ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к способам получения растений с увеличенной биомассой корней.
УРОВЕНЬ ТЕХНИКИ
Корни сосудистых растений имеют множество функций. Они включают: 1) всасывание воды и неорганических питательных веществ, 2) заякоривание растения в почве и поддержание его, 3) запасание питательных веществ и нутриентов и 4) вегетативное размножение. В зависимости от концентрации питательных веществ корни также синтезируют цитокинин, который выступает в качестве сигнала, влияющего на скорость роста побегов.
Таким образом, растения с увеличенной биомассой корней потенциально будут иметь ряд преимуществ, включающих улучшенное заякоривание, более эффективное поглощение воды, более эффективное поглощение питательных веществ и повышенную толерантность к засухе. Комбинация этих признаков также может приводить к увеличению урожая, включая увеличение биомассы зерен или плодов и/или увеличение биомассы листьев.
В настоящее время понимание генетических механизмов, контролирующих биомассу корней в растениях, ограничено.
Было бы полезным наличие доступных альтернативных способов контроля биомассы корней в растениях.
Таким образом, задачей изобретения является предоставление способов и материалов для повышения продукции биомассы корней в растениях и/или по меньшей мере предоставление обществу приемлемого выбора.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Ранее White (2006) выявил два соседних гомологичных гена в Arabidopsis (названных PEAPOD, PPD1 и PPD2), которые регулируют клеточную пролиферацию меристемоидов на последних стадиях развития листьев и семянок. Делеция этих генов в арабидопсисе приводила к увеличенным листьям и широким семянкам, в то время как сверхэкспрессия PPD1 приводила к уменьшению размера листьев и стручков (White, 2006).
Заявители в рамках изобретения неожиданно обнаружили, что сверхэкспрессия генов PEAPOD в растениях приводит к увеличению продукции биомассы корней.
Таким образом, изобретение заявителя относится к способу увеличения биомассы корней в растениях посредством эктопической экспрессии PEAPOD. В частности, изобретение относится к эктопической экспрессии белков PEAPOD, которые характеризуются присутствием консенсусных аминокислотных мотивов, общих для всех белков PEAPOD, описанных для широкого диапазона видов растений.
Способы
В первом аспекте изобретение относится к способу увеличения биомассы корней в растении, причем способ включает стадию увеличения экспрессии по меньшей мере одного белка PEAPOD в растении.
В одном варианте осуществления биомасса корней увеличивается относительно биомассы корней в контрольном растении того же вида или сорта.
В одном варианте осуществления увеличенная экспрессия по меньшей мере одного белка PEAPOD является следствием трансформации растения, или растения-предшественника или клетки растения-предшественника, полинуклеотидом, кодирующим белок PEAPOD.
В следующем варианте осуществления растение является трансгенным по меньшей мере по одному полинуклеотиду, кодирующему и экспрессирующему белок PEAPOD.
В следующем аспекте изобретение относится к способу продуцирования растения с увеличенной биомассой корней, причем способ включает стадию повышения экспрессии по меньшей мере одного белка PEAPOD в растении.
В одном варианте осуществления растение трансформируют по меньшей мере одним полинуклеотидом, кодирующим белок PEAPOD.
В следующем варианте осуществления способ включает стадию трансформации растения или трансформацию клетки растения, которая регенерирует в растение, полинуклеотидом, кодирующим белок PEAPOD.
В одном варианте осуществления способ включает дополнительную стадию тестирования или оценки растения в отношении увеличенной биомассы корней.
В следующем варианте осуществления способ включает стадию продуцирования дополнительных растений с увеличенной биомассой корней посредством бесполого или полового размножения растений, исследованных в отношении увеличенной биомассы.
Белки PEAPOD
В одном варианте осуществления белок PEAPOD представляет собой полипептид, содержащий последовательность согласно по меньшей мере одной из SEQ ID NO: 28, 29, 31, 32, 34 и 35.
В следующем варианте осуществления белок PEAPOD содержит последовательность SEQ ID NO: 28. В следующем варианте осуществления белок содержит PEAPOD последовательность SEQ ID NO: 29. В следующем варианте осуществления белок PEAPOD содержит последовательность SEQ ID NO: 31. В следующем варианте осуществления белок PEAPOD содержит последовательность SEQ ID NO: 32. В следующем варианте осуществления белок PEAPOD содержит последовательность SEQ ID NO: 34. В следующем варианте осуществления белок PEAPOD содержит последовательность SEQ ID NO: 35.
В следующем варианте осуществления белок PEAPOD представляет собой полипептид, содержащий последовательность с по меньшей мере 70% идентичностью с любой из SEQ ID NO: 1-26.
В следующем варианте осуществления белок PEAPOD представляет собой полипептид, содержащий последовательность, выбранную из любой из SEQ ID NO: 1-26.
В следующем варианте осуществления белок PEAPOD представляет собой полипептид, содержащий последовательность по меньшей мере с 70% идентичностью с SEQ ID NO: 1.
В следующем варианте осуществления белок PEAPOD представляет собой полипептид, содержащий последовательность SEQ ID NO: 1.
Экспрессия PEAPOD
Способы экспрессии белков в растениях хорошо известны специалистам в данной области и описаны в настоящем описании. Все такие способы входят в объем изобретения.
Повышение экспрессии PEAPOD путем введения полинуклеотида
В одном варианте осуществления экспрессию повышают путем введения по меньшей мере одного полинуклеотида в клетку растения или в растение.
В предпочтительном варианте осуществления полинуклеотид кодирует белок PEAPOD, как определено в настоящем описании.
В следующем варианте осуществления полинуклеотид содержит последовательность по меньшей мере с 70% идентичностью с кодирующей последовательностью согласно любой из SEQ ID NO: 83-107.
В следующем варианте осуществления полинуклеотид содержит последовательность по меньшей мере с 70% идентичностью с последовательностью согласно любой из SEQ ID NO: 83-107.
В следующем варианте осуществления полинуклеотид содержит кодирующую последовательность согласно любой из SEQ ID NO: 83-107.
В следующем варианте осуществления полинуклеотид содержит последовательность согласно любой из SEQ ID NO: 83-107.
В следующем варианте осуществления полинуклеотид содержит фрагмент последовательностей, описанных выше, который способен кодировать полипептид с той же функцией, что и у белка PEAPOD. В одном варианте осуществления фрагмент кодирует полипептид, способный увеличивать биомассу корней.
Экспрессия PEAPOD посредством экспрессирующей конструкции
В предпочтительном варианте осуществления полинуклеотид вводят в растение в качестве части экспрессирующей конструкции.
В предпочтительном варианте осуществления экспрессирующая конструкция содержит промотор, функционально связанный с полинуклеотидом.
Промотор для повышения экспрессии PEAPOD
В одном варианте осуществления промотор способен запускать или запускает конститутивную экспрессию функционально связанного полинуклеотида во всех тканях растения.
В следующем варианте осуществления промотор представляет собой тканепредпочтительный промотор.
В следующем варианте осуществления промотор способен запускать или запускает экспрессию функционально связанного полинуклеотида в подземных тканях растения.
В одном варианте осуществления промотор представляет собой тканепредпочтительный промотор для подземных тканей.
В следующем варианте осуществления промотор представляет собой тканеспецифический промотор для подземных тканей.
В одном варианте осуществления промотор представляет собой репрессируемый светом промотор.
В следующем варианте осуществления промотор способен запускать или запускает экспрессию функционально связанного полинуклеотида в корнях растения.
В одном варианте осуществления промотор представляет собой предпочтительный для корней промотор.
В следующем варианте осуществления промотор представляет собой специфический для корней промотор.
Источник полинуклеотидов и полипептидов
Полинуклеотиды и варианты полинуклеотидов по изобретению или используемые в способах по изобретению могут происходить из любого вида. Полинуклеотиды и варианты также могут быть продуцированными способами синтеза или рекомбинантными способами, а также они могут быть продуктами подходов "шаффлинга генов".
Полипептиды и варианты полипептидов по изобретению или используемые в способах по изобретению могут происходить из любого вида. Полипептиды и варианты также могут быть продуцированы рекомбинантными способами, а также могут экспрессироваться с продуктов подходов "шаффлинга генов".
В одном варианте осуществления полинуклеотид, полипептид или вариант происходит из вида растения.
В следующем варианте осуществления полинуклеотид, полипептид или вариант происходит из вида голосеменных растений.
В следующем варианте осуществления полинуклеотид, полипептид или вариант происходит из вида покрытосеменных растений.
В следующем варианте осуществления полинуклеотид, полипептид или вариант происходит из вида двудольного растения.
В следующем варианте осуществления полинуклеотид, полипептид или вариант происходит из вида двудольного растения.
В предпочтительном варианте осуществления полинуклеотид, полипептид или вариант происходит из вида эвдикот.
В следующем варианте осуществления полинуклеотид, полипептид или вариант происходит из вида растений группы эвдикот.
В следующем варианте осуществления полинуклеотид, полипептид или вариант происходит из вида однодольного растения. Предпочтительные однодольные растения включают: виды пальмы, банана, ряски и орхидей.
Клетки растений и растения, подлежащие трансформации
Клетки растений и растения по изобретению или используемые в способах по изобретению происходят из любого вида растений.
В одном варианте осуществления клетки растений или растения происходят из вида голосеменных растений.
В следующем варианте осуществления клетки растения или растения происходят из вида покрытосеменных растений.
В следующем варианте осуществления клетки растений или растения происходят из вида двудольных растений.
Предпочтительные роды однодольных растений включают: Agropyron, Allium, Alopecurus, Andropogon, Arrhenatherum, Asparagus, Avena, Bambusa, Bothrichloa, Bouteloua, Bromus, Calamovilfa, Cenchrus, Chloris, Cymbopogon, Cynodon, Dactylis, Dichanthium, Digitaria, Eleusine, Eragrostis, Fagopyrum, Festuca, Helianthus, Hordeum, Lolium, Miscanthis, Miscanthus x giganteus, Oryza, Panicum, Paspalum, Pennisetum, Phalaris, Phleum, Poa, Saccharum, Secale, Setaria, Sorgahastum, Sorghum, Triticum, Vanilla, X Triticosecale Triticale и Zea.
Предпочтительный вид однодольных растений включает: Agropyron cristatum, Agropyron desertorum, Agropyron elongatum, Agropyron intermedium, Agropyron smithii, Agropyron spicatum, Agropyron trachycaulum, Agropyron trichophorum, Allium ascalonicum, Allium cepa, Allium chinense, Allium porrum, Allium schoenoprasum, Allium fistulosum, Allium sativum, Alopecurus pratensis, Andropogon gerardi, Andropogon Gerardii, Andropogon scoparious, Arrhenatherum elatius, Asparagus officinalis, Avena nuda, Avena sativa, Bambusa vulgaris, Bothrichloa barbinodis, Bothrichloa ischaemum, Bothrichloa saccharoides, Bouteloua curipendula, Bouteloua eriopoda, Bouteloua gracilis, Bromus erectus, Bromus inermis, Bromus riparius, Calamovilfa longifilia, Cenchrus ciliaris, Chloris gayana, Cymbopogon nardus, Cynodon dactylon, Dactylis glomerata, Dichanthium annulatum, Dichanthium aristatum, Dichanthium sericeum, Digitaria decumbens, Digitaria smutsii, Dioscorea rotundata, Dicsorea alata, Dicscorea opposita, Dicscorea bulbifera, Dioscorea esculenta, Dioscorea trifida, Eleusine coracan, Elymus angustus, Elymus junceus, Eragrostis curvula, Eragrostis tef, Fagopyrum esculentum, Fagopyrum tataricum, Festuca arundinacea, Festuca ovina, Festuca pratensis, Festuca rubra, Helianthus annuus sunflower, Hordeum distichum, Hordeum vulgare, Lolium multiflorum, Lolium perenn, Miscanthis sinensis, Miscanthus x giganteus, Oryza sativa, Panicum italicium, Panicum maximum, Panicum miliaceum, Panicum purpurascens, Panicum virgatum, Panicum virgatum, Paspalum dilatatum, Paspalum notatum, Pennisetum clandestinum, Pennisetum glaucum, Pennisetum purpureum, Pennisetum spicatum, Phalaris arundinacea, Phleum bertolinii, Phleum pratense, Poa fendleriana, Poa pratensis, Poa. nemoralis, Saccharum officinarum, Saccharum robustum, Saccharum sinense, Saccharum spontaneum, Secale cereale, Setaria sphacelata, Sorgahastum nutans, Sorghastrum nutans, Sorghum dochna, Sorghum halepense, Sorghum sudanense, Sorghum vulgare, Sorghum vulgare, Triticum aestivum, Triticum dicoccum, Triticum durum, Triticum monococcum, Vanilla fragrans, X Triticosecale и Zea mays.
Предпочтительным семейством однодольных растений является семейство злаковых.
Предпочтительные подсемейства злаковых включают: Anomochlooideae, Pharoideae, Puelioideae, Bambusoideae, Pooideae, Ehrhartoideae, Aristidoideae, Arundinoideae, Chloridoideae, Panicoideae, Danthonioideae и Micrairoideae.
Предпочтительным подсемейством злаковых является подсемейство Pooideae. Предпочтительные растения Pooideae включают пшеницу, ячмень, овес, костер и вейник тростниковый.
Другим предпочтительным подсемейством злаковых является подсемейство Ehrhartoideae. Предпочтительные растения Ehrhartoideae включают рис.
Другим предпочтительным подсемейством злаковых является подсемейство Panicoideae. Предпочтительные растения Panicoideae включают петушье просо, кукурузу, сорго, сахарный тростник, энергетический тростник, миллет, фонио и бородатую траву.
Другим предпочтительным подсемейством злаковых является подсемейство Arundinoideae. Предпочтительные растения Arundinoideae включают Arundo donax.
Другим предпочтительным семейством злаковых является подсемейство Bambusoideae. Предпочтительные растения Bambusoideae включают бамбук.
Предпочтительные виды злаковых включают виды рода Lolium. Предпочтительные виды Lolium включают Lolium longiflorum, Lolium multiflorum, Lolium perenne, Lolium westerwoldicum, Lolium temulentum и Lolium hybridum.
Другие предпочтительные виды злаковых включают виды рода Festuca. Предпочтительные виды Festuca включают Festuca arundinacea, Festuca ovina, Festuca pratensis и Festuca rubra.
Предпочтительно клетки растений или растения происходят из видов двудольных растений.
Предпочтительные роды двудольных растений включают: Amygdalus, Anacardium, Arachis, Brassica, Cajanus, Cannabis, Carthamus, Carya, Ceiba, Cicer, Cocos, Coriandrum, Coronilla, Cossypium, Crotalaria, Dolichos, Elaeis, lycine, Gossypium, Helianthus, Lathyrus, Lens, Lespedeza, Linum, Lotus, Lupinus, Macadamia, Medicago, Melilotus, Mucuna, Olea, Onobrychis, Ornithopus, Papaver, Phaseolus, Phoenix, Pistacia, Pisum, Prunus, Pueraria, Ribes, Ricinus, Sesamum, Theobroma, Trifolium, Trigonella, Vicia и Vigna.
Предпочтительные виды двудольных растений включают: Amygdalus communis, Anacardium occidentale, Arachis hypogaea, Arachis hypogea, Brassica napus Rape, Brassica. nigra. Brassica campestris, Cajanus cajan, Cajanus indicus, Camelina sativa, Cannabis sativa, Carthamus tinctorius, Carya illinoinensis, Ceiba pentandra, Cicer arietinum, Cocos nucifera, Coriandrum sativum, Coronilla varia, Cossypium hirsutum, Crotalaria juncea, Dolichos lablab, Elaeis guineensis, Gossypium arboreum, Gossypium nanking, Gossypium barbadense, Gossypium herbaceum, Gossypium hirsutum, Glycine max, Glycine ussuriensis, Glycine gracilis, Helianthus annus, Jatropha cuneata, Jatropha curcas, Lupinus angustifolius, Lupinus luteus, Lupinus mutabilis, Lespedeza sericea, Lespedeza striata, Lotus uliginosus, Lathyrus sativus, Lens culinaris, Lespedeza stipulacea, Linum usitatissimum, Lotus corniculatus, Lupinus albus, Lupinus angustifolius, Lupinus luteus, Medicago arborea, Medicago falcate, Medicago hispida, Medicago officinalis, Medicago. sativa Alfalfa, Medicago tribuloides, Macadamia integrifolia, Medicago arabica, Melilotus albus, Millettia pinnata, Mucuna pruriens, Olea europaea, Onobrychis viciifolia, Ornithopus sativus, Phaseolus aureus, Prunus cerasifera, Prunus cerasus, Phaseolus coccineus, Prunus domestica, Phaseolus lunatus, Prunus. maheleb, Phaseolus mungo, Prunus. persica, Prunus. pseudocerasus, Phaseolus vulgaris, Papaver somniferum, Phaseolus acutifolius, Phoenix dactylifera, Pistacia vera, Pisum sativum, Prunus amygdalus, Prunus armeniaca, Pueraria thunbergiana, Ribes nigrum, Ribes rubrum, Ribes grossularia, Ricinus communis, Sesamum indicum, Solanum tuberosum, Trifolium augustifolium, Trifolium diffusum, Trifolium hybridum, Trifolium incarnatum, Trifolium ingrescens, Trifolium pratense, Trifolium repens, Trifolium resupinatum, Trifolium subterraneum, Theobroma cacao, Trifolium alexandrinum, Trigonella foenumgraecum, Vernicia fordii, Vicia angustifolia, Vicia atropurpurea, Vicia calcarata, Vicia dasycarpa, Vicia ervilia, Vaccinium oxycoccos, Vicia pannonica, Vigna sesquipedalis, Vigna sinensis, Vicia villosa, Vicia faba, Vicia sative и Vigna angularis.
Растения и части растений
В следующем аспекте изобретение относится к растению, которое имеет увеличенную биомассу корней в результате повышенной экспрессии белка PEAPOD или его фрагмента.
В одном варианте осуществления экспрессия белка PEAPOD или его фрагмента повышается вследствие трансформации растения, или его растения-предшественника или клетки растения-предшественника, полинуклеотидом, кодирующим белок PEAPOD или его фрагмент.
В следующем варианте осуществления растение является трансгенным по полинуклеотиду, экспрессирующему белок PEAPOD или его фрагмент.
В следующем варианте осуществления полинуклеотид или его фрагмент представляет собой полинуклеотид, функционально связанный с тканепредпочтительным промотором.
В следующем варианте осуществления промотор является предпочтительным для корней промотором.
В следующем варианте осуществления промотор является специфическим для корней промотором.
В следующем варианте осуществления белок PEAPOD является таким, как определено в настоящем описании.
В следующем варианте осуществления полинуклеотид, кодирующий белок PEAPOD, является таким, как определено в настоящем описании.
В следующем аспекте изобретение относится к клетке, части, сеянцу или потомству растения, которое является трансгенным по меньшей мере по одному из:
a) полинуклеотида, и
b) полинуклеотида и функционально связанного промотора.
ПОДРОБНОЕ ОПИСАНИЕ
В настоящем описании, когда упоминаются описания патентов, другие внешние документы или другие источники информации, это, как правило, предназначено для предоставления контекста обсуждения признаков изобретения. Если нет иных конкретных указаний, упоминание таких внешних документов не следует истолковывать как допущение того, что такие документы или такие источники информации, в любой юрисдикции, являются уровнем техники или формируют часть общей информации в данной области.
Термин "содержащий", как используют в настоящем описании, означает "по меньшей мере частично состоящий из". При интерпретации каждого утверждения в настоящем описании, который включает термин "содержащий", также могут присутствовать признаки, отличные от признаков, предшествующих термину. Родственные термины, такие как "содержать" и "содержат" следует интерпретировать аналогичным образом.
Увеличенная биомасса корней
Растение с "увеличенной биомассой корней" продуцирует больше биомассы, чем контрольное растение того же типа и возраста. Таким образом, "увеличенный" означает увеличенный относительно контрольного растения того же типа и возраста.
Предпочтительно растение с "увеличенной биомассой корней" продуцирует по меньшей мере на 10%, предпочтительно по меньшей мере на 20%, более предпочтительно по меньшей мере на 30%, более предпочтительно по меньшей мере на 40%, более предпочтительно по меньшей мере на 50%, более предпочтительно по меньшей мере на 60%, более предпочтительно по меньшей мере на 70%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90%, более предпочтительно по меньшей мере на 100%, более предпочтительно по меньшей мере на 150%, более предпочтительно по меньшей мере на 200%, более предпочтительно по меньшей мере на 300%, более предпочтительно по меньшей мере на 400% больше биомассы корней, чем контрольное растение того же типа и возраста.
В одном варианте осуществления растение с "увеличенной биомассой корней" имеет по меньшей мере одно из: более крупных корней, большего количества корней или более мощной корневой системы, чем контрольное растение.
Биомасса корней
Термин "биомасса корней" относится к общей массе ткани корней, продуцируемой растением. Ее можно оценивать посредством сухой массы или влажной массы.
Корень
Термин "корень", как используют в рамках изобретения, означает то же, что и стандартно употребляемый термин. Термин "корень" охватывает первичный корень, вторичные корни, адвентивные корни, ветви корней и корневые волоски. Корни обычно находятся под землей, однако термин также охватывает воздушные корни. В одном варианте осуществления термин "корень" охватывает части растения, не несущие листьев и узлов.
Увеличенная толерантность к засухе
В одном варианте осуществления растение с "увеличенной биомассой корней" также имеет увеличенную толерантность к засухе. Вновь, "увеличенный" означает увеличенный относительно контрольного растения того же типа и возраста.
Термин "увеличенная толерантность к засухе" описывает растение или растения, которые демонстрируют более благоприятные качества в любом аспекте их роста и развития в субоптиальных условиях увлажнения, чем подходящие контрольные растения в тех же условиях.
Контрольное растение
В одном варианте осуществления контрольное растение представляет собой растение дикого типа. В следующем варианте осуществления контрольное растение представляет собой нетрансформированное растение. В следующем варианте осуществления контрольное растение представляет собой растение, не трансформированное полинуклеотидом PEAPOD. В следующем варианте осуществления контрольное растение представляет собой растение, не трансформированное конструкцией. В следующем варианте осуществления контрольное растение представляет собой растение, трансформированное контрольной конструкцией. В одном варианте осуществления конструкция представляет собой пустую векторную конструкцию.
Эктопическая экспрессия
Термин "эктопическая экспрессия" следует интерпретировать широко. Термин относится к экспрессии полинуклеотида или полипептида в любом из:
- клетки, органа, ткани или растения, в которых они обычно не экспрессируются,
- клетки, органа, ткани или растения в момент времени или на стадии развития, когда они обычно не экспрессируются, и
- клетки, органа, ткани или растения на уровне, превышающем уровень, в котором они обычно экспрессируются в этой клетке, органе, ткани или растении.
Тканепредпочтительные промоторы
В определенных вариантах осуществления полинуклеотиды, кодирующие белок PEAPOD, экспрессируются под контролем тканепредпочтительных промоторов. Термин "предпочтительный" в отношении тканепредпочтительных промоторов означает, что промотор в основном запускает экспрессию в данной ткани. Таким образом, например, предпочтительный для корней промотор запускает более высокий уровень экспрессии функционально связанного полинуклеотида в ткани корней, чем в других тканях, или органах, или растении.
Предпочтительные для корней промоторы
Предпочтительный для корней промотор запускает более высокий уровень экспрессии функционально связанного полинуклеотида в ткани корней, чем в других тканях, или органах, или растении.
Предпочтительные для корней промоторы могут включать промоторы, предпочтительные для не фотосинтезирующих тканей, и репрессируемые светом регулируемые промоторы.
Промоторы, предпочтительные для не фотосинтезирующих тканей
Промоторы, предпочтительные для не фотосинтезирующих тканей, включают промоторы, предпочтительно экспрессируемые в не фотосинтезирующих тканях/органах растения.
Промоторы, предпочтительные для не фотосинтезирующих тканей, также могут включать репрессируемые светом промоторы.
Репрессируемые светом промоторы
Пример репрессируемого светом промотора находится в US 5639952 и в US 5656496.
Специфические для корней промоторы
Пример специфического для корней промотора находится в US 5837848; и US 2004/0067506 и US 2001/0047525.
Термин "предпочтительно экспрессируемый" в отношении промотора, предпочтительно экспрессируемого в определенной ткани, означает, что промотор экспрессируется на более высоком уровне в данной ткани, чем в других тканях растения.
Термин "тканеспецифический" в отношении промотора означает, что промотор экспрессируется по существу только в данной ткани, но не в других тканях растения.
В одном варианте осуществления предпочтительный для корней промотор представляет собой специфический для корней промотор.
Термин "ген", как используют в рамках изобретения, означает эндогенную геномную последовательность, которая включает кодирующую последовательность, которая кодирует полипептид или белок. Кодирующая последовательность может прерываться одним или несколькими интронами. Ген, как правило, также включает промоторную последовательность, 5'-нетранслируемую последовательность, 3'-нетранслируемую последовательность, и последовательность терминатора. Геномные последовательности, которые регулируют экспрессию белка, также можно считать частью гена.
Полинуклеотиды и фрагменты
Термин "полинуклеотид(ы)", как используют в рамках изобретения, означает одноцепочечный или двухцепочечный дезоксирибонуклеотидный или рибонуклеотидный полимер любой длины, но предпочтительно по меньшей мере 15 нуклеотидов, и он включает в качестве неограничивающих примеров кодирующие и некодирующие последовательности гена, смысловые и антисмысловые последовательности, комплементарные последовательности, экзоны, интроны, геномную ДНК, кДНК, пре-мРНК, мРНК, рРНК, миРНК, миРНК, тРНК, рибозимы, рекомбинантные полипептиды, выделенные и очищенные встречающиеся в природе последовательности ДНК или РНК, синтетические последовательности РНК и ДНК, зонды нуклеиновых кислот, праймеры и фрагменты.
"Фрагмент" полинуклеотида относится к непрерывной подпоследовательности более крупной полинуклеотидной последовательности. Предпочтительно фрагмент имеет по меньшей мере 15 нуклеотидов предпочтительно по меньшей мере 16 нуклеотидов, более предпочтительно по меньшей мере 17 нуклеотидов, более предпочтительно по меньшей мере 18 нуклеотидов, более предпочтительно по меньшей мере 19 нуклеотидов, более предпочтительно по меньшей мере 20 нуклеотидов, более предпочтительно по меньшей мере 21 нуклеотид, более предпочтительно по меньшей мере 22 нуклеотида, более предпочтительно по меньшей мере 23 нуклеотида, более предпочтительно по меньшей мере 24 нуклеотида, более предпочтительно по меньшей мере 25 нуклеотидов, более предпочтительно по меньшей мере 26 нуклеотидов, более предпочтительно по меньшей мере 27 нуклеотидов, более предпочтительно по меньшей мере 28 нуклеотидов, более предпочтительно по меньшей мере 29 нуклеотидов, более предпочтительно по меньшей мере 30 нуклеотидов, более предпочтительно по меньшей мере 31 нуклеотид, более предпочтительно по меньшей мере 32 нуклеотида, более предпочтительно по меньшей мере 33 нуклеотида, более предпочтительно по меньшей мере 34 нуклеотида, более предпочтительно по меньшей мере 35 нуклеотидов, более предпочтительно по меньшей мере 36 нуклеотидов, более предпочтительно по меньшей мере 37 нуклеотидов, более предпочтительно по меньшей мере 38 нуклеотидов, более предпочтительно по меньшей мере 39 нуклеотидов, более предпочтительно по меньшей мере 40 нуклеотидов, более предпочтительно по меньшей мере 41 нуклеотид, более предпочтительно по меньшей мере 42 нуклеотида, более предпочтительно по меньшей мере 43 нуклеотида, более предпочтительно по меньшей мере 44 нуклеотида, более предпочтительно по меньшей мере 45 нуклеотидов, более предпочтительно по меньшей мере 46 нуклеотидов, более предпочтительно по меньшей мере 47 нуклеотидов, более предпочтительно по меньшей мере 48 нуклеотидов, более предпочтительно по меньшей мере 49 нуклеотидов, более предпочтительно по меньшей мере 50 нуклеотидов, более предпочтительно по меньшей мере 51 нуклеотид, более предпочтительно по меньшей мере 52 нуклеотида, более предпочтительно по меньшей мере 53 нуклеотида, более предпочтительно по меньшей мере 54 нуклеотида, более предпочтительно по меньшей мере 55 нуклеотидов, более предпочтительно по меньшей мере 56 нуклеотидов, более предпочтительно по меньшей мере 57 нуклеотидов, более предпочтительно по меньшей мере 58 нуклеотидов, более предпочтительно по меньшей мере 59 нуклеотидов, более предпочтительно по меньшей мере 60 нуклеотидов, более предпочтительно по меньшей мере 61 нуклеотид, более предпочтительно по меньшей мере 62 нуклеотидов, более предпочтительно по меньшей мере 63 нуклеотида, более предпочтительно по меньшей мере 64 нуклеотида, более предпочтительно по меньшей мере 65 нуклеотидов, более предпочтительно по меньшей мере 66 нуклеотидов, более предпочтительно по меньшей мере 67 нуклеотидов, более предпочтительно по меньшей мере 68 нуклеотидов, более предпочтительно по меньшей мере 69 нуклеотидов, более предпочтительно по меньшей мере 70 нуклеотидов, более предпочтительно по меньшей мере 71 нуклеотид, более предпочтительно по меньшей мере 72 нуклеотида, более предпочтительно по меньшей мере 73 нуклеотида, более предпочтительно по меньшей мере 74 нуклеотида, более предпочтительно по меньшей мере 75 нуклеотидов, более предпочтительно по меньшей мере 76 нуклеотидов, более предпочтительно по меньшей мере 77 нуклеотидов, более предпочтительно по меньшей мере 78 нуклеотидов, более предпочтительно по меньшей мере 79 нуклеотидов, более предпочтительно по меньшей мере 80 нуклеотидов, более предпочтительно по меньшей мере 81 нуклеотид, более предпочтительно по меньшей мере 82 нуклеотида, более предпочтительно по меньшей мере 83 нуклеотида, более предпочтительно по меньшей мере 84 нуклеотида, более предпочтительно по меньшей мере 85 нуклеотидов, более предпочтительно по меньшей мере 86 нуклеотидов, более предпочтительно по меньшей мере 87 нуклеотидов, более предпочтительно по меньшей мере 88 нуклеотидов, более предпочтительно по меньшей мере 89 нуклеотидов, более предпочтительно по меньшей мере 90 нуклеотидов, более предпочтительно по меньшей мере 91 нуклеотид, более предпочтительно по меньшей мере 92 нуклеотида, более предпочтительно по меньшей мере 93 нуклеотида, более предпочтительно по меньшей мере 94 нуклеотида, более предпочтительно по меньшей мере 95 нуклеотидов, более предпочтительно по меньшей мере 96 нуклеотидов, более предпочтительно по меньшей мере 97 нуклеотидов, более предпочтительно по меньшей мере 98 нуклеотидов, более предпочтительно по меньшей мере 99 нуклеотида, более предпочтительно по меньшей мере 100 нуклеотидов, более предпочтительно по меньшей мере 150 нуклеотидов, более предпочтительно по меньшей мере 200 нуклеотидов, более предпочтительно по меньшей мере 250 нуклеотидов, более предпочтительно по меньшей мере 300 нуклеотидов, более предпочтительно по меньшей мере 350 нуклеотидов, более предпочтительно по меньшей мере 400 нуклеотидов, более предпочтительно по меньшей мере 450 нуклеотидов и наиболее предпочтительно по меньшей мере 500 нуклеотидов из непрерывных нуклеотидов описанного полинуклеотида. Фрагмент последовательности полинуклеотида можно использовать в антисмысловой технологии, РНК-интерференции (РНК-i), сайленсинге генов, трехспиральной или рибозимной технологии, или в качестве праймера, зонда, включенного в микрочип, или можно использовать в способах по изобретению для селекции на основе полинуклеотидов.
В одном варианте осуществления фрагмент кодирует полипептид, который выполняет или способен выполнять ту же функцию, что и полипептид, кодируемый более крупным полинуклеотидом, частью которого является фрагмент.
Термин "праймер" относится к короткому полинуклеотиду, обычно имеющему свободную группу 3'-OH, который гибридизуется или может гибридизоваться с матрицей и может использоваться в качестве затравки для полимеризации полинуклеотида, комплементарного мишени.
Термин "зонд" относится к короткому полинуклеотиду, который используют или может быть использован для обнаружения полинуклеотидной последовательности, которая комплементарна зонду, в анализе не основе гибридизации. Зонд может состоять из "фрагмента" полинуклеотида, как определено в настоящем описании.
Полипептиды и фрагменты
Термин "полипептид", как используют в рамках изобретения, охватывает цепи аминокислот любой длины, но предпочтительно по меньшей мере 5 аминокислот, включая полноразмерные белки, в которых аминокислотные остатки связаны ковалентными пептидными связями. Полипептиды по настоящему изобретению, или используемые в способах по изобретению, могут представлять собой очищенные природные продукты или их можно продуцировать частично или полностью с использованием рекомбинантных технологий или технологий синтеза. Термин может относиться к полипептиду, агрегату полипептидов, такому как димер или другой мультимер, слитый полипептид, фрагмент полипептида, вариант полипептида или их производное.
"Фрагмент" полинуклеотида относится к непрерывной подпоследовательности из более крупной полинуклеотидной последовательности. Предпочтительно фрагмент имеет длину по меньшей мере 5, более предпочтительно по меньшей мере 10, более предпочтительно по меньшей мере 20, более предпочтительно по меньшей мере 30, более предпочтительно по меньшей мере 40, более предпочтительно по меньшей мере 50, более предпочтительно по меньшей мере 100, более предпочтительно по меньшей мере 120, более предпочтительно по меньшей мере 150, более предпочтительно по меньшей мере 200, более предпочтительно по меньшей мере 250, более предпочтительно по меньшей мере 300, более предпочтительно по меньшей мере 350, более предпочтительно по меньшей мере 400.
В одном варианте осуществления фрагмент выполняет или способен выполнять ту же функцию, что и полипептид, частью которого фрагмент является.
Предпочтительно фрагмент выполняет функцию, которая требуется для биологической активности и/или обеспечивает трехмерную структуру полипептида.
Термин "выделенный", как используют в отношении полинуклеотидных или полипептидных последовательностей, описанных в настоящем описании, относится к последовательностям, которые удалены из их естественной клеточной среды. В одном варианте осуществления последовательность отделена от фланкирующих ее последовательностей, встречающихся в природе. Выделенную молекулу можно получать любым способом или комбинацией способов, включая биохимические, рекомбинантные способы и способы синтеза.
Термин "рекомбинантный" относится к полинуклеотидной последовательность, которую синтетически продуцируют или извлекают из последовательностей, которые окружают ее в ее природном контексте. Рекомбинантная последовательность может быть рекомбинирована с последовательностями, которые не присутствуют в ее природном контексте.
"Рекомбинантная" полипептидная последовательность продуцируется путем трансляции "рекомбинантной" полинуклеотидной последовательности.
Термин "происходящий из" в отношении полинуклеотидов или полипептидов по изобретению, происходящих из конкретного рода или вида, означает, что полинуклеотид или полипептид имеет ту же последовательность, что и полинуклеотид или полипептид, встречающийся в природе в данном роде или виде. Таким образом, полинуклеотид или полипептид, происходящий из конкретного рода или вида, можно продуцировать способами синтеза или рекомбинантными способами.
Варианты
Как используют в рамках изобретения, термин "вариант" относится к полинуклеотидным или полипептидным последовательностям, отличающимся от конкретно идентифицированных последовательностей, где один или несколько нуклеотидных или аминокислотных остатков удалены, замещены или вставлены. Варианты могут представлять собой встречающиеся в природе аллельные варианты или не встречающиеся в природе варианты. Варианты могут происходить из того же или из другого вида, и они могут охватывать гомологи, паралоги и ортологи. В определенных вариантах осуществления, варианты полипептидов и полинуклеотидов, описанных в настоящем описании, обладают видами биологической активности, которые являются такими же или сходными с видами биологической активности описанных полипептидов или полинуклеотидов. Термин "вариант" в отношении полипептидов и полинуклеотидов охватывает все формы полипептидов и полинуклеотидов, как определено в настоящем описании.
Варианты полинуклеотидов
Варианты полинуклеотидных последовательностей предпочтительно демонстрируют по меньшей мере 50%, более предпочтительно по меньшей мере 51%, более предпочтительно по меньшей мере 52%, более предпочтительно по меньшей мере 53%, более предпочтительно по меньшей мере 54%, более предпочтительно по меньшей мере 55%, более предпочтительно по меньшей мере 56%, более предпочтительно по меньшей мере 57%, более предпочтительно по меньшей мере 58%, более предпочтительно по меньшей мере 59%, более предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 61%, более предпочтительно по меньшей мере 62%, более предпочтительно по меньшей мере 63%, более предпочтительно по меньшей мере 64%, более предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 66%, более предпочтительно по меньшей мере 67%, более предпочтительно по меньшей мере 68%, более предпочтительно по меньшей мере 69%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 71%, более предпочтительно по меньшей мере 72%, более предпочтительно по меньшей мере 73%, более предпочтительно по меньшей мере 74%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 76%, более предпочтительно по меньшей мере 77%, более предпочтительно по меньшей мере 78%, более предпочтительно по меньшей мере 79%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 81%, более предпочтительно по меньшей мере 82%, более предпочтительно по меньшей мере 83%, более предпочтительно по меньшей мере 84%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 86%, более предпочтительно по меньшей мере 87%, более предпочтительно по меньшей мере 88%, более предпочтительно по меньшей мере 89%, более предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 91%, более предпочтительно по меньшей мере 92%, более предпочтительно по меньшей мере 93%, более предпочтительно по меньшей мере 94%, более предпочтительно по меньшей мере 95%, более предпочтительно по меньшей мере 96%, более предпочтительно по меньшей мере 97%, более предпочтительно по меньшей мере 98%, и наиболее предпочтительно по меньшей мере 99% идентичность с последовательностью по настоящему изобретению. Идентичность определяют на протяжении окна сравнения из по меньшей мере 20 нуклеотидных положений, предпочтительно по меньшей мере 50 нуклеотидных положений, более предпочтительно по меньшей мере 100 нуклеотидных положений и наиболее предпочтительно на протяжении всей длины полинуклеотида по изобретению.
Идентичность полинуклеотидных последовательностей можно определять следующим образом. Рассматриваемую полинуклеотидную последовательность сравнивают с полинуклеотидной последовательностью-кандидатом с использованием BLASTN (из пакета программ BLAST, версия 2.2.5 [ноябрь 2002 года]), в bl2seq (Tatiana A. Tatusova, Thomas L. Madden (1999), "Blast 2 sequences - a new tool for comparing protein and nucleotide sequences", FEMS Microbiol Lett. 174:247-250), который является общедоступным от NCBI (ftp://ftp.ncbi.nih.gov/blast/). В одном варианте осуществления используют параметры bl2seq по умолчанию. В следующем варианте осуществления используют параметры bl2seq по умолчанию, за исключением того, что фильтрация частей низкой комплексности должна быть выключена.
Идентичность полинуклеотидных последовательностей также можно вычислять по всей длине перекрывания между последовательностью-кандидатом и рассматриваемой полинуклеотидной последовательностью с использованием программ для глобального выравнивания последовательностей (например, Needleman, S. B. and Wunsch, C. D. (1970) J. Mol. Biol. 48, 443-453). Полное исполнение алгоритма глобального выравнивания Needleman-Wunsch может быть найдено в программе needle пакета EMBOSS (Rice, P. Longden, I. and Bleasby, A. EMBOSS: The European Molecular Biology Open Software Suite, Trends in Genetics June 2000, vol 16, No 6. pp.276-277), которая может быть найдена на http://www.hgmp.mrc.ac.uk/Software/EMBOSS/. Сервер European Bioinformatics Institute также обеспечивает возможность для выполнения глобального выравнивания EMBOSS-needle между двумя последовательностями через Интернет на http:/www.ebi.ac.uk/emboss/align/.
Альтернативно можно использовать программу GAP, которая проводит оптимальное глобальное выравнивание двух последовательностей без штрафа за концевые пропуски. GAP описана в следующей статье: Huang, X. (1994) On Global Sequence Alignment. Computer Applications in the Biosciences 10, 227-235.
Предпочтительный способ вычисления % идентичности полинуклеотидных последовательностей основан на выравнивании сравниваемых последовательностей с использованием Clustal X (Jeanmougin et al., 1998, Trends Biochem. Sci. 23, 403-5).
Полинуклеотидные варианты по настоящему изобретению также охватывают варианты, которые демонстрируют сходство с одной или несколькими из конкретно идентифицированных последовательностей, которые, вероятно, сохраняют функциональную эквивалентность с этими последовательностями и от которых нельзя ожидать, что они появятся случайным образом. Такое сходство последовательностей в отношении полипептидов можно определять с использованием общедоступной программы bl2seq из пакета программ BLAST (версии 2.2.5 [Nov 2002]) от NCBI (ftp://ftp.ncbi.nih.gov/blast/).
Альтернативно варианты полинуклеотидов по настоящему изобретению гибридизуются с определенными полинуклеотидными последовательностями или комплементарными им последовательностями в жестких условиях.
Термин "гибридизуются в жестких условиях" и его грамматические эквиваленты относится к способности полинуклеотидной молекулы гибридизоваться с молекулой полинуклеотида-мишени (такой как молекула полинуклеотида-мишени, иммобилизованная на ДНК- или РНК-блоте, таком как саузерн-блот или нозерн-блот) в определенных условиях температуры и концентрации соли. Способность гибридизоваться в жестких условиях гибридизации можно определять посредством первоначальной гибридизации в менее жестких условиях, а затем повышения жесткости до желаемой жесткости.
Что касается полинуклеотидных молекул длиной более чем приблизительно 100 оснований, типичные жесткие условия гибридизации представляют собой условия, не более чем на 25-30°C (например, 10°C) ниже температуры плавления (Tm) нативного дуплекса (см., главным образом, Sambrook et al., Eds, 1987, Molecular Cloning, A Laboratory Manual, 2nd Ed. Cold Spring Harbor Press; Ausubel et al., 1987, Current Protocols in Molecular Biology, Greene Publishing). Tm для полинуклеотидных молекул, имеющих размер более чем приблизительно 100 оснований, можно вычислять по формуле Tm=81,5+0,41%(G+C-log (Na+)). (Sambrook et al., Eds, 1987, Molecular Cloning, A Laboratory Manual, 2nd Ed. Cold Spring Harbor Press; Bolton and McCarthy, 1962, PNAS 84:1390). Типичные жесткие условия для полинуклеотида длиной более 100 оснований представляют собой такие условия гибридизации, как предварительное промывание в растворе 6× SSC, 0,2% SDS; гибридизация при 65°C, 6× SSC, 0,2% SDS, в течение ночи; а затем два промывания по 30 минут в каждом случае в 1× SSC, 0,1% SDS, при 65°C и два промывания по 30 минут в каждом случае в 0,2X SSC, 0,1% SDS, при 65°C.
Что касается полинуклеотидных молекул, имеющих длину менее 100 оснований, иллюстративные жесткие условия гибридизации представляют собой условия на 5-10°C ниже Tm. В среднем, Tm полинуклеотидной молекулы длиной менее 100 п.н. снижают приблизительно на (500/длина олигонуклеотида)°C.
Что касается миметиков ДНК, известных как пептидно-нуклеиновые кислоты (ПНК) (Nielsen et al., Science. 1991 Dec 6;254(5037):1497-500), их величины Tm являются более высокими, чем для гибридов ДНК-ДНК или ДНК-РНК, и их можно вычислять с использованием формулы, описанной в Giesen et al., Nucleic Acids Res. 1998 Nov 1; 26(21):5004-6. Иллюстративные жесткие условия гибридизации для гибрида ДНК-ПНК, имеющего длину менее 100 оснований, представляют собой условия на 5-10°C ниже Tm.
Варианты полинуклеотидов по настоящему изобретению также охватывают полинуклеотиды, которые отличаются от последовательностей по изобретению, но которые, вследствие вырожденности генетического кода, кодируют полипептид, обладающий сходной активностью с полипептидом, кодируемым полинуклеотидом по настоящему изобретению. Изменение последовательности, которое не изменяет аминокислотную последовательность полипептида, представляет собой "молчащее изменение". За исключением ATG (метионин) и TGG (триптофан), кодоны для одной и той же аминокислоты могут быть заменены общепринятыми способами, например, для оптимизации экспрессии кодонов в конкретном организме-хозяине.
Также изобретение включает изменения полинуклеотидной последовательности, которые приводят к консервативным заменам одной или нескольких аминокислот в кодируемой полипептидной последовательности без значительного изменения ее биологической активности. Квалифицированному специалисту известны способы получения фенотипически молчащих аминокислотных замен (см., например, Bowie et al., 1990, Science 247, 1306).
Варианты полинуклеотидов вследствие молчащих изменений и консервативных замен в кодируемой полипептидной последовательности можно определять с использованием общедоступной программы bl2seq из пакета программ BLAST (версия 2.2.5 [ноябрь 2002 года]) от NCBI (ftp://ftp.ncbi.nih.gov/blast/) через алгоритм tblastx, как описано ранее.
Варианты полипептидов
Термин "вариант" в отношении полипептидов охватывает встречающиеся в природе, продуцированные рекомбинантными способами и способами синтеза полипептиды. Варианты полипептидных последовательностей предпочтительно демонстрируют по меньшей мере 50%, более предпочтительно по меньшей мере 51%, более предпочтительно по меньшей мере 52%, более предпочтительно по меньшей мере 53%, более предпочтительно по меньшей мере 54%, более предпочтительно по меньшей мере 55%, более предпочтительно по меньшей мере 56%, более предпочтительно по меньшей мере 57%, более предпочтительно по меньшей мере 58%, более предпочтительно по меньшей мере 59%, более предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 61%, более предпочтительно по меньшей мере 62%, более предпочтительно по меньшей мере 63%, более предпочтительно по меньшей мере 64%, более предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 66%, более предпочтительно по меньшей мере 67%, более предпочтительно по меньшей мере 68%, более предпочтительно по меньшей мере 69%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 71%, более предпочтительно по меньшей мере 72%, более предпочтительно по меньшей мере 73%, более предпочтительно по меньшей мере 74%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 76%, более предпочтительно по меньшей мере 77%, более предпочтительно по меньшей мере 78%, более предпочтительно по меньшей мере 79%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 81%, более предпочтительно по меньшей мере 82%, более предпочтительно по меньшей мере 83%, более предпочтительно по меньшей мере 84%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 86%, более предпочтительно по меньшей мере 87%, более предпочтительно по меньшей мере 88%, более предпочтительно по меньшей мере 89%, более предпочтительно по меньшей мере 90%, более предпочтительно по меньшей мере 91%, более предпочтительно по меньшей мере 92%, более предпочтительно по меньшей мере 93%, более предпочтительно по меньшей мере 94%, более предпочтительно по меньшей мере 95%, более предпочтительно по меньшей мере 96%, более предпочтительно по меньшей мере 97%, более предпочтительно по меньшей мере 98%, и наиболее предпочтительно по меньшей мере 99% идентичность с последовательностями по настоящему изобретению. Идентичность определяют на протяжении окна сравнения по меньшей мере из 20 аминокислотных положений, предпочтительно по меньшей мере из 50 аминокислотных положений, более предпочтительно, по меньшей мере 100 аминокислотных положений и наиболее предпочтительно по всей длине полипептида по изобретению.
Идентичность полипептидных последовательностей можно определять следующим образом. Рассматриваемую полипептидную последовательность сравнивают с полипептидной последовательностью-кандидатом с использованием BLASTP (из пакета программ BLAST, версия 2.2.5 [ноябрь 2002 года]) в bl2seq, который является общедоступным от NCBI (ftp://ftp.ncbi.nih.gov/blast/). В одном варианте осуществления используют параметры bl2seq по умолчанию. В следующем варианте осуществления, используют параметры bl2seq по умолчанию, за исключением того, что фильтрация частей низкой комплексности должна быть выключена.
Идентичность полипептидных последовательностей также можно вычислять по всей длине перекрывания между последовательностью-кандидатом и рассматриваемой полинуклеотидной последовательностью с использованием программ для глобального выравнивания последовательностей. EMBOSS-needle (доступная http:/www.ebi.ac.uk/emboss/align/) и GAP (Huang, X. (1994) On Global Sequence Alignment. Computer Applications in the Biosciences 10, 227-235), как рассмотрено выше, также являются подходящими программами для выравнивания последовательностей для вычисления идентичности полипептидных последовательностей.
Предпочтительный способ вычисления % идентичности полипептидных последовательностей основан на выравнивании последовательностей, подлежащих сравнению, с использованием Clustal X (Jeanmougin et al., 1998, Trends Biochem. Sci. 23, 403-5).
Вариант полипептида включает полипептид, в котором аминокислотная последовательность отличается от полипептида, описанного в настоящем описании, одной или несколькими консервативными аминокислотными заменами, делециями, вставками или инсерциями, которые не влияют на биологическую активность пептида. Консервативные замены, как правило, включают замену одной аминокислоты другой аминокислотой со сходными характеристиками, например, замены в следующих группах: валин, глицин; глицин, аланин; валин, изолейцин, лейцин; аспарагиновая кислота, глутаминовая кислота; аспарагин, глутамин; серин, треонин; лизин, аргинин; и фенилаланин, тирозин.
Неконсервативные замены охватывают замену представителем одного из этих классов представителя другого класса.
Анализ эволюционировавших биологических последовательностей показал, что не все изменения последовательностей являются равновероятными, что отражает по меньшей мере частично отличия консервативных замен относительно неконсервативных замен на биологическом уровне. Например, определенные аминокислотные замены могут встречаться часто, в то время как другие являются очень редкими. Эволюционные изменения или замены аминокислотных остатков можно моделировать с помощью оценочной матрицы, также называемой матрицей замен. Такие матрицы используют в биоинформатическом анализе для идентификации взаимосвязи между последовательностями, одним из их примеров является матрица BLOSUM62, показанная ниже (таблица 1).
Таблица 1: Матрица BLOSUM62, содержащая все возможные показатели замен [Henikoff and Henikoff, 1992].

Представленную матрицу BLOSUM62 используют для получения показателя для каждой пары выровненных аминокислот, находящегося на пересечении соответствующего столбца и ряда. Например, показатель замены остатка глутаминовой кислоты (E) на остаток аспарагиновой кислоты (D) равен 2. По диагонали показаны показатели для аминокислот, которые не изменены. Большинство замен имеют отрицательный показатель. Матрица содержит только целые числа.
Полагают, что определение подходящей оценочной матрицы для осуществления наилучшего выравнивания для данного набора последовательностей входит в объем практических навыков в данной области. Матрицу BLOSUM62 в таблице 1 также используют в качестве матрицы по умолчанию в поиске BLAST, но не ограничиваясь этим.
Другие варианты включают пептиды с модификациями, которые влияют на стабильность пептидов. Такие аналоги могут содержать, например, одну или несколько непептидных связей (которые заменяют пептидные связи) в пептидной последовательности. Также включены аналоги, которые включают остатки, отличные от встречающихся в природе L-аминокислот, например D-аминокислоты или не встречающиеся в природе синтетические аминокислоты, например бета- или гамма-аминокислоты и циклические аналоги.
Конструкции, векторы и их компоненты
Термин "генетическая конструкция" относится к полинуклеотидной молекуле, обычно двухцепочечной ДНК, которая может быть встроена в другую полинуклеотидную молекулу (полинуклеотидная молекула-вставка), такую как, но не ограничиваясь ими, молекула кДНК. Генетическая конструкция может содержать необходимые элементы, которые позволяют транскрипцию полинуклеотидной молекулы-вставки, и, необязательно, трансляцию транскрипта в полипептид. Полинуклеотидная молекула-вставка может происходить из клетки-хозяина или может происходить из отличающейся клетки или организма, и/или она может представлять собой рекомбинантный полинуклеотид. После попадания внутрь клетки-хозяина генетическая конструкция может встраиваться в хромосомную ДНК хозяина. Генетическая конструкция может быть связана с вектором.
Термин "вектор" относится к полинуклеотидной молекуле, обычно двухцепочечной ДНК, которую используют для транспортировки генетической конструкции в клетку-хозяина. Вектор может быть способен к репликации по меньшей мере в одной дополнительной системе-хозяине, такой как E. coli.
Термин "экспрессирующая конструкция" относится к генетической конструкции, которая включает необходимые элементы, которые позволяют транскрипцию полинуклеотидной молекулы-вставки и необязательно трансляцию транскрипта в полипептид. Экспрессирующая конструкция, как правило, содержит в направлении 5'-3':
a) промотор, функциональный в клетке-хозяине, в которую будет трансформирована конструкция,
b) полинуклеотид, подлежащий экспрессии, и
c) терминатор, функциональный в клетке-хозяине, в которую конструкция будет трансформирована.
Термин "кодирующая область" или "открытая рамка считывания" (ORF) относится к смысловой цепи последовательности геномной ДНК или последовательности кДНК, которая способна продуцировать продукт транскрипции и/или полипептид под контролем соответствующих регуляторных последовательностей. Кодирующую последовательность идентифицируют по наличию 5'-кодона инициации трансляции и 3'-стоп-кодона трансляции. При встраивании в генетическую конструкцию "кодирующая последовательность" способна экспрессироваться, когда она функционально связана с последовательностями промотора и терминатора.
"Функционально связанный" означает, что последовательность, подлежащая экспрессии, помещена под контроль регуляторных элементов, которые включают промоторы, тканеспецифические регуляторные элементы, временные регуляторные элементы, энхансеры, репрессоры и терминаторы.
Термин "некодирующая область" относится к нетранслируемым последовательностям, находящимся выше участка начала трансляции и ниже участка остановки трансляции. Эти последовательности также обозначают, соответственно, как 5'-UTR и 3'-UTR. Эти области включают элементы, требуемые для инициации и терминации транскрипции и для регуляции эффективности трансляции.
Терминаторы представляют собой последовательности, которые терминируют транскрипцию и находятся на 3'-нетранслируемых концах генов ниже транслируемой последовательности. Терминаторы являются важными определяющими факторами стабильности мРНК и обнаружено, что в некоторых случаях они имеют пространственные регуляторные функции.
Термин "промотор" относится к нетранскрибируемым цис-регуляторным элементам выше кодирующей области, которые регулируют транскрипцию гена. Промоторы включают цис-инициирующие элементы, которые определяют участок инициации транскрипции, и консервативные боксы, такие как TATA-бокс, и мотивы, которые связываются факторами транскрипции.
Промотор может быть гомологичным в отношении экспрессируемого полинуклеотида. Это означает, что промотор и полинуклеотид встречаются в природе функционально связанными.
Альтернативно промотор может быть гетерологичным в отношении экспрессируемого полинуклеотида. Это означает, что промотор и полинуклеотид не встречаются в природе функционально связанными.
"Трансген" представляет собой полинуклеотид, который вводят в организм посредством трансформации. Трансген может происходить из того же вида, что и организм, в который вводят трансген, или из отличающегося вида. Трансген также может быть синтетическим и не встречающимся в природе ни в одном виде.
"Трансгенное растение" относится к растению, которое содержит новый генетический материал в результате генетического манипулирования или трансформации. Новый генетический материал может происходить из растения того же вида, что и конечное трансгенное растение, или из отличающегося вида, или он может быть синтетическим.
Предпочтительно "трансгенное растение" отличается от любого растения, встречающегося в природе, вследствие присутствия трансгена.
"Инвертированный повтор" представляет собой последовательность, которая является повторяющейся, где вторая половина повтора находится на комплементарной цепи, например,
(5')GATCTA…….TAGATC(3')
(3')CTAGAT…….ATCTAG(5')
Считывающая транскрипция приведет к транскрипту, который претерпевает комплементарное спаривание оснований с формированием шпилечной структуры при условии, что существует спейсер размером 3-5 п.н. между повторяющимися областями. Спейсер может представлять собой любую полинуклеотидную последовательность, однако, как правило, он имеет длину по меньшей мере 3 пары оснований.
Клетки-хозяева
Клетки-хозяева могут происходить, например, из организмов бактерий, грибов, насекомых, млекопитающих или растений.
Способы выделения или продуцирования полинуклеотидов
Молекулы полинуклеотидов по изобретению можно выделять с использованием различных способов, известных средним специалистам в данной области. В качестве примера, такие полипептиды можно выделять с использованием полимеразной цепной реакции (ПЦР) описанной в Mullis et al., Eds. 1994 The Polymerase Chain Reaction, Birkhauser, включенной в настоящее описание в качестве ссылки. Полипептиды по изобретению можно амплифицировать с использованием праймеров, как определено в настоящем описании, происходящих из полинуклеотидных последовательностей по изобретению.
Кроме того, способы выделения полинуклеотидов по изобретению включают применение целых полипептидов, имеющих последовательность, указанную в настоящем описании, или их частей в качестве зондов для гибридизации. Способ гибридизации меченных полинуклеотидных зондов с полинуклеотидами, иммобилизованными на твердых подложках, таких как нитроцеллюлозные фильтры или нейлоновые мембраны, можно использовать для скрининга геномных библиотек или библиотек кДНК. Иллюстративные условия гибридизации и промывания представляют собой: гибридизацию в течение 20 часов при 65°C в 5,0× SSC, 0,5% додецилсульфате натрия, 1x растворе Денхардта; промывание (три промывания по двадцать минут каждое при 55°C) в 1,0× SSC, 1% (масс./об.) додецилсульфате натрия, и необязательно одно промывание (в течение двадцати минут) в 0,5× SSC, 1% (масс./об.) додецилсульфате натрия, при 60°C. Можно проводить необязательное дополнительное промывание (в течение двадцати минут) в условиях 0,1× SSC, 1% (масс./об.) додецилсульфата натрия при 60°C.
Полинуклеотидные фрагменты по изобретению можно получать способами, хорошо известными в данной области, такими как расщепление эндонуклеазой рестрикции, олигонуклеотидный синтез и амплификация способом ПЦР.
Частичную полинуклеотидную последовательность можно использовать в способах, хорошо известных в данной области, для идентификации соответствующей полноразмерной полинуклеотидной последовательности. Такие способы включают способы на основе ПЦР, 5'RACE (Frohman MA, 1993, Methods Enzymol. 218: 340-56) и способ на основе гибридизации, компьютерные способы/способы на основе баз данных. Кроме того, в качестве примера, инвертированная ПЦР позволяет получение неизвестных последовательностей, фланкирующих полинуклеотидные последовательности, описанные в настоящем описании, начиная с праймеров, основанных на известной области (Triglia et al., 1998, Nucleic Acids Res 16, 8186, включенная в настоящее описание в качестве ссылки). В способе используется несколько ферментов рестрикции для получения подходящего фрагмента в известной области гена. Затем фрагмент замыкают посредством внутримолекулярного лигирования и используют в качестве матрицы для ПЦР. На основе известной области конструируют расходящиеся праймеры. Для физической сборки полноразмерных клонов можно использовать стандартные подходы молекулярной биологии (Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Ed. Cold Spring Harbor Press, 1987).
При получении трансгенного растения из конкретного вида может быть полезной трансформация такого растения последовательностью или последовательностями, происходящими из этого вида. Польза может состоять в уменьшении обеспокоенности общества в отношении перекрестно-видовой трансформации при получении трансгенных организмов. Кроме того, когда является желательным подавление гена, может быть необходимым использование последовательности, идентичной (или по меньшей мере в высокой степени сходной) последовательности в растении, для которого является желательной сниженная экспрессия. По этим причинам, среди прочего, является желательной возможность идентифицировать и выделить ортологи конкретного гена в нескольких различных видах растений.
Варианты (включая ортологи) можно идентифицировать описанными способами.
Способы идентификации вариантов
Физические способы
Варианты полипептидов можно идентифицировать с использованием способов на основе ПЦР (Mullis et al., Eds. 1994 The Polymerase Chain Reaction, Birkhauser). Как правило, полинуклеотидная последовательность праймера, пригодного для амплификации вариантов полинуклеотидных молекул по изобретению способом ПЦР, может быть основана на последовательности, кодирующей консервативную область соответствующей аминокислотной последовательности.
Альтернативно можно использовать способы скрининга библиотек, хорошо известные специалистам в данной области (Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Ed. Cold Spring Harbor Press, 1987). При идентификации вариантов последовательности зонда, жесткость гибридизации и/или промывания, как правило, являются сниженными относительно случая, когда ищут точно совпадающие последовательности.
Полипептидные варианты также можно идентифицировать физическими способами, например, посредством скрининга экспрессирующих библиотек с использованием антител, индуцированных против полипептидов по изобретению (Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Ed. Cold Spring Harbor Press, 1987), или посредством идентификации полипептидов из природных источников с помощью таких антител.
Компьютерные способы
Варианты последовательностей по изобретению, включая как полинуклеотидные, так и полипептидные, варианты также можно идентифицировать компьютерными способами, хорошо известными специалистам в данной области, с использованием общедоступных алгоритмов выравнивания последовательностей и инструментов поиска сходства последовательностей для поиска в базах данных последовательностей (общедоступные базы данных включают Genbank, EMBL, Swiss-Prot, PIR и прочие). См., например, Nucleic Acids Res. 29: 1-10 и 11-16, 2001 для примера Интернет-источников. Поиск сходства извлекает и выравнивает целевые последовательности для сравнения с последовательностью, подлежащей анализу (т.е. последовательностью запроса). Алогоритмы сравнения последовательности используют оценочные матрицы для присвоения итогового показателя каждому из выравниваний.
Иллюстративным семейством программ, пригодных для идентификации вариантов в базах данных последовательностей является пакет программ BLAST (версия 2.2.5 [ноябрь 2002 года]), включая BLASTN, BLASTP, BLASTX, tBLASTN и tBLASTX, который являются общедоступным от (ftp://ftp.ncbi.nih.gov/blast/) или от National Center for Biotechnology Information (NCBI), National Library of Medicine, Building 38A, Room 8N805, Bethesda, MD 20894 USA. Сервер NCBI также обеспечивает возможность использовать программы для скрининга ряда общедоступных баз данных последовательностей. BLASTN сравнивает нуклеотидную последовательность запроса против базы данных нуклеотидных последовательностей. BLASTP сравнивает аминокислотную последовательность запроса против базы данных белковых последовательностей. BLASTX сравнивает нуклеотидную последовательность запроса, транслируемую во всех рамках считывания, против базы данных белковых последовательностей. tBLASTN сравнивает белковую последовательность запроса против базы данных нуклеотидных последовательностей, динамично транслируемых во всех рамках считывания. tBLASTX сравнивает нуклеотидную последовательность запроса в шести рамках считывания против базы данных нуклеотидных последовательностей в шести рамках считывания. Программы BLAST можно использовать с параметрами по умолчанию или параметры можно изменять при необходимости для оптимизации скрининга.
Использование семейства алгоритмов BLAST, включая BLASTN, BLASTP и BLASTX, описано в публикации Altschul et al., Nucleic Acids Res. 25: 3389-3402, 1997.
"Совпадения" с одной или несколькими последовательностями базы данных с последовательностью запроса, выявленные с помощью BLASTN, BLASTP, BLASTX, tBLASTN, tBLASTX или сходного алгоритма, обеспечивают выравнивание и идентификацию сходных частей последовательностей. Совпадения ранжируют в порядке степени сходства и длины перекрывания последовательностей. Совпадения с последовательностью базы данных обычно отражают перекрывание на протяжении только части длины последовательности запроса.
Алгоритмы BLASTN, BLASTP, BLASTX, tBLASTN и tBLASTX также выдают величины "ожидания" для выравниваний. Величина ожидания (E) указывает на количество совпадений, которые можно "ожидать" случайным образом при поиске в базе данных того же размера, содержащей случайные непрерывные последовательности. Величину ожидания используют в качестве порога значимости для определения того, указывает ли совпадение с базой данных на истинное сходство. Например, величину E 0,1, присвоенную полинуклеотидному совпадению, интерпретируют как то, что в базе данных, имеющей размер подвергаемой скринингу базы данных, можно ожидать 0,1 соответствий на протяжении выровненной части последовательности со сходным показателем просто случайно. Для последовательностей, имеющих величину E 0,01 или менее на протяжении выровненных и приведенных в соответствие частей, вероятность нахождения соответствия случайным образом в этой базе данных составляет 1% или менее при использовании алгоритма BLASTN, BLASTP, BLASTX, tBLASTN или tBLASTX.
Множественное выравнивание последовательностей из группы родственных последовательностей можно проводить с помощью CLUSTALW (Thompson, J.D., Higgins, D.G. and Gibson, T.J. (1994) CLUSTALW: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, positions-specific gap penalties and weight matrix choice. Nucleic Acids Research, 22:4673-4680, http://www-igbmc.u-strasbg.fr/BioInfo/ClustalW/Top.html) или T-COFFEE (Cedric Notredame, Desmond G. Higgins, Jaap Heringa, T-Coffee: A novel method for fast and accurate multiple sequence alignment, J. Mol. Biol. (2000) 302: 205-217)) или PILEUP, который использует прогрессирующие попарные выравнивания (Feng and Doolittle, 1987, J. Mol. Evol. 25, 351).
Доступны паттерн-распознающие программные приложения для поиска мотивов или последовательностей сигнатур. Например, MEME (Multiple Em for Motif Elicitation) находит мотивы и последовательности сигнатур в наборе последовательностей и MAST (Motif Alignment and Search Tool) использует эти мотивы для идентификации сходных или одинаковых мотивов в последовательностях запроса. Результаты MAST предоставляют в качестве серии выравниваний с соответствующими статистическими данными и визуальным обзором найденных мотивов. MEME и MAST были разработаны в University of California, San Diego.
PROSITE (Bairoch and Bucher, 1994, Nucleic Acids Res. 22, 3583; Hofmann et al., 1999, Nucleic Acids Res. 27, 215) представляет собой способ идентификации функций неохарактеризованных белков, транслированных с геномных последовательностей или последовательностей кДНК. База данных PROSITE (www.expasy.org/prosite) содержит биологически значимые паттерны и профили и организована так, что ее можно использовать с соответствующими вычислительными инструментами для отнесения новой последовательности к известному семейству белков или для определения того, какой известный домен(ы) присутствует в последовательности (Falquet et al., 2002, Nucleic Acids Res. 30, 235). Prosearch представляет собой инструмент, который может осуществлять поиск в базах данных SWISS-PROT и EMBL с использованием данного паттерна или сигнатуры последовательности.
Способы выделения полипептидов
Полипептиды по изобретению или используемые в способах по изобретению, включая варианты полипептидов, можно получать с использованием способов синтеза пептидов, хорошо известных в данной области, таких как прямой синтез пептидов с использованием твердофазных способов (например Stewart et al., 1969, Solid-Phase Peptide Synthesis, WH Freeman Co, San Francisco California) или автоматизированного синтеза, например с использованием Applied Biosystems 431A Peptide Synthesizer (Foster City, California). Также в процессе такого синтеза можно получать мутантные формы полипептидов.
Полипептиды и варианты полипептидов по изобретению или используемые в способах по изобретению также можно очищать из природных источников с использованием различных способов, которые хорошо известны в данной области (например, Deutscher, 1990, Ed, Methods in Enzymology, Vol. 182, Guide to Protein Purification).
Альтернативно полипептиды и варианты полипептидов по изобретению или используемые в способах по изобретению можно экспрессировать рекомбинантными способами в подходящих клетках-хозяевах и отделять от клеток, как описано ниже.
Способы получения конструкций и векторов
Генетические конструкции по настоящему изобретению содержит одну или несколько полинуклеотидных последовательностей по изобретению и/или полинуклеотидов, кодирующих полипептиды по изобретению, и они могут быть пригодными для трансформации, например, организмов бактерий, грибов, насекомых, млекопитающих или растений. Генетические конструкции по изобретению включают экспрессирующие конструкции, как определено в настоящем описании.
Способы получения и применения генетических конструкций и векторов хорошо известны в данной области и описаны, главным образом, в Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Ed. Cold Spring Harbor Press, 1987; Ausubel et al., Current Protocols in Molecular Biology, Greene Publishing, 1987).
Способы получения клеток-хозяев, содержащих полинуклеотиды, конструкции или векторы
Изобретение относится к клетке-хозяину, которая содержит генетическую конструкцию или вектор по изобретению.
Клетки-хозяева, содержащие генетические конструкции, такие как экспрессирующие конструкции, по изобретению являются пригодными в способах, хорошо известных в данной области (например, Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Ed. Cold Spring Harbor Press, 1987; Ausubel et al., Current Protocols in Molecular Biology, Greene Publishing, 1987) для рекомбинантной продукции полипептидов по изобретению. Такие способы могут вовлекать культивирование клеток-хозяев в подходящей среде в условиях, подходящих для или способствующих экспрессии полипептида по изобретению. Затем экспрессированный рекомбинантный полипептид, который может необязательно секретироваться в культуру, можно отделять от среды, клеток-хозяев или культуральной среды способами, хорошо известными в данной области (например, Deutscher, Ed, 1990, Methods in Enzymology, Vol 182, Guide to Protein Purification).
Способы получения клеток растений и растений, содержащих конструкции и векторы
Кроме того, изобретение относится к клеткам растений, которые содержат генетическую конструкцию по изобретению, и к клеткам растений, модифицированным для изменения экспрессии полинуклеотида или полипептида по изобретению или используемых в способах по изобретению. Растения, содержащие такие клетки, также являются аспектом изобретения.
Способы трансформации клеток растений, растений и их частей полипептидами описаны в Draper et al., 1988, Plant Genetic Transformation and Gene Expression. A Laboratory Manual. Blackwell Sci. Pub. Oxford, p. 365; Potrykus and Spangenburg, 1995, Gene Transfer to Plants. Springer-Verlag, Berlin.; и Gelvin et al., 1993, Plant Molecular Biol. Manual. Kluwer Acad. Pub. Dordrecht. Обзор трансгенных растений, включая способы трансформации, предоставлен в Galun and Breiman, 1997, Transgenic Plants. Imperial College Press, London.
Способы генетического манипулирования растениями
Доступен ряд стратегий трансформации растений (например, Birch, 1997, Ann Rev Plant Phys Plant Mol Biol, 48, 297, Hellens RP, et al (2000) Plant Mol Biol 42: 819-32, Hellens R et al Plant Meth 1: 13). Например, стратегии можно разрабатывать для увеличения экспрессии полинуклеотида/полипептида в клетке растения, органе и/или на конкретной стадии развития где/когда он обычно экспрессируется, или для эктопической экспрессии полинуклеотида/полипептида в клетке, ткани, органе и/или на конкретной стадии развития, в которых/когда он в норме не экспрессируется. Экспрессируемый полинуклеотид/полипептид может происходить из вида растений, подлежащего трансформации, или он может происходить из отличающегося вида растений.
Генетические конструкции для экспрессии генов в трансгенных растениях обычно включают промоторы для запуска экспрессии одного или нескольких клонированных полинуклеотидов, терминаторы и последовательности селективных маркеров для обнаружения присутствия генетической конструкции в трансформированном растении.
Промоторы, пригодные для применения в генетических конструкциях, могут быть функциональными в клетке, ткани или органе однодольного или двудольного растения, и включают промоторы, специфические для клетки, ткани и органа, промоторы, специфические для клеточного цикла, временные промоторы, индуцибельные промоторы, конститутивные промоторы, которые являются активными в большинстве тканей растений, и рекомбинантные промоторы. Выбор промотора зависит от желаемой временной и пространственной экспрессии клонированного полинуклеотида. Промоторы могут представлять собой промоторы, обычно ассоциированные с представляющим интерес трансгеном, или промоторы, которые происходят из генов других растений, вирусов и патогенных для растений бактерий и грибов. Специалисты в данной области будут способны без излишнего экспериментирования выбрать промоторы, которые являются пригодными для применения для модификации и модулирования признаков растений с использованием генетических конструкций, содержащих полинуклеотидные последовательности по изобретению. Примеры конститутивных промоторов растений включают промотор CaMV 35S, промотор нопалинсинтазы и промотор октопинсинтазы и промотор Ubi 1 из кукурузы. Промоторы растений, которые являются активными в конкретных тканях, отвечают на внутренние сигналы развития или внешние абиотические или биотические стрессовые факторы, описаны в научной литературе. Иллюстративные промоторы описаны, например, в WO 02/00894, которая включена в настоящее описание в качестве ссылки.
Иллюстративные терминаторы, часто используемые в генетической конструкции для трансформации растений, включают, например, терминатор вируса мозаики цветной капусты (CaMV) 35S, терминаторы нопалинсинтазы или октопинсинтазы Agrobacterium tumefaciens, терминатор гена зеина Zea mays, терминатор пирофосфорилазы ADP-глюкозы Oryza sativa терминатор PI-II Solanum tuberosum.
Селективные маркеры, часто используемые для трансформации растений, включают ген неомицинфосфотрансферазы II (NPT II), который сообщает устойчивость к канамицину, ген aadA, который сообщает устойчивость к спектиномицину и стрептомицину, ген фосфинотрицинацетилтрансферазы (ген bar) для устойчивости к Ignite (AgrEvo) и Basta (Hoechst), и ген гигромицинфосфотрансферазы (hpt) для устойчивости к гигромицину.
Также предусматривается применение генетических конструкций, содержащих репортерные гены (кодирующие последовательности, которые проявляют активность, являющуюся чужеродной для хозяина, обычно ферментативную активность и/или видимый сигнал (например, люцифераза, GUS, GFP), которые можно использовать для анализа экспрессии с промотора в растениях и тканях растений. Репортерные гены рассмотрены в Herrera-Estrella et al., 1993, Nature 303, 209, и Schrott, 1995, Gene Transfer to Plants (Potrykus, T., Spangenberg. Eds) Springer Verlag. Berline, pp. 325-336.
Сайленсинг генов
Стратегии трансформации могут быть предназначены для снижения экспрессии полинуклеотида/полипептида в клетке, ткани, органе растения или на конкретной стадии развития, в которых/когда они обычно экспрессируются. Такие стратегии известны как стратегии сайленсинга генов.
Стратегии сайленсинга генов могут быть сфокусированы на самом гене или на регуляторных элементах, которые обеспечивают экспрессию кодируемого полипептида. "Регуляторные элементы" используют в настоящем описании в наиболее широком возможном значении, и они включают другие гены, которые взаимодействуют с представляющим интерес геном.
Генетические конструкции, сконструированные для снижения или сайленсинга экспрессии полинуклеотида/полипептида по изобретению, могут включать антисмысловую копию полинуклеотида по изобретению. В таких конструкциях полинуклеотид помещают в антисмысловой ориентации в отношении промотора и терминатора.
"Антисмысловой" полинуклеотид получают путем инвертирования полинуклеотида или сегмента полинуклеотида таким образом, чтобы продуцируемый транскрипт был комплементарен мРНК-транскрипту гена, например,
Генетические конструкции, сконструированные для сайленсинга генов, также могут включать инвертированный повтор. "Инвертированный повтор" представляет собой последовательность, которая повторяется, где вторая половина повтора находится на комплементарной цепи, например,
5'-GATCTA………TAGATC-3'
3'-CTAGAT………ATCTAG-5'
Образованный транскрипт может претерпевать образование комплементарных пар оснований с формированием шпилечной структуры. Обычно для возможности образования шпильки требуется спейсер размером по меньшей мере 3-5 п.н. между повторяющимися областями. Конструкции, включающие такие последовательности инвертированных повторов, можно использовать в РНК-интерференции (РНК-i) и, таким образом, они могут упоминаться как конструкции РНК-i.
Другой подход сайленсинга вовлекает применение малой антисмысловой РНК, нацеленной на транскрипт, эквивалентный миРНК (Llave et al., 2002, Science 297, 2053). Применение такой малой антисмысловой РНК, соответствующей полинуклеотиду по изобретению, прямо предусматривается.
Термин "генетическая" конструкция, как используют в рамках изобретения, также включает малые антисмысловые РНК и другие такие полипептиды, обеспечивающие сайленсинг генов.
Трансформация экспрессирующей конструкцией, как определено в настоящем описании, также может приводить к сайленсингу генов посредством процесса, известного как смысловое подавление (например, Napoli et al., 1990, Plant Cell 2, 279; de Carvalho Niebel et al., 1995, Plant Cell, 7, 347). В некоторых случаях смысловое подавление может вовлекать сверхэкспрессию целой или частичной кодирующей последовательности, но также может вовлекать экспрессию некодирующей области гена, такой как интрон или 5'- или 3'-нетранслируемая область (UTR). Химерные частичные смысловые конструкции можно использовать для координационного сайленсинга множества генов (Abbott et al., 2002, Plant Physiol. 128(3): 844-53; Jones et al., 1998, Planta 204: 499-505). Использование таких стратегий смыслового подавления для сайленсинга экспрессии полинуклеотида по изобретению также предусматривается.
Полинуклеотидные вставки в генетических конструкциях, предназначенных для сайленсинга генов, могут соответствовать кодирующей последовательности и/или некодирующей последовательности, такой как промотор, и/или интрон, и/или последовательность 5'- или 3'-UTR, соответствующего гена.
Другие стратегии сайленсинга генов включают доминантно-негативные подходы и использование конструкций рибозимов (McIntyre, 1996, Transgenic Res, 5, 257).
Претранскрипционный сайленсинг может быть достигнут посредством мутации самого гена или его регуляторных элементов. Такие мутации могут включать точковые мутации, сдвиг рамки считывания, инсерции, делеции и замены.
Протоколы трансформации
Ниже представлены репрезентативные публикации, в которых описаны протоколы генетической трансформации, которые можно использовать для генетической трансформации следующих видов растений: рис (Alam et al., 1999, Plant Cell Rep. 18, 572); яблоня (Yao et al., 1995, Plant Cell Reports 14, 407-412); кукуруза (патенты США с серийными номерами № 5177010 и 5981840); пшеница (Ortiz et al., 1996, Plant Cell Rep. 15, 1996, 877); томат (патент США с серийным номером № 5159135); картофель (Kumar et al., 1996 Plant J. 9: 821); маниока (Li et al., 1996 Nat. Biotechnology 14, 736); салат-латук (Michelmore et al., 1987, Plant Cell Rep. 6, 439); табак (Horsch et al., 1985, Science 227, 1229); хлопок (патенты США с серийными номерами № 5846797 и 5004863); травы (патенты США № 5187073 и 6020539); мята перечная (Niu et al., 1998, Plant Cell Rep. 17, 165); цитрусовые растения (Pena et al., 1995, Plant Sci.104, 183); тмин (Krens et al., 1997, Plant Cell Rep, 17, 39); банан (патент США с серийным номером № 5792935); соя (патенты США № 5416011; 5569834; 5824877; 556304455 и 5968830); ананас (патент США с серийным номером № 5952543); тополь (патент США № 4795855); однодольные растения в целом (патенты США № 5591616 и 6037522); капуста декоративная (патенты США № 5188958; 5463174 и 5750871); зерновые (патент США № 6074877); груша (Matsuda et al., 2005, Plant Cell Rep. 24(1):45-51); слива (Ramesh et al., 2006 Plant Cell Rep. 25(8):821-8; Song and Sink 2005 Plant Cell Rep. 2006; 25(2): 117-23; Gonzalez Padilla et al., 2003 Plant Cell Rep.22(1): 38-45); клубника (Oosumi et al., 2006 Planta. 223(6): 1219-30; Folta et al., 2006 Planta Apr 14; PMID: 16614818), роза (Li et al., 2003), рубус (Graham et al., 1995 Methods Mol Biol. 1995; 44: 129-33), томат (Dan et al., 2006, Plant Cell Reports V25: 432-441), яблоня (Yao et al., 1995, Plant Cell Rep. 14, 407-412) и Actinidia eriantha (Wang et al., 2006, Plant Cell Rep. 25.5: 425-31), белая береза (Keinonen-Mettala et al., 1998, Plant Cell Rep. 17: 356-361) и осина (Nilsson O, et al., 1992, Transgenic Research. 1: 209-220). Также изобретение предусматривает трансформацию других видов. Подходящие способы и протоколы доступны в научной литературе.
Для изменения выраженности активности нуклеотида и/или полипептида по изобретению можно использовать несколько других способов, известных в данной области. Такие способы включают, но не ограничиваются ими, Tilling (Till et al., 2003, Methods Mol Biol, 2%, 205), так называемую технологию "Deletagene" (Li et al., 2001, Plant Journal 27(3), 235) и использование искусственных факторов транскрипции, таких как синтетические факторы транскрипции с цинковыми пальцами (например, Jouvenot et al., 2003, Gene Therapy 10, 513). Кроме того, также для модулирования активности этого полипептида в растениях можно экспрессировать антитела или их фрагменты, нацеленные на конкретный полипептид, (Jobling et al., 2003, Nat. Biotechnol., 21(1), 35). Также можно использовать подходы маркирования транспозонов. Кроме того, пептиды, взаимодействующие с полипептидом по изобретению, можно идентифицировать способами, такими как фаговый дисплей (Dyax Corporation). Такие взаимодействующие пептиды можно экспрессировать в или наносить на растение для достижения активности полипептида по изобретению. Применение каждого из описанных выше подходов для изменения экспрессии нуклеотида и/или полипептида по изобретению прямо предусматривается.
Термины "для изменения экспрессии" и "измененная экспрессия" полинуклеотида или полипептида по изобретению или используемые в способах по изобретению, охватывают ситуацию, где геномная ДНК, соответствующая полинуклеотиду по изобретению, модифицирована таким образом, чтобы обеспечить измененную экспрессию полинуклеотида или полипептида по изобретению. Модификация геномной ДНК может быть осуществлена посредством генетической трансформации или других способов, известных в данной области, для индукции мутаций. "Измененная экспрессия" может относиться к увеличению или снижению количества матричной РНК и/или продуцируемого полипептида, а также может приводить к измененной активности полипептида вследствие изменений последовательности продуцируемого полинуклеотида и полипептида.
Способы селекции растений
Также предусматриваются способы селекции растений с увеличенной биомассой корней. Такие способы вовлекают исследование растений в отношении измененной экспрессии по меньшей мере одного полинуклеотида или полипептида PEAPOD, включая полинуклотиды или полипептиды, как определено или описано в настоящем описании. Такие способы можно использовать в раннем возрасте или на ранней стадии развития, когда характеристики увеличенной биомассы корней не обязательно могут быть легко поддающимися измерению.
Экспрессию полинуклеотида, такого как матричная РНК, часто используют в качестве индикатора экспрессии соответствующего полинуклеотида. Иллюстративные способы измерения экспрессии полинуклеотида включают, но не ограничиваются ими, нозерн-анализ, ОТ-ПЦР и дот-блоттинг (Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Ed. Cold Spring Harbor Press, 1987). Таким образом, полинуклеотиды или части полинуклеотидов по изобретению являются пригодными в качестве зондов или праймеров, как определено в настоящем описании, в способах идентификации растений с увеличенной биомассой корней. Полинуклеотиды по изобретению или описанные в настоящем описании можно использовать в качестве зондов в экспериментах по гибридизации или в качестве праймеров в экспериментах на основе ПЦР, сконструированных для идентификации таких растений.
Альтернативно антитела можно индуцировать против полипептидов PEAPOD, как описано или раскрыто в настоящем описании. Способы индукции и применения антител являются стандартными в данной области (см., например: Antibodies, A Laboratory Manual, Harlow A Lane, Eds, Cold Spring Harbour Laboratory, 1998). Такие антитела можно использовать в способах обнаружения измененной экспрессии таких полипептидов. Такие способы могут включать ELISA (Kemeny, 1991, A Practical Guide to ELISA, NY Pergamon Press) и вестерн-анализ (Towbin & Gordon, 1994, J Immunol Methods, 72, 313).
Эти подходы к анализу экспрессии полинуклеотидов или полипептидов и селекции растений с увеличенной биомассой корней являются пригодными в общепринятых программах разведения, предназначенных для получения сортов с такими измененными характеристиками.
Растения
Термин "растение" включает целое растение, любую часть растения, сеянцы и потомство растения.
Термин "сеянец" означает любую часть растения, которую можно использовать для воспроизведения или размножения, либо полового, либо бесполого, включая семена и черенки.
Растения по изобретению можно выращивать и либо подвергать самоопылению, либо скрещивать с другой линией растений, и можно идентифицировать полученные гибриды с желаемыми фенотипическими характеристиками. Можно выращивать два или более поколений, чтобы убедиться, что рассматриваемые фенотипические характеристики стабильно поддерживаются и наследуются. Растения, являющиеся результатом таких стандартных подходов к разведению, также составляют аспект настоящего изобретения.
Контроль роста и развития растений гиббереллинами (GA), брассиностероидами (BR) и другими гормонами растений
Гиббереллины (GA) и брассиностероиды (BR) представляют собой два класса гормонов растений; они вовлечены во многие аспекты морфогенеза и роста растений, включая: прорастание семян, удлинение клеток, развитие сосудов, размер семян, спрямленность листьев, цветение, старение листьев и плодов (Mathew et al 2009, NZJAR 52, 213-225; Hou et al 2010, Developmental Cell 19, 884-894; Jiang and Lin 2013, Plant Signaling and Behaviour 8:10, e25928).
Учитывая их роль в развитии растений, способность манипулировать уровнями либо GA, либо BR, или их нижеследующими мишенями является высокожелательной с точки зрения повышения как выхода, так и качества, во многих видах растений. Действительно существуют некоторые коммерческие примеры, когда экзогенные применения любого из этих гормонов используют для повышения агрономической ценности.
GA можно применять на пастбищах ржи для стимуляции несезонного роста, а также для ускорения цветения (Mathew et al 2009, NZJAR 52, 213-225), также ее можно использовать для противодействия неблагоприятным эффектам более холодных температур на сахарный тростник (тропическая трава C4). GA также используют для увеличения размера плодов бессемянного винограда и вишни, для стимуляции завязывания плода яблони и груши и для замедления старения кожуры конкретных цитрусовых культур (Sun 2011, Current Biology 21, R338-R345). Аналогично, препараты BR рекомендуют для повышения выхода культуры и улучшения качества томата, картофеля, огурца, перца и ячменя, риса, кукурузы, пшеницы, хлопка и табака (Prusakova et al 1999, Agrarian Russia, 41-44; Khripach et al 2000, Annals of Botany 86, 441-447; Anjum et al 2011, J. Agronomy Crop Sci. 197, 177-185; Vardhini 2012, J. Phytology 4, 1-3). Однако невысокое внедрение коммерчески применяемых брассиностероидов может отражать стоимость и тот факт, что растения не абсорбируют стероиды эффективно, когда их применяют экзогенно. Кроме того, необходимость жестко контролировать время и концентрацию экзогенно предоставляемых GA и BR ограничивает их применение.
По большей части биосинтез GA и BR и катаболитческие каскады покрытосеменных растений были охарактеризованы и включают отрицательные регуляторы и нижерасположенные мишени факторов транскрипции. При связывании GA или BR с их соответствующим рецептором возникает комплексный сигнальный каскад и в обоих случаях центральная точка регуляции вовлекает каскад убиквитин-протеасома, изменяющий уровень негативного регулятора DELLA (в случае GA) и регулятора транскрипции BZR1 (в случае BR).
Удаление белков DELLA приводит к устранению репрессии роста и стимуляции отвечающего на GA роста и развития. Напротив, обнаружение BR приводит к накоплению нефосфорилированного белка BZR1 в ядре. Дефосфорилирование BZR1 препятствует его деградации протеасомой и вместо этого позволяет связывание BZR1 с другими ДНК-связывающими факторами транскрипции и взаимодействие с кофакторами транскрипции. Это приводит к регуляции тысяч генов, вовлеченных в рост и другие клеточные процессы, включая ингибирование экспрессии генов биосинтеза BR (He et al 2005, Science 307, 1634-1638; Guo et al 2013, Current Opinion Plant Biol. 16, 545-553).
Существует ряд эндогенных сигналов и внешних стимулов, которые влияют на регуляторный модуль GA-GID1-DELLA, в котором DELLA интегрирует различные сигнальные виды активности посредством прямого белок-белкового взаимодействия с множеством ключевых регуляторных белков из других каскадов. По существу белки DELLA являются ведущими репрессорами роста, которые контролируют рост и развитие растений посредством встраивания внутренних сигналов из каскадов других гормонов (ауксин, абсцизовая кислота, жасмоновая кислота и этилен) и внешних биотических (патоген) и абиотических (условия освещения, холодовые и солевые стрессовые факторы) факторов (Sun 2011, Current Biology 21, R338-R345). Засуха является одним из наиболее важных средовых ограничений роста растений и сельскохозяйственной продуктивности. Неожиданно существует положительная корреляция между увеличенной толерантностью к засухе и более мощной корневой системой, включающей более глубокие корни и более латеральные корни, оба из которых обеспечивают освоение почвы и получение подземных ресурсов (Yu et al 2008, Plant Cell 20, 1134-1151; Werner et al 2010, Plant Cell 22, 3905-3920). Таким образом следует, что обычной сельскохозяйственной целью является оптимизация архитектуры корневой системы, чтобы помочь преодолеть ограничения урожая сельскохозяйственных растений, вызванные нехваткой воды или питательных веществ. Однако из всех абиотических стрессовых факторов, которые уменьшают продуктивность культуры, засуха является наиболее разрушительным и наиболее неподатливым усилиям селекционеров. Классические подходы разведения являются трудными, поскольку этот признак управляется многими генами и его трудно оценить (Werner et al 2010, Plant Cell 22, 3905-3920). В то время как селекция с помощью маркеров (MAS), локусы количественных признаков (QTL) и другие геномные подходы широко используют для облегчения усилий по разведению для получения устойчивых к засухе сортов (Tuberosa and Salvi, 2006, Trends in Plant Science, 11:405-412), система ограничена варьированием, присутствующим в подвергаемой скринингу популяции.
Интересно, что рис имеет только один белок DELLA (SLR1), кукуруза имеет два (d8 и d9) (Lawit et al 2010, Plant Cell Physiol 51, 1854-1868), в то время как арабидопсис имеет пять (GA1, RGA, RGL1, RGL2 и RGL3) (Achard and Genschik 2009, J. Exp. Bot. 60, 1085-1092). Более того, в последнем филогенетическом анализе Chen et al 2013 обнаружили, что пять из шести видов трав, которые они проанализировали, имели только один DELLA, в то время как 14 из 18 двудольных видов имели два или более белков DELLA. Напротив, существует 6 представителей семейства BZR в рисе, 10 в кукурузе (www.Grassius.org) и 6 в арабидопсисе (Wang et al 2002, Developmental Cell 2, 505-513).
Рост и развитие растений основаны на многочисленных связях между каскадами передачи сигнала, которые обеспечивают высокую пластичность при развитии, требуемую вследствие их сидячего образа жизни (Gallego-Bartolome et al 2012, PNAS 109, 13446-13451). Таким образом, вместо существования каждого каскада передачи сигнала гормонами в качестве обособленного модуля, современные данные указывают на то, что существует высокая степень взаимодействия между различными каскадами, и что данный гормон часто модулирует выход, обеспечиваемый остальными. В качестве примера, недавно было показано, что перекрестное взаимодействие между каскадами передачи сигнала GA и BR вовлекает прямое взаимодействие между DELLA и BZR1/BES1, посредством чего белки DELLA не только влияют на стабильность белка, но также ингибируют транскрипционную активность BZR1 (Li and He 2013, Plant Signaling and Behaviour 8:7, e24686 и ссылки в ней). Таким образом стимуляция удлинения клеток посредством GA частично происходит через устранение опосредуемого DELLA ингибирования BZR1.
Недавно было продемонстрировано, что рост и развитие растений можно модифицировать путем прямого манипулирования основными регуляторами роста DELLA (Lawit, Kundu, Rao and Tomes, 2007, ISOLATED POLYNUCLEOTIDE MOLECULES CORRESPONDING TO MUTANT AND WILD-TYPE ALLELES OF THE MAIZE D9 GENE AND METHODS OF USE, WO 2007124312 A2) и BZR1 (Chory and Wang, 2005, GENES INVOLVED IN BRASSINOSTEROID HORMONE ACTION IN PLANTS, US 6921848 B2).
Стероидные гормоны играют ключевую роль в координации широкого диапазона обусловленных развитием и физиологических процессов как в растениях, так и у животных (Thummel and Chory 2002, Genes Dev. 16, 3113-3129). В растениях стероидный гормон брассиностероид (BR) имеет обширные эффекты на рост, развитие и ответы как на биотические, так и на абиотические стрессовые факторы (Zhu et al 2013, Development 140, 1615-1620; Clouse 2011, Plant Cell 23, 1219-1230). В противоположность передаче сигнала стероидными гормонами животных, которые функционируют через ядерные рецепторы, в растениях BR связывается с внеклеточным доменом рецепторной киназы клеточной поверхности BRASSINOSTEROID INSENSITIVE 1 (BRI1) и активирует каскад внутриклеточной передачи сигнала, который регулирует экспрессию генов (Clouse 2011, Plant Cell 23, 1219-1230; Kinoshita et al 2005, Nature 433, 167-171). Существует множество стадий, вовлекающих активацию и инактивацию промежуточных соединений, приводящих к фосфорилированию двух факторов транскрипции BRASSINAZOLE RESISTANT 1 (BZR1) и BZR2 (также известный как BES1). Таким образом факторы транскрипции BZR передачи сигнала являются компонентами-мишенями, конвертирующими передачу сигнала в отвечающую на BR экспрессию генов.
Установлен паттерн в передаче сигнала гормонами растений, где факторы транскрипции-мишени, активируемые гормонами, также негативно регулируются конкретными репрессорными комплексами. Например, в каскадах передачи сигнала жасмоновой кислоты (JA), ауксина, абсцизовой кислоты (ABA) и стриголактона (SL) факторы транскрипции-мишени негативно регулируются репрессорными комплексами, использующими TOPLESS (TPL) в качестве общего корепрессора, привлекаемого специфическим для каскада гормонов репрессором (Pauwels et al 2010, Nature 464, 788-791). В каскаде передачи JA семейство репрессоров транскрипции JASMONATE ZIM DOMAIN (JAZ) как взаимодействует с отвечающим на JA активатором транскрипции MYC2, являющимся мишенью, так и привлекает TPL, либо прямо, либо через адаптерный белок NOVEL INTERACTOR OF JAZ (NINJA) (Pauwels et al 2010 Nature 464, 788-791).
Таким образом, способность регулировать каскады GA и BR для влияния на многие различные представляющие интерес сельскохозяйственные признаки имеет большое значение для коммерческого сельского хозяйства.
Изобретение заявителя
Как рассмотрено выше, настоящее изобретение относится к способу увеличения биомассы корней в растениях посредством эктопической экспрессии PEAPOD.
Без связи с теорией, заявители показали, что PEAPOD (PPD), по-видимому, вовлечен в модулирование каскадов как GA, так и BR, и посредством либо прямого, либо непрямого взаимодействия с основными регуляторами роста DELLA и BZR.
Анализ первичной аминокислотной структуры белков PPD указывает на присутствие высококонсервативного специфического для растений домена, присутствующего только в этих белках. Гомологи PPD существуют в широком диапазоне эвдикот, хвойных и некоторых однодольных растений (пальмы, банан, орхидеи, ряска), но не Poaceae (травы).
Гены PPD в Arabidopsis кодируют белки, которые являются представителями специфического для растений семейства TIFY, названного по мотиву TIF[F/Y]XG, находящемуся в домене, известном как ZIM (Vanholme et al 2007 Trends Plant Sci. 12, 239-244). Два белка PPD арабидопсиса, PPD1 и PPD2, включены в ту же группу класса II TIFY, что и двенадцать хорошо охарактеризованных белков JAZ, которые действуют в качестве репрессоров ответов на жасмонаты. Однако белки PPD и другой не являющийся JAZ белок в группе, по-видимому, не вовлечены в ответы на передачу сигнала гормонов на жасмонаты (Pauwels et al 2010 Nature 464, 788-791).
Вновь, без связи с теорией, заявители полагают, что увеличение биомассы корней согласно изобретению опосредуется новым механизмом регуляции каскадов как GA, так и BR, с использованием гена PPD. Примеры 3 и 4 ниже подтверждают это предположение.
Также можно широко сказать, что настоящее изобретение состоит в частях, элементах и признаках, упоминаемых или указанных в описании заявки, по отдельности или в совокупности, и в любых или всех комбинациях любых двух или более указанных частей, элементов или признаков, и где в настоящем описании упомянуты конкретные целые числа, которые имеют известные эквиваленты в области, к которой относится изобретение, такие эквиваленты считаются в включенными в настоящее описания, как если бы они были индивидуально указаны.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Настоящее изобретение станет более понятным с помощью прилагаемых чертежей, которые описывают следующее:
На фиг.1 представлены 46 аминокислотных остатков, содержащих область PEAPOD из ряда видов растений, идентичные остатки показаны звездочкой.
На фиг.2 представлены внутренние 27 аминокислотных остатков в области PEAPOD из ряда видов растений, идентичные остатки показаны звездочкой.
На фиг.3 представлены 6 аминокислотных остатков домена TIFY на белках PEAPOD из ряда видов растений, идентичные остатки показаны звездочкой.
На фиг.4 показано схематическое представление белка PPD и приблизительное положение консервативных областей PPD, TIFY и Jas*.
На фиг.5 представлена димеризация PPD и взаимодействие между TPL и NINJA в анализах Y2H.
На фиг.6 представлено взаимодействие между PPD и NINJA и взаимодействие между TPL и BZR1 в анализах Y2H.
На фиг.7 представлено взаимодействие PPD, NINJA, TPL и BZR1 в молодых (A и B) и старых (C) листьях с использованием анализов BiFC.
На фиг.8 показано схематическое представление комплекса PPD-NINJA-TPL-BZR1.
На фиг.9 показано взаимодействие между PPD и BZR1 в анализах Y2H.
На фиг.10 представлен ответ длины гипокотиля растений дикого типа, мутантов Δppd и сверхэкспрессирующих PEAPOD (PPD-OX) растений на экзогенное применение GA и PAC.
На фиг.11 показано увеличение роста корней растений плевела, сверхэкспрессирующих PPD (PPD-OX) из Arabidopsis thaliana или PEAPOD из Abroella trichopoda, по сравнению с диким типом и контролем вектора.
На фиг.12 представлено увеличение длины корней растений люцерны, сверхэкспрессирующих PPD (PPD-OX), по сравнению с контрольными растениями.
На фиг.13 показано, что белки PEAPOD из Arabidopsis thaliana; Picea sitchensis, Amborella trichopoda, Musa acuminate, Trifolium repens и Selaginella moellendorffii являются функционально эквивалентными. Оптимизированную кодирующую последовательность PEAPOD из каждого из них использовали для комплементации мутанта с делецией PEAPOD Δppd Arabidopsis thaliana (экотип Landsberg erecta). Изображения проростков получали на эквивалентных стадиях развития.
ПРИМЕРЫ
Далее изобретение проиллюстрировано с помощью следующих неограничивающих примеров.
Пример 1: Охарактеризация генов PEAPOD в множестве видов растений
Для идентификации ортологов гена PPD в других видах растений консервативную область PPD (46 аминокислот) из гена PPD1 арабидопсиса (SEQ ID NO: 27) использовали для поиска в общедоступных базах данных последовательностей генов растений с использованием программ поиска TBLASTN и BLASTP (Altschul et al 1990). Последовательности PEAPOD были идентифицированы в многообразном диапазоне видов растений, включая мхи, хвойные растения, все отряды исследованных двудольных растений и некоторые из отрядов однодольных растений, включая: пальмы, банан, орхидеи и ряску. Последовательности PEAPOD не обнаруживаются в травах. Репрезентативные последовательности белка и нуклеиновой кислоты PEAPOD представлены в SEQ ID NO: 1-26 и SEQ ID NO: 83-107, соответственно.
Область PEAPOD из 46 аминокислот из PPD1 Arabidopsis thaliana представлена в SEQ ID NO: 27. Эту область из полипептидов SEQ ID NO: 1-26 выравнивали с помощью Vector NTI (VNTI), как показано на фиг.1.
В SEQ ID NO: 28 показана консенсусная последовательность для этой области PPD из 46 аминокислот. В SEQ ID NO: 29 показана та же консенсусная область, однако показано, какие аминокислоты могут присутствовать в каждом из вариабельных положений.
Подпоследовательность из 27 аминокислот из области PEAPOD из 46 аминокислот из PPD1 Arabidopsis thaliana представлена в SEQ ID NO: 30.
Выравнивание этой подпоследовательности из 27 аминокислот для достижения тех же последовательностей, что и на фиг.1, представлено на фиг.2.
В SEQ ID NO: 30 представлена консенсусная последовательность для этой области PPD из 27 аминокислот. В SEQ ID NO: 32 представлена та же консенсусная область, однако показано, какие аминокислоты могут присутствовать в каждом из вариабельных положений.
В каждой из последовательностей пептидов PPD SEQ ID NO: 1-26 также присутствует консервативный мотив TIFY, который располагается после области PPD из 46 аминокислот. Количество аминокислотных остатков, разделяющих C-конец области PPD и N-конец мотива TIFY, зависит от источника PPD; например, количество варьируется от 46 до 140 аминокислот для SEQ ID NO: 1-26.
В SEQ ID NO: 33 показана последовательность PPD1 арабидопсиса на протяжении мотива TIFY. Выравнивание мотива TIFY (как описано Vanholme et al 2007 Trends Plant Sci. 12, 239-244) из SEQ ID NO: 1-26 представлено на фиг.3.
В SEQ ID NO: 34 представлена консенсусная последовательность для этого мотива TIFY из 6 аминокислот. В SEQ ID NO: 35 представлена та же консенсусная область, но показано, какие аминокислоты могут находиться в каждом из вариабельных положений.
Полностью консервативные остатки в доменах PPD и TIFY обозначены звездочками на фиг.1-3.
Заявители утверждают, что эти области и мотивы, описанные выше, обнаруживаются во всех идентифицированных белках PEAPOD и являются диагностическими для таких белков PEAPOD.
Пример 2: Демонстрация функциональности PEAPOD для последовательностей PEAPOD из множества видов растений
Функциональность любой последовательности PEAPOD можно подтверждать посредством комплементации мутантного фенотипа Δppd листьев арабидопсиса. Комплементацию мутантного фенотипа Δppd листьев арабидопсиса сначала использовали для идентификации гена PPD арабидопсиса (White 2006). Ее наблюдали по восстановлению фенотипа уплощенных листьев дикого типа и нормальной форме розетки в противоположность куполообразных листьев и скручиванию розетки в фенотип "пропеллера".
Последовательности PEAPOD, такие как последовательности SEQ IN NO: 1-26 (включая: пальму, хвойные растения, мох, орхидею и другие виды двудольных растений) или любую другую последовательность PEAPOD, подлежащую исследованию, можно трансформировать в мутантный арабидопсис Δppd способами, хорошо известными специалистам в данной области. Пример такого способа описан ниже.
Клонирование и генные конструкции
Получение конструкции CaMV35s::PPD1 Arabidopsis thaliana для сверхэкспрессии PPD1 арабидопсиса в мутантном арабидопсисе Δppd
Экспрессирующую конструкцию синтезировали для обеспечения сверхэкспрессии PPD1 Arabidopsis thaliana под контролем промотора CaMV35s (SEQ ID NO: 129) в мутантном арабидопсисе Δppd. ORF PPD оптимизировали для экспрессии в арабидопсисе; это включало модифицированную последовательность Joshi (Joshi 1997, Nucleic Acid Research 15, 6643-6653), оптимизацию кодонов, устранение нестабильности мРНК, удаление сигнальных последовательностей поли-A, удаление скрытых участков сплайсинга, добавление удаляемого посредством BamHI C-концевого эпитопа V5 и концевой части в виде His-метки (кодирующей SEQ ID NO: 37) и добавление двойного стоп-кодона. Затем конструкцию (с хвостовой частью и без нее) помещали между промотором CaMV35s и терминатором ocs посредством реакции LR GATEWAY®, которые кодировали SEQ ID NO: 105 и SEQ ID NO: 111, соответственно.
Получение конструкции CaMV35s::PPD Trifolium repens для сверхэкспрессии PPD1 Trifolium repens в мутантном арабидопсисе Δppd
Экспрессирующую конструкцию синтезировали для обеспечения сверхэкспрессии PPD1 Trifolium repens под контролем промотора CaMV35s (SEQ ID NO: 129) в мутантном арабидопсисе Δppd. ORF PPD оптимизировали для экспрессии в арабидопсисе; это включало модифицированную последовательность Joshi (Joshi 1997, Nucleic Acid Research 15, 6643-6653), оптимизацию кодонов, устранение нестабильности мРНК, удаление сигнальных последовательностей поли-A, удаление скрытых участков сплайсинга, добавление удаляемого посредством BamHI C-концевого эпитопа V5 и концевой части в виде His-метки (кодирующей SEQ ID NO: 37) и добавление двойного стоп-кодона. Затем конструкцию (с хвостовой частью и без нее) помещали между промотором CaMV35s и терминатором ocs посредством реакции LR GATEWAY®, которые кодировали SEQ ID NO: 106 и SEQ ID NO: 112, соответственно.
Получение конструкции CaMV35s::PPD Amborella trichopoda для сверхэкспрессии PPD Amborella trichopoda в мутантном арабидопсисе Δppd
Экспрессирующую конструкцию синтезировали для обеспечения сверхэкспрессии PPD1 Amborella trichopoda под контролем промотора CaMV35s (SEQ ID NO: 129) в мутантном арабидопсисе Δppd. ORF PPD оптимизировали для экспрессии в арабидопсисе; это включало модифицированную последовательность Joshi (Joshi 1997, Nucleic Acid Research 15, 6643-6653), оптимизацию кодонов, устранение нестабильности мРНК, удаление сигнальных последовательностей поли-A, удаление скрытых участков сплайсинга, добавление удаляемого посредством BamHI C-концевого эпитопа V5 и концевой части в виде His-метки (кодирующей SEQ ID NO: 37) и добавление двойного стоп-кодона. Затем конструкцию (с хвостовой частью и без нее) помещали между промотором CaMV35s и терминатором ocs посредством реакции LR GATEWAY®, которые кодировали SEQ ID NO: 107 и SEQ ID NO: 113, соответственно.
Получение конструкции CaMV35s::PPD Musa acuminate для сверхэкспрессии PPD Musa acuminate в мутантном арабидопсисе Δppd
Экспрессирующую конструкцию синтезировали для обеспечения сверхэкспрессии PPD1 Musa acuminate под контролем промотора CaMV35s (SEQ ID NO: 129) в мутантном арабидопсисе Δppd. ORF PPD оптимизировали для экспрессии в арабидопсисе; это включало модифицированную последовательность Joshi (Joshi 1997, Nucleic Acid Research 15, 6643-6653), оптимизацию кодонов, устранение нестабильности мРНК, удаление сигнальных последовательностей поли-A, удаление скрытых участков сплайсинга, добавление удаляемого посредством BamHI C-концевого эпитопа V5 и концевой части в виде His-метки (кодирующей SEQ ID NO: 37) и добавление двойного стоп-кодона. Затем конструкцию (с хвостовой частью и без нее) помещали между промотором CaMV35s и терминатором ocs посредством реакции LR GATEWAY®, которые кодировали SEQ ID NO: 108 и SEQ ID NO: 114, соответственно.
Получение конструкции CaMV35s::PPD1 Picea sitchensis для сверхэкспрессии PPD Picea sitchensis в мутантном арабидопсисе Δppd
Экспрессирующую конструкцию синтезировали для обеспечения сверхэкспрессии PPD1 Picea sitchensis под контролем промотора CaMV35s (SEQ ID NO: 129) в мутантном арабидопсисе Δppd. ORF PPD оптимизировали для экспрессии в арабидопсисе; это включало модифицированную последовательность Joshi (Joshi 1997, Nucleic Acid Research 15, 6643-6653), оптимизацию кодонов, устранение нестабильности мРНК, удаление сигнальных последовательностей поли-A, удаление скрытых участков сплайсинга, добавление удаляемого посредством BamHI C-концевого эпитопа V5 и концевой части в виде His-метки (кодирующей SEQ ID NO: 37) и добавление двойного стоп-кодона. Затем конструкцию (с хвостовой частью и без нее) помещали между промотором CaMV35s и терминатором ocs посредством реакции LR GATEWAY®, которые кодировали SEQ ID NO: 109 и SEQ ID NO: 115, соответственно.
Получение конструкции CaMV35s::PPD1 Selaginella moellendorffii для сверхэкспрессии PPD Selaginella moellendorffii у мутантном арабидопсисе Δppd
Экспрессирующую конструкцию синтезировали для обеспечения сверхэкспрессии PPD1 Selaginella moellendorffii под контролем промотора CaMV35s (SEQ ID NO: 129) в мутантном арабидопсисе Δppd. ORF PPD оптимизировали для экспрессии в арабидопсисе; это включало модифицированную последовательность Joshi (Joshi 1997, Nucleic Acid Research 15, 6643-6653), оптимизацию кодонов, устранение нестабильности мРНК, удаление сигнальных последовательностей поли-A, удаление скрытых участков сплайсинга, добавление удаляемого посредством BamHI C-концевого эпитопа V5 и концевой части в виде His-метки (кодирующей SEQ ID NO: 37) и добавление двойного стоп-кодона. Затем конструкцию (с хвостовой частью и без нее) помещали между промотором CaMV35s и терминатором ocs посредством реакции LR GATEWAY®, которые кодировали SEQ ID NO: 110 и SEQ ID NO: 116, соответственно.
Материалы растений и условия роста
Arabidopsis thaliana (L.) Heynh экотипа Ler можно использовать в качестве растения дикого типа (WT). Делеционный мутант с потерей функции Δppd (с делетированными PPD1 и PPD2) является таким, как описано ранее в White 2006 PNAS 103, 13238-13243.
Растения выращивают в теплице с контролируемой постоянной температурой 21°C или в помещении с контролируемыми условиями при 23°C с циклом 16 ч на свету 8 ч в темноте.
Трансформация Arabidopsis
Описанные выше конструкции можно трансформировать в Arabidopsis способом инфильтрации посредством погружения цветков (Clough and Bent, 1998 Plant J 16, 735-43). Линию Δppd трансформируют для экспрессии полипептидов PPD стандартными способами. Трансгенные растения подтверждают стандартными способами ПЦР-анализа с использованием комбинации специфических для трансгена праймеров и праймеров T-ДНК.
Комплементация линии Δppd с получением фенотипа листьев и фенотипа розеток дикого типа в проростках T1 (потомок растения после инфильтрации) подтверждает функциональность введенного гена PEAPOD, которая может быть показана на фотографиях.
Этот подход можно использовать для подтверждения функциональности любого гена PEAPOD, что по утверждению заявителя демонстрирует его пригодность для применения в рамках настоящего изобретения.
Посредством комплементации мутанта с делецией PEAPOD Δppd Arabidopsis thaliana экотипа Landsberg erecta было показано, что белки PEAPOD из Arabidopsis thaliana; Picea sitchensis, Amborella trichopoda, Musa acuminate и Selaginella moellendorffii являются функционально эквивалентными (фиг.13).
Пример 3: PEAPOD может быть вовлечен в регуляцию каскада передачи сигнала брассиностероидов
Заявители использовали дрожжевой двухгибридный (Y2H) анализ и бимолекулярную флуоресценцию (BiFC) для исследования взаимодействий между PPD, NINJA, TPL и BZR1.
Клонирование и конструкции
Конструкции для анализов Y2H и BiFC получали следующим образом. Последовательности ДНК арабидопсиса, кодирующие открытые рамки считывания для At4g14713 (PPD1) и укороченных и делеционных производных PPD1: PPD1; PPD1Δppd (N-концевое укорочение последовательностей, кодирующих а.к.1-61), PPD1Δtify (внутренняя делеция последовательностей, кодирующих а.к. 154-186), PPD1Δjas* (C-концевое укорочение последовательностей, кодирующих а.к. 229-313) (фиг.4), At4g28910 (NINJA), At1g15750 (TPL), At1g75080 (BZR1), синтетический промотор PUAS-35S и последовательности, кодирующие GAL4DBD и слитые белки c-myc, синтезировали и последовательность подтверждали в GeneArt. Большинство последовательностей было предоставлено в качестве клонов в pENTR221, готовой для клонирования Gateway в экспрессирующие векторы дрожжей и растений. В качестве исключения, промоторная последовательность для анализов активации транскрипции в растениях, включающая участки ферментов рестрикции 5' XhoI и 3' NcoI, была предоставлена клонированной в pMA-RQ. Плазмиды для временного репортерного анализа с LUC: синтетический промотор с 5× участками связывания ДНК UAS GAL4 выше промотора CaMV35S из 105 п.н. клонировали в участки XhoI-NcoI в двойной конструкции люциферазы pNWA62, которая содержит интрон-содержащий ген люциферазы светлячка (LUC) и 35Spro::люциферазу Renilla (REN) в качестве внутреннего стандарта для конструирования pAML7. Для сверхэкспрессии слитых белков GAL4DBD последовательности ДНК, кодирующие ДНК-связывающий домен GAL4 (а.к.1-147 GAL4DBD) и N-концевые слитые конструкции GAL4DBD (с использованием линкера, кодирующего GGGGS) с 2× доменом активатора VP16 (GAL4DBD-VP16) или PPD1 (GAL4DBD-PPD1), клонировали с использованием технологии Gateway в pRSh1 (Winichayakul et al 2008) для конструирования векторов pRSh1-GAL4DBD, pRSh1-GAL4DBD-VP16 и pRSh1-GAL4DBD-PPD1 для экспрессии слитых белков в растениях.
Плазмиды для дрожжевого двухгибридного анализа
Полноразмерные кодирующие последовательности BZR1, NINJA, TPL и PPD1, вместе с укороченными или делеционными производными PPD1 (PPD1Δppd, PPD1Δtify и PPD1Δjas*), подвергали субклонированию Gateway в pDEST32 (N-концевой GAL4DBD) или pDEST22 (N-концевой GAL4AD), для конструирования pDEST32-PPD1, pDEST32-PPD1Δppd, pDEST32-PPD1Δtify, pDEST32-PPD1Δjas*, pDEST32-TPL, в качестве векторов-приманок и pDEST22-PPD1, pDEST22-BZR1 и pDEST22-NINJA в качестве векторов-добычи. При экспрессии эти конструкции продуцировали белки, приведенные в SEQ IN NO: 56-67 и 70-72; включая: ДНК-связывающий домен (DBD), домен активации (AD), PPD1, слитый с DBD (PPD1-DBD), PPD1, слитый с AD (PPD1-AD), PPD1 без домена TIFY, слитого с AD (PPD1-tify-AD), PPD1 без домена jas, слитого с AD (PPD1-jas*-AD), TOPLESS (TPL), TPL, слитого с DBD (TPL-DBD), NINJA, NINJA, слитого с AD (NINJA-AD), BZR1, слитого с AD (BZR1-AD), PPD1 минус домен ppd, слитого с DBD (PPD1-Δppd-DBD), PPD1 минус домен TIFY, слитого с DBD (PPD1-tify-DBD), PPD1 минус домен jas, слитого с DBD (PPD1-jas*-DBD).
Плазмиды для бимолекулярной комплементации флуоресценции
Бинарные векторы BiFC-Gateway YFP: pDEST-VYNE(R)GW (Venus, а.к.1-173) и pDEST-VYCE(R)GW (Venus aa 156-239) с N-концевым слиянием использовали для конструирования следующих векторов: pDESTnYFP-BZR1, pDESTnYFP-NINJA, pDESTnYFP-PPD1, pDESTcYFP-BZR1, pDESTcYFP-PPD1, pDESTcYFP-PPD1Δppd, pDESTcYFP-PPD1Δtify и pDESTcYFP-PPD1Δjas*. Для временной экспрессии в растениях белков, взаимодействующих с PPD1 или BZR, NINJA и TPL подвергали субклонированию Gateway® в pRSh1 для конструирования pRSh1-NINJA и pRSh1-TPL. Плазмиды для коиммунопреципитации: синтезированную конструкцию ДНК, кодирующую PPD1, с C-концевым слиянием с 3× c-myc субклонировали в pRSh1 для получения pRSh1-PPD1-3xc-myc, в то время как последовательность кДНК NINJA субклонировали в pB7FWG2.0 (Karimi et al 2002, Trends Plant Sci. 7, 193-195) для конструирования pB7FWG2-NINJA-GFP. При экспрессии эти конструкции продуцировали белки, приведенные в SEQ ID NO: 63, 65, 73-82: включая TOPLESS (TPL), NINJA, Bimolecular Fluorescence (BiFC) nYFP, BiFC cYFP, BiFC nYFP-NINJA, BiFC nYFP-BZR1, BiFC cYFP-PPD1, BiFC cYFP-NINJA, BiFC cYFP-BZR1, BiFC cYFP-PPD1-ppd, BiFC cYFP-PPD1-tify, BiFC cYFP-PPD1-jas*.
Двухгибридную систему ProQuest (Invitrogen) использовали для анализа взаимодействий между PPD1, NINJA, TPL и BZR1. Комбинации конструкций приманок и добычи использовали для совместной трансформации штамма дрожжей MaV203 (Invitrogen) с селекцией на планшетах с агаром с синтетической средой с исключенными компонентами (SD) SD/-Leu/-Trp. Трансформированные штаммы исследовали в отношении взаимодействий с использованием 10-мкл капель в разведениях 1 к 10 и 1 к 100 на планшетах SD/-Leu/-Trp/-His с различными концентрациями 3-аминотриазола (3-AT) (Sigma).
Временные эксперименты BiFC проводили с использованием комбинаций плазмид pDESTnYFP и pDESTcYFP с плазмидами для экспрессии NINJA (pRSh1-NINJA) или TPL (pRSh1-TPL) или без них и инфильтрации агробактериями листьев Nicotiana benthamiana. Для инфильтрации штаммы Agrobacterium tumefaciens GV3101, содержавшие бинарные векторы, ресуспендировали из планшетов и подготавливали для трансформации, как описано для анализа LUC. Все штаммы YFP и экспрессирующие штаммы смешивали в соотношениях 1:1 (об./об.) с добавлением штамма P19 в 1/10 объема. Из каждого подвергнутого инфильтрации листа проводили взятие образцов пяти дисков листьев через 40 ч. За два часа до взятия образцов для микроскопического исследования флуоресценции листья подвергали инфильтрации посредством 1 мкг/мл раствора DAPI для окрашивания ядер. Флуоресценцию YFP и окрашивание DAPI обнаруживали с использованием конфокального лазерного сканирующего микроскопа Olympus Fluoview FV10i. Каждый эксперимент повторяли дважды.
Скрининг Y2H с использованием PPD1 в качестве белка-приманки идентифицировал NINJA как прямо взаимодействующий с PPD1. Результаты анализов BiFC показали, что PPD1 взаимодействовал с NINJA в растениях и что для этого взаимодействия также был необходимым мотив TIFY (фиг.7). Является возможным, что NINJA выполняет функцию мостика между TPL и PPD1. При использовании Y2H не наблюдали прямого взаимодействия между PPD1 и BZR1 (фиг.5). Однако недавние эксперименты по тандемной аффинной очистке (TAP) показали, что TPL может взаимодействовать с BZR1 (Wang et al 2013, Mol. Cell. Proteomics 12, 3653-3665) и здесь результаты Y2H подтвердили, что происходит прямое взаимодействие (фиг.6).
Для определения молекулярной функции белков PPD взаимодействия PPD1, NINJA, TPL и BZR1 исследовали в растениях. Бимолекулярную флуоресценцию (BiFC) использовали, чтобы показать, что в клетках мостовидного эпителия незрелых листьев Nicotiana benthamiana PPD1, по-видимому, взаимодействует с BZR1 в ядре (фиг.7A,B). Связывающий NINJA мотив TIFY в PPD1 был необходим для этого взаимодействия. Более того, взаимодействия не наблюдали, когда nYFP-PPD1 и cYFP-BZR1 коэкспрессировались в полностью расправленных листьях (фиг.7C). Интересно, что взаимодействие между PPD1 и BZR1 восстанавливалось при коэкспрессии NINJA, но не TPL отдельно, что указывает на отсутствие взаимодействия в зрелых листьях вследствие ограничения эндогенного NINJA. Что касается незрелых листьев взаимодействие между PPD1 и BZR1, даже в присутствии коэкспрессии NINJA и TPL, не наблюдали, когда NINJA-связывающий мотив TIFY PPD1 был делетирован (фиг.7C). Эти результаты указывают на то, что PPD1, NINJA, TPL и BZR1 существуют в качестве комплекса в растениях и что NINJA требуется для привлечения PPD1 для взаимодействия через TPL с BZR1.
Оказалось, что PPD1 не взаимодействует прямо с фактором транскрипции-мишенью BZR1. Вместо этого результаты экспериментов по взаимодействию белка PPD1 указывают на модель, в которой белки PPD привлекают корепрессоры транскрипции TPL, с использованием NINJA в качестве адаптера, и этот комплекс PPD-NINJA-TPL взаимодействует с мотивом EAR факторов транскрипции BZR (фиг.8). Таким образом, в этой модели белок PEAPOD1 (PPD1) Arabidopsis thaliana выступает в качестве репрессора каскада передачи сигнала BR и в комбинации с NINJA и TPL подавляет BZR1.
Пример 4: PEAPOD может быть вовлечен в регуляцию каскада передачи сигнала гиббереллина
Известно, что обработка гиббереллиновой кислотой (GA) снижает уровни белков DELLA (включая RGA1), которые являются репрессорами GA; для определения взаимосвязи между PPD, DELLA и каскадом передачи сигнала GA заявители провели дрожжевой двухгибридный (Y2H) анализ между PPD и DELLA (RGA1) и нанесли гормон гиббереллиновую кислоту (GA) и ингибитор биосинтеза GA (паклобутразол, PAC) на растение дикого типа, мутант Δppd и мутант Δppd, сверхэкспрессирующий PPD (PPD-OX).
Двухгибридную систему ProQuest (Invitrogen) использовали для анализа взаимодействий между PPD1 и RGA1. Полноразмерные кодирующие последовательности PPD1 вместе с укороченными или делетированными производными PPD1 (PPD1Δppd, PPD1Δtify и PPD1Δjas*) (фиг.4), подвергали субклонированию Gateway в pDEST32 (N-концевой GAL4DBD) или pDEST22 (N-концевой GAL4AD). При трансляции они давали следующие пептидные последовательности: SEQ ID NO: 58, 68, 69, 70, 71, 72, которые представляют собой PPD1-DBD, RGA1, RGA1-AD, PPD1-ppd-DBD, PPD1-tify-DBD, PP1-jas*-DBD, соответственно.
Комбинацию конструкций приманок и добычи использовали для котрансформации штамма дрожжей MaV203 (Invitrogen), с селекцией на планшетах с агаром с синтетической средой с исключенными компонентами (SD) SD/-Leu/-Trp. Трансформированные штаммы исследовали в отношении взаимодействий с использованием 10-мкл капель в разведениях 1 к 10 и 1 к 100 на планшетах SD/-Leu/-Trp/-His с различными концентрациями 3-аминотриазола (3-AT) (Sigma). Взаимодействие PPD1-RGA1 исследовали с помощью PPD1-DBD, использованного в качестве приманки. Трансформированные дрожжи наносили пятнами в качестве десятикратных разведений на контрольную среду (-2) или селективную среду (-3) с 15 мМ 3AT. Контроли представляли собой пустые векторы, DBD, ДНК-связывающий домен GAL4, AD, домен активации GAL4 (фиг.4). Результаты Y2H указывают на то, что PPD может прямо связываться с DELLA (фиг.9).
Для экзогенного применения GA или PAC семена подвергали поверхностной стерилизации 70% этанолом, 0,01% Triton X-100, в течение 10 мин, а затем 100% этанолом в течение 5 мин, сушили воздухом на стерильной фильтровальной бумаге и переносили на планшеты со средой, содержавшей половинные концентрации солей MS, 1% сахарозу и 0,8% агар. Планшеты инкубировали в течение 5 суток при 4°C в темноте, а затем переносили на 24°C с циклом 14 ч на свету/10 ч в темноте каждые сутки. Свет предоставляли посредством флуоресцентных трубок (Philips TLD 58W/865) с интенсивностью 100 мкм м-2 с-1. Проростки дикого типа (Col-0), мутанты по Δppd и трансгенные проростки PPD-OX выращивали в течение пяти суток на среде с различными концентрациями GA (фиг.10A) или PAC (фиг.10B). GA (ACROS organics) и PAC (Sigma-Aldrich) растворяли в этаноле и ацетоне, стерилизовали фильтрацией и добавляли в планшеты со средой. Этанол или ацетон (0,5%) использовали для имитирующих обработок. Проростки выращивали при 24°C при цикле 14 ч на свету/10 ч в темноте в течение 5 суток, а затем анализировали длину гипокотиля (n=35). Каждую обработку повторяли два раза; планки погрешности=стандартная ошибка среднего значения.
Уменьшение уровня DELLA приводит к увеличению транскрипции генов-мишеней DELLA, стимулирующих клеточную экспансию, и его можно количественно определять путем измерения удлинения гипокотиля проростков, растущих на среде, содержащей различные уровни GA. Наиболее низкая концентрация GA (1 мкМ) не стимулировала удлинение гипокотиля дикого типа (WT), в то время как проростки мутанта PPD с потерей функции (Δppd), так и трансгенные сверхэкспрессирующие PPD (PPD-OX) проростки продемонстрировали увеличенное удлинение гипокотиля (фиг.10A). При более высоких концентрациях GA (5-50 мкМ) удлинение гипокотиля WT происходило дозозависимым образом. Для сравнения проростки Δppd и PPD-OX продемонстрировали сверхчувствительное удлинение при вплоть до 5 и 10 мкМ GA, соответственно, где они оба достигли приблизительно одинаковой длины (фиг.10A).
Биосинтез GA ингибируется посредством применения экзогенного паклобутразола (PAC); это приводит к увеличению уровней белков-репрессоров DELLA и соответствующему снижению экспансии клеток. Проростки дикого типа продемонстрировали дозозависимое снижение удлинения гипокотиля от 0 до 10 мкМ PAC (фиг.10B). Вновь, проростки Δppd продемонстрировали сверхчувствительный ответ, который наблюдали в качестве более выраженного снижения удлинения гипокотиля на протяжении того же диапазона применений PAC. Однако проростки PPD-OX были относительно нечувствительными до тех пор, пока концентрацию PAC не увеличивали до уровня выше 0,1 мкМ, после чего они также продемонстрировали уменьшение длины гипокотиля (фиг.10B).
Сверхчувствительный ответ GA проростков Δppd потенциально отражает комбинацию увеличенного нацеливания DELLA на деградацию в отсутствие репрессии фактора транскрипции посредством PPD. Аналогично, добавление PAC на фоне Δppd возможно приводит к более высокому уменьшению удлинения гипокотиля по сравнению с WT, поскольку это происходит в отсутствие одного из природных антагонистов DELLA - PPD, что указывает на то, что PPD и GA конкурируют за связывание с DELLA.
Можно спрогнозировать, что сверхэкспрессия PPD приведет к более высокому уровню антагонизма DELLA, поскольку такое удлинение гипокотиля в этих растениях должно быть сверхчувствительным к GA; действительно, именно это наблюдалось в проростках PPD-OX. В обратной ситуации, когда уровень GA был снижен (вследствие применения PAC), проростки PPD-OX были не отвечающими до тех пор, пока концентрация PAC не превысила 0,1 мкМ. Это, вероятно, отражает точку, в которой происходит достаточное снижение уровней эндогенной GA, чтобы увидеть влияние белка DELLA без антагонизма сверхкспрессируемого PPD.
Пример 5: Увеличение биомассы корней посредством увеличения экспрессии PEAPOD в растениях
Конструкции
Ниже описано несколько конструкций для экспрессии последовательностей PEAPOD из различных видов под контролем различных промоторов для экспрессии в двудольных и однодольных растениях.
Получение конструкции pTobRB7 Δ1.3::PPD для двудольного растения для (предпочтительной для корней) сверхэкспрессии PPD арабидопсиса в двудольных растениях
Синтезировали конструкции, содержащие кодирующую последовательность PPD1, для экспрессии в корнях двудольных растений, последовательности нуклеиновых кислот представлены в SEQ ID NO: 40 и 126. ORF PPD первоначально происходила из кДНК PPD1 Arabidopsis thaliana (номер доступа № NM_202819), и она была модифицирована для включения модифицированной последовательности Joshi (Joshi 1997, Nucleic Acid Research 15, 6643-6653), удаления скрытых участков сплайсинга, в случае SEQ ID NO: 40 включения третьего интрона из DGAT1 Arabidopsis thaliana (SEQ ID NO: 39), добавления удаляемого посредством BamHI C-концевого эпитопа V5 и хвостовой части в виде His-метки (кодирующей SEQ ID NO: 37) и добавления двойного стоп-кодона. Положение интрона оптимизировали для прогнозирования участка сплайсинга, проводимого с помощью NetGene2 (http://www<dot>cbs<dot>dtu<dot>dk/services/NetGene2/).
Затем конструкцию SEQ ID NO: 40 (с хвостовой частью и без нее) помещали между промотором TobRB7 Δ1.3 (Yamamoto et al 1991, Plant Cell, 3:371-382) и терминатором NOS посредством реакции LR GATEWAY®, с получением SEQ ID NO: 42 и SEQ ID NO: 44, которые кодировали SEQ ID NO: 36 и SEQ ID NO: 38, соответственно. Сходную стадию субклонирования проводили для конструкции, показанной в SEQ ID NO: 126 (с C-концевой хвостовой частью и без нее), которая кодировала SEQ ID NO: 108 и SEQ ID NO: 114, соответственно.
Получение конструкции pTobRB7 Δ0.6::PPD для двудольных растений для (предпочтительной для корней) сверхэкспрессии PPD арабидопсиса в двудольных растениях
Синтезировали конструкции, содержащие кодирующую последовательность PPD1, для экспрессии в корнях двудольных растений, последовательности нуклеиновых кислот представлены в SEQ ID NO: 40 и 126. ORF PPD первоначально происходила из кДНК PPD1 Arabidopsis thaliana (Номер доступа № NM_202819), и она была модифицирована для включения модифицированной последовательности Joshi (Joshi 1997, Nucleic Acid Research 15, 6643-6653), удаления скрытых участков сплайсинга, в случае SEQ ID NO: 40 включения третьего интрона из DGAT1 Arabidopsis thaliana (SEQ ID NO: 39), добавления удаляемого посредством BamHI C-концевого эпитопа V5 и хвостовой части в виде His-метки (кодирующей SEQ ID NO: 37) и добавления двойного стоп-кодона. Положение интрона оптимизировали для прогнозирования участка сплайсинга, проводимого с помощью NetGene2 (http://www<dot>cbs<dot>dtu<dot>dk/services/NetGene2/).
Затем конструкцию (с хвостовой частью и без нее) помещали между промотором TobRB7 Δ0.6 (Yamamoto et al 1991, Plant Cell, 3:371-382) и терминатором NOS посредством реакции LR GATEWAY®, с получением SEQ ID NO: 43 и SEQ ID NO: 45, которые кодировали SEQ ID NO: 36 и SEQ ID NO: 38, соответственно. Сходную стадию субклонирования проводили для конструкции, показанной в SEQ ID NO: 126 (с C-концевой хвостовой частью и без нее), которая кодировала SEQ ID NO: 108 и SEQ ID NO: 114, соответственно.
Получение конструкции pAtWRKY6::PPD для двудольных растений для (предпочтительной для корней) сверхэкспрессии PPD арабидопсиса в двудольных растениях
Синтезировали конструкции, содержащие кодирующую последовательность PPD1, для экспрессии в корнях двудольных растений, последовательность нуклеиновой кислоты представлена в SEQ ID NO: 41 и 126. ORF PPD первоначально происходила из кДНК PPD1 Arabidopsis thaliana (Номер доступа № NM_202819), и она была модифицирована для включения модифицированной последовательности Joshi (Joshi 1997, Nucleic Acid Research 15, 6643-6653), удаления скрытых участков сплайсинга, в случае SEQ ID NO: 41 включения третьего интрона из DGAT1 Arabidopsis thaliana (SEQ ID NO: 39), добавления удаляемого посредством BamHI C-концевого эпитопа V5 и хвостовой части в виде His-метки (encoding SEQ ID NO: 37) и добавления двойного стоп-кодона. Положение интрона оптимизировали для прогнозирования участка сплайсинга, проводимого с помощью NetGene2 (http://www<dot>cbs<dot>dtu<dot>dk/services/NetGene2/).
Затем конструкцию (с хвостовой частью и без нее) помещали между промотором AtWRKY6 табака (Robatzek and Somssich 2001, The Plant Journal, 28: 123-133) и терминатором NOS посредством реакции LR GATEWAY®, с получением SEQ ID NO: 44 и SEQ ID NO: 47, которые кодировали SEQ ID NO: 36 и SEQ ID NO: 38, соответственно. Сходную стадию субклонирования проводили для конструкции, показанной в SEQ ID NO: 126 (с C-концевой хвостовой частью и без нее), которая кодировала SEQ ID NO: 108 и SEQ ID NO: 114, соответственно.
Получение конструкции pTobRB7 Δ1.3::PPD для однодольных растений для (предпочтительной для корней) сверхэкспрессии PPD арабидопсиса в однодольных растениях
Синтезировали конструкцию, содержащую кодирующую последовательность PPD1, для экспрессии в корнях однодольных растений, последовательности нуклеиновых кислот представлены в SEQ ID NO: 49 и SEQ ID NO: 120. ORF PPD оптимизировали для экспрессии в однодольных растениях; это включало модифицированную последовательность Joshi (Joshi 1997, Nucleic Acid Research 15, 6643-6653), оптимизацию кодонов, удаления последовательностей нестабильности мРНК, удаление сигнальных последовательностей поли-A, удаление скрытых участков сплайсинга, в случае SEQ ID NO: 49 включение третьего интрона из DGAT1 Lolium perenne (SEQ ID NO: 48), добавление удаляемого посредством BamHI C-концевого эпитопа V5 и хвостовой части в виде His-метки (кодирующей SEQ ID NO: 37) и добавление двойного стоп-кодона. Положение интрона оптимизировали для прогнозирования участка сплайсинга, проводимого с помощью deepc2 (http://www<dot>cbs<dot>dtu<dot>dk/services/NetGene2/).
Затем конструкцию (с хвостовой частью и без нее) помещали между промотором pTobRB7 Δ1.3 (Yamamoto et al 1991, Plant Cell, 3:371-382) и терминатором NOS посредством реакции LR GATEWAY®, с получением SEQ ID NO: 50 и SEQ ID NO: 53, которые кодировали SEQ ID NO: 36 и SEQ ID NO: 38, соответственно. Сходную стадию субклонирования проводили для конструкции, показанной в SEQ ID NO: 120 (с C-концевой хвостовой частью и без нее), которая кодировала SEQ ID NO: 108 и SEQ ID NO: 114, соответственно.
Получение конструкции pTobRB7 Δ0.6::PPD для однодольных растений для (предпочтительной для корней) сверхэкспрессии PPD арабидопсиса в двудольных растениях
Синтезировали конструкции, содержащие кодирующую последовательность PPD1, для экспрессии в корнях однодольных растений, последовательности нуклеиновых кислот представлены в SEQ ID NO: 49 и SEQ ID NO: 120. ORF PPD оптимизировали для экспрессии в однодольных растениях; это включало модифицированную последовательность Joshi (Joshi 1997, Nucleic Acid Research 15, 6643-6653), оптимизацию кодонов, удаление последовательностей нестабильности мРНК, удаление сигнальных последовательностей поли-A, удаление скрытых участков сплайсинга, в случае SEQ ID NO: 49 включение третьего интрона из DGAT1 Lolium perenne (SEQ ID NO: 48), добавление удаляемого посредством BamHI C-концевого эпитопа V5 и хвостовой части в виде His-метки (кодирующей SEQ ID NO: 37) и добавление двойного стоп-кодона. Положение интрона оптимизировали для прогнозирования участка сплайсинга, проводимого с помощью deepc2 (http://www<dot>cbs<dot>dtu<dot>dk/services/NetGene2/).
Затем конструкцию (с хвостовой частью и без нее) помещали между промотором pTobRB7 Δ0.6 (Yamamoto et al 1991, Plant Cell, 3:371-382) и терминатором NOS посредством реакции LR GATEWAY®, с получением SEQ ID NO: 51 и SEQ ID NO: 54, которые кодировали SEQ ID NO: 36 и SEQ ID NO: 38, соответственно. Сходную стадию субклонирования проводили для конструкции, показанной в SEQ ID NO: 120 (с C-концевой хвостовой частью и без нее), которая кодировала SEQ ID NO: 108 и SEQ ID NO: 114, соответственно.
Получение конструкции pAtWRKY6::PPD для однодольных растений для (предпочтительной для корней) сверхэкспрессии PPD арабидопсиса в однодольных растениях
Синтезировали конструкции, содержащие кодирующую последовательность PPD1, для экспрессии в корнях однодольных растений, последовательности нуклеиновых кислот представлены в SEQ ID NO: 49 и SEQ ID NO: 120. ORF PPD оптимизировали для экспрессии в однодольных растениях; это включало модифицированную последовательность Joshi (Joshi 1997, Nucleic Acid Research 15, 6643-6653), оптимизацию кодонов, удаление последовательностей нестабильности мРНК, удаление сигнальных последовательностей поли-A, удаление скрытых участков сплайсинга, в случае SEQ ID NO: 49 включение третьего интрона из DGAT1 Lolium perenne (SEQ ID NO: 48), добавление удаляемого посредством BamHI C-концевого эпитопа V5 и хвостовой части в виде His-метки (кодирующей SEQ ID NO: 37) и добавление двойного стоп-кодона. Положение интрона оптимизировали для прогнозирования участка сплайсинга, проводимого с помощью deepc2 (http://www<dot>cbs<dot>dtu<dot>dk/services/NetGene2/).
Затем конструкцию (с хвостовой частью и без нее) помещали между промотором pAtWRKY6 (Robatzek and Somssich 2001 The Plant Journal, 28: 123-133) и терминатором NOS посредством реакции LR GATEWAY®, с получением SEQ ID NO: 52 и SEQ ID NO: 55, которые кодировали SEQ ID NO: 36 и SEQ ID NO: 38, соответственно. Сходную стадию субклонирования проводили для конструкции, показанной в SEQ ID NO: 120 (с C-концевой хвостовой частью и без нее), которая кодировала SEQ ID NO: 108 и SEQ ID NO: 114, соответственно.
Получение экспрессирующих конструкции, содержащих PPD из других видов растений
Для демонстрации того, что можно использовать PPD из вида, отличного от арабидопсиса, авторы настоящего изобретения синтезировали конструкции, которые, когда их помещают под контроль подходящего промотора, экспрессируют PPD из Arabidopsis thaliana (контроль PPD), бобового растения Trifolium repens, примитивного покрытосеменного растения Amborella trichopoda, однодольного растения Musa acuminate, хвойных растений Picea abies и Picea sitchensis и плауновидного растения Selaginella moellendorffii. Их оптимизировали для экспрессии либо в рисе (включая интрон однодольного растения), либо в арабидопсисе (без интрона). Их модифицировали способами инженерии так, чтобы они содержали поддающуюся удалению хвостовую часть, которая состояла из двух участков BamHI (кодирующих глицин-серин) с каждой стороны последовательности нуклеиновой кислоты, кодирующей V5-His-метку. Пептидные последовательности (с C-концевой хвостовой частью V5-His-метка или без нее) представлены в последовательностях 108-119. Соответствующие последовательности нуклеиновых кислот, оптимизированные для экспрессии в рисе, представлены в последовательностях 120-125, и оптимизированные последовательности для экспрессии в арабидопсисе представлены в последовательностях 126-131.
Оптимизированные для арабидопсиса версии помещали ниже соответствующих промоторов в бинарных векторах посредством реакции GATEWAY LR и трансформировали в Δppd Arabidopsis thaliana способом погружения цветков.
Оптимизированные для риса версии помещали ниже соответствующих промоторов посредством реакции GATEWAY LR и трансформировали в плевел биолистическим способом.
Трансформация растений
Трансформация арабидопсиса
Арабидопсис трансформируют, как описано в примере 2.
Трансформация люцерны
Растения люцерны, сверхэкспрессирующие конструкцию Peapod, получали путем опосредуемой агробактериями трансформации с использованием адаптированного способа Samac et al. 2000 (Methods in Molecular Biology 343: 301-311).
Ткань растений
Идентифицировали генотипы Regen-SY (Bingham 1991 Crop Sci. 31, 1098) с высокой способностью к регенерации, которые были способны формировать целые растения из соматических зародышей в условиях in vitro. С использованием этого протокола был выбран один генотип для применения в экспериментах по трансформации растений.
Штаммы агробактерий и векторы
Во всех экспериментах по трансформации использовали бинарные векторы pRSh1::PPD-OP и pRSh1::PPD-OP-V5. Вектор pRSh1 является производным pART27 (Gleave 1992 Plant Mol Biol 20: 1203-1207) и содержит селективный ген bar, экспрессируемый с промотора CaMV, и оптимизированный для трав ген Peapod также под контролем экспрессии промотора CaMV 35S. Плазмиду переносили в штамм A. tumefaciens GV310 и селекцию трансформантов проводили на чашках с YM, содержавших 200 мг л-1 спектиномицина. Успешное перемещение плазмиды в агробактерии подтверждали рестрикционным картированием после получения плазмидной ДНК из бактериальной культуры.
Культуру бинарных плазмид добавляли к 25 мл бульона Луриа с маннитом и глутаматом (MGL), содержавшего 100 мг/л спектиномицина. Бактериальные культуры выращивали в течение ночи (16 часов) на ротационном шейкере (200 об/мин) при 28°C. Культуры собирали центрифугированием (3000× g), супернатант удаляли и клетки ресуспендировали в 5 мл раствора 10 мМ MgSO4 при подготовке для трансформации растений.
Трансформация растений
Второй и третий полностью развернутые листья из растения с отвечающим на тканевую культуру генотипом Regen-SY собирали и ополаскивали в течение 30 секунд в 50% растворе этанола, а затем проводили промывание в стерильной дистиллированной воде перед поверхностной стерилизацией в растворе гипохлорита натрия (3% доступный хлор). Затем стерильные листья разрезали на диски листьев, а затем опускали в раствор среды для индукции каллюса SH1.5D. Для трансформации дисков листьев добавляли приблизительно 3 мл бульона с агробактериями к 12 мл SH1.5D и инкубировали в течение 15 минут при нерегулярном перемешивании для тщательного покрытия листьев агробактериями. Затем раствор с бактериями удаляли и фрагменты листьев кратковременно промакивали стерильной фильтровальной бумагой, а затем их помещали адаксиальной стороной вверх на гель SH1.5D с 0,8% фитоагаром. Затем планшеты ингибировали в течение 5 суток в темноте при 25°C.
После инокуляции фрагменты листьев извлекали из чашек и ополаскивали 3-4 раза стерильной жидкой средой SH1.5D, промакивали стерильной фильтровальной бумагой и помещали на чашки SH1.5D, дополненные 5 мг/л глюфосината аммония и 100 мг/л цефотаксима и тиментина для селекции и инкубировали в течение 15 суток в темноте при 25°C. Затем каллюс, развившийся на фрагментах листьев, пассировали в свежую селективную среду SH1.5D на дополнительные 15 суток.
Затем трансформированный каллюс переносили в среду BL0, дополненную глюфосинатом аммония, цефотаксимом и тиментином, для формирования соматических зародышей. После развития зародышей их разрезали и переносили в среду MSN в половинной концентрации для селекции прорастания. Целые укоренившиеся растения переносили и выращивали в теплице.
Анализ способом ПЦР
Анализ способом ПЦР проводили для подтверждения стабильного встраивания T-ДНК в геном растений, восстановившихся после экспериментов по трансформации. Геномную ДНК экстрагировали приблизительно из 50 мг выращенных in vitro листьев с использованием набора Genomic DNA Mini Kit (Geneaid).
Пары праймеров, специфичные к сигналу полиаденилирования ocs3' (ocs3'-1f, 5'-GATATGCGAGACGCCTATGA-3' [SEQ ID NO: 132]; ocs3'-1r, 5'-GAGTTCCCTTCAGTGAACGT-3' [SEQ ID NO: 133]), использовали для получения продукта амплификации размером 439 п.н. Также были включены контрольные реакции, содержавшие плазмидную ДНК-матрицу, нетрансформированную ДНК растений или только воду. Протокол реакций ПЦР состоял из: первоначальной денатурации при 94°C в течение 5 минут, 30 циклов при 95°C 30 с, 55°C 15 с, 72°C 1 мин, и удлинения при 72°C в течение 10 мин. Продукты амплификации разделяли на 1,0% агарозных гелях посредством гель-электрофореза в буфере TAE и визуализировали на системе визуализации Bio-Rad Gel Doc.
Общий состав среды, использованной для трансформации люцерны
Трансформация плевела
Растения плевела, сверхэкспрессирующие конструкцию Peapod, получали посредством бомбардировки микрочастицами с использованием адаптированного способа Altpeter et al. 2000 (Molecular Breeding 6: 519-528).
Каллюсы для трансформации индуцировали из незрелых соцветий вплоть до 7 мм. Цветочные побеги собирали, подвергали поверхностной стерилизации в растворе гипохлорита натрия (4% доступный хлор), разрезали, а затем культивировали в темноте при 25°C в течение от четырех до шести недель перед трансформацией на базальной среде Murashige с макронутриентами, микнонутриентами и витаминами Skoog (MS) (1962 Physiol Plant. 15: 473-497), дополненной 30 г/л мальтозы, 5 мг/л 2,4-D, со значением pH, доведенном до 5,8, и отвержденной с помощью 6 г/л агарозы.
Плазмиды получали с использованием набора Invitrogen Pure Link Hi Pure Plasmid Maxiprep Kit с концентрацией, доведенной до 1 мкг/мкл. Плазмиду pAcH1, которая содержит экспрессирующую кассету, содержащую химерный ген гигромицинфосфотрансферазы (HPH) (Bilang et al. 1991 Gene 100: 247-250), экспрессируемый с промотора актина риса с первым интроном и завершающийся 3'-сигналом полиаденилирования nos, использовали для селекции. Плазмиды, содержавшие экспрессирующие кассеты PPD, смешивали в молярном соотношении 1:1 c pAcH1.
Плазмидные ДНК наносили на вольфрамовые частицы M17 (среднее распределение диаметра 1,4 мкМ) с использованием способа Sanford et al. 1993 (Meth. Enzymol. 217: 483-509.) и трансформировали в ткани-мишени с использованием DuPont PDS-1000/He Biolistic Particle Delivery System. Вплоть до 6 часов перед трансформацией каллюс субкультивировали на среде для инициации каллюса, содержавшей 64 г/л маннита. После трансформации (приблизительно 16 часов) трансформированные каллюсы переносили на свободную от маннита базальную среду MS, дополненную 2 мг/л 2,4-D. Через 2 суток каллюсы переносили на ту же среду, содержавшую 200 мг/л гигромицина, и культивировали в темноте в течение 4 недель для селекции трансгенных событий. Регенерация целых растений из соматических зародышей происходила при освещении на базальной среде MS, дополненной 0,2 мг/л кинетина, 30 г/л, сахарозы и 50 мг/л гигромицина, доведенной до pH 5,8 и отвержденной с помощью 8 г/л фитоагара. Трансформированные растения переносили в изолированные условия теплицы для анализа.
Анализ трансформантов способом ПЦР
Анализ способом ПЦР проводили для подтверждения стабильного встраивания трансгенов HPH и PPD в геном для получения растений, восстановленных после экспериментов по трансформации. Геномную ДНК экстрагировали из приблизительно 50 мг листьев, выращенных in vitro, с использованием набора Genomic DNA Mini Kit (Geneaid). Пары праймеров, специфичных к гену HPH (hpt-1, 5'-GCTGGGGCGTCGGTTTCCACTATCCG-3' [SEQ ID NO: 134]; hpt-2, 5'-CGCATAACAGCGGTCATTGACTGGAGC-3' [SEQ ID NO: 135]) и сигналу полиаденилирования nos3' (nos3'-1f, 5'-CTGTTGCCGGTCTTGCGATG-3' [SEQ ID NO: 136]; nos3'-1r, 5'-GTCACATAGATGACACCGCG-3' [SEQ ID NO: 137]), использовали для получения продуктов амплификации размером 375 п.н. и 202 п.н., соответственно. Также были включены контрольные реакции, содержавшие плазмидную ДНК-матрицу, нетрансформированную ДНК растений или только воду. Протокол для реакций ПЦР состоял из: первоначальной денатурации при 94°C в течение 5 минут, 30 циклов 95°C 30 с, 55°C 15 с, 72°C 1 мин, и удлинения при 72°C в течение 10 мин. Продукты амплификации разделяли на 1,0% агарозных гелях посредством гель-электрофореза в буфере TAE и визуализировали с помощью системы визуализации Bio-Rad Gel Doc.
Анализ с использованием саузерн-блоттинга трансформантов трав
Гибридизацию с использованием саузерн-блоттинга использовали для оценки количества копий трансгена на линию. Геномную ДНК экстрагировали из материала листьев растений, выращенных в теплице, для гибридизации с использованием саузерн-блоттинга способом Doyle J and Doyle J 1990 (Focus, 12:13-15). ДНК (20 мкг) расщепляли и разделяли на 0,8% агарозном геле и переносили на нейлоновую мембрану (Roche) с использованием капиллярного переноса посредством 0,4 Н NaOH. Геномную ДНК расщепляли посредством XbaI или HindIII при исследовании в отношении трансгенов HPH и PPD, соответственно. Зонды получали с использованием набора для ПЦР-синтеза DIG. Пары праймеров, специфичные к гену HPH (rgh1, 5'-CTCGTGCTTTCAGCTTCGATGTAG-3' [SEQ ID NO: 138]; rgh5, 5'-GCTGGGGCGTCGGTTTCCACTATCGG-3' [SEQ ID NO: 139]) и PPD (GrPPD1F, 5'-CACAGGATGGATTCTCCAAGG-3' [SEQ ID NO: 140]; GrPPD1R, 5'-TAAGGTCCACGGAGAGGTTC-3' [SEQ ID NO: 141]) использовали для получения продуктов амплификации размером 906 п.н. и 586 п.н. для зондов, соответственно. Предгибридизацию (1 час) и гибридизацию (12 часов) проводили при 45°C с использованием стандартных буферов (Roche). Обнаружение осуществляли с использованием нерадиоактивного способа в соответствии с протоколом изготовителя с CDP-Star в качестве хемилюминесцентного субстрата. Световые сигналы обнаруживали с использованием системы Bio-Rad ChemiDoc MP System и программного обеспечения.
Получение поликлональных антител против белка PPD1 и иммуноблоттинг
Специально изготовленные поликлональные антитела кролика против PPD1, подвергнутые аффинной очистки, изготавливали в GenScript с использованием полноразмерного белка PPD1 Arabidopsis thaliana. При разведении 1:5000 антитела были способны обнаруживать менее чем 10 нг очищенного белка PPD посредством иммуноблоттинга. Ткань растений замораживали в жидком азоте и растирали в тонкий порошок. Порошок замороженных тканей добавляли в буфер для экстракции, содержавший 50 мМ Tris pH 7,5, 150 мМ NaCl, 1 мМ EDTA, 10% (об./об.) глицерин, 5 мМ DTT, 1% (об./об.) полный коктейль ингибиторов протеаз (Sigma) и 1% (об./об.) Triton X-100 в соотношении 1,0/1,5 (масс./об.), гомогенизировали до размораживания, а затем центрифугировали в течение 12 мин при 16300 g и 4°C. Тотальный растворимый белок в супернатанте количественно определяли с использованием анализа Брэдфорд (Coomassie Plus, Thermo Scientific), доводили до эквивалентных концентраций тотального белка на образец (как правило, 10-40 мкг), денатурировали в 1× буфере для образца NuPAGE LDS (Invitrogen) и разделяли на 4-12% геле Bis-Tris SDS/PAGE (Novex). После блоттинга на мембране из PVDF с использованием устройства iBlot (Invitrogen) проводили обнаружение белка с использованием 10 поликлональных антител против PPD1 в разведении 1:5000, а затем 20 антител козы против антител кролика с HRP (Sigma) в разведении 1:5000, наносили реагент Western Bright ECL (Advansta) и получали изображение с использованием устройства ChemiDoc™ (BioRad).
Анализ биомассы корней трансформантов
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD арабидопсиса под контролем конститутивного промотора. Побеги сеяли в пластмассовые мешки для выращивания, содержавшие горшечную смесь, и подрезали до равной высоты. Растения выращивали в течение приблизительно 6 недель в теплице; увеличение биомассы/роста/длины/ветвления корней растений плевела, трансформированных PPD арабидопсиса под контролем конститутивного промотора, по сравнению с растениями WT можно было наблюдать путем изучения роста корней вне мешка для выращивания (фиг.11). Увеличение биомассы корней количественно определяли путем промывания корней проточной водой, затем удаления прикрепленной надземной части (листья и стебли) и высушивания корней при 65°C в течение 48 ч, а затем определения сухой массы (таблица 2).
Таблица 2
(n=12)
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD арабидопсиса под контролем одного из трех промоторов корней. Побеги сеяли в пластмассовые мешки для выращивания, содержавшие горшечную смесь, и подрезали до равной высоты. Растения выращивали в течение приблизительно 6 недель в теплице; увеличение биомассы/роста/длины/ветвления корней растений плевела, трансформированных PPD арабидопсиса под контролем конститутивного промотора, по сравнению с растениями WT можно было наблюдать путем изучения роста корней вне мешка для выращивания (фиг.12). Увеличение биомассы корней количественно определяли путем промывания корней проточной водой, затем удаления прикрепленной надземной части (листья и стебли) и высушивания корней при 65°C в течение 48 ч, а затем определения сухой массы (таблица 3 и таблица 4).
Таблица 3
(n=12)
Таблица 4
(n=12)
Черенки приблизительно равного размера получали из растений люцерны WT и растений люцерны (сорт USD5), трансформированных PPD арабидопсиса под контролем конститутивного промотора. Черенки сеяли в пластмассовые мешки для выращивания, содержавшие горшечную смесь. Растения выращивали в течение приблизительно 6 недель в помещении для выращивания с длительностью дня x часов; увеличение биомассы/роста/длины/ветвления корней растений люцерны, трансформированных PPD арабидопсиса под контролем конститутивного промотора, по сравнению с растениями WT можно было наблюдать путем изучения роста корней вне мешка для выращивания. Увеличение биомассы корней количественно определяли путем промывания корней проточной водой (их фотографировали, фиг.12), затем прикрепленную надземную часть (листья и стебли) удаляли и корни сушили при 65°C в течение 48 ч, а затем взвешивали, таблица 5.
Таблица 5
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD T. repens под контролем конститутивного промотора. Побеги сеяли в пластмассовые мешки для выращивания, содержавшие горшечную смесь, и подрезали до равной высоты. Растения выращивали в течение приблизительно 6 недель в теплице; увеличение биомассы/роста/длины/ветвления корней растений плевела, трансформированных PPD T. repens под контролем конститутивного промотора, по сравнению с растениями WT можно было наблюдать путем изучения роста корней вне мешка для выращивания. Увеличение биомассы корней можно количественно определять путем промывания корней под проточной водой, затем удаления прикрепленной надземной части (листья и стебли) и высушивания корней при 65°C в течение 48 ч, а затем определения сухой массы.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD T. repens под контролем одного из трех промоторов корней. Побеги сеяли в пластмассовые мешки для выращивания, содержавшие горшечную смесь, и подрезали до равной высоты. Растения выращивали в течение приблизительно 6 недель в теплице; увеличение биомассы/роста/длины/ветвления корней растений плевела, трансформированных PPD T. repens под контролем конститутивного промотора, по сравнению с растениями WT можно было наблюдать путем изучения роста корней вне мешка для выращивания. Увеличение биомассы корней можно количественно определять путем промывания корней под проточной водой, затем удаления прикрепленной надземной части (листья и стебли) и высушивания корней при 65°C в течение 48 ч, а затем определения сухой массы.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD A. trichopoda под контролем конститутивного промотора. Побеги сеяли в пластмассовые мешки для выращивания, содержавшие горшечную смесь, и подрезали до равной высоты. Растения выращивали в течение приблизительно 6 недель в теплице; увеличение биомассы/роста/длины/ветвления корней растений плевела, трансформированных PPD A. trichopoda под контролем конститутивного промотора, по сравнению с растениями WT можно было наблюдать путем изучения роста корней вне мешка для выращивания (фиг.11). Увеличение биомассы корней можно количественно определять путем промывания корней под проточной водой, затем удаления прикрепленной надземной части (листья и стебли) и высушивания корней при 65°C в течение 48 ч, а затем определения сухой массы.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD A. trichopoda под контролем одного из трех промоторов корней. Побеги сеяли в пластмассовые мешки для выращивания, содержавшие горшечную смесь, и подрезали до равной высоты. Растения выращивали в течение приблизительно 6 недель в теплице; увеличение биомассы/роста/длины/ветвления корней растений плевела, трансформированных PPD A. trichopoda под контролем конститутивного промотора, по сравнению с растениями WT можно было наблюдать путем изучения роста корней вне мешка для выращивания. Увеличение биомассы корней можно количественно определять путем промывания корней под проточной водой, затем удаления прикрепленной надземной части (листья и стебли) и высушивания корней при 65°C в течение 48 ч, а затем определения сухой массы.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD M. acuminate под контролем конститутивного промотора. Побеги сеяли в пластмассовые мешки для выращивания, содержавшие горшечную смесь, и подрезали до равной высоты. Растения выращивали в течение приблизительно 6 недель в теплице; увеличение биомассы/роста/длины/ветвления корней растений плевела, трансформированных PPD M. acuminate под контролем конститутивного промотора, по сравнению с растениями WT можно было наблюдать путем изучения роста корней вне мешка для выращивания. Увеличение биомассы корней можно количественно определять путем промывания корней под проточной водой, затем удаления прикрепленной надземной части (листья и стебли) и высушивания корней при 65°C в течение 48 ч, а затем определения сухой массы.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD M. acuminate под контролем одного из трех промоторов корней. Побеги сеяли в пластмассовые мешки для выращивания, содержавшие горшечную смесь, и подрезали до равной высоты. Растения выращивали в течение приблизительно 6 недель в теплице; увеличение биомассы/роста/длины/ветвления корней растений плевела, трансформированных PPD M. acuminate под контролем конститутивного промотора, по сравнению с растениями WT можно было наблюдать путем изучения роста корней вне мешка для выращивания. Увеличение биомассы корней можно количественно определять путем промывания корней под проточной водой, затем удаления прикрепленной надземной части (листья и стебли) и высушивания корней при 65°C в течение 48 ч, а затем определения сухой массы.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD P. sitchensis под контролем конститутивного промотора. Побеги сеяли в пластмассовые мешки для выращивания, содержавшие горшечную смесь, и подрезали до равной высоты. Растения выращивали в течение приблизительно 6 недель в теплице; увеличение биомассы/роста/длины/ветвления корней растений плевела, трансформированных PPD P. sitchensis под контролем конститутивного промотора, по сравнению с растениями WT можно было наблюдать путем изучения роста корней вне мешка для выращивания. Увеличение биомассы корней можно количественно определять путем промывания корней под проточной водой, затем удаления прикрепленной надземной части (листья и стебли) и высушивания корней при 65°C в течение 48 ч, а затем определения сухой массы.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD P. sitchensis под контролем одного из трех промоторов корней. Побеги сеяли в пластмассовые мешки для выращивания, содержавшие горшечную смесь, и подрезали до равной высоты. Растения выращивали в течение приблизительно 6 недель в теплице; увеличение биомассы/роста/длины/ветвления корней растений плевела, трансформированных PPD P. sitchensis под контролем конститутивного промотора, по сравнению с растениями WT можно было наблюдать путем изучения роста корней вне мешка для выращивания. Увеличение биомассы корней можно количественно определять путем промывания корней под проточной водой, затем удаления прикрепленной надземной части (листья и стебли) и высушивания корней при 65°C в течение 48 ч, а затем определения сухой массы.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD S. moellendorffii под контролем конститутивного промотора. Побеги сеяли в пластмассовые мешки для выращивания, содержавшие горшечную смесь, и подрезали до равной высоты. Растения выращивали в течение приблизительно 6 недель в теплице; увеличение биомассы/роста/длины/ветвления корней растений плевела, трансформированных PPD S. moellendorffii под контролем конститутивного промотора, по сравнению с растениями WT можно было наблюдать путем изучения роста корней вне мешка для выращивания. Увеличение биомассы корней можно количественно определять путем промывания корней под проточной водой, затем удаления прикрепленной надземной части (листья и стебли) и высушивания корней при 65°C в течение 48 ч, а затем определения сухой массы.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD S. moellendorffii под контролем одного из трех промоторов корней. Побеги сеяли в пластмассовые мешки для выращивания, содержавшие горшечную смесь, и подрезали до равной высоты. Растения выращивали в течение приблизительно 6 недель в теплице; увеличение биомассы/роста/длины/ветвления корней растений плевела, трансформированных PPD S. moellendorffii под контролем конститутивного промотора, по сравнению с растениями WT можно было наблюдать путем изучения роста корней вне мешка для выращивания. Увеличение биомассы корней можно количественно определять путем промывания корней под проточной водой, затем удаления прикрепленной надземной части (листья и стебли) и высушивания корней при 65°C в течение 48 ч, а затем определения сухой массы.
На семена растений арабидопсиса WT и растений арабидопсиса Δppd, трансформированных PPD T. repens под контролем конститутивного промотора наносили насечки, и их высевали в горшечную смесь. Растения выращивали в течение приблизительно 6 недель в помещении для роста с длиной дня x часов; увеличение биомассы/роста/длины/ветвления корней растений арабидопсиса, трансформированных PPD T. repens под контролем конститутивного промотора, по сравнению с растениями WT, можно было наблюдать путем изучения роста корней вне горшка. Увеличение биомассы корней количественно определяли путем промывания корней проточной водой, затем прикрепленную надземную часть (листья и стебли) удаляли и корни сушили при 65°C в течение 48 ч, а затем взвешивали.
На семена растений арабидопсиса WT и растений арабидопсиса Δppd, трансформированных PPD M. acuminate под контролем конститутивного промотора наносили насечки, и их высевали в горшечную смесь. Растения выращивали в течение приблизительно 6 недель в помещении для роста с длиной дня x часов; увеличение биомассы/роста/длины/ветвления корней растений арабидопсиса, трансформированных PPD M. acuminate под контролем конститутивного промотора, по сравнению с растениями WT можно наблюдать путем изучения роста корней вне горшка. Увеличение биомассы корней количественно определяли путем промывания корней проточной водой, затем прикрепленную надземную часть (листья и стебли) удаляли и корни сушили при 65°C в течение 48 ч, а затем взвешивали.
На семена растений арабидопсиса WT и растений арабидопсиса Δppd, трансформированных PPD A. trichopoda под контролем конститутивного промотора наносили насечки, и их высевали в горшечную смесь. Растения выращивали в течение приблизительно 6 недель в помещении для роста с длиной дня x часов; увеличение биомассы/роста/длины/ветвления корней растений арабидопсиса, трансформированных PPD A. trichopoda под контролем конститутивного промотора, по сравнению с растениями WT, можно было наблюдать путем изучения роста корней вне горшка. Увеличение биомассы корней количественно определяли путем промывания корней проточной водой, затем прикрепленную надземную часть (листья и стебли) удаляли и корни сушили при 65°C в течение 48 ч, а затем взвешивали.
На семена растений арабидопсиса WT и растений арабидопсиса Δppd, трансформированных PPD P. abies под контролем конститутивного промотора наносили насечки, и их высевали в горшечную смесь. Растения выращивали в течение приблизительно 6 недель в помещении для роста с длиной дня x часов; увеличение биомассы/роста/длины/ветвления корней растений арабидопсиса, трансформированных PPD P. abies под контролем конститутивного промотора, по сравнению с растениями WT, можно было наблюдать путем изучения роста корней вне горшка. Увеличение биомассы корней количественно определяли путем промывания корней проточной водой, затем прикрепленную надземную часть (листья и стебли) удаляли и корни сушили при 65°C в течение 48 ч, а затем взвешивали.
На семенах растений арабидопсиса WT и растений арабидопсиса Δppd, трансформированных PPD S. moellendorffii под контролем конститутивного промотора делали насечки, и их высевали в горшечную смесь. Растения выращивали в течение приблизительно 6 недель в помещении для роста с длиной дня x часов; увеличение биомассы/роста/длины/ветвления корней растений арабидопсиса, трансформированных PPD S. moellendorffii под контролем конститутивного промотора, по сравнению с растениями WT, можно было наблюдать путем изучения роста корней вне горшка. Увеличение биомассы корней количественно определяли путем промывания корней проточной водой, затем прикрепленную надземную часть (листья и стебли) удаляли и корни сушили при 65°C в течение 48 ч, а затем взвешивали.
Анализ толерантности трансформантов к засухе
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD арабидопсиса под контролем конститутивного промотора. Побеги высевали в большие горшки, содержавшие горшечную смесь и почву. Растениям позволяли акклиматизироваться в теплице перед тем, как половину растений каждого типа подвергали водяному стрессу (как правило, гравиметрическое содержание воды 12%, немного выше точки устойчивого завядания), в то время как другую половину оставляли поливаемой (как правило, гравиметрическое содержание воды 22%, приблизительная полевая влагоемкость). Увеличенную толерантность к стрессовому воздействию засухи у растений, сверхэкспрессирующих PPD, можно количественно определять путем сравнения биомассы корней и ростков с растениями WT.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD T. repens под контролем конститутивного промотора. Побеги высевали в большие горшки, содержавшие горшечную смесь и почву. Растениям позволяли акклиматизироваться в теплице перед тем, как половину растений каждого типа подвергали водяному стрессу (как правило, гравиметрическое содержание воды 12%, немного выше точки устойчивого завядания), в то время как другую половину оставляли поливаемой (как правило, гравиметрическое содержание воды 22%, приблизительная полевая влагоемкость). Увеличенную толерантность к стрессовому воздействию засухи у растений, сверхэкспрессирующих PPD, можно количественно определять путем сравнения биомассы корней и ростков с растениями WT.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD M. acuminate под контролем конститутивного промотора. Побеги высевали в большие горшки, содержавшие горшечную смесь и почву. Растениям позволяли акклиматизироваться в теплице перед тем, как половину растений каждого типа подвергали водяному стрессу (как правило, гравиметрическое содержание воды 12%, немного выше точки устойчивого завядания), в то время как другую половину оставляли поливаемой (как правило, гравиметрическое содержание воды 22%, приблизительная полевая влагоемкость). Увеличенную толерантность к стрессовому воздействию засухи у растений, сверхэкспрессирующих PPD, можно количественно определять путем сравнения биомассы корней и ростков с растениями WT.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD A. trichopoda под контролем конститутивного промотора. Побеги высевали в большие горшки, содержавшие горшечную смесь и почву. Растениям позволяли акклиматизироваться в теплице перед тем, как половину растений каждого типа подвергали водяному стрессу (как правило, гравиметрическое содержание воды 12%, немного выше точки устойчивого завядания), в то время как другую половину оставляли поливаемой (как правило, гравиметрическое содержание воды 22%, приблизительная полевая влагоемкость). Увеличенную толерантность к стрессовому воздействию засухи у растений, сверхэкспрессирующих PPD, можно количественно определять путем сравнения биомассы корней и ростков с растениями WT.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD P. sitchensis под контролем конститутивного промотора. Побеги высевали в большие горшки, содержавшие горшечную смесь и почву. Растениям позволяли акклиматизироваться в теплице перед тем, как половину растений каждого типа подвергали водяному стрессу (как правило, гравиметрическое содержание воды 12%, немного выше точки устойчивого завядания), в то время как другую половину оставляли поливаемой (как правило, гравиметрическое содержание воды 22%, приблизительная полевая влагоемкость). Увеличенную толерантность к стрессовому воздействию засухи у растений, сверхэкспрессирующих PPD, можно количественно определять путем сравнения биомассы корней и ростков с растениями WT.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD S. moellendorffii под контролем конститутивного промотора. Побеги высевали в большие горшки, содержавшие горшечную смесь и почву. Растениям позволяли акклиматизироваться в теплице перед тем, как половину растений каждого типа подвергали водяному стрессу (как правило, гравиметрическое содержание воды 12%, немного выше точки устойчивого завядания), в то время как другую половину оставляли поливаемой (как правило, гравиметрическое содержание воды 22%, приблизительная полевая влагоемкость). Увеличенную толерантность к стрессовому воздействию засухи у растений, сверхэкспрессирующих PPD, можно количественно определять путем сравнения биомассы корней и ростков с растениями WT.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD арабидопсиса под контролем одного из трех промоторов корней. Побеги высевали в большие горшки, содержавшие горшечную смесь и почву. Растениям позволяли акклиматизироваться в теплице перед тем, как половину растений каждого типа подвергали водяному стрессу (как правило, гравиметрическое содержание воды 12%, немного выше точки устойчивого завядания), в то время как другую половину оставляли поливаемой (как правило, гравиметрическое содержание воды 22%, приблизительная полевая влагоемкость). Увеличенную толерантность к стрессовому воздействию засухи у растений, сверхэкспрессирующих PPD, можно количественно определять путем сравнения биомассы корней и ростков с растениями WT.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD T. repens под контролем одного из трех промоторов корней. Побеги высевали в большие горшки, содержавшие горшечную смесь и почву. Растениям позволяли акклиматизироваться в теплице перед тем, как половину растений каждого типа подвергали водяному стрессу (как правило, гравиметрическое содержание воды 12%, немного выше точки устойчивого завядания), в то время как другую половину оставляли поливаемой (как правило, гравиметрическое содержание воды 22%, приблизительная полевая влагоемкость). Увеличенную толерантность к стрессовому воздействию засухи у растений, сверхэкспрессирующих PPD, можно количественно определять путем сравнения биомассы корней и ростков с растениями WT.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD M. acuminate под контролем одного из трех промоторов корней. Побеги высевали в большие горшки, содержавшие горшечную смесь и почву. Растениям позволяли акклиматизироваться в теплице перед тем, как половину растений каждого типа подвергали водяному стрессу (как правило, гравиметрическое содержание воды 12%, немного выше точки устойчивого завядания), в то время как другую половину оставляли поливаемой (как правило, гравиметрическое содержание воды 22%, приблизительная полевая влагоемкость). Увеличенную толерантность к стрессовому воздействию засухи у растений, сверхэкспрессирующих PPD, можно количественно определять путем сравнения биомассы корней и ростков с растениями WT.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD A. trichopoda под контролем одного из трех промоторов корней. Побеги высевали в большие горшки, содержавшие горшечную смесь и почву. Растениям позволяли акклиматизироваться в теплице перед тем, как половину растений каждого типа подвергали водяному стрессу (как правило, гравиметрическое содержание воды 12%, немного выше точки устойчивого завядания), в то время как другую половину оставляли поливаемой (как правило, гравиметрическое содержание воды 22%, приблизительная полевая влагоемкость). Увеличенную толерантность к стрессовому воздействию засухи у растений, сверхэкспрессирующих PPD, можно количественно определять путем сравнения биомассы корней и ростков с растениями WT.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD P. sitchensis под контролем одного из трех промоторов корней. Побеги высевали в большие горшки, содержавшие горшечную смесь и почву. Растениям позволяли акклиматизироваться в теплице перед тем, как половину растений каждого типа подвергали водяному стрессу (как правило, гравиметрическое содержание воды 12%, немного выше точки устойчивого завядания), в то время как другую половину оставляли поливаемой (как правило, гравиметрическое содержание воды 22%, приблизительная полевая влагоемкость). Увеличенную толерантность к стрессовому воздействию засухи у растений, сверхэкспрессирующих PPD, можно количественно определять путем сравнения биомассы корней и ростков с растениями WT.
Равное количество (как правило, 4-10) побегов получали из растений плевела WT и растений плевела, трансформированных PPD S. moellendorffii под контролем одного из трех промоторов корней. Побеги высевали в большие горшки, содержавшие горшечную смесь и почву. Растениям позволяли акклиматизироваться в теплице перед тем, как половину растений каждого типа подвергали водяному стрессу (как правило, гравиметрическое содержание воды 12%, немного выше точки устойчивого завядания), в то время как другую половину оставляли поливаемой (как правило, гравиметрическое содержание воды 22%, приблизительная полевая влагоемкость). Увеличенную толерантность к стрессовому воздействию засухи у растений, сверхэкспрессирующих PPD, можно количественно определять путем сравнения биомассы корней и ростков с растениями WT.
Равное количество (как правило, 4-10) проростков арабидопсиса WT и проростков арабидопсиса Δppd, трансформированных PPD арабидопсиса под контролем конститутивного промотора, высевали в большие горшки, содержавшие горшечную смесь и почву. Растениям позволяли акклиматизироваться в теплице перед тем, как половину растений каждого типа подвергали водяному стрессу (как правило, гравиметрическое содержание воды 12%, немного выше точки устойчивого завядания), в то время как другую половину оставляли поливаемой (как правило, гравиметрическое содержание воды 22%, приблизительная полевая влагоемкость). Увеличенную толерантность к стрессовому воздействию засухи у растений, сверхэкспрессирующих PPD, можно количественно определять путем сравнения биомассы корней и ростков с растениями WT.
Равное количество (как правило, 4-10) проростков арабидопсиса WT и проростков арабидопсиса Δppd, трансформированных PPD T. repens под контролем конститутивного промотора, весевали в большие горшки, содержавшие горшечную смесь и почву. Растениям позволяли акклиматизироваться в теплице перед тем, как половину растений каждого типа подвергали водяному стрессу (как правило, гравиметрическое содержание воды 12%, немного выше точки устойчивого завядания), в то время как другую половину оставляли поливаемой (как правило, гравиметрическое содержание воды 22%, приблизительная полевая влагоемкость). Увеличенную толерантность к стрессовому воздействию засухи у растений, сверхэкспрессирующих PPD, можно количественно определять путем сравнения биомассы корней и ростков с растениями WT.
Равное количество (как правило, 4-10) проростков арабидопсиса WT и проростков арабидопсиса Δppd, трансформированных PPD M. acuminate под контролем конститутивного промотора, высевали в большие горшки, содержавшие горшечную смесь и почву. Растениям позволяли акклиматизироваться в теплице перед тем, как половину растений каждого типа подвергали водяному стрессу (как правило, гравиметрическое содержание воды 12%, немного выше точки устойчивого завядания), в то время как другую половину оставляли поливаемой (как правило, гравиметрическое содержание воды 22%, приблизительная полевая влагоемкость). Увеличенную толерантность к стрессовому воздействию засухи у растений, сверхэкспрессирующих PPD, можно количественно определять путем сравнения биомассы корней и ростков с растениями WT.
Равное количество (как правило, 4-10) проростков арабидопсиса WT и проростков арабидопсиса Δppd, трансформированных PPD A. trichopoda под контролем конститутивного промотора, высевали в большие горшки, содержавшие горшечную смесь и почву. Растениям позволяли акклиматизироваться в теплице перед тем, как половину растений каждого типа подвергали водяному стрессу (как правило, гравиметрическое содержание воды 12%, немного выше точки устойчивого завядания), в то время как другую половину оставляли поливаемой (как правило, гравиметрическое содержание воды 22%, приблизительная полевая влагоемкость). Увеличенную толерантность к стрессовому воздействию засухи у растений, сверхэкспрессирующих PPD, можно количественно определять путем сравнения биомассы корней и ростков с растениями WT.
Равное количество (как правило, 4-10) проростков арабидопсиса WT и проростков арабидопсиса Δppd, трансформированных PPD P. abies под контролем конститутивного промотора, высевали в большие горшки, содержавшие горшечную смесь и почву. Растениям позволяли акклиматизироваться в теплице перед тем, как половину растений каждого типа подвергали водяному стрессу (как правило, гравиметрическое содержание воды 12%, немного выше точки устойчивого завядания), в то время как другую половину оставляли поливаемой (как правило, гравиметрическое содержание воды 22%, приблизительная полевая влагоемкость). Увеличенную толерантность к стрессовому воздействию засухи у растений, сверхэкспрессирующих PPD, можно количественно определять путем сравнения биомассы корней и ростков с растениями WT.
Равное количество (как правило, 4-10) проростков арабидопсиса WT и проростков арабидопсиса Δppd, трансформированных PPD S. moellendorffii под контролем конститутивного промотора, высевали в большие горшки, содержавшие горшечную смесь и почву. Растениям позволяли акклиматизироваться в теплице перед тем, как половину растений каждого типа подвергали водяному стрессу (как правило, гравиметрическое содержание воды 12%, немного выше точки устойчивого завядания), в то время как другую половину оставляли поливаемой (как правило, гравиметрическое содержание воды 22%, приблизительная полевая влагоемкость). Увеличенную толерантность к стрессовому воздействию засухи у растений, сверхэкспрессирующих PPD, можно количественно определять путем сравнения биомассы корней и ростков с растениями WT.
ССЫЛКИ
Achard, P. Genschik P. (2009). Releasing the brakes of plant growth: how Gas shutdown DELLA proteins. J. Exp. Bot., 60:1985-1092.
Altschul, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.J. (1990). Basic local alignment search tool. J. Mol. Biol. 215:403-410.
Anjum, SA., Wang, LC., Farooq, M., Hussain, M., Zou, CM. (2011) Brassinolide application improves the drought tolerance in maize through modulation of enzymatic antioxidants and leaf gas exchange. J. Agronomy and Crop Sci. 197:177-185.
Chory and Wang, 2005, Genes involved in brassinosteroid hormone action in plants, US 6,921,848 B2
Clough S.J. and Bent A.F. (1998) Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16: 735-743.
Clouse, S.D. Brassinosteroid signal transduction: from receptor kinase activation to transcriptional networks regulating plant development. Plant Cell 23, 1219-1230 (2011).
Clouse, S. D. & Sasse, J. M. BRASSINOSTEROIDS: Essential regulators of plant growth and development. Annu. Rev. Plant Physiol. Plant Mol. Biol. 16, 427-451 (1998).
Gallego-Bartolome, J., Minguet, EG., Grau-Enguix, F., Abbas, M., Locascio, A., Thomas, SG., Alabadi, D., Blazquez, MA. (2012). Molecular mechanism for the interaction between gibberellin and brassionosteroid signalling pathways in Arabidopsis. PNAS, 109:13446-13451.
Garcia, M.E., Lynch, T., Peeters, J., Snowden, C. & Finkelstein, R. A small plant-specific protein family of ABI five binding proteins (AFPs) regulates stress response in germinating Arabidopsis seeds and seedlings. Plant Mol. Biol. 67, 643-658 (2008).
Guo, H., Li, L., Aluru, M., Sluru, S., Yin, Y. (2013). Mechanisms and networkes for brassinosteroid regulated gene expression. Current Opinion in Plant Biology 16:545-553.
He, J-X., Gendron, JM., Sun, Y., Gampaia, SL., Gendron, N., Sun, CQ., Wang, Z-Y. (2005). BZR1 is a transcriptional repressor with dual roles in brassinosteroid homeostasis and growth responses. Science 307:1634-1638.
Hothorn, M., Belkhadir, Y., Dreux, M., Dabi, T., Noel, J. P., Wilson, I. A. & Chory, J. Structural basis of steroid hormone perception by the receptor kinase BRI1. Nature 474, 467-471 (2011).
Hou, X., Lee LYC, Xia, K., Yan, Y., Yu H. (2010). DELLAs modulate jasmonate signalling via competitive binding to JAZs. Devel. Cell 19:884-894.
Jiang, W-B., Lin, W-H. (2013). Brassinosteroid functions in Arabidopsis seed development. Plant Signaling & Behavior 8:10, e25928; October 2013.
Jiang, L., Liu, X., Xiong, G., Liu, H., Chen, F., Wang, L., Meng, X., Liu, G., Yu, H., Yuan, Y., Yi, W., Zhao, L., Ma, H., He, Y., Wu, Z., Melcher, K., Qian, Q., Xu, H.E., Wang, Y. & Li, J. DWARF 53 acts as a repressor of strigolactone signalling in rice. Nature 504, 401-405 (2013).
Karimi, M., Inzé, D. & Depicker, A. Gateway vectors for Agrobacterium-mediated plant transformation. Trends Plant Sci. 7, 193-195 (2002).
Khripach, V., Zhabinskii, V., Groot, AD. (2000). Twenty years of brassinosteroids: Steroidal plant hormones warrant better crops for the XXI century. Ann. Bot., 86:441-447.
Kinoshita, T., Caňo-Delgado, A., Seto, H., Hiranuma, S., Fujioka, S., Yoshida, S. & Chory, J. Binding of brassinosteroids to the extracellular domain of plant receptor kinase BRI1. Nature 433, 167-171 (2005).
Lawit, Kundu, Rao and Tomes, 2007, Isolated polynucleotide molecules corresponding to mutant and wild-type alleles of the maize D9 gene and methods of use, WO 2007124312 A2
Li, Q-F., He, J-X. (2013). Mechanisms of signalling crosstalk between brassinosteroids and gibberellins. Plant Signaling & Behavior 8:7,e24686; July 2013; © 2013 Landes Bioscience.
Mathew, C., Hofmann, WA., Osborne, MA. (2009). Pasture response to gibberellins: A review and recommendations. NZ. J. Agric. Res., 52:213-225.
Pauwels, L., Barbero, G.F., Geerink, J., Tilleman, S., Grunewald, W., Pérez, A.C., Chico, J.M., Bossche, R.V., Sewell, J., Gil, E., Garcia-Casado, G., Witters, E., Inzé, D., Long, J.A., Jaeger, G.D., Solano, R. & Goossens, A. NINJA connects the co-repressor TOPLESS to jasmonate signalling. Nature 464, 788-791 (2010).
Pérez, A.C., Durand, A.N., Bossche, R.V., De Clercq, R., Persiau, G., Van Wees, S.C.M., Pieterse, C.M.J., Gevaert, K., De Jaeger, G., Goossens, A. & Pauwels, L. The non-JAZ TIFY protein TIFY8 from Arabidopsis thaliana is a transcriptional repressor. PLos ONE 9, e84891 (2014).
Prusakova, LD., Ezhov, MN., Salnikov, AI. (1999). The use of emistim, epibrassinolide and uniconazole to overcome quality difference of buckwheat grains. Agrarian Russia: 41-44.
Robatzek and Somssich (2001). The Plant Journal, 28: 123-133
She, J., Han, Z., Kim, T. W., Wang, J., Cheng, W., Chang, J., Shi, S., Wang, J., Yang, M., Wang, Z-Y. & Chai, J. Structural insight into brassinosteroid perception by BRI1. Nature 474, 472-476 (2011).
Sun, T-P. (2011). The molecular mechanism and evolution of the GA-GID1-DELLA signalling module in plants. Review. Current Biology, 21:R338-R345.
Szemenyei, H., Hannon, M. & Long, J.A. TOPLESS mediates auxin-dependent transcriptional repression during Arabidopsis embryogenesis. Science 319, 1384-1386 (2008).
Thummel, C. S. & Chory, J. Steroid signalling in plants and insects-common themes, different pathways. Genes Dev. 16, 3113-3129 (2002).
Vanhaeren, H., Gonzalez, N, Coppens, F, De Milde, L, Van Daele, T, Vermeesch, M, Eloy, NB, Storme, V, Inez, D. (2014). Combining growth-promoting genes leads topositive epistasis in Arabidopsis thaliana. eLife 2014;3:e02252. DOI: 10.7554/eLife.02252
Vanholme, B., Grunewald, W., Bateman, A., Kohchi, T., Gheysen, G. (2007). The tify family previously known as ZIM. Trends in Plant Science, 12:239-244.
Vardhini, BV. (2012). Application of brassinolide mitigates saline stress of certain metabolites of sorghum grown in Karaikal. J. Phytology, 4:1-3.
Wang, Z-Y., Nakano T., Gendron, J., He, J., Chen, M., Vafeados, D., Yang, Y., Fujioka, S., Yoshida, S., Asami, T., Chory, J. (2002). Nuclear-localised BZR1 mediates brassinosteroid-induced growth and feedback suppression of brassinosteroid biosynthesis. Developmental Cell 2:505-513.
Wang, C-M., Shang, J-X., Chen, Q-X., Oses-Prieto, J.A., Bai, M-Y, Yang, Y., Yuan, M., Zhang, Y-L., Mu, C-C., Deng, Z., Wei C-Q., Burlingame, A.L., Wang, Z-Y. & Sun, Y. Identification of BZR1-interacting proteins as potential components of the brassinosteroid signalling pathway in Arabidopsis through tandem affinity purification. Mol. Cell. Proteomics 12, 3653-3665 (2013).
Winichayakul, S., Pernthaner, A., Scott, R., Vlaming, R. & Roberts, N. (2008) Head-to-tail fusions of camelid antibodies can be expressed in planta and bind in rumen fluid. Biotech. & Appl. Biochem. 53, 111-122.
White, D. (2006) PEAPOD regulates lamina size and curvature in Arabidopsis. PNAS, 103:13238-13243.
White, D. (2007). Novel plant genes and uses thereof, WO 2007105967A8.
Yamamoto et al (1991). Characterization of cis-acting sequences regulating root-specific gene expression in tobacco. Plant Cell, 3:371-382.
Zhu, J-Y., Sae-Seaw, J. & Wang, Z-Y. Brassinosteroid signalling. Development 140, 1615-1620 (2013).
ОБОБЩЕНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ:
<110> AGRESEARCH LIMITED
<120> СПОСОБЫ УСОВЕРШЕНСТВОВАНИЯ РАСТЕНИЙ
<130> 791261 HCF/mjw
<150> NZ 701643
<151> 2014-11-04
<160> 141
<170> PatentIn version 3.5
<210> 1
<211> 313
<212> БЕЛОК
<213> Arabidopsis thaliana
<400> 1
Met Asp Val Gly Val Ser Pro Ala Lys Ser Ile Leu Ala Lys Pro Leu
1 5 10 15
Lys Leu Leu Thr Glu Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys
20 25 30
Arg Lys Phe Leu Lys Asp Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
35 40 45
Ser Gln Ala Ile Gln Gln Val Leu Ser Leu Lys Ala Leu Tyr Glu Pro
50 55 60
Gly Asp Asp Ser Gly Ala Gly Ile Phe Arg Lys Ile Leu Val Ser Gln
65 70 75 80
Pro Val Asn Pro Pro Arg Val Thr Thr Thr Leu Ile Glu Pro Ser Asn
85 90 95
Glu Leu Glu Ala Cys Gly Arg Val Ser Tyr Pro Glu Asp Asn Gly Ala
100 105 110
Cys His Arg Met Asp Ser Pro Arg Ser Ala Glu Phe Ser Gly Gly Ser
115 120 125
Gly His Phe Val Ser Glu Lys Asp Gly His Lys Thr Thr Ile Ser Pro
130 135 140
Arg Ser Pro Ala Glu Thr Ser Glu Leu Val Gly Gln Met Thr Ile Phe
145 150 155 160
Tyr Ser Gly Lys Val Asn Val Tyr Asp Gly Ile Pro Pro Glu Lys Ala
165 170 175
Arg Ser Ile Met His Phe Ala Ala Asn Pro Ile Asp Leu Pro Glu Asn
180 185 190
Gly Ile Phe Ala Ser Ser Arg Met Ile Ser Lys Leu Ile Ser Lys Glu
195 200 205
Lys Met Met Glu Leu Pro Gln Lys Gly Leu Glu Lys Ala Asn Ser Ser
210 215 220
Arg Asp Ser Gly Met Glu Gly Gln Ala Asn Arg Lys Val Ser Leu Gln
225 230 235 240
Arg Tyr Arg Glu Lys Arg Lys Asp Arg Lys Phe Ser Lys Ala Lys Lys
245 250 255
Cys Pro Gly Val Ala Ser Ser Ser Leu Glu Met Phe Leu Asn Cys Gln
260 265 270
Pro Arg Met Lys Ala Ala Tyr Ser Gln Asn Leu Gly Cys Thr Gly Ser
275 280 285
Pro Leu His Ser Gln Ser Pro Glu Ser Gln Thr Lys Ser Pro Asn Leu
290 295 300
Ser Val Asp Leu Asn Ser Glu Gly Ile
305 310
<210> 2
<211> 315
<212> БЕЛОК
<213> Arabidopsis thaliana
<400> 2
Met Asp Val Gly Val Thr Thr Ala Lys Ser Ile Leu Glu Lys Pro Leu
1 5 10 15
Lys Leu Leu Thr Glu Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys
20 25 30
Arg Lys Phe Leu Lys Glu Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
35 40 45
Ser Gln Ala Ile Gln Gln Val Leu Ser Leu Lys Ala Leu Tyr Glu Pro
50 55 60
Gly Asp Asp Ser Gly Ala Gly Ile Leu Arg Lys Ile Leu Val Ser Gln
65 70 75 80
Pro Pro Asn Pro Pro Arg Val Thr Thr Thr Leu Ile Glu Pro Arg Asn
85 90 95
Glu Leu Glu Ala Cys Gly Arg Ile Pro Leu Gln Glu Asp Asp Gly Ala
100 105 110
Cys His Arg Arg Asp Ser Pro Arg Ser Ala Glu Phe Ser Gly Ser Ser
115 120 125
Gly Gln Phe Val Ala Asp Lys Asp Ser His Lys Thr Val Ser Val Ser
130 135 140
Pro Arg Ser Pro Ala Glu Thr Asn Ala Val Val Gly Gln Met Thr Ile
145 150 155 160
Phe Tyr Ser Gly Lys Val Asn Val Tyr Asp Gly Val Pro Pro Glu Lys
165 170 175
Ala Arg Ser Ile Met His Phe Ala Ala Asn Pro Ile Asp Leu Pro Glu
180 185 190
Asn Gly Ile Phe Ala Ser Ser Arg Met Ile Ser Lys Pro Met Ser Lys
195 200 205
Glu Lys Met Val Glu Leu Pro Gln Tyr Gly Leu Glu Lys Ala Pro Ala
210 215 220
Ser Arg Asp Ser Asp Val Glu Gly Gln Ala Asn Arg Lys Val Ser Leu
225 230 235 240
Gln Arg Tyr Leu Glu Lys Arg Lys Asp Arg Arg Phe Ser Lys Thr Lys
245 250 255
Lys Ala Pro Gly Val Ala Ser Ser Ser Leu Glu Met Phe Leu Asn Arg
260 265 270
Gln Pro Arg Met Asn Ala Ala Tyr Ser Gln Asn Leu Ser Gly Thr Gly
275 280 285
His Cys Glu Ser Pro Glu Asn Gln Thr Lys Ser Pro Asn Ile Ser Val
290 295 300
Asp Leu Asn Ser Asp Leu Asn Ser Glu Asp Asn
305 310 315
<210> 3
<211> 375
<212> БЕЛОК
<213> Populus trichocarpa
<400> 3
Met Gln Pro Gly Glu Thr Val Phe Arg Ser Ala Leu Asp Lys Pro Leu
1 5 10 15
His Gln Leu Thr Glu Asp Asp Ile Ser Gln Val Thr Arg Glu Asp Cys
20 25 30
Arg Arg Tyr Leu Lys Glu Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
35 40 45
Ser Gln Ala Ile Gln Gln Val Ile Ser Leu Lys Thr Leu Leu Glu Ala
50 55 60
Thr Pro Glu Thr Glu Ser Pro Arg Arg Arg Leu Tyr Ile Pro Arg Pro
65 70 75 80
Pro Pro His Pro Pro Asp Asn Thr Pro Arg Val Arg Phe Ser Ala Val
85 90 95
Pro Pro Asn Ser Ser Val Ser Glu Arg Gly Ala Ser Ala Glu Thr Pro
100 105 110
Ile Ser Val Pro Ala Glu Glu Pro Val Pro Cys Arg Gln His Asp Pro
115 120 125
Pro Asn Pro Asp Asp Pro Ala Asp Pro Leu Pro Pro Val His Ala Ala
130 135 140
Val Thr Glu Asn Ala Ser Val Ser Pro Arg Thr Thr Gly Met Ala Glu
145 150 155 160
Glu Ser Ala Gly Gln Met Thr Ile Phe Tyr Cys Gly Lys Val Asn Val
165 170 175
Tyr Asp Asp Val Pro Gly Asp Lys Ala Gln Ala Ile Met His Leu Ala
180 185 190
Ala Ser Pro Phe Ala Pro Pro Gln Asp Ala Ser Ser Asp Val Ile Pro
195 200 205
Thr Leu Arg Pro Leu Gln Cys Gln Leu Asp Thr Pro Gly Val Lys Ala
210 215 220
Ala Pro Asn Ser Ile Val Ala Asn Phe Pro Thr Leu Pro Thr Val Lys
225 230 235 240
Gly Ala Asp Ser Gly Gln Leu Leu Trp Glu Glu Ser Asn Ile Ala Arg
245 250 255
Glu Asp Asn Leu Glu Gly Ser Thr Ser Arg Lys Ala Ser Leu Gln Arg
260 265 270
Tyr Phe Glu Lys Lys Lys Asp Arg Phe Lys Asn Lys Arg Lys Val Ala
275 280 285
Val Pro Ser Ala Ser Leu Asp Val Phe Leu Ser His Leu Val Gly Asp
290 295 300
Gln Ile Ser Asn Asp His Trp Asn Leu Asn Asp Ala Cys Ser Pro Ser
305 310 315 320
Gln Pro Arg Pro Pro Gln Thr Pro Asn Arg Cys Asn Ser Val Asp Asn
325 330 335
Val Ala Lys Asn Gly Ile Leu Lys Ala Asp Leu Asn Asn Lys Gly Asp
340 345 350
Ala Asp Leu Ser Cys Cys Leu Asp Phe Ser Ser Lys Gln Ile Asn Ala
355 360 365
Trp Cys Leu Cys Leu Gly Cys
370 375
<210> 4
<211> 346
<212> БЕЛОК
<213> Picea abies
<400> 4
Met Arg Gly Gly Gly Gly Ala Asp Arg Leu Pro Ala Arg Ala Asn Leu
1 5 10 15
Glu Lys Pro Leu Glu Asp Leu Ser His Glu Asp Ile Met Gln Leu Thr
20 25 30
Arg Glu Asp Cys Arg Arg Tyr Leu Ile Glu Lys Gly Met Arg Arg Pro
35 40 45
Ser Trp Asn Lys Ser Gln Ala Ile Gln Gln Val Leu Ser Leu Lys Lys
50 55 60
Leu Phe Glu Ser Gly Pro Asn Asp Glu Lys Arg Ser Ala Ala Thr Asn
65 70 75 80
Arg Pro Asn Pro Asp Glu Asn Leu Lys Glu Ala Ala Ser Val Ser Leu
85 90 95
Leu Tyr Gly Ser Gln Pro Glu Ser Pro Ser Val Val Phe Ala Ser Lys
100 105 110
Asp Ser Asp Thr Phe Asn Leu Glu Trp Leu Ala Lys Thr Glu Leu Pro
115 120 125
Val Leu Ala Ser Gln Pro Arg His Ile Ala Gln Gln Asn Val Phe Leu
130 135 140
Ser Ser Leu Ser Ala Gln Gln Ser Gly Ala Gln Leu Thr Ile Phe Tyr
145 150 155 160
Ser Gly Asn Val Asn Val Tyr Asp Asp Val Pro Ala Glu Lys Ala Gln
165 170 175
Glu Ile Met Leu Leu Ala Gly Ser Gly Asn Tyr Pro Pro Ser Ser Thr
180 185 190
Cys Gln Ser Thr Arg Asn Thr Gln Gln Asn Ala Val Arg Ala Ala Tyr
195 200 205
Pro Ser Asn Pro Thr Asn Thr Pro Phe Ile His Gly Val Gly Pro Pro
210 215 220
Leu Ala Thr Val Ala Ser Ser Ser Val Met Ser Ser Pro Ile His Lys
225 230 235 240
Glu Ser Pro Ile Thr Arg Lys Ala Ser Leu Gln Arg Phe Leu Glu Lys
245 250 255
Arg Lys Asp Arg Ser Arg Gly Lys Leu Gly Ala Pro Thr Ile Ser Lys
260 265 270
Lys Pro Leu Leu Met Gly Met Phe Met His Pro Ser Ile Val His Arg
275 280 285
Gln Tyr Trp Thr Asp Thr Ala Lys Arg Lys Ser Gly Lys Pro Asp Ile
290 295 300
Pro Ala Ser Ile Ser Pro Thr Arg Pro Pro His Thr Pro Arg Arg Thr
305 310 315 320
Ser Ser Asp Glu Gln Leu Ser Ala Arg His Ala Arg Gly Asp Ile Ser
325 330 335
Ala Gln Gly Gly Ser Leu His Asn Ser Asn
340 345
<210> 5
<211> 222
<212> БЕЛОК
<213> Picea sitchensis
<400> 5
Met Arg Gly Gly Glu Arg Ala Pro Gly Ser Arg Pro Ser Leu Asp Lys
1 5 10 15
Pro Leu Glu Glu Leu Thr Glu Glu Asp Ile Phe Gln Leu Thr Arg Glu
20 25 30
Asp Cys Arg Arg Tyr Leu Lys Glu Lys Gly Met Arg Arg Pro Ser Trp
35 40 45
Asn Lys Ser Gln Ala Ile Gln Gln Val Leu Ser Leu Lys Ser Leu Phe
50 55 60
Glu Ser Lys Pro Asn Gln Gln Ser Lys Lys Pro Ser Lys His Lys Pro
65 70 75 80
Ala Thr Leu Gln Phe Glu Thr Ala Arg Asp Ser Thr Phe Ala Gln Ser
85 90 95
Ser Val Ser Gln Glu Gln Ser Leu Gly Phe Ser Trp Ser Lys Glu Val
100 105 110
Leu Asp Lys Gly Thr Ala Glu Arg Gln Arg Leu Cys Ser Asp Ser Gln
115 120 125
Glu Ala His Glu Ile Pro Arg Leu Gly Ser Lys Pro Pro Gln Ser Asn
130 135 140
Thr Glu Gly Lys Arg Cys Ala His Asp Gly His Gly Arg Lys Ser Ala
145 150 155 160
Gln Pro Leu Val Arg Leu Pro Ala Asn Phe Lys Asn Asp Cys Ser Asn
165 170 175
Arg Gln Ser Ser His Thr Ser Glu Ser Gln Pro Asp Thr Leu Leu Arg
180 185 190
Ser Asp Ser Phe Gln Gln Pro Thr Ala Gln Leu Thr Ile Phe Tyr Ala
195 200 205
Gly Met Val Asn Val Tyr Asp Asp Val Pro Leu Asp Lys Ala
210 215 220
<210> 6
<211> 365
<212> БЕЛОК
<213> Gossypium raimondii
<400> 6
Met Glu Ala Gly Val Thr Thr Thr Ala Thr Thr Thr Ala Ser Phe Ser
1 5 10 15
Ser Ile Leu Asp Lys Pro Leu Ser Gln Leu Thr Glu Glu Asp Ile Ser
20 25 30
Gln Leu Thr Arg Glu Asp Cys Arg Lys Phe Leu Lys Glu Lys Gly Met
35 40 45
Arg Arg Pro Ser Trp Asn Lys Ser Gln Ala Ile Gln Gln Val Ile Ser
50 55 60
Phe Lys Ala Leu Leu Glu Ser Asn Glu Asp Ser Gly Ala Gly Ala Arg
65 70 75 80
Arg Lys Ile Leu Val Cys Pro Pro Pro Ser His Phe Pro Pro Gln Asn
85 90 95
Ala Val Ala Ser Asn Ser Gly Glu Ser Val Lys Glu Ala Val Phe Gly
100 105 110
Glu Glu Glu Ser Leu Tyr Gly Gln Lys Asp Leu Ser Leu Lys Ala Ala
115 120 125
Pro Val Val Gln Met Asn Cys Gln Gly Gly Asp Thr Asp Asp Lys Thr
130 135 140
Leu Ser Pro Ser Leu Gly Ser Pro Arg Glu Tyr Ser Lys Leu Pro Gly
145 150 155 160
Arg Ser Gln Cys Glu Thr Asn Glu Leu Gly Gly Gln Met Thr Ile Phe
165 170 175
Tyr Cys Gly Lys Ile Asn Val Tyr Asp Gly Val Pro Leu Ala Lys Ala
180 185 190
Arg Ala Ile Met His Leu Ala Ala Ser Pro Ile Asp Phe Pro Gln Gly
195 200 205
Asn Leu Cys Asn Gln Asn Gly Ala Phe Arg Ser Phe Leu Gly His Val
210 215 220
Gln Glu Ala Glu Asp Lys Asn Asp Leu Thr Ser Ser Ile Ala Leu Asn
225 230 235 240
Leu Asn Ser His Thr Met His Thr Glu Lys Met Thr Glu Tyr Gln Gln
245 250 255
Gln Phe Arg Gly Lys Ala Asn Ile Ser Arg Asp Ser Asp Val Asp Gly
260 265 270
Gln Val Ser Arg Lys Glu Ser Leu Gln Arg Tyr Leu Glu Lys Arg Lys
275 280 285
Asp Arg Gly Arg Phe Phe Lys Gly Arg Lys Asn Ala Gly Gln Ala Leu
290 295 300
Ser Ser Ser Glu Met Tyr Leu Asn His Gln Ile Arg Ala His Tyr Leu
305 310 315 320
Asn Gly Gln Thr Asn Gln Ser Arg Thr Ser Ser Pro Pro Gln Ser Gly
325 330 335
Val Pro His Ala Phe Tyr Ser Ser Ala Asp Asn Gln Glu Leu Val Asn
340 345 350
Phe Ser Val Asp Leu Asn Asp Glu Gly Gly Gln Glu His
355 360 365
<210> 7
<211> 357
<212> БЕЛОК
<213> Aquilegia coerulea
<400> 7
Met Lys Pro Asp Glu Thr Val Ser Arg Ser Pro Leu Asp Lys Pro Leu
1 5 10 15
Phe Gln Leu Thr Asp Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys
20 25 30
Arg Lys Phe Leu Arg Asp Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
35 40 45
Ser Gln Ala Ile Glu Gln Val Ile Ser Leu Lys Thr Leu Leu Glu Pro
50 55 60
Arg Thr Glu Ser Asp Thr Asn Ala Thr Gly Ile Arg Gln Lys Leu Leu
65 70 75 80
Val Ser Arg Leu Glu Asn Ser Thr Gln Val Pro Leu Asn Asp Lys Thr
85 90 95
Asn Ala Ser Asn Leu Lys Thr Ser Val Gln Ala Ile Asn Ser Gly Lys
100 105 110
Ala Asp Ile His Gly Asp Arg Pro Cys Arg Val Pro Val Pro Val Pro
115 120 125
Asp Asp Asn Thr Ile Thr Val Pro Val Pro Asp Asn Asn Thr Ile Thr
130 135 140
Val Pro Val Pro Asp Asn Asn Ile Thr Ser Ser Arg Asn Leu Asn Ser
145 150 155 160
Thr Asn Gly Leu Val Gly Gln Met Thr Ile Phe Tyr Cys Gly Lys Val
165 170 175
Ile Val Tyr Asp Asp Met Pro Ala Glu Lys Ala His Ala Ile Met Lys
180 185 190
Phe Ala Gly Ser His Ile Asn Val Pro Glu Asp Ser Ser Pro Ala Gly
195 200 205
Ala Ala Val Ile Gln Ser Phe Ala Cys Gln Leu Gln Ala Ala Ser Ile
210 215 220
Arg His Gly Leu Ala Phe Pro Ser Ala Val Ser Pro Pro Leu His Asn
225 230 235 240
Val Val Ala Asp Thr Ser Gln His Cys Arg Glu Glu Val Thr Val Ser
245 250 255
Arg Glu Val Glu Pro Glu Gly Pro Val Ser Arg Lys Ala Ser Val Gln
260 265 270
Arg Tyr Leu Glu Lys Arg Lys Asp Arg Gly Arg Phe Lys Asn Lys Arg
275 280 285
Lys Ile Glu Ser Ser Ser Ser Leu Glu Ile Tyr Leu Asn His Gln Leu
290 295 300
Gly Asp Gln Tyr Leu Asn Glu Lys Ser Ser Gln Ser Arg Ala Cys Ser
305 310 315 320
Pro Pro Gln Pro Arg Ala Pro His Thr Pro Thr Arg Cys Ser Ser Val
325 330 335
Glu Asn Gln Val Thr Asn Val Val Phe Ser Ile Asp Leu Asn Asp Asn
340 345 350
Asp Val Arg Glu Gly
355
<210> 8
<211> 347
<212> БЕЛОК
<213> Aquilegia coerulea
<400> 8
Met Lys Pro Asp Glu Thr Val Ser Arg Ser Pro Leu Asp Lys Pro Leu
1 5 10 15
Phe Gln Leu Thr Asp Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys
20 25 30
Arg Lys Phe Leu Arg Asp Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
35 40 45
Ser Gln Ala Ile Glu Gln Val Ile Ser Leu Lys Thr Leu Leu Glu Pro
50 55 60
Arg Thr Glu Ser Asp Thr Asn Ala Thr Gly Ile Arg Gln Lys Leu Leu
65 70 75 80
Val Ser Arg Leu Glu Asn Ser Thr Gln Val Pro Leu Asn Asp Lys Thr
85 90 95
Asn Ala Ser Asn Leu Lys Thr Ser Val Gln Ala Ile Asn Ser Gly Glu
100 105 110
Ala Asp Ile His Gly Asp Arg Pro Cys Arg Val Pro Val Pro Val Pro
115 120 125
Asp Asp Asn Thr Ile Thr Val Pro Val Pro Asp Asn Asn Ile Thr Ser
130 135 140
Ser Arg Asn Leu Asn Ser Thr Asn Gly Leu Val Gly Gln Met Thr Ile
145 150 155 160
Phe Tyr Cys Gly Lys Val Ile Val Tyr Asp Gly Met Pro Ala Glu Lys
165 170 175
Ala His Ala Ile Met Lys Phe Ala Gly Ser His Ile Asn Val Pro Glu
180 185 190
Asp Ser Ser Pro Ala Gly Ala Ala Val Ile Gln Ser Phe Ala Cys Gln
195 200 205
Leu Gln Ala Ala Ser Ile Arg His Gly Leu Ala Phe Pro Ser Ala Val
210 215 220
Ser Pro Pro Leu His Asn Val Val Ala Asp Thr Ser Gln His Cys Arg
225 230 235 240
Glu Glu Val Thr Val Ser Arg Glu Val Glu Pro Glu Gly Pro Val Ser
245 250 255
Arg Lys Ala Ser Val Gln Arg Tyr Leu Glu Lys Arg Lys Asp Arg Gly
260 265 270
Arg Phe Lys Asn Lys Arg Lys Ile Glu Ser Ser Ser Ser Leu Glu Ile
275 280 285
Tyr Leu Asn His Gln Leu Gly Asp Gln Tyr Leu Asn Glu Lys Ser Ser
290 295 300
Gln Ser Arg Ala Cys Ser Pro Pro Gln Pro Arg Ala Pro His Thr Pro
305 310 315 320
Thr Arg Cys Ser Ser Val Glu Asn Gln Val Thr Asn Val Val Phe Ser
325 330 335
Ile Asp Leu Asn Asp Asn Asp Val Arg Glu Gly
340 345
<210> 9
<211> 328
<212> БЕЛОК
<213> Medicago truncatula
<400> 9
Met Asn Gly Gly Ser Thr Val Ser Phe Arg Ser Ile Leu Asp Arg Pro
1 5 10 15
Leu Asn Gln Leu Thr Glu Asp Asp Ile Ser Gln Leu Thr Arg Glu Asp
20 25 30
Cys Arg Arg Phe Leu Lys Asp Lys Gly Met Arg Arg Pro Ser Trp Asn
35 40 45
Lys Ser Gln Ala Ile Gln Gln Val Ile Ser Leu Lys Ala Leu Leu Glu
50 55 60
Pro Thr Asp Asp Asp Ile Pro Ala Thr Val Gly Val Gly Val Ser Ser
65 70 75 80
Ala Ile His His His His His His His Pro Pro Gln Pro Pro Pro Lys
85 90 95
Ala Leu Asp Pro Glu Asp Thr Ala Leu Glu Leu Gln Lys Ser Thr Ser
100 105 110
Pro Val Ala Glu Arg Pro Thr Glu Thr Asn Asp Ala Asn Val Val Asn
115 120 125
Asn Pro Gly Gly Cys Ala Pro Ser Gly Ser Phe Gly Gln Met Thr Ile
130 135 140
Phe Tyr Cys Gly Lys Val Asn Val Tyr Asp Gly Val Ser Pro Asp Lys
145 150 155 160
Ala Arg Ser Ile Met Gln Leu Ala Ala Ala Cys Pro Ser Ser Phe Pro
165 170 175
Gln Asp Asn Pro Ser Asn Lys Asn Ala Ala Val Trp Ala Ser Pro Cys
180 185 190
Asn Leu Pro Ile Asp Lys Glu Val Leu Phe Pro Thr Asp Thr Ala Ile
195 200 205
Leu Gln Val Ala Gln Thr Asp Lys Met Val Glu Tyr Pro Leu Gln Tyr
210 215 220
Arg Glu Lys Gly Ser Thr Ala Arg Asp Ala Glu Gly Gln Ala Ser Arg
225 230 235 240
Lys Val Ser Leu Gln Arg Tyr Leu Glu Lys Arg Lys Asp Arg Gly Arg
245 250 255
Ser Lys Gly Lys Lys Leu Thr Gly Ile Thr Ser Ser Asn Phe Glu Met
260 265 270
Tyr Leu Asn Leu Pro Val Lys Leu His Ala Ser Asn Gly Asn Ser Ser
275 280 285
Arg Ser Ser Thr Asp Ser Pro Pro Gln Pro Arg Leu Pro Leu Val Ser
290 295 300
Ser Gly Ser Ala Glu Asn Gln Pro Lys Val Thr Leu Pro Ile Asp Leu
305 310 315 320
Asn Asp Lys Asp Val Gln Glu Cys
325
<210> 10
<211> 339
<212> БЕЛОК
<213> Solanum lycopersicum
<400> 10
Met Ser Leu Glu Gln Thr Val Tyr Lys Ser Pro Leu Asp Lys Pro Leu
1 5 10 15
Tyr Leu Leu Thr Asp Asp Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys
20 25 30
Arg Arg Phe Leu Lys Ala Lys Gly Met Arg Lys Pro Ser Trp Asn Lys
35 40 45
Ser Gln Ala Ile Gln Gln Val Ile Ser Leu Lys Ala Leu Phe Glu Thr
50 55 60
Thr Pro Glu Ser Asp Thr Gly Gln Arg Lys Lys Arg His Ile Pro Arg
65 70 75 80
Pro Asp Thr Ser Leu Gln Arg Val Gln Lys Glu Thr Ser Ile Asp Ala
85 90 95
Glu Phe Ala Glu Ser Ala Glu Glu Thr Val Pro Tyr Gly Arg Lys Pro
100 105 110
Pro Asn Lys Pro Asp Leu Ser Gly Asp Lys Ala Ala Ser Ala Val Ala
115 120 125
Val Val Asn Asn Leu Ala Pro Ser Arg Thr Thr Asp Ser Gly Asn Ala
130 135 140
Ser Ser Gly Gln Leu Thr Ile Phe Tyr Cys Gly Lys Val Asn Val Tyr
145 150 155 160
Asp Asp Val Pro Ala Glu Lys Ala Glu Ala Ile Met His Leu Ala Ala
165 170 175
Ser Pro Leu Phe Val Pro Ser Glu Thr Pro Leu Asp Ala Asn Arg Ala
180 185 190
Ala Gln His Ser Glu Cys His Leu Gln Ala Ala Asn Val Lys Leu Gly
195 200 205
Gln Asp Ser Pro Met Val Phe Met Pro Thr Met Gln Thr Gly Lys Ile
210 215 220
Thr Glu Val Thr Arg Leu His Leu Glu Glu Ser Asn Thr Ser Tyr Glu
225 230 235 240
Asp Asn Pro Glu Ala Val Asn His Val Ser Arg Lys Ala Leu Leu Glu
245 250 255
Arg Tyr Arg Glu Lys Arg Lys Asp Arg Phe Lys Arg Lys Met Gly Met
260 265 270
Pro Ser Ser Ala Ser Leu Asp Ile Tyr Leu Asn His Arg Thr Ile Asn
275 280 285
His Thr Gln Ser Glu Leu Ser Ser Arg Ser Asn Thr Cys Ser Pro Pro
290 295 300
Ala Ile Arg Leu Ser Ala Ala Pro Ala Pro Ser Gly Ser Met Asp Asn
305 310 315 320
Ile Leu Gln Met Asp Ala Asn Ala Ser Gly Phe Leu Asp Asp Lys Asp
325 330 335
Gly Lys Glu
<210> 11
<211> 338
<212> БЕЛОК
<213> Trifolium repens
<400> 11
Met Asn Gly Gly Ser Thr Val Ser Phe Arg Ser Ile Leu Asp Lys Pro
1 5 10 15
Leu Asn Gln Leu Thr Glu Asp Asp Ile Ser Gln Leu Thr Arg Glu Asp
20 25 30
Cys Arg Arg Phe Leu Lys Asp Lys Gly Met Arg Arg Pro Ser Trp Asn
35 40 45
Lys Ser Gln Ala Ile Gln Gln Val Ile Ser Leu Lys Ala Leu Leu Glu
50 55 60
Pro Thr Asp Asp Asp Leu Pro Ala Pro Val Gly Val Ser Ser Ala Ile
65 70 75 80
His His His His His His His Pro Gln Pro Pro Gln Arg Asn Leu Asn
85 90 95
Glu Ala Pro Val Lys Gly Ser Asp Leu Asp Asp Thr Gly Phe His Thr
100 105 110
Ala Glu Asp Leu Asn Lys Ser Thr Ser Thr Ala Val Glu Ile Pro Thr
115 120 125
Glu Thr Asn Asp Ala Asn Val Val Lys Ser Ser Gly Gly Cys Val Ala
130 135 140
Ser Gly Ser Phe Gly Gln Met Thr Ile Phe Tyr Cys Gly Lys Val Asn
145 150 155 160
Val Tyr Asp Gly Val Ser Pro Asp Lys Ala Arg Ser Ile Met Gln Leu
165 170 175
Ala Ala Cys Pro Ser Ser Phe Pro Gln Asp Asn Leu Leu Asn Lys Asn
180 185 190
Ala Ala Val Trp Ala Ser Pro Cys Asn Ile Pro Ile Asp Lys Asp Val
195 200 205
Leu Phe Pro Asn Asp Thr Ala Ile Leu Gln Val Ala Gln Thr Asp Lys
210 215 220
Met Val Glu Tyr Pro Leu Gln Tyr Arg Glu Lys Gly Ser Ile Ala Arg
225 230 235 240
Asp Ala Asp Val Glu Gly Gln Ala Ser Arg Asn Ala Ser Leu Gln Arg
245 250 255
Tyr Arg Glu Lys Arg Lys Asp Arg Gly Arg Ser Lys Gly Asn Lys Leu
260 265 270
Thr Gly Ile Thr Ser Ser Asn Phe Glu Met Tyr Leu Asn Leu Pro Val
275 280 285
Lys Leu His Ala Ser Asn Gly Asn Ser Ser Arg Ser Ser Thr Asp Ser
290 295 300
Pro Pro Gln Pro Arg Leu Pro Leu Val Ser Gly Gly Ser Ala Glu Asn
305 310 315 320
Gln Pro Lys Val Thr Leu Pro Ile Asp Leu Asn Asp Lys Asp Val Gln
325 330 335
Glu Cys
<210> 12
<211> 311
<212> БЕЛОК
<213> Amborella trichopoda
<400> 12
Met Thr Ala Gly Asp Gly Ser Ile Arg Ser Ile Leu Asp Lys Pro Leu
1 5 10 15
Glu Glu Leu Thr Glu Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys
20 25 30
Arg Arg Tyr Leu Lys Glu Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
35 40 45
Tyr Gln Ala Ile Gln Gln Val Leu Ser Leu Lys Gly Leu Leu Glu Gly
50 55 60
Lys Pro Cys Asp Asp Asn Ser Asp Val Phe Ser His Arg Ser Pro Ile
65 70 75 80
Thr Val Ile Pro Asn Val Gly Ser Met Arg Glu Lys Glu Lys Ala Val
85 90 95
Asn Ile Ala Asp Pro Glu Ile Ser Gly Ser His Gln Pro Asn Phe Arg
100 105 110
Arg Glu Ile His Glu Thr Thr Arg Glu Arg Ala Leu Pro Ala Ser Asp
115 120 125
Trp Pro Pro Ser Gln Glu Pro Val Ser Gln Met Thr Ile Phe Tyr Ala
130 135 140
Gly Ala Val Asn Val Tyr Asn Asp Ile Pro Glu Asp Lys Val Gln Ala
145 150 155 160
Ile Ile Tyr Leu Ala Gly Lys Ser Asp Ser Leu Gln Gln Thr Asn Val
165 170 175
Ile Arg Thr Gly Pro Asp Gln Cys Ile Ala Ser Ala Ala Ser Pro Ser
180 185 190
Leu Asn Asp Leu His Ser Arg Arg Ile His Pro Thr Ser Asn Ile Thr
195 200 205
Thr Ser Gln Ser Leu Arg Val Ala Thr Ser Leu Pro Val Gly Pro His
210 215 220
Ser Glu Val Pro Lys Thr Arg Lys Thr Ser Val Gln Arg Phe Leu Glu
225 230 235 240
Lys Arg Lys Asp Arg Gly Arg Leu Lys Gly Thr Leu Ala Ser Gly Gly
245 250 255
Ser Ser Lys Arg Gly Ser Ser Cys Leu Glu Leu Tyr Ala Thr Ser Arg
260 265 270
Leu Lys Ser Glu Gly Val Ala Thr Thr Thr Thr Gln Ser Asn Met Asn
275 280 285
Asn Val Val Val Ser Pro Ser Asn Pro Arg Met Pro Leu Asn Pro Gly
290 295 300
Ser Cys Ser Trp Val Glu Asn
305 310
<210> 13
<211> 416
<212> БЕЛОК
<213> Selaginella moellendorffii
<400> 13
Met Ala Ala Ser Ile Leu Gly Cys Gly Ser Ser Asn Gly Val Ala Val
1 5 10 15
Thr Gly Asn Pro Ala Pro Ala Ala Ala Ala Glu Val Pro Ala Pro Leu
20 25 30
Arg Pro Leu Glu Glu Leu Thr Glu Leu Asp Ile Arg Gln Leu Thr Arg
35 40 45
Glu Asp Cys Arg Arg Tyr Leu Lys Glu Arg Gly Met Arg Arg Pro Ser
50 55 60
Trp Asn Lys Ala Gln Ala Ile Gln Gln Val Leu Ser Leu Arg Ser Leu
65 70 75 80
Leu Cys Pro Ser Asn Pro Val Gly Pro Ser Ser Lys Asn Pro Gly Ser
85 90 95
Ala Ala Asn Ala Pro Pro Ala Glu Ala Ala Ala Ala Gly His Thr Lys
100 105 110
Gln Leu Leu Asp Lys Val Ser Gln Gln Ser Met Pro Asp Ser Cys Pro
115 120 125
Ser Asn Asn Ala Ser Asp Pro Arg Pro Leu Ala Gly Cys Phe Gly Ser
130 135 140
Leu Ala Pro Thr Leu Ser Val Leu Asn Pro Asp Ala Lys Arg Asn Pro
145 150 155 160
Leu Ser Ser Lys Pro Ala Ser Thr Thr Lys Pro His Ser Ala Gln Leu
165 170 175
Thr Ile Phe Tyr Ser Gly Ile Val Asn Val Tyr Asp Asp Val Pro Leu
180 185 190
Asp Lys Ala Gln Ala Ile Met Leu Leu Ala Ala Ser Lys Thr Phe His
195 200 205
Val Pro Thr Ser Ser Val Pro Gly His Pro Pro Phe Thr Ser Ala Thr
210 215 220
Gln Gln Gln Gln Gln Gln Gln Arg Glu Leu Asn Gln Gln Thr Glu Ala
225 230 235 240
Thr Gln Lys Tyr Pro Met Gln His Gln Gln Ala Pro Gln Ile Tyr Leu
245 250 255
Ser Ser Gly Ser Ala Leu Pro Asp Glu Ser Cys Thr Glu Pro Gly Leu
260 265 270
Pro Gln Val Arg Ser Ala Ser Leu Gln Arg Phe Leu Ala Lys Arg Arg
275 280 285
Asp Arg Leu Ser Gly Asn Pro Ser Ser Ser Arg Arg Asn Asp Arg Ser
290 295 300
Lys Lys Arg Arg Phe Ser Pro Pro Pro Ser Pro Leu Thr Ser Ala Ser
305 310 315 320
Phe Gln Phe Pro Pro Ser Ala Arg Thr Ser Gln Val Leu Arg Tyr Ser
325 330 335
Thr Thr Ser Thr Thr Thr Ile Thr Thr Ala Thr Ala Thr Ala Ala Thr
340 345 350
Thr Thr Thr Thr Thr Gly Thr Thr Asn Gly Gly His Cys Ser Asn Ser
355 360 365
Asn Gln Ala Ser Glu Asn Ala Gly Ser Asp Thr Ser Gly Gly Ser Ser
370 375 380
Gly Thr Pro Asp Thr Ser Asp Thr Thr Arg Asp Asn Asp Asn Gly Arg
385 390 395 400
Val Ser Asn Glu Asn Gly Arg Val Ser Thr Thr Cys Leu Ala Ala Thr
405 410 415
<210> 14
<211> 234
<212> БЕЛОК
<213> Selaginella moellendorffii
<400> 14
Met Ser Ser Met Val Asp Phe Leu Gly Ile Glu Glu Lys Val Ser Thr
1 5 10 15
Ser Val Ser Ala Glu Arg Leu Lys Lys Leu Glu Glu Leu Thr Asp Glu
20 25 30
Asp Val Met Gln Leu Thr Arg Glu Asp Cys Arg Arg Tyr Leu Lys Glu
35 40 45
Lys Gly Met Arg Arg Pro Ser Trp Asn Lys Ala Gln Ala Val Gln Gln
50 55 60
Leu Leu Ser Leu Lys Ser Leu Cys Asp Pro Ser Pro Ala Ser Ser Gly
65 70 75 80
Ala Ala Lys Arg Ser Pro Ser Pro Pro Leu Asp Glu Ala Pro Ala Lys
85 90 95
Lys Pro Met Ala Met Thr Ser Ile Asp Leu Lys Ala Ala Ala Ala Val
100 105 110
Asp Ala Ala Asn Leu Thr Met Phe Tyr Asp Gly Ala Val Ser Val Phe
115 120 125
Asp Asp Val Ser Pro Asp Lys Ala Ser Leu Phe Pro Leu Ala Tyr Ala
130 135 140
Ile Met Leu Leu Ala Gly Asn Val Lys Ser Trp Pro Ser Ile Asn Val
145 150 155 160
Ala Ala Asn Thr Asn Lys Val Val Ile Ser Ser Tyr Glu Leu Pro Gln
165 170 175
Ala Arg Lys Ala Ser Leu Gln Arg Phe Leu Gln Arg Arg Arg Glu Lys
180 185 190
Thr Ala Lys Glu Ala Ala Ser Lys Gly Asn Ser Asn Lys Ser Pro Cys
195 200 205
His Gly Glu Ser Ser Gly Lys His Ala Ser Asp Ala Thr Asp Pro Ala
210 215 220
Thr Ser Pro Leu Leu Thr Glu Val Ser Ser
225 230
<210> 15
<211> 271
<212> БЕЛОК
<213> Nicotiana tabacum
<400> 15
Met Pro Pro Glu Glu Thr Val Ser Lys Ser Pro Leu Asp Lys Pro Leu
1 5 10 15
His Leu Leu Thr Asp Asp Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys
20 25 30
Arg Arg Tyr Leu Lys Glu Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
35 40 45
Ser Gln Ala Ile Gln Gln Val Ile Ser Leu Lys Ala Leu Leu Glu Thr
50 55 60
Thr Pro Asp Ser Asp Thr Gly Pro Arg Arg Lys Leu His Ile Pro Arg
65 70 75 80
Pro Asp Thr Arg Val Gln Gln Val Gln Lys Gly Thr Asp Thr Asp Ala
85 90 95
Glu Phe Ser Lys Ser Ala Glu Gly Met Val Pro Tyr Gly Arg Lys His
100 105 110
Ser Lys Lys Pro Asp Ile Pro Gly Asp Ile Ala Ala Gly Ser Val Ala
115 120 125
Val Ala Ala Gly Asn Asn Leu Ala Pro Ser Arg Thr Thr Asp Leu Gly
130 135 140
Asn Thr Pro Ala Ser Gln Leu Thr Ile Phe Tyr Cys Gly Lys Val Asn
145 150 155 160
Val Tyr Asp Asp Val Pro Ala Glu Lys Ala Gln Ala Ile Met His Leu
165 170 175
Ala Ala Thr Pro Leu Phe Val Pro Ser Glu Thr Pro Leu Gly Ala Thr
180 185 190
Leu Ala Ala Arg His Ser Glu Cys His Leu Gln Ala Ala Ser Val Lys
195 200 205
Gln Gly Pro Asp Ser Ala Met Val Leu Met Pro Thr Met Gln Thr Gly
210 215 220
Lys Met Ser Glu Val Thr Arg Leu Arg Leu Glu Glu Ser Asn Thr Phe
225 230 235 240
Tyr Glu Asp Asn Ser Ala Asn Tyr Ala Glu Ala Val Glu Gly His Pro
245 250 255
Ser Arg Lys Ala Ser Val Gln Arg Tyr Leu Glu Lys Arg Lys Asp
260 265 270
<210> 16
<211> 327
<212> БЕЛОК
<213> Solanum tuberosum
<400> 16
Met Pro Pro Glu Glu Thr Val Ser Lys Ser Pro Leu Asp Lys Pro Leu
1 5 10 15
Asn Gln Leu Thr Asp Asp Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys
20 25 30
Arg Arg Tyr Leu Lys Gln Lys Gly Met Arg Lys Pro Ser Trp Asn Lys
35 40 45
Ser Gln Ala Ile Gln Gln Val Ile Ser Leu Lys Ala Leu Leu Glu Pro
50 55 60
Asp Thr Asp Ala Gly Thr Arg Lys Lys Leu His Ile Pro Arg Ala Asp
65 70 75 80
Thr His Val Gln Ser Gly Lys Asn Thr Tyr Gly Glu Pro Ser Glu Pro
85 90 95
Val Pro Asp Arg Arg Asn Gln Gln Asp Arg Pro Asp Leu Ser Ser His
100 105 110
Ile Thr Ala Leu Pro Val Ala Val Val Asp Asn Ser Ala Pro Ser Arg
115 120 125
Thr Ile Gly Ser Ala Asp Lys Pro Val Gly Gln Met Thr Ile Phe Tyr
130 135 140
Arg Gly Lys Val Asn Val Tyr Asp Asp Val Pro Ala Asp Lys Ala Gln
145 150 155 160
Lys Ile Met Cys Leu Ala Ser Ser Pro Leu Cys Val Pro Ser Glu Thr
165 170 175
Pro Ser Asn Ala Thr Val Ala Ala Arg His Ser Ala Cys Cys Leu Gln
180 185 190
Ala Ala Asn Ser Lys Leu Arg Leu Asp Thr Asn Ile Val Pro Thr Ile
195 200 205
Gln Thr Val Lys Met Ser Glu Val Ser Arg Val Pro Ile Glu Glu Ser
210 215 220
Asn Arg Leu Tyr Asn Asp Asn Pro Glu Ala Val Glu Ser Pro Ala Ser
225 230 235 240
Arg Lys Ala Ser Val Gln Arg Tyr Leu Glu Lys Arg Lys Glu Arg Phe
245 250 255
Lys Trp Lys Arg Lys Val Glu Thr Thr Ser Ser Ala Ser Leu Asp Ile
260 265 270
Tyr Leu Ser Asp Arg Ile Gly Thr Arg Thr Pro Ser Asp Tyr Ala Ser
275 280 285
Gly Ala Asp Leu Cys Phe Thr Pro His Ile Thr Pro Thr Gly Ser Gly
290 295 300
Pro Ile Gln Asp Asn Ile Gln Met Asn Pro Thr Phe Ser Ser Asp Leu
305 310 315 320
Asn Asp Arg Asp Val Arg Glu
325
<210> 17
<211> 348
<212> БЕЛОК
<213> Glycine max
<400> 17
Met Asn Gly Gly Ala Thr Thr Ala Thr Phe Arg Ser Ile Leu Asp Lys
1 5 10 15
Pro Leu Asn Gln Leu Thr Glu Asp Asp Ile Ser Gln Leu Thr Arg Glu
20 25 30
Asp Cys Arg Arg Phe Leu Lys Glu Lys Gly Met Arg Arg Pro Ser Trp
35 40 45
Asn Lys Ser Gln Ala Ile Gln Gln Val Ile Ser Leu Lys Ala Leu Leu
50 55 60
Glu Pro Ser Asp Asp Asp Thr Pro Pro Pro Thr Ala Met His His Arg
65 70 75 80
Ser His Ala Pro Pro Pro Pro Pro Gln Pro Gln Ser Gln Val Asn Leu
85 90 95
Thr Glu Pro Pro Pro Pro Pro Lys Ala Pro Pro Pro Glu Glu Ser Ser
100 105 110
Phe His Ala Ala Glu Asp Ile Gln Lys Pro Ala Ser Ser Gly Glu Lys
115 120 125
Pro Ser Glu Thr Asn Asp Thr Asn Thr Asn Val Ala Ser Pro Lys Gly
130 135 140
Cys Ala Thr Ser Gly Ser Phe Gly Gln Met Thr Ile Phe Tyr Cys Gly
145 150 155 160
Lys Val Asn Val Tyr Asp Gly Val Ser Pro Asp Lys Ala Arg Ala Ile
165 170 175
Met Gln Leu Ala Val Ser Pro Val Gln Phe Thr Gln Asp Asp Pro Ser
180 185 190
Asn Gly Asn Ala Ala Val Trp Pro Ser Pro Cys His Leu Pro Met Asp
195 200 205
Lys Asp Val Leu Ile Pro Val Asp Thr Thr Ile Leu Gln Val Ala Gln
210 215 220
Ser Asp Lys Met Met Glu Tyr Pro Leu Gln Tyr Arg Glu Lys Gly Ser
225 230 235 240
Ile Ala Arg Asp Ala Glu Gly Gln Ala Ser Arg Lys Val Ser Leu Gln
245 250 255
Arg Tyr Leu Glu Lys Arg Lys Asp Arg Gly Arg Leu Lys Gly Lys Lys
260 265 270
Leu Thr Gly Ile Thr Ser Ser Asn Phe Glu Met Tyr Leu Asn Leu Pro
275 280 285
Val Lys Val His Ala Ser Asn Gly Asn Ser Ser Arg Ser Ser Thr Ser
290 295 300
Ser Pro Pro Gln Pro Arg Leu Pro Leu Val Ser Ser Gly Ser Ala Asp
305 310 315 320
Asn Gln Leu Lys Val Ala Leu Pro Ile Asp Leu Asn Asp Lys Val Ser
325 330 335
Leu Gln Met Phe Lys Asn Ala Lys Thr Leu Thr Arg
340 345
<210> 18
<211> 338
<212> БЕЛОК
<213> Citrus clementine
<400> 18
Met Asp Val Asp Gly Gly Val Thr Ser Cys Arg Ser Ile Leu Glu Lys
1 5 10 15
Pro Leu Ser Gln Leu Thr Glu Glu Asp Ile Thr Gln Leu Thr Arg Glu
20 25 30
Asp Cys Arg Lys Phe Leu Lys Glu Lys Gly Met Arg Arg Pro Ser Trp
35 40 45
Asn Lys Ser Gln Ala Ile Gln Gln Val Ile Ser Leu Lys Ala Leu Leu
50 55 60
Glu Ser Ser Gly Asp Ser Gly Ser Gly Val Leu Arg Arg Val Leu Val
65 70 75 80
Ser Pro Pro Glu Ser Met Pro Pro Arg Val Asn Val Thr Ser Asn Ser
85 90 95
Ala Asp Leu Val Lys Glu Pro Thr Ile Ser Val Ser Gly Asp Gln Asn
100 105 110
Ser Ala Tyr Arg Arg Lys Tyr Pro Arg Asn Cys Ala Val Asp Ala Asp
115 120 125
Asn Lys Thr Ile Ser Asn Arg Asn Pro Cys Glu Ala Asn Gly Ser Ile
130 135 140
Gly Gln Met Thr Ile Phe Tyr Cys Gly Lys Val Asn Val Tyr Glu Gly
145 150 155 160
Val Pro Thr Asp Lys Ala Gln Glu Ile Met His Leu Ala Ala Thr Pro
165 170 175
Ile Asp Phe Ser Gln Asn Gly Ser Phe Gly Gly Ile Thr Ala Tyr Arg
180 185 190
Ala Ile Pro Cys His Leu Gln Val Thr Ser Asn Arg His Val Ser Leu
195 200 205
Pro Leu Arg Pro Ala Ala Met Ile Ser Gln Phe Met Gln Thr Gly Lys
210 215 220
Ile Ala Asp Tyr Ser Gln Glu Tyr Arg Glu Lys Ala Ile Ser Thr His
225 230 235 240
Asp Ser Asp Val Asp Gly Gln Val Asn Arg Lys Val Ser Leu Gln Arg
245 250 255
Tyr Leu Glu Lys Arg Lys Asp Arg Gly Arg Phe Phe Lys Gly Lys Lys
260 265 270
Asn Thr Gly Pro Thr Pro Ser Leu Glu Met Tyr Leu Asn His Pro Gly
275 280 285
Lys Thr His Ala Ser Asn Gly Gln Gln Ser Gln Ser Asn Thr Ser Ser
290 295 300
Pro Thr Gln Pro Glu Leu Ser Asn Thr Leu Gly Thr Ser Pro Asp Asn
305 310 315 320
Gln Ala Lys Thr Val Met Leu Pro Val Asp Leu Asn Asn Glu Asp Ile
325 330 335
Gln Asp
<210> 19
<211> 335
<212> БЕЛОК
<213> Ricinus communis
<400> 19
Met Asp Ala Gly Val Thr Ser Phe Arg Ser Ile Leu Asp Lys Pro Leu
1 5 10 15
Thr Gln Leu Thr Glu Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys
20 25 30
Arg Lys Tyr Leu Lys Glu Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
35 40 45
Ser Gln Ala Ile Gln Gln Val Ile Ser Leu Lys Ala Leu Leu Glu Thr
50 55 60
Ser Glu Asp Ser Gly Ala Gly Ala Leu Arg Arg Ile Leu Val Ser Lys
65 70 75 80
Pro Pro Val Thr Ser Asn Ser Val Asp Ser Ala Lys Glu Pro Ser Asp
85 90 95
Ser Asn Asn Asn Asn Leu Leu Asp Glu Thr Ala Pro His Asp Ser Pro
100 105 110
Lys Ser Pro Pro Pro Ala Pro Ser Leu Asp Cys Pro Leu Glu Glu Ala
115 120 125
Asp Asn Lys Val Ile Ser Ser Arg Ser Pro Gly Ala Thr Asp Gly Leu
130 135 140
Val Gly Gln Met Thr Ile Phe Tyr Cys Gly Lys Val Asn Val Tyr Asp
145 150 155 160
Gly Val Pro Pro Asp Lys Ala Gln Ala Ile Met His Leu Ala Ala Thr
165 170 175
Pro Ile His Ser Pro Leu Asp Asp Pro Ile Arg Arg Pro Val Phe Ala
180 185 190
Phe Pro Tyr His Leu Gln Thr Pro Ser Asp Lys His Val Phe Val Pro
195 200 205
Ser Asn Ala Ala Ile Ser Pro Thr Thr Pro Thr Glu Lys Val Thr Glu
210 215 220
Tyr Ser Gln Gln Cys Arg Glu Lys Gly Asn Val Thr Tyr Asp His Asp
225 230 235 240
Val Glu Gly Gln Ala Asn Arg Lys Met Ser Leu Gln Arg Tyr Leu Glu
245 250 255
Lys Lys Lys Asp Arg Gly Arg Phe Lys Gly Arg Lys Asn Leu Gly Pro
260 265 270
Asn Ser Ser Ser Leu Asp Ala Tyr Leu Asn His Gln Met Arg Thr His
275 280 285
Ile Ser Asn Glu Gln Ser Thr Arg Ser Ser Thr Ser Ser Pro Thr Gln
290 295 300
Pro Gly Val Pro His Thr Ser Ser Asn Ser Ala Glu Asp Gln Leu Lys
305 310 315 320
Thr Ala Ser Phe Ala Val Asp Leu Asn Glu Asp Val Gln Glu Pro
325 330 335
<210> 20
<211> 393
<212> БЕЛОК
<213> Vitis vinifera
<400> 20
Met Asn Pro Gly Val Thr Thr Leu Arg Ser Ile Leu Asp Lys Pro Leu
1 5 10 15
His Glu Leu Thr Glu Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys
20 25 30
Arg Lys Tyr Leu Lys Glu Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
35 40 45
Ser Gln Ala Ile Gln Gln Val Ile Ser Leu Lys Ser Leu Leu Glu Thr
50 55 60
Ser Glu Gly Ser Gly Ala Gly Val Leu Arg Lys Ile Thr Asp Ser Pro
65 70 75 80
Pro Ala Glu Asn Leu Pro Pro Val Thr Ser Asn Ser Ala Asp Ser Gly
85 90 95
Lys Glu Leu Ser Ala Asp Ile Gln Ile Ser Val Ser Ala Asp Glu Leu
100 105 110
Val Pro Leu Pro Pro Lys Asp His His Pro Glu Ser Thr Pro Ser Gly
115 120 125
Glu Leu Ala Ser Arg Pro Pro Glu Ala Asp Thr Lys His Thr Cys Pro
130 135 140
Arg Ser Pro Gly Ala Thr Asn Cys Leu Val Gly Gln Met Thr Ile Phe
145 150 155 160
Tyr Cys Gly Lys Val Asn Val Tyr Asp Gly Val Pro Asp Asp Lys Ala
165 170 175
Gln Ala Ile Met His Leu Ala Ala Ser Pro Phe His Leu Pro Ser Asp
180 185 190
Asp Pro Phe Ser Gly Ala Ala Met Leu Cys Ser Ser Pro Cys His Leu
195 200 205
His Thr Ala Asn Val Lys His Gly His Ile Pro Pro Arg Ala Met Val
210 215 220
Ser Gln Thr Met Gln Thr Glu Lys Phe Thr Glu Tyr Ser Gln Gln Tyr
225 230 235 240
Arg Glu Glu Val Asn Phe Thr Arg Gly His Gly Ser Glu Ala Leu Ser
245 250 255
Gly Leu Arg Thr Val Gly Ser Pro Thr Ala Arg Pro Thr Glu Asp Met
260 265 270
Glu Gln Thr Thr Cys Leu Thr Ile Trp Gly Thr Phe Arg Tyr Lys Val
275 280 285
Met Pro Phe Glu Ile Tyr Glu Gly Ile Met Asp Val Glu Gly Gln Val
290 295 300
Asp Arg Lys Leu Ser Leu Gln Arg Tyr Phe Glu Lys Arg Lys Asp Arg
305 310 315 320
Phe Lys Ser Arg Lys Lys Ile Gly Leu Pro Ser Gly Ser Leu Glu Met
325 330 335
Tyr Val Asn His Gln Ala Arg Thr Gln Pro Ser Asn Gly Gln Ser Ser
340 345 350
Arg Ser Gly Thr Ser Ser Pro Pro Gln His Gly Leu Ser His Thr Leu
355 360 365
Cys Ser Ser Ala Asp Asn His Thr Lys Asn Phe Thr Pro Phe Val Asp
370 375 380
Leu Asn Ser Lys Asp Ile Gln Glu Ser
385 390
<210> 21
<211> 343
<212> БЕЛОК
<213> Morus notabilis
<400> 21
Met Ser Ala Gly Thr Thr Ala Phe Arg Ser Ile Leu Asp Lys Pro Leu
1 5 10 15
Asn Gln Leu Thr Glu Asp Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys
20 25 30
Arg Lys Tyr Leu Lys Glu Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
35 40 45
Ser Gln Ala Ile Gln Gln Val Ile Ser Leu Lys Ala Leu Leu Glu Pro
50 55 60
Cys Asp Asp Ser Gly Ala Gly Ala Leu Arg Arg Ile Val Ala Ser Thr
65 70 75 80
Pro Pro Pro Pro Pro Thr Gln Asn Ala Pro Arg Val Ser Thr Phe Ser
85 90 95
Val Thr Ser Asn Ser Ala Asp Ser Gly Lys Glu Ala Ser Val Asp Val
100 105 110
Gln Val Ser Ala Glu Glu Ser Gly Pro Cys Gln Arg Lys Glu Gln Ala
115 120 125
Lys Ser Ala Pro Glu Thr Glu Glu Arg Pro Ala Asp Ala Gly Glu Arg
130 135 140
Ala Ser Pro Arg Ser His Cys Ala Thr Asp Ala Leu Val Gly Gln Met
145 150 155 160
Thr Ile Phe Tyr Cys Gly Lys Val Asn Val Tyr Glu Gly Val Pro Pro
165 170 175
Glu Lys Ala Arg Ala Ile Met His Leu Ala Ala Ser Pro Ile Pro Leu
180 185 190
Ser Arg Glu Asn Ser Phe Gly Val Leu Ala Ala Pro Arg Ser Phe Pro
195 200 205
Trp His Leu His Ala Ala Ser Asp Lys Gly Gly Leu Leu Pro Pro Ser
210 215 220
Ala Thr Ile Ser Gln Pro Met Gln Thr Asp Lys Leu Ala Asp Tyr Ser
225 230 235 240
Gln Gln Cys Trp Glu Lys Glu Asn Asp Gly Gln Ala Ser Arg Lys Leu
245 250 255
Ser Leu Gln Lys Tyr Arg Glu Lys Lys Lys Asp Arg Gly Arg Leu Lys
260 265 270
Thr Lys Arg Ser Thr Gly Phe Asn Ser Ser Ser Met Glu Val Tyr Phe
275 280 285
Asn His Gln Val Lys Thr His Met Ser Asn Gly Asn Ser Ser Arg Ser
290 295 300
Ser Thr Ser Ser Pro Thr Gln Pro Gly Leu Pro Gln Thr Leu Cys Ser
305 310 315 320
Thr Val Asp Asn Gln Pro Lys Ile Pro Cys Leu Pro Val Asp Leu Asn
325 330 335
Glu Lys Leu Thr Ile Glu Met
340
<210> 22
<211> 364
<212> БЕЛОК
<213> Phoenix dactylifera
<400> 22
Met Tyr Trp Val Gly Ser Ala Gln Glu Arg Arg Arg Asp Gly Gly Arg
1 5 10 15
Ser Pro Leu Asp Lys Pro Leu Ser Leu Leu Thr Glu Glu Asp Ile Ala
20 25 30
Gln Leu Thr Arg Glu Asp Cys Arg Arg Phe Leu Lys Glu Lys Gly Met
35 40 45
Arg Arg Pro Ser Trp Asn Lys Ser Gln Ala Ile Gln Gln Val Ile Ser
50 55 60
Leu Lys Ala Leu Leu Glu Gly Arg Pro Glu Ser Gly Glu Leu Pro Val
65 70 75 80
Gly Ala Gly Tyr Arg Gln Lys Pro Pro Pro Arg Arg Pro Ala Ser Leu
85 90 95
Pro Ser Leu Gln Glu Ala Ala Gly Asp Ser Thr Ala Ala Ala Lys Glu
100 105 110
Pro Ser Pro Ser Ser Ser Leu Ser Pro Tyr Arg Arg Arg Asp Pro Ile
115 120 125
Pro Pro Ile Ile Ser Ala Gly Gly Pro Ser Cys Arg Phe Pro Val Ala
130 135 140
Gly Arg Asp Gln Gln Pro Pro Glu Thr Pro Ser Pro Ser Leu Arg Val
145 150 155 160
Thr Ala Glu Val Pro Ala Gly Gln Met Thr Ile Phe Tyr Asp Gly Lys
165 170 175
Val Asn Val Tyr Ser Asp Val Thr Val Asp Lys Ala Arg Ala Ile Leu
180 185 190
Leu Leu Ala Gly Arg Arg Asp Cys Tyr Gly Ala Ala Ala Leu Pro Gly
195 200 205
Pro Val His Ser Pro Gln Pro Ala Phe Leu Gly Pro Gly Gln Gly Pro
210 215 220
Val Pro Thr Ala Pro Pro Leu Ala Ala Ala Leu Pro Thr Ser Pro Ala
225 230 235 240
Gly Arg Leu Ala His Arg Phe Glu Gly Pro Ser Gly Val Pro Arg Gly
245 250 255
Lys Ser Ser Leu Val Arg Glu Arg Ser Thr Ser Pro Glu Gly Pro Thr
260 265 270
Ser Arg Lys Ala Ser Leu Gln Arg Tyr Leu Glu Lys Arg Lys Asp Arg
275 280 285
Leu Lys Gly Arg Lys Thr Leu Gly Gly Ala Ser Ser Ser Ser Met Glu
290 295 300
Ile Met Phe Leu Ser Gln Lys Phe Gly Gly Gln Ile Pro Asn Glu Gln
305 310 315 320
Leu Ser Arg Ser Asn Thr Ser Ser Pro Thr Gln Pro Arg Pro Pro Gly
325 330 335
Thr Pro Thr Arg Cys Ser Ser Ile Glu Asn Gln Ala Gln Lys Asn His
340 345 350
Leu Ser Val Asp Leu Asn Asp Asp Gly Cys Gly Asn
355 360
<210> 23
<211> 354
<212> БЕЛОК
<213> Theobroma cacao
<400> 23
Met Glu Ala Gly Val Ala Thr Thr Thr Thr Thr Thr Glu Ser Phe Arg
1 5 10 15
Ser Ile Leu Asp Lys Pro Leu Ser Gln Leu Thr Glu Glu Asp Ile Ser
20 25 30
Gln Leu Thr Arg Glu Asp Cys Arg Lys Phe Leu Lys Glu Lys Gly Met
35 40 45
Arg Arg Pro Ser Trp Asn Lys Ser Gln Ala Ile Gln Gln Val Ile Ser
50 55 60
Leu Lys Ala Leu Leu Glu Ser Asn Glu Asp Ser Gly Ala Gly Ala Ile
65 70 75 80
Arg Lys Ile Leu Val Ser Pro Pro Ser Pro Ser Val Pro Pro Gln Asn
85 90 95
Ala Ala Ala Arg Val Ala Ser Asn Ser Cys Asp Ser Val Lys Glu Ala
100 105 110
Val Val Gly Glu Glu Gly Ser Pro Tyr Arg Arg Lys Asp Pro Pro Leu
115 120 125
Lys Pro Ser Pro Val Gly Glu Ile Asn Cys Leu Gly Gly Asp Thr Asp
130 135 140
Asn Lys Asn Leu Ser Pro Arg Ser Pro Cys Glu Ser Asn Glu Leu Gly
145 150 155 160
Gly Gln Met Thr Ile Phe Tyr Cys Gly Lys Val Asn Val Tyr Asp Gly
165 170 175
Val Pro Leu Asp Lys Ala Arg Ala Ile Met His Leu Ala Ala Thr Pro
180 185 190
Ile Asp Phe Pro Gln Asp Asn Gln Cys Ser Gly Asn Ala Ala Leu Arg
195 200 205
Ser Phe Met Cys His Val Gln Ala Val Gly Asp Lys Asn Gly Leu Val
210 215 220
Ala Ser Thr Ala Leu Asn Ser His Thr Met Gln Thr Glu Lys Leu Thr
225 230 235 240
Glu Tyr Gln His Gln Phe Arg Glu Lys Gly Asn Ile Ala Arg Asp Ala
245 250 255
Asp Val Asp Gly Gln Val Asn Arg Lys Val Ser Leu Gln Arg Tyr Arg
260 265 270
Glu Lys Arg Lys Asp Arg Gly Arg Phe Phe Lys Gly Arg Lys Asn Thr
275 280 285
Gly Gln Ala Ser Ser Ser Leu Glu Met Tyr Leu Asn His Gln Ile Arg
290 295 300
Thr His Asn Ser Asn Gly Gln Ser Ser Arg Ser Ser Thr Gly Ser Pro
305 310 315 320
Pro Gln Ser Gly Leu Pro His Ala Phe Cys Ser Ser Ala Asp Asn Gln
325 330 335
Ala Lys Leu Val Asn Leu Ser Val Asp Leu Asn Asp Lys Ser Val Gln
340 345 350
Glu His
<210> 24
<211> 293
<212> БЕЛОК
<213> Spirodela polyrrhiza
<400> 24
Met Ala Gly Ser Glu Ala Ala Ala Pro Glu Glu Ala Gly Arg Ala Gly
1 5 10 15
Glu Glu Glu Val Arg Ala Ala Ala Gly Ala Ala Ala Val Lys Ser Pro
20 25 30
Leu Glu Lys Pro Leu Ser Glu Leu Thr Glu Glu Asp Ile Ala Gln Val
35 40 45
Thr Arg Glu Asp Cys Arg Arg Phe Leu Lys Glu Lys Gly Met Arg Arg
50 55 60
Pro Ser Trp Asn Lys Ser Gln Ala Val Gln Gln Val Ile Ser Leu Lys
65 70 75 80
Ala Leu Leu Glu Pro Cys His Asp Ala Asp Asp Asp Ala Pro Ser Ala
85 90 95
Gly Ala Val Pro Ser Ile Ser Ser Phe Phe Ser Lys Arg Pro Ser Asp
100 105 110
Ala Leu Leu Pro Ala Ala Ala Ala Gln Phe Pro Val Ser Ser Pro Met
115 120 125
Arg Gly Glu Pro Ala Gly Gly Ala Pro Gln Ile Val Ser Glu Arg Pro
130 135 140
His Gly Arg Asp Pro Leu Ala Asn Val Phe Thr Cys Ser Asp Ala Leu
145 150 155 160
Gly Arg Phe Pro Ala Thr Gly Asn Gly Ala Leu Pro Pro Asn Ser Ala
165 170 175
Thr Leu Pro Pro Arg Gly Val Ala Ser Ala Glu Thr Leu Glu Gly Gln
180 185 190
Leu Thr Ile Phe Tyr Asp Gly Lys Ile Asn Val Tyr Asp Gly Val Thr
195 200 205
Pro Glu Lys Val Arg Ser Gly Gln Lys Gly Pro Thr Ser Arg Ala Ala
210 215 220
Ser Leu Gln Arg Tyr Leu Glu Lys Arg Lys Asp Arg Arg Asp Pro Gly
225 230 235 240
Pro Ala Ala Val Ala Thr Leu Tyr Arg Lys Val Phe Leu Ser Ala Thr
245 250 255
Ala Leu Leu Ile Gly Cys Ser Ser Gly Ala Asn Val Val Leu Pro Arg
260 265 270
Ala Glu Gly Pro Thr Ser Arg Ala Ala Ser Leu Gln Arg Tyr Leu Glu
275 280 285
Lys Arg Lys Asp Arg
290
<210> 25
<211> 293
<212> БЕЛОК
<213> Musa acuminata
<400> 25
Met Asn Pro Gly Glu Thr Thr Pro Pro Ser Pro Leu Asp Lys Pro Leu
1 5 10 15
Ala Glu Leu Thr Glu Glu Asp Ile Ala Gln Leu Thr Arg Glu Asp Cys
20 25 30
Arg Arg Phe Leu Lys Ala Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
35 40 45
Ser Gln Ala Ile Gln Gln Val Ile Ser Leu Lys Ala Leu Leu Glu Gly
50 55 60
Arg Pro Gly Cys Asp Asp Cys Pro Ala Gly Gly Gly Ile Leu Gln Lys
65 70 75 80
Leu Leu Thr Ser Ser Pro Ser Glu Pro Leu Ser Pro Pro Gln Asp Ser
85 90 95
Pro Pro Pro Ala Pro Lys Glu Gly Gly Ser Gly Ser Gln Pro Leu Ala
100 105 110
Lys Glu Pro Ser Pro Tyr Arg Arg Arg Asp Pro Ile Pro Pro Pro Tyr
115 120 125
Ser Ala Gly Asn Pro Thr Cys Gln Thr Pro Ile Ala Gly Ala Asp Leu
130 135 140
Pro His Pro Pro Glu Lys Arg Cys Pro Ser Pro Arg Leu Thr Ala Glu
145 150 155 160
Val Pro Val Gly Gln Met Thr Ile Phe Tyr Asp Gly Met Val Asn Val
165 170 175
Tyr Asp Gly Val Ser Ala Asp Gln Ala Arg Ser Ile Met Glu Leu Ala
180 185 190
Ala Ser Pro Val Cys Phe Asp Asp Pro Thr Gly Ala Phe Ser Pro Ala
195 200 205
Arg Pro Pro Ala Phe Arg Phe Pro Pro Gly Leu Pro Arg Pro Ala Pro
210 215 220
Val Pro Thr Ala Pro Ser Phe Val Gly Thr Phe Pro Ile Ser Pro Ala
225 230 235 240
Gly Lys Arg Cys Tyr Ser Tyr Cys Ser Phe Arg Ser Ser Val Ser Leu
245 250 255
Leu Thr Thr Thr Glu Gly Pro Thr Ser Arg Lys Ala Ser Leu Gln Arg
260 265 270
Tyr Leu Glu Lys Arg Lys Asp Arg Tyr Gly His Leu Pro Thr Glu Ser
275 280 285
Ile Leu Leu Val Ser
290
<210> 26
<211> 171
<212> БЕЛОК
<213> Phalaenopsis aphrodite
<220>
<221> дополнительный признак
<222> (72)..(87)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<400> 26
Met Asn Ser Asp Ala Ile Thr Met Gly Lys Ser Leu Leu Glu Lys Pro
1 5 10 15
Leu Ser Leu Leu Thr Glu Asp Asp Ile Ala Gln Ile Thr Arg Glu Glu
20 25 30
Cys Arg Arg Phe Leu Lys Asp Arg Gly Met Arg Arg Pro Ser Trp Asn
35 40 45
Lys Ser Gln Ala Ile Gln Gln Val Ile Ser Leu Lys Ala Leu Phe Glu
50 55 60
Asn Arg Ser Asp Leu Glu Asp Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
65 70 75 80
Xaa Xaa Xaa Xaa Xaa Xaa Xaa Phe Pro Glu His Ala Asp Leu Ser Ser
85 90 95
Ile Ser Pro Thr Ala Glu Ala Lys Glu Pro Glu Lys Ala Gln Leu Thr
100 105 110
Ile Phe Tyr Gly Gly Lys Val Leu Val Phe Asp Asn Phe Pro Val Asn
115 120 125
Lys Ala Gln Asp Leu Met Gln Ile Ala Gly Lys Glu Gln Asn Gln Asn
130 135 140
Tyr Gly Thr Ala Asn Thr Val Ala Pro Ser Ala Pro Ala Ala Asp Leu
145 150 155 160
His Ser Leu Pro Leu Pro Ala Lys Pro Pro Ala
165 170
<210> 27
<211> 46
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 27
Leu Lys Leu Leu Thr Glu Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp
1 5 10 15
Cys Arg Lys Phe Leu Lys Asp Lys Gly Met Arg Arg Pro Ser Trp Asn
20 25 30
Lys Ser Gln Ala Ile Gln Gln Val Leu Ser Leu Lys Ala Leu
35 40 45
<210> 28
<211> 46
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<220>
<221> дополнительный признак
<222> (2)..(3)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (5)..(7)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (9)..(10)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (12)..(12)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (16)..(16)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (19)..(20)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (22)..(24)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (28)..(28)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (34)..(34)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (37)..(38)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (40)..(41)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (43)..(45)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<400> 28
Leu Xaa Xaa Leu Xaa Xaa Xaa Asp Xaa Xaa Gln Xaa Thr Arg Glu Xaa
1 5 10 15
Cys Arg Xaa Xaa Leu Xaa Xaa Xaa Gly Met Arg Xaa Pro Ser Trp Asn
20 25 30
Lys Xaa Gln Ala Xaa Xaa Gln Xaa Xaa Ser Xaa Xaa Xaa Leu
35 40 45
<210> 29
<211> 46
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (2)..(2)
<223> Xaa представляет собой Glu, Phe, His, Tyr, Asn, Trp, Ser, Lys или Thr
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (3)..(3)
<223> Xaa представляет собой Glu, Gln или Leu
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (5)..(5)
<223> Xaa представляет собой Thr или Ser
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (6)..(6)
<223> Xaa представляет собой Glu или Asp
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (7)..(7)
<223> Xaa представляет собой Glu, Leu или Asp
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (9)..(9)
<223> Xaa представляет собой Ile или Val
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (10)..(10)
<223> Xaa представляет собой Ser, Phe, Arg, Met или Thr
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (12)..(12)
<223> Xaa представляет собой Leu или Val
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (16)..(16)
<223> Xaa представляет собой Asp или Glu
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (19)..(19)
<223> Xaa представляет собой Arg или Lys
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (20)..(20)
<223> Xaa представляет собой Tyr или Phe
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (22)..(22)
<223> Xaa представляет собой Lys или Arg
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (23)..(23)
<223> Xaa представляет собой Glu, Asp, Gln или Ala
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (24)..(24)
<223> Xaa представляет собой Lys или Arg
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (28)..(28)
<223> Xaa представляет собой Lys или Arg
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (34)..(34)
<223> Xaa представляет собой Tyr, Ser или Ala
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (37)..(37)
<223> Xaa представляет собой Ile или Val
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (38)..(38)
<223> Xaa представляет собой Gln или Glu
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (40)..(40)
<223> Xaa представляет собой Val или Leu
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (41)..(41)
<223> Xaa представляет собой Leu или Ile
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (43)..(43)
<223> Xaa представляет собой Leu или Phe
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (44)..(44)
<223> Xaa представляет собой Lys или Arg
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (45)..(45)
<223> Xaa представляет собой Gly, Ser, Thr или Ala
<400> 29
Leu Xaa Xaa Leu Xaa Xaa Xaa Asp Xaa Xaa Gln Xaa Thr Arg Glu Xaa
1 5 10 15
Cys Arg Xaa Xaa Leu Xaa Xaa Xaa Gly Met Arg Xaa Pro Ser Trp Asn
20 25 30
Lys Xaa Gln Ala Xaa Xaa Gln Xaa Xaa Ser Xaa Xaa Xaa Leu
35 40 45
<210> 30
<211> 27
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 30
Thr Arg Glu Asp Cys Arg Lys Phe Leu Lys Asp Lys Gly Met Arg Arg
1 5 10 15
Pro Ser Trp Asn Lys Ser Gln Ala Ile Gln Gln
20 25
<210> 31
<211> 27
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<220>
<221> дополнительный признак
<222> (4)..(4)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (7)..(8)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (10)..(12)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (16)..(16)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (22)..(22)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (25)..(26)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<400> 31
Thr Arg Glu Xaa Cys Arg Xaa Xaa Leu Xaa Xaa Xaa Gly Met Arg Xaa
1 5 10 15
Pro Ser Trp Asn Lys Xaa Gln Ala Xaa Xaa Gln
20 25
<210> 32
<211> 27
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (4)..(4)
<223> Xaa представляет собой Asp или Glu
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (7)..(7)
<223> Xaa представляет собой Lys или Arg
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (8)..(8)
<223> Xaa представляет собой Phe или Tyr
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (10)..(10)
<223> Xaa представляет собой Lys или Arg
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (11)..(11)
<223> Xaa представляет собой Asp, Ala, Glu или Gln
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (12)..(12)
<223> Xaa представляет собой Lys или Arg
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (16)..(16)
<223> Xaa представляет собой Lys или Arg
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (22)..(22)
<223> Xaa представляет собой Tyr, Ser или Ala
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (25)..(25)
<223> Xaa представляет собой Ile или Val
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (26)..(26)
<223> Xaa представляет собой Gln или Glu
<400> 32
Thr Arg Glu Xaa Cys Arg Xaa Xaa Leu Xaa Xaa Xaa Gly Met Arg Xaa
1 5 10 15
Pro Ser Trp Asn Lys Xaa Gln Ala Xaa Xaa Gln
20 25
<210> 33
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 33
Thr Ile Phe Tyr Ser Gly
1 5
<210> 34
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<220>
<221> дополнительный признак
<222> (2)..(2)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<220>
<221> дополнительный признак
<222> (5)..(5)
<223> Xaa может представлять собой любую встречающуюся в природе аминокислоту
<400> 34
Thr Xaa Phe Tyr Xaa Gly
1 5
<210> 35
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (2)..(2)
<223> Xaa представляет собой Ile или Met
<220>
<221> ДОПОЛНИТЕЛЬНЫЙ ПРИЗНАК
<222> (5)..(5)
<223> Xaa представляет собой Ala, Ser, Asp, Cys, Arg или Gly
<400> 35
Thr Xaa Phe Tyr Xaa Gly
1 5
<210> 36
<211> 355
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 36
Met Asp Val Gly Val Ser Pro Ala Lys Ser Ile Leu Ala Lys Pro Leu
1 5 10 15
Lys Leu Leu Thr Glu Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys
20 25 30
Arg Lys Phe Leu Lys Asp Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
35 40 45
Ser Gln Ala Ile Gln Gln Val Leu Ser Leu Lys Ala Leu Tyr Glu Pro
50 55 60
Gly Asp Asp Ser Gly Ala Gly Ile Phe Arg Lys Ile Leu Val Ser Gln
65 70 75 80
Pro Val Asn Pro Pro Arg Val Thr Thr Thr Leu Ile Glu Pro Ser Asn
85 90 95
Glu Leu Glu Ala Cys Gly Arg Val Ser Tyr Pro Glu Asp Asn Gly Ala
100 105 110
Cys His Arg Met Asp Ser Pro Arg Ser Ala Glu Phe Ser Gly Gly Ser
115 120 125
Gly His Phe Val Ser Glu Lys Asp Gly His Lys Thr Thr Ile Ser Pro
130 135 140
Arg Ser Pro Ala Glu Thr Ser Glu Leu Val Gly Gln Met Thr Ile Phe
145 150 155 160
Tyr Ser Gly Lys Val Asn Val Tyr Asp Gly Ile Pro Pro Glu Lys Ala
165 170 175
Arg Ser Ile Met His Phe Ala Ala Asn Pro Ile Asp Leu Pro Glu Asn
180 185 190
Gly Ile Phe Ala Ser Ser Arg Met Ile Ser Lys Leu Ile Ser Lys Glu
195 200 205
Lys Met Met Glu Leu Pro Gln Lys Gly Leu Glu Lys Ala Asn Ser Ser
210 215 220
Arg Asp Ser Gly Met Glu Gly Gln Ala Asn Arg Lys Val Ser Leu Gln
225 230 235 240
Arg Tyr Arg Glu Lys Arg Lys Asp Arg Lys Phe Ser Lys Ala Lys Lys
245 250 255
Cys Pro Gly Val Ala Ser Ser Ser Leu Glu Met Phe Leu Asn Cys Gln
260 265 270
Pro Arg Met Lys Ala Ala Tyr Ser Gln Asn Leu Gly Cys Thr Gly Ser
275 280 285
Pro Leu His Ser Gln Ser Pro Glu Ser Gln Thr Lys Ser Pro Asn Leu
290 295 300
Ser Val Asp Leu Asn Ser Glu Gly Ile Gly Ser Gly Gly Gly Ser Ala
305 310 315 320
Lys Gly Glu Leu Arg Gly His Pro Phe Glu Gly Lys Pro Ile Pro Asn
325 330 335
Pro Leu Leu Gly Leu Asp Ser Thr Arg Thr Gly His His His His His
340 345 350
His Gly Ser
355
<210> 37
<211> 42
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 37
Gly Ser Gly Gly Gly Ser Ala Lys Gly Glu Leu Arg Gly His Pro Phe
1 5 10 15
Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp Ser Thr Arg
20 25 30
Thr Gly His His His His His His Gly Ser
35 40
<210> 38
<211> 315
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 38
Met Asp Val Gly Val Ser Pro Ala Lys Ser Ile Leu Ala Lys Pro Leu
1 5 10 15
Lys Leu Leu Thr Glu Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys
20 25 30
Arg Lys Phe Leu Lys Asp Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
35 40 45
Ser Gln Ala Ile Gln Gln Val Leu Ser Leu Lys Ala Leu Tyr Glu Pro
50 55 60
Gly Asp Asp Ser Gly Ala Gly Ile Phe Arg Lys Ile Leu Val Ser Gln
65 70 75 80
Pro Val Asn Pro Pro Arg Val Thr Thr Thr Leu Ile Glu Pro Ser Asn
85 90 95
Glu Leu Glu Ala Cys Gly Arg Val Ser Tyr Pro Glu Asp Asn Gly Ala
100 105 110
Cys His Arg Met Asp Ser Pro Arg Ser Ala Glu Phe Ser Gly Gly Ser
115 120 125
Gly His Phe Val Ser Glu Lys Asp Gly His Lys Thr Thr Ile Ser Pro
130 135 140
Arg Ser Pro Ala Glu Thr Ser Glu Leu Val Gly Gln Met Thr Ile Phe
145 150 155 160
Tyr Ser Gly Lys Val Asn Val Tyr Asp Gly Ile Pro Pro Glu Lys Ala
165 170 175
Arg Ser Ile Met His Phe Ala Ala Asn Pro Ile Asp Leu Pro Glu Asn
180 185 190
Gly Ile Phe Ala Ser Ser Arg Met Ile Ser Lys Leu Ile Ser Lys Glu
195 200 205
Lys Met Met Glu Leu Pro Gln Lys Gly Leu Glu Lys Ala Asn Ser Ser
210 215 220
Arg Asp Ser Gly Met Glu Gly Gln Ala Asn Arg Lys Val Ser Leu Gln
225 230 235 240
Arg Tyr Arg Glu Lys Arg Lys Asp Arg Lys Phe Ser Lys Ala Lys Lys
245 250 255
Cys Pro Gly Val Ala Ser Ser Ser Leu Glu Met Phe Leu Asn Cys Gln
260 265 270
Pro Arg Met Lys Ala Ala Tyr Ser Gln Asn Leu Gly Cys Thr Gly Ser
275 280 285
Pro Leu His Ser Gln Ser Pro Glu Ser Gln Thr Lys Ser Pro Asn Leu
290 295 300
Ser Val Asp Leu Asn Ser Glu Gly Ile Gly Ser
305 310 315
<210> 39
<211> 86
<212> ДНК
<213> Lolium perenne
<400> 39
gtaatgttca gctctgctat agtgtgtgcc accctgcttg tttaataatg cgttctcttc 60
gtttttatga tatcttattc ttccag 86
<210> 40
<211> 1157
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 40
atggacgtgg gcgtgtcccc ggccaagtct attctcgcca agccggtaat gttcagctct 60
gctatagtgt gtgccaccct gcttgtttaa taatgcgttc tcttcgtttt tatgatatct 120
tattcttcca gctcaagctc ctcaccgagg aggacatctc tcagctcaca agagaggact 180
gccgcaagtt cctgaaggac aaggggatga gaaggccttc ctggaacaag tcccaggcca 240
tccagcaagt gctcagcctc aaggcccttt acgagccagg cgacgactcc ggcgctggca 300
ttttcagaaa gatcctcgtg tcccagccgg tgaacccacc aagggtgacc accacactca 360
tcgagccgtc caatgagctt gaggcttgcg gcagagtgtc ctacccagag gataatggcg 420
cctgccacag gatggattct ccaaggtctg ctgagttctc tggcggctcc ggccatttcg 480
tgtctgagaa ggatggccac aagaccacca tctccccaag atccccagcc gagacatctg 540
agcttgtggg ccagatgacc atcttctact ccggcaaggt gaacgtgtac gacggcatcc 600
caccagagaa ggcccgctcc attatgcact tcgccgccaa cccaatcgac ctcccagaga 660
atggcatctt cgcctccagc cgcatgatct ccaagctcat ctccaaggag aagatgatgg 720
agctgccgca gaagggcctc gagaaggcta attcctctcg cgactccggc atggagggcc 780
aggctaatag aaaggtgtcc ctccaacgct accgcgagaa gaggaaggac cgcaagttct 840
ccaaggccaa gaagtgccca ggcgttgcct cttccagcct cgagatgttc ctcaactgcc 900
agccgagaat gaaggccgcc tactcccaaa atctcggctg cacaggctcc ccactccatt 960
ctcagtcccc agagtctcag accaagtccc cgaacctctc cgtggacctt aactccgagg 1020
gcatcggatc cggcggcggc tctgctaagg gcgagctgag gggccacccg ttcgagggca 1080
agccaattcc aaatccactc ctcggcctcg actctaccag gaccggccac catcaccatc 1140
accacggatc ctaatga 1157
<210> 41
<211> 2921
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 41
gcggccgctc tagaactagt ccccttattg tacttcaatt aattatcatt atatcagcat 60
aaacattata ataagtttct tgcgtgttgg aacgtcattt tagttattct aaagaggaaa 120
tagtttcttt tttgctcatg acatcagaca tctggactac tatactggag tttacctttt 180
cttctcctct ttttcttatt gttcctctaa aaaaaattat cactttttaa atgcattagt 240
taaacttatc tcaacaacgt ttaaaattca tttcttgaat gcccattaca atgtaatagt 300
ataacttaat tagtcgtctc catgaaccat taatacgtac ggagtaatat aaaacaccat 360
tggggagttc aatttgcaat aatttcttgc aaaaatgtaa agtacctttt tgttcttgca 420
aaattttaca aataaaaatt tgcagctctt ttttttctct ctctccaaat actagctcaa 480
aacccacaaa tatttttgaa tttatggcat acttttagaa tgcgtttgat gcaactattt 540
tcctttagga aatattcaca acaatctaag acaatcaaaa agtagaaaat agtttgtaaa 600
aagggatgtg gaggacatct taatcaaata ttttcagttt aaaacttgaa aatgaaaaaa 660
cacccgaaag gaaatgattc gttctttaat atgtcctaca caatgtgaat ttgaattagt 720
ttggtcatac ggtatatcat atgattataa ataaaaaaaa ttagcaaaag aatataattt 780
attaaatatt ttacaccata ccaaacacaa ccgcattata tataatctta attatcatta 840
tcaccagcat caacattata atgattcccc tatgcgttgg aacgtcatta tagttattct 900
aaacaagaaa gaaatttgtt cttgacatca gacatctagt attataactc tagtggagct 960
taccttttct tttccttctt ttttttcttc ttaaaaaaat tatcactttt taaatcttgt 1020
atattagtta agcttatcta aacaaagttt taaattcatt tcttaaacgt ccattacaat 1080
gtaatataac ttagtcgtct caattaaacc attaatgtga aatataaatc aaaaaaagcc 1140
aaagggcggt gggacggcgc caatcatttg tcctagtcca ctcaaataag gcccatggtc 1200
ggcaaaacca aacacaaaat gtgttatttt taattttttc ctcttttatt gttaaagttg 1260
caaaatgtgt tatttttggt aagaccctat ggatatataa agacaggtta tgtgaaactt 1320
ggaaaaccat caagttttaa gcaaaaccct cttaagaact taaattgagc ttcttttggg 1380
gcatttttct agtgagaact aaaaacaagt ttgtacaaaa aagcaggcta aaaaaaacca 1440
tggacgtggg cgtgtccccg gccaagtcta ttctcgccaa gccggtaatg ttcagctctg 1500
ctatagtgtg tgccaccctg cttgtttaat aatgcgttct cttcgttttt atgatatctt 1560
attcttccag ctcaagctcc tcaccgagga ggacatctct cagctcacaa gagaggactg 1620
ccgcaagttc ctgaaggaca aggggatgag aaggccttcc tggaacaagt cccaggccat 1680
ccagcaagtg ctcagcctca aggcccttta cgagccaggc gacgactccg gcgctggcat 1740
tttcagaaag atcctcgtgt cccagccggt gaacccacca agggtgacca ccacactcat 1800
cgagccgtcc aatgagcttg aggcttgcgg cagagtgtcc tacccagagg ataatggcgc 1860
ctgccacagg atggattctc caaggtctgc tgagttctct ggcggctccg gccatttcgt 1920
gtctgagaag gatggccaca agaccaccat ctccccaaga tccccagccg agacatctga 1980
gcttgtgggc cagatgacca tcttctactc cggcaaggtg aacgtgtacg acggcatccc 2040
accagagaag gcccgctcca ttatgcactt cgccgccaac ccaatcgacc tcccagagaa 2100
tggcatcttc gcctccagcc gcatgatctc caagctcatc tccaaggaga agatgatgga 2160
gctgccgcag aagggcctcg agaaggctaa ttcctctcgc gactccggca tggagggcca 2220
ggctaataga aaggtgtccc tccaacgcta ccgcgagaag aggaaggacc gcaagttctc 2280
caaggccaag aagtgcccag gcgttgcctc ttccagcctc gagatgttcc tcaactgcca 2340
gccgagaatg aaggccgcct actcccaaaa tctcggctgc acaggctccc cactccattc 2400
tcagtcccca gagtctcaga ccaagtcccc gaacctctcc gtggacctta actccgaggg 2460
catcggatcc ggcggcggct ctgctaaggg cgagctgagg ggccacccgt tcgagggcaa 2520
gccaattcca aatccactcc tcggcctcga ctctaccagg accggccacc atcaccatca 2580
ccacggatcc taatgaaccc agctttcttg tacaaagtgg tctagtgggt accgcgaatt 2640
tccccgatcg ttcaaacatt tggcaataaa gtttcttaag attgaatcct gttgccggtc 2700
ttgcgatgat tatcatataa tttctgttga attacgttaa gcatgtaata attaacatgt 2760
aatgcatgac gttatttatg agatgggttt ttatgattag agtcccgcaa ttatacattt 2820
aatacgcgat agaaaacaaa atatagcgcg caaactagga taaattatcg cgcgcggtgt 2880
catctatgtt actagatcga agcggccgct ctagaactag t 2921
<210> 42
<211> 2249
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 42
gcggccgctc tagaactagt gtcctacaca atgtgaattt gaattagttt ggtcatacgg 60
tatatcatat gattataaat aaaaaaaatt agcaaaagaa tataatttat taaatatttt 120
acaccatacc aaacacaacc gcattatata taatcttaat tatcattatc accagcatca 180
acattataat gattccccta tgcgttggaa cgtcattata gttattctaa acaagaaaga 240
aatttgttct tgacatcaga catctagtat tataactcta gtggagctta ccttttcttt 300
tccttctttt ttttcttctt aaaaaaatta tcacttttta aatcttgtat attagttaag 360
cttatctaaa caaagtttta aattcatttc ttaaacgtcc attacaatgt aatataactt 420
agtcgtctca attaaaccat taatgtgaaa tataaatcaa aaaaagccaa agggcggtgg 480
gacggcgcca atcatttgtc ctagtccact caaataaggc ccatggtcgg caaaaccaaa 540
cacaaaatgt gttattttta attttttcct cttttattgt taaagttgca aaatgtgtta 600
tttttggtaa gaccctatgg atatataaag acaggttatg tgaaacttgg aaaaccatca 660
agttttaagc aaaaccctct taagaactta aattgagctt cttttggggc atttttctag 720
tgagaactaa aaacaagttt gtacaaaaaa gcaggctaaa aaaaaccatg gacgtgggcg 780
tgtccccggc caagtctatt ctcgccaagc cggtaatgtt cagctctgct atagtgtgtg 840
ccaccctgct tgtttaataa tgcgttctct tcgtttttat gatatcttat tcttccagct 900
caagctcctc accgaggagg acatctctca gctcacaaga gaggactgcc gcaagttcct 960
gaaggacaag gggatgagaa ggccttcctg gaacaagtcc caggccatcc agcaagtgct 1020
cagcctcaag gccctttacg agccaggcga cgactccggc gctggcattt tcagaaagat 1080
cctcgtgtcc cagccggtga acccaccaag ggtgaccacc acactcatcg agccgtccaa 1140
tgagcttgag gcttgcggca gagtgtccta cccagaggat aatggcgcct gccacaggat 1200
ggattctcca aggtctgctg agttctctgg cggctccggc catttcgtgt ctgagaagga 1260
tggccacaag accaccatct ccccaagatc cccagccgag acatctgagc ttgtgggcca 1320
gatgaccatc ttctactccg gcaaggtgaa cgtgtacgac ggcatcccac cagagaaggc 1380
ccgctccatt atgcacttcg ccgccaaccc aatcgacctc ccagagaatg gcatcttcgc 1440
ctccagccgc atgatctcca agctcatctc caaggagaag atgatggagc tgccgcagaa 1500
gggcctcgag aaggctaatt cctctcgcga ctccggcatg gagggccagg ctaatagaaa 1560
ggtgtccctc caacgctacc gcgagaagag gaaggaccgc aagttctcca aggccaagaa 1620
gtgcccaggc gttgcctctt ccagcctcga gatgttcctc aactgccagc cgagaatgaa 1680
ggccgcctac tcccaaaatc tcggctgcac aggctcccca ctccattctc agtccccaga 1740
gtctcagacc aagtccccga acctctccgt ggaccttaac tccgagggca tcggatccgg 1800
cggcggctct gctaagggcg agctgagggg ccacccgttc gagggcaagc caattccaaa 1860
tccactcctc ggcctcgact ctaccaggac cggccaccat caccatcacc acggatccta 1920
atgaacccag ctttcttgta caaagtggtc tagtgggtac cgcgaatttc cccgatcgtt 1980
caaacatttg gcaataaagt ttcttaagat tgaatcctgt tgccggtctt gcgatgatta 2040
tcatataatt tctgttgaat tacgttaagc atgtaataat taacatgtaa tgcatgacgt 2100
tatttatgag atgggttttt atgattagag tcccgcaatt atacatttaa tacgcgatag 2160
aaaacaaaat atagcgcgca aactaggata aattatcgcg cgcggtgtca tctatgttac 2220
tagatcgaag cggccgctct agaactagt 2249
<210> 43
<211> 2852
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 43
gcggccgctc tagaactagt tcacgtggaa cgcgccgcag taaacggagc ggtggatcaa 60
acttttcgtc cgtttgatca aacagaagag aactagtcaa tgctctttct tcatatcaca 120
atttaatagt ctcaagacga ttacgccaca taaccatttt ctcgtgattt cgacatcaaa 180
atttaataaa aggaactgat tgattggtca tcatgttaca agtgtcaaat gagctaatcc 240
gttttacagt ggcatagttt acgatcaatt tacaaatttt tggttttata acatacttgt 300
agttaaaact atttataagc tatttatagt gagttagctt ataaaaccct attcttttat 360
ctaaaattat gttttgactc gtttcatgat aaaattttat ccttttcatc ggaataaaaa 420
actttattat ttatttggca aaataattgg tgtaaaaatt atgtatatgt taataacaaa 480
aaatattaat ctgattcata atcttaaaaa agaaaaattt cttgaaataa actttagaca 540
ttgtaaataa aaaaacatta tttttatata atgggatgtt tatatgtaat tttttataaa 600
aaaataaaag ttgtttacta gtaatccgat tggctttaac tatcgtcgcc aaaagaataa 660
tgtagaactg actttgaggt aaaactaaaa gaaatttgta agataatagt cacattaaat 720
gctaaaatta atacatactg atatatcgta taaaatttat gaaaactaca ccttaacctg 780
aatcatacac tgtaataaaa aaaacaaatt atatataaac cctaaaaact aatcataaat 840
cccaaacggt gtactctcta ttagctttga aaggattgcc caattgtttg ttaaaaattt 900
ctaataatag tacaatgttt tgtttcattt ttccttttcg tcaacctgtt acccaatagc 960
aaatgaagtt tttatgtgtg tgtgtgtgtg tgaatttcca tgaaaatgaa acgggcttag 1020
aatcccggtg tattatgggt cgggtcgtaa ccgggcaatg acgcaggatc tgacgtaaaa 1080
ctcccaagaa tttttttaaa aagtctccgg aaaataaaat caaagttcat taacttaaaa 1140
agaaaaaaca aaatcggtcc acgtcccaaa ccctttttat aggagagtct tatgttctgg 1200
cagaagactt cacagactct ttcttaatct ctctctcttt caaccaaacc cctaaacaaa 1260
aaaaaaatac attttctgat ctctctaaaa atctttctcc ttcgttaatc tcgtgatctc 1320
tttctttttc tatatacaag tttgtacaaa aaagcaggct aaaaaaaacc atggacgtgg 1380
gcgtgtcccc ggccaagtct attctcgcca agccggtaat gttcagctct gctatagtgt 1440
gtgccaccct gcttgtttaa taatgcgttc tcttcgtttt tatgatatct tattcttcca 1500
gctcaagctc ctcaccgagg aggacatctc tcagctcaca agagaggact gccgcaagtt 1560
cctgaaggac aaggggatga gaaggccttc ctggaacaag tcccaggcca tccagcaagt 1620
gctcagcctc aaggcccttt acgagccagg cgacgactcc ggcgctggca ttttcagaaa 1680
gatcctcgtg tcccagccgg tgaacccacc aagggtgacc accacactca tcgagccgtc 1740
caatgagctt gaggcttgcg gcagagtgtc ctacccagag gataatggcg cctgccacag 1800
gatggattct ccaaggtctg ctgagttctc tggcggctcc ggccatttcg tgtctgagaa 1860
ggatggccac aagaccacca tctccccaag atccccagcc gagacatctg agcttgtggg 1920
ccagatgacc atcttctact ccggcaaggt gaacgtgtac gacggcatcc caccagagaa 1980
ggcccgctcc attatgcact tcgccgccaa cccaatcgac ctcccagaga atggcatctt 2040
cgcctccagc cgcatgatct ccaagctcat ctccaaggag aagatgatgg agctgccgca 2100
gaagggcctc gagaaggcta attcctctcg cgactccggc atggagggcc aggctaatag 2160
aaaggtgtcc ctccaacgct accgcgagaa gaggaaggac cgcaagttct ccaaggccaa 2220
gaagtgccca ggcgttgcct cttccagcct cgagatgttc ctcaactgcc agccgagaat 2280
gaaggccgcc tactcccaaa atctcggctg cacaggctcc ccactccatt ctcagtcccc 2340
agagtctcag accaagtccc cgaacctctc cgtggacctt aactccgagg gcatcggatc 2400
cggcggcggc tctgctaagg gcgagctgag gggccacccg ttcgagggca agccaattcc 2460
aaatccactc ctcggcctcg actctaccag gaccggccac catcaccatc accacggatc 2520
ctaatgaacc cagctttctt gtacaaagtg gtctagtggg taccgcgaat ttccccgatc 2580
gttcaaacat ttggcaataa agtttcttaa gattgaatcc tgttgccggt cttgcgatga 2640
ttatcatata atttctgttg aattacgtta agcatgtaat aattaacatg taatgcatga 2700
cgttatttat gagatgggtt tttatgatta gagtcccgca attatacatt taatacgcga 2760
tagaaaacaa aatatagcgc gcaaactagg ataaattatc gcgcgcggtg tcatctatgt 2820
tactagatcg aagcggccgc tctagaacta gt 2852
<210> 44
<211> 2801
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 44
gcggccgctc tagaactagt ccccttattg tacttcaatt aattatcatt atatcagcat 60
aaacattata ataagtttct tgcgtgttgg aacgtcattt tagttattct aaagaggaaa 120
tagtttcttt tttgctcatg acatcagaca tctggactac tatactggag tttacctttt 180
cttctcctct ttttcttatt gttcctctaa aaaaaattat cactttttaa atgcattagt 240
taaacttatc tcaacaacgt ttaaaattca tttcttgaat gcccattaca atgtaatagt 300
ataacttaat tagtcgtctc catgaaccat taatacgtac ggagtaatat aaaacaccat 360
tggggagttc aatttgcaat aatttcttgc aaaaatgtaa agtacctttt tgttcttgca 420
aaattttaca aataaaaatt tgcagctctt ttttttctct ctctccaaat actagctcaa 480
aacccacaaa tatttttgaa tttatggcat acttttagaa tgcgtttgat gcaactattt 540
tcctttagga aatattcaca acaatctaag acaatcaaaa agtagaaaat agtttgtaaa 600
aagggatgtg gaggacatct taatcaaata ttttcagttt aaaacttgaa aatgaaaaaa 660
cacccgaaag gaaatgattc gttctttaat atgtcctaca caatgtgaat ttgaattagt 720
ttggtcatac ggtatatcat atgattataa ataaaaaaaa ttagcaaaag aatataattt 780
attaaatatt ttacaccata ccaaacacaa ccgcattata tataatctta attatcatta 840
tcaccagcat caacattata atgattcccc tatgcgttgg aacgtcatta tagttattct 900
aaacaagaaa gaaatttgtt cttgacatca gacatctagt attataactc tagtggagct 960
taccttttct tttccttctt ttttttcttc ttaaaaaaat tatcactttt taaatcttgt 1020
atattagtta agcttatcta aacaaagttt taaattcatt tcttaaacgt ccattacaat 1080
gtaatataac ttagtcgtct caattaaacc attaatgtga aatataaatc aaaaaaagcc 1140
aaagggcggt gggacggcgc caatcatttg tcctagtcca ctcaaataag gcccatggtc 1200
ggcaaaacca aacacaaaat gtgttatttt taattttttc ctcttttatt gttaaagttg 1260
caaaatgtgt tatttttggt aagaccctat ggatatataa agacaggtta tgtgaaactt 1320
ggaaaaccat caagttttaa gcaaaaccct cttaagaact taaattgagc ttcttttggg 1380
gcatttttct agtgagaact aaaaacaagt ttgtacaaaa aagcaggcta aaaaaaacca 1440
tggacgtggg cgtgtccccg gccaagtcta ttctcgccaa gccggtaatg ttcagctctg 1500
ctatagtgtg tgccaccctg cttgtttaat aatgcgttct cttcgttttt atgatatctt 1560
attcttccag ctcaagctcc tcaccgagga ggacatctct cagctcacaa gagaggactg 1620
ccgcaagttc ctgaaggaca aggggatgag aaggccttcc tggaacaagt cccaggccat 1680
ccagcaagtg ctcagcctca aggcccttta cgagccaggc gacgactccg gcgctggcat 1740
tttcagaaag atcctcgtgt cccagccggt gaacccacca agggtgacca ccacactcat 1800
cgagccgtcc aatgagcttg aggcttgcgg cagagtgtcc tacccagagg ataatggcgc 1860
ctgccacagg atggattctc caaggtctgc tgagttctct ggcggctccg gccatttcgt 1920
gtctgagaag gatggccaca agaccaccat ctccccaaga tccccagccg agacatctga 1980
gcttgtgggc cagatgacca tcttctactc cggcaaggtg aacgtgtacg acggcatccc 2040
accagagaag gcccgctcca ttatgcactt cgccgccaac ccaatcgacc tcccagagaa 2100
tggcatcttc gcctccagcc gcatgatctc caagctcatc tccaaggaga agatgatgga 2160
gctgccgcag aagggcctcg agaaggctaa ttcctctcgc gactccggca tggagggcca 2220
ggctaataga aaggtgtccc tccaacgcta ccgcgagaag aggaaggacc gcaagttctc 2280
caaggccaag aagtgcccag gcgttgcctc ttccagcctc gagatgttcc tcaactgcca 2340
gccgagaatg aaggccgcct actcccaaaa tctcggctgc acaggctccc cactccattc 2400
tcagtcccca gagtctcaga ccaagtcccc gaacctctcc gtggacctta actccgaggg 2460
catcggatcc taatgaaccc agctttcttg tacaaagtgg tctagtgggt accgcgaatt 2520
tccccgatcg ttcaaacatt tggcaataaa gtttcttaag attgaatcct gttgccggtc 2580
ttgcgatgat tatcatataa tttctgttga attacgttaa gcatgtaata attaacatgt 2640
aatgcatgac gttatttatg agatgggttt ttatgattag agtcccgcaa ttatacattt 2700
aatacgcgat agaaaacaaa atatagcgcg caaactagga taaattatcg cgcgcggtgt 2760
catctatgtt actagatcga agcggccgct ctagaactag t 2801
<210> 45
<211> 2129
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 45
gcggccgctc tagaactagt gtcctacaca atgtgaattt gaattagttt ggtcatacgg 60
tatatcatat gattataaat aaaaaaaatt agcaaaagaa tataatttat taaatatttt 120
acaccatacc aaacacaacc gcattatata taatcttaat tatcattatc accagcatca 180
acattataat gattccccta tgcgttggaa cgtcattata gttattctaa acaagaaaga 240
aatttgttct tgacatcaga catctagtat tataactcta gtggagctta ccttttcttt 300
tccttctttt ttttcttctt aaaaaaatta tcacttttta aatcttgtat attagttaag 360
cttatctaaa caaagtttta aattcatttc ttaaacgtcc attacaatgt aatataactt 420
agtcgtctca attaaaccat taatgtgaaa tataaatcaa aaaaagccaa agggcggtgg 480
gacggcgcca atcatttgtc ctagtccact caaataaggc ccatggtcgg caaaaccaaa 540
cacaaaatgt gttattttta attttttcct cttttattgt taaagttgca aaatgtgtta 600
tttttggtaa gaccctatgg atatataaag acaggttatg tgaaacttgg aaaaccatca 660
agttttaagc aaaaccctct taagaactta aattgagctt cttttggggc atttttctag 720
tgagaactaa aaacaagttt gtacaaaaaa gcaggctaaa aaaaaccatg gacgtgggcg 780
tgtccccggc caagtctatt ctcgccaagc cggtaatgtt cagctctgct atagtgtgtg 840
ccaccctgct tgtttaataa tgcgttctct tcgtttttat gatatcttat tcttccagct 900
caagctcctc accgaggagg acatctctca gctcacaaga gaggactgcc gcaagttcct 960
gaaggacaag gggatgagaa ggccttcctg gaacaagtcc caggccatcc agcaagtgct 1020
cagcctcaag gccctttacg agccaggcga cgactccggc gctggcattt tcagaaagat 1080
cctcgtgtcc cagccggtga acccaccaag ggtgaccacc acactcatcg agccgtccaa 1140
tgagcttgag gcttgcggca gagtgtccta cccagaggat aatggcgcct gccacaggat 1200
ggattctcca aggtctgctg agttctctgg cggctccggc catttcgtgt ctgagaagga 1260
tggccacaag accaccatct ccccaagatc cccagccgag acatctgagc ttgtgggcca 1320
gatgaccatc ttctactccg gcaaggtgaa cgtgtacgac ggcatcccac cagagaaggc 1380
ccgctccatt atgcacttcg ccgccaaccc aatcgacctc ccagagaatg gcatcttcgc 1440
ctccagccgc atgatctcca agctcatctc caaggagaag atgatggagc tgccgcagaa 1500
gggcctcgag aaggctaatt cctctcgcga ctccggcatg gagggccagg ctaatagaaa 1560
ggtgtccctc caacgctacc gcgagaagag gaaggaccgc aagttctcca aggccaagaa 1620
gtgcccaggc gttgcctctt ccagcctcga gatgttcctc aactgccagc cgagaatgaa 1680
ggccgcctac tcccaaaatc tcggctgcac aggctcccca ctccattctc agtccccaga 1740
gtctcagacc aagtccccga acctctccgt ggaccttaac tccgagggca tcggatccta 1800
atgaacccag ctttcttgta caaagtggtc tagtgggtac cgcgaatttc cccgatcgtt 1860
caaacatttg gcaataaagt ttcttaagat tgaatcctgt tgccggtctt gcgatgatta 1920
tcatataatt tctgttgaat tacgttaagc atgtaataat taacatgtaa tgcatgacgt 1980
tatttatgag atgggttttt atgattagag tcccgcaatt atacatttaa tacgcgatag 2040
aaaacaaaat atagcgcgca aactaggata aattatcgcg cgcggtgtca tctatgttac 2100
tagatcgaag cggccgctct agaactagt 2129
<210> 46
<211> 2732
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 46
gcggccgctc tagaactagt tcacgtggaa cgcgccgcag taaacggagc ggtggatcaa 60
acttttcgtc cgtttgatca aacagaagag aactagtcaa tgctctttct tcatatcaca 120
atttaatagt ctcaagacga ttacgccaca taaccatttt ctcgtgattt cgacatcaaa 180
atttaataaa aggaactgat tgattggtca tcatgttaca agtgtcaaat gagctaatcc 240
gttttacagt ggcatagttt acgatcaatt tacaaatttt tggttttata acatacttgt 300
agttaaaact atttataagc tatttatagt gagttagctt ataaaaccct attcttttat 360
ctaaaattat gttttgactc gtttcatgat aaaattttat ccttttcatc ggaataaaaa 420
actttattat ttatttggca aaataattgg tgtaaaaatt atgtatatgt taataacaaa 480
aaatattaat ctgattcata atcttaaaaa agaaaaattt cttgaaataa actttagaca 540
ttgtaaataa aaaaacatta tttttatata atgggatgtt tatatgtaat tttttataaa 600
aaaataaaag ttgtttacta gtaatccgat tggctttaac tatcgtcgcc aaaagaataa 660
tgtagaactg actttgaggt aaaactaaaa gaaatttgta agataatagt cacattaaat 720
gctaaaatta atacatactg atatatcgta taaaatttat gaaaactaca ccttaacctg 780
aatcatacac tgtaataaaa aaaacaaatt atatataaac cctaaaaact aatcataaat 840
cccaaacggt gtactctcta ttagctttga aaggattgcc caattgtttg ttaaaaattt 900
ctaataatag tacaatgttt tgtttcattt ttccttttcg tcaacctgtt acccaatagc 960
aaatgaagtt tttatgtgtg tgtgtgtgtg tgaatttcca tgaaaatgaa acgggcttag 1020
aatcccggtg tattatgggt cgggtcgtaa ccgggcaatg acgcaggatc tgacgtaaaa 1080
ctcccaagaa tttttttaaa aagtctccgg aaaataaaat caaagttcat taacttaaaa 1140
agaaaaaaca aaatcggtcc acgtcccaaa ccctttttat aggagagtct tatgttctgg 1200
cagaagactt cacagactct ttcttaatct ctctctcttt caaccaaacc cctaaacaaa 1260
aaaaaaatac attttctgat ctctctaaaa atctttctcc ttcgttaatc tcgtgatctc 1320
tttctttttc tatatacaag tttgtacaaa aaagcaggct aaaaaaaacc atggacgtgg 1380
gcgtgtcccc ggccaagtct attctcgcca agccggtaat gttcagctct gctatagtgt 1440
gtgccaccct gcttgtttaa taatgcgttc tcttcgtttt tatgatatct tattcttcca 1500
gctcaagctc ctcaccgagg aggacatctc tcagctcaca agagaggact gccgcaagtt 1560
cctgaaggac aaggggatga gaaggccttc ctggaacaag tcccaggcca tccagcaagt 1620
gctcagcctc aaggcccttt acgagccagg cgacgactcc ggcgctggca ttttcagaaa 1680
gatcctcgtg tcccagccgg tgaacccacc aagggtgacc accacactca tcgagccgtc 1740
caatgagctt gaggcttgcg gcagagtgtc ctacccagag gataatggcg cctgccacag 1800
gatggattct ccaaggtctg ctgagttctc tggcggctcc ggccatttcg tgtctgagaa 1860
ggatggccac aagaccacca tctccccaag atccccagcc gagacatctg agcttgtggg 1920
ccagatgacc atcttctact ccggcaaggt gaacgtgtac gacggcatcc caccagagaa 1980
ggcccgctcc attatgcact tcgccgccaa cccaatcgac ctcccagaga atggcatctt 2040
cgcctccagc cgcatgatct ccaagctcat ctccaaggag aagatgatgg agctgccgca 2100
gaagggcctc gagaaggcta attcctctcg cgactccggc atggagggcc aggctaatag 2160
aaaggtgtcc ctccaacgct accgcgagaa gaggaaggac cgcaagttct ccaaggccaa 2220
gaagtgccca ggcgttgcct cttccagcct cgagatgttc ctcaactgcc agccgagaat 2280
gaaggccgcc tactcccaaa atctcggctg cacaggctcc ccactccatt ctcagtcccc 2340
agagtctcag accaagtccc cgaacctctc cgtggacctt aactccgagg gcatcggatc 2400
ctaatgaacc cagctttctt gtacaaagtg gtctagtggg taccgcgaat ttccccgatc 2460
gttcaaacat ttggcaataa agtttcttaa gattgaatcc tgttgccggt cttgcgatga 2520
ttatcatata atttctgttg aattacgtta agcatgtaat aattaacatg taatgcatga 2580
cgttatttat gagatgggtt tttatgatta gagtcccgca attatacatt taatacgcga 2640
tagaaaacaa aatatagcgc gcaaactagg ataaattatc gcgcgcggtg tcatctatgt 2700
tactagatcg aagcggccgc tctagaacta gt 2732
<210> 47
<211> 102
<212> ДНК
<213> Arabidopsis thaliana
<400> 47
gtaaaagaag atgtttttta tttccagcaa tgttacattg ttatacgtat aatgatgagt 60
ttagtgatca agttcctctt tgattcttct ttcttgttgc ag 102
<210> 48
<211> 1173
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 48
atggatgtcg gagtttcacc ggcgaagtct atacttgcga aacctctgaa gctactcact 60
gaagaggaca tttctcagct cactcgcgaa gactgcgtaa aagaagatgt tttttatttc 120
cagcaatgtt acattgttat acgtataatg atgagtttag tgatcaagtt cctctttgat 180
tcttctttct tgttgcagcg caaattcctc aaagacaaag gaatgcgaag accgtcgtgg 240
aacaaatctc aggcgatcca gcaagtttta tctcttaaag ctctctatga gcctggagac 300
gattccggcg ccggtatctt ccgcaagatc ctcgtttctc agccagtaaa tccgcctcgc 360
gtcacaacaa cgttgattga gccaagcaac gagctggaag cttgtggccg ggtttcttat 420
ccggaagata acggcgcgtg ccatagaatg gattctccaa gatcagctga gttttccggt 480
gggtctggtc actttgtatc cgagaaagat ggccacaaga cgactatttc tcccagaagc 540
ccagctgaaa caagtgagct cgttgggcaa atgacgatat tctatagtgg aaaagtgaat 600
gtgtatgatg gaataccacc tgaaaaggcc cggtcaatca tgcactttgc agccaatcca 660
attgatttgc ctgaaaacgg tatttttgct tccagtagaa tgatttcaaa gctcataagt 720
aaagagaaga tgatggaact tccccaaaaa ggccttgaga aggcgaattc ttctcgtgat 780
tctggtatgg agggccaggc gaacagaaag gtatctttgc aaagatatcg tgaaaagcgg 840
aaagacagaa aattctcaaa ggccaaaaag tgtccaggag ttgcgtcctc tagcttggag 900
atgtttctga attgtcagcc acggatgaaa gctgcatatt cgcaaaacct aggctgcacc 960
ggatctccac tgcatagcca gtcacctgaa agccagacaa aaagtcccaa tctttcagtt 1020
gatctaaaca gtgaaggcat tggatccgga ggtggatctg ctaagggaga acttagagga 1080
catccatttg aaggaaagcc aattccaaat ccacttcttg gacttgattc tactagaact 1140
ggacatcacc atcaccatca tggatcctaa tga 1173
<210> 49
<211> 1173
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 49
atggatgtcg gagtttcacc ggtaaaagaa gatgtttttt atttccagca atgttacatt 60
gttatacgta taatgatgag tttagtgatc aagttcctct ttgattcttc tttcttgttg 120
caggcgaagt ctatacttgc gaaacctctg aagctactca ctgaagagga catttctcag 180
ctcactcgcg aagactgccg caaattcctc aaagacaaag gaatgcgaag accgtcgtgg 240
aacaaatctc aggcgatcca gcaagtttta tctcttaaag ctctctatga gcctggagac 300
gattccggcg ccggtatctt ccgcaagatc ctcgtttctc agccagtaaa tccgcctcgc 360
gtcacaacaa cgttgattga gccaagcaac gagctggaag cttgtggccg ggtttcttat 420
ccggaagata acggcgcgtg ccatagaatg gattctccaa gatcagctga gttttccggt 480
gggtctggtc actttgtatc cgagaaagat ggccacaaga cgactatttc tcccagaagc 540
ccagctgaaa caagtgagct cgttgggcaa atgacgatat tctatagtgg aaaagtgaat 600
gtgtatgatg gaataccacc tgaaaaggcc cggtcaatca tgcactttgc agccaatcca 660
attgatttgc ctgaaaacgg tatttttgct tccagtagaa tgatttcaaa gctcataagt 720
aaagagaaga tgatggaact tccccaaaaa ggccttgaga aggcgaattc ttctcgtgat 780
tctggtatgg agggccaggc gaacagaaag gtatctttgc aaagatatcg tgaaaagcgg 840
aaagacagaa aattctcaaa ggccaaaaag tgtccaggag ttgcgtcctc tagcttggag 900
atgtttctga attgtcagcc acggatgaaa gctgcatatt cgcaaaacct aggctgcacc 960
ggatctccac tgcatagcca gtcacctgaa agccagacaa aaagtcccaa tctttcagtt 1020
gatctaaaca gtgaaggcat tggatccgga ggtggatctg ctaagggaga acttagagga 1080
catccatttg aaggaaagcc aattccaaat ccacttcttg gacttgattc tactagaact 1140
ggacatcacc atcaccatca tggatcctaa tga 1173
<210> 50
<211> 2897
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 50
ccccttattg tacttcaatt aattatcatt atatcagcat aaacattata ataagtttct 60
tgcgtgttgg aacgtcattt tagttattct aaagaggaaa tagtttcttt tttgctcatg 120
acatcagaca tctggactac tatactggag tttacctttt cttctcctct ttttcttatt 180
gttcctctaa aaaaaattat cactttttaa atgcattagt taaacttatc tcaacaacgt 240
ttaaaattca tttcttgaat gcccattaca atgtaatagt ataacttaat tagtcgtctc 300
catgaaccat taatacgtac ggagtaatat aaaacaccat tggggagttc aatttgcaat 360
aatttcttgc aaaaatgtaa agtacctttt tgttcttgca aaattttaca aataaaaatt 420
tgcagctctt ttttttctct ctctccaaat actagctcaa aacccacaaa tatttttgaa 480
tttatggcat acttttagaa tgcgtttgat gcaactattt tcctttagga aatattcaca 540
acaatctaag acaatcaaaa agtagaaaat agtttgtaaa aagggatgtg gaggacatct 600
taatcaaata ttttcagttt aaaacttgaa aatgaaaaaa cacccgaaag gaaatgattc 660
gttctttaat atgtcctaca caatgtgaat ttgaattagt ttggtcatac ggtatatcat 720
atgattataa ataaaaaaaa ttagcaaaag aatataattt attaaatatt ttacaccata 780
ccaaacacaa ccgcattata tataatctta attatcatta tcaccagcat caacattata 840
atgattcccc tatgcgttgg aacgtcatta tagttattct aaacaagaaa gaaatttgtt 900
cttgacatca gacatctagt attataactc tagtggagct taccttttct tttccttctt 960
ttttttcttc ttaaaaaaat tatcactttt taaatcttgt atattagtta agcttatcta 1020
aacaaagttt taaattcatt tcttaaacgt ccattacaat gtaatataac ttagtcgtct 1080
caattaaacc attaatgtga aatataaatc aaaaaaagcc aaagggcggt gggacggcgc 1140
caatcatttg tcctagtcca ctcaaataag gcccatggtc ggcaaaacca aacacaaaat 1200
gtgttatttt taattttttc ctcttttatt gttaaagttg caaaatgtgt tatttttggt 1260
aagaccctat ggatatataa agacaggtta tgtgaaactt ggaaaaccat caagttttaa 1320
gcaaaaccct cttaagaact taaattgagc ttcttttggg gcatttttct agtgagaact 1380
aaaaacaagt ttgtacaaaa aagcaggcta aaaaaaacca tggatgtcgg agtttcaccg 1440
gcgaagtcta tacttgcgaa acctctgaag ctactcactg aagaggacat ttctcagctc 1500
actcgcgaag actgcgtaaa agaagatgtt ttttatttcc agcaatgtta cattgttata 1560
cgtataatga tgagtttagt gatcaagttc ctctttgatt cttctttctt gttgcagcgc 1620
aaattcctca aagacaaagg aatgcgaaga ccgtcgtgga acaaatctca ggcgatccag 1680
caagttttat ctcttaaagc tctctatgag cctggagacg attccggcgc cggtatcttc 1740
cgcaagatcc tcgtttctca gccagtaaat ccgcctcgcg tcacaacaac gttgattgag 1800
ccaagcaacg agctggaagc ttgtggccgg gtttcttatc cggaagataa cggcgcgtgc 1860
catagaatgg attctccaag atcagctgag ttttccggtg ggtctggtca ctttgtatcc 1920
gagaaagatg gccacaagac gactatttct cccagaagcc cagctgaaac aagtgagctc 1980
gttgggcaaa tgacgatatt ctatagtgga aaagtgaatg tgtatgatgg aataccacct 2040
gaaaaggccc ggtcaatcat gcactttgca gccaatccaa ttgatttgcc tgaaaacggt 2100
atttttgctt ccagtagaat gatttcaaag ctcataagta aagagaagat gatggaactt 2160
ccccaaaaag gccttgagaa ggcgaattct tctcgtgatt ctggtatgga gggccaggcg 2220
aacagaaagg tatctttgca aagatatcgt gaaaagcgga aagacagaaa attctcaaag 2280
gccaaaaagt gtccaggagt tgcgtcctct agcttggaga tgtttctgaa ttgtcagcca 2340
cggatgaaag ctgcatattc gcaaaaccta ggctgcaccg gatctccact gcatagccag 2400
tcacctgaaa gccagacaaa aagtcccaat ctttcagttg atctaaacag tgaaggcatt 2460
ggatccggag gtggatctgc taagggagaa cttagaggac atccatttga aggaaagcca 2520
attccaaatc cacttcttgg acttgattct actagaactg gacatcacca tcaccatcat 2580
ggatcctaat gaacccagct ttcttgtaca aagtggtcta gtgggtaccg cgaatttccc 2640
cgatcgttca aacatttggc aataaagttt cttaagattg aatcctgttg ccggtcttgc 2700
gatgattatc atataatttc tgttgaatta cgttaagcat gtaataatta acatgtaatg 2760
catgacgtta tttatgagat gggtttttat gattagagtc ccgcaattat acatttaata 2820
cgcgatagaa aacaaaatat agcgcgcaaa ctaggataaa ttatcgcgcg cggtgtcatc 2880
tatgttacta gatcgaa 2897
<210> 51
<211> 2225
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 51
gtcctacaca atgtgaattt gaattagttt ggtcatacgg tatatcatat gattataaat 60
aaaaaaaatt agcaaaagaa tataatttat taaatatttt acaccatacc aaacacaacc 120
gcattatata taatcttaat tatcattatc accagcatca acattataat gattccccta 180
tgcgttggaa cgtcattata gttattctaa acaagaaaga aatttgttct tgacatcaga 240
catctagtat tataactcta gtggagctta ccttttcttt tccttctttt ttttcttctt 300
aaaaaaatta tcacttttta aatcttgtat attagttaag cttatctaaa caaagtttta 360
aattcatttc ttaaacgtcc attacaatgt aatataactt agtcgtctca attaaaccat 420
taatgtgaaa tataaatcaa aaaaagccaa agggcggtgg gacggcgcca atcatttgtc 480
ctagtccact caaataaggc ccatggtcgg caaaaccaaa cacaaaatgt gttattttta 540
attttttcct cttttattgt taaagttgca aaatgtgtta tttttggtaa gaccctatgg 600
atatataaag acaggttatg tgaaacttgg aaaaccatca agttttaagc aaaaccctct 660
taagaactta aattgagctt cttttggggc atttttctag tgagaactaa aaacaagttt 720
gtacaaaaaa gcaggctaaa aaaaaccatg gatgtcggag tttcaccggc gaagtctata 780
cttgcgaaac ctctgaagct actcactgaa gaggacattt ctcagctcac tcgcgaagac 840
tgcgtaaaag aagatgtttt ttatttccag caatgttaca ttgttatacg tataatgatg 900
agtttagtga tcaagttcct ctttgattct tctttcttgt tgcagcgcaa attcctcaaa 960
gacaaaggaa tgcgaagacc gtcgtggaac aaatctcagg cgatccagca agttttatct 1020
cttaaagctc tctatgagcc tggagacgat tccggcgccg gtatcttccg caagatcctc 1080
gtttctcagc cagtaaatcc gcctcgcgtc acaacaacgt tgattgagcc aagcaacgag 1140
ctggaagctt gtggccgggt ttcttatccg gaagataacg gcgcgtgcca tagaatggat 1200
tctccaagat cagctgagtt ttccggtggg tctggtcact ttgtatccga gaaagatggc 1260
cacaagacga ctatttctcc cagaagccca gctgaaacaa gtgagctcgt tgggcaaatg 1320
acgatattct atagtggaaa agtgaatgtg tatgatggaa taccacctga aaaggcccgg 1380
tcaatcatgc actttgcagc caatccaatt gatttgcctg aaaacggtat ttttgcttcc 1440
agtagaatga tttcaaagct cataagtaaa gagaagatga tggaacttcc ccaaaaaggc 1500
cttgagaagg cgaattcttc tcgtgattct ggtatggagg gccaggcgaa cagaaaggta 1560
tctttgcaaa gatatcgtga aaagcggaaa gacagaaaat tctcaaaggc caaaaagtgt 1620
ccaggagttg cgtcctctag cttggagatg tttctgaatt gtcagccacg gatgaaagct 1680
gcatattcgc aaaacctagg ctgcaccgga tctccactgc atagccagtc acctgaaagc 1740
cagacaaaaa gtcccaatct ttcagttgat ctaaacagtg aaggcattgg atccggaggt 1800
ggatctgcta agggagaact tagaggacat ccatttgaag gaaagccaat tccaaatcca 1860
cttcttggac ttgattctac tagaactgga catcaccatc accatcatgg atcctaatga 1920
acccagcttt cttgtacaaa gtggtctagt gggtaccgcg aatttccccg atcgttcaaa 1980
catttggcaa taaagtttct taagattgaa tcctgttgcc ggtcttgcga tgattatcat 2040
ataatttctg ttgaattacg ttaagcatgt aataattaac atgtaatgca tgacgttatt 2100
tatgagatgg gtttttatga ttagagtccc gcaattatac atttaatacg cgatagaaaa 2160
caaaatatag cgcgcaaact aggataaatt atcgcgcgcg gtgtcatcta tgttactaga 2220
tcgaa 2225
<210> 52
<211> 2828
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 52
tcacgtggaa cgcgccgcag taaacggagc ggtggatcaa acttttcgtc cgtttgatca 60
aacagaagag aactagtcaa tgctctttct tcatatcaca atttaatagt ctcaagacga 120
ttacgccaca taaccatttt ctcgtgattt cgacatcaaa atttaataaa aggaactgat 180
tgattggtca tcatgttaca agtgtcaaat gagctaatcc gttttacagt ggcatagttt 240
acgatcaatt tacaaatttt tggttttata acatacttgt agttaaaact atttataagc 300
tatttatagt gagttagctt ataaaaccct attcttttat ctaaaattat gttttgactc 360
gtttcatgat aaaattttat ccttttcatc ggaataaaaa actttattat ttatttggca 420
aaataattgg tgtaaaaatt atgtatatgt taataacaaa aaatattaat ctgattcata 480
atcttaaaaa agaaaaattt cttgaaataa actttagaca ttgtaaataa aaaaacatta 540
tttttatata atgggatgtt tatatgtaat tttttataaa aaaataaaag ttgtttacta 600
gtaatccgat tggctttaac tatcgtcgcc aaaagaataa tgtagaactg actttgaggt 660
aaaactaaaa gaaatttgta agataatagt cacattaaat gctaaaatta atacatactg 720
atatatcgta taaaatttat gaaaactaca ccttaacctg aatcatacac tgtaataaaa 780
aaaacaaatt atatataaac cctaaaaact aatcataaat cccaaacggt gtactctcta 840
ttagctttga aaggattgcc caattgtttg ttaaaaattt ctaataatag tacaatgttt 900
tgtttcattt ttccttttcg tcaacctgtt acccaatagc aaatgaagtt tttatgtgtg 960
tgtgtgtgtg tgaatttcca tgaaaatgaa acgggcttag aatcccggtg tattatgggt 1020
cgggtcgtaa ccgggcaatg acgcaggatc tgacgtaaaa ctcccaagaa tttttttaaa 1080
aagtctccgg aaaataaaat caaagttcat taacttaaaa agaaaaaaca aaatcggtcc 1140
acgtcccaaa ccctttttat aggagagtct tatgttctgg cagaagactt cacagactct 1200
ttcttaatct ctctctcttt caaccaaacc cctaaacaaa aaaaaaatac attttctgat 1260
ctctctaaaa atctttctcc ttcgttaatc tcgtgatctc tttctttttc tatatacaag 1320
tttgtacaaa aaagcaggct aaaaaaaacc atggatgtcg gagtttcacc ggtaaaagaa 1380
gatgtttttt atttccagca atgttacatt gttatacgta taatgatgag tttagtgatc 1440
aagttcctct ttgattcttc tttcttgttg caggcgaagt ctatacttgc gaaacctctg 1500
aagctactca ctgaagagga catttctcag ctcactcgcg aagactgccg caaattcctc 1560
aaagacaaag gaatgcgaag accgtcgtgg aacaaatctc aggcgatcca gcaagtttta 1620
tctcttaaag ctctctatga gcctggagac gattccggcg ccggtatctt ccgcaagatc 1680
ctcgtttctc agccagtaaa tccgcctcgc gtcacaacaa cgttgattga gccaagcaac 1740
gagctggaag cttgtggccg ggtttcttat ccggaagata acggcgcgtg ccatagaatg 1800
gattctccaa gatcagctga gttttccggt gggtctggtc actttgtatc cgagaaagat 1860
ggccacaaga cgactatttc tcccagaagc ccagctgaaa caagtgagct cgttgggcaa 1920
atgacgatat tctatagtgg aaaagtgaat gtgtatgatg gaataccacc tgaaaaggcc 1980
cggtcaatca tgcactttgc agccaatcca attgatttgc ctgaaaacgg tatttttgct 2040
tccagtagaa tgatttcaaa gctcataagt aaagagaaga tgatggaact tccccaaaaa 2100
ggccttgaga aggcgaattc ttctcgtgat tctggtatgg agggccaggc gaacagaaag 2160
gtatctttgc aaagatatcg tgaaaagcgg aaagacagaa aattctcaaa ggccaaaaag 2220
tgtccaggag ttgcgtcctc tagcttggag atgtttctga attgtcagcc acggatgaaa 2280
gctgcatatt cgcaaaacct aggctgcacc ggatctccac tgcatagcca gtcacctgaa 2340
agccagacaa aaagtcccaa tctttcagtt gatctaaaca gtgaaggcat tggatccgga 2400
ggtggatctg ctaagggaga acttagagga catccatttg aaggaaagcc aattccaaat 2460
ccacttcttg gacttgattc tactagaact ggacatcacc atcaccatca tggatcctaa 2520
tgaacccagc tttcttgtac aaagtggtct agtgggtacc gcgaatttcc ccgatcgttc 2580
aaacatttgg caataaagtt tcttaagatt gaatcctgtt gccggtcttg cgatgattat 2640
catataattt ctgttgaatt acgttaagca tgtaataatt aacatgtaat gcatgacgtt 2700
atttatgaga tgggttttta tgattagagt cccgcaatta tacatttaat acgcgataga 2760
aaacaaaata tagcgcgcaa actaggataa attatcgcgc gcggtgtcat ctatgttact 2820
agatcgaa 2828
<210> 53
<211> 2777
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 53
ccccttattg tacttcaatt aattatcatt atatcagcat aaacattata ataagtttct 60
tgcgtgttgg aacgtcattt tagttattct aaagaggaaa tagtttcttt tttgctcatg 120
acatcagaca tctggactac tatactggag tttacctttt cttctcctct ttttcttatt 180
gttcctctaa aaaaaattat cactttttaa atgcattagt taaacttatc tcaacaacgt 240
ttaaaattca tttcttgaat gcccattaca atgtaatagt ataacttaat tagtcgtctc 300
catgaaccat taatacgtac ggagtaatat aaaacaccat tggggagttc aatttgcaat 360
aatttcttgc aaaaatgtaa agtacctttt tgttcttgca aaattttaca aataaaaatt 420
tgcagctctt ttttttctct ctctccaaat actagctcaa aacccacaaa tatttttgaa 480
tttatggcat acttttagaa tgcgtttgat gcaactattt tcctttagga aatattcaca 540
acaatctaag acaatcaaaa agtagaaaat agtttgtaaa aagggatgtg gaggacatct 600
taatcaaata ttttcagttt aaaacttgaa aatgaaaaaa cacccgaaag gaaatgattc 660
gttctttaat atgtcctaca caatgtgaat ttgaattagt ttggtcatac ggtatatcat 720
atgattataa ataaaaaaaa ttagcaaaag aatataattt attaaatatt ttacaccata 780
ccaaacacaa ccgcattata tataatctta attatcatta tcaccagcat caacattata 840
atgattcccc tatgcgttgg aacgtcatta tagttattct aaacaagaaa gaaatttgtt 900
cttgacatca gacatctagt attataactc tagtggagct taccttttct tttccttctt 960
ttttttcttc ttaaaaaaat tatcactttt taaatcttgt atattagtta agcttatcta 1020
aacaaagttt taaattcatt tcttaaacgt ccattacaat gtaatataac ttagtcgtct 1080
caattaaacc attaatgtga aatataaatc aaaaaaagcc aaagggcggt gggacggcgc 1140
caatcatttg tcctagtcca ctcaaataag gcccatggtc ggcaaaacca aacacaaaat 1200
gtgttatttt taattttttc ctcttttatt gttaaagttg caaaatgtgt tatttttggt 1260
aagaccctat ggatatataa agacaggtta tgtgaaactt ggaaaaccat caagttttaa 1320
gcaaaaccct cttaagaact taaattgagc ttcttttggg gcatttttct agtgagaact 1380
aaaaacaagt ttgtacaaaa aagcaggcta aaaaaaacca tggatgtcgg agtttcaccg 1440
gcgaagtcta tacttgcgaa acctctgaag ctactcactg aagaggacat ttctcagctc 1500
actcgcgaag actgcgtaaa agaagatgtt ttttatttcc agcaatgtta cattgttata 1560
cgtataatga tgagtttagt gatcaagttc ctctttgatt cttctttctt gttgcagcgc 1620
aaattcctca aagacaaagg aatgcgaaga ccgtcgtgga acaaatctca ggcgatccag 1680
caagttttat ctcttaaagc tctctatgag cctggagacg attccggcgc cggtatcttc 1740
cgcaagatcc tcgtttctca gccagtaaat ccgcctcgcg tcacaacaac gttgattgag 1800
ccaagcaacg agctggaagc ttgtggccgg gtttcttatc cggaagataa cggcgcgtgc 1860
catagaatgg attctccaag atcagctgag ttttccggtg ggtctggtca ctttgtatcc 1920
gagaaagatg gccacaagac gactatttct cccagaagcc cagctgaaac aagtgagctc 1980
gttgggcaaa tgacgatatt ctatagtgga aaagtgaatg tgtatgatgg aataccacct 2040
gaaaaggccc ggtcaatcat gcactttgca gccaatccaa ttgatttgcc tgaaaacggt 2100
atttttgctt ccagtagaat gatttcaaag ctcataagta aagagaagat gatggaactt 2160
ccccaaaaag gccttgagaa ggcgaattct tctcgtgatt ctggtatgga gggccaggcg 2220
aacagaaagg tatctttgca aagatatcgt gaaaagcgga aagacagaaa attctcaaag 2280
gccaaaaagt gtccaggagt tgcgtcctct agcttggaga tgtttctgaa ttgtcagcca 2340
cggatgaaag ctgcatattc gcaaaaccta ggctgcaccg gatctccact gcatagccag 2400
tcacctgaaa gccagacaaa aagtcccaat ctttcagttg atctaaacag tgaaggcatt 2460
ggatcctaat gaacccagct ttcttgtaca aagtggtcta gtgggtaccg cgaatttccc 2520
cgatcgttca aacatttggc aataaagttt cttaagattg aatcctgttg ccggtcttgc 2580
gatgattatc atataatttc tgttgaatta cgttaagcat gtaataatta acatgtaatg 2640
catgacgtta tttatgagat gggtttttat gattagagtc ccgcaattat acatttaata 2700
cgcgatagaa aacaaaatat agcgcgcaaa ctaggataaa ttatcgcgcg cggtgtcatc 2760
tatgttacta gatcgaa 2777
<210> 54
<211> 2105
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 54
gtcctacaca atgtgaattt gaattagttt ggtcatacgg tatatcatat gattataaat 60
aaaaaaaatt agcaaaagaa tataatttat taaatatttt acaccatacc aaacacaacc 120
gcattatata taatcttaat tatcattatc accagcatca acattataat gattccccta 180
tgcgttggaa cgtcattata gttattctaa acaagaaaga aatttgttct tgacatcaga 240
catctagtat tataactcta gtggagctta ccttttcttt tccttctttt ttttcttctt 300
aaaaaaatta tcacttttta aatcttgtat attagttaag cttatctaaa caaagtttta 360
aattcatttc ttaaacgtcc attacaatgt aatataactt agtcgtctca attaaaccat 420
taatgtgaaa tataaatcaa aaaaagccaa agggcggtgg gacggcgcca atcatttgtc 480
ctagtccact caaataaggc ccatggtcgg caaaaccaaa cacaaaatgt gttattttta 540
attttttcct cttttattgt taaagttgca aaatgtgtta tttttggtaa gaccctatgg 600
atatataaag acaggttatg tgaaacttgg aaaaccatca agttttaagc aaaaccctct 660
taagaactta aattgagctt cttttggggc atttttctag tgagaactaa aaacaagttt 720
gtacaaaaaa gcaggctaaa aaaaaccatg gatgtcggag tttcaccggc gaagtctata 780
cttgcgaaac ctctgaagct actcactgaa gaggacattt ctcagctcac tcgcgaagac 840
tgcgtaaaag aagatgtttt ttatttccag caatgttaca ttgttatacg tataatgatg 900
agtttagtga tcaagttcct ctttgattct tctttcttgt tgcagcgcaa attcctcaaa 960
gacaaaggaa tgcgaagacc gtcgtggaac aaatctcagg cgatccagca agttttatct 1020
cttaaagctc tctatgagcc tggagacgat tccggcgccg gtatcttccg caagatcctc 1080
gtttctcagc cagtaaatcc gcctcgcgtc acaacaacgt tgattgagcc aagcaacgag 1140
ctggaagctt gtggccgggt ttcttatccg gaagataacg gcgcgtgcca tagaatggat 1200
tctccaagat cagctgagtt ttccggtggg tctggtcact ttgtatccga gaaagatggc 1260
cacaagacga ctatttctcc cagaagccca gctgaaacaa gtgagctcgt tgggcaaatg 1320
acgatattct atagtggaaa agtgaatgtg tatgatggaa taccacctga aaaggcccgg 1380
tcaatcatgc actttgcagc caatccaatt gatttgcctg aaaacggtat ttttgcttcc 1440
agtagaatga tttcaaagct cataagtaaa gagaagatga tggaacttcc ccaaaaaggc 1500
cttgagaagg cgaattcttc tcgtgattct ggtatggagg gccaggcgaa cagaaaggta 1560
tctttgcaaa gatatcgtga aaagcggaaa gacagaaaat tctcaaaggc caaaaagtgt 1620
ccaggagttg cgtcctctag cttggagatg tttctgaatt gtcagccacg gatgaaagct 1680
gcatattcgc aaaacctagg ctgcaccgga tctccactgc atagccagtc acctgaaagc 1740
cagacaaaaa gtcccaatct ttcagttgat ctaaacagtg aaggcattgg atcctaatga 1800
acccagcttt cttgtacaaa gtggtctagt gggtaccgcg aatttccccg atcgttcaaa 1860
catttggcaa taaagtttct taagattgaa tcctgttgcc ggtcttgcga tgattatcat 1920
ataatttctg ttgaattacg ttaagcatgt aataattaac atgtaatgca tgacgttatt 1980
tatgagatgg gtttttatga ttagagtccc gcaattatac atttaatacg cgatagaaaa 2040
caaaatatag cgcgcaaact aggataaatt atcgcgcgcg gtgtcatcta tgttactaga 2100
tcgaa 2105
<210> 55
<211> 2708
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 55
tcacgtggaa cgcgccgcag taaacggagc ggtggatcaa acttttcgtc cgtttgatca 60
aacagaagag aactagtcaa tgctctttct tcatatcaca atttaatagt ctcaagacga 120
ttacgccaca taaccatttt ctcgtgattt cgacatcaaa atttaataaa aggaactgat 180
tgattggtca tcatgttaca agtgtcaaat gagctaatcc gttttacagt ggcatagttt 240
acgatcaatt tacaaatttt tggttttata acatacttgt agttaaaact atttataagc 300
tatttatagt gagttagctt ataaaaccct attcttttat ctaaaattat gttttgactc 360
gtttcatgat aaaattttat ccttttcatc ggaataaaaa actttattat ttatttggca 420
aaataattgg tgtaaaaatt atgtatatgt taataacaaa aaatattaat ctgattcata 480
atcttaaaaa agaaaaattt cttgaaataa actttagaca ttgtaaataa aaaaacatta 540
tttttatata atgggatgtt tatatgtaat tttttataaa aaaataaaag ttgtttacta 600
gtaatccgat tggctttaac tatcgtcgcc aaaagaataa tgtagaactg actttgaggt 660
aaaactaaaa gaaatttgta agataatagt cacattaaat gctaaaatta atacatactg 720
atatatcgta taaaatttat gaaaactaca ccttaacctg aatcatacac tgtaataaaa 780
aaaacaaatt atatataaac cctaaaaact aatcataaat cccaaacggt gtactctcta 840
ttagctttga aaggattgcc caattgtttg ttaaaaattt ctaataatag tacaatgttt 900
tgtttcattt ttccttttcg tcaacctgtt acccaatagc aaatgaagtt tttatgtgtg 960
tgtgtgtgtg tgaatttcca tgaaaatgaa acgggcttag aatcccggtg tattatgggt 1020
cgggtcgtaa ccgggcaatg acgcaggatc tgacgtaaaa ctcccaagaa tttttttaaa 1080
aagtctccgg aaaataaaat caaagttcat taacttaaaa agaaaaaaca aaatcggtcc 1140
acgtcccaaa ccctttttat aggagagtct tatgttctgg cagaagactt cacagactct 1200
ttcttaatct ctctctcttt caaccaaacc cctaaacaaa aaaaaaatac attttctgat 1260
ctctctaaaa atctttctcc ttcgttaatc tcgtgatctc tttctttttc tatatacaag 1320
tttgtacaaa aaagcaggct aaaaaaaacc atggatgtcg gagtttcacc ggtaaaagaa 1380
gatgtttttt atttccagca atgttacatt gttatacgta taatgatgag tttagtgatc 1440
aagttcctct ttgattcttc tttcttgttg caggcgaagt ctatacttgc gaaacctctg 1500
aagctactca ctgaagagga catttctcag ctcactcgcg aagactgccg caaattcctc 1560
aaagacaaag gaatgcgaag accgtcgtgg aacaaatctc aggcgatcca gcaagtttta 1620
tctcttaaag ctctctatga gcctggagac gattccggcg ccggtatctt ccgcaagatc 1680
ctcgtttctc agccagtaaa tccgcctcgc gtcacaacaa cgttgattga gccaagcaac 1740
gagctggaag cttgtggccg ggtttcttat ccggaagata acggcgcgtg ccatagaatg 1800
gattctccaa gatcagctga gttttccggt gggtctggtc actttgtatc cgagaaagat 1860
ggccacaaga cgactatttc tcccagaagc ccagctgaaa caagtgagct cgttgggcaa 1920
atgacgatat tctatagtgg aaaagtgaat gtgtatgatg gaataccacc tgaaaaggcc 1980
cggtcaatca tgcactttgc agccaatcca attgatttgc ctgaaaacgg tatttttgct 2040
tccagtagaa tgatttcaaa gctcataagt aaagagaaga tgatggaact tccccaaaaa 2100
ggccttgaga aggcgaattc ttctcgtgat tctggtatgg agggccaggc gaacagaaag 2160
gtatctttgc aaagatatcg tgaaaagcgg aaagacagaa aattctcaaa ggccaaaaag 2220
tgtccaggag ttgcgtcctc tagcttggag atgtttctga attgtcagcc acggatgaaa 2280
gctgcatatt cgcaaaacct aggctgcacc ggatctccac tgcatagcca gtcacctgaa 2340
agccagacaa aaagtcccaa tctttcagtt gatctaaaca gtgaaggcat tggatcctaa 2400
tgaacccagc tttcttgtac aaagtggtct agtgggtacc gcgaatttcc ccgatcgttc 2460
aaacatttgg caataaagtt tcttaagatt gaatcctgtt gccggtcttg cgatgattat 2520
catataattt ctgttgaatt acgttaagca tgtaataatt aacatgtaat gcatgacgtt 2580
atttatgaga tgggttttta tgattagagt cccgcaatta tacatttaat acgcgataga 2640
aaacaaaata tagcgcgcaa actaggataa attatcgcgc gcggtgtcat ctatgttact 2700
agatcgaa 2708
<210> 56
<211> 161
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 56
Met Lys Leu Leu Ser Ser Ile Glu Glu Ala Cys Asn Ile Cys Arg Leu
1 5 10 15
Lys Lys Leu Lys Cys Ser Lys Glu Lys Pro Lys Cys Ala Lys Cys Leu
20 25 30
Lys Asn Asn Trp Glu Cys Arg Tyr Ser Pro Lys Thr Lys Arg Ser Pro
35 40 45
Leu Thr Arg Ala His Leu Thr Glu Val Glu Ser Arg Leu Glu Arg Leu
50 55 60
Glu Gln Leu Phe Leu Leu Ile Phe Pro Arg Glu Asn Leu Asn Met Ile
65 70 75 80
Leu Lys Met Asp Ser Leu Gln Asp Ile Lys Ala Leu Leu Thr Gly Leu
85 90 95
Phe Val Gln Asp Asn Val Asn Lys Asp Ala Val Thr Asp Arg Leu Ala
100 105 110
Ser Val Glu Thr Asp Met Pro Leu Thr Leu Arg Gln His Arg Ile Ser
115 120 125
Ala Thr Ser Ser Ser Glu Glu Ser Ser Asn Lys Gly Gln Arg Gln Leu
130 135 140
Thr Val Ser Ser Arg Ser Asn Gln Thr Ser Leu Tyr Lys Lys Ala Gly
145 150 155 160
Ser
<210> 57
<211> 139
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 57
Met Pro Lys Lys Lys Arg Lys Val Ser Ser Gly Ala Asn Phe Asn Gln
1 5 10 15
Ser Gly Asn Ile Ala Asp Ser Ser Leu Ser Phe Thr Phe Thr Asn Ser
20 25 30
Ser Asn Gly Pro Asn Leu Ile Thr Thr Gln Thr Asn Ser Gln Ala Leu
35 40 45
Ser Gln Pro Ile Ala Ser Ser Asn Val His Asp Asn Phe Met Asn Asn
50 55 60
Glu Ile Thr Ala Ser Lys Ile Asp Asp Gly Asn Asn Ser Lys Pro Leu
65 70 75 80
Ser Pro Gly Trp Thr Asp Gln Thr Ala Tyr Asn Ala Phe Gly Ile Thr
85 90 95
Thr Gly Met Phe Asn Thr Thr Thr Met Asp Asp Val Tyr Asn Tyr Leu
100 105 110
Phe Asp Asp Glu Asp Thr Pro Pro Asn Pro Lys Lys Glu Gly Gly Ser
115 120 125
Asn Gln Thr Ser Leu Tyr Lys Lys Ala Gly Ser
130 135
<210> 58
<211> 474
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 58
Met Lys Leu Leu Ser Ser Ile Glu Glu Ala Cys Asn Ile Cys Arg Leu
1 5 10 15
Lys Lys Leu Lys Cys Ser Lys Glu Lys Pro Lys Cys Ala Lys Cys Leu
20 25 30
Lys Asn Asn Trp Glu Cys Arg Tyr Ser Pro Lys Thr Lys Arg Ser Pro
35 40 45
Leu Thr Arg Ala His Leu Thr Glu Val Glu Ser Arg Leu Glu Arg Leu
50 55 60
Glu Gln Leu Phe Leu Leu Ile Phe Pro Arg Glu Asn Leu Asn Met Ile
65 70 75 80
Leu Lys Met Asp Ser Leu Gln Asp Ile Lys Ala Leu Leu Thr Gly Leu
85 90 95
Phe Val Gln Asp Asn Val Asn Lys Asp Ala Val Thr Asp Arg Leu Ala
100 105 110
Ser Val Glu Thr Asp Met Pro Leu Thr Leu Arg Gln His Arg Ile Ser
115 120 125
Ala Thr Ser Ser Ser Glu Glu Ser Ser Asn Lys Gly Gln Arg Gln Leu
130 135 140
Thr Val Ser Ser Arg Ser Asn Gln Thr Ser Leu Tyr Lys Lys Ala Gly
145 150 155 160
Ser Met Asp Val Gly Val Ser Pro Ala Lys Ser Ile Leu Ala Lys Pro
165 170 175
Leu Lys Leu Leu Thr Glu Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp
180 185 190
Cys Arg Lys Phe Leu Lys Asp Lys Gly Met Arg Arg Pro Ser Trp Asn
195 200 205
Lys Ser Gln Ala Ile Gln Gln Val Leu Ser Leu Lys Ala Leu Tyr Glu
210 215 220
Pro Gly Asp Asp Ser Gly Ala Gly Ile Phe Arg Lys Ile Leu Val Ser
225 230 235 240
Gln Pro Val Asn Pro Pro Arg Val Thr Thr Thr Leu Ile Glu Pro Ser
245 250 255
Asn Glu Leu Glu Ala Cys Gly Arg Val Ser Tyr Pro Glu Asp Asn Gly
260 265 270
Ala Cys His Arg Met Asp Ser Pro Arg Ser Ala Glu Phe Ser Gly Gly
275 280 285
Ser Gly His Phe Val Ser Glu Lys Asp Gly His Lys Thr Thr Ile Ser
290 295 300
Pro Arg Ser Pro Ala Glu Thr Ser Glu Leu Val Gly Gln Met Thr Ile
305 310 315 320
Phe Tyr Ser Gly Lys Val Asn Val Tyr Asp Gly Ile Pro Pro Glu Lys
325 330 335
Ala Arg Ser Ile Met His Phe Ala Ala Asn Pro Ile Asp Leu Pro Glu
340 345 350
Asn Gly Ile Phe Ala Ser Ser Arg Met Ile Ser Lys Leu Ile Ser Lys
355 360 365
Glu Lys Met Met Glu Leu Pro Gln Lys Gly Leu Glu Lys Ala Asn Ser
370 375 380
Ser Arg Asp Ser Gly Met Glu Gly Gln Ala Asn Arg Lys Val Ser Leu
385 390 395 400
Gln Arg Tyr Arg Glu Lys Arg Lys Asp Arg Lys Phe Ser Lys Ala Lys
405 410 415
Lys Cys Pro Gly Val Ala Ser Ser Ser Leu Glu Met Phe Leu Asn Cys
420 425 430
Gln Pro Arg Met Lys Ala Ala Tyr Ser Gln Asn Leu Gly Cys Thr Gly
435 440 445
Ser Pro Leu His Ser Gln Ser Pro Glu Ser Gln Thr Lys Ser Pro Asn
450 455 460
Leu Ser Val Asp Leu Asn Ser Glu Gly Ile
465 470
<210> 59
<211> 452
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 59
Met Pro Lys Lys Lys Arg Lys Val Ser Ser Gly Ala Asn Phe Asn Gln
1 5 10 15
Ser Gly Asn Ile Ala Asp Ser Ser Leu Ser Phe Thr Phe Thr Asn Ser
20 25 30
Ser Asn Gly Pro Asn Leu Ile Thr Thr Gln Thr Asn Ser Gln Ala Leu
35 40 45
Ser Gln Pro Ile Ala Ser Ser Asn Val His Asp Asn Phe Met Asn Asn
50 55 60
Glu Ile Thr Ala Ser Lys Ile Asp Asp Gly Asn Asn Ser Lys Pro Leu
65 70 75 80
Ser Pro Gly Trp Thr Asp Gln Thr Ala Tyr Asn Ala Phe Gly Ile Thr
85 90 95
Thr Gly Met Phe Asn Thr Thr Thr Met Asp Asp Val Tyr Asn Tyr Leu
100 105 110
Phe Asp Asp Glu Asp Thr Pro Pro Asn Pro Lys Lys Glu Gly Gly Ser
115 120 125
Asn Gln Thr Ser Leu Tyr Lys Lys Ala Gly Ser Met Asp Val Gly Val
130 135 140
Ser Pro Ala Lys Ser Ile Leu Ala Lys Pro Leu Lys Leu Leu Thr Glu
145 150 155 160
Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys Arg Lys Phe Leu Lys
165 170 175
Asp Lys Gly Met Arg Arg Pro Ser Trp Asn Lys Ser Gln Ala Ile Gln
180 185 190
Gln Val Leu Ser Leu Lys Ala Leu Tyr Glu Pro Gly Asp Asp Ser Gly
195 200 205
Ala Gly Ile Phe Arg Lys Ile Leu Val Ser Gln Pro Val Asn Pro Pro
210 215 220
Arg Val Thr Thr Thr Leu Ile Glu Pro Ser Asn Glu Leu Glu Ala Cys
225 230 235 240
Gly Arg Val Ser Tyr Pro Glu Asp Asn Gly Ala Cys His Arg Met Asp
245 250 255
Ser Pro Arg Ser Ala Glu Phe Ser Gly Gly Ser Gly His Phe Val Ser
260 265 270
Glu Lys Asp Gly His Lys Thr Thr Ile Ser Pro Arg Ser Pro Ala Glu
275 280 285
Thr Ser Glu Leu Val Gly Gln Met Thr Ile Phe Tyr Ser Gly Lys Val
290 295 300
Asn Val Tyr Asp Gly Ile Pro Pro Glu Lys Ala Arg Ser Ile Met His
305 310 315 320
Phe Ala Ala Asn Pro Ile Asp Leu Pro Glu Asn Gly Ile Phe Ala Ser
325 330 335
Ser Arg Met Ile Ser Lys Leu Ile Ser Lys Glu Lys Met Met Glu Leu
340 345 350
Pro Gln Lys Gly Leu Glu Lys Ala Asn Ser Ser Arg Asp Ser Gly Met
355 360 365
Glu Gly Gln Ala Asn Arg Lys Val Ser Leu Gln Arg Tyr Arg Glu Lys
370 375 380
Arg Lys Asp Arg Lys Phe Ser Lys Ala Lys Lys Cys Pro Gly Val Ala
385 390 395 400
Ser Ser Ser Leu Glu Met Phe Leu Asn Cys Gln Pro Arg Met Lys Ala
405 410 415
Ala Tyr Ser Gln Asn Leu Gly Cys Thr Gly Ser Pro Leu His Ser Gln
420 425 430
Ser Pro Glu Ser Gln Thr Lys Ser Pro Asn Leu Ser Val Asp Leu Asn
435 440 445
Ser Glu Gly Ile
450
<210> 60
<211> 391
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 60
Met Pro Lys Lys Lys Arg Lys Val Ser Ser Gly Ala Asn Phe Asn Gln
1 5 10 15
Ser Gly Asn Ile Ala Asp Ser Ser Leu Ser Phe Thr Phe Thr Asn Ser
20 25 30
Ser Asn Gly Pro Asn Leu Ile Thr Thr Gln Thr Asn Ser Gln Ala Leu
35 40 45
Ser Gln Pro Ile Ala Ser Ser Asn Val His Asp Asn Phe Met Asn Asn
50 55 60
Glu Ile Thr Ala Ser Lys Ile Asp Asp Gly Asn Asn Ser Lys Pro Leu
65 70 75 80
Ser Pro Gly Trp Thr Asp Gln Thr Ala Tyr Asn Ala Phe Gly Ile Thr
85 90 95
Thr Gly Met Phe Asn Thr Thr Thr Met Asp Asp Val Tyr Asn Tyr Leu
100 105 110
Phe Asp Asp Glu Asp Thr Pro Pro Asn Pro Lys Lys Glu Gly Gly Ser
115 120 125
Asn Gln Thr Ser Leu Tyr Lys Lys Ala Gly Ser Tyr Glu Pro Gly Asp
130 135 140
Asp Ser Gly Ala Gly Ile Phe Arg Lys Ile Leu Val Ser Gln Pro Val
145 150 155 160
Asn Pro Pro Arg Val Thr Thr Thr Leu Ile Glu Pro Ser Asn Glu Leu
165 170 175
Glu Ala Cys Gly Arg Val Ser Tyr Pro Glu Asp Asn Gly Ala Cys His
180 185 190
Arg Met Asp Ser Pro Arg Ser Ala Glu Phe Ser Gly Gly Ser Gly His
195 200 205
Phe Val Ser Glu Lys Asp Gly His Lys Thr Thr Ile Ser Pro Arg Ser
210 215 220
Pro Ala Glu Thr Ser Glu Leu Val Gly Gln Met Thr Ile Phe Tyr Ser
225 230 235 240
Gly Lys Val Asn Val Tyr Asp Gly Ile Pro Pro Glu Lys Ala Arg Ser
245 250 255
Ile Met His Phe Ala Ala Asn Pro Ile Asp Leu Pro Glu Asn Gly Ile
260 265 270
Phe Ala Ser Ser Arg Met Ile Ser Lys Leu Ile Ser Lys Glu Lys Met
275 280 285
Met Glu Leu Pro Gln Lys Gly Leu Glu Lys Ala Asn Ser Ser Arg Asp
290 295 300
Ser Gly Met Glu Gly Gln Ala Asn Arg Lys Val Ser Leu Gln Arg Tyr
305 310 315 320
Arg Glu Lys Arg Lys Asp Arg Lys Phe Ser Lys Ala Lys Lys Cys Pro
325 330 335
Gly Val Ala Ser Ser Ser Leu Glu Met Phe Leu Asn Cys Gln Pro Arg
340 345 350
Met Lys Ala Ala Tyr Ser Gln Asn Leu Gly Cys Thr Gly Ser Pro Leu
355 360 365
His Ser Gln Ser Pro Glu Ser Gln Thr Lys Ser Pro Asn Leu Ser Val
370 375 380
Asp Leu Asn Ser Glu Gly Ile
385 390
<210> 61
<211> 427
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 61
Met Pro Lys Lys Lys Arg Lys Val Ser Ser Gly Ala Asn Phe Asn Gln
1 5 10 15
Ser Gly Asn Ile Ala Asp Ser Ser Leu Ser Phe Thr Phe Thr Asn Ser
20 25 30
Ser Asn Gly Pro Asn Leu Ile Thr Thr Gln Thr Asn Ser Gln Ala Leu
35 40 45
Ser Gln Pro Ile Ala Ser Ser Asn Val His Asp Asn Phe Met Asn Asn
50 55 60
Glu Ile Thr Ala Ser Lys Ile Asp Asp Gly Asn Asn Ser Lys Pro Leu
65 70 75 80
Ser Pro Gly Trp Thr Asp Gln Thr Ala Tyr Asn Ala Phe Gly Ile Thr
85 90 95
Thr Gly Met Phe Asn Thr Thr Thr Met Asp Asp Val Tyr Asn Tyr Leu
100 105 110
Phe Asp Asp Glu Asp Thr Pro Pro Asn Pro Lys Lys Glu Gly Gly Ser
115 120 125
Asn Gln Thr Ser Leu Tyr Lys Lys Ala Gly Ser Met Asp Val Gly Val
130 135 140
Ser Pro Ala Lys Ser Ile Leu Ala Lys Pro Leu Lys Leu Leu Thr Glu
145 150 155 160
Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys Arg Lys Phe Leu Lys
165 170 175
Asp Lys Gly Met Arg Arg Pro Ser Trp Asn Lys Ser Gln Ala Ile Gln
180 185 190
Gln Val Leu Ser Leu Lys Ala Leu Tyr Glu Pro Gly Asp Asp Ser Gly
195 200 205
Ala Gly Ile Phe Arg Lys Ile Leu Val Ser Gln Pro Val Asn Pro Pro
210 215 220
Arg Val Thr Thr Thr Leu Ile Glu Pro Ser Asn Glu Leu Glu Ala Cys
225 230 235 240
Gly Arg Val Ser Tyr Pro Glu Asp Asn Gly Ala Cys His Arg Met Asp
245 250 255
Ser Pro Arg Ser Ala Glu Phe Ser Gly Gly Ser Gly His Phe Val Ser
260 265 270
Glu Lys Asp Gly His Lys Thr Thr Ile Ser Pro Arg Ser Pro Ala Glu
275 280 285
Thr Ser Glu Leu Ile Met His Phe Ala Ala Asn Pro Ile Asp Leu Pro
290 295 300
Glu Asn Gly Ile Phe Ala Ser Ser Arg Met Ile Ser Lys Leu Ile Ser
305 310 315 320
Lys Glu Lys Met Met Glu Leu Pro Gln Lys Gly Leu Glu Lys Ala Asn
325 330 335
Ser Ser Arg Asp Ser Gly Met Glu Gly Gln Ala Asn Arg Lys Val Ser
340 345 350
Leu Gln Arg Tyr Arg Glu Lys Arg Lys Asp Arg Lys Phe Ser Lys Ala
355 360 365
Lys Lys Cys Pro Gly Val Ala Ser Ser Ser Leu Glu Met Phe Leu Asn
370 375 380
Cys Gln Pro Arg Met Lys Ala Ala Tyr Ser Gln Asn Leu Gly Cys Thr
385 390 395 400
Gly Ser Pro Leu His Ser Gln Ser Pro Glu Ser Gln Thr Lys Ser Pro
405 410 415
Asn Leu Ser Val Asp Leu Asn Ser Glu Gly Ile
420 425
<210> 62
<211> 368
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 62
Met Pro Lys Lys Lys Arg Lys Val Ser Ser Gly Ala Asn Phe Asn Gln
1 5 10 15
Ser Gly Asn Ile Ala Asp Ser Ser Leu Ser Phe Thr Phe Thr Asn Ser
20 25 30
Ser Asn Gly Pro Asn Leu Ile Thr Thr Gln Thr Asn Ser Gln Ala Leu
35 40 45
Ser Gln Pro Ile Ala Ser Ser Asn Val His Asp Asn Phe Met Asn Asn
50 55 60
Glu Ile Thr Ala Ser Lys Ile Asp Asp Gly Asn Asn Ser Lys Pro Leu
65 70 75 80
Ser Pro Gly Trp Thr Asp Gln Thr Ala Tyr Asn Ala Phe Gly Ile Thr
85 90 95
Thr Gly Met Phe Asn Thr Thr Thr Met Asp Asp Val Tyr Asn Tyr Leu
100 105 110
Phe Asp Asp Glu Asp Thr Pro Pro Asn Pro Lys Lys Glu Gly Gly Ser
115 120 125
Asn Gln Thr Ser Leu Tyr Lys Lys Ala Gly Ser Met Asp Val Gly Val
130 135 140
Ser Pro Ala Lys Ser Ile Leu Ala Lys Pro Leu Lys Leu Leu Thr Glu
145 150 155 160
Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys Arg Lys Phe Leu Lys
165 170 175
Asp Lys Gly Met Arg Arg Pro Ser Trp Asn Lys Ser Gln Ala Ile Gln
180 185 190
Gln Val Leu Ser Leu Lys Ala Leu Tyr Glu Pro Gly Asp Asp Ser Gly
195 200 205
Ala Gly Ile Phe Arg Lys Ile Leu Val Ser Gln Pro Val Asn Pro Pro
210 215 220
Arg Val Thr Thr Thr Leu Ile Glu Pro Ser Asn Glu Leu Glu Ala Cys
225 230 235 240
Gly Arg Val Ser Tyr Pro Glu Asp Asn Gly Ala Cys His Arg Met Asp
245 250 255
Ser Pro Arg Ser Ala Glu Phe Ser Gly Gly Ser Gly His Phe Val Ser
260 265 270
Glu Lys Asp Gly His Lys Thr Thr Ile Ser Pro Arg Ser Pro Ala Glu
275 280 285
Thr Ser Glu Leu Val Gly Gln Met Thr Ile Phe Tyr Ser Gly Lys Val
290 295 300
Asn Val Tyr Asp Gly Ile Pro Pro Glu Lys Ala Arg Ser Ile Met His
305 310 315 320
Phe Ala Ala Asn Pro Ile Asp Leu Pro Glu Asn Gly Ile Phe Ala Ser
325 330 335
Ser Arg Met Ile Ser Lys Leu Ile Ser Lys Glu Lys Met Met Glu Leu
340 345 350
Pro Gln Lys Gly Leu Glu Lys Ala Asn Ser Ser Arg Asp Ser Gly Met
355 360 365
<210> 63
<211> 1131
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 63
Met Ser Ser Leu Ser Arg Glu Leu Val Phe Leu Ile Leu Gln Phe Leu
1 5 10 15
Asp Glu Glu Lys Phe Lys Glu Thr Val His Lys Leu Glu Gln Glu Ser
20 25 30
Gly Phe Phe Phe Asn Met Lys Tyr Phe Glu Asp Glu Val His Asn Gly
35 40 45
Asn Trp Asp Glu Val Glu Lys Tyr Leu Ser Gly Phe Thr Lys Val Asp
50 55 60
Asp Asn Arg Tyr Ser Met Lys Ile Phe Phe Glu Ile Arg Lys Gln Lys
65 70 75 80
Tyr Leu Glu Ala Leu Asp Lys His Asp Arg Pro Lys Ala Val Asp Ile
85 90 95
Leu Val Lys Asp Leu Lys Val Phe Ser Thr Phe Asn Glu Glu Leu Phe
100 105 110
Lys Glu Ile Thr Gln Leu Leu Thr Leu Glu Asn Phe Arg Glu Asn Glu
115 120 125
Gln Leu Ser Lys Tyr Gly Asp Thr Lys Ser Ala Arg Ala Ile Met Leu
130 135 140
Val Glu Leu Lys Lys Leu Ile Glu Ala Asn Pro Leu Phe Arg Asp Lys
145 150 155 160
Leu Gln Phe Pro Thr Leu Arg Asn Ser Arg Leu Arg Thr Leu Ile Asn
165 170 175
Gln Ser Leu Asn Trp Gln His Gln Leu Cys Lys Asn Pro Arg Pro Asn
180 185 190
Pro Asp Ile Lys Thr Leu Phe Val Asp His Ser Cys Gly Pro Pro Asn
195 200 205
Gly Ala Arg Ala Pro Ser Pro Val Asn Asn Pro Leu Leu Gly Gly Ile
210 215 220
Pro Lys Ala Gly Gly Phe Pro Pro Leu Gly Ala His Gly Pro Phe Gln
225 230 235 240
Pro Thr Ala Ser Pro Val Pro Thr Pro Leu Ala Gly Trp Met Ser Ser
245 250 255
Pro Ser Ser Val Pro His Pro Ala Val Ser Ala Gly Ala Ile Ala Leu
260 265 270
Gly Gly Pro Ser Ile Pro Ala Ala Leu Lys His Pro Arg Thr Pro Pro
275 280 285
Thr Asn Ala Ser Leu Asp Tyr Pro Ser Ala Asp Ser Glu His Val Ser
290 295 300
Lys Arg Thr Arg Pro Met Gly Ile Ser Asp Glu Val Asn Leu Gly Val
305 310 315 320
Asn Met Leu Pro Met Ser Phe Ser Gly Gln Ala His Gly His Ser Pro
325 330 335
Ala Phe Lys Ala Pro Asp Asp Leu Pro Lys Thr Val Ala Arg Thr Leu
340 345 350
Ser Gln Gly Ser Ser Pro Met Ser Met Asp Phe His Pro Ile Lys Gln
355 360 365
Thr Leu Leu Leu Val Gly Thr Asn Val Gly Asp Ile Gly Leu Trp Glu
370 375 380
Val Gly Ser Arg Glu Arg Leu Val Gln Lys Thr Phe Lys Val Trp Asp
385 390 395 400
Leu Ser Lys Cys Ser Met Pro Leu Gln Ala Ala Leu Val Lys Glu Pro
405 410 415
Val Val Ser Val Asn Arg Val Ile Trp Ser Pro Asp Gly Ser Leu Phe
420 425 430
Gly Val Ala Tyr Ser Arg His Ile Val Gln Leu Tyr Ser Tyr His Gly
435 440 445
Gly Glu Asp Met Arg Gln His Leu Glu Ile Asp Ala His Val Gly Gly
450 455 460
Val Asn Asp Ile Ser Phe Ser Thr Pro Asn Lys Gln Leu Cys Val Ile
465 470 475 480
Thr Cys Gly Asp Asp Lys Thr Ile Lys Val Trp Asp Ala Ala Thr Gly
485 490 495
Val Lys Arg His Thr Phe Glu Gly His Glu Ala Pro Val Tyr Ser Val
500 505 510
Cys Pro His Tyr Lys Glu Asn Ile Gln Phe Ile Phe Ser Thr Ala Leu
515 520 525
Asp Gly Lys Ile Lys Ala Trp Leu Tyr Asp Asn Met Gly Ser Arg Val
530 535 540
Asp Tyr Asp Ala Pro Gly Arg Trp Cys Thr Thr Met Ala Tyr Ser Ala
545 550 555 560
Asp Gly Thr Arg Leu Phe Ser Cys Gly Thr Ser Lys Asp Gly Glu Ser
565 570 575
Phe Ile Val Glu Trp Asn Glu Ser Glu Gly Ala Val Lys Arg Thr Tyr
580 585 590
Gln Gly Phe His Lys Arg Ser Leu Gly Val Val Gln Phe Asp Thr Thr
595 600 605
Lys Asn Arg Tyr Leu Ala Ala Gly Asp Asp Phe Ser Ile Lys Phe Trp
610 615 620
Asp Met Asp Ala Val Gln Leu Leu Thr Ala Ile Asp Gly Asp Gly Gly
625 630 635 640
Leu Gln Ala Ser Pro Arg Ile Arg Phe Asn Lys Glu Gly Ser Leu Leu
645 650 655
Ala Val Ser Gly Asn Glu Asn Val Ile Lys Ile Met Ala Asn Ser Asp
660 665 670
Gly Leu Arg Leu Leu His Thr Phe Glu Asn Ile Ser Ser Glu Ser Ser
675 680 685
Lys Pro Ala Ile Asn Ser Ile Ala Ala Ala Ala Ala Ala Ala Ala Thr
690 695 700
Ser Ala Gly His Ala Asp Arg Ser Ala Asn Val Val Ser Ile Gln Gly
705 710 715 720
Met Asn Gly Asp Ser Arg Asn Met Val Asp Val Lys Pro Val Ile Thr
725 730 735
Glu Glu Ser Asn Asp Lys Ser Lys Ile Trp Lys Leu Thr Glu Val Ser
740 745 750
Glu Pro Ser Gln Cys Arg Ser Leu Arg Leu Pro Glu Asn Leu Arg Val
755 760 765
Ala Lys Ile Ser Arg Leu Ile Phe Thr Asn Ser Gly Asn Ala Ile Leu
770 775 780
Ala Leu Ala Ser Asn Ala Ile His Leu Leu Trp Lys Trp Gln Arg Asn
785 790 795 800
Glu Arg Asn Ala Thr Gly Lys Ala Thr Ala Ser Leu Pro Pro Gln Gln
805 810 815
Trp Gln Pro Ala Ser Gly Ile Leu Met Thr Asn Asp Val Ala Glu Thr
820 825 830
Asn Pro Glu Glu Ala Val Pro Cys Phe Ala Leu Ser Lys Asn Asp Ser
835 840 845
Tyr Val Met Ser Ala Ser Gly Gly Lys Ile Ser Leu Phe Asn Met Met
850 855 860
Thr Phe Lys Thr Met Ala Thr Phe Met Pro Pro Pro Pro Ala Ala Thr
865 870 875 880
Phe Leu Ala Phe His Pro Gln Asp Asn Asn Ile Ile Ala Ile Gly Met
885 890 895
Asp Asp Ser Thr Ile Gln Ile Tyr Asn Val Arg Val Asp Glu Val Lys
900 905 910
Ser Lys Leu Lys Gly His Ser Lys Arg Ile Thr Gly Leu Ala Phe Ser
915 920 925
Asn Val Leu Asn Val Leu Val Ser Ser Gly Ala Asp Ala Gln Leu Cys
930 935 940
Val Trp Asn Thr Asp Gly Trp Glu Lys Gln Arg Ser Lys Val Leu Pro
945 950 955 960
Leu Pro Gln Gly Arg Pro Asn Ser Ala Pro Ser Asp Thr Arg Val Gln
965 970 975
Phe His Gln Asp Gln Ala His Phe Leu Val Val His Glu Thr Gln Leu
980 985 990
Ala Ile Tyr Glu Thr Thr Lys Leu Glu Cys Met Lys Gln Trp Ala Val
995 1000 1005
Arg Glu Ser Leu Ala Pro Ile Thr His Ala Thr Phe Ser Cys Asp
1010 1015 1020
Ser Gln Leu Val Tyr Ala Ser Phe Met Asp Ala Thr Val Cys Val
1025 1030 1035
Phe Ser Ser Ala Asn Leu Arg Leu Arg Cys Arg Val Asn Pro Ser
1040 1045 1050
Ala Tyr Leu Pro Ala Ser Leu Ser Asn Ser Asn Val His Pro Leu
1055 1060 1065
Val Ile Ala Ala His Pro Gln Glu Pro Asn Met Phe Ala Val Gly
1070 1075 1080
Leu Ser Asp Gly Gly Val His Ile Phe Glu Pro Leu Glu Ser Glu
1085 1090 1095
Gly Lys Trp Gly Val Ala Pro Pro Ala Glu Asn Gly Ser Ala Ser
1100 1105 1110
Gly Ala Pro Thr Ala Pro Ser Val Gly Ala Ser Ala Ser Asp Gln
1115 1120 1125
Pro Gln Arg
1130
<210> 64
<211> 1292
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 64
Met Lys Leu Leu Ser Ser Ile Glu Glu Ala Cys Asn Ile Cys Arg Leu
1 5 10 15
Lys Lys Leu Lys Cys Ser Lys Glu Lys Pro Lys Cys Ala Lys Cys Leu
20 25 30
Lys Asn Asn Trp Glu Cys Arg Tyr Ser Pro Lys Thr Lys Arg Ser Pro
35 40 45
Leu Thr Arg Ala His Leu Thr Glu Val Glu Ser Arg Leu Glu Arg Leu
50 55 60
Glu Gln Leu Phe Leu Leu Ile Phe Pro Arg Glu Asn Leu Asn Met Ile
65 70 75 80
Leu Lys Met Asp Ser Leu Gln Asp Ile Lys Ala Leu Leu Thr Gly Leu
85 90 95
Phe Val Gln Asp Asn Val Asn Lys Asp Ala Val Thr Asp Arg Leu Ala
100 105 110
Ser Val Glu Thr Asp Met Pro Leu Thr Leu Arg Gln His Arg Ile Ser
115 120 125
Ala Thr Ser Ser Ser Glu Glu Ser Ser Asn Lys Gly Gln Arg Gln Leu
130 135 140
Thr Val Ser Ser Arg Ser Asn Gln Thr Ser Leu Tyr Lys Lys Ala Gly
145 150 155 160
Ser Met Ser Ser Leu Ser Arg Glu Leu Val Phe Leu Ile Leu Gln Phe
165 170 175
Leu Asp Glu Glu Lys Phe Lys Glu Thr Val His Lys Leu Glu Gln Glu
180 185 190
Ser Gly Phe Phe Phe Asn Met Lys Tyr Phe Glu Asp Glu Val His Asn
195 200 205
Gly Asn Trp Asp Glu Val Glu Lys Tyr Leu Ser Gly Phe Thr Lys Val
210 215 220
Asp Asp Asn Arg Tyr Ser Met Lys Ile Phe Phe Glu Ile Arg Lys Gln
225 230 235 240
Lys Tyr Leu Glu Ala Leu Asp Lys His Asp Arg Pro Lys Ala Val Asp
245 250 255
Ile Leu Val Lys Asp Leu Lys Val Phe Ser Thr Phe Asn Glu Glu Leu
260 265 270
Phe Lys Glu Ile Thr Gln Leu Leu Thr Leu Glu Asn Phe Arg Glu Asn
275 280 285
Glu Gln Leu Ser Lys Tyr Gly Asp Thr Lys Ser Ala Arg Ala Ile Met
290 295 300
Leu Val Glu Leu Lys Lys Leu Ile Glu Ala Asn Pro Leu Phe Arg Asp
305 310 315 320
Lys Leu Gln Phe Pro Thr Leu Arg Asn Ser Arg Leu Arg Thr Leu Ile
325 330 335
Asn Gln Ser Leu Asn Trp Gln His Gln Leu Cys Lys Asn Pro Arg Pro
340 345 350
Asn Pro Asp Ile Lys Thr Leu Phe Val Asp His Ser Cys Gly Pro Pro
355 360 365
Asn Gly Ala Arg Ala Pro Ser Pro Val Asn Asn Pro Leu Leu Gly Gly
370 375 380
Ile Pro Lys Ala Gly Gly Phe Pro Pro Leu Gly Ala His Gly Pro Phe
385 390 395 400
Gln Pro Thr Ala Ser Pro Val Pro Thr Pro Leu Ala Gly Trp Met Ser
405 410 415
Ser Pro Ser Ser Val Pro His Pro Ala Val Ser Ala Gly Ala Ile Ala
420 425 430
Leu Gly Gly Pro Ser Ile Pro Ala Ala Leu Lys His Pro Arg Thr Pro
435 440 445
Pro Thr Asn Ala Ser Leu Asp Tyr Pro Ser Ala Asp Ser Glu His Val
450 455 460
Ser Lys Arg Thr Arg Pro Met Gly Ile Ser Asp Glu Val Asn Leu Gly
465 470 475 480
Val Asn Met Leu Pro Met Ser Phe Ser Gly Gln Ala His Gly His Ser
485 490 495
Pro Ala Phe Lys Ala Pro Asp Asp Leu Pro Lys Thr Val Ala Arg Thr
500 505 510
Leu Ser Gln Gly Ser Ser Pro Met Ser Met Asp Phe His Pro Ile Lys
515 520 525
Gln Thr Leu Leu Leu Val Gly Thr Asn Val Gly Asp Ile Gly Leu Trp
530 535 540
Glu Val Gly Ser Arg Glu Arg Leu Val Gln Lys Thr Phe Lys Val Trp
545 550 555 560
Asp Leu Ser Lys Cys Ser Met Pro Leu Gln Ala Ala Leu Val Lys Glu
565 570 575
Pro Val Val Ser Val Asn Arg Val Ile Trp Ser Pro Asp Gly Ser Leu
580 585 590
Phe Gly Val Ala Tyr Ser Arg His Ile Val Gln Leu Tyr Ser Tyr His
595 600 605
Gly Gly Glu Asp Met Arg Gln His Leu Glu Ile Asp Ala His Val Gly
610 615 620
Gly Val Asn Asp Ile Ser Phe Ser Thr Pro Asn Lys Gln Leu Cys Val
625 630 635 640
Ile Thr Cys Gly Asp Asp Lys Thr Ile Lys Val Trp Asp Ala Ala Thr
645 650 655
Gly Val Lys Arg His Thr Phe Glu Gly His Glu Ala Pro Val Tyr Ser
660 665 670
Val Cys Pro His Tyr Lys Glu Asn Ile Gln Phe Ile Phe Ser Thr Ala
675 680 685
Leu Asp Gly Lys Ile Lys Ala Trp Leu Tyr Asp Asn Met Gly Ser Arg
690 695 700
Val Asp Tyr Asp Ala Pro Gly Arg Trp Cys Thr Thr Met Ala Tyr Ser
705 710 715 720
Ala Asp Gly Thr Arg Leu Phe Ser Cys Gly Thr Ser Lys Asp Gly Glu
725 730 735
Ser Phe Ile Val Glu Trp Asn Glu Ser Glu Gly Ala Val Lys Arg Thr
740 745 750
Tyr Gln Gly Phe His Lys Arg Ser Leu Gly Val Val Gln Phe Asp Thr
755 760 765
Thr Lys Asn Arg Tyr Leu Ala Ala Gly Asp Asp Phe Ser Ile Lys Phe
770 775 780
Trp Asp Met Asp Ala Val Gln Leu Leu Thr Ala Ile Asp Gly Asp Gly
785 790 795 800
Gly Leu Gln Ala Ser Pro Arg Ile Arg Phe Asn Lys Glu Gly Ser Leu
805 810 815
Leu Ala Val Ser Gly Asn Glu Asn Val Ile Lys Ile Met Ala Asn Ser
820 825 830
Asp Gly Leu Arg Leu Leu His Thr Phe Glu Asn Ile Ser Ser Glu Ser
835 840 845
Ser Lys Pro Ala Ile Asn Ser Ile Ala Ala Ala Ala Ala Ala Ala Ala
850 855 860
Thr Ser Ala Gly His Ala Asp Arg Ser Ala Asn Val Val Ser Ile Gln
865 870 875 880
Gly Met Asn Gly Asp Ser Arg Asn Met Val Asp Val Lys Pro Val Ile
885 890 895
Thr Glu Glu Ser Asn Asp Lys Ser Lys Ile Trp Lys Leu Thr Glu Val
900 905 910
Ser Glu Pro Ser Gln Cys Arg Ser Leu Arg Leu Pro Glu Asn Leu Arg
915 920 925
Val Ala Lys Ile Ser Arg Leu Ile Phe Thr Asn Ser Gly Asn Ala Ile
930 935 940
Leu Ala Leu Ala Ser Asn Ala Ile His Leu Leu Trp Lys Trp Gln Arg
945 950 955 960
Asn Glu Arg Asn Ala Thr Gly Lys Ala Thr Ala Ser Leu Pro Pro Gln
965 970 975
Gln Trp Gln Pro Ala Ser Gly Ile Leu Met Thr Asn Asp Val Ala Glu
980 985 990
Thr Asn Pro Glu Glu Ala Val Pro Cys Phe Ala Leu Ser Lys Asn Asp
995 1000 1005
Ser Tyr Val Met Ser Ala Ser Gly Gly Lys Ile Ser Leu Phe Asn
1010 1015 1020
Met Met Thr Phe Lys Thr Met Ala Thr Phe Met Pro Pro Pro Pro
1025 1030 1035
Ala Ala Thr Phe Leu Ala Phe His Pro Gln Asp Asn Asn Ile Ile
1040 1045 1050
Ala Ile Gly Met Asp Asp Ser Thr Ile Gln Ile Tyr Asn Val Arg
1055 1060 1065
Val Asp Glu Val Lys Ser Lys Leu Lys Gly His Ser Lys Arg Ile
1070 1075 1080
Thr Gly Leu Ala Phe Ser Asn Val Leu Asn Val Leu Val Ser Ser
1085 1090 1095
Gly Ala Asp Ala Gln Leu Cys Val Trp Asn Thr Asp Gly Trp Glu
1100 1105 1110
Lys Gln Arg Ser Lys Val Leu Pro Leu Pro Gln Gly Arg Pro Asn
1115 1120 1125
Ser Ala Pro Ser Asp Thr Arg Val Gln Phe His Gln Asp Gln Ala
1130 1135 1140
His Phe Leu Val Val His Glu Thr Gln Leu Ala Ile Tyr Glu Thr
1145 1150 1155
Thr Lys Leu Glu Cys Met Lys Gln Trp Ala Val Arg Glu Ser Leu
1160 1165 1170
Ala Pro Ile Thr His Ala Thr Phe Ser Cys Asp Ser Gln Leu Val
1175 1180 1185
Tyr Ala Ser Phe Met Asp Ala Thr Val Cys Val Phe Ser Ser Ala
1190 1195 1200
Asn Leu Arg Leu Arg Cys Arg Val Asn Pro Ser Ala Tyr Leu Pro
1205 1210 1215
Ala Ser Leu Ser Asn Ser Asn Val His Pro Leu Val Ile Ala Ala
1220 1225 1230
His Pro Gln Glu Pro Asn Met Phe Ala Val Gly Leu Ser Asp Gly
1235 1240 1245
Gly Val His Ile Phe Glu Pro Leu Glu Ser Glu Gly Lys Trp Gly
1250 1255 1260
Val Ala Pro Pro Ala Glu Asn Gly Ser Ala Ser Gly Ala Pro Thr
1265 1270 1275
Ala Pro Ser Val Gly Ala Ser Ala Ser Asp Gln Pro Gln Arg
1280 1285 1290
<210> 65
<211> 425
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 65
Met Asp Asp Asp Asn Gly Leu Glu Leu Ser Leu Gly Leu Ser Cys Gly
1 5 10 15
Gly Ser Thr Gly Lys Ala Lys Gly Asn Asn Asn Asn Asn Ala Gly Ser
20 25 30
Ser Ser Glu Asn Tyr Arg Ala Glu Gly Gly Asp Arg Ser Ala Lys Val
35 40 45
Ile Asp Asp Phe Lys Asn Phe Leu His Pro Thr Ser Gln Arg Pro Ala
50 55 60
Glu Pro Ser Ser Gly Ser Gln Arg Ser Asp Ser Gly Gln Gln Pro Pro
65 70 75 80
Gln Asn Phe Phe Asn Asp Leu Ser Lys Ala Pro Thr Thr Glu Ala Glu
85 90 95
Ala Ser Thr Lys Pro Leu Trp Val Glu Asp Glu Ser Arg Lys Glu Ala
100 105 110
Gly Asn Lys Arg Lys Phe Gly Phe Pro Gly Met Asn Asp Asp Lys Lys
115 120 125
Lys Glu Lys Asp Ser Ser His Val Asp Met His Glu Lys Lys Thr Lys
130 135 140
Ala Ser His Val Ser Thr Ala Thr Asp Glu Gly Ser Thr Ala Glu Asn
145 150 155 160
Glu Asp Val Ala Glu Ser Glu Val Gly Gly Gly Ser Ser Ser Asn His
165 170 175
Ala Lys Glu Val Val Arg Pro Pro Thr Asp Thr Asn Ile Val Asp Asn
180 185 190
Leu Thr Gly Gln Arg Arg Ser Asn His Gly Gly Ser Gly Thr Glu Glu
195 200 205
Phe Thr Met Arg Asn Met Ser Tyr Thr Val Pro Phe Thr Val His Pro
210 215 220
Gln Asn Val Val Thr Ser Met Pro Tyr Ser Leu Pro Thr Lys Glu Ser
225 230 235 240
Gly Gln His Ala Ala Ala Thr Ser Leu Leu Gln Pro Asn Ala Asn Ala
245 250 255
Gly Asn Leu Pro Ile Met Phe Gly Tyr Ser Pro Val Gln Leu Pro Met
260 265 270
Leu Asp Lys Asp Gly Ser Gly Gly Ile Val Ala Leu Ser Gln Ser Pro
275 280 285
Phe Ala Gly Arg Val Pro Ser Asn Ser Ala Thr Ala Lys Gly Glu Gly
290 295 300
Lys Gln Pro Val Ala Glu Glu Gly Ser Ser Glu Asp Ala Ser Glu Arg
305 310 315 320
Pro Thr Gly Asp Asn Ser Asn Leu Asn Thr Ala Phe Ser Phe Asp Phe
325 330 335
Ser Ala Ile Lys Pro Gly Met Ala Ala Asp Val Lys Phe Gly Gly Ser
340 345 350
Gly Ala Arg Pro Asn Leu Pro Trp Val Ser Thr Thr Gly Ser Gly Pro
355 360 365
His Gly Arg Thr Ile Ser Gly Val Thr Tyr Arg Tyr Asn Ala Asn Gln
370 375 380
Ile Lys Ile Val Cys Ala Cys His Gly Ser His Met Ser Pro Glu Glu
385 390 395 400
Phe Val Arg His Ala Ser Glu Glu Tyr Val Ser Pro Glu Ser Ser Met
405 410 415
Gly Met Thr Ala Ala Ser Ala His Thr
420 425
<210> 66
<211> 564
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 66
Met Pro Lys Lys Lys Arg Lys Val Ser Ser Gly Ala Asn Phe Asn Gln
1 5 10 15
Ser Gly Asn Ile Ala Asp Ser Ser Leu Ser Phe Thr Phe Thr Asn Ser
20 25 30
Ser Asn Gly Pro Asn Leu Ile Thr Thr Gln Thr Asn Ser Gln Ala Leu
35 40 45
Ser Gln Pro Ile Ala Ser Ser Asn Val His Asp Asn Phe Met Asn Asn
50 55 60
Glu Ile Thr Ala Ser Lys Ile Asp Asp Gly Asn Asn Ser Lys Pro Leu
65 70 75 80
Ser Pro Gly Trp Thr Asp Gln Thr Ala Tyr Asn Ala Phe Gly Ile Thr
85 90 95
Thr Gly Met Phe Asn Thr Thr Thr Met Asp Asp Val Tyr Asn Tyr Leu
100 105 110
Phe Asp Asp Glu Asp Thr Pro Pro Asn Pro Lys Lys Glu Gly Gly Ser
115 120 125
Asn Gln Thr Ser Leu Tyr Lys Lys Ala Gly Ser Met Asp Asp Asp Asn
130 135 140
Gly Leu Glu Leu Ser Leu Gly Leu Ser Cys Gly Gly Ser Thr Gly Lys
145 150 155 160
Ala Lys Gly Asn Asn Asn Asn Asn Ala Gly Ser Ser Ser Glu Asn Tyr
165 170 175
Arg Ala Glu Gly Gly Asp Arg Ser Ala Lys Val Ile Asp Asp Phe Lys
180 185 190
Asn Phe Leu His Pro Thr Ser Gln Arg Pro Ala Glu Pro Ser Ser Gly
195 200 205
Ser Gln Arg Ser Asp Ser Gly Gln Gln Pro Pro Gln Asn Phe Phe Asn
210 215 220
Asp Leu Ser Lys Ala Pro Thr Thr Glu Ala Glu Ala Ser Thr Lys Pro
225 230 235 240
Leu Trp Val Glu Asp Glu Ser Arg Lys Glu Ala Gly Asn Lys Arg Lys
245 250 255
Phe Gly Phe Pro Gly Met Asn Asp Asp Lys Lys Lys Glu Lys Asp Ser
260 265 270
Ser His Val Asp Met His Glu Lys Lys Thr Lys Ala Ser His Val Ser
275 280 285
Thr Ala Thr Asp Glu Gly Ser Thr Ala Glu Asn Glu Asp Val Ala Glu
290 295 300
Ser Glu Val Gly Gly Gly Ser Ser Ser Asn His Ala Lys Glu Val Val
305 310 315 320
Arg Pro Pro Thr Asp Thr Asn Ile Val Asp Asn Leu Thr Gly Gln Arg
325 330 335
Arg Ser Asn His Gly Gly Ser Gly Thr Glu Glu Phe Thr Met Arg Asn
340 345 350
Met Ser Tyr Thr Val Pro Phe Thr Val His Pro Gln Asn Val Val Thr
355 360 365
Ser Met Pro Tyr Ser Leu Pro Thr Lys Glu Ser Gly Gln His Ala Ala
370 375 380
Ala Thr Ser Leu Leu Gln Pro Asn Ala Asn Ala Gly Asn Leu Pro Ile
385 390 395 400
Met Phe Gly Tyr Ser Pro Val Gln Leu Pro Met Leu Asp Lys Asp Gly
405 410 415
Ser Gly Gly Ile Val Ala Leu Ser Gln Ser Pro Phe Ala Gly Arg Val
420 425 430
Pro Ser Asn Ser Ala Thr Ala Lys Gly Glu Gly Lys Gln Pro Val Ala
435 440 445
Glu Glu Gly Ser Ser Glu Asp Ala Ser Glu Arg Pro Thr Gly Asp Asn
450 455 460
Ser Asn Leu Asn Thr Ala Phe Ser Phe Asp Phe Ser Ala Ile Lys Pro
465 470 475 480
Gly Met Ala Ala Asp Val Lys Phe Gly Gly Ser Gly Ala Arg Pro Asn
485 490 495
Leu Pro Trp Val Ser Thr Thr Gly Ser Gly Pro His Gly Arg Thr Ile
500 505 510
Ser Gly Val Thr Tyr Arg Tyr Asn Ala Asn Gln Ile Lys Ile Val Cys
515 520 525
Ala Cys His Gly Ser His Met Ser Pro Glu Glu Phe Val Arg His Ala
530 535 540
Ser Glu Glu Tyr Val Ser Pro Glu Ser Ser Met Gly Met Thr Ala Ala
545 550 555 560
Ser Ala His Thr
<210> 67
<211> 475
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 67
Met Pro Lys Lys Lys Arg Lys Val Ser Ser Gly Ala Asn Phe Asn Gln
1 5 10 15
Ser Gly Asn Ile Ala Asp Ser Ser Leu Ser Phe Thr Phe Thr Asn Ser
20 25 30
Ser Asn Gly Pro Asn Leu Ile Thr Thr Gln Thr Asn Ser Gln Ala Leu
35 40 45
Ser Gln Pro Ile Ala Ser Ser Asn Val His Asp Asn Phe Met Asn Asn
50 55 60
Glu Ile Thr Ala Ser Lys Ile Asp Asp Gly Asn Asn Ser Lys Pro Leu
65 70 75 80
Ser Pro Gly Trp Thr Asp Gln Thr Ala Tyr Asn Ala Phe Gly Ile Thr
85 90 95
Thr Gly Met Phe Asn Thr Thr Thr Met Asp Asp Val Tyr Asn Tyr Leu
100 105 110
Phe Asp Asp Glu Asp Thr Pro Pro Asn Pro Lys Lys Glu Gly Gly Ser
115 120 125
Asn Gln Thr Ser Leu Tyr Lys Lys Ala Gly Ser Met Thr Ser Asp Gly
130 135 140
Ala Thr Ser Thr Ser Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala Ala
145 150 155 160
Arg Arg Lys Pro Ser Trp Arg Glu Arg Glu Asn Asn Arg Arg Arg Glu
165 170 175
Arg Arg Arg Arg Ala Val Ala Ala Lys Ile Tyr Thr Gly Leu Arg Ala
180 185 190
Gln Gly Asp Tyr Asn Leu Pro Lys His Cys Asp Asn Asn Glu Val Leu
195 200 205
Lys Ala Leu Cys Val Glu Ala Gly Trp Val Val Glu Glu Asp Gly Thr
210 215 220
Thr Tyr Arg Lys Gly Cys Lys Pro Leu Pro Gly Glu Ile Ala Gly Thr
225 230 235 240
Ser Ser Arg Val Thr Pro Tyr Ser Ser Gln Asn Gln Ser Pro Leu Ser
245 250 255
Ser Ala Phe Gln Ser Pro Ile Pro Ser Tyr Gln Val Ser Pro Ser Ser
260 265 270
Ser Ser Phe Pro Ser Pro Ser Arg Gly Glu Pro Asn Asn Asn Met Ser
275 280 285
Ser Thr Phe Phe Pro Phe Leu Arg Asn Gly Gly Ile Pro Ser Ser Leu
290 295 300
Pro Ser Leu Arg Ile Ser Asn Ser Cys Pro Val Thr Pro Pro Val Ser
305 310 315 320
Ser Pro Thr Ser Lys Asn Pro Lys Pro Leu Pro Asn Trp Glu Ser Ile
325 330 335
Ala Lys Gln Ser Met Ala Ile Ala Lys Gln Ser Met Ala Ser Phe Asn
340 345 350
Tyr Pro Phe Tyr Ala Val Ser Ala Pro Ala Ser Pro Thr His Arg His
355 360 365
Gln Phe His Thr Pro Ala Thr Ile Pro Glu Cys Asp Glu Ser Asp Ser
370 375 380
Ser Thr Val Asp Ser Gly His Trp Ile Ser Phe Gln Lys Phe Ala Gln
385 390 395 400
Gln Gln Pro Phe Ser Ala Ser Met Val Pro Thr Ser Pro Thr Phe Asn
405 410 415
Leu Val Lys Pro Ala Pro Gln Gln Met Ser Pro Asn Thr Ala Ala Phe
420 425 430
Gln Glu Ile Gly Gln Ser Ser Glu Phe Lys Phe Glu Asn Ser Gln Val
435 440 445
Lys Pro Trp Glu Gly Glu Arg Ile His Asp Val Gly Met Glu Asp Leu
450 455 460
Glu Leu Thr Leu Gly Asn Gly Lys Ala Arg Gly
465 470 475
<210> 68
<211> 587
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 68
Met Lys Arg Asp His His Gln Phe Gln Gly Arg Leu Ser Asn His Gly
1 5 10 15
Thr Ser Ser Ser Ser Ser Ser Ile Ser Lys Asp Lys Met Met Met Val
20 25 30
Lys Lys Glu Glu Asp Gly Gly Gly Asn Met Asp Asp Glu Leu Leu Ala
35 40 45
Val Leu Gly Tyr Lys Val Arg Ser Ser Glu Met Ala Glu Val Ala Leu
50 55 60
Lys Leu Glu Gln Leu Glu Thr Met Met Ser Asn Val Gln Glu Asp Gly
65 70 75 80
Leu Ser His Leu Ala Thr Asp Thr Val His Tyr Asn Pro Ser Glu Leu
85 90 95
Tyr Ser Trp Leu Asp Asn Met Leu Ser Glu Leu Asn Pro Pro Pro Leu
100 105 110
Pro Ala Ser Ser Asn Gly Leu Asp Pro Val Leu Pro Ser Pro Glu Ile
115 120 125
Cys Gly Phe Pro Ala Ser Asp Tyr Asp Leu Lys Val Ile Pro Gly Asn
130 135 140
Ala Ile Tyr Gln Phe Pro Ala Ile Asp Ser Ser Ser Ser Ser Asn Asn
145 150 155 160
Gln Asn Lys Arg Leu Lys Ser Cys Ser Ser Pro Asp Ser Met Val Thr
165 170 175
Ser Thr Ser Thr Gly Thr Gln Ile Gly Gly Val Ile Gly Thr Thr Val
180 185 190
Thr Thr Thr Thr Thr Thr Thr Thr Ala Ala Gly Glu Ser Thr Arg Ser
195 200 205
Val Ile Leu Val Asp Ser Gln Glu Asn Gly Val Arg Leu Val His Ala
210 215 220
Leu Met Ala Cys Ala Glu Ala Ile Gln Gln Asn Asn Leu Thr Leu Ala
225 230 235 240
Glu Ala Leu Val Lys Gln Ile Gly Cys Leu Ala Val Ser Gln Ala Gly
245 250 255
Ala Met Arg Lys Val Ala Thr Tyr Phe Ala Glu Ala Leu Ala Arg Arg
260 265 270
Ile Tyr Arg Leu Ser Pro Pro Gln Asn Gln Ile Asp His Cys Leu Ser
275 280 285
Asp Thr Leu Gln Met His Phe Tyr Glu Thr Cys Pro Tyr Leu Lys Phe
290 295 300
Ala His Phe Thr Ala Asn Gln Ala Ile Leu Glu Ala Phe Glu Gly Lys
305 310 315 320
Lys Arg Val His Val Ile Asp Phe Ser Met Asn Gln Gly Leu Gln Trp
325 330 335
Pro Ala Leu Met Gln Ala Leu Ala Leu Arg Glu Gly Gly Pro Pro Thr
340 345 350
Phe Arg Leu Thr Gly Ile Gly Pro Pro Ala Pro Asp Asn Ser Asp His
355 360 365
Leu His Glu Val Gly Cys Lys Leu Ala Gln Leu Ala Glu Ala Ile His
370 375 380
Val Glu Phe Glu Tyr Arg Gly Phe Val Ala Asn Ser Leu Ala Asp Leu
385 390 395 400
Asp Ala Ser Met Leu Glu Leu Arg Pro Ser Asp Thr Glu Ala Val Ala
405 410 415
Val Asn Ser Val Phe Glu Leu His Lys Leu Leu Gly Arg Pro Gly Gly
420 425 430
Ile Glu Lys Val Leu Gly Val Val Lys Gln Ile Lys Pro Val Ile Phe
435 440 445
Thr Val Val Glu Gln Glu Ser Asn His Asn Gly Pro Val Phe Leu Asp
450 455 460
Arg Phe Thr Glu Ser Leu His Tyr Tyr Ser Thr Leu Phe Asp Ser Leu
465 470 475 480
Glu Gly Val Pro Asn Ser Gln Asp Lys Val Met Ser Glu Val Tyr Leu
485 490 495
Gly Lys Gln Ile Cys Asn Leu Val Ala Cys Glu Gly Pro Asp Arg Val
500 505 510
Glu Arg His Glu Thr Leu Ser Gln Trp Gly Asn Arg Phe Gly Ser Ser
515 520 525
Gly Leu Ala Pro Ala His Leu Gly Ser Asn Ala Phe Lys Gln Ala Ser
530 535 540
Met Leu Leu Ser Val Phe Asn Ser Gly Gln Gly Tyr Arg Val Glu Glu
545 550 555 560
Ser Asn Gly Cys Leu Met Leu Gly Trp His Thr Arg Pro Leu Ile Thr
565 570 575
Thr Ser Ala Trp Lys Leu Ser Thr Ala Ala Tyr
580 585
<210> 69
<211> 726
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 69
Met Pro Lys Lys Lys Arg Lys Val Ser Ser Gly Ala Asn Phe Asn Gln
1 5 10 15
Ser Gly Asn Ile Ala Asp Ser Ser Leu Ser Phe Thr Phe Thr Asn Ser
20 25 30
Ser Asn Gly Pro Asn Leu Ile Thr Thr Gln Thr Asn Ser Gln Ala Leu
35 40 45
Ser Gln Pro Ile Ala Ser Ser Asn Val His Asp Asn Phe Met Asn Asn
50 55 60
Glu Ile Thr Ala Ser Lys Ile Asp Asp Gly Asn Asn Ser Lys Pro Leu
65 70 75 80
Ser Pro Gly Trp Thr Asp Gln Thr Ala Tyr Asn Ala Phe Gly Ile Thr
85 90 95
Thr Gly Met Phe Asn Thr Thr Thr Met Asp Asp Val Tyr Asn Tyr Leu
100 105 110
Phe Asp Asp Glu Asp Thr Pro Pro Asn Pro Lys Lys Glu Gly Gly Ser
115 120 125
Asn Gln Thr Ser Leu Tyr Lys Lys Ala Gly Ser Met Lys Arg Asp His
130 135 140
His Gln Phe Gln Gly Arg Leu Ser Asn His Gly Thr Ser Ser Ser Ser
145 150 155 160
Ser Ser Ile Ser Lys Asp Lys Met Met Met Val Lys Lys Glu Glu Asp
165 170 175
Gly Gly Gly Asn Met Asp Asp Glu Leu Leu Ala Val Leu Gly Tyr Lys
180 185 190
Val Arg Ser Ser Glu Met Ala Glu Val Ala Leu Lys Leu Glu Gln Leu
195 200 205
Glu Thr Met Met Ser Asn Val Gln Glu Asp Gly Leu Ser His Leu Ala
210 215 220
Thr Asp Thr Val His Tyr Asn Pro Ser Glu Leu Tyr Ser Trp Leu Asp
225 230 235 240
Asn Met Leu Ser Glu Leu Asn Pro Pro Pro Leu Pro Ala Ser Ser Asn
245 250 255
Gly Leu Asp Pro Val Leu Pro Ser Pro Glu Ile Cys Gly Phe Pro Ala
260 265 270
Ser Asp Tyr Asp Leu Lys Val Ile Pro Gly Asn Ala Ile Tyr Gln Phe
275 280 285
Pro Ala Ile Asp Ser Ser Ser Ser Ser Asn Asn Gln Asn Lys Arg Leu
290 295 300
Lys Ser Cys Ser Ser Pro Asp Ser Met Val Thr Ser Thr Ser Thr Gly
305 310 315 320
Thr Gln Ile Gly Gly Val Ile Gly Thr Thr Val Thr Thr Thr Thr Thr
325 330 335
Thr Thr Thr Ala Ala Gly Glu Ser Thr Arg Ser Val Ile Leu Val Asp
340 345 350
Ser Gln Glu Asn Gly Val Arg Leu Val His Ala Leu Met Ala Cys Ala
355 360 365
Glu Ala Ile Gln Gln Asn Asn Leu Thr Leu Ala Glu Ala Leu Val Lys
370 375 380
Gln Ile Gly Cys Leu Ala Val Ser Gln Ala Gly Ala Met Arg Lys Val
385 390 395 400
Ala Thr Tyr Phe Ala Glu Ala Leu Ala Arg Arg Ile Tyr Arg Leu Ser
405 410 415
Pro Pro Gln Asn Gln Ile Asp His Cys Leu Ser Asp Thr Leu Gln Met
420 425 430
His Phe Tyr Glu Thr Cys Pro Tyr Leu Lys Phe Ala His Phe Thr Ala
435 440 445
Asn Gln Ala Ile Leu Glu Ala Phe Glu Gly Lys Lys Arg Val His Val
450 455 460
Ile Asp Phe Ser Met Asn Gln Gly Leu Gln Trp Pro Ala Leu Met Gln
465 470 475 480
Ala Leu Ala Leu Arg Glu Gly Gly Pro Pro Thr Phe Arg Leu Thr Gly
485 490 495
Ile Gly Pro Pro Ala Pro Asp Asn Ser Asp His Leu His Glu Val Gly
500 505 510
Cys Lys Leu Ala Gln Leu Ala Glu Ala Ile His Val Glu Phe Glu Tyr
515 520 525
Arg Gly Phe Val Ala Asn Ser Leu Ala Asp Leu Asp Ala Ser Met Leu
530 535 540
Glu Leu Arg Pro Ser Asp Thr Glu Ala Val Ala Val Asn Ser Val Phe
545 550 555 560
Glu Leu His Lys Leu Leu Gly Arg Pro Gly Gly Ile Glu Lys Val Leu
565 570 575
Gly Val Val Lys Gln Ile Lys Pro Val Ile Phe Thr Val Val Glu Gln
580 585 590
Glu Ser Asn His Asn Gly Pro Val Phe Leu Asp Arg Phe Thr Glu Ser
595 600 605
Leu His Tyr Tyr Ser Thr Leu Phe Asp Ser Leu Glu Gly Val Pro Asn
610 615 620
Ser Gln Asp Lys Val Met Ser Glu Val Tyr Leu Gly Lys Gln Ile Cys
625 630 635 640
Asn Leu Val Ala Cys Glu Gly Pro Asp Arg Val Glu Arg His Glu Thr
645 650 655
Leu Ser Gln Trp Gly Asn Arg Phe Gly Ser Ser Gly Leu Ala Pro Ala
660 665 670
His Leu Gly Ser Asn Ala Phe Lys Gln Ala Ser Met Leu Leu Ser Val
675 680 685
Phe Asn Ser Gly Gln Gly Tyr Arg Val Glu Glu Ser Asn Gly Cys Leu
690 695 700
Met Leu Gly Trp His Thr Arg Pro Leu Ile Thr Thr Ser Ala Trp Lys
705 710 715 720
Leu Ser Thr Ala Ala Tyr
725
<210> 70
<211> 413
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 70
Met Lys Leu Leu Ser Ser Ile Glu Glu Ala Cys Asn Ile Cys Arg Leu
1 5 10 15
Lys Lys Leu Lys Cys Ser Lys Glu Lys Pro Lys Cys Ala Lys Cys Leu
20 25 30
Lys Asn Asn Trp Glu Cys Arg Tyr Ser Pro Lys Thr Lys Arg Ser Pro
35 40 45
Leu Thr Arg Ala His Leu Thr Glu Val Glu Ser Arg Leu Glu Arg Leu
50 55 60
Glu Gln Leu Phe Leu Leu Ile Phe Pro Arg Glu Asn Leu Asn Met Ile
65 70 75 80
Leu Lys Met Asp Ser Leu Gln Asp Ile Lys Ala Leu Leu Thr Gly Leu
85 90 95
Phe Val Gln Asp Asn Val Asn Lys Asp Ala Val Thr Asp Arg Leu Ala
100 105 110
Ser Val Glu Thr Asp Met Pro Leu Thr Leu Arg Gln His Arg Ile Ser
115 120 125
Ala Thr Ser Ser Ser Glu Glu Ser Ser Asn Lys Gly Gln Arg Gln Leu
130 135 140
Thr Val Ser Ser Arg Ser Asn Gln Thr Ser Leu Tyr Lys Lys Ala Gly
145 150 155 160
Ser Tyr Glu Pro Gly Asp Asp Ser Gly Ala Gly Ile Phe Arg Lys Ile
165 170 175
Leu Val Ser Gln Pro Val Asn Pro Pro Arg Val Thr Thr Thr Leu Ile
180 185 190
Glu Pro Ser Asn Glu Leu Glu Ala Cys Gly Arg Val Ser Tyr Pro Glu
195 200 205
Asp Asn Gly Ala Cys His Arg Met Asp Ser Pro Arg Ser Ala Glu Phe
210 215 220
Ser Gly Gly Ser Gly His Phe Val Ser Glu Lys Asp Gly His Lys Thr
225 230 235 240
Thr Ile Ser Pro Arg Ser Pro Ala Glu Thr Ser Glu Leu Val Gly Gln
245 250 255
Met Thr Ile Phe Tyr Ser Gly Lys Val Asn Val Tyr Asp Gly Ile Pro
260 265 270
Pro Glu Lys Ala Arg Ser Ile Met His Phe Ala Ala Asn Pro Ile Asp
275 280 285
Leu Pro Glu Asn Gly Ile Phe Ala Ser Ser Arg Met Ile Ser Lys Leu
290 295 300
Ile Ser Lys Glu Lys Met Met Glu Leu Pro Gln Lys Gly Leu Glu Lys
305 310 315 320
Ala Asn Ser Ser Arg Asp Ser Gly Met Glu Gly Gln Ala Asn Arg Lys
325 330 335
Val Ser Leu Gln Arg Tyr Arg Glu Lys Arg Lys Asp Arg Lys Phe Ser
340 345 350
Lys Ala Lys Lys Cys Pro Gly Val Ala Ser Ser Ser Leu Glu Met Phe
355 360 365
Leu Asn Cys Gln Pro Arg Met Lys Ala Ala Tyr Ser Gln Asn Leu Gly
370 375 380
Cys Thr Gly Ser Pro Leu His Ser Gln Ser Pro Glu Ser Gln Thr Lys
385 390 395 400
Ser Pro Asn Leu Ser Val Asp Leu Asn Ser Glu Gly Ile
405 410
<210> 71
<211> 449
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 71
Met Lys Leu Leu Ser Ser Ile Glu Glu Ala Cys Asn Ile Cys Arg Leu
1 5 10 15
Lys Lys Leu Lys Cys Ser Lys Glu Lys Pro Lys Cys Ala Lys Cys Leu
20 25 30
Lys Asn Asn Trp Glu Cys Arg Tyr Ser Pro Lys Thr Lys Arg Ser Pro
35 40 45
Leu Thr Arg Ala His Leu Thr Glu Val Glu Ser Arg Leu Glu Arg Leu
50 55 60
Glu Gln Leu Phe Leu Leu Ile Phe Pro Arg Glu Asn Leu Asn Met Ile
65 70 75 80
Leu Lys Met Asp Ser Leu Gln Asp Ile Lys Ala Leu Leu Thr Gly Leu
85 90 95
Phe Val Gln Asp Asn Val Asn Lys Asp Ala Val Thr Asp Arg Leu Ala
100 105 110
Ser Val Glu Thr Asp Met Pro Leu Thr Leu Arg Gln His Arg Ile Ser
115 120 125
Ala Thr Ser Ser Ser Glu Glu Ser Ser Asn Lys Gly Gln Arg Gln Leu
130 135 140
Thr Val Ser Ser Arg Ser Asn Gln Thr Ser Leu Tyr Lys Lys Ala Gly
145 150 155 160
Ser Met Asp Val Gly Val Ser Pro Ala Lys Ser Ile Leu Ala Lys Pro
165 170 175
Leu Lys Leu Leu Thr Glu Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp
180 185 190
Cys Arg Lys Phe Leu Lys Asp Lys Gly Met Arg Arg Pro Ser Trp Asn
195 200 205
Lys Ser Gln Ala Ile Gln Gln Val Leu Ser Leu Lys Ala Leu Tyr Glu
210 215 220
Pro Gly Asp Asp Ser Gly Ala Gly Ile Phe Arg Lys Ile Leu Val Ser
225 230 235 240
Gln Pro Val Asn Pro Pro Arg Val Thr Thr Thr Leu Ile Glu Pro Ser
245 250 255
Asn Glu Leu Glu Ala Cys Gly Arg Val Ser Tyr Pro Glu Asp Asn Gly
260 265 270
Ala Cys His Arg Met Asp Ser Pro Arg Ser Ala Glu Phe Ser Gly Gly
275 280 285
Ser Gly His Phe Val Ser Glu Lys Asp Gly His Lys Thr Thr Ile Ser
290 295 300
Pro Arg Ser Pro Ala Glu Thr Ser Glu Leu Ile Met His Phe Ala Ala
305 310 315 320
Asn Pro Ile Asp Leu Pro Glu Asn Gly Ile Phe Ala Ser Ser Arg Met
325 330 335
Ile Ser Lys Leu Ile Ser Lys Glu Lys Met Met Glu Leu Pro Gln Lys
340 345 350
Gly Leu Glu Lys Ala Asn Ser Ser Arg Asp Ser Gly Met Glu Gly Gln
355 360 365
Ala Asn Arg Lys Val Ser Leu Gln Arg Tyr Arg Glu Lys Arg Lys Asp
370 375 380
Arg Lys Phe Ser Lys Ala Lys Lys Cys Pro Gly Val Ala Ser Ser Ser
385 390 395 400
Leu Glu Met Phe Leu Asn Cys Gln Pro Arg Met Lys Ala Ala Tyr Ser
405 410 415
Gln Asn Leu Gly Cys Thr Gly Ser Pro Leu His Ser Gln Ser Pro Glu
420 425 430
Ser Gln Thr Lys Ser Pro Asn Leu Ser Val Asp Leu Asn Ser Glu Gly
435 440 445
Ile
<210> 72
<211> 390
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 72
Met Lys Leu Leu Ser Ser Ile Glu Glu Ala Cys Asn Ile Cys Arg Leu
1 5 10 15
Lys Lys Leu Lys Cys Ser Lys Glu Lys Pro Lys Cys Ala Lys Cys Leu
20 25 30
Lys Asn Asn Trp Glu Cys Arg Tyr Ser Pro Lys Thr Lys Arg Ser Pro
35 40 45
Leu Thr Arg Ala His Leu Thr Glu Val Glu Ser Arg Leu Glu Arg Leu
50 55 60
Glu Gln Leu Phe Leu Leu Ile Phe Pro Arg Glu Asn Leu Asn Met Ile
65 70 75 80
Leu Lys Met Asp Ser Leu Gln Asp Ile Lys Ala Leu Leu Thr Gly Leu
85 90 95
Phe Val Gln Asp Asn Val Asn Lys Asp Ala Val Thr Asp Arg Leu Ala
100 105 110
Ser Val Glu Thr Asp Met Pro Leu Thr Leu Arg Gln His Arg Ile Ser
115 120 125
Ala Thr Ser Ser Ser Glu Glu Ser Ser Asn Lys Gly Gln Arg Gln Leu
130 135 140
Thr Val Ser Ser Arg Ser Asn Gln Thr Ser Leu Tyr Lys Lys Ala Gly
145 150 155 160
Ser Met Asp Val Gly Val Ser Pro Ala Lys Ser Ile Leu Ala Lys Pro
165 170 175
Leu Lys Leu Leu Thr Glu Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp
180 185 190
Cys Arg Lys Phe Leu Lys Asp Lys Gly Met Arg Arg Pro Ser Trp Asn
195 200 205
Lys Ser Gln Ala Ile Gln Gln Val Leu Ser Leu Lys Ala Leu Tyr Glu
210 215 220
Pro Gly Asp Asp Ser Gly Ala Gly Ile Phe Arg Lys Ile Leu Val Ser
225 230 235 240
Gln Pro Val Asn Pro Pro Arg Val Thr Thr Thr Leu Ile Glu Pro Ser
245 250 255
Asn Glu Leu Glu Ala Cys Gly Arg Val Ser Tyr Pro Glu Asp Asn Gly
260 265 270
Ala Cys His Arg Met Asp Ser Pro Arg Ser Ala Glu Phe Ser Gly Gly
275 280 285
Ser Gly His Phe Val Ser Glu Lys Asp Gly His Lys Thr Thr Ile Ser
290 295 300
Pro Arg Ser Pro Ala Glu Thr Ser Glu Leu Val Gly Gln Met Thr Ile
305 310 315 320
Phe Tyr Ser Gly Lys Val Asn Val Tyr Asp Gly Ile Pro Pro Glu Lys
325 330 335
Ala Arg Ser Ile Met His Phe Ala Ala Asn Pro Ile Asp Leu Pro Glu
340 345 350
Asn Gly Ile Phe Ala Ser Ser Arg Met Ile Ser Lys Leu Ile Ser Lys
355 360 365
Glu Lys Met Met Glu Leu Pro Gln Lys Gly Leu Glu Lys Ala Asn Ser
370 375 380
Ser Arg Asp Ser Gly Met
385 390
<210> 73
<211> 142
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 73
Met Val Ser Lys Gly Glu Glu Leu Phe Thr Gly Val Val Pro Ile Leu
1 5 10 15
Val Glu Leu Asp Gly Asp Val Asn Gly His Lys Phe Ser Val Ser Gly
20 25 30
Glu Gly Glu Gly Asp Ala Thr Tyr Gly Lys Leu Thr Asp Asp Gly Asn
35 40 45
Tyr Lys Thr Arg Ala Glu Val Lys Phe Glu Gly Asp Thr Leu Val Asn
50 55 60
Arg Ile Glu Leu Lys Gly Ile Asp Phe Lys Glu Asp Gly Asn Ile Leu
65 70 75 80
Gly His Lys Leu Glu Tyr Asn Tyr Asn Ser His Asn Val Tyr Ile Thr
85 90 95
Ala Asp Lys Gln Lys Asn Gly Ile Lys Ala Asn Phe Lys Ile Arg His
100 105 110
Asn Ile Glu Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu Gly Pro Arg
115 120 125
Pro Thr Ser Gly Ser Val Asp Leu Glu Gly Thr Ala Pro Gly
130 135 140
<210> 74
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 74
Met Asp Lys Gln Lys Asn Gly Ile Lys Ala Asn Phe Lys Ile Arg His
1 5 10 15
Asn Ile Glu Asp Gly Gly Val Gln Leu Ala Asp His Tyr Gln Gln Asn
20 25 30
Thr Pro Ile Gly Asp Gly Pro Val Leu Leu Pro Asp Asn His Tyr Leu
35 40 45
Ser Tyr Gln Ser Ala Leu Ser Lys Asp Pro Asn Glu Lys Arg Asp His
50 55 60
Met Val Leu Leu Glu Phe Val Thr Ala Ala Gly Ile Thr Leu Gly Met
65 70 75 80
Asp Glu Leu Tyr Lys Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Gly Pro
85 90 95
Arg Pro Thr Ser Gly Ser Val Asp Leu Glu Gly Thr Ala Pro Gly
100 105 110
<210> 75
<211> 568
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 75
Met Val Ser Lys Gly Glu Glu Leu Phe Thr Gly Val Val Pro Ile Leu
1 5 10 15
Val Glu Leu Asp Gly Asp Val Asn Gly His Lys Phe Ser Val Ser Gly
20 25 30
Glu Gly Glu Gly Asp Ala Thr Tyr Gly Lys Leu Thr Asp Asp Gly Asn
35 40 45
Tyr Lys Thr Arg Ala Glu Val Lys Phe Glu Gly Asp Thr Leu Val Asn
50 55 60
Arg Ile Glu Leu Lys Gly Ile Asp Phe Lys Glu Asp Gly Asn Ile Leu
65 70 75 80
Gly His Lys Leu Glu Tyr Asn Tyr Asn Ser His Asn Val Tyr Ile Thr
85 90 95
Ala Asp Lys Gln Lys Asn Gly Ile Lys Ala Asn Phe Lys Ile Arg His
100 105 110
Asn Ile Glu Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu Gly Pro Arg
115 120 125
Pro Thr Ser Gly Ser Val Asp Leu Glu Gly Thr Ala Pro Gly Ser Met
130 135 140
Asp Asp Asp Asn Gly Leu Glu Leu Ser Leu Gly Leu Ser Cys Gly Gly
145 150 155 160
Ser Thr Gly Lys Ala Lys Gly Asn Asn Asn Asn Asn Ala Gly Ser Ser
165 170 175
Ser Glu Asn Tyr Arg Ala Glu Gly Gly Asp Arg Ser Ala Lys Val Ile
180 185 190
Asp Asp Phe Lys Asn Phe Leu His Pro Thr Ser Gln Arg Pro Ala Glu
195 200 205
Pro Ser Ser Gly Ser Gln Arg Ser Asp Ser Gly Gln Gln Pro Pro Gln
210 215 220
Asn Phe Phe Asn Asp Leu Ser Lys Ala Pro Thr Thr Glu Ala Glu Ala
225 230 235 240
Ser Thr Lys Pro Leu Trp Val Glu Asp Glu Ser Arg Lys Glu Ala Gly
245 250 255
Asn Lys Arg Lys Phe Gly Phe Pro Gly Met Asn Asp Asp Lys Lys Lys
260 265 270
Glu Lys Asp Ser Ser His Val Asp Met His Glu Lys Lys Thr Lys Ala
275 280 285
Ser His Val Ser Thr Ala Thr Asp Glu Gly Ser Thr Ala Glu Asn Glu
290 295 300
Asp Val Ala Glu Ser Glu Val Gly Gly Gly Ser Ser Ser Asn His Ala
305 310 315 320
Lys Glu Val Val Arg Pro Pro Thr Asp Thr Asn Ile Val Asp Asn Leu
325 330 335
Thr Gly Gln Arg Arg Ser Asn His Gly Gly Ser Gly Thr Glu Glu Phe
340 345 350
Thr Met Arg Asn Met Ser Tyr Thr Val Pro Phe Thr Val His Pro Gln
355 360 365
Asn Val Val Thr Ser Met Pro Tyr Ser Leu Pro Thr Lys Glu Ser Gly
370 375 380
Gln His Ala Ala Ala Thr Ser Leu Leu Gln Pro Asn Ala Asn Ala Gly
385 390 395 400
Asn Leu Pro Ile Met Phe Gly Tyr Ser Pro Val Gln Leu Pro Met Leu
405 410 415
Asp Lys Asp Gly Ser Gly Gly Ile Val Ala Leu Ser Gln Ser Pro Phe
420 425 430
Ala Gly Arg Val Pro Ser Asn Ser Ala Thr Ala Lys Gly Glu Gly Lys
435 440 445
Gln Pro Val Ala Glu Glu Gly Ser Ser Glu Asp Ala Ser Glu Arg Pro
450 455 460
Thr Gly Asp Asn Ser Asn Leu Asn Thr Ala Phe Ser Phe Asp Phe Ser
465 470 475 480
Ala Ile Lys Pro Gly Met Ala Ala Asp Val Lys Phe Gly Gly Ser Gly
485 490 495
Ala Arg Pro Asn Leu Pro Trp Val Ser Thr Thr Gly Ser Gly Pro His
500 505 510
Gly Arg Thr Ile Ser Gly Val Thr Tyr Arg Tyr Asn Ala Asn Gln Ile
515 520 525
Lys Ile Val Cys Ala Cys His Gly Ser His Met Ser Pro Glu Glu Phe
530 535 540
Val Arg His Ala Ser Glu Glu Tyr Val Ser Pro Glu Ser Ser Met Gly
545 550 555 560
Met Thr Ala Ala Ser Ala His Thr
565
<210> 76
<211> 479
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 76
Met Val Ser Lys Gly Glu Glu Leu Phe Thr Gly Val Val Pro Ile Leu
1 5 10 15
Val Glu Leu Asp Gly Asp Val Asn Gly His Lys Phe Ser Val Ser Gly
20 25 30
Glu Gly Glu Gly Asp Ala Thr Tyr Gly Lys Leu Thr Asp Asp Gly Asn
35 40 45
Tyr Lys Thr Arg Ala Glu Val Lys Phe Glu Gly Asp Thr Leu Val Asn
50 55 60
Arg Ile Glu Leu Lys Gly Ile Asp Phe Lys Glu Asp Gly Asn Ile Leu
65 70 75 80
Gly His Lys Leu Glu Tyr Asn Tyr Asn Ser His Asn Val Tyr Ile Thr
85 90 95
Ala Asp Lys Gln Lys Asn Gly Ile Lys Ala Asn Phe Lys Ile Arg His
100 105 110
Asn Ile Glu Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu Gly Pro Arg
115 120 125
Pro Thr Ser Gly Ser Val Asp Leu Glu Gly Thr Ala Pro Gly Ser Met
130 135 140
Thr Ser Asp Gly Ala Thr Ser Thr Ser Ala Ala Ala Ala Ala Ala Ala
145 150 155 160
Ala Ala Ala Ala Arg Arg Lys Pro Ser Trp Arg Glu Arg Glu Asn Asn
165 170 175
Arg Arg Arg Glu Arg Arg Arg Arg Ala Val Ala Ala Lys Ile Tyr Thr
180 185 190
Gly Leu Arg Ala Gln Gly Asp Tyr Asn Leu Pro Lys His Cys Asp Asn
195 200 205
Asn Glu Val Leu Lys Ala Leu Cys Val Glu Ala Gly Trp Val Val Glu
210 215 220
Glu Asp Gly Thr Thr Tyr Arg Lys Gly Cys Lys Pro Leu Pro Gly Glu
225 230 235 240
Ile Ala Gly Thr Ser Ser Arg Val Thr Pro Tyr Ser Ser Gln Asn Gln
245 250 255
Ser Pro Leu Ser Ser Ala Phe Gln Ser Pro Ile Pro Ser Tyr Gln Val
260 265 270
Ser Pro Ser Ser Ser Ser Phe Pro Ser Pro Ser Arg Gly Glu Pro Asn
275 280 285
Asn Asn Met Ser Ser Thr Phe Phe Pro Phe Leu Arg Asn Gly Gly Ile
290 295 300
Pro Ser Ser Leu Pro Ser Leu Arg Ile Ser Asn Ser Cys Pro Val Thr
305 310 315 320
Pro Pro Val Ser Ser Pro Thr Ser Lys Asn Pro Lys Pro Leu Pro Asn
325 330 335
Trp Glu Ser Ile Ala Lys Gln Ser Met Ala Ile Ala Lys Gln Ser Met
340 345 350
Ala Ser Phe Asn Tyr Pro Phe Tyr Ala Val Ser Ala Pro Ala Ser Pro
355 360 365
Thr His Arg His Gln Phe His Thr Pro Ala Thr Ile Pro Glu Cys Asp
370 375 380
Glu Ser Asp Ser Ser Thr Val Asp Ser Gly His Trp Ile Ser Phe Gln
385 390 395 400
Lys Phe Ala Gln Gln Gln Pro Phe Ser Ala Ser Met Val Pro Thr Ser
405 410 415
Pro Thr Phe Asn Leu Val Lys Pro Ala Pro Gln Gln Met Ser Pro Asn
420 425 430
Thr Ala Ala Phe Gln Glu Ile Gly Gln Ser Ser Glu Phe Lys Phe Glu
435 440 445
Asn Ser Gln Val Lys Pro Trp Glu Gly Glu Arg Ile His Asp Val Gly
450 455 460
Met Glu Asp Leu Glu Leu Thr Leu Gly Asn Gly Lys Ala Arg Gly
465 470 475
<210> 77
<211> 425
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 77
Met Asp Lys Gln Lys Asn Gly Ile Lys Ala Asn Phe Lys Ile Arg His
1 5 10 15
Asn Ile Glu Asp Gly Gly Val Gln Leu Ala Asp His Tyr Gln Gln Asn
20 25 30
Thr Pro Ile Gly Asp Gly Pro Val Leu Leu Pro Asp Asn His Tyr Leu
35 40 45
Ser Tyr Gln Ser Ala Leu Ser Lys Asp Pro Asn Glu Lys Arg Asp His
50 55 60
Met Val Leu Leu Glu Phe Val Thr Ala Ala Gly Ile Thr Leu Gly Met
65 70 75 80
Asp Glu Leu Tyr Lys Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Gly Pro
85 90 95
Arg Pro Thr Ser Gly Ser Val Asp Leu Glu Gly Thr Ala Pro Gly Ser
100 105 110
Met Asp Val Gly Val Ser Pro Ala Lys Ser Ile Leu Ala Lys Pro Leu
115 120 125
Lys Leu Leu Thr Glu Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys
130 135 140
Arg Lys Phe Leu Lys Asp Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
145 150 155 160
Ser Gln Ala Ile Gln Gln Val Leu Ser Leu Lys Ala Leu Tyr Glu Pro
165 170 175
Gly Asp Asp Ser Gly Ala Gly Ile Phe Arg Lys Ile Leu Val Ser Gln
180 185 190
Pro Val Asn Pro Pro Arg Val Thr Thr Thr Leu Ile Glu Pro Ser Asn
195 200 205
Glu Leu Glu Ala Cys Gly Arg Val Ser Tyr Pro Glu Asp Asn Gly Ala
210 215 220
Cys His Arg Met Asp Ser Pro Arg Ser Ala Glu Phe Ser Gly Gly Ser
225 230 235 240
Gly His Phe Val Ser Glu Lys Asp Gly His Lys Thr Thr Ile Ser Pro
245 250 255
Arg Ser Pro Ala Glu Thr Ser Glu Leu Val Gly Gln Met Thr Ile Phe
260 265 270
Tyr Ser Gly Lys Val Asn Val Tyr Asp Gly Ile Pro Pro Glu Lys Ala
275 280 285
Arg Ser Ile Met His Phe Ala Ala Asn Pro Ile Asp Leu Pro Glu Asn
290 295 300
Gly Ile Phe Ala Ser Ser Arg Met Ile Ser Lys Leu Ile Ser Lys Glu
305 310 315 320
Lys Met Met Glu Leu Pro Gln Lys Gly Leu Glu Lys Ala Asn Ser Ser
325 330 335
Arg Asp Ser Gly Met Glu Gly Gln Ala Asn Arg Lys Val Ser Leu Gln
340 345 350
Arg Tyr Arg Glu Lys Arg Lys Asp Arg Lys Phe Ser Lys Ala Lys Lys
355 360 365
Cys Pro Gly Val Ala Ser Ser Ser Leu Glu Met Phe Leu Asn Cys Gln
370 375 380
Pro Arg Met Lys Ala Ala Tyr Ser Gln Asn Leu Gly Cys Thr Gly Ser
385 390 395 400
Pro Leu His Ser Gln Ser Pro Glu Ser Gln Thr Lys Ser Pro Asn Leu
405 410 415
Ser Val Asp Leu Asn Ser Glu Gly Ile
420 425
<210> 78
<211> 537
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 78
Met Asp Lys Gln Lys Asn Gly Ile Lys Ala Asn Phe Lys Ile Arg His
1 5 10 15
Asn Ile Glu Asp Gly Gly Val Gln Leu Ala Asp His Tyr Gln Gln Asn
20 25 30
Thr Pro Ile Gly Asp Gly Pro Val Leu Leu Pro Asp Asn His Tyr Leu
35 40 45
Ser Tyr Gln Ser Ala Leu Ser Lys Asp Pro Asn Glu Lys Arg Asp His
50 55 60
Met Val Leu Leu Glu Phe Val Thr Ala Ala Gly Ile Thr Leu Gly Met
65 70 75 80
Asp Glu Leu Tyr Lys Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Gly Pro
85 90 95
Arg Pro Thr Ser Gly Ser Val Asp Leu Glu Gly Thr Ala Pro Gly Ser
100 105 110
Met Asp Asp Asp Asn Gly Leu Glu Leu Ser Leu Gly Leu Ser Cys Gly
115 120 125
Gly Ser Thr Gly Lys Ala Lys Gly Asn Asn Asn Asn Asn Ala Gly Ser
130 135 140
Ser Ser Glu Asn Tyr Arg Ala Glu Gly Gly Asp Arg Ser Ala Lys Val
145 150 155 160
Ile Asp Asp Phe Lys Asn Phe Leu His Pro Thr Ser Gln Arg Pro Ala
165 170 175
Glu Pro Ser Ser Gly Ser Gln Arg Ser Asp Ser Gly Gln Gln Pro Pro
180 185 190
Gln Asn Phe Phe Asn Asp Leu Ser Lys Ala Pro Thr Thr Glu Ala Glu
195 200 205
Ala Ser Thr Lys Pro Leu Trp Val Glu Asp Glu Ser Arg Lys Glu Ala
210 215 220
Gly Asn Lys Arg Lys Phe Gly Phe Pro Gly Met Asn Asp Asp Lys Lys
225 230 235 240
Lys Glu Lys Asp Ser Ser His Val Asp Met His Glu Lys Lys Thr Lys
245 250 255
Ala Ser His Val Ser Thr Ala Thr Asp Glu Gly Ser Thr Ala Glu Asn
260 265 270
Glu Asp Val Ala Glu Ser Glu Val Gly Gly Gly Ser Ser Ser Asn His
275 280 285
Ala Lys Glu Val Val Arg Pro Pro Thr Asp Thr Asn Ile Val Asp Asn
290 295 300
Leu Thr Gly Gln Arg Arg Ser Asn His Gly Gly Ser Gly Thr Glu Glu
305 310 315 320
Phe Thr Met Arg Asn Met Ser Tyr Thr Val Pro Phe Thr Val His Pro
325 330 335
Gln Asn Val Val Thr Ser Met Pro Tyr Ser Leu Pro Thr Lys Glu Ser
340 345 350
Gly Gln His Ala Ala Ala Thr Ser Leu Leu Gln Pro Asn Ala Asn Ala
355 360 365
Gly Asn Leu Pro Ile Met Phe Gly Tyr Ser Pro Val Gln Leu Pro Met
370 375 380
Leu Asp Lys Asp Gly Ser Gly Gly Ile Val Ala Leu Ser Gln Ser Pro
385 390 395 400
Phe Ala Gly Arg Val Pro Ser Asn Ser Ala Thr Ala Lys Gly Glu Gly
405 410 415
Lys Gln Pro Val Ala Glu Glu Gly Ser Ser Glu Asp Ala Ser Glu Arg
420 425 430
Pro Thr Gly Asp Asn Ser Asn Leu Asn Thr Ala Phe Ser Phe Asp Phe
435 440 445
Ser Ala Ile Lys Pro Gly Met Ala Ala Asp Val Lys Phe Gly Gly Ser
450 455 460
Gly Ala Arg Pro Asn Leu Pro Trp Val Ser Thr Thr Gly Ser Gly Pro
465 470 475 480
His Gly Arg Thr Ile Ser Gly Val Thr Tyr Arg Tyr Asn Ala Asn Gln
485 490 495
Ile Lys Ile Val Cys Ala Cys His Gly Ser His Met Ser Pro Glu Glu
500 505 510
Phe Val Arg His Ala Ser Glu Glu Tyr Val Ser Pro Glu Ser Ser Met
515 520 525
Gly Met Thr Ala Ala Ser Ala His Thr
530 535
<210> 79
<211> 448
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 79
Met Asp Lys Gln Lys Asn Gly Ile Lys Ala Asn Phe Lys Ile Arg His
1 5 10 15
Asn Ile Glu Asp Gly Gly Val Gln Leu Ala Asp His Tyr Gln Gln Asn
20 25 30
Thr Pro Ile Gly Asp Gly Pro Val Leu Leu Pro Asp Asn His Tyr Leu
35 40 45
Ser Tyr Gln Ser Ala Leu Ser Lys Asp Pro Asn Glu Lys Arg Asp His
50 55 60
Met Val Leu Leu Glu Phe Val Thr Ala Ala Gly Ile Thr Leu Gly Met
65 70 75 80
Asp Glu Leu Tyr Lys Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Gly Pro
85 90 95
Arg Pro Thr Ser Gly Ser Val Asp Leu Glu Gly Thr Ala Pro Gly Ser
100 105 110
Met Thr Ser Asp Gly Ala Thr Ser Thr Ser Ala Ala Ala Ala Ala Ala
115 120 125
Ala Ala Ala Ala Ala Arg Arg Lys Pro Ser Trp Arg Glu Arg Glu Asn
130 135 140
Asn Arg Arg Arg Glu Arg Arg Arg Arg Ala Val Ala Ala Lys Ile Tyr
145 150 155 160
Thr Gly Leu Arg Ala Gln Gly Asp Tyr Asn Leu Pro Lys His Cys Asp
165 170 175
Asn Asn Glu Val Leu Lys Ala Leu Cys Val Glu Ala Gly Trp Val Val
180 185 190
Glu Glu Asp Gly Thr Thr Tyr Arg Lys Gly Cys Lys Pro Leu Pro Gly
195 200 205
Glu Ile Ala Gly Thr Ser Ser Arg Val Thr Pro Tyr Ser Ser Gln Asn
210 215 220
Gln Ser Pro Leu Ser Ser Ala Phe Gln Ser Pro Ile Pro Ser Tyr Gln
225 230 235 240
Val Ser Pro Ser Ser Ser Ser Phe Pro Ser Pro Ser Arg Gly Glu Pro
245 250 255
Asn Asn Asn Met Ser Ser Thr Phe Phe Pro Phe Leu Arg Asn Gly Gly
260 265 270
Ile Pro Ser Ser Leu Pro Ser Leu Arg Ile Ser Asn Ser Cys Pro Val
275 280 285
Thr Pro Pro Val Ser Ser Pro Thr Ser Lys Asn Pro Lys Pro Leu Pro
290 295 300
Asn Trp Glu Ser Ile Ala Lys Gln Ser Met Ala Ile Ala Lys Gln Ser
305 310 315 320
Met Ala Ser Phe Asn Tyr Pro Phe Tyr Ala Val Ser Ala Pro Ala Ser
325 330 335
Pro Thr His Arg His Gln Phe His Thr Pro Ala Thr Ile Pro Glu Cys
340 345 350
Asp Glu Ser Asp Ser Ser Thr Val Asp Ser Gly His Trp Ile Ser Phe
355 360 365
Gln Lys Phe Ala Gln Gln Gln Pro Phe Ser Ala Ser Met Val Pro Thr
370 375 380
Ser Pro Thr Phe Asn Leu Val Lys Pro Ala Pro Gln Gln Met Ser Pro
385 390 395 400
Asn Thr Ala Ala Phe Gln Glu Ile Gly Gln Ser Ser Glu Phe Lys Phe
405 410 415
Glu Asn Ser Gln Val Lys Pro Trp Glu Gly Glu Arg Ile His Asp Val
420 425 430
Gly Met Glu Asp Leu Glu Leu Thr Leu Gly Asn Gly Lys Ala Arg Gly
435 440 445
<210> 80
<211> 364
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 80
Met Asp Lys Gln Lys Asn Gly Ile Lys Ala Asn Phe Lys Ile Arg His
1 5 10 15
Asn Ile Glu Asp Gly Gly Val Gln Leu Ala Asp His Tyr Gln Gln Asn
20 25 30
Thr Pro Ile Gly Asp Gly Pro Val Leu Leu Pro Asp Asn His Tyr Leu
35 40 45
Ser Tyr Gln Ser Ala Leu Ser Lys Asp Pro Asn Glu Lys Arg Asp His
50 55 60
Met Val Leu Leu Glu Phe Val Thr Ala Ala Gly Ile Thr Leu Gly Met
65 70 75 80
Asp Glu Leu Tyr Lys Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Gly Pro
85 90 95
Arg Pro Thr Ser Gly Ser Val Asp Leu Glu Gly Thr Ala Pro Gly Ser
100 105 110
Tyr Glu Pro Gly Asp Asp Ser Gly Ala Gly Ile Phe Arg Lys Ile Leu
115 120 125
Val Ser Gln Pro Val Asn Pro Pro Arg Val Thr Thr Thr Leu Ile Glu
130 135 140
Pro Ser Asn Glu Leu Glu Ala Cys Gly Arg Val Ser Tyr Pro Glu Asp
145 150 155 160
Asn Gly Ala Cys His Arg Met Asp Ser Pro Arg Ser Ala Glu Phe Ser
165 170 175
Gly Gly Ser Gly His Phe Val Ser Glu Lys Asp Gly His Lys Thr Thr
180 185 190
Ile Ser Pro Arg Ser Pro Ala Glu Thr Ser Glu Leu Val Gly Gln Met
195 200 205
Thr Ile Phe Tyr Ser Gly Lys Val Asn Val Tyr Asp Gly Ile Pro Pro
210 215 220
Glu Lys Ala Arg Ser Ile Met His Phe Ala Ala Asn Pro Ile Asp Leu
225 230 235 240
Pro Glu Asn Gly Ile Phe Ala Ser Ser Arg Met Ile Ser Lys Leu Ile
245 250 255
Ser Lys Glu Lys Met Met Glu Leu Pro Gln Lys Gly Leu Glu Lys Ala
260 265 270
Asn Ser Ser Arg Asp Ser Gly Met Glu Gly Gln Ala Asn Arg Lys Val
275 280 285
Ser Leu Gln Arg Tyr Arg Glu Lys Arg Lys Asp Arg Lys Phe Ser Lys
290 295 300
Ala Lys Lys Cys Pro Gly Val Ala Ser Ser Ser Leu Glu Met Phe Leu
305 310 315 320
Asn Cys Gln Pro Arg Met Lys Ala Ala Tyr Ser Gln Asn Leu Gly Cys
325 330 335
Thr Gly Ser Pro Leu His Ser Gln Ser Pro Glu Ser Gln Thr Lys Ser
340 345 350
Pro Asn Leu Ser Val Asp Leu Asn Ser Glu Gly Ile
355 360
<210> 81
<211> 400
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 81
Met Asp Lys Gln Lys Asn Gly Ile Lys Ala Asn Phe Lys Ile Arg His
1 5 10 15
Asn Ile Glu Asp Gly Gly Val Gln Leu Ala Asp His Tyr Gln Gln Asn
20 25 30
Thr Pro Ile Gly Asp Gly Pro Val Leu Leu Pro Asp Asn His Tyr Leu
35 40 45
Ser Tyr Gln Ser Ala Leu Ser Lys Asp Pro Asn Glu Lys Arg Asp His
50 55 60
Met Val Leu Leu Glu Phe Val Thr Ala Ala Gly Ile Thr Leu Gly Met
65 70 75 80
Asp Glu Leu Tyr Lys Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Gly Pro
85 90 95
Arg Pro Thr Ser Gly Ser Val Asp Leu Glu Gly Thr Ala Pro Gly Ser
100 105 110
Met Asp Val Gly Val Ser Pro Ala Lys Ser Ile Leu Ala Lys Pro Leu
115 120 125
Lys Leu Leu Thr Glu Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys
130 135 140
Arg Lys Phe Leu Lys Asp Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
145 150 155 160
Ser Gln Ala Ile Gln Gln Val Leu Ser Leu Lys Ala Leu Tyr Glu Pro
165 170 175
Gly Asp Asp Ser Gly Ala Gly Ile Phe Arg Lys Ile Leu Val Ser Gln
180 185 190
Pro Val Asn Pro Pro Arg Val Thr Thr Thr Leu Ile Glu Pro Ser Asn
195 200 205
Glu Leu Glu Ala Cys Gly Arg Val Ser Tyr Pro Glu Asp Asn Gly Ala
210 215 220
Cys His Arg Met Asp Ser Pro Arg Ser Ala Glu Phe Ser Gly Gly Ser
225 230 235 240
Gly His Phe Val Ser Glu Lys Asp Gly His Lys Thr Thr Ile Ser Pro
245 250 255
Arg Ser Pro Ala Glu Thr Ser Glu Leu Ile Met His Phe Ala Ala Asn
260 265 270
Pro Ile Asp Leu Pro Glu Asn Gly Ile Phe Ala Ser Ser Arg Met Ile
275 280 285
Ser Lys Leu Ile Ser Lys Glu Lys Met Met Glu Leu Pro Gln Lys Gly
290 295 300
Leu Glu Lys Ala Asn Ser Ser Arg Asp Ser Gly Met Glu Gly Gln Ala
305 310 315 320
Asn Arg Lys Val Ser Leu Gln Arg Tyr Arg Glu Lys Arg Lys Asp Arg
325 330 335
Lys Phe Ser Lys Ala Lys Lys Cys Pro Gly Val Ala Ser Ser Ser Leu
340 345 350
Glu Met Phe Leu Asn Cys Gln Pro Arg Met Lys Ala Ala Tyr Ser Gln
355 360 365
Asn Leu Gly Cys Thr Gly Ser Pro Leu His Ser Gln Ser Pro Glu Ser
370 375 380
Gln Thr Lys Ser Pro Asn Leu Ser Val Asp Leu Asn Ser Glu Gly Ile
385 390 395 400
<210> 82
<211> 341
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 82
Met Asp Lys Gln Lys Asn Gly Ile Lys Ala Asn Phe Lys Ile Arg His
1 5 10 15
Asn Ile Glu Asp Gly Gly Val Gln Leu Ala Asp His Tyr Gln Gln Asn
20 25 30
Thr Pro Ile Gly Asp Gly Pro Val Leu Leu Pro Asp Asn His Tyr Leu
35 40 45
Ser Tyr Gln Ser Ala Leu Ser Lys Asp Pro Asn Glu Lys Arg Asp His
50 55 60
Met Val Leu Leu Glu Phe Val Thr Ala Ala Gly Ile Thr Leu Gly Met
65 70 75 80
Asp Glu Leu Tyr Lys Tyr Pro Tyr Asp Val Pro Asp Tyr Ala Gly Pro
85 90 95
Arg Pro Thr Ser Gly Ser Val Asp Leu Glu Gly Thr Ala Pro Gly Ser
100 105 110
Met Asp Val Gly Val Ser Pro Ala Lys Ser Ile Leu Ala Lys Pro Leu
115 120 125
Lys Leu Leu Thr Glu Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys
130 135 140
Arg Lys Phe Leu Lys Asp Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
145 150 155 160
Ser Gln Ala Ile Gln Gln Val Leu Ser Leu Lys Ala Leu Tyr Glu Pro
165 170 175
Gly Asp Asp Ser Gly Ala Gly Ile Phe Arg Lys Ile Leu Val Ser Gln
180 185 190
Pro Val Asn Pro Pro Arg Val Thr Thr Thr Leu Ile Glu Pro Ser Asn
195 200 205
Glu Leu Glu Ala Cys Gly Arg Val Ser Tyr Pro Glu Asp Asn Gly Ala
210 215 220
Cys His Arg Met Asp Ser Pro Arg Ser Ala Glu Phe Ser Gly Gly Ser
225 230 235 240
Gly His Phe Val Ser Glu Lys Asp Gly His Lys Thr Thr Ile Ser Pro
245 250 255
Arg Ser Pro Ala Glu Thr Ser Glu Leu Val Gly Gln Met Thr Ile Phe
260 265 270
Tyr Ser Gly Lys Val Asn Val Tyr Asp Gly Ile Pro Pro Glu Lys Ala
275 280 285
Arg Ser Ile Met His Phe Ala Ala Asn Pro Ile Asp Leu Pro Glu Asn
290 295 300
Gly Ile Phe Ala Ser Ser Arg Met Ile Ser Lys Leu Ile Ser Lys Glu
305 310 315 320
Lys Met Met Glu Leu Pro Gln Lys Gly Leu Glu Lys Ala Asn Ser Ser
325 330 335
Arg Asp Ser Gly Met
340
<210> 83
<211> 942
<212> ДНК
<213> Arabidopsis thaliana
<400> 83
atggatgtcg gagtttcacc ggcgaagtct atacttgcga aacctctgaa gctactcact 60
gaagaggaca tttctcagct cactcgcgaa gactgccgca aattcctcaa agacaaagga 120
atgcgaagac cgtcgtggaa caaatctcag gcgatccagc aagttttatc tcttaaagct 180
ctctatgagc ctggagacga ttccggcgcc ggtatcttcc gcaagatcct cgtttctcag 240
ccagtaaatc cgcctcgcgt cacaacaacg ttgattgagc caagcaacga gctggaagct 300
tgtggccggg tttcttatcc ggaagataac ggcgcgtgcc atagaatgga ttctccaaga 360
tcagctgagt tttccggtgg gtctggtcac tttgtatccg agaaagatgg ccacaagacg 420
actatttctc ccagaagccc agctgaaaca agtgagctcg ttgggcaaat gacgatattc 480
tatagtggaa aagtgaatgt gtatgatgga ataccacctg aaaaggcccg gtcaatcatg 540
cactttgcag ccaatccaat tgatttgcct gaaaacggta tttttgcttc cagtagaatg 600
atttcaaagc tcataagtaa agagaagatg atggaacttc cccaaaaagg ccttgagaag 660
gcgaattctt ctcgtgattc tggtatggag ggccaggcga acagaaaggt atctttgcaa 720
agatatcgtg aaaagcggaa agacagaaaa ttctcaaagg ccaaaaagtg tccaggagtt 780
gcgtcctcta gcttggagat gtttctgaat tgtcagccac ggatgaaagc tgcatattcg 840
caaaacctag gctgcaccgg atctccactg catagccagt cacctgaaag ccagacaaaa 900
agtcccaatc tttcagttga tctaaacagt gaaggcattt aa 942
<210> 84
<211> 950
<212> ДНК
<213> Arabidopsis thaliana
<400> 84
agggatatgg atgtaggagt tactacggcg aagtctatac ttgagaagcc tctgaagctt 60
ctcactgaag aagacatttc tcagcttact cgcgaagatt gccgcaaatt cctcaaagag 120
aaaggaatgc gcaggccttc gtggaataaa tctcaggcga tccagcaagt tttatctctt 180
aaagctctct atgaacctgg agatgattcc ggcgccggaa tcctccgcaa gatccttgtt 240
tctcagccgc caaatccgcc tcgcgttaca acaacgttga ttgagccaag gaacgagctc 300
gaagcttgtg gaaggattcc tttacaggaa gatgatggtg cgtgccatag aagggattct 360
ccaagatcag ctgagttttc tggtagttct ggtcagtttg ttgcggataa agatagccac 420
aagactgttt ctgtttcccc cagaagccca gctgaaacaa atgcggtggt tgggcaaatg 480
acgatatttt atagtggcaa agtgaatgta tatgatggag taccacctga aaaggcccgg 540
tctatcatgc attttgcagc caatccaatt gatttgcctg aaaatggtat ttttgcttct 600
agtagaatga tttcgaaacc catgagtaaa gagaagatgg tggagcttcc ccaatatgga 660
cttgaaaagg cacctgcttc tcgtgattct gatgttgagg gtcaggcgaa cagaaaagta 720
tcgttgcaaa gatatcttga aaagcggaaa gacagaagat tttctaagac caagaaggct 780
ccaggagttg cgtcctctag cttggagatg tttctgaatc gtcagccacg gatgaacgct 840
gcatattcac aaaaccttag tggcacaggg cattgcgagt cacctgaaaa tcaaacaaaa 900
agtcccaata tctcagttga tctaaacagt gatctaaaca gcgaagataa 950
<210> 85
<211> 1128
<212> ДНК
<213> Populus trichocarpa
<400> 85
atgcagccgg gagagacagt tttccggtca gctctggaca aacccctaca ccagctaaca 60
gaagatgata tttctcaggt cactcgcgaa gattgccgcc gttacctcaa agaaaaaggt 120
atgagaaggc cgtcgtggaa caaatcgcag gcaatacagc aagtgatttc actcaaaaca 180
ctcctggaag cgacgccgga gactgaatct ccaaggcgac gactctacat tccccgccct 240
cctcctcatc ctcctgataa tactcctcgt gtgcgtttct ctgccgtccc tccaaattcc 300
tctgtttcag agaggggagc aagtgctgaa acgccgatct cggtgccagc cgaggagcca 360
gttccgtgcc ggcaacacga tcctccaaat cccgatgatc ctgccgatcc tctgcctcct 420
gtccatgccg ccgtcaccga gaatgcttcg gtttcaccaa gaactacagg catggcagaa 480
gaatcagcag gacagatgac aattttttac tgtgggaagg taaacgtcta tgatgatgta 540
ccgggagaca aggcgcaagc aataatgcat cttgctgcaa gcccatttgc tccacctcag 600
gatgcttctt cagatgtaat tcctacatta aggcctttac aatgccagtt agacactcca 660
ggtgtcaaag ctgctccaaa ttcaattgtg gcgaactttc caaccctgcc aacagtgaaa 720
ggggcagata gtggtcagct tctctgggaa gaaagcaaca tagctcgtga agacaaccta 780
gaaggctcta caagcagaaa agcatcctta caaagatatt ttgagaagaa gaaagacagg 840
ttcaagaaca agagaaaggt ggcagtgcct tctgctagct tggacgtctt cttaagccat 900
ctggttggag atcaaatctc aaatgatcat tggaacctaa atgatgcctg ctctccttcc 960
caacccaggc ctccccaaac gcctaaccgg tgcaactctg ttgacaatgt agcaaaaaat 1020
ggcatcctca aagctgacct taacaacaaa ggtgatgcag atttatcttg ttgtcttgac 1080
tttagttcca agcagattaa tgcgtggtgc ttatgcttgg gatgttga 1128
<210> 86
<211> 2114
<212> ДНК
<213> Picea abies
<400> 86
atgcgaggag gaggaggcgc ggacagactc cccgctagag ctaacctcga gaaacccttg 60
gaagatctca gccatgaaga cattatgcag ctcaccaggg aagactgccg gcgatacttg 120
atagaaaaag gtagcttcca aaatcttttg ctttctcctc aaataacgct ttgctctgag 180
taaatatatg aatataaatg aatgtaatct agtaatcgag ccctaagcgc gacatttaaa 240
gtatttgtaa aggtctgcgg ctgttttggc tttgtctacg gagacaaaaa tcttttttcc 300
gttgggttga gctcagaatc aatggcggtt cttctcgtgg atttgatttt gtttcgtttg 360
gcttgacttt tgccagtatt tacgccctgc cttgtcctaa aatgctttta cagaaaaaat 420
taaatcctct aaattttctt taaacctttc cagtgttaac ctctgaacat tgccaataaa 480
agcgtatagg aaaattttca attaaagctt tatataccgt aaggccatgt agaatctttt 540
aatttttgcc gtttcagaaa acggtttaaa gggataaatt tcttagaagc tcttaaaata 600
gaaaatagag cttgaaatac cactgatcca atgccagaat tgtatataat ttgacttcag 660
accttaaata ctgcttattc ggacggacat ttgctcagaa atcgttttaa tatttatgca 720
taacagattt taaatttctt tgtaggcatg cgacggcctt cgtggaacaa gtctcaggcg 780
attcagcagg tactctcgtt gaagaaattg tttgaatccg ggccgaacga tgaaaagagg 840
tcggcggcaa caaatcggcc gaatccggat gaaaacttaa atgaaaagag gtcggcggca 900
acaaatcggc cgaatccgga tgaaaactta aaggaagctg cgtccgtttc tttgctttac 960
ggttcacagc ctgaaagtcc ttcggtaggc tttttctttt aaacaatgtc gtctaatcga 1020
gcttaaacct gcagaaacgc tggcattgtc tttaatttgg ctaggtattt caaagttgaa 1080
catttctgtc tccattgtaa tgtttgttca ggttgtcttc gccagtaaag actcagacac 1140
ttttaatttg gagtggttgg cgaagacaga gttgccagta ttagcaagcc aaccccgaca 1200
catagcacag cagaatgttt tcttaagctc tttatctgct cagcaatccg gagctcagct 1260
caccattttt tactcgggaa atgttaatgt ctacgacgat gtgcctgcag aaaaggtatc 1320
tacagattta cagttcgatt ctcgtaaaga tgtgcttaaa atttccaatg atataggctt 1380
aaaattctca agatctatat ttgcaggcac aagaaataat gctgttggcc gggagcggaa 1440
attatcctcc gtcgtcgacg tgtcagtcca cacgaaatac acaacaaaac gcagtacgtg 1500
cggcgtatcc atcaaatcct acgaatacgc cgttcattca cggagtaggg ccgcctcttg 1560
caactgtggc gagctcttcc gtcatgagca gtccaataca taaaggtatc gtccattgta 1620
tcccaatgcg gaatgaaacg aaactaaaaa attgacccaa atttatcaaa atttgggcgt 1680
ccgaacataa tttggttgtt tccgatgcag agagtccgat tacaagaaaa gcatcgctgc 1740
aaagatttct ggagaaaaga aaggacaggt acagatagaa aaggttttac tccattatca 1800
tgagattcgt ggttaaaaat gcaatgaatg caattaaata tttttgattg caggagtcgt 1860
ggcaagttgg gggctcccac tatatcgaaa aagcctctgc tgatgggtat gtttatgcat 1920
ccctccattg ttcatcgtca gtattggact gatacggcca agaggaaatc cggaaaaccg 1980
gacatacctg cttctatttc tccgacccgg cctcctcaca cgccgcgtcg gacttcgtcg 2040
gacgaacaac ttagtgcacg ccatgctcgt ggtgatataa gtgcgcaggg ggggtcgcta 2100
cataattcca acta 2114
<210> 87
<211> 1098
<212> ДНК
<213> Gossypium raimondii
<400> 87
atggaggctg gggtaacgac gacggcgact acaacagcgt cgttcagttc gatacttgat 60
aaacccctca gccaactaac cgaagaagac atttctcaac tcactcgcga agactgtcgc 120
aaattcctca aagaaaaagg aatgcgtagg ccgtcatgga acaaatcgca ggcgatccag 180
caagtgattt cgttcaaggc gttgttggaa agcaacgaag attccggcgc cggagctcgc 240
cggaaaatcc ttgtttgtcc accaccgtca cattttcctc cgcaaaatgc ggtagcttca 300
aattctggtg agtcagtaaa agaagcagtc tttggagaag aagaaagcct gtacggccaa 360
aaagatcttt ctttgaaagc tgctccggtg gtgcagatga attgtcaggg cggtgacacg 420
gatgacaaga ctctttcgcc tagtttaggc tctccacggg agtattcaaa attgcctggc 480
agaagtcaat gtgaaacaaa tgagttgggt gggcaaatga caatttttta ctgtggaaag 540
atcaatgtgt acgatggtgt accacttgct aaggcacgag caatcatgca cctggcagct 600
tctcctattg attttcctca gggcaatcta tgtaatcaaa atggtgcctt taggtccttt 660
ctgggtcatg tacaagaagc cgaagacaaa aacgacctta cttcatctat tgctttgaac 720
ttgaattctc ataccatgca cactgagaag atgacagaat atcagcagca gtttagggga 780
aaagcaaaca tcagtcgtga ttctgatgta gatggacagg tgagcagaaa agaatcattg 840
cagcgatatc ttgaaaagcg aaaagacagg ggaagattct ttaaaggcag gaaaaatgca 900
ggacaagctt tgtctagctc ggagatgtac ctgaaccatc agataagagc tcactactta 960
aatggacaaa caaaccagag cagaacaagt tctccaccac agtctggagt gccacatgca 1020
ttttatagct cagctgacaa ccaagagctt gtgaattttt ctgtagatct caatgatgaa 1080
ggtggtcaag aacactga 1098
<210> 88
<211> 1074
<212> ДНК
<213> Aquilegia coerulea
<400> 88
atgaaacctg acgagacagt ttcccggtca ccacttgata aacctttgtt tcaacttact 60
gatgaagata tttcacagct cactcgtgaa gattgccgga aatttctcag agacaaaggt 120
atgagacgtc cttcatggaa caaatctcag gcgattgaac aagtgatctc acttaaaacg 180
ttgctagaac caagaacgga atctgataca aatgccaccg gaatccggca gaaattactt 240
gtttctcggc tagaaaattc tacccaagta cctttaaatg acaagacaaa tgcctcaaat 300
ttaaagacat ctgttcaggc aataaactcc gggaaagccg atattcatgg tgacaggccg 360
tgtcgggtcc ctgttccagt ccctgacgat aacacaatca ctgttccagt ccctgacaat 420
aacacaatca ctgttccagt ccctgacaat aacatcactt catccagaaa cctgaactcc 480
accaatggac tggttggtca gatgacaatt ttctactgcg gcaaggtgat cgtctacgat 540
gatatgcctg ctgagaaggc acatgcaatc atgaaatttg caggaagcca tatcaatgtg 600
cctgaggatt cttcaccagc tggagctgca gtaattcaat cctttgcatg ccaattacag 660
gcagcatcca tcagacatgg acttgctttc ccgtcagcgg tctctccacc cttgcacaat 720
gtggtagccg atacttctca gcattgcagg gaggaagtga cagtttctcg tgaagttgaa 780
cccgagggtc cagtgagtag aaaagcatct gtacaaagat atttggagaa gcgaaaagac 840
agggggcggt ttaagaacaa gcgaaagata gagtcatctt ctagcttaga gatatacttg 900
aaccatcaac tgggggatca gtaccttaat gagaaatcaa gtcagagcag ggcatgttcc 960
ccaccccaac ctagagcacc acacactccc actcgttgca gttcagttga gaaccaggtc 1020
acaaatgtcg tgttctccat tgatctcaat gataacgatg ttcgggaagg ctga 1074
<210> 89
<211> 1044
<212> ДНК
<213> Aquilegia coerulea
<400> 89
atgaaacctg acgagacagt ttcccggtca ccacttgata aacctttgtt tcaacttact 60
gatgaagata tttcacagct cactcgtgaa gattgccgga aatttctcag agacaaaggt 120
atgagacgtc cttcatggaa caaatctcag gcgattgaac aagtgatctc acttaaaacg 180
ttgctagaac caagaacgga atctgacaca aatgccaccg gaatccggca gaaattactt 240
gtttctcggc tagaaaattc tacccaagta cctttaaatg acaagacaaa tgcctcaaat 300
ttaaagacat ctgttcaggc aataaactcc ggggaagccg atattcatgg tgacaggccg 360
tgtcgggtcc ctgttccagt ccctgacgat aacacaatca ctgttccagt ccctgacaat 420
aacatcactt catccagaaa cctgaactcc accaatggac tggttggtca gatgacaatt 480
ttctactgcg gcaaggtgat cgtctacgat ggtatgcctg ctgagaaggc acatgcaatc 540
atgaaatttg caggaagcca tatcaatgtg cctgaggatt cttcaccagc tggagctgca 600
gtaattcaat cctttgcatg ccaattacag gcagcatcca tcagacatgg acttgctttc 660
ccgtcagcgg tctctccacc cttgcacaat gtggtagccg atacttctca gcattgcagg 720
gaggaagtga cagtttctcg tgaagttgaa cccgagggtc cagtgagtag aaaagcatct 780
gtacaaagat atttggagaa gcgaaaagac agggggcggt ttaagaacaa gcgaaagata 840
gagtcatctt ctagcttaga gatatacttg aaccatcaac tgggggatca gtaccttaat 900
gagaaatcaa gtcagagcag ggcatgttcc ccaccccaac ctagagcacc acacactccc 960
actcgttgca gttcagttga gaaccaggtc acaaatgtcg tgttctccat tgatctcaat 1020
gataacgatg ttcgggaagg ctga 1044
<210> 90
<211> 987
<212> ДНК
<213> Medicago truncatula
<400> 90
atgaacggcg gaagcaccgt ttccttccga tccatcctcg acagacctct taaccaactc 60
actgaagatg acatttctca actcactcgc gaagactgtc gcagattcct caaagataaa 120
gggatgcgca ggccttcctg gaacaaatca caggcgatcc agcaggtgat ttctctcaaa 180
gcgcttctag aacctaccga cgatgatatc ccggctaccg tcggcgttgg tgtctcctcc 240
gccattcacc accatcacca ccaccaccct cctcaacctc cgccgaaggc tttggatccc 300
gaagatactg ctttggaact acagaaatcc acttcacctg ttgctgagag acccacggaa 360
accaatgatg ccaatgttgt taacaatccc ggagggtgcg cacctagtgg gtcatttggg 420
caaatgacaa ttttctactg tggtaaggtg aatgtctatg atggagtctc gccggataag 480
gcacgatcaa tcatgcagct tgctgctgca tgtccgtcct cctttcctca ggataatcct 540
tcaaataaaa atgcagcagt ttgggcttct ccttgcaact tacctattga taaggaagtc 600
ctcttcccta ctgacacagc aatccttcaa gttgctcaaa cagataagat ggtggaatac 660
cctctgcaat acagggagaa aggaagcaca gctcgtgatg ctgagggtca ggcaagcaga 720
aaagtgtcac tgcagcgata tcttgaaaag cgaaaggaca ggggaagatc gaagggcaag 780
aaactgactg gcataacttc atctaacttt gagatgtatt tgaaccttcc agtgaagctc 840
catgcctcaa atgggaattc aagtcgtagt agcactgact ctccaccaca gcctagactg 900
cctttagttt ccagtggctc agctgaaaac cagccaaaag ttacccttcc cattgatttg 960
aatgataaag atgttcaaga atgctaa 987
<210> 91
<211> 1020
<212> ДНК
<213> Solanum lycopersicum
<400> 91
atgtcgctgg aacaaactgt ttacaagtct cctctggaca aaccgcttta cctacttacc 60
gatgacgaca tttctcagct cactcgcgaa gattgccgac gttttcttaa agctaaagga 120
atgagaaagc cgtcatggaa taaatcacag gcgattcagc aggtgatttc actgaaggcg 180
ttgtttgaga cgacgccgga atccgacacc ggtcagcgga aaaagcgtca cattcctcgc 240
ccggacacta gtttacagcg agtccagaaa gaaacgagta tcgatgcaga atttgctgaa 300
tcggctgaag aaacggtgcc gtacggtaga aaacctccca ataaacctga tctttccggc 360
gacaaagctg caagtgctgt tgccgttgtc aataacttag ctccttctag aaccacagat 420
tcaggaaatg catcatcagg tcaattgaca atcttctatt gtggcaaggt gaatgtgtat 480
gatgatgtac ctgctgaaaa ggcagaagca atcatgcatc ttgctgcaag cccactcttt 540
gtcccttcag aaactccatt ggatgctaac agagcagctc aacattccga atgccatttg 600
caagctgcaa atgttaaact gggtcaagat tctcctatgg tgttcatgcc aaccatgcaa 660
acagggaaaa taactgaagt tactcgcctg catttggagg aaagcaacac ttcctatgaa 720
gacaatcctg aagcagtgaa ccacgtaagc aggaaagcat tactggaaag atatcgtgag 780
aagcggaaag acaggttcaa gagaaagatg ggaatgcctt catctgctag cttggacatc 840
tatttgaacc atcgaaccat aaatcatacc caaagcgagc tctcaagtag gagcaacact 900
tgttccccgc ccgcaattag attatctgct gcgcctgctc caagtggttc aatggataac 960
attctccaaa tggatgccaa tgcttctggt tttctcgacg acaaagatgg taaagagtga 1020
<210> 92
<211> 1018
<212> ДНК
<213> Trifolium repens
<400> 92
atgaacggcg gaagcaccgt ttccttccga tccatcctcg acaaacccct taaccagctc 60
accgaagatg acatttctca actcactcgt gaagactgtc gcagattcct caaagataaa 120
gggatgcgca ggccttcctg gaacaaatct caggcgatcc agcaagtcat ttctctcaaa 180
gcacttctag aacctaccga cgatgatctc cctgctcccg tcggtgtctc ctccgccatt 240
caccaccatc atcaccacca ccctcaacct cctcagagga atttgaatga agctccggtg 300
aagggctccg atctcgatga taccggtttt catactgcgg aggatcttaa caaatctact 360
tcaactgctg tggaaattcc tactgaaacg aatgatgcca atgttgttaa atcctctggg 420
gggtgcgtag ctagtgggtc gtttgggcaa atgacaattt tctactgtgg taaggtgaat 480
gtctatgatg gagtctcacc ggataaggca cgatcaatca tgcagcttgc tgcatgtcca 540
tcctcgtttc ctcaggataa tcttttaaat aaaaatgcag cagtgtgggc ttctccttgc 600
aacataccaa ttgataagga tgtcctcttt cccaatgaca cagcaatcct tcaggttgct 660
caaacagata agatggtgga atatcctctg caatacaggg agaaagggag catagctcgt 720
gatgctgatg tagagggtca ggcaagcaga aatgcgtcgc tgcagcgata tcgtgaaaag 780
cgaaaggaca ggggaagatc gaaaggcaac aaactgactg gcataacttc atctaacttt 840
gagatgtatt tgaaccttcc agtgaagctc catgcctcaa atggtaattc aagtcgtagt 900
agcactgact ctccaccaca gcctagactg cctctagttt ccggtggctc agctgaaaac 960
cagccaaaag ttacccttcc cattgatttg aatgataaag atgttcaaga atgctaat 1018
<210> 93
<211> 936
<212> ДНК
<213> Amborella trichopoda
<400> 93
atgacggccg gtgatggctc catacgatca atattggaca agcccttgga agagctcacg 60
gaggaggaca tctcgcagct cactcgtgaa gactgtcgca ggtacctcaa agaaaaaggg 120
atgcgaaggc cttcgtggaa caagtatcag gcaattcagc aggttctctc tctaaaaggc 180
ctcttagagg ggaagccttg cgatgacaac agcgatgttt tcagtcaccg atcaccgatc 240
acggtcattc ccaatgttgg gagcatgaga gagaaagaaa aggccgtaaa tattgcggat 300
ccggagatat cggggtctca tcagccgaat tttcgccgag aaattcacga aaccacccgg 360
gaaagagctt taccggcttc cgactggcca ccttctcagg agccggtatc tcagatgacc 420
attttctatg ctggagccgt taacgtatac aacgacattc ctgaagataa ggtgcaagcc 480
atcatttatc ttgctgggaa gtcagactcc ttacagcaaa ctaatgttat cagaacggga 540
ccggaccaat gcatagcatc tgctgcaagc ccctcattga acgatctcca cagtagacga 600
atccacccaa cttcaaacat caccacttct cagtctcttc gtgttgcaac ttcccttcct 660
gttgggcctc attcagaggt tcctaagacg aggaaaacct cggtgcagcg attcttggag 720
aagcggaagg acagggggcg cttgaaggga acattggcga gtggtgggag ctctaagagg 780
ggttcctcat gcctagaatt gtatgcaact tcaagattaa agagtgaggg ggtggccacg 840
actacaactc aatccaatat gaacaatgtg gtcgtatcac cttctaaccc aagaatgcct 900
ctaaatcccg ggagttgcag ctgggttgag aactaa 936
<210> 94
<211> 1251
<212> ДНК
<213> Selaginella moellendorffii
<400> 94
atggcggcct cgattctagg ttgcggttct agcaatggcg tcgcggtcac cggtaatcct 60
gctccagccg cggcggccga ggtgcccgcg cctctcaggc cgctggagga gctcacggag 120
ctggatatca ggcagctcac gcgggaggac tgtagacgct atctcaagga acggggaatg 180
cgcaggccgt cttggaacaa ggcacaggcg atccagcaag tgctgtcttt gaggagtttg 240
ctttgtcctt ccaatccggt aggcccttcc tccaagaacc cgggaagtgc cgcgaacgcc 300
cctccggctg aagcagctgc tgctggtcac accaaacaat tactggacaa ggtctctcag 360
caaagcatgc cagattcttg tccatctaac aacgcctctg atcctaggcc gctcgccgga 420
tgctttggat ctcttgcccc gacgttatca gttctcaatc ccgatgcgaa acgtaacccg 480
ctgagttcta aacccgcgtc aacgacaaag cctcacagtg cccagctgac cattttctac 540
tccggtattg tgaacgtgta cgacgatgtc ccgcttgaca aggcacaagc tataatgctt 600
cttgccgcga gtaaaacgtt tcacgttccg acaagttcag tgcctggcca tccgccgttt 660
acgagtgcaa cccaacaaca acaacaacaa caacgagagc ttaaccaaca aaccgaagcc 720
acgcaaaagt acccgatgca gcaccaacaa gctcctcaaa tatatctaag ctcgggttca 780
gctctacccg acgaaagctg cacggaacct gggcttccac aggtcagaag tgcatcgcta 840
cagagattcc tggctaaacg acgagacagg ttgtcaggga atccttcctc gtctaggcgg 900
aacgacagat ccaaaaagcg gaggttctcc ccgccaccgt caccgttaac ttcggcttcg 960
ttccagtttc ctccaagtgc tagaacatcg caagttttaa gatactccac tacttctaca 1020
actacgatca ctactgccac tgctactgcc gctactacca ctactactac gggtaccacg 1080
aatggtggac actgttccaa ttccaatcaa gcaagcgaga atgcaggcag cgatacctcc 1140
ggtggaagtt ctggaacgcc ggacacaagc gacacaacaa gggacaacga caatggacga 1200
gtttccaacg aaaatggacg agtgtccacc acttgtctcg cagcaacgtg a 1251
<210> 95
<211> 705
<212> ДНК
<213> Selaginella moellendorffii
<400> 95
atgtctagca tggtcgattt cctggggatc gaggagaagg tgtccaccag cgtcagcgcc 60
gagaggttga agaagctgga ggagctcacg gacgaggacg tgatgcagct cactcgggag 120
gattgccggc gctacctcaa ggagaaggga atgcgtcgtc cgtcttggaa caaggcccag 180
gccgtgcagc aacttctctc gctcaagagc ctgtgcgatc cttccccggc ttccagtgga 240
gccgccaaga ggagcccatc tccgccgctc gacgaggctc cagcgaagaa acccatggca 300
atgacaagca tcgatctcaa ggctgccgct gctgtggacg ccgccaatct tacgatgttt 360
tatgatggag cagtgtccgt gtttgacgac gtttcgccag acaaggcatc tcttttccct 420
ttggcttatg cgatcatgct cctggccggg aatgtgaagt cgtggccttc gatcaacgtt 480
gctgctaaca ccaacaaagt tgtgatctct tcttatgagt tgccacaggc gcgaaaggca 540
tcactccagc gttttcttca gagacgccgt gagaaaactg cgaaagaggc agcatccaaa 600
gggaactcta ataagtcgcc ttgtcatggc gagagctcgg ggaagcacgc atcggatgct 660
actgatccag ccacttctcc cttgctcacg gaggtctctt cctag 705
<210> 96
<211> 813
<212> ДНК
<213> Nicotiana tabacum
<400> 96
atgccgccgg aagaaacagt ttccaaatca cctctggaca aaccgctcca cctacttacc 60
gatgacgaca tttctcagct tactcgcgaa gattgccgcc gttaccttaa agaaaaagga 120
atgagaaggc cgtcatggaa taaatcacag gcgattcagc aggtgatttc actgaaggcg 180
cttctcgaga cgacaccgga ttccgacacc ggccctcgga gaaaacttca cattcctcgc 240
ccagacacta gagtacaaca agtccagaaa ggaacggata ccgatgcaga attttcgaaa 300
tctgctgaag ggatggtgcc atacggaaga aaacattcga aaaaacctga tattcccggt 360
gatatagctg ccggttcagt tgccgttgcc gccggcaata acttagctcc ttctagaacc 420
acagatttgg gaaacacacc agcaagtcaa ttgacaatct tctattgtgg caaggtgaat 480
gtgtacgatg atgtgcccgc tgaaaaggca caagcaatca tgcatcttgc tgcaactcca 540
ctctttgtgc cttcagaaac tccattgggt gctaccttag cggctcgaca ttccgaatgc 600
catttgcaag ctgcaagtgt taaacagggt ccagattctg ctatggtgct catgccaacc 660
atgcaaacag ggaaaatgag tgaagtgact cgcctgcgtc tggaggaaag caataccttc 720
tatgaagaca actctgccaa ttatgcagaa gcagtggaag gccacccaag caggaaagca 780
tcagtacaaa gatatcttga gaagcggaaa gac 813
<210> 97
<211> 984
<212> ДНК
<213> Solanum tuberosum
<400> 97
atgccgccgg aagaaacagt ttccaagtca cctctcgata aacctctcaa tcaactcact 60
gacgatgaca tttctcagct cacacgcgaa gactgccgtc gttacctcaa acaaaaagga 120
atgagaaagc cgtcatggaa taaatcacag gcgattcagc aagttatatc gttgaaggct 180
ctcctcgagc cggatactga cgccggaact cggaagaaac ttcacattcc tcgtgcagat 240
actcatgtcc agagcgggaa aaatacctat ggcgaacctt ctgaaccagt gcctgataga 300
agaaatcagc aggacagacc tgatctttcc agtcatatta ctgcccttcc ggtcgctgtt 360
gtggataatt cagctccttc tagaacaata ggttcagcag ataaaccagt aggacaaatg 420
acaatcttct atagaggcaa ggtgaatgtc tatgatgatg tgcctgccga caaggcacaa 480
aaaatcatgt gtcttgcttc aagccctctt tgtgtgcctt cagaaactcc atcgaatgcc 540
actgtagcag ctcgacattc agcatgctgc ttacaagctg caaatagtaa actacgccta 600
gatactaata ttgtaccgac tattcaaaca gtgaaaatga gtgaggtttc tcgagttcct 660
atagaagaaa gcaaccgctt atacaatgat aatcctgaag cagtggagag ccccgcaagc 720
aggaaagcat cagtacaaag atatcttgag aagcgaaaag aaaggttcaa gtggaagaga 780
aaggtagaaa caacttcatc agctagcttg gatatctatt taagtgatcg aattgggact 840
cgtacgccaa gtgactatgc aagtggggct gatctttgct tcacacccca cattacacct 900
acaggaagtg gtcctataca agacaatatt cagatgaatc ccactttttc tagtgatctc 960
aatgacagag atgttagaga gtga 984
<210> 98
<211> 1047
<212> ДНК
<213> Glycine max
<400> 98
atgaacggcg gtgccaccac cgccaccttc cgatccatcc tcgacaagcc cctcaaccag 60
ctcaccgagg atgacatttc tcagctcact cgcgaagact gtcgcagatt cctcaaagaa 120
aaagggatgc gcaggccttc ctggaacaaa tcgcaggcga tccaacaggt catttccctg 180
aaagcgctgc tggaaccttc cgacgatgat actcctcctc ctaccgccat gcaccaccgt 240
agtcatgctc ctccccctcc acctcaacct caatctcaag tgaatttgac tgaacctcct 300
cctccgccca aggctccgcc acctgaagaa tcctcttttc atgctgctga agacattcag 360
aaacctgcgt cgtctgggga aaaaccttcg gaaactaatg acaccaacac caacgttgct 420
agccccaaag ggtgtgcaac tagtggatca tttgggcaaa tgacaatttt ctattgtggt 480
aaggtgaatg tctatgatgg agtctcgcct gataaggcac gagcaatcat gcagcttgcg 540
gtgagtcctg tccagtttac tcaagatgat ccttcaaatg gaaatgcagc tgtttggcct 600
tctccttgcc acttaccaat ggataaggat gtcctcattc ctgtagatac aacaatcctt 660
caggttgctc aatcagataa gatgatggaa tatcctctgc aatatagaga gaaaggtagc 720
atagctcgtg atgctgaggg tcaggcaagc agaaaagtgt cattgcagcg atatcttgaa 780
aagcgtaagg acagggggag attgaaaggc aagaaattga ctgggataac ttcatctaac 840
ttcgagatgt atttgaacct tccagtgaag gtccatgcct caaatgggaa ttcaagccgt 900
agcagtacta gctctccacc acagcctaga ctgcctctag tatctagtgg ttcagctgac 960
aaccagctaa aggttgccct tcccattgat ctcaatgaca aagtgtcatt gcagatgttc 1020
aagaatgcta aaactctaac tagatag 1047
<210> 99
<211> 1017
<212> ДНК
<213> Citrus clementine
<400> 99
atggacgtgg acggtggcgt gacgtcgtgc cggtcaatac tcgagaaacc tctcagtcag 60
ctcactgaag aggacattac gcagctcaca cgcgaagatt gccgcaaatt tctcaaggag 120
aaaggaatgc gcagaccatc gtggaacaaa tcgcaggcga tccagcaggt gatctctctc 180
aaagctttgc tcgagtccag cggcgattcc ggctcaggtg ttttacgcag agtactcgtc 240
tcgcctccgg aaagtatgcc gccgcgcgtg aatgtgactt caaattcagc tgatttagta 300
aaggaaccga ccatctcagt ttctggagac caaaacagtg cgtataggcg gaagtaccct 360
cgcaactgtg ctgttgatgc agataacaag accatctcta acagaaatcc ctgtgaagca 420
aatgggtcca tagggcagat gacgattttc tattgtggca aggtgaacgt gtacgaagga 480
gtgccaactg ataaggcaca ggagattatg caccttgcag caactccaat tgatttttcc 540
cagaacggtt catttggtgg aattacggca tatagggcca ttccatgcca tttacaagtg 600
acaagcaaca gacatgtgtc tctccctctt cgtcctgctg ccatgatctc tcagttcatg 660
caaacaggga agatagcaga ttattctcag gagtataggg agaaagcgat tagtactcat 720
gactctgatg tggatggtca ggttaaccga aaagtctcgt tgcagaggta tcttgaaaag 780
cggaaagaca ggggaaggtt tttcaaggga aagaaaaata caggaccaac tcctagtttg 840
gagatgtacc tgaaccatcc ggggaagaca catgcctcca atggacaaca gagccagagc 900
aacacaagct ctccgaccca gcctgagttg tccaacacat tggggacctc cccagacaac 960
caggcgaaga ctgtcatgct tccggttgat ctcaacaatg aagatattca agactga 1017
<210> 100
<211> 1008
<212> ДНК
<213> Ricinus communis
<400> 100
atggacgccg gagtgacgtc gttcaggtca atactagata aacccctaac tcagctaact 60
gaagaagaca tttctcaact cacacgcgaa gattgccgca aatacctcaa agaaaaagga 120
atgcgaagac cttcatggaa caaatcgcaa gcgatccagc aagtgatttc tctaaaagca 180
cttcttgaaa ctagtgaaga ttccggtgcc ggtgctctcc gtagaatctt agtttctaaa 240
cctccggtta cttcaaattc tgttgattca gctaaggaac caagtgatag caacaataat 300
aacttactag atgagacagc tcctcatgat tctcccaaat ctcctcctcc ggcgccatcg 360
ttggattgtc cactggaaga ggcagataat aaagtcattt cttcaagaag tcctggtgca 420
acagatgggt tggtcgggca aatgacgatt ttctattgtg gaaaggtgaa tgtttatgat 480
ggagtcccac ccgataaggc ccaggcgatc atgcatcttg cagcgactcc aattcactca 540
cctttagacg atccaattcg tagacctgta tttgcttttc cgtatcattt acagacccca 600
agtgacaaac atgtctttgt tccttctaat gctgcaattt ctccaaccac accaacagag 660
aaggtgacag aatattctca gcagtgtagg gagaaaggaa atgtaactta tgatcatgat 720
gtagagggtc aagcaaaccg aaaaatgtca ttgcagagat atctggagaa gaaaaaggat 780
aggggaagat tcaagggtag gaaaaattta gggcctaatt cgtctagctt ggatgcatat 840
ttgaaccatc aaatgaggac acatatctca aacgagcaat caaccaggag cagtacaagc 900
tctccaaccc agcctggagt gccacatact tcgagtaact cggccgaaga tcagctgaag 960
actgccagtt ttgctgttga tcttaatgaa gatgtccaag aaccttga 1008
<210> 101
<211> 1182
<212> ДНК
<213> Vitis vinifera
<400> 101
atgaatcccg gcgtcaccac tctccgctct atactggaca aaccccttca cgaactcacc 60
gaagaagaca tttctcagct cactcgtgaa gattgtcgca aatacctcaa agaaaaagga 120
atgcgtcgtc cttcctggaa caaatcgcag gcgatccagc aggttatttc gcttaaatcg 180
ttgctcgaaa ccagtgaggg cagcggtgcc ggagttttga ggaagatcac cgattcaccg 240
ccggcggaaa atctacctcc ggttacctcc aattcagctg attcaggcaa ggagctgagt 300
gctgatatcc agatctcagt atcagctgat gaactggttc cccttccgcc aaaagatcat 360
catccagaat ccaccccttc tggcgaatta gccagccggc ctccagaggc agacaccaag 420
catacttgtc ccagaagtcc aggtgcaaca aattgtttgg ttgggcagat gacaattttc 480
tactgtggaa aggtgaatgt gtatgatgga gttccagatg ataaggcaca agcaatcatg 540
catcttgcag caagcccatt ccatttgcct tcagatgacc cctttagtgg tgctgctatg 600
ctttgctcct ctccatgcca tttgcatact gccaatgtta aacatggcca tattcctcct 660
cgagccatgg tttctcagac tatgcaaaca gagaaattta ctgaatattc tcaacagtac 720
agagaagaag tgaactttac ccgtggacat ggatcggaag cactttctgg gcttaggacg 780
gtaggaagcc caacggccag gcctaccgaa gatatggaac agaccacttg tctcactata 840
tggggtacct tccgctacaa ggttatgcca ttcgagatat atgagggcat catggatgtt 900
gaaggtcagg ttgacagaaa attatcattg caaagatatt tcgaaaagcg aaaagacaga 960
tttaagagca ggaaaaaaat aggactacct tctggtagct tggagatgta tgtgaaccat 1020
caagcaagga cacaaccctc gaatgggcaa tcaagccgga gtggcacaag ctctccaccc 1080
cagcatggat tgtcgcacac cctgtgcagc tcagctgaca accatacaaa gaatttcact 1140
ccttttgttg atctaaacag taaagatatc caagaaagtt ga 1182
<210> 102
<211> 1029
<212> ДНК
<213> Morus notabilis
<400> 102
atgagcgccg gcacgacggc gtttcggtcc atactggaca agcccctgaa ccagctcacc 60
gaggatgaca tttctcagct cacccgtgaa gattgccgca aatacctcaa ggaaaaaggg 120
atgcgaaggc cgtcgtggaa caaatcgcag gcgatccagc aagtgatttc gctcaaggct 180
ttgttggagc cctgcgacga ttccggcgcc ggagccctaa ggaggatcgt cgcttcgacg 240
ccgccgccac cgccgacaca aaacgcgcca cgtgtctcca ctttcagtgt tacttcgaac 300
tcggcagatt cgggtaagga agcaagtgtt gatgtccagg tttcggcgga ggaatcggga 360
ccgtgtcaga ggaaggagca ggcgaaatct gctccggaga ctgaggaaag accggctgat 420
gcgggtgaga gggcaagtcc aagaagtcat tgtgcaactg atgcattggt cggacaaatg 480
acaattttct attgtggcaa agtgaatgtg tacgaagggg ttccacctga gaaggcacga 540
gcaatcatgc accttgctgc aagtccaatc cctttatctc gagaaaattc atttggggtc 600
cttgcagcac ctagatcttt tccatggcat ttacatgctg cgagtgacaa gggtggcctt 660
ctccctccta gtgccacaat atcacaaccc atgcagacag ataagctagc cgactacagc 720
caacagtgct gggagaaaga aaatgatggt caggcgagca gaaaactctc attgcagaaa 780
taccgtgaaa agaaaaaaga tagggggagg ttgaagacca agagaagcac gggatttaat 840
tcttctagca tggaggtcta ttttaaccac caggtaaaga cccacatgtc aaatggtaat 900
tcaagtcgaa gtagcacaag ctctccgacc cagcctggac taccacaaac attgtgtagc 960
acagtcgaca atcagccaaa gattccctgt cttcctgttg atctcaatga aaaactaact 1020
attgagatg 1029
<210> 103
<211> 1095
<212> ДНК
<213> Phoenix dactylifera
<400> 103
atgtattggg ttggatcggc tcaagaacgc cgccgagacg ggggccggtc gccgctcgac 60
aagccactca gcctgctcac agaggaggat atcgcccagc tcacccgcga ggactgccgc 120
cgattcctca aagagaaagg catgcgacgg ccgtcctgga ataagtcgca ggcgatccaa 180
caggtcatct ccctcaaggc cctcctcgag ggacgaccgg agtccggcga actccccgtc 240
ggcgccggct accgccagaa gcctccccct cggcggccgg cctctcttcc ttcgctgcag 300
gaggcggccg gcgactcgac ggcggcggcg aaggagccgt cgccgtcgtc gtcgctgtct 360
ccgtaccgga gaagagatcc gatcccgccg atcatctccg ccggcgggcc gtcttgccgg 420
ttcccggtcg ccggcaggga ccaacaaccg ccagagaccc cctccccctc gctcagggtg 480
acggcggaag taccggcggg tcagatgacg atcttctacg acggcaaggt gaacgtctac 540
agcgacgtga cggtcgataa ggcgcgggcg atcctgctgc tcgcggggag acgagactgc 600
tacggcgctg cggctctacc gggtccggtt cactcgcccc agccggcttt tctcggaccg 660
ggtcagggcc cggtccccac cgctcccccg ctggccgctg ctttacccac ctcgccagct 720
gggaggttag cccaccgttt cgagggaccg agtggagtgc cgcgcgggaa atcgagcctg 780
gtaagagagc ggagcacgtc accggagggt ccaacaagta gaaaagcatc attgcagcgg 840
tacctggaga aaaggaagga caggttaaaa ggtagaaaaa ctcttggagg ggcatcttct 900
tcaagcatgg aaataatgtt cttgagccaa aaatttgggg gtcagatacc aaatgagcag 960
ttaagtagga gcaacactag ctcccctacc caacccagac cacctggcac accaactaga 1020
tgcagttcaa tagagaacca ggctcagaaa aatcatctct cagttgatct caatgatgat 1080
ggttgcggca actga 1095
<210> 104
<211> 1065
<212> ДНК
<213> Theobroma cacao
<400> 104
atggaggcgg gggtagcgac gacgacgaca acgacggagt cgtttaggtc gatacttgat 60
aaacccctca gccaactaac agaagaagac atttctcagc tcactcggga agattgtcga 120
aaattcctca aggaaaaagg aatgcggagg ccgtcgtgga acaaatcgca ggcgatccag 180
caagtaattt cactcaaggc gttgttggag agcaacgaag attccggcgc cggagctatc 240
cggaagatcc tcgtttctcc accatcaccg tcagtgcctc cgcaaaatgc agcggcgcgt 300
gtggcttcca attcatgtga ttcagtaaaa gaagcggttg tcggagaaga aggaagcccg 360
tatcggcgaa aagatcctcc tttgaaacct tctccggtgg gggagataaa ttgccttggc 420
ggtgacacgg ataacaagaa tctctctcct agaagtccat gtgaatcaaa tgagttgggt 480
gggcaaatga caattttcta ctgtggaaag gtcaatgtgt atgatggagt accacttgat 540
aaggcacggg caatcatgca tctggcagcg actcctattg attttcctca ggacaatcaa 600
tgtagtggaa atgcagccct taggtccttt atgtgccatg tccaagcagt cggtgacaaa 660
aatggccttg ttgcttctac tgccttgaac tctcatacca tgcaaacaga gaagttgaca 720
gaatatcagc atcagtttag ggaaaaagga aatatcgctc gtgacgctga tgtagatggg 780
caggtgaaca gaaaagtatc attgcagaga tatcgtgaaa agcgaaaaga caggggaaga 840
ttttttaagg gcaggaagaa tacaggacaa gcttcctcta gcttggagat gtacctgaac 900
catcagataa gaactcacaa ctcaaatgga caatcaagcc ggagcagcac gggttctcca 960
ccacagtctg gattgccaca tgcattttgt agctcagctg acaaccaagc aaaacttgtg 1020
aatctttctg tagatctcaa tgacaaaagt gttcaagaac actga 1065
<210> 105
<211> 3789
<212> ДНК
<213> Spirodela polyrrhiza
<400> 105
atggccggga gcgaggcggc ggcgccggag gaggccggaa gggcggggga ggaggaggtg 60
agagcggcgg cgggggctgc ggcggtgaag tcgccgctgg agaagccgct gtcggagctc 120
acggaggagg acatcgcgca ggtcacgcgc gaggactgcc gtcggttcct caaggaaaaa 180
ggtgctctag tccttttcct ttggttttcc tccttgtttc tcttcttttc ctttggaggt 240
cgcggtggag ctgatctcga tatccacgtg gccgcccgcg gtgacggcgg ctcccgtttt 300
ctctgtgcaa acgaatgcag gtatgcgccg cccttcgtgg aacaagtcgc aggccgtcca 360
gcaggtcatc tccctcaagg cgctcctgga gccctgccac gatgcagacg acgacgcacc 420
ttccgccggt gctgttccct ccatctcctc cttcttctcg aaaaggccgt ccgacgccct 480
gcttccggcc gccgcggcgc aggtgaggag gggtatcctg ttccaaactt ccgtcgcctt 540
ctcttctagc tcgagctggc aacattttgt gaaattgttc cttctcgatt tagttggaga 600
aacgcgtgtg gtttattgtg ttctgtttac ttctcagttt cccgtctctt ctccaatgag 660
gggtgaacct gccggcggcg cgccccaaat tgtctccgaa cgtccccacg gaagggaccc 720
gctggcgaac gtcttcacct gctccgacgc cctcggtcga ttcccggcaa cggggaacgg 780
tgctcttccg ccaaacagtg ccaccctccc gcccaggtgc gtcccttctc agcacaacgc 840
cccctctatt caactctttt atttccccgg attgtccctc agatgttaac gggcgcccat 900
ggtgttgtag aggggttgct tccgctgaga cgctggaggg acagctgaca atcttctacg 960
atggcaagat taatgtctat gatggtgtga cgccggagaa ggtgaggtcc ggtcagagta 1020
agttggcggt gaccccttcc tgtggtagat ctaggcactc atggcagtgt atttcccgtc 1080
tttgaaggcg cgggtgatcg tacaatttgc ggggagcccg agctgctacg atatgccgcc 1140
gatgccttcg ccgtctttct acccaaaccg accccctaaa tgccacgacc tggctctgcc 1200
agcgttatct caagcgacag gtggtggttc cttcaatccg ccaccgcctc cgccgccgtt 1260
gcagccaccg ccgtctcatc ctgcccagcc tatcggctat tcccaggttc cccagaacgc 1320
tggtacgttt ttttctccgt catgactttc ctgcgggttc gcctatgctt gatcacgttc 1380
acgataacgc cctgcggtgg ccctcctgct cacgttgagt tctattgccc tggcaggaag 1440
gttcccgcag caatttcggg aaggcatgga agagtggaga agctcgcgag aagttgaacc 1500
aggtgagaga ccttggaccc ttttcttcag tatcagtcgt tattctcgtt tctctctgct 1560
taactgacct atgcacccgg cctgaacagg cagggtatgc tgggtgattg aggagattac 1620
gcctatatgt cagtctatta ccatgctact gtttaagggt gtccttctgt ctaagttaac 1680
tagagtgacc ttaagctcct ggatttctta acctaccaca ccctgatggt acatggtaca 1740
ctcacagttg acctcaaagc ctttctcatt cctgcggggg aagagggtgg gggtgggcgt 1800
tgactgagaa aattttgaaa gcaaattgct aatttctgtt gttctttata atgacatttt 1860
aggcattgcg cgcaagatag ctcttttttc ctttttaaga actgtgcttc ttgattccat 1920
tagcccggga tttaggttcc gttcattgct tagttagcct ccccaattgg cgctgtttat 1980
gaacgattgg cctcgaccct cttcagcatc cttatcaggg atcttgagat tgagcttaat 2040
cttatcgcct cttctcaaat gattatggct gccgcgtttt tttccaattt ccagttgatt 2100
tacctatttt ttggggagag gatgactggt acagtatggc aacttatcca gttttgaatt 2160
gatgctgata ctgttcttct ttataaggcc tagaaattga tggtaattaa agtaacgtgt 2220
cagtgattcc atggatcaca ttaatgccct aaacttctgt gtcttatgct attccaaatc 2280
tgaaaacctt atgaatcaga ccaacaatag tatgaagaaa atttatttca tgctagggtc 2340
catggtgttc tattcaagaa ctaccctttt tttggatcat gcagatgatc tattaagaga 2400
acaaaatgca gtagaaataa gttaacgaaa gacaacaagg tcatgtcatt tgacgaatga 2460
agaatttcag aatcaaattg aagcacatta catcttctca tgagttaaaa aaaatgctca 2520
tgcactgctt ttaatgctcc tttataatac ctgtcatagg ctgaattatt tcaaagtttc 2580
cttgtatggg aagttctact gggtggttct tttgtatgaa tcttcgtaaa taatcaaagc 2640
ctgtttattg ttcttcatga cgtggaaata attattttta tgtatcactc acgaaatgga 2700
tacagcacct atggttacat gatatttaac ttacgtccgc tcaaataaat gagcaataat 2760
aagataacct accatttctc tccttcattc ccattttgca aagcgtctgt tggtcgtatg 2820
ggaaaccaca tttctacaag aggaatccat catcttctat cacatctccc ttgagtgggc 2880
gcattttatg tggatccaaa tatctcaatt aataataatg tttactggat cattatatag 2940
ttttttttat gcattcctct gatgatctta gtggctctcg gaaatttttt attcatgaat 3000
agaaattttg tcatccctga aagctgctgc tcctttgcgt tcatcatttc tattctatta 3060
ttctcccttc ttcctgtatg gcctcctgaa gagttcttct tcatgcgaga gaaaaactat 3120
ggtcttaccc gaatagggtg ggatatttgt atgatcttcc caaggttttg tgtgggcccc 3180
atatgggggc ggtggcgaag cgggctttcg ggccggcgag cctagagaga cccgggcccg 3240
gcggctgttg ctaccctcta tagaaaagtt ttcctcagtg ctaccgcgct actgattggt 3300
tgctcttctg gtgccaatgt cgtcttgccg cgagcagagg gccccaccag cagagccgca 3360
tcgctccagc ggtacctgga gaaacggaag gataggtaaa gatggtgggc gtggggggtt 3420
gaccgactgg ttgactgccg ccgccgctgg ctgacttcgc cctctttccc tgtcggaagg 3480
ttcaagtgca agaagcacgc cggcgggggc tcttccggcg tggagctcct cctgagccag 3540
cggatcaggg accagattcc ggccgcccac ctctcatgcg gagagatgta cgcggcggcg 3600
ccgccggccc cgcggccccc tcccccgccg gcccgctgta gcgccgccgg ccacctccgc 3660
ttctccatcg acctcaacga cggcggtgag cgctctctct atctctctca cctctttttc 3720
atcgcctctg cttttgaaga gagtacccat gccaccgccg cctgcgtttc agatgtccgg 3780
gaagcttga 3789
<210> 106
<211> 882
<212> ДНК
<213> Musa acuminata
<400> 106
atgaaccccg gggagaccac gcccccgtcg ccgctcgaca agcccctcgc cgagctcacc 60
gaggaggaca tcgcccagct cacccgcgag gactgccgcc gcttcctcaa agcgaaaggc 120
atgcgacggc cgtcgtggaa caagtcgcag gcgatccagc aggtcatctc tctcaaggcc 180
cttctcgagg ggcggcccgg ctgtgacgac tgccctgctg gcggcggaat cctccaaaag 240
ctgctcactt cttctccttc ggagccgcta tcgccaccgc aggactcacc tcctcccgcg 300
ccgaaggagg gcggtagcgg atcacagccg ctggcgaagg agccgtcgcc gtatcgaagg 360
agggacccga tcccaccgcc ctattcagcc ggaaatccga cgtgccagac cccaattgcc 420
ggagctgacc ttccccaccc gccggagaag cgctgcccct cccccaggtt gacggcggaa 480
gtaccggtcg gccagatgac gatcttctac gacgggatgg tcaacgtata cgacggcgtc 540
tcggccgatc aggccaggtc gatcatggaa cttgcggcca gcccggtctg cttcgacgat 600
ccgaccggtg cattctctcc ggcccggccg ccggccttcc gcttccctcc gggtctcccc 660
cgaccggccc cggtccccac cgctccctcg ttcgtgggga ccttcccgat ctcgccggct 720
ggtaaacgtt gctactccta ctgttcgttc cggtcaagcg tcagcctttt aaccacaaca 780
gagggcccaa caagcagaaa agcatcattg cagagatact tggagaaaag gaaagacagg 840
tatggtcatt taccaacaga aagtatacta cttgttagct ga 882
<210> 107
<211> 498
<212> ДНК
<213> Phalaenopsis aphrodite
<220>
<221> дополнительный признак
<222> (214)..(243)
<223> n представляет собой a, c, g или t
<400> 107
atgaactccg atgcaataac catggggaaa tctctgcttg agaaacccct tagccttcta 60
accgaagacg atatcgcaca gattacaaga gaagaatgcc gtagattcct caaagataga 120
ggcatgcgtc gcccctcttg gaacaagtcg caggcgatcc agcaagtgat ttctctcaaa 180
gccctgttcg aaaaccgatc agatctagaa gatnnnnnnn nnnnnnnnnn nnnnnnnnnn 240
nnntttcccg aacacgcaga tctcagttcg atctcgccga ctgcggaggc caaggaacca 300
gagaaagctc agctcactat attctacggg gggaaggtgc ttgtgttcga caattttccg 360
gttaataagg cacaggattt gatgcagatt gcaggaaaag agcagaatca aaattacggg 420
acagcaaaca ctgtggctcc atctgcccct gcagcagacc tccatagttt acctctgccg 480
gctaagcctc cggcgtaa 498
<210> 108
<211> 238
<212> ДНК
<213> Arabidopsis thaliana
<220>
<221> дополнительный признак
<222> (31)..(31)
<223> n представляет собой a, c, g или t
<220>
<221> дополнительный признак
<222> (52)..(52)
<223> n представляет собой a, c, g или t
<220>
<221> дополнительный признак
<222> (59)..(59)
<223> n представляет собой a, c, g или t
<220>
<221> дополнительный признак
<222> (68)..(68)
<223> n представляет собой a, c, g или t
<220>
<221> дополнительный признак
<222> (110)..(110)
<223> n представляет собой a, c, g или t
<220>
<221> дополнительный признак
<222> (123)..(123)
<223> n представляет собой a, c, g или t
<220>
<221> дополнительный признак
<222> (125)..(125)
<223> n представляет собой a, c, g или t
<220>
<221> дополнительный признак
<222> (143)..(143)
<223> n представляет собой a, c, g или t
<220>
<221> дополнительный признак
<222> (153)..(153)
<223> n представляет собой a, c, g или t
<220>
<221> дополнительный признак
<222> (180)..(180)
<223> n представляет собой a, c, g или t
<220>
<221> дополнительный признак
<222> (189)..(189)
<223> n представляет собой a, c, g или t
<220>
<221> дополнительный признак
<222> (202)..(202)
<223> n представляет собой a, c, g или t
<220>
<221> дополнительный признак
<222> (206)..(206)
<223> n представляет собой a, c, g или t
<220>
<221> дополнительный признак
<222> (223)..(223)
<223> n представляет собой a, c, g или t
<400> 108
mdvgvsaksa kktdstrdcr kkdkgmrrsw nksavskayg ddsgagrkvs vnrvtttsna 60
cgrvsydnga chrmdsrsas ggsghvskdg hkttsrsats vgmtysgkvn vydgkarsmh 120
aandngassr mskskkmmkg kanssrdsgm ganrkvsryr krkdrkskak kcgvasssmn 180
crmkaaysng ctgshssstk snsvdnsggs gggsakgrgh gkngdstrtg hhhhhhgs 238
<210> 109
<211> 380
<212> БЕЛОК
<213> Trifolium repens
<400> 109
Met Asn Gly Gly Ser Thr Val Ser Phe Arg Ser Ile Leu Asp Lys Pro
1 5 10 15
Leu Asn Gln Leu Thr Glu Asp Asp Ile Ser Gln Leu Thr Arg Glu Asp
20 25 30
Cys Arg Arg Phe Leu Lys Asp Lys Gly Met Arg Arg Pro Ser Trp Asn
35 40 45
Lys Ser Gln Ala Ile Gln Gln Val Ile Ser Leu Lys Ala Leu Leu Glu
50 55 60
Pro Thr Asp Asp Asp Leu Pro Ala Pro Val Gly Val Ser Ser Ala Ile
65 70 75 80
His His His His His His His Pro Gln Pro Pro Gln Arg Asn Leu Asn
85 90 95
Glu Ala Pro Val Lys Gly Ser Asp Leu Asp Asp Thr Gly Phe His Thr
100 105 110
Ala Glu Asp Leu Asn Lys Ser Thr Ser Thr Ala Val Glu Ile Pro Thr
115 120 125
Glu Thr Asn Asp Ala Asn Val Val Lys Ser Ser Gly Gly Cys Val Ala
130 135 140
Ser Gly Ser Phe Gly Gln Met Thr Ile Phe Tyr Cys Gly Lys Val Asn
145 150 155 160
Val Tyr Asp Gly Val Ser Pro Asp Lys Ala Arg Ser Ile Met Gln Leu
165 170 175
Ala Ala Cys Pro Ser Ser Phe Pro Gln Asp Asn Leu Leu Asn Lys Asn
180 185 190
Ala Ala Val Trp Ala Ser Pro Cys Asn Ile Pro Ile Asp Lys Asp Val
195 200 205
Leu Phe Pro Asn Asp Thr Ala Ile Leu Gln Val Ala Gln Thr Asp Lys
210 215 220
Met Val Glu Tyr Pro Leu Gln Tyr Arg Glu Lys Gly Ser Ile Ala Arg
225 230 235 240
Asp Ala Asp Val Glu Gly Gln Ala Ser Arg Asn Ala Ser Leu Gln Arg
245 250 255
Tyr Arg Glu Lys Arg Lys Asp Arg Gly Arg Ser Lys Gly Asn Lys Leu
260 265 270
Thr Gly Ile Thr Ser Ser Asn Phe Glu Met Tyr Leu Asn Leu Pro Val
275 280 285
Lys Leu His Ala Ser Asn Gly Asn Ser Ser Arg Ser Ser Thr Asp Ser
290 295 300
Pro Pro Gln Pro Arg Leu Pro Leu Val Ser Gly Gly Ser Ala Glu Asn
305 310 315 320
Gln Pro Lys Val Thr Leu Pro Ile Asp Leu Asn Asp Lys Asp Val Gln
325 330 335
Glu Cys Gly Ser Gly Gly Gly Ser Ala Lys Gly Glu Leu Arg Gly His
340 345 350
Pro Phe Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp Ser
355 360 365
Thr Arg Thr Gly His His His His His His Gly Ser
370 375 380
<210> 110
<211> 353
<212> БЕЛОК
<213> Amborella trichopoda
<400> 110
Met Thr Ala Gly Asp Gly Ser Ile Arg Ser Ile Leu Asp Lys Pro Leu
1 5 10 15
Glu Glu Leu Thr Glu Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys
20 25 30
Arg Arg Tyr Leu Lys Glu Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
35 40 45
Tyr Gln Ala Ile Gln Gln Val Leu Ser Leu Lys Gly Leu Leu Glu Gly
50 55 60
Lys Pro Cys Asp Asp Asn Ser Asp Val Phe Ser His Arg Ser Pro Ile
65 70 75 80
Thr Val Ile Pro Asn Val Gly Ser Met Arg Glu Lys Glu Lys Ala Val
85 90 95
Asn Ile Ala Asp Pro Glu Ile Ser Gly Ser His Gln Pro Asn Phe Arg
100 105 110
Arg Glu Ile His Glu Thr Thr Arg Glu Arg Ala Leu Pro Ala Ser Asp
115 120 125
Trp Pro Pro Ser Gln Glu Pro Val Ser Gln Met Thr Ile Phe Tyr Ala
130 135 140
Gly Ala Val Asn Val Tyr Asn Asp Ile Pro Glu Asp Lys Val Gln Ala
145 150 155 160
Ile Ile Tyr Leu Ala Gly Lys Ser Asp Ser Leu Gln Gln Thr Asn Val
165 170 175
Ile Arg Thr Gly Pro Asp Gln Cys Ile Ala Ser Ala Ala Ser Pro Ser
180 185 190
Leu Asn Asp Leu His Ser Arg Arg Ile His Pro Thr Ser Asn Ile Thr
195 200 205
Thr Ser Gln Ser Leu Arg Val Ala Thr Ser Leu Pro Val Gly Pro His
210 215 220
Ser Glu Val Pro Lys Thr Arg Lys Thr Ser Val Gln Arg Phe Leu Glu
225 230 235 240
Lys Arg Lys Asp Arg Gly Arg Leu Lys Gly Thr Leu Ala Ser Gly Gly
245 250 255
Ser Ser Lys Arg Gly Ser Ser Cys Leu Glu Leu Tyr Ala Thr Ser Arg
260 265 270
Leu Lys Ser Glu Gly Val Ala Thr Thr Thr Thr Gln Ser Asn Met Asn
275 280 285
Asn Val Val Val Ser Pro Ser Asn Pro Arg Met Pro Leu Asn Pro Gly
290 295 300
Ser Cys Ser Trp Val Glu Asn Gly Ser Gly Gly Gly Ser Ala Lys Gly
305 310 315 320
Glu Leu Arg Gly His Pro Phe Glu Gly Lys Pro Ile Pro Asn Pro Leu
325 330 335
Leu Gly Leu Asp Ser Thr Arg Thr Gly His His His His His His Gly
340 345 350
Ser
<210> 111
<211> 335
<212> БЕЛОК
<213> Musa acuminata
<400> 111
Met Asn Pro Gly Glu Thr Thr Pro Pro Ser Pro Leu Asp Lys Pro Leu
1 5 10 15
Ala Glu Leu Thr Glu Glu Asp Ile Ala Gln Leu Thr Arg Glu Asp Cys
20 25 30
Arg Arg Phe Leu Lys Ala Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
35 40 45
Ser Gln Ala Ile Gln Gln Val Ile Ser Leu Lys Ala Leu Leu Glu Gly
50 55 60
Arg Pro Gly Cys Asp Asp Cys Pro Ala Gly Gly Gly Ile Leu Gln Lys
65 70 75 80
Leu Leu Thr Ser Ser Pro Ser Glu Pro Leu Ser Pro Pro Gln Asp Ser
85 90 95
Pro Pro Pro Ala Pro Lys Glu Gly Gly Ser Gly Ser Gln Pro Leu Ala
100 105 110
Lys Glu Pro Ser Pro Tyr Arg Arg Arg Asp Pro Ile Pro Pro Pro Tyr
115 120 125
Ser Ala Gly Asn Pro Thr Cys Gln Thr Pro Ile Ala Gly Ala Asp Leu
130 135 140
Pro His Pro Pro Glu Lys Arg Cys Pro Ser Pro Arg Leu Thr Ala Glu
145 150 155 160
Val Pro Val Gly Gln Met Thr Ile Phe Tyr Asp Gly Met Val Asn Val
165 170 175
Tyr Asp Gly Val Ser Ala Asp Gln Ala Arg Ser Ile Met Glu Leu Ala
180 185 190
Ala Ser Pro Val Cys Phe Asp Asp Pro Thr Gly Ala Phe Ser Pro Ala
195 200 205
Arg Pro Pro Ala Phe Arg Phe Pro Pro Gly Leu Pro Arg Pro Ala Pro
210 215 220
Val Pro Thr Ala Pro Ser Phe Val Gly Thr Phe Pro Ile Ser Pro Ala
225 230 235 240
Gly Lys Arg Cys Tyr Ser Tyr Cys Ser Phe Arg Ser Ser Val Ser Leu
245 250 255
Leu Thr Thr Thr Glu Gly Pro Thr Ser Arg Lys Ala Ser Leu Gln Arg
260 265 270
Tyr Leu Glu Lys Arg Lys Asp Arg Tyr Gly His Leu Pro Thr Glu Ser
275 280 285
Ile Leu Leu Val Ser Gly Ser Gly Gly Gly Ser Ala Lys Gly Glu Leu
290 295 300
Arg Gly His Pro Phe Glu Gly Lys Pro Ile Pro Asn Pro Leu Leu Gly
305 310 315 320
Leu Asp Ser Thr Arg Thr Gly His His His His His His Gly Ser
325 330 335
<210> 112
<211> 264
<212> БЕЛОК
<213> Picea sitchensis
<400> 112
Met Arg Gly Gly Glu Arg Ala Pro Gly Ser Arg Pro Ser Leu Asp Lys
1 5 10 15
Pro Leu Glu Glu Leu Thr Glu Glu Asp Ile Phe Gln Leu Thr Arg Glu
20 25 30
Asp Cys Arg Arg Tyr Leu Lys Glu Lys Gly Met Arg Arg Pro Ser Trp
35 40 45
Asn Lys Ser Gln Ala Ile Gln Gln Val Leu Ser Leu Lys Ser Leu Phe
50 55 60
Glu Ser Lys Pro Asn Gln Gln Ser Lys Lys Pro Ser Lys His Lys Pro
65 70 75 80
Ala Thr Leu Gln Phe Glu Thr Ala Arg Asp Ser Thr Phe Ala Gln Ser
85 90 95
Ser Val Ser Gln Glu Gln Ser Leu Gly Phe Ser Trp Ser Lys Glu Val
100 105 110
Leu Asp Lys Gly Thr Ala Glu Arg Gln Arg Leu Cys Ser Asp Ser Gln
115 120 125
Glu Ala His Glu Ile Pro Arg Leu Gly Ser Lys Pro Pro Gln Ser Asn
130 135 140
Thr Glu Gly Lys Arg Cys Ala His Asp Gly His Gly Arg Lys Ser Ala
145 150 155 160
Gln Pro Leu Val Arg Leu Pro Ala Asn Phe Lys Asn Asp Cys Ser Asn
165 170 175
Arg Gln Ser Ser His Thr Ser Glu Ser Gln Pro Asp Thr Leu Leu Arg
180 185 190
Ser Asp Ser Phe Gln Gln Pro Thr Ala Gln Leu Thr Ile Phe Tyr Ala
195 200 205
Gly Met Val Asn Val Tyr Asp Asp Val Pro Leu Asp Lys Ala Gly Ser
210 215 220
Gly Gly Gly Ser Ala Lys Gly Glu Leu Arg Gly His Pro Phe Glu Gly
225 230 235 240
Lys Pro Ile Pro Asn Pro Leu Leu Gly Leu Asp Ser Thr Arg Thr Gly
245 250 255
His His His His His His Gly Ser
260
<210> 113
<211> 276
<212> БЕЛОК
<213> Selaginella moellendorffii
<400> 113
Met Ser Ser Met Val Asp Phe Leu Gly Ile Glu Glu Lys Val Ser Thr
1 5 10 15
Ser Val Ser Ala Glu Arg Leu Lys Lys Leu Glu Glu Leu Thr Asp Glu
20 25 30
Asp Val Met Gln Leu Thr Arg Glu Asp Cys Arg Arg Tyr Leu Lys Glu
35 40 45
Lys Gly Met Arg Arg Pro Ser Trp Asn Lys Ala Gln Ala Val Gln Gln
50 55 60
Leu Leu Ser Leu Lys Ser Leu Cys Asp Pro Ser Pro Ala Ser Ser Gly
65 70 75 80
Ala Ala Lys Arg Ser Pro Ser Pro Pro Leu Asp Glu Ala Pro Ala Lys
85 90 95
Lys Pro Met Ala Met Thr Ser Ile Asp Leu Lys Ala Ala Ala Ala Val
100 105 110
Asp Ala Ala Asn Leu Thr Met Phe Tyr Asp Gly Ala Val Ser Val Phe
115 120 125
Asp Asp Val Ser Pro Asp Lys Ala Ser Leu Phe Pro Leu Ala Tyr Ala
130 135 140
Ile Met Leu Leu Ala Gly Asn Val Lys Ser Trp Pro Ser Ile Asn Val
145 150 155 160
Ala Ala Asn Thr Asn Lys Val Val Ile Ser Ser Tyr Glu Leu Pro Gln
165 170 175
Ala Arg Lys Ala Ser Leu Gln Arg Phe Leu Gln Arg Arg Arg Glu Lys
180 185 190
Thr Ala Lys Glu Ala Ala Ser Lys Gly Asn Ser Asn Lys Ser Pro Cys
195 200 205
His Gly Glu Ser Ser Gly Lys His Ala Ser Asp Ala Thr Asp Pro Ala
210 215 220
Thr Ser Pro Leu Leu Thr Glu Val Ser Ser Gly Ser Gly Gly Gly Ser
225 230 235 240
Ala Lys Gly Glu Leu Arg Gly His Pro Phe Glu Gly Lys Pro Ile Pro
245 250 255
Asn Pro Leu Leu Gly Leu Asp Ser Thr Arg Thr Gly His His His His
260 265 270
His His Gly Ser
275
<210> 114
<211> 315
<212> БЕЛОК
<213> Arabidopsis thaliana
<400> 114
Met Asp Val Gly Val Ser Pro Ala Lys Ser Ile Leu Ala Lys Pro Leu
1 5 10 15
Lys Leu Leu Thr Glu Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys
20 25 30
Arg Lys Phe Leu Lys Asp Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
35 40 45
Ser Gln Ala Ile Gln Gln Val Leu Ser Leu Lys Ala Leu Tyr Glu Pro
50 55 60
Gly Asp Asp Ser Gly Ala Gly Ile Phe Arg Lys Ile Leu Val Ser Gln
65 70 75 80
Pro Val Asn Pro Pro Arg Val Thr Thr Thr Leu Ile Glu Pro Ser Asn
85 90 95
Glu Leu Glu Ala Cys Gly Arg Val Ser Tyr Pro Glu Asp Asn Gly Ala
100 105 110
Cys His Arg Met Asp Ser Pro Arg Ser Ala Glu Phe Ser Gly Gly Ser
115 120 125
Gly His Phe Val Ser Glu Lys Asp Gly His Lys Thr Thr Ile Ser Pro
130 135 140
Arg Ser Pro Ala Glu Thr Ser Glu Leu Val Gly Gln Met Thr Ile Phe
145 150 155 160
Tyr Ser Gly Lys Val Asn Val Tyr Asp Gly Ile Pro Pro Glu Lys Ala
165 170 175
Arg Ser Ile Met His Phe Ala Ala Asn Pro Ile Asp Leu Pro Glu Asn
180 185 190
Gly Ile Phe Ala Ser Ser Arg Met Ile Ser Lys Leu Ile Ser Lys Glu
195 200 205
Lys Met Met Glu Leu Pro Gln Lys Gly Leu Glu Lys Ala Asn Ser Ser
210 215 220
Arg Asp Ser Gly Met Glu Gly Gln Ala Asn Arg Lys Val Ser Leu Gln
225 230 235 240
Arg Tyr Arg Glu Lys Arg Lys Asp Arg Lys Phe Ser Lys Ala Lys Lys
245 250 255
Cys Pro Gly Val Ala Ser Ser Ser Leu Glu Met Phe Leu Asn Cys Gln
260 265 270
Pro Arg Met Lys Ala Ala Tyr Ser Gln Asn Leu Gly Cys Thr Gly Ser
275 280 285
Pro Leu His Ser Gln Ser Pro Glu Ser Gln Thr Lys Ser Pro Asn Leu
290 295 300
Ser Val Asp Leu Asn Ser Glu Gly Ile Gly Ser
305 310 315
<210> 115
<211> 340
<212> БЕЛОК
<213> Trifolium repens
<400> 115
Met Asn Gly Gly Ser Thr Val Ser Phe Arg Ser Ile Leu Asp Lys Pro
1 5 10 15
Leu Asn Gln Leu Thr Glu Asp Asp Ile Ser Gln Leu Thr Arg Glu Asp
20 25 30
Cys Arg Arg Phe Leu Lys Asp Lys Gly Met Arg Arg Pro Ser Trp Asn
35 40 45
Lys Ser Gln Ala Ile Gln Gln Val Ile Ser Leu Lys Ala Leu Leu Glu
50 55 60
Pro Thr Asp Asp Asp Leu Pro Ala Pro Val Gly Val Ser Ser Ala Ile
65 70 75 80
His His His His His His His Pro Gln Pro Pro Gln Arg Asn Leu Asn
85 90 95
Glu Ala Pro Val Lys Gly Ser Asp Leu Asp Asp Thr Gly Phe His Thr
100 105 110
Ala Glu Asp Leu Asn Lys Ser Thr Ser Thr Ala Val Glu Ile Pro Thr
115 120 125
Glu Thr Asn Asp Ala Asn Val Val Lys Ser Ser Gly Gly Cys Val Ala
130 135 140
Ser Gly Ser Phe Gly Gln Met Thr Ile Phe Tyr Cys Gly Lys Val Asn
145 150 155 160
Val Tyr Asp Gly Val Ser Pro Asp Lys Ala Arg Ser Ile Met Gln Leu
165 170 175
Ala Ala Cys Pro Ser Ser Phe Pro Gln Asp Asn Leu Leu Asn Lys Asn
180 185 190
Ala Ala Val Trp Ala Ser Pro Cys Asn Ile Pro Ile Asp Lys Asp Val
195 200 205
Leu Phe Pro Asn Asp Thr Ala Ile Leu Gln Val Ala Gln Thr Asp Lys
210 215 220
Met Val Glu Tyr Pro Leu Gln Tyr Arg Glu Lys Gly Ser Ile Ala Arg
225 230 235 240
Asp Ala Asp Val Glu Gly Gln Ala Ser Arg Asn Ala Ser Leu Gln Arg
245 250 255
Tyr Arg Glu Lys Arg Lys Asp Arg Gly Arg Ser Lys Gly Asn Lys Leu
260 265 270
Thr Gly Ile Thr Ser Ser Asn Phe Glu Met Tyr Leu Asn Leu Pro Val
275 280 285
Lys Leu His Ala Ser Asn Gly Asn Ser Ser Arg Ser Ser Thr Asp Ser
290 295 300
Pro Pro Gln Pro Arg Leu Pro Leu Val Ser Gly Gly Ser Ala Glu Asn
305 310 315 320
Gln Pro Lys Val Thr Leu Pro Ile Asp Leu Asn Asp Lys Asp Val Gln
325 330 335
Glu Cys Gly Ser
340
<210> 116
<211> 313
<212> БЕЛОК
<213> Amborella trichopoda
<400> 116
Met Thr Ala Gly Asp Gly Ser Ile Arg Ser Ile Leu Asp Lys Pro Leu
1 5 10 15
Glu Glu Leu Thr Glu Glu Asp Ile Ser Gln Leu Thr Arg Glu Asp Cys
20 25 30
Arg Arg Tyr Leu Lys Glu Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
35 40 45
Tyr Gln Ala Ile Gln Gln Val Leu Ser Leu Lys Gly Leu Leu Glu Gly
50 55 60
Lys Pro Cys Asp Asp Asn Ser Asp Val Phe Ser His Arg Ser Pro Ile
65 70 75 80
Thr Val Ile Pro Asn Val Gly Ser Met Arg Glu Lys Glu Lys Ala Val
85 90 95
Asn Ile Ala Asp Pro Glu Ile Ser Gly Ser His Gln Pro Asn Phe Arg
100 105 110
Arg Glu Ile His Glu Thr Thr Arg Glu Arg Ala Leu Pro Ala Ser Asp
115 120 125
Trp Pro Pro Ser Gln Glu Pro Val Ser Gln Met Thr Ile Phe Tyr Ala
130 135 140
Gly Ala Val Asn Val Tyr Asn Asp Ile Pro Glu Asp Lys Val Gln Ala
145 150 155 160
Ile Ile Tyr Leu Ala Gly Lys Ser Asp Ser Leu Gln Gln Thr Asn Val
165 170 175
Ile Arg Thr Gly Pro Asp Gln Cys Ile Ala Ser Ala Ala Ser Pro Ser
180 185 190
Leu Asn Asp Leu His Ser Arg Arg Ile His Pro Thr Ser Asn Ile Thr
195 200 205
Thr Ser Gln Ser Leu Arg Val Ala Thr Ser Leu Pro Val Gly Pro His
210 215 220
Ser Glu Val Pro Lys Thr Arg Lys Thr Ser Val Gln Arg Phe Leu Glu
225 230 235 240
Lys Arg Lys Asp Arg Gly Arg Leu Lys Gly Thr Leu Ala Ser Gly Gly
245 250 255
Ser Ser Lys Arg Gly Ser Ser Cys Leu Glu Leu Tyr Ala Thr Ser Arg
260 265 270
Leu Lys Ser Glu Gly Val Ala Thr Thr Thr Thr Gln Ser Asn Met Asn
275 280 285
Asn Val Val Val Ser Pro Ser Asn Pro Arg Met Pro Leu Asn Pro Gly
290 295 300
Ser Cys Ser Trp Val Glu Asn Gly Ser
305 310
<210> 117
<211> 295
<212> БЕЛОК
<213> Musa acuminata
<400> 117
Met Asn Pro Gly Glu Thr Thr Pro Pro Ser Pro Leu Asp Lys Pro Leu
1 5 10 15
Ala Glu Leu Thr Glu Glu Asp Ile Ala Gln Leu Thr Arg Glu Asp Cys
20 25 30
Arg Arg Phe Leu Lys Ala Lys Gly Met Arg Arg Pro Ser Trp Asn Lys
35 40 45
Ser Gln Ala Ile Gln Gln Val Ile Ser Leu Lys Ala Leu Leu Glu Gly
50 55 60
Arg Pro Gly Cys Asp Asp Cys Pro Ala Gly Gly Gly Ile Leu Gln Lys
65 70 75 80
Leu Leu Thr Ser Ser Pro Ser Glu Pro Leu Ser Pro Pro Gln Asp Ser
85 90 95
Pro Pro Pro Ala Pro Lys Glu Gly Gly Ser Gly Ser Gln Pro Leu Ala
100 105 110
Lys Glu Pro Ser Pro Tyr Arg Arg Arg Asp Pro Ile Pro Pro Pro Tyr
115 120 125
Ser Ala Gly Asn Pro Thr Cys Gln Thr Pro Ile Ala Gly Ala Asp Leu
130 135 140
Pro His Pro Pro Glu Lys Arg Cys Pro Ser Pro Arg Leu Thr Ala Glu
145 150 155 160
Val Pro Val Gly Gln Met Thr Ile Phe Tyr Asp Gly Met Val Asn Val
165 170 175
Tyr Asp Gly Val Ser Ala Asp Gln Ala Arg Ser Ile Met Glu Leu Ala
180 185 190
Ala Ser Pro Val Cys Phe Asp Asp Pro Thr Gly Ala Phe Ser Pro Ala
195 200 205
Arg Pro Pro Ala Phe Arg Phe Pro Pro Gly Leu Pro Arg Pro Ala Pro
210 215 220
Val Pro Thr Ala Pro Ser Phe Val Gly Thr Phe Pro Ile Ser Pro Ala
225 230 235 240
Gly Lys Arg Cys Tyr Ser Tyr Cys Ser Phe Arg Ser Ser Val Ser Leu
245 250 255
Leu Thr Thr Thr Glu Gly Pro Thr Ser Arg Lys Ala Ser Leu Gln Arg
260 265 270
Tyr Leu Glu Lys Arg Lys Asp Arg Tyr Gly His Leu Pro Thr Glu Ser
275 280 285
Ile Leu Leu Val Ser Gly Ser
290 295
<210> 118
<211> 224
<212> БЕЛОК
<213> Picea abies
<400> 118
Met Arg Gly Gly Glu Arg Ala Pro Gly Ser Arg Pro Ser Leu Asp Lys
1 5 10 15
Pro Leu Glu Glu Leu Thr Glu Glu Asp Ile Phe Gln Leu Thr Arg Glu
20 25 30
Asp Cys Arg Arg Tyr Leu Lys Glu Lys Gly Met Arg Arg Pro Ser Trp
35 40 45
Asn Lys Ser Gln Ala Ile Gln Gln Val Leu Ser Leu Lys Ser Leu Phe
50 55 60
Glu Ser Lys Pro Asn Gln Gln Ser Lys Lys Pro Ser Lys His Lys Pro
65 70 75 80
Ala Thr Leu Gln Phe Glu Thr Ala Arg Asp Ser Thr Phe Ala Gln Ser
85 90 95
Ser Val Ser Gln Glu Gln Ser Leu Gly Phe Ser Trp Ser Lys Glu Val
100 105 110
Leu Asp Lys Gly Thr Ala Glu Arg Gln Arg Leu Cys Ser Asp Ser Gln
115 120 125
Glu Ala His Glu Ile Pro Arg Leu Gly Ser Lys Pro Pro Gln Ser Asn
130 135 140
Thr Glu Gly Lys Arg Cys Ala His Asp Gly His Gly Arg Lys Ser Ala
145 150 155 160
Gln Pro Leu Val Arg Leu Pro Ala Asn Phe Lys Asn Asp Cys Ser Asn
165 170 175
Arg Gln Ser Ser His Thr Ser Glu Ser Gln Pro Asp Thr Leu Leu Arg
180 185 190
Ser Asp Ser Phe Gln Gln Pro Thr Ala Gln Leu Thr Ile Phe Tyr Ala
195 200 205
Gly Met Val Asn Val Tyr Asp Asp Val Pro Leu Asp Lys Ala Gly Ser
210 215 220
<210> 119
<211> 236
<212> БЕЛОК
<213> Selaginella moellendorffii
<400> 119
Met Ser Ser Met Val Asp Phe Leu Gly Ile Glu Glu Lys Val Ser Thr
1 5 10 15
Ser Val Ser Ala Glu Arg Leu Lys Lys Leu Glu Glu Leu Thr Asp Glu
20 25 30
Asp Val Met Gln Leu Thr Arg Glu Asp Cys Arg Arg Tyr Leu Lys Glu
35 40 45
Lys Gly Met Arg Arg Pro Ser Trp Asn Lys Ala Gln Ala Val Gln Gln
50 55 60
Leu Leu Ser Leu Lys Ser Leu Cys Asp Pro Ser Pro Ala Ser Ser Gly
65 70 75 80
Ala Ala Lys Arg Ser Pro Ser Pro Pro Leu Asp Glu Ala Pro Ala Lys
85 90 95
Lys Pro Met Ala Met Thr Ser Ile Asp Leu Lys Ala Ala Ala Ala Val
100 105 110
Asp Ala Ala Asn Leu Thr Met Phe Tyr Asp Gly Ala Val Ser Val Phe
115 120 125
Asp Asp Val Ser Pro Asp Lys Ala Ser Leu Phe Pro Leu Ala Tyr Ala
130 135 140
Ile Met Leu Leu Ala Gly Asn Val Lys Ser Trp Pro Ser Ile Asn Val
145 150 155 160
Ala Ala Asn Thr Asn Lys Val Val Ile Ser Ser Tyr Glu Leu Pro Gln
165 170 175
Ala Arg Lys Ala Ser Leu Gln Arg Phe Leu Gln Arg Arg Arg Glu Lys
180 185 190
Thr Ala Lys Glu Ala Ala Ser Lys Gly Asn Ser Asn Lys Ser Pro Cys
195 200 205
His Gly Glu Ser Ser Gly Lys His Ala Ser Asp Ala Thr Asp Pro Ala
210 215 220
Thr Ser Pro Leu Leu Thr Glu Val Ser Ser Gly Ser
225 230 235
<210> 120
<211> 1226
<212> ДНК
<213> Arabidopsis thaliana
<400> 120
acaagtttgt acaaaaaagc aggctcaaaa aaaaccatgg acgtgggcgt gtccccggcc 60
aagtctattc tcgccaagcc ggtaatgttc agctctgcta tagtgtgtgc caccctgctt 120
gtttaataat gcgttctctt cgtttttatg atatcttatt cttccagctc aagctcctca 180
ccgaggagga catctctcag ctcacaagag aggactgccg caagttcctc aaggacaagg 240
gcatgagaag gccgtcctgg aacaagtccc aggccatcca acaagtgctc agcctcaagg 300
ccctctacga gccaggcgac gactccggcg ctggcatttt cagaaagatc ctcgtgtccc 360
agccggtgaa cccaccaagg gtgaccacca cactcatcga gccgtccaat gagcttgagg 420
cctgcggcag agtttcctac ccagaggata atggcgcctg ccacaggatg gattctccaa 480
ggtctgctga gttctctggc ggctccggcc atttcgtgtc tgagaaggat ggccacaaga 540
ccaccatctc cccaagatcc ccagccgaga catctgagct tgtgggccag atgaccatct 600
tctactccgg caaggtgaac gtgtacgacg gcatcccacc agagaaggcc cgctccatta 660
tgcacttcgc cgccaaccca atcgacctcc cagagaatgg catcttcgcc tccagccgca 720
tgatctccaa gctcatctcc aaggagaaga tgatggagct gccgcagaag ggcctcgaga 780
aggctaattc ctctcgcgac tccggcatgg agggccaggc taatagaaag gtgtccctcc 840
aacgctaccg cgagaagagg aaggaccgca agttctccaa ggccaagaag tgcccaggcg 900
ttgcctcttc cagcctcgag atgttcctca actgccagcc gagaatgaag gccgcctact 960
cccaaaatct cggctgcaca ggctccccac tccattctca gtccccagag tctcagacca 1020
agtccccgaa cctctccgtg gaccttaact ccgagggcat cggatccggc ggcggctctg 1080
ctaagggcga gctgaggggc cacccgttcg agggcaagcc aattccaaat ccactcctcg 1140
gcctcgactc taccaggacc ggccaccatc accatcacca cggatcctaa tgaagaccca 1200
gctttcttgt acaaagtggt caggct 1226
<210> 121
<211> 1295
<212> ДНК
<213> Trifolium repens
<400> 121
acaagtttgt acaaaaaagc aggctcaaaa aaaaccatga acggcggctc caccgtgtcc 60
ttcagatcca tcctcgataa gccggtaatg ttcagctctg ctatagtgtg tgccaccctg 120
cttgtttaat aatgcgttct cttcgttttt atgatatctt attcttccag ctcaaccagc 180
tcaccgagga cgacatctct cagctcacac gcgaggattg ccgccgcttc cttaaggaca 240
agggcatgag aaggccgtcc tggaacaagt cccaggccat ccagcaagtg atctccctca 300
aggctctcct cgagccgacc gacgatgatc tcccagcccc ggtgggcgtg tcatctgcca 360
tccaccatca ccaccaccac catcctcaac cgccacagag gaacctcaat gaggccccag 420
ttaagggctc cgacctcgac gataccggct tccatacagc cgaggacctc aacaagtcta 480
cctccaccgc cgtcgagatc ccgaccgaga caaacgatgc caacgtggtg aagtctagcg 540
gcggctgcgt ggcctccggc tccttcggcc agatgaccat tttctactgc ggcaaggtga 600
acgtgtacga cggcgtgtca ccagataagg cccgctccat tatgcaactc gccgcttgcc 660
catctagctt cccgcaggat aacctcctca acaagaacgc cgccgtttgg gcctccccat 720
gcaacatccc gatcgacaag gatgtcctct tcccgaacga caccgccatt ctccaggtgg 780
cccagaccga taagatggtc gagtacccac tccagtaccg cgagaagggc tctattgcca 840
gggatgccga tgttgagggc caggcctcca gaaatgcttc cctgcaacgc tatcgcgaga 900
agcgcaagga cagaggcaga tccaagggca acaagctgac cggcatcacc tcctccaact 960
tcgagatgta cctcaacctc ccggtgaagc tccatgcctc caacggcaac tcctctaggt 1020
cctccacaga ttccccaccg cagccaagac tcccactcgt gtccggcggc tctgccgaga 1080
accagccaaa ggtgaccctc ccgatcgacc tcaacgacaa ggacgtgcaa gagtgcggat 1140
ccggcggcgg ctctgctaag ggcgagctga ggggccaccc gttcgagggc aagccaattc 1200
caaatccact cctcggcctc gactctacca ggaccggcca ccatcaccat caccacggat 1260
cctaatgaag acccagcttt cttgtacaaa gtggt 1295
<210> 122
<211> 1214
<212> ДНК
<213> Amborella trichopoda
<400> 122
acaagtttgt acaaaaaagc aggctcaaaa aaaaccatga cagccggcga cggctccatc 60
cgctctatcc ttgacaagcc ggtaatgttc agctctgcta tagtgtgtgc caccctgctt 120
gtttaataat gcgttctctt cgtttttatg atatcttatt cttccagctc gaggagctga 180
ccgaggagga catctctcaa ctcacccgcg aggattgccg ccgctacctc aaggagaagg 240
ggatgcgcag gccatcctgg aacaagtacc aggccatcca gcaggtcctc agcctcaagg 300
gcctcctcga gggcaagcca tgcgacgata actccgacgt gttctcccac aggtccccga 360
tcaccgtgat cccaaatgtt ggctccatgc gcgagaagga gaaggccgtc aatatcgccg 420
acccagagat ctccggctcc caccagccga actttaggcg cgagatccat gagacaacac 480
gcgagagagc cctcccagct tctgattggc cgccgtcaca agagccagtg tcccagatga 540
ccatcttcta cgctggcgcc gtgaacgtgt acaacgacat cccagaggac aaggtgcagg 600
ccatcatcta cctcgccggc aagtctgatt ccctccagca gaccaacgtg atcaggaccg 660
gcccagatca gtgcattgct tctgctgctt ccccgtccct caacgacctc cattctaggc 720
gcatccaccc gacctccaac atcaccacat ctcagtctct ccgcgtggcc acatctctcc 780
cagtgggccc gcactccgag gtgccaaaga ccagaaagac aagcgtgcag cgcttcctcg 840
agaagaggaa ggataggggc aggctcaagg gcacactcgc ctccggcggc tcctccaaga 900
ggggctcctc ctgcctcgag ctttacgcta catcccgcct taagtctgag ggcgtggcca 960
ccacaaccac ccagtccaac atgaacaacg tggtggtgtc cccgtccaac ccgaggatgc 1020
cgctcaaccc gggctcctgc tcctgggtcg agaacggatc cggcggcggc tctgctaagg 1080
gcgagctgag gggccacccg ttcgagggca agccaattcc aaatccactc ctcggcctcg 1140
actctaccag gaccggccac catcaccatc accacggatc ctaatgaaga cccagctttc 1200
ttgtacaaag tggt 1214
<210> 123
<211> 1160
<212> ДНК
<213> Musa acuminata
<400> 123
acaagtttgt acaaaaaagc aggctcaaaa aaaaccatga acccgggcga gacaaccccg 60
ccatctccac ttgacaagcc ggtaatgttc agctctgcta tagtgtgtgc caccctgctt 120
gtttaataat gcgttctctt cgtttttatg atatcttatt cttccagctc gccgagctga 180
ccgaggagga tattgctcaa ctcacccgcg aggactgccg cagattcctt aaggctaagg 240
gcatgcgcag gccgtcctgg aacaagtctc aggccatcca gcaagtgatc tccctcaagg 300
ctctcctcga gggcaggcca ggttgcgatg actgcccggc cggcggcggc atcctccaga 360
agctcctcac ctccagcccg tctgagccgc tctccccgcc acaggactcc ccgccaccgg 420
ctccaaaaga gggcggctcc ggctcccagc ctctcgccaa ggagccgtcc ccgtacaggc 480
gcagggaccc gatcccgcca ccgtactccg ccggcaaccc gacctgccag accccgatcg 540
ctggcgccga cctcccgcac ccgccagaga agaggtgccc gtccccgagg ctcacagccg 600
aggtgccggt gggccagatg accattttct acgacggcat ggtgaacgtg tacgacggcg 660
tgtcagctga tcaggcccgc tccattatgg agcttgccgc ttctccggtg tgcttcgatg 720
atccaacagg cgcctttagc ccagccagac caccagcttt cagattccca ccaggcctcc 780
caaggccagc tccggtgccg accgccccgt ccttcgtggg caccttcccg atctccccag 840
ccggcaagag atgctactcc tactgctcct tccgctcctc cgtgtcactc ctcacaacaa 900
ccgagggccc aacatctagg aaggcctcac tccaacgcta cctcgagaag cgcaaggaca 960
ggtacggcca tctcccaacc gagtccattc tcctcgtgtc cggatccggc ggcggctctg 1020
ctaagggcga gctgaggggc cacccgttcg agggcaagcc aattccaaat ccactcctcg 1080
gcctcgactc taccaggacc ggccaccatc accatcacca cggatcctaa tgaagaccca 1140
gctttcttgt acaaagtggt 1160
<210> 124
<211> 947
<212> ДНК
<213> Picea abies
<400> 124
acaagtttgt acaaaaaagc aggctcaaaa aaaaccatga gaggcggcga gagagcccca 60
ggctccaggc cggtaatgtt cagctctgct atagtgtgtg ccaccctgct tgtttaataa 120
tgcgttctct tcgtttttat gatatcttat tcttccagtc cctcgacaag ccgctcgagg 180
agcttaccga ggaggacatc ttccagctca cccgcgagga ttgcaggcgc tacctcaagg 240
agaaggggat gagaaggccg tcctggaaca agtcccaggc catccaacaa gtgctcagcc 300
tcaagagcct cttcgagtcc aagccgaacc agcagtccaa gaagccgtcc aagcacaagc 360
cagccaccct ccaattcgag acagccaggg attctacctt cgcccagtcc tccgtgtccc 420
aagagcaatc tctcggcttc tcctggtcca aggaggtgct cgataagggc acagccgaga 480
gacaaaggct ctgctccgat tcccaagagg cccacgagat tccaaggctc ggctctaagc 540
caccgcagtc caacaccgag ggcaagagat gcgctcatga tggccatggg agaaagtccg 600
cccaaccact cgttaggctc ccggccaact tcaagaacga ctgctccaac aggcagtcct 660
cccacacatc tgagtcccag ccagataccc tcctccgctc cgattctttc cagcagccaa 720
cagcccagct caccatcttc tacgccggca tggtgaacgt gtacgacgac gtgccactcg 780
acaaggccgg atccggcggc ggctctgcta agggcgagct gaggggccac ccgttcgagg 840
gcaagccaat tccaaatcca ctcctcggcc tcgactctac caggaccggc caccatcacc 900
atcaccacgg atcctaatga agacccagct ttcttgtaca aagtggt 947
<210> 125
<211> 897
<212> ДНК
<213> Selaginella moellendorffii
<400> 125
acaagtttgt acaaaaaagc aggctcaaaa aaaaccatgt cctcgatggt ggacttcctc 60
ggcatcgagg agaaggtgtc cacctccgtg tccgccgaga ggctcaagaa gctcgaggag 120
ctgaccgacg aggacgtgat gcaactcaca cgcgaggatt gccgccgcta ccttaaggag 180
aaggggatga gaaggccgtc ctggaacaag gcccaagccg tgcaacaact cctcagcctc 240
aagtccctct gcgatccatc tccggcctcc agcggagctg ccaagaggtc cccgtccccg 300
ccactcgacg aggccccagc caagaagccg atggccatga cctccatcga tctcaaggcc 360
gctgccgccg ttgatgccgc caatctcacc atgttctacg acggcgccgt gtccgtgttc 420
gatgatgtgt ctccagacaa ggcctccctc ttcccactcg cctacgccat tatgctcctc 480
gccggcaatg tgaagtcctg gccgtctatc aacgtggccg ccaacaccaa caaggtggtg 540
atctccagct acgagctgcc gcaagctaga aaggcttccc tccagcgctt ccttcagaga 600
aggcgcgaga agacagccaa ggaggccgct tctaagggca actccaacaa gtccccatgc 660
cacggcgagt ctagcggcaa gcacgcctct gatgctaccg atccagctac ctctccactc 720
ctcacagagg tgtcatccgg atccggcggc ggctctgcta agggcgagct gaggggccac 780
ccgttcgagg gcaagccaat tccaaatcca ctcctcggcc tcgactctac caggaccggc 840
caccatcacc atcaccacgg atcctaatga agacccagct ttcttgtaca aagtggt 897
<210> 126
<211> 1134
<212> ДНК
<213> Arabidopsis thaliana
<400> 126
acaagtttgt acaaaaaagc aggctcaaaa aaaaccatgg atgtgggagt gtctccagct 60
aagtctatcc ttgctaagcc tctcaagctc ctcaccgaag aggatatctc tcagctcact 120
agagaggatt gcagaaagtt cctcaaggat aagggaatga gaaggccatc ttggaacaag 180
tctcaggcta tccagcaggt tctcagtctc aaggctcttt acgagcctgg tgatgattct 240
ggtgctggaa tcttcagaaa gatcctcgtg tctcagcctg tgaaccctcc tagagttact 300
actactctca tcgagccttc taacgagctt gaggcttgcg gaagagtttc ttaccctgag 360
gataacggtg cttgccacag gatggattct ccaagatctg ctgagttctc tggtggatct 420
ggacacttcg tgtctgagaa ggatggacac aagactacta tctctccaag aagtcctgct 480
gagacttctg agcttgtggg acagatgacc atcttctact ctggaaaggt gaacgtgtac 540
gatggaatcc ctcctgagaa ggctagatct atcatgcact tcgctgctaa ccctatcgat 600
ctccctgaga acggaatctt cgcttcttct aggatgatct ctaagctcat ctctaaagaa 660
aagatgatgg aactccctca gaagggactc gagaaggcta actcttctag ggattctgga 720
atggaaggac aggctaacag aaaggtgtca ctccagaggt acagagagaa gaggaaggat 780
aggaagttct ctaaggctaa gaaatgccct ggtgtggctt cttcatctct cgagatgttc 840
cttaactgcc agcctaggat gaaggctgct tactctcaga acctcggatg tactggatct 900
ccactccatt ctcagtctcc agagtctcag accaagtctc ctaacctctc tgtggatctc 960
aactctgagg gaatcggatc cggtggtgga tctgctaagg gtgagcttag aggtcatcct 1020
ttcgagggta agcctatccc taaccctctt ctcggtctcg attctactag aactggtcat 1080
catcatcacc atcacggatc ctaatgaaga cccagctttc ttgtacaaag tggt 1134
<210> 127
<211> 1209
<212> ДНК
<213> Trifolium repens
<400> 127
acaagtttgt acaaaaaagc aggctcaaaa aaaaccatga acggtggatc taccgtgtct 60
ttcagatcta tcctcgataa gcctctcaac cagctcaccg aggatgatat ctctcagctc 120
actagagagg attgcagaag attcctcaag gataagggaa tgagaaggcc atcttggaac 180
aagtctcagg ctatccagca ggttatctct ctcaaggctc tcctcgagcc taccgatgat 240
gatcttcctg ctcctgtggg agtgtcatct gctatccatc atcatcacca tcaccaccct 300
caacctccac agagaaatct taacgaggct cctgtgaagg gatctgatct cgatgatact 360
ggattccaca ccgctgagga tctcaacaag tctacttcta ccgctgttga gatccctacc 420
gagactaacg atgctaacgt ggtgaagtca tctggtggat gtgtggcttc tggatctttc 480
ggacagatga ccatcttcta ctgcggaaag gtgaacgtgt acgatggtgt gtctcctgat 540
aaggctagat ctatcatgca gctcgctgct tgcccttcta gtttccctca ggataacctc 600
ctcaacaaga acgctgctgt ttgggcttct ccttgcaaca tccctattga taaggatgtt 660
ctcttcccta acgataccgc tatcctccaa gtggctcaga ccgataagat ggttgagtac 720
cctctccagt acagagagaa gggatctatc gctagggatg ctgatgttga gggacaggct 780
tctagaaacg cttcactcca gaggtacagg gaaaagagga aggatagggg aaggtctaag 840
ggaaacaagc tcaccggaat cacctcttct aacttcgaga tgtacctcaa cctccctgtg 900
aagctccatg cttctaacgg aaactcttct aggtctagta ccgattcacc tcctcagcct 960
agactccctc ttgtttctgg tggatctgct gagaaccagc ctaaggttac actccctatc 1020
gatctcaacg ataaggatgt gcaagagtgc ggatccggtg gtggatctgc taagggtgag 1080
cttagaggtc atcctttcga gggtaagcct atccctaacc ctcttctcgg tctcgattct 1140
actagaactg gtcatcatca tcaccatcac ggatcctaat gaagacccag ctttcttgta 1200
caaagtggt 1209
<210> 128
<211> 1128
<212> ДНК
<213> Amborella trichopoda
<400> 128
acaagtttgt acaaaaaagc aggctcaaaa aaaaccatga ccgctggtga tggatctatc 60
agatctatcc tcgataagcc tctcgaggaa ctcaccgaag aggatatctc tcagctcacc 120
agagaggatt gcagaagata cctcaaagaa aagggtatga gaaggccatc ttggaacaag 180
taccaggcta tccagcaggt tctcagtctt aagggacttc tcgagggaaa gccttgtgat 240
gataactctg atgtgttctc tcacaggtca cctatcaccg tgatccctaa cgtgggatct 300
atgagagaga aagagaaggc tgttaacatt gctgatcctg agatctctgg ttctcaccag 360
cctaacttta gaagagagat ccacgagact accagagaga gagctttgcc tgcttctgat 420
tggcctccat ctcaagagcc tgtgtctcag atgaccatct tctacgctgg tgctgtgaac 480
gtgtacaacg atatccctga ggataaggtg caggctatca tctacctcgc tggaaagtct 540
gattctctcc agcagaccaa cgtgatcaga actggacctg atcagtgtat cgcttctgct 600
gcttctcctt ctctcaacga tctccactct agaagaatcc accctacctc taacatcacc 660
acctctcagt ctctcagagt ggctacttct cttcctgtgg gacctcattc tgaggtgcca 720
aagactagaa agacctctgt gcagagattc ctcgagaaga ggaaggatag aggtaggctc 780
aagggaactc ttgcttctgg tggatcttct aagaggggat cttcttgcct cgagctttac 840
gctacctcta ggcttaagtc tgagggtgtg gctactacta ccacccagtc taacatgaac 900
aacgtggtgg tgtctccatc taaccctagg atgcctctta accctggatc ttgctcttgg 960
gttgagaacg gatccggtgg tggatctgct aagggtgagc ttagaggtca tcctttcgag 1020
ggtaagccta tccctaaccc tcttctcggt ctcgattcta ctagaactgg tcatcatcat 1080
caccatcacg gatcctaatg aagacccagc tttcttgtac aaagtggt 1128
<210> 129
<211> 1074
<212> ДНК
<213> Musa acuminata
<400> 129
acaagtttgt acaaaaaagc aggctcaaaa aaaaccatga accctggtga gactacccct 60
ccatctccac ttgataagcc tctcgctgag cttaccgaag aggatatcgc tcagctcact 120
agagaggatt gcagaagatt cctcaaggct aagggaatga gaaggccatc ttggaacaag 180
tctcaggcta tccagcaggt tatctctctc aaggctctcc ttgaaggtag gcctggatgt 240
gatgattgtc ctgctggtgg tggaatcctc cagaagctcc ttacttctag tccttctgag 300
cctctcagtc ctcctcaaga ttctccacct cctgctccta aagagggagg atctggatct 360
cagcctcttg ctaaagagcc ttctccatac agaagaagag atcctatccc tcctccttac 420
tctgctggaa accctacttg tcagactcct atcgctggtg ctgatcttcc tcatcctcct 480
gagaagagat gcccatctcc tagacttact gctgaggttc cagtgggaca gatgaccatc 540
ttctacgatg gaatggtgaa cgtgtacgat ggtgtgtctg ctgatcaggc tagatctatt 600
atggaactcg ctgcttctcc tgtgtgcttc gatgatccta ctggtgcttt cagtcctgct 660
agacctcctg cttttagatt ccctccagga cttcctagac ctgctcctgt tcctactgct 720
ccttctttcg ttggaacctt ccctatctct cctgctggaa agaggtgcta ctcttactgc 780
tctttcaggt ctagtgtgtc tctcttgact accactgagg gacctacctc tagaaaggct 840
tcactccaga gatacctcga gaagaggaag gatagatacg gacacctccc taccgagtct 900
atccttctcg tttctggatc cggtggtgga tctgctaagg gtgagcttag aggtcatcct 960
ttcgagggta agcctatccc taaccctctt ctcggtctcg attctactag aactggtcat 1020
catcatcacc atcacggatc ctaatgaaga cccagctttc ttgtacaaag tggt 1074
<210> 130
<211> 1233
<212> ДНК
<213> Picea abies
<400> 130
acaagtttgt acaaaaaagc aggctcaaaa aaaaccatga ggggtggtgg tggtgctgat 60
agacttcctg ctagagctaa ccttgagaag cctctcgagg atctctctca cgaggatatc 120
atgcagctca ccagagagga ttgcagaaga tacctcatcg agaagggaat gagaaggcca 180
tcttggaaca agtctcaggc tatccagcag gttctcagtc tcaagaagct tttcgagtct 240
ggacctaacg atgagaagag gtctgctgct accaacaggc ctaatcctga tgagaacctc 300
aaagaggctg cttctgtttc tctcttgtac ggatctcagc ctgagtctcc ttctgtggtg 360
ttcgcttcta aggattctga taccttcaac ctcgagtggc tcgctaagac tgagcttcct 420
gttcttgctt ctcagcctag gcatatcgct cagcagaacg tgttcctctc ttcactttct 480
gctcagcagt ctggtgctca gctcaccatt ttctactctg gaaacgtgaa cgtgtacgat 540
gatgtgcctg ctgagaaggc tcaagagatc atgcttcttg ctggatctgg aaactaccct 600
ccttcatcta cttgccagtc taccagaaac acccagcaga acgctgttag agctgcttac 660
ccttctaacc ctaccaacac ccctttcatc catggtgttg gaccacctct tgctaccgtg 720
gcttcttctt ctgtgatgtc atctcctatc cacaaagagt ctccaatcac cagaaaggct 780
tcactccaga gattcctcga gaagaggaag gataggtcta ggggtaagct tggtgctcct 840
actatctcta agaaacctct cctcatggga atgttcatgc acccttctat cgtgcacaga 900
cagtactgga ctgataccgc taagaggaag tctggaaagc ctgatatccc tgcttctatc 960
tctcctacca gacctcctca cactcctaga aggacatctt ctgatgagca gctctctgct 1020
agacacgcta ggggagatat ttctgctcaa ggtggaagtc tccacaactc taacggatcc 1080
ggtggtggat ctgctaaggg tgagcttaga ggtcatcctt tcgagggtaa gcctatccct 1140
aaccctcttc tcggtctcga ttctactaga actggtcatc atcatcacca tcacggatcc 1200
taatgaagac ccagctttct tgtacaaagt ggt 1233
<210> 131
<211> 835
<212> ДНК
<213> Selaginella moellendorffii
<400> 131
acaagtttgt acaaaaaagc aggctcaaaa aaaaccatgt caagtatggt ggatttcctc 60
ggaatcgaag agaaggtttc aacctctgtg tctgctgaga ggcttaagaa gctcgaggaa 120
ctcactgatg aggatgtgat gcagctcacc agagaggatt gcagaagata cctcaaagaa 180
aagggtatga gaaggccatc ttggaacaag gctcaagctg ttcagcagct cctcagtctt 240
aagtctctct gcgatccttc accagcttca tctggtgctg ctaagagatc tccttcacct 300
cctttggatg aggctcctgc taagaaacct atggctatga cctctatcga tctcaaggct 360
gctgctgctg ttgatgctgc taacctcacc atgttctacg atggtgctgt gtctgtgttc 420
gatgatgtgt ctcctgataa ggcttctctc ttcccactcg cttacgctat catgcttctc 480
gctggaaacg tgaagtcttg gccttctatc aacgtggcag ctaacaccaa caaggtggtg 540
atctcttctt acgaactccc tcaggctagg aaggcttcac ttcagagatt cctccagaga 600
agaagggaaa agaccgctaa agaggctgct tctaagggaa actctaacaa gtctccttgc 660
cacggtgagt ctagtggaaa gcacgcttct gatgctactg atcctgctac ttctccactc 720
ctcactgagg tgtcatctgg atccggtggt ggatctgcta agggtgagct tagaggtcat 780
cctttcgagg gtaagcctat ccctaaccct cttctcggtc tcgattctac tagaa 835
<210> 132
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 132
gatatgcgag acgcctatga 20
<210> 133
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 133
gagttccctt cagtgaacgt 20
<210> 134
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 134
gctggggcgt cggtttccac tatccg 26
<210> 135
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 135
cgcataacag cggtcattga ctggagc 27
<210> 136
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 136
ctgttgccgg tcttgcgatg 20
<210> 137
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 137
gtcacataga tgacaccgcg 20
<210> 138
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 138
ctcgtgcttt cagcttcgat gtag 24
<210> 139
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 139
gctggggcgt cggtttccac tatcgg 26
<210> 140
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 140
cacaggatgg attctccaag g 21
<210> 141
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Консенсусная последовательность
<400> 141
taaggtccac ggagaggttc 20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ УСОВЕРШЕНСТВОВАНИЯ ОДНОДОЛЬНЫХ РАСТЕНИЙ | 2015 |
|
RU2727428C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ПРОМЫШЛЕННЫХ ПРОДУКТОВ ИЗ РАСТИТЕЛЬНЫХ ЛИПИДОВ | 2015 |
|
RU2743384C2 |
| Конструкции для направленного редактирования генов и способы с их применением | 2020 |
|
RU2832109C2 |
| РАСТЕНИЯ С МОДИФИЦИРОВАННЫМИ ПРИЗНАКАМИ | 2017 |
|
RU2809117C2 |
| Ферменты биосинтеза люциферина и их применение | 2018 |
|
RU2730038C2 |
| Гиспидин-синтазы и их применение | 2020 |
|
RU2809367C2 |
| Кофеилпируват-гидролазы и их применение | 2020 |
|
RU2812474C2 |
| Поликетидсинтазы III типа и их применение | 2020 |
|
RU2822532C2 |
| ПОЛИПЕПТИДЫ, МОДУЛИРУЮЩИЕ SIGLEC-ЗАВИСИМЫЕ ИММУННЫЕ ОТВЕТЫ | 2017 |
|
RU2776807C2 |
| HPV-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2018 |
|
RU2804664C2 |





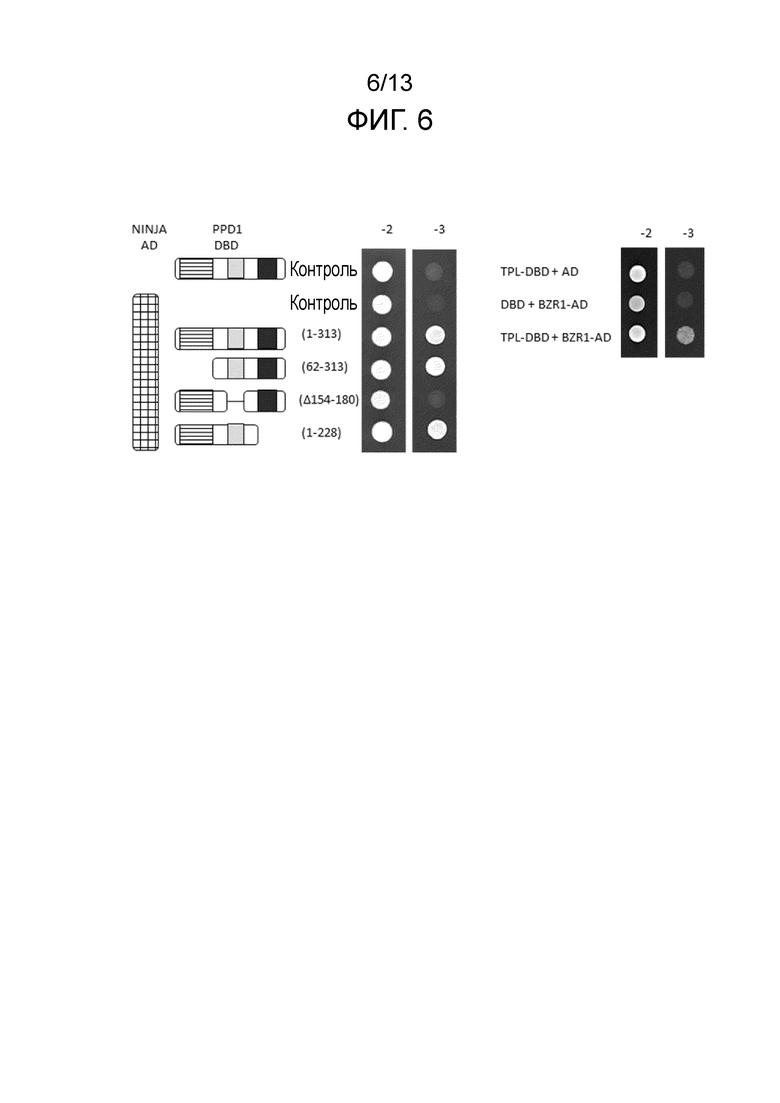

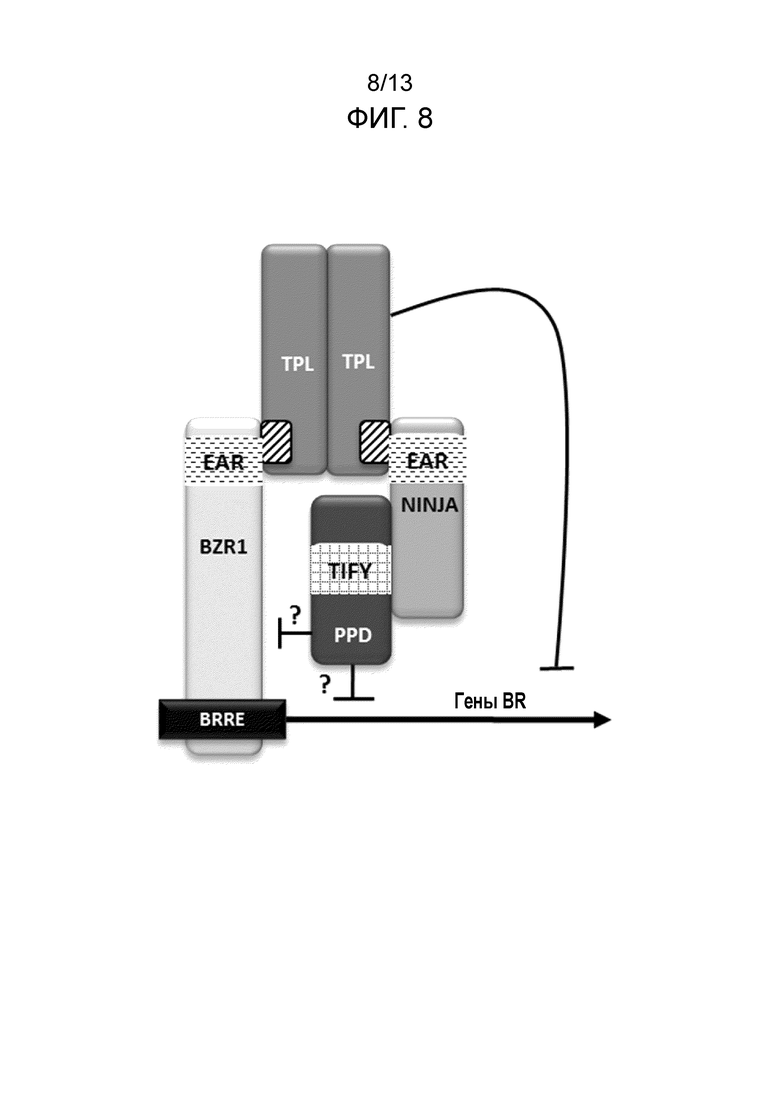
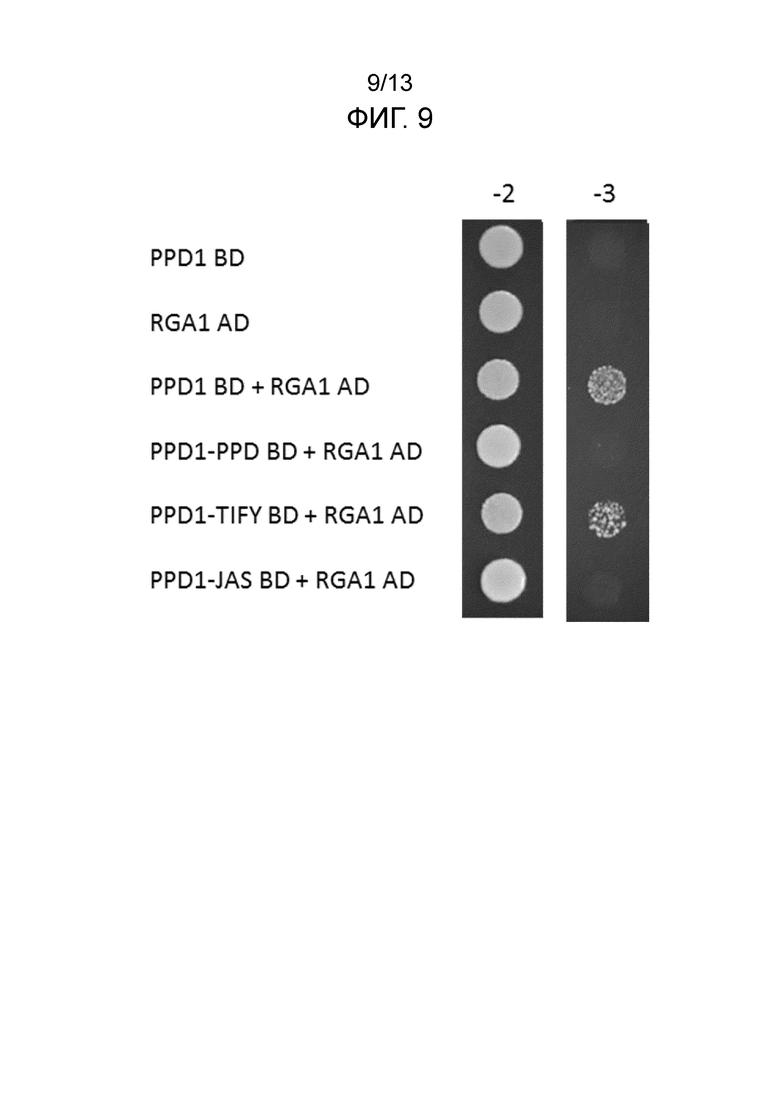




Изобретение относится к области биохимии, в частности к способу увеличения биомассы корней в растении, включающему стадию повышения экспрессии по меньшей мере одного белка PEAPOD в растении. Также раскрыт способ получения растения с увеличенной биомассой корней, который включает стадию трансформации растения или трансформации клетки растения, которая регенерирует в растение, полинуклеотидом, кодирующим белок PEAPOD. Изобретение также относится к растению, которое имеет увеличенную биомассу корней в результате повышения экспрессии белка PEAPOD вследствие трансформации растения полинуклеотидом, кодирующим белок PEAPOD. Изобретение позволяет эффективно увеличивать биомассу корней в растении. 3 н. и 20 з.п. ф-лы, 13 ил., 5 табл., 5 пр.
1. Способ увеличения биомассы корней в растении, причем способ включает стадию повышения экспрессии по меньшей мере одного белка PEAPOD в растении, в котором повышенная экспрессия по меньшей мере одного белка PEAPOD является следствием трансформации растения полинуклеотидом, кодирующим белок PEAPOD.
2. Способ по п.1, в котором растение является трансгенным по меньшей мере по одному полинуклеотиду, кодирующему и экспрессирующему белок PEAPOD.
3. Способ получения растения с увеличенной биомассой корней, причем способ включает стадию трансформации растения или трансформации клетки растения, которая регенерирует в растение, полинуклеотидом, кодирующим белок PEAPOD, тем самым повышая экспрессию по меньшей мере одного белка PEAPOD в растении, где биомасса корней увеличивается по сравнению с таковой в растении того же типа и возраста, которое не было трансформировано полинуклеотидом.
4. Способ по п.3, в котором растение трансформируют по меньшей мере одним полинуклеотидом, кодирующим белок PEAPOD.
5. Способ по п.3 или 4, который включает дополнительную стадию исследования или оценки растения в отношении увеличения биомассы корней.
6. Способ по любому из пп.1-5, в котором белок PEAPOD содержит последовательность согласно по меньшей мере одной из SEQ ID NO: 28, 29, 31, 32, 34 и 35.
7. Способ по любому из пп.1-6, в котором белок PEAPOD обладает полипептидной последовательностью с по меньшей мере 70% идентичностью с любой из SEQ ID NO: 1-26.
8. Способ по любому из пп.1-7, в котором полинуклеотид обладает последовательностью с по меньшей мере 70% идентичностью с кодирующей последовательностью согласно любой из SEQ ID NO: 83-107.
9. Способ по любому из пп.1-7, в котором полинуклеотид обладает последовательностью с по меньшей мере 70% идентичностью с последовательностью согласно любой из SEQ ID NO: 83-107.
10. Способ по любому из пп.8, 9, в котором полинуклеотид вводят в растение в качестве части экспрессирующей конструкции, которая содержит промотор, функционально связанный с полинуклеотидом.
11. Способ по п.10, в котором промотор способен запускать или запускает конститутивную экспрессию функционально связанного полинуклеотида во всех тканях растения.
12. Способ по п.10, в котором промотор представляет собой предпочтительный для тканей промотор.
13. Способ по п.10, в котором промотор способен запускать или запускает экспрессию функционально связанного полинуклеотида в подземных тканях растения.
14. Способ по п.10, в котором промотор представляет собой промотор, предпочтительный для подземных тканей растения.
15. Способ по п.10, в котором промотор представляет собой репрессируемый светом промотор.
16. Способ по п.10, в котором промотор способен запускать или запускает экспрессию функционально связанного полинуклеотида в корнях растения.
17. Способ по п.10, в котором промотор представляет собой предпочтительный для корней промотор.
18. Способ по п.10, в котором промотор представляет собой специфический для корней промотор.
19. Растение, которое имеет увеличенную биомассу корней в результате повышения экспрессии белка PEAPOD вследствие трансформации растения полинуклеотидом, кодирующим белок PEAPOD, где биомасса корней увеличивается по сравнению с таковой в растении того же типа и возраста, которое не было трансформировано полинуклеотидом.
20. Растение по п.19, которое является трансгенным в отношении полинуклеотида, экспрессирующего белок PEAPOD.
21. Растение по п.19 или 20, в котором полинуклеотид представляет собой полинуклеотид, функционально связанный с тканепредпочтительным промотором.
22. Растение по п.21, в котором промотор представляет собой предпочтительный для корней промотор.
23. Растение по п.21, в котором промотор представляет собой специфический для корней промотор.
| WO2007105967 A1, 20.09.2007 | |||
| US2007039067 A1, 15.02.2007 | |||
| ROJAS CA et al., Genetically modified crops for biomass increase | |||
| Genes and strategies, GM Crops, 2010, Vol.1, N.3, pp.137-142 | |||
| RU2012156243 A, 27.06.2014. |

