтетическим препаратом для лечения CF, таким как потенцирующие соединения, например, ивакафтор, или корректирующие соединения, например, лумакафтор и/или VX-661.
Для облегчения доступа к клеткам-мишеням можно применять бронхоальвеолярный лаваж (BAL) с целью очистить легкие перед введением олигонуклеотидных конструкций по изобретению.
В данном изобретении также предусматривается олигонуклеотидная конструкция, содержащая нуклеотидную последовательность согласно любой из SEQ ID NOs: с 1 до 3:
GFP SEQ ID NO: 1 
СЕР290 SEQ ID NO: 2 
CFTR SEQ ID NO: 3 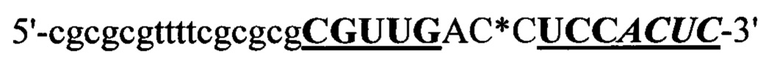
Соответствующая последовательность SEQ ID NO: 9 G551D мРНК 
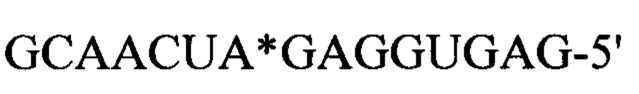
Строчные буквы = ДНК с предрасположенностью образовывать Z-ДНК структуру Жирный шрифт, подчеркнуто = 2'-O-метил
Курсив = фосфоротиоатные межнуклеозидные связи и 2'-O-метил
С* = основание, противостоящее целевому аденозину, подлежащему редактированию
Аналогично, в данном изобретении предусматривается также олигонуклеотидная конструкция, содержащая нуклеотидную последовательность в соответствии с любой из SEQ ID NOs: с 16 до 22 (и необязательно модификации нуклеиновых кислот, описанные в Примере 1 для таких нуклеотидных последовательностей) или с любой из SEQ ID NOs: 2, 3, 28, 38, 39, 40 и 41 (и необязательно модификации нуклеиновых кислот, описанные в Примерах 2-5 для таких нуклеотидных последовательностей).
Общие замечания
Термины "аденин", "гуанин", "цитозин", "тимин", "урацил" и гипоксантин (нуклеиновое основание в инозине) относятся к нуклеиновым основаниям как таковым.
Термины аденозин, гуанозин, цитидин, тимидин, уридин и инозин относятся к нуклеиновым основаниям, связанным с остатком сахара (дезокси)рибозы ((дезокси)рибозильным фрагментом).
Термин "нуклеозид" относится к нуклеиновому основанию, связанному с (дезокси)рибозильным фрагментом.
Термин нуклеотид относится к соответствующей структуре нуклеиновое основание - (дезокси)рибозил-фосфорсодержащий линкер, а также к любым химическим модификациям рибозного фрагмента или фосфорсодержащей группы. Таким образом термин включает нуклеотид, содержащий закрытый рибозильный фрагмент (содержащий 2'-4' мостик, содержащий метиленовую группу или любую другую группу, хорошо известную в уровне техники), нуклеотид, включающий линкер, представляющий собой фосфодиэфирный, фосфотриэфирный, фосфо(ди)тиоатный, метилфосфонатный, фосфорамидатные линкеры и т.п.
Иногда термины аденозин и аденин, гуанозин и гуанин, цитозин и цитидин, урацил и уридин, тимин и тимидин, инозин и гипоксантин применяются как синонимы по отношению к соответствующему нуклеиновому основанию, нуклеозиду или нуклеотиду.
Иногда термины нуклеиновое основание, нуклеозид и нуклеотид применяются как синонимы, если только в соответствии с контекстом явно не требуется иное.
Во всех случаях, когда дается ссылка на "олигонуклеотид", имеются в виду как олигорибонуклеотиды, так и дезоксиолигорибонуклеотиды, если контекст не требует иного. Во всех случаях, когда дается ссылка на олигорибонуклеотид, он может включать основания A, G, С, U или I. Во всех случаях, когда дается ссылка на дезоксиолигорибонуклеотид, он может включать основания A, G, С, Т или I.
Во всех случаях, когда дается ссылка на нуклеотиды в олигонуклеотидной конструкции, то имеются в виду такие нуклеотиды, как цитозин, 5-метилцитозин, 5-гидроксиметилцитозин и β-D-глюкозил-5-гидроксиметилцитозин; когда дается ссылка на аденин, то охватываются N6-метиладенин и 7-метиладенин; когда дается ссылка на урацил, то охватываются дигидроурацил, 4-тиоурацил и 5-гидроксиметилурацил; когда дается ссылка на гуанин, то она включает 1-метилгуанин.
Во всех случаях, когда дается ссылка на нуклеозиды или нуклеотиды, то они включают производные рибофуранозы, такие как 2'-дезокси, 2'-гидрокси и 2'-O - замещенные варианты, такие как 2'-O-метил, а также другие модификации, включая 2'-4' мостиковые варианты.
Во всех случаях, когда дается ссылка на олигонуклеотиды, связи между двумя мононуклеотидами могут представлять собой фосфодиэфирные связи, а также их модификации, включая фосфодиэфирные, фосфотриэфирные, фосфо(ди)тиоатные, метилфосфонатные, фосфорамидатные линкеры и т.п.
Термин "содержащий" является синонимом "включающий", а также "состоящий из", например, композиция, "содержащая" X, может состоять исключительно из X или может включать нечто еще, например, X+Y.
Термин "примерно", "около" по отношению к числовой величине x является необязательным и означает, например, x±10%.
Термин "по существу", "практически" не исключает "полностью", "совсем", например, композиция, которая "по существу не содержит" Y, может совсем не содержать Y. Где это уместно, слово "по существу", "практически" можно исключить из формулы изобретения.
Термин "ниже", "3"' применительно к последовательности нуклеиновой кислоты а означает "дальше по ходу последовательности в 3' направлении"; термин "выше", "5"' означает обратное. Таким образом, в любой последовательности, кодирующей полипептид, стартовый кодон находится выше стоп-кодона (терминирующего кодона) в смысловой цепи, но ниже стоп-кодона в антисмысловой цепи.
Ссылки на "гибридизацию" обычно относятся к специфической гибридизации и исключают неспецифическую гибридизацию. Специфическая гибридизация может происходить в выбранных условиях эксперимента с применением методов, хорошо известных в уровне техники, чтобы гарантировать, что большинство устойчивых взаимодействий между зондом и мишенью представляет собой такие взаимодействия, в которых идентичность последовательностей зонда и мишени составляет по меныпе70%, предпочтительно, по меньшей мере 80%, более предпочтительно, по меньшей мере 90%.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1: Показана целевая РНК-последовательность и олигонуклеотидная конструкция по изобретению; А: олигонуклеотидная конструкция изображена как единичная олигонуклеотидная "одноногая" конструкция с нацеливающим участком на ее 3' конце и рекрутирующим участком на ее 5' конце. В: олигонуклеотидная конструкция изображена как единичная олигонуклеотидная "одноногая" конструкция с нацеливающим участком на ее 5' конце и рекрутирующим участком на ее 3' конце; редактирующая структура покрашена в серый цвет, серым цветом показан домен узнавания на левой стороне и каталитический домен на правой стороне, указывающий на реакцию дезаминирования ('молния') в сайте-мишени.
Фигура 2: Показана целевая РНК структура с олигонуклеотидной конструкцией: на рисунке А показана олигонуклеотидная конструкция с линкером между нацеливающим участком и рекрутирующим участком; на рисунке В показан вариант, в котором олигонуклеотидная конструкция содержит две олигонуклеотидных последовательности - не связанные ковалентной связью-взаимодействующие посредством антисмыслового спаривания оснований по Уотсону-Крику по своим соответствующим 3' сегментам.
Фигура 3: Показана целевая РНК структура с олигонуклеотидной конструкцией в "двуногом" формате, причем нацеливающий участок разделен (или "расщеплен") рекрутирующим участком; на рисунке А показан сайт редактирования в целевой РНК-последовательности выше относительно рекрутирующего участка олигонуклеотидной конструкции; на рисунке В показан сайт редактирования ниже относительно рекрутирующего участка олигонуклеотидной конструкции.
Фигура 4: Флуоресцентная спектроскопия лунок, показывающая зеленую флуоресценцию в HeLa клетках после трансфекции с использованием липофектамина с pGFPstop57 и указанными олигонуклеотидами ("oligo"), за исключением: на верхнем левом рисунке представлены необработанные клетки; на четырех нижних рисунках представлены контрольные образцы, которые не получали по меньшей мере один из указанных компонентов. Фигуры 4а и 4b отличаются только контрастностью.
Фигура 5: Флуоресцентная микроскопия лунок, показывающая зеленую флуоресценцию в HeLa клетках после трансфекции с использованием липофектамина с pGFPstop57, pADAR2 и олигонуклеотидом #11 с различным соотношением двух плазмид. На верхнем левом рисунке представлены необработанные клетки; на нижних рисунках показаны клетки с плазмидой, кодирующей скорее немутантный GFP, нежели pGFPstop57.
Фигура 6: Флуоресцентная микроскопия лунок, показывающая зеленую флуоресценцию в HeLa клетках через 24 часа или через 48 часов после отсутствия обработки, или трансфекцию с использованием 50 нМ или 200 нМ олигонуклеотида #11 (или с 500 нМ контрольного олигонуклеотида) совместно с pGFPstop57.
Фигура 7: Спектры FACS необработанных (левый пик) или трансфицированных (правый пик) HeLa клеток. На рисунках 7A-7F клетки трансфицировали с использованием pGFPstop57 и олигонуклеотида #11. На рисунках 7B-7F (но не 7А) клетки также трансфицировали с использованием pADAR2. На рисунках 7G и 7Н экспрессировали немутантный GFP.
Все варианты, проиллюстрированные на Фигурах, можно объединить, как объясняется в подробном описании настоящего изобретения.
СПОСОБЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Пример 1: Реверсия (обращение) нонсенс-мутации в GFP целевой РНК посредством сайт-направленного редактирования А в I
Олигонуклеотидная конструкция для применения: 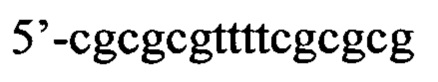
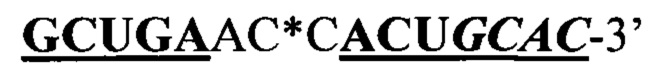 (SEQ ID NO: 1). Клетки HeLa (АТСС, CCL-2) культивировали в модифицированной по способу Дульбекко среде Игла (Life Technologies), дополненной 10% термоинактивированной фетальной бычьей сывороткой. Клетки хранили в атмосфере влажного воздуха с 5% СО2. Клетки засевали в 24-луночный планшет за день до трансфекции и по достижении конфлюентности 70-80%. Использовали GFP репортерную конструкцию, лишенную GFP активности ввиду того, что был применен терминирующий (стоп) кодон TGA. 100-200 нг плазмидной ДНК и 500 нг олигонуклеотидной (10-100 пмолей) конструкции смешивали в соответствующем количестве среды Opti-MEM I (Life Technologies) и реагента Lipofectamine 2000 и клетки трансфицировали согласно методике производителя. Клетки инкубировали при 37°C в СО2 термостате и среду заменяли через 4-6 часов. Через 48 часов клетки анализировали под флуоресцентным микроскопом с целью оценки эффективности олигонуклеотида для восстановления экспрессии GFP.
(SEQ ID NO: 1). Клетки HeLa (АТСС, CCL-2) культивировали в модифицированной по способу Дульбекко среде Игла (Life Technologies), дополненной 10% термоинактивированной фетальной бычьей сывороткой. Клетки хранили в атмосфере влажного воздуха с 5% СО2. Клетки засевали в 24-луночный планшет за день до трансфекции и по достижении конфлюентности 70-80%. Использовали GFP репортерную конструкцию, лишенную GFP активности ввиду того, что был применен терминирующий (стоп) кодон TGA. 100-200 нг плазмидной ДНК и 500 нг олигонуклеотидной (10-100 пмолей) конструкции смешивали в соответствующем количестве среды Opti-MEM I (Life Technologies) и реагента Lipofectamine 2000 и клетки трансфицировали согласно методике производителя. Клетки инкубировали при 37°C в СО2 термостате и среду заменяли через 4-6 часов. Через 48 часов клетки анализировали под флуоресцентным микроскопом с целью оценки эффективности олигонуклеотида для восстановления экспрессии GFP.
В дальнейших экспериментах снова применяли мутантный GFP, содержащий внутренний TAG стоп-кодон вследствие точковой мутации G→А, экспрессирующий при использовании плазмиды ('pGFPstop57'). Клетки дополнительно трансфицировали при использовании плазмиды, кодирующей ADAR2, с целью гарантировать, что клетки были способны осуществлять редактирование РНК. Различные олигонуклеотиды получали для восстановления экспрессии GFP посредством дезаминирования мутантного остатка А, на основе принципа нацеливания участка, специфического к GFP мутации, и рекрутирующего участка на основе GluR-B.
Было проведено испытание семи РНК олигонуклеотидов, и они нацеливались на короткие, средние или длинные формы GluR-B (S/M/L; различная длина рекрутирующего участка). Помимо этого, они содержали нацеливающие и рекрутирующие участки в любом порядке (выше (5')/ниже (3')), и в некоторых случаях олигонуклеотиды включали химически модифицированные области (рекрутирующий участок химически модифицировали таким образом, чтобы он включал 2'-ОМе сахара и фосфоротиоатные связи; нацеливающий участок модифицировали таким же образом, за исключением положения мутантного остатка А (дважды подчеркнут) и двух фланкирующих его нуклеотидов). Все олигонуклеотиды включают SEQ ID NO: 7 (подчеркнута), и участки, нацеленные на GFP, показаны жирным шрифтом:

Клетки HeLa культивировали в модифицированной по способу Дульбекко среде Игла, дополненной 10% термоинактивированной фетальной бычьей сывороткой. Клетки хранили в атмосфере влажного воздуха с 5% СО2. 8×104 клеток засевали в одну лунку 24-луночного планшета за один день до трансфекции, и когда они достигали конфлюентности 70-80%, их временно трансфицировали, используя (i) олигонуклеотид + плазмиду GFPstop57 или (ii) олигонуклеотид + GFPstop57 + ADAR2 плазмиды, применяя реагент Lipocetamine 2000 согласно методике производителя. Среду заменяли на свежую через 24 часа после трансфекции и экспрессию GFP проверяли под флуоресцентным микроскопом еще через 24 часа.
Также проводили анализ FACS. Клетки трипсинизировали и собирали в пробирки Эппендорфа, а затем ресуспендировали в окрашивающем буфере для проточной цитометрии. Интактные клетки отбирали на основании морфологических свойств, используя прямое светорассеяние и угловое светорассеяние, исключающие дебрис. Рассчитывали медианное и среднее значения GFP средней интенсивности флуоресценции (MFI) интактных клеток. Наложение гистограмм делали с помощью редактора форматов.
Результаты для клеток, трансфицированных в отсутствие ADAR плазмиды, показаны на Фигуре 4, а уровни зеленой флуоресценции, превышающие уровни, наблюдаемые в контрольных образцах (которые объясняются аутофлуоресценцией клеток и/или компонентов среды), четко видны для всех семи олигонуклеотидов. Олигонуклеотиды #10 и #11 показали наилучшие результаты (оба имеют длинный GluR-B рекрутирующий участок), и для олигонуклеотида с 5' рекрутирующим участком (#11) результаты были немного лучше.
Поэтому для последующих испытаний был выбран олигонуклеотид #11 в комбинации с плазмидой, кодирующей ADAR2. Опять же на Фигуре 5 видно, что уровни флуоресценции клеток, обработанных олигонуклеотидом #11, выше уровней флуоресценции отрицательного контроля. Для сравнения клетки трансфицировали при использовании плазмиды, кодирующей немутантный GFP, и наблюдали ожидаемую флуоресценцию (Фигура 5, нижний рисунок).
Проводили испытания с разными концентрациями олигонуклеотида #11 в диапазоне от 50 до 1500 нМ. На Фигуре 6 показаны результаты через 24 часа или через 48 часов при концентрации 50 и 200 нМ, наряду с необработанными клетками (слева) и контрольными клетками с концентрацией контрольного олигонуклеотида 500 нМ, имеющего ту же длину и те же химические модификации, что и олигонуклеотид #11. Для олигонуклеотида #11 наблюдается увеличение флуоресценции в зависимости от времени, которое не наблюдается для контрольного олигонуклеотида.
Влияние олигонуклеотида на экспрессию GFP также наблюдали методом FACS (Фигура 7). Клетки, обработанные олигонуклеотидом #11, в присутствии (7B-7F) или в отсутствие (7А) плазмиды ADAR2, показывали увеличение GFP флуоресценции по сравнению с необработанными клетками (левый пик). В качестве положительного контроля использовали немутантный GFP (7G-7H).
Пример 2: Введение скрытого сайта сплайсинга в целевую РНК для СЕР290 посредством сайт-направленного редактирования А в I
Применяли олигонуклеотидную конструкцию: 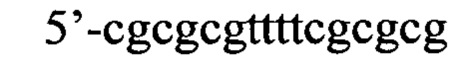
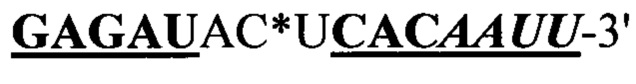 (SEQ ID NO: 2).
(SEQ ID NO: 2).
Все линии клеток являются линиями человеческих фибробластов, взятых при биопсии кожи. Клетки FBL1 (CL10-00008) и FBL2 (CL12-00027) являются клетками дикого типа и представляют собой клетки контрольных линий, как клетки FBL3 (CL12-00035), так и клетки FBL4 (CL12-00036) являются are гомозиготным мутантом по мутации в СЕР290 (c.2991+1655A>G). Все клеточные линии выращивали в среде DMEM (Life Technologies), дополненной 20% FBS, 1% Pen/strep и 1% пирувата натрия.
За день до трансфекции клетки засевали при плотности 2×105/лунка в 6-луночный планшет в общем объеме среды 2.5 мл. В день трансфекции в каждую лунку добавляли AON (антисмысловые олигонуклеотиды) для тестирования с конечной концентрацией 100 нМ, используя maxPEI (Poliscience) в качестве агента для трансфекции при соотношении масс олигонуклеотид:РЕ1 1:4. Через 24 ч клетки отмывали с помощью PBS, клеточный лизат собирали и замораживали при -80°С.
РНК выделяли из клеточных лизатов, которые хранили при -80°С, используя набор Promega ReliaPrep RNA Cell Miniprep System. Тотальную РНК количественно определяли на спектрофотометре Nanodrop 2000.
400 нг РНК использовали в качестве матрицы для синтеза кДНК с применением набора для синтеза кДНК Verso (Thermoscientific) согласно инструкциям производителя.
Для этой реакции кДНК разводили 2.5х и 2 мкл этой разведенной кДНК использовали в качестве матрицы. Для амплификации целевой последовательности использовали AmpliTaq Gold® 360 ДНК полимеразу от компании Life Technologies. Использовали праймеры ex26_Fw (SEQ ID NO: 10) и ex27_Rv (SEQ ID NO: 11) в следующих условиях ПЦР (PCR): выдерживание 5 мин при 95°С, денатурация 30 сек при 95°С, отжиг праймеров 30 сек при 58°С и элонгация (удлинение) праймеров 35 сек при 72°С, 35 циклов, конечное удлинение 7 мин при 72°С.
ПЦР фрагменты анализировали на приборе Agilent 2100 Bioanalyzer с применением набора DNA 1000 Kit от компании "Agilent technologies". Этот набор содержит чип, состоящий из связанных между собой микроканалов, с помощью которых фрагменты разделяются в соответствии с их размерами по мере их продвижения в процессе электрофореза. Для измерения уровня экспрессии СЕР290 мРНК транскрипт дикого типа и мутантный транскрипт амплифицировали в виде фрагментов 93 п.о. и 117 п.о., соответственно. Для нормализации использовали большую мРНК для человеческого Р0 рибосомного белка (RPLP0). Использовали праймеры wt_Fw (SEQ ID NO: 12), wt-Rv (SEQ ID NO: 13), mt_Fw (SEQ ID NO: 14) и mt_Rv (SEQ ID NO: 15). Для этой реакции в качестве матрицы использовали смесь SYBR Select Master Mix от компании "Life Technologies" вместе с кДНК в разведении 10х. Программа ПЦР: 50°С в течение 2 мин, 95°С в течение 2 мин, 50 циклов при 95°С в течение 15 сек, 62.5°С в течение 1 мин.
Пример 3: Реверсия аминокислотной замены в мутантной CFTR G551D целевой РНК посредством сайт-направленного редактирования А в I
Олигонуклеотидная конструкция для применения: 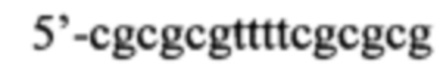
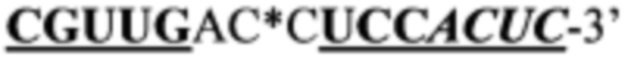 (SEQ ID NO: 3).
(SEQ ID NO: 3).
Линии клеток являются линиями человеческих фибробластов, взятых при биопсии кожи и перикарда (околосердечной сумки), GM00142 и GM03465, соответственно, (Coriell Institute Cell Repository). Обе они являются гетерозиготными: один аллель несет делеционную мутацию дельтаF508 (Phe508Del), а второй аллель несет транзицию G→A в нуклеотиде 1784 (1784G>A), которая превращает кодон gly-551 (GGT) в asp (GAT), в результате приводя к миссенс-мутации в экзоне 11 в CFTR гене [Gly551 Asp (G551D)]. Все клеточные линии выращивали в минимальной эссенциальной среде Игла (Life Technologies) с солями Эрла и заменимыми аминокислотами, дополненной 15% неактивированной FBS и 1% Pen/strep.
За день до трансфекции клетки засевали при плотности 2×105/лунка в 6-луночный планшет в общем объеме среды 2.5 мл. В день трансфекции в каждую лунку добавляли олигонуклеотид для тестирования с конечной концентрацией 100 нМ, используя maxPEI (Poliscience) в качестве агента для трансфекции при соотношении масс олигонуклеотид:РЕ1 1:4. Через 24 ч клетки отмывали с помощью PBS и клеточный лизат собирали и замораживали при -80°С.
РНК выделяли из клеточных лизатов, которые хранили при -80°С, используя набор Promega ReliaPrep RNA Cell Miniprep System. Тотальную РНК количественно определяли на спектрофотометре Nanodrop 2000. 400 нг РНК использовали в качестве матрицы для синтеза кДНК с применением набора для синтеза кДНК Verso (Thermoscientific) согласно инструкциям производителя. Сначала проводили ПЦР, используя 1 мкл кДНК с 0.4 мкМ прямого и обратного праймеров, объединяли 25 мкМ каждого из dNTP, 1× AmpliTaq Gold® 360 буфера, 3.125 30 мМ MgCl2 и 1.0 единиц AmpliTaq Gold® 360 полимеразы (все от Life Technologies). Применяли следующие праймеры: SEQ ID NOs: 26 (прямой) и 27 (обратный). Циклы ПЦР осуществляли в следующих условиях: начальная стадия денатурации при 95°С в течение 7 мин с последующими 30 циклами по 30 с при 95°С, 30 с при 55°С и 45 с при 72°С. ПЦР амплификацию заканчивали финальной элонгацией (удлинением) в течение 7 мин при 72°С. 1 мкл продуктов предыдущей ПЦР использовали в программе гнездовой (нестед) ПЦР, содержащей 0.4 мкМ прямых праймеров и уникальный MiSeq индексный праймер на один образец, 25 мкМ каждого dNTP, 1× AmpliTaq Gold® 360 буфера, 3.125 мМ MgCl2 и 1.0 единицу AmpliTaq Gold® 360 полимеразы. ПЦР циклы проводили в следующих условиях: начальная стадия денатурации при 95°С в течение 7 мин, а затем 25 циклов по 30 с при 95°С, 30 с при 60°С и 45 с при 72°С. Реакции заканчивали финальной элонгацией (удлинением) в течение 7 мин при 72°С. Перед загрузкой в секвенатор ПЦР-продуктов, содержащих MiSeq праймерные последовательности, определяли концентрацию очищенных ПЦР-продуктов на флуориметре Qubit® 2.0 (Life Technologies) в соответствии с протоколом производителя. Короче говоря, готовили две пробирки для анализа в качестве стандартов 20-кратным разведением 2 стандартов в рабочем растворе. Для каждого образца 200 мкл рабочего раствора готовили в 10 отдельных пробирках, 1 мкл ПЦР-продукта вводили в этот раствор и интенсивно перемешивали в течение пары секунд. Количественно определяли образцы по сравнению с двумя стандартными образцами и соответственно рассчитывали концентрацию в нг/мкл. Секвенирование ПЦР-продуктов осуществляли на приборе MiSeq™ от Illumina, который использует секвенирование методом синтеза (SBS) для быстрого получения высококачественных данных о последовательности.
Пример 4: Реверсия мутации замены аминокислоты в транскрипте α-1-антитрипсина (А1АТ) посредством целевого редактирования А в I для лечения недостаточности А1АТ
Олигонуклеотиды (SEQ ID NO: 28):
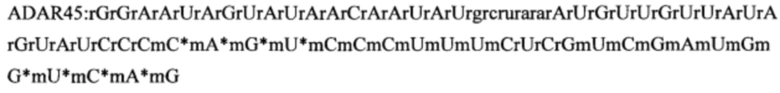

r = нет модификации
m = 2'O-Ме
* = фосфоротиоатная связь
Трансфекция фибробластов печени
Фибробласты печени, предоставленные Coriell Cell Repository (GM11423), получали от донора, гомозиготного по Z аллелю (ZZ), который является результатом транзиции G>A в нуклеотиде 9989 в экзоне 5 гена SERPINA1 [9989G>A], приводящей к замене лизина на глутаминовую кислоту в кодоне 342 [Glu342Lys (E342K)]. Клетки сохраняли в среде ЕМЕМ (Life Technologies) и дополняли 15% FBS. За день до трансфекции клетки засевали в 6-луночный планшет в общем объеме 2 мл среды. В день трансфекции олигонуклеотид добавляли к 1x PBS (Thermo Fisher Scientific) в микроцентрифужную пробирку на 1.5 мл, смешивали с MaxPei (Polysciences) и совместно инкубировали в течение 20 мин. Одновременно клеточную культуральную среду удаляли из клеток и добавляли соответствующее количество свежей среды ЕМЕМ с 15% FBS. Затем на клетки осторожно по каплям из пипетки добавляли ДНК/Олигонуклеотид-MaxPei разведенную смесь. Клетки инкубировали при 37°С и заменяли среду на свежую через 6-24 ч.
Выделение РНК
Выделение РНК проводили, используя систему Reliaprep RNA Cell Miniprep System (Promega) согласно методике производителя. Коротко говоря, культуральную среду удаляли и клетки отмывали холодным PBS. 250 мкл буфера для лизиса добавляли в каждую лунку 6-луночного планшета. Планшет осторожно перемешивали на качалке и клетки полностью лизировали многократным пипетированием по поверхности лунки. К лизату добавляли 85 мкл изопропанола в соответствии с рекомендациями и лизат переносили на миниколонку и центрифугировали в течение 30 сек при 12,000-14,000 g (RT), жидкости отбрасывали. Добавляли 500 мкл раствора для отмывки РНК и снова центрифугировали в течение 30 сек при 12,000-14,000g. В стерильной пробирке готовили мастер-микс для инкубации ДНКазы I, смешивая образец 24 мкл Yellow Core буфера, 3 мкл 0.09М MnCl2, 3 мкл ДНКазы I и 30 мкл свежеприготовленной смеси ДНКазы I (микс), помещали на мембране в колонке каждого образца, инкубировали в течение 15 мин при RT. Затем прибавляли 200 мкл промывного раствора для колонок и центрифугировали в течение 15 сек при 12,000-14,000g. После этого добавляли 500 мкл раствора для отмывки РНК, центрифугировали в течение 30 сек при 12,000-14,000g и жидкость отбрасывали. Миниколонку помещали в новую пробирку-сборник и добавляли 300 мкл раствора для отмывки РНК и центрифугировали при 14,000g в течение 2 мин. Каждую миниколонку помещали в пробирку для элюирования и на мембрану добавляли безнуклеазную воду и центрифугировали 1 мин. Под конец определяли концентрацию РНК на спектрофотометре Nanodrop.
Синтез кДНК
Синтез кДНК проводили, используя набор Verso cDNA synthesis kit (Thermo Fisher) с загрузкой по РНК 500 нг или 1000 нг. Смесь РНК (микс) готовили из заданного количества РНК, доводя водой до нужного объема 11 мкл. Эту смесь нагревали в течение 5 мин. при 70°С, затем охлаждали. Смесь кДНК готовили согласно схеме пипетирования, предусмотренной поставщиком. 9 мкл смеси кДНК помещали в реакционную пробирку и добавляли 11 мкл смеси РНК и выдерживали в амплификаторе (термоциклере) в течение 30 мин. при 42°С и 2 мин. при 95°С.
ПЦР
ПЦР проводили, используя набор AmpliTaq Gold 360 DNA Polymerase 1000U Kit (Applied Biosystems). Мастер-микс для ПЦР готовили, добавляя 1 мкл кДНК, 2.5 мкл 10х буфера, 3 мкл MgCl2, 0.5 мкл dNTPs, 1 мкл каждого праймера, 0.5 мкл Taq ДНК полимеразы, и дополняли водой до объема 25 мкл. Ниже дается программа ПЦР: 95°С в течение 5 мин, 95°С в течение 30 с, 55°С в течение 30 с, 72°С в течение 1 мин - 35 циклов и 72°С в течение 7 мин. После ПЦР образцы анализировали на Биоанализаторе, используя набор DNA 1000 kit (Agilent) и программу.
Очистка образца ПЦР
Образцы очищали, используя набор NucleoSpin PCR Clean-up kit (Macherey-Nagel), чтобы гарантировать удаление примесей из продуктов ПЦР. 1 объем образца ПЦР смешивали с 2 объемами NT1 и образцы нагружали на гель NucleoSpin® и PCR Clean- up колонку помещали в пробирку-сборник и центрифугировали в течение 30 с при 11000 x g. Жидкость отбрасывали и колонку снова помещали в пробирку-сборник. 700 мкл буфера NT3 добавляли к гелю NucleoSpin® и центрифугировали в течение 30 с при 11000 x g. Стадию отмывки повторяли и пробирки центрифугировали в течение 1 мин при 11000 x g для удаления буфера NT3.
Гель NucleoSpin® и колонку PCR Clean-up помещали в новую пробирку Эппендорфа на 1.5 мл, добавляли 15-30 мкл буфера NE и инкубировали в течение 1 мин. при комнатной температуре, затем центрифугировали в течение 1 мин. при 11000 x g. Концентрацию ДНК определяли методом спектрофотометрии на приборе Nanodrop.
Расщепление рестриктазами
После очистки продуктов ПЦР ампликон SERPINA1 подвергали расщеплению рестриктазой по сайту Hyp99I. Фермент распознавал последний G в последовательности CGACG в WT и разрезал ампликон на два фрагмента, но не мог разрезать последовательность CGACA в мутантном варианте вследствие отсутствия второго остатка G. Таким образом, если редактирование было успешным, должно быть некоторое количество WT ДНК в качестве субстрата для расщепления рестриктазой по Hyp99I. 1 единицу Hyp99I (NEB) использовали для расщепления 0.1 мкг ДНК. Загрузка ДНК составляла 0.5 мкг. Образцы разводили в безнуклеазной воде и инкубировали в течение 1-1.5 часов при 37°С, фермент инактивировали последующей инкубацией образцов при 65°С в течение 20 мин. После инкубации образцы загружали в Биоанализатор, используя набор DNA1000 kit (Agilent) и программу для визуализации результатов.
TaqMan ПЦР
SNP-генотипирование осуществляли, применяя обычное SERPINA1 SNP-генотипирование (ThermoFisher Scientific). Этот анализ проводили параллельно с расщеплением с помощью рестриктаз с целью выяснить, какой анализ обладает большей чувствительностью к обнаружению редактирования. Зонды, специфические к WT и мутантному гену SERPINA1, связаны с различными флуоресцентными группами, VIC и FAM, соответственно. Поэтому мы могли количественно определить увеличение или уменьшение числа транскриптов WT/мутанта. Реакционную смесь готовили, прибавляя 5 мкл мастер-микса и 0.5 мкл смеси зондов и праймеров. Использовали следующие олигонуклеотидные последовательности: WT зонд (SEQ ID NO: 29), "мутантный" зонд (SEQ ID NO: 30), прямой праймер (SEQ ID NO: 31) и обратный праймер (SEQ ID NO: 32). 5.5 мкл реакционной смеси добавляли в определенные лунки 96-луночного планшета. В каждую лунку добавляли 4.5 мкл кДНК.. Устанавливали следующую программу ПЦР: 95°С в течение 10 мин, 92°С в течение 15 сек и 60°С в течение 90 сек, 50 циклов. Результаты анализировали с применением программы CFX Manager.
Пример 5: Реверсия мутации замены аминокислоты G2019S в транскрипте LRRK2 посредством нацеленного редактирования А в I для лечения болезни Паркинсона
Мутации в каталитическом Roc-COR и киназном доменах гена богатой лейциновыми повторами киназы 2 (LRRK2 Gene ID: 120892) являются общей причиной семейной болезни Паркинсона (PD). Мы решили сделать мишенью G2019S мутацию в LRRK2 пре-мРНК транскрипте (Transcript RefSeq NM_l98578.3), используя AONs, способные рекрутировать ADAR1 и 2, посредством полноразмерного или укороченного GluRB участка в качестве рекрутирующего участка, связанного с нацеливающим участком с комплементарностью последовательности, прилегающей к мутации G в А в экзоне 41 в положении G6055 (см. нижеприведенную последовательность с подчеркнутым мутантным G). Эта последовательность также идентифицируется как Genbank dbSNP вариант rs34637584, обычно именуемый G2019S.
Последовательность LRRK2 Экзона 41 cwt G остатком, выделенным в положении 6055:
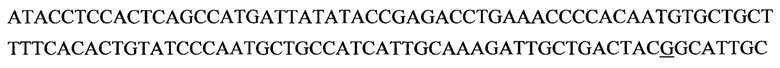
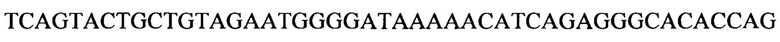 (SEQ ID NO: 33)
(SEQ ID NO: 33)
Мутантный аллель и трансляция:
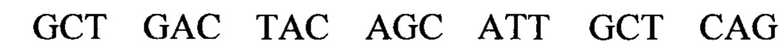 SEQ ID NO: 34
SEQ ID NO: 34
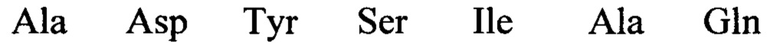 SEQ ID NO: 35
SEQ ID NO: 35
Нормальный аллель и трансляция (после редактирования А в I):
_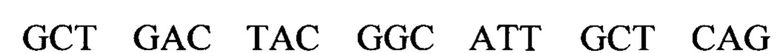 SEQ ID NO: 36
SEQ ID NO: 36
 SEQ ID NO: 37
SEQ ID NO: 37
Нижеприведенные последовательности предназначены для нацеливания на LRRK2 G2019S мутацию. Нуклеотиды, показанные жирным шрифтом, образуют нацеливающий участок; все нуклеотиды содержат 2'-ОМе группы, за исключением подчеркнутых нуклеотидов; *обозначает PS-связь. AONs меняют длину нацеливающего участка и содержат либо 25 (LRRK2-ADAR1 и LRRK2-ADAR3), либо 30 нуклеотидов (LRRK-ADAR2 и 4). AONs меняют длину рекрутирующего участка: укороченный GluRB рекрутирующий участок (LRRK2-ADAR3 и 4) и полноразмерный GluRB рекрутирующий участок (LRRK2-ADAR1 и 2).

Корректирование LRRK2 оценивают в LRRK2G6055A мутантных фибробластах методами секвенирования РНК, вестерн-блоттингом (ауто)фосфорилированного состояния LRRK2, и методом функциональных считываний хорошо известных LRRK2 -ассоциированных митохондриальных фенотипов (скорость поглощения кислорода и трансмембранного потенциала мембраны митохондрий). См.: Tatiana D. Papkovskaia et al. (2012) Hum. Mol. Genet. 21 (19): 4201-4213; Smiths al. (2015) Molecular Neurobiology, 1-17; и Griinewald et al. (2014) Antioxidants & Redox Signaling, 20(13), 1955-1960.
LRRK2G6055A гомозиготную линию фибробластов fff-028 (Telethon Network of Genetic Biobanks), гетерозиготные no G6055A линии ND29492, ND29542, ND29802 и контрольные линии клеток от здоровых доноров GM023074, GM08402 (Coriell Institute), трансфицировали с помощью LRRK2-ADAR AON, используя Lipofectamine 2000. После инкубации в течение 48-96 часов проводили следующие анализы:
1) Для детекции A-в-I редактируемого LRRK2 транскрипта клетки лизировали, РНК выделяли стандартными методами и проводили полуколичественное РНК-секвенирование с использованием следующих праймеров для секвенирования: SEQ ID NOs: 42 и 43. После трансфекции LRRK2-ADAR-AON появлялись A-в-I редактированные мРНК-последовательности.
2) Ранее было показано, что белок LRRK2G6055A (Smith et al, 2015) проявляет повышенное аутофосфорилирование по остатку серина 955 после стрессовой обработки валиномицином (10 мкМ за 24 ч), агентом, вызывающим деполяризацию митохондриальной мембраны вследствие повышенной каталитической активности киназного домена по сравнению с LRRK2wt. Обработка посредством LRRK2-ADAR-AON снижала фосфорилирование по серину-955 обработанных валиномицином LRRK2G6055A клеток, по меньшей мере частично, возможно, даже полностью до уровней дикого типа. Это можно определить вестерн-блоттингом (Smith et al), используя антитела к LRRK2 фосфо-8955 (Abeam клон MJF-R11-(75-l)) и тотальные антитела к LRRK2 (Abeam клон MJFF2 (с41-2)).
3) Фибробласты LRRK2G6055A продемонстрировали изменения митохондриального дыхания (OXPHOS), включая повышенную утечку протонов (Smith et al, 2015;  et al, 2014), которая может стать обратимой после LRRK2-ADAR-AON трансфекции. Скорость поглощения кислорода в условиях основного и интенсивного дыхания оценивали с помощью анализатора внеклеточного потока Seahorse XF24 (Seahorse Bioscience), прибора, который измеряет концентрацию растворенного кислорода в культуральной среде с интервалами 2 сек посредством твердотелых сенсорных зондов. Этот анализ можно проводить в сочетании с набором XF Xell Mito Stress Test kit (Seahorse Bioscience), который посредством последовательной обработки олигомицином, карбонил цианил-4-(трифторметокси)фенилгидразоном (FCCP), ротеноном и антимицином-А может определять метаболические показатели, например, такие как основное дыхание, продуцирование АТФ (АТР), утечку протонов и максимальное дыхание. Анализ проводили в соответствии с инструкциями производителя и утечку протонов определяли как поглощение остаточного кислорода после обработки олигомицином.
et al, 2014), которая может стать обратимой после LRRK2-ADAR-AON трансфекции. Скорость поглощения кислорода в условиях основного и интенсивного дыхания оценивали с помощью анализатора внеклеточного потока Seahorse XF24 (Seahorse Bioscience), прибора, который измеряет концентрацию растворенного кислорода в культуральной среде с интервалами 2 сек посредством твердотелых сенсорных зондов. Этот анализ можно проводить в сочетании с набором XF Xell Mito Stress Test kit (Seahorse Bioscience), который посредством последовательной обработки олигомицином, карбонил цианил-4-(трифторметокси)фенилгидразоном (FCCP), ротеноном и антимицином-А может определять метаболические показатели, например, такие как основное дыхание, продуцирование АТФ (АТР), утечку протонов и максимальное дыхание. Анализ проводили в соответствии с инструкциями производителя и утечку протонов определяли как поглощение остаточного кислорода после обработки олигомицином.
4) Фибробласты LRRK2G6055A продемонстрировали пониженный мембранный потенциал митохондрий по сравнению с контрольными клетками (Smith et al, 2015; Grunewald et al, 2014). Мембранный потенциал митохондрий определяли с помощью липофильного катионного красителя метилового эфира тетраметилродамина (TMRM), применяемого без разведения (0.5-30 нм, определяли экспериментально), вводили в культуральную среду без фенола красного в течение 20 минут. После загрузки красителя флуоресценцию TMRM в митохондриях в равновесии количественно определяли конфокальной микроскопией живых клеток и визуальным анализом. Трансфекция LRRK2-ADAR-AON может повышать мембранный потенциал митохондрий, по меньшей мере частично, возможно, даже полностью до контрольных уровней.
Выводы
Описанные выше примеры показывают, как осуществить нужные изменения в целевой РНК-последовательности посредством сайт-направленного редактирования нуклеотидов в целевой молекуле РНК, используя олигонуклеотидные конструкции по изобретению. В Примерах объясняется, как удалить терминирующий (стоп-) кодон, чтобы снова открыть рамку считывания экспрессионной конструкции GFP, создать сайт сплайсинга, чтобы изменить паттерн сплайсинга целевой РНК, кодирующей СЕР290, и как ввести нужную аминокислотную замену посредством изменения в кодоне в целевой РНК-последовательности, кодирующей мутантный G551D CFTR белок. Успешное редактирование РНК можно наглядно подтвердить, например, посредством наблюдения за флуоресцентными клетками в случае реверсивной (обратной) несмысловой мутации GFP, посредством наблюдения за сдвигом RT-PR полос в геле от дикого типа к мутантному в случае введения скрытого сайта сплайсинга в РНК, кодирующей СЕР290, и посредством секвенирования, или с помощью функционального анализа (например, анализа в камере Уссинга) в случае реверсии мутации G551D в РНК, кодирующей CFTR, и т.п. Аналогичную работу можно также провести на α-1-антитрипсине (A1AT) и LRRK2.
Понятно, что данное изобретение описано выше исключительно как пример, и можно делать модификации, не отходя от объема и сущности изобретения.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ




| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И ПРОДУКТЫ ДЛЯ ЭКСПРЕССИИ БЕЛКОВ В КЛЕТКАХ | 2013 |
|
RU2711249C2 |
| РЕДАКТИРОВАНИЕ ГЕНОВ С ИСПОЛЬЗОВАНИЕМ МОДИФИЦИРОВАННОЙ ДНК С ЗАМКНУТЫМИ КОНЦАМИ (зкДНК) | 2018 |
|
RU2811724C2 |
| ТЕРАПЕВТИЧЕСКИЕ СОЕДИНЕНИЯ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ХАНТИНГТОНА | 2015 |
|
RU2711147C2 |
| КОМПОНЕНТЫ СИСТЕМЫ CRISPR-CAS, СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ МАНИПУЛЯЦИИ С ПОСЛЕДОВАТЕЛЬНОСТЯМИ | 2013 |
|
RU2701662C2 |
| Способ редактирования гена GJB2 для исправления патогенного варианта c.del35G в клетках человека, культивируемых in vitro | 2021 |
|
RU2780677C1 |
| КОМПОЗИЦИИ И СПОСОБЫ, ПРЕДНАЗНАЧЕННЫЕ ДЛЯ ЛЕЧЕНИЯ ГЕМОГЛОБИНОПАТИЙ | 2018 |
|
RU2834231C2 |
| ЛЕЧЕНИЕ ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С САЙТ-1 МЕМБРАНОСВЯЗАННОЙ ПЕПТИДАЗОЙ ТРАНСКРИПЦИОННЫХ ФАКТОРОВ (MBTPS1), ПУТЕМ ИНГИБИРОВАНИЯ ПРИРОДНОГО АНТИСМЫСЛОВОГО ТРАНСКРИПТА К MBTPS1 | 2010 |
|
RU2639550C2 |
| РНК-НАПРАВЛЯЕМАЯ ИНЖЕНЕРИЯ ГЕНОМА ЧЕЛОВЕКА | 2013 |
|
RU2766685C2 |
| ОЛИГОНУКЛЕОТИДЫ ДЛЯ ВВЕДЕНИЯ ИЗМЕНЕНИЯ В ПОСЛЕДОВАТЕЛЬНОСТЬ МОЛЕКУЛЫ ЦЕЛЕВОЙ РНК В ЖИВОЙ КЛЕТКЕ | 2013 |
|
RU2663110C2 |
| ОПОСРЕДОВАННАЯ НУКЛЕАЗОЙ СБОРКА ДНК | 2015 |
|
RU2707911C2 |









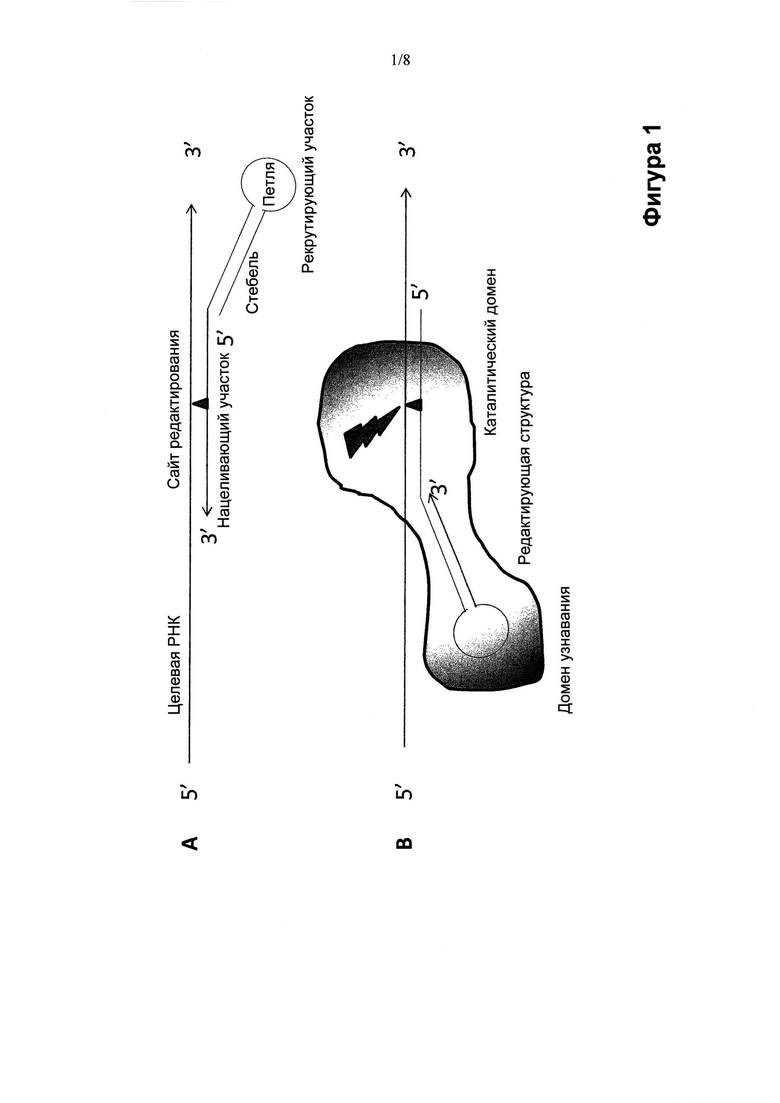
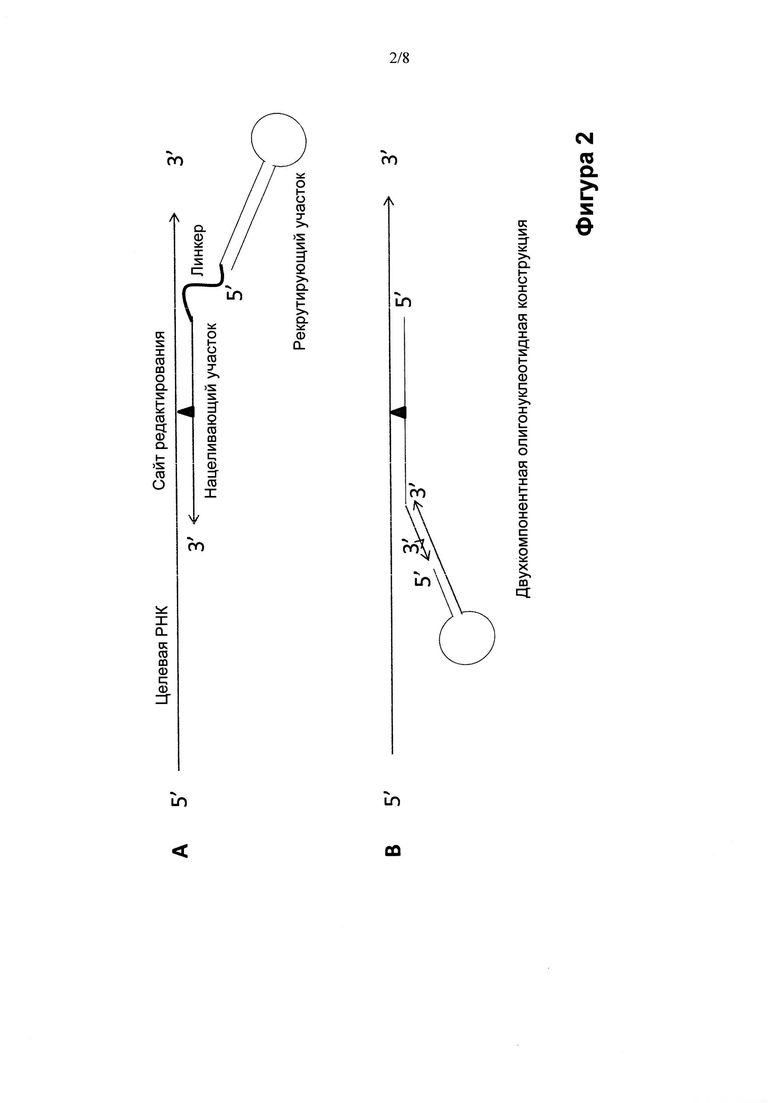
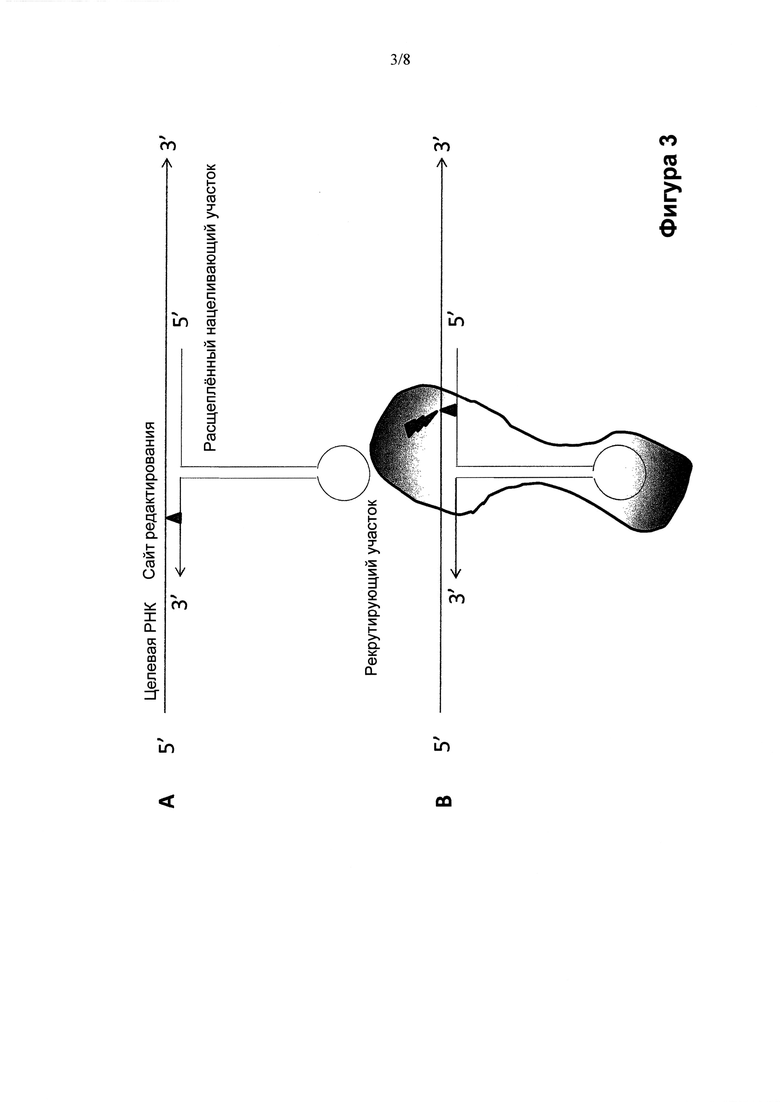



Изобретение относится к биотехнологии и представляет собой способ редактирования целевой РНК. Редактирование РНК осуществляют, используя олигонуклеотидные конструкции, содержащие (i) нацеливающий участок, специфический к целевой нуклеотидной последовательности, подлежащей редактированию; и (ii) рекрутирующий участок, способный связывать и рекрутировать структуру, редактирующую нуклеиновую кислоту, естественно присутствующую в клетке. Структура, редактирующая нуклеиновую кислоту, например, такая, как ADAR, перенацеливается на заданный сайт-мишень при посредстве нацеливающего участка, тем самым инициируя редактирование заданных нуклеотидных остатков в области целевой РНК, которая соответствует нацеливающему участку. 2 н. и 25 з.п. ф-лы, 7 ил., 3 табл., 5 пр.
1. Олигонуклеотидная конструкция для сайт-направленного редактирования нуклеотида в целевой РНК-последовательности в эукариотической клетке, содержащая:
(a) нацеливающий участок, содержащий антисмысловую последовательность, комплементарную части целевой РНК; и
(b) рекрутирующий участок, который способен образовывать внутримолекулярную структуру "стебель-петля"; способен связывать и рекрутировать фермент, редактирующий РНК, естественно присутствующий в указанной клетке; и способен осуществлять редактирование указанного нуклеотида.
2. Олигонуклеотидная конструкция по п. 1, в которой рекрутирующий участок содержит последовательность, полученную из любого из следующих фрагментов:
(i) РНК-последовательности, кодирующей В-домен человеческого белка GluR;
(ii) РНК-последовательности, кодирующей связывающий домен РНК-редактирующего фермента из GluR-C, GluR-D или 5-НТ2с серотонинового рецептора;
(iii) РНК- или ДНК-структуры стебель-петля, содержащей последовательность (RY или YR)nNm(RY или YR)n, где R обозначает аденозин или гуанозин, Y обозначает уридин или цитидин, N обозначает аденозин, гуанозин, цитидин, уридин или инозин, n обозначает 3 или более, m обозначает 4 или более, и где N образует петлю, а две последовательности (RY)n или (YR)n образуют структуру двухцепочечного стебля посредством комплементарного спаривания оснований;
(iv) ДНК-структуры стебель-петля, содержащей последовательность (CG)3N1-Nn(CG)3, где каждый с N1 по Nn могут быть одинаковыми или различными и могут быть выбраны из гуанозина, аденозина, тимидина, цитидина и инозина, и 'n' имеет значение от 2 до 20;
(v) пентануклеотида, имеющего последовательность: GCUMA, где G обозначает гуанозин, С обозначает цитидин, М обозначает аденозин или цитидин и U обозначает уридин; и
(vi) аптамера, выбранного для связывания с РНК-редактирующим ферментом.
3. Олигонуклеотидная конструкция по п. 1 или 2, в которой рекрутирующий участок не является комплементарным целевой РНК.
4. Олигонуклеотидная конструкция по любому из пп. 1-3, в которой нацеливающий участок содержит олигонуклеотидную последовательность, образующую дцРНК структуру посредством спаривания оснований с целевой РНК-последовательностью.
5. Олигонуклеотидная конструкция по любому из пп. 1-3, в которой нацеливающий участок содержит некомплементарный нуклеотид в положении, противостоящем нуклеотиду, подлежащему редактированию в целевой РНК-последовательности.
6. Олигонуклеотидная конструкция по любому из пп. 1-3, в которой нуклеотид, являющийся мишенью редактирования, представляет собой аденозин или цитидин, а фермент, редактирующий РНК, содержит дезаминазную активность.
7. Олигонуклеотидная конструкция по любому из пп. 1-3, в которой нуклеотид, являющийся мишенью редактирования, представляет собой аденозин, а редактирующий фермент представляет собой аденозиндезаминазу.
8. Олигонуклеотидная конструкция по любому из пп. 1-3, причем клетка является человеческой клеткой.
9. Олигонуклеотидная конструкция по любому из пп. 1-3, в которой редактирующий фермент обладает активностью hADAR1 или hADAR2.
10. Олигонуклеотидная конструкция по п. 5, в которой некомплементарный нуклеотид представляет собой цитидин или уридин, предпочтительно, цитидин.
11. Олигонуклеотидная конструкция по п. 3, в которой последовательность нуклеиновой кислоты, способная образовывать внутримолекулярную структуру стебель-петля, представляет собой олигорибонуклеотидную (РНК) последовательность.
12. Олигонуклеотидная конструкция по любому из пп. 1-3, в которой рекрутирующий участок содержит нуклеотидную последовательность:
5'-(AUANa)nUAUAACAAUAUgcuaaAUGUUGUUAUA(NbUAU)n-3',
где Na и Nb, каждый, обозначают единичные нуклеотиды, которые могут представлять собой A, G, С или U, при условии, что Na и Nb образуют пару неспаренных оснований (мисмэтчей) при образовании структуры стебель-петля и n обозначает 1 или 0.
13. Олигонуклеотидная конструкция по п. 11, в которой рекрутирующий участок содержит последовательность, выбранную из SEQ ID NOs: 6, 7, 20, 21, 22, 23 и 24.
14. Олигонуклеотидная конструкция по любому из пп. 1-3, в которой рекрутирующий участок содержит дезоксиолигонуклеотидную последовательность, включающую последовательность (CG)3T4(CG)3 (SEQ ID NO: 5), где С обозначает цитидин, G обозначает гуанозин и Т обозначает тимидин.
15. Олигонуклеотидная конструкция по п. 1, в которой один или более нуклеотидов содержат химическую модификацию.
16. Олигонуклеотидная конструкция по п. 15, в которой нацеливающий участок олигонуклеотидной конструкции содержит один или более 2'-O рибозил-замещенных уридинов, предпочтительно, 2'-ОМе замещенных уридинов.
17. Олигонуклеотидная конструкция по п. 16, в которой все уридины, противостоящие аденозинам в целевой РНК-последовательности, которые не являются мишенями редактирования, представляют собой 2'-метокси-(2'-ОМе) уридин.
18. Олигонуклеотидная конструкция по любому из пп. 1-3, в которой нацеливающий участок и рекрутирующий участок разделены линкерной последовательностью, содержащей олигонуклеотидную последовательность, олигопептидную последовательность или другую ковалентную химическую связь, например, такую как ПЭГ.
19. Олигонуклеотидная конструкция по любому из пп. 1-3, в которой нацеливающий участок по существу не прерывается и образует 5' часть олигонуклеотидной конструкции, а рекрутирующий участок образует 3' часть олигонуклеотидной конструкции.
20. Олигонуклеотидная конструкция по любому из пп. 1-3, в которой нацеливающий участок по существу не прерывается и образует 3' часть олигонуклеотидной конструкции, а рекрутирующий участок образует 5' часть олигонуклеотидной конструкции.
21. Олигонуклеотидная конструкция по любому из пп. 1-3, в которой нацеливающий участок прерывается рекрутирующим участком, так что нацеливающий участок образует 5' часть и 3' часть олигонуклеотидной конструкции, а рекрутирующий участок образует средний фрагмент олигонуклеотидной конструкции.
22. Олигонуклеотидная конструкция по п. 21, в которой рекрутирующий участок образует петлю, когда нацеливающий участок отжигается с целевой РНК-последовательностью посредством комплементарного спаривания оснований.
23. Олигонуклеотидная конструкция по любому из предыдущих пунктов, в которой длина олигонуклеотида составляет от 20 до 100 нуклеотидов, предпочтительно, от 24 до 60 нуклеотидов, более предпочтительно, от 30 до 50 нуклеотидов.
24. Олигонуклеотидная конструкция по любому из предыдущих пунктов, в которой целевая РНК-последовательность выбрана из группы, состоящей из предшественника матричной РНК (пре-мРНК), матричной РНК (мРНК), рибосомной РНК (рРНК), транспортной РНК (тРНК) и микроРНК.
25. Олигонуклеотидная конструкция по любому из пп. 1-3, 10, 11, 13, 15-17 или 22, где целевая РНК кодирует CFTR (например, для редактирования мутации 1784G>A), СЕР290 (например, для редактирования мутации c. 2991+1655A>G), А1АТ (например, для редактирования мутации 9989G>A) или LRRK2 (например, для редактирования мутации G6055).
26. Способ сайт-направленного редактирования нуклеотида в целевой РНК в эукариотической клетке, предпочтительно в клетке млекопитающего, более предпочтительно в человеческой клетке, с помощью РНК-редактирующего фермента, естественно присутствующего в указанной клетке и способного осуществлять редактирование указанного нуклеотида, включающий стадии: (i) предоставление указанной клетке олигонуклеотидной конструкции по любому предыдущему пункту; (ii) выдерживание в течение времени, достаточного для поглощения клеткой указанной олигонуклеотидной конструкции, отжиг указанной олигонуклеотидной конструкции с целевой РНК-последовательностью и предоставление РНК-редактирующего фермента для осуществления реакции редактирования нуклеотида в указанной целевой РНК-последовательности; и (iii) идентификация присутствия редактированного нуклеотида в РНК-последовательности.
27. Способ по п. 26, где целевая РНК кодирует CFTR (например, для редактирования мутации 1784G>A), СЕР290 (например, для редактирования мутации c.2991+1655A>G), А1АТ (например, для редактирования мутации 9989G>A) или LRRK2 (например, для редактирования мутации G6055).
| WO 2013142578 A1, 26.09.2013 | |||
| Wenyan Jiang, CRISPR-assisted editing of bacterial genomes, Nat Biotechnol., 2013 March, 31(3), 233-239 | |||
| Kazuko Nishikura, Functions and Regulation of RNA Editing by ADAR Deaminases, Annu Rev Biochem | |||
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| A | |||
| Киран, ИДЕНТИФИКАЦИЯ A→I РЕДАКТИРОВАНИЯ РНК: РАССТАВЛЕНИЕ ТОЧЕК НАД I В ТРАНСКРИПТОМЕ ЧЕЛОВЕКА, | |||

