Изобретение относится к биотехнологии микроводорослей и может быть использовано при промышленном культивировании зеленой микроводоросли Chromochloris zofingiensis 
 & Lewis (Chlorophyceae, Sphaeropleales) (штамм CALU-190) как сырья для получения липидов и кетокаротиноидов (ККар) группы астаксантина (Аст).
& Lewis (Chlorophyceae, Sphaeropleales) (штамм CALU-190) как сырья для получения липидов и кетокаротиноидов (ККар) группы астаксантина (Аст).
Chromochloris zofingiensis ранее Chlorella zofingiensis 
 Lewis, 2012) относится к группе высоко перспективных в коммерческом отношении зеленых микроводорослей, являющихся источником одновременно двух стабильно востребованных рынком продуктов: технических липидов, пригодных для получения качественного биодизеля, и высокоценных С40-кетокаротиноидов (ККар) группы астаксантина (Аст) (астаксантина, кантаксантина и адониксантина) с ярко выраженной антиоксидантной и регуляторной активностью, нашедших широкое применение в производстве кормов для аквакультуры и птицеводства, нутрицевтиков и элитной косметики [Mata et al. 2010; Surai, 2012; Han et al., 2013; Maoka et al., 2013]. Важными в биотехнологическом аспекте преимуществами вида по сравнению с основным источником Аст зеленой микроводорослью Haematococcus pluvialis, являются более высокая скорость роста, более легкое разрушение клеточных оболочек и более полное извлечение липидов и ККар из биомассы [Liu et al. 2014; Kim et al., 2015]. Вместе с тем, содержание Act в биомассе С. zofingiensis (0.05-0.2% сухого вещества) намного ниже, чем у H. pluvialis 2-5%) [Ip, Chen, 2005; Li et al., 2009; Sun et al., 200]. Поэтому данный вид следует расценивать, главным образом, как промышленно перспективный продуцент липидов для биодизеля, обладающий существенным преимуществом перед другими масличными культурами в виде возможности получения дополнительного дорогостоящего и высоко востребованного продукта - кетокаротиноидов группы астаксантина [Liu et al., 2010].
Lewis, 2012) относится к группе высоко перспективных в коммерческом отношении зеленых микроводорослей, являющихся источником одновременно двух стабильно востребованных рынком продуктов: технических липидов, пригодных для получения качественного биодизеля, и высокоценных С40-кетокаротиноидов (ККар) группы астаксантина (Аст) (астаксантина, кантаксантина и адониксантина) с ярко выраженной антиоксидантной и регуляторной активностью, нашедших широкое применение в производстве кормов для аквакультуры и птицеводства, нутрицевтиков и элитной косметики [Mata et al. 2010; Surai, 2012; Han et al., 2013; Maoka et al., 2013]. Важными в биотехнологическом аспекте преимуществами вида по сравнению с основным источником Аст зеленой микроводорослью Haematococcus pluvialis, являются более высокая скорость роста, более легкое разрушение клеточных оболочек и более полное извлечение липидов и ККар из биомассы [Liu et al. 2014; Kim et al., 2015]. Вместе с тем, содержание Act в биомассе С. zofingiensis (0.05-0.2% сухого вещества) намного ниже, чем у H. pluvialis 2-5%) [Ip, Chen, 2005; Li et al., 2009; Sun et al., 200]. Поэтому данный вид следует расценивать, главным образом, как промышленно перспективный продуцент липидов для биодизеля, обладающий существенным преимуществом перед другими масличными культурами в виде возможности получения дополнительного дорогостоящего и высоко востребованного продукта - кетокаротиноидов группы астаксантина [Liu et al., 2010].
Оценке продукционного потенциала С. zofingiensis при различных условиях культивирования посвящен ряд исследований, показавших, что этот вид можно успешно культивировать автотрофным, миксотрофным и гетеротрофным способами [Ip, Chen, 2005; Feng et al., 2012; Chen et al., 2017; Azaman et al, 2017; Zang et al., 2017; Mao et al., 2018 и др.]. В условиях гетеротрофного питания продуктивность водоросли по биомассе и липидам может быть в 2-3 раза выше, чем при автотрофном выращивании на минеральных средах [Li et al., 2009; Sun et al., 2008; Zang et al., 2017]. Однако высокая себестоимость конечных продуктов, получаемых миксотрофным и гетеротрофным методами в закрытых фотобиореакторах (ферментерах), является в настоящее время серьезным препятствием для внедрения этого метода в промышленную культуру, поэтому более дешевые автотрофные способы культивирования видов-продуцентов липидов и каротиноидов в открытых прудах или простых пленочных фотореакторах по-прежнему сохраняют свою актуальность [Hannon et al., 2010; Richardson et al., 2012; Liu et al., 2016].
В фототрофных системах по получению липидов из микроводорослей в качестве источника азота, как правило, используют соли азотной кислоты (KNO3 и NaNO3). Причем в структуре общих затрат на реагенты для питательных сред нитраты занимают второе место после расходов на СО2 - на производство 1 тонны биодизеля необходимо 250-500 кг нитратного азота [Markou, Monlau, 2019]. Выявление хорошо доступных для С. zofingiensis и вместе с тем более дешевых, чем нитраты, источников азота будет способствовать повышению рентабельности технологий автотрофного культивирования этого вида как сырья для получения липидов и ККар группы Аст.
Показано, что при автотрофном культивировании многих видов зеленых микроводорослей альтернативным источником азота может служить более дешевая и более богатая азотом мочевина (СО(NH2)2), в которой его массовая доля выше, чем в NaNO3 и KNO3 в 2.8 и 3.4 раза соответственно. В литературе имеются сведения о наличии в клетках Chlorophyceae эффективных путей метаболизма мочевины, а также данные, подтверждающие возможность полноценной замены нитратов на мочевину при культивировании отдельных видов [Bekheet, Syrett, 1977; Hsieh, Wu, 2009; Choochote et al., 2010]. Сведения о способах культивирования С. zofingiensis на мочевине для одновременного получения липидов и кетокаротинодов группы Аст в доступной нам литературе отсутствуют. Предлагаемый способ восполняет этот пробел.
Известен способ выращивания С. zofingiensis в колоночных культиваторах из оргстекла методом одностадийной накопительной культуры, на питательной среде BG11 [Rippka et al., 1979] с заменой нитратов на мочевину или ее смеси с бикарбонатом аммония (NH4HCO3) различного состава [Huo et al. 2018]. Наилучшие результаты получены при составе смеси 300 мг/л СО(NH2)2 и 200 мг/л NH4HCO3. К недостаткам способа относятся: а) отсутствие сведений об использованном штамме С. zofingiensis; б) слишком короткий период наблюдений (5 сут); в) регистрация роста водоросли только по оптической плотности клеточных суспензий; г) отсутствие в схеме культивирования условий для индукции биосинтеза кетокаротиноидов; д) отсутствие данных по продуктивности культур С. zofingiensis по сухой биомассе, липидам и их жирнокислотном составе - ключевом показателе пригодности липидов для производства биодизеля [Islam et al., 2013].
Известен способ миксотрофного культивирования С. zofingiensis для получения биотоплива, основанный на методе одностадийной накопительной культуры, использовании люминесцентных ламп белого, красного и синего света и смеси минеральных и органических удобрений, включающей мочевину [Sudibyo et al., 2010]. Наилучшие результаты по содержанию сухой биомассы в культуре (680 мг/л) и массовой доле суммарных липидов в сухом веществе (47%) получены при выращивании на лампах синего света и концентрации смеси питательных веществ в среде 19 мг/л. Основной недостаток способа состоит в том, что он не предполагает получение второго полезного продукта - ККар группы Аст. Кроме того, в описании способа отсутствуют конкретные сведения об использованном штамме С. zofingiensis, концентрации мочевины в питательной смеси, продуктивности культур по сухому веществу, суммарным липидам и их жирнокислотному составу. Отсутствие этих важных сведений не позволяет оценить перспективность данного способа для получения липидов и производства из них биотоплива.
Известен способ автотрофного культивирования Chlorella sp. на мочевине для получения технических масел [Hsieh, Wu, 2009]. В основе трех вариантов способа (периодической культуры, периодической культуры с добавлением субстрата и полупроточной культуры) лежит один принцип - поддержание подострого дефицита азота в среде на протяжении всего периода культивирования (21 сут) как главного фактора накопления липидов в клетках водоросли. В условиях периодической культуры при однократном внесении мочевины в среду в концентрации 0.100 г/л средняя продуктивность водоросли по липидам составляла 0.124 г/л сут, итоговое содержание сухой биомассы - 2.027 г/л, массовая доля липидов - 3.6%. При накопительном культивировании с добавлением субстрата (0.025 г/л мочевины ежедневно) средняя продуктивность по липидам была сходной. Культивирование в полупроточном режиме с добавлением 0.025 г/л мочевины на каждой ранней стационарной фазе позволило увеличить среднюю продуктивность по липидам до 0,139 г/л сут. Недостатком способа является отсутствие сведений, позволяющих оценить его пригодность для культивирования Chromochloris (Chlorella) zofingiensis, вследствие филогенетической удаленности родов Chlorella и Chromochloris [ , Lewis, 2012]. Кроме того, в описании данного способа так же, как и в предыдущих, отсутствуют сведения о жирнокислотном составе липидов, без которых невозможна оценка их пригодности для получения биодизеля [Islam et al., 2013]. Также отсутствуют сведения о содержании в биомассе кетокаротиноидов, что, по всей вероятности, обусловлено неспособностью Chlorella spp. синтезировать астаксантин
, Lewis, 2012]. Кроме того, в описании данного способа так же, как и в предыдущих, отсутствуют сведения о жирнокислотном составе липидов, без которых невозможна оценка их пригодности для получения биодизеля [Islam et al., 2013]. Также отсутствуют сведения о содержании в биомассе кетокаротиноидов, что, по всей вероятности, обусловлено неспособностью Chlorella spp. синтезировать астаксантин 
Наиболее близким к заявляемому способу по задаче одновременного получения из биомассы Chromochloris zofingiensis сразу двух полезных продуктов и использованному штамму (АТСС 30412) является способ, предложенный Wang и сотр. [Wang et al., 2013], основанный на миксотрофном одностадийном культивировании вида на минеральной среде [Kuhl and Lorenzen, 1964], содержащей в качестве источника азота 1.011 г KNO3, а в качестве источника органического углерода глюкозу. В первые 3-е суток концентрация глюкозы в среде составляла 50 мМ. На 4-е сутки в среду дополнительно внесли глюкозу и KNO3 до концентрации 200 и 5 мМ соответственно, а также соли двухвалентных металлов (Mn+2 Mg+2 и Fe+2) для активирования биосинтеза Аст. Интенсивность фотосинтетической радиации (ФАР), создаваемой люминесцентными лампами на поверхности колб (I), составляла 150 ммоль фотонов/м2 с), температура питательной среды (Т) - +26-27°С. Наилучшие результаты по содержанию Аст и жирных кислот (ЖК) в сухой биомассе были получены при концентрации Fe+2 в среде 0.2 мМ - 0.22% и 41.8% соответственно.
Основными недостатками известного способа являются: а) использование более дорогого, чем мочевина, источника азота; б) необходимость использования аксеничной культуры и закрытого фотобиореактора при культивировании на глюкозе; в) применение для индукции биосинтеза Аст соли Fe+2, приводящей к частичной потере биомассы и ухудшению качества биомассы как сырья для получения технических липидов. Использование эмпирических уравнений для прогнозной оценки технических характеристик биодизеля по жирнокислотному составу липидов, приведенному в работе [Islam et al., 2013], показало, что внесение в среду 0.2 мМ Fe+2 приводит к ухудшению ряда важных показателей качества по сравнению с контролем: увеличению йодного числа (IV) на 11%, снижению цетанового числа (CN) на 5% и окислительной стабильности биодизеля (Y) на 14%.
Задачей изобретения «Способ культивирования микроводоросли Chromochloris zofingiensis для получения липидов и каротиноидов» является создание условий автотрофного культивирования С. zofingiensis, обеспечивающих одновременное накопление в биомассе двух целевых продуктов: липидов, пригодных для получения высоко качественного биодизеля, и С40-кетокаротиноидов группы астаксантина, а также снижение материальных затрат на реагенты для питательной среды.
Техническим результатом изобретения является разработка нового варианта двухстадийного способа автотрофного культивирования С. zofingiensis на мочевине, обеспечивающего:
- снижение затрат на источник азота для питательной среды примерно в 3.3 раз, путем полной замены в питательной среде ВВМ 3N нитрата натрия на мочевину;
- коммерчески значимую, продуктивность культуры С. zofingiensis по суммарным липидам (347±21 мг/л сут) и каротиноидам (1.3±0.01 мг/л сут);
- коммерчески значимое содержание суммарных липидов (69.4±4.6%) и суммарных каротиноидов (0.3±0.01%) в сухой биомассе;
- специфический жирнокислотный профиль суммарных липидов, необходимый для изготовления высококачественного биодизеля, отвечающего требованиям европейского стандарта EN14214;
- преобладание кетокаротиноидов группы Аст в получаемой смеси каротиноидов (86.8% от суммы) при относительном содержании Аст - 46.3%; Кан - 25.5%, Адк - 14.9% от суммы Кар.
Поставленная задача достигается тем, что в Способе культивирования микроводоросли Chromochloris zofingiensis для получения липидов и каротиноидов, включающем культивирование на питательной среде, создание стрессовых условий внешней среды для индукции биосинтеза астаксантина и липидов, получение биомассы, предусмотрены следующие отличия. Водоросли культивируют методом двухстадийной накопительной культуры на модифицированной питательной среде ВВМ (МВВМ), модифицированной путем эквивалентной по азоту (8.86 ммоль N/л) замены нитрата натрия (NaNO3) на мочевину (СО(NH2)2) с соблюдением на I («зеленой») стадии условий внешней среды, способствующих активному делению клеток и накоплению биомассы в культуре, а на II («красной») стадии условий для интенсивного накопления липидов и С40-кетокаротиноидов. На I («зеленой») стадии С.zofingiensis культивируют в течение 17 суток при следующих условиях: начальная численность клеток в культуре (nнач) - 2.5×106 кл/мл, освещенность (I) - 120 мкмоль фотонов/⋅м2⋅с, фотопериод - 15 ч свет/9 ч темнота, температура среды (Т) - 22-23°С и скорость продувки культуры воздухом (v) - 0.3 л⋅/(мин л). В состав питательной среды МВВМ для «зеленой» стадии культивирования входят (г/л дистиллированной воды): CO(NH2)2 - 0.265; MgSO4⋅7H2O - 0.075; NaCl - 0.025; K2HPO4⋅3H2O - 0.0745; KH2PO4 - 0.1755; Na2-ЭДТА - 0.05; KOH - 0.031; Н3ВО3 - 0.011; ZnSO4⋅7H2O - 0.00882; MnCl2⋅4H2O - 0.00144; Na2MoO4⋅2H2O - 0.00242; CuSO4⋅5H2O - 0.00157; Co(NO3)2⋅6H2O - 0.00049; FeSO4⋅7H2O - 0.00498; CaCl2 - 0.0189; H2SO4 конц - 1 мкл.
Перевод культуры на II («красную») стадию осуществляют путем ее разведения до nнач=4×106 кл/л питательной средой МВВМ со сниженным в 10 раз содержанием азота и фосфора. На этой стадии С. ofingiensis выращивают в течение 15 суток при продолжительности светового периода 24 ч, I=240 мкмоль фотонов/⋅м2⋅с, Т=24-25°С, v=0.6 л⋅/(мин л). В состав питательной среды МВВМ для «красной» стадии культивирования входят (г/л дистиллированной воды): CO(NH2)2 - 0.0265; MgSO4⋅7H2O - 0.075; NaCl - 0.025; K2HPO4⋅3H2O - 0.0075; KH2PO4 - 0.01775; Na2-ЭДТА - 0.050; KOH - 0.031; Н3ВО3 - 0.011; ZnSO4⋅7H2O - 0.00882; MnCl2⋅4H2O - 0.00144; Na2MoO4⋅2H2O - 0.00242; CuSO4⋅5H2O - 0.00157; Co(NO3)2⋅6H2O - 0.00049; FeSO4⋅7H2O - 0.00498; CaCl2 - 0.0189; H2SO4 конц - 1 мкл.
Для достижения технического результата изобретения культуру водоросли С. ofingiensis выращивают оптимизированным двухстадийным методом в сочетании эффективных приемов по интенсификации роста и метаболизма водоросли, разработанных авторами, и направленных на достижение коммерчески значимого выхода целевых продуктов технических липидов и кетокаротиноидов группы астаксантина.
Заявляемый способ культивирования микроводоросли Chromochloris zofingiensis для получения липидов и каротиноидов отличается от прототипа следующими параметрами:
а) использованием мочевины вместо нитрата калия в качестве единственного источника азота;
б) использованием вместо миксотрофного метода культивирования на глюкозе автотрофного метода, при котором источником углерода является СО2, поступающий в культуру при ее барботаже воздухом, и СО2, образующийся при ферментативном гидролизе мочевины под действием карбамидлиазы:
CO(NH2)2+АТФ+H2O↔CO2+2NH3+АДФ+Pi [Bekheet, Syrett, 1977];
в) использованием двухстадийного метода культивирования, в ходе которого на I «зеленой» стадии создают условия для активного деления клеток и накопления биомассы, а на II «красной» стадии культуру подвергают резкому стресс-воздействию, индуцирующему биосинтез запасных липидов и кетокаротиноидов группы Аст.
г) использованием приема индукции биосинтеза запасных липидов и кетокаротиноидов группы Аст в клетках водоросли, не вызывающего потерь биомассы в результате стресса. Вместо внесения в среду ионов Fe2+, приводящих наряду с активизацией биосинтеза целевых продуктов к гибели значительной части клеток, эту задачу решают путем разведения культуры редуцированной по азоту и фосфору питательной средой в сочетании с увеличением интенсивности и продолжительности освещения. Такой прием сопровождается не снижением, а увеличением численности клеток в культуре и обеспечивает специфический состав жирных кислот, необходимый для получения биодизеля высокого качества, отвечающего требованиям европейских стандартов.
д) возможностью использования неаксеничной культуры Chromochloris zofingiensis, а также более простых и более дешевых, чем закрытые фотобиореакторы / ферментеры, пленочных культиваторов.
е) использованием расчетных методов предварительной оценки технических характеристик биодизеля, который может быть изготовлен из данного сырья, по результатам анализа жирнокислотного профиля общих липидов.
Сходство с прототипом заключается:
а) в использовании одного и того же штамма Chromochloris zofingiensis CALU 190 (= АТСС 30412);
б) возможности получения из биомассы С. zofingiensis в результате реализации способа сразу двух высокоценных продуктов: технических липидов и кетокаротиноидов группы астаксантина.
в) в общем принципе культивирования каротиногенных микроводорослей, состоящем в создании стрессовых условий внешней среды для индукции биосинтеза астаксантина и липидов.
Изобретение поясняется таблицами и иллюстрациями:
Таблица 1. Состав питательных сред, использованных на «зеленой» и «красной» стадиях культивирования Chromochloris zofingiensis (г/л Н2Одист.) (ВВМ - контроль, МВВМ - по изобретению).
Таблица 2. Жирнокислотный профиль суммарных липидов Chromochloris zofingiensis в конце «зеленой» и «красной» стадий культивирования в зависимости от источника азота в питательной среде.
Таблица 3. Прогнозные свойства биодизеля из Chromochloris zofingiensis, рассчитанные по профилю жирных кислот в конце «зеленой» и «красной» стадий в зависимости от источника азота в питательной среде.
Таблица 4. Состав вторичных каротиноидов в биомассе Chromochloris zofingiensis в конце «красной» стадии в зависимости от химической формы азота в питательной среде.
Таблица 5. Сравнительная характеристика некоторых технологических показателей мочевины и нитрата натрия как источников азота в питательных средах для культивирования С. zofingiensis.
Фиг. 1 - Динамика численности клеток (А) и сухой биомассы (Б) в культурах Chromochloris zofingiensis на протяжении "красной" стадии культивирования и валовый выход сухой биомассы за весь период культивирования (32 сут) из литра исходной культуры в зависимости от химической формы азота в среде. 1 - нитрат натрия. 2 - мочевина.
Фиг. 2 - Содержание суммарных липидов в клетках (А), культурах (Б) и сухой биомассе (В) Chromochloris zofingiensis в конце "красной" стадии и валовый выход липидов из литра исходной культуры за 32 сут (Д) в зависимости химической формы азота в питательной среде. 1 - нитрат натрия. 2 - мочевина.
Фиг. 3 - Продукционные характеристики культур Chromochloris zofingiensis по вторичным каротиноидам в зависимости от химической формы азота в питательной среде: А - динамика накопления вторичных каротиноидов в культурах на "красной" стадии; Б - средняя продуктивность культур по суммарным каротиноидам на "красной" стадии. В - содержание суммарных каротиноидов в полученной биомассе. Г - валовый выход вторичных каротиноидов из 1 литра исходной культуры за 32 сут. 1 - нитрат натрия. 2 - мочевина.
Для реализации предлагаемого способа получения биомассы Chromochloris zofingiensis используют штамм CALU-190, который культивируют методом двухстадийной накопительной культуры. На I («зеленой») стадии С. zofingiensis выращивают в условиях, благоприятных для роста на питательной среде, содержащей в качестве единственного источника азота мочевину. Продолжительность «зеленой» стадии определяется достижением культурой стационарной фазы роста, которую выявляют по результатам измерения динамики одного или нескольких показателей скорости деления клеток (оптической плотности культуры (OD), численности клеток (n), содержания сухого вещества (СВ) или содержания хлорофилла а (Хла) в мл культуры.
Перевод культуры на II («красную») стадию осуществляют при помощи резкого стресс-воздействия, вызывающего интенсификацию свободно-радикальных процессов в клетках водоросли и, как следствие, индукцию комплекса морфо-физиологических защитных реакций, направленных на сохранение жизнеспособности клеток путем их перехода в покоящееся состояние. В аспекте предлагаемого способа наиболее важными из них являются активация биосинтеза антиоксидантных кетокаротиноидов (астаксантина и его ближайших метаболических предшественников) и накопление в цитоплазме запасных липидов со специфическим составом жирных кислот (ЖК), пригодных для получения высококачественного биодизеля.
Главными стресс-факторами при переводе культуры на «красную» стадию являются острый дефицит питания, создаваемый разведением культуры редуцированной по азоту и фосфору питательной средой, в сочетании с резким увеличением облученности клеток, которое достигается путем снижения плотности культуры при ее разведении, 2-х кратным увеличением интенсивности внешнего освещения и продолжительности светового периода до 24 ч.
Пример реализации способа культивирования Chromochloris zofingiensis для получения липидов и каротиноидов
Получение инокулята (посевного материала)
Штамм Chromochloris zofingiensis CALU 190 (= ССАР-211/14 = UTEX 32 = SAG 211-14 = ATTC 30412) с момента получения в 2006 г из коллекции Музея живых культур микроорганизмов Санкт-Петербургского государственного университета до проведения исследования хранили на агаризованной питательной среде ВВМ (1.5%) при Т = 15-16°С и I=40 μЕ⋅м-2⋅с-1 с пересевом каждые 45 суток. Для получения инокулята биомассу с твердой среды перенесли в жидкую среду ВВМ и выращивали в течение 7 суток в стеклянных конических колбах объемом 250 мл при естественном рассеянном свете I=30 μЕ⋅м-2⋅с-1, Т = 20-22°С и продувке стерильным воздухом со скоростью 0.3 л⋅мин-1⋅л-1 культуры. Затем культуру сконцентрировали центрифугированием при 800-1000g в течение 5 мин, супернатант удалили, а осадок клеток перенесли в колбы объемом 1 л со свежей средой ВВМ и снова культивировали в течение 4 дней при I = 120 μЕ⋅м-2⋅с-1, Т = 20-22°С. Полученную культуру, состоящую из молодых, активно делящихся клеток, использовали в качестве инокулята для засева колб при проведении «зеленой» стадии культивирования.
Проведение «зеленой» стадии
Для культивирования С.zofingiensis на «зеленой» стадии использовали два варианта питательной среды ВВМ [Bischoff, Bold, 1963]. В варианте №1, служившем контролем, в качестве источника азота использовали нитрат натрия, а в варианте №2 - мочевину. Содержание азота в обеих средах было одинаковым и составляло 8.83 ммоль/л (таблица 1).
В состав питательной среды в варианте №1 входили (г/л дистиллированной воды): CO(NH2)2 - 0.265; MgSO4⋅7H2O - 0.075; NaCl - 0.025; K2HPO4⋅3H2O - 0.0745; KH2PO4 - 0.1755; Na2-ЭДТА - 0.05; KOH - 0.031; H3BO3 - 0.011; ZnSO4⋅7H2O - 0.00882; MnCl2⋅4H2O - 0.00144; Na2MoO4⋅2H2O - 0.00242; CuSO4⋅5H2O - 0.00157; Co(NO3)2⋅6H2O - 0.00049; FeSO4⋅7H2O - 0.00498; CaCl2 - 0.0189; H2SO4 конц - 1 мкл.
В состав питательной среды МВВМ в варианте №2 входили (г/л дистиллированной воды): CO(NH2)2 - 0.265; MgSO4⋅7H2O - 0.075; NaCl - 0.025; K2HPO4⋅3H2O - 0.0745; KH2PO4 - 0.1755; Na2-ЭДТА - 0.05; KOH - 0.031; H3BO3 - 0.011; ZnSO4⋅7H2O - 0.00882; MnCl2⋅4H2O - 0.00144; Na2MoO4⋅2H2O - 0.00242; CuSO4⋅5H2O - 0.00157; Co(NO3)2⋅6H2O - 0.00049; FeSO4⋅7H2O - 0.00498; CaCl2 - 0.0189; H2SO4 конц - 1 мкл.
В среде варианта №2 содержание мочевины составляло 0.265 г/л, K2HPO4⋅3H2O - 0.00745 г/л, KH2PO4 - 0.01755 г/л, содержание всех остальных компонентов было таким же, как и в варианте №1. В обоих вариантах водоросль выращивали в течение 17 суток в стеклянных конических колбах объемом 1 л при объеме культуры 0.45 л, начальной численности клеток 2.3×106 кл⋅мл-1, освещенности 120 мкмоль фотонов/⋅м2⋅с, фотопериоде 15 ч свет/9 ч темнота, температуре среды 22-23°С и скорости продувки культуры воздухом - 0.3 л /(мин⋅л).
Проведение «красной» стадии
Перевод культур на II («красную») стадию осуществляли путем сочетания двух ключевых факторов вторичного каротиногенеза: дефицита элементов питания и высокой освещенности. Для этого культуры, полученные в конце «зеленой» стадии, развели редуцированными в 10 раз по азоту и фосфору средами ВВМ и МВВМ (таблица 1) до nнач=4×106 кл/л и перевели на круглосуточное двухстороннее освещение при I=240 мкмоль фотонов/⋅м2⋅с.
Состав среды МВВМ для «красной» стадии культивирования (г/л дистиллированной воды): CO(NH2)2 - 0.0265; MgSO4⋅7H2O - 0.075; NaCl - 0.025; K2HPO4⋅3H2O - 0.0075; KH2PO4 - 0.01775; Na2-ЭДТА - 0.050; KOH - 0.031; Н3ВО3 - 0.011; ZnSO4⋅7H2O - 0.00882; MnCl2⋅4H2O - 0.00144; Na2MoO4⋅2H2O - 0.00242; CuSO4⋅5H2O - 0.00157; Co(NO3)2⋅6H2O - 0.00049; FeSO4⋅7H2O - 0.00498; CaCl2 - 0.0189; H2SO4 конц - 1 мкл. Молярные концентрации азота и фосфора в редуцированной среде составляли соответственно 0.88 и 0.16 ммоль/л. Т = 24-25°С, v = 0.6 л⋅/(мин л). В таком режиме культуры С.zofingiensis выращивали в течение 15 суток до появления первых признаков снижения плотности культуры. В ходе культивирования на обеих стадиях контролировали динамику плотности культур по численности клеток и сухому веществу, скорость поглощения клетками нитратного азота и мочевины, содержание суммарных пигментов и липидов в культурах и биомассе, удельные скорости роста и продуктивности культур по пигментам и суммарным липидам [Минюк и др., 2016; Лелевич и др., 2016; Minyuk et al., 2017]. В конце «зеленой» и «красной» стадий анализировали ВЭЖХ-профиль суммарных каротиноидов и ГЖХ-профиль жирных кислот [Минюк и др., 2016; Minyuk et al., 2017]. Прогнозные характеристики биодизеля, который можно изготовить из биомассы, полученной предлагаемым способом, рассчитывали по эмпирическим уравнениям, приведенным в [Islam et al., 2013].
Все измерения проводили в трех биологических и трех аналитических повторностях. На рисунках и в тексте приведены средние значения показателей и их стандартные отклонения. Достоверность различий между средними определяли по t-критерию Стьюдента при уровне значимости р<0.05.
На «зеленой» стадии средние скорости поглощения нитратного и амидного азота были сходны и составляли соответственно 0.626 и 0.631 ммоль/(л сут). На 13-е сутки содержание азота в обеих культурах не превышало следовых количеств. Максимальная удельная скорость роста (μмакс), рассчитанная по численности клеток, была отмечена в культуре, росшей на мочевине (1.15±0.04 сут-1 против 0.89±0.04 сут-1 на нитрате). Средние удельные скорости роста (μср) на нитратном и амидном азоте достоверно не различались и составляли соответственно 0.23±0.009 и 0.021±0.002 сут-1. Особенностью питательной среды на мочевине было сохранение оптимального для роста водоросли диапазона рН среды (7.01-7.6), в то время как в культуре, росшей на нитрате натрия, отмечено постепенное защелачивание среды - к концу «зеленой» стадии рН здесь вырос с 6.7 до 9.1. При этом индекс Кар/Хл на протяжении «зеленой» стадии при культивировании водоросли на мочевине был ниже (0.18-0.22), чем при культивировании на нитратах (0.20-0.27), что указывает на лучшее функциональное состояние клеток в первом случае [Минюк и др., 2010].
На «красной» стадии обе культуры, независимо от источника азота, на которой они были получены на «зеленой» стадии, проявили высокую устойчивость к стресс-воздействию, использованному для индукции липогенеза и биосинтеза Аст. Ни в одном из вариантов массовой смертности клеток, типичной в сходных условиях для многих других видов зеленых микроводорослей [Минюк и др. 2007; Чубчикова и др. 2009], не наблюдалось. Напротив, численность клеток неуклонно росла на протяжении всей «красной» стадии и к ее окончанию увеличилась в 3-3.5 раза, причем средняя продуктивность по сухому веществу культуры, выращенной на «зеленой» стадии на CO(NH2)2, была на 21% выше, чем выращенной на NaNO3 (Фиг. 1).
Значительную часть сухого вещества, накопленного клетками к концу «красной» стадии, составляли липиды. В варианте СО(NH2)2 их содержание в клетках (37.09±1.64 пг/кл), культуре (471.19±24.74 мг/л) и биомассе (69.36±4.60%), а также валовый выход из литра исходной культуры (11.10±0.67 г/л), и средняя за весь период культивирования (32 сут) продуктивность (347.19±20.80 мг/(л сут), рассчитанные с учетом разведения культуры, были выше, чем в варианте NaNO3, на 25.1, 27.7, 52.8, 30.0 и 30.0%% соответственно (Фиг. 2). Все указанные различия были существенны (р≤0.05). При этом в составе жирных кислот (ЖК) суммарных липидов произошли существенные изменения, приведшие к снижению индекса ненасыщенности ЖК в обоих вариантах примерно в 1.3 раза и обусловленные, главным образом, увеличением доли олеиновой кислоты в 1.8-1.9 раза и снижением доли полиненасыщенных кислот в 2.1-2.2 раза (таблица 2). Указанные изменения, в свою очередь, определили существенное улучшение качества липидов С. zofingiensis как технического сырья, на что указывают рассчитанные прогнозные характеристики биодизеля [Islam etal. 2013], который можно произвести из липидов, полученных при реализации предлагаемого способа культивирования С. zofingiensis (таблица 3). Все ключевые расчетные показатели качества потенциального биодизеля из липидов, полученных в конце «красной» стадии, соответствуют требованиям одного из самых строгих европейских стандартов на биодизель EN14214 [www.rob-oil.com - 08.04.2019 - дата посещения электронного документа удаленного доступа], что наряду с высокой продуктивностью культур по липидам, может служить необходимой предпосылкой для внедрения в данного способа в практику.
Интенсивное накопление липидов клетками С. zofingiensis, готовящимися к переходу в стадию покоя, происходило на фоне деструкции их фотосинтетического аппарата и активизации биосинтеза вторичных С40-кетокаротиноидов (Аст и его ближайших метаболических предшественников), выполняющих важнейшую в условиях стресса функцию – сдерживание свободнорадикального окисления ключевых макромолекул в субклеточных структурах [Han et al., 2013]. Более интенсивно этот процесс происходил в культуре, выращенной на «зеленой» стадии на мочевине. Средняя продуктивность по Кар здесь была на 16.3% выше, чем в культуре, полученной на NaNO3 (Фиг. 3а, б). Тем не менее, валовый выход суммарных Кар из литра исходной культуры, рассчитанный с учетом разведения при переходе на «красную» стадию (43.61±1.05 и 41.76±2.3 мг/л), и их массовая доля в сухом веществе (0.3% в обоих случаях) были сходны (Фиг. 3в, г).
Состав суммарных Кар в биомассе, собранной на заключительной стадии культивирования, мало зависел от условий азотного питания С. zofingiensis и на обеих формах азота был представлен смесью ККар (88.5-91%), небольшого количества первичных (6-8.5%) и неидентифицированных Кар (≈3%) (таблица 4). Ключевыми фракциями, независимо от источника азота, были астаксантин (Аст), кантаксантин (Кан) и адониксантин (Адк). Аст и Адк накапливались как в свободной форме, так и форме моно- и диацильных эфиров (главным образом эфиров олеиновой кислоты). Причем, в обоих вариантах культивирования Аст накапливался преимущественно в форме моноэфиров, а Адк - в виде диэфиров. Наиболее четко влияние химической формы азота в питательной среде на состав Кар проявилось в соотношении свободных и этерифицированных форм гидроксилированных ККар. При выращивании на мочевине доля свободных Аст и Адк была достоверно ниже (на 60 и 187% соответственно), а суммарная доля их моно- и диэфиров на 19 и 40% выше, чем при выращивании на NaNO3. Эти различия, наряду с более высоким содержанием липидов и сухого вещества в клетках, могут указывать на более быстрое завершение процессов подготовки клеток к переходу в состояние покоя при их выращивании на мочевине, служащей для клеток дополнительным источником СО2, образующегося при ее ферментативном гидролизе под действием карбамидлиазы [Bekheet, Syrett, 1977].
Результаты определения продукционных показателей С. zofingiensis по сухой массе, суммарным липидам и каротиноидам при выращивании на различных химических формах азота позволяют сделать заключение о возможности полной замены нитратов (как традиционного компонента питательных сред для Chlorophyceae) на карбамид в условиях двухстадийной накопительной культуры. Такая замена даст весомое снижение затрат на питательную среду (примерно в 3.3 раза), определяющееся более высокой массовой долей азота в мочевине и, как следствие, меньшим расходом соли на кубометр среды, а также более низкой оптовой ценой на мочевину, по сравнению с нитратом натрия (таблица 5).
В условиях предлагаемого способа двухстадийного культивирования на среде MBBN, содержащей в качестве источника азота мочевину, Chromochloris zofingiensis характеризуется высокой, коммерчески значимой продуктивностью по липидам - 347 мг/(л сут), что на 30% превышает данный показатель при выращивании С. zofingiensis на нитрате натрия - традиционном для зеленых микроводорослей источнике азота. С учетом разведения культур при переводе на «красную» стадию, валовый выход липидов из литра исходной культуры с начальной плотностью 2×106 клеток/мл за 32 суток составил 11.68 г против 9.14 г при выращивании на нитрате натрия.
Валовый выход суммарных Кар из литра исходной культуры и их массовая доля в сухом веществе при выращивании на мочевине и нитрате натрия достоверно не различались и составляли соответственно 41.8-43.6 мг/л и 0.3% сухого вещества.
Замена в питательной среде нитрата натрия на мочевину не снижала качественных показателей получаемой биомассы как потенциального сырья для производства биодизеля и кетокаротиноидов группы астаксантина:
- жирнокислотный состав суммарных липидов обеспечивает полное соответствие расчетных технических характеристик биодизелия, который можно изготовить из полученной биомассы, требованиям европейского стандарта EN14214.
- высокая доля кетокаротиноидов в суммарных каротиноидах (88.5%) представленных тремя наиболее сильными по антиоксидантной активности С40-кетокаротиноидами астаксантином (46.3%), кантаксантином (25.5%) и адониксантином (14.8%), определяет возможность получения дополнительного целевого продукта - экстрактов высоко ценных кетокаротиноидов медицинского и пищевого назначения.
Полученные результаты позволяют сделать заключение об эффективности полной замены нитрата натрия на мочевину при выращивании Cromochloris zofingiensis для получения липидов и каротиноидов. Такая замена обеспечивает снижение затрат на питательную среду примерно в 3.3 раза, что определяется более высокой массовой долей азота в мочевине, меньшим расходом соли на кубометр среды, а также более низкой оптовой ценой на мочевину, по сравнению с нитратом натрия.
Источники информации, принятые во внимание
1.  С., Lewis, L.E. Intersection of Chlorella, Muriella and Bracteacoccus: Resurrecting the genus Chromochloris Kol et Chodat Chlorophyceae, Chlorophyta) // Fottea. - 2012. - V. 12, No 1. - P. 83-93.
С., Lewis, L.E. Intersection of Chlorella, Muriella and Bracteacoccus: Resurrecting the genus Chromochloris Kol et Chodat Chlorophyceae, Chlorophyta) // Fottea. - 2012. - V. 12, No 1. - P. 83-93.
2. Mata T.M., Martins A.A., Caetano N.S. Microalgae for biodiesel production and other applications: A review // Renewable and Sustainable Energy Reviews. - 2010. - V. 14, N 1. - P. 217-232.
3. Surai P.F. The antioxidant properties of canthaxanthin and its potential effects in the poultry eggs and on embryonic development of the chick. Part 2. //  Poultry Sci. J. - 2012. - Vol. 68. - P. 717-726.
Poultry Sci. J. - 2012. - Vol. 68. - P. 717-726.
4. Han D., Li Y., Hu Q. Astaxanthin in microalgae: pathways, functions and biotechnological implications // Algae. - 2013. - V. 28, N 2. - . P. 131-147.
5. Maoka Т., Yasui H., Ohmori A., Tokuda H., Suzuki N., Osawa A., Shindo K., Ishibashi T. Anti-oxidative, anti-tumor-promoting, and anticarcinogenic activities of adonirubin and adonixanthin // J. Oleo Sci. - 2013. - V. 62. - P. 181-186.
6. Liu, J., Sun, Z., Gerken, H., Liu, Z., Jiang, Y., Chen, F. Chlorella zofingiensis as an Alternative Microalgal Producer of Astaxanthin: Biology and Industrial Potential // Mar. Drugs. - 2014. - V. - 12. - P. 3487-3515.
7. Kim D-Y., Vijayan D., Praveenkumar R., Han J-I., Lee K., Park J-Y., Chang W-S., Lee J-S., Oh Y-K. Cell-wall disruption and lipid/astaxanthin extraction from microalgae: Chlorella and Haematococcus II Электронный ресурс Biores. Technol. - 2015. dx.doi.org/10.1016/j.biortech.2015.08.107.
8. Ip P.F., Chen F. Production of astaxanthin by the green microalga Chlorella zofingiensis in the dark // Process Biochem. - 2005. - V. 40. - P. 733-738.
9. Li Y., Huang J., Sandmann G., Chen F. High-light and sodium chloride stress differentially regulate the biosynthesis of astaxanthin in Chlorella zofingiensis (Chlorophyceae) // J. Phycol. - 2009. - V. 5. - P. 635-641.
10. Sun N., Wang Y., Li Y.Т., Huang J.C., Chen, F. (). Sugar-based growth, astaxanthin accumulation and carotenogenic transcription of heterotrophic Chlorella zofingiensis (Chlorophyta) // Process Biochem. - 2008. - V. 43. - 1288-1292.
11. Liu J., Huang J., Fan K.W., Jiang Y., Zhong Y., Sun Z., Chen F. Production potential of Chlorella zofingienesis as a feedstock for biodiesel // Biores. Technol. - 2010. - V. 101. - P. 8658-8663.
12. Feng P., Deng Z., Fan L., Hu Z. Lipid accumulation and growth characteristics of Chlorella zofingiensis under different nitrate and phosphate concentrations // J. Biosci. Bioeng. - 2012. - V. 114, No. 4. - P. 405-410.
13. Chen J., Liu L., Wei D. Enhanced production of astaxanthin by Chromochloris zofingiensis in a microplate-based culture system under high light irradiation // Biores. Technol. - 2017. - V. 245. - P. 518-529.
14. Azaman S.N.A., Nagao N., Yusoff F.M., Tan S.W., Yeap S.K. A comparison of the morphological and biochemical characteristics of Chlorella sorokiniana and Chlorella zofingiensis cultured under photoautotrophic and mixotrophic conditions // PeerJ. 5: e3473; DOI 10.7717/peerj.3473.
15. Zhang Z., Huang J.J., Sun D., Lee Y., Chen F. Two-step cultivation for production of astaxanthin in Chlorella zofingiensis using a patented energy-free rotating floating photobioreactor (RFP) // Biores. Technol. - 2017. - N 224. - P. 515-522.
16. Мао X., Wu Т., Sun D., Zhang Z., Chen, F. Differential responses of the green microalga Chlorella zofingiensis to the starvation of various nutrients for oil and astaxanthin production // Biores. Technol. - 2018. - V. 249. - P. 791-798.
17. Hannon M., Gimpel J., Tran M., Rasala В., Mayfield S. Biofuels from algae: challenges and potential // Biofuels. - 2010. - V. 1, №5. - P. 763-784.
18. Richardson, J.W., Johnson, M.D., Outlaw, J.L. Economic comparison of open pond raceways to photo bio-reactors for profitable production of algae for transportation fuels in the Southwest // Algal Res. - 2012. - V. 1. - P. 93-100.
19. Liu J., Мао X., Zhou W., Guarnieri M.T. Simultaneous production of triacylglycerol and high-value carotenoids by the astaxanthin-producing oleaginous green microalga Chlorella zofingiensis // Biores. Technol. - 2016. - V. 214. - P. 319-327.
20. Markou G., Monlau F. 2019. Nutrient recycling for sustainable production of algal biofuels // Biofuels from Algae. - Elsevier, 2019. - P. 109-133.
21. Bekheet I.A., Syrett P.J. Urea-degrading enzymes in algae // British Phycol. J. - 1977. - V. 12, N 2. - P. 137-143.
22. Hsieh C-H., Wu W-T. Cultivation of microalgae for oil production with a cultivation strategy of urea limitation // Biores. Technol. - 2009. - V. 100. - P. 3921-3926.
23. Choochote W., Paiboonsin K., Ruangpan S., Pharuang A. Effects of Urea and Light Intensity on the Growth of Chlorella sp. // The 8th International Symposium on Biocontrol and Biotechnology. - 2010. - P. 127-134.
24. Rippka R., Deruelles J., Waterbury J., Herdman M., Stanier, R. Generic assignments, strain histories and properties of pure cultures of cyanobacteria // J. Gen. Microbiol. - 1979 - V. 111. - P. 1-61.
25. Huo S., Wang Z., Zhu S., Shu Q., Zhu L., Qin L., Zhou W., Feng P., Zhu F., Yuan Z., Dong R. Biomass accumulation of Chlorella zofingiensis G1 cultures grown outdoors in photobioreactors // Front. Energy Res. - 2018. - V. 6. art. 49. doi: 0.3389/fenrg.2018.00049.
26. Islam M.A., Magnusson M., Brown R.J., Ayoko G.A., Nabi Md. N., Heimann K. Microalgal species selection for biodiesel production based on fuel properties derived from fatty acid profiles // Energies. - 2013. - V. 6. - P. 5676-5702.
27. Sudibyo Н., Pradana Y.S., Samudra T.T., Budiman A., Suyono E.A. Study of cultivation under different colors of light and growth kinetic study of Chlorella zofingiensis Donz for biofuel production // Energy Procedia. - 2017. - V. 105. - P. 270-276.
28. Wang Y., Liu Z., Qin S. Effects of iron on fatty acid and astaxanthin accumulation in mixotrophic Chromochloris zofingiensis II Biotechnol. Lett. - 2013. - V. 35. - P. 351-357.
29. Kuhl A., Lorenzen Н. Handling and culturing of Chlorella // In: Prescott, D.M. (ed.), Methods in cell physiology. Acad Press: NY- London, 1964. - V. 1. - P. 159-187.
30. Bishoff Н., Bold Н.С. Phycological studies. IV. Some soil algae from enchanted rock and related algae species. University of Texas Publication, Austin, TX, 1963. P. 1-95.
31. Минюк Г.С., Челебиева Э.С., Чубчикова И.Н., Данцюк Н.В., Дробецкая И.В., Сахонь Е.Г., Чивкунова О.Б., Чеканов К.А., Лобакова Е.С., Сидоров Р.А., Соловченко А.Е. Влияние рН и СО2 на рост и метаболизм микроводоросли Coelastrella (Scotiellopsis) rubescens // Физиология растений. - 2016. - Т. 63. - С. 601-610.
32. Лелевич В.В., Климович В.В., Масловская А.А., Леднева И.О., Курбат М.Н. Биологическая химия: практикум для студентов педиатрического факультета / Гродно: ГрГМУ, 2016. - 108 с.
33. Minyuk G., Chelebieva E, ⋅Chubchikova I., ⋅Dantsyuk N., ⋅Drobetskaya I., ⋅Sakhon E., Chivkunova O., Chekanov K., Lobakova E., Solovchenko A. Stress-induced secondary carotenogenesis in Coelastrella rubescens, a producer of value-added keto-carotenoids // Algae 2017. - V. 32., N 3. P. - 245-259.
34. Минюк Г.С., Дробецкая И.В., Чубчикова И.Н., Данцюк Н.В., Челебиева Э.С. Скрининг зеленых микроводорослей как потенциальных источников природных кетокаротиноидов. Актуальность, стратегия и тактика исследований // Экология моря. - 2010. - Вып. 80. - С. 67-78.
35. Минюк Г.С., Терентъева Н.В., Дробецкая И.В. Сравнительная характеристика морфологических и физиолого-биохимических признаков трех штаммов Haematococcus pluvialis Flotow (Chlorophyta, Chlamydomonadales) // Альгология. - 2007. - Том. 17, N 2. - С. 148-159.
36. Чубчикова И.Н., Минюк Г.С. Дробецкая И.В., Терентъева Н.В. Хлорококковые микроводоросли как потенциальный источник природных кетокаротиноидов // Экология моря. - 2009. - Вып. 77. - С. 77-83.
Таблица 1. Состав питательный сред, использованных на «зеленой» и «красной" стадиях культивирования Chromochloris zofingiensis (г/л Н2Одист.)
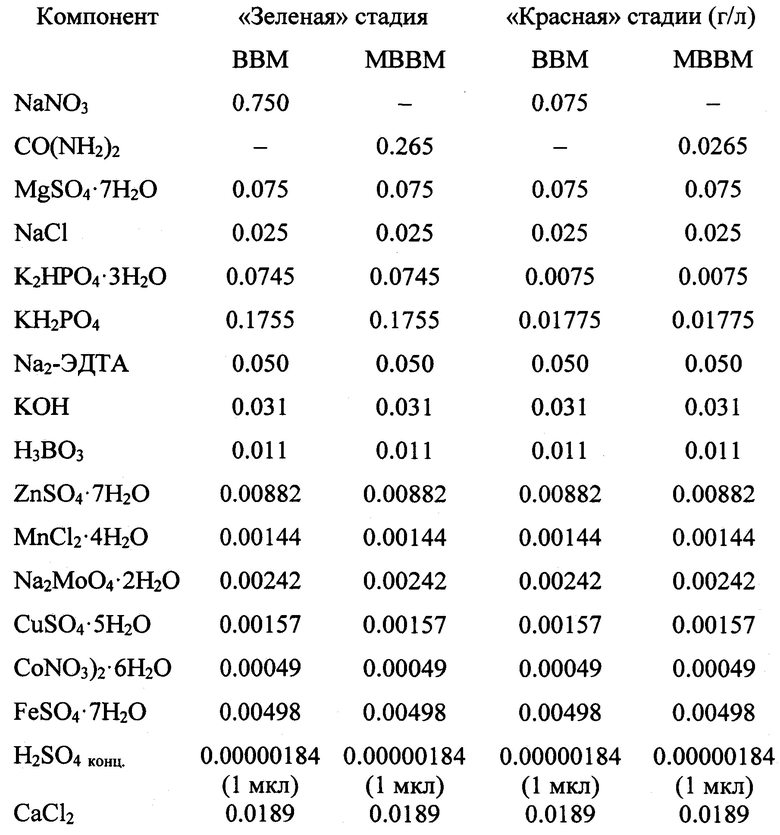
Таблица 2. Жирнокислотный профиль суммарных липидов Chromochloris zofingiensis в начале и конце «красной» стадии культивирования в зависимости от источника азота в питательной среде
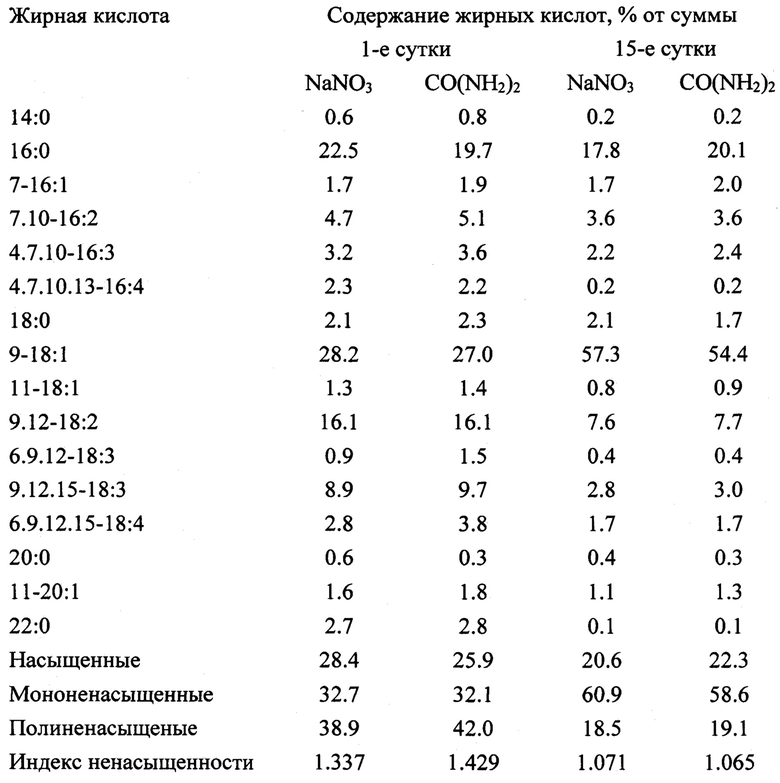
Таблица 3. Прогнозные характеристики биодизеля из Chromochloris zofingiensis, рассчитанные по профилю жирных кислот в начале и конце «красной» стадии
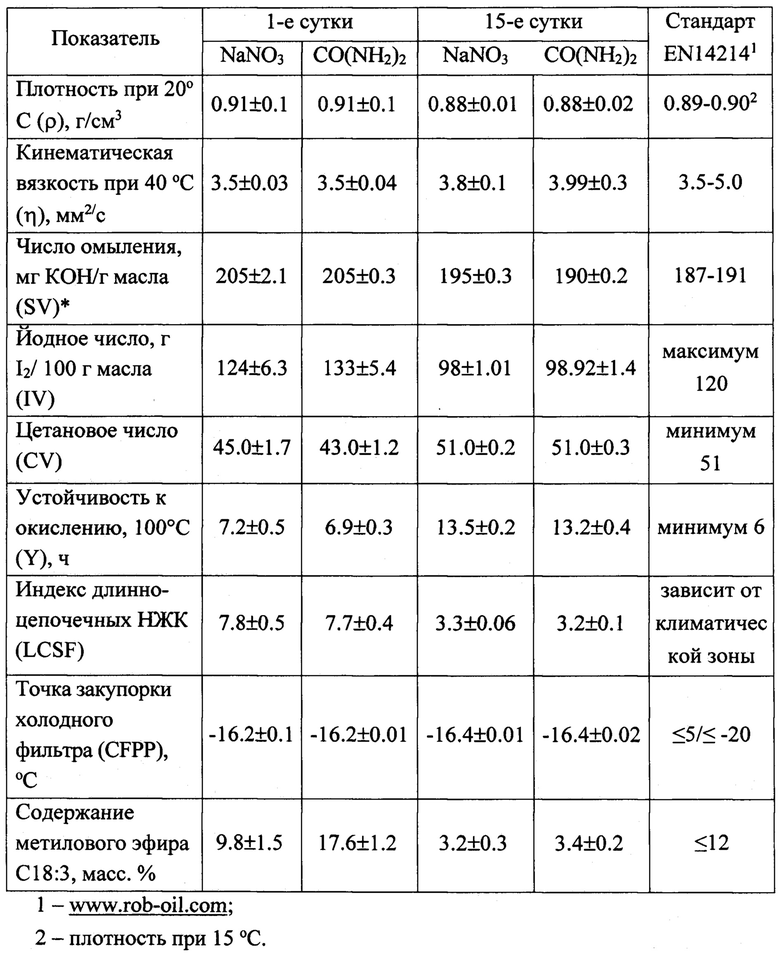
Таблица 4. Состав суммарных каротиноидов в биомассе Chromochloris zofingiensis в конце «красной» стадии в зависимости от химической формы азота в питательной среде

Таблица 5. Сравнительная характеристика некоторых технологических показателей мочевины и нитрата натрия как источников азота в питательных средах для культивирования С. zofingiensis

| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ КУЛЬТИВИРОВАНИЯ МИКРОВОДОРОСЛИ COELASTRELLA RUBESCENS ДЛЯ ПОЛУЧЕНИЯ КАРОТИНОИДОВ И ЛИПИДОВ | 2017 |
|
RU2661086C1 |
| ШТАММ МИКРОВОДОРОСЛИ Coelastrella sp. - ПРОДУЦЕНТ СМЕСИ НАТУРАЛЬНОГО БИОАНТИОКСИДАНТА АСТАКСАНТИНА И β-КАРОТИНА | 2018 |
|
RU2703420C1 |
| ШТАММ МИКРОВОДОРОСЛИ HAEMATOCOCCUS PLUVIALIS - ПРОДУЦЕНТ НАТУРАЛЬНОГО АСТАКСАНТИНА | 2014 |
|
RU2573944C1 |
| ШТАММ МИКРОВОДОРОСЛИ Bracteacoccus aggregatus - ПРОДУЦЕНТ СМЕСИ НАТУРАЛЬНОГО БИОАНТИОКСИДАНТА АСТАКСАНТИНА И ПРОВИТАМИНА А | 2019 |
|
RU2710131C1 |
| СПОСОБ КУЛЬТИВИРОВАНИЯ ОДНОКЛЕТОЧНОЙ ЗЕЛЕНОЙ ВОДОРОСЛИ HAEMATOCOCCUS PLUVIALIS ДЛЯ ПОЛУЧЕНИЯ АСТАКСАНТИНА | 2014 |
|
RU2541455C1 |
| СПОСОБ ПРОИЗВОДСТВА ПРИГОДНОГО ДЛЯ ИСПОЛЬЗОВАНИЯ В ДВИГАТЕЛЕ СЛОЖНОГО МЕТИЛОВОГО ЭФИРА ЖИРНЫХ КИСЛОТ | 2012 |
|
RU2603748C2 |
| ШТАММ МИКРОВОДОРОСЛИ Chlorella vulgaris, ПРЕДНАЗНАЧЕННЫЙ ДЛЯ ОЧИСТКИ СТОЧНЫХ ВОД СЕЛЬСКОХОЗЯЙСТВЕННЫХ И СПИРТОВЫХ ПРОИЗВОДСТВ | 2013 |
|
RU2555519C2 |
| СПОСОБ КУЛЬТИВИРОВАНИЯ ВИДА HAEMATOCOCCUS ДЛЯ ПРОИЗВОДСТВА АСТАКСАНТИНА | 2016 |
|
RU2730670C2 |
| СПОСОБ ХИМИЧЕСКОЙ МОДИФИКАЦИИ ЛИПИДОВ МИКРОВОДОРОСЛЕЙ, СПОСОБ ПОЛУЧЕНИЯ МЫЛА И МЫЛО, ВКЛЮЧАЮЩЕЕ СОЛИ ЖИРНЫХ КИСЛОТ ОМЫЛЕННЫХ ЛИПИДОВ МИКРОВОДОРОСЛЕЙ | 2009 |
|
RU2542374C2 |
| ШТАММ МИКРОВОДОРОСЛИ Desmodesmus sp. ДЛЯ КОНВЕРСИИ УГЛЕКИСЛОТЫ ИЗ ПРОМЫШЛЕННЫХ СБРОСНЫХ ГАЗОВ В СЫРЬЕ ДЛЯ ПРОИЗВОДСТВА БИОТОПЛИВА И КОРМОВЫХ ДОБАВОК | 2013 |
|
RU2555520C2 |

Изобретение относится к биотехнологии. Способ культивирования микроводоросли Chromochloris zofingiensis предусматривает культивирование микроводоросли Chromochloris zofingiensis на «зеленой стадии» на питательной среде МВВМ в течение 17 суток при температуре 22-23°С при заданном освещении и скорости продувки культуры воздухом. Осуществляют перевод культуры на «красную стадию» путем разведения культуры питательной средой со сниженным содержанием азота и фосфора и выращивают в течение 15 суток при продолжительности светового периода 24 ч, при заданной температуре и заданной освещенности. При этом питательная среда для «зеленой» и «красной» стадии культивирования содержит CO(NH2)2, MgSO4⋅7H2O, NaCl, K2HPO4⋅3H2O, KH2PO4, Na2-ЭДТА, KOH, H3BO3, ZnSO4⋅7H2O, MnCl2⋅4H2O, Na2MoO4⋅2H2O, CuSO4⋅5H2O, Co(NO3)2⋅6H2O, FeSO4⋅7H2O, CaCl2, H2SO4 конц и дистиллированную воду при заданном содержании компонентов. Изобретение позволяет повысить выход липидов и кетокаратиноидов. 2 з.п. ф-лы, 3 ил., 5 табл., 1 пр.
1. Способ культивирования микроводоросли Chromochloris zofingiensis для получения липидов и каротиноидов, включающий культивирование на питательной среде, создание стрессовых условий внешней среды для индукции биосинтеза астаксантина и липидов, получение биомассы, отличающийся тем, что водоросли культивируют методом двухстадийной накопительной культуры на модифицированной питательной среде ВВМ (МВВМ), модифицированной путем эквивалентной по азоту (8.86 ммоль N/л) замены нитрата натрия (NaNO3) на мочевину (CO(NH2)2) с соблюдением на I («зеленой») стадии условий внешней среды, способствующих активному делению клеток и накоплению биомассы в культуре, а на II («красной») стадии условий для интенсивного накопления липидов и С40-кетокаротиноидов, и для этого на I («зеленой») стадии С.Zofingiensis культивируют в течение 17 суток при следующих условиях: начальная численность клеток в культуре (nнач) - 2.5×106 кл/мл, освещенность (I) - 120 мкмоль фотонов/⋅м2⋅с, фотопериод - 15 ч свет/9 ч темнота, температура среды (Т) - 22-23°С и скорость продувки культуры воздухом (v) - 0.3 л⋅/(мин л), а перевод культуры на II («красную») стадию осуществляют путем ее разведения до nнач=4×106 кл/л питательной средой МВВМ со сниженным в 10 раз содержанием азота и фосфора и выращивают в течение 15 суток при продолжительности светового периода 24 ч, I=240 мкмоль фотонов/⋅м2⋅с, Т=24-25°С, v=0.6 л⋅/(мин л). (Т) - 22-23°С.
2. Способ по п. 1, отличающийся тем, что в состав питательной среды МВВМ для «зеленой» стадии культивирования входят (г/л дистиллированной воды): CO(NH2)2 - 0,265; MgSO4⋅7H2O - 0,075; NaCl - 0,025; K2HPO4⋅3H2O - 0,0745; KH2PO4 - 0,1755; Na2-ЭДТА - 0,05; KOH - 0,031; H3BO3 - 0,011; ZnSO4⋅7H2O - 0,00882; MnCl2⋅4H2O - 0,00144; Na2MoO4⋅2H2O - 0,00242; CuSO4⋅5H2O - 0,00157; Co(NO3)2⋅6H2O - 0,00049; FeSO4⋅7H2O - 0,00498; CaCl2 - 0,0189; H2SO4 конц - 1 мкл.
3. Способ по п. 1, отличающийся тем, что питательная среда МВВМ для «красной» стадии культивирования состоит (г/л дистиллированной воды): CO(NH2)2 - 0.0265; MgSO4⋅7H2O - 0.075; NaCl - 0.025; K2HPO4⋅3H2O - 0.0075; KH2PO4 - 0.01775; Na2-ЭДТА - 0.050; KOH - 0.031; H3BO3 - 0.011; ZnSO4⋅7H2O - 0.00882; MnCl2⋅4H2O - 0.00144; Na2MoO4⋅2H2O - 0.00242; CuSO4⋅5H2O - 0.00157; Co(NO3)2⋅6H2O - 0.00049; FeSO4⋅7H2O - 0.00498; CaCl2 - 0.0189; H2SO4 конц - 1 мкл.
| WANG Y., LIU Z., QIN S | |||
| Effects of iron on fatty acid and astaxanthin accumulation in mixotrophic Chromochloris zofingiensis II Biotechnol | |||
| Lett | |||
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| - V | |||
| Скоропечатный станок для печатания со стеклянных пластинок | 1922 |
|
SU35A1 |
| - P | |||
| Деревобетонный каток | 1916 |
|
SU351A1 |
| SUDIBYO Н., PRADANA Y.S., et | |||
| al | |||
| Study of cultivation under different colors of light and growth kinetic study of Chlorella zofingiensis Donz for biofuel production // | |||

