Настоящее изобретение относится к новому парвовирусу свиней, к белкам этого вируса и к вакцинам на основе этого вируса и его белков. Изобретение также относится к фрагментам ДНК, включающим ген вируса, и к ДНК вакцинам на основе генов вируса. Кроме того, изобретение относится к антителам, которые реагируют с новым вирусом, и к диагностическим тестам для определения вируса или антител к вирусу.
В течение последних десятилетий во всем мире наблюдают существенное увеличение потребления свиного мяса. Как следствие, наблюдают увеличение количества и размера ферм с целью соответствия увеличивающейся потребности на рынке. Как известно из животноводства в общем, большое количество животных, живущих тесно вместе, подвержены всем типам заболеваний, даже заболеваниям, плохо известным или наблюдаемым, или даже неизвестным до периода широкомасштабного коммерческого фермерского хозяйства.
Одним из заболеваний у свиней, которое известно в течение более 60 лет, в настоящее время является геморрагический кишечный синдром (HBS). Указанное заболевание называют синдромом из-за того факта, что причина заболевания неясна и согласованность различных клинических признаков не всегда полностью очевидна.
HBS представляет собой заболевание, которое развивается редкими, взрывными вспышками. Преимущественно поражаются быстро растущие свиньи в возрасте 4-6 месяцев. В большинстве случаев заболевание наблюдают у свиней на откормке. Свиньи внезапно умирают без проявлений диареи, хотя степень смертности варьирует(1-3). Шведские данные аутопсии на основании более чем 16000 свиней продемонстрировали частоту HBS 2,66%. В USA сообщают, что HBS вызывает 0,5%-7% всей смертности в течение фазы завершения роста(3).
Геморрагический кишечный синдром не рассматривается как единственное заболевание с единственной причиной.
Наиболее выраженным симптомом HBS являются кишечные кровоизлияния, часто в сочетании с заворотом кишечника (скручивание кишечника). Однако такие симптомы часто являются признаками желудочных язв и илеита, что осложняет диагностику HBS. Может развиваться ротация всего кишечника, вызывая скопление и застой крови. Заворот кишечника может наблюдаться в до 80% случаев HBS(1-3). Другими часто наблюдаемыми симптомами заболевания являются тонкие стенки кишечника и кровянистая жидкость в кишечнике.
Точная этиология HBS неясна. Как показано выше, скорее, чем одна причина, наиболее вероятно этиология синдрома является многофакторной. Могут играть роль стресс, некоторые аспекты окружающей среды и содержания. Предрасполагающие факторы могут включать интенсивную физическую нагрузку, погрузку, драки, складирование или нерегулярное кормление. Не существует убедительных доказательств, что инфекционный агент (бактериальный или вирусный) может вызывать HBS(1,3), хотя Clostridium sp. и E. coli выделяли от животных, страдающих от HBS. Попытки воспроизвести заболевание путем введения кишечного содержимого от животных, страдающих от HBS, внутривенно или перорально, здоровым животным, провалились. Попытки воспроизвести заболевание путем пероральной инокуляции E. coli и Clostridium perfringens типа A, выделенных от инфицированных свиней, также оказались безуспешными.
С другой стороны, известно, что частота заболевания может быть снижена в определенной степени путем введения в пищу антибиотиков. Это усиливает идею, что заболевание является действительно многофакторным: комбинированный эффект, например, стресса и одного или более патогенов.
Задачей настоящего изобретения является получение нового инфекционного агента, ассоциированного с таким заболеванием, а также вакцин, нацеленных на борьбу с заболеванием или по меньшей мере снижение смертности от заболевания. Более того, задачей настоящего изобретения является обеспечение средств для определения и идентификации инфекционного агента, ассоциированного с заболеванием.
Некоторое время назад свиней с диагнозом HBS собрали с нескольких ферм во время вспышки заболевания в Мексике.
Пораженные свиньи не имели предшествующих симптомов заболевания и умирали неожиданно, между 2 и 6 часами после первых признаков заболевания.
При аутопсии у свиней наблюдали патологию тонкой кишки, среди прочего, геморрагические симптомы, истончение стенки кишечника и кровянистую жидкость в кишечнике. Не обнаруживали патологии в других органах, за исключением увеличенных, отекших красных лимфатических узлов. Заболевание подтверждали как HBS.
Образцы от вскрытых пораженных свиней с различных ферм анализировали в отношении присутствия вирусов и неожиданно новый вирус обнаружили у 76% животных. Тот факт, что вирус не определяли у всех животных, может иметь место из-за количества времени, прошедшего между смертью животных, и моментом, когда их подвергали посмертному вскрытию. Такой вывод может быть сделан из того факта, что количества вируса, обнаруживаемые у животного, варьировались в большой степени. Авторы предположили, что у свиней, у которых вирус очевидно отсутствовал, это происходило вероятно из-за того факта, что количество присутствующего вируса у таких свиней было ниже уровня определения в момент анализа. Более того, место исходной репликации вируса неизвестно для указанного нового вируса и, следовательно, основное место репликации вируса после инфицирования может быть не определено.
Так как новый вирус определяли у свиней с диагнозом HBS, вирус далее называют как HBS-ассоциированный вирус. Геморрагический синдром кишечника в настоящее время может быть охарактеризован присутствием нового вируса в соответствии с изобретением на некоторых стадиях в течение заболевания в органах животных, страдающих от HBS, в комбинации со следующими клиническими симптомами: кишечные кровоизлияния, часто сопровождаемые заворотом кишечника, истончением стенок кишечника и кровянистой жидкостью в кишечнике.
Анализировали последовательность вирусного генома и выявили, что новый вирус несет некоторый, хотя и относительно низкий уровень сходства с в последнее время идентифицированным родом субсемейства Parvovirinae в Parvoviridae.
Парвовирусы являются линейными, несегментированными одноцепочечными ДНК вирусами со средним размером генома 5000 нуклеотидов и размером в диапазоне 18-26 нм в диаметре.
Почти полнодлинновая последовательность ДНК, характерная для парвовируса свиней, представлена в SEQ ID NO: 10.
Новый вирус включает две больших открытых рамки считывания (ORF): ORF1, кодирующая неструктурный белок 1 (NS1), состоящий из 662 аминокислот, обнаруживается в положении 0134-2122 SEQ ID NO: 10 и ORF2, кодирующая капсидный белок (CP), состоящий из 1189 аминокислот, обнаруживается в положении 2130-5699 SEQ ID NO: 10.
Пример последовательности ДНК ORF2, гена, кодирующего капсидный белок, представлен в SEQ ID NO: 1. SEQ ID NO: 2 представляет собой последовательность аминокислот капсидного белка.
Пример последовательности ДНК ORF1, кодирующей неструктурный белок NS1, представлен в SEQ ID NO: 3. SEQ ID NO: 4 представляет собой последовательность аминокислот неструктурного белка NS1.
Подсемейство Parvovirinae в настоящее время включает 7 родов(15):
1) PARV4-подобный вирус
2) эритровирус
3) бокавирус
4) депендовирус
5) амдовирус
6) парвовирус
7) заново предложенная парво филогенетическая ветвь
В настоящее время были идентифицированы шесть различных парвовирусов, которые инфицируют свиней:
а) классический парвовирус свиней тип 1 (PPV1), член рода Parvovirus
b) парвовирус свиней тип 2 (PPV2), член рода PARV4-подобных вирусов
с) парвовирус свиней тип 3 (PPV3, также известный как свиной PARV4, хоковирус или партетравирус), также член рода PARV4-подобного вируса
d) парвовирус свиней тип 4 (PPV4), член заново предложенной филогенетической ветви
e) парвовирус свиней тип 5 (PPV5), также член заново предложенной филогенетической ветви
f) бокавирус свиней (PBoV), член рода Bocavirus
PPV1 известен как этиологический агент SMEDI, синдрома, связанного с мертворождением, мумификацией, смертью эмбриона и бесплодием.(4,5)
Не подтверждено, что PPV2 вызывает заболевание, как таковое, но предположительно является кофактором развития заболевания, ассоциированного с цирковирусом свиней (PCVAD)(6,7).
Не подтверждено, что PPV3 вызывает заболевание, как таковое, но возможно он также является кофактором в развитии заболевания, ассоциированного с цирковирусом свиней (PCVAD)8-11).
PPV4 выделили исходно из ткани легких свиней. Ткань, вероятно, совместно инфицирована цирковирусом свиней. Неизвестно, вызывает ли он заболевание и также четко не был ассоциирован с заболеванием, вызванным другим патогеном(12-14).
Также неизвестно, вызывает ли PPV5 какие-то симптомы или повреждения, и он не ассоциирован с заболеванием, вызванным другим патогеном(15-16).
Бокавирус свиней является относительно новым типом парвовируса свиней, для которого до настоящего времени клиническая значимость и эпидемиология широко не исследованы(17).
Последовательности аминокислот ORF1 и ORF2 нового вируса использовали для получения филогенетических схем на основании метода максимальной вероятности, модели коррекции Пуассона и бутстрэп-анализа (500 репликатов).
Такие схемы получали с использованием программы MEGA, версия 5, с использованием стандартных настроек. (MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Koichiro Tamura, Daniel Peterson, Nicholas Peterson, Glen Stecher, Masatoshi Nei and Sudhir Kumar. Mol. Biol. Evol. 28(10): 2731-2739. 2011 doi:10.1093/molbev/msr121 Advance Access publication May 4, 2011).
Филогенетическая схема ORF1 представлена на фиг. 1, таковая ORF2 на фиг. 2. Процент бустрэп поддержки указан в узлах. Столбики расстояния показывают количество замен нуклеотидов на сайт.
Как следует из указанных филогенетических схем новый парвовирус свиней скорее относится к парвовирусу свиней 5 (PPV5) и парвовирусу свиней 4 (PPV4), чем к PPV1, 2 или 3, или бокавирусам. Было обнаружено, что и последовательность, кодирующая NS1, и последовательность, кодирующая капсидный белок, демонстрируют определенное соответствие в той части филогенетической схемы Parvovirinae, которая также включает несвязанные вирусы PPV4 и PPV5. По этой причине авторы изобретения решили условно поместить новый вирус в группу вирусов новой филогенетической ветви.
Однако, идентичность последовательности с существующими парвовирусами свиней, даже в группе вирусов новой филогенетической ветви является относительно низкой. По этой причине более вероятно, что новый вирус принадлежит к новому роду в семействе Parvovirinae.
SEQ ID NO: 1 и 3 демонстрируют типичные примеры последовательности нуклеотидов генов, кодирующих капсидный белок и неструктурный белок NS1 вируса по изобретению.
Понятно, что для указанных белков могут существовать натуральные варианты среди отдельных представителей HBS-ассоциированного вируса. Существуют генетические варианты, приводящие к минимальным изменениям в, например, последовательности капсидного белка. Это в равной степени верно для гена NS1. Прежде всего, существует так называемое ʺколебание во втором и третьем основанииʺ, объясняющее, что могут существовать изменения нуклеотидов, которые остаются незамеченными в последовательности аминокислот, они кодируют: например, триплеты TTA, TTG, TCA, TCT, TCG и TCC, все кодирующие лейцин. Кроме того, могут наблюдаться минимальные варианты в последовательности аминокислот характерных представителей нового парвовируса свиней в соответствии с изобретением. Такие варианты могут быть отражены различием(ями) аминокислот в целой последовательности или посредством делеций, замен, вставок, инверсий или добавлений аминокислот(ы) в указанной последовательности. Замены аминокислот, которые существенно не изменяют биологическую и иммунологическую активности, были описаны, например, Neurath et al в "The Proteins" Academic Press New York (1979). Замены аминокислот между связанными аминокислотами или замены, которые часто возникают в эволюции, представляют собой, между прочим, Ser/Ala, Ser/Gly, Asp/Gly, Asp/Asn, Ile/Val (см. Dayhof, M.D., Atlas of protein sequence and structure, Nat. Biomed. Res. Found., Washington D.C., 1978, vol. 5, suppl. 3). Другие замены аминокислот включают Asp/Glu, Thr/Ser, Ala/Gly, Ala/Thr, Ser/Asn, Ala/Val, Thr/Phe, Ala/Pro, Lys/Arg, Leu/Ile, Leu/Val и Ala/Glu. На основании указанной информации, Lipman и Pearson разработали метод быстрого и чувствительного сравнения белка (Science 227, 1435-1441, 1985) и определения функционального сходства между гомологичными белками. Такие замены аминокислот примерных вариантов осуществления изобретения, а также варианты, имеющие делеции и/или вставки, находятся в рамках изобретения.
Это объясняет, почему капсидный белок и неструктурный белок NS1 при выделении из различных представителей парвовируса свиней по изобретению, могут иметь уровень гомологии, который значительно ниже 100%, при этом все еще представляя собой капсидный белок и неструктурный белок NS1 парвовируса свиней по изобретению.
Это четко отражено, например, в филогенетической схеме на фиг. 1 статьи Xiao et al.,(6) где показано, что даже в одном единственном роде PARV4-подобного вируса, роде, состоящем из высоко связанных парвовирусов, тем не менее имеются различные общие геномные последовательности нуклеотидов, а также достоверно различающиеся последовательности нуклеотидов гена NS1.
Следовательно, вирус по изобретению описан, среди прочего, как выделенный вирус, который является членом подсемейства Parvovirinae семейства Parvoviridae, указанный вирус характеризуется тем, что
a) вирус является HBS-ассоциированным вирусом и
b) вирус имеет вирусный геном, включающий ген, кодирующий капсидный белок (CP), где последовательность нуклеотидов гена CP имеет уровень идентичности по меньшей мере 80% относительно последовательности нуклеотидов, как изображено в SEQ ID NO: 1.
Для целей настоящего изобретения уровень идентичности понимают как уровень идентичности последовательности SEQ ID NO: 1 и соответствующего участка, кодирующего капсидный белок парвовируса свиней, уровень идентичности которого определяют.
Подходящей программой для определения уровня идентичности является нуклеотидная бласт программа (blastn) NCBI's Basic Local Alignment Search Tool, с использованием опции ʺвыравнивание двух или более последовательностейʺ и стандартных установок (http://blast.ncbi.nlm.nih.gov/Blast.cgi).
Для целей настоящего изобретения изолированный обозначает: выделенный из ткани, с которой вирус ассоциирован в природе. Примером изолированного вируса является вирус, который присутствует в культуре клеток.
Предпочтительная форма варианта осуществления настоящего изобретения относится к вирусу, который имеет ген капсидного белка, который имеет уровень идентичности по меньшей мере 82%, более предпочтительно 84%, 86%, 88%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или даже 100%, в таком порядке предпочтительности, последовательности нуклеотидов капсидного белка, как изображено в SEQ ID NO: 1.
Альтернативный путь описания вируса по настоящему изобретению относится к последовательности гена NS1 вируса.
SEQ ID NO: 3 демонстрирует типичный пример последовательности нуклеотидов гена NS1 вируса по изобретению. Как объясняется выше, однако обнаруживают натуральные варианты, приводящие к минимальным изменениям последовательности NS1.
Следовательно, вирус по изобретению также может быть описан как выделенный вирус, который является членом субсемейства Parvovirinae семейства Parvoviridae, указанный вирус отличается тем, что
a) вирус является HBS-ассоциированным вирусом и
b) вирус имеет вирусный геном, включающий ген, кодирующий неструктурный белок 1 (NS1), где последовательность нуклеотидов гена NS1 имеет уровень иденичности по меньшей мере 80% относительно последовательности нуклеотидов, как изображено в SEQ ID NO: 3.
Предпочтительная форма варианта осуществления настоящего изобретения относится к такому вирусу, который имеет NS1 ген, который имеет уровень идентичности по меньшей мере 82%, более предпочтительно 84%, 86%, 88%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или даже 100%, в таком порядке предпочтительности относительно последовательности нуклеотидов гена NS1, как изображено в SEQ ID NO: 3.
Следовательно, в общем вирус по настоящему изобретению представляет собой выделенный вирус, который является членом субсемейства Parvovirinae семейства Parvoviridae, указанный вирус отличается тем, что
a) вирус представляет собой HBS-ассоциированный вирус и
b) вирус имеет вирусный геном, включающий ген, кодирующий капсидный белок (CP), и ген, кодирующий неструктурный белок 1 (NS1), где последовательность нуклеотидов гена CP имеет уровень идентичности по меньшей мере 80% относительно последовательности нуклеотидов, как изображено в SEQ ID NO: 1, или последовательность нуклеотидов гена NS1 имеет уровень идентичности по меньшей мере 80% относительно последовательности нуклеотидов, как изображено в SEQ ID NO: 3.
Предпочтительная форма варианта осуществления настоящего изобретения относится к изолированному вирусу, который является членом субсемейства Parvovirinae семейства Parvoviridae, указанный вирус отличается тем, что
a) вирусом является HBS-ассоциированный вирус и
b) вирус имеет вирусный геном, включающий ген, кодирующий капсидный белок (CP), и ген, кодирующий неструктурный белок 1 (NS1), где последовательность нуклеотидов гена CP имеет уровень идентичности по меньшей мере 80% относительно последовательности нуклеотидов, как изображено в SEQ ID NO: 1, и последовательность нуклеотидов гена NS1 имеет уровень идентичности по меньшей мере 80% относительно последовательности нуклеотидов, как изображено в SEQ ID NO: 3.
Еще один альтернативный вариант характеристики вируса по изобретению зависит от ПЦР-тестирования с использованием группы праймеров, которые являются специфичными для последовательности гена капсидного белка или последовательности гена NS1 вируса по изобретению. Две различные группы праймеров, последовательность которых изображена в SEQ ID NO: 5-6 и SEQ ID NO: 7-8, были выбраны из-за их специфичности к вирусу. В ПЦР-исследовании с использованием первой группы праймеров (SEQ ID NO: 5-6), которые специфически реагируют с геном капсидного белка вируса, используют два праймера Bowl_Q_ORF2_FW: CTACATCTGCGCCTGAC и Bowl_Q_ORF2_REV: GTGGTGAGAAGGCAAGAC,
Для количественного(К)-ПЦР анализа используют зонд Bowl_Q_ORF2_PROBE: 6FAM-CACGAGCTAGAGCGTGCTAAACAG-BHQ1 как изображено в SEQ DI NO.: 9, в добавление к указанным двум праймерам.
В ПЦР исследовании с использованием второй группы праймеров (SEQ ID NO: 7-8) специфически реагируют с геном NS1 вируса и применение двух праймеров Bowl_ORF1_774_F: TGTTGAGTGTGGTGGATTGG и Bowl_ORF1_1626_R: AAGGAAGCTGGACCGAGAG.
Тесты, которые описаны более подробно в разделе Примеры, являются стандартными ПЦР тестами.
Если член субсемейства Parvovirinae в рамках Parvoviridae анализируют с использованием группы праймеров, описанных выше, может быть сказано следующее: если анализ ПЦР-продукта первой группы праймеров выявляет ПЦР продукт приблизительно из 140 пар оснований, или если анализ ПЦР-продукта второй группы праймеров выявляет ПЦР продукт из приблизительно 853 пар оснований, это однозначно демонстрирует, что анализируемый вирус принадлежит к вирусам по изобретению.
Исключительно в качестве примера: ПЦР продукт приблизительно из 853 пар оснований представляет собой ПЦР продукт с длиной от 853+10 и 853-10 пар оснований. ПЦР продукт приблизительно из 140 пар оснований представляет собой ПЦР продукт с длиной от 140+10 до 140-10 пар оснований.
Следовательно, еще одна форма варианта осуществления настоящего изобретения относится к изолированному вирусу, который является членом субсемейства Parvovirinae subfamily в Parvoviridae, отличающемуся тем, что:
a) вирус является HBS-ассоциированным вирусом и
b) вирусная геномная ДНК реагирует в ПЦР реакции с группой праймеров, как изображено в SEQ ID NO: 5 и 6, с образованием ПЦР продукта из 140+/-10 пар оснований, или реагирует в ПЦР реакции с группой праймеров, как изображено в SEQ ID NO: 7 и 8, с образованием продукта ПЦР из 853+/-10 пар оснований.
Предпочтительная форма варианта осуществления настоящего изобретения относится к вирусу по изобретению, где вирусная геномная ДНК реагирует в ПЦР реакции с группой праймеров, как изображено в SEQ ID NO: 5 и 6, для получения ПЦР продукта из 140+/-10 пар оснований, и реагирует в ПЦР реакции с группой праймеров, как изображено в SEQ ID NO: 7 и 8, для получения ПЦР продукта из 853+/-10 пар оснований.
Более предпочтительная форма варианта осуществления настоящего изобретения относится к вирусу по изобретению, где вирус имеет вирусный геном, включающий ген, кодирующий капсидный белок (CP), и ген, кодирующий неструктурный белок 1 (NS1), где последовательность нуклеотидов гена CP имеет уровень идентичности по меньшей мере 80% относительно последовательности нуклеотидов, как изображено в SEQ ID NO: 1, или последовательность нуклеотидов гена NS1 имеет уровень идентичности по меньшей мере 80% относительно последовательности нуклеотидов, как изображено SEQ ID NO: 3, и где вирусная геномная ДНК реагирует в ПЦР реакции с группой праймеров, как изображено в SEQ ID NO: 5 и 6, с получением ПЦР продукта из 140+/-10 пар оснований и реагирует в ПЦР реакции с группой праймеров, как изображено в SEQ ID NO: 7 и 8, с образованием ПЦР продукта из 853+/-10 пар оснований.
Вирус по изобретению может быть в живой, живой ослабленной или инактивированной форме.
Как указано выше в настоящее время были охарактеризованы последовательности ДНК генов, кодирующих CP и NS1 вируса. Идентификация таких генов является крайне полезной, так как они могут использоваться в настоящее время между прочим как основа для ДНК вакцин, для использования субъединичных вакцин на основе указанных белков или для диагностических целей, как подробно объясняется ниже.
Другой вариант осуществления настоящего изобретения относится к фрагменту ДНК, включающему ген, кодирующий капсидный белок, отличающемуся тем, что ген имеет уровень идентичности по меньшей мере 80% относительно последовательности нуклеотидов гена CP, как изображено в SEQ ID NO: 1.
Предпочтительная форма варианта осуществления настоящего изобретения относится к такому фрагменту ДНК, включающему ген, имеющий уровень идентичности по меньшей мере 82%, более предпочтительно 84%, 86%, 88%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или даже 100%, в таком порядке предпочтительности, относительно последовательности нуклеотидов CP, как изображено в SEQ ID NO: 1.
Еще один вариант осуществления настоящего изобретения относится к фрагменту ДНК, включающему ген, кодирующий NS1, отличающемуся тем, что ген имеет уровень идентичности по меньшей мере 80% относительно последовательности нуклеотидов гена NS1, как изображено в SEQ ID NO: 3.
Предпочтительная форма варианта осуществления настоящего изобретения относится к такому фрагменту ДНК, включающему ген, имеющий уровень идентичности по меньшей мере 82%, более предпочтительно 84%, 86%, 88%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или даже 100%, в таком порядке предпочтения, относительно последовательности нуклеотидов NS1, как изображено в SEQ ID NO: 3.
Еще один вариант осуществления настоящего изобретения относится к CP, отличающемуся тем, что указанный CP кодируется фрагментом ДНК, кодирующим CP по изобретению.
Такие CP вируса по изобретению являются крайне подходящими, так как они подходят для применения в вакцинах, точнее в субъединичных вакцинах, они могут быть использованы для увеличения количества антител и делают возможными диагностические тесты, как объясняется ниже.
Предпочтительная форма варианта осуществления настоящего изобретения относится к CP, имеющему последовательность аминокислот, как изображено в SEQ ID NO: 2.
Еще один вариант осуществления настоящего изобретения относится к NS1, отличающемуся тем, что NS1 кодируется фрагментом ДНК, кодирующим NS1 по изобретению.
Такие NS1 вируса по изобретению являются очень подходящими, между прочим, так как они позволяют проводить диагностические тесты, как объяснено ниже.
Предпочтительная форма варианта осуществления настоящего изобретения относится к NS1, имеющему последовательность аминокислот, как изображено в SEQ ID NO: 4.
Одним из достоинств настоящего изобретения является то, что впервые возможно следовать течению вирусной инфекции и анализировать наличие или отсутствие нового вируса в различных органах и жидкостях тела свиней, страдающих от HBS. Это помогает получить больше информации о развитии заболевания.
Известно, что в течение недель или даже дней до того, как у определенной свиньи проявятся все клинические признаки HBS, патологических изменений не обнаруживают. Средний промежуток времени между первыми симптомами и смертью животных составляет около 2-6 часов. Незадолго до смерти животные, вероятно, страдают от растяжения живота и некоторые из них кричат перед смертью. Животных на фермах, у которых развивались клинические признаки, умерщвляли до того, как они умирали от заболевания.
На различных мексиканских фермах, с которых собирали свиней с HBS, частота HBS варьировалась между 1-2%.
Всего 33 умерщвленных животных с диагнозом HBS получили с различных ферм, одну группу из 17 в возрасте между 18-27 недель (Пример 1) и одну группу из 16 в возрасте между 12-26 неделями (Пример 2), анализировали. ПЦР реакции с группой праймеров, как описано выше, выявили, что в группе 1, 5 из 14 сывороток (3 сыворотки отсутствовали) оказались положительными в отношении вируса и один ректальный мазок оказался положительным. Всего 8 образцов цельной крови оказались положительными и 12 из 15 лимфатических узлов оказались положительными. В группе 2, 5 из 16 сывороток оказались в отношении вируса и один ректальный мазок оказались положительным. От характерного животного из каждой группы (животное 2 группа 1, животное 10 группа 2) органы анализировали в отношении присутствия вируса. Было обнаружено, что все взятые органы, включая лимфатические узлы, легкое, селезенку, кишечник, почки и печень, а также фекалии показали положительный результат теста на вирус.
Другим достоинством настоящего изобретения является то, что в настоящее время возможно инфицировать здоровых свиней новым вирусом и исследовать путь вирусной инфекции. С указанной целью, материал органов и фекалии от животных с HBS гомогенизировали в среде для культивирования тканей. Гомогенаты замораживали-оттаивали однократно (-70°C), центрифугировали и фильтровали на фильтрах 5 мкм, 0,45 мкм и 0,22 мкм для удаления оставшегося тканевого материала.
Инокулят A получали из следующего материала характерных животных: фекалии, лимфатические узлы, легкие, селезенка и кишечник животного 10 группы 2.
Инокулят B получали из следующего материала другого характерного животного: фекалии, лимфатические узлы, легкое, почка и печень животного 2 группа 1. Полные подробности проведенных экспериментов даны в разделе Примеры ниже (Пример 3).
Инокуляты (A или B) давали в виде 4×2 мл в/м дозу и пероральную дозу 20 мл всего 4 самцам и 8 самкам (Landrace/хорошее состояние здоровья/SPF) в возрасте 12-14 недель в момент инокуляции, как указано далее: Группа 1: пять животных, из которых трое животных получали инокулят A, двое служили в качестве особей-часовых. Группа 2: четверо животных, из которых трое животных получали инокулят B, один служил в качестве особи-часового. Группа 3: трое животных, все из которых получали инокулят B. Одно животное (самец) умерщвляли до инокуляции и он служил в качестве отрицательного контроля. Животных скринировали в отношении присутствия нового вируса в сыворотке, фекалиях, мазке из полости носа и мазке из глаз, до инокуляции.
Образцы крови, ректальные/назальные/глазные мазки получали в нескольких временных точках. Все подробности экспериментов на животных даны в разделе Примеры ниже.
Было обнаружено, что у 6 из 6 инокулированных животных вирус был определен в сыворотке, а также в ректальных, назальных и глазных мазках через 7 дней после инокуляции. У всех из 3 неинокулированных животных, особях-часовых, вирус был обнаружен в сыворотке через 14 дней после инокуляции других животных. Ректальные/назальные и глазные мазки всех особей-часовых были положительными на 7 день после инокуляции (Пример 3, группы 1,2). В 3 из 3 сывороток инокулированных животных на 3 день после инокуляции обнаруживали вирус (Пример 3, группа 3).
Следовательно, хотя сообщают об относительно низкой частоте HBS, при условии высококонтагиозной природы вируса в комбинации с его высокой скоростью инфицирования, можно ожидать, что, однозначно большинство, если не все, свиньи на фермах, где появлялся HBS, подвергаются инфицированию вирусом. Следовательно, крайне желательно вакцинировать всех животных на фермах, где появляется HBS, против инфекции HBS-ассоциированным свиным парвовирусом по изобретению. Такая вакцинация устранит по меньшей мере вирусный компонент многофакторного синдрома. И это в свою очередь будет предотвращать или по меньшей мере уменьшать тяжесть заболевания.
Также одним из достоинств настоящего изобретения является то, что новый парвовирус свиней был выделен и ассоциирован с HBS, вирус и/или защитные субъединицы вируса могут быть использованы в качестве исходного материала для целей вакцинации.
Следовательно, другой вариант осуществления настоящего изобретения относится к вакцинам для борьбы с HBS у свиней, где такие вакцины включают вирус по изобретению и фармацевтически приемлемый носитель.
Примеры фармацевтически приемлемых носителей, которые являются подходящими для применения в вакцине по изобретению, представляют собой стерильную воду, солевой раствор, водные буферы, такие как PBS, и подобные. Кроме того, вакцина по изобретению может включать другие добавки, такие как адъюванты, стабилизаторы, антиоксиданты и другие, как описано ниже.
Борьба в таком отношении должна интерпретироваться в широком смысле: предполагают, что борьба с HBS включает вакцинацию для предотвращения признаков заболевания, а также вакцинацию для облегчения признаков заболевания, как указано выше.
Терапевтическая вакцинация после диагностики вируса у инфицированного животного, которое еще не страдает от синдрома, является, конечно, также эффективной. Терапевтическая вакцинация после диагностики синдрома кажется неэффективной, с учетом очень короткого периода между первыми клиническими признаками и смертью.
Вакцина по изобретению может включать вирус по изобретению в ослабленной живой или инактивированной форме.
Ослабленные живые вирусные вакцины, т.е. вакцины, включающие вирус по изобретению в живой ослабленной форме, имеют преимущества над инактивированными вакцинами, так как они лучше имитируют естественный путь инфицирования. Кроме того, их способность к репликации позволяет осуществлять вакцинацию небольшим количеством вируса; их количество автоматически увеличивается до того, как оно достигает триггерного уровня иммунной системы. С указанного момента иммунная система запускается и, в конечном счете, устраняет вирусы.
Живой ослабленный вирус представляет собой вирус, который имеет сниженный уровень вирулентности при сравнении с вирусом, выделенным из среды. Вирус, имеющий сниженный уровень вирулентности, расценивается как вирус, который даже в комбинации с другими факторами, вовлеченными в HBS, не вызывает смертности у свиней.
Следовательно, одна предпочтительная форма варианта осуществления настоящего изобретения относится к вакцине, включающей вирус по изобретению, где указанный вирус находится в живой ослабленной форме.
Ослабленные вирусы могут, например, быть получены путем выращивания вирусов по изобретению в присутствии мутагенного агента, с последующей селекцией вируса, которая демонстрирует уменьшение количества потомства и/или скорости репликации. Множество таких агентов известны в области техники.
Другим очень часто используемым методом является серийный пассаж in vitro. Вирусы затем адаптируются к клеточной линии, используемой для серийного пассажа, так что они становятся ослабленными при переносе в естественный организм-хозяин снова в виде вакцины. Еще одним способом получения ослабленных вирусов является их выращивание при температуре, отличающейся от температуры их естественной среды обитания. Способы выбора мутантов, чувствительных к температуре (Ts-мутантов), хорошо известны в области техники. Такие методы включают выращивание вирусов в присутствии мутагена с последующим ростом при субоптимальной температуре, титрацию потомства вируса на клеточных слоях и визуальный отбор тех бляшек, которые растут медленнее при оптимальной температуре. Такие небольшие бляшки включают медленно растущие и, следовательно, желаемые живые ослабленные вирусы.
Живые ослабленные вакцины для борьбы со свиным парвовирусом типа PPV были описаны, между прочим, Paul & Mengeling(32), Paul & Mengeling(33) и Fujisaki e& Murakami(34).
Возможным недостатком использования живых ослабленных вирусов может быть по своей сути определенный уровень остаточной вирулентности. Это не является реальным недостатком, пока уровень вирулентности является приемлемым, т.е. пока вакцина по меньшей мере предотвращает смерть свиней. Конечно, чем меньше остаточная вирулентность ослабленной вакцины, тем меньшее влияние вакцинация оказывает на набор веса во время/после вакцинации.
Инактивированные вакцины представляют собой, в противоположность их живым ослабленным аналогам, по существу безопасные, так как они не имеют остаточного уровня вирулентности. Несмотря на тот факт, что они обычно включают несколько более высокую дозу вирусов по сравнению с живыми ослабленными вакцинами, они могут, например, быть предпочтительной формой вакцины у свиней, которые уже страдают от других заболеваний. Свиньи, которых содержат в субоптимальных условиях, таких как неполноценное питание или субоптимальное размещение, также получат пользу от инактивированных вакцин.
Следовательно, другая предпочтительная форма варианта осуществления настоящего изобретения относится к вакцине, включающей вирус по изобретению, где указанный вирус находится в инактивированной форме.
Известно, что полностью инактивированные парвовирусы в общем, являясь свиными или собачьими парвовирусами, являются очень эффективной и безопасной основой для вакцин. Исключительно в качестве примера: MSD AH (Boxmeer, The Netherlands) производит коммерчески доступную вакцину инактивированного парвовируса свиней тип PPV: Porcilis Parvo. Hipra (Испания) также производит коммерчески доступную вакцину инактивированного парвовируса свиней тип PPV: PARVOSUIN® MR/AD. Zoetis производит инактивированный собачий парвовирус: PARVAC и вакцину инактивированного парвовируса свиней типа PPV: Porcine PARVAC. Novartis обеспечивает способы инактивации парвовируса в патенте США US 4193991.
Такие инактивированные вакцины целого вируса могут равным образом быть получены для нового парвовируса свиней по изобретению. Как в случае для известных вакцин парвовируса, получение в основном включает стадии выращивания нового парвовируса на подходящих свиных клетках, сбор вируса, инактивацию вируса и смешивание инактивированного вируса с фармацевтически приемлемым носителем.
Стандартным способом инактивации является классическая обработка формальдегидом. Другими методами инактивации, хорошо известными в области техники, является УФ-излучение, гамма-излучение, обработка бинарным этиленимином, тимеросалом и подобными. Специалист в области техники знает, как применять указанные методы. Предпочтительно вирус инактивируют с помощью β-пропиолактона, глютаральдегида, этиленимина или формальдегида. Не говоря о том, что другие пути инактивации вируса также включены в настоящее изобретение.
Как указано выше, вирус может быть выращен в культуре клеток на предрасположенных свиных клетках или клеточных линиях.
Следовательно, другой вариант осуществления изобретения относится к культуре клеток, включающей HBS-ассоциированный парвовирус свиней по настоящему изобретению. Примеры клеток и клеточных линий представляют собой SK6, PK15, первичные или бессмертные свиные клетки почек, первичные или бессмертные свиные альвеолярные легочные макрофаги.
Практически полный вирусный геном нового парвовируса свиней в настоящее время был определен, и ДНК последовательность представителя нового вируса представлена в SEQ ID NO: 10. Инвертированные концевые повторы (ITR) генома не представлены в настоящем описании. Так как парвовирусы по определению принадлежат к наименьшим известным вирусам, полная ss-ДНК, кодирующая парвовирус по изобретению, может быть легко получена синтетически. По этой причине, парвовирус может быть легко получен in vivo с использованием вирусной ДНК в качестве исходного материала. Инвертированные концевые повторы (ITR) генома другого, известного, парвовируса, такого как, между прочим, описанный Qiu et al.(37) и Wang et al.(38), могут быть использованы для завершения вирусного генома, как представлено в SEQ ID NO: 10. ITR играют роль исключительно в репликации вирусного генома и, как таковые, они не важны с иммунологической точки зрения. Следовательно, для целей получения вируса по изобретению ITR являются взаимозаменяемыми.
Клонирование полнодлинновой парвовирусной ДНК в плазмиде, такой как, например, Bluescript II SK, и последующее получение целого парвовируса посредством трансфекции свиных клеток с помощью плазмиды экспрессии, кодирующей новый парвовирус свиней, описано, между прочим Qiu et al.(37) и Wang et al.(38). Пермиссивная клеточная линия, такая как SK6, PK15, первичные или бессмертные свиные клетки почек, первичные или бессмертные свиные альвеолярные легочные макрофаги будут предпочтительной клеточной линией для этой цели. Тем не менее, если желательно, также могут быть использованы непермиссивные клеточные линии: геном нового парвовируса может, между прочим, быть реплицирован в непермиссивных клетках с помощью аденовирусных генов, как описано Guan et al.(39).
Хотя целые инактивированные парвовирусы обеспечивают хорошую основу для вакцин, их продукция может быть дорогой, в зависимости от, между прочим, типа используемых клеток-хозяев, субстрата и используемой среды для культивирования клеток. В специфическом случае парвовирусов привлекательной альтернативой для использования целых вирусов является применение CP субъединиц парвовирусов, более предпочтительно субъединиц в форме так называемых пустых капсидов. Такие пустые капсиды являются в основном вирус-подобными частицами, которые однако не включают парвовирусного генома. Как следствие, нет необходимости инактивировать парвовирусные пустые капсидные частицы до использования в вакцине, и, следовательно, они имеют дополнительное преимущество в том, что они являются внутренне безопасными.
Пустые капсиды могут быть получены посредством обычной экспрессии ORF2, кодирующей капсидный белок, в подходящей системе экспрессии. Образующиеся таким образом капсидные белки самоорганизуются в пустые вирусные частицы. Парвовирусные пустые капсиды могут легко быть получены в больших количествах, и они являются высоко иммуногенными. Однозначно, большинство систем экспрессии, используемых в настоящее время для получения парвовирусных пустых капсидов, являются системы экспрессии на основе бакуловируса.
Способы получения высоко иммуногенных парвовирусных пустых капсидов в системах экспрессии бакуловирусов были, например, описаны для парвовируса свиней типа PPV Martinez(18), Casal(19), Zhou et al.(20) и Hao Feng(21). Для других парвовирусов, такие методы были описаны, например, Saliki(22) и Brown(23).
Более того, системы экспрессии бакуловирусов и векторы экспрессии бакуловирусов в общем были описаны подробно в руководствах, таких как O'Reilly at al.(24) и Murhammer(25).
Системы экспрессии на основе бакуловирусов также являются коммерчески доступными, например, от Invitrogen Corporation, 1600 Faraday Avenue, Carlsbad, California 92008, USA.
Альтернативой для систем экспрессии на основе бакуловирусов являются системы экспрессии на основе дрожжей. Системы экспрессии дрожжей, например, описаны Gellissen et al.(29).
Готовые к использованию системы экспрессии являются, между прочим, коммерчески доступными от Research Corp. Technologies, 5210 East Williams Circle, Suite 240, Tucson, AZ 85711-4410 USA. Системы экспрессии клеток дрожжей и насекомых являются также коммерчески доступными от Clontech Laboratories, Inc. 4030 Fabian Way, Palo Alto, California 94303-4607, USA.
Экспрессия капсидного белка, конечно, также возможна в системах экспрессии на основе клеток млекопитающих, как известно в области техники, но такие системы будут скорее всего более дорогими в использовании при сравнении с системами экспрессии на основе бакуловируса.
Следовательно, другая форма варианта осуществления настоящего изобретения относится к вакцине для борьбы с HBS-ассоциированным свиным парвовирусом, отличающейся тем, что указанная вакцина включает иммуногенно эффективное количество капсидного белка по изобретению и фармацевтически приемлемый носитель.
Предпочтительная форма варианта осуществления настоящего изобретения относится к вакцине для борьбы с HBS-ассоциированным свиным парвовирусом у свиней, отличающейся тем, что указанная вакцина включает иммуногенно эффективное количество капсидного белка по изобретению в форме пустых капсидов.
Количество пустых капсидов в вакцине и путь введения будут совместимыми с таковыми инактивированных целых частиц вируса, так как в отношении иммуногенности и сходства капсида они являются сравнимыми с инактивированными целыми частицами вируса.
Обычно количество от 1 до 100 мкг новых пустых капсидов парвовируса является подходящим в качестве дозы вакцины. С точки зрения затрат, предпочтительное количество находится в диапазоне 1-50 мкг пустых капсидов, более предпочтительно в диапазоне 1-25 мкг.
Casal(19) описывает, что дозы и собачьего парвовируса и парвовируса свиней, такие как 1-3 мкг, в присутствии обычных добавок, обеспечивают тотальную защиту соответствующего организма-хозяина от заболевания.
Вакцина по изобретению на основании инактивированного целого вируса или пустых капсидов предпочтительно включает добавку. Обычные добавки, хорошо известные в области техники, представляют собой полный и неполный адъювант Фрейнда, витамин E, неионные блокполимеры, мурамилдипептиды, Quill A(R), минеральное масло, например, Bayol(R) или Markol(R), растительное масло и Carbopol(R) (гомополимер), или Diluvac(R) Forte. Вакцина также может включать так называемый "носитель". Носителем является соединение, к которому полипептид прилипает, без ковалентной связи с ним. Часто используемыми соединениями носителями являются, например, гидроксид, фосфат или оксид алюминия, диоксид кремния, каолин и бентонит.
Casal(19) в своей вакцине парвовируса Quill A успешно использовали между прочим гидроксид алюминия.
В принципе вакцина по изобретению может быть введена даже однократно. Однако, особенно в случае инактивированных вакцин, будут ли это вакцины из целых вирусов или вакцины из пустых капсидов, проводят предпочтительно также первую и вероятно вторую бустерную вакцинацию. Первый бустер обычно вводят по меньшей мере через две недели после первой вакцинации. Особенно подходящим моментом для бустерной вакцинации является от 3 до 16 недель после первой вакцинации. Второй бустер, если необходимо, обычно вводят между 4 и 50 неделями после первого бустера.
Альтернативой подходу вакцины с инактивированным целым вирусом и подходу вакцины с пустыми капсидами является применение живых рекомбинатных непарвовирусных векторов, животными хозяевами которых являются свиньи, в качестве носителей гена капсидного белка нового парвовируса свиней.
Среди подходящих рекомбинантных непарвовирусных векторов, животными хозяевами которых являются свиньи, два вектора являются особенно подходящими в качестве носителей: вирус псевдобешенства (PRV) и вирус классической свиной лихорадки (CSFV).
Применение таких рекомбинантных вирусов в вакцинах имеет дополнительное преимущество в том, что вакцинированные животные одновременно вакцинируются против и PRV и PPV или и CSFV и PPV.
Chen et al.(27) описывают получение и применение живого ослабленного PRV рекомбинатного вектора, экспрессирующего капсидный белок парвовируса свиней тип PPV. Такой рекомбинант PRV вводили восьмидневным поросятам в количестве 5×105TCID50 и указанное количество вакцины оказалось безопасным и обеспечивающим превосходный иммунитет против и PRV и PPV.
Живые ослабленные векторы CSFV также являются крайне подходящими в качестве живых рекомбинатных векторов. Исключительно в качестве примера; живые ослабленные CSFV, у которых ген Npro был удален, были описаны Mayer et al.(28) Такой живой ослабленный вирус позволяет, между прочим, вставлять ген, кодирующий капсидный белок, в месте делеции гена Npro. Такой живой рекомбинатный CSFV вектор также образует подходящий носитель для гена капсидного белка нового парвовируса свиней.
Экспрессия гена капсидного белка может быть осуществлена под контролем любого подходящего гетерологичного промотора, который является функциональным в клетках млекопитающих (см. ниже). Гетерологичным промотором является промотор, который не является промотером, ответственным за транскрипцию гена CP в форме дикого типа нового парвовируса свиней по изобретению. Это может быть парвовирусный промотор, ответственный за транскрипцию CP или NS1 другого парвовируса, который не принадлежит к парвовирусам по изобретению, или это может быть непарвовирусный промотор.
Следовательно, другой вариант осуществления настоящего изобретения относится к фрагменту ДНК, включающему ген, кодирующий CP по изобретению, отличающийся тем, что указанный ген находится под контролем функционального гетерологичного промотора.
Chen et al(27) использовали CMV промотор для запуска экспрессии гена капсидного белка, но в области техники известны другие подходящие промоторы, которые являются функциональными в клетках млекопитающих. Промотор, который является функциональным в клетках млекопитающих, представляет собой промотор, который способен запускать транскрипцию гена, который расположен в клетках млекопитающих после промотора.
Примеры подходящих промоторов, которые являются функциональными в клетках млекопитающих, включают классические промоторы, такие как (человеческий) цитомегаловирусный немедленный ранний промотор (Seed, B. et al., Nature 329, 840-842, 1987; Fynan, E.F. et al., PNAS 90, 11478-11482,1993; Ulmer, J.B. et al., Science 259, 1745-1748, 1993), вирус саркомы Рауса LTR (RSV, Gorman, C.M. et al., PNAS 79, 6777-6781, 1982; Fynan et al., supra; Ulmer et al., выше), MPSV LTR (Stacey et al., J. Virology 50, 725-732, 1984), SV40 немедленный ранний промотор (Sprague J. et al., J. Virology 45, 773,1983), промотор SV-40 (Berman, P.W. et al., Science, 222, 524-527, 1983), металлотионеиновый промотор (Brinster, R.L. et al., Nature 296, 39-42, 1982), промотор теплового шока (Voellmy et al., Proc. Natl. Acad. Sci. USA, 82, 4949-53, 1985), крупный поздний промотор Ad2 и промотор β-актина (Tang et al., Nature 356, 152-154, 1992). Регуляторные последовательности также могут включать терминатор и последовательности полиаденилирования. Среди последовательностей, которые могут быть использованы, хорошо известны последовательность полиаденилирования бычьего гормона роста, последовательность полиаденилирования SV40, человеческий терминатор цитомегаловируса (hCMV) и последовательности полиаденилирования.
Следовательно, другая форма указанного варианта осуществления изобретения относится к вакцине для борьбы с HBS-ассоциированным свиным парвовирусом у свиней, отличающейся тем, что указанная вакцина включает живой рекомбинантный непарвовирусный вектор, включающий фрагмент ДНК, включающий ген, кодирующий CP по изобретению, под контролем функционального промотора и фармацевтически приемлемого носителя.
Не нужно говорить, что живой рекомбинатный непарвовирусный вектор должен экспрессировать иммуногенно эффективное количество капсидного белка.
Альтернативой вакцинации инактивированной вакциной с целым вирусом, вакциной с пустыми капсидами или живым рекомбинатным непарвовирусным вектором, является применение ДНК вакцинации.
Такая ДНК вакцинация основана на внесении фрагмента ДНК, несущего ген, кодирующий капсидный белок, под контролем подходящего промотора, в организм животного хозяина. Когда ДНК потребляется клеткой хозяином, ген, кодирующий капсидный белок, транскрибируется и транскрипт транслируется в капсидный белок в клетке хозяине. Это четко имитирует процесс натурального инфицирования парвовирусом.
Подходящими промоторами являются промоторы, которые являются функциональными в клетках млекопитающих, как проиллюстрировано выше.
Фрагмент ДНК, несущий ген, кодирующий капсидный белок, под контролем подходящего промотора, может, например, быть плазмидой. Указанная плазмида может быть в кольцевой или линейной форме.
Примеры успешной ДНК вакцинации свиней представляют собой между прочим успешную вакцинацию против болезни Ауески, как описано в Gerdts et al.(30), Journal of General Virology 78: 2139-2146 (1997). Они описывают ДНК вакцину, где используют фрагмент ДНК, который несет гликопротеин C, под контролем основного немедленного раннего промотора человеческого цитомегаловируса. Вакцинацию проводили четыре раза с интервалами в две недели с количеством 50 мкг ДНК. У вакцинированных животных образовывались антитела в сыворотке, которые распознавали соответствующий антиген в иммуноблоте и которые проявляли нейтрализующую активность.
Другой пример успешной ДНК вакцинации свиней дан Gorres et al.(31). Они описывают успешную ДНК вакцинацию свиней от пандемического и классического свиного гриппа H1N1. Их вакцинировали первичной вакцинацией и 2 гомологичными бустами через 3 и 6 недель после примирования ДНК вакциной, включающей НА ген гриппа H1N1 под контролем функционального промотора.
Следовательно, еще одна форма по настоящему изобретению относится к вакцине для борьбы с HBS-ассоциированным свиным парвовирусом у свиней, отличающейся тем, что указанная вакцина включает ДНК фрагмент, включающий ген, кодирующий капсидный белок по настоящему изобретению, под контролем функционального промотора, и фармацевтически приемлемый носитель.
Не нужно говорить, что ДНК фрагмент, включающий ген, кодирующий капсидный белок, должен экспрессировать иммуногенно эффективное количество капсидного белка.
"Иммуногенно эффективное количество" для вакцины по изобретению, которое основано на цельном парвовирусе по изобретению, пустом капсиде по изобретению, живом рекомбинантном векторе или ДНК вакцине по изобретению зависит от желаемого эффекта и от организма-мишени.
Термин "иммуногенно эффективное количество", как используется в настоящем описании, относится к количеству парвовируса, пустого капсида, живого рекомбинантного вектора или ДНК вакцины, которые необходимы для индукции иммунного ответа у свиней в такой степени, чтобы он уменьшал патологические эффекты, вызванные инфицированием диким типом HBS-ассоциированного парвовируса свиней при сравнении с патологическими эффектами, вызванными инфицированием диким типом HBS-ассоциированного парвовируса свиней у неиммунизированных свиней.
В рамках знаний специалиста определить, является ли лечение "иммунологически эффективным", например, путем проведения экспериментального инфицирования вакцинированных животных и затем определения клинических признаков заболевания животного-мишени, серологических параметров или путем измерения ре-изоляции патогена, с последующим сравнением полученных открытий с таковыми, наблюдаемыми у свиней, инфицированных в среде.
Количество вводимого вируса будет зависеть от пути введения, присутствия добавки и момента введения.
Предпочтительное количество живой вакцины, включающей вирус по изобретению, выражено, например, как инфекционная доза культуры ткани (TCID50). Например, для живого вируса преимущественно может быть использован диапазон доз от 10 до 109 TCID50 на животное, в зависимости от остаточной вирулентности вируса. Предпочтительно используют диапазон от 102 до 106 TCID50.
Как известно в области техники может применяться множество путей введения. Вакцины по изобретению предпочтительно вводят животному посредством инъекции (внутримышечной или посредством интраперитонеального пути) или перорально.
Протокол для введения может быть оптимизирован в соответствии со стандартной практикой вакцинации. Во всех случаях введение посредством внутрикожного инжектора (IDAL) является предпочтительным путем введения.
Если вакцина включает инактивированный вирус или пустые капсиды по изобретению, доза также выражается, как количество вирусных частиц для введения. Доза обычно будет несколько выше при сравнении с введением живых частиц вируса, так как живые вирусные частицы в определенной степени реплицируются у животного-мишени, до того, как они удаляются иммунной системой. Для вакцин на основе инактивированного вируса обычно подходящим является количество вирусных частиц в диапазоне от около 104 до 109 частиц, в зависимости от используемого адъюванта.
Если вакцина включает субъединицы, например, CP по изобретению, доза также может быть выражена в микрограммах белка. Для вакцин на основании субъединиц, подходящая доза обычно находится в диапазоне от 5 до 500 микрограмм белка, также в зависимости от используемого адъюванта.
Если вакцина включает фрагмент ДНК, включающий ген, кодирующий капсидный белок, доза выражается в микрограммах ДНК. Для вакцин на основе субъединиц, подходящая доза обычно находится в диапазоне от 5 и 500 микрограмм ДНК, т.е. в зависимости от эффективности используемой плазмиды экспрессии. Во множестве случаев количество от 20 до 50 микрограмм плазмиды на животное будет достаточным для эффективной вакцинации.
Вакцина по изобретению может принимать любую форму, которая является подходящей для введения в контексте разведения свиней, и которая соответствует желаемому пути применения и желаемому эффекту. Получение вакцины по изобретению осуществляют при помощи средств, обычных для специалиста.
Пероральные пути являются предпочтительными, когда они облегчают введение вакцины.
Для перорального введения вакцину предпочтительно смешивают с подходящим носителем для перорального введения, т.е. целлюлозой, пищей или метаболизируемыми веществами, такими как альфа-целлюлоза или различные масла растительного или животного происхождения.
На практике, свиней вакцинируют против ряда патогенетических вирусов или микроорганизмов.
Следовательно, является крайне привлекательным, по практическим и экономическим причинам, комбинировать вакцину для свиней по изобретению с, например, дополнительным иммуногеном вируса или микроорганизма, патогенного для свиней, или генетической информацией, кодирующей иммуноген указанного вируса или микроорганизма.
Следовательно, предпочтительная форма настоящего варианте осуществления изобретения относится к вакцине по изобретению, где вакцина включает по меньшей мере один другой микроорганизм, патогенный для свиней, или вирус, патогенный для свиней, и/или по меньшей мере один другой иммуногенный компонент и/или генетический материал, кодирующий указанный другой иммуногенный компонент, указанного микроорганизма, патогенного для свиней или вируса, патогенного для свиней. Иммуноген или иммуногенный компонент представляет собой соединение, которое индуцирует иммунный ответ у животного. Это может быть, например, целый вирус или бактерия, или белковый или сахарный фрагмент указанного вируса или бактерии.
Наиболее частыми патогенными вирусами и микроорганизмами, которые являются патогенными для свиней, являются Brachyspira hyodysenteriae, вирус Африканской свиной лихорадки, вирус Нипах, свиной цирковирус, свиной вирус гепатита TTV, вирус псевдобешенства, вирус свиного гриппа, парвовирус свиней, свиной вирус респираторного и репродуктивного синдрома (PRRS), свиной вирус эпидемической диареи (PEDV), вирус ящура, вирус переносимого гастроэнтерита, ротавирус, Escherichia coli, Erysipelo rhusiopathiae, Bordetella bronchiseptica, Salmonella cholerasuis, Haemophilus parasuis, Pasteurella multocida, Streptococcus suis, Mycoplasma hyopneumoniae и Actinobacillus pleuropneumoniae.
Следовательно, более предпочтительная форма изобретения относится к вакцине по изобретению, где вирус или микроорганизм, патогенный для свиней, выбирают из группы Brachyspira hyodysenteriae, вируса Африканской свиной лихорадки, вируса Нипах, свиного цирковируса, свиного вируса гепатита TTV, вируса псевдобешенства, вируса свиного гриппа, парвовируса свиней, свиного вируса респираторного и репродуктивного синдрома (PRRS), свиного вируса эпидемической диареи (PEDV), вируса ящура, вируса переносимого гастроэнтерита, ротавируса, Escherichia coli, Erysipelo rhusiopathiae, Bordetella bronchiseptica, Salmonella cholerasuis, Haemophilus parasuis, Pasteurella multocida, Streptococcus suis, Mycoplasma hyopneumoniae и Actinobacillus pleuropneumoniae.
Еще один вариант осуществления изобретения относится к способу получения вакцины по изобретению, где способ включает смешивание вируса по изобретению и/или пустого капсида и/или CP по изобретению и/или фрагмента ДНК, кодирующего CP по изобретению, и/или живого рекомбинантного непарвовирусного вектора, кодирующего CP по изобретению, и фармацевтически приемлемого носителя.
Еще один вариант осуществления настоящего изобретения относится к вирусу по изобретению и/или пустому капсиду и/или CP по изобретению и/или фрагменту ДНК, кодирующему CP по изобретению, и/или живому рекомбинантному непарвовирусному вектору, кодирующему CP по изобретению, для применения в вакцине.
Как упомянуто выше, геморрагический кишечный синдром представляет собой многофакторный синдром. Это сочетание факторов, которые в конечном счете запускают HBS. Это означает, что важно знать, присутствует ли HBS-ассоциированный парвовирус свиней в определенной популяции свиней задолго до проявления первых клинических признаков. Следовательно, для эффективной защиты от заболевания важным является быстрое и правильное определение присутствия HBS-ассоциированного парвовируса свиней.
Следовательно, другой задачей настоящего изобретения является обеспечение диагностических средств, подходящих для определения HBS-ассоциированного парвовируса свиней.
Указанные средства частично полагаются на доступность антител к вирусу. Такие антитела могут, например, быть использованы в диагностических тестах для HBS-ассоциированного парвовируса свиней.
Антитела или антисыворотка, включающие антитела против HBS-ассоциированного парвовируса свиней по изобретению, могут быстро и легко быть получены посредством вакцинации, например, свиней, птицы или, например, кроликов вирусом по изобретению с последующим, через около четырех недель, забором крови, центрифугированием коагулированной крови и отделением сыворотки. Такие методы хорошо известны в области техники.
Другие методы получения антител против HBS-ассоциированного парвовируса свиней, которые могут быть поликлональными, моноспецифическими или моноклональными (или их производными) также известны в области техники. Если желательны поликлональные антитела, методики получения и обработки поликлональной сыворотки хорошо известны в области техники в течение десятилетий, см., например, Mayer and Walter(35).
Моноклональные антитела, реактивные против вируса по изобретению, могут быть получены путем иммунизации разведенных при близкородственном скрещивании мышей, методиками, также длительно известными в области техники, см., например, Kohler and Milstein(36).
Следовательно, другой вариант осуществления настоящего изобретения относится к антителам или антисыворотке, которые реагируют с вирусом по изобретению.
Диагностический тестовый набор, основанный на определении вируса по изобретению или антигенного материала указанного вируса и, следовательно, подходящий для определения инфекции HBS-ассоциированного парвовируса свиней, может, например, включать стандартный тест ELISA. В одном примере такого теста стенки ячеек планшета ELISA покрывают антителами, направленными против вируса. После инкубации с тестируемым материалом, меченные антитела, реагирующие с вирусом, добавляют к ячейкам. Если тестируемый материал действительно включает новый парвовирус свиней по изобретению, такой вирус связывается с антителами, покрывающими ячейки ELISA. Меченные антитела, реагирующие с вирусом, которые впоследствии реагируют с вирусом, в свою очередь связываются с вирусом и цветовая реакция затем выявляет присутствие антигенного материала вируса.
Следовательно, еще один вариант осуществления настоящего изобретение относится к диагностическим тестовым наборам для определения вируса по изобретению или антигенного материала вируса, которые включают антитела, реагирующие с вирусом по изобретению или его антигенным материалом. Антигенный материал вируса интерпретируют в широком смысле. Он может быть, например, вирусом в разрушенной форме или материалом вирусной оболочки, включающим белки наружной мембраны вируса. Пока материал вируса реагирует с антисывороткой против вируса, материал расценивают как антигенный материал.
Диагностический тестовый набор, основанный на определении в сыворотке антител, реагирующих с вирусом по изобретению или антигенным материалом вируса и, следовательно, подходящий для определения инфицирования HBS-ассоциированным свиным парвовирусом, также может, например, включать стандартный тест ELISA. В таком тесте стенки ячеек планшета ELISA могут, например, быть покрыты вирусом по изобретению или его антигенным материалом. После инкубации с тестируемым материалом, например, сывороткой животного, предположительно инфицированного новым свиным парвовирусом по изобретению, меченные антитела, реагирующие с вирусом по изобретению, добавляют в ячейки. Если антитела к HBS-ассоциированному парвовирусу свиней присутствуют в тестируемой сыворотке, такие антитела связываются с вирусами, нанесенными на ячейки ELISA. Как следствие, последние добавленные меченные антитела, реагирующие с вирусом, не будут связываться и не будет обнаруживаться реакция окрашивания. Отсутствие реакции окрашивания, следовательно, выявит присутствие антител, реагирующих с вирусом по изобретению.
Следовательно, еще один вариант осуществления настоящего изобретения относится к диагностическим тестовым наборам для определения антител, реагирующих с вирусом по изобретению или с антигенным материалом вируса, который включает вирус по изобретению или его антигенный материал.
Дизайн иммуноанализа может варьироваться. Например, иммуноанализ может быть основан на конкуренции или прямой реакции. Более того, в протоколах можно использовать твердые подложки или можно использовать клеточный материал. Определение комплекса антитело-антиген может включать применение меченных антител; метками могут быть, например, ферментные, флуоресцентные, хемилюминесцентные, радиоактивные или красящие молекулы.
Подходящие методы для определения антител, реагирующих с вирусом по настоящему изобретению в образце, включают, в добавление к ELISA, упомянутому выше, иммунофлуоресцентный тест (IFT) и вестерн-блоттинг анализ.
Альтернативным, но быстрым и легким диагностическим тестом для диагностики присутствия или отсутствия вируса по изобретению, является ПЦР тест, как указано выше, включающий группу праймеров ПЦР, реагирующих со специфическим участком гена CP или NS1 HBS-ассоциированного парвовируса свиней. Специфический в таком контексте обозначает уникальный, для например, гена CP или NS1 HBS-ассоциированного парвовируса свиней, т.е. не присутствующий в других членах семейства Parvoviridae.
Предпочтительно в таком тесте используют группу праймеров (SEQ ID NO: 5-6), которые специфически реагируют с капсидным белком вируса, или группу праймеров (SEQ ID NO: 7-8), специфически реагирующих с NS1 вируса.
Не нужно говорить, что может быть использовано больше праймеров, чем праймеры, указанные выше. Настоящее изобретение впервые обеспечивает уникальную последовательность гена CP и NS1 HBS-ассоциированного парвовируса свиней. Это позволяет специалисту выбрать без каких-то дополнительных усилий другие селективные праймеры. Посредством простого компьютерного анализа последовательности гена CP или NS1 HBS-ассоциированного парвовируса свиней с известным геном CP или NS1 других, не-HBS-ассоциированных членов свиных парвовирусов семейства Parvoviridae, специалист в области техники способен разработать другие специфические ПЦР-праймеры для диагностических тестов для определения HBS-ассоциированного парвовируса свиней и/или различия между HBS-ассоциированным свиным парвовирусом и другими вирусными (свиными) патогенами.
PCR-праймерами, которые специфически реагируют с геном CP или NS1 HBS-ассоциированного парвовируса свиней, являются те праймеры, которые реагируют только с геном CP или NS1 HBS-ассоциированного парвовируса свиней, но не с геном CP или NS1 другого (свиного) патогенного вируса, или группы (свиных) патогенных вирусов.
Следовательно, другой вариант осуществления изобретения относится к диагностическому тестовому набору для определения вируса по изобретению, отличающемуся тем, что указанный тестовый набор включает группу ПЦР праймеров, которые специфически реагируют с участком гена CP или NS1 HBS-ассоциированного парвовируса свиней.
Предпочтительная форма настоящего изобретения относится к диагностическому тестовому набору для определения вируса по изобретению, отличающемуся тем, что указанный тест включает группу праймеров, как изображено в SEQ ID NO: 5-6, или группу праймеров, как изображено в SEQ ID NO: 7-8.
Примеры
Пример 1: Анализ заболевших животных в группе образцов 1.
Описание группы образцов 1
Группа образцов 1: 17 животных, 16 самцов/1 самка свиней, в возрасте 18-27 недель. 7 ферм. Получены 31 июля 2013.
Клинические симптомы: у животных внезапно развилось вздутие живота, некоторые из них кричали перед смертью. Не было отмечено симптомов в течение недель до смерти. Время от наблюдения первых симптомов и смерти составило 2-6 часов. После развития симптомов, животных умерщвляли путем электрооглушения и вскрывали.
Органные симптомы: патология тонкой кишки. Геморрагические симптомы, тонкая кишечная стенка, кровянистая жидкость в кишечнике. Отсутствие патологии в других органах, за исключением покрасневших, отечных лимфатических узлов. См. фиг.3.
Органы замораживали при -70°C. Сыворотку получали из свернувшейся крови посредством центрифугирования при 3000xg и впоследствии хранили при -70°C.
Протоколы ПЦР:
Изолированную ДНК скринировали посредством ПЦР с использованием праймеров, полученных из вирусных последовательностей (таблица 1). ПЦР проводили с использованием стандартных методов с температурой отжига 58°С для группы праймеров Bowl_ORF1_774_F/1626R, и 52°C для группы праймеров Bowl_Q_ORF2_FW/REV. Зонд создавали для К-ПЦР (таблица 1). К-ПЦР проводили с использованием стандартного метода с температурой отжига 50°C. Данные К-ПЦР анализировали с использованием Bio-Rad CFX Manager 2.0.
Таблица 1: Последовательности праймеров Bowl
Результаты ПЦР анализа:
Результаты ПЦР анализа изображены в таблице 2. Всего, 76% образцов оказались положительными.
Таблица 2: Анализ результатов образцов группы 1. (+): положительный в отношении нового парвовируса. (-) отрицательный в отношении нового парвовируса
Пример 2: Анализ пораженных животных в группе образцов 2.
Описание группы образцов 2
16 животных, 13 самцов/3 самок свиней, в возрасте 12-26 недель. 7 ферм, 4 дополнительные фермы сравнивали с группой образцов 1 (всего 11 ферм в группе образцов 1+2). Получили 22 авг 2013.
Клинические симптомы: У животных внезапно развивалось вздутие живота, некоторые из них кричали перед смертью. Не было отмечено симптомов в течение недель до смерти. Время между обнаружением первых симптомов и смертью составило 2-6 часов. После развития симптомов животных умерщвляли путем электрошока и вскрывали.
Органные симптомы: Патология тонкого кишечника. Геморрагические симптомы, тонкая стенка кишечника, кровянистая жидкость в кишечнике. Не было патологии в других органах, за исключением увеличенных, красных, отечных лимфатических узлов. См. фиг. 3. Не все лимфатические узлы и образцы крови тестировали.
Органы замораживали при -70°C. Сыворотку получали из свернувшейся крови путем центрифугирования при 3000xg и последующего хранения при -70°C.
Протоколы ПЦР:
Выделенную ДНК скринировали посредством ПЦР с использованием праймеров, полученных из вирусных последовательностей (таблица 1). PCR проводили с использованием стандартных методов с температурой отжига 58°С для группы праймеров Bowl_ORF1_774_F/1626R, и 52°C для группы праймеров Bowl_Q_ORF2_FW/REV. Зонд создавали для К-ПЦР (таблица 1). К-ПЦР проводили с использованием стандартного метода с температурой отжига 50°C. Данные К-ПЦР анализировали с использованием Bio-Rad CFX Manager 2.0.
Результаты анализа ПЦР:
Результаты анализа ПЦР изображены в таблице 3. Всего, 25% образцов оказались положительными. Кровь не анализировали. Анализировали только два лимфатических узла.
Таблица 3: Результаты анализа образцов группы 2. (+): положительные в отношении нового парвовируса. (-) отрицательные в отношении нового парвовируса
Пример 3: Репликация нового парвовируса у свиней
Получение материала животных:
Замороженный материал органов и фекалии групп образцов 1 и 2 хранили при -70°C до анализа. Все процедуры проводили на льду. Органы размораживали и впоследствии гомогенизировали (10% масс/об) в среде для культивирования тканей. Гомогенат замораживали-оттаивали однократно (-70°C). Впоследствии гомогенат центрифугировали и фильтровали на фильтрах 5 мкм, 0,45 мкм и 0,22 мкм для удаления оставшегося тканевого материала. Фильтрованный гомогенат хранили при -70°C до инокуляции.
Инокулят (A или B) давали как дозу 4×2мл в/м (4 различных органа, шея слева, шея справа, левая нога, правая нога) и пероральную дозу 20 мл (10 гомогенат фекалий+4×2,5 мл гомогената различных органов). Инокуляты вводили при комнатной температуре.
Инокулят A: Животное 10 группа образцов 2 (см. Пример 2)
Фекалии, лимфатические узлы, легкое, селезенка, кишечник
Инокулят B: Животное 2 группа образов 1 (см. пример 1)
Фекалии, лимфатические узлы, легкое, почка, печень
Животные
Тринадцать свиней (5 хряков/8 свинок/Ландрас/хорошее состояние здоровья/SPF/в возрасте 12-14 недель в момент инокуляции) кормили и выращивали на ферме MSD в Stevensbeek, Нидерланды, и размещали в соответствии с руководствами института. Животных скринировали в отношении присутствия нового парвовируса в сыворотке, фекалиях, назальном мазке и глазных мазках до инокуляции, как описано в примере 1. Одного самца умерщвляли как контрольное животное (не инфицированное). Другие двенадцать свиней размещали в 3 отдельных группах.
Лечение
Группа 1: Пять животных
Трое животных получали инокулят A, два служили как особи-часовые
Группа 2: Четыре животных
Трое животных получали инокулят B, два служили как особи-часовые
Группа 3: Три животных
Трое животных получали инокулят B
Забор образцов и вскрытие
Группа 1, 2:
Образцы крови, ректальные/назальные/глазные мазки в день -3, 0, 7, 14, 21, 28 после инокуляции (если не умерщвляли)
Ректальные/назальные/глазные мазки в день 3, 10, 17, 24 после инокуляции (если не умерщвляли)
Группа 3:
Образцы крови, ректальные/назальные/глазные мазки в день -4, 0, 3, 6 после инокуляции (если не умерщвляли)
На основании результатов ПЦР, животных распределяли для вскрытия:
Группа 1:
Инокулированные: день 10 п.и.; день 25 п.и.; день 31 п.и.
Особи-часовые: день 18 п.и.; день 31 п.и.
Группа 2:
Инокулированные: день 14 п.и.; день 29 п.и. (2 животных)
Особи-часовые: день 22 п.и.
Группа 3:
Инокулированные: день 4 п.и. (1 животное); день 7 п.и. (2 животных).
Результаты
ПЦР
Животное 10 группа образцов 2 использовали для инокулята:
Все органы оказались положительными в отношении нового парвовируса: фекалии, лимфатические узлы, легкое, селезенка, кишечник, почка, печень
Животное 2 группа образцов 1 использовали для инокулята:
Все органы оказались положительными в отношении нового парвовируса: фекалии, лимфатические узлы, легкое, почка, печень
Эксперимент на животных
Результаты ПЦР на мазках/сыворотке: Таблица 4A-B-C
Образцы органов получали для гистологии, для анализа ПЦР и для выделения вируса. Органы для выделения вируса хранили при -70°C. Печеночные лимфатические узлы (10% гомогенаты) анализировали с использованием ПЦР.
Группа 1: сыворотка всех инокулированных животных+(положительная) в день 7
Сыворотка особей-часовых- (отрицательная) в день 7
Мазки: см. таблицу 4A
Группа 2: сыворотка всех инокулированных животных+в день 7
Сыворотка особей-часовых -в день 7
Мазки: см. Таблицу 4B
Группа 3: сыворотка всех инокулированных животных+в день 4
Мазки: см. Таблицу 4C
Результаты ПЦР
Результаты лимфатических узлов
Печеночные лимфатические узлы всех 13 животных, полученные в момент вскрытия, гомогенизировали 10% (масс/об) в культуральной среде. ДНК выделяли из гомогената и присутствие вируса анализировали посредством ПЦР. Контрольный лимфатический узел был отрицательным в отношении нового парвовируса, все 12 инокулированных или особей-часовых свиней оказались положительными в отношении вируса.
Заключение:
На основании представленных данных может быть сделано заключение, что новый парвовирус реплицируется у свиней. Путем передачи наиболее вероятно является пероральный/назальный посредством прямого контакта, но не может быть исключена орально/фекальная передача и передача через воздух. Экскреция с фекалиями, однако, ограничена.
Вирус обнаруживается во множестве органов.
На основании комбинированных результатов в примере 1-3 ожидают, что в подгруппе животных у около 1-2% всех инфицированных животных, инфицирование новым парвовирусом вызывает заболевание примерно в момент появления вируса в крови (виремия). Сброс в фекалии является минимальным, но вирус остается присутствовать в крови >30 дней после инфицирования. Также назальные и глазные мазки остаются ПЦР положительными >30 дней после инфицирования. У инфицированных свиней, как описано в примере 3, не наблюдали геморрагического кишечного синдрома, но это было ожидаемо на основании низкой частоты заболевания; 1-2% в общей популяции и факта, что животные, используемые в примере 3, были относительно молодыми и в превосходном состоянии. Они, в отличие от свиней в условиях коммерческих ферм, не имели предрасполагающих факторов риска.
Таблица 4A: Результаты группы 1: 3 инфицированных животных и 2 особи-часовых, Инокулят A
Новый парвовирус свиней по изобретению называют как ʺBOWLʺ.
R: ректальный мазок
N: назальный мазок
E: глазной мазок
S: сыворотка
н/а: не анализировали (не получали образец)
Таблица 4B: Результаты группы 2: 3 инфицированных животных и 1 особь-часовой, Инокулят B
Новый парвовирус свиней по изобретению называют как ʺBOWLʺ.
R: ректальный мазок
N: назальный мазок
E: глазной мазок
S: сыворотка
н/а: не анализировали (не получали образец)
Таблица 4C: Результаты группы 3: 3 инфицированных животных, Инокулят B
Новый парвовирус свиней по изобретению называют как ʺBOWLʺ.
R: ректальный мазок
N: назальный мазок
E: глазной мазок
S: сыворотка
н/а: не анализировали (не получали образец)
Подписи к чертежам
Фиг. 1: филогенетическая схема, показывающая родство NS1 нового парвовируса свиней по изобретению с NS1 других парвовирусов. Новый парвовирус свиней по изобретению называют как ʺBOWLʺ.
Фиг. 2: филогенетическая схема, показывающая родство капсидного белка нового парвовируса свиней по изобретению с капсидным белком других парвовирусов. Новый парвовирус свиней по изобретению называют как ʺBOWLʺ.
Фиг. 3: пример геморрагического кишечного синдрома, как видно из группы образцов 1 и 2
Список литературы
1)http://www.merckmanuals.com/vet/digestive_system/intestinal_diseases_in_pigs/hemorrhagic_bowel_syndrome_in_pigs.html
2) Häni et al. (1993) Schweiz Arch Tierheilkd. 135:117-124
3)http://www.pork.org/filelibrary/Factsheets/PIGFactsheets/NEWfactSheets/04-01-01g.pdf
4) Streck et al., Journal of General Virology 92: 2628-2636 (2011)
5) Ren et al., Virus research 178: 392-397 (2013)
6) Xiao et al., Veterinary Microbiology 161: 325-330 (2013)
7) Opriessnig et al., Veterinary Microbiology 163: 177-183 (2013)
8) Chenbin Li et al., Archives of Virology 158: 1987-1991 (2013)
9) Xiao et al., Veterinary Microbiology 160: 290-296 (2012)
10) Cadar et al., Archives of Virology 156: 2233-2239 (2011)
11) Lau et al., Journal of General Virology 89: 1840-1848 (2008)
12) Cheung et al., Archives of Virology 155: 801-806(2010)
13) Huang et al., Virology Journal 7: 333-336 (2010)
14) Cheung et al., Archives of Virology 156: 2071-2078 (2011)
15) Xiao et al., PLoS One 8: e65312. (2013)
16) Xiao et al., Genome Announcements 1:e00021-12. (2013)
17) Cheng et al., PLoS One 5: e13583. (2010)
18) Martinez et al., Vaccine 10: 684-690 (1992)
19) Casal et al., Biotechnology and Applied Biochemistry 29: 141-150 (1999)
20) Zhou et al, Virology Journal 7: 366 (2010)
21) Hao Feng, PlosOne; January 17, 2014, DOI: 10.1371/journal.pone.007957
22) Saliki et al., Journ. Gen. Virol. 73: 369-74 (1992)
23) Brown et al., Journ. of Virol. 65: 2702 (1991)
24) Baculovirus Expression Vectors, A Laboratory Manual. By David R. O'Reilly, Lois K. Miller, and Verne A. Luckow. Publisher: Oxford University Press, USA ISBN-10: 0195091310 (September 23, 1993), ISBN-13: 978-0195091311 (May 1994).
25) Baculovirus and Insect Cell Expression Protocols. In: Methods in Molecular Biology™, Volume 388 (2007). Editors: David W. Murhammer. ISBN: 978-1-58829-537-8 (Print) 978-1-59745-457-5 (Online)
26) Fujisaki, Y., and Murikami, Y (1982). "Immunity to infection with porcine parvovirus in pigs inoculated with attenuated HT-strain". Natl Inst Anim Health (Tokyo) 22: 36-37
27) Chen et al., Virology Journal 8:307 (2011)
28) Mayer et al., Vaccine 22: 317-328 (2004)
29) Production of recombinant proteins: novel microbial and eukaryotic expression systems by Gerd Gellissen, ISBN: 3-527-31036-3
30) Gerdts et al, Journal of General Virology 78: 2139-2146 (1997)
31) Gorres et al., Clinical Vaccine Immunology 18: 1987-1995 (2011)
32) Paul & Mengeling, Am. J. Vet. Res.: 41: 2007-2011 (1980)
33) Paul & Mengeling, Am. J. Vet. Res.: 45: 2481-2485 (1984)
34) Fujisaki e& Murakami, Nat. Inst. of Animal Health quarterly (Tokyo) 22: 36-37 (1982)
35) Mayer and Walter, eds. Immunochemical Methods in Cell and Molecular Biology, Academic Press, London, 1987)
36) Kohler and Milstein, Nature, 256: 495-497 (1975)
37) Qiu et al., J. Virol. 79: 11035-11044 (2005)
38) Wang et al., J. Virol. Meth. 200: 41-46 (2014)
39) Guan et al., J. Virol. 83: 9541-9553 (2009)
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
--->
<110> Intervet International B.V.
<120> Новый свиной парвовирус
<130> 23755
<160> 10
<170> PatentIn version 3.5
<210> 1
<211> 3570
<212> ДНК
<213> Свиной парвовирус
<220>
<221> CDS
<222> (1)..(3570)
<400> 1
atg agc cgt tct act caa aga gat ctt tgg tct ttg tta agg gag aga 48
Met Ser Arg Ser Thr Gln Arg Asp Leu Trp Ser Leu Leu Arg Glu Arg
1 5 10 15
ctt gaa agg tat aag gat cga gtt aaa tat tat ggt att ttg gtg cca 96
Leu Glu Arg Tyr Lys Asp Arg Val Lys Tyr Tyr Gly Ile Leu Val Pro
20 25 30
gaa cgt cct tct acc tta gca tct tat ttt agt aaa gac cca cct cca 144
Glu Arg Pro Ser Thr Leu Ala Ser Tyr Phe Ser Lys Asp Pro Pro Pro
35 40 45
gat cct cca act gtt aaa ttt gat aaa ccc tat cag gat gta gat aga 192
Asp Pro Pro Thr Val Lys Phe Asp Lys Pro Tyr Gln Asp Val Asp Arg
50 55 60
ttc tgg ttt cca aca gac gat tat agt gac tgg tat gtc tgg cat gga 240
Phe Trp Phe Pro Thr Asp Asp Tyr Ser Asp Trp Tyr Val Trp His Gly
65 70 75 80
gag gac aga cct cct aga ttt acc gag cat tcc ggt gtt cag ggt ttt 288
Glu Asp Arg Pro Pro Arg Phe Thr Glu His Ser Gly Val Gln Gly Phe
85 90 95
aaa act agg tgt gag ggg ggg ctt cct tta atg ccc agt gat ccc aaa 336
Lys Thr Arg Cys Glu Gly Gly Leu Pro Leu Met Pro Ser Asp Pro Lys
100 105 110
ttt tgc ccc ata caa aat ttt tgg gat cag ttc gct aat ttt gat gag 384
Phe Cys Pro Ile Gln Asn Phe Trp Asp Gln Phe Ala Asn Phe Asp Glu
115 120 125
ggg tct ccg tcc acc cag atc ggt gag agt gtt tca tgg agt gat cat 432
Gly Ser Pro Ser Thr Gln Ile Gly Glu Ser Val Ser Trp Ser Asp His
130 135 140
ttt gga agt ccc gat ccc tct cat cat cag gag gtg agg gat gct gat 480
Phe Gly Ser Pro Asp Pro Ser His His Gln Glu Val Arg Asp Ala Asp
145 150 155 160
gaa gtt act tcc gag cgg cag caa tat aaa aat agg att gtc acc tta 528
Glu Val Thr Ser Glu Arg Gln Gln Tyr Lys Asn Arg Ile Val Thr Leu
165 170 175
tta aga aaa gtt tat tgg gct aag cag tgg tct ggg aaa tta caa att 576
Leu Arg Lys Val Tyr Trp Ala Lys Gln Trp Ser Gly Lys Leu Gln Ile
180 185 190
aat gtt cct tcc ctc gaa agc ctt tat gag cag atc ccc tat atg cta 624
Asn Val Pro Ser Leu Glu Ser Leu Tyr Glu Gln Ile Pro Tyr Met Leu
195 200 205
gcc tat atg gac gct gat aat tgg cgt cag aat ttg tta gct gct aaa 672
Ala Tyr Met Asp Ala Asp Asn Trp Arg Gln Asn Leu Leu Ala Ala Lys
210 215 220
act ctg cga acc act ttg gaa gca ttt tcc tgt gtt cca gat cct tcc 720
Thr Leu Arg Thr Thr Leu Glu Ala Phe Ser Cys Val Pro Asp Pro Ser
225 230 235 240
acg tgc gat gtc aca atc tcc aca ccc cta tct ggt gaa acg gat ccc 768
Thr Cys Asp Val Thr Ile Ser Thr Pro Leu Ser Gly Glu Thr Asp Pro
245 250 255
gcc tct ttt gca aaa tac tta tgt tcc ctt gta tgc aac agg tct caa 816
Ala Ser Phe Ala Lys Tyr Leu Cys Ser Leu Val Cys Asn Arg Ser Gln
260 265 270
gaa aaa gaa cag gcg cag acg cct tct ttg tct cca tct aaa caa gaa 864
Glu Lys Glu Gln Ala Gln Thr Pro Ser Leu Ser Pro Ser Lys Gln Glu
275 280 285
ggg cag atg tca agt cct gat tct gca tcg atc tcc caa cct ccc cct 912
Gly Gln Met Ser Ser Pro Asp Ser Ala Ser Ile Ser Gln Pro Pro Pro
290 295 300
gaa agt cac aag gat aga ctg ctt cct aaa act gat ccc ctt cag gaa 960
Glu Ser His Lys Asp Arg Leu Leu Pro Lys Thr Asp Pro Leu Gln Glu
305 310 315 320
gca ggg ccc att ccc gct ccc ccc aca gct cag aag cct att atc tct 1008
Ala Gly Pro Ile Pro Ala Pro Pro Thr Ala Gln Lys Pro Ile Ile Ser
325 330 335
aag ggt gca ggc ggt gga ggg tct tct ggc ttc ata atc cct cca aaa 1056
Lys Gly Ala Gly Gly Gly Gly Ser Ser Gly Phe Ile Ile Pro Pro Lys
340 345 350
cct cct agc ccc gat cac cct aaa gat ccc ccc cct cct cct cct ccc 1104
Pro Pro Ser Pro Asp His Pro Lys Asp Pro Pro Pro Pro Pro Pro Pro
355 360 365
tct cca att cct cct tct aca tct gcg cct gac gca gaa aag cac gag 1152
Ser Pro Ile Pro Pro Ser Thr Ser Ala Pro Asp Ala Glu Lys His Glu
370 375 380
cta gag cgt gct aaa cag gag aaa caa gag gaa gat gag ctc atg cac 1200
Leu Glu Arg Ala Lys Gln Glu Lys Gln Glu Glu Asp Glu Leu Met His
385 390 395 400
aga atc aaa tca gga gaa gga gaa gga gaa cga gga ggc ctc gtc ttg 1248
Arg Ile Lys Ser Gly Glu Gly Glu Gly Glu Arg Gly Gly Leu Val Leu
405 410 415
cct tct cac cac tac act ggt cct aga aat cct gtc cca gct ggc aag 1296
Pro Ser His His Tyr Thr Gly Pro Arg Asn Pro Val Pro Ala Gly Lys
420 425 430
cct gct gac ccc gtt gat gaa tct tct gcg aga cat gac atc agg tat 1344
Pro Ala Asp Pro Val Asp Glu Ser Ser Ala Arg His Asp Ile Arg Tyr
435 440 445
ggg caa cgt ctt aaa cat gga gac tgg cca tac ctg tgg ggg aag gac 1392
Gly Gln Arg Leu Lys His Gly Asp Trp Pro Tyr Leu Trp Gly Lys Asp
450 455 460
ttg gat aat gct cag cga gat gag att atc aaa gct ctt cat agt cat 1440
Leu Asp Asn Ala Gln Arg Asp Glu Ile Ile Lys Ala Leu His Ser His
465 470 475 480
gtc aaa gtg gga acc caa ttg gca ggg aat ata gtg agg agt att tgg 1488
Val Lys Val Gly Thr Gln Leu Ala Gly Asn Ile Val Arg Ser Ile Trp
485 490 495
aag gct aag gag ctc tta aca gaa cct gtg tat gag ctg tta aag tct 1536
Lys Ala Lys Glu Leu Leu Thr Glu Pro Val Tyr Glu Leu Leu Lys Ser
500 505 510
att ctc cct cct tca gat tta tct aaa gtt cct ctt cct cat tcc caa 1584
Ile Leu Pro Pro Ser Asp Leu Ser Lys Val Pro Leu Pro His Ser Gln
515 520 525
cag aca gac aga aca gaa gat cca gaa act cca ggg gag act aga gga 1632
Gln Thr Asp Arg Thr Glu Asp Pro Glu Thr Pro Gly Glu Thr Arg Gly
530 535 540
act gga tca gac agt cct cga tct cct cgg cct tct gga tca act gaa 1680
Thr Gly Ser Asp Ser Pro Arg Ser Pro Arg Pro Ser Gly Ser Thr Glu
545 550 555 560
gac ggg gga ggt cct tct tcc gag tcc aga tta cct ggg act aaa gtt 1728
Asp Gly Gly Gly Pro Ser Ser Glu Ser Arg Leu Pro Gly Thr Lys Val
565 570 575
cca gta gac cca tct gcc acc acg tcc gaa gca aag agg cag agg act 1776
Pro Val Asp Pro Ser Ala Thr Thr Ser Glu Ala Lys Arg Gln Arg Thr
580 585 590
gag gag ggg atg gac ata tct tct tgc tgt cca ggg ggg att tct gct 1824
Glu Glu Gly Met Asp Ile Ser Ser Cys Cys Pro Gly Gly Ile Ser Ala
595 600 605
tct ggg gct gct tca aat aac tct ggt ctt gct tgt ggg ggt ggg ggg 1872
Ser Gly Ala Ala Ser Asn Asn Ser Gly Leu Ala Cys Gly Gly Gly Gly
610 615 620
ggt act aat tta ggg aca gaa tct ctt gta tcc ggc tgt cag ttt ggt 1920
Gly Thr Asn Leu Gly Thr Glu Ser Leu Val Ser Gly Cys Gln Phe Gly
625 630 635 640
aaa aac tct gtg atc act tca tct ttt aga cga tgt ctc att tca ccc 1968
Lys Asn Ser Val Ile Thr Ser Ser Phe Arg Arg Cys Leu Ile Ser Pro
645 650 655
tgg cct gat aaa tac tgt tgt tct tct gct cac gat ctt att ccc gga 2016
Trp Pro Asp Lys Tyr Cys Cys Ser Ser Ala His Asp Leu Ile Pro Gly
660 665 670
gtg gtc tac gag acc cct tgg tgc tat tat gat ctg aac gtc atc tca 2064
Val Val Tyr Glu Thr Pro Trp Cys Tyr Tyr Asp Leu Asn Val Ile Ser
675 680 685
gca cat ttt tct cct tct gct tgg cag agg ctt ttg gag gat tat gat 2112
Ala His Phe Ser Pro Ser Ala Trp Gln Arg Leu Leu Glu Asp Tyr Asp
690 695 700
gcc ttt cga cct aaa tcc ctt aaa gtt acc atc cag tct tta gtt ttt 2160
Ala Phe Arg Pro Lys Ser Leu Lys Val Thr Ile Gln Ser Leu Val Phe
705 710 715 720
aaa gat gtc tgt caa ggt gca gaa aaa caa act aca gtt cag gat tcc 2208
Lys Asp Val Cys Gln Gly Ala Glu Lys Gln Thr Thr Val Gln Asp Ser
725 730 735
cag tca gcc acc att gct atc ttt gag gat aaa gac tat gac tac ccc 2256
Gln Ser Ala Thr Ile Ala Ile Phe Glu Asp Lys Asp Tyr Asp Tyr Pro
740 745 750
tat gtg atg gga ggg ggt caa aaa aca gtt ccg ggt cac ttg ccc ggt 2304
Tyr Val Met Gly Gly Gly Gln Lys Thr Val Pro Gly His Leu Pro Gly
755 760 765
caa cct tat aat ctt ccc aag tat tct tac aga acc ctt ggt tca gtc 2352
Gln Pro Tyr Asn Leu Pro Lys Tyr Ser Tyr Arg Thr Leu Gly Ser Val
770 775 780
aaa gaa agt aat agg gcc agt atg ggc ggt tca ggg tac act ttc aaa 2400
Lys Glu Ser Asn Arg Ala Ser Met Gly Gly Ser Gly Tyr Thr Phe Lys
785 790 795 800
tcc aat caa gat acg gaa ttg ttt ctg ctt gaa aca cat gat gcc act 2448
Ser Asn Gln Asp Thr Glu Leu Phe Leu Leu Glu Thr His Asp Ala Thr
805 810 815
ctt att cga ggc ggg ggt act ttt gag cag tac tat gaa ttt cca aac 2496
Leu Ile Arg Gly Gly Gly Thr Phe Glu Gln Tyr Tyr Glu Phe Pro Asn
820 825 830
gat ctt cct ttc gaa aat ttg act cag tat cct tgg gat atc cgc cgt 2544
Asp Leu Pro Phe Glu Asn Leu Thr Gln Tyr Pro Trp Asp Ile Arg Arg
835 840 845
cag gat aac ccc ctc tat cag cag agg atc act gtc atg tca ggt tct 2592
Gln Asp Asn Pro Leu Tyr Gln Gln Arg Ile Thr Val Met Ser Gly Ser
850 855 860
gac aga gat acg gta ggc att tta gat gga gat ttt tac tct cct ttt 2640
Asp Arg Asp Thr Val Gly Ile Leu Asp Gly Asp Phe Tyr Ser Pro Phe
865 870 875 880
cgg ttc aaa ggt cat gat aga ccc gcc atg tgg ctg cca gga cag agg 2688
Arg Phe Lys Gly His Asp Arg Pro Ala Met Trp Leu Pro Gly Gln Arg
885 890 895
ttg att cag ggc aaa ttc ata gat acg cac cca ata ccc aat aca ggg 2736
Leu Ile Gln Gly Lys Phe Ile Asp Thr His Pro Ile Pro Asn Thr Gly
900 905 910
agg agt ggg gtt cat cct aat gat ttt cac aca agg ggc gat ggt cat 2784
Arg Ser Gly Val His Pro Asn Asp Phe His Thr Arg Gly Asp Gly His
915 920 925
ggt gac acc cat aga aca cat gaa gag agg atc tac agt cta gat aca 2832
Gly Asp Thr His Arg Thr His Glu Glu Arg Ile Tyr Ser Leu Asp Thr
930 935 940
ggt ctt gct gct atg cca cgt gcc gct cat aga ccc acc ctt cag ccc 2880
Gly Leu Ala Ala Met Pro Arg Ala Ala His Arg Pro Thr Leu Gln Pro
945 950 955 960
gga cct agg act ctg tct cat gcc gta cgc aga ccc gat ggt tcc acc 2928
Gly Pro Arg Thr Leu Ser His Ala Val Arg Arg Pro Asp Gly Ser Thr
965 970 975
gtg gtc acg gct aat gcg tgt gct tac gct tac acc cag gag aat cct 2976
Val Val Thr Ala Asn Ala Cys Ala Tyr Ala Tyr Thr Gln Glu Asn Pro
980 985 990
cat cag gaa ccc tgg agt gat ctc aat gtc aga cat acc atg tat agg 3024
His Gln Glu Pro Trp Ser Asp Leu Asn Val Arg His Thr Met Tyr Arg
995 1000 1005
tta gcc tat caa cgt caa aaa ggt ttt cag caa ccc ggg gac cct 3069
Leu Ala Tyr Gln Arg Gln Lys Gly Phe Gln Gln Pro Gly Asp Pro
1010 1015 1020
ctt cat att cga acc cat gct tgt tat ggg gac ggg gat gtt acc 3114
Leu His Ile Arg Thr His Ala Cys Tyr Gly Asp Gly Asp Val Thr
1025 1030 1035
att cca aaa gaa gag tcc tta tgg cct act gtt ctg ggt agt tgc 3159
Ile Pro Lys Glu Glu Ser Leu Trp Pro Thr Val Leu Gly Ser Cys
1040 1045 1050
aca gaa aag tcc cct gcc tgt tta gag tcc cag att tgg tgt aaa 3204
Thr Glu Lys Ser Pro Ala Cys Leu Glu Ser Gln Ile Trp Cys Lys
1055 1060 1065
aca cca aat gtg gac atg gtc tat gga gaa cac aca ccc cct ctt 3249
Thr Pro Asn Val Asp Met Val Tyr Gly Glu His Thr Pro Pro Leu
1070 1075 1080
gct tta tgg ggt atg cat gct ccc cca ccc cat gta ttt ctc agg 3294
Ala Leu Trp Gly Met His Ala Pro Pro Pro His Val Phe Leu Arg
1085 1090 1095
atg ctt gct caa gag ggt cct cct aat gtc agt act tgc aga ccg 3339
Met Leu Ala Gln Glu Gly Pro Pro Asn Val Ser Thr Cys Arg Pro
1100 1105 1110
gct caa tct ggt cag acc ttc atc aat caa tat ggt cag ttt ctc 3384
Ala Gln Ser Gly Gln Thr Phe Ile Asn Gln Tyr Gly Gln Phe Leu
1115 1120 1125
ctc tgt ttt acc atg gta tgg gaa gtt aag cct aga ccc aag tcc 3429
Leu Cys Phe Thr Met Val Trp Glu Val Lys Pro Arg Pro Lys Ser
1130 1135 1140
atc aag caa tgg aat cca cgt ccg ccc atc agc att cct gtt ggt 3474
Ile Lys Gln Trp Asn Pro Arg Pro Pro Ile Ser Ile Pro Val Gly
1145 1150 1155
cag tct ggt cct gct ttc att ctc gat caa gat ggc tac tac cgt 3519
Gln Ser Gly Pro Ala Phe Ile Leu Asp Gln Asp Gly Tyr Tyr Arg
1160 1165 1170
ctc cca gaa cat gtc tgg tct gcc agg gaa cgt atc cgc agc aaa 3564
Leu Pro Glu His Val Trp Ser Ala Arg Glu Arg Ile Arg Ser Lys
1175 1180 1185
cgc tag 3570
Arg
<210> 2
<211> 1189
<212> Белок
<213> Свиной парвовирус
<400> 2
Met Ser Arg Ser Thr Gln Arg Asp Leu Trp Ser Leu Leu Arg Glu Arg
1 5 10 15
Leu Glu Arg Tyr Lys Asp Arg Val Lys Tyr Tyr Gly Ile Leu Val Pro
20 25 30
Glu Arg Pro Ser Thr Leu Ala Ser Tyr Phe Ser Lys Asp Pro Pro Pro
35 40 45
Asp Pro Pro Thr Val Lys Phe Asp Lys Pro Tyr Gln Asp Val Asp Arg
50 55 60
Phe Trp Phe Pro Thr Asp Asp Tyr Ser Asp Trp Tyr Val Trp His Gly
65 70 75 80
Glu Asp Arg Pro Pro Arg Phe Thr Glu His Ser Gly Val Gln Gly Phe
85 90 95
Lys Thr Arg Cys Glu Gly Gly Leu Pro Leu Met Pro Ser Asp Pro Lys
100 105 110
Phe Cys Pro Ile Gln Asn Phe Trp Asp Gln Phe Ala Asn Phe Asp Glu
115 120 125
Gly Ser Pro Ser Thr Gln Ile Gly Glu Ser Val Ser Trp Ser Asp His
130 135 140
Phe Gly Ser Pro Asp Pro Ser His His Gln Glu Val Arg Asp Ala Asp
145 150 155 160
Glu Val Thr Ser Glu Arg Gln Gln Tyr Lys Asn Arg Ile Val Thr Leu
165 170 175
Leu Arg Lys Val Tyr Trp Ala Lys Gln Trp Ser Gly Lys Leu Gln Ile
180 185 190
Asn Val Pro Ser Leu Glu Ser Leu Tyr Glu Gln Ile Pro Tyr Met Leu
195 200 205
Ala Tyr Met Asp Ala Asp Asn Trp Arg Gln Asn Leu Leu Ala Ala Lys
210 215 220
Thr Leu Arg Thr Thr Leu Glu Ala Phe Ser Cys Val Pro Asp Pro Ser
225 230 235 240
Thr Cys Asp Val Thr Ile Ser Thr Pro Leu Ser Gly Glu Thr Asp Pro
245 250 255
Ala Ser Phe Ala Lys Tyr Leu Cys Ser Leu Val Cys Asn Arg Ser Gln
260 265 270
Glu Lys Glu Gln Ala Gln Thr Pro Ser Leu Ser Pro Ser Lys Gln Glu
275 280 285
Gly Gln Met Ser Ser Pro Asp Ser Ala Ser Ile Ser Gln Pro Pro Pro
290 295 300
Glu Ser His Lys Asp Arg Leu Leu Pro Lys Thr Asp Pro Leu Gln Glu
305 310 315 320
Ala Gly Pro Ile Pro Ala Pro Pro Thr Ala Gln Lys Pro Ile Ile Ser
325 330 335
Lys Gly Ala Gly Gly Gly Gly Ser Ser Gly Phe Ile Ile Pro Pro Lys
340 345 350
Pro Pro Ser Pro Asp His Pro Lys Asp Pro Pro Pro Pro Pro Pro Pro
355 360 365
Ser Pro Ile Pro Pro Ser Thr Ser Ala Pro Asp Ala Glu Lys His Glu
370 375 380
Leu Glu Arg Ala Lys Gln Glu Lys Gln Glu Glu Asp Glu Leu Met His
385 390 395 400
Arg Ile Lys Ser Gly Glu Gly Glu Gly Glu Arg Gly Gly Leu Val Leu
405 410 415
Pro Ser His His Tyr Thr Gly Pro Arg Asn Pro Val Pro Ala Gly Lys
420 425 430
Pro Ala Asp Pro Val Asp Glu Ser Ser Ala Arg His Asp Ile Arg Tyr
435 440 445
Gly Gln Arg Leu Lys His Gly Asp Trp Pro Tyr Leu Trp Gly Lys Asp
450 455 460
Leu Asp Asn Ala Gln Arg Asp Glu Ile Ile Lys Ala Leu His Ser His
465 470 475 480
Val Lys Val Gly Thr Gln Leu Ala Gly Asn Ile Val Arg Ser Ile Trp
485 490 495
Lys Ala Lys Glu Leu Leu Thr Glu Pro Val Tyr Glu Leu Leu Lys Ser
500 505 510
Ile Leu Pro Pro Ser Asp Leu Ser Lys Val Pro Leu Pro His Ser Gln
515 520 525
Gln Thr Asp Arg Thr Glu Asp Pro Glu Thr Pro Gly Glu Thr Arg Gly
530 535 540
Thr Gly Ser Asp Ser Pro Arg Ser Pro Arg Pro Ser Gly Ser Thr Glu
545 550 555 560
Asp Gly Gly Gly Pro Ser Ser Glu Ser Arg Leu Pro Gly Thr Lys Val
565 570 575
Pro Val Asp Pro Ser Ala Thr Thr Ser Glu Ala Lys Arg Gln Arg Thr
580 585 590
Glu Glu Gly Met Asp Ile Ser Ser Cys Cys Pro Gly Gly Ile Ser Ala
595 600 605
Ser Gly Ala Ala Ser Asn Asn Ser Gly Leu Ala Cys Gly Gly Gly Gly
610 615 620
Gly Thr Asn Leu Gly Thr Glu Ser Leu Val Ser Gly Cys Gln Phe Gly
625 630 635 640
Lys Asn Ser Val Ile Thr Ser Ser Phe Arg Arg Cys Leu Ile Ser Pro
645 650 655
Trp Pro Asp Lys Tyr Cys Cys Ser Ser Ala His Asp Leu Ile Pro Gly
660 665 670
Val Val Tyr Glu Thr Pro Trp Cys Tyr Tyr Asp Leu Asn Val Ile Ser
675 680 685
Ala His Phe Ser Pro Ser Ala Trp Gln Arg Leu Leu Glu Asp Tyr Asp
690 695 700
Ala Phe Arg Pro Lys Ser Leu Lys Val Thr Ile Gln Ser Leu Val Phe
705 710 715 720
Lys Asp Val Cys Gln Gly Ala Glu Lys Gln Thr Thr Val Gln Asp Ser
725 730 735
Gln Ser Ala Thr Ile Ala Ile Phe Glu Asp Lys Asp Tyr Asp Tyr Pro
740 745 750
Tyr Val Met Gly Gly Gly Gln Lys Thr Val Pro Gly His Leu Pro Gly
755 760 765
Gln Pro Tyr Asn Leu Pro Lys Tyr Ser Tyr Arg Thr Leu Gly Ser Val
770 775 780
Lys Glu Ser Asn Arg Ala Ser Met Gly Gly Ser Gly Tyr Thr Phe Lys
785 790 795 800
Ser Asn Gln Asp Thr Glu Leu Phe Leu Leu Glu Thr His Asp Ala Thr
805 810 815
Leu Ile Arg Gly Gly Gly Thr Phe Glu Gln Tyr Tyr Glu Phe Pro Asn
820 825 830
Asp Leu Pro Phe Glu Asn Leu Thr Gln Tyr Pro Trp Asp Ile Arg Arg
835 840 845
Gln Asp Asn Pro Leu Tyr Gln Gln Arg Ile Thr Val Met Ser Gly Ser
850 855 860
Asp Arg Asp Thr Val Gly Ile Leu Asp Gly Asp Phe Tyr Ser Pro Phe
865 870 875 880
Arg Phe Lys Gly His Asp Arg Pro Ala Met Trp Leu Pro Gly Gln Arg
885 890 895
Leu Ile Gln Gly Lys Phe Ile Asp Thr His Pro Ile Pro Asn Thr Gly
900 905 910
Arg Ser Gly Val His Pro Asn Asp Phe His Thr Arg Gly Asp Gly His
915 920 925
Gly Asp Thr His Arg Thr His Glu Glu Arg Ile Tyr Ser Leu Asp Thr
930 935 940
Gly Leu Ala Ala Met Pro Arg Ala Ala His Arg Pro Thr Leu Gln Pro
945 950 955 960
Gly Pro Arg Thr Leu Ser His Ala Val Arg Arg Pro Asp Gly Ser Thr
965 970 975
Val Val Thr Ala Asn Ala Cys Ala Tyr Ala Tyr Thr Gln Glu Asn Pro
980 985 990
His Gln Glu Pro Trp Ser Asp Leu Asn Val Arg His Thr Met Tyr Arg
995 1000 1005
Leu Ala Tyr Gln Arg Gln Lys Gly Phe Gln Gln Pro Gly Asp Pro
1010 1015 1020
Leu His Ile Arg Thr His Ala Cys Tyr Gly Asp Gly Asp Val Thr
1025 1030 1035
Ile Pro Lys Glu Glu Ser Leu Trp Pro Thr Val Leu Gly Ser Cys
1040 1045 1050
Thr Glu Lys Ser Pro Ala Cys Leu Glu Ser Gln Ile Trp Cys Lys
1055 1060 1065
Thr Pro Asn Val Asp Met Val Tyr Gly Glu His Thr Pro Pro Leu
1070 1075 1080
Ala Leu Trp Gly Met His Ala Pro Pro Pro His Val Phe Leu Arg
1085 1090 1095
Met Leu Ala Gln Glu Gly Pro Pro Asn Val Ser Thr Cys Arg Pro
1100 1105 1110
Ala Gln Ser Gly Gln Thr Phe Ile Asn Gln Tyr Gly Gln Phe Leu
1115 1120 1125
Leu Cys Phe Thr Met Val Trp Glu Val Lys Pro Arg Pro Lys Ser
1130 1135 1140
Ile Lys Gln Trp Asn Pro Arg Pro Pro Ile Ser Ile Pro Val Gly
1145 1150 1155
Gln Ser Gly Pro Ala Phe Ile Leu Asp Gln Asp Gly Tyr Tyr Arg
1160 1165 1170
Leu Pro Glu His Val Trp Ser Ala Arg Glu Arg Ile Arg Ser Lys
1175 1180 1185
Arg
<210> 3
<211> 1989
<212> ДНК
<213> Свиной парвовирус
<220>
<221> CDS
<222> (1)..(1989)
<400> 3
atg tct cag acc ttc tgg acg ggt att tgc agg ctt ttc cct gat att 48
Met Ser Gln Thr Phe Trp Thr Gly Ile Cys Arg Leu Phe Pro Asp Ile
1 5 10 15
ggg aag att cct ggg gtg act cct gag cgg tat att tat gct gta aac 96
Gly Lys Ile Pro Gly Val Thr Pro Glu Arg Tyr Ile Tyr Ala Val Asn
20 25 30
gtg agt acc cgt aat ggg caa aag tgg ccg aaa gtt ggg acc gag ggg 144
Val Ser Thr Arg Asn Gly Gln Lys Trp Pro Lys Val Gly Thr Glu Gly
35 40 45
cca ttg gct gca ggt gtc tta cag ggg gag gcc ctt ttc cgt gag acc 192
Pro Leu Ala Ala Gly Val Leu Gln Gly Glu Ala Leu Phe Arg Glu Thr
50 55 60
ctt aaa gag gtc cgg aag gtg tgc cgg ctg ccg caa gac ccc cat ctt 240
Leu Lys Glu Val Arg Lys Val Cys Arg Leu Pro Gln Asp Pro His Leu
65 70 75 80
ttc ttt caa ttg gag aaa gtt gat tcc aaa ggg ggt ctt cac ctt cat 288
Phe Phe Gln Leu Glu Lys Val Asp Ser Lys Gly Gly Leu His Leu His
85 90 95
ttt tgt att agt gtt gct gct ggc act cct aga gat gtg tct tcg atg 336
Phe Cys Ile Ser Val Ala Ala Gly Thr Pro Arg Asp Val Ser Ser Met
100 105 110
ttt aaa gcc att gag cgt cga gtt tcc ttt tac tac ttt ggt gta gag 384
Phe Lys Ala Ile Glu Arg Arg Val Ser Phe Tyr Tyr Phe Gly Val Glu
115 120 125
ggc ttg act ttt ttt act cct cat aaa aat aag cat ggt gct tgg aag 432
Gly Leu Thr Phe Phe Thr Pro His Lys Asn Lys His Gly Ala Trp Lys
130 135 140
agt aat gat gag ggc ttt att gtg aat tat ctt ctt aaa aaa ttg cct 480
Ser Asn Asp Glu Gly Phe Ile Val Asn Tyr Leu Leu Lys Lys Leu Pro
145 150 155 160
ttg tct gag tgt gtg tat gcg tgg act aat atg gat ggg gtg att ggc 528
Leu Ser Glu Cys Val Tyr Ala Trp Thr Asn Met Asp Gly Val Ile Gly
165 170 175
gat gcc tgt ctg aat gaa gat aag cgc agg gaa ttg ttg tct gag cgt 576
Asp Ala Cys Leu Asn Glu Asp Lys Arg Arg Glu Leu Leu Ser Glu Arg
180 185 190
caa gat cag gga gtc att aaa gag ctc act gct cct act ttt aaa gct 624
Gln Asp Gln Gly Val Ile Lys Glu Leu Thr Ala Pro Thr Phe Lys Ala
195 200 205
gct acg ggt gat aaa atg ttg agt gtg gtg gat tgg atg tgt gat aat 672
Ala Thr Gly Asp Lys Met Leu Ser Val Val Asp Trp Met Cys Asp Asn
210 215 220
gat gtg act acg gag cgc cga tgg gag gaa ata tcg gct gcg tcc ctg 720
Asp Val Thr Thr Glu Arg Arg Trp Glu Glu Ile Ser Ala Ala Ser Leu
225 230 235 240
tac tct ttc ctt gct acc cca gct ggc act cat ttg gct aaa cag tgt 768
Tyr Ser Phe Leu Ala Thr Pro Ala Gly Thr His Leu Ala Lys Gln Cys
245 250 255
ctt aga gct gcg aat cag cgt att gtg agc act aaa cct ctt ggt ttg 816
Leu Arg Ala Ala Asn Gln Arg Ile Val Ser Thr Lys Pro Leu Gly Leu
260 265 270
agt ctc tgt aaa ttt gcc agt gaa aag gag ctt cgt gct ttt caa aat 864
Ser Leu Cys Lys Phe Ala Ser Glu Lys Glu Leu Arg Ala Phe Gln Asn
275 280 285
ggg gat cct gag ttg tct tca gac aac aat agg atc tat tac tta ttt 912
Gly Asp Pro Glu Leu Ser Ser Asp Asn Asn Arg Ile Tyr Tyr Leu Phe
290 295 300
gct atg aat aat tac tgc cct gat atg gcc agt atg att ttc ttt tgg 960
Ala Met Asn Asn Tyr Cys Pro Asp Met Ala Ser Met Ile Phe Phe Trp
305 310 315 320
tgg tct atg cgc caa acg ggt aag aga aac agt att tgg ctg ttt gga 1008
Trp Ser Met Arg Gln Thr Gly Lys Arg Asn Ser Ile Trp Leu Phe Gly
325 330 335
cct gct aca acc ggt aaa acc aat tta gca agt gct att gca cat act 1056
Pro Ala Thr Thr Gly Lys Thr Asn Leu Ala Ser Ala Ile Ala His Thr
340 345 350
gct gcc agt tat ggg tgt gtt aac tgg aat aac gca aat ttc ccg ttt 1104
Ala Ala Ser Tyr Gly Cys Val Asn Trp Asn Asn Ala Asn Phe Pro Phe
355 360 365
caa gac att gtg aat gtg caa ctg ggt tgg tgg gag gaa gga aag atg 1152
Gln Asp Ile Val Asn Val Gln Leu Gly Trp Trp Glu Glu Gly Lys Met
370 375 380
aca gag gat gtg gtt gaa tgt gct aag gca ttg cta ggg gga agt aat 1200
Thr Glu Asp Val Val Glu Cys Ala Lys Ala Leu Leu Gly Gly Ser Asn
385 390 395 400
gta aga gtt gac cgc aaa tgt atg caa tct gcg gaa gtg caa tct cct 1248
Val Arg Val Asp Arg Lys Cys Met Gln Ser Ala Glu Val Gln Ser Pro
405 410 415
cct ttt att att aca tcc aat acg gat atg tgc ctt gtt tct caa ggc 1296
Pro Phe Ile Ile Thr Ser Asn Thr Asp Met Cys Leu Val Ser Gln Gly
420 425 430
agt tac atc agt ttt gag cac cag cag cct ctc cag gat aga atg att 1344
Ser Tyr Ile Ser Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Ile
435 440 445
aaa ttt gag ttt aac cat gtt ctt cct ggc aat ttc ggc ctc att tct 1392
Lys Phe Glu Phe Asn His Val Leu Pro Gly Asn Phe Gly Leu Ile Ser
450 455 460
gaa gat gaa gtc gtg gcc ttc ttc aga aat ggt gct tat aat gtg ttg 1440
Glu Asp Glu Val Val Ala Phe Phe Arg Asn Gly Ala Tyr Asn Val Leu
465 470 475 480
aag cat gcg tac atg aga aag gcc cag ctt ttt gct ctc ggt cca gct 1488
Lys His Ala Tyr Met Arg Lys Ala Gln Leu Phe Ala Leu Gly Pro Ala
485 490 495
tcc ttg cca tat aag ccc cct acg ggt gaa tta gtg tgt ata gac caa 1536
Ser Leu Pro Tyr Lys Pro Pro Thr Gly Glu Leu Val Cys Ile Asp Gln
500 505 510
gct aag tct cct tct ccc tct gct tct cgc cgc agt gtg aga aat tgg 1584
Ala Lys Ser Pro Ser Pro Ser Ala Ser Arg Arg Ser Val Arg Asn Trp
515 520 525
atg acg tgt ccc cct gat cct gtc cag gac gat ccc gct gaa ctg gac 1632
Met Thr Cys Pro Pro Asp Pro Val Gln Asp Asp Pro Ala Glu Leu Asp
530 535 540
gag tat ttt cct cca gat act cct cca gtg gat tgt cct tgt cct att 1680
Glu Tyr Phe Pro Pro Asp Thr Pro Pro Val Asp Cys Pro Cys Pro Ile
545 550 555 560
tct cct gtg agg gag tct tgt ccc tcg cca ggg cct tgc cct acc cct 1728
Ser Pro Val Arg Glu Ser Cys Pro Ser Pro Gly Pro Cys Pro Thr Pro
565 570 575
cct cgc aaa aaa caa cgg aag agc aag cat tgc tcc ttg tct gtc tct 1776
Pro Arg Lys Lys Gln Arg Lys Ser Lys His Cys Ser Leu Ser Val Ser
580 585 590
gcg ggt aaa gtt cct gtg gtg gtt gtg ggt gat tct gac tca gtc ccc 1824
Ala Gly Lys Val Pro Val Val Val Val Gly Asp Ser Asp Ser Val Pro
595 600 605
ccc gaa gaa aaa gaa aaa gag gaa gtt tcc gtg ggg gaa tcc cag gat 1872
Pro Glu Glu Lys Glu Lys Glu Glu Val Ser Val Gly Glu Ser Gln Asp
610 615 620
cca caa ctg tac tgg gac cta act ctc agt caa tcc gac gtt cct gtg 1920
Pro Gln Leu Tyr Trp Asp Leu Thr Leu Ser Gln Ser Asp Val Pro Val
625 630 635 640
cct gaa gac gag agt acc cag ttt cct gac gac gct gtg gac gct tct 1968
Pro Glu Asp Glu Ser Thr Gln Phe Pro Asp Asp Ala Val Asp Ala Ser
645 650 655
gat ctc att gct gaa cag tag 1989
Asp Leu Ile Ala Glu Gln
660
<210> 4
<211> 662
<212> Белок
<213> Свиной парвовирус
<400> 4
Met Ser Gln Thr Phe Trp Thr Gly Ile Cys Arg Leu Phe Pro Asp Ile
1 5 10 15
Gly Lys Ile Pro Gly Val Thr Pro Glu Arg Tyr Ile Tyr Ala Val Asn
20 25 30
Val Ser Thr Arg Asn Gly Gln Lys Trp Pro Lys Val Gly Thr Glu Gly
35 40 45
Pro Leu Ala Ala Gly Val Leu Gln Gly Glu Ala Leu Phe Arg Glu Thr
50 55 60
Leu Lys Glu Val Arg Lys Val Cys Arg Leu Pro Gln Asp Pro His Leu
65 70 75 80
Phe Phe Gln Leu Glu Lys Val Asp Ser Lys Gly Gly Leu His Leu His
85 90 95
Phe Cys Ile Ser Val Ala Ala Gly Thr Pro Arg Asp Val Ser Ser Met
100 105 110
Phe Lys Ala Ile Glu Arg Arg Val Ser Phe Tyr Tyr Phe Gly Val Glu
115 120 125
Gly Leu Thr Phe Phe Thr Pro His Lys Asn Lys His Gly Ala Trp Lys
130 135 140
Ser Asn Asp Glu Gly Phe Ile Val Asn Tyr Leu Leu Lys Lys Leu Pro
145 150 155 160
Leu Ser Glu Cys Val Tyr Ala Trp Thr Asn Met Asp Gly Val Ile Gly
165 170 175
Asp Ala Cys Leu Asn Glu Asp Lys Arg Arg Glu Leu Leu Ser Glu Arg
180 185 190
Gln Asp Gln Gly Val Ile Lys Glu Leu Thr Ala Pro Thr Phe Lys Ala
195 200 205
Ala Thr Gly Asp Lys Met Leu Ser Val Val Asp Trp Met Cys Asp Asn
210 215 220
Asp Val Thr Thr Glu Arg Arg Trp Glu Glu Ile Ser Ala Ala Ser Leu
225 230 235 240
Tyr Ser Phe Leu Ala Thr Pro Ala Gly Thr His Leu Ala Lys Gln Cys
245 250 255
Leu Arg Ala Ala Asn Gln Arg Ile Val Ser Thr Lys Pro Leu Gly Leu
260 265 270
Ser Leu Cys Lys Phe Ala Ser Glu Lys Glu Leu Arg Ala Phe Gln Asn
275 280 285
Gly Asp Pro Glu Leu Ser Ser Asp Asn Asn Arg Ile Tyr Tyr Leu Phe
290 295 300
Ala Met Asn Asn Tyr Cys Pro Asp Met Ala Ser Met Ile Phe Phe Trp
305 310 315 320
Trp Ser Met Arg Gln Thr Gly Lys Arg Asn Ser Ile Trp Leu Phe Gly
325 330 335
Pro Ala Thr Thr Gly Lys Thr Asn Leu Ala Ser Ala Ile Ala His Thr
340 345 350
Ala Ala Ser Tyr Gly Cys Val Asn Trp Asn Asn Ala Asn Phe Pro Phe
355 360 365
Gln Asp Ile Val Asn Val Gln Leu Gly Trp Trp Glu Glu Gly Lys Met
370 375 380
Thr Glu Asp Val Val Glu Cys Ala Lys Ala Leu Leu Gly Gly Ser Asn
385 390 395 400
Val Arg Val Asp Arg Lys Cys Met Gln Ser Ala Glu Val Gln Ser Pro
405 410 415
Pro Phe Ile Ile Thr Ser Asn Thr Asp Met Cys Leu Val Ser Gln Gly
420 425 430
Ser Tyr Ile Ser Phe Glu His Gln Gln Pro Leu Gln Asp Arg Met Ile
435 440 445
Lys Phe Glu Phe Asn His Val Leu Pro Gly Asn Phe Gly Leu Ile Ser
450 455 460
Glu Asp Glu Val Val Ala Phe Phe Arg Asn Gly Ala Tyr Asn Val Leu
465 470 475 480
Lys His Ala Tyr Met Arg Lys Ala Gln Leu Phe Ala Leu Gly Pro Ala
485 490 495
Ser Leu Pro Tyr Lys Pro Pro Thr Gly Glu Leu Val Cys Ile Asp Gln
500 505 510
Ala Lys Ser Pro Ser Pro Ser Ala Ser Arg Arg Ser Val Arg Asn Trp
515 520 525
Met Thr Cys Pro Pro Asp Pro Val Gln Asp Asp Pro Ala Glu Leu Asp
530 535 540
Glu Tyr Phe Pro Pro Asp Thr Pro Pro Val Asp Cys Pro Cys Pro Ile
545 550 555 560
Ser Pro Val Arg Glu Ser Cys Pro Ser Pro Gly Pro Cys Pro Thr Pro
565 570 575
Pro Arg Lys Lys Gln Arg Lys Ser Lys His Cys Ser Leu Ser Val Ser
580 585 590
Ala Gly Lys Val Pro Val Val Val Val Gly Asp Ser Asp Ser Val Pro
595 600 605
Pro Glu Glu Lys Glu Lys Glu Glu Val Ser Val Gly Glu Ser Gln Asp
610 615 620
Pro Gln Leu Tyr Trp Asp Leu Thr Leu Ser Gln Ser Asp Val Pro Val
625 630 635 640
Pro Glu Asp Glu Ser Thr Gln Phe Pro Asp Asp Ala Val Asp Ala Ser
645 650 655
Asp Leu Ile Ala Glu Gln
660
<210> 5
<211> 17
<212> ДНК
<213> Свиной парвовирус
<400> 5
ctacatctgc gcctgac 17
<210> 6
<211> 18
<212> ДНК
<213> Свиной парвовирус
<400> 6
gtggtgagaa ggcaagac 18
<210> 7
<211> 20
<212> ДНК
<213> Свиной парвовирус
<400> 7
tgttgagtgt ggtggattgg 20
<210> 8
<211> 19
<212> ДНК
<213> Свиной парвовирус
<400> 8
aaggaagctg gaccgagag 19
<210> 9
<211> 24
<212> ДНК
<213> Свиной парвовирус
<400> 9
cacgagctag agcgtgctaa acag 24
<210> 10
<211> 5999
<212> ДНК
<213> Свиной парвовирус
<400> 10
ttaatggacc aataggattt acttctgctt tttagggtat aaaagttcgt acttctgctt 60
tttccctcat tctgtgcttg ggagctgatc tgtttagatc tttttgcttc tctgcgccga 120
aagcctctgt attatgtctc agaccttctg gacgggtatt tgcaggcttt tccctgatat 180
tgggaagatt cctggggtga ctcctgagcg gtatatttat gctgtaaacg tgagtacccg 240
taatgggcaa aagtggccga aagttgggac cgaggggcca ttggctgcag gtgtcttaca 300
gggggaggcc cttttccgtg agacccttaa agaggtccgg aaggtgtgcc ggctgccgca 360
agacccccat cttttctttc aattggagaa agttgattcc aaagggggtc ttcaccttca 420
tttttgtatt agtgttgctg ctggcactcc tagagatgtg tcttcgatgt ttaaagccat 480
tgagcgtcga gtttcctttt actactttgg tgtagagggc ttgacttttt ttactcctca 540
taaaaataag catggtgctt ggaagagtaa tgatgagggc tttattgtga attatcttct 600
taaaaaattg cctttgtctg agtgtgtgta tgcgtggact aatatggatg gggtgattgg 660
cgatgcctgt ctgaatgaag ataagcgcag ggaattgttg tctgagcgtc aagatcaggg 720
agtcattaaa gagctcactg ctcctacttt taaagctgct acgggtgata aaatgttgag 780
tgtggtggat tggatgtgtg ataatgatgt gactacggag cgccgatggg aggaaatatc 840
ggctgcgtcc ctgtactctt tccttgctac cccagctggc actcatttgg ctaaacagtg 900
tcttagagct gcgaatcagc gtattgtgag cactaaacct cttggtttga gtctctgtaa 960
atttgccagt gaaaaggagc ttcgtgcttt tcaaaatggg gatcctgagt tgtcttcaga 1020
caacaatagg atctattact tatttgctat gaataattac tgccctgata tggccagtat 1080
gattttcttt tggtggtcta tgcgccaaac gggtaagaga aacagtattt ggctgtttgg 1140
acctgctaca accggtaaaa ccaatttagc aagtgctatt gcacatactg ctgccagtta 1200
tgggtgtgtt aactggaata acgcaaattt cccgtttcaa gacattgtga atgtgcaact 1260
gggttggtgg gaggaaggaa agatgacaga ggatgtggtt gaatgtgcta aggcattgct 1320
agggggaagt aatgtaagag ttgaccgcaa atgtatgcaa tctgcggaag tgcaatctcc 1380
tccttttatt attacatcca atacggatat gtgccttgtt tctcaaggca gttacatcag 1440
ttttgagcac cagcagcctc tccaggatag aatgattaaa tttgagttta accatgttct 1500
tcctggcaat ttcggcctca tttctgaaga tgaagtcgtg gccttcttca gaaatggtgc 1560
ttataatgtg ttgaagcatg cgtacatgag aaaggcccag ctttttgctc tcggtccagc 1620
ttccttgcca tataagcccc ctacgggtga attagtgtgt atagaccaag ctaagtctcc 1680
ttctccctct gcttctcgcc gcagtgtgag aaattggatg acgtgtcccc ctgatcctgt 1740
ccaggacgat cccgctgaac tggacgagta ttttcctcca gatactcctc cagtggattg 1800
tccttgtcct atttctcctg tgagggagtc ttgtccctcg ccagggcctt gccctacccc 1860
tcctcgcaaa aaacaacgga agagcaagca ttgctccttg tctgtctctg cgggtaaagt 1920
tcctgtggtg gttgtgggtg attctgactc agtccccccc gaagaaaaag aaaaagagga 1980
agtttccgtg ggggaatccc aggatccaca actgtactgg gacctaactc tcagtcaatc 2040
cgacgttcct gtgcctgaag acgagagtac ccagtttcct gacgacgctg tggacgcttc 2100
tgatctcatt gctgaacagt agttaaggta tgagccgttc tactcaaaga gatctttggt 2160
ctttgttaag ggagagactt gaaaggtata aggatcgagt taaatattat ggtattttgg 2220
tgccagaacg tccttctacc ttagcatctt attttagtaa agacccacct ccagatcctc 2280
caactgttaa atttgataaa ccctatcagg atgtagatag attctggttt ccaacagacg 2340
attatagtga ctggtatgtc tggcatggag aggacagacc tcctagattt accgagcatt 2400
ccggtgttca gggttttaaa actaggtgtg agggggggct tcctttaatg cccagtgatc 2460
ccaaattttg ccccatacaa aatttttggg atcagttcgc taattttgat gaggggtctc 2520
cgtccaccca gatcggtgag agtgtttcat ggagtgatca ttttggaagt cccgatccct 2580
ctcatcatca ggaggtgagg gatgctgatg aagttacttc cgagcggcag caatataaaa 2640
ataggattgt caccttatta agaaaagttt attgggctaa gcagtggtct gggaaattac 2700
aaattaatgt tccttccctc gaaagccttt atgagcagat cccctatatg ctagcctata 2760
tggacgctga taattggcgt cagaatttgt tagctgctaa aactctgcga accactttgg 2820
aagcattttc ctgtgttcca gatccttcca cgtgcgatgt cacaatctcc acacccctat 2880
ctggtgaaac ggatcccgcc tcttttgcaa aatacttatg ttcccttgta tgcaacaggt 2940
ctcaagaaaa agaacaggcg cagacgcctt ctttgtctcc atctaaacaa gaagggcaga 3000
tgtcaagtcc tgattctgca tcgatctccc aacctccccc tgaaagtcac aaggatagac 3060
tgcttcctaa aactgatccc cttcaggaag cagggcccat tcccgctccc cccacagctc 3120
agaagcctat tatctctaag ggtgcaggcg gtggagggtc ttctggcttc ataatccctc 3180
caaaacctcc tagccccgat caccctaaag atcccccccc tcctcctcct ccctctccaa 3240
ttcctccttc tacatctgcg cctgacgcag aaaagcacga gctagagcgt gctaaacagg 3300
agaaacaaga ggaagatgag ctcatgcaca gaatcaaatc aggagaagga gaaggagaac 3360
gaggaggcct cgtcttgcct tctcaccact acactggtcc tagaaatcct gtcccagctg 3420
gcaagcctgc tgaccccgtt gatgaatctt ctgcgagaca tgacatcagg tatgggcaac 3480
gtcttaaaca tggagactgg ccatacctgt gggggaagga cttggataat gctcagcgag 3540
atgagattat caaagctctt catagtcatg tcaaagtggg aacccaattg gcagggaata 3600
tagtgaggag tatttggaag gctaaggagc tcttaacaga acctgtgtat gagctgttaa 3660
agtctattct ccctccttca gatttatcta aagttcctct tcctcattcc caacagacag 3720
acagaacaga agatccagaa actccagggg agactagagg aactggatca gacagtcctc 3780
gatctcctcg gccttctgga tcaactgaag acgggggagg tccttcttcc gagtccagat 3840
tacctgggac taaagttcca gtagacccat ctgccaccac gtccgaagca aagaggcaga 3900
ggactgagga ggggatggac atatcttctt gctgtccagg ggggatttct gcttctgggg 3960
ctgcttcaaa taactctggt cttgcttgtg ggggtggggg gggtactaat ttagggacag 4020
aatctcttgt atccggctgt cagtttggta aaaactctgt gatcacttca tcttttagac 4080
gatgtctcat ttcaccctgg cctgataaat actgttgttc ttctgctcac gatcttattc 4140
ccggagtggt ctacgagacc ccttggtgct attatgatct gaacgtcatc tcagcacatt 4200
tttctccttc tgcttggcag aggcttttgg aggattatga tgcctttcga cctaaatccc 4260
ttaaagttac catccagtct ttagttttta aagatgtctg tcaaggtgca gaaaaacaaa 4320
ctacagttca ggattcccag tcagccacca ttgctatctt tgaggataaa gactatgact 4380
acccctatgt gatgggaggg ggtcaaaaaa cagttccggg tcacttgccc ggtcaacctt 4440
ataatcttcc caagtattct tacagaaccc ttggttcagt caaagaaagt aatagggcca 4500
gtatgggcgg ttcagggtac actttcaaat ccaatcaaga tacggaattg tttctgcttg 4560
aaacacatga tgccactctt attcgaggcg ggggtacttt tgagcagtac tatgaatttc 4620
caaacgatct tcctttcgaa aatttgactc agtatccttg ggatatccgc cgtcaggata 4680
accccctcta tcagcagagg atcactgtca tgtcaggttc tgacagagat acggtaggca 4740
ttttagatgg agatttttac tctccttttc ggttcaaagg tcatgataga cccgccatgt 4800
ggctgccagg acagaggttg attcagggca aattcataga tacgcaccca atacccaata 4860
cagggaggag tggggttcat cctaatgatt ttcacacaag gggcgatggt catggtgaca 4920
cccatagaac acatgaagag aggatctaca gtctagatac aggtcttgct gctatgccac 4980
gtgccgctca tagacccacc cttcagcccg gacctaggac tctgtctcat gccgtacgca 5040
gacccgatgg ttccaccgtg gtcacggcta atgcgtgtgc ttacgcttac acccaggaga 5100
atcctcatca ggaaccctgg agtgatctca atgtcagaca taccatgtat aggttagcct 5160
atcaacgtca aaaaggtttt cagcaacccg gggaccctct tcatattcga acccatgctt 5220
gttatgggga cggggatgtt accattccaa aagaagagtc cttatggcct actgttctgg 5280
gtagttgcac agaaaagtcc cctgcctgtt tagagtccca gatttggtgt aaaacaccaa 5340
atgtggacat ggtctatgga gaacacacac cccctcttgc tttatggggt atgcatgctc 5400
ccccacccca tgtatttctc aggatgcttg ctcaagaggg tcctcctaat gtcagtactt 5460
gcagaccggc tcaatctggt cagaccttca tcaatcaata tggtcagttt ctcctctgtt 5520
ttaccatggt atgggaagtt aagcctagac ccaagtccat caagcaatgg aatccacgtc 5580
cgcccatcag cattcctgtt ggtcagtctg gtcctgcttt cattctcgat caagatggct 5640
actaccgtct cccagaacat gtctggtctg ccagggaacg tatccgcagc aaacgctagt 5700
gcccccagca atacacttac tacagtattg atgtgtcagg cattctggtt gattgtttta 5760
ttttggctcc gcctactgta tggcccatgt aaacgcatct attatgaaaa taaaatacgt 5820
caattgctga tgtaattcgt gttgtaattc ttgtttgaaa agcgcatatt ttcttgccgg 5880
tctgagtaac accacctatg acatcatata aatttgatta cgtaacttcc tctttttact 5940
tcctcttttt tgattggatt tacgcaatat cacaattcta gcagttatct gagcgggct 5999
1
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МАТЕРИАЛЫ И МЕТОДЫ, ИСПОЛЬЗУЕМЫЕ ДЛЯ ЛЕЧЕНИЯ РЕСПИРАТОРНЫХ ЗАБОЛЕВАНИЙ У СОБАК | 2020 |
|
RU2811752C2 |
| ВИРУС ГРИППА, СПОСОБНЫЙ ИНФИЦИРОВАТЬ СОБАЧЬИХ, И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2802222C2 |
| Вакцина против герпеса | 2019 |
|
RU2731073C1 |
| КОНСТРУКЦИИ НУКЛЕИНОВОЙ КИСЛОТЫ И ВЕКТОРЫ ДЛЯ ГЕНОТЕРАПИИ ДЛЯ ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ВИЛЬСОНА И ДРУГИХ СОСТОЯНИЙ | 2015 |
|
RU2745567C2 |
| ИММУНОГЕННЫЕ КОМПОЗИЦИИ ГЛИКОПРОТЕИНА G ВИРУСОВ HENDRA И NIPAH | 2012 |
|
RU2787820C2 |
| БИОСЕНСОРНАЯ СИСТЕМА ДЛЯ БЫСТРОГО ОБНАРУЖЕНИЯ ОПРЕДЕЛЯЕМЫХ КОМПОНЕНТОВ | 2016 |
|
RU2717658C2 |
| ГЕННАЯ ТЕРАПИЯ ГИПОФОСФАТЕМИЧЕСКИХ ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С ФАКТОРОМ РОСТА ФИБРОБЛАСТОВ 23 | 2020 |
|
RU2815545C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2019 |
|
RU2781542C2 |
| Антитела против белка р17 ВИЧ-1 субтипа А | 2019 |
|
RU2727673C1 |
| ИММУНОИНДУЦИРУЮЩЕЕ СРЕДСТВО | 2016 |
|
RU2744843C2 |
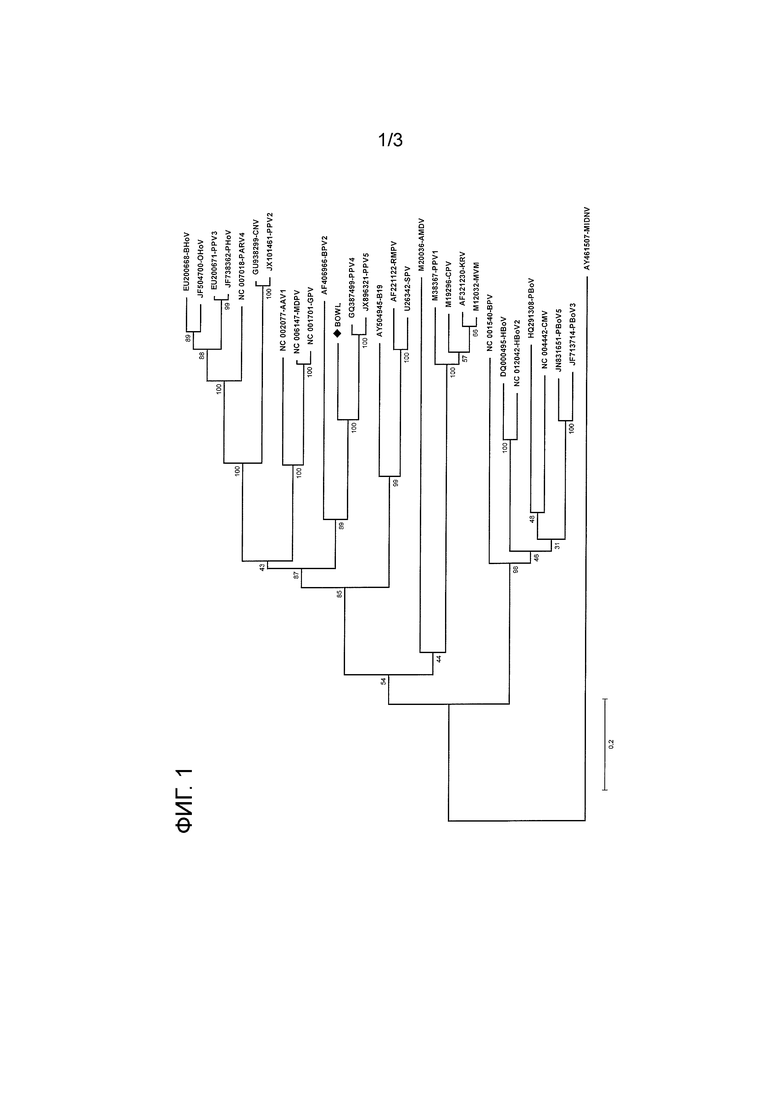

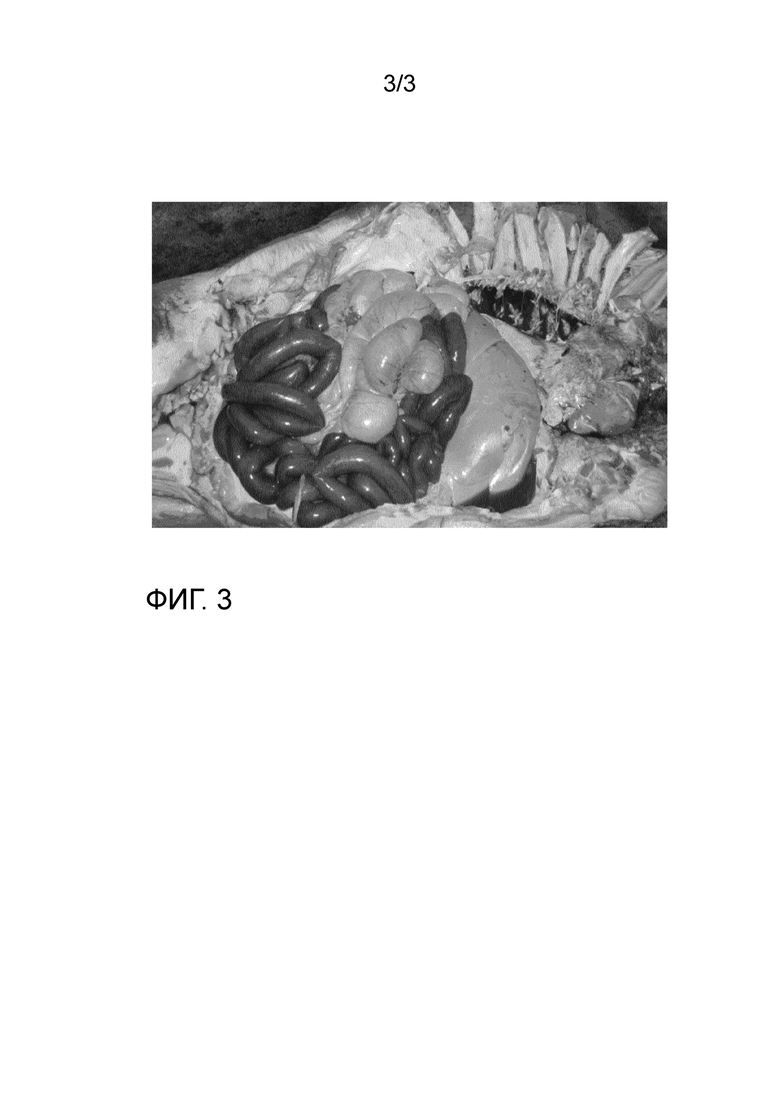
Изобретение относится к биотехнологии. Описана вакцина для борьбы с HBS-ассоциированным парвовирусом у свиней. Вакцина включает иммуногенно эффективное количество капсидного белка (CP), кодируемого фрагментом ДНК, содержащим ген, кодирующий CP. Причем нуклеотидная последовательность указанного гена имеет уровень идентичности по меньшей мере 80% относительно последовательности нуклеотидов гена CP, как показано в SEQ ID NO: 1, и фармацевтически приемлемый носитель. Также описан набор для диагностического тестирования, содержащий набор праймеров для ПЦР, как показано в SEQ ID NO: 5 и 6, который специфически реагирует с участком гена капсидного белка HBS-ассоциированного парвовируса свиней, или набор праймеров, как показано в SEQ ID NO: 7 и 8, который специфически реагирует с участком гена NS1 HBS-ассоциированного парвовируса свиней. Изобретение расширяет арсенал средств для борьбы с HBS-ассоциированным парвовирусом у свиней. 2 н.п. ф-лы, 3 ил., 4 табл., 3 пр.
1. Вакцина для борьбы с HBS-ассоциированным парвовирусом у свиней, отличающаяся тем, что указанная вакцина включает иммуногенно эффективное количество капсидного белка (CP), кодируемого фрагментом ДНК, содержащим ген, кодирующий CP, причем нуклеотидная последовательность указанного гена имеет уровень идентичности по меньшей мере 80% относительно последовательности нуклеотидов гена CP, как показано в SEQ ID NO: 1, и фармацевтически приемлемый носитель.
2. Набор для диагностического тестирования, отличающийся тем, что указанный набор для тестирования содержит набор праймеров для ПЦР, как показано в SEQ ID NO: 5 и 6, который специфически реагирует с участком гена капсидного белка HBS-ассоциированного парвовируса свиней, или набор праймеров, как показано в SEQ ID NO: 7 и 8, который специфически реагирует с участком гена NS1 HBS-ассоциированного парвовируса свиней.
| WO2011107534 A1, 09.09.2011 | |||
| Прибор для равномерного смешения зерна и одновременного отбирания нескольких одинаковых по объему проб | 1921 |
|
SU23A1 |
| EM_STD: KF999685 | |||
| Прибор для равномерного смешения зерна и одновременного отбирания нескольких одинаковых по объему проб | 1921 |
|
SU23A1 |
| EM_STD: KF999683 | |||
| ИММУНОГЕННАЯ КОМПОЗИЦИЯ, ВЫЗЫВАЮЩАЯ ИММУННЫЙ ОТВЕТ ПРОТИВ ПАРВОВИРУСА И ЦИРКОВИРУСА СВИНЬИ, ВАКЦИНА ПРОТИВ МНОГОСИСТЕМНОГО СИНДРОМА ИСТОЩЕНИЯ СВИНЕЙ (PMWS) (ВАРИАНТЫ), НАБОР ДЛЯ ВАКЦИНАЦИИ И СПОСОБ ЕЕ ОСУЩЕСТВЛЕНИЯ | 1999 |
|
RU2237492C2 |

