Область изобретения
Настоящее изобретение относится к микроорганизму, продуцирующему ксантозин-5'-монофосфат, и к способу получения ксантозин-5'-монофосфата с использованием этого микроорганизма.
Предшествующий уровень техники
Ксантозин-5'-монофосфат (ХМР) не только имеет физиологическое значение у животных и растений как промежуточное соединение в биосинтезе нуклеиновых кислот и метаболических путях, но также его используют для различных целей: в пищевых продуктах, лекарственных средствах и для различных медицинских применений. В частности, когда его используют вместе с однозамещенным глутаматом натрия (MSG), присутствует синергетический эффект в усилении ароматов. Соответственно, ХМР представляет собой один из усилителей аромата на основе нуклеиновой кислоты, который привлек внимание как приправа.
Кроме того, ксантозин-5'-монофосфат представляет собой промежуточное соединение в метаболических путях биосинтеза пуриновых нуклеотидов и представляет собой важное исходное вещество для получения гуанозин-5'-монофосфата (GMP). Хорошо известный способ получения гуанозинмонофосфата, который имеет большую промышленную ценность, и действие которого проявляется в увеличении интенсивности вкуса, представляет собой ферментацию микроорганизмом, который включает продуцирование ксантозин-5'-монофосфата и его ферментативное превращение в гуанозин-5'-монофосфат. Поскольку этот способ является наиболее экономически выгодным, ксантозин-5'-монофосфат является таким же необходимым как гуанозин-5'-монофосфат.
В качестве способов получения ксантозин-5'-монофосфата известны: (1) химический синтез, (2) дезаминирование полученного ксантозин-5'-монофосфата, (3) ферментативное получение путем добавления ксантина в качестве предшественника в культуральную среду, (4) прямая ферментация мутантным штаммом микроорганизма, продуцирующего ксантозин-5'-монофосфат, и так далее. Среди различных способов прямая ферментация ксантозин-5'-монофосфата штаммом мутантного микроорганизма является экономически наиболее выгодным способом. Тем не менее, все еще существует необходимость в исследованиях по поиску способов для получения ксантозин-5'-монофосфата с высоким выходом.
Подробное описание изобретения
Техническая задача
Авторы настоящего изобретения поставили задачу разработать микроорганизмы, способные продуцировать ксантозин-5'-монофосфат с высоким выходом, и в результате они обнаружили, что продукционный выход ксантозин-5'-монофосфата увеличивается при повышении активности определенного белка, тем самым создав настоящее изобретение.
Техническое решение
Задача настоящего изобретения заключается в том, чтобы предложить микроорганизм рода Corynebacterium, продуцирующий ксантозин-5'-монофосфат, у которого активность белка, содержащего аминокислотную последовательность SEQ ID NO: 2, повышена.
Другая задача настоящего изобретения заключается в том, чтобы предложить способ получения ксантозин-5'-монофосфата с использованием этого микроорганизма.
Полезные эффекты изобретения
Микроорганизм рода Corynebacterium по настоящему изобретению обладает повышенной активностью белка, который экспортирует ксантозин-5'-монофосфат, тем самым обеспечивая повышенное продуцирование ксантозин-5'-монофосфата, и это может вносить существенный вклад в снижение себестоимости получения ксантозин-5'-монофосфата.
Наилучшее воплощение изобретения
Далее настоящее изобретение описано более подробно. Между тем, каждое описание и пример воплощения, раскрытые в настоящем описании, могут быть применены к другим описаниям и примерам воплощений. То есть, объем настоящего изобретения охватывает все комбинации различных элементов, раскрытых в настоящем описании. Кроме того, объем настоящего изобретения не следует рассматривать как ограниченный конкретным описанием, представленным ниже.
В одном аспекте настоящего изобретения для решения вышеуказанных задач предложен микроорганизм рода Corynebacterium, продуцирующий ксантозин-5'-монофосфат, у которого активность белка, содержащего аминокислотную последовательность SEQ ID NO: 2, повышена.
Как его используют здесь, термин "ксантозин-5'-монофосфат" относится к соединению под названием 5'-ксантиловая кислота, ксантозин и так далее. В настоящем описании термин "ксантозин-5'-монофосфат" может быть использован взаимозаменяемо с "ХМР".
Как его используют здесь, термин "белок, содержащий аминокислотную последовательность SEQ ID NO: 2" относится к белку, эндогенно присутствующему в микроорганизме рода Corynebacterium, кодируемому геном транспортера суперсемейства "major facilitator" (MFS-транспортер), экспортирующему ксантозин-5'-монофосфат из этого микроорганизма. В частности, он может представлять собой белок-MFS-транспортер, содержащий аминокислотную последовательность SEQ ID NO: 2, который эндогенно присутствует в микроорганизме рода Corynebacterium. Кроме того, этот белок может представлять собой белок, состоящий из или составленный из аминокислотной последовательности SEQ ID NO: 2, но он не ограничен этим. Белок MFS-транспортер в настоящем описании может быть назван "xmpE" или "белок xmpE".
Как его используют здесь, термин "белок, содержащий аминокислотную последовательность определенной SEQ ID NO", при условии что он обладает активностью идентичной или соответствующей активности белка, включающего такую же SEQ ID NO, может включать в своей последовательности вставку, которая не изменяет функцию белка, в начале или в конце аминокислотной последовательности; природную мутацию; или молчащую мутацию. В случае белков, имеющих в последовательности такую вставку или мутацию, очевидно, что они также включены в объем настоящего изобретения.
Кроме того, белок по настоящему изобретению может содержать аминокислотную последовательность SEQ ID NO: 2, но также последовательность, имеющую по меньшей мере 60% гомологии или идентичности с SEQ ID NO: 2. Белок, содержащий аминокислотную последовательность, имеющую по меньшей мере 60% гомологии или идентичности с SEQ ID NO: 2, может представлять собой белок, включающий аминокислотную последовательность, имеющую по меньшей мере 60%, в частности 70%, более конкретно 80%, еще более конкретно 83%, 84%, 85%, 88%, 90%, 93%, 95% или 97% гомологии или идентичности с SEQ ID NO: 2. Аминокислотная последовательность, имеющая гомологию или идентичность, может исключать таковую, имеющую 100% идентичности, из указанного диапазона или представлять собой таковую, имеющую идентичность менее 100%. Очевидно, что при условии, когда аминокислотная последовательность, как и последовательность, имеющая гомологию или идентичность последовательности, по существу обладает биологической активностью идентичной или соответствующей таковой SEQ ID NO: 2, она включена в объем настоящего изобретения, даже если часть этой последовательности имеет делецию, модификацию, замену или вставку аминокислотной последовательности.
Дополнительно, нуклеотидная последовательность гена, кодирующего белок, содержащий аминокислотную последовательность SEQ ID NO: 2, может быть получена из известной базы данных, такой как NCBI GenBank, но не ограничена этим. В частности, белок, содержащий аминокислотную последовательность SEQ ID NO: 2, может кодироваться геном, включающим нуклеотидную последовательность SEQ ID NO: 1, а также геном, состоящим из нуклеотидной последовательности SEQ ID NO: 1 или составленным из таковой, но не ограничен этим.
Кроме того, нуклеотидная последовательность SEQ ID NO: 1 может включать не только саму нуклеотидную последовательность SEQ ID NO: 1, но также нуклеотидную последовательность, имеющую по меньшей мере 80% гомологии или идентичности с SEQ ID NO: 1.
В частности, любая нуклеотидная последовательность, способная кодировать аминокислотную последовательность, имеющую по меньшей мере 80% гомологии или идентичности с SEQ ID NO: 2, включена в объем настоящего изобретения, но белок может кодироваться геном, включающим нуклеотидную последовательность, имеющую по меньшей мере 80%, в частности 83%, 84%, 85%, 88%, 90%, 93%, 95% или 97% гомологии или идентичности с SEQ ID NO: 1. Однако очевидно, что нуклеотидная последовательность включена в объем настоящего изобретения без ограничения, при условии что она кодирует белок, обладающий активностью, соответствующей таковой белка, включающего аминокислотную последовательность SEQ ID NO: 2.
Кроме того, очевидно, что вследствие вырожденности генетического кода, полинуклеотид, который может быть транслирован в белок, включающий такую же аминокислотную последовательность, или белок, имеющий гомологию или идентичность с ним, может быть включен в объем настоящего изобретения. Далее, зонд, который может быть получен из известной последовательности гена, например любой последовательности, кодирующей белок, обладающий активностью белка, включающего аминокислотную последовательность SEQ ID NO: 2, путем гибридизации всей или части нуклеотидной последовательности с комплементарной ей последовательностью в строгих условиях, может быть включен без ограничения. Понятие "строгие условия" относится к условиям, обеспечивающим возможность специфической гибридизации между полинуклеотидами. Эти условия подробно описаны в литературе (J. Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989); например, условия, при которых гены, имеющие высокую степень гомологии или идентичности по меньшей мере 40%, в частности 85%, в частности 90%, более конкретно 95%, еще более конкретно 97% и в особенности 99%, гибридизуются, и те, которые имеют более низкую степень гомологии или идентичности, не гибридизуются; или условия традиционной гибридизации по Саузерну, то есть однократная промывка, в частности от двукратной до трехкратной, при температуре и концентрации соли, эквивалетным 60°С, 1×SSC (стандартный раствор хлорида и цитрата натрия), 0,1% SDS (додецилсульфат натрия), в частности 60°С, 0,1×SSC, 0,1% SDS, более точно 68°С, 0,1×SSC, 0,1% SDS. Хотя гибридизация может допускать несоответствие между основаниями в зависимости от степени строгости, для нее требуется, чтобы две нуклеиновые кислоты имели комплементарные друг другу последовательности. Как его используют здесь, термин "комплементарный" используют для описания соответствия между нуклеотидными основаниями, способными гибридизоваться друг с другом. В отношении ДНК, например, аденозин является комплементарным тимину, и цитозин является комплементарным гуанину. Соответственно, настоящее изобретение также может включать выделенный фрагмент нуклеиновой кислоты, комплементарный не только по существу подобной последовательности нуклеиновой кислоты, но также полной последовательности.
В частности, полинуклеотид, имеющий гомологию или идентичность, может быть выявлен с использованием условий гибридизации, включающих стадию гибридизации при значении Tm 55°С и в вышеописанных условиях. Кроме того, значение Tm может составлять 60°С, 63°С или 65°С, но оно не ограничено этим и может быть соответствующим образом скорректировано специалистом в данной области техники.
Подходящая степень строгости для гибридизации полинуклеотидов зависит от длины и степени комплементарности полинуклеотидов, и переменные хорошо известны в соответствующей области техники (см. Sambrook et al., supra, 9.50-9.51, 11.7-11.8).
Как его используют здесь, термин "гомология" относится к степени идентичности между двумя заданными аминокислотными последовательностями или нуклеотидными последовательностями и может быть выражена в процентах. В настоящем описании гомологичная последовательность, имеющая одинаковую или сходную активность с заданной аминокислотной последовательностью или полинуклеотидной последовательностью, может быть выражена в "% гомологии".
Как его используют здесь, термин "идентичность" относится к степени родства последовательностей между аминокислотными или нуклеотидными последовательностями, и в некоторых случаях ее можно определить по соответствию между цепями таких последовательностей.
Термины "гомология" и "идентичность" часто используют взаимозаменяемо друг с другом.
Гомологию или идентичность последовательностей консервативных полинуклеотидов или полипептидов можно определить с помощью стандартных алгоритмов выравнивания и использовать со штрафом за пропуск в последовательности по умолчанию, установленным используемой программой. По существу обычно ожидают, что гомологичные или идентичные полинуклеотиды или полипептиды гибридизуются в умеренно строгих или очень строгих условиях вдоль по меньшей мере примерно 50%, 60%, 70%, 80% или 90% всей длины всех или целевых полинуклеотидов или полипептидов. Рассматривают также полинуклеотиды, которые содержат вырожденные кодоны вместо кодонов в гибридизуемых полипетидах.
Имеют ли две полинуклеотидные или полипептидные последовательности гомологию или идентичность, например, по меньшей мере 50%, 55%, 60%, 65% 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99%, можно определить с помощью известного компьютерного алгоритма, такого как программа "FASTA" (например, Pearson et al., (1988) Proc. Natl. Acad. Sci. USA 85: 2444), используя параметры по умолчанию. Альтернативно, это можно определить с использованием алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48: 443-453), который выполнен в программе "Needleman" пакета "EMBOSS" (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16: 276-277) (предпочтительно версия 5.0.0 или более поздняя). (Включая пакет программ "GCG" (Devereux, J., et al., Nucleic Acids Research 12: 387 (1984)), BLASTP, BLASTN, FASTA (Atschul, [S.] [F.,] [ET AL., J MOLEC BIOL 215]: 403 (1990); Guide to Huge Computers, Martin J. Bishop, [ED.,] Academic Press, San Diego, 1994, и [CARILLO ETA/.](1988) SIAM J Applied Math 48: 1073). Например, гомологию или идентичность можно определить с использованием "BLAST" или "ClustalW" от Национального Центра Биотехнологической Информации (NCBI).
Гомологию или идентичность полинуклеотидов или полипептидов можно определить путем сравнения информации о последовательностях как она опубликована (например Smith and Waterman, Adv. Appl. Math (1981) 2:482), например с использованием компьютерной программы "GAP", как раскрыто в Needleman et al. (1970), J Mol Biol.48 : 443. Таким образом, программа "GAP" определяет гомологию или идентичность как величину, полученную путем деления числа одинаково выровненных символов (то есть нуклеотидов или аминокислот) на общее число символов в более короткой из двух последовательностей. Параметры по умолчанию для программы "GAP" могут включать: (1) одинарную матрицу сравнения (содержащую значение 1 для совпадений и 0 для несовпадений) и взвешенную матрицу сравнения от Gribskov et al. (1986), Nucl. Acids Res. 14:6745, как раскрыто в Schwartz and Dayhoff, eds., Atlas of Белок Sequence and Structure, National Biomedical Research Foundation, pp. 353-358, 1979; (2) штраф 3,0 за каждый пропуск и дополнительно штраф 0,10 за каждый символ в каждом пропуске (или штраф за внесение пропуска 10 и штраф за продление пропуска 0,5); и (3) отсутствие штрафа за окончание пропуска. Соответственно, как его используют здесь, термин "гомология" или "идентичность" относится к соответствию между полипептидами или полинуклеотидами.
Микроорганизм рода Corynebacterium, продуцирующий ксантозин-5'-монофосфат, в настоящем изобретении может обладать повышенной активностью белка, содержащего аминокислотную последовательность SEQ ID NO: 2.
Понятие "повышение активности белка, содержащего аминокислотную последовательность SEQ ID NO: 2" можно использовать взаимозаменяемо с понятием "повышение активности xmpE " или "повышение активности белка, кодируемого геном, содержащим нуклеотидную последовательность SEQ ID NO: 1".
Как его используют здесь, термин "повышенная активность белка, содержащего аминокислотную последовательность SEQ ID NO: 2" относится к увеличенной экспрессии белка в сравнении с родительским штаммом или к таковой для белка, включающего аминокислотную последовательность SEQ ID NO: 2, в сравнении с не модифицированным штаммом; его повышенной активности с эквивалентной экспрессией; или его повышенной активности и экспрессии. Также этот термин относится к повышенной активности белка по сравнению с его эндогенной активностью и повышенной экспрессии или активности xmpE в сравнении с его родительским штаммом или немодифицированным штаммом.
В настоящем изобретении повышение активности белка может быть достигнуто использованием различных способов, известных в данной области техники. Например, повышение активности может быть достигнуто путем увеличения числа копий полинуклеотидов, кодирующих этот белок, повышения активности промотора, замены стартового кодона, или их комбинации, в частности, путем: 1) увеличения числа копий полинуклеотида, кодирующего белок; 2) модификации последовательности регулятора экспрессии (промотора, оператора и так далее), с тем чтобы увеличить экспрессию полинуклеотида; 3) модификации последовательности гена (стартового кодона и так далее) в хромосоме, с тем чтобы повысить активность белка, или их комбинации, но не ограничиваясь этим.
В частности, в отношении повышения активности белка, способ модификации последовательности регулятора экспрессии может быть выполнен с использованием различных способов, известных в данной области техники. Например, может быть проведена модификация для повышения активности последовательности регулятора экспрессии посредством введения модификации путем делеции, вставки или неконсервативной или консервативной замены, или их комбинации в регуляторной последовательности; или замены нуклеотидной последовательности на нуклеотидную последовательность, имеющую повышенную активность. Регулятор экспрессии включает промотор, энхансер, оператор, сайт связывания с рибосомой и последовательность, которая регулирует терминацию транскрипции и трансляции, но не ограничиваясь этим.
Дополнительно, может быть выполнен способ модификации последовательности гена для повышения активности белка путем введения модификации в последовательности путем делеции, вставки, неконсервативной или консервативной замены, или их комбинации, или замены последовательности гена на модифицированную последовательность гена, имеющего повышенную активность, но не ограничиваясь этим.
В определенном примере воплощения повышение активности белка может быть достигнуто путем увеличения числа копий xmpE; замены промотора xmpE на другой промотор, имеющий повышенную активность; замены стартового кодона xmpE, или их комбинации.
Как его используют здесь, термин "вектор" относится к ДНК-конструкции, содержащей нуклеотидную последовательность полинуклеотида, кодирующего целевой белок, функционально связанную с подходящей регуляторной последовательностью для экспрессии целевого белка в подходящем хозяине. Регуляторная последовательность может включать промотор, который может инициировать транскрипцию, любую операторную последовательность для контроля транскрипции, последовательность для кодирования подходящего сайта связывания мРНК с рибосомой и последовательность контроля терминации транскрипции и трансляции. Вектором можно трансфицировать подходящего хозяина, и затем он может реплицироваться или функционировать независимо от генома хозяина и может сам интегрировать в геном.
Как его используют здесь, термин "трансформация" относится к введению вектора, содержащего полинуклеотид, кодирующий целевой белок, в клетку-хозяина, так что белок, кодируемый полинуклеотидом, может экспрессироваться в клетке-хозяине. При условии, что он может экспрессироваться в клетке-хозяине, трансформированный полинуклеотид может быть интегрирован в хромосому и локализоваться в хромосоме клетки-хозяина, или локализоваться вне хромосомы. Кроме того, полинуклеотид включает ДНК и РНК, кодирующие целевой белок. Полинуклеотид может быть введен в любой форме, при условии что он может быть введен в клетку-хозяина и экспрессироваться в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, которая представляет собой полинуклеотидную конструкцию, включающую все элементы, необходимые для ее автономной экспрессии. Экспрессионная кассета может включать промотор, функционально связанный с геном, сигнал терминации транскрипции, сайт связывания с рибосомой и сигнал терминации трансляции. Экспрессионная кассета может находиться в форме самореплицирующегося вектора экспрессии. Кроме того, полинуклеотид может быть введен в клетку-хозяина как есть и функционально связан с последовательностью, необходимой для экспрессии в клетке-хозяине.
Как его используют здесь, термин "микроорганизм, продуцирующий ксантозин-5'-монофосфат" или "микроорганизм, обладающий способностью продуцировать ксантозин-5'-монофосфат" относится к микроорганизму, от природы обладающему способностью продуцировать ксантозин-5'-монофосфат, или к микроорганизму, в котором способностью продуцировать ксантозин-5'-монофосфат наделен родительский штамм, не обладающий способностью продуцировать ксантозин-5'-монофосфат. В частности, он может представлять собой микроорганизм, обладающий способностью продуцировать ксантозин-5'-монофосфат, у которого повышена активность белка, содержащего аминокислотную последовательность SEQ ID NO: 2 или белка xmpE.
Кроме того, в отношении задачи настоящего изобретения, способность микроорганизма продуцировать ксантозин-5'-монофосфат может быть улучшена/повышена вследствие повышения ксантозин-5'-монофосфат-экспортирующей способности.
В частности, термины "способность продуцировать" и "способность экспортировать" могут быть использованы взаимозаменяемо. Кроме того, термин "микроорганизм" может быть использован взаимозаменяемо с термином "микроорганизм, экспортирующий ксантозин-5'-монофосфат" или "микроорганизм, обладающий ксантозин-5'-монофосфат-экспортирующей способностью". Он может относиться к микроорганизму, от природы обладающему ксантозин-5'-монофосфат-экспортирующей способностью, или к микроорганизму, в котором ксантозин-5'-монофосфат-экспортирующей способностью наделен родительский штамм, не обладающий ксантозин-5'-монофосфат-экспортирующей способностью.
В частности, микроорганизм рода Corynebacterium, продуцирующий ксантозин-5'-монофосфат, может представлять собой Corynebacterium stationis, или Corynebacterium glutamicum, или Corynebacterium ammoniagenes; и более конкретно, может представлять собой Corynebacterium stationis, но не ограничен этим.
В другом аспекте настоящего изобретения предложен способ получения ксантозин-5'-монофосфата, включающий культивирование микроорганизма рода Corynebacterium в культуральной среде и выделение ксантозин-5'-монофосфата из микроорганизма или из культуральной среды.
Микроорганизм и ксантозин-5'-монофосфат по настоящему изобретению являются такими же как описано выше.
Микроорганизм рода Corynebacterium по настоящему изобретению может представлять собой Corynebacterium stationis, но не ограничиваясь этим.
В отношении способа по настоящему изобретению микроорганизм рода Corynebacterium можно культивировать с использованием любых условий культивирования и способов, известных в данной области техники.
Как его используют здесь, термин "культура" относится к культивированию микроорганизма в умеренно искусственно контролируемых условиях окружающей среды. Процесс культивирования по настоящему изобретению можно проводить в соответствии с подходящей культуральной средой и условиями культивирования, известными в данной области техники. Кроме того, способ культивирования включает периодическое культивирование, непрерывное культивирование и периодическое культивирование с подпиткой; в частности, путем периодического процесса, или периодического процесса с подпиткой, или повторяющегося периодического процесса с подпиткой можно культивировать непрерывно, но способ культивирования не ограничен этим.
Среда, используемая для культивирования, должна надлежащим образом соответствовать требованиям конкретных штаммов. Культуральная среда для микроорганизма рода Corynebacterium известна в данной области техники (например, Manual of Methods for General Bacteriology by the American Society for Bacteriology, Washington D.C., USA, 1981).
В частности, источники сахара, которые можно использовать в составе среды, включают сахариды и углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, мальтоза, крахмал, целлюлоза и так далее; масла и жиры, такие как соевое масло, подсолнечное масло, касторовое масло, кокосовое масло и так далее; жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота, линолевая кислота и так далее; глицерин; спирты, такие как этанол, и органические кислоты, такие как уксусная кислота. Эти вещества можно использовать по отдельности или с составе смеси, но не ограничиваясь этим.
Источники углерода, которые можно использовать, могут представлять собой неочищенную сахарозу, или глюкозу, или мелассу, содержащую большое количество сахарозы, и в частности могут представлять собой очищенную глюкозу, но не ограничиваясь этим. Можно использовать другие различные источники углерода.
Источники азота, которые можно использовать, включают пептон, дрожжевой экстракт, мясной экстракт, солодовый экстракт, жидкий кукурузный экстракт, соевую муку и мочевину или неорганические соединения, например сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Источники азота также можно использовать по отдельности или в форме смеси, но не ограничиваясь этим.
Источники фосфата, которые можно использовать, включают дигидрофосфат калия, гидроортофосфат калия или соответствующие им натрийсодержащие соли.
Кроме того, культуральная среда может содержать соли металлов, такие как сульфат магния или сульфат железа. Дополнительно к вышеуказанным веществам можно использовать необходимые для роста вещества, такие как аминокислоты и витамины. Соответствующие предшественники также можно использовать в культуральной среде. Исходные вещества, описанные выше, можно добавлять в инкубатор партиями или непрерывно надлежащим образом.
рН культуральной среды может быть подведен с использованием основного соединения, такого как гидроксид натрия, гидроксид калия и аммиак; или кислого соединения, такого как фосфорная кислота и серная кислота, надлежащим образом. Кроме того, образование пены можно подавить путем использования пеногасителя, такого как полигликолевый сложный эфир жирной кислоты. Кислород или кислородсодержащий газ (например воздух) можно вводить в культуральную среду для поддержания аэробных условий.
В частности, температура культивирования обычно составляет от 30°С до 37°С, более конкретно от 32°С до 33°С. Культивирование можно продолжать до тех пор, когда будет получено желаемое количество ксантозин-5'-монофосфата, но в частности может быть осуществлено в течение от 40 часов до 120 часов.
Выделение ксантозин-5'-монофосфата из культуры можно проводить традиционным способом, известным в данной области техники, например центрифугированием, фильтрованием, ионообменной хроматографией, кристаллизацией и так далее. Например, культуру можно центрифугировать с низкой скоростью для удаления биомассы и затем полученный супернатант можно отделять путем ионообменной хроматографии. Однако выделение не ограничено этим.
Стадия выделения может дополнительно включать процесс очистки.
Примеры
Далее настоящее изобретение будет описано более подробно со ссылкой на следующие примеры воплощений. Однако эти примеры предназначены исключительно для иллюстративных целей и не предназначены каким-либо образом ограничивать объем настоящего изобретения.
Пример 1: Обнаружение ксантозин-5'-монофосфат (ХМР)-экспортирующего белка
Для идентификации мембранного белка Corynebacterium, вовлеченного в экспорт ХМР, была получена библиотека геномной ДНК Corynebacterium stationis (АТСС6872). Геномную ДНК штамма АТСС6872 (то есть, Corynebacterium stationis дикого типа) экстрагировали с использованием "G-spin Total DNA Extraction Minin Kit" от "Intron" (Кат. No. 17045) в соответствии с прилагаемым к нему протоколом. Библиотека мембранных белков была получена с использованием экстрагированной геномной ДНК в качестве матрицы.
Для исследования того, какой белок обладает экспортирующей функцией, полученную библиотеку геномной ДНК вводили в штамм Corynebacterium КССМ-10530, используемый в качестве родительского штамма в патенте Кореи No. 10-2011-0105662.
Затем ХМР дополнительно добавляли к минимальной культуральной среде, содержащей 1,7% агара, для создания условий скрининга, в которых штамм КССМ-10530 демонстрирует ингибирование роста. Штамм КССМ-10530 трансформировали путем электропорации плазмидой геномной библиотеки мембранного белка штамма АТСС6872 и отбирали колонии, которые нормально росли в условиях избыточного количества ХМР, добавленного в культуральную среду. Из выбранной колонии получали плазмиду и анализировали ее нуклеотидную последовательность методом анализа нуклеотидной последовательности. С помощью вышеописанного эксперимента был идентифицирован один тип мембранного белка, вовлеченный в устранение ингибирования роста в условиях избыточного количества добавленного ХМР.
Было подтверждено, что мембранный белок Corynebacterium кодируется нуклеотидной последовательностью SEQ ID NO: 1 и имеет аминокислотную последовательность SEQ ID NO: 2 (NCBI GenBank: NZ_CP014279.1, WP_066795121, MFS-транспортер). Этот мембранный белок известен как MFS-транспортер, но его функция не полностью изучена. Кроме того, известно, что он обладает функцией экспортирования ХМР. В настоящем описании этот мембранный белок обозначен как "xmpE"
Пример 2: Идентификация xmpE
Пример 2-1: Получение вектора, дефицитного по xmpE
"Дефицитный" вектор был получен для подтверждения того, уменьшится ли ХМР-экспортирующая способность, когда xmpE, белок, вовлеченный в устранение ингибирования роста, вызванного ХМР, идентифицированный в Примере 1, удален из ХМР-продуцирующего штамма.
Фрагмент гена для получения вектора был получен путем ПЦР с использованием геномной ДНК штамма АТСС6872, используемого в качестве родительского штамма. Более точно, ПЦР для хтрЕ проводили с использованием праймеров SEQ ID NO: 3 и 4. Праймеры, которые использовали, были получены на основе информации о генах Corynebacterium stationis (АТСС6872), зарегистрированной в "GenBank" Национального Института Здоровья (NIH) (NCBI Genbank: NZ_СР014279.1), и окружающей их нуклеотидной последовательности.
ПЦР проводили в следующих условиях: денатурация при 94°С в течение 5 минут; 25 циклов денатурации при 94°С в течение 30 секунд, отжига при 52°С в течение 30 секунд и полимеризации при 72°С в течение 1 минуты. Затем проводили заключительную полимеризацию при 72°С в течение 5 минут. Поскольку фрагмент гена xmpE амплифицировали с использованием праймеров SEQ ID NO: 3 и 4, был получен полинуклеотидный образец примерно 1,0 т.п.н. Полученный фрагмент гена клонировали с использованием вектора Т (Solgent) для получения вектора "ТОРО-ΔxmpE".
Пример 2-2: Получение штамма, дефицитного по xmpE
Штамм КССМ-10530 трансформировали путем электропорации вектором, полученным в примере 2-1 (используя метод биотрансформации согласно Appl. Microbiol. Biotechnol. (1999) 52:541-545), и штамм, имеющий вектор, вставленный в хромосому вследствие рекомбинации гомологичных последовательностей, отбирали в культуральной среде, содержащей 25 мг/л канамицина. Выбранный штамм, дефицитный по xmpE, был обозначен как "КССМ-10530_ΔxmpE", и была оценена его способность продуцировать ХМР.
Родительский штамм Corynebacterium stationis КССМ-10530 и штамм КССМ-10530_ΔxmpE инокулировали в пробирку на 14 мл, содержащую 3 мл посевной культуральной среды, описанной ниже, и культивировали при 30°С в течение 24 часов со встряхиванием при 170 об/мин. Затем посевные культуры добавляли в количестве 0,7 мл в 32 мл следующей продукционной культуральной среды (24 мл основной культуральной среды плюс 8 мл дополнительной стерильной культуральной среды) в соответствующие колбы с угловыми перегородками на 250 мл с последующим культивированием при 30°С в течение 72 часов со встряхиванием при 170 об/мин. После завершения культивирования выполняли анализ ВЭЖХ (высокоэффективная жидкостная хроматография) для измерения количества продуцированного ХМР. Состав культуральной среды и результаты по концентрациям ХМР в культуральной среде представлены в Таблице 1 ниже.
Минимальная культуральная среда ХМР
Глюкоза 2%, сульфат натрия 0,3%, дигидрофосфат калия 0,1%, гидроортофосфат калия 0,3%, сульфат магния 0,3%, хлорид кальция 10 мг/л, сульфат железа 10 мг/л, сульфат цинка 1 мг/л, хлорид марганца 3,6 мг/л, L-цистеин 20 мг/л, пантотенат кальция 10 мг/л, тиамина гидрохлорид 5 мг/л, биотин 30 мкг/л, аденин 20 мг/л, гуанин 20 мг/л, рН 7,3
Питательная культуральная среда ХМР
Пептон 1%, мясной экстракт 0,5%, хлорид натрия 0,25%, дрожжевой экстракт 1%, мочевина 0,3%, аденин 50 мг/л, гуанин 50 мг/л, агар 2%, рН 7,2
Посевная культуральная среда ХМР для колбы
Глюкоза 20 г/л, пептон 10 г/л, дрожжевой экстракт 10 г/л, хлорид натрия 2,5 г/л, мочевина 3 г/л, аденин 150 мг/л, гуанин 150 мг/л, рН 7,0
Продукционная культуральная среда ХМР для колбы (основная культуральная среда)
Глюкоза 50 г/л, сульфат магния 10 г/л, хлорид кальция 100 мг/л, сульфат железа 20 мг/л, сульфат марганца 10 мг/л, сульфат цинка 10 мг/л, сульфат меди 0,8 мг/л, гистидин 20 мг/л, цистеин 15 мг/л, бета-аланин 15 мг/л, биотин 100 мкг/л, тиамин 5 мг/л, аденин 50 мг/л, гуанин 25 мг/л, ниацин 5 мг/л, рН 7,0
Продукционная культуральная среда ХМР для колбы (дополнительная стерильная культуральная среда)
Дигидрофосфат калия 18 г/л, гидроортофосфат калия 42 г/л, мочевина 7 г/л, сульфат аммония 5 г/л
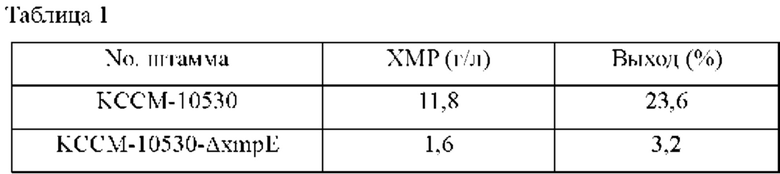
Сравнивали концентрации ХМР в культуральной среде родительского штамма КССМ-10530 и штамма КССМ-10530-ΔxmpE, дефицитного по xmpE; в результате, как показано в Таблице 1 выше, концентрация ХМР штамма КССМ-10530-ΔxmpE уменьшалась по меньшей мере примерно на 10 г/л по сравнению с родительским штаммом в тех же условиях.
Соответственно, было подтверждено, что xmpE представляет собой белок, вовлеченный в экспорт ХМР.
Пример 3: Повышение активности белка xmpE
Активность белка xmpE у штамма была повышена в соответствии со следующими примерами, и штамм с повышенной активностью xmpE был изучен в отношении того, была ли увеличена его способность продуцировать/экспортировать ХМР.
Пример 3-1: Увеличение числа копий xmpE
Пример 3-1-1: Получение вектора для увеличения числа копий xmpE
Для подтверждения того, увеличивается ли ХМР-экспортирующая способность при повышении активности xmpE, который, как ожидается, вовлечен в ХМР-экспортирующую способность, был получен вектор для усиления гена xmpE. С использованием способа увеличения числа копий в качестве способа усиления был проведен следующий эксперимент.
Фрагмент гена для получения вектора был получен путем ПЦР с использованием геномной ДНК АТСС6872 в качестве матрицы. В частности, ген xmpE был амплифицирован так, чтобы содержать 484 п.н. выше по ходу транскрипции от гена xmpE с использованием пары праймеров SEQ ID NO: 5 и 6. Амплифицированный фрагмент гена xmpE обрабатывали ферментами рестрикции XbaI и SpeI. Клонирование проводили по сайту XbaI вектора pDZ (патент Кореи No. 10-0924065 и международная публикация No. 2008-033001) для получения вектора pDZ-xmpE. Затем проводили ПЦР на основе гена xmpE с использованием пары праймеров SEQ ID NO: 6 и 7 для получения вектора, содержащего две копии. Каждый фрагмент ДНК, полученный таким образом, разрезали ферментом рестрикции ДНК SpeI и клонировали в вектор pDZ xmpE, расщепленный ферментом рестрикции ДНК XbaI, с тем чтобы получить вектор. Вектор, содержащий две копии гена xmpE, был обозначен как "pDZ-xmpEX2".
Пример 3-1-2: Оценка способности штамма, имеющего увеличенное число копий xmpE, продуцировать ХМР
Штамм КССМ-10530 трансформировали путем электропорации вектором pDZ-xmpEX2, полученным в Примере 3-1-1 (используя метод трансформации согласно Appl. Microbiol. Biotechnol. (1999) 52:541-545), и штамм, имеющий вектор, вставленный в хромосому вследствие рекомбинации гомологичных последовательностей, отбирали в культуральной среде, содержащей 25 мг/л канамицина. Штамм, в котором произошла вставка целевого гена, отбирали путем вторичного кроссинговера выбранной первичной цепи. Успешную вставку гена конечного трансформированного штамма подтверждали путем ПЦР с использованием пары праймеров SEQ ID NO: 8 и 9. Выбранный штамм с увеличенным числом копий xmpE был обозначен как "КССМ-10530-xmpEX2", и была оценена его способность продуцировать ХМР.
Для измерения способности штамма продуцировать ХМР использовали такой же способ как в Примере 2-2. После завершения культивирования проводили анализ ВЭЖХ для измерения количества продуцированного ХМР, и результат представлен в Таблице 2 ниже.
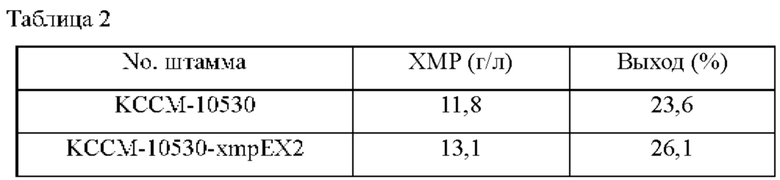
Сравнивали концентрации ХМР в культуральной среде родительского штамма Corynebacterium stationis КССМ-10530 и штамма КССМ-10530-xmpEX2, имеющего увеличенное число копий xmpE; в результате, как показано в Таблице 2 выше, концентрация ХМР штамма КССМ-10530-xmpEX2 составила 1,3 г/л, указывая на то, что концентрация увеличилась примерно на 11% по сравнению с родительским штаммом в таких же условиях.
Это можно понимать как очень значимый результат, подтверждающий, что такое увеличение количества продуцированного ХМР является следствием повышенной активности белка xmpE.
Пример 3-2: Повышение уровня экспрессии xmpE путем замены промотора
Пример 3-2-1: Получение вектора для замены промотора xmpE
Для подтверждения того, увеличивается ли ХМР-экспортирующая способность при повышении активности xmpE, который, как ожидается, вовлечен в ХМР-экспортирующую способность, был получен вектор, в котором промотор гена xmpE заменен на промотор, способный обеспечивать высокий уровень экспрессии. Фрагмент гена для получения вектора был получен путем ПЦР с использованием геномной ДНК штамма АТСС6872 в качестве матрицы.
Pcj7, который, как сообщается в патенте Кореи No. 10-0620092, экспрессируется на высоком уровне в Corynebacterium stationis, использовали в качестве промотора.
Для амплификации фрагмента гена Pcj7 применяли пару праймеров SEQ ID NO: 10 и 11 с использованием геномной ДНК штамма АТСС6872 в качестве матрицы. Каждый ген xmpE амплифицировали путем ПЦР с использованием пары праймеров SEQ ID NO: 12 и 13, и 14 и 15, соответственно. Реакцию ПЦР проводили в следующих условиях: денатурация при 94°С в течение 5 минут; 25 циклов денатурации при 94°С в течение 30 секунд, отжига при 52°С в течение 30 секунд и полимеризации при 72°С в течение 1 минуты. Затем проводили заключительную полимеризацию при 72°С в течение 5 минут. Полинуклеотидный образец 2,3 т.п.н. был получен путем проведения полимеразной цепной реакции с перекрывающимися праймерами с использованием трех амплифицированных генных фрагментов Pcj7, xmpE-1 и xmpE-2 в качестве матриц. Полученный фрагмент гена расщепляли ферментом рестрикции XbaI и клонировали в линейный вектор pDZ, расщепленный XbaI, используя лигазу Т4 для получения вектора "pDZ-Pcj7/xmpE".
Пример 3-2-2: Получение штамма с замененным промотором xmpE и оценка его способности продуцировать ХМР
Штамм КССМ10530 трансформировали путем электропорации вектором pDZ-Pcj7/xmpE, полученным в Примере 3-2-1 (используя метод трансформации согласно Appl. Microbiol. Biotechnol. (1999) 52:541-545), и штамм, имеющий вектор, вставленный в хромосому вследствие рекомбинации гомологичных последовательностей, отбирали в культуральной среде, содержащей 25 мг/л канамицина. Штамм, в котором целевой ген усилен, отбирали путем вторичного кроссинговера выбранной первичной цепи. Успешную вставку промотора гена конечного трансформированного штамма подтверждали путем ПЦР с использованием пары праймеров SEQ ID NO: 16 и 17. Штамм, в котором промотор заменен на более сильный промотор, был обозначен как "KCCM-10530-Pcj7/xmpE", и была оценена его способность продуцировать ХМР.
Для измерения способности штамма продуцировать ХМР использовали такой же способ, как в Примере 2-2. После завершения культивирования проводили анализ ВЭЖХ для измерения количества продуцированного ХМР, и результаты представлены в Таблице 3 ниже.
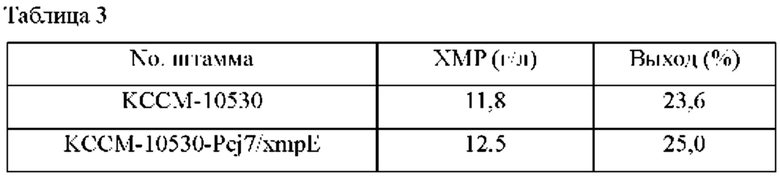
Сравнивали концентрации ХМР в культуральной среде родительского штамма Corynebacterium stationis КССМ-10530 и штамма KCCM-10530-Pcj7/xmpE, обладающего повышенным уровнем экспрессии xmpE благодаря более сильному промотору; в результате, как показано в Таблице 3 выше, концентрация ХМР штамма KCCM-10530-Pcj7/xmpE составила 0,7 г/л, указывая на увеличение концентрации примерно на 6% по сравнению с родительским штаммом в таких же условиях.
Это можно понимать как очень значимый результат, подтверждающий, что такое увеличение количества продуцированного ХМР является следствием повышенной активности белка xmpE.
Пример 3-3: Замена стартового кодона xmpE
Пример 3-3-1: Получение вектора для замены стартового кодона xmpE
Для подтверждения того, увеличивается ли способность экскретировать ХМР при повышении уровня экспрессии белка xmpE, который, как ожидается, вовлечен в способность экскретировать ХМР, был получен вектор, в котором стартовый кодон gtg заменен на atg. Фрагмент гена для получения вектора был получен путем ПЦР с использованием геномной ДНК штамма АТСС6872 в качестве матрицы. Для получения вектора, в котором стартовый кодон gtg заменен на atg, были получены два фрагмента гена А и В с использованием пар праймеров SEQ ID NO: 18 и 19, и 20 и 21, соответственно. Реакцию ПЦР проводили в следующих условиях: 25 циклов денатурации при 94°С в течение 5 минут; денатурации при 94°С в течение 30 секунд; отжига при 52°С в течение 30 секунд и полимеризации при 72°С в течение 1 минуты. Затем проводили заключительную полимеризацию при 72°С в течение 5 минут. В результате были получены полинуклеотиды примерно 0,7 т.п.н. и 1 т.п.н. для фрагментов А и В, соответственно. С использованием этих двух фрагментов в качестве матриц проводили ПЦР, используя пару праймеров SEQ ID NO: 18 и 21 для получения продукта ПЦР примерно 1,7 т.п.н. (далее "фрагмент gla"). Полимеризацию проводили в следующих условиях: 25 циклов денатурации при 94°С в течение 5 минут; денатурации при 94°С в течение 30 секунд; отжига при 55°С в течение 30 секунд и полимеризации при 72°С в течение 120 секунд. Затем проводили заключительную полимеризацию при 72°С в течение 7 минут.
Полученный фрагмент гена расщепляли ферментом рестрикции XbaI и клонировали в линейный вектор pDZ, расщепленный XbaI, с использованием лигазы Т4 для получения вектора "pDZ-xmpE (gla)".
Пример 3-3-2: Получение штамма с замененным стартовым кодоном xmpE и оценка его способности продуцировать ХМР
Штамм КССМ-10530 трансформировали путем электропорации вектором pDZ-xmpE(gla), полученным в Примере 3-3-1 (используя метод трансформации согласно Appl. Microbiol. Biotechnol. (1999) 52:541-545), и штамм, имеющий вектор, вставленный в хромосому вследствие рекомбинации гомологичных последовательностей, отбирали в культуральной среде, содержащей 25 мг/л канамицина. Штамм, в котором целевой ген усилен, отбирали путем вторичного кроссинговера выбранной первичной цепи. Успешную вставку промотора гена конечного трансформированного штамма подтверждали путем ПЦР с использованием пары праймеров SEQ ID NO: 18 и 21 с последующим анализом замененной нуклеотидной последовательности методом анализа нуклеотидной последовательности. Выбранный штамм, в котором стартовый кодон xmpE заменен на atg, был обозначен как CJX1662, и была оценена его способность продуцировать ХМР.
Для измерения способности штамма продуцировать ХМР использовали такой же способ, как в Примере 2-2. После завершения культивирования проводили анализ ВЭЖХ для измерения количества продуцированного ХМР, и результаты представлены в Таблице 4 ниже.
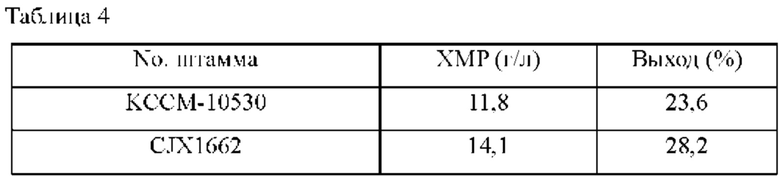
Сравнивали концентрации ХМР в культуральной среде родительского штамма Corynebacterium stationis KCCM-10530 и штамма CJX1662, имеющего замену стартового кодона xmpE; в результате, как показано в Таблице 4 выше, концентрация ХМР у штамма CJX1662 составила 2,3 г/л, показывая увеличение концентрации примерно на 20% по сравнению с родительским штаммом в таких же условиях.
Это можно понимать как очень значимый результат, подтверждающий, что такое увеличение количества продуцированного ХМР является следствием повышенной активности белка xmpE.
Далее, штамм CJX1662, полученный выше, был депонирован в Корейском Центре Культур Микроорганизмов (КССМ), международном органе депонирования, в соответствии с Будапештским договором 11 Апреля 2018 года с номером доступа КССМ12248Р.
Пример 3-4: Получение штамма с усиленным xmpE на основе ХМР-продуцирующей клеточной линии дикого типа
Пример 3-4-1: Получение штамма с усиленным xmpE на основе ХМР-продуцирующей клеточной линии дикого типа
Штамм, обладающий способностью эффективно продуцировать ХМР был получен путем ослабления активностей аденилосукцинатсинтетазы и ХМР-дегидрогеназы, которые действуют в пути расщепления ХМР у штамма АТСС6872 дикого типа. Для снижения активностей этих двух ферментов был получен штамм, у которого 'а', первый нуклеотид нуклеотидной последовательности purA и guaA, генов, кодирующих два этих фермента, соответственно, заменен на 't'. Более конкретно, штамм, полученный путем ослабления экспрессии этих двух генов у штамма АТСС6872, был обозначен как CJX1663.
Полученный штамм CJX1663 трансформировали путем электропорации вектором pDZ-xmpE(gla), полученным в Примере 3-3-2, и штамм, имеющий вектор, вставленный в хромосому вследствие рекомбинации гомологичных последовательностей, отбирали в культуральной среде, содержащей 25 мг/л канамицина. Штамм, в котором целевой ген усилен, отбирали путем вторичного кроссинговера выбранной первичной цепи. Успешную вставку промотора гена конечного трансформированного штамма подтверждали путем ПЦР с использованием пары праймеров SEQ ID NO: 18 и 21.
Штамм, в котором выбранный стартовый кодон xmpE заменен на atg, был обозначен как "CJX1663_xmpE(gla)", и была оценена его способность продуцировать ХМР.
После завершения культивирования проводили анализ ВЭЖХ для измерения количества продуцированного ХМР, и результаты представлены в Таблице 5 ниже.
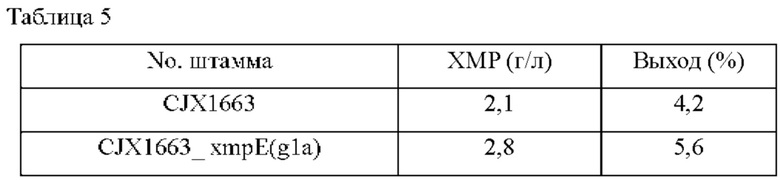
Сравнивали концентрации ХМР в культуральной среде родительского штамма Corynebacterium stationis CJX1663 и штамма CJX1663_xmpE(gla), имеющего замену стартового кодона xmpE; в результате, как показано в Таблице 5 выше, концентрация ХМР штамма CJX1663_xmpE(gla) составила 2,8 г/л, указывая на увеличение концентрации примерно на 33% по сравнению с родительским штаммом в таких же условиях.
Этот результат подтверждает, что путем повышения активности белка (xmpE) по настоящему изобретению, который экспортирует ксантозин-5'-монофосфат, может быть увеличена продуктивность по ХМР.
Следует отметить, что последовательности праймеров, использованные в настоящем изобретении, являются такими как показано в Таблице 6.


Специалисту в данной области техники понятно, что настоящее изобретение может быть воплощено в других конкретных формах без отступления от его идеи или существенных характеристик. В этом отношении описанные воплощения следует рассматривать во всех отношениях только как иллюстративные и не ограничивающие. Следовательно, объем настоящего изобретения определен прилагаемой формулой, а не вышеизложенным описанием. Все изменения, которые происходят в рамках значения и диапазона эквивалентности прилагаемой формулы попадают в объем настоящего изобретения.
--->
<110> CJ CheilJedang Corporation
<120> Microorganism of the genus Corynebacterium producing 5’-xanthosine monophosphate and method for preparing 5’-xanthosine monophosphate using the same
<130> OPA18214
<150> KR 10-2018-0065681
<151> 2018-06-07
<160> 21
<170> KoPatentIn 3.0
<210> 1
<211> 1242
<212> DNA
<213> Corynebacterium sp.
<400> 1
gtgcacacaa ccgaagaggt caaactcttt acgccgacgt ttatcatggg gtgggtcgcc 60
aacttcctgc agttcttggt gttctacttc ctcatcacca cgatggcgct ctacgcaacc 120
aaggaattca gcgcatcgga aaccgaagca ggctttgccg ccagtgcgat tgttatcggt 180
gcggtctttt cccgtttggt ttccggatac attattgacc gcttcggtcg ccgcaaaatt 240
gtggtggtct ccgtcatcgc gaccactatt gcgtgcgcgc tatatatccc gatcgattct 300
ttggggctgc tctacgctga ccgcttcttc cacggtgtag cttatgcctt tgcgtgcacc 360
gcgattatgg cgatggtcca ggaactcatt ccttctgcac gccgctccga aggcactggc 420
tacctggctt tgggtaccac cgtttcggct gctatcggac cagcgctagc gctatttttg 480
ctgggttctt tcaactacga agtcctcttc gttgtcgtcc tcggcatttc gattgtctct 540
ttgatcgctt cgctagtcat ctatttccgc acctccgacc cagagccaga gctggatgaa 600
aacggcaatg ctgctgagcc cattaagttc agcttcaagt ccatcattaa ccctaaagtc 660
ttgccgattg gcctcttcat gctgctggta gcctttgcct actccggcgt gatcgcacat 720
atcaacgctt ttgctgaaaa ccgcgacgtt gttactggcg caggcctatt ctttatcgct 780
tacgccatct ccatgttcgt gatgcgctcc taccttggta aattgcaaga ccgccggggc 840
gataacagcg ttatctactt tggtctcgta ttctttgtta tctcatttat cgtgctctcg 900
ctttctaccg ccaactggca tgtcgttgtc gctggcgtgc tagcaggtct gggctacggc 960
accttgatgc cagctgctca agcagtatct gttggtgttg tcgacaagag cgagttcggc 1020
tcagcatttt ccaccttgtt ccttttcgtt gacctcggct tcggcttcgg cccagtcatc 1080
cttggtgcag tggtttccgc gattggctac ggttcgatgt atgcagtgct cgtcggcgtc 1140
ggcgttattg ctggcatcta ctacctgttc acccacgcac gcaccgagcg cgcaaagcac 1200
ggcgtagtca agcatgtaga aaccatggtt ctggctcaat aa 1242
<210> 2
<211> 413
<212> PRT
<213> Corynebacterium sp.
<400> 2
Val His Thr Thr Glu Glu Val Lys Leu Phe Thr Pro Thr Phe Ile Met
1 5 10 15
Gly Trp Val Ala Asn Phe Leu Gln Phe Leu Val Phe Tyr Phe Leu Ile
20 25 30
Thr Thr Met Ala Leu Tyr Ala Thr Lys Glu Phe Ser Ala Ser Glu Thr
35 40 45
Glu Ala Gly Phe Ala Ala Ser Ala Ile Val Ile Gly Ala Val Phe Ser
50 55 60
Arg Leu Val Ser Gly Tyr Ile Ile Asp Arg Phe Gly Arg Arg Lys Ile
65 70 75 80
Val Val Val Ser Val Ile Ala Thr Thr Ile Ala Cys Ala Leu Tyr Ile
85 90 95
Pro Ile Asp Ser Leu Gly Leu Leu Tyr Ala Asp Arg Phe Phe His Gly
100 105 110
Val Ala Tyr Ala Phe Ala Cys Thr Ala Ile Met Ala Met Val Gln Glu
115 120 125
Leu Ile Pro Ser Ala Arg Arg Ser Glu Gly Thr Gly Tyr Leu Ala Leu
130 135 140
Gly Thr Thr Val Ser Ala Ala Ile Gly Pro Ala Leu Ala Leu Phe Leu
145 150 155 160
Leu Gly Ser Phe Asn Tyr Glu Val Leu Phe Val Val Val Leu Gly Ile
165 170 175
Ser Ile Val Ser Leu Ile Ala Ser Leu Val Ile Tyr Phe Arg Thr Ser
180 185 190
Asp Pro Glu Pro Glu Leu Asp Glu Asn Gly Asn Ala Ala Glu Pro Ile
195 200 205
Lys Phe Ser Phe Lys Ser Ile Ile Asn Pro Lys Val Leu Pro Ile Gly
210 215 220
Leu Phe Met Leu Leu Val Ala Phe Ala Tyr Ser Gly Val Ile Ala His
225 230 235 240
Ile Asn Ala Phe Ala Glu Asn Arg Asp Val Val Thr Gly Ala Gly Leu
245 250 255
Phe Phe Ile Ala Tyr Ala Ile Ser Met Phe Val Met Arg Ser Tyr Leu
260 265 270
Gly Lys Leu Gln Asp Arg Arg Gly Asp Asn Ser Val Ile Tyr Phe Gly
275 280 285
Leu Val Phe Phe Val Ile Ser Phe Ile Val Leu Ser Leu Ser Thr Ala
290 295 300
Asn Trp His Val Val Val Ala Gly Val Leu Ala Gly Leu Gly Tyr Gly
305 310 315 320
Thr Leu Met Pro Ala Ala Gln Ala Val Ser Val Gly Val Val Asp Lys
325 330 335
Ser Glu Phe Gly Ser Ala Phe Ser Thr Leu Phe Leu Phe Val Asp Leu
340 345 350
Gly Phe Gly Phe Gly Pro Val Ile Leu Gly Ala Val Val Ser Ala Ile
355 360 365
Gly Tyr Gly Ser Met Tyr Ala Val Leu Val Gly Val Gly Val Ile Ala
370 375 380
Gly Ile Tyr Tyr Leu Phe Thr His Ala Arg Thr Glu Arg Ala Lys His
385 390 395 400
Gly Val Val Lys His Val Glu Thr Met Val Leu Ala Gln
405 410
<210> 3
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> xmpE del F
<400> 3
gtcaaactct ttacgccgac g 21
<210> 4
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> xmpE del R
<400> 4
cgacaacacc aacagatact gc 22
<210> 5
<211> 31
<212> DNA
<213> Artificial Sequence
<220>
<223> xmpE 2copy F (xbaI)
<400> 5
atgctctaga ctagatcttc tcgacgggca g 31
<210> 6
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<223> xmpE 2copy R (speI)
<400> 6
atgatactag tccttgggga cttcgcgtgt cg 32
<210> 7
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<223> xmpE 2copy F (speI)
<400> 7
atgatactag tctagatctt ctcgacgggc ag 32
<210> 8
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> xmpE 2copy CF
<400> 8
ttcggctcag cattttccac 20
<210> 9
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> xmpE 2copy CR
<400> 9
caatagtggt cgcgatgacg 20
<210> 10
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> Pcj7 F
<400> 10
gcagtaagga gaatcagaaa catcccagcg ctacta 36
<210> 11
<211> 40
<212> DNA
<213> Artificial Sequence
<220>
<223> Pcj7 R
<400> 11
acctcttcgg ttgtgtgcac gagtgtttcc tttcgttggg 40
<210> 12
<211> 31
<212> DNA
<213> Artificial Sequence
<220>
<223> xmpE P 1
<400> 12
atgctctaga tccagtgtgg ttaagaagtc g 31
<210> 13
<211> 33
<212> DNA
<213> Artificial Sequence
<220>
<223> xmpE P 2
<400> 13
cgctgggatg tttctgattc tccttactgc agt 33
<210> 14
<211> 43
<212> DNA
<213> Artificial Sequence
<220>
<223> xmpE P 3
<400> 14
gtacccaacg aaaggaaaca ctcgtgcaca caaccgaaga ggt 43
<210> 15
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> xmpE P 4
<400> 15
atgctctaga ttattgagcc agaaccatgg 30
<210> 16
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Pcj7/xmpE CF
<400> 16
ctagatcttc tcgacgggca g 21
<210> 17
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Pcj7/xmpE CR
<400> 17
caatagtggt cgcgatgacg 20
<210> 18
<211> 31
<212> DNA
<213> Artificial Sequence
<220>
<223> xmpE atg 1
<400> 18
atgctctaga tccagtgtgg ttaagaagtc g 31
<210> 19
<211> 41
<212> DNA
<213> Artificial Sequence
<220>
<223> xmpE atg 2
<400> 19
gacctcttcg gttgtgtgca tgattctcct tactgcagtt a 41
<210> 20
<211> 41
<212> DNA
<213> Artificial Sequence
<220>
<223> xmpE atg 3
<400> 20
taactgcagt aaggagaatc atgcacacaa ccgaagaggt c 41
<210> 21
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> xmpE atg 4
<400> 21
atgctctaga ctcgctcttg tcgacaacac 3
<---
Настоящее изобретение относится к микроорганизму рода Corynebacterium, продуцирующему ксантозин-5'-монофосфат, и к способу получения ксантозин-5'-монофосфата с использованием этого микроорганизма. 2 н. и 4 з.п. ф-лы.
1. Микроорганизм рода Corynebacterium, продуцирующий ксантозин-5'-монофосфат, где активность белка, содержащего аминокислотную последовательность SEQ ID NO: 2, повышена.
2. Микроорганизм рода Corynebacterium по п. 1, где указанный белок кодируется геном, содержащим нуклеотидную последовательность SEQ ID NO: 1.
3. Микроорганизм рода Corynebacterium по п. 1, где повышение активности достигается путем увеличения числа копий полинуклеотидов, кодирующих указанный белок, повышения активности промотора, замены стартового кодона, или их комбинацией.
4. Микроорганизм рода Corynebacterium по п. 1, где микроорганизм рода Corynebacterium, продуцирующий ксантозин-5'-монофосфат, представляет собой Corynebacterium stationis.
5. Способ получения ксантозин-5'-монофосфата, включающий:
культивирование микроорганизма рода Corynebacterium по любому из пп. 1-4 в культуральной среде; и
выделение ксантозин-5'-монофосфата из микроорганизма или из культуральной среды.
6. Способ получения ксантозин-5'-монофосфата по п. 5, где микроорганизм рода Corynebacterium, продуцирующий ксантозин-5'-монофосфат, представляет собой Corynebacterium stationis.
| СПОСОБ ПОЛУЧЕНИЯ КСАНТОЗИН-5'-МОНОФОСФАТА, ШТАММ CORYNEBACTERIUM AMMONIAGENES - ПРОДУЦЕНТ КСАНТОЗИН-5'-МОНОФОСФАТА (ВАРИАНТЫ) | 2000 |
|
RU2209249C2 |
| RU 2000128988 A, 20.02.2003 | |||
| KR 100862172 B1, 09.10.2008 | |||
| US 8859236 B2, 14.10.2014. | |||

