Изобретение относится к области биохимии, а именно, к получению новой, модифицированной формы термостабильного белка GroEL микроорганизма Т. thermophilus. Может быть использовано в фармакологической промышленности в качестве белка-носителя для биосинтеза рекомбинантных пептидов, синтез которых не рентабелен существующими технологиями, в научно-исследовательских лабораториях для наработки различных пептидов в целях поиска обладающих необходимыми свойствами.
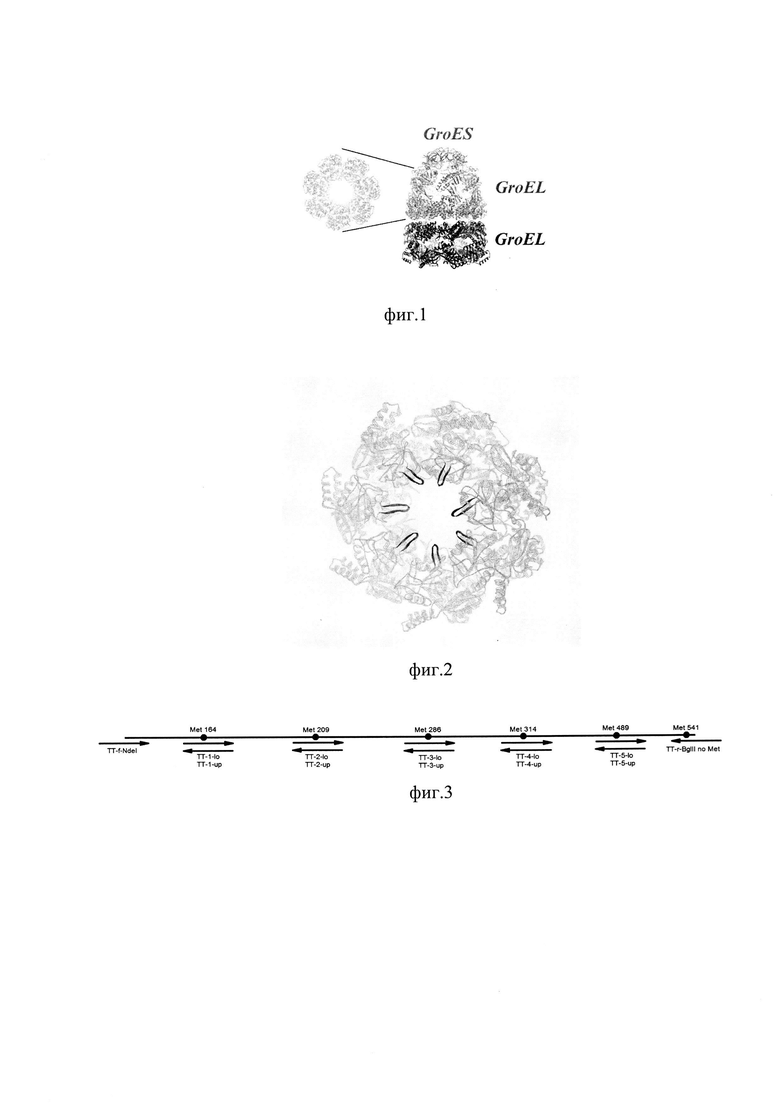
Семейство белков-шаперонов GroEL выглядит привлекательным как белок-носитель для системы слитых белков. Данный шаперон представляет собой большую частицу, состоящую из двух гептамеров. Полость, образованная протомером, вмещает белковый субстрат.GroEL является хорошо изученным шапероном, его структура и функционирование подробно описаны. Обычно одна молекула белкового субстрата связывается с одним протомером GroEL; что касается всей частицы, обычно между двумя протомерами при связывании субстратного белка наблюдается отрицательная кооперативность. Общая полость, образованная внутри гептамера, достаточна для размещения достаточно большого субстратного белка.
Одним из распространенных подходов для биосинтетического получения целевых рекомбинантных полипептидов и белков является создание слитых белков. В этом подходе используется белок-носитель, который обладает высоким уровнем экспрессии и также часто растворимостью. В качестве белков-носителей используются и чрезвычайно популярны такие белки, как тиоредоксин, мальтоза-связывющий белок, глютотион-S-трансфераза и многие другие [Kapust R.B., Waugh D.S. Escherichia coli maltose-binding protein is uncommonly effective at promoting the solubility of polypeptides to which it is fused // Protein Sci. Publ. Protein Soc. 1999. T. 8. №8. C. 1668-1674; Riggs P., La Vallie E.R., McCoy J.M. Introduction to expression by fusion protein vectors // Curr. Protoc. Mol. Biol. Ed. Frederick M Ausubel Al. 2001. T. Chapter 16. C. Unit16.4A; Terpe K. Overview of tag protein fusions: from molecular and biochemical fundamentals to commercial systems // Appl. Microbiol. Biotechnol. 2003. T. 60. №5. C. 523-533; Bell M.R. и др. To fuse or not to fuse: what is your purpose? // Protein Sci. Publ. Protein Soc. 2013. T. 22. №11. C. 1466-1477]. Наиболее широко используются системы с тиоредоксином, поскольку данный белок экспрессируется в клетках е coli с высоким уровнем, является хорошо растворимым и стабильным. Тем не менее, используемые системы слитых белков с указанными а также другими используемыми белками-носителями не позволяют решить проблемы получения многих целевых полипептидов, востребованных как в научных исследованиях, так и в биотехнологических и медицинских приложениях. Методами генной инженерии создается генный конструкт кодирующий белок-носитель и целевой полипептид, таким образом что они составляют единую полипептидную цепь. Экспрессия данного генетического конструкта в соответствующих микроорганизмах-продуцентах дает слитый белок, состоящий из белка-носителя и целевого пептида. Такой подход позволяет получить высокие уровни экспрессии и растворимости для многих целевых белков и пептидов, которые сами по себе плохо экспрессируются, либо являются нестабильными и/или плохо растворимыми. При необходимости использовать отдельно целевой полипептид требуется отделить его от белка-носителя. Это можно достичь, используя высокоспецифичные протеазы, например энтерокиназу человека или TEV-протеазу. Альтернативным методом является использование химического расщепления, специфичного к определенной аминокислоте. Одним из популярных реагентов для данного направления является бромциан натрия, специфичный к метионину. При наличии метионина между белком-носителем и целевым полипептидом, такая связь гидролизуется бромцианом, что позволяет выделять целевой полипептид. Как правило, в любом белке присутствуют остатки метионина внутри полипептидной цепи, поэтому при наличии в белке-носителе остатков метионина он также будет гидролизоваться бромцианом, давая набор полипептидов, которые будут затруднять выделение целевого продукта. Замена остатков метионина в белке-носителе на другие аминокислотные остатки, позволяет избежать действия на него бромциана натрия, и таким образом упростить процедуру очистки целевых пептидов.
Молекулярные шапероны это белки предназначенные для сворачивания и поддержания в растворимом состоянии вновь свернутых белков, таким образом основная функция шаперонов делает их привлекательными в качестве белков-носителей для создания слитых белковых систем [Alexey N. Fedorov*, and Maria S. Yurkova. Molecular Chaperone GroEL - toward a Nano Toolkit in Protein Engineering, Production and Pharmacy // NanoWorld J. 2018. T. 4. №1. C. 8-15]. Наиболее изученным классом молекулярных шаперонов является семейство GroEL. Это белок, мономер которого представляет собой полипептидную цепь весом 60 кДа, которые формируют частицу, состоящую из двух колец, каждое кольцо формируется из семи мономеров [Horwich A.L., Fair G.W., Fenton W.A. GroEL-GroES-mediated protein folding // Chem. Rev. 2006. T. 106. №5. C. 1917-1930; Horwich A.L. и др. Two families of chaperonin: physiology and mechanism // Annu Rev Cell Dev Biol. 2007. T. 23. C. 115-145]. Шапероны этого семейства взаимодействуют с большим кругом белков, стабилизируя их в растворимом виде и помогая их сворачиванию. Исходный GroEL испытывался в качестве белка носителя в слитых системах, однако система не нашла своего распространения в силу целого ряда причин. Одна из них -сложность в получении целевого полипептида из состава слитого белка.
Для успешного использования шаперонов в качестве белков-носителей должны быть разработаны их формы, которые могли бы адекватно функционировать как белки-носители. В состав полипептидной цепи мономера GroEL входит 6 остатков Метионина [Horwich A.L., Farr G.W., Fenton W.A. GroEL-GroES-mediated protein folding // Chem. Rev. 2006. T. 106. №5. C. 1917-1930], достаточно равномерно распределенных вдоль полипептидной цепи. Это делает затруднительным использование бромциана натрия, в силу того что пришлось бы выделять целевой полипептид среди большого числа пептидов, близких по размерам и физико-химическим свойствам. Привлекательным подходом является замена остатков метионина на другие аминокислотные остатки, что делает белок нечувствительным к указанному химическому реагенту. Необходимым условием использования такого подхода, т.е. замена остатков метионина, является сохранение структуры и функциональности белка-носителя, в данном случае частицы GroEL. Молекулярные шапероны семейства GroEL являются необходимыми для выживания любого организма, в том числе и микроорганизмов и присутствует также у микроорганизмов, адаптированных к экстремальным условиям функционирования, в частности к высоким температурам. GroEL таких микроорганизмов отличается чрезвычайно высокой термостабильностью структуры и функциональностью при повышенных температурах. Термостабильный GroEL может быть использован в качестве белка-носителя в слитых системах с целью упрощения очистки целевых пептидов от внутриклеточных белков
Присоединение дополнительных аминокислотных последовательностей к последовательности GroEL влияет на стабильность шаперона. И его N-конец, и С-конец глубоко погружены в белковую глобулу, так что присоединение любых дополнительных последовательностей к любым из концов может повлиять на третичную и четвертичную структуры. Некоторые результаты исследований показывают, что размещение 6His-tag дестабилизирует исходную структуру частиц GroEL и что очистка His-tagged GroEL происходит с низкой эффективностью и в результате дает только мономерную форму. Изучение трехмерной структуры GroEL предполагает, что присоединение любых дополнительных последовательностей к его концам должно дестабилизировать его структуру. Основной целью данного исследования было достижение способности GroEL являться белком-носителем для присоединения целевых пептидов, сохраняя при этом свою третичную и четвертичную структуры. Единственное исследование, которое ранее рассматривало этот вопрос, было основано на анализе случайных вставок транспозонов в геном GroEL. Было обнаружено, что транспозоновые вставки дестабилизировали частицу GroEL, все они приводили к дестабилизации колец гептамера, многие мутанты были нерастворимы. Естественное ограничение этого подхода и, как следствие, ограничение его выводов следует из анализа ограниченного числа вариантов транспозонов, которые не были предварительно выбраны для оптимального размещения вставки с учетом третичной структуры. Известно, что случайные вставки транспозона обычно инактивируют белки и влияют на их нативную структуру.
Задача изобретения состоит в том, что аминокислотная последовательность модифицированного белка GroEL не должна содержать остатков метионина, для обеспечения последующей возможности химического отщепления целевого пептида; должна быть стабильной, термостабильной и высокоэкспрессируемой в клетке-хозяине, не подвергаться протеолизу.
Технический результат изобретения достигается тем, что создается безметиониновый конструкт версии шаперона GroEL микроорганизма Т. thermophilus, с помощью изменения кодирующей последовательности ДНК GroEL. Все триплеты последовательности ATG, кодирующие метионин в положениях Met164, Met209, Met286, Met314, Met489, Met541, заменяли на последовательности CTG, кодирующие лейцин. Частица GroELS обладает высокой молекулярной массой (более 850 кДа), что технологически упрощает выделение конструкта из клеточного лизата методами гель-фильтрации и ультрафильтрации. Использование шаперона термофильного микроорганизма, помимо того, позволяет проводить первую стадию очистки с помощью тепловой денатурации эндогенных белков мезофильной бактерии.
Модифицированный белок после замены аминокислот не должен изменять нативную конформацию, должен оставаться стабильным и хорошо растворимым.
Существенными признаками, характеризующими изобретение, в отличие от исходной версии белка, является использование модифицированной термостабильной, безметиониновой версии шаперона GroEL микроорганизма Т. thermophilics.
Краткое описание фигур и чертежей:
фиг. 1 Структура комплекса GroELS
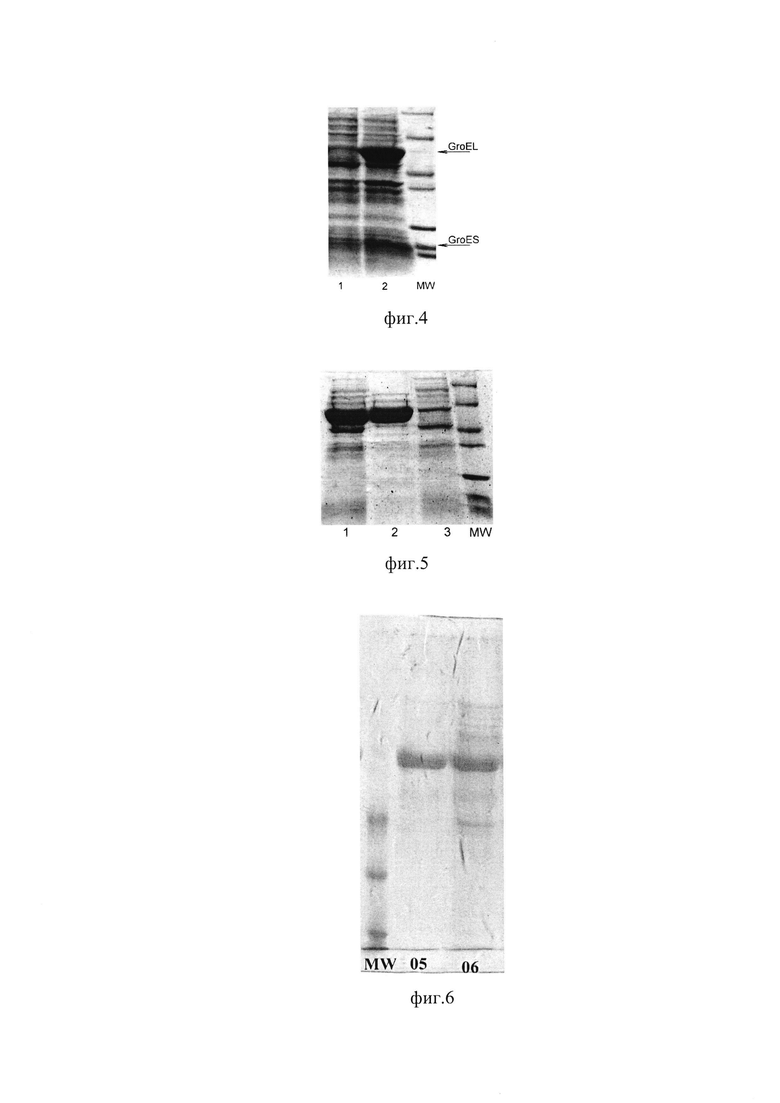
фиг. 2 Протомер GroEL. Выделены места введения полилинкера на субстрат-связывающей поверхности апикального домена
фиг. 3 Условные обозначения расположения метионинов в гене GroEL бактерии Т. thermophilus и используемые праймеры для синтеза модифицированного гена
фиг. 4 Экспрессия GroES и модифицированного безметионинового GroEL бактерии Т. thermophilus в экспрессионном бицистронном векторе. 1 - до индукции ИПТГ, 2 - после индукции ИПТГ, MW - белковый маркер молекулярного веса.
фиг. 5 Электрофорез белкового профиля GroEL. 1 - клеточный лизат до нагревания, 2 - супернатант после прогревания, 3 - осадок после прогревания, MW - белковый маркер молекулярного веса.
фиг. 6 Электрофорез GroELS. 06 - супернатант после прогревания при 62°С, 05 - дополнительная очистка супернатанта ультрафильтрацией, MW - белковый маркер молекулярного веса
Предложенный способ поясняется следующими материалами:
Все кодоны, кодирующие метионин, были заменены теми, которые кодируют лейцин. GroEL из Т. thermophilus (RefSeq assembly: GCF_000091545.1; GenBank: AP008226.1) имеет шесть остатков Met в своей последовательности в положениях Met164, Met209, Met286, Met314, Met489, Met541 (инициатор Met отщепляется во время полипептидного синтеза, как это происходит почти для всех белков); ген, кодирующий GroEL, со всеми кодонами ATG, кодирующими Met, замененными на кодоны CTG, кодирующие Leu, получали методом ПЦР с использованием пар праймеров SeqID №1-10.
Реакцию проводили в следующих условиях: Pfu-буфер 1×(20 mM Tris-HCl рН=8.8; 10 mM (NH4)SO4, 10 mM KCl, 0.1% Triton Х-100 v/v, 0.1 mg/ml BSA), 1.2u Pfu-полимеразы, 10 пкмоль каждого праймера, по следующей схеме ПЦР: 94°С 3 минуты, 40 циклов (94°С 40 сек, 60°С 40 сек, 72°С 2 минуты), 72°С 3 минуты.
Комбинационную ПЦР проводили со праймерами SeqID№11-12. Реакцию проводили в следующих условиях: Pfu-буфер 1×(20 mM Tris-HCl рН=8.8; 10 mM (NH4)SO4, 10 mM KCl, 0.1% Triton Х-100 v/v, 0.1 mg/ml BSA), 1.2u Pfu-полимеразы, 10 пкмоль каждого праймера, по следующей схеме ПЦР: 94°С 3 минуты, 40 циклов (94°С 40 сек, 60°С 40 сек, 72°С 2 минуты), 72°С 3 минуты. Схема метода представлена на фиг. 3. В результате получали ген безметионинового варианта GroEL, заканчивающийся стоп-кодоном ТАА и фланкированный рестрикционными сайтами NdeI и BglII.
Фрагмент очищали методом агарозного гель-электрофореза в 1% агарозном геле в 1х буфере ТБЕ, очищали фрагмент с помощью набора для очистки ДНК из геля «QIAEX II Gel Extraction Kit», и далее проводили расщепление рестриктазами NdeI и BglII в следующих условиях: 1х буфер О (Orange) (50 mM Tris-HCl рН=7.5, 10 mM MgCl2, 100 mM NaCl, 0.1 mg/ml BSA), по 5ед. а. каждого фермента BglII и NdeI, 50 нг ДНК и инкубировали при 37°С в течение 16 часов. Далее рестрикционный фрагмент клонировали во второй полилинкер экспрессирующего бицистронного вектора рЕТ Duet вместе с геном, кодирующим Т. thermophilus GroES, с получением вектора экспрессии pGroEL/ES. GroES амплифицировали с использованием праймеров SeqID№13-14 содержащие сайты рестрикции для NcoI и NotI, соответственно, и генома Т. thermophilus в качестве матрицы в следующих условиях: Pfu-буфер l×(20 mM Tris-HCl рН=8.8; 10 mM (NH4)SO4, 10 mM KCl, 0.1% Triton Х-100 v/v, 0.1 mg/ml BSA), 1.2u Pfu-полимеразы, 10 пкмоль каждого праймера, по следующей схеме ПЦР: 94°С 3 минуты, 40 циклов (94°С 40 сек, 60°С 40 сек, 72°С 2 минуты), 72°С 3 минуты. Полученный фрагмент ПЦР расщепляли ферментами NcoI и NotI в следующих условиях: Tango-буфер 2×(66 mM Tris-acetate рН=7.9, 20 mM magnesium acetate, 132 mM potassium acetate, 0.2 mg/ml BSA), 5 ед. а. каждого фермента NcoI и NotI; инкубировали при 37°С в течение 16 часов. Далее клонировали в первый полилинкер экспрессии бицистронного вектора рЕТ Duet вместе с геном, кодирующим GroEL Т. thermophilus, не содержащим метионин, и получали вектор экспрессии pGroEL/ES. Лигирование проводили в следующих условиях: Ligation-буфер 1×(40 mM Tris-HCl рН=7.9, 10 mM MgCl2, 10 mM DTT, 0.5 mM АТР), 20нг плазмиды, 1:5 молярном соотношении вектор-вставка, 2 ед. а. лигазы по Вейсу. Лигирование проводили при 22°С в течение 1 часа.
Химически компетентные клетки штамма E.coli pLysE в объеме 50 мкл размораживали на льду, затем добавляли 5 мкл лигазной смеси и инкубировали 30 минут на льду. Затем проводили термический шок при 42°С в течение 30 сек с последующим инкубированием на льду в течение 5 минут. Далее к трансформантам приливали 1 мл среды SOC предварительно нагретой до 37°С и инкубировали в течение 1 часа при постоянном покачивании в термостате-качалке. Далее 20 мкл проинкубированной смеси рассевали на чашки с твердой питательной средой LB, содержащей 100 мкг/мл ампициллина. Выращивали колонии в течение ночи при 37°С. Далее проводили аналитическую экспрессию по индивидуальным клонам, с индуктором 0.4 мМ ИПТГ.
Экспрессия генов в экспрессионном штамме E.coli pLysE осуществлялась в среде LB, при 37°С в течение 3 часов; селективный агент - ампициллин 100 мкг / мл. фиг. 4
Для очистки целевого белка проводили лизис с помощью френчпресса в буфере PBS(0.137M NaCl, 0.0027 М KCl, 0.01М Na2HPO4, 0.0018M KH2PO4) при 4°C. Далее разбавляли в 5 раз буфером PBS с последующим прогреванием клеточного лизата при 62°С в течение 5 мин на водяной бане, и далее центрифугировали при 13000g в течение 15 мин (фиг. 5).
Предварительно очищенный полипептид доочищали методом ультрафильтрации на кассете 300K Minimate capsule with Omega membrane, PALL, США в буфере PBS, в режиме диафильтрации с добавлением свежего буфера по мере уменьшения объема при давлении 2 бар и потоке 65 мл/мин и температуре окружающей среды 4°С (фиг. 6).
Пример 1 - получение модифицированного белка GroEL
Стадия 1
Провести ПЦР с использованием пар праймеров SeqID №1-10 в следующих условиях: Pfu-буфер 1×(20 mM Tris-HCl рН=8.8; 10 mM (NH4)SO4, 10 mM KCl, 0.1% Triton Х-100 v/v, 0.1 mg/ml BSA), 1.2u Pfu-полимеразы, 10 пкмоль каждого праймера, 10 нг геномной ДНК Т. thermophilus по следующей схеме ПЦР: 94°С 3 минуты, 40 циклов (94°С 40 сек, 60°С 40 сек, 72°С 2 минуты), 72°С 3 минуты.
Провести комбинационную ПЦР с праймерами SeqID №11-12 в следующих условиях: Pfu-буфер 1×(20 mM Tris-HCl рН=8.8; 10 mM (NH4)SO4, 10 mM KCl, 0.1% Triton Х-100 v/v, 0.1 mg/ml BSA), 1.2u Pfu-полимеразы, 10 пкмоль каждого праймера, по следующей схеме ПЦР: 94°С 3 минуты, 40 циклов (94°С 40 сек, 60°С 40 сек, 72°С 2 минуты), 72°С 3 минуты.
Стадия 2
Фрагмент очистить методом агарозного гель-электрофореза в 1% агарозном геле в 1х буфере ТБЕ с помощью набора для очистки ДНК из геля «QIAEX II Gel Extraction Kit», и далее провести расщепление рестриктазами NdeI и BglII в следующих условиях: 1х буфер О (Orange) (50 mM Tris-HCl рН=7.5, 10 mM MgCl2, 100 mM NaCl, 0.1 mg/ml BSA), по 5ед. а. каждого фермента BglII и NdeI, 50нг ДНК и инкубировать при 37°С в течении 16 часов. Далее рестрикционный фрагмент клонировать во второй полилинкер экспрессирующего бицистронного вектора рЕТ Duet вместе с геном, кодирующим Т. thermophilus GroES, с получением вектора экспрессии pGroEL/ES. GroES амплифицировать с использованием праймеров SeqID№13-14 в следующих условиях: Pfu-буфер 1×(20 mM Tris-HCl рН=8.8; 10 mM (NH4)SO4, 10 mM KCl, 0.1% Triton Х-100 v/v, 0.1 mg/ml BSA), 1.2u Pfu-polymerase, 10 пкмоль каждого праймера, 10 нг геномной ДНК Т. thermophilus по следующей схеме ПЦР: 94°С 3 минуты, 40 циклов (94°С 40 сек, 60°С 40 сек, 72°С 2 минуты), 72°С 3 минуты. Полученный фрагмент ПЦР расщепить ферментами NcoI и NotI в следующих условиях: Tango-буфер 2×(66 mM Tris-acetate рН=7.9, 20 mM magnesium acetate, 132 mM potassium acetate, 0.2 mg/ml BSA), 5 ед. а. каждого фермента NcoI и NotI; инкубировать при 37°С в течение 16 часов.
Далее клонировать в первый полилинкер экспрессии бицистронного вектора рЕТ Duet вместе с геном, кодирующим GroEL Т. thermophilus. Лигирование проводить в следующих условиях: Ligation-буфер 1×(40 mM Tris-HCl рН=7.9, 10 mM MgCl2, 10 mM DTT, 0.5 mM АТР), 20 нг плазмиды, 1:5 молярном соотношении вектор-вставка, 2ед. а. лигазы по Вейсу. Лигирование провести при 22°С в течение 1 часа.
Стадия 3
Химически компетентные клетки штамма E. coli pLysE в объеме 50 мкл разморозить на льду, затем добавить 5 мкл лигазной смеси и инкубировать 30 минут на льду. Затем провести термический шок при 42°С в течение 30 сек с последующим инкубированием на льду в течение 5 минут. Далее к трансформантам прилить 1 мл среды SOC предварительно нагретой до 37°С и инкубировать в течение 1 часа при постоянном покачивании в термостате-качалке. Далее 20 мкл проинкубированной смеси рассеить на чашки с твердой питательной средой LB, содержащей 100 мкг/мл ампициллина. Выращивать колонии в течение ночи при 37°С. Провести ПЦР колоний на определение наличия желаемой конструкции.

Изобретение относится к области биохимии, а именно к получению новой, модифицированной формы термостабильного белка GroEL микроорганизма Т. thermophilus. Модифицированный белок GroEL Thermus thermopiles имеет аминокислотную последовательность термостабильного шаперона GroEL микроорганизма Т. thermopilus, в которой остатки метионинов Met164, Met209, Met286, Met314, Met489, Met541 заменены на остатки лейцина. Также предложен фрагмент ДНК, кодирующий модифицированный белок GroEL. Полученный модифицированный белок GroEL является стабильным, термостабильным и высокоэкспрессируемым в клетке-хозяине, не подвергается протеолизу. Может быть использован в фармакологической промышленности в качестве белка-носителя целевых пептидов в слитых белковых конструкциях путем встраивания последовательности ДНК, кодирующей целевой пептид, в кодирующую белок-носитель последовательность, для биосинтеза рекомбинантных пептидов, синтез которых не рентабелен существующими технологиями, в научно-исследовательских лабораториях для наработки различных, обладающих необходимыми свойствами. 2 н.п. ф-лы, 6 ил., 1 пр.
1. Модифицированный белок GroEL микроорганизма Thermus thermopilus, способный выступать в качестве белка-носителя целевых пептидов в слитых белковых конструкциях путем встраивания последовательности ДНК, кодирующей целевой пептид, в кодирующую белок-носитель последовательность, имеющий аминокислотную последовательность термостабильного шаперона GroEL микроорганизма Т. thermopilus, в которой остатки метионинов Met164, Met209, Met286, Met314, Met489, Met541 заменены на остатки лейцина.
2. Фрагмент ДНК, кодирующий модифицированный белок GroEL по п. 1, способный выступать в качестве белка-носителя целевых пептидов в слитых белковых конструкциях путем встраивания последовательности ДНК, кодирующей целевой пептид, в кодирующую белок-носитель последовательность.
| HORWICH A.L | |||
| et al | |||
| "GroEL-GroES-mediated protein folding" | |||
| Chem | |||
| Rev | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Светоэлектрический измеритель длин и площадей | 1919 |
|
SU106A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| HIDEKI TAGUCH et al | |||
| "A Chaperonin from a Thermophilic Bacterium, Thermus thermophilus, That Controls Refoldings of Several Thermophilic Enzyme." The Journal of Biological Chemistry, 1991, v | |||
| Способ нагрева эквипотенциального катода в электронных вакуумных реле | 1921 |
|
SU266A1 |
| CN 104593400 B, 04.07.2017 | |||

