ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
[0001] Настоящее раскрытие относится к композиции против инфекции Mycoplasma hyorhinis, в частности, к субъединичной вакцине против инфекции Mycoplasma hyorhinis.
ОПИСАНИЕ УРОВНЯ ТЕХНИКИ
[0002] Mycoplasma hyopneumoniae и Mycoplasma hyorhinis представляют собой основные патогены, вызывающие энзоотическую пневмонию свиней, SEP. Mycoplasma hyorhinis также участвует во вспышках полисерозита или артрита. Инфекция или Mycoplasma hyopneumoniae или Mycoplasma hyorhinis снижает эффективность кормления и вызывает замедление роста свиней и также делает свиней более уязвимыми для вторичной инфекции других вирусных или бактериальных патогенов, что, следовательно, ведет к экономическим потерям в свиноводстве. В мире производителями вакцин для животных разработаны инактивированные (убитые) вакцины против Mycoplasma hyopneumoniae, которые широко используются в свиноводстве. Однако ни одна вакцина против Mycoplasma hyorhinis не появилась на рынке до сих пор. Введение инактивированных вакцин против Mycoplasma hyopneumoniae может лишь предотвращать инфекцию Mycoplasma hyopneumoniae, но не может защищать свиней от инфекции Mycoplasma hyorhinis.
[0003] Для того чтобы решить проблему недостаточной защиты у существующих вакцин, необходимо разработать вакцину против Mycoplasma hyorhinis. Стандартные вакцины в этой области преимущественно представляют собой инактивированные (убитые) вакцины. Поскольку Mycoplasma spp. нелегко поддаются культивированию, среда для их культивирования является дорогостоящей, и концентрация Mycoplasma spp., получаемых из нее, не достаточно высока, стоимость изготовления инактивированной (убитой) вакцин против Mycoplasma hyorhinis остается высокой. Следовательно, субъединичную вакцину, которую легко получать и которая обладает высокой безопасностью, рассматривают в качестве другой возможности для разработки вакцины. До настоящего времени, антигены, пригодные для использования в вакцинах против Mycoplasma hyorhinis, не предложены ни в каком хорошо обоснованном отчете об исследованиях в этой области. Ввиду вышесказанного, основная цель настоящего раскрытия состоит в том, чтобы разработать дешевую и эффективную субъединичную вакцину против Mycoplasma hyorhinis, тем самым усовершенствуя общую работу по предотвращению эпидемий в свиноводстве.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0004] Одна цель настоящего раскрытия состоит в получении субъединичной вакцины против инфекции Mycoplasma hyorhinis с улучшением, тем самым, работу по предотвращению эпидемий в свиноводстве.
[0005] Другая цель настоящего раскрытия состоит в получении экспрессирующего антиген вектора и разработке способа получения субъединичной вакцины, где ген антигена в указанном экспрессирующем векторе экспрессируют в прокариотической экспрессирующей системе, тем самым снижая стоимость производства указанной субъединичной вакцины.
[0006] В отношении достижения указанных целей, настоящее раскрытие относится к композиции для предотвращения заболеваний, обусловленных инфекцией Mycoplasma hyorhinis, которая содержит активный ингредиент, содержащий XylF, DnaK, P72 или их комбинацию, и адъювант; где указанный XylF содержит SEQ ID № 01, указанный DnaK содержит SEQ ID № 02 и указанный P72 содержит SEQ ID № 03; где указанное заболевание представляет собой по меньшей мере одно заболевание, выбранное из перитонита, плеврита, перикардита и отека суставов.
[0007] Предпочтительно, указанный активный ингредиент содержит по меньшей мере два ингредиента, выбранных из группы, состоящей из XylF, DnaK, P72 или их комбинацию; более предпочтительно, указанный активный ингредиент содержит комбинацию XylF, DnaK и P72.
[0008] Предпочтительно, указанный активный ингредиент имеет концентрацию от 50 до 300 мкг/мл, на основе общего объема указанной композиции.
[0009] Предпочтительно, указанный адъювант включает полный адъювант Фрейнда, неполный адъювант Фрейнда, гель оксида алюминия, поверхностно-активное средство, анионный полимер, пептид, масляную эмульсию или их комбинацию.
[0010] Предпочтительно, указанное заболевание представляет собой перитонит, плеврит или их комбинацию, при условии, что указанным активным ингредиентом является XylF.
[0011] Предпочтительно, указанное заболевание представляет собой перитонит, при условии, что указанным активным ингредиентом является DnaK.
[0012] Предпочтительно, указанное заболевание представляет собой плеврит, при условии, что указанным активным ингредиентом является P72.
[0013] Предпочтительно, указанное заболевание представляет собой перитонит, плеврит, перикардит и отек суставов, при условии, что указанным активным ингредиентом является комбинация XylF, DnaK и P72.
[0014] Кроме того, настоящее раскрытие относится к экспрессирующему вектору для получения активного ингредиента указанной композиции в прокариотической экспрессирующей системе, где указанный экспрессирующий вектор содержит:
экспрессионный элемент, который содержит промотор и участок связывания рибосомы;
нуклеотидную последовательность, кодирующую указанный XylF, указанный DnaK, указанный P72 или их комбинацию; и
последовательность, кодирующую партнера слияния;
где указанная нуклеотидная последовательность содержит SEQ ID № 04, SEQ ID № 05 или SEQ ID № 06 или их комбинацию.
[0015] Предпочтительно, партнер слияния представляет собой DsbC из E. coli, MsyB из E. coli, FklB из E. coli или их комбинацию. Более предпочтительно, для экспрессирующего вектора, указанным партнером слияния является DsbC из E. coli, при условии, что указанная нуклеотидная последовательность кодирует XylF; указанным партнером слияния является MsyB из E. coli, при условии, что указанная нуклеотидная последовательность кодирует DnaK; или указанным партнером слияния является FklB из E. coli, при условии, что указанная нуклеотидная последовательность кодирует P72.
[0016] Предпочтительно, указанный экспрессирующий вектор дополнительно содержит последовательность, кодирующую гистидиновую метку, глутатион-S-трансферазу (метку GST) или их комбинацию.
[0017] Предпочтительно, указанный экспрессирующий вектор содержит SEQ ID № 07, SEQ ID № 08 или SEQ ID № 09.
[0018] Предпочтительно, указанная прокариотическая экспрессирующая система представляет собой экспрессирующую систему E. coli.
[0019] Настоящее раскрытие, кроме того, относится к способу получения растворимого белка; где указанный белок представляет собой XylF, DnaK, P72 или их комбинацию; где указанный способ включает
(1) получение прокариотической экспрессирующей системы; и
(2) экспрессию указанной нуклеотидной последовательности экспрессирующего вектора в указанной прокариотической экспрессирующей системе.
[0020] Предпочтительно, указанный способ дополнительно включает стадию пропускания продукта, полученного в вышеуказанной стадии (2), через аффинную колонку с ионами никеля или глутатионовую аффинную колонку для получения указанного растворимого белка.
[0021] В целом, настоящее раскрытие относится к композиции против инфекции Mycoplasma hyorhinis, с помощью которой можно обеспечить предотвращение заболеваний, обусловленных инфекцией Mycoplasma hyorhinis. В настоящем раскрытии также раскрыт экспрессирующий антиген вектор и способ получения указанного активного ингредиента указанной композиции в прокариотической экспрессирующей системе.
КРАТКОЕ ОПИСАНИЕ ФИГУР
[0022] На фиг. 1 представлен результат электрофореза белков, который проводили в примере 3 для наблюдения растворимости рекомбинантного антигена по настоящему раскрытию (где T представляет общие клеточные лизаты и S представляет растворимую фракцию в общих клеточных лизатах.
[0023] На фиг. 2 представлен результат электрофореза белков, который проводили в примере 3 для наблюдения очистки рекомбинантного антигена по настоящему раскрытию.
ПОДРОБНОЕ ОПИСАНИЕ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
[0024] Ввиду отсутствия композиций против инфекции Mycoplasma hyorhinis, авторы настоящего изобретения выполнили сложное исследование и доказали, что XylF, DnaK и P72, отдельно или в комбинации, можно использовать в качестве активных ингредиентов композиции для предотвращения заболевания, обусловленного инфекцией Mycoplasma hyorhinis. Термин «заболевание, обусловленное инфекцией Mycoplasma hyorhinis», в этом тексте означает по меньшей мере одно заболевание, выбранное из перитонита, плеврита, перикардита и отека суставов. В одном из вариантов осуществления «указанное заболевание, обусловленное инфекцией Mycoplasma hyorhinis», можно оценивать посредством способа, предложенного в Magnusson et al. (Vet. Immunol. Immunopathol., 61:83-96, 1998).
[0025] В одном из примеров результат эксперимента по настоящему раскрытию показывал, что XylF, когда его используют в качестве активного ингредиента композиции согласно настоящему раскрытию, в частности, можно использовать в облегчении перитонита, плеврита или их комбинации. В другом примере согласно настоящему раскрытию результат эксперимента показывал, что DnaK, когда его используют в качестве активного ингредиента композиции согласно настоящему раскрытию, в частности, можно использовать в облегчении перитонита. В дополнительном примере согласно настоящему раскрытию, результат эксперимента показывал, что P72, когда его используют в качестве активного ингредиента композиции согласно настоящему раскрытию, в частности, можно использовать в облегчении плеврита. В другом примере согласно настоящему раскрытию, результат эксперимента показывал, что комбинацию XylF, DnaK и P72, когда ее используют в качестве активных ингредиентов, в частности, можно использовать при условии, что заболевания, подлежащие облегчению, представляют собой перитонит, плеврит, перикардит и отек суставов.
[0026] В одном из аспектов, настоящее раскрытие относится к композиции для предотвращения заболеваний, обусловленных инфекцией Mycoplasma hyorhinis. Указанная композиция содержит активный ингредиент и адъювант. Термин «активный ингредиент» обозначает ингредиент, который позволяет достигать цели использования указанной композиции. Указанный активный ингредиент содержит XylF, DnaK, P72 или их комбинацию; где указанный XylF содержит SEQ ID № 01, указанный DnaK содержит SEQ ID № 02 и указанный P72 содержит SEQ ID № 03.
[0027] В одном из вариантов осуществления указанный активный ингредиент представляет собой ингредиент, выбранный из группы, состоящей из XylF, DnaK и P72. В другом варианте осуществления указанный активный ингредиент содержит два ингредиента, выбранных из группа, состоящей из XylF, DnaK и P72. В предпочтительном варианте осуществления указанный активный ингредиент представляет собой комбинацию XylF, DnaK и P72. Специалисту в данной области будет понятно, что указанный активный ингредиент может представлять собой рекомбинантный белок, который может содержать аминокислотные последовательности по меньшей мере двух белков, выбранных из группы, состоящей из XylF, DnaK и P72, при условии, что структура эпитопа каждого из XylF, DnaK и P72 не изменена. В альтернативном варианте осуществления указанная композиция может содержать смесь указанных активных ингредиентов, получаемую посредством смешивания по меньшей мере двух белков, выбранных из группы, состоящей из XylF, DnaK и P72.
[0028] Специалисту в данной области будет понятно, что если вакцина содержит два или более активных ингредиентов, ее эффект является непредсказуемым, в частности, когда указанные два или более активных ингредиентов направлены против одной и той же патогенной инфекции, поскольку нежелательный результат может возникать из-за взаимного влияния между указанными двумя или больше активными ингредиентами друг с другом. В другом аспекте, даже несмотря на то, что указанные два или более активных ингредиентов не взаимодействуют друг с другом и не возникает какой-либо нежелательный результат, все так же нет мотивации с экономической точки зрения для того, чтобы комбинировать указанные два или более активных ингредиентов в одной единственной вакцине, если объединение указанных двух или более активных ингредиентов не вызывает более хороший эффект (например, индукция более сильного иммунного ответ). Ввиду вышесказанного, объединение двух или более активных ингредиентов в одной единственной вакцине может иметь промышленные эффекты, только когда комбинация указанных двух или более активных ингредиентов может вызывать более хороший эффект. Однако без проведения экспериментов нельзя сказать, какой тип рассматриваемых ингредиентов или композиций может вызывать более хороший эффект.
[0029] В одном из вариантов осуществления указанный активный ингредиент указанной композиции имеет концентрацию от 50 до 300 мкг/мл, на основе общего объема указанной композиции. В предпочтительном варианте осуществления указанный активный ингредиент указанной композиции представляет собой комбинацию XylF, DnaK и P72, в которой каждый из XylF, DnaK и P72 имеет концентрацию 100 мкг/мл, на основе общего объема указанной композиции. Специалисту будет понятно, что указанную концентрацию можно корректировать в зависимости от цели применения композиции. Например, для того, чтобы сделать транспортировку и хранение композиции более удобными, специалист в данной области может получить указанную композицию с более высокой концентрацией указанного активного ингредиента(ов) и развести указанную композицию непосредственно перед использованием.
[0030] Термин «адъювант» имеет то же значение, которое общеизвестно в области лекарственных средств/вакцин. Например, указанный адъювант используют для усовершенствования иммуногенного эффекта указанных активных ингредиентов и/или стабилизации указанных активных ингредиентов. Указанный адъювант включает, но не ограничиваясь этим, полный адъювант Фрейнда, неполный адъювант Фрейнда, гель оксида алюминия, поверхностно-активное средство, анионный полимер, пептид, масляную эмульсию или их комбинацию. В одном из вариантов осуществления указанный адъювант представляет собой гель оксида алюминия.
[0031] В другом аспекте настоящего раскрытия, настоящее раскрытие относится к экспрессирующему антиген вектору. Цель конструкции указанного экспрессирующего антиген вектора состоит в том, чтобы экспрессировать активный ингредиент (т. е. рекомбинантный антиген) указанной композиции в прокариотической экспрессирующей системе с тем, чтобы обеспечить массовую продукцию указанного рекомбинантного антигена. Несмотря на то, что проведено множество опытов по экспрессии белков в прокариотической экспрессирующей системе, различные условия экспрессии могут быть необходимы для различных белков в силу биоразнообразия. Следовательно, все еще необходимо проводить большое количество экспериментов для того, чтобы тестировать условия экспрессии любого конкретного антигена. Путем исследований согласно настоящему раскрытию, экспрессирующий вектор, который может экспрессировать рекомбинантные антигены в прокариотической экспрессирующей системе, успешно сконструирован, предпочтительно для экспрессирующей системы E. coli. Экспрессирующий вектор согласно настоящему раскрытию, который может экспрессировать рекомбинантные антигены в экспрессирующей системе E. coli, может быть легко модифицирован специалистом, тем самым позволяя экспрессирующему вектору согласно настоящему раскрытию экспрессировать рекомбинантные антигены в других прокариотических экспрессирующих системах.
[0032] В другом аспекте одной из существующих технических проблем при получении указанных активных ингредиентов является очистка указанных активных ингредиентов, экспрессируемых в указанной прокариотической экспрессирующей системе. Рекомбинантные белки, экспрессируемые в прокариотической экспрессирующей системе, обычно имеют низкую растворимость, что повышает сложность и стоимость стадии выделения/очистки. Ввиду вышесказанного, экспрессирующий антиген вектор согласно настоящему раскрытию разрабатывают для того, чтобы он мог экспрессировать рекомбинантный белок, который имеет хорошую растворимость, тем самым упрощая стадию выделения/очистки рекомбинантного белка и снижая его стоимость.
[0033] Экспрессирующий вектор согласно настоящему раскрытию содержит экспрессионный элемент; нуклеотидную последовательность, кодирующую указанный XylF, указанный DnaK, указанный P72 или их комбинацию; и последовательность, кодирующую партнер слияния. В одном из вариантов осуществления, в котором указанная нуклеотидная последовательность содержит SEQ ID № 04, SEQ ID № 05 или SEQ ID № 06 или их комбинацию. Специалист может модифицировать указанную нуклеотидную последовательность, в зависимости от кодонных предпочтение выбранной прокариотической экспрессирующей системы, при условии, что указанная нуклеотидная последовательность может кодировать указанный XylF, указанный DnaK, указанный P72 или их комбинацию.
[0034] В предпочтительном варианте осуществления исследование согласно настоящему раскрытию доказывает, что DsbC из E. coli, MsyB из E. coli, FklB из E. coli или их комбинацию предпочтительно использовать в качестве партнера слияния для экспрессии XylF, DnaK, P72 или их комбинации, тем самым придавая рекомбинантным белкам, получаемым в прокариотической экспрессирующей системе, желаемую растворимость. В другом предпочтительном варианте осуществления, для облегчения стадии очистки указанный экспрессирующий вектор дополнительно может содержать последовательность, кодирующую гистидиновую метку, метку GST или их комбинацию, тем самым получаемые рекомбинантные белки можно очищать посредством пропускания через аффинную колонку с ионами никеля или глутатионовую аффинную колонку.
[0035] В одном из вариантов осуществления указанный экспрессионный элемент по меньшей мере содержит промотор и участок связывания рибосомы для осуществления транскрипции и /или трансляции. В другом варианте осуществления, чтобы содействовать операции генетической инженерии, указанный экспрессирующий вектор дополнительно может содержать сайт множественного клонирования, состоящий из сайтов разрезания рестрикционных ферментов, селективный маркер или их комбинацию. Указанный селективный маркер может представлять собой ген устойчивости к антибиотику или ауксотрофный ген.
[0036] В другом аспекте настоящее раскрытие относится к способу получения растворимого белка, в котором указанный растворимый белок представляет собой XylF, DnaK, P72 или их комбинацию. Способ согласно настоящему раскрытию включает (1) получение прокариотической экспрессирующей системы; и (2) экспрессию гена антигена в экспрессирующем векторе в указанной прокариотической экспрессирующей системе. Термин «растворимый» в этом тексте относится к такому свойству, что указанный белок стремится растворяться в водном растворе. Термин «экспрессия» в этом тексте указывает, что экспрессирующий вектор индуцируют к транскрипции и трансляции целевого гена в вышеуказанной прокариотической экспрессирующей системе с помощью любых средств. Средства могут представлять собой, но не ограничиваясь этим, добавление изопропил-β-D-тиогалактозида (IPTG) в указанную прокариотическую экспрессирующую систему.
Пример 1. Исследование белков, подходящих в качестве активных ингредиентов композиции согласно настоящему раскрытию
(1) Получение инактивированной вакцины против Mycoplasma hyorhinis
[0037] Mycoplasma hyorhinis (ATIT-7) культивировали в общеизвестной среде Фриза и затем использовали для того, чтобы получать инактивированную вакцину против Mycoplasma hyorhinis в соответствии со способом, как раскрыто в тайваньском патенте на изобретение № 1238721.
(2) Получение антисыворотки, направленной на Mycoplasma hyorhinis
[0038] Трех свиней SPF в возрасте 4 недель приобретали на свиноферме второго поколения SPF в Animal Technology Laboratories of Agricultural Technology Research Institute. Всех свиней кормили в одних и тех же условиях индуцирования в постройке для экспериментальных свиней SPF. Свиней индуцировали к возрасту 32 суток, 46 суток и 60 суток, соответственно, и в каждый из вышеуказанных моментов времени 2 мл инактивированной вакцины против Mycoplasma hyorhinis вводили свиньям посредством внутримышечной инъекции. Когда свиней индуцировали к возрасту 74 суток, у свиней из яремной вены брали образцы крови и держали их при комнатной температуре (приблизительно 25°C) в течение 1 часа. Затем образцы крови оставляли при 4°C. На следующий день образцы крови центрифугировали при 1107×g в течение 30 мин. Супернатант (а именно антисыворотку) переносили в чистую пробирку для центрифуги и хранили при -20°C для использования на последующих стадиях.
(3) Экстрагирование общего белка Mycoplasma hyorhinis
[0039] Общий белок экстрагировали из Mycoplasma hyorhinis с использованием набора для экстрагирования белка (набор для экстрагирования белка Ready Prep™; Bio-Rad, USA). Сначала Mycoplasma hyorhinis, культивированную в среде Фриза, центрифугировали (10,000×g, 20 мин) для того, чтобы собирать бактериальные клетки. Затем бактериальные клетки промывали 3 раза в низкосолевом буферном растворе (100 мМ основания Tris, 250 мМ сахарозы, pH 8,0), суспендировали в 1 мл буфера для образцов (полный 2-D буфер для регидратации/образцов) с добавлением 10 мкл восстанавливающего средства TBF (восстанавливающее средство ReadyPrep™ TBP), подходящего количества амфолита Bio-Lyte 3/10 (конечная концентрация амфолита 0,2%) и подходящего количества протеазного ингибитора. Бактериальные клетки разрушали обработкой ультразвуком. Разрушенные клетки удаляли посредством центрифугирования, и супернатант, который содержал общий белок Mycoplasma hyorhinis, сохраняли и использовали на последующих стадиях.
[0040] Концентрацию общего белка определяли с помощью набора для анализа белка (RC DC™ Protein Assay Kit; Bio-Rad, USA). 100 мкл указанного супернатанта, содержащего общий белок, смешивали тщательно с 500 мкл реактива RC I, и смесь помещали в комнатную температуру (приблизительно 25°C) и позволяли реакции протекать в течение 1 мин. Затем добавляли 500 мкл реактива RC II, тщательно смешивали и центрифугировали (15000×g, 5 мин). Осадители собирали и затем тщательно смешивали с 510 мкл реактива A'. Смесь помещали в комнатную температуру (приблизительно 25°C) и позволяли реакции протекать в течение 5 мин или до растворения всех осадителей. После этого 4 мл реактива B добавляли и смесь снова помещали в комнатную температуру (приблизительно 25°C) и позволяли реакции протекать в течение 15 мин. Поглощение раствора при длине волны 750 нм определяли посредством спектрофотометра. Бычий сывороточный альбумин (BSA) использовали в качестве стандартного образца для создания калибровочной кривой зависимости концентрации белка от поглощения. Концентрацию общих белков в указанном супернатанте можно получать посредством интерполяции поглощения образца супернатанта на калибровочной кривой и вычисления концентрации общего белка в нем. Образец с известной концентрацией общего белка можно использовать на последующей стадии электрофореза белков.
(4) Двухмерный электрофорез белков
[0041] Двухмерный электрофорез белков включает две стадии: электрофорез с изоэлектрическим фокусированием (IEF) и электрофорез в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE), как указано в следующих абзацах.
Электрофорез с изоэлектрическим фокусированием (IEF)
[0042] Электрофорез с изоэлектрическим фокусированием (IEF) представляет собой прием для разделения различных белков по различиям в их изоэлектрических точках. Сначала брали 1 мг супернатанта, содержащего общий белок Mycoplasma hyorhinis, и смешивали с подходящим количеством буфера для регидратации таким образом, что общий объем получаемой смеси составлял 400 мкл. Затем смесь добавляли в щели для образцов в планшете для фокусирования (Bio-Rad, USA). Два куска фильтровальной бумаги, которую увлажняли дистиллированной деионизированной водой, помещали, соответственно, на положительный электрод и отрицательный электрод. Указанная фильтровальная бумага может адсорбировать примеси и соли в образце, тем самым избегая влияния указанных примесей и слей на последующий эксперимент и повреждение электродов. Затем IPG гелевую полоску, а именно ReadyStrip™ (pH 5-8/17 см) медленно помещали в планшет для фокусирования. Брали 2,5 мл минерального масла и распределяли его равномерно по IPG гелевой полоске для того, чтобы избегать испарения образца, которое могло влиять на последующий эксперимент. Планшет для фокусирования накрывали крышкой и помещали на прибор с ячейкой для электрофореза с изоэлектрическим фокусированием PROTEAN IEF (Bio-Rad, USA). После того, как задавали программу для ячейки PROTEAN IEF, одномерный электрофорез проводили в 5 этапов. Первый этап представляет собой регидратацию при 50 В в течение 12 часов, чтобы заставлять IPG гелевую полоску абсорбировать образец. Второй этап представляет собой удаление ионов солей и примесей при 250 В в течение 15 мин. Третий этап представляет собой повышение напряжения, в котором напряжение линейно повышают до фокусирующего напряжения 10000 В за 4 часа. Четвертый этап представлял собой стадию изоэлектрического фокусирования, проводимую при 50000 В×ч. Пятый этап представлял собой поддержание напряжения 500 В для того, чтобы избегать избыточной реакции. После завершения одномерного электрофореза, IPG гелевую полоску можно хранить при -80°C, или после уравновешивания быстро использовать на последующей стадии SDS-PAGE.
Электрофорез в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE)
[0043] Электрофорез в полиакриламидном геле с додецилсульфатом натрия представляет собой прием для разделения различных белков по различиям в их молекулярной массе. Сначала, после промывания IPG гелевой полоски деионизированной водой, минеральное масло и воду на обратной стороне геля абсорбировали с помощью фильтровальной бумаги. Затем IPG гелевую полоску помещали в одноразовый регидратационный планшет. Туда добавляли 6 мл уравновешивающего буфера I (6 M мочевина, 2% SDS, 0,375 M Tris, 20% глицерин, 130 мМ DTT, pH 8,8) и получаемую смесь встряхивали в течение 20 мин. Затем вынимали IPG гелевую полоску и уравновешивающий буфер I, оставшийся на обратной стороне геля, абсорбировали фильтровальной бумагой. IPG гелевую полоску снова помещали в одноразовый регидратационный планшет. Туда добавляли 6 мл уравновешивающего буфера II (6 M мочевина, 2% SDS, 0,375 M Tris, 20% глицерин, 135 мМ йодацетамид, pH 8,8) и получаемую смесь встряхивали в течение 20 мин. После завершения указанной уравновешивающей обработки проводили SDS-PAGE.
[0044] Сначала получали 12,5% разделяющий гель. IPG гелевую полоску помещали на верхнюю поверхность разделяющего геля. Затем туда добавляли подходящее количество растворенной агарозы (ReadyPrep™ Overlay Agarose, Bio-Rad, USA). Чтобы сделать определение молекулярной массы более удобным, фильтровальную бумагу с точками стандартов молекулярных масс белков на ней помещали рядом IPG гелевой полоской. После коагуляции агарозы, гелевую полоску и фильтровальную бумагу со стандартами молекулярных масс белков фиксировали на разделяющем геле. Кусок электрофоретического геля помещали в электрофоретический сосуд (Bio-Rad, USA) и электрофоретический буфер (25 мМ Tris, 192 мМ глицин, 0,1% SDS, pH 8,3) вводили в сосуд. Электрофорез проводили при электрическом токе 26 мА в течение 15 часов, тем самым разделяя белки с различными молекулярными массами.
(5) Способ вестерн-блоттинга
[0045] После электрофореза белков, вышеуказанный кусок геля погружали в буфер для блоттинга [25 мМ основания Tris, 192 мМ глицина, 10% (об./об.), метанол, pH 8,3]. PVDF пленку обрезали до надлежащего размера и погружали в метанол на несколько секунд, затем промывали 1 раз деионизированной водой и погружали в буфер для блоттинга. После погружения куска геля и PVDF пленки в буфер для блоттинга на 15 мин, кусок фильтровальной бумаги, кусок геля, PVDF пленку и еще один кусок фильтровальной бумаги помещали последовательно в аппарат для переноса блоттингом и проводили перенос при электрическом токе 1300 мА в течение 1,5 часа.
[0046] После завершения переноса, указанную PVDF пленку погружали в блокирующий буфер [20 мМ основание Tris, 150 мМ NaCl, 5% (масс./об.) снятое молоко, pH 7,4] при комнатной температуре в течение 1 часа. Подходящее количество антисыворотки (разведение в 1000 раз) против Mycoplasma hyorhinis, полученной в вышеуказанном эксперименте, добавляли и встряхивали при комнатной температуре в течение 1 часа. После того, как блокирующий раствор выливали и указанную PVDF пленку промывали подходящим количеством буфера TBST [20 мМ основание Tris, 150 мМ NaCl, 0,05% (об./об.) Tween-20, pH 7,4] 3 раза (5 мин/раз), добавляли блокирующий буфер, содержащий конъюгированное с щелочной фосфатазой IgG козы против свиньи (H+L) (разведение 2000 раз). После встряхивания в темноте в течение 1 часа, указанную PVDF пленку промывали буфером TBST 3 раза и затем раствор NBT/BCIP (Thermo Fisher Scientific, USA) добавляли для того, чтобы индуцировать реакцию развития окраски.
(6) Идентификация белков
[0047] Окрашенные точки (всего 17 точек, не показано на фиг.), как показано в результатах реакции развития окраски в указанном способе вестерн-блоттинга, показывали белки Mycoplasma hyorhinis, способные реагировать с антисывороткой против Mycoplasma hyorhinis. Сравнивая вышеуказанный результат реакции развития окраски с куском геля, который подвергали электрофорезу, полосы куска геля, которые находились в положении, соответствующем положению окрашенных точек, брали с помощью микропипетки и использовали в масс-спектрометрическом анализе. Идентификацию белков, полученных выше, проводили посредством сравнения их аминокислотных последовательностей с базой данных о последовательностях белков. Как результат, идентифицировали 3 белка, а именно XylF, DnaK и P72, которые, соответственно, имеют последовательности, как показано в SEQ ID № 01, SEQ ID № 02 и SEQ ID № 03. Настоящее раскрытие продолжает последующее исследование этих 3 белков.
Пример 2. Конструкция экспрессирующего вектора согласно настоящему раскрытию
(1) Точечная мутация и клонирование генов антигенов
[0048] В соответствии с данными из National Center for Biotechnology Information (NCBI) of America, последовательности указанных генов XylF, DnaK и P72, соответственно, имели 4, 1 и 8 кодонов TGA. Кодон TGA считали терминирующим кодоном в экспрессирующей системе E. coli. Для того чтобы избегать неспособности экспрессирующей системы E. coli продуцировать полноразмерные вышеуказанные белки, кодоны TGA в последовательностях генов антигенов мутировали в кодоны TGG посредством полимеразной цепной реакции.
Экстрагирование геномной ДНК Mycoplasma hyorhinis
[0049] Геномную ДНК Mycoplasma hyorhinis экстрагировали с использованием набора для очистки ДНК (Tissue & Cell Genomic DNA Purification Kit; GMbiolab, Taiwan). Сначала 4,5 мл жидкой культуры Mycoplasma hyorhinis брали и помещали в пробирку для центрифуги. После центрифугирования (5870×g, 5 мин), выливали супернатант и собирали осадок. Затем добавляли 20 мкл протеазы K (10 мг/мл) и 200 мкл реактива для экстрагирования и оставляли реагировать при 56°C в течение 3 часов. Добавляли 200 мкл связывающего раствора и оставляли реагировать при 70°C в течение 10 мин. После завершения реакции 200 мкл абсолютного спирта добавляли и переносили в микропробирку для центрифуги и тщательно смешивали, получаемый раствор (включая преципитат) пипетировали в центрифужную колонку, и затем помещали центрифужную колонку в собирающую пробирку. После центрифугирования (17970×g) в течение 2 мин, супернатант выливали и в центрифужную колонку снова добавляли 300 мкл связывающего реактива. После центрифугирования (17970×g) в течение 2 мин, супернатант выливали. 700 мкл промывающего раствора добавляли в центрифужную колонку и смесь центрифугировали (17970×g) в течение 2 мин и затем выливали супернатант. Вышеуказанную стадию промывания повторяли один раз. В конце смесь центрифугировали при 17970×g в течение 5 мин для того, чтобы удалять оставшийся спирт. Центрифужную колонку помещали в стерилизованную микропробирку для центрифуги и добавляли подходящее количество стерильной деионизированной воды, чтобы смывать геномную ДНК.
Точечная мутация гена антигена XylF
[0050] Разрабатывали праймеры XylF/XylR и мутационные праймеры от XylM1 до XylM8 для гена антигена XylF, последовательности этих праймеров перечислены в таблице 1.
Таблица 1
[0051] Используя геномную ДНК Mycoplasma hyorhinis в качестве матрицы, фрагменты ДНК амплифицировали с использованием пар праймеров XylF/XylM2, XylM1/XylM4, XylM3/XylM6, XylM5/XylM8 и XylM7/XylR и т. д., соответственно. 50 мкл реакционной смеси ПЦР содержали 1× GDP-HiFi ПЦР буфер B, 200 мкМ dNTP (dATP, dTTP, dGTP и dCTP), 1 мкМ праймеров для амплификации, 200 нг геномной ДНК Mycoplasma hyorhinis и 1 Ед. GDP-HiFi ДНК полимеразы. Условия реакции ПЦР представляют собой 96°C в течение 5 мин (одна стадия); 94°C в течение 30 с, 55°C в течение 30 с, 68°C в течение 30 с (35 циклов); и 68°C в течение 5 мин (одна стадия).
[0052] После завершения ПЦР, электрофорез в агарозном геле проводили для того, чтобы подтверждать, имеют ли место фрагменты ДНК с предсказанными размерами. ПЦР продукты извлекали с помощью набора для системы экстрагирования гелей Gel-MTM. Амплификацию гена проводили с использованием 5 извлеченных ПЦР продуктов в качестве матриц и пары праймеров XylF/XylR. Условия реакции ПЦР представляют собой 96°C в течение 2 мин (одна стадия); 94°C в течение 30 с, 55°C в течение 30 с, 68°C в течение 45 с (35 циклов); и 68°C в течение 5 мин (одна стадия). Ген XylF с точечной мутацией получали посредством указанной ПЦР. Продукты ПЦР извлекали с помощью PCR-MTM Clean Up System Kit. В соответствии с результатом определения последовательности, ген XylF согласно настоящему раскрытию имеет нуклеотидную последовательность, как показано в SEQ ID № 4.
Точечная мутация гена антигена DnaK
[0053] Разрабатывали праймеры DnaKF/DnaKR и мутационные праймеры от DnaKM1 до DnaKM2 для гена антигена DnaK, последовательности этих праймеров перечислены в таблице 2.
Таблица 2
[0054] Используя геномную ДНК Mycoplasma hyorhinis в качестве матриц, фрагменты ДНК амплифицировали посредством соответствующего использования пар праймеров DnaKF/DnaKM2 и DnaKM1/DnaKR. 50 мкл реакционной смеси ПЦР содержали 1× GDP-HiFi ПЦР буфер B, 200 мкМ dNTP (dATP, dTTP, dGTP, и dCTP), 1 мкМ праймеров для амплификации, 200 нг геномной ДНК Mycoplasma hyorhinis и 1 Ед. GDP-HiFi ДНК полимеразы. Условия реакции ПЦР представляют собой 96°C в течение 5 мин (одна стадия); 94°C в течение 30 с, 55°C в течение 30 с, 68°C в течение 30 с (35 циклов); и 68°C в течение 5 мин (одна стадия).
[0055] После завершения ПЦР, электрофорез в агарозном геле проводили для того, чтобы подтверждать, имеют ли место фрагменты ДНК с предсказанными размерами. ПЦР продукты извлекали с помощью набора для системы экстрагирования гелей Gel-M™. Амплификацию гена проводили с использованием 2 извлеченных ПЦР продуктов в качестве матриц и пары праймеров DnaKF/DnaK. Условия реакции ПЦР представляют собой 96°C в течение 2 мин (одна стадия); 94°C в течение 30 с, 55°C в течение 30 с, 68°C в течение 45 с (35 циклов); и 68°C в течение 5 мин (одна стадия). Ген DnaK с точечной мутацией получали посредством указанной ПЦР. Наконец, продукты ПЦР извлекали посредством PCR-M™ Clean Up System Kit. В соответствии с результатом определения последовательности, ген DnaK согласно настоящему раскрытию имел нуклеотидную последовательность, как показано в SEQ ID № 5.
Точечная мутация гена P72
[0056] Разрабатывали праймеры P72F/P72R и мутационные праймеры от P72M1 до P72M8 для гена антигена P72, последовательности этих праймеров перечислены в таблице 3.
Таблица 3
[0057] Используя геномную ДНК Mycoplasma hyorhinis в качестве матриц, фрагменты ДНК амплифицировали посредством соответствующего использования пар праймеров P72F/P72M2, P72M1/P72M4, P72M3/P72M6, P72M5/P72M8, P72M7/P72M10, P72M9/P72M12, P72M11/P72M14, P72M13/P72M16 и P72M15/P72R. 50 мкл реакционной смеси ПЦР содержали 1× GDP-HiFi ПЦР буфер B, 200 мкМ dNTP (dATP, dTTP, dGTP, и dCTP), 1 мкМ праймеров для амплификации, 200 нг геномной ДНК Mycoplasma hyorhinis и 1 Ед. GDP-HiFi ДНК полимеразы. Условия реакции ПЦР представляют собой 96°C в течение 5 мин (одна стадия); 94°C в течение 30 с, 55°C в течение 30 с, 68°C в течение 30 с (35 циклов); и 68°C в течение 5 мин (одна стадия).
[0058] После завершения ПЦР, электрофорез в агарозном геле проводили для того, чтобы подтверждать, имеют ли место фрагменты ДНК с предсказанными размерами. ПЦР продукты извлекали с помощью набора для системы экстрагирования гелей Gel-M™. Амплификацию гена проводили с использованием 9 извлеченных ПЦР продуктов в качестве матриц и пары праймеров P72F/P72R. Условия реакции ПЦР представляют собой 96°C в течение 2 мин (одна стадия); 94°C в течение 30 с, 55°C в течение 30 с, 68°C в течение 1 мин (35 циклов); и 68°C в течение 5 мин (одна стадия). Ген P72 с точечной мутацией получали посредством указанной ПЦР. Наконец, продукты ПЦР извлекали посредством PCR-M™ Clean Up System Kit. В соответствии с результатом определения последовательности, ген P72 согласно настоящему раскрытию имел нуклеотидную последовательность, как показано в SEQ ID № 6.
(2) Конструкция экспрессирующих векторов для антигенов Mycoplasma hyorhinis
[0059] Экспрессирующие векторы для антигенов Mycoplasma hyorhinis конструировали с использованием векторов, содержащих различные гены партнеров слияния, в качестве остова. Гены партнеров слияния, соответственно, представляют собой последовательности ДНК DsbC, MsyB, FklB из E. coli. Схема создания указанных экспрессирующих векторов представляет собой следующее.
[0060] Ген XylF, ген DnaK и ген P72 согласно настоящему раскрытию, полученные в вышеприведенных экспериментах, соответственно, резали с помощью BamHI и SalI и затем, с использованием T4 ДНК лигазы, фрагменты ДНК, полученные выше, соответствующим образом лигировали в слитый с геном DsbC экспрессирующий вектор, слитый с геном MsyB экспрессирующий вектор или слитый с геном FklB экспрессирующий вектор, которые резали с использованием тех же рестрикционных ферментов. После этого лигированные продукты трансформировали в E. coli ECOS 9-5. Трансформированные штаммы отбирали посредством ПЦР колоний. Электрофорез ДНК проводили для того, чтобы подтверждать, если имели место фрагменты ДНК с предсказанными размерами. После подтверждения того, что рекомбинантные векторы из трансформированных штаммов несли вставленную ДНК, векторы экстрагировали из трансформированных штаммов и определяли их последовательности ДНК. Векторы с правильными последовательностями ДНК, соответственно, называли pET-DsbC-XylF (SEQ ID № 07), pET-MysB-DnaK (SEQ ID № 08) и pET-FklB-P72 (SEQ ID № 09).
Пример 3. Получение и применение субъединичной вакцины согласно настоящему раскрытию
(1) Экспрессия рекомбинантных антигенов посредством экспрессирующих векторов согласно настоящему раскрытию
[0061] Вектор для экспрессии антигенов Mycoplasma hyorhinis трансформировали в E. coli BL21 (DE3). Одну колонию штамма, экспрессирующего антигены, выбирали и инокулировали в 12 мл среды LB, содержащей канамицин (конечная концентрация: 30 мкг/мл), затем культивировали при 37°C и в условиях 180 об./мин в течение ночи. После этого, 10 мл жидкой культуры E. coli брали и добавляли в 1 л среды LB, содержащей канамицин (конечная концентрация: 30 мкг/мл), и культивировали при встряхивании (37°C, 180 об./мин) до тех пор, пока OD600 не достигала приблизительно 0,4-0,6. Затем добавляли 0,1 мМ IPTG при 28°C для того, чтобы индуцировать экспрессию белка. После индуцирования в течение 4 часов, культуру центрифугировали (10000×g, 10 мин, 4°C) для того, чтобы собирать бактериальные клетки. После бактериальные клетки ресуспендировали в 10 мл фосфатного буфера (20 мМ фосфат натрия, 500 мМ NaCl, pH 7,4) и разрушали посредством обработки ультразвуком, суспензию центрифугировали (30966×g, 30 мин) для того, чтобы собирать супернатант. Наконец, супернатант фильтровали через фильтровальную мембрану с размером пор 0,22 мкМ и собирали фильтрат. Электрофорез белков проводили для того, чтобы наблюдать статус экспрессии и растворимость рекомбинантных антигенов. Результат показан на фиг. 1. На фиг. 1 можно наблюдать, что E. coli хорошо экспрессировали рекомбинантные антигены согласно настоящему раскрытию. Кроме того, рекомбинантные антигены согласно настоящему раскрытию имели превосходную растворимость, что указывает на то, что партнеры слияния, выбранные в настоящем раскрытии, являются подходящими.
[0062] Затем использовали аффинную хроматографию с иммобилизованными ионами металлов для очистки белка через ковалентное связывание между гистидиновой меткой на рекомбинантных антигенах и ионами никеля или ионами кобальта. Рекомбинантные антигены очищали с помощью 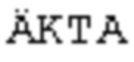 Prime Plus (GE Healthcare, Sweden), оборудованного 5 мл колонкой HiTrap™ Ni excel (GE Healthcare, Sweden). Сначала вышеуказанный супернатант вводили в колонку HiTrap™ Ni excel, затем колонку уравновешивали с использованием 25 мл фосфатного буферного раствора. После завершения введения образца, 100 мл буферного раствора для промывания, содержащего 30 мМ имидазола (20 мМ фосфата натрия, 500 мМ NaCl, 30 мМ имидазола, pH 7,4), использовали для промывания колонки с тем, чтобы можно было удалять белки, не специфически прилипшие к ней. Наконец, 150 мл элюирующего буферного раствора, содержащего 250 мМ имидазола (20 мМ фосфата натрия, 500 мМ NaCl, 250 мМ имидазола, pH 7,4), использовали для вымывания рекомбинантных антигенов из смолы, где имидазол в высокой концентрации может конкурировать за сайт связывания на смоле с рекомбинантными антигенами и, тем самым, вызывать вымывание рекомбинантных антигенов. Раствор очищенного антигена помещали в пробирку для центрифуги Amicon™ ultra-15 ultracel-30K (Merck Millipore, USA), центрифугировали (2600×g) ниже 4°C, корректировали до подходящего объема и затем хранили при 4°C для использования на последующей стадии. Результат очистки показан на фиг. 2. На фиг. 2 можно видеть, что рекомбинантные антигены согласно настоящему раскрытию, полученные в этом эксперименте, имели высокую чистоту.
Prime Plus (GE Healthcare, Sweden), оборудованного 5 мл колонкой HiTrap™ Ni excel (GE Healthcare, Sweden). Сначала вышеуказанный супернатант вводили в колонку HiTrap™ Ni excel, затем колонку уравновешивали с использованием 25 мл фосфатного буферного раствора. После завершения введения образца, 100 мл буферного раствора для промывания, содержащего 30 мМ имидазола (20 мМ фосфата натрия, 500 мМ NaCl, 30 мМ имидазола, pH 7,4), использовали для промывания колонки с тем, чтобы можно было удалять белки, не специфически прилипшие к ней. Наконец, 150 мл элюирующего буферного раствора, содержащего 250 мМ имидазола (20 мМ фосфата натрия, 500 мМ NaCl, 250 мМ имидазола, pH 7,4), использовали для вымывания рекомбинантных антигенов из смолы, где имидазол в высокой концентрации может конкурировать за сайт связывания на смоле с рекомбинантными антигенами и, тем самым, вызывать вымывание рекомбинантных антигенов. Раствор очищенного антигена помещали в пробирку для центрифуги Amicon™ ultra-15 ultracel-30K (Merck Millipore, USA), центрифугировали (2600×g) ниже 4°C, корректировали до подходящего объема и затем хранили при 4°C для использования на последующей стадии. Результат очистки показан на фиг. 2. На фиг. 2 можно видеть, что рекомбинантные антигены согласно настоящему раскрытию, полученные в этом эксперименте, имели высокую чистоту.
(2) Получение субъединичной вакцины согласно настоящему раскрытию и тест на протективный иммунный ответ на нее
[0063] При условиях, как показано в таблицах в следующих абзацах, рекомбинантные антигены согласно настоящему раскрытию, полученные в вышеуказанном эксперименте, соответствующим образом тщательно смешивали с адъювантом (гель оксида алюминия), тем самым получая различные субъединичные вакцины, содержащие один антиген, и вакцинные коктейли, содержащие несколько антигенов. Доза вакцины составляла 2 мл/доза, где содержание каждого рекомбинантного антигена, содержащегося в вакцине, составляло 200 мкг.
Эксперимент 1 по определению протективного иммунного ответа на вакцину из одного антигена
[0064] Этот эксперимент проводили в здании для генетически модифицированных организмов (GMO) в Animal Drugs Inspection Branch of Animal Health Research Institute. 12 свиней в возрасте 3 недель, у которых тесты показали отсутствие антител к Mycoplasma hyorhinis, случайно разделяли на группы с A до D. В каждой группе было по 3 свиньи, где группы с A до C представляли собой экспериментальные группы и группа D представляла собой контрольную группу. Каждую из свиней из групп с A до C иммунизировали вакциной из этого эксперимента через внутримышечную инъекцию (2 мл/доза) один раз, соответственно, в возрасте 3 недель и в возрасте 5 недель. Свиней группы D не иммунизировали. Компоненты вакцин как показаны в таблице 4.
Таблица 4
[0065] Когда свиньи были в возрасте 7 недель (то есть, через 2 недели после иммунизации), тест абдоминальной стимуляции проводили с использованием среды для культивирования, содержащей выделенный дикий штамм ATIT-2 Mycoplasma hyorhinis. Когда свиньи были в возрасте 10 недель (то есть, через 3 недели после теста стимуляции), проводили патологоанатомическое исследование. Вычисляли процентные доли патологических изменений, таких как перитонит, плеврит, перикардит и отек суставов и т. д. Видимые патологические изменения оценивали в соответствии со способом из Magnusson et al. (Vet. Immunol. Immunopathol., 61:83-96, 1998).
[0066] Результат эксперимента показан в таблице 5. Субъединичная вакцина, полученная из XylF, может снижать заболеваемость перитонитом и плевритом свиней, кроме того, усредненные оценки патологических изменений у иммунизированных свиней были ниже таковых у не иммунизированных свиней (контрольная группа), что указывает на то, что XylF может индуцировать значимый протективный иммунный ответ. Субъединичные вакцины, полученные из DnaK и P72, соответственно, могут снижать заболеваемость перитонитом и плевритом.
[0067] Следует отметить, что данные в таблице 5 не следует интерпретировать как то, что субъединичная вакцина, полученная из XylF, благоприятна только в облегчении перитонита и плеврита, они лишь указывают на то, что субъединичная вакцина, полученная из XylF, имела менее заметный эффект в облегчении других состоянии в течение эксперимента. По той же причине данные в таблице 5 не следует интерпретировать как то, что субъединичные вакцины, полученные из DnaK и из P72, благоприятны, соответственно, в облегчении только перитонита и плеврита, они лишь указывают на то, что субъединичная вакцина, полученная из DnaK или из P72, оказывает менее заметный эффект в облегчении других состояний в течение эксперимента.
Таблица 5
Эксперимент 2 по определению протективного иммунного ответа вакцинного коктейля
[0068] Этот эксперимент проводили в здании для генетически модифицированных организмов (GMO) в Animal Drugs Inspection Branch of Animal Health Research Institute. 24 свиньи в возрасте 4 недель, у которых тесты показывали отсутствие антител к Mycoplasma hyorhinis, отбирали и случайно разделяли на 2 группы, а именно группу иммунизации (группа E) и контрольную группу (группа F). В каждой группе было 12 свиней. Каждую из свиней группы иммунизации иммунизировали вакциной из этого эксперимента через внутримышечную инъекцию (2 мл/доза) один раз, соответственно, в возрасте 4 недель и в возрасте 6 недель. Свиней из контрольной группы не иммунизировали. Компоненты вакцины показаны в таблице 6.
Таблица 6
[0069] Результат эксперимента показан в таблице 7. Вакцинный коктейль согласно настоящему раскрытию (содержащий смесь 3 рекомбинантных антигенов согласно настоящему раскрытию в этом эксперименте) может значительно облегчать клинические симптомы перитонита, плеврита, перикардита и отека суставов и т. д., которые обусловлены инфекцией Mycoplasma hyorhinis, и он демонстрировал более хороший эффект по сравнению с эффектом субъединичной вакцины, содержащей один антиген (как показано в таблице 5).
Таблица 7
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Agricultural Technology Research Institute
<120> КОМПОЗИЦИЯ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ ИНФЕКЦИИ MYCOPLASMA HYORHINIS И СПОСОБ ПОЛУЧЕНИЯ УКАЗАННОЙ КОМПОЗИЦИИ
<140> PCT/CN2016/094104
<141> 2016-08-09
<160> 41
<170> PatentIn версии 3.5
<210> 1
<211> 447
<212> Белок
<213> Mycoplasma hyorhinis
<400> 1
Met Lys Lys Leu Ile Thr Lys Lys Phe Leu Tyr Leu Ser Thr Ile Ser
1 5 10 15
Thr Ala Ser Phe Ile Ala Phe Ala Ser Phe Val Ala Cys Gly Thr Thr
20 25 30
Ala Thr Gly Leu Ser Gln Thr Lys Asp His Ala Val Thr Asn Glu Ser
35 40 45
Ile Arg Val Ala Leu Thr Asp Pro Asp Asn Pro Arg Trp Ile Asn Ala
50 55 60
Gln Lys Asp Ile Leu Asn Tyr Ile Asp Lys Thr Glu Gly Ala Ile Ser
65 70 75 80
Thr Ile Thr Lys Asp Gln Asn Ala Gln Asn Asn Trp Leu Thr Gln Gln
85 90 95
Ala Asn Leu Asn Pro Ala Pro Lys Gly Phe Ile Ile Ala Pro Glu Asn
100 105 110
Gly Gly Gly Val Gly Thr Ala Val Asn Ser Ile Ala Glu Lys Asn Ile
115 120 125
Pro Ile Val Ala Tyr Asp Arg Leu Ile Thr Gly Ser Thr Ser Tyr Asp
130 135 140
Trp Tyr Val Ser Tyr Asp Asn Glu Lys Val Gly Glu Leu Gln Gly Leu
145 150 155 160
Ser Leu Ala Ala Gly Leu Leu Gly Lys Thr Asp Gly Ala Phe Lys Asp
165 170 175
Glu Thr Glu Met Leu Asn Tyr Leu Lys Asp His Met Pro Gln Glu Thr
180 185 190
Val Ser Phe Tyr Ala Val Ala Gly Ser Gln Asp Asp Asn Asn Ser Gln
195 200 205
Tyr Phe Tyr Asn Gly Ala Met Lys Ile Leu Lys Lys Leu Met Glu Asn
210 215 220
Ser Asn Gly Lys Val Val Asp Leu Ser Pro Arg Gly Asn Ala Val Tyr
225 230 235 240
Val Pro Gly Trp Asn Tyr Gly Asp Ala Gly Gln Arg Ile Gln Gln Phe
245 250 255
Phe Thr Gln Tyr Arg Asp Ser Ser Val Pro Asn Gly Val Leu Pro Val
260 265 270
Thr Tyr Asp Asn Val Thr Ile Pro Lys Ser Val Leu Lys Gly Phe Leu
275 280 285
Ala Pro Asn Asp Gly Met Ala Glu Gln Ala Val Asp Lys Leu Lys Ala
290 295 300
Thr Asn Tyr Asp Val Gln Lys Val Phe Ile Thr Gly Gln Asp Tyr Asn
305 310 315 320
Asp Thr Ala Lys Lys Leu Ile Lys Asn Gly Glu Gln Asn Met Thr Ile
325 330 335
Tyr Lys Pro Asp Val Ile Leu Gly Lys Val Ala Val Glu Val Leu Lys
340 345 350
Thr Leu Ile Asn Lys Lys Lys Ala Asn Ala Thr Ala Thr Lys Glu Asp
355 360 365
Val Glu Asn Ala Leu Lys Asn Asn Glu Asn Thr Lys Asp Ile His Phe
370 375 380
Arg Tyr Asp Asp Thr Thr Tyr Lys Ala Gly Ser Val Gly Ala Tyr Lys
385 390 395 400
Asn Ile Lys Ala Ile Leu Val Asn Pro Val Val Val Thr Lys Gln Asn
405 410 415
Val Asp Asn Pro Leu Ala Asn Ala Gln Gln Ser Thr Ser Ala Asn Val
420 425 430
Ser Val Ser Ala Ser Gly Thr Val Pro Thr Asn Gly Val Thr Gln
435 440 445
<210> 2
<211> 597
<212> Белок
<213> Mycoplasma hyorhinis
<400> 2
Met Ala Lys Glu Ile Ile Leu Gly Ile Asp Leu Gly Thr Thr Asn Ser
1 5 10 15
Val Val Ser Ile Ile Glu Asn Gly Lys Pro Val Val Leu Glu Asn Pro
20 25 30
Asn Gly Lys Asn Thr Thr Pro Ser Val Val Ala Phe Lys Asn Gly Glu
35 40 45
Glu Ile Val Gly Asp Ala Ala Lys Arg Gln Leu Glu Thr Asn Pro Asp
50 55 60
Ala Ile Ala Ser Ile Lys Arg Leu Met Gly Thr Asp Lys Thr Val Lys
65 70 75 80
Ala Asn Gly Lys Asp Tyr Lys Pro Glu Glu Ile Ser Ala Lys Ile Leu
85 90 95
Ser Tyr Leu Lys Gln Tyr Ala Glu Lys Lys Ile Gly Asn Lys Val Ser
100 105 110
Lys Ala Val Ile Thr Val Pro Ala Tyr Phe Asn Asn Ala Gln Arg Glu
115 120 125
Ala Thr Lys Asn Ala Gly Lys Ile Ala Gly Leu Glu Val Glu Arg Ile
130 135 140
Ile Asn Glu Pro Thr Ala Ala Ala Leu Ala Phe Gly Leu Glu Lys Thr
145 150 155 160
Glu Lys Glu Met Lys Val Leu Val Tyr Asp Leu Gly Gly Gly Thr Phe
165 170 175
Asp Val Ser Val Leu Glu Leu Ser Gly Gly Thr Phe Glu Val Leu Ser
180 185 190
Thr Ser Gly Asp Asn Lys Leu Gly Gly Asp Asp Cys Asp His Val Ile
195 200 205
Val Asp Trp Leu Val Asp Lys Ile Lys Lys Glu Tyr Glu Phe Asp Pro
210 215 220
Ser Lys Asp Lys Met Ala Leu Ser Arg Leu Lys Glu Glu Ala Glu Lys
225 230 235 240
Thr Lys Ile Ser Leu Ser Asn Gln Ser Val Ala Thr Ile Ser Leu Pro
245 250 255
Phe Leu Gly Ile Gly Pro Lys Gly Pro Ile Asn Val Glu Leu Glu Leu
260 265 270
Lys Arg Ser Glu Phe Glu Lys Met Thr Ala His Leu Val Asp Lys Thr
275 280 285
Arg Lys Pro Ile Met Asp Ala Leu Lys Glu Ala Lys Leu Glu Ala Lys
290 295 300
Asp Leu Asp Glu Val Leu Leu Val Gly Gly Ser Thr Arg Ile Pro Ala
305 310 315 320
Val Gln Thr Met Ile Glu His Thr Leu Gly Lys Lys Pro Asn Arg Ser
325 330 335
Ile Asn Pro Asp Glu Val Val Ala Ile Gly Ala Ala Ile Gln Gly Gly
340 345 350
Val Leu Ala Gly Glu Ile Asn Asp Val Leu Leu Leu Asp Val Thr Pro
355 360 365
Leu Thr Leu Gly Ile Glu Thr Leu Gly Gly Ile Ala Thr Pro Leu Ile
370 375 380
Pro Arg Asn Thr Thr Ile Pro Val Thr Lys Ser Gln Ile Phe Ser Thr
385 390 395 400
Ala Glu Asn Asn Gln Thr Glu Val Thr Ile Ser Val Val Gln Gly Glu
405 410 415
Arg Gln Leu Ala Ala Asp Asn Lys Leu Leu Gly Arg Phe Asn Leu Ser
420 425 430
Gly Ile Glu Gln Ala Pro Arg Gly Val Pro Gln Ile Glu Val Ser Phe
435 440 445
Ser Ile Asp Val Asn Gly Ile Thr Thr Val Ser Ala Lys Asp Lys Lys
450 455 460
Thr Asn Lys Glu Gln Thr Ile Thr Ile Lys Asn Thr Thr Ser Leu Ser
465 470 475 480
Glu Glu Glu Ile Glu Arg Met Val Lys Glu Ala Glu Glu Asn Arg Glu
485 490 495
Ala Asp Ala Lys Lys Lys Glu Lys Ile Glu Val Thr Val Arg Ala Glu
500 505 510
Ala Leu Ile Asn Gln Leu Glu Lys Ser Leu Glu Asp Gln Gly Asp Lys
515 520 525
Val Asp Ala Lys Gln Lys Glu Thr Leu Glu Lys Gln Ile Gln Glu Leu
530 535 540
Lys Asp Leu Val Lys Glu Glu Lys Ile Glu Glu Leu Lys Thr Lys Leu
545 550 555 560
Asp Gln Ile Glu Gln Ala Ala Gln Ala Phe Ala Gln Ala Ala Ala Gln
565 570 575
Gln Ala Asn Thr Ser Asp Thr Ser Ser Asp Asp Gln Pro Ile Glu Ala
580 585 590
Glu Val Lys Glu Asn
595
<210> 3
<211> 511
<212> Белок
<213> Mycoplasma hyorhinis
<400> 3
Met Asn Lys Lys His Ile Lys Thr Leu Ile Ser Ser Val Ser Ile Leu
1 5 10 15
Thr Pro Val Ala Ile Leu Ala Ser Cys Gly Gln Pro Thr Thr Ile Lys
20 25 30
Phe Ala Thr Ser Gln Gly Ser Tyr Trp Pro Met Met Leu Gly Met Lys
35 40 45
Glu Ile Ile Lys Ile Tyr Asn Glu Gln His Lys Asn Asp Ala Asp Phe
50 55 60
Ile Pro Val Glu Leu Leu Thr Ser Asp Val Thr His Lys Asn Ser Glu
65 70 75 80
Gly Gln Leu Leu Ser Ser Leu Asp Ser Asp Leu Ser Thr Lys Gln Ser
85 90 95
Ser Asp Trp Asn Leu Ile Leu Gly Asn Lys Ala Thr Ala Tyr Val Ala
100 105 110
Asn Ser Tyr Asp Lys Leu Leu Asp Val Gly Thr Ser Thr Val Asn Pro
115 120 125
Asn Ser Phe Pro Thr Lys Ile Ile Asn Asn Tyr Asn Lys Leu Leu Gly
130 135 140
Val Glu Gly Gln Thr Thr Leu Lys Ser Leu Pro Tyr Asn Ile Asn Asp
145 150 155 160
Thr Asp Gly Ile Val Phe Asn Leu Asp Ile Met Lys Val Leu Phe Asp
165 170 175
Ile Ile Lys Gln Gly Gly Gly Thr Ile Asp Glu Asn Ser Ile Ile Ala
180 185 190
Lys Lys Val Lys Glu Ala Glu Gly Lys Gly Asn His Ile Pro Ser Ser
195 200 205
Ser Met Phe Ser Ala Ile Lys Ile Lys Glu Ser Ser Lys Asn Thr Gly
210 215 220
Phe Ser Gly Tyr Thr Val Asn Asp Ser Thr Phe Ser Asp Ile Lys Lys
225 230 235 240
Ala Phe Glu Phe Ala Gln Lys Ile Tyr Asp Asn Thr Glu Ile Asp Thr
245 250 255
Thr Lys Leu Ser Lys Asp Val Lys Asp Ser Glu Ile Phe Ala Ile Asp
260 265 270
Tyr Ala Ser Asp Val Phe Arg Lys Glu Ile Leu Ser Lys Thr Asn Lys
275 280 285
Thr Phe Trp Ser Glu Lys Ser Leu Gln Asn Asn Lys Ile Thr Leu Asp
290 295 300
Val Asn Leu Asn Thr Asn Gln Ala Leu Lys Thr Glu Val Glu Asn Gln
305 310 315 320
Tyr Gln Glu Trp Glu Asn Thr Leu Lys Gln Thr Gln Phe Ile Pro Thr
325 330 335
Thr Thr Thr Gln Ala Asn Thr Glu Thr Thr Ser Trp Thr Lys Lys Asp
340 345 350
Ile Gln Thr Lys Ser Ala Thr Asp Asp Ser Gln Gln Ser Ile Asn Ser
355 360 365
Lys Thr Phe Tyr Ser Val Lys Phe Thr Glu Tyr Phe Lys Pro Ile Asn
370 375 380
Gln Trp Gly Ser Phe Glu Ile Arg Gln Tyr Leu Thr Ala Phe Thr Tyr
385 390 395 400
Ala Pro Leu Val Gly Thr Asn Tyr Ser Val Asp Ser Pro Trp Ser Arg
405 410 415
Ala Phe Phe Lys Lys Asp Leu Glu Glu Gly Lys Gln Lys Ala Ser Glu
420 425 430
Trp Thr Thr Arg Glu Asp Val Tyr Ala Thr Asn Gln Ala Met Lys Ala
435 440 445
Asp Asp Asn Ala Gln Tyr Leu Ala Tyr Asn Ala Gly Gly Phe Ser Leu
450 455 460
Ile Ala Ile Lys Ser Asn Ser Asp Ile Ile Asn Lys Asn Ile Ile Lys
465 470 475 480
Phe Val Asp Phe Leu Tyr Asn Gly Thr Gly Leu Thr Asp Leu Thr Gly
485 490 495
Ala Lys Ile Ser Ala Ala Asp Phe Met Ala Glu Lys Ser Ala Ala
500 505 510
<210> 4
<211> 1344
<212> ДНК
<213> Искусственная последовательность
<220>
<223> антиген
<400> 4
atgaaaaagt taattactaa aaaattttta tatttatcaa caatttcaac agcttcattt 60
atagcttttg cttcatttgt agcttgtgga acaacagcaa caggtttatc acaaacaaaa 120
gatcatgctg ttactaatga atccattaga gttgcattaa cagatccaga taatccaaga 180
tggattaatg cacaaaaaga tatattaaat tatatagaca aaacagaagg tgcaatttct 240
actattacta aagaccaaaa tgcacaaaac aactggttaa ctcaacaagc aaacttaaat 300
ccagcaccta aaggatttat tattgctcct gaaaatggag gaggagttgg aaccgctgtt 360
aattcaatag ctgaaaaaaa tattcctatc gttgcttatg atagattaat tactggttca 420
acttcatatg attggtatgt ttcttacgat aatgaaaaag tgggtgaact acaaggtctt 480
tcattagcag ctggattatt aggaaaaact gatggtgcat ttaaagatga aactgaaatg 540
ttaaactatc taaaagatca catgccacaa gaaactgttt cattttatgc agtagctgga 600
tcacaagatg acaacaattc tcaatacttt tacaacggag caatgaaaat tttaaaaaaa 660
ttaatggaaa actccaatgg taaagtagta gatttgtctc caagaggtaa tgcagtttat 720
gttccaggat ggaattacgg agacgctgga caaagaattc aacaattctt cacacaatat 780
cgtgattcat cagtgccaaa tggagtatta ccagttacat atgataatgt aactatccct 840
aaatctgttt taaaaggatt tttagcacca aatgatggaa tggcagaaca agcagtagac 900
aaactaaaag ctactaatta tgatgtacaa aaagttttca ttactggtca agactacaat 960
gatacagcaa aaaaacttat taaaaatggt gaacaaaata tgaccatcta caaaccagat 1020
gtaattttag gtaaagtagc tgtagaagta ttaaaaacat taataaacaa aaagaaagca 1080
aatgcaacag caactaaaga agatgttgaa aatgctttaa aaaacaacga aaatacaaaa 1140
gatattcact ttagatatga tgatacaaca tataaagctg gttcagtagg tgcttacaaa 1200
aacattaaag caattttagt taatccagtt gtagtaacta aacaaaatgt agataaccca 1260
ttagctaatg cacaacaatc tacaagtgca aatgtttctg tttctgcatc aggtacagta 1320
ccaactaatg gtgtaacaca ataa 1344
<210> 5
<211> 1794
<212> ДНК
<213> Искусственная последовательность
<220>
<223> антиген
<400> 5
atggcaaaag aaattatttt aggaatagat ttaggaacaa caaattctgt tgtttctata 60
atagaaaatg gtaagccagt tgtattagaa aatccaaatg gaaaaaatac aacaccttct 120
gttgttgctt ttaaaaacgg agaagaaatc gtaggagatg ctgctaaaag acaattagaa 180
acaaatcctg acgcaattgc ttcaattaaa agattaatgg gaacagataa aacagtaaaa 240
gcaaatggaa aagactataa accagaagaa atttctgcaa aaattctttc ttatttaaaa 300
caatatgcag agaaaaaaat tggtaacaaa gtttcaaaag cagttattac tgttccagct 360
tacttcaata atgcacaaag agaagcaaca aaaaatgcag gaaaaattgc tggcttagaa 420
gtagaaagaa ttattaatga accaactgca gctgctttag cttttggact agaaaaaact 480
gaaaaagaaa tgaaagtatt agtttacgac ttaggtggag gaacatttga cgtttctgta 540
ttagaacttt caggtggaac tttcgaagtt ttatcaacta gtggagataa caaattagga 600
ggagatgact gtgatcatgt tattgttgat tggttagttg acaaaattaa aaaagaatat 660
gaatttgatc cttctaaaga caaaatggct ttatctcgtt taaaagaaga agcagaaaaa 720
acaaaaattt ctttatccaa ccaatcagtt gctacaattt cattaccatt tttgggtatt 780
ggaccaaaag gacctattaa tgttgagtta gaactaaaaa gatcagaatt tgaaaaaatg 840
acagctcatt tagtagataa aacaagaaaa ccaattatgg atgctttaaa agaagcaaaa 900
ttagaagcaa aagatttaga tgaagtttta ttagttggtg gatcaacaag aattcctgct 960
gttcaaacaa tgattgagca cacattaggt aaaaaaccaa atagatccat taatccagat 1020
gaagttgttg ctattggagc tgcaattcaa ggtggtgttt tagctggaga aattaatgat 1080
gttttattac ttgacgttac acctttaact ttaggtattg aaacattagg tggaattgct 1140
actccattaa ttccaagaaa cacaacaatt cctgttacta aatcacaaat tttctcaact 1200
gctgaaaata accaaacaga agttacaatt tcagttgttc aaggagaaag acaattagct 1260
gctgataata aattattagg tagatttaat ctctcaggaa ttgaacaagc acctagagga 1320
gttcctcaaa ttgaagttag tttctcaatt gatgttaacg gaataacaac tgtatcagca 1380
aaagataaaa aaacaaacaa agaacaaact attactatta aaaatacaac cagcttatct 1440
gaagaagaaa ttgaaagaat ggtaaaagaa gctgaagaaa atcgtgaagc agatgctaag 1500
aaaaaagaaa aaattgaagt aacagttaga gctgaagctt taattaatca attagaaaaa 1560
tctctagaag accaaggtga taaagtagat gctaaacaaa aagaaacttt agaaaaacaa 1620
attcaagaat taaaagactt ggtaaaagaa gaaaaaattg aagaattaaa aacaaaatta 1680
gaccaaattg aacaagctgc acaagctttt gctcaagctg cagcacaaca agcaaatact 1740
tctgatactt cttcagatga tcaaccaatt gaagctgaag ttaaagaaaa ttaa 1794
<210> 6
<211> 1536
<212> ДНК
<213> Искусственная последовательность
<220>
<223> антиген
<400> 6
atgaataaaa aacatataaa aacacttatt tcaagtgtaa gtattttaac tcctgttgcc 60
attttagctt cttgtggaca accaaccaca attaaatttg ctacttctca aggttcatat 120
tggccaatga tgctaggaat gaaagaaatt attaagattt ataatgaaca acacaaaaac 180
gatgctgatt tcattcctgt tgaactttta acatcagatg taactcataa aaatagtgaa 240
ggtcaacttt taagtagttt agatagtgat ttatctacta agcaaagctc tgattggaat 300
cttattttag gaaacaaagc tactgcttat gttgcaaatt cttatgataa attattagac 360
gttggtactt ctacagtaaa tcccaattca ttccctacaa aaataattaa taattacaat 420
aaattattag gtgttgaagg acaaacaaca ttaaaaagtt taccttacaa catcaacgat 480
acagatggta ttgtttttaa tttagacatc atgaaagttc tttttgatat cataaaacaa 540
ggtggcggaa ctattgatga aaactcaata attgctaaaa aagtaaaaga agcagaagga 600
aaaggtaatc atattccatc aagttcaatg ttttctgcaa ttaaaattaa agagtcttca 660
aaaaacactg gattttctgg atatacagta aatgattcta cttttagtga tattaaaaaa 720
gcttttgagt ttgctcaaaa aatttatgat aacacagaaa ttgatacgac caaattatca 780
aaagatgtaa aagattcaga gatttttgca atagattatg cttcagatgt ttttagaaaa 840
gaaattcttt ctaaaactaa taaaactttt tggagtgaaa aaagtttaca aaataacaaa 900
attacattag atgtaaatct caatacaaat caagctctaa aaacagaagt tgaaaaccaa 960
tatcaagaat gggaaaacac tttaaaacaa acacaattta taccaactac aacaactcaa 1020
gcaaatactg aaactacttc atggacaaaa aaagatattc aaactaaatc agcaactgat 1080
gatagtcaac aatccattaa ttctaaaaca ttttattctg ttaaatttac tgaatacttt 1140
aaacctataa atcaatgggg atcttttgaa attagacaat atttaactgc atttacttat 1200
gctcctctag taggaacaaa ctattctgta gattctcctt ggtcacgtgc attttttaaa 1260
aaagatttag aagaaggaaa acaaaaagct tcagaatgga caacaagaga agatgtttat 1320
gctacaaatc aagcaatgaa agcagatgat aatgcacaat atttagcata taacgctggt 1380
ggtttttctt taattgctat caaatccaac agtgatataa taaataaaaa tatcattaaa 1440
tttgtagact ttttatacaa tggaacagga ctaacagatt taactggtgc taaaatttca 1500
gctgctgatt ttatggccga aaaatctgca gcttaa 1536
<210> 7
<211> 7245
<212> ДНК
<213> Искусственная последовательность
<220>
<223> вектор
<400> 7
atccggatat agttcctcct ttcagcaaaa aacccctcaa gacccgttta gaggccccaa 60
ggggttatgc tagttattgc tcagcggtgg cagcagccaa ctcagcttcc tttcgggctt 120
tgttagcagc cggatctcag tggtggtggt ggtggtgctc gagtgcggcc gcaagcttgt 180
cgacttgtgt tacaccatta gttggtactg tacctgatgc agaaacagaa acatttgcac 240
ttgtagattg ttgtgcatta gctaatgggt tatctacatt ttgtttagtt actacaactg 300
gattaactaa aattgcttta atgtttttgt aagcacctac tgaaccagct ttatatgttg 360
tatcatcata tctaaagtga atatcttttg tattttcgtt gttttttaaa gcattttcaa 420
catcttcttt agttgctgtt gcatttgctt tctttttgtt tattaatgtt tttaatactt 480
ctacagctac tttacctaaa attacatctg gtttgtagat ggtcatattt tgttcaccat 540
ttttaataag tttttttgct gtatcattgt agtcttgacc agtaatgaaa actttttgta 600
catcataatt agtagctttt agtttgtcta ctgcttgttc tgccattcca tcatttggtg 660
ctaaaaatcc ttttaaaaca gatttaggga tagttacatt atcatatgta actggtaata 720
ctccatttgg cactgatgaa tcacgatatt gtgtgaagaa ttgttgaatt ctttgtccag 780
cgtctccgta attccatcct ggaacataaa ctgcattacc tcttggagac aaatctacta 840
ctttaccatt ggagttttcc attaattttt ttaaaatttt cattgctccg ttgtaaaagt 900
attgagaatt gttgtcatct tgtgatccag ctactgcata aaatgaaaca gtttcttgtg 960
gcatgtgatc ttttagatag tttaacattt cagtttcatc tttaaatgca ccatcagttt 1020
ttcctaataa tccagctgct aatgaaagac cttgtagttc acccactttt tcattatcgt 1080
aagaaacata ccaatcatat gaagttgaac cagtaattaa tctatcataa gcaacgatag 1140
gaatattttt ttcagctatt gaattaacag cggttccaac tcctcctcca ttttcaggag 1200
caataataaa tcctttaggt gctggattta agtttgcttg ttgagttaac cagttgtttt 1260
gtgcattttg gtctttagta atagtagaaa ttgcaccttc tgttttgtct atataattta 1320
atatatcttt ttgtgcatta atccatcttg gattatctgg atctgttaat gcaactctaa 1380
tggattcatt agtaacagca tgatcttttg tttgtgataa acctgttgct gttgttccac 1440
aagctacaaa tgaggatcct ttaccgctgg tcattttttg gtgttcgtcg aggaattctt 1500
tcatctcttt cggcggctgg taacccggaa caagtgtgcc attgctcagc acaactgccg 1560
gagtaccgct aacgccaagc tggacgccaa gtgcgtaatg gtcggcaata tccacgtcgc 1620
aactggctgg tgcgacgctt ttacctgcca tcacatcatc aaacgctttg tttttatctt 1680
tcgcacacca gatagctttc atttctttct ctgcatcgct gtccagcccc tggcgcggga 1740
aagcaagata acgcacggtg atccccagcg cgttgtagtc tgccatttgc tcatgcagtt 1800
tgtggcagta accacaggta atatcagtaa acacggtgat gacgtgtttt tcctgcggcg 1860
ctttataaac gatcatctct ttttcaagcg cattcaactg ctttaacagc atcttattgg 1920
tgacattgac cggagccgtg ccactaacgt catacattgg cccctgaatg atatgtttac 1980
catcatcggt gatgtacaac acgccgctgt tagtcagaac tgtcttcatg ccagctacag 2040
gcgcgggctg aatatcgctg cttttgatgc ccattttggc taacgtttgt tgaattgccg 2100
cgtcatcggt acccagatct gggctgtcca tgtgctggcg ttcgaattta gcagcagcgg 2160
tttctttcat atgtatatct ccttcttaaa gttaaacaaa attatttcta gaggggaatt 2220
gttatccgct cacaattccc ctatagtgag tcgtattaat ttcgcgggat cgagatcgat 2280
ctcgatcctc tacgccggac gcatcgtggc cggcatcacc ggcgccacag gtgcggttgc 2340
tggcgcctat atcgccgaca tcaccgatgg ggaagatcgg gctcgccact tcgggctcat 2400
gagcgcttgt ttcggcgtgg gtatggtggc aggccccgtg gccgggggac tgttgggcgc 2460
catctccttg catgcaccat tccttgcggc ggcggtgctc aacggcctca acctactact 2520
gggctgcttc ctaatgcagg agtcgcataa gggagagcgt cgagatcccg gacaccatcg 2580
aatggcgcaa aacctttcgc ggtatggcat gatagcgccc ggaagagagt caattcaggg 2640
tggtgaatgt gaaaccagta acgttatacg atgtcgcaga gtatgccggt gtctcttatc 2700
agaccgtttc ccgcgtggtg aaccaggcca gccacgtttc tgcgaaaacg cgggaaaaag 2760
tggaagcggc gatggcggag ctgaattaca ttcccaaccg cgtggcacaa caactggcgg 2820
gcaaacagtc gttgctgatt ggcgttgcca cctccagtct ggccctgcac gcgccgtcgc 2880
aaattgtcgc ggcgattaaa tctcgcgccg atcaactggg tgccagcgtg gtggtgtcga 2940
tggtagaacg aagcggcgtc gaagcctgta aagcggcggt gcacaatctt ctcgcgcaac 3000
gcgtcagtgg gctgatcatt aactatccgc tggatgacca ggatgccatt gctgtggaag 3060
ctgcctgcac taatgttccg gcgttatttc ttgatgtctc tgaccagaca cccatcaaca 3120
gtattatttt ctcccatgaa gacggtacgc gactgggcgt ggagcatctg gtcgcattgg 3180
gtcaccagca aatcgcgctg ttagcgggcc cattaagttc tgtctcggcg cgtctgcgtc 3240
tggctggctg gcataaatat ctcactcgca atcaaattca gccgatagcg gaacgggaag 3300
gcgactggag tgccatgtcc ggttttcaac aaaccatgca aatgctgaat gagggcatcg 3360
ttcccactgc gatgctggtt gccaacgatc agatggcgct gggcgcaatg cgcgccatta 3420
ccgagtccgg gctgcgcgtt ggtgcggaca tctcggtagt gggatacgac gataccgaag 3480
acagctcatg ttatatcccg ccgttaacca ccatcaaaca ggattttcgc ctgctggggc 3540
aaaccagcgt ggaccgcttg ctgcaactct ctcagggcca ggcggtgaag ggcaatcagc 3600
tgttgcccgt ctcactggtg aaaagaaaaa ccaccctggc gcccaatacg caaaccgcct 3660
ctccccgcgc gttggccgat tcattaatgc agctggcacg acaggtttcc cgactggaaa 3720
gcgggcagtg agcgcaacgc aattaatgta agttagctca ctcattaggc accgggatct 3780
cgaccgatgc ccttgagagc cttcaaccca gtcagctcct tccggtgggc gcggggcatg 3840
actatcgtcg ccgcacttat gactgtcttc tttatcatgc aactcgtagg acaggtgccg 3900
gcagcgctct gggtcatttt cggcgaggac cgctttcgct ggagcgcgac gatgatcggc 3960
ctgtcgcttg cggtattcgg aatcttgcac gccctcgctc aagccttcgt cactggtccc 4020
gccaccaaac gtttcggcga gaagcaggcc attatcgccg gcatggcggc cccacgggtg 4080
cgcatgatcg tgctcctgtc gttgaggacc cggctaggct ggcggggttg ccttactggt 4140
tagcagaatg aatcaccgat acgcgagcga acgtgaagcg actgctgctg caaaacgtct 4200
gcgacctgag caacaacatg aatggtcttc ggtttccgtg tttcgtaaag tctggaaacg 4260
cggaagtcag cgccctgcac cattatgttc cggatctgca tcgcaggatg ctgctggcta 4320
ccctgtggaa cacctacatc tgtattaacg aagcgctggc attgaccctg agtgattttt 4380
ctctggtccc gccgcatcca taccgccagt tgtttaccct cacaacgttc cagtaaccgg 4440
gcatgttcat catcagtaac ccgtatcgtg agcatcctct ctcgtttcat cggtatcatt 4500
acccccatga acagaaatcc cccttacacg gaggcatcag tgaccaaaca ggaaaaaacc 4560
gcccttaaca tggcccgctt tatcagaagc cagacattaa cgcttctgga gaaactcaac 4620
gagctggacg cggatgaaca ggcagacatc tgtgaatcgc ttcacgacca cgctgatgag 4680
ctttaccgca gctgcctcgc gcgtttcggt gatgacggtg aaaacctctg acacatgcag 4740
ctcccggaga cggtcacagc ttgtctgtaa gcggatgccg ggagcagaca agcccgtcag 4800
ggcgcgtcag cgggtgttgg cgggtgtcgg ggcgcagcca tgacccagtc acgtagcgat 4860
agcggagtgt atactggctt aactatgcgg catcagagca gattgtactg agagtgcacc 4920
atatatgcgg tgtgaaatac cgcacagatg cgtaaggaga aaataccgca tcaggcgctc 4980
ttccgcttcc tcgctcactg actcgctgcg ctcggtcgtt cggctgcggc gagcggtatc 5040
agctcactca aaggcggtaa tacggttatc cacagaatca ggggataacg caggaaagaa 5100
catgtgagca aaaggccagc aaaaggccag gaaccgtaaa aaggccgcgt tgctggcgtt 5160
tttccatagg ctccgccccc ctgacgagca tcacaaaaat cgacgctcaa gtcagaggtg 5220
gcgaaacccg acaggactat aaagatacca ggcgtttccc cctggaagct ccctcgtgcg 5280
ctctcctgtt ccgaccctgc cgcttaccgg atacctgtcc gcctttctcc cttcgggaag 5340
cgtggcgctt tctcatagct cacgctgtag gtatctcagt tcggtgtagg tcgttcgctc 5400
caagctgggc tgtgtgcacg aaccccccgt tcagcccgac cgctgcgcct tatccggtaa 5460
ctatcgtctt gagtccaacc cggtaagaca cgacttatcg ccactggcag cagccactgg 5520
taacaggatt agcagagcga ggtatgtagg cggtgctaca gagttcttga agtggtggcc 5580
taactacggc tacactagaa ggacagtatt tggtatctgc gctctgctga agccagttac 5640
cttcggaaaa agagttggta gctcttgatc cggcaaacaa accaccgctg gtagcggtgg 5700
tttttttgtt tgcaagcagc agattacgcg cagaaaaaaa ggatctcaag aagatccttt 5760
gatcttttct acggggtctg acgctcagtg gaacgaaaac tcacgttaag ggattttggt 5820
catgaacaat aaaactgtct gcttacataa acagtaatac aaggggtgtt atgagccata 5880
ttcaacggga aacgtcttgc tctaggccgc gattaaattc caacatggat gctgatttat 5940
atgggtataa atgggctcgc gataatgtcg ggcaatcagg tgcgacaatc tatcgattgt 6000
atgggaagcc cgatgcgcca gagttgtttc tgaaacatgg caaaggtagc gttgccaatg 6060
atgttacaga tgagatggtc agactaaact ggctgacgga atttatgcct cttccgacca 6120
tcaagcattt tatccgtact cctgatgatg catggttact caccactgcg atccccggga 6180
aaacagcatt ccaggtatta gaagaatatc ctgattcagg tgaaaatatt gttgatgcgc 6240
tggcagtgtt cctgcgccgg ttgcattcga ttcctgtttg taattgtcct tttaacagcg 6300
atcgcgtatt tcgtctcgct caggcgcaat cacgaatgaa taacggtttg gttgatgcga 6360
gtgattttga tgacgagcgt aatggctggc ctgttgaaca agtctggaaa gaaatgcata 6420
aacttttgcc attctcaccg gattcagtcg tcactcatgg tgatttctca cttgataacc 6480
ttatttttga cgaggggaaa ttaataggtt gtattgatgt tggacgagtc ggaatcgcag 6540
accgatacca ggatcttgcc atcctatgga actgcctcgg tgagttttct ccttcattac 6600
agaaacggct ttttcaaaaa tatggtattg ataatcctga tatgaataaa ttgcagtttc 6660
atttgatgct cgatgagttt ttctaagaat taattcatga gcggatacat atttgaatgt 6720
atttagaaaa ataaacaaat aggggttccg cgcacatttc cccgaaaagt gccacctgaa 6780
attgtaaacg ttaatatttt gttaaaattc gcgttaaatt tttgttaaat cagctcattt 6840
tttaaccaat aggccgaaat cggcaaaatc ccttataaat caaaagaata gaccgagata 6900
gggttgagtg ttgttccagt ttggaacaag agtccactat taaagaacgt ggactccaac 6960
gtcaaagggc gaaaaaccgt ctatcagggc gatggcccac tacgtgaacc atcaccctaa 7020
tcaagttttt tggggtcgag gtgccgtaaa gcactaaatc ggaaccctaa agggagcccc 7080
cgatttagag cttgacgggg aaagccggcg aacgtggcga gaaaggaagg gaagaaagcg 7140
aaaggagcgg gcgctagggc gctggcaagt gtagcggtca cgctgcgcgt aaccaccaca 7200
cccgccgcgc ttaatgcgcc gctacagggc gcgtcccatt cgcca 7245
<210> 8
<211> 7500
<212> ДНК
<213> Искусственная последовательность
<220>
<223> вектор
<400> 8
atccggatat agttcctcct ttcagcaaaa aacccctcaa gacccgttta gaggccccaa 60
ggggttatgc tagttattgc tcagcggtgg cagcagccaa ctcagcttcc tttcgggctt 120
tgttagcagc cggatctcag tggtggtggt ggtggtgctc gagtgcggcc gcaagcttgt 180
cgacttaatt ttctttaact tcagcttcaa ttggttgatc atctgaagaa gtatcagaag 240
tatttgcttg ttgtgctgca gcttgagcaa aagcttgtgc agcttgttca atttggtcta 300
attttgtttt taattcttca attttttctt cttttaccaa gtcttttaat tcttgaattt 360
gtttttctaa agtttctttt tgtttagcat ctactttatc accttggtct tctagagatt 420
tttctaattg attaattaaa gcttcagctc taactgttac ttcaattttt tcttttttct 480
tagcatctgc ttcacgattt tcttcagctt cttttaccat tctttcaatt tcttcttcag 540
ataagctggt tgtattttta atagtaatag tttgttcttt gtttgttttt ttatcttttg 600
ctgatacagt tgttattccg ttaacatcaa ttgagaaact aacttcaatt tgaggaactc 660
ctctaggtgc ttgttcaatt cctgagagat taaatctacc taataattta ttatcagcag 720
ctaattgtct ttctccttga acaactgaaa ttgtaacttc tgtttggtta ttttcagcag 780
ttgagaaaat ttgtgattta gtaacaggaa ttgttgtgtt tcttggaatt aatggagtag 840
caattccacc taatgtttca atacctaaag ttaaaggtgt aacgtcaagt aataaaacat 900
cattaatttc tccagctaaa acaccacctt gaattgcagc tccaatagca acaacttcat 960
ctggattaat ggatctattt ggttttttac ctaatgtgtg ctcaatcatt gtttgaacag 1020
caggaattct tgttgatcca ccaactaata aaacttcatc taaatctttt gcttctaatt 1080
ttgcttcttt taaagcatcc ataattggtt ttcttgtttt atctactaaa tgagctgtca 1140
ttttttcaaa ttctgatctt tttagttcta actcaacatt aataggtcct tttggtccaa 1200
tacccaaaaa tggtaatgaa attgtagcaa ctgattggtt ggataaagaa atttttgttt 1260
tttctgcttc ttcttttaaa cgagataaag ccattttgtc tttagaagga tcaaattcat 1320
attctttttt aattttgtca actaaccaat caacaataac atgatcacag tcatctcctc 1380
ctaatttgtt atctccacta gttgataaaa cttcgaaagt tccacctgaa agttctaata 1440
cagaaacgtc aaatgttcct ccacctaagt cgtaaactaa tactttcatt tctttttcag 1500
ttttttctag tccaaaagct aaagcagctg cagttggttc attaataatt ctttctactt 1560
ctaagccagc aatttttcct gcattttttg ttgcttctct ttgtgcatta ttgaagtaag 1620
ctggaacagt aataactgct tttgaaactt tgttaccaat ttttttctct gcatattgtt 1680
ttaaataaga aagaattttt gcagaaattt cttctggttt atagtctttt ccatttgctt 1740
ttactgtttt atctgttccc attaatcttt taattgaagc aattgcgtca ggatttgttt 1800
ctaattgtct tttagcagca tctcctacga tttcttctcc gtttttaaaa gcaacaacag 1860
aaggtgttgt attttttcca tttggatttt ctaatacaac tggcttacca ttttctatta 1920
tagaaacaac agaatttgtt gttcctaaat ctattcctaa aataatttct tttgccatgg 1980
atccggtgcc gccaccagaa cgttcatccc actcatcagc tgctgcgcgt ccttcctcgg 2040
tatccagcgg tggctcaagc tgaaattccc cctcgtccca ttcatgtaat gtattctctt 2100
cctgccactc ctggcgtatc tctatctcat catagtcgcc atcaaaaaca ctttgcgcgg 2160
cttcaccgct aagcataggt aaacattcac cttcttcccc ttcgtcggca aaaaactcaa 2220
cttgccacat gatgtcgccg tcctgcaaaa cgtatttttg ggcattgaac tgttgcacat 2280
tcgcatcttc ggcgtcgatg ccggggttgt ctgcaagaaa ttcttcgcgt gcagcgtcaa 2340
tggcttcttc aagcgttgcg tacatggtca tggtaccacc accgctgtga tggtgatggt 2400
gatgcgatac gaattttttt ttcatatgta tatctccttc ttaaagttaa acaaaattat 2460
ttctagaggg gaattgttat ccgctcacaa ttcccctata gtgagtcgta ttaatttcgc 2520
gggatcgaga tcgatctcga tcctctacgc cggacgcatc gtggccggca tcaccggcgc 2580
cacaggtgcg gttgctggcg cctatatcgc cgacatcacc gatggggaag atcgggctcg 2640
ccacttcggg ctcatgagcg cttgtttcgg cgtgggtatg gtggcaggcc ccgtggccgg 2700
gggactgttg ggcgccatct ccttgcatgc accattcctt gcggcggcgg tgctcaacgg 2760
cctcaaccta ctactgggct gcttcctaat gcaggagtcg cataagggag agcgtcgaga 2820
tcccggacac catcgaatgg cgcaaaacct ttcgcggtat ggcatgatag cgcccggaag 2880
agagtcaatt cagggtggtg aatgtgaaac cagtaacgtt atacgatgtc gcagagtatg 2940
ccggtgtctc ttatcagacc gtttcccgcg tggtgaacca ggccagccac gtttctgcga 3000
aaacgcggga aaaagtggaa gcggcgatgg cggagctgaa ttacattccc aaccgcgtgg 3060
cacaacaact ggcgggcaaa cagtcgttgc tgattggcgt tgccacctcc agtctggccc 3120
tgcacgcgcc gtcgcaaatt gtcgcggcga ttaaatctcg cgccgatcaa ctgggtgcca 3180
gcgtggtggt gtcgatggta gaacgaagcg gcgtcgaagc ctgtaaagcg gcggtgcaca 3240
atcttctcgc gcaacgcgtc agtgggctga tcattaacta tccgctggat gaccaggatg 3300
ccattgctgt ggaagctgcc tgcactaatg ttccggcgtt atttcttgat gtctctgacc 3360
agacacccat caacagtatt attttctccc atgaagacgg tacgcgactg ggcgtggagc 3420
atctggtcgc attgggtcac cagcaaatcg cgctgttagc gggcccatta agttctgtct 3480
cggcgcgtct gcgtctggct ggctggcata aatatctcac tcgcaatcaa attcagccga 3540
tagcggaacg ggaaggcgac tggagtgcca tgtccggttt tcaacaaacc atgcaaatgc 3600
tgaatgaggg catcgttccc actgcgatgc tggttgccaa cgatcagatg gcgctgggcg 3660
caatgcgcgc cattaccgag tccgggctgc gcgttggtgc ggacatctcg gtagtgggat 3720
acgacgatac cgaagacagc tcatgttata tcccgccgtt aaccaccatc aaacaggatt 3780
ttcgcctgct ggggcaaacc agcgtggacc gcttgctgca actctctcag ggccaggcgg 3840
tgaagggcaa tcagctgttg cccgtctcac tggtgaaaag aaaaaccacc ctggcgccca 3900
atacgcaaac cgcctctccc cgcgcgttgg ccgattcatt aatgcagctg gcacgacagg 3960
tttcccgact ggaaagcggg cagtgagcgc aacgcaatta atgtaagtta gctcactcat 4020
taggcaccgg gatctcgacc gatgcccttg agagccttca acccagtcag ctccttccgg 4080
tgggcgcggg gcatgactat cgtcgccgca cttatgactg tcttctttat catgcaactc 4140
gtaggacagg tgccggcagc gctctgggtc attttcggcg aggaccgctt tcgctggagc 4200
gcgacgatga tcggcctgtc gcttgcggta ttcggaatct tgcacgccct cgctcaagcc 4260
ttcgtcactg gtcccgccac caaacgtttc ggcgagaagc aggccattat cgccggcatg 4320
gcggccccac gggtgcgcat gatcgtgctc ctgtcgttga ggacccggct aggctggcgg 4380
ggttgcctta ctggttagca gaatgaatca ccgatacgcg agcgaacgtg aagcgactgc 4440
tgctgcaaaa cgtctgcgac ctgagcaaca acatgaatgg tcttcggttt ccgtgtttcg 4500
taaagtctgg aaacgcggaa gtcagcgccc tgcaccatta tgttccggat ctgcatcgca 4560
ggatgctgct ggctaccctg tggaacacct acatctgtat taacgaagcg ctggcattga 4620
ccctgagtga tttttctctg gtcccgccgc atccataccg ccagttgttt accctcacaa 4680
cgttccagta accgggcatg ttcatcatca gtaacccgta tcgtgagcat cctctctcgt 4740
ttcatcggta tcattacccc catgaacaga aatccccctt acacggaggc atcagtgacc 4800
aaacaggaaa aaaccgccct taacatggcc cgctttatca gaagccagac attaacgctt 4860
ctggagaaac tcaacgagct ggacgcggat gaacaggcag acatctgtga atcgcttcac 4920
gaccacgctg atgagcttta ccgcagctgc ctcgcgcgtt tcggtgatga cggtgaaaac 4980
ctctgacaca tgcagctccc ggagacggtc acagcttgtc tgtaagcgga tgccgggagc 5040
agacaagccc gtcagggcgc gtcagcgggt gttggcgggt gtcggggcgc agccatgacc 5100
cagtcacgta gcgatagcgg agtgtatact ggcttaacta tgcggcatca gagcagattg 5160
tactgagagt gcaccatata tgcggtgtga aataccgcac agatgcgtaa ggagaaaata 5220
ccgcatcagg cgctcttccg cttcctcgct cactgactcg ctgcgctcgg tcgttcggct 5280
gcggcgagcg gtatcagctc actcaaaggc ggtaatacgg ttatccacag aatcagggga 5340
taacgcagga aagaacatgt gagcaaaagg ccagcaaaag gccaggaacc gtaaaaaggc 5400
cgcgttgctg gcgtttttcc ataggctccg cccccctgac gagcatcaca aaaatcgacg 5460
ctcaagtcag aggtggcgaa acccgacagg actataaaga taccaggcgt ttccccctgg 5520
aagctccctc gtgcgctctc ctgttccgac cctgccgctt accggatacc tgtccgcctt 5580
tctcccttcg ggaagcgtgg cgctttctca tagctcacgc tgtaggtatc tcagttcggt 5640
gtaggtcgtt cgctccaagc tgggctgtgt gcacgaaccc cccgttcagc ccgaccgctg 5700
cgccttatcc ggtaactatc gtcttgagtc caacccggta agacacgact tatcgccact 5760
ggcagcagcc actggtaaca ggattagcag agcgaggtat gtaggcggtg ctacagagtt 5820
cttgaagtgg tggcctaact acggctacac tagaaggaca gtatttggta tctgcgctct 5880
gctgaagcca gttaccttcg gaaaaagagt tggtagctct tgatccggca aacaaaccac 5940
cgctggtagc ggtggttttt ttgtttgcaa gcagcagatt acgcgcagaa aaaaaggatc 6000
tcaagaagat cctttgatct tttctacggg gtctgacgct cagtggaacg aaaactcacg 6060
ttaagggatt ttggtcatga acaataaaac tgtctgctta cataaacagt aatacaaggg 6120
gtgttatgag ccatattcaa cgggaaacgt cttgctctag gccgcgatta aattccaaca 6180
tggatgctga tttatatggg tataaatggg ctcgcgataa tgtcgggcaa tcaggtgcga 6240
caatctatcg attgtatggg aagcccgatg cgccagagtt gtttctgaaa catggcaaag 6300
gtagcgttgc caatgatgtt acagatgaga tggtcagact aaactggctg acggaattta 6360
tgcctcttcc gaccatcaag cattttatcc gtactcctga tgatgcatgg ttactcacca 6420
ctgcgatccc cgggaaaaca gcattccagg tattagaaga atatcctgat tcaggtgaaa 6480
atattgttga tgcgctggca gtgttcctgc gccggttgca ttcgattcct gtttgtaatt 6540
gtccttttaa cagcgatcgc gtatttcgtc tcgctcaggc gcaatcacga atgaataacg 6600
gtttggttga tgcgagtgat tttgatgacg agcgtaatgg ctggcctgtt gaacaagtct 6660
ggaaagaaat gcataaactt ttgccattct caccggattc agtcgtcact catggtgatt 6720
tctcacttga taaccttatt tttgacgagg ggaaattaat aggttgtatt gatgttggac 6780
gagtcggaat cgcagaccga taccaggatc ttgccatcct atggaactgc ctcggtgagt 6840
tttctccttc attacagaaa cggctttttc aaaaatatgg tattgataat cctgatatga 6900
ataaattgca gtttcatttg atgctcgatg agtttttcta agaattaatt catgagcgga 6960
tacatatttg aatgtattta gaaaaataaa caaatagggg ttccgcgcac atttccccga 7020
aaagtgccac ctgaaattgt aaacgttaat attttgttaa aattcgcgtt aaatttttgt 7080
taaatcagct cattttttaa ccaataggcc gaaatcggca aaatccctta taaatcaaaa 7140
gaatagaccg agatagggtt gagtgttgtt ccagtttgga acaagagtcc actattaaag 7200
aacgtggact ccaacgtcaa agggcgaaaa accgtctatc agggcgatgg cccactacgt 7260
gaaccatcac cctaatcaag ttttttgggg tcgaggtgcc gtaaagcact aaatcggaac 7320
cctaaaggga gcccccgatt tagagcttga cggggaaagc cggcgaacgt ggcgagaaag 7380
gaagggaaga aagcgaaagg agcgggcgct agggcgctgg caagtgtagc ggtcacgctg 7440
cgcgtaacca ccacacccgc cgcgcttaat gcgccgctac agggcgcgtc ccattcgcca 7500
<210> 9
<211> 7431
<212> ДНК
<213> Искусственная последовательность
<220>
<223> вектор
<400> 9
atccggatat agttcctcct ttcagcaaaa aacccctcaa gacccgttta gaggccccaa 60
ggggttatgc tagttattgc tcagcggtgg cagcagccaa ctcagcttcc tttcgggctt 120
tgttagcagc cggatctcag tggtggtggt ggtggtgctc gagtgcggcc gcaagcttgt 180
cgacttagtg atggtgatgg tgatgagctg cagatttttc ggccataaaa tcagcagctg 240
aaattttagc accagttaaa tctgttagtc ctgttccatt gtataaaaag tctacaaatt 300
taatgatatt tttatttatt atatcactgt tggatttgat agcaattaaa gaaaaaccac 360
cagcgttata tgctaaatat tgtgcattat catctgcttt cattgcttga tttgtagcat 420
aaacatcttc tcttgttgtc cattctgaag ctttttgttt tccttcttct aaatcttttt 480
taaaaaatgc acgtgaccaa ggagaatcta cagaatagtt tgttcctact agaggagcat 540
aagtaaatgc agttaaatat tgtctaattt caaaagatcc ccattgattt ataggtttaa 600
agtattcagt aaatttaaca gaataaaatg ttttagaatt aatggattgt tgactatcat 660
cagttgctga tttagtttga atatcttttt ttgtccatga agtagtttca gtatttgctt 720
gagttgttgt agttggtata aattgtgttt gttttaaagt gttttcccat tcttgatatt 780
ggttttcaac ttctgttttt agagcttgat ttgtattgag atttacatct aatgtaattt 840
tgttattttg taaacttttt tcactccaaa aagttttatt agttttagaa agaatttctt 900
ttctaaaaac atctgaagca taatctattg caaaaatctc tgaatctttt acatcttttg 960
ataatttggt cgtatcaatt tctgtgttat cataaatttt ttgagcaaac tcaaaagctt 1020
ttttaatatc actaaaagta gaatcattta ctgtatatcc agaaaatcca gtgttttttg 1080
aagactcttt aattttaatt gcagaaaaca ttgaacttga tggaatatga ttaccttttc 1140
cttctgcttc ttttactttt ttagcaatta ttgagttttc atcaatagtt ccgccacctt 1200
gttttatgat atcaaaaaga actttcatga tgtctaaatt aaaaacaata ccatctgtat 1260
cgttgatgtt gtaaggtaaa ctttttaatg ttgtttgtcc ttcaacacct aataatttat 1320
tgtaattatt aattattttt gtagggaatg aattgggatt tactgtagaa gtaccaacgt 1380
ctaataattt atcataagaa tttgcaacat aagcagtagc tttgtttcct aaaataagat 1440
tccaatcaga gctttgctta gtagataaat cactatctaa actacttaaa agttgacctt 1500
cactattttt atgagttaca tctgatgtta aaagttcaac aggaatgaaa tcagcatcgt 1560
ttttgtgttg ttcattataa atcttaataa tttctttcat tcctagcatc attggccaat 1620
atgaaccttg agaagtagca aatttaattg tggttggttg tccacaagag gatccgagga 1680
tttccagcag ttcgacttca aacaccaggg tgctgaacgg agggatggat gcgcctgcgc 1740
cgcgctcgcc atatgccagt tcctgcggga tagtcagttc ccatttagaa cctaccggca 1800
tcagagtcag tgcttcaatc cagccaggga tcacgccatt aaccgggaat tcagcgggtt 1860
caccacgagc aacggagctg tcaaacacgg tgccgtcgat cagtttaccg gtgtaatgaa 1920
cacgaacgcg gtcggtacgt gccggaattg cgccttcacc ctggttgatc acgcggaatt 1980
gcaggccaga ttcggtgcta ttcacacctt cttttttggc gttttcttcc aggtatttca 2040
caccttcagc agccatcgcc tggaaacgct gacgacgaac ggcatcggcg cgctcgtgga 2100
tttcacgcag cgcgcgatgc accacatcaa ccggaacagc cggatgtttg ccttccagcg 2160
catcggcaat acctgcaacc agtgcttctg gcagcagccc ttccaggcca gattcactca 2220
gttgttgccc tacctgcaaa ccaatgccgt agcttgcttg cgcttcgatg gtgtcaaaag 2280
ttggggtggt catggtaccc agatctgggc tgtccatgtg ctggcgttcg aatttagcag 2340
cagcggtttc tttcatatgt atatctcctt cttaaagtta aacaaaatta tttctagagg 2400
ggaattgtta tccgctcaca attcccctat agtgagtcgt attaatttcg cgggatcgag 2460
atcgatctcg atcctctacg ccggacgcat cgtggccggc atcaccggcg ccacaggtgc 2520
ggttgctggc gcctatatcg ccgacatcac cgatggggaa gatcgggctc gccacttcgg 2580
gctcatgagc gcttgtttcg gcgtgggtat ggtggcaggc cccgtggccg ggggactgtt 2640
gggcgccatc tccttgcatg caccattcct tgcggcggcg gtgctcaacg gcctcaacct 2700
actactgggc tgcttcctaa tgcaggagtc gcataaggga gagcgtcgag atcccggaca 2760
ccatcgaatg gcgcaaaacc tttcgcggta tggcatgata gcgcccggaa gagagtcaat 2820
tcagggtggt gaatgtgaaa ccagtaacgt tatacgatgt cgcagagtat gccggtgtct 2880
cttatcagac cgtttcccgc gtggtgaacc aggccagcca cgtttctgcg aaaacgcggg 2940
aaaaagtgga agcggcgatg gcggagctga attacattcc caaccgcgtg gcacaacaac 3000
tggcgggcaa acagtcgttg ctgattggcg ttgccacctc cagtctggcc ctgcacgcgc 3060
cgtcgcaaat tgtcgcggcg attaaatctc gcgccgatca actgggtgcc agcgtggtgg 3120
tgtcgatggt agaacgaagc ggcgtcgaag cctgtaaagc ggcggtgcac aatcttctcg 3180
cgcaacgcgt cagtgggctg atcattaact atccgctgga tgaccaggat gccattgctg 3240
tggaagctgc ctgcactaat gttccggcgt tatttcttga tgtctctgac cagacaccca 3300
tcaacagtat tattttctcc catgaagacg gtacgcgact gggcgtggag catctggtcg 3360
cattgggtca ccagcaaatc gcgctgttag cgggcccatt aagttctgtc tcggcgcgtc 3420
tgcgtctggc tggctggcat aaatatctca ctcgcaatca aattcagccg atagcggaac 3480
gggaaggcga ctggagtgcc atgtccggtt ttcaacaaac catgcaaatg ctgaatgagg 3540
gcatcgttcc cactgcgatg ctggttgcca acgatcagat ggcgctgggc gcaatgcgcg 3600
ccattaccga gtccgggctg cgcgttggtg cggacatctc ggtagtggga tacgacgata 3660
ccgaagacag ctcatgttat atcccgccgt taaccaccat caaacaggat tttcgcctgc 3720
tggggcaaac cagcgtggac cgcttgctgc aactctctca gggccaggcg gtgaagggca 3780
atcagctgtt gcccgtctca ctggtgaaaa gaaaaaccac cctggcgccc aatacgcaaa 3840
ccgcctctcc ccgcgcgttg gccgattcat taatgcagct ggcacgacag gtttcccgac 3900
tggaaagcgg gcagtgagcg caacgcaatt aatgtaagtt agctcactca ttaggcaccg 3960
ggatctcgac cgatgccctt gagagccttc aacccagtca gctccttccg gtgggcgcgg 4020
ggcatgacta tcgtcgccgc acttatgact gtcttcttta tcatgcaact cgtaggacag 4080
gtgccggcag cgctctgggt cattttcggc gaggaccgct ttcgctggag cgcgacgatg 4140
atcggcctgt cgcttgcggt attcggaatc ttgcacgccc tcgctcaagc cttcgtcact 4200
ggtcccgcca ccaaacgttt cggcgagaag caggccatta tcgccggcat ggcggcccca 4260
cgggtgcgca tgatcgtgct cctgtcgttg aggacccggc taggctggcg gggttgcctt 4320
actggttagc agaatgaatc accgatacgc gagcgaacgt gaagcgactg ctgctgcaaa 4380
acgtctgcga cctgagcaac aacatgaatg gtcttcggtt tccgtgtttc gtaaagtctg 4440
gaaacgcgga agtcagcgcc ctgcaccatt atgttccgga tctgcatcgc aggatgctgc 4500
tggctaccct gtggaacacc tacatctgta ttaacgaagc gctggcattg accctgagtg 4560
atttttctct ggtcccgccg catccatacc gccagttgtt taccctcaca acgttccagt 4620
aaccgggcat gttcatcatc agtaacccgt atcgtgagca tcctctctcg tttcatcggt 4680
atcattaccc ccatgaacag aaatccccct tacacggagg catcagtgac caaacaggaa 4740
aaaaccgccc ttaacatggc ccgctttatc agaagccaga cattaacgct tctggagaaa 4800
ctcaacgagc tggacgcgga tgaacaggca gacatctgtg aatcgcttca cgaccacgct 4860
gatgagcttt accgcagctg cctcgcgcgt ttcggtgatg acggtgaaaa cctctgacac 4920
atgcagctcc cggagacggt cacagcttgt ctgtaagcgg atgccgggag cagacaagcc 4980
cgtcagggcg cgtcagcggg tgttggcggg tgtcggggcg cagccatgac ccagtcacgt 5040
agcgatagcg gagtgtatac tggcttaact atgcggcatc agagcagatt gtactgagag 5100
tgcaccatat atgcggtgtg aaataccgca cagatgcgta aggagaaaat accgcatcag 5160
gcgctcttcc gcttcctcgc tcactgactc gctgcgctcg gtcgttcggc tgcggcgagc 5220
ggtatcagct cactcaaagg cggtaatacg gttatccaca gaatcagggg ataacgcagg 5280
aaagaacatg tgagcaaaag gccagcaaaa ggccaggaac cgtaaaaagg ccgcgttgct 5340
ggcgtttttc cataggctcc gcccccctga cgagcatcac aaaaatcgac gctcaagtca 5400
gaggtggcga aacccgacag gactataaag ataccaggcg tttccccctg gaagctccct 5460
cgtgcgctct cctgttccga ccctgccgct taccggatac ctgtccgcct ttctcccttc 5520
gggaagcgtg gcgctttctc atagctcacg ctgtaggtat ctcagttcgg tgtaggtcgt 5580
tcgctccaag ctgggctgtg tgcacgaacc ccccgttcag cccgaccgct gcgccttatc 5640
cggtaactat cgtcttgagt ccaacccggt aagacacgac ttatcgccac tggcagcagc 5700
cactggtaac aggattagca gagcgaggta tgtaggcggt gctacagagt tcttgaagtg 5760
gtggcctaac tacggctaca ctagaaggac agtatttggt atctgcgctc tgctgaagcc 5820
agttaccttc ggaaaaagag ttggtagctc ttgatccggc aaacaaacca ccgctggtag 5880
cggtggtttt tttgtttgca agcagcagat tacgcgcaga aaaaaaggat ctcaagaaga 5940
tcctttgatc ttttctacgg ggtctgacgc tcagtggaac gaaaactcac gttaagggat 6000
tttggtcatg aacaataaaa ctgtctgctt acataaacag taatacaagg ggtgttatga 6060
gccatattca acgggaaacg tcttgctcta ggccgcgatt aaattccaac atggatgctg 6120
atttatatgg gtataaatgg gctcgcgata atgtcgggca atcaggtgcg acaatctatc 6180
gattgtatgg gaagcccgat gcgccagagt tgtttctgaa acatggcaaa ggtagcgttg 6240
ccaatgatgt tacagatgag atggtcagac taaactggct gacggaattt atgcctcttc 6300
cgaccatcaa gcattttatc cgtactcctg atgatgcatg gttactcacc actgcgatcc 6360
ccgggaaaac agcattccag gtattagaag aatatcctga ttcaggtgaa aatattgttg 6420
atgcgctggc agtgttcctg cgccggttgc attcgattcc tgtttgtaat tgtcctttta 6480
acagcgatcg cgtatttcgt ctcgctcagg cgcaatcacg aatgaataac ggtttggttg 6540
atgcgagtga ttttgatgac gagcgtaatg gctggcctgt tgaacaagtc tggaaagaaa 6600
tgcataaact tttgccattc tcaccggatt cagtcgtcac tcatggtgat ttctcacttg 6660
ataaccttat ttttgacgag gggaaattaa taggttgtat tgatgttgga cgagtcggaa 6720
tcgcagaccg ataccaggat cttgccatcc tatggaactg cctcggtgag ttttctcctt 6780
cattacagaa acggcttttt caaaaatatg gtattgataa tcctgatatg aataaattgc 6840
agtttcattt gatgctcgat gagtttttct aagaattaat tcatgagcgg atacatattt 6900
gaatgtattt agaaaaataa acaaataggg gttccgcgca catttccccg aaaagtgcca 6960
cctgaaattg taaacgttaa tattttgtta aaattcgcgt taaatttttg ttaaatcagc 7020
tcatttttta accaataggc cgaaatcggc aaaatccctt ataaatcaaa agaatagacc 7080
gagatagggt tgagtgttgt tccagtttgg aacaagagtc cactattaaa gaacgtggac 7140
tccaacgtca aagggcgaaa aaccgtctat cagggcgatg gcccactacg tgaaccatca 7200
ccctaatcaa gttttttggg gtcgaggtgc cgtaaagcac taaatcggaa ccctaaaggg 7260
agcccccgat ttagagcttg acggggaaag ccggcgaacg tggcgagaaa ggaagggaag 7320
aaagcgaaag gagcgggcgc tagggcgctg gcaagtgtag cggtcacgct gcgcgtaacc 7380
accacacccg ccgcgcttaa tgcgccgcta cagggcgcgt cccattcgcc a 7431
<210> 10
<211> 37
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 10
gatataggat cctcatttgt agcttgtgga acaacag 37
<210> 11
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 11
caatatgtcg acttgtgtta caccattagt tggtactgta cc 42
<210> 12
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 12
gatccagata atccaagatg gattaatgca caaaaagata 40
<210> 13
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 13
tatctttttg tgcattaatc catcttggat tatctggatc 40
<210> 14
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 14
caaaatgcac aaaacaactg gttaactcaa caagcaaact 40
<210> 15
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 15
agtttgcttg ttgagttaac cagttgtttt gtgcattttg 40
<210> 16
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 16
ggttcaactt catatgattg gtatgtttct tacgataatg 40
<210> 17
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 17
cattatcgta agaaacatac caatcatatg aagttgaacc 40
<210> 18
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 18
ttatgttcca ggatggaatt acggagacgc tg 32
<210> 19
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 19
cagcgtctcc gtaattccat cctggaacat aa 32
<210> 20
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 20
gatataggat ccatggcaaa agaaattatt ttaggaatag atttag 46
<210> 21
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 21
caatatgtcg acttaatttt ctttaacttc agcttcaatt ggttg 45
<210> 22
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 22
gatgactgtg atcatgttat tgttgattgg ttagttgaca aaattaaaaa agaatatg 58
<210> 23
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 23
catattcttt tttaattttg tcaactaacc aatcaacaat aacatgatca cagtcatc 58
<210> 24
<211> 43
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 24
caatatggat cctcttgtgg acaaccaacc acaattaaat ttg 43
<210> 25
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 25
gtataagtcg acttagtgat ggtgatggtg atgagctgca gatttttcgg ccataaaatc 60
<210> 26
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 26
cttctcaagg ttcatattgg ccaatgatgc taggaatg 38
<210> 27
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 27
cattcctagc atcattggcc aatatgaacc ttgagaag 38
<210> 28
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 28
actaagcaaa gctctgattg gaatcttatt ttaggaaaca 40
<210> 29
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 29
tgtttcctaa aataagattc caatcagagc tttgcttagt 40
<210> 30
<211> 53
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 30
ctttctaaaa ctaataaaac tttttggagt gaaaaaagtt tacaaaataa caa 53
<210> 31
<211> 53
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 31
ttgttatttt gtaaactttt ttcactccaa aaagttttat tagttttaga aag 53
<210> 32
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 32
gttgaaaacc aatatcaaga atgggaaaac actttaaaac aaacac 46
<210> 33
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 33
gtgtttgttt taaagtgttt tcccattctt gatattggtt ttcaac 46
<210> 34
<211> 53
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 34
caagcaaata ctgaaactac ttcatggaca aaaaaagata ttcaaactaa atc 53
<210> 35
<211> 53
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 35
gatttagttt gaatatcttt ttttgtccat gaagtagttt cagtatttgc ttg 53
<210> 36
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 36
ctgaatactt taaacctata aatcaatggg gatcttttga aattagacaa tatttaac 58
<210> 37
<211> 58
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 37
gttaaatatt gtctaatttc aaaagatccc cattgattta taggtttaaa gtattcag 58
<210> 38
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 38
ctattctgta gattctcctt ggtcacgtgc attttttaaa aaag 44
<210> 39
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 39
cttttttaaa aaatgcacgt gaccaaggag aatctacaga atag 44
<210> 40
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 40
gaaaacaaaa agcttcagaa tggacaacaa gagaagatgt ttatg 45
<210> 41
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 41
cataaacatc ttctcttgtt gtccattctg aagctttttg ttttc 45
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИЯ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ ИНФЕКЦИИ MYCOPLASMA HYORHINIS И СПОСОБ ПОЛУЧЕНИЯ УКАЗАННОЙ КОМПОЗИЦИИ | 2020 |
|
RU2737971C1 |
| СУБЪЕДИНИЧНАЯ ВАКЦИНА ПРОТИВ MYCOPLASMA SPP | 2019 |
|
RU2759427C2 |
| КОМПОЗИЦИЯ ПО ПРЕДУПРЕЖДЕНИЮ ИНФЕКЦИИ Mycoplasma spp. | 2013 |
|
RU2646137C2 |
| КОМПОЗИЦИИ NEISSERIA MENINGITIDIS И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2739504C2 |
| РЕКОМБИНАНТНЫЕ ВЕКТОРЫ, ЭКСПРЕССИРУЮЩИЕ АНТИГЕНЫ ВИРУСА ПТИЧЬЕГО ГРИППА, И ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2761869C2 |
| ПОЛИВАЛЕНТНАЯ ИММУНОГЕННАЯ КОМПОЗИЦИЯ ПРОТИВ ПАПИЛЛОМАВИРУСА ЧЕЛОВЕКА | 2020 |
|
RU2806424C2 |
| КОМПОЗИЦИЯ МИКОБАКТЕРИАЛЬНОГО АНТИГЕНА | 2015 |
|
RU2702194C2 |
| Способ создания живого штамма энтерококка L3-SARS на основе биологически активного штамма Е. faecium L3 | 2021 |
|
RU2782529C1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРЕДОТВРАЩЕНИЯ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2016 |
|
RU2714205C2 |
| ПАРВОВИРУС СВИНЕЙ | 2015 |
|
RU2817872C2 |
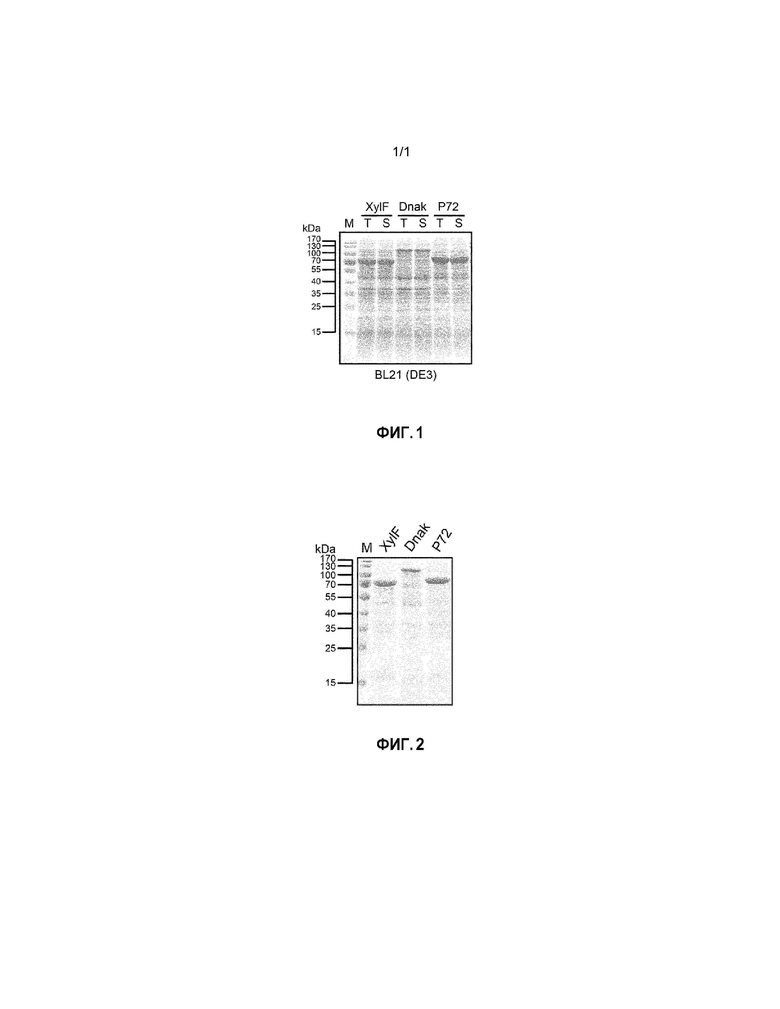
Изобретение относится к области биотехнологии, конкретно к получению субъединичных вакцин против Mycoplasma hyorhinis, и может быть использовано в медицине для профилактики инфекции Mycoplasma hyorhinis. Предложена вакцинная композиция на основе белка XylF с SEQ ID NO:1 в сочетании с адъювантом. Для рекомбинантного получения активного начала вакцины предложен экспрессирующий вектор для получения XylF в прокариотической экспрессирующей системе, содержащий экспрессионный элемент, который содержит промотор и участок связывания рибосомы; нуклеотидную последовательность, кодирующую указанный XylF; и последовательность, кодирующую партнер слияния, выбранный из группы, состоящей из DsbC из E. coli, MsyB из E. coli, FklB из E. coli или их комбинации. Изобретение обеспечивает достижение сильного иммунного ответа против Mycoplasma hyorhinis. 4 н. и 7 з.п. ф-лы, 7 табл., 3 пр., 2 ил.
1. Композиция для предотвращения заболевания, обусловленного инфекцией Mycoplasma hyorhinis, содержащая:
эффективное количество активного ингредиента, представляющего собой XylF; и
адъювант;
где указанный XylF представляет собой SEQ ID NO: 01.
2. Композиция по п.1, в которой указанный активный ингредиент имеет концентрацию от 50 до 300 мкг/мл на основе общего объема указанной композиции.
3. Композиция по любому из пп. 1-2, в которой указанный адъювант включает полный адъювант Фрейнда, неполный адъювант Фрейнда, гель оксида алюминия, поверхностно-активное средство, анионный полимер, пептид, масляную эмульсию или их комбинацию.
4. Композиция по п.1, где указанное заболевание представляет собой перитонит, плеврит или их комбинацию при условии, что указанный активный ингредиент представляет собой XylF.
5. Экспрессирующий вектор для получения XylF в прокариотической экспрессирующей системе, содержащий:
экспрессионный элемент, который содержит промотор и участок связывания рибосомы;
нуклеотидную последовательность, кодирующую указанный XylF; и
последовательность, кодирующую партнер слияния, выбранный из группы, состоящей из DsbC из E. coli, MsyB из E. coli, FklB из E. coli или их комбинации;
где указанная нуклеотидная последовательность представляет собой SEQ ID NO: 04.
6. Экспрессирующий вектор по п.5, дополнительно содержащий последовательность, кодирующую гистидиновую метку, метку глутатион-S-трансферазы или их комбинацию.
7. Экспрессирующий вектор по п.5, представляющий собой нуклеотидную последовательность, как показано в SEQ ID NO: 07.
8. Экспрессирующий вектор по п.5, в котором указанная прокариотическая экспрессирующая система представляет собой экспрессирующую систему E. coli.
9. Способ получения растворимого белка, где указанный растворимый белок представляет собой XylF; где указанный способ включает:
получение прокариотической экспрессирующей системы;
экспрессию указанной нуклеотидной последовательности экспрессирующего вектора по любому из пп. 5-8 в указанной прокариотической экспрессирующей системе; и
пропускание продукта, полученного на вышеуказанной стадии экспрессии, через аффинную колонку с ионами никеля или глутатионовую аффинную колонку для получения указанного растворимого белка.
10. Применение композиции по любому из пп. 1-4 для получения вакцины для предотвращения заболевания, обусловленного инфекцией Mycoplasma hyorhinis,
где указанное заболевание представляет собой по меньшей мере одно заболевание, выбранное из перитонита, плеврита, перикардита и отека суставов.
11. Применение по п.10, где указанное заболевание представляет собой перитонит, плеврит или их комбинацию, при условии, что указанный активный ингредиент представляет собой XylF.
| WO 2014121433 A1, 14.08.2014 | |||
| LIU M | |||
| et al., Mycoplasma hyopneumoniae DnaK Gene and ELISA Method, Journal of Jinling Institute of Technology, 2011, v | |||
| Прибор с двумя призмами | 1917 |
|
SU27A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| FENG Z | |||
| et al., The Progress of Prevention for Mycoplasma Pneumonia of Swine, Proceedings of the Third Symposium on Prevention and Control of Swine Disease of Chinese | |||

