Область техники
Изобретение относится к генной инженерии и может быть использовано в биотехнологии, медицине и сельском хозяйстве для создания препаратов генной терапии.
Уровень техники
На сегодняшний день одним из перспективных методов лечения наследственных и приобретенных заболеваний связан с использованием современных методов генной терапии, с помощью которых проводится доставка генетического материала в клетки пациента для коррекции дефекта/мутации, компенсации овер- или гипоэкспрессии гена(ов). В результате такой терапии в клетках запускается экспрессия или молекул РНК или белка, которые и проявляют терапевтический эффект, посредством регуляции белковой активности или активации/подавления экспрессии таргетных протеинов. Так или иначе конечной целью генетической терапии является регуляция на уровне трансляции или транскрипции метаболических или физиологических нарушений, связанных с повреждением или ненормальной активностью генетического аппарата клетки. В предлагаемом изобретении под экспрессией гена подразумевается продукция молекулы белка с определенной аминокислотной последовательностью, определяемой нуклеотидной последовательностью этого гена.
Ряд белков, являющихся модуляторами иммунной системы - IFNB1, IFNA14, IL12A, IL12B, IFNA2, имеют общее название – цитокины. Гены, кодирующие данные белки, играют важную роль во множестве как нормальных, так и патологических процессов у человека и животных. На сегодняшний день продемонстрирована корреляция между различными патологическими состояниями у человека с нарушением уровня экспрессии данных цитокинов, что делает их потенциальными мишенями для генетической терапии с целью коррекции.
Одни из цитокинов - интерфероны I типа проявляют противовирусную и противопухолевую активность и являются важнейшим компонентом врожденного иммунитета, а также вовлечены в дифференцировку клеток. Контакт с патогенами приводит к секреции интерферона I типа, который связывается с соотвествующим рецепторным комплексом, что приводит к активации транскрипции генов хемокинов и провоспалительных цитокинов. У нокаутных мышей по этому интерферону наблюдается нарушение созревание B клеточного звена иммунитета и повышается восприимчивость к вирусной инфекции.
К интерферонам I типа относятся две группы белков - IFN-α и β, которые обладают множеством иммунорегуляторных функций, обеспечивающих связь между врожденными и адаптивными иммунными ответами. Показано участие IFN-α / β в индукции экспрессии MHC класса I и активации NK клеток, а также регуляции экспрессии других цитокинов.
Исходя из данных современной мировой литературы, известен следующий механизм действия интерферонов I типа: при контролируемой вирусной инфекции, не сильно выраженных и опосредованных индукторов функционирование интерферонов I типа носит не системный, а локальный эффект. В случае активации другого провоспалительного цитокина - IL-12, наряду с IFN-α / β, продуцируется IFN-γ CD4 Т клетками, в том числе и, при экспозиции антигенов в составе MHC I CD8 T-клеткок. При распространенной вирусной инфекции и высоких уровнях секреции IFN-α / β значительно повышается цитотоксичность NK-клеток и с превалированием функционирования CD8-Т-клеточного звена.
Исходя из данных литературы, противовирусные свойства IFNB1 позволяют контролировать репликацию ретровирусов, включая ВИЧ. Использование иммунодефицитных животных моделей подтверждает данное положение. Так было показано, что инфекция ВИЧ мононуклеарных клеток пациентов с уже экспрессирующимся геном IFNB1, приводила к подавлению репликации ВИЧ по сравнению с контрольными клетками. При этом, трансплантация данных клеток мышам супрессировала репликацию ВИЧ до недетектируемого стандартными методами уровня у 40% животных (Vieillard V et al., 1999). Подобные данные были получены в работе на приматах (Gay W et al., 2004).
Противоопухолевая активность IFNB1 была показана в экспериментах с помощью трансфекции клеток глиомы человека IFNB1 экспрессирующим плазмидным вектором, что приводило эффективному подавлению пролиферативной активности клеток в культуре (Mizuno M et al., 1990). В другой работе с помощью трансфекции клеток меланомы вирусным вектором, экспрессирующим ген IFNB1, было показано значительное повышение эффективности цитостатика блеомицина in vitro, приводящего к почти полной элиминации опухолевых клеток (Fondello C et al., 2018). Противопухолевая активность IFNB1 модифицированных клеток также была показана на мышиной модели метастазирующей хориокарциномы (Kim GS et al., 2018).
Другие патологии, в которые вовлечен IFNB1 – это аутоимуннные заболевания, среди которых особое внимание уделяется рассеянному склерозу (РС). В настоящий момент терапия РС IFNB1 является основной (Rebif, Betaseron, Avonex и др.). Использование геннотерапевтических конструкций с геном IFNB1 представляется перспективным подходом в терапии РС (Hamana A et al., 2018).
Экспрессия IFNB1 в нервной ткани также играет роль и в патогенезе болезни Паркинсона. Результаты эксперимента на мышах с экспериментальной моделью болезни Паркинсона, которым вводили лентивирусный вектор с геном IFNB1, тормозило развитие патологии (Ejlerskov P et al., 2015).
Ряд генно-инженерных продуктов уже находятся на начальных фазах клинических испытаний. Так, уже на первой стадии КИ аденоассоциированный вирусный вектор, экспрессирующий ген IFNB1, который вводится внутрь сустава пациентам с ревматоидным артритом (NCT02727764).
Другие гены IFNA14, IFNA2 отвечают за экспрессию цитокинов IFN-α подтипов 14 и 2, относящиеся также к интерферонам I типа, которые связываются с одним и тем же рецептором интерферона I типа (IFNAR), при этом, афинность к нему внутри этой группы интереферонов различается, что связывают с индукцией различных сигнальных путей. Эффективность разных подтипов интерферонов при инфекции HSV, MCMV и вируса гриппа у мышей различается. Было показано, что в случае заражения ВИЧ-1, IFN-α2 может играть роль самого патогенетически значимого подтипа. Хотя в данном исследовании анализировали всего шесть подтипов IFN-α с использованием имплементированного к лабораторным исследованиям штамма ВИЧ-1 (Sperber SJ et al., 1992). В другой работе было показано, что существует обратная зависимость между экспрессией подтипов IFN-α и эффективностью подавления in vitro репликации ВИЧ-1. Обнаружено, что под влиянием ВИЧ-инфекции наиболее распространенные подтипы, IFN-α2 и -α1 / 13 были менее эффективны по сравнению с IFN-α6, -α14 и -α8 (Harper et al., 2015). С помощью плазмидных векторов, экспрессирующих различные подтипы IFN-I, продемонстрирован наибольший по длительности подавляющий репликацию ВИЧ эффект у мышей, получавших плазмиды, экспрессирующие гены IFNB1, IFNA14 (Abraham S et al., 2016).
Таким образом, несмотря на противоречивые данные о сравнительной эффективности различных интерферонов I типа, каждый из подтипов может быть использован для терапии различных патологий иммунной системы.
Другой цитокин - IL-12, являющийся провоспалительным, экспрессируется дендритными клетками, макрофагами и В-клетками в ответ на инфекционные агенты или опухолевые антигены. Гены IL12A, IL12B кодируют соотвествующие субъединицы – p35 и p40. Его индукция стимулирует продукцию IFN-g Т-клетками и приводит к потенциированию презентации антигена антиген-презентирующими клетками, стимулируя, в свою очередь, дифференцировку TH1 клеток. Кроме этого, он может индуцировать экспрессию IFN-g NK-клетками. Клиническое проявление многих инфекций зависит от способности агента индуцировать продукцию IL-12. В частности, заражение Candida albicans активирует синтез IL-12, что способствует дальнейшей эффективной защите клеток от возбудителя. ВИЧ, наоборот подавляет экспрессию IL-I2, вызывая многочисленные нарушения клеточного иммуного звена. Аналогично ведут себы лейшмании, подавляя синтез IL-12, что способствует хронизации инфекции. Выборочное подавление продукции IL-12, при сохранении экспрессии других провоспалительных цитокинов, таких как IL-1, TNF-a, приводит к длительному персистированию возбудителей в организме хозяина (Okwor I, Uzonna JE., 2016). В этой связи, исследователи предложили генотерапевтический метод с использованием экспрессирующих IL-12 плазмидных векторов, для получения протективного эффекта при поражении лейшманиями и трипаносомами (Sakai T et al., 2000). IL-12 опосредованная стимуляция Th1 позволила использовать его в качестве вакцины в составе геннотерапевтических конструкций для иммуномодулирующего эффекта (Tapia E et al., 2003).
При ряде патологий – таких как рассеянный склероз, системная красная волчанка, первичный билиарный цирроз, болезнь Бехчета, целиакия, синдром Шегрена найдены ассоциированые с генами IL12A, IL12B мутации. Противоопухолевое действие IL-12 при геннотерапевтическом подходе стимулирующее дендритные клетки и, как следствие, восстанавливающее противоопухолевый иммунитет, изучено в ряде работ (Razi Soofiyani S et al., 2016). Denies и савт. показали противоопухолевое действие с использованием плазмидного вектора, экспрессирующего IL-12, в комбинации с противоопухолевой адьювантной вакцинацией (Denies S et al., 2014).
Благодаря тому, что субъединица р40 входит в состав ряда других цитокинов из семейства IL-12, модуляция экспрессии гена IL12B с помощью генотерапевтического подхода дает потенциальные возможности влиять на патологии, связанные с недостаточной экспрессией таких цитокинов как IL-23, IL-27 (Waldner MJ, Neurath MF, 2009).
Таким образом предшествующий уровень техники свидетельствует о том, гены IFNB1, IFNA14, IFNA2, IL12A, IL12B обладают потенциалом для коррекции ряда отклонений, включающих в себя, но не ограничивающихся, инфекционными, аутоиммунными заболеваниями, раком, наследственными и приобретенными патологическими состояниями, а также необходимостью модуляции иммунного ответа. Этим обусловлено объединение генов IFNB1, IFNA14, IFNA2, IL12A, IL12B в рамках данного патента в группу генов. Генетические конструкции, обеспечивающие экспрессию белков, кодируемых генами из группы IFNB1, IFNA14, IFNA2, IL12A, IL12B могут быть использованы для разработки лекарственных препаратов для предотвращения и терапии различных заболеваний и патологических состояний.
Более того, приведенные данные свидетельствуют о том, что недостаточная экспрессия белков, кодируемых генами IFNB1, IFNA14, IFNA2, IL12A, IL12B входящими в группу генов, связана не только с патологическими состояниями, но и с предрасположенность к их развитию. Также приведенные данные свидетельствуют о том, что недостаточная экспрессия данных белков может не проявляться в явном виде в форме патологии, которая может быть однозначно описана в рамках существующих стандартов клинической практики (например, с применением кода МКБ), однако при этом вызывать состояния, которые неблагоприятны для человека и животных и связанны с ухудшением качества жизни.
Анализ подходов для повышения экспрессии целевых генов подразумевает возможность использования различных генотерапевтических векторов.
Генотерапевтические векторы разделяют на вирусные, клеточные и ДНК-векторы (Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal Products EMA/CAT/80183/2014). В последнее время в генной терапии всё большее внимание уделяется разработке невирусных систем доставки генетического материала, среди которых лидируют плазмидные векторы. Плазмидные векторы лишены недостатков, присущих клеточным и вирусным векторам. В клетке-мишени они существуют в эписомальной форме, не интегрируют в геном, производство их достаточно дешево, отсутствие иммунного ответа и побочных реакций на введение плазмидного вектора делают их удобным инструментом генной терапии и генетической профилактики (ДНК-вакцины) (Li L, Petrovsky N. // Expert Rev Vaccines. 2016;15(3):313-29).
Тем не менее, ограничениями для использования плазмидных векторов для генной терапии являются: 1) наличие генов устойчивости к антибиотикам для наработки в бактериальных штаммах, 2) наличие различных регуляторных элементов, представленных последовательностями вирусных геномов 3) размер терапевтического плазмидного вектора, определяющий эффективность проникновения вектора в клетку-мишень.
Известно, что Европейское агентство по лекарственным средствам считает необходимым избегать введения маркеров антибиотикорезистентности в разрабатываемые плазмидные векторы для генной терапии (Reflection paper on design modifications of gene therapy medicinal products during development / 14 December 2011 EMA/CAT/GTWP/44236/2009 Committee for advanced therapies). Данная рекомендация связана, в первую очередь, с потенциальной опасностью проникновения ДНК-вектора или горизонтального переноса генов антибиотикорезистентности в клетки бактерий, представленных в организме в составе нормальной или оппортунистической микрофлоры. Помимо этого, наличие генов антибиотикорезистентности значительно увеличивает размер ДНК-вектора, что приводит к снижению эффективности его проникновения в эукариотические клетки.
Необходимо отметить, что гены антибиотикорезистентности также вносят принципиальный вклад в способ получения ДНК-векторов. В случае наличия генов антибиотикорезистентности штаммы для наработки ДНК-векторов обычно культивируются в среде, содержащей селективный антибиотик, что создает риск наличия следовых количеств антибиотика в недостаточно очищенных препаратах ДНК-векторов. Таким образом, получение ДНК-векторов для генной терапии, в которых отсутствуют гены антибиотикорезистентности, связано с получением штаммов, обладающих такой отличительной особенностью как способность к стабильной амплификации целевых ДНК-векторов в среде без содержания антибиотиков.
Кроме того, Европейское Медицинское Агентство рекомендует избегать наличия в составе терапевтических плазмидных векторов регуляторных элементов для повышения экспрессии целевых генов (промоторов, энхансеров, посттрансляционных регуляторных элементов), являющихся нуклеотидными последовательностями геномов различных вирусов (Draft Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal products,http://www.ema.europa.eu/docs/en_GB/document_library/Scientific_guideline/2015/05/WC500187020.pdf). Данные последовательности, хотя и могут увеличивать уровень экспрессии целевого трансгена, однако создают риск рекомбинации с генетическим материалом вирусов дикого типа и интеграции в геном эукариотической клетки. Более того, целесообразность гиперэкспрессии того или иного гена в целях терапии остается нерешенным вопросом.
Также, существенным моментом является размер терапевтического вектора. Известно, что современные плазмидные векторы зачастую перегружены нефункциональными участками, серьезно увеличивающими размер вектора (Mairhofer J, Grabherr R. // Mol Biotechnol. 2008.39(2):97-104). Например, ген устойчивости к ампициллину в векторах серии pBR322, как правило, состоит из не менее чем 1000 п.н., что составляет более 20% от размера самого вектора. При этом наблюдается обратная зависимость между размером вектора и его способностью проникать в эукариотические клетки – ДНК-векторы с небольшим размером эффективней проникаю в клетки человека и животных. Так, например, в серии экспериментов по трансфекции клеток HELA ДНК-векторами с размером от 383 до 4548 п.н. было показано, что разница в эффективности проникновения может достигать двух порядков (отличаться в 100 раз) (Hornstein BD et al. // PLoS ONE. 2016;11(12): e0167537.).
Таким образом при выборе ДНК-вектора в целях безопасности и наибольшей эффективности следует отдавать предпочтение тем конструкциям, в которых не содержатся гены устойчивости к антибиотикам, последовательности вирусного происхождения и размер которых позволяет эффективно проникать в эукариотические клетки. Штамм для получения такого ДНК-вектора в количествах, достаточных для целей генной терапии, должен обеспечивать возможность стабильной амплификации ДНК-вектора с использованием питательных сред, не содержащих антибиотики.
Примером использования рекомбинантных ДНК-векторов для генной терапии является способ получения рекомбинантного вектора для генетической иммунизации по патенту US 9550998 В2. Вектор представляет собой суперскрученный плазмидный ДНК-вектор и предназначен для экспрессии клонированных генов в клетках животных и человека. Вектор состоит из ориджина репликации, регуляторных элементов, включающих промотор и энхансер цитомегаловируса человека, регуляторные элементы из Т-лимфотропного вируса человека.
Накопление вектора проводят в специальном штамме E. coli без использования антибиотиков за счет антисенс-комплементации гена sacB, введенного в штамм посредством бактериофага. Недостатком данного изобретения является наличие в составе ДНК-вектора регуляторных элементов, представляющих собой последовательности вирусных геномов.
Прототипами настоящего изобретения в части использования генотерапевтических подходов для повышения уровня экспрессии генов из группы IFNB1, IFNA14, IFNA2, IL12A, IL12B являются следующие заявки.
В патенте US4808523A описан плазмидный вектор, несущий ген, кодирующий IFNB1. Данный вектор позволяет повышать экспрессию IFNB1 в клетках млекопитающих. Недостатком данного изобретения является способ использования, ограниченный продукцией белка IFNB1 в условиях in vitro и отличный от генотерапевтического использования данного вектора. Также недостатком данного изобретения является наличие в составе вектора последовательностей вирусного происхождения.
В патенте RU2678756 описан генотерапевтический ДНК-вектор VTvaf17, способ его получения, штамм Escherichia coli SCS110-AF, способ его получения, штамм Escherichia coli SCS110-AF/VTvaf17, несущий генотерапевтический ДНК-вектор VTvaf17, способ его получения. Недостатком данного изобретения является значительный размер векторной части, что может негативно отразиться на эффективности доставки ДНК-вектора в клетки.
В заявке US20040203118A1 описаны полинуклеотиды, кодирующие различные варианты IFNA14, а также терапевтическое средство, представляющее собой вектор, экспрессирующий различные варианты IFNA14. Недостатком данного изобретения является неопределенность целей использования данного изобретения и неопределенные требования к безопасности используемых векторов.
В заявке CN101304758A описана нуклеотидная последовательность, кодирующая IFNA2, а также вектор, экспрессирующий данную последовательность для терапии различных заболеваний, связанных с нарушением экспрессии этого гена. Недостатком данного изобретения является неопределенность требований к безопасности используемого вектора.
В патенте US5723127A описан способ использования белка IL12 для терапевтической иммуномодуляции, в частности в качестве адъюванта. Недостатком данного изобретения является использование белка IL12 вместо генотерапевтического вектора, экспрессирующего ген, кодирующий белок IL12.
Раскрытие изобретения
Задачей изобретения является конструирование генотерапевтических ДНК-векторов для повышения уровня экспрессии группы генов IFNB1, IFNA14, IFNA2, IL12A, IL12B в организме человека и животных, сочетающих в себе следующие свойства:
I) Эффективность генотерапевтического ДНК-вектора для повышения уровня экспрессии целевых генов в эукариотических клетках.
II) Возможность безопасного применения для генетической терапии человека и животных за счет отсутствия в составе генотерапевтического ДНК-вектора регуляторных элементов, представляющих собой нуклеотидные последовательности вирусных геномов.
III) Возможность безопасного применения для генетической терапии человека и животных за счет отсутствия в составе генотерапевтического ДНК-вектора генов антибиотикорезистентности.
IV) Технологичность получения и возможность наработки генотерапевтического ДНК-вектора в промышленных масштабах.
Пункты II и III предусмотрены в данном техническом решении в соответствии с рекомендациями государственных регуляторов к лекарственным средствам для генной терапии, в частности, Европейского Агентства по лекарственным средствам касательно отказа от введения маркеров антибиотикорезистентности в разрабатываемые плазмидные векторы для генной терапии (Reflection paper on design modifications of gene therapy medicinal products during development / 14 December 2011, EMA/CAT/GTWP/44236/2009 Committee for advanced therapies) и касательно отказа от введения в разрабатываемые плазмидные векторы для генной терапии элементов вирусных геномов (Guideline on the quality, non-clinical and clinical aspects of gene therapy medicinal products / 23 March 2015, EMA/CAT/80183/2014, Committee for Advanced Therapies).
Задачей изобретения также является конструирование штаммов, несущих эти генотерапевтические ДНК-вектора, для наработки и производства в промышленных масштабах генотерапевтических ДНК-векторов.
Поставленная задача решается за счет того, что создан генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 для лечения заболеваний, характеризующихся нарушением врожденного и адаптивного иммунитета, для терапии аутоиммунных, онкологических, вирусных заболеваний, связанных с дисбалансом иммунной системы, для иммуномодуляции в том числе при противоопухолевой вакцинации в качестве генотерапевтического адъюванта, при этом генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1 содержит кодирующую часть целевого гена IFNB1, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с нуклеотидной последовательностью SEQ ID №1, генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNA14 содержит кодирующую часть целевого гена IFNA14, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с нуклеотидной последовательностью SEQ ID №2, генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNA2 содержит кодирующую часть целевого гена IFNA2, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с нуклеотидной последовательностью SEQ ID №3, генотерапевтический ДНК-вектор GDTT1.8NAS12-IL12A содержит кодирующую часть целевого гена IL12A, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с нуклеотидной последовательностью SEQ ID №4, генотерапевтический ДНК-вектор GDTT1.8NAS12-IL12B содержит кодирующую часть целевого гена IL12B, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с нуклеотидной последовательностью SEQ ID №5.
Каждый из созданных генотерапевтических ДНК-векторов: GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-IFNA14, или GDTT1.8NAS12-IFNA2, или GDTT1.8NAS12-IL12A, или GDTT1.8NAS12-IL12B за счет ограниченного размера векторной части GDTT1.8NAS12, не превышающей 2600 п.н., обладает способностью эффективно проникать в клетки человека и животных и экспрессировать клонированный в него целевой ген IFNB1, или IFNA14, или IFNA2, или IL12A, или IL12B.
В составе каждого из созданных генотерапевтических ДНК-векторов: GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-IFNA14, или GDTT1.8NAS12-IFNA2, или GDTT1.8NAS12-IL12A, или GDTT1.8NAS12-IL12B в качестве структурных элементов используются нуклеотидные последовательности, которые не являются генами антибиотикорезистентности, вирусными генами и регуляторными элементами вирусных геномов, обеспечивая возможность его безопасного применения для генетической терапии человека и животных.
Создан также способ получения генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IFNB1, IFNA14, IFNA2, IL12A, IL12B, заключающийся в том, что каждый из генотерапевтических ДНК-векторов: GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-IFNA14, или GDTT1.8NAS12-IFNA2, или GDTT1.8NAS12-IL12A, или GDTT1.8NAS12-IL12B получают следующим образом: кодирующую часть целевого гена IFNB1, или IFNA14, или IFNA2, или IL12A, или IL12B клонируют в генотерапевтический ДНК-вектор GDTT1.8NAS12 и получают генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1, SEQ ID №1, или GDTT1.8NAS12-IFNA14, SEQ ID №2 или GDTT1.8NAS12-IFNA2, SEQ ID №3, или GDTT1.8NAS12-IL12A, SEQ ID №4, или GDTT1.8NAS12-IL12B, SEQ ID №5 соответственно, при этом кодирующую часть целевого гена IFNB1, или IFNA14, или IFNA2, или IL12A, или IL12B получают путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции и ПЦР-амплификации с использованием созданных олигонуклеотидов и расщеплением продукта амплификации соответствующими эндонуклеазами рестрикции, причем клонирование в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят по сайтам рестрикции SalI и KpnI, или по сайтам рестрикции BamHI и SalI, причем селекцию проводят без антибиотиков,
при этом при получении генотерапевтического ДНК-вектора GDTT1.8NAS12- IFNB1, SEQID №1 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды
IFNB1-Fw AAAGGATCCACCATGACCAACAAGTGTCTCCTCCAAA
IFNB1-Rv TTTGTCGACTCAGTTTCGGAGGTAACCTGTAAGTCTG,
а расщепление продукта амплификации и клонирование кодирующей части гена IFNB1 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14, SEQID №2 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды
IFNA14-Fw TTTGGATCCACCATGGCATTGCCCTTTGCTTTAAT
IFNA14-Rv TTTGTCGACTCAATCCTTCCTCCTTAATCTTTTTTGC,
а расщепление продукта амплификации и клонирование кодирующей части гена IFNA14 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12- IFNA2, SEQID №3 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды
IFNA2-Fw GGGGGATCCACCATGGCCTTGACCTTTGCTTTACTGG
IFNA2-Rv TTTGTCGACTCATTCCTTACTTCTTAAACTTTCTTGC,
а расщепление продукта амплификации и клонирование кодирующей части гена IFNA2 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12A, SEQID №4 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды
IL12A-up TTTGTCGACCACCATGTGGCCCCCTGGGTCAGCCTCC
IL12A-lo AATGGTACCTTAGGAAGCATTCAGATAGCTCATCACTCTATC,
а расщепление продукта амплификации и клонирование кодирующей части гена IL12A в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции SalI и KpnI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12- IL12B, SEQID №5 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды
IL12B-up TTTGTCGACCACCATGTGTCACCAGCAGTTGGTCATCTC
IL12B-lo AATGGTACCTAACTGCAGGGCACAGATGCCCA,
а расщепление продукта амплификации и клонирование кодирующей части гена IL12B в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции SalI-KpnI.
Создан способ использования генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IFNB1, IFNA14, IFNA2, IL12A, IL12B для лечения заболеваний, характеризующихся нарушением врожденного и адаптивного иммунитета, для терапии аутоиммунных, онкологических, вирусных заболеваний, связанных с дисбалансом иммунной системы, для иммуномодуляции в том числе при противоопухолевой вакцинации в качестве генотерапевтического адъюванта, заключающийся в трансфекции выбранным генотерапевтическим ДНК-вектором, несущим целевой ген на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, или несколькими выбранными генотерапевтическими ДНК-векторами, несущими целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, из созданных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, клеток органов и тканей пациента или животного, и/или во введении в органы и ткани пациента или животного аутологичных клеток этого пациента или животного, трансфицированных выбранным генотерапевтическим ДНК-вектором, несущим целевой ген на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 или несколькими выбранными генотерапевтическими ДНК-векторами, несущими целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, из созданных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 и/или во введении в органы и ткани пациента или животного выбранного генотерапевтического ДНК-вектора, несущего целевой ген на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 или нескольких выбранных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, из созданных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, или в сочетании обозначенных способов.
Создан способ получения штамма для производства генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся нарушением врожденного и адаптивного иммунитета, для терапии аутоиммунных, онкологических, вирусных заболеваний, связанных с дисбалансом иммунной системы, для иммуномодуляции в том числе при противоопухолевой вакцинации в качестве генотерапевтического адъюванта, заключающийся в получении электрокомпетентных клеток штамма Escherichia coli JM 110-NAS с последующим проведением электропорации этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNB1, или ДНК-вектором GDTT1.8NAS12-IFNA14, или ДНК-вектором GDTT1.8NAS12- IFNA2, или ДНК-вектором GDTT1.8NAS12- IL12A, или ДНК-вектором GDTT1.8NAS12- IL12B, после чего клетки высеивают на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола с получением в результате штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1 или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA14, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA2, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12A, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12B.
Заявлен штамм Escherichiacoli JM110-NAS/GDTT1.8NAS12-IFNB1, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора или штамм Escherichiacoli JM110-NAS/GDTT1.8NAS12-IFNA14, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNA14, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA2, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNA2, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12A, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-IL12A, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12B, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12-IL12B, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся нарушением врожденного и адаптивного иммунитета, для терапии аутоиммунных, онкологических, вирусных заболеваний, связанных с дисбалансом иммунной системы, для иммуномодуляции в том числе при противоопухолевой вакцинации в качестве генотерапевтического адъюванта.
Создан способ производства в промышленных масштабах генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IFNB1 или IFNA14 или IFNA2 или IL12A или IL12B для лечения заболеваний, характеризующихся нарушением врожденного и адаптивного иммунитета, для терапии аутоиммунных, онкологических, вирусных заболеваний, связанных с дисбалансом иммунной системы, для иммуномодуляции в том числе при противоопухолевой вакцинации в качестве генотерапевтического адъюванта, заключающийся в том, что генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1, или генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNA14, или генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNA2, или генотерапевтический ДНК-вектор GDTT1.8NAS12-IL12A, или генотерапевтический ДНК-вектора GDTT1.8NAS12-IL12B получают путем того, что засевают в колбу с приготовленной средой затравочную культуру, выбранную из штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA14, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA2, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12A, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12B, затем инкубируют затравочную среду в шейкере-инкубаторе и переносят ее в промышленный ферментер, после чего растят до достижения стационарной фазы, затем выделяют фракцию, содержащую целевой ДНК-продукт, многостадийно фильтруют и очищают хроматографическими методами.
Краткое описание чертежей
На фиг.1 приведена схема генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген, выбранный из группы генов IFNB1, IFNA14, IFNA2, IL12A, IL12B, который представляет собой кольцевую двуцепочечную молекулу ДНК, способную к автономной репликации в клетках бактерии Escherichia coli.
На фиг.1 приведены схемы, соответствующие:
1A - генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1,
1B - генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNA14,
1C - генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNA2,
1D - генотерапевтический ДНК-вектор GDTT1.8NAS12-IL12A,
1E - генотерапевтический ДНК-вектор GDTT1.8NAS12-IL12B.
На схемах отмечены следующие структурные элементы вектора:
EF1a pr - промоторная область гена человеческого фактора элонгации EF1A с собственным энхансером, содержащимся в первом интроне гена. Служит для обеспечения высокого уровня транскрипции рекомбинантного гена в большинстве тканей человека;
Рамка считывания целевого гена, соответствующая кодирующей части гена IFNB1 (фиг. 1A), или IFNA14 (фиг. 1B), или IFNA2 (фиг. 1C), или IL12A (фиг. 1D) или IL12B (фиг. 1E) соответственно;
hGH TA – терминатор транскрипции;
pUCori – ориджин репликации, служащий для автономной репликации с однонуклеотидной заменой для повышения копийности плазмиды в клетках большинства штаммов Escherichia coli;
RNAout – регуляторный элемент РНК-out транспозона Tn 10, обеспечивающий возможность положительной селекции без использования антибиотиков при использовании штамма Eshcerichia coli JM110NAS.
Отмечены уникальные сайты рестрикции.
На фиг.2 показаны графики накопления ампликонов кДНК целевого гена, а именно, гена IFNB1, в первичной культуре клеток фибробластов легких человека (ATCС PCS-201-020) до их трансфекции и через 48 часов после трансфекции этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNB1 с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.2 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 – кДНК гена IFNB1 в первичной культуре клеток фибробластов легких человека до трансфекции ДНК-вектором GDTT1.8NAS12-IFNB1,
2 – кДНК гена IFNB1 в первичной культуре клеток фибробластов легких человека после трансфекции ДНК-вектором GDTT1.8NAS12-IFNB1,
3 – кДНК гена B2M в первичной культуре клеток фибробластов легких человека до трансфекции ДНК-вектором GDTT1.8NAS12-IFNB1,
4 – кДНК гена B2M в первичной культуре клеток фибробластов легких человека после трансфекции ДНК-вектором GDTT1.8NAS12-IFNB1.
В качестве референтного гена использовали ген B2M человека (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг. 3 показаны графики накопления ампликонов кДНК целевого гена, а именно гена IFNA14, в культуре клеток аденокарциномы легкого человека SK-LU-1 (ATCC® HTB-57™) до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IFNA14 с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.3 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена IFNA14 в культуре клеток SK-LU-1 до трансфекции ДНК-вектором GDTT1.8NAS12-IFNA14
2 – кДНК гена IFNA14 в культуре клеток SK-LU-1 после трансфекции ДНК-вектором GDTT1.8NAS12-IFNA14,
3 - кДНК гена B2M в культуре клеток SK-LU-1 до трансфекции ДНК-вектором GDTT1.8NAS12-IFNA14,
4 - кДНК гена B2M в культуре клеток SK-LU-1 после трансфекции ДНК-вектором GDTT1.8NAS12-IFNA14.
В качестве референтного гена использовали ген B2M человека (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.4 показаны графики накопления ампликонов кДНК целевого гена, а именно гена IFNA2 в клетках эпителия бронхов человека BZR (ATCC® CRL-9483™) до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IFNA2 с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.4 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена IFNA2 в клетках эпителия бронхов человека BZR до трансфекции ДНК-вектором GDTT1.8NAS12-IFNA2,
2 – кДНК гена IFNA2 в клетках эпителия бронхов человека BZR после трансфекции ДНК-вектором GDTT1.8NAS12-IFNA2,
3 - кДНК гена B2M в клетках эпителия бронхов человека BZR до трансфекции ДНК-вектором GDTT1.8NAS12-IFNA2,
4 - кДНК гена B2M в клетках эпителия бронхов человека BZR после трансфекции ДНК-вектором GDTT1.8NAS12-IFNA2.
В качестве референтного гена использовали ген B2M человека (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.5 показаны графики накопления ампликонов кДНК целевого гена, а именно гена IL12A, в культуре клеток эндотелия пупочной вены человека HUVEC (ATCC® PCS-100-010™) до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IL12A с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.5 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена IL12A в культуре клеток эндотелия пупочной вены человека HUVEC до трансфекции ДНК-вектором GDTT1.8NAS12-IL12A,
2 – кДНК гена IL12A в культуре клеток эндотелия пупочной вены человека HUVEC после трансфекции ДНК-вектором GDTT1.8NAS12-IL12A,
3 - кДНК гена B2M в культуре клеток эндотелия пупочной вены человека HUVEC до трансфекции ДНК-вектором GDTT1.8NAS12-IL12A,
4 - кДНК гена B2M в культуре клеток эндотелия пупочной вены человека HUVEC после трансфекции ДНК-вектором GDTT1.8NAS12-IL12A.
В качестве референтного гена использовали ген B2M человека (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.6 показаны графики накопления ампликонов кДНК целевого гена, а именно гена IL12B, в культуре клеток скелетных миобластов человека HSkM до их трансфекции и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IL12B с целью оценки способности проникать в эукариотические клетки и функциональной активности, то есть экспрессии целевого гена на уровне мРНК.
На фиг.6 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена IL12B в культуре клеток скелетных миобластов человека HSkM до трансфекции ДНК-вектором GDTT1.8NAS12-IL12B,
2 – кДНК гена IL12B в культуре клеток скелетных миобластов человека HSkM после трансфекции ДНК-вектором GDTT1.8NAS12-IL12B,
3 - кДНК гена B2M в культуре клеток скелетных миобластов человека HSkM до трансфекции ДНК-вектором GDTT1.8NAS12-IL12B,
4 - кДНК гена B2M в культуре клеток скелетных миобластов человека HSkM после трансфекции ДНК-вектором GDTT1.8NAS12-IL12B,
В качестве референтного гена использовали ген B2M человека (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2.
На фиг.7 показана диаграмма концентрации белка IFNB1 в клеточном лизате первичной культуры клеток фибробластов легких человека (ATCС PCS-201-020) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12- IFNB1 с целью оценки функциональной активности, то есть экспрессии на уровне белка, по изменению количества белка IFNB1 в лизате клеток.
На фиг.7 отмечены следующие элементы:
культура А – первичная культура клеток фибробластов легких человека, трансфицированных водным раствором дендримеров без плазмидной ДНК (контроль),
культура B - первичная культура клеток фибробластов легких человека, трансфицированных ДНК-вектором GDTT1.8NAS12,
культура C – первичная культура клеток фибробластов легких человека, трансфицированных ДНК-вектором GDTT1.8NAS12-IFNB1.
На фиг. 8 показана диаграмма концентрации белка IFNA14 в лизате клеток аденокарциномы легкого человека SK-LU-1 (ATCC® HTB-57™), после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IFNA14 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IFNA14.
На фиг.8 отмечены следующие элементы:
культура А - клетки аденокарциномы легкого человека SK-LU-1, трансфицированные водным раствором дендримеров без плазмидной ДНК (контроль),
культура B - клетки аденокарциномы легкого человека SK-LU-1, трансфицированные ДНК-вектором GDTT1.8NAS12,
культура C - клетки аденокарциномы легкого человека SK-LU-1, трансфицированные ДНК-вектором GDTT1.8NAS12-IFNA14.
На фиг.9 показана диаграмма концентрации белка IFNA2 в лизате культуры клеток эпителия бронхов человека BZR (ATCC® CRL-9483™) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IFNA2 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IFNA2.
На фиг.9 отмечены следующие элементы:
культура А - культура клеток эпителия бронхов человека BZR, трансфицированных водным раствором дендримеров без плазмидной ДНК (контроль),
культура B - культура клеток эпителия бронхов человека BZR, трансфицированных ДНК-вектором GDTT1.8NAS12,
культура C - культура клеток эпителия бронхов человека BZR, трансфицированных ДНК-вектором GDTT1.8NAS12-IFNA2.
На фиг. 10 показана диаграмма концентрации белка IL12A в лизате культуры клеток эндотелия пупочной вены человека HUVEC (ATCC® PCS-100-010™) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IL12A с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IL12A.
На фиг.10 отмечены следующие элементы:
культура А - культура клеток эндотелия пупочной вены человека HUVEC, трансфицированных водным раствором дендримеров без плазмидной ДНК (контроль),
культура B - культура клеток эндотелия пупочной вены человека HUVEC, трансфицированных ДНК-вектором GDTT1.8NAS12,
культура C - культура клеток эндотелия пупочной вены человека HUVEC, трансфицированных ДНК-вектором GDTT1.8NAS12-IL12A.
На фиг. 11 показана диаграмма концентрации белка IL12B в лизате культуры клеток скелетных миобластов человека HSkM (Gibco, Кат., № A12555) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IL12B с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IL12B.
На фиг.10 отмечены следующие элементы:
культура А - культура клеток скелетных миобластов человека HSkM, трансфицированных водным раствором дендримеров без плазмидной ДНК (контроль),
культура B - культура клеток скелетных миобластов человека HSkM, трансфицированных ДНК-вектором GDTT1.8NAS12,
культура C - культура клеток скелетных миобластов человека HSkM, трансфицированных ДНК-вектором GDTT1.8NAS12-IL12B.
На фиг. 12 показана диаграмма концентрации белка IL12A в биоптатах кожи трех пациентов после введения в кожу этих пациентов генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12A с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IL12A.
На фиг.12 отмечены следующие элементы:
П1I – биоптат кожи пациента П1 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IL12A,
П1II – биоптат кожи пациента П1 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо),
П1III – биоптат кожи пациента П1 из интактного участка,
П2I – биоптат кожи пациента П2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IL12A
П2II – биоптат кожи пациента П2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо),
П2III – биоптат кожи пациента П2 из интактного участка,
П3I – биоптат кожи пациента П3 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IL12A,
П3II – биоптат кожи пациента П3 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо),
П3III – биоптат кожи пациента П3 из интактного участка.
На фиг. 13 показана диаграмма концентрации белка IFNA2 в биоптатах икроножной мышцы трех пациентов после введения в икроножную мышцу этих пациентов генотерапевтического ДНК-вектора GDTT1.8NAS12- IFNA2, с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IFNA2.
На фиг.13 отмечены следующие элементы:
П1I – биоптат икроножной мышцы пациента П1 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12- IFNA2,
П1II – биоптат икроножной мышцы пациента П1 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо),
П1III – биоптат интактного участка икроножной мышцы пациента П1,
П2I – биоптат икроножной мышцы пациента П2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12- IFNA2,
П2II – биоптат икроножной мышцы пациента П2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо),
П2III – биоптат интактного участка икроножной мышцы пациента П2,
П3I – биоптат икроножной мышцы пациента П3 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12- IFNA2,
П3II – биоптат икроножной мышцы пациента П3 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо),
П3III – биоптат интактного участка икроножной мышцы пациента П3.
На фиг.14 показана диаграмма концентрации белка IFNA14 в биоптатах кожи трех пациентов после введения в кожу этих пациентов генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14 с целью оценки функциональной активности, то есть экспрессии целевого гена на уровне белка, и возможности повышения уровня экспрессии белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IFNA14.
На фиг.14 отмечены следующие элементы:
П1I – биоптат кожи пациента П1 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IFNA14,
П1II – биоптат кожи пациента П1 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо)
П1III – биоптат кожи пациента П1 из интактного участка,
П2I – биоптат кожи пациента П2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IFNA14,
П2II – биоптат кожи пациента П2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо),
П2III – биоптат кожи пациента П2 из интактного участка,
П3I – биоптат кожи пациента П3 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IFNA14,
П3II – биоптат кожи пациента П3 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо),
П3III – биоптат кожи пациента П3 из интактного участка.
На фиг. 15 показана диаграмма концентрации белка IFNA14 в биоптатах кожи человека после введения в кожу культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA14 с целью демонстрации способа применения путем введения аутологичных клеток, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA14
На фиг.15 отмечены следующие элементы:
П1С – биоптат кожи пациента П1 в зоне введения культуры аутологичных фибробластов пациента, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA14,
П1B – биоптат кожи пациента П1 в зоне введения аутологичных фибробластов пациента, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12,
П1А – биоптат кожи пациента П1 из интактного участка.
На фиг. 16 показана диаграмма концентраций белков: белка IFNB1 человека, белка IFNA14 человека, белка IFNA2 человека, белка IL12A человека, белка IL12B человека в мышечной ткани трех крыс линии Wistar после инъекционного введения смеси генотерапевтических векторов: генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, генотерапевтического ДНК-вектора, GDTT1.8NAS12-IFNA14, генотерапевтического ДНК-вектора, GDTT1.8NAS12-IFNA2, генотерапевтического ДНК-вектора, GDTT1.8NAS12-IL12A, генотерапевтического ДНК-вектора, GDTT1.8NAS12-IL12B с целью демонстрации способа применения, смеси генотерапевтических ДНК-векторов.
На фиг.16 отмечены следующие элементы:
К1I – фрагмент мышечной ткани крысы К1 в зоне введения смеси генотерапевтических ДНК векторов: GDTT1.8NAS12- IFNB1, GDTT1.8NAS12- IFNA14, GDTT1.8NAS12- IFNA2, GDTT1.8NAS12- IL12A и GDTT1.8NAS12- IL12B,
К1II – фрагмент мышечной ткани крысы К1 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо),
К1III – фрагмент мышечной ткани контрольного интактного участка крысы К1,
К2I – фрагмент мышечной ткани крысы К2 в зоне введения смеси генотерапевтических ДНК векторов: GDTT1.8NAS12- IFNB1, GDTT1.8NAS12- IFNA14, GDTT1.8NAS12- IFNA2, GDTT1.8NAS12- IL12A и GDTT1.8NAS12- IL12B,
К2II – фрагмент мышечной ткани крысы К2 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо),
К2III – фрагмент мышечной ткани контрольного интактного участка крысы К2,
К3I – фрагмент мышечной ткани крысы К3 в зоне введения смеси генотерапевтических ДНК векторов: GDTT1.8NAS12- IFNB1, GDTT1.8NAS12- IFNA14, GDTT1.8NAS12- IFNA2, GDTT1.8NAS12- IL12A и GDTT1.8NAS12- IL12B,
К3II – фрагмент мышечной ткани крысы К3 в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12 (плацебо),
К3III – фрагмент мышечной ткани контрольного интактного участка крысы К3.
На фиг. 17 показаны графики накопления ампликонов кДНК целевого гена IFNA2 в клетках эндотелия легочной артерии быка CPAE (ATCC® CCL-209) до и через 48 часов после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IFNA2 с целью демонстрации способа применения путем введения генотерапевтического ДНК-вектора животным
На фиг.17 отмечены кривые накопления ампликонов в ходе реакции, соответствующие:
1 - кДНК гена IFNA2 в клетках эндотелия легочной артерии быка CPAE до трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12- IFNA2,
2 – кДНК гена IFNA2 в клетках эндотелия легочной артерии быка CPAE после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12- IFNA2
3 - кДНК гена ACT в клетках эндотелия легочной артерии быка CPAE до трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12- IFNA2
4 - кДНК гена ACT в клетках эндотелия легочной артерии быка CPAE после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12- IFNA2
В качестве референтного гена использовали ген актина быка/ коровы (АСТ), приведенного в базе данных GenBank под номером AH001130.2.
Реализация изобретения
На основе ДНК-вектора GDTT1.8NAS12 размером 2591 п.н. созданы генотерапевтические ДНК-векторы, несущие целевые гены человека, предназначенные для повышения уровня экспрессии этих целевых генов в тканях человека и животных. При этом способ получения каждого генотерапевтического ДНК-вектора, несущего целевые гены заключается в том, что в полилинкер генотерапевтического ДНК-вектора GDTT1.8NAS12 клонируют белок-кодирующую последовательность целевого гена, выбранного из группы генов: ген IFNB1 (кодирует белок IFNB1), ген IFNA14 (кодирует белок IFNA14), ген IFNA2 (кодирует белок IFNA2), ген IL12A (кодирует белок IL12A), ген IL12B (кодирует белок IL12B) человека. Известно, что способность ДНК-векторов проникать в эукариотические клетки обусловлена, главным образом, размером вектора. При этом ДНК-вектора с наименьшим размером обладают более высокой проникающей способностью. Таким образом, предпочтительным является отсутствие в составе вектора элементов, которые не несут функциональной нагрузки, но при этом увеличивают размер ДНК-вектора. Данные особенности ДНК-векторов были учтены при получении генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген, выбранный из группы генов IFNB1, IFNA14, IFNA2, IL12A, IL12B путем отсутствия в составе вектора крупных нефункциональных последовательностей и генов антибиотикорезистентности, что позволило, помимо технологических преимуществ и преимуществ в плане безопасности применения, значительно уменьшить размер полученного генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген, выбранный из группы генов IFNB1, IFNA14, IFNA2, IL12A, IL12B. Таким образом, способность проникать в эукариотические клетки полученного генотерапевтического ДНК-вектора обусловлена его небольшими размерами.
Каждый из генотерапевтических ДНК-векторов: ДНК-вектор GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-IFNA14, или GDTT1.8NAS12-IFNA2, или GDTT1.8NAS12-IL12A, или GDTT1.8NAS12-IL12B получали следующим образом: кодирующую часть целевого гена IFNB1, или IFNA14, или IFNA2, или IL12A, или IL12B клонировали в генотерапевтический ДНК-вектор GDTT1.8NAS12 и получали генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1, SEQ ID №1, или GDTT1.8NAS12-IFNA14, SEQ ID №2 или GDTT1.8NAS12-IFNA2, SEQ ID №3, или GDTT1.8NAS12-IL12A, SEQ ID №4, или GDTT1.8NAS12-IL12B, SEQ ID №5 соответственно. Кодирующую часть гена IFNB1 размером 564 п.н., или гена IFNA14 размером 570 п.н., или гена IFNA2 размером 567 п.н., или гена IL12A размером 762 п.н., или гена IL12B размером 987 п.н. получали путем выделения суммарной РНК из биологического образца ткани здорового человека. Для получения первой цепи кДНК генов IFNB1, IFNA14, IFNA2, IL12A, IL12B человека использовали реакцию обратной транскрипции. Амплификацию проводили с использованием созданных для этого методом химического синтеза олигонуклеотидов. Расщепление продукта амплификации специфическими эндонуклеазами рестрикции проводили с учетом оптимальной процедуры дальнейшего клонирования, причем клонирование в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводили по сайтам рестрикции BamHI, EcoRI, HindIII, KpnI, SalI, расположенными в полилинкере вектора GDTT1.8NAS12. Выбор сайтов рестрикции проводили таким образом, чтобы клонированный фрагмент попадал в рамку считывания экспрессионной кассеты вектора GDTT1.8NAS12, при этом белок-кодирующая последовательность не содержала сайты рестрикции для выбранных эндонуклеаз. При этом специалистам в данной области техники понятно, что методическая реализация получения генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-IFNA14, или GDTT1.8NAS12-IFNA2, или GDTT1.8NAS12-IL12A, или GDTT1.8NAS12-IL12B может варьировать в рамках выбора известных методов молекулярного клонирования генов, при этом эти способы подпадают под объем настоящего изобретения. Так, например, могут быть использованы различные последовательности олигонуклеотидов для амплификации гена IFNB1, или IFNA14, или IFNA2, или IL12A, или IL12B различные эндонуклеазы рестрикции или такие лабораторные техники как безлигазное клонирование генов.
Генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-IFNA14, или GDTT1.8NAS12-IFNA2, или GDTT1.8NAS12-IL12A, или GDTT1.8NAS12-IL12B обладает нуклеотидной последовательностью SEQ ID №1, или SEQ ID №2, или SEQ ID №3, или SEQ ID №4, или SEQ ID №5, соответственно. При этом специалистам в данной области техники известно свойство вырожденности генетического кода, из которого следует, что под объем настоящего изобретения также подпадают варианты нуклеотидных последовательностей, отличающихся инсерцией, делецией или заменой нуклеотидов, которые не приводят к изменению полипептидной последовательности, кодируемой целевым геном, и/или не приводят к потере функциональной активности регуляторных элементов вектора GDTT1.8NAS12. При этом специалистам в данной области техники известно явление генетического полиморфизма, из которого следует, что под объем настоящего изобретения также подпадают варианты нуклеотидных последовательностей генов из группы генов IFNB1, IFNA14, IFNA2, IL12A, IL12B, которые при этом кодируют различные варианты аминокислотных последовательностей белков IFNB1, IFNA14, IFNA2, IL12A, IL12B не отличающихся от приведенных по своей функциональной активности при физиологических условиях.
Способность проникать в эукариотические клетки и функциональную активность, то есть способность экспрессировать целевой ген, полученного генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-IFNA14, или GDTT1.8NAS12-IFNA2, или GDTT1.8NAS12-IL12A, или GDTT1.8NAS12-IL12B подтверждают путем введения в эукариотические клетки полученного вектора и последующим анализом экспрессии специфической мРНК и/или белкового продукта целевого гена. Наличие специфической мРНК в клетках, в которые был введен генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-IFNA14, или GDTT1.8NAS12-IFNA2, или GDTT1.8NAS12-IL12A, или GDTT1.8NAS12-IL12B свидетельствует как о способности полученного вектора проникать в эукариотические клетки, так и о его способности экспрессировать мРНК целевого гена. При этом, как известно специалистам в данной области техники, наличие мРНК гена является обязательным условием, но не доказательством трансляции белка, кодируемого целевым геном. Поэтому для подтверждения свойства генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-IFNA14, или GDTT1.8NAS12-IFNA2, или GDTT1.8NAS12-IL12A, или GDTT1.8NAS12-IL12B экспрессировать целевой ген на уровне белка в эукариотических клетках, в которые был введен генотерапевтический ДНК-вектор, проводят анализ концентрации белков, кодируемых целевыми генами, с использованием иммунологических методов. Наличие белка IFNB1, или IFNA14, или IFNA2, или IL12A, или IL12B подтверждает эффективность экспрессии целевых генов в эукариотических клетках и возможность повышения уровня концентрации белка с помощью генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген, выбранный из группы генов IFNB1, IFNA14, IFNA2, IL12A, IL12B. Таким образом для подтверждения эффективности экспрессии созданного генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, несущего целевой ген, а именно, ген IFNB1, генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14, несущего целевой ген, а именно, ген IFNA14, генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA2, несущего целевой ген, а именно, ген IFNA2, генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12A, несущего целевой ген, а именно, ген IL12A, генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12B, несущего целевой ген, а именно, ген IL12B, использовали следующие методы:
А) ПЦР в реальном времени - изменение накопления ампликонов кДНК целевых генов в лизате клеток человека и животного, после трансфекции различных клеточных линий человека и животного генотерапевтическим ДНК-векторами;
B) Иммуноферментный анализ - изменение количественного уровня целевых белков в лизате клеток человека, после трансфекции различных клеточных линий человека генотерапевтическими ДНК-векторами;
C) Иммуноферментный анализ - изменение количественного уровня целевых белков в супернатанте биоптатов тканей человека и животного, после введения в эти ткани генотерапевтических ДНК-векторов;
D) Иммуноферментный анализ - изменение количественного уровня целевых белков в супернатанте биоптатов тканей человека, после введения в эти ткани аутологичных клеток этого человека, трансфицированных генотерапевтическими ДНК-векторами.
Для подтверждения реализуемости способа применения созданного генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, несущего целевой ген, а именно, ген IFNB1, генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14, несущего целевой ген, а именно, ген IFNA14, генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA2, несущего целевой ген, а именно, ген IFNA2, генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12A, несущего целевой ген, а именно, ген IL12A, генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12B, несущего целевой ген, а именно, ген IL12B, выполняли:
А) трансфекцию генотерапевтическими ДНК-векторами различных клеточных линий человека и животного;
B) введение генотерапевтических ДНК-векторов в различные ткани человека и животного;
С) введение в ткани животного смеси генотерапевтических ДНК-векторов;
D) введение в ткани человека аутологичных клеток, трансфицированных генотерапевтическими ДНК-векторами.
Указанные способы применения характеризуются отсутствием потенциальных рисков для генетической терапии человека и животных за счет отсутствия в составе генотерапевтического ДНК-вектора регуляторных элементов, представляющих собой нуклеотидные последовательности вирусных геномов, и за счет отсутствия в составе генотерапевтического ДНК-вектора генов устойчивости к антибиотикам, что подтверждается отсутствием участков, гомологичных вирусным геномам и генам антибиотикорезистентности в нуклеотидных последовательностях генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, или генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14, или генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA2, или генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12A, или генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12B (SEQ ID №1 или SEQ ID №2 или SEQ ID №3 или SEQ ID №4 или SEQ ID №5 соответственно).
Как известно специалистам в данной области техники, гены антибиотикорезистентности в составе генотерапевтических ДНК-векторов используются с целью получения этих векторов в препаративных количествах путем наращивания бактериальной биомассы в питательной среде, содержащей селективный антибиотик. В рамках настоящего изобретения в целях возможности безопасного применения генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IFNB1, или IFNA14, или IFNA2, или IL12A, или IL12B, использование селективных питательных сред, содержащих антибиотик, не представляется возможным. В качестве технологического решения для получения генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген, выбранный из группы генов IFNB1, или IFNA14, или IFNA2, или IL12A, или IL12B для возможности масштабирования до промышленных масштабов получения генотерапевтических векторов предлагается способ получения штаммов для наработки указанных генотерапевтических векторов на основе бактерии Escherichia coli JM110-NAS. Способ получения штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA14, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA2, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12A, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12B, заключается в получении компетентных клеток штамма Escherichia coli JM110-NAS с введением в эти клетки генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, или ДНК-вектора GDTT1.8NAS12-IFNA14, или ДНК-вектора GDTT1.8NAS12-IFNA2, или ДНК-вектора GDTT1.8NAS12-IL12A, или ДНК-вектора GDTT1.8NAS12-IL12B соответственно с помощью методов трансформации (электропорации), общеизвестных специалистам в данной области техники. Полученный штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA14, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA2, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12A, или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12B используется для наработки генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-IFNA14, или GDTT1.8NAS12-IFNA2, или GDTT1.8NAS12-IL12A, или GDTT1.8NAS12-IL12B соответственно с возможностью использования сред без содержания антибиотиков.
Для подтверждения получения штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA14, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA2, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12A, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12B проводили трансформацию, селекцию и последующее наращивание с выделением плазмидной ДНК.
Для подтверждения технологичности получения и возможности масштабирования до промышленного производства генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, несущего целевой ген, а именно, ген IFNB1, генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14, несущего целевой ген, а именно, ген IFNA14, генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA2, несущего целевой ген, а именно, ген IFNA2, генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12A, несущего целевой ген, а именно, ген IL12A, генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12B, несущего целевой ген, а именно, ген IL12B, выполняли ферментацию в промышленном масштабе штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1 или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA14 или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA2, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12A, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12B, каждый из которых содержит генотерапевтический ДНК-вектор GDTT1.8NAS12, несущий целевой ген, а именно IFNB1, или IFNA14, или IFNA2, или IL12A, или IL12B.
Способ масштабирования получения бактериальной массы до промышленных масштабов для выделения генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген, выбранный из группы генов IFNB1, IFNA14, IFNA2, IL12A, IL12B заключается в том, что затравочную культуру штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA14, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA2, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12A, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12B инкубируют в объеме питательной среды без содержания антибиотика обеспечивающим подходящую динамику накопления биомассы, по достижению достаточного количества биомассы в логарифмической фазе роста, бактериальную культуру переносят в промышленный ферментер, после чего растят до достижения стационарной фазы роста, затем выделяют фракцию, содержащую целевой ДНК-продукт - генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1, или генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNA14, или генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNA2, или генотерапевтический ДНК-вектор GDTT1.8NAS12-IL12A или генотерапевтический ДНК-вектор GDTT1.8NAS12-IL12B многостадийно фильтруют и очищают хроматографическими методами. При этом специалистам в данной области техники понятно, что условия культивирования штаммов, состав питательных сред (за исключением содержания антибиотиков), используемое оборудование, методы очистки ДНК могут варьировать в рамках стандартных операционных процедур в зависимости от отдельно взятой производственной линии, но известные подходы к масштабированию, промышленному получению и очистке ДНК-векторов с использованием штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA14, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA2, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12A, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12B или подпадают под объем настоящего изобретения.
Описанное раскрытие изобретения подтверждается примерами реализации настоящего изобретения.
Изобретение поясняется следующими примерами.
Пример 1.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, несущего целевой ген, а именно, гена IFNB1.
Генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1 конструировали клонированием кодирующей части гена IFNB1 размером 564 п.н. в ДНК-вектор GDTT1.8NAS12 размером 2591 п.н. по сайтам рестрикции BamHI и Sal. Кодирующую часть гена IFNB1 размером 564 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов:
IFNB1-Fw AAAGGATCCACCATGACCAACAAGTGTCTCCTCCAAA
IFNB1-Rv TTTGTCGACTCAGTTTCGGAGGTAACCTGTAAGTCTG
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США).
Генотерапевтический ДНК-вектор GDTT1.8NAS12 конструировали объединением шести фрагментов ДНК, полученных из разных источников:
(а) ориджин репликации получали путем ПЦР-амплификации участка коммерческой плазмиды pUC19 с использованием олигонуклеотидов UCori-Bam, UCori-Nco (перечень последовательностей, (1) -(2));
(б) терминатор транскрипции hGH-TA получали путем ПЦР-амплификации участка геномной ДНК человека с использованием олигонуклеотидов hGH-F и hGH-R (перечень последовательностей, (3) и (4));
(в) регуляторный участок транспозона Tn10 РНК-out получали путем синтеза из олигонуклеотидов RO-F, RO-R, RO-1, RO-2, RO-3 (перечень последовательностей, (5) -(9));
(г) ген устойчивости к канамицину получали путем ПЦР-амплификации участка коммерческой плазмиды pET-28 человека с использованием олигонуклеотидов Kan-F и Kan-R (перечень последовательностей, (10) и (11));
(д) полилинкер получали кинированием и отжигом четырех синтетических олигонуклеотидов MCS1, MCS2, MCS3 и MCS4 (перечень последовательностей, (12) - (15));
(е) промоторно-регуляторный участок человеческого гена фактора элонгации EF1а с собственным энхансером получали путем ПЦР-амплификации участка геномной ДНК человека с использованием олигонуклеотидов EF1-Xho и EF1-R (перечень последовательностей, (16) - (17)).
ПЦР-амплификацию проводили с использованием коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs) в соответствии с инструкцией производителя. Фрагменты (а), (б), (в) и (г) имели перекрывающиеся области для возможности их объединения с последующей ПЦР-амплификацией. Объединяли фрагменты (а), (б), (в) и (г) с использованием олигонуклеотидов hGH-F и Kan-R (список последовательностей, (3) и (11)). Далее, полученные фрагменты ДНК объединялись путем рестрикции с последующим лигированием по сайтам BamHI и NcoI. В результате получали вектор, пока еще не содержащий полилинкер. Для его введения проводили расщепление плазмиды эндонуклеазами рестрикции по сайтам BamHI и EcoRI с последующим лигированием с фрагментом (д). В результате получали промежуточный вектор размером 2408 п.н, несущий ген устойчивости к канамицину, пока еще не содержащий промоторно-регуляторный участок гена фактора элонгации EF1а с собственным энхансером. Полученный вектор расщепляли эндонуклеазами рестрикции по сайтам SalI и BamHI с последующим лигированием с фрагментом (е). В результате получали вектор размером 3608 п.н., несущий ген устойчивости к канамицину и промоторно-регуляторный участок гена гена фактора элонгации EF1а с собственным энхансером. Далее ген устойчивости к канамицину выщепляли по сайтам рестрикции SpeI, после чего оставшийся фрагмент лигировали сам на себя. Таким образом, получали генотерапевтический ДНК-вектор GDTT1.8NAS12 размером 2591 п.н., который является рекомбинантным, с возможностью селекции без антибиотиков. Расщепление продукта амплификации кодирующей части гена IFNB1 и ДНК-вектора GDTT1.8NAS12 проводили эндонуклеазами рестрикции BamHI и SalI (New England Biolabs, США).
В результате получали ДНК-вектор GDTT1.8NAS12-IFNB1 размером 3120 п.н. с нуклеотидной последовательностью SEQ ID №1 и общей структурой изображенной на фиг.1A.
Пример 2.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14, несущего целевой ген, а именно, гена IFNA14.
Генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNA14 конструировали клонированием кодирующей части гена IFNA14 размером 570 п.н. в ДНК-вектор GDTT1.8NAS12 размером 2591 п.н. по сайтам рестрикции BamHI и SalI. Кодирующую часть гена IFNA14 размером 570 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов:
IFNA14-Fw TTTGGATCCACCATGGCATTGCCCTTTGCTTTAAT
IFNA14-Rv TTTGTCGACTCAATCCTTCCTCCTTAATCTTTTTTGC
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США), расщепление продукта амплификации и ДНК-вектора GDTT1.8NAS12 проводили эндонуклеазами рестрикции BamHI и SalI (New England Biolabs, США).
В результате получали ДНК-вектор GDTT1.8NAS12-IFNA14 размером 3126 п.н. с нуклеотидной последовательностью SEQ ID №2 и общей структурой изображенной на фиг.1B.
При этом генотерапевтический ДНК-вектор GDTT1.8NAS12 конструировали по Примеру 1.
Пример 3
Получение ДНК-вектора GDTT1.8NAS12-IFNA2, несущего целевой ген, а именно, гена IFNA2 человека.
Генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNA2 конструировали клонированием кодирующей части гена IFNA2 размером 567 п.н. в ДНК-вектор GDTT1.8NAS12 размером 2591 п.н. по сайтам рестрикции BamHI и SalI. Кодирующую часть гена IFNA2 размером 567 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген, Россия) и ПЦР-амплификации с использованием олигонуклеотидов:
IFNA2-Fw GGGGGATCCACCATGGCCTTGACCTTTGCTTTACTGG
IFNA2-Rv TTTGTCGACTCATTCCTTACTTCTTAAACTTTCTTGC
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США), расщепление продукта амплификации и ДНК-вектора GDTT1.8NAS12 проводили эндонуклеазами рестрикции BamHI и SalI (New England Biolabs, США).
В результате получали ДНК-вектор GDTT1.8NAS12-IFNA2 размером 3123 п.н. с нуклеотидной последовательностью SEQ ID №3 и общей структурой изображенной на фиг.1C.
При этом генотерапевтический ДНК-вектор GDTT1.8NAS12 конструировали по Примеру 1.
Пример 4.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12A, несущего целевой ген, а именно, гена IL12A.
Генотерапевтический ДНК-вектор GDTT1.8NAS12-IL12A конструировали клонированием кодирующей части гена IL12A размером 762 п.н. в ДНК-вектор GDTT1.8NAS12 размером 2591 п.н. по сайтам рестрикции SalI и KpnI. Кодирующую часть гена IL12A размером 762 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген) и ПЦР-амплификации с использованием олигонуклеотидов:
IL12A-up TTTGTCGACCACCATGTGGCCCCCTGGGTCAGCCTCC
IL12A-lo AATGGTACCTTAGGAAGCATTCAGATAGCTCATCACTCTATC
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США), расщепление продукта амплификации и ДНК-вектора GDTT1.8NAS12 проводили эндонуклеазами рестрикции SalI и KpnI (New England Biolabs, США).
В результате получали ДНК-вектор GDTT1.8NAS12-IL12A размером 3307 п.н. с нуклеотидной последовательностью SEQ ID №4 и общей структурой изображенной на фиг.1D.
При этом генотерапевтический ДНК-вектор GDTT1.8NAS12 конструировали по Примеру 1.
Пример 5.
Получение генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12B, несущего целевой ген, а именно, гена IL12B.
Генотерапевтический ДНК-вектор GDTT1.8NAS12-IL12B конструировали клонированием кодирующей части гена IL12B размером 987 п.н. в ДНК-вектор GDTT1.8NAS12 размером 2591 п.н. по сайтам рестрикции SalI и KpnI. Кодирующую часть гена IL12B размером 987 п.н. получали путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции с использованием коммерческого набора Mint-2 (Евроген) и ПЦР-амплификации с использованием олигонуклеотидов:
IL12B-up TTTGTCGACCACCATGTGTCACCAGCAGTTGGTCATCTC
IL12B-lo AATGGTACCTAACTGCAGGGCACAGATGCCCA
и коммерческого набора Phusion® High-Fidelity DNA Polymerase (New England Biolabs, США), расщепление продукта амплификации и ДНК-вектора GDTT1.8NAS12 проводили эндонуклеазами рестрикции SalI и KpnI (New England Biolabs, США).
В результате получали ДНК-вектор GDTT1.8NAS12-IL12B размером 3543 п.н. с нуклеотидной последовательностью SEQ ID №5 и общей структурой изображенной на фиг.1E
При этом генотерапевтический ДНК-вектор GDTT1.8NAS12 конструировали по Примеру 1.
Пример 6.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, несущего целевой ген, а именно, ген IFNB1, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена IFNB1, в первичной культуре клеток фибробластов легких человека (ATCС PCS-201-020) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNB1, несущим ген IFNB1 человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Первичную культуру клеток фибробластов легких человека выращивали в стандартных условиях (37 ˚С, 5% СО2) с использованием набора для приготовления питательной среды Fibroblast Growth Kit-Low serum ATCC® PCS-201-041 и среды Fibroblast Basal Medium ATCC® PCS-201-030. В процессе культивирования каждые 48 ч происходила смена ростовой среды.
Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNB1, экспрессирующим ген IFNB1 человека, проводили с использованием Lipofectamine 3000 (ThermoFisher Scientific, США) согласно рекомендациям производителя. В пробирке №1 к 25 мкл среды Opti-MEM (Gibco, США) добавляли 1 мкл раствора ДНК-вектора GDTT1.8NAS12-IFNB1 (концентрация 500 нг/мкл) и 1 мкл реагента Р3000. Аккуратно перемешивали легким встряхиванием. В пробирке №2 к 25 мкл среды Opti-MEM (Gibco, США) добавляли 1 мкл раствора Lipofectamin 3000. Аккуратно перемешивали легким встряхиванием. Добавляли содержимое пробирки №1 к содержимому пробирки №2, инкубировали 5 мин при комнатной температуре. Полученный раствор по каплям добавляли к клеткам в объеме 40 мкл.
В качестве контроля использовали первичную культуру клеток фибробластов легких человека трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, не содержащим вставку целевого гена (кДНК гена IFNB1 до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12, не содержащим вставку целевого гена на фигурах не показано). Подготовку контрольного вектора GDTT1.8NAS12 для трансфекции проводили как описано выше.
Суммарную РНК из первичной культуры клеток фибробластов легких человека выделяли с использованием Trizol Reagent (Invitrogen, США) согласно рекомендациям производителя. В лунку с клетками добавляли 1 мл Trizol Reagent и гомогенизировали с последующим прогреванием в течении 5 мин при 65 0С. Далее образец центрифугировали при 14 000g в течении 10 мин и снова прогревали в течении 10 мин при 65 0С. Далее добавляли 200 мкл хлороформа, плавно перемешивали и центрифугировали при 14 000g в течении 10 мин. Затем отбирали водную фазу, добавляли к ней 1/10 объема 3М ацетата натрия, рН 5.2 и равный объем изопропилового спирта. Инкубировали образец при -20 0С в течении 10 мин с последующим центрифугированием при 14 000g в течении 10 мин. Осадок промывали 1 мл 70% этилового спирта, высушивали на воздухе и растворяли в 10 мкл воды, свободной от РНКаз. Определение уровня экспрессии мРНК гена IFNB1 после трансфекции проводили путем оценки динамики накопления ампликонов кДНК методом ПЦР в режиме реального времени. Для получения и амплификации кДНК, специфичной для гена IFNB1 человека, использовали олигонуклеотиды:
IFNB1_SF GTGGCAATTGAATGGGAGGC
IFND1_RW ATAGATGGTCAATGCGGCGT
дина продукта амплификации - 118 п.н.
Реакцию обратной транскрипции и ПЦР-амплификацию проводили с помощью набора реагентов SYBR GreenQuantitect RT–PCR Kit (Qiagen, США) для ПЦР в режиме реального времени. Реакцию проводили в объеме 20 мкл, содержащих: 25 мкл QuantiTect SYBR Green RT-PCR MasterMix, 2,5 мM хлорида магния, по 0,5 мкМ каждого праймера, 5 мкл РНК. Реакцию осуществляли на амплификаторе CFX96 (Bio-Rad, США) при следующих условиях: 1 цикл обратной транскрипции при 42оС - 30 минут, денатурация 98оС - 15 мин, затем 40 циклов, включающих денатурацию 94оС - 15 сек, отжиг праймеров 60оC - 30 сек и элонгацию 72оС - 30 сек. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов IFNB1 и B2M. В качестве отрицательного контроля использовали деионизированную воду. Количество динамику накопления ампликонов кДНК генов IFNB1 и B2M оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа представлены на фиг. 2.
Из фигуры 2 следует, что в результате трансфекции первичной культуры клеток фибробластов легких человека генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNB1, уровень специфической мРНК гена IFNB1 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IFNB1 на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1 для повышения уровня экспрессии гена IFNB1 в эукариотических клетках.
Пример 7.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14, несущего целевой ген, а именно, ген IFNA14, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена IFNA14, в культуре клеток аденокарциномы легкого человека SK-LU-1 (ATCC® HTB-57™) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA14, несущим ген IFNA14 человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Клетки аденокарциномы легкого человека SK-LU-1 культивировали в среде DMEM (GIBCO) с добавлением 10 % сыворотки эмбрионов коров в стандартных условиях (37˚С, 5% СО2). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA14, экспрессирующим ген IFNA14 человека, проводили как описано в примере 6. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве контроля использовали клетки аденокарциномы легкого человека SK-LU-1 , трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, не несущим целевой ген (кДНК гена IFNA14 до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12, не содержащим вставку целевого гена на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили как описано в примере 6, за исключением олигонуклеотидов с отличающимися от примера 6 последовательностями. Для амплификации кДНК, специфичной для гена IFNA14 человека, использовали олигонуклеотиды
IFN14_SF GGCAACCAGTTCCAGAAAGC
IFN14_SR CACACAGGCTTCCAGGTCAT
длина продукта амплификации – 168 п.н.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов IFNA14 и B2M. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов – кДНК генов IFNA14 и B2M, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа представлены на фиг. 3.
Из фигуры 3 следует, что в результате трансфекции клетки аденокарциномы легкого человека SK-LU-1 генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA14, уровень специфической мРНК гена IFNA14 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IFNA14 на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14 для повышения уровня экспрессии гена IFNA14 в эукариотических клетках.
Пример 8.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA2, несущего целевой ген, а именно, ген IFNA2, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена IFNA2 в культуре клеток эпителия бронхов человека BZR (ATCC® CRL-9483™) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA2, несущим ген IFNA2 человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Культуру клеток эпителия бронхов человека BZR выращивали с использованием набора BEGM Kit (Lonza, Catalog No. CC-3170) в стандартных условиях (37˚С, 5% СО2). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA2, экспрессирующим ген IFNA2 человека, проводили как описано в примере 6. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве контроля использовали культуру клеток эпителия бронхов человека BZR трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, не несущим целевой ген (кДНК гена IFNA2 до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12, не содержащим вставку целевого гена на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили как описано в примере 5, за исключением олигонуклеотидов с отличающимися от примера 6 последовательностями. Для амплификации кДНК, специфичной для гена IFNA2 человека, использовали олигонуклеотиды
IFNA2_SF AGCTGAATGACCTGGAAGCC
IFNA2_SR CTGCTCTGACAACCTCCCAG
длина продукта амплификации – 165 п.н.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов IFNA2 и B2M. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов – кДНК генов IFNA2 и B2M, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа представлены на фиг. 4.
Из фигуры 4 следует, что в результате трансфекции культуры клеток эпителия бронхов человека BZR генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA2 уровень специфической мРНК гена IFNA2 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IFNA2 на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA2 для повышения уровня экспрессии гена IFNA2 в эукариотических клетках.
Пример 9.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12A, несущего целевой ген, а именно, ген IL12A, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена IL12A, в культуре клеток эндотелия пупочной вены человека HUVEC (ATCC® PCS-100-010™) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL12A, несущим ген IL12A человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Культуру клеток эндотелия пупочной вены человека HUVEC выращивали с использованием среды Vascular Cell Basal Medium и набора Endothelial Cell Growth Kit-BBE (ATCC PCS-100-040) в стандартных условиях (37˚С, 5% СО2). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL12A, экспрессирующим ген IL12A человека, проводили как описано в примере 6. В качестве контроля использовали культуру клеток клеток эндотелия пупочной вены человека HUVEC, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, не несущим целевой ген (кДНК гена IL12A до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12, не содержащим вставку целевого гена на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили как описано в примере 6, за исключением олигонуклеотидов с отличающимися от примера 6 последовательностями. Для амплификации кДНК, специфичной для гена IL12A человека, использовали олигонуклеотиды
IL12A_SF GCTCCAGAAGGCCAGACAAA
IL12A_SR GCCAGGCAACTCCCATTAGT
длина продукта амплификации – 183 п.н.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов IL12A и B2M. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов – кДНК генов IL12A и B2M, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа представлены на фиг. 5.
Из фигуры 5 следует, что в результате трансфекции культуры клеток эндотелия пупочной вены человека HUVEC генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL12A, уровень специфической мРНК гена IL12A человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IL12A на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL12A для повышения уровня экспрессии гена IL12A в эукариотических клетках.
Пример 10.
Подтверждение способности генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12B, несущего целевой ген, а именно, ген IL12B, проникать в эукариотические клетки и подтверждение его функциональной активности на уровне экспрессии мРНК целевого гена. Данный пример также демонстрирует реализуемость способа применения генотерапевтического ДНК-вектора, несущего целевой ген.
Оценивали изменения накопления мРНК целевого гена IL12B, в культуре клеток скелетных миобластов человека HSkM (Gibco, Кат., № A12555) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL12B, несущим ген IL12B человека. Количество мРНК определяли по динамике накопления ампликонов кДНК в реакции ПРЦ в реальном времени.
Культуру клеток скелетных миобластов человека HSkM выращивали согласно инструкции фирмы-производителя в стандартных условиях (37˚С, 5% СО2). Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент Lipofectamine 3000 (ThermoFisher Scientific, США). Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL12B, экспрессирующим ген IL12B человека, проводили как описано в примере 6. В качестве контроля использовали культуру клеток скелетных миобластов человека HSkM, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, не несущим целевой ген (кДНК гена IL12B до и после трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12, не содержащим вставку целевого гена на фигурах не показано). Выделение РНК, реакцию обратной транскрипции и ПЦР в реальном времени проводили как описано в примере 6, за исключением олигонуклеотидов с отличающимися от примера 6 последовательностями. Для амплификации кДНК, специфичной для гена IL12B человека, использовали олигонуклеотиды
IL12B_SF ATGCCCCTGGAGAAATGGTG
IL12B_SR GGCCAGCATCTCCAAACTCT
длина продукта амплификации – 141 п.н.
В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов IL12B и B2M. В качестве референтного гена использовали ген B2M (Бета-2-микроглобулин) приведенный в базе данных GenBank под номером NM 004048.2. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов – кДНК генов IL12B и B2M, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1 (Bio-Rad, США). Графики, полученные в результате анализа представлены на фиг. 5.
Из фигуры 5 следует, что в результате трансфекции культуры клеток скелетных миобластов человека HSkM генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL12B, уровень специфической мРНК гена IL12B человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IL12B на уровне мРНК. Представленные результаты также подтверждают реализуемость способа применения генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL12B для повышения уровня экспрессии гена IL12B в эукариотических клетках.
Пример 11.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, несущего ген IFNB1, для повышения экспрессии белка IFNB1 в клетках млекопитающих.
Оценивали изменение количества белка IFNB1 в лизате первичной культуре клеток фибробластов легких человека после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IFNB1, несущим ген IFNB1 человека.
Клеточную культуру фибробластов легких человека выращивали как это описано в примере 6.
Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевали в 24-луночный планшет из расчета 5×104 клеток/лунку. Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена IFNB1 (В), в качестве трансфицируемых агентов - ДНК-вектор GDTT1.8NAS12-IFNB1, несущий ген IFNB1 человека (С). Приготовление комплекса ДНК/дендример проводили по методике производителя (QIAGEN, SuperFect Transfection Reagent Handbook, 2002) с некоторыми изменениями. Для трансфекции клеток в одной лунке 24-луночного планшета к 1 мкг ДНК-вектора, растворенного в буфере ТЕ, добавляли культуральную среду до конечного объема 60 мкл, затем добавляли 5 мкл SuperFect Transfection Reagent и осторожно перемешивали пятикратным пипетированием. Комплекс инкубировали при комнатной температуре в течении 10-15 мин. Далее отбирали из лунок культуральную среду, лунки промывали 1 мл буфера PBS. К образовавшемуся комплексу добавляли 350 мкл среды, содержащей 10 мкг/мл гентамицина, осторожно перемешивали и добавляли к клеткам. Клетки инкубировали с комплексом 2-3 часа при 37°С в атмосфере, содержащей 5% СО2.
Затем осторожно удаляли среду, промывали клеточный монослой 1 мл буфера PBS. Затем добавляли среду, содержащую 10 мкг/мл гентамицина, и инкубировали 24-48 часов при 37°С в атмосфере 5% СО2.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка. Количественное определение белка IFNB1 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Human IFN-β (Interferon Beta) ELISA Kit (ElabBioscience E-EL-H0085) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка IFNB1, входящим в состав набора. Чувствительность метода составляла не менее 18,75 пг/мл, диапазон измерения - от 31,25 пг/мл до 2000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа представлены на фиг. 7.
Из фигуры 7 следует, что трансфекция первичной культуры клеток фибробластов легких человека генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNB1 приводит к увеличению количества белка IFNB1 по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IFNB1 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1 для повышения уровня экспрессии IFNB1 в эукариотических клетках.
Пример 12.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14, несущего ген IFNA14, для повышения экспрессии белка IFNA14 в клетках млекопитающих.
Оценивали изменение количества белка в культуре клеток аденокарциномы легкого человека SK-LU-1 (ATCC® HTB-57™) после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IFNA14, несущим ген IFNA14 человека. Клетки выращивали как описано в примере 7.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена IFNA14 (В), в качестве трансфицируемых агентов - ДНК-вектор GDTT1.8NAS12-IFNA14, несущий ген IFNA14 человека (С). Приготовление комплекса ДНК-дендример и трансфекцию клеток SK-LU-1 проводили как описано в примере 11.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка. Количественное определение белка IFNA14 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Human IFNA14 / Interferon Alpha 14 ELISA Kit (Sandwich ELISA) (LifeSpan BioSciences LS-F13512-1) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка IFNA14, входящим в состав набора. Чувствительность метода составляла не менее 15,6 пг/мл, диапазон измерения - от 15,6 пг/мл до 1000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа представлены на фиг. 8.
Из фигуры 8 следует, что трансфекция культуры клеток аденокарциномы легкого человека SK-LU-1 генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA14 приводит к увеличению количества белка IFNA14 по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IFNA14 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14 для повышения уровня экспрессии IFNA14 в эукариотических клетках.
Пример 13.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA2, несущего ген IFNA2, для повышения экспрессии белка IFNA2 в клетках млекопитающих.
Оценивали изменение количества белка IFNA2 в лизате клеток эпителия бронхов человека BZR после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IFNA2, несущим ген IFNA2 человека. Клетки культивировали как описано в примере 8.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена IFNA2 (В), в качестве трансфицируемых агентов - ДНК-вектор GDTT1.8NAS12-IFNA2, несущий ген IFNA2 человека (С). Приготовление комплекса ДНК-дендример и трансфекцию клеток эпителия бронхов человека BZR проводили как описано в примере 11.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка. Количественное определение белка IFNA2 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Human Interferon Alpha 2 ELISA Kit (Abcam, ab233622) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка IFNA2, входящим в состав набора. Чувствительность метода составляла не менее 56.04 пг/мл, диапазон измерения - от 187.5 пг/мл до 12000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа представлены на фиг. 9.
Из фигуры 9 следует, что трансфекция клеток эпителия бронхов человека BZR генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA2 приводит к увеличению количества белка IFNA2 по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IFNA2 на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA2 для повышения уровня экспрессии IFNA2 в эукариотических клетках.
Пример 14.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12A, несущего ген IL12A, для повышения экспрессии белка IL12A в клетках млекопитающих.
Оценивали изменение количества белка IL12A в лизате культуры клеток эндотелия пупочной вены человека HUVEC после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IL12A, несущим ген IL12A человека. Клетки культивировали как описано в примере 9.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена IL12A (В), в качестве трансфицируемых агентов - ДНК-вектор GDTT1.8NAS12-IL12A, несущий ген IL12A человека (С). Приготовление комплекса ДНК-дендримеры и трансфекцию клеток HUVEC проводили как описано в примере 11.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка.
Количественное определение белка IL12A проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Human IL12A / p35 ELISA Kit (Sandwich ELISA) (LifeSpan BioSciences LS-F24142-1) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка IL12A, входящим в состав набора. Чувствительность метода составляла не менее 4.688 пг/мл, диапазон измерения - от 7.813 пг/мл до 500 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа, представлены на фиг. 10.
Из фигуры 10 следует, что трансфекция культуры клеток HUVEC генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL12A приводит к увеличению количества белка IL12A по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IL12A на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12A для повышения уровня экспрессии IL12A в эукариотических клетках.
Пример 15.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12B, несущего ген IL12B, для повышения экспрессии белка IL12B в клетках млекопитающих.
Оценивали изменение количества белка IL12B в лизате культуры клеток скелетных миобластов человека HSkM после трансфекции этих клеток ДНК-вектором GDTT1.8NAS12-IL12B, несущим ген IL12B человека. Клетки культивировали как описано в примере 10.
Для трансфекции использовали реагент SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия). В качестве контроля использовали водный раствор дендримеров без ДНК-вектора (А), ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена IL12B (В), в качестве трансфицируемых агентов - ДНК-вектор GDTT1.8NAS12-IL12B, несущий ген IL12B человека (С). Приготовление комплекса ДНК-дендримеры и трансфекцию клеток HSkM проводили как описано в примере 11.
После трансфекции к 0,5 мл культуральной жидкости добавляли 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовали смесь, добавляя 0,1 мл 1.2М NaOH/0.5M HEPES (pH 7-7,6) и тщательно перемешивали. Отбирали супернатант и использовали его для количественного определения целевого белка.
Количественное определение белка IL12B проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Human IL12B / IL12 p40 ELISA Kit (Sandwich ELISA) (LifeSpan BioSciences LS-F24549-1) согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США).
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка IL12B, входящим в состав набора. Чувствительность метода составляла не менее 2 пг/мл, диапазон измерения - от 31,2 пг/мл до 2000 пг/мл. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа, представлены на фиг. 11.
Из фигуры 11 следует, что трансфекция культуры клеток HSkM генотерапевтическим ДНК-вектором GDTT1.8NAS12-IL12B приводит к увеличению количества белка IL12B по сравнению с контрольными образцами, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IL12B на уровне белка. Представленные результаты также подтверждают реализуемость способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12B для повышения уровня экспрессии IL12B в эукариотических клетках.
Пример 16.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12A, несущего ген IL12A, для повышения экспрессии белка IL12A в тканях человека.
Для подтверждения эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12A, несущего целевой ген, а именно, ген IL12A и реализуемости способа его применения оценивали изменения количества белка IL12A в коже человека при введении в кожу человека генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12A, несущего ген человека IL12A.
С целью анализа изменения количества белка IL12A трем пациентам вводили в кожу предплечья генотерапевтический ДНК-вектор GDTT1.8NAS12-IL12A, несущий ген IL12A и параллельно вводили плацебо, представляющее собой генотерапевтический ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена IL12A.
Пациент 1, мужчина 66 лет, (П1); пациент 2, женщина 65 лет, П2); пациент 3, мужчина, 52 года, (П3). В качестве транспортной системы использовали полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, Франция). Генотерапевтический ДНК-вектор GDTT1.8NAS12-IL12A, который содержит кДНК гена IL12A и генотерапевтический ДНК-вектор GDTT1.8NAS12, используемый в качестве плацебо, который не содержит кДНК гена IL12A, каждый из которых растворяли в стерильной воде степени очистки Nuclease-Free. Для получения генетической конструкции готовили комплексы ДНК- cGMP grade in-vivo-jetPEI в соответствии с рекомендациями производителя.
Генотерапевтический ДНК-вектор GDTT1.8NAS12 (плацебо) и генотерапевтический ДНК вектор GDTT1.8NAS12-IL12A, несущий ген IL12A вводили в количестве 1 мг для каждой генетической конструкции тоннельным методом иглой 30G на глубину 3 мм. Объем вводимого раствора генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо) и генотерапевтического ДНК вектора GDTT1.8NAS12-IL12A, несущего ген IL12A - 0,3 мл для каждой генетической конструкции. Очаги введения каждой генетической конструкции располагались на участке предплечья на расстоянии 8-10 см друг от друга.
Биопсийные образцы отбирали на 2-е сутки после введения генетических конструкций генотерапевтических ДНК-векторов. Взятие биопсии осуществляли из участков кожи пациентов в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IL12A, несущего ген IL12A (I), генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо) (II), а также из интактных участков кожи (III), используя устройство для взятия биопсии Epitheasy 3.5 (Medax SRL, Италия). Кожу пациентов в зоне отбора биоптата предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер биопсийного образца составлял около 10 куб. мм, масса - около 11 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000g. Отбирали супернатант и использовали его для количественного определения целевого белка методом твердофазного иммуноферментного анализа (ELISA). Количественное определение белка IL12A проводили методом твердофазного иммуноферментного анализа как это описано в примере 14.
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка IL12A, входящим в состав набора. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). согласно методике производителя с детекцией оптической плотности при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness Technology Inc., США). Графики, полученные в результате анализа, представлены на фиг. 12.
Из фигуры 12 следует, что в коже всех трех пациентов в области введения генотерапевтического ДНК вектора GDTT1.8NAS12- IL12A, несущего целевой ген IL12A человека, произошло увеличение количества белка IL12A в сравнении с количеством белка IL12A в области введения генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо), не содержащего ген IL12A человека, что свидетельствует об эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12A и подтверждает реализуемость способа его применения, в частности при внутрикожном введении генотерапевтического ДНК-вектора в ткани человека.
Пример 17.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA2, несущего ген IFNA2, для повышения экспрессии белка IFNA2 в тканях человека.
Для подтверждения эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA2, несущего целевой ген IFNA2 и реализуемости способа его применения оценивали изменение количества белка IFNA2 в мышечной ткани человека при введении генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA2, несущего целевой ген, а именно, ген IFNA2 человека.
С целью анализа изменения количества белка IFNA2 трем пациентам вводили в ткань икроножной мышцы генотерапевтический ДНК-вектор GDTT1.8NAS12- IFNA2, несущий ген IFNA2 с транспортной молекулой, и параллельно вводили плацебо, представляющее собой генотерапевтический ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена IFNA2 с транспортной молекулой.
Пациент 1, женщина 48 лет, (П1); пациент 2, мужчина 32 года (П2); пациент 3, мужчина 54 года, (П3). В качестве транспортной системы использовали полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, Франция), пробоподготовку проводили в соответствии с рекомендациями производителя.
Генотерапевтический ДНК-вектор GDTT1.8NAS12 (плацебо) и генотерапевтический ДНК вектор GDTT1.8NAS12-IFNA2, несущий ген IFNA2 вводили в количестве 1 мг для каждой генетической конструкции тоннельным методом иглой 30G на глубину около 10 мм. Объем вводимого раствора генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо) и генотерапевтического ДНК вектора GDTT1.8NAS12-IFNA2, несущего ген IFNA2 - 0,3 мл для каждой генетической конструкции. Очаги введения каждой генетической конструкции располагали медиально на расстоянии 8-10 см друг от друга.
Биопсийные образцы отбирали на 2-е сутки после введения генетических конструкций генотерапевтических ДНК-векторов. Взятие биопсии осуществляли из участков мышечной ткани пациентов в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IFNA2, несущего ген IFNA2 (I), генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо) (II), а также из интактного участка икроножной мышцы (III), используя устройство для взятия биопсии MAGNUM (BARD, США). Кожу пациентов в зоне отбора биоптата предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер биопсийного образца составлял около 20 куб. мм, масса - около 22 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000g. Отбирали супернатант и использовали его для количественного определения целевого белка.
Количественное определение белка IFNA2 проводили методом твердофазного иммуноферментного анализа как это описано в примере 13.
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка IFNA2, входящим в состав набора. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа представлены на фиг. 13.
Из фигуры 13 следует, что в икроножной мышце всех трех пациентов в области введения генотерапевтического ДНК вектора GDTT1.8NAS12-IFNA2, несущего целевой ген, а именно, ген IFNA2 человека, произошло увеличение количества белка IFNA2 в сравнении с количеством белка IFNA2 в области введения генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо), не содержащего ген IFNA2 человека, что свидетельствует об эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA2 и подтверждает реализуемость способа его применения, в частности при внутримышечном введении генотерапевтического ДНК-вектора в ткани человека.
Пример 18.
Подтверждение эффективности и реализуемости способа применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14, несущего ген IFNA14, для повышения экспрессии белка IFNA14 в тканях человека.
Для подтверждения эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14, несущего целевой ген, а именно, ген IFNA14 и реализуемости способа его применения оценивали изменения количества белка IFNA14 в коже человека при введении в кожу человека генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14, несущего ген человека IFNA14.
С целью анализа изменения количества белка IFNA14 трем пациентам вводили в кожу предплечья генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNA14, несущий ген IFNA14 и параллельно вводили плацебо, представляющее собой генотерапевтический ДНК-вектор GDTT1.8NAS12, не содержащий кДНК гена IFNA14.
Пациент 1, женщина 55 лет (П1); пациент 2, мужчина 47 лет (П2); пациент 3, мужчина, 60 лет (П3). В качестве транспортной системы использовали полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, Франция). Генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNA14, который содержит кДНК гена IFNA14 и генотерапевтический ДНК-вектор GDTT1.8NAS12, используемый в качестве плацебо, который не содержит кДНК гена IFNA14, каждый из которых растворяли в стерильной воде степени очистки Nuclease-Free. Для получения генетической конструкции готовили комплексы ДНК- cGMP grade in-vivo-jetPEI в соответствии с рекомендациями производителя.
Генотерапевтический ДНК-вектор GDTT1.8NAS12 (плацебо) и генотерапевтический ДНК вектор GDTT1.8NAS12-IFNA14, несущий ген IFNA14 вводили в количестве 1 мг для каждой генетической конструкции тоннельным методом иглой 30G на глубину 3 мм. Объем вводимого раствора генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо) и генотерапевтического ДНК вектора GDTT1.8NAS12-IFNA14, несущего ген IFNA14 - 0,3 мл для каждой генетической конструкции. Очаги введения каждой генетической конструкции располагались на участке предплечья на расстоянии 8-10 см друг от друга.
Биопсийные образцы отбирали на 2-е сутки после введения генетических конструкций генотерапевтических ДНК-векторов. Взятие биопсии осуществляли из участков кожи пациентов в зоне введения генотерапевтического ДНК вектора GDTT1.8NAS12-IFNA14, несущего ген IFNA14 (I), генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо) (II), а также из интактных участков кожи (III), используя устройство для взятия биопсии Epitheasy 3.5 (Medax SRL, Италия). Кожу пациентов в зоне отбора биоптата предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер биопсийного образца составлял около 10 куб. мм, масса - около 11 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000g. Отбирали супернатант и использовали его для количественного определения целевого белка методом твердофазного иммуноферментного анализа как это описано в примере 12.
Численное значение концентрации определяли с помощью калибровочной кривой, построенной по стандартным образцам с известной концентрацией белка IFNA14, входящим в состав набора. Статистическую обработку полученных результатов осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Графики, полученные в результате анализа, представлены на фиг. 14.
Из фигуры 14 следует, что в коже всех трех пациентов в области введения генотерапевтического ДНК вектора GDTT1.8NAS12-IFNA14, несущего целевой ген IFNA14 человека, произошло увеличение количества белка IFNA14 в сравнении с количеством белка IFNA14 в области введения генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо), не содержащего ген IFNA14 человека, что свидетельствует об эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14 и подтверждает реализуемость способа его применения, в частности при внутрикожном введении генотерапевтического ДНК-вектора в ткани человека.
Пример 19.
Подтверждение эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14, несущего ген IFNA14 и реализуемости способа его применения для повышения уровня экспрессии белка IFNA14 в тканях человека путем введения аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA14.
Для подтверждения эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14, несущего ген IFNA14 и реализуемости способа его применения оценивали изменения количества белка IFNA14 в коже человека при введении в кожу пациента культуры аутологичных фибробластов этого пациента, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA14.
Пациенту вводили в кожу предплечья соответствующую культуру аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA14, несущим ген IFNA14, и параллельно вводили плацебо, представляющее собой культуру аутологичных фибробластов пациента, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, не несущим ген IFNA14.
Первичную культуру фибробластов человека выделяли из биоптатов кожи пациента. Используя устройство для взятия биопсии кожи Epitheasy 3.5 (Medax SRL, Италия) брали образец биоптата кожи из зоны, защищенной от действия ультрафиолета, а именно за ушной раковиной или с боковой внутренней поверхности в зоне локтевого сустава, размером около 10 кв. мм, массой - около 11 мг. Кожу пациента предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Культивирование первичной культуры клеток осуществляли при 370С в атмосфере, содержащей 5 % СО2 в среде DMEM с 10 % фетальной телячьей сывороткой и ампициллином 100 Ед/мл. Пересев культуры и смену культуральной среды осуществляли каждые 2 дня. Общая продолжительность роста культуры не превышала 25-30 дней. Из культуры клеток отбирали аликвоту, содержащую 5×104 клеток. Культуру фибробластов пациента трансфицировали генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA14, несущим ген IFNA14 или плацебо – вектор GDTT1.8NAS12, не несущий целевой ген IFNA14.
Трансфекцию осуществляли с помощью катионного полимера полиэтиленимина JETPEI (Polyplus Transfection, Франция), согласно инструкции фирмы-производителя. Клетки культивировали 72 часа и затем вводили пациенту. Введение пациенту культуры аутологичных фибробластов этого пациента, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA14, и плацебо, представляющее собой культуру аутологичных фибробластов пациента, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, осуществляли туннельным методом в область предплечья иглой 30G длиной 13 мм на глубину около 3 мм. Концентрация модифицированных аутологичных фибробластов в вводимой суспензии составляла примерно 5 млн клеток на 1 мл суспензии, количество вводимых клеток не превышало 15 млн. Очаги введения культуры аутологичных фибробластов располагались на расстоянии 8-10 см друг от друга.
Биопсийные образцы отбирали на 4-е сутки после введения культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA14, несущим целевой ген, а именно, ген IFNA14 и плацебо. Взятие биопсии осуществляли из участков кожи пациента в зоне введения культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA14, несущим целевой ген, а именно, ген IFNA14 (C), культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, не несущим целевой ген IFNA14 (плацебо) (B), а также из участков интактной кожи (A), используя устройство для взятия биопсии Epitheasy 3.5 (Medax SRL, Италия). Кожу в зоне отбора биоптата пациентов предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер биопсийного образца составлял около 10 куб. мм, масса - около 11 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000g. Отбирали супернатант и использовали его для количественного определения целевого белка как это описано в примере 12.
Графики, полученные в результате анализа, представлены на фиг. 15.
Из фигуры 15 следует, что в коже пациента в области введения культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA14, несущим ген IFNA14, произошло увеличение количества белка IFNA14 в сравнении с количеством белка IFNA14 в области введения культуры аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12, не несущим ген IFNA14 (плацебо), что свидетельствует об эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14 и подтверждает реализуемость способа его применения для повышения уровня экспрессии IFNA14 в тканях человека, в частности при введении аутологичных фибробластов, трансфицированных генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA14.
Пример 20.
Подтверждение эффективности и реализуемости способа сочетанного применения генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, несущего ген IFNB1, генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14, несущего ген IFNA14, генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA2, несущего ген IFNA2, генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12A, несущего ген IL12A, генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12B, несущего ген IL12B, для повышения уровня экспрессии белков IFNB1, IFNA14, IFNA2, IL12A и IL12B в тканях млекопитающих.
Оценивали изменение количества белков IFNB1, IFNA14, IFNA2, IL12A и IL12B в зоне мышечной ткани при сочетанном введении в эту зону смеси генотерапевтических векторов. Исследование проводили на 3 лабораторных животных – самцах крысы линии Wistar 8 месячного возраста массой 240–290 г. В качестве транспортной системы использовали полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, Франция). Эквимолярную смесь генотерапевтических ДНК-векторов растворяли в стерильной воде степени очистки Nuclease-Free. Для получения генетической конструкции готовили комплексы ДНК- cGMP grade in-vivo-jetPEI в соответствии с рекомендациями производителя.
Объем внутримышечно вводимого раствора составлял 0,1 мл с суммарным количеством ДНК 100 мкг. Введение раствора осуществляли с помощью инсулинового шприца. Через 2 сутки после процедуры крыс декапитировали.
Образцы отбирали на 2 сутки после введения генотерапевтических ДНК-векторов. Взятие биопсийного материала осуществляли из участков правой мышцы бедра в зоне введения смеси пяти генотерапевтических ДНК-векторов, несущих гены IFNB1, IFNA14, IFNA2, IL12A, IL12B (зона I), из участков левой мышцы бедра в зоне введения генотерапевтического ДНК-вектора GDTT1.8NAS12 (плацебо) (зона II), а также из участков мышечной ткани, не подвергавшихся никаким манипуляциям ( зона III). Каждый образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторида, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 мин при 14000g. Отбирали супернатант и использовали его для количественного определения целевых белков как это описано в примере 11 (количественное определение белка IFNB1), примере 12 (количественное определение белка IFNA14), примере 13 (количественное определение белка IFNA2), примере 14 (количественное определение белка IL12A), примере 15 (количественное определение белка IL12B). Графики, полученные в результате анализа, представлены на фиг. 16.
Из фигур 16 следует, что в зоне I, в которую вводилась смесь генотерапевтических векторов: генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, несущего целевой ген IFNB1, генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14, несущего целевой ген IFNA14, генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA2, несущего целевой ген IFNA2, генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12A, несущего целевой ген IL12A, генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12B, несущего целевой ген IL12B, произошло увеличение количества белков IFNB1, IFNA14, IFNA2, IL12A и IL12B по сравнению с зоной II (зона плацебо) и зоной III (интактная зона). Полученные результаты свидетельствуют об эффективности сочетанного применения генотерапевтических ДНК-векторов и подтверждает реализуемость способа применения для повышения уровня экспрессии целевых белков в тканях млекопитающих.
Пример 21.
Подтверждение эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA2, несущего ген IFNA2 и реализуемости способа его применения для повышения уровня экспрессии белка IFNA2 в клетках млекопитающих.
Для подтверждения эффективности генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA2, несущего ген IFNA2 оценивали изменение накопления мРНК целевого гена IFNA2 в клетках эндотелия легочной артерии быка CPAE (ATCC® CCL-209) через 48 часов после их трансфекции генотерапевтическим ДНК-вектором GDTT1.8NAS12- IFNA2, несущим ген IFNA2 человека.
Клетки СРАЕ культивировали в среде DMEM (Gibco) с добавлением 10% сыворотки эмбрионов коров (Gibco) в стандартных условиях. Трансфекцию генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA2, несущим ген IFNA2 человека и ДНК-вектором GDTT1.8NAS12, не несущим ген IFNA2 человека (контроль), выделение РНК, реакцию обратной транскрипции, ПЦР амплификации и анализ данных проводили как описано в примере 8. В качестве референтного гена использовали ген актина быка/ коровы (АСТ), приведенного в базе данных GenBank под номером AH001130.2. В качестве положительного контроля использовали ампликоны, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности генов IFNA2 и АСТ. В качестве отрицательного контроля использовали деионизированную воду. Количество ПЦР продуктов – кДНК генов IFNA2 и ACT, полученных в результате амплификации, оценивали в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1. (Bio-Rad, USA).
Графики, полученные в результате анализа представлены на фиг. 17.
Из фигуры 17 следует, что в результате трансфекции клеток эндотелия легочной артерии быка CPAE генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA2, уровень специфической мРНК гена IFNA2 человека вырос многократно, что подтверждает способность вектора проникать в эукариотические клетки и экспрессировать ген IFNA2 на уровне мРНК. Представленные результаты подтверждают реализуемость способа применения генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNA2 для повышения уровня экспрессии гена IFNA2 в клетках млекопитающих.
Пример 22.
Штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1 или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA14 или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA2 или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12A или штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12B, несущий генотерапевтический ДНК-вектор, и способ его получения.
Конструирование штамма для наработки в промышленных масштабах генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген, выбранный из группы генов: IFNB1, IFNA14, IFNA2, IL12A, IL12B а именно штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA14, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA2, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12A или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12B, несущего генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-IFNA14, или GDTT1.8NAS12-IFNA2, или GDTT1.8NAS12-IL12A или GDTT1.8NAS12-IL12B соответственно для его наработки с возможностью селекции без использования антибиотиков заключается в получении электрокомпетентных клеток штамма Escherichia coli JM110-NAS с проведением электропорации этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNB1, или ДНК-вектором GDTT1.8NAS12-IFNA14, или ДНК-вектором GDTT1.8NAS12-IFNA2, или ДНК-вектором GDTT1.8NAS12-IL12A, или ДНК-вектором GDTT1.8NAS12-IL12B после чего клетки высеваются на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола. При этом штамм Escherichia coli JM110-NAS для наработки генотерапевтического ДНК-вектора GDTT1.8NAS12 или генотерапевтических ДНК-векторов на его основе с возможностью положительной селекции без использования антибиотиков получали путем конструирования линейного фрагмента ДНК, содержащего регуляторный элемент RNA-in транспозона Tn10 для селекции без применения антибиотиков размером 64 п.н., ген левансахаразы sacB, продукт которого обеспечивает селекцию на сахарозо-содержащей среде размером 1422 п.н., ген устойчивости к хлорамфениколу catR, необходимый для отбора клонов штамма, в которых прошла гомологичная рекомбинация размером 763 п.н. и две гомологичные последовательности, обеспечивающие процесс гомологичной рекомбинации в области гена recA с одновременной его инактивацией размером 329 п.н. и 233 п.н., после чего проводили трансформацию клеток Escherichia coli путем электропорации и отбирали клоны, выжившие на среде, содержащей 10 мкг/мл хлорамфеникола.
Полученные штаммы для наработки были внесены в коллекции Национального биоресурсного центра - Всероссийская коллекция промышленных микроорганизмов (НБЦ ВКПМ), РФ и NCIMB Patent Deposit Service, UK с присвоением регистрационных номеров:
штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1 – регистрационный № ВКПМ В-13534, дата депонирования 27.11.2019; INTERNATIONAL DEPOSITARY AUTHORITY № NCIMB - 43525, дата депонирования 28.11.2019;
штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA14 - регистрационный № ВКПМ В-13539, дата депонирования 27.11.2019; INTERNATIONAL DEPOSITARY AUTHORITY № NCIMB - 43520 дата депонирования 28.11.2019;
штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA2 - регистрационный № ВКПМ В-13536, дата депонирования 27.11.2019; INTERNATIONAL DEPOSITARY AUTHORITY № NCIMB - 43521, дата депонирования 28.11.2019;
штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12A - регистрационный № ВКПМ В-13537, дата депонирования 27.11.2019; INTERNATIONAL DEPOSITARY AUTHORITY № NCIMB - 43524, дата депонирования 28.11.2019;
штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12B - регистрационный № ВКПМ В-13535, дата депонирования 27.11.2019; INTERNATIONAL DEPOSITARY AUTHORITY № NCIMB - 43523, дата депонирования 28.11.2019.
Пример 23.
Способ масштабирования получения генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген, выбранный из группы генов: IFNB1, IFNA14, IFNA2, IL12A, IL12B до промышленного масштаба.
Для подтверждения технологичности получения и возможности производства в промышленных масштабах генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1 (SEQ ID №1), или GDTT1.8NAS12-IFNA14 (SEQ ID №2), или GDTT1.8NAS12-IFNA2 (SEQ ID №3), или GDTT1.8NAS12-IL12A (SEQ ID №4), или GDTT1.8NAS12-IL12B (SEQ ID №5) проводили масштабную ферментацию штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA14, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA2, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12A, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12B, каждый из которых содержит генотерапевтический ДНК-вектор GDTT1.8NAS12, несущий целевой ген, а именно IFNB1, или IFNA14, или IFNA2, или IL12A, или IL12B. Каждый штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA14, или Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA2, или Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12A, или Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12B получали на базе штамма Escherichia coli JM110-NAS (Genetic Diagnostics and Therapy 21 Ltd, Великобритания) по примеру 22 путем электропорации компетентных клеток этого штамма генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-IFNA14, или GDTT1.8NAS12-IFNA2, или GDTT1.8NAS12-IL12A, или GDTT1.8NAS12-IL12B, несущим целевой ген, а именно, IFNB1, или IFNA14, или IFNA2, или IL12A или IL12B с последующим высевом трансформированных клеток на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы и отбором отдельных клонов.
Ферментацию полученного штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, несущего генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1 проводили в ферментере объемом 10 л с последующим выделением генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1.
Для ферментации штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1 готовили среду, содержащую на 10 л: 100 г триптон, 50 г дрожжевой экстракт (Becton Dickinson, США), доводили водой до 8800 мл и автоклавировали при 121 0С 20 мин, затем добавляли 1200 мл 50% (вес/объем) сахарозы. Далее засевали в колбу затравочную культуру штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1 в объеме 100 мл. Инкубировали в шейкере-инкубаторе 16 ч при 30°С. Переносили затравочную культуру в ферментер Techfors S (Infors HT, Швейцария), растили до достижения стационарной фазы. Контроль осуществляли измерением оптической плотности культуры при длине волны 600 нм. Клетки осаждали центрифугированием 30 мин при 5000-10000 g. Супернатант удаляли, клеточную массу ресуспендировали в 10% по объему фосфатно-солевого буфера. Повторно центрифугировали 30 мин при 5000-10000g. Супернатант удаляли, к клеточной массе добавляли раствор 20 мМ TrisCl, 1 мМ ЭДТА, 200 г/л сахароза, рН 8,0 в объеме 1000 мл, тщательно перемешивали до образования гомогенной суспензии. Добавляли раствор яичного лизоцима до конечной концентрации 100мкг/мл. Инкубировали 20 мин на льду при бережном перемешивании. Далее, добавляли 2500 мл раствора 0,2 М NaOH, 10 г/л додецилсульфат натрия, инкубировали 10 мин на льду при бережном перемешивании, затем добавляли 3500 мл раствора 3M ацетат натрия, 2М уксусная кислота, рН 5-5,5, инкубировали 10 мин на льду при бережном перемешивании. Полученный образец центрифугировали 20-30 мин при 15000 g или более. Раствор аккуратно декантировали, от остатков осадка избавлялись фильтрацией через грубый фильтр (фильтровальную бумагу). Добавляли РНКазу А (Sigma, США) до конечной концентрации 20 нг/мл, инкубировали ночь 16 ч при комнатной температуре. Раствор центрифугировали 20-30 мин при 15000g, затем фильтровали через мембранный фильтр с порами 0,45 мкм (Millipore, США). Далее проводили ультрафильтрацию через мембрану размером отсечения 100кДа (Millipore, США) и разбавляли до начального объема буфером 25 мМ TrisCl, pH 7.0. Операцию повторяли три – четыре раза. Наносили раствор, на колонку с 250 мл сорбента DEAE sepharose HP (GE, США), уравновешенную раствором 25 мМ TrisCl, pH 7.0. После нанесения образца колонку промывали тремя объемами этого же раствора, а затем проводили элюцию генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1 линейным градиентом от раствора 25 мМ TrisCl, pH 7.0 до раствора 25 мМ TrisCl, pH 7.0, 1М NaCl в объеме пяти объемов колонки. Контроль элюции осуществляли по оптической плотности сходящего раствора при 260 нм. Хроматографические фракции, содержащие генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1, объединяли и проводили гель-фильтрацию на сорбенте Superdex 200 (GE, США). Колонку уравновешивали фосфатно-солевым буфером. Контроль элюции осуществляли по оптической плотности сходящего раствора при 260 нм, фракции анализировали электрофорезом в агарозном геле. Фракции, содержащие генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1 объединяли и хранили при -20°С. Для оценки воспроизводимости техпроцесса обозначенные технологические операции повторяли пятикратно. Все технологические операции для штаммов Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA14, или Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA2, или Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12A или Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12B проводили аналогичным образом.
Воспроизводимость техпроцесса и количественные характеристики выхода конечного продукта подтверждают технологичность получения и возможность производства в промышленных масштабах генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-IFNA14, или GDTT1.8NAS12-IFNA2, или GDTT1.8NAS12-IL12A, или GDTT1.8NAS12-IL12B.
Таким образом, созданный генотерапевтический ДНК-вектор с целевым геном может быть использован для введения в клетки организма животных и человека, характеризующихся сниженной или недостаточной экспрессией белка, кодируемого данным геном, обеспечивая таким образом достижение терапевтического эффекта.
Задача, поставленная в данном изобретении, решена, а именно: конструирование генотерапевтических ДНК-векторов для повышения уровня экспрессии генов IFNB1, IFNA14, IFNA2, IL12A, IL12B, сочетающих в себе следующие свойства:
I) Эффективность повышения экспрессии целевых генов в эукариотических клетках за счет полученных генотерапевтических векторов с минимальным размером;
II) Возможность безопасного применения для генетической терапии человека и животных за счет отсутствия в составе генотерапевтического ДНК-вектора регуляторных элементов, представляющих собой нуклеотидные последовательности вирусных геномов и генов антибиотикорезистентности;
III) Технологичность получения и возможность наработки в штаммах в промышленных масштабах;
IV) Также решена задача по конструированию штаммов, несущих эти генотерапевтические ДНК-вектора для производства этих генотерапевтических ДНК-векторов.
Это подтверждается примерами:
для п. I – пример 1, 2, 3, 4, 5; 6; 7; 8; 9; 10; 11; 12; 13; 14; 15; 16; 17; 18, 19, 20, 21;
для п. II – пример 1, 2, 3, 4, 5;
для п. III и п. IV – пример 22, 23.
Промышленная применимость:
Все представленные выше примеры подтверждают промышленную применимость предлагаемого генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген, выбранный из группы генов: ген IFNB1, ген IFNA14, ген IFNA2, ген IL12A, ген IL12B для повышения уровня экспрессии этих целевых генов, штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1 или Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA14 или Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA2 или Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12A или Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12B, несущего генотерапевтический ДНК-вектор, способа получения генотерапевтического ДНК-вектора, способа производства в промышленных масштабах генотерапевтического ДНК-вектора.
Перечень сокращений
GDTT1.8NAS12 – вектор генотерапевтический, не содержащий последовательностей вирусных геномов и маркеров антибиотико-резистентности (vector therapeutic virus-antibiotic-free)
ДНК - дезоксирибонуклеиновая кислота
кДНК - комплементарная дезоксирибонуклеиновая кислота
РНК - рибонуклеиновая кислота
мРНК - матричная рибонуклеиновая кислота
п.н. – пар нуклеотидов
ПЦР - полимеразная цепная реакция
мл – миллилитр, мкл – микролитр
куб. мм – кубический миллиметр
л - литр
мкг - микрограмм
мг - миллиграмм
г - грамм
мкМ - микромоль
мМ – миллимоль
мин – минута
сек - секунда
об/мин - обороты в минуту
нм - нанометр
см - сантиметр
мВт - милливатт
о.е. ф-относительная единица флуоресценции
РВС – фосфатно-солевой буфер
PBMC – мононуклеарные клетки периферической крови
МНС – главный комплекс гистосовместимости (major histocompatibility complex)
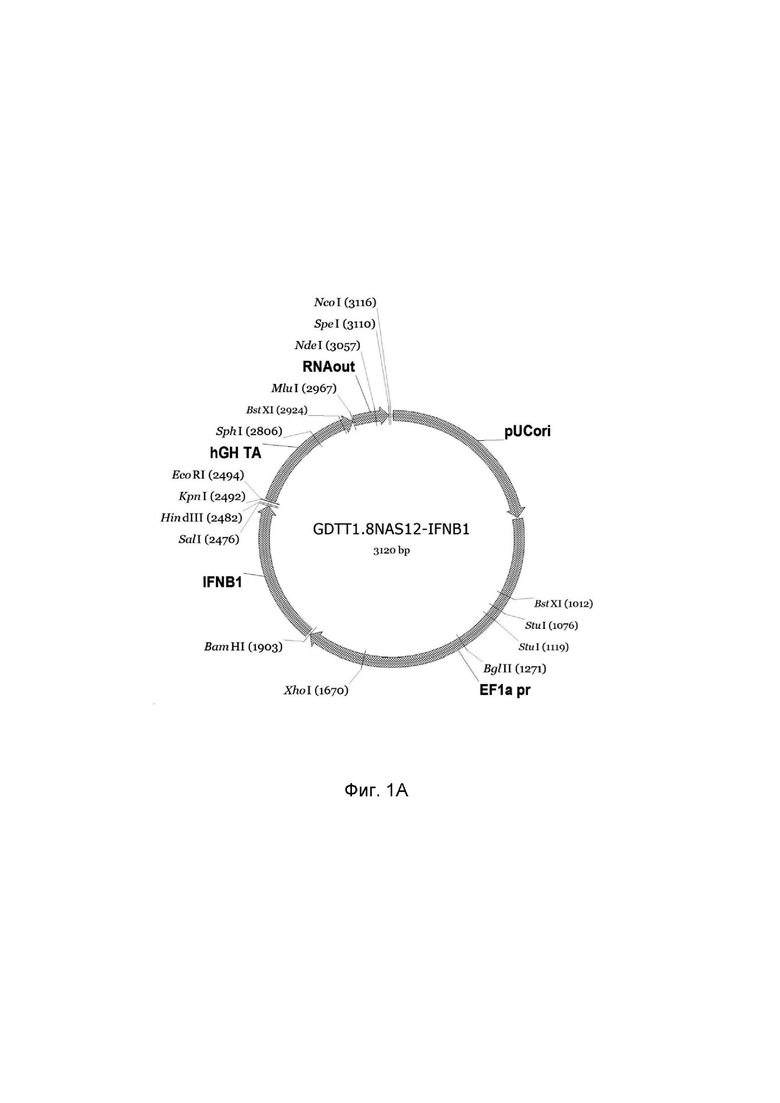
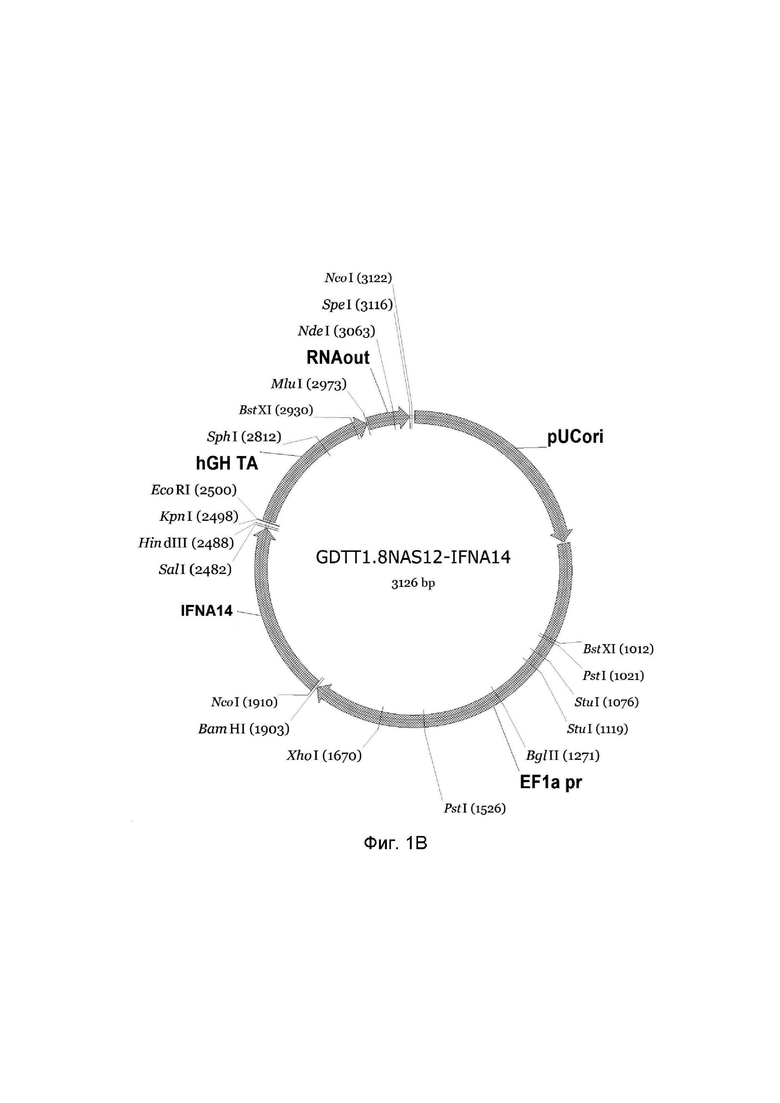

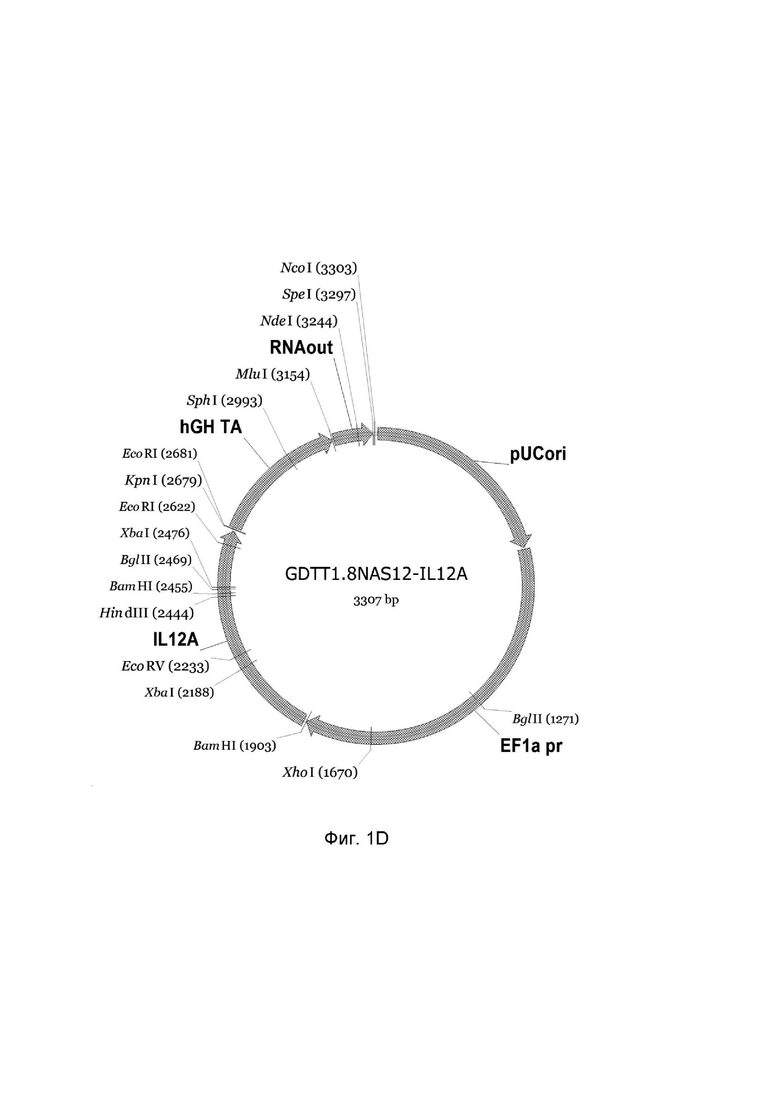
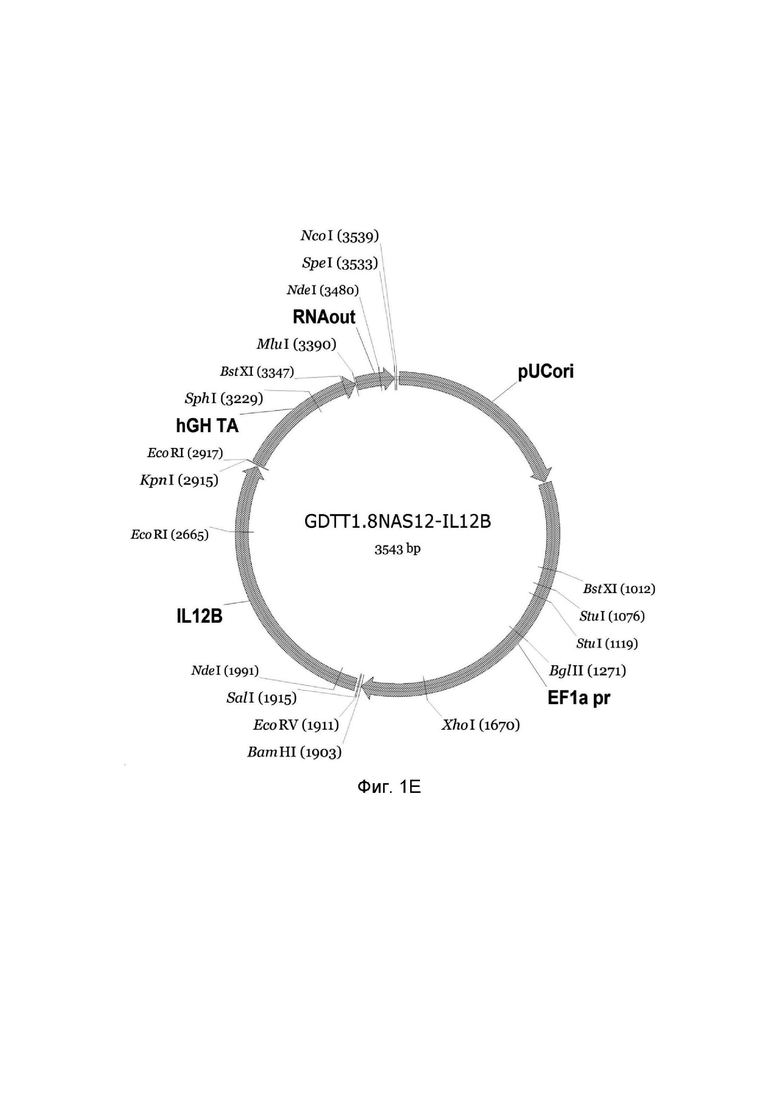
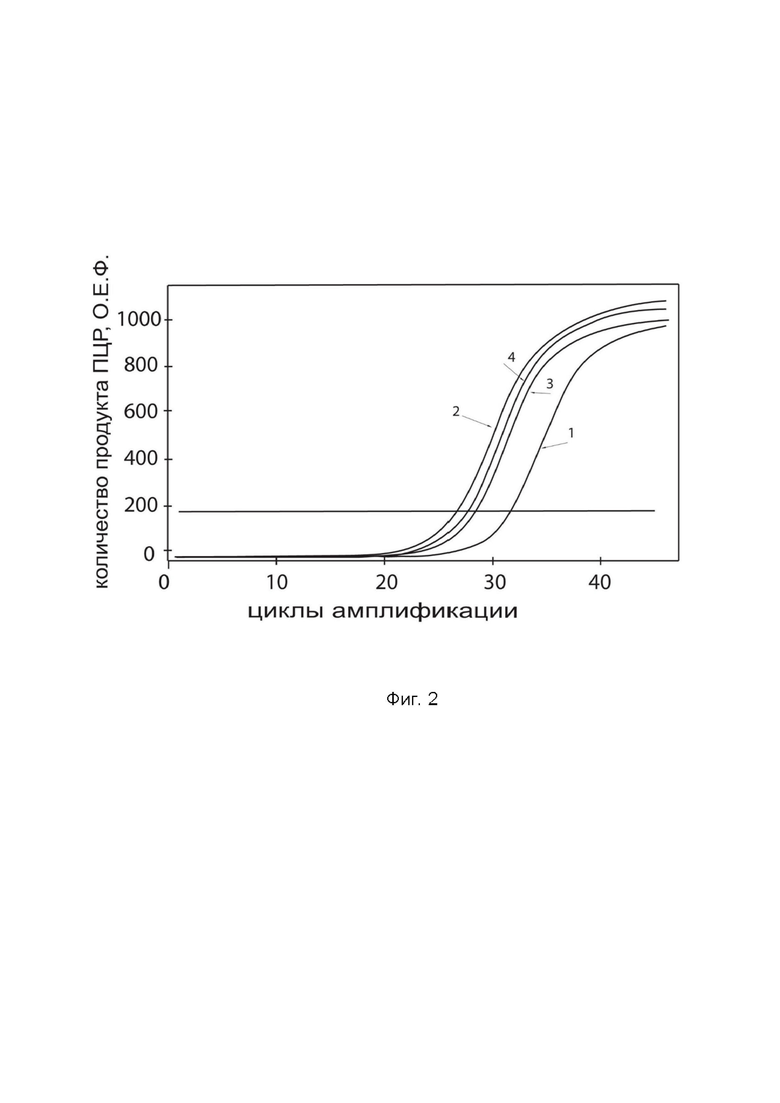


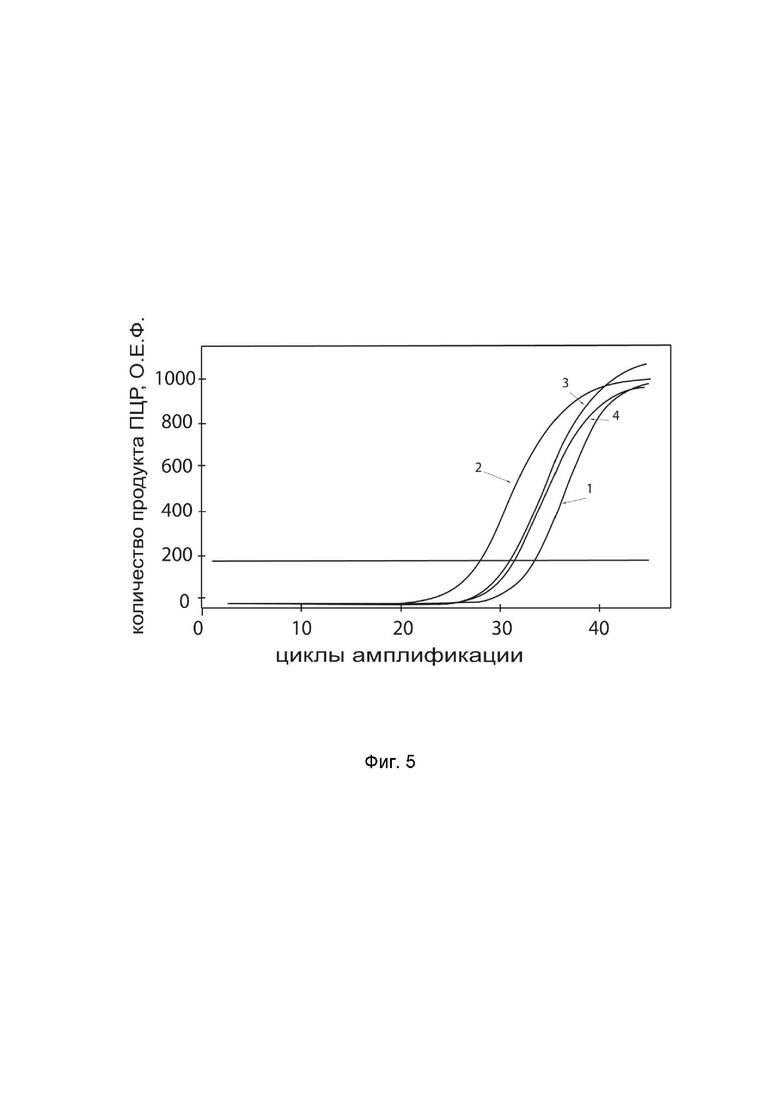
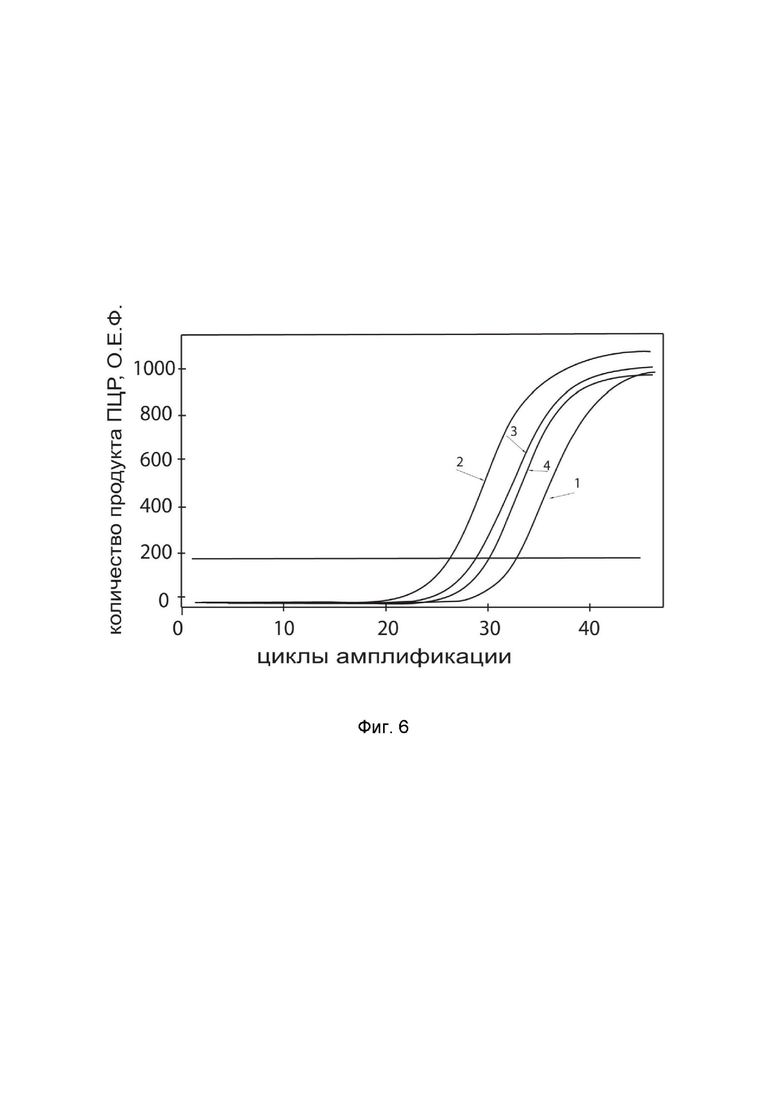




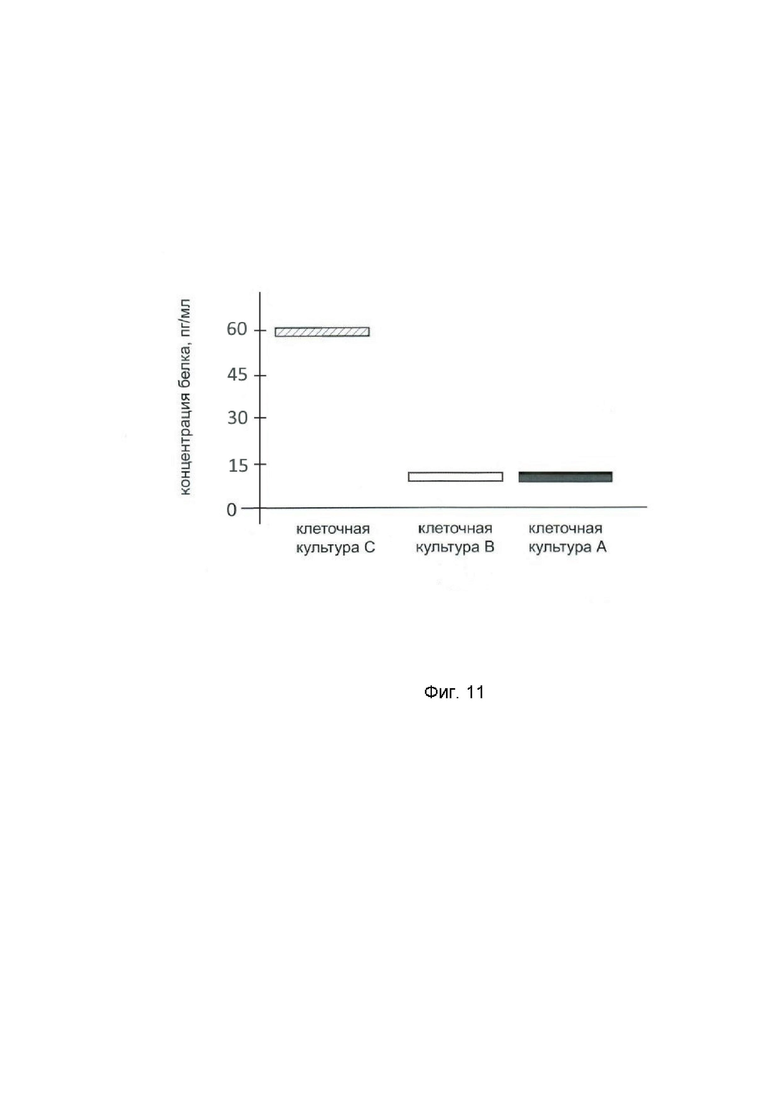
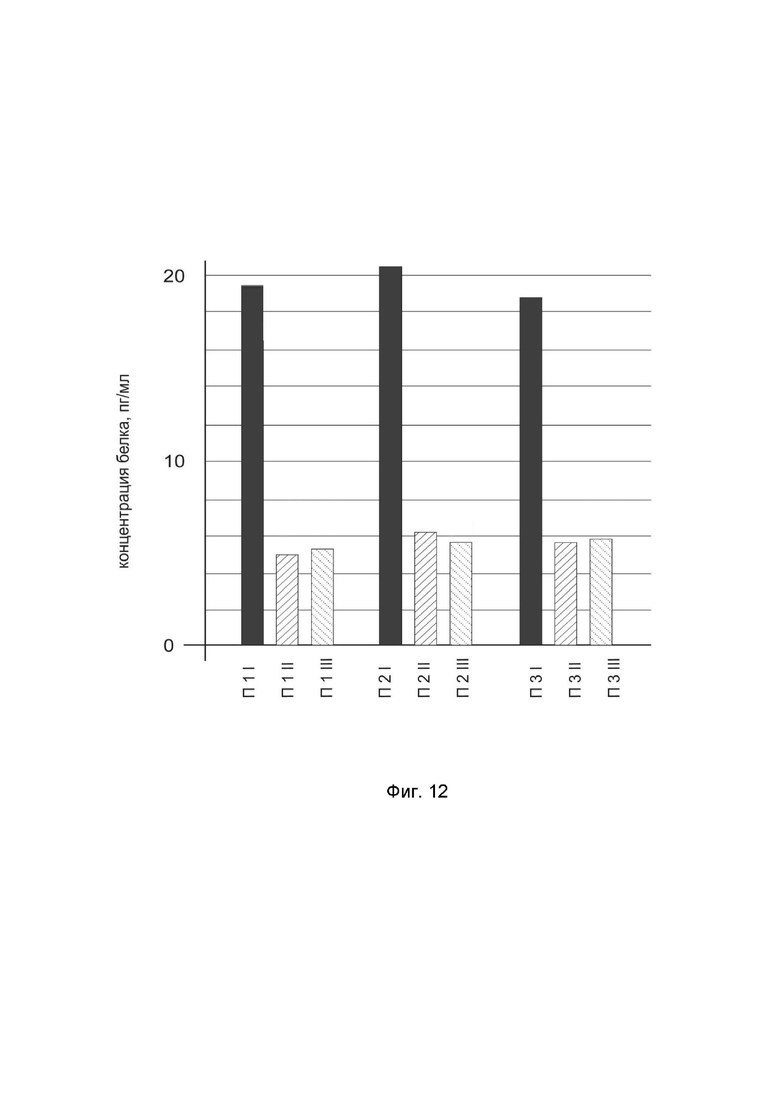


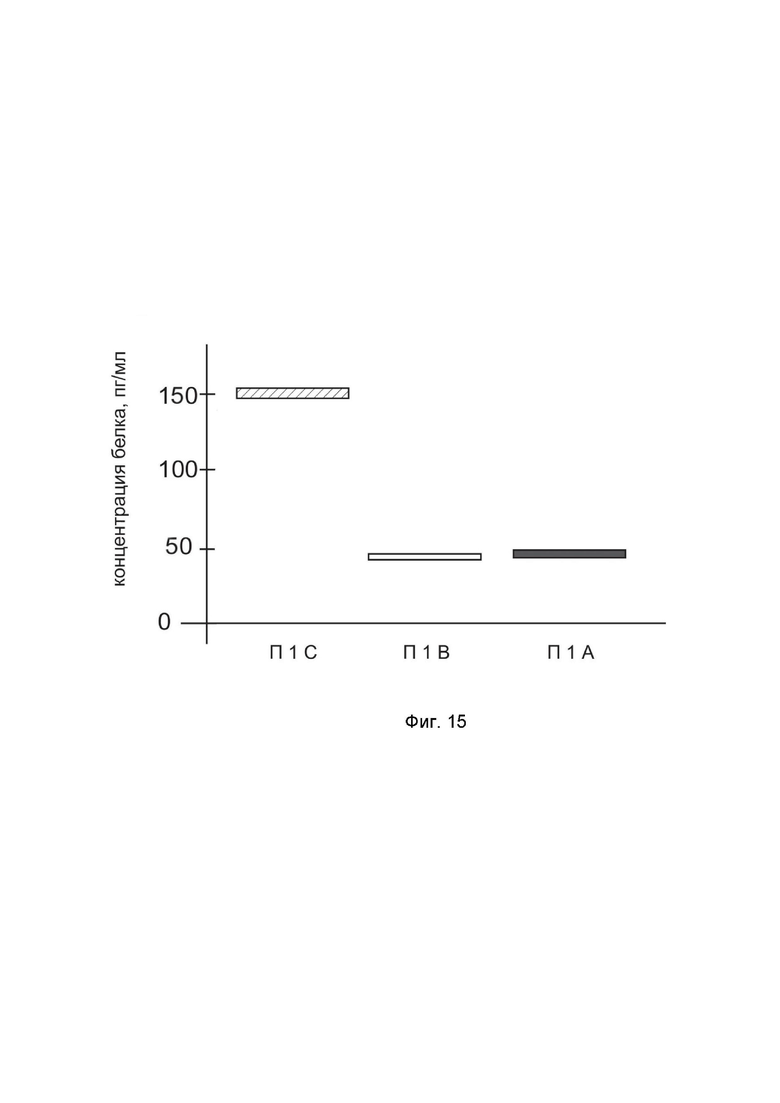

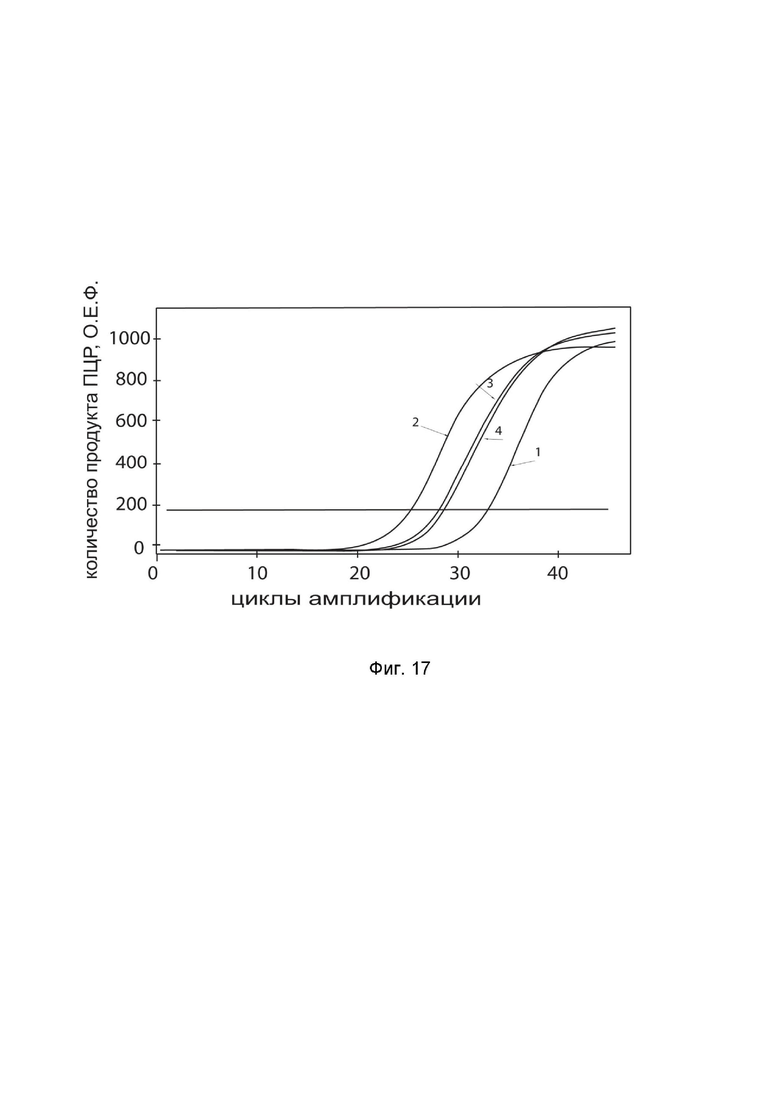
Изобретение относится к генной инженерии и может быть использовано в биотехнологии, медицине и сельском хозяйстве для создания препаратов генной терапии. Создан генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 для лечения заболеваний, характеризующихся нарушением врожденного и адаптивного иммунитета, для терапии аутоиммунных, онкологических, вирусных заболеваний, связанных с дисбалансом иммунной системы, для иммуномодуляции в том числе при противоопухолевой вакцинации в качестве генотерапевтического адъюванта, при этом генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1 содержит кодирующую часть целевого гена IFNB1, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с нуклеотидной последовательностью SEQ ID №1, генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNA14 содержит кодирующую часть целевого гена IFNA14, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с нуклеотидной последовательностью SEQ ID №2, генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNA2 содержит кодирующую часть целевого гена IFNA2, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с нуклеотидной последовательностью SEQ ID №3, генотерапевтический ДНК-вектор GDTT1.8NAS12-IL12A содержит кодирующую часть целевого гена IL12A, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с нуклеотидной последовательностью SEQ ID №4, генотерапевтический ДНК-вектор GDTT1.8NAS12-IL12B содержит кодирующую часть целевого гена IL12B, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с нуклеотидной последовательностью SEQ ID №5. 16 н.п. ф-лы, 17 ил., 23 пр.
1. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 для лечения заболеваний, характеризующихся нарушением врожденного и адаптивного иммунитета, для терапии аутоиммунных, онкологических, вирусных заболеваний, связанных с дисбалансом иммунной системы, для иммуномодуляции в том числе при противоопухолевой вакцинации в качестве генотерапевтического адъюванта, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена IFNB1, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNB1, с нуклеотидной последовательностью SEQ ID №1.
2. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 для лечения заболеваний, характеризующихся нарушением врожденного и адаптивного иммунитета, для терапии аутоиммунных, онкологических, вирусных заболеваний, связанных с дисбалансом иммунной системы, для иммуномодуляции в том числе при противоопухолевой вакцинации в качестве генотерапевтического адъюванта, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена IFNA14, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14, с нуклеотидной последовательностью SEQ ID №2.
3. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 для лечения заболеваний, характеризующихся нарушением врожденного и адаптивного иммунитета, для терапии аутоиммунных, онкологических, вирусных заболеваний, связанных с дисбалансом иммунной системы, для иммуномодуляции в том числе при противоопухолевой вакцинации в качестве генотерапевтического адъюванта, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена IFNA2, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA2, с нуклеотидной последовательностью SEQ ID №3.
4. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 для лечения заболеваний, характеризующихся нарушением врожденного и адаптивного иммунитета, для терапии аутоиммунных, онкологических, вирусных заболеваний, связанных с дисбалансом иммунной системы, для иммуномодуляции в том числе при противоопухолевой вакцинации в качестве генотерапевтического адъюванта, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена IL12A, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12A, с нуклеотидной последовательностью SEQ ID №4.
5. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 для лечения заболеваний, характеризующихся нарушением врожденного и адаптивного иммунитета, для терапии аутоиммунных, онкологических, вирусных заболеваний, связанных с дисбалансом иммунной системы, для иммуномодуляции в том числе при противоопухолевой вакцинации в качестве генотерапевтического адъюванта, при этом генотерапевтический ДНК-вектор содержит кодирующую часть целевого гена IL12B, клонированную в генотерапевтический ДНК-вектор GDTT1.8NAS12, с получением генотерапевтического ДНК-вектора GDTT1.8NAS12-IL12B, с нуклеотидной последовательностью SEQ ID №5.
6. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущий целевой ген IFNB1, IFNA14, IFNA2, IL12A, IL12B по пп. 1-4 или 5, отличающийся тем, что каждый из созданных генотерапевтических ДНК-векторов: GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-IFNA14, или GDTT1.8NAS12-IFNA2, или GDTT1.8NAS12-IL12A, или GDTT1.8NAS12-IL12B по пп. 1-4 или 5, за счет ограниченного размера векторной части GDTT1.8NAS12, не превышающей 2600 п.н., обладает способностью эффективно проникать в клетки человека и животных и экспрессировать клонированный в него целевой ген IFNB1, или IFNA14, или IFNA2, или IL12A, или IL12B.
7. Генотерапевтический ДНК-вектор на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущий целевой ген IFNB1, или IFNA14, или IFNA2, или IL12A, или IL12B по пп. 1-4 или 5, отличающийся тем, что в составе каждого из созданных генотерапевтических ДНК-векторов: GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-IFNA14, или GDTT1.8NAS12-IFNA2, или GDTT1.8NAS12-IL12A, или GDTT1.8NAS12-IL12B по пп. 1-4 или 5, в качестве структурных элементов используют нуклеотидные последовательности, которые не являются генами антибиотикорезистентности, вирусными генами и регуляторными элементами вирусных геномов, обеспечивая возможность его безопасного применения для генетической терапии человека и животных.
8. Способ получения генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IFNB1, или IFNA14, или IFNA2, или IL12A, или IL12B по пп. 1-4 или 5, заключающийся в том, что каждый из генотерапевтических ДНК-векторов: GDTT1.8NAS12-IFNB1, или GDTT1.8NAS12-IFNA14, или GDTT1.8NAS12-IFNA2, или GDTT1.8NAS12-IL12A, или GDTT1.8NAS12- получают следующим образом: кодирующую часть целевого гена IFNB1, или IFNA14, или IFNA2, или IL12A, или IL12B клонируют в генотерапевтический ДНК-вектор GDTT1.8NAS12 и получают генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNB1, SEQ ID №1, или GDTT1.8NAS12-IFNA14, SEQ ID №2 или GDTT1.8NAS12-IFNA2, SEQ ID №3, или GDTT1.8NAS12-IL12A, SEQ ID №4, или GDTT1.8NAS12-IL12B, SEQ ID №5 соответственно, при этом кодирующую часть целевого гена IFNB1, или IFNA14, или IFNA2, или IL12A, или IL12B получают путем выделения суммарной РНК из биологического образца ткани человека с последующим проведением реакции обратной транскрипции и ПЦР-амплификации с использованием созданных олигонуклеотидов и расщеплением продукта амплификации соответствующими эндонуклеазами рестрикции, причем клонирование в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят по сайтам рестрикции SalI и KpnI, или по сайтам рестрикции BamHI и SalI, причем селекцию проводят без антибиотиков, при этом при получении генотерапевтического ДНК-вектора GDTT1.8NAS12 IFNB1, SEQID №1 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
IFNB1-Fw AAAGGATCCACCATGACCAACAAGTGTCTCCTCCAAA,
IFNB1-Rv TTTGTCGACTCAGTTTCGGAGGTAACCTGTAAGTCTG,
а расщепление продукта амплификации и клонирование кодирующей части гена IFNB1 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12-IFNA14, SEQID №2 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
IFNA14-Fw TTTGGATCCACCATGGCATTGCCCTTTGCTTTAAT,
IFNA14-Rv TTTGTCGACTCAATCCTTCCTCCTTAATCTTTTTTGC,
а расщепление продукта амплификации и клонирование кодирующей части гена IFNA14 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12- IFNA2, SEQID №3 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
IFNA2-Fw GGGGGATCCACCATGGCCTTGACCTTTGCTTTACTGG,
IFNA2-Rv TTTGTCGACTCATTCCTTACTTCTTAAACTTTCTTGC,
а расщепление продукта амплификации и клонирование кодирующей части гена IFNA2 в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции BamHI и SalI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12- IL12A, SEQID №4 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
IL12A-up TTTGTCGACCACCATGTGGCCCCCTGGGTCAGCCTCC
IL12A-lo, AATGGTACCTTAGGAAGCATTCAGATAGCTCATCACTCTATC,
а расщепление продукта амплификации и клонирование кодирующей части гена IL12A в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции SalI и KpnI,
причем при получении генотерапевтического ДНК-вектора GDTT1.8NAS12- IL12B, SEQID №5 для проведения реакции обратной транскрипции и ПЦР-амплификации, в качестве созданных для этого олигонуклеотидов используют олигонуклеотиды:
IL12B-up TTTGTCGACCACCATGTGTCACCAGCAGTTGGTCATCTC,
IL12B-lo AATGGTACCTAACTGCAGGGCACAGATGCCCA,
а расщепление продукта амплификации и клонирование кодирующей части гена IL12B в генотерапевтический ДНК-вектор GDTT1.8NAS12 проводят с использованием эндонуклеаз рестрикции SalI и KpnI.
9. Способ использования генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IFNB1, IFNA14, IFNA2, IL12A, IL12B по пп. 1-4 и/или 5, для лечения заболеваний, характеризующихся нарушением врожденного и адаптивного иммунитета, для терапии аутоиммунных, онкологических, вирусных заболеваний, связанных с дисбалансом иммунной системы, для иммуномодуляции в том числе при противоопухолевой вакцинации в качестве генотерапевтического адъюванта, заключающийся в трансфекции выбранным генотерапевтическим ДНК-вектором, несущим целевой ген на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, или несколькими выбранными генотерапевтическими ДНК-векторами, несущими целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, из созданных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, клеток органов и тканей пациента или животного, и/или во введении в органы и ткани пациента или животного аутологичных клеток этого пациента или животного, трансфицированных выбранным генотерапевтическим ДНК-вектором, несущим целевой ген на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 или несколькими выбранными генотерапевтическими ДНК-векторами, несущими целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, из созданных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 и/или во введении в органы и ткани пациента или животного выбранного генотерапевтического ДНК-вектора, несущего целевой ген на основе генотерапевтического ДНК-вектора GDTT1.8NAS12 или нескольких выбранных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, из созданных генотерапевтических ДНК-векторов, несущих целевые гены на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, или в сочетании обозначенных способов.
10. Способ получения штамма для производства генотерапевтического ДНК-вектора по пп. 1-4 или 5 для лечения заболеваний, характеризующихся нарушением врожденного и адаптивного иммунитета, для терапии аутоиммунных, онкологических, вирусных заболеваний, связанных с дисбалансом иммунной системы, для иммуномодуляции в том числе при противоопухолевой вакцинации в качестве генотерапевтического адъюванта, заключающийся в получении электрокомпетентных клеток штамма Escherichia coli JM110-NAS с последующим проведением электропорации этих клеток генотерапевтическим ДНК-вектором GDTT1.8NAS12-IFNB1, или ДНК-вектором GDTT1.8NAS12-IFNA14, или ДНК-вектором GDTT1.8NAS12- IFNA2, или ДНК-вектором GDTT1.8NAS12- IL12A, или ДНК-вектором GDTT1.8NAS12- IL12B, после чего клетки высеивают на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола с получением в результате штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1 или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12- IFNA14, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12- IFNA2, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12- IL12A, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12- IL12B.
11. Штамм Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, полученный способом по п. 10, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12- IFNB1, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся нарушением врожденного и адаптивного иммунитета, для терапии аутоиммунных, онкологических, вирусных заболеваний, связанных с дисбалансом иммунной системы, для иммуномодуляции в том числе при противоопухолевой вакцинации в качестве генотерапевтического адъюванта.
12. Штамм Escherichia coli JM110-NAS/GDTT1.8NAS12- IFNA14, полученный способом по п. 10, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12- IFNA14, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся нарушением врожденного и адаптивного иммунитета, для терапии аутоиммунных, онкологических, вирусных заболеваний, связанных с дисбалансом иммунной системы, для иммуномодуляции в том числе при противоопухолевой вакцинации в качестве генотерапевтического адъюванта.
13. Штамм Escherichia coli JM110-NAS/GDTT1.8NAS12- IFNA2, полученный способом по п. 10, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12- IFNA2, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся нарушением врожденного и адаптивного иммунитета, для терапии аутоиммунных, онкологических, вирусных заболеваний, связанных с дисбалансом иммунной системы, для иммуномодуляции в том числе при противоопухолевой вакцинации в качестве генотерапевтического адъюванта.
14. Штамм Escherichia coli JM110-NAS/GDTT1.8NAS12- IL12A, полученный способом по п. 10, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12- IL12A, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся нарушением врожденного и адаптивного иммунитета, для терапии аутоиммунных, онкологических, вирусных заболеваний, связанных с дисбалансом иммунной системы, для иммуномодуляции в том числе при противоопухолевой вакцинации в качестве генотерапевтического адъюванта.
15. Штамм Escherichia coli JM110-NAS/GDTT1.8NAS12- IL12B, полученный способом по п. 10, несущий генотерапевтический ДНК-вектор GDTT1.8NAS12- IL12B, для его наработки с возможностью селекции без использования антибиотиков при получении генотерапевтического ДНК-вектора для лечения заболеваний, характеризующихся нарушением врожденного и адаптивного иммунитета, для терапии аутоиммунных, онкологических, вирусных заболеваний, связанных с дисбалансом иммунной системы, для иммуномодуляции в том числе при противоопухолевой вакцинации в качестве генотерапевтического адъюванта.
16. Способ производства в промышленных масштабах генотерапевтического ДНК-вектора на основе генотерапевтического ДНК-вектора GDTT1.8NAS12, несущего целевой ген IFNB1 или IFNA14 или IFNA2 или IL12A или IL12B, по пп. 1-4 или 5, для лечения заболеваний, характеризующихся нарушением врожденного и адаптивного иммунитета, для терапии аутоиммунных, онкологических, вирусных заболеваний, связанных с дисбалансом иммунной системы, для иммуномодуляции в том числе при противоопухолевой вакцинации в качестве генотерапевтического адъюванта, заключающийся в том, что генотерапевтический ДНК-вектор GDTT1.8NAS12- IFNB1, или генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNA14, или генотерапевтический ДНК-вектор GDTT1.8NAS12-IFNA2, или генотерапевтический ДНК-вектор GDTT1.8NAS12-IL12A, или генотерапевтический ДНК-вектора GDTT1.8NAS12-IL12B получают путем того, что засевают в колбу с приготовленной средой затравочную культуру, выбранную из штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNB1, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA14, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IFNA2, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12A, или штамма Escherichia coli JM110-NAS/GDTT1.8NAS12-IL12B, затем инкубируют затравочную среду в шейкере-инкубаторе и переносят ее в промышленный ферментер, после чего растят до достижения стационарной фазы, затем выделяют фракцию, содержащую целевой ДНК-продукт, многостадийно фильтруют и очищают хроматографическими методами.
| Генотерапевтический ДНК-вектор VTvaf17, способ его получения, штамм Escherichia coli SCS110-AF, способ его получения, штамм Escherichia coli SCS110-AF/VTvaf17, несущий генотерапевтический ДНК-вектор VTvaf17, способ его получения | 2017 |
|
RU2678756C1 |
| US 5723127 A1, 03.03.1998 | |||
| CN 101304758 B, 21.08.2013. | |||

