Изобретение относится к области биотехнологии и может быть использовано при промышленном производстве эффективной, безопасной и удобной в применении антирабической инактивированной эмульсионной культуральной вакцины для профилактической иммунизации домашних плотоядных и сельскохозяйственных животных.
Бешенство (Rabies) занимает первоочередное место в ряду вирусных болезней человека и животных, является одним из самых опасных зоонозов, вызывая поражение центральной нервной системы, энцефаломиелиты, параличи с неизбежным летальным исходом. Данное заболевание представляет собой мировую проблему, которой уделяют особое внимание международные организации (ВОЗ, МЭБ, ФАО, GARC) и ветеринарные службы многих стран мира [1, 2, 3].
Вирус бешенства принадлежит к порядку Mononegavirales, семейству Rhabdoviridae, роду Lyssavirus, виду Rabies lyssavirus [4]. Вирионы вируса бешенства имеют пулевидную форму, длиной ≈180 нм (100-430 нм), диаметром ≈75 нм (45-100 нм). На наружной поверхности вирусной частицы имеются выступы в виде шипов длиной 10 нм, которые прикреплены к двуслойной липидной оболочке [1]. Геном вируса бешенства представлен несегментированной одноцепочечной негативной спиральной РНК длиной около 12000 н.о., который кодирует 5 основных белков: нуклеопротеин (N-белок), фосфопротеин (P-белок), матриксный белок (М-белок), гликопротеин (G-белок), РНК-зависимую РНК-полимеразу (L-белок) [1]. При этом L-ген занимает более половины вирусного генома. Между G- и L-цистронами располагается псевдоген (ψ-фрагмент) [1]. За процесс связывания вируса бешенства с рецепторами клетки-мишени и последующего индуцирования иммунного ответа в организме хозяина ответственен G-белок [5].
Род Lyssavirus включает классический вирус бешенства (филогенетическая группа 1, генетическая линия RABV), распространенный среди различных видов животных по всему миру, а также 13 родственных генотипов, но встречающихся преимущественно у летучих мышей [6]. Род Lyssavirus включает в себя 4 филогенетические группы. К первой филогруппе относятся следующие виды вирусов: Rabies virus, Duvenhage virus, European bat lissavirus 1, European bat lissavirus 2, Australian batlissa virus, Aravan virus, Khujand virus, Irkut virus, ко второй филогруппе - Mokola virus, Logos bat virus, Shimoni bat, к третьей филогруппе - West Caucasian virus, к четвертой филогруппе - Ikoma [1]. Между представленными видами существуют значительные нуклеотидные отличия (количество замен нуклеотидов составляет 0,449-0,543 при стандартной ошибке 0,006-0,008) [1]. Подавляющее большинство случаев бешенства у человека и животных вызывается классическим вирусом бешенства [1, 7]. По результатам исследований Y.T. Arai и соавт. (1997), эффект накопления несинонимических мутаций в ходе эволюции вируса бешенства не приводит к существенным изменениям эпитопов внутри генетической линии, что подтверждает строгое торможение процесса эволюции N-гена. По всей видимости, именно данный ген характеризуется наибольшей консервативностью [8].
Важную роль в эпизоотологии бешенства играют дикие и домашние животные следующих семейств: псовые (Canidae), скунсовые (Mephitidae), куньи (Mustelidae), енотовые (Procyonidae), кошачьи (Felidae), мангустовые (Herpestidae), виверровые (Viverridae), летучие мыши (Microchiroptera), грызуны (Rodentia), которые являются резервуарами и переносчиками вируса [1, 9].
Заболевание приводит к значительным затратам, которые связаны с гибелью животных, ликвидацией последствий вспышек болезни, проведением профилактических и карантинных мероприятий, регулированием численности диких плотоядных животных, отловом бродячих кошек и собак и осуществлением лабораторных исследований по постановке диагноза [10, 11]. По оценкам специалистов, ежегодный экономический ущерб от бешенства составляет более 8,6 млрд. долларов США [7].
Вирус бешенства легко распространяется, по этой причине данное заболевание может приобретать размах эпизоотий [9]. В настоящее время сложная эпидемиологическая и эпизоотическая ситуация по бешенству отмечается более чем в 110 странах мира за исключением некоторых островных государств и на всех континентах, кроме Антарктиды [12]. Так, в Российской Федерации имеется большое количество природных очагов заболевания, которые поддерживаются преимущественно за счет диких плотоядных животных, являющихся основным резервуаром и источником вируса бешенства. Возбудитель заболевания передается домашним и сельскохозяйственным животным, что требует повышенного внимания в свете их непосредственной близости к человеку [13]. При этом считается, что наибольший вклад в процесс передачи вируса бешенства человеку вносят заболевшие собаки.
Наиболее важную роль в борьбе с бешенством на сегодняшний день играет профилактика заболевания у восприимчивых животных, которая заключается в их специфической иммунизации, и оценка их иммунного статуса [1, 14]. Учитывая, что в эпизоотический процесс все более интенсивно стали вовлекаться домашние плотоядные и сельскохозяйственные животные, их специфическая профилактика становится особенно актуальной [15].
Для профилактики бешенства у животных в настоящее время используют живые оральные и инактивированные культуральные антирабические вакцины [16]. Используемые антирабические вакцины обладают протективными свойствами против лиссавирусов филогенетической группы 1 (классический вирус бешенства генетической линии RABV) [1].
В соответствии с международными требованиями МЭБ (OIE), антирабические вакцины должны быть безопасными, иметь индекс иммуногенности не менее 1,0 ME в наименьшей предписанной дозе, формировать надежный иммунитет против бешенства у целевых видов животных, вызывать выработку вируснейтрализующих антител (ВНА) против вируса бешенства с защитным титром ≥0,5 МЕ/см3 [2, 14].
Для практического применения разработано и апробировано множество различных антирабических вакцин для животных (мозговые, эмбриональные, культуральные и другие). Процесс усовершенствования технологии изготовления позволяет отказываться от устаревших образцов в пользу более прогрессивных и качественных препаратов. Антирабические вакцины, изготовленные на основе культурального вируса бешенства, являются более безопасными, ареактогенными, относительно дешевыми по себестоимости и эффективными [1].
В настоящее время для профилактики бешенства у домашних плотоядных и сельскохозяйственных животных в России применяют антирабические инактивированные культуральные вакцины из различных фиксированных штаммов вируса бешенства, в том числе «Щелково-51» и «ВНИИЗЖ» [16, 17, 18].
Штамм «Щелково-51» вируса бешенства был получен на Щелковском биокомбинате (РФ) в результате адаптации варианта Пастеровского штамма «Овечий-ВГНКИ» к перевиваемой клеточной сублинии из почки новорожденного сирийского хомячка (ВНК-21/13) [1]. Данный штамм вируса бешенства относится к филогенетической группе 1, генетической линии RABV [19]. На основе штамма «Щелково-51» вируса бешенства производят следующие вакцинные препараты: вакцину антирабическую инактивированную сухую культуральную из штамма «Щелково-51» для собак и кошек «Рабикан»; вакцину антирабическую из штамма «Щелково-51» инактивированную жидкую культуральную «Рабиков»; вакцину антирабическую для животных «УНИРЭВ»; вакцину антирабическую сухую для крупного и мелкого рогатого скота.
Вакцина «Рабикан» предназначена для профилактической и вынужденной иммунизации домашних плотоядных животных против бешенства. Производителем данной вакцины является ФКП Щелковский биокомбинат (РФ). Препарат представляет собой лиофилизированный, инактивированный β-пропиолактоном фиксированный вирус бешенства штамма «Щелково-51», выращенный в культуре клеток ВНК-21. У первично вакцинированных животных иммунитет начинает формироваться с 5-7 суток после однократного введения вакцины, достигает своего максимума к 30-40 суткам и сохраняется до года. После вторичной вакцинации, проведенной через 30-50 суток после первой, защитная реакция организма усиливается и напряженный иммунитет сохраняется в течение 2 лет. Иммуногенная активность препарата составляет не менее 1,0 МЕ/см3 [16, 17]. Недостатком аналога является то, что при однократной вакцинации домашних плотоядных животных поствакцинальные антирабические антитела практически полностью исчезают спустя 12 месяцев после иммунизации и, следовательно, требуется повторная инокуляция. Также не предусмотрено применение данного препарата для вакцинации сельскохозяйственных животных.
Вакцина «Рабиков» предназначена для профилактики и вынужденной иммунизации крупного рогатого скота, овец и коз, северных оленей и лошадей против бешенства. Препарат представляет собой гомогенную суспензию светло-серого цвета. Вакцина изготовлена из фиксированного вируса бешенства штамма «Щелково-51», репродуцированного в клеточной линии ВНК-21, инактивированного β-пропиолактоном и адсорбированного на гидрате окиси алюминия (Al2O3⋅nH2O). У первично вакцинированных животных иммунитет начинает формироваться с 5-7 суток после введения вакцины, достигает своего максимума к 30-40 суткам и сохраняется до года. После вторичной вакцинации, проведенной через 30-50 суток после первой, защитная реакция организма усиливается и напряженный иммунитет может сохраняться в течение 2-х лет. Иммуногеная активность вакцины составляет не менее 1,0 МЕ/см3 [16, 17]. Недостатком аналога является то, что при непродолжительном хранении на дне формируется рыхлый осадок светло-серого цвета и прозрачная надосадочная жидкость с желтоватым оттенком. Кроме того, в вакцине используется гидрат окиси алюминия, иммуностимулирующая активность которого не во всех случаях [20] проявляется на уровне, обеспечивающим после первичной и однократной вакцинации индуцирование антирабических антител с титром ≥0,5 МЕ/см3, признанным OIE достаточным для защиты животных от бешенства [2].
Вакцина антирабическая для животных «УНИРЭВ» предназначена для профилактики кошек, собак и крупного рогатого скота от бешенства. Изготовителем препарата является ГНУ Всероссийский научно-исследовательский и технологический институт биологической промышленности Россельхозакадемии (ФГБНУ «ВНИТИБП», РФ). Данная вакцина является инактивированной сорбированной. Она включает в себя инактивированный с помощью УФ-света и этанола вирус бешенства штамма «Щелково-51» (мас. 70-80%) и в качестве адъюванта гидроокись алюминия (мас. 9-21%). Дополнительно препарат содержит в своем составе этанол (мас. 9-11%), который надежно защищает вакцину от бактериального пророста и повышает антителоиндуцирующую активность для целевых животных. Вакцина содержит не менее 2,0 ME вирусного материала в 1,0 см3 препарата. Формирование иммунного ответа против вируса бешенства наступает на 14-21 сутки после вакцинации животных. При использовании вакцины, хранящейся в течение 14 месяцев при температуре 6-8°С, количество вируснейтрализующих антител в сыворотках крови собак (щенят), иммунизированных первично и однократно, на 27 сутки после вакцинации равно 2,5-3,4 МЕ/см3, у котят - 1,7-2,2 МЕ/см3. Продолжительность иммунитета домашних плотоядных животных составляет 12 месяцев. Однако, сведения о напряженности иммунитета животных против бешенства в течение года не представлены [20].
Недостатками аналога является то, что спустя 12 месяцев после первичной и однократной иммунизации антирабической сорбированной вакциной количество антител против вируса бешенства ниже защитного уровня (0,5 МЕ/см3), а также не предусмотрена возможность применения данного препарата для профилактики сельскохозяйственных животных.
Вакцина антирабическая сухая культуральная для крупного и мелкого рогатого скота (производитель - ФГБУ «ВНИИЗЖ», РФ) включает антирабический антиген, среду культивирования, защитную среду для высушивания на основе пептона, сахарозы и желатина, а также стимулятор иммунитета сапонин. Среда высушивания дополнительно содержит гидрат окиси алюминия при следующем соотношении компонентов в конечном продукте (мас. %): гидрат окиси алюминия с адсорбированном на нем инактивированным вирусом бешенства - 6,0-9,0, сапонин - 0,75-1,50, пептон - 30,0-33,0, сахароза - 35,0-37,0, желатин - 9,0-10,0, среда культивирования - остальное. Сухая вакцина обладает высоким уровнем иммуногенной активности. Антителоиндуцирующая активность антирабической вакцины в опыте на крупном рогатом скоте через 45 суток после иммунизации составляет 3,5-5,1 МЕ/см3, через 350 суток - 0,38-0,65 МЕ/см3. Иммуногенность данной антирабической вакцины в опыте на овцах и козах через 35 суток после инокуляции равна 2,2-2,8 МЕ/см3 и 3,1-4,2 МЕ/см3, соответственно, через 14 суток после повторной вакцинации у овец и коз - 65-87 МЕ/см3 и 40-48 МЕ/см3, соответственно [21].
Недостатком аналога является то, что спустя 350 дней после иммунизации данной вакциной количество антител против вируса бешенства у некоторых животных было ниже защитного уровня (0,5 МЕ/см3), а также не предусмотрена возможность применения данного препарата для профилактики бешенства у домашних плотоядных животных. Вакцина обладает высокой иммуногенной активностью в течение 11-12 месяцев, однако, содержит сапонин, который может вызывать токсический эффект у мелкого рогатого скота, проявляющийся в нарушении половой функции и расстройствах нервной системы [22].
В 1997 году путем адаптации вакцинного штамма «Щелково-51» вируса бешенства к суспензионной клеточной сублинии ВНК-21/2-17b был получен производственный штамм «ВНИИЗЖ» вируса бешенства.
Известна вакцина антирабическая инактивированная культуральная жидкая (ФГБУ «ВНИИЗЖ») [1, 23, 24, 25], которая предназначена для проведения профилактической и вынужденной иммунизации крупного и мелкого рогатого скота, свиней, лошадей, собак и кошек против бешенства. Данная вакцина является инактивированной сорбированной. Она содержит активное вещество в виде авирулентного и очищенного антигена вируса бешенства штамма «ВНИИЗЖ», полученного в чувствительной биологической системе, и целевые добавки в виде минерального адъюванта гидроокиси алюминия (Al(OH)3) и консерванта мертиолята в соотношении, мас. %:
В качестве чувствительной биологической системы для репродукции вируса бешенства используют перевиваемую суспензионную клеточную линию из почки новорожденного сирийского хомячка ВНК-21. Поддерживающая среда представляет собой раствор Эрла без внесения сыворотки, с добавлением ферментативного гидролизата мышц сухого (ФГМС), гидролизата белков крови сухого (ГБКС) и антибиотиков (рН среды 7,7-7,9). Для инактивации вируса используют аминоэтилэтиленимин (АЭЭИ). Очистку вируссодержащей суспензии от балластных примесей осуществляют методом простой седиментации частиц.
Вакцина вызывает формирование иммунного ответа к вирусу бешенства через 21 сутки после однократного применения, который сохраняется в течение 12 месяцев.
Недостатком данной вакцины является то, что при использовании сорбированной вакцины спустя 12 месяцев для формирования напряженного иммунитета против бешенства требуется повторная инокуляция животных.
Существенные недостатки известных сухих и сорбированных вакцин против бешенства заключаются в их недостаточной иммуногенной активности и необходимости случаев дополнительной иммунизации животных, что объясняется отсутствием более длительного действия и лучшей сохранности антигена, а также невозможностью введения более концентрированной антигенной части.
Наиболее близким аналогом заявляемой вакцины по совокупности существенных признаков является культуральная инактивированная вакцина против бешенства из штамма «ВНИИЗЖ» (вакцина-аналог), которая предназначена для проведения профилактической иммунизации крупного и мелкого рогатого скота, лошадей и собак против бешенства [26]. Данная вакцина содержит активное вещество в виде инактивированного антигена вируса бешенства штамма «ВНИИЗЖ», полученного в чувствительной биологической системе, и целевую добавку в виде масляного адъюванта ISA-70 в соотношении, мас. %:
В качестве чувствительной биологической системы для репродукции вируса бешенства используют перевиваемую суспензионную клеточную линию из почки новорожденного сирийского хомячка ВНК-21-2-17b. Поддерживающая среда состоит из раствора Эрла без внесения сыворотки, с добавлением ферментативного гидролизата мышц сухого (ФГМС), гидролизата белков крови сухого (ГБКС) и антибиотиков (рН среды 7,4-7,6). Для инактивации вируса используют аминоэтилэтиленимин (АЭЭИ) в количестве 0,02% (по объему). При изготовлении вакцины в качестве масляного адъюванта применяют ISA-70. Очистку вируссодержащей суспензии от балластных примесей осуществляют методом простой седиментации частиц.
Иммунизация данной вакциной в однократной дозе индуцирует выработку антирабических антител у КРС в количестве 5,2; 4,7; 5,1; 5,0 log2 или 2,30; 1,62; 2,14; 2,00 МЕ/см3 через 1, 2, 3 и 4 месяца после вакцинации, соответственно. Титр антител против вируса бешенства у лошадей составлял 5,0; 5,3; 5,3; 5,0 log2 или 2,00; 2,46; 2,46; 2,00 МЕ/см3 через 1, 2, 3 и 4 месяца после однократной иммунизации данным препаратом, соответственно. Однократная инокуляция данной вакциной овец вызывает выработку антирабических антител в титрах 5,50; 5,53; 5,00; 5,00 log2 или 2,83; 2,89; 2,00; 2,00 МЕ/см3 через 1, 2, 3 и 4 месяца после введения данной вакцины, соответственно. При этом введение указанной вакцины, включающей в свой состав адъювант ISA-70, активирует только гуморальный иммунитет. Сведения об уровне напряженности поствакцинального иммунитета спустя 5 и более месяцев после вакцинации предложенной экспериментальной вакциной не представлены [26].
Индекс иммуногенности данной вакцины после изготовления составляет 1,30-1,45 МЕ/см3, а через 12 месяцев хранения при температуре 4-8°С - 1,30-1,40 МЕ/см3 [26]. Таким образом, культуральная инактивированная вакцина против бешенства из штамма «ВНИИЗЖ» в течение года при соблюдении условий хранения сохраняет необходимые показатели иммуногенности.
Существенный недостаток антирабической вакцины-аналога заключается в отсутствии более пролонгированного действия инактивированного антигена вируса бешенства и возможности введения концентрированной антигенной части вакцины [27] в количестве более 30%, что, в свою очередь, не позволяет сформировать более продолжительный напряженный иммунитет против рабической инфекции. Кроме того, использование вакцины-аналога не обеспечивает активацию клеточного иммунитета, который играет существенную роль в формировании защиты против возбудителя бешенства [1].
Для решения этой проблемы было создано настоящее изобретение, в которое входила разработка вакцины антирабической инактивированной эмульсионной культуральной для профилактической иммунизации домашних плотоядных и сельскохозяйственных животных.
Технический результат от использования предлагаемого изобретения заключается в повышении иммуногенности антирабической эмульсионной вакцины, обеспечивающей активацию долгосрочного напряженного гуморального и клеточного иммунитета домашних плотоядных и сельскохозяйственных животных за счет введения в состав вакцинного препарата большего количества антигенной части (40% по весу) и применения масляного адъюванта Montanide ISA-61 VG [28].
Указанный технический результат достигнут созданием вакцины антирабической инактивированной эмульсионной культуральной для профилактической иммунизации домашних плотоядных и сельскохозяйственных животных, охарактеризованной следующей совокупностью признаков.
Разработанная вакцина в 1,0 см3 препарата содержит следующие компоненты: 1) активное вещество в виде авирулентного и очищенного антигенного материала из штамма «ВНИИЗЖ» вируса бешенства, репродуцированного предпочтительно в перевиваемой суспензионной клеточной сублинии ВНК-21/SUSP/ARRIAH, с количествами гликопротеина вирусной оболочки не менее 1,5 мкг и нуклеопротеина вириона не менее 1,5 мкг, что соответствует индексу иммуногенности (JJ) не ниже 4,5 ME; 2) масляный адъювант Montanide ISA-61 VG («SEPPIC») предпочтительно в количестве 575000,0-700000,0 мкг.
Производственный штамм «Щелково-51» вируса бешенства, послуживший источником для получения штамма «ВНИИЗЖ», был создан путем проведения более 50 последовательных пассажей в культуре клеток ВНК-21/2 в 1997 году с последующей адаптацией в 2016-2018 гг. к суспензионному культивированию в клеточной сублинии ВНК-21/SUSP/ARRIAH.
Штамм «ВНИИЗЖ» вируса бешенства депонирован в коллекцию штаммов микроорганизмов Федерального государственного бюджетного учреждения «Федеральный центр охраны здоровья животных» (ФГБУ «ВНИИЗЖ»), под регистрационным номером: «ВНИИЗЖ»-ДЕП фиксированного вируса бешенства от 20.12.2018 г.
Штамм «ВНИИЗЖ» вируса бешенства адаптирован к первичной культуре клеток почка свиньи (СП) (Abelseth M.K., 1964), первичной клеточной линии почки хомяка (Kissling R.Е., 1958), монослойной клеточной линии мышиной нейробластомы (NA-2), монослойной культуре клеток из почки зеленой мартышки (Vero) (Montagnon B.J. et al., 1985), монослойной культуре клеток из почки новорожденного сирийского хомячка (ВНК-21/2-17) (ФГБУ «ВНИИЗЖ), суспензионной клеточной сублинии из почки новорожденного сирийского хомячка (ВНК-21/SUSP/ARRIAH) (ФГБУ «ВНИИЗЖ) и другим.
Для изготовления вакцины в качестве чувствительной биологической системы используется предпочтительно перевиваемая суспензионная культура клеток ВНК-21/SUSP/ARRIAH. В качестве поддерживающей среды применяется среду Игла MEM без внесения сыворотки крови КРС с добавлением ФГМС и жидкого гидролизата белков крови (рН среды 7,4-7,6). Инактивацию вируса бешенства проводят с использованием АЭЭИ с концентрацией до 0,025% (от объема). После окончания инактивации проводят охлаждение суспензии до температуры 8-10°С, вносят мертиолят до 0,002% (по весу) и на 48 часов оставляют для процесса седиментации. Необходимую концентрацию антигена в вакцине обеспечивают благодаря его концентрированию методом проточной ультрафильтрации [29, 30].
Для приготовления вакцины используют авирулентный и очищенный антигенный материал из штамма «ВНИИЗЖ» вируса бешенства в виде суспензии с количеством гликопротеина не менее 3,75 мкг/см3 и нуклеопротеина не менее 3,75 мкг/см3, что соответствует индексу иммуногенности не менее 4,5 ME в 1,0 см3 вакцины. Данные концентрации иммуногенных компонентов обеспечивают содержание гликопротеина и нуклеопротеина в количествах каждого не менее 1,5 ME в 1,0 см3 препарата, учитывая, что при изготовлении вакцины антигенная часть составляет 40% (по весу). Определение концентрации гликопротеина и нуклеопротеина вируса бешенства проводят с применением твердофазного иммуноферментного анализа с применением поликлональных антител, специфичных против указанных антигенов [19]. Оценку иммуногенной потенции осуществляют в соответствии с рекомендациями Manual of diagnostic tests and vaccines for terrestrial animals [2].
Антирабическую инактивированную эмульсионную культуральную вакцину получают путем диспергирования при помощи гомогенизатора концентрата антигена вируса бешенства штамма «ВНИИЗЖ» и масляного адъюванта Montanide ISA-61 VG производства фирмы «SEPPIC» в соотношении 40/60 по массе, соответственно. Полученная вакцина представляет собой молокоподобную жидкость, которая не растворима в воде.
Предлагаемое изобретение включает следующую совокупность существенных признаков, обеспечивающих получение технического результата во всех случаях, на которые спрашивается правовая охрана:
1. Антирабическая инактивированная эмульсионная культуральная вакцина для профилактической иммунизации домашних плотоядных и сельскохозяйственных животных.
2. Активное вещество в виде авирулентного и очищенного антигенного материала вируса бешенства из штамма «ВНИИЗЖ» (классический вариант филогенетической группы 1, генетической линии RABV) в эффективном количестве.
3. Целевые добавки.
Существенные отличительные признаки предлагаемой вакцины заключаются в том, что в качестве активного вещества применяется авирулентный очищенный антигенный материал вируса бешенства штамма «ВНИИЗЖ» в эффективном количестве (антигенный концентрат в количестве 40% от массы препарата), а для усиления гуморального и клеточного иммунного ответа используется масляный адъювант Montanide ISA-61 VG.
Предлагаемое изобретение характеризуется также другими отличительными признаками, выражающими конкретные формы выполнения или особые условия его использования:
1. Авирулентный и очищенный антигенный материал из штамма «ВНИИЗЖ» вируса бешенства генетической линии RABV, полученный предпочтительно в суспензионной перевиваемой клеточной линии ВНK-21/SUSP/ARRIAH и представляющий собой суспензию, содержащую преимущественно иммуногенные компоненты вируса бешенства в эффективных количествах.
2. Авирулентный и очищенный антигенный материал из штамма «ВНИИЗЖ» вируса бешенства генетической линии RABV, полученный предпочтительно в суспензионной перевиваемой клеточной линии ВНК-21/SUSP/ARRIAH и представляющий собой суспензию, содержащую иммуногенные компоненты вируса бешенства в виде гликопротеина в количестве не менее 3,75 мкг/см3 и нуклеопротеина не менее 3,75 мкг/см3 (40% от массы препарата), что соответствует индексу иммуногенности вакцины не менее 4,5 ME в 1,0 см3 готового препарата.
3. Из целевых добавок вакцина содержит масляный адъювант.
4. Из целевых добавок вакцина содержит масляный адъювант Montanide ISA-61 VG.
5. Вакцина содержит масляный адъювант Montanide ISA-61 VG в количестве 575000,0-700000,0 мкг (60% от массы препарата) в 1,0 см3 готового препарата.
6. Вакцина содержит консервант мертиолят в количестве 0,002% (по весу).
7. Авирулентный и очищенный антигенный материал из штамма «ВНИИЗЖ» вируса бешенства генетической линии RABV, полученный предпочтительно в суспензионной перевиваемой клеточной линии ВНK-21/SUSP/ARRIAH, масляный адъювант Montanide ISA-61 VG и консервант мертиолят в следующих количествах готового препарата (1,0 см3):

Предлагаемая вакцина обладает высокой иммуногенной активностью и обеспечивает надежную продолжительную защиту против вируса бешенства генетической линии RABV, циркулирующего во всем мире [1].
Достижение технического результата от использования изобретения обеспечивается тем, что предлагаемая антирабическая вакцина изготовлена из авирулентного антигена вируса бешенства штамма «ВНИИЗЖ» в эффективном количестве в сочетании с масляным адъюваном Montanide ISA-61 VG и консерватном мертиолятом. Данное сочетание обуславливает высокую иммуногенную активностью вакцины, что создает эффективную раннюю и длительную защиту домашних плотоядных и сельскохозяйственных животных против бешенства, вызванного лиссавирусами генетической линии RABV, а также активирует у целевых животных не только гуморальный, но и клеточный иммунитет против рабической инфекции.
Сущность изобретения отражена на графических изображениях:
Фиг. 1 - Дендрограмма, отражающая филогенетическое родство штамма «ВНИИЗЖ» с другими штаммами вируса бешенства (при сравнении полных нуклеотидных последовательностей гена N (1350 н.о.)). Маркером ♦ обозначен штамм «ВНИИЗЖ» вируса бешенства. Аббревиатурой обозначены представители различных генотипов рода Lyssavirus: EBLV-1 - European bat lyssavims 1; EBLV-2 - European bat lyssavims 2; KHUV - Khujand virus; BBLV - Bokeloh bat lyssavims; ARAV - Aravan virus; IRKV - Irkut virus; DUVV - Duvenhage virus; ABLV - Australian bat lyssavims; RABV - Rabies virus (вирус бешенства, лиссавирус 1-го генотипа); IKOV - Ikoma virus; WCBV - West Caucasian bat virus; SHIBV - Shimon bat virus; LBV - Lagos bat virus; MOKV - Mokola virus.
Фиг. 2 - Антигенное родство штамма «ВНИИЗЖ» с производственными штаммами вируса бешенства «RV-97», «SAD В19», «Щелково-51» и полевыми изолятами RV 257 Omsk, 24/2010 Tuva, 1634/2008 Saratov (Европейская группа), 1352/2008 Krasnodar, RV 308 Georgia, 18/2005 Krasnodar (Кавказская группа), 33/2005 Bryansk, 552/2010 Kaliningrad (Северо-Европейская группа), 211/2010 Vladimir, 110/2009 NNovgorod, 705/2009 Moscow (Центральная группа), 304c Chita, SG 21 Yakutia, 1410/2008 Komi (Арктическая группа) в реакции нейтрализации.
Фиг. 3 - Результаты культивирования штамма «ВНИИЗЖ» вируса бешенства в первичных культурах клеток почки свиньи и почки хомяка, в гетероплоидных клеточных линиях из почки новорожденного сирийского хомячка ВНК-21/2-17 и ВНК-21/SUSP/ARRIAH, в клетках нейробластомы мыши (NA-2) и почки зеленой мартышки (Vero) (n=3).
Фиг. 4 - Сравнительный анализ биологических свойств штамма «ВНИИЗЖ» с производственными штаммами вируса бешенства генетической линии RABV («RV-97», «Щелково-51», «CVS-27») (n=3).
Фиг. 5 - Электрофореграмма фрагментов ДНК штамма «ВНИИЗЖ» вируса бешенства, положительного и отрицательного контролей с системами праймеров D. Oligo-R721 (а) (на ген N), G0326-G0327 (б) (на ген G), G0341-G0342 (в) (на ген N).
Фиг. 6 - Графики амплификации фрагментов комплементарной ДНК производственных штаммов «ВНИИЗЖ» (A), «RV-97» (Б), «SAD В19» (В), «Щелково-51» (Г) вируса бешенства в ПЦР-РВ с разными системами праймеров (F4924-R5063, CF17-R230, F11-SR220) (на ген N) и зондов (Су5 и FAM).
Фиг. 7 - Динамика уровня субпопуляций лимфоцитов CD4+ и CD8+ в крови КРС в разные сроки после введения одной дозы антирабической инактивированной эмульсионной культуральной вакцины (№1), культуральной инактивированной вакцины против бешенства из штамма «ВНИИЗЖ» (№2). Контроль без вакцинации - №3.
Фиг. 8 - Динамика уровня CD4+/CD8+ -клеток в крови КРС в разные сроки после введения одной дозы антирабической инактивированной эмульсионной культуральной вакцины (№1), культуральной инактивированной вакцины против бешенства из штамма «ВНИИЗЖ» (№2). Контроль без вакцинации - №3.
Фиг. 9 - Динамика уровня субпопуляций CD4+ и CD8+ -клеток в крови коз в разные сроки после введения одной дозы антирабической инактивированной эмульсионной культуральной вакцины (№1), культуральной инактивированной вакцины против бешенства из штамма «ВНИИЗЖ» (№2). Контроль без вакцинации - №3.
Фиг. 10 - Динамика уровня CD4+/CD8+ -клеток в крови коз в разные сроки после введения одной дозы антирабической инактивированной эмульсионной культуральной вакцины (№1), культуральной инактивированной вакцины против бешенства из штамма «ВНИИЗЖ» (№2). Контроль без вакцинации - №3.
Фиг. 11 - Динамика уровня субпопуляций CD4+ и CD8+ -клеток в крови кошек в разные сроки после введения одной дозы антирабической инактивированной эмульсионной культуральной вакцины (№1), культуральной инактивированной вакцины против бешенства из штамма «ВНИИЗЖ» (№2). Контроль без вакцинации - №3.
Фиг. 12 - Динамика уровня CD4+/CD8+ -клеток в крови кошек в разные сроки после введения одной дозы антирабической инактивированной эмульсионной культуральной вакцины (№1), культуральной инактивированной вакцины против бешенства из штамма «ВНИИЗЖ» (№2). Контроль без вакцинации - №3.
Фиг. 13 - Динамика уровня субпопуляций CD4+ и CD8+ -клеток в крови собак в разные сроки после введения одной дозы антирабической инактивированной эмульсионной культуральной вакцины (№1), культуральной инактивированной вакцины против бешенства из штамма «ВНИИЗЖ» (№2). Контроль без вакцинации - №3.
Фиг. 14 - Динамика уровня CD4+/CD8+ -клеток в крови собак в разные сроки после введения одной дозы антирабической инактивированной эмульсионной культуральной вакцины (№1), культуральной инактивированной вакцины против бешенства из штамма «ВНИИЗЖ» (№2). Контроль без вакцинации - №3.
Фиг. 15 - Динамика уровня субпопуляций CD4+ и CD8+ -клеток в крови свиней в разные сроки после введения одной дозы антирабической инактивированной эмульсионной культуральной вакцины (№1), культуральной инактивированной вакцины против бешенства из штамма «ВНИИЗЖ» (№2). Контроль без вакцинации - №3.
Фиг. 16 - Динамика уровня CD4+/CD8+ -клеток в крови свиней в разные сроки после введения одной дозы антирабической инактивированной эмульсионной культуральной вакцины (№1), культуральной инактивированной вакцины против бешенства из штамма «ВНИИЗЖ» (№2). Контроль без вакцинации - №3.
Сущность изобретения изложена в перечне последовательностей, в котором:
SEQ ID NO: 1 представляет последовательность нуклеотидов гена N (1350 н.о.) штамма «ВНИИЗЖ» вируса бешенства филогенетической группы 1, генетической линии RABV;
SEQ ID NO: 2 отражает последовательность аминокислот (450 а.о.), соответствующих нуклеотидной последовательности гена N штамма «ВНИИЗЖ» вируса бешенства филогенетической группы 1, генетической линии RABV.
SEQ ID NO: 3 представляет фрагмент последовательности нуклеотидов псевдогена (ψ-фрагмента) (540 н.о.) штамма «ВНИИЗЖ» филогенетической группы 1, генетической линии RABV;
SEQ ID NO: 4 отражает последовательность аминокислот (180 а.о.), соответствующих фрагменту нуклеотидной последовательности псевдогена (ψ-фрагмента) штамма «ВНИИЗЖ» филогенетической группы 1 генетической линии RABV.
Штамм «ВНИИЗЖ» вируса бешенства филогенетической группы 1 генетической линии RABV характеризуется следующими признаками и свойствами.
Морфологические признаки
Штамм «ВНИИЗЖ» вируса бешенства относится к порядку Mononegavirales, семейству Rhabdoviridae, роду Lyssavirus, виду Rabies lyssavirus и обладает морфологическими признаками, характерными для возбудителя бешенства: форма вириона пулевидная, длина ≈180 нм (100-430 нм), диаметр ≈75 нм (45-100 нм). На наружной поверхности вирусной частицы имеются выступы в виде шипов длиной 10 нм, которые прикреплены к двуслойной липидной оболочке. Вирион состоит из несегментированного генома, представленного одной молекулой спиралеобразно скрученной негативной РНК, длиной приблизительно 12000 н.о. [1].
Антигенные свойства
Вирион вируса бешенства штамма «ВНИИЗЖ» содержит 2 основных антигена: гликопротеин (G-белок) вирусной оболочки и внутренний нуклеопротеин (N-белок) вириона [1].
G-белок индуцирует формирование вируснейтрализующих антител и определяет развитие гуморального иммунитета у животных против вируса бешенства.
Штамм «ВНИИЗЖ» и большинство других штаммов и полевых изолятов вируса бешенства, выделенных на различных континентах, содержат общий нуклеопротеиновый антиген, который обнаруживается с помощью реакции иммунофлуоресценции (РИФ), или методом флуоресцирующих антител (МФА), а также в реакции диффузной преципитации (РДП) [19]. Нуклеопротеиновый антиген обладает способностью стимулировать перекрестные реакции между изолятами вируса бешенства. N-белок играет важную роль в формировании клеточного иммунитета, в частности, взаимодействует с Т-хелперами, активирует CD4+ и CD8+ -клетки [31]. Нуклеопротеин способен стимулировать иммунную систему без осуществления внутриклеточного процессинга и сплайсинга, при этом формируется потенциально протективный иммунный ответ на высококонсервативные эпитопы N-белка (позиции 404-418) [32].
Методом нуклеотидного секвенирования была определена первичная структура гена N штамма «ВНИИЗЖ» вируса бешенства. Сравнительный анализ нуклеотидных последовательностей гена N показал, что штамм «ВНИИЗЖ» вируса бешенства принадлежит к генетической линии RABV первой филогенетической группы (фиг. 1).
Антигенное родство штамма «ВНИИЗЖ» с производственными штаммами «RV-97», «SAD В19», «Щелково-51» вируса бешенства, а также с полевыми изолятами из разных географических групп на территории РФ: RV 257 Omsk, 24/2010 Tuva, 1634/2008 Saratov (Европейская группа), 1352/2008 Krasnodar, RV 308 Georgia, 18/2005 Krasnodar (Кавказская группа), 33/2005 Bryansk, 552/2010 Kaliningrad (Северо-Европейская группа), 211/2010 Vladimir, 110/2009 NNovgorod, 705/2009 Moscow (Центральная группа), 304c Chita, SG 21 Yakutia, 1410/2008 Komi (Арктическая группа) изучено в реакции нейтрализации. В качестве исходного контрольного штамма при постановке реакции нейтрализации использовали референтный штамм «CVS-27», адаптированный к культуре клеток (ANSES, Nancy, France). Референтной сывороткой служила сыворотка против штамма «ВНИИЗЖ» вируса бешенства с титром 2,5 МЕ/см3 (ФГБУ «ВНИИЗЖ»). Изучение антигенного родства (матчинг) в реакции нейтрализации выполняли в соответствии с рекомендациями МЭБ [2]. Значение коэффициента антигенного родства (r1) вычисляли по формуле:
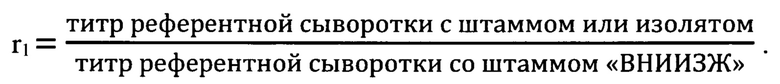
При значении r1>0,30 исследуемые штаммы являются близкородственными, и вакцина из производственного штамма будет защищать от эпизоотического вируса, при значении r1<0,30 полевой изолят отличается от производственного штамма, и вакцина из данного штамма не обеспечит защиту от полевого изолята. Результаты исследований представлены в таблице 1 и на фиг. 2.
Из данных таблицы 1 и фиг. 2 следует, что коэффициент r1 для штамма «CVS-27» составил 0,41; «RV-97» - 0,65; «SAD В19» - 0,42; «Щелково-51» - 0,71; RV 257 Omsk - 0,40; 24/2010 Tuva - 0,45; 1634/2008 Saratov - 0,51; 1352/2008 Krasnodar - 0,48; RV 308 Georgia - 0,58; 18/2005 Krasnodar - 0,51; 33/2005 Bryansk - 0,39; 552/2010 Kaliningrad - 0,46; 211/2010 Vladimir - 0,65; 110/2009 NNovgorod - 0,62; 705/2009 Moscow - 0,53; 304c Chita - 0,61; SG 21 Yakutia - 0,64; 1410/2008 Komi - 0,62. Таким образом, представленный штамм «ВНИИЗЖ» вируса бешенства показывает перекрестные серологические реакции с представителями первой филогенетической линии генетической группы RABV и позволяет обеспечить защиту от заражения изолятами вирусом бешенства данной группы.
Биотехнологические характеристики
Штамм «ВНИИЗЖ» вируса бешенства репродуцируется в первичных культурах клеток: почка свиньи (Abelseth M.K., 1964), почка хомяка (Kissling R.Е., 1958); в гетероплоидных клеточных линиях: монослойная культура клеток из почки новорожденного сирийского хомячка (ВНК-21/2-17) (ФГБУ «ВНИИЗЖ», г. Владимир); суспензионная клеточная сублиния из почки новорожденного сирийского хомячка (ВНК-21/SUSP/ARRIAH) (ФГБУ «ВНИИЗЖ», г. Владимир); клетки нейробластомы мыши (NA-2); клетки из почки зеленой мартышки (Vero) (Montagnon B.J. et al., 1985) и др. Доза заражения составляла 0,005 ККИД50/клетка. Для определения биологической активности проводили титрование вируса каждого пассажа в перевиваемой монослойной культуре клеток ВНК-21/2-17b [33]. В течение 48-96 часов инкубирования количество вируса в указанных культурах клеток достигает высоких значений от 5,60±0,25 до 7,75±0,20 lg ККИД50/см3 (таблица 2). Наиболее высокие показатели увеличения количества штамма «ВНИИЗЖ» вируса бешенства достигаются при репродукции в суспензионной клеточной линии ВНК-21/SUSP/ARRIAH. Данный штамм сохраняет исходные характеристики при пассировании в указанных клеточных культурах на протяжении 10 последовательных пассажей (срок наблюдения).
Гено- и хемотаксономические характеристики
Штамм «ВНИИЗЖ» вируса бешенства принадлежит генетической группе RABV филогенетической линии 1.
Возбудитель является РНК-содержащим вирусом со средним молекулярным весом 0,011926 Mb [32]. Нуклеиновая кислота представлена одноцепочной негативной несегментированной спиральной молекулой длиной приблизительно 12000 н.о. Вирион вируса бешенства имеет пулевидную форму, длиной ≈180 нм (100-430 нм), диаметром ≈75 нм (45-100 нм). На наружной поверхности вирусной частицы имеются выступы в виде шипов длиной 10 нм, которые прикреплены к двуслойной липидной оболочке [1].
Основными антигенными белками являются гликопротеин (G-белок) вирусной оболочки и внутренний нуклеопротеин (N-белок) вириона. В очищенных препаратах штамма «ВНИИЗЖ» вируса бешенства содержится приблизительно 1% РНК, 72% белков, 22% липидов и 3% углеводов.
РНК вируса бешенства штамма «ВНИИЗЖ» кодирует 5 основных белков: нуклеопротеин (N-белок), фосфопротеин (P-белок), матриксный белок (М-белок), гликопротеин (G-белок), РНК-зависимую РНК-полимеразу (L-белок) [1]. Представленные белки и связанная с ними РНК формирует рибонуклеопротеин, который контролирует процессы транскрипции [34].
Устойчивость к внешним факторам
Штамм «ВНИИЗЖ» вируса бешенства чувствителен к эфиру, хлороформу, ацетону, метанолу, трипсину, фосфолипазе. Дезинфицирующие вещества, такие как 1-5% раствор формальдегида, 1% раствор хлорной извести, 1% раствор гипохлорита натрия, 3-5% раствор хлороводородной кислоты, 45-70% раствор этанола, β-пропиолактон и другие, инактивируют вирус бешенства.
При температуре 70°С штамм «ВНИИЗЖ» вируса бешенства мгновенно инактивируется, при температуре 60°С - в течение 5-10 минут, при 50°С - в течение 60 минут, при 35°С - в течение 20 суток, при 23°С - в течение 35 суток. Низкие температуры консервируют данный вирус. В замороженном состоянии данный штамм сохраняет свою активность в течение около 10 лет. Вирус хорошо сохраняется в лиофилизированном состоянии и в смеси с 50% глицерина.
УФ-свет разрушает штамм «ВНИИЗЖ» вируса бешенства в течение 5-10 минут. Вирус относительно стабилен при значениях водородного показателя 5-10. Наиболее стабилен при рН 7,4-7,6. Вирус бешенства высокочувствителен к очень кислой (рН<3) и очень щелочной среде (рН>11).
Дополнительные признаки и свойства
Реактогенность - реактогенными свойствами не обладает.
Патогенность - патогенен для человека, лисицы, волка, койота, шакала, собаки, кошки, крысы, полевки, хомяка, кролика, овцы, козы, лошади, КРС, белки, приматов и других.
Вирулентность - вирулентен для естественно-восприимчивых животных при контактном заражении (преимущественно через укус, при ослюнении поврежденных поверхностей) и аэрогенным путем, а также при трансплантации органов и тканей.
Стабильность - сохраняет исходные биологические свойства при пассировании в чувствительных биологических системах в течение 10 пассажей (срок наблюдения) в первичных клеточных линиях из почки свиньи и почки хомяка, а также в гетероплоидных культурах клеток ВНК-21/2-17 (монослойная), ВНК-21/SUSP/ARRIAH (суспензионная), NA-2, Vero.
Исходя из полученных данных анализа морфологических, биотехнологических, антигенных и дополнительных характеристик, можно утверждать, что штамм «ВНИИЗЖ» вируса бешенства обладает высокой стабильностью при репродукции в чувствительных клеточных линиях, характеризуется высокими показателями титра инфекционной активности, показывает перекрестные серологические реакции с различными представителями первой филогенетической группы генетической линии RABV и позволяет обеспечить защиту от заражения изолятами вирусом бешенства данной генлинии.
Для снижения эпизоотической опасности вируса бешенства генетической линии RABV и предотвращения возникновения новых очагов болезни важна своевременная вакцинопрофилактика, что требует разработки безопасной, удобной в применении антирабической вакцины с высокой эффективностью.
Получена антирабическая инактивированная эмульсионная культуральная вакцина для профилактической иммунизации домашних плотоядных и сельскохозяйственных животных.
Сущность предлагаемого изобретения пояснена примерами его использования, которые не ограничивают объем изобретения.
Пример 1. Исследование биологических свойств штамма «ВНИИЗЖ» вируса бешенства.
Для исследования биологических свойств штамма «ВНИИЗЖ» вируса бешенства использовали вирусный материал, изготовленный в виде 10%-ой суспензия из пораженных вирусом бешенства тканей мозга овцы.
Культивирование штамма «ВНИИЗЖ» вируса бешенства в первичных культурах клеток: почка свиньи (Abelseth M.K., 1964), почка хомяка (Kissling R.E., 1958); в гетероплоидных клеточных линиях: монослойная культура клеток из почки новорожденного сирийского хомячка (ВНК-21/2-17) (ФГБУ «ВНИИЗЖ», г. Владимир); суспензионная клеточная сублиния из почки новорожденного сирийского хомячка (ВНК-21/SUSP/ ARRIAH) (ФГБУ «ВНИИЗЖ», г. Владимир); клетки нейробластомы мыши (NA-2); клетки из почки зеленой мартышки (Vero) (Montagnon B.J. et al., 1985).
Инфекционную активность штамма «ВНИИЗЖ» вируса бешенства определяли титрованием с 10-кратным шагом в монослойной культуре клеток ВНК-21/2-17b. Планшеты с клеточным монослоем и необходимыми количествами вируса инкубировали при температуре 37°С в атмосфере 5% СО2 в течение 48 часов, затем монослой фиксировали 80% раствором ацетона с последующим отмыванием лунок и внесением ФИТЦ-коньюгата. Учет результатов реакции проводят с использованием люминесцентного микроскопа. Антиген вируса бешенства в инфицированных клетках, обработанных антирабическими ФИТЦ-иммунолобулинами, обнаруживали по изумрудному или ярко-зеленому диффузному свечению цитоплазмы и гранул-включений, как в единичных пораженных клетках, так и в клетках, располагающихся группами (флуоресцирующие фокусы) [2].
При репродукции штамма «ВНИИЗЖ» вируса бешенства в первичной культуре клеток почки свиньи с 1 по 10 пассажей наблюдалось увеличение титра инфекционной активности с 5,60±0,25 до 6,50±0,20 lg ККИД50/см3, в первичной клеточной линии почки хомяка - с 5,60±0,20 до 6,50±0,25 lg ККИД50/см3, в монослойной культуре клеток из почки сирийского хомячка (ВНК-21/2-17) - с 5,85±0,15 до 7,25±0,20 lg ККИД50/см3, в суспензионной сублинии клеток из почки сирийского хомячка (ВНК-21/SUSP/ARRIAH) - с 6,00±0,10 до 7,75±0,15 lg ККИД50/см3, в клетках нейробластомы мыши (NA-2) - с 5,70±0,15 до 6,75±0,15 lg ККИД50/см3, в клетках почки зеленой мартышки (Vero) - с 5,60±0,15 до 6,50±0,20 lg ККИД50/см3 (таблица 2, фиг. 3).
Результаты исследований, представленные в таблице 2 и на фиг. 3, свидетельствуют о высокой адаптационной активности штамма «ВНИИЗЖ» вируса бешенства к указанным клеточным культурам. Вирус, репродуцированный в суспензионной культуре клеток ВНК-21/SUSP/ARRIAH, был использован для получения антигена с целью изготовления экспериментальной серии вакцины.
Пример 2. Сравнительный анализ биологических свойств штамма «ВНИИЗЖ» с производственными штаммами вируса бешенства генетической линии RABV.
При изучении свойств штамма «ВНИИЗЖ» вируса бешенства выявлены значительные преимущества в сравнении с другими используемыми производственными штаммами «RV-97», «Щелково-51», «CVS-27» (контрольный). В качестве тест-системы для заражения использовали суспензию клеток сублинии ВНК-21/SUSP/ARRIAH с концентрацией 3,0-3,5 млн клеток/см3. Доза заражения составляла 0,005 ККИД50/ клетка.
В таблице 3 и на фиг. 4 приведены результаты культивирования представленных штаммов вируса бешенства. Данные по репродукции вируса свидетельствуют о преимуществах штамма «ВНИИЗЖ», поскольку для него по итогам 10 последовательных пассажей в чувствительной клеточной сублинии ВНК-21/SUSP/ARRIAH характерны более высокие значения титра инфекционной активности (7,75±0,15 lg ККИД50/см3) по сравнению со штаммами «RV-97», «Щелково-51», «CVS-27» для которых значения титра инфекционности соответствовали показаниям 7,00±0,10, 7,25±0,11, 7,31±0,16 lg ККИД50/см3. По итогам сравнительного анализа выявлено, что накопление гликопротеина вируса бешенства штамма «ВНИИЗЖ» составляло 2,11±0,11 мкг/см3, что на 67,4, 44,5, 41,6% выше по сравнению с репродукцией штаммов «RV-97», «Щелково-51», «CVS-27», соответственно. Количество репродуцированного нуклеопротеина вируса бешенства для штамма «ВНИИЗЖ» составляло 1,90±0,12 мкг/см3, что на 140,5; 40,7; 53,2% выше по сравнению с культивированием штаммов «RV-97», «Щелково-51», «CVS-27», соответственно. Таким образом, штамм «ВНИИЗЖ» вируса бешенства по сравнению с представленными производственными штаммами при репродукции в чувствительной клеточной сублинии ВНК-21/SUSP/ARRIAH характеризуется более высокими показателями накопления иммуногенных компонентов для производства антирабической инактивированной эмульсионной вакцины.
Пример 3. Генетическая идентификация штамма «ВНИИЗЖ» вируса бешенства.
На первом этапе анализа подтверждали принадлежность штамма «ВНИИЗЖ» вируса бешенства к генетической линии RABV. Выделение РНК штамма «ВНИИЗЖ» вируса бешенства проводили с использованием набора для выделения РНК «РИБО-сорб» (ФГУН ЦНИИЭ, г. Москва) в соответствии с инструкцией производителя. Полученный элюат РНК использовали в качестве матрицы для постановки реакции обратной транскрипции, которую осуществляли в термоциклере С 1000 ТМ Thermo Cycler (или аналог) при следующих параметрах: 42°С в течение 15 минут, 99°С - 5 минут. В результате реакции получали ДНК, комплементарную РНК вируса бешенства, которую использовали для проведения полимеразной цепной реакции (ПЦР). Постановку реакции проводили в амплификаторе Master Cycler GSX1 при следующих параметрах термоциклирования: 1) предварительная денатурация - 95°С в течение 10 минут; 2) ПЦР: 35 циклов, каждый из которых включает в себя денатурацию, которую проводили при температуре 94°С в течение 1 минуты, отжиг праймеров - 55°С в течение 1 минуты, элонгацию - 72°С в течение 1 минуты. Реакцию проводили с применением системы праймеров, специфичных для гена N (D. Oligo-R721, G0341-G0342) и для гена G (G0326-G0327). Размеры фрагментов составляли 705, 280, 384 п.н., соответственно. Положительным контролем служила суспензия штамма «CVS-27» вируса бешенства. В качестве отрицательного контроля применяли суспензию клеток ВНК-21/SUSP/ARRIAH, не инфицированную вирусом бешенства. После получения продуктов ПЦР проводили горизонтальный электрофорез в 2% агарозном геле при напряжении 170 V в течение 12 минут с последующей детекцией в УФ-потоке трансиллюминатора и документированием в видеосистеме «Взгляд». В качестве маркера использовали pUC Mix Marker, 8. Результаты исследования представлены на фиг. 5, из которой видно, что штамм «ВНИИЗЖ» принадлежит к вирусу бешенства генетической линии RABV.
На следующем этапе исследования проводили дифференциацию штамма «ВНИИЗЖ» и других производственных штаммов вируса бешенства. ДНК, комплементарную РНК штаммов «ВНИИЗЖ», «RV-97», «SAD В19», «Щелково-51» вируса бешенства, использовали для проведения полимеразной цепной реакции в режиме реального времени (ПЦР-РВ). Постановку реакции проводили в амплификаторе iCycler iQ5 при следующих параметрах термоциклирования: 1) предварительная денатурация - 95°С в течение 10 минут; 2) ПЦР: 40 циклов, каждый из которых включает в себя денатурацию, проводимую при температуре 95°С в течение 10 секунд, отжиг праймеров и зонда, элонгацию - 60°С в течение 60 секунд.
Учитывая, что штаммы вируса бешенства принадлежат к разным геногруппам, отличающимся друг от друга по последовательности нуклеотидов, при постановке ПЦР-РВ применяли 2 системы праймеров (CF 17 - R 230, F 11 - SR 220), специфичных для вируса бешенства разных геногрупп, за исключением штамма «ВНИИЗЖ». Для амплификации фрагмента генома штамма «ВНИИЗЖ» использовали специально разработанную систему праймеров (F4924-R5063), что позволяет с высокой степенью достоверности отличать вакцинный штамм «ВНИИЗЖ» от других производственных штаммов вируса бешенства. Данные системы праймеров комплементарны участкам генома вируса бешенства на границе G- и псевдогенов.
Для детекции накопления амплифицируемых фрагментов комплементарной ДНК вируса бешенства применяли 2 зонда «TaqMan»: ISO, специфичный для множества производственных штаммов и полевых изолятов вируса бешенства (исключение составляет вакцинный штамм «ВНИИЗЖ»); VAC, который высокоспецифичен для штамма «ВНИИЗЖ» вируса бешенства. Учитывая, что зонды имеют флуоресцентные красители с разной длиной волны, по наличию сигнала цианинового флуоресцентного красителя Су5 судили о детекции генома вирусного штамма «ВНИИЗЖ», красителя FAM (карбоксифлуоресцеина) - о наличии множества полевых изолятов и вакцинных штаммов вируса, за исключением штамма «ВНИИЗЖ». Результаты исследования представлены на фиг.6, из которой видно, что при использовании системы праймеров F4924-R5063 наблюдалось накопление сигнала флуоресценции, что свидетельствует о наличии ДНК штамма «ВНИИЗЖ» вируса бешенства. При этом тестирование данного материала с системами праймеров CF 17 - R 230, F 11 - SR 220 детекции сигнала не отмечали (фиг. 6А). Анализ производственных штаммов «ВНИИЗЖ»,
«RV-97», «SAD В19», «Щелково-51» показал, что при использовании систем праймеров CF 17 - R 230, F 11 - SR 220 идентифицированы штаммы «RV-97», «SAD В19», «Щелково-51» вируса бешенства и не обнаружены с использованием системы олигонуклеотидов F4924-R5063 (фиг. 6). В то же время система праймеров F4924-R5063 позволила доказать, что штамм «ВНИИЗЖ» относится к вирусу бешенства генетической линии RABV, но идентифицировала штаммы «RV-97», «SAD В19», «Щелково-51». Полученные результаты свидетельствуют о том, что штамм «ВНИИЗЖ» отличается от других производственных штаммов вируса бешенства по последовательности нуклеотидов псевдогена (ψ-фрагмент).
Пример 4. Получение инактивированного антигена, исследование влияния инактивации с помощью АЭЭИ и очистки антигенного материала на количество иммуногенных компонентов штамма «ВНИИЗЖ» вируса бешенства.
Антирабическую инактивированную эмульсионную культуральную вакцину получали на основе штамма «ВНИИЗЖ» вируса бешенства, выращенного в суспензионной перевиваемой клеточной линии ВНК-21/SUSP/ARRIAH. В качестве поддерживающей среды применяли среду Игла MEM без внесения сыворотки крови КРС с добавлением ФГМС, жидкого гидролизата белков крови (рН среды 7,4-7,6). Культуру клеток заражали данным штаммом вируса бешенства из расчета 0,005 ККИД50/клетка. Через каждые 12 ч культивирования отбирали пробы для определения водородного показателя и корректировки значений рН в указанных пределах.
По окончании репродукции штамм «ВНИИЗЖ» вируса бешенства в суспензию добавляли раствор мертиолята до конечной концентрации 0,002% (по весу) и раствор АЭЭИ до концентрации 0,025% (по весу). Инактивацию вируса бешенства проводили при температуре 36-37°С в течение 24 ч с периодическим перемешиванием. После процесса инактивации суспензию инактивированного вируса охлаждали до температуры 4-6°С. Очистку вирусной суспензии от клеточного детрита осуществляли путем простой седиментации частиц.
Результаты исследований по изучению влияния инактивации и очистки антигенного материала с помощью АЭЭИ на количество иммуногенных компонентов штамма «ВНИИЗЖ» вируса бешенства на примере 10-ти образцов отражены в таблице 4. Определение концентрации гликопротеина и нуклеопротеина вируса бешенства проводили с применением твердофазного иммуноферментного анализа с использованием поликлональных антител, специфичных против указанных антигенов [19]. Из данных таблицы 4 видно, что до инактивации средние значения концентрации гликопротеина и нуклеопротеина штамма «ВНИИЗЖ» вируса бешенства составили 2,20±0,06 и 2,02±0,03 мкг/см3, соответственно. После процесса инактивации средние показатели для гликопротеина и нуклеопротеина снизились на 8,02-10,83 и 7,61-10,44%, и были равны 2,01±0,06 и 1,84±0,03 мкг/см3, соответственно. Таким образом, иммуногенные компоненты штамма «ВНИИЗЖ» вируса бешенства устойчивы к процессам инактивации с помощью 0,025% АЭЭИ и очистки методом простой седиментации.
Пример 5. Концентрирование антигена штамма «ВНИИЗЖ» вируса бешенства для получения вакцины.
Для достижения необходимой концентрации иммуногенных компонентов штамма «ВНИИЗЖ» вируса бешенства в 1,0 см3 вакцины (гликопротеин - не менее 1,5 мкг, нуклеопротеин - не менее 1,5 мкг) требуется проводить концентрирование антигенов до количеств каждого не менее 3,75 мкг в 1,0 см3 вирусной суспензии, поскольку на этапе диспергирования соотношение антигена и адъюванта составляет 40/60. Необходимую концентрацию иммуногенных компонентов штамма «ВНИИЗЖ» вируса бешенства в прививной дозе эмульсионной вакцины получали путем концентрирования антигена проточной ультрафильтрацией с применением ультрафильтров при давлением 1,5 атм. Полученный концентрат антигена хранили при температуре 4-6°С до момента использования. Получаемую вакцину расфасовывают в стеклянные или пластиковые флаконы и проводят контроль стерильности продукции в соответствии с ГОСТ 28085-89.
Пример 6. Процесс диспергирования антигена и адъюванта для получения антирабической инактивированной эмульсионной культуральной вакцины из штамма «ВНИИЗЖ».
Антирабическую инактивированную эмульсионную культуральную вакцину получали путем диспергирования при помощи гомогенизатора концентрата антигена вируса бешенства штамма «ВНИИЗЖ» и масляного адъюванта Montanide ISA-61 VG в соотношении 40/60 по массе, соответственно.
В результате получали эмульсионную вакцину, которая представляла собой молокоподобную жидкость, не растворимую в воде. Вакцина имела жидкую и удобную в применении консистенцию, рассасывалась в месте введения, не вызывала образования абсцессов, общей реакции в виде повышения температуры. Кинематическая вязкость полученного препарата составляла 3,5-4,0 сСт, что является средней степенью вязкости и находится в пределах от 3,0 до 7,0 сСт [35].
Оптимальный компонентный состав полученной вакцины представлен на странице 15.
Пример 7. Оценка индекса иммуногенности антирабической инактивированной эмульсионной культуральной вакцины из штамма «ВНИИЗЖ».
Оценку иммуногенной потенции осуществляли в соответствии с рекомендациями Manual of diagnostic tests and vaccines for terrestrial animals [2]. Тестировали разработанную антирабическую вакцину (с применением адъюванта Montanide ISA-61 VG) и вакцину-аналог (с применением адъюванта ISA-70). В качестве контроля использовали референтную вакцину против бешенства из штамма «CVS-27» с индексом иммуногенности (JJреф), равным 2,0 МЕ/см3 (изготовитель - ФГБУ «ВНИИЗЖ»). Для анализа каждого препарата использовали по пять мышей массой 18-20 г. Каждую мышь вакцинировали внутримышечно с использованием 1/5 объема наименьшей дозы, что составило 0,1 см3. Через 14 дней после инъекции отбирали образцы крови. Сыворотки тестировали на наличие антитела к вирусу бешенства с использованием теста FAVN [2]. Результаты оценки индекса иммуногенности испытуемых вакцин (JJисп) отражены в таблице 5.
В 1,0 см3 готового препарата содержалось не менее 3,0 мкг иммуногенных компонентов вируса бешенства (гликопротеина не менее 1,5 мкг и нуклеопротеина не менее 1,5 мкг), что соответствовало индексу иммуногенности вакцины не менее 4,5 МЕ/см3. Из данных, отраженных в таблице 5, следует, что антирабическая инактивированная эмульсионная культуральная вакцина из штамма «ВНИИЗЖ», репродуцированного в суспензионной клеточной линии ВНК-21/SUSP/ARRIAH и масляного адъюванта Montanide ISA-61 VG, характеризовалась индексом иммуногенности 5,76±0,10 ME в 1,0 см3 препарата, что выше по сравнению с инактивированной жидкой вакциной против бешенства из штамма «ВНИИЗЖ», репродуцированного в культуре клеток ВНК-21/2-17b, и ISA-70 на 4,02 ME в 1,0 см3 препарата. Иными словами, разработанная вакцина отличается высокими значениями индекса иммуногенности и удовлетворяла заявленным выше требованиям к данному препарату (JJ не менее 4,50 ME в 1,0 см3 вакцины).
Пример 8. Оценка ареактогенности и безвредности антирабической инактивированной эмульсионной культуральной вакцины.
Оценку ареактогенности и безвредности антирабической инактивированной эмульсионной культуральной вакцины проводили на КРС, МРС, свиньях, собаках, кошках и кроликах.
Вакцину вводили всем видам животных внутримышечно в дозе, рекомендованной для каждого вида животных. Прививная доза 2,0 см3 предназначена для КРС, 1,0 см3 - для овец и коз, взрослых собак крупных и средних пород, 0,5 см3 - для кошек, щенков с 2-х месячного возраста всех пород, взрослых собак мелких пород (болонки, таксы, шпицы и др.). За животными осуществляли клиническое наблюдение. Температуру тела измеряли в течение 7 суток после инъекции, затем через 10 и 14 суток после вакцинации. Критерием оценки клинического наблюдения было развитие или отсутствие общей и местной реакции на введение испытуемого препарата. Результаты исследования представлены в таблице 6, из которой следует, что все испытуемые животные были клинически здоровы, общей и местной реакции на введение препарата не было выявлено. Повышение температуры тела у животных было незначительным (0,3-0,7°С) в 1-2 сутки после ведения вакцины. В течение этого времени наблюдения разница температур (ΔT=Tmax-Т на 0 день после вакцинации) у КРС составила 0,3-0,6°С, у МРС - 0,3-0,4°С, у свиней - 0,3-0,7°С, у собак - 0,3-0,6°С, у кошек - 0,3-0,7°С, у кроликов - 0,3-0,5°С. Начиная с 3 суток температура животных была в норме. Местная реакция (отек, болезненность, хромота) у целевых видов животных не отмечена. Таким образом, по результатам исследований антирабическая инактивированная эмульсионная культуральная вакцина является безвредным и аректогенным препаратом для целевых видов животных.
Пример 9. Оценка гуморального и клеточного иммунитета у КРС при использовании антирабической инактивированной эмульсионной культуральной вакцины в сравнении с вакциной-аналогом. Исследование протективных свойств разработанной вакцины на примере коров.
Проведена оценка гуморального иммунитета у КРС, привитых антирабической инактивированной эмульсионной культуральной вакциной, в сравнении с вакциной-аналогом. В качестве наиболее близкого прототипа использовали культуральную инактивированную вакцину против бешенства из штамма «ВНИИЗЖ», которая предназначена для проведения профилактической вакцинации крупного и мелкого рогатого скота, лошадей и собак против бешенства [26]. Данная вакцина содержит активное вещество в виде инактивированного антигена вируса бешенства штамма «ВНИИЗЖ», полученного в культуре клеток ВНК-21/2-17 (30% по весу), и масляный адъювант ISA-70 (70% по весу).
Испытания двух представленных вакцин проводили на десяти головах КРС черно-пестрой породы в возрасте 1 года со средней массой 200 кг. Животных прививали вакцинами в дозе 2,0 см3 внутримышечно однократно. Для оценки напряженности поствакцинального антирабического иммунитета у животных, в соответствии с рекомендациями МЭБ (OIE), применяли реакцию нейтрализации в монослойной культуре клеток ВНК-21 (модификация реакции - FAVN (Fluorescent antibody virus neutralization)), выражая значения титра антирабических антител в МЕ/см3 [2]. Для анализа использовали положительный сывороточный контрольный стандарт, разведенный до иммуногенности 0,5 МЕ/см3, и отрицательный сывороточный контрольный стандарт с иммуногенностью <0,1 МЕ/см3 [2].
В соответствии с международными требованиями, защитным является уровень ВНА против вируса бешенства ≥0,50 МЕ/см3 [2]. В реакции нейтрализации оценивали сыворотки крови от КРС до и на 21 сутки, а также через 2, 3, 6, 12, 15, 18 месяцев после вакцинации.
До иммунизации в крови животных антирабические антитела не были выявлены. Уровень антирабических вируснейтрализующих антител у КРС после введения одной рекомендованной дозы для этого вида животных (2,0 см3) антирабической инактивированной эмульсионной культуральной вакцины на 21 сутки составлял 25,90±0,42 МЕ/см3, через 2 месяца - 42,50±0,65 МЕ/см3, через 3 месяца - 38,51±0,68 МЕ/см3, через 6 месяцев - 24,80±0,46 МЕ/см3, через 12 месяцев - 13,42±0,84 МЕ/см3, через 15 месяцев - 4,83±0,42 МЕ/см3, через 18 месяцев - 0,41±0,11 МЕ/см3 (таблица 7). Иными словами, введение одной дозы (2,0 см3) антирабической инактивированной эмульсионной культуральной вакцины обеспечивает накопление высокого уровня антител (до 42,50±0,65 МЕ/см3) в крови коров и сохранение защитного уровня антител против вируса бешенства (≥0,50 МЕ/см3) до 15 месяцев.
Уровень антирабических вируснейтрализующих антител у КРС после введения рекомендованной для него одной дозы (2,0 см3) инактивированной жидкой вакцины-аналога против бешенства из штамма «ВНИИЗЖ» на 21 сутки составлял 2,30±0,43 МЕ/см3, через 2 месяца - 1,62±0,56 МЕ/см3, через 3 месяца - 2,14±0,33 МЕ/см3, через 6 месяцев - 1,95±0,39 МЕ/см3, через 12 месяцев - 0,67±0,19 МЕ/см3, через 15 месяцев - 0,15±0,09 МЕ/см3, через 18 месяцев - 0,07±0,02 МЕ/см3. Иными словами, введение одной дозы (2,0 см3) культуральной инактивированной вакцины против бешенства из штамма «ВНИИЗЖ» обеспечивает накопление защитного уровня антител против вируса бешенства до 12 месяцев. При этом вакцина-аналог индуцирует выработку антирабических антител в 18,48 раз ниже по сравнению с разработанной вакциной.
На следующем этапе исследования проводили оценку клеточного иммунитета у КРС после введения одной дозы (2,0 см3) разработанной вакцины и вакцины-аналога.
Иммунизацию животных проводили так же, как указано выше в примере 9. Кровь от КРС отбирали через 1, 2, 3 недели после вакцинации. Из крови КРС проводили выделение лимфоцитов. Для этого кровь обрабатывали 2% раствором этилендиаминтетраацетатом (ЭДТА) и центрифугировали в стандартном градиенте плотности Ficoll-Paque PLUS. На 3 см3 Ficoll-Paque наслаивали 2,0 см3 крови, разведенной 1:1 раствором среды Игла MEM, и центрифугировали 30 минут при 400 g. Выделенную фракцию лимфоцитов отмывали 1/15 М фосфатным буферным раствором (ФБР) и полученный осадок ресуспендировали в 200 мкл ФБР. Подсчет клеток проводили в счетчике клеток Countess Automated Cell Counter. Концентрацию лимфоцитов доводили раствором ФБР до 106-107 клеток/см3.
Анализ субпопуляции лимфоцитов CD4+ и CD8+ и определение их относительного количества в крови КРС проводили цитофлуориметрическим методом. Динамика уровня субпопуляций CD4+ и CD8+-клеток в крови КРС в разные сроки после введения одной дозы (2,0 см3) антирабической инактивированной эмульсионной культуральной вакцины, культуральной инактивированной вакцины против бешенства из штамма «ВНИИЗЖ» в сравнении с контролем без вакцинации представлена на фиг. 7. Из данных фиг. 7А и 7Б следует, что относительное количество CD4+ и CD8+-клеток в организме животных после введения разработанной вакцины увеличивалось и составляло 12-14 и 6-10%, соответственно. Содержание лимфоцитов CD4+ и CD8+ после введения вакцины аналога находилось на уровне значений контрольных животных без вакцинации и составляло 3-5%.
Анализируя значения отношения CD4+/CD8+-клеток после введения 2,0 см3 разработанной вакцины, можно отметить тенденцию к его снижению у КРС с 2,0 до 1,2, иными словами отмечалось увеличение относительного количества клеток CD8+ (фиг. 8). Таким образом, введение в организм КРС антирабической инактивированной эмульсионной культуральной вакцины с применением масляного адъюванта Montanide ISA-61 VG вызывало изменение фенотипа Т-клеток, что свидетельствовало об активации клеточного иммунитета. После иммунизации КРС инактивированной жидкой вакциной-аналогом против бешенства из штамма «ВНИИЗЖ» с использованием адъюванта ISA-70 индуцирования клеточного иммунитета не отмечали.
Для определения протективных свойств разработанной антирабической инактивированной эмульсионной культуральной вакцины проводили контрольное заражение штаммом «CVS-27» вируса бешенства в дозе 104 ЛД50/0,20 см3 десяти голов КРС на 21 сутки, а также через 6, 12 и 15 месяцев после иммунизации и пяти голов невакцинированных животных. Животные опытных групп оставались живыми и клинически здоровыми, в контрольной группе была отмечена гибель всех голов КРС. Таким образом, разработанная антирабическая инактивированная эмульсионная культуральная вакцина является эффективной для иммунизации КРС.
Пример 10. Оценка гуморального и клеточного иммунитета у MP при использовании антирабической инактивированной эмульсионной культуральной вакцины в сравнении с вакциной-аналогом. Исследование протективных свойств разработанной вакцины на примере коз.
Оценку гуморального иммунитета у коз, привитых антирабической инактивированной эмульсионной культуральной вакцины, проводили в сравнении с инактивированной жидкой вакциной-аналогом против бешенства из штамма «ВНИИЗЖ».
Испытания данных вакцин проводили на десяти головах коз Зааненской породы в возрасте 1 года. Животных привили вакциной в дозе 1,0 см3 внутримышечно однократно. До иммунизации и спустя 21 сутки, а также через 2, 3, 6, 12, 15, 18 месяцев после вакцинации у коз отбирали кровь и исследовали полученные сыворотки на наличие антирабических антител в реакции нейтрализации (модификация FAVN) [2].
До иммунизации в крови животных антирабические антитела не были выявлены. Уровень антирабических вируснейтрализующих антител у коз после введения одной рекомендованной для данного вида животных дозы (1,0 см3) антирабической инактивированной эмульсионной культуральной вакцины на 21 сутки составлял 52,70±0,99 МЕ/см3, через 2 месяца - 84,65±0,85 МЕ/см3, через 3 месяца - 79,65±0,74 МЕ/см3, через 6 месяцев - 45,16±0,89 МЕ/см3, через 12 месяцев - 25,42±0,74 МЕ/см3, через 15 месяцев - 7,56±0,40 МЕ/см3, через 18 месяцев - 0,49±0,06 МЕ/см3 (таблица 8). Таким образом, введение одной дозы (1,0 см3) антирабической инактивированной эмульсионной культуральной вакцины обеспечивает накопление высокого уровня антител (до 84,65±0,85 МЕ/см3) в крови коз и сохранение защитного уровня антител против вируса бешенства (≥0,50 МЕ/см3) до 15-18 месяцев.
Титр антирабических вируснейтрализующих антител у коз после введения рекомендованной для них одной дозы (1,0 см3) культуральной инактивированной вакцины-аналога против бешенства из штамма «ВНИИЗЖ» на 21 сутки составлял 2,98±0,51 МЕ/см3, через 2 месяца - 3,15±0,64 МЕ/см3, через 3 месяца - 3,09±0,41 МЕ/см3, через 6 месяцев - 2,43±0,53 МЕ/см3, через 12 месяцев - 0,85±0,22 МЕ/см3, через 15 месяцев - 0,38±0,11 МЕ/см3, через 18 месяцев - 0,11±0,03 МЕ/см3. Иными словами, использование одной дозы (1,0 см3) культуральной инактивированной вакцины против бешенства из штамма «ВНИИЗЖ» обеспечивает накопление защитного уровня антител против вируса бешенства до 12 месяцев. При этом данная вакцина-аналог индуцирует выработку антирабических антител в 26,87 раз ниже по сравнению с разработанной вакциной.
Следующий этап исследования был посвящен оценке клеточного иммунитета у коз после введения одной дозы (1,0 см3) представленной вакцины и вакцины-аналога.
Иммунизацию животных, отбор крови, выделение и анализ лимфоцитов проводили так же, как указано выше в примере 9. Динамика уровня субпопуляций лимфоцитов CD4+ и CD8+ в крови коз через 1, 2 и 3 недели после введения одной дозы (1,0 см3) антирабической инактивированной эмульсионной культуральной вакцины, культуральной инактивированной вакцины против бешенства из штамма «ВНИИЗЖ» в сравнении с контролем без вакцинации представлена на фиг. 9. Из данных фиг. 9А и 9Б видно, что относительное количество CD4+ и CD8+ -клеток в организме животных после введения разработанной вакцины увеличивалось и составляло 19-24 и 10-13%, соответственно. Содержание CD4+ и CD8+ лимфоцитов после введения вакцины аналога находилось на уровне значений контрольных животных без вакцинации и составляло 3-6%.
Анализ значений отношения клеток CD4+/CD8+ после введения разработанной вакцины показал, что отмечалась незначительная тенденция к его снижению у коз с 1,90 до 1,83 (фиг. 10). Максимальное относительное количество лимфоцитов CD4+ и CD8+ наблюдалось через 2 недели после вакцинации. В целом определено, что введение в организм коз антирабической инактивированной эмульсионной культуральной вакцины с применением масляного адъюванта Montanide ISA-61 VG активировало клеточный иммунитет, что проявлялось в изменении фенотипа Т-клеток. После иммунизации коз культуральной инактивированной вакцины против бешенства из штамма «ВНИИЗЖ» с использованием адъюванта ISA-70 активации клеточного иммунитета не наблюдали.
Протективные свойства разработанной антирабической инактивированной эмульсионной культуральной вакцины оценивали так, как отражено в примере 9. После контрольного заражения все 10 коз, привитых одной дозой (1,0 см3) вакцины, оставались живыми и клинически здоровыми. В контрольной группе была отмечена гибель всех коз. Таким образом, разработанная антирабическая инактивированная эмульсионная культуральная вакцина является эффективной для иммунизации коз.
Пример 11. Оценка гуморального и клеточного иммунитета у кошек при использовании антирабической инактивированной эмульсионной культуральной вакцины в сравнении с вакциной-аналогом. Исследование протективных свойств разработанной вакцины на примере кошек.
Определение напряженности гуморального иммунитета у кошек, привитых антирабической инактивированной эмульсионной культуральной вакцины, проводили в сравнении с инактивированной жидкой вакциной-аналогом против бешенства из штамма «ВНИИЗЖ».
Тестирование данных вакцин проводили на десяти головах кошек беспородных в возрасте от 2 до 6 месяцев массой 0,5 кг. Всем животным вводили вакцину внутримышечно в объеме 0,5 см3 однократно. До вакцинации и спустя 21 сутки, а также через 2, 3, 6, 12, 15, 18 месяцев после иммунизации у кошек отбирали кровь и полученные сыворотки исследовали на наличие антител против вируса бешенства в реакции нейтрализации (модификация FAVN) [2].
До иммунизации в крови животных антирабические антитела не были детектированы. Количество антител против вируса бешенства у кошек после введения одной рекомендованной для них дозы (0,5 см3) антирабической инактивированной эмульсионной культуральной вакцины на 21 сутки составлял 82,65±1,03 МЕ/см3, через 2 месяца - 92,14±1,21 МЕ/см3, через 3 месяца - 85,12±0,85 МЕ/см3, через 6 месяцев - 49,16±0,98 МЕ/см3, через 12 месяцев - 28,31±0,82 МЕ/см3, через 15 месяцев - 8,12±0,52 МЕ/см3, через 18 месяцев - 0,76±0,14 МЕ/см3 (таблица 9). Таким образом, введение одной дозы (0,5 см3) антирабической инактивированной эмульсионной культуральной вакцины обеспечивает накопление высокого уровня антител (до 92,14±1,21 МЕ/см3) в крови кошек и сохранение защитного уровня антител против вируса бешенства (≥0,50 МЕ/см3) до 15-18 месяцев.
Уровень антирабических вируснейтрализующих антител в крови кошек после введения рекомендованной для них одной дозы (0,5 см3) инактивированной вакцины-аналога против бешенства из штамма «ВНИИЗЖ» на 21 сутки составлял 3,15±0,68 МЕ/см3, через 2 месяца - 3,35±0,74 МЕ/см3, через 3 месяца - 3,14±0,50 МЕ/см3, через 6 месяцев - 2,75±0,62 МЕ/см3, через 12 месяцев - 0,91±0,31 МЕ/см3, через 15 месяцев - 0,44±0,13 МЕ/см3, через 18 месяцев - 0,21±0,05 МЕ/см3. Иными словами, введение одной дозы (0,5 см3) инактивированной вакцины-аналога против бешенства из штамма «ВНИИЗЖ» обеспечивает накопление защитного уровня антител в крови кошек против вируса бешенства до 12 месяцев. При этом данная вакцина-аналог индуцирует выработку антирабических антител в 27,50 раз ниже по сравнению с разработанной вакциной.
На следующем этапе работы проводили исследование клеточного иммунитета у кошек после введения одной дозы (0,5 см3) представленной вакцины и вакцины-аналога.
Вакцинацию животных, отбор крови, выделение и анализ лимфоцитов проводили так же, как указано выше в примере 9. Динамика уровня субпопуляций CD4+ и CD8+ -клеток в крови кошек через 1, 2 и 3 недели после введения одной дозы (0,5 см3) антирабической инактивированной эмульсионной культуральной вакцины, инактивированной вакцины-аналога против бешенства из штамма «ВНИИЗЖ» в сравнении с контролем без вакцинации представлена на фиг. 11. Из данных фиг. 11А и 11Б видно, что относительное содержание лимфоцитов CD4+ и CD8+ в организме животных после введения разработанной вакцины увеличивалось и составляло 21-28 и 12-19%, соответственно. Количество CD4+ и CD8+ лимфоцитов после введения вакцины-аналога находилось на уровне значений контрольных животных без вакцинации и составляло 4-6%.
Анализ значений отношения клеток CD4+/CD8+ после введения разработанной вакцины показал, что со 2 недели после вакцинации отмечалась тенденция к его снижению данного показателя у кошек с 1,75 до 1,42 (фиг. 12). Максимальное относительное количество лимфоцитов CD4+ и CD8+ зафиксировано через 1-2 недели после инокуляции разработанной вакцины. Относительное количество лимфоцитов CD8+ возрастало с 1 по 3 недели тестирования. Выявлено, что введение в организм кошек антирабической инактивированной эмульсионной культуральной вакцины с применением масляного адъюванта Montanide ISA-61 VG активировало клеточный иммунитет, вызывая количественные изменения фенотипа Т-клеток. После иммунизации кошек культуральной инактивированной вакциной-аналогом против бешенства из штамма «ВНИИЗЖ» с использованием адъюванта ISA-70 активации клеточного иммунитета не отмечали.
Протективные свойства разработанной антирабической инактивированной эмульсионной культуральной вакцины оценивали так, как отражено в примере 9. После контрольного заражения все 10 кошек, привитых одной дозой (0,5 см3) вакцины, оставались живыми и клинически здоровыми. В контрольной группе была отмечена гибель всех животных. Таким образом, разработанная антирабическая инактивированная эмульсионная культуральная вакцина является эффективной для иммунизации кошек.
Пример 12. Оценка гуморального и клеточного иммунитета у собак при использовании антирабической инактивированной эмульсионной культуральной вакцины в сравнении с вакциной-аналогом. Исследование протективных свойств разработанной вакцины на примере собак.
Исследование напряженности гуморального иммунитета у собак, привитых антирабической инактивированной эмульсионной культуральной вакцины, проводили в сравнении с инактивированной жидкой вакциной-аналогом против бешенства из штамма «ВНИИЗЖ».
Тестирование указанных вакцин проводили на десяти головах собак беспородных в возрасте 1-3 года с массой тела от 10-25 кг. Всем животным вводили препарат внутримышечно в объеме 1,0 см3 однократно. До иммунизации и спустя 21 сутки, а также через 2, 3, 6, 12, 15, 18 месяцев после иммунизации у собак отбирали кровь и полученные сыворотки исследовали на наличие антирабических антител в реакции нейтрализации (модификация FAVN) [2].
До вакцинации в крови собак антитела против вируса бешенства не были выявлены. Количество вируснейтрализующих антител у собак после введения одной рекомендованной для данного вида животных дозы (1,0 см3) антирабической инактивированной эмульсионной культуральной вакцины на 21 сутки составлял 24,12±0,56 МЕ/см3, через 2 месяца - 29,65±0,65 МЕ/см3, через 3 месяца - 25,63±0,74 МЕ/см3, через 6 месяцев - 18,16±0,51 МЕ/см3, через 12 месяцев - 6,45±0,13 МЕ/см3, через 15 месяцев - 0,95±0,12 МЕ/см3, через 18 месяцев - 0,32±0,12 МЕ/см3 (таблица 10). Таким образом, введение одной дозы (1,0 см3) антирабической инактивированной эмульсионной культуральной вакцины обеспечивает накопление высокого уровня антител (до 29,65±0,65 МЕ/см3) в крови собак и сохранение защитного уровня антител против вируса бешенства (≥0,50 МЕ/см3) до 15 месяцев.
Титр антирабических вируснейтрализующих антител в крови собак после введения одной рекомендованной для них одной дозы (1,0 см3) культуральной инактивированной вакцины-аналога против бешенства из штамма «ВНИИЗЖ» на 21 сутки составлял 1,84±0,21 МЕ/см3, через 2 месяца - 2,17±0,34 МЕ/см3, через 3 месяца - 2,75±0,42 МЕ/см3, через 6 месяцев - 2,01±0,46 МЕ/см3, через 12 месяцев - 0,79±0,42 МЕ/см3, через 15 месяцев - 0,36±0,10 МЕ/см3, через 18 месяцев - 0,09±0,02 МЕ/см3. Таким образом, введение одной дозы культуральной инактивированной вакцины-аналога против бешенства из штамма «ВНИИЗЖ» обеспечивает накопление защитного уровня антител в крови собак против вируса бешенства до 12 месяцев. При этом данная вакцина-аналог индуцирует выработку антирабических антител в 10,78 раз ниже по сравнению с разработанной вакциной.
На следующем этапе тестирования проводили оценку клеточного иммунитета у собак после введения одной дозы (1,0 см3) представленной вакцины и вакцины-аналога.
Иммунизацию животных, отбор крови, выделение и анализ лимфоцитов проводили так же, как указано выше в примере 9. Динамика уровня субпопуляций CD4+ и CD8+ -клеток в крови собак через 1, 2 и 3 недели после введения одной дозы (1,0 см3) антирабической инактивированной эмульсионной культуральной вакцины, культуральной инактивированной вакцины-аналога против бешенства из штамма «ВНИИЗЖ» в сравнении с контролем без вакцинации представлена на фиг. 13. Из данных фиг. 13 видно, что относительное содержание лимфоцитов CD4+ и CD8+ в крови собак после введения предложенной вакцины увеличивалось и составляло 13-19 и 8-13%, соответственно. Количество клеток CD4+ и CD8+ после введения вакцины-аналога находилось на уровне значений контрольных животных без вакцинации и было равно 3-6%.
Уровень CD4+/CD8+ -клеток после введения разработанной вакцины показал, что с 1 недели после вакцинации наблюдалось снижение данного показателя у собак с 1,63 до 1,31 и, как следствие, увеличение относительного количества лимфоцитов CD8+ с 8 до 13 (фиг. 14). Максимальное относительное количество лимфоцитов CD4+ и CD8+ отмечалось через 1 неделю после вакцинации. Выявлено, что введение в организм собак антирабической инактивированной эмульсионной культуральной вакцины с применением масляного адъюванта Montanide ISA-61 VG активировало клеточный иммунитет, что выражалось в количественных изменениях фенотипа Т-клеток. После иммунизации собак культуральной инактивированной вакцины-аналога против бешенства из штамма «ВНИИЗЖ» с использованием адъюванта ISA-70 активации клеточного иммунитета не наблюдали.
Протективные свойства предложенной антирабической инактивированной эмульсионной культуральной вакцины оценивали так, как отражено в примере 9. После контрольного заражения все 10 собак, привитых одной дозой (1,0 см3) вакцины, оставались живыми и клинически здоровыми. В контрольной группе была отмечена гибель всех животных. Таким образом, разработанная антирабическая инактивированная эмульсионная культуральная вакцина является эффективной для иммунизации собак.
Пример 12. Оценка гуморального и клеточного иммунитета у свиней при использовании антирабической инактивированной эмульсионной культуральной вакцины в сравнении с вакциной-аналогом. Исследование протективных свойств разработанной вакцины на примере свиней.
Исследование напряженности гуморального иммунитета у свиней, привитых антирабической инактивированной эмульсионной культуральной вакцины, проводили в сравнении с инактивированной жидкой вакциной-аналогом против бешенства из штамма «ВНИИЗЖ».
Тестирование указанных вакцин проводили на десяти головах в возрасте 2 месяцев массой около 30 кг. Всем животным вводили вакцину внутримышечно однократно в объеме 2,0 см3. До иммунизации и спустя 21 сутки, а также через 2, 3, 6, 12, 15, 18 месяцев после иммунизации у собак отбирали кровь и полученные сыворотки исследовали на наличие антирабических антител в реакции нейтрализации (модификация FAVN) [2].
До вакцинации в крови свиней антитела против вируса бешенства не были выявлены. Количество вируснейтрализующих антител у свиней после введения одной рекомендованной для данного вида животных дозы (2,0 см3) антирабической инактивированной эмульсионной культуральной вакцины на 21 сутки составлял 4,50±0,31 МЕ/см3, через 2 месяца - 5,12±0,37 МЕ/см3, через 3 месяца - 5,08±0,44 МЕ/см3, через 6 месяцев - 3,98±0,45 МЕ/см3, через 12 месяцев - 1,85±0,29 МЕ/см3, через 15 месяцев - 0,63±0,10 МЕ/см3, через 18 месяцев - 0,08±0,01 МЕ/см3 (таблица 11). Таким образом, введение одной дозы (2,0 см3) антирабической инактивированной эмульсионной культуральной вакцины обеспечивает накопление достаточно высокого уровня антител (до 5,12±0,37 МЕ/см3) в крови собак и сохранение защитного уровня антител против вируса бешенства (≥0,50 МЕ/см3) до 15 месяцев.
Титр антирабических вируснейтрализующих антител в крови свиней после введения одной рекомендованной для них одной дозы (2,0 см3) культуральной инактивированной вакцины-аналога против бешенства из штамма «ВНИИЗЖ» на 21 сутки составлял 1,45±0,28 МЕ/см3, через 2 месяца - 1,86±0,29 МЕ/см3, через 3 месяца - 1,95±0,31 МЕ/см3, через 6 месяцев - 1,25±0,27 МЕ/см3, через 12 месяцев - 0,79±0,19 МЕ/см3, через 15 месяцев - 0,14±0,07 МЕ/см3, через 18 месяцев - 0,03±0,01 МЕ/см3. Таким образом, введение одной дозы (2,0 см3) культуральной инактивированной вакцины против бешенства из штамма «ВНИИЗЖ» (с применением адъюванта ISA-70) обеспечивает накопление защитного уровня антител в крови собак против вируса бешенства до 12 месяцев. При этом данная вакцина-аналог индуцирует выработку антирабических антител в 2,63 раза ниже по сравнению с разработанной вакциной.
На следующем этапе тестирования проводили оценку клеточного иммунитета у свиней после введения одной дозы (2,0 см3) представленной вакцины и вакцины-аналога.
Иммунизацию животных, отбор крови, выделение и анализ лимфоцитов проводили так же, как указано выше в примере 9. Динамика уровня субпопуляций CD4+ и CD8+ -клеток в крови свиней через 1, 2 и 3 недели после введения одной дозы (2,0 см3) антирабической инактивированной эмульсионной культуральной вакцины, культуральной инактивированной вакцины-аналога против бешенства из штамма «ВНИИЗЖ» в сравнении с контролем без вакцинации представлена на фиг. 15. Из данных фиг. 15 видно, что относительное содержание лимфоцитов CD4+ и CD8+ в крови свиней после введения предложенной вакцины увеличивалось и составляло 7-15 и 6-10%, соответственно. Количество клеток CD4+ и CD8+ после введения вакцины-аналога находилось на уровне значений контрольных животных без вакцинации и было равно 3-4%.
Уровень лимфоцитов CD4+/CD8+ после введения разработанной вакцины через 1, 2, 3 недели у свиней составлял 1,17, 1,67, 1,50, соответственно (фиг. 16). Данный факт свидетельствовал о том, что содержание клеток CD4+ увеличивалось с 1 до 2 недели и сохранялось на уровне 15% в течение 3 недели. Относительное количество лимфоцитов CD8+ с 1 по 3 недели оценки увеличивалось с 4 до 10% (фиг. 15). Выявлено, что введение в организм свиней антирабической инактивированной эмульсионной культуральной вакцины с применением масляного адъюванта Montanide ISA-61 VG активировало клеточный иммунитет, что выражалось в количественных изменениях фенотипа Т-клеток. После иммунизации свиней культуральной инактивированной вакциной-аналогом против бешенства из штамма «ВНИИЗЖ» с использованием адъюванта ISA-70 активации клеточного иммунитета не отмечали.
Протективные свойства предложенной антирабической инактивированной эмульсионной культуральной вакцины оценивали так, как отражено в примере 9. После контрольного заражения все 10 свиней, привитых одной дозой (2,0 см3) вакцины, оставались живыми и клинически здоровыми. В контрольной группе была отмечена гибель всех животных. Таким образом, разработанная антирабическая инактивированная эмульсионная культуральная вакцина является эффективной для иммунизации свиней.
Пример 14. Определение индекса иммуногенности и стабильности антирабической инактивированной эмульсионной культуральной вакцины при хранении в течение 30 месяцев в сравнении с вакциной-аналогом.
Срок сохранности иммуногенной потенции антирабической инактивированной эмульсионной культуральной вакцины и инактивированной жидкой вакцины-аналога против бешенства из штамма «ВНИИЗЖ» определяли в испытаниях с 5 экспериментальными сериями данных вакцин, хранившихся в течение 30 месяцев после изготовления при температуре 2,0-8,0°С. Тестирование вакцин проводились каждые 6 месяцев после изготовления. Определение индекса иммуногенности препаратов проводили так, как указано в примере 7 [2]. Результаты исследований разработанной вакцины и вакцины-аналога представлены в таблице 12, из которой видно, что через 12, 18, 24, 30 месяцев хранения в указанных условиях индекс иммуногенности разработанной вакцины снижался на 1,19; 2,00; 2,61; 3,44%, а вакцины-аналога - на 2,94; 4,80; 6,06; 7,36%, соответственно. Иными словами, предложенная вакцина позволяла в течение всего наблюдения сохранять высокий индекс иммуногенности.
При этом следует отметить, что все серии исследуемых вакцин в течение 24 месяцев сохраняли стабильность эмульсии. Отделение водной фазы отмечали только через 30 месяцев хранения при температуре 2,0-8,0°С.
Полученные результаты позволили сделать вывод, что антирабическая инактивированная эмульсионная культуральная вакцина гарантированно сохраняла свои физические и биологические свойства в течение 24 месяцев при условии хранения при температуре 2,0-8,0°С.
Таким образом, приведенная выше информация свидетельствует о том, что антирабическая инактивированная эмульсионная культуральная вакцина, воплощающая предлагаемое изобретение, предназначена для использования в сельском хозяйстве, а именно в ветеринарной вирусологии и биотехнологии; подтверждена возможность осуществления представленного; вакцина, изготовленная из штамма «ВНИИЗЖ», масляного адъюванта Montanide ISA-61 VG и мертиолята в соответствии с предлагаемым изобретением, обладает высокой иммуногенной активностью, способна активировать гуморальный и клеточный иммунитет у домашних плотоядных и сельскохозяйственных животных, обеспечивая их раннюю и длительную защиту против вируса бешенства генетической линии RABV.
Источники информации, принятые во внимание при составлении описания изобретения к заявке на выдачу патента РФ на изобретение «Вакцина антирабическая инактивированная эмульсионная культуральная для профилактической иммунизации домашних плотоядных и сельскохозяйственных животных»:
1. Груздев К.Н., Метлин А.Е. Бешенство животных. - Владимир: ФГБУ «ВНИИЗЖ», 2019. - 394 с.
2. OIE. Manual of diagnostic tests and vaccines for terrestrial animals. - Paris, 2018. - Chapter 3.1.17. - P. 578-612.
3. WHO Rabies vaccines: WHO position paper-recommendations. Vaccine. - 2010. - Vol. 28. - P. 7140-7142.
4. Метлин A.E. Современные аспекты классификации лиссавирусов // Ветеринария сегодня. - 2017. - №3 (22). - С. 52-57.
5. A single aminoacide change in rabies virus glycoprotein increasesvirus spread and enhances virus pathogenicity / M. Faber, M.L. Faber, A. Papanery [et al.] // J. Virology. - 2005. - Vol. 79 (22). - P. 14141-14148.
6. Greene, С.E. Rabies and other Lyssavirus infections / С.E. Greene // Infectious Diseases of the Dog and Cat // ed. C.E. Greene. - 4th ed. - Saunders, 2012. - Chap. 20. - P. 179-197.
7. Estimating the global burden of endemic canine rabies / K. Hampson, L. Coudelive, T. Lembo [et al.] // PLoS Negl. Trop. Dis. - 2015. - Vol. 9 (4):e0003709; DOI: 10/1371/jornal.pntd.0003709.
8. Nucleoprotein gene analysis of fixed and street rabies virus variants using RT-PCR / Y.T. Arai, K. Yamada, Y. Kameoka [et al.] // Arch. Virol. - 1997. - Vol. 142. - P. 1787-1796.
9. Груздев Л.К., Уласов В.И., Груздев К.Н. Экология вируса бешенства и проблемы контроля заболевания // Актуальные проблемы биологии и ветеринарной медицины мелких домашних животных: материалы Междунар. науч.-практ. конф. - Троицк, 2000. - С. 155.
10. Недосеков В.В. Современные вакцины против бешенства животных // Ветеринария. - 2003. - С. 23-25.
11. Barrat, J. Rabies diagnosis / J. Barrat, E. Picard-Meyer, F. Cliquet // Developments in Biologicals. - 2006. - Vol. 125. - P. 71-77.
12. Борисов A.B. Разработка технологии изготовления вирусвакцин против бешенства диких плотоядных: Автореф. дис. канд. вет. наук. - Владимир, 2003.
13. Сидорова Д.Г. Современные экологические особенности проявления эпизоотического процесса бешенства в природных очагах: автореф. дис. … канд. биол. наук. - Новосибирск, 2009. - 25 с.
14. Основные принципы специфической профилактики бешенства, обеспечивающие эффективность антирабических мероприятий / А.В. Саввин, Ю.В. Пашкина, К.Н. Груздев, В.В. Сочнев // Ветеринарная патология. - 2005. - №4. - С. 102-106.
15. Полюшкина Г.С., Груздев Л.К. Бешенство сельскохозяйственных и диких животных. // Тез. Докладов 1-й региональной конф. по болезням мелких домашних животных. Новосибирск, 1996. - С. 62.
16. Елаков А.Л. Антирабические вакцины для животных, применяемые в России // VetPharma. - 2013. - №4 (15). - С. 32-34.
17. Мовсесянц А.А., Агеенко Г.Б. Препараты против бешенства, применяемые в Российской Федерации // РЭТ-Инфо. - 2006. - №1. - С. 44-45.
18. Иванов, И.В. Антирабическая вакцина для крупного рогатого скота «РАБАВАК» и ее иммуногенность / И.В. Иванов // Научные основы производства вет. биол. препаратов: материалы Междунар. научно-практ. конф. - Щелково, 2005. - С. 94-96.
19. Сухарьков А.Ю. Разработка методов оценки оральной антирабической вакцинации животных: Автореф. дис. канд. биол. наук. - Владимир, 2014. - 25 с.
20. Патент 2366457 Российская Федерация, МПК A61K 39/205 (2006.01). Вакцина антирабическая для животных (УНИРЭВ) / Иванов B.C., Иванов И.В., Самуйленко А.Я. [и др.] - №2008107039/13; заявл. 27.02.2008; опубл. 10.09.2009, Бюл. №25.
21. Патент 2402348 Российская Федерация, МПК A61K 39/205 (2006.01). Вакцина антирабическая сухая для крупного и мелкого рогатого скота / Иванов B.C., Иванов И.В., Самуйленко А.Я [и др.]. - №2009117725/13; заявл. 13.05.2009; опубл. 27.10.2010, Бюл. №30.
22. Vinken J.P., Heng L., de Groot A., Gruppen H. Saponins, classification and occurance in plant kingdom // Phytochemistry. - 2007. - T. 68. - C. 275-297. - DOI: 10.1016/j.phytochem.2006.10.008.
23. Прохорова M.E. Бешенство не дремлет // Ветеринария и жизнь. - 2017. №3. - С. 2.
24. Вакцина антирабическая инактивированная культуральная жидкая (ВНИИЗЖ) [Электронный ресурс]. - Режим доступа: http://www.arriah.ru/main/production/price-vaccines/113/vakcina-antirabicheskaya-inaktivirovannaya-kulturalnaya-zhidkaya (Дата обращения: 23 сентября 2019 г.).
25. Влияние географических и антропогенных факторов на эпизоотический процесс бешенства. Профилактика и меры борьбы [Электронный ресурс]. - Режим доступа: https://studwood.ru/1992944/meditsina/profilaktika_mery_borby (Дата обращения: 23 сентября 2019 г. ).
26. Цеденхуу Пуревхуу. Культуральная инактивированная вакцина против бешенства из штаммов ВНИИЗЖ и ERA: Автореф. дис. канд. ветер, наук. - Владимир, 2005. - 27 с.
27. Сумм Б.Д. Основы коллоидной химии. М., - 2007. - 240 с.
28. MONTANIDE™ ISA for W/O emulsions - Application [Электронный ресурс]. - Режим доступа: https://www.seppic.com/montanide-isa-w-o (Дата обращения: 14 сентября 2019 г.).
29. Бирюков В.В. Основы промышленной биотехнологии. М.: КолосС. - 2004. - 296 с.
30. Комиссаров А.В. Применение ультрафильтрации для концентрирования и очистки антигенов / А.В. Комиссаров, Ю.А. Алешина, О.В. Громова [и др.] // Проблемы особо опасных инфекций. - 2015. - №1. - С. 79-84.
31. Perry, L.L., Lodmell, D.L. (1991). Role of CD4+ and CD8+ T-cells in murine resistance to Street Rabies Virus // J. Virol. - 1991. - Vol. 65(7). - P. 3429-3434.
32. Rabies lyssavirus. Links from Taxonomy [Электронный ресурс]. - Режим доступа: https://www.ncbi.nlm.nih.gov/genome?LinkName=taxonomy_genome&from_uid=11292 (Дата обращения: 07 сентября 2019 г. ).
33. Юрков С.Г., Зуев В.В. Каталог коллекции клеточных культур ВНИИВВиМ - Покров. - 2000. - С. 62.
34. Rabies / ed. А.С. Jackson, W.Н. Wunner. - 2nd ed. - London: Academic Press, 2007. - 660 p.
35. Таблицы вязкости [Электронный ресурс]. - Режим доступа: http://uodn.ru/docs/vyzkos (Дата обращения: 10 сентября 2019 г.).
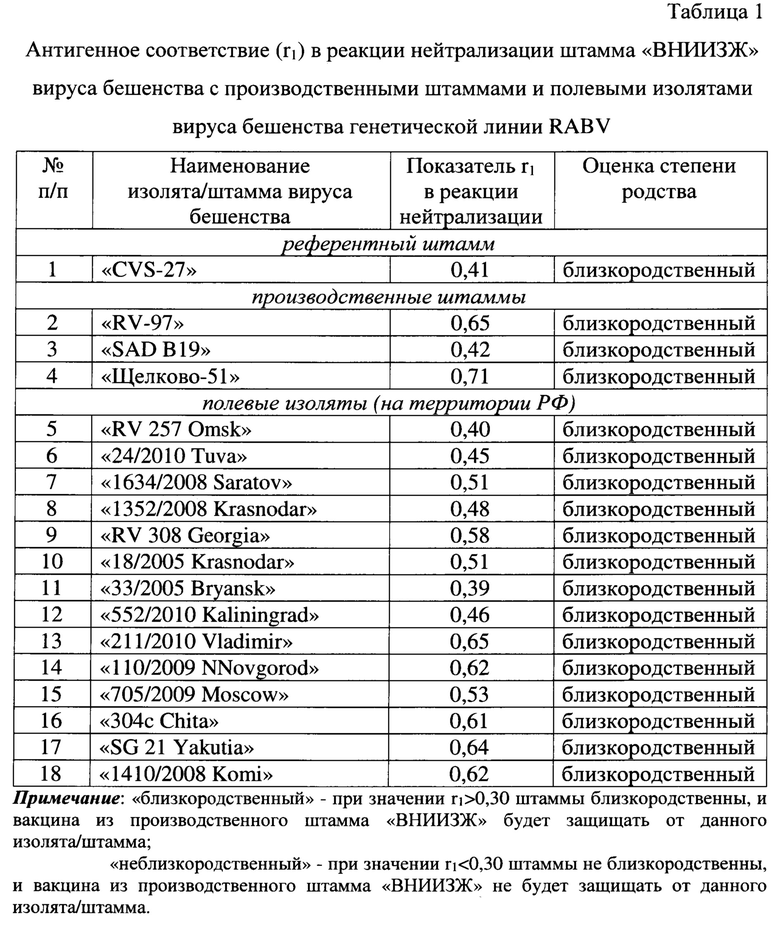
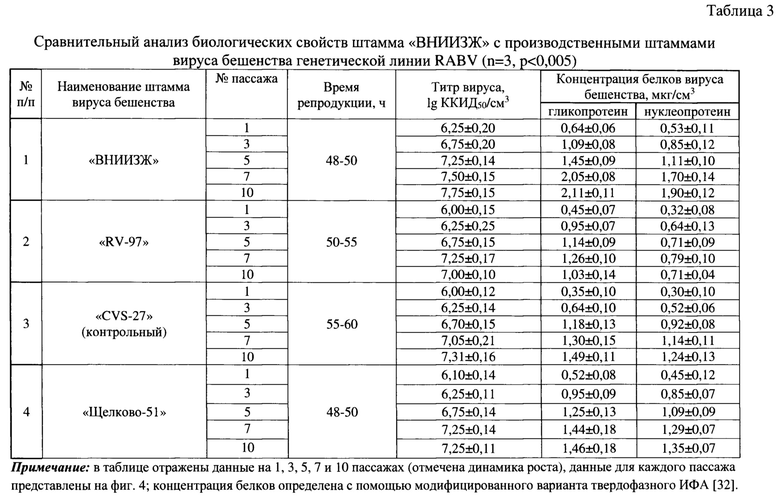
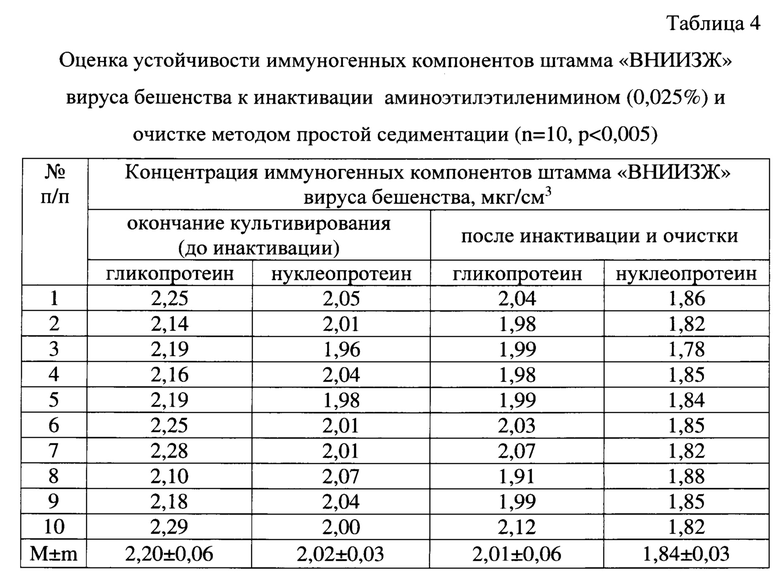
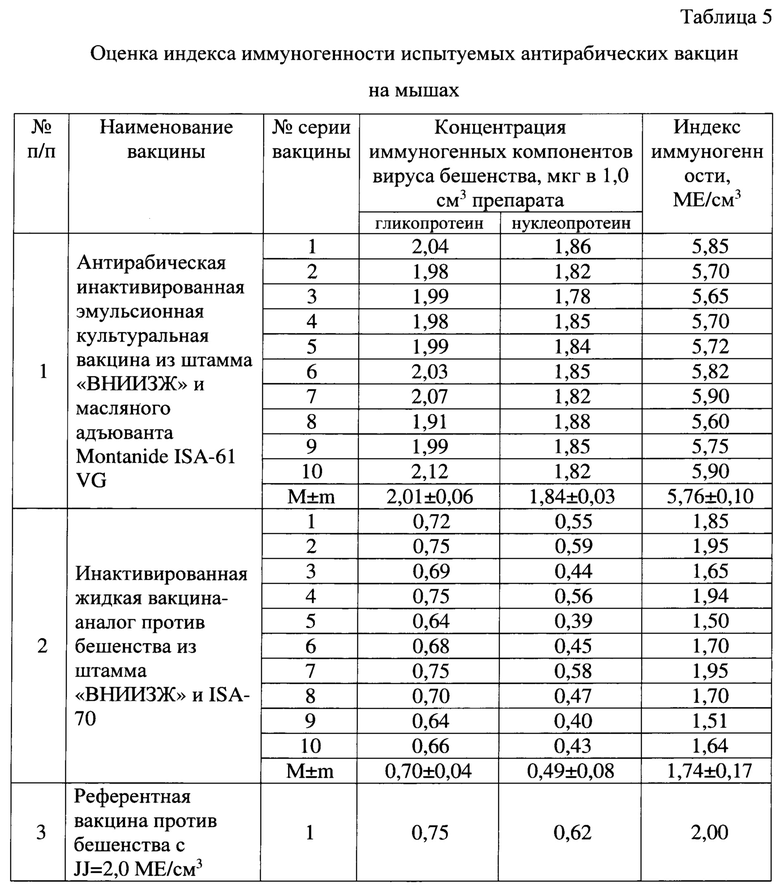
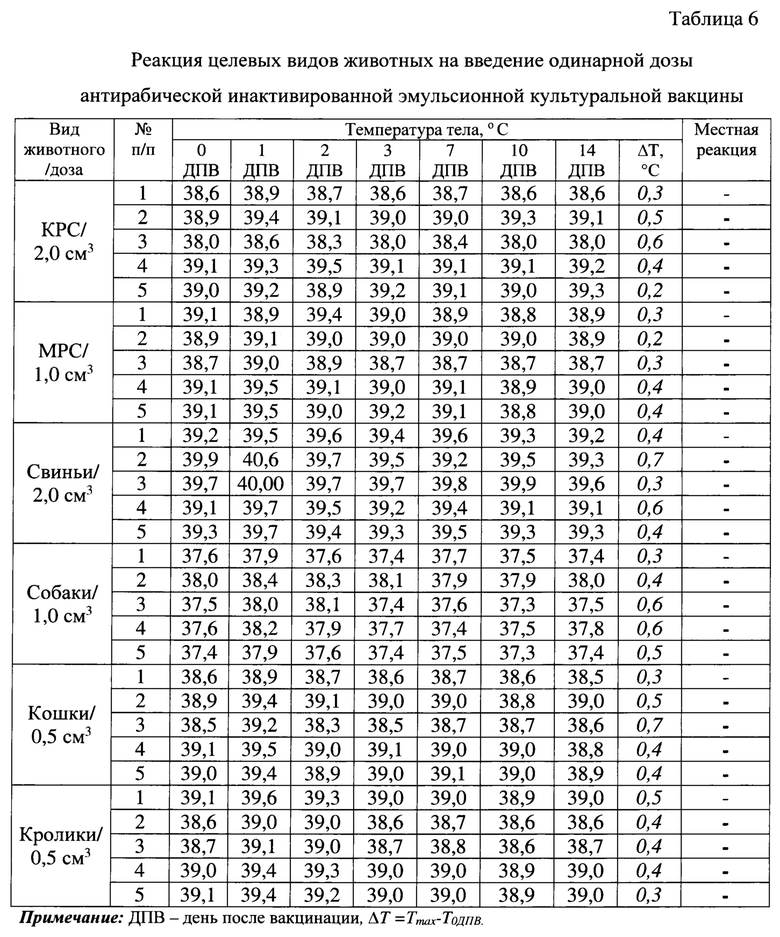
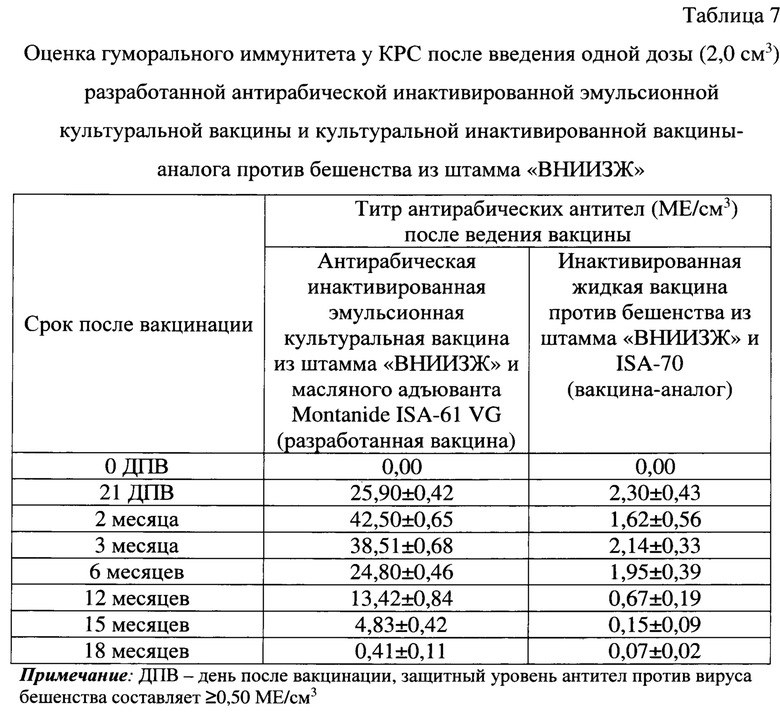
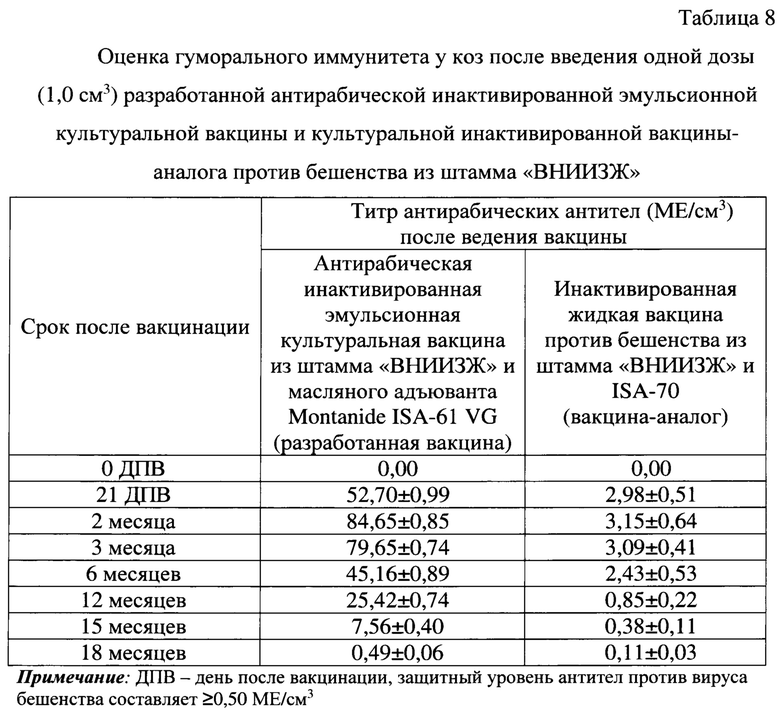
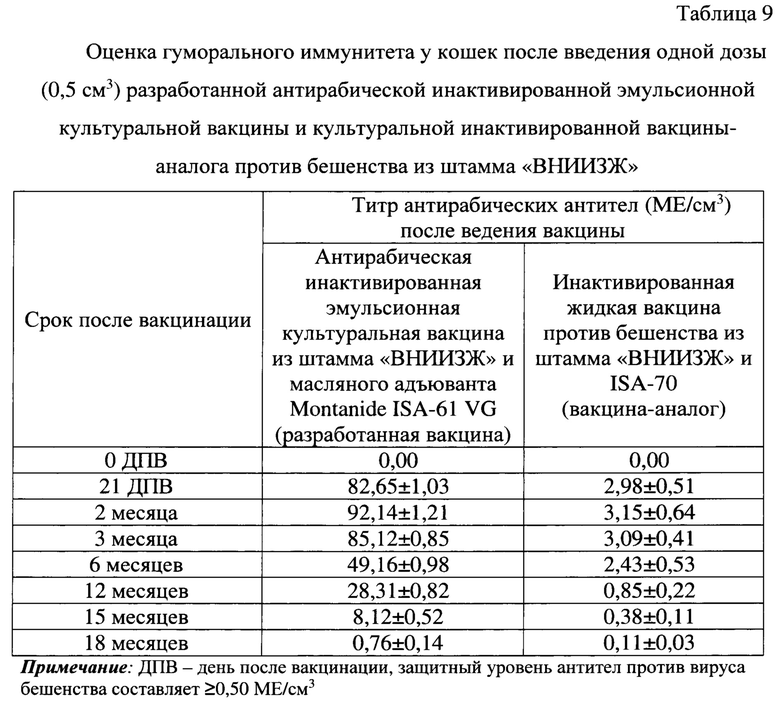
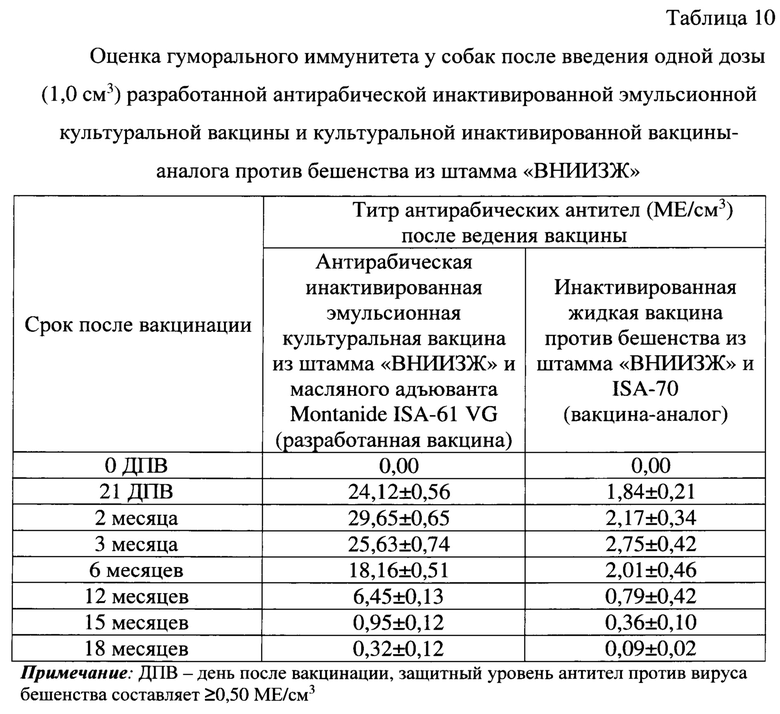
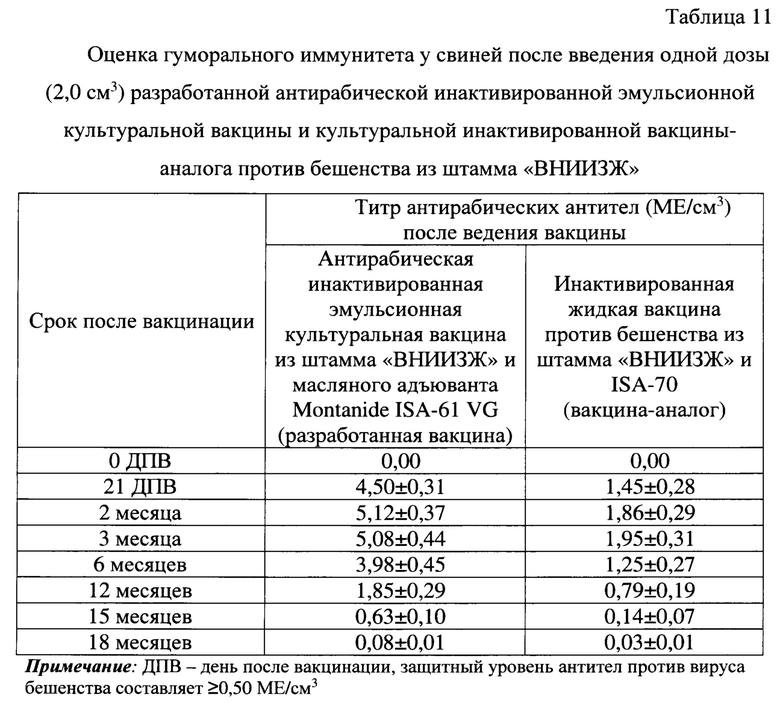
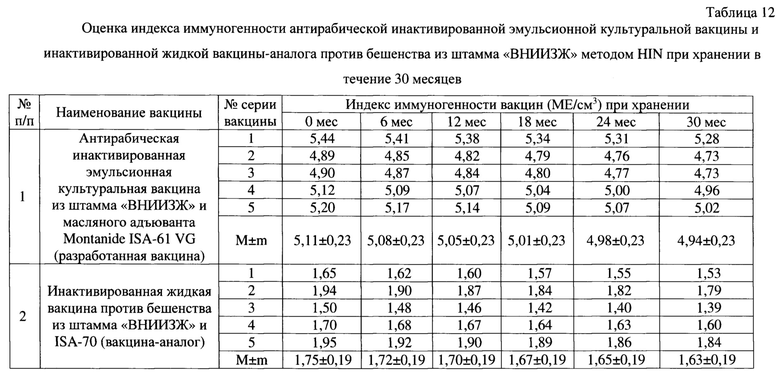
--->
Перечень последовательностей
<110> Федеральное государственное бюджетное учреждение «Федеральный центр охраны здоровья животных» (ФГБУ «ВНИИЗЖ») Federal Governmental Budgetary Institution «Federal Centre for Animal Health» (FGBI «ARRIAH»)
<120> Вакцина антирабическая инактивированная эмульсионная культуральная для профилактической иммунизации домашних плотоядных и сельскохозяйственных животных
<160> 4
<210> 1
<211> 1350
<212> ДНК
<213> вирус бешенства
<400> 1
1 atggatgccg acaagattgt cttcaaagtc aataatcagg tggtctcttt gaagcctgag 60
61 atcatagtgg atcaatatga gtacaagtac cctgctatca aagatttgaa aaagccctgt 120
121 ataaccctag ggaaagcccc cgacttgaac aaagcataca agtcaatttt atcaggcatg 180
181 aatgctgcca agctagatcc tgatgatgta tgttcctact tggcagcggc aatgcagttc 240
241 tttgagggga cgtgtccgga agactggacc agctatggaa tcttgattgc acgaaaagga 300
301 aacaagatca ccccggattc tctggtggaa ataaagcgta ctgatgtaga aggaaattgg 360
361 gctctgacag gaggcatgga attgactagg gaccccactg tctctgaaca tgcgtctttg 420
421 gtcggtcttc tcttgagtct gtataggttg agcaaaatat caggacaaaa cactggtaac 480
481 tataaaacaa acatcgcaga taggatagag cagatctttg agaccgcccc ttttgttaaa 540
541 atcgtggagc accacactct aatgacgact cacaaaatgt gtgctaattg gagtactata 600
601 ccgaatttca gatttttggc cgggacctac gacatgtttt tctcccggat tgagcatcta 660
661 tattcagcaa tcagagtggg cacggttgtt actgcttatg aagactgttc aggtctggta 720
721 tcatttacag ggttcataaa gcagatcaat ctcaccgcaa gagaagcaat actatacttc 780
781 ttccacaaga actttgagga agagataaga agaatgtttg agccagggca ggagacagct 840
841 gttcctcact cctatttcat ccacttccgt tcactaggct tgagcgggaa gtctccttat 900
901 tcatcaaatg ctgttggtca tgtgttcaat ctcattcact ttgttggatg ctatatgggt 960
961 caagtcagat ccctaaacgc aacagttatt gccgcatgtg ctcctcatga gatgtctgtt 1020
1021 ttagggggct atctgggaga agagttcttc gggaaaggaa cattcgagag aagattcttc 1080
1081 agagatgaga aagaacttca agaatacgag gcagctgaac tgacaaagac tgacgtggct 1140
1141 ttggcagatg atggaactgt ccactcggac gatgaggact acttttccgg agagaccaga 1200
1201 agtccagaag ccgtttatac tcgaatcatg atgaatggag gtcgactaaa gagattgcat 1260
1261 atacggagat atgtttcagt cagttcaaat catcaagctc gtcctaactc atttgccgag 1320
1321 tttctaaaca agacgtattc gaacgactcс 1350
<210> 2
<211> 450
<212> ПРТ
<213> вирус бешенства
<400> 2
1 MetAspAlaAspLysIleValPheLysValAsnAsnGlnValValSerLeuLysProGlu 20
21 IleIleValAspGlnTyrGluTyrLysTyrProAlaIleLysAspLeuLysLysProCys 40
41 IleThrLeuGlyLysAlaProAspLeuAsnLysAlaTyrLysSerIleLeuSerGlyMet 60
61 AsnAlaAlaLysLeuAspProAspAspValCysSerTyrLeuAlaAlaAlaMetGlnPhe 80
81 PheGluGlyThrCysProGluAspTrpThrSerTyrGlyIleLeuIleAlaArgLysGly 100
101 AsnLysIleThrProAspSerLeuValGluIleLysArgThrAspValGluGlyAsnTrp 120
121 AlaLeuThrGlyGlyMetGluLeuThrArgAspProThrValSerGluHisAlaSerLeu 140
141 ValGlyLeuLeuLeuSerLeuTyrArgLeuSerLysIleSerGlyGlnAsnThrGlyAsn 160
161 TyrLysThrAsnIleAlaAspArgIleGluGlnIlePheGluThrAlaProPheValLys 180
181 IleValGluHisHisThrLeuMetThrThrHisLysMetCysAlaAsnTrpSerThrIle 200
201 ProAsnPheArgPheLeuAlaGlyThrTyrAspMetPhePheSerArgIleGluHisLeu 220
221 TyrSerAlaIleArgValGlyThrValValThrAlaTyrGluAspCysSerGlyLeuVal 240
241 SerPheThrGlyPheIleLysGlnIleAsnLeuThrAlaArgGluAlaIleLeuTyrPhe 260
261 PheHisLysAsnPheGluGluGluIleArgArgMetPheGluProGlyGlnGluThrAla 280
281 ValProHisSerTyrPheIleHisPheArgSerLeuGlyLeuSerGlyLysSerProTyr 300
301 SerSerAsnAlaValGlyHisValPheAsnLeuIleHisPheValGlyCysTyrMetGly 320
321 GlnValArgSerLeuAsnAlaThrValIleAlaAlaCysAlaProHisGluMetSerVal 340
341 LeuGlyGlyTyrLeuGlyGluGluPhePheGlyLysGlyThrPheGluArgArgPhePhe 360
361 ArgAspGluLysGluLeuGlnGluTyrGluAlaAlaGluLeuThrLysThrAspValAla 380
381 LeuAlaAspAspGlyThrValHisSerAspAspGluAspTyrPheSerGlyGluThrArg 400
401 SerProGluAlaValTyrThrArgIleMetMetAsnGlyGlyArgLeuLysArgLeuHis 420
421 IleArgArgTyrValSerValSerSerAsnHisGlnAlaArgProAsnSerPheAlaGlu 440
441 PheLeuAsnLysThrTyrSerAsnAspSer 450
<210> 3
<211> 540
<212> ДНК
<213> вирус бешенства
<400> 1
1 ccgaagatca cctccccttt aagtttgctt tattgggggg aatctccggg ttcaagagtc 60
61 ctccttgaac tccatgcgac agagtatatt caagagtcat gtgactctca ttattcctct 120
121 cagttgatca aatcaagtca tgtagattct cataacgcgg gagatcttct agcagtttca 180
181 gtgcccaact gtgctttcat tctccaggaa ctaatactaa gggttgtgga cggaccaaga 240
241 ggtatctccg atgatcccgt gcttgggcac gggtataggt catattacgt cccacgatat 300
301 cggactccac atgaaccgat tgagaaaggc aatctgcctc ccatgaagga cgcaagtaat 360
361 agctcacaat cgtcttgcat ctcagcgaag tatgtatatt tatatagagc tgggtcctct 420
421 aagcttttca gtcaagaaaa aaactgtaga tcaaaagagc aactggcaac acttctcatc 480
481 ctaagaccta catcaagatg ctatatccag gagaggtcta tgatgacccc attgatccaa 540
<210> 4
<211> 180
<212> ПРТ
<213> вирус бешенства
<400> 2
1 ProLysIleThrSerProLeuSerLeuLeuTyrTrpGlyGluSerProGlySerArgVal 20
21 LeuLeuGluLeuHisAlaThrGluTyrIleGlnGluSerCysAspSerHisTyrSerSer 40
41 GlnLeuIleLysSerSerHisValAspSerHisAsnAlaGlyAspLeuLeuAlaValSer 60
61 ValProAsnCysAlaPheIleLeuGlnGluLeuIleLeuArgValValAspGlyProArg 80
81 GlyIleSerAspAspProValLeuGlyHisGlyTyrArgSerTyrTyrValProArgTyr 100
101 ArgThrProHisGluProIleGluLysGlyAsnLeuProProMetLysAspAlaSerAsn 120
121 SerSerGlnSerSerCysIleSerAlaLysTyrValTyrLeuTyrArgAlaGlySerSer 140
141 LysLeuPheSerGlnGluLysAsnCysArgSerLysGluGlnLeuAlaThrLeuLeuIle 160
161 LeuArgProThrSerArgCysTyrIleGlnGluArgSerMetMetThrProLeuIleGln 180
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Бессывороточная среда "ВНИИЗЖ" для культивирования клеток почки сирийского хомячка ВНК-21 и получения иммуногенных компонентов культуральных вирусов ящура и бешенства для изготовления вакцин | 2021 |
|
RU2770814C1 |
| Ассоциированная вакцина против панлейкопении, калицивироза, вирусного ринотрахеита и бешенства кошек | 2024 |
|
RU2827230C1 |
| Ассоциированная вакцина против чумы плотоядных, парвовирусного и коронавирусного энтеритов, аденовирусной инфекции и бешенства собак | 2023 |
|
RU2817255C1 |
| Вакцина для ранней защиты против ящура типа О инактивированная эмульсионная | 2019 |
|
RU2699671C1 |
| Вакцина для ранней защиты против ящура из штамма А 2205/G IV культуральная инактивированная эмульсионная | 2021 |
|
RU2772713C1 |
| Вакцина инактивированная эмульсионная против ящура типа А | 2017 |
|
RU2665850C1 |
| Вакцина инактивированная эмульсионная против ящура типа О | 2017 |
|
RU2682876C1 |
| Вакцина для ранней защиты против ящура типа Азия-1 инактивированная эмульсионная | 2020 |
|
RU2741639C1 |
| Вакцина против ящура генотипа O/ME-SA/Ind-2001e из штамма "О N2620/Оренбургский/2021" культуральная инактивированная эмульсионная | 2023 |
|
RU2815537C1 |
| Вакцина против ящура серотипа О из штамма «O/ARRIAH/Mya-98» культуральная инактивированная эмульсионная | 2023 |
|
RU2816944C1 |
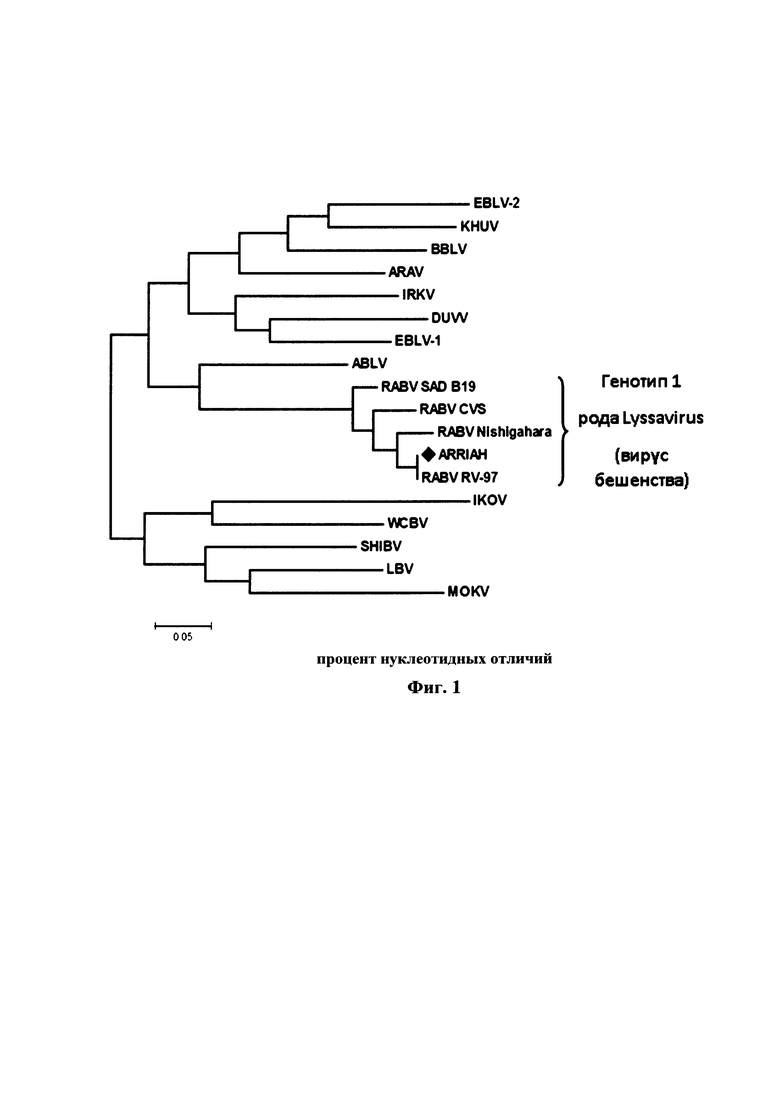
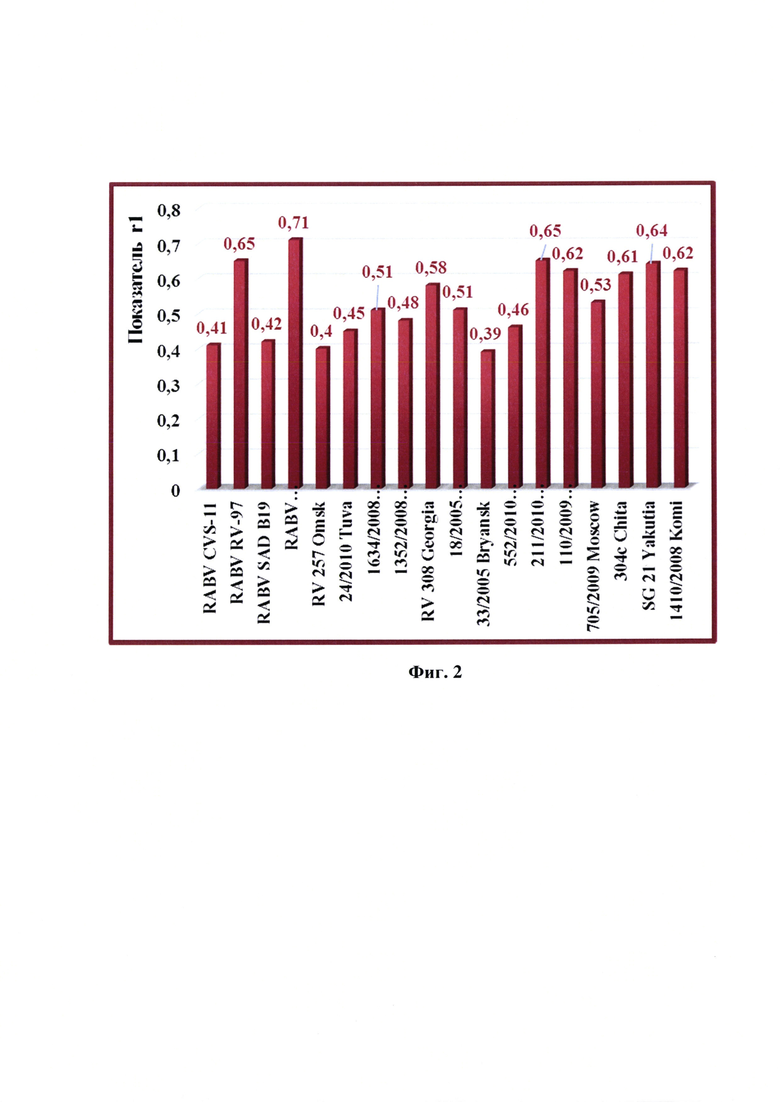
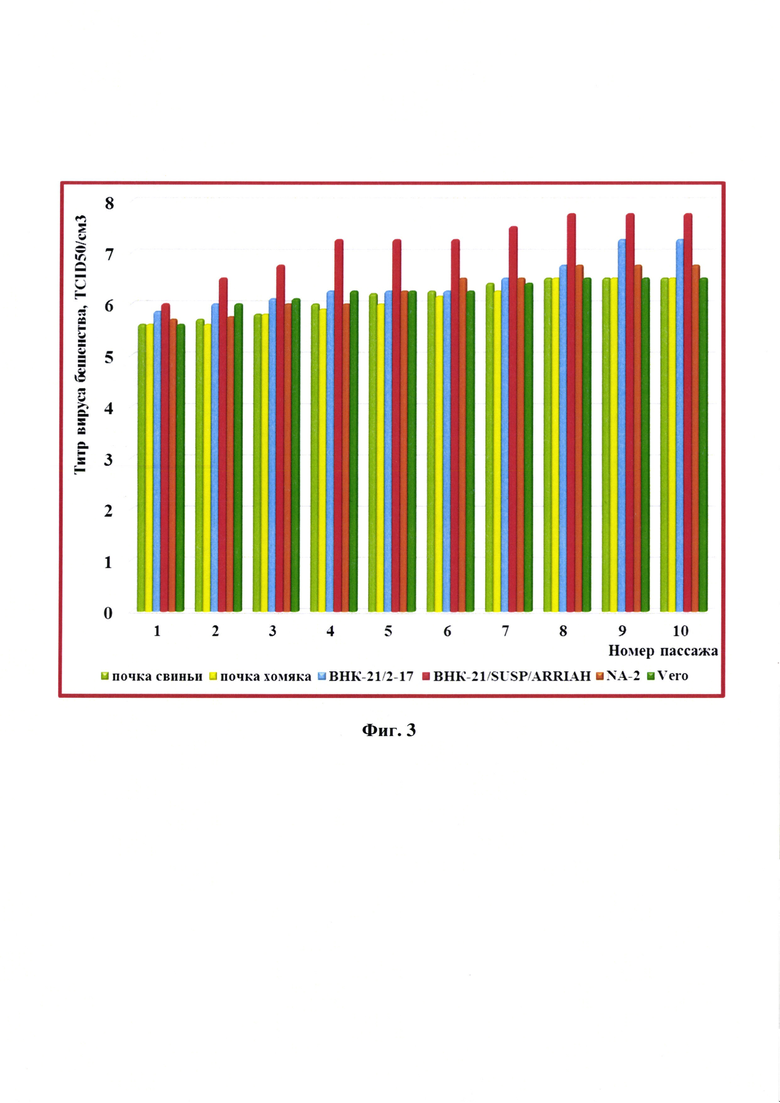


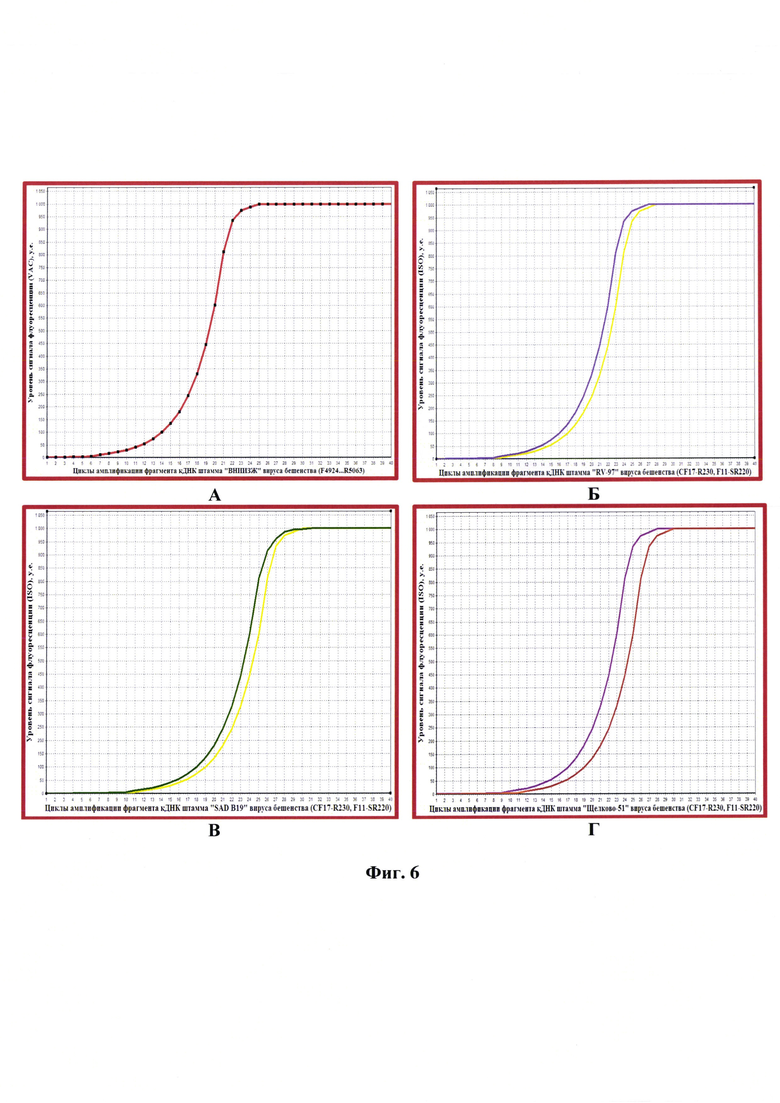
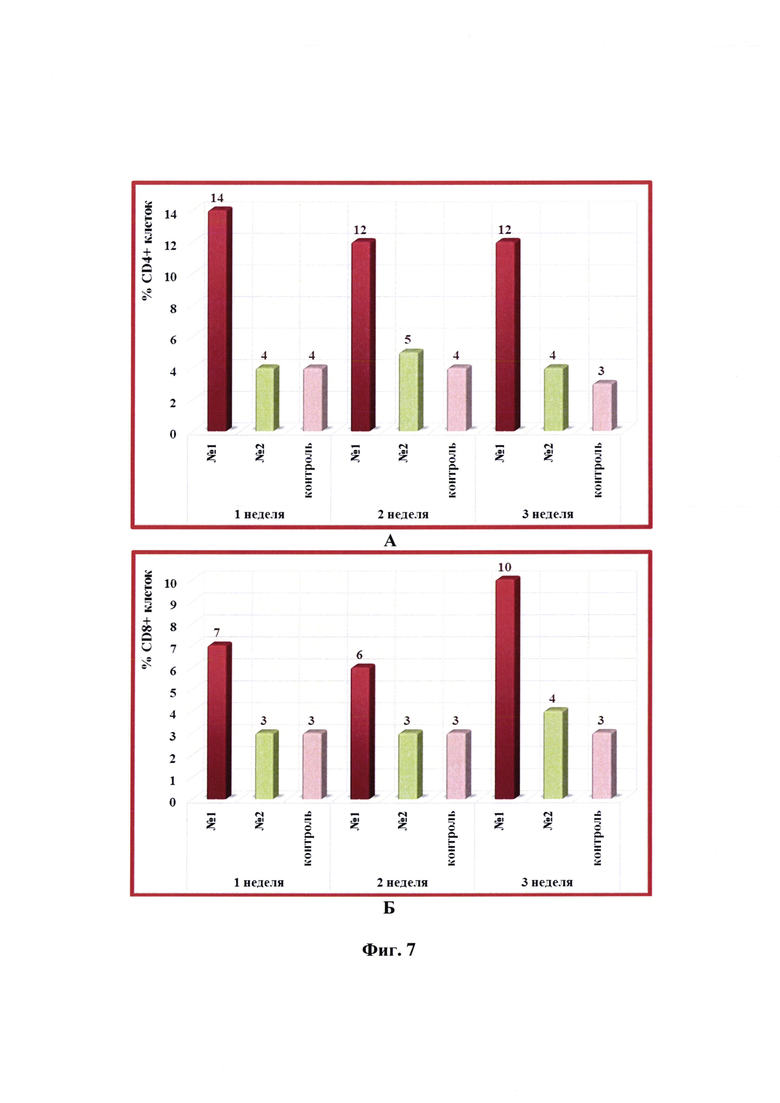

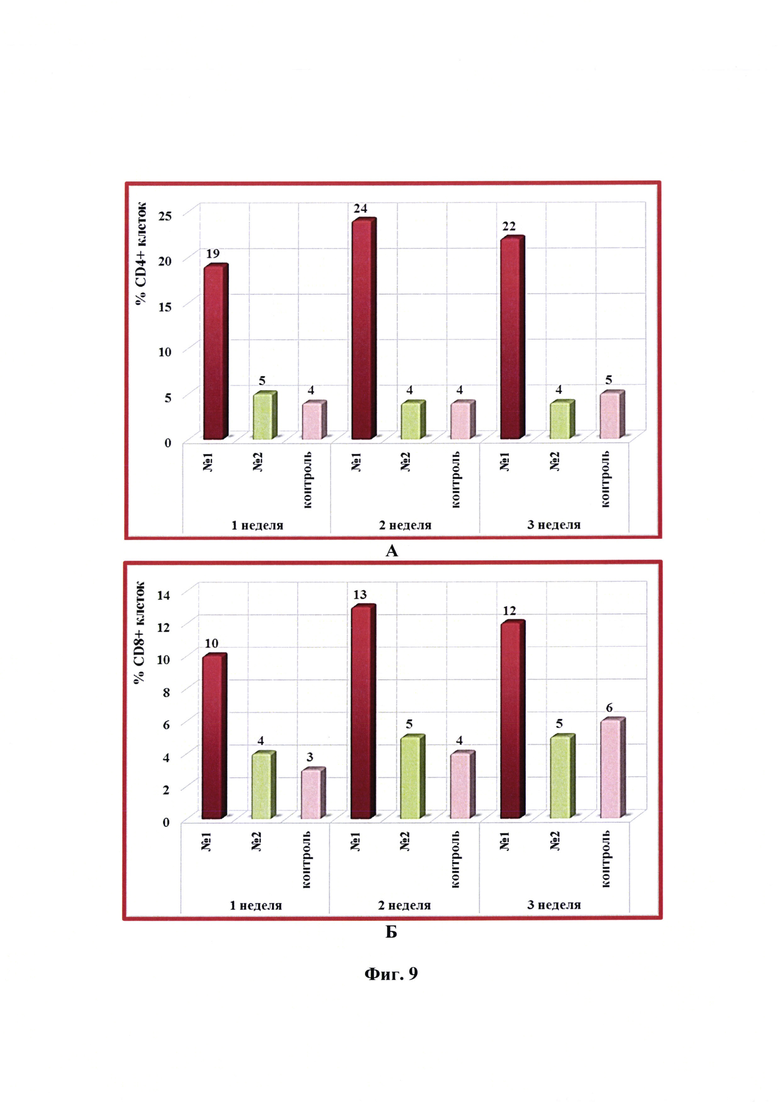
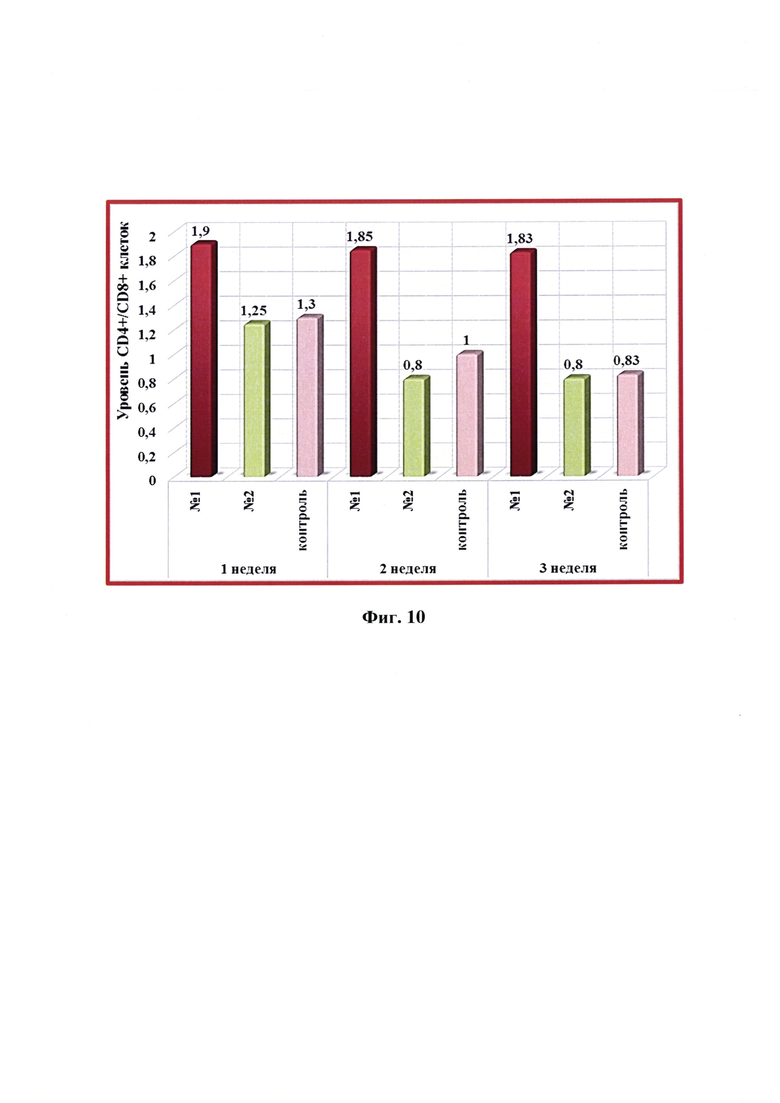
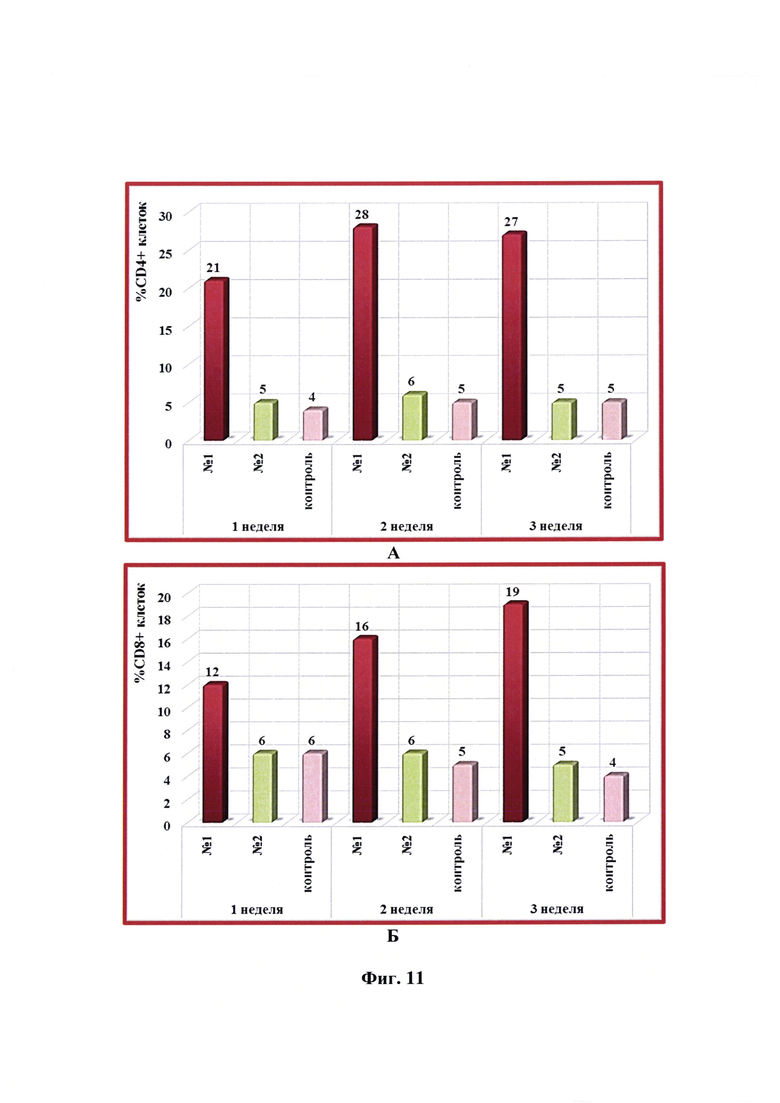
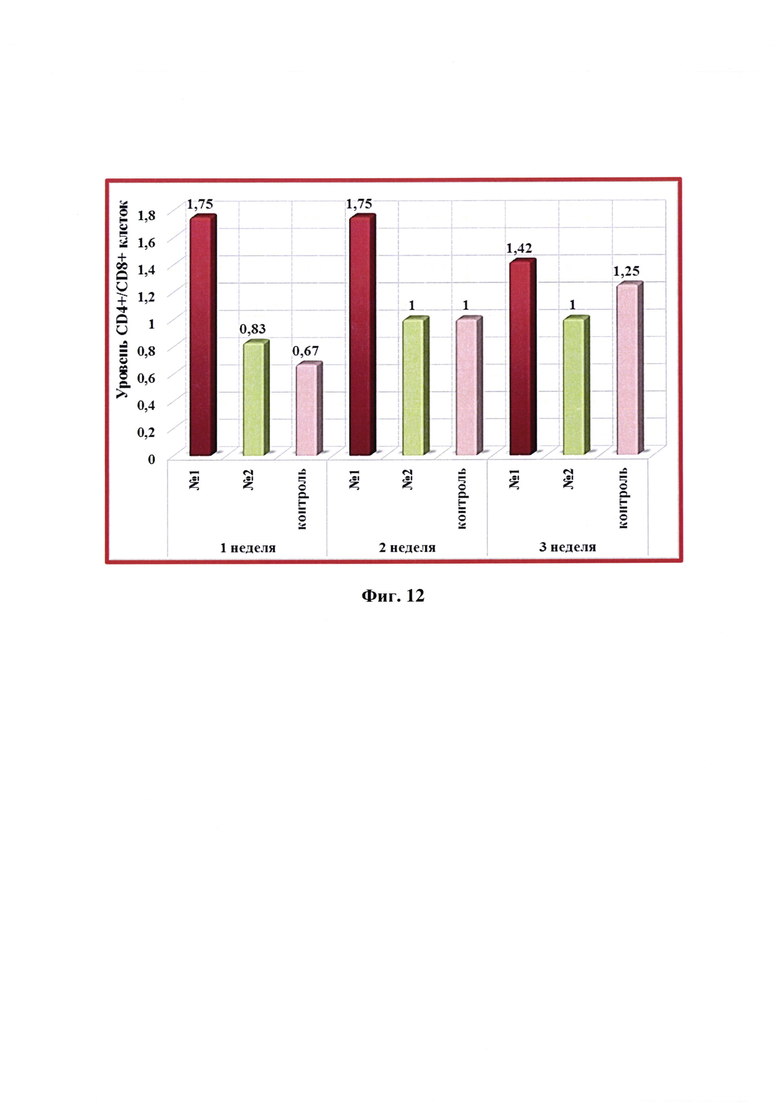
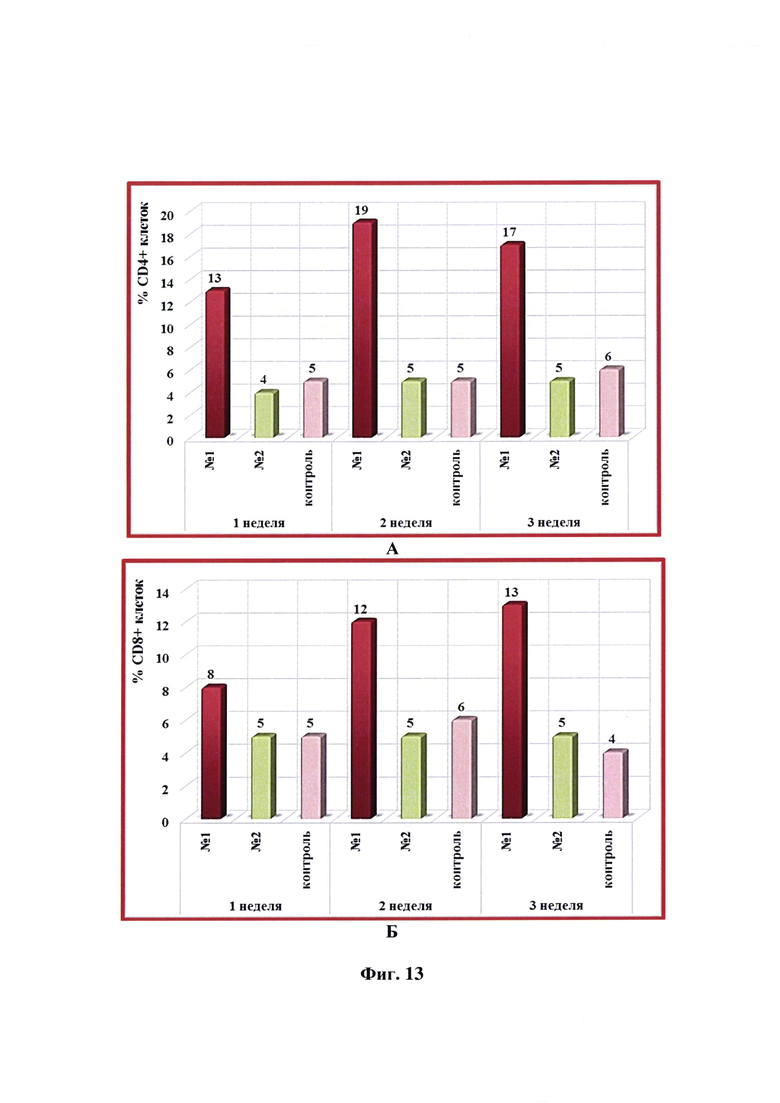
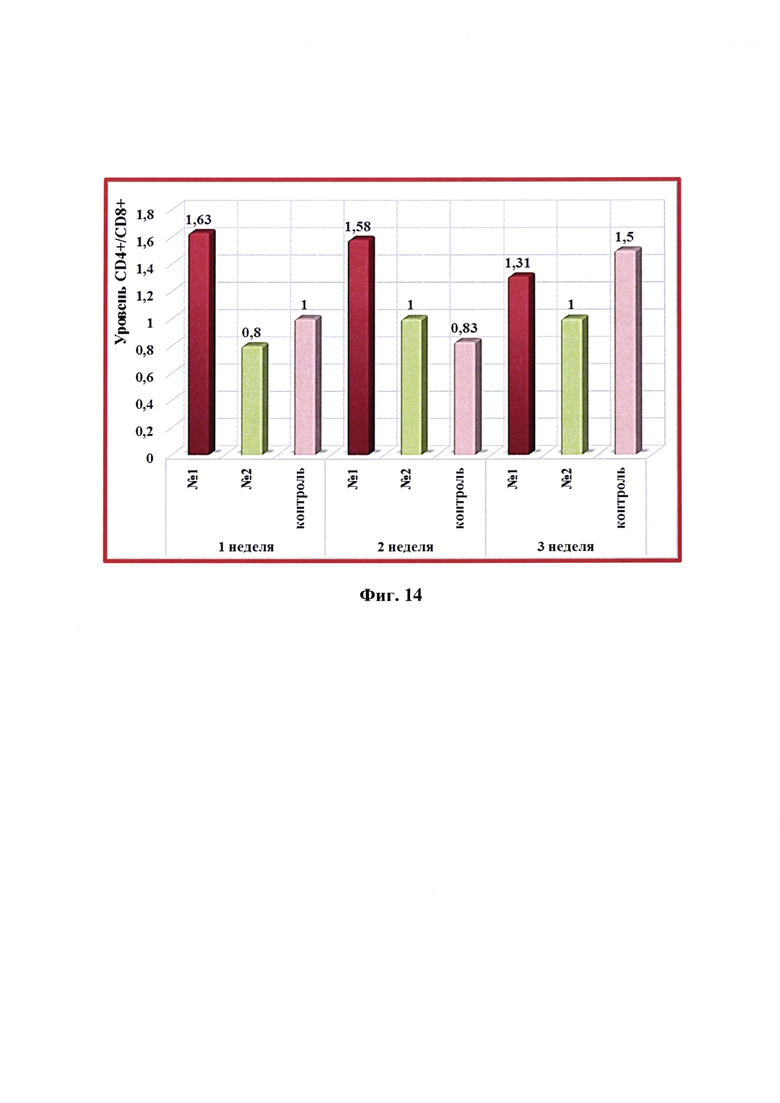
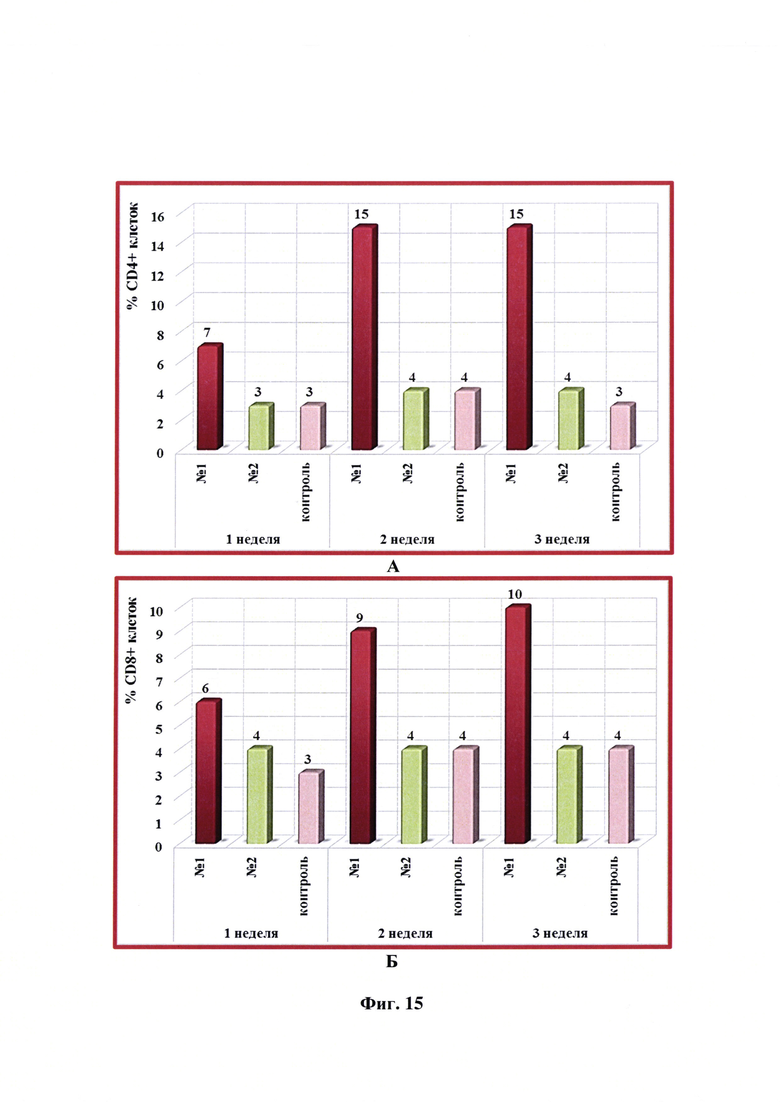
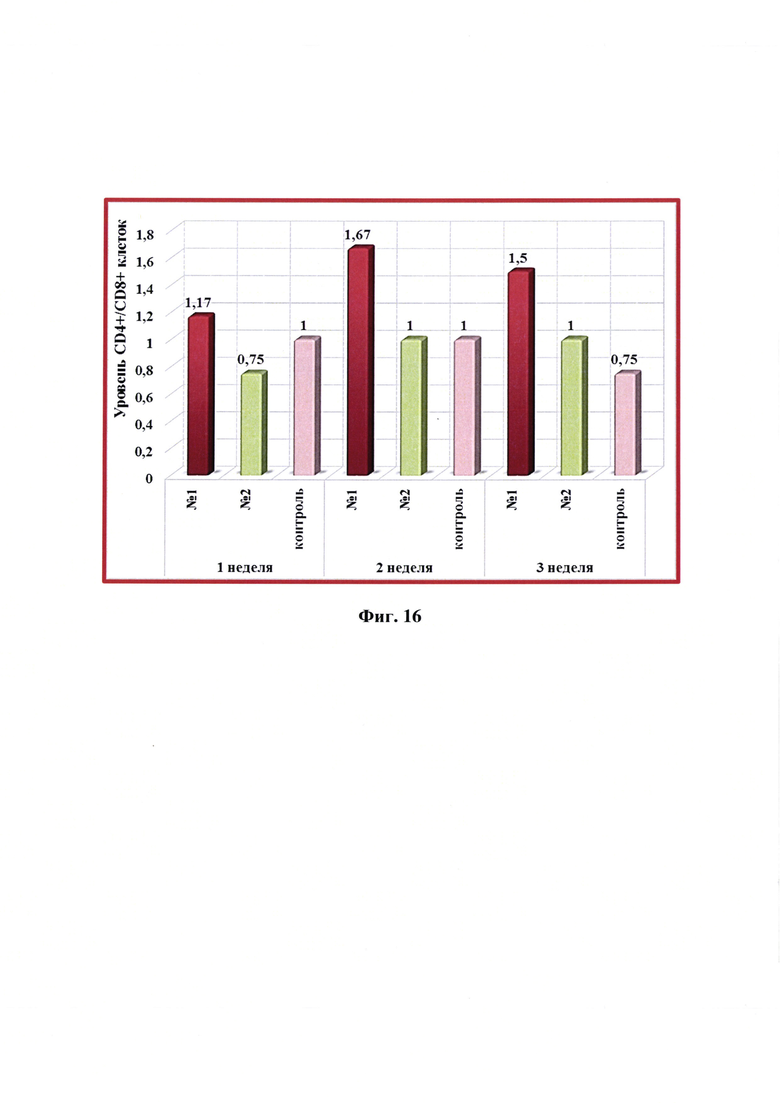
Изобретение относится к области биотехнологии, а именно к разработке вакцины антирабической инактивированной эмульсионной культуральной для профилактической иммунизации домашних плотоядных и сельскохозяйственных животных. Вакцина содержит авирулентный и очищенный антигенный материал из штамма «ВНИИЗЖ» вируса бешенства, полученный в перевиваемой суспензионной клеточной линии из почки новорожденного сирийского хомячка (BHK-21/SUSP/ARRIAH), представляющий собой суспензию, содержащую преимущественно иммуногенные компоненты вируса бешенства (гликопротеин и нуклеопротеин), масляный адъювант Montanide ISA-61 VG, консервант мертиолят в эффективных соотношениях. Вакцина обладает высокой иммуногенностью и способна обеспечить длительную и надежную защиту от вируса бешенства генетической линии RABV. 9 з.п. ф-лы, 16 ил., 12 табл., 14 пр.
1. Вакцина антирабическая инактивированная эмульсионная культуральная для профилактической иммунизации домашних плотоядных и сельскохозяйственных животных, содержащая активное вещество, адъювант, консервант, отличающаяся тем, что в качестве активного вещества она содержит авирулентный и очищенный антигенный материал из штамма «ВНИИЗЖ» вируса бешенства, семейства Rhabdoviridae, рода Lyssavirus, вида Rabies lyssavirus, депонированого в Коллекцию штаммов микроорганизмов ФГБУ «Федеральный центр охраны здоровья животных» (ФГБУ «ВНИИЗЖ»), под регистрационным номером «ВНИИЗЖ»-ДЕП фиксированного вируса бешенства от 20.12.2018 г., в эффективном количестве.
2. Вакцина по п. 1, отличающаяся тем, что она содержит авирулентный и очищенный антигенный материал из гомологичного возбудителю инфекции штамма «ВНИИЗЖ» вируса бешенства, полученный в чувствительной биологической системе и представляющий собой суспензию, содержащую преимущественно иммуногенные компоненты (гликопротеин и нуклеопротеин) вируса бешенства генетической линии RABV.
3. Вакцина по п. 2, отличающаяся тем, что она содержит авирулентный и очищенный антигенный материал из гомологичного возбудителю инфекции штамма «ВНИИЗЖ» вируса бешенства, полученный предпочтительно в перевиваемой суспензионной клеточной сублинии ВНК-21/SUSP/ARRIAH и представляющий собой суспензию, содержащую преимущественно иммуногенные компоненты (гликопротеин и нуклеопротеин) вируса бешенства генетической линии RABV.
4. Вакцина по п. 3, отличающаяся тем, что она содержит авирулентный и очищенный антигенный материал из гомологичного возбудителю инфекции штамма «ВНИИЗЖ» вируса бешенства, полученный предпочтительно в перевиваемой суспензионной клеточной сублинии из почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH и представляющий собой суспензию, содержащую преимущественно иммуногенные компоненты вируса бешенства генетической линии RABV: гликопротеин в количестве не менее 1,5 мкг и нуклеопротеин в количестве не менее 1,5 мкг в 1,0 см3 готового препарата, что соответствует индексу иммуногенности вакцины не менее 4,5 МЕ/см.
5. Вакцина по п. 1, отличающаяся тем, что в качестве модулятора иммунного ответа она содержит масляный адъювант Montanide ISA-61 VG.
6. Вакцина по п. 5, отличающаяся тем, что содержит масляный адъювант Montanide ISA-61 VG предпочтительно в количестве 575000,0-700000,0 мкг в 1,0 см3 готового препарата.
7. Вакцина по п. 5, отличающаяся тем, что содержит масляный адъювант Montanide ISA-61 VG предпочтительно в количестве 575000,0-700000,0 мкг в 1,0 см3 готового препарата и активирует гуморальный и клеточный иммунитет у домашних плотоядных и сельскохозяйственных животных.
8. Вакцина по п. 1, отличающаяся тем, что в качестве консерванта содержит мертиолят.
9. Вакцина по п. 8, отличающаяся тем, что в качестве консерванта содержит мертиолят в количестве 0,002% (по весу).
10. Вакцина по любому из пп. 1-9, отличающаяся тем, что она содержит авирулентный и очищенный антигенный материал из гомологичного возбудителю инфекции штамма «ВНИИЗЖ» вируса бешенства, полученный предпочтительно в перевиваемой суспензионной клеточной сублинии из почки новорожденного сирийского хомячка ВНК-21/SUSP/ARRIAH и представляющий собой суспензию, содержащую преимущественно иммуногенные компоненты вируса бешенства генетической линии RABV, масляный адъювант Montanide ISA-61 VG, консервант мертиолят в соотношении, мкг в 1,0 см3 готового препарата:

| ЦЕДЕНХУУ ПУРЕВХУУ, Культуральная инактивированная вакцина против бешенства из штаммов "ВНИИЗЖ" и "ERA", автореферат диссертации, Владимир, 2005 | |||
| ВАКЦИНА АНТИРАБИЧЕСКАЯ ДЛЯ ЖИВОТНЫХ (УНИРЭВ) | 2008 |
|
RU2366457C1 |
| FABER M., et al, A single amino acid change in rabies virus glycoprotein increases virus spread and enhances virus pathogenicity, Journal of virology, Nov.2005, p.14141-14148, | |||

