Область техники
Изобретение относится к генной инженерии и может быть использовано в биотехнологии, медицине и сельском хозяйстве для создания препаратов генной терапии.
Уровень техники
Генная терапия - это современный медицинский подход, направленный на лечение наследственных и приобретенных заболеваний путем введения нового генетического материала в клетки пациента с целью компенсации или подавления функции мутантного гена и/или исправления генетического дефекта.
Переносчики генетического материала (генотерапевтические векторы) разделяют на вирусные и невирусные. В качестве вирусных векторов с успехом применяют ретровирусы, лентивирусы, аденоассоциированный вирус, герпесвирусы, поксвирусы и аденовирусы (Lukashev AN, Zamyatnin АА Jr. Viral Vectors for Gene Therapy: Current State and Clinical Perspectives. Biochemistry (Mosc). 2016. 81:700-708.). В невирусной доставке генетического материала лидируют плазмидные векторы, несущие терапевтический ген в комплексе с различными носителями - липидами, катионными полимерами, дендримерами, полипептидами, наночастицами (Mintzer MA, Simanek ЕЕ. Nonviral vectors for gene delivery. Chem Rev. 2009. 109:259-302).
Несмотря на то что вирус, в силу своей природы, является почти идеальным агентом по скорости и эффективности доставки рекомбинантной ДНК в клетку, существуют практические ограничения для широкого использования вирусной системы доставки. Это трудности, связанные с промышленным производством, отсутствием селективности, возникновением иммунного ответа, потенциальной угрозой канцерогенеза, а также воспалением, возникающим после трансдукции клеток. Часть этих проблем остается до сих пор нерешенной. Поэтому в последнее время в генной терапии все большее внимание уделяется разработке невирусных систем доставки генетического материала.
Плазмидный вектор - автономно реплицирующаяся внехромосомная кольцевая ДНК. Плазмидные векторы могут иметь в своем составе гены устойчивости к антибиотикам, ионам тяжелых металлов, а также гены, контролирующие катаболизм некоторых органических соединений (Lipps G. (editor). (2008). Plasmids: Current Research and Future Trends. Caister Academic Press. ISBN 978-1-904455-35-6). Плазмидные вектора - своего рода мобильные генетические элементы, могут передаваться от одной бактериальной клетки к другой путем конъюгации, являясь средством горизонтального переноса генов.
Плазмидные векторы лишены недостатков, присущих вирусным векторам. В клетке-мишени они существуют в эписомальной форме, не интегрируясь в геном, производство их достаточно дешево, отсутствие иммунного ответа и побочных реакций на введение плазмидного вектора делают их удобным инструментом генетической терапии (перенос терапевтических генов) и генетической профилактики (ДНК-вакцины) (Li L, Petrovsky N. Molecular mechanisms for enhanced DNA vaccine immunogenicity. Expert Rev Vaccines. 2016; 15(3):313-29).
Помимо того, что плазмидные векторы являются весьма перспективным средством доставки в генетической терапии, уже долгое время они служат важным инструментом в молекулярно-биологических и биотехнологических научных лабораториях и с успехом применяются в области молекулярного клонирования и получения рекомбинантных белков (Russell, David W.; Sambrook, Joseph (2001), Molecular cloning: a laboratory manual. Cold Spring Harbor, N.Y; Cold Spring Harbor Laboratory).
Несмотря на очевидную перспективность для генной терапии, критическим ограничением для использования плазмидных векторов в качестве терапевтических средств является наличие в их составе: i) генов устойчивости к антибиотикам для наработки в штаммах-носителях, ii) различных регуляторных элементов, представленных последовательностями вирусных геномов. Еще одним ограничением является размер терапевтического плазмидного вектора, определяющий эффективность проникновения вектора в клетку-мишень.
Как известно, на протяжении последних лет во всем мире отмечается значительный рост устойчивости возбудителей инфекционных заболеваний к антимикробным препаратам. Возникновение антимикробной резистентности является естественным биологическим ответом на использование антибиотиков, которые создают селективное давление, способствующее отбору, выживанию и размножению резистентных штаммов микроорганизмов. Резистентность к антибиотикам имеет большое социально-экономическое значение и рассматривается как угроза национальной безопасности (MacPherson D.W., Gushulak B.D., Baine W.B., Bala S., Gubbins P.O., Holtom P., Segarra-Newnham M. 2009. Population mobility, globalization, and antimicrobial drug resistance. Emerg Infect Dis 15:1727-1732). Именно с помощью плазмидных векторов обеспечивается механизм горизонтального переноса генов, в том числе и антибиотикорезистентности, внутри популяции микроорганизмов, что обеспечивает им селективное преимущество. Таким образом, с горизонтальным переносом генов связывают распространение устойчивых к современным антибиотикам возбудителей инфекционных заболеваний человека (Ramirez MS, Traglia GM, Lin DL, Tran T, Tolmasky ME. Plasmid-Mediated Antibiotic Resistance and Virulence in Gram-Negatives: the Klebsiella pneumoniae Paradigm. Microbiol Spectr. 2014 (5).
В связи с этим, Европейское агентство по лекарственным средствам считает необходимым избегать введения маркеров антибиотикорезистентности в разрабатываемые плазмидные векторы для генной терапии (Reflection paper on design modifications of gene therapy medicinal products during development/14 December 2011 EMA/CAT/GTWP/44236/2009 Committee for advanced therapies).
Другим существенным ограничением использования терапевтических плазмидных векторов является наличие в их составе регуляторных элементов для повышения экспрессии целевых генов (промоторы, энхансеры, посттрансляционные регуляторные элементы), которые в большинстве случаев представляют собой нуклеотидные последовательности геномов различных вирусов (Draft Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal products, http://www.ema.europa.eu/docs/en_GB/document_library/Scientific_guideline/2015/05/WC500187020.pdf).
Еще одним недостатком существующих плазмидных векторов для генетической терапии является их размер (длина). Известно, что чем больше длина плазмидного вектора, тем менее эффективно он проникает в клетку-мишень. Существующие плазмидные векторы зачастую перегружены нефункциональными участками, серьезно увеличивающими размер вектора (Mairhofer J, Grabherr R. Rational vector design for efficient non-viral gene delivery: challenges facing the use of plasmid DNA. Mol Biotechnol. 2008.39(2):97-104).
Еще одним важным ограничением применения терапевтических плазмидных векторов является отсутствие тканеспецифичной экспрессии целевых генов. Использование в терапии плазмидных векторов, не обладающих функцией тканеспецифичной экспрессии целевых генов в целевых клетках (целевой ткани, органе) в ряде случаев может привести к побочным эффектам, таким как токсичность продукта вследствие гиперэкспрессии, ингибирование экспрессии других генов по принципу обратной связи, риск от которых может превышать пользу. Известно, что экспрессия генов обеспечивается регуляторными элементами в геноме, такими как промоторы, энхансеры и сайленсеры различной структуры. Эти регуляторные элементы содержат последовательности для связывания регуляторных белков, в том числе факторов транскрипции. Именно это обеспечивает контроль экспрессии генов на уровне транскрипции. Определенные гены могут транскрибироваться во всех тканях организма, другие - в нескольких тканях, часто гены экспрессируются в клетках только одной ткани. Такие регуляторные участки генов, характеризующихся тканеспецифической транскрипцией, можно использовать для создания генотерапевтических векторов с целью специфической экспрессии целевых генов в строго определенной ткани (или в группе тканей) организма, что, в свою очередь повышает эффективность и безопасность этих генотерапевтических векторов (Gill DR, Pringle IA, Hyde SC. Progress and prospects: the design and production of plasmid vectors. Gene Ther. 2009. 16:165-171) и является подходом таргетной генной терапии. Кроме того, вводя в подобные векторы различные нуклеазы, например, Cas9, можно реализовать подход терапевтического геномного редактирования, что также является таргетной генной терапией.
Известен способ накопления плазмидных векторов в штаммах Escherichia coli без использования антибиотиков (Cranenburgh RM, Hanak JA, Williams SG, Sherratt DJ. Escherichia coli strains that allow antibiotic-free plasmid selection and maintenance by repressor titration. Nucleic Acids Res. 2001. 29(5):E26). Были созданы штаммы Escherichia coli DH1lacdapD и DH1lacP2dapD, в которых ген dapD, кодирующий фермент 2,3,4,5-тетрагидропиридин-2,6-дикарбоксилат-N-сукцинилтрансферазу, участвующий в биосинтезе L-лизина, находится под контролем lac-промотора. В отсутствие индуктора IPTG (изопропил-β-D-1-тиогалактопиранозид) эти штаммы подвержены лизису. При введении мультикопийного вектора pORT, содержащего lac-оператор, индуцируется экспрессия гена dapD и, таким образом, трансформированные клоны могут быть отобраны и размножены. Однако, эти штаммы характеризуются низким уровнем трансформации и ее нестабильностью.
Известен также способ получения штаммов Escherichia coli для наработки плазмидных векторов без использования селекции с антибиотиками (Mairhofer J, Pfaffenzeller I, Merz D, Grabherr R.A novel antibiotic free plasmid selection system: advances in safe and efficient DNA therapy. Biotechnol J. 2008. 3(1):83-89). Выбранные бактериальные штаммы (например DG5α, JM109, MG1655) были модифицированы таким образом, что ингибитор репликации плазмидных векторов RNA I мог подавлять трансляцию генов, необходимых для жизнедеятельности бактерий (например, murA, кодирующего фермент UDP-N-ацетилглюкозамин 1-карбоксивинил-трансферазу, участвующего в биосинтезе пептидогликана клеточной стенки бактерий) путем образования дуплекса РНК/антисенс-РНК. Ген murA находился под контролем белка репрессора tetR и мог экспрессироваться только в присутствии сконструированного плазмидного вектора, несущего RNA I. Однако было обнаружено, что добавление индуктора IPTG приводило к появлению колоний Escherichia coli, не содержащих целевого плазмидного вектора. Механизм ингибирования селекции остается неизвестным.
Известен способ получения минимального по длине вектора. Была получена небольшая, имеющая суперскрученную форму молекула ДНК, лишенная всех прокариотических нуклеотидных последовательностей, содержащая только ориджин репликации и ген устойчивости к антибиотикам (так называемое «миникольцо»). Вектор был получен путем интеграза-опосредованной внутримолекулярной интеграции с использованием фага ϕС31 (Chen ZY, Не CY, Ehrhardt A, Kay MA. Minicircle DNA vectors devoid of bacterial DNA result in persistent and high-level transgene expression in vivo. Mol Ther. 2003. 8(3):495-500). Недостатком такого рода плазмидных векторов является трудоемкость их получения и невозможность наработки в промышленных масштабах.
Известно решение по патентной заявке US 2011152377/10, описывающей получение экспрессионного плазмидного вектора без устойчивости к антибиотику, которая содержит полинуклеотид, кодирующий белок-репрессор. Экспрессия указанного белка-репрессора регулирует экспрессию токсичного генного продукта, встроенного в участок генома E.coli. Однако, как и все методы селекции, основанные на использовании белков-репрессоров, данный метод отличается нестабильностью трансформации и ее низкой эффективностью.
Известен патент (US 9,644,211), описывающий получение минимального по размеру вектора («миникольца»). Данный вектор не содержит прокариотических последовательностей и продуцируется путем parA-опосредованной рекомбинации, проходящей в специально полученном штамме E.coli. Недостатком данного метода получения минимального по размеру вектора является невозможность его использования при масштабировании производства.
Известен патент (US 7,341,847), описывающий использование промотора бета-цепи тромбоцитарного фактора роста PDGF в сочетании с энхансером немедленно-ранних генов цитомегаловируса человека в составе вектора на основе плазмиды или вируса для экспрессии трансгенов в нейрональной ткани. Однако наличие элементов вирусных геномов в составе плазмидного вектора ограничивает его применение для генной терапии человека.
Прототипом настоящего изобретения в части использования рекомбинантных ДНК-векторов для генной терапии является способ получения рекомбинантного вектора для генетической иммунизации (US 9,550,998). Плазмидный вектор представляет собой суперскученный плазмидный ДНК-вектор и предназначен для экспрессии клонированных генов в клетках животных и человека. Вектор состоит из ориджина репликации, регуляторных элементов, включающих промотор и энхансер цитомегаловируса человека, регуляторные элементы из Т-лимфотропного вируса человека.
Накопление вектора производят в специальном штамме E.coli без использования антибиотиков за счет антисенс-комплементации гена sacB, введенного в штамм посредством бактериофага. Ограничением использования данного ДНК - вектора для генетической терапии является наличие в его составе регуляторных элементов, представляющих собой последовательности вирусных геномов.
Раскрытие изобретения
Задачей изобретения является конструирование группы генотерапевтических ДНК-векторов для таргетной генной терапии, сочетающих в себе:
I) возможность безопасного использования для генетической терапии человека и животных за счет отсутствия в составе генотерапевтических ДНК-векторов генов антибиотикорезистентности.
II) длину, обеспечивающую эффективное проникновение в клетку-мишень;
III) наличие регуляторных элементов, обеспечивающих эффективную экспрессию целевых генов и, в то же время, не представляющих собой нуклеотидные последовательности вирусных геномов;
IV) возможность выбора для таргетной генной терапии генотерапевтического ДНК-вектора из группы генотерапевтических ДНК-векторов, несущих различные промоторно-регуляторные участки, для конструирования генотерапевтического ДНК-вектора, содержащего целевой ген, исходя из критериев обеспечения его тканеспецифичной экспрессии и максимальной эффективности в целевой клеточной линии, целевой ткани или органе человека.
V) технологичность получения и возможность наработки в промышленных масштабах.
Пункты I и III являются обязательными, и предусмотрены в данном техническом решении в соответствии с требованиями государственных регуляторов к лекарственным средствам для генной терапии, в частности, Европейского Агентства по лекарственным средствам касательно отказа от введения маркеров антибиотикорезистентности в разрабатываемые плазмидные векторы для генной терапии (Reflection paper on design modifications of gene therapy medicinal products during development / 14 December 2011, EMA/CAT/GTWP/44236/2009 Committee for advanced therapies) и касательно отказа от введения в разрабатываемые плазмидные векторы для генной терапии элементов вирусных геномов (Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal products / 23 March 2015, EMA/CAT/80183/2014, Committee for Advanced Therapies).
Задачей изобретения также является конструирование штаммов, несущих эти генотерапевтические ДНК-вектора для производства в промышленных масштабах этих генотерапевтических ДНК-векторов.
Указанная задача решается за счет того, что создан генотерапевтический ДНК-вектор для таргетной генной терапии, выбранный из GDTT1.8NAS1 размером 1736 п.н., или GDTT1.8NAS2 1654 п.н., или GDTT1.8NAS3 1783 п.н., или GDTT1.8NAS4 1953 п.н., или GDTT1.8NAS5 2019 п.н., или GDTT1.8NAS6 1940 п.н., или GDTT1.8NAS7 2620 п.н., или GDTT1.8NAS8 2240 п.н., или GDTT1.8NAS9 2604 п.н., или GDTT1.8NAS10 2048 п.н., или GDTT1.8NAS11 1978 п.н. с нуклеотидной последовательностью соответственно: SEQ ID №1, SEQ ID №2, SEQ ID №3, SEQ ID №4, SEQ ID №5, SEQ ID №6, SEQ ID №7, SEQ ID №8, SEQ ID №9, SEQ ID №10, SEQ ID №11.
При этом каждый из выбранных генотерапевтических ДНК-векторов содержит различные тканеспецифичные промоторно-регуляторные участки гена человека: гена миоглобина человека, или гена эластазы человека, или гена молекулы межклеточной адгезии 2 человека, или гена остеокальцина 2 человека, или гена белка сурфактанта В, или гена синапсина I человека, или гена нефрина человека, или гена общего лейкоцитарного антигена CD45 человека, или гена белка В29 человека, или гена белка CD68 человека, или гена инсулина человека соответственно.
Способ получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS1 заключается в том, что сначала конструируют вектор размером 2408 п.н., содержащий ориджин репликации размером 688 п.н., терминатор транскрипции hGH-TA размером 467 п.н., регуляторный участок транспозона Tn10 РНК-out размером 137 п.н., ген устойчивости к канамицину размером 1018 п.н. и полилинкер размером 68 п.н., затем его расщепляют с использованием эндонуклеаз рестрикции SalI и BamHI и лигируют с промоторно-регуляторным участком, содержащим промоторную область гена миоглобина человека размером 351 п.н., а затем ген устойчивости к канамицину выщепляют по сайтам рестрикции Spel (вариант 1).
Способ получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS2 заключается в том, что сначала конструируют вектор размером 2408 п.н., содержащий ориджин репликации размером 688 п.н., терминатор транскрипции hGH-TA размером 467 п.н., регуляторный участок транспозона Tn10 РНК-out размером 137 п.н., ген устойчивости к канамицину размером 1018 п.н. и полилинкер размером 68 п.н., затем его расщепляют с использованием эндонуклеаз рестрикции SalI и BamHI и лигируют с промоторно-регуляторным участком, содержащим промоторную область гена эластазы человека размером 269 п.н., а затем ген устойчивости к канамицину выщепляют по сайтам рестрикции Spel (вариант 2).
Способ получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS3, заключается в том, что сначала конструируют вектор размером 2408 п.н., содержащий ориджин репликации размером 688 п.н., терминатор транскрипции hGH-TA размером 467 п.н., регуляторный участок транспозона Tn10 РНК-out размером 137 п.н., ген устойчивости к канамицину размером 1018 п.н. и полилинкер размером 68 п.н., затем его расщепляют с использованием эндонуклеаз рестрикции SalI и BamHI и лигируют с промоторно-регуляторным участком, содержащим промоторную область гена молекулы межклеточной адгезии 2 человека размером 398 п.н., а затем ген устойчивости к канамицину выщепляют по сайтам рестрикции Spel (вариант 3).
Способ получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS4 по, заключается в том, что сначала конструируют вектор размером 2408 п.н., содержащий ориджин репликации размером 688 п.н., терминатор транскрипции hGH-TA размером 467 п.н., регуляторный участок транспозона Tn10 РНК-out размером 137 п.н., ген устойчивости к канамицину размером 1018 п.н. и полилинкер размером 68 п.н., затем его расщепляют с использованием эндонуклеаз рестрикции SalI и BamHI и лигируют с промоторно-регуляторным участком, содержащим промоторную область гена остеокальцина 2 человека размером 568 п.н., а затем ген устойчивости к канамицину выщепляют по сайтам рестрикции Spel (вариант 4).
Способ получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS5, заключается в том, что сначала конструируют вектор размером 2408 п.н., содержащий ориджин репликации размером 688 п.н., терминатор транскрипции hGH-TA размером 467 п.н., регуляторный участок транспозона Tn10 РНК-out размером 137 п.н., ген устойчивости к канамицину размером 1018 п.н. и полилинкер размером 68 п.н., затем его расщепляют с использованием эндонуклеаз рестрикции SalI и BamHI и лигируют с промоторно-регуляторным участком, содержащим промоторную область гена белка сурфактанта В человека размером 634 п.н., а затем ген устойчивости к канамицину выщепляют по сайтам рестрикции Spel (вариант 5).
Способ получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS6 заключается в том, что сначала конструируют вектор размером 2408 п.н., содержащий ориджин репликации размером 688 п.н., терминатор транскрипции hGH-TA размером 467 п.н., регуляторный участок транспозона Tn10 РНК-out размером 137 п.н., ген устойчивости к канамицину размером 1018 п.н. и полилинкер размером 68 п.н., затем его расщепляют с использованием эндонуклеаз рестрикции SalI и BamHI и лигируют с промоторно-регуляторным участком, содержащим промоторную область гена белка синапсина I человека размером 555 п.н., а затем ген устойчивости к канамицину выщепляют по сайтам рестрикции Spel (вариант 6).
Способ получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS7 заключается в том, что сначала конструируют вектор размером 2408 п.н., содержащий ориджин репликации размером 688 п.н., терминатор транскрипции hGH-TA размером 467 п.н., регуляторный участок транспозона Tn10 РНК-out размером 137 п.н., ген устойчивости к канамицину размером 1018 п.н. и полилинкер размером 68 п.н., затем его расщепляют с использованием эндонуклеаз рестрикции SalI и BamHI и лигируют с промоторно-регуляторным участком, содержащим промоторную область гена нефрина человека размером 1235 п.н., а затем ген устойчивости к канамицину выщепляют по сайтам рестрикции Spel (вариант 7).
Способ получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS8 заключается в том, что сначала конструируют вектор размером 2408 п.н., содержащий ориджин репликации размером 688 п.н., терминатор транскрипции hGH-TA размером 467 п.н., регуляторный участок транспозона Tn10 РНК-out размером 137 п.н., ген устойчивости к канамицину размером 1018 п.н. и полилинкер размером 68 п.н., затем его расщепляют с использованием эндонуклеаз рестрикции SalI и BamHI и лигируют с промоторно-регуляторным участком, содержащим промоторную область гена общего лейкоцитарного антигена CD45 человека размером 855 п.н., а затем ген устойчивости к канамицину выщепляют по сайтам рестрикции Spel (вариант 8).
Способ получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS9 заключается в том, что сначала конструируют вектор размером 2408 п.н., содержащий ориджин репликации размером 688 п.н., терминатор транскрипции hGH-TA размером 467 п.н., регуляторный участок транспозона Tn10 РНК-out размером 137 п.н., ген устойчивости к канамицину размером 1018 п.н. и полилинкер размером 68 п.н., затем его расщепляют с использованием эндонуклеаз рестрикции SalI и BamHI и лигируют с промоторно-регуляторным участком, содержащим промоторную область гена белка В29 человека размером 1219 п.н., а затем ген устойчивости к канамицину выщепляют по сайтам рестрикции Spel (вариант 9).
Способ получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS10 заключается в том, что сначала конструируют вектор размером 2408 п.н., содержащий ориджин репликации размером 688 п.н., терминатор транскрипции hGH-TA размером 467 п.н., регуляторный участок транспозона Tn10 РНК-out размером 137 п.н., ген устойчивости к канамицину размером 1018 п.н. и полилинкер размером 68 п.н., затем его расщепляют с использованием эндонуклеаз рестрикции SalI и BamHI и лигируют с промоторно-регуляторным участком, содержащим промоторную область гена белка CD68 человека размером 657 п.н., а затем ген устойчивости к канамицину выщепляют по сайтам рестрикции Spel (вариант 10).
Способ получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS11 заключается в том, что сначала конструируют вектор размером 2408 п.н., содержащий ориджин репликации размером 688 п.н., терминатор транскрипции hGH-TA размером 467 п.н., регуляторный участок транспозона Tn10 РНК-out размером 137 п.н., ген устойчивости к канамицину размером 1018 п.н. и полилинкер размером 68 п.н., затем его расщепляют с использованием эндонуклеаз рестрикции SalI и BamHI и лигируют с промоторно-регуляторным участком, содержащим промоторную область гена инсулина человека размером 593 п.н., а затем ген устойчивости к канамицину выщепляют по сайтам рестрикции Spel (вариант 11).
Способ получения штамма Escherichia coli JM110-NAS для наработки генотерапевтического ДНК-вектора, выбранного из: GDTT1.8NAS1, или GDTT1.8NAS2, или GDTT1.8NAS3, или GDTT1.8NAS4, или GDTT1.8NAS5, или GDTT1.8NAS6, или GDTT1.8NAS7, или GDTT1.8NAS8, или GDTT1.8NAS9, или GDTT1.8NAS10, или GDTT1.8NAS11, заключается в том, что конструируют линейный фрагмент ДНК, содержащий регуляторный элемент RNA-in транспозона Tn10 для селекции без применения антибиотиков размером 64 п.н., ген левансахаразы sacB, продукт которого обеспечивает селекцию на сахарозо-содержащей среде размером 1422 п.н., ген устойчивости к хлорамфениколу catR, необходимый для отбора клонов штамма, в которых прошла гомологичная рекомбинация размером 763 п.н. и две гомологичные последовательности, обеспечивающие процесс гомологичной рекомбинации в области гена recA с одновременной его инактивацией размером 329 п.н. и 233 п.н., причем указанные гомологичные последовательности являются последовательностями, полученными путем проведения ПЦР амплификации фрагментов гена recA с использованием геномной ДНК Eshcerichia coli JM110-NAS в качестве матрицы и пары праймеров (84) и (85) и пары праймеров (86) и 87), после чего проводят трансформацию клеток Escherichia coli путем электропорации и отбирают клоны, выжившие на среде, содержащей 10 мкг/мл хлорамфеникола.
Получен штамм Escherichia coli JM110-NAS для наработки генотерапевтического ДНК-вектора, выбранного из GDTT1.8NAS1, или GDTT1.8NAS2, или GDTT1.8NAS3, или GDTT1.8NAS4, или GDTT1.8NAS5, или GDTT1.8NAS6, или GDTT1.8NAS7, или GDTT1.8NAS8, или GDTT1.8NAS9, или GDTT1.8NAS10, или GDTT1.8NAS11 с возможностью положительной селекции без использования антибиотиков, и содержащий в хромосоме в области гена recA линейный фрагмент, состоящий из регуляторного элемента RNA-in транспозона Tn10, гена левансахарозы sacB и гена устойчивости к хлорамфениколу catR.
Разработан способ получения штамма, несущего генотерапевтический ДНК-вектор, выбранный из GDTT1.8NAS1, или GDTT1.8NAS2, или GDTT1.8NAS3, или GDTT1.8NAS4, или GDTT1.8NAS5, или GDTT1.8NAS6, или GDTT1.8NAS7, или GDTT1.8NAS8, или GDTT1.8NAS9, или GDTT1.8NAS10, или GDTT1.8NAS11, а именно: Escherichia coli JM-110-NAS-GDTT1.8NAS1, или Escherichia coli JM-110-NAS- GDTT1.8NAS2, или Escherichia coli JM-110-NAS - GDTT1.8NAS3, или Escherichia coli JM-110-NAS- GDTT1.8NAS4, или Escherichia coli JM-110-NAS- GDTT1.8NAS5, или Escherichia coli JM-110-NAS - GDTT1.8NAS6, или Escherichia coli JM-110-NAS- GDTT1.8NAS7, или Escherichia coli JM-110-NAS- GDTT1.8NAS8, или Escherichia coli JM-110-NAS- GDTT1.8NAS9, или Escherichia coli JM-110-NAS- GDTT1.8NAS10, или Escherichia coli JM-110-NAS- GDTT1.8NAS11, заключающийся в том, что получают электрокомпетентные клетки штамма Escherichia coli JM110-NAS и проводят электропорацию этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS1, или GDTT1.8NAS2, или GDTT1.8NAS3, или GDTT1.8NAS4, или GDTT1.8NAS5, или GDTT1.8NAS6, или GDTT1.8NAS7, или GDTT1.8NAS8, или GDTT1.8NAS9, или GDTT1.8NAS10, или GDTT1.8NAS11, после чего клетки высеивают на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола.
Получен штамм Escherichia coli JM-110-NAS-GDTT1.8NAS1, или Escherichia coli JM-110-NAS-GDTT1.8NAS2, или Escherichia coli JM-110-NAS- GDTT1.8NAS3, или Escherichia coli JM-110-NAS- GDTT1.8NAS4, или Escherichia coli JM-110-NAS - GDTT1.8NAS5, или Escherichia coli JM-110-NAS- GDTT1.8NAS6, или Escherichia coli JM-110-NAS- GDTT1.8NAS7, или Escherichia coli JM-110-NAS- GDTT1.8NAS8, или Escherichia coli JM-110-NAS- GDTT1.8NAS9, или Escherichia coli JM-110-NAS- GDTT1.8NAS10, или Escherichia coli JM-110-NAS- GDTT1.8NAS11, несущий генотерапевтический ДНК-вектор GDTT1.8NAS1, или GDTT1.8NAS2, или GDTT1.8NAS3, или GDTT1.8NAS4, или GDTT1.8NAS5, или GDTT1.8NAS6, или GDTT1.8NAS7, или GDTT1.8NAS8, или GDTT1.8NAS9, или GDTT1.8NAS10, или GDTT1.8NAS11 для наработки генотерапевтического ДНК-вектора GDTT1.8NAS1, или GDTT1.8NAS2, или GDTT1.8NAS3, или GDTT1.8NAS4, или GDTT1.8NAS5, или GDTT1.8NAS6, или GDTT1.8NAS7, или GDTT1.8NAS8, или GDTT1.8NAS9, или GDTT1.8NAS10, или GDTT1.8NAS11 с возможностью селекции без использования антибиотиков.
Создан способ производства в промышленных масштабах генотерапевтического ДНК-вектора, заключающийся в масштабировании бактериальной культуры штамма до количеств, необходимых для наращивания бактериальной биомассы в промышленном ферментере, после чего биомассу используют для выделения фракции, содержащей целевой ДНК-продукт - генотерапевтический ДНК-вектор GDTT1.8NAS1, или GDTT1.8NAS2, или GDTT1.8NAS3, или GDTT1.8NAS4, или GDTT1.8NAS5, или GDTT1.8NAS6, или GDTT1.8NAS7, или, GDTT1.8NAS8, или GDTT1.8NAS9, или GDTT1.8NAS10, или GDTT1.8NAS11, многостадийно фильтруют и очищают хроматографическими методами.
Разработан способ выбора для тагетной генной терапии генотерапевтического ДНК-вектора GDTT1.8NAS1, или GDTT1.8NAS2, или GDTT1.8NAS3, или GDTT1.8NAS4, или GDTT1.8NAS5, или GDTT1.8NAS6, или GDTT1.8NAS7, или GDTT1.8NAS8, или GDTT1.8NAS9, или GDTT1.8NAS10, или GDTT1.8NAS11, каждый из которых содержит различные тканеспецифичные промоторно-регуляторные участки гена человека, для конструирования генотерапевтического ДНК-вектора, содержащего целевой ген, исходя из критериев обеспечения его тканеспецифичной экспрессии и максимальной эффективности в определенной целевой клеточной линии, целевой ткани или органе человека, заключающийся в том, что для конструирования генотерапевтического ДНК-вектора, содержащего целевой ген для тагетной генной терапии, направленной на повышение экспрессии этого целевого гена в клетках мышечной ткани, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS1, для конструирования генотерапевтического ДНК-вектора, содержащего целевой ген для тагетной генной терапии, направленной на повышение экспрессии этого целевого гена в клетках кожи, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS2, для конструирования генотерапевтического ДНК-вектора, содержащего целевой ген для тагетной генной терапии, направленной на повышение экспрессии этого целевого гена в клетках эндотелия сосудов, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS3, для конструирования генотерапевтического ДНК-вектора, содержащего целевой ген для тагетной генной терапии, направленной на повышение экспрессии этого целевого гена в остеобластах и одонтобластах, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS4, для конструирования генотерапевтического ДНК-вектора, содержащего целевой ген для тагетной генной терапии, направленной на повышение экспрессии этого целевого гена в эпителиальных клетках бронхов и альвеол, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS5, для конструирования генотерапевтического ДНК-вектора, содержащего целевой ген для тагетной генной терапии, направленной на повышение экспрессии этого целевого гена в нейронах, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS6, для конструирования генотерапевтического ДНК-вектора, содержащего целевой ген для тагетной генной терапии, направленной на повышение экспрессии этого целевого гена в подоцитах почек, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS7, для конструирования генотерапевтического ДНК-вектора, содержащего целевой ген для тагетной генной терапии, направленной на повышение экспрессии этого целевого гена в гематопоэтических клетках, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS8, для конструирования генотерапевтического ДНК-вектора, содержащего целевой ген для тагетной генной терапии, направленной на повышение экспрессии этого целевого гена в лимфоцитах, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS9, для конструирования генотерапевтического ДНК-вектора, содержащего целевой ген для тагетной генной терапии, направленной на повышение экспрессии этого целевого гена в макрофагах, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS10, для конструирования генотерапевтического ДНК-вектора, содержащего целевой ген для тагетной генной терапии, направленной на повышение экспрессии этого целевого гена в бета-клетках поджелудочной железы, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS11.
Изобретение поясняется чертежами, где
на фиг. 1
приведена схема генотерапевтического ДНК-вектора GDTT1.8NAS1, который представляет собой кольцевую двуцепочечную молекулу ДНК размером 1736 п.н., способную к автономной репликации в клетках бактерии Eshcerichia coli при отсутствии селективного антибиотика в среде культивирования.
На фиг. 1 отмечены следующие структурные элементы вектора:
(1) pUCori (позиция 1 - 688 п.н.) - ориджин репликации с однонуклеотидной заменой для повышения копийности вектора, служащий для автономной репликации в клетках Eshcerichia coli.
(2) prom (позиция 695 - 1046 п.н.) - промоторная область гена миоглобина человека. Служит для обеспечения высокого специфического уровня транскрипции целевого терапевтического гена в клетках мышечной ткани различных органов человека.
(3) MCS (позиция 1047 - 1114 п.н.) - полилинкер (сайт множественного клонирования), содержащий сайты рестрикции BamHI, EcoRV, SalI, KpnI, EcoRI, XbaI и NotI и предназначенный для клонирования целевых терапевтических генов.
(4) hGH-TA (позиция 1115 - 1581 п.н.) - терминатор транскрипции и сайт полиаденилирования гена фактора роста человека;
(5) RNA-out (позиция 1588 - 1724 п.н.) - регуляторный элемент РНК-out транспозона Tn10, обеспечивающий возможность положительной селекции без использования антибиотиков при использовании штамма Escherichia coli JM110-NAS.
На фиг. 2
приведена схема генотерапевтического ДНК-вектора GDTT1.8NAS2, который представляет собой кольцевую двуцепочечную молекулу ДНК размером 1654 п.н., способную к автономной репликации в клетках бактерии Eshcerichia coli при отсутствии селективного антибиотика в среде культивирования.
На фиг. 2 отмечены следующие структурные элементы вектора:
(1) pUCori (позиция 1 - 688 п.н.) - ориджин репликации с однонуклеотидной заменой для повышения копийности вектора, служащий для автономной репликации в клетках Eshcerichia coli.
(2) prom (позиция 695-964 п.н.) - промоторная область гена эластазы человека. Служит для обеспечения высокого специфического уровня транскрипции целевого терапевтического гена в клетках в клетках кожи человека.
(3) MCS (позиция 965-1032 п.н.) - полилинкер (сайт множественного клонирования), содержащий сайты рестрикции BamHI, EcoRV, SalI, KpnI, EcoRI, XbaI и NotI и предназначенный для клонирования целевых терапевтических генов.
(4) hGH-TA (позиция 1033-1499 п.н.) - терминатор транскрипции и сайт полиаденилирования гена фактора роста человека;
(5) RNA-out (позиция 1506-1642 п.н.) - регуляторный элемент РНК-out транспозона Tn10, обеспечивающий возможность положительной селекции без использования антибиотиков при использовании штамма Escherichia coli JM110-NAS.
На фиг. 3
приведена схема генотерапевтического ДНК-вектора GDTT1.8NAS3, который представляет собой кольцевую двуцепочечную молекулу ДНК размером 1783 п.н., способную к автономной репликации в клетках бактерии Eshcerichia coli при отсутствии селективного антибиотика в среде культивирования.
На фиг. 3 отмечены следующие структурные элементы вектора:
(1) pUCori (позиция 1 - 688 п.н.) - ориджин репликации с однонуклеотидной заменой для повышения копийности вектора, служащий для автономной репликации в клетках Eshcerichia coli.
(2) prom (позиция 695-1093 п.н.) - промоторная область гена молекулы межклеточной адгезии 2 человека. Служит для обеспечения высокого специфического уровня транскрипции целевого терапевтического гена в клетках в клетках эндотелия сосудов.
(3) MCS (позиция 1094-1161 п.н.) - полилинкер (сайт множественного клонирования), содержащий сайты рестрикции BamHI, EcoRV, SalI, KpnI, EcoRI, XbaI и NotI и предназначенный для клонирования целевых терапевтических генов.
(4) hGH-TA (позиция 1162-1628 п.н.) - терминатор транскрипции и сайт полиаденилирования гена фактора роста человека;
(5) RNA-out (позиция 1635-1771 п.н.) - регуляторный элемент РНК-out транспозона Tn10, обеспечивающий возможность положительной селекции без использования антибиотиков при использовании штамма Escherichia coli JM110-NAS.
На фиг. 4
приведена схема генотерапевтического ДНК-вектора GDTT1.8NAS4, который представляет собой кольцевую двуцепочечную молекулу ДНК размером 1953 п.н., способную к автономной репликации в клетках бактерии Eshcerichia coli при отсутствии селективного антибиотика в среде культивирования.
На фиг. 4 отмечены следующие структурные элементы вектора:
(1) pUCori (позиция 1 - 688 п.н.) - ориджин репликации с однонуклеотидной заменой для повышения копийности вектора, служащий для автономной репликации в клетках Eshcerichia coli.
(2) prom (позиция 695-1263 п.н.) - промоторная область гена остеокальцина 2 человека. Служит для обеспечения высокого специфического уровня транскрипции целевого терапевтического гена в остеобластах и одонтобластах.
(3) MCS (позиция 1264-1331 п.н.) - полилинкер (сайт множественного клонирования), содержащий сайты рестрикции BamHI, EcoRV, SalI, KpnI, EcoRI, XbaI и NotI и предназначенный для клонирования целевых терапевтических генов.
(4) hGH-TA (позиция 1332-1798 п.н.) - терминатор транскрипции и сайт полиаденилирования гена фактора роста человека;
(5) RNA-out (позиция 1805-1941 п.н.) - регуляторный элемент РНК-out транспозона Tn10, обеспечивающий возможность положительной селекции без использования антибиотиков при использовании штамма Escherichia coli JM110-NAS.
На фиг. 5
приведена схема генотерапевтического ДНК-вектора GDTT1.8NAS5, который представляет собой кольцевую двуцепочечную молекулу ДНК размером 2019 п.н., способную к автономной репликации в клетках бактерии Eshcerichia coli при отсутствии селективного антибиотика в среде культивирования.
На фиг. 5 отмечены следующие структурные элементы вектора:
(1) pUCori (позиция 1 - 688 п.н.) - ориджин репликации с однонуклеотидной заменой для повышения копийности вектора, служащий для автономной репликации в клетках Eshcerichia coli.
(2) prom (позиция 695-1329 п.н) - промоторная область гена белка сурфактанта В человека. Служит для обеспечения высокого специфического уровня транскрипции целевого терапевтического гена в клетках в эпителиальных клетках бронхов и альвеол.
(3) MCS (позиция 1330-1397 п.н.) - полилинкер (сайт множественного клонирования), содержащий сайты рестрикции BamHI, EcoRV, SalI, KpnI, EcoRI, XbaI и NotI и предназначенный для клонирования целевых терапевтических генов.
(4) hGH-TA (позиция 1398-1864 п.н.) - терминатор транскрипции и сайт полиаденилирования гена фактора роста человека;
(5) RNA-out (позиция 1871-2007 п.н.) - регуляторный элемент РНК-out транспозона Tn10, обеспечивающий возможность положительной селекции без использования антибиотиков при использовании штамма Escherichia coli JM110-NAS.
На фиг. 6
приведена схема генотерапевтического ДНК-вектора GDTT1.8NAS6, который представляет собой кольцевую двуцепочечную молекулу ДНК размером 1940 п.н., способную к автономной репликации в клетках бактерии Eshcerichia coli при отсутствии селективного антибиотика в среде культивирования.
На фиг. 6 отмечены следующие структурные элементы вектора:
(1) pUCori (позиция 1 - 688 п.н.) - ориджин репликации с однонуклеотидной заменой для повышения копийности вектора, служащий для автономной репликации в клетках Eshcerichia coli.
(2) prom (позиция 695-1250 п.н.) - промоторная область гена синапсина I человека. Служит для обеспечения высокого специфического уровня транскрипции целевого терапевтического гена в клетках в нейронах.
(3) MCS (позиция 1251-1318 п.н.) - полилинкер (сайт множественного клонирования), содержащий сайты рестрикции BamHI, EcoRV, SalI, KpnI, EcoRI, XbaI и NotI и предназначенный для клонирования целевых терапевтических генов.
(4) hGH-TA (позиция 1319-1785 п.н.) - терминатор транскрипции и сайт полиаденилирования гена фактора роста человека;
(5) RNA-out (позиция 1792-1928 п.н.) - регуляторный элемент РНК-out транспозона Tn10, обеспечивающий возможность положительной селекции без использования антибиотиков при использовании штамма Escherichia coli JM110-NAS.
На фиг. 7
приведена схема генотерапевтического ДНК-вектора GDTT1.8NAS7, который представляет собой кольцевую двуцепочечную молекулу ДНК размером 2620 п.н., способную к автономной репликации в клетках бактерии Eshcerichia coli при отсутствии селективного антибиотика в среде культивирования.
На фиг. 7
отмечены следующие структурные элементы вектора:
(1) pUCori (позиция 1 - 688 п.н.) - ориджин репликации с однонуклеотидной заменой для повышения копийности вектора, служащий для автономной репликации в клетках Eshcerichia coli.
(2) prom (позиция 695-1930 п.н.) - промоторная область гена нефрина человека. Служит для обеспечения высокого специфического уровня транскрипции целевого терапевтического гена в подоцитах почек.
(3) MCS (позиция 1937-1998 п.н.) - полилинкер (сайт множественного клонирования), содержащий сайты рестрикции BamHI, EcoRV, SalI, KpnI, EcoRI, XbaI и NotI и предназначенный для клонирования целевых терапевтических генов.
(4) hGH-TA (позиция 1999-2465 п.н.) - терминатор транскрипции и сайт полиаденилирования гена фактора роста человека;
(5) RNA-out (позиция 2472-2608 п.н.) - регуляторный элемент РНК-out транспозона Tn10, обеспечивающий возможность положительной селекции без использования антибиотиков при использовании штамма Escherichia coli JM110-NAS.
На фиг. 8
приведена схема генотерапевтического ДНК-вектора GDTT1.8NAS8, который представляет собой кольцевую двуцепочечную молекулу ДНК размером 2240 п.н., способную к автономной репликации в клетках бактерии Eshcerichia coli при отсутствии селективного антибиотика в среде культивирования.
На фиг. 8 отмечены следующие структурные элементы вектора:
(1) pUCori (позиция 1 - 688 п.н.) - ориджин репликации с однонуклеотидной заменой для повышения копийности вектора, служащий для автономной репликации в клетках Eshcerichia coli.
(2) prom (позиция 695-1550 п.н.) - промоторная область гена общего лейкоцитарного антигена CD45 человека. Служит для обеспечения высокого специфического уровня транскрипции целевого терапевтического гена в гематопоэтических клетках.
(3) MCS (позиция 1551-1618 п.н.) - полилинкер (сайт множественного клонирования), содержащий сайты рестрикции BamHI, EcoRV, SalI, KpnI, EcoRI, XbaI и NotI и предназначенный для клонирования целевых терапевтических генов.
(4) hGH-TA (позиция 1619-2085 п.н.) - терминатор транскрипции и сайт полиаденилирования гена фактора роста человека;
(5) RNA-out (позиция 2092-2228 п.н.) - регуляторный элемент РНК-out транспозона Tn10, обеспечивающий возможность положительной селекции без использования антибиотиков при использовании штамма Escherichia coli JM110-NAS.
На фиг. 9
приведена схема генотерапевтического ДНК-вектора GDTT1.8NAS9, который представляет собой кольцевую двуцепочечную молекулу ДНК размером 2604 п.н., способную к автономной репликации в клетках бактерии Eshcerichia coli при отсутствии селективного антибиотика в среде культивирования.
На фиг. 9 отмечены следующие структурные элементы вектора:
(1) pUCori (позиция 1 - 688 п.н.) - ориджин репликации с однонуклеотидной заменой для повышения копийности вектора, служащий для автономной репликации в клетках Eshcerichia coli.
(2) prom (позиция 695-1914 п.н.) - промоторная область гена белка В29 человека. Служит для обеспечения высокого специфического уровня транскрипции целевого терапевтического гена в лимфоцитах.
(3) MCS (позиция 1915-1982 п.н.) - полилинкер (сайт множественного клонирования), содержащий сайты рестрикции BamHI, EcoRV, SalI, KpnI, EcoRI, XbaI и NotI и предназначенный для клонирования целевых терапевтических генов.
(4) hGH-TA (позиция 1983-2449 п.н.) - терминатор транскрипции и сайт полиаденилирования гена фактора роста человека;
(5) RNA-out (позиция 2456-2592 п.н.) - регуляторный элемент РНК-out транспозона Tn10, обеспечивающий возможность положительной селекции без использования антибиотиков при использовании штамма Escherichia coli JM110-NAS.
На фиг. 10
приведена схема генотерапевтического ДНК-вектора GDTT1.8NAS10, который представляет собой кольцевую двуцепочечную молекулу ДНК размером 2048 п.н., способную к автономной репликации в клетках бактерии Eshcerichia coli при отсутствии селективного антибиотика в среде культивирования.
На фиг. 10 отмечены следующие структурные элементы вектора:
(1) pUCori (позиция 1 - 688 п.н.) - ориджин репликации с однонуклеотидной заменой для повышения копийности вектора, служащий для автономной репликации в клетках Eshcerichia coli.
(2) prom (позиция 695-1352 п.н.) - промоторная область гена белка CD68 человека. Служит для обеспечения высокого специфического уровня транскрипции целевого терапевтического гена в макрофагах.
(3) MCS (позиция 1359-1426 п.н.) - полилинкер (сайт множественного клонирования), содержащий сайты рестрикции BamHI, EcoRV, SalI, KpnI, EcoRI, XbaI и NotI и предназначенный для клонирования целевых терапевтических генов.
(4) hGH-TA (позиция 1427-1893 п.н.) - терминатор транскрипции и сайт полиаденилирования гена фактора роста человека;
(5) RNA-out (позиция 1900-2036 п.н.) - регуляторный элемент РНК-out транспозона Tn10, обеспечивающий возможность положительной селекции без использования антибиотиков при использовании штамма Escherichia coli JM110-NAS.
На фиг. 11
приведена схема генотерапевтического ДНК-вектора GDTT1.8NAS11, который представляет собой кольцевую двуцепочечную молекулу ДНК размером 1978 п.н., способную к автономной репликации в клетках бактерии Eshcerichia coli при отсутствии селективного антибиотика в среде культивирования.
На фиг. 11 отмечены следующие структурные элементы вектора:
(1) pUCori (позиция 1 - 688 п.н.) - ориджин репликации с однонуклеотидной заменой для повышения копийности вектора, служащий для автономной репликации в клетках Eshcerichia coli.
(2) prom (позиция 695-1288 п.н.) - промоторная область гена инсулина человека. Служит для обеспечения высокого специфического уровня транскрипции целевого терапевтического гена в бета-клетках поджелудочной железы.
(3) MCS (позиция 1289-1356 п.н.) - полилинкер (сайт множественного клонирования), содержащий сайты рестрикции BamHI, EcoRV, SalI, KpnI, EcoRI, XbaI и NotI и предназначенный для клонирования целевых терапевтических генов.
(4) hGH-TA (позиция 1357-1823 п.н.) - терминатор транскрипции и сайт полиаденилирования гена фактора роста человека;
(5) RNA-out (позиция 1830-1966 п.н.) - регуляторный элемент РНК-out транспозона Tn10, обеспечивающий возможность положительной селекции без использования антибиотиков при использовании штамма Escherichia coli JM110-NAS.
На фиг. 12
изображена структура фрагмента ДНК для гомологичной рекомбинации в области гена recA Eshcerichia coli для получения штамма Eshcerichia coli JM 110, где:
RNA-in - линейный фрагмент, состоящий из кассеты, несущей в себе регуляторный элемент RNA-in транспозона Tn10 для селекции без применения антибиотиков (64 п.н.),
sacB - ген левансахаразы sacB, продукт которого обеспечивает селекцию на сахарозо-содержащей среде (1422 п.н.),
catR - ген устойчивости к хлорамфениколу catR, необходимый для отбора клонов штамма, в которых прошла гомологичная рекомбинация (763 п.н.).
LHA и RHA - кассета фланкирована двумя гомологичными плечами, обеспечивающими процесс рекомбинации в области гена recA с одновременной его инактивацией (329 п.н. и 233 п.н. для левого и правого плеча, соответственно).
На фиг. 13
показаны графики накопления мРНК гена VEGF человека в скелетных миобластах человека HSkM до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS1- VEGF, несущим ген VEGF человека с целью оценки изменения накопления мРНК целевого гена, например, гена VEGF, в скелетных миобластах человека HSkM до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS1-VEGF, несущим ген VEGF человека, где:
1 - кДНК гена VEGF после трансфекции генотерапевтическим вектором GDTT1.8NAS1;
2 - кДНК гена VEGF после трансфекции генотерапевтическим вектором GDTT1.8NAS1-VEGF, несущим ген VEGF человека;
3 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS1;
4 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS1-VEGF, несущим ген VEGF человека.
На фиг. 14
показана диаграмма концентрации белка VEGF в клеточном лизате скелетных миобластов человека HSkM при трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS1-VEGF, несущим ген VEGF человека и генотерапевтическим ДНК-вектором GDTT1.8NAS1, не несущим ген VEGF человека с целью сравнения количества целевого белка, например белка VEGF, где:
культура А - клеточная культура скелетных миобластов человека HSkM, трансфицированных
без плазмидной ДНК (контроль)
культура В - клеточная культура скелетных миобластов человека HSkM, трансфицированных ДНК-вектором GDTT1.8NAS1
культура С - клеточная культура скелетных миобластов человека HSkM, трансфицированных ДНК-вектором GDTT1.8NAS1-VEGF.
На фиг. 15
показаны графики накопления мРНК гена CAT человека в первичной культуре фибробластов кожи человека HDFa до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS2-CAT, несущим ген CAT человека с целью оценки изменения накопления мРНК целевого гена, например, гена CAT, в первичной культуре фибробластов кожи человека HDFa до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS2-CAT, несущим ген CAT человека, где:
1 - кДНК гена CAT после трансфекции генотерапевтическим вектором GDTT1.8NAS2;
2 - кДНК гена CAT после трансфекции генотерапевтическим вектором GDTT1.8NAS2-CAT, несущим ген CAT человека;
3 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS2;
4 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS2-CAT, несущим ген CAT человека.
На фиг. 16
показана диаграмма концентрации белка CAT в клеточном лизате первичной культуры фибробластов кожи человека HDFa при трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS2-CAT, несущим ген CAT человека и генотерапевтическим ДНК-вектором GDTT1.8NAS2, не несущим ген CAT человека с целью сравнения количества целевого белка, например белка CAT, где:
культура А - клеточная культура фибробластов кожи человека HDFa, трансфицированных раствором реагента Lipofectamine 3000 без плазмидной ДНК (контроль)
культура В - клеточная культура фибробластов кожи человека HDFa, трансфицированных ДНК-вектором GDTT1.8NAS2
культура С - клеточная культура фибробластов кожи человека HDFa, трансфицированных ДНК-вектором GDTT1.8NAS2-CAT
На фиг. 17
показаны графики накопления мРНК гена HIF1a человека в клетках эндотелия пупочной вены человека HUVEC до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS3-HIF1a, несущим ген HIF1a человека с целью оценки изменения накопления мРНК целевого гена, например, гена HIF1a, в клетках эндотелия пупочной вены человека HUVEC до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS3- HIF1a, несущим ген HIF1a человека, где:
1 - кДНК гена HIF1a после трансфекции генотерапевтическим вектором GDTT1.8NAS3;
2 - кДНК гена HIF1a после трансфекции генотерапевтическим вектором GDTT1.8NAS3-HIF1a, несущим ген HIF1a человека;
3 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS3;
4 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS3-HIF1a, несущим ген HIF1a человека.
На фиг. 18
показана диаграмма концентрации белка HIF1a в клеточном лизате клеток эндотелия пупочной вены человека HUVEC при трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS3-HIF1a, несущим ген HIF1a человека и генотерапевтическим ДНК-вектором GDTT1.8NAS3, не несущим ген HIF1a человека с целью сравнения количества целевого белка, например белка HIF1a, где:
культура А - клеточная культура эндотелия пупочной вены человека HUVEC, трансфицированных раствором реагента Lipofectamine 3000 без плазмидной ДНК (контроль)
культура В - клеточная культура эндотелия пупочной вены человека HUVEC, трансфицированных ДНК-вектором GDTT1.8NAS3
культура С - клеточная культура эндотелия пупочной вены человека HUVEC, трансфицированных ДНК-вектором GDTT1.8NAS3-HIF1a.
На фиг. 19
показаны графики накопления мРНК гена ВМР2 человека в клетках остеосаркомы человека MG-63 до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS4-BMP2, несущим ген ВМР2 человека с целью оценки изменения накопления мРНК целевого гена, например, гена ВМР2, в клетках остеосаркомы человека MG-63 до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS4-BMP2, несущим ген ВМР2 человека, где:
1 - кДНК гена ВМР2 после трансфекции генотерапевтическим вектором GDTT1.8NAS4;
2 - кДНК гена ВМР2 после трансфекции генотерапевтическим вектором GDTT1.8NAS4-ВМР2, несущим ген ВМР2 человека;
3 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS4;
4 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS4-ВМР2, несущим ген ВМР2 человека.
На фиг. 20
показана диаграмма концентрации белка ВМР2 в клеточном лизате остеосаркомы человека MG-63 при трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS4-BMP2, несущим ген ВМР2 человека и генотерапевтическим ДНК-вектором GDTT1.8NAS4, не несущим ген ВМР2 человека с целью сравнения количества целевого белка, например белка ВМР2, где:
культура А - клеточная культура остеосаркомы человека MG-63, трансфицированных раствором реагента Lipofectamine 3000 без плазмидной ДНК (контроль)
культура В - клеточная культура остеосаркомы человека MG-63, трансфицированных ДНК-вектором GDTT1.8NAS4
культура С - клеточная культура остеосаркомы человека MG-63, трансфицированных ДНК-вектором GDTT1.8NAS4-ВМР2.
На фиг. 21
показаны графики накопления мРНК гена CFTR человека в клетках трахеального эпителия человека CFTE29o- до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS5-CFTR, несущим ген CFTR человека с целью оценки изменения накопления мРНК целевого гена, например, гена CFTR, в клетках трахеального эпителия человека CFTE29o- до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS5-CFTR, несущим ген CFTR человека, где:
1 - кДНК гена CFTR после трансфекции генотерапевтическим вектором GDTT1.8NAS5;
2 - кДНК гена CFTR после трансфекции генотерапевтическим вектором GDTT1.8NAS5-CFTR, несущим ген CFTR человека;
3 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS5;
4 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS5-CFTR, несущим ген CFTR человека.
На фиг. 22
показана диаграмма концентрации белка CFTR в клеточном лизате трахеального эпителия человека CFTE29o- при трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS5-CFTR, несущим ген CFTR человека и генотерапевтическим ДНК-вектором GDTT1.8NAS5, не несущим ген CFTR человека с целью сравнения количества целевого белка, например белка CFTR, где:
культура А - клеточная культура трахеального эпителия человека CFTE29o-, трансфицированных раствором реагента Lipofectamine 3000 без плазмидной ДНК (контроль)
культура В - клеточная культура трахеального эпителия человека CFTE29o-, трансфицированных ДНК-вектором GDTT1.8NAS5
культура С - клеточная культура трахеального эпителия человека CFTE29o-, трансфицированных ДНК-вектором GDTT1.8NAS5- CFTR.
На фиг. 23
показаны графики накопления мРНК гена BDNF человека в клетках нейробластомы человека SH-SY5Y до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS6-BDNF, несущим ген BDNF человека с целью оценки изменения накопления мРНК целевого гена, например, гена BDNF, в клетках нейробластомы человека SH-SY5Y до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS6-BDNF, несущим ген BDNF человека, где:
1 - кДНК гена BDNF после трансфекции генотерапевтическим вектором GDTT1.8NAS6;
2 - кДНК гена BDNF после трансфекции генотерапевтическим вектором GDTT1.8NAS6-BDNF, несущим ген BDNF человека;
3 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS6;
4 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS6-BDNF, несущим ген BDNF человека.
На фиг. 24
показана диаграмма концентрации белка BDNF в клеточном лизате нейробластомы человека SH-SY5Y при трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS6-BDNF, несущим ген BDNF человека и генотерапевтическим ДНК-вектором GDTT1.8NAS6, не несущим ген BDNF человека с целью сравнения количества целевого белка, например белка BDNF, где:
культура А - клеточная культура нейробластомы человека SH-SY5Y, трансфицированных раствором реагента Lipofectamine 3000 без плазмидной ДНК (контроль)
культура В - клеточная культура нейробластомы человека SH-SY5Y, трансфицированных ДНК-вектором GDTT1.8NAS6
культура С - клеточная культура нейробластомы человека SH-SY5Y, трансфицированных ДНК-вектором GDTT1.8NAS6- BDNF.
На фиг. 25
показаны графики накопления мРНК гена ATGL человека в первичной смешанной культуре эпителия почки человека HREC до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS7-ATGL, несущим ген ATGL человека с целью оценки изменения накопления мРНК целевого гена, например, гена ATGL, в первичной смешанной культуре эпителия почки человека HREC до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS7-ATGL, несущим ген ATGL человека, где:
1 - кДНК гена ATGL после трансфекции генотерапевтическим вектором GDTT1.8NAS7;
2 - кДНК гена ATGL после трансфекции генотерапевтическим вектором GDTT1.8NAS7-ATGL, несущим ген ATGL человека;
3 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS7;
4 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS7-ATGL, несущим ген ATGL человека.
На фиг. 26
показана диаграмма концентрации белка ATGL в клеточном лизате первичной смешанной культуры эпителия почки человека HREC при трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS7-ATGL, несущим ген ATGL человека и генотерапевтическим ДНК-вектором GDTT1.8NAS7, не несущим ген ATGL человека с целью сравнения количества целевого белка, например белка ATGL, где:
культура А - первичная смешанная культура эпителия почки человека HREC, трансфицированных раствором реагента Lipofectamine 3000 без плазмидной ДНК (контроль)
культура В - первичная смешанная культура эпителия почки человека HREC, трансфицированных ДНК-вектором GDTT1.8NAS7
культура С - первичная смешанная культура эпителия почки человека HREC, трансфицированных ДНК-вектором GDTT1.8NAS7-ATGL.
На фиг. 27
показаны графики накопления мРНК гена CAS9 человека в первичной культуре костного мозга человека до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS8-CAS9, несущим ген CAS9 человека с целью оценки изменения накопления мРНК целевого гена, например, гена CAS9, в первичной культуре костного мозга человека до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS8-CAS9, несущим ген CAS9 человека, где:
1 - кДНК гена CAS9 после трансфекции генотерапевтическим вектором GDTT1.8NAS8;
2 - кДНК гена CAS9 после трансфекции генотерапевтическим вектором GDTT1.8NAS8-CAS9, несущим ген CAS9 человека;
3 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS8;
4 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS8-CAS9, несущим ген CAS9 человека.
На фиг. 28
показана диаграмма концентрации белка CAS9 в клеточном лизате первичной культуры костного мозга человека при трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS8-CAS9, несущим ген CAS9 человека и генотерапевтическим ДНК-вектором GDTT1.8NAS8, не несущим ген CAS9 человека с целью сравнения количества целевого белка, например белка CAS9, где:
культура А - первичная культура костного мозга человека, трансфицированных раствором реагента Lipofectamine 3000 без плазмидной ДНК (контроль)
культура В - первичная культура костного мозга человека, трансфицированных ДНК-вектором GDTT1.8NAS8
культура С - первичная культура костного мозга человека, трансфицированных ДНК-вектором GDTT1.8NAS8-CAS9.
На фиг. 29
показаны графики накопления мРНК гена BTK человека в первичной культуре периферических мононуклеаров крови человека РВМС до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS9-BTK, несущим ген ВТК человека с целью оценки изменения накопления мРНК целевого гена, например, гена BTK, в первичной культуре периферических мононуклеаров крови человека РВМС до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS9-VEGF, несущим ген ВТК человека, где:
1 - кДНК гена BTK после трансфекции генотерапевтическим вектором GDTT1.8NAS9;
2 - кДНК гена BTK после трансфекции генотерапевтическим вектором GDTT1.8NAS9-ВТК, несущим ген ВТК человека;
3 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS9;
4 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS9-BTK, несущим ген ВТК человека.
На фиг. 30
показана диаграмма концентрации белка BTK в клеточном лизате первичной культуры периферических мононуклеаров крови человека РВМС при трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS9-BTK, несущим ген BTK человека и генотерапевтическим ДНК-вектором GDTT1.8NAS9, не несущим ген BTK человека с целью сравнения количества целевого белка, например белка ВТК, где:
культура А - первичная культура периферических мононуклеаров крови человека РВМС, трансфицированных раствором реагента Lipofectamine 3000 без плазмидной ДНК (контроль)
культура В - первичная культура периферических мононуклеаров крови человека РВМС, трансфицированных ДНК-вектором GDTT1.8NAS9
культура С - первичная культура периферических мононуклеаров крови человека РВМС, трансфицированных ДНК-вектором GDTT1.8NAS9- BTK.
На фиг. 31
показаны графики накопления мРНК гена GBA человека в макрофагах периферической крови человека SC до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS10-GBA, несущим ген GBA человека с целью оценки изменения накопления мРНК целевого гена, например, гена GBA, в макрофагах периферической крови человека SC до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS10- GBA, несущим ген GBA человека, где:
1 - кДНК гена GBA после трансфекции генотерапевтическим вектором GDTT1.8NAS10;
2 - кДНК гена GBA после трансфекции генотерапевтическим вектором GDTT1.8NAS10- GBA, несущим ген GBA человека;
3 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS10;
4 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS10- GBA, несущим ген GBA человека.
На фиг. 32
показана диаграмма концентрации белка GBA в клеточном лизате макрофагов периферической крови человека SC при трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS10-GBA, несущим ген GBA человека и генотерапевтическим ДНК-вектором GDTT1.8NAS10, не несущим ген GBA человека с целью сравнения количества целевого белка, например белка GBA, где:
культура А - клеточная культура макрофагов периферической крови человека SC, трансфицированных раствором реагента Lipofectamine 3000 без плазмидной ДНК (контроль)
культура В - клеточная культура макрофагов периферической крови человека SC, трансфицированных ДНК-вектором GDTT1.8NAS10
культура С - клеточная культура макрофагов периферической крови человека SC, трансфицированных ДНК-вектором GDTT1.8NAS10-GBA.
На фиг. 33
показаны графики накопления мРНК гена PDX1 человека в клетках аденокарциномы поджелудочной железы Panc 10.05 до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS11-PDX1, несущим ген PDX1 человека с целью оценки изменения накопления мРНК целевого гена, например, гена PDX1, в клетках аденокарциномы поджелудочной железы Panc 10.05 до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS11-PDX1, несущим ген PDX1 человека, где:
1 - кДНК гена PDX1 после трансфекции генотерапевтическим вектором GDTT1.8NAS11;
2 - кДНК гена PDX1 после трансфекции генотерапевтическим вектором GDTT1.8NAS11- PDX1, несущим ген PDX1 человека;
3 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS11;
4 - кДНК гена В2М после трансфекции генотерапевтическим вектором GDTT1.8NAS11- PDX1, несущим ген PDX1 человека.
На фиг. 34
показана диаграмма концентрации белка PDX1 в клеточном лизате аденокарциномы поджелудочной железы Panc 10.05 при трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS11- PDX1, несущим ген PDX1 человека и генотерапевтическим ДНК-вектором GDTT1.8NAS11, не несущим ген PDX1 человека с целью сравнения количества целевого белка, например белка PDX1, где:
культура А - клеточная культура аденокарциномы поджелудочной железы Panc 10.05, трансфицированных раствором реагента Lipofectamine 3000 без плазмидной ДНК (контроль)
культура В - клеточная культура аденокарциномы поджелудочной железы Panc 10.05, трансфицированных ДНК-вектором GDTT1.8NAS11
культура С - клеточная культура аденокарциномы поджелудочной железы Panc 10.05, трансфицированных ДНК-вектором GDTT1.8NAS11- PDX1
На фиг. 35
показана диаграмма концентрации белка VEGF в биоптатах икроножной мышцы трех пациентов при введении в икроножную мышцу этих пациентов генотерапевтического ДНК-вектора GDTT1.8NAS1- VEGF, несущего ген VEGF человека и параллельном введении генотерапевтического ДНК-вектора GDTT1.8NAS1, не несущего ген VEGF человека с целью сравнения при этом количества целевого белка, например белка VEGF, где:
П1I - биоптат икроножной мышцы пациента П1 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS1- VEGF
П1II - биоптат икроножной мышцы пациента П1 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS1 (плацебо)
П1III - биоптат интактного участка икроножной мышцы пациента П1
П2I - биоптат икроножной мышцы пациента П2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS1- VEGF
П2II - биоптат икроножной мышцы пациента П2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS1 (плацебо)
П2III - биоптат интактного участка икроножной мышцы пациента П2
П3I - биоптат икроножной мышцы пациента П3 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS1- VEGF
П3II - биоптат икроножной мышцы пациента П3 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS1 (плацебо)
П3III - биоптат интактного участка икроножной мышцы пациента П3
На фиг. 36
показана диаграмма концентрации белка CAT в биоптатах кожи трех пациентов при введении в кожу этих пациентов генотерапевтического ДНК-вектора GDTT1.8NAS2-CAT, несущего ген CAT человека и параллельном введении генотерапевтического ДНК-вектора GDTT1.8NAS2, не несущего ген CAT человека с целью сравнения при этом количества целевого белка, например белка CAT, где:
П1I - биоптат кожи пациента П1 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS2-CAT
П1II - биоптат кожи пациента П1 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS2 (плацебо)
П1III - биоптат интактного участка кожи пациента П1
П2I - биоптат кожи пациента П2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS2-CAT
П2II - биоптат кожи пациента П2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS2 (плацебо)
П2III - биоптат интактного участка кожи пациента П2
П3I - биоптат кожи пациента П3 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS2-CAT
П3II - биоптат кожи пациента П3 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS2 (плацебо)
П3III - биоптат интактного участка кожи пациента П3
На фиг. 37
показаны графики накопления мРНК гена ATGL человека в клетках почки быка MDBK до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS7-ATGL, несущим ген ATGL человека с целью оценки изменения накопления мРНК целевого гена, например, гена ATGL, в клетках почки быка MDBK до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS1-ATGL, несущим ген ATGL человека, где:
1 - кДНК гена ATGL после трансфекции генотерапевтическим вектором GDTT1.8NAS7;
2 - кДНК гена ATGL после трансфекции генотерапевтическим вектором GDTT1.8NAS7- ATGL, несущим ген ATGL человека;
3 - кДНК гена ACT после трансфекции генотерапевтическим вектором GDTT1.8NAS7;
4 - кДНК гена ACT после трансфекции генотерапевтическим вектором GDTT1.8NAS7- ATGL, несущим ген ATGL человека.
На фиг. 38
показана диаграмма концентрации белка ATGL в лизате клеток почки быка MDBK при трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS7-ATGL, несущим ген ATGL человека и генотерапевтическим ДНК-вектором GDTT1.8NAS7, не несущим ген ATGL человека с целью сравнения количества целевого белка, например белка ATGL, где:
культура А - культура клеток почки быка MDBK, трансфицированных раствором реагента Lipofectamine 3000 без плазмидной ДНК (контроль)
культура В - культура клеток почки быка MDBK, трансфицированных ДНК-вектором GDTT1.8NAS7
культура С - культура клеток почки быка MDBK, трансфицированных ДНК-вектором GDTT1.8NAS7-ATGL
Реализация изобретения
Для подтверждения эффективности созданного генотерапевтического ДНК-вектора GDTT1.8NAS1 или GDTT1.8NAS2 или GDTT1.8NAS3 или GDTT1.8NAS4 или GDTT1.8NAS5 или GDTT1.8NAS6 или GDTT1.8NAS7 или GDTT1.8NAS8 или GDTT1.8NAS9 или GDTT1.8NAS10 или GDTT1.8NAS11 оценивали:
A) изменение накопления мРНК целевых /терапевтических генов в лизате клеток человека и животного, после трансфекции различных клеточных линий человека и животного генотерапевтическими ДНК-векторами;
B) изменение количественного уровня целевых /терапевтических белков в лизате клеток человека и животного, после трансфекции различных клеточных линий человека и животного генотерапевтическими ДНК-векторами;
C) изменение количественного уровня целевых /терапевтических белков в супернатанте биоптатов тканей человека, после введения в эти ткани генотерапевтических ДНК-векторов;
Для подтверждения реализуемости способа использования созданного генотерапевтического ДНК-вектора GDTT1.8NAS1 или GDTT1.8NAS2 или GDTT1.8NAS3 или GDTT1.8NAS4 или GDTT1.8NAS5 или GDTT1.8NAS6 или GDTT1.8NAS7 или GDTT1.8NAS8 или GDTT1.8NAS9 или GDTT1.8NAS10 или GDTT1.8NAS11 выполняли:
D) трансфекцию генотерапевтическими ДНК-векторами различных клеточных линий человека и животного;
Е) введение генотерапевтических ДНК-векторов в различные ткани человека;
Для подтверждения тканевой специфичности созданного генотерапевтического ДНК-вектора GDTT1.8NAS1 или GDTT1.8NAS2 или GDTT1.8NAS3 или GDTT1.8NAS4 или GDTT1.8NAS5 или GDTT1.8NAS6 или GDTT1.8NAS7 или GDTT1.8NAS8 или GDTT1.8NAS9 или GDTT1.8NAS10 или GDTT1.8NAS11 выполняли:
F) трансфекцию несколькими генотерапевтическими ДНК-векторами нескольких клеточных линий человека;
Для подтверждения технологичности получения и возможности производства в промышленных масштабах генотерапевтического ДНК-вектора GDTT1.8NAS1 или GDTT1.8NAS2 или GDTT1.8NAS3 или GDTT1.8NAS4 или GDTT1.8NAS5 или GDTT1.8NAS6 или GDTT1.8NAS7 или GDTT1.8NAS8 или GDTT1.8NAS9 или GDTT1.8NAS10 или GDTT1.8NAS11 выполняли:
Н) ферментацию в промышленном масштабе штамма Escherichia coli JM-110-NAS- GDTT1.8NAS1 или Escherichia coli JM-110-NAS- GDTT1.8NAS2 или Escherichia coli JM-110-NAS-GDTT1.8NAS3 или Escherichia coli JM-110-NAS- GDTT1.8NAS4 или Escherichia coli JM-110-NAS- GDTT1.8NAS5 или Escherichia coli JM-110-NAS- GDTT1.8NAS6 или Escherichia coli JM-110-NAS-GDTT1.8NAS7 или Escherichia coli JM-110-NAS- GDTT1.8NAS8 или Escherichia coli JM-110-NAS- GDTT1.8NAS9 или Escherichia coli JM-110-NAS- GDTT1.8NAS10 или Escherichia coli JM-110-NAS- GDTT1.8NAS11 каждый из которых содержит генотерапевтический ДНК-вектор GDTT1.8NAS1 или GDTT1.8NAS2 или GDTT1.8NAS3 или GDTT1.8NAS4 или GDTT1.8NAS5 или GDTT1.8NAS6 или GDTT1.8NAS7 или GDTT1.8NAS8 или GDTT1.8NAS9 или GDTT1.8NAS10 или GDTT1.8NAS11.
Изобретение поясняется следующими примерами.
Пример 1.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS1, содержащего промоторно-регуляторный участок гена миоглобина человека, для повышения экспрессии целевых генов в клетках мышечной ткани.
Генотерапевтический ДНК-вектор GDTT1.8NAS1 конструировали объединением шести фрагментов ДНК, полученных из разных источников:
(а) ориджин репликации получали путем ПЦР-амплификации участка коммерческой плазмиды pUC19 с использованием олигонуклеотидов UCori-Bam, UCori-Nco (перечень последовательностей, (1)-(2));
(б) терминатор транскрипции hGH-TA получали путем ПЦР-амплификации участка геномной ДНК человека с использованием олигонуклеотидов hGH-F и hGH-R (перечень последовательностей, (3) и (4));
(в) регуляторный участок транспозона Tn10 PHK-out получали путем синтеза из олигонуклеотидов RO-F, RO-R, RO-1, RO-2, RO-3 (перечень последовательностей, (5) -(9));
(г) ген устойчивости к канамицину получали путем ПЦР-амплификации участка коммерческой плазмиды рЕТ-28 человека с использованием олигонуклеотидов Kan-F и Kan-R (перечень последовательностей, (10) и (11));
(д) полилинкер получали кинированием и отжигом четырех синтетических олигонуклеотидов MCS1, MCS2, MCS3 и MCS4 (перечень последовательностей, (12) - (15));
(е) промоторно-регуляторный участок гена миоглобина человека получали путем ПЦР-амплификации участка геномной ДНК человека с использованием олигонуклеотидов Myo-F и Myo-R (перечень последовательностей, (16) - (17)).
ПЦР-амплификацию проводили с использованием коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs) в соответствии с инструкцией производителя. Фрагменты (б), (в) и (г) имели перекрывающиеся области для возможности их объединения с последующей ПЦР-амплификацией. Объединяли фрагменты (б), (в) и (г) с использованием олигонуклеотидов hGH-F и Kan-R (список последовательностей, (3) и (11)). Далее, полученные фрагменты ДНК объединялись путем рестрикции с последующим лигированием по сайтам BamHI и NcoI. В результате получали плазмиду, пока еще не содержащую полилинкер. Для его введения проводили расщепление плазмиды эндонуклеазами рестрикции по сайтам BamHI и EcoRI с последующим лигированием с фрагментом (д). В результате получали промежуточный вектор размером 2408 п.н, несущий ген устойчивости к канамицину, пока еще не содержащий промоторно-регуляторный участок гена миоглобина человека. Полученный вектор расщепляли эндонуклеазами рестрикции по сайтам XhoI и BamHI с последующим лигированием с фрагментом (е). В результате получали вектор размером 2753 п.н., несущий ген устойчивости к канамицину и промоторно-регуляторный участок гена миоглобина человека. Далее ген устойчивости к канамицину выщепляли по сайтам рестрикции SpeI, после чего оставшийся фрагмент лигировали сам на себя. Таким образом получали генотерапевтический ДНК-вектор GDTT1.8NAS1 размером 1736 п.н., который является рекомбинантным, с возможностью селекции без антибиотиков и возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в клетках мышечной ткани (SEQ ID №1).
Пример 2.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS2, содержащего промоторно-регуляторный участок гена эластазы человека, для повышения экспрессии целевых генов в клетках кожи.
Генотерапевтический ДНК-вектор GDTT1.8NAS2 конструировали, как описано в примере 1, за исключением того, что фрагмент (е), представляющий собой промоторно-регуляторный участок гена эластазы человека получали путем ПЦР-амплификации участка геномной ДНК человека с использованием олигонуклеотидов Els-F и Els-R (перечень последовательностей, (18)-(19)).
Далее, промежуточный вектор размером 2408 п.н., несущий ген устойчивости к канамицину, пока еще не содержащий промоторно-регуляторный участок гена эластазы человека расщепляли эндонуклеазами рестрикции по сайтам XhoI и BamHI с последующим лигированием с фрагментом (е). В результате получали вектор размером 2671 п.н., несущий ген устойчивости к канамицину и промоторно-регуляторный участок гена эластазы человека. Далее ген устойчивости к канамицину выщепляли по сайтам рестрикции SpeI, после чего оставшийся фрагмент лигировали сам на себя. Таким образом получали генотерапевтический ДНК-вектор GDTT1.8NAS2 размером 1654 п.н., который является рекомбинантным, с возможностью селекции без антибиотиков и возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в клетках кожи (SEQ ID №2).
Пример 3.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS3, содержащего промоторно-регуляторный участок гена молекулы межклеточной адгезии 2 человека, для повышения экспрессии целевых генов в клетках эндотелия сосудов.
Генотерапевтический ДНК-вектор GDTT1.8NAS3 конструировали, как описано в примере 1, за исключением того, что фрагмент (е), представляющий собой промоторно-регуляторный участок гена молекулы межклеточной адгезии 2 человека получали путем ПЦР-амплификации участка геномной ДНК человека с использованием олигонуклеотидов ICAM-F и ICAM-R (перечень последовательностей, (20) - (21)).
Далее, промежуточный вектор размером 2408 п.н., несущий ген устойчивости к канамицину, пока еще не содержащий промоторно-регуляторный участок гена молекулы межклеточной адгезии 2 человека расщепляли эндонуклеазами рестрикции по сайтам XhoI и BamHI с последующим лигированием с фрагментом (е). В результате получали вектор размером 2800 п.н., несущий ген устойчивости к канамицину и промоторно-регуляторный участок гена молекулы межклеточной адгезии 2 человека. Далее ген устойчивости к канамицину выщепляли по сайтам рестрикции SpeI, после чего оставшийся фрагмент лигировали сам на себя. Таким образом получали генотерапевтический ДНК-вектор GDTT1.8NAS3 размером 1783 п.н., который является рекомбинантным, с возможностью селекции без антибиотиков и возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в клетках эндотелия сосудов (SEQ ID №3).
Пример 4.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS4, содержащего промоторно-регуляторный участок гена остеокальцина 2 человека, для повышения экспрессии целевых генов в остеобластах и одонтобластах.
Генотерапевтический ДНК-вектор GDTT1.8NAS4 конструировали, как описано в примере 1, за исключением того, что фрагмент (е), представляющий собой промоторно-регуляторный участок гена остеокальцина 2 человека получали путем ПЦР-амплификации участка геномной ДНК человека с использованием олигонуклеотидов OS2-F и OS2-R (перечень последовательностей, (22) - (23)).
Далее, промежуточный вектор размером 2408 п.н., несущий ген устойчивости к канамицину, пока еще не содержащий промоторно-регуляторный участок гена остеокальцина 2 человека расщепляли эндонуклеазами рестрикции по сайтам XhoI и BamHI с последующим лигированием с фрагментом (е). В результате получали вектор размером 2970 п.н., несущий ген устойчивости к канамицину и промоторно-регуляторный участок гена остеокальцина 2 человека. Далее ген устойчивости к канамицину выщепляли по сайтам рестрикции SpeI, после чего оставшийся фрагмент лигировали сам на себя. Таким образом получали генотерапевтический ДНК-вектор GDTT1.8NAS4 размером 1953 п.н., который является рекомбинантным, с возможностью селекции без антибиотиков и возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в остеобластах и одонтобластах (SEQ ID №4).
Пример 5.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS5, содержащего промоторно-регуляторный участок гена белка сурфактанта В человека, для повышения экспрессии целевых генов в эпителиальных клетках бронхов и альвеол.
Генотерапевтический ДНК-вектор GDTT1.8NAS5 конструировали как описано в примере 1, за исключением того, что фрагмент (е), представляющий собой промоторно-регуляторный участок гена белка сурфактанта В человека получали путем ПЦР-амплификации участка геномной ДНК человека с использованием олигонуклеотидов Spb-F и Spb-R (перечень последовательностей, (24) - (25)).
Далее, промежуточный вектор размером 2408 п.н, несущий ген устойчивости к канамицину, пока еще не содержащий промоторно-регуляторный участок гена белка сурфактанта В человека расщепляли эндонуклеазами рестрикции по сайтам XhoI и BamHI с последующим лигированием с фрагментом (е). В результате получали вектор размером 3036 п. н., несущий ген устойчивости к канамицину и промоторно-регуляторный участок гена белка сурфактанта В человека. Далее ген устойчивости к канамицину выщепляли по сайтам рестрикции SpeI, после чего оставшийся фрагмент лигировали сам на себя. Таким образом получали генотерапевтический ДНК-вектор GDTT1.8NAS5 размером 2019 п.н., который является рекомбинантным, с возможностью селекции без антибиотиков и возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в эпителиальных клетках бронхов и альвеол (SEQ ID №5).
Пример 6.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS6, содержащего промоторно-регуляторный участок гена белка синапсина I человека, для повышения экспрессии целевых генов в нейронах.
Генотерапевтический ДНК-вектор GDTT1.8NAS6 конструировали, как описано в примере 1, за исключением того, что фрагмент (е), представляющий собой промоторно-регуляторный участок гена синапсина I человека получали путем ПЦР-амплификации участка геномной ДНК человека с использованием олигонуклеотидов Syn-F и Syn-R (перечень последовательностей, (26) - (27)).
Далее, промежуточный вектор размером 2408 п.н., несущий ген устойчивости к канамицину, пока еще не содержащий промоторно-регуляторный участок гена синапсина I человека расщепляли эндонуклеазами рестрикции по сайтам XhoI и BamHI с последующим лигированием с фрагментом (е). В результате получали вектор размером 2957 п.н., несущий ген устойчивости к канамицину и промоторно-регуляторный участок гена синапсина I человека. Далее ген устойчивости к канамицину выщепляли по сайтам рестрикции SpeI, после чего оставшийся фрагмент лигировали сам на себя. Таким образом получали генотерапевтический ДНК-вектор GDTT1.8NAS6 размером 1940 п.н., который является рекомбинантным, с возможностью селекции без антибиотиков и возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в нейронах (SEQ ID №6).
Пример 7.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS7, содержащего промоторно-регуляторный участок гена нефрина человека, для повышения экспрессии целевых генов в подоцитах почек.
Генотерапевтический ДНК-вектор GDTT1.8NAS7 конструировали, как описано в примере 1, за исключением того, что фрагмент (е), представляющий собой промоторно-регуляторный участок гена нефрина человека получали путем ПЦР-амплификации участка геномной ДНК человека с использованием олигонуклеотидов NPHS-F и NPHS-R (перечень последовательностей, (28) - (29)).
Далее, промежуточный вектор размером 2408 п.н., несущий ген устойчивости к канамицину, пока еще не содержащий промоторно-регуляторный участок гена нефрина человека расщепляли эндонуклеазами рестрикции по сайтам XhoI и BamHI с последующим лигированием с фрагментом (е). В результате получали вектор размером 3637 п.н., несущий ген устойчивости к канамицину и промоторно-регуляторный участок гена нефрина человека. Далее ген устойчивости к канамицину выщепляли по сайтам рестрикции SpeI, после чего оставшийся фрагмент лигировали сам на себя. Таким образом получали генотерапевтический ДНК-вектор GDTT1.8NAS7 размером 2620 п.н., который является рекомбинантным, с возможностью селекции без антибиотиков и возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в подоцитах почек (SEQ ID №7).
Пример 8.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS8, содержащего промоторно-регуляторный участок гена общего лейкоцитарного антигена CD45 человека, для повышения экспрессии целевых генов в гематопоэтических клетках.
Генотерапевтический ДНК-вектор GDTT1.8NAS8 конструировали, как описано в примере 1, за исключением того, что фрагмент (е), представляющий собой промоторно-регуляторный участок гена общего лейкоцитарного антигена CD45 человека получали путем ПЦР-амплификации участка геномной ДНК человека с использованием олигонуклеотидов cd45-F и cd45-R (перечень последовательностей, (30) - (31)).
Далее, промежуточный вектор размером 2408 п.н., несущий ген устойчивости к канамицину, пока еще не содержащий промоторно-регуляторный участок гена общего лейкоцитарного антигена CD45 человека расщепляли эндонуклеазами рестрикции по сайтам XhoI и BamHI с последующим лигированием с фрагментом (е). В результате получали вектор размером 3257 п.н., несущий ген устойчивости к канамицину и промоторно-регуляторный участок гена общего лейкоцитарного антигена CD45 человека. Далее ген устойчивости к канамицину выщепляли по сайтам рестрикции SpeI, после чего оставшийся фрагмент лигировали сам на себя. Таким образом получали генотерапевтический ДНК-вектор GDTT1.8NAS8 размером 2240 п.н., который является рекомбинантным, с возможностью селекции без антибиотиков и возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в гематопоэтических клетках (SEQ ID №8).
Пример 9.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS9, содержащего промоторно-регуляторный участок гена белка В29 человека, для повышения экспрессии целевых генов в лимфоцитах.
Генотерапевтический ДНК-вектор GDTT1.8NAS9 конструировали, как описано в примере 1, за исключением того, что фрагмент (е), представляющий собой промоторно-регуляторный участок гена белка В29 человека получали путем ПЦР-амплификации участка геномной ДНК человека с использованием олигонуклеотидов b29-F и b29-R (перечень последовательностей, (32) - (33)).
Далее, промежуточный вектор размером 2408 п.н., несущий ген устойчивости к канамицину, пока еще не содержащий промоторно-регуляторный участок гена белка В29 человека расщепляли эндонуклеазами рестрикции по сайтам XhoI и BamHI с последующим лигированием с фрагментом (е). В результате получали вектор размером 3621 п.н., несущий ген устойчивости к канамицину и промоторно-регуляторный участок гена белка В29 человека. Далее ген устойчивости к канамицину выщепляли по сайтам рестрикции SpeI, после чего оставшийся фрагмент лигировали сам на себя. Таким образом получали генотерапевтический ДНК-вектор GDTT1.8NAS9 размером 2604 п.н., который является рекомбинантным, с возможностью селекции без антибиотиков и возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в лимфоцитах (SEQ ID №9).
Пример 10.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS10, содержащего промоторно-регуляторный участок гена белка CD68 человека, для повышения экспрессии целевых генов в макрофагах.
Генотерапевтический ДНК-вектор GDTT1.8NAS10 конструировали, как описано в примере 1, за исключением того, что фрагмент (е), представляющий собой промоторно-регуляторный участок гена белка CD68 человека получали путем ПЦР-амплификации участка геномной ДНК человека с использованием олигонуклеотидов cd68-F и cd68-R (перечень последовательностей, (34) - (35)).
Далее, промежуточный вектор размером 2408 п.н., несущий ген устойчивости к канамицину, пока еще не содержащий промоторно-регуляторный участок гена CD68 человека расщепляли эндонуклеазами рестрикции по сайтам XhoI и BgIII с последующим лигированием с фрагментом (е). В результате получали вектор размером 3065 п.н., несущий ген устойчивости к канамицину и промоторно-регуляторный участок гена CD68 человека. Далее ген устойчивости к канамицину выщепляли по сайтам рестрикции SpeI, после чего оставшийся фрагмент лигировали сам на себя. Таким образом получали генотерапевтический ДНК-вектор GDTT1.8NAS10 размером 2048 п.н., который является рекомбинантным, с возможностью селекции без антибиотиков и возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в макрофагах (SEQ ID №10).
Пример 11.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS11, содержащего промоторно-регуляторный участок гена инсулина человека, для повышения экспрессии целевых генов в бета-клетках поджелудочной железы.
Генотерапевтический ДНК-вектор GDTT1.8NAS11 конструировали, как описано в примере 1, за исключением того, что фрагмент (е), представляющий собой промоторно-регуляторный участок гена инсулина человека получали путем ПЦР-амплификации участка геномной ДНК человека с использованием олигонуклеотидов Ins-F и Ins-R (перечень последовательностей, (36) - (37)).
Далее, промежуточный вектор размером 2408 п.н., несущий ген устойчивости к канамицину, пока еще не содержащий промоторно-регуляторный участок гена инсулина человека расщепляли эндонуклеазами рестрикции по сайтам XhoI и BamHI с последующим лигированием с фрагментом (е). В результате получали вектор размером 2995 п.н., несущий ген устойчивости к канамицину и промоторно-регуляторный участок гена инсулина человека. Далее ген устойчивости к канамицину выщепляли по сайтам рестрикции SpeI, после чего оставшийся фрагмент лигировали сам на себя. Таким образом получали генотерапевтический ДНК-вектор GDTT1.8NAS11 размером 1978 п.н., который является рекомбинантным, с возможностью селекции без антибиотиков и возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в макрофагах (SEQ ID №11).
Пример 12.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS1 проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена и уровне целевого белка. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS1.
Для подтверждения функциональной активности ДНК-вектора GDTT1.8NAS1 и реализуемости способа его использования клонировали в полилинкер целевой ген, например, ген VEGF, приведенный в базе данных GenBank под номером NM_001025368. Получение генотерапевтического ДНК-вектора GDTT1.8NAS1-VEGF, несущего участок, кодирующий целевой ген, например, ген, VEGF. Кодирующую часть гена VEGF получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов VEGF165_F и VEGF165_R (перечень последовательностей, (38) и (39)). Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции BamHI и EcoRI и лигировали с ДНК-вектором размером 2753 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS1-VEGF размером 2269 п.н. с возможностью селекции без антибиотиков.
Оценивали изменения накопления мРНК целевого гена VEGF, в клетках первичной культуры скелетных миобластов человека HSkM (ThermoFisher Scientific # A12555) до и через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS1-VEGF, несущим ген VEGF человека, а также сравнивали количественное содержание целевого белка VEGF после трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS1-VEGF, несущим ген VEGF человека и ДНК-вектором GDTT1.8NAS1, не несущим ген VEGF человека.
Клеточную культуру HSkM выращивали в стандартных условиях (37°С, 5% CO2) с использованием питательной среды DMEM. В процессе культивирования каждые 48 ч происходила смена ростовой среды.
Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS1-VEGF, экспрессирующим ген VEGF человека, проводили с использованием Lipofectamine 3000 (ThermoFisher Scientific, США) согласно рекомендациям производителя. В пробирке №1 к 25 мкл среды Opti-MEM (Gibco, США) добавляли 1 мкл раствора ДНК-вектора GDTT1.8NAS1-VEGF (концентрация 500 нг/мкл) и 1 мкл реагента Р3000. Аккуратно перемешивали легким встряхиванием. В пробирке №2 к 25 мкл среды Opti-MEM (Gibco, США) добавляли 1 мкл раствора Lipofectamin 3000. Аккуратно перемешивали легким встряхиванием. Добавляли содержимое пробирки №1 к содержимому пробирки №2, инкубировали 5 мин при комнатной температуре. Полученный раствор по каплям добавляли к клеткам в объеме 40 мкл.
В качестве контроля использовали клетки HSkM, трансфицированные генотерапевтическим ДНК-вектором GDTT1.8NAS1, не содержащим вставку целевого гена VEGF (кДНК гена VEGF до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS1, не содержащим вставку целевого гена на фигурах не показано). Подготовку контрольного вектора GDTT1.8NAS1 для трансфекции проводили, как описано выше.
Суммарную РНК из клеток HSkM выделяли с использованием Trizol Reagent (Invitrogen, США) согласно рекомендациям производителя. В лунку с клетками добавляли 1 мл Trizol Reagent и гомогенизировали с последующим прогреванием в течение 5 мин при 65°С. Далее образец центрифугировали при 14000 g в течение 10 мин и снова прогревали в течение 10 мин при 65°С. Далее добавляли 200 мкл хлороформа, плавно перемешивали и центрифугировали при 14000 g в течение 10 мин. Затем отбирали водную фазу, добавляли к ней 1/10 объема 3М ацетата натрия, рН 5.2 и равный объем изопропилового спирта. Инкубировали образец при -20°С в течение 10 мин с последующим центрифугированием при 14000 g в течение 10 мин. Осадок промывали 1 мл 70% этилового спирта, высушивали на воздухе и растворяли в 10 мкл воды, свободной от РНКаз. Определение уровня экспрессии мРНК гена VEGF после трансфекции проводили путем оценки динамики накопления ампликонов кДНК методом ПЦР в режиме реального времени. Для получения и амплификации кДНК, специфичной для гена VEGF человека, использовали олигонуклеотиды VEGF165_SF и VEGF165_SК (перечень последовательностей (40) и (41)). Длина продукта амплификации - 576 п.н.
Реакцию обратной транскрипции и ПЦР-амплификацию проводили с помощью набора реагентов Quantitect SYBR Green RT-PCR Kit (Qiagen, США) для ПЦР в режиме реального времени. Реакцию проводили в объеме 20 мкл, содержащих: 25 мкл QuantiTect SYBR Green RT-PCR MasterMix, 2,5 мМ хлорида магния, по 0,5 мкМ каждого праймера, 5 мкл РНК. Реакцию осуществляли на амплификаторе CFX96 (Bio-Rad, США) при следующих условиях: 1 цикл обратной транскрипции при 42°С - 30 минут, денатурация 98°С - 15 мин, затем 40 циклов, включающих денатурацию 94°С - 15 сек, отжиг праймеров 60°С - 30 сек и элонгацию 72°С - 30 сек. В качестве референтного гена использовали ген В2М (Бета-2-микроглобулин человека) приведенный в базе данных GenBank под номером NM 004048.2. В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов VEGF и В2М. В качестве отрицательного контроля использовали деионизированную воду. Количество динамику накопления ампликонов кДНК генов VEGF и В2М оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа, представлены на фиг. 13.
Количественное определение белка VEGF проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор ELISA Kit for Vascular Endothelial Growth Factor A (VEGFA), Cloud-Clone Corp., USA, согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка VEGF, входящим в состав набора. Чувствительность метода составляла не менее 6,2 пг/мл, диапазон измерения - от 15,6 пг/мл до 1000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графические материалы, полученные в результате анализа, представлены на фиг. 14.
Из фигуры 13 следует, что в результате трансфекции первичной культуры скелетных миобластов человека HSkM генотерапевтическим ДНК-вектором GDTT1.8NAS1-VEGF, уровень специфической мРНК гена VEGF человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген VEGF на уровне мРНК. Из фигуры 14 следует, что трансфекция клеток первичной культуры скелетных миобластов человека HSkM генотерапевтическим ДНК-вектором GDTT1.8NAS1- VEGF приводит к увеличению количества белка VEGF по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген VEGF на уровне белка.
Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS1-VEGF для повышения уровня экспрессии гена VEGF в эукариотических клетках.
Пример 13.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS2 проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена и уровне целевого белка. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS2.
Для подтверждения функциональной активности ДНК-вектора GDTT1.8NAS2 и реализуемости способа его использования клонировали в полилинкер целевой ген, например, ген CAT, приведенный в базе данных GenBank под номером NM_001752 Получение генотерапевтического ДНК-вектора GDTT1.8NAS2-CAT, несущего участок, кодирующий целевой ген. например, ген, CAT. Кодирующую часть гена CAT получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов CAT_F и CAT_R (перечень последовательностей, (42) и (43)). Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции EcoRV и XbaI и лигировали с ДНК-вектором размером 2671 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS2-CAT размером 3237 п.н. с возможностью селекции без антибиотиков.
Оценивали изменения накопления мРНК целевого гена CAT, в клетках первичной культуры фибробластов кожи человека HDFa (АТСС® PCS-201-012™) до и через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS2-CAT, несущим ген CAT человека, а также сравнивали количественное содержание целевого белка CAT после трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS2-CAT, несущим ген CAT человека и ДНК-вектором GDTT1.8NAS2, не несущим ген CAT человека.
Клеточную культуру HDFa выращивали в стандартных условиях (37°С, 5% CO2) с использованием питательной среды Fibroblast Growth Kit-Serum-Free (ATCC® PCS-201-040). В процессе культивирования каждые 48 ч происходила смена ростовой среды.
Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS2-CAT, экспрессирующим ген CAT человека, проводили с использованием Lipofectamine 3000 (ThermoFisher Scientific, США) как описано в примере 12. В качестве контроля использовали клетки HDFa трансфицированные генотерапевтическим ДНК-вектором GDTT1.8NAS2, не содержащим вставку целевого гена CAT (кДНК гена CAT до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS2, не содержащим вставку целевого гена на фигурах не показано). Подготовку контрольного вектора GDTT1.8NAS2 для трансфекции проводили, как описано выше.
Суммарную РНК из клеток HDFa выделяли с использованием Trizol Reagent (Invitrogen, США) как описано в примере 12. Определение уровня экспрессии мРНК гена CAT после трансфекции проводили путем оценки динамики накопления ампликонов кДНК методом ПЦР в режиме реального времени, как описано в примере 12. Для получения и амплификации кДНК, специфичной для гена CAT человека, использовали олигонуклеотиды CAT_SF и CAT_SR (перечень последовательностей (44) и (45)). Длина продукта амплификации - 272 п.н.
Графики, полученные в результате анализа, представлены на фиг. 15.
Количественное определение белка CAT проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор ELISA Kit for Catalase (CAT) Cloud-Clone Corp.согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка CAT, входящим в состав набора. Чувствительность метода составляла 0.124 нг/мл, диапазон измерения - от 0,312 нг/мл до 20 нг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графические материалы, полученные в результате анализа, представлены на фиг. 16.
Из фигуры 15 следует, что в результате трансфекции первичной культуры фибробластов кожи человека HDFa генотерапевтическим ДНК-вектором GDTT1.8NAS2- CAT, уровень специфической мРНК гена CAT человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген CAT на уровне мРНК. Из фигуры 16 следует, что трансфекция клеток первичной культуры фибробластов кожи человека HDFa генотерапевтическим ДНК-вектором GDTT1.8NAS2-CAT приводит к увеличению количества белка CAT по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген CAT на уровне белка.
Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS2-CAT для повышения уровня экспрессии гена CAT в эукариотических клетках.
Пример 14.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS3 проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена и уровне целевого белка. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS3.
Для подтверждения функциональной активности ДНК-вектора GDTT1.8NAS3 и реализуемости способа его использования клонировали в полилинкер целевой ген, например, ген HIF1a, приведенный в базе данных GenBank под номером NM_001530. Получение генотерапевтического ДНК-вектора GDTT1.8NA3-HIF1a, несущего участок, кодирующий целевой ген, например, ген, HIF1a. Кодирующую часть гена HIF1a получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов HIF1a_F и HIF1a_R (перечень последовательностей, (46) и (47)). Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции BamHI и XbaI и лигировали с ДНК-вектором размером 2800 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS3-HIF1a размером 4257 п.н. с возможностью селекции без антибиотиков.
Оценивали изменения накопления мРНК целевого гена HIF1a, в клетках эндотелия пупочной вены человека HUVEC (ATCC® PCS-100-013™) до и через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS3-HIF1a, несущим ген HIF1a человека, а также сравнивали количественное содержание целевого белка HIF1a после трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS3-HIF1a, несущим ген HIF1a человека и ДНК-вектором GDTT1.8NAS3, не несущим ген HIF1a человека.
Клеточную культуру HUVEC выращивали в стандартных условиях (37°С, 5% CO2) с использованием питательной среды Endothelial Cell Growth Kit-BBE (ATCC® PCS-100-040). В процессе культивирования каждые 48 ч происходила смена ростовой среды.
Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS3-HIF1a, экспрессирующим ген HIF1a человека, проводили с использованием Lipofectamine 3000 (ThermoFisher Scientific, США) как описано в примере 12.
В качестве контроля использовали клетки HUVEC, трансфицированные генотерапевтическим ДНК-вектором GDTT1.8NAS3, не содержащим вставку целевого гена HIF1a (кДНК гена HIF1a до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS3, не содержащим вставку целевого гена на фигурах не показано). Подготовку контрольного вектора GDTT1.8NAS3 для трансфекции проводили, как описано выше.
Суммарную РНК из клеток HUVEC выделяли с использованием Trizol Reagent (Invitrogen, США) как описано в примере 12. Определение уровня экспрессии мРНК гена HIF1a после трансфекции проводили путем оценки динамики накопления ампликонов кДНК методом ПЦР в режиме реального времени как описано в примере 12. Для получения и амплификации кДНК, специфичной для гена HIF1a человека, использовали олигонуклеотиды HIF1a _SF и HIF1a _SR (перечень последовательностей, (48) и (49)). Длина продукта амплификации - 379 п.н.
Графики, полученные в результате анализа, представлены на фиг. 17.
Количественное определение белка HIF1a проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор ELISA Kit for Hypoxia Inducible Factor 1 Alpha (HIF1a), Cloud-Clone Corp., USA, согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка HIF1a, входящим в состав набора. Чувствительность метода составляла не менее 0,054 нг/мл, диапазон измерения - от 0,156 нг/мл до 10 нг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графические материалы, полученные в результате анализа, представлены на фиг. 18.
Из фигуры 17 следует, что в результате трансфекции культуры эндотелия пупочной вены человека HUVEC генотерапевтическим ДНК-вектором GDTT1.8NAS3-HIF1a, уровень специфической мРНК гена HIF1a человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген HIF1a на уровне мРНК. Из фигуры 18 следует, что трансфекция клеток культуры эндотелия пупочной вены человека HUVEC генотерапевтическим ДНК-вектором GDTT1.8NAS3-HIF1a приводит к увеличению количества белка HIF1a по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген HIF1a на уровне белка.
Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS3-HIF1a для повышения уровня экспрессии гена HIF1a в эукариотических клетках.
Пример 15.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS4 проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена и уровне целевого белка. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS4.
Для подтверждения функциональной активности ДНК-вектора GDTT1.8NAS4 и реализуемости способа его использования клонировали в полилинкер целевой ген, например, ген ВМР2, приведенный в базе данных GenBank под номером NM_001200. Получение генотерапевтического ДНК-вектора GDTT1.8NAS4-ВМР2, несущего участок, кодирующий целевой ген, например, ген, ВМР2. Кодирующую часть гена ВМР2 получали выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов BMP2_F и BMP2_R (перечень последовательностей, (50) и (51)). Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции BamHI и EcoRI и лигировали с ДНК-вектором размером 2970 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS4-BMP2 размером 3104 п.н. с возможностью селекции без антибиотиков.
Оценивали изменения накопления мРНК целевого гена ВМР2, в клетках остеосаркомы человека MG-63 (ATCC® CRL-1427™) до и через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS4-BMP2, несущим ген ВМР2 человека, а также сравнивали количественное содержание целевого белка ВМР2 после трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS4-BMP2, несущим ген ВМР2 человека и ДНК-вектором GDTT1.8NAS4, не несущим ген ВМР2 человека.
Клеточную культуру MG-63 выращивали в стандартных условиях (37°С, 5% CO2) с использованием питательной среды DMEM (Gibco) с добавлением 10% сыворотки эмбрионов коров (Gibco). В процессе культивирования каждые 48 ч происходила смена ростовой среды.
Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS4-BMP2, экспрессирующим ген ВМР2 человека, проводили с использованием Lipofectamine 3000 (ThermoFisher Scientific, США) как описано в примере 12.
В качестве контроля использовали клетки MG-63, трансфицированные генотерапевтическим ДНК-вектором GDTT1.8NAS4, не содержащим вставку целевого гена ВМР2 (кДНК гена ВМР2 до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS4, не содержащим вставку целевого гена на фигурах не показано). Подготовку контрольного вектора GDTT1.8NAS4 для трансфекции проводили, как описано выше.
Суммарную РНК из клеток MG-63 выделяли с использованием Trizol Reagent (Invitrogen, США) как описано в примере 12. Определение уровня экспрессии мРНК гена ВМР2 после трансфекции проводили путем оценки динамики накопления ампликонов кДНК методом ПЦР в режиме реального времени как описано в примере 12. Для получения и амплификации кДНК, специфичной для гена ВМР2 человека, использовали олигонуклеотиды ВМР2 _SF и ВМР2 _SR (перечень последовательностей, (52) и (53)). Длина продукта амплификации - 353 п.н.
Графики, полученные в результате анализа, представлены на фиг. 19.
Количественное определение белка ВМР2 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор ELISA Kit for Bone Morphogenetic Protein 2 (ВМР2), Cloud-Clone Corp.USA, согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка ВМР2, входящим в состав набора. Чувствительность метода составляла не менее 6,1 пг/мл, диапазон измерения - от 15,6 пг/мл до 1000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графические материалы, полученные в результате анализа, представлены на фиг.20.
Из фигуры 19 следует, что в результате трансфекции клеточной культуры остеосаркомы человека MG-63 генотерапевтическим ДНК-вектором GDTT1.8NAS4-BMP2, уровень специфической мРНК гена ВМР2 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген ВМР2 на уровне мРНК. Из фигуры 20 следует, что трансфекция клеток клеточной культуры остеосаркомы человека MG-63 генотерапевтическим ДНК-вектором GDTT1.8NAS4- ВМР2 приводит к увеличению количества белка ВМР2 по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген ВМР2 на уровне белка.
Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS4-BMP2 для повышения уровня экспрессии гена ВМР2 в эукариотических клетках.
Пример 16.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS5 проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена и уровне целевого белка. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS5.
Для подтверждения функциональной активности ДНК-вектора GDTT1.8NAS5 и реализуемости способа его использования клонировали в полилинкер целевой ген, например, ген CFTR, приведенный в базе данных GenBank под номером NM_000492. Получение генотерапевтического ДНК-вектора GDTT1.8NAS5-CFTR, несущего участок, кодирующий целевой ген, например, ген, CFTR. Кодирующую часть гена CFTR получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов CFTR_F и CFTR_R (перечень последовательностей, (54) и (55)). Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции SalI и NotI и лигировали с ДНК-вектором размером 3036 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS5-CFTR размером 6448 п.н. с возможностью селекции без антибиотиков.
Оценивали изменения накопления мРНК целевого гена CFTR, в клетках трахеального эпителия человека CFTE29o- (коллекция Института Цитологии РАН) до и через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NA5-CFTR, несущим ген CFTR человека, а также сравнивали количественное содержание целевого белка CFTR после трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS5- CFTR, несущим ген CFTR человека и ДНК-вектором GDTT1.8NAS5, не несущим ген CFTR человека.
Клеточную культуру CFTE29o- выращивали в стандартных условиях (37°С, 5% CO2) с использованием питательной среды DMEM (Gibco) с добавлением 10% сыворотки эмбрионов коров (Gibco). В процессе культивирования каждые 48 ч происходила смена ростовой среды.
Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS5-CFTR, экспрессирующим ген CFTR человека, проводили с использованием Lipofectamine 3000 (ThermoFisher Scientific, США) как описано в примере 12.
В качестве контроля использовали клетки CFTE29o-, трансфицированные генотерапевтическим ДНК-вектором GDTT1.8NAS5, не содержащим вставку целевого гена CFTR (кДНК гена CFTR до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS5, не содержащим вставку целевого гена на фигурах не показано). Подготовку контрольного вектора GDTT1.8NAS5 для трансфекции проводили, как описано выше.
Суммарную РНК из клеток CFTE29o- выделяли с использованием Trizol Reagent (Invitrogen, США) как описано в примере 12. Определение уровня экспрессии мРНК гена CFTR после трансфекции проводили путем оценки динамики накопления ампликонов кДНК методом ПЦР в режиме реального времени как описано в примере 12. Для получения и амплификации кДНК, специфичной для гена CFTR человека, использовали олигонуклеотиды CFTR _SF и CFTR _SR (перечень последовательностей, (56) и (57)). Длина продукта амплификации - 328 п.н.
Графики, полученные в результате анализа, представлены на фиг. 21.
Количественное определение белка CFTR проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор ELISA Kit for Cystic Fibrosis Transmembrane Conductance Regulator (CFTR), Cloud-Clone Corp, USA, согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка CFTR, входящим в состав набора. Чувствительность метода составляла 0.059 нг/мл, диапазон измерения - 0.156-10 нг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графические материалы, полученные в результате анализа, представлены на фиг. 22.
Из фигуры 21 следует, что в результате трансфекции клеточной культуры трахеального эпителия человека CFTE29o-генотерапевтическим ДНК-вектором GDTT1.8NAS5-CFTR, уровень специфической мРНК гена CFTR человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген CFTR на уровне мРНК. Из фигуры 22 следует, что трансфекция клеток клеточной культуры трахеального эпителия человека CFTE29o- генотерапевтическим ДНК-вектором GDTT1.8NAS5-CFTR приводит к увеличению количества белка CFTR по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген CFTR на уровне белка.
Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS5-CFTR для повышения уровня экспрессии гена CFTR в эукариотических клетках.
Пример 17.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS6 проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена и уровне целевого белка. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS6.
Для подтверждения функциональной активности ДНК-вектора GDTT1.8NAS6 и реализуемости способа его использования клонировали в полилинкер целевой ген, например, ген BDNF, приведенный в базе данных GenBank под номером NM_170735. Получение генотерапевтического ДНК-вектора GDTT1.8NAS6-BDNF, несущего участок, кодирующий целевой ген, например, ген, BDNF. Кодирующую часть гена BDNF получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов BDNF_F и BDNF_R (перечень последовательностей, (58) и (59)). Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции BamHI и EcoRI и лигировали с ДНК-вектором размером 2957 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS6-BDNF размером 2637 п.н. с возможностью селекции без антибиотиков.
Оценивали изменения накопления мРНК целевого гена BDNF, в клеточной культуре нейробластомы человека SH-SY5Y (АТСС® CRL-2266™) до и через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS6-BDNF, несущим ген BDNF человека, а также сравнивали количественное содержание целевого белка BDNF после трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS6-BDNF, несущим ген BDNF человека и ДНК-вектором GDTT1.8NAS6, не несущим ген BDNF человека.
Клеточную культуру SH-SY5Y выращивали в стандартных условиях (37°С, 5% CO2) с использованием смеси питательных сред (1:1) ATCC-formulated Eagle's Minimum Essential Medium (АТСС) и F12 Medium (Gibco). В процессе культивирования каждые 48 ч происходила смена ростовой среды.
Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS6-BDNF, экспрессирующим ген BDNF человека, проводили с использованием Lipofectamine 3000 (ThermoFisher Scientific, США) как описано в примере 12. В качестве контроля использовали клетки SH-SY5Y, трансфицированные генотерапевтическим ДНК-вектором GDTT1.8NAS6, не содержащим вставку целевого гена BDNF (кДНК гена BDNF до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS6, не содержащим вставку целевого гена на фигурах не показано). Подготовку контрольного вектора GDTT1.8NAS6 для трансфекции проводили, как описано выше.
Суммарную РНК из клеток SH-SY5Y выделяли с использованием Trizol Reagent (Invitrogen, США) как описано в примере 12. Определение уровня экспрессии мРНК гена BDNF после трансфекции проводили путем оценки динамики накопления ампликонов кДНК методом ПЦР в режиме реального времени как описано в примере 12. Для получения и амплификации кДНК, специфичной для гена BDNF человека, использовали олигонуклеотиды BDNF_SF и BDNF_SR (перечень последовательностей, (60) и (61)). Длина продукта амплификации - 199 п.н.
Графики, полученные в результате анализа, представлены на фиг. 23.
Количественное определение белка BDNF проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор ELISA Kit for Brain Derived Neurotrophic Factor (BDNF), Cloud-Clone Corp, USA, согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка BDNF, входящим в состав набора. Чувствительность метода составляла не менее 12,6 пг/мл, диапазон измерения - от 31,2 пг/мл до 2000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графические материалы, полученные в результате анализа, представлены на фиг. 24.
Из фигуры 23 следует, что в результате трансфекции клеточной культуры нейробластомы человека SH-SY5Y генотерапевтическим ДНК-вектором GDTT1.8NAS6-BDNF, уровень специфической мРНК гена BDNF человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген BDNF на уровне мРНК. Из фигуры 24 следует, что трансфекция клеток клеточной культуры нейробластомы человека SH-SY5Y генотерапевтическим ДНК-вектором GDTT1.8NAS6- BDNF приводит к увеличению количества белка BDNF по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген BDNF на уровне белка.
Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS6-BDNF для повышения уровня экспрессии гена BDNF в эукариотических клетках.
Пример 18.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS7 проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена и уровне целевого белка. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS7.
Для подтверждения функциональной активности ДНК-вектора GDTT1.8NAS7 и реализуемости способа его использования клонировали в полилинкер целевой ген, например, ген ATGL, приведенный в базе данных GenBank под номером NM_020376. Получение генотерапевтического ДНК-вектора GDTT1.8NAS7-ATGL, несущего участок, кодирующий целевой ген, например, ген, ATGL. Кодирующую часть гена ATGL получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов ATGL_F и ATGL_R (перечень последовательностей, (62) и (63)). Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции BamHI и EcoRI и лигировали с ДНК-вектором размером 3637 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS7-ATGL размером 4088 п.н. с возможностью селекции без антибиотиков.
Оценивали изменения накопления мРНК целевого гена ATGL, в клетках первичной смешанной культуры эпителия почки человека HREC (АТСС® PCS-400-012™) до и через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS7-ATGL, несущим ген ATGL человека, а также сравнивали количественное содержание целевого белка ATGL после трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS7- ATGL, несущим ген ATGL человека и ДНК-вектором GDTT1.8NAS7, не несущим ген ATGL человека.
Клеточную культуру HREC выращивали в стандартных условиях (37°С, 5% CO2) с использованием набора реагентов Renal Epithelial Cell Growth Kit (АТСС). В процессе культивирования каждые 48 ч происходила смена ростовой среды.
Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS7-ATGL, экспрессирующим ген ATGL человека, проводили с использованием Lipofectamine 3000 (ThermoFisher Scientific, США) как описано в примере 12.
В качестве контроля использовали клетки HREC, трансфицированные генотерапевтическим ДНК-вектором GDTT1.8NAS7, не содержащим вставку целевого гена ATGL (кДНК гена ATGL до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS7, не содержащим вставку целевого гена на фигурах не показано). Подготовку контрольного вектора GDTT1.8NAS7 для трансфекции проводили, как описано выше.
Суммарную РНК из клеток HREC выделяли с использованием Trizol Reagent (Invitrogen, США) как описано в примере 12. Определение уровня экспрессии мРНК гена ATGL после трансфекции проводили путем оценки динамики накопления ампликонов кДНК методом ПЦР в режиме реального времени как описано в примере 12. Для получения и амплификации кДНК, специфичной для гена ATGL человека, использовали олигонуклеотиды ATGL_SF и ATGL_SR (перечень последовательностей, (64) и (65)). Длина продукта амплификации - 417 п.н.
Графики, полученные в результате анализа, представлены на фиг. 25.
Количественное определение белка ATGL проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор ELISA Kit for Patatin Like Phospholipase Domain Containing Protein 2 (PNPLA2), Cloud-Clone Corp, USA, согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка ATGL, входящим в состав набора. Чувствительность метода составляла не менее 0,128 нг/мл, диапазон измерения - от 0,312 нг/мл до 20 нг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графические материалы, полученные в результате анализа, представлены на фиг. 26.
Из фигуры 25 следует, что в результате трансфекции первичной смешанной культуры эпителия почки человека HREC генотерапевтическим ДНК-вектором GDTT1.8NAS7-ATGL, уровень специфической мРНК гена ATGL человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген ATGL на уровне мРНК. Из фигуры 26 следует, что трансфекция клеток первичной смешанной культуры эпителия почки человека HREC генотерапевтическим ДНК-вектором GDTT1.8NAS7-ATGL приводит к увеличению количества белка ATGL по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген ATGL на уровне белка.
Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS7-ATGL для повышения уровня экспрессии гена ATGL в эукариотических клетках.
Пример 19.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS8 проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена и уровне целевого белка. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS8.
Для подтверждения функциональной активности ДНК-вектора GDTT1.8NAS8 и реализуемости способа его использования клонировали в полилинкер целевой ген, например, ген CAS9, приведенный в базе данных GenBank под номером EZM83080.1.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS8-CAS9, несущего участок, кодирующий целевой ген, например, ген, CAS9. Кодирующую часть гена CAS9 получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов CAS9_F и CAS9_R (перечень последовательностей, (66) и (67)). Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции BamHI и EcoRI и лигировали с ДНК-вектором размером 3257 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS8-CAS9 размером 6461 п.н. с возможностью селекции без антибиотиков.
Оценивали изменения накопления мРНК целевого гена CAS9, в клетках первичной культуры костного мозга человека (АТСС® PCS-800-012™) до и через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS8-CAS9, несущим ген CAS9 человека, а также сравнивали количественное содержание целевого белка CAS9 после трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS8-CAS9, несущим ген CAS9 человека и ДНК-вектором GDTT1.8NAS8, не несущим ген CAS9 человека.
Первичную клеточную культуру клеток CD34+ костного мозга человека (АТСС® PCS-800-012™) за 24 часа до постановки трансфекции размораживали и переносили в 24-луночный планшет в растворе Хенкса с добавлением 10% сыворотки эмбрионов коров (Gibco) из расчета 5×104 клеток/лунку. Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS8-CAS9, экспрессирующим ген CAS9 человека, проводили с использованием Lipofectamine 3000 (ThermoFisher Scientific, США) как описано в примере 12.
В качестве контроля использовали клетки костного мозга человека, трансфицированные генотерапевтическим ДНК-вектором GDTT1.8NAS8, не содержащим вставку целевого гена CAS9 (кДНК гена CAS9 до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS8, не содержащим вставку целевого гена на фигурах не показано). Подготовку контрольного вектора GDTT1.8NAS8 для трансфекции проводили, как описано выше.
Суммарную РНК из клеток костного мозга человека выделяли с использованием Trizol Reagent (Invitrogen, США) как описано в примере 12. Определение уровня экспрессии мРНК гена CAS9 после трансфекции проводили путем оценки динамики накопления ампликонов кДНК методом ПЦР в режиме реального времени как описано в примере 12. Для получения и амплификации кДНК, специфичной для гена CAS9 человека, использовали олигонуклеотиды CAS9_SF и CAS9_SR (перечень последовательностей, (68) и (69)).
Длина продукта амплификации - 275 п.н.
Графики, полученные в результате анализа, представлены на фиг. 27.
Количественное определение белка CAS9 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Cas9 (CRISPR Associated Protein 9) ELISA Kit, Cell Biolabs Inc, согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка CAS9, входящим в состав набора. Чувствительность метода составляла не менее 1,5 нг/мл, диапазон измерения - от 1.56 нг/мл до 100 нг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графические материалы, полученные в результате анализа, представлены на фиг. 28.
Из фигуры 27 следует, что в результате трансфекции первичной культуры костного мозга человека генотерапевтическим ДНК-вектором GDTT1.8NAS8-CAS9, уровень специфической мРНК гена CAS9 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген CAS9 на уровне мРНК. Из фигуры 28 следует, что трансфекция клеток первичной культуры костного мозга человека генотерапевтическим ДНК-вектором GDTT1.8NAS8-CAS9 приводит к увеличению количества белка CAS9 по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген CAS9 на уровне белка.
Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS8-CAS9 для повышения уровня экспрессии гена CAS9 в эукариотических клетках.
Пример 20.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS9 проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена и уровне целевого белка. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS9.
Для подтверждения функциональной активности ДНК-вектора GDTT1.8NAS9 и реализуемости способа его использования клонировали в полилинкер целевой ген, например, ген BTK, приведенный в базе данных GenBank под номером NM_000061. Получение генотерапевтического ДНК-вектора GDTT1.8NAS9-BTK, несущего участок, кодирующий целевой ген, например, ген, BTK. Кодирующую часть гена BTK получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов BTK_F и BTK_R (перечень последовательностей, (70) и (71)). Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции BamHI и EcoRI и лигировали с ДНК-вектором размером 3621 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS9-BTK размером 2637 п.н. с возможностью селекции без антибиотиков.
Оценивали изменения накопления мРНК целевого гена BTK, в клетках первичной культуры периферических мононуклеаров крови человека РВМС (АТСС® PCS-800-011 ™) до и через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS9-BTK, несущим ген BTK человека, а также сравнивали количественное содержание целевого белка BTK после трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS9-BTK, несущим ген BTK человека и ДНК-вектором GDTT1.8NAS9, не несущим ген BTK человека.
Клеточную культуру РВМС за 24 часа до постановки трансфекции размораживали и переносили в 24-луночный планшет в растворе Хенкса с добавлением 10% сыворотки эмбрионов коров (Gibco) из расчета 5×104 клеток/лунку. Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS9-BTK, экспрессирующим ген BTK человека, проводили с использованием Lipofectamine 3000 (ThermoFisher Scientific, США) как описано в примере 12. В качестве контроля использовали клетки РВМС, трансфицированные генотерапевтическим ДНК-вектором GDTT1.8NAS9, не содержащим вставку целевого гена BTK (кДНК гена BTK до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS9, не содержащим вставку целевого гена на фигурах не показано). Подготовку контрольного вектора GDTT1.8NAS9 для трансфекции проводили, как описано выше.
Суммарную РНК из клеток РВМС выделяли с использованием Trizol Reagent (Invitrogen, США) как описано в примере 12. Определение уровня экспрессии мРНК гена BTK после трансфекции проводили путем оценки динамики накопления ампликонов кДНК методом ПЦР в режиме реального времени как описано в примере 12. Для получения и амплификации кДНК, специфичной для гена BTK человека, использовали олигонуклеотиды BTK_SF и BTK_SR (перечень последовательностей, (72) и (73)). Длина продукта амплификации - 579 п.н.
Графики, полученные в результате анализа, представлены на фиг. 29.
Количественное определение белка BTK проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор ELISA Kit for Bruton'S Tyrosine Kinase (Btk), Cloud-Clone Corp, USA, согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка BTK, входящим в состав набора. Чувствительность метода составляла не менее 0,055 нг/мл, диапазон измерения - от 0,156 нг/мл до 10 нг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графические материалы, полученные в результате анализа, представлены на фиг. 30.
Из фигуры 29 следует, что в результате трансфекции первичной культуры периферических мононуклеаров крови человека РВМС генотерапевтическим ДНК-вектором GDTT1.8NAS9-BTK, уровень специфической мРНК гена BTK человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген BTK на уровне мРНК. Из фигуры 30 следует, что трансфекция первичной культуры периферических мононуклеаров крови человека РВМС генотерапевтическим ДНК-вектором GDTT1.8NAS9-BTK приводит к увеличению количества белка BTK по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген BTK на уровне белка.
Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS9-BTK для повышения уровня экспрессии гена BTK в эукариотических клетках.
Пример 21.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS10 проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена и уровне целевого белка. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS10.
Для подтверждения функциональной активности ДНК-вектора GDTT1.8NAS10 и реализуемости способа его использования клонировали в полилинкер целевой ген, например, ген GBA, приведенный в базе данных GenBank под номером NM_000157. Получение генотерапевтического ДНК-вектора GDTT1.8NAS10-GBA, несущего участок, кодирующий целевой ген, например, ген, GBA. Кодирующую часть гена GBA получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов GBA_F и GBA_R (перечень последовательностей, (74) и (75)). Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции EcoRV и EcoRI и лигировали с ДНК-вектором размером 3065 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS10-GBA размером 3617 п.н. с возможностью селекции без антибиотиков.
Оценивали изменения накопления мРНК целевого гена GBA, в макрофагах периферической крови человека SC (АТСС® CRL-9855™) до и через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS10-GBA, несущим ген GBA человека, а также сравнивали количественное содержание целевого белка GBA после трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS10-GBA, несущим ген GBA человека и ДНК-вектором GDTT1.8NAS10, не несущим ген GBA человека.
Клеточную культуру SC выращивали в соответствии с рекомендациями поставщика (https://www.lgcstandards-atcc.org/Products/AII/CRL-9855.aspx#culturemethod). В процессе культивирования каждые 48 ч происходила смена ростовой среды.
Для получения 90% конфлюэнтности за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS10-GBA, экспрессирующим ген GBA человека, проводили с использованием Lipofectamine 3000 (ThermoFisher Scientific, США) как описано в примере 12. В качестве контроля использовали клетки SC, трансфицированные генотерапевтическим ДНК-вектором GDTT1.8NAS10, не содержащим вставку целевого гена GBA (кДНК гена GBA до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS10, не содержащим вставку целевого гена на фигурах не показано). Подготовку контрольного вектора GDTT1.8NAS10 для трансфекции проводили, как описано выше.
Суммарную РНК из клеток SC выделяли с использованием Trizol Reagent (Invitrogen, США) как описано в примере 12. Определение уровня экспрессии мРНК гена GBA после трансфекции проводили путем оценки динамики накопления ампликонов кДНК методом ПЦР в режиме реального времени как описано в примере 12. Для получения и амплификации кДНК, специфичной для гена GBA человека, использовали олигонуклеотиды GBA_SF и GBA_SR (перечень последовательностей, (76) и (77)). Длина продукта амплификации - 397 п.н.
Графики, полученные в результате анализа, представлены на фиг. 31.
Количественное определение белка GBA проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор ELISA Kit for Glucosidase Beta, Acid (GbA), Cloud-Clone Corp, USA, согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка GBA, входящим в состав набора. Чувствительность метода составляла не менее 12,8 пг/мл, диапазон измерения - от 31,2 пг/мл до 2000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графические материалы, полученные в результате анализа, представлены на фиг. 32.
Из фигуры 31 следует, что в результате трансфекции клеточной культуры макрофагов периферической крови человека SC генотерапевтическим ДНК-вектором GDTT1.8NAS10-GBA, уровень специфической мРНК гена GBA человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген GBA на уровне мРНК. Из фигуры 32 следует, что трансфекция клеток клеточной культуры макрофагов периферической крови человека SC генотерапевтическим ДНК-вектором GDTT1.8NAS10-GBA приводит к увеличению количества белка GBA по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген GBA на уровне белка.
Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS10-GBA для повышения уровня экспрессии гена GBA в эукариотических клетках.
Пример 22.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS11 проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена и уровне целевого белка. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS11.
Для подтверждения функциональной активности ДНК-вектора GDTT1.8NAS11 и реализуемости способа его использования клонировали в полилинкер целевой ген, например, ген PDX1, приведенный в базе данных GenBank под номером NM_000209. Получение генотерапевтического ДНК-вектора GDTT1.8NAS11-PDX1, несущего участок, кодирующий целевой ген, например, ген, PDX1. Кодирующую часть гена PDX1 получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов PDX1_F и PDX1_R (перечень последовательностей, (78) и (79)). Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции BamHI и EcoRI и лигировали с ДНК-вектором размером 2995 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS11-PDX1 размером 2784 п.н. с возможностью селекции без антибиотиков.
Оценивали изменения накопления мРНК целевого гена PDX1, в клетках аденокарциномы поджелудочной железы Panc 10.05 (АТСС® CRL-2547™) до и через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS11-PDX1, несущим ген PDX1 человека, а также сравнивали количественное содержание целевого белка PDX1 после трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS11-PDX1, несущим ген PDX1 человека и ДНК-вектором GDTT1.8NAS11, не несущим ген PDX1 человека.
Клеточную культуру Panc 10.05 выращивали в стандартных условиях (37°С, 5% CO2) с использованием питательной среды ATCC-formulated RPMI-1640 Medium (АТСС) с добавлением 10 Ед/мл рекомбинантного инсулина человека и 15% сыворотки эмбрионов коров (Gibco). В процессе культивирования каждые 48 ч происходила смена ростовой среды.
Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS11-PDX1, экспрессирующим ген PDX1 человека, проводили с использованием Lipofectamine 3000 (ThermoFisher Scientific, США) как описано в примере 12. В качестве контроля использовали клетки Panc 10.05, трансфицированные генотерапевтическим ДНК-вектором GDTT1.8NAS11, не содержащим вставку целевого гена PDX1 (кДНК гена PDX1 до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS11, не содержащим вставку целевого гена на фигурах не показано). Подготовку контрольного вектора GDTT1.8NAS11 для трансфекции проводили, как описано выше.
Суммарную РНК из клеток Panc 10.05 выделяли с использованием Trizol Reagent (Invitrogen, США) как описано в примере 12. Определение уровня экспрессии мРНК гена PDX1 после трансфекции проводили путем оценки динамики накопления ампликонов кДНК методом ПЦР в режиме реального времени как описано в примере 12. Для получения и амплификации кДНК, специфичной для гена PDX1 человека, использовали олигонуклеотиды PDX1_SF и PDX1_SR (перечень последовательностей, (80) и (81)). Длина продукта амплификации - 365 п.н.
Графики, полученные в результате анализа, представлены на фиг. 33.
Количественное определение белка PDX1 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Human PDX1 (Pancreas/duodenum homeobox protein 1) ELISA Kit, Wuhan Fine Biological Technology Co., Ltd., China, согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка PDX1, входящим в состав набора. Чувствительность метода составляла не менее 9,375 пг/мл, диапазон измерения - от 15,6 пг/мл до 1000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графические материалы, полученные в результате анализа, представлены на фиг. 34.
Из фигуры 33 следует, что в результате трансфекции клеточной культуры аденокарциномы поджелудочной железы Panc 10.05 генотерапевтическим ДНК-вектором GDTT1.8NAS11-PDX1, уровень специфической мРНК гена PDX1 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген PDX1 на уровне мРНК. Из фигуры 34 следует, что трансфекция клеток клеточной культуры аденокарциномы поджелудочной железы Panc 10.05 генотерапевтическим ДНК-вектором GDTT1.8NAS11-PDX1 приводит к увеличению количества белка PDX1 по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген PDX1 на уровне белка.
Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS11-PDX1 для повышения уровня экспрессии гена PDX1 в эукариотических клетках.
Пример 23.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS1 проникать в эукариотические клетки и подтверждение его функциональной активности на уровне целевого белка. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS1.
Для подтверждения функциональной активности ДНК-вектора GDTT1.8NAS1 и реализуемости способа его использования клонировали в полилинкер целевой ген, например, ген VEGF, приведенный в базе данных GenBank под номером NM_001025368. Получение генотерапевтического ДНК-вектора GDTT1.8NAS1-VEGF, несущего участок, кодирующий целевой ген, например, ген VEGF проводили, как это описано в примере 12.
Для подтверждения эффективности генотерапевтического ДНК-вектора GDTT1.8NAS1-VEGF, несущего целевой ген, а именно, ген VEGF и реализуемости способа его применения оценивали изменения количества белка VEGF в мышечной ткани человека при введении в мышечную ткань человека генотерапевтического ДНК-вектора GDTT1.8NAS1-VEGF, несущего ген человека VEGF.
С целью анализа изменения количества белка VEGF трем пациентам вводили в ткань икроножной мышцы генотерапевтический ДНК-вектор GDTT1.8NAS1-VEGF, несущий ген VEGF с транспортной молекулой, и параллельно вводили плацебо, представляющее собой генотерапевтический ДНК-вектор GDTT1.8NAS1, не содержащий кДНК гена VEGF с транспортной молекулой.
Пациент 1, мужчина 60 лет, (П1); пациент 2, женщина 53 года (П2); пациент 3, мужчина, 49 лет, (П3). В качестве транспортной системы использовали полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, Франция), пробоподготовку проводили в соответствии с рекомендациями производителя.
Генотерапевтический ДНК-вектор GDTT1.8NAS1 (плацебо) и генотерапевтический ДНК вектор GDTT1.8NAS1-VEGF, несущий ген VEGF вводили в количестве 1 мг для каждой генетической конструкции тоннельным методом иглой 30G на глубину около 10 мм. Объем вводимого раствора генотерапевтического ДНК-вектора GDTT1.8NAS1 (плацебо) и генотерапевтического ДНК вектора GDTT1.8NAS1-VEGF, несущего ген VEGF - 0,3 мл для каждой генетической конструкции. Очаги введения каждой генетической конструкции располагали медиально на расстоянии 8-10 см друг от друга.
Биопсийные образцы отбирали на 2-е сутки после введения генетических конструкций генотерапевтических ДНК-векторов. Взятие биопсии осуществляли из участков мышечной ткани пациентов в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS1-VEGF, несущего ген VEGF (I), генотерапевтического ДНК-вектора GDTT1.8NAS1 (плацебо) (II), а также из интактного участка икроножной мышцы (III), используя устройство для взятия биопсии MAGNUM (BARD, США). Кожу пациентов в зоне отбора биоптата предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер биопсийного образца составлял около 10 куб. мм, масса - около 11 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000 g. Отбирали супернатант и использовали его для количественного определения целевого белка.
Количественное определение белка VEGF проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор ELISA Kit for Vascular Endothelial Growth Factor A (VEGFA), Cloud-Clone Corp., USA, согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка VEGF, входящим в состав набора. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графические материалы, полученные в результате анализа, представлены на фиг. 35.
Из фигуры 35 следует, что в икроножной мышце всех трех пациентов в области введения генотерапевтического ДНК вектора GDTT1.8NAS1-VEGF, несущего целевой ген, а именно, ген VEGF человека, произошло увеличение количества белка VEGF в сравнении с количеством белка VEGF в области введения генотерапевтического ДНК-вектора GDTT1.8NAS1 (плацебо), не содержащего ген VEGF человека, что свидетельствует об эффективности генотерапевтического ДНК-вектора GDTT1.8NAS1-VEGF и подтверждает реализуемость способа его применения, в частности при внутримышечном введении генотерапевтического ДНК-вектора в ткани человека.
Пример 24.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS2 проникать в эукариотические клетки и подтверждение его функциональной активности на уровне целевого белка. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS2.
Для подтверждения функциональной активности ДНК-вектора GDTT1.8NAS2 и реализуемости способа его использования клонировали в полилинкер целевой ген, например, ген CAT, приведенный в базе данных GenBank под номером NM_001752. Получение генотерапевтического ДНК-вектора GDTT1.8NAS2-CAT, несущего участок, кодирующий целевой ген, например, ген CAT проводили, как это описано в примере 13. В результате получали ДНК-вектор GDTT1.8NAS2-CAT размером 3237 п.н. с возможностью селекции без антибиотиков.
Для подтверждения эффективности генотерапевтического ДНК-вектора GDTT1.8NAS2-CAT, несущего целевой ген, а именно, ген CAT и реализуемости способа его применения оценивали изменения количества белка CAT в коже человека при введении в кожу человека генотерапевтического ДНК-вектора GDTT1.8NAS2-CAT, несущего ген человека CAT.
С целью анализа изменения количества белка CAT трем пациентам вводили в кожу предплечья генотерапевтический ДНК-вектор GDTT1.8NAS2-CAT, несущий ген CAT и параллельно вводили плацебо, представляющее собой генотерапевтический ДНК-вектор GDTT1.8NAS2, не содержащий кДНК гена CAT.
Пациент 1, женщина 38 лет, (П1); пациент 2, женщина 66 лет, П2); пациент 3, мужчина, 49 лет, (П3). В качестве транспортной системы использовали полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, Франция). Генотерапевтический ДНК-вектор GDTT1.8NAS2-CAT, который содержит кДНК гена CAT и генотерапевтический ДНК-вектор GDTT1.8NAS2, используемый в качестве плацебо, который не содержит кДНК гена CAT, каждый из которых растворяли в стерильной воде степени очистки Nuclease-Free. Для получения генетической конструкции готовили комплексы ДНК-cGMP grade in-vivo-jetPEI в соответствии с рекомендациями производителя.
Генотерапевтический ДНК-вектор GDTT1.8NAS2 (плацебо) и генотерапевтический ДНК вектор GDTT1.8NAS2-CAT, несущий ген CAT вводили в количестве 1 мг для каждой генетической конструкции тоннельным методом иглой 30G на глубину 3 мм. Объем вводимого раствора генотерапевтического ДНК-вектора GDTT1.8NAS2 (плацебо) и генотерапевтического ДНК вектора GDTT1.8NAS2-CAT, несущего ген CAT - 0,3 мл для каждой генетической конструкции. Очаги введения каждой генетической конструкции располагались на участке предплечья на расстоянии 8-10 см друг от друга.
Биопсийные образцы отбирали на 2-е сутки после введения генетических конструкций генотерапевтических ДНК-векторов. Взятие биопсии осуществляли из участков кожи пациентов в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS2-CAT, несущего ген CAT (I), генотерапевтического ДНК-вектора GDTT1.8NAS2 (плацебо) (II), а также из интактных участков кожи (III), используя устройство для взятия биопсии Epitheasy 3.5 (Medax SRL, Италия). Кожу пациентов в зоне отбора биоптата предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер биопсийного образца составлял около 10 куб. мм, масса - около 11 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000 g. Отбирали супернатант и использовали его для количественного определения целевого белка методом твердофазного иммуноферментного анализа (ELISA), используя набор ELISA Kit for Catalase (CAT) Cloud-Clone Corp. согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка CAT, входящим в состав набора. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графические материалы, полученные в результате анализа, представлены на фиг. 36.
Из фигуры 36 следует, что в коже всех трех пациентов в области введения генотерапевтического ДНК вектора GDTT1.8NAS2-CAT, несущего целевой ген CAT человека, произошло увеличение количества белка CAT в сравнении с количеством белка CAT в области введения генотерапевтического ДНК-вектора GDTT1.8NAS2 (плацебо), не содержащего ген CAT человека, что свидетельствует об эффективности генотерапевтического ДНК-вектора GDTT1.8NAS2-CAT и подтверждает реализуемость способа его применения, в частности при внутрикожном введении генотерапевтического ДНК-вектора в ткани человека.
Пример 25.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS7 проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена и уровне целевого белка. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS7 в клетках животного.
Для подтверждения функциональной активности ДНК-вектора GDTT1.8NAS7 и реализуемости способа его использования клонировали в полилинкер целевой ген, например, ген ATGL, приведенный в базе данных GenBank под номером NM_020376. Получение генотерапевтического ДНК-вектора GDTT1.8NAS7-ATGL, несущего участок, кодирующий целевой ген, например, ген ATGL проводили, как это описано в примере 18. В результате получали ДНК-вектор GDTT1.8NAS7-ATGL размером 4088 п.н. с возможностью селекции без антибиотиков.
Оценивали изменения накопления мРНК целевого гена ATGL, в клетках почки быка MDBK (АТСС CLL-22) до и через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS7-ATGL, несущим ген ATGL человека, а также сравнивали количественное содержание целевого белка ATGL после трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS7-ATGL, несущим ген ATGL человека и ДНК-вектором GDTT1.8NAS7, не несущим ген ATGL человека.
Клеточную культуру почки быка MDBK выращивали в среде MEM (ThermoFisher Scientific, США), содержащей 10% сыворотки эмбрионов коров (ThermoFisher Scientific, США), 1 г/л глюкозы и 2 мМ глутамина в атмосфере 5% CO2 при 37°С. В процессе культивирования каждые 48 ч происходила смена ростовой среды.
Для получения 90% конфлюэнтности за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS7-ATGL, экспрессирующим ген ATGL человека, проводили с использованием Lipofectamine 3000 (ThermoFisher Scientific, США) согласно рекомендациям производителя. В пробирке №1 к 25 мкл среды Opti-MEM (Gibco, США) добавляли 1 мкл раствора ДНК-вектора GDTT1.8NAS7-ATGL (концентрация 500 нг/мкл) и 1 мкл реагента Р3000. Аккуратно перемешивали легким встряхиванием. В пробирке №2 к 25 мкл среды Opti-MEM (Gibco, США) добавляли 1 мкл раствора Lipofectamin 3000. Аккуратно перемешивали легким встряхиванием. Добавляли содержимое пробирки №1 к содержимому пробирки №2, инкубировали 5 мин при комнатной температуре. Полученный раствор по каплям добавляли к клеткам в объеме 40 мкл.
В качестве контроля использовали клетки почки быка MDBK, трансфицированные генотерапевтическим ДНК-вектором GDTT1.8NAS7, не содержащим вставку целевого гена ATGL (кДНК гена ATGL до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS7, не содержащим вставку целевого гена на фигурах не показано). Подготовку контрольного вектора GDTT1.8NAS7 для трансфекции проводили, как описано выше.
Суммарную РНК из клеток почки быка MDBK выделяли с использованием Trizol Reagent (Invitrogen, США) согласно рекомендациям производителя. В лунку с клетками добавляли 1 мл Trizol Reagent и гомогенизировали с последующим прогреванием в течении 5 мин при 65°С. Далее образец центрифугировали при 14000 g в течении 10 мин и снова прогревали в течении 10 мин при 65°С. Далее добавляли 200 мкл хлороформа, плавно перемешивали и центрифугировали при 14000 g в течении 10 мин. Затем отбирали водную фазу, добавляли к ней 1/10 объема 3М ацетата натрия, рН 5.2 и равный объем изопропилового спирта. Инкубировали образец при -20°С в течении 10 мин с последующим центрифугированием при 14000 g в течении 10 мин. Осадок промывали 1 мл 70% этилового спирта, высушивали на воздухе и растворяли в 10 мкл воды, свободной от РНКаз. Определение уровня экспрессии мРНК гена ATGL после трансфекции проводили путем оценки динамики накопления ампликонов кДНК методом ПЦР в режиме реального времени. Для получения и амплификации кДНК, специфичной для гена ATGL человека, использовали олигонуклеотиды ATGL_SF и ATGL_SR (перечень последовательностей (64) и (65)). Длина продукта амплификации - 417 п.н.
Реакцию обратной транскрипции и ПЦР-амплификацию проводили с помощью набора реагентов SYBR GreenQuantitect RT-PCR Kit (Qiagen, США) для ПЦР в режиме реального времени. Реакцию проводили в объеме 20 мкл, содержащих: 25 мкл QuantiTect SYBR Green RT-PCR MasterMix, 2,5 мМ хлорида магния, по 0,5 мкМ каждого праймера, 5 мкл РНК. Реакцию осуществляли на амплификаторе CFX96 (Bio-Rad, США) при следующих условиях: 1 цикл обратной транскрипции при 42°С - 30 минут, денатурация 98°С - 15 мин, затем 40 циклов, включающих денатурацию 94°С - 15 сек, отжиг праймеров 60°С - 30 сек и элонгацию 72°С - 30 сек. В качестве референтного гена использовали ген актина быка/коровы (ACT), приведенного в базе данных GenBank под номером АН001130.2. В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов ATGL и ACT. В качестве отрицательного контроля использовали деионизированную воду. Количество динамику накопления ампликонов кДНК генов ATGL и ACT оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа, представлены на фиг. 37.
Количественное определение белка ATGL проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор ELISA Kit for Patatin Like Phospholipase Domain Containing Protein 2 (PNPLA2), Cloud-Clone Corp, USA, согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка ATGL, входящим в состав набора. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графические материалы, полученные в результате анализа, представлены на фиг. 38.
Из фигуры 37 следует, что в результате трансфекции клеточной культуры почки быка MDBK генотерапевтическим ДНК-вектором GDTT1.8NAS7-ATGL, уровень специфической мРНК гена ATGL человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген ATGL на уровне мРНК. Из фигуры 38 следует, что трансфекция клеток клеточной культуры почки быка MDBK генотерапевтическим ДНК-вектором GDTT1.8NAS7-ATGL приводит к увеличению количества белка ATGL по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген ATGL на уровне белка.
Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS7-ATGL для повышения уровня экспрессии гена ATGL в эукариотических клетках.
Пример 26.
Подтверждение функциональной активности на уровне целевого белка генотерапевтического ДНК-вектора GDTT1.8NAS1, обладающего возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в клетках мышечной ткани, генотерапевтического ДНК-вектора GDTT1.8NAS2, обладающего возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в клетках кожи, генотерапевтического ДНК-вектора GDTT1.8NAS3, обладающего возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в клетках эндотелия сосудов, генотерапевтического ДНК-вектора GDTT1.8NAS4, обладающего возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в остеобластах и одонтобластах, генотерапевтического ДНК-вектора GDTT1.8NAS5, обладающего возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в эпителиальных клетках бронхов и альвеол, генотерапевтического ДНК-вектора GDTT1.8NAS6, обладающего возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в нейронах, генотерапевтического ДНК-вектора GDTT1.8NAS7, обладающего возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в подоцитах почек, генотерапевтического ДНК-вектора GDTT1.8NAS8, обладающего возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в гематопоэтических клетках, генотерапевтического ДНК-вектора GDTT1.8NAS9, обладающего возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в лимфоцитах, генотерапевтического ДНК-вектора GDTT1.8NAS10, обладающего возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в макрофагах, генотерапевтического ДНК-вектора GDTT1.8NAS11, обладающего возможностью тканеспецифичной экспрессии клонированных в него целевых генов преимущественно в бета-клетках поджелудочной железы, с учетом тканевой специфичности этих генотерапевтических ДНК-векторов, т.е. возможностью экспрессировать клонированные в него целевые гены только в определенной целевой клеточной линии, целевой ткани или органе человека. Данный пример также демонстрирует реализуемость способа применения для таргетной генной терапии генотерапевтических ДНК-векторов GDTT1.8NAS1, GDTT1.8NAS2, GDTT1.8NAS3, GDTT1.8NAS4, GDTT1.8NAS5, GDTT1.8NAS6, GDTT1.8NAS7, GDTT1.8NAS8, GDTT1.8NAS9, GDTT1.8NAS10, GDTT1.8NAS11 с учетом их тканевой специфичности.
Для подтверждения функциональной активности генотерапевтических ДНК-векторов GDTT1.8NAS1, GDTT1.8NAS2, GDTT1.8NAS3, GDTT1.8NAS4, GDTT1.8NAS5, GDTT1.8NAS6, GDTT1.8NAS7, GDTT1.8NAS8, GDTT1.8NAS9, GDTT1.8NAS10, GDTT1.8NAS11 и реализуемости способа их использования клонировали в полилинкер каждого генотерапевтического ДНК-вектора целевой ген, например, ген GFP.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS1-eGFP, несущего целевой ген, например, ген, кодирующий зеленый флуоресцирующий белок (GFP).
Кодирующую часть гена зеленого флуоресцентного белка получали путем ПЦР-амплификации коммерческой плазмиды pEGFP-N1 (Clontech) с использованием олигонуклеотидов MVGFP-F и MVGFP-R (перечень последовательностей, (82) и (83)). Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции BamHI и EcoRI и лигировали с ДНК-вектором размером 2753 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS1-eGFP размером 2413 п.н. с возможностью селекции без антибиотиков.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS2-eGFP, несущего целевой ген, например, ген, кодирующий зеленый флуоресцирующий белок (GFP).
Кодирующую часть гена зеленого флуоресцентного белка получали путем ПЦР-амплификации коммерческой плазмиды pEGFP-N1 (Clontech), как описано выше. Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции BamHI и EcoRI и лигировали с ДНК-вектором размером 2671 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS2-eGFP размером 2331 п.н. с возможностью селекции без антибиотиков.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS3-eGFP, несущего целевой ген, например, ген, кодирующий зеленый флуоресцирующий белок (GFP).
Кодирующую часть гена зеленого флуоресцентного белка получали путем ПЦР-амплификации коммерческой плазмиды pEGFP-N1 (Clontech), как описано выше. Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции BamHI и EcoRI и лигировали с ДНК-вектором размером 2800 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS3-eGFP размером 2460 п.н. с возможностью селекции без антибиотиков.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS4-eGFP, несущего целевой ген, например, ген, кодирующий зеленый флуоресцирующий белок (GFP).
Кодирующую часть гена зеленого флуоресцентного белка получали путем ПЦР-амплификации коммерческой плазмиды pEGFP-N1 (Clontech) как это описано выше. Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции BamHI и EcoRI и лигировали с ДНК-вектором размером 2970 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS4-eGFP размером 2630 п.н. с возможностью селекции без антибиотиков.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS5-eGFP, несущего целевой ген, например, ген, кодирующий зеленый флуоресцирующий белок (GFP).
Кодирующую часть гена зеленого флуоресцентного белка получали путем ПЦР-амплификации коммерческой плазмиды pEGFP-N1 (Clontech) как это описано выше. Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции BamHI и EcoRI и лигировали с ДНК-вектором размером 3036 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS5-eGFP размером 2696 п.н. с возможностью селекции без антибиотиков.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS6-eGFP, несущего целевой ген, например, ген, кодирующий зеленый флуоресцирующий белок (GFP).
Кодирующую часть гена зеленого флуоресцентного белка получали путем ПЦР-амплификации коммерческой плазмиды pEGFP-N1 (Clontech) как это описано выше. Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции BamHI и EcoRI и лигировали с ДНК-вектором размером 2957 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS6-eGFP размером 2617 п.н. с возможностью селекции без антибиотиков.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS7-eGFP, несущего целевой ген, например, ген, кодирующий зеленый флуоресцирующий белок (GFP).
Кодирующую часть гена зеленого флуоресцентного белка получали путем ПЦР-амплификации коммерческой плазмиды pEGFP-N1 (Clontech) как это описано выше. Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции BamHI и EcoRI и лигировали с ДНК-вектором размером 3637 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS7-eGFP размером 3297 п.н. с возможностью селекции без антибиотиков.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS8-eGFP, несущего целевой ген, например, ген, кодирующий зеленый флуоресцирующий белок (GFP).
Кодирующую часть гена зеленого флуоресцентного белка получали путем ПЦР-амплификации коммерческой плазмиды pEGFP-N1 (Clontech) как это описано выше. Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции BamHI и EcoRI и лигировали с ДНК-вектором размером 3257 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS8-eGFP размером 2917 п.н. с возможностью селекции без антибиотиков.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS9-eGFP, несущего целевой ген, например, ген, кодирующий зеленый флуоресцирующий белок (GFP).
Кодирующую часть гена зеленого флуоресцентного белка получали путем ПЦР-амплификации коммерческой плазмиды pEGFP-N1 (Clontech) как это описано выше. Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции BamHI и EcoRI и лигировали с ДНК-вектором размером 3621 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS9-eGFP размером 3281 п.н. с возможностью селекции без антибиотиков.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS10-eGFP, несущего целевой ген, например, ген, кодирующий зеленый флуоресцирующий белок (GFP).
Кодирующую часть гена зеленого флуоресцентного белка получали путем ПЦР-амплификации коммерческой плазмиды pEGFP-N1 (Clontech) как это выше. Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции BamHI и EcoRI и лигировали с ДНК-вектором размером 3065 п.н., несущим ген устойчивости к канамицину, расщепленным эндонуклеазами рестрикции BgIII и EcoRI. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS10-eGFP размером 2719 п.н. с возможностью селекции без антибиотиков.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS11-eGFP, несущего целевой ген, например, ген, кодирующий зеленый флуоресцирующий белок (GFP).
Кодирующую часть гена зеленого флуоресцентного белка получали путем ПЦР-амплификации коммерческой плазмиды pEGFP-N1 как это описано выше. Полученный ПЦР-фрагмент расщепляли эндонуклеазами рестрикции BamHI и EcoRI и лигировали с ДНК-вектором размером 2995 п.н., несущим ген устойчивости к канамицину, расщепленным теми же ферментами. Далее, из полученного вектора удаляли ген устойчивости к канамицину, как это описано в примере 1. В результате получали ДНК-вектор GDTT1.8NAS11-eGFP размером 2655 п.н. с возможностью селекции без антибиотиков.
Сравнение уровня накопления продукта целевого гена, например, зеленого флуоресцентного белка (GFP) в каждой из клеточных линий: в клетках первичной культуры скелетных миобластов человека HSkM (ThermoFisher Scientific # А12555), в клетках первичной культуры фибробластов кожи человека HDFa (АТСС® PCS-201-012™), в клетках эндотелия пупочной вены человека HUVEC (АТСС® PCS-100-013™), в клетках остеосаркомы человека MG-63 (АТСС® CRL-1427™), в клетках трахеального эпителия человека CFTE29o- (коллекция Института Цитологии РАН РФ), в клеточной культуре нейробластомы человека SH-SY5Y (АТСС® CRL-2266™), в клетках первичной смешанной культуры эпителия почки человека HREC (АТСС® PCS-400-012™), в клетках первичной культуры костного мозга человека (АТСС® PCS-800-012™), в клетках первичной культуры периферических мононуклеаров крови человека РВМС (АТСС® PCS-800-011™), в макрофагах периферической крови человека SC (АТСС® CRL-9855™), в клетках аденокарциномы поджелудочной железы Panc 10.05 (АТСС® CRL-2547™) через 48 часов после их трансфекции каждым генотерапевтическим ДНК-вектором: генотерапевтическим ДНК-вектором GDTT1.8NAS1-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS2-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS3-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS4-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS5-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS6-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS7-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS8-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS9-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS10eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS11-eGFP.
Для оценки уровня накопления зеленого флуоресцентного белка GFP в клетках первичной культуры скелетных миобластов человека HSkM, в клетках первичной культуры фибробластов кожи человека HDFa, в клетках эндотелия пупочной вены человека HUVEC, в клетках остеосаркомы человека MG-63, в клетках трахеального эпителия человека CFTE29o-, в клеточной культуре нейробластомы человека SH-SY5Y, в клетках первичной смешанной культуры эпителия почки человека HREC, в клетках первичной культуры костного мозга человека, в клетках первичной культуры периферических мононуклеаров крови человека РВМС, в макрофагах периферической крови человека SC, в клетках аденокарциномы поджелудочной железы Panc 10.05 проводили трансфекцию каждой клеточной линии каждым генотерапевтическим ДНК-вектором: генотерапевтическим ДНК-вектором GDTT1.8NAS1-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS2-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS3-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS4-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS5-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS6-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS7-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS8-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS9-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS10eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS11-eGFP. В качестве контроля осуществляли аналогичную трансфекцию этих клеточных линий плазмидным вектором pEFGP-N1 (Clontech).
Клетки первичной культуры скелетных миобластов человека HSkM, клетки первичной культуры фибробластов кожи человека HDFa, клетки эндотелия пупочной вены человека HUVEC, клетки остеосаркомы человека MG-63, клетки трахеального эпителия человека CFTE29o-, клеточную культуру нейробластомы человека SH-SY5Y, клетки первичной смешанной культуры эпителия почки человека HREC, клетки первичной культуры костного мозга человека, клетки первичной культуры периферических мононуклеаров крови человека РВМС, макрофаги периферической крови человека SC, клетки аденокарциномы поджелудочной железы Panc 10.05 выращивали, как описано в примерах 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22. Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки каждой линии высевали в 24-луночный планшет из расчета 4×104 клеток/лунку. Для трансфекции каждой клеточной линии использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Для трансфекции каждой клеточной линии каждым генотерапевтическим ДНК-вектором в каждой пробирке №1 к 25 мкл среды Opti-MEM (Gibco) добавляли 1 мкл раствора каждого генотерапевтического ДНК-вектора: генотерапевтического ДНК-вектора GDTT1.8NAS2-eGFP, генотерапевтического ДНК-вектора GDTT1.8NAS3-eGFP, генотерапевтического ДНК-вектора GDTT1.8NAS4-eGFP, генотерапевтического ДНК-вектора GDTT1.8NAS5-eGFP, генотерапевтического ДНК-вектора GDTT1.8NAS6-eGFP, генотерапевтического ДНК-вектора GDTT1.8NAS7-eGFP, генотерапевтического ДНК-вектора GDTT1.8NAS8-eGFP, генотерапевтического ДНК-вектора GDTT1.8NAS9-eGFP, генотерапевтического ДНК-вектора GDTT1.8NAS10eGFP, генотерапевтического ДНК-вектора GDTT1.8NAS11-eGFP (концентрация каждого 500 нг/мкл) и 1 мкл реагента Р3000. Аккуратно перемешивали легким встряхиванием. В каждой пробирке №2 к 25 мкл среды Opti-MEM (Gibco) добавляли 1 мкл раствора Lipofectamine 3000. Аккуратно перемешивали легким встряхиванием. Добавляли содержимое каждой пробирки №1 к содержимому соответствующей пробирки №2, инкубировали 5 мин при комнатной температуре. Полученный раствор по каплям добавляли к каждой клеточной линии в объеме 40 мкл.
Учет результатов проводили путем измерения флуоресценции белка, выделенного из каждой трансфицированной линии клеток. Для этого клетки каждой трансфицированной линии из лунки смывали пипетированием и осаждали центрифугированием на 6000 об/мин в течение 10 мин, дважды промывали и затем ресуспендировали осадки в 1 мл натрий-фосфатного буфера. Клетки разрушали с помощью трехкратного замораживания-оттаивания при -70°С. Далее гомогенат разрушенных клеток осаждали центрифугированием при 13000 g в течение 15 мин. Полученные супернатанты переносили в 96-луночный планшет (Grainer Bio-one), в четырех технических повторах для каждого образца, после чего оценивали относительную флуоресценцию белка GFP в пробе (поглощение 455 нм / испускание 538 нм) с помощью флуориметра Fluoroskan Ascent (Labsystems). Нормировку полученных значений осуществляли в зависимости от суммарной концентрации белка в пробе, которую определяли методом Бредфорд. Для этого в качестве красителя использовали Coomassie Brilliant Blue R-250 (BioRad). Каждый технический повтор в ячейках 96-луночного планшета (4 повтора для каждого образца) разводили водой в 100 раз, после чего добавляли краситель. Затем с помощью сканера Multiskan Ascent (Thermo) проводили оценку оптической плотности всех проб на длине волны 620 нм. Полученные значения сравнивали с калибровочной кривой, построенной для бычьего сывороточного альбумина (Bio-Rad) с использованием серии разведений от 20 до 2,5 мкг/мл. Расчет осуществляли по формуле:
Σколичество белка (мкг) = {[х] - σ} ÷ k * М,
где [х] - среднее значение OD620 из четырех повторов для каждого образца, σ - среднее отклонение, k - коэффициент наклона калибровочной кривой для БСА, М - число раз, в которое был разведен образец.
Исходя из полученных значений суммарного количества белка, выделенного из клеток, нормировку флуоресценции GFP в образцах осуществляли по формуле:
OEn = [ОЕ] ÷ Σколичество белка (мг)
где
[ОЕ] - среднее значение из четырех повторов для каждого образца в относительных единицах флуоресценции.
Ниже приведена таблица, отражающая уровень флуоресценции (в отн. ед. флуоресценции), выделенного суммарного белка из каждой клеточной линии: из клеток первичной культуры скелетных миобластов человека HSkM, из клеток первичной культуры фибробластов кожи человека HDFa, из клеток эндотелия пупочной вены человека HUVEC, из клеток остеосаркомы человека MG-63, из клеток трахеального эпителия человека CFTE29o-, из клеточной культуры нейробластомы человека SH-SY5Y, из клеток первичной смешанной культуры эпителия почки человека HREC, из клеток первичной культуры костного мозга человека, из клеток первичной культуры периферических мононуклеаров крови человека РВМС, из макрофагов периферической крови человека SC, из клеток аденокарциномы поджелудочной железы Panc 10.05 HSkM через 48 часов после трансфекции каждой из них каждым из генотерапевтических ДНК-векторов: генотерапевтическим ДНК-вектором GDTT1.8NAS1-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS2-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS3-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS4-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS5-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS6-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS7-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS8-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS9-eGFP, генотерапевтическим ДНК-вектоома GDTT1.8NAS10eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS11-eGFP с целью сравнения уровня накопления продукта целевого гена, например, зеленого флуоресцентного белка (GFP) в каждой клеточной линии: в клетках первичной культуры скелетных миобластов человека HSkM, в клетках первичной культуры фибробластов кожи человека HDFa, в клетках эндотелия пупочной вены человека HUVEC, в клетках остеосаркомы человека MG-63, в клетках трахеального эпителия человека CFTE29o-, в клеточной культуре нейробластомы человека SH-SY5Y, в клетках первичной смешанной культуры эпителия почки человека HREC, в клетках первичной культуры костного мозга человека, в клетках первичной культуры периферических мононуклеаров крови человека РВМС, в макрофагах периферической крови человека SC, в клетках аденокарциномы поджелудочной железы Panc 10.05 HSkM через 48 часов после трансфекции каждой из них каждым из генотерапевтических ДНК-векторов: генотерапевтическим ДНК-вектором GDTT1.8NAS1-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS2-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS3-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS4-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS5-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS6-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS7-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS8-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS9-eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS10eGFP, генотерапевтическим ДНК-вектором GDTT1.8NAS11-eGFP для оценки тканеспецифичности данных генотерапевтических ДНК-векторов.
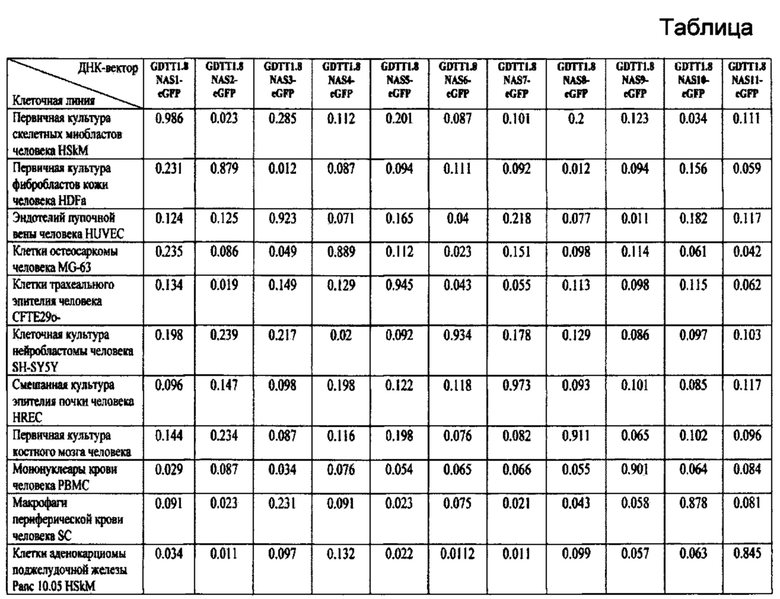
Из таблицы следует, что трансфекция клеток скелетных миобластов человека HSkM генотерапевтическим ДНК-вектором GDTT1.8NAS1-eGFP, обладающим возможностью тканеспецифичной экспрессии клонированных в него целевых генов в клетках мышечной ткани, приводит к достоверному увеличению накопления зеленого флуоресцентного белка по сравнению с накоплением зеленого флуоресцентного белка при трансфекции этих клеток остальными генотерапевтическими ДНК-векторами, не обладающими возможностью тканеспецифичной экспрессии клонированных в него целевых генов в клетках мышечной ткани; трансфекция клеток первичной культуры фибробластов кожи человека HDFa генотерапевтическим ДНК-вектором GDTT1.8NAS2-eGFP, обладающим возможностью тканеспецифичной экспрессии клонированных в него целевых генов в клетках кожи, приводит к достоверному увеличению накопления зеленого флуоресцентного белка по сравнению с накоплением зеленого флуоресцентного белка при трансфекции этих клеток остальными генотерапевтическими ДНК-векторами, не обладающими возможностью тканеспецифичной экспрессии клонированных в него целевых генов в клетках кожи; трансфекция клеток эндотелия пупочной вены человека HUVEC генотерапевтическим ДНК-вектором GDTT1.8NAS3-eGFP, обладающим возможностью тканеспецифичной экспрессии клонированных в него целевых генов в клетках эндотелия сосудов, приводит к достоверному увеличению накопления зеленого флуоресцентного белка по сравнению с накоплением зеленого флуоресцентного белка при трансфекции этих клеток остальными генотерапевтическими ДНК-векторами, не обладающими возможностью тканеспецифичной экспрессии клонированных в него целевых генов в клетках эндотелия сосудов; трансфекция клеток остеосаркомы человека MG-63 генотерапевтическим ДНК-вектором GDTT1.8NAS4-eGFP, обладающим возможностью тканеспецифичной экспрессии клонированных в него целевых генов в остеобластах и одонтобластах, приводит к достоверному увеличению накопления зеленого флуоресцентного белка по сравнению с накоплением зеленого флуоресцентного белка при трансфекции этих клеток остальными генотерапевтическими ДНК-векторами, не обладающими возможностью тканеспецифичной экспрессии клонированных в него целевых генов в остеобластах и одонтобластах; трансфекция клеток трахеального эпителия человека CFTE29o- генотерапевтическим ДНК-вектором GDTT1.8NAS5-eGFP, обладающим возможностью тканеспецифичной экспрессии клонированных в него целевых генов в эпителиальных клетках бронхов и альвеол, приводит к достоверному увеличению накопления зеленого флуоресцентного белка по сравнению с накоплением зеленого флуоресцентного белка при трансфекции этих клеток остальными генотерапевтическими ДНК-векторами, не обладающими возможностью тканеспецифичной экспрессии клонированных в него целевых генов в эпителиальных клетках бронхов и альвеол; трансфекция клеточной культуры нейробластомы человека SH-SY5Y генотерапевтическим ДНК-вектором GDTT1.8NAS6-eGFP, обладающим возможностью тканеспецифичной экспрессии клонированных в него целевых генов в нейронах, приводит к достоверному увеличению накопления зеленого флуоресцентного белка по сравнению с накоплением зеленого флуоресцентного белка при трансфекции этих клеток остальными генотерапевтическими ДНК-векторами, не обладающими возможностью тканеспецифичной экспрессии клонированных в него целевых генов в нейронах; трансфекция клеток первичной смешанной культуры эпителия почки человека HREC генотерапевтическим ДНК-вектором GDTT1.8NAS7-eGFP, обладающим возможностью тканеспецифичной экспрессии клонированных в него целевых генов в подоцитах почек, приводит к достоверному увеличению накопления зеленого флуоресцентного белка по сравнению с накоплением зеленого флуоресцентного белка при трансфекции этих клеток остальными генотерапевтическими ДНК-векторами, не обладающими возможностью тканеспецифичной экспрессии клонированных в него целевых генов в подоцитах почек; трансфекция клеток первичной культуры костного мозга человека генотерапевтическим ДНК-вектором GDTT1.8NAS8-eGFP, обладающим возможностью тканеспецифичной экспрессии клонированных в него целевых генов в гематопоэтических клетках, приводит к достоверному увеличению накопления зеленого флуоресцентного белка по сравнению с накоплением зеленого флуоресцентного белка при трансфекции этих клеток остальными генотерапевтическими ДНК-векторами, не обладающими возможностью тканеспецифичной экспрессии клонированных в него целевых генов в гематопоэтических клетках; трансфекция клеток первичной культуры периферических мононуклеаров крови человека РВМС генотерапевтическим ДНК-вектором GDTT1.8NAS9-eGFP, обладающим возможностью тканеспецифичной экспрессии клонированных в него целевых генов в лимфоцитах, приводит к достоверному увеличению накопления зеленого флуоресцентного белка по сравнению с накоплением зеленого флуоресцентного белка при трансфекции этих клеток остальными генотерапевтическими ДНК-векторами, не обладающими возможностью тканеспецифичной экспрессии клонированных в него целевых генов в лимфоцитах; трансфекция макрофагов периферической крови человека SC генотерапевтическим ДНК-вектором GDTT1.8NAS10-eGFP, обладающим возможностью тканеспецифичной экспрессии клонированных в него целевых генов в макрофагах, приводит к достоверному увеличению накопления зеленого флуоресцентного белка по сравнению с накоплением зеленого флуоресцентного белка при трансфекции этих клеток остальными генотерапевтическими ДНК-векторами, не обладающими возможностью тканеспецифичной экспрессии клонированных в него целевых генов в макрофагах; трансфекция клеток аденокарциномы поджелудочной железы Рапс 10.05 генотерапевтическим ДНК-вектором GDTT1.8NAS11-eGFP, обладающим возможностью тканеспецифичной экспрессии клонированных в него целевых генов в бета-клетках поджелудочной железы, приводит к достоверному увеличению накопления зеленого флуоресцентного белка по сравнению с накоплением зеленого флуоресцентного белка при трансфекции этих клеток остальными генотерапевтическими ДНК-векторами, не обладающими возможностью тканеспецифичной экспрессии клонированных в него целевых генов в бета-клетках поджелудочной железы.
Результаты примера 26 подтверждает способность генотерапевтических ДНК-векторов: генотерапевтического ДНК-вектора GDTT1.8NAS1-eGFP, генотерапевтического ДНК-вектора GDTT1.8NAS2-eGFP, генотерапевтического ДНК-вектора GDTT1.8NAS3-eGFP, генотерапевтического ДНК-вектора GDTT1.8NAS4-eGFP, генотерапевтического ДНК-вектора GDTT1.8NAS5-eGFP, генотерапевтического ДНК-вектора GDTT1.8NAS6-eGFP, генотерапевтического ДНК-вектора GDTT1.8NAS7-eGFP, генотерапевтического ДНК-вектора GDTT1.8NAS8-eGFP, генотерапевтического ДНК-вектора GDTT1.8NAS9-eGFP, генотерапевтического ДНК-вектора GDTT1.8NAS10eGFP, генотерапевтического ДНК-вектора GDTT1.8NAS11-eGFP проникать в эукариотические клетки, их функциональную активность, а также тканевую специфичность этих генотерапевтических ДНК-векторов в виде возможности достоверного увеличения уровня экспрессии целевого гена, клонированного в эти векторы, в определенной клеточной линии, ткани или органе человека.
Представленные результаты также подтверждают реализуемость способа выбора генотерапевтического ДНК-вектора для конструирования генотерапевтического ДНК-вектора, содержащего целевой ген для таргетной генной терапии.
Пример 27.
Получение штамма Escherichia coli JM-110-NAS для продукции генотерапевтического ДНК-вектора GDTT1.8NAS1 или GDTT1.8NAS2 или GDTT1.8NAS3 или GDTT1.8NAS4 или GDTT1.8NAS5 или GDTT1.8NAS6 или GDTT1.8NAS7 или GDTT1.8NAS8 или GDTT1.8NAS9 или GDTT1.8NAS10 или GDTT1.8NAS11 и/или генотерапевтических векторов, несущих целевые гены на его основе.
Штамм Escherichia coli JM-110-NAS для продукции генотерапевтического ДНК-вектора GDTT1.8NAS1 или GDTT1.8NAS2 или GDTT1.8NAS3 или GDTT1.8NAS4 или GDTT1.8NAS5 или GDTT1.8NAS6 или GDTT1.8NAS7 или GDTT1.8NAS8 или GDTT1.8NAS9 или GDTT1.8NAS10 или GDTT1.8NAS11 и/или генотерапевтических векторов, несущих целевые гены на его основе получали методом гомологичной рекомбинации путем введения в его хромосому в область гена recA линейного фрагмента, содержащего регуляторный элемент RNA-in транспозона Tn10 для селекции без применения антибиотиков (64 п.н.), ген левансахаразы sacB, продукт которого обеспечивает селекцию на сахарозо-содержащей среде (1422 п.н.), ген устойчивости к хлорамфениколу catR, необходимый для отбора клонов штамма, в которых прошла гомологичная рекомбинация (763 п.н.) и две гомологичные последовательности (плечи гомологии), обеспечивающих процесс гомологичной рекомбинации в области гена recA с одновременной его инактивацией (329 п.н. и 233 п.н. для левого и правого плеча, соответственно).
Для получения левого и правого плеч гомологии проводили ПЦР амплификацию фрагментов гена recA с использованием геномной ДНК Eshcerichia coli JM110 (Agilent Technologies, кат. №200239) в качестве матрицы. Для получения левого плеча гомологии использовали праймеры LHA-F и LHA-R (перечень последовательностей, (84) и (85)), для получения правого плеча гомологии используют праймеры RHA-F и RHA-R (перечень последовательностей, (86) и (87)). Фрагмент RNA-in достраивали из синтетических олигонуклеотидов IN-F, IN-1, IN-2, IN-R (перечень последовательностей, (88), (89), (90), (91)). Ген sacB получали ПЦР-амплификацией с использованием геномной ДНК B.subtilis 168НТ в качестве матрицы и SacB-F и SacB-R в качестве праймеров (перечень последовательностей, (92) и (93)). Для получения гена catR проводили ПЦР-амплификацию с использованием штамма Eshcerichia coli BL21 pLysS в качестве матрицы и CatR-F и CatR-R (перечень последовательностей, (94) и (95)) в качестве праймеров. ПЦР продукты LHA (левое плечо гомологии), SacB, RHA (правое плечо) амплифицировали при условиях 94°С - 3 мин; 30 циклов: 94°С - 20 сек, 60°С - 20 сек и 72°С - 60 сек, с финальной элонгацией 72°С - 5 мин. ПЦР продукт RNA-in получали при условиях 94°С - 3 мин; 30 циклов: 94°С - 10 сек, 60°С - 10 сек и 72°С - 10 сек использованием IN-F, IN-1, IN-2, IN-R олигонуклеотидов (перечень последовательностей, (88), (89), (90), (91)) для сборки фрагмента, при этом использовали 10 мкМ праймеров IN-F и IN-R, и 5 мкМ праймеров IN-1 и IN-2. ПЦР-амплификацию проводили с использованием коммерческого набора Phusion® High-Fidelity DNA Polymerase (Thermo Fisher Scientific) в соответствии с инструкцией производителя.
Линейный фрагмент для гомологичной рекомбинации получали путем объединения пяти продуктов ПЦР амплификации. Все пять продуктов имели перекрывающиеся области для дальнейшей сборки в единый фрагмент. Все фрагменты смешивали в количестве 10 нг в объеме 50 мкл. ПЦР продукт получали при условиях 94°С - 3 мин; 10 циклов: 94°С - 30 сек, 60°С - 30 сек и 72°С - 2 мин без добавления праймеров, затем добавляли праймеры LHA-F, RHA-R (перечень последовательностей, (84), (85)), после чего проводили дополнительно 25 циклов ПЦР: 94°С - 30 сек, 60°С - 30 сек и 72°С - 2 мин с финальной элонгацией 72°С - 5 мин. В результате получали ПЦР фрагмент длиной 2811 п.н. следующей структуры: LHA-RNA-IN-SacB-CatR-RHA. Данный фрагмент выделяли препаративно из агарозного геля с помощью набора для элюции ДНК из агарозного геля (BioSilica, Россия) согласно рекомендациям производителя. Структура фрагмента ДНК для гомологичной рекомбинации в области гена recA Eshcerichia coli для получения штамма Eshcerichia coli JM 110 приведена на фиг. 12.
Для получения штамма Escherichia coli JM110-NAS готовили элетрокомпетентные клетки. Для этого одиночной колонией штамма Eshcerichia coli JM 110 (Agilent Technologies) инокулировали 10 мл среды LB и культивировали ночь в орбитальном шейкере при 150 об/мин и 37°С. На следующий день пересеивали 1/20 в 100 мл среды LB и культивировали в орбитальном шейкере при 150 об/мин и 37°С до достижения OD600=0,5. По достижении необходимой оптической плотности, клетки охлаждали до 0°С и центрифугировали 10 мин при 4000g. Далее, удаляли среду, клетки отмывали от остатков среды охлажденной во льду 100 мл бидистилированной водой дважды, затем промывали 20 мл 10% глицерина. После этого клетки ресуспендировали в 1 мл 10% глицерина и использовали для трансформации.
Трансформацию полученным линейным фрагментом проводили методом электропорации в кюветах 1 мм при 2 кВ, 200 Ом, 25 мкФ на приборе Gene Pulser Xcell (Bio-Rad, США). Время импульса составляло от 4,9 до 5,1 мсек. После чего клетки культивировали в течение 2,5 часов в среде SOC в шейкере-инкубаторе при 30°С. Затем, клетки высеивали на чашки Петри с агаризованной средой LB, содержащей 10 мкг/мл хлорамфеникола. Клетки культивировали 48 часов при 30°С. Отобранные клоны проверяли на выживаемость на селективной среде, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола. Полученный штамм имеет генотип -recA rpsL (Strr) thr leu endA thi-1 lacY galK galT ara tonA tsx dam dcm supE44 Δ(lac-proAB) [F' traD36 proAB laclq ZΔM15]CmR sacB+
Пример 28.
Получение штамма Escherichia coli JM-110-NAS-GDTT1.8NAS1 несущего генотерапевтический ДНК-вектор GDTT1.8NAS1 и/или генотерапевтические ДНК-вектора, содержащие в своем составе целевые гены на его основе для его наработки.
Для приготовления электрокомпетентных клеток штамма Escherichia coli JM110-NAS, одиночной колонией заражали 10 мл среды LB и культивировали ночь в орбитальном шейкере при 150 об/мин и 37°С. На следующий день пересеивали 1/20 в 100 мл среды LB и культивировали в орбитальном шейкере при 150 об/мин и 37°С до достижения OD600=0,5. По достижении необходимой оптической плотности, клетки охлаждали до 0°С и центрифугировали 10 мин при 4000g. Далее удаляли среду, клетки отмывали от остатков среды охлажденной во льду 100 мл бидистилированной водой дважды, затем промывали 20 мл 10% глицерина. После этого, клетки ресуспендировали в 1 мл 10% глицерина и использовали для трансформации путем электропорации генотерапевтическим ДНК-вектором GDTT1.8NAS1. Электропорацию проводили в кюветах 1 мм при 2 кВ, 200 Ом, 25 мкФ на приборе Gene Pulser Xcell (Bio-Rad, США). Время импульса составляло от 4,9 до 5,1 мсек, с использованием 1-10 нг вектора. После чего клетки культивировали в течение 2,5 часов в среде SOC в шейкере-инкубаторе при 30°С. Затем, клетки высеивали на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола. Таким образом получали штамм Escherichia coli JM110-NAS/GDTT1.8NAS1, несущий генотерапевтический ДНК-вектор GDTT1.8NAS1. Через 48 часов одиночной колонией инокулировали 10 мл жидкой селективной среды, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола, и культивировали ночь в орбитальном шейкере при 150 об/мин и 37°С. На следующий день клетки осаждали центрифугированием и выделяли ДНК-вектор методом щелочного лизиса с помощью набора GeneJET Plasmid Miniprep Kit (Thermo Fisher Scientific) согласно рекомендациям производителя.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS1 задепонирован в коллекциях Национального биоресурсного центра - Всероссийская коллекция промышленных микроорганизмов (НБЦ ВКПМ), РФ и INTERNATIONAL DEPOSITARY AUTHORITY NCIMB Patent Deposit Service, UK (регистрационный № ВКПМ B-13223, дата депонирования 21.08.2018; accession № NCIMB 43108, дата депонирования 19.07.2018).
Пример 29.
Получение штамма Escherichia coli JM-110-NAS-GDTT1.8NAS2 несущего генотерапевтический ДНК-вектор GDTT1.8NAS2 и/или генотерапевтические ДНК-вектора, содержащие в своем составе целевые гены на его основе для его наработки.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS2, несущий генотерапевтический ДНК-вектор GDTT1.8NAS2 получали по примеру 28 с отличием в том, что трансформацию электрокомпетентных клеток штамма Escherichia coli JM110-NAS путем электропорации проводили генотерапевтическим ДНК-вектором GDTT1.8NAS2.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS2 задепонирован в коллекциях Национального биоресурсного центра - Всероссийская коллекция промышленных микроорганизмов (НБЦ ВКПМ), РФ и INTERNATIONAL DEPOSITARY AUTHORITY NCIMB Patent Deposit Service, UK (регистрационный № ВКПМ B-13224, дата депонирования 21.08.2018; accession № NCIMB 43109, дата депонирования 19.07.2018).
Пример 30.
Получение штамма Escherichia coli JM-110-NAS-GDTT1.8NAS3 несущего генотерапевтический ДНК-вектор GDTT1.8NAS3 и/или генотерапевтические ДНК-вектора, содержащие в своем составе целевые гены на его основе для его наработки.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS3, несущий генотерапевтический ДНК-вектор GDTT1.8NAS3 получали по примеру 28 с отличием в том, что трансформацию электрокомпетентных клеток штамма Escherichia coli JM110-NAS путем электропорации проводили генотерапевтическим ДНК-вектором GDTT1.8NAS3.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS3 задепонирован в коллекциях Национального биоресурсного центра - Всероссийская коллекция промышленных микроорганизмов (НБЦ ВКПМ), РФ и INTERNATIONAL DEPOSITARY AUTHORITY NCIMB Patent Deposit Service, UK (регистрационный № ВКПМ B-13225, дата депонирования 21.08.2018; accession № NCIMB 43110, дата депонирования 19.07.2018).
Пример 31.
Получение штамма Escherichia coli JM-110-NAS-GDTT1.8NAS4 несущего генотерапевтический ДНК-вектор GDTT1.8NAS4 и/или генотерапевтические ДНК-вектора, содержащие в своем составе целевые гены на его основе для его наработки.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS4, несущий генотерапевтический ДНК-вектор GDTT1.8NAS4 получали по примеру 28 с отличием в том, что трансформацию электрокомпетентных клеток штамма Escherichia coli JM110-NAS путем электропорации проводили генотерапевтическим ДНК-вектором GDTT1.8NAS4.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS4 задепонирован в коллекциях Национального биоресурсного центра - Всероссийская коллекция промышленных микроорганизмов (НБЦ ВКПМ), РФ и INTERNATIONAL DEPOSITARY AUTHORITY NCIMB Patent Deposit Service, UK (регистрационный № ВКПМ B-13226, дата депонирования 21.08.2018; accession № NCIMB 43111, дата депонирования 19.07.2018).
Пример 32.
Получение штамма Escherichia coli JM-110-NAS-GDTT1.8NAS5 несущего генотерапевтический ДНК-вектор GDTT1.8NAS5 и/или генотерапевтические ДНК-вектора, содержащие в своем составе целевые гены на его основе для его наработки.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS5, несущий генотерапевтический ДНК-вектор GDTT1.8NAS5 получали по примеру 28 с отличием в том, что трансформацию электрокомпетентных клеток штамма Escherichia coli JM110-NAS путем электропорации проводили генотерапевтическим ДНК-вектором GDTT1.8NAS5.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS5 задепонирован в коллекциях Национального биоресурсного центра - Всероссийская коллекция промышленных микроорганизмов (НБЦ ВКПМ), РФ и INTERNATIONAL DEPOSITARY AUTHORITY NCIMB Patent Deposit Service, UK (регистрационный № ВКПМ B-13227, дата депонирования 21.08.2018; accession № NCIMB 43112, дата депонирования 19.07.2018).
Пример 33.
Получение штамма Escherichia coli JM-110-NAS-GDTT1.8NAS6 несущего генотерапевтический ДНК-вектор GDTT1.8NAS6 и/или генотерапевтические ДНК-вектора, содержащие в своем составе целевые гены на его основе для его наработки.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS6, несущий генотерапевтический ДНК-вектор GDTT1.8NAS6 получали по примеру 28 с отличием в том, что трансформацию электрокомпетентных клеток штамма Escherichia coli JM110-NAS путем электропорации проводили генотерапевтическим ДНК-вектором GDTT1.8NAS6.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS6 задепонирован в коллекциях Национального биоресурсного центра - Всероссийская коллекция промышленных микроорганизмов (НБЦ ВКПМ), РФ и INTERNATIONAL DEPOSITARY AUTHORITY NCIMB Patent Deposit Service, UK (регистрационный № ВКПМ B-13228, дата депонирования 21.08.2018; accession № NCIMB 43113, дата депонирования 19.07.2018).
Пример 34.
Получение штамма Escherichia coli JM-110-NAS-GDTT1.8NAS7 несущего генотерапевтический ДНК-вектор GDTT1.8NAS7 и/или генотерапевтические ДНК-вектора, содержащие в своем составе целевые гены на его основе для его наработки.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS7, несущий генотерапевтический ДНК-вектор GDTT1.8NAS7 получали по примеру 28 с отличием в том, что трансформацию электрокомпетентных клеток штамма Escherichia coli JM110-NAS путем электропорации проводили генотерапевтическим ДНК-вектором GDTT1.8NAS7.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS7 задепонирован в коллекциях Национального биоресурсного центра - Всероссийская коллекция промышленных микроорганизмов (НБЦ ВКПМ), РФ и INTERNATIONAL DEPOSITARY AUTHORITY NCIMB Patent Deposit Service, UK (регистрационный № ВКПМ B-13229, дата депонирования 21.08.2018; accession № NCIMB 43114, дата депонирования 19.07.2018).
Пример 35.
Получение штамма Escherichia coli JM-110-NAS-GDTT1.8NAS8 несущего генотерапевтический ДНК-вектор GDTT1.8NAS8 и/или генотерапевтические ДНК-вектора, содержащие в своем составе целевые гены на его основе для его наработки.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS8, несущий генотерапевтический ДНК-вектор GDTT1.8NAS8 получали по примеру 28 с отличием в том, что трансформацию электрокомпетентных клеток штамма Escherichia coli JM110-NAS путем электропорации проводили генотерапевтическим ДНК-вектором GDTT1.8NAS8.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS8 задепонирован в коллекциях Национального биоресурсного центра - Всероссийская коллекция промышленных микроорганизмов (НБЦ ВКПМ), РФ и INTERNATIONAL DEPOSITARY AUTHORITY NCIMB Patent Deposit Service, UK (регистрационный № ВКПМ B-13230, дата депонирования 21.08.2018; accession № NCIMB 43115, дата депонирования 19.07.2018).
Пример 36.
Получение штамма Escherichia coli JM-110-NAS-GDTT1.8NAS9 несущего генотерапевтический ДНК-вектор GDTT1.8NAS9 и/или генотерапевтические ДНК-вектора, содержащие в своем составе целевые гены на его основе для его наработки.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS9, несущий генотерапевтический ДНК-вектор GDTT1.8NAS9 получали по примеру 28 с отличием в том, что трансформацию электрокомпетентных клеток штамма Escherichia coli JM110-NAS путем электропорации проводили генотерапевтическим ДНК-вектором GDTT1.8NAS9.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS9 задепонирован в коллекциях Национального биоресурсного центра - Всероссийская коллекция промышленных микроорганизмов (НБЦ ВКПМ), РФ и INTERNATIONAL DEPOSITARY AUTHORITY NCIMB Patent Deposit Service, UK (регистрационный № ВКПМ B-13231, дата депонирования 21.08.2018; accession № NCIMB 43116, дата депонирования 19.07.2018).
Пример 37.
Получение штамма Escherichia coli JM-110-NAS GDTT1.8NAS10 несущего генотерапевтический ДНК-вектор GDTT1.8NAS10 и/или генотерапевтические ДНК-вектора, содержащие в своем составе целевые гены на его основе для его наработки.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS10, несущий генотерапевтический ДНК-вектор GDTT1.8NAS10 получали по примеру 28 с отличием в том, что трансформацию электрокомпетентных клеток штамма Escherichia coli JM110-NAS путем электропорации проводили генотерапевтическим ДНК-вектором GDTT1.8NAS10.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS10 задепонирован в коллекциях Национального биоресурсного центра - Всероссийская коллекция промышленных микроорганизмов (НБЦ ВКПМ), РФ и INTERNATIONAL DEPOSITARY AUTHORITY NCIMB Patent Deposit Service, UK (регистрационный № ВКПМ B-13232, дата депонирования 21.08.2018; accession № NCIMB 43117, дата депонирования 19.07.2018).
Пример 38.
Получение штамма Escherichia coli JM-110-NAS - GDTT1.8NAS11 несущего генотерапевтический ДНК-вектор GDTT1.8NAS11 и/или генотерапевтические ДНК-вектора, содержащие в своем составе целевые гены на его основе для его наработки.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS11, несущий генотерапевтический ДНК-вектор GDTT1.8NAS11 получали по примеру 28 с отличием в том, что трансформацию электрокомпетентных клеток штамма Escherichia coli JM110-NAS путем электропорации проводили генотерапевтическим ДНК-вектором GDTT1.8NAS11.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS11 задепонирован в коллекциях Национального биоресурсного центра - Всероссийская коллекция промышленных микроорганизмов (НБЦ ВКПМ), РФ и INTERNATIONAL DEPOSITARY AUTHORITY NCIMB Patent Deposit Service, UK (регистрационный № ВКПМ B-13233, дата депонирования 21.08.2018; accession № NCIMB 43118, дата депонирования 19.07.2018).
Пример 39.
Способ производства в промышленных масштабах генотерапевтического ДНК-вектора GDTT1.8NAS1 или GDTT1.8NAS2 или GDTT1.8NAS3 или GDTT1.8NAS4 или GDTT1.8NAS5 или GDTT1.8NAS6 или GDTT1.8NAS7 или GDTT1.8NAS8 или GDTT1.8NAS9 или GDTT1.8NAS10 или GDTT1.8NAS11 и/или генотерапевтических ДНК-векторов, несущих целевые гены на его основе.
Для подтверждения технологичности получения и возможности производства в промышленных масштабах генотерапевтического ДНК-вектора GDTT1.8NAS1 или GDTT1.8NAS2 или GDTT1.8NAS3 или GDTT1.8NAS4 или GDTT1.8NAS5 или GDTT1.8NAS6 или GDTT1.8NAS7 или GDTT1.8NAS8 или GDTT1.8NAS9 или GDTT1.8NAS10 или GDTT1.8NAS11 и/или генотерапевтических ДНК-векторов, несущих целевые гены на его основе проводили масштабную ферментацию штаммов Escherichia coli JM-110-NAS-GDTT1.8NAS1 или Escherichia coli JM-110-NAS-GDTT1. или Escherichia coli JM-110-NAS-GDTT1.8NAS3 или Escherichia coli JM-110-NAS-GDTT1.8NAS4 или Escherichia coli JM-110-NAS-GDTT1.8NAS5 или Escherichia coli JM-110-NAS-GDTT1.8NAS6 или Escherichia coli JM-110-NAS-GDTT1.8NAS7 или Escherichia coli JM-110-NAS-GDTT1.8NAS8 или Escherichia coli JM-110-NAS-GDTT1.8NAS9 или Escherichia coli JM-110-NAS-GDTT1.8NAS10 или Escherichia coli JM-110-NAS-GDTT1.8NAS11, каждый из которых несет генотерапевтический ДНК-вектор GDTT1.8NAS1 или GDTT1.8NAS2 или GDTT1.8NAS3 или GDTT1.8NAS4 или GDTT1.8NAS5 или GDTT1.8NAS6 или GDTT1.8NAS7 или GDTT1.8NAS8 или GDTT1.8NAS9 или GDTT1.8NAS10 или GDTT1.8NAS11.
Каждый штамм Escherichia coli JM-110-NAS-GDTT1.8NAS1 или Escherichia coli JM-110-NAS-GDTT1.8NAS2 или Escherichia coli JM-110-NAS-GDTT1.8NAS3 или Escherichia coli JM-110-NAS-GDTT1.8NAS4 или Escherichia coli JM-110-NAS-GDTT1.8NAS5 или Escherichia coli JM-110-NAS-GDTT1.8NAS6 или Escherichia coli JM-110-NAS-GDTT1.8NAS7 или Escherichia coli JM-110-NAS-GDTT1.8NAS8 или Escherichia coli JM-110-NAS-GDTT1.8NAS9 или Escherichia coli JM-110-NAS-GDTT1.8NAS10 или Escherichia coli JM-110-NAS-GDTT1.8NAS11 получали по Примеру 28 или 29 или 30 или 31 или 32 или 33 или 34 или 35 или 36 или 37 или 38.
Проводили ферментацию штамма Escherichia coli JM110-NAS/GDTT1.8NAS1 несущего генотерапевтический ДНК-вектор GDTT1.8NAS1 в ферментере объемом 10 л с последующим выделением генотерапевтического ДНК-вектора GDTT1.8NAS1.
Для ферментации штамма Escherichia coli JM110-NAS/GDTT1.8NAS1 готовили среду, содержащую на 10 л: 100 г триптон, 50 г дрожжевой экстракт (Becton Dickinson), доводили водой до 8800 мл и автокпавировали при 121 0С 20 мин, затем добавляли 1200 мл 50% (вес/объем) сахарозы. Далее засевали в колбу затравочную культуру штамма Escherichia coli JM110-NAS/GDTT1.8NAS1 в объеме 100 мл. Инкубировали в шейкере-инкубаторе 16 ч при 30°С. Переносили затравочную культуру в ферментер Techfors S (Infors HT, Швейцария), растили до достижения стационарной фазы. Контроль осуществляли измерением оптической плотности культуры при длине волны 600 нм. Клетки осаждали центрифугированием 30 мин при 5000-10000 g. Супернатант удаляли, клеточную массу ресуспендировали в 10% по объему фосфатно-солевого буфера. Повторно центрифугировали 30 мин при 5000-10000g. Супернатант удаляли, к клеточной массе добавляли раствор 20 мМ TrisCl, 1 мМ ЭДТА, 200 г/л сахароза, рН 8,0 в объеме 1000 мл, тщательно перемешивали до образования гомогенной суспензии. Добавляли раствор яичного лизоцима до конечной концентрации 100 мкг/мл. Инкубировали 20 мин на льду при бережном перемешивании. Далее, добавляли 2500 мл раствора 0,2 М NaOH, 10 г/л додецилсульфат натрия, инкубировали 10 мин на льду при бережном перемешивании, затем добавляли 3500 мл раствора 3М ацетат натрия, 2М уксусная кислота, рН 5-5,5, инкубировали 10 мин на льду при бережном перемешивании. Полученный образец центрифугировали 20-30 мин при 15000 g или более. Раствор аккуратно декантировали, от остатков осадка избавляли фильтрацией через грубый фильтр (фильтровальную бумагу). Добавляли РНКазу A (Sigma) до конечной концентрации 20 нг/мл, инкубировали ночь 16 ч при комнатной температуре. Раствор центрифугировали 20-30 мин при 15000g, затем фильтровали через мембранный фильтр с порами 0,45 мкм (Millipore). Далее проводили ультрафильтрацию через мембрану размером отсечения 100 кДа (Millipore) и разбавляли до начального объема буфером 25 мМ TrisCl, рН 7.0. Операцию повторяли три - четыре раза. Наносили раствор, на колонку с 250 мл сорбента DEAE Sepharose HP (GE, США), уравновешенную раствором 25 мМ TrisCl, pH 7.0. После нанесения образца колонку промывали тремя объемами этого же раствора, а затем проводили элюцию генотерапевтического ДНК-вектора GDTT1.8NAS1 линейным градиентом от раствора 25 мМ TrisCl, рН 7.0 до раствора 25 мМ TrisCl, рН 7.0, 1М NaCl в объеме пяти объемов колонки. Контроль элюции осуществляли по оптической плотности сходящего раствора при 260 нм. Хроматографические фракции, содержащие генотерапевтический ДНК-вектор GDTT1.8NAS1, объединяли и проводили гель-фильтрацию на сорбенте Superdex 200 (GE, США). Колонку уравновешивали фосфатно-солевым буфером. Контроль элюции осуществляли по оптической плотности сходящего раствора при 260 нм, фракции анализировали электрофорезом в агарозном геле. Фракции, содержащие генотерапевтический ДНК-вектор GDTT1.8NAS1 объединяли и хранили при -20°С. Для оценки воспроизводимости техпроцесса обозначенные технологические операции повторяли четырехкратно. Выход конечного продукта был достаточен для организации масштабного промышленного производства генотерапевтического ДНК-вектора GDTT1.8NAS1.
Все технологические операции для штаммов Escherichia coli JM-110-NAS-GDTT1.8NAS2 или Escherichia coli JM-110-NAS-GDTT1.8NAS3 или Escherichia coli JM-110-NAS-GDTT1.8NAS4 или Escherichia coli JM-110-NAS-GDTT1.8NAS5 или Escherichia coli JM-110-NAS-GDTT1.8NAS6 или Escherichia coli JM-110-NAS-GDTT1.8NAS7 или Escherichia coli JM-110-NAS-GDTT1.8NAS8 или Escherichia coli JM-110-NAS-GDTT1.8NAS9 или Escherichia coli JM-110-NAS-GDTT1.8NAS10 или Escherichia coli JM-110-NAS-GDTT1.8NAS11 проводили аналогичным образом. Воспроизводимость техпроцесса и количественные характеристики выхода конечного продукта подтверждают технологичность получения и возможность производства в промышленных масштабах генотерапевтического ДНК-вектора GDTT1.8NAS1 или GDTT1.8NAS2 или GDTT1.8NAS3 или GDTT1.8NAS4 или GDTT1.8NAS5 или GDTT1.8NAS6 или GDTT1.8NAS7 или GDTT1.8NAS8 или GDTT1.8NAS9 или GDTT1.8NAS10 или GDTT1.8NAS11 и/или генотерапевтических ДНК-векторов, несущих целевые гены на его основе.
Созданные генотерапевтические ДНК-вектора с целевыми генами могут быть использованы для введения в клетки организма животных и человека, характеризующихся сниженной или недостаточной экспрессией данного гена, обеспечивая таким образом достижение терапевтического эффекта, а также возможность редактирования последовательности генома человека. При этом тканевая специфичность экспрессии целевого гена определяется наличием в генотерапевтическом ДНК-векторе специальных регуляторных элементов.
Таким образом, задача, поставленная в данном изобретении, а именно: конструирование генотерапевтического ДНК-вектора для генетической модификации клеток животных и человека, оптимально сочетающего в себе:
I) возможность безопасного использования для генетической терапии человека и животных за счет отсутствия в составе генотерапевтического ДНК-вектора генов антибиотикорезистентности;
II) длину, обеспечивающую эффективное проникновение в клетку-мишень;
III) наличие регуляторных элементов, обеспечивающих эффективную экспрессию целевых генов и, в то же время, не представляющих собой нуклеотидные последовательности вирусных геномов;
IV) возможность выбора для тагетной генной терапии генотерапевтического ДНК-вектора из группы генотерапевтических ДНК-векторов, несущих различные промоторно-регуляторные участки, для конструирования генотерапевтического ДНК-вектора, содержащего целевой ген, исходя из критериев обеспечения его тканеспецифичной экспрессии и максимальной эффективности в целевой клеточной линии, целевой ткани или органе человека.
V) технологичность получения и возможность наработки в промышленных масштабах, а также - конструирование штаммов, несущих эти генотерапевтические ДНК-вектора, для производства в промышленных масштабах этих генотерапевтических ДНК-векторов - решена, что подтверждается примерами: для п. I -пример 1, 2, 3, 4, 5; 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26; для п.II - пример 1, 2, 3, 4, 5; 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21,22, 23, 24, 25, 26; для п. III - пример 1, 2, 3, 4, 5; 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26; для п. IV - пример 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26; для п. V - пример 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39.
Промышленная применимость
Все представленные выше примеры подтверждают промышленную применимость предлагаемых генотерапевтического ДНК-вектора GDTT1.8NAS1 или GDTT1.8NAS2 или GDTT1.8NAS3 или GDTT1.8NAS4 или GDTT1.8NAS5 или GDTT1.8NAS6 или GDTT1.8NAS7 или GDTT1.8NAS8 или GDTT1.8NAS9 или GDTT1.8NAS10 или GDTT1.8NAS11, способа его получения, штамма Escherichia coli JM-110-NAS, способа его получения, штамма Escherichia coli JM-110-NAS-GDTT1.8NAS1 или Escherichia coli JM-110-NAS-GDTT1.8NAS2 или Escherichia coli JM-110-NAS-GDTT1.8NAS3 или Escherichia coli JM-110-NAS-GDTT1.8NAS4 или Escherichia coli JM-110-NAS-GDTT1.8NAS5 или Escherichia coli JM-110-NAS-GDTT1.8NAS6 или Escherichia coli JM-110-NAS-GDTT1.8NAS7 или Escherichia coli JM-110-NAS-GDTT1.8NAS8 или Escherichia coli JM-110-NAS-GDTT1.8NAS9 или Escherichia coli JM-110-NAS-GDTT1.8NAS10 или Escherichia coli JM-110-NAS-GDTT1.8NAS11, несущего генотерапевтический ДНК-вектор GDTT1.8NAS1 или GDTT1.8NAS2 или GDTT1.8NAS3 или GDTT1.8NAS4 или GDTT1.8NAS5 или GDTT1.8NAS6 или GDTT1.8NAS7 или GDTT1.8NAS8 или GDTT1.8NAS9 или GDTT1.8NAS10 или GDTT1.8NAS11, способа его получения, способа производства в промышленных масштабах генотерапевтического ДНК-вектора
Перечень последовательностей олигонулеотидов:


Перечень сокращений
GDTT1.8NAS_ - вектор генотерапевтический, не содержащий последовательностей вирусных геномов и маркеров антибиотико-резистентности (GDT - аббревиатура производителя, Т - vector therapeutic, 1.8 - базовая длинна, NA - no antibiotic)
ДНК - дезоксирибонуклеиновая кислота
кДНК - комплементарная дезоксирибонуклеиновая кислота
РНК - рибонуклеиновая кислота
мРНК - матричная рибонуклеиновая кислота
п.н. - пар оснований
ПЦР - полимеразная цепная реакция
мл - миллилитр, мкл - микролитр
л - литр
мкг - микрограмм
мг - миллиграмм
г - грамм
мкМ - микромоль
мМ - миллимоль
мин - минута
сек - секунда
об/мин - обороты в минуту
нм - нанометр
см - сантиметр
мВт - милливатт
о.е.ф - относительная единица флуоресценции
РВС - фосфатно-солевой буфер.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110>ГЕНЕТИК ДИАГНОСТИКС ЭНД ТЕРАПИ 21 ЛТД (GENETIC DIAGNOSTICS AND THERAPY 21 LTD), Общество с ограниченной ответственностью «РЕКОМБИТЕХ»
<120>Генотерапевтический ДНК-вектор для таргетной генной терапии, способ его получения (варианты), штамм для его производства, способ его получения.
<160>11
<170> BiSSAP 1.3.6
<210> 1
<211>1736
<212> DNA
<213> Homo Sapiens
<400> 1
ataacgcagg aaagaacatg tgagcaaaag gccagcaaaa ggccagggac cgtaaaaagg 60
ccgcgttgct ggcgtttttc cataggctcc gcccccctga cgagcatcac aaaaatcgac 120
gctcaagtca gaggtggcga aacccgacag gactataaag ataccaggcg tttccccctg 180
gaagctccct cgtgcgctct cctgttccga ccctgccgct taccggatac ctgtccgcct 240
ttctcccttc gggaagcgtg gcgctttctc atagctcacg ctgtaggtat ctcagttcgg 300
tgtaggtcgt tcgctccaag ctgggctgtg tgcacgaacc ccccgttcag cccgaccgct 360
gcgccttatc cggtaactat cgtcttgagt ccaacccggt aagacacgac ttatcgccac 420
tggcagcagc cactggtaac aggattagca gagcgaggta tgtaggcggt gctacagagt 480
tcttgaagtg gtggcctaac tacggctaca ctagaagaac agtatttggt atctgcgctc 540
tgctgaagcc agttaccttc ggaaaaagag ttggtagctc ttgatccggc aaacaaacca 600
ccgctggtag cggtggtttt tttgtttgca agcagcagat tacgcgcaga aaaaaaggat 660
ctcaagaaga tcctttgatc ttttctacct cgaggtggcc aagcttagaa acatgacagg 720
tcctcttggg agggctgacc gcagggagcg ttgggtttca ggctgctggc gtcggcttct 780
gtggtgccct ttctgtcggc tatgagagtc cagacagtgc ccaacctcct ccccttcttt 840
ccacacgcac aaccacccca ccccctgtgg cctgagctgt cctgcctcgc cacaatggca 900
cctgccctaa aatagcttcc catgtgaggg ctagagaaag gaaaagatta gaccctccct 960
ggatgagaga gagaaagtga aggagggcag gggaggggga cagcgagcca ttgagcgatc 1020
tttgtcaagc atcccagaag gtataaggat ccgatatcgt cgacaagctt ggtacctccg 1080
gagcggccgc tctagagcta gcgacgtcga attccctgtg acccctcccc agtgcctctc 1140
ctggccctgg aagttgccac tccagtgccc accagccttg tcctaataaa attaagttgc 1200
atcattttgt ctgactaggt gtccttctat aatattatgg ggtggagggg ggtggtatgg 1260
agcaaggggc aagttgggaa gacaacctgt agggcctgcg gggtctattg ggaaccaagc 1320
tggagtgcag tggcacaatc ttggctcact gcaatctccg cctcctgggt tcaagcgatt 1380
ctcctgcctc agcctcccga gttgttggga ttccaggcat gcatgaccag gctcagctaa 1440
tttttgtttt tttggtagag acggggtttc accatattgg ccaggctggt ctccaactcc 1500
taatctcagg tgatctaccc accttggcct cccaaattgc tgggattaca ggcgtgaacc 1560
actgctccct tccctgtcct tacgcgtaga attggtaaag agagtcgtgt aaaatatcga 1620
gttcgcacat cttgttgtct gattattgat ttttggcgaa accatttgat catatgacaa 1680
gatgtgtatc taccttaact taatgatttt gataaaaatc attaactagt ccatgg 1736
<210> 2
<211>1654
<212>DNA
<213>HomoSapiens
<400> 2
ataacgcagg aaagaacatg tgagcaaaag gccagcaaaa ggccagggac cgtaaaaagg 60
ccgcgttgct ggcgtttttc cataggctcc gcccccctga cgagcatcac aaaaatcgac 120
gctcaagtca gaggtggcga aacccgacag gactataaag ataccaggcg tttccccctg 180
gaagctccct cgtgcgctct cctgttccga ccctgccgct taccggatac ctgtccgcct 240
ttctcccttc gggaagcgtg gcgctttctc atagctcacg ctgtaggtat ctcagttcgg 300
tgtaggtcgt tcgctccaag ctgggctgtg tgcacgaacc ccccgttcag cccgaccgct 360
gcgccttatc cggtaactat cgtcttgagt ccaacccggt aagacacgac ttatcgccac 420
tggcagcagc cactggtaac aggattagca gagcgaggta tgtaggcggt gctacagagt 480
tcttgaagtg gtggcctaac tacggctaca ctagaagaac agtatttggt atctgcgctc 540
tgctgaagcc agttaccttc ggaaaaagag ttggtagctc ttgatccggc aaacaaacca 600
ccgctggtag cggtggtttt tttgtttgca agcagcagat tacgcgcaga aaaaaaggat 660
ctcaagaaga tcctttgatc ttttctacct cgagcatctt atttctgaaa aagagtttgt 720
tggatgcttt tcaattaaaa gcagatgatt aattcagccc tggccttgtt ctgtctttga 780
ataacagata cattagttta cttgaaaaga aatattttca tttgtatttc catgtcactt 840
gtgattttcc ctgccttgca ccctcagccc tgccagttgg caagaagaca gtcagctttg 900
ctgctaagag gagtataaag agggcttggt ccaagcaaga aggcagtggt ctactccatc 960
ggcaggatcc gatatcgtcg acaagcttgg tacctccgga gcggccgctc tagagctagc 1020
gacgtcgaat tccctgtgac ccctccccag tgcctctcct ggccctggaa gttgccactc 1080
cagtgcccac cagccttgtc ctaataaaat taagttgcat cattttgtct gactaggtgt 1140
ccttctataa tattatgggg tggagggggg tggtatggag caaggggcaa gttgggaaga 1200
caacctgtag ggcctgcggg gtctattggg aaccaagctg gagtgcagtg gcacaatctt 1260
ggctcactgc aatctccgcc tcctgggttc aagcgattct cctgcctcag cctcccgagt 1320
tgttgggatt ccaggcatgc atgaccaggc tcagctaatt tttgtttttt tggtagagac 1380
ggggtttcac catattggcc aggctggtct ccaactccta atctcaggtg atctacccac 1440
cttggcctcc caaattgctg ggattacagg cgtgaaccac tgctcccttc cctgtcctta 1500
cgcgtagaat tggtaaagag agtcgtgtaa aatatcgagt tcgcacatct tgttgtctga 1560
ttattgattt ttggcgaaac catttgatca tatgacaaga tgtgtatcta ccttaactta 1620
atgattttga taaaaatcat taactagtcc atgg 1654
<210>3
<211>1783
<212>DNA
<213>HomoSapiens
<400>3
ataacgcagg aaagaacatg tgagcaaaag gccagcaaaa ggccagggac cgtaaaaagg 60
ccgcgttgct ggcgtttttc cataggctcc gcccccctga cgagcatcac aaaaatcgac 120
gctcaagtca gaggtggcga aacccgacag gactataaag ataccaggcg tttccccctg 180
gaagctccct cgtgcgctct cctgttccga ccctgccgct taccggatac ctgtccgcct 240
ttctcccttc gggaagcgtg gcgctttctc atagctcacg ctgtaggtat ctcagttcgg 300
tgtaggtcgt tcgctccaag ctgggctgtg tgcacgaacc ccccgttcag cccgaccgct 360
gcgccttatc cggtaactat cgtcttgagt ccaacccggt aagacacgac ttatcgccac 420
tggcagcagc cactggtaac aggattagca gagcgaggta tgtaggcggt gctacagagt 480
tcttgaagtg gtggcctaac tacggctaca ctagaagaac agtatttggt atctgcgctc 540
tgctgaagcc agttaccttc ggaaaaagag ttggtagctc ttgatccggc aaacaaacca 600
ccgctggtag cggtggtttt tttgtttgca agcagcagat tacgcgcaga aaaaaaggat 660
ctcaagaaga tcctttgatc ttttctacct cgaggtctcc caggcatgac tccaacaatg 720
catcccatgg gatttggggt tccccagatc tggggcttgt aggcctgact ctcccctgtg 780
cacacgtctc atacacgcat gcgtgcaccc attgcctgcc ccgccccttg cacagggagt 840
cagcagggag gactgggtta tgccctgctt atcagcagct tcccagcttc ctctgcctgg 900
attcttagag gcctggggtc ctagaacgag ctggtgcacg tggcttccca aagatctctc 960
agataatgag aggaaatgca gtcatcagtt tgcagaaggc tagggattct gggccatagc 1020
tcagacctgc gcccaccatc tccctccagg cagcccttgg ctggtccctg cgagcccgtg 1080
gagactgcca gtcggatccg atatcgtcga caagcttggt acctccggag cggccgctct 1140
agagctagcg acgtcgaatt ccctgtgacc cctccccagt gcctctcctg gccctggaag 1200
ttgccactcc agtgcccacc agccttgtcc taataaaatt aagttgcatc attttgtctg 1260
actaggtgtc cttctataat attatggggt ggaggggggt ggtatggagc aaggggcaag 1320
ttgggaagac aacctgtagg gcctgcgggg tctattggga accaagctgg agtgcagtgg 1380
cacaatcttg gctcactgca atctccgcct cctgggttca agcgattctc ctgcctcagc 1440
ctcccgagtt gttgggattc caggcatgca tgaccaggct cagctaattt ttgttttttt 1500
ggtagagacg gggtttcacc atattggcca ggctggtctc caactcctaa tctcaggtga 1560
tctacccacc ttggcctccc aaattgctgg gattacaggc gtgaaccact gctcccttcc 1620
ctgtccttac gcgtagaatt ggtaaagaga gtcgtgtaaa atatcgagtt cgcacatctt 1680
gttgtctgat tattgatttt tggcgaaacc atttgatcat atgacaagat gtgtatctac 1740
cttaacttaa tgattttgat aaaaatcatt aactagtcca tgg 1783
<210>4
<211>1953
<212>DNA
<213>HomoSapiens
<400>4
ataacgcagg aaagaacatg tgagcaaaag gccagcaaaa ggccagggac cgtaaaaagg 60
ccgcgttgct ggcgtttttc cataggctcc gcccccctga cgagcatcac aaaaatcgac 120
gctcaagtca gaggtggcga aacccgacag gactataaag ataccaggcg tttccccctg 180
gaagctccct cgtgcgctct cctgttccga ccctgccgct taccggatac ctgtccgcct 240
ttctcccttc gggaagcgtg gcgctttctc atagctcacg ctgtaggtat ctcagttcgg 300
tgtaggtcgt tcgctccaag ctgggctgtg tgcacgaacc ccccgttcag cccgaccgct 360
gcgccttatc cggtaactat cgtcttgagt ccaacccggt aagacacgac ttatcgccac 420
tggcagcagc cactggtaac aggattagca gagcgaggta tgtaggcggt gctacagagt 480
tcttgaagtg gtggcctaac tacggctaca ctagaagaac agtatttggt atctgcgctc 540
tgctgaagcc agttaccttc ggaaaaagag ttggtagctc ttgatccggc aaacaaacca 600
ccgctggtag cggtggtttt tttgtttgca agcagcagat tacgcgcaga aaaaaaggat 660
ctcaagaaga tcctttgatc ttttctacct cgagcaccca cagcaggctg cctttggtga 720
ctcaccgggt gaacgggggc attgcgaggc atcccctccc tgggtttggc tcctgcccac 780
ggggctgaca gtagaaatca caggctgtga gacagctgga gcccagctct gcttgaacct 840
attttaggtc tctgatcccc gcttcctctt tagactcccc tagagctcag ccagtgctca 900
acctgaggct gggggtctct gaggaagagt gagttggagc tgaggggtct ggggctgtcc 960
cctgagagag gggccagagg cagtgtcaag agccgggcag tctgattgtg gctcaccctc 1020
catcactccc aggggcccct ggcccagcag ccgcagctcc caaccacaat atcctttggg 1080
gtttggccta cggagctggg gcggatgacc cccaaatagc cctggcagat tccccctaga 1140
cccgcccgca ccatggtcag gcatgcccct cctcatcgct gggcacagcc cagagggtat 1200
aaacagtgct ggaggctggc ggggcaggcc agctgagtcc tgagcagcag cccagcgcag 1260
ccaggatccg atatcgtcga caagcttggt acctccggag cggccgctct agagctagcg 1320
acgtcgaatt ccctgtgacc cctccccagt gcctctcctg gccctggaag ttgccactcc 1380
agtgcccacc agccttgtcc taataaaatt aagttgcatc attttgtctg actaggtgtc 1440
cttctataat attatggggt ggaggggggt ggtatggagc aaggggcaag ttgggaagac 1500
aacctgtagg gcctgcgggg tctattggga accaagctgg agtgcagtgg cacaatcttg 1560
gctcactgca atctccgcct cctgggttca agcgattctc ctgcctcagc ctcccgagtt 1620
gttgggattc caggcatgca tgaccaggct cagctaattt ttgttttttt ggtagagacg 1680
gggtttcacc atattggcca ggctggtctc caactcctaa tctcaggtga tctacccacc 1740
ttggcctccc aaattgctgg gattacaggc gtgaaccact gctcccttcc ctgtccttac 1800
gcgtagaatt ggtaaagaga gtcgtgtaaa atatcgagtt cgcacatctt gttgtctgat 1860
tattgatttt tggcgaaacc atttgatcat atgacaagat gtgtatctac cttaacttaa 1920
tgattttgat aaaaatcatt aactagtcca tgg 1953
<210> 5
<211> 2019
<212>DNA
<213>HomoSapiens
<400> 5
ataacgcagg aaagaacatg tgagcaaaag gccagcaaaa ggccagggac cgtaaaaagg 60
ccgcgttgct ggcgtttttc cataggctcc gcccccctga cgagcatcac aaaaatcgac 120
gctcaagtca gaggtggcga aacccgacag gactataaag ataccaggcg tttccccctg 180
gaagctccct cgtgcgctct cctgttccga ccctgccgct taccggatac ctgtccgcct 240
ttctcccttc gggaagcgtg gcgctttctc atagctcacg ctgtaggtat ctcagttcgg 300
tgtaggtcgt tcgctccaag ctgggctgtg tgcacgaacc ccccgttcag cccgaccgct 360
gcgccttatc cggtaactat cgtcttgagt ccaacccggt aagacacgac ttatcgccac 420
tggcagcagc cactggtaac aggattagca gagcgaggta tgtaggcggt gctacagagt 480
tcttgaagtg gtggcctaac tacggctaca ctagaagaac agtatttggt atctgcgctc 540
tgctgaagcc agttaccttc ggaaaaagag ttggtagctc ttgatccggc aaacaaacca 600
ccgctggtag cggtggtttt tttgtttgca agcagcagat tacgcgcaga aaaaaaggat 660
ctcaagaaga tcctttgatc ttttctacct cgaggtatag ggctgtctgg gagccactcc 720
agggccacag aaatcttgtc tctgactcag ggtattttgt tttctgtttt gtgtaaatgc 780
tcttctgact aatgcaaacc atgtgtccat agaaccagaa gatttttcca ggggaaaagg 840
taaggaggtg gtgagagtgt cctgggtctg cccttccagg gcttgccctg ggttaagagc 900
caggcaggaa gctctcaaga gcattgctca agagtagagg gggcctggga ggcccaggga 960
ggggatggga ggggaacacc caggctgccc ccaaccagat gccctccacc ctcctcaacc 1020
tccctcccac ggcctggaga ggtgggacca ggtatggagg cttgagagcc cctggttgga 1080
ggaagccaca agtccaggaa catgggagtc tgggcagggg gcaaaggagg caggaacagg 1140
ccatcagcca ggacaggtgg taaggcaggc aggagtgttc ctgctgggaa aaggtgggat 1200
caagcacctg gagggctctt cagagcaaag acaaacactg aggtcgctgc cactcctaca 1260
gagcccccac gccccgccca gctataaggg gccatgcccc aagcagggta cccaggctgc 1320
agaggtgccg gatccgatat cgtcgacaag cttggtacct ccggagcggc cgctctagag 1380
ctagcgacgt cgaattccct gtgacccctc cccagtgcct ctcctggccc tggaagttgc 1440
cactccagtg cccaccagcc ttgtcctaat aaaattaagt tgcatcattt tgtctgacta 1500
ggtgtccttc tataatatta tggggtggag gggggtggta tggagcaagg ggcaagttgg 1560
gaagacaacc tgtagggcct gcggggtcta ttgggaacca agctggagtg cagtggcaca 1620
atcttggctc actgcaatct ccgcctcctg ggttcaagcg attctcctgc ctcagcctcc 1680
cgagttgttg ggattccagg catgcatgac caggctcagc taatttttgt ttttttggta 1740
gagacggggt ttcaccatat tggccaggct ggtctccaac tcctaatctc aggtgatcta 1800
cccaccttgg cctcccaaat tgctgggatt acaggcgtga accactgctc ccttccctgt 1860
ccttacgcgt agaattggta aagagagtcg tgtaaaatat cgagttcgca catcttgttg 1920
tctgattatt gatttttggc gaaaccattt gatcatatga caagatgtgt atctacctta 1980
acttaatgat tttgataaaa atcattaact agtccatgg 2019
<210> 6
<211> 1940
<212>DNA
<213>HomoSapiens
<400> 6
ataacgcagg aaagaacatg tgagcaaaag gccagcaaaa ggccagggac cgtaaaaagg 60
ccgcgttgct ggcgtttttc cataggctcc gcccccctga cgagcatcac aaaaatcgac 120
gctcaagtca gaggtggcga aacccgacag gactataaag ataccaggcg tttccccctg 180
gaagctccct cgtgcgctct cctgttccga ccctgccgct taccggatac ctgtccgcct 240
ttctcccttc gggaagcgtg gcgctttctc atagctcacg ctgtaggtat ctcagttcgg 300
tgtaggtcgt tcgctccaag ctgggctgtg tgcacgaacc ccccgttcag cccgaccgct 360
gcgccttatc cggtaactat cgtcttgagt ccaacccggt aagacacgac ttatcgccac 420
tggcagcagc cactggtaac aggattagca gagcgaggta tgtaggcggt gctacagagt 480
tcttgaagtg gtggcctaac tacggctaca ctagaagaac agtatttggt atctgcgctc 540
tgctgaagcc agttaccttc ggaaaaagag ttggtagctc ttgatccggc aaacaaacca 600
ccgctggtag cggtggtttt tttgtttgca agcagcagat tacgcgcaga aaaaaaggat 660
ctcaagaaga tcctttgatc ttttctacct cgagagtatc tgcagagggc cctgcgtatg 720
agtgcaagtg ggttttagga ccaggatgag gcggggtggg ggtgcctacc tgacgaccga 780
ccccgaccca ctggacaagc acccaacccc cattccccaa attgcgcatc ccctatcaga 840
gagggggagg ggaaacagga tgcggcgagg cgcgtgcgca ctgccagctt cagcaccgcg 900
gacagtgcct tcgcccccgc ctggcggcgc gcgccaccgc cgcctcagca ctgaaggcgc 960
gctgacgtca ctcgccggtc ccccgcaaac tccccttccc ggccaccttg gtcgcgtccg 1020
cgccgccgcc ggcccagccg gaccgcacca cgcgaggcgc gagatagggg ggcacgggcg 1080
cgaccatctg cgctgcggcg ccggcgactc agcgctgcct cagtctgcgg tgggcagcgg 1140
aggagtcgtg tcgtgcctga gagcgcagct gtgctcctgg gcaccgcgca gtccgccccc 1200
gcggctcctg gccagaccac ccctaggacc ccctgcccca agtcgcagcc ggatccgata 1260
tcgtcgacaa gcttggtacc tccggagcgg ccgctctaga gctagcgacg tcgaattccc 1320
tgtgacccct ccccagtgcc tctcctggcc ctggaagttg ccactccagt gcccaccagc 1380
cttgtcctaa taaaattaag ttgcatcatt ttgtctgact aggtgtcctt ctataatatt 1440
atggggtgga ggggggtggt atggagcaag gggcaagttg ggaagacaac ctgtagggcc 1500
tgcggggtct attgggaacc aagctggagt gcagtggcac aatcttggct cactgcaatc 1560
tccgcctcct gggttcaagc gattctcctg cctcagcctc ccgagttgtt gggattccag 1620
gcatgcatga ccaggctcag ctaatttttg tttttttggt agagacgggg tttcaccata 1680
ttggccaggc tggtctccaa ctcctaatct caggtgatct acccaccttg gcctcccaaa 1740
ttgctgggat tacaggcgtg aaccactgct cccttccctg tccttacgcg tagaattggt 1800
aaagagagtc gtgtaaaata tcgagttcgc acatcttgtt gtctgattat tgatttttgg 1860
cgaaaccatt tgatcatatg acaagatgtg tatctacctt aacttaatga ttttgataaa 1920
aatcattaac tagtccatgg 1940
<210> 7
<211>2620
<212>DNA
<213>HomoSapiens
<400> 7
ataacgcagg aaagaacatg tgagcaaaag gccagcaaaa ggccagggac cgtaaaaagg 60
ccgcgttgct ggcgtttttc cataggctcc gcccccctga cgagcatcac aaaaatcgac 120
gctcaagtca gaggtggcga aacccgacag gactataaag ataccaggcg tttccccctg 180
gaagctccct cgtgcgctct cctgttccga ccctgccgct taccggatac ctgtccgcct 240
ttctcccttc gggaagcgtg gcgctttctc atagctcacg ctgtaggtat ctcagttcgg 300
tgtaggtcgt tcgctccaag ctgggctgtg tgcacgaacc ccccgttcag cccgaccgct 360
gcgccttatc cggtaactat cgtcttgagt ccaacccggt aagacacgac ttatcgccac 420
tggcagcagc cactggtaac aggattagca gagcgaggta tgtaggcggt gctacagagt 480
tcttgaagtg gtggcctaac tacggctaca ctagaagaac agtatttggt atctgcgctc 540
tgctgaagcc agttaccttc ggaaaaagag ttggtagctc ttgatccggc aaacaaacca 600
ccgctggtag cggtggtttt tttgtttgca agcagcagat tacgcgcaga aaaaaaggat 660
ctcaagaaga tcctttgatc ttttctacct cgaggtctgt aatcccagca ttttgggagg 720
ctgaggcaga tggatcacct gaggtcagga gttcgagacc agcgtggcca acatgatgaa 780
accccgtctc tagtaaaaat acaaaaatta gccaggcatg gtgctatata cctgtagcac 840
cagctacttg ggagacagag gtgggagaat tacttgaacc tgggaggttc aagccatggg 900
aggtggaagt tgcagtgagc cgagatgcca ctgcactcca gcctgagcaa cagagcaaga 960
ctatctcaag aaaagaaaga aagaaagaaa gagacttgcc aaggtcatgt atcagggcaa 1020
ggaagagctg ggggcccagc tggctgctcc cctgctgagc tgggagacca ccttgatctg 1080
acttctccca tcttcccagc ctaagccagg ccctggggtc acggaggctg gggaggcacc 1140
gaggaacgcg cctggcatgt gctgacaggg gattttatgc tccagctggg ccagctggga 1200
ggagcctgct gggcagaggc cagagctggg ggctctggaa ggtacctggg ggaggttgca 1260
ctgtgagaat gagctcaagc tgggtcagag agcagggctg actctgccag tgcctgcatc 1320
agcctcatcg ctctcctagg ctcctggcct gctggactct gggctgcagg tccttcttga 1380
aaggctgtga gtagtgagac aaggagcagg agtgaggggt ggcaggagag aagatagaga 1440
ttgagagaga gagagagaga gagacagaga gagaggaaga gacagagaca aaaggagaga 1500
gaacggctta gacaaggaga gaaagatgga aagataaaga gactgggcgc agtggctcac 1560
gcctgtaatc ccaacacttg gggaggccaa ggtgggagga tggcttgaag gaaagagtct 1620
gagatcaacc tggccaacat agtgagaccc cgtctctaaa aaaaaaaaaa agaaaaaaaa 1680
aagaaaaaag aaaaaaaagt ttttttaaag agacagagaa agagactcag agattgagac 1740
tgagagcaag acagagagag atactcacag ggaagagggg aagaggaaaa cgagaaaggg 1800
aggagagtaa cggaaagaga taaaaaagaa aagcaggtgg cagagacaca cagagaggga 1860
cccagagaaa gccagacaga cgcaggtggc tggcagcggg cgctgtgggg gtcacagtag 1920
ggggacctgt ggatccgata tcgtcgacaa gcttggtacc tccggagcgg ccgctctaga 1980
gctagcgacg tcgaattccc tgtgacccct ccccagtgcc tctcctggcc ctggaagttg 2040
ccactccagt gcccaccagc cttgtcctaa taaaattaag ttgcatcatt ttgtctgact 2100
aggtgtcctt ctataatatt atggggtgga ggggggtggt atggagcaag gggcaagttg 2160
ggaagacaac ctgtagggcc tgcggggtct attgggaacc aagctggagt gcagtggcac 2220
aatcttggct cactgcaatc tccgcctcct gggttcaagc gattctcctg cctcagcctc 2280
ccgagttgtt gggattccag gcatgcatga ccaggctcag ctaatttttg tttttttggt 2340
agagacgggg tttcaccata ttggccaggc tggtctccaa ctcctaatct caggtgatct 2400
acccaccttg gcctcccaaa ttgctgggat tacaggcgtg aaccactgct cccttccctg 2460
tccttacgcg tagaattggt aaagagagtc gtgtaaaata tcgagttcgc acatcttgtt 2520
gtctgattat tgatttttgg cgaaaccatt tgatcatatg acaagatgtg tatctacctt 2580
aacttaatga ttttgataaa aatcattaac tagtccatgg 2620
<210>8
<211>2240
<212>DNA
<213>HomoSapiens
<400> 8
ataacgcagg aaagaacatg tgagcaaaag gccagcaaaa ggccagggac cgtaaaaagg 60
ccgcgttgct ggcgtttttc cataggctcc gcccccctga cgagcatcac aaaaatcgac 120
gctcaagtca gaggtggcga aacccgacag gactataaag ataccaggcg tttccccctg 180
gaagctccct cgtgcgctct cctgttccga ccctgccgct taccggatac ctgtccgcct 240
ttctcccttc gggaagcgtg gcgctttctc atagctcacg ctgtaggtat ctcagttcgg 300
tgtaggtcgt tcgctccaag ctgggctgtg tgcacgaacc ccccgttcag cccgaccgct 360
gcgccttatc cggtaactat cgtcttgagt ccaacccggt aagacacgac ttatcgccac 420
tggcagcagc cactggtaac aggattagca gagcgaggta tgtaggcggt gctacagagt 480
tcttgaagtg gtggcctaac tacggctaca ctagaagaac agtatttggt atctgcgctc 540
tgctgaagcc agttaccttc ggaaaaagag ttggtagctc ttgatccggc aaacaaacca 600
ccgctggtag cggtggtttt tttgtttgca agcagcagat tacgcgcaga aaaaaaggat 660
ctcaagaaga tcctttgatc ttttctacct cgaggccaag aacatcttaa gtcacagaaa 720
cattagtttt tggaagcagg gtttgctgta actatagtag aaatgacatt ctgattccac 780
tcctagcttc acaaggatat ctgtgaaaga tttggggcaa aactgttaag ctgtctgaaa 840
gtgcttttgc ataagaaatg ggttttactg ctaaaactgt catattgctg agttttgaat 900
gccctaatgg taaatgatac tgggttgcca aaaataacca gattagtagt tttttcattc 960
atttggccgt ctcagtaagt caaatattga tactttctac taagtcatct tgccaacacc 1020
cattttgtta tacttatgct gaatctgttt gtcatctctt aagtaagaaa attattgatt 1080
attttgtggg gatttaattt aaaaaaaatg gtaatggata ctgtaaagga gcattatttg 1140
gatggtttaa aaacatcttc cttgatggga aaatctttta aaaggctttc taacttggtg 1200
taattacttg aattaaggaa gtgcaatgcc attctactga cttagaacaa cttttttgac 1260
ttcctgcaaa gaggaccctt acagtatttt tggagaagtt agtaaaaccg aatctgacat 1320
catcacctag cagttcatgc agctagcaag tggtttgttc ttagggtaac agaggaggaa 1380
attgttcctc gtctgataag acaacagtgg agagtatgca tttatttatt tacttttaca 1440
tttttgattc gtttttacag agaaaaactt ctacagagat aacaattatt ttgcttttca 1500
gaaggacgca tgctgtttct tagggacacg gctgacttcc agatatgacc ggatccgata 1560
tcgtcgacaa gcttggtacc tccggagcgg ccgctctaga gctagcgacg tcgaattccc 1620
tgtgacccct ccccagtgcc tctcctggcc ctggaagttg ccactccagt gcccaccagc 1680
cttgtcctaa taaaattaag ttgcatcatt ttgtctgact aggtgtcctt ctataatatt 1740
atggggtgga ggggggtggt atggagcaag gggcaagttg ggaagacaac ctgtagggcc 1800
tgcggggtct attgggaacc aagctggagt gcagtggcac aatcttggct cactgcaatc 1860
tccgcctcct gggttcaagc gattctcctg cctcagcctc ccgagttgtt gggattccag 1920
gcatgcatga ccaggctcag ctaatttttg tttttttggt agagacgggg tttcaccata 1980
ttggccaggc tggtctccaa ctcctaatct caggtgatct acccaccttg gcctcccaaa 2040
ttgctgggat tacaggcgtg aaccactgct cccttccctg tccttacgcg tagaattggt 2100
aaagagagtc gtgtaaaata tcgagttcgc acatcttgtt gtctgattat tgatttttgg 2160
cgaaaccatt tgatcatatg acaagatgtg tatctacctt aacttaatga ttttgataaa 2220
aatcattaac tagtccatgg 2240
<210> 9
<211> 2604
<212>DNA
<213>HomoSapiens
<400> 9
ataacgcagg aaagaacatg tgagcaaaag gccagcaaaa ggccagggac cgtaaaaagg 60
ccgcgttgct ggcgtttttc cataggctcc gcccccctga cgagcatcac aaaaatcgac 120
gctcaagtca gaggtggcga aacccgacag gactataaag ataccaggcg tttccccctg 180
gaagctccct cgtgcgctct cctgttccga ccctgccgct taccggatac ctgtccgcct 240
ttctcccttc gggaagcgtg gcgctttctc atagctcacg ctgtaggtat ctcagttcgg 300
tgtaggtcgt tcgctccaag ctgggctgtg tgcacgaacc ccccgttcag cccgaccgct 360
gcgccttatc cggtaactat cgtcttgagt ccaacccggt aagacacgac ttatcgccac 420
tggcagcagc cactggtaac aggattagca gagcgaggta tgtaggcggt gctacagagt 480
tcttgaagtg gtggcctaac tacggctaca ctagaagaac agtatttggt atctgcgctc 540
tgctgaagcc agttaccttc ggaaaaagag ttggtagctc ttgatccggc aaacaaacca 600
ccgctggtag cggtggtttt tttgtttgca agcagcagat tacgcgcaga aaaaaaggat 660
ctcaagaaga tcctttgatc ttttctacct cgagaaacgg agggttgtga ggagagtgag 720
aggtggacag agggcaccga cgatttagca tctcttcctc tcctgggggt cgaggatgag 780
agacaaaaaa gaagctgcca ggaaacataa aattcagagg gctcagctgc agggctgagg 840
tctgcaagca tgctgtgtac acttgtgcat gttgtgccct gcacaagggc atctctgaag 900
gggctgcatt ggacccaggg gcaggggcgc aaaggtgagt ttatatcagt tcctgagcac 960
tgtggctcca tccagcactc tgaggacagg caggatacag ctggaggacc tgagggctcc 1020
cccacaccag cttctgttcc ctgcccaaga ccccctggac ctgcagacaa caattcaacg 1080
cactcagagt cccacagtta agaactccct gaagaagccc ccagtggctg cgtggtggat 1140
tttcgcaaag ctgtctccac ctacatccac cctgtttggc agcccctaca tactctttca 1200
cagcatgagg aagggaggcc tctcaccaag acctggactg aatcttctcc cagtggctgc 1260
cacacctgac ctgctcttgc tccagaacct ctgtggctcc catactccac agggtcaact 1320
tccaacatgg ctgcctgcac tccagccaag aggctctgct ctgggcccct ccagatgcct 1380
gacctgggtc tgtggctgcc ctgtccttct tcagtgctcc tcttcccgct gggtgaggaa 1440
tagttcagga cagaggagct aagttcaggt tcattcatag gacaggtgcc tatttcgctc 1500
acggcccagg aatagagact tgccgggctc ggcccttcgg ggagctggca gacggcagag 1560
gggaggctgg ctggcccagg ggatgaccac cggtggggta agcacagaca gaggggagca 1620
caggcttccc ccagaagact gagaggcccc ccagaggcat ccacagagga ccccagctgt 1680
gctgcccaag ctgggcgacc gccaaacctt agcggcccag ctgacaaaag cctgccctcc 1740
cccagggtcc ccggagagct ggtgcctccc ctgggtccca atttgcatgg caggaagggg 1800
cctggtgagg aagaggcggg gaggggacag gctgcagccg gtgcagttac acgttttcct 1860
ccaaggagcc tcggacgttg tcacgggttt ggggtcgggg acagagcggt gaccggatcc 1920
gatatcgtcg acaagcttgg tacctccgga gcggccgctc tagagctagc gacgtcgaat 1980
tccctgtgac ccctccccag tgcctctcct ggccctggaa gttgccactc cagtgcccac 2040
cagccttgtc ctaataaaat taagttgcat cattttgtct gactaggtgt ccttctataa 2100
tattatgggg tggagggggg tggtatggag caaggggcaa gttgggaaga caacctgtag 2160
ggcctgcggg gtctattggg aaccaagctg gagtgcagtg gcacaatctt ggctcactgc 2220
aatctccgcc tcctgggttc aagcgattct cctgcctcag cctcccgagt tgttgggatt 2280
ccaggcatgc atgaccaggc tcagctaatt tttgtttttt tggtagagac ggggtttcac 2340
catattggcc aggctggtct ccaactccta atctcaggtg atctacccac cttggcctcc 2400
caaattgctg ggattacagg cgtgaaccac tgctcccttc cctgtcctta cgcgtagaat 2460
tggtaaagag agtcgtgtaa aatatcgagt tcgcacatct tgttgtctga ttattgattt 2520
ttggcgaaac catttgatca tatgacaaga tgtgtatcta ccttaactta atgattttga 2580
taaaaatcat taactagtcc atgg 2604
<210> 10
<211> 2048
<212>DNA
<213>HomoSapiens
<400> 10
ataacgcagg aaagaacatg tgagcaaaag gccagcaaaa ggccagggac cgtaaaaagg 60
ccgcgttgct ggcgtttttc cataggctcc gcccccctga cgagcatcac aaaaatcgac 120
gctcaagtca gaggtggcga aacccgacag gactataaag ataccaggcg tttccccctg 180
gaagctccct cgtgcgctct cctgttccga ccctgccgct taccggatac ctgtccgcct 240
ttctcccttc gggaagcgtg gcgctttctc atagctcacg ctgtaggtat ctcagttcgg 300
tgtaggtcgt tcgctccaag ctgggctgtg tgcacgaacc ccccgttcag cccgaccgct 360
gcgccttatc cggtaactat cgtcttgagt ccaacccggt aagacacgac ttatcgccac 420
tggcagcagc cactggtaac aggattagca gagcgaggta tgtaggcggt gctacagagt 480
tcttgaagtg gtggcctaac tacggctaca ctagaagaac agtatttggt atctgcgctc 540
tgctgaagcc agttaccttc ggaaaaagag ttggtagctc ttgatccggc aaacaaacca 600
ccgctggtag cggtggtttt tttgtttgca agcagcagat tacgcgcaga aaaaaaggat 660
ctcaagaaga tcctttgatc ttttctacct cgagtcattt cttacctccc cttccctctc 720
ccacctgcta ctgggtgcat ctctgctccc cccttcccca gcagatggtt acctttgggc 780
tgttgctttc ttgtcaccat ctgagttctc agacgctgga aagccatgtt ctcggctctg 840
tgaatgacaa tgctgactgg agtgctgccc ctctgtaaag ggctgggtgt ggatggtcac 900
aagcccccca catgcctcag ccaagaggaa gtagtacagg ggtcagccca gaggtccagg 960
ggaaaggagt ggaaaccgat ttccccacca agggaggggc ctgtacctca gctgttccca 1020
tagctacttg ccacaactgc caagcaagtt tcgctgagtt tgacacatgg atccctgtgg 1080
atcaactgcc ctaggactcc gtttgcaccc atgtgacact gttgactttg ccctgatgaa 1140
gcagggccaa cagtccccta acttaattac aaaaactaat gactaagaga gaggtggcta 1200
gagctgaggc ccctgagtca ggctgtgggt gggatcatct ccagtacagg aagtgagact 1260
ttcatttcct cctttccaag agagggctga gggagcaggg ttgagcaact ggtgcagaca 1320
gcctagctgg actttgggtg aggcggttca gcagatctgg atccgatatc gtcgacaagc 1380
ttggtacctc cggagcggcc gctctagagc tagcgacgtc gaattccctg tgacccctcc 1440
ccagtgcctc tcctggccct ggaagttgcc actccagtgc ccaccagcct tgtcctaata 1500
aaattaagtt gcatcatttt gtctgactag gtgtccttct ataatattat ggggtggagg 1560
ggggtggtat ggagcaaggg gcaagttggg aagacaacct gtagggcctg cggggtctat 1620
tgggaaccaa gctggagtgc agtggcacaa tcttggctca ctgcaatctc cgcctcctgg 1680
gttcaagcga ttctcctgcc tcagcctccc gagttgttgg gattccaggc atgcatgacc 1740
aggctcagct aatttttgtt tttttggtag agacggggtt tcaccatatt ggccaggctg 1800
gtctccaact cctaatctca ggtgatctac ccaccttggc ctcccaaatt gctgggatta 1860
caggcgtgaa ccactgctcc cttccctgtc cttacgcgta gaattggtaa agagagtcgt 1920
gtaaaatatc gagttcgcac atcttgttgt ctgattattg atttttggcg aaaccatttg 1980
atcatatgac aagatgtgta tctaccttaa cttaatgatt ttgataaaaa tcattaacta 2040
gtccatgg 2048
<210> 11
<211> 1978
<212>DNA
<213>HomoSapiens
<400> 11
ataacgcagg aaagaacatg tgagcaaaag gccagcaaaa ggccagggac cgtaaaaagg 60
ccgcgttgct ggcgtttttc cataggctcc gcccccctga cgagcatcac aaaaatcgac 120
gctcaagtca gaggtggcga aacccgacag gactataaag ataccaggcg tttccccctg 180
gaagctccct cgtgcgctct cctgttccga ccctgccgct taccggatac ctgtccgcct 240
ttctcccttc gggaagcgtg gcgctttctc atagctcacg ctgtaggtat ctcagttcgg 300
tgtaggtcgt tcgctccaag ctgggctgtg tgcacgaacc ccccgttcag cccgaccgct 360
gcgccttatc cggtaactat cgtcttgagt ccaacccggt aagacacgac ttatcgccac 420
tggcagcagc cactggtaac aggattagca gagcgaggta tgtaggcggt gctacagagt 480
tcttgaagtg gtggcctaac tacggctaca ctagaagaac agtatttggt atctgcgctc 540
tgctgaagcc agttaccttc ggaaaaagag ttggtagctc ttgatccggc aaacaaacca 600
ccgctggtag cggtggtttt tttgtttgca agcagcagat tacgcgcaga aaaaaaggat 660
ctcaagaaga tcctttgatc ttttctacct cgaggacagc agcgcaaaga gccccgccct 720
gcagcctcca gctctcctgg tctaatgtgg aaagtggccc aggtgagggc tttgctctcc 780
tggagacatt tgcccccagc tgtgagcagg gacaggtctg gccaccgggc ccctggttaa 840
gactctaatg acccgctggt cctgaggaag aggtgctgac gaccaaggag atcttcccac 900
agacccagca ccagggaaat ggtccggaaa ttgcagcctc agcccccagc catctgccga 960
ccccctcacc ccaggcccta atgggccagg cggcaggggt tgacaggtag gggagatggg 1020
ctctgagact ataaagccag cgggggccca gcagccctca gccctccagg acaggctgca 1080
tcagaagagg ccatcaagca ggtctgttcc aagggccttt gcgtcaggtg ggctcagggt 1140
tccagggtgg ctggacccca ggccccagct ctgcagcagg gaggacgtgg ctgggctcgt 1200
gaagcatgtg ggggtgagcc caggggcccc aaggcagggc acctggcctt cagcctgcct 1260
cagccctgcc tgtctcccaa tcactgtcgg atccgatatc gtcgacaagc ttggtacctc 1320
cggagcggcc gctctagagc tagcgacgtc gaattccctg tgacccctcc ccagtgcctc 1380
tcctggccct ggaagttgcc actccagtgc ccaccagcct tgtcctaata aaattaagtt 1440
gcatcatttt gtctgactag gtgtccttct ataatattat ggggtggagg ggggtggtat 1500
ggagcaaggg gcaagttggg aagacaacct gtagggcctg cggggtctat tgggaaccaa 1560
gctggagtgc agtggcacaa tcttggctca ctgcaatctc cgcctcctgg gttcaagcga 1620
ttctcctgcc tcagcctccc gagttgttgg gattccaggc atgcatgacc aggctcagct 1680
aatttttgtt tttttggtag agacggggtt tcaccatatt ggccaggctg gtctccaact 1740
cctaatctca ggtgatctac ccaccttggc ctcccaaatt gctgggatta caggcgtgaa 1800
ccactgctcc cttccctgtc cttacgcgta gaattggtaa agagagtcgt gtaaaatatc 1860
gagttcgcac atcttgttgt ctgattattg atttttggcg aaaccatttg atcatatgac 1920
aagatgtgta tctaccttaa cttaatgatt ttgataaaaa tcattaacta gtccatgg 1978
<---

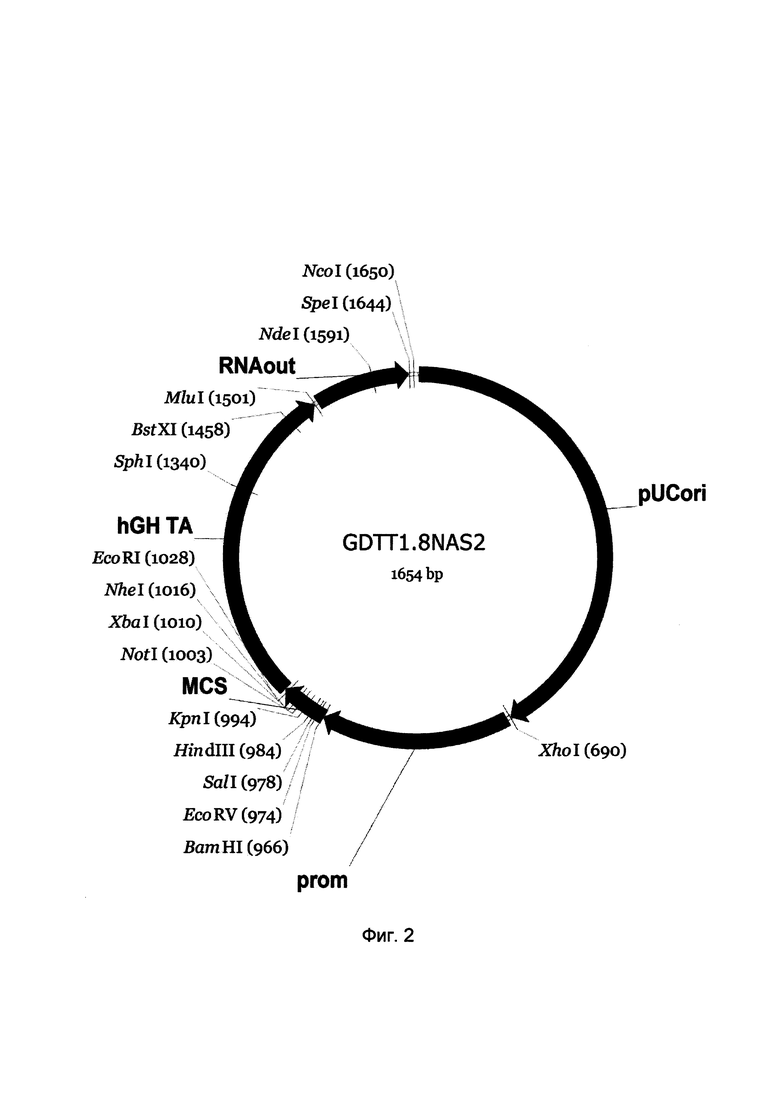
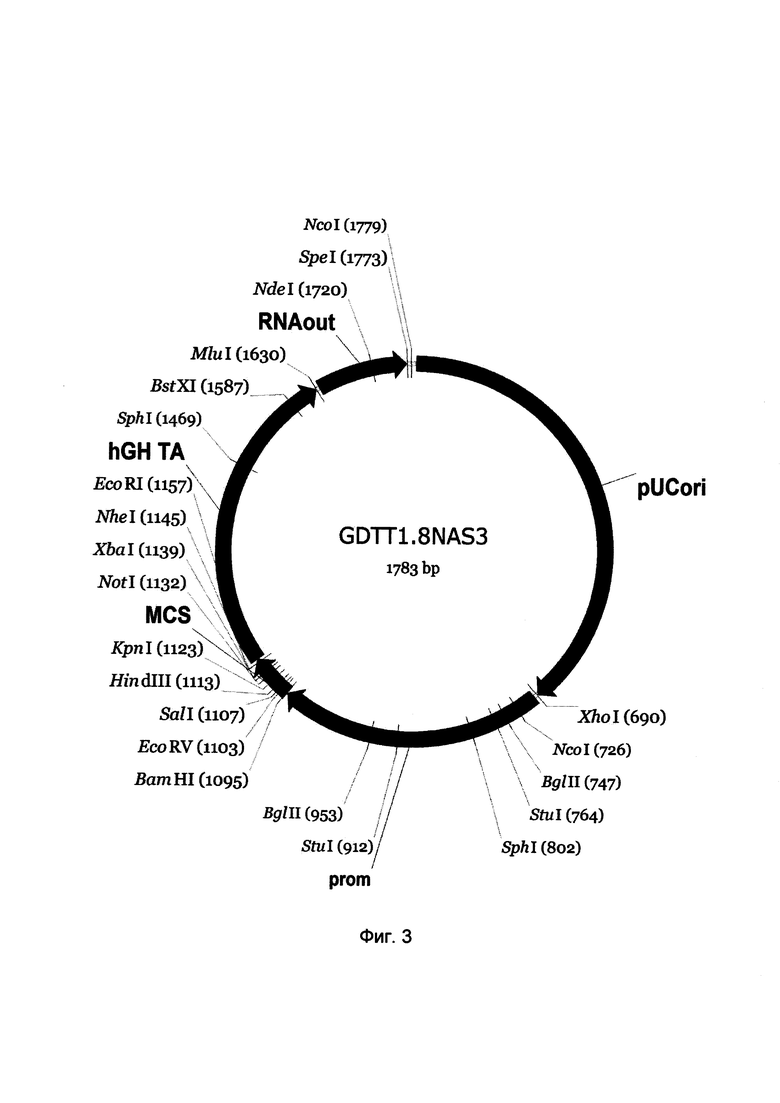
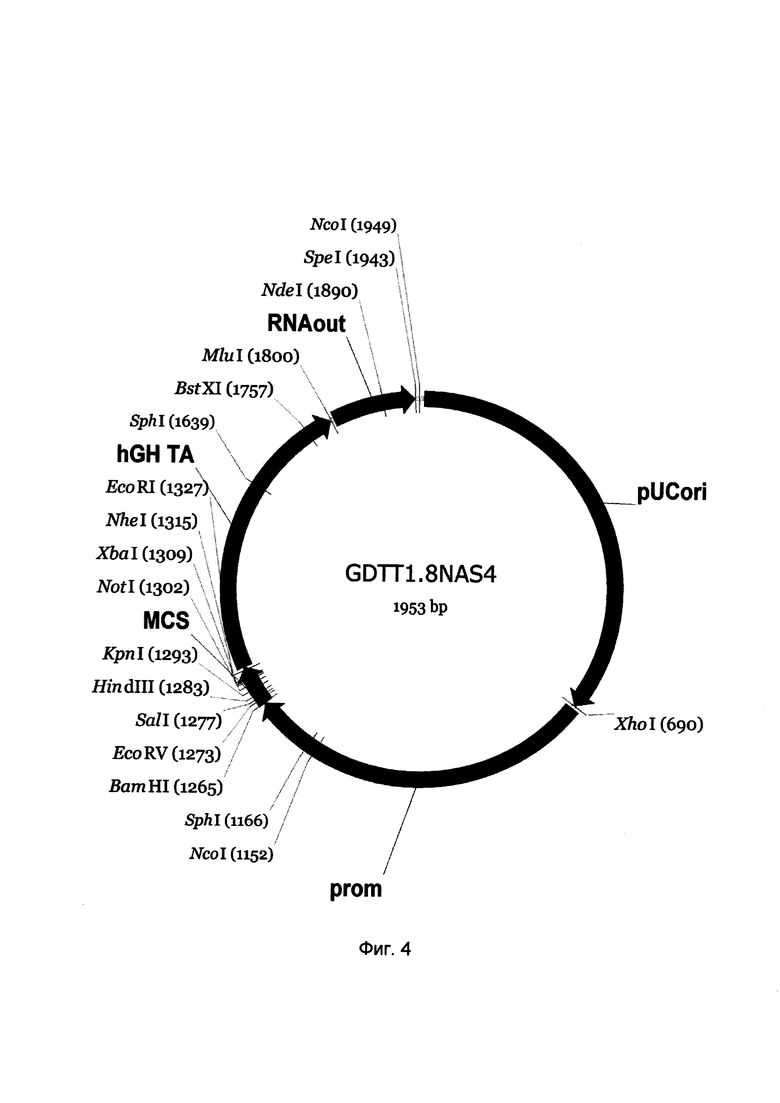
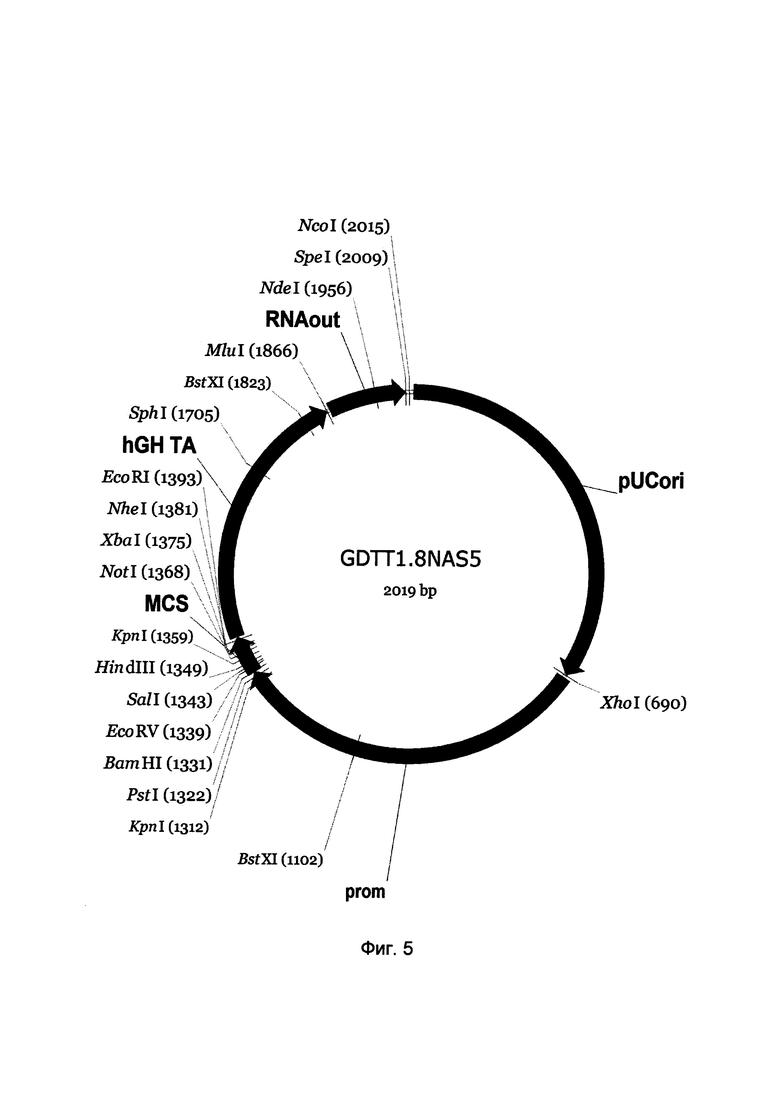

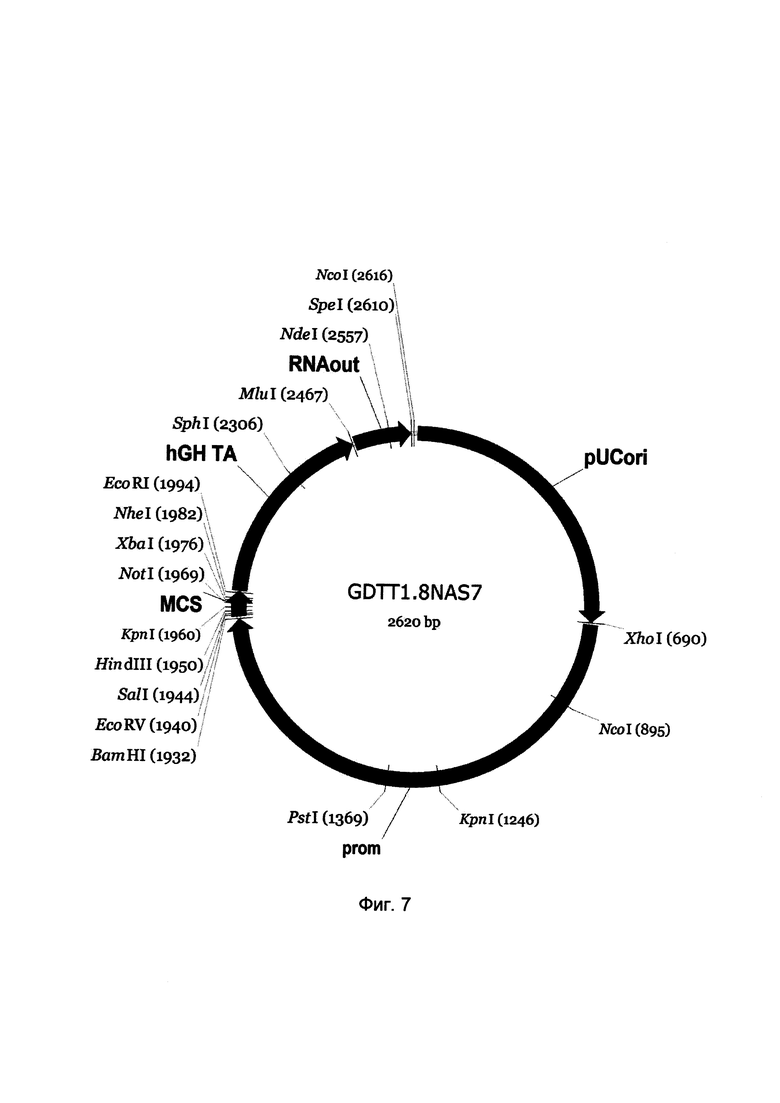
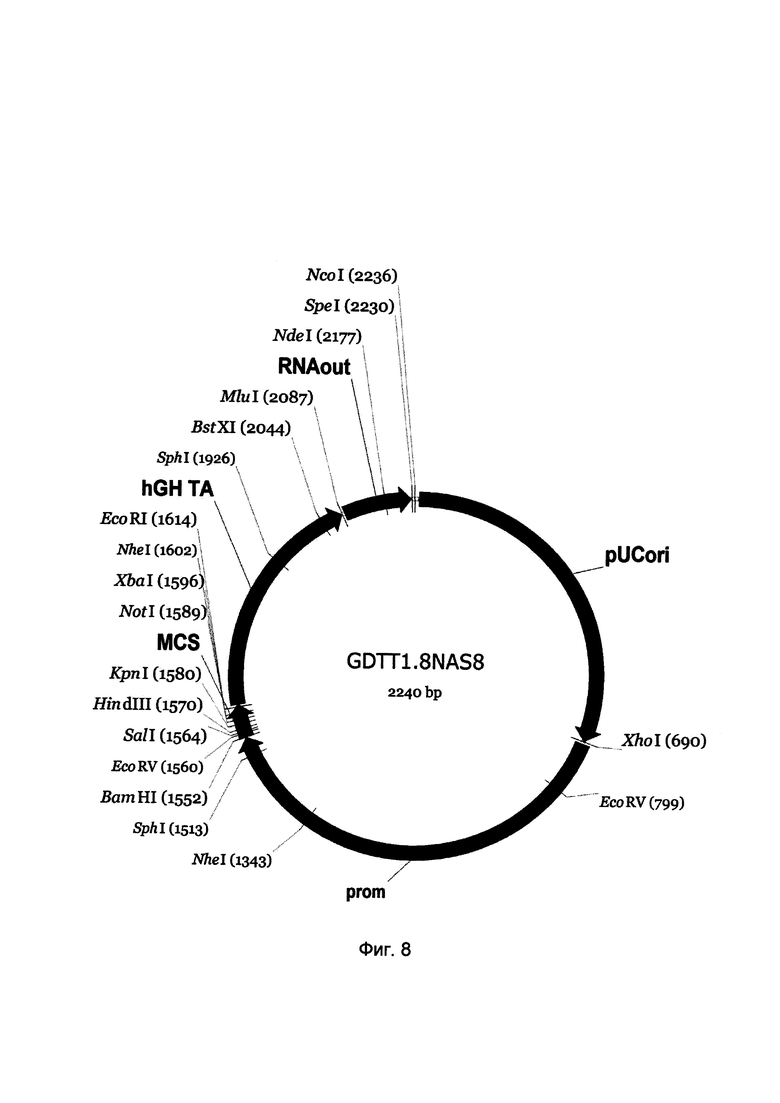
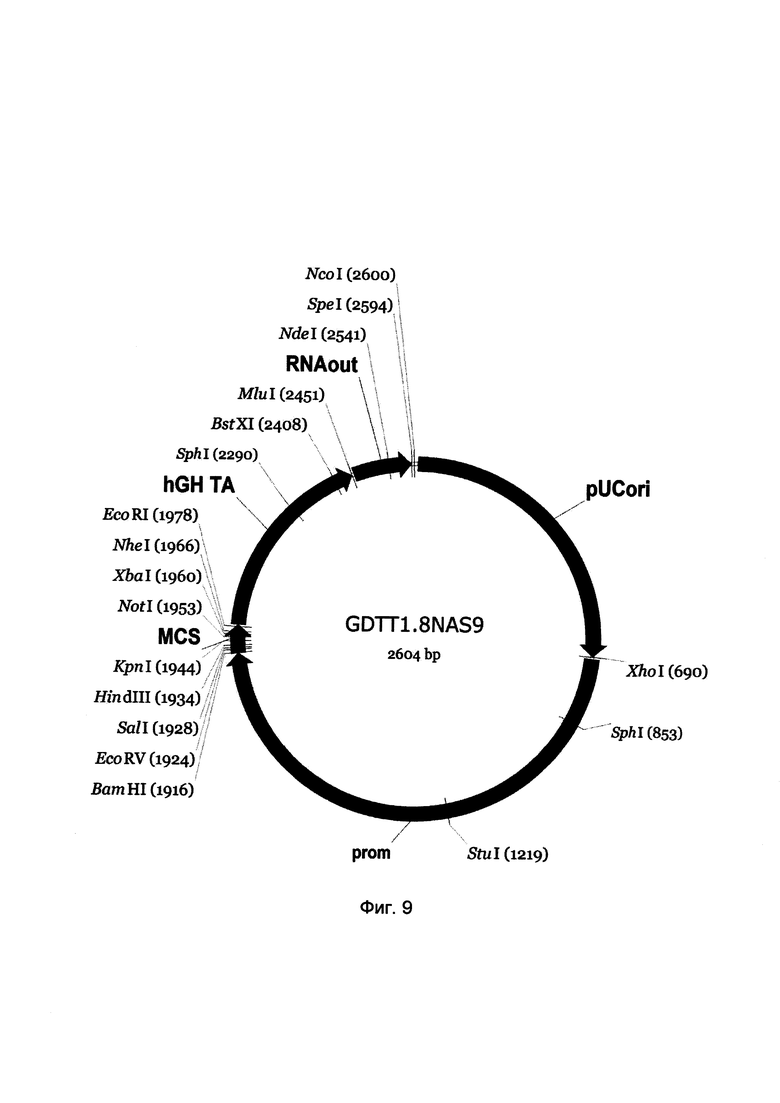
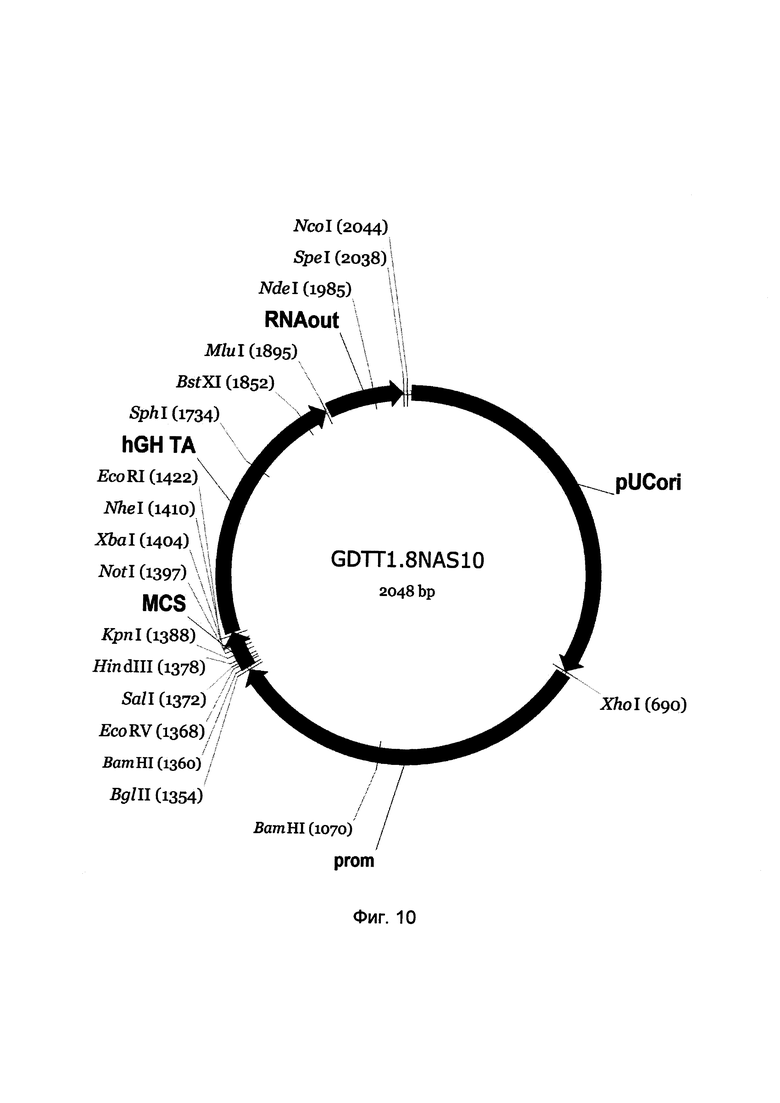
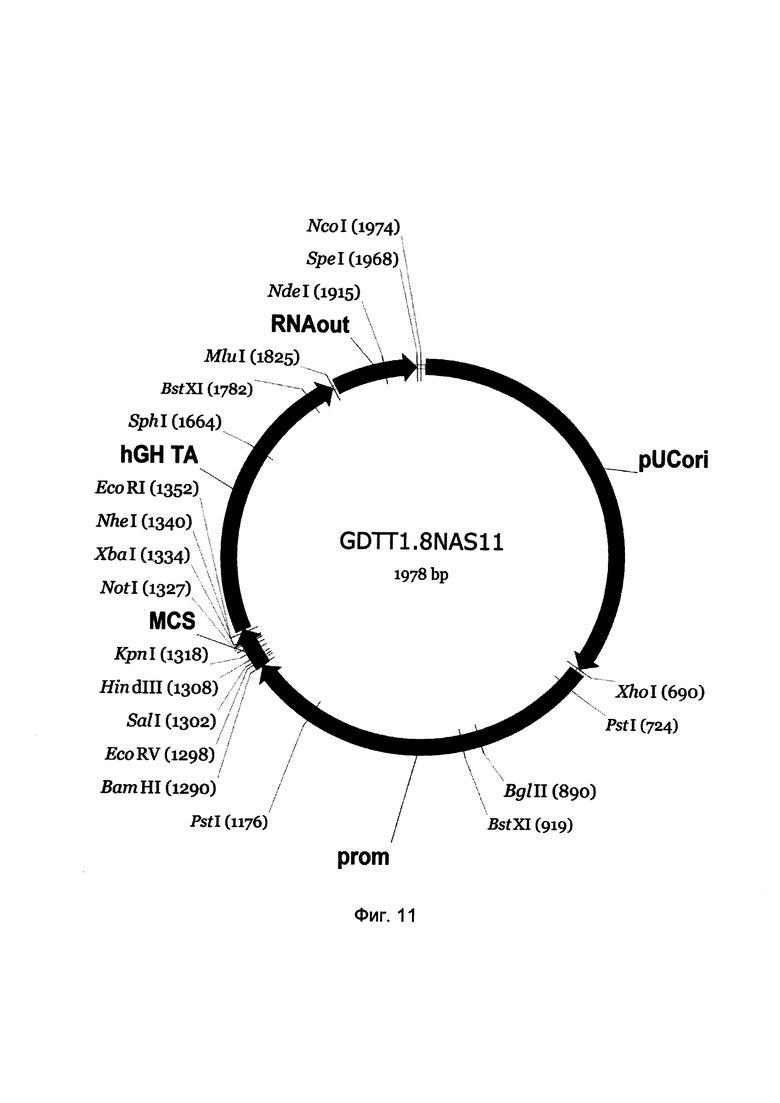
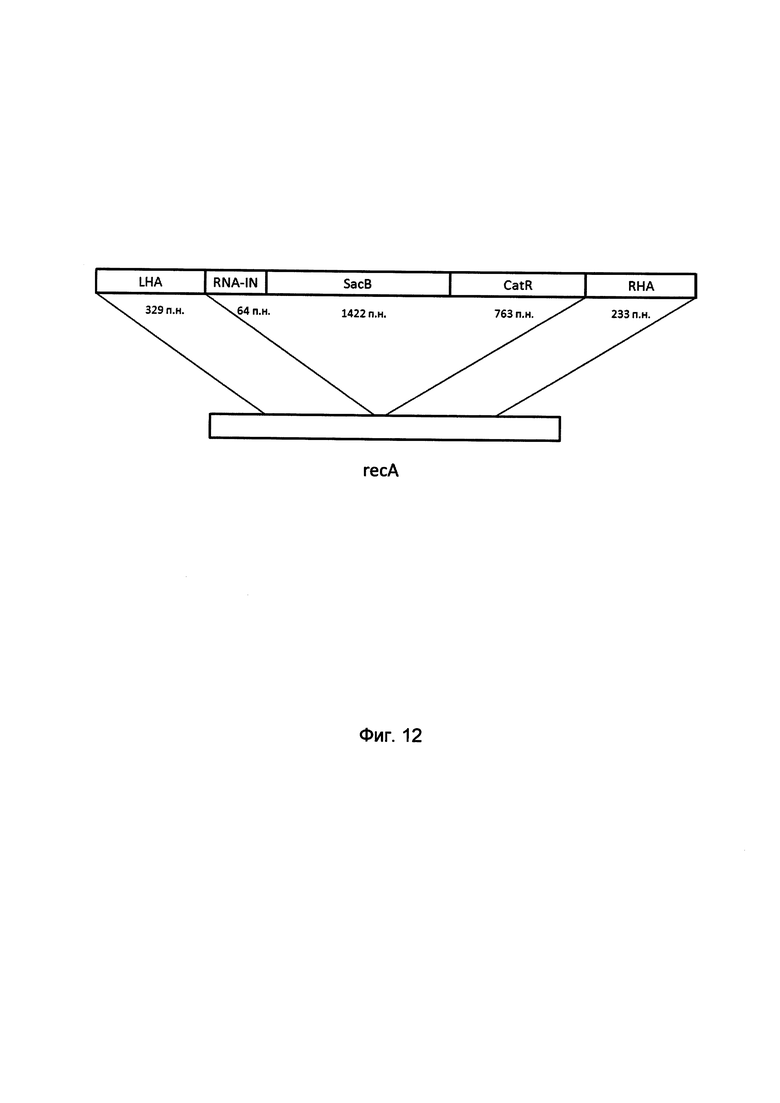
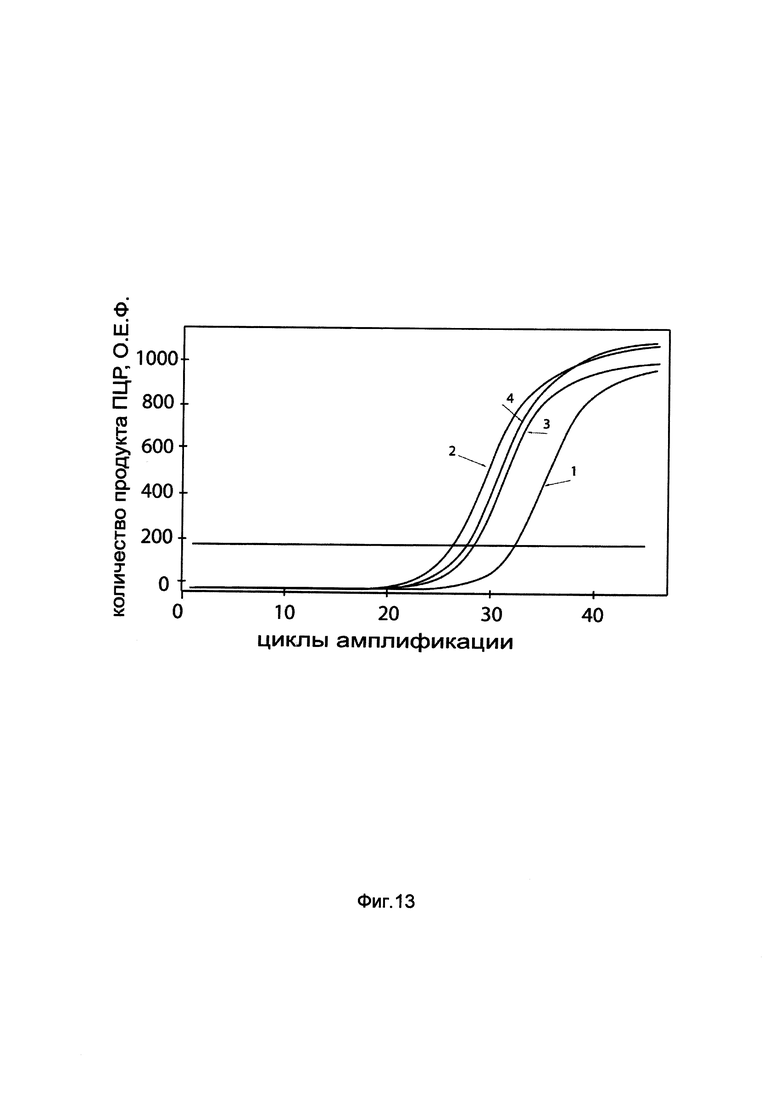
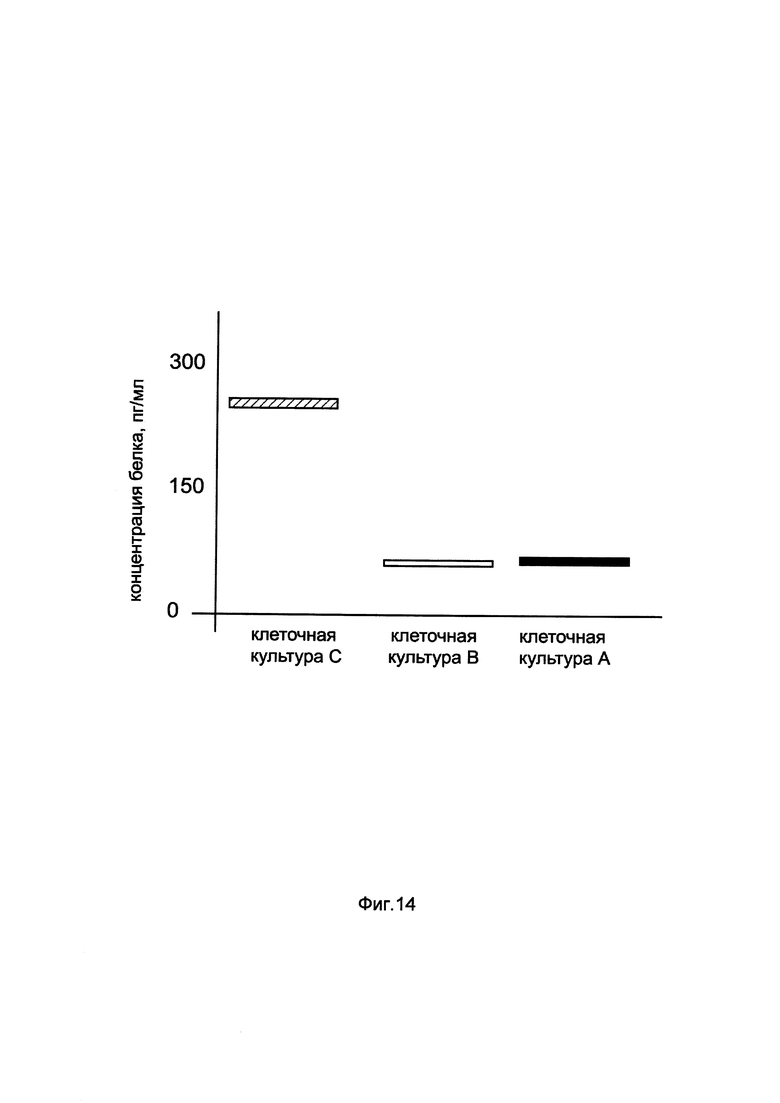
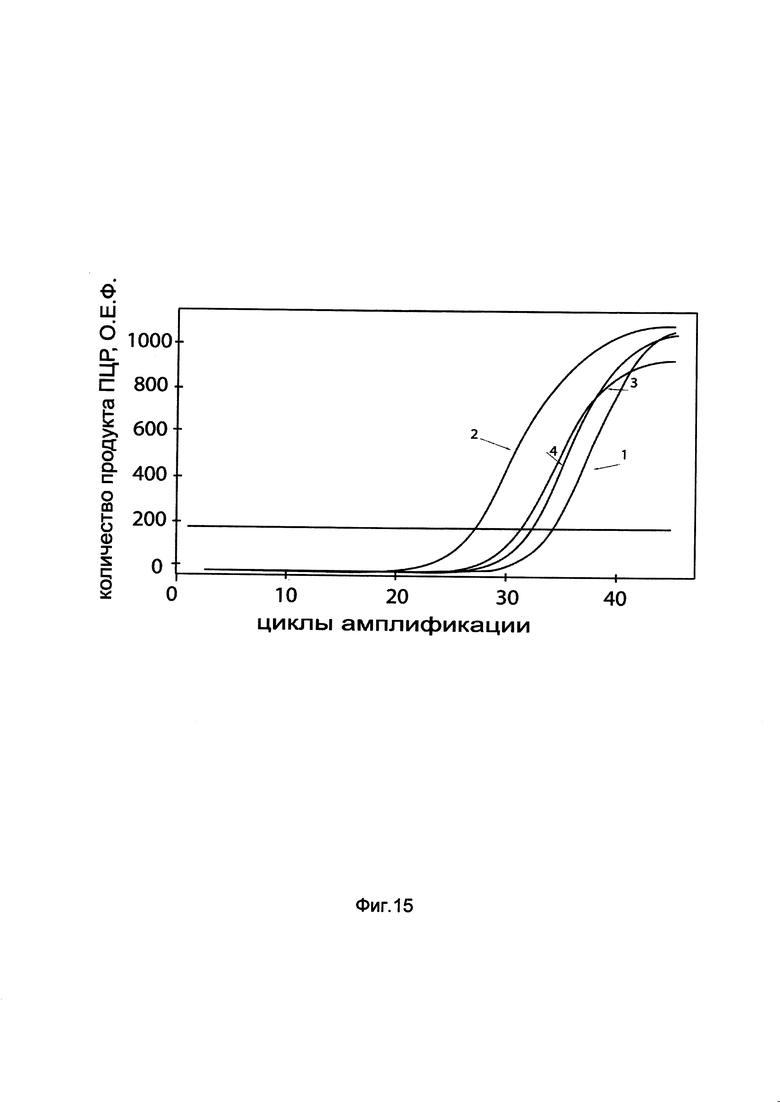
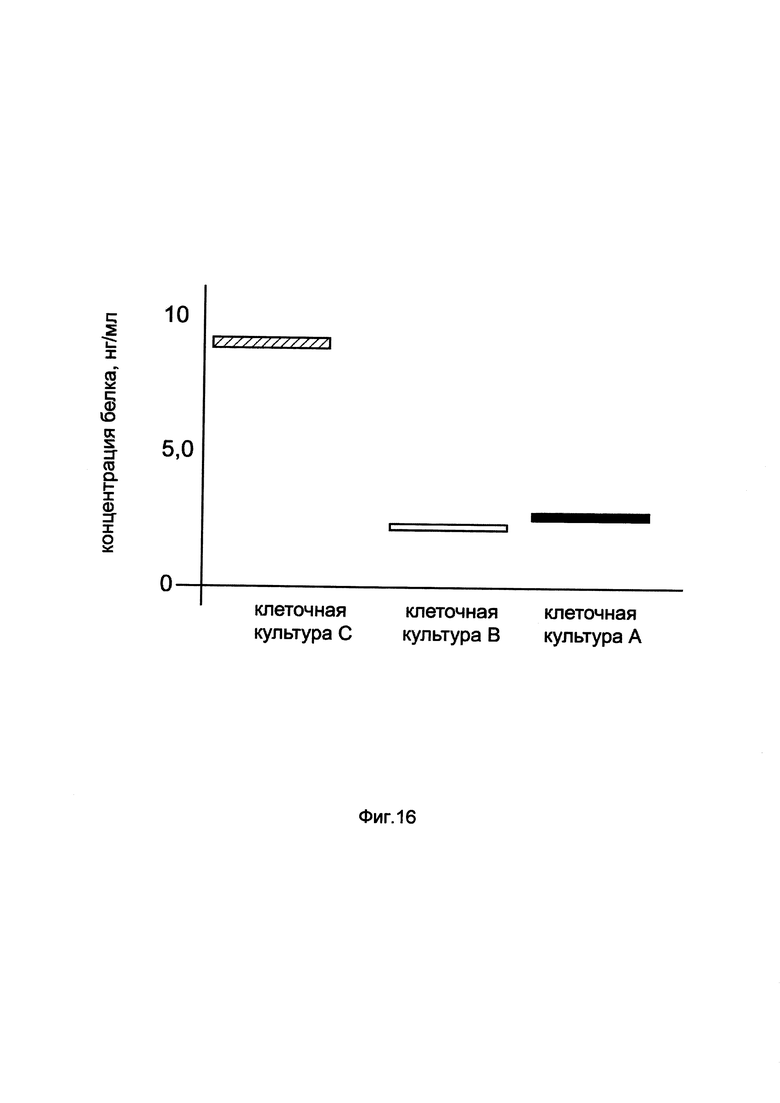
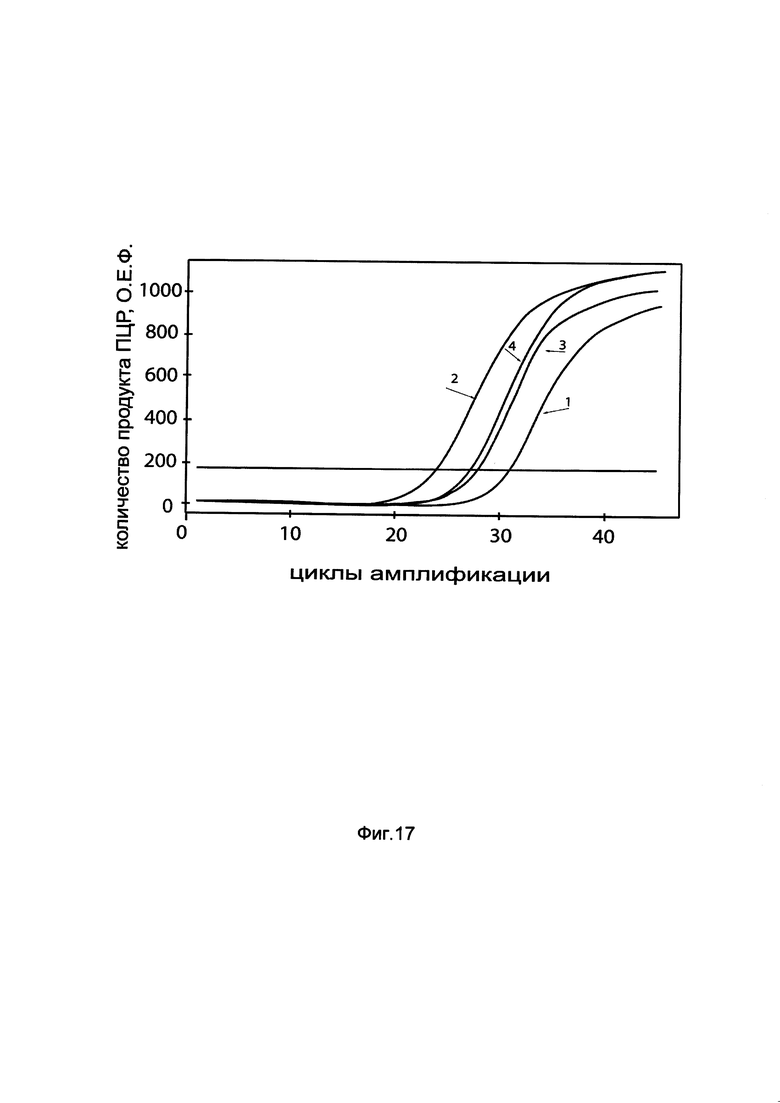
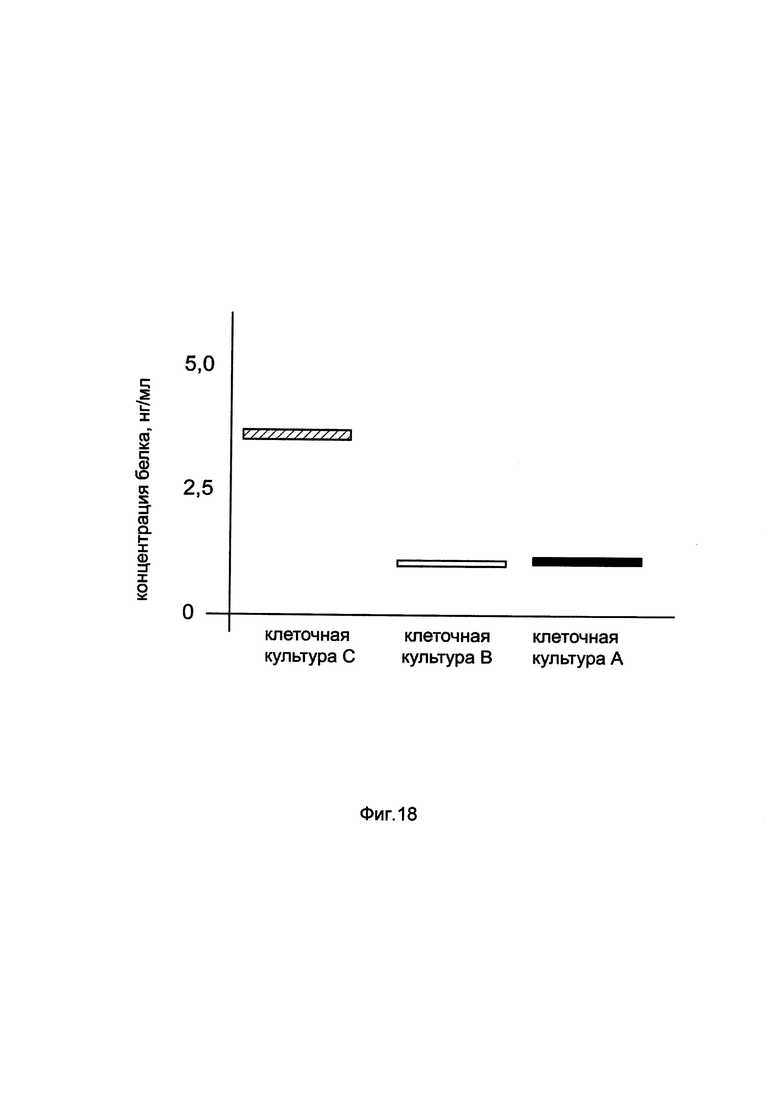
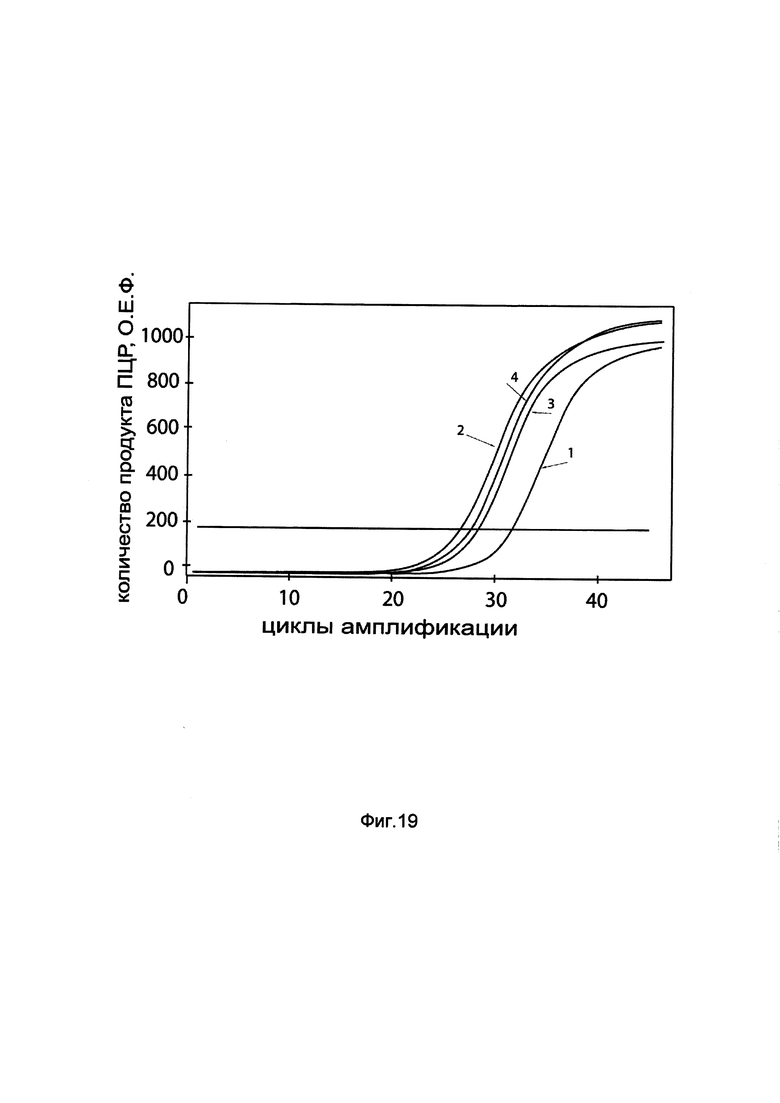
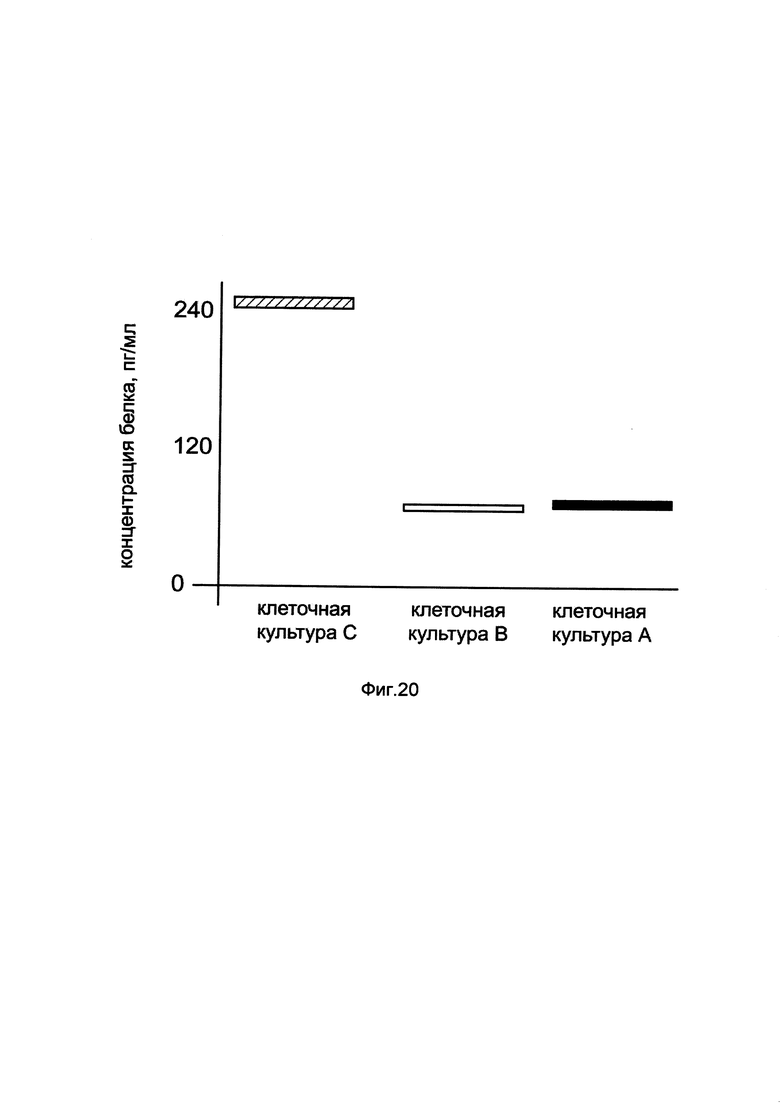
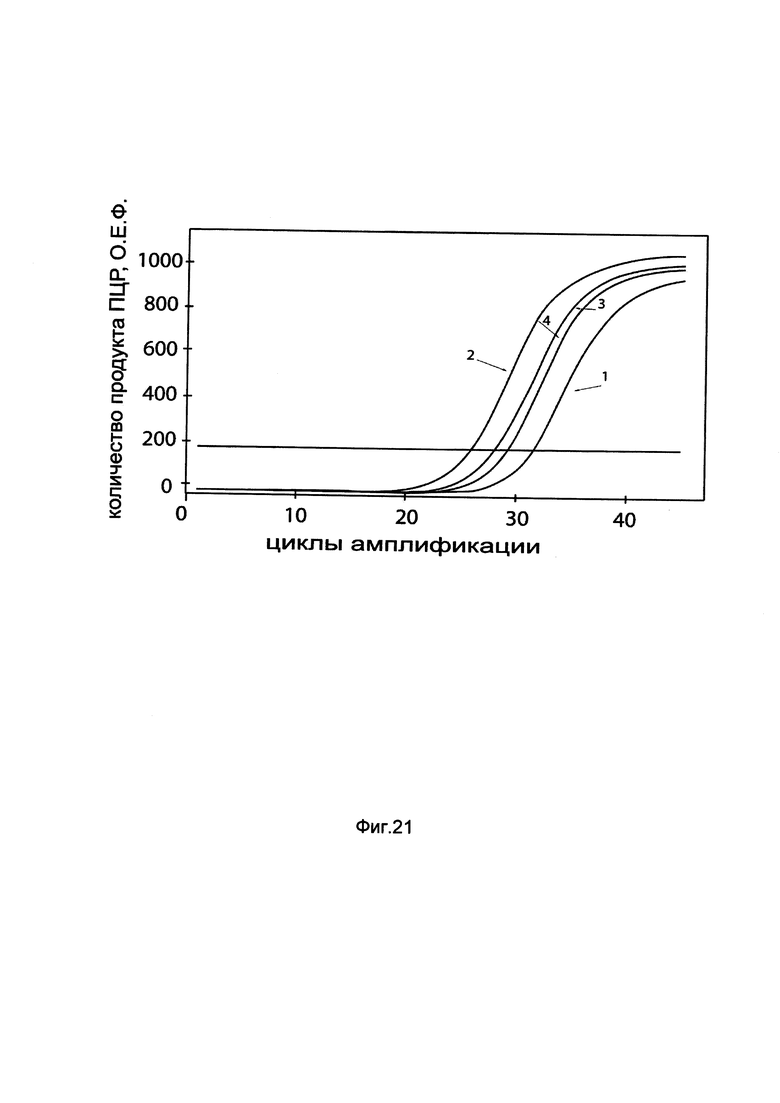
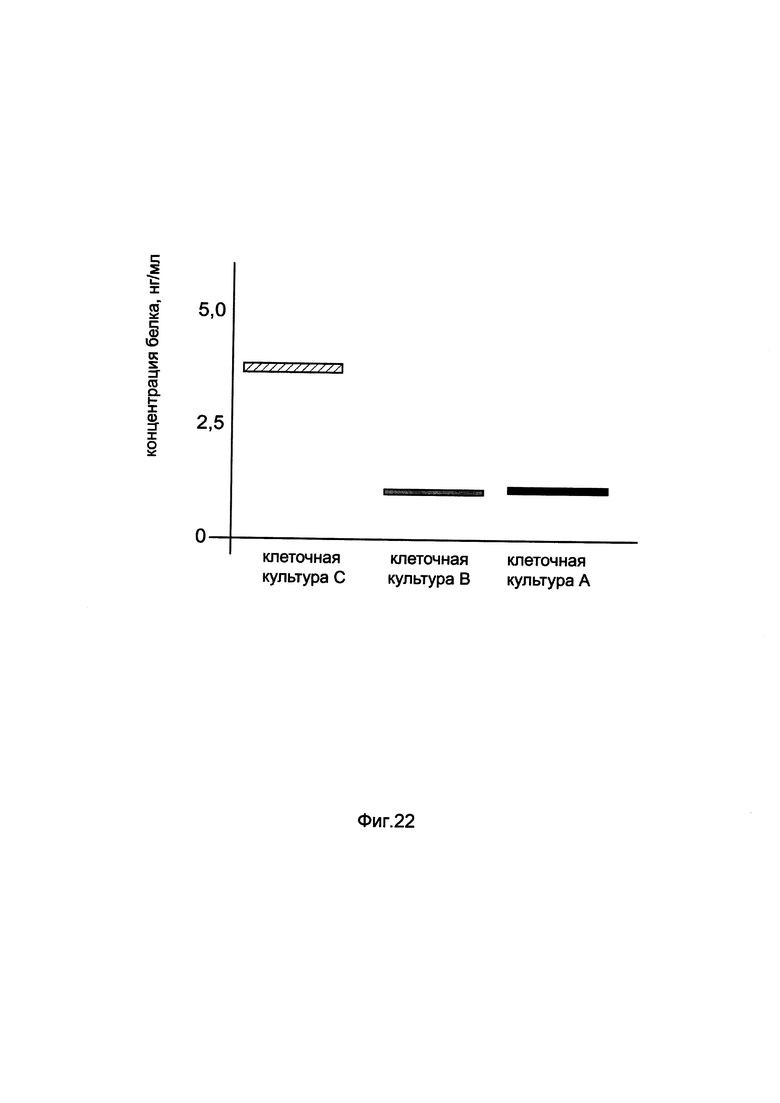
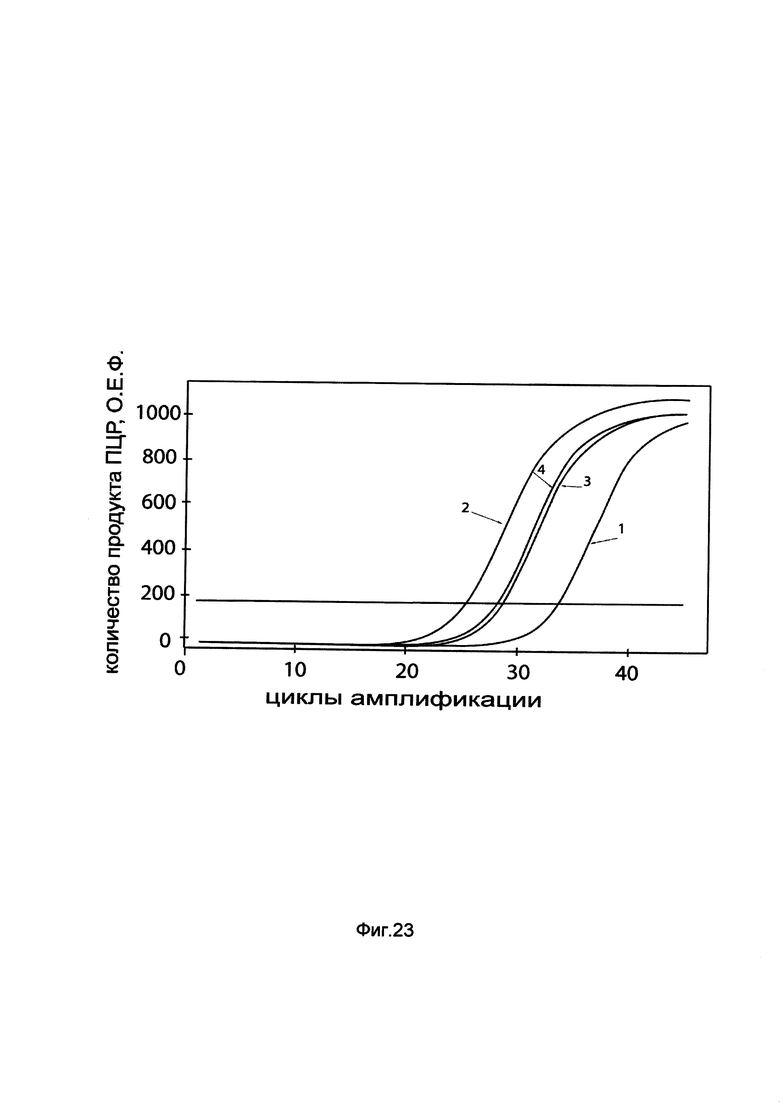
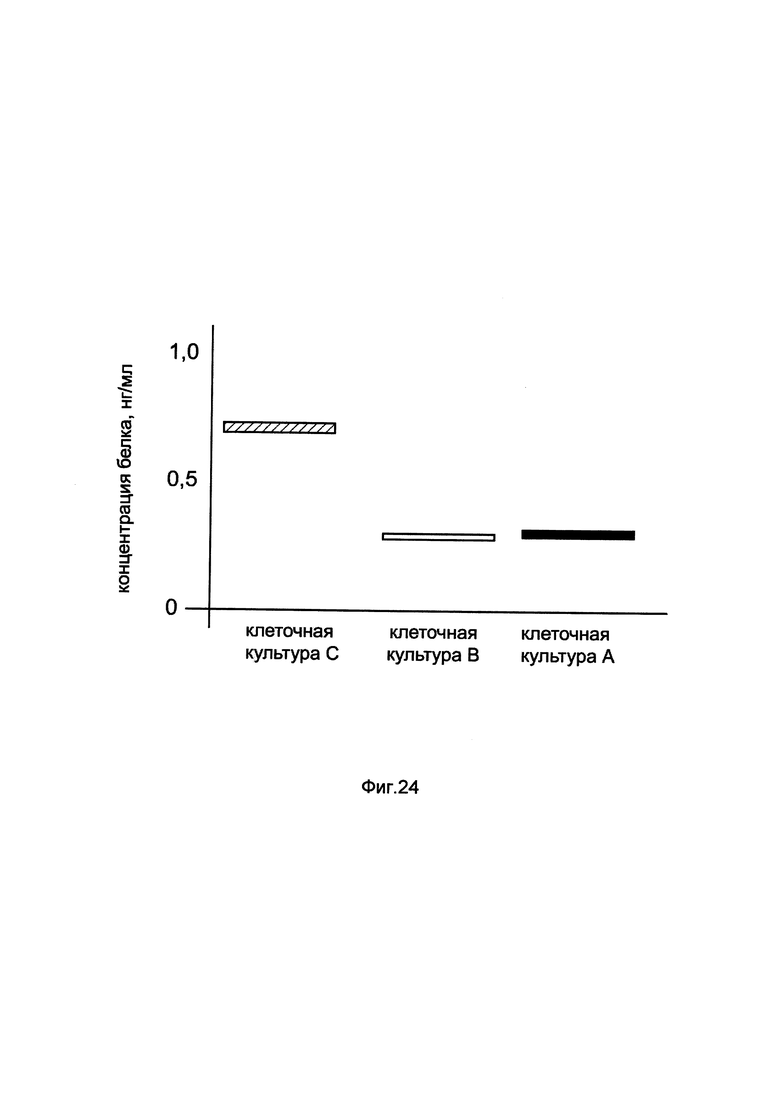
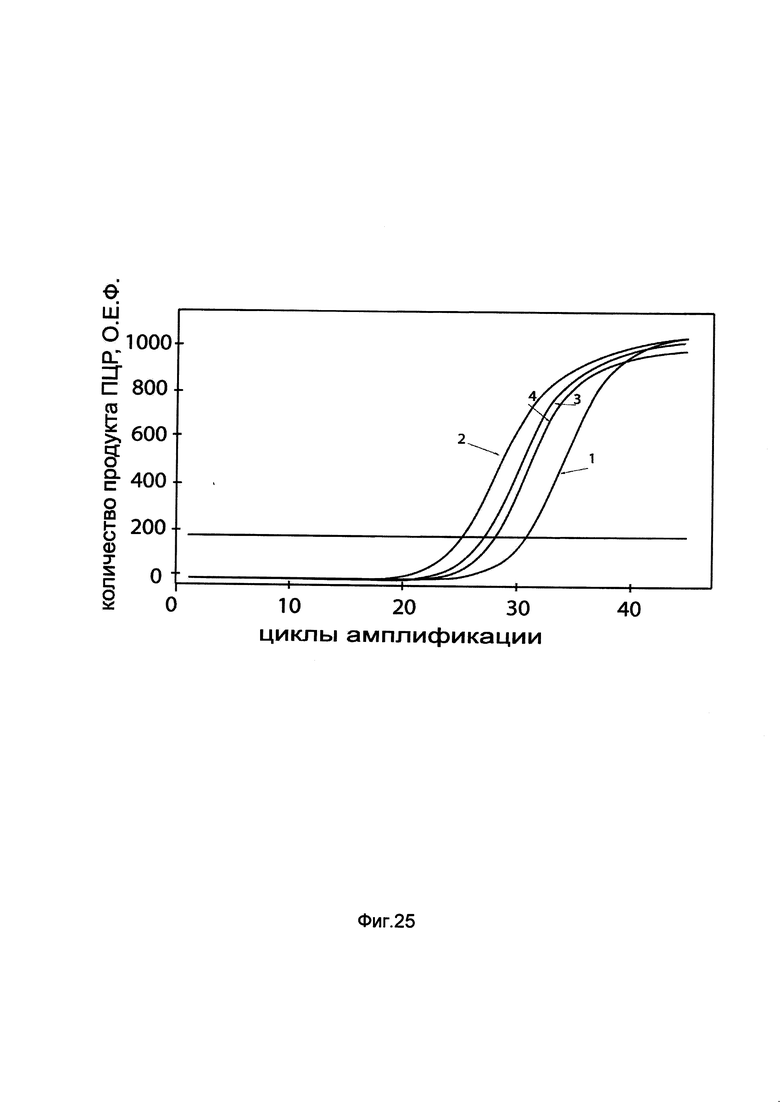
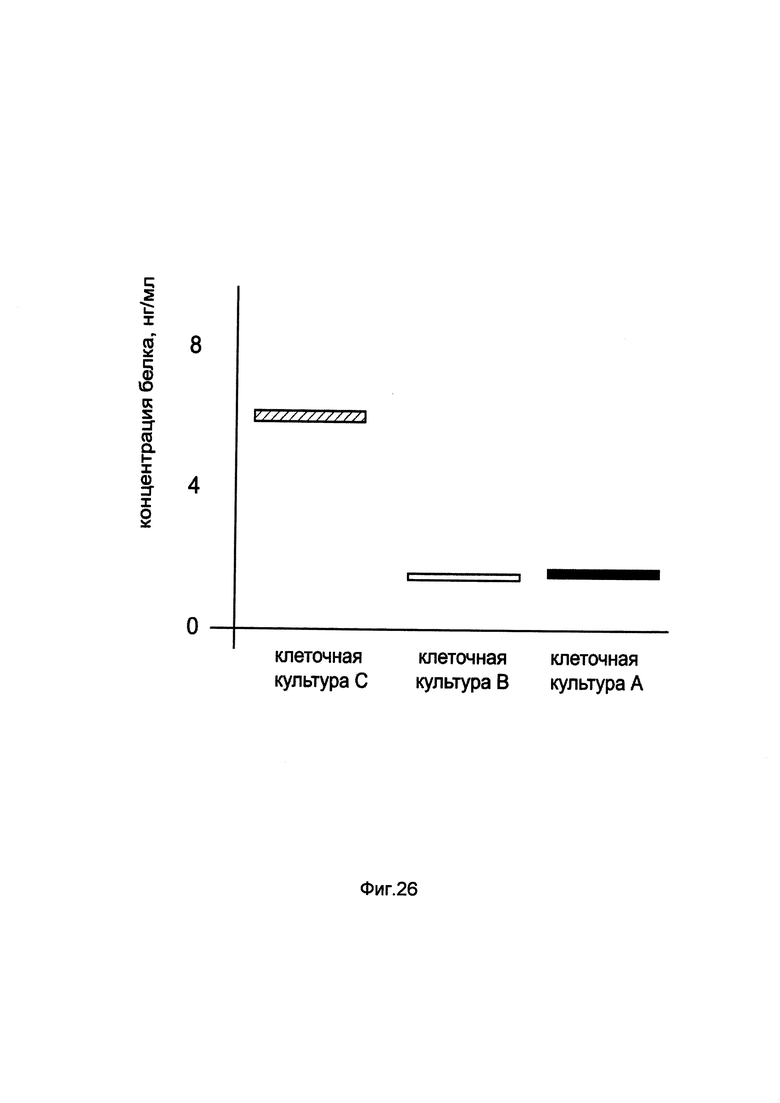
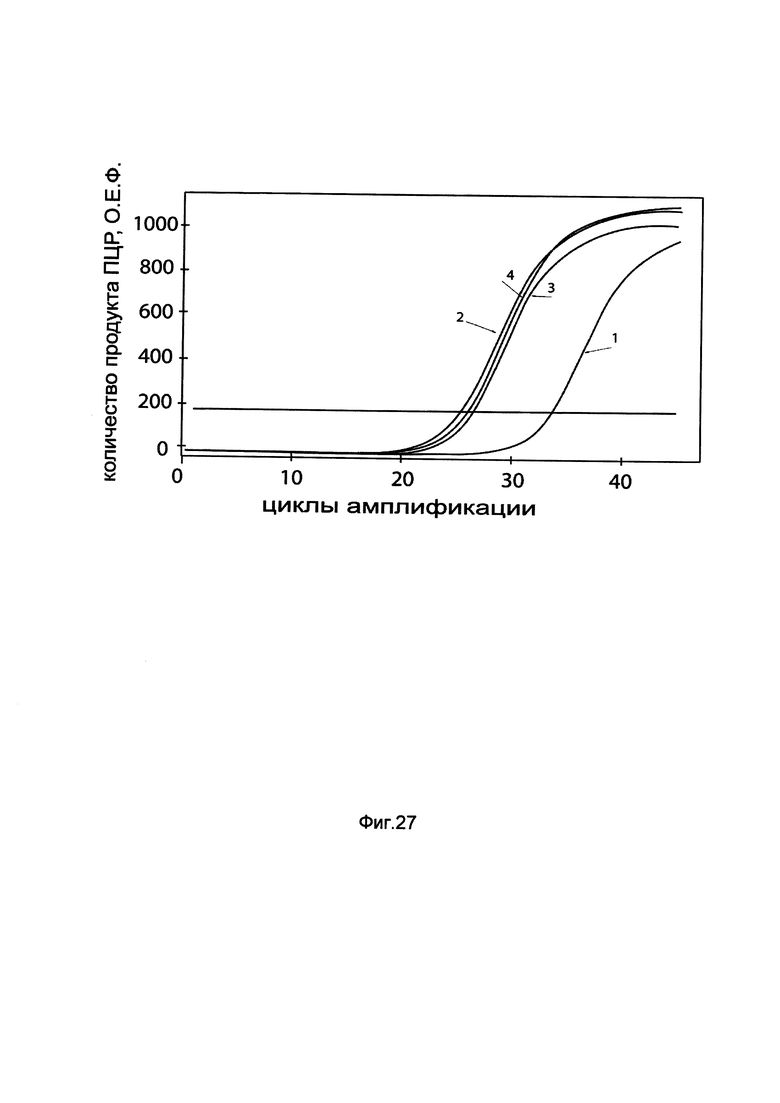
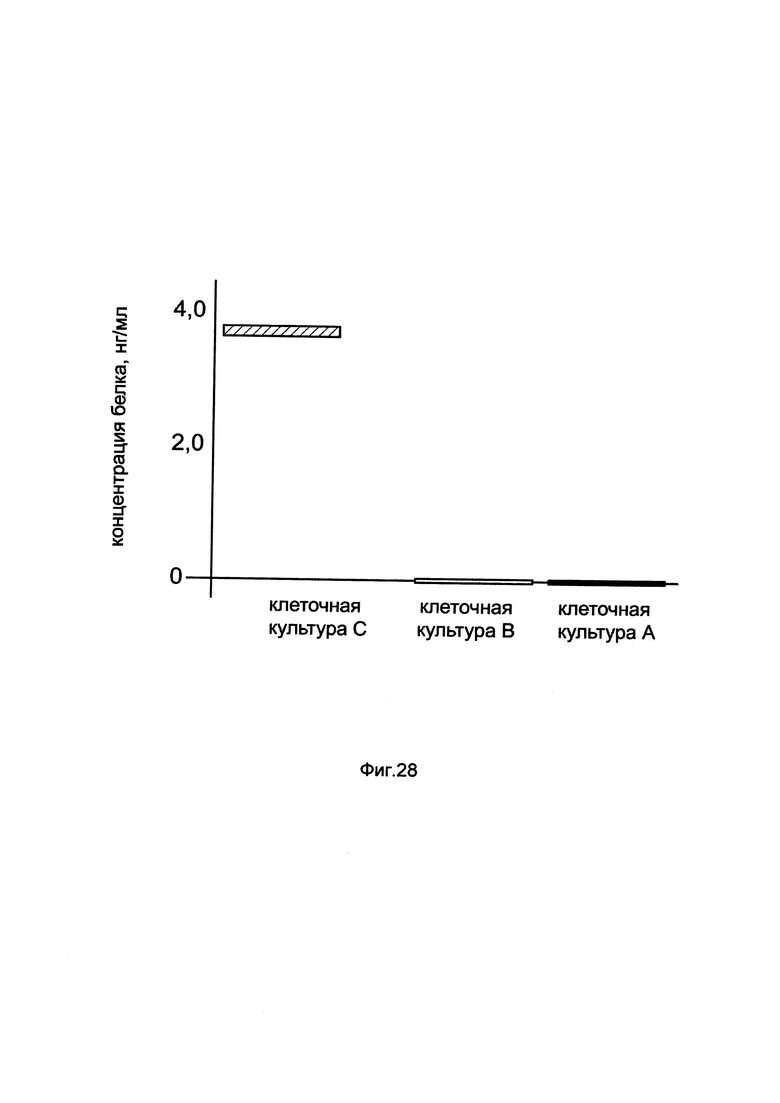
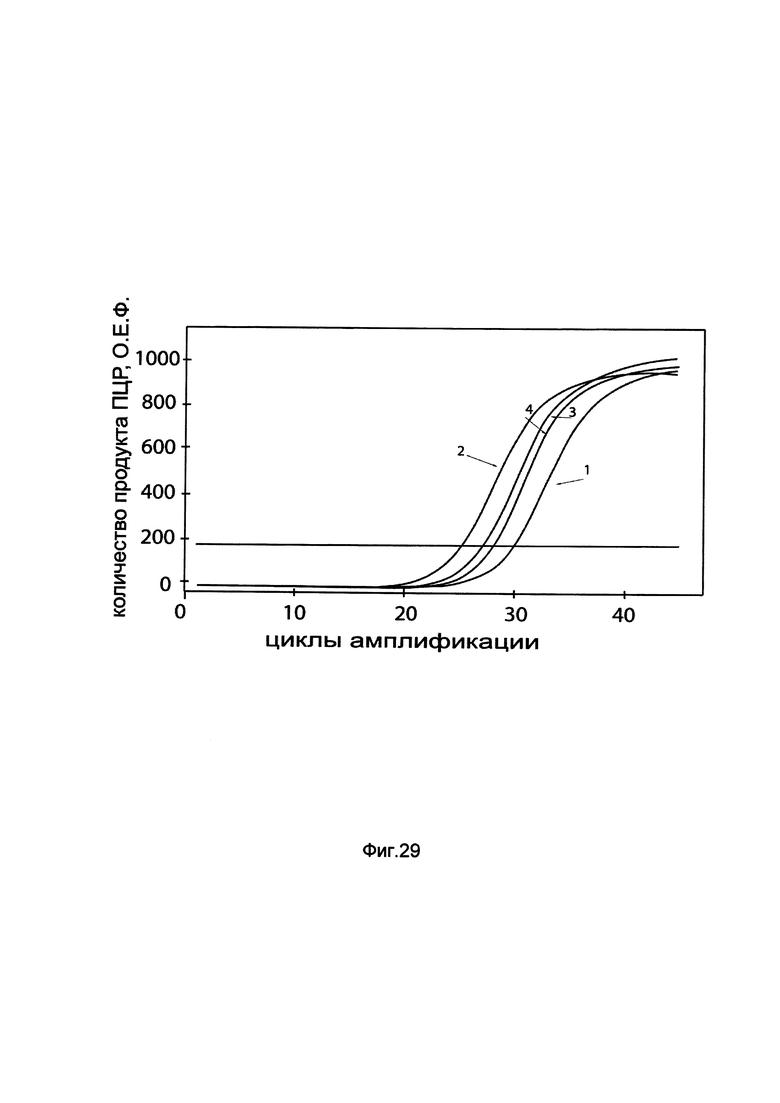
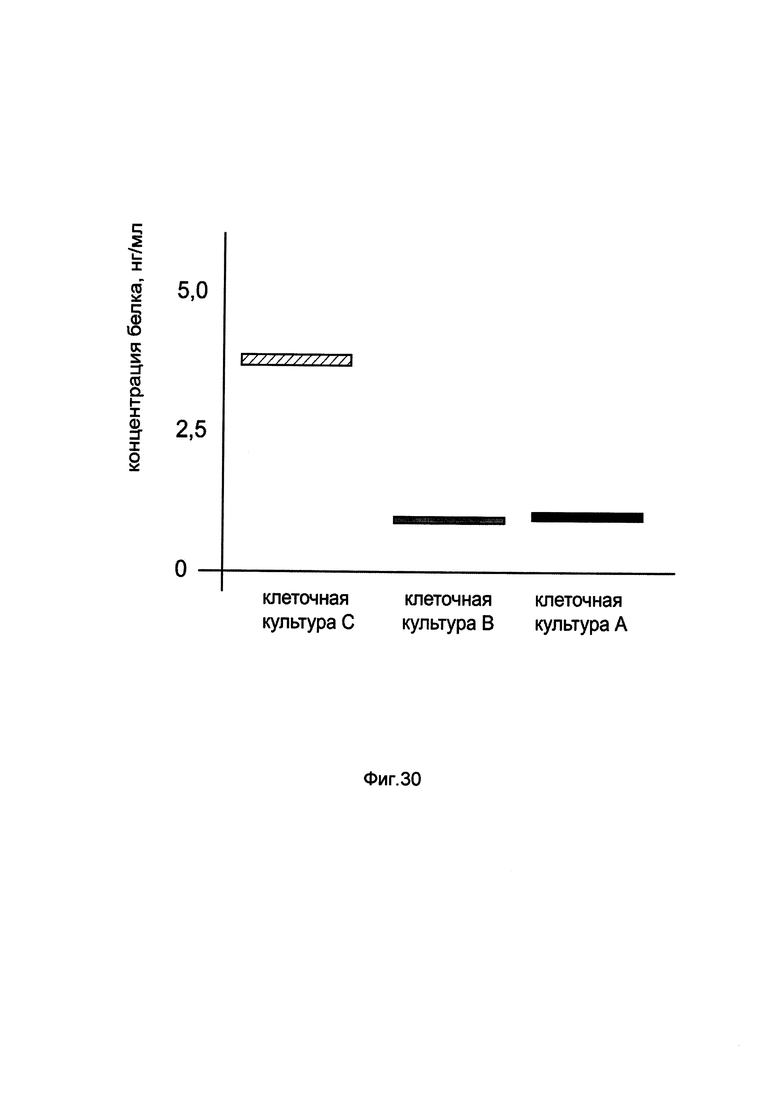
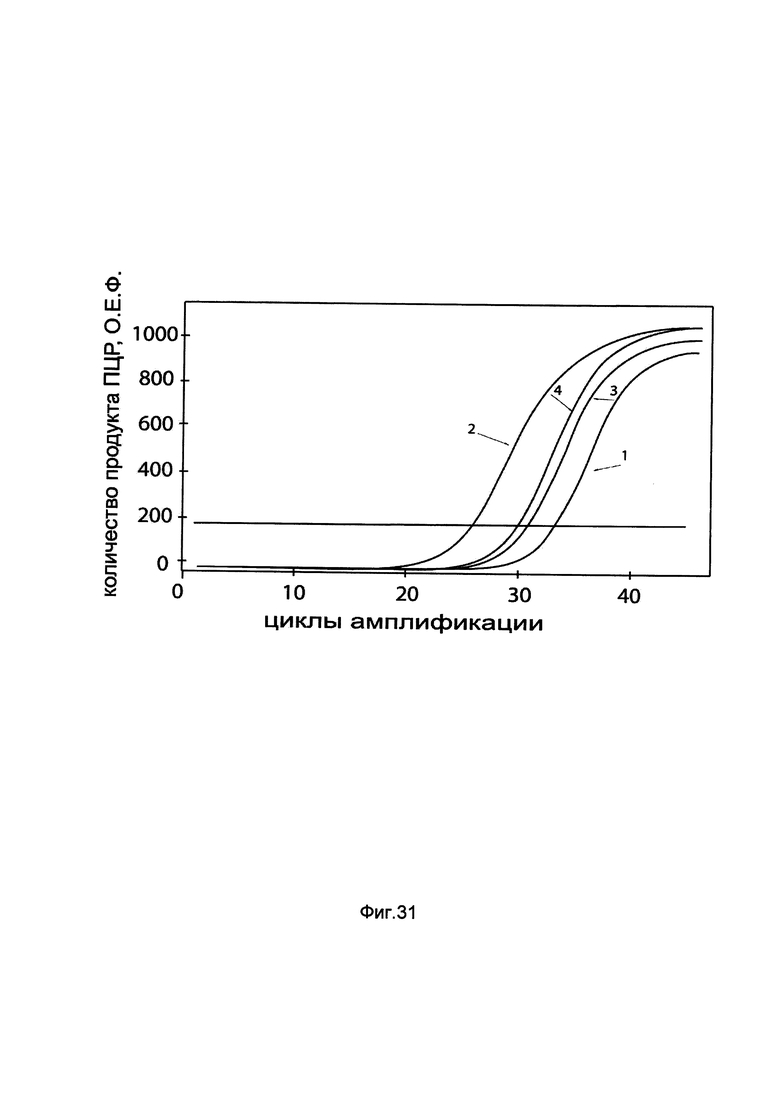
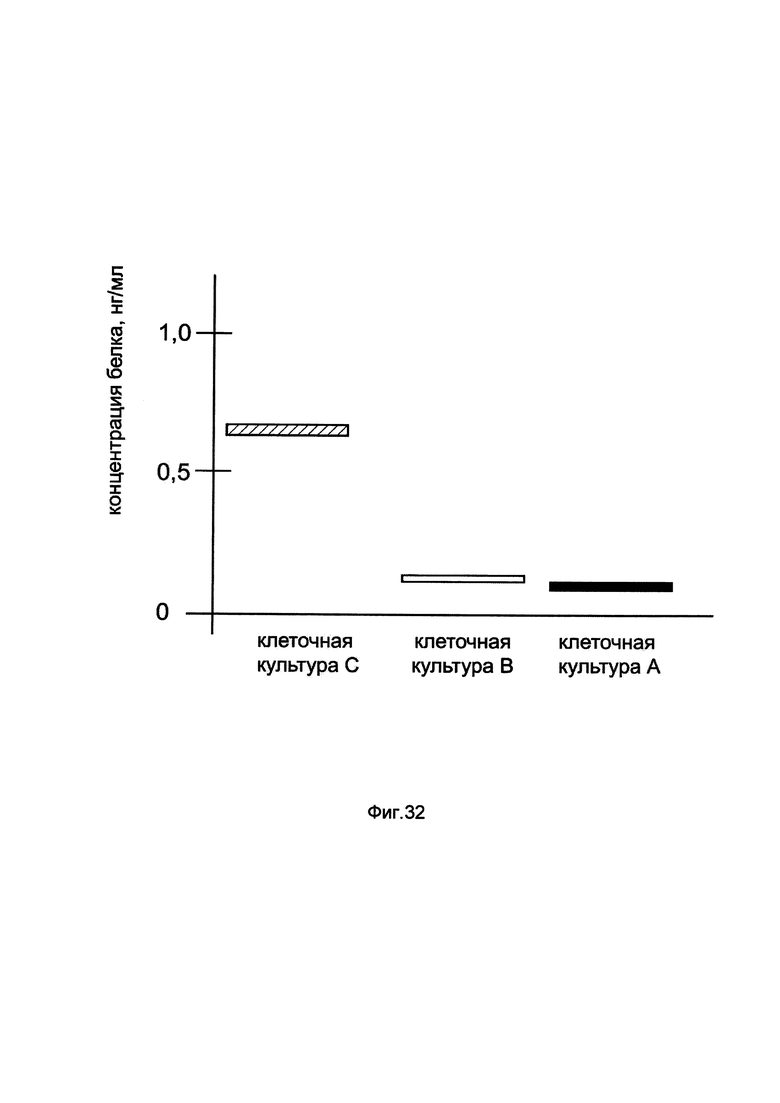
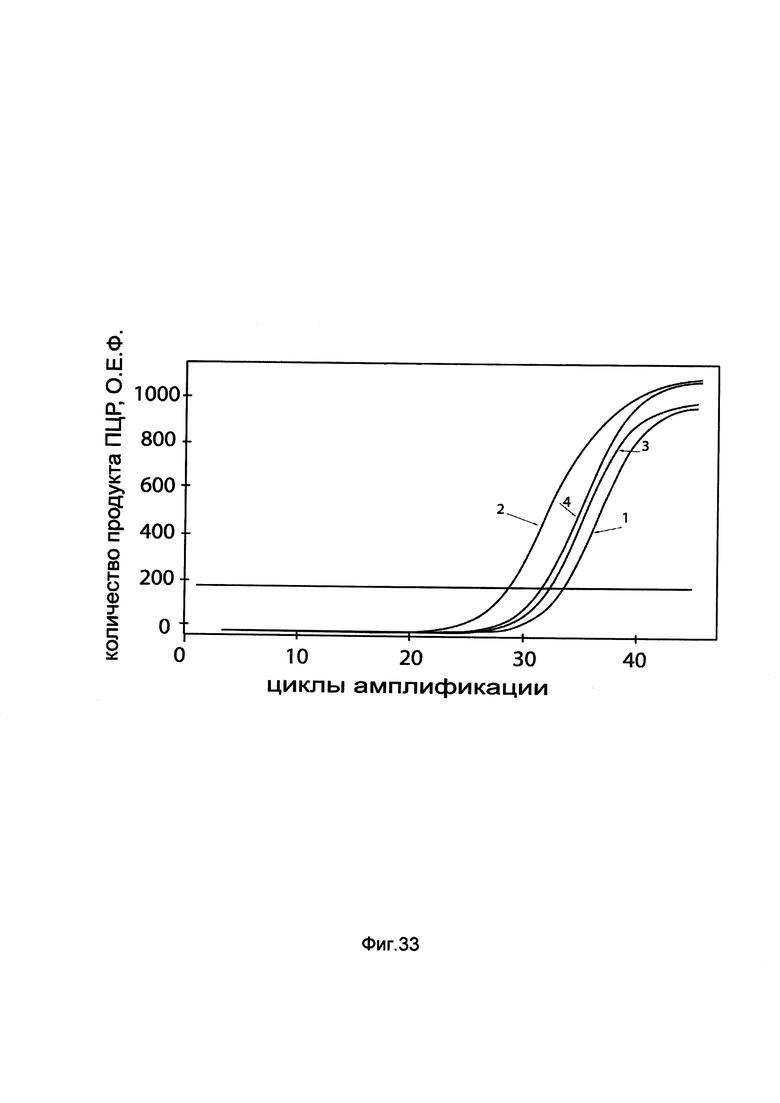
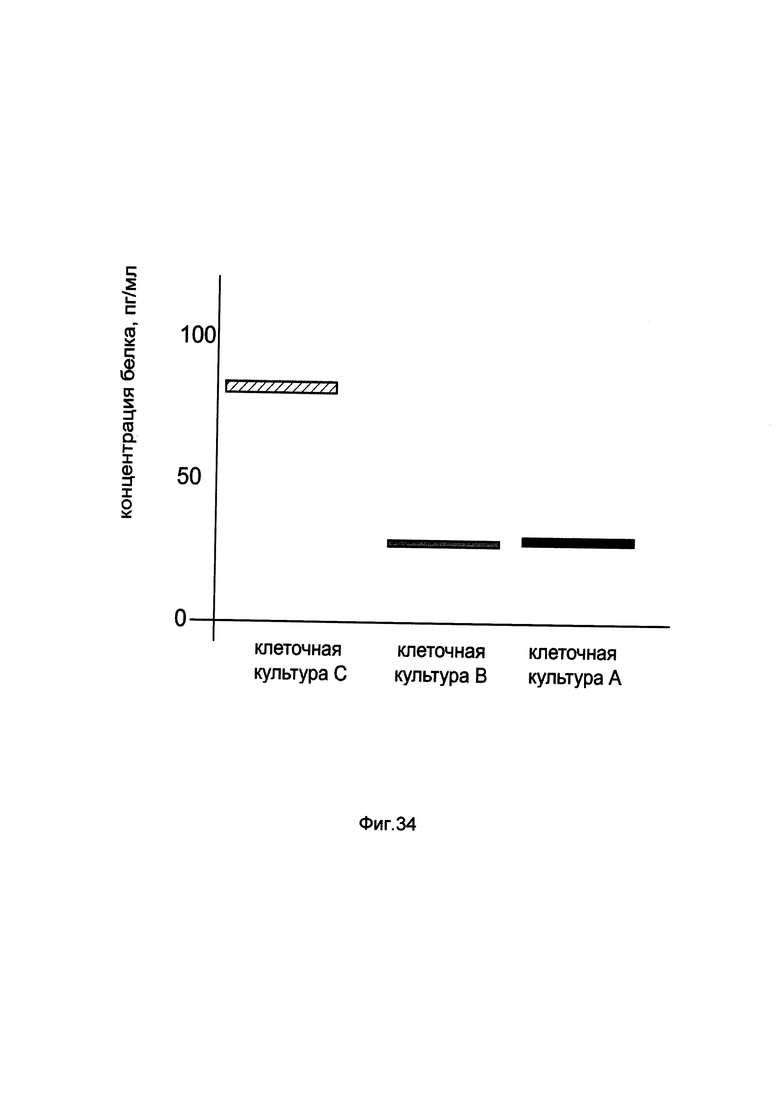
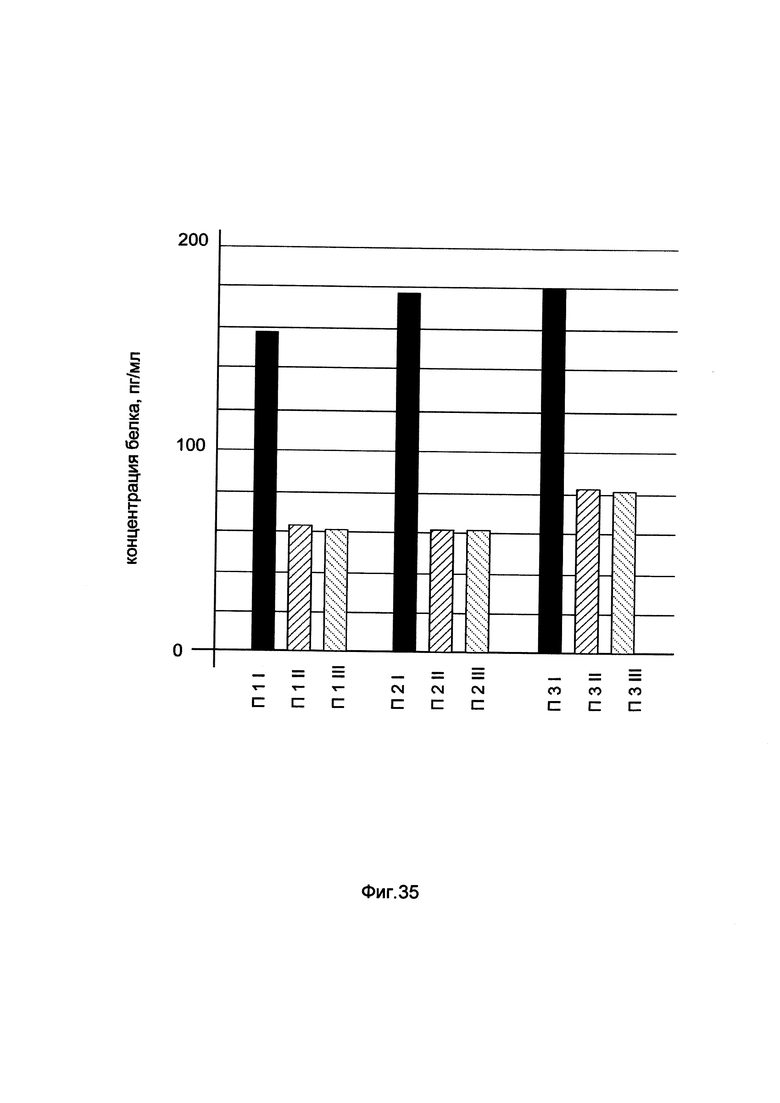
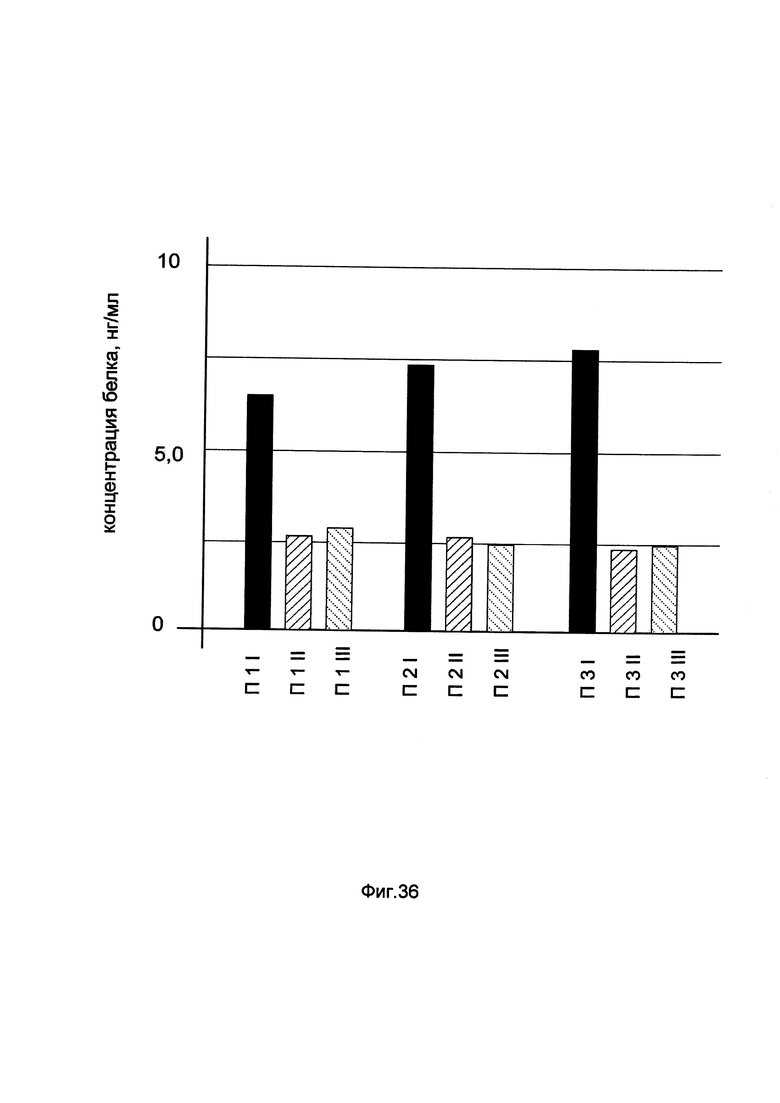
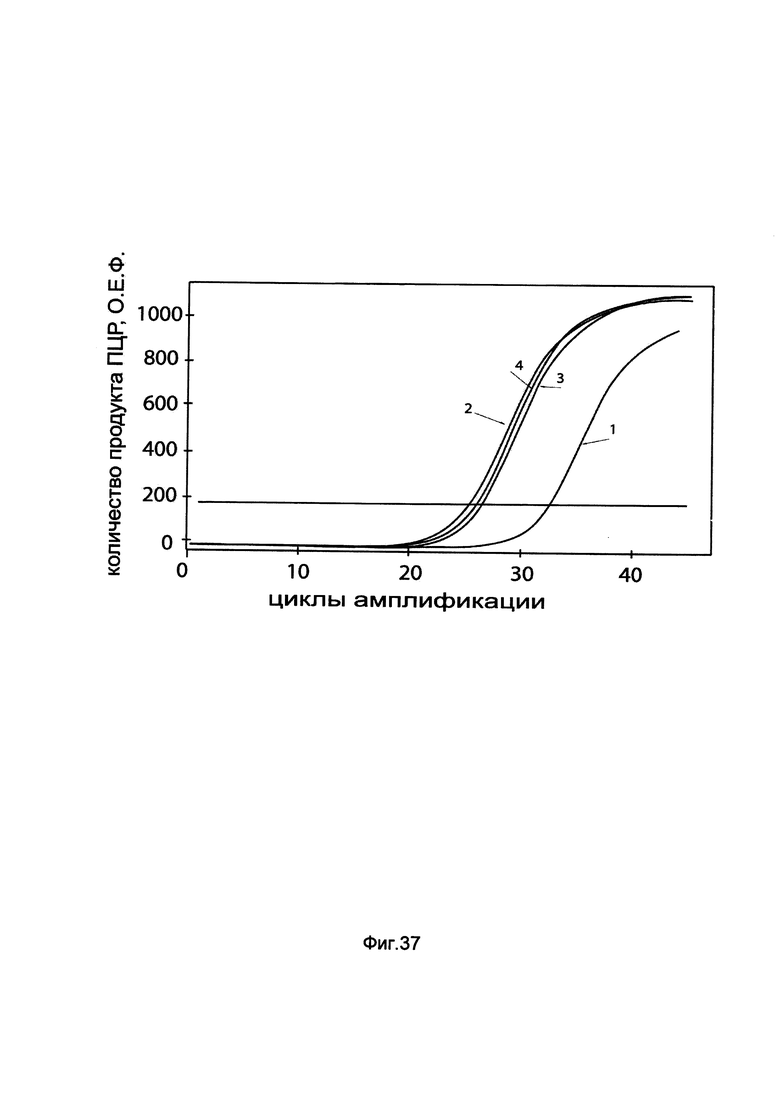
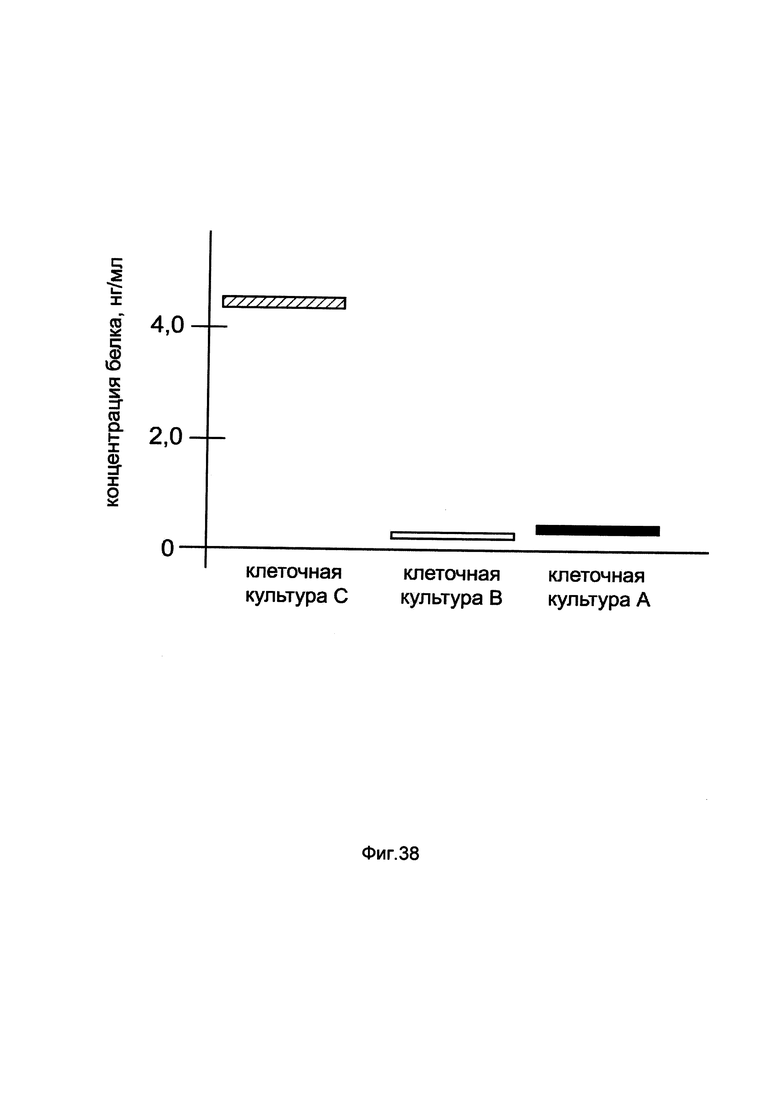
Изобретение относится к области биотехнологии, а именно к генотерапевтическим ДНК-векторам для таргетной терапии, штамму Escherichia coli JM110-NAS и выбору для таргетной генной терапии генотерапевтического ДНК-вектора. Способ генотерапевтического ДНК-вектора для таргетной генной терапии включает конструирование вектора размером 2408 п.н., содержащего ориджин репликации размером 688 п.н., терминатор транскрипции hGH-TA размером 467 п.н., регуляторный участок транспозона Tn10 PHK-out размером 137 п.н., ген устойчивости к канамицину размером 1018 п.н. и полилинкер размером 68 п.н. Затем его расщепляют с использованием эндонуклеаз рестрикции XhoI и BamHI и лигируют с промоторно-регуляторным участком, причем для получения интересующего генотерапевтического ДНК-вектора для таргетной генной терапии используют участок, содержащий промоторную область интересующего человека, а затем ген устойчивости к канамицину выщепляют по сайтам рестрикции SpeI, после чего оставшийся фрагмент лигируют сам на себя. Способ получения штамма Escherichia coli JM110-NAS для наработки генотерапевтического ДНК-вектора включает конструирование линейного фрагмента ДНК, содержащего регуляторный элемент RNA-in транспозона Tn11 для селекции без применения антибиотиков размером 64 п.н., ген левансахаразы sacB, продукт которого обеспечивает селекцию на сахарозо-содержащей среде размером 1422 п.н., ген устойчивости к хлорамфениколу catR, необходимый для отбора клонов штамма, в которых прошла гомологичная рекомбинация размером 763 п.н. и две гомологичные последовательности, обеспечивающие процесс гомологичной рекомбинации в области гена гесА с одновременной его инактивацией размером 329 п.н. и 233 п.н., причем указанные гомологичные последовательности являются последовательностями, полученными путем проведения ПЦР амплификации фрагментов гена гесА с использованием геномной ДНК Eshcerichia coli JM110-NAS в качестве матрицы и пары праймеров LHA-F (5'-GCTGACGCTGCAGGTGATC) и LHA-R (5-GACAAGATGTGTGTCTACCGCTTCAGGTTACCCGCCAG), и пары праймеров RHA-F (5'-TGGCAGGGCGGGGCGTAACTACGCCTCTGTTCGTCTCGA) и RHA-R (5'-CTCAGCAGCAACTCACGTAC). Затем проводят трансформацию клеток Escherichia coli путем электропорации и отбирают клоны, выжившие на среде, содержащей 10 мкг/мл хлорамфеникола. Изобретение позволяет расширить арсенал технических средств. 28 н.п. ф-лы, 1 табл., 38 ил., 39 пр.
1. Генотерапевтический ДНК-вектор GDTT1.8NAS1 для таргетной генной терапии, направленной на повышение экспрессии терапевтического гена в клетках целевой ткани человека, характеризующийся избирательностью, с обеспечением экспрессии терапевтического гена в клетках именно этой целевой ткани за счет наличия в составе генотерапевтического ДНК-вектора промоторно-регуляторного участка тканеспецифичного для клеток этой целевой ткани, при этом генотерапевтический ДНК-вектор GDTT1.8NAS1 имеет размер 1736 п.н. с нуклеотидной последовательностью SEQ ID №1 и содержит промоторно-регуляторный участок гена миоглобина человека тканеспецифичный для клеток мышечной ткани.
2. Генотерапевтический ДНК-вектор GDTT1.8NAS2 для таргетной генной терапии, направленной на повышение экспрессии терапевтического гена в клетках целевой ткани человека, характеризующийся избирательностью, с обеспечением экспрессии терапевтического гена в клетках именно этой целевой ткани за счет наличия в составе генотерапевтического ДНК-вектора промоторно-регуляторного участка тканеспецифичного для клеток этой целевой ткани, при этом генотерапевтический ДНК-вектор GDTT1.8NAS2 имеет размер 1654 п.н. с нуклеотидной последовательностью SEQ ID №2 и содержит промоторно-регуляторный участок гена эластазы человека тканеспецифичный для клеток кожи.
3. Генотерапевтический ДНК-вектор GDTT1.8NAS3 для таргетной генной терапии, направленной на повышение экспрессии терапевтического гена в клетках целевой ткани человека, характеризующийся избирательностью, с обеспечением экспрессии терапевтического гена в клетках именно этой целевой ткани за счет наличия в составе генотерапевтического ДНК-вектора промоторно-регуляторного участка тканеспецифичного для клеток этой целевой ткани, при этом генотерапевтический ДНК-вектор GDTT1.8NAS3 имеет размер 1783 п.н. с нуклеотидной последовательностью SEQ ID №3 и содержит промоторно-регуляторный участок гена молекулы межклеточной адгезии 2 человека тканеспецифичный для клеток эндотелия сосудов.
4. Генотерапевтический ДНК-вектор GDTT1.8NAS4 для таргетной генной терапии, направленной на повышение экспрессии терапевтического гена в клетках целевой ткани человека, характеризующийся избирательностью, с обеспечением экспрессии терапевтического гена в клетках именно этой целевой ткани за счет наличия в составе генотерапевтического ДНК-вектора промоторно-регуляторного участка тканеспецифичного для клеток этой целевой ткани, при этом генотерапевтический ДНК-вектор GDTT1.8NAS4 имеет размер 1953 п.н. с нуклеотидной последовательностью SEQ ID №4 и содержит промоторно-регуляторный участок гена остеокальцина 2 человека тканеспецифичный для клеток костной ткани, а именно остеобластов и одонтобластов.
5. Генотерапевтический ДНК-вектор GDTT1.8NAS5 для таргетной генной терапии, направленной на повышение экспрессии терапевтического гена в клетках целевой ткани человека, характеризующийся избирательностью, с обеспечением экспрессии терапевтического гена в клетках именно этой целевой ткани за счет наличия в составе генотерапевтического ДНК-вектора промоторно-регуляторного участка тканеспецифичного для клеток этой целевой ткани, при этом генотерапевтический ДНК-вектор GDTT1.8NAS5 имеет размер 2019 п.н. с нуклеотидной последовательностью SEQ ID №5 и содержит промоторно-регуляторный участок гена сурфактанта В человека тканеспецифичный для эпителиальных клеток бронхов и альвеол легких.
6. Генотерапевтический ДНК-вектор GDTT1.8NAS6 для таргетной генной терапии, направленной на повышение экспрессии терапевтического гена в клетках целевой ткани человека, характеризующийся избирательностью, с обеспечением экспрессии терапевтического гена в клетках именно этой целевой ткани за счет наличия в составе генотерапевтического ДНК-вектора промоторно-регуляторного участка тканеспецифичного для клеток этой целевой ткани, при этом генотерапевтический ДНК-вектор GDTT1.8NAS6 имеет размер 1940 п.н. с нуклеотидной последовательностью SEQ ID №6 и содержит промоторно-регуляторный участок гена синапсина I человека тканеспецифичный для клеток нервной ткани, а именно нейронов.
7. Генотерапевтический ДНК-вектор GDTT1.8NAS7 для таргетной генной терапии, направленной на повышение экспрессии терапевтического гена в клетках целевой ткани человека, характеризующийся избирательностью, с обеспечением экспрессии терапевтического гена в клетках именно этой целевой ткани за счет наличия в составе генотерапевтического ДНК-вектора промоторно-регуляторного участка тканеспецифичного для клеток этой целевой ткани, при этом генотерапевтический ДНК-вектор GDTT1.8NAS7 имеет размер 2620 п.н. с нуклеотидной последовательностью SEQ ID №7 и содержит промоторно-регуляторный участок гена нефрина человека тканеспецифичный для клеток почек, а именно подоцитов.
8. Генотерапевтический ДНК-вектор GDTT1.8NAS8 для таргетной генной терапии, направленной на повышение экспрессии терапевтического гена в клетках целевой ткани человека, характеризующийся избирательностью, с обеспечением экспрессии терапевтического гена в клетках именно этой целевой ткани за счет наличия в составе генотерапевтического ДНК-вектора промоторно-регуляторного участка тканеспецифичного для клеток этой целевой ткани, при этом генотерапевтический ДНК-вектор GDTT1.8NAS8 имеет размер 2240 п.н. с нуклеотидной последовательностью SEQ ID №8 и содержит промоторно-регуляторный участок гена общего лейкоцитарного антигена CD45 человека тканеспецифичный для клеток крови, а именно гематопоэтических клеток крови.
9. Генотерапевтический ДНК-вектор GDTT1.8NAS9 для таргетной генной терапии, направленной на повышение экспрессии терапевтического гена в клетках целевой ткани человека, характеризующийся избирательностью, с обеспечением экспрессии терапевтического гена в клетках именно этой целевой ткани за счет наличия в составе генотерапевтического ДНК-вектора промоторно-регуляторного участка тканеспецифичного для клеток этой целевой ткани, при этом генотерапевтический ДНК-вектор GDTT1.8NAS9 имеет размер 2604 п.н. с нуклеотидной последовательностью SEQ ID №9 и содержит промоторно-регуляторный участок гена белка В29 человека тканеспецифичный для клеток крови, а именно лимфоцитов.
10. Генотерапевтический ДНК-вектор GDTT1.8NAS10 для таргетной генной терапии, направленной на повышение экспрессии терапевтического гена в клетках целевой ткани человека, характеризующийся избирательностью, с обеспечением экспрессии терапевтического гена в клетках именно этой целевой ткани за счет наличия в составе генотерапевтического ДНК-вектора промоторно-регуляторного участка тканеспецифичного для клеток этой целевой ткани, при этом генотерапевтический ДНК-вектор GDTT1.8NAS10 имеет размер 2048 п.н. с нуклеотидной последовательностью SEQ ID №10 и содержит промоторно-регуляторный участок гена белка CD68 человека тканеспецифичный для клеток крови, а именно макрофагов.
11. Генотерапевтический ДНК-вектор GDTT1.8NAS11 для таргетной генной терапии, направленной на повышение экспрессии терапевтического гена в клетках целевой ткани человека, характеризующийся избирательностью, с обеспечением экспрессии терапевтического гена в клетках именно этой целевой ткани за счет наличия в составе генотерапевтического ДНК-вектора промоторно-регуляторного участка тканеспецифичного для клеток этой целевой ткани, при этом генотерапевтический ДНК-вектор GDTT1.8NAS11 имеет размер 1978 п.н. с нуклеотидной последовательностью SEQ ID №11 и содержит промоторно-регуляторный участок гена инсулина человека тканеспецифичный для клеток поджелудочной железы, а именно бета-клеток поджелудочной железы.
12. Способ получения генотерапевтического ДНК-вектора для таргетной генной терапии по пп. 1-11, заключающийся в том, что сначала конструируют вектор размером 2408 п.н., содержащий ориджин репликации размером 688 п.н., терминатор транскрипции hGH-TA размером 467 п.н., регуляторный участок транспозона Tn10 PHK-out размером 137 п.н., ген устойчивости к канамицину размером 1018 п.н. и полилинкер размером 68 п.н., затем его расщепляют с использованием эндонуклеаз рестрикции XhoI и BamHI и лигируют с промоторно-регуляторным участком, причем для получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS1 по п. 1 используют участок, содержащий промоторную область гена миоглобина человека размером 352 п.н.; для получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS2 по п. 2 используют участок, содержащий промоторную область гена эластазы человека размером 270 п.н.; для получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS3 по п. 3 используют участок, содержащий промоторную область гена молекулы межклеточной адгезии 2 человека размером 399 п.н.; для получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS4 по п. 4 используют участок, содержащий промоторную область гена остеокальцина 2 человека размером 569 п.н.; для получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS5 по п. 5 используют участок, содержащий промоторную область гена белка сурфактанта В человека размером 635 п.н.; для получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS6 по п. 6 используют участок, содержащий промоторную область гена белка синапсина I человека размером 556 п.н.; для получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS7 по п. 7 используют участок, содержащий промоторную область гена нефрина человека размером 1236 п.н.; для получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS8 по п. 8 используют участок, содержащий промоторную область гена общего лейкоцитарного антигена CD45 человека размером 856 п.н.; для получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS9 по п. 9 используют участок, содержащий промоторную область гена белка В29 человека размером 1220 п.н.; для получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS10 по п. 10 используют участок, содержащий промоторную область гена белка CD68 человека размером 658 п.н.; для получения генотерапевтического ДНК-вектора для таргетной генной терапии GDTT1.8NAS11 по п. 11 используют участок, содержащий промоторную область гена инсулина человека размером 594 п.н., а затем ген устойчивости к канамицину выщепляют по сайтам рестрикции SpeI, после чего оставшийся фрагмент лигируют сам на себя.
13. Способ получения штамма Escherichia coli JM110-NAS для наработки генотерапевтического ДНК-вектора по пп. 1-11, выбранного из: GDTT1.8NAS1, или GDTT1.8NAS2, или GDTT1.8NAS3, или GDTT1.8NAS4, или GDTT1.8NAS5, или GDTT1.8NAS6, или GDTT1.8NAS7, или GDTT1.8NAS8, или GDTT1.8NAS9, или GDTT1.8NAS10, или GDTT1.8NAS11, заключающийся в том, что конструируют линейный фрагмент ДНК, содержащий регуляторный элемент RNA-in транспозона Tn11 для селекции без применения антибиотиков размером 64 п.н., ген левансахаразы sacB, продукт которого обеспечивает селекцию на сахарозо-содержащей среде размером 1422 п.н., ген устойчивости к хлорамфениколу catR, необходимый для отбора клонов штамма, в которых прошла гомологичная рекомбинация размером 763 п.н. и две гомологичные последовательности, обеспечивающие процесс гомологичной рекомбинации в области гена гесА с одновременной его инактивацией размером 329 п.н. и 233 п.н., причем указанные гомологичные последовательности являются последовательностями, полученными путем проведения ПЦР амплификации фрагментов гена гесА с использованием геномной ДНК Eshcerichia coli JM110-NAS в качестве матрицы и пары праймеров LHA-F (5'-GCTGACGCTGCAGGTGATC) и LHA-R (5-GACAAGATGTGTGTCTACCGCTTCAGGTTACCCGCCAG), и пары праймеров RHA-F (5'-TGGCAGGGCGGGGCGTAACTACGCCTCTGTTCGTCTCGA) и RHA-R (5'-CTCAGCAGCAACTCACGTAC), после чего проводят трансформацию клеток Escherichia coli путем электропорации и отбирают клоны, выжившие на среде, содержащей 10 мкг/мл хлорамфеникола.
14. Штамм Escherichia coli JM110-NAS, полученный способом по п. 13, для наработки генотерапевтического ДНК-вектора по пп. 1-11, выбранного из GDTT1.8NAS1, или GDTT1.8NAS2, или GDTT1.8NAS3, или GDTT1.8NAS4, или GDTT1.8NAS5, или GDTT1.8NAS6, или GDTT1.8NAS7, или GDTT1.8NAS8, или GDTT1.8NAS9, или GDTT1.8NAS10, или GDTT1.8NAS11 с возможностью положительной селекции без использования антибиотиков, и содержащий в хромосоме в области гена гесА линейный фрагмент, состоящий из регуляторного элемента RNA-in транспозона Tn10, гена левансахарозы sacB и гена устойчивости к хлорамфениколу catR.
15. Способ получения штамма, несущего генотерапевтический ДНК-вектор по пп. 1-11, выбранный из GDTT1.8NAS1, или GDTT1.8NAS2, или GDTT1.8NAS3, или GDTT1.8NAS4, или GDTT1.8NAS5, или GDTT1.8NAS6, или GDTT1.8NAS7, или GDTT1.8NAS8, или GDTT1.8NAS9, или GDTT1.8NAS10, или GDTT1.8NAS11, а именно: Escherichia coli JM-110-NAS-GDTT1.8NAS1, или Escherichia coli JM-110-NAS-GDTT1.8NAS2, или Escherichia coli JM-110-NAS-GDTT1.8NAS3, или Escherichia coli JM-110-NAS-GDTT1.8NAS4, или Escherichia coli JM-110-NAS-GDTT1.8NAS5, или Escherichia coli JM-110-NAS-GDTT1.8NAS6, или Escherichia coli JM-110-NAS-GDTT1.8NAS7, или Escherichia coli JM-110-NAS- GDTT1.8NAS8, или Escherichia coli JM-110-NAS-GDTT1. 8NAS9, или Escherichia coli JM-110-NAS-GDTT1.8NAS10, или Escherichia coli JM-110-NAS-GDTT1.8NAS11, заключающийся в том, что получают электрокомпетентные клетки штамма Escherichia coli JM110-NAS по п. 13 и проводят электропорацию этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS1, или GDTT1.8NAS2, или GDTT1.8NAS3, или GDTT1.8NAS4, или GDTT1.8NAS5, или GDTT1.8NAS6, или GDTT1.8NAS7, или GDTT1.8NAS8, или GDTT1.8NAS9, или GDTT1.8NAS10, или GDTT1.8NAS11, после чего клетки высевают на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола для отбора стабильных клонов штамма.
16. Штамм Escherichia coli JM-110-NAS-GDTT1.8NAS1, полученный по п. 15, несущий генотерапевтический ДНК-вектор GDTT1.8NAS1, для наработки генотерапевтического ДНК-вектора GDTT1.8NAS1, с возможностью селекции без использования антибиотиков.
17. Штамм Escherichia coli JM-110-NAS-GDTT1.8NAS2, полученный по п. 15, несущий генотерапевтический ДНК-вектор GDTT1.8NAS2, для наработки генотерапевтического ДНК-вектора GDTT1.8NAS2, с возможностью селекции без использования антибиотиков.
18. Штамм Escherichia coli JM-110-NAS-GDTT1.8NAS3, полученный по п. 15, несущий генотерапевтический ДНК-вектор GDTT1.8NAS3, для наработки генотерапевтического ДНК-вектора GDTT1.8NAS3, с возможностью селекции без использования антибиотиков.
19. Штамм Escherichia coli JM-110-NAS-GDTT1.8NAS4, полученный по п. 15, несущий генотерапевтический ДНК-вектор GDTT1.8NAS4, для наработки генотерапевтического ДНК-вектора GDTT1.8NAS4, с возможностью селекции без использования антибиотиков.
20. Штамм Escherichia coli JM-110-NAS-GDTT1.8NAS5, полученный по п. 15, несущий генотерапевтический ДНК-вектор GDTT1.8NAS5, для наработки генотерапевтического ДНК-вектора GDTT1.8NAS5, с возможностью селекции без использования антибиотиков.
21. Штамм Escherichia coli JM-110-NAS-GDTT1.8NAS6, полученный по п. 15, несущий генотерапевтический ДНК-вектор GDTT1.8NAS6, для наработки генотерапевтического ДНК-вектора GDTT1.8NAS6, с возможностью селекции без использования антибиотиков.
22. Штамм Escherichia coli JM-110-NAS-GDTT1.8NAS7, полученный по п. 15, несущий генотерапевтический ДНК-вектор GDTT1.8NAS7, для наработки генотерапевтического ДНК-вектора GDTT1.8NAS7, с возможностью селекции без использования антибиотиков.
23. Штамм Escherichia coli JM-110-NAS-GDTT1.8NAS8, полученный по п. 15, несущий генотерапевтический ДНК-вектор GDTT1.8NAS8, для наработки генотерапевтического ДНК-вектора GDTT1.8NAS8, с возможностью селекции без использования антибиотиков.
24. Штамм Escherichia coli JM-110-NAS-GDTT1.8NAS9, полученный по п. 15, несущий генотерапевтический ДНК-вектор GDTT1.8NAS9, для наработки генотерапевтического ДНК-вектора GDTT1.8NAS9, с возможностью селекции без использования антибиотиков.
25. Штамм Escherichia coli JM-110-NAS-GDTT1.8 NAS 10, полученный по п. 15, несущий генотерапевтический ДНК-вектор GDTT1.8NAS10, для наработки генотерапевтического ДНК-вектора GDTT1.8NAS10, с возможностью селекции без использования антибиотиков.
26. Штамм Escherichia coli JM-110-NAS-GDTT1.8NAS11, полученный по п. 15, несущий генотерапевтический ДНК-вектор GDTT1.8NAS11, для наработки генотерапевтического ДНК-вектора GDTT1.8NAS11, с возможностью селекции без использования антибиотиков.
27. Способ производства в промышленных масштабах генотерапевтического ДНК-вектора по пп. 1-11, заключающийся в масштабировании бактериальной культуры штамма по пп. 16-26 до количеств, необходимых для наращивания бактериальной биомассы в промышленном ферментере, после чего биомассу используют для выделения фракции, содержащей целевой ДНК-продукт - генотерапевтический ДНК-вектор GDTT1.8NAS1, или GDTT1.8NAS2, или GDTT1.8NAS3, или GDTT1.8NAS4, или GDTT1.8NAS5, или GDTT1.8NAS6, или GDTT1.8NAS7, или, GDTT1.8NAS8, или GDTT1.8NAS9, или GDTT1.8NAS10, или GDTT1.8NAS11 по пп. 1-11, многостадийно фильтруют и очищают хроматографическими методами.
28. Способ выбора для таргетной генной терапии генотерапевтического ДНК-вектора GDTT1.8NAS1, или GDTT1.8NAS2, или GDTT1.8NAS3, или GDTT1.8NAS4, или GDTT1.8NAS5, или GDTT1.8NAS6, или GDTT1.8NAS7, или GDTT1.8NAS8, или GDTT1.8NAS9, или GDTT1.8NAS10, или GDTT1.8NAS11 по пп. 1-11, каждый из которых содержит различные тканеспецифичные промоторно-регуляторные участки гена человека, для конструирования тканеспецифичного генотерапевтического ДНК-вектора, содержащего терапевтический ген, для обеспечения эффективной экспрессии этого терапевтического гена исключительно в целевой клеточной линии, целевой ткани человека за счет наличия в составе генотерапевтического ДНК-вектора промоторно-регуляторного участка тканеспецифичного только для этой целевой клеточной линии, целевой ткани человека, заключающегося в том, что для конструирования генотерапевтического ДНК-вектора, содержащего терапевтический ген для таргетной генной терапии, направленной на повышение экспрессии этого терапевтического гена в клетках мышечной ткани, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS1, для конструирования генотерапевтического ДНК-вектора, содержащего терапевтический ген для таргетной генной терапии, направленной на повышение экспрессии этого терапевтического гена в клетках кожи, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS2, для конструирования генотерапевтического ДНК-вектора, содержащего терапевтический ген для таргетной генной терапии, направленной на повышение экспрессии этого терапевтического гена в клетках эндотелия сосудов, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS3, для конструирования генотерапевтического ДНК-вектора, содержащего терапевтический ген для таргетной генной терапии, направленной на повышение экспрессии этого терапевтического гена в клетках костной ткани, а именно в остеобластах и одонтобластах, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS4, для конструирования генотерапевтического ДНК-вектора, содержащего терапевтический ген для таргетной генной терапии, направленной на повышение экспрессии этого терапевтического гена в эпителиальных клетках бронхов и альвеол легких, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS5, для конструирования генотерапевтического ДНК-вектора, содержащего терапевтический ген для таргетной генной терапии, направленной на повышение экспрессии этого терапевтического гена в клетках нервной ткани, а именно в нейронах, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS6, для конструирования генотерапевтического ДНК-вектора, содержащего терапевтический ген для таргетной генной терапии, направленной на повышение экспрессии этого терапевтического гена в клетках почек, а именно в подоцитах, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS7, для конструирования генотерапевтического ДНК-вектора, содержащего терапевтический ген для таргетной генной терапии, направленной на повышение экспрессии этого терапевтического гена в клетках крови, а именно в гематопоэтических клетках крови, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS8, для конструирования генотерапевтического ДНК-вектора, содержащего терапевтический ген для таргетной генной терапии, направленной на повышение экспрессии этого терапевтического гена в клетках крови, а именно в лимфоцитах, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS9, для конструирования генотерапевтического ДНК-вектора, содержащего терапевтический ген для таргетной генной терапии, направленной на повышение экспрессии этого терапевтического гена в клетках крови, а именно в макрофагах, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS10, для конструирования генотерапевтического ДНК-вектора, содержащего терапевтический ген для таргетной генной терапии, направленной на повышение экспрессии этого терапевтического гена в клетках поджелудочной железы, а именно в бета-клетках поджелудочной железы, выбирают генотерапевтический ДНК-вектор GDTT1.8NAS11.
| WO2017121831 A1, 20.07.2017 | |||
| СУПОТНИЦКИЙ М.В | |||
| "Генотерапевтические векторные системы на основе вирусов", Биопрепараты | |||
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| CHEN Z.Y., et al | |||
| "Minicircle DNA vectors devoid of bacterial DNA result in persistent and high-level transgene expression in vivo", Mol Ther | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |

