Изобретение относится к области молекулярной биологии и может быть для увеличения содержания антоцианов в растениях для повышения устойчивости к абиотическим стрессам, а также для их выделения и дальнейшего использования в биотехнологии или пищевой промышленности.
Антоцианы – группа природных пигментов, раскрашивающие фрукты и овощи в яркие цвета. Соединения благотворно влияют на человеческий организм, поскольку проявляют антиоксидантные, бактерицидные, противоспалительные, адаптогенные и спазмолитические свойства. Природные источники пигментов: черника, бузина, чёрная смородина, ежевика, голубика черноплодная рябина. Природные красители используют в составе комплексной терапии сахарного диабета, сезонных инфекций (гриппа, ОРВИ), онкологии, дегенеративных расстройств, офтальмологических патологий (дистрофии сетчатки, близорукости, диабетической ретинопатии, катаракты, глаукомы). Кроме того, антоцианы применяются в пищевой индустрии (при изготовлении кондитерских изделий, йогуртов, напитков), косметологии (как коллаген), электротехнической отрасли (для краски солнечных батарей). В связи с тем, что антоцианы не могут образовываться в организме человека, они не способны накапливаться в организме, поэтому быстро выводятся из него. В организм человека они должны поступать с пищей. Следовательно, разработка способов повышения антоцианов в растениях на сегодняшний день является актуальным вопросом.
Известны подходы, в которых для получения растений способных синтезировать большое количество антоцианов, используют гены, ассоциированные с биосинтезов данных соединений. В частности, описаны растения, трансформированные генами, кодирующими белки, вовлеченные в биосинтез антоцианов Arabidopsis thaliana, для обеспечения высокого уровня продукции антоцианов у трансформированных растений.
Так, известен способ, в котором увеличение содержания антоцианов осуществлялось за счет генетической трансформации растений геном, кодирующим фактор транскрипции генов участвующих в работе финилпропаноидного пути. Высокое содержание антоцианов достигалась за счет работы генов кодирующих транскрипционные факторы, которые активируют транскрипцию генов, непосредственно участвующих в биосинтеза антоцианов. В представленном изобретении были использованы гены кодирующие факторы транскрипции PAP1 и PAP2 активирующие производство антоцианового пигмента (патент US № 6573432, МПК C07K14/415; C12N15/82; C12Q1/68, опубл. 03.06.2003).
Известно также использование генов халкон-синтазы (CHS), халкон-изомеразы (CHI), флаванон-3-гидроксилазы (F3H) и флавонол-синтазы (FLS), в частности, для увеличении содержания флаваноидов, в том числе антоцианов (патент US № 7208659, МПК C12N15/82; C12N9/02; C12N9/10; опубл. 29.05.2003).
Главный недостаток вышеприведенных решений заключается в том, что регуляция работы генов достигается путем генетической модификации генома растений. Данная процедура является сложной и долговременной технологией, возможные последствия которой до конца не изучены. Также во многих странах существуют законодательные запреты на выращивание геномодицицированных растений.
Наиболее близким техническим решением к заявляемому является способ увеличения содержания антоцианов с помощью воздействия на растения двумя генами факторов транскрипции (ROS1 / DEL), выделенными из львиного зева (Antirrhinum) В известном способе используются гены факторов транскрипции Rosea1 (ROS1) и Delila (DEL), выделенные из цветков Antirrhinum majus для генетической модификации томата (Solanum lycopersicum) с целью повышения содержания антоцианов (Nikolay S. Outchkourov et al. Transcription Factor-Mediated Control of Anthocyanin Biosynthesis in Vegetative Tissues // Plant Physiology. 2018. Т. 176 (2). С. 1862-1878).
Основным недостатком данного технического решения следует отнести использование генетической трансформации. Генетическая трансформация осуществляется путем введения в геном томатов генов факторов транскрипции Rosea1 (ROS1) и Delila (DEL), выделенные из цветков Antirrhinum majus. Это приводит к получению геномодифицированных томатов, имеющих физиологические и морфологические изменения в вегетативных тканях, которые отрицательно влияют на рост и развитие томатов. Кроме того, процедура генной модификации является сложной и долговременной технологией, возможные последствия которой до конца не изучены. Также во многих странах существуют законодательные запреты на выращивание геномодицицированных растений, в том числе и в Российской Федерации.
Технической проблемой, решаемой настоящим изобретением, является разработка эффективного метода увеличения общего содержания антоцианов в растениях без генетической модификации генома, а также упрощение и сокращение времени, необходимого для достижения высокой концентрации антоцианов в растениях.
Поставленная техническая проблема решается тем, что в известном способе увеличения содержания антоцианов в растениях, включающим воздействие полинуклеотидами на растения, согласно изобретению, в качестве полинуклеотидов используют двухцепочечную РНК комплементарную участку генов ANAC032 или MYBL2, при этом двухцепочечная РНК содержит одну из последовательностей SEQ ID NO: 1-6, имеющую 100% комплементрарность с последовательностью матричной РНК SEQ ID NO: 7-12, а воздействие на растение осуществляют путем нанесения их на поверхность растения.
По сравнению с другими генами, участвующими в биосинтезе антоцианов, использование в качестве мишени матричной РНК гена MYBL2 позволяет добиться эффективного синтеза антоцианов, за счет того, что MYBL2 один из ключевых клеточных элементов регуляции биосинтеза антоцианов в ответ на свет и другие факторы окружающей среды. Ингибирование гена MYBL2 с помощью внешней обработки двухцепочечными РНК позволит повысить содержание антоцианов без генетической модификации растения, тем самым данный подход является более быстрым и простым способом повышения уровня антоцианов.
Использование в качестве мишени для двухцепочечной РНК матричной РНК гена ANAC032 позволяет добиться эффективного синтеза антоцианов, за счет того, что данный ген подавляет биосинтез антоцианов в ответ на повышения уровня сахара, HL и окислительный стресс. Подавляя двухцепочечными РНК, которые наносятся на поверхность растения, данный ген позволяет повысить уровень антоцианов в растениях в короткие сроки и с минимальными усилиями.
Использование последовательностей SEQ ID NO: 1-6, имеющую 100% комплементарность с последовательностью матричной РНК SEQ ID NO: 7-12 позволяет достичь максимального уровня и скорости накопления антоцианов растениями, за счет того, что данные последовательности обеспечивают максимальную комплементарность.
Воздействие на растение может осуществляться путем нанесения двухцепочечной РНК на поверхность растения методом растирания, распыления, инфильтрации и инокуляции.
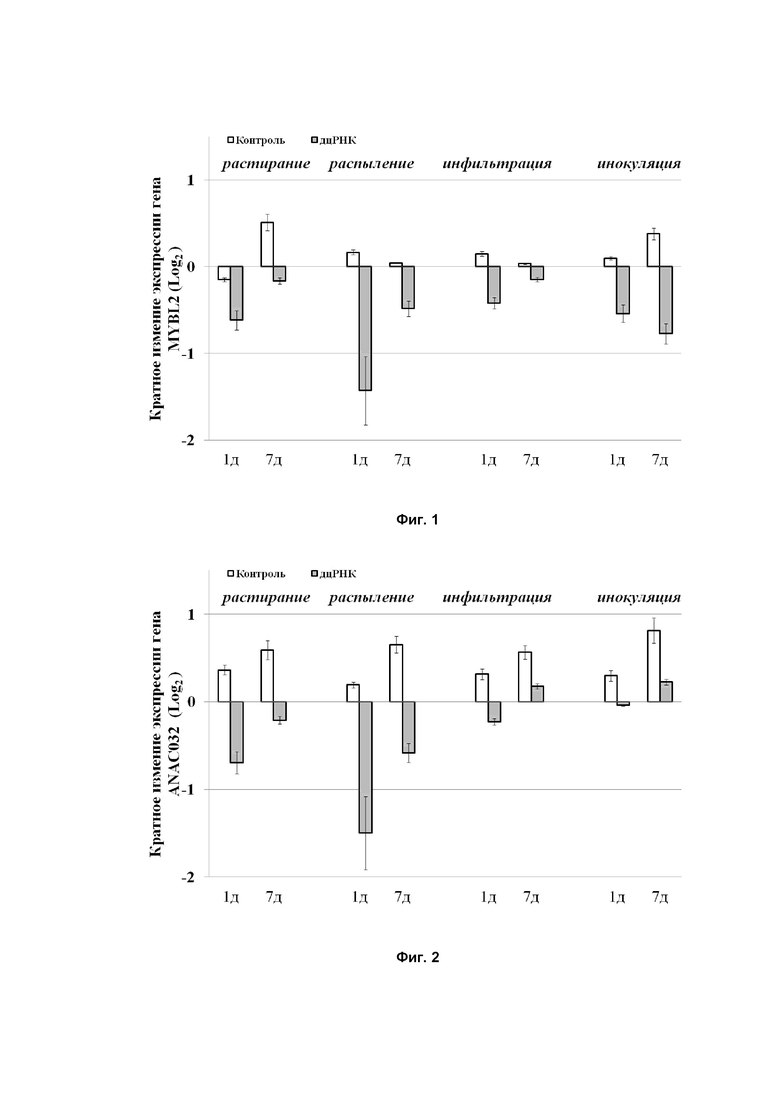
Показано, что максимальное влияние на уровень матричной РНК генов ANAC032 или MYBL2 и как следствие на содержание антоцианов было достигнуто при воздействии на растение двухцепочечными РНК комплементарными участку генов ANAC032 или MYBL2, которые наносятся на поверхность растения методом растирания и распыления. При этом, данный метод существенно упрощает нанесение двухцепочечной РНК на поверхность растения за счет минимальных манипуляций и возможности воздействовать одновременно на несколько растений. Поэтому для ускорения и упрощения обработки растения двухцепочечной РНК комплементарной участку генов ANAC032 или MYBL2 целесообразно вести методом распыления пульверизатором или растиранием по поверхности листа с помощью мягких кисточек (Фиг.1,2).
При инокуляции двухцепочечная РНК комплементарную участку генов ANAC032 или MYBL2 наносится на поверхность растения с использованием абразива, которым посыпается поверхность листа перед инокуляцией. В качестве абразива в данном способе применяется карборунд, но возможно использование любого другого абразива.
При инфильтрации двухцепочечная РНК комплементарная участку генов ANAC032 или MYBL2 наносится на поверхность растения путем процеживания шприцом через поверхность листа без попадания в среднюю жилку.
Способ иллюстрируется фиг.1, на которой приведено влияние различных методов воздействия воды в качестве контроля и двухцепочечной РНК(дцРНК) на подавление экспрессии MYBL2 Arabidopsis thaliana на 1 и 7 день после воздействия;
на фиг. 2 – приведено влияние различных методов воздействия воды в качестве контроля и двухцепочечной РНК(дцРНК) на подавление экспрессии ANAC032 Arabidopsis thaliana на 1 и 7 день после воздействия;
в табл. приведено содержание антоцианов в контрольных (Kont) и обработанных двухцепочечной РНК комплементарной участку генов ANAC032 или MYBL2 растениях Arabidopsis thaliana, Solanum lycopersicum, Solanum tuberosum.
Способ осуществляют следующим образом.
Перед реализацией заявленного способа предварительно РНК выделяется из растения, конкретного вида, который будет подвержен обработке двух цепочечными РНК, далее получаем комплементарную ДНК генов ANAC032 или MYBL2 методом полимеразной цепной реакции с обратной транскрипцией (ОТ-ПЦР), ПЦР продукт используется в качестве матриц для транскрипции in vitro и синтеза двухепочечных РНК с использованием набора MEGAscript RNAi (ThermoFisher Scientific, Уолтем, Массачусетс, США) или иного другого коммерческого набора, далее полученные двухцепочечные РНК анализируются методом гель-электрофорез и спектрофотометрически для оценки количества, чистоты и целостности двухцепочечных РНК, затем двухцепочечные РНК наносятся на поверхность листьев методом распыления пульверизатором или втиранием в поверхность листа механическим способом.
Пример 1
Растения Arabidopsis thaliana экотип Columbia L. в количестве 100 штук выращивали в горшках, заполненных коммерчески доступной богатой почвой, в контролируемой климатической камере при температуре +22˚C (Sanyo MLR-352, Panasonic, Япония), выдерживали цикл 16/8 часов день/ночь при интенсивности света ~120 мкмоль м-2 с-1.
Для выделения РНК из взрослых листьев A . thaliana использовали протокол на основе цетилтриметиламмонийбромида (CTAB). Образцы тканей A . thaliana (0,05 г листьев) гомогенизировали в 1,4 мл буфера, содержащего 2% меркаптоэтанола (AppliChem, Германия); 100 мМ Трис (Biomedicals, Франция), pH 8,0; 2 мМ натрия хлорид (NaCl, Мосреактив, Россия); 25 мМ дигидрат динатриевой соли этилендиаминтетрауксусной кислоты (EDTA, Biomedicals, Германия), pH 8,0; 2% N-цетил-N, N, N-триметиламмоний бромид (CTAB, Panreac, Испания); и 2% поливинилпирролидон K30 (PVP, AppliChem, Германия). Смесь инкубировали 5 мин при 65˚C на водяной бане. После указанной выше инкубации добавляли 700 мкл хлороформа. Затем образцы центрифугировали на скорости 10000g в течение 15 минут (5415R, Эппендорф, Германия). Примерно 1 мл водной фазы смешивали с 250 мкл 10 М хлорида лития. (LiCl, Panreac, Испания) и инкубировали при ~ 4˚C в течение ночи. После инкубации образцы центрифугировали на скорости 10000g в течение 20 мин. Осадок сушили при -37 ° C в течение 40 мин, а затем растворяют в 100 мкл дистиллированной воды и добавляли 300 мкл этанола, и затем образцы инкубировали при -20°C в течение ночи. После указанной выше инкубации образцы центрифугировали на полной скорости в течение 20 мин. Осадок сушили при -37°C в течение 20 мин и окончательно растворить в 50 мкл дистиллированной воды.
Для синтеза двухцепочечных РНК использовали коммерческий набор MEGAscript RNAi (ThermoFisher Scientific, Уолтем, Массачусетс, США). Промоторная последовательность T7 была введена как в 5', так и в 3' концы ANAC032 и MYBL2 с помощью ПЦР. Полученные продукты ПЦР использовались в качестве матриц для транскрипции in vitro и синтеза двухцепочечных РНК (SEQ ID NO: 1-2) в соответствии с протоколом производителя. Полученные двухцепочечные РНК анализировали гель-электрофорезом и подвергали спектрофотометрическому анализу для оценки количества, чистоты и целостности двухцепочечных РНК.
Для наружного применения синтезированные дцРНК разводили в воде до конечной концентрации 0,35 мкг/мкл. ДцРНК (100 мкл каждой дцРНК на отдельное растение, т.е. 35 мкг) наносили на поверхность листа (как с адаксиальной, так и с абаксиальной сторон) четырехнедельных A. thaliana методом распыления пульверизатором. После обработки A. thaliana инкубировали в течение семи дней в контролируемой климатической камере при температуре +22˚C (Sanyo MLR-352, Panasonic, Япония), выдерживали цикл 16/8 часов день/ночь при интенсивности света ~120 мкмоль м-2 с-1.
Для анализа содержания стильбенов проводят аналитическую ВЭЖХ с использованием аналитической ВЭЖХ-системы LC-20AD XR (Shimadzu, Киото, Япония). Данные DAD регистрировались в диапазоне 200–800 нм, а хроматограммы для количественного определения получали при 530 нм. Хроматографическое разделение проводили на колонке Shim-pack GIST C18 (150 мм, внутренний диаметр 2,1 нм, размер части 3 мкм; Shimadzu, Япония). Антоцианы разделяли с использованием 0,1% муравьиной кислоты и ацетонитрила в качестве подвижных фаз A и B, соответственно, со следующим профилем элюирования: от 0 до 35 мин 0% B; От 35 до 40 мин 40% B; 40-50 мин. 50% B; От 50 до 65 мин. 100% B. 5 мкл экстракта образца вводили при постоянной температуре колонки, поддерживаемой на уровне 40°C, и скорости потока, поддерживаемой на уровне 0,2 мл/мин. Содержание антоцианов определяли методом внешнего стандарта с использованием калибровочных кривых четырехточечной регрессии, построенных с использованием имеющихся стандартов.
В итоге получают растения, которые содержат до 1 мкг/г от свежей массы, что в 2,5 раза больше по сравнению с растениями, не подвергаемыми воздействию двухцепочечной РНК (см. таблицу).
Пример 2
Проводят аналогично примеру 1, с той лишь разницей, что воздействие двухцепочечной РНК идентичной последовательности (SEQ ID NO: 2-4), которая комплементарна участку генов ANAC032 или MYBL2 осуществляется на растения томата (Solanum lycopersicum).
В результате воздействия двухцепочечной РНК на растения Solanum lycopersicum получают растения, содержащие до 1,3 мкг/г от свежей массы, что в 5,4 раз больше по сравнению с растениями, не подвергаемыми воздействию двухцепочечной РНК. Результаты представлены в таблице.
Пример 3
Проводят аналогично примеру 1, с той лишь разницей, что воздействие двухцепочечной РНК идентичной последовательности (SEQ ID NO: 5-6), которая комплементарна участку генов ANAC032 или MYBL2 осуществляется на растения паслёна клубненосного (Solanum tuberosum).
В результате воздействия двухцепочечной РНК на растения Solanum tuberosum получают растения, содержащие до 1,5 мкг/г от свежей массы, что в 5,5 раз больше по сравнению с растениями, не подвергаемыми воздействию двухцепочечной РНК. Результаты представлены в таблице.
Как следует из результатов, приведенных в примерах и представленных в таблице воздействие синтетической двухцепочечной РНК, нацеленными на гены ANAC032 или MYBL2 различных растений, способны существенно увеличивать содержание антоцианов. При этом, высокое содержание антоцианов в растениях достигается за более короткий промежуток времени по сравнению с традиционными методами, такими как генетическая модификация. Более того, данная технология существенно упрощает достижение высоких концентраций антоцианов в растении за счет минимальных манипуляций и возможности воздействовать одновременно на несколько растений, а также отсутствия необходимости модификации генома растения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Супрун Андрей Романович
<120> Способ увеличения содержания антоцианов в растениях
<160> 12
<210> 1
<211> 762
<212> РНК
<213> Arabidopsis thaliana
<400> 1
augaugaaaucuggggcugauuugcaauuuccaccaggauuuagauuucauccuacggaugaggagcuaguc
cucauguaucucugucguaaaugcgcgucgcagccgaucccugcuccgauuaucaccgaacucgauuuguac
cgauaugauccuugggaccuucccgacauggcuuuguacggugaaaaggagugguauuuuuucucaccaaga
gaucgaaaguauccaaacgguucaagacccaaccgugcagcugguacuggauauuggaaagcuaccggagcu
gauaaaccaauaggucguccuaaaccgguugguauuaagaaggcucuaguguuuuacucgggaaaaccucca
aauggagagaaaaccaauuggauuaugcacgaauaccggcucgcugacguugaccggucgguucguaagaaa
aacagucuaagauuggacgauuggguauugugucguauauauaacaagaaaggugucaucgagaagcgacga
agcgauaucgaggacggguuaaagccugugacugacacguguccaccggaaucuguggcgagauugaucucc
ggcucggagcaagcggugucaccggaauucacguguagcaacggucgguugaguaaugcccuugauuuuccg
uuuaauuacguagaugccaucgccgauaacgagauugugucacggcuauugggcgggaaucagauguggucg
acgacgcuugauccacuugugguuaggcagggaacuuucuga
<210> 2
<211> 588
<212> РНК
<213> Arabidopsis thaliana
<400> 2
augaacaaaacccgccuucgugcucucuccccaccuuccgguaugcaacaccguaagagaugucgauugaga
ggucgaaacuacguaaggccagaaguuaaacaacgcaacuucucaaaagaugaagacgaucucauccucaag
cuucaugcacuucuuggcaauagauggucauugauagcgggaagauugccaggacgaaccgacaacgaaguu
aggauccauugggaaacuuaccuaaaaaggaagcucguaaaaaugggaaucgacccaaccaaucaucgucuc
caccaucacaccaacuacauuucuagacgucaccuccauucuucacauaaggaacaugaaaccaagauuauu
agugaucaaucuucuucgguauccgaaucaugugguguaacaauuuugcccauuccaaguaccaauugcucg
gaggauaguacuaguaccggacgaagucauuugccugaccuaaacauuggucucaucccggccgugacuucu
uugccagcucuuugccuucaggacucuagcgaauccucuaccaaugguucaacaggucaagaaacgcuucuu
cuauuccgauga
<210> 3
<211> 519
<212> РНК
<213> Solanum lycopersicum
<400> 3
auggcagauuggaauagaucaagcacaucagauaaugccucaguggucucaccugauucgacccgagugguu
gcauuagagacuaccaacgaagaaaccucuaaacuugaauuuucagaagaugaagaaaugcucauugcuaaa
auguucagcuugguuagagagagguggucauuaauugcuggaagaaucccaggaagaaaugcugaugagauu
gaaaaauauuggaaaucaaaauacuccaaaagccaggaggaaauacaggcccaaucucaagaugaagcucau
ggcauuagauugauagagaaaacugggccuucaacggggcacccucagcaguguguuuuuccagaaaaaaag
cccauaucugucugucaaccuucagauauugggccucaacuauugggcccaacaacgaaugggccucaacag
cuauugggcccuaaaacaacugggcccaauaacaucacuguagaccaaaaguuaguuaacccagcccaagac
caaauaaaccaauaa
<210> 4
<211> 285
<212> РНК
<213> Solanum lycopersicum
<400> 4
auggaucaaaaucuccaucaccgacacaaacucaugcaccaccgauguugcagccaugaagaagagguuaau
aguauggagugggaauucauuagaaugagcaagcaagaagaagaucuuauuuacaggaugcacaaacuuguu
ggagacaggugggggcuuauagcagguagaauaccagggagaacagcagaagaaauagaaagguuuuggaua
augagacacagugauggguuugcacacaagagaagacaaacaauuaagaaaagucuaccaccuacauaa
<210> 5
<211> 955
<212> РНК
<213> Solanum tuberosum
<400> 5
ucaaggagaugagaagguugauaugcucaugaagucggagaagaaauccaagaaaaggaaaaagaaugcugu
caccgaugauucgaaagugacaaagggagauguuaaagcuguugagcaugauucauguuucaaggaagagag
ucucucuuuuacagggaucgggaaagaaagggauaagacaaagauggaaacugaagcaccucgagaaaguga
agauagugacaucaagagaaagaaagaacgagguaauggaaaacagccugggaaauccaguaaagacgggug
cgaggaugcuguuaaaaauguuuuaaauuagaaaaagaaguugaagaaugagcaggagauacugauaaugca
cgaugcuaguuuagacaccaaaacacuugcucaugaaagugcaagcggaacacaggaaaucaaggcaguuga
gaugcucaaugagggcucugguggaaauuuuacagaugaggugaagaggaagaaaaagaaaaaagauaagag
ggaaaaagacgauggacagguagauauuguggcuggugucauucaagguguuguuucagcugacgaggugaa
gaggaagaaaaggaaaaaggauaagaggaauaaagaggauggacagguagauaucguggcuggcgucauuca
aggugauguuucagcuauagaagagacgaaagauauacaaauagaugaugcuaauaucagaaagaugaaaga
gacaaaauuagggaacaauucgaaagaucuuucuaaugaaaagacugaaaaaagagugagauuuucugauga
uguacaguuuuuccauccaaucagugaaccuagcaaugagaggcaugaaaauaacaaacaagaauuacuguu
uggcaaaaaauucacacaggaagaagaugaaaucgccaaagaugcuguuuguagauacauagagguauauaa
uuugggcgaugaaggguug
<210> 6
<211> 844
<212> РНК
<213> Solanum tuberosum
<400> 6
cuggauuaaacagaugcagaaaaaguuguagacugagguggcuaaauuaucuaaggccacauaucaagagag
gugacuuugaaccagaugaaguggaucucaucuugagacuucauaagcucuuaggcaaccgauggucacuua
uugcugguagacuuccaggaaggacagcuaacgaugugaaaaacuauuggaacacuaaccuucuaaggaagc
uaaauacuaguacuaaauuugcuccucaaccacaagaaggaauuaauacuaguacuauugcuccucaaccac
aagaaggaauuaaguaugggcaagccaaugccauaauaagaccucaaccucagaaauucacaagcuccauga
agauuaaugucucuuggugcaacaacaauaguaugguaaauaaugaagaagcaucgaaagacaacaacgaua
ugcaauggugggcaaauauacuggaaaauugcaaugacauuggagaaggagaagcugaaagaacacuaccuu
cauguaaggaaauuaauugcaaugaaauugauaaaauaccaaguuuguuacaugauggaggcaacuccaugc
aacaaggacaaagugauggugguugggaugaauuuucucuagaugauuuauggaaucuacuuaauuagcggg
ugauuuacacgaaugaccugagaggugaauggucuugaacuuuugauuucgcuugucuuauccuacugauaa
aacuucaaguuuuacaaguccuuauuuuuauucuugaagauuugccuauggaguaaauugagguaaaaaauu
cgagaccacucauauaauuaggugcauggucuuuguauuaucuacgugugua
<210> 7
<211> 1730
<212> РНК
<213> Arabidopsis thaliana
<400> 7
aaccauuuuucuauuuuuguuugagauuccuuuccaccaacacaaagccccugucuauccgccaugugucca
cgaccuuucuuauauauauagucaucuuucuucuuugcucgagagaaguuucaacuuucaaacauaucuuua
caguucuuucuugcuaaacaacaauaaaggggaaaauagguuaauuaugaugaaaucuggggcugauuugca
auuuccaccaggauuuagauuucauccuacggaugaggagcuaguccucauguaucucugucguaaaugcgc
gucgcagccgaucccugcuccgauuaucaccgaacucgauuuguaccgauaugauccuugggaccuucccga
cauggcuuuguacggugaaaaggagugguauuuuuucucaccaagagaucgaaaguauccaaacgguucaag
acccaaccgugcagcugguacuggauauuggaaagcuaccggagcugauaaaccaauaggucguccuaaacc
gguugguauuaagaaggcucuaguguuuuacucgggaaaaccuccaaauggagagaaaaccaauuggauuau
gcacgaauaccggcucgcugacguugaccggucgguucguaagaaaaacagucuaagauuggacgauugggu
auugugucguauauauaacaagaaaggugucaucgagaagcgacgaagcgauaucgaggacggguuaaagcc
ugugacugacacguguccaccggaaucuguggcgagauugaucuccggcucggagcaagcggugucaccgga
auucacguguagcaacggucgguugaguaaugcccuugauuuuccguuuaauuacguagaugccaucgccga
uaacgagauugugucacggcuauugggcgggaaucagauguggucgacgacgcuugauccacuugugguuag
gcagggaacuuucugaguugucacgugcgauuagaguuaguggaaaguggaaacuaucacugucuguuuucg
cacgugucggguagccgucuugaaacccgcuagcugauccagaagugaccucauugggcuuuucaacaacaa
uugggccuuagccuuucuucucccuuaguaagacaagcuacuguauuguguuguuauuuucaggaagcaaua
uaaauucauuuuaaucacuuauauauucuuuacauuuuuuuuucacucaaaacuacaaucuagcuucuucuu
uuuuuuuuuuuuuuuuuguuaugguuguacuacaaucuagcuuuguuauauauauauauaaguuaaaaauag
uaacuaaauuacaaacuaaaauaaaauaaagaaauuggaagauaucacuaguguuugggguauuagggaggg
aggggagcucgagauccgauuuuacugacugcaugguaucgaaaaagugucgguaaugguagggucugacag
auaacagucuuaucguugcaacuugcauucaaacaaaucucauuguucgcuuuaauuucuaauuaauuuguu
uuugucuuuuguauauuuauuugauuuugacacgaguuugucuuuccuaauaagaccgaggcaugauuaguu
uaaccgguuugaauuguuuagguuaaaucggaguccaagucaaauacaucgcuaucuaacgaaaacgacgac
ugaaauuucguaaaguuccuuacuauaaaacugaucuaacuaucuucuacaagaacaauauaauaaauuugaug
<210> 8
<211> 1222
<212> РНК
<213> Arabidopsis thaliana
<400> 8
cacacuuccacauucaugauuccacaaauucuaauggauuugaacaaauaugggaccaauuuuuauuuguuu
caacuccuaaaugugggaccaauuaacaagguugauguauucuucuuguucuuccucguucauuaauaguuu
gauugagcaugcacgcuucucucaacccaccaguccaagucaaaccuccucaagccauccccuaauccuaua
uaucaguuucuucucgcaaaccaccucacaaucuaucaacaguuuuaaaaaaagacauaacucaaccaaucu
cacuuugaaaaagacaugaacaaaacccgccuucgugcucucuccccaccuuccgguaugcaacaccguaag
agaugucgauugagaggucgaaacuacguaaggccagaaguuaaacaacgcaacuucucaaaagaugaagac
gaucucauccucaagcuucaugcacuucuuggcaauagauggucauugauagcgggaagauugccaggacga
accgacaacgaaguuaggauccauugggaaacuuaccuaaaaaggaagcucguaaaaaugggaaucgaccca
accaaucaucgucuccaccaucacaccaacuacauuucuagacgucaccuccauucuucacauaaggaacau
gaaaccaagauuauuagugaucaaucuucuucgguauccgaaucaugugguguaacaauuuugcccauucca
aguaccaauugcucggaggauaguacuaguaccggacgaagucauuugccugaccuaaacauuggucucauc
ccggccgugacuucuuugccagcucuuugccuucaggacucuagcgaauccucuaccaaugguucaacaggu
caagaaacgcuucuucuauuccgaugagaaaugguguauuuguuuguacacauuuguuauauuacaaaugca
cauauuauaauauucacuagagagaugguauaacgauguuugaaauauguuaauccugaauuggaccggcug
gucauugaacaacuauucucuuaaauguccgguucuguuuucagugauuaguauuauaguagaucguuaaua
acuaaccgacgauaaccgcugcuuauuugaucaaucaagcuguaagaaauuauagcuaauacagcaauguua
acggugcacauugacgggaaguauuaauuaacucucuuaaaauggacaaguauguucauuuuccuccaaa
<210> 9
<211> 859
<212> РНК
<213> Solanum lycopersicum
<400> 9
gaccacauucaaaucuuuguucuccacauauacucuuccuuaauuaguacuccaauucauuuaauuaaaguu
uuauuucucuugauggcagauuggaauagaucaagcacaucagauaaugccucaguggucucaccugauucg
acccgagugguugcauuagagacuaccaacgaagaaaccucuaaacuugaauuuucagaagaugaagaaaug
cucauugcuaaaauguucagcuugguuagagagagguggucauuaauugcuggaagaaucccaggaagaaau
gcugaugagauugaaaaauauuggaaaucaaaauacuccaaaagccaggaggaaauacaggcccaaucucaa
gaugaagcucauggcauuagauugauagagaaaacugggccuucaacggggcacccucagcaguguguuuuu
ccagaaaaaaagcccauaucugucugucaaccuucagauauugggccucaacuauugggcccaacaacgaau
gggccucaacagcuauugggcccuaaaacaacugggcccaauaacaucacuguagaccaaaaguuaguuaac
ccagcccaagaccaaauaaaccaauaaccuaugcccagcccagcccaagauuggucaaacaugacacuagac
aaaccuacaauauauuuucuauuuacauauuuucgcaaauaccuacuacucaauuaugcuauguauagcuau
aguuugcauauuuacuaguuguaguuacaguuuaaguuaucucguuuguauaauuuccauauauguuuaaau
aagugaguuauauuuguauaaacucaaaauaaugaauuuauacaaacacaaacauuuaaacuuuaaa
<210> 10
<211> 1067
<212> РНК
<213> Solanum lycopersicum
<400> 10
uucuuuuuguaacuaccugaaauuguaugucaauuggaccuaccaaucaaauucauaaggcaaauaaugauc
aaugaagacccccuucuuuucuuuuuuuuaagggcucagguuaaucagucuacuaaguuuugcuuuucuugu
agucgauccugaauguucuuugcuccuuucagcuuaaaaugccuaauguggagaaacaaguaacaacagcuu
auauuauuggggagaaauuaagauuugaacuuuaacuauccacuggauauaaaugaaagaaacguauguaaa
aauguaaguagauaaugggaaggaaugacaacugcaagucaaagggaaagaaagaguaguuuguuugguuag
gaccagccguuguuugauguuugaugauaaacaaaugcagacaaaucugucgaaccauugaggacaaguaua
aaaaaacagaagcugucccuccuaaucaacagcaacucucugcuaguccucuuuucuugauccucauuauuc
uuaucuuucuauuucucuacucuuuuucuuuuguaccuaaaaauggaucaaaaucuccaucaccgacacaaa
cucaugcaccaccgauguugcagccaugaagaagagguuaauaguauggagugggaauucauuagaaugagc
aagcaagaagaagaucuuauuuacaggaugcacaaacuuguuggagacaggugggggcuuauagcagguaga
auaccagggagaacagcagaagaaauagaaagguuuuggauaaugagacacagugauggguuugcacacaag
agaagacaaacaauuaagaaaagucuaccaccuacauaauuauuauuauccuucuuuuucuuuaaaccaaaa
aaaaaaaaaacuuuuugaggaaagcagauucucuuuuuccuccucauccuuucuuuuccauuuuguuugggg
acccaagcuauugaucauuuuaucauuuucuugucuguguacuuuuauuuuuuagauuuauaucuuaugccu
uuuugguuuacuuguuauguuuaugugggcaauauauauguaaucauauuaucuucuua
<210> 11
<211> 2368
<212> РНК
<213> Solanum tuberosum
<400> 11
cccuaauuuuuuuuucaguacuugaaacugagaauucacaaaucaauuacguuuggauuugauuggauuuuu
ugcauuuauugcugcucuccagugauuuuggcauucagagagcccacuuuuggugcuuugcgagaggacaca
uagguggcauuuucuaaaagaggcaugguucaaggagaugagaagguugauaugcucaugaagucggagaag
aaauccaagaaaaggaaaaagaaugcugucaccgaugauucgaaagugacaaagggagauguuaaagcuguu
gagcaugauucauguuucaaggaagagagucucucuuuuacagggaucgggaaagaaagggauaagacaaag
auggaaacugaagcaccucgagaaagugaagauagugacaucaagagaaagaaagaacgagguaauggaaaa
cagccugggaaauccaguaaagacgggugcgaggaugcuguuaaaaauguuuuaaauuagaaaaagaaguug
aagaaugagcaggagauacugauaaugcacgaugcuaguuuagacaccaaaacacuugcucaugaaagugca
agcggaacacaggaaaucaaggcaguugagaugcucaaugagggcucugguggaaauuuuacagaugaggug
aagaggaagaaaaagaaaaaagauaagagggaaaaagacgauggacagguagauauuguggcuggugucauu
caagguguuguuucagcugacgaggugaagaggaagaaaaggaaaaaggauaagaggaauaaagaggaugga
cagguagauaucguggcuggcgucauucaaggugauguuucagcuauagaagagacgaaagauauacaaaua
gaugaugcuaauaucagaaagaugaaagagacaaaauuagggaacaauucgaaagaucuuucuaaugaaaag
acugaaaaaagagugagauuuucugaugauguacaguuuuuccauccaaucagugaaccuagcaaugagagg
caugaaaauaacaaacaagaauuacuguuuggcaaaaaauucacacaggaagaagaugaaaucgccaaagau
gcuguuuguagauacauagagguauauaauuugggcgaugaaggguugcaaaagguuuugaauugcaagucu
uauccugaaguaaggggcugcuggaaagagauagggaaggcuauaccauacaggccuuacagagcaguuuau
uaucgugcacaacguuuguuucgaaugggagagaagcguaaauggacugaagaagagcaugggaugcuacgg
aaguuccagggacaacaugggaauaaguggaccgucuuggcggaugaacuugggaaacauccgguucaugua
ggaaaugcauggcauaggauaaaacuggaaaaucagaagaggggacagugggaucaggaggaaguacagaaa
uuguuugauuuaguaaacaccgaucugcaacuuaagcucucugacgaaaggaaaucuaagcaugggauguua
cgggauaauauuuguuggaguucaauuagugacaauuuguccaccaggauuucccaccauugcugcaauaaa
ugguacagacaauuaacauccucgauggugguugcaggugaaugggcagauacugaugacuaucgcuuaauu
gcugcccuuuuugaacuggacgcaagcugcauagaggacguggauugggacaaucuucucucccacaggcau
ggagauuuaugucgaaaaagauggaaagagauggugcgucaaauaagucaacaugaaaacaagucauuugau
gcacaaguacaaguguuagcaaagagauaccgcccugauuuaguuggagcaagggaggucugggauaguaaa
ccacuuguuccaugacauguucaacacucacauggguguuucuguacagagaugcuuaaggcacuaguauac
acuucugucuacaacuuagcgcgucacuucuguacuuuauuucuuucguucuuuuaauuuuugagagguuua
uaaaacaucugcaaauauuagucacucgguacauaugcuuguaggagguauuagguacucgugaaacaguag
aggugcuuguuggcuugggcaccaaacauuauccaaaaagaaaauguucacagauguuuuggaagagauucu
uucuugucugguggaaagaaaaggaaaaaaagcuuucacaauuugaugucauuaaggauugaaagagaugag
ccgagggucuaucugacacaaaugcucuaccucgccaagguagggacauaauuuuuccccaucccaaacccc
acuugugagauuaugguguguauguuguguaccugccuaaagaaaagauuacaggauauuauua
<210> 12
<211> 1120
<212> РНК
<213> Solanum tuberosum
<400> 12
aaagguaguaaaauauaugacuucacauguaaugaucaugaguacuccuaugauguguacauuuuugggagu
aauaaggaaagguucauggacugaagaagaagauauucuuuugaggaaauguauugauaaguauggagaagg
aaaguggcaucuuguuccaacuagagcuggauuaaacagaugcagaaaaaguuguagacugagguggcuaaa
uuaucuaaggccacauaucaagagaggugacuuugaaccagaugaaguggaucucaucuugagacuucauaa
gcucuuaggcaaccgauggucacuuauugcugguagacuuccaggaaggacagcuaacgaugugaaaaacua
uuggaacacuaaccuucuaaggaagcuaaauacuaguacuaaauuugcuccucaaccacaagaaggaauuaa
uacuaguacuauugcuccucaaccacaagaaggaauuaaguaugggcaagccaaugccauaauaagaccuca
accucagaaauucacaagcuccaugaagauuaaugucucuuggugcaacaacaauaguaugguaaauaauga
agaagcaucgaaagacaacaacgauaugcaauggugggcaaauauacuggaaaauugcaaugacauuggaga
aggagaagcugaaagaacacuaccuucauguaaggaaauuaauugcaaugaaauugauaaaauaccaaguuu
guuacaugauggaggcaacuccaugcaacaaggacaaagugauggugguugggaugaauuuucucuagauga
uuuauggaaucuacuuaauuagcgggugauuuacacgaaugaccugagaggugaauggucuugaacuuuuga
uuucgcuugucuuauccuacugauaaaacuucaaguuuuacaaguccuuauuuuuauucuugaagauuugcc
uauggaguaaauugagguaaaaaauucgagaccacucauauaauuaggugcauggucuuuguauuaucuacg
uguguacuuuuguguuuuuaaggugaucaaguuuauaguggaaugugaaaguauuccaagugugaaguaugu
aauaugauauaugaaaugaagugaguauuauauuauauua
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МОЛЕКУЛЫ РНК | 2018 |
|
RU2819868C2 |
| МОЛЕКУЛЫ РНК, ВКЛЮЧАЮЩИЕ НЕКАНОНИЧЕСКИЕ ПАРЫ ОСНОВАНИЙ | 2019 |
|
RU2812710C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ИЗМЕНЕНИЯ ЦВЕТЕНИЯ И АРХИТЕКТУРЫ РАСТЕНИЙ ДЛЯ УЛУЧШЕНИЯ ПОТЕНЦИАЛЬНОЙ УРОЖАЙНОСТИ | 2017 |
|
RU2775653C2 |
| Композиция на основе интерферирующей дцРНК для защиты растений от фитофтороза | 2020 |
|
RU2808216C2 |
| Способ увеличения устойчивости растений к воздействию ультрафиолета | 2020 |
|
RU2733119C1 |
| ПОЛИНУКЛЕОТИД И СПОСОБ БОРЬБЫ С НАШЕСТВИЕМ НАСЕКОМЫХ | 2019 |
|
RU2781829C2 |
| РАСТЕНИЕ ТОМАТА, ОБРАЗУЮЩЕЕ ПЛОДЫ С МОДИФИЦИРОВАННЫМ СОДЕРЖАНИЕМ САХАРОВ | 2019 |
|
RU2817600C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ПРОМЫШЛЕННЫХ ПРОДУКТОВ ИЗ РАСТИТЕЛЬНЫХ ЛИПИДОВ | 2015 |
|
RU2743384C2 |
| ЭНХАНСЕР ПАЛОЧКОВИДНОГО ВИРУСА САХАРНОГО ТРОСТНИКА (SCBV) И ЕГО ПРИМЕНЕНИЕ В ФУНКЦИОНАЛЬНОЙ ГЕНОМИКЕ РАСТЕНИЙ | 2013 |
|
RU2639517C2 |
| ПОЛИНУКЛЕОТИД И СПОСОБ, ИСПОЛЬЗУЕМЫЙ ДЛЯ ОСУЩЕСТВЛЕНИЯ КОНТРОЛЯ НАД НАШЕСТВИЕМ НАСЕКОМЫХ | 2019 |
|
RU2775717C1 |

Изобретение относится к области биохимии, в частности к способу увеличения содержания антоцианов в растениях Arabidopsis thaliana, Solanum lycopersicum, Solanum tuberosum. Изобретение позволяет эффективно увеличивать общее содержание антоцианов в растениях без генетической модификации генома, а также упрощает и сокращает время, необходимое для достижения высокой концентрации антоцианов в растениях. 2 з.п. ф-лы, 2 ил., 3 пр.
1. Способ увеличения содержания антоцианов в растениях Arabidopsis thaliana, Solanum lycopersicum, Solanum tuberosum, включающий воздействие полинуклеотидами на растения, отличающийся тем, что в качестве полинуклеотидов используют двухцепочечную РНК, смысловая цепь которой комплементарна участку матричной РНК генов ANAC032 или MYBL2 (MYB-LIKE), при этом:
для растения Arabidopsis thaliana смысловая цепь двухцепочечной РНК SEQ ID NO: 1, комплементарна с участком последовательности матричной РНК SEQ ID NO: 7, SEQ ID NO: 2 комплементарна с участком последовательности матричной SEQ ID NO: 8,
для растения Solanum lycopersicum SEQ ID NO: 3 комплементарна с участком последовательности матричной SEQ ID NO: 9, SEQ ID NO: 4 комплементарна SEQ ID NO: 10, SEQ ID NO: 5 комплементарна с участком последовательности матричной SEQ ID NO:10,
для растения Solanum tuberosum SEQ ID NO: 6 комплементарна с участком последовательности матричной SEQ ID NO: 11, SEQ ID NO: 6 комплементарна с участком последовательности матричной SEQ ID NO: 12, а воздействие на растения осуществляют путем нанесения двухцепочечной РНК на поверхность указанного растения.
2. Способ по п. 1, отличающийся тем, что нанесение на поверхность растения двухцепочечной РНК ведут методом распыления пульверизатором.
3. Способ по п. 1, отличающийся тем, что нанесение на поверхность растения двухцепочечной РНК ведут растиранием по поверхности листа механическим способом.
| WO 2016016208 A1, 04.02.2016 | |||
| US 20180127769 A1, 10.05.2018 | |||
| СТАБИЛЬНЫЕ КОМПОЗИЦИИ С ИНОКУЛЯНТОМ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2016 |
|
RU2730863C2 |
| H.MAKI and et.al., ANAC032 regulates root growth through the MYB30 gene regulatory network, Scientific Reports, 2019, pp.1-13 | |||
| K | |||
| MAHMOOD and et.al., The Arabidopsis Transcription Factor ANAC032 Represses Anthocyanin Biosynthesis in Response to High Sucrose | |||

