Область изобретения
Настоящее изобретение связано с диагностикой и лечением опухолей, в частности, с новым опухолевым биомаркером, а также со способами и наборами для детектирования возникновения злокачественной опухоли и метастазов. Настоящее изобретение связано также со способами и лекарственными средствами для лечения злокачественной опухоли и/или ее метастазов.
Предшествующий уровень техники
В настоящее время ежегодно во всем мире приблизительно у 11 миллионов людей диагностируют опухоли, и предполагается, что к 2020 году это количество возрастет более чем до 16 миллионов. В 2005 году из 58 миллионов смертей 7,6 миллионов было вызвано злокачественной опухолью (что составляет приблизительно 13%). Это количество возрастет, и предполагается, что к 2015 и 2030 годам от злокачественных опухолей погибнет, соответственно, 9 и 11,4 миллиона человек. (Всемирная Организация Здравоохранения, 2006).
Опухолевые маркеры являются веществами, количество которых уменьшается или увеличивается при генных мутациях или в результате вариации экспрессии генов в процессе клеточного канцерогенеза. Обычно опухолевые маркеры включают в себя антигены и другие биологически активные вещества, которые могут быть использованы для раннего детектирования злокачественных опухолей, а также для мониторинга прогрессии заболевания и реакции на лечение (ASCO, 1996). Это создает огромные преимущества для клинического лечения злокачественных опухолей, в особенности, когда злокачественную опухоль удается обнаружить еще до появления какого-либо заметного клинического признака, или когда опухолевые маркеры могут быть использованы для мониторинга реакции пациентов на конкретный вид лечения. В настоящее время на развитие опухолевых маркеров затрачиваются бóльшие усилия, с тем, чтобы полнее удовлетворить клинические потребности.
Применимость большей части опухолевых биомаркеров, используемых в настоящее время в клинике, более или менее ограничена в силу недостаточной чувствительности и специфичности. Например, уровень AFP [альфа-фетопротеин] и ультразвуковые исследования широко используются для детектирования рака печени. Хотя их чувствительность и не очень высока, они действительно продлевают срок выживаемости пациентов при диагностировании у людей наличия высокой степени риска. Опухолевый антиген CA-125 имеет повышенную чувствительность, но лишен специфичности. Сходным образом, опухолевый биомаркер CA15-3 в крови, показатель злокачественной опухоли молочной железы, трудно использовать для ранней диагностики ввиду его низкой чувствительности. Следовательно, в настоящее время способы раннего детектирования рака и определение злокачественности опухоли в клинике недоступны. Необходимо развитие новых технологий, а также новых способов.
Развитие опухолевой протеомики дает основание надеяться на возможность идентификации новых опухолевых биомаркеров и скрининга, ранней диагностики и прогнозирования опухолей. Злокачественная трансформация опухолей всегда приводит к изменению экспрессии белков, которая может быть идентифицирована и количественно оценена на белковом уровне. Таким образом, можно получить большое количество информации и данных, посредством которых можно будет открывать потенциальные биомаркеры и оценивать их в плане дальнейшего развития и клинических применений.
Hsp90α (белок теплового шока 90α, Hsp90α) является молекулярным шапероном, функционирование которого связано со стабилизацией зависимых от него белков в их активных состояниях. Hsp90α является одним из наиболее изобилующих белков в эукариотических клетках и составляет примерно 1-2% от общего количества клеточных белков. Внутриклеточный Hsp90α главным образом функционирует, стабилизируя связанные с ним белки (то есть рецептор эстрогена), и способствует их созреванию (то есть некоторые киназы и сигнальные белки). Кроме того, Hsp90α участвует также и в других физиологических явлениях, таких как эволюция мутировавших белков, реорганизация цитоскелета, транслокация ядерных белков, клеточная пролиферация и апоптоз, деградация белков, процессинг антигенов и распознавание ЛПС и т.д. Hsp90α связан также со многими заболеваниями, такими как злокачественные опухоли, аутоиммунные заболевания и сердечнососудистые заболевания. Например, моноклональное антитело против антигена последовательности LKVIRK, полученной из Hsp90α, может быть использовано для лечения инфекции, сопутствующей грибковой, и такое клиническое испытание проводится в настоящее время компанией Neutec (под коммерческим названием: Mycogrip).
Сообщалось также, что Hsp90α может быть секретирован под действием некоторых стимулов (Liao et al. (2000) J. Biol. Chem. 275, 189-96). Поскольку Hsp90α является классическим внутриклеточным белком, мало сообщений относительно функции внеклеточного Hsp90α. В известных ранее литературных источниках Hsp90α был идентифицирован как белок, облегчающий процессинг антигенов в клетках, в которых антиген подвергается процессингу (АПС, антиген-представляющие клетки), и было обнаружено, что он является одним из четырех белков в липополисахарид (ЛПС)-активирующихся кластерах, которые могут взаимодействовать с ЛПС таким образом, чтобы происходил запуск ответа внутриклеточных белков (Triantafilou et al. (2002) Trends in Immunology 23, 301-4).
Было обнаружено также, что Hsp90α в большом количестве экспрессируется на поверхности некоторых опухолевых клеток, включая клетки немелкоклеточного рака легкого, клетки меланомы и клетки рака печени (Ferrarini et al. (1992) Int. J. Cancer 51, 613-19). Предполагалось, что высокий уровень экспрессии Hsp90α на клеточной поверхности указанных клеток связан с процессингом антигена, тогда как прямых доказательств этого представлено не было.
Сообщалось также, что Hsp90α может способствовать транслокации трансмембранных белков (Schlatter et al. (2002) Biochem. J. 362, 675-84), а также то, что он связан с выходом из клеток некоторых лекарственных средств против лейкемии, рака легкого и рака шейки матки (Rappa et al. (2002) Oncol. Res. 12, 113-9 и Rappa et al. (2000) Anticancer Drug Des 15,127-34).
Внутриклеточный Hsp90α является важной мишенью для разработки противораковых лекарственных средств, поскольку он участвует в регуляции многих путей передачи сигналов, которые являются критичными в клеточном канцерогенезе. Ингибирование внутриклеточного Hsp90α может приводить к селективной деградации белков, связанных с клеточной пролиферацией, с контролем клеточного цикла, а также с апоптозом. В настоящее время считается, что некоторые известные антибиотики, такие как гелданамицин, радицикол и кумермицин A1 являются природными ингибиторами Hsp90α. В патенте WO 00/53169 описывается указанный механизм и высказывается предположение, что предотвращение взаимодействия между шаперонами и связанными с ними белками является способом ингибирования активности шаперонов. Считается, что в ряду указанных антибиотиков указанной активностью обладает кумарин и его производные. Однако ингибиторы, описанные в патенте WO 00/53169, в основном нацелены на внутриклеточный Hsp90α.
Аналог гелданамицина 17-AAG также является ингибитором Hsp90α, и в настоящее время он проходит проверку в клинических испытаниях. Однако в некоторых сообщениях показано, что 17-AAG может вызывать неспецифические ингибирующие эффекты и клеточную токсичность в результате взаимодействия со многими другими клеточными компонентами. Кроме того, в силу ограниченности познаний в отношении многообразия процессов передачи клеточных сигналов с участием Hsp90α и связанных с ним белков, непосредственное ингибирование внутриклеточного Hsp90α является рискованным.
В другом патенте (EP 1457499 A1) описана функция внеклеточного Hsp90α по облегчению инвазии опухолевой клетки путем содействия секреции или активации MMP-2. На основе указанных механизмов в данном патенте предполагается, что ингибирование внеклеточного Hsp90α может препятствовать опухолевой инвазии и метастазированию, и что путем детектирования ответа опухолевых клеток на лечение под действием ингибитора Hsp90α можно сделать заключение об инвазивной способности клеток и их связи с Hsp90α.
Авторы изобретения по заявке на патент WO/2008/070472 предлагают осуществлять мониторинг противоопухолевой эффективности терапии, нацеленной на Hsp90α, путем детектирования в плазме Hsp90α и других связанных с ним факторов. В указанной заявке ими на мышиных моделях определена взаимосвязь между уровнем Hsp90α в плазме и эффективностью нацеленной на Hsp90α терапии с помощью ингибиторов этого белка, включая 17-AAG и 17-DMAG, а также взаимосвязь между уровнем Hsp90α в плазме и объемом опухоли. Однако ими не было представлено никакого доказательства относительно точной формы плазматического Hsp90α и не было приведено соотношения между уровнем Hsp90α в плазме и злокачественностью опухоли, в частности, опухолевыми метастазами. Ими не было предложено также и применение плазматического Hsp90α в качестве независимого опухолевого биомаркера в диагностике и прогнозировании опухолей.
Одна из групп исследователей сообщала, что сывороточный уровень Hsp90α связан со стадиями немелкоклеточного рака легких (Xu et al. (2007) J. Cancer Mol. 3, 107-112). Сывороточный уровень Hsp90α у этих пациентов с раком легких был значительно выше, чем у нормальных людей или у пациентов с доброкачественной опухолью. Однако в указанной публикации опять же не была идентифицирована ни точная форма сывороточного Hsp90α, ни его взаимосвязь с опухолевыми метастазами. Кроме того, в ней исследовался только немелкоклеточный рак легкого, тогда как взаимосвязь и специфичность сывороточного уровня Hsp90α в отношении рака молочной железы, рака печени и рака поджелудочной железы оставались неизвестными. Кроме того, в указанной публикации описано только количественное изменение сывороточного Hsp90α у опухолевых пациентов, там не представлено никакой информации относительно точного содержания Hsp90α в сыворотке, а также относительно порогового значения нормального или аномального уровня, что является необходимым для диагностики и прогнозирования опухолей.
Краткое содержание изобретения
Настоящее изобретение основано на обнаружении того, что уровень Hsp90α в крови коррелирует с развитием, злокачественностью и метастазированием многих типов злокачественных опухолей. Соответственно, Hsp90α в крови может быть использован в качестве нового опухолевого биомаркера. Авторы изобретения обнаружили, что Hsp90α в крови отличается от внутриклеточного Hsp90α тем, что он лишен четырех аминокислотных остатков на своем C-конце, по сравнению с внутриклеточным Hsp90α.
Следовательно, в одном из аспектов настоящее изобретение связано с изолированным полипептидом, содержащим аминокислотную последовательность SEQ ID NO: 1 или состоящим из нее.
Полипептид согласно изобретению может быть фосфорилирован. В частности, в полипептиде согласно настоящему изобретению один или более аминокислотных остатков в аминокислотной последовательности SEQ ID NO: 1 в положениях, выбранных из группы, состоящей из Thr90, Ser231, Ser263, Tyr309 и их сочетания, фосфорилированы. Предпочтительно, чтобы в полипептиде согласно настоящему изобретению был фосфорилирован Thr90.
Полипептид согласно настоящему изобретению может служить в качестве опухолевого биомаркера. С помощью агента, специфически связывающегося с полипептидом согласно настоящему изобретению, можно осуществить детектирование уровня указанного полипептида в крови и, следовательно, определить наличие, степень злокачественности, а также метастазирование злокачественных опухолей.
Соответственно, в другом аспекте настоящее изобретение связано с применением агента, способного специфически связываться с полипептидом согласно настоящему изобретению, при изготовлении набора для определения наличия, стадии и/или метастазирования злокачественной опухоли у субъекта путем определения уровня полипептида согласно изобретению в плазме, для скрининга наличия злокачественной опухоли среди популяции высокого риска путем детектирования уровня полипептида согласно изобретению в плазме, для определения прогноза в отношении пациента, имеющего злокачественную опухоль, путем детектирования уровня полипептида согласно изобретению в плазме, для определения эффективности хирургического вмешательства, радиационной терапии или химиотерапии в отношении пациента, имеющего опухоль, путем детектирования уровня полипептида согласно настоящему изобретению в плазме.
Предпочтительно, чтобы указанный агент, способный к специфическому связыванию с полипептидом согласно изобретению, являлся антителом, специфичным в отношении данного полипептида. Предпочтительно, чтобы указанное антитело было моноклональным антителом или его антиген-связывающим фрагментом, таким как scFv, Fab, Fab' и F(ab')2. В одном из специфических воплощений указанное антитело является MAb E9 или D10, которое продуцируется клеточными линиями, депонированными, соответственно, под регистрационными номерами CGMCC No. 2903 или 2904.
Согласно настоящему изобретению, указанное антитело специфически связывается с полипептидом согласно изобретению и, предпочтительно, специфически связывается с полипептидом согласно изобретению, присутствующим в плазме. Предпочтительно, чтобы указанное антитело специфически связывалось с фосфорилированной формой полипептида согласно изобретению, где указанная фосфорилированная форма полипептида содержит один или более фосфорилированных аминокислотных остатков в аминокислотной последовательности SEQ ID NO: 1 в положениях, выбранных из группы, состоящей из Thr90, Ser231, Ser263, Tyr309 и их сочетания. Предпочтительно, чтобы Thr90 в полипептиде согласно настоящему изобретению был фосфорилирован.
В другом аспекте настоящее изобретение связано с применением ингибитора полипептида согласно изобретению при получении лекарственного средства для предотвращения или лечения метастазов злокачественной опухоли у субъекта. В одном из воплощений данного аспекта указанный ингибитор представляет собой антитело, специфичное в отношении полипептида согласно изобретению. Предпочтительно, чтобы указанное антитело было гуманизированным антителом или его антигенсвязывающим фрагментом. В одном из воплощений указанное антитело специфически связывается с фосфорилированной формой полипептида согласно изобретению, где указанная фосфорилированная форма полипептида содержит один или более фосфорилированных аминокислотных остатков в аминокислотной последовательности SEQ ID NO: 1 в положениях, выбранных из группы, состоящей из Thr90, Ser231, Ser263, Tyr309 и их сочетания. В предпочтительном воплощении указанное антитело специфически связывается с полипептидом, который фосфорилирован в положении Thr90. В одном из специфических воплощений указанное антитело представляет собой MAb E9 или D10, продуцируемые клеточными линиями, депонированными, соответственно, под регистрационными номерами CGMCC No. 2903 или 2904.
В указанных выше аспектах настоящего изобретения указанной злокачественной опухолью может быть, например, рак легкого, рак печени, рак желудка, рак желудочно-кишечного тракта, рак пищевода, остеосаркома, рак поджелудочной железы, лимфома, рак толстой и прямой кишок, рак молочной железы, рак предстательной железы, рак ротовой полости, рак носоглотки, рак шейки матки, рак яичника, лейкемия, злокачественная меланома, саркома, почечноклеточный рак и холангиокарцинома.
Настоящее изобретение включает в себя также антитела, которые могут специфически связываться с полипептидом согласно настоящему изобретению в плазме. В одном из специфических аспектов указанное антитело представляет собой MAb E9 или D10, продуцируемые клеточными линиями, депонированными, соответственно, под регистрационными номерами CGMCC No. 2903 или 2904. Предпочтительно, чтобы указанное антитело было гуманизированным антителом или его антиген-связывающим фрагментом. В одном из воплощений указанное антитело специфически связывается с фосфорилированной формой указанного полипептида, где указанная фосфорилированная форма полипептида содержит один или более фосфорилированных аминокислотных остатков в аминокислотной последовательности SEQ ID NO: 1 в положениях, выбранных из группы, состоящей из Thr90, Ser231, Ser263, Tyr309 и их сочетания. В предпочтительном воплощении указанное антитело специфически связывается с полипептидом, который фосфорилирован в положении Thr90.
В другом аспекте настоящее изобретение связано со способом ингибирования инвазии и метастазирования злокачественной опухоли, включающим в себя стадию ингибирования фосфорилирования внутриклеточного Hsp90α в злокачественных клетках. В одном из воплощений настоящее изобретение связано со способом ингибирования инвазии и метастазирования злокачественной опухоли, включающим в себя стадию ингибирования фосфорилирования Hsp90α по Thr90 в злокачественных клетках. В одном из специфических воплощений данного аспекта указанный способ включает в себя стадию повышения экспрессии молекулы нуклеиновой кислоты, кодирующей протеинфосфатазу 5 (PP5) в злокачественных клетках, например, путем внедрения гена.
Краткое описание графического материала
Фиг. 1: Уровень Hsp90α в плазме мышей-опухоленосителей значительно выше, чем таковой у нормальных мышей.
Фиг. 2: Уровень Hsp90α в плазме пациентов значительно выше, чем таковой у нормальных людей.
Фиг. 3: Исследование титра мышиных моноклональных антител E9 и D10.
Фиг. 4: Стандартная кривая плазматического Hsp90α, полученная с помощью мышиного моноклонального антитела E9 и кроличьего поликлонального антитела S2 (сэндвич-ELISA).
Фиг. 5: Количественное определение уровня Hsp90α в плазме у пациентов с раком легкого, печени, поджелудочной железы, а также у пациентов с доброкачественной ретенционной кистой молочной железы и миомой матки, полученное с помощью сэндвич-ELISA:
A: Уровень Hsp90α в плазме у пациентов с раком печени, измеряемый методом сэндвич-ELISA. Уровень Hsp90α в плазме у пациентов с доброкачественной опухолью печени варьирует от 2 до 10 нг/мл, преимущественно от 2 до 5 нг/мл, тогда как уровень Hsp90α в плазме у 69% (20/29) пациентов с раком печени составляет выше 50 нг/мл, что в среднем более чем в 10 раз выше, чем у пациентов с доброкачественной опухолью, и это согласуется с результатами вестерн-блоттинга. Это свидетельствует о том, что уровень Hsp90α в плазме положительно коррелирует со злокачественностью опухоли.
B: Уровень Hsp90α в плазме у пациентов с раком легкого, измеряемый методом сэндвич-ELISA. Уровень Hsp90α в плазме у 64% (9/14) пациентов с раком легкого составляет выше 50 нг/мл, что в среднем более чем в 10 раз выше, чем у пациентов с доброкачественной опухолью. Это свидетельствует о том, что уровень Hsp90α в плазме положительно коррелирует со злокачественностью опухоли.
C: Уровень Hsp90α в плазме у пациентов со злокачественной опухолью молочной железы, измеряемый методом сэндвич-ELISA. По сравнению с пациентами с доброкачественной опухолью, наиболее высокий уровень Hsp90α в плазме составляет более чем 5-кратное повышение. В целом повышение среднего уровня Hsp90α в плазме пациентов со злокачественной опухолью молочной железы является статистически значимым по сравнению с таковым у пациентов с доброкачественной опухолью.
D: Уровень Hsp90α в плазме у пациентов со злокачественной опухолью поджелудочной железы, измеряемый методом сэндвич-ELISA. Уровень Hsp90α в плазме у 100% (10/10) пациентов со злокачественной опухолью поджелудочной железы составляет более 50 нг/мл, что в среднем более чем в 10 раз выше, чем у пациентов с доброкачественной опухолью. Это свидетельствует о том, что уровень Hsp90α в плазме положительно коррелирует со злокачественностью опухоли.
Фиг. 6: Количественное определение уровня Hsp90α в плазме пациентов со злокачественной опухолью при наличии или отсутствии у них метастазов, предпринятое с помощью метода сэндвич-ELISA:
A: Пациентов со злокачественной опухолью печени делили на две группы, в одной из которых пациенты имели метастазы, а в другой не имели метастазов, и затем в указанных двух группах сравнивали уровни Hsp90α. Уровни Hsp90α в плазме пациентов со злокачественной опухолью и метастазами всегда были выше 200 нг/мл, тогда как у пациентов без метастазов они колебались от 50 до 200 нг/мл.
B: Пациентов со злокачественной опухолью легкого делили на две группы, в одной из которых пациенты имели метастазы, а в другой не имели метастазов, и затем в указанных двух группах сравнивали уровни Hsp90α. Уровни Hsp90α в плазме пациентов со злокачественной опухолью и метастазами всегда были выше 200 нг/мл, тогда как у пациентов без метастазов они колебались от 50 до 200 нг/мл.
C: Пациентов со злокачественной опухолью молочной железы делили на две группы, в одной из которых пациенты имели метастазы, а в другой не имели метастазов, а затем в указанных двух группах сравнивали уровни Hsp90α. Уровни Hsp90α в плазме пациентов со злокачественной опухолью и метастазами всегда были статистически значимо выше таковых у пациентов без метастазов.
Фиг. 7: Количественное определение уровня Hsp90α в плазме пациентов с воспалением (пневмония и гепатит), здоровых людей и опухолевых пациентов, предпринятое с помощью метода сэндвич-ELISA:
A: Для того, чтобы убедиться, что повышенный уровень Hsp90α в плазме пациентов со злокачественной опухолью является опухолеспецифическим, авторы изобретения сравнивали уровень Hsp90α в плазме пациентов с пневмонией, в плазме здоровых людей и опухолевых пациентов, и обнаружили, что уровень Hsp90α в плазме пациентов с пневмонией изменяется в интервале от 2 до 10 нг/мл, что указывает на статистически незначимые изменения по сравнению с таковыми у нормальных людей.
B: Уровень Hsp90α в плазме пациентов с гепатитом (гепатит A и B) колеблется от 2 до 10 нг/мл, что указывает на отсутствие статистически значимых изменений по сравнению с таковыми у нормальных людей.
Фиг. 8: Hsp90α в плазме, секретируется опухолевыми клетками.
Фиг. 9: Hsp90α, секретируемый опухолевыми клетками, представляет собой форму, укороченную на C-конце.
Фиг. 10: Плазматический Hsp90α лишен четырех C-концевых аминокислотных остатков.
Фиг. 11: Плазматический Hsp90α является фосфорилированным.
Фиг. 12: Уровень Hsp90α, фосфорилированного по остатку Thr90, в плазме пациентов-опухоленосителей повышен.
Фиг. 13: В плазме пациентов-опухоленосителей повышение уровня Hsp90α, фосфорилированного по остатку Thr90, сопоставимо с уровнем Hsp90α.
Фиг. 14: Фосфорилирование Hsp90α по остатку Thr90 является необходимым условием, предшествующим секреции Hsp90α.
Фиг. 15: Протеинфосфатаза 5 (PP5) осуществляет дефосфорилирование Hsp90α, фосфорилированного по остатку Thr90.
A: Очищенную PP5 инкубировали совместно с Hsp90α, фосфорилированным по остатку Thr90 (pT90-Hsp90α), и количественно оценивали выделяющуюся фосфатную группу. Пептид является положительным контролем. В результате было показано, что PP5 может напрямую осуществлять дефосфорилирование Hsp90α, фосфорилированного по остатку Thr90.
B: В клетках линии MCF-7 злокачественной опухоли молочной железы человека повышенная экспрессия человеческой PP5 приводит к ингибированию Thr90-фосфорилирования Hsp90α (составляя 0,55 такового в контрольной группе), а РНК-интерференция эндогенной человеческой PP5 приводит к повышению Thr90-фосфорилирования Hsp90α (составляя 1,58 такового в контрольной группе).
Фиг. 16: PP5 регулирует секрецию Hsp90α.
A: В линии клеток MCF-7 злокачественной опухоли молочной железы человека повышенная экспрессия человеческой PP5 приводит к ингибированию секреции Hsp90α.
B: В линии клеток MCF-7 злокачественной опухоли молочной железы человека РНК-интерференция эндогенной человеческой PP5 приводит к повышению секреции Hsp90α.
Фиг. 17: Взаимозависимость уровня экспрессии PP5 и уровня секреции Hsp90α.
Фиг. 18: Взаимозависимость уровня экспрессии PP5 и инвазивной способности опухолевых клеток.
Фиг. 19: Специфическое антитело против Hsp90α может ингибировать миграцию опухолевых клеток.
Фиг. 20: Специфическое антитело против Hsp90α может ингибировать метастазирование опухоли.
Информация, связанная с депонированием биологических материалов
Линию клеток мышиной гибридомы SP2/0-Agl4, которая продуцирует моноклональное антитело E9, депонировали в Китайском Центре Коллекционирования Общих Микробиологических Культур (CGMCC, Chinese Academy of Sciences Institute of Microbiology, Datun Road, Chaoyang District, Beijing) 24 февраля 2009 года под регистрационным номером No. CGMCC No.2903.
Линию клеток мышиной гибридомы SP2/0-Agl4, которая продуцирует моноклональное антитело E9, депонировали в Китайском Центре Коллекционирования Общих Микробиологических Культур (CGMCC, Chinese Academy of Sciences Institute of Microbiology, Datun Road, Chaoyang District, Beijing) 24 февраля 2009 года под регистрационным номером No. CGMCC No.2904.
Подробное описание изобретения
Канцерогенез вызывает изменения определенных путей передачи внутриклеточных сигналов, сопровождаемые изменениями экспрессии, модификации и распределения белков. Указанные изменения могут быть использованы для мониторинга развития и прогрессии опухоли, а указанные белки называются опухолевыми маркерами. С развитием технологий протеомики становится возможным осуществление мониторинга изменений опухолевого протеома качественно или количественно. Обнаружено много новых опухолевых маркеров, обеспечивающих более точные и достоверные данные для реализации клинической диагностики и прогноза опухолей.
Настоящее изобретение основано на открытии нового опухолевого маркера в плазме, то есть плазматического Hsp90α. По сравнению с внутриклеточным Hsp90α (аминокислотной последовательностью которого является последовательность SEQ ID NO: 3, а кодирующей его последовательностью нуклеиновой кислоты является последовательность SEQ ID NO: 4), плазматический Hsp90α имеет делецию 4 аминокислот на С-конце.
Соответственно, в одном из аспектов настоящее изобретение относится к изолированному полипептиду, Hsp90α, обнаруживаемому в плазме или в сыворотке, который содержит аминокислотную последовательность SEQ ID NO: 1 или состоит из нее. Используемый здесь термин "полипептид согласно изобретению" относится к Hsp90α, присутствующему в плазме или в сыворотке, который содержит аминокислотную последовательность SEQ ID NO: 1 или состоит из нее. Предпочтительно, чтобы используемый здесь термин "полипептид согласно изобретению" относился к полипептиду, состоящему из аминокислотной последовательности SEQ ID NO: 1. В настоящей заявке термин "Hsp90α в плазме" или "Hsp90α в сыворотке" может быть использован взаимозаменяемым образом, указывая на свободную форму белка Hsp90α, присутствующего в крови, в отличие от внутриклеточного или связанного с клеточной поверхностью белка Hsp90α. В таком применении термин "полипептид" и "белок" может быть использован взаимозаменяемым образом.
Настоящее изобретение относится также к полинуклеотиду, кодирующему полипептид, содержащий аминокислотную последовательность SEQ ID NO: 1 или состоящий из нее. В специфическом воплощении указанный полинуклеотид содержит или состоит из нуклеиновокислотной последовательности SEQ ID NO: 2.
Авторы изобретения обнаружили также, что в плазме полипептид согласно изобретению представлен в фосфорилированной форме, в которой один или более аминокислотных остатков в аминокислотной последовательности, соответствующей SEQ ID NO: 1, выбранных из группы, состоящей из Thr90, Ser231, Ser263, Tyr309 и их сочетаний, являются фосфорилированными. Предпочтительно, чтобы в полипептиде согласно настоящему изобретению фосфорилированным был остаток Thr90.
Такая специальная форма присутствующего в плазме Hsp90α (фосфорилированная и усеченная на C-конце) еще не описана. При этом никогда еще не было описано также и то, что плазматический белок Hsp90α связан с развитием и прогрессией опухоли. В документе EP 1457499 A1 описана внеклеточная форма Hsp90α, а также высказано предположение, что ингибиторы Hsp90α могут быть использованы для лечения опухолевых метастазов, для детекции инвазии опухоли и для определения зависимости инвазивности опухоли от Hsp90α. Однако способ, описанный в EP 1457499 A1, используется для детекции Hsp90α клеточной поверхности, и он не годится для определения Hsp90α в плазме. В патенте EP 1457499 A1 не содержится ни описание, ни предположение о том, что плазматический Hsp90α может быть использован для определения степени и стадии злокачественности или для мониторинга терапевтического ответа и прогнозирования злокачественной опухоли путем измерения уровня Hsp90α. В документе W0/2008/070472 сообщается о том, что эффекты противораковой терапии, нацеленной на Hsp90α, могут быть определены путем детектирования Hsp90α и связанных с ним факторов в плазме, но там нет указания на то, что Hsp90α является независимым опухолевым маркером в диагностике и прогнозировании злокачественной опухоли.
Авторы настоящего изобретения изучали образцы крови приблизительно одной сотни пациентов (с раком молочной железы, раком печени, раком поджелудочной железы, раком легкого и т.д.) и обнаружили, что уровень Hsp90α в плазме коррелирует со злокачественностью опухоли, а в особенности с метастазированием опухоли; в то же время воспалительный ответ не влияет на уровень Hsp90α в плазме. Следовательно, плазматический Hsp90α может служить в качестве опухолевого маркера, который может быть использован при диагностике и прогнозировании опухолей и метастазов.
Соответственно, в другом аспекте настоящее изобретение связано с набором для определения плазматических уровней полипептида согласно изобретению. Набор согласно изобретению содержит средство, способное специфически связываться с полипептидом согласно изобретению. Набор согласно изобретению может быть использован для детектирования уровня Hsp90α в плазме.
Настоящее изобретение связано также с применением средства, способного специфически связываться с полипептидом согласно изобретению, для изготовления набора для детектирования уровня Hsp90α в образце плазмы субъекта. Набор согласно изобретению может быть использован для определения наличия, злокачественности и метастазирования опухоли; для скрининга злокачественной опухоли в популяции высокого риска; для определения прогноза у пациентов со злокачественной опухолью; а также для определения эффективности лечения путем хирургического вмешательства, радиационной терапии или химиотерапии и/или для определения того, надо ли - или когда именно надо - прекращать указанное терапевтическое воздействие.
Используемый здесь термин "средство, способное специфически связываться с полипептидом согласно изобретению" относится к молекулам, которые могут с высокой степенью аффинности связываться с полипептидом согласно изобретению. Указанные средства включают в себя также молекулы, которые могут связываться с внутриклеточным Hsp90α и с Hsp90α клеточной поверхности. Средство, способное специфически связываться с полипептидом согласно изобретению, может быть представлено белками, в частности, антителами, специфичными в отношении Hsp90α. В предпочтительных примерах указанное выше антитело является моноклональным антителом или его антигенсвязывающим фрагментом, таким как scFv, Fab, Fab' и F(ab')2. В специфическом воплощении указанные антитела являются моноклональным антителом E9 или D10, которые продуцируются клеточной линией с регистрационным номером депозита CGMCC No. 2903 или 2904, соответственно.
Моноклональные антитела получают путем скрининга клеток, которые могут секретировать указанные антитела, а затем путем культивирования таких клеток in vitro. Указанный способ хорошо известен специалистам в данной области (Kohler G & Milstein C. (1975) Nature. 256, 495-7). Способ получения Hsp90α-специфического моноклонального антитела является следующим: в качестве первой иммунизации, рекомбинантный человеческий Hsp90α (rhHsp90α, 100 мкг) с полным адъювантом Фрейнда подкожно инъецируют мышам BALB/c в несколько участков на бедре; через 3 недели такую же дозу rhHsp90α инъецируют внутрибрюшинно (в/б) с неполным адъювантом Фрейнда; третью иммунизацию производят через 3 недели путем в/б инъекции той же самой дозы (через 5 - 7 дней проверяют титр антител в крови); еще через 3 недели производят бустер-иммунизацию путем в/б введения 200 мкг rhHsp90α. Через 3 дня клетки селезенки сливают с гибридомой SP2/0-Ag14 (SP2/0) (источник: ATCC CRL-1581), используя HAT для скрининга. Затем осуществляют предельное разведение клеток гибридомы. Для идентификации и, в конечном счете, для селекции клеточных линий, которые могут секретировать специфичные в отношении Hsp90α антитела, были использованы методы иммуноблоттинга и ELISA.
Согласно настоящему изобретению, антитела, используемые для изготовления набора, могут специфически связываться с Hsp90α, и предпочтительно, с Hsp90α плазмы крови. В одном из воплощений указанные антитела специфически связываются с Hsp90α, в котором один или более из следующих аминокислотных остатков: Thr90, Ser231, Ser263, Tyr309 и их сочетание - являются фосфорилированными. В предпочтительном воплощении описанные здесь антитела могут специфически связываться с Hsp90α, содержащим фосфорилированный Thr90.
Настоящее изобретение связано также со способом детектирования уровня полипептида согласно изобретению в плазме. Уровень Hsp90α в плазме может быть детектирован любым из подходящих способов. Описанные здесь способы включают в себя прямой и непрямой способы детектирования полипептида согласно изобретению, который может быть использован для диагностики развития, злокачественности и метастазирования опухоли.
Прямые измерения включают в себя способы детектирования описанного полипептида согласно изобретению с помощью средства, способного специфически связываться с указанным полипептидом, например, с помощью антител, специфичных в отношении указанных полипептидов, методом вестерн-блоттинга или ELISA.
Концентрация Hsp90α может быть определена также и непрямым способом путем детектирования активности Hsp90α. Примером является анализ тепловой денатурации люциферазы, который может быть использован для детектирования активности шаперона Hsp90α (Johnson et al. (2000) J. Biol. Chem., 275, 32499-32507).
Предпочтительно, чтобы уровень Hsp90α в плазме можно было детектировать путем ELISA или вестерн-блоттинга, включающих в себя следующие стадии:
а) сбор цельной крови субъекта, например, пациента со злокачественной опухолью, и получение из нее плазмы или сыворотки путем центрифугирования;
b) детектирование с помощью методов ELISA или вестерн-блоттинга уровня Hsp90α в плазме или сыворотке, полученных на стадии (a), при этом образец плазмы здорового человека используется в качестве отрицательного контроля, а образец плазмы пациентов со злокачественными опухолями используется в качестве положительного контроля, и, необязательно, получение стандартной кривой концентрации Hsp90α; и
c) определение злокачественности и стадии опухоли в соответствии с уровнем Hsp90α в плазме, и, следовательно, определение диагноза, прогноза или эффективности лечения для данного субъекта.
На стадии (b) могут быть использованы также и другие методы, например, другие способы на основе реакций антиген-антитело, а также способы, основанные на других принципах прямого или непрямого отражения концентраций Hsp90α, например, способы определения концентрации Hsp90α путем детектирования активности Hsp90α.
Стандартный образец Hsp90α, используемый для получения стандартной кривой ELISA, может быть очищен из плазмы пациентов со злокачественной опухолью и может быть получен также с помощью рекомбинантных технологий. Стандартный образец Hsp90α может быть полноразмерным Hsp90α, его фрагментами, а также другими рекомбинантными белками или конъюгатами, содержащими последовательность Hsp90α. "Стандартная кривая концентрации Hsp90α" относится к корреляционной кривой между концентрацией Hsp90α и значениями поглощения, определяемыми при ELISA с использованием стандартных образцов Hsp90α. "Стандартный образец Hsp90α" относится к плазматическому белку Hsp90α, рекомбинантному белку Hsp90α, его фрагментам или производным со степенью чистоты выше 95%.
"Определение злокачественности опухоли" относится к вынесению заключения относительно злокачественности опухоли в результате изучения концентрации Hsp90α в образцах плазмы пациентов и сравнения полученной величины с отрицательным и положительным контролями.
Для определения уровня Hsp90α в плазме могут быть использованы как метод сэндвич-ELISA, так и конкурентный метод ELISA. Конкурентный метод ELISA с более высокой чувствительностью является предпочтительным.
Обычно метод сэндвич-ELISA включает в себя следующие стадии: a) иммобилизацию специфического антитела на твердой подложке, и удаление не связавшихся антител и примесей с помощью одной или нескольких стадий очистки; b) добавление тестируемых образцов и инкубацию в течение периода, позволяющего антигену в указанных образцах связаться с иммобилизованным антителом и на твердой подложке образовать иммунный комплекс антиген-антитело, а затем удаление не связавшихся веществ; c) добавление конъюгированного с ферментом антитела, которое может связываться с указанным антигеном в твердофазных иммунных комплексах, а затем удаление не связавшегося конъюгированного с ферментом антитела путем тщательного промывания (количество конъюгированного с ферментом антитела, связавшегося с твердой подложкой, положительно коррелирует с количеством тестируемого антигена); и d) добавление субстрата и качественное или количественное определение количества антигена в соответствии с цветной реакцией.
Обычно конкурентный метод ELISA включает в себя следующие стадии: a) иммобилизацию специфического антитела на твердой подложке и тщательное промывание; b) добавление смеси тестируемого образца и определенного количества конъюгированного с ферментом антигена, и инкубацию в течение достаточного временнóго периода, а затем тщательное промывание. (Если в образце антигена нет, конъюгированный с ферментом антиген будет связываться с твердофазным антителом неконкурентным образом. Если в образце антиген есть, указанный антиген в образце будет конкурировать с антигеном, меченным ферментом, за связывание с иммобилизованным антителом, и количество меченного ферментом антигена, связанного с иммобилизованным антителом, будет уменьшаться. В качестве контроля, только эталонная пробирка содержит меченный ферментом антиген); c) добавление субстрата и получение значения поглощения для каждой пробирки. Значение в эталонной пробирке является наиболее высоким, и разница между эталонной пробиркой и пробиркой с образцом представляет собой количество антигенов в образце. Чем бледнее цвет, тем больше антигенов содержится в образце.
При использовании сэндвич-ELISA антитела представляют собой два разных типа антител, специфичных в отношении Hsp90α плазмы. Иммобилизованное антитело может быть кроличьим поликлональным антителом со способностью к прочному связыванию, тогда как детектируемое антитело является мышиным моноклональным антителом с высокой специфичностью. Два этих антитела не должны перекрестно реагировать.
При использовании конкурентной ELISA антитела должны быть специфическими антителами против плазматического Hsp90α, который должен обладать способностью к прочному связыванию и иметь высокую специфичность, как в отношении конкурентного вещества, так и в отношении Hsp90α.
При использовании конкурентной ELISA конкурентным веществом может быть меченый стандартный белок плазмы Hsp90α. И метка не мешает связыванию стандартного белка Hsp90α с его антителом.
При использовании метода вестерн-блоттинга первичным используемым антителом является специфическое антитело против плазматического Hsp90α; вторичное антитело может быть конъюгировано с пероксидазой хрена или щелочной фосфатазой; субстратом может быть DAB или флуоресцентный субстрат. Предпочтительным является флуоресцентный субстрат с высокой чувствительностью.
Чувствительность указанных методов должна составлять 10 нг/мл, или она должна быть еще более чувствительной.
Используемое в настоящем изобретении антитело, специфичное в отношении Hsp90α плазмы, может быть полноразмерным антителом или его фрагментами и производными.
Антитела согласно изобретению могут быть также заменены другими средствами, способными к специфическому связыванию с Hsp90α, при этом указанные средства включают в себя низкомолекулярные соединения, пептиды и их производные.
Конкурентные стандартные белки плазмы Hsp90α могут быть мечены биотином и различными флуоресцентно меченными реагентами. Биотин является предпочтительным.
Злокачественные опухоли, которые могут быть детектированы посредством описанного в настоящем изобретении набора или способа, включают в себя, но не ограничены перечисленным, рак легкого, рак печени, рак желудка, рак пищевода, остеосаркому, рак поджелудочной железы, лимфому, рак толстой и прямой кишок, рак молочной железы, рак предстательной железы, рак ротовой полости, рак носоглотки, рак шейки матки, лейкемию, злокачественную меланому, саркому, почечноклеточный рак, холангиокарциному. "Опухоль и ее злокачественность" относится к тому, является ли опухоль доброкачественной, злокачественной или метастазирующей.
Авторы изобретения показали, что уровень Hsp90α в плазме у человека-неопухоленосителя составляет от 2 до 50 нг/мл, в основном в интервале от 2 до 10 нг/мл. Уровень Hsp90α в плазме у пациентов, у которых диагностирована злокачественная опухоль, выше нормального уровня, тогда как уровень Hsp90α в плазме у пациентов с метастазами злокачественной опухоли составляет выше 50 нг/мл, в основном выше чем 200 нг/мл. Это делает плазматический Hsp90α новым опухолевым маркером, который будет полезен для диагностики злокачественных опухолей, особенно метастазирующих.
Таким образом, в одном из воплощений набор или способ согласно изобретению может быть использован для определения существования опухоли, в особенности злокачественных опухолей и опухолевых метастазов. Для этих целей набор или способ согласно изобретению может быть использован для измерения уровня Hsp90α в образцах плазмы пациентов, у которых подозревают наличие опухоли, или же пациентов, у которых подозревают наличие опухолевых метастазов, и, необязательно, сравнения с нормальными контролями, а затем определения вероятности того, что у пациента есть опухоль или опухолевые метастазы, в соответствии с уровнем Hsp90α в образце. Повышенный уровень Hsp90α в плазме предполагает повышенную вероятность того, что пациент страдает от злокачественной опухоли, а в случае пациентов с известным диагнозом злокачественной опухоли повышенный уровень Hsp90α строго свидетельствует о возможности опухолевых метастазов.
В другом воплощении набор или способ согласно изобретению может быть использован для скрининга популяций высокого риска в отношении опухолей путем детектирования уровня Hsp90α в плазме. Для этих целей набор или способ согласно изобретению может быть использован для детектирования уровня Hsp90α в образцах плазмы в популяциях повышенного риска, и, необязательно, детектируемый уровень может быть сравнен с нормальными контролями, и затем может быть определена вероятность того, что пациент входит в состав популяции, имеющей опухоль, в соответствии с указанным уровнем в образце. Повышенный уровень Hsp90α в плазме указывает на повышенную вероятность наличия злокачественной опухоли. Специалистам в данной области хорошо известно, как определить различные группы высокого риска, в зависимости от типа злокачественной опухоли, которую подвергают скринингу, от индивидуальных факторов, таких как возраст, семейный анамнез, образ жизни, производственная среда, история подверженности действию вредных соединений и т.д. Например, пациенты с хроническим гепатитом В или гепатитом С имеют высокий риск развития HCC.
В другом воплощении набор или способ согласно изобретению может быть использован для предсказания прогнозов у пациентов со злокачественными опухолями путем детектирования уровня Hsp90α в плазме. Для этих целей набор или способ согласно изобретению может быть использован для детектирования уровня Hsp90α в образцах плазмы пациентов со злокачественной опухолью, и, необязательно, детектируемый уровень может быть сравнен с нормальными контролями или с прежними уровнями Hsp90α в плазме у данного пациента, и затем можно сделать прогнозирование у пациента со злокачественной опухолью в соответствии с уровнем Hsp90α в образцах плазмы. Поддержание Hsp90α на высоком уровне или дальнейшее повышение уровня Hsp90α может быть связано с плохим прогнозом. Следовательно, клиницисты смогут быть готовы к тому, чтобы обеспечить более тщательное наблюдение за пациентом и, если это необходимо, изменить курс лечения.
В другом воплощении набор или способ согласно изобретению может быть использован для детектирования уровня Hsp90α в образцах плазмы, который, таким образом, используется для определения того, является ли лечение, такое как хирургическое вмешательство, радиационная терапия или химиотерапия эффективной для пациента со злокачественной опухолью. Для этих целей набор или способ согласно изобретению может быть использован для детектирования уровня Hsp90α в образцах плазмы пациентов со злокачественной опухолью, и, необязательно, детектируемый уровень может быть сравнен с нормальными контролями или с прежними уровнями Hsp90α в плазме у данного пациента, и затем можно определить, будет ли хирургическое вмешательство, радиационная терапия или химиотерапия эффективной для пациента со злокачественной опухолью, или же следует прервать лечение.
Специалистам в данной области будет понятно, что, подобно всем другим опухолевым маркерам или способам скрининга и диагностики на основе таких опухолевых маркеров, способ согласно изобретению, предназначенный для определения наличия опухоли или ее метастазов путем детектирования уровня Hsp90α в образце плазмы, является дополнительным способом, полезным при диагностике опухолей. Обычно результаты, получаемые способами согласно изобретению, как таковые не являются достаточными ни для того, чтобы специалист в данной области непосредственно вынес моментальное заключение о том, страдает ли субъект злокачественной опухолью или имеет ли метастазы, ни для того, чтобы их можно было непосредственно использовать для определения гистологической природы и патологического признака злокачественной опухоли или участков расположения или количества метастазов. Следовательно, установление клинического диагноза все еще зависит от результатов клинической технологии визуализации и патологического обследования, также как и от заключения клинициста-онколога. Как таковой целью способов согласно изобретению является обеспечение клинических врачей или исследователей некоторой дополнительной информацией для дополнительного контроля. Тем не менее, как здесь показано, уровень Hsp90α в плазме обеспечит полезную информацию для определения наличия злокачественной опухоли, в частности, метастазов злокачественной опухоли. Сходным образом, способы согласно изобретению, предназначенные для скрининга, прогностической оценки и оценки эффективности терапии опухоли, также могут предоставить полезную информацию для практикующих клиницистов или исследователей.
Авторы изобретения дополнительно показали, что в плазму Hsp90α секретируется опухолевыми клетками, и что он отличается от такового, присутствующего внутри опухолевых клеток. Следовательно, делается заключение, что можно вызвать супрессию развития опухоли и метастазов путем ингибирования секреции Hsp90α в плазму. Это было показано на мышиной модели опухолевых метастазов с помощью специфического антитела против плазматического Hsp90α. Следовательно, секреция Hsp90α может быть использована в качестве новой мишени для скрининга новых противоопухолевых лекарственных средств.
В другом аспекте настоящее изобретение связано с применением ингибитора полипептида согласно изобретению при изготовлении фармацевтической композиции для предотвращения или лечения опухолевых метастазов.
В соответствии с одним из аспектов настоящего изобретения, указанный выше ингибитор является специфическим антителом против Hsp90α. Предпочтительно, чтобы указанное антитело было гуманизированным антителом или его фрагментом. В одном из воплощений указанные антитела специфически связываются с фосфорилированной формой Hsp90α, которая содержит одну или более фосфорилированных аминокислотных остатков в аминокислотной последовательности, соответствующей SEQ ID NO: 1, выбранных из группы, состоящей из Thr90, Ser231, Ser263, Tyr309 и их сочетания. В предпочтительном воплощении указанное антитело может связываться с Hsp90α, который фосфорилирован по Thr90. В одном из специфических воплощений указанное антитело представляет собой MAb E9 или D10, которое продуцируется клеточной линией, депонированной, соответственно, под регистрационным номером CGMCC No. 2903 или 2904. Как здесь показано, такие антитела могут у мышиной модели полностью ингибировать опухолевые метастазы в лимфатический узел, и могут вплоть до 56% ингибировать метастазы в легкое.
Соответственно, в другом аспекте настоящее изобретение связано также с антителом, которое специфически связывается с полипептидом согласно изобретению, и в частности, с Hsp90α плазмы. В специфическом воплощении указанным антителом является MAb E9 или D10, которое продуцируются, соответственно, клеточной линией, депонированной под регистрационным номером CGMCC No. 2903 или 2904. Предпочтительно, чтобы указанное антитело было гуманизированным антителом или его антигенсвязывающим фрагментом. В специфическом воплощении указанное антитело специфически связывается с фосфорилированной формой Hsp90α, которая содержит один или более аминокислотных остатков, выбранных из группы, состоящей из Thr90, Ser231, Ser263, Tyr309 и их сочетания. В одном из специфических воплощений указанное антитело специфически связывается с Hsp90α, фосфорилированным по Thr90. Предпочтительно, чтобы указанное антитело ингибировало опухолевый рост, в частности, метастазирование. Настоящее изобретение связано также с конъюгатом, содержащим указанное антитело, и с частичной диагностикой или лечением. Например, диагностической частью является флюорофор, тогда как лечебным фрагментом является химиотерапевтическое средство.
Автором изобретения обнаружено также, что Hsp90α, секретируемый опухолевыми клетками, связан с уровнем протеинфосфатазы 5 (PP5) в опухолевых клетках. В доброкачественных опухолях секретируемого Hsp90α мало, но уровень экспрессии PP5 является высоким. В злокачественной опухоли секретируемого Hsp90α много, но уровень экспрессии PP5 является низким. Следовательно, уровень секретируемого Hsp90α отрицательно взаимосвязан с уровнем PP5. В результате, путем детектирования уровня экспрессии PP5 в образце опухолевой ткани можно предсказать количество секретируемого Hsp90α.
Авторами изобретения обнаружено также, что секреция Hsp90α может быть ингибирована внутриклеточным PP5. Повышенная экспрессия молекулы нуклеиновой кислоты, кодирующей PP5 в клетке, ингибирует секрецию Hsp90α, тогда как снижение уровня экспрессии PP5 приведет к повышению секреции Hsp90α. Таким образом, путем повышения экспрессии PP5 можно ингибировать секрецию Hsp90α, и можно ингибировать прогрессию и метастазирование опухоли. На основе экспериментов авторов изобретения им удалось показать, что повышенная экспрессия PP5 ингибирует миграцию клеток MCF-7 злокачественной опухоли молочной железы. Следовательно, PP5 может быть новой терапевтической мишенью для лечения опухолей.
Соответственно, в следующем аспекте настоящее изобретение связано со способом ингибирования опухолевой инвазии и метастазирования, предусматривающим стадию ингибирования фосфорилирования Hsp90α внутри опухолевых клеток. В одном из воплощений указанный способ включает в себя стадию ингибирования фосфорилирования Hsp90α в опухолевых клетках. В одном из специфических воплощений указанный способ включает в себя стадию повышенной экспрессии молекулы нуклеиновой кислоты, кодирующей PP5 в опухолевых клетках. Предпочтительно, чтобы повышенная экспрессия PP5 достигалась путем введения гена. В одном из воплощений указанный способ включает в себя стадию повышенной экспрессии молекулы нуклеиновой кислоты, кодирующей PP5, имеющей аминокислотную последовательность SEQ ID NO: 5. В одном из специфических воплощений указанная молекула нуклеиновой кислоты содержит нуклеотидную последовательность SEQ ID NO: 6. Настоящее изобретение связано также с применением вектора, несущего полинуклеотид, кодирующий PP5, оперативно связанного с промотором, при получении лекарственного средства для ингибирования фосфорилирования Hsp90α в опухолевых клетках путем повышения экспрессии PP5 в опухолевых клетках. Такое лекарственное средство может быть использовано для ингибирования инвазивности и метастазирования опухолей.
Настоящее изобретение связано также со способами и моделями скрининга противоопухолевых лекарственных средств с применением Hsp90α плазмы или его производных. Такие противоопухолевые лекарственные средства включают в себя, но не ограничиваются перечисленным, белки, связывающие плазматический Hsp90α, низкомолекулярные пептиды и низкомолекулярные соединения, а также ингибиторы, которые могут вызывать супрессию активности плазматического Hsp90α.
Примеры
Пример 1: Сбор и получение образцов мышиной плазмы, и детектирование Hsp90α в плазме Мышей Balb/c со средним весом тела в 20 граммов (приобретенных в корпорации Beijing Vital River Laboratory Animal Technology Co., Ltd.) случайным образом делили на две группы, по 3 животных в каждую. Мышам экспериментальной группы инокулировали по 106 клеток H22 (CCTCC ID: GDC091), тогда как мышам контрольной группы инокуляцию не производили. Когда диаметр опухоли достигал приблизительно 2 см (примерно через 20 дней), кровь отбирали из нижнего венозного сплетения глаза. Для предотвращения гемолиза добавляли антикоагулянт. В случае возникновении гемолиза кровь нужно было брать повторно. Образцы крови дважды центрифугировали при 4°С и при 6000×g, и собирали супернатант. Количество белка Hsp90α в плазме детектировали методом вестерн-блоттинга с использованием кроличьего поликлонального антитела pAb против человеческого Hsp90α (Labvasion). Метод BCA (Pierce) был использован для определения общего количества суммарного белка в образцах, для гарантии того, что нанесенные количества разных образов равны между собой. Результаты представлены на Фиг. 1. По сравнению с контрольными мышами, уровень Hsp90α в плазме у мышей-опухоленосителей повышался.
Пример 2: Отбор и получение образцов плазмы, полученных у здоровых людей и у пациентов-опухоленосителей, и детектирование Hsp90α в плазме
Отбирали образцы крови здоровых людей и пациентов-опухоленосителей, и в течение 24 часов доставляли их в лабораторию, храня их в течение указанного периода при 4°С. При возникновении гемолиза кровь приходилось брать повторно. Образцы крови дважды центрифугировали при 4°С и при 6000×g, и собирали супернатант. Количество белка Hsp90α определяли методом вестерн-блоттинга. В случае если образцы не удавалось исследовать сразу же, их хранили при -80°С. Путем сравнения результатов с клиническим диагнозом устанавливали корреляцию между Hsp90α и злокачественностью опухоли.
Методика проведения вестерн-блоттинга: образцы плазмы смешивали с загрузочным буфером в соотношении 1:1. 1-2 мкл образца наносили на ДСН-ПААГ. Первичное антитело является тем самым антителом, которое может специфически распознавать Hsp90α в плазме (крысиное моноклональное антитело mAb SPA-840, Stressgen). Вторичным антителом является козье антитело против крысиного антитела, конъюгированного с пероксидазой хрена (HRP) (приобретено в Zhongshanjinqiao). На фиг. 2 представлены результаты, согласно которым количество Hsp90α у пациентов с раком печени в 10 раз превышает его количество у здоровых людей (A), тогда как у пациентов с доброкачественной ретенционной кистой молочной железы (галактоцеле) и с миомой матки оно повышено в 2 раза по сравнению со здоровыми людьми.
Пример 3: Получение кроличьего pAb и мышиного mAb против Hsp90α
Для клонирования гена Hsp90α были использованы праймеры со следующими последовательностями:
Hsp90α-Sal1-Re: ACGCGTCGACTTAGTCTACTTCTTCCATGC (SEQ ID NO:8) и Hsp90α-Sph1-For: ACATGCATGCATGCCTGAGGAAACCCAGACC (SEQ ID NO:9). Праймеры были синтезированы компанией Invitrogen. Pfu-ДНК-полимеразы (NEB) были синтезированы для амплификации Hsp90α из библиотек кДНК печени человека (Stratagene). Sph1 и Sal1 (NEB) были использованы для расщепления амплифицированных продуктов ПЦР и вектора pQE80L (Qiagen). Затем для лигирования были использованы T4-лигазы. Указанные продукты были транпортированы в клетки Top 10 (Transgen) с целью амплификации и подтверждения правильности выборки. Затем подтвержденные плазмиды были дополнительно транпортированы в клетки BL21DE3 (Transgen) для экспрессии. Рекомбинантные белки Hsp90α очищали путем ионообменной хроматографии SP HP, pH 6,8, собирая пик элюции в 10 мсек/мл, и Q HP, pH 7,8, собирая пик элюции в 19 мсек/мл.
Рекомбинантный человеческий Hsp90α со степенью чистоты, превышающей 95%, использовали для иммунизации самцов новозеландских кроликов путем подкожных дорсальных инъекций в несколько участков суммарным количеством в 100 мкг. Через две недели проводили вторичную иммунизацию таким же способом, за исключением того, что количество инъецируемого Hsp90α уменьшали до 50 мкг. Усиливающие буст-инъекции двукратно производили каждые 2 недели после вторичной иммунизации. Титр антитела в сыворотке определяли через 7-10 дней после каждой буст-иммунизации. Через восемь дней после последней иммунизации сыворотку собирали и хранили при -20°С. Аффинную хроматографию конъюгированного антигена использовали для очистки антитела от сыворотки. Очищенное кроличье pAb было названо S2.
Мышей BALB/C иммунизировали рекомбинантным человеческим Hsp90α. Первичная иммунизация: 100 мкг антигена и полного адъюванта Фрейнда вводили в виде подкожных дорсальных инъекций в несколько участков. Вторую иммунизацию проводили через 3 недели, с неполным адъювантом Фрейнда, используя ту же самую дозу, путем внутрибрюшинной инъекции. Третью иммунизацию проводили еще через 3 недели, без адъюванта, используя ту же самую дозу (кровь на анализ брали через 5-7 дней). Еще через 3 недели 200 мкг антигена вводили в виде в/б инъекции в качестве бустер-иммунизации. Через 3 дня получали клетки селезенки, и сливали их с гибридомами SP2/0-Ag14(SP2/0) (ATCC: CRL-1581). Для скрининга использовали HAT. Для образования гибридомных колоний использовали ограниченное разведение. Для идентификации использовали методы вестерн-блоттинга и ELISA. В конечном итоге были получены E9 и D10, секретирующие специфическое антитело против Hsp90α, которые 24 февраля 2009 года были задепонированы в банке [клеточных линий] под регистрационными номерами CGMCC No.2903 и 2904.
Непрямой метод ELISA использовали для определения титров E9 и D10. Как показано на фиг. 3, E9 и D10, с титрами в 500000, могут быть использованы для детектирования Hsp90α в плазме. Непрямой метод ELISA: планшет покрывали рекомбинантным человеческим Hsp90α в концентрации 10 мкг/мл и выдерживали при 4°С в течение ночи. Затем указанный планшет блокировали в течение 1 часа при 37°С. Потом добавляли E9 или D10 в разведениях 1:400, 1:1600, 1:6400, 1:25600, 1:102400, 1:509600 и инкубировали в течение 2 часов при комнатной температуре. После этого добавляли вторичное антитело, козье противомышиное антитело, конъюгированное с HRP (приобретенной в компании Zhongshanjinqiao), и инкубировали в течение 1 часа при комнатной температуре. В качестве субстрата добавляли O-фенилендиамин, и измеряли поглощение при ОП 490 нм.
Пример 4: Измерение концентрации Hsp90α с помощью сэндвич-ELISA с использованием E9 и S2
В методе сэндвич-ELISA, предпринятом для измерения концентрации Hsp90α в плазме, были использованы два антитела из разных источников. S2 с высокой связывающей способностью (получение S2 было описано в примере 3) было использовано в качестве покрывающего антитела, а E9 (получение E9 было описано в примере 3) с высокой специфичностью связывания было использовано в качестве детектирующего антитела. Перекрестной реакции между двумя указанными антителами выявлено не было. Указанный метод является воспроизводимым и чувствительным. На фиг. 4 показано, что чувствительность указанного метода составляет 5 нг/мл.
Пример 5: Отбор и получение плазмы человека, детекция Hsp90α в плазме и определение злокачественности опухоли (сэндвич-ELISA)
Образцы крови здоровых людей, пациентов со злокачественной опухолью и пациентов с воспалением доставляли в лабораторию при низкой температуре в течение 24 часов. В случае возникновения гемолиза кровь нужно было брать повторно. Образцы крови дважды центрифугировали при 4°С и при 6000×g, и собирали супернатант. Количество белка Hsp90α в плазме детектировали методом ELISA. Указанные образцы делили на аликвоты и хранили при -80°С, если их не удавалось исследовать сразу же. Путем сравнения результатов с клиническим диагнозом устанавливали корреляцию между Hsp90α и злокачественностью опухоли.
В методе сэндвич-ELISA были использованы два антитела против Hsp90α. Антителом S2 покрывали планшет и инкубировали при 4°С в течение ночи. Затем указанный планшет блокировали в течение 1 часа при 37°С. Потом образцы плазмы разбавляли в 10 раз и добавляли в указанный планшет (по 100 мкл/лунку). После 2-часовой инкубации при 37°С добавляли E9 и инкубировали при 37°С еще в течение 2 часов. К инкубату добавляли козье противомышиное антитело, конъюгированное с HRP, и инкубировали в течение еще 1 часа. В качестве субстрата добавляли O-фенилендиамин, и измеряли поглощение при ОП 490 нм. Результаты представлены на фиг. 5, 6 и 7. Стандартная кривая включает в себя серию образцов, содержащих градиент концентрации стандартных белков Hsp90α и 10% отрицательной плазмы в каждом образце, которые были использованы для исключения фона плазмы.
Как показано на фиг. 5, количество Hsp90α в плазме у пациентов с доброкачественной опухолью (включая пациентов с доброкачественной галактоцеле и миомой матки, семь случаев) составляет от 2 до 10 нг/мл, в основном от 2 до 5 нг/мл. Уровень Hsp90α в плазме у 69% (20/29) пациентов с раком печени составляет более 50 нг/мл, что в среднем в 10 раз выше такового у пациентов с доброкачественной опухолью, p=0,00263, t-тест Стьюдента (A); Уровень Hsp90α в плазме у 64% (9/14) пациентов с раком легкого составляет более 50 нг/мл. Средняя концентрация в 10 раз выше таковой у пациентов с доброкачественной опухолью, p=0,0497, t-тест Стьюдента (B). Уровень Hsp90α в плазме у 78% (25/32) пациентов со злокачественной опухолью молочной железы составляет более 50 нг/мл. Среднее значение в 10 раз выше такового у пациентов с доброкачественной опухолью, p<0,001, t-тест Стьюдента (C). Уровень Hsp90α в плазме у 100% (10/10) пациентов с раком предстательной железы составляет более 50 нг/мл. Среднее значение в 10 раз выше такового у пациентов с доброкачественной опухолью, p<0,05, t-тест Стьюдента (D).
Как показано на фиг. 6, у пациентов с раком печени (17 случаев, 7 случаев с метастазами) (A), раком легкого (10 случаев, 2 случая с метастазами) (B) и раком молочной железы (21 случай, 10 случаев с метастазами) (С) уровни Hsp90α при наличии у пациентов метастазов были значительно выше, чем таковые у пациентов без метастазов. При раке печени p=0,003, при раке молочной железы p=0,002, t-тест Стьюдента.
Как показано на фиг. 7, у пациентов с воспалением, включая 10 случаев пневмонии (A), 10 случаев гепатита (типа A, 5 случаев, типа В, 5 случаев) (B) уровни Hsp90α в плазме составляют от 2 до 10 нг/мл, что не дает значимых различий по сравнению с таковыми, наблюдаемыми у здоровых персон (3 случая), соответственно, p=0,2988, 0,5177, 0,138, согласно t-тесту Стьюдента.
Образцы [плазмы] здоровых людей отбирали у здоровых добровольцев, тогда как образцы, полученные у пациентов-опухоленосителей и у пациентов с воспалением, получали из госпиталей Beijing Cancer Hospital и Xiamen First Hospital.
Пример 6: Hsp90α плазмы секретируется опухолевыми клетками
Мышей nude (приобретенных в компании Beijing Vital River Laboratory Aimal Technology Co., Ltd.) со средним весом тела в 20 г разделили на две группы, по 6 мышей в каждой группе. Каждой мыши инъецировали по 106 клеток Hela (ATCC: CCL-2). Контрольную группу составляли нормальные мыши без опухолей. Когда диаметр опухоли достигал 2 см (приблизительно через 20 дней), кровь отбирали из венозного сплетения глаза. Антитело против Hsp90α (крысиное mAb, Stressgen), которое специфически распознает человеческий Hsp90α, но не распознает мышиный Hsp90α, использовали для детекции Hsp90α в плазме. Как показано на фиг. 8, Hsp90α в плазме мышей-опухоленосителей специфически распознается человеческим, но не мышиным анти-Hsp90α-антителом, что указывает на то, что Hsp90α плазмы секретируется ксенотрансплантатом опухолевых клеток.
Пример 7: Hsp90α, секретируемый опухолевыми клетками, усечен на C-конце
Используя Hsp90α-pc3.1-Nhe1-For-Myc: GCTAGCTAGCGCCACCA TGGAACAAAAACTCATCTCAGAAGAGGATCTGCCTGAGGAAACCCAGACCCAAGAC (SEQ ID NO: 10) и Hsp90α-pc3.1-Xhol-Re-nostop: CCCGCTCGA GTGTCTACTTCTTCCATGCGTGATG (SEQ ID NO: 12) в качестве праймеров (синтезированных компанией Invitrogen), а также Pfu-ДНК-полимеразу (NEB), Hsp90α амплифицировали из матрицы pQE80L-Hsp90α (полученной в примере 3). Продукты расщепляли и встраивали в pcDNA3.1/Myc-His(-) (Invitrogen), с получением Hsp90α с меткой Мус на N-конце, которая была названа Myc-H. Сходным образом, His-Myc-H с N-концевыми тандемными метками His-Myc амплифицировали посредством Hsp90α-pc3.1-Nhe1-For-His-Myc: GCTAGCTAGCGCCACCATGCATCATCATCATCATCATGAACAAAAACTCATCTCAGAAGAGGATCTGCCTGAGGAAACCCAGACCCAAGAC (SEQ ID NO: 11) и SEQ ID NO: 12 в качестве праймеров. Эти две плазмиды были транзитно трансфицированы в MCF-7 для обнаружения секреции экзогенного Hsp90α. Антитела против Hsp90α, Мус и His использовали для детекции секретируемого Hsp90α. Результаты показали, что секретируемый Hsp90α был усечен на C-конце (Фиг. 9A).
С использованием Myc-His-H в качестве матрицы, были созданы мутации последних четырех аминокислот (EEVD) с С-конца Hsp90α. EE-> AA представляют собой мутации двух EE на два Ala (с использованием SEQ ID NO: 11 и Hsp90α-EE-AA: GGCCGCTCGAGTGTCTACTGCTGCCATGCGTGATGTG (SEQ ID NO: 13) в качестве праймеров), VD-> AA означает, что VD были заменены на два Ala (с использованием SEQ ID NO: 11 и Hsp90α-VD-AA: GGCCGCTCGA GTTGCTGCTTCTTCCATGCGTGATGTG (SEQ ID NO: 14) в качестве праймеров). Все Ala представляют собой мутацию EEVD на четыре Ala (с использованием SEQ ID NO: 11 и Hsp90α-EE VD-AAAA: GGCCGCTCGAGTTGCTGCTGCTGCCATGCGTGATGTG (SEQ ID NO: 15) в качестве праймеров), СΔ4 представляют собой делецию последних четырех аминокислот EEVD (с использованием SEQ ID NO: 11 и Hsp90α-CΔ4-Xho: CCGCTCGAGTCATGCGTGATGTGTCGTCATCTC (SEQ ID NO: 16) в качестве праймеров). Линию клеток злокачественной опухоли молочной железы человека MCF-7 транзитно трансфицировали полученными мутантами для обнаружения секреции экзогенного Hsp90α (Hsp90α, экспрессируемого в повышенных количествах, который отличается от эндогенного). Антитело против Hsp90α было использовано для детекции изменений секретируемого во внеклеточную среду Hsp90α. Результаты показали, что последние четыре C-концевых аминокислотных остатка регулируют секрецию Hsp90α. Любая сайт-направленная мутация или делеция указанных четырех аминокислотных остатков может приводить к секреции Hsp90α без C-концевого укорочения, а это и предполагает, что C-концевые аминокислотные остатки EEVD в секретируемом внеклеточном Hsp90α подвергаются делеции (фиг. 9B).
Пример 8: Изучение существующей формы Hsp90α в плазме человека
Были отобраны образцы цельной крови пациентов с раком печени, которые в течение 24 часов после взятия подвергали двукратному центрифугированию, а затем определению Hsp90α в плазме с использованием метода иммунопреципитации и иммуноблоттинга. Сначала для иммунопреципитации было использовано специфическое кроличье поликлональное антитело против Hsp90α (источник: Labvision), а затем кроличье поликлональное антитело, которое способно специфически распознавать четыре C-концевых аминокислотных остатка EEVD белка Hsp90α (лабораторная база, антиген, используемый для иммунизации, является белком-носителем, соединенным с трижды повторяемыми пептидами EEVD, синтезированными в компании SBS Genetech Co., Ltd.), использовали для детекции Hsp90α в плазме. Как показано на фиг. 10, анти-EEVD-антитело специфически распознает Hsp90α из лизата цельных клеток, но не распознает Hsp90α плазмы, что свидетельствует о том, что Hsp90α плазмы лишен четырех C-концевых аминокислотных остатков EEVD и отличается от внутриклеточного белка Hsp90α. (Фиг. 10).
Пример 9: Детекция фосфорилированной формы Hsp90α в плазме
Образцы цельной крови пациентов с раком печени в течение 24 часов после взятия крови подвергали двукратному центрифугированию, с получением плазмы. Затем определяли Hsp90α в плазме с использованием методов иммунопреципитации и иммуноблоттинга. Сначала для иммунопреципитации белка Hsp90α плазмы было использовано специфическое кроличье поликлональное антитело против Hsp90α (источник: Labvision), а затем антитело (кроличье поликлональное антитело pAb против фосфорилированного субстрата протеинкиназы PKA, анти-фосфо-(Ser/Thr), Cell signaling), которое способно специфически распознавать фосфорилированный по Thr90 белок Hsp90, использовали для детекции фосфорилированного статуса Hsp90 плазмы. Как показано на фиг. 11, белок Hsp90α в плазме является фосфорилированным по Thr90.
Пример 10: Детектирование концентрации фосфорилированного по Thr90 белка Hsp90α в плазме
Образцы цельной крови пациентов с раком печени и здоровых людей в течение 24 часов после взятия крови подвергали двукратному центрифугированию, с получением плазмы. Относительный уровень Hsp90α в плазме определяли с использованием метода сэндвич-ELISA. Протокол: сначала полученным своими силами поликлональным кроличьим антителом S2 покрывали планшет, который выдерживали в течение ночи при 4°С, затем в каждую лунку добавляли по 100 мкл 10-кратно разведенных образцов плазмы. После инкубации в течение 2 часов при 37°С добавляли антитело, которое способно специфически распознавать фосфорилированный по Thr90 белок Hsp90α (cell signal). Планшет инкубировали в течение 2 часов при 37°С; затем добавляли вторичные антитела, конъюгированные с пероксидазой хрена, и инкубировали в течение 1 часа. В конце добавляли O-фенилендиамин, чтобы измерить поглощение при ОП 490 нм. Результаты показывают, что уровни Hsp90α у пациентов, больных раком печени, выше таковых у здоровых людей, P=0,003, t-тест Стьюдента, что свидетельствует о том, что уровень фосфорилированного по Thr90 белка Hsp90 у пациентов с раком печени является повышенным (фиг. 12).
Пример 11: Детектирование постоянства уровня фосфорилированного по Thr90 белка Hsp90 и суммарный уровень Hsp90α в плазме
Были определены уровни фосфорилированного по Thr90 белка Hsp90 и суммарный уровень Hsp90α в плазме пациентов с раком печени (8 случаев). Способ детектирования суммарного количества Hsp90α в плазме был таким же, который использовали и в примере 5; способ детектирования уровня фосфорилированного по Thr90 белка Hsp90 был таким же, который использовали в примере 10. Полученные результаты показывают, что уровень фосфорилированного по Thr90 белка Hsp90 в плазме соответствует суммарному количеству Hsp90α в плазме, что дополнительно указывает на то, что фосфорилирование содержащегося в плазме Hsp90α происходит по Thr90, и приращение суммарного количества Hsp90α может представлять собой приращение фосфорилированного по Thr90 белка Hsp90α (фиг. 13).
Пример 12: Фосфорилирование Thr90 является необходимым для секреции Hsp90α
Используя плазмиду pcDNA3.1/Myc-His-Hsp90α (известную также как Hsp90α дикого типа (WT Hsp90α)) в качестве матрицы, создали мутантный Hsp90α (T90A, в котором произвели замену треонина на аланин) посредством quick-change-ПЦР с использованием следующих праймеров: Hsp90α-T89A-смысловой: GATCGAACTCTTGCAATTGTGGATACTGG AATTGGAATG (SEQ ID NO: 17) и Hsp90α-T89-антисмысловой: CATTCCAATTCCAGTATCCACAATTGCAAGAGTTCGATC (SEQ ID NO: 18). T90A-мутант белка Hsp90α не может быть фосфорилирован по Thr90. Линию клеток злокачественной опухоли молочной железы человека MCF-7 (приобретенной в ATCC, No. HTB-22) трансфицировали Hsp90α дикого типа (WT) или мутантным Hsp90α (T90A). Затем собирали среду, и в ней с помощью анти-Hsp90α-антитела определяли секрецию экзогенного Hsp90α.
Полученные результаты показывают, что во внеклеточной среде можно обнаружить повышенную экспрессию экзогенного Hsp90α дикого типа, тогда как T90A-мутант не обнаруживается, что указывает на то, что фосфорилирование по остатку Thr90 является необходимым условием для секреции Hsp90α (фиг. 14).
Пример 13: PP5 ответственна за дефосфорилирование pT90-Hsp90α
Получение фосфорилированного по Thr90 белка Hsp90α (pT90-Hsp90α): Рекомбинантный человеческий белок Hsp90α и рекомбинантную протеинкиназу A (Promega Corporation, USA) инкубировали в реакционном буфере (NEB UK company) при 30°С в течение 1 часа, затем производили очистку белка pT90-Hsp90α. После удаления свободного фосфата путем диализа очищенный белок pT90-Hsp90α, смешанный с рекомбинантным человеческим белком PP5, инкубировали при 30°С, свободный фосфат, высвобождаемый из pT90-Hsp90α, определяли с помощью набора для нерадиоактивного анализа серин/треонин-фосфатазы (Promega Corporation USA). Пептидный субстрат входит в состав данного набора, и его использовали в качестве положительного контроля. Как показано на фиг. 15A, когда PP5 инкубировали с пептидным субстратом, высвобождение свободного фосфата повышалось значимым образом, значение P <0,005, t-тест Стьюдента, что указывает на то, что PP5 может напрямую осуществлять дефосфорилирование данного пептидного субстрата (положительный контроль). Когда PP5 инкубировали с белком pT90-Hsp90α, высвобождение свободного фосфата также повышалось значимым образом, значение P <0,005, t-тест Стьюдента, что указывает на то, что PP5 может напрямую осуществлять дефосфорилирование pT90-Hsp90α.
Нуклеотидную последовательность протеинфосфатазы 5 (PP5) (SEQ ID NO:6) амплифицировали из библиотек кДНК печени человека и встраивали в вектор pcDNA3.1/Myc-His(-) (источник вектора: Invitrogen). Вектор PP5 трансфицировали в линии клеток MCF-7 злокачественной опухоли молочной железы человека и добились его повышенной экспрессии. С другой стороны, с использованием технологии интерферирующей РНК можно было также «выключить» повышенную экспрессию PP5 с помощью ки-РНК, имеющей последовательность 5'-ACTCGAACACCTCGCTAAAGAGCTC-3'(SEQ ID NO:7) (синтезированную компанией Invitrogen). Затем изучали статус Thr90-фосфорилирования Hsp90α на уровне повышенной экспрессии или «выключения» PP5. Как показано на фиг. 15B, при повышенной экспрессии человеческой PP5 уровень Thr90-фосфорилированного Hsp90α (pT90-Hsp90α) был в значительной степени понижен (составляя 0,55 такового в контрольной группе). Когда экспрессия человеческой PP5 была заторможена, уровень Thr90-фосфорилированного Hsp90α (pT90-Hsp90α) был в значительной степени повышен (составляя 1,58 такового в контрольной группе).
Пример 14: Регуляция секреции Hsp90α в результате промоции или ингибирования экспрессии PP5
PP5 может осуществлять дефосфорилирование Thr90-фосфорилированного Hsp90α. Для амплификации PP5 из библиотеки кДНК печени человека были использованы следующие праймеры: PP5-NheI-For: CTAGCTAGCATGTACCCATACGACGTCCCAGACTACGCT (SEQ ID NO: 19) и PP5-XhoI-Re: CCGCTCGAGTTAATGATGATGATGATG ATGCACGTGTACC (SEQ ID NO: 20). Полноразмерную PP5 клонировали в вектор pcDNA3.1/Myc-His(-) (источник вектора: Invitrogen). Линию клеток MCF-7 злокачественной опухоли молочной железы человека трансфицировали вектором PP5, затем изучали секрецию Hsp90α из указанных клеток. Полученные результаты показали, что после повышения экспрессии человеческой PP5 секреция Hsp90α значительно снижалась (фиг. 16А).
С другой стороны, когда экспрессию человеческой PP5 в клетках MCF-7 «выключали» посредством интерферирующей РНК (против человеческой PP5, Invitrogen), секреция Hsp90α значительно повышалась (фиг. 16B).
Пример 15: Уровень PP5 и злокачественность опухоли
Взаимосвязь между уровнем экспрессии PP5 и секрецией Hsp90α изучали в линиях клеток злокачественной опухоли молочной железы человека MCF-7, SKBR3, MDA-MB-453, 435s и 231 (ATCC, номера, соответственно, HTB-22, -30, -131, -129 и HTB-26) с использованием метода вестерн-блоттинга. Линии клеток MCF-7, SKBR3 злокачественной опухоли молочной железы являются менее злокачественными клеточными линиями. У мышей nude эти две клеточные линии могут образовывать первичные опухоли, но не вызывают метастазирование. Линии MDA-MB-453, 435s и 231 являются более злокачественными. Они могут образовывать не только первичные опухоли, но также и вызывать образование метастазов в отдаленные органы мышей nude. Таким образом, MDA-MB-435s и 231 часто используют для получения моделей метастазирующих опухолей.
На фиг. 17 показано, что клетки, которые экспрессируют высокий уровень PP5, секретируют низкие количества Hsp90α; тогда как клетки с низким уровнем экспрессии PP5 могут секретировать больше Hsp90α. В то же время результаты также показывают, что уровень секретируемого Hsp90α положительно коррелирует, а уровень экспрессии PP5 отрицательно коррелирует со злокачественностью опухоли (фиг. 17), свидетельствуя о том, что уровень секретируемого Hsp90α и его регуляторные факторы, такие как PP5, могут быть использованы для определения степени злокачественности опухоли.
Пример 16: Уровень экспрессии PP5 связан с подвижностью опухоли
Модель ранозаживления использовали для изучения взаимосвязи между уровнем экспрессии PP5 и миграцией опухолевых клеток.
Линию клеток злокачественной опухоли молочной железы человека MCF-7 трансфицировали человеческим вектором PP5 или ки-РНК PP5. Затем клетки инокулировали в 12-луночный планшет. Когда рост клеток достиг конфлуэнтности, кончик пипетки использовали для нанесения царапины, соскребя слой клеток, с образованием "раны". Соскребенные клетки удалили, а оставшиеся клетки культивировали в свежей среде DMEM (GIBCO) при 37°С в инкубаторе с 5% CO2. Изображения такой "раны" были сделаны во времена 0 ч, 12 ч и 24 ч (фиг. 18A). Влияние экспрессии PP5 на миграцию клеток изучали, анализируя скорость заживления "раны". Результаты показывают, что повышенная экспрессия PP5 ингибирует миграцию клеток MCF-7, тогда как PP5-ки-РНК может стимулировать миграцию клеток MCF-7 (фиг. 18B).
Пример 17: Влияние специфических антител против Hsp90α плазмы на миграцию опухолевых клеток
Модель ранозаживления использовали для детектирования влияния антитела, специфичного в отношении Hsp90α плазмы, на миграцию опухолевых клеток.
Клетки MCF-7 или MDA-MB-231 (ATCC, No. HTB-22 и HTB-26,соответственно) инокулировали в 12-луночный планшет. Когда рост клеток достиг конфлуэнтности, кончики пипеток использовали для нанесения царапин, соскребая слой клеток, с образованием "раны". Соскребенные клетки удаляли, а оставшиеся клетки культивировали в свежей среде DMEM (GIBCO). При этом добавляли E9 (20 мкг/мл), мышиное моноклональное антитело, специфичное в отношении Hsp90α, или контрольный IgG (20 нг/мл). После этого планшеты инкубировали при 37°С в инкубаторе с 5% CO2. Изображения полученной "раны" были сделаны во времена 0 ч, 6 ч, 12 ч, 24 ч, 48 ч и 72 ч после начала инкубации (фиг. 18A). Влияние антител, специфичных в отношении Hsp90 плазмы, на миграцию опухолевых клеток изучали, осуществляя мониторинг заживления "раны". Как показано на фиг. 19, антитело, специфичное в отношении Hsp90α плазмы, способно существенно ингибировать миграцию как MDA-MB-231 (фиг. 19A), так и MCF-7 (фиг. 19B) (степень ингибирования >40%).
Пример 18: Определение влияния антитела, специфичного в отношении Hsp90α плазмы, на опухолевые метастазы
Мышей nude со средним весом тела в 20 г приобретали в компании Beijing Vital River Laboratory Animal Technology Co., Ltd. Клетки меланомы мышей B16/F10 (ATCC, регистрационный номер CRL-6475) (2×105) инъецировали мышам в хвостовую вену. На следующий день мышей произвольно разделили на две группы (n=8): контрольную группу (вводили IgG) и опытную группу, которую обрабатывали Hsp90α Ab (мышиное моноклональное антитело E9). Антитела вводили через день в дозе 40 мкг/мышь. Метастазы изучали через 15 дней после инокуляции. Как показано на фиг. 20, антитело, специфичное в отношении Hsp90α плазмы, может полностью затормозить метастазирование клеток B16/F10 в лимфатические узлы (A), а метастазирование в легкие может ингибироваться на 56% (B).
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Tsinghua University
Protgen Ltd
<120> НОВЫЙ ОПУХОЛЕВЫЙ БИОМАРКЕР
<130> P2010653C
<160> 20
<170> PatentIn version 3.3
<210> 1
<211> 728
<212> Белок
<213> Homo sapiens
<400> 1
Met Pro Glu Glu Thr Gln Thr Gln Asp Gln Pro Met Glu Glu Glu Glu
1 5 10 15
Val Glu Thr Phe Ala Phe Gln Ala Glu Ile Ala Gln Leu Met Ser Leu
20 25 30
Ile Ile Asn Thr Phe Tyr Ser Asn Lys Glu Ile Phe Leu Arg Glu Leu
35 40 45
Ile Ser Asn Ser Ser Asp Ala Leu Asp Lys Ile Arg Tyr Glu Thr Leu
50 55 60
Thr Asp Pro Ser Lys Leu Asp Ser Gly Lys Glu Leu His Ile Asn Leu
65 70 75 80
Ile Pro Asn Lys Gln Asp Arg Thr Leu Thr Ile Val Asp Thr Gly Ile
85 90 95
Gly Met Thr Lys Ala Asp Leu Ile Asn Asn Leu Gly Thr Ile Ala Lys
100 105 110
Ser Gly Thr Lys Ala Phe Met Glu Ala Leu Gln Ala Gly Ala Asp Ile
115 120 125
Ser Met Ile Gly Gln Phe Gly Val Gly Phe Tyr Ser Ala Tyr Leu Val
130 135 140
Ala Glu Lys Val Thr Val Ile Thr Lys His Asn Asp Asp Glu Gln Tyr
145 150 155 160
Ala Trp Glu Ser Ser Ala Gly Gly Ser Phe Thr Val Arg Thr Asp Thr
165 170 175
Gly Glu Pro Met Gly Arg Gly Thr Lys Val Ile Leu His Leu Lys Glu
180 185 190
Asp Gln Thr Glu Tyr Leu Glu Glu Arg Arg Ile Lys Glu Ile Val Lys
195 200 205
Lys His Ser Gln Phe Ile Gly Tyr Pro Ile Thr Leu Phe Val Glu Lys
210 215 220
Glu Arg Asp Lys Glu Val Ser Asp Asp Glu Ala Glu Glu Lys Glu Asp
225 230 235 240
Lys Glu Glu Glu Lys Glu Lys Glu Glu Lys Glu Ser Glu Asp Lys Pro
245 250 255
Glu Ile Glu Asp Val Gly Ser Asp Glu Glu Glu Glu Lys Lys Asp Gly
260 265 270
Asp Lys Lys Lys Lys Lys Lys Ile Lys Glu Lys Tyr Ile Asp Gln Glu
275 280 285
Glu Leu Asn Lys Thr Lys Pro Ile Trp Thr Arg Asn Pro Asp Asp Ile
290 295 300
Thr Asn Glu Glu Tyr Gly Glu Phe Tyr Lys Ser Leu Thr Asn Asp Trp
305 310 315 320
Glu Asp His Leu Ala Val Lys His Phe Ser Val Glu Gly Gln Leu Glu
325 330 335
Phe Arg Ala Leu Leu Phe Val Pro Arg Arg Ala Pro Phe Asp Leu Phe
340 345 350
Glu Asn Arg Lys Lys Lys Asn Asn Ile Lys Leu Tyr Val Arg Arg Val
355 360 365
Phe Ile Met Asp Asn Cys Glu Glu Leu Ile Pro Glu Tyr Leu Asn Phe
370 375 380
Ile Arg Gly Val Val Asp Ser Glu Asp Leu Pro Leu Asn Ile Ser Arg
385 390 395 400
Glu Met Leu Gln Gln Ser Lys Ile Leu Lys Val Ile Arg Lys Asn Leu
405 410 415
Val Lys Lys Cys Leu Glu Leu Phe Thr Glu Leu Ala Glu Asp Lys Glu
420 425 430
Asn Tyr Lys Lys Phe Tyr Glu Gln Phe Ser Lys Asn Ile Lys Leu Gly
435 440 445
Ile His Glu Asp Ser Gln Asn Arg Lys Lys Leu Ser Glu Leu Leu Arg
450 455 460
Tyr Tyr Thr Ser Ala Ser Gly Asp Glu Met Val Ser Leu Lys Asp Tyr
465 470 475 480
Cys Thr Arg Met Lys Glu Asn Gln Lys His Ile Tyr Tyr Ile Thr Gly
485 490 495
Glu Thr Lys Asp Gln Val Ala Asn Ser Ala Phe Val Glu Arg Leu Arg
500 505 510
Lys His Gly Leu Glu Val Ile Tyr Met Ile Glu Pro Ile Asp Glu Tyr
515 520 525
Cys Val Gln Gln Leu Lys Glu Phe Glu Gly Lys Thr Leu Val Ser Val
530 535 540
Thr Lys Glu Gly Leu Glu Leu Pro Glu Asp Glu Glu Glu Lys Lys Lys
545 550 555 560
Gln Glu Glu Lys Lys Thr Lys Phe Glu Asn Leu Cys Lys Ile Met Lys
565 570 575
Asp Ile Leu Glu Lys Lys Val Glu Lys Val Val Val Ser Asn Arg Leu
580 585 590
Val Thr Ser Pro Cys Cys Ile Val Thr Ser Thr Tyr Gly Trp Thr Ala
595 600 605
Asn Met Glu Arg Ile Met Lys Ala Gln Ala Leu Arg Asp Asn Ser Thr
610 615 620
Met Gly Tyr Met Ala Ala Lys Lys His Leu Glu Ile Asn Pro Asp His
625 630 635 640
Ser Ile Ile Glu Thr Leu Arg Gln Lys Ala Glu Ala Asp Lys Asn Asp
645 650 655
Lys Ser Val Lys Asp Leu Val Ile Leu Leu Tyr Glu Thr Ala Leu Leu
660 665 670
Ser Ser Gly Phe Ser Leu Glu Asp Pro Gln Thr His Ala Asn Arg Ile
675 680 685
Tyr Arg Met Ile Lys Leu Gly Leu Gly Ile Asp Glu Asp Asp Pro Thr
690 695 700
Ala Asp Asp Thr Ser Ala Ala Val Thr Glu Glu Met Pro Pro Leu Glu
705 710 715 720
Gly Asp Asp Asp Thr Ser Arg Met
725
<210> 2
<211> 2184
<212> ДНК
<213> Homo sapiens
<400> 2
atgcctgagg aaacccagac ccaagaccaa ccgatggagg aggaggaggt tgagacgttc 60
gcctttcagg cagaaattgc ccagttgatg tcattgatca tcaatacttt ctactcgaac 120
aaagagatct ttctgagaga gctcatttca aattcatcag atgcattgga caaaatccgg 180
tatgaaactt tgacagatcc cagtaaatta gactctggga aagagctgca tattaacctt 240
ataccgaaca aacaagatcg aactctcact attgtggata ctggaattgg aatgaccaag 300
gctgacttga tcaataacct tggtactatc gccaagtctg ggaccaaagc gttcatggaa 360
gctttgcagg ctggtgcaga tatctctatg attggccagt tcggtgttgg tttttattct 420
gcttatttgg ttgctgagaa agtaactgtg atcaccaaac ataacgatga tgagcagtac 480
gcttgggagt cctcagcagg gggatcattc acagtgagga cagacacagg tgaacctatg 540
ggtcgtggaa caaaagttat cctacacctg aaagaagacc aaactgagta cttggaggaa 600
cgaagaataa aggagattgt gaagaaacat tctcagttta ttggatatcc cattactctt 660
tttgtggaga aggaacgtga taaagaagta agcgatgatg aggctgaaga aaaggaagac 720
aaagaagaag aaaaagaaaa agaagagaaa gagtcggaag acaaacctga aattgaagat 780
gttggttctg atgaggaaga agaaaagaag gatggtgaca agaagaagaa gaagaagatt 840
aaggaaaagt acatcgatca agaagagctc aacaaaacaa agcccatctg gaccagaaat 900
cccgacgata ttactaatga ggagtacgga gaattctata agagcttgac caatgactgg 960
gaagatcact tggcagtgaa gcatttttca gttgaaggac agttggaatt cagagccctt 1020
ctatttgtcc cacgacgtgc tccttttgat ctgtttgaaa acagaaagaa aaagaacaat 1080
atcaaattgt atgtacgcag agttttcatc atggataact gtgaggagct aatccctgaa 1140
tatctgaact tcattagagg ggtggtagac tcggaggatc tccctctaaa catatcccgt 1200
gagatgttgc aacaaagcaa aattttgaaa gttatcagga agaatttggt caaaaaatgc 1260
ttagaactct ttactgaact ggcggaagat aaagagaact acaagaaatt ctatgagcag 1320
ttctctaaaa acataaagct tggaatacac gaagactctc aaaatcggaa gaagctttca 1380
gagctgttaa ggtactacac atctgcctct ggtgatgaga tggtttctct caaggactac 1440
tgcaccagaa tgaaggagaa ccagaaacat atctattata tcacaggtga gaccaaggac 1500
caggtagcta actcagcctt tgtggaacgt cttcggaaac atggcttaga agtgatctat 1560
atgattgagc ccattgatga gtactgtgtc caacagctga aggaatttga ggggaagact 1620
ttagtgtcag tcaccaaaga aggcctggaa cttccagagg atgaagaaga gaaaaagaag 1680
caggaagaga aaaaaacaaa gtttgagaac ctctgcaaaa tcatgaaaga catattggag 1740
aaaaaagttg aaaaggtggt tgtgtcaaac cgattggtga catctccatg ctgtattgtc 1800
acaagcacat atggctggac agcaaacatg gagagaatca tgaaagctca agccctaaga 1860
gacaactcaa caatgggtta catggcagca aagaaacacc tggagataaa ccctgaccat 1920
tccattattg agaccttaag gcaaaaggca gaggctgata agaacgacaa gtctgtgaag 1980
gatctggtca tcttgcttta tgaaactgcg ctcctgtctt ctggcttcag tctggaagat 2040
ccccagacac atgctaacag gatctacagg atgatcaaac ttggtctggg tattgatgaa 2100
gatgacccta ctgctgatga taccagtgct gctgtaactg aagaaatgcc accccttgaa 2160
ggagatgacg acacatcacg catg 2184
<210> 3
<211> 732
<212> Белок
<213> Homo sapiens
<400> 3
Met Pro Glu Glu Thr Gln Thr Gln Asp Gln Pro Met Glu Glu Glu Glu
1 5 10 15
Val Glu Thr Phe Ala Phe Gln Ala Glu Ile Ala Gln Leu Met Ser Leu
20 25 30
Ile Ile Asn Thr Phe Tyr Ser Asn Lys Glu Ile Phe Leu Arg Glu Leu
35 40 45
Ile Ser Asn Ser Ser Asp Ala Leu Asp Lys Ile Arg Tyr Glu Thr Leu
50 55 60
Thr Asp Pro Ser Lys Leu Asp Ser Gly Lys Glu Leu His Ile Asn Leu
65 70 75 80
Ile Pro Asn Lys Gln Asp Arg Thr Leu Thr Ile Val Asp Thr Gly Ile
85 90 95
Gly Met Thr Lys Ala Asp Leu Ile Asn Asn Leu Gly Thr Ile Ala Lys
100 105 110
Ser Gly Thr Lys Ala Phe Met Glu Ala Leu Gln Ala Gly Ala Asp Ile
115 120 125
Ser Met Ile Gly Gln Phe Gly Val Gly Phe Tyr Ser Ala Tyr Leu Val
130 135 140
Ala Glu Lys Val Thr Val Ile Thr Lys His Asn Asp Asp Glu Gln Tyr
145 150 155 160
Ala Trp Glu Ser Ser Ala Gly Gly Ser Phe Thr Val Arg Thr Asp Thr
165 170 175
Gly Glu Pro Met Gly Arg Gly Thr Lys Val Ile Leu His Leu Lys Glu
180 185 190
Asp Gln Thr Glu Tyr Leu Glu Glu Arg Arg Ile Lys Glu Ile Val Lys
195 200 205
Lys His Ser Gln Phe Ile Gly Tyr Pro Ile Thr Leu Phe Val Glu Lys
210 215 220
Glu Arg Asp Lys Glu Val Ser Asp Asp Glu Ala Glu Glu Lys Glu Asp
225 230 235 240
Lys Glu Glu Glu Lys Glu Lys Glu Glu Lys Glu Ser Glu Asp Lys Pro
245 250 255
Glu Ile Glu Asp Val Gly Ser Asp Glu Glu Glu Glu Lys Lys Asp Gly
260 265 270
Asp Lys Lys Lys Lys Lys Lys Ile Lys Glu Lys Tyr Ile Asp Gln Glu
275 280 285
Glu Leu Asn Lys Thr Lys Pro Ile Trp Thr Arg Asn Pro Asp Asp Ile
290 295 300
Thr Asn Glu Glu Tyr Gly Glu Phe Tyr Lys Ser Leu Thr Asn Asp Trp
305 310 315 320
Glu Asp His Leu Ala Val Lys His Phe Ser Val Glu Gly Gln Leu Glu
325 330 335
Phe Arg Ala Leu Leu Phe Val Pro Arg Arg Ala Pro Phe Asp Leu Phe
340 345 350
Glu Asn Arg Lys Lys Lys Asn Asn Ile Lys Leu Tyr Val Arg Arg Val
355 360 365
Phe Ile Met Asp Asn Cys Glu Glu Leu Ile Pro Glu Tyr Leu Asn Phe
370 375 380
Ile Arg Gly Val Val Asp Ser Glu Asp Leu Pro Leu Asn Ile Ser Arg
385 390 395 400
Glu Met Leu Gln Gln Ser Lys Ile Leu Lys Val Ile Arg Lys Asn Leu
405 410 415
Val Lys Lys Cys Leu Glu Leu Phe Thr Glu Leu Ala Glu Asp Lys Glu
420 425 430
Asn Tyr Lys Lys Phe Tyr Glu Gln Phe Ser Lys Asn Ile Lys Leu Gly
435 440 445
Ile His Glu Asp Ser Gln Asn Arg Lys Lys Leu Ser Glu Leu Leu Arg
450 455 460
Tyr Tyr Thr Ser Ala Ser Gly Asp Glu Met Val Ser Leu Lys Asp Tyr
465 470 475 480
Cys Thr Arg Met Lys Glu Asn Gln Lys His Ile Tyr Tyr Ile Thr Gly
485 490 495
Glu Thr Lys Asp Gln Val Ala Asn Ser Ala Phe Val Glu Arg Leu Arg
500 505 510
Lys His Gly Leu Glu Val Ile Tyr Met Ile Glu Pro Ile Asp Glu Tyr
515 520 525
Cys Val Gln Gln Leu Lys Glu Phe Glu Gly Lys Thr Leu Val Ser Val
530 535 540
Thr Lys Glu Gly Leu Glu Leu Pro Glu Asp Glu Glu Glu Lys Lys Lys
545 550 555 560
Gln Glu Glu Lys Lys Thr Lys Phe Glu Asn Leu Cys Lys Ile Met Lys
565 570 575
Asp Ile Leu Glu Lys Lys Val Glu Lys Val Val Val Ser Asn Arg Leu
580 585 590
Val Thr Ser Pro Cys Cys Ile Val Thr Ser Thr Tyr Gly Trp Thr Ala
595 600 605
Asn Met Glu Arg Ile Met Lys Ala Gln Ala Leu Arg Asp Asn Ser Thr
610 615 620
Met Gly Tyr Met Ala Ala Lys Lys His Leu Glu Ile Asn Pro Asp His
625 630 635 640
Ser Ile Ile Glu Thr Leu Arg Gln Lys Ala Glu Ala Asp Lys Asn Asp
645 650 655
Lys Ser Val Lys Asp Leu Val Ile Leu Leu Tyr Glu Thr Ala Leu Leu
660 665 670
Ser Ser Gly Phe Ser Leu Glu Asp Pro Gln Thr His Ala Asn Arg Ile
675 680 685
Tyr Arg Met Ile Lys Leu Gly Leu Gly Ile Asp Glu Asp Asp Pro Thr
690 695 700
Ala Asp Asp Thr Ser Ala Ala Val Thr Glu Glu Met Pro Pro Leu Glu
705 710 715 720
Gly Asp Asp Asp Thr Ser Arg Met Glu Glu Val Asp
725 730
<210> 4
<211> 2199
<212> ДНК
<213> Homo sapiens
<400> 4
atgcctgagg aaacccagac ccaagaccaa ccgatggagg aggaggaggt tgagacgttc 60
gcctttcagg cagaaattgc ccagttgatg tcattgatca tcaatacttt ctactcgaac 120
aaagagatct ttctgagaga gctcatttca aattcatcag atgcattgga caaaatccgg 180
tatgaaactt tgacagatcc cagtaaatta gactctggga aagagctgca tattaacctt 240
ataccgaaca aacaagatcg aactctcact attgtggata ctggaattgg aatgaccaag 300
gctgacttga tcaataacct tggtactatc gccaagtctg ggaccaaagc gttcatggaa 360
gctttgcagg ctggtgcaga tatctctatg attggccagt tcggtgttgg tttttattct 420
gcttatttgg ttgctgagaa agtaactgtg atcaccaaac ataacgatga tgagcagtac 480
gcttgggagt cctcagcagg gggatcattc acagtgagga cagacacagg tgaacctatg 540
ggtcgtggaa caaaagttat cctacacctg aaagaagacc aaactgagta cttggaggaa 600
cgaagaataa aggagattgt gaagaaacat tctcagttta ttggatatcc cattactctt 660
tttgtggaga aggaacgtga taaagaagta agcgatgatg aggctgaaga aaaggaagac 720
aaagaagaag aaaaagaaaa agaagagaaa gagtcggaag acaaacctga aattgaagat 780
gttggttctg atgaggaaga agaaaagaag gatggtgaca agaagaagaa gaagaagatt 840
aaggaaaagt acatcgatca agaagagctc aacaaaacaa agcccatctg gaccagaaat 900
cccgacgata ttactaatga ggagtacgga gaattctata agagcttgac caatgactgg 960
gaagatcact tggcagtgaa gcatttttca gttgaaggac agttggaatt cagagccctt 1020
ctatttgtcc cacgacgtgc tccttttgat ctgtttgaaa acagaaagaa aaagaacaat 1080
atcaaattgt atgtacgcag agttttcatc atggataact gtgaggagct aatccctgaa 1140
tatctgaact tcattagagg ggtggtagac tcggaggatc tccctctaaa catatcccgt 1200
gagatgttgc aacaaagcaa aattttgaaa gttatcagga agaatttggt caaaaaatgc 1260
ttagaactct ttactgaact ggcggaagat aaagagaact acaagaaatt ctatgagcag 1320
ttctctaaaa acataaagct tggaatacac gaagactctc aaaatcggaa gaagctttca 1380
gagctgttaa ggtactacac atctgcctct ggtgatgaga tggtttctct caaggactac 1440
tgcaccagaa tgaaggagaa ccagaaacat atctattata tcacaggtga gaccaaggac 1500
caggtagcta actcagcctt tgtggaacgt cttcggaaac atggcttaga agtgatctat 1560
atgattgagc ccattgatga gtactgtgtc caacagctga aggaatttga ggggaagact 1620
ttagtgtcag tcaccaaaga aggcctggaa cttccagagg atgaagaaga gaaaaagaag 1680
caggaagaga aaaaaacaaa gtttgagaac ctctgcaaaa tcatgaaaga catattggag 1740
aaaaaagttg aaaaggtggt tgtgtcaaac cgattggtga catctccatg ctgtattgtc 1800
acaagcacat atggctggac agcaaacatg gagagaatca tgaaagctca agccctaaga 1860
gacaactcaa caatgggtta catggcagca aagaaacacc tggagataaa ccctgaccat 1920
tccattattg agaccttaag gcaaaaggca gaggctgata agaacgacaa gtctgtgaag 1980
gatctggtca tcttgcttta tgaaactgcg ctcctgtctt ctggcttcag tctggaagat 2040
ccccagacac atgctaacag gatctacagg atgatcaaac ttggtctggg tattgatgaa 2100
gatgacccta ctgctgatga taccagtgct gctgtaactg aagaaatgcc accccttgaa 2160
ggagatgacg acacatcacg catggaagaa gtagactaa 2199
<210> 5
<211> 499
<212> Белок
<213> Homo sapiens
<400> 5
Met Ala Met Ala Glu Gly Glu Arg Thr Glu Cys Ala Glu Pro Pro Arg
1 5 10 15
Asp Glu Pro Pro Ala Asp Gly Ala Leu Lys Arg Ala Glu Glu Leu Lys
20 25 30
Thr Gln Ala Asn Asp Tyr Phe Lys Ala Lys Asp Tyr Glu Asn Ala Ile
35 40 45
Lys Phe Tyr Ser Gln Ala Ile Glu Leu Asn Pro Ser Asn Ala Ile Tyr
50 55 60
Tyr Gly Asn Arg Ser Leu Ala Tyr Leu Arg Thr Glu Cys Tyr Gly Tyr
65 70 75 80
Ala Leu Gly Asp Ala Thr Arg Ala Ile Glu Leu Asp Lys Lys Tyr Ile
85 90 95
Lys Gly Tyr Tyr Arg Arg Ala Ala Ser Asn Met Ala Leu Gly Lys Phe
100 105 110
Arg Ala Ala Leu Arg Asp Tyr Glu Thr Val Val Lys Val Lys Pro His
115 120 125
Asp Lys Asp Ala Lys Met Lys Tyr Gln Glu Cys Asn Lys Ile Val Lys
130 135 140
Gln Lys Ala Phe Glu Arg Ala Ile Ala Gly Asp Glu His Lys Arg Ser
145 150 155 160
Val Val Asp Ser Leu Asp Ile Glu Ser Met Thr Ile Glu Asp Glu Tyr
165 170 175
Ser Gly Pro Lys Leu Glu Asp Gly Lys Val Thr Ile Ser Phe Met Lys
180 185 190
Glu Leu Met Gln Trp Tyr Lys Asp Gln Lys Lys Leu His Arg Lys Cys
195 200 205
Ala Tyr Gln Ile Leu Val Gln Val Lys Glu Val Leu Ser Lys Leu Ser
210 215 220
Thr Leu Val Glu Thr Thr Leu Lys Glu Thr Glu Lys Ile Thr Val Cys
225 230 235 240
Gly Asp Thr His Gly Gln Phe Tyr Asp Leu Leu Asn Ile Phe Glu Leu
245 250 255
Asn Gly Leu Pro Ser Glu Thr Asn Pro Tyr Ile Phe Asn Gly Asp Phe
260 265 270
Val Asp Arg Gly Ser Phe Ser Val Glu Val Ile Leu Thr Leu Phe Gly
275 280 285
Phe Lys Leu Leu Tyr Pro Asp His Phe His Leu Leu Arg Gly Asn His
290 295 300
Glu Thr Asp Asn Met Asn Gln Ile Tyr Gly Phe Glu Gly Glu Val Lys
305 310 315 320
Ala Lys Tyr Thr Ala Gln Met Tyr Glu Leu Phe Ser Glu Val Phe Glu
325 330 335
Trp Leu Pro Leu Ala Gln Cys Ile Asn Gly Lys Val Leu Ile Met His
340 345 350
Gly Gly Leu Phe Ser Glu Asp Gly Val Thr Leu Asp Asp Ile Arg Lys
355 360 365
Ile Glu Arg Asn Arg Gln Pro Pro Asp Ser Gly Pro Met Cys Asp Leu
370 375 380
Leu Trp Ser Asp Pro Gln Pro Gln Asn Gly Arg Ser Ile Ser Lys Arg
385 390 395 400
Gly Val Ser Cys Gln Phe Gly Pro Asp Val Thr Lys Ala Phe Leu Glu
405 410 415
Glu Asn Asn Leu Asp Tyr Ile Ile Arg Ser His Glu Val Lys Ala Glu
420 425 430
Gly Tyr Glu Val Ala His Gly Gly Arg Cys Val Thr Val Phe Ser Ala
435 440 445
Pro Asn Tyr Cys Asp Gln Met Gly Asn Lys Ala Ser Tyr Ile His Leu
450 455 460
Gln Gly Ser Asp Leu Arg Pro Gln Phe His Gln Phe Thr Ala Val Pro
465 470 475 480
His Pro Asn Val Lys Pro Met Ala Tyr Ala Asn Thr Leu Leu Gln Leu
485 490 495
Gly Met Met
<210> 6
<211> 1500
<212> ДНК
<213> Homo sapiens
<400> 6
atggcgatgg cggagggcga gaggactgag tgtgctgagc ccccccggga cgaacccccg 60
gctgatggag ctctgaagcg ggcagaggag ctcaagactc aggccaatga ctacttcaaa 120
gccaaggact acgagaacgc catcaagttc tacagccagg ccatcgagct gaaccccagc 180
aatgccatct actatggcaa ccgcagcctg gcctacctgc gcactgagtg ctatggctac 240
gcgctgggag acgccacgcg ggccattgag ctggacaaga agtacatcaa gggttattac 300
cgccgggctg ccagcaacat ggcactgggc aagttccggg ccgcgctgcg agactacgag 360
acggtggtca aggtgaagcc ccatgacaag gatgccaaaa tgaaatacca ggagtgcaac 420
aagatcgtga agcagaaggc ctttgagcgg gccatcgcgg gcgacgagca caagcgctcc 480
gtggtggact cgctggacat cgagagcatg accattgagg atgagtacag cggacccaag 540
cttgaagacg gcaaagtgac aatcagtttc atgaaggagc tcatgcagtg gtacaaggac 600
cagaagaaac tgcaccggaa atgtgcctac cagattctgg tacaggtcaa agaggtcctc 660
tccaagctga gcacgctcgt ggaaaccaca ctcaaagaga cagagaagat tacagtatgt 720
ggggacaccc atggccagtt ctatgacctc ctcaacatat tcgagctcaa cggtttaccc 780
tcggagacca acccctatat atttaatggt gactttgtgg accgaggctc cttctctgta 840
gaagtgatcc tcaccctttt cggcttcaag ctcctgtacc cagatcactt tcacctcctt 900
cgaggcaacc acgagacaga caacatgaac cagatctacg gtttcgaggg tgaggtgaag 960
gccaagtaca cagcccagat gtacgagctc tttagcgagg tgttcgagtg gctcccgttg 1020
gcccagtgca tcaacggcaa agtgctgatc atgcacggag gcctgttcag tgaagacggt 1080
gtcaccctgg atgacatccg gaaaattgag cggaatcgac aacccccaga ttcagggccc 1140
atgtgtgacc tgctctggtc agatccacag ccacagaacg ggcgctcgat cagcaagcgg 1200
ggcgtgagct gtcagtttgg gcctgacgtc accaaggcct tcttggaaga gaacaacctg 1260
gactatatca tccgcagcca cgaagtcaag gccgagggct acgaggtggc tcacggaggc 1320
cgctgtgtca ccgtcttctc tgcccccaac tactgcgacc agatggggaa caaagcctcc 1380
tacatccacc tccagggctc tgacctacgg cctcagttcc accagttcac agcagtgcct 1440
catcccaacg tcaagcccat ggcctatgcc aacacgctgc tgcagctagg aatgatgtga 1500
<210> 7
<211> 25
<212> ДНК
<213> Искусственная
<220>
<223> Последовательность-мишень PP5-ки-РНК
<400> 7
actcgaacac ctcgctaaag agctc 25
<210> 8
<211> 30
<212> ДНК
<213> Искусственная
<220>
<223> Hsp90α-Sal1-Re
<400> 8
acgcgtcgac ttagtctact tcttccatgc 30
<210> 9
<211> 31
<212> ДНК
<213> Искусственная
<220>
<223> Hsp90α-Sph1-For
<400> 9
acatgcatgc atgcctgagg aaacccagac c 31
<210> 10
<211> 73
<212> ДНК
<213> Искусственная
<220>
<223> Hsp90α-pc3.1-Nhe1-For-Myc:
<400> 10
gctagctagc gccaccatgg aacaaaaact catctcagaa gaggatctgc ctgaggaaac 60
ccagacccaa gac 73
<210> 11
<211> 91
<212> ДНК
<213> Искусственная
<220>
<223> Hsp90α-pc3.1-Nhe1-For-His-Myc:
<400> 11
gctagctagc gccaccatgc atcatcatca tcatcatgaa caaaaactca tctcagaaga 60
ggatctgcct gaggaaaccc agacccaaga c 91
<210> 12
<211> 34
<212> ДНК
<213> Искусственная
<220>
<223> Hsp90α-pc3.1-Xho1-Re-nostop
<400> 12
cccgctcgag tgtctacttc ttccatgcgt gatg 34
<210> 13
<211> 37
<212> ДНК
<213> Искусственная
<220>
<223> Hsp90α-EE-AA
<400> 13
ggccgctcga gtgtctactg ctgccatgcg tgatgtg 37
<210> 14
<211> 37
<212> ДНК
<213> Искусственная
<220>
<223> Hsp90α-VD-AA
<400> 14
ggccgctcga gttgctgctt cttccatgcg tgatgtg 37
<210> 15
<211> 37
<212> ДНК
<213> Искусственная
<220>
<223> Hsp90α-EEVD-AAAA
<400> 15
ggccgctcga gttgctgctg ctgccatgcg tgatgtg 37
<210> 16
<211> 33
<212> ДНК
<213> Искусственная
<220>
<223> Hsp90α-CΔ4-Xho
<400> 16
ccgctcgagt catgcgtgat gtgtcgtcat ctc 33
<210> 17
<211> 39
<212> ДНК
<213> Искусственная
<220>
<223> Hsp90α-T89A-смысловой
<400> 17
gatcgaactc ttgcaattgt ggatactgga attggaatg 39
<210> 18
<211> 39
<212> ДНК
<213> Искусственная
<220>
<223> Hsp90α-T89-антисмысловой
<400> 18
cattccaatt ccagtatcca caattgcaag agttcgatc 39
<210> 19
<211> 39
<212> ДНК
<213> Искусственная
<220>
<223> PP5-NheI-For
<400> 19
ctagctagca tgtacccata cgacgtccca gactacgct 39
<210> 20
<211> 40
<212> ДНК
<213> Искусственная
<220>
<223> PP5-XhoI-Re
<400> 20
ccgctcgagt taatgatgat gatgatgatg cacgtgtacc 40
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ПРОГНОЗИРОВАНИЯ ТЕРАПЕВТИЧЕСКОЙ ЭФФЕКТИВНОСТИ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ И ПРОГНОЗА ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2016 |
|
RU2785291C2 |
| ЭКЗОСОМАЛЬНЫЕ БИОМАРКЕРЫ | 2015 |
|
RU2712223C2 |
| КОМПОЗИЦИИ ИММУНОГЕННЫХ/ТЕРАПЕВТИЧЕСКИХ ГЛИКОКОНЪЮГАТОВ И ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2720295C2 |
| НОВАЯ КОМБИНАЦИЯ ДЛЯ ПРИМЕНЕНИЯ ПРИ ЛЕЧЕНИИ ЗЛОКАЧЕСТВЕННОГО НОВООБРАЗОВАНИЯ | 2016 |
|
RU2742494C2 |
| КОНЪЮГАТЫ ЛЕКАРСТВЕННЫХ СРЕДСТВ, СОДЕРЖАЩИЕ АНТИТЕЛА ПРОТИВ КЛАУДИНА 18.2 | 2016 |
|
RU2841168C2 |
| АДЕНОВИРУСЫ И СПОСОБЫ ПРИМЕНЕНИЯ АДЕНОВИРУСОВ | 2019 |
|
RU2782528C1 |
| АНТИ-СЕАСАМ6 АНТИТЕЛА И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2739163C2 |
| Белки, связывающие NKG2D, CD16 и опухолеассоциированный антиген | 2018 |
|
RU2816716C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ С АТИПИЧНЫМИ МУТАЦИЯМИ BRAF | 2018 |
|
RU2812706C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2019 |
|
RU2781542C2 |
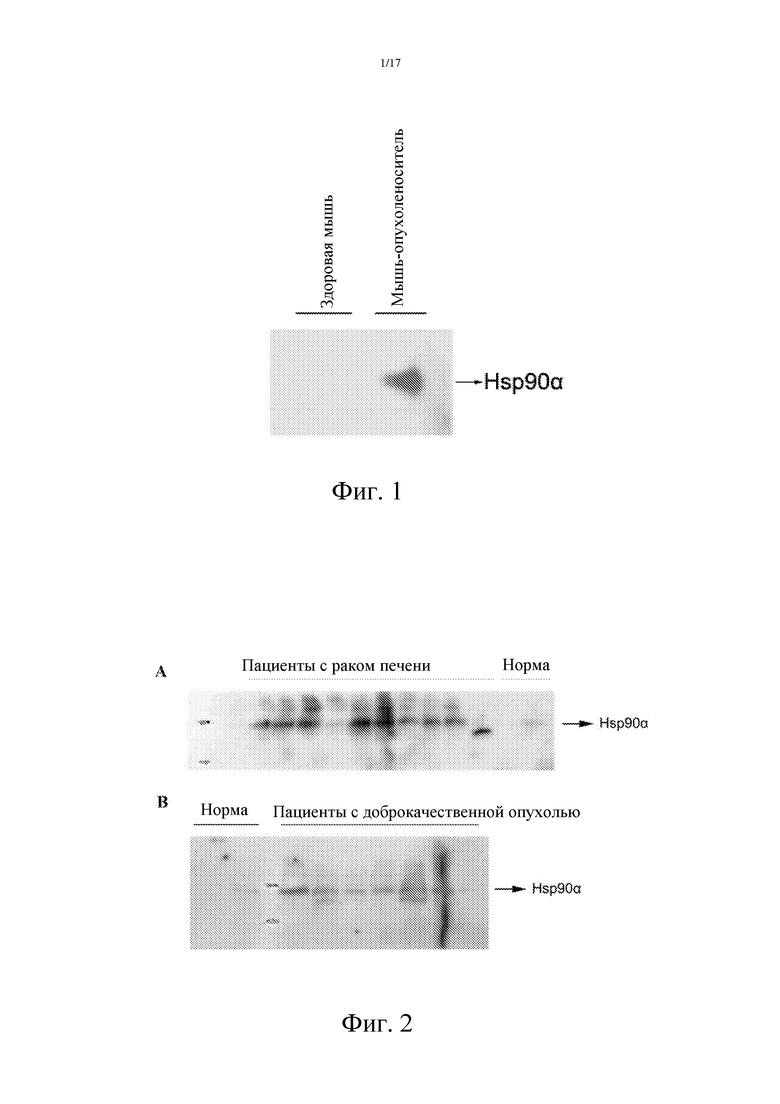
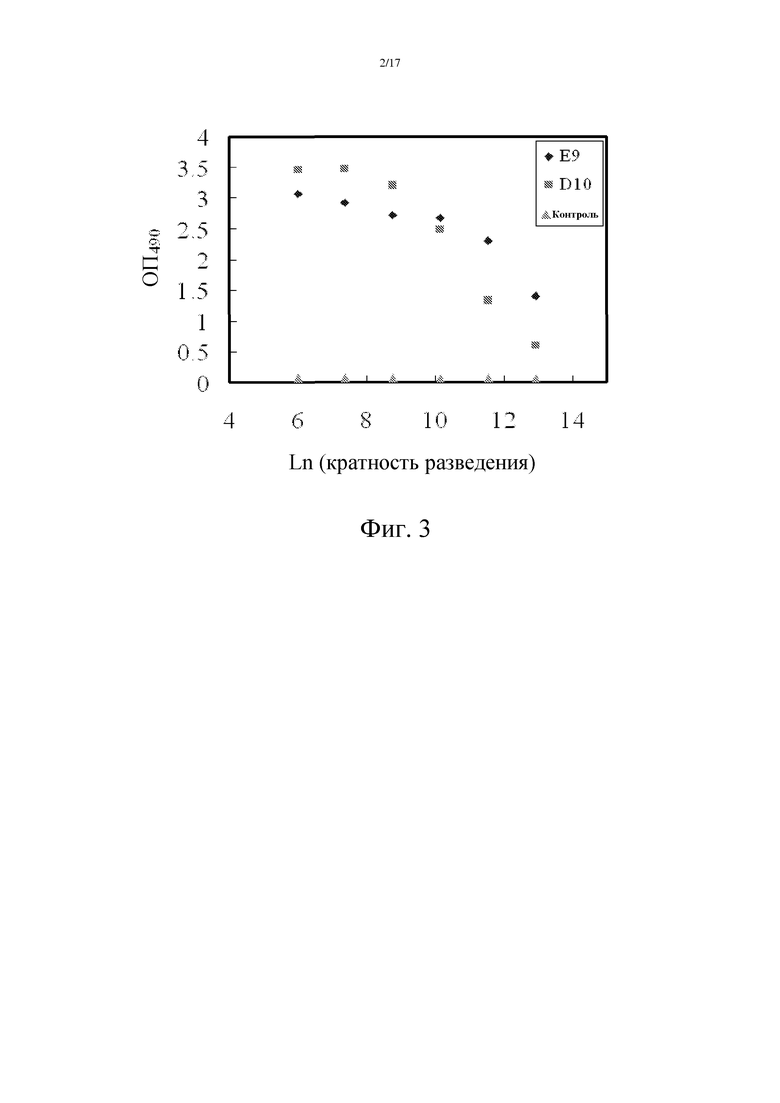
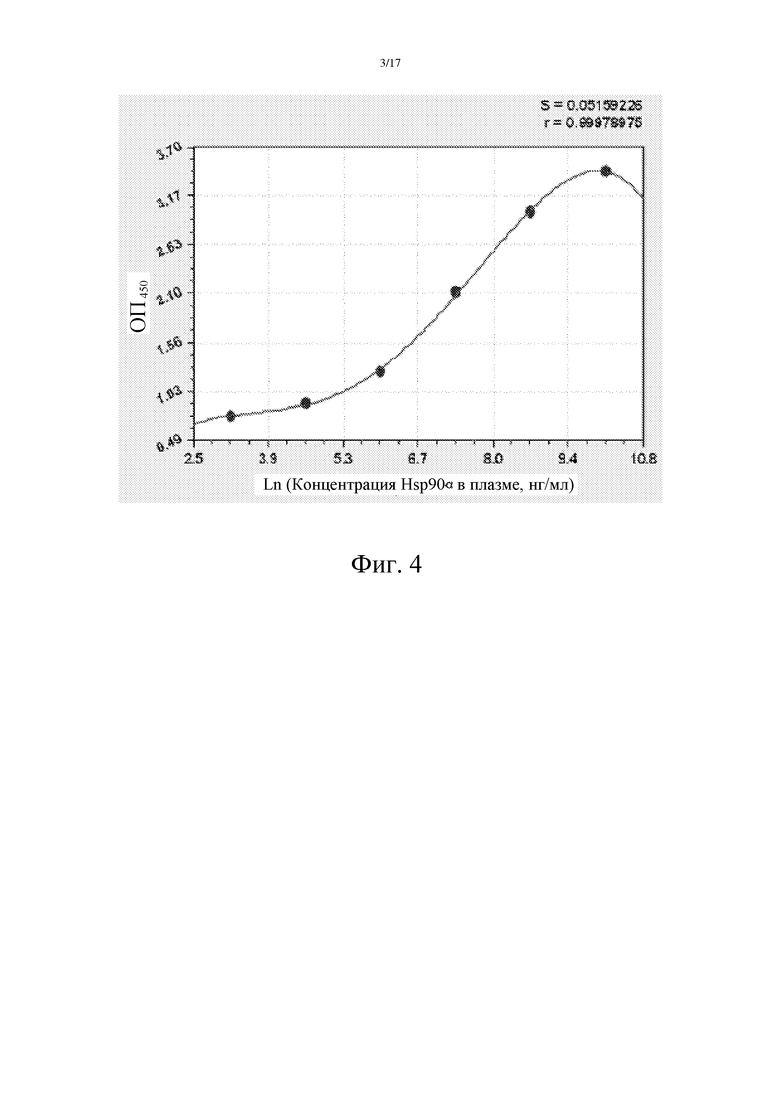
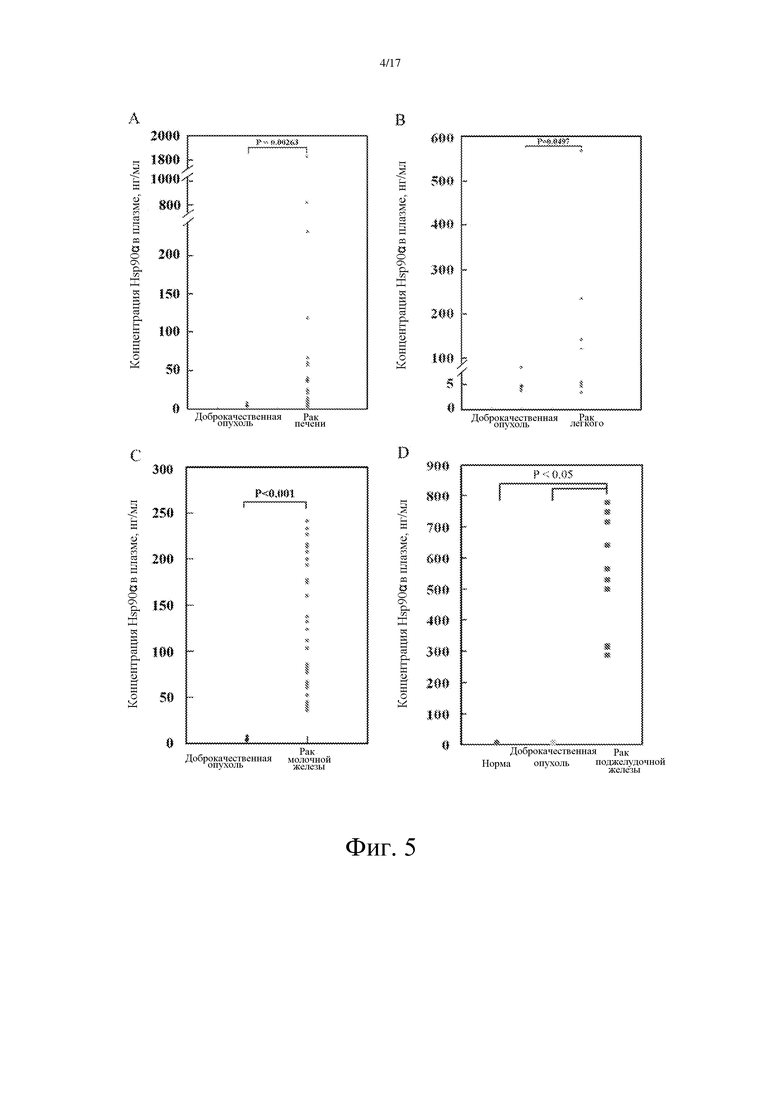
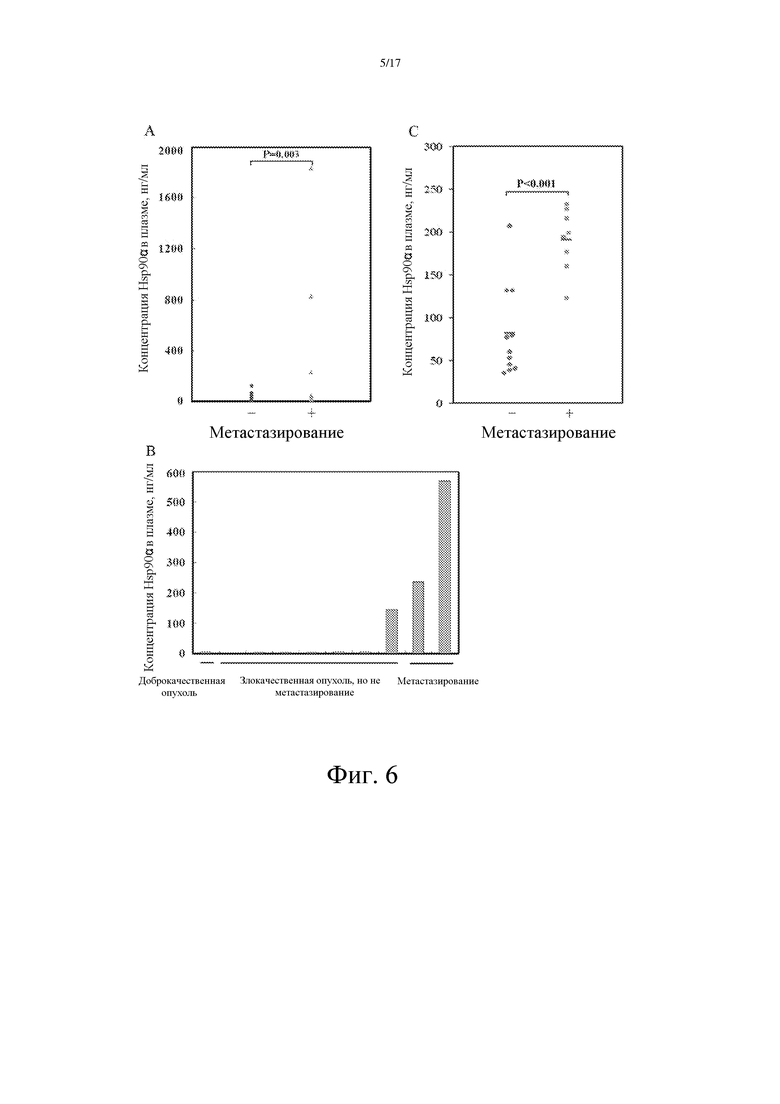
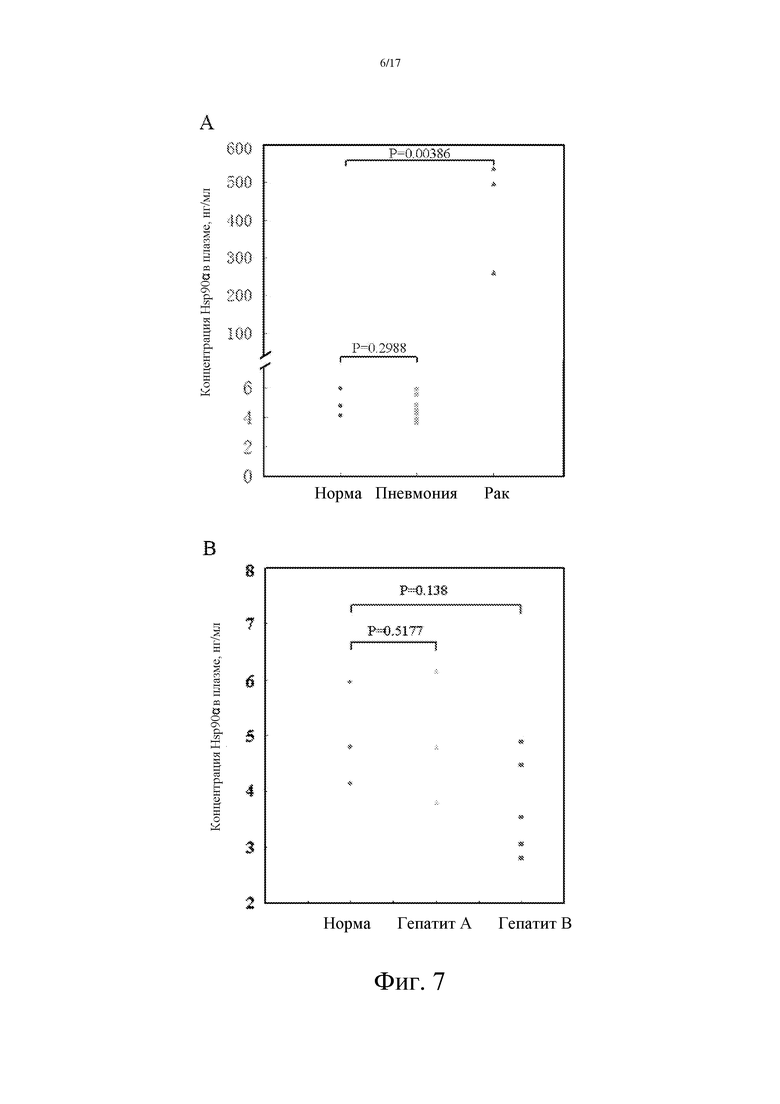

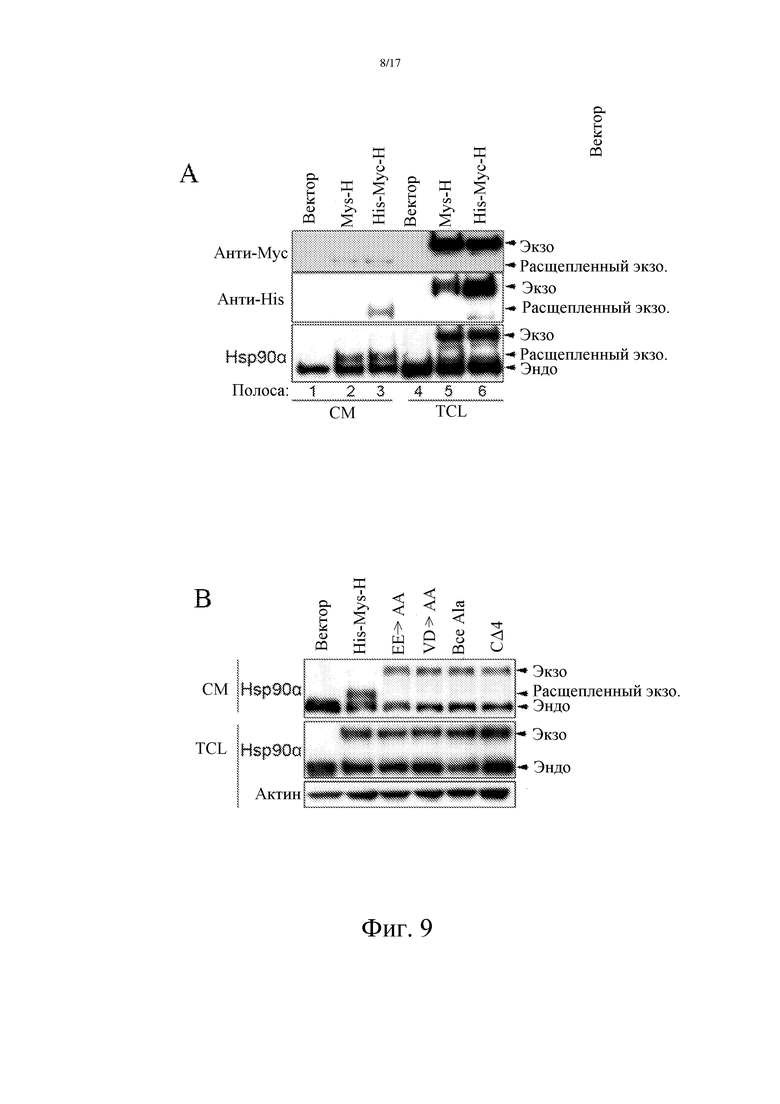
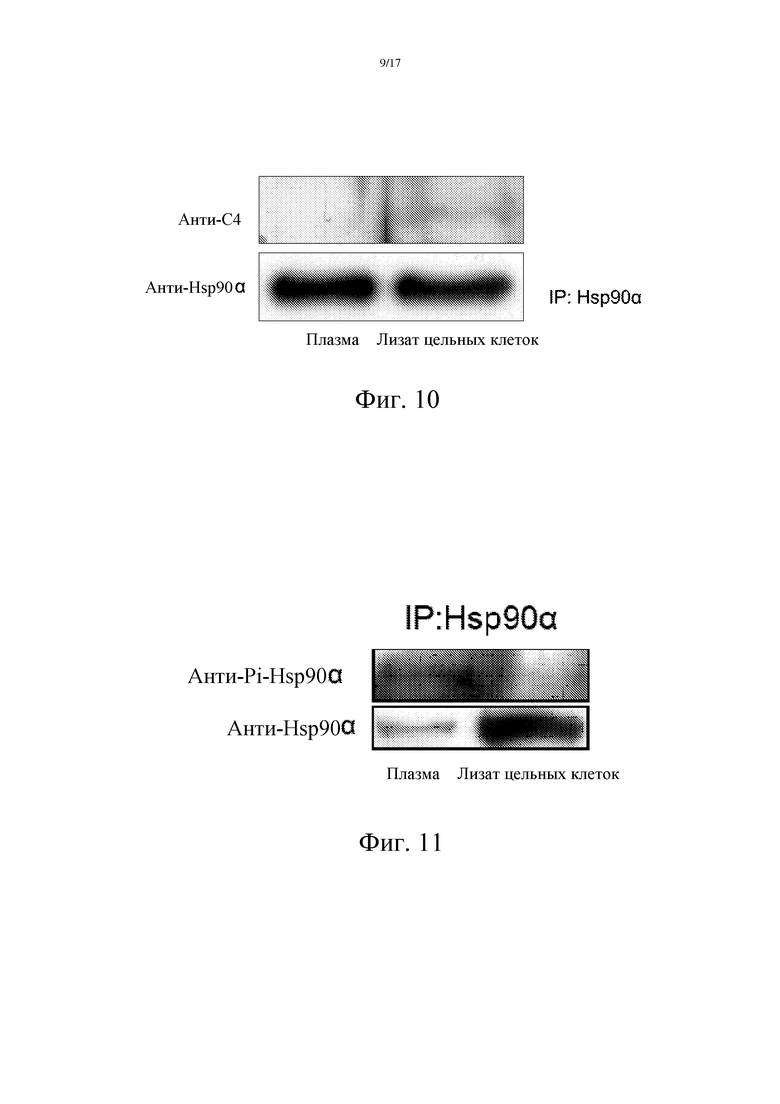
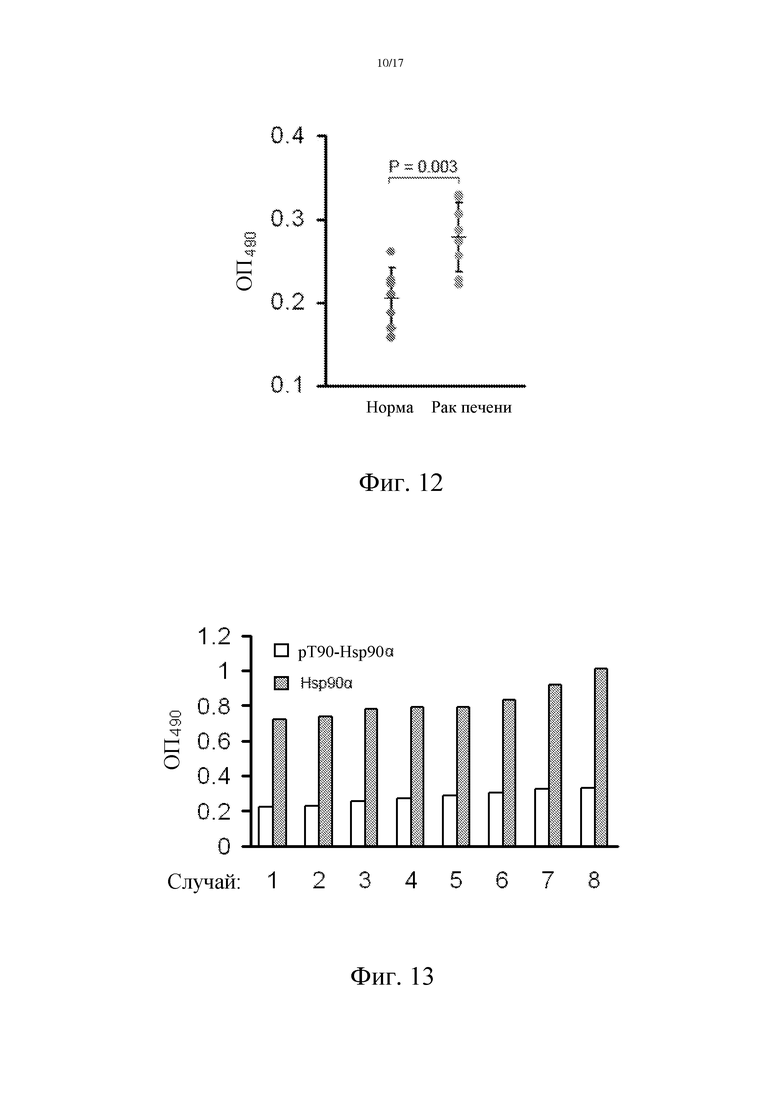

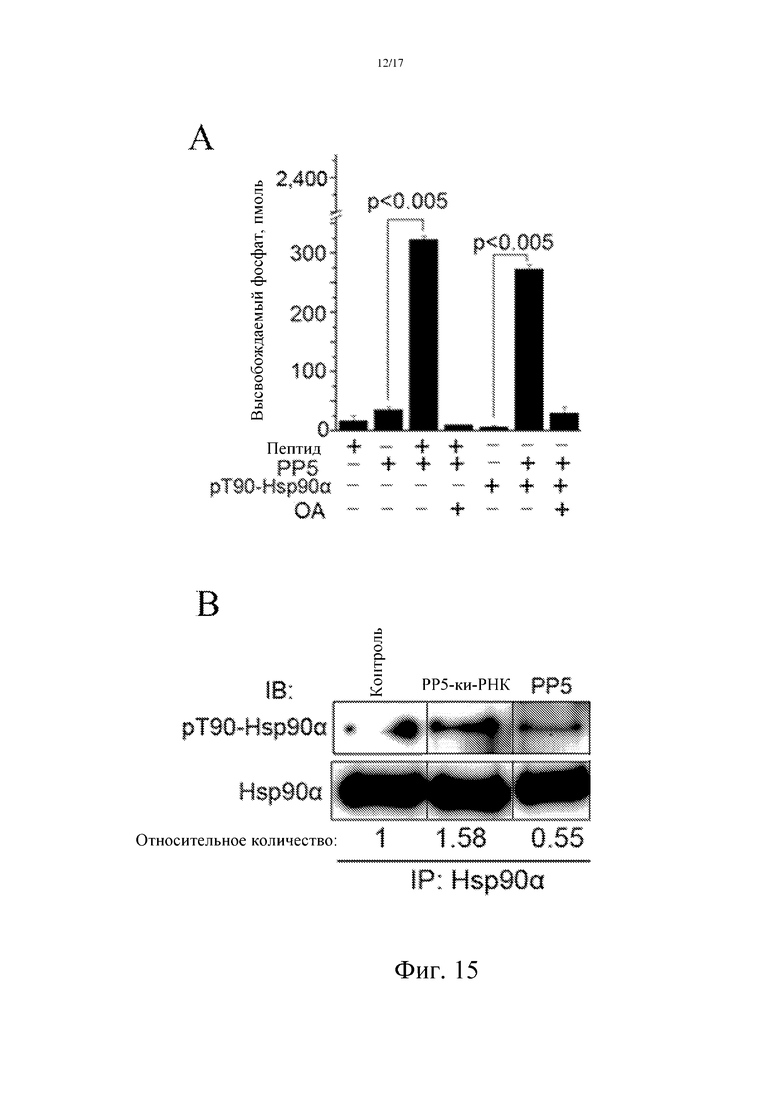
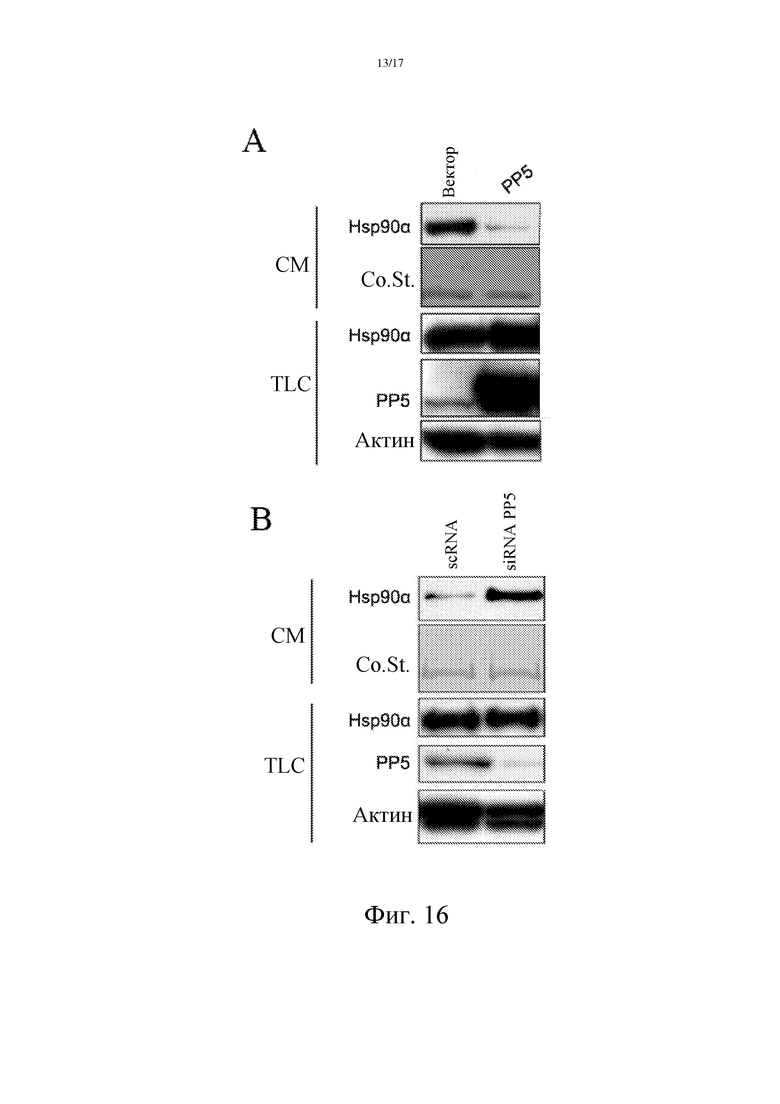

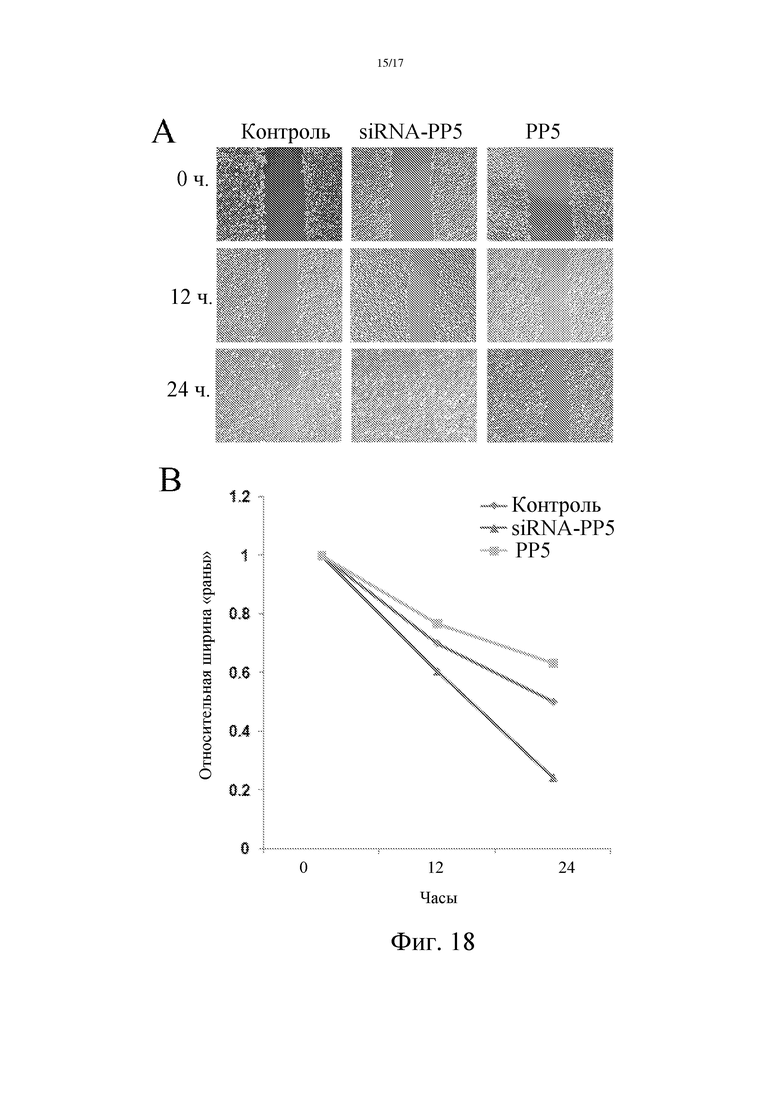
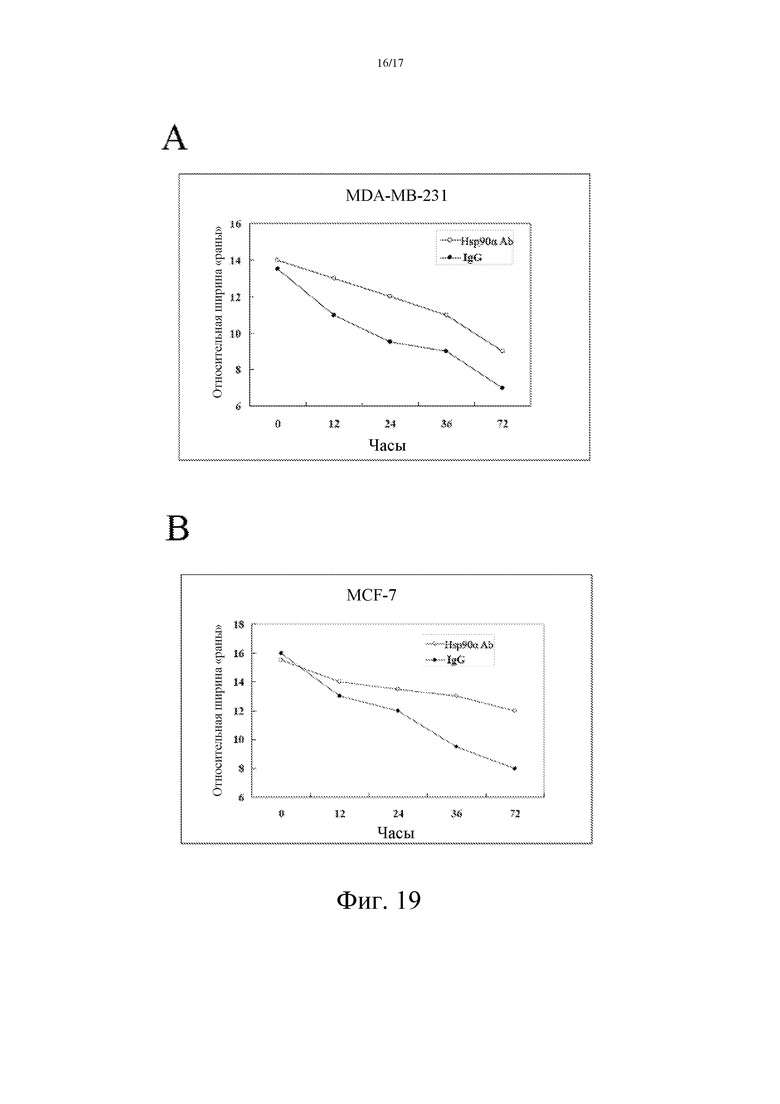
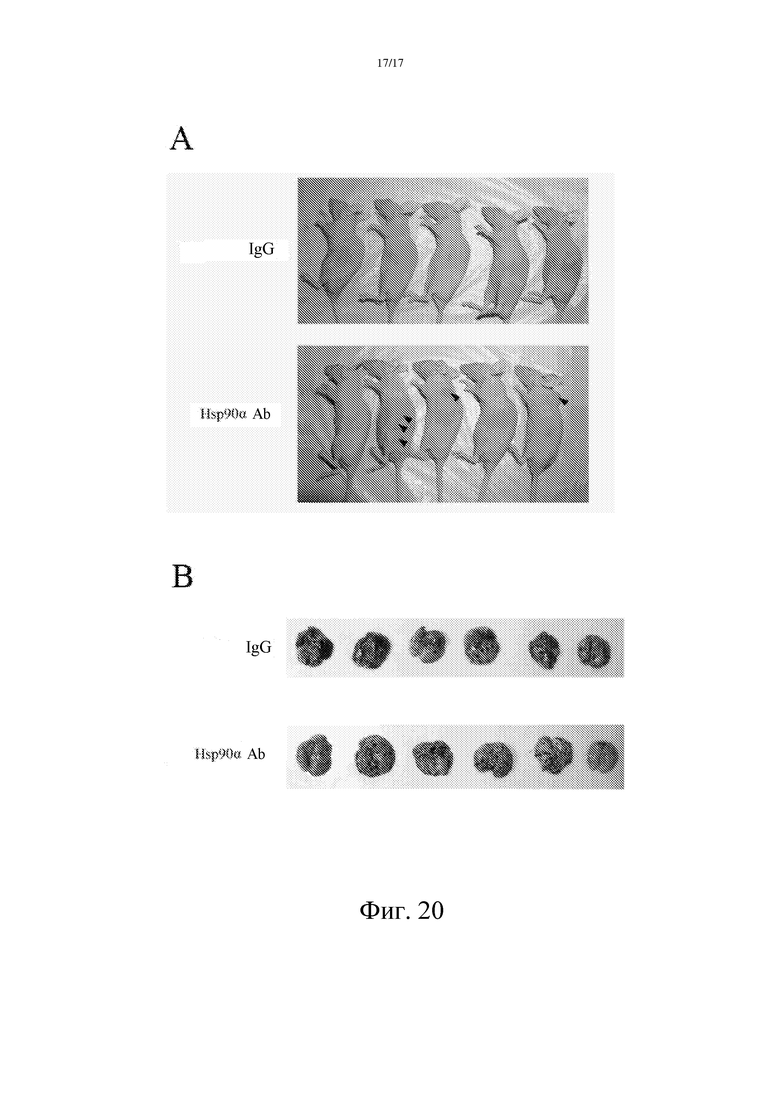
Изобретение относится к биотехнологии и связано с диагностикой и лечением злокачественных опухолей, а конкретно, со способом диагностики наличия метастазов злокачественной опухоли путем детектирования в плазме Hsp90α, имеющего аминокислотную последовательность SEQ ID NO: 1, в качестве опухолевого маркера. Кроме того, настоящее изобретение связано также со способом лечения злокачественной опухоли и метастазов. 4 н. и 15 з.п. ф-лы, 20 ил., 18 пр.
1. Применение антитела, способного специфично связываться с полипептидом, состоящим из аминокислотной последовательности SEQ ID NO:1, в производстве фармацевтической композиции для предотвращения или лечения метастазов злокачественной опухоли, где Thr90 указанного полипептида является фосфорилированным.
2. Применение по п.1, где метастазы злокачественной опухоли представляют собой метастазы в лимфатический узел или метастазы в легкое.
3. Применение по п.1, где указанное антитело представляет собой моноклональное антитело или его антиген-связывающий фрагмент.
4. Применение по п.1, где указанное антитело представляет собой гуманизированное антитело или его антиген-связывающий фрагмент.
5. Применение по п.1, где указанное антитело представляет собой MAb E9 или D10, продуцируемые клеточной линией, депонированной под регистрационным номером CGMCC No. 2903 или 2904, соответственно.
6. Применение по п.1, где указанная злокачественная опухоль выбрана из группы, состоящей из рака легкого, рака печени, рака желудка, рака пищевода, остеосаркомы, рака поджелудочной железы, лимфомы, рака толстой и прямой кишок, рака молочной железы, рака предстательной железы, рака яичника, рака ротовой полости, карциномы носоглотки, карциномы шейки матки, лейкоза, злокачественной меланомы, саркомы, почечноклеточной карциномы и холангиокарциномы.
7. Фармацевтическая композиция для предотвращения или лечения метастазов злокачественной опухоли, содержащая эффективное количество антитела, способного специфично связываться с полипептидом, состоящим из аминокислотной последовательности SEQ ID NO:1, где Thr90 указанного полипептида является фосфорилированным.
8. Фармацевтическая композиция по п.7, где метастазы злокачественной опухоли представляют собой метастазы в лимфатический узел или метастазы в легкое.
9. Фармацевтическая композиция по п.7, где указанное антитело представляет собой моноклональное антитело или его антиген-связывающий фрагмент.
10. Фармацевтическая композиция по п.7, где указанное антитело представляет собой гуманизированное антитело или его антиген-связывающий фрагмент.
11. Фармацевтическая композиция по п.7, где указанное антитело представляет собой MAb E9 или D10, продуцируемые клеточной линией, депонированной под регистрационным номером CGMCC No. 2903 или 2904, соответственно.
12. Фармацевтическая композиция по п.7, где указанная злокачественная опухоль выбрана из группы, состоящей из рака легкого, рака печени, рака желудка, рака пищевода, остеосаркомы, рака поджелудочной железы, лимфомы, рака толстой и прямой кишок, рака молочной железы, рака предстательной железы, рака яичника, рака ротовой полости, карциномы носоглотки, карциномы шейки матки, лейкоза, злокачественной меланомы, саркомы, почечноклеточной карциномы и холангиокарциномы.
13. Способ предотвращения или лечения метастазов злокачественной опухоли у субъекта, включающий введение субъекту антитела, способного специфично связываться с полипептидом, состоящим из аминокислотной последовательности SEQ ID NO:1, где Thr90 указанного полипептида является фосфорилированным.
14. Способ по п.13, где метастазы злокачественной опухоли представляют собой метастазы в лимфатический узел или метастазы в легкое.
15. Способ по п.13, где указанное антитело представляет собой моноклональное антитело или его антиген-связывающий фрагмент.
16. Способ по п.13, где указанное антитело представляет собой гуманизированное антитело или его антиген-связывающий фрагмент.
17. Способ по п.13, где указанное антитело представляет собой MAb E9 или D10, продуцируемые клеточной линией, депонированной под регистрационным номером CGMCC No. 2903 или 2904, соответственно.
18. Способ по п.13, где указанная злокачественная опухоль выбрана из группы, состоящей из рака легкого, рака печени, рака желудка, рака пищевода, остеосаркомы, рака поджелудочной железы, лимфомы, рака толстой и прямой кишок, рака молочной железы, рака предстательной железы, рака яичника, рака ротовой полости, карциномы носоглотки, карциномы шейки матки, лейкоза, злокачественной меланомы, саркомы, почечноклеточной карциномы и холангиокарциномы.
19. Полипептид-маркер метастазов злокачественной опухоли, состоящий из аминокислотной последовательности SEQ ID NO:1, где Thr90 указанного полипептида является фосфорилированным.
| Xiaofeng Wang et al., The regulatory mechanism of Hsp90α secretion and its function in tumor malignancy, PNAS, vol | |||
| Светоэлектрический измеритель длин и площадей | 1919 |
|
SU106A1 |
| Устройство для выпрямления многофазного тока | 1923 |
|
SU50A1 |
| Никитин К | |||
| Д., Белки теплового шока: биологические функции и перспективы применения, Клиническая онкогематология | |||
| Фундаментальные исследования и клиническая практика, том 1, номер 2, стр | |||

