Область техники
[0001] Настоящее изобретение относится к способу продукции N-ацетилгепаросана (далее в настоящем описании обозначенного как гепаросан), представляющего собой капсульный полисахарид бактерии из рода Escherichia, посредством ферментной продукции с использованием этой бактерии.
Уровень техники
[0002] Гепарин, представляющий собой сульфатированный полисахарид, представляет собой антикоагулянтное средство, и его используют для лечения тромбоэмболии и синдрома диссеминированного внутрисосудистого свертывания, и для предотвращения свертывания во время искусственного диализа и при экстракорпоральном кровообращении, и т.п. В промышленности, большинство используемого гепарина выделено и очищено из слизистой оболочки кишечника свиней.
[0003] Поскольку случай со смертельным исходом произошел в 2008 г. из-за контаминации гепарина свиного происхождения примесями, потребовались исследования и разработка гепарина, не из животных источников, с контролем производства и с контролем качества. В качестве конкретного примера, способ, в котором ферментно полученный и очищенный N-ацетилгепаросан, представляющий собой капсульный полисахарид из грамотрицательных микроорганизмов, химически N-деацетилируют и N-сульфатируют, с последующим ферментными эпимеризацией и сульфатированием для получения гепарина, имеющего такую же структуру и антикоагулянтную активность, как происходящие из свиней (NPL 1 и 2).
[0004] В вышеуказанном способе, гепаросан, в качестве основной структуры представляет собой сахарную цепь, состоящую из повторяющейся дисахаридной структуры из глюкуроновой кислоты (GlcA) и N-ацетил-D-глюкозамина (GlcNAc). Для продукции гепаросана, опубликованы способ применения Escherichia coli штамма K5, исходно имеющей способность к продукции гепаросана (PTL 1 и NPL 1), способ применения Escherichia coli штамма Nissle 1917, также имеющей способность к продукции гепаросана (NPL 2), и способ применения Escherichia coli, исходно не имеющей способность к продукции гепаросана (PTL 4 и NPL 3 и 4).
[0005] Гепаросан классифицируют как капсульные полисахариды группы 2, и известно, что группы генов из области I, области II и области III, которые формируют кластер в геноме, вовлечены в синтез и транспорт гепаросана в Escherichia coli штамма K5 (NPL 4).
[0006] Утверждают, что среди белков, кодированных этими группами генов, KpsS и KpsC обеспечивают перенос множества остатков 3-дезокси-D-маннооктулозоновой кислоты (Kdo) к фосфатидилглицерину на внутренней мембране, и гликозилтрансферазы KfiA и KfiC добавляют предшественник сахара нуклеотида, таким образом, продолжается синтез гепаросана (NPL 5 и 6).
[0007] Кроме того, KfiD вовлечен в синтез предшественника UDP-GlcA; KpsF и KpsU вовлечены в синтез CMP-Kdo, который является субстратом для синтеза линкера Kdo; и KpsM, KpsT, KpsE и KpsD вовлечены в транспорт гепаросана, синтезированного на внутренней мембране, во внешнее окружение бактериальных клеток (NPL 6).
[0008] В качестве способа осуществления ферментации гепаросана с использованием штамма BL21 и штамма K-12 Escherichia coli, которые не имеют способность к продукции гепаросана, в качестве хозяев, известен способ введения кластера генов KfiABCD из области II, полученного из Escherichia coli штамма K5, в хозяина. Кроме того, опубликованы гены, улучшающие продукцию гепаросана, такие как rfaH, nusG и rpoE, (PTL 4, и NPL 3 и 4).
Список литературы
Патентные документы
[0009] [PTL 1] Японский патент No. 5830464
[PTL 2] Патент США No. 8771995
[PTL 3] WO2018/048973
[PTL 4] Патент США No. 9975928
Непатентные документы
[0010] [NPL 1] Biotechnology and Bioengineering 107 (2010) 964-973
[NPL 2] Applied Microbiology and Biotechnology 103 (2019) 6771-6782
[NPL 3] Metabolic Engineering 14 (2012) 521-527
[NPL 4] Carbohydrate Research 360 (2012) 19-24
[NPL 5] Proceedings of the National Academy of Sciences of USA 110 (2013) 20753-20758
[NPL 6] Carbohydrate Research 378 (2013) 35-44
Сущность изобретения
Техническая проблема
[0011] Как описано выше, проводили исследование и разработку продукции гепарина, происходящего не из животных источников, с контролем производства и с контролем качества, однако, общепринятые способы продукции имели недостаточную эффективность. Тем не менее, каким образом каждый из генов, кодируемых областями I, II и III, влияет на продукцию гепаросана бактериями из рода Escherichia, имеющими способность к продукции гепаросана, неизвестно до сих пор.
[0012] Соответственно, целью настоящего изобретения является предоставление способа эффективной продукции гепаросана посредством улучшения эффективности продукции гепаросана посредством генетической модификации бактерии из рода Escherichia, имеющей способность к продукции гепаросана.
Решение проблемы
[0013] Авторы настоящего изобретения обнаружили, что эффективность продукции гепаросана улучшается посредством использования бактерии из рода Escherichia, которая имеет специфическую генетическую модификацию и имеет способность к продукции гепаросана, и таким образом, завершили настоящее изобретение.
[0014] Таким образом, настоящее изобретение представляет собой следующее.
1. Способ продукции гепаросана, включающий: культивирование бактерии из рода Escherichia, которая имеет следующую генетическую модификацию (1) и имеет способность к продукции гепаросана, в среде для продукции гепаросана:
(1) генетическую модификацию, увеличивающую экспрессию гена kpsS.
2. Способ продукции гепаросана по 1, где бактерия из рода Escherichia дополнительно имеет по меньшей мере одну из следующих генетических модификаций (2) и (3):
(2) генетическую модификацию, увеличивающую экспрессию по меньшей мере одного гена, выбранного из гена kfiA, гена kfiB, гена kfiC и гена kfiD, и
(3) генетическую модификацию, вызывающую потерю функции гена yhbJ.
3. Способ продукции гепаросана по 1 или 2, где генетическая модификация (1) представляет собой по меньшей мере одну из модификации области контроля экспрессии гена kpsS и увеличения количества копий гена kpsS.
4. Способ продукции гепаросана по 2 или 3, где генетическая модификация (2) представляет собой по меньшей мере одну из модификации области контроля экспрессии по меньшей мере одного гена, выбранного из гена kfiA, гена kfiB, гена kfiC и гена kfiD, и увеличения количества копий по меньшей мере одного гена, выбранного из гена kfiA, гена kfiB, гена kfiC и гена kfiD.
5. Способ продукции гепаросана по любому из 2-4, где генетическая модификация (3) представляет собой делецию гена yhbJ.
6. Способ продукции гепаросана по любому из 1-5, где бактерия из рода Escherichia представляет собой Escherichia coli.
7. Способ продукции гепаросана по любому из 1-6, где ген kpsS представляет собой ДНК, содержащую нуклеотидную последовательность, показанную в SEQ ID NO: 33, или ДНК, содержащую нуклеотидную последовательность, имеющую 90% или более идентичность с нуклеотидной последовательностью, показанной в SEQ ID NO: 33, и имеющую свойство увеличения способности к продукции гепаросана бактерии из рода Escherichia, имеющей способность к продукции гепаросана, когда уровень экспрессии увеличен в бактерии.
8. Способ продукции гепаросана по любому из 2-7, где ген kfiA представляет собой ДНК, содержащую нуклеотидную последовательность, показанную в SEQ ID NO: 34, или ДНК, содержащую нуклеотидную последовательность, имеющую 90% или более идентичность с нуклеотидной последовательностью, показанной в SEQ ID NO: 34, и имеющую свойство увеличения способности к продукции гепаросана бактерии из рода Escherichia, имеющей способность к продукции гепаросана, когда уровень экспрессии увеличен в бактерии, ген kfiB представляет собой ДНК, содержащую нуклеотидную последовательность, показанную в SEQ ID NO: 35, или ДНК, содержащую нуклеотидную последовательность, имеющую 90% или более идентичность с нуклеотидной последовательностью, показанной в SEQ ID NO: 35, и имеющую свойство увеличения способности к продукции гепаросана бактерии из рода Escherichia, имеющей способность к продукции гепаросана, когда уровень экспрессии увеличен в бактерии, ген kfiC представляет собой ДНК, содержащую нуклеотидную последовательность, показанную в SEQ ID NO: 36, или ДНК, содержащую нуклеотидную последовательность, имеющую 90% или более идентичность с нуклеотидной последовательностью, показанной в SEQ ID NO: 36, и имеющую свойство увеличения способности к продукции гепаросана бактерии из рода Escherichia, имеющей способность к продукции гепаросана, когда уровень экспрессии увеличен в бактерии, и ген kfiD представляет собой ДНК, содержащую нуклеотидную последовательность, показанную в SEQ ID NO: 37, или ДНК, содержащую нуклеотидную последовательность, имеющую 90% или более идентичность с нуклеотидной последовательностью, показанной в SEQ ID NO: 37, и имеющую свойство увеличения способности к продукции гепаросана бактерии из рода Escherichia, имеющей способность к продукции гепаросана, когда уровень экспрессии увеличен в бактерии.
9. Способ продукции гепаросана по любому из 2-8, где ген yhbJ представляет собой ДНК, содержащую нуклеотидную последовательность, показанную в SEQ ID NO: 38, или ДНК, содержащую нуклеотидную последовательность, имеющую 90% или более идентичность с нуклеотидной последовательностью, показанной в SEQ ID NO: 38, и имеющую свойство увеличения способности к продукции гепаросана бактерии из рода Escherichia, имеющей способность к продукции гепаросана, когда уровень экспрессии уменьшен в бактерии.
10. Способ продукции гепаросана по любому из 1-9, где бактерия из рода Escherichia не имеет следующую генетическую модификацию (4):
(4) генетическую модификацию, увеличивающую экспрессию гена kpsC.
11. Бактерия из рода Escherichia, имеющая способность к продукции гепаросана и имеющая следующую генетическую модификацию (1):
(1) генетическую модификацию, увеличивающую экспрессию гена kpsS.
12. Бактерия из рода Escherichia по 11, дополнительно имеющая по меньшей мере одну из следующих генетических модификаций (2) и (3):
(2) генетической модификации, увеличивающей экспрессию по меньшей мере одного гена, выбранного из гена kfiA, гена kfiB, гена kfiC и гена kfiD, и
(3) генетической модификации, вызывающей потерю функции гена yhbJ.
13. Бактерия из рода Escherichia по 11 или 12, не имеющая следующую генетическую модификацию (4):
(4) генетическую модификацию, увеличивающую экспрессию гена kpsC.
Обеспечивающие преимущество эффекты изобретения
[0015] В соответствии со способом продукции гепаросана по настоящему изобретению, гепаросан, происходящий не из животных источников, можно получать с отличной эффективностью продукции с использованием бактерии из рода Escherichia, которая имеет специфическую генетическую модификацию и имеет способность к продукции гепаросана.
Краткое описание чертежей
[0016] [Фиг. 1] На ФИГ. 1 показана схематическая диаграмма кластера генов синтеза гепаросана на хромосоме Escherichia coli штамма K5.
[Фиг. 2] На ФИГ. 2 показана схематическая диаграмма ферментов, вовлеченных в продукцию гепаросана.
[Фиг. 3] На ФИГ. 3 показана схематическая диаграмма биосинтетического пути гепаросана.
Описание вариантов осуществления
[0017] Далее в настоящем описании, термины, используемые в настоящем описании, имеют значения, общеупотребительные в данной области, если не указано иное.
[0018] Бактерия по настоящему изобретению
В способе продукции гепаросана по настоящему изобретению, используют бактерию из рода Escherichia, которая имеет следующую генетическую модификацию (1) и имеет способность к продукции гепаросана (далее в настоящем описании обозначенного как бактерия по настоящему изобретению):
(1) генетическую модификацию, увеличивающую экспрессию гена kpsS.
[0019] Бактерия по настоящему изобретению, предпочтительно, дополнительно имеет по меньшей мере одну из следующих генетических модификаций (2) и (3):
(2) генетической модификации, увеличивающей экспрессию по меньшей мере одного гена, выбранного из гена kfiA, гена kfiB, гена kfiC и гена kfiD, и
(3) генетической модификации, вызывающей потерю функции гена yhbJ. Соответственно, примеры генетических модификаций в бактерии из рода Escherichia включают (1) и (2), описанные выше, (1) и (3) описанные выше, и (1) - (3), описанные выше.
[0020] Бактерия по настоящему изобретению, предпочтительно, не имеет следующую генетическую модификацию (4):
(4) генетическую модификацию, увеличивающую экспрессию гена kpsC.
[0021] В качестве примера бактерии из рода Escherichia, на ФИГ. 1 показана схематическая диаграмма кластера генов синтеза гепаросана на хромосоме Escherichia coli штамма K5 [J. Nzakizwanayo et al., PLOS ONE, (2015)]. Кроме того, на ФИГ. 2 показана схематическая диаграмма ферментов, вовлеченных в продукцию гепаросана.
[0022] Как показано на ФИГ. 1, kpsS и kpsC представляют собой гены, кодируемые областью I, среди группы генов из области I, области II и области III. Как показано на ФИГ. 2, kpsS и kpsC вовлечены в инициацию синтеза гепаросана. В продукции гепаросана, kpsS, вместе с kpsC, играет роль в добавлении множества линкеров Kdo к фосфатидилглицерину на внутренней мембране.
[0023] В качестве гена kpsS, ген kpsS, происходящий из рода Escherichia, является предпочтительным. Его конкретные примеры включают ген kpsS из Escherichia coli штамма K5. Нуклеотидная последовательность гена kpsS из Escherichia coli штамма K5 и аминокислотная последовательность белка, кодируемого геном, могут быть получены из публичных баз данных. Ген kpsS Escherichia coli штамма K5 зарегистрирован как номер доступа в GenBank CAA52659.1.
[0024] Примеры генов kpsS включают ДНК, содержащую нуклеотидную последовательность, показанную в SEQ ID NO: 33, или ДНК, содержащую нуклеотидную последовательность, имеющую 90% или более идентичность с нуклеотидной последовательностью, показанной в SEQ ID NO: 33, и имеющую свойство увеличения способности к продукции гепаросана бактерии из рода Escherichia, имеющей способность к продукции гепаросана, когда уровень экспрессии увеличен в бактерии.
[0025] Как показано на ФИГ. 1, kfiA, kfiB, kfiC и kfiD представляют собой гены, кодированные областью II, среди группы генов из области I, области II и области III. Как показано на ФИГ. 2, kfiA, kfiB, kfiC и kfiD вовлечены в синтез гепаросана, и играют роль добавления сахарида и таким образом, синтеза гепаросана.
[0026] В качестве гена kfiA, kfiB, kfiC или kfiD, ген kfiA, kfiB, kfiC или kfiD, происходящий из рода Escherichia, является предпочтительным. Его конкретные примеры включают ген kfiA, kfiB, kfiC или kfiD из Escherichia coli штамма K5. Нуклеотидная последовательность гена kfiA, kfiB, kfiC или kfiD из Escherichia coli штамма K5 и аминокислотная последовательность белка, кодированного геном, могут быть получены из публичных баз данных. kfiA зарегистрирован как номер доступа в GenBank CAA54711.1; kfiB зарегистрирован как номер доступа в GenBank CAE55824.1; kfiC зарегистрирован как номер доступа в GenBank CAA54709.1; и kfiD зарегистрирован как номер доступа в GenBank CAA54708.1.
[0027] Примеры генов kfiA включают ДНК, содержащую нуклеотидную последовательность, показанную в SEQ ID NO: 34, или ДНК, содержащую нуклеотидную последовательность, имеющую 90% или более идентичность с нуклеотидной последовательностью, показанной в SEQ ID NO: 34, и имеющую свойство увеличения способности к продукции гепаросана бактерии из рода Escherichia, имеющей способность к продукции гепаросана, когда уровень экспрессии увеличен в бактерии.
[0028] Примеры генов kfiB включают ДНК, содержащую нуклеотидную последовательность, показанную в SEQ ID NO: 35, или ДНК, содержащую нуклеотидную последовательность, имеющую 90% или более идентичность с нуклеотидной последовательностью, показанной в SEQ ID NO: 35, и имеющую свойство увеличения способности к продукции гепаросана бактерии из рода Escherichia, имеющей способность к продукции гепаросана, когда уровень экспрессии увеличен в бактерии.
[0029] Примеры генов kfiC включают ДНК, содержащую нуклеотидную последовательность, показанную в SEQ ID NO: 36, или ДНК, содержащую нуклеотидную последовательность, имеющую 90% или более идентичность с нуклеотидной последовательностью, показанной в SEQ ID NO: 36, и имеющую свойство увеличения способности к продукции гепаросана бактерии из рода Escherichia, имеющей способность к продукции гепаросана, когда уровень экспрессии увеличен в бактерии.
[0030] Примеры генов kfiD включают ДНК, содержащую нуклеотидную последовательность, показанную в SEQ ID NO: 37, или ДНК, содержащую нуклеотидную последовательность, имеющую 90% или более идентичность с нуклеотидной последовательностью, показанной в SEQ ID NO: 37, и имеющую свойство увеличения способности к продукции гепаросана бактерии из рода Escherichia, имеющей способность к продукции гепаросана, когда уровень экспрессии увеличен в бактерии.
[0031] На ФИГ. 3 показана схематическая диаграмма биосинтетического пути гепаросана. Как показано на ФИГ. 3, GlmS представляет собой первый фермент в пути снабжения UDP-N-ацетилглюкозамином, являющимся предшественником гепаросана, и представляет собой фермент, который катализирует реакцию из фруктоза-6-фосфата до глюкозамин-6-фосфата. YhbJ представляет собой фермент, отрицательно контролирующий GlmS.
[0032] В качестве гена yhbJ, ген yhbJ, происходящий из рода Escherichia, является предпочтительным. Его конкретные примеры включают ген yhbJ из Escherichia coli штамма K-12. Нуклеотидная последовательность гена yhbJ из Escherichia coli штамма K-12 и аминокислотная последовательность белка, кодированного геном, могут быть получены из публичных баз данных. Ген yhbJ из Escherichia coli штамма K-12 зарегистрирован как номер доступа в GenBank BAE77249.1.
[0033] Примеры генов yhbJ включают ДНК, содержащую нуклеотидную последовательность, показанную в SEQ ID NO: 38, или ДНК, содержащую нуклеотидную последовательность, имеющую 90% или более идентичность с нуклеотидной последовательностью, показанной в SEQ ID NO: 38, и имеющую свойство увеличения способности к продукции гепаросана бактерии из рода Escherichia, имеющей способность к продукции гепаросана, когда уровень экспрессии уменьшен в бактерии.
[0034] Каждый из генов в (1) - (3) выше может быть легко получен из публичных баз данных посредством, например, поиска BLAST или поиска FASTA с использованием нуклеотидной последовательности каждого гена, описанной выше. Кроме того, гомолог каждого гена может быть получен, например, посредством ПЦР с использованием хромосомы микроорганизма, такого как бактерия, в качестве матрицы и с использованием олигонуклеотида, полученного на основе этих известных последовательностей генов, в качестве праймера.
[0035] Каждый из генов в (1) - (3) выше могут представлять собой варианты генов, при условии, что исходные функции (например, активность или свойство) белка, кодированного генами, сохраняется. Можно проверять, сохраняет или нет белок, кодированный вариантами генов, свою исходную функцию; конкретно, например, когда исходной функцией является улучшение способности к продукции гепаросана, посредством введения варианта гена в микроорганизм, принадлежащий к прокариотам, имеющим способность к продукции гепаросана.
[0036] Варианты каждого из генов в (1) - (3) выше могут быть получены в соответствии со способом сайт-направленного мутагенеза, посредством модификации кодирующей области гена таким образом, что аминокислотные остатки в специфических положениях кодированного белка заменены, делетированы, вставлены или добавлены. Кроме того, варианты каждого из генов в (1) - (3) выше могут быть также получены, например, посредством обработки для мутагенеза.
[0037] При условии, что их исходные функции сохраняются, каждый из генов в (1) - (3) выше могут представлять собой гены, кодирующие белок, имеющий аминокислотную последовательность, в которой одна или несколько аминокислот в одном или нескольких положениях заменены, делетированы, вставлены или добавлены. Например, в кодированном белке, его N-конец и/или C-конец может быть удлинен или укорочен. Фраза «одна или несколько» отличается, в зависимости от положения и типа аминокислотного остатка в трехмерной структуре белка. Конкретные примеры этого включают 1-50, 1-40, 1-30, и это составляет, предпочтительно, 1-20, более предпочтительно, 1-10, даже более предпочтительно, 1-5, и особенно предпочтительно, 1-3.
[0038] Замена, делеция, вставка или добавление одной или нескольких аминокислот, как описано выше, представляет собой консервативную мутацию, которая нормально сохраняет функцию белка. Репрезентативными консервативными мутациями являются консервативные замены. Консервативная замена представляет собой мутацию, в которой замена возникает между Phe, Trp и Tyr в случае, когда участок замены представляет собой ароматическую аминокислоту, замена возникает между Leu, Ile и Val, в случае, когда участок замены представляет собой гидрофобную аминокислоту, замена возникает между Gln и Asn, в случае, когда участок замены представляет собой полярную аминокислоту, замена возникает между Lys, Arg и His, в случае, когда участок замены представляет собой основную аминокислоту, замена возникает между Asp и Glu, в случае, когда участок замены представляет собой кислую аминокислоту, и замена возникает между Ser и Thr, в случае, когда участок замены представляет собой аминокислоту, имеющую гидроксильные группы. Конкретные примеры замен, рассматриваемых как консервативная замена, включают замену Ala на Ser или Thr; замену Arg на Gln, His или Lys; замену Asn на Glu, Gln, Lys, His или Asp; замену Asp на Asn, Glu или Gln; замену Cys на Ser или Ala; замену Gln на Asn, Glu, Lys, His, Asp или Arg; замену Glu на Gly, Asn, Gln, Lys или Asp; замену Gly на Pro; замену His на Asn, Lys, Gln, Arg или Tyr; замену Ile на Leu, Met, Val или Phe; замену Leu на Ile, Met, Val или Phe; замену Lys на Asn, Glu, Gln, His или Arg; замену Met на Ile, Leu, Val или Phe; замену Phe на Trp, Tyr, Met, Ile или Leu; замену Ser на Thr или Ala; замену Thr на Ser или Ala; замену Trp на Phe или Tyr; замену Tyr на His, Phe или Trp; и замену Val на Met, Ile или Leu. Кроме того, замены, делеции, вставки, добавления аминокислот, как описано выше, их инверсии и т.п. включают замены, делеции, вставки и добавления, инверсии, и т.п., которые вызваны мутациями (мутанты или варианты), возникающими естественным образом, такими как мутации, основанные на индивидуальных различиях или видовых различиях в организме, из которого происходит ген.
[0039] Кроме того, при условии, что их исходные функции сохраняются, каждый из генов в (1)-(3) выше могут представлять собой гены, кодирующие белок, имеющий 80% или более, предпочтительно, 90% или более, более предпочтительно, 95% или более, даже более предпочтительно, 97% или более, и особенно предпочтительно, 99% или более идентичность с полной аминокислотной последовательностью.
[0040] Кроме того, при условии, что их исходные функции сохраняются, каждый из генов в (1) - (3) выше может представлять собой ДНК, которая гибридизуется, в строгих условиях, с зондом, который может быть получен из известных последовательностей гена, таких как последовательность, комплементарная полной или части нуклеотидной последовательности. Термин «строгие условия» относится к условиям, в которых образуются так называемые специфические гибриды, и неспецифические гибриды не образуются. Их примеры включают условия, в которых ДНК, имеющие идентичность друг с другом на более высоком уровне, например, ДНК, имеющие 80% или более, предпочтительно 90% или более, более предпочтительно 95% или более, даже более предпочтительно, 97% или более, и особенно предпочтительно 99% или более идентичность друг с другом, гибридизуются друг с другом, и ДНК, имеющие идентичность друг с другом на более низком уровне, не гибридизуются друг с другом; или условия, представляющие собой условия для отмывки при нормальной Саузерн-гибридизации, в которых отмывку проводят один раз, предпочтительно, от двух до трех раз, при концентрации соли и температуре, соответствующих 60 градусам C, 1 x SSC и 0,1% SDS, предпочтительно, 60 градусам C, 0,1 x SSC и 0,1% SDS, и более предпочтительно, 68 градусам C, 0,1 x SSC и 0,1% SDS.
[0041] Зонд, используемый для вышеуказанной гибридизации, может являться частью последовательности, комплементарной каждому гену. Такой зонд может быть получен посредством ПЦР с использованием олигонуклеотида, полученного на основе известной последовательности гена, в качестве праймера и с использованием фрагмента ДНК, содержащего каждый из генов в (1) - (3), выше, в качестве матрицы. Например, фрагмент ДНК, имеющий длину приблизительно 300 п.о., можно использовать в качестве зонда. В случае, когда фрагмент ДНК, имеющий длину приблизительно 300 п.о., используют в качестве зонда, примеры условий для отмывки при гибридизации включают условия 50 градусов C, 2 x SSC и 0,1% SDS.
[0042] Кроме того, поскольку вырожденность кодонов отличается, в зависимости от хозяев, каждый из генов в (1) - (3) выше, могут представлять собой гены, полученные посредством замены любого кодона на эквивалентный кодон, при условии, что их исходные функции сохраняются. Например, гены в таблицах 1-3 можно модифицировать таким образом, чтобы они имели оптимальные кодоны, в зависимости от частоты использования кодонов для используемого хозяина.
[0043] Примеры обработок для мутагенеза включают способ обработки in vitro молекулы ДНК, имеющей нуклеотидную последовательность каждого из генов в (1) - (3) выше, с использованием гидроксиламина или т.п.; способ обработки микроорганизмов, несущих каждый из генов в (1) - (3) выше, с использованием рентгеновского излучения, ультрафиолетового излучения или мутагена, такого как N-метил-N’-нитро-N-нитрозогуанидин (NTG), этилметансульфонат (EMS) и метилметансульфонат (MMS), и т.п.
[0044] Генетическая модификация для увеличения уровня экспрессии гена
Фраза «увеличение экспрессии гена» означает, что экспрессия гена увеличена, по сравнению с экспрессией для немодифицированного штамма. Примеры одного аспекта увеличения экспрессии гена включают аспект, в котором экспрессия гена предпочтительно, увеличена в 1,5 раза или более, более предпочтительно, увеличена в 2 раза или более, и даже более предпочтительно, увеличена в 3 раза или более, по сравнению с экспрессией для немодифицированного штамма.
[0045] Кроме того, фраза «экспрессия гена увеличена» означает не только увеличение экспрессии целевого гена в штамме, в котором целевой ген исходно экспрессируется, но также означает, что целевой ген экспрессируется в штамме, в котором целевой ген исходно не экспрессируется. То есть, фраза «экспрессия гена увеличена» включает, например, случай, в котором целевой ген вводят в штамм, не имеющий целевого гена, и целевой ген экспрессируется в нем. Кроме того, фраза «экспрессия гена увеличена» также обозначена как фразы «экспрессия гена усилена» и «экспрессия гена повышена».
[0046] Увеличения экспрессии гена можно достигать, например, посредством увеличения количества копий гена. Увеличения количества копий гена можно достигать посредством введения гена в хромосому хозяина. Введение гена в хромосому можно осуществлять с использованием, например, гомологичной рекомбинации (Miller I, J.H. Experiments in Molecular Genetics, 1972, Cold Spring Harbor Laboratory). Можно вводить только одну копию гена, или можно вводить две или более его копий.
[0047] Например, множество копий гена можно вводить в хромосому посредством проведения гомологичной рекомбинации с нацеливанием в то же время на последовательность, имеющую множество копий на хромосоме. Примеры последовательности, имеющей множество копий на хромосоме, включают повторяющуюся последовательность ДНК (повторяющуюся ДНК) и инвертированные повторы, присутствующие на обоих концах транспозона.
[0048] Альтернативно, гомологичную рекомбинацию можно проводить с нацеливанием в то же время на соответствующую последовательность на хромосоме, такую как ген, не являющийся необходимым для продукции целевого вещества. Гомологичную рекомбинацию можно проводить, например, посредством способа с использованием линейной ДНК, способа с использованием плазмиды, содержащей термочувствительную точку начала репликации, способа с использованием плазмиды, способной к конъюгативному переносу, способа с использованием суицидного вектора, не имеющего точку начала репликации, которая функционирует в хозяине, или способа трансдукции с использованием фага. Кроме того, ген можно также случайным образом вводить в хромосому с использованием транспозона или Mini-Mu (JP-A-H2-109985).
[0049] Введен ли целевой ген в хромосому, можно проверять посредством Саузерн-гибридизации с использованием зонда, имеющего последовательность, комплементарную всей или части гена, ПЦР с использованием праймера, сконструированного на основе последовательности гена, или т.п.
[0050] Кроме того, количество копий гена также можно увеличивать посредством введения вектора, содержащего ген, в хозяина. Например, является возможным увеличивать количество копий гена посредством конструирования экспрессирующего вектора для гена, посредством лигирования фрагмента ДНК, содержащего целевой ген, с вектором, который функционирует в хозяине, и трансформации хозяина с использованием экспрессирующего вектора. Фрагмент ДНК, содержащий целевой ген, можно получать, например, посредством ПЦР с использованием геномной ДНК микроорганизма, имеющего целевой ген, в качестве матрицы. Способ трансформации не является конкретно ограниченным, и можно использовать общеизвестный способ.
[0051] В качестве вектора, можно использовать вектор, который может автономно реплицироваться в клетке-хозяине. Вектор, предпочтительно, представляет собой мультикопийный вектор. Кроме того, вектор, предпочтительно, имеет маркер, такой как ген устойчивости к антибиотику или другой ген, описанный в литературе [Karl Friehs, Plasmid Copy Number and Plasmid Stability, Adv Biochem Engin/Biotechnol 86: 47-82 (2004)], для отбора трансформанта. Кроме того, вектор может иметь промотор или терминатор для экспрессии вставленного гена. Примеры вектора включают вектор, происходящий из бактериальной плазмиды, вектор, происходящий из дрожжевой плазмиды, вектор, происходящий из бактериофага, космиды, фагмиды и т.п.
[0052] Конкретные примеры векторов, способных к автономной репликации в бактериях из числа Enterobacteriaceae, таких как Escherichia coli, включают pUC19, pUC18, pHSG299, pHSG399, pHSG398, pBR322 и pSTV29 (все из Takara Bio Inc.), pACYC184, pMW219, pMW118 и pMW119 (все из Nippon Gene), pTrc99A (Pharmacia), вектор pPROK (Clontech), pKK233-2 (Clontech), вектор pET (Novagen), вектор pQE (Qiagen) и вектор с широким спектром хозяев RSF1010.
[0053] В случае введения гена, является достаточным, чтобы ген сохранялся в бактерии из рода Escherichia, имеющей генетическую модификацию по настоящему изобретению. Конкретно, является достаточным, чтобы ген был введен таким образом, чтобы он экспрессировался под контролем промоторной последовательности, которая функционирует в бактерии по настоящему изобретению. Промотор может представлять собой промотор, происходящий из хозяина, или гетерологичный промотор. Промотор может представлять собой собственный промотор гена, подлежащего введению, или промотор других генов. В качестве промотора, например, можно использовать более сильный промотор, который будет описан позже.
[0054] Терминатор для терминации транскрипции может быть расположен ниже гена. Терминатор не является конкретно ограниченным, при условии, что он функционирует в бактерии по настоящему изобретению. Терминатор может представлять собой терминатор, происходящий из хозяина, или гетерологичный терминатор. Терминатор может представлять собой терминатор, специфический для гена, подлежащего введению, или может представлять собой терминатор из других генов. Конкретные примеры терминаторов включают терминатор T7, терминатор T4, терминатор фага fd, терминатор tet и терминатор trpA.
[0055] Векторы, промоторы и терминаторы, которые можно использовать в различных микроорганизмах, подробно описаны, например, в «Basic Lecture 8 on Microbiology, Gene Engineering, KYORITSU SHUPPAN, 1987», и их можно использовать.
[0056] Кроме того, в случае, когда вводят два или более генов, является достаточным, чтобы каждый ген сохранялся в бактерии по настоящему изобретению подходящим для экспрессии образом. Например, каждый ген из всех может сохраняться в отдельном экспрессирующем векторе, или все могут сохраняться на хромосоме. Кроме того, каждый ген может отдельно сохраняться на множестве экспрессирующих векторов, или может отдельно сохраняться на одном или множестве экспрессирующих векторов и на хромосоме. Кроме того, два или более генов могут формировать оперон и быть введены. Примеры «случаев, когда вводят два или более генов», включают случай введения генов, соответственно, кодирующих два или более ферментов, случай введения генов, соответственно, кодирующих две или более субъединиц, которые формируют один фермент, и их комбинации.
[0057] Ген, подлежащий введению, не является конкретно ограниченным, при условии, что он кодирует белок, который функционирует в хозяине. Ген, подлежащий введению, может представлять собой ген, происходящий из хозяина, или гетерологичный ген. Ген, подлежащий введению, может быть получен, например, посредством ПЦР с использованием праймеров, сконструированных на основе нуклеотидной последовательности гена, и с использованием геномной ДНК из организма, имеющего ген, или плазмиды или т.п., несущей ген, в качестве матрицы. Кроме того, ген, подлежащий введению, можно полностью синтезировать, например, на основании нуклеотидной последовательности гена [Gene, 60 (1), 115-127 (1987)].
[0058] Кроме того, увеличения экспрессии гена можно достигать посредством улучшения эффективности транскрипции гена. Улучшения эффективности транскрипции гена можно достигать, например, посредством замены промотора гена на хромосоме на более сильный промотор. «Более сильный промотор» относится к промотору, который усиливает транскрипцию гена, по сравнению с природным промотором дикого типа.
[0059] Примеры «более сильных промоторов» включают известные промоторы с высокой экспрессией, такие как промотор uspA, промотор T7, промотор trp, промотор lac, промотор thr, промотор tac, промотор trc, промотор tet, промотор araBAD, промотор rpoH, промотор PR и промотор PL.
[0060] Кроме того, в качестве более сильного промотора, общепринятый промотор высоко активного типа может быть получен с использованием различных репортерных генов. Например, активность промотора можно увеличивать посредством переноса областей -35 и -10 в промоторной области ближе к консенсусной последовательности (WO2000/18935).
[0061] Примеры промоторов высоко активного типа включают различные tac-подобные промоторы (Katashkina JI et al. Патентная заявка Российской Федерации 2006134574) и промотор pnlp8 (WO2010/027045). Способы оценки силы промотора и примеры сильных промоторов описаны в известных литературных источниках [Prokaryotic promoters in biotechnology. Biotechnol. Annu. Rev., 1, 105-128 (1995) и т.п.].
[0062] Кроме того, увеличения уровня экспрессии гена можно достигать посредством улучшения эффективности трансляции гена. Улучшения эффективности трансляции гена можно достигать, например, посредством замены последовательности Шайна-Дальгарно (SD) (также называемой участок связывания рибосомы (RBS)) гена на хромосоме на более сильную последовательность SD.
[0063] «Более сильная последовательность SD» относится к последовательности SD, для которой трансляция мРНК является улучшенной, по сравнением с природной последовательностью SD дикого типа. Примеры более сильной последовательности SD включают RBS гена 10 из фага T7 (Olins P.O. et al, Gene, 1988, 73, 227-235). Кроме того, известно, что замена, вставка или делеция нескольких нуклеотидов в спейсерной области между RBS и инициирующим кодоном, в частности, в последовательности, непосредственно выше инициирующего кодона (5’-UTR), значительно влияет на стабильность и эффективность трансляции мРНК, и таким образом, эффективность трансляции гена можно улучшать посредством их модификации.
[0064] По настоящему изобретению, участки, влияющие на экспрессию гена, такие как промотор, последовательность SD и спейсерная область между RBS и инициирующим кодоном, также в совокупности обозначены как «области контроля экспрессии». Области контроля экспрессии можно определять с использованием программного обеспечения для поиска генов, такого как vector или GENETYX для поиска промоторов. Модификацию этих областей контроля экспрессии можно проводить, например, посредством способа с использованием термочувствительного вектора или способа направляемой Red интеграции (WO2005/010175).
[0065] Улучшения эффективности трансляции генов можно также достигать, например, посредством модификации кодонов. Конкретно, например, в случае, когда проводят гетерологичную экспрессию гена, и т.п., эффективность трансляции гена можно улучшать посредством замены редких кодонов, присутствующих в гене, на синонимические кодоны, используемые более часто.
[0066] Замену кодонов можно проводить, например, посредством способа сайт-направленного мутагенеза, в котором целевую мутацию вводят в целевой участок в ДНК. Примеры способов сайт-направленного мутагенеза включают способ с использованием ПЦР [Higuchi, R., 61, in PCR technology, Erlich, H.A. Eds., Stockton press (1989); Carter, P., Meth. In Enzymol., 154, 382 (1987)], и способ с использованием фага [Kramer, W. and Frits, H.J., Meth. In Enzymol., 154, 350 (1987); Kunkel, T.A. et al., Meth. In Enzymol., 154, 367 (1987)]. Альтернативно, фрагмент гена, в котором кодоны заменены, можно полностью синтезировать. Частота использования кодонов в различных организмах описана в «базе данных использования кодонов» [http://www.kazusa.or.jp/codon; Nakamura, Y. et al, Nucl. Acids Res., 28, 292 (2000)].
[0067] Кроме того, экспрессию гена можно также увеличивать посредством амплификации регулятора, который увеличивает экспрессию гена, или посредством делеции или ослабления регулятора, который уменьшает экспрессию гена. Такие способы увеличения экспрессии гена, как описано выше, можно использовать отдельно или можно использовать в любой комбинации.
[0068] Увеличение экспрессии гена можно проверять, например, посредством проверки увеличения уровня транскрипции гена или посредством проверки увеличения количества белка, экспрессированного с гена. Кроме того, увеличение экспрессии гена можно проверять, например, посредством проверки увеличения активности белка, экспрессированного с гена.
[0069] Проверку увеличения уровня транскрипции гена можно проводить посредством сравнения количества мРНК, транскрибированной с гена, с немодифицированным штаммом, таким как штамм дикого типа или родительский штамм. Примеры способов оценки количества мРНК включают Нозерн-гибридизацию, RT-ПЦР и т.п. [Sambrook, J., et al., Molecular Cloning A Laboratory Manual/Third Edition, Cold spring Harbor Laboratory Press, Cold spring Harbor (USA), 2001]. Увеличение количества мРНК относится, например, к случаю, в котором количество мРНК, предпочтительно, увеличено в 1,5 раза или более, более предпочтительно, увеличено в 2 раза или более, и даже более предпочтительно, увеличено в 3 раза или более, по сравнению с количеством в немодифицированном штамме.
[0070] Увеличение количества белка можно проверять, например, посредством Вестерн-блоттинга с использованием антитела. Увеличение количества белка относится, например, к случаю, в котором количество белка, предпочтительно, увеличено в 1,5 раза или более, более предпочтительно, увеличено в 2 раза или более, и даже более предпочтительно, увеличено в 3 раза или более, по сравнению с количеством в немодифицированном штамме.
[0071] Увеличение активности белка можно проверять, например, посредством измерения активности белка. Увеличение активности белка относится, например, к случаю, в котором активность белка, предпочтительно, увеличена в 1,5 раза или более, более предпочтительно, увеличена в 2 раза или более, и даже более предпочтительно, увеличена в 3 раза или более, по сравнению с активностью в немодифицированном штамме.
[0072] Вышеописанные способы увеличения экспрессии гена можно использовать для усиления экспрессии каждого из генов (1) и (2) выше.
[0073] Генетическая модификация, которая увеличивает экспрессию гена kpsS, предпочтительно, представляет собой по меньшей мере одну из модификации областей контроля экспрессии гена kpsS и генетической модификации, увеличивающей количество копий. Ген kpsFEDUCS присутствует в качестве группы генов продукции гепаросана, но как будет описано позднее в примерах, авторы настоящего изобретения обнаружили, что эффект особенного улучшения продукции гепаросана получен посредством увеличения экспрессии только гена kpsS среди них. Соответственно, в качестве генетической модификации, которая увеличивает экспрессию гена kpsS, генетическая модификация для увеличения количества копий гена kpsS является особенно предпочтительной.
[0074] В то время как бактерия по настоящему изобретению имеет генетическую модификацию, которая увеличивает экспрессию гена kpsS, бактерия по настоящему изобретению, предпочтительно, не имеет генетическую модификацию, которая увеличивает экспрессию гена kpsC, более предпочтительно, не имеет генетическую модификацию, которая увеличивает экспрессию гена kpsC, и по меньшей мере одного гена, выбранного из генов kpsF, kpsE, kpsD и kpsU, и наиболее предпочтительно, не имеет генетическую модификацию, которая увеличивает экспрессию всех генов kpsC, kpsF, kpsE, kpsD и kpsU.
[0075] Генетическая модификация, которая увеличивает экспрессию по меньшей мере одного гена, выбранного из генов kfiA, kfiB, kfiC и kfiD, предпочтительно, представляет собой по меньшей мере одну из модификации областей контроля экспрессии по меньшей мере одного гена, выбранного из генов kfiA, kfiB, kfiC и kfiD, и увеличения количества копий генов. Гены kfiA, kfiB, kfiC и kfiD составляют, оперон, как показано на ФИГ. 1. Генетическая модификация, которая усиливает весь оперон, составляемый генами kfiA, kfiB, kfiC и kfiD, является предпочтительной, и модификация области контроля экспрессии генов kfiA, kfiB, kfiC и kfiD, является более предпочтительной.
[0076] Генетическая модификация, которая вызывает потерю функции гена
Примеры генетических модификаций, которые вызывают потерю функции гена yhbJ из (3) выше, включают генетическую модификацию, при которой функция белка, кодированного частью, соответствующей yhbJ, уменьшена или полностью остановлена посредством модификации ДНК, кодирующей часть, соответствующую yhbJ, в геномной ДНК бактерии из рода Escherichia в качестве хозяина.
[0077] В способе по настоящему изобретению, форма модификации, подлежащей добавлению к ДНК, кодирующей часть, соответствующую yhbJ, не является конкретно ограниченной, при условии, что функция белка, кодированного частью, соответствующей yhbJ, уменьшена или полностью остановлена, и можно соответствующим образом использовать известный способ.
[0078] Примеры форм, которые уменьшают или полностью останавливают функцию белка, кодированного частью, соответствующей yhbJ, включают любую из следующих модификаций (a) - (c):
(a) Вся или часть ДНК, кодирующей часть, соответствующую yhbJ, удалена,
(b) Одна или несколько замен, делеций или добавлений внесены в ДНК, кодирующую часть, соответствующую yhbJ, и
(c) ДНК, кодирующая часть, соответствующую yhbJ, заменена на последовательность ДНК, имеющую менее чем 80% идентичность с последовательностью ДНК до модификации.
[0079] Примеры потери функции гена yhbJ включают случай, в котором активность гена yhbJ составляет, предпочтительно, 20% или менее, более предпочтительно, 10% или менее, и даже более предпочтительно, 5% или менее, по сравнению с активностью немодифицированного штамма. Активность yhbJ можно проверять посредством проверки уровня экспрессии glmS посредством способа Нозерн-блоттинга, способа Вестерн-блоттинга или т.п. [Kalamorz F. et al, (2007) «Feedback control of glucosamine-6-phosphate synthase GlmS expression depends on the small RNA GlmZ and involves the novel protein YhbJ in Escherichia coli». Mol Microbiol. 65 (6):1518-33].
[0080] Бактерия из рода Escherichia
Бактерия, принадлежащая к роду Escherichia, не является конкретно ограниченной, и ее примеры включают бактерию, классифицированную в род Escherichia, по классификации, известной экспертам в микробиологии. Примеры бактерий, принадлежащих к роду Escherichia, включают бактерии, описанные в литературе [Backmann, B.J. 1996. Derivations and Genotypes of some mutant derivatives of Escherichia coli K-12, p. 2460-2488. Таблица 1. In F.D. Neidhardt (ed.), Escherichia coli and Salmonella Cellular and Molecular Biology/Second Edition, American Society for Microbiology Press, Washington, DC].
[0081] Примеры бактерий, принадлежащих к роду Escherichia, включают Escherichia coli. Примеры Escherichia coli включают Escherichia coli штаммов K-12, таких как штамм W3110 (ATCC 27325) и штамм MG1655 (ATCC 47076); Escherichia coli штамма K5 (ATCC 23506); Escherichia coli штаммов B, таких как штамм BL21 (DE3); Escherichia coli штамма Nissle 1917 (DSM 6601); и их производные штаммы.
[0082] Эти штаммы могут быть заказаны, например, из Американской коллекции типовых культур (адрес: 12301 Parklawn Drive, Rockville, Maryland 20852, P.O. Box 1549, Manassas, VA 20108, United States of America). То есть, регистрационный номер присвоен каждому штамму, и штаммы могут быть заказаны с использованием этого регистрационного номера (ссылка https://www.atcc.org/). Регистрационный номер, соответствующий каждому штамму, описан в каталоге Американской коллекции типовых культур. Кроме того, штамм BL21 (DE3) является доступным, например, из Life Technologies (номер продукта C6000-03).
[0083] Бактерия по настоящему изобретению может исходно иметь способность к продукции гепаросана, или может являться модифицированной, чтобы иметь способность к продукции гепаросана. Бактерия, имеющая способность к продукции гепаросана, может быть получена, например, посредством придания способности к продукции гепаросана вышеупомянутой бактерии.
[0084] Способность к продукции гепаросана можно придавать посредством введения гена, кодирующего белок, вовлеченный в продукцию гепаросана, со ссылкой на Metabolic Engineering 14 (2012) 521-527, Carbohydrate Research 360 (2012) 19-24, Патент США No. 9975928 и т.п. Примеры белков, вовлеченных в продукцию гепаросана, включают гликозилтрансферазу и белок-носитель для эффлюкса гепаросана. По настоящему изобретению, можно вводить один вид гена, или можно вводить два или более видов генов. Введение гена можно проводить таким же образом, как в вышеописанном способе увеличения количества копий гена.
[0085] Способ продукции гепаросана
Способ продукции гепаросана по настоящему изобретению, включающий культивирование бактерии по настоящему изобретению в среде для продукции и накопления гепаросана в среде. Способ продукции гепаросана по настоящему изобретению может дополнительно включать, при необходимости, сбор гепаросана из среды.
[0086] Используемая среда не является конкретно ограниченной, при условии, что бактерия по настоящему изобретению может расти, и гепаросан продуцируется и накапливается. В качестве среды, например, можно использовать нормальную среду, используемую для культивирования бактерий. Примеры сред включают среду LB (среду Луриа-Бертани), однако, примеры не являются ограниченными этим. В качестве среды, является возможным использовать, например, среду, содержащую компонент, выбранный из источника углерода, источника азота, источника фосфата, источника серы, и других различных органических компонентов и неорганических компонентов, по необходимости. Специалист в данной области может подходящим образом устанавливать тип и концентрацию компонентов среды.
[0087] Источник углерода не является конкретно ограниченным, при условии, что он может быть использован бактерией по настоящему изобретению таким образом, что может продуцироваться гепаросан. Примеры источников углерода включают сахара, такие как глюкоза, фруктоза, сахароза, лактоза, галактоза, ксилоза, арабиноза, меласса, гидролизат крахмала и гидролизат биомассы, органические кислоты, такие как уксусная кислота, фумаровая кислота, лимонная кислота, янтарная кислота и яблочная кислота, спирты, такие как глицерин, неочищенный глицерин и этанол, и жирные кислоты. В качестве источника углерода, можно использовать один вид источника углерода, или два или более видов источников углерода можно комбинировать.
[0088] Примеры источников азота включают соли аммония, такие как сульфат аммония, хлорид аммония и фосфат аммония, органические источники азота, такие как пептон, дрожжевой экстракт, мясной экстракт и продукты разложения соевого белка, аммиак и мочевину. В качестве источника азота, можно использовать один вид источника азота, или два или более видов источников азота можно комбинировать.
[0089] Примеры источников фосфата включают фосфаты, такие как дигидрофосфат калия и гидрофосфат дикалия, и полимеры фосфорной кислоты, такие как пирофосфорная кислота. В качестве источника фосфата, можно использовать один вид источника фосфата, или два или более видов источников фосфата можно комбинировать.
[0090] Примеры источников серы включают неорганические соединения серы, такие как сульфаты, тиосульфаты и сульфиты, и содержащие серу аминокислоты, такие как цистеин, цистин и глутатион. В качестве источника серы, можно использовать один вид источника серы, или два или более видов источников серы можно комбинировать.
[0091] Конкретные примеры различных других органических компонентов и неорганических компонентов включают неорганические соли, такие как хлорид натрия и хлорид калия; металлические микроэлементы, такие как железо, марганец, магний и кальций; витамины, такие как витамин B1, витамин B2, витамин B6, никотиновая кислота, никотинамид и витамин B12; аминокислоты; нуклеиновые кислоты; содержащие их органические компоненты, такие как пептон, казаминовая кислота, дрожжевой экстракт и продукты разложения соевого белка. В качестве различных других органических компонентов и неорганических компонентов, можно использовать один вид компонента, или два или более видов компонентов можно использовать в комбинации.
[0092] Кроме того, в случае, когда используют ауксотрофный мутант, требующий аминокислоту или т.п. для роста, является предпочтительным дополнение питательного вещества, необходимого для среды. Кроме того, когда ген вводят с использованием вектора, несущего ген устойчивости к антибиотику, является предпочтительным добавление соответствующего антибиотика в среду.
[0093] Условия культивирования не являются конкретно ограниченными, при условии, что бактерия по настоящему изобретению может расти, и гепаросан продуцируется и накапливается. Культивирование можно проводить, например, в нормальных условиях, используемых для культивирования бактерий. Специалист в данной области может подходящим образом устанавливать условия культивирования.
[0094] Культивирование можно проводить аэробно, например, посредством аэрационной культуры или встряхиваемой культуры с использованием жидкой среды. Температура культивирования может составлять, например, 30 градусов C - 37 градусов C. Период культивирования может составлять, например, 16-72 часов. Культивирование можно проводить посредством периодического культивирования, периодического культивирования с подпиткой, непрерывного культивирования или их комбинации. Кроме того, культивирование можно разделять на предварительное культивирование и основное культивирование. Предварительное культивирование можно проводить с использованием, например, среды на чашках или жидкой среды.
[0095] Посредством культивирования бактерии по настоящему изобретению, как описано выше, гепаросан накапливается в среде.
[0096] Способ сбора гепаросана из культурального раствора не является конкретно ограниченным, при условии, что гепаросан можно собирать. Примеры способов сбора гепаросана из культурального раствора включают способы, описанные в примерах. Конкретно, например, культуральный супернатант можно отделять от культурального раствора, и затем гепаросан в супернатанте можно преципитировать посредством преципитации этанолом. Количество добавленного этанола может составлять, например, в 2,5-3,5 раза больше количества жидкого супернатанта. Для преципитации гепаросана, можно использовать не только этанол, но также органический растворитель, необязательно способный смешиваться с водой.
[0097] Примеры органических растворителей включают этанол, метанол, н-пропанол, изопропанол, n-бутанол, т-бутанол, втор-бутанол, пропиленгликоль, ацетонитрил, ацетон, DMF, DMSO, N-метилпирролидон, пиридин, 1,2-диметоксиэтан, 1,4-диоксан и THF.
[0098]
Дополнительные примеры способа сбора гепаросана из культурального раствора включают способ, включающий очистку гепаросана посредством нацеливания на Kdo, присутствующий на конце гепаросана.
[0099] Преципитатированный гепаросан можно растворять, например, в воде, составляющей двойное количество исходного супернатанта. Собранный гепаросан может содержать такие компоненты, как бактериальные клетки, компоненты среды, вода и побочные продукты метаболизма бактерий, в дополнение к гепаросану. Гепаросан можно очищать до желательной степени. Степень чистоты гепаросана может составлять, например, 30% (масс./масс.) или более, 50% (масс./масс.) или более, 70% (масс./масс.) или более, 80% (масс./масс.) или более, 90% (масс./масс.) или более, или 95% (масс./масс.) или более.
[0100] Детекцию и количественную оценку гепаросана можно проводить посредством известного способа. Конкретно, например, гепаросан можно детектировать и количественно оценивать посредством способа с использованием карбазола, как будет описано позже в примерах. Способ с использованием карбазола представляет собой способ, широко используемый в качестве способа количественной оценки уроновой кислоты, и является возможным детектировать и количественно оценивать гепаросан посредством термической реакции гепаросана с карбазолом в присутствии серной кислоты, и измерения поглощения при 530 нм посредством образования окрашенного вещества [Bitter T. and Muir H.M., (1962) «A modified uronic acid carbazole reaction». Analytical Biochemistry, 4 (4):330-334]. Кроме того, например, гепаросан можно детектировать и количественно оценивать посредством обработки гепаросана с использованием гепариназы III, которая представляет собой осуществляющий деградацию гепаросана фермент, и проведения анализа состава дисахаридов.
[0101] ПРИМЕРЫ
Примеры показаны ниже, но настоящее изобретение не является ограниченным следующими примерами.
Пример 1
[0102] Конструирование штамма с делецией гена
(a) Конструирование фрагмента маркерного гена для делеции гена
Фрагмент ДНК, содержащий ген устойчивости к хлорамфениколу (cat) и ген левансахаразы (sacB), используемые в качестве маркерных генов для делеции генов Escherichia coli, с использованием гомологичной рекомбинации, получали следующим образом.
[0103] ПЦР проводили с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 1 и 2, в качестве группы праймеров, и с использованием плазмиды pHSG396 (Takara Bio Inc.), в качестве матрицы, и таким образом, получали фрагмент ДНК, содержащий ген cat. ПЦР проводили с использованием ДНК-полимеразы PrimeSTAR Max (Takara Bio Inc.), в соответствии с описанием в руководстве пользователя. Кроме того, ПЦР проводили с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 3 и 4, в качестве группы праймеров, и с использованием pMOB3 (происходящей из ATCC 77282), в качестве матрицы, и таким образом, получали фрагмент ДНК, содержащий ген sacB.
[0104] После очистки фрагмента ДНК, содержащего ген cat, и фрагмента ДНК, содержащего ген sacB, фрагменты ДНК разрезали с использованием SalI. Проводили обработку фенолом/хлороформом и преципитацию этанолом, и два фрагмента смешивали в эквимолярном соотношении и лигировали с использованием набора для лигирования ДНК вер.2 (Takara Bio Inc.). Реакционный раствор для лигирования подвергали обработке фенолом/хлороформом и очищали посредством преципитации этанолом, и ПЦР проводили с использованием полученного таким образом продукта в качестве матрицы, и с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 5 и 6, в качестве группы праймеров. Полученную амплифицированную ДНК очищали с использованием набора для очистки продуктов ПЦР Qiaquick (Qiagen), и таким образом, получали фрагмент ДНК (фрагмент cat-sacB), содержащий ген cat и ген sacB.
[0105] (b) Конструирование дефектного по yhbJ штамма
Первую ПЦР проводили с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 7 и 8, и синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 9 и 10, в качестве групп праймеров, и с использованием геномной ДНК Escherichia coli штамма Nissle 1917 [DSM 6601, Mutaflor (Pharma-Zentrale GmbH), далее в настоящем описании обозначенного как штамм Nissle], полученной общепринятым способом, в качестве матрицы. В результате, получали фрагменты ДНК, соответственно, содержащие 1000 п.о. выше из области, близкой к инициирующему кодону гена yhbJ и приблизительно 1000 п.о. ниже из области, близкой к стоп-кодону гена yhbJ. ПЦР проводили с использованием ДНК-полимеразы PrimeSTAR Max (Takara Bio Inc.), в соответствии с описанием в руководстве пользователя.
[0106] Вторую ПЦР проводили с использованием, в качестве матрицы, смеси, полученной посредством смешивания амплифицированного продукта, очищенного с использованием набора для очистки продуктов ПЦР QIAquick (изготовленного в Qiagen), и фрагмента cat-sacB в эквимолярном соотношении. ПЦР проводили с использованием ДНК-полимеразы PrimeSTAR GXL (Takara Bio Inc.), в соответствии с описанием в руководстве пользователя. Для группы праймеров, использовали синтетические ДНК, состоящие из нуклеотидных последовательностей, показанных в SEQ ID NO: 7 и 10. Амплифицированный продукт подвергали электрофорезу в агарозном геле для отделения фрагмента ДНК приблизительно 4,6 т.п.о., и таким образом, получали фрагмент ДНК, содержащий периферическую область yhbJ, в который вставлен фрагмент cat-sacB.
[0107] Кроме того, первую ПЦР проводили с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 7 и 11, и синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 12 и 10, в качестве групп праймеров, и с использованием геномной ДНК Escherichia coli штамма Nissle, в качестве матрицы. В результате, получали фрагменты ДНК, соответственно, содержащие 1000 п.о. выше из области, близкой к инициирующему кодону гена yhbJ, и приблизительно 1000 п.о. ниже из области, близкой к стоп-кодону гена yhbJ.
[0108] Вторую ПЦР проводили с использованием смеси, полученной посредством смешивания очищенного амплифицированного продукта в эквимолярном соотношении, в качестве матрицы. Для группы праймеров, использовали синтетическую ДНК, состоящую из нуклеотидных последовательностей, показанных в 7 и 10. Амплифицированный продукт подвергали электрофорезу в агарозном геле для отделения фрагмента ДНК приблизительно 2,0 т.п.о., и таким образом, получали фрагмент ДНК, содержащий периферическую область yhbJ дефектного гена yhbJ.
[0109] Затем, Escherichia coli штамма Nissle (штамма Nissle/pKD46), сохраняющего pKD46, культивировали в среде LB [10 г/л бактотриптона (изготовленного в Difco), 5 г/л дрожжевого экстракта (изготовленного в Difco) и 5 г/л хлорида натрия] в присутствии 15 г/л L-арабинозы и 100 мг/л ампициллина. Плазмида pKD46 имеет ген рекомбиназы лямбда-Red, и таким образом, экспрессию гена можно индуцировать посредством L-арабинозы. Соответственно, когда Escherichia coli, сохраняющую pKD46, которую выращивали в присутствии L-арабинозы, трансформируют с использованием линейной ДНК, гомологичная рекомбинация происходит с высокой частотой. Кроме того, поскольку pKD46 имеет термочувствительную точку начала репликации, плазмиду можно легко удалять посредством выращивания pKD46 при 42 градусах C. Получали компетентные клетки штамма Nissle/pKD46, и фрагмент ДНК, содержащий периферическую область yhbJ, в которую вставлен фрагмент cat-sacB, полученный выше, вводили в них посредством электропорации.
[0110] Полученный трансформант наносили на среду LB с агаром, содержащую 15 мг/л хлорамфеникола и 100 мг/л ампициллина (LB+хлорамфеникол+ампициллин), и культивировали, и отбирали устойчивые к хлорамфениколу колонии. Поскольку штамм, в котором произошла гомологичная рекомбинация, является устойчивым к хлорамфениколу и чувствительным к сахарозе, отобранные колонии подвергали репликации в среде LB с агаром, содержащей 10% сахарозы и 100 мг/л ампициллина, и LB+хлорамфеникол+ампициллин (LB+сахароза+ампициллин), и таким образом, отбирали штамм, имеющий устойчивость к хлорамфениколу и чувствительность к сахарозе.
[0111] ПЦР колоний проводили для отобранного штамма с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 13 и 10, в качестве группы праймеров, и проверяли, что фрагмент cat-sacB вставлен в положении гена yhbJ. Штамм, в котором фрагмент cat-sacB вставлен в положении гена yhbJ, культивировали таким же способом, как описано выше, для получения компетентных клеток, и фрагмент ДНК, полученный выше и содержащий периферическую область yhbJ дефектного гена yhbJ, вводили в них посредством электропорации.
[0112] Полученный трансформант культивировали с среде LB+сахароза с агаром, и обирали устойчивые к сахарозе колонии. Поскольку штамм, подвергнутый гомологичной рекомбинации, не содержит фрагмент cat-sacB и таким образом, является чувствительным к хлорамфениколу и устойчивым к сахарозе, отобранные колонии подвергали репликации в среде LB+хлорамфеникол с агаром и в среде LB+сахароза с агаром, и таким образом, получали штамм, имеющий чувствительность к хлорамфениколу и устойчивость к сахарозе.
[0113] ПЦР колоний проводили для отобранного штамма с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 13 и 10, в качестве группы праймеров, и проверяли, что ген yhbJ делетирован. Штамм, в котором ген yhbJ, как проверено, был делетирован, наносили на среду LB с агаром и культивировали при 42 градусах C, и затем, отбирали штамм, имеющий чувствительность к ампициллину, то есть, штамм, из которого pKD46 удалена.
[0114] Штамм с недостаточностью гена yhbJ получен, как описано выше, и обозначен как Escherichia coli штамма NY.
Пример 2
[0115] Конструирование штамма с усиленным геном
Штамм с замененной промоторной областью гена kfiA конструировали посредством следующего способа. Первую ПЦР проводили с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 14 и 15, и синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 16 и 17, в качестве групп праймеров, и с использованием геномной ДНК из Escherichia coli штамма Nissle, полученной посредством обычного способа, в качестве матрицы. В результате, получали фрагменты ДНК, соответственно, содержащие 1000 п.о. выше от приблизительно 100 п.о., вышележащих от инициирующего кодона гена kfiA, и приблизительно 1000 п.о. ниже из области, близкой к инициирующему кодону гена kfiA.
[0116] Вторую ПЦР проводили с использованием, в качестве матрицы, смеси, полученной посредством смешивания амплифицированного продукта, очищенного с использованием набора для очистки продуктов ПЦР QIAquick (изготовленного в Qiagen), и фрагмента cat-sacB в эквимолярном соотношении. Для группы праймеров, использовали синтетические ДНК, состоящие из нуклеотидных последовательностей, показанных в SEQ ID NO: 14 и 17. Амплифицированный продукт подвергали электрофорезу в агарозном геле для отделения фрагмента ДНК приблизительно 4,6 т.п.о., и таким образом, получали фрагмент ДНК, содержащий периферическую промоторную область гена kfiA, в которую вставлен фрагмент cat-sacB.
[0117] Первую ПЦР проводили с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 14 и 18, и синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 19 и 17, в качестве групп праймеров, и с использованием геномной ДНК Escherichia coli штамма Nissle, полученной посредством обычного способа, в качестве матрицы. В результате, получали фрагменты ДНК, соответственно, содержащие 1000 п.о. выше от приблизительно 100 п.о., вышележащих от инициирующего кодона гена kfiA, и приблизительно 1000 п.о. ниже из области, близкой к инициирующему кодону гена kfiA. Кроме того, ПЦР проводили с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 20 и 21, в качестве группы праймеров, и с использованием геномной ДНК Escherichia coli штамма W (ATCC 9637), полученной посредством обычного способа, в качестве матрицы, и таким образом, получали фрагмент ДНК приблизительно 300 п.о., содержащий промотор uspA.
[0118] Вторую ПЦР проводили с использованием смеси, полученной посредством смешивания очищенных амплифицированных продуктов в эквимолярном соотношении, в качестве матрицы. Для группы праймеров, использовали синтетические ДНК, состоящие из нуклеотидных последовательностей, показанных в 14 и 17. Амплифицированный продукт подвергали электрофорезу в агарозном геле для отделения фрагмента ДНК приблизительно 2,3 т.п.о., и таким образом, получали фрагмент ДНК с недостаточностью промоторной области kfiA из периферической промоторной области kfiA, но вместо этого содержащий промотор uspA.
[0119] Затем, Escherichia coli штамма Nissle (штамма Nissle/pKD46), сохраняющего плазмиду pKD46, содержащую ген, кодирующий рекомбиназу гамма-Red, и Escherichia coli штамма NY (штамма NY/pKD46), сконструированного в примере 1, культивировали в присутствии 15 г/л L-арабинозы и 100 мг/л ампициллина. Получали компетентные клетки обоих штаммов, и фрагмент ДНК, содержащий периферическую промоторную область kfiA, в которую вставлен фрагмент cat-sacB, полученный выше, вводили в них посредством электропорации.
[0120] Полученный трансформант культивировали в среде LB+хлорамфеникол+ампициллин с агаром, и отбирали устойчивые к хлорамфениколу колонии. Штамм, имеющий устойчивость к хлорамфениколу и чувствительность к сахарозе, далее отбирали из отобранных колоний.
[0121] ПЦР колоний проводили для отобранного штамма с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 22 и 17, в качестве группы праймеров, и проверяли, что фрагмент cat-sacB вставлен в промоторную область kfiA.
[0122] Штамм, в котором фрагмент cat-sacB вставлен в промоторную область kfiA, культивировали таким же способом, как описано выше, для получения компетентных клеток, и фрагмент ДНК, полученный выше и с недостаточностью промоторной области kfiA из периферической промоторной области kfiA, но содержащий вместо этого промотор uspA, вводили в них посредством электропорации.
[0123] Полученный трансформант культивировали в среде LB+сахароза с агаром, и отбирали колонии с устойчивостью к сахарозе. Штамм, имеющий чувствительность к хлорамфениколу и устойчивость к сахарозе, далее отбирали из отобранных колоний.
[0124] ПЦР колоний проводили для отобранного штамма с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 22 и 17, в качестве группы праймеров, и проверяли, что промотор uspA вставлен в промоторную область kfiA. Штамм, в котором, как проверено, промотор uspA вставлен в промоторную область kfiA, наносили на среду LB с агаром и культивировали при 42 градусах C, и затем отбирали штамм, имеющий чувствительность к ампициллину, то есть, штамм, из которого удалена pKD46.
[0125] Штаммы, в которых промотор uspA вставлен в промоторную область kfiA, как описано выше, получены, и соответственно, обозначены как штамм NA и штамм NYA Escherichia coli.
Пример 3
[0126] Тестирование продукции гепаросана с использованием штамма NY, штамма NA и штамма NYA
(a) Культивирование продуцирующего гепаросан штамма
Мутант с недостаточностью yhbJ штамма NY, полученный в примере 1, штамм NA с заменой промотора kfiA и штамм NYA с заменой промотора kfiA, с недостаточностью yhbJ, полученный в примере 2, и штамм Nissle, представляющий собой родительский штамм, культивировали при 30 градусах C в течение 24 часов на среде LB с агаром, и, соответственно, инокулировали в 2 л колбу с отражателями, содержащую 330 мл среды для предварительного культивирования [10 г/л соевого пептида (Hinute AM; изготовленного в Fuji Oil Co., Ltd.), 5 г/л хлорида натрия, 5 г/л порошка дрожжевого экстракта (AY-80; изготовленного в Asahi Food and Healthcare Co., Ltd.), доводили с использованием гидроксида натрия, таким образом, что pH становился pH 7,2], и культивировали при 30 градусах C в течение 18 часов.
[0127] В емкостный ферментер, содержащий 760 мл среды для основного культивирования [в которой 20 г/л глюкозы, 13,5 г/л дигидрофосфата калия, 4 г/л фосфата диаммония, 1,7 г/л лимонной кислоты, 1,7 г/л гептагидрата сульфата магния, 10 мг/л гидрохлорида тиамина и 10 мл/л раствора микроэлементов доводили с использованием 5 моль/л гидроксида натрия, таким образом, что pH становился pH 6,7, и глюкозу и гептагидрат сульфата магния добавляли по отдельности после стерилизации автоклавированием (при 120 градусах C в течение 20 минут)], 40 мл полученной предварительной культуры инокулировали и культивировали при 37 градусах C в течение 72 часов при скорости перемешивания 800 об./мин и скорости аэрации 1,5 л/мин.
[0128] Раствор микроэлементов относится к раствору, в котором 10 г/л гептагидрата сульфата железа, 2 г/л хлорида кальция, 2,2 г/л гептагидрата сульфата цинка, 0,5 г/л тетрагидрата сульфата марганца, 1 г/л пентагидрата сульфата меди, 0,1 г/л тетрагидрата гептамолибдата гексааммония и 0,02 г/л декагидрата тетрабората натрия растворяли в 5 моль/л соляной кислоты.
[0129] От времени, когда концентрация глюкозы в культуральном растворе становилась 0 г/л (0 часов), до 72 часов, подпитывающий раствор [500 г/л глюкозы, 33,6 г/л дигидрофосфата калия, 14,3 г/л сульфата магния, 0,4 г/л гидрохлорида тиамина и 14,3 мл/л раствора микроэлементов] добавляли при 7,0 мл/час. Количество подпитывающего раствора, добавленного к концу культивирования, составляло приблизительно 450 мл.
[0130] (b) Частичная очистка гепаросана из культурального растворе продуцирующего гепаросан штамма После инкубации культурального раствора, разведенного в 10 раз, с дистиллированной водой при 100 градусах C в течение 30 минут, бактериальные клетки удаляли из культурального раствора посредством центрифугирования, и 200 микролитров полученного супернатанта переносили в 1,5 мл пробирку Eppendorf. После добавления и перемешивания 40 микролитров 0,5 моль/л водного раствора сульфата натрия, 400 микролитров 10 г/л водного раствора хлорида гексадецилпиридиния добавляли и перемешивали посредством переворачивания, и смеси позволяли стоять при 37 градусах C в течение 1 часа.
[0131] Раствор центрифугировали для формирования преципитата, и супернатант удаляли. После промывки преципитата дистиллированной водой, 100 микролитров раствора [водного раствора, содержащего 0,5 моль/л хлорида натрия и 4% (об./об.) этанола] добавляли для растворения преципитата. После дополнительного обеспечения возможности стоять при 4 градусах C в течение ночи, 900 микролитров 0,25 моль/л водного раствора хлорида натрия добавляли и перемешивали, и таким образом, получали раствор неочищенного гепаросана. Получали также нулевой контроль посредством обработки 200 микролитров основной культуральной среды таким же способом.
[0132] (c) Измерение количества накопленного гепаросана посредством способа с карбазолом-серной кислотой
В то время как 20 микролитров неочищенного раствора гепаросана охлаждали на льду, 100 микролитров раствора серной кислоты [раствора, в котором 9,5 г/л декагидрата тетрабората натрия растворяли в концентрированной серной кислоте] добавляли и перемешивали, с последующей инкубацией при 100 градусах C в течение 10 минут. В то время как раствор снова охлаждали на льду, 4 микролитров раствора карбазола [раствора, в котором 1,25 г/л карбазола растворяли в 100% этанола] добавляли к нему и перемешивали, и инкубировали при 100 градусах C в течение 15 минут.
[0133] После повторного охлаждения раствора на льду, температуру возвращали к комнатной температуре, и оптическую плотность при 530 нм измеряли с использованием считывателя для микропланшетов. С использованием 0, 0,1 и 0,2 г/л моногидрата глюкуроната натрия для калибровочных кривых, получали калибровочные кривые с концентрацией (г/л) глюкуроновой кислоты, в качестве горизонтальной оси, и оптической плотности при 530 нм, в качестве вертикальной оси.
[0134] Концентрацию гепаросана рассчитывали, в соответствии с выражением для расчета, описанным ниже. Когда желательная концентрация гепаросана представляет собой H (г/л), полученная калибровочная кривая представляет собой y=ax+b, значение измерения A530 для неочищенного образца гепаросана представляет собой h, нулевое значение измерения A530 представляет собой k, и конечная кратность разведения представляет собой n. 0,5387 обозначает содержание глюкуроновой кислоты в гепаросане, и 216/234 обозначает содержание глюкуроната натрия в моногидрате глюкуроната натрия.
[0135]
[Математическая формула 1]

[0136] (d) Результаты измерения количества накопленного гепаросана
В таблице 1 показано количество накопленного гепаросана, когда его измеряют посредством вышеуказанного способа.
[0137]
[0138] Как показано в таблице 1, количество накопленного гепаросана в штамме NY с недостаточностью yhbJ было более чем в два раза выше, чем в родительском штамме Nissle. Кроме того, даже в штамме NA, в котором промотор kfiA заменен на промотор uspA, количество накопленного гепаросана было увеличено, по сравнению с количеством в родительском штамме Nissle. Для штамма NYA, в котором эти две мутации скомбинированы, показана даже более высокий выход продукции гепаросана.
Пример 4
[0139] Конструирование плазмиды, экспрессирующей ген, вовлеченный в продукцию гепаросана
Реакцию ПЦР проводили с использованием хромосомной ДНК Escherichia coli штамма Nissle в качестве матрицы, и с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 23 и 24, в качестве группы праймеров, и таким образом, получали фрагмент ДНК приблизительно 3,3 т.п.о., содержащий области генов kpsC и kpsS (далее в настоящем описании обозначенный как амплифицированный фрагмент гена kpsCS).
[0140] Реакцию ПЦР проводили с использованием вектора pMW118 в качестве матрицы, и с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 25 и 26, в качестве группы праймеров, и таким образом, получали линейный фрагмент ДНК pMW118 приблизительно 4 т.п.о. Кроме того, реакцию ПЦР проводили с использованием хромосомной ДНК Escherichia coli штамма W (ATCC 9637) в качестве матрицы, и с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 20 и 21, в качестве группы праймеров, и таким образом, получали фрагмент ДНК приблизительно 300 п.о., содержащий промоторную область uspA.
[0141] Линейный фрагмент ДНК pMW118, фрагмент ДНК, содержащий промоторную область uspA, и амплифицированный фрагмент гена kpsCS, полученные, как выше, смешивали и лигировали с использованием набора для клонирования In-Fusion HD (Takara Bio Inc.).
[0142] Escherichia coli штамма DH5 альфа трансформировали с использованием полученной лигированной ДНК, и трансформанты отбирали с использованием устойчивость к ампициллину в качестве показателя. Плазмиду выделяли из трансформанта, в соответствии с известным способом, и реакцию ПЦР проводили с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 27 и 24, в качестве группы праймеров, и таким образом, проверяли, что получена плазмида для экспрессии гена, которая обозначена как pMW118-kpsCS.
[0143] Кроме того, реакцию ПЦР проводили с использованием хромосомной ДНК Escherichia coli штамма Nissle в качестве матрицы, и с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 28 и 24, и синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 29 и 24, в качестве групп праймеров, и таким образом, соответственно, получали фрагмент ДНК приблизительно 1,2 т.п.о., содержащий область гена kpsS (далее в настоящем описании обозначенный как амплифицированный фрагмент гена kpsS), и фрагмент ДНК приблизительно 7,9 т.п.о., содержащий области генов kpsF, kpsE, kpsD, kpsU, kpsC и kpsS (далее в настоящем описании обозначенный как амплифицированный фрагмент гена kpsFEDUCS).
[0144] Кроме того, реакцию ПЦР проводили с использованием плазмиды pMW118-kpsCS, полученной выше, в качестве матрицы, и с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 25 и 21, в качестве группы праймеров, и таким образом, получали линейный фрагмент ДНК pMW118 приблизительно 4,3 т.п.о., содержащий промоторную последовательность uspA.
[0145] Линейный фрагмент ДНК pMW118, содержащий промоторную последовательность uspA, и амплифицированный фрагмент гена kpsS, полученный выше, смешивали и лигировали с использованием набора для клонирования In-Fusion HD (Takara Bio Inc.). Таким же способом, линейный фрагмент ДНК pMW118, содержащий промоторную последовательность uspA, и амплифицированный фрагмент гена kpsFEDUCS смешивали и лигировали.
[0146] Escherichia coli штамма DH5 альфа трансформировали с использованием каждой из полученных лигированных ДНК, и трансформанты отбирали с использованием устойчивости к ампициллину в качестве показателя. Плазмиду выделяли из трансформанта, в соответствии с известным способом, и реакцию ПЦР проводили с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 20 и 24, в качестве группы праймеров для плазмиды, в которую лигирован амплифицированный фрагмент гена kpsS, и с использованием синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 27 и 30, и синтетических ДНК, состоящих из нуклеотидных последовательностей, показанных в SEQ ID NO: 31 и 32, в качестве группы праймеров для плазмиды, в которую лигирован амплифицированный фрагмент гена kpsFEDUCS. Соответственно, проверяли, что, соответственно, получены плазмиды, экспрессирующие гены, которые, соответственно обозначены как pMW118-kpsS и pMW118-kpsFEDUCS.
Пример 5
[0147] Тестирование продукции гепаросана с использованием штамма, сохраняющего плазмиду для экспрессии гена - 1
Плазмидами pMW118 и pMW118-PuspA-kpsS, pMW118-PuspA-kpsCS, и pMW118-PuspA-kpsFEDUCS, полученными в примере 4, соответственно, трансформировали штамм NY, полученный в примере 1. Полученные трансформанты, соответственно, обозначены как штамм NY/pMW118, штамм NY/pMW118-PuspA-kpsS, штамм NY/pMW118-PuspA-kpsCS и штамм NY/pMW118-PuspA-kpsFEDUCS.
[0148] Трансформанты, полученные выше, инокулировали в большую пробирку, содержащую 5 мл среды LB, содержащей 100 мг/л ампициллина, и культивировали при 37 градусах C в течение 15 часов. 1% культуральный раствор инокулировали в пробирку, содержащую 5 мл среды R, содержащей 100 мг/л ампициллина [в которой 20 г/л глюкозы, 13,5 г/л дигидрофосфата калия, 4 г/л фосфата диаммония, 1,7 г/л лимонной кислоты, 1 г/л гептагидрата сульфата магния, 10 мг/л гидрохлорида тиамина и 10 мл/л раствора микроэлементов, доводили с использованием 5 моль/л гидроксида натрия, таким образом, что pH становился pH 6,8, и глюкозу и гептагидрат сульфата магния добавляли по отдельности после стерилизации автоклавированием (при 120 градусах C в течение 20 минут)], и культивировали при 37 градусах C в течение 24 часов. Кроме того, такую же операцию проводили для штамма NY в условиях без содержания ампициллина.
[0149] Полученный культуральный раствор обрабатывали посредством способа, описанного в примере 3, и измеряли количество накопленного гепаросана в культуральном растворе. Результаты показаны в таблице 2.
[0150]
[0151] Как показано в таблице 2, в штамме NY/pMW118-PuspA-kpsS, экспрессирующем kpsS, количество накопленного гепаросана было явно увеличено, по сравнению с количеством в родительском штамме NY и штамме NY/pMW118, сохраняющем только вектор pMW118.
[0152] Тем не менее, на основании того результата, что для штамма NY/pMW118-PuspA-kpsCS, экспрессирующего kpsC и kpsS, и штамма NY/pMW118-PuspA-kpsFEDUCS, экспрессирующего kpsF, kpsE, kpsD, kpsU, kpsC и kpsS, показан почти такой же уровень продукции гепаросана, что и для штамма NY/pMW118-PuspA-kpsS, обнаружено, что эффект улучшения способности к продукции гепаросана был в достаточной степени получен посредством усиления экспрессия только kpsS в kpsFEDUCS, представляющей собой группу генов продукции гепаросана.
Пример 6
[0153]
Тестирование продукции гепаросана с использованием штамма, сохраняющего плазмиду для экспрессии гена - 2
Плазмидами pMW118 и pMW118-PuspA-kpsS, pMW118-PuspA-kpsCS, и pMW118-PuspA-kpsFEDUCS, полученными в примере 4, соответственно, трансформировали штамм дикого типа Nissle Escherichia coli. Полученные трансформанты, соответственно, обозначены как штамм N/pMW118, штамм N/pMW118-PuspA-kpsS, штамм N/pMW118-PuspA-kpsCS и штамм N/pMW118-PuspA-kpsFEDUCS.
[0154]
Трансформанты, полученные выше, инокулировали в большую пробирку, содержащую 5 мл среды LB, содержащей 100 мг/л ампициллина, и культивировали при 37 градусах C в течение 15 часов. 1% культуральном растворе инокулировали в пробирку, содержащую 5 мл среды R, содержащей 100 мг/л ампициллина [в которой 20 г/л глюкозы, 13,5 г/л дигидрофосфата калия, 4 г/л фосфата диаммония, 1,7 г/л лимонной кислоты, 1 г/л гептагидрата сульфата магния, 10 мг/л гидрохлорида тиамина и 10 мл/л раствора микроэлементов доводили с использованием 5 моль/л гидроксида натрия, таким образом, что pH становился pH 6,8, и глюкозу и гептагидрат сульфата магния добавляли по отдельности после стерилизации автоклавированием (при 120 градусах C в течение 20 минут)], и культивировали при 37 градусах C в течение 24 часов. Кроме того, такую же операцию проводили для штамма дикого типа Nissle в условиях без содержания ампициллина.
[0155]
Полученный культуральный раствор обрабатывали посредством способа, описанного в примере 3, и измеряли количество накопленного гепаросана в культуральном растворе. Результаты показаны в таблице 3.
[0156]
[0157]
Как показано в таблице 3, в штамме N/pMW118-PuspA-kpsS, экспрессирующем kpsS, количество накопленного гепаросана было увеличено в три раза или более, по сравнению с количеством в родительском штамме дикого типа Nissle и штамме N/pMW118, сохраняющем только вектор pMW118.
[0158] Тем не менее, в штамме N/pMW118-PuspA-kpsCS, экспрессирующем kpsC и kpsS, и в штамме N/pMW118-PuspA-kpsFEDUCS, экспрессирующем kpsF, kpsE, kpsD, kpsU, kpsC и kpsS, титр улучшался, самое большее, в 1,8 раз, по сравнению с родительским штаммом. В этом отношении, обнаружено, что эффект улучшения способности к продукции гепаросана был в достаточной степени получен посредством усиления экспрессии только kpsS, среди группы генов продукции гепаросана kpsFEDUCS, и что способность к продукции гепаросана становилась выше, когда экспрессия kpsC и kpsFEDU не была усилена, вместо того, чтобы являться усиленной.
[0159] В то время как изобретение описано подробно и со ссылкой на конкретные варианты его осуществления, специалисту в данной области очевидно, что можно осуществлять его различные изменения и модификации, без отклонения от его содержания и объема. Полное содержание всех ссылок, процитированных в настоящем описании, приведено в настоящем описании в качестве ссылки. Эта заявка основана на Международной заявке No. PCT/JP2020/015384, поданной 3 апреля 2020 г., полное содержание которой приведено в настоящем описании в качестве ссылки.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Rensselaer Polytechnic Institute
Otsuka Pharmaceutical Factory, Inc
KYOWA HAKKO BIO CO.,LTD.
<120> Способ продукции гепаросана и бактерия из рода Escherichia, имеющая
способность к продукции гепаросана
<130> W528319
<160> 38
<170> PatentIn версии 3.5
<210> 1
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 1
aatcgggatc cgcggccgca gaggcggttt gcgtattgga gc 42
<210> 2
<211> 51
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 2
ggatttgact acgggcctaa agtcgacaga ataaataaat cctggtgtcc c 51
<210> 3
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 3
acggagcatg cgcggccgct caaaatcggt ggagctgcat ga 42
<210> 4
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 4
gggacaccag gatttattta ttctgtcgac tttaggcccg tagtctgcaa atcc 54
<210> 5
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 5
cacttattca ggcgtagcac c 21
<210> 6
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 6
atcggcattt tctttgcgtt ttta 24
<210> 7
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 7
gaagtaccga gagtctttat cc 22
<210> 8
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 8
ctacgcctga ataagtggta ctgtttctcc tcacaacg 38
<210> 9
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 9
gcaaagaaaa tgccgatccg tcaagcaaac tgttgaaatc 40
<210> 10
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 10
tcgtttccat aaccgctctt gc 22
<210> 11
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 11
caacagtttg cttgacgggt actgtttctc ctcacaacg 39
<210> 12
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 12
gtgaggagaa acagtacccg tcaagcaaac tgttgaaatc 40
<210> 13
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 13
gaaattaatc gcggcggaaa accc 24
<210> 14
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 14
gtatagatga gtataaaatg tatc 24
<210> 15
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 15
ctacgcctga ataagtggtt gattagaatg actccgcac 39
<210> 16
<211> 37
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 16
gcaaagaaaa tgccgatatg attgttgcaa atatgtc 37
<210> 17
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 17
ggtggcgtta tgtgataata c 21
<210> 18
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 18
caccacaaaa gcggttgttg attagaatga ctccgcac 38
<210> 19
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 19
tggaaggagt aacactatga ttgttgcaaa tatgtc 36
<210> 20
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 20
aaccgctttt gtggtgacca 20
<210> 21
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 21
agtgttactc cttccataaa g 21
<210> 22
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 22
gaaagagttt tgtgatgtgg cg 22
<210> 23
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 23
ctttatggaa ggagtaacac tatgattggc atttactcgc ctgg 44
<210> 24
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 24
gtaaaacgac ggccagtgcc ttaataataa accgcattaa cctg 44
<210> 25
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 25
ggcactggcc gtcgttttac aacgtc 26
<210> 26
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 26
gcgctttctc atagctcacg ctg 23
<210> 27
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 27
caccgaactg agatacctac agc 23
<210> 28
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 28
ctttatggaa ggagtaacac tatgcaaggt aatgcactaa ccg 43
<210> 29
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 29
caactttatg gaaggagtaa cactatgtct gaaagacatt tacc 44
<210> 30
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 30
gttgcgcgtc atgatggctt tcc 23
<210> 31
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 31
gaagtttggg tggcaacaga cgatc 25
<210> 32
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтезированный олигонуклеотид для праймера
<400> 32
cagggttttc ccagtcacga cgttg 25
<210> 33
<211> 1170
<212> ДНК
<213> Escherichia coli
<400> 33
atgcaaggta atgcactaac cgttttatta tccggtaaaa aatatctgct attgcagggg 60
ccaatgggac cctttttcag tgatgttgcc gagtggctag agtcattagg tcgtaacgct 120
gtgaatgttg tattcaacgg tggggatcgt ttttactgcc gccatcgaca atacctagct 180
tactaccaga caccgaaaga gtttcccgga tggttacggg atctccaccg gcaatatgac 240
tttgacacaa tcctctgctt tggcgactgc cgcccattgc ataaagaagc aaaacgctgg 300
gcaaagtcga aagggatccg cttcctggca tttgaagaag gatatttacg cccgcaattt 360
attaccgttg aagaaggcgg agtgaacgca tattcatcgc taccgcgcga tccggatttt 420
tatcgtaagt taccagatat gcctacgccg cacgttgaga acttaaaacc ttcaacgatg 480
aaacgtatag gccatgctat gtggtattac ctgatgggct ggcattaccg tcatgagttt 540
cctcgctacc gccaccacaa atcattttcc ccctggtatg aagcacgttg ctgggttcgt 600
gcatactggc gcaagcaact ttacaaggta acacagcgta aggtattacc gaggttaatg 660
aacgaactgg accagcgtta ttatcttgct gttttgcagg tgtataacga tagccagatt 720
cgtaaccaca gcagttataa cgatgtgcgt gactatatta atgaagtcat gtactcattt 780
tcgcgtaaag cgccgaaaga aagttatttg gtgatcaaac atcatccgat ggatcgtggt 840
cacagactct atcgaccatt aattaaacgg ttgagtaagg aatatggctt aggtgagcga 900
atcctttatg tgcacgatct cccgatgccg gaattattac gccatgcaaa agcggtggtg 960
acgattaaca gtacggcggg gatctctgcg ctgattcata acaaaccact caaagtgatg 1020
ggcaatgccc tgtacgacat caagggcttg acgtatcaag ggcatttgca ccagttctgg 1080
caggctgatt ttaaaccaga tatgaaactg tttaagaagt ttcgtgggta tttattggtg 1140
aagacgcagg ttaatgcggt ttattattaa 1170
<210> 34
<211> 717
<212> ДНК
<213> Escherichia coli
<400> 34
atgattgttg caaatatgtc atcataccca cctcgaaaaa aagagttggt gcattctata 60
caaagtttac atgctcaagt agataaaatt aatctttgcc tgaatgagtt tgaagaaatt 120
cctgaggaat tagatggttt ttcaaaatta aatccagtta ttccagataa agattataag 180
gatgtgggca aatttatatt tccttgcgct aaaaatgata tgatcgtact tacagatgat 240
gatattattt accctcccga ttatgtagaa aaaatgctca atttttataa ttcctttgca 300
atattcaatt gcattgttgg gattcatggc tgtatataca tagatgcatt tgatggagat 360
cagtctaaaa gaaaagtatt ttcatttact caagggctat tgcgaccgag agttgtaaat 420
caattaggta cagggactgt ttttcttaag gcagatcaat taccatcttt aaaatatatg 480
gatggttctc aacgattcgt cgatgttaga ttttctcgct atatgttaga gaatgaaatt 540
ggtatgatat gtgttcccag agaaaaaaac tggctaagag aggtctcatc aggttcaatg 600
gaaggacttt ggaacacatt tacaaaaaaa tggcctttag acatcataaa agaaacacaa 660
gcaatcgcag gatattcaaa acttaacctc gaattagtgt ataatgtgga agggtaa 717
<210> 35
<211> 1689
<212> ДНК
<213> Escherichia coli
<400> 35
atgaataaat tagtgctagt cggacatcct ggctcaaagt atcagatagt tgaacatttt 60
ttgaaagaaa ttggcatgaa ctcaccaaat tattctacaa gtaataaaat ttccccagaa 120
tatatcaccg cttcattatg tcaattttat caaacaccag aagttaatga tgtagtagat 180
gagagagaat tctcagctgt tcaagtctca accatgtggg atagcatggt tcttgaacta 240
atgatgaaca atctaaataa caaactttgg gggtgggcag atccatctat aatatttttt 300
cttgattttt ggaaaaatat agataaaagc ataaaattca tcatgatata tgatcaccct 360
aaatataatt taatgcgttc agtaaataat gcccctctct ctttaaatat aaataatagt 420
gtagataact ggattgcata taataaaaga ttgcttgatt tttttttgga gaataaagaa 480
cgatgtgtgt tgattaattt tgaggcgttt caaagcaata agaaaaatat tataaagcca 540
ttgagtaata ttataaaaat agataatcta atgtctgcgc attacaaaaa ttcaatattg 600
tttgatgtgg ttgagaataa tgattataca aaatcaaatg aaattgccct gcttgaaaaa 660
tatacaactt tattttcttt aagtgcaaat gagactgaaa ttacatttaa tgatacaaag 720
gttagtgagt acttagtatc tgaattaata aaagaaagaa ccgaggttct gaagctttat 780
aatgagttac aagcctatgc aaacctacct tatatagaaa catcgaaaga taacgtttcg 840
gctgaggctg cattatggga ggtagtcgaa gagagaaatt ctatcttcaa tattgtatct 900
catttggtgc aagagtcaaa aaagaaggat gcagatattg aattgactaa atctatattt 960
aagaaaagac aatttttatt attgaacagg attaatgagc taaaaaaaga aaaggaagag 1020
gtaattaaac tttcaaaaat aaatcacaac gatgttgtga gacaagaaaa atatccagat 1080
gatattgaaa aaaaaataaa tgacatacag aaatatgaag aagagataag cgaaaaagaa 1140
tcaaaactca ctcaggcaat atcagaaaaa gaacagattt taaaacaatt gcataaatat 1200
gaagaagaga taagcgaaaa agaatcaaaa ctcactcagg caatatcaga aaaagaacag 1260
attttaaaac aattgcatat agtgcaagag cagttggaac actattttat agaaaatcag 1320
gaaattaaaa agaaacttcc acctgtgcta tatggagcag ctgagcagat aaaacaagag 1380
ttaggttatc gacttggtta tattatagtc tcgtattcta aatccctcaa ggggattatt 1440
accatgccat ttgcacttat ccgtgagtgt gtttttgaaa aaaaacgtaa gaagagttat 1500
ggcgttgatg tgccactcta tttatatgct gatgctgata aggctgaaag agttaagaaa 1560
catttatctt atcaattagg gcaggctatt atctccagtg ctaattcgat atttggattc 1620
attacccttc catttaagtt aattgttgtt gtttataaat ataggagagc taaaatcaag 1680
ggctgttaa 1689
<210> 36
<211> 1563
<212> ДНК
<213> Escherichia coli
<400> 36
atgaacgcag aatatataaa tttagttgaa cgtaaaaaga aattagggac aaatattggt 60
gctcttgatt ttttattatc aattcataag gagaaagttg atcttcaaca taaaaactcg 120
cctttaaaag gtaacgataa ccttattcac aaaagaataa acgaatacga caatgtactt 180
gaactatcta agaatgtatc agctcagaat tctggcaatg agttttctta tttattggga 240
tatgcagatt ctcttagaaa agttggtatg ttggatactt atattaaaat tgtttgttat 300
ctaacaattc aatctcgtta ttttaaaaat ggcgaacgag ttaagctttt tgaacatata 360
agtaacgctc tacggtattc aaggagtgat tttctcatta atcttatttt tgaacgatat 420
atcgaatata taaaccatct aaaattgtcg cccaaacaaa aagattttta tttttgtacg 480
aagttttcaa aatttcatga ttatactaaa aatggatata aatatttagc atttgataat 540
caagccgatg cagggtatgg cctgacttta ttattaaatg caaacgatga tatgcaagat 600
agttataatc tactccctga gcaagaactt tttatttgta atgctgtaat agataatatg 660
aatatttata ggagtcaatt taacaaatgt ctacgaaaat acgatttatc agaaataact 720
gatatatacc caaataaaat tatattgcaa ggaattaagt tcgataagaa aaaaaatgtt 780
tatggaaaag atcttgttag tataataatg tcagtattca attcagaaga tactattgca 840
tactcattac attcattgtt gaatcaaaca tatgaaaata ttgaaattct cgtgtgcgat 900
gattgttcat cggacaaaag ccttgaaata attaagagca tagcttattc tgattcaaga 960
gtgaaagtat atagctcacg aaaaaaccaa ggcccttata atataagaaa tgagctaata 1020
aaaaaagcac acggtaattt catcaccttt caagatgcag atgatctttc tcatccggag 1080
agaatacaaa gacaagttga ggttcttcgc aataataagg ctgtaatctg tatggctaac 1140
tggatccgtg ttgcgtcaaa tggaaaaatt caattcttct atgatgataa agccacaaga 1200
atgtctgttg tatcgtcaat gataaaaaaa gatatttttg cgacagttgg tggctataga 1260
caatctttaa ttggtgcaga tacggagttt tatgaaacag taataatgcg ttatgggcga 1320
gaaagtattg taagattact gcagccattg atattggggt tatggggaga ctccggactt 1380
accaggaata aaggaacaga agctctacct gatggatata tatcacaatc tcgaagagaa 1440
tatagtgata tcgcggcaag acaacgagtg ttagggaaaa gtatcgtaag tgataaagat 1500
gtacgtggtt tattatctcg ctatggtttg tttaaagatg tatcaggaat aattgaacaa 1560
tag 1563
<210> 37
<211> 1179
<212> ДНК
<213> Escherichia coli
<400> 37
atgttcggaa cactaaaaat aactgtttca ggcgctggtt acgttgggct ttcaaatgga 60
attctaatgg ctcaaaatca tgaagtggtt gcatttgata cccatcaaaa aaaagttgac 120
ttacttaatg ataaactctc tcctatagag gataaggaaa ttgaaaatta tctttcaact 180
aaaatactta attttcgcgc aactactaac aaatatgaag cctataaaaa tgccaattac 240
gttattattg ctacaccaac gaattatgac ccaggttcaa attactttga tacatcaagc 300
gttgaagctg tcattcgtga cgtaacggaa atcaacccaa acgcaattat ggtggttaaa 360
tctacggtcc cagtaggttt cacaaaaaca attaaagaac atttaggtat taataatatt 420
atcttctctc cagaattttt acgagaagga agagccctat acgataatct ccatccatct 480
cgcattatta tcggtgaatg ttctgaacgg gcagaacgtt tggcagtgtt atttcaggaa 540
ggagcgatta aacaaaatat acccgtttta tttacagatt ctacggaagc ggaagcgatt 600
aagttatttt caaatactta tttggctatg cgagttgcat tttttaatga attggatagt 660
tacgcagaaa gttttggtct gaatacgcgt cagattattg acggtgtttg tttggatccg 720
cgcattggta attactacaa taatccttct tttggttatg gtggctactg tttgccaaaa 780
gataccaagc aattattagc caactatcag tctgttccga ataaacttat atctgcaatt 840
gttgatgcta accgtacacg taaggacttt atcactaatg ttattttgaa acatagacca 900
caagttgtgg gggtttatcg tttgattatg aaaagtggtt cagataattt tagagattct 960
tctattcttg gtattataaa gcgtatcaag aaaaaaggcg tgaaagtaat tatttatgag 1020
ccgcttattt ctggagatac attctttaac tcacctttgg aacgggagct ggcgatcttt 1080
aaagggaaag ctgatattat tatcactaac cgaatgtcag aggagttgaa cgatgtggtc 1140
gacaaagtct atagtcgcga tttgtttaaa tgtgactaa 1179
<210> 38
<211> 855
<212> ДНК
<213> Escherichia coli
<400> 38
atggtactga tgatcgtcag cggacgttca ggttcaggta aatctgtcgc cctgcgtgcg 60
ctggaagata tgggttttta ctgcgtggat aaccttcccg tagtgttgtt acccgatctg 120
gctcgaactc tggccgatcg agagatttct gccgccgtca gcattgatgt tcgtaatatg 180
ccggagtcac cagaaatatt cgaacaggcg atgagtaacc tgcctgacgc tttctcaccg 240
caactactgt tcctggatgc cgaccgtaat accttaattc gtcgttacag tgacacgcgc 300
cgactgcatc cgctttccag caaaaacctg tcgctggaaa gtgctatcga caaagaaagc 360
gatttgctgg agcctctgcg ttcgcgagcg gatctgattg tcgacacctc agaaatgtcc 420
gttcacgagc tggcagaaat gctgcgtacc cgtctgctgg gtaaacgtga acgtgaactg 480
accatggtct ttgagtcttt cggcttcaaa cacggtatcc ctatcgatgc agattacgtc 540
tttgacgtgc gcttcttgcc gaacccgcac tgggatccga aactgcgtcc aatgacaggt 600
cttgataaac ctgtcgccgc gttcctcgac cgccacacag aagtacacaa ttttatctac 660
cagacgcgaa gctatcttga gctatggtta cctatgctgg aaaccaacaa ccgtagctac 720
ctgacggtcg ccattggttg taccggcggg aagcaccgtt cggtgtatat tgcagagcaa 780
ctggcagact acttccgctc gcgcggtaaa aacgtccagt cacgccatcg gacgctggaa 840
aaacgtaaac catga 855
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПРОДУКЦИИ СУЛЬФАТИРОВАННОГО ПОЛИСАХАРИДА И СПОСОБ ПРОДУКЦИИ PAPS | 2021 |
|
RU2811941C1 |
| БАКТЕРИЯ - ПРОДУЦЕНТ ПРОДУКТА РЕАКЦИИ, КАТАЛИЗИРУЕМОЙ БЕЛКОМ, ОБЛАДАЮЩИМ АКТИВНОСТЬЮ 2-ОКСОГЛУТАРАТ-ЗАВИСИМОГО ФЕРМЕНТА, И СПОСОБ ПРОДУКЦИИ УКАЗАННОГО ПРОДУКТА | 2008 |
|
RU2444568C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ЦИСТЕИНА, L-ЦИСТИНА, S-СУЛЬФОЦИСТЕИНА ИЛИ ТИАЗОЛИДИНОВОГО ПРОИЗВОДНОГО L-ЦИСТЕИНА, ИЛИ ИХ СМЕСИ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА ENTEROBACTERIACEAE | 2010 |
|
RU2458982C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ЦИСТЕИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА ENTEROBACTERIACEAE | 2010 |
|
RU2458981C2 |
| МУТАНТНАЯ АЦЕТОЛАКТАТСИНТАЗА И СПОСОБ ПРОДУКЦИИ РАЗВЕТВЛЕННЫХ L-АМИНОКИСЛОТ | 2006 |
|
RU2355763C2 |
| СПОСОБ КОНСТРУИРОВАНИЯ РЕКОМБИНАНТНЫХ БАКТЕРИЙ, ПРИНАДЛЕЖАЩИХ К РОДУ PANTOEA, И СПОСОБ ПРОДУКЦИИ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ПРИНАДЛЕЖАЩИХ К РОДУ PANTOEA | 2006 |
|
RU2418069C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2009 |
|
RU2518677C2 |
| СПОСОБ ПРОДУЦИРОВАНИЯ L-ЛИЗИНА ИЛИ L-ТРЕОНИНА С ПОМОЩЬЮ БАКТЕРИЙ ESCHERICHIA, ИМЕЮЩИХ АТТЕНУИРОВАННУЮ АКТИВНОСТЬ МАЛЕИНОВОГО ФЕРМЕНТА | 2004 |
|
RU2337140C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА И L-АРГИНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН ybiV | 2005 |
|
RU2320719C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА И L-АРГИНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН cpxR | 2005 |
|
RU2320723C2 |

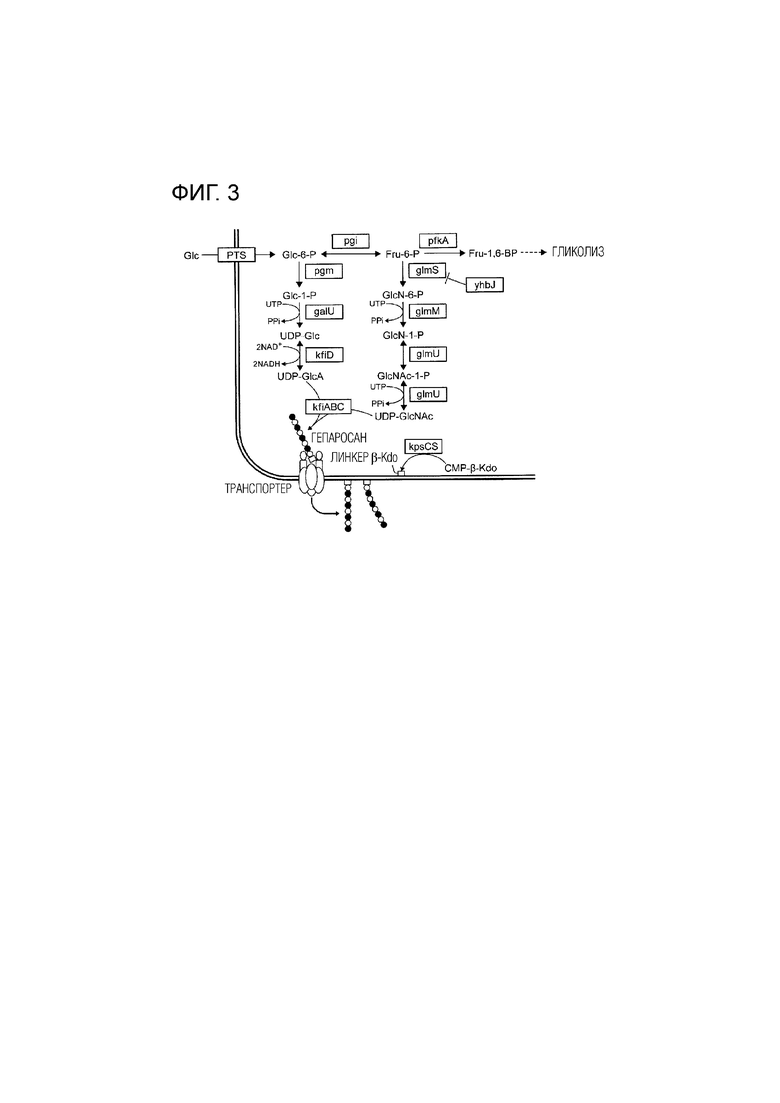
Изобретение относится к биотехнологии. Предложена бактерия из рода Escherichia, которая продуцирует гепаросан и имеет генетическую модификацию, увеличивающую экспрессию гена kpsS по сравнению с экспрессией для немодифицированной бактерией, и не имеет генетическую модификацию, увеличивающую экспрессию гена kpsC по сравнению с экспрессией для немодифицированной бактерией. Также предложен способ продукции гепаросана с использованием указанной бактерии. Изобретение обеспечивает эффективную продукцию гепаросана. 2 н. и 9 з.п. ф-лы, 3 ил., 3 табл., 6 пр.
1. Способ продукции гепаросана, включающий: культивирование бактерии из рода Escherichia, которая:
имеет следующую генетическую модификацию (1) и продуцирует гепаросан, в среде для продукции гепаросана:
(1) генетическую модификацию, увеличивающую экспрессию гена kpsS, по сравнению с экспрессией для немодифицированной бактерией;
и не имеет следующую генетическую модификацию (4):
(4) генетическую модификацию, увеличивающую экспрессию гена kpsC по сравнению с экспрессией для немодифицированной бактерией.
2. Способ продукции гепаросана по п.1, где бактерия из рода Escherichia дополнительно имеет по меньшей мере одну из следующих генетических модификаций (2) и (3):
(2) генетической модификации, увеличивающей экспрессию по меньшей мере одного гена, выбранного из гена kfiA, гена kfiB, гена kfiC и гена kfiD, по сравнению с экспрессией для немодифицированной бактерией, и
(3) генетической модификации, вызывающей потерю функции гена yhbJ.
3. Способ продукции гепаросана по п.1 или 2, где генетическая модификация (1) представляет собой по меньшей мере одну из модификации области контроля экспрессии гена kpsS и увеличения количества копий гена kpsS.
4. Способ продукции гепаросана по п.2 или 3, где генетическая модификация (2) представляет собой по меньшей мере одну из модификации области контроля экспрессии по меньшей мере одного гена, выбранного из гена kfiA, гена kfiB, гена kfiC и гена kfiD, и увеличения количества копий по меньшей мере одного гена, выбранного из гена kfiA, гена kfiB, гена kfiC и гена kfiD.
5. Способ продукции гепаросана по любому из пп. 2-4, где генетическая модификация (3) представляет собой делецию гена yhbJ.
6. Способ продукции гепаросана по любому из пп. 1-5, где бактерия из рода Escherichia представляет собой Escherichia coli.
7. Способ продукции гепаросана по любому из пп. 1-6, где ген kpsS представляет собой ДНК, содержащую нуклеотидную последовательность, показанную в SEQ ID NO: 33, или ДНК, содержащую нуклеотидную последовательность, имеющую 90% или более идентичность с нуклеотидной последовательностью, показанной в SEQ ID NO: 33, и увеличивающую продукцию гепаросана бактерии из рода Escherichia, которая продуцирует гепаросан, когда уровень экспрессии увеличен в бактерии.
8. Способ продукции гепаросана по любому из пп. 2-7, где ген kfiA представляет собой ДНК, содержащую нуклеотидную последовательность, показанную в SEQ ID NO: 34, или ДНК, содержащую нуклеотидную последовательность, имеющую 90% или более идентичность с нуклеотидной последовательностью, показанной в SEQ ID NO: 34, и увеличивающую продукцию гепаросана бактерии из рода Escherichia, которая продуцирует гепаросан, когда уровень экспрессии увеличен в бактерии, ген kfiB представляет собой ДНК, содержащую нуклеотидную последовательность, показанную в SEQ ID NO: 35, или ДНК, содержащую нуклеотидную последовательность, имеющую 90% или более идентичность с нуклеотидной последовательностью, показанной в SEQ ID NO: 35, и увеличивающую продукцию гепаросана бактерии из рода Escherichia, которая продуцирует гепаросан, когда уровень экспрессии увеличен в бактерии, ген kfiC представляет собой ДНК, содержащую нуклеотидную последовательность, показанную в SEQ ID NO: 36, или ДНК, содержащую нуклеотидную последовательность, имеющую 90% или более идентичность с нуклеотидной последовательностью, показанной в SEQ ID NO: 36, и увеличивающую продукцию гепаросана бактерии из рода Escherichia, которая продуцирует гепаросан, когда уровень экспрессии увеличен в бактерии, и ген kfiD представляет собой ДНК, содержащую нуклеотидную последовательность, показанную в SEQ ID NO: 37, или ДНК, содержащую нуклеотидную последовательность, имеющую 90% или более идентичность с нуклеотидной последовательностью, показанной в SEQ ID NO: 37, и увеличивающую продукцию гепаросана бактерии из рода Escherichia, которая продуцирует гепаросан, когда уровень экспрессии увеличен в бактерии.
9. Способ продукции гепаросана по любому из пп. 2-8, где ген yhbJ представляет собой ДНК, содержащую нуклеотидную последовательность, показанную в SEQ ID NO: 38, или ДНК, содержащую нуклеотидную последовательность, имеющую 90% или более идентичность с нуклеотидной последовательностью, показанной в SEQ ID NO: 38, и увеличивающую продукцию гепаросана бактерии из рода Escherichia, которая продуцирует гепаросан, когда уровень экспрессии уменьшен в бактерии.
10. Бактерия из рода Escherichia, которая продуцирует гепаросан и
имеет следующую генетическую модификацию (1):
(1) генетическую модификацию, увеличивающую экспрессию гена kpsS по сравнению с экспрессией для немодифицированной бактерией;
и не имеет следующую генетическую модификацию (4):
(4) генетическую модификацию, увеличивающую экспрессию гена kpsC по сравнению с экспрессией для немодифицированной бактерией.
11. Бактерия из рода Escherichia по п.10, дополнительно имеющая по меньшей мере одну из следующих генетических модификаций (2) и (3):
(2) генетической модификации, увеличивающей экспрессию по меньшей мере одного гена, выбранного из гена kfiA, гена kfiB, гена kfiC и гена kfiD по сравнению с экспрессией для немодифицированной бактерией, и
(3) генетической модификации, вызывающую потерю функции гена yhbJ.
| WILLIS L | |||
| M | |||
| et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| ZHANG C | |||
| et al | |||
| Выбрасывающий ячеистый аппарат для рядовых сеялок | 1922 |
|
SU21A1 |

