Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущий целевой ген, выбранный из группы генов DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2 для повышения уровня экспрессии этих целевых генов, способ его получения и применения, штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC или Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10 или Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13 или Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1 или Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4 или Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10 или Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2 или Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF или Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2, несущий генотерапевтический ДНК-вектор, способ его получения, способ производства в промышленных масштабах генотерапевтического ДНК-вектора.
Область техники
Изобретение относится к генной инженерии и может быть использовано в биотехнологии, медицине и сельском хозяйстве для создания препаратов генной терапии.
Уровень техники
Генная терапия — это современный медицинский подход, направленный на лечение наследственных и приобретенных заболеваний путем введения нового генетического материала в клетки пациента с целью компенсации или подавления функции мутантного гена и/или исправления генетического дефекта. Конечным продуктом экспрессии гена может являться молекула РНК или белка. Однако осуществление большей части физиологических процессов в организме связано с функциональной активностью белковых молекул, тогда как молекулы РНК являются либо промежуточным продуктом в синтезе белков, либо осуществляют регуляторные функции. Таким образом, целью генной терапии является, в большинстве случаев, введение в организм генов, обеспечивающих транскрипцию и последующую трансляцию белковых молекул, кодируемых этими генами. В рамках описания настоящего изобретения под экспрессией гена подразумевается продукция белковой молекулы, аминокислотная последовательность которой кодируется этим геном.
Гены DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2, входящие в группу генов, играют ключевую роль в ряде процессов в организме человека и животных. Показана связь низких/недостаточных концентраций белков, кодируемых этими генами с различными неблагоприятными состояниями человека, которая, в ряде случаев, подтверждена нарушениями в нормальной экспрессии генов, кодирующих эти белки. Таким образом, генотерапевтическое повышение экспрессии гена, выбранного из группы генов DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2, обладает потенциалом для коррекции различных состояний человека и животных.
Ген DCC (Dopa decarboxylase, Gene ID: 1644) кодирует фермент декарбоксилазу ароматических L-аминокислот (ДАА), который катализирует реакцию декарбоксилирования, обеспечивая биосинтез дофамина из L-3,4-дигидроксифенилаланина (L-DOPA), серотонина из L-5-гидрокситриптофана и триптамина из L-триптофана. L-DOPA получается из тирозина в реакции, катализируемой ферментом тирозин гидроксилазой. Поскольку дофамин не проникает сквозь гематоэнцефалический барьер L-DOPA применяется как фармакологический препарат для компенсации дефицита дофамина у пациентов с Болезнью Паркинсона, купируя, таким образом, симптомы Болезни Паркинсона. Применение L-DOPA дает заметный положительный эффект только на начальных стадиях развития Болезни Паркинсона. Это связано с тем, что, несмотря на значительное снижение числа дофаминергических окончаний в стриатуме, в них все еще содержится достаточное количество ДАА для поддержания необходимого уровня дофамина (Marsden and Parkes, 1977). Однако по мере прогрессирования болезни уровень ДАА падает, поэтому необходимо увеличивать дозу и частоту применения L-DOPA для достижения желаемого клинического эффекта (Nagatsu et al., 1979). В этой связи, генная терапия путем доставки гена, кодирующего ДАА, в стриатум рассматривается как подход для улучшения клинического ответа на терапию L-DOPA при прогрессировании Болезни Паркинсона. Так на модели БП, вызванной введением MPTP, у обезьян была проведена комплексная терапия, включающая доставку рекомбинантного гена DCC человека в стриатум с помощью адено-ассоциированного вируса и систематическое применение L-DOPA (Bankiewicz et al., 2000, 2006). Полученные результаты подтвердили экспрессию целевого гена в течение по крайней мере 6-ти лет после введения и показали восстановление способности клеток в стриатуме превращать L-DOPA в дофамин. Это приводило к тому, что желаемые терапевтические эффекты у животных, которым был введен трансген достигались при существенно меньших дозах L-DOPA. Уменьшение дозы L-DOPA также снижало побочные эффекты его применения. В похожем исследовании положительные эффекты доставки гена DCC на модели Болезни Паркинсона у обезьян удалось наблюдать на протяжении 15 лет (Sehara et al., 2017). В настоящее время несколько исследовательских центров проводят клинические испытания генной терапии конструкцией, кодирующей ген DCC, на основе адено-ассоциированного вируса серотипа 2 (Eberling et al., 2008; Muramatsu et al., 2010; Mittermeyer et al., 2012; Valles et al., 2010).
Interleukin 10 (IL-10) (Gene ID: 3586). Интерлейкин 10 относится к иммуномодулирующим цитокинам и продуцируется преимущественно моноцитами и в меньшей степени лимфоцитами. Он оказывает разнообразные воздействия на регуляцию иммунитета и воспаления. На модели экспериментального энцефаломиелита (ЭЭМ), вызванного введением крысиного гликопротеина миелина (MOG1-125), конструкцию с геном интерлейкина 10 с точечной мутацией IL-10F129S вводили интратекально (Sloane et al., 2008). Результаты анализа экспериментальных и контрольных животных показали торможение развития паралича, замедление потери массы тела, снижение проникновения клеток воспалительного ряда в паренхиму мозга, подавление активации астроцитов ЦНС и подавление аллодении (болевой чувствительности к неболевым стимулам). Помимо экспериментального энцефаломиелита, генная терапия с использованием гена, кодирующего IL-10, была использована на моделях хронической боли, вызванной перевязкой седалищного нерва (Dengler et al., 2014; Milligan et al., 2006) или введением таксола (Ledeboer et al., 2007). При этом в этих работах доставка осуществлялась интратекально с помощью плазмидного вектора. Анализ эффекта генной терапии на индукцию хронической боли проводили с помощью теста на механическую аллодению. Во всех исследованных случаях, доставка гена, кодирующего IL-10, приводила пороговые значения чувствительности к механическому раздражению к уровню нормальных животных. В качестве основного механизма действия такой генной терапии на моделях хронической боли рассматривается подавление интерлейкином 10 активации астроцитов и клеток микроглии в спинном мозге (Lau et al., 2012).
Interleukin 13 (IL-13) (Gene ID: 3596). Интерлейкин 13 относится к иммуномодулирующим цитокинам и продуцируется преимущественно активированными Т-хелперными клетками 2-го типа. Он регулирует некоторые стадии созревания и дифференцировки антителопродуцирующих B-клеток, обеспечивает переключение B-клеток на продукцию антител IgE изотипа и подавляет активность макрофагов, ингибируя синтез провоспалительных цитокинов и хемокинов (Mao et al., 2018). IL-13, как показывают последние исследования, играет определенную роль в патогенезе аутоиммунных заболеваний, в том числе и рассеянного склероза (Mao et al., 2018). На модели индуцированной купризоном демиелинизации ген, кодирующий IL-13, доставляли с помощью лентивирусного вектора в утолщение мозолистого тела в головном мозге (Guglielmetti et al., 2016). Это приводило к изменению ответа микроглии и макрофагов на демиелинизирующее повреждение. В частности, IL-13 меняет фенотип микроглии и инфильтрующих ткани мозга макрофагов, микроокружение, способствующее развитию воспалительной реакции. Генная терапия с использованием конструкции, кодирующей IL-13, таким образом, способствует уменьшению последствий демиелинизации, вызванной купризоном.
Interferon beta 1 (INFB1) (Gene ID: 3456). Этот ген кодирует цитокин, который принадлежит к семейству интерферонов, принимающих участите в активации ответа врожденного иммунитета на проникновение внутрь организма патогена. IFNB1 важен для защиты преимущественно от вирусных инфекций. В случае с рассеянным склерозом рекомбинантный IFNB1 применяется как препарат первой линии для облегчения симптомов хронического воспаления и демиелинизации при рецидивирующей форме заболевания (Moreno et al., 2010). На модели экспериментального энцефаломиелита (ЭЭМ), вызванного введением пептида гликопротеина миелина (MOG35-55) (Hamana et al., 2017), через 7 дней после индукции вводили плазмидный вектор, кодирующий мышиный INFB, в хвостовую вену экспериментальных животных. Введение конструкции приводило к существенному снижению выраженности симптомов ЭЭМ спустя месяц. Кроме того, анализ проницаемости гематоэнцефалического барьера (ГЭБ) показал, что введение конструкции подавляет нарушения ГЭБа характерные для модели ЭЭМ. Гистологический анализ позволил установить, что в спинной мозг мышей, подвергнутых генной терапии, проникает гораздо меньше клеток воспалительного ряда чем в контрольных животных. Авторы работы пришли к заключению, что однократного введения конструкции, кодирующей IFNB, достаточно для долгосрочного противовоспалительного эффекта в модели ЭЭМ.
TNF receptor superfamily member 4 (TNFRSF4, OX40) (Gene ID: 7293) и TNF superfamily member 10 (TNFSF10, TRAIL) (Gene ID: 8743). Ген ОХ40 кодирует один из белков, относящихся к суперсемейству рецепторов к факторам некроза опухоли. Нокаут этого гена подавляет апоптоз в трансформированных и опухолевых клетках. Белок, кодируемый этим геном принимает участие в регуляции пролиферации и дифференцировки В клеток. Ген TRAIL кодирует цитокин, который принадлежит к семейству апоптоз-индуцирующих лигандов, зависимых от фактора некроза опухоли. Он индуцирует апоптоз в трансформированных и опухолевых клетках, связываясь со своими рецепторами и активируя MAPK8/JNK, каспазы 8 и 3. На модели экспериментального энцефаломиелита (ЭЭМ), вызванного введением гликопротеина миелина (MOG) (Yellayi et al., 2011), на 8-ой день после индукции (т.е. до появления симптомов) интратекально (в большую цистерну) вводили плазмиду в составе ДНК-липидного комплекса (lipoplex). Генетическая конструкция кодировала слитый белок OX40-TRAIL. Экспрессия генетической конструкции приводила к синтезу слитого белка, который обладает иммуномодулирующим свойством. Анализ животных, в котором оценивали формирование симптомов (слабость хвоста, слабость и паралич задних конечностей и т.д.), проводили в течение 30 дней после введения плазмиды. Результаты анализа показали достоверное снижение клинических проявлений ЭЭМ, а гистологический анализ подтвердил экспрессию самой конструкции в субэпиндимальной области 3-го желудочка и заметное снижение числа клеток воспалительного ряда, инфильтрующих белое вещество спинного мозга.
B-Cell Lymphoma protein-2 (BCL2 apoptosis regulator, BCL-2) (Gene ID: 596). Ген BCL-2 кодирует белок, встраивающийся во внешнюю мембрану митохондрий и блокирующий апоптоз в некоторых типах клеток. Он регулирует клеточную гибель в условиях низкого содержания кислорода (Snyder and Chandel, 2009). Так, крысам с церебральной ишемией, вызванной временной (1.5 часа) перевязкой срединной церебральной артерии, интратекально (в большую цистерну) сразу после индукции повреждения вводили плазмиды pCMV-bcl2 или pHRE-bcl2 в составе ДНК-липосомного комплекса (lipoplex) (Cao et al., 2002). Белок BCL-2 локализуется во внешней мембране митохондрий, где он ингибирует действие различных проапоптотических факторов, обеспечивая, таким образом, выживание клетки. В данном случае генная терапия с использованием гена BCL-2 была направлена на повышение выживаемости нейронов в области мозга, подвергнутой ишемии. В работе было проведено сравнение векторов c промотерами CMV (цитомегаловирусный промотер) и HRE (индуцируемый гипоксией промотер фактора роста эндотелия человека). В первом случае трансген должен экспрессироваться повсеместно в мозге, а во-втором – преимущественно в области затронутой ишемией. Результаты морфологического анализа показали, что обе конструкции приводили к значительному снижению числа апоптотических клеток, причем в случае с CMV промотером это снижение было более существенным. Объем же инфарктной зоны достоверно снижался только в случае с CMV промотером. Полученные результаты позволили авторам заключить, что интратекальная доставка векторов с генами антиапоптотических факторов оказывает нейропротекторное действие при ишемическом повреждении.
Hepatocyte growth factor (HGF) (Gene ID: 3082). Фактор роста гепатоцитов – плазминогензависимый плеотропный ростовой фактор, происходящий из мезенхимных клеток (Nakamura et al., 2011). Он регулирует рост клеток, клеточную подвижность и морфогенез различных типов клеток, включая эпителиальные и эндотелиальные. Помимо этого, он играет роль в ангиогенезе, регенерации некоторых органов и тканей, воспалительных реакциях и усилении роста отростков нервных клеток ганглиев дорзальных корешков (Maina and Klein, 1999). Также он выполняет несколько функций в центральной (ЦНС) и периферической (ПНС) нервных системах: защищает от гибели затронутые повреждением нейроны и обеспечивает регенерацию нервных отростков в ЦНС и ПНС. Плазмидные векторы, кодирующие HGF, вводили либо внутримышечно (Tsuchihara et al., 2009), либо интратекально (Hu et al., 2017) на модели перевязки седалищного нерва или его ветвей. Результаты теста на механочувствительность показали восстановление порога, при котором животные отдергивают лапу, после проведения генной терапии. Основным механизмом действия HGF на моделях хронической боли, как и в случае с интерлейкином 10, рассматривается подавление активации астроцитов и клеток микроглии в спинном мозге.
Interleukin 2 (IL-2) (Gene ID: 3558). Интерлейкин 2 – это цитокин, который усиливает пролиферацию Т и В лимфоцитов после стимулирования антигеном. Помимо иммуномодуляции, IL-2 действует и как регуляторная молекула в ЦНС. Он и его рецептор продуцируется в различных областях мозга, включая гиппокамп, гипоталамус, мозжечок, неостриатум и ганглии дорзальных корешков (Hanisch and Quirion, 1995). Накопленные данные показывают, что IL-2 играет важную роль в нервной и нейроэндокринной регуляции организма. Также одним из проявлений IL-2 в нервной системе является его антиноцицептивная функция, как в периферических нервах, так и в ЦНС. Он способен связываться с опиоидными рецепторами (Wang et al., 1996). На модели перевязки седалищного нерва плазмидный вектор, кодирующий IL-2, вводили интратекально спустя 30 дней после индукции хронической боли крысам (Yao et al., 2002). Анализ чувствительности к нагреванию показал, что однократное введение конструкции вместе с липофектамином приводило к увеличению латентного периода в ответ на нагревание, и этот эффект длился в течение нескольких дней. Тот факт, что антиноцицептивный эффект трансгена блокировался налаксоном, говорит о прямом действии IL-2 на опиоидные рецепторы. Авторы пришли к заключению, что генная терапия с использованием гена, кодирующего IL-2, может быть использована для облегчения невропатической боли.
Таким образом, предшествующий уровень техники свидетельствует о том, гены DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2 обладают потенциалом для коррекции ряда отклонений и состояний организма, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе, с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте, или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз.
Этим обусловлено объединение генов DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2 в рамках данного патента в группу генов. Генетические конструкции, обеспечивающие экспрессию белков, кодируемых генами из группы DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2 могут быть использованы для разработки лекарственных препаратов для предотвращения и терапии различных заболеваний и патологических состояний.
Более того, приведенные данные свидетельствуют о том, что недостаточная экспрессия белков, кодируемых генами DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2, входящими в группу генов, связана не только с патологическими состояниями, но и с предрасположенность к их развитию. Также приведенные данные свидетельствуют о том, что недостаточная экспрессия данных белков может не проявляться в явном виде в форме патологии, которая может быть однозначно описана в рамках существующих стандартов клинической практики (например, с применением кода МКБ), однако при этом вызывать состояния, которые неблагоприятны для человека и животных и связанны с ухудшением качества жизни.
Анализ подходов для повышения экспрессии целевых генов подразумевает возможность использования различных генотерапевтических векторов.
Генотерапевтические векторы разделяют на вирусные, клеточные и ДНК-векторы (Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal Products EMA/CAT/80183/2014). В последнее время в генной терапии всё большее внимание уделяется разработке невирусных систем доставки генетического материала, среди которых лидируют плазмидные векторы. Плазмидные векторы лишены недостатков, присущих клеточным и вирусным векторам. В клетке-мишени они существуют в эписомальной форме, не интегрируют в геном, производство их достаточно дешево, отсутствие иммунного ответа и побочных реакций на введение плазмидного вектора делают их удобным инструментом генной терапии и генетической профилактики (ДНК-вакцины) (Li L, Petrovsky N. // Expert Rev Vaccines. 2016;15(3):313-29).
Тем не менее, ограничениями для использования плазмидных векторов для генной терапии являются: 1) наличие генов устойчивости к антибиотикам для наработки в бактериальных штаммах, 2) наличие различных регуляторных элементов, представленных последовательностями вирусных геномов 3) размер терапевтического плазмидного вектора, определяющий эффективность проникновения вектора в клетку-мишень.
Известно, что Европейское агентство по лекарственным средствам считает необходимым избегать введения маркеров антибиотикорезистентности в разрабатываемые плазмидные векторы для генной терапии (Reflection paper on design modifications of gene therapy medicinal products during development / 14 December 2011 EMA/CAT/GTWP/44236/2009 Committee for advanced therapies). Данная рекомендация связана, в первую очередь, с потенциальной опасностью проникновения ДНК-вектора или горизонтального переноса генов антибиотикорезистентности в клетки бактерий, представленных в организме в составе нормальной или оппортунистической микрофлоры. Помимо этого, наличие генов антибиотикорезистентности значительно увеличивает размер ДНК-вектора, что приводит к снижению эффективности его проникновения в эукариотические клетки.
Необходимо отметить, что гены антибиотикорезистентности также вносят принципиальный вклад в способ получения ДНК-векторов. В случае наличия генов антибиотикорезистентности штаммы для наработки ДНК-векторов обычно культивируются в среде, содержащей селективный антибиотик, что создает риск наличия следовых количеств антибиотика в недостаточно очищенных препаратах ДНК-векторов. Таким образом, получение ДНК-векторов для генной терапии, в которых отсутствуют гены антибиотикорезистентности, связано с получением штаммов, обладающих такой отличительной особенностью как способность к стабильной амплификации целевых ДНК-векторов в среде без содержания антибиотиков.
Кроме того, Европейское Медицинское Агентство рекомендует избегать наличия в составе терапевтических плазмидных векторов регуляторных элементов для повышения экспрессии целевых генов (промоторов, энхансеров, посттрансляционных регуляторных элементов), являющихся нуклеотидными последовательностями геномов различных вирусов (Draft Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal products,http://www.ema.europa.eu/docs/en_GB/document_library/Scientific_guideline/2015/05/WC500187020.pdf). Данные последовательности, хотя и могут увеличивать уровень экспрессии целевого трансгена, однако создают риск рекомбинации с генетическим материалом вирусов дикого типа и интеграции в геном эукариотической клетки. Более того, целесообразность гиперэкспрессии того или иного гена в целях терапии остается нерешенным вопросом.
Также, существенным моментом является размер терапевтического вектора. Известно, что современные плазмидные векторы зачастую перегружены нефункциональными участками, серьезно увеличивающими размер вектора (Mairhofer J, Grabherr R. // Mol Biotechnol. 2008.39(2):97-104). Например, ген устойчивости к ампициллину в векторах серии pBR322, как правило, состоит из не менее чем 1000 п.н., что составляет более 20% от размера самого вектора. При этом наблюдается обратная зависимость между размером вектора и его способностью проникать в эукариотические клетки – ДНК-векторы с небольшим размером эффективней проникают в клетки человека и животных. Так, например, в серии экспериментов по трансфекции клеток HELA ДНК-векторами с размером от 383 до 4548 п.н. было показано, что разница в эффективности проникновения может достигать двух порядков (отличаться в 100 раз) (Hornstein BD et al. // PLoS ONE. 2016;11(12): e0167537.).
Таким образом, при выборе ДНК-вектора в целях безопасности и наибольшей эффективности следует отдавать предпочтение тем конструкциям, в которых не содержатся гены устойчивости к антибиотикам, последовательности вирусного происхождения и размер которых позволяет эффективно проникать в эукариотические клетки., Штамм для получения такого ДНК-вектора в количествах, достаточных для целей генной терапии, должен обеспечивать возможность стабильной амплификации ДНК-вектора с использованием питательных сред, не содержащих антибиотики.
Примером использования рекомбинантных ДНК-векторов для генной терапии является способ получения рекомбинантного вектора для генетической иммунизации по патенту US 9550998 В2. Вектор представляет собой суперскрученный плазмидный ДНК-вектор и предназначен для экспрессии клонированных генов в клетках животных и человека. Вектор состоит из ориджина репликации, регуляторных элементов, включающих промотор и энхансер цитомегаловируса человека, регуляторные элементы из Т-лимфотропного вируса человека.
Накопление вектора проводят в специальном штамме E. coli без использования антибиотиков за счет антисенс-комплементации гена sacB, введенного в штамм посредством бактериофага. Недостатком данного изобретения является наличие в составе ДНК-вектора регуляторных элементов, представляющих собой последовательности вирусных геномов.
Известен патент на изобретение US 10316074, описывающий способ получения белка IL2 для использования в медицинских целях. Ген, кодирующий IL2, был включен в состав рекомбинантной конструкции для экспрессии в дрожжах. Недостатком данного изобретения является использование белка IL2 вместо генотерапевтического вектора, экспрессирующего ген, кодирующий белок IL2.
Известен патент на изобретение US 6090791, в котором описывается способ повышения концентрации терапевтического белка, в частности, IL10, путем введения в целевые клетки плазмидного вектора, содержащего последовательность с неметелированными нуклеотидами цитозином и гуанином. Недостатком данного метода является ограниченное число клеток, способных индуцировать выработку целевого белка и неопределенность требований к безопасности используемого вектора.
Известен патент на изобретение US 7598058, где описаны способы получения модифицированного белка IL13, включая синтез полипептида, последовательностей ДНК, кодирующих этот ген, а также клонирование этих последовательностей в плазмидные векторы. Недостатком данного изобретения является наличие в составе ДНК-вектора регуляторных элементов, представляющих собой последовательности вирусных геномов, а также наличие последовательностей генов антибиотикорезистентности.
Прототипами предлагаемого решения, а именно, использования генотерапевтических подходов для повышения уровня экспрессии генов из группы DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2, являются следующие патенты и заявки на получение патента:
Патент на изобретение US 4808523A, где описан плазмидный вектор, несущий ген, кодирующий IFNB1. Данный вектор позволяет повышать экспрессию IFNB1 в клетках млекопитающих. Недостатком данного изобретения является способ использования, ограниченный продукцией белка IFNB1 в условиях in vitro и отличный от генотерапевтического использования данного вектора. Также недостатком данного изобретения является наличие в составе вектора последовательностей вирусного происхождения.
Патент на изобретение RU 2678756, где описан генотерапевтический ДНК-вектор VTvaf17, способ его получения, штамм Escherichia coli SCS110-AF, способ его получения, штамм Escherichia coli SCS110-AF/VTvaf17, несущий генотерапевтический ДНК-вектор VTvaf17, способ его получения. Недостатком данного изобретения является значительный размер векторной части, что может негативно отразиться на эффективности доставки ДНК-вектора в клетки.
Патент на изобретение US 8647618, где описан способ доставки генотерапевтического вектора, несущего последовательность гена IL2 в составе клеток аттенуированного штамма Salmonella typhimurium для лечения онкологических заболеваний. Недостатком данного способа является индукция иммунного ответа на повторные введения бактериальных клеток, и, соответственно, критическое снижение терапевтического эффекта, а также неопределенность требований к безопасности используемого вектора.
Патент на изобретение US 8389492, где описан экспрессирующий плазмидный вектор, в составе которого находится ген, кодирующий фактор роста гепатоцитов (HGF) для лечения ишемических заболеваний и заболеваний печени. В патенте описаны способы введения вектора, в том числе, в виде «голой» ДНК, а также в составе липосомальных комплексов. Недостатком данного изобретения является наличие в составе вектора гена резистентности к антибиотику канамицину.
Раскрытие изобретения
Задачей изобретения является конструирование генотерапевтических ДНК-векторов для повышения уровня экспрессии группы генов DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2 в организме человека и животных, сочетающих в себе следующие свойства:
I) Эффективность генотерапевтического ДНК-вектора для повышения уровня экспрессии целевых генов в эукариотических клетках.
II) Возможность безопасного применения для генетической терапии человека и животных за счет отсутствия в составе генотерапевтического ДНК-вектора регуляторных элементов, представляющих собой нуклеотидные последовательности вирусных геномов.
III) Возможность безопасного применения для генетической терапии человека и животных за счет отсутствия в составе генотерапевтического ДНК-вектора генов антибиотикорезистентности.
IV) Технологичность получения и возможность наработки генотерапевтического ДНК-вектора в промышленных масштабах.
Пункты II и III предусмотрены в данном техническом решении в соответствии с рекомендациями государственных регуляторов к лекарственным средствам для генной терапии, в частности, Европейского Агентства по лекарственным средствам касательно отказа от введения маркеров антибиотикорезистентности в разрабатываемые плазмидные векторы для генной терапии (Reflection paper on design modifications of gene therapy medicinal products during development / 14 December 2011, EMA/CAT/GTWP/44236/2009 Committee for advanced therapies) и касательно отказа от введения в разрабатываемые плазмидные векторы для генной терапии элементов вирусных геномов (Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal products / 23 March 2015, EMA/CAT/80183/2014, Committee for Advanced Therapies).
Задачей изобретения также является конструирование штаммов, несущих эти генотерапевтические ДНК-вектора, для наработки и производства в промышленных масштабах генотерапевтических ДНК-векторов.
Поставленная задача решается за счет того, что создан генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе, с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте, или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз, при этом генотерапевтический ДНК-вектор GDTT1.8NAS12-DDC содержит кодирующую часть целевого гена DDC, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с нуклеотидной последовательностью SEQ ID №1, генотерапевтический ДНК-вектор GDTT1.8NAS12-IL10 содержит кодирующую часть целевого гена IL10, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с нуклеотидной последовательностью SEQ ID №2, генотерапевтический ДНК-вектор GDTT1.8NAS12-IL13 содержит кодирующую часть целевого гена IL13, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с нуклеотидной последовательностью SEQ ID №3, генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1 содержит кодирующую часть целевого гена IFNB1, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с нуклеотидной последовательностью SEQ ID №4, генотерапевтический ДНК-вектор GDTT1.8NAS12-TNFRSF4 содержит кодирующую часть целевого гена TNFRSF4, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с нуклеотидной последовательностью SEQ ID №5, генотерапевтический ДНК-вектор GDTT1.8NAS12-TNFSF10 содержит кодирующую часть целевого гена TNFSF10, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с нуклеотидной последовательностью SEQ ID №6, генотерапевтический ДНК-вектор GDTT1.8NAS12-BCL2 содержит кодирующую часть целевого гена BCL2, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с нуклеотидной последовательностью SEQ ID №7, генотерапевтический ДНК-вектор GDTT1.8NAS12-HGF содержит кодирующую часть целевого гена HGF, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с нуклеотидной последовательностью SEQ ID №8, генотерапевтический ДНК-вектор GDTT1.8NAS12-IL2 содержит кодирующую часть целевого гена IL2, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с нуклеотидной последовательностью SEQ ID №9.
Каждый из созданных генотерапевтических ДНК-векторов: GDTT1.8NAS12-DDC или GDTT1.8NAS12-IL10 или GDTT1.8NAS12-IL13 или GDTT1.8NAS12-IFNB1 или GDTT1.8NAS12-TNFRSF4 или GDTT1.8NAS12-TNFSF10 или GDTT1.8NAS12-BCL2 или GDTT1.8NAS12-HGF или GDTT1.8NAS12-IL2 за счет ограниченного размера векторной части GDTT1.8NAS12, не превышающей 2600 п.н., обладает способностью эффективно проникать в клетки человека и животных и экспрессировать клонированный в него целевой ген DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2.
В составе каждого из созданных генотерапевтических ДНК-векторов: GDTT1.8NAS12-DDC или GDTT1.8NAS12-IL10 или GDTT1.8NAS12-IL13 или GDTT1.8NAS12-IFNB1 или GDTT1.8NAS12-TNFRSF4 или GDTT1.8NAS12-TNFSF10 или GDTT1.8NAS12-BCL2 или GDTT1.8NAS12-HGF или GDTT1.8NAS12-IL2 в качестве структурных элементов используются нуклеотидные последовательности, которые не являются генами антибиотикорезистентности, вирусными генами и регуляторными элементами вирусных геномов, обеспечивая возможность его безопасного применения для генетической терапии человека и животных.
Создан также способ получения генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген DDC, или IL10, или IL13, или IFNB1, или TNFRSF4, или TNFSF10, или BCL2, или HGF, или IL2, заключающийся в том, что каждый из генотерапевтических ДНК-векторов: GDTT1.8NAS12-DDC или GDTT1.8NAS12-IL10 или GDTT1.8NAS12-IL13 или GDTT1.8NAS12-IFNB1 или GDTT1.8NAS12-TNFRSF4 или GDTT1.8NAS12-TNFSF10 или GDTT1.8NAS12-BCL2 или GDTT1.8NAS12-HGF или GDTT1.8NAS12-IL2 получают следующим образом: кодирующую часть целевого гена DDC или IL10 или IL13 или IFNB1 или TNFRSF4 или TNFSF10 или BCL2 или HGF или IL2 клонируют в генотерапевтический ДНК-вектор GDTT1.8NAS12 и получают генотерапевтический ДНК-вектор GDTT1.8NAS12-DDC, SEQ ID №1, или GDTT1.8NAS12-IL10, SEQ ID №2 или GDTT1.8NAS12-IL13, SEQ ID №3, или GDTT1.8NAS12-IFNB1, SEQ ID №4, или GDTT1.8NAS12-TNFRSF4, SEQ ID №5 или GDTT1.8NAS12-TNFSF10, SEQ ID №6 или GDTT1.8NAS12-BCL2, SEQ ID №7, или GDTT1.8NAS12-HGF, SEQ ID №8, или GDTT1.8NAS12-IL2, SEQ ID №9 соответственно, при этом кодирующую часть целевого гена DDC или IL10 или IL13 или IFNB1 или TNFRSF4 или TNFSF10 или BCL2 или HGF или IL2 получают путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции и ПЦР-амплификации с использованием созданных олигонуклеотидов и расщеплением продукта амплификации соответствующими эндонуклеазами рестрикции, причем клонирование в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят по сайтам рестрикции SalI и HindIII, или SalI и KpnI, или BamHI и SalI, причем селекцию проводят без антибиотиков,
при этом при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, SEQ ID №1 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
DDC-F ATCGTCGACCACCATGAACGCAAGTGAGTTCCGA,
DDC-R ACCAAGCTTCTACTCCCTCTCTG,
а расщепление продукта амплификации и клонирование кодирующей части гена DDC в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции SalI и HindIII,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-IL10, SEQ ID №2 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
IL10-F AGGATCCACCATGCACAGCTCAGCACTGC,
IL10-R CTTGTCGACTCAGTTTCGTATCTTCATTGTC,
а расщепление продукта амплификации и клонирование кодирующей части гена IL10 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-IL13, SEQ ID №3 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
IL13-F AGGATCCACCATGCATCCGCTCCTCAATC,
IL13-R CTTGTCGACTCAGTTGAACTGTCCCTCG,
а расщепление продукта амплификации и клонирование кодирующей части гена IL13 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, SEQ ID №4 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
IFNB1-F AAAGGATCCACCATGACCAACAAGTGTCTCCTCCAAA,
IFNB1-R TTTGTCGACTCAGTTTCGGAGGTAACCTGTAAGTCTG,
а расщепление продукта амплификации и клонирование кодирующей части гена IFNB1 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFRSF4, SEQ ID №5 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
TNFRSF4-F AGGATCCACCATGTGCGTGGGGGCTCGG,
TNFRSF4-R CTTGTCGACTCAGATCTTGGCCAGGGTG,
а расщепление продукта амплификации и клонирование кодирующей части гена TNFRSF4 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFSF10, SEQ ID №6 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
TNFSF10-F AGGATCCACCATGGCTATGATGGAGGTCCA,
TNFSF10-R TGTCGACTCAGCCAACTAAAAAGGCCCCGA,
а расщепление продукта амплификации и клонирование кодирующей части гена TNFSF10 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-BCL2, SEQ ID №7 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
BCL-F ATCGTCGACCACCATGGCGCACGCTGGGAGA,
BCL-R TTCGGTACCTCACTTGTGGCCCA,
а расщепление продукта амплификации и клонирование кодирующей части гена BCL2 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции SalI и KpnI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-HGF, SEQ ID №8 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
HGF_F TTTGGATCCACCATGTGGGTGACCAAACTCCTGCCA,
HGF_R AATGTCGACCTATGACTGTGGTACCTTATATGTTAAAAT,
а расщепление продукта амплификации и клонирование кодирующей части гена HGF в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-IL2, SEQ ID №9 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
IL2-F AGGATCCACCATGTACAGGATGCAACTCCT,
IL2-R TGTCGACTCAAGTCAGTGTTGAGATGATG,
а расщепление продукта амплификации и клонирование кодирующей части гена IL2 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI.
Создан способ использования генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2 для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе, с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте, или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз, заключающийся в трансфекции выбранным генотерапевтическим ДНК-вектором, несущим целевой ген на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, или несколькими выбранными генотерапевтическими ДНК-векторами, несущими целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, из созданных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, клеток органов и тканей пациента или животного, и/или во введении в органы и ткани пациента или животного аутологичных клеток этого пациента или животного, трансфицированных выбранным генотерапевтическим ДНК-вектором, несущим целевой ген на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 или несколькими выбранными генотерапевтическими ДНК-векторами, несущими целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, из созданных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 и/или во введении в органы и ткани пациента или животного выбранного генотерапевтического ДНК-вектора, несущего целевой ген на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 или нескольких выбранных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, из созданных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, или в сочетании обозначенных способов.
Создан способ получения штамма для производства генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе, с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте, или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз, заключающийся в получении электрокомпетентных клеток штамма Escherichia coli JM110-NAS с последующим проведением электропорации этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS12-DDC или GDTT1.8NAS12-IL10 или GDTT1.8NAS12-IL13 или GDTT1.8NAS12-IFNB1 или GDTT1.8NAS12-TNFRSF4 или GDTT1.8NAS12-TNFSF10 или GDTT1.8NAS12-BCL2 или GDTT1.8NAS12-HGF или GDTT1.8NAS12-IL2, после чего клетки высеивают на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола с получением в результате штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10 или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13 или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1 или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4 или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10 или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2 или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2.
Заявлен штамм Escherichiacoli JM110-NAS/GDTT1.8NAS12-DDC, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-DDC, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-IL10, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-IL13, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-TNFRSF4, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-TNFSF10, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-BCL2, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-HGF, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12- IL2, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе, с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте, или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз.
Создан способ производства в промышленных масштабах генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген DDC, или IL10, или IL13, или IFNB1, или TNFRSF4, или TNFSF10, или BCL2, или HGF, или IL2 для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе, с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте, или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз, заключающийся в том, что генотерапевтический ДНК-вектор GDTT1.8NAS12-DDC или генотерапевтический ДНК-вектор GDTT1.8NAS12-IL10 или генотерапевтический ДНК-вектор GDTT1.8NAS12-IL13 или генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1 или генотерапевтический ДНК-вектора GDTT1.8NAS12-TNFRSF4 или генотерапевтический ДНК-вектор GDTT1.8NAS12-TNFSF10 или генотерапевтический ДНК-вектор GDTT1.8NAS12-BCL2 или генотерапевтический ДНК-вектор GDTT1.8NAS12-HGF или генотерапевтический ДНК-вектор GDTT1.8NAS12-IL2 получают путем того, что засевают в колбу с приготовленной средой затравочную культуру, выбранную из штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10 или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13 или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1 или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4 или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10 или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2 или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2, затем инкубируют затравочную среду в шейкере-инкубаторе и переносят ее в промышленный ферментер, после чего растят до достижения стационарной фазы, затем выделяют фракцию, содержащую целевой ДНК-продукт, многостадийно фильтруют и очищают хроматографическими методами.
Краткое описание чертежей
На фиг.1
приведена схема генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген, выбранный из группы генов: DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2, который представляет собой кольцевую двуцепочечную молекулу ДНК, способную к автономной репликации в клетках бактерии Escherichia coli.
На фиг.1 приведены схемы, соответствующие:
Фиг. 1A - генотерапевтический ДНК-вектор GDTT1.8NAS12-DDC,
Фиг. 1B - генотерапевтический ДНК-вектор GDTT1.8NAS12-IL10,
Фиг. 1C - генотерапевтический ДНК-вектор GDTT1.8NAS12-IL13,
Фиг. 1D - генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1,
Фиг. 1E - генотерапевтический ДНК-вектор GDTT1.8NAS12-TNFRSF4,
Фиг. 1F - генотерапевтический ДНК-вектор GDTT1.8NAS12-TNFSF10,
Фиг. 1G - генотерапевтический ДНК-вектор GDTT1.8NAS12-BCL2,
Фиг. 1H - генотерапевтический ДНК-вектор GDTT1.8NAS12-HGF,
Фиг. 1I - генотерапевтический ДНК-вектор GDTT1.8NAS12-IL2.
На схемах отмечены следующие структурные элементы вектора:
EF1a pr - промоторная область гена человеческого фактора элонгации EF1A с собственным энхансером, содержащимся в первом интроне гена. Служит для обеспечения высокого уровня транскрипции рекомбинантного гена в большинстве тканей человека;
Рамка считывания целевого гена, соответствующая кодирующей части гена DDC (фиг. 1A), или IL10 (фиг. 1B), или IL13 (фиг. 1C), или IFNB1 (фиг. 1D) или TNFRSF4 (фиг. 1E), или TNFSF10 (фиг. 1F), или BCL2 (фиг. 1G), или HGF (фиг. 1H), или IL2 (фиг. 1I) соответственно;
hGH TA – терминатор транскрипции;
pUCori – ориджин репликации, служащий для автономной репликации с однонуклеотидной заменой для повышения копийности плазмиды в клетках большинства штаммов Escherichia coli;
RNA out – регуляторный элемент РНК-out транспозона Tn 10, обеспечивающий возможность положительной селекции без использования антибиотиков при использовании штамма Eshcerichia coli JM110-NAS.
Отмечены уникальные сайты рестрикции.
На фиг.2
показаны графики накопления ампликонов кДНК целевого гена, а именно, гена DDC, в культуре клеток кортикальных нейронов человека HCN-2 (ATCC CRL-10742) до их трансфекции и через 48 часов после трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS12-DDC с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.2 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 – кДНК гена DDC в культуре клеток HCN-2 до трансфекции ДНК-вектором GDTT1.8NAS12-DDC;
2 – кДНК гена DDC в культуре клеток HCN-2 после трансфекции ДНК-вектором GDTT1.8NAS12-DDC;
3 – кДНК гена B2M в культуре клеток HCN-2 до трансфекции ДНК-вектором GDTT1.8NAS12-DDC;
4 – кДНК гена B2M в культуре клеток HCN-2 после трансфекции ДНК-вектором GDTT1.8NAS12-DDC.
В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.3
показаны графики накопления ампликонов кДНК целевого гена, а именно гена IL10, в культуре клеток нейробластомы человека IMR-32 (ATCC CCL-127) человека до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IL10 с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.3 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена IL10 в культуре IMR-32 до трансфекции ДНК-вектором GDTT1.8NAS12-IL10;
2 – кДНК гена IL10 в культуре IMR-32 после трансфекции ДНК-вектором GDTT1.8NAS12-IL10;
3 - кДНК гена B2M в культуре IMR-32 до трансфекции ДНК-вектором GDTT1.8NAS12-IL10;
4 - кДНК гена B2M в культуре IMR-32 после трансфекции ДНК-вектором GDTT1.8NAS12-IL10.
В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.4
показаны графики накопления ампликонов кДНК целевого гена, а именно гена IL13 в культуре клеток глиобластомы человека U-118 MG (ATCC HTB-15) до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IL13 с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.4 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена IL13 в клетках U-118 MG до трансфекции ДНК-вектором GDTT1.8NAS12-IL13;
2 – кДНК гена IL13 в клетках U-118 MG после трансфекции ДНК-вектором GDTT1.8NAS12-IL13;
3 - кДНК гена B2M в клетках U-118 MG до трансфекции ДНК-вектором GDTT1.8NAS12-IL13;
4 - кДНК гена B2M в клетках U-118 MG после трансфекции ДНК-вектором GDTT1.8NAS12-IL13.
В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.5
показаны графики накопления ампликонов кДНК целевого гена, а именно гена IFNB1, в культуре клеток нейробластомы человека SH-SY5Y (ATCC CRL-2266) до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IFNB1 с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.5 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена IFNB1 в культуре клеток SH-SY5Y до трансфекции ДНК-вектором GDTT1.8NAS12-IFNB1;
2 – кДНК гена IFNB1 в культуре клеток SH-SY5Y после трансфекции ДНК-вектором GDTT1.8NAS12-IFNB1;
3 - кДНК гена B2M в культуре клеток SH-SY5Y до трансфекции ДНК-вектором GDTT1.8NAS12-IFNB1;
4 - кДНК гена B2M в культуре клеток SH-SY5Y после трансфекции ДНК-вектором GDTT1.8NAS12-IFNB1.
В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.6
показаны графики накопления ампликонов кДНК целевого гена, а именно гена TNFRSF4, в культуре клеток астроглии человека SVG p12 (ATCC CRL-8621) до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-TNFRSF4 с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.6 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена TNFRSF4 в культуре клеток SVG p12 до трансфекции ДНК-вектором GDTT1.8NAS12-TNFRSF4;
2 – кДНК гена TNFRSF4 в культуре клеток SVG p12 после трансфекции ДНК-вектором GDTT1.8NAS12-TNFRSF4;
3 - кДНК гена B2M в культуре клеток SVG p12 до трансфекции ДНК-вектором GDTT1.8NAS12-TNFRSF4;
4 - кДНК гена B2M в культуре клеток SVG p12 после трансфекции ДНК-вектором GDTT1.8NAS12-TNFRSF4.
В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.7
показаны графики накопления ампликонов кДНК целевого гена, а именно гена TNFSF10, в культуре нейроэктодермальных клеток опухоли головного мозга человека PFSK-1 (ATCC number CRL-2060) человека до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-TNFSF10 с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.7 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена TNFSF10 в культуре PFSK-1 до трансфекции ДНК-вектором GDTT1.8NAS12-TNFSF10;
2 – кДНК гена TNFSF10 в культуре PFSK-1 после трансфекции ДНК-вектором GDTT1.8NAS12-TNFSF10;
3 - кДНК гена B2M в культуре PFSK-1 до трансфекции ДНК-вектором GDTT1.8NAS12-TNFSF10;
4 - кДНК гена B2M в культуре PFSK-1 после трансфекции ДНК-вектором GDTT1.8NAS12-TNFSF10.
В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.8
показаны графики накопления ампликонов кДНК целевого гена, а именно гена BCL2 в культуре клеток медуллобластомы человека D341 Med (ATCC HTB-187) до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-BCL2 с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.8 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена BCL2 в клетках D341 Med до трансфекции ДНК-вектором GDTT1.8NAS12-BCL2;
2 – кДНК гена BCL2 в клетках D341 Med после трансфекции ДНК-вектором GDTT1.8NAS12- BCL2;
3 - кДНК гена B2M в клетках D341 Med до трансфекции ДНК-вектором GDTT1.8NAS12-BCL2;
4 - кДНК гена B2M в клетках D341 Med после трансфекции ДНК-вектором GDTT1.8NAS12- BCL2.
В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.9
показаны графики накопления ампликонов кДНК целевого гена, а именно гена HGF, в культуре клеток астроцитомы головного мозга человека A-172 (ATCC CRL-1620) до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-HGF с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.9 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена HGF в культуре клеток A-172 до трансфекции ДНК-вектором GDTT1.8NAS12-HGF;
2 – кДНК гена HGF в культуре клеток A-172 после трансфекции ДНК-вектором GDTT1.8NAS12-HGF;
3 - кДНК гена B2M в культуре клеток A-172 до трансфекции ДНК-вектором GDTT1.8NAS12-HGF;
4 - кДНК гена B2M в культуре клеток A-172 после трансфекции ДНК-вектором GDTT1.8NAS12-HGF.
В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.10
показаны графики накопления ампликонов кДНК целевого гена, а именно гена IL2, в культуре нейрональных прогениторных клеток человека ATCC-BXS0117 (ATCC ACS-5003) до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IL2 с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.10 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена IL2 в культуре клеток ATCC-BXS0117 до трансфекции ДНК-вектором GDTT1.8NAS12-IL2;
2 – кДНК гена IL2 в культуре клеток ATCC-BXS0117 после трансфекции ДНК-вектором GDTT1.8NAS12-IL2;
3 - кДНК гена B2M в культуре клеток ATCC-BXS0117 до трансфекции ДНК-вектором GDTT1.8NAS12-IL2;
4 - кДНК гена B2M в культуре клеток ATCC-BXS0117 после трансфекции ДНК-вектором GDTT1.8NAS12-IL2.
В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.11
показана диаграмма концентрации белка DDC в клеточном лизате кортикальных нейронов человека HCN-2 (ATCC CRL-10742) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-DDC с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген DDC.
На фиг.11 отмечены следующие элементы:
культура А – культура HCN-2, трансфицированная водным раствором дендримеров без плазмидной ДНК (контроль);
культура B - культура HCN-2, трансфицированных ДНК-вектором GDTT1.8NAS12;
культура C - культура HCN-2, трансфицированных ДНК-вектором GDTT1.8NAS12-DDC.
На фиг. 12
показана диаграмма концентрации белка IL10 в лизате культура клеток нейробластомы человека IMR-32 (ATCC CCL-127), после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IL10 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IL10.
На фиг.12 отмечены следующие элементы:
культура А - культура IMR-32, трансфицированная водным раствором дендримеров без плазмидной ДНК (контроль);
культура B - культура IMR-32, трансфицированная ДНК-вектором GDTT1.8NAS12;
культура C - культура IMR-32, трансфицированных ДНК-вектором GDTT1.8NAS12-IL10.
На фиг.13
показана диаграмма концентрации белка IL13 в лизате культуры клеток глиобластомы человека U-118 MG (ATCC HTB-15) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IL13 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IL13.
На фиг 13 отмечены следующие элементы:
культура А - культура клеток U-118 MG, трансфицированных водным раствором дендримеров без плазмидной ДНК (контроль);
культура B - культура клеток U-118 MG, трансфицированных ДНК-вектором GDTT1.8NAS12;
культура C - культура клеток U-118 MG, трансфицированных ДНК-вектором GDTT1.8NAS12-IL13.
На фиг. 14
показана диаграмма концентрации белка IFNB1 в лизате культуры клеток нейробластомы человека SH-SY5Y (ATCC CRL-2266) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IFNB1 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IFNB1.
На фиг.14 отмечены следующие элементы:
культура А - культура клеток SH-SY5Y, трансфицированных водным раствором дендримеров без плазмидной ДНК (контроль);
культура B - культура клеток SH-SY5Y, трансфицированных ДНК-вектором GDTT1.8NAS12;
культура C - культура клеток SH-SY5Y, трансфицированных ДНК-вектором GDTT1.8NAS12-IFNB1.
На фиг. 15
показана диаграмма концентрации белка TNFRSF4 в лизате клеток астроглии человека SVG p12 (ATCC CRL-8621после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-TNFRSF4 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген TNFRSF4.
На фиг.15 отмечены следующие элементы:
культура А - культура клеток SVG p12, трансфицированных водным раствором дендримеров без плазмидной ДНК (контроль);
культура B - культура клеток SVG p12, трансфицированных ДНК-вектором GDTT1.8NAS12;
культура C - культура клеток SVG p12, трансфицированных ДНК-вектором GDTT1.8NAS12-TNFRSF4.
На фиг. 16
показана диаграмма концентрации белка TNFSF10 в лизате нейроэктодермальных клеток опухоли головного мозга человека PFSK-1 (ATCC number CRL-2060) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-TNFSF10 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген TNFSF10.
На фиг.16 отмечены следующие элементы:
культура А - культура PFSK-1, трансфицированная водным раствором дендримеров без плазмидной ДНК (контроль);
культура B - культура PFSK-1, трансфицированная ДНК-вектором GDTT1.8NAS12;
культура C - культура PFSK-1, трансфицированных ДНК-вектором GDTT1.8NAS12-TNFSF10.
На фиг.17
показана диаграмма концентрации белка BCL2 в лизате культуры клеток медуллобластомы человека D341 Med (ATCC HTB-187) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-BCL2 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген BCL2.
На фиг.17 отмечены следующие элементы:
культура А - культура клеток D341 Med, трансфицированных водным раствором дендримеров без плазмидной ДНК (контроль);
культура B - культура клеток D341 Med, трансфицированных ДНК-вектором GDTT1.8NAS12;
культура C - культура клеток D341 Med, трансфицированных ДНК-вектором GDTT1.8NAS12-BCL2.
На фиг.18
показана диаграмма концентрации белка HGF в лизате культуры клеток астроцитомы головного мозга человека A-172 (ATCC CRL-1620) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-HGF с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген HGF.
На фиг.18 отмечены следующие элементы:
культура А - культура клеток A-172, трансфицированных водным раствором дендримеров без плазмидной ДНК (контроль);
культура B - культура клеток A-172, трансфицированных ДНК-вектором GDTT1.8NAS12;
культура C - культура клеток A-172, трансфицированных ДНК-вектором GDTT1.8NAS12-HGF.
На фиг. 19
показана диаграмма концентрации белка IL2 в лизате культуры нейрональных прогениторных клеток человека ATCC-BXS0117 (ATCC ACS-5003) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IL2 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IL2.
На фиг.19 отмечены следующие элементы:
культура А - культура клеток ATCC-BXS0117, трансфицированных водным раствором дендримеров без плазмидной ДНК (контроль);
культура B - культура клеток ATCC-BXS0117, трансфицированных ДНК-вектором GDTT1.8NAS12;
культура C - культура клеток ATCC-BXS0117, трансфицированных ДНК-вектором GDTT1.8NAS12-IL2.
На фиг. 20
показана диаграмма концентрации белка IFNB1 в биоптатах кожи трех пациентов после введения в кожу этих пациентов генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IFNB1.
На фиг.20 отмечены следующие элементы:
П1I – биоптат кожи пациента П1 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IFNB1;
П1II – биоптат кожи пациента П1 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо);
П1III – биоптат кожи пациента П1 из интактного участка;
П2I – биоптат кожи пациента П2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IFNB1;
П2II – биоптат кожи пациента П2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо);
П2III – биоптат кожи пациента П2 из интактного участка;
П3I – биоптат кожи пациента П3 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IFNB1;
П3II – биоптат кожи пациента П3 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо);
П3III – биоптат кожи пациента П3 из интактного участка.
На фиг. 21
показана диаграмма концентрации белка IL13 в биоптатах икроножной мышцы трех пациентов после введения в икроножную мышцу этих пациентов генотерапевтического ДНК-вектора GDTT1.8NAS12- IL13, с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IL13.
На фиг.21 отмечены следующие элементы:
П1I – биоптат икроножной мышцы пациента П1 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IL13;
П1II – биоптат икроножной мышцы пациента П1 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо);
П1III – биоптат интактного участка икроножной мышцы пациента П1;
П2I – биоптат икроножной мышцы пациента П2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IL13;
П2II – биоптат икроножной мышцы пациента П2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо);
П2III – биоптат интактного участка икроножной мышцы пациента П2;
П3I – биоптат икроножной мышцы пациента П3 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IL13;
П3II – биоптат икроножной мышцы пациента П3 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо);
П3III – биоптат интактного участка икроножной мышцы пациента П3.
На фиг.22
показана диаграмма концентрации белка IL2 в биоптатах кожи трех пациентов после введения в кожу этих пациентов генотерапевтического ДНК-вектора GDTT1.8NAS12-IL2 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IL2.
На фиг.22 отмечены следующие элементы:
П1I – биоптат кожи пациента П1 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IL2;
П1II – биоптат кожи пациента П1 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо);
П1III – биоптат кожи пациента П1 из интактного участка;
П2I – биоптат кожи пациента П2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IL2;
П2II – биоптат кожи пациента П2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо);
П2III – биоптат кожи пациента П2 из интактного участка;
П3I – биоптат кожи пациента П3 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IL2;
П3II – биоптат кожи пациента П3 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо);
П3III – биоптат кожи пациента П3 из интактного участка.
На фиг. 23
показана диаграмма концентрации белка IL10 в биоптатах кожи человека после введения в кожу культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL10 с целью демонстрации способа применения путем введения аутологичных клеток, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL10
На фиг. 23 отмечены следующие элементы:
П1С – биоптат кожи пациента П1 в зоне введения культуры аутологичных фибробластов пациента, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL10;
П1B – биоптат кожи пациента П1 в зоне введения аутологичных фибробластов пациента, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12;
П1А – биоптат кожи пациента П1 из интактного участка.
На фиг. 24
показана диаграмма концентраций белков: белка DDC человека, белка BCL2 человека, белка HGF человека, белка TNFSF10 человека, белка TNFRSF4 человека в мышечной ткани трех групп крыс линии Wistar после инъекционного введения смеси генотерапевтических векторов: генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, генотерапевтического ДНК-вектора GDTT1.8NAS12-BCL2, генотерапевтического ДНК-вектора GDTT1.8NAS12-HGF, генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFSF10, генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFRSF4 с целью демонстрации способа применения смеси генотерапевтических ДНК-векторов.
На фиг. 24 отмечены следующие элементы:
К1I – фрагмент мышечной ткани крысы I группы К1 в зоне введения смеси генотерапевтических ДНК векторов: GDTT1.8NAS12- DDC, GDTT1.8NAS12-BCL2, GDTT1.8NAS12-HGF, GDTT1.8NAS12-TNFSF10 и GDTT1.8NAS12-TNFRSF4;
К1II – фрагмент мышечной ткани крысы II группы К1 в зоне введения смеси генотерапевтических ДНК векторов: GDTT1.8NAS12- DDC, GDTT1.8NAS12-BCL2, GDTT1.8NAS12-HGF, GDTT1.8NAS12-TNFSF10 и GDTT1.8NAS12-TNFRSF4;
К1III – фрагмент мышечной ткани крысы III группы К1 в зоне введения смеси генотерапевтических ДНК векторов: GDTT1.8NAS12- DDC, GDTT1.8NAS12-BCL2, GDTT1.8NAS12-HGF, GDTT1.8NAS12-TNFSF10 и GDTT1.8NAS12-TNFRSF4;
К1IV – фрагмент мышечной ткани крысы IV группы К1 в зоне введения смеси генотерапевтических ДНК векторов: GDTT1.8NAS12- DDC, GDTT1.8NAS12-BCL2, GDTT1.8NAS12-HGF, GDTT1.8NAS12-TNFSF10 и GDTT1.8NAS12-TNFRSF4;
К1V – фрагмент мышечной ткани крысы V группы К1 в зоне введения смеси генотерапевтических ДНК векторов: GDTT1.8NAS12- DDC, GDTT1.8NAS12-BCL2, GDTT1.8NAS12-HGF, GDTT1.8NAS12-TNFSF10 и GDTT1.8NAS12-TNFRSF4;
К2I – фрагмент мышечной ткани крысы I группы К2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо);
К2II – фрагмент мышечной ткани крысы II группы К2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо);
К2III – фрагмент мышечной ткани крысы III группы К2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо);
К2IV– фрагмент мышечной ткани крысы IV группы К2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо);
К2V – фрагмент мышечной ткани крысы V группы К2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо);
К3I – фрагмент мышечной ткани контрольного интактного участка крысы I группы К3;
К3II – фрагмент мышечной ткани контрольного интактного участка крысы II группы К3;
К3III – фрагмент мышечной ткани контрольного интактного участка крысы III группы К3;
К3IV – фрагмент мышечной ткани контрольного интактного участка крысы IV группыК3;
К3V – фрагмент мышечной ткани контрольного интактного участка крысы V группы К3.
На фиг. 25
показаны графики накопления ампликонов кДНК целевого гена IL13 в мононуклеарных клетках периферической крови (PBMC) быка до и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IL13 с целью демонстрации способа применения путем введения генотерапевтического ДНК-вектора животным
На фиг.25 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена IL13 в клетках PBMC быка до трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL13;
2 – кДНК гена IL13 в клетках PBMC быка после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL13;
3 - кДНК гена ACT в клетках PBMC быка до трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL13;
4 - кДНК гена ACT в клетках PBMC быка после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL13.
В качестве референтного гена использовали ген актина быка/ коровы (АСТ), приведенного в базе данных GenBank под номером AH001130.2.
Реализация изобретения
На основе ДНК-вектора GDTT1.8NAS12 размером 2591 п.н. созданы генотерапевтические ДНК-векторы, несущие целевые гены человека, предназначенные для повышения уровня экспрессии этих целевых генов в тканях человека и животных. При этом способ получения каждого генотерапевтического ДНК-вектора, несущего целевые гены заключается в том, что в полилинкер генотерапевтического ДНК-вектора GDTT1.8NAS12 клонируют белок-кодирующую последовательность целевого гена, выбранного из группы генов: ген DDC (кодирует белок DDC), ген IL10 (кодирует белок IL10), ген IL13 (кодирует белок IL13), ген IFNB1 (кодирует белок IFNB1), ген TNFRSF4 (кодирует белок TNFRSF4), ген TNFSF10 (кодирует белок TNFSF10), ген BCL2 (кодирует белок BCL2), ген HGF (кодирует белок HGF), ген IL2 (кодирует белок IL2) человека. Известно, что способность ДНК-векторов проникать в эукариотические клетки обусловлена, главным образом, размером вектора. При этом ДНК-вектора с наименьшим размером обладают более высокой проникающей способностью. Таким образом, предпочтительным является отсутствие в составе вектора элементов, которые не несут функциональной нагрузки, но при этом увеличивают размер ДНК-вектора. Данные особенности ДНК-векторов были учтены при получении генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген, выбранный из группы генов: DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2, путем отсутствия в составе вектора крупных нефункциональных последовательностей и генов антибиотикорезистентности, что позволило, помимо технологических преимуществ и преимуществ в плане безопасности применения, значительно уменьшить размер полученного генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген, выбранный из группы генов: DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2. Таким образом, способность проникать в эукариотические клетки полученного генотерапевтического ДНК-вектора обусловлена его небольшими размерами.
Каждый из генотерапевтических ДНК-векторов: ДНК-вектор GDTT1.8NAS12-DDC, или GDTT1.8NAS12-IL10, или GDTT1.8NAS12-IL13, или GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-TNFRSF4, или GDTT1.8NAS12-TNFSF10, или GDTT1.8NAS12-BCL2, или GDTT1.8NAS12-HGF, или GDTT1.8NAS12-IL2 получали следующим образом: кодирующую часть целевого гена DDC, или IL10, или IL13, или IFNB1, или TNFRSF4, или TNFSF10, или BCL2, или HGF, или IL2 клонировали в генотерапевтический ДНК-вектор GDTT1.8NAS12 и получали генотерапевтический ДНК-вектор GDTT1.8NAS12-DDC, SEQ ID №1, или GDTT1.8NAS12-IL10, SEQ ID №2 или GDTT1.8NAS12-IL13, SEQ ID №3, или GDTT1.8NAS12-IFNB1, SEQ ID №4, или GDTT1.8NAS12-TNFRSF4, SEQ ID №5, или GDTT1.8NAS12-TNFSF10, SEQ ID №6, или GDTT1.8NAS12-BCL2, SEQ ID №7, или GDTT1.8NAS12-HGF, SEQ ID №8, или GDTT1.8NAS12-IL2, SEQ ID №9 соответственно. Кодирующую часть гена DDC размером 1447 п.н., или гена IL10 размером 540 п.н., или гена IL13 размером 444 п.н., или гена IFNB1 размером 567 п.н., или гена TNFRSF4 размером 837 п.н., или гена TNFSF10 размером 849 п.н., или гена BCL2 размером 724 п.н., или гена HGF размером 2190 п.н., или гена IL2 размером 465 п.н. получали путем выделения суммарной РНК из биологического образца ткани здорового человека. Для получения первой цепи кДНК генов DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2 человека использовали реакцию обратной транскрипции. Амплификацию проводили с использованием созданных для этого методом химического синтеза олигонуклеотидов. Расщепление продукта амплификации специфическими эндонуклеазами рестрикции проводили с учетом оптимальной процедуры дальнейшего клонирования, причем клонирование в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводили по сайтам рестрикции BamHI, EcoRI, HindIII, KpnI, SalI, расположенными в полилинкере вектора GDTT1.8NAS12. Выбор сайтов рестрикции проводили таким образом, чтобы клонированный фрагмент попадал в рамку считывания экспрессионной кассеты вектора GDTT1.8NAS12, при этом белок-кодирующая последовательность не содержала сайты рестрикции для выбранных эндонуклеаз. При этом специалистам в данной области техники понятно, что методическая реализация получения генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, или GDTT1.8NAS12-IL10, или GDTT1.8NAS12-IL13, или GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-TNFRSF4, или GDTT1.8NAS12-TNFSF10, или GDTT1.8NAS12-BCL2, или GDTT1.8NAS12-HGF, или GDTT1.8NAS12-IL2 может варьировать в рамках выбора известных методов молекулярного клонирования генов, при этом эти способы подпадают под объем настоящего изобретения. Так, например, могут быть использованы различные последовательности олигонуклеотидов для амплификации гена DDC, или IL10, или IL13, или IFNB1, или TNFRSF4, или TNFSF10, или BCL2, или HGF, или IL2 различные эндонуклеазы рестрикции или такие лабораторные техники, как безлигазное клонирование генов.
Генотерапевтический ДНК-вектор GDTT1.8NAS12-DDC, или GDTT1.8NAS12-IL10, или GDTT1.8NAS12-IL13, или GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-TNFRSF4, или GDTT1.8NAS12-TNFSF10, или GDTT1.8NAS12-BCL2, или GDTT1.8NAS12-HGF, или GDTT1.8NAS12-IL2 обладает нуклеотидной последовательностью SEQ ID №1 или SEQ ID №2 или SEQ ID №3 или SEQ ID №4 или SEQ ID №5 или SEQ ID №6 или SEQ ID №7 или SEQ ID №8 или SEQ ID №9 соответственно. При этом специалистам в данной области техники известно свойство вырожденности генетического кода, из которого следует, что под объем настоящего изобретения также подпадают варианты нуклеотидных последовательностей, отличающихся инсерцией, делецией или заменой нуклеотидов, которые не приводят к изменению полипептидной последовательности, кодируемой целевым геном, и/или не приводят к потере функциональной активности регуляторных элементов вектора GDTT1.8NAS12. При этом специалистам в данной области техники известно явление генетического полиморфизма, из которого следует, что под объем настоящего изобретения также подпадают варианты нуклеотидных последовательностей генов из группы генов: DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2, которые при этом кодируют различные варианты аминокислотных последовательностей белков DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2, не отличающихся от приведенных по своей функциональной активности при физиологических условиях.
Способность проникать в эукариотические клетки и функциональную активность, то есть способность экспрессировать целевой ген, полученного генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, или GDTT1.8NAS12-IL10, или GDTT1.8NAS12-IL13, или GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-TNFRSF4, или GDTT1.8NAS12-TNFSF10, или GDTT1.8NAS12-BCL2, или GDTT1.8NAS12-HGF, или GDTT1.8NAS12-IL2 подтверждают путем введения в эукариотические клетки полученного вектора и последующим анализом экспрессии специфической мРНК и/или белкового продукта целевого гена. Наличие специфической мРНК в клетках, в которые был введен генотерапевтический ДНК-вектор GDTT1.8NAS12-DDC, или GDTT1.8NAS12-IL10, или GDTT1.8NAS12-IL13, или GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-TNFRSF4, или GDTT1.8NAS12-TNFSF10, или GDTT1.8NAS12-BCL2, или GDTT1.8NAS12-HGF, или GDTT1.8NAS12-IL2 свидетельствует как о способности полученного вектора проникать в эукариотические клетки, так и о его способности экспрессировать мРНК целевого гена. При этом, как известно специалистам в данной области техники, наличие мРНК гена является обязательным условием, но не доказательством трансляции белка, кодируемого целевым геном. Поэтому для подтверждения свойства генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, или GDTT1.8NAS12-IL10, или GDTT1.8NAS12-IL13, или GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-TNFRSF4, или GDTT1.8NAS12-TNFSF10, или GDTT1.8NAS12-BCL2, или GDTT1.8NAS12-HGF, или GDTT1.8NAS12-IL2 экспрессировать целевой ген на уровне белка в эукариотических клетках, в которые был введен генотерапевтический ДНК-вектор, проводят анализ концентрации белков, кодируемых целевыми генами, с использованием иммунологических методов. Наличие белка DDC, или IL10, или IL13, или IFNB1, или TNFRSF4, или TNFSF10, или BCL2, или HGF, или IL2 подтверждает эффективность экспрессии целевых генов в эукариотических клетках и возможность повышения уровня концентрации белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген, выбранный из группы генов: DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2. Таким образом для подтверждения эффективности экспрессии созданного генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, несущего целевой ген, а именно, ген DDC, генотерапевтического ДНК-вектора GDTT1.8NAS12-IL10, несущего целевой ген, а именно, ген IL10, генотерапевтического ДНК-вектора GDTT1.8NAS12-IL13, несущего целевой ген, а именно, ген IL13, генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, несущего целевой ген, а именно, ген IFNB1, генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFRSF4, несущего целевой ген, а именно, ген TNFRSF4, генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFSF10, несущего целевой ген, а именно, ген TNFSF10, генотерапевтического ДНК-вектора GDTT1.8NAS12-BCL2, несущего целевой ген, а именно, ген BCL2, генотерапевтического ДНК-вектора GDTT1.8NAS12-HGF, несущего целевой ген, а именно, ген HGF, генотерапевтического ДНК-вектора GDTT1.8NAS12-IL2, несущего целевой ген, а именно, ген IL2 использовали следующие методы:
А) ПЦР в реальном времени - изменение накопления ампликонов кДНК целевых генов в лизате клеток человека и животного, после трансфекции различных клеточных линий человека и животного генотерапевтическим ДНК-векторами;
B) Иммуноферментный анализ - изменение количественного уровня целевых белков в лизате клеток человека, после трансфекции различных клеточных линий человека генотерапевтическими ДНК-векторами;
C) Иммуноферментный анализ - изменение количественного уровня целевых белков в супернатанте биоптатов тканей человека и животного, после введения в эти ткани генотерапевтических ДНК-векторов;
D) Иммуноферментный анализ - изменение количественного уровня целевых белков в супернатанте биоптатов тканей человека, после введения в эти ткани аутологичных клеток этого человека, трансфицированных генотерапевтическими ДНК-векторами.
Для подтверждения реализуемости способа применения созданного генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, несущего целевой ген, а именно, ген DDC, генотерапевтического ДНК-вектора GDTT1.8NAS12-IL10, несущего целевой ген, а именно, ген IL10, генотерапевтического ДНК-вектора GDTT1.8NAS12-IL13, несущего целевой ген, а именно, ген IL13, генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, несущего целевой ген, а именно, ген IFNB1, генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFRSF4, несущего целевой ген, а именно, ген TNFRSF4, генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFSF10, несущего целевой ген, а именно, ген TNFSF10, генотерапевтического ДНК-вектора GDTT1.8NAS12-BCL2, несущего целевой ген, а именно, ген BCL2, генотерапевтического ДНК-вектора GDTT1.8NAS12-HGF, несущего целевой ген, а именно, ген HGF, генотерапевтического ДНК-вектора GDTT1.8NAS12-IL2, несущего целевой ген, а именно, ген IL2 выполняли:
А) трансфекцию генотерапевтическими ДНК-векторами различных клеточных линий человека и животного;
B) введение генотерапевтических ДНК-векторов в различные ткани человека и животного;
С) введение в ткани животного смеси генотерапевтических ДНК-векторов;
D) введение в ткани человека аутологичных клеток, трансфицированных генотерапевтическими ДНК-векторами.
Указанные способы применения характеризуются отсутствием потенциальных рисков для генетической терапии человека и животных за счет отсутствия в составе генотерапевтического ДНК-вектора регуляторных элементов, представляющих собой нуклеотидные последовательности вирусных геномов, и за счет отсутствия в составе генотерапевтического ДНК-вектора генов устойчивости к антибиотикам, что подтверждается отсутствием участков, гомологичных вирусным геномам и генам антибиотикорезистентности в нуклеотидных последовательностях генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, или генотерапевтического ДНК-вектора GDTT1.8NAS12-IL10, или генотерапевтического ДНК-вектора GDTT1.8NAS12-IL13, или генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, или генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFRSF4, или генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFSF10, или генотерапевтического ДНК-вектора GDTT1.8NAS12-BCL2, или генотерапевтического ДНК-вектора GDTT1.8NAS12-HGF, или генотерапевтического ДНК-вектора GDTT1.8NAS12-IL2 (SEQ ID №1 или SEQ ID №2 или SEQ ID №3 или SEQ ID №4 или SEQ ID №5 или SEQ ID №6 или SEQ ID №7 или SEQ ID №8 или SEQ ID №9 соответственно).
Как известно специалистам в данной области техники, гены антибиотикорезистентности в составе генотерапевтических ДНК-векторов используются с целью получения этих векторов в препаративных количествах путем наращивания бактериальной биомассы в питательной среде, содержащей селективный антибиотик. В рамках настоящего изобретения в целях возможности безопасного применения генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген DDC, или IL10, или IL13, или IFNB1, или TNFRSF4, или TNFSF10, или BCL2, или HGF, или IL2 использование селективных питательных сред, содержащих антибиотик, не представляется возможным. В качестве технологического решения для получения генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген, выбранный из группы генов: DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2 для возможности масштабирования до промышленных масштабов получения генотерапевтических векторов предлагается способ получения штаммов для наработки указанных генотерапевтических векторов на основе бактерии Escherichia coli JM110-NAS. Способ получения штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1 или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2 заключается в получении компетентных клеток штамма Escherichia coli JM110-NAS с введением в эти клетки генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, или ДНК-вектора GDTT1.8NAS12-IL10, или ДНК-вектора GDTT1.8NAS12-IL13, или ДНК-вектора GDTT1.8NAS12-IFNB1, или ДНК-вектора GDTT1.8NAS12-TNFRSF4, или ДНК-вектора GDTT1.8NAS12-TNFSF10, или ДНК-вектора GDTT1.8NAS12-BCL2, или ДНК-вектора GDTT1.8NAS12-HGF, или ДНК-вектора GDTT1.8NAS12-IL2 соответственно с помощью методов трансформации (электропорации), общеизвестных специалистам в данной области техники. Полученный штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2 используется для наработки генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, или GDTT1.8NAS12-IL10, или GDTT1.8NAS12-IL13, или GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-TNFRSF4, или GDTT1.8NAS12-TNFSF10, или GDTT1.8NAS12-BCL2, или GDTT1.8NAS12-HGF, или GDTT1.8NAS12-IL2 соответственно с возможностью использования сред без содержания антибиотиков.
Для подтверждения получения штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2 проводили трансформацию, селекцию и последующее наращивание с выделением плазмидной ДНК.
Для подтверждения технологичности получения и возможности масштабирования до промышленного производства генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, несущего целевой ген, а именно, ген DDC, генотерапевтического ДНК-вектора GDTT1.8NAS12-IL10, несущего целевой ген, а именно, ген IL10, генотерапевтического ДНК-вектора GDTT1.8NAS12-IL13, несущего целевой ген, а именно, ген IL13, генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, несущего целевой ген, а именно, ген IFNB1, генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFRSF4, несущего целевой ген, а именно, ген TNFRSF4, генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFSF10, несущего целевой ген, а именно, ген TNFSF10, генотерапевтического ДНК-вектора GDTT1.8NAS12-BCL2, несущего целевой ген, а именно, ген BCL2, генотерапевтического ДНК-вектора GDTT1.8NAS12-HGF, несущего целевой ген, а именно, ген HGF, генотерапевтического ДНК-вектора GDTT1.8NAS12-IL2, несущего целевой ген, а именно, ген IL2, выполняли ферментацию в промышленном масштабе штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2, каждый из которых содержит генотерапевтический ДНК-вектор GDTT1.8NAS12, несущий целевой ген, а именно DDC, или IL10, или IL13, или IFNB1, или TNFRSF4, или TNFSF10, или BCL2, или HGF, или IL2.
Способ масштабирования получения бактериальной массы до промышленных масштабов для выделения генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген, выбранный из группы генов DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2 заключается в том, что затравочную культуру штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2 инкубируют в объеме питательной среды без содержания антибиотика обеспечивающим подходящую динамику накопления биомассы, по достижению достаточного количества биомассы в логарифмической фазе роста, бактериальную культуру переносят в промышленный ферментер, после чего растят до достижения стационарной фазы роста, затем выделяют фракцию, содержащую целевой ДНК-продукт - генотерапевтический ДНК-вектор GDTT1.8NAS12-DDC, или генотерапевтический ДНК-вектор GDTT1.8NAS12-IL10, или генотерапевтический ДНК-вектор GDTT1.8NAS12-IL13, или генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1, или генотерапевтический ДНК-вектор GDTT1.8NAS12-TNFRSF4, или GDTT1.8NAS12-TNFSF10, или GDTT1.8NAS12-BCL2, или GDTT1.8NAS12-HGF, или GDTT1.8NAS12-IL2 многостадийно фильтруют и очищают хроматографическими методами. При этом специалистам в данной области техники понятно, что условия культивирования штаммов, состав питательных сред (за исключением содержания антибиотиков), используемое оборудование, методы очистки ДНК могут варьировать в рамках стандартных операционных процедур в зависимости от отдельно взятой производственной линии, но известные подходы к масштабированию, промышленному получению и очистке ДНК-векторов с использованием штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2 подпадают под объем настоящего изобретения.
Описанное раскрытие изобретения подтверждается примерами реализации настоящего изобретения.
Изобретение поясняется следующими примерами.
Пример 1.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, несущего целевой ген, а именно, ген DDC.
Генотерапевтический ДНК-вектор GDTT1.8NAS12-DDC конструировали клонированием кодирующей части гена DDC размером 1447 п.н. в ДНК-вектор GDTT1.8NAS12 размером 2591 п.н. по сайтам рестрикции SalI-HindIII. Кодирующую часть гена DDC размером 1447 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов:
DDC-F ATCGTCGACCACCATGAACGCAAGTGAGTTCCGA,
DDC-R ACCAAGCTTCTACTCCCTCTCTG
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США).
Генотерапевтический ДНК-вектор GDTT1.8NAS12 конструировали объединением шести фрагментов ДНК, полученных из разных источников:
(а) ориджин репликации получали путем ПЦР-амплификации участка коммерческой плазмиды pUC19 с использованием олигонуклеотидов UCori-Bam, UCori-Nco (перечень последовательностей олигонуклеотидов, (1) -(2));
(б) терминатор транскрипции hGH-TA получали путем ПЦР-амплификации участка геномной ДНК человека с использованием олигонуклеотидов hGH-F и hGH-R (перечень последовательностей олигонуклеотидов, (3) и (4));
(в) регуляторный участок транспозона Tn10 РНК-out получали путем синтеза из олигонуклеотидов RO-F, RO-R, RO-1, RO-2, RO-3 (перечень последовательностей олигонуклеотидов, (5) -(9));
(г) ген устойчивости к канамицину получали путем ПЦР-амплификации участка коммерческой плазмиды pET-28 человека с использованием олигонуклеотидов Kan-F и Kan-R (перечень последовательностей олигонуклеотидов, (10) и (11));
(д) полилинкер получали кинированием и отжигом четырех синтетических олигонуклеотидов MCS1, MCS2, MCS3 и MCS4 (перечень последовательностей олигонуклеотидов, (12) - (15));
(е) промоторно-регуляторный участок человеческого гена фактора элонгации EF1а с собственным энхансером получали путем ПЦР-амплификации участка геномной ДНК человека с использованием олигонуклеотидов EF1-Xho и EF1-R (перечень последовательностей олигонуклеотидов, (16) - (17)).
ПЦР-амплификацию проводили с использованием коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs) в соответствии с инструкцией производителя. Фрагменты (б), (в) и (г) имели перекрывающиеся области для возможности их объединения с последующей ПЦР-амплификацией. Объединяли фрагменты (б), (в) и (г) с использованием олигонуклеотидов hGH-F и Kan-R (перечень последовательностей олигонуклеотидов, (3) и (11)). Далее, полученные фрагменты ДНК объединялись путем рестрикции с последующим лигированием по сайтам BamHI и NcoI. В результате получали вектор, пока еще не содержащий полилинкер. Для его введения проводили расщепление плазмиды эндонуклеазами рестрикции по сайтам BamHI и EcoRI с последующим лигированием с фрагментом (д). В результате получали промежуточный вектор размером 2408 п.н, несущий ген устойчивости к канамицину, пока еще не содержащий промоторно-регуляторный участок гена фактора элонгации EF1а с собственным энхансером. Полученный вектор расщепляли эндонуклеазами рестрикции по сайтам SalI и BamHI с последующим лигированием с фрагментом (е). В результате получали вектор размером 3608 п.н., несущий ген устойчивости к канамицину и промоторно-регуляторный участок гена фактора элонгации EF1а с собственным энхансером. Далее ген устойчивости к канамицину выщепляли по сайтам рестрикции SpeI, после чего оставшийся фрагмент лигировали сам на себя. Таким образом получали генотерапевтический ДНК-вектор GDTT1.8NAS12 размером 2591 п.н., который является рекомбинантным, с возможностью селекции без антибиотиков и возможностью экспрессии клонированных в него целевых генов в большинстве типов тканей человека и животных.
Расщепление продукта амплификации кодирующей части гена DDC и ДНК-вектора GDTT1.8NAS12 проводили эндонуклеазами рестрикции SalI и HindIII.
В результате получали ДНК-вектор GDTT1.8NAS12-DDC размером 4006 п.н. с нуклеотидной последовательностью SEQ ID №1 и общей структурой изображенной на фиг.1A.
Пример 2.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS12-IL10, несущего целевой ген, а именно, ген IL10.
Генотерапевтический ДНК-вектор GDTT1.8NAS12-IL10 конструировали клонированием кодирующей части гена IL10 размером 540 п.н. в ДНК-вектор GDTT1.8NAS12 размером 2591 п.н. по сайтам рестрикции BamHI и SalI. Кодирующую часть гена IL10 размером 540 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов:
IL10-F AGGATCCACCATGCACAGCTCAGCACTGC,
IL10-R CTTGTCGACTCAGTTTCGTATCTTCATTGTC
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США), расщепление продукта амплификации и ДНК-вектора GDTT1.8NAS12 проводили эндонуклеазами рестрикции BamHI и SalI.
В результате получали ДНК-вектор GDTT1.8NAS12-IL10 размером 3093 п.н. с нуклеотидной последовательностью SEQ ID №2 и общей структурой изображенной на фиг.1B.
При этом генотерапевтический ДНК-вектор GDTT1.8NAS12 конструировали по Примеру 1.
Пример 3
Получение ДНК-вектора GDTT1.8NAS12-IL13, несущего целевой ген, а именно, ген IL13 человека.
Генотерапевтический ДНК-вектор GDTT1.8NAS12-IL13 конструировали клонированием кодирующей части гена IL13 размером 444 п.н. в ДНК-вектор GDTT1.8NAS12 размером 2591 п.н. по сайтам рестрикции BamHI и SalI. Кодирующую часть гена IL13 размером 444 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов:
IL13-F AGGATCCACCATGCATCCGCTCCTCAATC,
IL13-R CTTGTCGACTCAGTTGAACTGTCCCTCG
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США), расщепление продукта амплификации и ДНК-вектора GDTT1.8NAS12 проводили эндонуклеазами рестрикции BamHI и SalI.
В результате получали ДНК-вектор GDTT1.8NAS12-IL13 размером 2997 п.н. с нуклеотидной последовательностью SEQ ID №3 и общей структурой изображенной на фиг.1C.
При этом генотерапевтический ДНК-вектор GDTT1.8NAS12 конструировали по Примеру 1.
Пример 4.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, несущего целевой ген, а именно, ген IFNB1.
Генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1 конструировали клонированием кодирующей части гена IFNB1 размером 567 п.н. в ДНК-вектор GDTT1.8NAS12 размером 2591 п.н. по сайтам рестрикции BamHI и SalI. Кодирующую часть гена IFNB1 размером 567 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген) и ПЦР-амплификации с использованием олигонуклеотидов:
IFNB1-F AAAGGATCCACCATGACCAACAAGTGTCTCCTCCAAA,
IFNB1-R TTTGTCGACTCAGTTTCGGAGGTAACCTGTAAGTCTG
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США), расщепление продукта амплификации и ДНК-вектора GDTT1.8NAS12 проводили эндонуклеазами рестрикции BamHI и SalI.
В результате получали ДНК-вектор GDTT1.8NAS12-IFNB1 размером 3120 п.н. с нуклеотидной последовательностью SEQ ID №4 и общей структурой изображенной на фиг.1D.
При этом генотерапевтический ДНК-вектор GDTT1.8NAS12 конструировали по Примеру 1.
Пример 5.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFRSF4, несущего целевой ген, а именно, ген TNFRSF4.
Генотерапевтический ДНК-вектор GDTT1.8NAS12-TNFRSF4 конструировали клонированием кодирующей части гена TNFRSF4 размером 837 п.н. в ДНК-вектор GDTT1.8NAS12 размером 2591 п.н. по сайтам рестрикции BamHI и SalI. Кодирующую часть гена TNFRSF4 размером 837 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген) и ПЦР-амплификации с использованием олигонуклеотидов:
TNFRSF4-F AGGATCCACCATGTGCGTGGGGGCTCGG,
TNFRSF4-R CTTGTCGACTCAGATCTTGGCCAGGGTG
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США), расщепление продукта амплификации и ДНК-вектора GDTT1.8NAS12 проводили эндонуклеазами рестрикции BamHI и SalI.
В результате получали ДНК-вектор GDTT1.8NAS12-TNFRSF4 размером 3390 п.н. с нуклеотидной последовательностью SEQ ID №5 и общей структурой изображенной на фиг.1E.
При этом генотерапевтический ДНК-вектор GDTT1.8NAS12 конструировали по Примеру 1.
Пример 6.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS12- TNFSF10, несущего целевой ген, а именно, ген TNFSF10.
Генотерапевтический ДНК-вектор GDTT1.8NAS12-TNFSF10 конструировали клонированием кодирующей части гена TNFSF10 размером 849 п.н. в ДНК-вектор GDTT1.8NAS12 размером 2591 п.н. по сайтам рестрикции BamHI, HindIII. Кодирующую часть гена TNFSF10 размером 849 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген) и ПЦР-амплификации с использованием олигонуклеотидов:
TNFSF10-F AGGATCCACCATGGCTATGATGGAGGTCCA,
TNFSF10-R TGTCGACTCAGCCAACTAAAAAGGCCCCGA
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США), расщепление продукта амплификации и ДНК-вектора GDTT1.8NAS12 проводили эндонуклеазами рестрикции BamHI и SalI.
В результате получали ДНК-вектор GDTT1.8NAS12-TNFSF10 размером 3402 п.н. с нуклеотидной последовательностью SEQ ID №6 и общей структурой изображенной на фиг.1F.
При этом генотерапевтический ДНК-вектор GDTT1.8NAS12 конструировали по Примеру 1.
Пример 7.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS12- BCL2, несущего целевой ген, а именно, ген BCL2.
Генотерапевтический ДНК-вектор GDTT1.8NAS12-BCL2 конструировали клонированием кодирующей части гена BCL2 размером 724 п.н. в ДНК-вектор GDTT1.8NAS12 размером 2591 п.н. по сайтам рестрикции SalI и KpnI. Кодирующую часть гена BCL2 размером 724 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген) и ПЦР-амплификации с использованием олигонуклеотидов:
BCL-F ATCGTCGACCACCATGGCGCACGCTGGGAGA,
BCL-R TTCGGTACCTCACTTGTGGCCCA
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США), расщепление продукта амплификации и ДНК-вектора GDTT1.8NAS12 проводили эндонуклеазами рестрикции SalI и KpnI.
В результате получали ДНК-вектор GDTT1.8NAS12-BCL2 размером 3277 п.н. с нуклеотидной последовательностью SEQ ID №7 и общей структурой изображенной на фиг.1G.
При этом генотерапевтический ДНК-вектор GDTT1.8NAS12 конструировали по Примеру 1.
Пример 8.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS12- HGF, несущего целевой ген, а именно, ген HGF.
Генотерапевтический ДНК-вектор GDTT1.8NAS12-HGF конструировали клонированием кодирующей части гена HGF размером 2190 п.н. в ДНК-вектор GDTT1.8NAS12 размером 2591 п.н. по сайтам рестрикции BamHI и SalI. Кодирующую часть гена HGF размером 2190 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген) и ПЦР-амплификации с использованием олигонуклеотидов:
HGF_F TTTGGATCCACCATGTGGGTGACCAAACTCCTGCCA,
HGF_R AATGTCGACCTATGACTGTGGTACCTTATATGTTAAAAT
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США), расщепление продукта амплификации и ДНК-вектора GDTT1.8NAS12 проводили эндонуклеазами рестрикции BamHI и SalI.
В результате получали ДНК-вектор GDTT1.8NAS12-HGF размером 4775 п.н. с нуклеотидной последовательностью SEQ ID №8 и общей структурой изображенной на фиг.1H.
При этом генотерапевтический ДНК-вектор GDTT1.8NAS12 конструировали по Примеру 1.
Пример 9.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS12- IL2, несущего целевой ген, а именно, ген IL2.
Генотерапевтический ДНК-вектор GDTT1.8NAS12- IL2 конструировали клонированием кодирующей части гена IL2 размером 465 п.н. в ДНК-вектор GDTT1.8NAS12 размером 2591 п.н. по сайтам рестрикции BamHI и SalI. Кодирующую часть гена IL2 размером 465 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген) и ПЦР-амплификации с использованием олигонуклеотидов:
IL2-F AGGATCCACCATGTACAGGATGCAACTCCT,
IL2-R TGTCGACTCAAGTCAGTGTTGAGATGATG
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США), расщепление продукта амплификации и ДНК-вектора GDTT1.8NAS12 проводили эндонуклеазами рестрикции BamHI и SalI.
В результате получали ДНК-вектор GDTT1.8NAS12-IL2 размером 3018 п.н. с нуклеотидной последовательностью SEQ ID №9 и общей структурой изображенной на фиг.1I.
При этом генотерапевтический ДНК-вектор GDTT1.8NAS12 конструировали по Примеру 1.
Пример 10.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, несущего целевой ген, а именно, ген DDC, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена DDC, в культуре клеток кортикальных нейронов человека HCN-2 (ATCC CRL-10742) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12-DDC, несущим ген DDC человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Клеточную культуру HCN-2 выращивали в стандартных условиях (37 ˚С, 5% СО2) в культуральной среде DMEM с добавлением 10% сыворотки эмбрионов коров (Gibco). В процессе культивирования каждые 48 ч происходила смена ростовой среды.
Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS12-DDC, экспрессирующим ген DDC человека, проводили с использованием Lipofectamine 3000 (ThermoFisher Scientific, США) согласно рекомендациям производителя. В пробирке №1 к 25 мкл среды Opti-MEM (Gibco, США) добавляли 1 мкл раствора ДНК-вектора GDTT1.8NAS12-DDC (концентрация 500 нг/мкл) и 1 мкл реагента Р3000. Аккуратно перемешивали легким встряхиванием. В пробирке №2 к 25 мкл среды Opti-MEM (Gibco, США) добавляли 1 мкл раствора Lipofectamin 3000. Аккуратно перемешивали легким встряхиванием. Добавляли содержимое пробирки №1 к содержимому пробирки №2, инкубировали 5 мин при комнатной температуре. Полученный раствор по каплям добавляли к клеткам в объеме 40 мкл.
В качестве контроля использовали клетки HCN-2, трансфицированные генотерапевтическим ДНК-вектором GDTT1.8NAS12, не содержащим вставку целевого гена (кДНК гена DDC до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12, не содержащим вставку целевого гена на фигурах не показано). Подготовку контрольного вектора GDTT1.8NAS12 для трансфекции проводили, как описано выше.
Суммарную РНК из клеток HCN-2 выделяли с использованием Trizol Reagent (Invitrogen, США) согласно рекомендациям производителя. В лунку с клетками добавляли 1 мл Trizol Reagent и гомогенизировали с последующим прогреванием в течение 5 мин при 65 0С. Далее образец центрифугировали при 14 000g в течение 10 мин и снова прогревали в течение 10 мин при 65 0С. Далее добавляли 200 мкл хлороформа, плавно перемешивали и центрифугировали при 14 000g в течение 10 мин. Затем отбирали водную фазу, добавляли к ней 1/10 объема 3М ацетата натрия, рН 5.2 и равный объем изопропилового спирта. Инкубировали образец при -20 0С в течение 10 мин с последующим центрифугированием при 14 000g в течение 10 мин. Осадок промывали 1 мл 70% этилового спирта, высушивали на воздухе и растворяли в 10 мкл воды, свободной от РНКаз. Определение уровня экспрессии мРНК гена DDC после трансфекции проводили путем оценки динамики накопления ампликонов кДНК методом ПЦР в режиме реального времени. Для получения и амплификации кДНК, специфичной для гена DDC человека, использовали олигонуклеотиды DDC_SF и DDC_SR:
DDC_SF GGAGAAGGGGGAGGAGTGAT,
DDC_SR CCCAGCTCTTTCCACTGAGG.
Длина продукта амплификации - 186 п.н.
Реакцию обратной транскрипции и ПЦР-амплификацию проводили с помощью набора реагентов SYBR GreenQuantitect RT–PCR Kit (Qiagen, США) для ПЦР в режиме реального времени. Реакцию проводили в объеме 20 мкл, содержащих: 25 мкл QuantiTect SYBR Green RT-PCR MasterMix, 2,5 мM хлорида магния, по 0,5 мкМ каждого праймера, 5 мкл РНК. Реакцию осуществляли на амплификаторе CFX96 (Bio-Rad, США) при следующих условиях: 1 цикл обратной транскрипции при 42оС - 30 минут, денатурация 98оС - 15 мин, затем 40 циклов, включающих денатурацию 94оС - 15 сек, отжиг праймеров 60оC - 30 сек и элонгацию 72оС - 30 сек. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов DDC и B2M. В качестве отрицательного контроля использовали деионизированную воду. Количество динамику накопления ампликонов кДНК генов DDC и B2M оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа, представлены на фиг. 2.
Из фигуры 2 следует, что в результате трансфекции культуры клеток кортикальных нейронов человека HCN-2 генотерапевтическим ДНК-вектором GDTT1.8NAS12-DDC, уровень специфической мРНК гена DDC человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген DDC на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC для повышения уровня экспрессии гена DDC в эукариотических клетках.
Пример 11.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS12-IL10, несущего целевой ген, а именно, ген IL10, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена IL10, в культуре клеток нейробластомы человека IMR-32 (ATCC CCL-127) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL10, несущим ген IL10 человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Клеточную культуру IMR-32 выращивали в стандартных условиях (37 ˚С, 5% СО2) в культуральной среде EMEM с добавлением 10% сыворотки эмбрионов коров (Gibco). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL10, экспрессирующим ген IL10 человека, проводили, как описано в примере 10. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве контроля использовали культуру клеток нейробластомы человека IMR-32, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, не несущим целевой ген (кДНК гена IL10 до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12, не содержащим вставку целевого гена на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили, как описано в примере 10, за исключением олигонуклеотидов с отличающимися от примера 10 последовательностями. Для амплификации кДНК, специфичной для гена IL10 человека, использовали олигонуклеотиды IL10_SF и IL10_SR:
IL10_SF AAGACCCAGACATCAAGGCG,
IL10_SR AGGCATTCTTCACCTGCTCC.
Длина продукта амплификации – 135 п.н.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов IL10 и B2M. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов – кДНК генов IL10 и B2M, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа, представлены на фиг. 3.
Из фигуры 3 следует, что в результате трансфекции культуры клеток нейробластомы человека IMR-32 генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL10, уровень специфической мРНК гена IL10 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IL10 на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IL10 для повышения уровня экспрессии гена IL10 в эукариотических клетках.
Пример 12.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS12-IL13, несущего целевой ген, а именно, ген IL13, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена IL13 в культуре клеток глиобластомы человека U-118 MG (ATCC HTB-15) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL13, несущим ген IL13 человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Клеточную культуру U-118 MG выращивали в стандартных условиях (37 ˚С, 5% СО2) в культуральной среде MEM с добавлением 10% сыворотки эмбрионов коров (Gibco). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL13, экспрессирующим ген IL13 человека, проводили, как описано в примере 10. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве контроля использовали культуру клеток глиобластомы человека U-118 MG, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, не несущим целевой ген (кДНК гена IL13 до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12, не содержащим вставку целевого гена на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили, как описано в примере 10, за исключением олигонуклеотидов с отличающимися от примера 10 последовательностями. Для амплификации кДНК, специфичной для гена IL13 человека, использовали олигонуклеотиды IL13_SF и IL13_SR:
IL13_SF CATGGCGCTTTTGTTGACCA,
IL13_SR AGCTGTCAGGTTGATGCTCC.
Длина продукта амплификации – 181 п.н.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов IL13 и B2M. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов – кДНК генов IL13 и B2M, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа, представлены на фиг. 4.
Из фигуры 4 следует, что в результате трансфекции культуры клеток глиобластомы человека U-118 MG генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL13, уровень специфической мРНК гена IL13 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IL13 на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IL13 для повышения уровня экспрессии гена IL13 в эукариотических клетках.
Пример 13.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, несущего целевой ген, а именно, ген IFNB1, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена IFNB1, в культуре клеток нейробластомы человека SH-SY5Y (ATCC CRL-2266) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNB1, несущим ген IFNB1 человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Клеточную культуру SH-SY5Y выращивали в стандартных условиях (37 ˚С, 5% СО2) в культуральной среде DMEM:F12 (Gibco) 1:1 с добавлением 10% сыворотки эмбрионов коров (Gibco). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNB1, экспрессирующим ген IFNB1 человека, проводили, как описано в примере 10. В качестве контроля использовали культуру клеток нейробластомы человека SH-SY5Y, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, не несущим целевой ген (кДНК гена IFNB1 до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12, не содержащим вставку целевого гена на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили, как описано в примере 10, за исключением олигонуклеотидов с отличающимися от примера 10 последовательностями. Для амплификации кДНК, специфичной для гена IFNB1 человека, использовали олигонуклеотиды IFNB1_SF и IFNB1_SR:
IFNB1_SF GTGGCAATTGAATGGGAGGC,
IFNB1_SR ATAGATGGTCAATGCGGCGT.
Длина продукта амплификации – 118 п.н.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов IFNB1 и B2M. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов – кДНК генов IFNB1 и B2M, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа, представлены на фиг. 5.
Из фигуры 5 следует, что в результате трансфекции культуры клеток нейробластомы человека SH-SY5Y генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNB1, уровень специфической мРНК гена IFNB1 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IFNB1 на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNB1 для повышения уровня экспрессии гена IFNB1 в эукариотических клетках.
Пример 14.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFRSF4, несущего целевой ген, а именно, ген TNFRSF4, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена TNFRSF4, в культуре клеток астроглии человека SVG p12 (ATCC CRL-8621) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12-TNFRSF4, несущим ген TNFRSF4 человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Клеточную культуру SVG p12 выращивали в стандартных условиях (37 ˚С, 5% СО2) в культуральной среде MEM с добавлением 10% сыворотки эмбрионов коров (Gibco). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS12-TNFRSF4, экспрессирующим ген TNFRSF4 человека, проводили, как описано в примере 10. В качестве контроля использовали культуру клеток астроглии человека SVG p12, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, не несущим целевой ген (кДНК гена TNFRSF4 до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12, не содержащим вставку целевого гена на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили, как описано в примере 10, за исключением олигонуклеотидов с отличающимися от примера 10 последовательностями. Для амплификации кДНК, специфичной для гена TNFRSF4 человека, использовали олигонуклеотиды TNFRSF4_SF и TNFRSF4_SR:
TNFRSF4_SF CTGGACAGCTACAAGCCTGG,
TNFRSF4_SR CCTGTCCTCACAGATTGCGT.
Длина продукта амплификации – 162 п.н.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов TNFRSF4 и B2M. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов – кДНК генов TNFRSF4 и B2M, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа, представлены на фиг. 6.
Из фигуры 6 следует, что в результате трансфекции культуры клеток астроглии человека SVG p12 генотерапевтическим ДНК-вектором GDTT1.8NAS12-TNFRSF4, уровень специфической мРНК гена TNFRSF4 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген TNFRSF4 на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтическим ДНК-вектором GDTT1.8NAS12-TNFRSF4 для повышения уровня экспрессии гена TNFRSF4 в эукариотических клетках.
Пример 15.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFSF10, несущего целевой ген, а именно, ген TNFSF10, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена TNFSF10, в культуре нейроэктодермальных клеток опухоли головного мозга человека PFSK-1 (ATCC number CRL-2060) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12-TNFSF10, несущим ген TNFSF10 человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Клеточную культуру PFSK-1 выращивали в стандартных условиях (37 ˚С, 5% СО2) в культуральной среде RPMI-1640 с добавлением 10% сыворотки эмбрионов коров (Gibco). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS12-TNFSF10, экспрессирующим ген TNFSF10 человека, проводили, как описано в примере 10. В качестве контроля использовали культуру нейроэктодермальных клеток опухоли головного мозга человека PFSK-1, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, не несущим целевой ген (кДНК гена TNFSF10 до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12, не содержащим вставку целевого гена на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили, как описано в примере 10, за исключением олигонуклеотидов с отличающимися от примера 10 последовательностями. Для амплификации кДНК, специфичной для гена TNFSF10 человека, использовали олигонуклеотиды TNFSF10_SF и TNFSF10_SR:
TNFSF10_SF CCTGCAGTCTCTCTGTGTGG,
TNFSF10_SR ACGGAGTTGCCACTTGACTT.
Длина продукта амплификации – 181 п.н.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов TNFSF10 и B2M. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов – кДНК генов TNFSF10 и B2M, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа, представлены на фиг. 7.
Из фигуры 7 следует, что в результате трансфекции культуры нейроэктодермальных клеток опухоли головного мозга человека PFSK-1 генотерапевтическим ДНК-вектором GDTT1.8NAS12-TNFSF10, уровень специфической мРНК гена TNFSF10 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген TNFSF10 на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтическим ДНК-вектором GDTT1.8NAS12-TNFSF10 для повышения уровня экспрессии гена TNFSF10 в эукариотических клетках.
Пример 16.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS12-BCL2, несущего целевой ген, а именно, ген BCL2, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена BCL2, в культуре клеток медуллобластомы человека D341 Med (ATCC HTB-187) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12-BCL2, несущим ген BCL2 человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Клеточную культуру D341 Med выращивали в стандартных условиях (37 ˚С, 5% СО2) в культуральной среде MEM с добавлением 10% сыворотки эмбрионов коров (Gibco). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS12-BCL2, экспрессирующим ген BCL2 человека, проводили, как описано в примере 10. В качестве контроля использовали культуру клеток медуллобластомы человека D341 Med, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, не несущим целевой ген (кДНК гена BCL2 до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12, не содержащим вставку целевого гена на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили, как описано в примере 10, за исключением олигонуклеотидов с отличающимися от примера 10 последовательностями. Для амплификации кДНК, специфичной для гена BCL2 человека, использовали олигонуклеотиды BCL2_SF и BCL2_SR:
BCL_SF GAACTGGGGGAGGATTGTGG,
BCL_SR CATCCCAGCCTCCGTTATCC.
Длина продукта амплификации – 164 п.н.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов BCL2 и B2M. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов – кДНК генов BCL2 и B2M, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа, представлены на фиг. 8.
Из фигуры 8 следует, что в результате трансфекции культуры клеток медуллобластомы человека D341 Med генотерапевтическим ДНК-вектором GDTT1.8NAS12-BCL2, уровень специфической мРНК гена BCL2 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген BCL2 на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтическим ДНК-вектором GDTT1.8NAS12-BCL2 для повышения уровня экспрессии гена BCL2 в эукариотических клетках.
Пример 17.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS12-HGF, несущего целевой ген, а именно, ген HGF, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена HGF, в культуре клеток астроцитомы головного мозга человека A-172 (ATCC CRL-1620) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12-HGF, несущим ген HGF человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Клеточную культуру A-172 выращивали в стандартных условиях (37 ˚С, 5% СО2 в культуральной среде DMEM с добавлением 10% сыворотки эмбрионов коров (Gibco). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS12-HGF, экспрессирующим ген HGF человека, проводили, как описано в примере 10. В качестве контроля использовали культуру клеток астроцитомы головного мозга человека A-172, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, не несущим целевой ген (кДНК гена HGF до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12, не содержащим вставку целевого гена на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили, как описано в примере 10, за исключением олигонуклеотидов с отличающимися от примера 10 последовательностями. Для амплификации кДНК, специфичной для гена HGF человека, использовали олигонуклеотиды HGF_SF и HGF_SR:
HGF_SF ACCCTGGTGTTTCACAAGCA,
HGF_FR GCAAGAATTTGTGCCGGTGT.
Длина продукта амплификации – 182 п.н.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов HGF и B2M. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов – кДНК генов HGF и B2M, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа, представлены на фиг. 9.
Из фигуры 9 следует, что в результате трансфекции культуры клеток астроцитомы головного мозга человека A-172 генотерапевтическим ДНК-вектором GDTT1.8NAS12-HGF, уровень специфической мРНК гена HGF человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген HGF на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтическим ДНК-вектором GDTT1.8NAS12-HGF для повышения уровня экспрессии гена HGF в эукариотических клетках.
Пример 18.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS12-IL2, несущего целевой ген, а именно, ген IL2, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена IL2, в культуре нейрональных прогениторных клеток человека ATCC-BXS0117 (ATCC ACS-5003) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL2, несущим ген IL2 человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Клеточную культуру ATCC-BXS0117 выращивали в стандартных условиях (37 ˚С, 5% СО2) в культуральной среде DMEM:F12 (Gibco) 1:1 с добавлением 10% сыворотки эмбрионов коров (Gibco) и с использованием набора реагентов Growth Kit for Neural Progenitor Cell Expansion (ATCC ACS-3003). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL2, экспрессирующим ген IL2 человека, проводили, как описано в примере 10. В качестве контроля использовали культуру нейрональных прогениторных клеток человека ATCC-BXS0117, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, не несущим целевой ген (кДНК гена IL2 до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12, не содержащим вставку целевого гена на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили, как описано в примере 10, за исключением олигонуклеотидов с отличающимися от примера 10 последовательностями. Для амплификации кДНК, специфичной для гена IL2 человека, использовали олигонуклеотиды IL2_SF и IL2_SR:
IL2_SF ACAGGATGCAACTCCTGTCT,
IL2_SR GCATCCTGGTGAGTTTGGGA.
Длина продукта амплификации – 192 п.н.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов IL2 и B2M. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов – кДНК генов IL2 и B2M, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа, представлены на фиг. 10.
Из фигуры 10 следует, что в результате трансфекции культуры нейрональных прогениторных клеток человека ATCC-BXS0117 генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL2, уровень специфической мРНК гена IL2 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IL2 на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL2 для повышения уровня экспрессии гена IL2 в эукариотических клетках.
Пример 19.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, несущего ген DDC, для повышения экспрессии белка DDC в клетках млекопитающих.
Оценивали изменение количества белка DDC в лизате культуры клеток кортикальных нейронов человека HCN-2 (ATCC CRL-10742) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-DDC, несущим ген DDC человека.
Клеточную культуру кортикальных нейронов человека HCN-2 выращивали, как это описано в примере 10.
Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена DDC (В), в качестве трансфицируемых агентов - ДНК-вектор GDTT1.8NAS12-DDC, несущий ген DDC человека (С). Приготовление комплекса ДНК/дендример проводили по методике производителя (QIAGEN, SuperFect Transfection Reagent Handbook, 2002) с некоторыми изменениями. Для трансфекции клеток в одной лунке 24-луночного планшета к 1 мкг ДНК-вектора, растворенного в буфере ТЕ, добавляли культуральную среду до конечного объема 60 мкл, затем добавляли 5 мкл SuperFect Transfection Reagent и осторожно перемешивали пятикратным пипетированием. Комплекс инкубировали при комнатной температуре в течение 10-15 мин. Далее отбирали из лунок культуральную среду, лунки промывали 1 мл буфера PBS. К образовавшемуся комплексу добавляли 350 мкл среды, содержащей 10 мкг/мл гентамицина, осторожно перемешивали и добавляли к клеткам. Клетки инкубировали с комплексом 2-3 часа при 37°С в атмосфере, содержащей 5% СО2.
Затем осторожно удаляли среду, промывали клеточный монослой 1 мл буфера PBS. Затем добавляли среду, содержащую 10 мкг/мл гентамицина, и инкубировали 24-48 часов при 37°С в атмосфере 5% СО2.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка. Количественное определение белка DDC проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор ELISA Kit for Dopa Decarboxylase (DDC) (SEG474Hu Cloud-Clone Corp., США) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка DDC, входящим в состав набора. Чувствительность метода составляла не менее 0,61нг/мл, диапазон измерения - от 1,56 нг/мл до 100 нг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа, представлены на фиг.11.
Из фигуры 11 следует, что трансфекция кортикальных нейронов человека HCN-2 генотерапевтическим ДНК-вектором GDTT1.8NAS12-DDC приводит к увеличению количества белка DDC по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген DDC на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC для повышения уровня экспрессии DDC в эукариотических клетках.
Пример 20.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IL10, несущего ген IL10, для повышения экспрессии белка IL10 в клетках млекопитающих.
Оценивали изменение количества белка в культуре клеток нейробластомы человека IMR-32 (ATCC CCL-127) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IL10, несущим ген IL10 человека. Клетки выделяли и выращивали, как описано в примере 11.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена IL10 (В), в качестве трансфицируемых агентов - ДНК-вектор GDTT1.8NAS12-IL10, несущий ген IL10 человека (С). Приготовление комплекса ДНК-дендример и трансфекцию культуры клеток нейробластомы человека IMR-32 проводили, как описано в примере 19.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка. Количественное определение белка IL10 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор ELISA Kit for Interleukin 10 (IL10) (SEA056Hu, Cloud-Clone Corp., США), согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка IL10, входящим в состав набора. Чувствительность метода составляла не менее 2,8 пг/мл, диапазон измерения - от 7,8 пг/мл до 500 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа, представлены на фиг. 12.
Из фигуры 12 следует, что трансфекция культуры клеток нейробластомы человека IMR-32 генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL10 приводит к увеличению количества белка IL10 по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IL10 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IL10 для повышения уровня экспрессии IL10 в эукариотических клетках.
Пример 21.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IL13, несущего ген IL13, для повышения экспрессии белка IL13 в клетках млекопитающих.
Оценивали изменение количества белка IL13 в лизате культуры клеток глиобластомы человека U-118 MG (ATCC HTB-15) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IL13, несущим ген IL13 человека. Клетки культивировали, как описано в примере 12.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена IL13 (В), в качестве трансфицируемых агентов - ДНК-вектор GDTT1.8NAS12-IL13, несущий ген IL13 человека (С). Приготовление комплекса ДНК-дендример и трансфекцию культуры клеток глиобластомы человека U-118 MG проводили, как описано в примере 19.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка. Количественное определение белка IL13 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор ELISA Kit for Interleukin 13 (IL13) (SEA060Hu, Cloud-Clone Corp., США), согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка IL13, входящим в состав набора. Чувствительность метода составляла не менее 6,7 пг/мл, диапазон измерения - от 15,6 пг/мл до 1000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа, представлены на фиг. 13.
Из фигуры 13 следует, что трансфекция культуры клеток глиобластомы человека U-118 MG генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL13 приводит к увеличению количества белка IL13 по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IL13 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IL13 для повышения уровня экспрессии IL13 в эукариотических клетках.
Пример 22.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, несущего ген IFNB1, для повышения экспрессии белка IFNB1 в клетках млекопитающих.
Оценивали изменение количества белка IFNB1 в лизате культуры клеток нейробластомы человека SH-SY5Y (ATCC CRL-2266) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IFNB1, несущим ген IFNB1 человека. Клетки культивировали, как описано в примере 13.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена IFNB1 (В), в качестве трансфицируемых агентов - ДНК-вектор GDTT1.8NAS12-IFNB1, несущий ген IFNB1 человека (С). Приготовление комплекса ДНК-дендримеры и трансфекцию культуры клеток нейробластомы человека SH-SY5Y проводили, как описано в примере 19.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка.
Количественное определение белка IFNB1 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор ELISA Kit for Interferon Beta (IFNb) (SEA222Hu, Cloud-Clone Corp., США), согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка IFNB1, входящим в состав набора. Чувствительность метода составляла не менее 2,7 пг/мл, диапазон измерения - от 7,8 пг/мл до 500 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа, представлены на фиг. 14.
Из фигуры 14 следует, что трансфекция культуры клеток нейробластомы человека SH-SY5Y генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNB1 приводит к увеличению количества белка IFNB1 по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IFNB1 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1 для повышения уровня экспрессии IFNB1 в эукариотических клетках.
Пример 23.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFRSF4, несущего ген TNFRSF4, для повышения экспрессии белка TNFRSF4 в клетках млекопитающих.
Оценивали изменение количества белка TNFRSF4 в лизате культуры клеток астроглии человека SVG p12 (ATCC CRL-8621) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-TNFRSF4, несущим ген TNFRSF4 человека. Клетки культивировали, как описано в примере 14.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена TNFRSF4 (В), в качестве трансфицируемых агентов - ДНК-вектор GDTT1.8NAS12-TNFRSF4, несущий ген TNFRSF4 человека (С). Приготовление комплекса ДНК-дендримеры и трансфекцию культуры клеток астроглии человека SVG p12 проводили, как описано в примере 19.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка.
Количественное определение белка TNFRSF4 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Human OX40 ELISA Kit (ab231926, Abcam, США), согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка TNFRSF4, входящим в состав набора. Чувствительность метода составляла не менее 23 пг/мл, диапазон измерения - от 46,9 пг/мл до 3000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа, представлены на фиг. 15.
Из фигуры 15 следует, что трансфекция культуры клеток астроглии человека SVG p12 генотерапевтическим ДНК-вектором GDTT1.8NAS12-TNFRSF4 приводит к увеличению количества белка TNFRSF4 по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген TNFRSF4 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFRSF4 для повышения уровня экспрессии TNFRSF4 в эукариотических клетках.
Пример 24.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFSF10, несущего ген TNFSF10, для повышения экспрессии белка TNFSF10 в клетках млекопитающих.
Оценивали изменение количества белка TNFSF10 в лизате нейроэктодермальных клеток опухоли головного мозга человека PFSK-1 (ATCC number CRL-2060) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-TNFSF10, несущим ген TNFSF10 человека. Клетки культивировали, как описано в примере 15.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена TNFSF10 (В), в качестве трансфицируемых агентов - ДНК-вектор GDTT1.8NAS12- TNFSF10, несущий ген TNFSF10 человека (С). Приготовление комплекса ДНК-дендримеры и трансфекцию нейроэктодермальных клеток опухоли головного мозга человека PFSK-1 проводили, как описано в примере 19.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка.
Количественное определение белка TNFSF10 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Human TRAIL/TNFSF10 Quantikine ELISA Kit (DTRL00, R&D Systems, Inc., США) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка TNFSF10, входящим в состав набора. Чувствительность метода составляла не менее 7,87 пг/мл, диапазон измерения - от 15,6 пг/мл до 1000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа, представлены на фиг. 16.
Из фигуры 16 следует, что трансфекция культуры нейроэктодермальных клеток опухоли головного мозга человека PFSK-1 генотерапевтическим ДНК-вектором GDTT1.8NAS12-TNFSF10 приводит к увеличению количества белка TNFSF10 по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген TNFSF10 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFSF10 для повышения уровня экспрессии TNFSF10 в эукариотических клетках.
Пример 25.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-BCL2, несущего ген BCL2, для повышения экспрессии белка BCL2 в клетках млекопитающих.
Оценивали изменение количества белка BCL2 в лизате культуры клеток медуллобластомы человека D341 Med (ATCC HTB-187) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-BCL2, несущим ген BCL2 человека. Клетки культивировали, как описано в примере 16.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена BCL2 (В), в качестве трансфицируемых агентов - ДНК-вектор GDTT1.8NAS12-BCL2, несущий ген BCL2 человека (С). Приготовление комплекса ДНК-дендримеры и трансфекцию культуры клеток медуллобластомы человека D341 Med проводили, как описано в примере 19.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка.
Количественное определение белка BCL2 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор ELISA Kit for B-Cell Leukemia/Lymphoma 2 (Bcl2) (SEA778Hu, Cloud-Clone Corp., США) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка BCL2, входящим в состав набора. Чувствительность метода составляла не менее 0,061 нг/мл, диапазон измерения - от 0,156 нг/мл до 10 нг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа, представлены на фиг. 17.
Из фигуры 17 следует, что трансфекция культуры клеток медуллобластомы человека D341 Med генотерапевтическим ДНК-вектором GDTT1.8NAS12-BCL2 приводит к увеличению количества белка BCL2 по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген BCL2 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-BCL2 для повышения уровня экспрессии BCL2 в эукариотических клетках.
Пример 26.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-HGF, несущего ген HGF, для повышения экспрессии белка HGF в клетках млекопитающих.
Оценивали изменение количества белка HGF в лизате культуры клеток астроцитомы головного мозга человека A-172 (ATCC CRL-1620) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-HGF, несущим ген HGF человека. Клетки культивировали, как описано в примере 17.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена HGF (В), в качестве трансфицируемых агентов - ДНК-вектор GDTT1.8NAS12-HGF, несущий ген HGF человека (С). Приготовление комплекса ДНК-дендримеры и трансфекцию клеток астроцитомы головного мозга человека A-172 проводили, как описано в примере 19.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка.
Количественное определение белка HGF проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Human HGF ELISA Kit (ab100534, Abcam, США) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка HGF, входящим в состав набора. Чувствительность метода составляла не менее 3 пг/мл, диапазон измерения - от 2,74 пг/мл до 2000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа, представлены на фиг. 18.
Из фигуры 18 следует, что трансфекция культуры клеток астроцитомы головного мозга человека A-172 генотерапевтическим ДНК-вектором GDTT1.8NAS12-HGF приводит к увеличению количества белка HGF по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген HGF на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-HGF для повышения уровня экспрессии HGF в эукариотических клетках.
Пример 27.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IL2, несущего ген IL2, для повышения экспрессии белка IL2 в клетках млекопитающих.
Оценивали изменение количества белка IL2 в лизате культуры нейрональных прогениторных клеток человека ATCC-BXS0117 (ATCC ACS-5003) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IL2, несущим ген IL2 человека. Клетки культивировали, как описано в примере 18.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена IL2 (В), в качестве трансфицируемых агентов - ДНК-вектор GDTT1.8NAS12-IL2, несущий ген IL2 человека (С). Приготовление комплекса ДНК-дендримеры и трансфекцию нейрональных прогениторных клеток человека ATCC-BXS0117 проводили, как описано в примере 19.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка.
Количественное определение белка IL2 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор ELISA Kit for Interleukin 2 (IL2) (SEA073Hu, Cloud-Clone Corp., США) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка IL2, входящим в состав набора. Чувствительность метода составляла не менее 5,9 пг/мл, диапазон измерения - от 15,6 пг/мл до 1000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа, представлены на фиг. 19.
Из фигуры 19 следует, что трансфекция культуры нейрональных прогениторных клеток человека ATCC-BXS0117 генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL2 приводит к увеличению количества белка IL2 по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IL2 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IL2 для повышения уровня экспрессии IL2 в эукариотических клетках.
Пример 28.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, несущего ген IFNB1, для повышения экспрессии белка IFNB1 в тканях человека.
Для подтверждения эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, несущего целевой ген, а именно, ген IFNB1 и реализуемости способа его применения оценивали изменения количества белка IFNB1 в коже человека при введении в кожу человека генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, несущего ген человека IFNB1.
С целью анализа изменения количества белка IFNB1 трем пациентам вводили в кожу предплечья генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1, несущий ген IFNB1 и параллельно вводили плацебо, представляющее собой генотерапевтический ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена IFNB1.
Пациент 1, женщина 48 лет, (П1); пациент 2, женщина 66 лет, П2); пациент 3, мужчина, 44 года, (П3). В качестве транспортной системы использовали полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, Франция). Генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1, который содержит кДНК гена IFNB1 и генотерапевтический ДНК-вектор GDTT1.8NAS12, используемый в качестве плацебо, который не содержит кДНК гена IFNB1, каждый из которых растворяли в стерильной воде степени очистки Nuclease-Free. Для получения генетической конструкции готовили комплексы ДНК- cGMP grade in-vivo-jetPEI в соответствии с рекомендациями производителя.
Генотерапевтический ДНК-вектор GDTT1.8NAS12 (плацебо) и генотерапевтический ДНК вектор GDTT1.8NAS12-IFNB1, несущий ген IFNB1 вводили в количестве 1 мг для каждой генетической конструкции тоннельным методом иглой 30G на глубину 3 мм. Объем вводимого раствора генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо) и генотерапевтического ДНК вектора GDTT1.8NAS12-IFNB1, несущего ген IFNB1 – 0,5 мл для каждой генетической конструкции. Очаги введения каждой генетической конструкции располагались на участке предплечья на расстоянии 8-10 см друг от друга.
Биопсийные образцы отбирали на 2-е сутки после введения генетических конструкций генотерапевтических ДНК-векторов. Взятие биопсии осуществляли из участков кожи пациентов в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IFNB1, несущего ген IFNB1 (I), генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо) (II), а также из интактных участков кожи (III), используя устройство для взятия биопсии Epitheasy 3.5 (Medax SRL, Италия). Кожу пациентов в зоне отбора биоптата предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер биопсийного образца составлял около 10 куб. мм, масса - около 11 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000g. Отбирали супернатант и использовали его для количественного определения целевого белка методом твердофазного иммуноферментного анализа (ELISA). Количественное определение белка IFNB1 проводили методом твердофазного иммуноферментного анализа, как это описано в примере 22.
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка IFNB1, входящим в состав набора. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США). Графики, полученные в результате анализа, представлены на фиг. 20.
Из фигуры 20 следует, что в коже всех трех пациентов в области введения генотерапевтического ДНК вектора GDTT1.8NAS12- IFNB1, несущего целевой ген IFNB1 человека, произошло увеличение количества белка IFNB1 в сравнении с количеством белка IFNB1 в области введения генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо), не содержащего ген IFNB1 человека, что свидетельствует об эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1 и подтверждает реализуемость способа его применения, в частности при внутрикожном введении генотерапевтического ДНК-вектора в ткани человека.
Пример 29.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IL13, несущего ген IL13, для повышения экспрессии белка IL13 в тканях человека.
Для подтверждения эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IL13, несущего целевой ген IL13 и реализуемости способа его применения оценивали изменение количества белка IL13 в мышечной ткани человека при введении генотерапевтического ДНК-вектора GDTT1.8NAS12-IL13, несущего целевой ген, а именно, ген IL13 человека.
С целью анализа изменения количества белка IL13 трем пациентам вводили в ткань икроножной мышцы генотерапевтический ДНК-вектор GDTT1.8NAS12- IL13, несущий ген IL13 с транспортной молекулой, и параллельно вводили плацебо, представляющее собой генотерапевтический ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена IL13 с транспортной молекулой.
Пациент 1, женщина 49 лет, (П1); пациент 2, мужчина 54 года (П2); пациент 3, мужчина 53 года, (П3). В качестве транспортной системы использовали полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, Франция), пробоподготовку проводили в соответствии с рекомендациями производителя.
Генотерапевтический ДНК-вектор GDTT1.8NAS12 (плацебо) и генотерапевтический ДНК вектор GDTT1.8NAS12-IL13, несущий ген IL13 вводили в количестве 1 мг для каждой генетической конструкции тоннельным методом иглой 30G на глубину около 10 -12 мм. Объем вводимого раствора генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо) и генотерапевтического ДНК вектора GDTT1.8NAS12-IL13, несущего ген IL13 – 1мл для каждой генетической конструкции. Очаги введения каждой генетической конструкции располагали медиально на расстоянии 8-10 см друг от друга.
Биопсийные образцы отбирали на 2-е сутки после введения генетических конструкций генотерапевтических ДНК-векторов. Взятие биопсии осуществляли из участков мышечной ткани пациентов в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IL13, несущего ген IL13 (I), генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо) (II), а также из интактного участка икроножной мышцы (III), используя устройство для взятия биопсии MAGNUM (BARD, США). Кожу пациентов в зоне отбора биоптата предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер биопсийного образца составлял около 20 куб. мм, масса - около 22 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000g. Отбирали супернатант и использовали его для количественного определения целевого белка.
Количественное определение белка IL13 проводили методом твердофазного иммуноферментного анализа, как это описано в примере 21.
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка IL13, входящим в состав набора. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа, представлены на фиг. 21.
Из фигуры 21 следует, что в икроножной мышце всех трех пациентов в области введения генотерапевтического ДНК вектора GDTT1.8NAS12-IL13, несущего целевой ген, а именно, ген IL13 человека, произошло увеличение количества белка IL13 в сравнении с количеством белка IL13 в области введения генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо), не содержащего ген IL13 человека, что свидетельствует об эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IL13 и подтверждает реализуемость способа его применения, в частности при внутримышечном введении генотерапевтического ДНК-вектора в ткани человека.
Пример 30.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IL2, несущего ген IL2, для повышения экспрессии белка IL2 в тканях человека.
Для подтверждения эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IL2, несущего целевой ген, а именно, ген IL2 и реализуемости способа его применения оценивали изменения количества белка IL2 в коже человека при введении в кожу человека генотерапевтического ДНК-вектора GDTT1.8NAS12-IL2, несущего ген человека IL2.
С целью анализа изменения количества белка IL2 трем пациентам вводили в кожу предплечья генотерапевтический ДНК-вектор GDTT1.8NAS12-IL2, несущий ген IL2 и параллельно вводили плацебо, представляющее собой генотерапевтический ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена IL2.
Пациент 1, женщина 52 года (П1); пациент 2, мужчина 39 лет (П2); пациент 3, мужчина, 40 лет (П3). В качестве транспортной системы использовали полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, Франция). Генотерапевтический ДНК-вектор GDTT1.8NAS12-IL2, который содержит кДНК гена IL2 и генотерапевтический ДНК-вектор GDTT1.8NAS12, используемый в качестве плацебо, который не содержит кДНК гена IL2, каждый из которых растворяли в стерильной воде степени очистки Nuclease-Free. Для получения генетической конструкции готовили комплексы ДНК- cGMP grade in-vivo-jetPEI в соответствии с рекомендациями производителя.
Генотерапевтический ДНК-вектор GDTT1.8NAS12 (плацебо) и генотерапевтический ДНК вектор GDTT1.8NAS12-IL2, несущий ген IL2 вводили в количестве 1 мг для каждой генетической конструкции тоннельным методом иглой 30G на глубину 3 мм. Объем вводимого раствора генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо) и генотерапевтического ДНК вектора GDTT1.8NAS12-IL2, несущего ген IL2 – 0,5 мл для каждой генетической конструкции. Очаги введения каждой генетической конструкции располагались на участке предплечья на расстоянии 8-10 см друг от друга.
Биопсийные образцы отбирали на 2-е сутки после введения генетических конструкций генотерапевтических ДНК-векторов. Взятие биопсии осуществляли из участков кожи пациентов в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IL2, несущего ген IL2 (I), генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо) (II), а также из интактных участков кожи (III), используя устройство для взятия биопсии Epitheasy 3.5 (Medax SRL, Италия). Кожу пациентов в зоне отбора биоптата предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер биопсийного образца составлял около 10 куб. мм, масса - около 11 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000g. Отбирали супернатант и использовали его для количественного определения целевого белка методом твердофазного иммуноферментного анализа, как это описано в примере 27.
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка IL2, входящим в состав набора. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа, представлены на фиг. 22.
Из фигуры 22 следует, что в коже всех трех пациентов в области введения генотерапевтического ДНК вектора GDTT1.8NAS12-IL2, несущего целевой ген IL2 человека, произошло увеличение количества белка IL2 в сравнении с количеством белка IL2 в области введения генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо), не содержащего ген IL2 человека, что свидетельствует об эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-I2 и подтверждает реализуемость способа его применения, в частности при внутрикожном введении генотерапевтического ДНК-вектора в ткани человека.
Пример 31.
Подтверждение эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IL10, несущего ген IL10 и реализуемости способа его применения для повышения уровня экспрессии белка IL10 в тканях человека путем введения аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL10.
Для подтверждения эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IL10, несущего ген IL10 и реализуемости способа его применения оценивали изменения количества белка IL10 в коже человека при введении в кожу пациента культуры аутологичных фибробластов этого пациента, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL10.
Пациенту вводили в кожу предплечья соответствующую культуру аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL10, несущим ген IL10, и параллельно вводили плацебо, представляющее собой культуру аутологичных фибробластов пациента, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, не несущим ген IL10.
Первичную культуру фибробластов человека выделяли из биоптатов кожи пациента. Используя устройство для взятия биопсии кожи Epitheasy 3.5 (Medax SRL, Италия) брали образец биоптата кожи из зоны, защищенной от действия ультрафиолета, а именно за ушной раковиной или с боковой внутренней поверхности в зоне локтевого сустава, размером около 10 кв. мм, массой - около 11 мг. Кожу пациента предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Культивирование первичной культуры клеток осуществляли при 370С в атмосфере, содержащей 5 % СО2 в среде DMEM с 10 % фетальной телячьей сывороткой и ампициллином 100 Ед/мл. Пересев культуры и смену культуральной среды осуществляли каждые 2 дня. Общая продолжительность роста культуры не превышала 25-30 дней. Из культуры клеток отбирали аликвоту, содержащую 5×104 клеток. Культуру фибробластов пациента трансфицировали генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL10, несущим ген IL10 или плацебо – вектор GDTT1.8NAS12, не несущий целевой ген IL10.
Трансфекцию осуществляли с помощью катионного полимера полиэтиленимина JETPEI (Polyplus Transfection, Франция), согласно инструкции фирмы-производителя. Клетки культивировали 72 часа и затем вводили пациенту. Введение пациенту культуры аутологичных фибробластов этого пациента, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL10, и плацебо, представляющее собой культуру аутологичных фибробластов пациента, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, осуществляли туннельным методом в область предплечья иглой 30G длиной 13 мм на глубину около 3 мм. Концентрация модифицированных аутологичных фибробластов в вводимой суспензии составляла примерно 5 млн клеток на 1 мл суспензии, количество вводимых клеток не превышало 15 млн. Очаги введения культуры аутологичных фибробластов располагались на расстоянии 8-10 см друг от друга.
Биопсийные образцы отбирали на 4-е сутки после введения культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL10, несущим целевой ген, а именно, ген IL10 и плацебо. Взятие биопсии осуществляли из участков кожи пациента в зоне введения культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL10, несущим целевой ген, а именно, ген IL10 (C), культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, не несущим целевой ген IL10 (плацебо) (B), а также из участков интактной кожи (A), используя устройство для взятия биопсии Epitheasy 3.5 (Medax SRL, Италия). Кожу в зоне отбора биоптата пациентов предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер биопсийного образца составлял около 10 куб. мм, масса - около 11 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000g. Отбирали супернатант и использовали его для количественного определения целевого белка, как это описано в примере 12.
Графики, полученные в результате анализа, представлены на фиг. 23.
Из фигуры 23 следует, что в коже пациента в области введения культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL10, несущим ген IL10, произошло увеличение количества белка IL10 в сравнении с количеством белка IL10 в области введения культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, не несущим ген IL10 (плацебо), что свидетельствует об эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IL10 и подтверждает реализуемость способа его применения для повышения уровня экспрессии IL10 в тканях человека, в частности при введении аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL10.
Пример 32.
Подтверждение эффективности и реализуемости способа сочетанного применения генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, несущего ген DDC, генотерапевтического ДНК-вектора GDTT1.8NAS12-BCL2, несущего ген BCL2, генотерапевтического ДНК-вектора GDTT1.8NAS12-HGF, несущего ген HGF, генотерапевтического ДНК-вектора GDTT1.8NAS12- TNFSF10, несущего ген TNFSF10, генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFRSF4, несущего ген TNFRSF4, для повышения уровня экспрессии белков DDC, BCL2, HGF, TNFSF10 и TNFRSF4 в тканях млекопитающих.
Оценивали изменение количества белков DDC, BCL2, HGF, TNFSF10 и TNFRSF4 в мышечной ткани крысы при сочетанном введении в зону этой ткани смеси генотерапевтических векторов: генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, генотерапевтического ДНК-вектора GDTT1.8NAS12-BCL2, генотерапевтического ДНК-вектора GDTT1.8NAS12-HGF, генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFSF10, генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFRSF4. Исследование проводили на 15 лабораторных животных – самцах крысы линии Wistar 8-ми месячного возраста массой 240–290 г., разбитых на три группы: группа К1 (5 животных) - введение смеси пяти генотерапевтических ДНК-векторов, несущих гены DDC, BCL2, HGF, TNFSF10, TNFRSF4; группа К2 (5 животных) - введения генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо); группа К3 (5 животных) – интактные животные. В качестве транспортной системы использовали полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, Франция). Эквимолярную смесь генотерапевтических ДНК-векторов GDTT1.8NAS12-DDC, GDTT1.8NAS12-BCL2, GDTT1.8NAS12-HGF, GDTT1.8NAS12-TNFSF10, GDTT1.8NAS12-TNFRSF4 в пропорции 30 мкг каждого вектора растворяли в стерильной воде степени очистки Nuclease-Free. Для получения генетической конструкции готовили комплексы ДНК- cGMP grade in-vivo-jetPEI в соответствии с рекомендациями производителя.
Объем внутримышечно вводимого раствора составлял 0,15 мл с суммарным количеством ДНК 150 мкг. Введение раствора осуществляли с помощью инсулинового шприца. Через 2 сутки после процедуры крыс декапитировали.
Образцы отбирали на 2 сутки после введения генотерапевтических ДНК-векторов. Взятие биопсийного материала осуществляли из участков правой мышцы бедра всех животных: в зоне введения смеси пяти генотерапевтических ДНК-векторов, несущих гены DDC, BCL2, HGF, TNFSF10, TNFRSF4 (группа 1), в зоне введения плацебо (группа 2), а также из участков мышечной ткани правой мышцы бедра животных, не подвергавшихся никаким манипуляциям (группа 3). Каждый образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторида, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000g. Отбирали супернатант и использовали его для количественного определения целевых белков, как это описано в примере 19 (количественное определение белка DDC), примере 25 (количественное определение белка BCL2), примере 26 (количественное определение белка HGF), примере 24 (количественное определение белка TNFSF10), примере 23 (количественное определение белка TNFRSF4). Графики, полученные в результате анализа, представлены на фиг. 24.
Из фигуры 24 следует, что в зоне мышечной ткани животных группы К1, которым вводилась смесь генотерапевтических векторов: генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, несущего целевой ген DDC, генотерапевтического ДНК-вектора GDTT1.8NAS12-BCL2, несущего целевой ген BCL2, генотерапевтического ДНК-вектора GDTT1.8NAS12-HGF, несущего целевой ген HGF, генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFSF10, несущего целевой ген TNFSF10, генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFRSF4, несущего целевой ген TNFRSF4, произошло увеличение количества белков DDC, BCL2, HGF, TNFSF10 и TNFRSF4 по сравнению с зоной мышечной ткани группы животных К2 (плацебо) и зоной мышечной ткани животных группы К3 (интактная группа). Полученные результаты свидетельствуют об эффективности сочетанного применения генотерапевтических ДНК-векторов и подтверждают реализуемость способа применения для повышения уровня экспрессии целевых белков в тканях млекопитающих.
Пример 33.
Подтверждение эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IL13, несущего ген IL13 и реализуемости способа его применения для повышения уровня экспрессии белка IL13 в клетках млекопитающих.
Для подтверждения эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IL13, несущего ген IL13 оценивали изменение накопления мРНК целевого гена IL13 в мононуклеарных клетках периферической крови быка через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12- IL13, несущим ген IL13 человека.
Мононуклеарные клетки периферической крови быка выделяли из 50 мл венозной крови путем центрифугирования в градиенте раствора фиколла 1,077 (Панэко, Р052п) и культивировали в среде RPMI Media 1640 (GIBCO®, 11875-085) с добавлением 10 % лошадиной сыворотки Horse Serum (ATCC® 30-2040™) в стандартных условиях. Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL13, несущим ген IL13 человека и ДНК-вектором GDTT1.8NAS12, не несущим ген IL13 человека (контроль), выделение РНК, реакцию обратной транскрипции, ПЦР амплификации и анализ данных проводили как описано в примере 12. В качестве референтного гена использовали ген актина быка/ коровы (АСТ), приведенного в базе данных GenBank под номером AH001130.2. В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности генов IL13 и АСТ. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов – кДНК генов IL13 и ACT, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1. (Bio-Rad, USA).
Графики, полученные в результате анализа, представлены на фиг. 25.
Из фигуры 25 следует, что в результате трансфекции мононуклеарных клеток периферической крови быка генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL13, уровень специфической мРНК гена IL13 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IL13 на уровне мРНК. Представленные результаты подтверждают реализуемость способа применения генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL13 для повышения уровня экспрессии гена IL13 в клетках млекопитающих.
Пример 34.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2, несущий генотерапевтический ДНК-вектор, и способ его получения.
Конструирование штамма для наработки в промышленных масштабах генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген, выбранный из группы генов: DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2, а именно штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2, несущего генотерапевтический ДНК-вектор GDTT1.8NAS12-DDC или GDTT1.8NAS12-IL10 или GDTT1.8NAS12-IL13 или GDTT1.8NAS12-IFNB1 или GDTT1.8NAS12-TNFRSF4 или GDTT1.8NAS12-TNFSF10 или GDTT1.8NAS12-BCL2 или GDTT1.8NAS12-HGF или GDTT1.8NAS12-IL2 соответственно для его наработки с возможностью селекции без использования антибиотиков заключается в получении электрокомпетентных клеток штамма Escherichia coli JM110-NAS с проведением электропорации этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS12-DDC, или ДНК-вектором GDTT1.8NAS12-IL10, или ДНК-вектором GDTT1.8NAS12-IL13, или ДНК-вектором GDTT1.8NAS12-IFNB1, или ДНК-вектором GDTT1.8NAS12-TNFRSF4, или ДНК-вектором GDTT1.8NAS12-TNFSF10, или ДНК-вектором GDTT1.8NAS12-BCL2, или ДНК-вектором GDTT1.8NAS12-HGF, или ДНК-вектором GDTT1.8NAS12-IL2, после чего клетки высеваются на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола. При этом штамм Escherichia coli JM110-NAS для наработки генотерапевтического ДНК-вектора GDTT1.8NAS12 или генотерапевтических ДНК-векторов на его основе с возможностью положительной селекции без использования антибиотиков получали путем конструирования линейного фрагмента ДНК, содержащего регуляторный элемент RNA-in транспозона Tn10 для селекции без применения антибиотиков размером 64 п.н., ген левансахаразы sacB, продукт которого обеспечивает селекцию на сахарозо-содержащей среде размером 1422 п.н., ген устойчивости к хлорамфениколу catR, необходимый для отбора клонов штамма, в которых прошла гомологичная рекомбинация размером 763 п.н. и две гомологичные последовательности, обеспечивающие процесс гомологичной рекомбинации в области гена recA с одновременной его инактивацией размером 329 п.н. и 233 п.н., после чего проводили трансформацию клеток Escherichia coli путем электропорации и отбирали клоны, выжившие на среде, содержащей 10 мкг/мл хлорамфеникола.
Полученные штаммы для наработки были внесены в коллекции Национального биоресурсного центра - Всероссийская коллекция промышленных микроорганизмов (НБЦ ВКПМ), РФ и NCIMB Patent Deposit Service, UK с присвоением регистрационных номеров:
штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC – регистрационный № ВКПМ В-_____, дата депонирования 14.01.2020; INTERNATIONAL DEPOSITARY AUTHORITY № NCIMB 43543, дата депонирования 06.01.2020;
штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10 - регистрационный № ВКПМ В-______, дата депонирования 14.01.2020; INTERNATIONAL DEPOSITARY AUTHORITY № NCIMB 43540, дата депонирования 06.01.2020;
штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13 - регистрационный № ВКПМ В-______, дата депонирования 14.01.2020; INTERNATIONAL DEPOSITARY AUTHORITY № NCIMB 43542, дата депонирования 06.01.2020;
штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1 - регистрационный № ВКПМ В-13534, дата депонирования 27.11.2019; INTERNATIONAL DEPOSITARY AUTHORITY № NCIMB 43525, дата депонирования 28.11.2019;
штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4 - регистрационный № ВКПМ В-______, дата депонирования 14.01.2020; INTERNATIONAL DEPOSITARY AUTHORITY № NCIMB 43545, дата депонирования 06.01.2020;
штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10 - регистрационный № ВКПМ В-______, дата депонирования 14.01.2020; INTERNATIONAL DEPOSITARY AUTHORITY № NCIMB 43539, дата депонирования 06.01.2020;
штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2 - регистрационный № ВКПМ В-______, дата депонирования 14.01.2020; INTERNATIONAL DEPOSITARY AUTHORITY № NCIMB 43546, дата депонирования 06.01.2020;
штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF - регистрационный № ВКПМ В-______, дата депонирования 14.01.2020; INTERNATIONAL DEPOSITARY AUTHORITY № NCIMB 43541, дата депонирования 06.01.2020;
штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2 - регистрационный № ВКПМ В-______, дата депонирования 14.01.2020; INTERNATIONAL DEPOSITARY AUTHORITY № NCIMB 43544, дата депонирования 06.01.2020.
Пример 35.
Способ масштабирования получения генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген, выбранный из группы генов: DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2 до промышленного масштаба.
Для подтверждения технологичности получения и возможности производства в промышленных масштабах генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC (SEQ ID №1), или GDTT1.8NAS12-IL10 (SEQ ID №2), или GDTT1.8NAS12-IL13 (SEQ ID №3), или GDTT1.8NAS12-IFNB1 (SEQ ID №4), или GDTT1.8NAS12-TNFRSF4 (SEQ ID №5), или GDTT1.8NAS12-TNFSF10 (SEQ ID №6), или GDTT1.8NAS12-BCL2 (SEQ ID №7), или GDTT1.8NAS12-HGF (SEQ ID №8), или GDTT1.8NAS12-IL2 (SEQ ID №9) проводили масштабную ферментацию штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2, каждый из которых содержит генотерапевтический ДНК-вектор GDTT1.8NAS12, несущий целевой ген, а именно DDC, или IL10, или IL13, или IFNB1, или TNFRSF4, или TNFSF10, или BCL2, или HGF, или IL2. Каждый штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC, или Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10, или Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13, или Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4, или Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10, или Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2, или Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF, или Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2, получали на базе штамма Escherichia coli JM110-NAS (Genetic Diagnostics and Therapy 21 Ltd, Великобритания) по примеру 34 путем электропорации компетентных клеток этого штамма генотерапевтическим ДНК-вектором GDTT1.8NAS12-DDC, или GDTT1.8NAS12-IL10, или GDTT1.8NAS12-IL13, или GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-TNFRSF4, или GDTT1.8NAS12-TNFSF10, или GDTT1.8NAS12-BCL2, или GDTT1.8NAS12-HGF, или GDTT1.8NAS12-IL2, несущим целевой ген, а именно DDC, или IL10, или IL13, или IFNB1, или TNFRSF4, или TNFSF10, или BCL2, или HGF, или IL2 с последующим высевом трансформированных клеток на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы и отбором отдельных клонов.
Ферментацию полученного штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC, несущего генотерапевтический ДНК-вектор GDTT1.8NAS12-DDC проводили в ферментере объемом 10 л с последующим выделением генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC.
Для ферментации штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC готовили среду, содержащую на 10 л: 100 г триптон, 50 г дрожжевой экстракт (Becton Dickinson, США), доводили водой до 8800 мл и автоклавировали при 121 0С 20 мин, затем добавляли 1200 мл 50% (вес/объем) сахарозы. Далее засевали в колбу затравочную культуру штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC в объеме 100 мл. Инкубировали в шейкере-инкубаторе 16 ч при 30°С. Переносили затравочную культуру в ферментер Techfors S (Infors HT, Швейцария), растили до достижения стационарной фазы. Контроль осуществляли измерением оптической плотности культуры при длине волны 600 нм. Клетки осаждали центрифугированием 30 мин при 5000-10000 g. Супернатант удаляли, клеточную массу ресуспендировали в 10% по объему фосфатно-солевого буфера. Повторно центрифугировали 30 мин при 5000-10000g. Супернатант удаляли, к клеточной массе добавляли раствор 20 мМ TrisCl, 1 мМ ЭДТА, 200 г/л сахароза, рН 8,0 в объеме 1000 мл, тщательно перемешивали до образования гомогенной суспензии. Добавляли раствор яичного лизоцима до конечной концентрации 100мкг/мл. Инкубировали 20 мин на льду при бережном перемешивании. Далее, добавляли 2500 мл раствора 0,2 М NaOH, 10 г/л додецилсульфат натрия, инкубировали 10 мин на льду при бережном перемешивании, затем добавляли 3500 мл раствора 3M ацетат натрия, 2М уксусная кислота, рН 5-5,5, инкубировали 10 мин на льду при бережном перемешивании. Полученный образец центрифугировали 20-30 мин при 15000 g или более. Раствор аккуратно декантировали, от остатков осадка избавлялись фильтрацией через грубый фильтр (фильтровальную бумагу). Добавляли РНКазу А (Sigma, США) до конечной концентрации 20 нг/мл, инкубировали ночь 16 ч при комнатной температуре. Раствор центрифугировали 20-30 мин при 15000g, затем фильтровали через мембранный фильтр с порами 0,45 мкм (Millipore, США). Далее проводили ультрафильтрацию через мембрану размером отсечения 100кДа (Millipore, США) и разбавляли до начального объема буфером 25 мМ TrisCl, pH 7.0. Операцию повторяли три – четыре раза. Наносили раствор, на колонку с 250 мл сорбента DEAE sepharose HP (GE, США), уравновешенную раствором 25 мМ TrisCl, pH 7.0. После нанесения образца колонку промывали тремя объемами этого же раствора, а затем проводили элюцию генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC линейным градиентом от раствора 25 мМ TrisCl, pH 7.0 до раствора 25 мМ TrisCl, pH 7.0, 1М NaCl в объеме пяти объемов колонки. Контроль элюции осуществляли по оптической плотности сходящего раствора при 260 нм. Хроматографические фракции, содержащие генотерапевтический ДНК-вектор GDTT1.8NAS12-DDC, объединяли и проводили гель-фильтрацию на сорбенте Superdex 200 (GE, США). Колонку уравновешивали фосфатно-солевым буфером. Контроль элюции осуществляли по оптической плотности сходящего раствора при 260 нм, фракции анализировали электрофорезом в агарозном геле. Фракции, содержащие генотерапевтический ДНК-вектор GDTT1.8NAS12-DDC объединяли и хранили при -20°С. Для оценки воспроизводимости техпроцесса обозначенные технологические операции повторяли пятикратно. Все технологические операции для штаммов Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10, или Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13, или Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4, или Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10, или Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2, или Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF, или Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2 проводили аналогичным образом.
Воспроизводимость техпроцесса и количественные характеристики выхода конечного продукта подтверждают технологичность получения и возможность производства в промышленных масштабах генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC или GDTT1.8NAS12-IL10 или GDTT1.8NAS12-IL13 или GDTT1.8NAS12-IFNB1 или GDTT1.8NAS12-TNFRSF4 или GDTT1.8NAS12-TNFSF10 или GDTT1.8NAS12-BCL2 или GDTT1.8NAS12-HGF или GDTT1.8NAS12-IL2.
Таким образом, созданный генотерапевтический ДНК-вектор с целевым геном может быть использован для введения в клетки организма животных и человека, характеризующихся сниженной или недостаточной экспрессией белка, кодируемого данным геном, обеспечивая, таким образом, достижение терапевтического эффекта.
Задача, поставленная в данном изобретении, решена, а именно: конструирование генотерапевтических ДНК-векторов для повышения уровня экспрессии генов DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2, сочетающих в себе следующие свойства:
I) Эффективность повышения экспрессии целевых генов в эукариотических клетках за счет полученных генотерапевтических векторов с минимальным размером;
II) Возможность безопасного применения для генетической терапии человека и животных за счет отсутствия в составе генотерапевтического ДНК-вектора регуляторных элементов, представляющих собой нуклеотидные последовательности вирусных геномов и генов антибиотикорезистентности;
III) Технологичность получения и возможность наработки в штаммах в промышленных масштабах;
IV) Также решена задача по конструированию штаммов, несущих эти генотерапевтические ДНК-вектора для производства этих генотерапевтических ДНК-векторов.
Это подтверждается примерами:
для п. I – пример 1, 2, 3, 4, 5; 6; 7; 8; 9; 10; 11; 12; 13; 14; 15; 16; 17; 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33;
для п. II – пример 1, 2, 3, 4, 5; 6; 7; 8; 9; 10; 11; 12; 13; 14; 15; 16; 17; 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33;
для п. III и п. IV – пример 34, 35.
Промышленная применимость:
Все представленные выше примеры подтверждают промышленную применимость предлагаемого генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген, выбранный из группы генов: ген DDC, ген IL10, ген IL13, ген IFNB1, ген TNFRSF4, ген TNFSF10, ген BCL2, ген HGF, ген IL2 для повышения уровня экспрессии этих целевых генов, штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC, или Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10, или Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13, или Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4, или Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10, или Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2, или Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF, или Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2, несущего генотерапевтический ДНК-вектор, способа получения генотерапевтического ДНК-вектора, способа производства в промышленных масштабах генотерапевтического ДНК-вектора.
Перечень сокращений
GDTT1.8NAS12 – вектор генотерапевтический, не содержащий последовательностей вирусных геномов и маркеров антибиотико-резистентности (vector therapeutic virus-antibiotic-free)
L-DOPA - l-3,4-dihydroxyphenylalanine
ДНК - дезоксирибонуклеиновая кислота
ДАА - декарбоксилаза ароматических L-аминокислот
кДНК - комплементарная дезоксирибонуклеиновая кислота
РНК - рибонуклеиновая кислота
мРНК - матричная рибонуклеиновая кислота
п.н. – пар нуклеотидов
ПЦР - полимеразная цепная реакция
мл – миллилитр, мкл – микролитр
куб. мм – кубический миллиметр
л - литр
мкг - микрограмм
мг - миллиграмм
г - грамм
мкМ - микромоль
мМ – миллимоль
MPTP - 1-метил-4-фенил-1,2,3,6-тетрагидропиридин
мин – минута
сек - секунда
об/мин - обороты в минуту
нм - нанометр
см - сантиметр
мВт - милливатт
о.е. ф - относительная единица флуоресценции
РВС – фосфатно-солевой буфер
PBMC – мононуклеарные клетки периферической крови
ЦНС – центральная нервная система
ЭЭМ - экспериментальный энцефаломиелит.
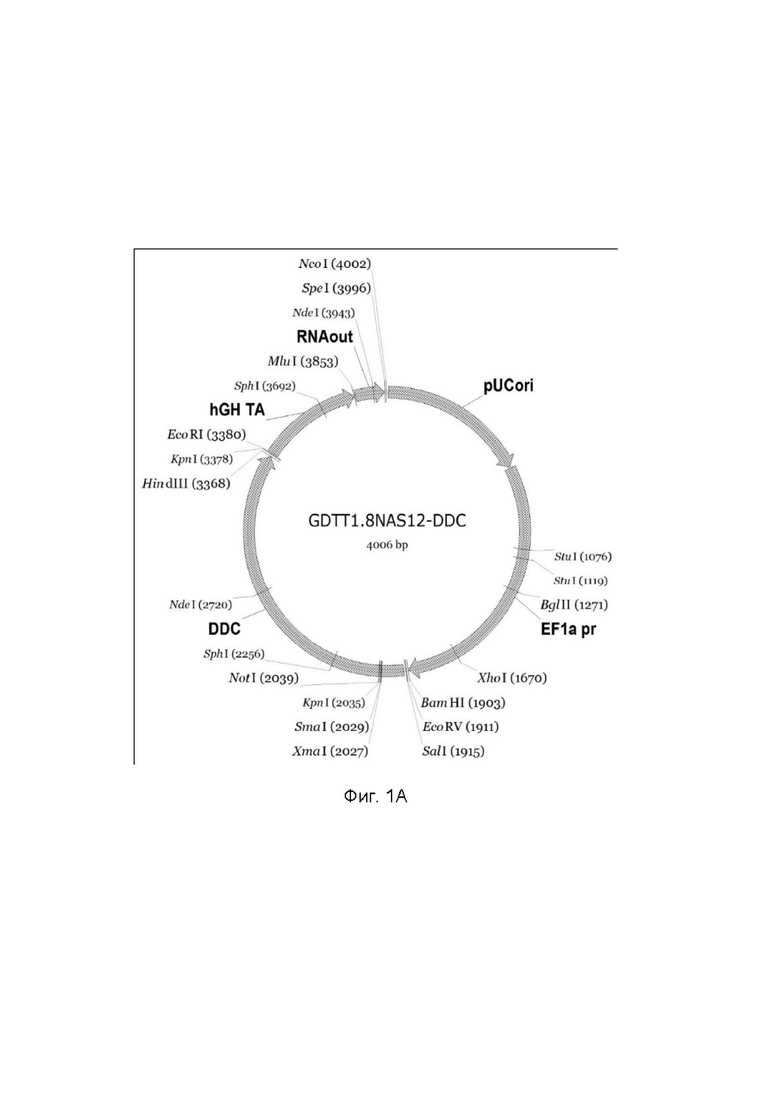

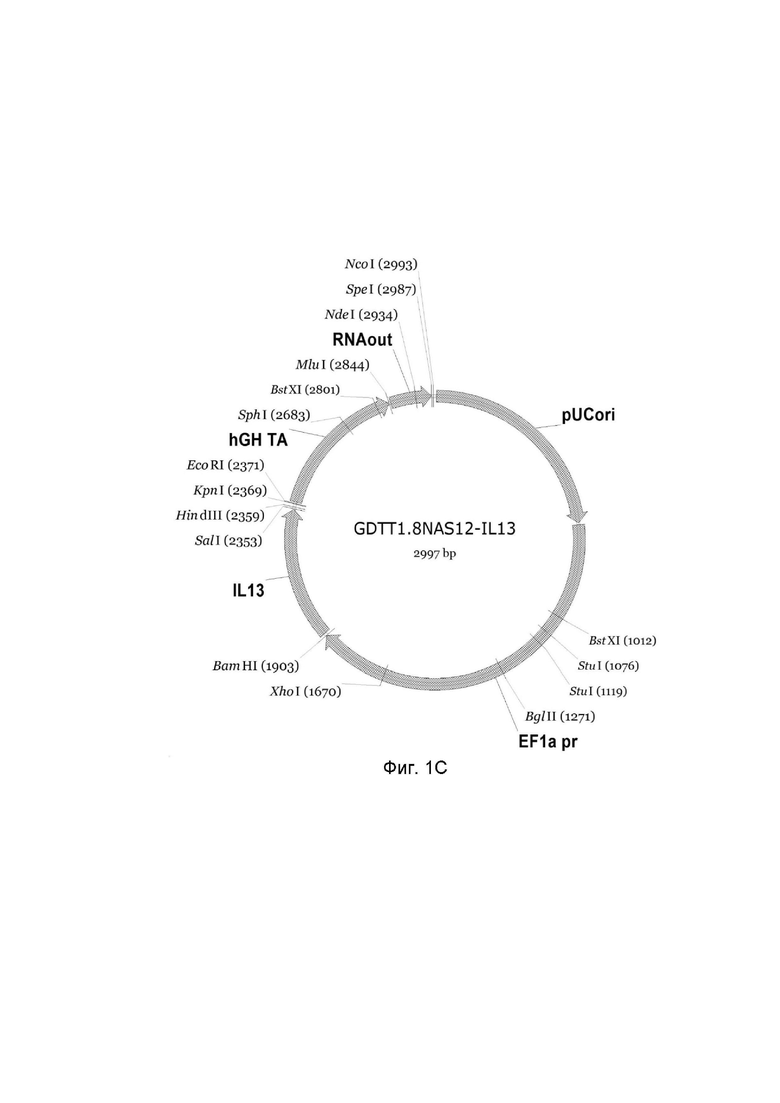
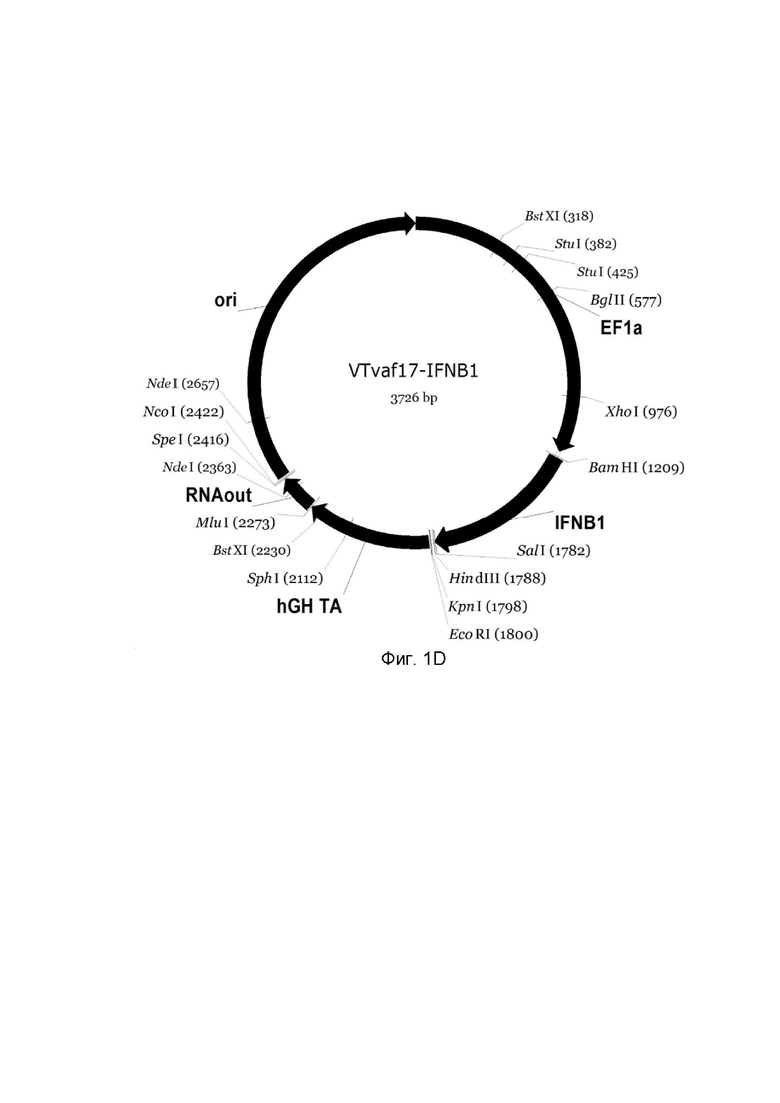
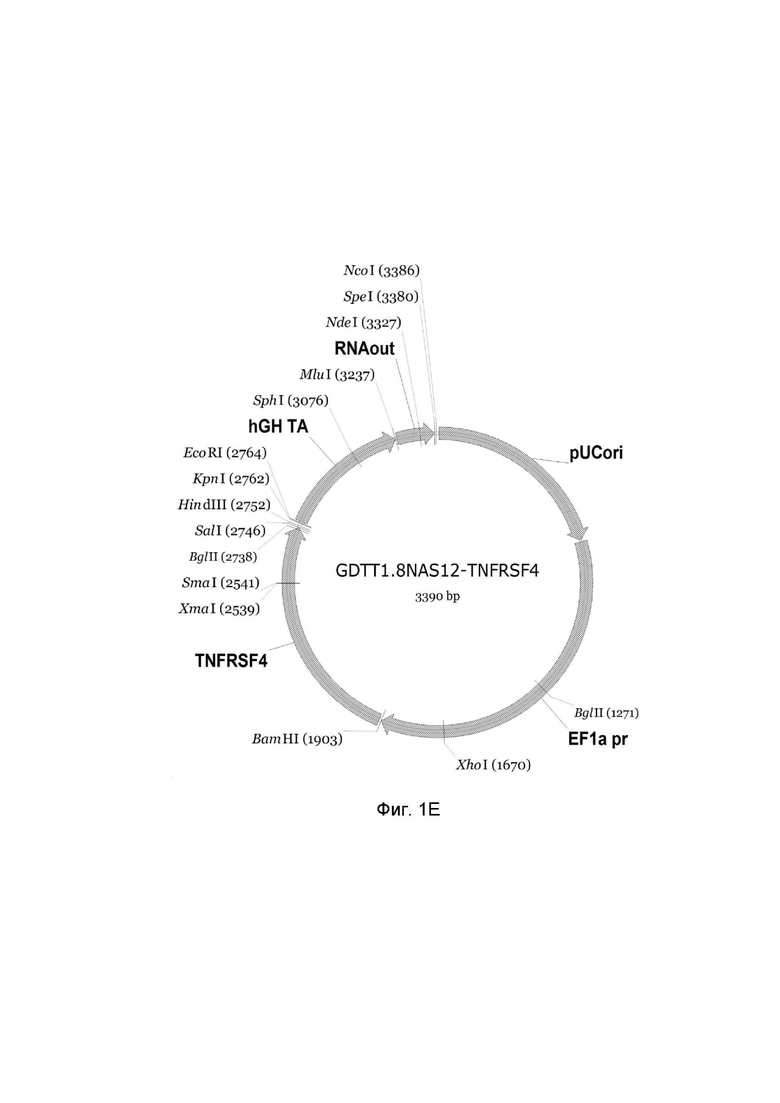


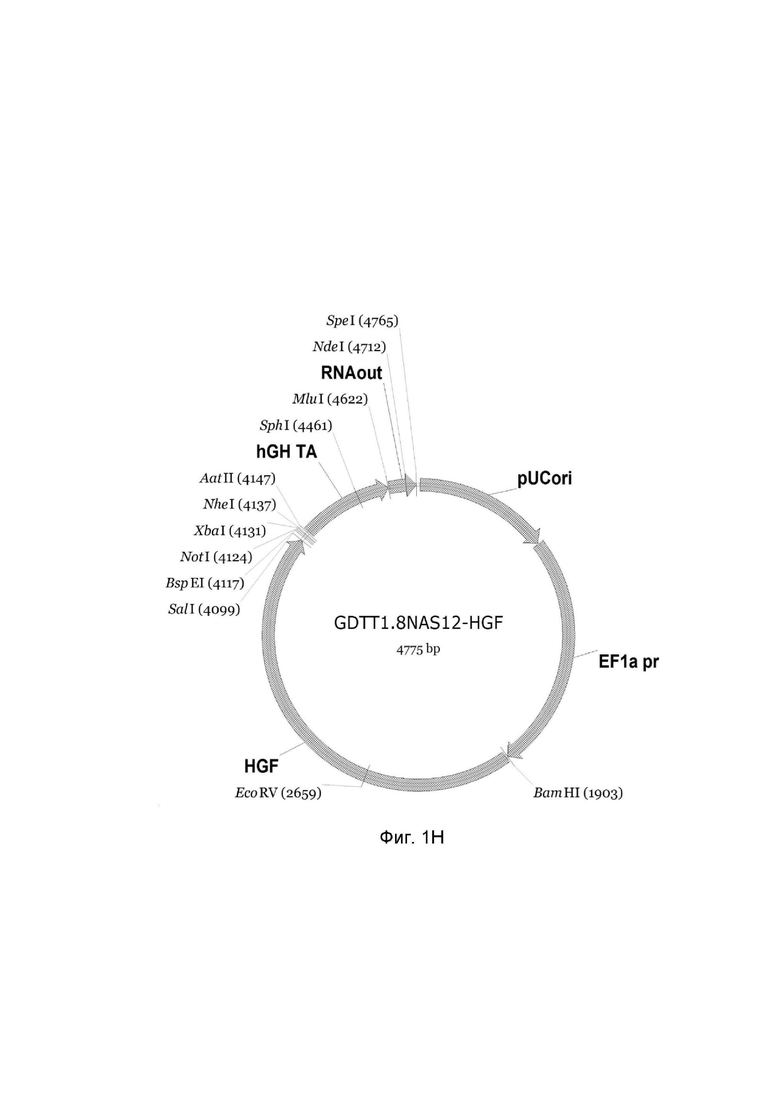
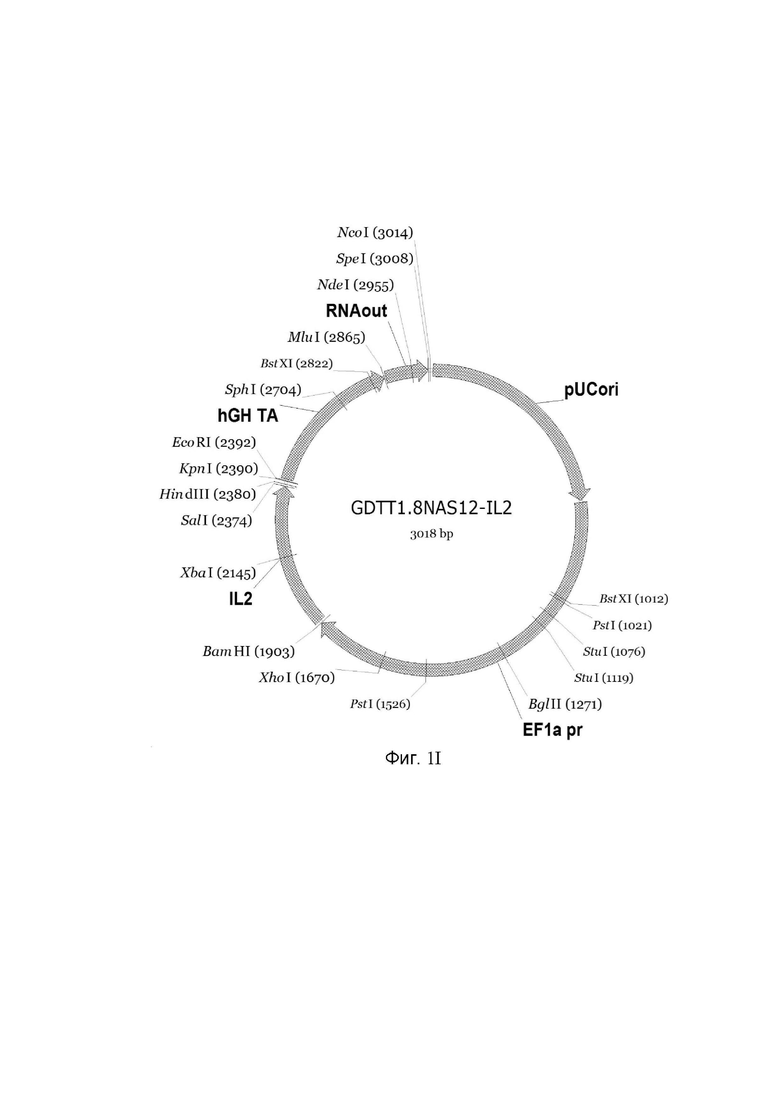
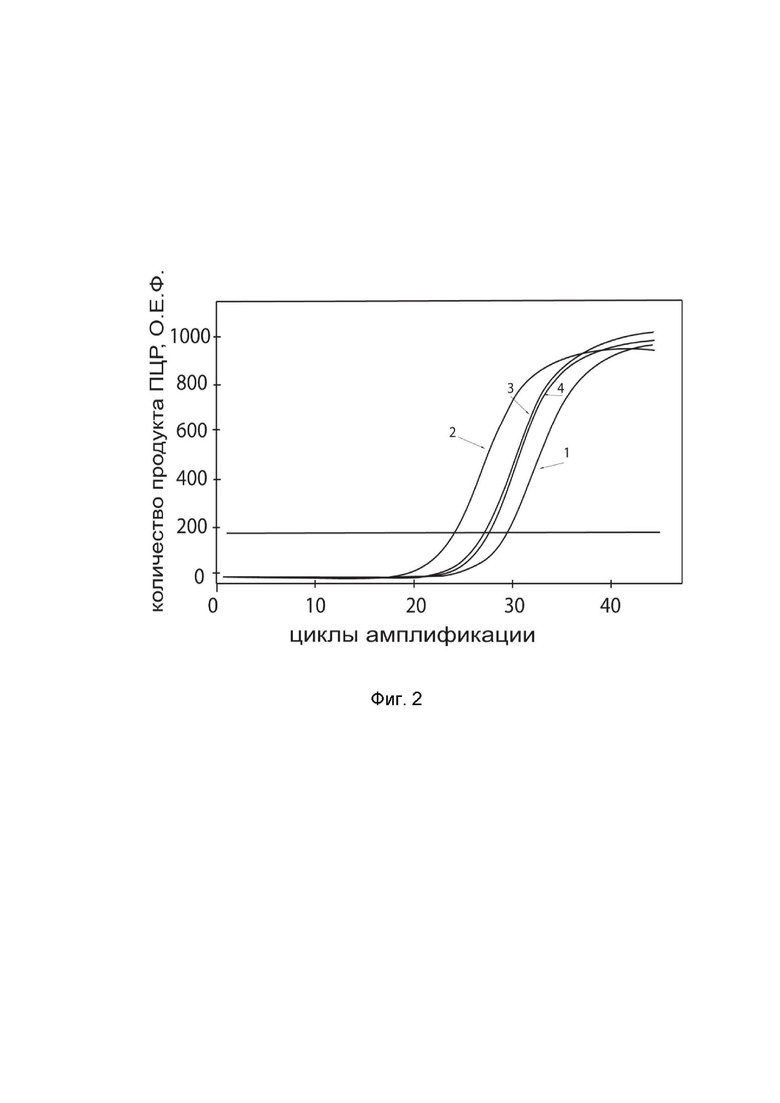
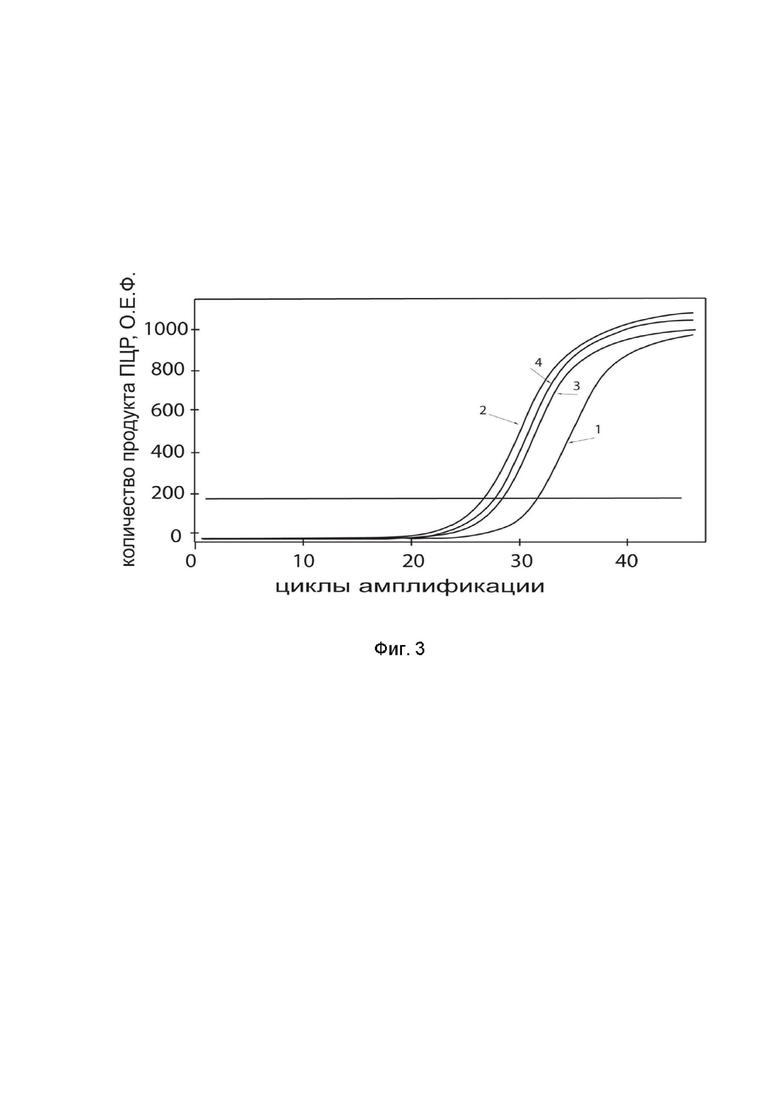
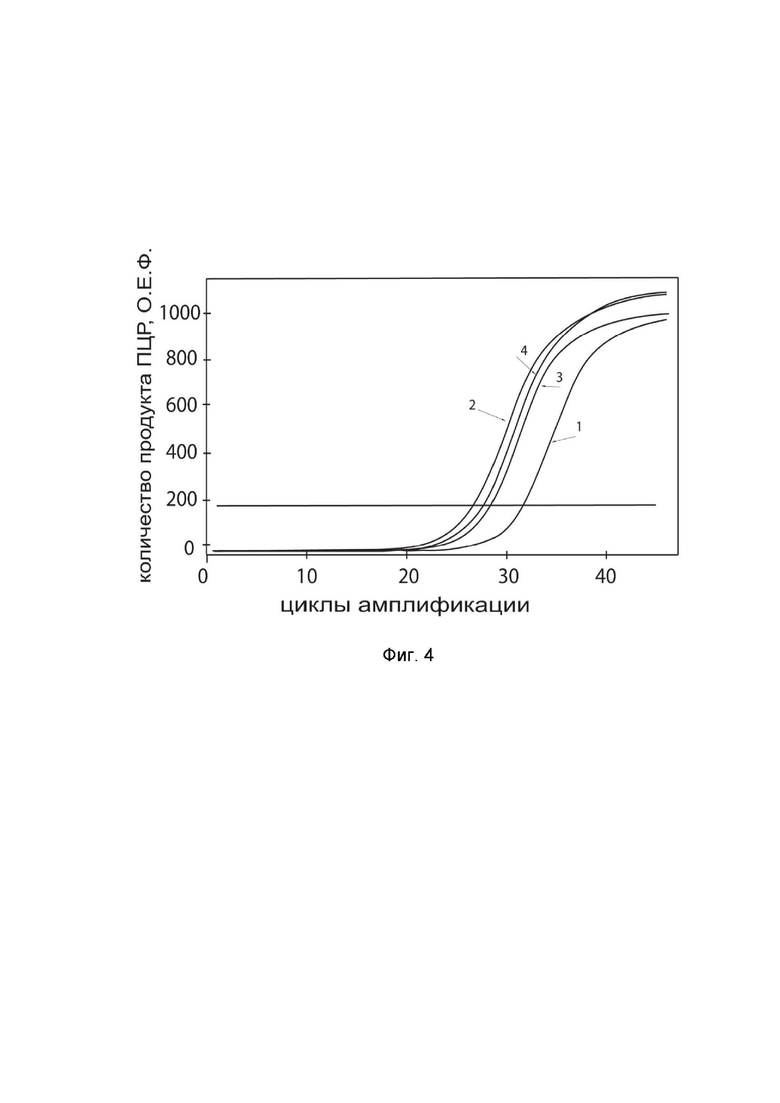
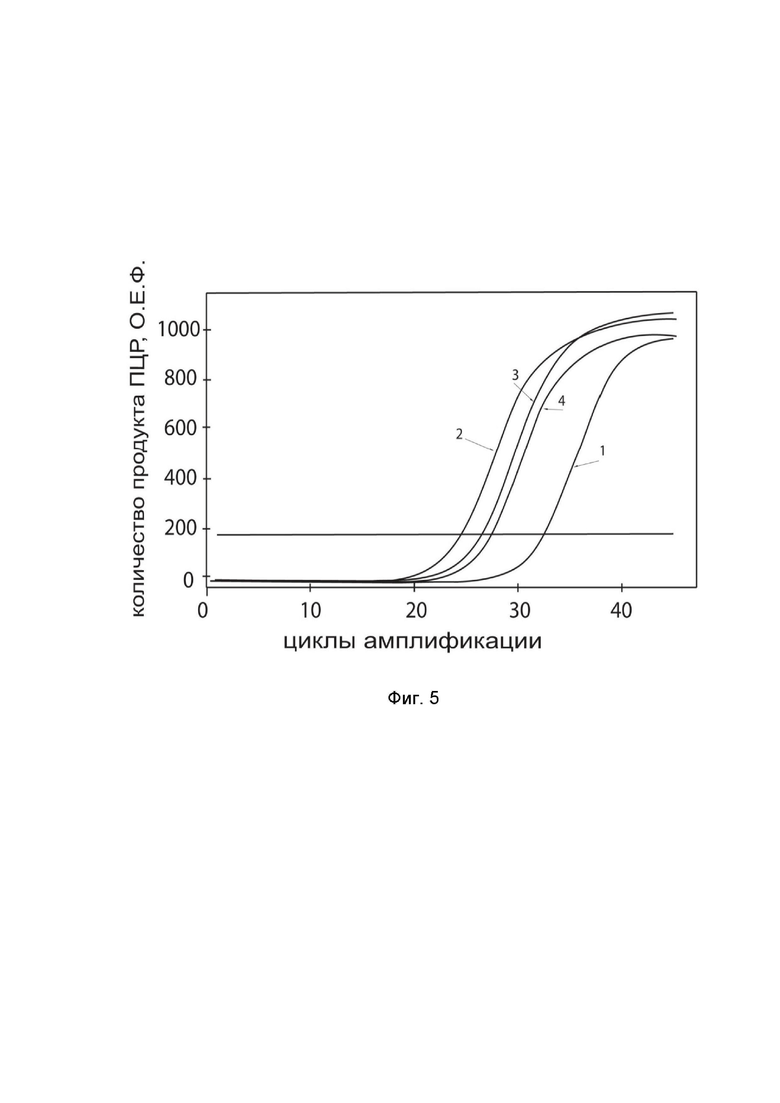
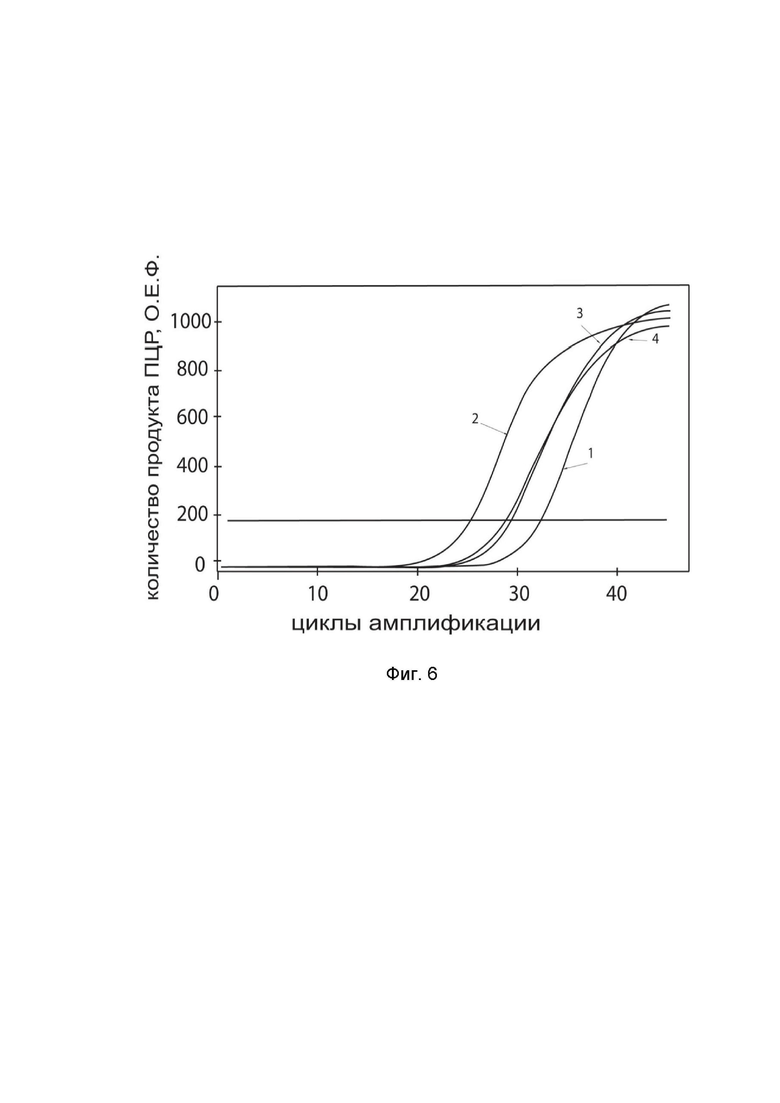
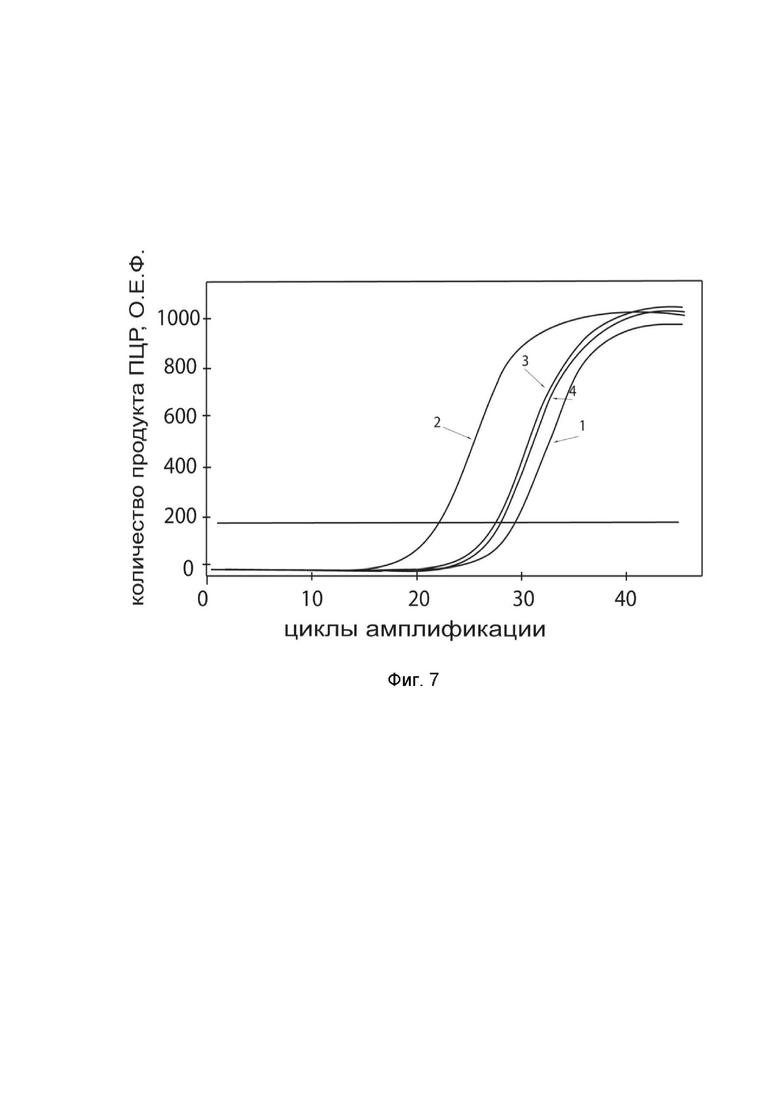
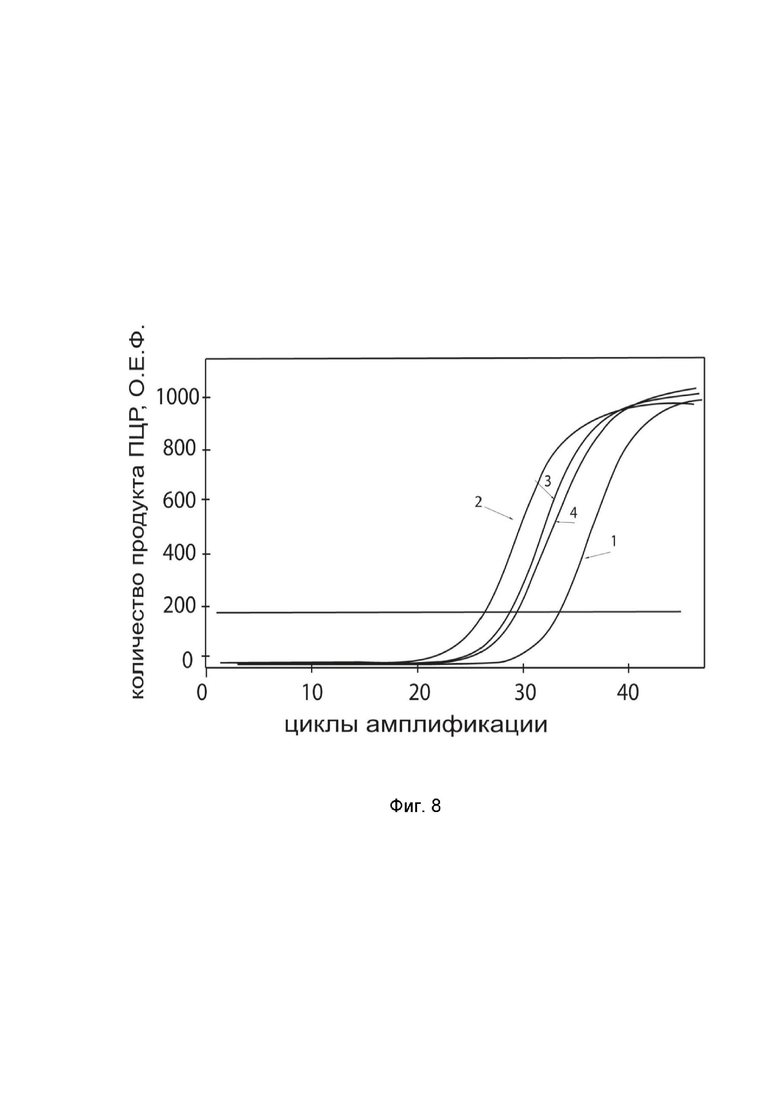
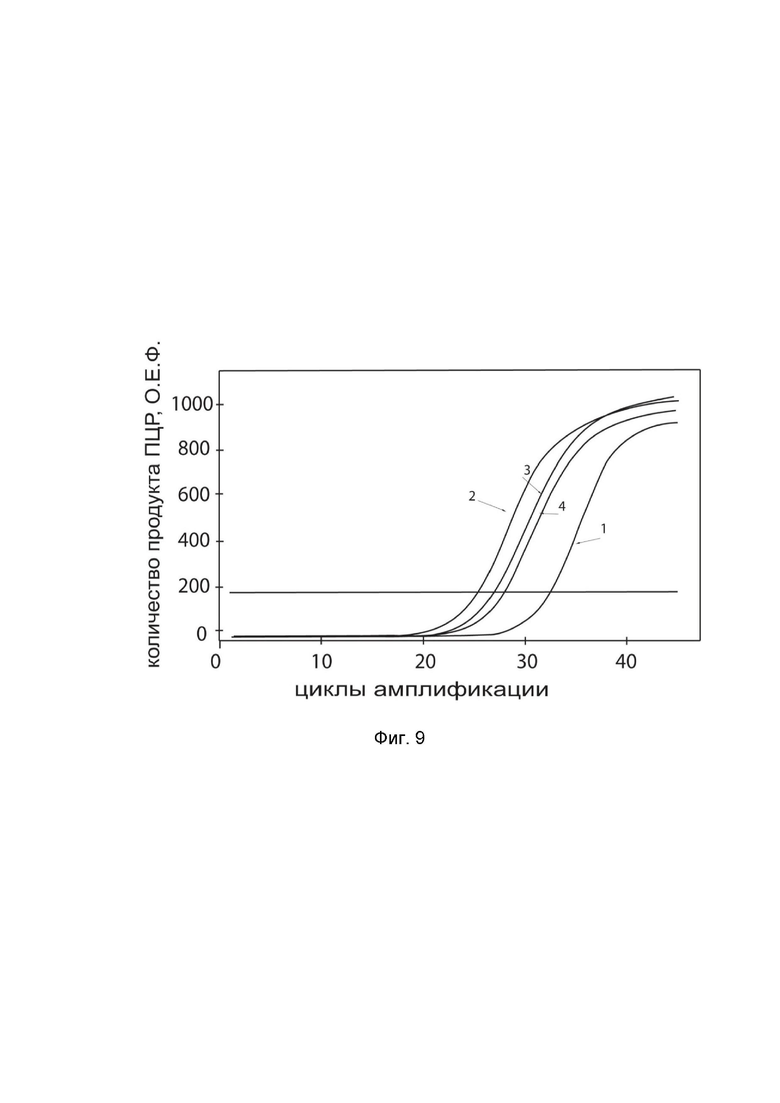
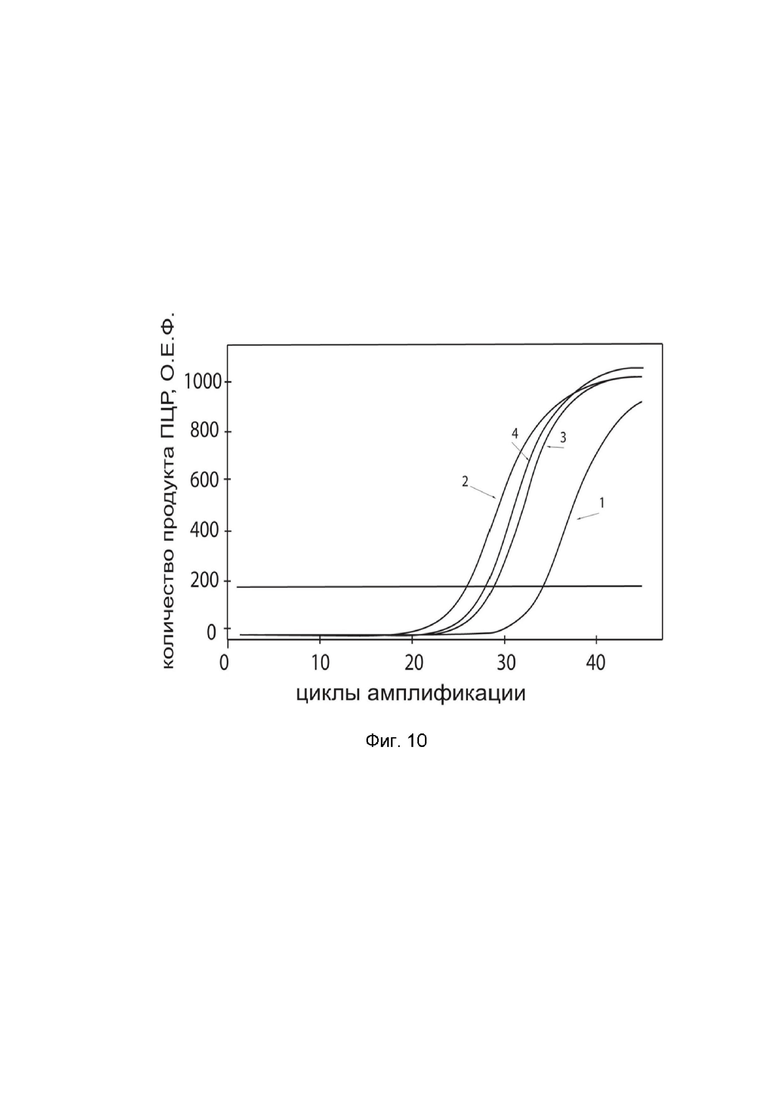
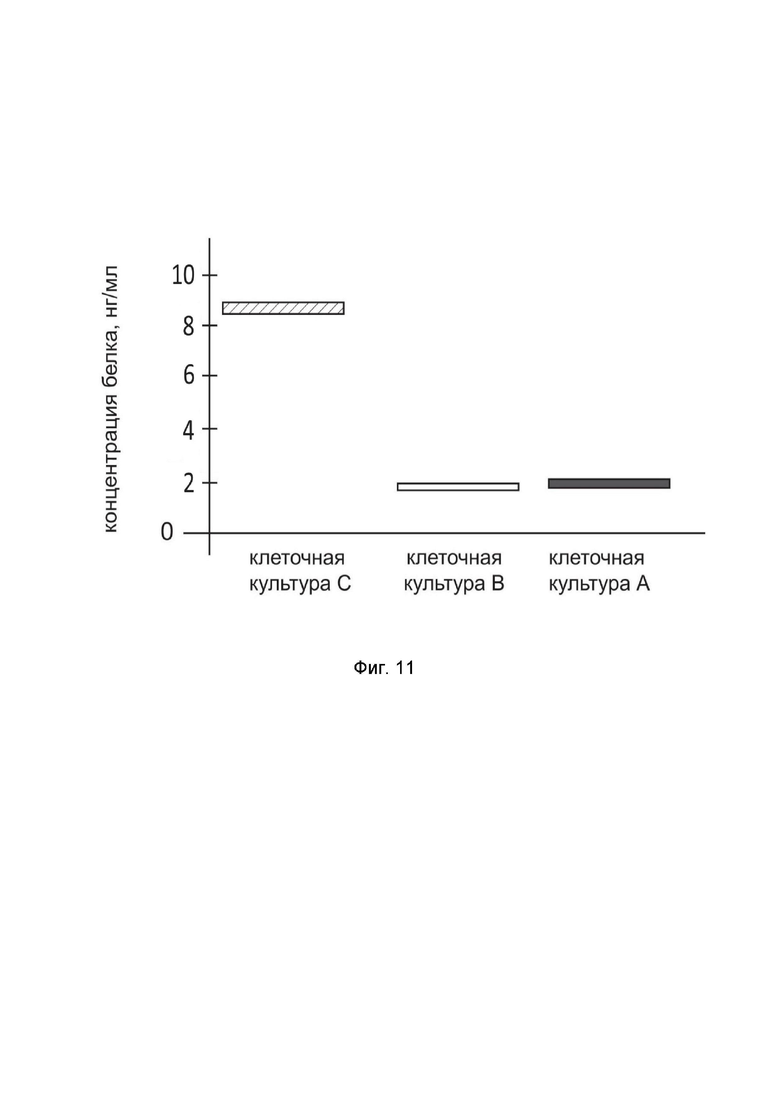
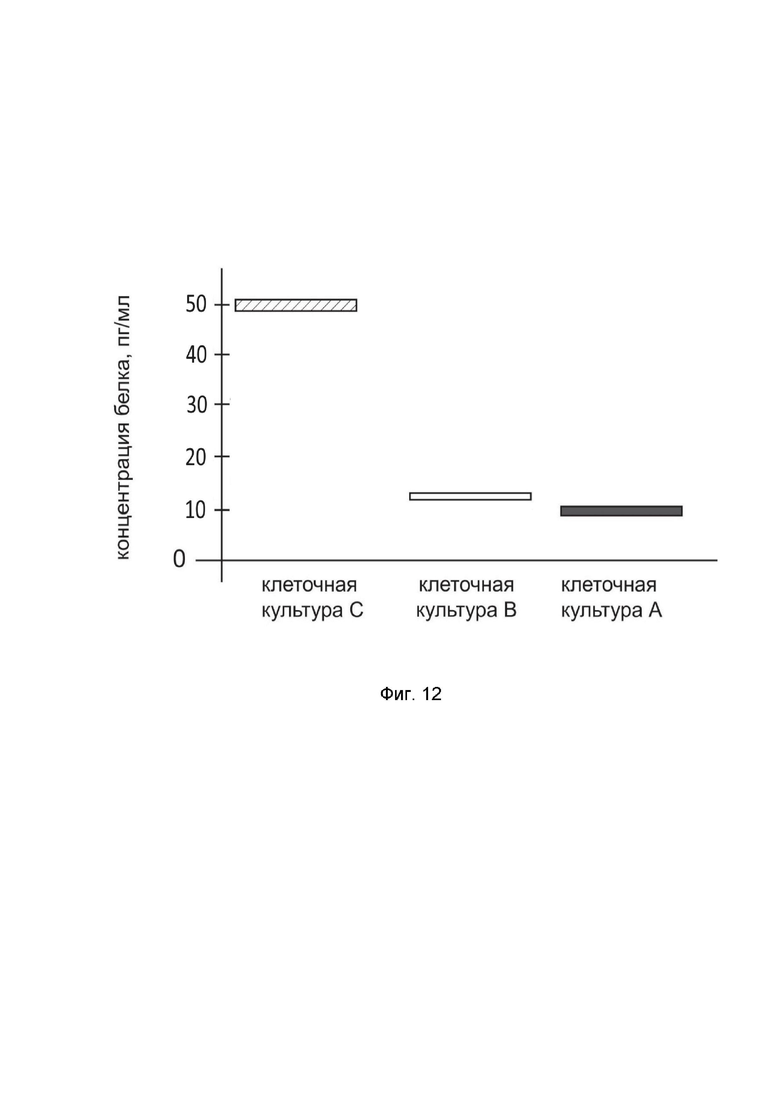
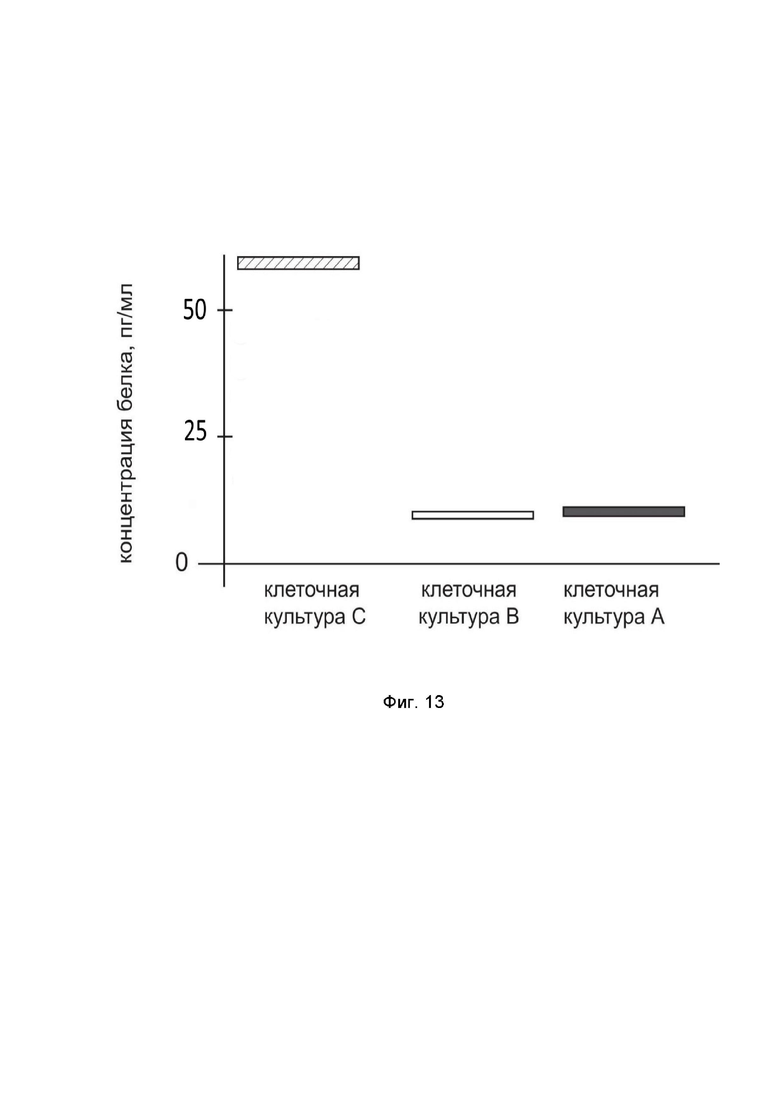
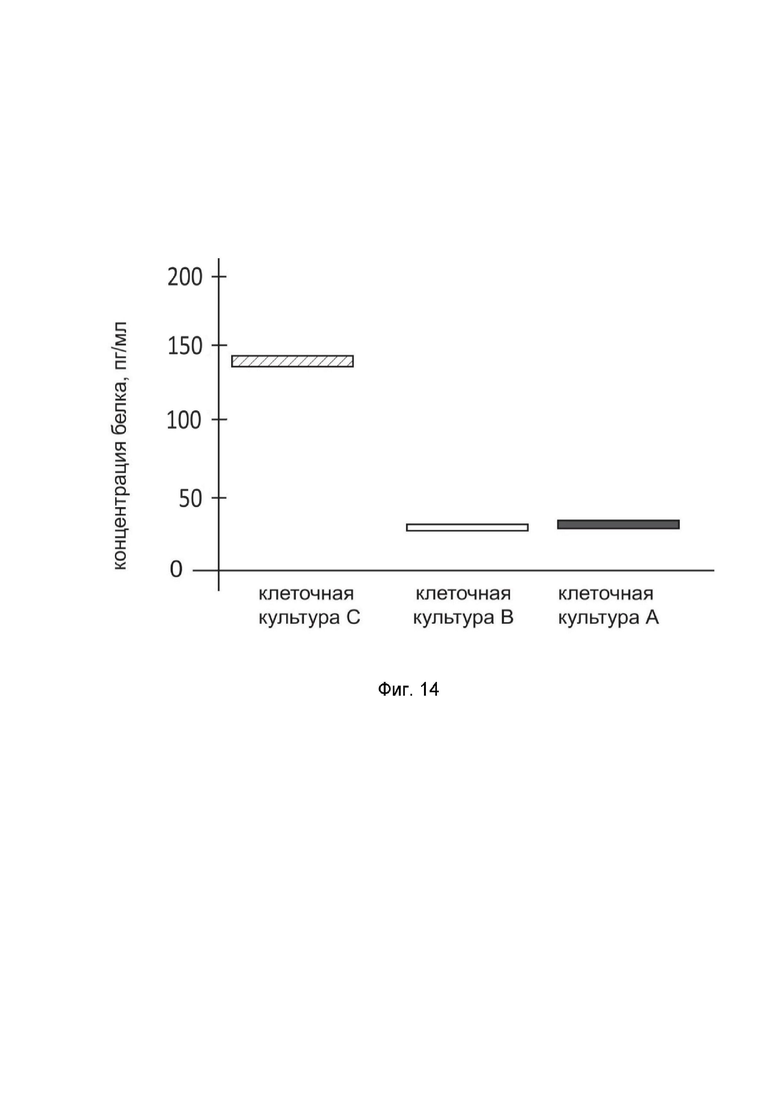
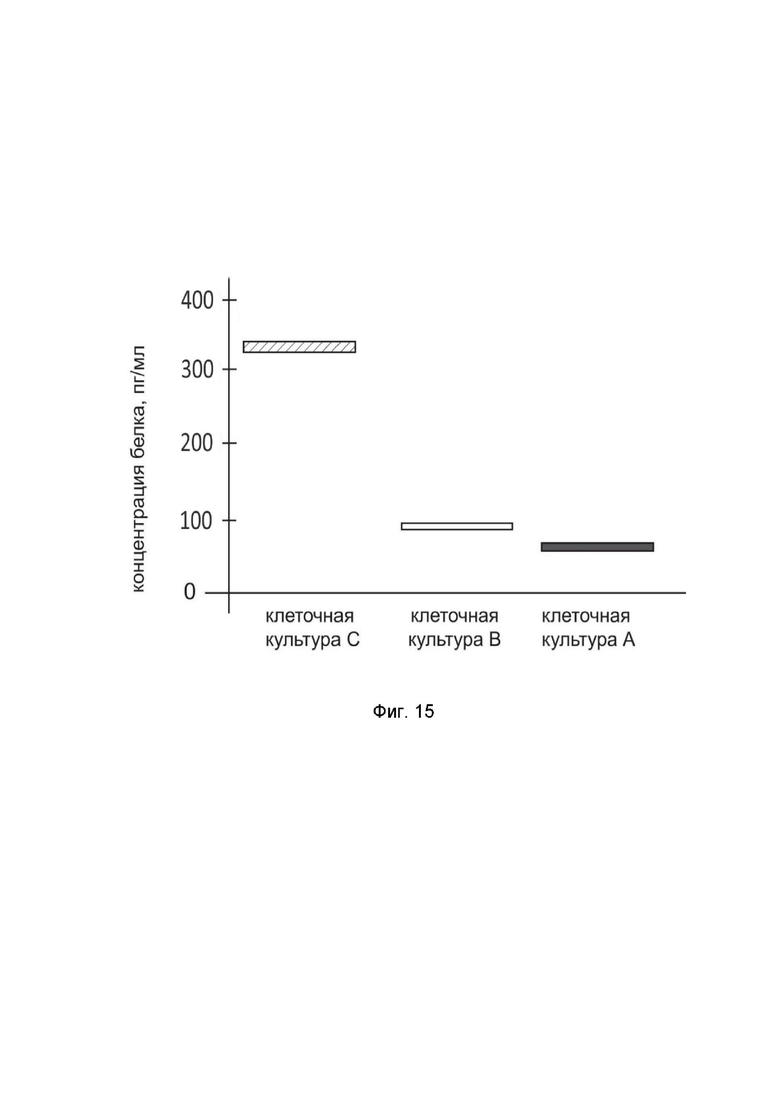
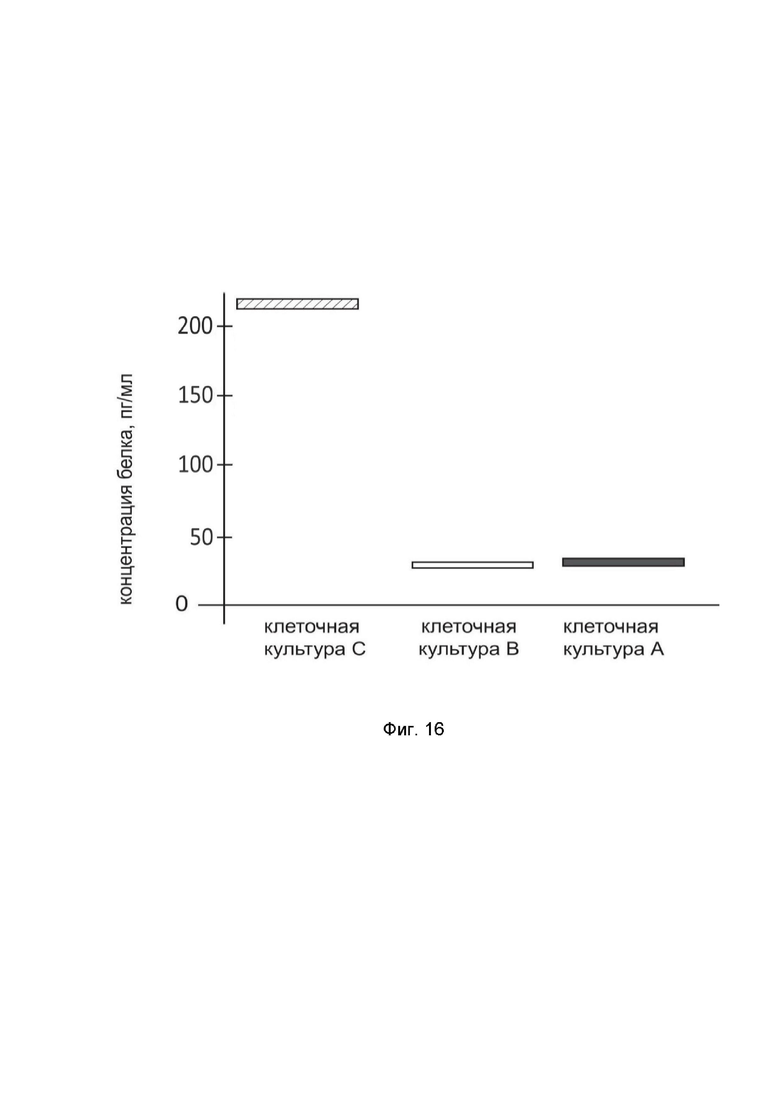
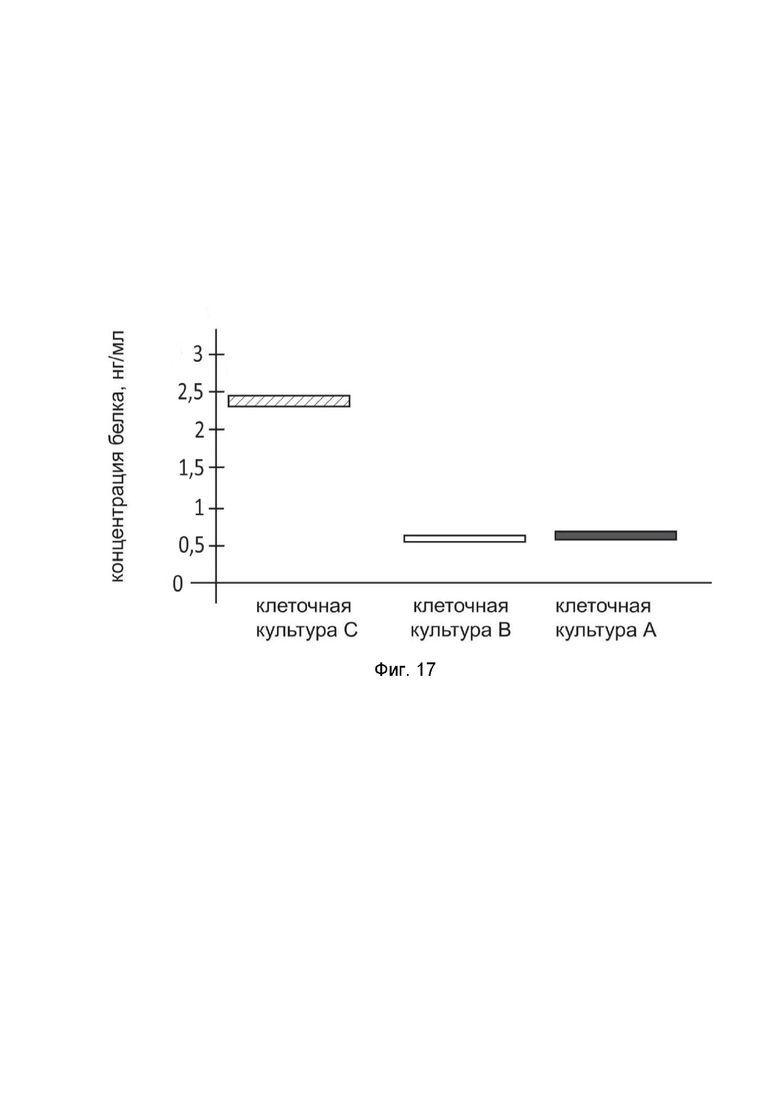
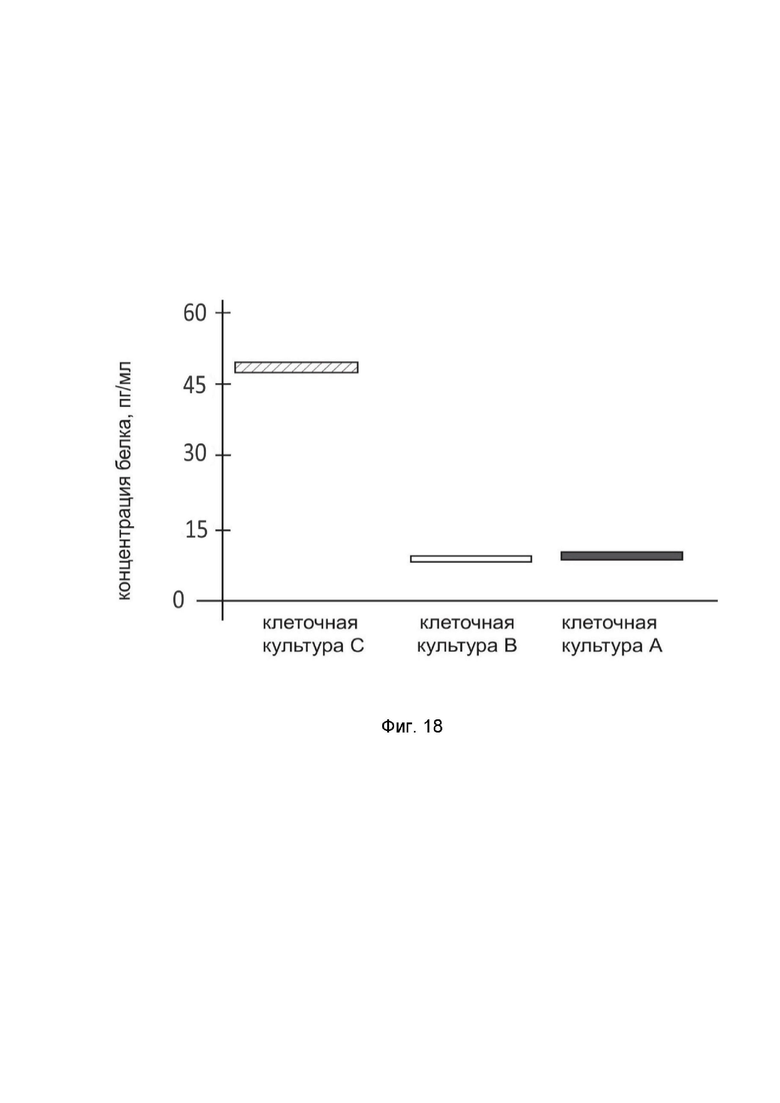
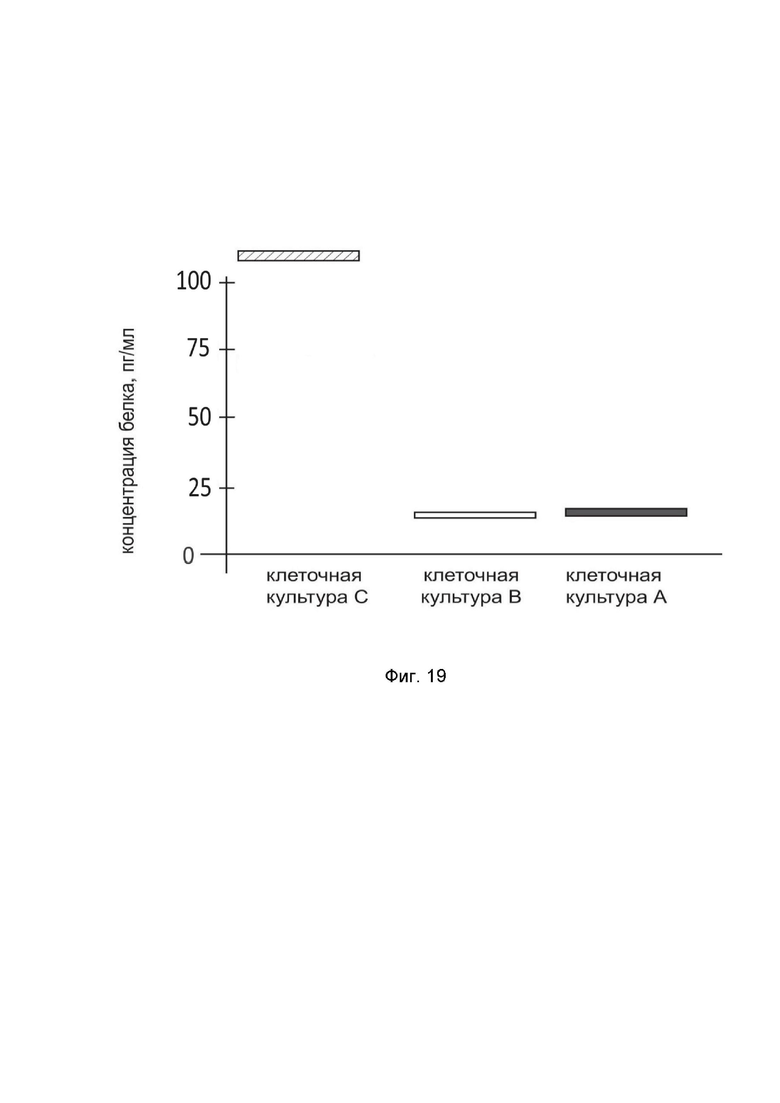
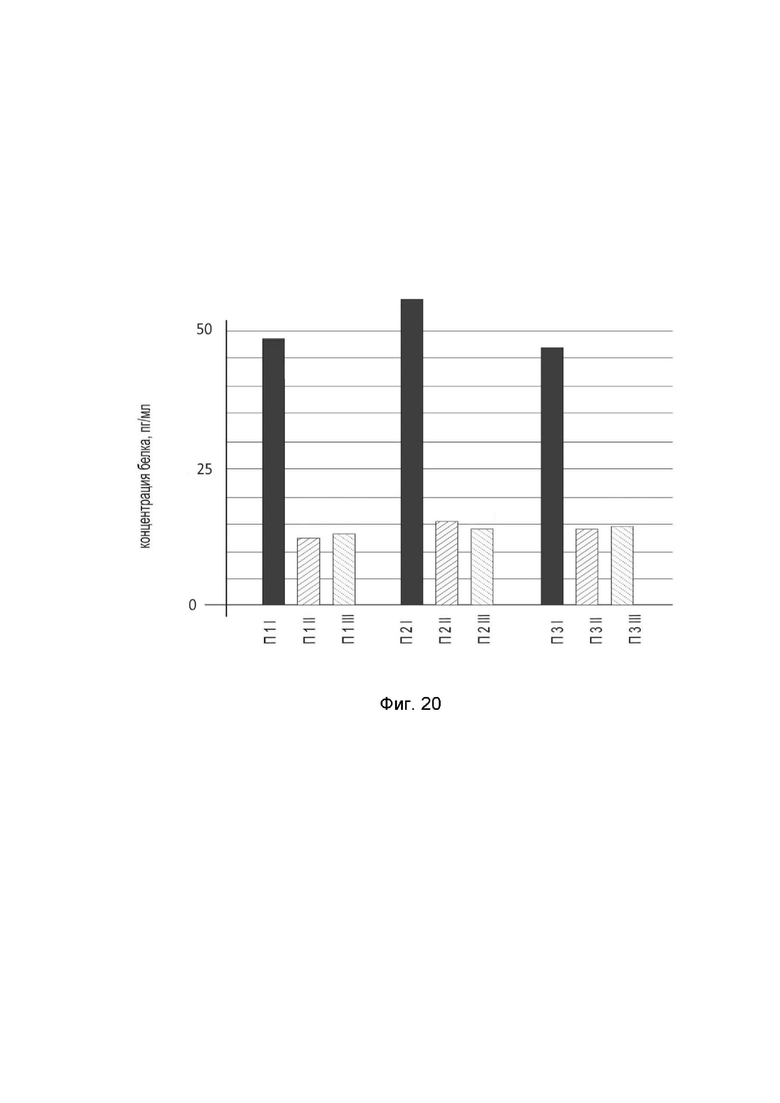
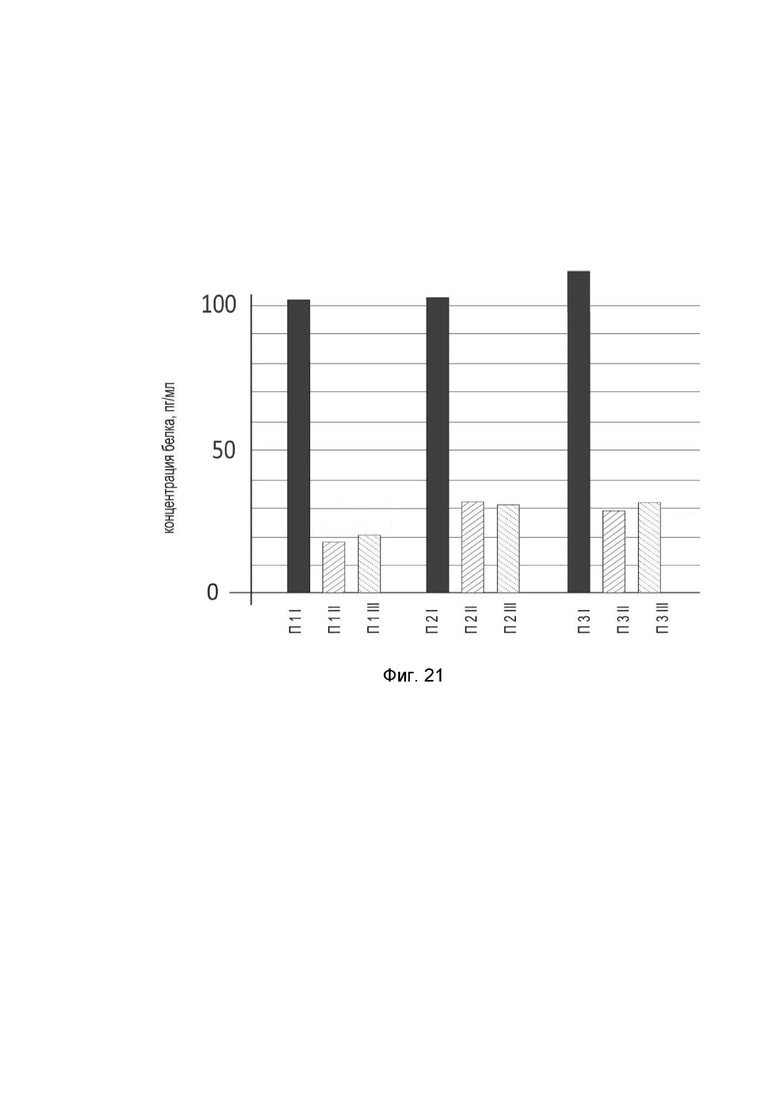
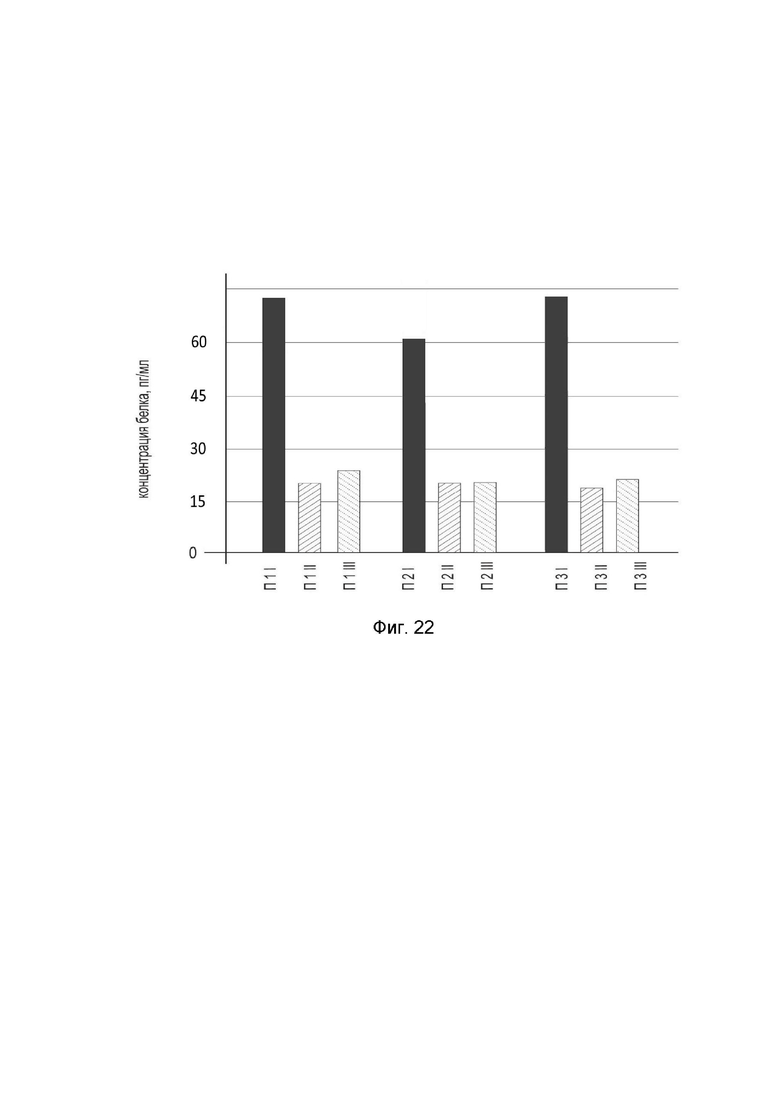
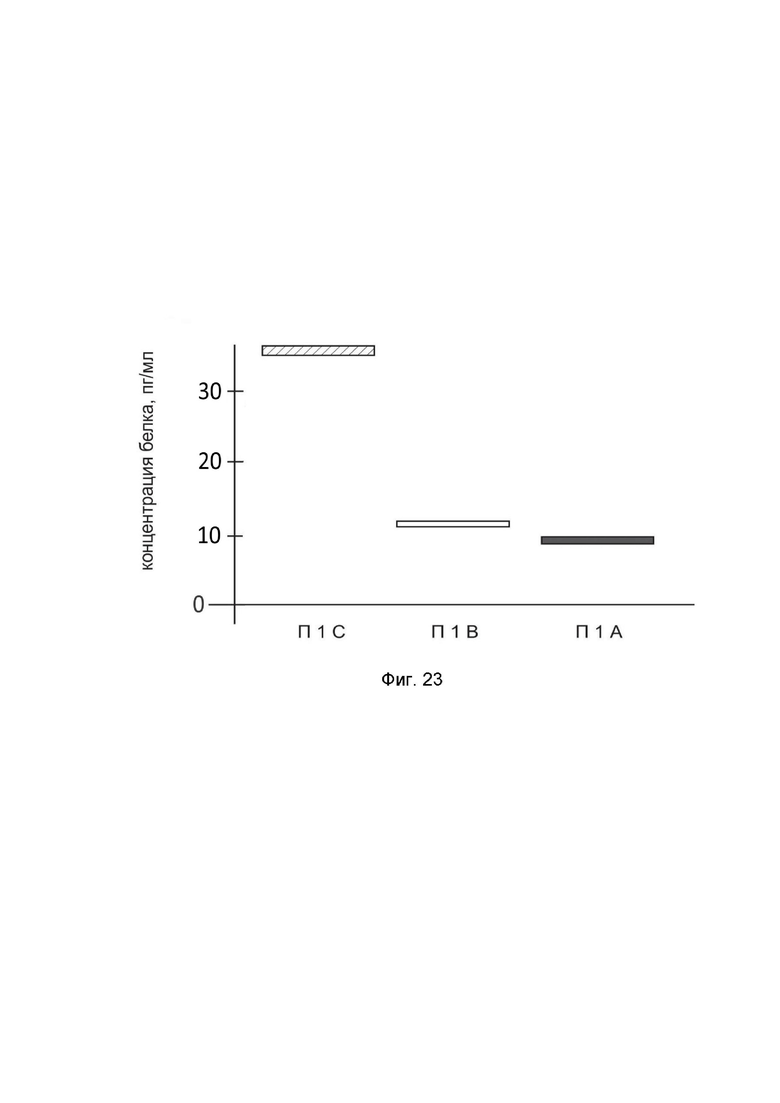
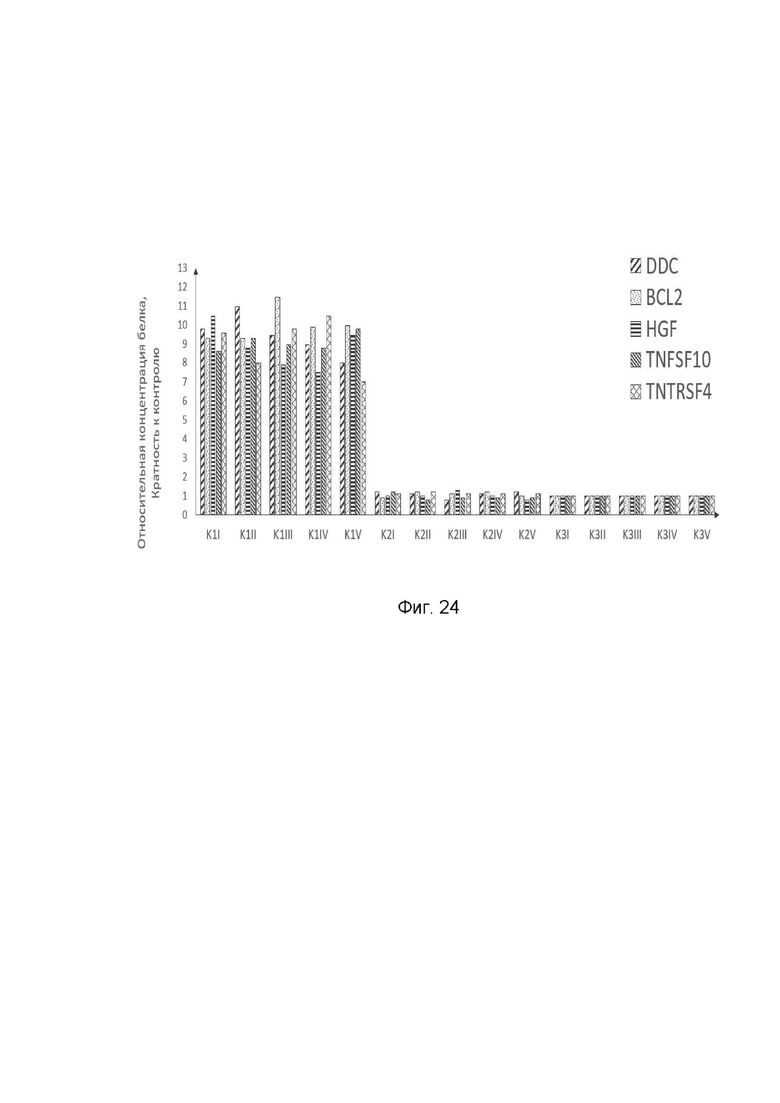
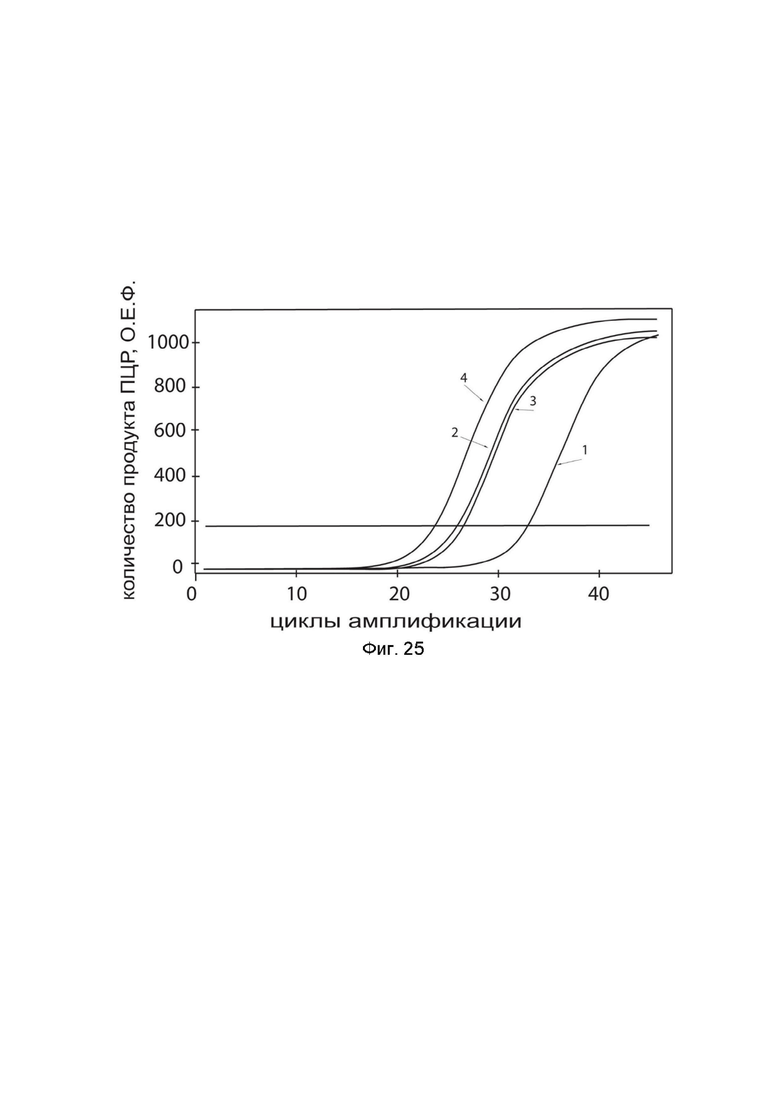
Изобретение относится к генной инженерии и может быть использовано в биотехнологии, медицине и сельском хозяйстве для создания препаратов генной терапии. Создан генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена DDC, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, с нуклеотидной последовательностью SEQ ID №1, или IL10, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-IL10, с нуклеотидной последовательностью SEQ ID №2, или IL13, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-IL13, с нуклеотидной последовательностью SEQ ID №3, или IFNB1, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, с нуклеотидной последовательностью SEQ ID №4, или TNFRSF4, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFRSF4, с нуклеотидной последовательностью SEQ ID №5, или TNFSF10, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFSF10, с нуклеотидной последовательностью SEQ ID №6, или BCL2, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-BCL2, с нуклеотидной последовательностью SEQ ID №7, или HGF, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-HGF, с нуклеотидной последовательностью SEQ ID №8, или IL2, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-IL2, с нуклеотидной последовательностью SEQ ID №9. 22 н. и 2 з.п. ф-лы, 25 ил., 35 пр.
1. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена DDC, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, с нуклеотидной последовательностью SEQ ID №1.
2. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена IL10, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-IL10, с нуклеотидной последовательностью SEQ ID №2.
3. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена IL13, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-IL13, с нуклеотидной последовательностью SEQ ID №3.
4. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена IFNB1, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, с нуклеотидной последовательностью SEQ ID №4.
5. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена TNFRSF4, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFRSF4, с нуклеотидной последовательностью SEQ ID №5.
6. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена TNFSF10, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFSF10, с нуклеотидной последовательностью SEQ ID №6.
7. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена BCL2, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-BCL2, с нуклеотидной последовательностью SEQ ID №7.
8. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена HGF, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-HGF, с нуклеотидной последовательностью SEQ ID №8.
9. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена IL2, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-IL2, с нуклеотидной последовательностью SEQ ID №9.
10. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущий целевой ген DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2 по пп.1-9 соответственно, отличающийся тем, что каждый из созданных генотерапевтических ДНК-векторов: GDTT1.8NAS12-DDC, или GDTT1.8NAS12-IL10, или GDTT1.8NAS12-IL13, или GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-TNFRSF4, или GDTT1.8NAS12-TNFSF10, или GDTT1.8NAS12-BCL2, или GDTT1.8NAS12-HGF, или GDTT1.8NAS12-IL2 по пп.1-9 соответственно за счет ограниченного размера векторной части GDTT1.8NAS12, не превышающей 2600 п.н., обладает способностью эффективно проникать в клетки человека и животных и экспрессировать клонированный в него целевой ген DDC, или IL10, или IL13, или IFNB1, или TNFRSF4, или TNFSF10, или BCL2, или HGF, или IL2.
11. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущий целевой ген DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2 по пп.1-9 соответственно, отличающийся тем, что в составе каждого из созданных генотерапевтических ДНК-векторов: GDTT1.8NAS12-DDC, или GDTT1.8NAS12-IL10, или GDTT1.8NAS12-IL13, или GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-TNFRSF4, или GDTT1.8NAS12-TNFSF10, или GDTT1.8NAS12-BCL2, или GDTT1.8NAS12-HGF, или GDTT1.8NAS12-IL2 по пп.1-9 соответственно в качестве структурных элементов используются нуклеотидные последовательности, которые не являются генами антибиотикорезистентности, вирусными генами и регуляторными элементами вирусных геномов, обеспечивая возможность его безопасного применения для генетической терапии человека и животных.
12. Способ получения генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2 по пп.1-9 соответственно, заключающийся в том, что каждый из генотерапевтических ДНК-векторов: GDTT1.8NAS12-DDC, или GDTT1.8NAS12-IL10, или GDTT1.8NAS12-IL13, или GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-TNFRSF4, или GDTT1.8NAS12-TNFSF10, или GDTT1.8NAS12-BCL2, или GDTT1.8NAS12-HGF, или GDTT1.8NAS12-IL2 получают следующим образом: кодирующую часть целевого гена DDC, или IL10, или IL13, или IFNB1, или TNFRSF4, или TNFSF10, или BCL2, или HGF, или IL2 клонируют в генотерапевтический ДНК-вектор GDTT1.8NAS12 и получают генотерапевтический ДНК-вектор GDTT1.8NAS12-DDC, SEQ ID №1, или GDTT1.8NAS12-IL10, SEQ ID №2, или GDTT1.8NAS12-IL13, SEQ ID №3, или GDTT1.8NAS12-IFNB1, SEQ ID №4, или GDTT1.8NAS12-TNFRSF4, SEQ ID №5, или GDTT1.8NAS12-TNFSF10, SEQ ID №6, или GDTT1.8NAS12-BCL2, SEQ ID №7, или GDTT1.8NAS12-HGF, SEQ ID №8, или GDTT1.8NAS12-IL2, SEQ ID №9, соответственно, при этом кодирующую часть целевого гена DDC, или IL10, или IL13, или IFNB1, или TNFRSF4, или TNFSF10, или BCL2, или HGF, или IL2 получают путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции и ПЦР-амплификации с использованием созданных олигонуклеотидов и расщеплением продукта амплификации соответствующими эндонуклеазами рестрикции, причем клонирование в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят по сайтам рестрикции SalI и HindIII, или SalI и KpnI, или BamHI и SalI, причем селекцию проводят без антибиотиков, при этом при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-DDC, SEQ ID №1, для проведения реакции обратной транскрипции и ПЦР-амплификации в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
DDC-F ATCGTCGACCACCATGAACGCAAGTGAGTTCCGA,
DDC-R ACCAAGCTTCTACTCCCTCTCTG,
а расщепление продукта амплификации и клонирование кодирующей части гена DDC в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции SalI и HindIII,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-IL10, SEQ ID №2, для проведения реакции обратной транскрипции и ПЦР-амплификации в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
IL10-F AGGATCCACCATGCACAGCTCAGCACTGC,
IL10-R CTTGTCGACTCAGTTTCGTATCTTCATTGTC,
а расщепление продукта амплификации и клонирование кодирующей части гена IL10 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-IL13, SEQ ID №3, для проведения реакции обратной транскрипции и ПЦР-амплификации в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
IL13-F AGGATCCACCATGCATCCGCTCCTCAATC,
IL13-R CTTGTCGACTCAGTTGAACTGTCCCTCG,
а расщепление продукта амплификации и клонирование кодирующей части гена IL13 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, SEQ ID №4, для проведения реакции обратной транскрипции и ПЦР-амплификации в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
IFNB1-F AAAGGATCCACCATGACCAACAAGTGTCTCCTCCAAA,
IFNB1-R TTTGTCGACTCAGTTTCGGAGGTAACCTGTAAGTCTG,
а расщепление продукта амплификации и клонирование кодирующей части гена IFNB1 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFRSF4, SEQ ID №5, для проведения реакции обратной транскрипции и ПЦР-амплификации в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
TNFRSF4-F AGGATCCACCATGTGCGTGGGGGCTCGG,
TNFRSF4-R CTTGTCGACTCAGATCTTGGCCAGGGTG,
а расщепление продукта амплификации и клонирование кодирующей части гена TNFRSF4 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-TNFSF10, SEQ ID №6, для проведения реакции обратной транскрипции и ПЦР-амплификации в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
TNFSF10-F AGGATCCACCATGGCTATGATGGAGGTCCA,
TNFSF10-R TGTCGACTCAGCCAACTAAAAAGGCCCCGA,
а расщепление продукта амплификации и клонирование кодирующей части гена TNFSF10 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-BCL2, SEQ ID №7, для проведения реакции обратной транскрипции и ПЦР-амплификации в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
BCL-F ATCGTCGACCACCATGGCGCACGCTGGGAGA,
BCL-R TTCGGTACCTCACTTGTGGCCCA,
а расщепление продукта амплификации и клонирование кодирующей части гена BCL2 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции SalI и KpnI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-HGF, SEQ ID №8, для проведения реакции обратной транскрипции и ПЦР-амплификации в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
HGF_F TTTGGATCCACCATGTGGGTGACCAAACTCCTGCCA,
HGF_R AATGTCGACCTATGACTGTGGTACCTTATATGTTAAAAT,
а расщепление продукта амплификации и клонирование кодирующей части гена HGF в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-IL2, SEQ ID №9, для проведения реакции обратной транскрипции и ПЦР-амплификации в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
IL2-F AGGATCCACCATGTACAGGATGCAACTCCT,
IL2-R TGTCGACTCAAGTCAGTGTTGAGATGATG,
а расщепление продукта амплификации и клонирование кодирующей части гена IL2 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI.
13. Способ использования генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген DDC, IL10, IL13, IFNB1, TNFRSF4, TNFSF10, BCL2, HGF, IL2 по пп.1-9 соответственно, для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз, заключающийся в трансфекции выбранным генотерапевтическим ДНК-вектором, несущим целевой ген на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, или несколькими выбранными генотерапевтическими ДНК-векторами, несущими целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, из созданных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, клеток органов и тканей пациента или животного, и/или во введении в органы и ткани пациента или животного аутологичных клеток этого пациента или животного, трансфицированных выбранным генотерапевтическим ДНК-вектором, несущим целевой ген на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, или несколькими выбранными генотерапевтическими ДНК-векторами, несущими целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, из созданных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, и/или во введении в органы и ткани пациента или животного выбранного генотерапевтического ДНК-вектора, несущего целевой ген на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, или нескольких выбранных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, из созданных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, или в сочетании обозначенных способов.
14. Способ получения штамма для производства генотерапевтического ДНК-вектора по любому из пп.1-9 для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз, заключающийся в получении электрокомпетентных клеток штамма Escherichia coli JM110-NAS с последующим проведением электропорации этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS12-DDC, или генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL10, или генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL13, или генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNB1, или генотерапевтическим ДНК-вектором GDTT1.8NAS12-TNFRSF4, или генотерапевтическим ДНК-вектором GDTT1.8NAS12-TNFSF10, или генотерапевтическим ДНК-вектором GDTT1.8NAS12-BCL2, или генотерапевтическим ДНК-вектором GDTT1.8NAS12-HGF, или генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL2, после чего клетки высеивают на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола с получением в результате штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2.
15. Штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC, полученный способом по п.14, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-DDC, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз.
16. Штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10, полученный способом по п.14, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-IL10, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз.
17. Штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13, полученный способом по п.14, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-IL13, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз.
18. Штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, полученный способом по п.14, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз.
19. Штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4, полученный способом по п.14, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-TNFRSF4, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз.
20. Штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10, полученный способом по п.14, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-TNFSF10, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз.
21. Штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2, полученный способом по п.14, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-BCL2, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз.
22. Штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF, полученный способом по п.14, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-HGF, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз.
23. Штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2, полученный способом по п.14, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-IL2, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз.
24. Способ производства в промышленных масштабах генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген DDC, или IL10, или IL13, или IFNB1, или TNFRSF4, или TNFSF10, или BCL2, или HGF, или IL2 по пп.1-9 соответственно, для лечения заболеваний, характеризующихся прогрессирующим патологическим изменением структуры нервной ткани и функции нейронов, в том числе их гибелью, ассоциированным с генетическими факторами, в том числе с мутациями в генах, кодирующих критически значимые для нормального функционирования нейронов белки, включая болезнь Гентингтона, наследуемые формы бокового амиотрофического склероза, а также с нарушением сворачивания третичной структуры белков, включая болезнь Паркинсона, болезнь Альцгеймера, с травмами центральной нервной системы, с нарушением кислородного снабжения головного или спинного мозга, с отклонениями в энергетическом метаболизме нейронов и аксональном транспорте или с аутоиммунными демиелинизирующими процессами, включая рассеянный склероз, заключающийся в том, что генотерапевтический ДНК-вектор GDTT1.8NAS12-DDC, или генотерапевтический ДНК-вектор GDTT1.8NAS12-IL10, или генотерапевтический ДНК-вектор GDTT1.8NAS12-IL13, или генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1, или генотерапевтический ДНК-вектор GDTT1.8NAS12-TNFRSF4, или генотерапевтический ДНК-вектор GDTT1.8NAS12-TNFSF10, или генотерапевтический ДНК-вектор GDTT1.8NAS12-BCL2, или генотерапевтический ДНК-вектор GDTT1.8NAS12-HGF, или генотерапевтический ДНК-вектор GDTT1.8NAS12-IL2 получают путем того, что засевают в колбу с приготовленной средой затравочную культуру, выбранную из штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-DDC, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL10, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL13, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFRSF4, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-TNFSF10, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-BCL2, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-HGF, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL2, затем инкубируют затравочную среду в шейкере-инкубаторе и переносят ее в промышленный ферментер, после чего растят до достижения стационарной фазы, затем выделяют фракцию, содержащую целевой ДНК-продукт, многостадийно фильтруют и очищают хроматографическими методами.
| Генотерапевтический ДНК-вектор VTvaf17, способ его получения, штамм Escherichia coli SCS110-AF, способ его получения, штамм Escherichia coli SCS110-AF/VTvaf17, несущий генотерапевтический ДНК-вектор VTvaf17, способ его получения | 2017 |
|
RU2678756C1 |
| US 7598058 B2, 06.10.2009 | |||
| US 4808523 A1, 28.02.1989. | |||

