Изобретение относится к биотехнологии и сельскому хозяйству и может быть использовано для увеличения устойчивости растений против абиотических стрессов, главным образом к жесткому солнечному облучению (ультрафиолету B, УФ-В).
Известны решения, в которых для получения устойчивых к стрессу культур используют гены, ассоциированные с толерантностью к стрессу. В частности, описаны растения, трансформированные генами, кодирующими белки, вовлеченные в адаптацию к абиотическим стрессам Arabidopsis thaliana, для обеспечения адаптации к холоду, засухе и засолению у трансформированных растений.
Так, известен способ устойчивость растений к абиотическим стрессам, а именно обезвоживание, низкие температуры и засоление, приобреталось за счет генетической трансформации геном, кодирующим фактор транскрипции. Толерантность достигалась за счет работы генов, кодирующих транскрипционные факторы, которые при воздействии неблагоприятных условий активируют транскрипцию генов, которые участвуют в резистентности растений конкретному стрессу. В данном изобретении использовались гены промоутеров rd29A, rd17, DERIAN, erb 1 и kin 1. (патент США № 6670528, МПК A01H 5/00, C12N 15/29, опубл. 30.12.2002 г.).
Известно также использование генов транскрипционных факторов из Physcomitrella patens: фосфолипаза С-1 (PLC-1), фосфолипаза С-2 (PLC-2), 14-3-3 белок-1 (14-3-3P-1), 14-3-3 белок-2 (14-3-3P-2) и Ca2 – связывающий белок-1 (CPB-1), в частности, для устойчивости против засухи и пониженной температуры (заявка США № 2002069432, МПК A01H 5/00, C12N 15/29, опубл. 26.06.2002).
Основным недостатком описанных выше технических решений является отсутствие устойчивости растений к действию ультрафиолетового излучения, которое в свою очередь является одним из самых агрессивных абиотических факторов действующих на растения.
Сильнейшим стресс-фактором для растений, как всего живого, является ультрафиолетовое излучение (УФ). Обычно УФ - излучение разделяют на:
- коротковолновое, УФ-С, с длиной волны от 200 до 280 нм;
- средневолновое, УФ-В, с длиной волны 280-315 нм;
- длинноволновое, УФ-А, с длиной волны 315-380 нм.
Под воздействием УФ - излучения изменяются физиологические и биохимические процессы растительной клетки. Изменения зависят от строения ткани растения, стадии его развития и генотипа. Сказывается на изменениях в растении длительность его облучения и длина волны УФ - излучения. Так под влиянием коротковолнового излучения в растительной клетке поражается ДНК, средневолновое излучение разрушает белки (но в незначительных количествах оно необходимо растениям), длинноволновое же излучение опасно для клеток растений только в больших дозах.
Как правило, земная атмосфера задерживает УФ - С и УФ - В, но в ситуации истончения озонового слоя поверхности земли достигают и эти виды УФ–излучения.
Растения выработали биохимические защитные механизмы от воздействия больших доз УФ-излучения: они вырабатывают флавоноидные пигменты и другие фенольные соединения, являющиеся его протекторами: флавоноиды, антоцианы и др. Флавоноиды – это крупнейший класс растительных полифенолов. С химической точки зрения, флавоноиды представляют собой гидроксипроизводные флавона (собственно флавоноиды). Антоцианы - являются растительными пигментами, обуславливающими окраску цветов и плодов растений, и также относятся к классу флавоноидов. Эти вещества накапливаются в эпидермисе клетки и блокируют до 99% УФ-излучения.
Однако простое увеличение концентрации флавоноидов в листе полностью не исключает отрицательный эффект УФ-В. Так, фотосинтетический аппарат некоторых видов растений, произрастающих в тропических регионах на большой высоте над уровнем моря, достаточно устойчив к повышенному уровню ультрафиолетовой радиации. Однако эта устойчивость далеко не всегда связана с повышенной концентрацией флавоноидов. Поэтому считается, что накопление флавоноидов может вносить определенный вклад в устойчивость, однако простые причинно-следственные связи при этом не всегда могут существовать. (Найдено 22.01.2020, http://industrial-wood.ru/fiziologiya-ustoychivosti/10721-vliyanie-ultrafioletovogo-izlucheniya-na-fiziologicheskie-processy-v-rastenii.html)
Стильбены — это природные фенольные соединения, которые встречаются у некоторых видов различных семейств растений, таких как Pinaceae (ель, сосна), Fabaceae (ложная акация, арахис), Polygonaceae (полигонум) и Vitaceae (виноград). (Dubrovina A.S., Kiselev K. V. Regulation of stilbene biosynthesis in plants // Planta. 2017. Т. 246. № 4. С. 597–623.)
В данной работе описываются все известные виды растений, которые содержат стильбены, но отсутствует какая-либо информация об устойчивости растений к действию ультрафиолета.
Наиболее близким техническим решением к заявляемому является способ повышения устойчивости к засухе и засолению путем использования стильбен синтазы (He X. et al. Overexpressing fusion proteins of 4-coumaroyl-CoA ligase (4CL) and stilbene synthase (STS) in tobacco plants leading to resveratrol accumulation and improved stress tolerance // Plant Biotechnol. Rep. 2018. Т. 12. № 5. С. 295–302).
В известном способе используются ген 4-кумароил-КоА-лигаза стильбен синтазы для генетической модификации табака (Nicotiana benthamiana) с целью повышения устойчивости к различным патогенам, таким как бурая гниль персика ( Monilinia fructicola ), возбудитель яблочной гнили ( Valsa mali ), ( Fusicoccum aesculi ), ( Pythium aphanidermatum ) и ( Corynespora cassiicola ), а также улучшение толерантности к солевым и осмотическим стрессам.
Известно, что растения с повышенным содержанием стильбенов, обладают более высокой устойчивостью к засухе и засолению (He X. et al. Overexpressing fusion proteins of 4-coumaroyl-CoA ligase (4CL) and stilbene synthase (STS) in tobacco plants leading to resveratrol accumulation and improved stress tolerance // Plant Biotechnol. Rep. 2018. Т. 12. № 5. С. 295–302). Однако в данной работе отсутствует любое упоминание об улучшении устойчивости растений против действия ультрафиолета, вследствие чего, можно сделать вывод, что использование ген 4-кумароил-КоА-лигаза стильбен синтазы не приводит к повышению устойчивости растений против действия ультрафиолета.
Технической проблемой, решаемой настоящим изобретением, является разработка эффективного способа повышения устойчивости растений к действию ультрафиолетового излучения.
Поставленная техническая проблема решается тем, что в известном способе повышения устойчивости растений к действию ультрафиолетового излучения, включающим трансформацию растений геном стильбенсинтазы, согласно изобретению, в качестве гена стильбенсинтазы применяют ген VaSTS1 винограда амурского, путем трансформации растений с использованием плазмиды pZP-RCS2-nptII под контролем двойного промотора вируса цветной капусты 35s (CaMV35S). Затем проводят селекционный отбор по устойчивости растений к канамицину, отобранные растения используют для культивирования.
По сравнению с другими генами стильбенсинтаз, использование гена VaSTS1 позволяет добиться эффективного синтеза стильбенов и, как следствие, увеличение устойчивости растений к действию ультрафиолета, за счет наиболее оптимальной аминокислотной последовательности и высокой экспрессии данного гена в растении под влиянием ультрафиолетового облучения.
Плазмида pZP-RCS2-nptII это наиболее распространенная и эффективная векторная система для генетической трансформации растений, которая позволяет наиболее быстро и качественно получить генетическую конструкцию. В свою очередь, применение промоутера вируса цветной капусты 35s (CaMV35S) позволяет добиться высокой и постоянной экспрессии необходимого гена, а именно VaSTS1.
Селекционный отбор по устойчивости растений к антибиотику канамицину, который возможен благодаря гену неомицинфосфотрансферазе 2 (nptII), поскольку он заключен в используемую генетическую конструкцию. Селекционный отбор позволяет выделить только растения, которые были трансформированы генетическим вектором, что, в конечном результате, приводит к повышению устойчивости растений к действию ультрафиолетового излучения.
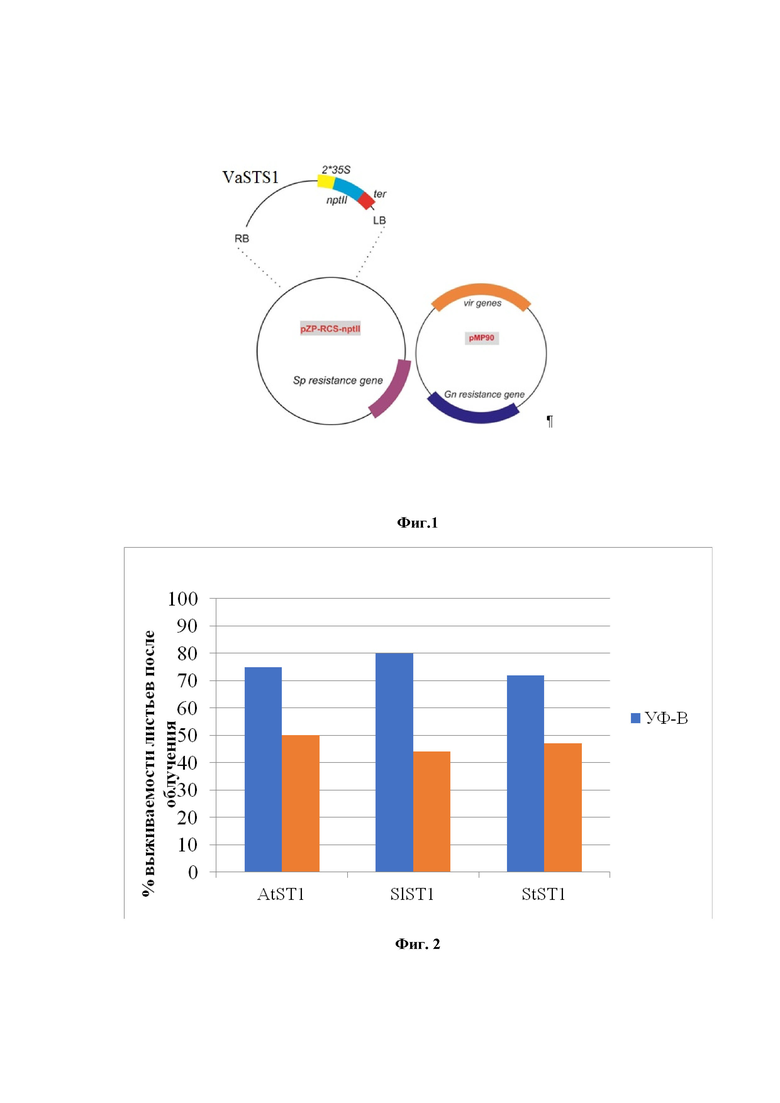
Способ иллюстрируется схемой бинарного вектора, представленного на фиг.1, где: в состав вектора входит плазмида pZP-RCS2-nptII, содержащая RB и LB – правый и левый бордер, 2*35S – конститутивный промотор вируса мозаики цветной капусты, VaSTS1 – целевой ген, nptII – ген неомицин-фосфотрансферазы II, ter – терминатор, Sp resistance gene – ген устойчивости к спектиномицину; плазмида pMP90, содержащая vir-гены и ген устойчивости к гентамицину (Gn resistance gene);
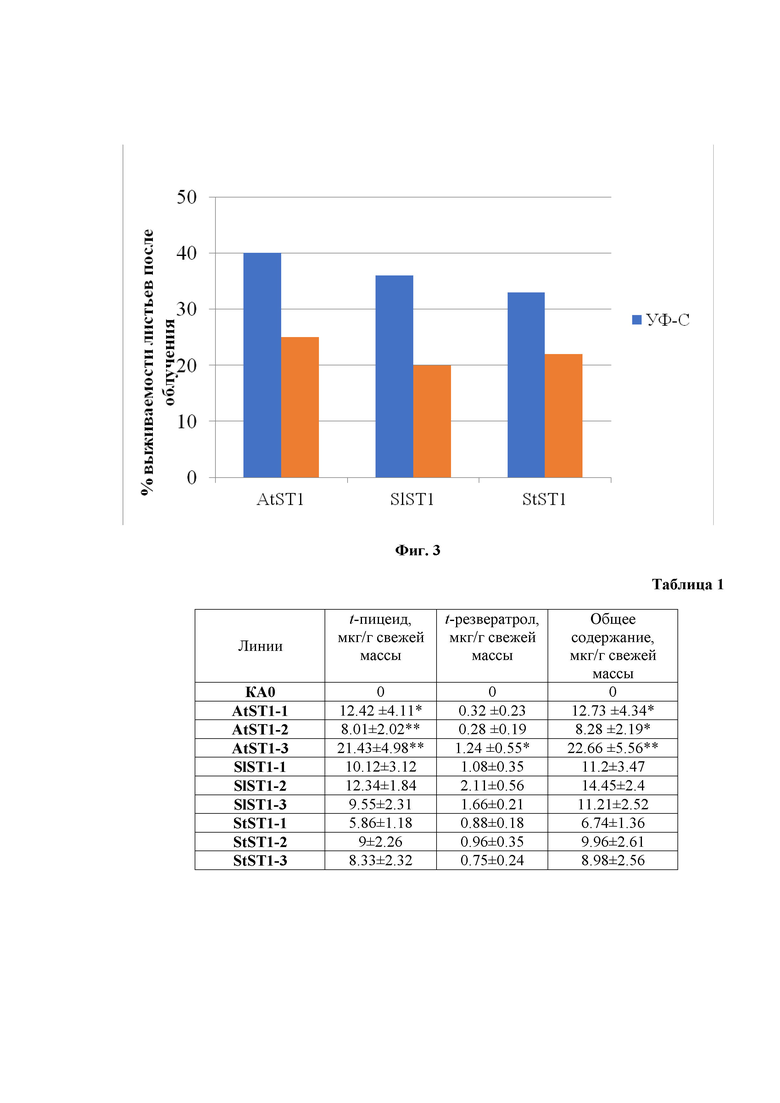
на фиг. 2 - приведена схема устойчивость контрольных и трансгенных растений Arabidopsis thaliana (AtST1), Solanum lycopersicum (SlST1), Solanum tuberosum (StST1) к действию ультрафиолета-В;
на фиг. 3 - приведена схема устойчивость контрольных и трансгенных растений Arabidopsis thaliana (AtST1), Solanum lycopersicum (SlST1), Solanum tuberosum (StST1) к действию ультрафиолета-С;
в табл. 1 приведено содержание транс-пицеида и транс-ресвератрола в контрольной и трансгенной линии;
в табл. 2 приведены праймеры, используемые для проверки наличие гена VaSTS1 в геноме растения.
Способ осуществляется следующим образом.
Перед реализацией заявленного способа предварительнго ген стильбенсинтазы VaSTS1 выделяется из амурского винограда (Vítis amurensis) и вставляется в бинарный вектор, содержащий плазмиду pZP-RCS2-nptII, несущую ген неомицинфосфотрансферазы 2 (nptII) и двойной 35S – конститутивный промотор вируса мозаики цветной капусты, полученной генетической конструкцией, методом цветочной трансформации, трансформируем растения, проводим селекционный отбор на обедненной в два раза среде Murashige и Skoog, дополненной 50 мкг / мл канамицина (Km), наличие встройки гена VaSTS1 в ДНК растения проверяется методом полимеразой цепной реакции, трансгенные растения высаживаем в землю и выращивают в течении 30-35 суток, после указанного периода растения проверяются на устойчивость к ультрафиолету, наиболее устойчивые линии отбираются для дальнейшей селекции.
Пример 1.
Растения Arabidopsis thaliana экотип Columbia L. в количестве 100 штук выращивали в горшках, заполненных коммерчески доступной богатой почвой, в контролируемой климатической камере при температуре +22˚C (Sanyo MLR-352, Panasonic, Япония), выдерживали цикл 16/8 часов день/ночь при интенсивности света ~120 мкмоль м-2 с-1.
Для создания трансгенных линий растений арабидопсиса (Arabidopsis thaliana), сверхэкспрессирующих ген VaSTS1, мы использовали плазмидную конструкцию pzp-RCS2-(VaSTS1) - nptII (Kiselev K.V., Aleynova O.A. Influence of overexpression of stilbene synthase VaSTS7 gene on resveratrol production in transgenic cell cultures of grape Vitis amurensis Rupr. // Appl. Biochem. Microbiol. 2016. Т. 52. № 1. С. 56–60; Aleynova O.A. et al. Stilbene accumulation in cell cultures of Vitis amurensis Rupr. overexpressing VaSTS1, VaSTS2, and VaSTS7 genes // Plant Cell. Tissue Organ Cult. 2016. Т. 125. № 2. С. 329–339). Сверхэкспрессионная конструкция pZP-RCS2-(VaSTS1)-nptII или пустой вектор pZP-RCS2-nptII была введена в штамм Agrobacterium tumefaciens GV3101::pMP90 и преобразована методом цветочной трансформации, в растения дикого типа A. Thaliana в количестве 100 единиц. Обработанные семена через 35-40 дней дают трансгенные семена, которые проращивают на обедненной в два раза среде Murashige и Skoog (Murashige T., Skoog F. Murashige1962Revised.Pdf // Physiol. Plant. 1962. Т. 15. С. 474–497), с добавлением 50 мкг/мл канамицина (Km), в течении 7 дней. Таким путем отбор трансгенных гомозиготных растений осуществляют с помощью селекции в присутствии канамицина в концентрации 50 мкг/мл в течение 3 поколений. Устойчивые к антибиотику растения проверяют на наличие в геноме гена VaSTS1 методом полимеразой цепной реакции (ПЦР) с использованием праймеров, перечисленных в таблице 2. Продукты ПЦР подвергают верификации с помощью секвенирования ДНК.
Ультрафиолетовая обработка проводится облучением растений в течение 10 мин на расстоянии 15 см над горшками, пиковая мощность составляла 254 нм, как описано (Tyunin A.P., Kiselev K. V. Alternations in VaSTS gene cytosine methylation and t-resveratrol production in response to UV-C irradiation in Vitis amurensis Rupr. cells // Plant Cell. Tissue Organ Cult. 2016. Т. 124. № 1. С. 33–45). Выживаемость листьев определяют как количество видимых зеленых листьев через 1 день после окончания стрессовых воздействий.
Для анализа содержания стильбенов проводят аналитическую ВЭЖХ с использованием аналитической ВЭЖХ-системы 1260 Infinity (Agilent Technologies, Санта-Клара, Калифорния, США), оснащенной детектором фотодиодной матрицы G1315D, четвертичным насосом G1311C, колонной g1316a и автоматическим пробоотборником G1329B. Экстракты разделяются на колонке Zorbax C18 (150 мм, 2,1 мм i. d., размер детали 3,5 мкм, Agilent Technologies, США); температура колонки составляет 40°C. подвижная фаза состоит из градиентного элюирования 0,1%-ной водной уксусной кислоты (а) и ацетонитрила (Б). Градиентный профиль со скоростью потока 0,2 мл / мин составлял: 0 мин 0% B; 35 мин 40% B; 40 мин 50% B. 50 мин 100% B, затем элюент B до 60 мин. Вводимый объем составлял 1 мкл. УФ-спектры регистрировали в диапазоне 310 нм. Все определяемые компоненты экстрактов рассчитываются на основе четырехточечных регрессионных кривых, построенных по внешним стандартам.
В итоге получают 3 трансгенные линии растений, которые обладают способностью синтезировать стильбены в количестве до 22 мкг/г от свежей массы (Табл.1). Более того растения, несущие ген VaSTS1, обладают в 1,5 раза лучшей устойчивостью к действию ультрафиолета-В по сравнению с растениями не несущими ген VaSTS1 и не синтезирующими стильбены (Фиг. 2).
Пример 2
Проводят аналогично примеру 1, с той лишь разницей, что в качестве трансформируемого объекта используются растения томата Solanum lycopersicum.
В результате получают трансгенные растения томата, несущие ген VaSTS1, которые обладают способностью синтезировать стильбены в количестве до 15 мкг/г от свежей массы (Табл.1). Более того, вследствие способности синтезировать стильбены, полученные растения томата обладают лучшей устойчивостью к действию ультрафиолета-В от 1,8 до 2 раз по сравнению с растениями не несущими ген VaSTS1 и не синтезирующими стильбены (Фиг. 2).
Пример 3
Проводят аналогично примеру 1, с той лишь разницей, что в качестве трансформируемого объекта используются растения паслёна клубненосного (Solanum tuberosum).
В результате генетической трансформации растений Solanum tuberosum получают трансгенные растения, несущие ген VaSTS1, которые обладают способностью синтезировать стильбены в количестве до 10 мкг/г от свежей массы (Табл. 1). Более того, вследствие способности синтезировать стильбены, полученные растения паслёна клубненосного обладают лучшей устойчивостью к действию ультрафиолета-В от 1,2–1,3 раз по сравнению с растениями не несущими ген VaSTS1 и не синтезирующими стильбены (Фиг. 2).
Результаты, представленные в примерах 1-3, иллюстрируют то, что выделенный ген VaSTS1, способен увеличивать толерантность растений к действию ультрафиолетового стресса. Кроме того, данный ген, способствует накоплению транс-ресвератрола и транс-пицеида, в норме не содержащихся в арабидопсисе. (Таблица 1). Следовательно, результаты четко указывают на то, что описанный ген стильбенсинтазы играет роль в устойчивости к абиотическому стрессу можно легко выделять и применять для значительного увеличения толерантности к абиотическому стрессу растений.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ РЕЗВЕРАТРОЛА | 2006 |
|
RU2326165C1 |
| СПОСОБЫ ПОВЫШЕНИЯ ТОЛЕРАНТНОСТИ К АБИОТИЧЕСКОМУ СТРЕССУ И/ИЛИ БИОМАССЫ У РАСТЕНИЙ И ПОЛУЧАЕМЫЕ ТАКИМ ОБРАЗОМ РАСТЕНИЯ | 2004 |
|
RU2350653C2 |
| Способ увеличения содержания антоцианов в растениях | 2021 |
|
RU2779748C1 |
| Промотор pro-SmAMP-D1 из растения звездчатка средняя (Stellaria media L.) для биотехнологии растений | 2022 |
|
RU2799014C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ ТАБАКА С ПОВЫШЕННЫМ СОДЕРЖАНИЕМ ПРОЛИНА | 2006 |
|
RU2324737C1 |
| Промотор pro-SmAMP-X из растения звездчатка белая (Stellaria media L.) для экспрессии рекомбинантных генов в клетках растений | 2020 |
|
RU2766095C1 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ С ПОНИЖЕННОЙ ВОСПРИИМЧИВОСТЬЮ К РАСТИТЕЛЬНЫМ ПАРАЗИТИЧЕСКИМ НЕМАТОДАМ (ВАРИАНТЫ), РЕКОМБИНАНТНАЯ ДНК (ВАРИАНТЫ), ТРАНСФОРМИРУЮЩИЙ РАСТЕНИЕ ВЕКТОР, ШТАММ AGROBACTERIUM И СПОСОБ СНИЖЕНИЯ УЩЕРБА УРОЖАЮ | 1992 |
|
RU2143000C1 |
| НОВЫЕ ГЕНЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2000 |
|
RU2241749C2 |
| Способ получения безмаркерных трансгенных растений каланхоэ перистого, экспрессирующих ген цекропина Р1 | 2015 |
|
RU2624042C2 |
| СПОСОБ ПОЛУЧЕНИЯ ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫХ РАСТЕНИЙ КАЛАНХОЭ, ЭКСПРЕССИРУЮЩИХ ГЕН ЦЕКРОПИНА P1 | 2010 |
|
RU2445768C1 |

Изобретение относится к области биотехнологии. Изобретение представляет собой способ повышения устойчивости растений к действию ультрафиолетового излучения, включающий трансформацию растений геном стильбенсинтазы, при этом в качестве гена стильбенсинтазы применяют ген VaSTS1 винограда амурского, путем трансформации растений с использованием плазмиды pZP-RCS2-nptII под контролем двойного промотора вируса цветной капусты 35s (CaMV35S). Затем проводят селекционный отбор по устойчивости растений к канамицину, отобранные растения используют для культивирования. Разработанный способ увеличивает толерантность растений к действию ультрафиолетового стресса. Кроме того, данный ген способствует накоплению транс-ресвератрола и транс-пицеида, в норме не содержащихся в арабидопсисе. 3 ил., 2 табл., 3 пр.
Способ увеличения устойчивости растений к воздействию ультрафиолета, включающий трансформацию растений геном стильбенсинтазы, отличающийся тем, что в качестве гена стильбенсинтазы применяют ген VaSTS1 винограда амурского, которым трансформируют растения с использованием плазмиды pZP-RCS2-nptII под контролем двойного промотора вируса цветной капусты 35s (CaMV35S), затем проводят селекционный отбор по устойчивости растений к канамицину, отобранные растения используют для культивирования.
| He X | |||
| et al, Overexpressing fusion proteins of 4-coumaroyl-CoA ligase (4CL) and stilbene synthase (STS) in tobacco plants leading to resveratrol accumulation and improved stress tolerance, Plant Biotechnol | |||
| Rep | |||
| Способ получения цианистых соединений | 1924 |
|
SU2018A1 |
| Т | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| С | |||
| УСТРОЙСТВО ПАРОПЕРЕГРЕВАТЕЛЯ | 1920 |
|
SU295A1 |
| KISELEV K.V., et al, Influence of overexpression of stilbene synthase VaSTS7 gene on resveratrol | |||

