Изобретение относится к медицине, а именно к иммунологии, и может быть использовано для получения высокоэффективного гипосенсибилизирующего аллергоида для проведения аллерген-специфической иммунотерапии больных, страдающих аллергией к яду пчел.
Инсектная аллергия - раздел аллергологии, объединяющий реакции гиперчувствительности, возникающие при ужалениях, укусах насекомыми, при соприкосновении с ними, вдыхании частиц тел насекомых и/или продуктов их жизнедеятельности. Аллергические реакции на ужаление пчелой, осой, шершнем -насекомыми, относящимися к отряду "Перепончатокрылые" (Hymenoptera) характеризуются тяжестью симптомов, бурным течением и возможностью летального исхода [1].
По способу сенсибилизации различают следующие пути попадания инсектного аллергена в организм:
- с ядом при ужалении, что характерно для отряда перепончатокрылых насекомых (пчел, ос и других жалящих насекомых);
- со слюной при укусах насекомыми отряда двукрылых (например, комаров);
- ингаляционным и контактным способом с чешуйками и дебрисом насекомых нескольких отрядов (например, жуков, тараканов) [1].
Распространенность инсектной аллергии с одной стороны определяется географическим нахождением мест обитания насекомых, с другой - интенсивностью иммунного ответа у лиц, проживающих в эндемичных регионах. Характер аллергической реакции зависит от вида насекомого, источника аллергенного материала (секрет, слюна, частица тела и пр.), путей введения (попадания в организм), от продолжительности и интенсивности воздействия, а также от индивидуальной чувствительности и реактивности человека к аллергенам насекомого.
Распространенность инсектной аллергии к перепончатокрылым в России составляет 0,4-8% [2]. В США выраженные местные реакции на ужаления жалящих насекомых встречаются в 2,4-26,4% общей популяции населения. Системные реакции выявлены у 0,5-3,3% обследованных жителей США. Среди взрослого населения Европы сенсибилизацию к жалящим насекомым выявляют в 9,2-28,7% случаев, а статистическое наблюдение системных реакций колеблется от 0,3 до 7,5% [3, 4]. В России системные реакции отмечены у 6,4% обследованных пациентов на гиперчувствительность к ужалению перепончатокрылыми насекомыми [5]. Смертность от анафилактических реакций на яд жалящих насекомых составляет 0,03-0,48% случая на миллион населения ежегодно [6]. К группе риска относятся пчеловоды, так как у 15-43% возникает аллергия к яду пчел. Атопические заболевания зарегистрированы у 31-35% больных аллергией к ужалению перепончатокрылым насекомым [1].
АСИТ является методом патогенетической терапии, позволяющим снизить повышенную чувствительность организма к аллергенам жалящих насекомых. АСИТ проводится в осенне-зимний период, вне сезона лета насекомых. Данный метод терапии осуществляется высококвалифицированными аллергологами в специализированных стационарах при наличии отделения интенсивной терапии, поскольку существует угроза развития анафилаксии [1, 7, 8]. В зарубежной литературе принято данный метод, применительно к лечению пациентов с гиперчувствительностью к яду жалящих несекомых, называть специфической иммунотерапией ядом (venom-specific immunotherapy - VIT) [6, 9].
Современными терапевтическими продуктами для специфической иммунотерапии аллергии к яду перепончатокрылых является экстракты ядов, которые очищаются по стандартной процедуре. Производственный процесс этих продуктов регулируется территориально-зависимыми руководствами. Вследствие конфиденциальности и специфичности производственного процесса, производимые экстракты ядов могут отличатся по компонентам состава и аллергенной активности. Продукты для специфической иммунотерапии ядом доступны в качестве водных (или лиофилизированных) экстрактов или, как депонированных препаратов адсорбированных на гидроксиде алюминия [10, 11, 12]. В Европе и Америке для проведения VIT используют следующие препараты: 1) Venomil (Allergy Therapeutics, Worthing, UK); 2) Reless (также известный как Pharmalgen; ALK-Abello, Hamburg, Germany); 3) ALK lyophylisiert SQ (также известный как Aquagen SQ; ALK-Abello), 4) Venomenhal (HAL Allergy, Leiden, Netherlands).
Несмотря на исследования показавшие уменьшение симптомных показателей после специфической иммунотерапии ядом у пациентов аллергией на ужаление пчелами, существует необходимость усовершенствовать препараты для клинического применения [13].
Аллерген-специфическая иммунотерапия пчелиным ядом имеет значительный риск системных реакций до 14,2% [14]. К факторам риска для тяжелых побочных эффектов в течение специфической иммунотерапии пчелиным ядом является повышенный уровень базальной сывороточной триптазы или мастоцитоза [15]. Возможные побочные эффекты, длительная продолжительность терапии (3-5 лет) и высокая стоимость приводит к низкой приверженности среди пациентов [87, 88].
Следовательно, существует необходимость поиска новых адъювантных систем, чтобы улучшить эффективность и увеличить безопасность аллерген-специфической иммунотерапии. Распространенным адъювантом для аллерген-специфической иммунотерапии является гидроксид алюминия (75%) [16, 17]. В продуктах для VIT гидроксид алюминия в избыточном количестве смешивается с экстрактом яда и смесь вводят подкожно. К сожалению, для профилактических вакцин, производимых для аллерген-специфической терапии, нет необходимости устанавливать количество гидроксида алюминия в описании характеристик продукта или самой упаковке [17]. В Европе установлено, что количество гидроксида алюминия 1,25 мг на инъекцию является максимально допустимым [18]. Основным аргументом против использования адъювантов на алюминиевой основе, является их способность стимулировать иммунную систему в направлении активации Th2 (Т хелперов второго типа) и как следствие индуцирование продукции IgE [19, 20].
В качестве альтернативных адъювантов, которые одобрены для иммунотерапии, могут использоваться L-тирозин, для депонирующего эффекта, и агонист Toll-подобного рецептора монофосфорил липид А. В настоящие время исследуются следующие соединения, в качестве будущих адъювантов: липополисахариды, CpG-олигодеоксинуклеотиды и имиквимод/резимиквимод (R 837, R 848) [21]. В качестве примера, подобной разработки следует указать патент WO 2005049647 А2 "Композиции, содержащие мелиттин-производные пептиды и методы для потенцирования иммунных ответов против мишеней антигенов" (Rinaldo Zurbriggen, 2005). Настоящим изобретением предусматриваются новые адъювантные системы, способные уменьшить риск токсических побочных эффектов связанных с использованием известных адъювантов, а также обеспечить сильную стимуляцию иммунных ответов против мишеней антигенов. Эти новые адъювантные системы, основанные на использовании мелиттина из яда медоносных пчел или специфических катионных пептидов производных от мелиттина, способны вызвать сильные иммунные ответы против мишеней антигенов. В будущих иммуногенетических разработках эти адъювантные системы могут выборочно комбинированы с синтетическими олигодеоксинуклеотидами (ODNs) подобно любым иммуностимуляторным последовательностям таким как CpG мотивы [22].
Предполагается использование новых систем доставки: липосомы, вирус-подобные частицы или биорасщепленные полимерные носители [21]. К подобным разработкам относится работа по двойной модификации пчелиного яда посредством алкилирования и сукцинилирования, с целью ингибирования мелиттина и фосфолипазы А2, с последующим инкапсулированием модифицированного яда в липосомы [23].
В недавних исследованиях были продемонстрированы новые системы доставки такие как микросферы на основе poly-lactide-co-glycolide acid и микропузырьки, которые могут быть эффективно нагружены ядом или аллергенами яда [24, 25].
Указанный, новой подход к VIT заключался в инкапсулировании пчелиного яда (bee venom - BV) внутри микросфер (MS) состоящего из poly-lactide-co-glycolide acid (PLGA), представляющий собой контролируемую систему доставки. Микросферы из PLGA, содержащие пчелиный яд (BV-MS-PLGA) были приготовлены с помощью метода испарения двойной эмульсии растворителя (вода/масло/вода) и протестированы на биологическую эффективность, посредством исследования ex vivo поглощения макрофагами пустых микросфер и нагруженных пчелиным ядом, а также исследований in vivo по стимулированию у мышей продукции IgG1 и IgG2a антител после проведения VIT. BV-MS-PLGA предотвращал воспалительную реакцию, препятствуя прямому контакту между пчелиным ядом и тканью организма. Повышения уровня фагоцитоза BV-MS-PLGA макрофагами способствовало доставке антигена в направлении внутриклеточной среды. Более того, при использовании BV-MS-PLGA, антительная продукция была сопоставима с традиционной VIT (свободный пчелиный разведенный в фосфатносолевом буфере), с преимуществом защиты организма против вредных воздействий токсинов пчелиного яда и уменьшением количества инъекций необходимого для достижения такого же защитного эффекта [24].
Микропузырьки нагруженные аллергеном пчелиного яда были использованы для предотвращения сенсибилизации ядом медоносных пчел у мышей [25]. В этом исследовании авторами была оценена, на мышиной модели аллергии к пчелиному яду, индуцированная защита, посредством профилактического использования, главного аллергена фосфолипазы А2 (ФЛА2) ассоциированного с микропузырьками (МП). Методы: Антитела и Т-клеточные ответы, определяемые с помощью ELISA и пролиферативных методов, основанных на использовании carboxyfluorescein diacetate succinimidyl ester (CFSE), сперва были обследованы после профилактической иммунизации CBA/J мышей с ФЛА2-МП, повторно после сенсибилизации с нативным ФЛА2. В конечном итоге, мышам было проведено испытание (протестированы) с летальной дозой ФЛА2, чтобы оценить защиту против анафилаксии. Результаты: Профилактическая иммунизация с ФЛА2-МП индуцировала ФЛА2-специфические IgG и IgA антитела, запускало продукцию IFN-γ и IL-10 и дифференциацию ФЛА2-специфических Foxp3(+)Treg.
Иммунизированные/сенсибилизированные мыши показали следующее: 1) титров потенциально блокирующих IgG1, IgG2a и IgG3 антител, 2) снижет аллерген-специфическую Т клеточную пролиферацию и Тh2-типа цитокиновую продукцию и 3) повышает экспрессию специфических Foxp3(+) в Treg клетках и увеличивает продукцию TGF-β, при сравнении наивных и сенсибилизированных животных. Иммуномодуляция коррелирует с уменьшением сигналов анафилаксии после аллергенной провакации. Данные полученные авторами, демонстрируют возможность ФЛА2-МП обеспечивать профилактическую защиту на срок до 4-х месяцев против последующей сенсибилизацией и вызывающей смерть животных провокации ФЛА, таким образом, показывая иммуномодулирующие свойства МП. Эти данные, в сочетании с безопасным использованием микропузырьков, в качестве контрастного средства для in situ визуализации у людей, делает их иммунотерапевтическим средством, представляющим интерес для последующего исследования [25].
Однако модифицированные препараты имели определенные недостатки, которые негативно проявлялись при АСИТ. Например, аллергены, эмульгированные в минеральном масле, были эффективны, но их действие приводило к образованию узелков, сохранявшихся несколько лет. Сорбированные формы аллергенов - комплекс аллергена с сорбентом, созданный на основе ковалентных химических связей, обладал стабильностью, был эффективен, позволял сократить курс инъекций с 36 до 8, но подкожное введение этих препаратов вызывало формирование на коже окрашенных пятен.
Для уменьшения побочных эффектов после инъекций аллергенов, связанных с неблагоприятными IgE-опосредованными реакциями, и увеличения безопасности иммунотерапии, аллергены могут быть химически модифицированы. Для этого предполагается использовать рекомбинантные гипоаллергенные молекулы, димеры, тримеры, сплав протеинов или пептидов [21].
К настоящему времени есть запатентованные рекомбинантные аллергены яда медоносных пчел:
- Клонированный полипептид Api m 3 (кислая фосфатаза) для диагностического и фармацевтического использования [26];
- Клонированный полипептид Api m 5 (аллерген С) для диагностического и фармацевтического использования [27];
- Аллерген-пептидные фрагменты для АСИТ, композиция, состоящая из множества смежных частично совпадающих пептидных фрагментов, которые вместе составляют полную аминокислотную последовательность аллергена (фосфолипаза А2) яда пчелы. Фрагменты способны вызывать Т-клеточный ответ у пациентов с гиперчувствительностью к этому аллергену [28];
- Идентифицирован белок Api m 6, существующий 4 изоформах по 8 kD каждая, очищенный Api m 6 может быть использован для модуляции иммунного ответа у людей чувствительных к яду пчел [29];
- Клонирование и экспрессия икарапина, нового IgE-связывающего протеина пчелиного яда [30].
В контексте перечисленных работ особого внимания заслуживают следующая работа. Исследовательская работа коллектива авторов (Karamloo F., Schmid-Grendelmeier P., Kussebi F., Akdis M., Salagianni M., von Beust B.R., Reimers A., Zumkehr J., Soldatova L., Housley-Markovic Z.,  David M. Kemeny D., Spangfort M. D., Blaser K. and Akdis C. A., 2005) "Предупреждение аллергии рекомбинантной мульти-аллергенной вакциной с уменьшением IgE связывания и предохранением Т клеточных эпитопов". В этой работе представлен, рекомбинантный химерный протеин, который включает в себя полные аминокислотные последовательности трех главных аллергенов пчелиного яда сконструированный и используемый для предотвращения чувствительности к пчелиному яду у мышей. Фрагменты фосфолипазы А2 (Api m1), гиалуронидазы (Api m2) и мелиттина (Api m3) с совпадающими аминокислотами были собраны в несхожем порядке в химерный протеин, который предохраняет в целом Т клеточные эпитопы, тогда как В клеточные эпитопы всех трех аллергенов были упразднены. Соответственно, IgE перекрестное связывание приводящие высвобождению медиаторов из тучных клеток и базофилов у людей было полностью уменьшено. В подтверждение этих данных, Api m (1/2/3) вызывает от 100 до 1000 раз меньше реактивность по кожному тесту первого типа у аллергических пациентов. Лечение мышей с Api m (1/2/3) приводило к значительному снижению специфического IgE проявления к нативному аллергену, отображающие защитный эффект вакцины in vivo. Эти результаты демонстрируют новый прототип превентивной аллерго-вакцины, которая предохраняет в целом Т клеточный эпитопный репертуар, но обходит индукцию IgE по отношению нативному аллергену, и побочные эффекты имеющие отношение к перекрестному связыванию с рецепторами IgE FceRI тучной клетки/базофила у чувствительных индивидуумов [31].
David M. Kemeny D., Spangfort M. D., Blaser K. and Akdis C. A., 2005) "Предупреждение аллергии рекомбинантной мульти-аллергенной вакциной с уменьшением IgE связывания и предохранением Т клеточных эпитопов". В этой работе представлен, рекомбинантный химерный протеин, который включает в себя полные аминокислотные последовательности трех главных аллергенов пчелиного яда сконструированный и используемый для предотвращения чувствительности к пчелиному яду у мышей. Фрагменты фосфолипазы А2 (Api m1), гиалуронидазы (Api m2) и мелиттина (Api m3) с совпадающими аминокислотами были собраны в несхожем порядке в химерный протеин, который предохраняет в целом Т клеточные эпитопы, тогда как В клеточные эпитопы всех трех аллергенов были упразднены. Соответственно, IgE перекрестное связывание приводящие высвобождению медиаторов из тучных клеток и базофилов у людей было полностью уменьшено. В подтверждение этих данных, Api m (1/2/3) вызывает от 100 до 1000 раз меньше реактивность по кожному тесту первого типа у аллергических пациентов. Лечение мышей с Api m (1/2/3) приводило к значительному снижению специфического IgE проявления к нативному аллергену, отображающие защитный эффект вакцины in vivo. Эти результаты демонстрируют новый прототип превентивной аллерго-вакцины, которая предохраняет в целом Т клеточный эпитопный репертуар, но обходит индукцию IgE по отношению нативному аллергену, и побочные эффекты имеющие отношение к перекрестному связыванию с рецепторами IgE FceRI тучной клетки/базофила у чувствительных индивидуумов [31].
Алергоиды представляют собой модифицированные аллергены путем обработки формальдегидом (карбоксилирование), янтарной кислотой (сукцинилирование), цианатом калия (карбамилирование). Полученные таким образом соединения должны характеризоваться низкой аллергенной активностью при сохранении хороших иммунизирующих свойств, обеспечивающих продукцию специфических блокирующих антинел.
Известен патент US 20100297188 А1 // Allergens and allergoids from bee venom // дата публикации: 25.11.2010, в котором раскрыты аллергены для иммунотерапии полученные диализом нативного пчелиного яда с использованием мембраны с границей пропускания 10 кДа и представляют собой пчелиный яд, по существу не содержащий мелиттин. Аллергоиды для иммунотерапии на основе пчелиного яда получены путем карбамилирования или тиокарбамилирования или обработки с образованием групп гуанидинового типа первичных аминогрупп белковых частей мелиттин-истощенного пчелиного яда. [32, 33].
В описании работы приводится методика получения пчелиного яда, не содержащего мелиттина: 50 мг лиофилизированного пчелиного яда (Latoxan, Valence, France) растворяют в 10 мл изотонического физиологического раствора (0,9% NaCl в дистиллированной воде) и оставляют перемешиваться в течение десяти минут или около того. Этот раствор затем пропускают через фильтр 0,45 мкм (Puradisc 25 AS, Whatman, Biomap, Agrate Brianza, Milan, Italy) и разделяют на две аликвоты по 25 мл каждая, затем подвергают диализу, одну аликвоту с мембраной с границей пропускания 3,5 кДа, а другую с границей пропускания 10 кДа (Spectra/Por 7, Spectrumlabs, Prodotti Gianni, Italy). Диализ проводят исчерпывающе против изотонического солевого раствора при 4°С, с помощью магнитной мешалки, контролируя его протекание с помощью SDS-PAGE, с тем, чтобы установить исчезновение полосы, связанной с мелиттином, для образца, подвергнутого диализу с мембраной, имеющей границу пропускания 10 кДа. Указанный мониторинг можно даже проводить, используя другие способы, например такие как масс-спектрометрия. Диализ против изотонического солевого раствора в течение 72 часов может быть эффективным, как продемонстрировано профилем SDS-PAGE.
При проведении диализа пчелиного яда в течение 72 ч истощается содержание мелиттина в пчелином яде. Однако, за счет эффекта разбавления при диализе резко сокращается содержание высокомолекулярных аллергенов, которые в исходном пчелином яде содержатся в минимальных количествах. В итоге остается только фосфолипаза А2.
Химическую модификацию мелиттин-истощенного пчелиного яда (в концентрации 2 мг/мл), проводят путем карбамилирования с использованием цианата калия и тетрабората натрия декагидрата, в твердой форме и в таких количествах, чтобы получить молярность 0,05 М или 0,5 М цианата и 0,1 М тетрабората. Добавленные соли растворяют перемешиванием и рН доводят до 9,3 добавлением 1 М NaOH. Всю смесь затем выдерживают при медленном перемешивании в течение 18 часов на термостатичесокй бане при 40°С в контейнере, герметично закрытом соответствующим образом, контролируя рН и поддерживая постоянным. По завершении реакции полученный в результате раствор подвергают гель-фильтрации через колонку Sephadex G-25 (Pharmacia, Milan, Italy) для удаления избытка цианата и уравновешивают с изотоническим солевым раствором, перед фильтрованием через 0,22 мкм мембрану и хранят при 4°С для последующих тестов.
Авторы проверяли с помощью ELISA специфический IgG ответ в отношении иммуногена (модифицированного мелиттин-истощенного пчелиного яда), также оценивали способность распознавать белок дикого типа (нативный мелиттин-истощенный пчелиный яд) [32, 33].
Однако, определение уровней специфических IgE не выявлено преимущества иммунизирующего действия модифицированного мелиттин-истощенного пчелиного яда перед нативным мелиттин-истощенным пчелиным ядом.
В отношении модифицированного мелиттин-истощенного пчелиного яда можно сказать, следующие процесс карбамилирования, как способ снижения аллергенных свойств следует рассматривать в качестве перспективного направления. При этом, также следует учитывать, что препарат должен обладать иммуногенными свойствами для стимулирования синтеза блокирующих антител, в конечном итоге теми свойствами на которые рассчитано проведение АСИТ.
Наиболее близким аналогом изобретения является аллергоид для аллерген-специфической иммунотерапии больных с аллергическими реакциями на ужаления пчелами, содержащий аллергенные фракции с молекулярной массой 3-12; 16-31; 38-46, а также 20 и 60 kD [34]. Способ получения данного аллергоида характеризуется тем, что яд пчел экстрагируют, при этом яд растворяют в физрастворе, далее стабилизируют сывороточным альбумином человека, осветляют, затем очищают диализом не более 3 ч и выделяют суммарную аллергенную фракцию с молекулярной массой 3-12; 16-31; 38-46, а также 20 и 60 kD, которую затем обрабатывают формальдегидом для полимеризации белков при смене температуры от 4 до 22°С, по окончании полимеризации проводят диализ, центрифугируют при 6000 об/мин в течение 30 мин, полученный аллергоид стабилизируют NaBH4 и лиофильно высушивают.
Задачей изобретения была разработка технологии получения аллергоида, действующего на аллерген-распознающий рецептор, для лечения аллергии к пчелиному яду.
Разработан способ получения аллергоида на основе пчелиного яда для проведения аллерген-специфической иммунотерапии, включающий экстрагирование яда пчелы, где пчелиный яд-сырец растворяют в дистиллированной воде до конечной концентрация 10 мг/мл, инкубируют, полученный экстракт центрифугируют при 15000 g и фильтруют супернатант через мембранный фильтр с диаметром пор 0,22 мкм, затем профильтрованный экстракт дополнительно пропускают через колонку Sephadex G25 и проводят концентрирование фракций методом ультрафильтрации, получая очищенный аллерген пчелиного яда, который для получения аллергоида обрабатывают формальдегидом и инкубируют 20-30 дней при комнатной температуре, далее полученный раствор аллергоида стабилизируют боргидридом натрия и проводят диализ аллергоида, по окончании которого аллергоид центрифугируют 20 минут при 8000 об/мин и фильтруют на мембранных фильтрах 0,22 мкм.
Колонку Sephadex G25, сорбирующую пигменты, заполняют из расчета 10 мл матрицы на каждые 100 мл экстракта. Концентрирование фракций проводят методом ультрафильтрации на фильтрах с диаметром пор 6-8 кДа. Очищенный аллерген пчелиного яда доводят до концентрации 6 мг аллергена на 1 мл буфера 0,01 М PBS, рН 7,5. Для получения аллергоида обрабатывают формальдегидом, где к прозрачному раствору аллергена по каплям при перемешивании добавляют формальдегид так, чтобы концентрация формальдегида в растворе аллергена составляла 1%. Для проведения диализа аллергоида используют диализную ленту с диаметром пор 6-8 кДа.
Техническими результатами заявленного изобретения является модификация главных аллергенов пчелиного яда, очищенных от низкомолекулярных примесей аллергенов, входящих в суммарную аллергенную фракцию, в аллергоид, путем химической модификации в присутствии формальдегида, которое приводит к взаимодействию высоко- и низкомолекулярных компонентов и способствует сохранению свойств аллергоида индуцировать синтез антител. Положительное влияние на иммуногенность оказывает увеличение молекулярной массы и создание более жестких молекулярных связей. Это обеспечивает увеличение стимулирования продукции IgG-антител при иммунизации аллергоидом в сравнении с исходными водно-солевыми экстрактами аллергенов в модельных экспериментах на мышах.
Способность аллергоида к индукции синтеза антител обеспечивается за счет жесткой, стабильной структуры молекулы аллергоида, устойчивой к влиянию денатурирующих агентов в отличие от исходного аллергена. Снижение аллергенности в полимеризованных аллергенах связано, с одной стороны с уменьшением числа антигенных детерминант в результате их реакции с альдегидными группами, а с другой стороны - недоступностью для IgE-антител остальных детерминант, скрытых внутри высокомолекулярного соединения.
Данный способ выделения основных аллергенных фракций из яда-сырцам методом гель-фильтрации позволяет получить аллерген, не содержащий мелиттин.
В тоже время разработанная технология позволяет уменьшить сроки полимеризации аллергена без потери свойств аллергоида.
Дополнительная стадия диализа аллергоида удаляет остатки NaBH4, формалина и метанола, т.е. токсических веществ, что уменьшает нежелательные побочные реакции у пациентов.
Краткое описание чертежей.
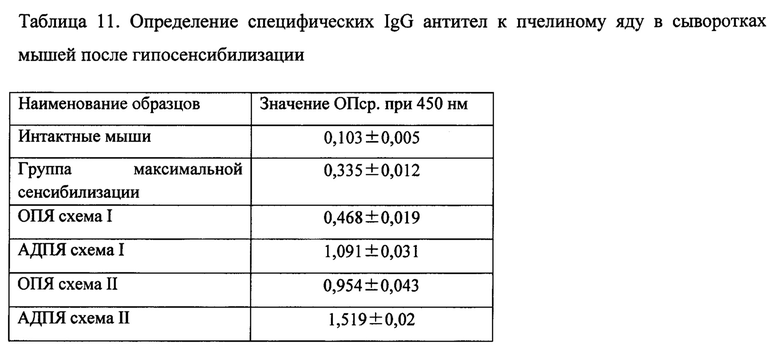
Фиг. 1. Хроматография образца на Superdex 75, 16x60, 3,6 мл, 10,0 мг/мл, PBS, 1,5 мл/мин 5,0 мл/см 2,0 OUFS.
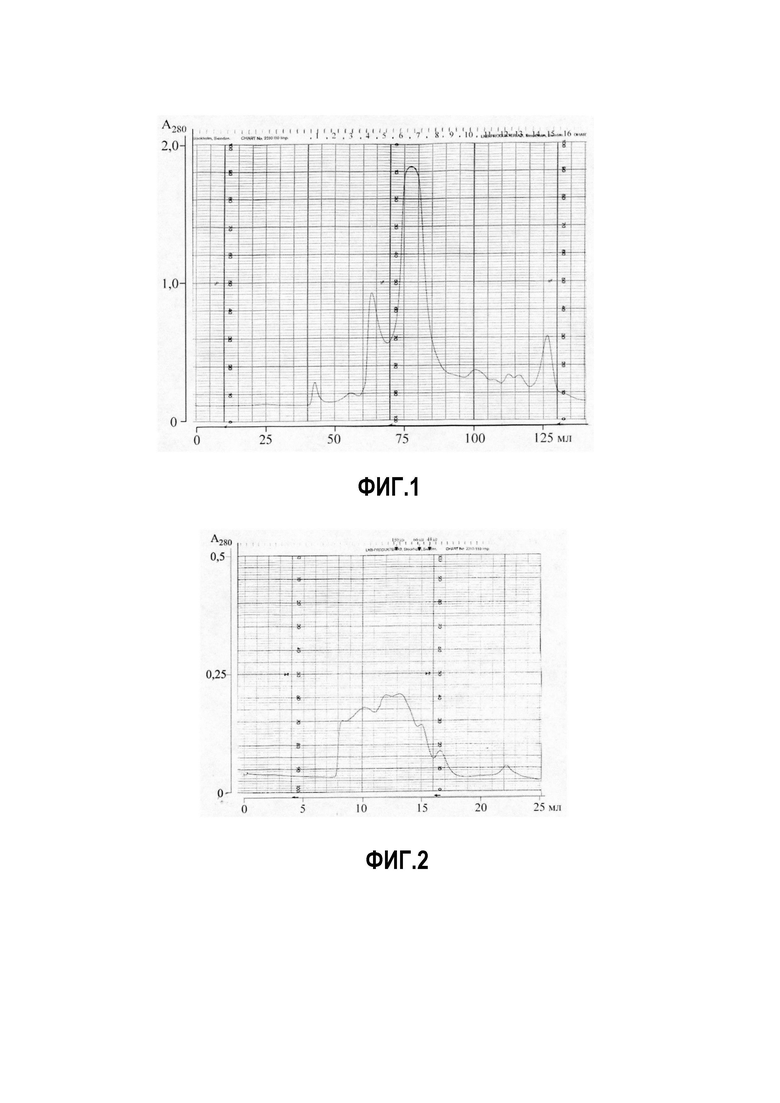
Фиг. 2. Аналитическая хроматография Superdex 200, 10x30 Аллергоид из пчелиного яда, 0,2 мл, PBS, 0,5 мл/мин 1,0 мл/см 0,5 OUFS.
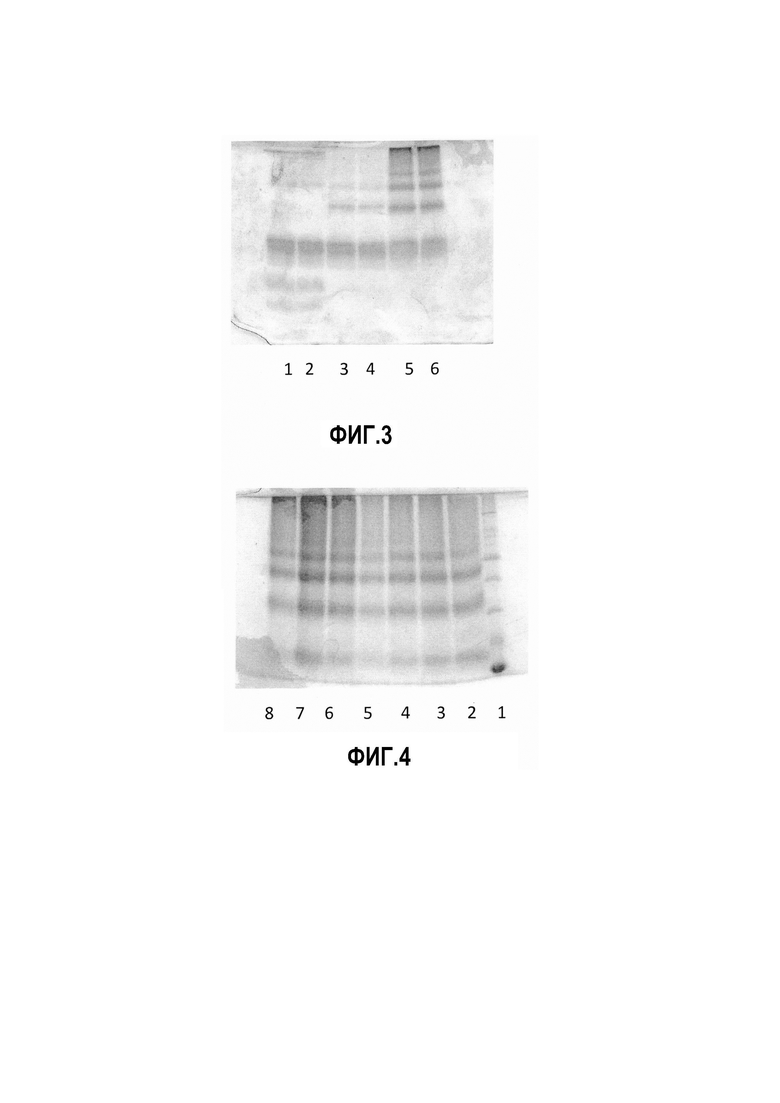
Фиг. 3. Вертикальный электрофорез в 15% ПААГ аллергоидов и их исходного материала. 1- очищенный пчелиный яд фирмы Sigma (USA), 2- ОПЯ, 3- АНПЯ из очищенного пчелиного яда фирмы Sigma (USA), 4 -АНПЯ из ОПЯ, 5- АДПЯ из очищенного пчелиного яда фирмы Sigma (USA), 6- АДПЯ из ОПЯ.
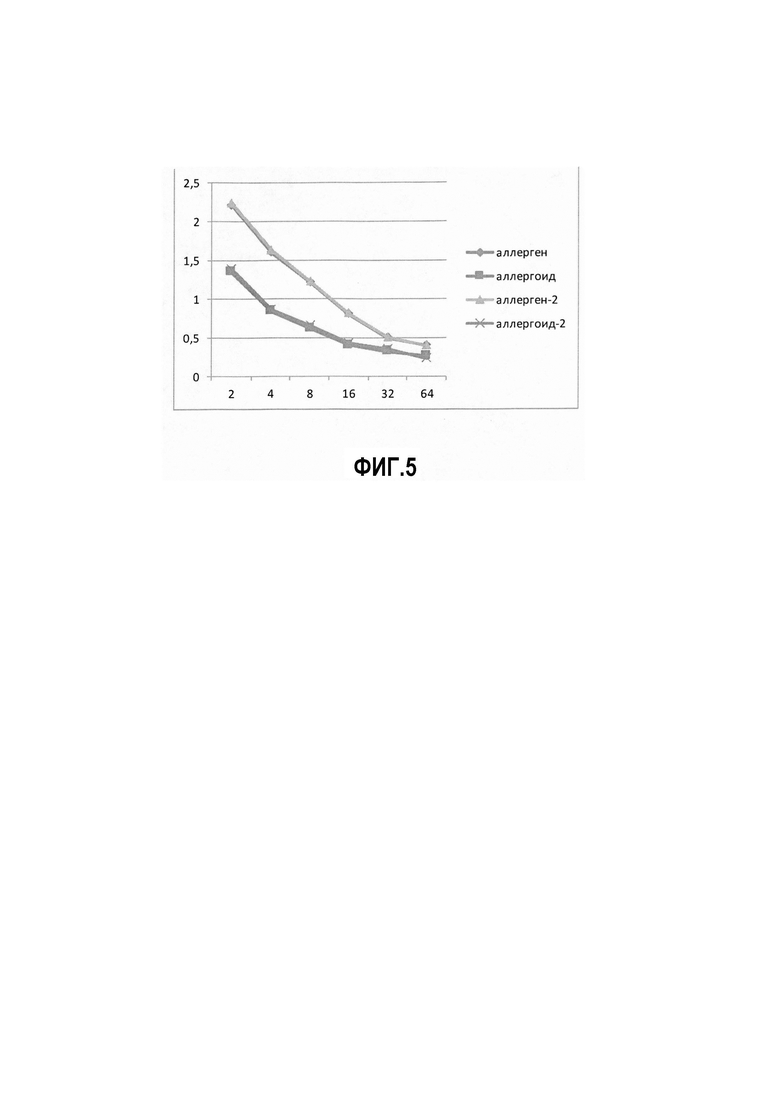
Фиг. 4. Вертикальный электрофорез в 12% ПААГ, модельные АДПЯ в зависимости от времени полимеризации 1- маркеры молекулярной массы (Fermentas) 14-110 КДа, 2-АДПЯ 1 сутки, 3- АДПЯ 5 суток, 4- АДПЯ 10 суток, 5 - АДПЯ 15 суток, 6- АДПЯ 20 суток, 7- АДПЯ 25 суток, 8- АДПЯ 30 суток
Фиг. 5. Сравнительный анализ калибровочных кривых для аллергенов (АНПЯ) и аллергоидов (АДПЯ) двух разных серий.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Обозначения и сокращения:
АНПЯ - аллерген пчелиного яда, не содержащий мелиттин
АДПЯ - аллергоид из пчелиного яда
ОПЯ - очищенный пчелиный яд
ПЯс - пчелиного яда-сырца
У пациентов с аллергическими реакциями IgE ответ преимущественно направлен только на определенные гликопротеины. Из них фосфолипаза А2 является наиболее значительным аллергеном с клинической точки зрения, поскольку она распознается сывороточным IgE почти у всех индивидуумов с аллергическими реакциями. Другие, менее важные, чем фосфолипаза, аллергены - гиалуронидаза и кислая фосфатаза. Все эти компоненты находятся в диапазоне молекулярной массы 60-16 кДа. Высокомолекулярные протеины пчелиного яда практически не способны связываться с IgE и, следовательно, считаются менее клинически релевантными или даже полностью нерелевантными. Низкомолекулярные компоненты необходимо удалить в связи с их токсическими свойствами. В этой связи, представлялось оптимальным использование гельфильтрации на носителе Superdex 75, диапазон фракционирования которого 3-70 кДа.
ПРИМЕР 1.
Получение аллергоида на основе пчелиного пчелиного яда-сырца.
Подготовку препарата пчелиного яда-сырца (ПЯс) для гель-хроматографии проводили следующим образом: 1 г ПЯс растворяли в 100 мл дистиллированной воды (конечная концентрация 10 мг/мл) и инкубировали при комнатной температуре в течение 2 часов при периодическом перемешивании. Полученный экстракт центрифугировали при 15000 g и супернатант профильтровали через мембранный фильтр с диаметром пор 0,22 мкм.
Поскольку экстракт ПЯс содержал заметное количество пигментов (возможно растительного происхождения из содержащихся в ПЯс примесей пыльцы), которые активно сорбируются на используемый для гель-хроматографии супердекс, то для предотвращения загрязнения основной разделяющей колонки Superdex 75 16x60 профильтрованный экстракт ПЯс дополнительно пропускали через сорбирующую пигменты колонку, заполненную сефадексом G25 из расчета 10 мл матрицы на каждые 100 мл экстракта. Не связавшийся с сефадексом материал собирали под визуальным контролем и лиофилизировали. Результаты взвешивания количественного выхода ОПЯ (очищенный пчелиный яд) из 1 г ПЯс для 6 серий полупродукта представлены в таблице 1.
На фиг. 1 представлена хроматография образца на Superdex 75, 16x60, суммируются фракции 1, 2, 3, 4, 5.
Полученная в результате препаративного разделения на Superdex 75 очищенного пчелиного яда фракция 6 (рисунок 13.3), наряду со значительным, количеством фосфолипазы А2 (мажорный аллерген) 16 кДа, имеет значительную примесь мелиттина, 3 кДа (дорожка 1 на рисунке 13.4). Фракции 7-8 состоят более чем на 90% из мелиттина. Для получения АДПЯ используются фракции 1-5. Данный способ выделения основных аллергенных фракций из яда-сырцам методом гель-фильтрации позволяет получить аллерген, не содержащий мелиттин.
Таким образом, взвешивание полученного после лиофилизации препарата показало, что количественный выход ОПЯ из ПЯс каждый раз составлял не менее 80%.
Концентрирование фракций проводят методом ультрафильтрации на фильтрах с диаметром пор 6-8 кДа. Таким образом, был получен очищенный аллерген пчелиного яда, оптимальный по белковому составу, для последующего изготовления аллергоида методом полимеризации формальдегидом. Проводится вертикальный электрофорез в ПААГ для контроля белкового профиля полупродукта.
Очищенный аллерген доводили до концентрации 6 мг аллергена на 1 мл буфера 0,01 М PBS Sigma (USA), рН 7,5. Для получения аллергоида к прозрачному раствору аллергена по каплям при перемешивании добавляли формальдегид (36-40%) так, чтобы концентрация формальдегида в растворе аллергена составляла 1%. При концентрации формальдегида 40% на 50 мл раствора аллергена добавляли 1,292 мл формальдегида. Полученную смесь фильтровали через фильтр с пористостью 22 мкм. Реакционную смесь инкубировали 30 дней при комнатной температуре, периодически перемешивая.
Полученный раствор аллергоида стабилизировали боргидридом натрия. Раствор боргидрида натрия (NaBH4) готовили перед употреблением. Навеску NaBHi в количестве 0,02 г растворяли в 5 мл дистиллированной воды (4 мг/мл). На 50 мл раствора аллергоида добавляли 1 мл раствора боргидрида натрия и инкубировали 2 часа при комнатной температуре, периодически помешивая.
Диализ аллергоида
Используют диализную ленту с диаметром пор 6-8 кДа. Диализ проводят против 10 л апирогенной воды 48 часов. Диализ проводят в холодильнике при 8°С. По окончании диализа аллергоид центрифугируют 20 минут при 8000 об/мин и фильтруют на мембранных фильтрах 0,22 мкм. Затем определяют концентрацию белка.
На фигуре 2 представлена аналитическая хроматография Superdex 200, 10x30 Аллергоид из пчелиного яда, 0,2 мл, PBS, 0,5 мл/мин 1,0 мл/см 0,5 OUFS.
Полученный аллергоид, как видно на фиг. 2, представляет собой многокомпонентный пик, с молекулярной массой от 16-600 кДа. Наибольшее количество белка в аллергоиде, находится в диапазоне молекулярных масс 120-220 кДа, что характерно для полимеризации формальдегидом. Дополнительная стадия диализа аллергоида удаляет остатки NaBH4, формалина и метанола, т.е. токсических веществ, что уменьшает нежелательные побочные реакции у пациентов.
ПРИМЕР 2.
Выполнены эксперименты по оценке качества получаемого полупродукта - ОПЯ. Качество получаемого препарата соответствовало качеству очищенного пчелиного яда фирмы Sigma (USA), как референс-образца. Из очищенного пчелиного яда фирмы Sigma (USA) были изготовлены аллерген, не содержащий мелиттин (АНПЯ) и аллергоид (АДПЯ). Параллельно из ОПЯ, в соответствии с лабораторной технологией, были получены АНПЯ и АДПЯ. Полученные препараты сравнивали в вертикальном электрофорезе в 15% ПААГ. Результаты представлены на Фиг. 3 (Вертикальный электрофорез в 15% ПААГ аллергоидов и их исходного материала. 1- очищенный пчелиный яд фирмы Sigma (USA), 2- ОПЯ, 3- АНПЯ из очищенного пчелиного яда фирмы Sigma (USA), 4 -АНПЯ из ОПЯ, 5- АДПЯ из очищенного пчелиного яда фирмы Sigma (USA), 6- АДПЯ из ОПЯ).
Видно, что различий по белковому составу очищенного пчелиного яда фирмы Sigma (USA) и ОПЯ, а также полученных из них аллергенов и аллергоидов не обнаружено. Таким образом, полученный нами очищенный пчелиный яд соответствует по качеству признанным мировым стандартам. В рамках оптимизации лабораторной технологии разработана простая и дешевая методика очистки яда-сырца, позволяющая получить дорогостоящий продукт - очищенный пчелиный яд. ОПЯ можно использовать как основу для изготовления других лекарственных средств на основе пчелиного яда, для исследовательских целей и т.д.
Кроме того, использование данного способа очистки не требует частой очистки основной колонки, что сокращает вынужденные простои и увеличивает срок службы оборудования.
Полимеризация аллергоида формальдегидом, по общепринятой технологии, осуществляется в течение 30 суток, при комнатной температуре. Это длительный срок.
Уменьшение срока, без потери свойств аллергоида, было бы экономически эффективно. Для этого проведено исследование по изучению влияния сроков полимеризации АДПЯ в 1% формальдегиде на свойства аллергоида. АНПЯ доводили до концентрации 6 мг аллергена на 1 мл буфера 0,01 М PBS Sigma (USA), рН 7,5. Для получения аллергоида к прозрачному раствору аллергена по каплям при перемешивании добавляли формальдегид (36-40%) так, чтобы концентрация формальдегида в растворе аллергена составляла 1%. При концентрации формальдегида 40% на 50 мл раствора аллергена добавляли 1,292 мл формальдегида. Полученную смесь фильтровали через фильтр с пористостью 22 мкм. Реакционную смесь инкубировали дней при комнатной температуре, периодически перемешивая. Через определенные временные интервалы времени - 1, 5, 10, 15, 20, 25 и 30 суток, из емкости, где проводили полимеризацию, отбирали пробы по 7 мл. Полученный раствор аллергоида стабилизировали боргидридом натрия. Раствор боргидрида натрия (NaBH4) готовили перед употреблением. Навеску NaBH4 в количестве 0,02 г растворяли в 5 мл дистиллированной воды (4 мг/мл). На 7 мл раствора аллергоида добавляли 0,14 мл раствора боргидрида натрия и инкубировали 2 часа при комнатной температуре, периодически помешивая. Реакционную смесь затем помещали в диализные мешки и ставили на диализ против апирогенной воды, при 4°С в течение 2 дней. После завершения диализа раствор аллергоида и лиофильно высушивали. Полученные экспериментальные аллергоиды исследовали по способности растворяться в стандартной разводящей жидкости (Биомед). Результаты представлены в таблице 2.
Как видно из представленных данных, растворимость препарата оказалась зависимой от времени полимеризации. Оптимальные значения растворимости достигались только на 20 сутки полимеризации формальдегидом (препарат должен растворятся в течение 5 минут).
Учитывая, что во флаконе содержится 1 мг - 10 ТД (10×100 мкг) и флакон будут разводить не менее чем 10 мл разводящей жидкости, растворимость 2,5 мг/мл в течение 5 минут является вполне достаточной.
Вместе с тем, видимых изменений белкового профиля АДПЯ в зависимости от времени полимеризации формальдегидом не наблюдалось. Белковый профиль модельных аллергоидов исследовали в вертикальном электрофорезе в ПААГ. Результаты представлены на Фиг. 4 (Вертикальный электрофорез в 12% ПААГ, модельные АДПЯ в зависимости от времени полимеризации 1- маркеры молекулярной массы (Fermentas) 14- 110 КДа, 2- АДПЯ 1 сутки, 3- АДПЯ 5 суток, 4- АДПЯ 10 суток, 5 - АДПЯ 15 суток, 6-АДПЯ 20 суток, 7- АДПЯ 25 суток, 8- АДПЯ 30 суток).
Из представленных данных видно, что независимо от времени полимеризации АДПЯ, набор белковых фракций у модельных аллергоидов одинаков по молекулярной массе.
Таким образом, разработанный способ позволяет уменьшить сроки полимеризации аллергена без потери свойств аллергоида.
ПРИМЕР 3.
ГФХIV (действие с 1.12.2018) требует для оценки подлинности аллергенов и аллергоидов использования конкурентного иммуноанализа. В этой связи, возникла необходимость в разработке конкурентного ИФА для оценки специфической активности аллергоида. Был модифицирован набор реагентов «АллергоИФА-специфические IgE» предназначенный для количественного определения специфических Ig Е в сыворотке крови человека (Алкор-Био). Контрольная - высокотитражная сыворотка. Для определение специфических IgE-антител человека был использован набор реагентов «АллергоИФА-специфические IgE» предназначенный для количественного определения специфических Ig Е в сыворотке крови человека (Алкор-Био).
В наборе «АллергоИФА-специфические IgE» реализован «сарturе» - вариант иммуноферментного анализа. Это новый метод для количественного определения специфических IgE, в котором используются твердая фаза с адсорбированными специфичными к IgE антителами и жидкие биотинилированные аллергены. Создание иммуносорбента на основе антител к IgE (МАТ), а не фиксированных на твердой фазе аллергенов, обеспечивает ряд преимуществ, в частности - исключаются перекрестные неспецифические реакции с иммуноглобулинами других классов: A, G, М и D, что положительно влияет на специфичность.
Использовались два вида жидких биотинилированных аллергенов:
Пчела медоносная (Apis mellifera) - i1
фосфолипаза А2 (nApi m1 Фосфолипаза A2 пчелы), как мажорный аллерген - 94% специфических IgE антител в сыворотках.
Исследование выполнялось в соответствии с инструкцией к набору. Единственным изменением было добавление исследуемого препарата и препаратов сравнения на стадии внесения биотинилированных аллергенов, для создания конкурентного связывания. Исследуемые препараты добавляли в количествах: 20, 10, 5, 2,5, 1,25 и 0,625 мкг/лунку. В качестве положительного контроля использовали добавление PBS (Sigma), на котором готовили разведение препаратов. Оптическую плотность определяли на многоканальном спектрофотометре, при длине волны 450 нм. Результаты оценивали по величине оптической плотности.
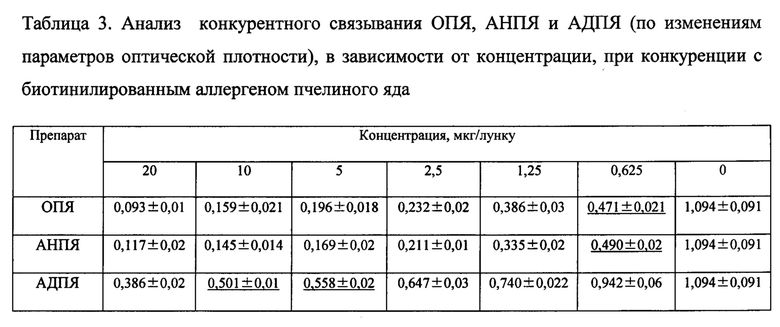
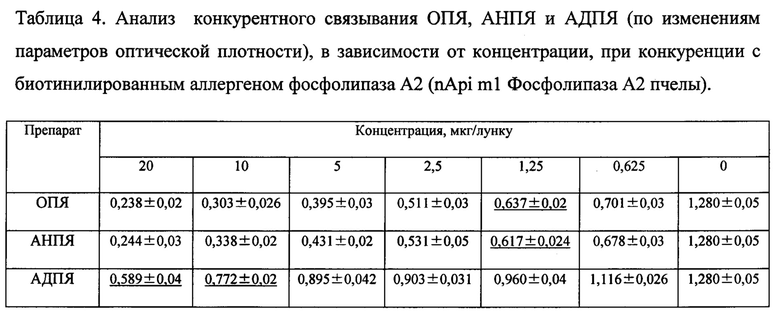
Известно, что аллергоиды, после полимеризации формальдегидом, характеризуются снижением способности специфического IgE связываться со аллергоидом, в сравнении с исходным аллергеном, что подтверждает снижение аллергенности. В конкурентном реверсивном ИФА сравнивали IgE-связывающую способность ОПЯ, АНПЯ и конечного продукта - аллергоида АДПЯ. В качестве конкурентов использовали коммерческие жидкие биотинилированные аллергены как к яду пчелы, так и мажорному аллергену фосфолипазе А2. Результаты оценивали по точке 50% снижения ОП исследуемого образца по отношению к положительному контролю (точка 50% ингибиции). Результаты представлены в таблицах 3 и 4.
Как видно из данных, приведенных в таблице 3, точка 50% ингибиции в конкурентном ИФА для АДПЯ находиться в пределах концентраций препарата 5-10 мкг/лунку при конкуренции с биотинилированным аллергеном пчелиного яда, тогда как для ОПЯ и АНПЯ эта точка соответствует 0,625 мкг/лунку.
Как видно из данных, приведенных в таблице 4, точка 50% ингибиции в конкурентном ИФА для АДПЯ находиться в пределах концентраций препарата 10-20 мкг/лунку при конкуренции с биотинилированным аллергеном фосфолипаза А2 (nApi m1 Фосфолипаза А2 пчелы), тогда как для ОПЯ и АНПЯ эта точка приходится на концентрацию 1,25 мкг/лунку.
Следовательно, в результате полимеризации формальдегидом, АДПЯ утратил большее количество сайтов связывания для специфических IgE антител к белкам пчелиного яда. Таким образом, средство, действующее на аллерген-распознающий рецептор, для лечения аллергии к пчелиному яду, АДПЯ соответствует требованиям предъявляемым к аллергоидам по критерию снижения аллергенности.
ПРИМЕР 4.
Для определения подлинности (выявления специфических аллергенных компонентов), в соответствии с ГФХIII, ОФС.1.7.2.0034.15 «Определение подлинности препаратов аллергенов», использовали определение специфических IgE твердофазным иммуноферментным анализом (ИФА). Используется тест-система «IgE-AT-ИФТС» (Микроген НПО ФГУП МЗ РФ, Россия). Данная система рекомендована Минздравом и приведена в качестве примера в ОФС «Определение подлинности препаратов аллергенов». На предыдущем этапе работы, нами была разработана собственная иммуноферментная тест система для определения специфических антител, с использованием в качестве антигена либо аллергена пчелиного яда (АНПЯ) либо полученного из него аллергоида (АДПЯ). Используя нашу тест-систему, мы провели сравнительный анализ калибровочных кривых для аллергенов и аллергоидов из наработанных серий.
Микропланшет сенсибилизировали раствором аллергена или аллергоида 10 мкг/мл в бикарбонатно-карбонатном буфере рН 9.6-9.8 в течение 16-18 часов при 4°С. После троекратной отмывки в лунку вносили по 100 мкл образца или контрольных сывороток. После инкубации в течение 1 часа при комнатной температуре и троекратной отмывки, в во все лунки вносили по 100 мкл конъюгата МАТ с ПХ против IgE в разведении 1:20000. После инкубации в течение 1 часа при комнатной температуре и троекратной отмывки реакцию проявляли ТМБ. В качестве контроля неспецифического связывания использовали результаты реакции в лунках без внесенной сыворотки. При регистрации значений ОП на многоканальном спектрофотометре результаты контроля автоматически вычитались. Контрольная положительная сыворотка была раститрована с разведения 1:2, с двойным шагом, до 1:64. Результаты представлены на Фиг. 5 (Сравнительный анализ калибровочных кривых для аллергенов (АНПЯ) и аллергоидов (АДПЯ) двух разных серий).
Из представленного анализа видно, что значения ОП для аллергенов и аллергоидов из разных серий, в одинаковых разведениях, очень близки, и достоверно не отличаются.
Показано отчетливое снижение способности специфического IgE связываться с АДПЯ- аллергоидом, в сравнении с аллергеном, АНПЯ, что подтверждает снижение аллергенности. Снижение связывания, оцениваемое по изменению ОП, отмечено для всех точек калибровочной кривой. Снижение связывания для аллергоида составило не менее 30%, по сравнению с исходным аллергеном. Результаты представлены в таблице 5.
Полученные данные свидетельствуют о хорошей воспроизводимости, от серии к серии, разработанной технологии получения аллергоида.
ПРИМЕР 5.
Изучение специфической активности лекарственного средства in vivo и in vitro.
Целью исследования являлось изучение специфической активности аллергоида действующего на аллерген-распознающий рецептор, для лечения аллергии к пчелиному яду, для определения возможности изучения данного препарата в клинических исследованиях.
В задачи исследования входило:
- изучение специфической активности in vivo аллергоида
- изучение специфической активности in vitro аллергоида
Тестируемый объект - средство, действующее на аллерген-распознающий рецептор, для лечения аллергии к пчелиному яду. Действующее средство - алергоид (АДПЯ), представляет собой модифицированный аллерген, путем обработки формальдегидом (карбоксилирование). Полученные таким образом соединения характеризуются низкой аллергенной активностью при сохранении хороших иммунизирующих свойств, обеспечивающих продукцию специфических блокирующих антител. Средство представляет собой флакон с лиофильно высушенным АДПЯ, 1 мг во флаконе. Препарат предназначен для подкожного введения. Эффективная терапевтическая доза (ТД) ЛС - 100 мкг/человека (1,43 мкг/кг) в объеме 1 мл, приравнена к дозам очищенных экстрактов пчелиного яда («Фармалген», «Веномил»), используемых для проведения аллерген-специфической иммунной терапии в Евросоюзе.
В связи с отсутствием, разрешенных к применению в РФ, аналогичных препаратов, в качестве препаратов сравнения использовали промежуточные продукты изготовления -очищенный пчелиный яд (ОПЯ) и аллерген пчелиного яда, не содержащий мелиттин и другие низкомолекулярные токсичные компоненты (АНПЯ). ОПЯ по составу аналогичен Фармалгену и Веномилу (эти препараты представляют собой очищенный пчелиный яд). Однако, учитывая, что в ОПЯ не менее 50% примеси мелиттина, а аллергоид получен полимеризацией белковых фракций яда, за исключением мелиттина и других низкомолекулярных компонентов, то для корректности сравнения мы использовали дополнительный препарат сравнения - АНПЯ, одинаковый по исходному белковому составу с аллергоидом.
5.1 Изучение специфической активности in vivo аллергоида.
Разрабатываемое средство должно удовлетворять следующим требованиям: структура средства - аллергоид, обладающий сниженной аллергенной активностью (по сравнению с исходным аллергеном), но обладающий большей иммуногенностью.
5.1.1. Изучение аллергенной активности тестируемого аллергоида.
Аллергенную активность оценивали в соответствии с методическими указаниями по изучению специфической активности аллергенов, изложенными в Руководстве по проведению доклинических исследований лекарственных средств. Использована модель сенсибилизации животных, с последующим сравнением уровней специфических IgE антител, поскольку, аллергия на пчелиный яд развивается именно по I типу, и обусловлена наличием специфического IgE. В Институте иммунологии отработана оптимальная модель сенсибилизации мышей различными аллергенами. Более того, протокол АСИТ предусматривает первоначальное введение 0,01 ТД с последующим плавным увеличением дозы препарата, под перманентным наблюдением за состоянием пациента. Поэтому изучение аллергенной активности препарата проводили с использованием одной дозы, для оценки снижения аллергенности у тестируемого аллергоида (средства) в сравнении с очищенным пчелиным ядом и безмелиттиновым аллергеном. В исследовании использовали самцов, поскольку, в отличие от самок, их гормональный фон более стабильный. Известно, что колебания гормонального фона могут оказать влияния на развитие иммунного ответа.
Проводили трехкратную (х3) подкожную сенсибилизацию мышей с интервалом 7 дней (1-ое введение 1 дн., 2-ое введение 7 дн., 3-ие введение 14 дн.). Доза ОПЯ, АНПЯ или АДПЯ, вводимая подкожно одной особи, составила 20 мкг. В качестве растворителя использована разводящая жидкость для аллергенов. Для создания депо сенсибилизирующего вещества в организме животного использовали адъювант - гидроксид алюминия (Аl(ОН)3) в дозе 2 мг/особь. Соотношение объемов вводимого вещества и адъюванта 1: 1, на одну особь 20 мкг ОПЯ, АНПЯ или АДПЯ, в 0,1 мл жидкости для разведения аллергенов и 2 мг Аl(ОН)3 в 0,1 мл забуференного физиологического раствора рН 7,2-7,4. Общий объем смеси вводимой подкожно на одно животное составил 0,2 мл. На 21 день от начала сенсибилизации проводили взятие крови у мышей для определения уровня специфических IgE.
Препарат сравнения 1 - ОПЯ
Препарат сравнения 2 - АНПЯ
Отрицательный контроль - разводящая жидкость для аллергенов (Биомед).
Сенсибилизацию исследовали на мышах СВА х Вl 57 (F1) самцы, количество 40 особей.
Экспериментальные группы представлены в таблице 6.
Количество циркулирующих специфических IgE-антител в сыворотках мышей определяли с использованием модифицированных коммерческих наборов Mouse IgE ELISA Kit (B&D). Вместо планшета из набора, с сорбированными антителами против IgE мыши, мы использовали планшеты Grainer high binding, с сорбированным на них ОПЯ. Сорбцию раствором ОПЯ, в концентрации 10 мкг/мл в карбонатно-бикарбонатном буфере рН9,5-9,7 (таблетированный, ПанЭко), проводили в течение 24 часов при 8°С. Далее исследование выполняли согласно инструкции к набору Mouse IgE ELISA Kit (B&D). Оптическую плотность определяли на многоканальном спектрофотометре, при длине волны 450 нм.
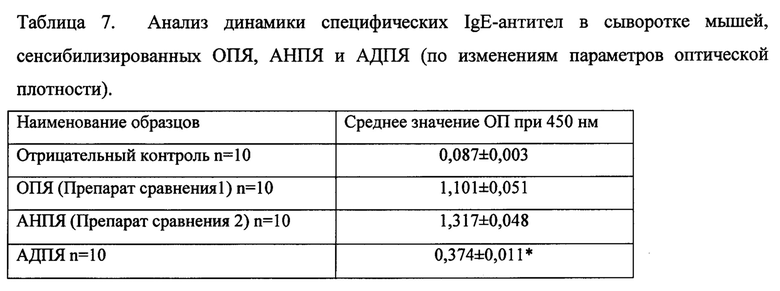
Результаты изучения аллергенной активности тестируемого аллергоида представлены в таблице 7 (Анализ динамики специфических IgE-антител в сыворотке мышей, сенсибилизированных ОПЯ, АНПЯ и АДПЯ (по изменениям параметров оптической плотности).
Из представленных данных видно, что тестируемое средство, действующее на аллерген-распознающий рецептор, для лечения аллергии к пчелиному яду, на модели сенсибилизации животных, демонстрирует снижение аллергенности. Способность аллергоида индуцировать синтез специфического IgE, после полимеризации формальдегидом, достоверно снижается, в сравнении, как с очищенным пчелиным ядом, так и с аллергеном пчелиного яда, который был использован для полимеризации.
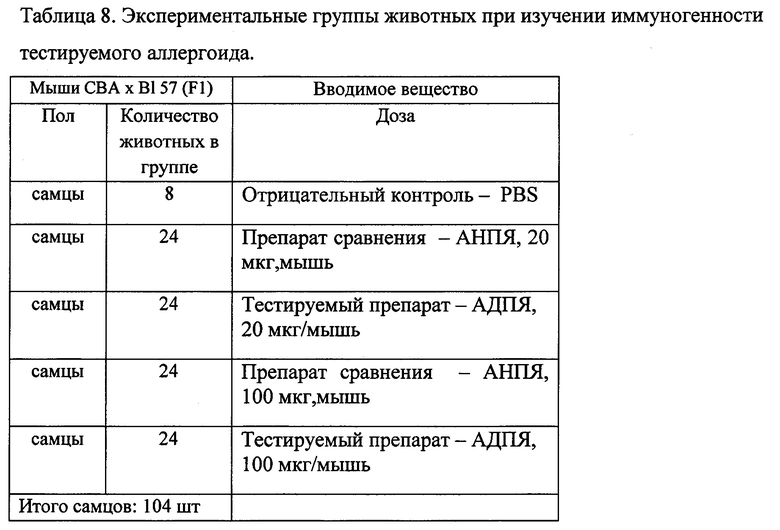
5.1.2. Изучение иммуногенности тестируемого аллергоида.
Иммуногенную активность исследовали на мышах (104 особи, самцы, весом 18-22 г линии (СВА х C57Bl/6)F1)), разделенных на 4 опытных группы по 24 мыши, и группу из 8 мышей - интактные. Доза АДПЯ или АНПЯ, вводимая внутрибрюшинно одной особи составила 20 мкг/мышь (группы 1 и 2) и 100 мкг/мышь (группы 3 и 4). В качестве растворителя использован PBS (Sigma) рН 7,2-7,4. Для создания депо препарата в организме животного использовали адъювант - гидроксид алюминия (Аl(ОН)3) в дозе 2 мг/особь. Соотношение объемов препарата и адъюванта 1:1, на одну особь 100 мкг АДПЯ или АНПЯ в 0,1 мл PBS и 2 мг Аl(ОН)3 в 0,1 мл PBS рН 7,2-7,4. Общий объем смеси, вводимой внутрибрюшинно, на одно животное составил 0,2 мл. Через 1, 2, 3, и 4 недели после начала иммунизации животных забивали и в их сыворотке, методом ИФА, определяли количество циркулирующих специфических IgG-антител. ОПЯ в эксперименте не использовали, т.к. для мыши расчетное LD50=660,8 мкг/мышь, и при введении ОПЯ с адъювантом не исключается токсическое действие (100 мкг - 1/6 LD). АНПЯ не содержит мелиттин, а токсическое действие пчелиного яда обусловлено одновременным наличием фосфолипазы А2 и мелиттина. В исследовании использовали самцов, поскольку, в отличие от самок, их гормональный фон более стабильный. Известно, что колебания гормонального фона могут оказать влияния на развитие иммунного ответа.
Экспериментальные группы представлены в таблице 8.
Количество циркулирующих специфических IgG-антител в сыворотках мышей определяли с использованием модифицированных коммерческих наборов Mouse IgG ELISA Kit (Sigma). Вместо планшета из набора, с сорбированными антителами против IgG мыши, мы использовали планшеты Grainer high binding, с сорбированным на них ОПЯ. Сорбцию раствором ОПЯ, в концентрации 10 мкг/мл в карбонатно-бикарбонатном буфере рН 9,5-9,7 (таблетированный, ПанЭко), проводили в течение 24 часов при 8°С. Далее исследование выполняли согласно инструкции к набору Mouse IgG ELISA Kit (Sigma). Оптическую плотность определяли на многоканальном спектрофотометре, при длине волны 450 нм.
Результаты исследования иммуногенности тестируемого аллергоида в дозах 20 и 100 мкг/мышь, представлены в таблице 9.
ПРИМЕР 6.
Изучение механизма действия аллергоида in vivo на модели гипосенсибилизации животных.
Средство представляет собой флакон с лиофильно высушенным АДПЯ, 1 мг во флаконе. Препарат предназначен для подкожного введения. Эффективная терапевтическая доза (ТД) ЛС - 100 мкг/человека (1,43 мкг/кг) в объеме 1 мл, приравнена к дозам очищенных экстрактов пчелиного яда («Фармалген», «Веномил»), используемых для проведения аллерген-специфической иммунной терапии в Евросоюзе
Разрабатываемое средство должно удовлетворять следующим требованиям: механизм действия лекарственного средства - введение по определенной схеме повышающихся доз аллергоида приводит к выработке против него значительного количества антител IgG класса, которые связывают поступающий в организм аллерген, не допуская, таким образом, связывания его с IgE и запуска каскада аллергопатологических реакций.
В этой связи в задачи исследования входило:
- Отработка оптимальной схемы и курсовой дозы препарата на модели гипосенсибилизации животных
- Определение специфических IgE антител к пчелиному яду в сыворотках мышей после гипосенсибилизации
- Определение специфических IgG антител к пчелиному яду в сыворотках мышей после гипосенсибилизации
6.1 Отработка оптимальной схемы и курсовой дозы препарата на модели гипосенсибилизации животных.
Гипосенсибилизацию АДПЯ исследовали на мышах, СВА х Вl 57 (F1) самцы, количество 50 особей. Проводили взятие крови у мышей для определения уровня IgE, IgG антител до проведения сенсибилизации. Осуществлен забой выборочно 9 мышей. Сбор крови в коническую пластиковую пробирку. Центрифугировали кровь 2000 об/мин в течение 10 мин. Полученную сыворотку крови аликвотировали по 0,2 мл (200 мкл) и использовали для определения уровня специфических IgE, IgG - группа интактные мыши.
Сенсибилизация:
Проводили трехкратную (х3) подкожную сенсибилизацию ОПЯ мышей с интервалом 7 дней (1-ое введение 1 дн., 2-ое введение 7 дн., 3-ие введение 14 дн.). Доза очищенного пчелиного яда, вводимая подкожно одной особи составила 20 мкг. В качестве растворителя использован PBS (Sigma) рН 7,2-7,4. Для создания депо сенсибилизирующего вещества в организме животного использовали адъювант - гидроксид алюминия (Аl(ОН)3) в дозе 2 мг/особь. Соотношение объемов очищенного пчелиного яда и адъюванта 1:1, на одну особь 20 мкг очищенного пчелиного яда в 0,1 мл PBS и 2 мг Аl(ОН)3 в 0,1 мл PBS рН 7,2-7,4. Общий объем смеси вводимой подкожно на одно животное составил 0,2 мл.
Общие количество ОПЯ, требующегося для трехкратной сенсибилизации 50-ти мышей составляет 3000 мкг (3,0 мг). 20 мкг х 50 особей=1000 мкг; 1000 мкг х 3 крат, сенсибил.=3000 мкг (3,0 мг).
На 21 день от начала сенсибилизации проводили взятие крови выборочно у 9 мышей для определения уровня специфических IgE, IgG - группа максимальной сенсибилизации.
Гипосенсибилизация мышей
Оставшихся 32 мышей гипосенсибилизировали подкожно ОПЯ (контрольная группа) и лекарственным средством АДПЯ по двум схемам: схема I, предложенная нами, (16 особей), и для сравнения схема II, разработанная ранее в Институте иммунологии (патент RU 2004133424 A от 17.11.2004), для ОПЯ, (16 особей). Схемы принципиально различаются по количеству вводимого за курс гипосенсибилизацию препарата, однако длительность курса гипосенсибилизации и количество инъекций на данном этапе исследования не изменяли.
Проведение гипосенсибилизации мышей по схеме I;
- Контрольная группа- гипосенсибилизация мышей очищенным пчелиным ядом - ОПЯ (8 особей);
- Экспериментальная группа - гипосенсибилизация АДПЯ (8 особей);
Гипосенсибилизацию проводили путем подкожного введения препаратов. Курс гипосенсибилизации составил 12 инъекций, по одной инъекции через день с увеличением дозы препарата. 1 инъекция 0,2 мл п/к 0,04 мкг/особь; 2 инъекция - 0,04 мкг/особь; 3 инъекция - 0,075 мкг/особь; 4 инъекция - 0,075 мкг/особь; 5 инъекция - 0,075 мкг/особь; 6 инъекция - 0,15 мкг/особь; 7 инъекция - 0,15 мкг/особь; 8 инъекция - 0,15 мкг/особь; 9 инъекция - 0,15 мкг/особь; 10 инъекция - 0,3 мкг/особь; 11 инъекция - 0,3 мкг/особь; 12 инъекция - 0,3 мкг/особь. Количество введенного п/к препарата составило 1,8 мкг/особь.
Через 3 дня по завершению гипосенсибилизации сбор крови, получение сыворотки для определения уровней специфических IgE и IgG.
Проведение гипосенсибилизации мышей по схеме II
- Контрольная группа - гипосенсибилизация мышей ОПЯ (8 особей);
- Экспериментальная группа - гипосенсибилизация АДПЯ (8 особей);
Гипосенсибилизацию проводили путем подкожного введения препаратов. Курс гипосенсибилизации составил 12 инъекций, по одной инъекции через день с увеличением дозы препарата. 1 инъекция 0,2 мл вв п/к 0,05 мкг/особь; 2 инъекция - 0,1 мкг/особь; 3 инъекция - 0,2 мкг/особь; 4 инъекция - 0,2 мкг/особь; 5 инъекция - 0,2 мкг/особь; 6 инъекция - 0,5 мкг/особь; 7 инъекция - 0,5 мкг/особь; 8 инъекция - 1,0 мкг/особь; 9 инъекция - 1,0 мкг/особь; 10 инъекция - 2,0 мкг/особь; 11 инъекция - 2,0 мкг/особь; 12 инъекция - 2,0 мкг/особь. Количество введенного п/к препарата составило 9,75 мкг/особь.
Через 3 дня по завершении гипосенсибилизации сбор крови, получение сыворотки для определения уровней специфических IgE и IgG.
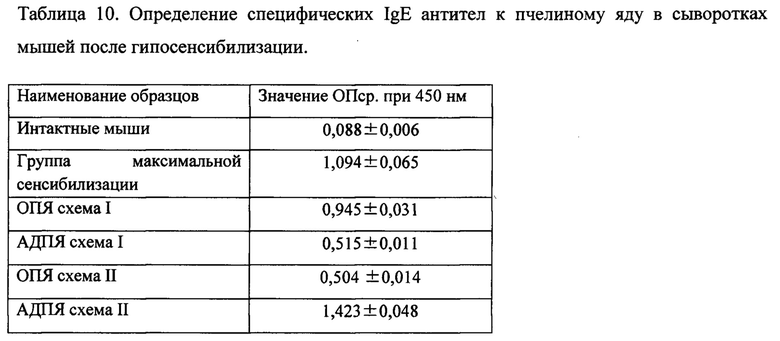
6.2. Определение специфических IgE антител к пчелиному яду в сыворотках мышей после гипосенсибилизации.
Количество циркулирующих специфических IgE-антител в сыворотках мышей определяли с использованием модифицированных коммерческих наборов Mouse IgE ELISA Kit (B&D). Вместо планшета из набора, с сорбированными антителами против IgE мыши, мы использовали планшеты Grainer high binding, с сорбированным на них ОПЯ. Сорбцию раствором ОПЯ, в концентрации 10 мкг/мл в карбонатно-бикарбонатном буфере рН9,5-9,7 (таблетированный, ПанЭко), проводили в течение 24 часов при 8°С. Далее исследование выполняли согласно инструкции к набору Mouse IgE ELISA Kit (B&D).
Оптическую плотность определяли на многоканальном спектрофотометре, при длине волны 450 нм. Результаты исследования представлены в таблице 10.
Как видно из таблицы 10, содержание специфических IgE антител к пчелиному яду в сыворотках мышей после гипосенсибилизации ощутимо снижается при терапии АДПЯ по схеме I и ОПЯ по схеме II, группы практически не различаются между собой. Однако, для достижения одинакового эффекта гипосенсибилизации АДПЯ по схеме I потребовалось в 5,4 раза меньше, чем для ОПЯ по схеме II (1,8 мкг/особь и 9,75 мкг/особь на курс, соответственно). При гипосенсибилизации ОПЯ по схеме I снижение специфических IgE практически не происходит. Поскольку в ОПЯ 50% приходится на мелиттин, то при использовании 1,8 мкг/особь на курс, количества препарата не хватает для полноценной гипосенсибилизации. При гипосенсибилизации мышей АДПЯ по схеме II происходит увеличение уровня специфических IgE антител к пчелиному яду. Подобное явление можно объяснить тем, что доза 9,75 мкг/особь слишком велика. Полученные результаты будут использованы для дальнейшей работы с целью оптимизации схем и используемых доз препаратов в эксперименте.
6.3. Определение специфических IgG антител к пчелиному яду в сыворотках мышей после гипосенсибилизации.
Количество циркулирующих специфических IgG-антител в сыворотках мышей определяли с использованием модифицированных коммерческих наборов Mouse IgG ELISA Kit (Sigma). Вместо планшета из набора, с сорбированными антителами против IgG мыши, мы использовали планшеты Grainer high binding, с сорбированным на них ОПЯ. Сорбцию раствором ОПЯ, в концентрации 10 мкг/мл в карбонатно-бикарбонатном буфере рН9,5-9,7 (таблетированный, ПанЭко), проводили в течение 24 часов при 8°С. Далее исследование выполняли согласно инструкции к набору Mouse IgG ELISA Kit (Sigma). Оптическую плотность определяли на многоканальном спектрофотометре, при длине волны 450 нм. Результаты представлены в таблице 11.
Как видно из таблицы 11, содержание специфических IgG антител к пчелиному яду в сыворотках мышей после гипосенсибилизации увеличивается, в сравнении группой максимальной сенсибилизации, при терапии АДПЯ по схеме I и ОПЯ по схеме II, при этом группы не различаются между собой. Однако, для достижения одинакового эффекта АДПЯ по схеме I потребовалось в 5,4 раза меньше, чем для ОПЯ по схеме II (1,8 мкг/особь и 9,75 мкг/особь на курс, соответственно). При гипосенсибилизации ОПЯ по схеме I увеличение специфических IgG антител практически не происходит. При гипосенсибилизации мышей АДПЯ по схеме II происходит увеличение уровня специфических IgG антител антител к пчелиному яду в наибольшей степени, что связано с большей дозой препарата на курс, однако это приводит к параллельному увеличению специфических IgE антител в сыворотках мышей, что недопустимо при АСИТ. Критерием успешности проведения гипосенсибилизации является увеличение количества специфических IgG при уменьшении IgE.
Таким образом, создана оптимальная модель гипосенсибилизации на животных, для изучения механизма действия средства. Показано, что в результате проведения гипосенсибилизации мышей аллергоидом пчелиного яда АДПЯ, уменьшается количество специфического IgE и увеличивается количество специфического IgG, что свидетельствует об эффективности терапии. Причем, для достижения одинакового эффекта, АДПЯ потребовалось в 5 раз меньше чем препарата сравнения - ОПЯ, при одинаковом количестве инъекций.
Таким образом, модификация главных аллергенов пчелиного яда, очищенных от низкомолекулярных примесей аллергенов, входящих в суммарную аллергенную фракцию, в аллергоид, путем химической модификации в присутствии формальдегида, приводит к взаимодействию высоко- и низкомолекулярных компонентов и способствует сохранению свойств аллергоида индуцировать синтез антител. Повышение иммуногенности за счет увеличения молекулярной массы и создания более жестких молекулярных связей обеспечивает увеличение стимулирования продукции IgG-антител при иммунизации аллергоидом. Разработанный способ позволяет уменьшить сроки полимеризации аллергена без потери свойств аллергоида.
Список литературы
1. Гущин И.С., Читаева В.Г. // Аллергия к насекомым - М. - 2003. - С. 7-48.
2. Швец С.М. Аллергические реакции на яд жалящих насекомых // Российский аллергологический журнал - 2004. - №3. С. 9-18.
3. Antonicelli L., Bilo М.В., Bonifazi F. Epidemiology of hyme-noptera allergy // Curr. Opin. Allergy Clin. Immunol. -2002 - V. 2 - P. 341-346.
4. Bilo M.B., Bonifazi F. Epidemiology of insect-venom anaphylaxis // Curr. Opin. Allergy Clin. Immunol. - 2008 - V. 8 - P. 330-337.
5. Шабанов Д.В., Мартынов А.И., Федоскова Т.Г., Федосеева В.Н., Гришина Т.И. Проблема аллергии к жалящим насекомым в условиях мегаполиса: Распространенность, Патогенетическое лечение // Сборник материалов IХ-ого Международного симпозиума "Экология человека и медико-биологическая безопасность населения" - Франтишковы Лазне, 25 октября-01 ноября 2014 г. - С. 141-144.
6. Bilo М., Bonifazi F. The natural history and epidemiology of insect venom allergy // J. Clin. Exp. Allergy - 2009 - V. 39(10) - P. 1467-1476.
7. Боброва E.E. Инсектная аллергия. Частная алергология / под ред. Г.Б. Федосеева -СПб.: Нордмедиздат - 2001.
8. Аллергология и иммунология. Национальное руководство / под ред. P.M. Хаитова, Н.И. Ильиной - М.: ГЭОТАР-Медиа - 2009.
9. Golden D.B.K., Demain J., Freeman Т., Graft D., Tankersley M., Tracy J., Blessing-Moore J., Bernstein D., Dinakar C, Greenhawt M., et al. Stinging insect hypersensitivity // Ann. Allergy Asthma Immunol. - 2017 - V. 118 - P. 28-54.
10. Schiner M., Graessel A., Ollert M., Schmidt-Weber C.B., Blank S. Allergen-specific immunotherapy of Hymenoptera venom allergy - also a matter of diagnosis // Human Vaccines and Immunotherapeutics - 2017 - V. 13 - №10 - P. 2467-2481.
11.  Francese S., Turillazzi S., Moneti G., Clench M., Barber D. In situ imaging of honeybee (Apis mellifera) venom components from aqueous and aluminum hydroxide-adsorbed venom immunotherapy preparations // J. Allergy Clin. Immunol. - 2012 May; - V. 129(5) - P. 1314-1320.
Francese S., Turillazzi S., Moneti G., Clench M., Barber D. In situ imaging of honeybee (Apis mellifera) venom components from aqueous and aluminum hydroxide-adsorbed venom immunotherapy preparations // J. Allergy Clin. Immunol. - 2012 May; - V. 129(5) - P. 1314-1320.
12. Carballada F., Boquete M.,  Lombardero M., de la Torre F. Follow-up of venom immunotherapy (VIT) based on conventional techniques and monitoring of immunoglobulin E to individual venom allergens // J. Investig. Allergol. Clin. Immunol. 2010-V. 20(6) - P. 506-513.
Lombardero M., de la Torre F. Follow-up of venom immunotherapy (VIT) based on conventional techniques and monitoring of immunoglobulin E to individual venom allergens // J. Investig. Allergol. Clin. Immunol. 2010-V. 20(6) - P. 506-513.
13. Calderon M.A., Alves В., Jacobson M., Hurwitz В., Sheikh A., Durham S. Allergen injection immunotherapy for seasonal allergic rhinitis // Cochrane Database Syst. Rev. - 2007-V. 5:CD001936.
14. Boyle R.J., Elremeli M., Hockenhull J., Cherry M.G., Bulsara M.K., Daniels M., Oude Elberink J.N. Venom immunotherapy for preventing allergic reactions to insect stings // Cochrane Database Syst. Rev. - 2012 - V. 10:CD008838; PMID:23076950.
15. Rueff F., Przybilla В., Bilo M.B.,  Scheipl F., Aberer W., Birnbaum J, Bodzenta-Lukaszyk A., Bonifazi F., Bucher C, et al. Predictors of side effects during the buildup phase of venom immunotherapy for Hymenoptera venom allergy: The importance of baseline serum tryptasey // J. Allergy Clin. Immunol. - 2010 - V. 126 - P. 105-111.e5.
Scheipl F., Aberer W., Birnbaum J, Bodzenta-Lukaszyk A., Bonifazi F., Bucher C, et al. Predictors of side effects during the buildup phase of venom immunotherapy for Hymenoptera venom allergy: The importance of baseline serum tryptasey // J. Allergy Clin. Immunol. - 2010 - V. 126 - P. 105-111.e5.
16. Exley C. Aluminium adjuvants and adverse events in sub-cutaneous allergy immunotherapy // Allergy Asthma Clin. Immunol. - 2014 - V. 10 - P. 4.
17. Kramer M.F., Heath M.D. Aluminium in allergen-specific subcutaneous immunotherapy // A German perspective Vaccine - 2014 -V. 32 - P. 4140-4148.
18. Council of Europe. European pharmacopoeia. Guideline on the clinical development of products for specific immunotherapy for the treatment of allergic diseases // Strasbourg: Council of Europe - 2013; 3971.
19. Lindblad E.B. Aluminium compounds for use in vaccines // Immunol. Cell. Biol. - 2004 - V. 82 - P. 497-505.
20. Brewer J.M., Conacher M., Satoskar A., Bluethmann H., Alexander J. In interleukin-4-deficient mice, alum not only generates T helper 1 responses equivalent to Freund's complete adjuvant, but continues to induce T helper 2 cytokine production // Eur. J. Immunol. - 1996 - V. 26 - P. 2062-2066.
21. Chesne J., Schmidt-Weber C.B., Esser von-Bieren J. The Use of Adjuvants for Enhancing Allergen Immunotherapy Efficacy // Immunol. Allergy Clin. North Am. - 2016 - V. 36 - 125-145.
22. Rinaldo Zurbriggen Compositions comprising melittin-derved peptides and methods for the potentiation of immune responses against target antigens // WO 2005049647 A2 - дата публикации: 02.06.2005 - дата приоритета: 05.11.2003.
23. Silva ТС, Moura Sde P., Ramos H.R., de Araujo P.S., da Costa M.H. Design of a modern liposome and bee venom formulation for the traditional VIT-venom immunotherapy // J. Liposome Res. - 2008 - vol. 18(4) - p. 353-368.
24. da Trindade R.A., Esteves Sant'Anna O.A.B., Rescia V.C, Bruni F.M., dos Anjos Lopes-Ferreira M.V., dos Santos D.F., Nicolete R., Faccioli L.H., de Araujo P.S., Bueno da Costa M.H. PLGA Microspheres as New Strategy to Improve the Efficiency of Venom Immunotherapy // J. Chem. and Pharm. Research - 2017 - V. 9(3) - P. 197-208.
25. Bioley G., Lassus A., Terrettaz J., Tranquart F., Corthesy B. Prophylactic immunization of mice with phospholipase A2-loaded gas-filled microbubbles is protective against Th2-mediated honeybee venom allergy // Clin. Exp. Allergy - 2016 - V. 46 - P. 153-162.
26. Thomas Grunwald Cloning of honey bee allergen // EP1846556 A1 - дата публикации: 24.10.2007 - дата приоритета: 14.12.2004.
27. Simon Blank, Benjamin Bockisch, Thomas Grunwald Cloning of honey bee allergen С // US 7888068 B2 - дата публикации: 15.02.2011 - дата приоритета: 26.06.2006.
28. Francois Spertini, Blaise Corthesy Allergen peptide fragments and use thereof // US 8075897 B2 - дата публикации: 13.12.2011 - дата приоритета: 14.03.2003.
29. Francois Spertini Bee venom polypeptides and methods of use thereof // US6780416 B1 - дата публикации: 24.08.2004 - дата приоритета: 18.02.2000.
30. Peirena N., de Graaf D.C., Brunaina M., Bridts C.H., Ebo D.G., Stevens W.J., Jacobs F.J. Molecular cloning and expression of icarapin, a novel IgE-binding bee venom protein // FEBS Letters - 2006 - V. 580 - P. 4895-4899.
31. Karamloo F., Schmid-Grendelmeier P., Kussebi 1 F., Akdis M., Salagianni M., von Beust B.R., Reimers A., Zumkehr J., Soldatova L., Housley-Markovic Z., , Thomas  David M. Kemeny D., Spangfort M. D., Blaser K. and Akdis C. A., Prevention of allergy by a recombinant multi-allergen vaccine with reduced IgE binding and preserved T cell epitopes // Eur. J. Immunol. - 2005 - V. 35 - P. 3268-3276.
David M. Kemeny D., Spangfort M. D., Blaser K. and Akdis C. A., Prevention of allergy by a recombinant multi-allergen vaccine with reduced IgE binding and preserved T cell epitopes // Eur. J. Immunol. - 2005 - V. 35 - P. 3268-3276.
32. Giovanni Mistrello, Daniela Roncarolo, Dario Zanoni, Paolo Falagiani Allergens and allergoids from bee venom //US 20100297188 A1 - дата публикации: 25.11. 2010. - дата приоритета: 21.12. 2007.
33. Мистрелло Джованни, Ронкароло Данеэла, Дзанони Дарио, Фаланджи Паоло Аллергены и аллергоиды из пчелиного яда // RU 2481122 - дата публикации: 10.05.2013 - дата приоритета: 16.12.2008.
34. Федосеева В.Н., Орлова И.А., Мартынов А.И., Федоскова Т.Г. Аллергоид из яда пчел для аллерген-специфической иммунотерапии больных с аллергическими реакции на ужаление пчелами и способ его получения // RU 2279888 - дата публикации: 20.07.2006 - дата приоритета: 17.11.2004.





| название | год | авторы | номер документа |
|---|---|---|---|
| АЛЛЕРГОИД ИЗ ЯДА ПЧЕЛ ДЛЯ АЛЛЕРГЕН-СПЕЦИФИЧЕСКОЙ ИММУНОТЕРАПИИ БОЛЬНЫХ С АЛЛЕРГИЧЕСКИМИ РЕАКЦИЯМИ НА УЖАЛЕНИЕ ПЧЕЛАМИ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2004 |
|
RU2279888C1 |
| КОМПЛЕКСНЫЙ ПРЕПАРАТ НА ОСНОВЕ АЛЛЕРГЕНОВ ПЫЛЬЦЫ БЕРЕЗЫ И МУРАМИЛПЕПТИДОВ | 2012 |
|
RU2537225C2 |
| АЛЛЕРГЕНЫ И АЛЛЕРГОИДЫ ИЗ ПЧЕЛИНОГО ЯДА | 2008 |
|
RU2481122C2 |
| АЛЛЕРГОТРОПИН ДЛЯ ЛЕЧЕНИЯ ПОЛИНОЗОВ И СПОСОБ ЛЕЧЕНИЯ ПОЛИНОЗОВ | 2001 |
|
RU2205661C2 |
| СПОСОБ ДИФФЕРЕНЦИАЛЬНОЙ ДИАГНОСТИКИ ГИПЕРЧУВСТВИТЕЛЬНОСТИ К ЯДУ ПЧЕЛЫ (APIS MELLIFERA) | 2016 |
|
RU2609839C1 |
| АЛЛЕРГЕН ОСИНОГО ЯДА С ПОНИЖЕННОЙ РЕАКЦИОННОЙ СПОСОБНОСТЬЮ IgE И СПОСОБ ЕГО ПОЛУЧЕНИЯ (ВАРИАНТЫ) | 2000 |
|
RU2264462C2 |
| Рекомбинантный полипептид на основе аллергена пыльцы березы и аллергена яблока в качестве вакцины от аллергии | 2020 |
|
RU2761431C1 |
| АЛЛЕРГОТРОПИНЫ ДЛЯ ЛЕЧЕНИЯ АЛЛЕРГИИ, ВЫЗЫВАЕМОЙ КЛЕЩАМИ ДОМАШНЕЙ ПЫЛИ | 2013 |
|
RU2548727C2 |
| СОПОЛИМЕРЫ ГЕТЕРОЦЕПНЫХ АЛИФАТИЧЕСКИХ ПОЛИ-N-ОКСИДОВ, ВАКЦИНИРУЮЩИЕ И ЛЕКАРСТВЕННЫЕ СРЕДСТВА НА ИХ ОСНОВЕ | 2010 |
|
RU2428991C9 |
| СПОСОБ ПОЛУЧЕНИЯ ЭКСТРАКТА АЛЛЕРГЕНА | 2011 |
|
RU2572230C2 |
Изобретение относится к медицине, а именно к иммунологии. Предложен способ получения аллергоида на основе пчелиного яда. Способ включает растворение пчелиного яда-сырца в дистиллированной воде до конечной концентрация 10 мг/мл, инкубирование, центрифугирование и фильтрацию супернатанта через мембранный фильтр с диаметром пор 0,22 мкм. Профильтрованный экстракт пропускают через колонку Sephadex G25 и концентрируют фракции методом ультрафильтрации на фильтрах с диаметром пор 6-8 кДа. Очищенный аллерген пчелиного яда обрабатывают формальдегидом и инкубируют 20-30 дней при комнатной температуре, полученный раствор аллергоида стабилизируют боргидридом натрия, проводят диализ, центрифугируют 20 мин при 8000 об/мин и фильтруют на мембранных фильтрах 0,22 мкм. Изобретение обеспечивает получение гипосенсибилизирующего аллергоида для проведения аллерген-специфической иммунотерапии больных, страдающих аллергией к яду пчел. 4 з.п. ф-лы, 5 ил., 11 табл., 6 пр.
1. Способ получения аллергоида на основе пчелиного яда для проведения аллерген-специфической иммунотерапии, включающий экстрагирование яда пчелы, полимеризацию формальдегидом, диализ и стабилизацию боргидридом натрия, отличающийся тем, что пчелиный яд-сырец растворяют в дистиллированной воде до конечной концентрация 10 мг/мл, инкубируют, полученный экстракт центрифугируют при 15000 g и фильтруют супернатант через мембранный фильтр с диаметром пор 0,22 мкм, затем профильтрованный экстракт дополнительно пропускают через колонку Sephadex G25 и проводят концентрирование фракций методом ультрафильтрации на фильтрах с диаметром пор 6-8 кДа, получая очищенный аллерген пчелиного яда, который для получения аллергоида обрабатывают формальдегидом и инкубируют 20-30 дней при комнатной температуре, далее полученный раствор аллергоида стабилизируют боргидридом натрия и проводят диализ аллергоида, по окончании которого аллергоид центрифугируют 20 мин при 8000 об/мин и фильтруют на мембранных фильтрах 0,22 мкм.
2. Способ получения аллергоида по п. 1, где колонку Sephadex G25, сорбирующую пигменты, заполняют из расчета 10 мл матрицы на каждые 100 мл экстракта.
3. Способ получения аллергоида по п. 1, где очищенный аллерген пчелиного яда доводят до концентрации 6 мг аллергена на 1 мл буфера 0,01 М PBS, рН 7,5.
4. Способ получения аллергоида по п. 1, где к прозрачному раствору аллергена по каплям при перемешивании добавляют формальдегид так, чтобы концентрация формальдегида в растворе аллергена составляла 1%.
5. Способ получения аллергоида по п. 1, где для проведения диализа аллергоида используют диализную ленту с диаметром пор 6-8 кДа.
| АЛЛЕРГОИД ИЗ ЯДА ПЧЕЛ ДЛЯ АЛЛЕРГЕН-СПЕЦИФИЧЕСКОЙ ИММУНОТЕРАПИИ БОЛЬНЫХ С АЛЛЕРГИЧЕСКИМИ РЕАКЦИЯМИ НА УЖАЛЕНИЕ ПЧЕЛАМИ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2004 |
|
RU2279888C1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| SU 1524226 A1, 27.05.1996. | |||

